
アポトーシスの制御
細胞中の選択したアポトーシス関連遺伝子の発現を抑制するための方法であって、(1)ゲノム中に存在する、選択した遺伝子にある部位、または該遺伝子と関係がある部位と結合する核酸結合部分、および(2)修飾部分を含む分子を、細胞中に導入することを含み、前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記抑制因子部分がポリペプチドまたはペプチド模倣体を含む方法。本発明の方法において使用するための分子を提供する。抑制因子または修飾部分は、ヒストン脱アセチル化酵素またはDNAメチラーゼを補うことができる、ヒストン脱アセチル化酵素またはDNAメチラーゼまたはポリペプチドの一部分であってよい。核酸結合部分は、三重鎖形成オリゴヌクレオチド(TFO)であってよい。アポトーシス関連遺伝子は、Bel-2であってよい。本発明の方法および分子は、癌の治療において有用である可能性がある。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、プログラムされた細胞死、アポトーシス、および関連遺伝子発現の制御に関するものであり、詳細には、本発明は、細胞中でのアポトーシスを制御するため、特定の選択したアポトーシス関連遺伝子の発現を調節、好ましくは抑制するための方法および手段に関する。
【背景技術】
【0002】
遺伝子の発現を選択的に抑制する能力は、生物学の多くの分野、例えば遺伝子の発現が望ましくない可能性がある治療法;特定の遺伝子の発現の欠如がその疾患と関係がある疾患モデルの作製;望ましい特性を生み出すための表現型の変更において有用である。したがって、遺伝子の発現を選択的に抑制する能力は、発生および分化の研究において、(野生型であれ突然変異型であれ)ヒト細胞中のヒト遺伝子を「ノックアウト」し、真核生物の遺伝子をノックアウトすることができる。
【0003】
細胞死を選択的に誘導する能力は、癌中の腫瘍細胞をターゲッティングし破壊する際に、ならびに他の状況、例えば炎症の消散において重要である。この能力は、発癌性状態になっている細胞、または腫瘍中の他の細胞(白血球など、例えばマクロファージ)を直接殺傷する際に、ならびに任意の他の療法を向上させる際に有用である。なぜなら、癌療法に対する腫瘍の耐性は、疾患を治療する際の主要な問題だからである。アポトーシスは、細胞死を誘導するための主要な機構の1つであり、アポトーシスに対する耐性は、腫瘍の進行中に自然に得られる特性である。これは通常、アポトーシスシグナル分子の制御の変更を伴い、腫瘍細胞をアポトーシス刺激に対して鈍感にする。
【0004】
特定の遺伝子の発現を抑制することを試みる方法は、主に3つの主要な範疇、すなわちアンチセンス技術、リボザイム技術、および相同的組換えによってもたらされる標的遺伝子の欠失に分類される。
【0005】
アンチセンス技法は、核酸分子の細胞中への導入に基づくものであり、核酸分子は典型的には、選択した遺伝子によって発現されるmRNAと相補的である。アンチセンス分子は、mRNA分子と結合したままの間は、典型的にはmRNA分子の翻訳を抑制し、遺伝子によってコードされるポリペプチドの発現を妨げる。アンチセンス技法に修正を加えれば、三重らせんを形成するために遺伝子のDNAと結合するアンチセンス分子(三重鎖形成オリゴヌクレオチド;TFO)により、選択した遺伝子の転写を防止できるであろう。この方法では、第三の鎖の存在がDNA転写を妨げ、その間、その鎖は結合したままの状態である。
【0006】
化学修飾基、例えばソラーレン架橋基は、TFO中に含まれているが、これらは不可逆的なDNAの損傷および突然変異をもたらす可能性がある。細胞中でのこのような化学修飾基の調節は困難でもある。これらは、分子の細胞内送達に関する欠点を有する可能性もある。
【0007】
リボザイム技法は、核酸分子の細胞中への導入に基づくものであり、核酸分子はRNA分子を発現し、標的RNA分子と結合し、標的RNA分子の選択的切断を触媒する。標的RNA分子は典型的にはmRNA分子であるが、それは例えば、レトロウイルスRNA分子であってよい。
【0008】
アンチセンスおよびリボザイムに基づく技法は、実施が困難であることが明らかになっており、これらの技法は、標的遺伝子の抑制または不活性化において、成功の程度にバラツキがある。さらに、これら2つの技法は、遺伝子不活性化物質の持続的な発現または投与を必要とする。
【0009】
TFOとVP16ウイルス活性化ドメインの結合(Kusnetsova他、(1999) Nucleic Acids Res20、3995〜4000)を使用して、TFOの適用を広げ、遺伝子活性化を含めている(遺伝子の抑制または不活性化における以前の使用とは対照的)。
【0010】
相同的組換えによる標的遺伝子の欠失は、2つの遺伝子不活性化事象(それぞれの対立遺伝子に関する事象)を必要とし、初代細胞、特に例えば、数回の継代の培養においてのみ維持することができる、初代ヒト乳房細胞には容易に適用できない。標的遺伝子の欠失は、植物において行うのは依然として困難である。cre-lox仲介の部位特異的組み込みが優れた方法となっているが、特異的組み込み事象の効率は低い(Alberts他、(1995) Plant J.7、649〜659; Vergunst & Hooykass (1998) Plant Mol.Biol.38、393〜406; Vergunst他、(1998) Nucl.Acids Res.26、2729〜2734)。
【0011】
WO01/02019は、真核生物ゲノム中に存在する部位に結合する核酸結合部分、およびクロマチン不活性化部分、例えばHDACまたはHDAC補充物質、PLZFなどを含むポリペプチドを使用して、選択した遺伝子を不活性化させるための方法を記載している。
【0012】
細胞死を誘導するための現在の方法は、満足のいくものではない。本発明者らは、これが、これらの方法が、細胞を殺傷するのに充分持続的または完全な、遺伝子発現に対する作用を生み出すことができないためであると考える。細胞のアポトーシス機構が素早く、有効に、充分に長く作用しない場合、通常、細胞は非常に早く耐性を発現する。既存の技術および方法にはこれらの重大な欠点があるので、別の戦略が求められる。
【非特許文献1】Kusnetsova他、(1999) Nucleic Acids Res20、3995〜4000
【非特許文献2】Alberts他、(1995) Plant J.7、649〜659
【非特許文献3】Vergunst & Hooykass (1998) Plant Mol.Biol.38、393〜406
【非特許文献4】Vergunst他、(1998) Nucl.Acids Res.26、2729〜2734
【特許文献1】WO01/02019
【非特許文献5】Sellers & Fisher (1999) The Journal of Clinical investigation、p.1655; Apoptosis and cancer Drug Targeting
【非特許文献6】Cory & Adams (2002) Nature Reviews Cancer、p489; The Bcl-2 family : regulators of the ceullular life-or-death switch
【非特許文献7】Chen & Li (1998) Crit.Rev.Eukaryotic Gene Expression 8、169〜190
【非特許文献8】Workman & Kingston (1998) Ann.Rev.Biochem.67、545〜579
【非特許文献9】Perlmann & Vennstrom (1995) Nature 377、387-
【非特許文献10】Wolfe (1997) Nature 387、16〜17
【非特許文献11】Grunstein (1997) Nature 389、349〜352
【非特許文献12】Pazin & Kadonaga (1997) Cell 89、325〜328
【非特許文献13】DePinho (1998) Nature 391、533〜536
【非特許文献14】Bestor (1998) Nature 393、311〜312
【非特許文献15】Grunstein (1998) Cell 93、325〜328
【非特許文献16】Taunton他、(1996) Nature 272、408〜441
【非特許文献17】Yang他、(1996) Proc.Natl.Acad.Sci.USA 93、12845〜12850
【非特許文献18】Dangond他、(1998) Biochem.Biophys.Res.Comm.242、648〜652
【非特許文献19】Horlein他、(1995) Nature 377、397〜404
【非特許文献20】Chen & Evans (1995) Nature 377、454〜457
【非特許文献21】Zhang他、(1998) Molecular Cell1、1021〜1031
【非特許文献22】Ayer他、(1995) Cell 80、767〜776
【非特許文献23】Schreiber-Agus他、(1995) Cell 80、777〜786
【非特許文献24】Zhang他、(1997) Cell 89、357〜364
【非特許文献25】Qian他、(1993) Nature 364、648〜652
【非特許文献26】Lin他、(1998) Nature 391、811〜814
【非特許文献27】Grignani他、(1998) Nature 391、815〜818
【非特許文献28】David他、(1998) Oncogene 16、2549〜2556
【非特許文献29】Hassig他、(1997) Cell 89、341〜347
【非特許文献30】Laherty他、(1997) Cell 89、349〜356
【非特許文献31】Zhang他、(1997) Cell 89, 357-364
【非特許文献32】Heinzel他、(1997) Nature 387、43〜48
【非特許文献33】Alland他、(1997) Nature 387、49〜55
【非特許文献34】Kadosh & Struhl (1997) Cell 89、365〜371
【非特許文献35】Nagy他、(1997) Cell 89、373〜380
【非特許文献36】Laherty他、(1998) Molecular Cell 2、33〜42
【非特許文献37】Grozinger他、(1999) Proc.Natl.Acad.Sci.USA 96、4868〜4873
【非特許文献38】Lussen他、(1997) Science 277、88〜91
【非特許文献39】Lechner他、(1996) Biochim.Biophys.Acta 1296、181〜188
【非特許文献40】Chen & Pickard (1997) Genes Dev.11、2124〜2136
【非特許文献41】Brosch他、(1995) Plant Cell 7、1941〜1950
【非特許文献42】Okano M、Xie S、LiE.(1998) Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases.Nat Genet19:219〜220
【非特許文献43】Adrian P.Bird and Alan P.Wolffe (1999) Methylation-Induced Repression:Belts、Braces、and Chromatin.Cell 99、451〜454
【非特許文献44】Soukchareun他、(1998)Bioconjugate Chem9、466〜475
【非特許文献45】Soukchareun他、(1995) Bioconjugate Chem6、43〜53
【非特許文献46】Eritja他、(1991) Tetrahedron 47、4113〜4120
【特許文献2】WO99/13719
【非特許文献47】Prochiantz (2000) Curr Opin Cell Biol9、420〜429
【特許文献3】WO01/14737
【非特許文献48】Kusnetsova他、(1999) Nucleic Acids Res 27、3995〜4000
【特許文献4】WO90/06934
【特許文献5】EP0375408
【特許文献6】WO91/06626
【特許文献7】EP0266099
【特許文献8】W094/17086
【特許文献9】W096/35706
【非特許文献49】Debin他、(1999) Nucl Acids Res 27 (13)、2699〜2707
【非特許文献50】Xodo他、(2001) Eur J Biochem 268、656〜664
【非特許文献51】Blume他、(1999) Nucl Acids Res 27、695〜702
【非特許文献52】Faria他、(2001) JMol Biol 306、15〜24
【非特許文献53】Cheng他、(2000) Biotech and Bioeng 70、467〜472
【非特許文献54】Vasquez & Wilson (1998) Trends Biochern Sci1、4〜9
【非特許文献55】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFFACTOR
【非特許文献56】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fum+pagelibinfo+-info+TFSITE
【非特許文献57】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFCELL
【非特許文献58】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFCLASS
【非特許文献59】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFMATRIX
【非特許文献60】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFGENE
【非特許文献61】Hill MおよびHemmings B (2002): Inhibition of protein kinase B/AKT.Implications for cancer therapy.Pharmacol Ther、93、p.243
【非特許文献62】Kanaseki T他、(2002) Identification of germline mutation of PTEN gene and analysis of apoptosis resistance of the lymphocytes in a patient with Cowden Disease.Pathobiolog y70、p.34
【非特許文献63】Basu他、(1995) Tetrahedron Lett36、4943
【非特許文献64】Ivanovskaya他、(1995) Nucl Nucl6、931〜934
【非特許文献65】Ivanovskaya他、(1987) Dokl Acad Nauk SSSR 293、477〜481
【非特許文献66】Kuznetsova他、(1999) Nuc Acids Res 27、3995〜4000
【非特許文献67】Tung & Stein (2000) Bioconjugate Chem 11 (5)、605〜618
【特許文献10】WO01/15737
【非特許文献68】Stetsenko & Gait (2000) Organic Chem 65 (16)、4900〜4908
【非特許文献69】Chen他、(1993) EMBO J.12、1161〜1167
【非特許文献70】Tommasino他、(1995) Bioessays 17、509〜518
【非特許文献71】Zhang他、(1997) Cell 89、357〜364
【非特許文献72】Ayer他、(1993) Cell 72、211〜222
【非特許文献73】Weinberg (1995) Cell 81、323〜330
【非特許文献74】Mu他、(1997) Plant Mol.Biol.34、357〜362
【非特許文献75】Cohen他、(1972) Proc.Natl.Acad.Sci.USA 69、2110
【非特許文献76】Sambrook他、(1989) Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Laboratory、Cold Spring Harbor、NY
【非特許文献77】Sherman他、(1986) Methods In Yeast Genetics、A Laboratory Manual、Cold Spring Harbor、NY
【非特許文献78】Beggs (1978) Nature 275、104〜109
【非特許文献79】J.DraperおよびR.Scott、D.Grierson(ed.)、「Plant Genetic Engineering」、Blackie、Glasgow and London、1991、vol.1、pp38〜81
【非特許文献80】J.Draper、R.Scott、A.KumarおよびG.Dury、ibid.、pp161〜198
【非特許文献81】Neuhaus & Spangenberg (1990) Physiol.Plant 79、213〜217
【非特許文献82】Gad他、(1990) Physiol.Plant 79、177〜183
【非特許文献83】Mathur & Koncz (1998) Method Mol.Biol.82、267〜276
【非特許文献84】D.McCabeおよびP.Christou、Plant Cell Tiss.Org.Cult.、3、227〜236 (1993)
【非特許文献85】P.Christou、Plant J.、3、275〜281(1992)
【非特許文献86】Bechtold他、(1993) Life Sciences、C.R.Acad.Sci.Paris 316、1194〜1199
【非特許文献87】J.DraperおよびR.Scott、D.Grierson(ed.)、「Plant Genetic Engineering」、Blackie、Glasgow and London、1991、vol.1 pp38〜81
【非特許文献88】Lazzeri (1995) Methods Mol.Biol.49、95〜106
【非特許文献89】Luchansky他、(1988) Mol.Microbiol.2、637〜646
【非特許文献90】Becker & Guarente (1990) Methods Enzymol.194、182
【非特許文献91】Vivanco & Sawyers (2002) The phosphatidylinositol 3-kinase-AKT pathway in human cancer.Nature Reviews Cancer、p489
【非特許文献92】Cory & Adams (2002) The Bcl-2family:regulators of the ceullular life-or-death switch
【非特許文献93】Mittler R.他、(1996) Inhibition of Programmed Cell Death in Tobacco Plants during a Pathogen-Induced Hypersensitive Response at Low Oxygen Pressure.Plant Cell、8、p.1991
【非特許文献94】Kuznetsova他、(1999) Nucl Acids Res 27 (20)、3995〜4000
【非特許文献95】Felgner他、(1997) Hum Gene Ther 8、511〜512
【非特許文献96】Madzak他、(1992) J.Gen.Viral.73、1533〜1536
【非特許文献97】Berkner (1992) Curr.Top.Microbiol.Immunol.158、39〜61
【非特許文献98】Berkner他、(1988) BioTechniques 6、616〜629
【非特許文献99】GorzigliaおよびKapikian (1992) J.Virol.66、4407〜4412
【非特許文献100】Quantin他、(1992) Proc.Natl. Acad. Sci.USA89、2581〜2584
【非特許文献101】Rosenfeld他、(1992) Cell 68、143〜155
【非特許文献102】Wilkinson他、(1992) Nucleic Acids Res.20、2233〜2239
【非特許文献103】Stratford-Perricaudet他、(1990) Hum.Gene Ther.1、241〜256
【非特許文献104】Moss (1992) Curr.Top.Microbiol.Immunol.158、25〜38
【非特許文献105】Muzyczka (1992) Curr.Top.Microbial.Immunol.158、97-123
【非特許文献106】Ohi他、(1990) Gene89、279〜282
【非特許文献107】Margolskee (1992) Curr.Top.Microbiol.Immunol.158、67〜90
【非特許文献108】Johnson他、(1992) J.Virol.66、2952〜2965
【非特許文献109】Fink他、(1992) Hum.Gene Ther.3、11〜19
【非特許文献110】BreakfieldおよびGeller (1987) Mol.Neurobiol.1、337〜371
【非特許文献111】Freese他、(1990) Biochem.Pharmacol.40、2189〜2199
【非特許文献112】BrandyopadhyayおよびTemin (1984) Mol.Cell.Biol.4、749〜754
【非特許文献113】Petropoulos他、(1992) J.Virol.66、3391〜3397
【非特許文献114】Miller (1992) Curt.Top.Microbiol.Immunol.158、1〜24
【非特許文献115】Miller他、(1985) Mol.Cell.Biol.5、431〜437
【非特許文献116】Sorge他、(1984) Mot.Cell.Biol.4、1730〜1737
【非特許文献117】Mann and Baltimore (1985) J.Virol.54、401〜407
【非特許文献118】Miller他、(1988) J.Virol.62、4337〜4345
【非特許文献119】Shimada他、(1991) J.Clin.Invest.88、1043〜1047
【非特許文献120】Helseth他、(1990) J.Virol.64、2416〜2420
【非特許文献121】Page他、(1990)J.Virol.64、5370〜5276
【非特許文献122】BuchschacherおよびPanganiban (1992) J.Virol.66、2731〜2739
【非特許文献123】Grahamおよびvan der Eb (1973) Virology52、456〜467
【非特許文献124】Pellicer他、(1980) Science209、1414〜1422)
【非特許文献125】Anderson他、(1980) Proc.Natl.Acad.Sci.USA77、5399〜5403
【非特許文献126】Brinster他、(1981)Cell27、223〜231
【非特許文献127】ConstantiniおよびLacy (1981) Nature 294、92〜94
【非特許文献128】Felgner他、(1987) Proc.Natl.Acad.Sci.USA 84、7413〜7417
【非特許文献129】WangおよびHuang (1989) Biochemistry 28、9508〜9514
【非特許文献130】Kaneda他、(1989) J.Biol.Chem.264、12126〜12129
【非特許文献131】Stewart他、(1992) Hum.Gene Ther.3、267〜275
【非特許文献132】Lim他、(1992) Circulation 83、2007〜2011
【非特許文献133】Wolff他、(1990) Science 247、1465〜1468
【非特許文献134】Wu他、(1991) J.Biol.Chem.266、14338〜14342
【非特許文献135】Zenke他、(1990) Proc.Natl.Acad.Sci.USA 87、3655〜3659
【非特許文献136】Wolff他、(1991) BioTecliniques 11、474〜485
【非特許文献137】Wagner他、(1991) Proc.Natl.Acad.Sci.USA 88、4255〜4259
【非特許文献138】Cotten他、(1990) Proc.Natl.Acad.Sci.USA 87、4033〜4037
【非特許文献139】Curiel他、(1991a) Proc.Natl.Acad.Sci.USA88、8850〜8854
【非特許文献140】Curiel他、(1991b) Hum.Gene Ther.3、147〜154
【非特許文献141】Feng他、(1997) Nature Biotechnology 15、866〜870
【非特許文献142】Ebbinghaus他、(1996) Gene Ther 3(4)、287〜297
【非特許文献143】Pichon他、(2000) Nucl Acids Res 28(2)
【非特許文献144】Active Motif、Morris他、(2001) Nature Biotech 19、1173〜1176
【非特許文献145】Nabel (1992) Hum.Gene Ther.3、399〜410
【非特許文献146】Vasquez KMおよびWilson JH、Trends Biochem Sci、1:4〜9、1998
【非特許文献147】Stetsenko & Gait (2000) Organic Chem 65(16)、4900〜4908
【非特許文献148】Perkins他、(1998) Biochemistry 37、11315〜11322
【発明の開示】
【課題を解決するための手段】
【0013】
本発明の第一の態様は、細胞中のアポトーシスを(誘導または回復を含めて)促進するための方法であって、(1)ゲノム中に存在する、選択したアポトーシス関連遺伝子にある部位、または該遺伝子と関係がある部位と結合する核酸結合部分、および(2)修飾部分を含む分子を、細胞中に導入するステップを含み、前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記修飾部分がポリペプチドまたはペプチド模倣体を含む方法を提供する。
【発明を実施するための最良の形態】
【0014】
一実施形態では、修飾部分は発現抑制因子部分である。修飾部分は、核酸またはクロマチンの共有結合修飾を調節することができる。
【0015】
以下でさらに論じるように、アポトーシス関連遺伝子は、細胞中のアポトーシスの制御と関係がある遺伝子である。アポトーシス関連遺伝子は、アポトーシス刺激に曝された細胞中で起こるアポトーシスを防ぐことができることを含めて(開始した場合でも、アポトーシスの終了を防ぐことができることを含む)、その産物がアポトーシスから細胞を救済することができる遺伝子であってよい。
【0016】
本発明の第二の態様は、(1)ゲノム中に存在する、選択したアポトーシス関連遺伝子にある部位、または該遺伝子と関係がある部位と結合する核酸結合部分、および(2)修飾部分を含み、前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記ポリペプチドまたはペプチド模倣体を含む分子を提供する。本発明の第一の態様と同様に、修飾部分は発現抑制因子部分であってよい。修飾部分は、核酸またはクロマチンの共有結合修飾を調節することができる。
【0017】
細胞またはゲノムは、真核生物の細胞またはゲノム、例えば真菌、動物または植物細胞であることが好ましい。
【0018】
選択した遺伝子標的(アポトーシス関連遺伝子)は、任意の様式の細胞増殖または細胞死の制御に関与するものである。アポトーシス関連遺伝子は、(少なくとも特定の条件下において)遺伝子が過剰発現されていない(あるいはその過小発現が、アポトーシスを受ける細胞の割合を低下させるのに、それ自体で充分である)以外は同一である細胞と比較した場合に、その過剰発現が、(少なくとも特定の条件下において)アポトーシスを受ける細胞の割合を増大させるのにそれ自体で充分である遺伝子であってよい。この遺伝子は、他の正常細胞または非改変型細胞において、アポトーシスを促進するのに充分である(あるいはその過小発現がアポトーシスを減少させるのに充分である)ものとすることができる。あるいは、この遺伝子は、例えば人工または天然の突然変異/淘汰(例えば腫瘍細胞中)の結果として他の遺伝子への改変、または幾つか他の方法で、例えばDNA結合部分を有する分子を使用し、前に記載したように分子を改変して、発現の改変がなされた細胞中においてのみ、アポトーシスを促進する(あるいは過小発現がアポトーシスを減少させる)ことができる。
【0019】
このような場合、遺伝子の発現を増大させてアポトーシスを促進することが望ましい。
【0020】
あるいは、アポトーシス関連遺伝子は、遺伝子が過剰発現されていない(あるいはその過小発現が、(少なくとも特定の条件下において)アポトーシスを受ける細胞の割合を増大させるのに、それ自体で充分である)こと以外は同一である細胞と比較した場合、その過剰発現が、(少なくとも特定の条件下において)アポトーシスを受ける細胞の割合を減少させるのに、それ自体で充分である遺伝子とすることができる。この遺伝子は、他の正常細胞または非改変型細胞において、アポトーシスを減少させるのに充分である(あるいはその過小発現は、アポトーシスを増大させるのに充分である)ものとすることができる。あるいは、この遺伝子は、例えば人為的または天然の突然変異/淘汰(例えば腫瘍細胞中)の結果として他の遺伝子への改変、または幾つか他の方法で、例えばDNA結合部分を有する分子を使用し、前に記載したように分子を改変して、発現の改変がなされた細胞中においてのみ、(あるいは過小発現はアポトーシスを増大させる)アポトーシスを減少させることができる。
【0021】
このような場合、遺伝子の発現を減少させてアポトーシスを促進することが望ましい。このような遺伝子(および遺伝子の発現の抑制)は、本発明の方法および分子における、非常に適した標的であると考えられる。
【0022】
分子の改変は、その破壊が「プログラムされた」細胞死(アポトーシスおよび任意の他の形の制御された細胞死、これらは用語アポトーシス中に一般に含めることができる)と相対する、壊死性細胞死に至ると予想される、中心的細胞活性と関係がある遺伝子ではなく、その産物が細胞増殖または細胞死の調節と関係がある遺伝子を標的とすることが好ましい。適切な標的遺伝子の例は、以下でさらに論じる。適切な標的遺伝子は、例えばSellers & Fisher (1999) The Journal of Clinical investigation、p.1655; Apoptosis and cancer Drug Targeting中、およびCory & Adams (2002) Nature Reviews Cancer、p489; The Bcl-2 family : regulators of the ceullular life-or-death switch中に述べられている。
【0023】
幾つかの実施形態では、本発明の分子の存在下において、細胞の生存能力は、同等の条件下での本発明の分子の不在下より、1.2倍、1.4倍、1.6倍、2倍、3倍、5倍、10倍、20倍、50倍、100倍、または1000倍低い。細胞の生存能力または細胞死は、細胞計数、生存能力アッセイまたは顕微鏡調査を含めた、任意の適切な技法を使用して測定することができる。アポトーシスは、ターミナルデオキシトランスフェラーゼ仲介dUTPニック末端標識(TUNEL)を使用することによって、壊死性細胞死と区別することができる。充分なニック末端標識によって、アポトーシスで見られるが壊死性細胞死では見られない、DNA断片化の型が示される。アポトーシスは、アネクシンVなどの細胞表面上の初期のアポトーシスマーカーを、測定することによって検出することもできる。これは例えば、染色およびFACS選別によって行うことができる。
【0024】
(影響を受けやすい細胞において)アポトーシスの誘導物質は、陽性対照として使用することができ、かつ/あるいは、(阻害剤の影響を受けやすい細胞において)アポトーシス阻害剤は、陰性対照として使用することができる。例えば、スタウロスポリンはアポトーシスの誘導物質として、Zvadまたは他のカスパーゼ阻害剤はアポトーシスの阻害剤として、使用することができる。修飾部分が(以下でさらに論じるような)ヒストン脱アセチル化酵素またはヒストン脱アセチル化酵素補充物質であるとき、脱アセチル化酵素阻害剤は、本発明の分子の影響を打ち消すことがある。
【0025】
抑制因子部分または修飾部分は、核酸またはクロマチンの共有結合修飾を調節することができることが好ましい。抑制因子または修飾部分は、クロマチン不活性化部分であることが好ましい。クロマチン不活性化部分は、クロマチン不活性化を直接的または間接的にもたらす任意のポリペプチドまたはその一部分であってよい。クロマチン不活性化を「直接的に」もたらすことによって、ポリペプチドまたはその一部自体がクロマチンに作用して、それを不活性化させることを意味する。クロマチン不活性化を「間接的に」もたらすことによって、ポリペプチドまたはその一部自体はクロマチンに作用しないが、細胞成分がそうすることを補うか促進することを意味する。
【0026】
クロマチン不活性化は一般に、遺伝子発現の抑制または不活性化をもたらす。典型的にはクロマチン不活性化は、遺伝子発現の抑制または不活性化が典型的には1個または数個の遺伝子に制限されるような、局所的事象である。したがって、クロマチン不活性化部分は任意の適切なポリペプチドであり、本発明の分子の一部分であるとき、および核酸結合部分近くの選択した遺伝子を標的とするとき、遺伝子の発現が不活性化または抑制されるように、選択した遺伝子と関係があるクロマチンを局所的に不活性化させる。ヒストン脱アセチル化はクロマチン不活性化と関係があり、したがって、クロマチン不活性化部分がヒストン脱アセチル化を容易にする場合、そのことは非常に好ましい。所与の遺伝子と関係があるヒストンの標的脱アセチル化は、ほぼ不可逆的に遺伝子の不活性化をもたらす。遺伝子発現の「抑制」または「不活性化」によって、本発明者らは、本発明の分子の存在下において、選択した標的遺伝子の発現は、均等な条件下での本発明の分子の不在下より、1.2倍、1.4倍、1.6倍、2倍、3倍、5倍、10倍、20倍、50倍、100倍、または1000倍低いことを意味する。遺伝子発現は、逆転写酵素-ポリメラーゼ連鎖反応(RT-PCR)、RNAハイブリダイゼーション、RNAse保護アッセイ、核ランオフアッセイ、およびDNAse1過敏性によって判断されるクロマチンの改変の使用を含めた、任意の適切な方法を使用して測定することができる。
【0027】
動物および植物細胞では、ヒストン脱アセチル化は、いわゆるヒストン脱アセチル化酵素複合体(HDAC)によってもたらされ、これは1つまたは複数のヒストン脱アセチル化酵素に加えて、複合体の形成および機能と関係がある補助タンパク質を含む。さらに、他のタンパク質成分が存在し、これらがHDACの一部分である可能性はないが、これらはHDACと結合し、あるいは他の場合はHDACと相互作用し、ヒストン脱アセチル化を容易にするのを助ける。
【0028】
ヒストンの脱アセチル化およびアセチル化は、よく知られている現象であり、以下の: Chen & Li (1998) Crit.Rev.Eukaryotic Gene Expression 8、169〜190; Workman & Kingston (1998) Ann.Rev.Biochem.67、545〜579; Perlmann & Vennstrom (1995) Nature 377、387-; Wolfe (1997) Nature 387、16〜17; Grunstein (1997) Nature 389、349〜352; Pazin & Kadonaga (1997) Cell 89、325〜328; DePinho (1998) Nature 391、533〜536; Bestor (1998) Nature 393、311〜312;およびGrunstein (1998) Cell 93、325〜328中に総説されている。
【0029】
HDAC複合体のポリペプチド組成は、現在調査中である。幾つかのHDAC複合体の一部分を形成することができる、あるいはそれらと結合するポリペプチドには、ヒストン脱アセチル化酵素1(HDAC1) Taunton他、(1996) Nature 272、408〜441);ヒストン脱アセチル化酵素2(HDAC2)(Yang他、(1996) Proc.Natl.Acad.Sci.USA 93、12845〜12850);ヒストン脱アセチル化酵素3(HDAC3)(Dangond他、(1998) Biochem.Biophys.Res.Comm.242、648〜652); N-CoR(Horlein他、(1995) Nature 377、397〜404); SMRT(Chen & Evans (1995) Nature 377、454〜457); SAP30(Zhang他、(1998) Molecular Cell1、1021〜1031).Sin3(Ayer他、(1995) Cell 80、767〜776; Schreiber-Agus他、(1995) Cell 80、777〜786)、SAP18(Zhang他、(1997) Cell 89、357〜364);およびRbAp48(Qian他、(1993) Nature 364、648〜652)がある。これらの論文はすべて、参照により本明細書に組み込まれている。HDAC複合体の他の成分が存在する可能性があり、あるいはそれらはHDAC複合体と相互作用し、それらは依然として発見されていないと考えられる。これらも本発明を実施する際に有用であると想定される。
【0030】
PLZFはN-CoRおよびSMRTと相互作用することが示されてきており、したがってHDAC複合体を補う。PLZFは、HDACとも直接相互作用する(Lin他、(1998) Nature 391、811〜814; Grignani他、(1998) Nature 391、815〜818; David他、(1998) Oncogene 16、2549〜2556)。
【0031】
Mad1はMadファミリーのメンバーであり、転写抑制因子として作用する能力を有する。Mad1はSin3と相互作用することができ、したがってクラスIのヒストン脱アセチル化酵素(HDAC1およびHDAC2)と相互作用することが示されてきている。Mad/Sin3は、多数のタンパク質-タンパク質相互作用が可能な、大きなタンパク質骨格として機能する(Hassig他、(1997) Cell 89、341〜347; Laherty他、(1997) Cell 89、349〜356; Zhang他、(1997) Cell 89、357〜364))。
【0032】
N-CoR、SMRT、Sin3、SAP18、SAP30およびヒストン脱アセチル化酵素のいずれかを含む、形成される複合体は、Heinzel他、(1997) Nature 387、43〜48; Alland他、(1997) Nature 387、49〜55; Hassig他、(1997) Cell 89、341〜347; Laherty他、(1997) Cell 89、349〜356; Zhang他、(1997) Cell 89、357〜364; Kadosh & Struhl (1997) Cell 89、365〜371; Nagy他、(1997) Cell 89、373〜380;およびLaherty他、(1998) Molecular Cell 2、33〜42中に記載されている。これらの論文はすべて、参照により本明細書に組み込まれている。
【0033】
したがって、HDAC複合体の成分、またはHDAC複合体と結合するか、あるいはHDAC複合体の補充を容易にするポリペプチドが、MAD1、E7、PLZF、SMRT、Sin3、SAP18、SAP30またはN-CoR、あるいはHDAC1、BDAC2またはHDAC3を含むHDAC、あるいはNuRD、MAD2、MAD3、MAD4またはRbのいずれか1つである場合、それは非常に好ましい。その機能的部分が存在する限り、すべてのポリペプチドが存在することが必要とされないこともあることは理解されよう。例えば、ヒストン脱アセチル化酵素(例えば、HDAC1、HDAC2またはHDAC3)に関しては、機能的部分はヒストン脱アセチル化酵素活性を保持する部分であってよく、あるいは機能的部分は、HDAC複合体の他の成分用の結合部位を含む部分、または他の場合はHDAC複合体を補い、ヒストン脱アセチル化を促進する部分であってよい。同様に、HDAC複合体の他の成分、またはHDAC複合体と結合するポリペプチドに関しては、機能的部分は、HDAC複合体の他の成分用の結合部位を含む部分であってよい。今日まで、6個の哺乳動物HDAC遺伝子が記載されてきており(Grozinger他、(1999) Proc.Natl.Acad.Sci.USA 96、4868〜4873)、これらの遺伝子の任意の1つまたは複数が、本発明を実施する際に有用である可能性があると考えられる。
【0034】
修飾部分はVP16またはKRABであってよいが、これは好ましくない。したがって、一実施形態では、VP16またはKRABは、用語「修飾部分」または「クロマチン不活性化部分」の意味の中には含まれない。VP16は、その作用形式がDNAまたはクロマチンの共有結合修飾と関係があるとは考えられない、転写活性因子である。KRABは、その作用形式がクロマチン不活性化以外の機構と関係があると考えられる、転写抑制因子である。好ましくはないが、KRABの任意の断片は、前に定義した分子/ポリペプチドの一部分であるとき、核酸結合部分付近の選択した遺伝子を標的化して、選択した遺伝子と関係があるクロマチンを局所的に不活性化させて、その結果遺伝子の発現を不活性化させるか抑制するとき、用語「クロマチン不活性化部分」内に含まれる。例えば、HDAC複合体と結合するか、HDAC複合体の補充を容易にすることができるKRABの任意の断片は、用語「クロマチン不活性化部分」内に含まれる。しかしながら、いかなるこのような断片も好ましくない。
【0035】
結合モチーフは、HDAC複合体の成分内、あるいはHDAC複合体と結合するポリペプチド内に存在し、これらのモチーフは、本発明のポリペプチド中においてクロマチン不活性化部分として作用するのに、充分である可能性があると考えられる。何故ならこれらは、HDAC複合体を補充することによってヒストン脱アセチル化を容易にすることができるからである。
【0036】
さらに、HDAC複合体の成分の変異体、またはHDAC複合体と結合するポリペプチドの変異体を使用できることは理解されよう。適切な変異体は、前に記載した機能的部分だけでなく、アミノ酸残基が欠失または置換または挿入されている変異体も含む。ただし変異体は、ヒストン脱アセチル化を容易にすることが依然としてできるものとする。したがって、適切な変異体は、そのアミノ酸配列が野生型と比較して修飾されているが、ヒストンを脱アセチル化するその能力を保持している、ヒストン脱アセチル化酵素の変異体を含む。同様に、適切な変異体は、例えばそのアミノ酸配列が野生型と比較して修飾されているが、HDAC複合体と、あるいはその中で相互作用するその能力を保持している、Sin3またはPLZFの変異体を含む。同様に、HDAC複合体の成分と相互作用する他のタンパク質の変異体、およびHDAC活性によって遺伝子の不活性化をもたらすことができる他の転写因子を、使用することができる。
【0037】
Rb、MADおよびMeCpG2タンパク質の全体または一部分は、HDAC複合体と相互作用することができる。
【0038】
哺乳動物系における、HDAC複合体、およびHDAC複合体の補充と関係があるポリペプチドに関して、多くの研究がなされてきているが、この系の根本的性質は、機能的に同等なポリペプチドが、他の真核細胞中、特に他の動物細胞または植物細胞中で見られると予想されることである。例えば、図5は、ヒトHDAC1と非常に関係があるポリペプチドが、シロイヌナズナ中および酵母菌中に存在することを示す。植物のHDAC複合体はトウモロコシから単離されており(Lussen他、(1997) Science 277、88〜91)、植物、真菌および脊椎動物細胞由来のヒストン脱アセチル化酵素の比較試験が着手されてきている(Lechner他、(1996) Biochim.Biophys.Acta 1296、181〜188)。ヒストン脱アセチル化酵素阻害剤は、アブラナ科植物中の不活性状態のrRNA遺伝子を活性化させることが示されてきており(Chen & Pickard (1997) Genes Dev.11、2124〜2136)、Cochliobolus carbonum由来の天然に存在する宿主選択的毒素(HC毒素)は、植物、真菌および哺乳動物のヒストン脱アセチル化酵素を阻害する(Brosch他、(1995) Plant Cell 7、1941〜1950)。
【0039】
クロマチン不活性化部分が、分子が導入される細胞と同じ細胞型または種に由来することは必要ではないが、このようなクロマチン不活性化部分が、その細胞中においてより効率よくクロマチンを不活性化することができる可能性がある場合、それが望ましい。
【0040】
分子のクロマチン不活性化部分が、ヒストン脱アセチル化を容易にすることができる、PLZF、E7、MAD1、RbまたはSAP18、あるいはPLZFまたはE7またはMAD1またはRbまたはSAP18の一部分、あるいはヒストン脱アセチル化によって遺伝子の活性化を引き起こすことが知られている、ポリペプチドまたはポリペプチドの一部分である場合、それが特に好ましい。例えば、APLと関係があるPLZF-RAR中のPLZFの部分は、N-CoRおよびSMRTと相互作用すると考えられる。
【0041】
好ましいクロマチン不活性化部分は実施例中に記載し、アミノ酸配列XXXMAVESRVTQEEIKKEPEKPIDREKTCPLLLRVF(前式でXXXは、例えばAAAまたはDDD連結基、または他の親水性の、好ましくは帯電した連結基である)を有するSAP18由来のポリペプチド/ポリペプチド模倣体または類似体、およびアミノ酸配列XXXMNIQMLLEAADYLERREREAEHGYASMLP(前式でXXXは、例えばAAAまたはDDD連結基、または他の親水性の、好ましくは帯電した連結基である)を有するMAD1由来のポリペプチドを含む。
【0042】
クロマチン不活性化部分が、HDAC1、HDAC2またはHDAC3中に含まれるようなヒストン脱アセチル化酵素活性を有するポリペプチドである場合、それも特に好ましい。
【0043】
あるいは、修飾部分が、共有結合修飾、例えば核酸、好ましくはDNAのメチル化を調節することができる部分であってよい。したがって、修飾部分はDNA修飾酵素であるか、あるいはこれを含んでよく、あるいはこのような酵素を補うことができてよい。この調節には、選択した遺伝子を抑制する効果があることが好ましい。
【0044】
修飾部分は、核酸の配列を変えないことが好ましい。修飾部分は、核酸骨格を切断しないことが好ましい。修飾部分は、リコンビナーゼまたは制限エンドヌクレアーゼではないことが好ましい。
【0045】
例えば、修飾部分は、例えばOkano M、Xie S、LiE.(1998) Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases.Nat Genet19:219〜220; Adrian P.Bird and Alan P.Wolffe (1999) Methylation-Induced Repression:Belts、Braces、and Chromatin.Cell 99、451〜454中に論じられたように、メチルトランスフェラーゼまたはメチルトランスフェラーゼ複合体の成分の全体または一部分を含むことができる(あるいは補充することができる)。
【0046】
抑制因子または修飾部分は、DNA鎖の持続的な破壊を生じるエンドヌクレアーゼまたは他の分子ではないことが好ましい。
【0047】
分子のポリペプチド/ポリペプチド模倣体または類似体部分(例えば修飾部分)は、11kDa未満、好ましくは8kDa未満、さらにより好ましくは6kDa未満の分子質量を有することが好ましい。例えば、ポリペプチド/ポリペプチド模倣体または類似体部分は、100個未満、さらにより好ましくは90、80、70、60、50、45、40、35、30、25または20個未満のアミノ酸(またはその模倣体または類似体)、最も好ましくは約60個と25個の間のアミノ酸(またはその模倣体または類似体)を有することが好ましい。
【0048】
修飾部分は、SAP18またはMAD1またはRb由来のペプチドおよび適切な連結基、例えば前および実施例1に記載した、SAP18またはMAD1由来のペプチドおよび連結基からなることが特に好ましい。
【0049】
本発明の分子は、細胞進入および/または核局在を容易にする部分(局在部分)をさらに含むことができる。この部分は、ポリペプチドまたはポリペプチド模倣体/類似体であってもよい。例えば、局在部分は、例えばSoukchareun他、(1998)Bioconjugate Chem9、466〜475およびその中に引用された参照文献、例えばSoukchareun他、(1995) Bioconjugate Cherry6、43〜53(ウイルス融合ペプチド)またはEritja他、(1991) Tetrahedron 47、4113〜4120(核輸送シグナル配列)中に論じられた、膜親和性活性を有するペプチドを含むか、あるいはこのペプチドからなっていてよい。局在部分は、核局在シグナルペプチド(例えばDDDPKKKRKV-NH2)、またはWO99/13719中に述べられたエンドソーム溶解ペプチド(エンドソーム部分からの分子の放出を容易にすることができる)であってよい。この部分は、3kDa未満、好ましくは2.5kDa未満であることが好ましい。分子の全ポリペプチド/模倣体/類似体含量は、11kDa未満であることが好ましい。典型的には、局在部分は、約7〜16個のアミノ酸を有することができる。
【0050】
局在部分の他の例には、修飾型アンテナペディアホメオドメイン系ペネトラチン(例えばRQIKIWFQNRRMKWKK)、またはTAT(例えばC(Acm)GRKKRRQRRRPPQC、前式でC(Acm)はCys-アセトアミドメチルである)またはVP22系分子(Prochiantz (2000) Curr Opin Cell Biol9、420〜429).)または塩基性HIV TAT内在ペプチドがある。
【0051】
例えば以下でより詳細に論じるように、本発明の分子は、本発明の態様によって与えられる方法および使用において、有用である可能性がある。特に、本発明のポリペプチドは、本発明の第一の態様の方法において、有用である可能性がある。
【0052】
本発明の分子が、天然に存在しないハイブリッド分子である場合、それが好ましい。例えば、核酸結合部分および修飾部分が、天然に存在する複合体または分子に由来しない場合、それが好ましい。核酸結合部分およびクロマチン不活性化部分が由来する分子は(存在する場合)、同種に由来してよく(例えば、以下でより詳細に記載するように、核酸結合部分は、ヒト核酸中に見られる配列を有するオリゴヌクレオチドであってよく、クロマチン不活性化部分はヒトPLZFの一部分であってよい)、あるいはそれらは異種に由来してよい(例えば、ヒト核酸中では見られない配列を有し、例えば細菌DNA配列と結合することができるオリゴヌクレオチドを、ヒトPLZFの一部分と融合させることができる)。
【0053】
したがって、特に好ましい実施形態では、本発明の分子は、化学合成法によって生成される分子であり、核酸結合部分および修飾部分またはクロマチン不活性化部分は、以下でより詳細に記載するように選択する。
【0054】
合成および結合技法は、WO01/14737およびその中の参照文献(参照により本明細書に組み込まれている)中に論じられている。WO01/147373の方法が好ましい。あるいは、Kusnetsova他、(1999) Nucleic Acids Res 27、3995〜4000中に記載された技法も使用することができる。
【0055】
真核生物ゲノム中に存在する部位は、その発現を調節、好ましくは抑制または不活性化するのが望ましい、選択した1つまたは複数の遺伝子の部位、あるいはその遺伝子と結合した部位である。その部位が真核生物ゲノム中に本来存在する部位である場合、それが好ましい。しかしながら、以下でより詳細に論じるように、部位はゲノム中で遺伝子工学処理された部位であってよく、あるいは部位は、挿入されるウイルス配列と関係がある部位であってよい。その発現が抑制される遺伝子の付近に存在するゲノム中に遺伝子工学処理された部位は、同種(ただしゲノム中の他の場所に存在する)由来の部位であってよく、あるいは部位は、異種に存在する部位であってよい。「ゲノム」によって、本発明者らは、染色体DNAだけでなく、真核生物の細胞中に存在する他のDNA、細胞中に導入されたDNA、例えばプラスミドまたはウイルスDNAなどを含むものとする。核酸結合部分が染色体DNAと、あるいは以下でより詳細に記載するように、染色体DNAから転写されたRNAと結合することができる場合、それが好ましい。
【0056】
一実施形態では、遺伝子は内因性遺伝子であることが好ましい。用語「内因性遺伝子」は、その細胞本来の、すなわちその細胞と異種ではなく、その本来のゲノム概念中の遺伝子を指す。この概念では、真核生物ゲノム中に存在する部位は、その発現を抑制または不活性化するのが望ましい、選択した1つまたは複数の内因性遺伝子の、あるいはその遺伝子と結合した部位である。その部位は真核生物ゲノム中に本来存在する部位であり、その本来のゲノム概念中に存在する。
【0057】
前述のように、遺伝子は(その産物を含めて)、細胞の死または生存能力の制御と関係がある。これは直接的または間接的である可能性があり、その効果は、物質による処理後の任意の時間で生じる可能性がある。
【0058】
当業者に知られているように、上記の部位は三重らせんを形成するためのオリゴヌクレオチドとの結合を促進する特定の配列特性を有することが望ましいことがある。しかしながら、本発明に関しては、このような配列特性は、ポリペプチド部分の不在下では、オリゴヌクレオチドほど重要ではない可能性があると考えられる。何故なら、本発明の分子の抑制または調節効果は、分子が標的部位とはもはや結合せず、したがって、結合の親和性がそれほど重要ではない可能性があるときでさえも持続する可能性があるからである。オリゴヌクレオチドの配列は一層重要であり、したがって特異的な認識が得られる。しかしながら、オリゴと標的配列の間に形成される結合は、ポリペプチド/またはペプチド模倣体部分が存在する場合、強力でなくでもよいこともある。
【0059】
その転写が抑制または調節される遺伝子に対する、オリゴヌクレオチド結合部位の位置も、オリゴヌクレオチド、例えば修飾部分を含まないTFOほど重要ではない可能性がある、なぜなら、本発明の分子の調節または抑制効果は(例えば、修飾ドメインがメチルトランスフェラーゼまたはヒストン脱アセチル化酵素であるか、あるいはこれらを補うことができるとき)、オリゴヌクレオチド結合部位のいずれかの側に広がる可能性があるからである。核酸結合部位は遺伝子プロモーターと結合することができるが、あるいは当該の遺伝子内またはその付近の他の配列と結合することができる。
【0060】
WO90/06934/EP0375408およびWO91/06626は、TFOの配列要件を論じている。三重らせんを形成するための2つのモチーフは、「CT」モチーフおよび「GT」モチーフと呼ばれる。これらの第一の文献は、TFOとしてのポリピリミジンオリゴヌクレオチドの使用に関する。それぞれのGC塩基対に関して、CはTFO中に存在し、それぞれのAT塩基対に関して、T(またはキサンチンまたはイノシンまたはハロゲン化誘導体)はTFO中に存在する。TFOは、二本鎖のプリン多量鎖に対して、平行方向を向いていると考えられる。あるいは、「GT」モチーフを使用すると、G(またはハロゲン化誘導体)がそれぞれのGC塩基対に関して存在し、T(またはキサンチンまたはイノシンまたはハロゲン化誘導体)がそれぞれのAT塩基対に関して存在し、TFOは、二本鎖のプリン多量鎖に対して、逆平行方向を向いていると考えられる。標的配列は、少なくとも約65%のプリン塩基、または少なくとも約65%のピリミジン塩基を有していなければならない。EP0266099も、どのようにして適切な標的配列を選択することができるかを論じている。
【0061】
W094/17086は、一本鎖の立体配座をとることができると考えられるDNA配列と結合させることを目的とする、オリゴヌクレオチドを論じている。このような配列はプリンが多量であり、相当な鏡面対称性を有する可能性がある。オリゴヌクレオチドはプリン鎖と相当に相補的であってよく、一本鎖標的DNAを有するワトソン-クリックおよびフーグスティン結合を形成することができる、環状またはステム-ループ機能構造を有していてよい。
【0062】
W096/35706は、特異的で安定した標的核酸(ピリミジン一本鎖核酸)との複合体形成を促進すると考えられ、標的核酸不在下における平行鎖ヘアピン構造の形成ために、高い安定性を有し得る構造および配列特性を有するオリゴヌクレオチドを記載している。
【0063】
Debin他、(1999) Nucl Acids Res 27 (13)、2699〜2707は、G、A三重らせんの安定性、およびTFO設計の結果に影響を与える因子を述べている。Xodo他、(2001) Eur J Biochem 268、656〜664も、標的部位とTFOの結合、例えば短いオリゴヌクレオチドと近辺部位の結合に影響を与える因子を調べている。
【0064】
Blume他、(1999) Nucl Acids Res 27、695〜702は、三重らせん形成における二価カチオンの関与、および形成をどのようにして、正または負に調節することができるかを調べている。Faria他、(2001) JMol Biol 306、15〜24は、細胞中のTFOおよびさまざまなオリゴヌクレオチドに関する結果を評価するためのアッセイを記載している。Cheng他、(2000) Biotech and Bioeng 70、467〜472は、TFO結合の数学的モデル化の結果、および結合部位およびTFO配列の選択に関する結果を示している。
【0065】
Demidov & Frank-Kamenetskiiは、ペプチド核酸(PNA)、特にカチオン性ピリミジンPNA(cpyPNA)と二本鎖DNAの結合を総説している。
【0066】
可能性のあるTFOを設計するためのルールは、Vasquez & Wilson (1998) Trends Biochem Sci1、4〜9中に総説されている。3つの型のTFO:ピリミジン多量(CT);プリン多量(GA)および混合型(GTまたはGAT)が有効であることが示されている。CTTFOは平行モチーフ中で結合し、その中では第三の鎖が、二本鎖のプリン鎖と同じ5'〜3'方向を有する。GATFOは逆平行モチーフ中で結合する。混合型TFOは、標的配列に応じて、いずれかの方式で結合することができる。他の性質も幾つかの型のTFOの間で異なり;例えばCTTFOはpH依存性である。それぞれの型のTFOが、本発明に関して適切である可能性がある。
【0067】
オリゴヌクレオチドまたは模倣部分は、約10〜80、好ましくは15〜40塩基長、さらにより好ましくは約20〜40塩基長であることが好ましい。20塩基未満のオリゴヌクレオチドは、弱いおよび/または特異性の低い結合を示す可能性があるが、それにもかかわらず、前述のように、例えばごく一時的な結合が必要とされるので、本発明の実施において有用である可能性がある。
【0068】
「DNA」または「オリゴ(デオキシ)ヌクレオチド」によって、本発明者らは、糖-連結基-糖の骨格を有する分子であって、糖残基が2'-デオキシリボース(および、2',3'ジデオキシリボース部分を含むヌクレオシドが末端であるDNA鎖をしたがって含む)を含み、1位置において糖残基と結合しているのが、アデニン(A)、シトシン(C)、グアニン(G)、チミジン(T)、イノシン(I)、ウリジン(U)およびその他などの塩基である分子を意味する。正常なDNAでは、糖残基間の結合(「糖-糖結合」)はリン酸部分であり、これが前記糖残基とジエステルを形成する。しかしながら、本発明者らは、用語「核酸」中に(およびより詳細には用語DNA中に)、非リン酸結合を有する分子を含むものとする。
【0069】
したがって、ホスホロチオエート結合およびホスホロセレノエート結合を含める。これらの結合が、正常なDNAよりも細胞のヌクレアーゼによる攻撃に対して耐性があることが、好ましい可能性がある。このような結合は、リン酸メチル、ホスホトリエステル、および天然に存在するホスホジエステルの鏡像異性体も含むことができる。
【0070】
用語「核酸」または「オリゴヌクレオチド」とは、非天然塩基類似体を含む分子;ペントース糖の2'および3'位置が独立に、-H、-OHまたは-NH2のいずれかである分子;および、ホスホジエステル結合中ではなくリン原子と結合している酸素が、-SH、SeH、-BH2、-NH2、-PH3、-F、-Cl、-CH3、-OCH3、-CNおよび-Hによって置換されている分子も含む。
【0071】
オリゴヌクレオチドは、オリゴリボヌクレオチドまたはオリゴデオキシリボヌクレオチドであってよい。オリゴデオキシリボヌクレオチドが好ましい、何故なら、オリゴリボヌクレオチドは、オリゴデオキシリボヌクレオチドよりも酵素による攻撃を受けやすい可能性があるからである。
【0072】
オリゴヌクレオチドまたは模倣体または類似体は、当業者に知られるように、例えばWO99/13719およびその中の参照文献中、およびDemidov & Frank-Kamenetskii(2001)上記中に記載されたように、ペプチド核酸であってよい。PNAは、DNAのデオキシリボース-リン酸骨格の代わりに2-アミノエチルグリシン単位を含む、ポリアミド(ペプチド)骨格を有する核酸類似体である。PNA骨格は中性であり(DNA骨格と異なり、それは負に帯電している)、したがって、対応するDNA分子よりも、帯電した核酸分子とより安定的に結合することができる。
【0073】
オリゴヌクレオチドまたは類似体または模倣体は、DNAオリゴヌクレオチド
であることが好ましい。
【0074】
オリゴヌクレオチドに対する言及は(それが適切な場合)、オリゴヌクレオチド模倣体または類似体、例えばPNAに対する言及を含む。
【0075】
オリゴヌクレオチドは、オリゴヌクレオチドの5'または3'末端と結合することができる、連結基を含むことができる。適切な連結基の例は、例えばWO90/06934中に記載されている。
【0076】
核酸結合部分は、ペプチド核酸(PNA)であるか、あるいはこれを含むことが、好ましい可能性がある。
【0077】
核酸結合部分は、植物または動物などの真核生物、ゲノム中に存在する部位と結合する、定義する任意の適切な結合部分であってよい。核酸結合部分は、本発明の分子のクロマチン不活性化部分の存在によって、その発現が変更、特に抑制される、選択した遺伝子の部位またはそれと関係がある部位と結合することができることが特に好ましい。核酸結合部分は、所望の部位と選択的に結合することが好ましい。核酸結合部分が結合することができる、1つまたは複数の所望の部位が存在する可能性がある。典型的には、標的ゲノム中には、1つの目的とする標的部位が存在する。誤解を避けるために、真核生物中に存在する部位は、天然に存在する部位であることが最も有用である可能性があり、あるいはそれは、真核生物中に存在させるように遺伝子工学処理した部位であってよい。部位は、同じまたは任意の他の真核生物に元来由来するものである必要はない。例えば部位は、TFOが事前に特徴付けられており、真核生物細胞、例えば植物細胞のDNA中に存在させるように遺伝子工学処理した、細菌またはウイルスの配列、あるいは人工配列であってよい。これらの例は、本明細書の実施例中に記載するEREおよびIREなどの応答エレメント、または他の特徴付けされた結合部位を含むことができる。部位の内因性調節因子を含まない細胞において、このような部位を使用することが望ましい可能性がある。あるいは、部位は、本来存在する応答エレメントの改変型であってよく、この改変型はTFOの結合部位として働くことができるが、本来存在する応答エレメントの本来存在する調節因子によって調節することができない。しかしながら、核酸結合部分が結合する部位が真核生物細胞中に本来存在し、それがゲノム中のその本来の位置に存在する場合、それが好ましい。
【0078】
核酸結合部分は、DNA結合部分またはRNA結合部分であってよい。したがって、核酸結合部分は、二本鎖核酸(例えばDNA)または一本鎖核酸(例えばRNAまたは一本鎖DNA)と結合することができる。RNA結合部分の場合、RNA結合部分と結合する、真核生物ゲノム中に存在する部位は、典型的には、不活性化用に選択した部位においてDNAから転写される新生RNAである。そのRNAは、その発現が抑制される遺伝子により転写されるRNAであってよく、あるいはそのRNAは、その発現が抑制される遺伝子と隣接しているか、あるいは少なくともそれと近い、遺伝子により転写されるRNAであってよい。RNA結合部分は、転写産物の5'端の、あるいはその近くのRNA配列と結合することが好ましい。転写される一方で、新生RNAは、その転写部位あるいはその近くに留まり、転写部位が、その発現が抑制される遺伝子、あるいはその近くに存在する場合、本発明の分子中にRNA結合部分を使用することは、所望の部位へのクロマチン不活性化部分の局在化を容易にすることは理解されよう。
【0079】
DNA結合部分が転写因子結合部位と結合し、例えばその結果、転写因子が結合する2つ以上の遺伝子の発現を調節することができる場合、それは有用である可能性がある。アポトーシス遺伝子と関係がある転写因子には、CREB、WT1、NF-kappaBおよびStat3がある。転写因子およびそれらの結合部位を列挙するデータベースを、以下に列挙する:
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFFACTOR
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fum+pagelibinfo+-info+TFSITE
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFCELL
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-
info+TFCLASS
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-
info+TFMATRIX
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-
info+TFGENE
【0080】
DNA結合部分が、転写開始部位のすぐ上流の、プロモーター領域または他の調節領域または配列と結合する場合、それが有用である可能性がある。幾つかの適用例では、遺伝子内の配列を標的化して、スプライス変異体を区別することが、好ましい可能性がある。
【0081】
前述のように、オリゴヌクレオチドを設計/遺伝子工学処理して、選択した遺伝子の、あるいはそれと結合した、特定の選択した標的DNA配列と結合させることができる。本発明の一実施形態では、オリゴヌクレオチドは、植物または動物細胞中には存在するが、同等な野生型配列中には存在しない、突然変異遺伝子配列中に存在する部位と結合させるために、遺伝子工学処理したオリゴヌクレオチドである。例えば、以下でより詳細に論じるように、オリゴヌクレオチドは、突然変異アポトーシス関連癌遺伝子などの、優性ネガティブ突然変異遺伝子と選択的に結合することができ、結合によって、DNAメチル化またはクロマチン不活性化が起こり、突然変異アポトーシス関連癌遺伝子の発現を抑制する。突然変異するアポトーシス関連癌遺伝子の例には、タンパク質キナーゼB/AKTおよびPTENがある(Hill MおよびHemmings B (2002): Inhibition of protein kinase B/AKT.Implications for cancer therapy.Pharmacol Ther、93、p.243; Kanaseki T他、(2002) Identification of germline mutation of PTEN gene and analysis of apoptosis resistance of the lymphocytes in a patient with Cowden Disease.Pathobiolog y70、p.34)。
【0082】
RAS(H-ras)およびBcl-10は、アポトーシス関連遺伝子の定義から除外される。
【0083】
典型的には、核酸結合部分と、修飾部分またはクロマチン不活性化部分を融合させる。例えばSoukchareun他、(1998)上記、およびその中に引用された参照文献中、およびBasu他、(1995) Tetrahedron Lett36、4943中に記載されたのと同様に、核酸結合部分と、修飾部分またはクロマチン不活性化部分は、例えば固相支持体上でのペプチド、および次いでオリゴヌクレオチドの連続的構築によって、1つの分子として合成することができる(全体的合成手法)。好ましくは、自動化された手順を使用する。
【0084】
あるいは、核酸結合部分と、修飾部分またはクロマチン不活性化部分は、当業者によく知られている技法を使用して別々に合成し、次いで結合する。ペプチド核酸とペプチドの結合に適した技法には、SPDP(N-スクシンイミジル3-(2-ピリジルジチオ)プロピオネート)およびSMCC(スクシンイミジル4-(N-マレイミドメチル)シクロヘキサン-カルボキシレート)などの、ヘテロ二官能性結合試薬の使用があり、例えばWO99/13719中、特に実施例12〜15中に記載されている。オリゴデオキシヌクレオチドとペプチドの結合に適した技法には、Soukehareun他、(1998)上記中に記載されたのと同様の、N_-Fmoc-システイン(S-チオブチル)誘導型オリゴデオキシヌクレオチドの使用がある。他の技法としては、ペプチド中の求核基(-NH2基など)を介して非保護ペプチドを結合させる前に、リン酸オリゴヌクレオチドの3'または5'端の、N-ヒドロキシベンゾトリアゾール(HOBT)エステル活性化を使用することがある(Ivanovskaya他、(1995) Nucl Nucl6、931〜934; Ivanovskaya他、(1987) Dokl Acad Nauk SSSR 293、477〜481; Kuznetsova他、(1999) Nuc Acids Res 27、3995〜4000を参照)。ペプチド-オリゴヌクレオチド結合技法は、例えばTung & Stein (2000) Bioconjugate Chem 11 (5)、605〜618中に総説されている。
【0085】
WO01/15737およびStetsenko & Gait (2000) Organic Chem 65 (16)、4900〜4908中に記載されたのと同様に、「本来の連結」技法を使用することが好ましい。N-末端チオエステル-官能化ペプチドを、5'-システイニルオリゴヌクレオチド誘導体と連結させる。
【0086】
適切には、核酸結合部分、および抑制因子、修飾部分またはクロマチン不活性化部分が結合し、その結果両方の部分が、例えば核酸結合部分は、植物または動物ゲノム中に存在する部位と結合することができ、結合によって、修飾部分が、核酸またはクロマチンの共有結合修飾を調節することが依然としてできる、例えばクロマチン不活性化部分が、クロマチンを不活性化させることが依然としてできるように、そのそれぞれの活性を保つ。この2つの部分は直接結合することができるが、これらは連結基ペプチドまたはオリゴヌクレオチドによって結合することができる。適切な連結基ペプチドは、ランダムコイルの立体配座を典型的にとるペプチドであり、例えばポリペプチドは、アラニンまたはプロリン、あるいはアラニン残基とプロリン残基の混合物を含むことができる。アミノ酸は溶解度を増大させることが好ましい;したがって連結基は、例えばアスパラギン酸残基などの、帯電または親水性アミノ酸を含むことができる。連結基はアスパラギン酸残基を含むか、あるいはそれらからなることが好ましい。アミノ酸は、フェニルアラニンまたはトリプトファンなどの、親水性アミノ酸ではないことが好ましい。連結基は、10個と100個の間、より好ましくは10個と50個の間、さらにより好ましくは10個と20個の間の、アミノ酸残基を含むことが好ましい。例えば、3個と9個の間のアミノ酸の短い連結基も、有用である可能性がある。任意の事象において、分子の部分間に連結基が存在しようとしまいと、分子はその標的核酸と結合することができ、発現を抑制するか、あるいは核酸またはクロマチンの共有結合修飾を調節する、例えばクロマチンを不活性化させ、それによって遺伝子発現を選択的に抑制するか不活性化させることができる。
【0087】
適切な抑制因子、修飾またはクロマチン不活性化部分をコードするポリヌクレオチドは、当分野において知られており、知られている配列から設計し、作製することが容易にできる。さまざまな適切なクロマチン不活性化部分をコードする、ポリヌクレオチド配列は、ポリペプチドを言及する参照文献中に前に与えており、あるいはGenBankまたはEMBLまたはdbESTから入手可能である。PLZFに関する参照文献は、Chen他、(1993) EMBO J.12、1161〜1167である。E7に関する参照文献は、Tommasino他、(1995) Bioessays 17、509〜518である。SAP18、MAD1およびRbに関する参照文献は、それぞれZhang他、(1997) Cell 89、357〜364; Ayer他、(1993) Cell 72、211〜222およびWeinberg (1995) Cell 81、323〜330である。
【0088】
適切な連結基ペプチドをコードするポリヌクレオチドは、連結基ペプチドの配列から設計し、作製することが容易にできる。
【0089】
したがって、本発明の分子の抑制因子または修飾部分をコードするポリヌクレオチドは、よく知られている遺伝子工学技法を使用して、容易に構築することができる。したがって、抑制因子または修飾部分は、当業者によく知られている分子生物学の技法を使用して、発現によって合成することができる。しかしながら、当業者に知られており本明細書で論じるように、有機化学の技法によって、本発明の分子のポリペプチド/類似体/模倣体部分を合成することが、好ましい可能性がある。
【0090】
本発明は、本発明の分子を用いて形質転換した、宿主細胞にも関する。宿主細胞は原核生物または真核生物であってよい。細菌細胞は、好ましい原核生物宿主細胞であり、典型的には大腸菌の菌株、例えばBethesda Research Laboratories Inc.、Bethesda、MD、USAから入手可能な大腸菌菌株DH5、およびAmerican Type Culture Collection (ATCC)of Rockville、MD、USAから入手可能なRR1などである(NoATCC31343)。好ましい真核生物細胞には、植物、酵母菌、昆虫および哺乳動物細胞、好ましくは脊椎動物細胞、マウス、ラット、サルまたはヒト線維芽細胞および腎臓細胞系由来の細胞などがある。酵母菌宿主細胞には、YPH499、YPH500およびYPH501があり、これらは一般にStratagene Cloning Systems、La Jolla、CA92037、USAから入手可能である。好ましい哺乳動物宿主細胞には、CCL61としてATCCから入手可能なチャイニーズハムスター卵巣(CHO)細胞、CRL1658としてATCCから入手可能なNIHスイスマウス胚細胞NIH/3T3、CRL1650としてATCCから入手可能なサル腎臓由来COS-1細胞;ヒト胚腎臓細胞である293細胞、およびHT1080ヒト骨線維肉腫細胞がある。
【0091】
形質転換用のプロトプラストは、典型的には、当分野で知られている方法によって、必要に応じて作製される。植物細胞系は、一般には利用可能ではない。しかしながら、一般的に使用される1つの細胞系は、タバコ由来のBright Yellow2細胞系である(BY2;Mu他、(1997) Plant Mol.Biol.34、357〜362)。
【0092】
本発明の分子を用いた適切な細胞宿主の形質転換は、よく知られている方法によって行い、これは典型的には、使用する分子の型に依存する。原核生物宿主細胞の形質転換に関しては、例えばCohen他、(1972) Proc.Natl.Acad.Sci.USA 69、2110、およびSambrook他、(1989) Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Laboratory、Cold Spring Harbor、NYを参照のこと。酵母菌細胞の形質転換は、Sherman他、(1986) Methods In Yeast Genetics、A Laboratory Manual、Cold Spring Harbor、NY中に記載されている。Beggs (1978) Nature 275、104〜109の方法も有用である。脊椎動物細胞に関しては、このような細胞をトランスフェクトする際に有用な試薬、例えばリン酸カルシウムおよびDEAE-デキストランまたはリポソーム配合物は、Stratagene Cloning Systems、またはLife Technologies Inc.、Gaithersburg、MD20877、USAから入手可能である。植物細胞および全植物に関しては、以下の植物形質転換手法(J.DraperおよびR.Scott、D.Grierson(ed.)、「Plant Genetic Engineering」、Blackie、Glasgow and London、1991、vol.1、pp38〜81中)を使用することができる:
i)DNA仲介の遺伝子の移送、ポリエチレングリコール刺激型の、プロトプラストへのDNAの取り込みによる、エレクトロポレーションによる、あるいはプロトプラストまたは植物細胞のマイクロインジェクション(J.Draper、R.Scott、A.KumarおよびG.Dury、ibid.、pp161〜198)。プロトプラストへの直接的な遺伝子の移送は、Neuhaus & Spangenberg (1990) Physiol.Plant 79、213〜217; Gad他、(1990) Physiol.Plant 79、177〜183;およびMathur & Koncz (1998) Method Mol.Biol.82、267〜276中にも記載されている;
ii)粒子の衝撃を使用する形質転換(D.McCabeおよびP.Christou、Plant Cell Tiss.Org.Cult.、3、227〜236 (1993); P.Christou、Plant J.、3、275〜281(1992))。
【0093】
好ましい技法には、エレクトロポレーション、マイクロインジェクションおよびリポソーム配合がある。
【0094】
幾つかの種は、直接的な形質転換の影響を受けやすく、組織または細胞培養の必要性が回避される(Bechtold他、(1993) Life Sciences、C.R.Acad.Sci.Paris316、1194〜1199)。
【0095】
全ての手法において、カナマイシンまたは除草剤耐性剤などの、適切な選択マーカー、あるいは代替的にスクリーニング可能なマーカー(「レポーター」)遺伝子、β-グルクロニダーゼまたはルシフェラーゼなどが好ましい(J.DraperおよびR.Scott、D.Grierson(ed.)、「Plant Genetic Engineering」、Blackie、Glasgow and London、1991、vol.1 pp38〜81中を参照)。
【0096】
エレクトロポレーションは、細胞を形質転換および/またはトランスフェクトするため、酵母菌細胞、細菌細胞、昆虫細胞、脊椎動物細胞および幾つかの植物細胞(例えばオオムギ細胞、Lazzeri (1995) Methods Mol.Biol.49、95〜106を参照)を形質転換するためにも有用であり、当分野でよく知られている。
【0097】
例えば、多くの細菌種は、参照により本明細書に組み込まれているLuchansky他、(1988) Mol.Microbiol.2、637〜646中に記載された方法によって、形質転換することができる。最大数の形質転換体が、25μFDで1cm当たり6250Vを使用する、2.5×PEBに懸濁させたDNA-細胞混合物のエレクトロポレーション後に、一貫して回収される。
【0098】
エレクトロポレーションによって酵母菌を形質転換するための方法は、Becker & Guarente (1990) Methods Enzymol.194、182中に開示されている。
【0099】
首尾よく形質転換された細胞、すなわち本発明の分子を含む細胞は、よく知られている技法によって同定することができる。例えば、標識したオリゴおよび/またはGFPマーカーを使用することができる。
【0100】
したがって、形質転換された宿主細胞そのもの以外にも、本発明は、これらの細胞の培養物、好ましくはモノクローナル(クローン的に均一な)培養物、または栄養培地中のモノクローナル培養物由来の培養物も企図する。
【0101】
細胞の性質のため(例えば、細胞は組換え細胞であり、例えばその細胞中に、本発明の分子が組換えコピーと結合しない点で標的遺伝子と異なる、標的遺伝子の組換えコピーが存在するので)、あるいは細胞/培養物を増殖または維持する条件が、細胞中のアポトーシスを誘導する条件ではないため、形質転換された細胞または培養物は、本発明の分子がアポトーシスを促進しない、細胞または培養物であってよいことは理解されよう。
【0102】
植物に関して、本発明は、単細胞由来の細胞の懸濁液培養物、単離プロトプラストまたは安定した形質転換型植物を含むことが想定される。
【0103】
本発明の分子は任意の適切な宿主細胞中に導入することができるが、適切な動物または植物細胞、特に本発明の分子が結合することができるそれらのDNA内に1つまたは複数の部位を有する細胞中において有効であるように、本発明の分子が本来設計されていることは理解されよう。
【0104】
したがって、その存在が特定の遺伝子の発現を抑制する、本発明の分子を含む動物または植物細胞、あるいはこれらの細胞を含む動物または植物は、遺伝子がもはや発現することができないという意味で「ノックアウトされた」遺伝子を有すると考えることができる。ヒストン脱アセチル化によるクロマチン不活性化は、他の介入がないほぼ不可逆的なものとし得る。当業者に知られているように、ヒストン脱アセチル化による抑制は、ヒストン脱アセチル化酵素の阻害剤、例えばトリコスタチンA(TSA)、トラポキシンンまたは酪酸ナトリウム(NaB)などを使用して逆行させることができる。同様にメチル化は、他の介入、例えばメチル化阻害剤/逆行物質の投与がないほぼ不可逆的なものである可能性があり、メチル化阻害剤/逆行物質は当分野では知られており、化合物アザシチジンを含む。他のメチル化阻害剤には、5デオキシアザシチジン、または例えば、DNAメチルトランスフェラーゼを対象とするアンチセンスオリゴ(または本明細書に記載する遺伝子発現抑制因子)がある。
【0105】
本発明の分子を動物または植物細胞中に導入することによって、核酸、例えばDNA内の適切な結合部位(ポリペプチドのDNA結合部分によって結合する)への分子の標的化を可能にし、遺伝子発現の抑制または不活性化、または他の調節を、例えば標的結合部位のクロマチン、または標的結合部位と結合したクロマチンを不活性化させることによって、可能にするであろうことは容易に理解されよう。典型的には、選択した遺伝子を標的化するように、本発明の分子を選択する。したがって、適切には、標的遺伝子は、それと関係がある分子のDNA結合部分によって結合した部位を有する。このように結合する部位は、遺伝子自体の中、例えば遺伝子のイントロンまたはエクソン内に存在してよく、あるいは部位は、遺伝子の転写された部分の5'領域中、例えばプロモーターまたはエンハンサー領域内あるいはその近辺に存在してよく;あるいは部位は、遺伝子の転写された部分の3'領域中に存在してよい。
【0106】
選択したアポトーシス関連遺伝子の発現を調節、特に抑制する能力は、生物学の多くの領域において有用である。
【0107】
典型的には、その発現が抑制される遺伝子が動物細胞中に存在するとき、その動物細胞は動物内の細胞であり、本発明の方法を使用して、動物(それはヒトまたは非ヒト動物であってよい)中において選択したアポトーシス関連遺伝子の発現を調節、例えば抑制し、それによってアポトーシスを調節、例えば促進する。動物細胞における個々の使用の例には、Bcl-2、Bcl-XI(Bcl-2ファミリーのメンバー)またはAKT/PKB(タンパク質キナーゼB;さまざまな対立遺伝子)の不活性化がある。これらの遺伝子は、例えばVivanco & Sawyers (2002) The phosphatidylinositol 3-kinase-AKT pathway in human cancer.Nature Reviews Cancer、p489; Cory & Adams (2002) The Bcl-2family:regulators of the ceullular life-or-death switch中に記載されたように、アポトーシスの制御と関係がある。
【0108】
Bcl2、AKTおよびBCI-XI遺伝子に関する寄託番号には、以下のものがある:Bcl2:NM_000657およびNM_000633、AKT:NM_005163、Bcl-XL:NM_138578。
【0109】
さらに典型的には、植物細胞は植物内の細胞であり、本発明の方法を使用して、植物中のアポトーシス関連の選択した遺伝子の発現を抑制する。これは例えば、侵入する病原体に対する防御機構において有用である可能性がある(Mittler R.他、(1996) Inhibition of Programmed Cell Death in Tobacco Plants during a Pathogen-Induced Hypersensitive Response at Low Oxygen Pressure.Plant Cell、8、p.1991)。
【0110】
適切には、本発明の方法を使用して、その発現を調節、抑制または不活性化することが望ましい、アポトーシス関連遺伝子の発現を調節、抑制または不活性化する。その発現を抑制または不活性化することが望ましい遺伝子には、アポトーシス救済遺伝子、例えばAKT/PKB、Bcl-XIまたはBcl-2、またはプロウイルスゲノム中に存在する遺伝子を含めたウイルス遺伝子があり、したがって、動物に関するこの方法は、医学的治療の方法を構成することができる。さらに、腫瘍遺伝子は幾つかの癌では過剰発現する可能性があり、他のアポトーシス促進調節と組み合わせて、それらの発現を抑制することが望ましい可能性がある。幾つかの腫瘍遺伝子は、活性化突然変異を有するために腫瘍原性である。
【0111】
突然変異腫瘍遺伝子の発現の選択的抑制は、突然変異腫瘍遺伝子の配列と選択的に結合する、DNA結合部分を使用して得ることができ、抑制因子または修飾部分、例えばクロマチン不活性化部分は、例えば腫瘍遺伝子がその中に存在するかあるいはそれと関係があるクロマチンを不活性化することによって、突然変異腫瘍遺伝子の発現を抑制する。腫瘍遺伝子の過剰発現、または突然変異(非常に活性化した)腫瘍遺伝子の発現を抑制することが、腫瘍遺伝子が役割を果たす癌の治療において、一般に望ましい。標的化することができる突然変異腫瘍遺伝子にはRasおよびBcl-10がある。これらは、配列特異的な方式で突然変異した遺伝子を認識することができるDNA結合部分によって、標的化することができる。
【0112】
アポトーシスはプログラムされた細胞死であり、これはさまざまな刺激によって誘導することができる。例えば、多くの化学療法剤は、アポトーシスの誘導によって癌細胞を除去する。腫瘍の発達および進行には、充分な増殖およびアポトーシスの阻害が必要であると考えられる。現在の治療プロトコルに対する原発性または後天性の耐性は、臨床腫瘍学における主な関心事のままであり、アポトーシスプログラムの欠陥によって引き起こされる可能性があり、化学療法に対する耐性は、部分的には低いアポトーシス率によることが知られている。幾つかの遺伝子は、アポトーシスの制御と関係があることが報告されてきており、例えば、bcl-2原-腫瘍遺伝子の発現は、さまざまなヒト血液悪性腫瘍および固形腫瘍中で見られる。Bcl-2タンパク質は、腫瘍細胞が放射線、化学療法、およびホルモン療法によって誘導されるアポトーシスを受けるのを防ぐことによって、その腫瘍原的役割を発揮する。したがって、アポトーシスの防止と関係がある、遺伝子の発現を妨げることを目的とする、療法を開発することが望ましい。
【0113】
本発明のこれらの方法は、本発明の分子を動物細胞または植物細胞に移送させることを、典型的には含む。
【0114】
オリゴヌクレオチドまたはオリゴヌクレオチド-ペプチド融合体を用いる有用な移送系が当業者に知られ、本発明の分子を動物の身体中または外の細胞に導入する、本発明の方法を実施する際に、有用である可能性がある。例えば、リポソームまたはウイルス系の方法を使用することができる。エレクトロポレーション(例えばKuznetsova他、(1999) Nucl Acids Res 27 (20)、3995〜4000を参照)、衝撃法、カチオン性脂質(例えばFelgner他、(1997) Hum Gene Ther 8、511〜512またはWO99/13719中に記載されたのと同様の)、または分子のオリゴヌクレオチドまたはポリペプチド部分と結合した特異的リガンド、または担体を、例えばWO99/13719中に記載されたのと同様に使用することができる。
【0115】
ウイルスまたは非ウイルス移送法を使用することができる。パラボウイルス、例えばSV40(Madzak他、(1992) J.Gen.Virol.73、1533〜1536)、アデノウイルス(Berkner (1992) Curr.Top.Microbiol.Immunol.158、39〜61; Berkner他、(1988) BioTechniques 6、616〜629; GorzigliaおよびKapikian (1992) J.Virol.66、4407〜4412; Quantin他、(1992) Proc.Natl. Acad. Sci.USA89、2581〜2584; Rosenfeld他、(1992) Cell 68、143〜155; Wilkinson他、(1992) Nucleic Acids Res.20、2233〜2239; Stratford-Perricaudet他、(1990) Hum.Gene Ther.1、241〜256)、ワクシニアウイルス(Moss (1992) Curr.Top.Microbiol.Immunol.158、25〜38)、アデノ随伴ウイルス(Muzyczka (1992) Curr.Top.Microbial.Immunol.158、97-123; Ohi他、(1990) Gene89、279〜282)、HSVおよびEBVを含めたヘルペスウイルス(Margolskee (1992) Curr.Top.Microbial.Immunol.158、67〜90; Johnson他、(1992) J.Virol.66、2952〜2965; Fink他、(1992) Hum.Gene Ther.3、11〜19; BreakfieldおよびGeller (1987) Mol.Neurobiol.1、337〜371; Freese他、(1990) Biochem.Pharmacol.40、2189〜2199)、ならびに鳥類(BrandyopadhyayおよびTemin (1984) Mol.Cell.Biol.4、749〜754; Petropoulos他、(1992) J.Virol.66、3391〜3397)、ネズミ(Miller (1992) Curr.Top.Microbiol.Immunol.158、1〜24; Miller
他、(1985) Mol.Cell.Biol.5、431〜437; Sorge他、(1984) Mot.Cell.Biol.4、1730〜1737; Mann and Baltimore (1985) J.Virol.54、401〜407; Miller他、(1988) J.Virol.62、4337〜4345)、およびヒト起源(Shimada他、(1991) J.Clin.Invest.88、1043〜1047; Helseth他、(1990) J.Virol.64、2416〜2420; Page他、(1990)J.Virol.64、5370〜5276; BuchschacherおよびPanganiban (1992) J.Virol.66、2731〜2739)のレトロウイルスを含めた、幾つかのウイルスが、遺伝子移送ベクターとして使用されてきている。今日まで、大部分のヒト遺伝子療法プロトコルは、欠陥のあるネズミレトロウイルスに基づいてきている。
【0116】
当分野で知られている非ウイルス性の遺伝子の移送法には、リン酸カルシウム共沈殿などの化学技法(Grahamおよびvan der Eb (1973) Virology52、456〜467;Pellicer他、(1980) Science209、1414〜1422);機械的技法、例えばマイクロインジェクション(Anderson他、(1980) Proc.Natl.Acad.Sci.USA77、5399〜5403; Gordon他、1980; Brinster他、(1981)Cell27、223〜231; ConstantiniおよびLacy (1981) Nature 294、92〜94);リポソームによる膜融合仲介型移送(Felgner他、(1987) Proc.Natl.Acad.Sci.USA 84、7413〜7417; WangおよびHuang (1989) Biochemistry 28、9508〜9514; Kaneda他、(1989) J.Biol.Chem.264、12126〜12129; Stewart他、(1992) Hum.Gene Ther.3、267〜275; Nabel他、1990; Lim他、(1992) Circulation 83、2007〜2011);および直接的なDNAの取り込みおよび受容体仲介型DNA移送(Wolff他、(1990) Science 247、1465〜1468; Wu他、(1991) J.Biol.Chem.266、14338〜14342; Zenke他、(1990) Proc.Natl.Acad.Sci.USA 87、3655〜3659; Wu他、1989b; Wolff他、(1991) BioTecliniques 11、474〜485; Wagner他、1990; Wagner他、(1991) Proc.Natl.Acad.Sci.USA 88、4255〜4259; Cotten他、(1990) Proc.Natl.Acad.Sci.USA 87、4033〜4037; Curiel他、(1991a) Proc.Natl.Acad.Sci.USA88、8850〜8854; Curiel他、(1991b) Hum.Gene Ther.3、147〜154)がある。ウイルス仲介型の遺伝子の移送は、リポソーム送達を使用
する直接的なin vivoの遺伝子の移送送達と組み合わせることができ、ウイルスベクターを、周囲の非分裂細胞ではなく腫瘍細胞に向けることが可能である。
【0117】
他の適切な系には、Feng他、(1997) Nature Biotechnology 15、866〜870によって記載されたレトロウイルス-アデノウイルスハイブリッド系、または適切な一本鎖Fv断片などの標的リガンドを用いるウイルス系がある。
【0118】
生物学的な遺伝子の移送法と物理学的な遺伝子の移送法を組み合わせる手法では、任意の大きさのプラスミドDNA(または例えばオリゴヌクレオチド/ペプチド融合体)を、アデノウイルスヘキソンタンパク質に特異的なポリリシン結合抗体と組み合わせ、生成した複合体をアデノウイルスベクターと結合させる。三分子の複合体を次いで使用して、細胞を感染させる。アデノウイルスベクターは、結合したDNAが損傷する前に、エンドソームの効率の良い結合、内在化、および分解を可能にする。
【0119】
Ebbinghaus他、(1996) Gene Ther 3(4)、287〜297は、アデノウイルス-ポリリシン複合体を使用して、TFOを細胞に送達することができる方法を記載している。Pichon他、(2000) Nucl Acids Res 28(2)は、ヒスチジル化オリゴリシンを使用して、オリゴヌクレオチドの取り込み、サイトゾル送達および核内蓄積を改善することができる方法を記載している。
【0120】
「Chariot」系(Active Motif、Morris他、(2001) Nature Biotech 19、1173〜1176)を使用することができる。非共有結合複合体が送達させる分子を用いて形成され、この複合体は細胞中に効率良く内在化される。
【0121】
リポソーム/DNA複合体は、直接的なin vivoの遺伝子の移送を仲介することができることが示されてきている。標準的なリポソーム調製では、遺伝子の移送プロセスは非特異的であり、例えば直接的なin situ投与の後の腫瘍堆積において、局所的なin vivoの取り込みおよび発現が報告されてきている (Nabel (1992) Hum.Gene Ther.3、399〜410)。
【0122】
分子を直接標的細胞または組織に誘導する、遺伝子の移送技法が好ましい。受容体仲介の遺伝子移送は、例えば、ポリリシンを介したDNAとタンパク質リガンドの結合によって行われる。標的細胞/組織型の細胞表面上の、対応するリガンド受容体の存在に基づいて、リガンドが選択される。これらのリガンド-DNA結合体は、望むならば血液中に直接注射することができ、受容体結合およびDNA-タンパク質複合体の内在化が起こる標的組織に向けられる。DNAの細胞内破壊の問題を克服するために、アデノウイルスによる混合感染を、エンドソーム機能を害するために含めることができる。
【0123】
選択した遺伝子の発現を抑制または調節する方法を使用して、ヒト細胞中の遺伝子の発現を抑制する(または調節する)ことが好ましく、1つの特に好ましい実施形態では、ヒト細胞はヒトの身体中に存在する。
【0124】
しかしながら、本発明の方法は、動物の身体外の(ヒト細胞を含めた)動物細胞の改変(すなわち細胞のex vivo処理)を含むことができ、そのように改変した細胞を、動物の身体中に再導入することができる。
【0125】
本発明の方法は、例えば潜在的な薬剤標的を同定する際、あるいはおそらく薬剤として有用な活性または性質に関して候補化合物をスクリーニングする際の、改変された細胞のin vitroの調査または特徴付けも含むことができる。
【0126】
本発明の分子、本発明の方法を使用して、特に化学療法剤耐性、または腫瘍発達の他の態様、または炎症の消散、または(他の形の制御された細胞死を含めた)アポトーシスが関与すると考えられる、他のプロセスにおける、遺伝子の役割を分析することができる。
【0127】
本発明の他の態様は、(好ましくは真核生物)細胞中における、選択したアポトーシス関連遺伝子の発現を抑制(または調節)するための物質の製造における、本発明の分子の使用を提供する。選択する遺伝子は、内因性遺伝子であることが好ましい。本発明の前の態様に関して前に示した、他の好ましい事項も当てはまる。
【0128】
動物中における選択したアポトーシス関連遺伝子の発現を抑制(または調節)するための、薬剤の調製において分子を使用する場合、それが非常に好ましいことは理解されよう。誤解を避けるために、「動物」によって本発明者らは、ヒトおよび非ヒト動物を含むものとする。
【0129】
本発明の他の態様は、選択したアポトーシス関連遺伝子の発現の抑制(または調節)を必要とする患者を治療する方法を提供し、この方法は、本発明の分子を有効量、患者に投与することを含む。
【0130】
本発明の他の態様は、アポトーシスの促進を必要とする患者を治療するための方法を提供し、この患者は組合せ法で治療し、この方法は、細胞死誘導物質および本発明の分子を有効量、患者に投与することを含む。腫瘍学の分野の当業者によく知られているように、細胞死誘導物質は、化学療法剤、または治療剤、例えば放射線治療剤であってよい。
【0131】
選択した遺伝子の発現または過剰発現が望ましくなく、患者の病状に貢献する場合、その選択した遺伝子の発現を抑制することが有用であることは理解されよう。遺伝子の望ましくない発現の例には、癌中の幾つかの活性化した腫瘍遺伝子の発現、およびアポトーシス阻害と関係がある遺伝子の発現がある。選択した遺伝子の不充分な発現の欠如が望ましくなく、患者の病状に貢献する場合、選択した遺伝子の発現の増大が有用である可能性がある。例えば、腫瘍抑制遺伝子またはアポトーシス促進遺伝子の不充分な発現は、癌に貢献する可能性があり;したがって、腫瘍抑制遺伝子またはアポトーシス促進遺伝子の、発現を増大させることが有用である可能性がある。
【0132】
Bcl-2ファミリーの遺伝子の発現の抑制は、さまざまな癌を治療する際に望ましい。同様に、プログラムされた細胞死を調節するタンパク質をコードする他の遺伝子、例えばBcl-XIまたはAktの発現の抑制は、癌を治療する際に望ましい。
【0133】
本発明の他の態様は、抑制(または調節)を必要とする患者の中での、選択した遺伝子を抑制(または調節)するための薬剤の製造における、本発明の分子の使用を提供する。
【0134】
本発明のさらに他の態様は、医学において使用するための本発明の分子を提供する。したがって、本発明の分子をパッケージ化して、医薬における使用を示す。
【0135】
本発明のさらに他の態様は、本発明の分子、および薬学的に許容される担体を含む薬剤組成物を提供する。
【0136】
「薬学的に許容される」とは、配合物が滅菌状態で発熱物質を含まないことが含まれる。適切な薬剤担体は、製薬の分野においてよく知られている。
【0137】
ここで本発明を、以下の図面および実施例を参照しながら、より詳細に説明する。
【実施例】
【0138】
(実施例1)
オリゴ-調節因子ペプチド融合分子の構築および使用
遺伝子調節分子として有用な、一連のオリゴヌクレオチド結合体を作製した。これらは、少なくとも2つの特異的な部分または単位、すなわち、選択した二本鎖標的配列とDNA三重らせんを形成することができるオリゴヌクレオチド(三重鎖形成オリゴヌクレオチド、すなわちTFO;単位1);および遺伝子抑制因子または活性化因子由来の別のペプチド配列(単位2)からなる。TFOは、抑制因子または活性化因子ペプチドと融合させる。
【0139】
一例として、TFOを設計して、Bcl-2プロモーター領域との三重鎖を形成する(受託番号:NM_000657およびNM_0006333)。選択する領域は1つのDNA鎖上に非常にプリンが豊富であり、したがって、フーグスティーン塩基対によってDNA三重鎖を形成するための候補配列である。可能なTFOを設計するためのルールは、Vasquez KMおよびWilson JH、Trends Biochem Sci、1:4〜9、1998中に要約されている。配列(5'GGGTGTGGGGTUTGTGTGTGGT3'(Bc1P)または5'TUGTGTGGGTGTGGTGUGGG3'または5'GGTGTUTTGGTTGGGTGT3'(BcIU))を、単位2ペプチドとの化学結合用の活性化5'端を有する、オリゴヌクレオチド(単位1)として生成させた。この試験で調べた単位2ペプチドは、ヒトMAD1転写抑制ドメイン(例えばアミノ酸XXXMNIQMLLEAADYLERREREAEHGYASMLP(前式でXXXは、例えばAAAまたはDDD連結基である))を含む。後者は、ヒストン脱アセチル化酵素複合体タンパク質Sin3aと、相互作用することが知られている領域である。さらに本発明者らは、Sin3aタンパク質と結合することも知られているヒトSap18タンパク質の、アミノ酸XXXMAVESRVTQEEIKKEPEKPIDREKTCPLLLRVF(前式でXXXは、例えばAAAまたはDDD連結基である)の使用を調べている。この領域は充分に進化保存された配列に対応し、遺伝子抑制を仲介することができる領域と重複する。単位2ペプチドは活性形で合成して、N末端チオエステル官能化ペプチドを5'-システイニルオリゴヌクレオチドと結合させる、「本来の結合」化学(WO01/15737およびStetsenko & Gait (2000) Organic Chem 65(16)、4900〜4908を参照のこと)による活性単位1オリゴヌクレオチドとのその後の結合を可能にする。
【0140】
トランスフェクション作業用の細胞系には、内因性Bcl-2遺伝子を発現する、前立腺癌LnCap細胞および膵臓癌PT45細胞があった。さまざまな量のオリゴヌクレオチド結合体を用いる、標準的なリポソーム系トランスフェクション法(あるいは代替として他の送達法、例えばエレクトロポレーションまたはマイクロインジェクション)を使用して、細胞を一時的にトランスフェクトした。特異性の対照として、細胞はオリゴヌクレオチド、または非特異的オリゴペプチド結合体でも処理した。適切な時間、例えば0.5、1、2、4、8、12、24および/または36時間後、細胞計数または以下に記載する他の方法を使用して、細胞の生存能力を測定した。
【0141】
細胞の生存能力を低下させる、さまざまなオリゴペプチド結合体の能力を調べた。高濃度の融合分子オリゴ-MAD1の送達は、濃度依存式に細胞死を増大させる。TFO(すなわちペプチドドメイン含まず)およびオリゴ-ペプチド融合分子を細胞に送達した後では、細胞死は、融合分子のみを用いて見られた細胞死より少なくなった、すなわち低濃度の融合分子を用いて見られる細胞死と同等であった。この結果は、抑制因子ペプチドを含む分子は、ペプチドドメインを含まないTFOより有効な遺伝子活性の調節因子であり、TFOは標的配列との結合に関して、オリゴ-ペプチド融合分子と競合する可能性があることを示す。オリゴおよび非特異的結合体分子には細胞殺傷効作用がなく、Bcl-2制御型細胞の生存能力におけるオリゴペプチド結合体の特異性が実証された。
【0142】
この実施例は、DNA結合オリゴヌクレオチドおよび機能性ペプチドからなる融合分子の設計および構築、ならびに細胞中へのそれらの送達を実証する。オリゴペプチド結合体は、標的方式で特異的な生物学的応答を誘導することができる。したがって、生物活性の強力な調節因子である、オリゴペプチド結合体を設計することができる。
【0143】
(実施例2)
オリゴ-調節因子ペプチド融合分子による染色体遺伝子の抑制
このような遺伝子をゲノム中に組み込むと、融合分子は遺伝子活性を調節することができる。MAD1と融合したDNA結合オリゴペプチド(TFO)を含む融合分子を、実施例1に記載したのと同様に設計し構築した。
【0144】
融合分子を細胞に送達し、Bcl-2遺伝子活性を測定するために実験を行った。
【0145】
Bcl-2遺伝子活性を抑制する、さまざまなオリゴペプチド結合体の能力を調べた。高濃度の融合分子オリゴ-MAD1の送達は、mRNAにより測定して、濃度依存式に、Bcl-2遺伝子活性を抑制した。TFO(すなわちペプチドドメイン含まず)、およびオリゴ-ペプチド融合分子を細胞に送達した後、遺伝子の抑制は、融合分子のみを用いて見られた抑制よりも低くなった、すなわち低濃度の融合分子を用いて見られる抑制と同等であった。この結果から、抑制因子ペプチドを含む分子が、ペプチドドメインを含まないTFOより有効な遺伝子活性の調節因子であることが示される。オリゴおよび非特異的結合体分子には細胞殺傷効果がなく、遺伝子調節におけるオリゴペプチド結合体の特異性が実証された。
【0146】
さらに、抑制を異なる時間で測定して、抑制因子効果の時間行程を確定した。融合分子は、TFO単独より有効な抑制因子であった。この効果も特異的であった(例えば、非融合オリゴヌクレオチドおよび非特異的分子には、オリゴ-ペプチド融合分子と同じ効果はなかった)。さらに、融合分子による抑制は、TFO単独によって見られた任意の抑制より後の時間地点で見られ、より持続的な効果が示唆された。
【0147】
したがって、実施例1に記載した融合分子は、染色体遺伝子の活性を調節することができる。DNA結合オリゴヌクレオチド標的部分を有する融合分子は、特定の染色体遺伝子を標的化することができる。DNA結合オリゴヌクレオチドと融合したGeneICE抑制因子ペプチドは、所定の染色体遺伝子の活性を抑制することができる。したがって、記載した融合分子は、染色体遺伝子の活性の強力な調節因子である。
【0148】
例えば遺伝子の転写を評価することによって(例えばPCRを使用して)、あるいはコードされているポリペプチドの量または活性を評価することによって、内因性遺伝子の調節を測定する。一例では、オリゴヌクレオチドは、Bcl-2遺伝子調節部位を対象とする。適切なRT-PCRプライマーの例は、以下のものを含む。
5'TCCGGTATTCGCAGAAGTCC3'
5'ATCAGAAGAGGATTCCTGCC3'
(Bc1Pを評価するために使用)および
5'TGATGGAGCTCAGAATTCC3'
5'TGCCTCTCCTCACGTTCC3'
(Bc1Uを評価するために使用)。
【0149】
一例では、前立腺癌細胞系LnCapおよび膵臓癌細胞系PT45を、実施例1に記載したMAD1を含むオリゴヌクレオチド-ペプチド融合体で処理する。トランスフェクトした細胞を、場合によっては同定および/または単離し(例えばGFPマーカーおよびFACS技法を使用する)、Bcl-2遺伝子の発現に関してアッセイする。
【0150】
例えば、GeneICE Bcl-2を用いて細胞をトランスフェクトした。12、24、48および72時間後、逆転写酵素PCRによってmRNAレベルを測定した。
【0151】
(実施例3)
標的配列およびヒストン脱アセチル化酵素複合体と結合する融合分子
この実施例は、融合分子が特定の標的配列、ヒストン脱アセチル化酵素複合体の成分と結合することを実証する。オリゴヌクレオチド(TFO)および抑制因子ペプチドを含む融合分子を、実施例1に記載したのと同様に作製した。
【0152】
異なる融合分子を、融合体のオリゴ部分と相補的にした、標識したオリゴヌクレオチドと共にインキュベートした。同じ融合分子も、ヒストン脱アセチル化酵素複合体の成分であるSin3と共にインキュベートした。さらに、これらの融合体を、相補的オリゴヌクレオチドおよびSin3タンパク質ともインキュベートした。
【0153】
次いでこれらの複合体を、標準的なバンド移送分析法によって分析した。融合分子は、標識した相補的オリゴヌクレオチドおよびSin3タンパク質と、別々かつ同時に結合することができた。非特異的な融合体は、標識したオリゴまたはSin3と結合することができず、したがって抑制因子融合体を用いた影響の特異性が実証された。
【0154】
抑制因子融合体は、それらの標的配列と特異的に結合することができると結論付けすることができる。抑制因子融合体は、この複合体の一部分であるタンパク質と結合し、それらの標的配列と結合し、ヒストン脱アセチル化酵素複合体を同時に補うことによって、ヒストン脱アセチル化酵素複合体を補うことができ、記載する融合分子は、遺伝子活性の非常に強力で特異的な抑制因子である。
【0155】
(実施例4)
標的確認プロトコル
当該の遺伝子の利用可能なDNA配列(側面配列含む)を分析して、オリゴ/ペプチドを向けるのに適切な部位を選択する。オリゴ/ペプチドを合成し、例えばレポーター遺伝子系を使用して、目的とする細胞または動物またはヒトにおいて使用する前に、オリゴ/ペプチドを試験することができる。オリゴ/ペプチドを細胞中、in vitroまたは動物またはヒト中において使用し、さらに試験することができる。
【0156】
ひとたび遺伝子配列を提供した後、プロセスは以下のことを含む。
当該の遺伝子(必要な場合、側面配列含む)を、生物情報科学データマイニングツール(例えば、Perkins他、(1998) Biochemistry 37、11315〜11322中で使用されたthe Genetics Computer Group(GCG)プログラム)を使用して、ヒトゲノム中の他の場所では見られないユニークな配列エレメントに関して調べる。(例えばTFOとして)同定した独特の配列と結合すると予想される、核酸系DNA結合分子を、設計し合成する。
【0157】
DNA結合分子はオリゴヌクレオチドである可能性があり、以下の特徴を有することは好ましい:
(a)少なくとも16ヌクレオチド長
(b)遺伝子のプロモーター、あるいは遺伝子の転写開始部位またはその近傍を標的化する。
【0158】
TFOと結合するための標的部位は、1本の鎖中にプリンが多量であることが好ましい。
【0159】
TFOはピリミジン多量(主にCまたはT);プリン多量(主にGまたはA)または混合型(主にGまたはT、またはG、AまたはT)であってよい。CTTFOは平行モチーフ中で結合すると考えられ、その中では第三の鎖(TFO)が、二本鎖のプリン鎖と同じ5'〜3'方向を有する。GATFOは逆平行モチーフ中で結合すると考えられ、その中ではTFOはプリン鎖に対して反対を向いている。混合型TFOは、標的配列に応じて、平行モチーフまたは逆平行モチーフ中で結合することができる。塩基対は、平行三重鎖(T:AT、C+:GCおよびG:GC)中のフーグスティン水素結合、および逆平行三重鎖(G:GC、A:ATおよびT:AT)中の逆フーグスティン水素結合の形成から生じる。
【0160】
オリゴヌクレオチドは、安定した化学修飾基をおそらく有するDNAオリゴヌクレオチドであると考えられる。他の塩基、例えばN6-メチル-8-オキソ-2-デオキシアデニンをシトシンの代わりに、2-デオキシ-6-チオグアニンをグアニンの代わりに、あるいは7-デアザ-2-デオキシキサンチンをチミンの代わりに使用することができる。
【0161】
抑制因子ペプチドまたはペプチドは、ペプチド合成装置を使用して大量に生成することができ、使用するまで凍結保存することができる。
【0162】
抑制因子ペプチド-DNA結合分子構築体を、調製し精製する。使用する化学物質は、WO01/15737中に記載されたものであってよい。キットはLink Technologiesから入手可能である。
【0163】
構築体は、質量分光法によって、かつ/あるいは(例えば蛍光、化学発光または酵素標識を使用して)標識した相補的オリゴヌクレオチド、または標識した抗体成分を使用することによって、品質管理することができる。典型的には、このような方法では、構築体を固形担体に加え、その上で、構築体のペプチド部分と結合する抗体を固定し、構築体のオリゴヌクレオチド部分と結合する標識したオリゴヌクレオチドを加える。この方法では、固形担体と結合した標識を検出することによって、構築体が完全であることが実証される。他の典型的な形式では、オリゴヌクレオチドは固形担体と結合することができ、抗体を標識することができる。
【0164】
レポーター遺伝子構築体を、当該の遺伝子用に作製することができる(しかしながら、これは一般に必要ではない)。
・候補DNA結合オリゴヌクレオチドまたはオリゴ/ペプチドは、以下の事項に関して試験することができる:
・標的配列に対する結合の親和性;
・全ゲノムDNAチップに陳列させることによる結合の特異性。
・オリゴ/ペプチドは、レポーター遺伝子系を使用して、有効性に関して試験することができる。
【0165】
したがってオリゴ/ペプチドは、当該の細胞または動物中の当該の遺伝子を、調節または抑制するために使用することができる。
【0166】
(実施例5)
標的確認
オリゴ/ペプチド融合分子を使用して、アポトーシス関連薬剤標的を確認することができる。これは以下のことを含むことができる:
・実施例1で述べたプロトコルを実施する。
・構築体の細胞または組織への送達。これらは、当該の分子の研究に適切な、正常または疾患組織、細胞系または初代細胞であってよい。
・任意の発現分析法による表現型の分析;または細胞の運動性の評価などの任意の機能分析、増殖またはアポトーシス分析。
・個々の疾患の任意の入手可能なデータとの比較、および細胞死または運動性などの望ましい効果の分析。
【0167】
得られたデータを使用して、薬剤開発プログラムの所定の薬剤標的を確認する。
【0168】
(実施例6)
患者治療の実施例
アポトーシス関連遺伝子、例えばBcl-2を標的とする、オリゴ/ペプチド融合体を、記載するように作製する。融合分子を滅菌環境で調製し、リポソームに配合する。融合体含有リポソームを、腫瘍、例えば乳または前立腺腫瘍の近くに向ける。リポソームは癌細胞によって採取し、アポトーシスをそれぞれの細胞において選択的に促進させる。
【0169】
(実施例7)
標的同定スクリーニング
オリゴ/ペプチド融合分子を使用して、アポトーシス関連薬剤標的を同定することができる。これは以下のことを含むであろう:
・実施例1で述べたのと同様の、融合分子の調製。
・融合分子の細胞または組織への送達。これらは、正常または疾患組織、細胞系または初代細胞であってよい。
・DNAアレイを使用する、遺伝子抑制から生じる遺伝子発現概略の分析。これは、モデル中全体の遺伝子発現に対する構築態の影響を示す。
・任意の発現分析法による表現型の分析、または細胞の運動性などの任意の機能分析、増殖またはアポトーシス分析。
【0170】
得られたデータを使用して、乳または前立腺癌などの疾患の、可能性のある薬剤標的を見出す。任意の他の同様のスクリーニング、in vitro法および細胞および動物モデルを含めた適切な方法によって、これらの標的をさらに確認することができる。
【0171】
(実施例8)
オリゴ/ペプチド融合分子による遺伝子発現の下方制御は、クロマチンヒストン脱アセチル化と関係がある
前に示したように、オリゴ/ペプチド融合分子は、遺伝子発現を抑制する際に有効である。これは、オリゴが結合するかあるいはその近くのDNAの、ヒストンアセチル化状態のMAD1仲介の変化によるものであると、本発明者らは考える。
【0172】
低下した遺伝子発現が、クロマチンヒストン脱アセチル化と関係があることを示すために、クロマチン免疫沈降(ChIP)法を使用することができる。使用製造元の指示書(Upstate Biotechnology、Bucks、UK)に従いChIPアッセイキットを使用して、クロマチン免疫沈降を行う。
【0173】
PT45またはBxPC-3膵臓癌細胞を増殖および繁殖させ、アポトーシス関連遺伝子の発現を抑制する際に有用な、オリゴ/ペプチド融合分子と共にインキュベートした。未処理細胞は対照として使用する。
【0174】
35cm組織培養プレート上、フェノールレッドを含まない、5%のDSS、P/S/GおよびG418(100μg/ml)を補ったDMEM中で、細胞を95%コンフルエントになるまで増殖させる。ヒグロマイシンB(80μg/ml)およびドキシサイクリン(1μg/mnl)を適切に加える。固定の30分前に、E2(10〜8M)またはエタノールを(対照として)、細胞に加える。37%ホルムアルデヒドを、1%の最終濃度まで培地に直接一滴ずつ加える。細胞は37℃で10分間インキュベートする。
【0175】
氷上で、培地をプレートから吸引し、1×プロテアーゼ阻害剤(PI)(Sigma、Dorset、UK)を含む氷冷PBSで二回、細胞を洗浄する。採取用に、1×PIを含む1mlの氷冷PBSをプレートに加え、ラバーポリスマンを使用して、予め冷却したミクロ遠心分離チューブに細胞をそぎ落とす。4℃における4分間2000rpmでの遠心分離によって、細胞をペレット状にする。PIを含む400μlの暖めたChIPSDS-溶解バッファー(1%のSDS;10mMのNaEDTA、pH 8.0;50mMのTris-HCl、pH8.1)中に、ペレットを再懸濁させ、氷上で10分間インキュベートする。
【0176】
溶解物を超音波処理して、DNAを200〜1000bpの長さに切る。超音波処理中、サンプルは氷-水ビーカー中に置いて、サンプルを冷たい状態に保ち、それらが分解するのを防ぐ。30秒間隔離して10秒の衝撃4回で、付属のSoniprep150エクスポネンシャルチタン製プローブ(Sanyo-Gallenkamp、Leics、UK)を有するSoniprep150超音波装置を使用して、超音波処理を行う。
【0177】
サンプルを、4℃において10分間13,000rpmの遠心分離にかける。上清は15mlの滅菌ファルコンチューブ中に回収し、3600μlのChIP希釈バッファー(0.01%のSDS;1.1%のTriton-X-100;1.2mMのNaEDTA、pH 8.0;16.7mMのTris-HCI、pH 8.1;167mMのNaCI)を用いて、10倍に希釈する。次いでサンプルを、2.5mlチューブ中の2つの2ml等分試料、1つは抗アセチル化ヒストンH4抗体、ChIPグレード(Upstate Biotechnology、Bucks、UK)とのインキュベーション用、もう1つは抗体を含まない対照としての使用するために分ける。
【0178】
非特異的な基底状態を低下させるために、それぞれの2ml等分試料を、80μlのサケ精子DNA/タンパク質Aアガロース50%スラリー(10mMのTris-HCl、pH 8.0;1mMのNaEDTA、pH 8.0に懸濁させたもの)を、垂直回転プラットホーム(Stuart Scientific、Staffs、UK)上において30分間4℃で加えることによって予め調製する。次いでアガロースビーズを、1000rpmで30秒間の遠心分離によってペレット状にし、上清分画を新たな2.5mlチューブに回収する。免疫沈降抗体は1:500の希釈で最初のサンプルに加えるが、抗体を含まない対照サンプルには加えない。両方のチューブを、垂直回転プラットホーム(Stuart Scientific、Staffs、UK)上において4℃で一晩インキュベートする。
【0179】
60μlのサケ精子DNA/タンパク質Aアガロース-50%スラリーを、回転させながら4℃で1時間、それぞれのチューブと共にインキュベートして、抗体/ヒストン複合体、または抗体を含まない対照の場合は非特異的に結合したタンパク質を回収する。アガロースビーズは、1分間800rpmでの簡単な遠心分離によって、ペレット状にする。上清は新たな2.5mlチューブに慎重に移し、-20℃で保存する。
【0180】
タンパク質Aアガロースビーズ/抗体/ヒストン複合体は、以下に列挙する順序で以下のバッファーそれぞれ1mlを用いて、4℃において垂直回転プラットホーム上で5分間洗浄する。
(a)低塩濃度免疫複合体洗浄バッファー、1回洗浄
(0.1%のSDS;1%のTriton-X-100;2mMのNaEDTA、pH 8.0;20mMのTris-HCl、pH 8.1;150mMのNaCl)
(b)高塩濃度免疫複合体洗浄バッファー、1回洗浄
(0.1%のSDS;1%のTriton-X-100;2mMのNaEDTA、pH 8.0;20mMのTris-HCl、pH 8.1;150mMのNaCl)
(c)LiCl免疫複合体洗浄バッファー、1回洗浄
(0.25MのLiCl;1%のNP40(nonidet);1%のデオキシコレート;1mMのNaEDTA、pH 8.0;10mMのTris-HCl、pH8.1)
(d)1×TE、2回洗浄
(10mMのTris-HCl、1mMのNaEDTA、pH 8.0)
【0181】
ヒストン/免疫複合体は、250μlの新たに調製した溶出バッファー(1%のSDS;O.1MのNaHCO3)を加えることによって、アガロースビーズから溶出させる。サンプルを軽く攪拌して混合させ、垂直回転プラットホーム上において15分間室温でインキュベートする。ビーズは室温において2分間1000rpmの遠心分離にかけ、溶出物は新たなミクロ遠心分離チューブに移す。他の250μlの溶出バッファーを用いて溶出ステップを繰り返し、溶出物を合わせる。
【0182】
5MのNaCl20μlを溶出物に加え、ヒストン-DNA架橋は、少なくとも4時間65℃に加熱することによって元の状態にする。10μlの0.5MNaEDTA、pH 8.0、20μlの1M Tris-HC1、pH 6.5および2μlの10mg/mlプロテイナーゼKを溶出サンプルに加える。IP由来の上清分画において、架橋を元の状態にする。4Oμlの0.5MNaEDTA、pH 8.0、80μlの1M Tris-HC1、pH 6.5および8μlの10mg/mlプロテイナーゼKを上清サンプルに加える。サンプルは45℃で1時間インキュベートして、サンプル中のタンパク質を分解する。
【0183】
20μgのグリコーゲンを不活性担体としてサンプルに加え、次いでサンプルDNAは、フェノール/クロロホルム/イソアミルアルコール抽出およびエタノール沈殿によって回収する。DNAペレットは、PCR反応用に50μlの滅菌水に再懸濁させる。1μlのサンプルおよび35〜40サイクルを、それぞれのPCR増幅用に使用する。コンピュータ系画像分析(NIH画像分析プログラム)を使用して、β-アクチンおよび抗体を含まない対照と比較した、Bcl2PCR産物の相対的レベルを評価する。これによって、他のサンプル内に含まれるそれと比較した、サンプル内に含まれる遺伝子転写産物の相対量を計算することができる。
【0184】
得ることができる結果の型の一例を、図6に示す。
【0185】
沈殿したBcl-2DNAの量、したがって、Bcl2と関係があるクロマチンヒストンタンパク質のアセチル化の程度は、未処理細胞と比較して、Bcl2遺伝子発現が記載する方法により阻害される細胞中では、約75%低下させることができる。
【0186】
(実施例9)
ゲルシフトアッセイ
前の実施例によって、オリゴ/ペプチド融合分子は、標的とする遺伝子の活性を調節することができることが実証された。これは、オリゴが結合するかあるいはその近くのDNAの、ヒストンアセチル化状態のMAD1仲介の変化によるものであると考えられる。
【0187】
ゲルシフトアッセイを使用して、オリゴ/ペプチド融合分子が、標的プロモーターを含むDNA断片と結合することができることを実証する。
【0188】
標的遺伝子配列を含むDNA断片を、オリゴヌクレオチドまたはオリゴ/ペプチド融合体と共にインキュベートする。サンプルに非変性ゲル電気泳動を施して、三重鎖仲介型の光付加体を特徴付けする。オリゴヌクレオチドまたはオリゴ/ペプチド融合体をそれぞれ量を増やして含むサンプル中の、付加体を検出する。
【0189】
このようにして、オリゴ/ペプチド融合分子が、標的プロモーターを含むDNA断片と結合することができることを示すことができる。
【0190】
(実施例10)
核局在シグナルを有するオリゴ/ペプチド融合分子
分子のペプチド部分は、分子を核に向けるための核局在シグナル(NLS)を有する可能性がある。この実施例で使用するペプチドは以下のものである。
a)DDD-MAD1-DDD-NLS、
これは以下のアミノ酸配列を有する:
(連結基)DDDMNIQMLLEAADYLERREREAEHGYASMLPDDDPKKKRK
V(カルボキサミド)
および
b)DDD-NLS-DDD-MAD1、
これは以下のアミノ酸配列を有する。
(連結基)DDDPKKKRKVDDDMNMQMLLEAADYLERREREAEHGYASML
P(カルボキサミド)
【0191】
このNLSは、SV40T-抗原由来の、7アミノ酸(配列PKKKRKV)機能的核局在シグナルである。
【0192】
DDD連結配列、および29アミノ酸のMADIアミノ酸配列は、前の実施例で論じた配列と同じである。
【0193】
DDD-MAD1-DDD-NLS、およびDDD-NLS-DDD-MAD1ペプチド配列が、核を標的化することを実証するために、PT45ヒト膵臓癌細胞を、標準リポフェクタミン2000プロトコルによって、Cy3標識オリゴヌクレオチドと連結した、DDD-MAD1-DDD-NLSおよびDDD-NLS-DDD-MAD1ペプチド配列からなる、GeneICEオリゴペプチドを使用してトランスフェクトした。次いで、製造元の推奨する手順を使用して、細胞をホルムアルデヒド中に固定し、DAPI核染色物質を用いて染色した。蛍光顕微鏡により細胞を調べることによって、Cy3標識GeneICEオリゴペプチド分子が、DAPI核染色物質と共に局在したことが示された。この生じた実験データから、ペプチドを核に向ける際に、NLSは非常に有効であると結論付けた。
【0194】
DDD-MAD1-DDD-NLS、およびDDD-NLS-DDD-MAD1ペプチド配列は、本発明のオリゴ/ペプチド分子中に取り込ませると、標的遺伝子の発現を仲介することができる。実施例8および9に概略した実験手法を使用して、これを実証することができる。
【0195】
例えば、DDD-MAD1-DDD-NLS、およびDDD-NLS-DDD-MAD1ペプチド配列は、以下のオリゴ/ペプチド分子中に取り込ませることができる:
Bcl2:-DDD-MAD1-DDD-NLS、および
Bcl2:-DDD-NLS-DDD-MAD1
【0196】
Bcl2は実施例8に示すTFOオリゴ配列である(5'GGGTGTGGGGTUTGTGTGTGGT3'または5'TUGTGTGGGTGTGGTGUGGG3'または5'GGTGTUTTGGTTGGGTGT3')。
【0197】
Bcl2:-DDD-MAD1-DDD-NLS、およびBcl2:-DDD-NLS-DDD-MAD1分子を、次いで試験細胞、例えばLNCap細胞にトランスフェクトする。前に概略した実験手順を使用して、Bcl2:-DDD-MAD1-DDD-NLS、およびBcl2:-DDD-NLS-DDD-MAD1分子の、標的遺伝子の発現に対する影響を示すことができる。
【0198】
(実施例11)
アポトーシス関連遺伝子の発現の調節に関する構築体の評価
結合:
TFOおよびGeneICE(商標)分子のそれらの標的DNA配列への結合を、ゲルシフトアッセイを使用することによって評価する。これを行うために、知られている量のTFOを、TFO結合バッファー(20mMのTRIS(pH 7.6)、10%v/vのグリセロール、10mMのMgCl2)中で、1時間37℃において、PCRおよびT4キナーゼを使用する末端標識を使用してゲノムDNAから前に増幅させた、微量の32P標識した標的DNAと共にインキュベートする。反応混合物は、非変性ローディングバッファー(10×ストック濃度:250mMのTRIS(pH 7.5)、40%v/vのグリセロール、10mMのMgCl2、0.2%w/vのブロモフェノールブルー)を使用して、12%非変性ポリアクリルアミドゲルに載せ、TRIS-ホウ酸マグネシウムランニングバッファー(0.5×TBE、10mMのMgCl2)中で3〜4時間、電気泳動に施す。バンドを目に見える状態にするために、オートラジオグラフィーフィルムに露光する前に、ゲルを乾燥させる。
【0199】
3つの異なる大きさのバンドが、ゲル上に予想される。TFOまたは構築体の濃度が低い場合、小さいDNAバンドのみが見られる。TFOの濃度が上昇して三重鎖DNAの形成が可能になるとき、中程度の大きさの三重鎖のバンドが現れ、GeneICE(商標)分子の濃度が上昇して三重鎖DNAの形成が可能になるとき、大きな三重鎖のバンドが見られる。TFOまたはGeneICE(商標)分子のKdは、等量の二量体DNAバンドと三重鎖DNAバンドが存在する濃度によって測定する。10-6〜10-9Mの範囲のKd値が予想され、GeneICE(商標)分子のKdは、TFOより1logまで高い。
【0200】
mRNA発現:
mRNA発現は、2つの方法によって調べることができる。最初に、全RNAを処理細胞から抽出し、RT-PCRまたはノーザンブロット分析用に使用する。RT-PCR分析では、逆転写酵素およびオリゴdTプライマーを使用することによって、mRNAがcDNAに逆転写される。このcDNAを、Bcl-2コード配列を増幅させるように設計したプライマーを使用する半定量PCR用に使用する。β-アクチンPCRも行い、相対量を使用して、GeneICE(商標)処理で見られた下方制御を定量化する。RT-PCR分析から得られる結果は、ノーザンブロット分析によって確認する。処理細胞から抽出した全RNAをTBEゲル上で電気泳動にかけ、ナイロン膜に移す。PCRにより32Pで前に標識した、一本鎖DNAプローブを使用して、Bcl-2mRNA配列にハイブリダイズさせる。非特異的結合は膜の厳密な洗浄によって除去し、オートラジオグラフィーを使用して、存在するmRNAの量を目に見える状態にする。β-アクチンmRNA用のプローブを使用することによって、RNA充填を調節する。
【0201】
mRNA発現の実験によって、Bcl-2標的GeneICE(商標)分子を用いた処理後の、Bcl-2遺伝子の特異的な下方制御が示される。下方制御の中間レベルは、TFOのみまたは送達ペプチドと結合したTFOを用いた処理によって上昇する。TSAなどの脱アセチル化酵素阻害剤を使用することによって、遺伝子発現のTFO仲介の下方制御ではなく、GeneICE(商標)分子による下方制御が元の状態になる。これらの実験は、GeneICE(商標)構築体の用量依存効果および時間依存効果も示し、用量の増大と共に、下方制御が非特異的になるまで、さらなる下方制御が示される。用量依存効果は1回量から長期の下方制御を示し、これは数時間後にピークに達し、数日間続く。
【0202】
タンパク質発現:
タンパク質発現は、イムノブロット分析によって評価する。細胞は氷上、プロテアーゼ阻害剤の存在下において溶解バッファーを含むSDS中で、溶解させる。細胞溶解物は遠心分離によって取り除き、タンパク質の量は、タンパク質アッセイを使用することによって測定する。それぞれのサンプルからの等量のタンパク質を、15%SDSポリアクリルアミドゲルのウエルに充填し、25mMのTRIS、192mMのグリシン、0.1%w/vのSDSランニングバッファーを使用して、150Vで2時間電気泳動にかける。分離後、48mMのTRIS、39mMのグリシン、0.0375%w/vのSDS、20%v/vのメタノール移送バッファーを使用して、5.5mA/cm2での半乾燥状態での電気泳動による移送によって、タンパク質をPVDF膜上に移す。非特異的な抗体結合は、5%w/vの無脂肪乳、PBS中/0.1%w/vのTween20(PBST)中でのインキュベーションによって阻害する。室温で1時間、2%乳中に1:2000に希釈した抗Bcl-2一次抗体と共に、膜をインキュベートする。過剰な抗体は2%乳を使用して洗浄除去し、西洋ワサビペルオキシダーゼ結合二次抗体を、2%のミルク中に1:10000で使用する。PBST中での2回の洗浄、次にPBS中での1回の洗浄を使用して、過剰な抗体を除去する。充分な化学発光およびオートラジオグラフィーを使用することによって、タンパク質を検出する。同じ手順を使用して、ただし1:500の希釈で抗アクチン一次抗体を抗Bcl-2抗体の代わりに使用して、アクチンを調べることによって、充填量を補正する。
【0203】
タンパク質発現は、製造元によって与えられる指示書に従って、Bcl-2ELISAキットを使用することによって確認する。
【0204】
mRNA発現と同様の実験を、タンパク質を分析するために行う。GeneICE(商標)構築体で処理した後の、タンパク質発現の時間および用量依存性の下方制御が予想され、これはmRNA発現の概略で見られるものに従う。タンパク質レベルは、mRNAレベルの約15〜20時間後に低下し、数日間は低い状態のままである。脱アセチル化酵素阻害剤は、この特異的なBcl-2タンパク質下方制御を妨げ、一方TFOによる処理によって、中間の表現型が表される。
【0205】
細胞死:
タンパク質発現の低下は、幾つかの細胞系では、アポトーシスによる細胞死を引き起こすのに充分である可能性がある。このことが当てはまる場合、非内因性プロモーター(SV40プロモーターなど)の制御下の、Bcl-2を有する発現プラスミドを、細胞系にトランスフェクトし、内因性mRNAのレベルは、内因性遺伝子に特異的なプライマーを使用して、記載するように評価する。
【0206】
細胞のトリパンブルー染色、および計測チャンバーを使用する計数によって、細胞死を評価する。細胞はGeneICE(商標)構築体で適切な長さの時間処理し、知られている体積のトリプシンを使用して、ウエルからトリプシン処理した。20μlの細胞懸濁液を20μlのトリパンブルーと混合させ、計測チャンバーに充填する。30個と300個の間の細胞を、計測チャンバーのグリッドを使用して、体積と所与数/mlの指標として計数し、細胞の生存能力は、非生存細胞を示す、トリパンブルー染色によって評価する。
【0207】
MTTアッセイを使用して、細胞計数によって与えられたデータを確認する。96ウエルプレート中の実験値を確定し、処理後、MTTを1時間までウエルに加える。培地を吸引し、細胞を洗浄する。残っている沈殿をDMSOに溶かし、吸光度を520nmで読み取る。
【0208】
GeneICE(商標)処理によって、培養物中の細胞数の減少、および培養物の生存能力の低下がもたらされる。脱アセチル化酵素阻害剤を使用することによって、アポトーシスの阻害剤と同様に、これらの影響が元の状態になる。TFO対照は、中間の表現型を示す。
【0209】
アポトーシス:
観察された細胞死の方法は、製造元により与えられる指示書に従い、末端デオキシトランスフェラーゼ仲介dUTPニック末端標識(TUNEL)キットを使用することによって調べる。実験は異なる時間地点の範囲で行って、最も多くの死細胞が見られる地点を決定する。GeneICE(商標)を用いた処理後に見られる細胞死は、アポトーシス機構を介したものであると予想される。アポトーシスの誘導物質を、陽性対照として使用して細胞を比較し、一方アポトーシス阻害剤は、TUNELアッセイにおいて細胞死、したがってアポトーシス細胞の出現を防ぐことができる。脱アセチル化酵素阻害剤は、GeneICE(商標)処理培養物において、アポトーシスが起こることも防ぐ。
【図面の簡単な説明】
【0210】
【図1】融合分子および標的部位との結合の概略図である。 Aでは、「1」は実施例1のオリゴヌクレオチド単位1を表し;「2」は、ポリペプチド単位2を表す。「L」は連結基領域を示す。Bでは、Dは、他の送達ペプチドを表す。ペプチドはオリゴヌクレオチドの一端または両端に連結することができ;例えば、抑制因子/修飾ポリペプチドは一端に連結することができ、送達ペプチドは他端に連結することができる。 Cは、オリゴヌクレオチド部分が二本鎖DNAを有する三重らせんを形成し、抑制因子ペプチドがSin3-HDAC複合体をその部位に補充した状況を表す。
【図2】Bcl-2のmRNAの定量化、およびGeneICEの内因性Bcl-2のmRNAレベルに対する影響を示す図である。 縦列は、対照mRNA値に対する比としての、示したBxPC-3膵臓癌細胞の3日間の処理後の、Bcl-2のmRNAの発現を表す。それぞれの縦列は、4つの独立したポリメラーゼ連鎖反応の、平均ODおよび平均の標準誤差を示すものである。平均光学濃度を測定した。 Bc1PおよびBc1Uに関するTFO配列である(5'から3'): Bc1P GGGTGTGGGGTUTGTGTGTGGT Bc1U GGTGTUTTGGTTGGGTGT L231はTFO-MAD-NLSに対応するペプチドであり、L232はTFO-NLS-MADに対応するペプチドである。
【0211】
【表1】
【図3】GeneICEbcl-2の細胞の生存能力に対する影響を示す図である。 示した3日間の処理後に合計mRNAによって測定した、PT45膵臓癌細胞の生存能力。縦列は合計mRNAのμg/mlを表す。
【0212】
【表2】
【図4】50pmol用量での一晩のインキュベーション後の、非結合TFOおよびBcIPL231結合体の、PT45細胞の生存能力に対する影響の図である。 特に「Chariot」送達系(Active Motif;Morris他、(2001) Nature Biotech 19、1173〜1176)を用いると、非結合TFOによってではなく、結合体によって、対照細胞と比べて、細胞数は減少する(すなわち、細胞死が誘導/促進される)。
【技術分野】
【0001】
本発明は、プログラムされた細胞死、アポトーシス、および関連遺伝子発現の制御に関するものであり、詳細には、本発明は、細胞中でのアポトーシスを制御するため、特定の選択したアポトーシス関連遺伝子の発現を調節、好ましくは抑制するための方法および手段に関する。
【背景技術】
【0002】
遺伝子の発現を選択的に抑制する能力は、生物学の多くの分野、例えば遺伝子の発現が望ましくない可能性がある治療法;特定の遺伝子の発現の欠如がその疾患と関係がある疾患モデルの作製;望ましい特性を生み出すための表現型の変更において有用である。したがって、遺伝子の発現を選択的に抑制する能力は、発生および分化の研究において、(野生型であれ突然変異型であれ)ヒト細胞中のヒト遺伝子を「ノックアウト」し、真核生物の遺伝子をノックアウトすることができる。
【0003】
細胞死を選択的に誘導する能力は、癌中の腫瘍細胞をターゲッティングし破壊する際に、ならびに他の状況、例えば炎症の消散において重要である。この能力は、発癌性状態になっている細胞、または腫瘍中の他の細胞(白血球など、例えばマクロファージ)を直接殺傷する際に、ならびに任意の他の療法を向上させる際に有用である。なぜなら、癌療法に対する腫瘍の耐性は、疾患を治療する際の主要な問題だからである。アポトーシスは、細胞死を誘導するための主要な機構の1つであり、アポトーシスに対する耐性は、腫瘍の進行中に自然に得られる特性である。これは通常、アポトーシスシグナル分子の制御の変更を伴い、腫瘍細胞をアポトーシス刺激に対して鈍感にする。
【0004】
特定の遺伝子の発現を抑制することを試みる方法は、主に3つの主要な範疇、すなわちアンチセンス技術、リボザイム技術、および相同的組換えによってもたらされる標的遺伝子の欠失に分類される。
【0005】
アンチセンス技法は、核酸分子の細胞中への導入に基づくものであり、核酸分子は典型的には、選択した遺伝子によって発現されるmRNAと相補的である。アンチセンス分子は、mRNA分子と結合したままの間は、典型的にはmRNA分子の翻訳を抑制し、遺伝子によってコードされるポリペプチドの発現を妨げる。アンチセンス技法に修正を加えれば、三重らせんを形成するために遺伝子のDNAと結合するアンチセンス分子(三重鎖形成オリゴヌクレオチド;TFO)により、選択した遺伝子の転写を防止できるであろう。この方法では、第三の鎖の存在がDNA転写を妨げ、その間、その鎖は結合したままの状態である。
【0006】
化学修飾基、例えばソラーレン架橋基は、TFO中に含まれているが、これらは不可逆的なDNAの損傷および突然変異をもたらす可能性がある。細胞中でのこのような化学修飾基の調節は困難でもある。これらは、分子の細胞内送達に関する欠点を有する可能性もある。
【0007】
リボザイム技法は、核酸分子の細胞中への導入に基づくものであり、核酸分子はRNA分子を発現し、標的RNA分子と結合し、標的RNA分子の選択的切断を触媒する。標的RNA分子は典型的にはmRNA分子であるが、それは例えば、レトロウイルスRNA分子であってよい。
【0008】
アンチセンスおよびリボザイムに基づく技法は、実施が困難であることが明らかになっており、これらの技法は、標的遺伝子の抑制または不活性化において、成功の程度にバラツキがある。さらに、これら2つの技法は、遺伝子不活性化物質の持続的な発現または投与を必要とする。
【0009】
TFOとVP16ウイルス活性化ドメインの結合(Kusnetsova他、(1999) Nucleic Acids Res20、3995〜4000)を使用して、TFOの適用を広げ、遺伝子活性化を含めている(遺伝子の抑制または不活性化における以前の使用とは対照的)。
【0010】
相同的組換えによる標的遺伝子の欠失は、2つの遺伝子不活性化事象(それぞれの対立遺伝子に関する事象)を必要とし、初代細胞、特に例えば、数回の継代の培養においてのみ維持することができる、初代ヒト乳房細胞には容易に適用できない。標的遺伝子の欠失は、植物において行うのは依然として困難である。cre-lox仲介の部位特異的組み込みが優れた方法となっているが、特異的組み込み事象の効率は低い(Alberts他、(1995) Plant J.7、649〜659; Vergunst & Hooykass (1998) Plant Mol.Biol.38、393〜406; Vergunst他、(1998) Nucl.Acids Res.26、2729〜2734)。
【0011】
WO01/02019は、真核生物ゲノム中に存在する部位に結合する核酸結合部分、およびクロマチン不活性化部分、例えばHDACまたはHDAC補充物質、PLZFなどを含むポリペプチドを使用して、選択した遺伝子を不活性化させるための方法を記載している。
【0012】
細胞死を誘導するための現在の方法は、満足のいくものではない。本発明者らは、これが、これらの方法が、細胞を殺傷するのに充分持続的または完全な、遺伝子発現に対する作用を生み出すことができないためであると考える。細胞のアポトーシス機構が素早く、有効に、充分に長く作用しない場合、通常、細胞は非常に早く耐性を発現する。既存の技術および方法にはこれらの重大な欠点があるので、別の戦略が求められる。
【非特許文献1】Kusnetsova他、(1999) Nucleic Acids Res20、3995〜4000
【非特許文献2】Alberts他、(1995) Plant J.7、649〜659
【非特許文献3】Vergunst & Hooykass (1998) Plant Mol.Biol.38、393〜406
【非特許文献4】Vergunst他、(1998) Nucl.Acids Res.26、2729〜2734
【特許文献1】WO01/02019
【非特許文献5】Sellers & Fisher (1999) The Journal of Clinical investigation、p.1655; Apoptosis and cancer Drug Targeting
【非特許文献6】Cory & Adams (2002) Nature Reviews Cancer、p489; The Bcl-2 family : regulators of the ceullular life-or-death switch
【非特許文献7】Chen & Li (1998) Crit.Rev.Eukaryotic Gene Expression 8、169〜190
【非特許文献8】Workman & Kingston (1998) Ann.Rev.Biochem.67、545〜579
【非特許文献9】Perlmann & Vennstrom (1995) Nature 377、387-
【非特許文献10】Wolfe (1997) Nature 387、16〜17
【非特許文献11】Grunstein (1997) Nature 389、349〜352
【非特許文献12】Pazin & Kadonaga (1997) Cell 89、325〜328
【非特許文献13】DePinho (1998) Nature 391、533〜536
【非特許文献14】Bestor (1998) Nature 393、311〜312
【非特許文献15】Grunstein (1998) Cell 93、325〜328
【非特許文献16】Taunton他、(1996) Nature 272、408〜441
【非特許文献17】Yang他、(1996) Proc.Natl.Acad.Sci.USA 93、12845〜12850
【非特許文献18】Dangond他、(1998) Biochem.Biophys.Res.Comm.242、648〜652
【非特許文献19】Horlein他、(1995) Nature 377、397〜404
【非特許文献20】Chen & Evans (1995) Nature 377、454〜457
【非特許文献21】Zhang他、(1998) Molecular Cell1、1021〜1031
【非特許文献22】Ayer他、(1995) Cell 80、767〜776
【非特許文献23】Schreiber-Agus他、(1995) Cell 80、777〜786
【非特許文献24】Zhang他、(1997) Cell 89、357〜364
【非特許文献25】Qian他、(1993) Nature 364、648〜652
【非特許文献26】Lin他、(1998) Nature 391、811〜814
【非特許文献27】Grignani他、(1998) Nature 391、815〜818
【非特許文献28】David他、(1998) Oncogene 16、2549〜2556
【非特許文献29】Hassig他、(1997) Cell 89、341〜347
【非特許文献30】Laherty他、(1997) Cell 89、349〜356
【非特許文献31】Zhang他、(1997) Cell 89, 357-364
【非特許文献32】Heinzel他、(1997) Nature 387、43〜48
【非特許文献33】Alland他、(1997) Nature 387、49〜55
【非特許文献34】Kadosh & Struhl (1997) Cell 89、365〜371
【非特許文献35】Nagy他、(1997) Cell 89、373〜380
【非特許文献36】Laherty他、(1998) Molecular Cell 2、33〜42
【非特許文献37】Grozinger他、(1999) Proc.Natl.Acad.Sci.USA 96、4868〜4873
【非特許文献38】Lussen他、(1997) Science 277、88〜91
【非特許文献39】Lechner他、(1996) Biochim.Biophys.Acta 1296、181〜188
【非特許文献40】Chen & Pickard (1997) Genes Dev.11、2124〜2136
【非特許文献41】Brosch他、(1995) Plant Cell 7、1941〜1950
【非特許文献42】Okano M、Xie S、LiE.(1998) Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases.Nat Genet19:219〜220
【非特許文献43】Adrian P.Bird and Alan P.Wolffe (1999) Methylation-Induced Repression:Belts、Braces、and Chromatin.Cell 99、451〜454
【非特許文献44】Soukchareun他、(1998)Bioconjugate Chem9、466〜475
【非特許文献45】Soukchareun他、(1995) Bioconjugate Chem6、43〜53
【非特許文献46】Eritja他、(1991) Tetrahedron 47、4113〜4120
【特許文献2】WO99/13719
【非特許文献47】Prochiantz (2000) Curr Opin Cell Biol9、420〜429
【特許文献3】WO01/14737
【非特許文献48】Kusnetsova他、(1999) Nucleic Acids Res 27、3995〜4000
【特許文献4】WO90/06934
【特許文献5】EP0375408
【特許文献6】WO91/06626
【特許文献7】EP0266099
【特許文献8】W094/17086
【特許文献9】W096/35706
【非特許文献49】Debin他、(1999) Nucl Acids Res 27 (13)、2699〜2707
【非特許文献50】Xodo他、(2001) Eur J Biochem 268、656〜664
【非特許文献51】Blume他、(1999) Nucl Acids Res 27、695〜702
【非特許文献52】Faria他、(2001) JMol Biol 306、15〜24
【非特許文献53】Cheng他、(2000) Biotech and Bioeng 70、467〜472
【非特許文献54】Vasquez & Wilson (1998) Trends Biochern Sci1、4〜9
【非特許文献55】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFFACTOR
【非特許文献56】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fum+pagelibinfo+-info+TFSITE
【非特許文献57】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFCELL
【非特許文献58】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFCLASS
【非特許文献59】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFMATRIX
【非特許文献60】http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFGENE
【非特許文献61】Hill MおよびHemmings B (2002): Inhibition of protein kinase B/AKT.Implications for cancer therapy.Pharmacol Ther、93、p.243
【非特許文献62】Kanaseki T他、(2002) Identification of germline mutation of PTEN gene and analysis of apoptosis resistance of the lymphocytes in a patient with Cowden Disease.Pathobiolog y70、p.34
【非特許文献63】Basu他、(1995) Tetrahedron Lett36、4943
【非特許文献64】Ivanovskaya他、(1995) Nucl Nucl6、931〜934
【非特許文献65】Ivanovskaya他、(1987) Dokl Acad Nauk SSSR 293、477〜481
【非特許文献66】Kuznetsova他、(1999) Nuc Acids Res 27、3995〜4000
【非特許文献67】Tung & Stein (2000) Bioconjugate Chem 11 (5)、605〜618
【特許文献10】WO01/15737
【非特許文献68】Stetsenko & Gait (2000) Organic Chem 65 (16)、4900〜4908
【非特許文献69】Chen他、(1993) EMBO J.12、1161〜1167
【非特許文献70】Tommasino他、(1995) Bioessays 17、509〜518
【非特許文献71】Zhang他、(1997) Cell 89、357〜364
【非特許文献72】Ayer他、(1993) Cell 72、211〜222
【非特許文献73】Weinberg (1995) Cell 81、323〜330
【非特許文献74】Mu他、(1997) Plant Mol.Biol.34、357〜362
【非特許文献75】Cohen他、(1972) Proc.Natl.Acad.Sci.USA 69、2110
【非特許文献76】Sambrook他、(1989) Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Laboratory、Cold Spring Harbor、NY
【非特許文献77】Sherman他、(1986) Methods In Yeast Genetics、A Laboratory Manual、Cold Spring Harbor、NY
【非特許文献78】Beggs (1978) Nature 275、104〜109
【非特許文献79】J.DraperおよびR.Scott、D.Grierson(ed.)、「Plant Genetic Engineering」、Blackie、Glasgow and London、1991、vol.1、pp38〜81
【非特許文献80】J.Draper、R.Scott、A.KumarおよびG.Dury、ibid.、pp161〜198
【非特許文献81】Neuhaus & Spangenberg (1990) Physiol.Plant 79、213〜217
【非特許文献82】Gad他、(1990) Physiol.Plant 79、177〜183
【非特許文献83】Mathur & Koncz (1998) Method Mol.Biol.82、267〜276
【非特許文献84】D.McCabeおよびP.Christou、Plant Cell Tiss.Org.Cult.、3、227〜236 (1993)
【非特許文献85】P.Christou、Plant J.、3、275〜281(1992)
【非特許文献86】Bechtold他、(1993) Life Sciences、C.R.Acad.Sci.Paris 316、1194〜1199
【非特許文献87】J.DraperおよびR.Scott、D.Grierson(ed.)、「Plant Genetic Engineering」、Blackie、Glasgow and London、1991、vol.1 pp38〜81
【非特許文献88】Lazzeri (1995) Methods Mol.Biol.49、95〜106
【非特許文献89】Luchansky他、(1988) Mol.Microbiol.2、637〜646
【非特許文献90】Becker & Guarente (1990) Methods Enzymol.194、182
【非特許文献91】Vivanco & Sawyers (2002) The phosphatidylinositol 3-kinase-AKT pathway in human cancer.Nature Reviews Cancer、p489
【非特許文献92】Cory & Adams (2002) The Bcl-2family:regulators of the ceullular life-or-death switch
【非特許文献93】Mittler R.他、(1996) Inhibition of Programmed Cell Death in Tobacco Plants during a Pathogen-Induced Hypersensitive Response at Low Oxygen Pressure.Plant Cell、8、p.1991
【非特許文献94】Kuznetsova他、(1999) Nucl Acids Res 27 (20)、3995〜4000
【非特許文献95】Felgner他、(1997) Hum Gene Ther 8、511〜512
【非特許文献96】Madzak他、(1992) J.Gen.Viral.73、1533〜1536
【非特許文献97】Berkner (1992) Curr.Top.Microbiol.Immunol.158、39〜61
【非特許文献98】Berkner他、(1988) BioTechniques 6、616〜629
【非特許文献99】GorzigliaおよびKapikian (1992) J.Virol.66、4407〜4412
【非特許文献100】Quantin他、(1992) Proc.Natl. Acad. Sci.USA89、2581〜2584
【非特許文献101】Rosenfeld他、(1992) Cell 68、143〜155
【非特許文献102】Wilkinson他、(1992) Nucleic Acids Res.20、2233〜2239
【非特許文献103】Stratford-Perricaudet他、(1990) Hum.Gene Ther.1、241〜256
【非特許文献104】Moss (1992) Curr.Top.Microbiol.Immunol.158、25〜38
【非特許文献105】Muzyczka (1992) Curr.Top.Microbial.Immunol.158、97-123
【非特許文献106】Ohi他、(1990) Gene89、279〜282
【非特許文献107】Margolskee (1992) Curr.Top.Microbiol.Immunol.158、67〜90
【非特許文献108】Johnson他、(1992) J.Virol.66、2952〜2965
【非特許文献109】Fink他、(1992) Hum.Gene Ther.3、11〜19
【非特許文献110】BreakfieldおよびGeller (1987) Mol.Neurobiol.1、337〜371
【非特許文献111】Freese他、(1990) Biochem.Pharmacol.40、2189〜2199
【非特許文献112】BrandyopadhyayおよびTemin (1984) Mol.Cell.Biol.4、749〜754
【非特許文献113】Petropoulos他、(1992) J.Virol.66、3391〜3397
【非特許文献114】Miller (1992) Curt.Top.Microbiol.Immunol.158、1〜24
【非特許文献115】Miller他、(1985) Mol.Cell.Biol.5、431〜437
【非特許文献116】Sorge他、(1984) Mot.Cell.Biol.4、1730〜1737
【非特許文献117】Mann and Baltimore (1985) J.Virol.54、401〜407
【非特許文献118】Miller他、(1988) J.Virol.62、4337〜4345
【非特許文献119】Shimada他、(1991) J.Clin.Invest.88、1043〜1047
【非特許文献120】Helseth他、(1990) J.Virol.64、2416〜2420
【非特許文献121】Page他、(1990)J.Virol.64、5370〜5276
【非特許文献122】BuchschacherおよびPanganiban (1992) J.Virol.66、2731〜2739
【非特許文献123】Grahamおよびvan der Eb (1973) Virology52、456〜467
【非特許文献124】Pellicer他、(1980) Science209、1414〜1422)
【非特許文献125】Anderson他、(1980) Proc.Natl.Acad.Sci.USA77、5399〜5403
【非特許文献126】Brinster他、(1981)Cell27、223〜231
【非特許文献127】ConstantiniおよびLacy (1981) Nature 294、92〜94
【非特許文献128】Felgner他、(1987) Proc.Natl.Acad.Sci.USA 84、7413〜7417
【非特許文献129】WangおよびHuang (1989) Biochemistry 28、9508〜9514
【非特許文献130】Kaneda他、(1989) J.Biol.Chem.264、12126〜12129
【非特許文献131】Stewart他、(1992) Hum.Gene Ther.3、267〜275
【非特許文献132】Lim他、(1992) Circulation 83、2007〜2011
【非特許文献133】Wolff他、(1990) Science 247、1465〜1468
【非特許文献134】Wu他、(1991) J.Biol.Chem.266、14338〜14342
【非特許文献135】Zenke他、(1990) Proc.Natl.Acad.Sci.USA 87、3655〜3659
【非特許文献136】Wolff他、(1991) BioTecliniques 11、474〜485
【非特許文献137】Wagner他、(1991) Proc.Natl.Acad.Sci.USA 88、4255〜4259
【非特許文献138】Cotten他、(1990) Proc.Natl.Acad.Sci.USA 87、4033〜4037
【非特許文献139】Curiel他、(1991a) Proc.Natl.Acad.Sci.USA88、8850〜8854
【非特許文献140】Curiel他、(1991b) Hum.Gene Ther.3、147〜154
【非特許文献141】Feng他、(1997) Nature Biotechnology 15、866〜870
【非特許文献142】Ebbinghaus他、(1996) Gene Ther 3(4)、287〜297
【非特許文献143】Pichon他、(2000) Nucl Acids Res 28(2)
【非特許文献144】Active Motif、Morris他、(2001) Nature Biotech 19、1173〜1176
【非特許文献145】Nabel (1992) Hum.Gene Ther.3、399〜410
【非特許文献146】Vasquez KMおよびWilson JH、Trends Biochem Sci、1:4〜9、1998
【非特許文献147】Stetsenko & Gait (2000) Organic Chem 65(16)、4900〜4908
【非特許文献148】Perkins他、(1998) Biochemistry 37、11315〜11322
【発明の開示】
【課題を解決するための手段】
【0013】
本発明の第一の態様は、細胞中のアポトーシスを(誘導または回復を含めて)促進するための方法であって、(1)ゲノム中に存在する、選択したアポトーシス関連遺伝子にある部位、または該遺伝子と関係がある部位と結合する核酸結合部分、および(2)修飾部分を含む分子を、細胞中に導入するステップを含み、前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記修飾部分がポリペプチドまたはペプチド模倣体を含む方法を提供する。
【発明を実施するための最良の形態】
【0014】
一実施形態では、修飾部分は発現抑制因子部分である。修飾部分は、核酸またはクロマチンの共有結合修飾を調節することができる。
【0015】
以下でさらに論じるように、アポトーシス関連遺伝子は、細胞中のアポトーシスの制御と関係がある遺伝子である。アポトーシス関連遺伝子は、アポトーシス刺激に曝された細胞中で起こるアポトーシスを防ぐことができることを含めて(開始した場合でも、アポトーシスの終了を防ぐことができることを含む)、その産物がアポトーシスから細胞を救済することができる遺伝子であってよい。
【0016】
本発明の第二の態様は、(1)ゲノム中に存在する、選択したアポトーシス関連遺伝子にある部位、または該遺伝子と関係がある部位と結合する核酸結合部分、および(2)修飾部分を含み、前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記ポリペプチドまたはペプチド模倣体を含む分子を提供する。本発明の第一の態様と同様に、修飾部分は発現抑制因子部分であってよい。修飾部分は、核酸またはクロマチンの共有結合修飾を調節することができる。
【0017】
細胞またはゲノムは、真核生物の細胞またはゲノム、例えば真菌、動物または植物細胞であることが好ましい。
【0018】
選択した遺伝子標的(アポトーシス関連遺伝子)は、任意の様式の細胞増殖または細胞死の制御に関与するものである。アポトーシス関連遺伝子は、(少なくとも特定の条件下において)遺伝子が過剰発現されていない(あるいはその過小発現が、アポトーシスを受ける細胞の割合を低下させるのに、それ自体で充分である)以外は同一である細胞と比較した場合に、その過剰発現が、(少なくとも特定の条件下において)アポトーシスを受ける細胞の割合を増大させるのにそれ自体で充分である遺伝子であってよい。この遺伝子は、他の正常細胞または非改変型細胞において、アポトーシスを促進するのに充分である(あるいはその過小発現がアポトーシスを減少させるのに充分である)ものとすることができる。あるいは、この遺伝子は、例えば人工または天然の突然変異/淘汰(例えば腫瘍細胞中)の結果として他の遺伝子への改変、または幾つか他の方法で、例えばDNA結合部分を有する分子を使用し、前に記載したように分子を改変して、発現の改変がなされた細胞中においてのみ、アポトーシスを促進する(あるいは過小発現がアポトーシスを減少させる)ことができる。
【0019】
このような場合、遺伝子の発現を増大させてアポトーシスを促進することが望ましい。
【0020】
あるいは、アポトーシス関連遺伝子は、遺伝子が過剰発現されていない(あるいはその過小発現が、(少なくとも特定の条件下において)アポトーシスを受ける細胞の割合を増大させるのに、それ自体で充分である)こと以外は同一である細胞と比較した場合、その過剰発現が、(少なくとも特定の条件下において)アポトーシスを受ける細胞の割合を減少させるのに、それ自体で充分である遺伝子とすることができる。この遺伝子は、他の正常細胞または非改変型細胞において、アポトーシスを減少させるのに充分である(あるいはその過小発現は、アポトーシスを増大させるのに充分である)ものとすることができる。あるいは、この遺伝子は、例えば人為的または天然の突然変異/淘汰(例えば腫瘍細胞中)の結果として他の遺伝子への改変、または幾つか他の方法で、例えばDNA結合部分を有する分子を使用し、前に記載したように分子を改変して、発現の改変がなされた細胞中においてのみ、(あるいは過小発現はアポトーシスを増大させる)アポトーシスを減少させることができる。
【0021】
このような場合、遺伝子の発現を減少させてアポトーシスを促進することが望ましい。このような遺伝子(および遺伝子の発現の抑制)は、本発明の方法および分子における、非常に適した標的であると考えられる。
【0022】
分子の改変は、その破壊が「プログラムされた」細胞死(アポトーシスおよび任意の他の形の制御された細胞死、これらは用語アポトーシス中に一般に含めることができる)と相対する、壊死性細胞死に至ると予想される、中心的細胞活性と関係がある遺伝子ではなく、その産物が細胞増殖または細胞死の調節と関係がある遺伝子を標的とすることが好ましい。適切な標的遺伝子の例は、以下でさらに論じる。適切な標的遺伝子は、例えばSellers & Fisher (1999) The Journal of Clinical investigation、p.1655; Apoptosis and cancer Drug Targeting中、およびCory & Adams (2002) Nature Reviews Cancer、p489; The Bcl-2 family : regulators of the ceullular life-or-death switch中に述べられている。
【0023】
幾つかの実施形態では、本発明の分子の存在下において、細胞の生存能力は、同等の条件下での本発明の分子の不在下より、1.2倍、1.4倍、1.6倍、2倍、3倍、5倍、10倍、20倍、50倍、100倍、または1000倍低い。細胞の生存能力または細胞死は、細胞計数、生存能力アッセイまたは顕微鏡調査を含めた、任意の適切な技法を使用して測定することができる。アポトーシスは、ターミナルデオキシトランスフェラーゼ仲介dUTPニック末端標識(TUNEL)を使用することによって、壊死性細胞死と区別することができる。充分なニック末端標識によって、アポトーシスで見られるが壊死性細胞死では見られない、DNA断片化の型が示される。アポトーシスは、アネクシンVなどの細胞表面上の初期のアポトーシスマーカーを、測定することによって検出することもできる。これは例えば、染色およびFACS選別によって行うことができる。
【0024】
(影響を受けやすい細胞において)アポトーシスの誘導物質は、陽性対照として使用することができ、かつ/あるいは、(阻害剤の影響を受けやすい細胞において)アポトーシス阻害剤は、陰性対照として使用することができる。例えば、スタウロスポリンはアポトーシスの誘導物質として、Zvadまたは他のカスパーゼ阻害剤はアポトーシスの阻害剤として、使用することができる。修飾部分が(以下でさらに論じるような)ヒストン脱アセチル化酵素またはヒストン脱アセチル化酵素補充物質であるとき、脱アセチル化酵素阻害剤は、本発明の分子の影響を打ち消すことがある。
【0025】
抑制因子部分または修飾部分は、核酸またはクロマチンの共有結合修飾を調節することができることが好ましい。抑制因子または修飾部分は、クロマチン不活性化部分であることが好ましい。クロマチン不活性化部分は、クロマチン不活性化を直接的または間接的にもたらす任意のポリペプチドまたはその一部分であってよい。クロマチン不活性化を「直接的に」もたらすことによって、ポリペプチドまたはその一部自体がクロマチンに作用して、それを不活性化させることを意味する。クロマチン不活性化を「間接的に」もたらすことによって、ポリペプチドまたはその一部自体はクロマチンに作用しないが、細胞成分がそうすることを補うか促進することを意味する。
【0026】
クロマチン不活性化は一般に、遺伝子発現の抑制または不活性化をもたらす。典型的にはクロマチン不活性化は、遺伝子発現の抑制または不活性化が典型的には1個または数個の遺伝子に制限されるような、局所的事象である。したがって、クロマチン不活性化部分は任意の適切なポリペプチドであり、本発明の分子の一部分であるとき、および核酸結合部分近くの選択した遺伝子を標的とするとき、遺伝子の発現が不活性化または抑制されるように、選択した遺伝子と関係があるクロマチンを局所的に不活性化させる。ヒストン脱アセチル化はクロマチン不活性化と関係があり、したがって、クロマチン不活性化部分がヒストン脱アセチル化を容易にする場合、そのことは非常に好ましい。所与の遺伝子と関係があるヒストンの標的脱アセチル化は、ほぼ不可逆的に遺伝子の不活性化をもたらす。遺伝子発現の「抑制」または「不活性化」によって、本発明者らは、本発明の分子の存在下において、選択した標的遺伝子の発現は、均等な条件下での本発明の分子の不在下より、1.2倍、1.4倍、1.6倍、2倍、3倍、5倍、10倍、20倍、50倍、100倍、または1000倍低いことを意味する。遺伝子発現は、逆転写酵素-ポリメラーゼ連鎖反応(RT-PCR)、RNAハイブリダイゼーション、RNAse保護アッセイ、核ランオフアッセイ、およびDNAse1過敏性によって判断されるクロマチンの改変の使用を含めた、任意の適切な方法を使用して測定することができる。
【0027】
動物および植物細胞では、ヒストン脱アセチル化は、いわゆるヒストン脱アセチル化酵素複合体(HDAC)によってもたらされ、これは1つまたは複数のヒストン脱アセチル化酵素に加えて、複合体の形成および機能と関係がある補助タンパク質を含む。さらに、他のタンパク質成分が存在し、これらがHDACの一部分である可能性はないが、これらはHDACと結合し、あるいは他の場合はHDACと相互作用し、ヒストン脱アセチル化を容易にするのを助ける。
【0028】
ヒストンの脱アセチル化およびアセチル化は、よく知られている現象であり、以下の: Chen & Li (1998) Crit.Rev.Eukaryotic Gene Expression 8、169〜190; Workman & Kingston (1998) Ann.Rev.Biochem.67、545〜579; Perlmann & Vennstrom (1995) Nature 377、387-; Wolfe (1997) Nature 387、16〜17; Grunstein (1997) Nature 389、349〜352; Pazin & Kadonaga (1997) Cell 89、325〜328; DePinho (1998) Nature 391、533〜536; Bestor (1998) Nature 393、311〜312;およびGrunstein (1998) Cell 93、325〜328中に総説されている。
【0029】
HDAC複合体のポリペプチド組成は、現在調査中である。幾つかのHDAC複合体の一部分を形成することができる、あるいはそれらと結合するポリペプチドには、ヒストン脱アセチル化酵素1(HDAC1) Taunton他、(1996) Nature 272、408〜441);ヒストン脱アセチル化酵素2(HDAC2)(Yang他、(1996) Proc.Natl.Acad.Sci.USA 93、12845〜12850);ヒストン脱アセチル化酵素3(HDAC3)(Dangond他、(1998) Biochem.Biophys.Res.Comm.242、648〜652); N-CoR(Horlein他、(1995) Nature 377、397〜404); SMRT(Chen & Evans (1995) Nature 377、454〜457); SAP30(Zhang他、(1998) Molecular Cell1、1021〜1031).Sin3(Ayer他、(1995) Cell 80、767〜776; Schreiber-Agus他、(1995) Cell 80、777〜786)、SAP18(Zhang他、(1997) Cell 89、357〜364);およびRbAp48(Qian他、(1993) Nature 364、648〜652)がある。これらの論文はすべて、参照により本明細書に組み込まれている。HDAC複合体の他の成分が存在する可能性があり、あるいはそれらはHDAC複合体と相互作用し、それらは依然として発見されていないと考えられる。これらも本発明を実施する際に有用であると想定される。
【0030】
PLZFはN-CoRおよびSMRTと相互作用することが示されてきており、したがってHDAC複合体を補う。PLZFは、HDACとも直接相互作用する(Lin他、(1998) Nature 391、811〜814; Grignani他、(1998) Nature 391、815〜818; David他、(1998) Oncogene 16、2549〜2556)。
【0031】
Mad1はMadファミリーのメンバーであり、転写抑制因子として作用する能力を有する。Mad1はSin3と相互作用することができ、したがってクラスIのヒストン脱アセチル化酵素(HDAC1およびHDAC2)と相互作用することが示されてきている。Mad/Sin3は、多数のタンパク質-タンパク質相互作用が可能な、大きなタンパク質骨格として機能する(Hassig他、(1997) Cell 89、341〜347; Laherty他、(1997) Cell 89、349〜356; Zhang他、(1997) Cell 89、357〜364))。
【0032】
N-CoR、SMRT、Sin3、SAP18、SAP30およびヒストン脱アセチル化酵素のいずれかを含む、形成される複合体は、Heinzel他、(1997) Nature 387、43〜48; Alland他、(1997) Nature 387、49〜55; Hassig他、(1997) Cell 89、341〜347; Laherty他、(1997) Cell 89、349〜356; Zhang他、(1997) Cell 89、357〜364; Kadosh & Struhl (1997) Cell 89、365〜371; Nagy他、(1997) Cell 89、373〜380;およびLaherty他、(1998) Molecular Cell 2、33〜42中に記載されている。これらの論文はすべて、参照により本明細書に組み込まれている。
【0033】
したがって、HDAC複合体の成分、またはHDAC複合体と結合するか、あるいはHDAC複合体の補充を容易にするポリペプチドが、MAD1、E7、PLZF、SMRT、Sin3、SAP18、SAP30またはN-CoR、あるいはHDAC1、BDAC2またはHDAC3を含むHDAC、あるいはNuRD、MAD2、MAD3、MAD4またはRbのいずれか1つである場合、それは非常に好ましい。その機能的部分が存在する限り、すべてのポリペプチドが存在することが必要とされないこともあることは理解されよう。例えば、ヒストン脱アセチル化酵素(例えば、HDAC1、HDAC2またはHDAC3)に関しては、機能的部分はヒストン脱アセチル化酵素活性を保持する部分であってよく、あるいは機能的部分は、HDAC複合体の他の成分用の結合部位を含む部分、または他の場合はHDAC複合体を補い、ヒストン脱アセチル化を促進する部分であってよい。同様に、HDAC複合体の他の成分、またはHDAC複合体と結合するポリペプチドに関しては、機能的部分は、HDAC複合体の他の成分用の結合部位を含む部分であってよい。今日まで、6個の哺乳動物HDAC遺伝子が記載されてきており(Grozinger他、(1999) Proc.Natl.Acad.Sci.USA 96、4868〜4873)、これらの遺伝子の任意の1つまたは複数が、本発明を実施する際に有用である可能性があると考えられる。
【0034】
修飾部分はVP16またはKRABであってよいが、これは好ましくない。したがって、一実施形態では、VP16またはKRABは、用語「修飾部分」または「クロマチン不活性化部分」の意味の中には含まれない。VP16は、その作用形式がDNAまたはクロマチンの共有結合修飾と関係があるとは考えられない、転写活性因子である。KRABは、その作用形式がクロマチン不活性化以外の機構と関係があると考えられる、転写抑制因子である。好ましくはないが、KRABの任意の断片は、前に定義した分子/ポリペプチドの一部分であるとき、核酸結合部分付近の選択した遺伝子を標的化して、選択した遺伝子と関係があるクロマチンを局所的に不活性化させて、その結果遺伝子の発現を不活性化させるか抑制するとき、用語「クロマチン不活性化部分」内に含まれる。例えば、HDAC複合体と結合するか、HDAC複合体の補充を容易にすることができるKRABの任意の断片は、用語「クロマチン不活性化部分」内に含まれる。しかしながら、いかなるこのような断片も好ましくない。
【0035】
結合モチーフは、HDAC複合体の成分内、あるいはHDAC複合体と結合するポリペプチド内に存在し、これらのモチーフは、本発明のポリペプチド中においてクロマチン不活性化部分として作用するのに、充分である可能性があると考えられる。何故ならこれらは、HDAC複合体を補充することによってヒストン脱アセチル化を容易にすることができるからである。
【0036】
さらに、HDAC複合体の成分の変異体、またはHDAC複合体と結合するポリペプチドの変異体を使用できることは理解されよう。適切な変異体は、前に記載した機能的部分だけでなく、アミノ酸残基が欠失または置換または挿入されている変異体も含む。ただし変異体は、ヒストン脱アセチル化を容易にすることが依然としてできるものとする。したがって、適切な変異体は、そのアミノ酸配列が野生型と比較して修飾されているが、ヒストンを脱アセチル化するその能力を保持している、ヒストン脱アセチル化酵素の変異体を含む。同様に、適切な変異体は、例えばそのアミノ酸配列が野生型と比較して修飾されているが、HDAC複合体と、あるいはその中で相互作用するその能力を保持している、Sin3またはPLZFの変異体を含む。同様に、HDAC複合体の成分と相互作用する他のタンパク質の変異体、およびHDAC活性によって遺伝子の不活性化をもたらすことができる他の転写因子を、使用することができる。
【0037】
Rb、MADおよびMeCpG2タンパク質の全体または一部分は、HDAC複合体と相互作用することができる。
【0038】
哺乳動物系における、HDAC複合体、およびHDAC複合体の補充と関係があるポリペプチドに関して、多くの研究がなされてきているが、この系の根本的性質は、機能的に同等なポリペプチドが、他の真核細胞中、特に他の動物細胞または植物細胞中で見られると予想されることである。例えば、図5は、ヒトHDAC1と非常に関係があるポリペプチドが、シロイヌナズナ中および酵母菌中に存在することを示す。植物のHDAC複合体はトウモロコシから単離されており(Lussen他、(1997) Science 277、88〜91)、植物、真菌および脊椎動物細胞由来のヒストン脱アセチル化酵素の比較試験が着手されてきている(Lechner他、(1996) Biochim.Biophys.Acta 1296、181〜188)。ヒストン脱アセチル化酵素阻害剤は、アブラナ科植物中の不活性状態のrRNA遺伝子を活性化させることが示されてきており(Chen & Pickard (1997) Genes Dev.11、2124〜2136)、Cochliobolus carbonum由来の天然に存在する宿主選択的毒素(HC毒素)は、植物、真菌および哺乳動物のヒストン脱アセチル化酵素を阻害する(Brosch他、(1995) Plant Cell 7、1941〜1950)。
【0039】
クロマチン不活性化部分が、分子が導入される細胞と同じ細胞型または種に由来することは必要ではないが、このようなクロマチン不活性化部分が、その細胞中においてより効率よくクロマチンを不活性化することができる可能性がある場合、それが望ましい。
【0040】
分子のクロマチン不活性化部分が、ヒストン脱アセチル化を容易にすることができる、PLZF、E7、MAD1、RbまたはSAP18、あるいはPLZFまたはE7またはMAD1またはRbまたはSAP18の一部分、あるいはヒストン脱アセチル化によって遺伝子の活性化を引き起こすことが知られている、ポリペプチドまたはポリペプチドの一部分である場合、それが特に好ましい。例えば、APLと関係があるPLZF-RAR中のPLZFの部分は、N-CoRおよびSMRTと相互作用すると考えられる。
【0041】
好ましいクロマチン不活性化部分は実施例中に記載し、アミノ酸配列XXXMAVESRVTQEEIKKEPEKPIDREKTCPLLLRVF(前式でXXXは、例えばAAAまたはDDD連結基、または他の親水性の、好ましくは帯電した連結基である)を有するSAP18由来のポリペプチド/ポリペプチド模倣体または類似体、およびアミノ酸配列XXXMNIQMLLEAADYLERREREAEHGYASMLP(前式でXXXは、例えばAAAまたはDDD連結基、または他の親水性の、好ましくは帯電した連結基である)を有するMAD1由来のポリペプチドを含む。
【0042】
クロマチン不活性化部分が、HDAC1、HDAC2またはHDAC3中に含まれるようなヒストン脱アセチル化酵素活性を有するポリペプチドである場合、それも特に好ましい。
【0043】
あるいは、修飾部分が、共有結合修飾、例えば核酸、好ましくはDNAのメチル化を調節することができる部分であってよい。したがって、修飾部分はDNA修飾酵素であるか、あるいはこれを含んでよく、あるいはこのような酵素を補うことができてよい。この調節には、選択した遺伝子を抑制する効果があることが好ましい。
【0044】
修飾部分は、核酸の配列を変えないことが好ましい。修飾部分は、核酸骨格を切断しないことが好ましい。修飾部分は、リコンビナーゼまたは制限エンドヌクレアーゼではないことが好ましい。
【0045】
例えば、修飾部分は、例えばOkano M、Xie S、LiE.(1998) Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases.Nat Genet19:219〜220; Adrian P.Bird and Alan P.Wolffe (1999) Methylation-Induced Repression:Belts、Braces、and Chromatin.Cell 99、451〜454中に論じられたように、メチルトランスフェラーゼまたはメチルトランスフェラーゼ複合体の成分の全体または一部分を含むことができる(あるいは補充することができる)。
【0046】
抑制因子または修飾部分は、DNA鎖の持続的な破壊を生じるエンドヌクレアーゼまたは他の分子ではないことが好ましい。
【0047】
分子のポリペプチド/ポリペプチド模倣体または類似体部分(例えば修飾部分)は、11kDa未満、好ましくは8kDa未満、さらにより好ましくは6kDa未満の分子質量を有することが好ましい。例えば、ポリペプチド/ポリペプチド模倣体または類似体部分は、100個未満、さらにより好ましくは90、80、70、60、50、45、40、35、30、25または20個未満のアミノ酸(またはその模倣体または類似体)、最も好ましくは約60個と25個の間のアミノ酸(またはその模倣体または類似体)を有することが好ましい。
【0048】
修飾部分は、SAP18またはMAD1またはRb由来のペプチドおよび適切な連結基、例えば前および実施例1に記載した、SAP18またはMAD1由来のペプチドおよび連結基からなることが特に好ましい。
【0049】
本発明の分子は、細胞進入および/または核局在を容易にする部分(局在部分)をさらに含むことができる。この部分は、ポリペプチドまたはポリペプチド模倣体/類似体であってもよい。例えば、局在部分は、例えばSoukchareun他、(1998)Bioconjugate Chem9、466〜475およびその中に引用された参照文献、例えばSoukchareun他、(1995) Bioconjugate Cherry6、43〜53(ウイルス融合ペプチド)またはEritja他、(1991) Tetrahedron 47、4113〜4120(核輸送シグナル配列)中に論じられた、膜親和性活性を有するペプチドを含むか、あるいはこのペプチドからなっていてよい。局在部分は、核局在シグナルペプチド(例えばDDDPKKKRKV-NH2)、またはWO99/13719中に述べられたエンドソーム溶解ペプチド(エンドソーム部分からの分子の放出を容易にすることができる)であってよい。この部分は、3kDa未満、好ましくは2.5kDa未満であることが好ましい。分子の全ポリペプチド/模倣体/類似体含量は、11kDa未満であることが好ましい。典型的には、局在部分は、約7〜16個のアミノ酸を有することができる。
【0050】
局在部分の他の例には、修飾型アンテナペディアホメオドメイン系ペネトラチン(例えばRQIKIWFQNRRMKWKK)、またはTAT(例えばC(Acm)GRKKRRQRRRPPQC、前式でC(Acm)はCys-アセトアミドメチルである)またはVP22系分子(Prochiantz (2000) Curr Opin Cell Biol9、420〜429).)または塩基性HIV TAT内在ペプチドがある。
【0051】
例えば以下でより詳細に論じるように、本発明の分子は、本発明の態様によって与えられる方法および使用において、有用である可能性がある。特に、本発明のポリペプチドは、本発明の第一の態様の方法において、有用である可能性がある。
【0052】
本発明の分子が、天然に存在しないハイブリッド分子である場合、それが好ましい。例えば、核酸結合部分および修飾部分が、天然に存在する複合体または分子に由来しない場合、それが好ましい。核酸結合部分およびクロマチン不活性化部分が由来する分子は(存在する場合)、同種に由来してよく(例えば、以下でより詳細に記載するように、核酸結合部分は、ヒト核酸中に見られる配列を有するオリゴヌクレオチドであってよく、クロマチン不活性化部分はヒトPLZFの一部分であってよい)、あるいはそれらは異種に由来してよい(例えば、ヒト核酸中では見られない配列を有し、例えば細菌DNA配列と結合することができるオリゴヌクレオチドを、ヒトPLZFの一部分と融合させることができる)。
【0053】
したがって、特に好ましい実施形態では、本発明の分子は、化学合成法によって生成される分子であり、核酸結合部分および修飾部分またはクロマチン不活性化部分は、以下でより詳細に記載するように選択する。
【0054】
合成および結合技法は、WO01/14737およびその中の参照文献(参照により本明細書に組み込まれている)中に論じられている。WO01/147373の方法が好ましい。あるいは、Kusnetsova他、(1999) Nucleic Acids Res 27、3995〜4000中に記載された技法も使用することができる。
【0055】
真核生物ゲノム中に存在する部位は、その発現を調節、好ましくは抑制または不活性化するのが望ましい、選択した1つまたは複数の遺伝子の部位、あるいはその遺伝子と結合した部位である。その部位が真核生物ゲノム中に本来存在する部位である場合、それが好ましい。しかしながら、以下でより詳細に論じるように、部位はゲノム中で遺伝子工学処理された部位であってよく、あるいは部位は、挿入されるウイルス配列と関係がある部位であってよい。その発現が抑制される遺伝子の付近に存在するゲノム中に遺伝子工学処理された部位は、同種(ただしゲノム中の他の場所に存在する)由来の部位であってよく、あるいは部位は、異種に存在する部位であってよい。「ゲノム」によって、本発明者らは、染色体DNAだけでなく、真核生物の細胞中に存在する他のDNA、細胞中に導入されたDNA、例えばプラスミドまたはウイルスDNAなどを含むものとする。核酸結合部分が染色体DNAと、あるいは以下でより詳細に記載するように、染色体DNAから転写されたRNAと結合することができる場合、それが好ましい。
【0056】
一実施形態では、遺伝子は内因性遺伝子であることが好ましい。用語「内因性遺伝子」は、その細胞本来の、すなわちその細胞と異種ではなく、その本来のゲノム概念中の遺伝子を指す。この概念では、真核生物ゲノム中に存在する部位は、その発現を抑制または不活性化するのが望ましい、選択した1つまたは複数の内因性遺伝子の、あるいはその遺伝子と結合した部位である。その部位は真核生物ゲノム中に本来存在する部位であり、その本来のゲノム概念中に存在する。
【0057】
前述のように、遺伝子は(その産物を含めて)、細胞の死または生存能力の制御と関係がある。これは直接的または間接的である可能性があり、その効果は、物質による処理後の任意の時間で生じる可能性がある。
【0058】
当業者に知られているように、上記の部位は三重らせんを形成するためのオリゴヌクレオチドとの結合を促進する特定の配列特性を有することが望ましいことがある。しかしながら、本発明に関しては、このような配列特性は、ポリペプチド部分の不在下では、オリゴヌクレオチドほど重要ではない可能性があると考えられる。何故なら、本発明の分子の抑制または調節効果は、分子が標的部位とはもはや結合せず、したがって、結合の親和性がそれほど重要ではない可能性があるときでさえも持続する可能性があるからである。オリゴヌクレオチドの配列は一層重要であり、したがって特異的な認識が得られる。しかしながら、オリゴと標的配列の間に形成される結合は、ポリペプチド/またはペプチド模倣体部分が存在する場合、強力でなくでもよいこともある。
【0059】
その転写が抑制または調節される遺伝子に対する、オリゴヌクレオチド結合部位の位置も、オリゴヌクレオチド、例えば修飾部分を含まないTFOほど重要ではない可能性がある、なぜなら、本発明の分子の調節または抑制効果は(例えば、修飾ドメインがメチルトランスフェラーゼまたはヒストン脱アセチル化酵素であるか、あるいはこれらを補うことができるとき)、オリゴヌクレオチド結合部位のいずれかの側に広がる可能性があるからである。核酸結合部位は遺伝子プロモーターと結合することができるが、あるいは当該の遺伝子内またはその付近の他の配列と結合することができる。
【0060】
WO90/06934/EP0375408およびWO91/06626は、TFOの配列要件を論じている。三重らせんを形成するための2つのモチーフは、「CT」モチーフおよび「GT」モチーフと呼ばれる。これらの第一の文献は、TFOとしてのポリピリミジンオリゴヌクレオチドの使用に関する。それぞれのGC塩基対に関して、CはTFO中に存在し、それぞれのAT塩基対に関して、T(またはキサンチンまたはイノシンまたはハロゲン化誘導体)はTFO中に存在する。TFOは、二本鎖のプリン多量鎖に対して、平行方向を向いていると考えられる。あるいは、「GT」モチーフを使用すると、G(またはハロゲン化誘導体)がそれぞれのGC塩基対に関して存在し、T(またはキサンチンまたはイノシンまたはハロゲン化誘導体)がそれぞれのAT塩基対に関して存在し、TFOは、二本鎖のプリン多量鎖に対して、逆平行方向を向いていると考えられる。標的配列は、少なくとも約65%のプリン塩基、または少なくとも約65%のピリミジン塩基を有していなければならない。EP0266099も、どのようにして適切な標的配列を選択することができるかを論じている。
【0061】
W094/17086は、一本鎖の立体配座をとることができると考えられるDNA配列と結合させることを目的とする、オリゴヌクレオチドを論じている。このような配列はプリンが多量であり、相当な鏡面対称性を有する可能性がある。オリゴヌクレオチドはプリン鎖と相当に相補的であってよく、一本鎖標的DNAを有するワトソン-クリックおよびフーグスティン結合を形成することができる、環状またはステム-ループ機能構造を有していてよい。
【0062】
W096/35706は、特異的で安定した標的核酸(ピリミジン一本鎖核酸)との複合体形成を促進すると考えられ、標的核酸不在下における平行鎖ヘアピン構造の形成ために、高い安定性を有し得る構造および配列特性を有するオリゴヌクレオチドを記載している。
【0063】
Debin他、(1999) Nucl Acids Res 27 (13)、2699〜2707は、G、A三重らせんの安定性、およびTFO設計の結果に影響を与える因子を述べている。Xodo他、(2001) Eur J Biochem 268、656〜664も、標的部位とTFOの結合、例えば短いオリゴヌクレオチドと近辺部位の結合に影響を与える因子を調べている。
【0064】
Blume他、(1999) Nucl Acids Res 27、695〜702は、三重らせん形成における二価カチオンの関与、および形成をどのようにして、正または負に調節することができるかを調べている。Faria他、(2001) JMol Biol 306、15〜24は、細胞中のTFOおよびさまざまなオリゴヌクレオチドに関する結果を評価するためのアッセイを記載している。Cheng他、(2000) Biotech and Bioeng 70、467〜472は、TFO結合の数学的モデル化の結果、および結合部位およびTFO配列の選択に関する結果を示している。
【0065】
Demidov & Frank-Kamenetskiiは、ペプチド核酸(PNA)、特にカチオン性ピリミジンPNA(cpyPNA)と二本鎖DNAの結合を総説している。
【0066】
可能性のあるTFOを設計するためのルールは、Vasquez & Wilson (1998) Trends Biochem Sci1、4〜9中に総説されている。3つの型のTFO:ピリミジン多量(CT);プリン多量(GA)および混合型(GTまたはGAT)が有効であることが示されている。CTTFOは平行モチーフ中で結合し、その中では第三の鎖が、二本鎖のプリン鎖と同じ5'〜3'方向を有する。GATFOは逆平行モチーフ中で結合する。混合型TFOは、標的配列に応じて、いずれかの方式で結合することができる。他の性質も幾つかの型のTFOの間で異なり;例えばCTTFOはpH依存性である。それぞれの型のTFOが、本発明に関して適切である可能性がある。
【0067】
オリゴヌクレオチドまたは模倣部分は、約10〜80、好ましくは15〜40塩基長、さらにより好ましくは約20〜40塩基長であることが好ましい。20塩基未満のオリゴヌクレオチドは、弱いおよび/または特異性の低い結合を示す可能性があるが、それにもかかわらず、前述のように、例えばごく一時的な結合が必要とされるので、本発明の実施において有用である可能性がある。
【0068】
「DNA」または「オリゴ(デオキシ)ヌクレオチド」によって、本発明者らは、糖-連結基-糖の骨格を有する分子であって、糖残基が2'-デオキシリボース(および、2',3'ジデオキシリボース部分を含むヌクレオシドが末端であるDNA鎖をしたがって含む)を含み、1位置において糖残基と結合しているのが、アデニン(A)、シトシン(C)、グアニン(G)、チミジン(T)、イノシン(I)、ウリジン(U)およびその他などの塩基である分子を意味する。正常なDNAでは、糖残基間の結合(「糖-糖結合」)はリン酸部分であり、これが前記糖残基とジエステルを形成する。しかしながら、本発明者らは、用語「核酸」中に(およびより詳細には用語DNA中に)、非リン酸結合を有する分子を含むものとする。
【0069】
したがって、ホスホロチオエート結合およびホスホロセレノエート結合を含める。これらの結合が、正常なDNAよりも細胞のヌクレアーゼによる攻撃に対して耐性があることが、好ましい可能性がある。このような結合は、リン酸メチル、ホスホトリエステル、および天然に存在するホスホジエステルの鏡像異性体も含むことができる。
【0070】
用語「核酸」または「オリゴヌクレオチド」とは、非天然塩基類似体を含む分子;ペントース糖の2'および3'位置が独立に、-H、-OHまたは-NH2のいずれかである分子;および、ホスホジエステル結合中ではなくリン原子と結合している酸素が、-SH、SeH、-BH2、-NH2、-PH3、-F、-Cl、-CH3、-OCH3、-CNおよび-Hによって置換されている分子も含む。
【0071】
オリゴヌクレオチドは、オリゴリボヌクレオチドまたはオリゴデオキシリボヌクレオチドであってよい。オリゴデオキシリボヌクレオチドが好ましい、何故なら、オリゴリボヌクレオチドは、オリゴデオキシリボヌクレオチドよりも酵素による攻撃を受けやすい可能性があるからである。
【0072】
オリゴヌクレオチドまたは模倣体または類似体は、当業者に知られるように、例えばWO99/13719およびその中の参照文献中、およびDemidov & Frank-Kamenetskii(2001)上記中に記載されたように、ペプチド核酸であってよい。PNAは、DNAのデオキシリボース-リン酸骨格の代わりに2-アミノエチルグリシン単位を含む、ポリアミド(ペプチド)骨格を有する核酸類似体である。PNA骨格は中性であり(DNA骨格と異なり、それは負に帯電している)、したがって、対応するDNA分子よりも、帯電した核酸分子とより安定的に結合することができる。
【0073】
オリゴヌクレオチドまたは類似体または模倣体は、DNAオリゴヌクレオチド
であることが好ましい。
【0074】
オリゴヌクレオチドに対する言及は(それが適切な場合)、オリゴヌクレオチド模倣体または類似体、例えばPNAに対する言及を含む。
【0075】
オリゴヌクレオチドは、オリゴヌクレオチドの5'または3'末端と結合することができる、連結基を含むことができる。適切な連結基の例は、例えばWO90/06934中に記載されている。
【0076】
核酸結合部分は、ペプチド核酸(PNA)であるか、あるいはこれを含むことが、好ましい可能性がある。
【0077】
核酸結合部分は、植物または動物などの真核生物、ゲノム中に存在する部位と結合する、定義する任意の適切な結合部分であってよい。核酸結合部分は、本発明の分子のクロマチン不活性化部分の存在によって、その発現が変更、特に抑制される、選択した遺伝子の部位またはそれと関係がある部位と結合することができることが特に好ましい。核酸結合部分は、所望の部位と選択的に結合することが好ましい。核酸結合部分が結合することができる、1つまたは複数の所望の部位が存在する可能性がある。典型的には、標的ゲノム中には、1つの目的とする標的部位が存在する。誤解を避けるために、真核生物中に存在する部位は、天然に存在する部位であることが最も有用である可能性があり、あるいはそれは、真核生物中に存在させるように遺伝子工学処理した部位であってよい。部位は、同じまたは任意の他の真核生物に元来由来するものである必要はない。例えば部位は、TFOが事前に特徴付けられており、真核生物細胞、例えば植物細胞のDNA中に存在させるように遺伝子工学処理した、細菌またはウイルスの配列、あるいは人工配列であってよい。これらの例は、本明細書の実施例中に記載するEREおよびIREなどの応答エレメント、または他の特徴付けされた結合部位を含むことができる。部位の内因性調節因子を含まない細胞において、このような部位を使用することが望ましい可能性がある。あるいは、部位は、本来存在する応答エレメントの改変型であってよく、この改変型はTFOの結合部位として働くことができるが、本来存在する応答エレメントの本来存在する調節因子によって調節することができない。しかしながら、核酸結合部分が結合する部位が真核生物細胞中に本来存在し、それがゲノム中のその本来の位置に存在する場合、それが好ましい。
【0078】
核酸結合部分は、DNA結合部分またはRNA結合部分であってよい。したがって、核酸結合部分は、二本鎖核酸(例えばDNA)または一本鎖核酸(例えばRNAまたは一本鎖DNA)と結合することができる。RNA結合部分の場合、RNA結合部分と結合する、真核生物ゲノム中に存在する部位は、典型的には、不活性化用に選択した部位においてDNAから転写される新生RNAである。そのRNAは、その発現が抑制される遺伝子により転写されるRNAであってよく、あるいはそのRNAは、その発現が抑制される遺伝子と隣接しているか、あるいは少なくともそれと近い、遺伝子により転写されるRNAであってよい。RNA結合部分は、転写産物の5'端の、あるいはその近くのRNA配列と結合することが好ましい。転写される一方で、新生RNAは、その転写部位あるいはその近くに留まり、転写部位が、その発現が抑制される遺伝子、あるいはその近くに存在する場合、本発明の分子中にRNA結合部分を使用することは、所望の部位へのクロマチン不活性化部分の局在化を容易にすることは理解されよう。
【0079】
DNA結合部分が転写因子結合部位と結合し、例えばその結果、転写因子が結合する2つ以上の遺伝子の発現を調節することができる場合、それは有用である可能性がある。アポトーシス遺伝子と関係がある転写因子には、CREB、WT1、NF-kappaBおよびStat3がある。転写因子およびそれらの結合部位を列挙するデータベースを、以下に列挙する:
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFFACTOR
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fum+pagelibinfo+-info+TFSITE
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-info+TFCELL
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-
info+TFCLASS
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-
info+TFMATRIX
http://www.embl-heidelberg.de/srs5bin/cgi-bin/wgetz?-fun+pagelibinfo+-
info+TFGENE
【0080】
DNA結合部分が、転写開始部位のすぐ上流の、プロモーター領域または他の調節領域または配列と結合する場合、それが有用である可能性がある。幾つかの適用例では、遺伝子内の配列を標的化して、スプライス変異体を区別することが、好ましい可能性がある。
【0081】
前述のように、オリゴヌクレオチドを設計/遺伝子工学処理して、選択した遺伝子の、あるいはそれと結合した、特定の選択した標的DNA配列と結合させることができる。本発明の一実施形態では、オリゴヌクレオチドは、植物または動物細胞中には存在するが、同等な野生型配列中には存在しない、突然変異遺伝子配列中に存在する部位と結合させるために、遺伝子工学処理したオリゴヌクレオチドである。例えば、以下でより詳細に論じるように、オリゴヌクレオチドは、突然変異アポトーシス関連癌遺伝子などの、優性ネガティブ突然変異遺伝子と選択的に結合することができ、結合によって、DNAメチル化またはクロマチン不活性化が起こり、突然変異アポトーシス関連癌遺伝子の発現を抑制する。突然変異するアポトーシス関連癌遺伝子の例には、タンパク質キナーゼB/AKTおよびPTENがある(Hill MおよびHemmings B (2002): Inhibition of protein kinase B/AKT.Implications for cancer therapy.Pharmacol Ther、93、p.243; Kanaseki T他、(2002) Identification of germline mutation of PTEN gene and analysis of apoptosis resistance of the lymphocytes in a patient with Cowden Disease.Pathobiolog y70、p.34)。
【0082】
RAS(H-ras)およびBcl-10は、アポトーシス関連遺伝子の定義から除外される。
【0083】
典型的には、核酸結合部分と、修飾部分またはクロマチン不活性化部分を融合させる。例えばSoukchareun他、(1998)上記、およびその中に引用された参照文献中、およびBasu他、(1995) Tetrahedron Lett36、4943中に記載されたのと同様に、核酸結合部分と、修飾部分またはクロマチン不活性化部分は、例えば固相支持体上でのペプチド、および次いでオリゴヌクレオチドの連続的構築によって、1つの分子として合成することができる(全体的合成手法)。好ましくは、自動化された手順を使用する。
【0084】
あるいは、核酸結合部分と、修飾部分またはクロマチン不活性化部分は、当業者によく知られている技法を使用して別々に合成し、次いで結合する。ペプチド核酸とペプチドの結合に適した技法には、SPDP(N-スクシンイミジル3-(2-ピリジルジチオ)プロピオネート)およびSMCC(スクシンイミジル4-(N-マレイミドメチル)シクロヘキサン-カルボキシレート)などの、ヘテロ二官能性結合試薬の使用があり、例えばWO99/13719中、特に実施例12〜15中に記載されている。オリゴデオキシヌクレオチドとペプチドの結合に適した技法には、Soukehareun他、(1998)上記中に記載されたのと同様の、N_-Fmoc-システイン(S-チオブチル)誘導型オリゴデオキシヌクレオチドの使用がある。他の技法としては、ペプチド中の求核基(-NH2基など)を介して非保護ペプチドを結合させる前に、リン酸オリゴヌクレオチドの3'または5'端の、N-ヒドロキシベンゾトリアゾール(HOBT)エステル活性化を使用することがある(Ivanovskaya他、(1995) Nucl Nucl6、931〜934; Ivanovskaya他、(1987) Dokl Acad Nauk SSSR 293、477〜481; Kuznetsova他、(1999) Nuc Acids Res 27、3995〜4000を参照)。ペプチド-オリゴヌクレオチド結合技法は、例えばTung & Stein (2000) Bioconjugate Chem 11 (5)、605〜618中に総説されている。
【0085】
WO01/15737およびStetsenko & Gait (2000) Organic Chem 65 (16)、4900〜4908中に記載されたのと同様に、「本来の連結」技法を使用することが好ましい。N-末端チオエステル-官能化ペプチドを、5'-システイニルオリゴヌクレオチド誘導体と連結させる。
【0086】
適切には、核酸結合部分、および抑制因子、修飾部分またはクロマチン不活性化部分が結合し、その結果両方の部分が、例えば核酸結合部分は、植物または動物ゲノム中に存在する部位と結合することができ、結合によって、修飾部分が、核酸またはクロマチンの共有結合修飾を調節することが依然としてできる、例えばクロマチン不活性化部分が、クロマチンを不活性化させることが依然としてできるように、そのそれぞれの活性を保つ。この2つの部分は直接結合することができるが、これらは連結基ペプチドまたはオリゴヌクレオチドによって結合することができる。適切な連結基ペプチドは、ランダムコイルの立体配座を典型的にとるペプチドであり、例えばポリペプチドは、アラニンまたはプロリン、あるいはアラニン残基とプロリン残基の混合物を含むことができる。アミノ酸は溶解度を増大させることが好ましい;したがって連結基は、例えばアスパラギン酸残基などの、帯電または親水性アミノ酸を含むことができる。連結基はアスパラギン酸残基を含むか、あるいはそれらからなることが好ましい。アミノ酸は、フェニルアラニンまたはトリプトファンなどの、親水性アミノ酸ではないことが好ましい。連結基は、10個と100個の間、より好ましくは10個と50個の間、さらにより好ましくは10個と20個の間の、アミノ酸残基を含むことが好ましい。例えば、3個と9個の間のアミノ酸の短い連結基も、有用である可能性がある。任意の事象において、分子の部分間に連結基が存在しようとしまいと、分子はその標的核酸と結合することができ、発現を抑制するか、あるいは核酸またはクロマチンの共有結合修飾を調節する、例えばクロマチンを不活性化させ、それによって遺伝子発現を選択的に抑制するか不活性化させることができる。
【0087】
適切な抑制因子、修飾またはクロマチン不活性化部分をコードするポリヌクレオチドは、当分野において知られており、知られている配列から設計し、作製することが容易にできる。さまざまな適切なクロマチン不活性化部分をコードする、ポリヌクレオチド配列は、ポリペプチドを言及する参照文献中に前に与えており、あるいはGenBankまたはEMBLまたはdbESTから入手可能である。PLZFに関する参照文献は、Chen他、(1993) EMBO J.12、1161〜1167である。E7に関する参照文献は、Tommasino他、(1995) Bioessays 17、509〜518である。SAP18、MAD1およびRbに関する参照文献は、それぞれZhang他、(1997) Cell 89、357〜364; Ayer他、(1993) Cell 72、211〜222およびWeinberg (1995) Cell 81、323〜330である。
【0088】
適切な連結基ペプチドをコードするポリヌクレオチドは、連結基ペプチドの配列から設計し、作製することが容易にできる。
【0089】
したがって、本発明の分子の抑制因子または修飾部分をコードするポリヌクレオチドは、よく知られている遺伝子工学技法を使用して、容易に構築することができる。したがって、抑制因子または修飾部分は、当業者によく知られている分子生物学の技法を使用して、発現によって合成することができる。しかしながら、当業者に知られており本明細書で論じるように、有機化学の技法によって、本発明の分子のポリペプチド/類似体/模倣体部分を合成することが、好ましい可能性がある。
【0090】
本発明は、本発明の分子を用いて形質転換した、宿主細胞にも関する。宿主細胞は原核生物または真核生物であってよい。細菌細胞は、好ましい原核生物宿主細胞であり、典型的には大腸菌の菌株、例えばBethesda Research Laboratories Inc.、Bethesda、MD、USAから入手可能な大腸菌菌株DH5、およびAmerican Type Culture Collection (ATCC)of Rockville、MD、USAから入手可能なRR1などである(NoATCC31343)。好ましい真核生物細胞には、植物、酵母菌、昆虫および哺乳動物細胞、好ましくは脊椎動物細胞、マウス、ラット、サルまたはヒト線維芽細胞および腎臓細胞系由来の細胞などがある。酵母菌宿主細胞には、YPH499、YPH500およびYPH501があり、これらは一般にStratagene Cloning Systems、La Jolla、CA92037、USAから入手可能である。好ましい哺乳動物宿主細胞には、CCL61としてATCCから入手可能なチャイニーズハムスター卵巣(CHO)細胞、CRL1658としてATCCから入手可能なNIHスイスマウス胚細胞NIH/3T3、CRL1650としてATCCから入手可能なサル腎臓由来COS-1細胞;ヒト胚腎臓細胞である293細胞、およびHT1080ヒト骨線維肉腫細胞がある。
【0091】
形質転換用のプロトプラストは、典型的には、当分野で知られている方法によって、必要に応じて作製される。植物細胞系は、一般には利用可能ではない。しかしながら、一般的に使用される1つの細胞系は、タバコ由来のBright Yellow2細胞系である(BY2;Mu他、(1997) Plant Mol.Biol.34、357〜362)。
【0092】
本発明の分子を用いた適切な細胞宿主の形質転換は、よく知られている方法によって行い、これは典型的には、使用する分子の型に依存する。原核生物宿主細胞の形質転換に関しては、例えばCohen他、(1972) Proc.Natl.Acad.Sci.USA 69、2110、およびSambrook他、(1989) Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Laboratory、Cold Spring Harbor、NYを参照のこと。酵母菌細胞の形質転換は、Sherman他、(1986) Methods In Yeast Genetics、A Laboratory Manual、Cold Spring Harbor、NY中に記載されている。Beggs (1978) Nature 275、104〜109の方法も有用である。脊椎動物細胞に関しては、このような細胞をトランスフェクトする際に有用な試薬、例えばリン酸カルシウムおよびDEAE-デキストランまたはリポソーム配合物は、Stratagene Cloning Systems、またはLife Technologies Inc.、Gaithersburg、MD20877、USAから入手可能である。植物細胞および全植物に関しては、以下の植物形質転換手法(J.DraperおよびR.Scott、D.Grierson(ed.)、「Plant Genetic Engineering」、Blackie、Glasgow and London、1991、vol.1、pp38〜81中)を使用することができる:
i)DNA仲介の遺伝子の移送、ポリエチレングリコール刺激型の、プロトプラストへのDNAの取り込みによる、エレクトロポレーションによる、あるいはプロトプラストまたは植物細胞のマイクロインジェクション(J.Draper、R.Scott、A.KumarおよびG.Dury、ibid.、pp161〜198)。プロトプラストへの直接的な遺伝子の移送は、Neuhaus & Spangenberg (1990) Physiol.Plant 79、213〜217; Gad他、(1990) Physiol.Plant 79、177〜183;およびMathur & Koncz (1998) Method Mol.Biol.82、267〜276中にも記載されている;
ii)粒子の衝撃を使用する形質転換(D.McCabeおよびP.Christou、Plant Cell Tiss.Org.Cult.、3、227〜236 (1993); P.Christou、Plant J.、3、275〜281(1992))。
【0093】
好ましい技法には、エレクトロポレーション、マイクロインジェクションおよびリポソーム配合がある。
【0094】
幾つかの種は、直接的な形質転換の影響を受けやすく、組織または細胞培養の必要性が回避される(Bechtold他、(1993) Life Sciences、C.R.Acad.Sci.Paris316、1194〜1199)。
【0095】
全ての手法において、カナマイシンまたは除草剤耐性剤などの、適切な選択マーカー、あるいは代替的にスクリーニング可能なマーカー(「レポーター」)遺伝子、β-グルクロニダーゼまたはルシフェラーゼなどが好ましい(J.DraperおよびR.Scott、D.Grierson(ed.)、「Plant Genetic Engineering」、Blackie、Glasgow and London、1991、vol.1 pp38〜81中を参照)。
【0096】
エレクトロポレーションは、細胞を形質転換および/またはトランスフェクトするため、酵母菌細胞、細菌細胞、昆虫細胞、脊椎動物細胞および幾つかの植物細胞(例えばオオムギ細胞、Lazzeri (1995) Methods Mol.Biol.49、95〜106を参照)を形質転換するためにも有用であり、当分野でよく知られている。
【0097】
例えば、多くの細菌種は、参照により本明細書に組み込まれているLuchansky他、(1988) Mol.Microbiol.2、637〜646中に記載された方法によって、形質転換することができる。最大数の形質転換体が、25μFDで1cm当たり6250Vを使用する、2.5×PEBに懸濁させたDNA-細胞混合物のエレクトロポレーション後に、一貫して回収される。
【0098】
エレクトロポレーションによって酵母菌を形質転換するための方法は、Becker & Guarente (1990) Methods Enzymol.194、182中に開示されている。
【0099】
首尾よく形質転換された細胞、すなわち本発明の分子を含む細胞は、よく知られている技法によって同定することができる。例えば、標識したオリゴおよび/またはGFPマーカーを使用することができる。
【0100】
したがって、形質転換された宿主細胞そのもの以外にも、本発明は、これらの細胞の培養物、好ましくはモノクローナル(クローン的に均一な)培養物、または栄養培地中のモノクローナル培養物由来の培養物も企図する。
【0101】
細胞の性質のため(例えば、細胞は組換え細胞であり、例えばその細胞中に、本発明の分子が組換えコピーと結合しない点で標的遺伝子と異なる、標的遺伝子の組換えコピーが存在するので)、あるいは細胞/培養物を増殖または維持する条件が、細胞中のアポトーシスを誘導する条件ではないため、形質転換された細胞または培養物は、本発明の分子がアポトーシスを促進しない、細胞または培養物であってよいことは理解されよう。
【0102】
植物に関して、本発明は、単細胞由来の細胞の懸濁液培養物、単離プロトプラストまたは安定した形質転換型植物を含むことが想定される。
【0103】
本発明の分子は任意の適切な宿主細胞中に導入することができるが、適切な動物または植物細胞、特に本発明の分子が結合することができるそれらのDNA内に1つまたは複数の部位を有する細胞中において有効であるように、本発明の分子が本来設計されていることは理解されよう。
【0104】
したがって、その存在が特定の遺伝子の発現を抑制する、本発明の分子を含む動物または植物細胞、あるいはこれらの細胞を含む動物または植物は、遺伝子がもはや発現することができないという意味で「ノックアウトされた」遺伝子を有すると考えることができる。ヒストン脱アセチル化によるクロマチン不活性化は、他の介入がないほぼ不可逆的なものとし得る。当業者に知られているように、ヒストン脱アセチル化による抑制は、ヒストン脱アセチル化酵素の阻害剤、例えばトリコスタチンA(TSA)、トラポキシンンまたは酪酸ナトリウム(NaB)などを使用して逆行させることができる。同様にメチル化は、他の介入、例えばメチル化阻害剤/逆行物質の投与がないほぼ不可逆的なものである可能性があり、メチル化阻害剤/逆行物質は当分野では知られており、化合物アザシチジンを含む。他のメチル化阻害剤には、5デオキシアザシチジン、または例えば、DNAメチルトランスフェラーゼを対象とするアンチセンスオリゴ(または本明細書に記載する遺伝子発現抑制因子)がある。
【0105】
本発明の分子を動物または植物細胞中に導入することによって、核酸、例えばDNA内の適切な結合部位(ポリペプチドのDNA結合部分によって結合する)への分子の標的化を可能にし、遺伝子発現の抑制または不活性化、または他の調節を、例えば標的結合部位のクロマチン、または標的結合部位と結合したクロマチンを不活性化させることによって、可能にするであろうことは容易に理解されよう。典型的には、選択した遺伝子を標的化するように、本発明の分子を選択する。したがって、適切には、標的遺伝子は、それと関係がある分子のDNA結合部分によって結合した部位を有する。このように結合する部位は、遺伝子自体の中、例えば遺伝子のイントロンまたはエクソン内に存在してよく、あるいは部位は、遺伝子の転写された部分の5'領域中、例えばプロモーターまたはエンハンサー領域内あるいはその近辺に存在してよく;あるいは部位は、遺伝子の転写された部分の3'領域中に存在してよい。
【0106】
選択したアポトーシス関連遺伝子の発現を調節、特に抑制する能力は、生物学の多くの領域において有用である。
【0107】
典型的には、その発現が抑制される遺伝子が動物細胞中に存在するとき、その動物細胞は動物内の細胞であり、本発明の方法を使用して、動物(それはヒトまたは非ヒト動物であってよい)中において選択したアポトーシス関連遺伝子の発現を調節、例えば抑制し、それによってアポトーシスを調節、例えば促進する。動物細胞における個々の使用の例には、Bcl-2、Bcl-XI(Bcl-2ファミリーのメンバー)またはAKT/PKB(タンパク質キナーゼB;さまざまな対立遺伝子)の不活性化がある。これらの遺伝子は、例えばVivanco & Sawyers (2002) The phosphatidylinositol 3-kinase-AKT pathway in human cancer.Nature Reviews Cancer、p489; Cory & Adams (2002) The Bcl-2family:regulators of the ceullular life-or-death switch中に記載されたように、アポトーシスの制御と関係がある。
【0108】
Bcl2、AKTおよびBCI-XI遺伝子に関する寄託番号には、以下のものがある:Bcl2:NM_000657およびNM_000633、AKT:NM_005163、Bcl-XL:NM_138578。
【0109】
さらに典型的には、植物細胞は植物内の細胞であり、本発明の方法を使用して、植物中のアポトーシス関連の選択した遺伝子の発現を抑制する。これは例えば、侵入する病原体に対する防御機構において有用である可能性がある(Mittler R.他、(1996) Inhibition of Programmed Cell Death in Tobacco Plants during a Pathogen-Induced Hypersensitive Response at Low Oxygen Pressure.Plant Cell、8、p.1991)。
【0110】
適切には、本発明の方法を使用して、その発現を調節、抑制または不活性化することが望ましい、アポトーシス関連遺伝子の発現を調節、抑制または不活性化する。その発現を抑制または不活性化することが望ましい遺伝子には、アポトーシス救済遺伝子、例えばAKT/PKB、Bcl-XIまたはBcl-2、またはプロウイルスゲノム中に存在する遺伝子を含めたウイルス遺伝子があり、したがって、動物に関するこの方法は、医学的治療の方法を構成することができる。さらに、腫瘍遺伝子は幾つかの癌では過剰発現する可能性があり、他のアポトーシス促進調節と組み合わせて、それらの発現を抑制することが望ましい可能性がある。幾つかの腫瘍遺伝子は、活性化突然変異を有するために腫瘍原性である。
【0111】
突然変異腫瘍遺伝子の発現の選択的抑制は、突然変異腫瘍遺伝子の配列と選択的に結合する、DNA結合部分を使用して得ることができ、抑制因子または修飾部分、例えばクロマチン不活性化部分は、例えば腫瘍遺伝子がその中に存在するかあるいはそれと関係があるクロマチンを不活性化することによって、突然変異腫瘍遺伝子の発現を抑制する。腫瘍遺伝子の過剰発現、または突然変異(非常に活性化した)腫瘍遺伝子の発現を抑制することが、腫瘍遺伝子が役割を果たす癌の治療において、一般に望ましい。標的化することができる突然変異腫瘍遺伝子にはRasおよびBcl-10がある。これらは、配列特異的な方式で突然変異した遺伝子を認識することができるDNA結合部分によって、標的化することができる。
【0112】
アポトーシスはプログラムされた細胞死であり、これはさまざまな刺激によって誘導することができる。例えば、多くの化学療法剤は、アポトーシスの誘導によって癌細胞を除去する。腫瘍の発達および進行には、充分な増殖およびアポトーシスの阻害が必要であると考えられる。現在の治療プロトコルに対する原発性または後天性の耐性は、臨床腫瘍学における主な関心事のままであり、アポトーシスプログラムの欠陥によって引き起こされる可能性があり、化学療法に対する耐性は、部分的には低いアポトーシス率によることが知られている。幾つかの遺伝子は、アポトーシスの制御と関係があることが報告されてきており、例えば、bcl-2原-腫瘍遺伝子の発現は、さまざまなヒト血液悪性腫瘍および固形腫瘍中で見られる。Bcl-2タンパク質は、腫瘍細胞が放射線、化学療法、およびホルモン療法によって誘導されるアポトーシスを受けるのを防ぐことによって、その腫瘍原的役割を発揮する。したがって、アポトーシスの防止と関係がある、遺伝子の発現を妨げることを目的とする、療法を開発することが望ましい。
【0113】
本発明のこれらの方法は、本発明の分子を動物細胞または植物細胞に移送させることを、典型的には含む。
【0114】
オリゴヌクレオチドまたはオリゴヌクレオチド-ペプチド融合体を用いる有用な移送系が当業者に知られ、本発明の分子を動物の身体中または外の細胞に導入する、本発明の方法を実施する際に、有用である可能性がある。例えば、リポソームまたはウイルス系の方法を使用することができる。エレクトロポレーション(例えばKuznetsova他、(1999) Nucl Acids Res 27 (20)、3995〜4000を参照)、衝撃法、カチオン性脂質(例えばFelgner他、(1997) Hum Gene Ther 8、511〜512またはWO99/13719中に記載されたのと同様の)、または分子のオリゴヌクレオチドまたはポリペプチド部分と結合した特異的リガンド、または担体を、例えばWO99/13719中に記載されたのと同様に使用することができる。
【0115】
ウイルスまたは非ウイルス移送法を使用することができる。パラボウイルス、例えばSV40(Madzak他、(1992) J.Gen.Virol.73、1533〜1536)、アデノウイルス(Berkner (1992) Curr.Top.Microbiol.Immunol.158、39〜61; Berkner他、(1988) BioTechniques 6、616〜629; GorzigliaおよびKapikian (1992) J.Virol.66、4407〜4412; Quantin他、(1992) Proc.Natl. Acad. Sci.USA89、2581〜2584; Rosenfeld他、(1992) Cell 68、143〜155; Wilkinson他、(1992) Nucleic Acids Res.20、2233〜2239; Stratford-Perricaudet他、(1990) Hum.Gene Ther.1、241〜256)、ワクシニアウイルス(Moss (1992) Curr.Top.Microbiol.Immunol.158、25〜38)、アデノ随伴ウイルス(Muzyczka (1992) Curr.Top.Microbial.Immunol.158、97-123; Ohi他、(1990) Gene89、279〜282)、HSVおよびEBVを含めたヘルペスウイルス(Margolskee (1992) Curr.Top.Microbial.Immunol.158、67〜90; Johnson他、(1992) J.Virol.66、2952〜2965; Fink他、(1992) Hum.Gene Ther.3、11〜19; BreakfieldおよびGeller (1987) Mol.Neurobiol.1、337〜371; Freese他、(1990) Biochem.Pharmacol.40、2189〜2199)、ならびに鳥類(BrandyopadhyayおよびTemin (1984) Mol.Cell.Biol.4、749〜754; Petropoulos他、(1992) J.Virol.66、3391〜3397)、ネズミ(Miller (1992) Curr.Top.Microbiol.Immunol.158、1〜24; Miller
他、(1985) Mol.Cell.Biol.5、431〜437; Sorge他、(1984) Mot.Cell.Biol.4、1730〜1737; Mann and Baltimore (1985) J.Virol.54、401〜407; Miller他、(1988) J.Virol.62、4337〜4345)、およびヒト起源(Shimada他、(1991) J.Clin.Invest.88、1043〜1047; Helseth他、(1990) J.Virol.64、2416〜2420; Page他、(1990)J.Virol.64、5370〜5276; BuchschacherおよびPanganiban (1992) J.Virol.66、2731〜2739)のレトロウイルスを含めた、幾つかのウイルスが、遺伝子移送ベクターとして使用されてきている。今日まで、大部分のヒト遺伝子療法プロトコルは、欠陥のあるネズミレトロウイルスに基づいてきている。
【0116】
当分野で知られている非ウイルス性の遺伝子の移送法には、リン酸カルシウム共沈殿などの化学技法(Grahamおよびvan der Eb (1973) Virology52、456〜467;Pellicer他、(1980) Science209、1414〜1422);機械的技法、例えばマイクロインジェクション(Anderson他、(1980) Proc.Natl.Acad.Sci.USA77、5399〜5403; Gordon他、1980; Brinster他、(1981)Cell27、223〜231; ConstantiniおよびLacy (1981) Nature 294、92〜94);リポソームによる膜融合仲介型移送(Felgner他、(1987) Proc.Natl.Acad.Sci.USA 84、7413〜7417; WangおよびHuang (1989) Biochemistry 28、9508〜9514; Kaneda他、(1989) J.Biol.Chem.264、12126〜12129; Stewart他、(1992) Hum.Gene Ther.3、267〜275; Nabel他、1990; Lim他、(1992) Circulation 83、2007〜2011);および直接的なDNAの取り込みおよび受容体仲介型DNA移送(Wolff他、(1990) Science 247、1465〜1468; Wu他、(1991) J.Biol.Chem.266、14338〜14342; Zenke他、(1990) Proc.Natl.Acad.Sci.USA 87、3655〜3659; Wu他、1989b; Wolff他、(1991) BioTecliniques 11、474〜485; Wagner他、1990; Wagner他、(1991) Proc.Natl.Acad.Sci.USA 88、4255〜4259; Cotten他、(1990) Proc.Natl.Acad.Sci.USA 87、4033〜4037; Curiel他、(1991a) Proc.Natl.Acad.Sci.USA88、8850〜8854; Curiel他、(1991b) Hum.Gene Ther.3、147〜154)がある。ウイルス仲介型の遺伝子の移送は、リポソーム送達を使用
する直接的なin vivoの遺伝子の移送送達と組み合わせることができ、ウイルスベクターを、周囲の非分裂細胞ではなく腫瘍細胞に向けることが可能である。
【0117】
他の適切な系には、Feng他、(1997) Nature Biotechnology 15、866〜870によって記載されたレトロウイルス-アデノウイルスハイブリッド系、または適切な一本鎖Fv断片などの標的リガンドを用いるウイルス系がある。
【0118】
生物学的な遺伝子の移送法と物理学的な遺伝子の移送法を組み合わせる手法では、任意の大きさのプラスミドDNA(または例えばオリゴヌクレオチド/ペプチド融合体)を、アデノウイルスヘキソンタンパク質に特異的なポリリシン結合抗体と組み合わせ、生成した複合体をアデノウイルスベクターと結合させる。三分子の複合体を次いで使用して、細胞を感染させる。アデノウイルスベクターは、結合したDNAが損傷する前に、エンドソームの効率の良い結合、内在化、および分解を可能にする。
【0119】
Ebbinghaus他、(1996) Gene Ther 3(4)、287〜297は、アデノウイルス-ポリリシン複合体を使用して、TFOを細胞に送達することができる方法を記載している。Pichon他、(2000) Nucl Acids Res 28(2)は、ヒスチジル化オリゴリシンを使用して、オリゴヌクレオチドの取り込み、サイトゾル送達および核内蓄積を改善することができる方法を記載している。
【0120】
「Chariot」系(Active Motif、Morris他、(2001) Nature Biotech 19、1173〜1176)を使用することができる。非共有結合複合体が送達させる分子を用いて形成され、この複合体は細胞中に効率良く内在化される。
【0121】
リポソーム/DNA複合体は、直接的なin vivoの遺伝子の移送を仲介することができることが示されてきている。標準的なリポソーム調製では、遺伝子の移送プロセスは非特異的であり、例えば直接的なin situ投与の後の腫瘍堆積において、局所的なin vivoの取り込みおよび発現が報告されてきている (Nabel (1992) Hum.Gene Ther.3、399〜410)。
【0122】
分子を直接標的細胞または組織に誘導する、遺伝子の移送技法が好ましい。受容体仲介の遺伝子移送は、例えば、ポリリシンを介したDNAとタンパク質リガンドの結合によって行われる。標的細胞/組織型の細胞表面上の、対応するリガンド受容体の存在に基づいて、リガンドが選択される。これらのリガンド-DNA結合体は、望むならば血液中に直接注射することができ、受容体結合およびDNA-タンパク質複合体の内在化が起こる標的組織に向けられる。DNAの細胞内破壊の問題を克服するために、アデノウイルスによる混合感染を、エンドソーム機能を害するために含めることができる。
【0123】
選択した遺伝子の発現を抑制または調節する方法を使用して、ヒト細胞中の遺伝子の発現を抑制する(または調節する)ことが好ましく、1つの特に好ましい実施形態では、ヒト細胞はヒトの身体中に存在する。
【0124】
しかしながら、本発明の方法は、動物の身体外の(ヒト細胞を含めた)動物細胞の改変(すなわち細胞のex vivo処理)を含むことができ、そのように改変した細胞を、動物の身体中に再導入することができる。
【0125】
本発明の方法は、例えば潜在的な薬剤標的を同定する際、あるいはおそらく薬剤として有用な活性または性質に関して候補化合物をスクリーニングする際の、改変された細胞のin vitroの調査または特徴付けも含むことができる。
【0126】
本発明の分子、本発明の方法を使用して、特に化学療法剤耐性、または腫瘍発達の他の態様、または炎症の消散、または(他の形の制御された細胞死を含めた)アポトーシスが関与すると考えられる、他のプロセスにおける、遺伝子の役割を分析することができる。
【0127】
本発明の他の態様は、(好ましくは真核生物)細胞中における、選択したアポトーシス関連遺伝子の発現を抑制(または調節)するための物質の製造における、本発明の分子の使用を提供する。選択する遺伝子は、内因性遺伝子であることが好ましい。本発明の前の態様に関して前に示した、他の好ましい事項も当てはまる。
【0128】
動物中における選択したアポトーシス関連遺伝子の発現を抑制(または調節)するための、薬剤の調製において分子を使用する場合、それが非常に好ましいことは理解されよう。誤解を避けるために、「動物」によって本発明者らは、ヒトおよび非ヒト動物を含むものとする。
【0129】
本発明の他の態様は、選択したアポトーシス関連遺伝子の発現の抑制(または調節)を必要とする患者を治療する方法を提供し、この方法は、本発明の分子を有効量、患者に投与することを含む。
【0130】
本発明の他の態様は、アポトーシスの促進を必要とする患者を治療するための方法を提供し、この患者は組合せ法で治療し、この方法は、細胞死誘導物質および本発明の分子を有効量、患者に投与することを含む。腫瘍学の分野の当業者によく知られているように、細胞死誘導物質は、化学療法剤、または治療剤、例えば放射線治療剤であってよい。
【0131】
選択した遺伝子の発現または過剰発現が望ましくなく、患者の病状に貢献する場合、その選択した遺伝子の発現を抑制することが有用であることは理解されよう。遺伝子の望ましくない発現の例には、癌中の幾つかの活性化した腫瘍遺伝子の発現、およびアポトーシス阻害と関係がある遺伝子の発現がある。選択した遺伝子の不充分な発現の欠如が望ましくなく、患者の病状に貢献する場合、選択した遺伝子の発現の増大が有用である可能性がある。例えば、腫瘍抑制遺伝子またはアポトーシス促進遺伝子の不充分な発現は、癌に貢献する可能性があり;したがって、腫瘍抑制遺伝子またはアポトーシス促進遺伝子の、発現を増大させることが有用である可能性がある。
【0132】
Bcl-2ファミリーの遺伝子の発現の抑制は、さまざまな癌を治療する際に望ましい。同様に、プログラムされた細胞死を調節するタンパク質をコードする他の遺伝子、例えばBcl-XIまたはAktの発現の抑制は、癌を治療する際に望ましい。
【0133】
本発明の他の態様は、抑制(または調節)を必要とする患者の中での、選択した遺伝子を抑制(または調節)するための薬剤の製造における、本発明の分子の使用を提供する。
【0134】
本発明のさらに他の態様は、医学において使用するための本発明の分子を提供する。したがって、本発明の分子をパッケージ化して、医薬における使用を示す。
【0135】
本発明のさらに他の態様は、本発明の分子、および薬学的に許容される担体を含む薬剤組成物を提供する。
【0136】
「薬学的に許容される」とは、配合物が滅菌状態で発熱物質を含まないことが含まれる。適切な薬剤担体は、製薬の分野においてよく知られている。
【0137】
ここで本発明を、以下の図面および実施例を参照しながら、より詳細に説明する。
【実施例】
【0138】
(実施例1)
オリゴ-調節因子ペプチド融合分子の構築および使用
遺伝子調節分子として有用な、一連のオリゴヌクレオチド結合体を作製した。これらは、少なくとも2つの特異的な部分または単位、すなわち、選択した二本鎖標的配列とDNA三重らせんを形成することができるオリゴヌクレオチド(三重鎖形成オリゴヌクレオチド、すなわちTFO;単位1);および遺伝子抑制因子または活性化因子由来の別のペプチド配列(単位2)からなる。TFOは、抑制因子または活性化因子ペプチドと融合させる。
【0139】
一例として、TFOを設計して、Bcl-2プロモーター領域との三重鎖を形成する(受託番号:NM_000657およびNM_0006333)。選択する領域は1つのDNA鎖上に非常にプリンが豊富であり、したがって、フーグスティーン塩基対によってDNA三重鎖を形成するための候補配列である。可能なTFOを設計するためのルールは、Vasquez KMおよびWilson JH、Trends Biochem Sci、1:4〜9、1998中に要約されている。配列(5'GGGTGTGGGGTUTGTGTGTGGT3'(Bc1P)または5'TUGTGTGGGTGTGGTGUGGG3'または5'GGTGTUTTGGTTGGGTGT3'(BcIU))を、単位2ペプチドとの化学結合用の活性化5'端を有する、オリゴヌクレオチド(単位1)として生成させた。この試験で調べた単位2ペプチドは、ヒトMAD1転写抑制ドメイン(例えばアミノ酸XXXMNIQMLLEAADYLERREREAEHGYASMLP(前式でXXXは、例えばAAAまたはDDD連結基である))を含む。後者は、ヒストン脱アセチル化酵素複合体タンパク質Sin3aと、相互作用することが知られている領域である。さらに本発明者らは、Sin3aタンパク質と結合することも知られているヒトSap18タンパク質の、アミノ酸XXXMAVESRVTQEEIKKEPEKPIDREKTCPLLLRVF(前式でXXXは、例えばAAAまたはDDD連結基である)の使用を調べている。この領域は充分に進化保存された配列に対応し、遺伝子抑制を仲介することができる領域と重複する。単位2ペプチドは活性形で合成して、N末端チオエステル官能化ペプチドを5'-システイニルオリゴヌクレオチドと結合させる、「本来の結合」化学(WO01/15737およびStetsenko & Gait (2000) Organic Chem 65(16)、4900〜4908を参照のこと)による活性単位1オリゴヌクレオチドとのその後の結合を可能にする。
【0140】
トランスフェクション作業用の細胞系には、内因性Bcl-2遺伝子を発現する、前立腺癌LnCap細胞および膵臓癌PT45細胞があった。さまざまな量のオリゴヌクレオチド結合体を用いる、標準的なリポソーム系トランスフェクション法(あるいは代替として他の送達法、例えばエレクトロポレーションまたはマイクロインジェクション)を使用して、細胞を一時的にトランスフェクトした。特異性の対照として、細胞はオリゴヌクレオチド、または非特異的オリゴペプチド結合体でも処理した。適切な時間、例えば0.5、1、2、4、8、12、24および/または36時間後、細胞計数または以下に記載する他の方法を使用して、細胞の生存能力を測定した。
【0141】
細胞の生存能力を低下させる、さまざまなオリゴペプチド結合体の能力を調べた。高濃度の融合分子オリゴ-MAD1の送達は、濃度依存式に細胞死を増大させる。TFO(すなわちペプチドドメイン含まず)およびオリゴ-ペプチド融合分子を細胞に送達した後では、細胞死は、融合分子のみを用いて見られた細胞死より少なくなった、すなわち低濃度の融合分子を用いて見られる細胞死と同等であった。この結果は、抑制因子ペプチドを含む分子は、ペプチドドメインを含まないTFOより有効な遺伝子活性の調節因子であり、TFOは標的配列との結合に関して、オリゴ-ペプチド融合分子と競合する可能性があることを示す。オリゴおよび非特異的結合体分子には細胞殺傷効作用がなく、Bcl-2制御型細胞の生存能力におけるオリゴペプチド結合体の特異性が実証された。
【0142】
この実施例は、DNA結合オリゴヌクレオチドおよび機能性ペプチドからなる融合分子の設計および構築、ならびに細胞中へのそれらの送達を実証する。オリゴペプチド結合体は、標的方式で特異的な生物学的応答を誘導することができる。したがって、生物活性の強力な調節因子である、オリゴペプチド結合体を設計することができる。
【0143】
(実施例2)
オリゴ-調節因子ペプチド融合分子による染色体遺伝子の抑制
このような遺伝子をゲノム中に組み込むと、融合分子は遺伝子活性を調節することができる。MAD1と融合したDNA結合オリゴペプチド(TFO)を含む融合分子を、実施例1に記載したのと同様に設計し構築した。
【0144】
融合分子を細胞に送達し、Bcl-2遺伝子活性を測定するために実験を行った。
【0145】
Bcl-2遺伝子活性を抑制する、さまざまなオリゴペプチド結合体の能力を調べた。高濃度の融合分子オリゴ-MAD1の送達は、mRNAにより測定して、濃度依存式に、Bcl-2遺伝子活性を抑制した。TFO(すなわちペプチドドメイン含まず)、およびオリゴ-ペプチド融合分子を細胞に送達した後、遺伝子の抑制は、融合分子のみを用いて見られた抑制よりも低くなった、すなわち低濃度の融合分子を用いて見られる抑制と同等であった。この結果から、抑制因子ペプチドを含む分子が、ペプチドドメインを含まないTFOより有効な遺伝子活性の調節因子であることが示される。オリゴおよび非特異的結合体分子には細胞殺傷効果がなく、遺伝子調節におけるオリゴペプチド結合体の特異性が実証された。
【0146】
さらに、抑制を異なる時間で測定して、抑制因子効果の時間行程を確定した。融合分子は、TFO単独より有効な抑制因子であった。この効果も特異的であった(例えば、非融合オリゴヌクレオチドおよび非特異的分子には、オリゴ-ペプチド融合分子と同じ効果はなかった)。さらに、融合分子による抑制は、TFO単独によって見られた任意の抑制より後の時間地点で見られ、より持続的な効果が示唆された。
【0147】
したがって、実施例1に記載した融合分子は、染色体遺伝子の活性を調節することができる。DNA結合オリゴヌクレオチド標的部分を有する融合分子は、特定の染色体遺伝子を標的化することができる。DNA結合オリゴヌクレオチドと融合したGeneICE抑制因子ペプチドは、所定の染色体遺伝子の活性を抑制することができる。したがって、記載した融合分子は、染色体遺伝子の活性の強力な調節因子である。
【0148】
例えば遺伝子の転写を評価することによって(例えばPCRを使用して)、あるいはコードされているポリペプチドの量または活性を評価することによって、内因性遺伝子の調節を測定する。一例では、オリゴヌクレオチドは、Bcl-2遺伝子調節部位を対象とする。適切なRT-PCRプライマーの例は、以下のものを含む。
5'TCCGGTATTCGCAGAAGTCC3'
5'ATCAGAAGAGGATTCCTGCC3'
(Bc1Pを評価するために使用)および
5'TGATGGAGCTCAGAATTCC3'
5'TGCCTCTCCTCACGTTCC3'
(Bc1Uを評価するために使用)。
【0149】
一例では、前立腺癌細胞系LnCapおよび膵臓癌細胞系PT45を、実施例1に記載したMAD1を含むオリゴヌクレオチド-ペプチド融合体で処理する。トランスフェクトした細胞を、場合によっては同定および/または単離し(例えばGFPマーカーおよびFACS技法を使用する)、Bcl-2遺伝子の発現に関してアッセイする。
【0150】
例えば、GeneICE Bcl-2を用いて細胞をトランスフェクトした。12、24、48および72時間後、逆転写酵素PCRによってmRNAレベルを測定した。
【0151】
(実施例3)
標的配列およびヒストン脱アセチル化酵素複合体と結合する融合分子
この実施例は、融合分子が特定の標的配列、ヒストン脱アセチル化酵素複合体の成分と結合することを実証する。オリゴヌクレオチド(TFO)および抑制因子ペプチドを含む融合分子を、実施例1に記載したのと同様に作製した。
【0152】
異なる融合分子を、融合体のオリゴ部分と相補的にした、標識したオリゴヌクレオチドと共にインキュベートした。同じ融合分子も、ヒストン脱アセチル化酵素複合体の成分であるSin3と共にインキュベートした。さらに、これらの融合体を、相補的オリゴヌクレオチドおよびSin3タンパク質ともインキュベートした。
【0153】
次いでこれらの複合体を、標準的なバンド移送分析法によって分析した。融合分子は、標識した相補的オリゴヌクレオチドおよびSin3タンパク質と、別々かつ同時に結合することができた。非特異的な融合体は、標識したオリゴまたはSin3と結合することができず、したがって抑制因子融合体を用いた影響の特異性が実証された。
【0154】
抑制因子融合体は、それらの標的配列と特異的に結合することができると結論付けすることができる。抑制因子融合体は、この複合体の一部分であるタンパク質と結合し、それらの標的配列と結合し、ヒストン脱アセチル化酵素複合体を同時に補うことによって、ヒストン脱アセチル化酵素複合体を補うことができ、記載する融合分子は、遺伝子活性の非常に強力で特異的な抑制因子である。
【0155】
(実施例4)
標的確認プロトコル
当該の遺伝子の利用可能なDNA配列(側面配列含む)を分析して、オリゴ/ペプチドを向けるのに適切な部位を選択する。オリゴ/ペプチドを合成し、例えばレポーター遺伝子系を使用して、目的とする細胞または動物またはヒトにおいて使用する前に、オリゴ/ペプチドを試験することができる。オリゴ/ペプチドを細胞中、in vitroまたは動物またはヒト中において使用し、さらに試験することができる。
【0156】
ひとたび遺伝子配列を提供した後、プロセスは以下のことを含む。
当該の遺伝子(必要な場合、側面配列含む)を、生物情報科学データマイニングツール(例えば、Perkins他、(1998) Biochemistry 37、11315〜11322中で使用されたthe Genetics Computer Group(GCG)プログラム)を使用して、ヒトゲノム中の他の場所では見られないユニークな配列エレメントに関して調べる。(例えばTFOとして)同定した独特の配列と結合すると予想される、核酸系DNA結合分子を、設計し合成する。
【0157】
DNA結合分子はオリゴヌクレオチドである可能性があり、以下の特徴を有することは好ましい:
(a)少なくとも16ヌクレオチド長
(b)遺伝子のプロモーター、あるいは遺伝子の転写開始部位またはその近傍を標的化する。
【0158】
TFOと結合するための標的部位は、1本の鎖中にプリンが多量であることが好ましい。
【0159】
TFOはピリミジン多量(主にCまたはT);プリン多量(主にGまたはA)または混合型(主にGまたはT、またはG、AまたはT)であってよい。CTTFOは平行モチーフ中で結合すると考えられ、その中では第三の鎖(TFO)が、二本鎖のプリン鎖と同じ5'〜3'方向を有する。GATFOは逆平行モチーフ中で結合すると考えられ、その中ではTFOはプリン鎖に対して反対を向いている。混合型TFOは、標的配列に応じて、平行モチーフまたは逆平行モチーフ中で結合することができる。塩基対は、平行三重鎖(T:AT、C+:GCおよびG:GC)中のフーグスティン水素結合、および逆平行三重鎖(G:GC、A:ATおよびT:AT)中の逆フーグスティン水素結合の形成から生じる。
【0160】
オリゴヌクレオチドは、安定した化学修飾基をおそらく有するDNAオリゴヌクレオチドであると考えられる。他の塩基、例えばN6-メチル-8-オキソ-2-デオキシアデニンをシトシンの代わりに、2-デオキシ-6-チオグアニンをグアニンの代わりに、あるいは7-デアザ-2-デオキシキサンチンをチミンの代わりに使用することができる。
【0161】
抑制因子ペプチドまたはペプチドは、ペプチド合成装置を使用して大量に生成することができ、使用するまで凍結保存することができる。
【0162】
抑制因子ペプチド-DNA結合分子構築体を、調製し精製する。使用する化学物質は、WO01/15737中に記載されたものであってよい。キットはLink Technologiesから入手可能である。
【0163】
構築体は、質量分光法によって、かつ/あるいは(例えば蛍光、化学発光または酵素標識を使用して)標識した相補的オリゴヌクレオチド、または標識した抗体成分を使用することによって、品質管理することができる。典型的には、このような方法では、構築体を固形担体に加え、その上で、構築体のペプチド部分と結合する抗体を固定し、構築体のオリゴヌクレオチド部分と結合する標識したオリゴヌクレオチドを加える。この方法では、固形担体と結合した標識を検出することによって、構築体が完全であることが実証される。他の典型的な形式では、オリゴヌクレオチドは固形担体と結合することができ、抗体を標識することができる。
【0164】
レポーター遺伝子構築体を、当該の遺伝子用に作製することができる(しかしながら、これは一般に必要ではない)。
・候補DNA結合オリゴヌクレオチドまたはオリゴ/ペプチドは、以下の事項に関して試験することができる:
・標的配列に対する結合の親和性;
・全ゲノムDNAチップに陳列させることによる結合の特異性。
・オリゴ/ペプチドは、レポーター遺伝子系を使用して、有効性に関して試験することができる。
【0165】
したがってオリゴ/ペプチドは、当該の細胞または動物中の当該の遺伝子を、調節または抑制するために使用することができる。
【0166】
(実施例5)
標的確認
オリゴ/ペプチド融合分子を使用して、アポトーシス関連薬剤標的を確認することができる。これは以下のことを含むことができる:
・実施例1で述べたプロトコルを実施する。
・構築体の細胞または組織への送達。これらは、当該の分子の研究に適切な、正常または疾患組織、細胞系または初代細胞であってよい。
・任意の発現分析法による表現型の分析;または細胞の運動性の評価などの任意の機能分析、増殖またはアポトーシス分析。
・個々の疾患の任意の入手可能なデータとの比較、および細胞死または運動性などの望ましい効果の分析。
【0167】
得られたデータを使用して、薬剤開発プログラムの所定の薬剤標的を確認する。
【0168】
(実施例6)
患者治療の実施例
アポトーシス関連遺伝子、例えばBcl-2を標的とする、オリゴ/ペプチド融合体を、記載するように作製する。融合分子を滅菌環境で調製し、リポソームに配合する。融合体含有リポソームを、腫瘍、例えば乳または前立腺腫瘍の近くに向ける。リポソームは癌細胞によって採取し、アポトーシスをそれぞれの細胞において選択的に促進させる。
【0169】
(実施例7)
標的同定スクリーニング
オリゴ/ペプチド融合分子を使用して、アポトーシス関連薬剤標的を同定することができる。これは以下のことを含むであろう:
・実施例1で述べたのと同様の、融合分子の調製。
・融合分子の細胞または組織への送達。これらは、正常または疾患組織、細胞系または初代細胞であってよい。
・DNAアレイを使用する、遺伝子抑制から生じる遺伝子発現概略の分析。これは、モデル中全体の遺伝子発現に対する構築態の影響を示す。
・任意の発現分析法による表現型の分析、または細胞の運動性などの任意の機能分析、増殖またはアポトーシス分析。
【0170】
得られたデータを使用して、乳または前立腺癌などの疾患の、可能性のある薬剤標的を見出す。任意の他の同様のスクリーニング、in vitro法および細胞および動物モデルを含めた適切な方法によって、これらの標的をさらに確認することができる。
【0171】
(実施例8)
オリゴ/ペプチド融合分子による遺伝子発現の下方制御は、クロマチンヒストン脱アセチル化と関係がある
前に示したように、オリゴ/ペプチド融合分子は、遺伝子発現を抑制する際に有効である。これは、オリゴが結合するかあるいはその近くのDNAの、ヒストンアセチル化状態のMAD1仲介の変化によるものであると、本発明者らは考える。
【0172】
低下した遺伝子発現が、クロマチンヒストン脱アセチル化と関係があることを示すために、クロマチン免疫沈降(ChIP)法を使用することができる。使用製造元の指示書(Upstate Biotechnology、Bucks、UK)に従いChIPアッセイキットを使用して、クロマチン免疫沈降を行う。
【0173】
PT45またはBxPC-3膵臓癌細胞を増殖および繁殖させ、アポトーシス関連遺伝子の発現を抑制する際に有用な、オリゴ/ペプチド融合分子と共にインキュベートした。未処理細胞は対照として使用する。
【0174】
35cm組織培養プレート上、フェノールレッドを含まない、5%のDSS、P/S/GおよびG418(100μg/ml)を補ったDMEM中で、細胞を95%コンフルエントになるまで増殖させる。ヒグロマイシンB(80μg/ml)およびドキシサイクリン(1μg/mnl)を適切に加える。固定の30分前に、E2(10〜8M)またはエタノールを(対照として)、細胞に加える。37%ホルムアルデヒドを、1%の最終濃度まで培地に直接一滴ずつ加える。細胞は37℃で10分間インキュベートする。
【0175】
氷上で、培地をプレートから吸引し、1×プロテアーゼ阻害剤(PI)(Sigma、Dorset、UK)を含む氷冷PBSで二回、細胞を洗浄する。採取用に、1×PIを含む1mlの氷冷PBSをプレートに加え、ラバーポリスマンを使用して、予め冷却したミクロ遠心分離チューブに細胞をそぎ落とす。4℃における4分間2000rpmでの遠心分離によって、細胞をペレット状にする。PIを含む400μlの暖めたChIPSDS-溶解バッファー(1%のSDS;10mMのNaEDTA、pH 8.0;50mMのTris-HCl、pH8.1)中に、ペレットを再懸濁させ、氷上で10分間インキュベートする。
【0176】
溶解物を超音波処理して、DNAを200〜1000bpの長さに切る。超音波処理中、サンプルは氷-水ビーカー中に置いて、サンプルを冷たい状態に保ち、それらが分解するのを防ぐ。30秒間隔離して10秒の衝撃4回で、付属のSoniprep150エクスポネンシャルチタン製プローブ(Sanyo-Gallenkamp、Leics、UK)を有するSoniprep150超音波装置を使用して、超音波処理を行う。
【0177】
サンプルを、4℃において10分間13,000rpmの遠心分離にかける。上清は15mlの滅菌ファルコンチューブ中に回収し、3600μlのChIP希釈バッファー(0.01%のSDS;1.1%のTriton-X-100;1.2mMのNaEDTA、pH 8.0;16.7mMのTris-HCI、pH 8.1;167mMのNaCI)を用いて、10倍に希釈する。次いでサンプルを、2.5mlチューブ中の2つの2ml等分試料、1つは抗アセチル化ヒストンH4抗体、ChIPグレード(Upstate Biotechnology、Bucks、UK)とのインキュベーション用、もう1つは抗体を含まない対照としての使用するために分ける。
【0178】
非特異的な基底状態を低下させるために、それぞれの2ml等分試料を、80μlのサケ精子DNA/タンパク質Aアガロース50%スラリー(10mMのTris-HCl、pH 8.0;1mMのNaEDTA、pH 8.0に懸濁させたもの)を、垂直回転プラットホーム(Stuart Scientific、Staffs、UK)上において30分間4℃で加えることによって予め調製する。次いでアガロースビーズを、1000rpmで30秒間の遠心分離によってペレット状にし、上清分画を新たな2.5mlチューブに回収する。免疫沈降抗体は1:500の希釈で最初のサンプルに加えるが、抗体を含まない対照サンプルには加えない。両方のチューブを、垂直回転プラットホーム(Stuart Scientific、Staffs、UK)上において4℃で一晩インキュベートする。
【0179】
60μlのサケ精子DNA/タンパク質Aアガロース-50%スラリーを、回転させながら4℃で1時間、それぞれのチューブと共にインキュベートして、抗体/ヒストン複合体、または抗体を含まない対照の場合は非特異的に結合したタンパク質を回収する。アガロースビーズは、1分間800rpmでの簡単な遠心分離によって、ペレット状にする。上清は新たな2.5mlチューブに慎重に移し、-20℃で保存する。
【0180】
タンパク質Aアガロースビーズ/抗体/ヒストン複合体は、以下に列挙する順序で以下のバッファーそれぞれ1mlを用いて、4℃において垂直回転プラットホーム上で5分間洗浄する。
(a)低塩濃度免疫複合体洗浄バッファー、1回洗浄
(0.1%のSDS;1%のTriton-X-100;2mMのNaEDTA、pH 8.0;20mMのTris-HCl、pH 8.1;150mMのNaCl)
(b)高塩濃度免疫複合体洗浄バッファー、1回洗浄
(0.1%のSDS;1%のTriton-X-100;2mMのNaEDTA、pH 8.0;20mMのTris-HCl、pH 8.1;150mMのNaCl)
(c)LiCl免疫複合体洗浄バッファー、1回洗浄
(0.25MのLiCl;1%のNP40(nonidet);1%のデオキシコレート;1mMのNaEDTA、pH 8.0;10mMのTris-HCl、pH8.1)
(d)1×TE、2回洗浄
(10mMのTris-HCl、1mMのNaEDTA、pH 8.0)
【0181】
ヒストン/免疫複合体は、250μlの新たに調製した溶出バッファー(1%のSDS;O.1MのNaHCO3)を加えることによって、アガロースビーズから溶出させる。サンプルを軽く攪拌して混合させ、垂直回転プラットホーム上において15分間室温でインキュベートする。ビーズは室温において2分間1000rpmの遠心分離にかけ、溶出物は新たなミクロ遠心分離チューブに移す。他の250μlの溶出バッファーを用いて溶出ステップを繰り返し、溶出物を合わせる。
【0182】
5MのNaCl20μlを溶出物に加え、ヒストン-DNA架橋は、少なくとも4時間65℃に加熱することによって元の状態にする。10μlの0.5MNaEDTA、pH 8.0、20μlの1M Tris-HC1、pH 6.5および2μlの10mg/mlプロテイナーゼKを溶出サンプルに加える。IP由来の上清分画において、架橋を元の状態にする。4Oμlの0.5MNaEDTA、pH 8.0、80μlの1M Tris-HC1、pH 6.5および8μlの10mg/mlプロテイナーゼKを上清サンプルに加える。サンプルは45℃で1時間インキュベートして、サンプル中のタンパク質を分解する。
【0183】
20μgのグリコーゲンを不活性担体としてサンプルに加え、次いでサンプルDNAは、フェノール/クロロホルム/イソアミルアルコール抽出およびエタノール沈殿によって回収する。DNAペレットは、PCR反応用に50μlの滅菌水に再懸濁させる。1μlのサンプルおよび35〜40サイクルを、それぞれのPCR増幅用に使用する。コンピュータ系画像分析(NIH画像分析プログラム)を使用して、β-アクチンおよび抗体を含まない対照と比較した、Bcl2PCR産物の相対的レベルを評価する。これによって、他のサンプル内に含まれるそれと比較した、サンプル内に含まれる遺伝子転写産物の相対量を計算することができる。
【0184】
得ることができる結果の型の一例を、図6に示す。
【0185】
沈殿したBcl-2DNAの量、したがって、Bcl2と関係があるクロマチンヒストンタンパク質のアセチル化の程度は、未処理細胞と比較して、Bcl2遺伝子発現が記載する方法により阻害される細胞中では、約75%低下させることができる。
【0186】
(実施例9)
ゲルシフトアッセイ
前の実施例によって、オリゴ/ペプチド融合分子は、標的とする遺伝子の活性を調節することができることが実証された。これは、オリゴが結合するかあるいはその近くのDNAの、ヒストンアセチル化状態のMAD1仲介の変化によるものであると考えられる。
【0187】
ゲルシフトアッセイを使用して、オリゴ/ペプチド融合分子が、標的プロモーターを含むDNA断片と結合することができることを実証する。
【0188】
標的遺伝子配列を含むDNA断片を、オリゴヌクレオチドまたはオリゴ/ペプチド融合体と共にインキュベートする。サンプルに非変性ゲル電気泳動を施して、三重鎖仲介型の光付加体を特徴付けする。オリゴヌクレオチドまたはオリゴ/ペプチド融合体をそれぞれ量を増やして含むサンプル中の、付加体を検出する。
【0189】
このようにして、オリゴ/ペプチド融合分子が、標的プロモーターを含むDNA断片と結合することができることを示すことができる。
【0190】
(実施例10)
核局在シグナルを有するオリゴ/ペプチド融合分子
分子のペプチド部分は、分子を核に向けるための核局在シグナル(NLS)を有する可能性がある。この実施例で使用するペプチドは以下のものである。
a)DDD-MAD1-DDD-NLS、
これは以下のアミノ酸配列を有する:
(連結基)DDDMNIQMLLEAADYLERREREAEHGYASMLPDDDPKKKRK
V(カルボキサミド)
および
b)DDD-NLS-DDD-MAD1、
これは以下のアミノ酸配列を有する。
(連結基)DDDPKKKRKVDDDMNMQMLLEAADYLERREREAEHGYASML
P(カルボキサミド)
【0191】
このNLSは、SV40T-抗原由来の、7アミノ酸(配列PKKKRKV)機能的核局在シグナルである。
【0192】
DDD連結配列、および29アミノ酸のMADIアミノ酸配列は、前の実施例で論じた配列と同じである。
【0193】
DDD-MAD1-DDD-NLS、およびDDD-NLS-DDD-MAD1ペプチド配列が、核を標的化することを実証するために、PT45ヒト膵臓癌細胞を、標準リポフェクタミン2000プロトコルによって、Cy3標識オリゴヌクレオチドと連結した、DDD-MAD1-DDD-NLSおよびDDD-NLS-DDD-MAD1ペプチド配列からなる、GeneICEオリゴペプチドを使用してトランスフェクトした。次いで、製造元の推奨する手順を使用して、細胞をホルムアルデヒド中に固定し、DAPI核染色物質を用いて染色した。蛍光顕微鏡により細胞を調べることによって、Cy3標識GeneICEオリゴペプチド分子が、DAPI核染色物質と共に局在したことが示された。この生じた実験データから、ペプチドを核に向ける際に、NLSは非常に有効であると結論付けた。
【0194】
DDD-MAD1-DDD-NLS、およびDDD-NLS-DDD-MAD1ペプチド配列は、本発明のオリゴ/ペプチド分子中に取り込ませると、標的遺伝子の発現を仲介することができる。実施例8および9に概略した実験手法を使用して、これを実証することができる。
【0195】
例えば、DDD-MAD1-DDD-NLS、およびDDD-NLS-DDD-MAD1ペプチド配列は、以下のオリゴ/ペプチド分子中に取り込ませることができる:
Bcl2:-DDD-MAD1-DDD-NLS、および
Bcl2:-DDD-NLS-DDD-MAD1
【0196】
Bcl2は実施例8に示すTFOオリゴ配列である(5'GGGTGTGGGGTUTGTGTGTGGT3'または5'TUGTGTGGGTGTGGTGUGGG3'または5'GGTGTUTTGGTTGGGTGT3')。
【0197】
Bcl2:-DDD-MAD1-DDD-NLS、およびBcl2:-DDD-NLS-DDD-MAD1分子を、次いで試験細胞、例えばLNCap細胞にトランスフェクトする。前に概略した実験手順を使用して、Bcl2:-DDD-MAD1-DDD-NLS、およびBcl2:-DDD-NLS-DDD-MAD1分子の、標的遺伝子の発現に対する影響を示すことができる。
【0198】
(実施例11)
アポトーシス関連遺伝子の発現の調節に関する構築体の評価
結合:
TFOおよびGeneICE(商標)分子のそれらの標的DNA配列への結合を、ゲルシフトアッセイを使用することによって評価する。これを行うために、知られている量のTFOを、TFO結合バッファー(20mMのTRIS(pH 7.6)、10%v/vのグリセロール、10mMのMgCl2)中で、1時間37℃において、PCRおよびT4キナーゼを使用する末端標識を使用してゲノムDNAから前に増幅させた、微量の32P標識した標的DNAと共にインキュベートする。反応混合物は、非変性ローディングバッファー(10×ストック濃度:250mMのTRIS(pH 7.5)、40%v/vのグリセロール、10mMのMgCl2、0.2%w/vのブロモフェノールブルー)を使用して、12%非変性ポリアクリルアミドゲルに載せ、TRIS-ホウ酸マグネシウムランニングバッファー(0.5×TBE、10mMのMgCl2)中で3〜4時間、電気泳動に施す。バンドを目に見える状態にするために、オートラジオグラフィーフィルムに露光する前に、ゲルを乾燥させる。
【0199】
3つの異なる大きさのバンドが、ゲル上に予想される。TFOまたは構築体の濃度が低い場合、小さいDNAバンドのみが見られる。TFOの濃度が上昇して三重鎖DNAの形成が可能になるとき、中程度の大きさの三重鎖のバンドが現れ、GeneICE(商標)分子の濃度が上昇して三重鎖DNAの形成が可能になるとき、大きな三重鎖のバンドが見られる。TFOまたはGeneICE(商標)分子のKdは、等量の二量体DNAバンドと三重鎖DNAバンドが存在する濃度によって測定する。10-6〜10-9Mの範囲のKd値が予想され、GeneICE(商標)分子のKdは、TFOより1logまで高い。
【0200】
mRNA発現:
mRNA発現は、2つの方法によって調べることができる。最初に、全RNAを処理細胞から抽出し、RT-PCRまたはノーザンブロット分析用に使用する。RT-PCR分析では、逆転写酵素およびオリゴdTプライマーを使用することによって、mRNAがcDNAに逆転写される。このcDNAを、Bcl-2コード配列を増幅させるように設計したプライマーを使用する半定量PCR用に使用する。β-アクチンPCRも行い、相対量を使用して、GeneICE(商標)処理で見られた下方制御を定量化する。RT-PCR分析から得られる結果は、ノーザンブロット分析によって確認する。処理細胞から抽出した全RNAをTBEゲル上で電気泳動にかけ、ナイロン膜に移す。PCRにより32Pで前に標識した、一本鎖DNAプローブを使用して、Bcl-2mRNA配列にハイブリダイズさせる。非特異的結合は膜の厳密な洗浄によって除去し、オートラジオグラフィーを使用して、存在するmRNAの量を目に見える状態にする。β-アクチンmRNA用のプローブを使用することによって、RNA充填を調節する。
【0201】
mRNA発現の実験によって、Bcl-2標的GeneICE(商標)分子を用いた処理後の、Bcl-2遺伝子の特異的な下方制御が示される。下方制御の中間レベルは、TFOのみまたは送達ペプチドと結合したTFOを用いた処理によって上昇する。TSAなどの脱アセチル化酵素阻害剤を使用することによって、遺伝子発現のTFO仲介の下方制御ではなく、GeneICE(商標)分子による下方制御が元の状態になる。これらの実験は、GeneICE(商標)構築体の用量依存効果および時間依存効果も示し、用量の増大と共に、下方制御が非特異的になるまで、さらなる下方制御が示される。用量依存効果は1回量から長期の下方制御を示し、これは数時間後にピークに達し、数日間続く。
【0202】
タンパク質発現:
タンパク質発現は、イムノブロット分析によって評価する。細胞は氷上、プロテアーゼ阻害剤の存在下において溶解バッファーを含むSDS中で、溶解させる。細胞溶解物は遠心分離によって取り除き、タンパク質の量は、タンパク質アッセイを使用することによって測定する。それぞれのサンプルからの等量のタンパク質を、15%SDSポリアクリルアミドゲルのウエルに充填し、25mMのTRIS、192mMのグリシン、0.1%w/vのSDSランニングバッファーを使用して、150Vで2時間電気泳動にかける。分離後、48mMのTRIS、39mMのグリシン、0.0375%w/vのSDS、20%v/vのメタノール移送バッファーを使用して、5.5mA/cm2での半乾燥状態での電気泳動による移送によって、タンパク質をPVDF膜上に移す。非特異的な抗体結合は、5%w/vの無脂肪乳、PBS中/0.1%w/vのTween20(PBST)中でのインキュベーションによって阻害する。室温で1時間、2%乳中に1:2000に希釈した抗Bcl-2一次抗体と共に、膜をインキュベートする。過剰な抗体は2%乳を使用して洗浄除去し、西洋ワサビペルオキシダーゼ結合二次抗体を、2%のミルク中に1:10000で使用する。PBST中での2回の洗浄、次にPBS中での1回の洗浄を使用して、過剰な抗体を除去する。充分な化学発光およびオートラジオグラフィーを使用することによって、タンパク質を検出する。同じ手順を使用して、ただし1:500の希釈で抗アクチン一次抗体を抗Bcl-2抗体の代わりに使用して、アクチンを調べることによって、充填量を補正する。
【0203】
タンパク質発現は、製造元によって与えられる指示書に従って、Bcl-2ELISAキットを使用することによって確認する。
【0204】
mRNA発現と同様の実験を、タンパク質を分析するために行う。GeneICE(商標)構築体で処理した後の、タンパク質発現の時間および用量依存性の下方制御が予想され、これはmRNA発現の概略で見られるものに従う。タンパク質レベルは、mRNAレベルの約15〜20時間後に低下し、数日間は低い状態のままである。脱アセチル化酵素阻害剤は、この特異的なBcl-2タンパク質下方制御を妨げ、一方TFOによる処理によって、中間の表現型が表される。
【0205】
細胞死:
タンパク質発現の低下は、幾つかの細胞系では、アポトーシスによる細胞死を引き起こすのに充分である可能性がある。このことが当てはまる場合、非内因性プロモーター(SV40プロモーターなど)の制御下の、Bcl-2を有する発現プラスミドを、細胞系にトランスフェクトし、内因性mRNAのレベルは、内因性遺伝子に特異的なプライマーを使用して、記載するように評価する。
【0206】
細胞のトリパンブルー染色、および計測チャンバーを使用する計数によって、細胞死を評価する。細胞はGeneICE(商標)構築体で適切な長さの時間処理し、知られている体積のトリプシンを使用して、ウエルからトリプシン処理した。20μlの細胞懸濁液を20μlのトリパンブルーと混合させ、計測チャンバーに充填する。30個と300個の間の細胞を、計測チャンバーのグリッドを使用して、体積と所与数/mlの指標として計数し、細胞の生存能力は、非生存細胞を示す、トリパンブルー染色によって評価する。
【0207】
MTTアッセイを使用して、細胞計数によって与えられたデータを確認する。96ウエルプレート中の実験値を確定し、処理後、MTTを1時間までウエルに加える。培地を吸引し、細胞を洗浄する。残っている沈殿をDMSOに溶かし、吸光度を520nmで読み取る。
【0208】
GeneICE(商標)処理によって、培養物中の細胞数の減少、および培養物の生存能力の低下がもたらされる。脱アセチル化酵素阻害剤を使用することによって、アポトーシスの阻害剤と同様に、これらの影響が元の状態になる。TFO対照は、中間の表現型を示す。
【0209】
アポトーシス:
観察された細胞死の方法は、製造元により与えられる指示書に従い、末端デオキシトランスフェラーゼ仲介dUTPニック末端標識(TUNEL)キットを使用することによって調べる。実験は異なる時間地点の範囲で行って、最も多くの死細胞が見られる地点を決定する。GeneICE(商標)を用いた処理後に見られる細胞死は、アポトーシス機構を介したものであると予想される。アポトーシスの誘導物質を、陽性対照として使用して細胞を比較し、一方アポトーシス阻害剤は、TUNELアッセイにおいて細胞死、したがってアポトーシス細胞の出現を防ぐことができる。脱アセチル化酵素阻害剤は、GeneICE(商標)処理培養物において、アポトーシスが起こることも防ぐ。
【図面の簡単な説明】
【0210】
【図1】融合分子および標的部位との結合の概略図である。 Aでは、「1」は実施例1のオリゴヌクレオチド単位1を表し;「2」は、ポリペプチド単位2を表す。「L」は連結基領域を示す。Bでは、Dは、他の送達ペプチドを表す。ペプチドはオリゴヌクレオチドの一端または両端に連結することができ;例えば、抑制因子/修飾ポリペプチドは一端に連結することができ、送達ペプチドは他端に連結することができる。 Cは、オリゴヌクレオチド部分が二本鎖DNAを有する三重らせんを形成し、抑制因子ペプチドがSin3-HDAC複合体をその部位に補充した状況を表す。
【図2】Bcl-2のmRNAの定量化、およびGeneICEの内因性Bcl-2のmRNAレベルに対する影響を示す図である。 縦列は、対照mRNA値に対する比としての、示したBxPC-3膵臓癌細胞の3日間の処理後の、Bcl-2のmRNAの発現を表す。それぞれの縦列は、4つの独立したポリメラーゼ連鎖反応の、平均ODおよび平均の標準誤差を示すものである。平均光学濃度を測定した。 Bc1PおよびBc1Uに関するTFO配列である(5'から3'): Bc1P GGGTGTGGGGTUTGTGTGTGGT Bc1U GGTGTUTTGGTTGGGTGT L231はTFO-MAD-NLSに対応するペプチドであり、L232はTFO-NLS-MADに対応するペプチドである。
【0211】
【表1】
【図3】GeneICEbcl-2の細胞の生存能力に対する影響を示す図である。 示した3日間の処理後に合計mRNAによって測定した、PT45膵臓癌細胞の生存能力。縦列は合計mRNAのμg/mlを表す。
【0212】
【表2】
【図4】50pmol用量での一晩のインキュベーション後の、非結合TFOおよびBcIPL231結合体の、PT45細胞の生存能力に対する影響の図である。 特に「Chariot」送達系(Active Motif;Morris他、(2001) Nature Biotech 19、1173〜1176)を用いると、非結合TFOによってではなく、結合体によって、対照細胞と比べて、細胞数は減少する(すなわち、細胞死が誘導/促進される)。
【特許請求の範囲】
【請求項1】
細胞中のアポトーシスを促進するための方法であって、(1)ゲノム中に存在する、選択したアポトーシス関連遺伝子にある部位、または該遺伝子と関係がある部位と結合する核酸結合部分、および(2)修飾部分を含む分子を、細胞中に導入するステップを含み、前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記修飾部分がポリペプチドまたはペプチド模倣体を含む方法。
【請求項2】
前記修飾部分が発現抑制因子部分である、請求項2に記載の方法。
【請求項3】
前記修飾部分が、核酸またはクロマチンの共有結合修飾を調節することができる、請求項1または2に記載の方法。
【請求項4】
前記抑制因子または修飾部分がクロマチン不活性化部分である、請求項1から3のいずれか一項に記載の方法。
【請求項5】
前記抑制因子または修飾部分が、DNAメチラーゼ複合体の一成分の全体または一部分、あるいはDNAメチラーゼ複合体と結合するかまたはDNAメチラーゼ複合体の補充を容易にするポリペプチドの全体または一部分である、請求項1から4のいずれか一項に記載の方法。
【請求項6】
前記抑制因子または修飾部分が、ヒストンアセチルトランスフェラーゼの一成分の全体または一部分、あるいはヒストンアセチルトランスフェラーゼ複合体と結合するかまたはヒストンアセチルトランスフェラーゼ複合体の補充を容易にするポリペプチドの全体または一部分である、請求項1から4のいずれか一項に記載の方法。
【請求項7】
前記分子のポリペプチドまたはペプチド模倣体部分が、11kDa未満の分子量を有する、請求項1から6のいずれか一項に記載の方法。
【請求項8】
前記核酸結合部分がDNA結合部分である、請求項1から7のいずれか一項に記載の方法。
【請求項9】
前記核酸結合部分がRNA結合部分であり、ゲノム中に存在する前記部位がDNAから転写される新生RNAである、請求項1から7のいずれか一項に記載の方法。
【請求項10】
前記オリゴヌクレオチドまたはオリゴヌクレオチド類似体または模倣体が、三重鎖形成オリゴヌクレオチド(TFO)である、請求項1から9のいずれか一項に記載の方法。
【請求項11】
前記オリゴヌクレオチド類似体または模倣体がペプチド核酸(PNA)である、請求項1から10のいずれかに記載の方法。
【請求項12】
前記クロマチン不活性化部分がヒストン脱アセチル化を容易にする、請求項4から11のいずれかに記載の方法。
【請求項13】
前記クロマチン不活性化部分が、ヒストン脱アセチル化(HDAC)複合体の一成分の全体または一部分、あるいはHDAC複合体と結合するかまたはHDAC複合体の補充を容易にするポリペプチドの全体または一部分である、請求項4から12のいずれかに記載の方法。
【請求項14】
前記HDAC複合体の成分、またはHDAC複合体と結合するかあるいはHDAC複合体の補充を容易にする前記ポリペプチドが、PLZF、N-CoR、SMRT、Sin3、SAP18、SAP30、HDAC、NuRD、MAD1、MAD2、MAD3、MAD4、RbまたはE7のいずれか1つである、請求項13に記載の方法。
【請求項15】
前記クロマチン不活性化部分がPLZF全体、あるいはPLZFのN-CoR-またはSMRT-結合部分である、請求項14に記載の方法。
【請求項16】
前記クロマチン不活性化部分がHDAC全体、あるいはHDACの酵素活性部分である、請求項14に記載の方法。
【請求項17】
前記クロマチン不活性化部分がSap18またはE7またはMAD1の全体、あるいはSap18またはE7またはMAD1のヒストン脱アセチル化酵素複合体-結合部分である、請求項14に記載の方法。
【請求項18】
前記分子が、細胞進入および/または核局在を容易にする部分をさらに含む、請求項1から17のいずれかに記載の方法。
【請求項19】
細胞進入および/または核局在を容易にする前記部分が、7〜16アミノ酸の小ペプチド、例えば修飾型アンテナペディアホメオドメイン(RQIKIWFQNRRMKWKK)、または塩基性HIV TAT内在ペプチド(C(Acm)GRKKRRQRRRPQC)であって、そこにおいて、C(Acm)がCys-アセトアミドメチルまたはSV40核局在シグナル(PKKKRKV-NH2)である、請求項18に記載の方法。
【請求項20】
前記核酸結合部分と前記抑制因子または修飾部分を融合させる、請求項1から19のいずれか一項に記載の方法。
【請求項21】
前記細胞が真核細胞である、請求項1から20のいずれかに記載の方法。
【請求項22】
前記アポトーシス関連遺伝子がBcl-2、Bcl-XIまたはAktである、請求項1から21のいずれか一項に記載の方法。
【請求項23】
前記細胞が動物細胞であり動物中に含まれているか、あるいは植物細胞であり植物中に含まれている、請求項1から22のいずれかに記載の方法。
【請求項24】
ヒト中における選択した遺伝子の発現が抑制される、請求項1から23のいずれかに記載の方法。
【請求項25】
複数の選択した遺伝子の発現が抑制される、請求項1から24のいずれかに記載の方法。
【請求項26】
細胞中での選択したアポトーシス関連遺伝子の発現を調節するための物質の製造における、請求項1から25のいずれかに関連して定義された分子の使用。
【請求項27】
前記物質が選択した遺伝子の発現を抑制するためのものである、請求項26に記載の使用。
【請求項28】
前記物質が、動物中での選択したアポトーシス関連遺伝子の発現を調節または抑制するための薬剤である、請求項26または27に記載の使用。
【請求項29】
選択したアポトーシス関連遺伝子の発現の抑制または調節を必要とする患者を治療する方法であって、請求項1から28のいずれかで定義した分子の有効量を、前記患者に投与することを含む方法。
【請求項30】
発現の抑制を必要とする患者中で選択したアポトーシス関連遺伝子の発現を抑制するための薬剤の製造における、請求項1から29のいずれかで定義した分子の使用。
【請求項31】
請求項1から30のいずれかで定義した分子。
【請求項32】
医薬において使用するための、請求項1から31のいずれかで定義した分子。
【請求項33】
請求項1から32のいずれかで定義した分子、および薬学的に許容される担体を含む薬剤組成物。
【請求項34】
分子の細胞への取り込みを促進するための手段、例えばリポソームまたはウイルス担体を含む、請求項33に記載の組成物。
【請求項35】
請求項1から34のいずれか一項で定義した分子を含む宿主細胞。
【請求項36】
細菌細胞である、請求項35に記載の宿主細胞。
【請求項37】
動物細胞である、請求項35に記載の宿主細胞。
【請求項38】
植物細胞である、請求項35に記載の宿主細胞。
【請求項39】
請求項37に記載の宿主細胞を含む動物。
【請求項40】
請求項38に記載の宿主細胞を含む植物。
【請求項41】
細胞中での選択したアポトーシス関連遺伝子の発現を抑制するための分子を設計するための方法であって、
(1)選択したアポトーシス関連遺伝子にある部位、または前記選択したアポトーシス関連遺伝子と関係がある部位を同定すること、
(2)前記部位(または前記部位のヌクレオチド配列を有するかあるいは含むポリヌクレオチド)と結合するか、あるいは結合すると予想される核酸結合部分を同定または設計すること、
(3)前記核酸結合部分および発現抑制因子部分を含む分子を調製することを含み、
前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記抑制因子部分がポリペプチドまたはペプチド模倣体を含む方法。
【請求項42】
細胞中での選択したアポトーシス関連遺伝子の発現を調節するための分子を設計するための方法であって、
(1)選択した遺伝子にある部位、または前記選択した遺伝子と関係がある部位を同定すること、
(2)前記部位(または前記部位のヌクレオチド配列を有するかあるいは含むポリヌクレオチド)と結合するか、あるいは結合すると予想される核酸結合部分を同定または設計すること、
(3)前記核酸結合部分および修飾部分を含む分子を調製することを含み、
前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記修飾部分が、核酸またはクロマチンの共有結合修飾を調節することができるポリペプチドまたはペプチド模倣体を含む方法。
【請求項43】
(4)分子調製について品質管理評価を行って、前記核酸結合部分と抑制因子または修飾部分が互いに結合していることを判定するステップ、および/または
(5)前記核酸結合部分と、前記部位および/または前記部位のヌクレオチド配列を有するかもしくは含むポリヌクレオチドとの結合の親和性および/または特異性を試験するステップ、および/または
(6)前記分子と前記部位および/または前記部位のヌクレオチド配列を有するかもしくは含むポリヌクレオチドとの結合の親和性および/または特異性を試験するステップ、および/または
(7)前記遺伝子および/または前記部位のヌクレオチド配列を含むレポーター遺伝子の発現の調節または抑制における、前記分子またはポリヌクレオチドの有効性を試験するステップをさらに含む、請求項41または42に記載の方法。
【請求項44】
本明細書に記載する、細胞、例えば植物または動物細胞中での選択したアポトーシス関連遺伝子の活性を調節する、例えば抑制する、任意の新規な方法。
【請求項45】
本明細書に記載する、細胞、例えば植物または動物細胞中での選択したアポトーシス関連遺伝子の活性を調節する、例えば抑制する、任意の新規な分子。
【請求項46】
アポトーシスの促進を必要とする患者を治療するための方法であって、細胞死誘導物質および請求項1から45のいずれかで定義した分子の有効量を、前記患者に投与することを含む方法。
【請求項47】
前記細胞死誘導物質が、化学療法剤および/または放射線治療物質を含む、請求項46に記載の方法。
【請求項1】
細胞中のアポトーシスを促進するための方法であって、(1)ゲノム中に存在する、選択したアポトーシス関連遺伝子にある部位、または該遺伝子と関係がある部位と結合する核酸結合部分、および(2)修飾部分を含む分子を、細胞中に導入するステップを含み、前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記修飾部分がポリペプチドまたはペプチド模倣体を含む方法。
【請求項2】
前記修飾部分が発現抑制因子部分である、請求項2に記載の方法。
【請求項3】
前記修飾部分が、核酸またはクロマチンの共有結合修飾を調節することができる、請求項1または2に記載の方法。
【請求項4】
前記抑制因子または修飾部分がクロマチン不活性化部分である、請求項1から3のいずれか一項に記載の方法。
【請求項5】
前記抑制因子または修飾部分が、DNAメチラーゼ複合体の一成分の全体または一部分、あるいはDNAメチラーゼ複合体と結合するかまたはDNAメチラーゼ複合体の補充を容易にするポリペプチドの全体または一部分である、請求項1から4のいずれか一項に記載の方法。
【請求項6】
前記抑制因子または修飾部分が、ヒストンアセチルトランスフェラーゼの一成分の全体または一部分、あるいはヒストンアセチルトランスフェラーゼ複合体と結合するかまたはヒストンアセチルトランスフェラーゼ複合体の補充を容易にするポリペプチドの全体または一部分である、請求項1から4のいずれか一項に記載の方法。
【請求項7】
前記分子のポリペプチドまたはペプチド模倣体部分が、11kDa未満の分子量を有する、請求項1から6のいずれか一項に記載の方法。
【請求項8】
前記核酸結合部分がDNA結合部分である、請求項1から7のいずれか一項に記載の方法。
【請求項9】
前記核酸結合部分がRNA結合部分であり、ゲノム中に存在する前記部位がDNAから転写される新生RNAである、請求項1から7のいずれか一項に記載の方法。
【請求項10】
前記オリゴヌクレオチドまたはオリゴヌクレオチド類似体または模倣体が、三重鎖形成オリゴヌクレオチド(TFO)である、請求項1から9のいずれか一項に記載の方法。
【請求項11】
前記オリゴヌクレオチド類似体または模倣体がペプチド核酸(PNA)である、請求項1から10のいずれかに記載の方法。
【請求項12】
前記クロマチン不活性化部分がヒストン脱アセチル化を容易にする、請求項4から11のいずれかに記載の方法。
【請求項13】
前記クロマチン不活性化部分が、ヒストン脱アセチル化(HDAC)複合体の一成分の全体または一部分、あるいはHDAC複合体と結合するかまたはHDAC複合体の補充を容易にするポリペプチドの全体または一部分である、請求項4から12のいずれかに記載の方法。
【請求項14】
前記HDAC複合体の成分、またはHDAC複合体と結合するかあるいはHDAC複合体の補充を容易にする前記ポリペプチドが、PLZF、N-CoR、SMRT、Sin3、SAP18、SAP30、HDAC、NuRD、MAD1、MAD2、MAD3、MAD4、RbまたはE7のいずれか1つである、請求項13に記載の方法。
【請求項15】
前記クロマチン不活性化部分がPLZF全体、あるいはPLZFのN-CoR-またはSMRT-結合部分である、請求項14に記載の方法。
【請求項16】
前記クロマチン不活性化部分がHDAC全体、あるいはHDACの酵素活性部分である、請求項14に記載の方法。
【請求項17】
前記クロマチン不活性化部分がSap18またはE7またはMAD1の全体、あるいはSap18またはE7またはMAD1のヒストン脱アセチル化酵素複合体-結合部分である、請求項14に記載の方法。
【請求項18】
前記分子が、細胞進入および/または核局在を容易にする部分をさらに含む、請求項1から17のいずれかに記載の方法。
【請求項19】
細胞進入および/または核局在を容易にする前記部分が、7〜16アミノ酸の小ペプチド、例えば修飾型アンテナペディアホメオドメイン(RQIKIWFQNRRMKWKK)、または塩基性HIV TAT内在ペプチド(C(Acm)GRKKRRQRRRPQC)であって、そこにおいて、C(Acm)がCys-アセトアミドメチルまたはSV40核局在シグナル(PKKKRKV-NH2)である、請求項18に記載の方法。
【請求項20】
前記核酸結合部分と前記抑制因子または修飾部分を融合させる、請求項1から19のいずれか一項に記載の方法。
【請求項21】
前記細胞が真核細胞である、請求項1から20のいずれかに記載の方法。
【請求項22】
前記アポトーシス関連遺伝子がBcl-2、Bcl-XIまたはAktである、請求項1から21のいずれか一項に記載の方法。
【請求項23】
前記細胞が動物細胞であり動物中に含まれているか、あるいは植物細胞であり植物中に含まれている、請求項1から22のいずれかに記載の方法。
【請求項24】
ヒト中における選択した遺伝子の発現が抑制される、請求項1から23のいずれかに記載の方法。
【請求項25】
複数の選択した遺伝子の発現が抑制される、請求項1から24のいずれかに記載の方法。
【請求項26】
細胞中での選択したアポトーシス関連遺伝子の発現を調節するための物質の製造における、請求項1から25のいずれかに関連して定義された分子の使用。
【請求項27】
前記物質が選択した遺伝子の発現を抑制するためのものである、請求項26に記載の使用。
【請求項28】
前記物質が、動物中での選択したアポトーシス関連遺伝子の発現を調節または抑制するための薬剤である、請求項26または27に記載の使用。
【請求項29】
選択したアポトーシス関連遺伝子の発現の抑制または調節を必要とする患者を治療する方法であって、請求項1から28のいずれかで定義した分子の有効量を、前記患者に投与することを含む方法。
【請求項30】
発現の抑制を必要とする患者中で選択したアポトーシス関連遺伝子の発現を抑制するための薬剤の製造における、請求項1から29のいずれかで定義した分子の使用。
【請求項31】
請求項1から30のいずれかで定義した分子。
【請求項32】
医薬において使用するための、請求項1から31のいずれかで定義した分子。
【請求項33】
請求項1から32のいずれかで定義した分子、および薬学的に許容される担体を含む薬剤組成物。
【請求項34】
分子の細胞への取り込みを促進するための手段、例えばリポソームまたはウイルス担体を含む、請求項33に記載の組成物。
【請求項35】
請求項1から34のいずれか一項で定義した分子を含む宿主細胞。
【請求項36】
細菌細胞である、請求項35に記載の宿主細胞。
【請求項37】
動物細胞である、請求項35に記載の宿主細胞。
【請求項38】
植物細胞である、請求項35に記載の宿主細胞。
【請求項39】
請求項37に記載の宿主細胞を含む動物。
【請求項40】
請求項38に記載の宿主細胞を含む植物。
【請求項41】
細胞中での選択したアポトーシス関連遺伝子の発現を抑制するための分子を設計するための方法であって、
(1)選択したアポトーシス関連遺伝子にある部位、または前記選択したアポトーシス関連遺伝子と関係がある部位を同定すること、
(2)前記部位(または前記部位のヌクレオチド配列を有するかあるいは含むポリヌクレオチド)と結合するか、あるいは結合すると予想される核酸結合部分を同定または設計すること、
(3)前記核酸結合部分および発現抑制因子部分を含む分子を調製することを含み、
前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記抑制因子部分がポリペプチドまたはペプチド模倣体を含む方法。
【請求項42】
細胞中での選択したアポトーシス関連遺伝子の発現を調節するための分子を設計するための方法であって、
(1)選択した遺伝子にある部位、または前記選択した遺伝子と関係がある部位を同定すること、
(2)前記部位(または前記部位のヌクレオチド配列を有するかあるいは含むポリヌクレオチド)と結合するか、あるいは結合すると予想される核酸結合部分を同定または設計すること、
(3)前記核酸結合部分および修飾部分を含む分子を調製することを含み、
前記核酸結合部分がオリゴヌクレオチドまたはオリゴヌクレオチド模倣体または類似体を含み、前記修飾部分が、核酸またはクロマチンの共有結合修飾を調節することができるポリペプチドまたはペプチド模倣体を含む方法。
【請求項43】
(4)分子調製について品質管理評価を行って、前記核酸結合部分と抑制因子または修飾部分が互いに結合していることを判定するステップ、および/または
(5)前記核酸結合部分と、前記部位および/または前記部位のヌクレオチド配列を有するかもしくは含むポリヌクレオチドとの結合の親和性および/または特異性を試験するステップ、および/または
(6)前記分子と前記部位および/または前記部位のヌクレオチド配列を有するかもしくは含むポリヌクレオチドとの結合の親和性および/または特異性を試験するステップ、および/または
(7)前記遺伝子および/または前記部位のヌクレオチド配列を含むレポーター遺伝子の発現の調節または抑制における、前記分子またはポリヌクレオチドの有効性を試験するステップをさらに含む、請求項41または42に記載の方法。
【請求項44】
本明細書に記載する、細胞、例えば植物または動物細胞中での選択したアポトーシス関連遺伝子の活性を調節する、例えば抑制する、任意の新規な方法。
【請求項45】
本明細書に記載する、細胞、例えば植物または動物細胞中での選択したアポトーシス関連遺伝子の活性を調節する、例えば抑制する、任意の新規な分子。
【請求項46】
アポトーシスの促進を必要とする患者を治療するための方法であって、細胞死誘導物質および請求項1から45のいずれかで定義した分子の有効量を、前記患者に投与することを含む方法。
【請求項47】
前記細胞死誘導物質が、化学療法剤および/または放射線治療物質を含む、請求項46に記載の方法。
【図1】

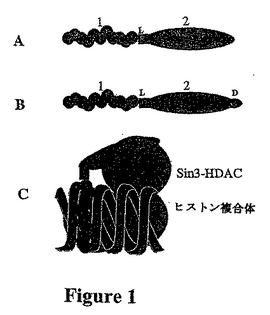
【図2】
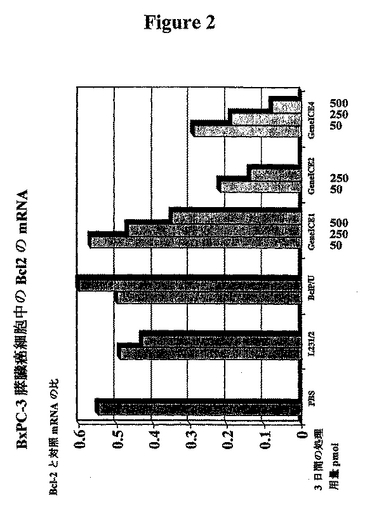
【図3】
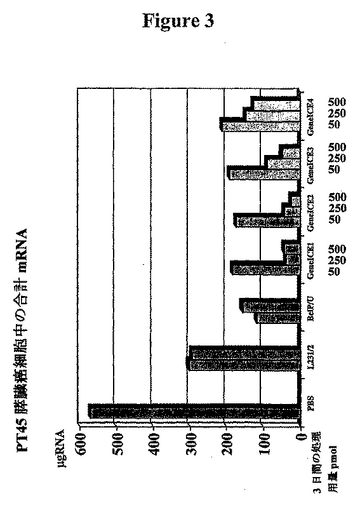
【図4】
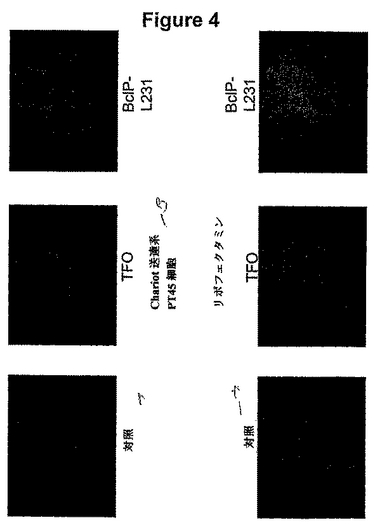
【図2】
【図3】
【図4】
【公表番号】特表2006−509010(P2006−509010A)
【公表日】平成18年3月16日(2006.3.16)
【国際特許分類】
【出願番号】特願2004−556552(P2004−556552)
【出願日】平成15年12月5日(2003.12.5)
【国際出願番号】PCT/GB2003/005321
【国際公開番号】WO2004/050885
【国際公開日】平成16年6月17日(2004.6.17)
【出願人】(500165809)インペリアル・カレッジ・イノベイションズ・リミテッド (32)
【Fターム(参考)】
【公表日】平成18年3月16日(2006.3.16)
【国際特許分類】
【出願日】平成15年12月5日(2003.12.5)
【国際出願番号】PCT/GB2003/005321
【国際公開番号】WO2004/050885
【国際公開日】平成16年6月17日(2004.6.17)
【出願人】(500165809)インペリアル・カレッジ・イノベイションズ・リミテッド (32)
【Fターム(参考)】
[ Back to top ]