
アミダーゼの保存剤、活性賦活剤及び失活予防剤
【課題】α−アミノ酸やα−ヒドロキシ酸等を製造するために用いるアミダーゼの保存剤、活性賦活剤及び失活予防剤を提供する。
【解決手段】Thermus sp.属、Thermus sp.0-3-1株、由来耐熱性アミダーゼ遺伝子を大腸菌JM109株にクローニングしてアミダーゼを生産することにより、アミダーゼ酵素溶液を調製した。該酵素溶液にニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を共存させることにより、アミダーゼ活性を安定化し、保存、活性賦活及び失活予防に有効である。
【解決手段】Thermus sp.属、Thermus sp.0-3-1株、由来耐熱性アミダーゼ遺伝子を大腸菌JM109株にクローニングしてアミダーゼを生産することにより、アミダーゼ酵素溶液を調製した。該酵素溶液にニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を共存させることにより、アミダーゼ活性を安定化し、保存、活性賦活及び失活予防に有効である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アミダーゼの保存剤、活性賦活剤及び失活予防剤、アミダーゼの保存用キット、活性賦活用キット及び失活予防用キット、並びにそれらの使用方法に関する。
【背景技術】
【0002】
光学活性なα−アミノ酸やα−ヒドロキシ酸は、各種工業製品等の中間体、食品添加物及び医薬品等として重要である。微生物あるいは微生物由来の酵素をα−アミノ酸アミド及びα−ヒドロキシ酸アミドの加水分解触媒として用いることにより、光学活性なα−アミノ酸(特許文献1及び2参照)やα−ヒドロキシ酸(特許文献3参照)を製造できることが知られている。近年、アミダーゼ遺伝子が導入された組換え体を用いた反応も報告されてきている(特許文献4及び5並びに非特許文献1参照)。
【0003】
このような加水分解反応において、アミダーゼ活性を効率的に発現させるため、培養液中や反応液中に亜鉛塩などの金属塩を添加する試みがなされている。
例えば、Enterobacter Cloacae N-7901株由来のアミダーゼについて、培養液中に亜鉛塩を添加する試みがなされている(特許文献4参照)。また、Ochrobactrum anthropi NCIMB40321株由来のアミダーゼについて、反応液中への亜鉛塩の添加効果、及び培養液中への亜鉛塩の添加効果が示されている(特許文献5参照)。さらに、同菌株ついて、亜鉛、マグネシウム、マンガンの金属塩の反応液中への添加効果が示されている(非特許文献1参照)。
【0004】
一方、上述のような、アミダーゼによる加水分解反応時の反応促進効果を目的とした金属塩の添加ではく、アミダーゼの保存性や活性の向上を目的として金属塩を利用する方法として、各種亜鉛塩を用いる方法が知られている(特許文献6参照)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開昭61−293394号公報
【特許文献2】特開昭62−55097号公報
【特許文献3】特開平2−84198号公報
【特許文献4】国際公開第00/63354号パンフレット
【特許文献5】特表2004−536608号公報
【特許文献6】特開2008−212027号公報
【非特許文献】
【0006】
【非特許文献1】Appl. Environ. Microbiol., vol. 71, p. 7961-7973, 2005
【発明の概要】
【発明が解決しようとする課題】
【0007】
このような状況下において、アミダーゼの保存性や活性の向上及び失活防止をより効果的に実現することができる新たな金属塩の利用が求められていた。
【課題を解決するための手段】
【0008】
本発明者は、上記課題を解決するため鋭意研究を行った結果、ニッケル、コバルト若しくはマンガン又はそれらの塩を酵素液に添加する又は添加して加熱処理すると、効果的にアミダーゼを保存し、活性を賦活し、失活を予防し得ること見出し、本発明を完成するに至った。
【0009】
(1)ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼの保存剤、活性賦活剤又は失活予防剤。
(2)ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼ保存用、アミダーゼ活性賦活用又はアミダーゼ失活予防用キット。
(3)アミダーゼを、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を用いて処理することを含む、アミダーゼの保存方法、活性賦活方法又は失活予防方法。
上記(3)の方法は、例えば、前記処理に際し保温又は加熱することを含んでいてもよい。
【発明の効果】
【0010】
本発明により、ニッケル、コバルト若しくはマンガン又はそれらの塩を含む、アミダーゼの保存剤、活性賦活剤及び失活防止剤や、アミダーゼの保存用キット、活性賦活用キット及び失活防止用キット、並びにそれらの使用方法を提供することができる。
本発明の保存剤等は、アミダーゼ活性の低下を抑制して、アミダーゼ酵素液を安定的に保存し、酵素活性を賦活し、使用することができる。従って、本発明の保存剤等は極めて有用なものである。
【図面の簡単な説明】
【0011】
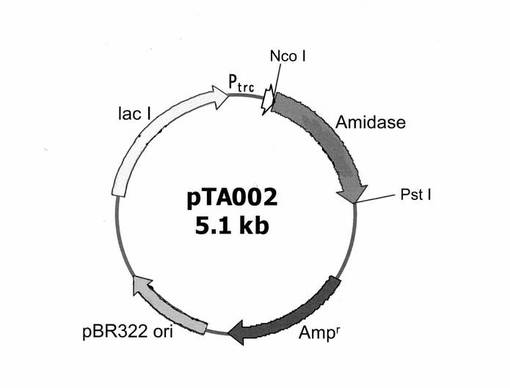
【図1】プラスミドpTA002の構造を示した概略図である。
【発明を実施するための形態】
【0012】
以下、本発明を詳細に説明する。本発明の範囲はこれらの説明に拘束されることはなく、以下の例示以外についても、本発明の趣旨を損なわない範囲で適宜変更し実施することができる。なお、本明細書において引用された全ての刊行物、例えば先行技術文献、及び公開公報、特許公報その他の特許文献は、参照として本明細書に組み込まれる。
【0013】
本発明は、ニッケル、コバルト若しくはマンガン又はそれらの塩を含む、アミダーゼの保存剤、活性賦活剤及び失活予防剤、アミダーゼの保存用キット、活性賦活用キット及び失活予防用キット、並びにそれらの使用方法である。
アミダーゼ酵素液に、ニッケル、コバルト若しくはマンガン又はそれらの塩を添加する、あるいは添加して熱処理する(ニッケル、コバルト若しくはマンガン又はそれらの塩の存在下で熱処理する)ことにより、それらを添加しない又は添加しないで熱処理した場合に比べて、活性が数倍から数十倍も向上することが示された。本発明は、このような知見に基づいて完成されたものである。
【0014】
1.アミダーゼの調製
本発明において、アミダーゼは、好熱菌由来、常温菌由来、好冷菌由来及び低温菌由来のいずれのものでもよいが、中でも好熱菌由来のものが好ましい。
ここで、「好熱菌」とは、一般には55℃以上で生育可能な細菌をいう(極限環境微生物ハンドブック、大島泰郎監修、株式会社サイエンスフォーラム)。好熱菌の例としては、限定されるわけではないが、サーマス(Thermus)属に属する細菌等(例えば、Thermus sp.、Thermus thermophilus、Thermus aquaticus等)を挙げることができる。
また、一般に「好冷菌」とは最適生育温度が15℃以下であって生育限界温度が20℃前後の菌を、「低温菌」とは最適生育温度が15℃以下であって生育限界温度が20℃以上のものをいう。
「常温菌」とは、上記好熱菌、好冷菌及び低温菌の定義に当てはまらないものを言い、従って、最適生育温度が15〜45℃であって55℃以上で生育不可能な細菌を指す。現在単離されている微生物の多くは常温菌であり、常温菌の例としては、限定されるわけではないが、エンテロバクター(Enterobacter属)に属する細菌、エッシェリヒア(Escherichia)属に属する細菌、クレブシエラ(Krebsiella)属に属する細菌、オクロバクトラム(Ochrobactrum)属に属する細菌、ロドコッカス(Rhodococcus)属に属する細菌、マイコバクテリウム(Mycobacterium)属に属する細菌等を挙げることができる。
【0015】
本発明において、アミダーゼは、例えば、配列番号2で示されるアミノ酸配列を含むタンパク質であって、かつアミダーゼ活性を有するタンパク質を含む。
【0016】
配列番号2で示されるアミノ酸配列からなるアミダーゼは、サーマス・エスピー(Thermus sp.)株由来のアミダーゼであり、配列番号1で示されるアミダーゼ遺伝子の塩基配列によりコードされる。
【0017】
また、本発明で用いられるアミダーゼには、配列番号2で示されるアミノ酸配列と65%以上、好ましくは70%以上、より好ましくは75%以上、さらに好ましくは85%以上、最も好ましくは90%、95%、97%、98%又は99%の相同性(同一性)を有するアミノ酸配列を含み、かつアミダーゼ活性を有するタンパク質も含まれる。
【0018】
さらに、本発明におけるアミダーゼには、配列番号2で示されるアミノ酸配列において、1又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列を含み、かつ、アミダーゼ活性を有するタンパク質も含まれる。
配列番号2で示されるアミノ酸配列において、1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列としては、例えば、(i) 配列番号2で示されるアミノ酸配列において、1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)のアミノ酸が欠失したアミノ酸配列、(ii)配列番号2で示されるアミノ酸配列の1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、(iii)配列番号2で示されるアミノ酸配列に1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)のアミノ酸が付加したアミノ酸配列、(iv)上記(i)〜(iii)を組み合わせたアミノ酸配列が挙げられる。
【0019】
本発明で用いられるアミダーゼは、酸アミド基を加水分解してカルボン酸とアミン又はアンモニアを生ずる反応(RCONHR’+H2O→RCOOH+NH2R’)を触媒する加水分解酵素であり、好ましくはα−アミノ酸アミド又はα−ヒドロキシ酸アミドを立体選択的に加水分解する活性を有する酵素である。したがって、本発明において、「アミダーゼ活性」とは、酸アミド基を加水分解してカルボン酸とアミン又はアンモニアとを生ずる反応を触媒する活性、好ましくはα−アミノ酸アミド又はα−ヒドロキシ酸アミドを立体選択的に加水分解して、L−α−アミノ酸又はL−α−ヒドロキシ酸と、アンモニアとを生ずる反応を触媒する活性を意味する。アミダーゼ活性の測定方法については、後述する。
【0020】
また、本発明で用いられるアミダーゼは、形質転換体、形質転換体懸濁液、形質転換体破砕物、粗精製アミダーゼ酵素液又はアミダーゼ酵素液を70℃で1時間処理した後も、処理前の活性と比較して、例えば、70%、好ましくは少なくとも80%、より好ましくは少なくとも90%、さらに好ましくは少なくとも95%の活性を保持していることを特徴とする。
【0021】
本発明のアミダーゼ遺伝子は、本発明で用いられるアミダーゼをコードする遺伝子である。本発明のアミダーゼ遺伝子は、例えば、配列番号1で示される塩基配列からなる遺伝子が挙げられるが、これに限定されるものではない。
【0022】
本発明のアミダーゼ遺伝子には、配列番号1で示される塩基配列に相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNAであって、かつアミダーゼ活性を有するタンパク質をコードする遺伝子も含まれる。
このような遺伝子の例として、配列番号1で示される塩基配列と65%以上、好ましくは70%以上、より好ましくは75%以上、さらに好ましくは85%以上、最も好ましくは90%、95%、97%、98%又は99%の相同性(同一性)を有する塩基配列を含み、かつアミダーゼ活性を有するタンパク質をコードする遺伝子が挙げられる。
【0023】
上記遺伝子は、配列番号1で示される塩基配列からなるDNAの一部をプライマーとして、公知の核酸増幅方法又はプライマーウォーキング法により取得することができる。また、本発明のアミダーゼ遺伝子は、配列番号1で示される塩基配列からなるDNAの一部をプローブとして、コロニーハイブリダーゼーション、プラークハイブリダイゼーション、サザンブロットなどの公知のハイブリダイゼーション法により取得することもできる。
上記方法には、アミダーゼ遺伝子を含むベクター、mRNA、total RNA、cDNA、ゲノムDNA、又はそれらのライブラリーを用いることができる。また、市販のライブラリーを使用しても良い。
本明細書において、ストリンジェントな条件は、ハイブリダイゼーション後の洗浄条件として、例えば、「2×SSC、0.1%SDS、42℃」、「1×SSC、0.1%SDS、37℃」、よりストリンジェントな条件としては、例えば、「1×SSC、0.1%SDS、65℃」、「0.5×SSC、0.1%SDS、50℃」等の条件を挙げることができる。
ハイブリダイゼーションは、公知の方法によって行うことができる。ハイブリダイゼーションの方法は、例えば、「Molecular Cloning, A Laboratory Manual 2nd ed.」(Cold Spring Harbor Laboratory Press(1989))、「Current Protocols in Molecular Biology」(John Wiley & Sons(1987-1997))等を参照することができる。
【0024】
さらに、本発明のアミダーゼ遺伝子には、配列番号1で示される塩基配列において、1又は数個の塩基配列が欠失、置換又は付加された塩基配列を含み、かつ、アミダーゼ活性を有するタンパク質をコードする遺伝子も含まれる。
配列番号1で示される塩基配列において、1個又は数個の塩基が欠失、置換又は付加された塩基配列としては、例えば、(i) 配列番号1で示される塩基配列において、1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)の塩基が欠失した塩基配列、(ii)配列番号1で示される塩基配列の1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)の塩基が他の塩基で置換された塩基配列、(iii)配列番号1で示される塩基配列に1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)の塩基が付加した塩基配列、(iv)上記(i)〜(iii)を組み合わせた塩基配列が挙げられる。
【0025】
配列番号1で示される塩基配列において1又は数個の核酸に欠失、置換又は付加などの変異の生じた塩基配列を含むDNAは、「Molecular Cloning, A Laboratory Manual 2nd ed.」(Cold Spring Harbor Press (1989))、「Current Protocols in Molecular Biology」(John Wiley & Sons (1987-1997))、Kunkel (1985) Proc. Natl. Acad. Sci. USA 82: 488-92、Kunkel (1988) Method. Enzymol. 85: 2763-6等に記載の部位特異的変異誘発法等の方法に従って調製することができる。
【0026】
また、DNAに変異を導入するには、Kunkel法やGapped duplex法等の公知手法により、部位特異的突然変異誘発法を利用した変異導入用キット、例えばQuikChangeTM Site-Directed Mutagenesis Kit(ストラタジーン社製)、GeneTailorTM Site-Directed Mutagenesis System(インビトロジェン社製)、TaKaRa Site-Directed Mutagenesis System(Mutan-K、Mutan-Super Express Km等:タカラバイオ社製)等を用いて行うことができる。
【0027】
本発明のアミダーゼ遺伝子の塩基配列の確認は、慣用の方法により配列決定することにより行うことができる。例えば、ジデオキシヌクレオチドチェーンターミネーション法(Sanger et al. (1977) Proc. Natl. Acad. Sci. USA 74: 5463)等により行うことができる。また、適当なDNAシークエンサーを利用して配列を解析することもできる。
【0028】
上述のアミダーゼを発現させるための宿主−ベクター系の選択は下記のようにして行われる。
【0029】
宿主に導入するベクターとしては、上記の酵素遺伝子を保持し、且つ複製可能であれば、特段の制約を受けるものではなく、それぞれの宿主に適したベクターを使用することができる。例えば、プラスミドDNA、バクテリオファージ等が挙げられる。
プラスミドDNAとしては、例えば、大腸菌由来のプラスミド(pBR322、pUC18、pUC19、pUC118、pUC119、pBluescriptなどのColE系プラスミド等)、放線菌由来のプラスミド(pIJ486等)、酵母由来のプラスミド(YEp13、YEp 24、Ycp50等)が挙げられる。ファージDNAとしては、λファージ(Charon4A、Charon21A、EMBL3、EMBL4、λgt10、λgt11等)、レトロトランスポゾンDNA、人工染色体DNA等が挙げられる。
【0030】
宿主としては、上記の酵素遺伝子が発現するものであれば良い。例えば、哺乳動物細胞、昆虫細胞、大腸菌、枯草菌、酵母、カビ、ロドコッカス属細菌、植物等を挙げることができる。好ましくは、大腸菌及びロドコッカス属細菌である。
大腸菌宿主としては、例えば大腸菌K12株やB株、あるいはそれら野生株由来の派生株であるJM109株、XL1-Blue株、C600株、W3110株等を挙げることができる。その他、これら菌株の変異体、組換え体及び遺伝子工学的手法による誘導体等も用いられ得る。
酵母宿主としては、例えばサッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)、ピヒア・パストリス(Pichia pastoris)等を挙げることができる。
ロドコッカス属細菌としては、例えばロドコッカス・ロドクロウス(Rhodococcus rhodochrous)ATCC 12674株やロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株(FERM BP-1478)等を挙げることができる。
【0031】
以下、より詳細にアミダーゼ遺伝子が導入された形質転換体(あるいは形質導入体)(以下、単に「形質転換体」と称する)の作製方法について説明する。
【0032】
プラスミドDNAは、宿主細胞中にてプラスミドが増殖するために必要なDNA配列、プロモーター、リボソーム結合配列、転写ターミネーター(転写終結配列)、さらに好ましくは形質転換体の選択マーカーとなる遺伝子を含む。
プロモーター配列としては、大腸菌由来のトリプトファンオペロンのtrpプロモーター、ラクトースオペロンのlacプロモーター、ラムダファージ由来のPLプロモーター及びPRプロモーター、枯草菌由来のグルコン酸合成酵素プロモーター(gnt)、アルカリプロテアーゼプロモーター(apr)、中性プロテアーゼプロモーター(npr)、α−アミラーゼプロモーター(amy)等を挙げることができる。また、tacプロモーター、trcプロモーターのように独自に改変、設計された配列も利用できる。
リボソーム結合配列としては、SD配列やKozak配列が知られており、これらの配列を変異遺伝子の上流に挿入することができる。原核生物を宿主に用いるときにはSD配列を、真核細胞を宿主に用いるときにはKozak配列をPCR法等により付加してもよい。SD配列としては、大腸菌由来、ロドコッカス属細菌又は枯草菌由来の配列等を挙げることができるが、所望の宿主内で機能する配列であれば特に限定されるものではない。例えば、16SリボゾームRNAの3’末端領域に相補的な配列が4塩基以上連続したコンセンサス配列をDNA合成により作製してこれを利用してもよい。
転写終結配列は必ずしも必要ではないが、ρ因子非依存性のもの、例えばリポプロテインターミネーター、trpオペロンターミネーター等が利用できる。
選択マーカーとしては、例えばジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子等を挙げることができる。
【0033】
大腸菌を宿主に用いた場合には、特に有用なベクターとしては、pTrc99A、pKK233-2、pFY529、pET-12、pET-26b等が例示される。
これらのベクターにアミダーゼをコードする遺伝子断片を組み込むには、アミダーゼ遺伝子を含むDNAを適当な制限酵素で切断し、必要であれば適当なリンカーを付加した後、適当な制限酵素で切断したベクターと結合させることにより行うことができる。
【0034】
このように作製した発現ベクターを宿主細胞に導入すれば、アミダーゼを高度に発現する形質転換体が得られる。当該形質転換体を培養することにより、これらの酵素を発現させることができる。
【0035】
発現ベクターの宿主への導入方法としては、DNAを導入する方法であれば特に限定されるものではない。例えばカルシウムイオンを用いる方法、エレクトロポレーション法等を挙げることができる。
酵母への発現プラスミドの導入方法としては、酵母にDNAを導入する方法であれば特に限定されず、例えばエレクトロポレーション法、スフェロプラスト法、酢酸リチウム法等を挙げることができる。
【0036】
次に、上記のように作製した形質転換体を培養する方法を説明する。培養に際し使用する培地には、特に制限は無く、宿主菌が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行うことができる培地であれば、天然培地、合成培地のいずれを用いてもよい。また培養条件に関しても、形質転換体が生育、増殖可能で且つ酵素産生が良好に行える条件を選択し、培養すればよい。
培養培地に用いる炭素源としては、グルコース、ガラクトース、フラクトース、スクロース、ラフィノース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール、グリセリン等のアルコール類を挙げることができる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸若しくは有機酸のアンモニウム塩又はその他の含窒素化合物を挙げることができる。その他、ペプトン、肉エキス、コーンスティープリカー、酵母エキス、各種アミノ酸等を用いてもよい。無機物としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム、硫酸亜鉛等を挙げることができる。その他、ビタミン等を必要に応じて適宜添加してもよい。培養中は必要に応じてアンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。
【0037】
プロモーターとして誘導性のプロモーターを用いた発現プラスミドを導入した形質転換体を培養する場合は、必要に応じてインデューサーを培地に添加してもよい。例えば、イソプロピル−β−D−チオガラクトシド(IPTG)で誘導可能なプロモーターを有する発現プラスミドを導入した形質転換体を培養するときは、IPTG等を培地に添加することができる。また、インドール酢酸(IAA)で誘導可能なtrpプロモーターを用いた発現プラスミドを導入した形質転換体を培養するときは、IAA等を培地に添加することができる。
【0038】
大腸菌の培養に際し、通常の固体培養法で培養してもよいが、液体培養法を採用して培養することが好ましい。培地の初発pHは7〜9に調整することが適当である。また、培養は、5℃〜40℃、好ましくは10℃〜37℃で5〜100時間行う。通気攪拌深部培養、振盪培養、静置培養、流加培養等により実施することが好ましい。
ロドコッカス属細菌の培養は、4〜36℃、好ましくは20〜30℃で18〜96時間行う。
【0039】
(破砕処理方法)
こうして得られた培養物から菌体や細胞を回収するには、遠心分離法や膜ろ過法を用いることができる。回収した形質転換体を、必要に応じて、リン酸−ナトリウム緩衝液、リン酸緩衝液等で洗浄し、懸濁することができる。
菌体又は細胞の破砕方法としては、超音波処理、フレンチプレスやホモジナイザーによる高圧処理、ガラスビーズなどによる磨砕処理、リゾチーム、セルラーゼ、ペクチナーゼ等を用いる酵素処理、凍結融解処理、低張液処理、ファージによる溶菌誘導処理等を利用することができる。破砕処理は、必要に応じて氷冷下で行う。破砕処理を行ったサンプルを「形質転換体破砕物」、「破砕物」又は「細胞抽出液全画分」とも称する。
破砕後、形質転換体破砕物から、必要に応じて菌体又は細胞の破砕残渣(細胞抽出液不溶性画分を含む)を除くことができる。残渣を除去する方法としては、例えば、遠心分離やろ過などが上げられ、必要に応じて、凝集剤やろ過助剤等を使用して残渣除去効率を上げることもできる。残渣を除去した後に得られた上清は、「細胞抽出液可溶性画分」であり、「粗精製したアミダーゼ溶液」とすることができる。
【0040】
2.ニッケル、コバルト若しくはマンガン又はそれらの塩によるアミダーゼ液の処理
(1)ニッケル、コバルト若しくはマンガン又はそれらの塩とアミダーゼとの接触処理
本発明において、「接触」とは、ニッケル、コバルト若しくはマンガン又はそれらの塩とアミダーゼとを共存させること、すなわちニッケル、コバルト若しくはマンガン又はそれらの塩とアミダーゼとを同一の反応系に存在させることを意味し、例えば、アミダーゼ酵素液を含む容器にニッケル、コバルト若しくはマンガン又はそれらの塩を添加すること、ニッケル、コバルト若しくはマンガン又はそれらの塩を含む培養液中でアミダーゼ遺伝子を含む菌体(形質転換体)を培養すること、アミダーゼ遺伝子を含む菌体(形質転換体)又はその破砕物を含有する溶液中にニッケル、コバルト若しくはマンガン又はそれらの塩を添加することなどが含まれる。ニッケル塩、コバルト塩及びマンガン塩の濃度は、それぞれ、好ましくは0.01 mM〜30 mM、好ましくは0.1 mM〜15 mMであり、より好ましくは2 mM〜15 mMであり、さらに好ましくは5 mM〜12 mMである。
ニッケル塩、コバルト塩及びマンガン塩としては、特に限定はされないが、例えば、ニッケル、コバルト及びマンガン各々の、塩酸塩、硫酸塩、硝酸塩、水酸化物塩、炭酸塩及びリン酸塩等が好ましく挙げられる。
このように、アミダーゼと、ニッケル、コバルト若しくはマンガン又はそれらの塩とを接触させることにより、アミダーゼの活性を増加(賦活化)させることができる。このことは、アミダーゼの活性低下(失活)を防止することを意味するものであり、これによりアミダーゼを有効に保存することができる。
なお、本発明において用いられる、ニッケル、コバルト若しくはマンガン又はそれらの塩は、1種のみには限定されず、2種以上を併用してもよく、限定はされない。すなわち、本発明において、アミダーゼと接触させるものとしては、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種が用いられる。
【0041】
(2)活性の測定
アミダーゼ活性は、DL−α−アミノ酸アミドやDL−α−ヒドロキシ酸アミドを基質として一定時間反応させた後、生成するα−アミノ酸やα−ヒドロキシ酸を高速液体クロマトグラフィー等により定量し、その定量値を活性の指標として測定することができる。
【0042】
(3)加熱処理
本発明においては、アミダーゼを、ニッケル、コバルト若しくはマンガン又はそれらの塩で処理する際は、常温下(20±15℃、例えば25℃程度)でも保温下(30〜40℃、例えば37℃程度)でもよいし、加熱処理を行ってもよい。適度な保温又は加熱処理を行うことにより、高活性アミダーゼを得ることができる。ここで、加熱処理とは、アミダーゼと、ニッケル、コバルト若しくはマンガン又はそれらの塩との混合物を、60℃〜80℃、好ましくは65℃〜75℃の温度範囲内の環境下に曝すことを意味し、例えば、アミダーゼ溶液に上記塩を添加した後、上記温度条件で5〜360分、好ましくは30〜60分処理する態様が挙げられる。
アミダーゼを上記塩による処理、又は上記塩による処理と加熱処理とを施すことにより、アミダーゼを24時間〜300日、好ましくは10〜100日保存することができる。また、アミダーゼ活性は、上記処理を施さないアミダーゼと比較して、好ましくは110〜5,000%、の活性に上昇させることが可能となる。その結果、アミダーゼの失活予防効果をより高くすることができる。
【0043】
3.キット
本発明において、ニッケル、コバルト若しくはマンガン又はそれらの塩は、アミダーゼの保存用キット、アミダーゼの活性賦活用キットあるいはアミダーゼの失活予防用キットの構成成分として使用される。
本発明のキットは、上記ニッケル、コバルト若しくはマンガン又はそれらの塩に加えて、一般の測定において慣用的な成分、例えば緩衝液、生理食塩水、pH調整剤等を含んでいてもよい。また、本発明のキットは、アミダーゼを保存する方法及び/又はアミダーゼの活性の測定方法等を記載した取扱説明書などを含んでいてもよい。
【0044】
以下に、実施例を挙げて本発明をより具体的に説明するが、本発明はこれらに限定されるものではない。
【実施例1】
【0045】
<プラスミドpTA002の作製>
(1)Thermus sp.属(Thermus sp.0-3-1株)由来耐熱性アミダーゼ遺伝子断片の調製
Thermus sp.0-3-1株由来のアミダーゼ遺伝子を含むプラスミドpM501KN(WO 03/020929参照)を鋳型として、下記の配列番号3に示される塩基配列からなる合成DNA(TAM-01)及び配列番号4に示される塩基配列からなる合成DNA(TAM-02)をプライマーとして用い、ポリメラーゼ伸長鎖反応(PCR)により、アミダーゼ遺伝子を含むDNA断片を増幅した。
【0046】
プライマー:
TAM-01: GCGGATCCATGGAGGGCTACAGGACCATTC (配列番号3)
TAM-02: GGACCTGCAGGTCCATCAGGCGAAAATATC (配列番号4)
【0047】
PCR用の反応混合物は、5μLの10×Pfu Turboバッファー、4μLのdNTP mix(2.5 mM each)、1μLのPfu Turbo DNAポリメラーゼ、37μLの蒸留水、1μLの各プライマー、および鋳型としてpLA205を1μL添加したものを用いた。PCRは、95℃で1分の変性を行った後、94℃で1分、60℃で1分、72℃で5分の反応を、計30サイクル行った。
反応終了後、1μLの反応終了液を1.5 %アガロースゲル電気泳動により分析し、増幅産物を確認した。増幅産物をGFX PCR DNA and Gel Band Purification Kit(GEヘルスケア バイオサイエンス社)を用いて精製し、約50μLのDNA溶液を得た。
次に、43μLのDNA溶液に、10×Kバッファー(タカラバイオ社)を5μL、制限酵素NcoIおよびSse8783Iを各1μL加え、37℃で2時間反応後、エタノール沈殿によりDNA断片を回収した。反応終了後1.5 %アガロースゲル電気泳動を行い、約0.93kbの目的バンド(アミダーゼ遺伝子断片)を切り出し、GFX PCR DNA and Gel Band Purification Kitを用いて精製した。
【0048】
(2)ベクター断片の調製とライゲーション
一方、5μLのベクターpTrc99A(GEバイオサイエンス社)に10×Kバッファー(タカラバイオ社)を3μL、滅菌水を23μL、制限酵素NcoIおよびPstIを各1μL加え、37℃で2時間反応後、アルカリフォスファターゼ処理(Shrimp Alkaline Phosphatase、プロメガ株式会社)を行い、エタノール沈殿によりDNA断片を回収した。
NcoIとPstIとにより切断されたこれらのDNA断片、すなわち上記(1)で得られたアミダーゼ遺伝子断片および本項(2)で得られたベクターDNA断片を各2μLと、solution I(DNA Ligation Kit ver.2(タカラバイオ社))(10μL)とを混合して、ライゲーション混合物を調製した。該混合物を、16℃で2時間インキュベートすることでアミダーゼ遺伝子断片とベクターpTrc99Aを結合させた。
【0049】
(3)大腸菌 JM109株のコンピテントセルの作製
大腸菌JM109株をLB培地(1%バクトトリプトン、0.5%バクトイーストエキス、0.5% NaCl) 1 mLに接種し37℃、5時間好気的に前培養した。得られた前培養液 0.4 mLをSOB培地 40 mL(2%バクトトリプトン、0.5%バクトイーストエキス、10mM NaCl 、2.5mM KCl 、1mM MgSO4 、1mM MgCl2) に加え、18℃で20時間培養した。当該培養物を遠心分離(3,700 G、10分間、4℃)により集菌した後、冷TF溶液 (20 mM PIPES−KOH (pH 6.0)、200 mM KCl 、10 mM CaCl2 、40mM MnCl2)を13 mL加え、0℃で10分間放置した。その後、再度遠心分離(3,700 G、10分間、4℃)し、上清を除いた。沈殿した大腸菌を冷TF溶液 3.2 mLに懸濁し、0.22 mLのジメチルスルホキシドを加え0℃で10分間放置した。その後、液体窒素下で冷凍した後、-80℃で保存した。
【0050】
(4)組換えプラスミドの調製
上記(3)で作製したコンピテントセル 200μLを氷上で解凍し、上記(2)で作製したライゲーション産物10μLに加え、0℃で30分放置した。続いて、当該コンピテントセルに42℃で30秒間ヒートショックを与え、0℃で2分間冷却した。その後、SOC 培地 (20 mM グルコース、2%バクトトリプトン、0.5%バクトイーストエキス、10 mM NaCl、2.5 mM KCl 、1 mM MgSO4、1mM MgCl2) 1 mLを添加し、37℃にて1時間振盪培養した。培養後の培養液を各200μLずつ、LB Amp寒天培地(アンピシリン 100 mg/L 、1.5%寒天を含有するLB培地)にまき、37℃で一晩培養した。寒天培地上に生育した形質転換体コロニー複数個を 1.5 mLのLB Amp培地(アンピシリン 100 mg/Lを含有するLB培地)にて37℃で一晩培養した。得られた培養液を各々集菌後、Flexi Prep(GEヘルスケア バイオサイエンス社製)を用いて組換えプラスミドを回収した。得られたプラスミドの塩基配列をCEQ DTCS Quick Start Kitおよび蛍光シーケンサCEQ 2000XL DNA Analysis system(いずれもBECKMAN COULTER、米国)を用いて解析した。目的とするプラスミドをpTA002と命名した(図1参照)。
【実施例2】
【0051】
<アミダーゼ酵素溶液の調製>
(1)形質転換体の作製及び培養
実施例1と同様にして、大腸菌JM109株のコンピテントセルに対しプラスミドpTA002による形質転換を行った。得られた形質転換体JM109/pTA002のコロニーを、5 mL 2×LB培地(20 g/Lポリペプトン、10 g/L乾燥酵母エキス、20 g/L NaCl、50μg/mLアンピシリン)に植菌し、37℃ 、250rpmで12時間振盪培養を行い前培養とした。前培養液を集菌(8,000rpm 10分間)し菌体を沈殿させ、約100μLとなるよう上清を除去した。菌体ペレットをピペットマンで懸濁し、2 L modified TB培地(12 g/Lポリペプトン、24 g/L乾燥酵母エキス、40 g/Lグリセロール、12.5 g/L K2HPO4、2.3 g/L KH2PO4、0.5 g/L ポリプロピレングリコール12000、50μg/mLアンピシリン)に植菌し、37℃、180 rpmで培養し、これを本培養とした。培養開始より4時間後にIPTGを1 mMとなるよう添加し14時間の誘導を行った。
菌体懸濁液の調製は、サンプリングして得られた培養液から遠心分離(3,700 G、10分間、4℃)により菌体を回収し、100 mM トリス塩酸緩衝液(pH 8.0)で洗浄した後、同緩衝液に懸濁することにより行った。
【0052】
(2)アミダーゼ酵素溶液の調製
(i) 無細胞抽出液の調製
本培養を行った培養液に対して6,000 rpmで20分間、遠心分離を行い、菌体の洗浄として50 mMトリス塩酸(pH 8.0)溶液200 mLで懸濁した後、再度6,000rpmで20分間、遠心分離した。集菌した菌体ペレットを50 mMトリス塩酸(pH 8.0)で湿菌体重の2倍量に懸濁し、150Wで20分間、超音波破砕を行った。この超音波破砕液を無細胞抽出液とした。
【0053】
(ii) 粗酵素溶液の調製
無細胞抽出液に対して、破砕液を14,000 rpmで15分間、遠心分離を行い、沈殿物を除去した。70℃で40分間のヒートショックを行った後、14,000 rpmで20分間、遠心分離を行った。沈殿したタンパク質を除去し、上清を採取した。
【0054】
(iii) 精製酵素溶液の調製
上記(ii)で得られた無細胞抽出液熱処理物を、硫酸アンモニウム沈殿法により4℃で分別した。硫酸アンモニウムを30%の飽和濃度になるまで攪拌しながらゆっくりと加え、次いで、この溶液を4℃、12,000 Gで、20分間で遠心分離し、得られたペレットを200 mLの50 mMトリス塩酸緩衝液(pH 8.0)に懸濁した。
得られた硫安沈殿画分のタンパク質量5 mg当たり1 mLのDEAEトヨパール650H(TOSOH)をカラムに充填し、担体の5倍量の50 mMトリス塩酸緩衝液(pH 8.0)で平衡化を行った。その後、タンパク質溶液をアプライし、平衡化で用いたものと同組成のパッファーを担体の15倍量添加し洗浄を行った。洗浄後、溶出用バッファー(50 mM トリス塩酸緩衝液(pH 8.0)、100 mM KCl)を7.5倍量アプライし、ステップワイズで本酵素の溶出を行った。最後に、押し出しとして溶出用緩衝液(50 mMトリス塩酸緩衝液(pH 8.0)、500 mM KCl)を添加した。溶出液は約7 mL(200ドロップ)ずつ試験管に分取し、タンパク質の溶出は各画分のO.D.280 nmを測定することにより決定した。
アミダーゼ活性は、50 mMトリス塩酸緩衝液(pH 8.0)、100 mM KClにより溶出を開始し、アミダーゼ活性を有する画分を集めてフェニルトヨパールによる精製を行った。
【0055】
タンパク質量5 mg当たり1 mLのフェニルトヨパール650H(TOSOH)をカラムに充填し、担体の5倍量の洗浄緩衝液(50 mMトリス塩酸緩衝液(pH 8.0)、10%硫酸アンモニウム)で平衡化を行った。次に、タンパク質溶液に10%となるように硫酸アンモニウムを添加し、懸濁した。その後、硫酸アンモニウムを添加したタンパク質溶液をカラムにアプライし、平衡化で用いたものと同組成の緩衝液を担体の15倍量添加し洗浄を行った。洗浄後、溶出用緩衝液(50 mMトリス塩酸緩衝液(pH 8.0))を7.5倍量アプライし、ステップワイズで本酵素の溶出を行った。溶出液は約7 mL(200ドロップ)ずつ試験管に分取し、タンパク質の溶出は各画分のO.D.280 mmを測定することにより決定した。
アミダーゼ活性は、50 mMトリス塩酸緩衝液(pH 8.0)、10%硫酸アンモニウムにより溶出を開始し、アミダーゼ活性を有する画分を集めてSephacryI S-200 HRによる精製を行った。
約700 mLのSephacryI S-200 High Resolutiomをカラムに充填し、担体の1〜4%までサンプルを濃縮した。担体の2倍量の50 mMトリス塩酸緩衝液(pH 8.0)で、1 mL/分の流速で平衡化を行った。
その後、マーカー(5 mg Blue Dextran 2000/700μL平衡化緩衝液)、タンパク質溶液を順にアプライし、平衡化で用いたものと同組成の緩衝液を担体の2倍量添加し、流速は一定のまま溶出を行った。溶出液は約7 mL(200ドロップ)ずつ試験管に分取し、タンパク質の溶出は各画分のO.D.280 mmを測定することにより決定した。また、それぞれのタンパク質のピークに合わせて活性測定を行い、活性の高い画分を濃縮して、精製酵素溶液とした。
【0056】
(3)酵素活性の測定方法
アミダーゼの活性測定は、0.32 mM ロイシン-p-ニトロアニリン、100 mM トリス塩酸緩衝液(pH 9.0)に適当量のアミダーゼ溶液を加え、80℃にて30分間反応させた。氷中に入れることにより反応を停止させ、反応液の405nmの吸光度を測定して、生成したp-ニトロアニリン量を決定し、アミダーゼ活性とした。
【0057】
(4)タンパク質濃度(酵素濃度)の定量方法
Protein Assayキット(Bio-Rad)を用いた。適当な濃度に希釈した0.8 mLの酵素溶液に0.2 mLのアッセイ試薬を添加し、10〜15分室温に静置して、O.D.595 nmを測定することにより試料のタンパク質濃度を決定した。検量線の作製には、BSA(1.25, 2.5, 5.0, 7.5, 10μg/mL)を用いた。
【実施例3】
【0058】
実施例2で調製したアミダーゼ精製酵素画分を用いて、金属塩の添加効果を検討した。
すなわち、アミダーゼ精製酵素画分150μLに、塩化コバルト、塩化マンガン、塩化ニッケルを、それぞれ、終濃度5 mMとなるように添加し、37℃及び80℃で、それぞれ、1時間インキュベートし、次いで、10,000 rpmで5分間遠心処理を行い、沈殿物を除去した。その後、各々150 mLのトリス塩酸緩衝液(pH 8.0)に対して透析を2時間実施し、緩衝液を交換して同様にもう一度透析を行った。その後、各処理酵素液のタンパク質濃度及び活性を測定した。塩化コバルト、塩化マンガン及び塩化ニッケルの添加により、それぞれ、700%、740%及び640%(37℃)、並びに860%、1230%及び4800%(80℃)の活性向上が認められた。
【実施例4】
【0059】
実施例2で調製したアミダーゼ精製酵素画分を用いて、酵素溶液をEDTAで処理した後、金属塩の添加効果を検討した。
すなわち、1.8 mLのアミダーゼ精製酵素画分に100 mM EDTA (pH 8.0)を終濃度 5 mMとなるように添加した後、80℃で60分間静置し、その後、500 mLのトリス塩酸緩衝液(pH8.0)に対して透析を2時間実施し、緩衝液を交換して同様にもう一度透析を行なうことによりEDTAを除去した。EDTA処理した、アミダーゼ精製酵素画分150μLに塩化コバルト、塩化マンガン、塩化ニッケル、塩化カルシウム、塩化マグネシウム、塩化亜鉛をそれぞれ、終濃度5mMとなるように添加し、37℃及び80℃で、それぞれ、1時間インキュベートし、次いで、10000 rpmで5分間遠心処理を行い、沈殿物を除去した。その後、各々150 mLのトリス塩酸緩衝液(pH 8.0)に対して透析を2時間実施し、緩衝液を交換して同様にもう一度透析を行った。その後、各処理酵素液のタンパク質濃度及び活性を測定した。
【0060】
EDTA処理を行なうことにより、アミダーゼ活性は、処理前の6.6%に減少した。
このEDTA処理したアミダーゼ精製酵素画分に金属塩を添加することにより、塩化コバルト、塩化マンガン、塩化ニッケル、塩化カルシウム、塩化マグネシウム及び塩化亜鉛の添加により、無処理アミダーゼ精製酵素画分の活性に対して、それぞれ、196%、77%、115%、13%、16%及び37%(37℃)、並びに397%、287%、236%、44%、28%及び94%(80℃)の活性回復が認められた。
【実施例5】
【0061】
実施例2で調製したアミダーゼ精製酵素画分を用いて、実施例4に記載した方法と同様の方法にほり酵素溶液をEDTAで処理した後、塩化コバルト、塩化マンガン、塩化ニッケルの添加濃度の影響を検討した。
すなわち、アミダーゼ精製酵素画分150μLに塩化コバルト、塩化マンガン、塩化ニッケルをそれぞれ、終濃度1 mM、5 mMあるいは10 mMとなるように添加し、80℃で、1時間インキュベートし、次いで、10,000 rpmで5分間遠心処理を行い、沈殿物を除去した。その後、各々150 mLのトリス塩酸緩衝液(pH 8.0)に対して透析を2時間実施し、緩衝液を交換して同様にもう一度透析を行った。その後、各処理酵素液のタンパク質濃度及び活性を測定した。塩化コバルトの添加により、それぞれ、100%(1 mM)、490%(5 mM)及び980%(10 mM)、塩化ニッケルの添加により、それぞれ、50%(1mM)、280%(5 mM)及び305%(10 mM)、並びに塩化マンガンの添加により、それぞれ、100%(1 mM)、220%(5 mM)及び205%(10 mM)の活性向上が認められた。
【配列表フリーテキスト】
【0062】
配列番号3:合成DNA
配列番号4:合成DNA
【技術分野】
【0001】
本発明は、アミダーゼの保存剤、活性賦活剤及び失活予防剤、アミダーゼの保存用キット、活性賦活用キット及び失活予防用キット、並びにそれらの使用方法に関する。
【背景技術】
【0002】
光学活性なα−アミノ酸やα−ヒドロキシ酸は、各種工業製品等の中間体、食品添加物及び医薬品等として重要である。微生物あるいは微生物由来の酵素をα−アミノ酸アミド及びα−ヒドロキシ酸アミドの加水分解触媒として用いることにより、光学活性なα−アミノ酸(特許文献1及び2参照)やα−ヒドロキシ酸(特許文献3参照)を製造できることが知られている。近年、アミダーゼ遺伝子が導入された組換え体を用いた反応も報告されてきている(特許文献4及び5並びに非特許文献1参照)。
【0003】
このような加水分解反応において、アミダーゼ活性を効率的に発現させるため、培養液中や反応液中に亜鉛塩などの金属塩を添加する試みがなされている。
例えば、Enterobacter Cloacae N-7901株由来のアミダーゼについて、培養液中に亜鉛塩を添加する試みがなされている(特許文献4参照)。また、Ochrobactrum anthropi NCIMB40321株由来のアミダーゼについて、反応液中への亜鉛塩の添加効果、及び培養液中への亜鉛塩の添加効果が示されている(特許文献5参照)。さらに、同菌株ついて、亜鉛、マグネシウム、マンガンの金属塩の反応液中への添加効果が示されている(非特許文献1参照)。
【0004】
一方、上述のような、アミダーゼによる加水分解反応時の反応促進効果を目的とした金属塩の添加ではく、アミダーゼの保存性や活性の向上を目的として金属塩を利用する方法として、各種亜鉛塩を用いる方法が知られている(特許文献6参照)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開昭61−293394号公報
【特許文献2】特開昭62−55097号公報
【特許文献3】特開平2−84198号公報
【特許文献4】国際公開第00/63354号パンフレット
【特許文献5】特表2004−536608号公報
【特許文献6】特開2008−212027号公報
【非特許文献】
【0006】
【非特許文献1】Appl. Environ. Microbiol., vol. 71, p. 7961-7973, 2005
【発明の概要】
【発明が解決しようとする課題】
【0007】
このような状況下において、アミダーゼの保存性や活性の向上及び失活防止をより効果的に実現することができる新たな金属塩の利用が求められていた。
【課題を解決するための手段】
【0008】
本発明者は、上記課題を解決するため鋭意研究を行った結果、ニッケル、コバルト若しくはマンガン又はそれらの塩を酵素液に添加する又は添加して加熱処理すると、効果的にアミダーゼを保存し、活性を賦活し、失活を予防し得ること見出し、本発明を完成するに至った。
【0009】
(1)ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼの保存剤、活性賦活剤又は失活予防剤。
(2)ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼ保存用、アミダーゼ活性賦活用又はアミダーゼ失活予防用キット。
(3)アミダーゼを、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を用いて処理することを含む、アミダーゼの保存方法、活性賦活方法又は失活予防方法。
上記(3)の方法は、例えば、前記処理に際し保温又は加熱することを含んでいてもよい。
【発明の効果】
【0010】
本発明により、ニッケル、コバルト若しくはマンガン又はそれらの塩を含む、アミダーゼの保存剤、活性賦活剤及び失活防止剤や、アミダーゼの保存用キット、活性賦活用キット及び失活防止用キット、並びにそれらの使用方法を提供することができる。
本発明の保存剤等は、アミダーゼ活性の低下を抑制して、アミダーゼ酵素液を安定的に保存し、酵素活性を賦活し、使用することができる。従って、本発明の保存剤等は極めて有用なものである。
【図面の簡単な説明】
【0011】
【図1】プラスミドpTA002の構造を示した概略図である。
【発明を実施するための形態】
【0012】
以下、本発明を詳細に説明する。本発明の範囲はこれらの説明に拘束されることはなく、以下の例示以外についても、本発明の趣旨を損なわない範囲で適宜変更し実施することができる。なお、本明細書において引用された全ての刊行物、例えば先行技術文献、及び公開公報、特許公報その他の特許文献は、参照として本明細書に組み込まれる。
【0013】
本発明は、ニッケル、コバルト若しくはマンガン又はそれらの塩を含む、アミダーゼの保存剤、活性賦活剤及び失活予防剤、アミダーゼの保存用キット、活性賦活用キット及び失活予防用キット、並びにそれらの使用方法である。
アミダーゼ酵素液に、ニッケル、コバルト若しくはマンガン又はそれらの塩を添加する、あるいは添加して熱処理する(ニッケル、コバルト若しくはマンガン又はそれらの塩の存在下で熱処理する)ことにより、それらを添加しない又は添加しないで熱処理した場合に比べて、活性が数倍から数十倍も向上することが示された。本発明は、このような知見に基づいて完成されたものである。
【0014】
1.アミダーゼの調製
本発明において、アミダーゼは、好熱菌由来、常温菌由来、好冷菌由来及び低温菌由来のいずれのものでもよいが、中でも好熱菌由来のものが好ましい。
ここで、「好熱菌」とは、一般には55℃以上で生育可能な細菌をいう(極限環境微生物ハンドブック、大島泰郎監修、株式会社サイエンスフォーラム)。好熱菌の例としては、限定されるわけではないが、サーマス(Thermus)属に属する細菌等(例えば、Thermus sp.、Thermus thermophilus、Thermus aquaticus等)を挙げることができる。
また、一般に「好冷菌」とは最適生育温度が15℃以下であって生育限界温度が20℃前後の菌を、「低温菌」とは最適生育温度が15℃以下であって生育限界温度が20℃以上のものをいう。
「常温菌」とは、上記好熱菌、好冷菌及び低温菌の定義に当てはまらないものを言い、従って、最適生育温度が15〜45℃であって55℃以上で生育不可能な細菌を指す。現在単離されている微生物の多くは常温菌であり、常温菌の例としては、限定されるわけではないが、エンテロバクター(Enterobacter属)に属する細菌、エッシェリヒア(Escherichia)属に属する細菌、クレブシエラ(Krebsiella)属に属する細菌、オクロバクトラム(Ochrobactrum)属に属する細菌、ロドコッカス(Rhodococcus)属に属する細菌、マイコバクテリウム(Mycobacterium)属に属する細菌等を挙げることができる。
【0015】
本発明において、アミダーゼは、例えば、配列番号2で示されるアミノ酸配列を含むタンパク質であって、かつアミダーゼ活性を有するタンパク質を含む。
【0016】
配列番号2で示されるアミノ酸配列からなるアミダーゼは、サーマス・エスピー(Thermus sp.)株由来のアミダーゼであり、配列番号1で示されるアミダーゼ遺伝子の塩基配列によりコードされる。
【0017】
また、本発明で用いられるアミダーゼには、配列番号2で示されるアミノ酸配列と65%以上、好ましくは70%以上、より好ましくは75%以上、さらに好ましくは85%以上、最も好ましくは90%、95%、97%、98%又は99%の相同性(同一性)を有するアミノ酸配列を含み、かつアミダーゼ活性を有するタンパク質も含まれる。
【0018】
さらに、本発明におけるアミダーゼには、配列番号2で示されるアミノ酸配列において、1又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列を含み、かつ、アミダーゼ活性を有するタンパク質も含まれる。
配列番号2で示されるアミノ酸配列において、1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列としては、例えば、(i) 配列番号2で示されるアミノ酸配列において、1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)のアミノ酸が欠失したアミノ酸配列、(ii)配列番号2で示されるアミノ酸配列の1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、(iii)配列番号2で示されるアミノ酸配列に1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)のアミノ酸が付加したアミノ酸配列、(iv)上記(i)〜(iii)を組み合わせたアミノ酸配列が挙げられる。
【0019】
本発明で用いられるアミダーゼは、酸アミド基を加水分解してカルボン酸とアミン又はアンモニアを生ずる反応(RCONHR’+H2O→RCOOH+NH2R’)を触媒する加水分解酵素であり、好ましくはα−アミノ酸アミド又はα−ヒドロキシ酸アミドを立体選択的に加水分解する活性を有する酵素である。したがって、本発明において、「アミダーゼ活性」とは、酸アミド基を加水分解してカルボン酸とアミン又はアンモニアとを生ずる反応を触媒する活性、好ましくはα−アミノ酸アミド又はα−ヒドロキシ酸アミドを立体選択的に加水分解して、L−α−アミノ酸又はL−α−ヒドロキシ酸と、アンモニアとを生ずる反応を触媒する活性を意味する。アミダーゼ活性の測定方法については、後述する。
【0020】
また、本発明で用いられるアミダーゼは、形質転換体、形質転換体懸濁液、形質転換体破砕物、粗精製アミダーゼ酵素液又はアミダーゼ酵素液を70℃で1時間処理した後も、処理前の活性と比較して、例えば、70%、好ましくは少なくとも80%、より好ましくは少なくとも90%、さらに好ましくは少なくとも95%の活性を保持していることを特徴とする。
【0021】
本発明のアミダーゼ遺伝子は、本発明で用いられるアミダーゼをコードする遺伝子である。本発明のアミダーゼ遺伝子は、例えば、配列番号1で示される塩基配列からなる遺伝子が挙げられるが、これに限定されるものではない。
【0022】
本発明のアミダーゼ遺伝子には、配列番号1で示される塩基配列に相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNAであって、かつアミダーゼ活性を有するタンパク質をコードする遺伝子も含まれる。
このような遺伝子の例として、配列番号1で示される塩基配列と65%以上、好ましくは70%以上、より好ましくは75%以上、さらに好ましくは85%以上、最も好ましくは90%、95%、97%、98%又は99%の相同性(同一性)を有する塩基配列を含み、かつアミダーゼ活性を有するタンパク質をコードする遺伝子が挙げられる。
【0023】
上記遺伝子は、配列番号1で示される塩基配列からなるDNAの一部をプライマーとして、公知の核酸増幅方法又はプライマーウォーキング法により取得することができる。また、本発明のアミダーゼ遺伝子は、配列番号1で示される塩基配列からなるDNAの一部をプローブとして、コロニーハイブリダーゼーション、プラークハイブリダイゼーション、サザンブロットなどの公知のハイブリダイゼーション法により取得することもできる。
上記方法には、アミダーゼ遺伝子を含むベクター、mRNA、total RNA、cDNA、ゲノムDNA、又はそれらのライブラリーを用いることができる。また、市販のライブラリーを使用しても良い。
本明細書において、ストリンジェントな条件は、ハイブリダイゼーション後の洗浄条件として、例えば、「2×SSC、0.1%SDS、42℃」、「1×SSC、0.1%SDS、37℃」、よりストリンジェントな条件としては、例えば、「1×SSC、0.1%SDS、65℃」、「0.5×SSC、0.1%SDS、50℃」等の条件を挙げることができる。
ハイブリダイゼーションは、公知の方法によって行うことができる。ハイブリダイゼーションの方法は、例えば、「Molecular Cloning, A Laboratory Manual 2nd ed.」(Cold Spring Harbor Laboratory Press(1989))、「Current Protocols in Molecular Biology」(John Wiley & Sons(1987-1997))等を参照することができる。
【0024】
さらに、本発明のアミダーゼ遺伝子には、配列番号1で示される塩基配列において、1又は数個の塩基配列が欠失、置換又は付加された塩基配列を含み、かつ、アミダーゼ活性を有するタンパク質をコードする遺伝子も含まれる。
配列番号1で示される塩基配列において、1個又は数個の塩基が欠失、置換又は付加された塩基配列としては、例えば、(i) 配列番号1で示される塩基配列において、1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)の塩基が欠失した塩基配列、(ii)配列番号1で示される塩基配列の1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)の塩基が他の塩基で置換された塩基配列、(iii)配列番号1で示される塩基配列に1〜10個(例えば、1〜5個、好ましくは1〜3個、さらに好ましくは1〜2個)の塩基が付加した塩基配列、(iv)上記(i)〜(iii)を組み合わせた塩基配列が挙げられる。
【0025】
配列番号1で示される塩基配列において1又は数個の核酸に欠失、置換又は付加などの変異の生じた塩基配列を含むDNAは、「Molecular Cloning, A Laboratory Manual 2nd ed.」(Cold Spring Harbor Press (1989))、「Current Protocols in Molecular Biology」(John Wiley & Sons (1987-1997))、Kunkel (1985) Proc. Natl. Acad. Sci. USA 82: 488-92、Kunkel (1988) Method. Enzymol. 85: 2763-6等に記載の部位特異的変異誘発法等の方法に従って調製することができる。
【0026】
また、DNAに変異を導入するには、Kunkel法やGapped duplex法等の公知手法により、部位特異的突然変異誘発法を利用した変異導入用キット、例えばQuikChangeTM Site-Directed Mutagenesis Kit(ストラタジーン社製)、GeneTailorTM Site-Directed Mutagenesis System(インビトロジェン社製)、TaKaRa Site-Directed Mutagenesis System(Mutan-K、Mutan-Super Express Km等:タカラバイオ社製)等を用いて行うことができる。
【0027】
本発明のアミダーゼ遺伝子の塩基配列の確認は、慣用の方法により配列決定することにより行うことができる。例えば、ジデオキシヌクレオチドチェーンターミネーション法(Sanger et al. (1977) Proc. Natl. Acad. Sci. USA 74: 5463)等により行うことができる。また、適当なDNAシークエンサーを利用して配列を解析することもできる。
【0028】
上述のアミダーゼを発現させるための宿主−ベクター系の選択は下記のようにして行われる。
【0029】
宿主に導入するベクターとしては、上記の酵素遺伝子を保持し、且つ複製可能であれば、特段の制約を受けるものではなく、それぞれの宿主に適したベクターを使用することができる。例えば、プラスミドDNA、バクテリオファージ等が挙げられる。
プラスミドDNAとしては、例えば、大腸菌由来のプラスミド(pBR322、pUC18、pUC19、pUC118、pUC119、pBluescriptなどのColE系プラスミド等)、放線菌由来のプラスミド(pIJ486等)、酵母由来のプラスミド(YEp13、YEp 24、Ycp50等)が挙げられる。ファージDNAとしては、λファージ(Charon4A、Charon21A、EMBL3、EMBL4、λgt10、λgt11等)、レトロトランスポゾンDNA、人工染色体DNA等が挙げられる。
【0030】
宿主としては、上記の酵素遺伝子が発現するものであれば良い。例えば、哺乳動物細胞、昆虫細胞、大腸菌、枯草菌、酵母、カビ、ロドコッカス属細菌、植物等を挙げることができる。好ましくは、大腸菌及びロドコッカス属細菌である。
大腸菌宿主としては、例えば大腸菌K12株やB株、あるいはそれら野生株由来の派生株であるJM109株、XL1-Blue株、C600株、W3110株等を挙げることができる。その他、これら菌株の変異体、組換え体及び遺伝子工学的手法による誘導体等も用いられ得る。
酵母宿主としては、例えばサッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)、ピヒア・パストリス(Pichia pastoris)等を挙げることができる。
ロドコッカス属細菌としては、例えばロドコッカス・ロドクロウス(Rhodococcus rhodochrous)ATCC 12674株やロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株(FERM BP-1478)等を挙げることができる。
【0031】
以下、より詳細にアミダーゼ遺伝子が導入された形質転換体(あるいは形質導入体)(以下、単に「形質転換体」と称する)の作製方法について説明する。
【0032】
プラスミドDNAは、宿主細胞中にてプラスミドが増殖するために必要なDNA配列、プロモーター、リボソーム結合配列、転写ターミネーター(転写終結配列)、さらに好ましくは形質転換体の選択マーカーとなる遺伝子を含む。
プロモーター配列としては、大腸菌由来のトリプトファンオペロンのtrpプロモーター、ラクトースオペロンのlacプロモーター、ラムダファージ由来のPLプロモーター及びPRプロモーター、枯草菌由来のグルコン酸合成酵素プロモーター(gnt)、アルカリプロテアーゼプロモーター(apr)、中性プロテアーゼプロモーター(npr)、α−アミラーゼプロモーター(amy)等を挙げることができる。また、tacプロモーター、trcプロモーターのように独自に改変、設計された配列も利用できる。
リボソーム結合配列としては、SD配列やKozak配列が知られており、これらの配列を変異遺伝子の上流に挿入することができる。原核生物を宿主に用いるときにはSD配列を、真核細胞を宿主に用いるときにはKozak配列をPCR法等により付加してもよい。SD配列としては、大腸菌由来、ロドコッカス属細菌又は枯草菌由来の配列等を挙げることができるが、所望の宿主内で機能する配列であれば特に限定されるものではない。例えば、16SリボゾームRNAの3’末端領域に相補的な配列が4塩基以上連続したコンセンサス配列をDNA合成により作製してこれを利用してもよい。
転写終結配列は必ずしも必要ではないが、ρ因子非依存性のもの、例えばリポプロテインターミネーター、trpオペロンターミネーター等が利用できる。
選択マーカーとしては、例えばジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子等を挙げることができる。
【0033】
大腸菌を宿主に用いた場合には、特に有用なベクターとしては、pTrc99A、pKK233-2、pFY529、pET-12、pET-26b等が例示される。
これらのベクターにアミダーゼをコードする遺伝子断片を組み込むには、アミダーゼ遺伝子を含むDNAを適当な制限酵素で切断し、必要であれば適当なリンカーを付加した後、適当な制限酵素で切断したベクターと結合させることにより行うことができる。
【0034】
このように作製した発現ベクターを宿主細胞に導入すれば、アミダーゼを高度に発現する形質転換体が得られる。当該形質転換体を培養することにより、これらの酵素を発現させることができる。
【0035】
発現ベクターの宿主への導入方法としては、DNAを導入する方法であれば特に限定されるものではない。例えばカルシウムイオンを用いる方法、エレクトロポレーション法等を挙げることができる。
酵母への発現プラスミドの導入方法としては、酵母にDNAを導入する方法であれば特に限定されず、例えばエレクトロポレーション法、スフェロプラスト法、酢酸リチウム法等を挙げることができる。
【0036】
次に、上記のように作製した形質転換体を培養する方法を説明する。培養に際し使用する培地には、特に制限は無く、宿主菌が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行うことができる培地であれば、天然培地、合成培地のいずれを用いてもよい。また培養条件に関しても、形質転換体が生育、増殖可能で且つ酵素産生が良好に行える条件を選択し、培養すればよい。
培養培地に用いる炭素源としては、グルコース、ガラクトース、フラクトース、スクロース、ラフィノース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール、グリセリン等のアルコール類を挙げることができる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸若しくは有機酸のアンモニウム塩又はその他の含窒素化合物を挙げることができる。その他、ペプトン、肉エキス、コーンスティープリカー、酵母エキス、各種アミノ酸等を用いてもよい。無機物としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム、硫酸亜鉛等を挙げることができる。その他、ビタミン等を必要に応じて適宜添加してもよい。培養中は必要に応じてアンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。
【0037】
プロモーターとして誘導性のプロモーターを用いた発現プラスミドを導入した形質転換体を培養する場合は、必要に応じてインデューサーを培地に添加してもよい。例えば、イソプロピル−β−D−チオガラクトシド(IPTG)で誘導可能なプロモーターを有する発現プラスミドを導入した形質転換体を培養するときは、IPTG等を培地に添加することができる。また、インドール酢酸(IAA)で誘導可能なtrpプロモーターを用いた発現プラスミドを導入した形質転換体を培養するときは、IAA等を培地に添加することができる。
【0038】
大腸菌の培養に際し、通常の固体培養法で培養してもよいが、液体培養法を採用して培養することが好ましい。培地の初発pHは7〜9に調整することが適当である。また、培養は、5℃〜40℃、好ましくは10℃〜37℃で5〜100時間行う。通気攪拌深部培養、振盪培養、静置培養、流加培養等により実施することが好ましい。
ロドコッカス属細菌の培養は、4〜36℃、好ましくは20〜30℃で18〜96時間行う。
【0039】
(破砕処理方法)
こうして得られた培養物から菌体や細胞を回収するには、遠心分離法や膜ろ過法を用いることができる。回収した形質転換体を、必要に応じて、リン酸−ナトリウム緩衝液、リン酸緩衝液等で洗浄し、懸濁することができる。
菌体又は細胞の破砕方法としては、超音波処理、フレンチプレスやホモジナイザーによる高圧処理、ガラスビーズなどによる磨砕処理、リゾチーム、セルラーゼ、ペクチナーゼ等を用いる酵素処理、凍結融解処理、低張液処理、ファージによる溶菌誘導処理等を利用することができる。破砕処理は、必要に応じて氷冷下で行う。破砕処理を行ったサンプルを「形質転換体破砕物」、「破砕物」又は「細胞抽出液全画分」とも称する。
破砕後、形質転換体破砕物から、必要に応じて菌体又は細胞の破砕残渣(細胞抽出液不溶性画分を含む)を除くことができる。残渣を除去する方法としては、例えば、遠心分離やろ過などが上げられ、必要に応じて、凝集剤やろ過助剤等を使用して残渣除去効率を上げることもできる。残渣を除去した後に得られた上清は、「細胞抽出液可溶性画分」であり、「粗精製したアミダーゼ溶液」とすることができる。
【0040】
2.ニッケル、コバルト若しくはマンガン又はそれらの塩によるアミダーゼ液の処理
(1)ニッケル、コバルト若しくはマンガン又はそれらの塩とアミダーゼとの接触処理
本発明において、「接触」とは、ニッケル、コバルト若しくはマンガン又はそれらの塩とアミダーゼとを共存させること、すなわちニッケル、コバルト若しくはマンガン又はそれらの塩とアミダーゼとを同一の反応系に存在させることを意味し、例えば、アミダーゼ酵素液を含む容器にニッケル、コバルト若しくはマンガン又はそれらの塩を添加すること、ニッケル、コバルト若しくはマンガン又はそれらの塩を含む培養液中でアミダーゼ遺伝子を含む菌体(形質転換体)を培養すること、アミダーゼ遺伝子を含む菌体(形質転換体)又はその破砕物を含有する溶液中にニッケル、コバルト若しくはマンガン又はそれらの塩を添加することなどが含まれる。ニッケル塩、コバルト塩及びマンガン塩の濃度は、それぞれ、好ましくは0.01 mM〜30 mM、好ましくは0.1 mM〜15 mMであり、より好ましくは2 mM〜15 mMであり、さらに好ましくは5 mM〜12 mMである。
ニッケル塩、コバルト塩及びマンガン塩としては、特に限定はされないが、例えば、ニッケル、コバルト及びマンガン各々の、塩酸塩、硫酸塩、硝酸塩、水酸化物塩、炭酸塩及びリン酸塩等が好ましく挙げられる。
このように、アミダーゼと、ニッケル、コバルト若しくはマンガン又はそれらの塩とを接触させることにより、アミダーゼの活性を増加(賦活化)させることができる。このことは、アミダーゼの活性低下(失活)を防止することを意味するものであり、これによりアミダーゼを有効に保存することができる。
なお、本発明において用いられる、ニッケル、コバルト若しくはマンガン又はそれらの塩は、1種のみには限定されず、2種以上を併用してもよく、限定はされない。すなわち、本発明において、アミダーゼと接触させるものとしては、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種が用いられる。
【0041】
(2)活性の測定
アミダーゼ活性は、DL−α−アミノ酸アミドやDL−α−ヒドロキシ酸アミドを基質として一定時間反応させた後、生成するα−アミノ酸やα−ヒドロキシ酸を高速液体クロマトグラフィー等により定量し、その定量値を活性の指標として測定することができる。
【0042】
(3)加熱処理
本発明においては、アミダーゼを、ニッケル、コバルト若しくはマンガン又はそれらの塩で処理する際は、常温下(20±15℃、例えば25℃程度)でも保温下(30〜40℃、例えば37℃程度)でもよいし、加熱処理を行ってもよい。適度な保温又は加熱処理を行うことにより、高活性アミダーゼを得ることができる。ここで、加熱処理とは、アミダーゼと、ニッケル、コバルト若しくはマンガン又はそれらの塩との混合物を、60℃〜80℃、好ましくは65℃〜75℃の温度範囲内の環境下に曝すことを意味し、例えば、アミダーゼ溶液に上記塩を添加した後、上記温度条件で5〜360分、好ましくは30〜60分処理する態様が挙げられる。
アミダーゼを上記塩による処理、又は上記塩による処理と加熱処理とを施すことにより、アミダーゼを24時間〜300日、好ましくは10〜100日保存することができる。また、アミダーゼ活性は、上記処理を施さないアミダーゼと比較して、好ましくは110〜5,000%、の活性に上昇させることが可能となる。その結果、アミダーゼの失活予防効果をより高くすることができる。
【0043】
3.キット
本発明において、ニッケル、コバルト若しくはマンガン又はそれらの塩は、アミダーゼの保存用キット、アミダーゼの活性賦活用キットあるいはアミダーゼの失活予防用キットの構成成分として使用される。
本発明のキットは、上記ニッケル、コバルト若しくはマンガン又はそれらの塩に加えて、一般の測定において慣用的な成分、例えば緩衝液、生理食塩水、pH調整剤等を含んでいてもよい。また、本発明のキットは、アミダーゼを保存する方法及び/又はアミダーゼの活性の測定方法等を記載した取扱説明書などを含んでいてもよい。
【0044】
以下に、実施例を挙げて本発明をより具体的に説明するが、本発明はこれらに限定されるものではない。
【実施例1】
【0045】
<プラスミドpTA002の作製>
(1)Thermus sp.属(Thermus sp.0-3-1株)由来耐熱性アミダーゼ遺伝子断片の調製
Thermus sp.0-3-1株由来のアミダーゼ遺伝子を含むプラスミドpM501KN(WO 03/020929参照)を鋳型として、下記の配列番号3に示される塩基配列からなる合成DNA(TAM-01)及び配列番号4に示される塩基配列からなる合成DNA(TAM-02)をプライマーとして用い、ポリメラーゼ伸長鎖反応(PCR)により、アミダーゼ遺伝子を含むDNA断片を増幅した。
【0046】
プライマー:
TAM-01: GCGGATCCATGGAGGGCTACAGGACCATTC (配列番号3)
TAM-02: GGACCTGCAGGTCCATCAGGCGAAAATATC (配列番号4)
【0047】
PCR用の反応混合物は、5μLの10×Pfu Turboバッファー、4μLのdNTP mix(2.5 mM each)、1μLのPfu Turbo DNAポリメラーゼ、37μLの蒸留水、1μLの各プライマー、および鋳型としてpLA205を1μL添加したものを用いた。PCRは、95℃で1分の変性を行った後、94℃で1分、60℃で1分、72℃で5分の反応を、計30サイクル行った。
反応終了後、1μLの反応終了液を1.5 %アガロースゲル電気泳動により分析し、増幅産物を確認した。増幅産物をGFX PCR DNA and Gel Band Purification Kit(GEヘルスケア バイオサイエンス社)を用いて精製し、約50μLのDNA溶液を得た。
次に、43μLのDNA溶液に、10×Kバッファー(タカラバイオ社)を5μL、制限酵素NcoIおよびSse8783Iを各1μL加え、37℃で2時間反応後、エタノール沈殿によりDNA断片を回収した。反応終了後1.5 %アガロースゲル電気泳動を行い、約0.93kbの目的バンド(アミダーゼ遺伝子断片)を切り出し、GFX PCR DNA and Gel Band Purification Kitを用いて精製した。
【0048】
(2)ベクター断片の調製とライゲーション
一方、5μLのベクターpTrc99A(GEバイオサイエンス社)に10×Kバッファー(タカラバイオ社)を3μL、滅菌水を23μL、制限酵素NcoIおよびPstIを各1μL加え、37℃で2時間反応後、アルカリフォスファターゼ処理(Shrimp Alkaline Phosphatase、プロメガ株式会社)を行い、エタノール沈殿によりDNA断片を回収した。
NcoIとPstIとにより切断されたこれらのDNA断片、すなわち上記(1)で得られたアミダーゼ遺伝子断片および本項(2)で得られたベクターDNA断片を各2μLと、solution I(DNA Ligation Kit ver.2(タカラバイオ社))(10μL)とを混合して、ライゲーション混合物を調製した。該混合物を、16℃で2時間インキュベートすることでアミダーゼ遺伝子断片とベクターpTrc99Aを結合させた。
【0049】
(3)大腸菌 JM109株のコンピテントセルの作製
大腸菌JM109株をLB培地(1%バクトトリプトン、0.5%バクトイーストエキス、0.5% NaCl) 1 mLに接種し37℃、5時間好気的に前培養した。得られた前培養液 0.4 mLをSOB培地 40 mL(2%バクトトリプトン、0.5%バクトイーストエキス、10mM NaCl 、2.5mM KCl 、1mM MgSO4 、1mM MgCl2) に加え、18℃で20時間培養した。当該培養物を遠心分離(3,700 G、10分間、4℃)により集菌した後、冷TF溶液 (20 mM PIPES−KOH (pH 6.0)、200 mM KCl 、10 mM CaCl2 、40mM MnCl2)を13 mL加え、0℃で10分間放置した。その後、再度遠心分離(3,700 G、10分間、4℃)し、上清を除いた。沈殿した大腸菌を冷TF溶液 3.2 mLに懸濁し、0.22 mLのジメチルスルホキシドを加え0℃で10分間放置した。その後、液体窒素下で冷凍した後、-80℃で保存した。
【0050】
(4)組換えプラスミドの調製
上記(3)で作製したコンピテントセル 200μLを氷上で解凍し、上記(2)で作製したライゲーション産物10μLに加え、0℃で30分放置した。続いて、当該コンピテントセルに42℃で30秒間ヒートショックを与え、0℃で2分間冷却した。その後、SOC 培地 (20 mM グルコース、2%バクトトリプトン、0.5%バクトイーストエキス、10 mM NaCl、2.5 mM KCl 、1 mM MgSO4、1mM MgCl2) 1 mLを添加し、37℃にて1時間振盪培養した。培養後の培養液を各200μLずつ、LB Amp寒天培地(アンピシリン 100 mg/L 、1.5%寒天を含有するLB培地)にまき、37℃で一晩培養した。寒天培地上に生育した形質転換体コロニー複数個を 1.5 mLのLB Amp培地(アンピシリン 100 mg/Lを含有するLB培地)にて37℃で一晩培養した。得られた培養液を各々集菌後、Flexi Prep(GEヘルスケア バイオサイエンス社製)を用いて組換えプラスミドを回収した。得られたプラスミドの塩基配列をCEQ DTCS Quick Start Kitおよび蛍光シーケンサCEQ 2000XL DNA Analysis system(いずれもBECKMAN COULTER、米国)を用いて解析した。目的とするプラスミドをpTA002と命名した(図1参照)。
【実施例2】
【0051】
<アミダーゼ酵素溶液の調製>
(1)形質転換体の作製及び培養
実施例1と同様にして、大腸菌JM109株のコンピテントセルに対しプラスミドpTA002による形質転換を行った。得られた形質転換体JM109/pTA002のコロニーを、5 mL 2×LB培地(20 g/Lポリペプトン、10 g/L乾燥酵母エキス、20 g/L NaCl、50μg/mLアンピシリン)に植菌し、37℃ 、250rpmで12時間振盪培養を行い前培養とした。前培養液を集菌(8,000rpm 10分間)し菌体を沈殿させ、約100μLとなるよう上清を除去した。菌体ペレットをピペットマンで懸濁し、2 L modified TB培地(12 g/Lポリペプトン、24 g/L乾燥酵母エキス、40 g/Lグリセロール、12.5 g/L K2HPO4、2.3 g/L KH2PO4、0.5 g/L ポリプロピレングリコール12000、50μg/mLアンピシリン)に植菌し、37℃、180 rpmで培養し、これを本培養とした。培養開始より4時間後にIPTGを1 mMとなるよう添加し14時間の誘導を行った。
菌体懸濁液の調製は、サンプリングして得られた培養液から遠心分離(3,700 G、10分間、4℃)により菌体を回収し、100 mM トリス塩酸緩衝液(pH 8.0)で洗浄した後、同緩衝液に懸濁することにより行った。
【0052】
(2)アミダーゼ酵素溶液の調製
(i) 無細胞抽出液の調製
本培養を行った培養液に対して6,000 rpmで20分間、遠心分離を行い、菌体の洗浄として50 mMトリス塩酸(pH 8.0)溶液200 mLで懸濁した後、再度6,000rpmで20分間、遠心分離した。集菌した菌体ペレットを50 mMトリス塩酸(pH 8.0)で湿菌体重の2倍量に懸濁し、150Wで20分間、超音波破砕を行った。この超音波破砕液を無細胞抽出液とした。
【0053】
(ii) 粗酵素溶液の調製
無細胞抽出液に対して、破砕液を14,000 rpmで15分間、遠心分離を行い、沈殿物を除去した。70℃で40分間のヒートショックを行った後、14,000 rpmで20分間、遠心分離を行った。沈殿したタンパク質を除去し、上清を採取した。
【0054】
(iii) 精製酵素溶液の調製
上記(ii)で得られた無細胞抽出液熱処理物を、硫酸アンモニウム沈殿法により4℃で分別した。硫酸アンモニウムを30%の飽和濃度になるまで攪拌しながらゆっくりと加え、次いで、この溶液を4℃、12,000 Gで、20分間で遠心分離し、得られたペレットを200 mLの50 mMトリス塩酸緩衝液(pH 8.0)に懸濁した。
得られた硫安沈殿画分のタンパク質量5 mg当たり1 mLのDEAEトヨパール650H(TOSOH)をカラムに充填し、担体の5倍量の50 mMトリス塩酸緩衝液(pH 8.0)で平衡化を行った。その後、タンパク質溶液をアプライし、平衡化で用いたものと同組成のパッファーを担体の15倍量添加し洗浄を行った。洗浄後、溶出用バッファー(50 mM トリス塩酸緩衝液(pH 8.0)、100 mM KCl)を7.5倍量アプライし、ステップワイズで本酵素の溶出を行った。最後に、押し出しとして溶出用緩衝液(50 mMトリス塩酸緩衝液(pH 8.0)、500 mM KCl)を添加した。溶出液は約7 mL(200ドロップ)ずつ試験管に分取し、タンパク質の溶出は各画分のO.D.280 nmを測定することにより決定した。
アミダーゼ活性は、50 mMトリス塩酸緩衝液(pH 8.0)、100 mM KClにより溶出を開始し、アミダーゼ活性を有する画分を集めてフェニルトヨパールによる精製を行った。
【0055】
タンパク質量5 mg当たり1 mLのフェニルトヨパール650H(TOSOH)をカラムに充填し、担体の5倍量の洗浄緩衝液(50 mMトリス塩酸緩衝液(pH 8.0)、10%硫酸アンモニウム)で平衡化を行った。次に、タンパク質溶液に10%となるように硫酸アンモニウムを添加し、懸濁した。その後、硫酸アンモニウムを添加したタンパク質溶液をカラムにアプライし、平衡化で用いたものと同組成の緩衝液を担体の15倍量添加し洗浄を行った。洗浄後、溶出用緩衝液(50 mMトリス塩酸緩衝液(pH 8.0))を7.5倍量アプライし、ステップワイズで本酵素の溶出を行った。溶出液は約7 mL(200ドロップ)ずつ試験管に分取し、タンパク質の溶出は各画分のO.D.280 mmを測定することにより決定した。
アミダーゼ活性は、50 mMトリス塩酸緩衝液(pH 8.0)、10%硫酸アンモニウムにより溶出を開始し、アミダーゼ活性を有する画分を集めてSephacryI S-200 HRによる精製を行った。
約700 mLのSephacryI S-200 High Resolutiomをカラムに充填し、担体の1〜4%までサンプルを濃縮した。担体の2倍量の50 mMトリス塩酸緩衝液(pH 8.0)で、1 mL/分の流速で平衡化を行った。
その後、マーカー(5 mg Blue Dextran 2000/700μL平衡化緩衝液)、タンパク質溶液を順にアプライし、平衡化で用いたものと同組成の緩衝液を担体の2倍量添加し、流速は一定のまま溶出を行った。溶出液は約7 mL(200ドロップ)ずつ試験管に分取し、タンパク質の溶出は各画分のO.D.280 mmを測定することにより決定した。また、それぞれのタンパク質のピークに合わせて活性測定を行い、活性の高い画分を濃縮して、精製酵素溶液とした。
【0056】
(3)酵素活性の測定方法
アミダーゼの活性測定は、0.32 mM ロイシン-p-ニトロアニリン、100 mM トリス塩酸緩衝液(pH 9.0)に適当量のアミダーゼ溶液を加え、80℃にて30分間反応させた。氷中に入れることにより反応を停止させ、反応液の405nmの吸光度を測定して、生成したp-ニトロアニリン量を決定し、アミダーゼ活性とした。
【0057】
(4)タンパク質濃度(酵素濃度)の定量方法
Protein Assayキット(Bio-Rad)を用いた。適当な濃度に希釈した0.8 mLの酵素溶液に0.2 mLのアッセイ試薬を添加し、10〜15分室温に静置して、O.D.595 nmを測定することにより試料のタンパク質濃度を決定した。検量線の作製には、BSA(1.25, 2.5, 5.0, 7.5, 10μg/mL)を用いた。
【実施例3】
【0058】
実施例2で調製したアミダーゼ精製酵素画分を用いて、金属塩の添加効果を検討した。
すなわち、アミダーゼ精製酵素画分150μLに、塩化コバルト、塩化マンガン、塩化ニッケルを、それぞれ、終濃度5 mMとなるように添加し、37℃及び80℃で、それぞれ、1時間インキュベートし、次いで、10,000 rpmで5分間遠心処理を行い、沈殿物を除去した。その後、各々150 mLのトリス塩酸緩衝液(pH 8.0)に対して透析を2時間実施し、緩衝液を交換して同様にもう一度透析を行った。その後、各処理酵素液のタンパク質濃度及び活性を測定した。塩化コバルト、塩化マンガン及び塩化ニッケルの添加により、それぞれ、700%、740%及び640%(37℃)、並びに860%、1230%及び4800%(80℃)の活性向上が認められた。
【実施例4】
【0059】
実施例2で調製したアミダーゼ精製酵素画分を用いて、酵素溶液をEDTAで処理した後、金属塩の添加効果を検討した。
すなわち、1.8 mLのアミダーゼ精製酵素画分に100 mM EDTA (pH 8.0)を終濃度 5 mMとなるように添加した後、80℃で60分間静置し、その後、500 mLのトリス塩酸緩衝液(pH8.0)に対して透析を2時間実施し、緩衝液を交換して同様にもう一度透析を行なうことによりEDTAを除去した。EDTA処理した、アミダーゼ精製酵素画分150μLに塩化コバルト、塩化マンガン、塩化ニッケル、塩化カルシウム、塩化マグネシウム、塩化亜鉛をそれぞれ、終濃度5mMとなるように添加し、37℃及び80℃で、それぞれ、1時間インキュベートし、次いで、10000 rpmで5分間遠心処理を行い、沈殿物を除去した。その後、各々150 mLのトリス塩酸緩衝液(pH 8.0)に対して透析を2時間実施し、緩衝液を交換して同様にもう一度透析を行った。その後、各処理酵素液のタンパク質濃度及び活性を測定した。
【0060】
EDTA処理を行なうことにより、アミダーゼ活性は、処理前の6.6%に減少した。
このEDTA処理したアミダーゼ精製酵素画分に金属塩を添加することにより、塩化コバルト、塩化マンガン、塩化ニッケル、塩化カルシウム、塩化マグネシウム及び塩化亜鉛の添加により、無処理アミダーゼ精製酵素画分の活性に対して、それぞれ、196%、77%、115%、13%、16%及び37%(37℃)、並びに397%、287%、236%、44%、28%及び94%(80℃)の活性回復が認められた。
【実施例5】
【0061】
実施例2で調製したアミダーゼ精製酵素画分を用いて、実施例4に記載した方法と同様の方法にほり酵素溶液をEDTAで処理した後、塩化コバルト、塩化マンガン、塩化ニッケルの添加濃度の影響を検討した。
すなわち、アミダーゼ精製酵素画分150μLに塩化コバルト、塩化マンガン、塩化ニッケルをそれぞれ、終濃度1 mM、5 mMあるいは10 mMとなるように添加し、80℃で、1時間インキュベートし、次いで、10,000 rpmで5分間遠心処理を行い、沈殿物を除去した。その後、各々150 mLのトリス塩酸緩衝液(pH 8.0)に対して透析を2時間実施し、緩衝液を交換して同様にもう一度透析を行った。その後、各処理酵素液のタンパク質濃度及び活性を測定した。塩化コバルトの添加により、それぞれ、100%(1 mM)、490%(5 mM)及び980%(10 mM)、塩化ニッケルの添加により、それぞれ、50%(1mM)、280%(5 mM)及び305%(10 mM)、並びに塩化マンガンの添加により、それぞれ、100%(1 mM)、220%(5 mM)及び205%(10 mM)の活性向上が認められた。
【配列表フリーテキスト】
【0062】
配列番号3:合成DNA
配列番号4:合成DNA
【特許請求の範囲】
【請求項1】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼの保存剤。
【請求項2】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼの活性賦活剤。
【請求項3】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼの失活予防剤。
【請求項4】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼ保存用キット。
【請求項5】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼ活性賦活用キット。
【請求項6】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼ失活予防用キット。
【請求項7】
アミダーゼを、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を用いて処理することを含む、アミダーゼの保存方法。
【請求項8】
アミダーゼを、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を用いて処理することを含む、アミダーゼの活性賦活方法。
【請求項9】
アミダーゼを、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を用いて処理することを含む、アミダーゼの失活予防方法。
【請求項10】
前記処理に際し保温又は加熱することを含む、請求項7〜9のいずれか1項に記載の方法。
【請求項1】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼの保存剤。
【請求項2】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼの活性賦活剤。
【請求項3】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼの失活予防剤。
【請求項4】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼ保存用キット。
【請求項5】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼ活性賦活用キット。
【請求項6】
ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を含む、アミダーゼ失活予防用キット。
【請求項7】
アミダーゼを、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を用いて処理することを含む、アミダーゼの保存方法。
【請求項8】
アミダーゼを、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を用いて処理することを含む、アミダーゼの活性賦活方法。
【請求項9】
アミダーゼを、ニッケル、コバルト及びマンガン並びにそれらの塩からなる群より選ばれる少なくとも1種を用いて処理することを含む、アミダーゼの失活予防方法。
【請求項10】
前記処理に際し保温又は加熱することを含む、請求項7〜9のいずれか1項に記載の方法。
【図1】


【公開番号】特開2011−41550(P2011−41550A)
【公開日】平成23年3月3日(2011.3.3)
【国際特許分類】
【出願番号】特願2009−193689(P2009−193689)
【出願日】平成21年8月24日(2009.8.24)
【出願人】(000006035)三菱レイヨン株式会社 (2,875)
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
【公開日】平成23年3月3日(2011.3.3)
【国際特許分類】
【出願日】平成21年8月24日(2009.8.24)
【出願人】(000006035)三菱レイヨン株式会社 (2,875)
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
[ Back to top ]