
アミロース比率を増加させたデンプンを有する米およびその製品
【課題】アミロース比率を増加させたデンプンを有するイネを提供する。
【解決手段】デンプン枝作り酵素のレベルを低下させたイネは、胚乳中の相対的アミロース含量が高い穀粒を産生する。本発明の米粒は、アミロペクチン合成経路に損傷があるにもかかわらずnon−shrunken表現型とすることができ、またこの穀粒は、遺伝子導入によるものでも、遺伝子導入によらないものでもよい。
【解決手段】デンプン枝作り酵素のレベルを低下させたイネは、胚乳中の相対的アミロース含量が高い穀粒を産生する。本発明の米粒は、アミロペクチン合成経路に損傷があるにもかかわらずnon−shrunken表現型とすることができ、またこの穀粒は、遺伝子導入によるものでも、遺伝子導入によらないものでもよい。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、相対アミロース含量が高い穀粒デンプンを有するイネに関する。本発明はまた、胚乳中のデンプン枝作り酵素(starch branching enzyme)IIa(SBEIIa)活性を低下させた米に関する。本発明はまた、穀粒およびデンプン、ならびにそれらから得られる食品および非食品製品に関する。
【背景技術】
【0002】
穀類デンプンは、2種類の分子、アミロースおよびアミロペクチンを含む。アミロースは、α−1,4結合したグルコシド単位からなるほぼ直鎖状の分子であるが、アミロペクチンは、α−1,6グルコシド結合で繋がった直鎖で高度に分岐している。
【0003】
高等植物の胚乳中でのデンプンの合成は、4つの重要な段階を触媒する1揃いの酵素によって行われる。第1に、ADP−グルコースピロホスホリラーゼ(ADGP)により、G−1−PおよびATPからのADP−グルコースの合成を介して、デンプンのモノマー前駆体が活性化される。第2に、デンプン合成酵素により、この活性化されたグリコシルドナーであるADP−グルコースが、既存のα−1,4結合した鎖の非還元末端に転移する。第3に、デンプン枝作り酵素(SBE)により、α−1,4結合したグルカンのある領域の切断を介して枝分かれ部位が導入された後、この切断された鎖が受容鎖に転移し、したがって新たなα−1,6結合が形成される。SBEは、α−1,6結合をα−ポリグルカン中に導入することができる唯一の酵素であり、したがって、アミロペクチンの形成において必須の一役割を果たしている。最後に、デンプン枝切り酵素(starch debranching enzyme)により、分岐結合の一部が切り離されるが、この酵素の作用機序はまだ解明されていない(Myersら、2000)。少なくともこれら4つの活性が高等植物における通常のデンプン顆粒合成に必要であることは明らかであるが、高等植物の胚乳中に、これら4つの活性それぞれのアイソフォームが複数存在しており、突然変異解析に基づいて(Wangら、1998a;Buleonら、1998)、あるいは遺伝子導入手法を使用して遺伝子発現レベルを改変することにより(Abelら、1996;Joblingら、1999;Schwallら、2000)、個々のアイソフォームに特異的な役割が提唱されている。しかし、各活性のそれぞれのアイソフォームのデンプン生合成への正確な寄与はまだ分かっておらず、またこうした寄与が種間で著しく異なるかどうかも分かっていない。
【0004】
穀類の胚乳中に、ADP−グルコースピロホスホリラーゼの2種のアイソフォームが存在しており、1種はアミロプラスト内にあり、また1種は細胞質中にある(Denyerら、1996;Thorbjornsenら、1996)。それぞれのアイソフォームは2つのサブユニット型からなる。トウモロコシのshrunken(sh2)変異体およびbrittle(bt2)変異体は、それぞれラージサブユニットおよびスモールサブユニットにおいて損傷を表す(GirouxおよびHannah、1994)。穀類の胚乳中で4つのクラスのデンプン合成酵素が見つかっており、1つのアイソフォームは、デンプン顆粒内に排他的に局在する顆粒結合型デンプン合成酵素(granule−bound starch synthase)(GBSS)であり、2つの型は、顆粒画分と可溶性画分に分配されており(SSI:Liら、1999a;SSII:Liら、1999b)、また第4の型は、可溶性画分全体に位置しているSSIIIである(Caoら、2000;Liら、1999b;Liら、2000)。GBSSは、アミロース合成に必須であることが示されており(Shureら、1983)、SSIIおよびSSIIIにおける変異によりアミロペクチン構造が変化することが示されている(Gaoら、1998;Craigら、1998)。イネGBSS(waxy)遺伝子の配列は記載されており(Wangら、1990)、アンチセンス法により発現が阻害される(Teradaら、2000)。waxy遺伝子は、胚乳および花粉中で発現するが、イネの他の器官では発現しない(HiranoおよびSano、2000)。
【発明の概要】
【発明が解決しようとする課題】
【0005】
植物では、2つの主要なクラスのSBE、すなわちSBEIおよびSBEIIが知られている。穀類では、SBEIIはさらに、SBEIIaおよびSBEIIbの2つのタイプに分類することができる(BoyerおよびPreiss、1978;Gaoら、1996;Fisherら、1996;HedmanおよびBoyer、1982;Mizunoら、1992;Sunら、1997;Sunら、1998)。また、一部の穀類では、SBEの追加の型、すなわちコムギからの推定149kDaのSBEI(Bagaら、2000)およびオオムギからの50/51kDaのSBEが報告されている(Sunら、1996)。イネ(NakamuraおよびYamanouchi、1992;Mizunoら、1992;Mizunoら、1993;Mizunoら、2001)、トウモロコシ(Babaら、1991;Fisherら、1993;Gaoら、1997)、コムギ(Repellinら、1997;Nairら、1997;Rahmanら、1997)、および他の穀類ついては、ゲノム配列およびcDNA配列が特徴付けられている。配列アラインメントにより、ヌクレオチドおよびアミノ酸のレベルでの高度の配列類似性が明らかになっており、SBEI、SBEIIa、およびSBEIIbのクラスに分類することが可能となる。SBEIIaおよびSBEIIbは一般に、特にその遺伝子の中央の領域においてお互いに約80%の配列同一性を示す。
【0006】
SBEIIa、SBEIIb、およびSBEIを、胚乳および他の組織におけるその時間的および空間的発現パターンによって区別することもできる。SBEIは、コムギおよびトウモロコシでは胚乳発達中期以降に発現される(Morellら、1997)。それとは対照的に、SBEIIaおよびSBEIIbは、胚乳発達の初期段階から発現される。トウモロコシでは、SBEIIbは、胚乳中で主役の型であるが、SBEIIaは、葉中に高い発現レベルで存在する(Gaoら、1997)。イネでは、SBEIIaおよびSBEIIbは、胚乳中におよそ等しい量で存在する(YamanouchiおよびNakamura、1992)。しかし、発現するタイミングおよび組織に違いがあった。SBEIIaは、種子発達のより初期の段階で発現され、したがって開花後日数(DAF)3日で検出され、また葉で発現されるが、SBEIIbは、DAF3日では検出不可能であり、DAF7〜10日の発達中の種子の中に最も豊富にあり、葉で発現されなかった(Mizunoら、2001)。コムギの胚乳中では、SBEI(Morellら、1997)は、その可溶性画分に排他的に存在するが、SBEIIaおよびSBEIIbは、可溶性画分およびデンプン顆粒を伴う画分のどちらにも存在する(Rahmanら、1995)。
【0007】
高等植物には2つの型の枝切り酵素が存在し、この酵素はその基質特異性に基づいて定義されており、すなわちそれはイソアミラーゼ型枝切り酵素およびプルラナーゼ型枝切り酵素である(Myersら、2000)。トウモロコシおよびイネのSugary−1変異は、どちらの枝切り酵素の欠損も伴うが(Jamesら、1995;Kuboら、1999)、その原因となる変異は、イソアミラーゼ型枝切り酵素遺伝子と同じ場所に位置する。イネでは、イソアミラーゼのアンチセンス阻害によりアミロペクチンの構造およびデンプンの特性が変化し(Fujitaら、2003)、したがってイソアミラーゼがアミロペクチン生合成に必要であることが示された。
【0008】
穀類からクローニングされた代表的なデンプン枝作り酵素遺伝子を表1に記載する。
【0009】
【表1】

【0010】
トウモロコシおよびイネでは、高アミロースの表現型は、amylose extender(ae)遺伝子としても知られるSBEIIb遺伝子の損傷に起因することが示されている(BoyerおよびPreiss、1981;Mizunoら、1993;Nishiら、2001)。こうしたSBEIIb変異体では、胚乳デンプン粒が異常な形態を示し、アミロース含量が有意に増大し、残存するアミロペクチンの分岐頻度が減少し、短鎖の割合(<DP17、特にDP8−12)が低下した。さらに、このデンプンの糊化温度が上昇した。さらに、アミロースとアミロペクチンとの「中間」と定義される、材料の重要なプールがあった(Boyerら、1980;Takedaら、1993b)。イネでは、SBEIIbの不活性化により、約18%のアミロースを有する野生型イネに比べて、約25%のアミロース含量がもたらされた(Nishiら、2001)。
【0011】
それとは対照的に、変異誘発(Mu)挿入配列、およびその結果としてのSBEIIaタンパク質発現の欠損による、SBEIIa遺伝子におけるトウモロコシ変異体では、葉のデンプンは変化したが、胚乳のデンプンの分岐は野生型植物と区別がつかなかった(Blauthら、2001)。同様に、SBEIIa活性を欠くイネ植物は、胚乳中のアミロペクチン鎖のプロファイルの有意な変化を示さなかった(Nakamura、2002)。トウモロコシおよびイネのどちらにおいても、SBEIIa遺伝子およびSBEIIb遺伝子はゲノム中で連鎖していない。
【0012】
超高アミロースのトウモロコシ品種がしばらく前から知られている。超高アミロース含量(>90%)のLAPS(低アミロペクチンデンプン)トウモロコシは、SBEII活性のほぼ完全な不活性化と共に、SBEI活性の相当な低下によって得られた(Sidebottomら、1998)。
【0013】
ジャガイモでは、塊茎における主要なSBE(SBE B、SBEIに相当)がアンチセンス法によってdown regulationされると、デンプンの新規の特徴がいくつか得られたが、アミロース含量は変化しなかった(Saffordら、1998)。量が豊富でない型のSBE(SBE A、穀類のSBEIIに類似)がアンチセンスによって阻害されると、アミロース含量が穏やかに38%にまで増大した(Joblingら、1999)。しかし、SBEIIおよびSBEIの両方がdown regulationされると、相対アミロース含量がSBEII単独がdown regulationされるよりもずっと大幅に60〜89%にまで増大した(Schwallら、2000)。
【0014】
コムギでは、SGP−1(SSII)タンパク質を完全に欠くある変異体が、アミロペクチン構造について改変されており、デンプン顆粒が変形し、アミロース含量がデンプンの約30〜37%にまで上昇したが、それは野生型のレベルを約8%超える増加であった(Yamamoriら、2000)。比色測定、電流滴定(どちらもヨウ素結合用)およびコンカナバリンA法により、アミロースが測定された。SSIIの無発現変異体からのデンプンは、対応する非変異体植物のデンプンに比べて糊化温度が低下した。この穀粒のデンプン含量は、野生型の60%から50%未満に低下した。
【0015】
トウモロコシでは、dull1変異が、変化の程度が遺伝的背景に依存した、胚乳中のデンプン含量の低下およびアミロースレベルの増大をもたらし、残存するアミロペクチンの分岐の程度を増大させる(ShannonおよびGarwood、1984)。この変異に相当する遺伝子は、トランスポゾン変異誘発因子(Mu)を使用したトランスポゾンタギング戦略によって同定および単離され、デンプン合成酵素II(SSII)と呼ばれる酵素をコードしていることが示された(Gaoら、1998)。この酵素は現在、穀類におけるSSIIIファミリーのメンバーとして認められている(Liら、2003)。変異体の胚乳は、dull1変異に伴いSBEIIaの活性レベルが低下した。こうした発見が他の穀類、例えばイネに関連しているかどうかは分かっていない。
【0016】
穀粒のデンプン中のアミロース比率を増加させたオオムギの系統が同定されている。こうした系統には、相対アミロース含量が約45%であり、かつ穀粒デンプン中のアミロースレベルが約65〜70%にまで上昇する、オオムギのSSIIa遺伝子の化学的に誘発される突然変異を有するHigh Amylose Glacier(AC38)が含まれる(国際公開第02/37955A1号;Morellら、2003)。このデンプンは糊化温度の低下を示した。
【課題を解決するための手段】
【0017】
イネ(Oryza sativa L.)は、発展途上の世界において最も重要な穀類収穫物であり、広く、特にアジアで栽培されており、そこでの産出量は全世界の約90%である。
【0018】
デンプンは、食物、紙、および化学工業において広く使用されている。デンプンの物理的構造は、食品もしくは非食品、または工業製品にとってのデンプンの栄養特性および取扱適性に重要な影響を与えることができる。いくつかの特徴を、アミロペクチン鎖長分布、結晶化の程度およびタイプ、ならびに糊化温度、粘度、膨潤容積などの特性を含めたデンプン構造の指標とみなしてよい。アミロペクチン鎖長の変化は、アミロペクチンの変化した結晶化、糊化、または老化の指標としてよい。
【0019】
デンプン組成物、特に高アミロース含量に関連する可能性があるレジスタントスターチと呼ばれる型は、腸の健康、特に大腸の健康のために重要な意味を有する。したがって、トウモロコシやオオムギのようなある種の穀粒において、腸の健康を促進させる手段として食品に使用するために高アミロースデンプンが開発されている。レジスタントスターチの有益な効果は、大腸への栄養分の供給に起因しており、それにより腸のミクロフローラはエネルギー源を受け取り、このエネルギー源が発酵してとりわけ短鎖の脂肪酸が形成される。こうした短鎖の脂肪酸は、結腸細胞に栄養分を提供し、大腸全域でいくつかの栄養分の摂取を増強し、結腸の生理的活性を促進する。一般に、レジスタントスターチまたは他の食物繊維が供給されない場合、結腸は代謝が比較的不活発である。
【0020】
化学的に、または別の方法で改変されたデンプンは食品に利用することができ、こうした食品は、未改変源からは通常もたらされない機能性を提供するが、このような加工は、改変に伴うプロセスに起因して、価値のある他の成分を変化させるか、あるいは望ましくない感覚をもたらす傾向がある。したがって、改変されていない形で食品に使用することができる構成成分の供給源を提供することが好ましい。
【0021】
したがって、アミロースの割合が40%を超えるデンプンを有する米は知られていない。高アミロースのトウモロコシおよびオオムギの品種は知られているが、米作地域にとっては超高アミロース米の方が好ましい。このような米由来のデンプンは、比較的難消化性であり、したがって超高アミロース米が世界人口の相当な割合に健康上重要な利益をもたらすと期待されている。
【図面の簡単な説明】
【0022】
【図1】米デンプン枝作り酵素I遺伝子(SBEI)−Genbankアクセッション番号D11082(配列番号1)をコードしているcDNAの配列を示す図である。
【図2】米デンプン枝作り酵素IIa遺伝子(SBEIIa)−Genbankアクセッション番号AB023498(配列番号2)をコードしているcDNAの配列を示す図である。
【図3】米デンプン枝作り酵素IIb遺伝子(SBEIIb)−Genbankアクセッション番号D16201(配列番号3)をコードしているcDNAの配列を示す図である。
【図4】pBC SK−中に挿入された、イネSBEI遺伝子のイントロン9の500bp断片を含むプラスミドpRint9_BCの略図である。
【図5】2重鎖RNA構築体の略図である。A.使用した遺伝子エレメントの順序は、プロモーター、センス方向のSBEIIaまたはSBEIIbのcDNA配列、イントロン(Rint9)、アンチセンス方向のSBEIIaまたはSBEIIbのcDNA配列、および転写ターミネーター/ポリアデニル化配列であった。ds−SBEIIaおよびds−SBEIIb遺伝子の転写物は、センス配列とアンチセンス配列の間のハイブリダイゼーションによって形成される2本鎖領域を有する「へアピン」RNA構造を形成する。GTヌクレオチドおよびAGヌクレオチドと境を接するイントロン配列は、スプライシングによって切り取られる。また、pRBEI.IR(pBC SK−(センス/Rint9/アンチセンス))の略図を示す。対応する構築体、pRBEIIa.IRおよびpRBEIIb.IRも作製した。
【図6】プラスミドベクターpBx17casNOTの略図である。
【図7−1】イネSBEIIa(配列番号2)およびイネSBEIIb(配列番号3)のcDNA配列の比較を示す図である。上側の配列(大文字)はSBEIIbのものであり、下側の配列(小文字)はSBEIIaのものである。5'末端および3'末端の配列は、十分な同一性がないので示さない。
【図7−2】イネSBEIIa(配列番号2)およびイネSBEIIb(配列番号3)のcDNA配列の比較を示す図である。上側の配列(大文字)はSBEIIbのものであり、下側の配列(小文字)はSBEIIaのものである。5'末端および3'末端の配列は、十分な同一性がないので示さない。
【図7−3】イネSBEIIa(配列番号2)およびイネSBEIIb(配列番号3)のcDNA配列の比較を示す図である。上側の配列(大文字)はSBEIIbのものであり、下側の配列(小文字)はSBEIIaのものである。5'末端および3'末端の配列は、十分な同一性がないので示さない。
【図7−4】イネSBEIIa(配列番号2)およびイネSBEIIb(配列番号3)のcDNA配列の比較を示す図である。上側の配列(大文字)はSBEIIbのものであり、下側の配列(小文字)はSBEIIaのものである。5'末端および3'末端の配列は、十分な同一性がないので示さない。
【図8】遺伝子発現抑制構築体に用いられたSBEIIa、SBEIIb、およびSBEIの3'配列を使用した遺伝子発現抑制プログラム(Gene Silencing program)からのBLAST出力を示す図である。
【発明を実施するための形態】
【0023】
<総論>
当分野の技術者であれば、本明細書に記載した本発明が、具体的に記載したもの以外の変更および改変を受けることに気付くであろう。本明細書に記載した本発明はそのようなすべての変更および改変を含むことを理解されたい。また、本発明は、個々に、またはひとまとめにして本明細書において言及し、あるいは示すこのようなすべての工程、特徴、組成物、および化合物、ならびに前記工程または特徴の任意の2つ以上の任意のあらゆる組合せを含む。
【0024】
本明細書を通じて、「含む(comprise)」という用語、ならびに「含む(comprises)」および「含む(comprising)」などの変形は、文脈上他の意味に解すべき場合を除き、記載した1つの完全体(integer)もしくは段階、または完全体(integer)もしくは段階の群を包含することを意味し、他のいかなる完全体(integer)もしくは段階、または完全体(integer)もしくは段階の群も除外することを意味するわけではないことが理解されよう。本発明は、本明細書に記載した特定の実施形態による範囲に限定するものではなく、例示のみを目的とするものである。機能的に同等な生成物、組成物、および方法は、明らかに、本明細書に記載した本発明の範囲内にある。
【0025】
本明細書において著者名で示した刊行物の書誌詳細を本記載の最後にまとめる。本明細書に記載する参考文献の全体を参照により本明細書に組み込む。本明細書における先行技術に関する参考文献は、1種または複数の任意の先行技術の文献を含めて、前記先行技術がオーストラリアにおける共通の一般知識であり、またはオーストラリアにおける共通の一般知識の一部を形成するものであるという認識あるいは示唆と解釈すべきではない。
【0026】
本明細書では、「に由来する(derived from)」という用語は、特定の完全体(integer)または完全体(integer)の群が、指定の種を起源としたものであって、それを必ずしも指定の供給源から直接得たものではないことを指すと解釈すべきである。
【0027】
本明細書で表すヌクレオチド残基の名称は、IUPAC−IUBの生化学命名法委員会によって推奨されるものであり、Aはアデニン、Cはシトシン、Gはグアニン、Tはチミジンを指す。
【0028】
<発明の概要>
第1の態様では、本発明は、イネ植物から得られる穀粒にあるものとしてもよく、この穀粒は、そのデンプン中のアミロースの割合が少なくとも40%であるデンプンを含む。この穀粒は、SBEIIaおよびSBEIIbの活性またはレベルを低下させたものであることが好ましく、ある形態では、これを2つ以上の遺伝的変異によって実施してよく、1つの遺伝的変異は、a)SBEIIaの発現および/または活性を阻害するSBEIIa遺伝子の突然変異、ならびにb)SBEIIaの発現および/または活性を阻害する導入核酸からなる群から選択され、第2の遺伝的変異は、a)SBEIIbの発現および/または活性を阻害するSBEIIb遺伝子の変異、ならびにb)SBEIIbの発現および/または活性を阻害する導入核酸からなる群から選択される。
【0029】
ある形態では、この穀粒はある導入遺伝子を含み、この導入遺伝子は、アンチセンス、コサプレッション、リボザイム、または2重鎖RNA分子をコードするものでよい。あるいは、この穀粒は、その阻害が染色体突然変異または染色体再配列に起因する、導入遺伝子によらないものでもよい。この穀粒は、SBEIIaまたはSBEIIb遺伝子の無発現変異を含むものでもよい。
【0030】
第1の態様で請求した穀粒は、SBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたものを含むものでよい。ある特定の形態では、この穀粒はさらに、SBEIのタンパク質レベルおよび/または活性レベルを低下させたものを含むものでもよく、それに加えて、またはその代わりに、ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型の枝切り酵素、およびプルラナーゼ型の枝切り酵素からなる群から選択される、タンパク質レベルおよび/または酵素活性レベルを変更したものも含むことができる。代替の特定の形態では、この穀粒は、GBSSのタンパク質および/または酵素活性のレベルを変更したものを含む。さらなる任意の形態では、この穀粒は、インディカ品種のものであるか、または対立遺伝子Wxaを含む。
【0031】
好ましいある形態では、穀粒のデンプン中のアミロースの割合が少なくとも50%である。
【0032】
この穀粒は、non−shrunkenであることが好ましく、特定のあるフォーム、すなわち玄米のフォームでは、平均重量が少なくとも約25mgであり、また好ましくは、デンプン含量が、同等であるが未改変の穀粒の少なくとも90%である。また、好ましくは、穀粒内のデンプン顆粒の少なくとも50%は、偏光下で観察すると非複屈折性に見える。
【0033】
第2の態様では、本発明はまた、本発明の第1の態様の穀粒を産生することができるイネ植物を包含する。
【0034】
本発明の第3および第4の態様は、本発明の第1の態様の穀粒から抽出されたデンプンおよびデンプン顆粒に関する。
【0035】
第5の態様では、本発明は、本発明の第1の態様の穀粒から生成した穀粉またはデンプンを含む製品に関する。この生成物は、穀粉またはデンプンと別の供給源からの穀粉またはデンプンとの混合物を含むものでもよい。この製品は食品でも非食品製品でもよい。
【0036】
本発明の第6の態様は、第3の態様のデンプン、および別の食品成分または水を含む組成物を包含する。
【0037】
本発明の第7の態様は、少なくとも40%のアミロースを含むデンプンを有する穀粒を産生することができるイネ植物を作出する方法にあると言ってもよく、この方法は、a)遺伝的変異を親イネ植物または種子中に導入するステップと、b)親イネ植物または種子親の子孫植物を同定するステップとを含み、この子孫植物の穀粒のデンプンが少なくとも40%のアミロースを含む。好ましくは、この遺伝的変異により、米植物の胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または活性のレベル低下がもたらされる。
【0038】
この方法の子孫イネ植物は、2つ以上の遺伝的変異を含むことが好ましく、1つの遺伝的変異が、a)SBEIIaの発現および/または活性を阻害するSBEIIa遺伝子の突然変異、ならびにb)SBEIIaの発現および/または活性を阻害する導入核酸からなる群から選択され、また第2の遺伝的変異が、c)SBEIIbの発現および/または活性を阻害するSBEIIb遺伝子の突然変異、ならびにd)SBEIIbの発現および/または活性を阻害する導入核酸からなる群から選択される。
【0039】
遺伝的変異を導入するステップは、外因性の核酸を導入することを含むものでよい。外因性の核酸をイネ細胞中に導入し、次いでこの細胞をイネ植物に再生させてもよい。この外因性の核酸は、SBEIIaおよび/またはSBEIIbの発現および/または活性の阻害物質をコードすることが好ましく、この阻害物質は、アンチセンス、コサプレッション、リボザイム、または2重鎖RNA分子であってよい。
【0040】
あるいは、遺伝的変異を導入するステップは、化学薬剤または放射線を用いた親イネ植物または種子の突然変異誘発を含むものでもよい。
【0041】
この子孫イネ植物は、SBEIIaおよび/またはSBEIIbにおける無発現変異を含むものでもよい。
【0042】
遺伝的変異を導入するステップはさらに、SBEIのタンパク質レベルおよび/または活性のレベルの低下に導くものでもよい。
【0043】
この子孫植物を、穀粒デンプン中のアミロースレベルに基づいて、あるいは子孫植物の胚乳中のSBEIIaおよび/またはSBEIIbのタンパク質レベルおよび/または活性のレベルの低下に基づいて同定してもよい。
【0044】
この方法はさらに、対立遺伝子Wxaのイネへの導入を含むものでもよく、これを交配によって導入してもよい。
【0045】
本発明の第8の態様は、胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または酵素活性のレベルを低下させたイネ植物を作出する方法にあるものとしてもよく、この方法は、a)SBEIIaのタンパク質レベルおよび/または酵素活性レベルを低下させた種子に突然変異を起こさせること、あるいはb)SBEIIbのタンパク質レベルおよび/または酵素活性レベルを低下させた種子に突然変異を起こさせること、あるいはc)SBEIIaのタンパク質レベルおよび/または酵素活性のレベルを低下させた植物を、SBEIIbのタンパク質レベルおよび/または酵素活性のレベルを低下させた植物と交配させること、ならびにd)胚乳中のSBEIIaおよびSBEIIbのタンパク質活性および/または酵素活性を低下させたイネ植物を同定することを含む。
【0046】
このイネ植物を同定するステップは、イネのSBEIIa遺伝子およびSBEIIb遺伝子に連鎖する分子マーカーを有するイネ植物の集団をスクリーニングすること、ならびに連鎖した分子マーカーを用いたスクリーニングからのシグナルの存在または非存在に基づいて植物を同定することを含むものでよい。
【0047】
このイネ植物を同定するステップは、イネのSBEIIaタンパク質およびSBEIIbタンパク質に結合する抗体を用いてイネ植物の集団からの種子をスクリーニングするステップ、ならびに結合した抗体の存在または非存在に基づいて植物を同定するステップを含むものでもよい。
【0048】
本発明はまた、改変した米デンプンを産生する方法を包含してもよく、この方法は、本発明の第1の態様の穀粒からデンプンを抽出するステップを含む。
【0049】
本発明はさらに、2つ以上の外因性の核酸分子の使用を包含してもよく、SBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたイネ植物を作出するために、その少なくとも1つがイネのSBEIIaの発現および/または活性の阻害物質をコードしており、またその少なくとも別の1つがイネのSBEIIbの発現および/または活性の阻害物質をコードしている。この阻害物質は、アンチセンス分子、コサプレッション分子、リボザイム、2重鎖RNA分子、およびそれらの任意の組合せからなる群から選択することができる。
【0050】
本発明はまた、イネSBEIIaの阻害物質およびイネSBEIIbの阻害物質をコードしている単離された核酸分子も包含しており、これらの阻害物質は、同一の分子であっても、異なる分子であってもよい。この単離されたベクターはベクターでよい。本発明が、単離された核酸分子を含む細胞を包含し、この細胞がイネ細胞であることが好ましいことを理解されたい。本発明はまた、単離された核酸分子を含むトランスジェニックイネ植物を包含することを理解されたい。
【0051】
<発明の詳細な説明>
イネ植物を作出する方法
一態様では、本発明は、穀粒中に改変したデンプンを有するイネ植物を作出する方法、特にデンプン中のアミロースの相対的な割合を少なくとも40%まで増加させる方法を提供する。本明細書で定義するデンプン中のアミロースの割合は、重量/重量(w/w)に基づき、すなわちデンプンの重量に対する百分率で示すアミロースの重量である。デンプン中のアミロースの割合は、少なくとも45%、50%、55%、または60%であることが好ましく、また少なくとも65%、70%、または75%であることがさらにより好ましい。米では通常、デンプン中のアミロースの割合は、約0〜35%の範囲にわたる。この方法には、米胚乳中のデンプン枝作り酵素IIa(SBEIIa)およびデンプン枝作り酵素IIb(SBEIIb)のタンパク質レベルまたは酵素活性レベルを低下させることが含まれ得る。このタンパク質または活性の低下は、改変していない米胚乳中のタンパク質または活性の相当するレベルに比べて、少なくとも40%、または好ましくは少なくとも60%、より好ましくは少なくとも75%、さらにより好ましくは少なくとも90%または95%でよい。米胚乳中でこのタンパク質のうちの一方または両方を検出できない可能性がある。この方法は、米のSBEIIaおよびSBEIIb遺伝子の発現の改変を含むものでよく、あるいは米におけるSBEIIaおよびSBEIIb遺伝子の突然変異またはこれらの組合せを含むものでよく、これにより胚乳中のSBEIIaおよびSBEIIbの活性がどちらも低下する。この遺伝子のうちのどちらかまたは両方の発現を、核酸の導入、例えば導入遺伝子によって阻害してよい。
【0052】
本明細書では、「増大した(increased)」、「減少した(decreased)」、「低下した(reduced)」、および「改変した(altered)」などの用語は、本発明に従って改変されていない対応する野生型の植物または生成物に比べての、本発明の植物または生成物の違いを指す比較用語であることは容易に明らかであろう。
【0053】
本明細書では、アミロースを、α−1,4結合グリコシド分子、およびアミロース様長鎖アミロペクチン(「中間物質」と呼ぶことがある、Takedaら、1993b;Fergason、1994)からなるほぼ直鎖状の分子を含むものと定義する。アミロース含量を、例えば90%(w/v)DMSO中でのサイズ排除HPLC、コンカナバリンA法(Megazyme社、アイルランド)、または好ましくは例えば実施例1に記載するヨウ素滴定法を含めた当技術分野で既知のいずれかの方法によって求めることができる。HPLC法は、デンプンの枝切りを伴うものでも(BateyおよびCurtin、1996)、枝切りを伴わないものでもよい。穀粒重量およびアミロース含量から、トランスジェニック系統および対照系統について、穀粒あたりの蓄積されたアミロースの量を計算し比較することができる。
【0054】
この方法は、米胚乳中のSBEIIaおよび/またはSBEIIbの活性を、好ましくは両方求めるステップを含むものでよい。これを、例えば免疫検出によりタンパク質のレベルを測定すること、あるいは、ノーザンブロットハイブリダイゼーション分析、スロットブロットハイブリダイゼーション、RNAseプロテクションアッセイ、マイクロアレイ分析、または逆転写ポリメラーゼ連鎖反応(RT−PCR)など当技術分野で既知の方法により、それらに対応するmRNAのレベルを測定することによって行ってもよい。この方法はさらに、胚乳中のSBEIIaおよび/またはSBEIIbの活性を低下させたイネ植物または穀粒をスクリーニングするステップ、あるいはこのような植物もしくは穀粒を選択または同定するステップを含むものでもよい。スクリーニング/選択のステップは、SBEIIaおよび/またはSBEIIbの活性レベルまたはタンパク質レベルを低下させることに基づくものでもよく、あるいは、増大させたアミロースの割合、または低下させたアミロペクチンの割合、または視覚的表現型、例えばShrunken穀粒などイネ植物の穀粒の表現型に基づくものでもよい。
【0055】
酵素アッセイ、例えばホスホリラーゼ刺激アッセイ(Phosphorylase stimulation assay)(BoyerおよびPreiss、1978)により、SBE活性を測定してよい。このアッセイでは、ホスホリラーゼaによるグルコース1−リン酸のメタノール不溶性ポリマー(α−D−グルカン)中への組込みの、SBEによる刺激が測定される。SBE活性を、グルカンポリマーの枝切りによって生じるグルカン−ポリヨウ素複合体の吸光度の減少を測定するヨード染色検査によって測定することができる。SBE活性を、イソアミラーゼ消化の後、基質である還元アミロースからの還元末端の形成を測定する分岐結合アッセイ(branch linkage assay)によって分析することもできる(Takedaら、1993a)。好ましくは、この活性をSBEI活性なしで測定する。SBEのアイソフォームは様々な基質特異性を示し、例えばSBEIは、アミロースを分岐させる際により高い活性を示すが、SBEIIaおよびSBEIIbは、アミロペクチン基質での分岐をより高い割合で示す。また、このアイソフォームを、転移するグルカン鎖の長さに基づいて区別してもよい。また、SBEタンパク質を、本明細書に記載するものなど特異的抗体を使用することによって測定してもよい。好ましい一実施形態では、イネSBEIIaおよびSBEIIbのN末端アミノ酸配列に相当するポリペプチド断片に対して産生された特異的抗体を使用した、ウエスタンブロット法またはELISA検査などの免疫学的方法により、SBEIIaおよびSBEIIbのタンパク質レベルを測定する。SBEII活性を、穀粒発達中の発達している胚乳中で、あるいはこのタンパク質が同等であるが未改変の穀粒中にまだ存在しており、それを免疫学的方法によって検査することができる成熟した穀粒中で測定してもよい。
【0056】
さらなる一態様では、本発明は、穀粒のデンプン中のアミロースの割合が少なくとも40%となるように、SBEIIaおよびSBEIIbの活性の低下と組み合わせて、イネにおける第3のデンプン生合成酵素の活性を改変する、好ましくは低下させる方法を提供する。好ましくは、胚乳中のSBEI活性も低下させる。SBEIIaおよびSBEIIbと組み合わせて改変してよい他のデンプン生合成の酵素活性は、SSI、SSII、およびSSIIIである。また、デンプン枝切り酵素、例えばイソアミラーゼまたはプルラナーゼ活性を改変してもよい。第3のデンプン生合成の酵素活性を、改変していないイネにおける活性に比べて、少なくとも40%、好ましくは60%または80%、より好ましくは少なくとも90%までに上昇または低下、好ましくは低下させてよい。
【0057】
さらなる一実施形態では、この植物のデンプン生合成酵素の活性を胚乳以外の植物組織中で改変してよく、例えばSBEIまたはSBEII、好ましくはSBEIIaの活性を葉において増大させて、胚乳中のSBEIIa活性の喪失に導く、この植物における遺伝的変異によって引き起こされる活性喪失の一部を補ってもよい。これは、遺伝的変異が、胚乳中だけでなく他の組織、特に葉においてSBEIIa活性の低下に導く場合に特に好ましい。胚乳以外の組織中でのこのような活性の補償は、例えば、胚乳中で発現しないプロモーターの制御下にある酵素コード領域に起因するであろうと理解されよう。これは、rbcSなど光合成に関係する遺伝子からのプロモーターでよい。あるいは、胚乳中のデンプン合成を、胚乳中のSBEIIaおよびSBEIIbの活性の低下と組み合わせた、1種または複数のデンプン生合成の酵素の過剰発現によってさらに改善してもよい。このような酵素をコードしている遺伝子は、様々な任意の供給源からのもの、例えば細菌や、イネ以外の他の供給源からのものでもよく、またこの遺伝子を改変して、触媒活性を変更、例えば酵素の温度依存を変更(WO94/09144)してもよい。
【0058】
高アミロース表現型は、SBEIIa遺伝子およびSBEIIb遺伝子の発現に対する部分的な、または十分な撹乱によって得てよい。本発明の方法は、SBEIIaおよび/またはSBEIIb遺伝子における無発現変異を有するイネ植物または穀粒をスクリーニングまたは同定または選択するステップを含むものでよい。本明細書では、「無発現変異」は、対象の植物組織中、好ましくは胚乳中の検出可能なタンパク質または酵素活性の欠損を起こす突然変異であると定義する。したがって、スクリーニング/同定するステップは、遺伝子レベルでの、例えばSBEIIaおよび/またはSBEIIbをコードしている遺伝子における欠失のためのスクリーニング、あるいは対象の遺伝子の発現のレベルでのスクリーニングを含むものでよい。この遺伝子が阻害される程度により、米粒中で形成されるデンプンの特徴がある程度決定される。好都合には、欠失のスクリーニングは、増幅産物の少なくとも一部が対象の遺伝子の少なくとも一部に及ぶように設計されたプライマーを使用したPCR増幅法によって好都合に実施してよい。改変した米胚乳から抽出されたタンパク質について実施される一連のゲル電気泳動法のいずれかにより、SBEIIaおよびSBEIIbの活性に対する改変の性質および程度が明らかになるであろう。改変は、SBEIIaおよび/またはSBEIIb活性の低下、または胚乳内の酵素活性の完全な消滅として起こるものでよい。こうした試験を実施するために、デンプンを米胚乳から抽出し、そのタンパク質を分析してよい。SDS−PAGEおよびイムノブロット法などの当技術分野で周知の技法を、可溶性画分およびデンプン顆粒画分について実施してよく、改変がSBEIIaおよびSBEIIb酵素に対して起こった植物または穀粒が同定される。
【0059】
本発明の方法は、イネ植物またはイネ祖先植物または種子への遺伝的変異の導入を含むものでよい。この遺伝的変異は、以下に記載するように導入遺伝子を含むものでよく、あるいは突然変異誘発によって、例えば化学的突然変異誘発によって、または放射線によって導入してもよい。
【0060】
イネ植物
さらなる一態様では、本発明は、デンプン中のアミロースの割合が少なくとも40%の穀粒を産生することができるイネ植物を提供する。本明細書では、イネ植物を、種Oryza sativa L.のいずれかであると定義する。このイネ植物は、O. sativa L.、すなわちJaponica(またはsinica)、Indica、およびJavanicaの認識されている3つの品種のいずれかのものでよく、Indica品種であることが好ましい。ぞれぞれの品種には栽培品種または変種が多数あり、すべて本発明の植物に含まれる。好ましい栽培品種は、例えば、栽培品種Amaroo、Ali Combo、Basmati、Bogan、Bombia、Doongara、Goolarah、Illabong、Jarrah、Koshihikari、Kyeema、Langi、Millin、Namage、Opus、Peldeを含めて、オーストラリアで栽培されているものである。アミロースの割合は、少なくとも45%、50%、55%、60%、65%、70%、または75%であることが好ましい。このイネ植物は、胚乳中のSBEIIaおよび/またはSBEIIbの発現および/または活性を阻害する少なくとも1種の遺伝的変異を含む。この遺伝的変異は、SBEIIaおよび/またはSBEIIbの発現または活性、ならびに上記のものの組合せを阻害する、SBEIIaおよびSBEIIb遺伝子における突然変異や、アンチセンス、増強されたアンチセンス、コサプレッション、リボザイム、2重鎖RNA、または類似の分子をコードしている遺伝子などの導入核酸など、米胚乳中のSBEIIaおよびSBEIIbの活性および/またはタンパク質の低減に導く任意の遺伝的変異でも、遺伝的変異の組合せでもよい。この遺伝的変異は、無発現変異であることが好ましい。SBEIIaおよびSBEIIbの活性を低下させた植物は、SBEIIaを低減させた植物を、SBEIIbを低減させた植物と交配させることによって、あるいはSBEIIaおよびSBEIIb遺伝子のどちらの発現も阻害する分子をコードしている導入遺伝子を導入することによって作出してよい。好ましい一実施形態では、このイネ植物は、SBEIIaおよびSBEIIbのどちらにおいても無発現変異を有する。
【0061】
本発明はまた、穀粒の発達の少なくとも一部の期間に胚乳中のSBEIIaおよびSBEIIb活性のレベルをどちらも低下させたイネ植物を提供し、このイネ植物は、同等であるが未改変の植物から抽出されたデンプンに比べて、高率のアミロースを含むデンプンを有する穀粒を生み出すことができる。好ましくは、SBEIIaおよびSBEIIbのレベルを、野生型に比べて、胚乳中で少なくとも50%、より好ましくは少なくとも75%、最も好ましくは少なくとも90%または95%低下させる。「野生型」という用語は、遺伝学の分野におけるその通常の意味を有し、それには、本明細書で教示するよう改変していないイネの栽培品種または遺伝子型が含まれる。
【0062】
本発明はまた、遺伝型および/または表現型において親イネ植物の所望の特徴を有する子孫植物および穀粒を提供する。本発明はまた、培養した組織または細胞など所望の特徴を有する植物を作出するのに使用できるイネ植物の任意の繁殖用材料にまで及ぶ。
【0063】
本発明の改変したイネ植物は、より望ましい遺伝的背景を含む植物と交配してよく、したがって、本発明は、他の遺伝的背景における1種(複数)の遺伝的変異を含む。最初の交配の後、戻し交配を適当な回数実施して、望ましくない背景を除去してもよい。所望の遺伝的背景には、商業収益、および農業生産力や非生物ストレス耐性など他の特徴を提供する遺伝子の適当な組合せが含まれ得る。また、この遺伝的背景には、改変した他のデンプン生合成、または修飾遺伝子、例えば、その原因となる遺伝子が分かっていないshrunken胚乳を有する他のイネ系統からの遺伝子が含まれ得る。
【0064】
好ましい一実施形態では、このイネ植物は、waxy遺伝子の対立遺伝子Wxaを含む。この対立遺伝子は大部分がイネのインディカ品種にみられるが、対立遺伝子Wxbは大部分がジャポニカ品種にみられる。対立遺伝子Wxbは、waxy遺伝子の第1イントロンの5'スプライス部位での置換突然変異(GTからTTに)を有し、その結果、対立遺伝子Wxaを含む相当する植物中よりも、waxy遺伝子発現が低く、したがってGBSS活性が低く、またアミロースレベルが低くなる(Isshikiら、1998;Hiranoら、1998;Francesら、1998)。
【0065】
この植物またはその穀粒は、遺伝子導入によるものでも、遺伝子導入によらないものでもよい。
【0066】
穀粒
本発明はまた、同等であるが未改変のイネから抽出されたデンプンに比べて、改変されたデンプンを含む米穀粒を提供する。本明細書では、穀粒をほぼ成熟した穀粒と定義する。この穀粒には、商業的環境で収穫された穀粒が含まれる。収穫時、米穀粒は、外皮を含む籾米の形でも、外皮が除去された「玄米」の形でもよい。玄米部分のみが食用である。玄米は、果皮、種皮および珠心の外層、胚芽(胚)、ならびに胚乳からなる。本明細書で定義する胚乳は、アリューロン層、および、サブアリューロン層とデンプン質の胚乳または内胚乳からなる本来の胚乳とからなる。玄米を研磨精米または摩擦精米にかけて、果皮、種皮、外種皮、アリューロン層、および胚を除去して、それにより本質的にデンプン質の胚乳を含む白米を得ることができる。精米の結果、脂肪、タンパク質、繊維、無機質、ならびにチアミン、リボフラビン、ナイアシン、a−トコフェロールなどのビタミンを含めた、種皮およびアリューロンの成分が一部失われる。炭水化物含量、主にデンプンは、玄米中よりも精米された米の中の方が多い。野生型の精米された米の穀粒は、炭水化物を約77〜89%、タンパク質を6.3〜7.1%、デンプンと結合した形のものもデンプンなしの形のものも含めた脂質を1.5〜1.7%、無機質を0.3〜0.8%、粗繊維を0.3〜0.5%、および中性デタージェント繊維を0.7〜2.3%含有しており、水分を約10〜15%含有するしていてよい。本明細書で定義する「米穀粒」または単に「米」には、籾米、玄米、および精米された米が含まれ、精米された米であることが好ましい。
【0067】
本発明の米穀粒は、この穀粒が由来するイネ植物について本明細書で定義する少なくとも1つの遺伝的変異を含む。1種(複数)の遺伝的変異は、米穀粒の胚乳の発達中に、SBEIIaおよびSBEIIbの活性および/またはタンパク質の低下をもたらす。この穀粒は、同等であるが未改変の植物の穀粒に比べて、(全デンプンに対する百分率で示す)アミロースが高率のもの、およびアミロペクチンの割合を低下させたものを含む。デンプンは、約90%の乾物を含む白米の主要成分である。本発明に従って改変されていない米胚乳デンプンのアミロース含量は、0〜37%の範囲にあるが、遺伝子型に依存する。「見掛けのアミロース含量」を測定するためのヨウ素滴定(比色)分析に基づき、精米された米は、waxy(1〜2%アミロース)、超低アミロース(2〜12%)、低アミロース(12〜20%)、中アミロース(20〜25%)、および高アミロース(25〜33%)に分類される(Juliano、1979, 1985)。HPLC分析を使用した最近の研究では、真のアミロース含量の最大値が約20%であり、追加のヨウ素結合がアミロペクチンの長い直鎖によるものであることが示された(Takedaら、1987)。「アミロース含量」または本明細書で定義する「見掛けのアミロース含量」を、当分野の技術者に既知のヨウ素滴定法、例えばMorrisonおよびLaignelet(1983)によって記載されている分光光度法によって求める。「真のアミロース」のみを分析する高速液体クロマトグラフィー(例えばHPLC、BateyおよびCurtin、1996)法などの他の方法は、本明細書で定義するアミロース含量を低く評価する可能性があることを理解されたい。
【0068】
本発明の穀粒は、少なくとも40%(w/w)のアミロースを含むデンプンを有する。アミロースの割合は、好ましくは全デンプンの少なくとも45%、50%、または55%であり、より好ましくは少なくとも60%であり、さらにより好ましくは少なくとも65%、70%、または75%である。好ましい一実施形態では、この穀粒は、遺伝子導入によるものではなく、このデンプンは、少なくとも40%のアミロースを含む。あるいは、この穀粒は、SBEIIaとSBEIIbのタンパク質レベルを低下させたものであり、このデンプンは少なくとも40%のアミロースを有する。増大したアミロースレベルを、光学顕微鏡下で観察した場合の異常なデンプン顆粒の形態、またはこの顆粒の複屈折性の喪失、あるいは当技術分野で既知の他の方法によって証明してもよい。
【0069】
好ましい一実施形態では、このイネは、インディカ品種のものであるか、またはwaxy遺伝子の対立遺伝子Wxaを含む。
【0070】
この穀粒は、物理的特徴を改変した、例えば、糊化温度を上昇もしくは低下させた、かつ/または糊化中と糊化後の膨潤特性を改変したデンプンを含むものでよい。
【0071】
この穀粒は、shrunkenでもnon−shrunkenでもよく、好ましくはnon−shrunken表現型を有する。本明細書では、「non−shrunken」を、大多数の穀粒、好ましくは個々の穀粒の少なくとも90%が丸々とした(plump)または実入りの十分な(fully−filled)表現型を示す場合と定義する。これは通常、正常な、もしくは正常に近いレベルのデンプン蓄積を伴う。それとは対照的に、本明細書では、表現型「shrunken」は、デンプン蓄積が低下した大多数の穀粒、特に少なくとも90%の穀粒を指す。slightly shrunken穀粒は、少なくとも30%の平均デンプン含量の低下を指し、moderately shrunken穀粒は、少なくとも50%の平均デンプン含量の低下を指し、highly shrunken穀粒は、少なくとも70%の平均デンプン含量の低下を指す。また、収縮性(shrunkenness)は、成熟穀粒重量に対する百分率で示す相対デンプン含量によって測定してよい。野生型玄米の穀粒のサイズおよび形のパラメータは、極長(extra long)>7.50mm、長(long)6.61〜7.50mm、並(medium)5.51〜6.60mm、短(short)<5.50mmと定義してよい。穀粒の形は、長さと幅の比に基づいて特徴付けてよく、細長(slender)>3.0、並(medium)2.1〜3.0、太(bold)1.1〜2.0、丸(round)<1.0と定義する。本発明の米粒ではこうした特徴をそれぞれ改変してもよい。
【0072】
本発明はまた、この穀粒から生成される穀粉、荒粉、または他の生成物を提供する。これらは、例えば分別または漂白によって加工されたものでも、加工されていないものでもよい。本発明はさらに、本発明のイネ植物から得られる、食糧生産に有用な米穀粒を提供する。さらに、本発明は、この穀粒を精米し、挽き、圧延し、精白し、粗挽きもしくは挽割りにし、またはゆでるといった、他の方法で加工された穀粒を包含する。
【0073】
デンプン
別の態様では、本発明は、アミロースの割合を増大させ、アミロペクチンの割合を低下させた、上記のイネ植物の穀粒から得られるデンプン顆粒またはデンプンを提供する。この穀粒が得られる植物は、胚乳中のSBEIIaおよびSBEIIbの活性レベルを低下させたものであり、より好ましくは、SBEIの活性も低下させる。別の態様では、本発明は、このイネ植物の穀粒から得られるデンプン顆粒またはデンプンを提供し、これらは、少なくとも40%のアミロース、好ましくは少なくとも45%、50%、55%、または60%のアミロース、さらにより好ましくは少なくとも65%、70%、75%のアミロースを含む。精製されたデンプンを、タンパク質、油、および繊維からのデンプンの分離を含む製粉加工、例えば湿式製粉加工により穀粒から得てよい。製粉加工の最初の生成物は、デンプン顆粒の混合物または組成物であり、したがって本発明はこのような顆粒を包含する。
【0074】
野生型米デンプン顆粒は、形が多角体であり、その大部分はサイズが3〜9μmで、サイズの単峰性分布で平均して約5μmである。タンパク質は主に、胚乳全体にわたってサイズが0.5〜4μmの球状のタンパク粒の形で生じる。
【0075】
このデンプンは、糊化温度を上昇させたものでも、低下させたものでもよく、糊化温度を上昇させたものであることが好ましい。この糊化温度、特に最初のピークの開始温度、または最初のピークの最高点の温度を、DSCによって測定して、類似のものであるが改変されていない穀粒に比べて、少なくとも3℃、好ましくは少なくとも5℃、またはより好ましくは少なくとも7℃上昇させてもよい。このデンプンは、レジスタントスターチのレベルを上昇させたものを含むものでもよく、改変された構造は、変化したデンプン顆粒形態、相当量のデンプン結合脂質の存在、変化した結晶化、および変化したアミロペクチン鎖長分布に起因する可能性がある、消化酵素への物理的接近の困難さからなる1種または複数の群を含めた特定の物理的特徴によって示される。アミロースの割合が高いこともレジスタントスターチのレベルに寄与している。
【0076】
本発明はまた、好ましくはレベルを上昇させたレジスタントスターチと組み合わせて、食物繊維の量を増加させたものを含む例示されたイネ植物の穀粒から得られるデンプンを提供する。また、この増加は少なくとも部分的に、アミロースの相対的レベルが高い結果である。
【0077】
遺伝子活性を低下させる方法:導入遺伝子
SBEIIa、SBEIIb、または他のデンプン生合成遺伝子もしくは修飾遺伝子の活性を、遺伝的変異をイネ植物に導入することによって改変することが好ましい。これは、導入遺伝子のイネ植物への導入によるものでよい。「遺伝的変異」は、この文脈においてSBEIIaおよびSBEIIb、ならびに場合によっては他のデンプン生合成遺伝子もしくは修飾遺伝子の活性の低下をもたらすゲノムの任意の改変を意味し、この変異には、点突然変異、置換、逆位、重複、転座、好ましくは欠失、ならびに遺伝子または制御エレメント中への導入遺伝子の導入などの突然変異が含まれる。好ましい一実施形態では、この遺伝的変異は、例えば逆位、重複、転座、欠失、フレームシフト、またはRNAスプライシング変異の結果である無発現変異である。本明細書では、「導入遺伝子」は、バイオテクノロジーの分野における通常の意味を有し、これには、組換えDNA技術またはRNA技術によって生成または改変され、対象の生物または細胞中に導入される遺伝子配列が含まれる。この導入遺伝子には、生物または細胞に由来する遺伝子配列、例えばアンチセンス配列が含まれ得る。好ましい一実施形態では、この導入遺伝子は、本明細書で定義するイネSBEIIa配列または本明細書で定義するイネSBEIIb配列の相補体である少なくとも19の連続したヌクレオチドと少なくとも94%の同一性を有する、少なくとも19の連続したヌクレオチドを有するヌクレオチド配列を含む。この導入遺伝子には通常、イネに由来してない外因性の核酸が含まれる。「トランスジェニック」とは、導入遺伝子を含むイネ植物または穀粒または細胞を指す。「非トランスジェニック」とは、イネ植物のゲノム、穀粒、または細胞中に全く導入遺伝子が存在しないことを指す。安定した遺伝のために、導入遺伝子をイネ植物のゲノム、穀粒、または細胞中に組み込むことが好ましい。
【0078】
本明細書で言及する「遺伝子」は、SBEIIa、SBEIIb、または他のデンプン生合成遺伝子、あるいはアンチセンス、増強されたアンチセンス、コサプレッション、リボザイム、2重鎖RNAなどをコードしている遺伝子を含めて、その最も広範な文脈において解釈すべきであり、この遺伝子には、ゲノムの遺伝子、ならびに存在するなら遺伝子のコード領域(すなわちエキソン)に対応するmRNAまたはcDNA、転写されるが翻訳されない配列、およびプロモーターと、転写ターミネーター/ポリアデニル化配列とを含めた調節領域が含まれる。また、「遺伝子」という用語は、機能的生成物のすべて、または一部をコードしている合成分子または融合分子を説明するのにも使用される。好ましい遺伝子は、標準の組換え技術による、自然に存在するSBEIIa、SBEIIb、またはデンプン生合成遺伝子に由来する。通常、遺伝子に、突然変異誘発を受けさせて、単数または複数のヌクレオチドの置換、欠失および/または付加を生じさせてよい。このような遺伝子のヌクレオチド挿入による派生物には、5'末端融合体および3'末端融合体、ならびに単数または複数のヌクレオチドの配列内挿入体が含まれる。ヌクレオチド配列の挿入による変異体は、1種または複数のヌクレオチドがヌクレオチド配列中の所定の部位に導入されているものであるが、得られた生成物の適当なスクリーニングによりランダム挿入も可能である。欠失による変異体は、1種または複数のヌクレオチドのその配列からの除去によって特徴付けられる。置換によるヌクレオチド変異体は、配列中の少なくとも1種のヌクレオチドが除去され、その場所に別のヌクレオチドが挿入されているものである。このような置換は、置換が、コドンによって定義されるアミノ酸を変化させないものである「サイレント」でもよい。あるいは、置換基は、1種のアミノ酸が類似する代理の別のアミノ酸に改変されるように設計される。典型的な保存的置換は、下記に従って作製されたものである。
【0079】
アミノ酸置換に適した残基
元の残基 典型的な置換
Ala Ser
Arg Lys
Asn Gln、His
Asp Glu
Cys Ser
Gln Asn
Glu Asp
Gly Ala
His Asn、Gln
Ile Leu、Val
Leu Ile、Val
Lys Arg、Gln、Glu
Met Leu、Ile
Phe Met、Leu、Tyr
Ser Thr
Thr Ser
Trp Tyr
Tyr Trp、Phe
Val Ile、Leu
【0080】
当分野の技術者であれば、細胞内での遺伝子またはそれに相補的な配列の発現には、前記遺伝子がプロモーター配列と作動可能に連結して位置する必要があることに気付くであろう。本目的のためのプロモーター、特に胚乳特異的プロモーターの選択は、必要な発現レベル、および/または発現が起こるべき組織、器官もしくは細胞によって変わり得る。
【0081】
プロモーター配列の調節制御下にある核酸分子の場所は、発現がプロモーター配列によって制御されるように前記分子を配置することを意味する。プロモーターは通常、必ずしもそうとは限らないが、それが調節する核酸分子の上流、すなわち5'末端に位置している。さらに、プロモーターを含む調節エレメントは通常、この遺伝子の転写開始部位から2kb以内に位置している。異種プロモーター/構造遺伝子の組合せの構築では、そのプロモーターと、プロモーターがその自然な状況で制御する遺伝子(すなわち、そのプロモーターが由来する遺伝子)との距離とほぼ同じである、遺伝子転写開始部位からの距離にプロモーターを配置することが通常好ましい。当技術分野で知られているように、プロモーター機能を喪失することなく、この距離においてある変異を受け入れることができる。同様に、その制御下に置くべき異種遺伝子に対する調節配列エレメントの好ましい配置は、その自然な状況でのエレメント(すなわち、そのプロモーターが由来する遺伝子)の配置によって定義される。さらに、当技術分野で知られているように、この距離においてある変異が起こり得る。
【0082】
本発明の遺伝子構築体での使用に適したプロモーターの例には、ウイルス、酵母、カビ、細菌、昆虫、鳥類、哺乳動物、および植物の遺伝子に由来するプロモーター、好ましくは植物細胞で機能することができるもの、より好ましくは米胚乳中で発現することができるものが含まれる。このプロモーターは、発現が起こる組織に関して、発現を構成的に、または示差的に調節してもよい。あるいは、発現は、発現が起こる発達段階に関して、または生理学的ストレスまたは温度など外部刺激に応じて示差的であってもよい。
【0083】
SBEIIaまたは他のデンプン生合成遺伝子の活性を低下させる方法は、導入遺伝子をイネ植物の再生可能な細胞中に導入するステップと、形質転換した細胞からトランスジェニックイネ植物を再生させるステップとを含むものでよい。アミロペクチンの合成に関与する枝作り酵素には、SBEI、SBEIIa、およびSBEIIbが含まれ、この1種(複数)の導入遺伝子は、こうした遺伝子を複数不活性化してもよい。さらに、SBEIIbおよび/またはSBEIの不活性化は、導入遺伝子(例えば、2重鎖RNA、アンチセンス、またはリボザイムRNAをコードするもの、以下参照)がSBEIIbまたはSBEI遺伝子の発現を直接標的とするという点で、直接的なものでもよく、この不活性化は、SBEIIbまたはSBEIの発現の低下を間接的にもたらすものでもよい。例えば、導入遺伝子RNAは、配列の同一性または塩基対形成という点で、SBEIIa遺伝子/RNAのみを標的としてもよいが、また胚乳中のタンパク質の安定性または分布を改変することによりSBEIIbまたはSBEIの活性の低下をもたらしてもよい。本発明のさらなる形態は、SBEIIaおよびSBEIIbの活性の低下と、SSI、SSII、SSIII、およびイソアミラーゼやプルラナーゼなどの枝切り酵素が含まれ得る他の1種または複数のアミロペクチン合成酵素の改変との組合せにある。こうした酵素のいずれかの、またはすべての発現を導入遺伝子の導入によって改変してもよい。特定の一実施形態では、イネ植物においてADP−グルコースピロホスホリラーゼ(ADGP)を過剰発現させており、これは収量および成長を増強させることが示されている(Smidanskyら、2003)。
【0084】
いくつかのDNA配列がイネにおけるアミロペクチン合成遺伝子で知られており、その任意の配列を、イネにおいて遺伝子を不活性化する導入遺伝子を設計するための基礎とすることができる。こうした配列には、イネcDNA SBEIIa(GenBankアクセッション番号E14723、特願平10−004970)、SBEIIb(D16201、Mizunoら、1993)、およびSBEI(D11082、Mizunoら、1992;D10752、NakamuraおよびYamanouchi、1992)が含まれる。イネのSBEI遺伝子は、Rahmanら、(1997)およびRahmanら、(1999)、またはアクセッション番号D10838、Kawasakiら、(1993)に記載されている。さらなる遺伝子配列を、以下のウェブサイト、http://www.ncbi.nlm.nih.gov/、http://www.tigr.org、およびhttp://www.gramene.org/about/index.htmlから得てもよい。
【0085】
コムギ、オオムギ、トウモロコシ、または他の近縁種由来のSBEIIa、SBEIIb、または他のアミロペクチン合成遺伝子の相同体を使用して、イネにおける遺伝子発現レベルを改変することもできる。このような遺伝子またはその断片は、PCR増幅または標識プローブに対するハイブリダイゼーションを含めた当技術分野で周知の方法によって得ることができる。導入遺伝子構築体を調製するのに使用する、相同体の1種(複数)の領域は、対応するイネ遺伝子と、その適切な領域において少なくとも85%の同一性、好ましくは少なくとも90%、さらにより好ましくは95〜100%の同一性を有するべきである。導入遺伝子はまた、米胚乳中で発現されるアミロペクチン合成遺伝子を特異的に標的とし、この植物の他の場所においてアミロペクチン合成に対して影響がないか、または最小限であることが好ましい。これを、導入遺伝子内の胚乳特異的プロモーターなど適当な調節配列を使用して行ってもよい。
【0086】
本明細書では、「ストリンジェントなハイブリダイゼーション条件」は、プローブと標的配列との間に少なくとも90%、好ましくは少なくとも95%の同一性がある場合にハイブリダイゼーションが通常生じることを意味する。ストリンジェントなハイブリダイゼーション条件の例は、50%ホルムアミド、5×SSC(1×SSC=150mM NaCl、15mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハルト溶液、10%硫酸デキストラン、およびサケ精子DNAなどを変性させ断片化した20μg/mlの担体DNAを含む溶液中で一晩インキュベートした後、ハイブリダイゼーション支持体を0.1×SSC中でおよそ65℃で洗浄することである。他のハイブリダイゼーション条件および洗浄条件は、周知であり、Sambrookら、Molecular Cloning:A Laboratory Manual,Second Edition,Cold Spring Harbor,NY(1989)、特に11章で例示されている。
【0087】
アンチセンス
植物において遺伝子活性を改変する、特に特異的に低下させるための既知の遺伝子工学または遺伝子導入手法は、当技術分野で周知である。遺伝的変異をイネ植物に導入するこうした方法には、標的遺伝子のRNAに相補的であり、そのRNAにハイブリッド形成することができる適当なアンチセンス分子の発現が含まれる。アンチセンス分子は、標的遺伝子のmRNAの翻訳、プロセッシングまたは安定性を妨げ、それによりmRNAの発現を不活性化すると考えられている。アンチセンス配列を考案する方法は当技術分野で周知であり、これらの例は、参照により本明細書に組み込む、米国特許第5190131号、欧州特許出願公開第0467349号明細書、欧州特許出願公開第0223399号明細書、および欧州特許出願公開第0240208号明細書で見られる。植物におけるアンチセンス技術の使用は、Bourque(1995)およびSenior(1998)にその総説がある。Bourqueは、遺伝子不活性化の方法としてのアンチセンス配列の植物系における使用例を数多く挙げている。彼女は、部分的阻害がおそらくその系の測定可能な変化をもたらすので、どんな酵素活性も必ずしも100%の阻害に達する必要はないと述べている。Senior(1998)は、アンチセンス法が現在、遺伝子発現を操作するための非常によく確立された技法であると述べている。
【0088】
イネのSBEIIa、SBEIIb、SBEI、または他のアミロペクチン生合成遺伝子に対するアンチセンス分子は、イネmRNA配列に基づくものとし、あるいは他の種、例えばオオムギに由来するDNAまたはmRNA配列との相同性に基づくものとすることができる。こうしたアンチセンス配列は、構造遺伝子、または遺伝子発現またはスプライシング事象の制御に影響を与える配列に対応するものでよい。例えば、このアンチセンス配列は、イネのSBEIIaまたは他の遺伝子の、標的とするコード領域、5'非翻訳領域(UTR)もしくは3'UTR、またはそれらの組合せに対応するものでよい。このアンチセンス配列は、転写中、または転写後にスプライシングによって切り取られるイントロン配列に一部が相補的であってもよく、好ましくは標的遺伝子のエキソン配列のみに相補的であってもよい。UTRは通常より大きな多様性があるという点から、こうした領域を標的とすることは、遺伝子阻害のより大きな特異性をもたらす。アンチセンス配列の長さは、少なくとも19の連続するヌクレオチド、好ましくは少なくとも50ヌクレオチド、およびより好ましくは少なくとも100、200、500、または1000ヌクレオチドであるべきである。遺伝子転写物全体に相補的な完全長配列を使用してよい。この長さは100〜2000ヌクレオチドであることが最も好ましい。標的転写物に対するアンチセンス配列の相同性の程度は、少なくとも85%、好ましくは少なくとも90%、より好ましくは95〜100%であるべきである。アンチセンスRNA分子はもちろん、分子を安定化させるために機能する可能性のある無関係の配列を含む。
【0089】
コサプレッション
使用してよい別の分子生物学的手法は、コサプレッションである。コサプレッションの機序は、十分に理解されていないが、それに転写後遺伝子発現抑制(PTGS)が関与していると考えられており、その点でアンチセンス抑制の多くの例に非常に類似している可能性がある。コサプレッションは、遺伝子の余分なコピーまたはその断片をある植物に、その発現用プロモーターに対してセンス方向に導入することを伴う。センス断片のサイズ、標的遺伝子領域に対するその対応性、および標的遺伝子との相同性の程度は、上記のアンチセンス配列の場合と同様である。場合によっては、遺伝子配列の追加のコピーが植物の標的遺伝子の発現を妨害する。コサプレッション手法を実施する方法については、国際公開第97/20936号、および欧州特許出願公開第0465572号明細書を参照されたい。
【0090】
2本鎖RNAによって媒介される遺伝子発現抑制
遺伝的変異をイネ植物に導入するのに使用してよいさらなる方法は、2重鎖もしくは2本鎖のRNAによって媒介される遺伝子発現抑制である。この方法にもPTGSが関与する。この方法では、不活性化すべき標的遺伝子との相同性を有する少なくとも部分的に2本鎖の1種(複数)のRNA産物を合成させるDNAが導入される。したがって、このDNAは、センス配列およびアンチセンス配列をどちらも含んでおり、このDNAがRNAに転写されると、ハイブリッド形成して2本鎖RNA領域を形成することができる。好ましい一実施形態では、このセンス配列およびアンチセンス配列は、RNAに転写される際にスプライシングによって切り取られるイントロンを含むスペーサー領域によって分離されている。この配置が、より効率の高い遺伝子発現抑制をもたらすことが示されている(Smithら、2000)。この2本鎖領域は、1種または2種のDNA領域のいずれかから転写された1種または2種のRNA分子を含むものでよい。2本鎖分子の存在は、標的遺伝子の活性を効率的に低下または消去する、2本鎖RNA、および植物の標的遺伝子からの相同なRNA転写物をも破壊する、内生植物系からの応答を誘発する。この技法を実施するための方法については、豪州特許出願公開第99/292514−A号明細書、および国際公開第99/53050号明細書を参照されたい。ハイブリッド形成するこのセンス配列およびアンチセンス配列の長さは、それぞれ少なくとも19の連続するヌクレオチド、好ましくは少なくとも30もしくは50ヌクレオチド、より好ましくは少なくとも100、200、500、または1000ヌクレオチドであるべきである。遺伝子転写物全体に対応する完全長配列を使用してよい。この長さは100〜2000ヌクレオチドであることが最も好ましい。標的転写物に対するセンス配列およびアンチセンス配列の相同性の程度は、少なくとも85%、好ましくは少なくとも90%、より好ましくは95〜100%であるべきである。このRNA分子はもちろん、この分子を安定化させるために機能する可能性がある無関係の配列を含む。このRNA分子を、RNAポリメラーゼIIまたはRNAポリメラーゼIIIプロモーターの制御下で発現させてよい。後者の例には、tRNAまたはsnRNAプロモーターが含まれる。
【0091】
このアンチセンス、コサプレッション、または2本鎖RNA分子は、国際公開第03/00292号に記載の通り、好ましくは核局在化シグナルを含む、大部分が2本鎖のRNA領域を含むものでもよい。好ましい一実施形態では、この大部分が2本鎖の領域は、PSTVd型ウイロイドに由来するか、または少なくとも35のCUGトリヌクレオチドの反復を含む。
【0092】
リボザイム
リボザイムを使用して、イネにおける所望の遺伝子発現の不活性化に関与する遺伝的変異を導入してよい。リボザイムは、酵素機能または触媒機能を有するRNA分子であり、この分子は、他のRNA分子を、1種または多くの場合2種のハイブリッド形成する配列によって定義される特異的部位で切断することができる。このRNAの切断により、標的遺伝子の発現が不活性化される。このリボザイムは、遺伝子不活性化に寄与することができるアンチセンス分子として作用することもできる。このリボザイムは、ハイブリッド形成する配列の間で、好ましくはハンマーヘッド型またはヘアピン型の1種または複数の触媒ドメインを含む。RNAseP、グループIもしくはIIイントロン、および肝炎デルタウイルス型を含めて、他のリボザイムのモチーフを使用してもよい。欧州特許出願公開第0321201号明細書および米国特許第6221661号を参照されたい。トランスジェニック植物の遺伝子を不活性化するためのリボザイムの使用は、例えばWegenerら(1994)によって実証されている。
【0093】
遺伝子構築体/ベクター
本発明はまた、遺伝子を阻害する分子をコードしているRNAおよび好ましくはDNAを含む単離された核酸分子を提供する。この核酸分子は、イネSBEIIaおよび/またはSBEIIb遺伝子の配列を標的とし、米穀粒の胚乳中でのそれらの発現を不活性化するのに効果的である、アンチセンス、センス(コサプレッション)、2本鎖RNA、またはリボザイム分子をコードしていることが好ましい。本発明はまた、単離された核酸分子を含む遺伝子構築体を提供するものであり、この構築体は、プロモーター、エンハンサー、および転写終結配列もしくはポリアデニル化配列などの1種または複数の調節エレメントを含む。このようなエレメントは当技術分野で周知である。この遺伝子構築体は、植物、特にイネなどの単子葉植物での導入遺伝子の発現を助けるイントロン配列を含むものでもよい。「イントロン」という用語は、その通常の意味で使用され、転写されるがタンパク質をコードしておらず、翻訳前にRNAからスプライシングによって切り取られる遺伝子断片を意味する。イントロンを、導入遺伝子が翻訳産物をコードしている場合は5'UTRまたはコード領域中に、あるいはそうでない場合は転写領域のどこにでも取り込んでよい。特定の一実施形態では、オオムギSBEII遺伝子のイントロン(Ahlandsbergら、2002)など胚乳特異的に発現させるイントロンを使用する。
【0094】
本発明はさらに、遺伝子構築体を含むベクター、例えばプラスミドベクターを提供する。「ベクター」という用語には、in vitroまたはin vivoで発現することができる発現ベクター、および1種の細胞または生物から別のものに移動することができる形質転換ベクターが含まれる。このベクターは、細胞、例えば大腸菌(E.coli)またはAgrobacteriumなどの原核細胞内での複製をもたらす配列を含む。好ましくは、このベクターは、イネ細胞中に導入することができる、少なくとも1種のT−DNA境界配列によって定義されるT−DNA配列を含むバイナリーベクターである。本発明はさらに、このベクターを含む細胞、例えばAgrobacteriumまたはイネ細胞を提供するものであり、この細胞は、未成熟胚の胚盤または胚形成カルスの細胞など再生可能な細胞であってもよい。あるいは、この細胞は、導入遺伝子を含む形質転換されたイネ細胞であってもよい。
【0095】
プロモーター/ターミネーター
本発明の導入遺伝子または他の遺伝子構築体は、米胚乳中で調節性の、または構成的な発現をもたらすことができる転写開始領域(プロモーター)を含むものでよい。このプロモーターは、組織特異的であってよく、したがって胚乳中で選択的または排他的発現を与える。このプロモーターを、胚乳特異的プロモーター(高分子量グルテニンプロモーター、イネSSIプロモーター、イネSBEIIプロモーター、イネGBSSプロモーターなど)、または胚乳特異的でないプロモーター(ユビキチンプロモーター、CaMV35Sまたは増強された35Sプロモーターなど)のどちらかから選択してよい。このプロモーターは、温度、光、ストレスなどの要因によって調節され得る。通常、プロモーターは、発現されるべき遺伝子配列の5'側に備えられる。この構築体は、nos3'もしくはocs3'ポリアデニル化領域、転写ターミネーターなど、転写を増強する他のエレメントを含むものでもよい。例示するDNAのこの領域を、適当な選択マーカー遺伝子配列および他のエレメントを含むベクター中か、あるいはこうした配列を含むベクターと同時形質転換されるベクター中に取り込む。
【0096】
イネの形質転換法
イネの形質転換の方法は、外因性の核酸の導入によりこの植物に遺伝的変異を導入するためのものであり、当技術分野で周知である。例えば、Chanら、1993;Hieiら、1994;Zhangら、1997;Buchholzら、1998を参照されたい。形質転換を、当技術分野で知られている、適当なAgrobacteriu菌株、微粒子銃による方法、ポリエチレングリコールの媒介によるイネプロトプラスト中への取り込みなどによって媒介してよい。所望のヌクレオチド配列または遺伝子構築体、および選択マーカーを載せたベクターを、組織培養された植物もしくは外植片、例えば、プロトプラスト、未成熟胚、またはカルスのような再生可能なイネ細胞中に導入してよい。この選択マーカー遺伝子は、イネ細胞に抗生物質耐性または除草剤耐性をもたらすことができ、あるいは基質、例えばマンノースの利用を成長のために可能にする。この選択マーカーにより、イネ細胞にジェネチシン耐性、ハイグロマイシン耐性またはフォスフィノスリシン耐性を与えることが好ましい。この再生可能なイネ細胞は、未成熟胚、成熟胚の胚盤、これらに由来するカルス、または分裂組織に由来するものであることが好ましい。形質転換する細胞を選択し、次いで実施例2に記載するものなど、当技術分野で周知の方法によって再生して、形質転換イネを作出する。
【0097】
形質転換植物は、選択マーカー遺伝子を含むものでもよく、またこのような遺伝子を、再生中、または再生後に、例えば選択マーカー遺伝子をゲノムの外に切り出すか、あるいは選択マーカー遺伝子を、SBEIIaおよび/またはSBEIIbの阻害をもたらす導入遺伝子から分離することによって除去してもよい。
【0098】
例えば、導入遺伝子に特異的な適当な核酸プローブ、または表現型観察を使用することにより、導入遺伝子または突然変異が染色体中に組み込まれた植物をスクリーニングすることができる。いくつかの任意の方法を使用して、形質転換植物であるかどうか判定することができる。例えば、ポリメラーゼ連鎖反応(PCR)を使用して形質転換植物に特有の配列を増幅し、ゲル電気泳動または他の方法により、この増幅した産物を検出することができる。この植物からDNAを、通常の方法を使用して抽出し、またプライマーを使用してPCRを実施してもよく、これにより形質転換植物と非形質転換植物とが区別される。例えば、この構築体を読み取って、形質転換用ベクター由来のDNAの領域を増幅するプライマーを設計してもよく、また対象の遺伝子からリバースプライマーを設計してもよい。この植物の形質転換が成功していたとしても、これらのプライマーはある断片を増幅するだけである。陽性の形質転換体を確認する代替方法は、当技術分野で周知のサザンブロットハイブリダイゼーションによるものである。また、形質転換植物または突然変異体植物を同定、すなわちその表現型により非形質転換植物または野生型植物と区別することができる。例えば、この表現型は、選択マーカー遺伝子の存在、または免疫学的方法による特定のタンパク質の存在によって、あるいはあるタンパク質の非存在、例えば、ELISAまたはウエスタンブロット分析によって検出される、胚乳中のSBEIIaタンパク質の非存在によって与えられる。また、このような植物をスクリーニングするのに使用する指標は、穀粒の表現形質の観察、例えばshrunken穀粒の目視検査または目測によるもの、アミロース含量が上昇したかどうか試験すること、あるいはデンプン顆粒の複屈折の存在を顕微鏡によって検査することによるものでよい。
【0099】
突然変異
米胚乳中のSBEIIaおよびSBEIIb酵素または他のデンプン生合成酵素の活性の低下をもたらす遺伝的変異の導入を、それぞれの遺伝子内または遺伝子の調節配列内の適切な突然変異によって行ってもよい。遺伝子が阻害される程度により、生成されるデンプンの特性がある程度まで決まる。この突然変異は切断(truncation)でも、無発現変異体でもよく、これらは、デンプンの性質に大きな影響を及ぼすことが知られているが、また、改変したデンプン構造は、アミロペクチン合成酵素活性を十分低下させてデンプンまたは米穀粒において対象の特性をもたらす漏出変異体に起因する。他に染色体再配列も効果的であり、これらには、欠失、逆位、重複、または点突然変異が含まれ得る。
【0100】
突然変異誘発は、化学的手段または放射線による手段、例えばEMSもしくはアジ化ナトリウム(ZwarおよびChandler、1995)による種子の処理、またはγ線照射によって行うことができる。γ線誘発変異については、種子を、60Co供給源からの20〜50kRの放射線量で照射してよい(ZikiryaevaおよびKasimov、1972)。EMSによる突然変異誘発を、Mullinsら、(1999)の通り、種子をEMS(0.03%、v/v)で処理することによって実施してよい。突然変異体の単離は、変異誘発された植物または種子をスクリーニングすることによって行うことができる。例えば、変異誘発したイネ集団を、穀粒中の高アミロース含量、かつ/または通常より長いアミロペクチン鎖長の分布、またはELISAによるSBEIIaおよび/またはSBEIIbタンパク質の喪失について、あるいは改変した穀粒形態についてスクリーニングしてよい(Greenら、1997)。スクリーニングは、SBE活性の1種をすでに欠いているイネの遺伝子型において、例えばSBEIIaまたはSBEIIb陰性の背景で行うことが好ましい。次いで、この突然変異体と所望の遺伝的背景をもつ植物とを交配し、戻し交配を適当な回数実施して本来所望でない親の背景を除去することにより、こうした変異を望ましい遺伝的背景に導入することができる。好ましい突然変異体は、イネにおいてSBEIIaおよびSBEIIbの発現または活性に影響を及ぼすものである。
【0101】
それにより、本発明は、高アミロースで、遺伝子導入によらない米粒およびそれらから得られる生成物を提供する。
【0102】
例えばGBSSのレベルを増大させるなど、アミロペクチンの合成に関与するSBEIIa、SBEIIb、または他の酵素をコードしている遺伝子における突然変異は、米胚乳のデンプン中のアミロースの割合の増大をもたらす。個々の穀粒あたりのアミロースの量は、アミロペクチンからアミロースへ炭素の流れが転換した結果として増大することもあり、あるいは、穀粒あたりのデンプン産生が著しく減少した場合、このアミロースの量は減少することもある。いずれの場合も、デンプンに対する百分率で示すアミロースの相対的レベルが増大する。
【0103】
デンプン顆粒を有する、形がゆがんだ種子が、高アミロースオオムギ(Morellら、2003)、およびデンプン中に約90%アミロースを有する低アミロペクチン(LAPS)トウモロコシ(Sidebottomら、1998)で報告されている。この表現型を、変異誘発したイネ集団をスクリーニングする際に使用することができる。複屈折もこれに使用することができる。複屈折は、2方向に光を屈折させる物質の能力であり、これは、偏光顕微鏡で見ると、各デンプン顆粒に「マルタクロス」と呼ばれる十字の影を形成する。複屈折は、顆粒内のポリマーの並んだ構造機構の程度の指標である(ThomasおよびAtwell、1999)。デンプン顆粒の複屈折の喪失は通常、アミロース含量の増大と関連している。
【0104】
食糧生産に適したもの
別の態様では、本発明は、食糧生産に有用なイネを提供するものであり、この穀粒は、高い相対アミロース含量、および低下したアミロペクチン含量を含むデンプンを有する。この穀粒が得らえるイネ植物は、発達中に胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたものであることが好ましい。本発明のイネ植物は、食糧生産、特に商業上の食糧生産に有用である。
【0105】
イネの所望の遺伝的背景には、農業収量および他の特性への考慮が含まれる。このような特性には、農業生産力、耐病性、および非生物ストレス耐性が含まれ得る。オーストラリアでは、改変したデンプン形質をAmaroo、Ali Combo、Basmati、Bogan、Bombia、Doongara、Goolarah、Illabong、Jarrah、Koshihikari、Kyeema、Langi、Millin、Namage、Opus、Pelde、他の一般に栽培されている品種などのイネ栽培品種中に入れることが望まれている。提供した例は、オーストラリアの生産地域に適したものであるが、その他の栽培地域に適した品種も他にある。本発明のイネ品種は、少なくとも一部の栽培地域で、対応する野生型品種の少なくとも80%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも95%の収量をもたらすことが好ましい。この収量は、管理された圃場試験において容易に測定することができる。
【0106】
穀粒のデンプン含量は、少なくとも約25%、好ましくは少なくとも35%もしくは45%、より好ましくは野生型のレベルに近い55〜65%(w/w)であるべきである。最も好ましくは、この穀粒は、デンプン含量が、同等であるが未改変のイネから得られる穀粒の少なくとも90%である。野生型よりもデンプン含量が低いのは、アミロペクチンレベルが低下した結果である可能性が高い。デンプン含量が低くなっても、高アミロース製品が比較的高価であるため、この穀粒は、商業上の食糧生産にいまだ有用である。他の望ましい特性には、穀粒を製粉する能力、特に穀粒硬度が含まれる。イネ植物の価値を高める可能性がある別の態様は、穀粒からのデンプン抽出の程度であり、より高い抽出率がより有用である。また、穀粒の形は、植物の商業上の有用性に影響を与える恐れがある別の特徴であり、したがって穀粒の形は、穀粒を製粉する際の容易さまたは別の点に影響を与える恐れがある。例えば、穀粒の細長い形態は、製粉および加工を困難にする可能性がある。
【0107】
デンプンは、標準の方法を使用して、例えばアルカリ溶液(水酸化ナトリウム)を用いた、brokersの湿式製粉を行ってタンパク質を除去することにより、米穀粒から容易に単離することができる。砕け米をアルカリ溶液中で24時間浸し、次いでアルカリ溶液を用いた、ピンミル、ハンマーミル、またはストーンミル粉砕機で湿式製粉を行う。このバッターを10〜24時間保存した後、それをスクリーンに通すことにより繊維を除去し、このデンプンを遠心分離により回収し、水で徹底的に洗浄し、乾燥させた。
【0108】
改変したデンプンの物理的特徴
本発明の別の態様では、米デンプンは、糊化温度が改変されたものでよく、これは、示差走査熱量測定(DSC)によって容易に測定することができる。糊化は、過剰水中でのデンプン顆粒内の分子整列の、熱駆動による崩壊(破壊)であり、顆粒の膨潤、結晶融解、複屈折の喪失、粘度の増大、およびデンプンの可溶化などの特性の付随する不可逆的変化を伴う。この糊化温度は、残りのアミロペクチンの鎖長によって、野生型植物から得られるデンプンに比べて、上昇させても、低下させてもよい。トウモロコシのae(amylose extender)変異体から得られる高アミロースデンプンは、正常なトウモロコシよりも高い糊化温度を示した(Fuwaら、1999;Kruegerら、1987)。一方、デンプン合成酵素IIaの活性を欠いているオオムギsex6変異体から得られるデンプンは、対照植物から得られるデンプンに比べた場合、糊化温度が低くなり、その糊化のピークのエンタルピーが低下した(Morellら、2003)。
【0109】
この改変した糊化温度は、比較的高いアミロース含量に伴うものである可能性がある。野生型米デンプンの糊化温度は通常、示差走査熱量測定で測定して、開始温度と定義される第1ピークの温度で約61〜67℃である(Rahmanら、2000)。
【0110】
このデンプンはまた、野生型デンプンに比べて、加熱した過剰水中でのその膨潤率によって特徴付けられる。通常、デンプンまたは穀粉のどちらかを過剰水と混合し、加熱して通常90℃以上の温度にまで上昇させることにより、膨潤容積を測定する。次いで、この試料を遠心分離によって回収し、沈降材料の質量をこの試料の乾燥重量で割ったものにより膨潤容積を表す。低い膨潤特性は、食品調製物、特に含水食品調製物のデンプン含量を増大させたい場合に有用である。
【0111】
本発明の選択される形態の米デンプン構造は、イネから単離された正常デンプンに比べて結晶化度が低下するという点から異なり得る。デンプンの結晶化度の低下は、感覚特性の向上に関係し、より滑らかな食感に寄与すると考えられる。したがって、このデンプンは、1種または複数のアミロペクチン合成酵素の活性レベルの低下に起因する結晶化度の低下をさらに示す可能性がある。結晶化度は通常、X線結晶解析によって調査する。
【0112】
改変したアミロペクチン構造の1つの尺度は、デンプンの鎖長の分布、または重合度である。この鎖長分布は、イソアミラーゼによる枝切りの後、FACE(fluorophore−assisted carbohydrate electrophoresis蛍光ラベル糖鎖電気泳動法)を使用して測定することができる。本発明のデンプンのアミロペクチンは、鎖長の分布が、枝切り後の野生型植物から得られるデンプンのその分布よりも大きい5〜60の範囲にあってよい。また、鎖長が長いデンプンは、分岐の頻度がそれに比例して低下する。したがって、このデンプンも、アミロペクチン中により長いアミロペクチン鎖長の分布を尚ももち得る。
【0113】
食品特性
米デンプンは、特にアジアでヒトの食物中の炭水化物の主要な供給源であり、本発明の穀粒およびそれから得られる生成物は、食品を調製するのに使用することができる。この食品は、ヒトまたは動物によって、例えば畜産またはペットフードにおいて消費され得る。改変したイネ植物から得られる穀粒は、食品加工方法に容易に使用することができ、したがって本発明には、精米され、挽かれ、粗挽きにされ、挽割りにされ、圧延され、煮沸された、またはパーボイルドの穀粒、あるいは穀粉、砕け米、米ぬか、および米ぬか油を含めた、米の加工された穀粒もしくは全粒から得られる製品が含まれる。この製品は、α米(precooked rice)もしくは迅速調理米(quick−cooking rice)、インスタント米、粉末米(granulated rice)、糊化米(gelatinized rice)、缶詰の米、またはライスプディングであってよい。この穀粒またはデンプンを、めん類、もち、ライスペーパー、または春巻の皮を含めて、加工された米製品の製造に、あるいは発酵めんなどの発酵製品または酒などの飲料に使用してもよい。この穀粒またはそれに由来するデンプンを、米の穀粉と、コムギもしくは他の穀粉、または増粘剤、結合剤などの食品添加物とを混合する場合を含めて、例えば、パン、ケーキ、クラッカー、ビスケットなどに、あるいは、飲料、めん類、パスタ、またはクイックスープを製造するのに使用してもよい。この米製品は、小麦を含まないダイエット食品での使用に適している。この穀粒または本発明の穀粒に由来する製品は、特に、パフライス、ライスフレークなどの朝食用シリアルに、あるいは押出製品として望ましい。本発明の高アミロースデンプンは、製菓業において有用であり、または成形および硬化時間が短縮される高強度ゲルを形成するのに使用することもできる。これらのデンプンは、例えば、十分に揚げたポテトまたは他の食品で油吸着を減少させる被覆剤として使用してもよい。
【0114】
食物繊維
本明細書における食物繊維は、炭水化物、および健康なヒトの小腸で吸収されないが大腸に入る、炭水化物の消化による産物である。この食物繊維には、レジスタントスターチ、ならびに他の可溶性および不溶性の炭水化物ポリマーが含まれる。この食物繊維は、大腸で常在ミクロフローラにより少なくとも一部が発酵性となる炭水化物の部分を含むものとする。
【0115】
本発明のデンプンは、比較的高レベルの食物繊維を含むことが好ましく、アミロースを含むことがより好ましい。本発明の穀粒の食物繊維含量は単に、胚乳の相対アミロース含量が増大したことに起因するものであっても、そうでなくてもよい。
【0116】
本発明の態様はまた、高レベルの食物繊維と組みあわせて、アリューロン層と胚芽の組合せから生じるものでもよい。特に、これは、より高い相対レベルのアリューロン層または胚芽が穀粒中に存在する場合に生じるものでよい。米穀粒が軽度に収縮している(slightly shrunken)場合、この胚乳の量が減少し、このアリューロン層および胚芽の量が比較的増大する。したがって、この米は、増大したレジスタントスターチと併せて、比較的高レベルのある種の有益な要素またはビタミンを有し、このような要素には、二価の陽イオン、生物利用可能なCa++、葉酸などのビタミン、またはトコフェロール、トコトリエノールなどの抗酸化剤が含まれる。製粉生成物の特定の一形態は、このアリューロン層が製粉生成物中に含まれる場合のものであってよい。特殊な製粉加工を行って、製粉生成物中のアリューロン層の量を高めてもよい。したがって、このように、糊粉層および胚芽を含むように製粉されたか、または別の方法で加工された穀粒由来の任意の生成物は、異なる供給源からこうした要素を添加する必要なしに、栄養面で追加の恩恵を受けている。
【0117】
レジスタントスターチ
レジスタントスターチを、デンプン、および健康なヒトの小腸に吸収されないが大腸に入るデンプンの消化産物の総量と定義する。したがって、レジスタントスターチには、小腸で消化吸収された産物が含まれない。レジスタントスターチには、物理的に接近しにくいデンプン(RS1型)、難消化性顆粒(RS2)、老化デンプン(RS3)、および化学修飾デンプン(RS4)が含まれる。本発明の改変したデンプン構造、特に高アミロースレベルのデンプンは、それを食品に費やすと、レジスタントスターチの増加をもたらす。このデンプンは、いくぶん消化されにくいRS1型でもよい。また、V錯体の結晶化度によって測定される、デンプンと脂質の結合は、レジスタントスターチのレベルに寄与している可能性が高い。
【0118】
本発明の1つの利点が、本発明が特に栄養面での特定の恩恵を受けた生成物を提供し、さらに、米穀粒のデンプンまたは他の成分を、収穫後に改変する必要なく役立つことを理解されたい。しかし、穀粒のデンプンまたは他の成分を改変することが望ましく、本発明は、改変されたそのような成分を包含する。改変の方法は周知であり、この方法には、従来の方法によりデンプンまたは他の成分を抽出すること、およびレジスタント型が増加するようにデンプンを改変することが含まれる。このデンプンを、熱および/または水分での処理により、物理的に(例えばボールミル)、酵素的に(例えばα−もしくはβ−アミラーゼ、プルラナーゼなどを使用して)、化学的加水分解(液体試薬もしくは気体試薬を使用した湿式または乾式)、酸化、二官能性試薬(例えばトリメタリン酸ナトリウム、オキシ塩化リン)での架橋、またはカルボキシメチル化によって改変してよい。
【0119】
血糖指数
血糖指数(GI)は、デンプンを含む食品の消化の速度に関するものであり、また血中グルコース濃度の変移に対する、試験食品の影響と白パンまたはグルコースの影響との比較である。この血糖指数は、食事後の血清中グルコース濃度に対する当該食品の影響可能性、および血中グルコースのホメオスタシスに対するインスリンの需要の程度である。本発明の食品によってもたらされるある重要な特性は、低下した血糖指数である。高アミロース米生成物のヒトボランティアによる消化の30分後の血清中グルコースレベルは、低アミロース米に比べて低くなった(Goddardら、1984)。さらに、この食品は、最終消化が低レベルであり、結果的に比較的低カロリーである可能性がある。低いカロリー製品は、製粉された米穀粒から生成される穀粉の含有物に基づく可能性がある。このような食品は、満腹にし、腸の健康を増進し、食事後の血清中グルコースおよび脂質の濃度を低下させ、また低カロリー食品を提供する効果がある可能性がある。
【0120】
非食品への応用
本発明は、アミロースレベルを上昇させ、その特性が様々な工業上の任意の要求を満たすアミロペクチンレベルの低下させる、改変または改良されたデンプンを提供する。デンプンは、フィルム、紙、繊維、波形加工(corrugating)、および例えば、のり剤などの接着剤の業界を含めた非食品業界で広く使用されている(Young、1984)。米デンプンを、グルコースシロップの製造またはエタノールの製造のための基質として使用してよい。改変されていないデンプンの物理的特性により、いくつかの用途でのその有用性が限定され、高価な、または他の欠点がある恐れがある化学的改変を強いられることが多い。本発明は、他の物理的特性と併せて、特にアミロペクチン含量を低下させることにより、収穫後の改変が少なくて済むデンプンを提供する。例えば、デンプンおよび本発明の穀粒から作製される生成物のペースティング温度(pasting temperature)、剪断応力に対する耐性、皮膜強度、および/または耐水性を改変してよい。このデンプンを、ポリスチレンまたは他の梱包材料の代替物として使用することができる、緩い充填の生分解性の梱包材料を調製するのに使用してもよい。
【0121】
本発明の態様に関して様々な示唆が与えられているが、本発明は、本発明の2種以上の態様の組合せにあってもよいことを理解されたい。
【0122】
<実施例>
(実施例1.材料および方法)
材料および培地
N6マクロ−エレメント(20×stock solution)
g/l
(NH4)2SO4 9.3
KNO3 56.6
KH2PO4 8
MgSO4.7H2O 3.7
CaCl2.2H2O 3.3
MSマクロ−エレメント(20×stock solution)
g/l
NH4NO3 33.0
KNO3 38.0
KH2PO4 3.4
MgSO4.7H2O 7.4
CaCl2.2H2O 8.8
N6マイクロ−エレメント(1000×stock solution)
mg/100ml
MnSO4.4H2O 440
ZnSO4.7H2O 150
H3BO3 160
KI 80
MSマイクロ−エレメント(1000×stock solution)
mg/l
MnSO4.4H2O 22300
Na2MoO4.2H2O 250
H3BO3 6220
ZnSO4.7H2O 8600
CuSO4.5H2O 25
CoCl2.6H2O 25
KI 830
B5マイクロ−エレメント(100×stock solution)
mg/l
MnSO4.4H2O 1000
Na2MoO4.2H2O 25
H3BO3 300
ZnSO4.7H2O 200
CuSO4.5H2O 3.87
CoCl2.6H2O 2.5
KI 75
N6ビタミン(100×stock solution)
mg/100ml
グリシン 20
チアミンHCl 10
ピリドキシンHCl 5
ニコチン酸 5
MSビタミン(100×stock solution)
mg/100ml
ミオイノシトール 1000
チアミンHCl 1
ピリドキシンHCl 5
ニコチン酸 5
B5ビタミン(100×stock solution)
mg/100ml
グリシン 1000
チアミンHCl 100
ピリドキシンHCl 10
ニコチン酸 10
MS鉄(200×stock solution)
mg/500ml
FeCl3(60%w/v) 2.7
MS Na2.EDTA(200×stock solution)
g/500ml
Na2.EDTA 3.7
【0123】
2,4−ジクロロ−フェノキシ酢酸(2,4−D)(1mg/ml、Sigma社製、番号D−6679)stock solutionを、100mgの2,4−Dを1ml無水エタノール中で溶解し、3mlの1N KOHを添加し、1N HClでpHを6に調整することによって調製した。
【0124】
6−ベンジルアミノプリン(1mg/ml BAP、Sigma社製、番号B−3408)およびナフタレン酢酸(1mg/ml NAA、Sigma社製、番号N−0640)の溶液を調製した。
【0125】
アブシジン酸(ABA、2.5mg/ml、Sigma社製、番号A−1049)を、250mgのABAを2mlの1M NaOH中で溶解し、滅菌水で100mlにすることによって調製した。
【0126】
チメンチン(timentin)(150mg/ml、Smith−Kline Beecham社製、6571−30)を、3.1gを20.66mlの滅菌水中で溶解することによって調製した。
【0127】
ハイグロマイシン(50mg/ml)をRoche社(番号843555)から、他の試薬をSigma社から入手した。
【0128】
カルス誘導用N6D培地
量/リットル
N6マクロ(20×) 50ml
N6マイクロ(1000×) 1ml
N6ビタミン(100×) 10ml
MS鉄(200×) 5ml
MS Na2 EDTA(200×) 5ml
ミオイノシトール 100mg
カサミノ酸 300mg
プロリン 2.9g
2,4−D(1mg/ml) 2ml
スクロース 30g
【0129】
1M KOHでpHを5.8に調整し、1リットルあたり3gのフィトゲル(phytogel)を添加し、この混合物をオートクレーブした。
【0130】
継代培養用NB培地
量/リットル
N6マクロ−エレメント(20×) 50ml
B5マイクロ−エレメント(100×)10ml
B5ビタミン(100×) 10ml
MS鉄(200×) 5ml
MS Na2 EDTA(200×) 5ml
2,4−D(1mg/ml) 2ml
スクロース 30g
プロリン 500mg
グルタミン 500mg
カゼイン酵素加水分解物(CEH) 300mg
【0131】
1M KOHでpHを5.8〜5.85に調整し、1リットルあたり3gのフィトゲルを添加し、この混合物をオートクレーブした。
【0132】
継代培養用MS培地
量/リットル
MSマクロ−エレメント(20×) 25ml
MSマイクロ−エレメント(1000×) 1ml
MSビタミン(100×) 10ml
MS鉄(200×) 5ml
MS Na2EDTA(200×) 5ml
スクロース 10g
【0133】
1M KOHでpHを5.8〜5.85に調整し、1リットルあたり2.5gのフィトゲルを添加し、この混合物をオートクレーブした。
NBO:NB培地に30g/lのマンニトールおよび30g/lのソルビトールを添加した後、pH調整する。
【0134】
NBHT30:添加前にNB培地をオートクレーブし、直後に30mg/lのハイグロマイシンおよび150mg/lのチメンチンを注ぐ。
【0135】
NBHT50:添加前にNB培地をオートクレーブし、直後に50mg/lのハイグロマイシンおよび150mg/lのチメンチンを注ぐ。
【0136】
PRHT50:NB培地(2,4−Dを含まない)をオートクレーブした後、以下のものを添加し、最終濃度をBAP(2mg/l)、NAA(1mg/l)、ABA(5mg/l)、ハイグロマイシン(50mg/l)、およびチメンチン(150mg/l)にする。
【0137】
RHT50:NB培地(2,4−Dを含まない)をオートクレーブした後、以下のものを添加し、最終濃度をBAP(3mg/l)、NAA(0.5mg/l)、ハイグロマイシン(50mg/l)、およびチメンチン(150mg/l)にする。
【0138】
MST培地:MS培地をオートクレーブした後、0.05mg/lのNAAおよび150mg/lのチメンチンを添加する。
【0139】
イネの形質転換
成熟した穀粒を、脱穀し、70%エタノールに1分間浸し、滅菌水で3回洗浄した後、50%漂白剤に30分間浸した。滅菌されたこの穀粒を、無菌条件下で滅菌水で徹底的に洗浄し、次いでN6D培地上に蒔いた。プレートをマイクロポアテープで密封し、カルス形成のため照明下、26〜28℃で6〜8週間インキュベートした。カルスを、穀粒から胚を切り出すことなく、おそらく胚の胚盤から形成した。継代培養を必要とする場合は、カルスをNB培地上に移し、このプレートをパラフィルムで密封した。このプレートを暗下、つまりアルミホイルで覆った箱の中で28℃で放置した。継代培養を4週間ごとに実施した。カルスを最高5回継代培養した後、形質転換に使用した。
【0140】
Agrobacteriumの媒介による形質転換のために、移入すべき遺伝子構築体を含むAgrobacterium菌株を、適切な抗生物質を含むプレート上で28℃で増殖させ、2日間の増殖後、このプレートから細胞を掻き取り、100μMアセトシリンゴンを含有するNB液体培地中で再懸濁させた。健常に見えるカルスを細菌懸濁液に10分間浸し、次いでこのカルスを簡単に排出し、100μMアセトシリンゴンを含有するNBOプレート上で暗下、25℃で2日間放置した。この期間を、遺伝子構築体を含むAgrobacteriumの存在下での「共培養」と呼ぶ。共培養の後、このカルスを、150mg/lのチメンチンを含む滅菌水中で洗浄し、簡単に乾燥し、150mg/lのチメンチンを含むNBHT30(選択薬剤ハイグロマイシンを含む)プレート上に蒔いた。26〜28℃で3〜4週間経過した後、増殖域を示した任意のカルスを、同じ培地上でさらに10〜24日間継代培養した。持続的に増殖させることにより、カルスはハイグロマイシン耐性、すなわち形質転換されたことを表した。こうしたカルスを、チメンチンを含むNBHT50プレート上に移し、暗下、26〜28℃でさらに14〜21日間インキュベートした。健常に見えるカルスをPRHT50プレートに移し、暗下でさらに8〜12日間インキュベートした。最後に、シュートをRHT50培地上で、照明下、28℃で30日間以上再生させた。根を形成したシュートを1/2MST培地に移し、十分大きくなると温室内の土壌に移植した。この方法は、ジャポニカ型およびインディカ型を含めて様々なイネ栽培品種で成功することが判明している。
【0141】
(実施例2.遺伝子のdown regulationのための構築体の調製)
遺伝子発現のdown regulationのための遺伝子構築体を調製するのに使用するために、イネSBEI、SBEIIa、およびSBEIIb遺伝子の断片をPCRによって増幅した。この遺伝子の3'末端に近いエキソン領域同士がより異なっているので、選択されたこの断片はこの遺伝子のこうした領域に由来したものであり、これにより、形質転換されたイネでの構築体によるこの遺伝子の交差発現抑制(cross−silencing)の可能性が低下すると考えられている。増幅された断片は、SBEI:GenBankアクセッション番号D11082のヌクレオチド1982〜2527、SBEIIa:アクセッション番号AB023498のヌクレオチド2458〜2997、およびSBEIIb:アクセッション番号D16201のヌクレオチド2414〜2912である(配列は図1〜3に示す)。次のクローニングの工程での便宜上、制限エンドヌクレアーゼ部位をその末端に含む追加の配列を含めた増幅断片をプラスミドベクターpGEM(登録商標)−T中にクローニングした。また、イネ由来のSBEIイントロン9配列を増幅させることにより、イントロン配列を獲得した。この断片に、ゲノム配列GenBankアクセッション番号D10838のヌクレオチド9112〜9606由来の配列、ならびに両端にあるSpeIおよびEcoRI制限部位を含めて、この断片をpBCSK(Stratagene社製)中に挿入してpRint9_BCを形成した(図4)。次いで、SBEI、SBEIIa、およびSBEIIb遺伝子由来のエキソン断片をpRint9 BC中に、そのSpeI/XbaIおよびXhoI/EcoRI部位を使用してそれぞれアンチセンス方向およびセンス方向にクローニングした。これにより、イントロン配列によって分離しているこれらの配列がそれぞれ逆方向反復を形成するように働いた。得られたプラスミドをpRBEI.IR、pRBEIIa.IR、およびpRBEIIb.IRと呼んだ(図5)。このキメラ断片を、BamHIおよびKpnIで切り取り、pBx17casNOTの同じ部位に挿入した(図6)。これにより、アンチセンス/イントロン/センスのキメラ断片がBx17プロモーター領域およびnos3'終結領域に、発現のための正しい方向に連結させた。次いで、各発現カセットを、HindIIIおよびNotIで消化することによって切り取り、植物細胞中での選択のための、植物で発現可能なハイグロマイシン遺伝子、ならびに細菌中での選択のためのスペクチノマイシン耐性遺伝子を含むバイナリーベクターpWBvec8中に挿入した(Wangら、Acta Hort 461:401−407、1998)。この構築体をdsSBEI、dsSBEIIa、およびdsSBEIIbと呼んだ。次いで、これらの構築体を、電気穿孔法によりAgrobacterium tumefaciens菌株(AGL1)の細胞に移入した(Lazoら(1991))。
【0142】
イネのSBEIIa遺伝子および場合によってはSBEIIb遺伝子の発現を低下させるために、コムギの対応するSBEIIa遺伝子由来の配列を使用して、さらなる2重鎖RNA(dsRNA)構築体を作製した。上記の他の構築体に関しては、SBEIIa遺伝子の部分に対応する所望の核酸配列が、プロモーターに対してセンス方向およびアンチセンス方向の両方で生じ、その結果、発現されたRNAが、塩基対合し、2重鎖もしくは2本鎖のRNAを形成することができる相補領域を含んでいた。センス配列およびアンチセンス配列の間のスペーサー領域は、イントロン配列を含んでおり、この配列は、形質転換された植物におけるRNAの部分として転写されるとスプライシングによって切り取られて、引き締まった「ヘアピン」2重鎖構造を形成するはずである。イントロンを含めると、2重鎖RNA構築体によって与えられる遺伝子発現抑制の効率が上昇することが分かっている(Smithら、2000)。所望の核酸を、コムギ由来の高分子量グルテニン(HMWG)プロモーター配列、およびAgrobacterium由来のノパリン合成遺伝子のターミネーター配列(nos3')に連結させた。これにより、dsRNA配列の胚乳特異的発現がもたらされた。
【0143】
このSBEIIaの2重鎖RNA構築体に、コムギSBEIIa遺伝子(GenBankアクセッション番号AF338431)からPCRによって増幅された1536bpのヌクレオチド配列を含めた。これに、どちらかの側にEcoRIおよびKpnI制限部位を有する、エキソン1および2の全体ならびにエキソン3の一部を含んだ468bpの配列(断片1)と、どちらかの側にKpnIおよびSacI部位を有する、SBEIIaのエキソン3および4の一部ならびにイントロン3の全体からなる512bpの配列(断片2)と、どちらかの側にBamHIおよびSacI部位を有する、SBEIIaの完全なエキソン1、2および3からなる528bp断片(断片3)とを含めた。使用する配列は、より狭い領域にわたるより高い相同性(50ヌクレオチドにわたり87%、27ヌクレオチドにわたり92%)を含めて、イネSBEIIa遺伝子(SBE4)と217ヌクレオチドにわたり80%の同一性を有し、したがって米胚乳におけるこの配列の発現により、イネSBEIIaの発現の相当な低下がもたらされることが期待された。また、コムギの配列は、SBEIIbの同等物であるイネ枝作り酵素3と113ヌクレオチドにわたり76%同一であり、これは、同様にこの転写物のレベルに影響を及ぼすことが予想された。
【0144】
次いで、断片3の配列が、断片1に対してアンチセンス方向に断片2と連結するように、断片1、2および3を連結させた。最初に、2重鎖RNA構築体を、HMWGプロモーター配列およびnos3'ターミネーターを含めたベクターpBx17casNOT(図6)において生成した。このベクターにおける遺伝子構築体をpBx17ds−wSBEIIaと呼び、この2重鎖RNA遺伝子をds−wSBEIIaと呼んだ。ds−wSBEIIa遺伝子を含むこのカセットを、pWBvec8中に挿入し、Agrobacterium菌株AGL1に導入し、これを使用して実施例1に記載の通りイネを形質転換させた。
(実施例3.SBE活性を低下させたイネの作出)
【0145】
AGL1細胞中の構築体dsSBEI、dsSBEIIa、dsSBEIIb、およびds−wSBEIIaを使用して、実施例1に記載の方法に従って、形質転換イネ植物(品種、ニッポンバレ)を作出した。各構築体につき500個のイネカルスを使用し、形質転換したカルスを選抜し、イネ植物を再生させた。この植物を土壌に移植した後、使用するSBEI、SBEIIa、もしくはSBEIIb遺伝子断片に特異的なプライマーまたはプローブを使用した、PCRあるいはサザンブロットハイブリダイゼーション分析により、この植物の形質転換を実証した。ds−wSBEIIaを用いた形質転換からの再生させた24の植物体のうち、21の植物体が、導入されたSBEIIa配列について陽性であることを示した。
【0146】
アクリルアミドゲルでの胚乳タンパク質のゲル電気泳動の後、それぞれのタンパク質に対する特異的抗体を使用したウエスタンブロット分析により、形質転換された植物から得られた穀粒(T1種子)をSBEタンパク質について検査する。SBEの活性は、形質転換された系統の大多数において低下する。穀粒のデンプン中のアミロースの割合を求めた。SBEIIaについて形質転換された系統には、少なくとも40%の相対アミロースレベルを示すものがあり、またそれらの中には50%以上のものもある。SBEIIaとSBEIIbの活性をどちらも低下させた場合、アミロースの割合はさらに増大する。
(実施例4.デンプンおよびタンパク質の分析)
【0147】
炭水化物の測定および分析
Takedaら,(1986)、Lumdubwongら,(2000)、Chiouら(2002)、またはSchulmanら,(1991)の方法を使用して、デンプンを発達中の胚乳から、または成熟した穀粒から単離する。Megazyme社(アイルランド共和国Wicklow州Bray)によって提供される総デンプン量分析キット(total starch analysis kit)を使用して、デンプン含量を求める。次いで、このデンプン含量を対照植物と比較する。総穀粒重量からデンプン重量を引いて、穀粒の総非デンプン含量を求めることにより、総重量の減少がデンプン含量の低下に起因するものであるかどうかが決まる。
【0148】
デンプン試料のアミロース含量を、以下の通り若干変更した、MorrisonおよびLaignelet(1983)の比色(ヨウ素滴定)法によって求める。蓋にゴム座金が取り付けられた2mlねじ込みキャップ式チューブ中に、デンプンをおよそ2mg正確に量り取る(0.1mgまで正確に)。脂質を除去するために、1mlの85%(v/v)メタノールをこのデンプンと混合し、チューブを時々ボルテックスしながら65℃のウォーターバス中で1時間加熱する。13,000gで5分間遠心分離した後、その上清を注意深く除去し、この抽出工程を繰り返す。次いで、このデンプンを65℃で1時間乾燥させ、2mgのデンプンにつき1mlのUDMSO(上記のように量り取る)を使用して、尿素−ジメチルスルホキシド溶液(UDMSO、6M尿素の容量1に対してジメチルスルホキシドの容量9)に溶解する。すぐにこの混合物を激しくボルテックスし、デンプンを完全に溶解するために断続的にボルテックスしながら95℃のウォーターバス中で1時間インキュベートする。デンプン−UDMSO溶液のアリコート(50μl)を、水1mlあたり2mgのヨウ素および20mgのヨウ化カリウムを含むI2−KI試薬20μlで処理する。この混合物を水で1mlにする。この混合物200μlをマイクロプレートに移し、Emax Precision Microplate Reader(Molecular Devices社製、米国)を使用して吸光度を読み取ることにより、その650nmでの吸光度を測定する。ジャガイモのアミロース、およびトウモロコシ(またはジャガイモ)のアミロペクチン(Sigma社製)から、0〜100%のアミロースおよび100%〜0%のアミロペクチンを含む標準試料を作製し、これを検査試料用のものとして扱う。この標準試料の吸光度から導き出した回帰方程式を使用して、その吸光度の値からアミロース含量(%アミロース)を求める。また、枝切りされなかったデンプンのアミロース/アミロペクチン比の分析を、Caseら,(1998)に従って、またはBateyおよびCurtin(1996)による記載の通り枝切りされたデンプンを分離するHPLC法によって実施してよい。
【0149】
デンプン試料の枝切りの後、デンプンにおける鎖長の分布を、Morellら(1998)に従って、キャピラリー電気泳動ユニットを使用したfluorophore assisted carbohydrate electrophoresis(FACE)によって分析してよい。デンプン試料の糊化温度のプロファイルをPyris 1 differential scanning calorimeter(Perkin Elmer社製、米国コネチカット州Norwalk)で測定してよい。デンプン溶液の粘度を、例えばBateyら、1997によって報告されている条件を使用して、Rapid−Visco−Analyser(RVA、Newport Scientific社製、シドニーWarriewood)で測定してよい。測定され得るパラメータには、最高粘度(peak viscosity)(最高ホットペースト粘度(hot paste viscosity))、保持強度(holding strength)、最終粘度、およびペースティング温度(pasting temperature)が含まれる。この穀粉またはデンプンの膨潤容積をKonik−Roseら(2001)の方法に従って求めてよい。一定の温度で水中で穀粉またはデンプン試料を混合する前、またその糊化した材料を回収した後で試料を量ることにより、水の吸収を測定する。
【0150】
β−グルカンレベルを、Megazyme社(アイルランド共和国Wicklow州Bray)によって提供されるキットを使用して求めてよい。
【0151】
胚乳中でのタンパク質発現の分析
胚乳中での特異的タンパク質の発現を、ウエスタンブロットの手順によって分析する。すべての母系組織から胚乳を切り出し、およそ0.2mgの試料を、5mM EDTA、20%グリセロール、5mM DTT、および1mM Pefablocを含むpH7.5の50mM KPi緩衝液(42mM K2HPO4および8mM KH2PO4)600μl中でホモジナイズする。この細かくした試料を13,000gで10分間遠心分離し、その上清を分取し、使用するまで−80℃で凍結させる。全タンパク質を測定するために、BSA標準曲線を、アリコート0μl、20μl、40μl、60μl、80μlおよび100μlの0.25mg/ml BSA標準液を使用して設定する。この試料(3μl)を蒸留水で100μlにし、それぞれに1mlのクーマシープラスプロテイン(Coomassie Plus Protein)試薬を添加する。5分後、その吸光度を595nmで読み取り、ブランクとして標準曲線からゼロのBSA試料を使用して、試料中のタンパク質レベルを求める。各胚乳から得られる20μgの全タンパク質を含む試料を、0.34MトリスHCl(pH8.8)、アクリルアミド(8.0%)、過硫酸アンモニウム(0.06%)、およびTEMED(0.1%)を含む8%非変性ポリアクリルアミドゲルで泳動する。電気泳動の後、このタンパク質を、Morellら、(1997)に従ってニトロセルロース膜に移し、SBEIIaまたはSBEIIbに特異的な抗体との免疫反応を行う。
(実施例5.イネゲノムに特有の配列を同定することによる標的遺伝子の最適化された遺伝子発現抑制)
【0152】
例えば、2重鎖RNA、アンチセンス、またはコサプレッション構築体を使用するなどの方法により標的遺伝子の発現を低減する(遺伝子発現抑制)のに使用する遺伝子配列は、標的遺伝子に高度に特異的であることが好ましい。すなわち、この発現抑制分子は、標的遺伝子またはその相補体の少なくとも19の連続するヌクレオチドの配列と少なくとも約95%同一である少なくとも19の連続するヌクレオチドの配列を含む。最高の特異性をもたらすためには、標的とする配列が、標的遺伝子に特有であり、植物ゲノムにおいて発現される遺伝子として他のどこにも存在しないことが理想的である。これは「オフ遺伝子効果(off−gene effect)」を最小限にするはずである。完全に近いイネゲノム配列の知識を使用して、SBE標的遺伝子配列をイネゲノムの残りの配列と比較して、それによりこうした遺伝子内の最適化された標的配列が同定されている。
【0153】
イネゲノムDNA配列データベース(OSA1.seq)をTIGRのウェブサイト(http://www.tigr.org/tdb/e2k1/osa1/)からFASTAフォーマットでダウンロードした。このデータベースをフォーマットし、これは、「formatdb」を使用してBLASTに、また「dbifasta」を使用してEMBOSS function seqretに使用可能となった。クエリー配列をFATSAフォーマットで作製し、1組のプリセットパラメータ(比較のためのオプション:ワード19およびストリンジェンシー18)を用いて、BLASTベースの遺伝子発現抑制プログラム(Gene Silencing program)(P.Waterhouseら、CSIRO Plant Industry,personal communication)を実行することにより、イネゲノムで相同配列を検索するのに使用した。
【0154】
実施例2に記載の通り遺伝子発現抑制構築体を調製するのに使用するSBEIIa、SBEIIb、およびSBEIの配列を、イネゲノムに対するクエリー配列として使用した。この出力を図8に示す。出力配列における多数の「NNNN...」により、これらの領域におけるクエリー配列が、少なくとも19の連続するヌクレオチドの領域において、イネゲノムにおける他の配列と相同性を有することが示唆された。使用したSBEIIa配列は、固有のものであり、使用したSBEIIb配列は、固有のものでない配列を一部含むが、使用したSBEI配列は、明らかであるように思われる末端の57ヌクレオチドを除いて、イネゲノムの他の場所で重複しているようであることが分かる(図8)。現行のイネゲノム配列の検査により、SBEI遺伝子領域を含むことができる重複する2種のBACクローン間のゲノムDNA配列が重複するようであることが明らかとなった。明らかなこの重複は、本当であるか、またはその領域におけるイネゲノムの組立てのエラーを表すものである恐れがある。
【0155】
<参考文献>
Abel et al., (1996). The Plant Journal 10, 981-991.
Ahlandsberg et al., (2002). Plant Cell Rep 20, 864-868.
Anderson et al., (1989). Nucl Acids Res 17, 461-462.
Baba et al., (1991). Biochem Biophys Res Commun 181: 87-94.
Baga et al., (2000). Plant Physiol. 124, 253-263.
Batey and Curtin. (1996). Starch 48, 338-344.
Batey et al., (1997). Cereal Chemistry 74, 497-501.
Blauth et al., (2001). Plant Physiology 125, 1396-1405.
Bourque. (1995). Plant Science 105, 125-149.
Boyer and Preiss, (1978). Carbohydrate Research 61, 321-334.
Boyer and Preiss, (1981). Plant Physiology 67, 1141-1145.
Boyer et al., (1980). Starch 32, 217-222.
Buchholz et al., (1998). Methods Mol Biol 81, 383-396.
Buleon et al., (1998). International Journal of Biological Macromolecules 23, 85-112.
Cao et al., (2000). Archives. of Biochemistry and Biophysics. 373, 135-146.
Case et al., (1998). Journal of Cereal Science 27, 301-314.
Chan et al., (1993). Plant Mol Biol 22, 491-506.
Chiou et al., (2002). Starch 54, 415-420.
Craig et al., (1998). Plant Cell 10, 413-426.
Denyer et al., (1996). Plant Physiology 112, 779-785.
Fergason. 1994. pp 55-77 in "Speciality Corns" eds , CRC Press Inc.
Filpse et al.,(1996). Planta 198, 340.
Fisher et al., (1993). Plant Physiol 102:1045-1046.
Fisher et al., (1996). Plant Physiol 110: 611-619.
Frances et al., (1998). Plant Mol Biol 38, 407-415.
Fujita et al., (2003). Plant Cell Physiol 44, 607-618.
Fuwa et al., (1999). Starch/Starke. 51, 147-151.
Gao et al., (1996) Plant Mol Biol 30, 1223-1232.
Gao et al., (1997). Plant Physiol 114: 69-78.
Gao et al., (1998). Plant Cell 10, 399-412.
Giroux and Hannah. (1994). Molecular and General Genetics 243, 400-408.
Goddard et al., (1984) Am J Clin Nutr 39, 388-392.
Green et al., (1997). Plant Physiology 114, 203-212.
Hedman and Boyer, (1982). Biochemical Genetics 20, 483-492.
Hiei et al., (1994). Plant J 6, 271-282.
Hirano et al., (1998). Mol Biol Evol 15, 978-987.
Hirano and Sano, (2000). Genes Genet Syst 75, 245-249.
Isshiki et al., (1998). Plant J 15, 133-138.
James et al., (1995). Plant Cell 7, 417-429.
Jobling et al., (1999). Plant Journal 18, 163-171.
Juliano, B.O. (1979). in Proceedings, Workshop on Chemical Aspects of Rice Grain Quality, p. 69-90. Los Banos, Laguna, the Philippines, IRRI.
Juliano, B.O., (1985). Rice: chemistry and technology, 2nd ed. St Paul, MN, USA, Am. Assoc. Cereal Chem. 774 pp.
Kawasaki et al., (1993). Mol Gen Genet 237, 10-16.
Konik-Rose et al (2001) Starch 53, 14-20.
Krueger et al., (1987). Cereal Chemistry 64, 187-190.
Kubo et al., (1999). Plant physiology. 121, 399-409.
Lazo et al., (1991). Bio/Technology 9, 963-967.
Li et al., (1999a). Plant physiology. 120, 1147-1155.
Li et al., (1999b). Theoretical and Applied Genetics 98, 1208-1216.
Li et al., (2000). Plant Physiology 123, 613-624.
Li et al., (2003). Funct Integr Genomics 3:76-85.
Lumdubwong et al., (2000). J Cereal Sci 31, 63-74.
Maniatis et al., (1982). Molecular cloning: a laboratory manual. Cold Spring Harbour Laboratory Press New York.
McCreery and Helentjaris (1994). Methods in Molecular Biology, Vol. 28: Protocols for nucleic acid analysis by non-radioactive probes, 67-71, Humana Press Inc., Totawa, NJ.
Mizuno et al., (1993). Journal of Biological Chemistry 268, 19084-19091.
Mizuno et al., (1992). Journal of Biochemistry 112, 643-651.
Mizuno et al., (2001). Plant Cell Physiol 42, 349-357.
Morell et al., (1997). Plant Physiology 113, 201-208.
Morell et al., (1998). Electrophoresis 19, 2603-2611.
Morell et al., (2003). Plant J. 34: 173-185.
Morrison and Laignelet (1983). Journal of Cereal Science 1:9-20.
Mullins et al., (1999). European Journal of Plant Pathology 105: 465-475.
Myers et al., (2000). Plant Physiology 122, 989-997.
Nakamura (2002). Plant Cell Physiology 43, 718-725.
Nakamura and Yamanouchi (1992). Plant Physiol 99: 1265-1266.
Nair et al., (1997). Plant Sci 122: 153-163.
Nishi et al., (2001). Plant Physiology 127, 459-472.
Rahman et al., (1995). Australian Journal of Plant Physiology 22, 793-803.
Rahman et al., (1997). Genome 40: 465-474.
Rahman et al., (1999). Theor Appl Genet. 98: 156-163.
Rahman et al., (2000). J Cereal Sci 31: 91-110.
Rahman et al., (2001). Plant Physiol 125: 1314-1324.
Repellin et al., (1997). Plant Gene Reg 97-094
Safford et al., (1998). Carbohydrate Polymers 35, 155-168.
Schulman and Kammiovirta, (1991). Starch 43, 387-389.
Schwall et al., (2000). Nature Biotechnology 18, 551-554.
Senior (1998). Biotechnology and Genetic Engineering Reviews 15, 79-119.
Shannon and Garwood, (1984). In Starch: Chemistry and Technology, Whistler et al., eds, Academic Press, Orlando, FL, pp25-86.
Shure et al., (1983). Cell 35, 225-233.
Sidebottom et al., (1998). Journal of Cereal Science 27, 279-287.
Smidansky et al., 2003) Planta 216, 656-664.
Smith et al., (2000) Nature 407, 319-320.
Sun et al., (1996). Physiol Plantarum 96, 474-483.
Sun et al., (1997). The New Phytologist 137 , 215-215.
Sun et al., (1998). Plant Physiol 118, 37-49.
Takeda et al., (1986). Carbohydr Res 148, 299-308.
Takeda et al., (1987). Carbohydr Res 168, 79-88.
Takeda et al., (1993a). Carbohydrate Research 240, 253-262.
Takeda et al., (1993b). Carbohydrate Research 246, 273-281.
Terada et al., (2000). Plant Cell Physiol 41, 881-888.
Thomas and Atwell 1999 Starches Eagen Press, St Paul, Minnesota, USA pp: 13-24.
Thorbjornsen et al., (1996). Plant Journal 10, 243-250.
Wang et al., (1990). Nucl Acids Res 18, 5898.
Wang et al., (1998a). Journal of Experimental Botany 49, 481-502.
Wang et al., (1998b). Acta Hort 461, 401-407.
Wegener et al., 1994. Mol. Gen Genet. 245, 465-470.
Yamamori et al., (2000). Theor.Appl.Genet. 101, 21-29
Yamanouchi and Nakamura (1992). Plant Cell Physiol 33, 985-991.
Young. (1984). in Whistler et al. (eds), Academic Press, Orlando, FL, chap 8.
Zhang et al., (1997). Mol Biotechnol 8, 223-231.
Zikiryaeva and Kasimov, (1972). Uzbekskii Biologicheskii Zhurnal 6, 18-20.
Zwar and Chandler, (1995). Planta 197, 39-48.
【技術分野】
【0001】
本発明は、相対アミロース含量が高い穀粒デンプンを有するイネに関する。本発明はまた、胚乳中のデンプン枝作り酵素(starch branching enzyme)IIa(SBEIIa)活性を低下させた米に関する。本発明はまた、穀粒およびデンプン、ならびにそれらから得られる食品および非食品製品に関する。
【背景技術】
【0002】
穀類デンプンは、2種類の分子、アミロースおよびアミロペクチンを含む。アミロースは、α−1,4結合したグルコシド単位からなるほぼ直鎖状の分子であるが、アミロペクチンは、α−1,6グルコシド結合で繋がった直鎖で高度に分岐している。
【0003】
高等植物の胚乳中でのデンプンの合成は、4つの重要な段階を触媒する1揃いの酵素によって行われる。第1に、ADP−グルコースピロホスホリラーゼ(ADGP)により、G−1−PおよびATPからのADP−グルコースの合成を介して、デンプンのモノマー前駆体が活性化される。第2に、デンプン合成酵素により、この活性化されたグリコシルドナーであるADP−グルコースが、既存のα−1,4結合した鎖の非還元末端に転移する。第3に、デンプン枝作り酵素(SBE)により、α−1,4結合したグルカンのある領域の切断を介して枝分かれ部位が導入された後、この切断された鎖が受容鎖に転移し、したがって新たなα−1,6結合が形成される。SBEは、α−1,6結合をα−ポリグルカン中に導入することができる唯一の酵素であり、したがって、アミロペクチンの形成において必須の一役割を果たしている。最後に、デンプン枝切り酵素(starch debranching enzyme)により、分岐結合の一部が切り離されるが、この酵素の作用機序はまだ解明されていない(Myersら、2000)。少なくともこれら4つの活性が高等植物における通常のデンプン顆粒合成に必要であることは明らかであるが、高等植物の胚乳中に、これら4つの活性それぞれのアイソフォームが複数存在しており、突然変異解析に基づいて(Wangら、1998a;Buleonら、1998)、あるいは遺伝子導入手法を使用して遺伝子発現レベルを改変することにより(Abelら、1996;Joblingら、1999;Schwallら、2000)、個々のアイソフォームに特異的な役割が提唱されている。しかし、各活性のそれぞれのアイソフォームのデンプン生合成への正確な寄与はまだ分かっておらず、またこうした寄与が種間で著しく異なるかどうかも分かっていない。
【0004】
穀類の胚乳中に、ADP−グルコースピロホスホリラーゼの2種のアイソフォームが存在しており、1種はアミロプラスト内にあり、また1種は細胞質中にある(Denyerら、1996;Thorbjornsenら、1996)。それぞれのアイソフォームは2つのサブユニット型からなる。トウモロコシのshrunken(sh2)変異体およびbrittle(bt2)変異体は、それぞれラージサブユニットおよびスモールサブユニットにおいて損傷を表す(GirouxおよびHannah、1994)。穀類の胚乳中で4つのクラスのデンプン合成酵素が見つかっており、1つのアイソフォームは、デンプン顆粒内に排他的に局在する顆粒結合型デンプン合成酵素(granule−bound starch synthase)(GBSS)であり、2つの型は、顆粒画分と可溶性画分に分配されており(SSI:Liら、1999a;SSII:Liら、1999b)、また第4の型は、可溶性画分全体に位置しているSSIIIである(Caoら、2000;Liら、1999b;Liら、2000)。GBSSは、アミロース合成に必須であることが示されており(Shureら、1983)、SSIIおよびSSIIIにおける変異によりアミロペクチン構造が変化することが示されている(Gaoら、1998;Craigら、1998)。イネGBSS(waxy)遺伝子の配列は記載されており(Wangら、1990)、アンチセンス法により発現が阻害される(Teradaら、2000)。waxy遺伝子は、胚乳および花粉中で発現するが、イネの他の器官では発現しない(HiranoおよびSano、2000)。
【発明の概要】
【発明が解決しようとする課題】
【0005】
植物では、2つの主要なクラスのSBE、すなわちSBEIおよびSBEIIが知られている。穀類では、SBEIIはさらに、SBEIIaおよびSBEIIbの2つのタイプに分類することができる(BoyerおよびPreiss、1978;Gaoら、1996;Fisherら、1996;HedmanおよびBoyer、1982;Mizunoら、1992;Sunら、1997;Sunら、1998)。また、一部の穀類では、SBEの追加の型、すなわちコムギからの推定149kDaのSBEI(Bagaら、2000)およびオオムギからの50/51kDaのSBEが報告されている(Sunら、1996)。イネ(NakamuraおよびYamanouchi、1992;Mizunoら、1992;Mizunoら、1993;Mizunoら、2001)、トウモロコシ(Babaら、1991;Fisherら、1993;Gaoら、1997)、コムギ(Repellinら、1997;Nairら、1997;Rahmanら、1997)、および他の穀類ついては、ゲノム配列およびcDNA配列が特徴付けられている。配列アラインメントにより、ヌクレオチドおよびアミノ酸のレベルでの高度の配列類似性が明らかになっており、SBEI、SBEIIa、およびSBEIIbのクラスに分類することが可能となる。SBEIIaおよびSBEIIbは一般に、特にその遺伝子の中央の領域においてお互いに約80%の配列同一性を示す。
【0006】
SBEIIa、SBEIIb、およびSBEIを、胚乳および他の組織におけるその時間的および空間的発現パターンによって区別することもできる。SBEIは、コムギおよびトウモロコシでは胚乳発達中期以降に発現される(Morellら、1997)。それとは対照的に、SBEIIaおよびSBEIIbは、胚乳発達の初期段階から発現される。トウモロコシでは、SBEIIbは、胚乳中で主役の型であるが、SBEIIaは、葉中に高い発現レベルで存在する(Gaoら、1997)。イネでは、SBEIIaおよびSBEIIbは、胚乳中におよそ等しい量で存在する(YamanouchiおよびNakamura、1992)。しかし、発現するタイミングおよび組織に違いがあった。SBEIIaは、種子発達のより初期の段階で発現され、したがって開花後日数(DAF)3日で検出され、また葉で発現されるが、SBEIIbは、DAF3日では検出不可能であり、DAF7〜10日の発達中の種子の中に最も豊富にあり、葉で発現されなかった(Mizunoら、2001)。コムギの胚乳中では、SBEI(Morellら、1997)は、その可溶性画分に排他的に存在するが、SBEIIaおよびSBEIIbは、可溶性画分およびデンプン顆粒を伴う画分のどちらにも存在する(Rahmanら、1995)。
【0007】
高等植物には2つの型の枝切り酵素が存在し、この酵素はその基質特異性に基づいて定義されており、すなわちそれはイソアミラーゼ型枝切り酵素およびプルラナーゼ型枝切り酵素である(Myersら、2000)。トウモロコシおよびイネのSugary−1変異は、どちらの枝切り酵素の欠損も伴うが(Jamesら、1995;Kuboら、1999)、その原因となる変異は、イソアミラーゼ型枝切り酵素遺伝子と同じ場所に位置する。イネでは、イソアミラーゼのアンチセンス阻害によりアミロペクチンの構造およびデンプンの特性が変化し(Fujitaら、2003)、したがってイソアミラーゼがアミロペクチン生合成に必要であることが示された。
【0008】
穀類からクローニングされた代表的なデンプン枝作り酵素遺伝子を表1に記載する。
【0009】
【表1】
【0010】
トウモロコシおよびイネでは、高アミロースの表現型は、amylose extender(ae)遺伝子としても知られるSBEIIb遺伝子の損傷に起因することが示されている(BoyerおよびPreiss、1981;Mizunoら、1993;Nishiら、2001)。こうしたSBEIIb変異体では、胚乳デンプン粒が異常な形態を示し、アミロース含量が有意に増大し、残存するアミロペクチンの分岐頻度が減少し、短鎖の割合(<DP17、特にDP8−12)が低下した。さらに、このデンプンの糊化温度が上昇した。さらに、アミロースとアミロペクチンとの「中間」と定義される、材料の重要なプールがあった(Boyerら、1980;Takedaら、1993b)。イネでは、SBEIIbの不活性化により、約18%のアミロースを有する野生型イネに比べて、約25%のアミロース含量がもたらされた(Nishiら、2001)。
【0011】
それとは対照的に、変異誘発(Mu)挿入配列、およびその結果としてのSBEIIaタンパク質発現の欠損による、SBEIIa遺伝子におけるトウモロコシ変異体では、葉のデンプンは変化したが、胚乳のデンプンの分岐は野生型植物と区別がつかなかった(Blauthら、2001)。同様に、SBEIIa活性を欠くイネ植物は、胚乳中のアミロペクチン鎖のプロファイルの有意な変化を示さなかった(Nakamura、2002)。トウモロコシおよびイネのどちらにおいても、SBEIIa遺伝子およびSBEIIb遺伝子はゲノム中で連鎖していない。
【0012】
超高アミロースのトウモロコシ品種がしばらく前から知られている。超高アミロース含量(>90%)のLAPS(低アミロペクチンデンプン)トウモロコシは、SBEII活性のほぼ完全な不活性化と共に、SBEI活性の相当な低下によって得られた(Sidebottomら、1998)。
【0013】
ジャガイモでは、塊茎における主要なSBE(SBE B、SBEIに相当)がアンチセンス法によってdown regulationされると、デンプンの新規の特徴がいくつか得られたが、アミロース含量は変化しなかった(Saffordら、1998)。量が豊富でない型のSBE(SBE A、穀類のSBEIIに類似)がアンチセンスによって阻害されると、アミロース含量が穏やかに38%にまで増大した(Joblingら、1999)。しかし、SBEIIおよびSBEIの両方がdown regulationされると、相対アミロース含量がSBEII単独がdown regulationされるよりもずっと大幅に60〜89%にまで増大した(Schwallら、2000)。
【0014】
コムギでは、SGP−1(SSII)タンパク質を完全に欠くある変異体が、アミロペクチン構造について改変されており、デンプン顆粒が変形し、アミロース含量がデンプンの約30〜37%にまで上昇したが、それは野生型のレベルを約8%超える増加であった(Yamamoriら、2000)。比色測定、電流滴定(どちらもヨウ素結合用)およびコンカナバリンA法により、アミロースが測定された。SSIIの無発現変異体からのデンプンは、対応する非変異体植物のデンプンに比べて糊化温度が低下した。この穀粒のデンプン含量は、野生型の60%から50%未満に低下した。
【0015】
トウモロコシでは、dull1変異が、変化の程度が遺伝的背景に依存した、胚乳中のデンプン含量の低下およびアミロースレベルの増大をもたらし、残存するアミロペクチンの分岐の程度を増大させる(ShannonおよびGarwood、1984)。この変異に相当する遺伝子は、トランスポゾン変異誘発因子(Mu)を使用したトランスポゾンタギング戦略によって同定および単離され、デンプン合成酵素II(SSII)と呼ばれる酵素をコードしていることが示された(Gaoら、1998)。この酵素は現在、穀類におけるSSIIIファミリーのメンバーとして認められている(Liら、2003)。変異体の胚乳は、dull1変異に伴いSBEIIaの活性レベルが低下した。こうした発見が他の穀類、例えばイネに関連しているかどうかは分かっていない。
【0016】
穀粒のデンプン中のアミロース比率を増加させたオオムギの系統が同定されている。こうした系統には、相対アミロース含量が約45%であり、かつ穀粒デンプン中のアミロースレベルが約65〜70%にまで上昇する、オオムギのSSIIa遺伝子の化学的に誘発される突然変異を有するHigh Amylose Glacier(AC38)が含まれる(国際公開第02/37955A1号;Morellら、2003)。このデンプンは糊化温度の低下を示した。
【課題を解決するための手段】
【0017】
イネ(Oryza sativa L.)は、発展途上の世界において最も重要な穀類収穫物であり、広く、特にアジアで栽培されており、そこでの産出量は全世界の約90%である。
【0018】
デンプンは、食物、紙、および化学工業において広く使用されている。デンプンの物理的構造は、食品もしくは非食品、または工業製品にとってのデンプンの栄養特性および取扱適性に重要な影響を与えることができる。いくつかの特徴を、アミロペクチン鎖長分布、結晶化の程度およびタイプ、ならびに糊化温度、粘度、膨潤容積などの特性を含めたデンプン構造の指標とみなしてよい。アミロペクチン鎖長の変化は、アミロペクチンの変化した結晶化、糊化、または老化の指標としてよい。
【0019】
デンプン組成物、特に高アミロース含量に関連する可能性があるレジスタントスターチと呼ばれる型は、腸の健康、特に大腸の健康のために重要な意味を有する。したがって、トウモロコシやオオムギのようなある種の穀粒において、腸の健康を促進させる手段として食品に使用するために高アミロースデンプンが開発されている。レジスタントスターチの有益な効果は、大腸への栄養分の供給に起因しており、それにより腸のミクロフローラはエネルギー源を受け取り、このエネルギー源が発酵してとりわけ短鎖の脂肪酸が形成される。こうした短鎖の脂肪酸は、結腸細胞に栄養分を提供し、大腸全域でいくつかの栄養分の摂取を増強し、結腸の生理的活性を促進する。一般に、レジスタントスターチまたは他の食物繊維が供給されない場合、結腸は代謝が比較的不活発である。
【0020】
化学的に、または別の方法で改変されたデンプンは食品に利用することができ、こうした食品は、未改変源からは通常もたらされない機能性を提供するが、このような加工は、改変に伴うプロセスに起因して、価値のある他の成分を変化させるか、あるいは望ましくない感覚をもたらす傾向がある。したがって、改変されていない形で食品に使用することができる構成成分の供給源を提供することが好ましい。
【0021】
したがって、アミロースの割合が40%を超えるデンプンを有する米は知られていない。高アミロースのトウモロコシおよびオオムギの品種は知られているが、米作地域にとっては超高アミロース米の方が好ましい。このような米由来のデンプンは、比較的難消化性であり、したがって超高アミロース米が世界人口の相当な割合に健康上重要な利益をもたらすと期待されている。
【図面の簡単な説明】
【0022】
【図1】米デンプン枝作り酵素I遺伝子(SBEI)−Genbankアクセッション番号D11082(配列番号1)をコードしているcDNAの配列を示す図である。
【図2】米デンプン枝作り酵素IIa遺伝子(SBEIIa)−Genbankアクセッション番号AB023498(配列番号2)をコードしているcDNAの配列を示す図である。
【図3】米デンプン枝作り酵素IIb遺伝子(SBEIIb)−Genbankアクセッション番号D16201(配列番号3)をコードしているcDNAの配列を示す図である。
【図4】pBC SK−中に挿入された、イネSBEI遺伝子のイントロン9の500bp断片を含むプラスミドpRint9_BCの略図である。
【図5】2重鎖RNA構築体の略図である。A.使用した遺伝子エレメントの順序は、プロモーター、センス方向のSBEIIaまたはSBEIIbのcDNA配列、イントロン(Rint9)、アンチセンス方向のSBEIIaまたはSBEIIbのcDNA配列、および転写ターミネーター/ポリアデニル化配列であった。ds−SBEIIaおよびds−SBEIIb遺伝子の転写物は、センス配列とアンチセンス配列の間のハイブリダイゼーションによって形成される2本鎖領域を有する「へアピン」RNA構造を形成する。GTヌクレオチドおよびAGヌクレオチドと境を接するイントロン配列は、スプライシングによって切り取られる。また、pRBEI.IR(pBC SK−(センス/Rint9/アンチセンス))の略図を示す。対応する構築体、pRBEIIa.IRおよびpRBEIIb.IRも作製した。
【図6】プラスミドベクターpBx17casNOTの略図である。
【図7−1】イネSBEIIa(配列番号2)およびイネSBEIIb(配列番号3)のcDNA配列の比較を示す図である。上側の配列(大文字)はSBEIIbのものであり、下側の配列(小文字)はSBEIIaのものである。5'末端および3'末端の配列は、十分な同一性がないので示さない。
【図7−2】イネSBEIIa(配列番号2)およびイネSBEIIb(配列番号3)のcDNA配列の比較を示す図である。上側の配列(大文字)はSBEIIbのものであり、下側の配列(小文字)はSBEIIaのものである。5'末端および3'末端の配列は、十分な同一性がないので示さない。
【図7−3】イネSBEIIa(配列番号2)およびイネSBEIIb(配列番号3)のcDNA配列の比較を示す図である。上側の配列(大文字)はSBEIIbのものであり、下側の配列(小文字)はSBEIIaのものである。5'末端および3'末端の配列は、十分な同一性がないので示さない。
【図7−4】イネSBEIIa(配列番号2)およびイネSBEIIb(配列番号3)のcDNA配列の比較を示す図である。上側の配列(大文字)はSBEIIbのものであり、下側の配列(小文字)はSBEIIaのものである。5'末端および3'末端の配列は、十分な同一性がないので示さない。
【図8】遺伝子発現抑制構築体に用いられたSBEIIa、SBEIIb、およびSBEIの3'配列を使用した遺伝子発現抑制プログラム(Gene Silencing program)からのBLAST出力を示す図である。
【発明を実施するための形態】
【0023】
<総論>
当分野の技術者であれば、本明細書に記載した本発明が、具体的に記載したもの以外の変更および改変を受けることに気付くであろう。本明細書に記載した本発明はそのようなすべての変更および改変を含むことを理解されたい。また、本発明は、個々に、またはひとまとめにして本明細書において言及し、あるいは示すこのようなすべての工程、特徴、組成物、および化合物、ならびに前記工程または特徴の任意の2つ以上の任意のあらゆる組合せを含む。
【0024】
本明細書を通じて、「含む(comprise)」という用語、ならびに「含む(comprises)」および「含む(comprising)」などの変形は、文脈上他の意味に解すべき場合を除き、記載した1つの完全体(integer)もしくは段階、または完全体(integer)もしくは段階の群を包含することを意味し、他のいかなる完全体(integer)もしくは段階、または完全体(integer)もしくは段階の群も除外することを意味するわけではないことが理解されよう。本発明は、本明細書に記載した特定の実施形態による範囲に限定するものではなく、例示のみを目的とするものである。機能的に同等な生成物、組成物、および方法は、明らかに、本明細書に記載した本発明の範囲内にある。
【0025】
本明細書において著者名で示した刊行物の書誌詳細を本記載の最後にまとめる。本明細書に記載する参考文献の全体を参照により本明細書に組み込む。本明細書における先行技術に関する参考文献は、1種または複数の任意の先行技術の文献を含めて、前記先行技術がオーストラリアにおける共通の一般知識であり、またはオーストラリアにおける共通の一般知識の一部を形成するものであるという認識あるいは示唆と解釈すべきではない。
【0026】
本明細書では、「に由来する(derived from)」という用語は、特定の完全体(integer)または完全体(integer)の群が、指定の種を起源としたものであって、それを必ずしも指定の供給源から直接得たものではないことを指すと解釈すべきである。
【0027】
本明細書で表すヌクレオチド残基の名称は、IUPAC−IUBの生化学命名法委員会によって推奨されるものであり、Aはアデニン、Cはシトシン、Gはグアニン、Tはチミジンを指す。
【0028】
<発明の概要>
第1の態様では、本発明は、イネ植物から得られる穀粒にあるものとしてもよく、この穀粒は、そのデンプン中のアミロースの割合が少なくとも40%であるデンプンを含む。この穀粒は、SBEIIaおよびSBEIIbの活性またはレベルを低下させたものであることが好ましく、ある形態では、これを2つ以上の遺伝的変異によって実施してよく、1つの遺伝的変異は、a)SBEIIaの発現および/または活性を阻害するSBEIIa遺伝子の突然変異、ならびにb)SBEIIaの発現および/または活性を阻害する導入核酸からなる群から選択され、第2の遺伝的変異は、a)SBEIIbの発現および/または活性を阻害するSBEIIb遺伝子の変異、ならびにb)SBEIIbの発現および/または活性を阻害する導入核酸からなる群から選択される。
【0029】
ある形態では、この穀粒はある導入遺伝子を含み、この導入遺伝子は、アンチセンス、コサプレッション、リボザイム、または2重鎖RNA分子をコードするものでよい。あるいは、この穀粒は、その阻害が染色体突然変異または染色体再配列に起因する、導入遺伝子によらないものでもよい。この穀粒は、SBEIIaまたはSBEIIb遺伝子の無発現変異を含むものでもよい。
【0030】
第1の態様で請求した穀粒は、SBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたものを含むものでよい。ある特定の形態では、この穀粒はさらに、SBEIのタンパク質レベルおよび/または活性レベルを低下させたものを含むものでもよく、それに加えて、またはその代わりに、ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型の枝切り酵素、およびプルラナーゼ型の枝切り酵素からなる群から選択される、タンパク質レベルおよび/または酵素活性レベルを変更したものも含むことができる。代替の特定の形態では、この穀粒は、GBSSのタンパク質および/または酵素活性のレベルを変更したものを含む。さらなる任意の形態では、この穀粒は、インディカ品種のものであるか、または対立遺伝子Wxaを含む。
【0031】
好ましいある形態では、穀粒のデンプン中のアミロースの割合が少なくとも50%である。
【0032】
この穀粒は、non−shrunkenであることが好ましく、特定のあるフォーム、すなわち玄米のフォームでは、平均重量が少なくとも約25mgであり、また好ましくは、デンプン含量が、同等であるが未改変の穀粒の少なくとも90%である。また、好ましくは、穀粒内のデンプン顆粒の少なくとも50%は、偏光下で観察すると非複屈折性に見える。
【0033】
第2の態様では、本発明はまた、本発明の第1の態様の穀粒を産生することができるイネ植物を包含する。
【0034】
本発明の第3および第4の態様は、本発明の第1の態様の穀粒から抽出されたデンプンおよびデンプン顆粒に関する。
【0035】
第5の態様では、本発明は、本発明の第1の態様の穀粒から生成した穀粉またはデンプンを含む製品に関する。この生成物は、穀粉またはデンプンと別の供給源からの穀粉またはデンプンとの混合物を含むものでもよい。この製品は食品でも非食品製品でもよい。
【0036】
本発明の第6の態様は、第3の態様のデンプン、および別の食品成分または水を含む組成物を包含する。
【0037】
本発明の第7の態様は、少なくとも40%のアミロースを含むデンプンを有する穀粒を産生することができるイネ植物を作出する方法にあると言ってもよく、この方法は、a)遺伝的変異を親イネ植物または種子中に導入するステップと、b)親イネ植物または種子親の子孫植物を同定するステップとを含み、この子孫植物の穀粒のデンプンが少なくとも40%のアミロースを含む。好ましくは、この遺伝的変異により、米植物の胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または活性のレベル低下がもたらされる。
【0038】
この方法の子孫イネ植物は、2つ以上の遺伝的変異を含むことが好ましく、1つの遺伝的変異が、a)SBEIIaの発現および/または活性を阻害するSBEIIa遺伝子の突然変異、ならびにb)SBEIIaの発現および/または活性を阻害する導入核酸からなる群から選択され、また第2の遺伝的変異が、c)SBEIIbの発現および/または活性を阻害するSBEIIb遺伝子の突然変異、ならびにd)SBEIIbの発現および/または活性を阻害する導入核酸からなる群から選択される。
【0039】
遺伝的変異を導入するステップは、外因性の核酸を導入することを含むものでよい。外因性の核酸をイネ細胞中に導入し、次いでこの細胞をイネ植物に再生させてもよい。この外因性の核酸は、SBEIIaおよび/またはSBEIIbの発現および/または活性の阻害物質をコードすることが好ましく、この阻害物質は、アンチセンス、コサプレッション、リボザイム、または2重鎖RNA分子であってよい。
【0040】
あるいは、遺伝的変異を導入するステップは、化学薬剤または放射線を用いた親イネ植物または種子の突然変異誘発を含むものでもよい。
【0041】
この子孫イネ植物は、SBEIIaおよび/またはSBEIIbにおける無発現変異を含むものでもよい。
【0042】
遺伝的変異を導入するステップはさらに、SBEIのタンパク質レベルおよび/または活性のレベルの低下に導くものでもよい。
【0043】
この子孫植物を、穀粒デンプン中のアミロースレベルに基づいて、あるいは子孫植物の胚乳中のSBEIIaおよび/またはSBEIIbのタンパク質レベルおよび/または活性のレベルの低下に基づいて同定してもよい。
【0044】
この方法はさらに、対立遺伝子Wxaのイネへの導入を含むものでもよく、これを交配によって導入してもよい。
【0045】
本発明の第8の態様は、胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または酵素活性のレベルを低下させたイネ植物を作出する方法にあるものとしてもよく、この方法は、a)SBEIIaのタンパク質レベルおよび/または酵素活性レベルを低下させた種子に突然変異を起こさせること、あるいはb)SBEIIbのタンパク質レベルおよび/または酵素活性レベルを低下させた種子に突然変異を起こさせること、あるいはc)SBEIIaのタンパク質レベルおよび/または酵素活性のレベルを低下させた植物を、SBEIIbのタンパク質レベルおよび/または酵素活性のレベルを低下させた植物と交配させること、ならびにd)胚乳中のSBEIIaおよびSBEIIbのタンパク質活性および/または酵素活性を低下させたイネ植物を同定することを含む。
【0046】
このイネ植物を同定するステップは、イネのSBEIIa遺伝子およびSBEIIb遺伝子に連鎖する分子マーカーを有するイネ植物の集団をスクリーニングすること、ならびに連鎖した分子マーカーを用いたスクリーニングからのシグナルの存在または非存在に基づいて植物を同定することを含むものでよい。
【0047】
このイネ植物を同定するステップは、イネのSBEIIaタンパク質およびSBEIIbタンパク質に結合する抗体を用いてイネ植物の集団からの種子をスクリーニングするステップ、ならびに結合した抗体の存在または非存在に基づいて植物を同定するステップを含むものでもよい。
【0048】
本発明はまた、改変した米デンプンを産生する方法を包含してもよく、この方法は、本発明の第1の態様の穀粒からデンプンを抽出するステップを含む。
【0049】
本発明はさらに、2つ以上の外因性の核酸分子の使用を包含してもよく、SBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたイネ植物を作出するために、その少なくとも1つがイネのSBEIIaの発現および/または活性の阻害物質をコードしており、またその少なくとも別の1つがイネのSBEIIbの発現および/または活性の阻害物質をコードしている。この阻害物質は、アンチセンス分子、コサプレッション分子、リボザイム、2重鎖RNA分子、およびそれらの任意の組合せからなる群から選択することができる。
【0050】
本発明はまた、イネSBEIIaの阻害物質およびイネSBEIIbの阻害物質をコードしている単離された核酸分子も包含しており、これらの阻害物質は、同一の分子であっても、異なる分子であってもよい。この単離されたベクターはベクターでよい。本発明が、単離された核酸分子を含む細胞を包含し、この細胞がイネ細胞であることが好ましいことを理解されたい。本発明はまた、単離された核酸分子を含むトランスジェニックイネ植物を包含することを理解されたい。
【0051】
<発明の詳細な説明>
イネ植物を作出する方法
一態様では、本発明は、穀粒中に改変したデンプンを有するイネ植物を作出する方法、特にデンプン中のアミロースの相対的な割合を少なくとも40%まで増加させる方法を提供する。本明細書で定義するデンプン中のアミロースの割合は、重量/重量(w/w)に基づき、すなわちデンプンの重量に対する百分率で示すアミロースの重量である。デンプン中のアミロースの割合は、少なくとも45%、50%、55%、または60%であることが好ましく、また少なくとも65%、70%、または75%であることがさらにより好ましい。米では通常、デンプン中のアミロースの割合は、約0〜35%の範囲にわたる。この方法には、米胚乳中のデンプン枝作り酵素IIa(SBEIIa)およびデンプン枝作り酵素IIb(SBEIIb)のタンパク質レベルまたは酵素活性レベルを低下させることが含まれ得る。このタンパク質または活性の低下は、改変していない米胚乳中のタンパク質または活性の相当するレベルに比べて、少なくとも40%、または好ましくは少なくとも60%、より好ましくは少なくとも75%、さらにより好ましくは少なくとも90%または95%でよい。米胚乳中でこのタンパク質のうちの一方または両方を検出できない可能性がある。この方法は、米のSBEIIaおよびSBEIIb遺伝子の発現の改変を含むものでよく、あるいは米におけるSBEIIaおよびSBEIIb遺伝子の突然変異またはこれらの組合せを含むものでよく、これにより胚乳中のSBEIIaおよびSBEIIbの活性がどちらも低下する。この遺伝子のうちのどちらかまたは両方の発現を、核酸の導入、例えば導入遺伝子によって阻害してよい。
【0052】
本明細書では、「増大した(increased)」、「減少した(decreased)」、「低下した(reduced)」、および「改変した(altered)」などの用語は、本発明に従って改変されていない対応する野生型の植物または生成物に比べての、本発明の植物または生成物の違いを指す比較用語であることは容易に明らかであろう。
【0053】
本明細書では、アミロースを、α−1,4結合グリコシド分子、およびアミロース様長鎖アミロペクチン(「中間物質」と呼ぶことがある、Takedaら、1993b;Fergason、1994)からなるほぼ直鎖状の分子を含むものと定義する。アミロース含量を、例えば90%(w/v)DMSO中でのサイズ排除HPLC、コンカナバリンA法(Megazyme社、アイルランド)、または好ましくは例えば実施例1に記載するヨウ素滴定法を含めた当技術分野で既知のいずれかの方法によって求めることができる。HPLC法は、デンプンの枝切りを伴うものでも(BateyおよびCurtin、1996)、枝切りを伴わないものでもよい。穀粒重量およびアミロース含量から、トランスジェニック系統および対照系統について、穀粒あたりの蓄積されたアミロースの量を計算し比較することができる。
【0054】
この方法は、米胚乳中のSBEIIaおよび/またはSBEIIbの活性を、好ましくは両方求めるステップを含むものでよい。これを、例えば免疫検出によりタンパク質のレベルを測定すること、あるいは、ノーザンブロットハイブリダイゼーション分析、スロットブロットハイブリダイゼーション、RNAseプロテクションアッセイ、マイクロアレイ分析、または逆転写ポリメラーゼ連鎖反応(RT−PCR)など当技術分野で既知の方法により、それらに対応するmRNAのレベルを測定することによって行ってもよい。この方法はさらに、胚乳中のSBEIIaおよび/またはSBEIIbの活性を低下させたイネ植物または穀粒をスクリーニングするステップ、あるいはこのような植物もしくは穀粒を選択または同定するステップを含むものでもよい。スクリーニング/選択のステップは、SBEIIaおよび/またはSBEIIbの活性レベルまたはタンパク質レベルを低下させることに基づくものでもよく、あるいは、増大させたアミロースの割合、または低下させたアミロペクチンの割合、または視覚的表現型、例えばShrunken穀粒などイネ植物の穀粒の表現型に基づくものでもよい。
【0055】
酵素アッセイ、例えばホスホリラーゼ刺激アッセイ(Phosphorylase stimulation assay)(BoyerおよびPreiss、1978)により、SBE活性を測定してよい。このアッセイでは、ホスホリラーゼaによるグルコース1−リン酸のメタノール不溶性ポリマー(α−D−グルカン)中への組込みの、SBEによる刺激が測定される。SBE活性を、グルカンポリマーの枝切りによって生じるグルカン−ポリヨウ素複合体の吸光度の減少を測定するヨード染色検査によって測定することができる。SBE活性を、イソアミラーゼ消化の後、基質である還元アミロースからの還元末端の形成を測定する分岐結合アッセイ(branch linkage assay)によって分析することもできる(Takedaら、1993a)。好ましくは、この活性をSBEI活性なしで測定する。SBEのアイソフォームは様々な基質特異性を示し、例えばSBEIは、アミロースを分岐させる際により高い活性を示すが、SBEIIaおよびSBEIIbは、アミロペクチン基質での分岐をより高い割合で示す。また、このアイソフォームを、転移するグルカン鎖の長さに基づいて区別してもよい。また、SBEタンパク質を、本明細書に記載するものなど特異的抗体を使用することによって測定してもよい。好ましい一実施形態では、イネSBEIIaおよびSBEIIbのN末端アミノ酸配列に相当するポリペプチド断片に対して産生された特異的抗体を使用した、ウエスタンブロット法またはELISA検査などの免疫学的方法により、SBEIIaおよびSBEIIbのタンパク質レベルを測定する。SBEII活性を、穀粒発達中の発達している胚乳中で、あるいはこのタンパク質が同等であるが未改変の穀粒中にまだ存在しており、それを免疫学的方法によって検査することができる成熟した穀粒中で測定してもよい。
【0056】
さらなる一態様では、本発明は、穀粒のデンプン中のアミロースの割合が少なくとも40%となるように、SBEIIaおよびSBEIIbの活性の低下と組み合わせて、イネにおける第3のデンプン生合成酵素の活性を改変する、好ましくは低下させる方法を提供する。好ましくは、胚乳中のSBEI活性も低下させる。SBEIIaおよびSBEIIbと組み合わせて改変してよい他のデンプン生合成の酵素活性は、SSI、SSII、およびSSIIIである。また、デンプン枝切り酵素、例えばイソアミラーゼまたはプルラナーゼ活性を改変してもよい。第3のデンプン生合成の酵素活性を、改変していないイネにおける活性に比べて、少なくとも40%、好ましくは60%または80%、より好ましくは少なくとも90%までに上昇または低下、好ましくは低下させてよい。
【0057】
さらなる一実施形態では、この植物のデンプン生合成酵素の活性を胚乳以外の植物組織中で改変してよく、例えばSBEIまたはSBEII、好ましくはSBEIIaの活性を葉において増大させて、胚乳中のSBEIIa活性の喪失に導く、この植物における遺伝的変異によって引き起こされる活性喪失の一部を補ってもよい。これは、遺伝的変異が、胚乳中だけでなく他の組織、特に葉においてSBEIIa活性の低下に導く場合に特に好ましい。胚乳以外の組織中でのこのような活性の補償は、例えば、胚乳中で発現しないプロモーターの制御下にある酵素コード領域に起因するであろうと理解されよう。これは、rbcSなど光合成に関係する遺伝子からのプロモーターでよい。あるいは、胚乳中のデンプン合成を、胚乳中のSBEIIaおよびSBEIIbの活性の低下と組み合わせた、1種または複数のデンプン生合成の酵素の過剰発現によってさらに改善してもよい。このような酵素をコードしている遺伝子は、様々な任意の供給源からのもの、例えば細菌や、イネ以外の他の供給源からのものでもよく、またこの遺伝子を改変して、触媒活性を変更、例えば酵素の温度依存を変更(WO94/09144)してもよい。
【0058】
高アミロース表現型は、SBEIIa遺伝子およびSBEIIb遺伝子の発現に対する部分的な、または十分な撹乱によって得てよい。本発明の方法は、SBEIIaおよび/またはSBEIIb遺伝子における無発現変異を有するイネ植物または穀粒をスクリーニングまたは同定または選択するステップを含むものでよい。本明細書では、「無発現変異」は、対象の植物組織中、好ましくは胚乳中の検出可能なタンパク質または酵素活性の欠損を起こす突然変異であると定義する。したがって、スクリーニング/同定するステップは、遺伝子レベルでの、例えばSBEIIaおよび/またはSBEIIbをコードしている遺伝子における欠失のためのスクリーニング、あるいは対象の遺伝子の発現のレベルでのスクリーニングを含むものでよい。この遺伝子が阻害される程度により、米粒中で形成されるデンプンの特徴がある程度決定される。好都合には、欠失のスクリーニングは、増幅産物の少なくとも一部が対象の遺伝子の少なくとも一部に及ぶように設計されたプライマーを使用したPCR増幅法によって好都合に実施してよい。改変した米胚乳から抽出されたタンパク質について実施される一連のゲル電気泳動法のいずれかにより、SBEIIaおよびSBEIIbの活性に対する改変の性質および程度が明らかになるであろう。改変は、SBEIIaおよび/またはSBEIIb活性の低下、または胚乳内の酵素活性の完全な消滅として起こるものでよい。こうした試験を実施するために、デンプンを米胚乳から抽出し、そのタンパク質を分析してよい。SDS−PAGEおよびイムノブロット法などの当技術分野で周知の技法を、可溶性画分およびデンプン顆粒画分について実施してよく、改変がSBEIIaおよびSBEIIb酵素に対して起こった植物または穀粒が同定される。
【0059】
本発明の方法は、イネ植物またはイネ祖先植物または種子への遺伝的変異の導入を含むものでよい。この遺伝的変異は、以下に記載するように導入遺伝子を含むものでよく、あるいは突然変異誘発によって、例えば化学的突然変異誘発によって、または放射線によって導入してもよい。
【0060】
イネ植物
さらなる一態様では、本発明は、デンプン中のアミロースの割合が少なくとも40%の穀粒を産生することができるイネ植物を提供する。本明細書では、イネ植物を、種Oryza sativa L.のいずれかであると定義する。このイネ植物は、O. sativa L.、すなわちJaponica(またはsinica)、Indica、およびJavanicaの認識されている3つの品種のいずれかのものでよく、Indica品種であることが好ましい。ぞれぞれの品種には栽培品種または変種が多数あり、すべて本発明の植物に含まれる。好ましい栽培品種は、例えば、栽培品種Amaroo、Ali Combo、Basmati、Bogan、Bombia、Doongara、Goolarah、Illabong、Jarrah、Koshihikari、Kyeema、Langi、Millin、Namage、Opus、Peldeを含めて、オーストラリアで栽培されているものである。アミロースの割合は、少なくとも45%、50%、55%、60%、65%、70%、または75%であることが好ましい。このイネ植物は、胚乳中のSBEIIaおよび/またはSBEIIbの発現および/または活性を阻害する少なくとも1種の遺伝的変異を含む。この遺伝的変異は、SBEIIaおよび/またはSBEIIbの発現または活性、ならびに上記のものの組合せを阻害する、SBEIIaおよびSBEIIb遺伝子における突然変異や、アンチセンス、増強されたアンチセンス、コサプレッション、リボザイム、2重鎖RNA、または類似の分子をコードしている遺伝子などの導入核酸など、米胚乳中のSBEIIaおよびSBEIIbの活性および/またはタンパク質の低減に導く任意の遺伝的変異でも、遺伝的変異の組合せでもよい。この遺伝的変異は、無発現変異であることが好ましい。SBEIIaおよびSBEIIbの活性を低下させた植物は、SBEIIaを低減させた植物を、SBEIIbを低減させた植物と交配させることによって、あるいはSBEIIaおよびSBEIIb遺伝子のどちらの発現も阻害する分子をコードしている導入遺伝子を導入することによって作出してよい。好ましい一実施形態では、このイネ植物は、SBEIIaおよびSBEIIbのどちらにおいても無発現変異を有する。
【0061】
本発明はまた、穀粒の発達の少なくとも一部の期間に胚乳中のSBEIIaおよびSBEIIb活性のレベルをどちらも低下させたイネ植物を提供し、このイネ植物は、同等であるが未改変の植物から抽出されたデンプンに比べて、高率のアミロースを含むデンプンを有する穀粒を生み出すことができる。好ましくは、SBEIIaおよびSBEIIbのレベルを、野生型に比べて、胚乳中で少なくとも50%、より好ましくは少なくとも75%、最も好ましくは少なくとも90%または95%低下させる。「野生型」という用語は、遺伝学の分野におけるその通常の意味を有し、それには、本明細書で教示するよう改変していないイネの栽培品種または遺伝子型が含まれる。
【0062】
本発明はまた、遺伝型および/または表現型において親イネ植物の所望の特徴を有する子孫植物および穀粒を提供する。本発明はまた、培養した組織または細胞など所望の特徴を有する植物を作出するのに使用できるイネ植物の任意の繁殖用材料にまで及ぶ。
【0063】
本発明の改変したイネ植物は、より望ましい遺伝的背景を含む植物と交配してよく、したがって、本発明は、他の遺伝的背景における1種(複数)の遺伝的変異を含む。最初の交配の後、戻し交配を適当な回数実施して、望ましくない背景を除去してもよい。所望の遺伝的背景には、商業収益、および農業生産力や非生物ストレス耐性など他の特徴を提供する遺伝子の適当な組合せが含まれ得る。また、この遺伝的背景には、改変した他のデンプン生合成、または修飾遺伝子、例えば、その原因となる遺伝子が分かっていないshrunken胚乳を有する他のイネ系統からの遺伝子が含まれ得る。
【0064】
好ましい一実施形態では、このイネ植物は、waxy遺伝子の対立遺伝子Wxaを含む。この対立遺伝子は大部分がイネのインディカ品種にみられるが、対立遺伝子Wxbは大部分がジャポニカ品種にみられる。対立遺伝子Wxbは、waxy遺伝子の第1イントロンの5'スプライス部位での置換突然変異(GTからTTに)を有し、その結果、対立遺伝子Wxaを含む相当する植物中よりも、waxy遺伝子発現が低く、したがってGBSS活性が低く、またアミロースレベルが低くなる(Isshikiら、1998;Hiranoら、1998;Francesら、1998)。
【0065】
この植物またはその穀粒は、遺伝子導入によるものでも、遺伝子導入によらないものでもよい。
【0066】
穀粒
本発明はまた、同等であるが未改変のイネから抽出されたデンプンに比べて、改変されたデンプンを含む米穀粒を提供する。本明細書では、穀粒をほぼ成熟した穀粒と定義する。この穀粒には、商業的環境で収穫された穀粒が含まれる。収穫時、米穀粒は、外皮を含む籾米の形でも、外皮が除去された「玄米」の形でもよい。玄米部分のみが食用である。玄米は、果皮、種皮および珠心の外層、胚芽(胚)、ならびに胚乳からなる。本明細書で定義する胚乳は、アリューロン層、および、サブアリューロン層とデンプン質の胚乳または内胚乳からなる本来の胚乳とからなる。玄米を研磨精米または摩擦精米にかけて、果皮、種皮、外種皮、アリューロン層、および胚を除去して、それにより本質的にデンプン質の胚乳を含む白米を得ることができる。精米の結果、脂肪、タンパク質、繊維、無機質、ならびにチアミン、リボフラビン、ナイアシン、a−トコフェロールなどのビタミンを含めた、種皮およびアリューロンの成分が一部失われる。炭水化物含量、主にデンプンは、玄米中よりも精米された米の中の方が多い。野生型の精米された米の穀粒は、炭水化物を約77〜89%、タンパク質を6.3〜7.1%、デンプンと結合した形のものもデンプンなしの形のものも含めた脂質を1.5〜1.7%、無機質を0.3〜0.8%、粗繊維を0.3〜0.5%、および中性デタージェント繊維を0.7〜2.3%含有しており、水分を約10〜15%含有するしていてよい。本明細書で定義する「米穀粒」または単に「米」には、籾米、玄米、および精米された米が含まれ、精米された米であることが好ましい。
【0067】
本発明の米穀粒は、この穀粒が由来するイネ植物について本明細書で定義する少なくとも1つの遺伝的変異を含む。1種(複数)の遺伝的変異は、米穀粒の胚乳の発達中に、SBEIIaおよびSBEIIbの活性および/またはタンパク質の低下をもたらす。この穀粒は、同等であるが未改変の植物の穀粒に比べて、(全デンプンに対する百分率で示す)アミロースが高率のもの、およびアミロペクチンの割合を低下させたものを含む。デンプンは、約90%の乾物を含む白米の主要成分である。本発明に従って改変されていない米胚乳デンプンのアミロース含量は、0〜37%の範囲にあるが、遺伝子型に依存する。「見掛けのアミロース含量」を測定するためのヨウ素滴定(比色)分析に基づき、精米された米は、waxy(1〜2%アミロース)、超低アミロース(2〜12%)、低アミロース(12〜20%)、中アミロース(20〜25%)、および高アミロース(25〜33%)に分類される(Juliano、1979, 1985)。HPLC分析を使用した最近の研究では、真のアミロース含量の最大値が約20%であり、追加のヨウ素結合がアミロペクチンの長い直鎖によるものであることが示された(Takedaら、1987)。「アミロース含量」または本明細書で定義する「見掛けのアミロース含量」を、当分野の技術者に既知のヨウ素滴定法、例えばMorrisonおよびLaignelet(1983)によって記載されている分光光度法によって求める。「真のアミロース」のみを分析する高速液体クロマトグラフィー(例えばHPLC、BateyおよびCurtin、1996)法などの他の方法は、本明細書で定義するアミロース含量を低く評価する可能性があることを理解されたい。
【0068】
本発明の穀粒は、少なくとも40%(w/w)のアミロースを含むデンプンを有する。アミロースの割合は、好ましくは全デンプンの少なくとも45%、50%、または55%であり、より好ましくは少なくとも60%であり、さらにより好ましくは少なくとも65%、70%、または75%である。好ましい一実施形態では、この穀粒は、遺伝子導入によるものではなく、このデンプンは、少なくとも40%のアミロースを含む。あるいは、この穀粒は、SBEIIaとSBEIIbのタンパク質レベルを低下させたものであり、このデンプンは少なくとも40%のアミロースを有する。増大したアミロースレベルを、光学顕微鏡下で観察した場合の異常なデンプン顆粒の形態、またはこの顆粒の複屈折性の喪失、あるいは当技術分野で既知の他の方法によって証明してもよい。
【0069】
好ましい一実施形態では、このイネは、インディカ品種のものであるか、またはwaxy遺伝子の対立遺伝子Wxaを含む。
【0070】
この穀粒は、物理的特徴を改変した、例えば、糊化温度を上昇もしくは低下させた、かつ/または糊化中と糊化後の膨潤特性を改変したデンプンを含むものでよい。
【0071】
この穀粒は、shrunkenでもnon−shrunkenでもよく、好ましくはnon−shrunken表現型を有する。本明細書では、「non−shrunken」を、大多数の穀粒、好ましくは個々の穀粒の少なくとも90%が丸々とした(plump)または実入りの十分な(fully−filled)表現型を示す場合と定義する。これは通常、正常な、もしくは正常に近いレベルのデンプン蓄積を伴う。それとは対照的に、本明細書では、表現型「shrunken」は、デンプン蓄積が低下した大多数の穀粒、特に少なくとも90%の穀粒を指す。slightly shrunken穀粒は、少なくとも30%の平均デンプン含量の低下を指し、moderately shrunken穀粒は、少なくとも50%の平均デンプン含量の低下を指し、highly shrunken穀粒は、少なくとも70%の平均デンプン含量の低下を指す。また、収縮性(shrunkenness)は、成熟穀粒重量に対する百分率で示す相対デンプン含量によって測定してよい。野生型玄米の穀粒のサイズおよび形のパラメータは、極長(extra long)>7.50mm、長(long)6.61〜7.50mm、並(medium)5.51〜6.60mm、短(short)<5.50mmと定義してよい。穀粒の形は、長さと幅の比に基づいて特徴付けてよく、細長(slender)>3.0、並(medium)2.1〜3.0、太(bold)1.1〜2.0、丸(round)<1.0と定義する。本発明の米粒ではこうした特徴をそれぞれ改変してもよい。
【0072】
本発明はまた、この穀粒から生成される穀粉、荒粉、または他の生成物を提供する。これらは、例えば分別または漂白によって加工されたものでも、加工されていないものでもよい。本発明はさらに、本発明のイネ植物から得られる、食糧生産に有用な米穀粒を提供する。さらに、本発明は、この穀粒を精米し、挽き、圧延し、精白し、粗挽きもしくは挽割りにし、またはゆでるといった、他の方法で加工された穀粒を包含する。
【0073】
デンプン
別の態様では、本発明は、アミロースの割合を増大させ、アミロペクチンの割合を低下させた、上記のイネ植物の穀粒から得られるデンプン顆粒またはデンプンを提供する。この穀粒が得られる植物は、胚乳中のSBEIIaおよびSBEIIbの活性レベルを低下させたものであり、より好ましくは、SBEIの活性も低下させる。別の態様では、本発明は、このイネ植物の穀粒から得られるデンプン顆粒またはデンプンを提供し、これらは、少なくとも40%のアミロース、好ましくは少なくとも45%、50%、55%、または60%のアミロース、さらにより好ましくは少なくとも65%、70%、75%のアミロースを含む。精製されたデンプンを、タンパク質、油、および繊維からのデンプンの分離を含む製粉加工、例えば湿式製粉加工により穀粒から得てよい。製粉加工の最初の生成物は、デンプン顆粒の混合物または組成物であり、したがって本発明はこのような顆粒を包含する。
【0074】
野生型米デンプン顆粒は、形が多角体であり、その大部分はサイズが3〜9μmで、サイズの単峰性分布で平均して約5μmである。タンパク質は主に、胚乳全体にわたってサイズが0.5〜4μmの球状のタンパク粒の形で生じる。
【0075】
このデンプンは、糊化温度を上昇させたものでも、低下させたものでもよく、糊化温度を上昇させたものであることが好ましい。この糊化温度、特に最初のピークの開始温度、または最初のピークの最高点の温度を、DSCによって測定して、類似のものであるが改変されていない穀粒に比べて、少なくとも3℃、好ましくは少なくとも5℃、またはより好ましくは少なくとも7℃上昇させてもよい。このデンプンは、レジスタントスターチのレベルを上昇させたものを含むものでもよく、改変された構造は、変化したデンプン顆粒形態、相当量のデンプン結合脂質の存在、変化した結晶化、および変化したアミロペクチン鎖長分布に起因する可能性がある、消化酵素への物理的接近の困難さからなる1種または複数の群を含めた特定の物理的特徴によって示される。アミロースの割合が高いこともレジスタントスターチのレベルに寄与している。
【0076】
本発明はまた、好ましくはレベルを上昇させたレジスタントスターチと組み合わせて、食物繊維の量を増加させたものを含む例示されたイネ植物の穀粒から得られるデンプンを提供する。また、この増加は少なくとも部分的に、アミロースの相対的レベルが高い結果である。
【0077】
遺伝子活性を低下させる方法:導入遺伝子
SBEIIa、SBEIIb、または他のデンプン生合成遺伝子もしくは修飾遺伝子の活性を、遺伝的変異をイネ植物に導入することによって改変することが好ましい。これは、導入遺伝子のイネ植物への導入によるものでよい。「遺伝的変異」は、この文脈においてSBEIIaおよびSBEIIb、ならびに場合によっては他のデンプン生合成遺伝子もしくは修飾遺伝子の活性の低下をもたらすゲノムの任意の改変を意味し、この変異には、点突然変異、置換、逆位、重複、転座、好ましくは欠失、ならびに遺伝子または制御エレメント中への導入遺伝子の導入などの突然変異が含まれる。好ましい一実施形態では、この遺伝的変異は、例えば逆位、重複、転座、欠失、フレームシフト、またはRNAスプライシング変異の結果である無発現変異である。本明細書では、「導入遺伝子」は、バイオテクノロジーの分野における通常の意味を有し、これには、組換えDNA技術またはRNA技術によって生成または改変され、対象の生物または細胞中に導入される遺伝子配列が含まれる。この導入遺伝子には、生物または細胞に由来する遺伝子配列、例えばアンチセンス配列が含まれ得る。好ましい一実施形態では、この導入遺伝子は、本明細書で定義するイネSBEIIa配列または本明細書で定義するイネSBEIIb配列の相補体である少なくとも19の連続したヌクレオチドと少なくとも94%の同一性を有する、少なくとも19の連続したヌクレオチドを有するヌクレオチド配列を含む。この導入遺伝子には通常、イネに由来してない外因性の核酸が含まれる。「トランスジェニック」とは、導入遺伝子を含むイネ植物または穀粒または細胞を指す。「非トランスジェニック」とは、イネ植物のゲノム、穀粒、または細胞中に全く導入遺伝子が存在しないことを指す。安定した遺伝のために、導入遺伝子をイネ植物のゲノム、穀粒、または細胞中に組み込むことが好ましい。
【0078】
本明細書で言及する「遺伝子」は、SBEIIa、SBEIIb、または他のデンプン生合成遺伝子、あるいはアンチセンス、増強されたアンチセンス、コサプレッション、リボザイム、2重鎖RNAなどをコードしている遺伝子を含めて、その最も広範な文脈において解釈すべきであり、この遺伝子には、ゲノムの遺伝子、ならびに存在するなら遺伝子のコード領域(すなわちエキソン)に対応するmRNAまたはcDNA、転写されるが翻訳されない配列、およびプロモーターと、転写ターミネーター/ポリアデニル化配列とを含めた調節領域が含まれる。また、「遺伝子」という用語は、機能的生成物のすべて、または一部をコードしている合成分子または融合分子を説明するのにも使用される。好ましい遺伝子は、標準の組換え技術による、自然に存在するSBEIIa、SBEIIb、またはデンプン生合成遺伝子に由来する。通常、遺伝子に、突然変異誘発を受けさせて、単数または複数のヌクレオチドの置換、欠失および/または付加を生じさせてよい。このような遺伝子のヌクレオチド挿入による派生物には、5'末端融合体および3'末端融合体、ならびに単数または複数のヌクレオチドの配列内挿入体が含まれる。ヌクレオチド配列の挿入による変異体は、1種または複数のヌクレオチドがヌクレオチド配列中の所定の部位に導入されているものであるが、得られた生成物の適当なスクリーニングによりランダム挿入も可能である。欠失による変異体は、1種または複数のヌクレオチドのその配列からの除去によって特徴付けられる。置換によるヌクレオチド変異体は、配列中の少なくとも1種のヌクレオチドが除去され、その場所に別のヌクレオチドが挿入されているものである。このような置換は、置換が、コドンによって定義されるアミノ酸を変化させないものである「サイレント」でもよい。あるいは、置換基は、1種のアミノ酸が類似する代理の別のアミノ酸に改変されるように設計される。典型的な保存的置換は、下記に従って作製されたものである。
【0079】
アミノ酸置換に適した残基
元の残基 典型的な置換
Ala Ser
Arg Lys
Asn Gln、His
Asp Glu
Cys Ser
Gln Asn
Glu Asp
Gly Ala
His Asn、Gln
Ile Leu、Val
Leu Ile、Val
Lys Arg、Gln、Glu
Met Leu、Ile
Phe Met、Leu、Tyr
Ser Thr
Thr Ser
Trp Tyr
Tyr Trp、Phe
Val Ile、Leu
【0080】
当分野の技術者であれば、細胞内での遺伝子またはそれに相補的な配列の発現には、前記遺伝子がプロモーター配列と作動可能に連結して位置する必要があることに気付くであろう。本目的のためのプロモーター、特に胚乳特異的プロモーターの選択は、必要な発現レベル、および/または発現が起こるべき組織、器官もしくは細胞によって変わり得る。
【0081】
プロモーター配列の調節制御下にある核酸分子の場所は、発現がプロモーター配列によって制御されるように前記分子を配置することを意味する。プロモーターは通常、必ずしもそうとは限らないが、それが調節する核酸分子の上流、すなわち5'末端に位置している。さらに、プロモーターを含む調節エレメントは通常、この遺伝子の転写開始部位から2kb以内に位置している。異種プロモーター/構造遺伝子の組合せの構築では、そのプロモーターと、プロモーターがその自然な状況で制御する遺伝子(すなわち、そのプロモーターが由来する遺伝子)との距離とほぼ同じである、遺伝子転写開始部位からの距離にプロモーターを配置することが通常好ましい。当技術分野で知られているように、プロモーター機能を喪失することなく、この距離においてある変異を受け入れることができる。同様に、その制御下に置くべき異種遺伝子に対する調節配列エレメントの好ましい配置は、その自然な状況でのエレメント(すなわち、そのプロモーターが由来する遺伝子)の配置によって定義される。さらに、当技術分野で知られているように、この距離においてある変異が起こり得る。
【0082】
本発明の遺伝子構築体での使用に適したプロモーターの例には、ウイルス、酵母、カビ、細菌、昆虫、鳥類、哺乳動物、および植物の遺伝子に由来するプロモーター、好ましくは植物細胞で機能することができるもの、より好ましくは米胚乳中で発現することができるものが含まれる。このプロモーターは、発現が起こる組織に関して、発現を構成的に、または示差的に調節してもよい。あるいは、発現は、発現が起こる発達段階に関して、または生理学的ストレスまたは温度など外部刺激に応じて示差的であってもよい。
【0083】
SBEIIaまたは他のデンプン生合成遺伝子の活性を低下させる方法は、導入遺伝子をイネ植物の再生可能な細胞中に導入するステップと、形質転換した細胞からトランスジェニックイネ植物を再生させるステップとを含むものでよい。アミロペクチンの合成に関与する枝作り酵素には、SBEI、SBEIIa、およびSBEIIbが含まれ、この1種(複数)の導入遺伝子は、こうした遺伝子を複数不活性化してもよい。さらに、SBEIIbおよび/またはSBEIの不活性化は、導入遺伝子(例えば、2重鎖RNA、アンチセンス、またはリボザイムRNAをコードするもの、以下参照)がSBEIIbまたはSBEI遺伝子の発現を直接標的とするという点で、直接的なものでもよく、この不活性化は、SBEIIbまたはSBEIの発現の低下を間接的にもたらすものでもよい。例えば、導入遺伝子RNAは、配列の同一性または塩基対形成という点で、SBEIIa遺伝子/RNAのみを標的としてもよいが、また胚乳中のタンパク質の安定性または分布を改変することによりSBEIIbまたはSBEIの活性の低下をもたらしてもよい。本発明のさらなる形態は、SBEIIaおよびSBEIIbの活性の低下と、SSI、SSII、SSIII、およびイソアミラーゼやプルラナーゼなどの枝切り酵素が含まれ得る他の1種または複数のアミロペクチン合成酵素の改変との組合せにある。こうした酵素のいずれかの、またはすべての発現を導入遺伝子の導入によって改変してもよい。特定の一実施形態では、イネ植物においてADP−グルコースピロホスホリラーゼ(ADGP)を過剰発現させており、これは収量および成長を増強させることが示されている(Smidanskyら、2003)。
【0084】
いくつかのDNA配列がイネにおけるアミロペクチン合成遺伝子で知られており、その任意の配列を、イネにおいて遺伝子を不活性化する導入遺伝子を設計するための基礎とすることができる。こうした配列には、イネcDNA SBEIIa(GenBankアクセッション番号E14723、特願平10−004970)、SBEIIb(D16201、Mizunoら、1993)、およびSBEI(D11082、Mizunoら、1992;D10752、NakamuraおよびYamanouchi、1992)が含まれる。イネのSBEI遺伝子は、Rahmanら、(1997)およびRahmanら、(1999)、またはアクセッション番号D10838、Kawasakiら、(1993)に記載されている。さらなる遺伝子配列を、以下のウェブサイト、http://www.ncbi.nlm.nih.gov/、http://www.tigr.org、およびhttp://www.gramene.org/about/index.htmlから得てもよい。
【0085】
コムギ、オオムギ、トウモロコシ、または他の近縁種由来のSBEIIa、SBEIIb、または他のアミロペクチン合成遺伝子の相同体を使用して、イネにおける遺伝子発現レベルを改変することもできる。このような遺伝子またはその断片は、PCR増幅または標識プローブに対するハイブリダイゼーションを含めた当技術分野で周知の方法によって得ることができる。導入遺伝子構築体を調製するのに使用する、相同体の1種(複数)の領域は、対応するイネ遺伝子と、その適切な領域において少なくとも85%の同一性、好ましくは少なくとも90%、さらにより好ましくは95〜100%の同一性を有するべきである。導入遺伝子はまた、米胚乳中で発現されるアミロペクチン合成遺伝子を特異的に標的とし、この植物の他の場所においてアミロペクチン合成に対して影響がないか、または最小限であることが好ましい。これを、導入遺伝子内の胚乳特異的プロモーターなど適当な調節配列を使用して行ってもよい。
【0086】
本明細書では、「ストリンジェントなハイブリダイゼーション条件」は、プローブと標的配列との間に少なくとも90%、好ましくは少なくとも95%の同一性がある場合にハイブリダイゼーションが通常生じることを意味する。ストリンジェントなハイブリダイゼーション条件の例は、50%ホルムアミド、5×SSC(1×SSC=150mM NaCl、15mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハルト溶液、10%硫酸デキストラン、およびサケ精子DNAなどを変性させ断片化した20μg/mlの担体DNAを含む溶液中で一晩インキュベートした後、ハイブリダイゼーション支持体を0.1×SSC中でおよそ65℃で洗浄することである。他のハイブリダイゼーション条件および洗浄条件は、周知であり、Sambrookら、Molecular Cloning:A Laboratory Manual,Second Edition,Cold Spring Harbor,NY(1989)、特に11章で例示されている。
【0087】
アンチセンス
植物において遺伝子活性を改変する、特に特異的に低下させるための既知の遺伝子工学または遺伝子導入手法は、当技術分野で周知である。遺伝的変異をイネ植物に導入するこうした方法には、標的遺伝子のRNAに相補的であり、そのRNAにハイブリッド形成することができる適当なアンチセンス分子の発現が含まれる。アンチセンス分子は、標的遺伝子のmRNAの翻訳、プロセッシングまたは安定性を妨げ、それによりmRNAの発現を不活性化すると考えられている。アンチセンス配列を考案する方法は当技術分野で周知であり、これらの例は、参照により本明細書に組み込む、米国特許第5190131号、欧州特許出願公開第0467349号明細書、欧州特許出願公開第0223399号明細書、および欧州特許出願公開第0240208号明細書で見られる。植物におけるアンチセンス技術の使用は、Bourque(1995)およびSenior(1998)にその総説がある。Bourqueは、遺伝子不活性化の方法としてのアンチセンス配列の植物系における使用例を数多く挙げている。彼女は、部分的阻害がおそらくその系の測定可能な変化をもたらすので、どんな酵素活性も必ずしも100%の阻害に達する必要はないと述べている。Senior(1998)は、アンチセンス法が現在、遺伝子発現を操作するための非常によく確立された技法であると述べている。
【0088】
イネのSBEIIa、SBEIIb、SBEI、または他のアミロペクチン生合成遺伝子に対するアンチセンス分子は、イネmRNA配列に基づくものとし、あるいは他の種、例えばオオムギに由来するDNAまたはmRNA配列との相同性に基づくものとすることができる。こうしたアンチセンス配列は、構造遺伝子、または遺伝子発現またはスプライシング事象の制御に影響を与える配列に対応するものでよい。例えば、このアンチセンス配列は、イネのSBEIIaまたは他の遺伝子の、標的とするコード領域、5'非翻訳領域(UTR)もしくは3'UTR、またはそれらの組合せに対応するものでよい。このアンチセンス配列は、転写中、または転写後にスプライシングによって切り取られるイントロン配列に一部が相補的であってもよく、好ましくは標的遺伝子のエキソン配列のみに相補的であってもよい。UTRは通常より大きな多様性があるという点から、こうした領域を標的とすることは、遺伝子阻害のより大きな特異性をもたらす。アンチセンス配列の長さは、少なくとも19の連続するヌクレオチド、好ましくは少なくとも50ヌクレオチド、およびより好ましくは少なくとも100、200、500、または1000ヌクレオチドであるべきである。遺伝子転写物全体に相補的な完全長配列を使用してよい。この長さは100〜2000ヌクレオチドであることが最も好ましい。標的転写物に対するアンチセンス配列の相同性の程度は、少なくとも85%、好ましくは少なくとも90%、より好ましくは95〜100%であるべきである。アンチセンスRNA分子はもちろん、分子を安定化させるために機能する可能性のある無関係の配列を含む。
【0089】
コサプレッション
使用してよい別の分子生物学的手法は、コサプレッションである。コサプレッションの機序は、十分に理解されていないが、それに転写後遺伝子発現抑制(PTGS)が関与していると考えられており、その点でアンチセンス抑制の多くの例に非常に類似している可能性がある。コサプレッションは、遺伝子の余分なコピーまたはその断片をある植物に、その発現用プロモーターに対してセンス方向に導入することを伴う。センス断片のサイズ、標的遺伝子領域に対するその対応性、および標的遺伝子との相同性の程度は、上記のアンチセンス配列の場合と同様である。場合によっては、遺伝子配列の追加のコピーが植物の標的遺伝子の発現を妨害する。コサプレッション手法を実施する方法については、国際公開第97/20936号、および欧州特許出願公開第0465572号明細書を参照されたい。
【0090】
2本鎖RNAによって媒介される遺伝子発現抑制
遺伝的変異をイネ植物に導入するのに使用してよいさらなる方法は、2重鎖もしくは2本鎖のRNAによって媒介される遺伝子発現抑制である。この方法にもPTGSが関与する。この方法では、不活性化すべき標的遺伝子との相同性を有する少なくとも部分的に2本鎖の1種(複数)のRNA産物を合成させるDNAが導入される。したがって、このDNAは、センス配列およびアンチセンス配列をどちらも含んでおり、このDNAがRNAに転写されると、ハイブリッド形成して2本鎖RNA領域を形成することができる。好ましい一実施形態では、このセンス配列およびアンチセンス配列は、RNAに転写される際にスプライシングによって切り取られるイントロンを含むスペーサー領域によって分離されている。この配置が、より効率の高い遺伝子発現抑制をもたらすことが示されている(Smithら、2000)。この2本鎖領域は、1種または2種のDNA領域のいずれかから転写された1種または2種のRNA分子を含むものでよい。2本鎖分子の存在は、標的遺伝子の活性を効率的に低下または消去する、2本鎖RNA、および植物の標的遺伝子からの相同なRNA転写物をも破壊する、内生植物系からの応答を誘発する。この技法を実施するための方法については、豪州特許出願公開第99/292514−A号明細書、および国際公開第99/53050号明細書を参照されたい。ハイブリッド形成するこのセンス配列およびアンチセンス配列の長さは、それぞれ少なくとも19の連続するヌクレオチド、好ましくは少なくとも30もしくは50ヌクレオチド、より好ましくは少なくとも100、200、500、または1000ヌクレオチドであるべきである。遺伝子転写物全体に対応する完全長配列を使用してよい。この長さは100〜2000ヌクレオチドであることが最も好ましい。標的転写物に対するセンス配列およびアンチセンス配列の相同性の程度は、少なくとも85%、好ましくは少なくとも90%、より好ましくは95〜100%であるべきである。このRNA分子はもちろん、この分子を安定化させるために機能する可能性がある無関係の配列を含む。このRNA分子を、RNAポリメラーゼIIまたはRNAポリメラーゼIIIプロモーターの制御下で発現させてよい。後者の例には、tRNAまたはsnRNAプロモーターが含まれる。
【0091】
このアンチセンス、コサプレッション、または2本鎖RNA分子は、国際公開第03/00292号に記載の通り、好ましくは核局在化シグナルを含む、大部分が2本鎖のRNA領域を含むものでもよい。好ましい一実施形態では、この大部分が2本鎖の領域は、PSTVd型ウイロイドに由来するか、または少なくとも35のCUGトリヌクレオチドの反復を含む。
【0092】
リボザイム
リボザイムを使用して、イネにおける所望の遺伝子発現の不活性化に関与する遺伝的変異を導入してよい。リボザイムは、酵素機能または触媒機能を有するRNA分子であり、この分子は、他のRNA分子を、1種または多くの場合2種のハイブリッド形成する配列によって定義される特異的部位で切断することができる。このRNAの切断により、標的遺伝子の発現が不活性化される。このリボザイムは、遺伝子不活性化に寄与することができるアンチセンス分子として作用することもできる。このリボザイムは、ハイブリッド形成する配列の間で、好ましくはハンマーヘッド型またはヘアピン型の1種または複数の触媒ドメインを含む。RNAseP、グループIもしくはIIイントロン、および肝炎デルタウイルス型を含めて、他のリボザイムのモチーフを使用してもよい。欧州特許出願公開第0321201号明細書および米国特許第6221661号を参照されたい。トランスジェニック植物の遺伝子を不活性化するためのリボザイムの使用は、例えばWegenerら(1994)によって実証されている。
【0093】
遺伝子構築体/ベクター
本発明はまた、遺伝子を阻害する分子をコードしているRNAおよび好ましくはDNAを含む単離された核酸分子を提供する。この核酸分子は、イネSBEIIaおよび/またはSBEIIb遺伝子の配列を標的とし、米穀粒の胚乳中でのそれらの発現を不活性化するのに効果的である、アンチセンス、センス(コサプレッション)、2本鎖RNA、またはリボザイム分子をコードしていることが好ましい。本発明はまた、単離された核酸分子を含む遺伝子構築体を提供するものであり、この構築体は、プロモーター、エンハンサー、および転写終結配列もしくはポリアデニル化配列などの1種または複数の調節エレメントを含む。このようなエレメントは当技術分野で周知である。この遺伝子構築体は、植物、特にイネなどの単子葉植物での導入遺伝子の発現を助けるイントロン配列を含むものでもよい。「イントロン」という用語は、その通常の意味で使用され、転写されるがタンパク質をコードしておらず、翻訳前にRNAからスプライシングによって切り取られる遺伝子断片を意味する。イントロンを、導入遺伝子が翻訳産物をコードしている場合は5'UTRまたはコード領域中に、あるいはそうでない場合は転写領域のどこにでも取り込んでよい。特定の一実施形態では、オオムギSBEII遺伝子のイントロン(Ahlandsbergら、2002)など胚乳特異的に発現させるイントロンを使用する。
【0094】
本発明はさらに、遺伝子構築体を含むベクター、例えばプラスミドベクターを提供する。「ベクター」という用語には、in vitroまたはin vivoで発現することができる発現ベクター、および1種の細胞または生物から別のものに移動することができる形質転換ベクターが含まれる。このベクターは、細胞、例えば大腸菌(E.coli)またはAgrobacteriumなどの原核細胞内での複製をもたらす配列を含む。好ましくは、このベクターは、イネ細胞中に導入することができる、少なくとも1種のT−DNA境界配列によって定義されるT−DNA配列を含むバイナリーベクターである。本発明はさらに、このベクターを含む細胞、例えばAgrobacteriumまたはイネ細胞を提供するものであり、この細胞は、未成熟胚の胚盤または胚形成カルスの細胞など再生可能な細胞であってもよい。あるいは、この細胞は、導入遺伝子を含む形質転換されたイネ細胞であってもよい。
【0095】
プロモーター/ターミネーター
本発明の導入遺伝子または他の遺伝子構築体は、米胚乳中で調節性の、または構成的な発現をもたらすことができる転写開始領域(プロモーター)を含むものでよい。このプロモーターは、組織特異的であってよく、したがって胚乳中で選択的または排他的発現を与える。このプロモーターを、胚乳特異的プロモーター(高分子量グルテニンプロモーター、イネSSIプロモーター、イネSBEIIプロモーター、イネGBSSプロモーターなど)、または胚乳特異的でないプロモーター(ユビキチンプロモーター、CaMV35Sまたは増強された35Sプロモーターなど)のどちらかから選択してよい。このプロモーターは、温度、光、ストレスなどの要因によって調節され得る。通常、プロモーターは、発現されるべき遺伝子配列の5'側に備えられる。この構築体は、nos3'もしくはocs3'ポリアデニル化領域、転写ターミネーターなど、転写を増強する他のエレメントを含むものでもよい。例示するDNAのこの領域を、適当な選択マーカー遺伝子配列および他のエレメントを含むベクター中か、あるいはこうした配列を含むベクターと同時形質転換されるベクター中に取り込む。
【0096】
イネの形質転換法
イネの形質転換の方法は、外因性の核酸の導入によりこの植物に遺伝的変異を導入するためのものであり、当技術分野で周知である。例えば、Chanら、1993;Hieiら、1994;Zhangら、1997;Buchholzら、1998を参照されたい。形質転換を、当技術分野で知られている、適当なAgrobacteriu菌株、微粒子銃による方法、ポリエチレングリコールの媒介によるイネプロトプラスト中への取り込みなどによって媒介してよい。所望のヌクレオチド配列または遺伝子構築体、および選択マーカーを載せたベクターを、組織培養された植物もしくは外植片、例えば、プロトプラスト、未成熟胚、またはカルスのような再生可能なイネ細胞中に導入してよい。この選択マーカー遺伝子は、イネ細胞に抗生物質耐性または除草剤耐性をもたらすことができ、あるいは基質、例えばマンノースの利用を成長のために可能にする。この選択マーカーにより、イネ細胞にジェネチシン耐性、ハイグロマイシン耐性またはフォスフィノスリシン耐性を与えることが好ましい。この再生可能なイネ細胞は、未成熟胚、成熟胚の胚盤、これらに由来するカルス、または分裂組織に由来するものであることが好ましい。形質転換する細胞を選択し、次いで実施例2に記載するものなど、当技術分野で周知の方法によって再生して、形質転換イネを作出する。
【0097】
形質転換植物は、選択マーカー遺伝子を含むものでもよく、またこのような遺伝子を、再生中、または再生後に、例えば選択マーカー遺伝子をゲノムの外に切り出すか、あるいは選択マーカー遺伝子を、SBEIIaおよび/またはSBEIIbの阻害をもたらす導入遺伝子から分離することによって除去してもよい。
【0098】
例えば、導入遺伝子に特異的な適当な核酸プローブ、または表現型観察を使用することにより、導入遺伝子または突然変異が染色体中に組み込まれた植物をスクリーニングすることができる。いくつかの任意の方法を使用して、形質転換植物であるかどうか判定することができる。例えば、ポリメラーゼ連鎖反応(PCR)を使用して形質転換植物に特有の配列を増幅し、ゲル電気泳動または他の方法により、この増幅した産物を検出することができる。この植物からDNAを、通常の方法を使用して抽出し、またプライマーを使用してPCRを実施してもよく、これにより形質転換植物と非形質転換植物とが区別される。例えば、この構築体を読み取って、形質転換用ベクター由来のDNAの領域を増幅するプライマーを設計してもよく、また対象の遺伝子からリバースプライマーを設計してもよい。この植物の形質転換が成功していたとしても、これらのプライマーはある断片を増幅するだけである。陽性の形質転換体を確認する代替方法は、当技術分野で周知のサザンブロットハイブリダイゼーションによるものである。また、形質転換植物または突然変異体植物を同定、すなわちその表現型により非形質転換植物または野生型植物と区別することができる。例えば、この表現型は、選択マーカー遺伝子の存在、または免疫学的方法による特定のタンパク質の存在によって、あるいはあるタンパク質の非存在、例えば、ELISAまたはウエスタンブロット分析によって検出される、胚乳中のSBEIIaタンパク質の非存在によって与えられる。また、このような植物をスクリーニングするのに使用する指標は、穀粒の表現形質の観察、例えばshrunken穀粒の目視検査または目測によるもの、アミロース含量が上昇したかどうか試験すること、あるいはデンプン顆粒の複屈折の存在を顕微鏡によって検査することによるものでよい。
【0099】
突然変異
米胚乳中のSBEIIaおよびSBEIIb酵素または他のデンプン生合成酵素の活性の低下をもたらす遺伝的変異の導入を、それぞれの遺伝子内または遺伝子の調節配列内の適切な突然変異によって行ってもよい。遺伝子が阻害される程度により、生成されるデンプンの特性がある程度まで決まる。この突然変異は切断(truncation)でも、無発現変異体でもよく、これらは、デンプンの性質に大きな影響を及ぼすことが知られているが、また、改変したデンプン構造は、アミロペクチン合成酵素活性を十分低下させてデンプンまたは米穀粒において対象の特性をもたらす漏出変異体に起因する。他に染色体再配列も効果的であり、これらには、欠失、逆位、重複、または点突然変異が含まれ得る。
【0100】
突然変異誘発は、化学的手段または放射線による手段、例えばEMSもしくはアジ化ナトリウム(ZwarおよびChandler、1995)による種子の処理、またはγ線照射によって行うことができる。γ線誘発変異については、種子を、60Co供給源からの20〜50kRの放射線量で照射してよい(ZikiryaevaおよびKasimov、1972)。EMSによる突然変異誘発を、Mullinsら、(1999)の通り、種子をEMS(0.03%、v/v)で処理することによって実施してよい。突然変異体の単離は、変異誘発された植物または種子をスクリーニングすることによって行うことができる。例えば、変異誘発したイネ集団を、穀粒中の高アミロース含量、かつ/または通常より長いアミロペクチン鎖長の分布、またはELISAによるSBEIIaおよび/またはSBEIIbタンパク質の喪失について、あるいは改変した穀粒形態についてスクリーニングしてよい(Greenら、1997)。スクリーニングは、SBE活性の1種をすでに欠いているイネの遺伝子型において、例えばSBEIIaまたはSBEIIb陰性の背景で行うことが好ましい。次いで、この突然変異体と所望の遺伝的背景をもつ植物とを交配し、戻し交配を適当な回数実施して本来所望でない親の背景を除去することにより、こうした変異を望ましい遺伝的背景に導入することができる。好ましい突然変異体は、イネにおいてSBEIIaおよびSBEIIbの発現または活性に影響を及ぼすものである。
【0101】
それにより、本発明は、高アミロースで、遺伝子導入によらない米粒およびそれらから得られる生成物を提供する。
【0102】
例えばGBSSのレベルを増大させるなど、アミロペクチンの合成に関与するSBEIIa、SBEIIb、または他の酵素をコードしている遺伝子における突然変異は、米胚乳のデンプン中のアミロースの割合の増大をもたらす。個々の穀粒あたりのアミロースの量は、アミロペクチンからアミロースへ炭素の流れが転換した結果として増大することもあり、あるいは、穀粒あたりのデンプン産生が著しく減少した場合、このアミロースの量は減少することもある。いずれの場合も、デンプンに対する百分率で示すアミロースの相対的レベルが増大する。
【0103】
デンプン顆粒を有する、形がゆがんだ種子が、高アミロースオオムギ(Morellら、2003)、およびデンプン中に約90%アミロースを有する低アミロペクチン(LAPS)トウモロコシ(Sidebottomら、1998)で報告されている。この表現型を、変異誘発したイネ集団をスクリーニングする際に使用することができる。複屈折もこれに使用することができる。複屈折は、2方向に光を屈折させる物質の能力であり、これは、偏光顕微鏡で見ると、各デンプン顆粒に「マルタクロス」と呼ばれる十字の影を形成する。複屈折は、顆粒内のポリマーの並んだ構造機構の程度の指標である(ThomasおよびAtwell、1999)。デンプン顆粒の複屈折の喪失は通常、アミロース含量の増大と関連している。
【0104】
食糧生産に適したもの
別の態様では、本発明は、食糧生産に有用なイネを提供するものであり、この穀粒は、高い相対アミロース含量、および低下したアミロペクチン含量を含むデンプンを有する。この穀粒が得らえるイネ植物は、発達中に胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたものであることが好ましい。本発明のイネ植物は、食糧生産、特に商業上の食糧生産に有用である。
【0105】
イネの所望の遺伝的背景には、農業収量および他の特性への考慮が含まれる。このような特性には、農業生産力、耐病性、および非生物ストレス耐性が含まれ得る。オーストラリアでは、改変したデンプン形質をAmaroo、Ali Combo、Basmati、Bogan、Bombia、Doongara、Goolarah、Illabong、Jarrah、Koshihikari、Kyeema、Langi、Millin、Namage、Opus、Pelde、他の一般に栽培されている品種などのイネ栽培品種中に入れることが望まれている。提供した例は、オーストラリアの生産地域に適したものであるが、その他の栽培地域に適した品種も他にある。本発明のイネ品種は、少なくとも一部の栽培地域で、対応する野生型品種の少なくとも80%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも95%の収量をもたらすことが好ましい。この収量は、管理された圃場試験において容易に測定することができる。
【0106】
穀粒のデンプン含量は、少なくとも約25%、好ましくは少なくとも35%もしくは45%、より好ましくは野生型のレベルに近い55〜65%(w/w)であるべきである。最も好ましくは、この穀粒は、デンプン含量が、同等であるが未改変のイネから得られる穀粒の少なくとも90%である。野生型よりもデンプン含量が低いのは、アミロペクチンレベルが低下した結果である可能性が高い。デンプン含量が低くなっても、高アミロース製品が比較的高価であるため、この穀粒は、商業上の食糧生産にいまだ有用である。他の望ましい特性には、穀粒を製粉する能力、特に穀粒硬度が含まれる。イネ植物の価値を高める可能性がある別の態様は、穀粒からのデンプン抽出の程度であり、より高い抽出率がより有用である。また、穀粒の形は、植物の商業上の有用性に影響を与える恐れがある別の特徴であり、したがって穀粒の形は、穀粒を製粉する際の容易さまたは別の点に影響を与える恐れがある。例えば、穀粒の細長い形態は、製粉および加工を困難にする可能性がある。
【0107】
デンプンは、標準の方法を使用して、例えばアルカリ溶液(水酸化ナトリウム)を用いた、brokersの湿式製粉を行ってタンパク質を除去することにより、米穀粒から容易に単離することができる。砕け米をアルカリ溶液中で24時間浸し、次いでアルカリ溶液を用いた、ピンミル、ハンマーミル、またはストーンミル粉砕機で湿式製粉を行う。このバッターを10〜24時間保存した後、それをスクリーンに通すことにより繊維を除去し、このデンプンを遠心分離により回収し、水で徹底的に洗浄し、乾燥させた。
【0108】
改変したデンプンの物理的特徴
本発明の別の態様では、米デンプンは、糊化温度が改変されたものでよく、これは、示差走査熱量測定(DSC)によって容易に測定することができる。糊化は、過剰水中でのデンプン顆粒内の分子整列の、熱駆動による崩壊(破壊)であり、顆粒の膨潤、結晶融解、複屈折の喪失、粘度の増大、およびデンプンの可溶化などの特性の付随する不可逆的変化を伴う。この糊化温度は、残りのアミロペクチンの鎖長によって、野生型植物から得られるデンプンに比べて、上昇させても、低下させてもよい。トウモロコシのae(amylose extender)変異体から得られる高アミロースデンプンは、正常なトウモロコシよりも高い糊化温度を示した(Fuwaら、1999;Kruegerら、1987)。一方、デンプン合成酵素IIaの活性を欠いているオオムギsex6変異体から得られるデンプンは、対照植物から得られるデンプンに比べた場合、糊化温度が低くなり、その糊化のピークのエンタルピーが低下した(Morellら、2003)。
【0109】
この改変した糊化温度は、比較的高いアミロース含量に伴うものである可能性がある。野生型米デンプンの糊化温度は通常、示差走査熱量測定で測定して、開始温度と定義される第1ピークの温度で約61〜67℃である(Rahmanら、2000)。
【0110】
このデンプンはまた、野生型デンプンに比べて、加熱した過剰水中でのその膨潤率によって特徴付けられる。通常、デンプンまたは穀粉のどちらかを過剰水と混合し、加熱して通常90℃以上の温度にまで上昇させることにより、膨潤容積を測定する。次いで、この試料を遠心分離によって回収し、沈降材料の質量をこの試料の乾燥重量で割ったものにより膨潤容積を表す。低い膨潤特性は、食品調製物、特に含水食品調製物のデンプン含量を増大させたい場合に有用である。
【0111】
本発明の選択される形態の米デンプン構造は、イネから単離された正常デンプンに比べて結晶化度が低下するという点から異なり得る。デンプンの結晶化度の低下は、感覚特性の向上に関係し、より滑らかな食感に寄与すると考えられる。したがって、このデンプンは、1種または複数のアミロペクチン合成酵素の活性レベルの低下に起因する結晶化度の低下をさらに示す可能性がある。結晶化度は通常、X線結晶解析によって調査する。
【0112】
改変したアミロペクチン構造の1つの尺度は、デンプンの鎖長の分布、または重合度である。この鎖長分布は、イソアミラーゼによる枝切りの後、FACE(fluorophore−assisted carbohydrate electrophoresis蛍光ラベル糖鎖電気泳動法)を使用して測定することができる。本発明のデンプンのアミロペクチンは、鎖長の分布が、枝切り後の野生型植物から得られるデンプンのその分布よりも大きい5〜60の範囲にあってよい。また、鎖長が長いデンプンは、分岐の頻度がそれに比例して低下する。したがって、このデンプンも、アミロペクチン中により長いアミロペクチン鎖長の分布を尚ももち得る。
【0113】
食品特性
米デンプンは、特にアジアでヒトの食物中の炭水化物の主要な供給源であり、本発明の穀粒およびそれから得られる生成物は、食品を調製するのに使用することができる。この食品は、ヒトまたは動物によって、例えば畜産またはペットフードにおいて消費され得る。改変したイネ植物から得られる穀粒は、食品加工方法に容易に使用することができ、したがって本発明には、精米され、挽かれ、粗挽きにされ、挽割りにされ、圧延され、煮沸された、またはパーボイルドの穀粒、あるいは穀粉、砕け米、米ぬか、および米ぬか油を含めた、米の加工された穀粒もしくは全粒から得られる製品が含まれる。この製品は、α米(precooked rice)もしくは迅速調理米(quick−cooking rice)、インスタント米、粉末米(granulated rice)、糊化米(gelatinized rice)、缶詰の米、またはライスプディングであってよい。この穀粒またはデンプンを、めん類、もち、ライスペーパー、または春巻の皮を含めて、加工された米製品の製造に、あるいは発酵めんなどの発酵製品または酒などの飲料に使用してもよい。この穀粒またはそれに由来するデンプンを、米の穀粉と、コムギもしくは他の穀粉、または増粘剤、結合剤などの食品添加物とを混合する場合を含めて、例えば、パン、ケーキ、クラッカー、ビスケットなどに、あるいは、飲料、めん類、パスタ、またはクイックスープを製造するのに使用してもよい。この米製品は、小麦を含まないダイエット食品での使用に適している。この穀粒または本発明の穀粒に由来する製品は、特に、パフライス、ライスフレークなどの朝食用シリアルに、あるいは押出製品として望ましい。本発明の高アミロースデンプンは、製菓業において有用であり、または成形および硬化時間が短縮される高強度ゲルを形成するのに使用することもできる。これらのデンプンは、例えば、十分に揚げたポテトまたは他の食品で油吸着を減少させる被覆剤として使用してもよい。
【0114】
食物繊維
本明細書における食物繊維は、炭水化物、および健康なヒトの小腸で吸収されないが大腸に入る、炭水化物の消化による産物である。この食物繊維には、レジスタントスターチ、ならびに他の可溶性および不溶性の炭水化物ポリマーが含まれる。この食物繊維は、大腸で常在ミクロフローラにより少なくとも一部が発酵性となる炭水化物の部分を含むものとする。
【0115】
本発明のデンプンは、比較的高レベルの食物繊維を含むことが好ましく、アミロースを含むことがより好ましい。本発明の穀粒の食物繊維含量は単に、胚乳の相対アミロース含量が増大したことに起因するものであっても、そうでなくてもよい。
【0116】
本発明の態様はまた、高レベルの食物繊維と組みあわせて、アリューロン層と胚芽の組合せから生じるものでもよい。特に、これは、より高い相対レベルのアリューロン層または胚芽が穀粒中に存在する場合に生じるものでよい。米穀粒が軽度に収縮している(slightly shrunken)場合、この胚乳の量が減少し、このアリューロン層および胚芽の量が比較的増大する。したがって、この米は、増大したレジスタントスターチと併せて、比較的高レベルのある種の有益な要素またはビタミンを有し、このような要素には、二価の陽イオン、生物利用可能なCa++、葉酸などのビタミン、またはトコフェロール、トコトリエノールなどの抗酸化剤が含まれる。製粉生成物の特定の一形態は、このアリューロン層が製粉生成物中に含まれる場合のものであってよい。特殊な製粉加工を行って、製粉生成物中のアリューロン層の量を高めてもよい。したがって、このように、糊粉層および胚芽を含むように製粉されたか、または別の方法で加工された穀粒由来の任意の生成物は、異なる供給源からこうした要素を添加する必要なしに、栄養面で追加の恩恵を受けている。
【0117】
レジスタントスターチ
レジスタントスターチを、デンプン、および健康なヒトの小腸に吸収されないが大腸に入るデンプンの消化産物の総量と定義する。したがって、レジスタントスターチには、小腸で消化吸収された産物が含まれない。レジスタントスターチには、物理的に接近しにくいデンプン(RS1型)、難消化性顆粒(RS2)、老化デンプン(RS3)、および化学修飾デンプン(RS4)が含まれる。本発明の改変したデンプン構造、特に高アミロースレベルのデンプンは、それを食品に費やすと、レジスタントスターチの増加をもたらす。このデンプンは、いくぶん消化されにくいRS1型でもよい。また、V錯体の結晶化度によって測定される、デンプンと脂質の結合は、レジスタントスターチのレベルに寄与している可能性が高い。
【0118】
本発明の1つの利点が、本発明が特に栄養面での特定の恩恵を受けた生成物を提供し、さらに、米穀粒のデンプンまたは他の成分を、収穫後に改変する必要なく役立つことを理解されたい。しかし、穀粒のデンプンまたは他の成分を改変することが望ましく、本発明は、改変されたそのような成分を包含する。改変の方法は周知であり、この方法には、従来の方法によりデンプンまたは他の成分を抽出すること、およびレジスタント型が増加するようにデンプンを改変することが含まれる。このデンプンを、熱および/または水分での処理により、物理的に(例えばボールミル)、酵素的に(例えばα−もしくはβ−アミラーゼ、プルラナーゼなどを使用して)、化学的加水分解(液体試薬もしくは気体試薬を使用した湿式または乾式)、酸化、二官能性試薬(例えばトリメタリン酸ナトリウム、オキシ塩化リン)での架橋、またはカルボキシメチル化によって改変してよい。
【0119】
血糖指数
血糖指数(GI)は、デンプンを含む食品の消化の速度に関するものであり、また血中グルコース濃度の変移に対する、試験食品の影響と白パンまたはグルコースの影響との比較である。この血糖指数は、食事後の血清中グルコース濃度に対する当該食品の影響可能性、および血中グルコースのホメオスタシスに対するインスリンの需要の程度である。本発明の食品によってもたらされるある重要な特性は、低下した血糖指数である。高アミロース米生成物のヒトボランティアによる消化の30分後の血清中グルコースレベルは、低アミロース米に比べて低くなった(Goddardら、1984)。さらに、この食品は、最終消化が低レベルであり、結果的に比較的低カロリーである可能性がある。低いカロリー製品は、製粉された米穀粒から生成される穀粉の含有物に基づく可能性がある。このような食品は、満腹にし、腸の健康を増進し、食事後の血清中グルコースおよび脂質の濃度を低下させ、また低カロリー食品を提供する効果がある可能性がある。
【0120】
非食品への応用
本発明は、アミロースレベルを上昇させ、その特性が様々な工業上の任意の要求を満たすアミロペクチンレベルの低下させる、改変または改良されたデンプンを提供する。デンプンは、フィルム、紙、繊維、波形加工(corrugating)、および例えば、のり剤などの接着剤の業界を含めた非食品業界で広く使用されている(Young、1984)。米デンプンを、グルコースシロップの製造またはエタノールの製造のための基質として使用してよい。改変されていないデンプンの物理的特性により、いくつかの用途でのその有用性が限定され、高価な、または他の欠点がある恐れがある化学的改変を強いられることが多い。本発明は、他の物理的特性と併せて、特にアミロペクチン含量を低下させることにより、収穫後の改変が少なくて済むデンプンを提供する。例えば、デンプンおよび本発明の穀粒から作製される生成物のペースティング温度(pasting temperature)、剪断応力に対する耐性、皮膜強度、および/または耐水性を改変してよい。このデンプンを、ポリスチレンまたは他の梱包材料の代替物として使用することができる、緩い充填の生分解性の梱包材料を調製するのに使用してもよい。
【0121】
本発明の態様に関して様々な示唆が与えられているが、本発明は、本発明の2種以上の態様の組合せにあってもよいことを理解されたい。
【0122】
<実施例>
(実施例1.材料および方法)
材料および培地
N6マクロ−エレメント(20×stock solution)
g/l
(NH4)2SO4 9.3
KNO3 56.6
KH2PO4 8
MgSO4.7H2O 3.7
CaCl2.2H2O 3.3
MSマクロ−エレメント(20×stock solution)
g/l
NH4NO3 33.0
KNO3 38.0
KH2PO4 3.4
MgSO4.7H2O 7.4
CaCl2.2H2O 8.8
N6マイクロ−エレメント(1000×stock solution)
mg/100ml
MnSO4.4H2O 440
ZnSO4.7H2O 150
H3BO3 160
KI 80
MSマイクロ−エレメント(1000×stock solution)
mg/l
MnSO4.4H2O 22300
Na2MoO4.2H2O 250
H3BO3 6220
ZnSO4.7H2O 8600
CuSO4.5H2O 25
CoCl2.6H2O 25
KI 830
B5マイクロ−エレメント(100×stock solution)
mg/l
MnSO4.4H2O 1000
Na2MoO4.2H2O 25
H3BO3 300
ZnSO4.7H2O 200
CuSO4.5H2O 3.87
CoCl2.6H2O 2.5
KI 75
N6ビタミン(100×stock solution)
mg/100ml
グリシン 20
チアミンHCl 10
ピリドキシンHCl 5
ニコチン酸 5
MSビタミン(100×stock solution)
mg/100ml
ミオイノシトール 1000
チアミンHCl 1
ピリドキシンHCl 5
ニコチン酸 5
B5ビタミン(100×stock solution)
mg/100ml
グリシン 1000
チアミンHCl 100
ピリドキシンHCl 10
ニコチン酸 10
MS鉄(200×stock solution)
mg/500ml
FeCl3(60%w/v) 2.7
MS Na2.EDTA(200×stock solution)
g/500ml
Na2.EDTA 3.7
【0123】
2,4−ジクロロ−フェノキシ酢酸(2,4−D)(1mg/ml、Sigma社製、番号D−6679)stock solutionを、100mgの2,4−Dを1ml無水エタノール中で溶解し、3mlの1N KOHを添加し、1N HClでpHを6に調整することによって調製した。
【0124】
6−ベンジルアミノプリン(1mg/ml BAP、Sigma社製、番号B−3408)およびナフタレン酢酸(1mg/ml NAA、Sigma社製、番号N−0640)の溶液を調製した。
【0125】
アブシジン酸(ABA、2.5mg/ml、Sigma社製、番号A−1049)を、250mgのABAを2mlの1M NaOH中で溶解し、滅菌水で100mlにすることによって調製した。
【0126】
チメンチン(timentin)(150mg/ml、Smith−Kline Beecham社製、6571−30)を、3.1gを20.66mlの滅菌水中で溶解することによって調製した。
【0127】
ハイグロマイシン(50mg/ml)をRoche社(番号843555)から、他の試薬をSigma社から入手した。
【0128】
カルス誘導用N6D培地
量/リットル
N6マクロ(20×) 50ml
N6マイクロ(1000×) 1ml
N6ビタミン(100×) 10ml
MS鉄(200×) 5ml
MS Na2 EDTA(200×) 5ml
ミオイノシトール 100mg
カサミノ酸 300mg
プロリン 2.9g
2,4−D(1mg/ml) 2ml
スクロース 30g
【0129】
1M KOHでpHを5.8に調整し、1リットルあたり3gのフィトゲル(phytogel)を添加し、この混合物をオートクレーブした。
【0130】
継代培養用NB培地
量/リットル
N6マクロ−エレメント(20×) 50ml
B5マイクロ−エレメント(100×)10ml
B5ビタミン(100×) 10ml
MS鉄(200×) 5ml
MS Na2 EDTA(200×) 5ml
2,4−D(1mg/ml) 2ml
スクロース 30g
プロリン 500mg
グルタミン 500mg
カゼイン酵素加水分解物(CEH) 300mg
【0131】
1M KOHでpHを5.8〜5.85に調整し、1リットルあたり3gのフィトゲルを添加し、この混合物をオートクレーブした。
【0132】
継代培養用MS培地
量/リットル
MSマクロ−エレメント(20×) 25ml
MSマイクロ−エレメント(1000×) 1ml
MSビタミン(100×) 10ml
MS鉄(200×) 5ml
MS Na2EDTA(200×) 5ml
スクロース 10g
【0133】
1M KOHでpHを5.8〜5.85に調整し、1リットルあたり2.5gのフィトゲルを添加し、この混合物をオートクレーブした。
NBO:NB培地に30g/lのマンニトールおよび30g/lのソルビトールを添加した後、pH調整する。
【0134】
NBHT30:添加前にNB培地をオートクレーブし、直後に30mg/lのハイグロマイシンおよび150mg/lのチメンチンを注ぐ。
【0135】
NBHT50:添加前にNB培地をオートクレーブし、直後に50mg/lのハイグロマイシンおよび150mg/lのチメンチンを注ぐ。
【0136】
PRHT50:NB培地(2,4−Dを含まない)をオートクレーブした後、以下のものを添加し、最終濃度をBAP(2mg/l)、NAA(1mg/l)、ABA(5mg/l)、ハイグロマイシン(50mg/l)、およびチメンチン(150mg/l)にする。
【0137】
RHT50:NB培地(2,4−Dを含まない)をオートクレーブした後、以下のものを添加し、最終濃度をBAP(3mg/l)、NAA(0.5mg/l)、ハイグロマイシン(50mg/l)、およびチメンチン(150mg/l)にする。
【0138】
MST培地:MS培地をオートクレーブした後、0.05mg/lのNAAおよび150mg/lのチメンチンを添加する。
【0139】
イネの形質転換
成熟した穀粒を、脱穀し、70%エタノールに1分間浸し、滅菌水で3回洗浄した後、50%漂白剤に30分間浸した。滅菌されたこの穀粒を、無菌条件下で滅菌水で徹底的に洗浄し、次いでN6D培地上に蒔いた。プレートをマイクロポアテープで密封し、カルス形成のため照明下、26〜28℃で6〜8週間インキュベートした。カルスを、穀粒から胚を切り出すことなく、おそらく胚の胚盤から形成した。継代培養を必要とする場合は、カルスをNB培地上に移し、このプレートをパラフィルムで密封した。このプレートを暗下、つまりアルミホイルで覆った箱の中で28℃で放置した。継代培養を4週間ごとに実施した。カルスを最高5回継代培養した後、形質転換に使用した。
【0140】
Agrobacteriumの媒介による形質転換のために、移入すべき遺伝子構築体を含むAgrobacterium菌株を、適切な抗生物質を含むプレート上で28℃で増殖させ、2日間の増殖後、このプレートから細胞を掻き取り、100μMアセトシリンゴンを含有するNB液体培地中で再懸濁させた。健常に見えるカルスを細菌懸濁液に10分間浸し、次いでこのカルスを簡単に排出し、100μMアセトシリンゴンを含有するNBOプレート上で暗下、25℃で2日間放置した。この期間を、遺伝子構築体を含むAgrobacteriumの存在下での「共培養」と呼ぶ。共培養の後、このカルスを、150mg/lのチメンチンを含む滅菌水中で洗浄し、簡単に乾燥し、150mg/lのチメンチンを含むNBHT30(選択薬剤ハイグロマイシンを含む)プレート上に蒔いた。26〜28℃で3〜4週間経過した後、増殖域を示した任意のカルスを、同じ培地上でさらに10〜24日間継代培養した。持続的に増殖させることにより、カルスはハイグロマイシン耐性、すなわち形質転換されたことを表した。こうしたカルスを、チメンチンを含むNBHT50プレート上に移し、暗下、26〜28℃でさらに14〜21日間インキュベートした。健常に見えるカルスをPRHT50プレートに移し、暗下でさらに8〜12日間インキュベートした。最後に、シュートをRHT50培地上で、照明下、28℃で30日間以上再生させた。根を形成したシュートを1/2MST培地に移し、十分大きくなると温室内の土壌に移植した。この方法は、ジャポニカ型およびインディカ型を含めて様々なイネ栽培品種で成功することが判明している。
【0141】
(実施例2.遺伝子のdown regulationのための構築体の調製)
遺伝子発現のdown regulationのための遺伝子構築体を調製するのに使用するために、イネSBEI、SBEIIa、およびSBEIIb遺伝子の断片をPCRによって増幅した。この遺伝子の3'末端に近いエキソン領域同士がより異なっているので、選択されたこの断片はこの遺伝子のこうした領域に由来したものであり、これにより、形質転換されたイネでの構築体によるこの遺伝子の交差発現抑制(cross−silencing)の可能性が低下すると考えられている。増幅された断片は、SBEI:GenBankアクセッション番号D11082のヌクレオチド1982〜2527、SBEIIa:アクセッション番号AB023498のヌクレオチド2458〜2997、およびSBEIIb:アクセッション番号D16201のヌクレオチド2414〜2912である(配列は図1〜3に示す)。次のクローニングの工程での便宜上、制限エンドヌクレアーゼ部位をその末端に含む追加の配列を含めた増幅断片をプラスミドベクターpGEM(登録商標)−T中にクローニングした。また、イネ由来のSBEIイントロン9配列を増幅させることにより、イントロン配列を獲得した。この断片に、ゲノム配列GenBankアクセッション番号D10838のヌクレオチド9112〜9606由来の配列、ならびに両端にあるSpeIおよびEcoRI制限部位を含めて、この断片をpBCSK(Stratagene社製)中に挿入してpRint9_BCを形成した(図4)。次いで、SBEI、SBEIIa、およびSBEIIb遺伝子由来のエキソン断片をpRint9 BC中に、そのSpeI/XbaIおよびXhoI/EcoRI部位を使用してそれぞれアンチセンス方向およびセンス方向にクローニングした。これにより、イントロン配列によって分離しているこれらの配列がそれぞれ逆方向反復を形成するように働いた。得られたプラスミドをpRBEI.IR、pRBEIIa.IR、およびpRBEIIb.IRと呼んだ(図5)。このキメラ断片を、BamHIおよびKpnIで切り取り、pBx17casNOTの同じ部位に挿入した(図6)。これにより、アンチセンス/イントロン/センスのキメラ断片がBx17プロモーター領域およびnos3'終結領域に、発現のための正しい方向に連結させた。次いで、各発現カセットを、HindIIIおよびNotIで消化することによって切り取り、植物細胞中での選択のための、植物で発現可能なハイグロマイシン遺伝子、ならびに細菌中での選択のためのスペクチノマイシン耐性遺伝子を含むバイナリーベクターpWBvec8中に挿入した(Wangら、Acta Hort 461:401−407、1998)。この構築体をdsSBEI、dsSBEIIa、およびdsSBEIIbと呼んだ。次いで、これらの構築体を、電気穿孔法によりAgrobacterium tumefaciens菌株(AGL1)の細胞に移入した(Lazoら(1991))。
【0142】
イネのSBEIIa遺伝子および場合によってはSBEIIb遺伝子の発現を低下させるために、コムギの対応するSBEIIa遺伝子由来の配列を使用して、さらなる2重鎖RNA(dsRNA)構築体を作製した。上記の他の構築体に関しては、SBEIIa遺伝子の部分に対応する所望の核酸配列が、プロモーターに対してセンス方向およびアンチセンス方向の両方で生じ、その結果、発現されたRNAが、塩基対合し、2重鎖もしくは2本鎖のRNAを形成することができる相補領域を含んでいた。センス配列およびアンチセンス配列の間のスペーサー領域は、イントロン配列を含んでおり、この配列は、形質転換された植物におけるRNAの部分として転写されるとスプライシングによって切り取られて、引き締まった「ヘアピン」2重鎖構造を形成するはずである。イントロンを含めると、2重鎖RNA構築体によって与えられる遺伝子発現抑制の効率が上昇することが分かっている(Smithら、2000)。所望の核酸を、コムギ由来の高分子量グルテニン(HMWG)プロモーター配列、およびAgrobacterium由来のノパリン合成遺伝子のターミネーター配列(nos3')に連結させた。これにより、dsRNA配列の胚乳特異的発現がもたらされた。
【0143】
このSBEIIaの2重鎖RNA構築体に、コムギSBEIIa遺伝子(GenBankアクセッション番号AF338431)からPCRによって増幅された1536bpのヌクレオチド配列を含めた。これに、どちらかの側にEcoRIおよびKpnI制限部位を有する、エキソン1および2の全体ならびにエキソン3の一部を含んだ468bpの配列(断片1)と、どちらかの側にKpnIおよびSacI部位を有する、SBEIIaのエキソン3および4の一部ならびにイントロン3の全体からなる512bpの配列(断片2)と、どちらかの側にBamHIおよびSacI部位を有する、SBEIIaの完全なエキソン1、2および3からなる528bp断片(断片3)とを含めた。使用する配列は、より狭い領域にわたるより高い相同性(50ヌクレオチドにわたり87%、27ヌクレオチドにわたり92%)を含めて、イネSBEIIa遺伝子(SBE4)と217ヌクレオチドにわたり80%の同一性を有し、したがって米胚乳におけるこの配列の発現により、イネSBEIIaの発現の相当な低下がもたらされることが期待された。また、コムギの配列は、SBEIIbの同等物であるイネ枝作り酵素3と113ヌクレオチドにわたり76%同一であり、これは、同様にこの転写物のレベルに影響を及ぼすことが予想された。
【0144】
次いで、断片3の配列が、断片1に対してアンチセンス方向に断片2と連結するように、断片1、2および3を連結させた。最初に、2重鎖RNA構築体を、HMWGプロモーター配列およびnos3'ターミネーターを含めたベクターpBx17casNOT(図6)において生成した。このベクターにおける遺伝子構築体をpBx17ds−wSBEIIaと呼び、この2重鎖RNA遺伝子をds−wSBEIIaと呼んだ。ds−wSBEIIa遺伝子を含むこのカセットを、pWBvec8中に挿入し、Agrobacterium菌株AGL1に導入し、これを使用して実施例1に記載の通りイネを形質転換させた。
(実施例3.SBE活性を低下させたイネの作出)
【0145】
AGL1細胞中の構築体dsSBEI、dsSBEIIa、dsSBEIIb、およびds−wSBEIIaを使用して、実施例1に記載の方法に従って、形質転換イネ植物(品種、ニッポンバレ)を作出した。各構築体につき500個のイネカルスを使用し、形質転換したカルスを選抜し、イネ植物を再生させた。この植物を土壌に移植した後、使用するSBEI、SBEIIa、もしくはSBEIIb遺伝子断片に特異的なプライマーまたはプローブを使用した、PCRあるいはサザンブロットハイブリダイゼーション分析により、この植物の形質転換を実証した。ds−wSBEIIaを用いた形質転換からの再生させた24の植物体のうち、21の植物体が、導入されたSBEIIa配列について陽性であることを示した。
【0146】
アクリルアミドゲルでの胚乳タンパク質のゲル電気泳動の後、それぞれのタンパク質に対する特異的抗体を使用したウエスタンブロット分析により、形質転換された植物から得られた穀粒(T1種子)をSBEタンパク質について検査する。SBEの活性は、形質転換された系統の大多数において低下する。穀粒のデンプン中のアミロースの割合を求めた。SBEIIaについて形質転換された系統には、少なくとも40%の相対アミロースレベルを示すものがあり、またそれらの中には50%以上のものもある。SBEIIaとSBEIIbの活性をどちらも低下させた場合、アミロースの割合はさらに増大する。
(実施例4.デンプンおよびタンパク質の分析)
【0147】
炭水化物の測定および分析
Takedaら,(1986)、Lumdubwongら,(2000)、Chiouら(2002)、またはSchulmanら,(1991)の方法を使用して、デンプンを発達中の胚乳から、または成熟した穀粒から単離する。Megazyme社(アイルランド共和国Wicklow州Bray)によって提供される総デンプン量分析キット(total starch analysis kit)を使用して、デンプン含量を求める。次いで、このデンプン含量を対照植物と比較する。総穀粒重量からデンプン重量を引いて、穀粒の総非デンプン含量を求めることにより、総重量の減少がデンプン含量の低下に起因するものであるかどうかが決まる。
【0148】
デンプン試料のアミロース含量を、以下の通り若干変更した、MorrisonおよびLaignelet(1983)の比色(ヨウ素滴定)法によって求める。蓋にゴム座金が取り付けられた2mlねじ込みキャップ式チューブ中に、デンプンをおよそ2mg正確に量り取る(0.1mgまで正確に)。脂質を除去するために、1mlの85%(v/v)メタノールをこのデンプンと混合し、チューブを時々ボルテックスしながら65℃のウォーターバス中で1時間加熱する。13,000gで5分間遠心分離した後、その上清を注意深く除去し、この抽出工程を繰り返す。次いで、このデンプンを65℃で1時間乾燥させ、2mgのデンプンにつき1mlのUDMSO(上記のように量り取る)を使用して、尿素−ジメチルスルホキシド溶液(UDMSO、6M尿素の容量1に対してジメチルスルホキシドの容量9)に溶解する。すぐにこの混合物を激しくボルテックスし、デンプンを完全に溶解するために断続的にボルテックスしながら95℃のウォーターバス中で1時間インキュベートする。デンプン−UDMSO溶液のアリコート(50μl)を、水1mlあたり2mgのヨウ素および20mgのヨウ化カリウムを含むI2−KI試薬20μlで処理する。この混合物を水で1mlにする。この混合物200μlをマイクロプレートに移し、Emax Precision Microplate Reader(Molecular Devices社製、米国)を使用して吸光度を読み取ることにより、その650nmでの吸光度を測定する。ジャガイモのアミロース、およびトウモロコシ(またはジャガイモ)のアミロペクチン(Sigma社製)から、0〜100%のアミロースおよび100%〜0%のアミロペクチンを含む標準試料を作製し、これを検査試料用のものとして扱う。この標準試料の吸光度から導き出した回帰方程式を使用して、その吸光度の値からアミロース含量(%アミロース)を求める。また、枝切りされなかったデンプンのアミロース/アミロペクチン比の分析を、Caseら,(1998)に従って、またはBateyおよびCurtin(1996)による記載の通り枝切りされたデンプンを分離するHPLC法によって実施してよい。
【0149】
デンプン試料の枝切りの後、デンプンにおける鎖長の分布を、Morellら(1998)に従って、キャピラリー電気泳動ユニットを使用したfluorophore assisted carbohydrate electrophoresis(FACE)によって分析してよい。デンプン試料の糊化温度のプロファイルをPyris 1 differential scanning calorimeter(Perkin Elmer社製、米国コネチカット州Norwalk)で測定してよい。デンプン溶液の粘度を、例えばBateyら、1997によって報告されている条件を使用して、Rapid−Visco−Analyser(RVA、Newport Scientific社製、シドニーWarriewood)で測定してよい。測定され得るパラメータには、最高粘度(peak viscosity)(最高ホットペースト粘度(hot paste viscosity))、保持強度(holding strength)、最終粘度、およびペースティング温度(pasting temperature)が含まれる。この穀粉またはデンプンの膨潤容積をKonik−Roseら(2001)の方法に従って求めてよい。一定の温度で水中で穀粉またはデンプン試料を混合する前、またその糊化した材料を回収した後で試料を量ることにより、水の吸収を測定する。
【0150】
β−グルカンレベルを、Megazyme社(アイルランド共和国Wicklow州Bray)によって提供されるキットを使用して求めてよい。
【0151】
胚乳中でのタンパク質発現の分析
胚乳中での特異的タンパク質の発現を、ウエスタンブロットの手順によって分析する。すべての母系組織から胚乳を切り出し、およそ0.2mgの試料を、5mM EDTA、20%グリセロール、5mM DTT、および1mM Pefablocを含むpH7.5の50mM KPi緩衝液(42mM K2HPO4および8mM KH2PO4)600μl中でホモジナイズする。この細かくした試料を13,000gで10分間遠心分離し、その上清を分取し、使用するまで−80℃で凍結させる。全タンパク質を測定するために、BSA標準曲線を、アリコート0μl、20μl、40μl、60μl、80μlおよび100μlの0.25mg/ml BSA標準液を使用して設定する。この試料(3μl)を蒸留水で100μlにし、それぞれに1mlのクーマシープラスプロテイン(Coomassie Plus Protein)試薬を添加する。5分後、その吸光度を595nmで読み取り、ブランクとして標準曲線からゼロのBSA試料を使用して、試料中のタンパク質レベルを求める。各胚乳から得られる20μgの全タンパク質を含む試料を、0.34MトリスHCl(pH8.8)、アクリルアミド(8.0%)、過硫酸アンモニウム(0.06%)、およびTEMED(0.1%)を含む8%非変性ポリアクリルアミドゲルで泳動する。電気泳動の後、このタンパク質を、Morellら、(1997)に従ってニトロセルロース膜に移し、SBEIIaまたはSBEIIbに特異的な抗体との免疫反応を行う。
(実施例5.イネゲノムに特有の配列を同定することによる標的遺伝子の最適化された遺伝子発現抑制)
【0152】
例えば、2重鎖RNA、アンチセンス、またはコサプレッション構築体を使用するなどの方法により標的遺伝子の発現を低減する(遺伝子発現抑制)のに使用する遺伝子配列は、標的遺伝子に高度に特異的であることが好ましい。すなわち、この発現抑制分子は、標的遺伝子またはその相補体の少なくとも19の連続するヌクレオチドの配列と少なくとも約95%同一である少なくとも19の連続するヌクレオチドの配列を含む。最高の特異性をもたらすためには、標的とする配列が、標的遺伝子に特有であり、植物ゲノムにおいて発現される遺伝子として他のどこにも存在しないことが理想的である。これは「オフ遺伝子効果(off−gene effect)」を最小限にするはずである。完全に近いイネゲノム配列の知識を使用して、SBE標的遺伝子配列をイネゲノムの残りの配列と比較して、それによりこうした遺伝子内の最適化された標的配列が同定されている。
【0153】
イネゲノムDNA配列データベース(OSA1.seq)をTIGRのウェブサイト(http://www.tigr.org/tdb/e2k1/osa1/)からFASTAフォーマットでダウンロードした。このデータベースをフォーマットし、これは、「formatdb」を使用してBLASTに、また「dbifasta」を使用してEMBOSS function seqretに使用可能となった。クエリー配列をFATSAフォーマットで作製し、1組のプリセットパラメータ(比較のためのオプション:ワード19およびストリンジェンシー18)を用いて、BLASTベースの遺伝子発現抑制プログラム(Gene Silencing program)(P.Waterhouseら、CSIRO Plant Industry,personal communication)を実行することにより、イネゲノムで相同配列を検索するのに使用した。
【0154】
実施例2に記載の通り遺伝子発現抑制構築体を調製するのに使用するSBEIIa、SBEIIb、およびSBEIの配列を、イネゲノムに対するクエリー配列として使用した。この出力を図8に示す。出力配列における多数の「NNNN...」により、これらの領域におけるクエリー配列が、少なくとも19の連続するヌクレオチドの領域において、イネゲノムにおける他の配列と相同性を有することが示唆された。使用したSBEIIa配列は、固有のものであり、使用したSBEIIb配列は、固有のものでない配列を一部含むが、使用したSBEI配列は、明らかであるように思われる末端の57ヌクレオチドを除いて、イネゲノムの他の場所で重複しているようであることが分かる(図8)。現行のイネゲノム配列の検査により、SBEI遺伝子領域を含むことができる重複する2種のBACクローン間のゲノムDNA配列が重複するようであることが明らかとなった。明らかなこの重複は、本当であるか、またはその領域におけるイネゲノムの組立てのエラーを表すものである恐れがある。
【0155】
<参考文献>
Abel et al., (1996). The Plant Journal 10, 981-991.
Ahlandsberg et al., (2002). Plant Cell Rep 20, 864-868.
Anderson et al., (1989). Nucl Acids Res 17, 461-462.
Baba et al., (1991). Biochem Biophys Res Commun 181: 87-94.
Baga et al., (2000). Plant Physiol. 124, 253-263.
Batey and Curtin. (1996). Starch 48, 338-344.
Batey et al., (1997). Cereal Chemistry 74, 497-501.
Blauth et al., (2001). Plant Physiology 125, 1396-1405.
Bourque. (1995). Plant Science 105, 125-149.
Boyer and Preiss, (1978). Carbohydrate Research 61, 321-334.
Boyer and Preiss, (1981). Plant Physiology 67, 1141-1145.
Boyer et al., (1980). Starch 32, 217-222.
Buchholz et al., (1998). Methods Mol Biol 81, 383-396.
Buleon et al., (1998). International Journal of Biological Macromolecules 23, 85-112.
Cao et al., (2000). Archives. of Biochemistry and Biophysics. 373, 135-146.
Case et al., (1998). Journal of Cereal Science 27, 301-314.
Chan et al., (1993). Plant Mol Biol 22, 491-506.
Chiou et al., (2002). Starch 54, 415-420.
Craig et al., (1998). Plant Cell 10, 413-426.
Denyer et al., (1996). Plant Physiology 112, 779-785.
Fergason. 1994. pp 55-77 in "Speciality Corns" eds , CRC Press Inc.
Filpse et al.,(1996). Planta 198, 340.
Fisher et al., (1993). Plant Physiol 102:1045-1046.
Fisher et al., (1996). Plant Physiol 110: 611-619.
Frances et al., (1998). Plant Mol Biol 38, 407-415.
Fujita et al., (2003). Plant Cell Physiol 44, 607-618.
Fuwa et al., (1999). Starch/Starke. 51, 147-151.
Gao et al., (1996) Plant Mol Biol 30, 1223-1232.
Gao et al., (1997). Plant Physiol 114: 69-78.
Gao et al., (1998). Plant Cell 10, 399-412.
Giroux and Hannah. (1994). Molecular and General Genetics 243, 400-408.
Goddard et al., (1984) Am J Clin Nutr 39, 388-392.
Green et al., (1997). Plant Physiology 114, 203-212.
Hedman and Boyer, (1982). Biochemical Genetics 20, 483-492.
Hiei et al., (1994). Plant J 6, 271-282.
Hirano et al., (1998). Mol Biol Evol 15, 978-987.
Hirano and Sano, (2000). Genes Genet Syst 75, 245-249.
Isshiki et al., (1998). Plant J 15, 133-138.
James et al., (1995). Plant Cell 7, 417-429.
Jobling et al., (1999). Plant Journal 18, 163-171.
Juliano, B.O. (1979). in Proceedings, Workshop on Chemical Aspects of Rice Grain Quality, p. 69-90. Los Banos, Laguna, the Philippines, IRRI.
Juliano, B.O., (1985). Rice: chemistry and technology, 2nd ed. St Paul, MN, USA, Am. Assoc. Cereal Chem. 774 pp.
Kawasaki et al., (1993). Mol Gen Genet 237, 10-16.
Konik-Rose et al (2001) Starch 53, 14-20.
Krueger et al., (1987). Cereal Chemistry 64, 187-190.
Kubo et al., (1999). Plant physiology. 121, 399-409.
Lazo et al., (1991). Bio/Technology 9, 963-967.
Li et al., (1999a). Plant physiology. 120, 1147-1155.
Li et al., (1999b). Theoretical and Applied Genetics 98, 1208-1216.
Li et al., (2000). Plant Physiology 123, 613-624.
Li et al., (2003). Funct Integr Genomics 3:76-85.
Lumdubwong et al., (2000). J Cereal Sci 31, 63-74.
Maniatis et al., (1982). Molecular cloning: a laboratory manual. Cold Spring Harbour Laboratory Press New York.
McCreery and Helentjaris (1994). Methods in Molecular Biology, Vol. 28: Protocols for nucleic acid analysis by non-radioactive probes, 67-71, Humana Press Inc., Totawa, NJ.
Mizuno et al., (1993). Journal of Biological Chemistry 268, 19084-19091.
Mizuno et al., (1992). Journal of Biochemistry 112, 643-651.
Mizuno et al., (2001). Plant Cell Physiol 42, 349-357.
Morell et al., (1997). Plant Physiology 113, 201-208.
Morell et al., (1998). Electrophoresis 19, 2603-2611.
Morell et al., (2003). Plant J. 34: 173-185.
Morrison and Laignelet (1983). Journal of Cereal Science 1:9-20.
Mullins et al., (1999). European Journal of Plant Pathology 105: 465-475.
Myers et al., (2000). Plant Physiology 122, 989-997.
Nakamura (2002). Plant Cell Physiology 43, 718-725.
Nakamura and Yamanouchi (1992). Plant Physiol 99: 1265-1266.
Nair et al., (1997). Plant Sci 122: 153-163.
Nishi et al., (2001). Plant Physiology 127, 459-472.
Rahman et al., (1995). Australian Journal of Plant Physiology 22, 793-803.
Rahman et al., (1997). Genome 40: 465-474.
Rahman et al., (1999). Theor Appl Genet. 98: 156-163.
Rahman et al., (2000). J Cereal Sci 31: 91-110.
Rahman et al., (2001). Plant Physiol 125: 1314-1324.
Repellin et al., (1997). Plant Gene Reg 97-094
Safford et al., (1998). Carbohydrate Polymers 35, 155-168.
Schulman and Kammiovirta, (1991). Starch 43, 387-389.
Schwall et al., (2000). Nature Biotechnology 18, 551-554.
Senior (1998). Biotechnology and Genetic Engineering Reviews 15, 79-119.
Shannon and Garwood, (1984). In Starch: Chemistry and Technology, Whistler et al., eds, Academic Press, Orlando, FL, pp25-86.
Shure et al., (1983). Cell 35, 225-233.
Sidebottom et al., (1998). Journal of Cereal Science 27, 279-287.
Smidansky et al., 2003) Planta 216, 656-664.
Smith et al., (2000) Nature 407, 319-320.
Sun et al., (1996). Physiol Plantarum 96, 474-483.
Sun et al., (1997). The New Phytologist 137 , 215-215.
Sun et al., (1998). Plant Physiol 118, 37-49.
Takeda et al., (1986). Carbohydr Res 148, 299-308.
Takeda et al., (1987). Carbohydr Res 168, 79-88.
Takeda et al., (1993a). Carbohydrate Research 240, 253-262.
Takeda et al., (1993b). Carbohydrate Research 246, 273-281.
Terada et al., (2000). Plant Cell Physiol 41, 881-888.
Thomas and Atwell 1999 Starches Eagen Press, St Paul, Minnesota, USA pp: 13-24.
Thorbjornsen et al., (1996). Plant Journal 10, 243-250.
Wang et al., (1990). Nucl Acids Res 18, 5898.
Wang et al., (1998a). Journal of Experimental Botany 49, 481-502.
Wang et al., (1998b). Acta Hort 461, 401-407.
Wegener et al., 1994. Mol. Gen Genet. 245, 465-470.
Yamamori et al., (2000). Theor.Appl.Genet. 101, 21-29
Yamanouchi and Nakamura (1992). Plant Cell Physiol 33, 985-991.
Young. (1984). in Whistler et al. (eds), Academic Press, Orlando, FL, chap 8.
Zhang et al., (1997). Mol Biotechnol 8, 223-231.
Zikiryaeva and Kasimov, (1972). Uzbekskii Biologicheskii Zhurnal 6, 18-20.
Zwar and Chandler, (1995). Planta 197, 39-48.
【特許請求の範囲】
【請求項1】
イネ植物から得られる穀粒であって、そのデンプン中のアミロースの割合が少なくとも40%であるデンプンを含む穀粒。
【請求項2】
2つ以上の遺伝的変異を含み、
1つの遺伝的変異が、
a)SBEIIaの発現および/または活性を阻害するSBEIIa遺伝子の突然変異、ならびに
b)SBEIIaの発現および/または活性を阻害する導入核酸
からなる群から選択され、
第2の遺伝的変異が、
c)SBEIIbの発現および/または活性を阻害するSBEIIb遺伝子の突然変異、ならびに
d)SBEIIbの発現および/または活性を阻害する導入核酸
からなる群から選択される請求項1に記載の穀粒。
【請求項3】
SBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたものを含む請求項1または2に記載の穀粒。
【請求項4】
前記穀粒のデンプン中のアミロースの割合が少なくとも50%である請求項1から3のいずれか一項に記載の穀粒。
【請求項5】
導入遺伝子を含む請求項1から4のいずれか一項に記載の穀粒。
【請求項6】
前記導入遺伝子が、アンチセンス、コサプレッション、リボザイム、または2重鎖RNA分子をコードしている請求項5に記載の穀粒。
【請求項7】
導入遺伝子によらないものである、請求項1から4のいずれか一項に記載の穀粒。
【請求項8】
SBEIのタンパク質レベルおよび/または活性レベルを低下させたものをさらに含む請求項2から7のいずれか一項に記載の穀粒。
【請求項9】
ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型の枝切り酵素、およびプルラナーゼ型の枝切り酵素からなる群から選択されるタンパク質レベルおよび/または酵素活性レベルを変更したものを含む請求項1から8のいずれか一項に記載の穀粒。
【請求項10】
GBSSのタンパク質レベルおよび/または酵素活性レベルを変更したものを含む請求項9に記載の穀粒。
【請求項11】
non−shrunkenである請求項1から10のいずれか一項に記載の穀粒。
【請求項12】
平均重量が少なくとも約25mgの玄米である請求項1から11のいずれか一項に記載の穀粒。
【請求項13】
穀粒内のデンプン顆粒の少なくとも50%が、偏光下で観察すると非複屈折性に見える請求項1から12のいずれか一項に記載の穀粒。
【請求項14】
デンプン含量が、同等のものであるが未改変の穀粒の少なくとも90%である請求項1から13のいずれか一項に記載の穀粒。
【請求項15】
SBEIIaまたはSBEIIb遺伝子の無発現変異を含む請求項2から14のいずれか一項に記載の穀粒。
【請求項16】
インディカ品種のものであるか、またはWxa対立遺伝子を含む、請求項1から15のいずれか一項に記載の穀粒。
【請求項17】
請求項1から16のいずれか一項に記載の穀粒を産生することができるイネ植物。
【請求項18】
請求項1から16のいずれか一項に記載の穀粒から抽出されたデンプン顆粒。
【請求項19】
請求項1から16のいずれか一項に記載の穀粒から抽出されたデンプン。
【請求項20】
請求項1から16のいずれか一項に記載の穀粒から生成される穀粉またはデンプンを含む生成物。
【請求項21】
前記穀粉またはデンプンを別の供給源からの穀粉またはデンプンと混合する、請求項20に記載の生成物。
【請求項22】
非食品である、請求項20に記載の生成物。
【請求項23】
請求項19に記載のデンプンと、別の食品成分または水を含む組成物。
【請求項24】
少なくとも40%のアミロースを含むデンプンを有する穀粒を産生することができるイネ植物を作出する方法であって、
a)遺伝的変異を親イネ植物または種子中に導入するステップと、
b)親イネ植物または種子の子孫植物を同定するステップとを含み、前記子孫植物の穀粒のデンプンが少なくとも40%のアミロースを含む方法。
【請求項25】
前記子孫イネ植物が2つ以上の遺伝的変異を含む方法であって、1つの遺伝的変異が、
e)SBEIIaの発現および/または活性を阻害するSBEIIa遺伝子の突然変異、ならびに
f)SBEIIaの発現および/または活性を阻害する導入核酸
からなる群から選択され、
第2の遺伝的変異が
g)SBEIIbの発現および/または活性を阻害するSBEIIb遺伝子の突然変異、ならびに
h)SBEIIbの発現および/または活性を阻害する導入核酸
からなる群から選択される、請求項24に記載の方法。
【請求項26】
前記遺伝的変異により、イネ植物の胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルの低下がもたらされる、請求項24または25に記載の方法。
【請求項27】
前記遺伝的変異を導入するステップが外因性の核酸を導入することを含む請求項24から27のいずれか一項に記載の方法。
【請求項28】
前記外因性の核酸をイネ細胞中に導入し、次いでこの細胞をイネ植物に再生させる請求項27に記載の方法。
【請求項29】
前記外因性の核酸が、SBEIIaおよび/またはSBEIIbの発現および/または活性の阻害物質をコードしている請求項28に記載の方法。
【請求項30】
前記阻害物質が、アンチセンス、コサプレッション、リボザイム、または2重鎖RNA分子である請求項29に記載の方法。
【請求項31】
前記遺伝的変異を導入するステップが、化学薬剤または放射線を用いた親イネ植物または種子の突然変異誘発を含む請求項24または25に記載の方法。
【請求項32】
前記子孫イネ植物が、SBEIIaおよび/またはSBEIIbにおける無発現変異を含む請求項25から31のいずれか一項に記載の方法。
【請求項33】
SBEIのタンパク質レベルおよび/または活性レベルの低下をもたらす遺伝的変異を導入するステップをさらに含む、請求項25から32のいずれか一項に記載の方法。
【請求項34】
前記子孫植物を、穀粒デンプン中のアミロースレベルに基づいて、あるいは子孫植物の胚乳中のSBEIIaおよび/またはSBEIIbのタンパク質レベルおよび/または活性レベルの低下について同定する、請求項24または25に記載の方法。
【請求項35】
対立遺伝子Wxaのイネ植物への導入をさらに含む請求項24から34のいずれか一項に記載の方法。
【請求項36】
前記対立遺伝子Wxaを交配によって導入する請求項35に記載の方法。
【請求項37】
胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または酵素活性レベルを低下させたイネ植物を作出する方法であって、
a)SBEIIaのタンパク質レベルおよび/または酵素活性レベルを低下させた種子に突然変異を起こさせること、あるいは
b)SBEIIbのタンパク質レベルおよび/または酵素活性レベルを低下させた種子に突然変異を起こさせること、あるいは
c)SBEIIaのタンパク質レベルおよび/または酵素活性レベルを低下させた植物を、SBEIIbのタンパク質および/または酵素活性のレベルを低下させた植物と交配させること、ならびに
d)胚乳中のSBEIIaとSBEIIbのタンパク質活性および/または酵素活性を低下させたイネ植物を同定することを含む方法。
【請求項38】
前記イネ植物を同定するステップが、イネのSBEIIa遺伝子またはSBEIIb遺伝子に連鎖する分子マーカーを有するイネ植物の集団をスクリーニングすること、ならびに連鎖した分子マーカーを用いたスクリーニングからのシグナルの存在または非存在に基づいて植物を同定することを含む、請求項37に記載の方法。
【請求項39】
前記イネ植物を同定するステップが、イネのSBEIIaタンパク質またはSBEIIbタンパク質に結合する抗体を用いてイネ植物の集団から種子をスクリーニングするステップ、ならびに結合した抗体の存在または非存在に基づいて植物を同定するステップを含む、請求項37に記載の方法。
【請求項40】
改変した米デンプンを産生する方法であって、請求項1から16のいずれか一項に記載の穀粒からのデンプンを抽出するステップを含む方法。
【請求項41】
2つ以上の外因性の核酸分子の使用であって、SBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたイネ植物を作出するために、その少なくとも1つがイネのSBEIIaの発現および/または活性の阻害物質をコードしており、またその少なくとも別の1つがイネのSBEIIbの発現および/または活性の阻害物質をコードしている使用。
【請求項42】
前記阻害物質が、アンチセンス分子、コサプレッション分子、リボザイム、2重鎖RNA分子、およびそれらの任意の組合せからなる群から選択される、請求項41に記載の使用。
【請求項43】
イネSBEIIaの阻害物質およびイネSBEIIbの阻害物質をコードしている単離された核酸分子であって、これらの阻害物質が同一の分子または異なる分子でもよい核酸分子。
【請求項44】
請求項43に記載の単離された核酸分子を含むベクター。
【請求項45】
請求項43に記載の単離された核酸分子を含む細胞。
【請求項46】
イネ細胞である、請求項45に記載の細胞。
【請求項47】
請求項43に記載の単離された核酸分子を含むトランスジェニックイネ植物。
【請求項1】
イネ植物から得られる穀粒であって、そのデンプン中のアミロースの割合が少なくとも40%であるデンプンを含む穀粒。
【請求項2】
2つ以上の遺伝的変異を含み、
1つの遺伝的変異が、
a)SBEIIaの発現および/または活性を阻害するSBEIIa遺伝子の突然変異、ならびに
b)SBEIIaの発現および/または活性を阻害する導入核酸
からなる群から選択され、
第2の遺伝的変異が、
c)SBEIIbの発現および/または活性を阻害するSBEIIb遺伝子の突然変異、ならびに
d)SBEIIbの発現および/または活性を阻害する導入核酸
からなる群から選択される請求項1に記載の穀粒。
【請求項3】
SBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたものを含む請求項1または2に記載の穀粒。
【請求項4】
前記穀粒のデンプン中のアミロースの割合が少なくとも50%である請求項1から3のいずれか一項に記載の穀粒。
【請求項5】
導入遺伝子を含む請求項1から4のいずれか一項に記載の穀粒。
【請求項6】
前記導入遺伝子が、アンチセンス、コサプレッション、リボザイム、または2重鎖RNA分子をコードしている請求項5に記載の穀粒。
【請求項7】
導入遺伝子によらないものである、請求項1から4のいずれか一項に記載の穀粒。
【請求項8】
SBEIのタンパク質レベルおよび/または活性レベルを低下させたものをさらに含む請求項2から7のいずれか一項に記載の穀粒。
【請求項9】
ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型の枝切り酵素、およびプルラナーゼ型の枝切り酵素からなる群から選択されるタンパク質レベルおよび/または酵素活性レベルを変更したものを含む請求項1から8のいずれか一項に記載の穀粒。
【請求項10】
GBSSのタンパク質レベルおよび/または酵素活性レベルを変更したものを含む請求項9に記載の穀粒。
【請求項11】
non−shrunkenである請求項1から10のいずれか一項に記載の穀粒。
【請求項12】
平均重量が少なくとも約25mgの玄米である請求項1から11のいずれか一項に記載の穀粒。
【請求項13】
穀粒内のデンプン顆粒の少なくとも50%が、偏光下で観察すると非複屈折性に見える請求項1から12のいずれか一項に記載の穀粒。
【請求項14】
デンプン含量が、同等のものであるが未改変の穀粒の少なくとも90%である請求項1から13のいずれか一項に記載の穀粒。
【請求項15】
SBEIIaまたはSBEIIb遺伝子の無発現変異を含む請求項2から14のいずれか一項に記載の穀粒。
【請求項16】
インディカ品種のものであるか、またはWxa対立遺伝子を含む、請求項1から15のいずれか一項に記載の穀粒。
【請求項17】
請求項1から16のいずれか一項に記載の穀粒を産生することができるイネ植物。
【請求項18】
請求項1から16のいずれか一項に記載の穀粒から抽出されたデンプン顆粒。
【請求項19】
請求項1から16のいずれか一項に記載の穀粒から抽出されたデンプン。
【請求項20】
請求項1から16のいずれか一項に記載の穀粒から生成される穀粉またはデンプンを含む生成物。
【請求項21】
前記穀粉またはデンプンを別の供給源からの穀粉またはデンプンと混合する、請求項20に記載の生成物。
【請求項22】
非食品である、請求項20に記載の生成物。
【請求項23】
請求項19に記載のデンプンと、別の食品成分または水を含む組成物。
【請求項24】
少なくとも40%のアミロースを含むデンプンを有する穀粒を産生することができるイネ植物を作出する方法であって、
a)遺伝的変異を親イネ植物または種子中に導入するステップと、
b)親イネ植物または種子の子孫植物を同定するステップとを含み、前記子孫植物の穀粒のデンプンが少なくとも40%のアミロースを含む方法。
【請求項25】
前記子孫イネ植物が2つ以上の遺伝的変異を含む方法であって、1つの遺伝的変異が、
e)SBEIIaの発現および/または活性を阻害するSBEIIa遺伝子の突然変異、ならびに
f)SBEIIaの発現および/または活性を阻害する導入核酸
からなる群から選択され、
第2の遺伝的変異が
g)SBEIIbの発現および/または活性を阻害するSBEIIb遺伝子の突然変異、ならびに
h)SBEIIbの発現および/または活性を阻害する導入核酸
からなる群から選択される、請求項24に記載の方法。
【請求項26】
前記遺伝的変異により、イネ植物の胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルの低下がもたらされる、請求項24または25に記載の方法。
【請求項27】
前記遺伝的変異を導入するステップが外因性の核酸を導入することを含む請求項24から27のいずれか一項に記載の方法。
【請求項28】
前記外因性の核酸をイネ細胞中に導入し、次いでこの細胞をイネ植物に再生させる請求項27に記載の方法。
【請求項29】
前記外因性の核酸が、SBEIIaおよび/またはSBEIIbの発現および/または活性の阻害物質をコードしている請求項28に記載の方法。
【請求項30】
前記阻害物質が、アンチセンス、コサプレッション、リボザイム、または2重鎖RNA分子である請求項29に記載の方法。
【請求項31】
前記遺伝的変異を導入するステップが、化学薬剤または放射線を用いた親イネ植物または種子の突然変異誘発を含む請求項24または25に記載の方法。
【請求項32】
前記子孫イネ植物が、SBEIIaおよび/またはSBEIIbにおける無発現変異を含む請求項25から31のいずれか一項に記載の方法。
【請求項33】
SBEIのタンパク質レベルおよび/または活性レベルの低下をもたらす遺伝的変異を導入するステップをさらに含む、請求項25から32のいずれか一項に記載の方法。
【請求項34】
前記子孫植物を、穀粒デンプン中のアミロースレベルに基づいて、あるいは子孫植物の胚乳中のSBEIIaおよび/またはSBEIIbのタンパク質レベルおよび/または活性レベルの低下について同定する、請求項24または25に記載の方法。
【請求項35】
対立遺伝子Wxaのイネ植物への導入をさらに含む請求項24から34のいずれか一項に記載の方法。
【請求項36】
前記対立遺伝子Wxaを交配によって導入する請求項35に記載の方法。
【請求項37】
胚乳中のSBEIIaとSBEIIbのタンパク質レベルおよび/または酵素活性レベルを低下させたイネ植物を作出する方法であって、
a)SBEIIaのタンパク質レベルおよび/または酵素活性レベルを低下させた種子に突然変異を起こさせること、あるいは
b)SBEIIbのタンパク質レベルおよび/または酵素活性レベルを低下させた種子に突然変異を起こさせること、あるいは
c)SBEIIaのタンパク質レベルおよび/または酵素活性レベルを低下させた植物を、SBEIIbのタンパク質および/または酵素活性のレベルを低下させた植物と交配させること、ならびに
d)胚乳中のSBEIIaとSBEIIbのタンパク質活性および/または酵素活性を低下させたイネ植物を同定することを含む方法。
【請求項38】
前記イネ植物を同定するステップが、イネのSBEIIa遺伝子またはSBEIIb遺伝子に連鎖する分子マーカーを有するイネ植物の集団をスクリーニングすること、ならびに連鎖した分子マーカーを用いたスクリーニングからのシグナルの存在または非存在に基づいて植物を同定することを含む、請求項37に記載の方法。
【請求項39】
前記イネ植物を同定するステップが、イネのSBEIIaタンパク質またはSBEIIbタンパク質に結合する抗体を用いてイネ植物の集団から種子をスクリーニングするステップ、ならびに結合した抗体の存在または非存在に基づいて植物を同定するステップを含む、請求項37に記載の方法。
【請求項40】
改変した米デンプンを産生する方法であって、請求項1から16のいずれか一項に記載の穀粒からのデンプンを抽出するステップを含む方法。
【請求項41】
2つ以上の外因性の核酸分子の使用であって、SBEIIaとSBEIIbのタンパク質レベルおよび/または活性レベルを低下させたイネ植物を作出するために、その少なくとも1つがイネのSBEIIaの発現および/または活性の阻害物質をコードしており、またその少なくとも別の1つがイネのSBEIIbの発現および/または活性の阻害物質をコードしている使用。
【請求項42】
前記阻害物質が、アンチセンス分子、コサプレッション分子、リボザイム、2重鎖RNA分子、およびそれらの任意の組合せからなる群から選択される、請求項41に記載の使用。
【請求項43】
イネSBEIIaの阻害物質およびイネSBEIIbの阻害物質をコードしている単離された核酸分子であって、これらの阻害物質が同一の分子または異なる分子でもよい核酸分子。
【請求項44】
請求項43に記載の単離された核酸分子を含むベクター。
【請求項45】
請求項43に記載の単離された核酸分子を含む細胞。
【請求項46】
イネ細胞である、請求項45に記載の細胞。
【請求項47】
請求項43に記載の単離された核酸分子を含むトランスジェニックイネ植物。
【図1】


【図2】
【図3】

【図4】

【図5】

【図6】
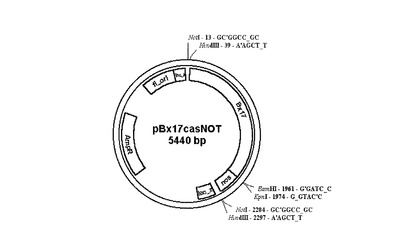
【図7−1】

【図7−2】

【図7−3】

【図7−4】

【図8】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7−1】
【図7−2】
【図7−3】
【図7−4】
【図8】
【公開番号】特開2012−179055(P2012−179055A)
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願番号】特願2012−106666(P2012−106666)
【出願日】平成24年5月8日(2012.5.8)
【分割の表示】特願2006−537005(P2006−537005)の分割
【原出願日】平成16年10月27日(2004.10.27)
【出願人】(305039998)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガニゼイション (92)
【Fターム(参考)】
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願日】平成24年5月8日(2012.5.8)
【分割の表示】特願2006−537005(P2006−537005)の分割
【原出願日】平成16年10月27日(2004.10.27)
【出願人】(305039998)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガニゼイション (92)
【Fターム(参考)】
[ Back to top ]