
アルカロイド含量を調節する機能を有する遺伝子、およびそれを利用した形質転換タバコ属植物
【課題】アルカロイド含量を大きく変化させることなく、ノルニコチン含量を有意に低減させた形質転換タバコ属植物を提供すること。
【解決手段】ノルニコチン型タバコの葉由来のアルカロイド含量調節機能を有するNor#35遺伝子、該遺伝子の発現を抑制することによってノルニコチン含量またはアルカロイド含量が低減された形質転換タバコ属植物、該形質転換タバコ属植物の作製方法。
【解決手段】ノルニコチン型タバコの葉由来のアルカロイド含量調節機能を有するNor#35遺伝子、該遺伝子の発現を抑制することによってノルニコチン含量またはアルカロイド含量が低減された形質転換タバコ属植物、該形質転換タバコ属植物の作製方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はアルカロイド含量を調節する機能を有する遺伝子、および当該遺伝子の発現を抑制することによってノルニコチン含量またはアルカロイド含量が低減された形質転換タバコ属植物等に関する。
【背景技術】
【0002】
タバコ栽培種に含まれている主要なアルカロイドはニコチンとノルニコチンである(Saitoh, F. et al.(1985),Phytochmistry 24, 477-480)。ノルニコチンは根で生合成されたニコチンが葉に移動し、そこで脱メチル化酵素(nicotine N-demethylase)の作用でノルニコチンに変換される。ノルニコチンのタバコに与える影響としては、タバコ黄色種において紅葉(Cherry-red leaf)をもたらす(岡ら、1961, 岡山たばこ試験場報告21,65-71)、あるいは、タバコバーレー種において香りの特性が変わる(生沼ら、1972, 盛岡たばこ試験場報告8,1-10)などが報告されており、いずれもタバコの原料価値を下げる原因となる。また、ノルニコチンはタバコ特異的ニトロソアミン(TSNA)の一種であるN−ニトロソノルニコチン(NNN)の前駆体となるため、タバコ原料中のノルニコチン含量を低減させることが望まれている。
【0003】
本来タバコ栽培種はノルニコチンをほとんど含まないニコチン型タバコである。ニコチン型タバコは2対のノルニコチン劣性遺伝子を持つが、その自殖後代でそのうち1対が優性遺伝子に突然変異し、前述の問題を有するノルニコチン型個体が出現すると考えられている。また、このニコチン型タバコからノルニコチン型タバコへの変異率は0.8%と極めて高いこともわかっている(Wernsman, E. et al.(1970), Tabaccc Science 12, 226-228)。
【0004】
タバコ葉中のノルニコチン含量を低減する試みとして、まずノルニコチン型に変異した個体を可能な限り採種用個体から排除する方法が知られている(非特許文献1)。しかしながら、本方法では多数の個体のノルニコチン解析という煩雑な作業を要する。一方、遺伝子工学的手法を用いてニコチンからノルニコチンへの変換を抑制することも試みられている。例えば、ある種のチトクロームP450遺伝子の発現を抑制することによって、ノルニコチン型タバコのノルニコチン含量を90%以上低減したタバコが作出されている(非特許文献2)。また、quinolate phosphoribosyl transferase (QPTase)遺伝子(NtQPT1)の発現を抑制することによって、非組換え(野生型)タバコと比較してノルニコチン含量を約1/3に低減したという報告がある(非特許文献3)が、この方法ではニコチン含量も1/10以下に低減し、その結果アルカロイド含量が大幅に低減している。アルカロイド含量はタバコの喫味に大きな影響を及ぼすことから、このようなアルカロイド含量の大幅な低減は望ましくない。
【0005】
その他、遺伝子工学的手法を用いてニコチン含量の増減を試みた例もある。例えば、ニコチン含量を増加させる報告としては、Nicotiana sylvestrisにPMTase遺伝子を導入することによってニコチン含量を約40%増加させた例(非特許文献4)、Nicotiana tabacumにVitreoscillaヘモグロンビン(VHb)をコードする遺伝子を導入することによってニコチン含量を34%増加させた例(特許文献1)、チトクロームP450遺伝子の発現を抑制することによってニコチン含量を増加させた例(非特許文献5)、一方、ニコチン含量を低減させる報告としては、ニコチン生合成遺伝子を用いてニコチン含量を低減させる方法(特許文献2)、ADC, ODC, SAMS, PMT, NADHといったニコチン生合成に関与する酵素遺伝子を用いてニコチン含量を低減させる方法(特許文献3、特許文献4)などが挙げられる。
【0006】
【特許文献1】WO 98/12913A
【特許文献2】WO 2002/100199A
【特許文献3】WO 00/67558A
【特許文献4】WO 2005/018307
【非特許文献1】佐藤ら、1982, 磐田たばこ試験場報告14,17-21
【非特許文献2】Siminszky,B. et al. (2005) Proc. Natl. Acad. Sci USA 102, 41,14919-14924
【非特許文献3】Biotechnology:A Tool for Reduced Risk Tobacco Products-The Nicotine Experience from Test Tube to Cigarette Pack, Jiahua Xie et al., Recent Advances in Tobacco Science Vol. 30: Biotechnology: A Tool for Reduced Risk Tobacco Products Symposium Proceedings 58th Meeting Tobacco Science Research Conference, 2004, 17-37
【非特許文献4】Proc. Natl. Acad. Sci. USA. January 2, 2001, Vol.98, No.1, 362-372
【非特許文献5】Gavilano, L. et al. (2006) J.Agric.Food Chem. 54, 9071-9078
【発明の開示】
【発明が解決しようとする課題】
【0007】
従って、本発明の課題は、アルカロイド含量を大きく変化させることなく、ノルニコチン含量を有意に低減させた形質転換タバコ属植物を提供することにある。また本発明のさらなる課題は、アルカロイド含量が調節された形質転換タバコ属植物を提供することにある。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決すべく鋭意研究を重ねた結果、ニコチン型タバコとノルニコチン型タバコの葉において発現量に違いがある遺伝子群の中から、幼苗個体であるか鉢上げ個体であるかを問わず、ノルニコチン型タバコの葉において発現量が高い遺伝子を見出した。また、その遺伝子の発現を抑制するため、RNA干渉 (以下、「RNAi」という)用のベクターを構築してタバコに導入することによって、ノルニコチン含量が低減した形質転換タバコを得ることに成功し、本発明を完成させるに至った。
【0009】
即ち、本発明は以下の発明を包含する。
(1) 以下の(a)〜(d)にポリヌクレオチドからなるNor#35遺伝子。
(a) 配列番号1に示す塩基配列からなるポリヌクレオチド
(b) 配列番号1に示す塩基配列において1若しくは数個の塩基が欠失、置換、若しくは付加された塩基配列からなり、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(c) 配列番号1に示す塩基配列からなるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(d) 配列番号1に示す塩基配列に対して80%以上の相同性を有する塩基配列からなり、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(2) 配列番号3に示す塩基配列を有する(1)に記載のNor#35遺伝子。
(3) (1)または(2)に記載のNor#35遺伝子を含む組換えベクター。
(4) (1)若しくは(2)に記載のNor#35遺伝子または(3)に記載の組換えベクターを導入した形質転換タバコ属植物。
(5) 植物が、植物体、植物器官、植物組織、または植物細胞である、(4)に記載の形質転換タバコ属植物。
(6) 配列番号1に示す塩基配列中の連続した100塩基以上からなる塩基配列をアンチセンス方向に含む、Nor#35遺伝子発現抑制用ベクター。
(7) 配列番号1に示す塩基配列中の連続した21塩基以上の塩基配列と該塩基配列の相補配列とを含む、Nor#35遺伝子発現抑制用ベクター。
(8) 配列番号1に示す塩基配列中の連続した21塩基以上の塩基配列と該塩基配列の相補配列との間にスペーサー配列を挿入した連続配列を含む、Nor#35遺伝子発現抑制用ベクター。
(9) (6)〜(8)のいずれかに記載のNor#35遺伝子発現抑制用ベクターを導入した形質転換タバコ属植物。
(10) 植物が、植物体、植物器官、植物組織、または植物細胞である、(9)に記載の形質転換タバコ属植物。
(11) (1)または(2)に記載のNor#35遺伝子の発現を抑制した植物細胞を作製する工程、および、該植物細胞から植物体を再生させる工程を含む、アルカロイド含量が低減した形質転換タバコ属植物の作製方法。
(12) (1)または(2)に記載のNor#35遺伝子の発現抑制が、RNAi法、アンチセンス法、遺伝子破壊法、および共抑制法から選ばれるいずれかの手法により行なわれることを特徴とする、(11)に記載の方法。
(13) 植物細胞内において(1)または(2)に記載のNor#35遺伝子の発現量を調節することを特徴とする、植物中のアルカロイド量を改変する方法。
【発明の効果】
【0010】
本発明によれば、アルカロイド含量を調節する機能を有する遺伝子が提供される。タバコ属植物において本遺伝子の発現を抑制させることにより、タバコの喫味に影響を与えるアルカロイド含量を大きく変化させることなく、ノルニコチン含量を低減させることが可能となる。従って、本発明は、ノルニコチンに起因するタバコの原料価値を下げる紅葉などの問題が少なく、しかも喫味のよいタバコを製造するのに有用である。
【発明を実施するための最良の形態】
【0011】
1.本発明の遺伝子
本発明の遺伝子であるNor#35遺伝子は、アルカロイド含量を調節する機能を有する遺伝子である。Nor#35遺伝子はノルニコチン型タバコとニコチン型タバコにおいて発現量の異なる遺伝子の1つであり、Amplified fragment length polymorphism Differential Display (以下、「AFLP-DD法」という)によって同定されたものである。
【0012】
Nor#35遺伝子は、配列番号1に示す塩基配列(cDNA配列)を有し、配列番号3に示すゲノムDNA配列を有する。Nor#35遺伝子のオープンリーディングフレーム(ORF)は、配列番号1に示す塩基配列の第107から第1153位までの領域(1047塩基)であり、配列番号2に示すアミノ酸配列(348アミノ酸残基)からなるタンパク質をコードする。従って、本発明の遺伝子には、配列番号1に示す塩基配列の第107から第1153位までの領域を含む限り、いずれの遺伝子も含まれる。
【0013】
本発明の遺伝子は、好ましくは、配列番号1に示す塩基配列からなるポリヌクレオチドからなる遺伝子であるが、当該遺伝子には限定はされず、アルカロイド含量を調節する機能を有する限り、配列番号1に示す塩基配列において1若しくは数個の塩基が欠失、置換、若しくは付加された塩基配列からなるポリヌクレオチド、配列番号1に示す塩基配列からなるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズするポリヌクレオチドであってもよい。
【0014】
ここで、欠失、置換、若しくは付加されてもよい塩基の数としては、部位特異的突然変異誘発法等の公知の変異導入法により欠失、置換、若しくは付加できる程度の数の塩基をいい、好ましくは、1個から数個である。例えば、配列番号1に示す塩基配列の1〜10個、好ましくは1〜5個の塩基が欠失してもよく、配列番号1に示す塩基配列に1〜10個、好ましくは1〜5個の塩基が付加してもよく、あるいは、配列番号1に示す塩基配列の1〜10個、好ましくは1〜5個の塩基が他の塩基に置換してもよい。また、ここにいう「変異」は、主には部位特異的突然変異誘発法等により人為的に導入された変異を意味するが、天然に存在する同様の変異であってもよい。
【0015】
また、ここで、ストリンジェントな条件とは、いわゆる塩基配列に特異的な2本鎖のポリヌクレオチドが形成され、非特異的な2本鎖のポリヌクレオチドが形成されない条件をいう。より具体的には、0.25M Na2HPO4, pH7.2, 7%SDS, 1mM EDTA, 1×デンハルト溶液からなる緩衝液中で温度が60〜68℃、好ましく65℃、さらに好ましくは68℃の条件下で16〜24時間ハイブリダイズさせ、さらに20mM Na2HPO4, pH7.2, 1%SDS, 1mM EDTAからなる緩衝液中で温度が60〜68℃、好ましく65℃、さらに好ましくは68℃の条件下で15分間の洗浄を2回を行う条件をいう。当業者であれば、Molecular Cloning(Sambrook, J. et al., Molecular Cloning :a Laboratory Manual 2nd ed., Cold Spring Harbor Laboratory Press, 10 Skyline Drive Plainview, NY (1989))等を参照することにより、こうしたホモログ遺伝子を容易に取得することができ、また、配列番号1に示す塩基配列との相同性は、FASTA検索やBLAST検索により決定することができる。
【0016】
また、本発明の遺伝子には、配列番号1に示す塩基配列に対して80%以上の相同性を有する塩基配列からなり、かつアルカロイド含量を調節する機能を有するポリヌクレオチドも含まれる。上記80%以上の相同性は、好ましくは85%以上、より好ましくは90%以上、最も好ましくは95%以上の相同性をいう。配列の同一性は、FASTA検索やBLAST検索により決定することができる。
【0017】
ここで、「アルカロイド含量」とは、タバコの主要アルカロイドであるニコチンとノルニコチンの総量をいい、「アルカロイド含量を調節する機能」には、ニコチンまたはノルニコチンのいずれかの含量を調節(増加または低減)する活性、および、ニコチンとノルニコチン両方の含量を調節(増加または低減)する活性を含むものとする。
【0018】
上記のホモログ遺伝子は、配列番号1に示す塩基配列からなるポリヌクレオチドを含む遺伝子を当該技術分野で公知の手法によって改変することによって得ることができる。遺伝子に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法またはこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-K(TAKARA-BIO社)やMutant-G(TAKARA-BIO社))などを用いて、あるいは、TAKARA-BIO社のLA PCR in vitro Mutagenesis シリーズキットを用いて変異が導入される。
【0019】
本発明の遺伝子を取得する方法は特に限定されるものではなく、一般的な方法が採用される。例えば、本発明の遺伝子を、それを有する生物のゲノムDNA、cDNAライブラリーなどから適切な制限酵素で切り出し、精製すればよい。本発明の遺伝子のゲノムDNAは、例えば、植物細胞または組織からゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作製し、このライブラリーから、配列番号1に示す塩基配列を基に設計したプローブあるいはプライマーを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションあるいは3次元PCRを行うことにより取得することができる。
【0020】
また、一端、本発明の遺伝子の塩基配列が確定されると、その後は化学合成によって、または本遺伝子のcDNAないしゲノムDNAを鋳型としたPCRによって、あるいは該塩基配列を有するDNA断片をプローブとしてハイブリダイズさせることにより本発明の遺伝子を得ることができる。
【0021】
2.植物形質転換用組換えベクター
本発明における植物形質転換用組換えベクターには、Nor#35遺伝子を植物細胞内で発現させるためのベクターとNor#35遺伝子の植物細胞内での発現を抑制させるためのベクターが含まれ、植物細胞内のアルカロイド含量を調節(増減)する上で、目的に応じて使用すればよい。
(1) Nor#35遺伝子発現ベクター
植物形質転換に用いる本発明の組換えベクターは、上記の本発明の遺伝子(以下、「目的遺伝子」ともいう)を適当なベクターに導入することにより構築することができる。ここで、ベクターとしては、アグロバクテリウムを介して植物細胞に目的遺伝子を導入することができる、pBI系、pPZP系、pSMA系のベクターなどが好適に用いられる。特に、バイナリーベクター系(pBI121、pBI101、pBI2113、pBI101.2等)、または中間ベクター系(pLGV23Neo、pNCAT等)のプラスミドが好ましい。バイナリーベクターとは大腸菌(Escherichia coli)およびアグロバクテリウムにおいて複製可能なシャトルベクターで、バイナリーベクターを保持するアグロバクテリムを植物細胞に感染させると、ベクター上にあるT-DNA領域と呼ばれているLB配列およびRB配列ではさまれた部分のDNAを植物の核DNAに組み込むことが可能である(EMBO Journal, 10(3), 697-704 (1991))。一方、植物細胞に遺伝子を直接導入も可能であり、例えばpUC系ベクター(pUC18、pUC19、pUC9等)が利用されている。また、カリフラワーモザイクウイルス(CaMV)、インゲンマメモザイクウイルス(BGMV)、タバコモザイクウイルス(TMV)等の植物ウイルスベクターも用いることができる。
【0022】
バイナリーベクター系プラスミドを用いる場合、上記のバイナリーベクターのT-DNA領域内に、目的遺伝子を含む遺伝子カセットを挿入し、この組換えベクターを大腸菌中で増幅する。次いで、増幅した組換えベクターをアグロバクテリウム・ツメファシエンスの菌株C58、LBA4404、EHA101、EHA105等に、凍結融解法、エレクトロポレーション法等用いて導入し、該アグロバクテリウムを植物細胞の形質転換に用いる。
【0023】
また、上記の方法以外にも、三者接合法(Nucleic Acids Research, 12:8711(1984))によって、植物細胞形質転換用アグロバクテリウムを調製することができる。すなわち、目的遺伝子を含むプラスミドを保有する大腸菌、ヘルパープラスミド(例えば、pRK2013等)を保有する大腸菌、およびアグロバクテリウムを混合培養し、リファンピシリンおよびカナマイシンを含む培地上で培養することにより植物細胞形質転換用のアグロバクテリウムを得ることができる。
【0024】
ベクターに目的遺伝子を挿入するには、まず、精製されたDNAを適当な制限酵素で消化し、適当なベクター DNAの制限酵素部位またはマルチクローニングサイトに挿入する方法などが採用される。
【0025】
また、組換えベクターには、目的遺伝子以外にプロモーター配列、エンハンサー配列、ターミネーター配列、ポリA付加シグナル、5'-UTR配列、選抜マーカー遺伝子、レポーター遺伝子、必要に応じて、アグロバクテリウム細菌中で増幅するための複製開始点などを配置することができる。
【0026】
「プロモーター」としては、植物細胞において機能し、植物の特定の組織内あるいは特定の発育段階において発現を導くことのできるDNAであれば、植物由来のものでなくてもよい。具体例としては、カリフラワーモザイクウイルス(CaMV)由来の35SRNAプロモーター、アグロバクテリウム由来のノパリン合成酵素遺伝子のプロモーター(Pnos)、トウモロコシ由来のユビキチン遺伝子プロモーター、イネ由来のアクチン遺伝子プロモーター、タバコ由来のPathogenesis related (PR)タンパク質遺伝子のプロモーター等が挙げられる。
【0027】
エンハンサー配列としては、例えば、目的遺伝子の発現効率を高めるために用いられ、CaMV35Sプロモーター内の上流側の配列を含むエンハンサー領域などが挙げられる。
【0028】
ターミネーター配列としては、プロモーター配列により転写された遺伝子のmRNA合成を終結できる配列であればよく、例えば、ノパリン合成酵素(NOS)遺伝子のターミネーター、オクトピン合成酵素(OCS)遺伝子のターミネーター、CaMV 35S RNA遺伝子のターミネーター等が挙げられる。
【0029】
選抜マーカー遺伝子としては、例えば、アンピシリン耐性遺伝子(Amp)、ネオマイシン耐性遺伝子(NTPII)、ハイグロマイシン耐性遺伝子(htp)、ビアラフォス耐性遺伝子(Bar)遺伝子、クロラムフェニコールアセチルトランスフェラーゼ(CAT)などが挙げられる。これらの選抜マーカー遺伝子を利用して例えば、アンピシリン、ネオマイシン、ハイグロマイシ、ビアラフォス、クロラムフェニコール等の選抜薬剤を含む培地上で目的遺伝子が導入された組換え体を簡単に選抜できるようになる。
【0030】
また、選抜マーカー遺伝子は上記のように目的遺伝子とともに同一の組換えベクター上に配置してもよい。あるいは、選抜マーカー遺伝子を目的遺伝子とそれぞれ別々の組換えベクター上に配置してもよい。別々に配置した場合は、各ベクターを宿主に同時導入(コトランスフェクト)する。
【0031】
(2) Nor#35遺伝子発現抑制用ベクター
本発明にかかる、Nor#35遺伝子発現抑制用ベクターは、Nor#35遺伝子の発現抑制を引き起こすための核酸分子を適当なベクターに挿入したもので、たとえば、アンチセンスベクターまたはRNAiベクターなどをいう。「Nor#35遺伝子の発現抑制」には、Nor#35遺伝子の転写物量の減少およびNor#35遺伝子がコードするタンパク質(翻訳産物)の減少が含まれる。Nor#35遺伝子発現抑制用ベクターは、目的遺伝子がNor#35遺伝子の発現抑制を引き起こす核酸分子である以外、基本的に、プロモーター配列等のその他の要素は前項のNor#35遺伝子発現ベクターと同じである。
【0032】
アンチセンスベクターとは、アンチセンスRNA法にてNor#35遺伝子の発現抑制を引き起こす核酸分子を含むベクターをいう。アンチセンスRNA法とは、目的遺伝子の転写産物と相補的なRNA(アンチセンスRNA)を発現させて、該アンチセンスRNAを目的遺伝子の転写産物と相補的に結合させることによって該遺伝子の発現を抑制する方法をいう。具体的には、植物細胞で機能する前記のプロモーター配列の下流に、Nor#35遺伝子のアンチセンス配列をつないで植物細胞に導入すると、上記mRNAと相補的なアンチセンスRNAを産生することができる。
【0033】
アンチセンスベクターに導入するNor#35遺伝子の発現を抑制する核酸分子は、例えば、Nor#35遺伝子の塩基配列(配列番号1)の全長のアンチセンス核酸分子であってもよいが、当該塩基配列中の連続した100塩基以上、好ましくは500塩基以上、より好ましくは1000塩基以上からなる塩基配列を有するアンチセンス核酸分子でもよい(特開昭60-232062号、特開平2000-23685号)。
【0034】
一方、RNAiベクター(特開2002-51602号、特開2004-52642号)は、RNAiを引き起こす二本鎖RNA(double-strand RNA、dsRNA)を発現するベクターである。前記のRNAiを引き起こすdsRNAは、二本鎖RNA特異的なRNase(Dicer)により21〜25塩基のsiRNAに切断され、その後RNA-induced silencing complex(RISC)と呼ばれる複合体を形成し、最終的にRISCは標的mRNAを塩基配列相同性により認識し分解する。植物のRNAiにはRNAiを引き起こすdsRNAをヘアピン型dsRNAとして発現するベクターが好適に利用される。これは数塩基以上のリンカー(スペーサー)配列の両端にIR(inverted repeat:逆位反復)となるようにdsRNA形成部分に対応したDNA配列を配置し、植物体内で高発現するプロモーターによりヘアピン型dsRNAを転写し、細胞内でsiRNAを産生するシステムである。また、siRNA発現システムには上記のようなヘアピンタイプのほか、タンデムタイプもある。タンデムタイプでは、2つのプロモーターからセンスRNAとアンチセンスRNAが転写され、細胞内でハイブリダイズしてsiRNAを産生する。RNAiを引き起こす二本鎖RNAの配列は、例えば、配列番号1および配列番号3の塩基配列中の連続した21塩基、好ましくは100塩基以上、さらに好ましくは300塩基以上を含む塩基配列およびその相補配列が用いられる。例えばNor#35遺伝子の転写産物の標的となる特定配列は、配列番号4に示す塩基配列に由来するRNAおよびその相補配列からなる。
【0035】
本発明において用いるRNAiベクターとしては、配列番号1に示す塩基配列に相同なDNA配列を、スペーサーをはさんで21塩基以上のIR(inverted repeat:逆位反復)となるように同一のベクター上に含むものが好ましい。
【0036】
このようなRNAiベクターとして、例えば、COMMONWEALTH SCIENTIFIC AND INDUSTRIAL RESEARCH ORGANIZATION 社のベクターが知られている(WO 99/53050、特表2002-511258)。
【0037】
3.形質転換タバコ属植物およびその製造方法
本発明の形質転換タバコ属植物は、上記の組換えベクターを用いて、対象のタバコ属植物を形質転換することによって作出することができる。形質転換タバコ属植物を作出する際には、既に報告され、確立されている種々の方法を適宜利用することができ、その好ましい例として、アグロバクテリウム法、PEG−リン酸カルシウム法、エレクトロポレーション法、リポソーム法、パーティクルガン法、マイクロインジェクション法等が挙げられるが、アグロバクテリウム法が好ましい。アグロバクテリウム法を用いる場合は、プロトプラストを用いる場合、培養細胞を用いる場合、組織を用いる場合がある。
【0038】
プロトプラストを用いる場合は、Tiプラスミドと組換えベクターをもつアグロバクテリウムと共存培養する方法、スフェロプラスト化したアグロバクテリウムと融合する方法(スフェロプラスト法)、培養細胞を用いる場合は、Tiプラスミドと組換えベクターをもつアグロバクテリウムと共存培養する方法、組織を用いる場合は、対象植物の滅菌した葉片に同様にTiプラスミドと組換えベクターをもつアグロバクテリウムを感染させる方法等により行うことができる。
【0039】
植物体が形質転換されたか否かの確認は、PCR法、サザンハイブリダイゼーション法、ノーザンハイブリダイゼーション法、ウェスタンブロッティング法等により行うことができる。例えば、導入した遺伝子配列に特異的なプライマーを設計し、植物体から調製したゲノムDNAを鋳型としてPCRを行う。特異的プライマーとは、導入した遺伝子の全長あるいはその一部を増幅可能なプライマーであって、対照由来のDNA、例えば非組換え体由来のDNAを鋳型としてPCRを行った場合、DNA断片を増幅しないあるいは鎖長の異なるDNA断片を増幅するプライマーである。PCRを行った後は、増幅産物をアガロースゲル電気泳動、ポリアクリルアミドゲル電気泳動またはキャピラリー電気泳動等で分離し、臭化エチジウム、SYBR Green液等により染色して検出することにより、形質転換されたことを確認することができる。また、予め蛍光色素等により標識したプライマーを用いてPCRを行い、増幅産物を検出することもできる。さらに、マイクロプレート等の固相に増幅産物を結合させ、蛍光または酵素反応等により増幅産物を確認する方法でもよい。
【0040】
あるいは、種々のレポーター遺伝子、例えばベータグルクロニダーゼ(GUS)、ルシフェラーゼ(LUC)、緑色蛍光タンパク質(GFP)、ベータガラクトシダーゼ(LacZ)等の遺伝子の発現によっても植物細胞が形質転換されたか否かを確認することができる。該レポーター遺伝子を含む発現カセットを目的遺伝子を含む発現カセットと同一ベクター上に配置したベクターを作製し、該ベクターを導入したアグロバクテリムを用いて上記と同様の方法で植物細胞を形質転換させ、該レポーター遺伝子の機能発現より確認できる。
【0041】
また、レポーター遺伝子を含む発現カセットを目的遺伝子とそれぞれ別々の組換えベクター上に配置してもよい。別々に配置した場合は、各ベクターを宿主に同時導入(コトランスフェクト)する。
【0042】
本発明においてタバコ属植物とは、タバコ(Nicotiana)属植物であれば特に限定はされず、例えば、ニコチアナ・シルベストリス(Nicotiana sylvestris)、ニコチアナ・トメントシフォルミス(Nicotiana tomentosiformis)、ニコチアナ・タバカム(Nicotiana tabacum)、ニコチアナ・プランバギニフォリア(Nicotiana plumbaginifolia)、ハナタバコ(Nicotiana affinis)、マルバタバコ(Nicotiana rustica)などが挙げられる。またニコチアナ・タバカム(Nicotiana tabacum)については、紙巻製品の原料となる黄色種、バーレー種、オリエント種、在来種、葉巻タバコの原料に用いられる葉巻種などを含む。
【0043】
本発明において、形質転換の対象とする植物材料としては、例えば、根、茎、葉、種子、胚、胚珠、子房、茎頂、葯、花粉等の植物組織やその切片、細胞、カルス、それを酵素処理して細胞壁を除いたプロプラスト等の植物細胞が挙げられる。
【0044】
また、本発明において形質転換植物体とは、植物体全体、植物器官(例えば根、茎、葉、花弁、種子、果実等)、植物組織(例えば表皮、篩部、柔組織、木部、維管束等)、植物細胞、カルス等のいずれをも意味するものである。
【0045】
得られた形質転換細胞から形質転換体を再生させるためには既知の組織培養法により器官または個体を再生させればよい。このような操作は、植物細胞から植物体への再生方法として一般的に知られている方法により、当業者であれば容易に行うことができる。植物細胞から植物体への再生については、例えば、以下のように行うことができる。
【0046】
形質転換の対象とする植物材料として植物組織、例えばリーフディスクを用いた場合、アグロバクテリウム感染後、これらを無機塩類、ビタミン類、炭素源(エネルギー源としての糖類など)、植物生長調節物質(オーキシン、サイトカイニン等の植物ホルモン)やカナマイシン等の選抜薬剤等を加えて滅菌した再分化固型培地上で適当な光および温度条件の下、培養することによって茎葉を形成させることができる。次に、上記固型培地より植物生長調節物質を除いた培地(発根培地)上で茎葉を培養することにより不定根を誘導し、完全な植物体へと再生することができる。
【0047】
本発明の形質転換植物体は、形質転換処理を施した再分化当代である「T0世代」やT0世代の植物の自殖種子である「T1世代」などの後代植物や、それらを片親にして交配した雑種植物やその後代植物を含む。
【0048】
本発明において、ノルニコチン含量を低減させた形質転換タパコ属植物を得る場合、上記のような通常の植物形質転換体作製の工程に、Nor#35遺伝子の発現を抑制した植物細胞を作製する工程を含む。Nor#35遺伝子の発現抑制する手段としては、例えば、アンチセンスRNA法、RNAi法、遺伝子破壊法、および共抑制法(Napoli, C. et al.(1990), Plant Cell 2, 279-289、Van Der Krol, A. R. et al (1990), Plant Cell 2, 291-299参照)等の方法を挙げることができる。
【0049】
アンチセンスRNA法、およびRNAi法は、それぞれ前項に記載したアンチセンスベクター、およびRNAiベクターをタバコ属植物細胞に導入することにより行うことができる。タバコ属植物へのベクターの導入は、前述したとおり、アグロバクテリウム法、ポリエチレングリコール法、エレクトロポレーション法等を用いることができ、形質転換されたタバコ属植物は、ベクター内に連結されたレポーター遺伝子や選抜マーカー遺伝子により容易に選抜される。
【0050】
Nor#35遺伝子の発現の抑制する他の手段として、触媒活性を有するRNA分子であるリボザイムを用いてもよい。標的のmRNAを切断できるよう設計されたリボザイムは、植物細胞中で転写されるように前述の組換えベクター内のカリフラワーモザイクウイルス(CaMV)35Sプロモーターなどのプロモーターの下流に連結して用いればよい。
【0051】
4.植物中のアルカロイド量を改変する方法
本発明はまた、植物細胞内においてNor#35遺伝子の発現量を調節することによって植物中のアルカロイド量を改変する方法を提供する。具体的には、Nor#35遺伝子の発現量の調節は、Nor#35遺伝子の遺伝子配列を利用して行ってもよいが、Nor#35遺伝子の転写調節因子の遺伝子配列を利用して行ってもよい。Nor#35遺伝子の発現量を調節する転写調節因子としては、Nor#35遺伝子がMH(マレイン酸ヒドラジド)および重炭酸ナトリウム処理によって発現誘導されることから、本薬剤処理によってNor#35遺伝子の発現に先立って発現誘導される転写調節因子等が挙げられる。
【実施例】
【0052】
以下、実施例によって本発明を更に具体的に説明するが、これらの実施例は本発明を限定するものでない。
(実施例1)ノルニコチン型タバコにおけるノルニコチンの薬剤誘導条件の決定
(1) 薬剤処理によるノルニコチン誘導
準同質遺伝子系統であるニコチン型タバコNS17(黄色種の品種Bright Yellow 103の半数体倍加系統)とノルニコチン型タバコB301(NS17のノルニコチン型に変異した系統)を試験材料として用いた。薬剤誘導処理にはマレイン酸ヒドラジド液剤(商品名OMH-K、大塚化学株式会社:以下、「MH」という)および重炭酸ナトリウムを、それぞれ0.2%(V/V)、0.8%(W/V)含む水溶液を用いた。
【0053】
B301(ノルニコチン型)、NS17(ニコチン型)の播種および植物体の栽培は温室内(昼夜温25℃、自然日長)で行った。播種後3週間目の芽生えは、仮植肥土を入れた25穴連結ビニールポット(信和株式会社、28.5cm×28.5cm)に1個体ずつ植え替え、2週間育苗した後、直径約12.5cmの素焼き鉢(4寸鉢)に各個体を移植した。移植後3週間目にノルニコチンを誘導するために、上記のMHおよび重炭酸ナトリウムを含む水溶液を植物体の地上部全体に均一に噴霧した(以下、本処理を「薬剤誘導処理」という)。薬剤誘導処理後5日間に渡り、毎日処理葉のサンプリングを行い、ガスクロマトグラフィーによるノルニコチンおよびニコチンの定量分析をアルカロイド分析法(Hybrid Method)にて行った(Chen, C. et al.(2005),Beitrage Tabakforschung 21, 369-379)。
【0054】
(2) 薬剤誘導後のノルニコチン蓄積パターンの経時的解析
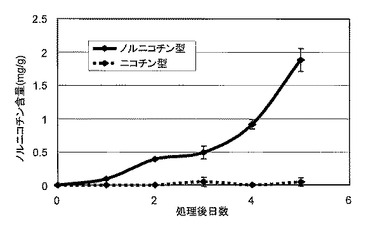
本試験にはB301(ノルニコチン型)、NS17(ニコチン型)それぞれ21個体を供試し、毎日、各3個体から展開した緑葉をサンプリングした。収穫葉(全葉)は熱風乾燥(70℃、約1日間)した後、直ぐにコーヒーミルで粉砕し、デシケーター内で室温保存した。本粉砕試料をノルニコチン分析に供試した。B301(ノルニコチン型)とNS17(ニコチン型)の薬剤誘導処理後の葉中ノルニコチン含量の経時的変化を図1に示す。ノルニコチン型では薬剤誘導処理後2日後には明らかなノルニコチンの蓄積が観察され始め、以降ノルニコチン含量の増加が確認されたが、ニコチン型では薬剤誘導処理後5日間経過してもノルニコチンの蓄積は認められなかった。本結果から、ノルニコチン蓄積の初期段階にある薬剤誘導処理後2-4日目の植物体から下記AFLP-DD解析に供試するmRNAを調製することとした。
【0055】
(実施例2)AFLP-DD解析
(1) AFLP-DD解析用試料の調製およびmRNAの単離
実施例1に記載した方法に従ってタバコ植物体を栽培し、薬剤誘導処理2-4日目の葉のラミナ部分を収穫し、速やかに液体窒素中で凍結後、マイナス80℃フリーザーで保存した。この凍結試料を全RNAおよびmRNAの単離に供した。全RNAについてはRNA WIZ (Ambion社) とPURE SCRIPT(Gentra社)を、mRNAについてはGene Elute mRNA Miniprep kit(SIGMA社)を用いてキットの指示書に従って単離を行った。その結果B301より45μg、NS17より42.5μgのmRNAを精製した。
【0056】
(2) AFLP-DD解析
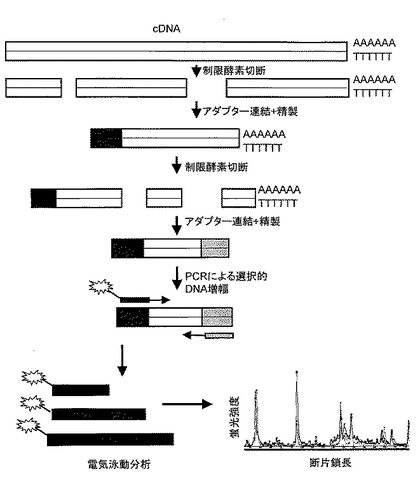
Messenger Scape社(東京都渋谷区幡ヶ谷3-20-2 TSビル101)にB301より調製したmRNA18μg、NS17より調製したmRNA 17μgを送付し、AFLP-DD解析を行った。AFLP-DD解析(Fukumura, R. et al. (2003) Nucleic. Acids. Res. 31, 16)の概要を図2に示す。AFLP-DD解析は、1)poly(A)+RNAからcDNAの合成、2)制限酵素Msp Iによる消化、3)Msp Iアダプター配列の付加、4)制限酵素Mse Iによる消化、5)Mse Iアダプター配列の付加、6)蛍光標識したプライマーを用いたPCRによるDNAの増幅、7)ABI PRISM 3100キャピラリー電気泳動装置(アプライドバイオシステムズ社)を用いた電気泳動によるPCR産物の分離およびピークの検出、8)フラグメントアナリシスソフトウェアGeneScan(アプライドバイオシステムズジャパン社)を用いた遺伝子発現プロファイル(横軸にDNA断片の推定鎖長を、縦軸に遺伝子発現量の指標となる各遺伝子断片の蛍光強度をピークとして表示したもの)および各DNA断片の蛍光強度の2サンプル間での比較データの自動作成、といった一連の作業から構成される。
【0057】
(3) ノルニコチン型タバコとニコチン型タバコで発現量の異なる遺伝子の一次選抜
AFLP-DD解析の結果、B301(ノルニコチン型)で発現量が高い275個の遺伝子断片、NS17(ニコチン型)で発現量が高い221個の遺伝子断片を見出した。合計496個の遺伝子断片のうち、蛍光強度比が2倍以上であった19個の遺伝子断片(下記表1記載のA1〜A9、A11〜A20)を選抜し、これらの塩基配列をMessenger Scape社にて決定した。
【0058】
【表1】

【0059】
Messenger Scape社によるフラグメントアナリシスソフトウェアGeneScanによる自動解析結果に加え、葉たばこ研究所においても遺伝子発現プロファイルの目視による解析を行った。何故なら前者の自動解析ではB301(ノルニコチン型)あるいはNS17(ニコチン型)のどちらか片方においてほとんどピークが検出されない場合、そうしたピークは自動選抜されてこないからである。前記の遺伝子発現プロファイルを目視で再解析した結果、ニコチン型でのみピークが検出される遺伝子断片(ニコチン型特異的断片)を15個(下記表2記載のB16〜B30)、ノルニコチン型でのみピークが検出される遺伝子断片(ノルニコチン型特異的断片)を8個(下記表3記載のC31〜C38)の合計23個を見出した。これら遺伝子断片の塩基配列解析をMessenger Scape社にて決定した。
【0060】
【表2】
【0061】
【表3】
【0062】
塩基配列解析はダイレクトシーケンス法にて行った。すなわち、AFLP-DD解析の過程で得られたPCR産物をポリアクリルアミドゲル電気泳動で分離し、選抜した各ピークの鎖長に対応する遺伝子断片を切り取り、ゲル中のDNAを溶出してPCRにて増幅後、シーケンス反応、シーケンス解析をし、塩基配列を決定した。決定した塩基配列をもとに、下記実施例3で用いる定量PCR解析用のプライマーを設計した。
【0063】
(実施例3)定量PCRによる遺伝子発現の再確認
(1) 薬剤誘導処理によりノルニコチン蓄積を誘導したタバコ(幼苗個体および鉢上げ個体)の調製
B301(ノルニコチン型)とNS17(ニコチン型)の1組み合わせ以外に、タバコ品種である、Bright Yellow103 (BY103)、Bright Yellow4 (BY4)、Burley21 (B21) のそれぞれのノルニコチン型とニコチン型を加えた合計4組の遺伝資源を温室内(昼夜温25℃、自然日長)で栽培した。幼苗個体のノルニコチン解析には、播種後3週間目の芽生えを36穴連結ビニールポット(信和株式会社製、28.5cm×28.5cm)に1個体ずつ植え替えた後1週間栽培し、薬剤誘導処理(MHおよび重炭酸ナトリウムを含む水溶液を約200μl/個体ずつ噴霧)後5日目にサンプリングした葉を供試した。鉢上げ個体のノルニコチン解析には、実施例1に記載の方法に従って植物体を4寸鉢で栽培し、薬剤誘導処理(MHおよび重炭酸ナトリウムを含む水溶液を植物体の地上部全体に均一に噴霧した)後5日目にサンプリングした葉を供試した。これらサンプリングした葉は実施例1に記載の方法に従い熱風乾燥および粉砕後、アルカロイド分析法(Hybrid Method)によりノルニコチン含量を解析した。その結果、いずれの遺伝資源においても薬剤誘導処理によってノルニコチン型においてのみノルニコチン蓄積が認められた。よって、本実験において用いた材料および方法(遺伝資源、栽培方法、薬剤誘導処理)を用いて、以下に述べる定量PCR解析用のサンプルを調製した。但し、薬剤誘導処理後5日目の葉はサンプリング後、熱風乾燥は行わず、速やかに液体窒素中で凍結後マイナス80℃のフリーザーで保存した。
【0064】
(2) 定量PCR解析による候補遺伝子の二次選抜
上記4組のノルニコチン型とニコチン型の幼苗個体および鉢上げ個体から調製した薬剤誘導処理後5日目の葉(マイナス80℃のフリーザーで保存した葉)から実施例2に記載の方法に従って全RNAを単離した。これら全RNAを鋳型として、B301(ノルニコチン型)およびNS17(ニコチン型)で発現量の異なった42個の遺伝子断片(表1、表2、表3)の定量PCR解析を行なった。
【0065】
定量PCR解析は、SYBR Green PCR Core Reagents (Applied Biosystems社)を用い、キット添付の指示書に従い、7500リアルタイムPCRシステム(Applied Biosystems社)上で、95℃で10分間処理後、94℃で15秒間および60℃で1分間の処理を40サイクル反応させることによって行った。幼苗個体の解析には、上記4組の遺伝資源に関し、それぞれ1組当りノルニコチン型24個体、ニコチン型24個体を供試し、また鉢上げ個体の解析には、それぞれ1組当りノルニコチン型6個体、ニコチン型6個体を供試した。また幼苗個体の解析では、12個体由来の全ての葉を混合したものを1サンプルとし、薬剤誘導処理直前(0日目)および薬剤誘導処理後5日目の植物体から各1サンプルずつ調製し、全RNA抽出に供試した。また鉢上げ個体の解析では、3個体由来の全ての葉を混合したものを1サンプルとし、薬剤誘導処理直前(0日目)および薬剤誘導処理後5日目の植物体から各1サンプルずつ調製し、全RNA抽出に供試した。一次選抜した42遺伝子断片の定量PCR解析を行った結果、4組全ての遺伝資源において、幼苗個体および鉢上げ個体を問わず、薬剤誘導処理5日目においてニコチン型と比較してノルニコチン型で5倍以上のmRNAの発現量が高い遺伝子断片4個を二次選抜した(下記表4)。これら4個の遺伝子断片のうち、3個は表1記載のピーク番号A2、A3およびA4、残りの1個は表3記載のピーク番号C35に対応する遺伝子断片であった。なお、薬剤誘導処理直前のノルニコチン型およびニコチン型では、これら4個の遺伝子断片の発現量は非常に低く、かつニコチン型とノルニコチン型で発現量の差は認められなかった。
【0066】
【表4】
【0067】
二次選抜した4個の遺伝子断片のうち、ピーク番号C35に対応する遺伝子断片(以下、「C35遺伝子断片」という)について更に詳細な解析を以降の実施例にて行った。C35遺伝子断片の塩基配列を配列番号7に示す。なお、C35遺伝子断片の定量PCR解析には下記の配列番号5のプライマーと配列番号6のプライマーを用いた。
配列番号5:Nor#35_ F1
GGTTCAGTCGGTAAATAGTGAACGGT
配列番号6:Nor#35_ R1
TCAGCGCAAGATTTCCAATTTGTCCGA
【0068】
(実施例4)Nor#35遺伝子のcDNAおよびゲノムDNAクローンの単離および構造解析
(1) Nor#35遺伝子の完全長cDNAの単離と塩基配列の決定
B301(ノルニコチン型)の全RNAより5'/3'RACE法(Loh,Y. et al. (1989) Science 243, 217;Ohara et al. (1989) Proc. Natl. Acad. Sci USA 86, 5673;Frohmann, M. (1994) PCR Methods and Applications, 4, 540-558)を用いて、C35遺伝子断片の塩基配列をその一部に有するcDNAの全長配列(以下、「Nor#35遺伝子」という)を明らかにした。5'/3'RACE法に用いたオリゴプライマーはC35遺伝子断片の塩基配列をもとに化学合成した(配列番号8〜13)。
【0069】
配列番号8:GSP1(3') primer
GGTTCAGTCGGTAAATAGTGAACGGT
配列番号9:GSP1(5') primer
TCAGCGCAAGATTTCCAATTTGTCCGA
配列番号10:GSP2(5') primer
CCGTTCACTATTTACCGACTGAACCCAT
配列番号11:GeneRacerTM 3' Primer(Invitrogen社)
GCTGTCAACGATACGCTACGTAACG
配列番号12:GeneRacerTM 5’Primer(Invitrogen社)
CGACTGGAGCACGAGGACACTGA
配列番号13:GeneRacerTM 5’Nested Primer(Invitrogen社)
GGACACTGACATGGACTGAAGGAGTA
【0070】
5'/3'RACEは、以下の手順で行った。まず、実施例1に記載の方法に従って温室栽培したノルニコチン誘導処理後3,4,5日目のB301(ノルニコチン型)の鉢上げ個体から全RNAを抽出した。次に、各全RNAを5μgずつ混合し、そのうち2μgを下記の1本鎖cDNA合成に用いた。1本鎖cDNA の合成はGeneRacerTMキット(Invitrogen社)を使用し、キットの指示書に従って行った。最終的に20μlの鋳型DNA溶液を得た。続いて、以下の条件で1st-PCRを行った。配列番号8のオリゴプライマー(15pmol)および配列番号11のオリゴプライマー(45pmol)を用いて、上記で得た鋳型DNA溶液1μlからcDNAの3’末端までの領域を含む1st-PCR産物(以下、「3’側1st-PCR産物」という)を増幅した。増幅反応(50μlの反応系)は、PfuUltraTM High-Fidelity DNA Polymerase (Stratagene社)およびGeneAmpTM PCR System 9700(Applied Biosystems社)を用い、以下のプログラムで行った。すなわち、95℃で2分間処理した後、95℃で30秒間および72℃で2分間の処理を5サイクル実施、95℃で30秒間および70℃で2分間の処理を5サイクル実施、95℃で30秒間および68℃で2分間の処理を25サイクル実施、72℃でさらに10分間の処理、からなるプログラムである。同様にして、配列番号9のオリゴプライマー(15pmol)および配列番号12のオリゴプライマー(45pmol)を用いて、cDNAの5’ 末端まで領域を含む1st-PCR産物(以下、「5’側1st-PCR産物」という)を増幅した。さらに,5'RACE では、5’ 側1st-PCR産物1μl、配列番号10のオリゴプライマー(15pmol)および配列番号13のオリゴプライマー(15pmol)を用いて、cDNAの5’側末端まで領域を含むnested-PCR産物(以下「5’側nested-PCR産物」という)を、以下のプログラムで増幅した。すなわち、95℃で2分間処理した後、95℃で30秒間および65℃で2分間の処理を25サイクル実施、72℃でさらに10分間の処理、からなるプログラムである。3’側1st-PCR産物および5’側nested-PCR産物を1.5%アガロースゲルで電気泳動したところ、それぞれ約500bp、約1200bpのバンドを確認した。
【0071】
次に、前述の3’側1st-PCR産物(約500bp)および5’側nested-PCR産物(約1200bp)を、おのおのクローニングベクターpCR 4Blunt-TOPO(Invitrogene社)に挿入した後、大腸菌TOP10株にエレクトロポレーション(Gene Puiser Xcell エレクトロポレーションシステムBIO-RAD社)法を用いて導入した。導入後、大腸菌は50mg/lアンピシリンを含むLuria Bertani (以下、「LB」という)固形培地(寒天1.5%)上で培養し、得られたアンピシリン耐性のコロニー約10個をLB(50mg/lアンピシリンを含む)液体培地中で終夜37℃で振とう培養した。本培養物からQIAprep Spin Miniprep Kit (QIAGEN社)を用いて指示書に従いプラスミドDNAを精製し、以下に述べるシークエンス解析の鋳型DNAとして用いた。BigDyeTMTerminator v3.1 Cycle Sequencing Kit(Applied Biosystems社)および3700 DNA Analyzer(Applied Biosystems社) を用いて、キットおよび装置の指示書に従ってシークエンス解析を実施した結果、3’側1st-PCR産物および5’側nested-PCR産物の塩基配列を決定することができた。また、Nor#35遺伝子の5’側末端までの領域を含む約1200bpのDNAが挿入されたプラスミド(以下、「プラスミド#35_5Race_05」という)は後にRNAiコンストラクトの構築に使用した。
【0072】
(2) Nor#35遺伝子がコードするアミノ酸配列の推定
3’側1st-PCR産物および5’側nested-PCR産物の塩基配列、および配列番号7記載の塩基配列を塩基配列解析ソフトウェアATGC, GENETYX(ゼネティックス社)を用いて結合処理した結果、1552bpからなるNor#35遺伝子の全長cDNA配列(配列番号1、 Nor#35_cDNA)を決定することができた。また、Nor#35遺伝子がコードするアミノ酸配列(配列番号2 、Nor#35_protein)も推定した。Nor#35遺伝子のアミノ酸配列(配列番号2)は、BLASTによる配列比較解析により、Phytophthora infestansの感染により誘導されるSolanum tuberosumのjasmonic acid 2翻訳産物(ACCESSION No.AAU12055)およびjasmonic acidに応答するLycopersicon esculentumのacid 2翻訳産物(ACCESSION No.AAF04915)と約70%の相同性があることが判明した。しかしながら、これら翻訳産物やこれら翻訳産物をコードする遺伝子の機能は解明されていない。
【0073】
(3) Nor#35遺伝子のゲノムDNAクローンの単離と塩基配列の決定
Nor#35遺伝子のゲノムクローン配列を葉たばこ研究所保有のBAC(Bacterial Artificial Chromosome)ライブラリーから単離した。該研究所保有のBACライブラリーは、タバコ品種BY4号のゲノムDNA由来で、平均約150Kbのインサートを有す約10万個のBACプラスミドから構成されている(Noguchi et al. (1999) Mol Gen Genet.262,822-829)。配列番号1記載のNor#35遺伝子配列を基に下記の配列番号14 (#35_F2)および配列番号15 (#35_R1)のプライマーを用いて3次元PCR法により上記BACライブラリーをスクリーニングした。その結果、2個のBACクローン(289F6および902D10)を選抜した。
【0074】
配列番号14:Nor#35_F2
TGGGTTCAGTCGGTAAATAGTGAACGGT
配列番号15:Nor#35_R1
TCAGCGCAAGATTTCCAATTTGTCCGA
【0075】
これら2個のBACクローンからBACプラスミドをQIAGEN Large-Construct Kit(QIAGEN社)を用いて各々精製し、そのうち各1μgのプラスミドをシークエンス解析の鋳型として用いた。BACプラスミドのインサートDNAのシークエンス解析は、BigDyeTMTerminator V3.1 Cycle Sequencing Kit(Applied Biosystems社)、3700 DNA Analyzer(Applied Biosystems社)、および下記に示す9種のプライマー(配列番号16〜24)を用い、キットおよび装置の指示書に従って行った。
【0076】
配列番号16:Nor#35_F6
CTACTTAGGATGACAAACAGCAGGAT
配列番号17:Nor#35_F7
CAAGATTTTGTTCATGTAACTGAGCAC
配列番号18:Nor#35_F8
TGCTGCCAAGAAGAATAATGGGTGTTCA
配列番号19:Nor#35_F9
AGTGCTCTTGTATAGCTACGGGTT
配列番号20:Nor#35_R2
CCGTTCACTATTTACCGACTGAACCCAT
配列番号21:Nor#35_R4
ATCTAGAAAATGACTACACGTTGCCGA
配列番号22:Nor#35_R5
GCCAGCAACTTTGCGACACAAGT
配列番号23:Nor#35_R6
AACCCGTAGCTATACAAGAGCACT
配列番号24:Nor#35_R7
TCCGTCAATTTCATTGTACTCTGCAT
【0077】
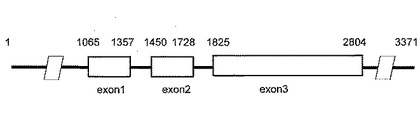
シークエンス解析により得た配列データを塩基配列結合ソフトウェアATGC, GENETYX(ゼネティックス社)を用いて結合処理することにより、最終的に3371bpのゲノムクローン配列(配列番号3)を得た。Nor#35遺伝子のゲノムクローン配列(配列番号3)と全長cDNA配列(配列番号1)を塩基配列解析ソフトウェアATGC, GENETYX(ゼネティックス社) (ATGC, GENETYX; ゼネティックス社)を用いて解析した結果、Nor#35遺伝子のゲノムクローン配列は,3つのエキソンと2つのイントロンを含む構造を持つことが分かった(図3)。
【0078】
(実施例5)Nor#35遺伝子の機能解析(遺伝子発現の抑制)
(1) RNAiベクターpSP107_Nor#35の構築
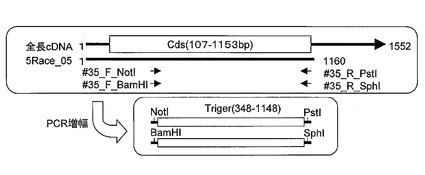
RNAiにおいては植物細胞内で導入遺伝子が転写されdsRNAを形成する必要性がある。dsRNAを形成する配列として配列番号4に示したDNA配列を選択し、本配列の5’および3’末端に異なる制限酵素部位を導入した2種類のDNA断片をPCR法で調製した。プラスミド#35_5Race_05を鋳型DNAとして配列番号25および26に示した#35_F_NotIと#35_R_PstIをプライマーとして、同様にプラスミド#35_5Race_05を鋳型DNAとして配列番号27および28に示した#35_F_BamHIと#35_R_SphIをプライマーとして、図4に示した2種のDNA断片をPCR法で増幅した。
【0079】
配列番号25:Nor#35_F_NotI
ACATAGCGGCCGCGAATGGATCACGACCAAACAGAGTAGCTG
配列番号26:Nor#35_R_PstI
ACTACTGCAGTTACCGACTGAACCCATATTTACTGTTTGGGT
配列番号27:Nor#35_ F_BamHI
CTATGGATCCGAATGGATCACGACCAAACAGAGTAGCTG
配列番号28:Nor#35_ R_SphI
TGTAGCATGCTTACCGACTGAACCCATATTTACTGTTTGGGT
【0080】
増幅した2種のDNA断片は,それぞれ5'/3'末端に、プライマーにより付与されたNotI/PstI、または、BamHI/SphIの制限酵素認識配列を持つ(図4)。増幅した2種のDNA断片は、それぞれこれら制限酵素で二重消化した後MinElute kit(QIAGEN社)で精製し、以下の実験に供試した。
【0081】
逆向反復配列(即ち、特定の配列に関し、正方向配列とその逆方向配列をタンデムに結合)を構築するためのベクターとして、pDNR-1(Clontech社)を以下に示すように改変したpSP148(図5)を利用した。pSP148(図5)の2個のloxP部位の間には、35Sプロモーター,マルチクローニングサイト1(MCS1),スペーサー配列,マルチクローニングサイト2(MCS2),NOSターミネーターがこの順番で存在する。このベクターのMCS1のPstI/NotI制限酵素認識部位およびMCS2のBamHI/SphI制限酵素認識部位に上記の制限酵素で二重消化後精製したDNA断片を挿入することによりpSP148_Nor#35FRを作製した(図6)。
【0082】
次に、pSP148_Nor#35FR上の2個のloxP部位にはさまれた領域を、Cre-loxP組換え(Saue, 1994, Curr. Opin. Biotechnol. 5:521-527;Abremski et al., 1984, J. Biol. Chem. 259:1509-1514)を利用してバイナリーベクターpSP107(図7)上のloxP部位に導入して、RNAiベクターpSP107_Nor#35を作製した(図8)。バイナリーベクターpSP107(図7)はpBI121(Clontech社)のHindIII-Sse8387I 部位にproK-loxP断片(Creator Acceptor Vector Construction Kit, Clontech社) を導入して作製したものである。また、Cre-loxP組換えはCre Recombinase kit(Clontech社)を用い、その指示書に従って行った。本RNAiベクター、pSP107_Nor#35をAgrobacterium tumefaciens LBA4404に導入し、以下の形質転換実験に使用した。
【0083】
(2)形質転換用アグロバクテリウムの調製
RNAiベクターpSP107_Nor#35を、Agrobacterium tumefaciens の菌株LBA4404(Hoekeman,A. et al. (1983) Nature 303)に凍結融解法(Ann,G. et al.(1988)Plant Molecular Biology Manual:1-19)により導入し、カナマイシン耐性であるコロニーを選抜し、本コロニーをタバコ植物の形質転換に用いた。また、形質転換当代(T0世代)植物の解析の際に対照として使用する植物を作出するため、カナマイシン耐性遺伝子およびGUS遺伝子のみからなるバイナリーベクターpBI121を導入したAgrobacterium tumefaciens 菌株LBA4404も調製した。
【0084】
(3)タバコの形質転換
温室内(昼夜温25℃、自然日長)で栽培したBY103(ノルニコチン型)の葉組織を使用して形質転換を行った(Komari,T. et al. (1989) ,Thor. Appl. Genet 77,547-552)。実施例1に記載の方法で栽培した移植後3週間目の鉢上げ植物より上位展開葉を採取し、70%(V/V)エチルアルコ−ルおよび1.0%(V/V)次亜塩素酸ナトリウム溶液で表面殺菌し、続いて十分の滅菌水で2回洗浄した後、剃刀を用いて約1cm角の葉片を調製した。この葉片20枚と上記カナマイシン耐性を示したAgrobacterium tumefaciens約108細胞とをLinsmaier and Skoog(Linsmaier, EM. et al.(1965),Physiol Plant 18, 100-127)の無機塩類と30 g/lのスクロースより成る液体培地中で共存培養(28℃で48時間)を行った。その後、葉片をセフォタキシム250 mg/lを含む滅菌水で1回洗浄し、細菌を洗い落とした後、Linsmaier and Skoog(1965)の無機塩類、インド−ル酢酸0.3 mg/l、6-(γ,γ-Dimethylallylamino)purine 10 mg/l、カナマイシン100 mg/l、セフォタキシム250 mg/lおよび寒天0.9%を含む再分化固型培地に置床し、約1ヶ月後、再分化した茎葉をLinsmaier and Skoog(1965)の無機塩類、30g/lのスクロ−ス、カナマイシン100 mg/l、セフォタキシム250 mg/lおよび寒天0.9%を含む発根培地に置床した。さらに1ヶ月後、発根した植物体(T0世代)を閉鎖系温室内で4寸鉢に移植し、以下の実験に供試した。
【0085】
(4)GUS染色による形質転換体の選抜
レポーター遺伝子であるGUS遺伝子の発現を指標として導入遺伝子が安定して発現していると思われる形質転換体を次のようにして選抜した。閉鎖系温室(昼夜温25℃、自然日長)に移植後3週間栽培した形質転換タバコ植物(T0世代)の展開緑葉から、鋏で約1cm角の葉片を切り出した。これら葉片を1.0%(V/V)Ttiton X-100 を含む50mMリン酸緩衝液(pH7.0)中で37℃で1時間インキュベートした後、50mMリン酸緩衝液(pH7.0)で2回洗浄し、1.0mM 5-bromo-4-chloro-3-indolyl-β-D-glucuronide(X-Gluc)、20%(V/V)メタノールを含む50mMリン酸緩衝液(pH7.0)中で37℃で一晩インキュベートした。葉片は99%(V/V)メタノールで洗浄後、発色の有無を最終確認した。その結果、pSP107_Nor#35のT-DNAを導入した60個体(以下「Nor#35 RNAi個体」という)、pBI121のT-DNAを導入した15個体(以下「対照個体」という)に関しGUS遺伝子の発現を確認した。GUS染色の詳細については樋江井ら(Hiei,Y. et al. (1994),Plant. J 6,271-282)の方法に従った。
【0086】
(5) Nor#35遺伝子の発現抑制の確認(T0世代)
次に、Nor#35 RNAi個体(T0世代)において内生のNor#35遺伝子の発現が低減したか否かを定量PCR法で調査した。Nor#35 RNAi個体から24個体、対照個体から4個体を無作為に選び、その展開緑葉から全RNAを抽出した。まず、実施例1に記載の方法に従って薬剤誘導処理した薬剤誘導処理後7日目の展開緑葉から直径約10mmのリーフディスクを採取した。1個体当り展開緑葉1枚からリーフディスク5枚を金属パンチで打ち抜き、RNAlater (Ambion社)に浸漬および保存したものを全RNA抽出用試料とした。全RNAはRNeasy Plant Mini Kit (QIAGEN社)を用いて抽出し、さらに、Omniscript RT Kit(QIAGEN社)を用いて指示書に従い逆転写反応を行い、その反応液の一部を定量PCR解析に供試した。定量PCR に用いるNor#35遺伝子およびβ-actin遺伝子(ハウスキーピング遺伝子)のTaqManTM(Roche Molecular Systems社)プローブとプライマーは、PrimerExpress (Applied Biosystems社)を用いて設計し(配列番号29〜34)、シグマ アルドリッチ ジャパン 株式会社 ライフサイエンス事業部で合成した。なお、リーフディスクを打ち抜いた葉の上位葉2枚のラミナ部分を、リーフディスクを打ち抜いた時に同時に収穫し、実施例1に記載の方法に従い、熱風乾燥後粉砕した試料をノルニコチン含量およびニコチン含量の分析に供試した。
【0087】
配列番号29:Nor#35_TMF_ver3
TTGGGAAAGGGTAAAATGTGAATG
配列番号30:Nor#35_TMR_ver3
TCCCCAACCAAAGAATATTTGC
配列番号31:Nor#35_TMP_ver3(FAM)
TGTTCGGACAAATTGGAAATCTTGCGC
配列番号32:actb_TMF
TCTGGATTGGAGGYTCTATCTTG
配列番号33:actb_TMR
TCGTCATACTCTGCCTTTGCA
配列番号34:actb_TMP(Cy5)
CCTCAGCACCTTCCAGCAGATGTGG
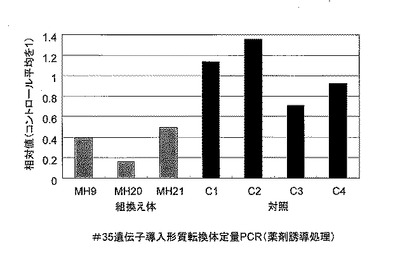
定量PCR解析は実施例3に記載の方法に従って行い、その結果を図9に示した。Nor#35 RNAi個体3個体 (MH9, MH20, MH21)において、内生のNor#35遺伝子の発現が対照個体(平均値)と比較して16-49%のレベルまで低減していた。
【0088】
(6) 薬剤誘導処理したNor#35遺伝子発現抑制個体(T0世代)のアルカロイド分析
Nor#35遺伝子の発現抑制を確認した3個体 (MH9, MH20, MH21)および対照個体3個体のアルカロイド分析を実施した。
【0089】
前述の熱風乾燥後粉砕した試料を用いてアルカロイド含量(ノルニコチン含量とニコチン含量の総量)、ノルニコチン含量およびノルニコチン指数(ノルニコチン含量/全アルカロイド含量×100)を解析した結果を下記表5に示した。対照個体(平均値)と比較してMH9ではアルカロイド含量は53%、ノルニコチン含量は80%、MH20ではアルカロイド含量は23%、ノルニコチン含量は69%、MH21ではアルカロイド含量は35%、ノルニコチン含量は76%、低減していた。
【0090】
【表5】
【0091】
(7) 黄色乾燥したNor#35遺伝子発現抑制個体(T0世代)のアルカロイド分析
Nor#35 RNAi個体のうち薬剤誘導処理を行わなかった20個体、対照個体のうち薬剤誘導処理を行わなかった6個体、を黄色乾燥実験に供試した。閉鎖系温室内で栽培開始後9週間目に芯止めし、その約1週間後に下位葉および中位葉を収穫し、黄色乾燥後、実施例1に記載のアルカロイド分析法(Hybrid Method)によりノルニコチンおよびニコチン含量を解析した。黄色乾燥は下記の13のステップから構成される。
【0092】
ステップ1:乾燥開始後3時間かけて昇温/加湿し、温度38℃/相対湿度100%に到達後、本条件で8時間処理、
ステップ2:次に1時間かけて昇温/排湿し、温度40℃/相対湿度87%に到達後、本条件で15時間処理、
ステップ3:次に1時間かけて昇温/排湿し、温度43℃/相対湿度63%に到達後、本条件で15時間処理、
ステップ4:次に1時間かけて昇温/排湿し、温度45℃/相対湿度52%に到達後、本条件で15時間処理、
ステップ5:次に1時間かけて昇温/排湿し、温度48℃/相対湿度43%に到達後、本条件で15時間処理、
ステップ6:次に1時間かけて昇温/排湿し、温度50℃/相対湿度37%に到達後、本条件で10時間処理、
ステップ7:次に1時間かけて昇温/排湿し、温度53℃/相対湿度31%に到達後、本条件で8時間処理、
ステップ8:次に1時間かけて昇温/排湿し、温度56℃/相対湿度26%に到達後、本条件で6時間処理、
ステップ9:次に1時間かけて昇温/排湿し、温度60℃/相対湿度20%に到達後、本条件で6時間処理、
ステップ10:次に1時間かけて昇温/排湿し、温度63℃/相対湿度16%に到達後、本条件で6時間処理、
ステップ11:次に2時間かけて昇温し、温度65℃/相対湿度16%に到達後、本条件で2時間処理、
ステップ12:次に1時間で昇温し、温度68℃/相対湿度16%に到達後、本条件で8時間処理、及び
ステップ13:5時間かけて、温度30℃/相対湿度60%まで降温/加湿し乾燥を終了。
【0093】
ノルニコチンおよびニコチン含量の分析結果を下記表6に示した。対照個体(平均値)と比較してYC3ではアルカロイド含量は21%、ノルニコチン含量は45%、YC4ではアルカロイド含量は25%、ノルニコチン含量は52%、YC10ではアルカロイド含量は43%、ノルニコチン含量は60%、低減していた。
【0094】
【表6】
【0095】
(8) 薬剤誘導処理したNor#35遺伝子発現抑制個体(自殖T1世代)のアルカロイド分析
T0世代においてアルカロイド含量およびノルニコチン含量の低減を確認した4個体(MH21、YC3、YC4、YC10)につき自殖T1種子を採種後、閉鎖系温室で栽培し、アルカロイド含量およびノルニコチン含量の解析を再度行った。
【0096】
上記選抜した4個体(MH21、YC3、YC4、YC10)のT1世代を実施例3に記載の方法に従って36穴連結ビニールポット2枚に栽培した。MH21のT1世代を72個体、YC3のT1世代を 72個体、YC4のT1世代を72個体、YC10のT1世代を72個体栽培した。36穴連結ビニールポットに仮植後1週間目に各個体から葉の一部をとって前述の方法に従ってGUS染色を行い、導入されたGUS遺伝子のT1世代における分離比検定を行った。検定の結果、YC10には2遺伝子座以上にGUS遺伝子が導入され、残り3個体(MH21、YC3、YC4)にはGUS遺伝子が1遺伝子座に導入されたものと推定された。以後の解析には、1遺伝子座に遺伝子が導入されたと推定されたMH21、YC3およびYC4のT1世代(3系統群)を供試した。
【0097】
次に、上記3系統群(MH21、YC3およびYC4のT1世代)の各個体は、36穴連結ビニールポットに仮植後3週間目に定量PCR解析用サンプルとして葉片を採取した、また葉片採取後同日内に薬剤誘導処理(MHおよび重炭酸ナトリウムを含む水溶液を約200μlずつ噴霧)を行った。植物体地上部に散布された薬剤が乾燥した後、鋏で各ビニールポットを切り離し、各個体間で葉が重ならないように間隔を空けて栽培した。薬剤誘導処理後7日目に、T1世代の各個体からラミナを採取し(全ての緑葉から採取)、実施例1に記載の方法に従って、熱風乾燥および粉砕後ニコチンおよびノルニコチン含量の解析を行った。なお上記3系統群(T1世代)の中で、GUS染色が認められずかつGUS遺伝子発現も定量PCR(配列番号35および配列番号36に示したプライマー及び配列番号37に示したTaqManプローブを使用)で認められなかった個体を対照個体として用いた。以下、YC3由来の対照個体をYC3-cont、YC4由来の対照個体をYC4-cont、MH21由来の対照個体をMH21-contと称する。一方、GUS染色が認められた個体を#35遺伝子発現抑制個体(T1世代)とした。以下、YC3個体由来の#35遺伝子発現抑制個体(T1世代)をYC3-T1、YC4個体由来の#35遺伝子発現抑制個体(T1世代)をYC4-T1、MH21個体由来の#35遺伝子発現抑制個体(T1世代)をMH21-T1と称する。ニコチンおよびノルニコチン含量の解析には5個体由来のラミナをまとめたものを1サンプルとして、MH21-T1、YC3-T1およびYC4-T1については5サンプル、YC3-contは3サンプル、YC4-contは5サンプル、MH21-contは2サンプルを解析に供試した。
【0098】
配列番号35:GUS_TM_F_ver2
GTTGCAACTGGACAAGGCACTA
配列番号36:GUS_TM_R_ver2
TGTCTGGCTTTTGGCTGTGA
配列番号37:GUS_TM_P_ver2_FAM
CTGGCAACCGGGTGAAGGTTATCTCTATGA
【0099】
薬剤誘導処理後7日目のYC3-T1、YC4-T1、MH21-T1、YC3-cont、YC4-cont、MH21-contのアルカロイド含量、ノルニコチン含量およびノルニコチン指数を下記表7に示した。YC3-T1はその対照である YC3-contに比較してアルカロイド含量は12%、ノルニコチン含量46%、YC4-T1はその対照であるYC4-cont に比較してアルカロイド含量は19%、ノルニコチン含量41%、MH21-T1はその対照であるMH21-cont に比較してアルカロイド含量は27%、ノルニコチン含量41%の低減が認められた。以上のように、Nor#35遺伝子の発現抑制により葉中アルカロイド含量を大きく変化させることなくノルニコチン含量を低減させたタバコ属植物を作出することができた。
【0100】
【表7】
【図面の簡単な説明】
【0101】
【図1】B301(ノルニコチン型)、NS17(ニコチン型)における薬剤処理によるノルニコチン含量の経時変化を示す。
【図2】AFLP-DD法の概要を示す。
【図3】Nor#35遺伝子のゲノム遺伝子の構造を示す。
【図4】Nor#35遺伝子の5’側末端領域を含むプラスミド(Nor #35_5Race_05)を鋳型としてPCR増幅したRNAiのトリガーとなる2種のDNA断片を示す。
【図5】Nor#35遺伝子の逆向反復配列を構築するためのベクターpSP148の構造を示す。
【図6】ヘアピン型dsRNAを発現するカセットを含むpSP148_Nor#35FRの構造を示す。
【図7】バイナリーベクターpSP107の構造を示す。
【図8】dsRNAを発現するカセットを含む断片をCre-lox組換えにより導入したバイナリーベクターpSP107_Nor#35の構造を示す。
【図9】#35組換え体T0世代におけるNor#35遺伝子発現の定量PCRによる解析結果を示す。
【技術分野】
【0001】
本発明はアルカロイド含量を調節する機能を有する遺伝子、および当該遺伝子の発現を抑制することによってノルニコチン含量またはアルカロイド含量が低減された形質転換タバコ属植物等に関する。
【背景技術】
【0002】
タバコ栽培種に含まれている主要なアルカロイドはニコチンとノルニコチンである(Saitoh, F. et al.(1985),Phytochmistry 24, 477-480)。ノルニコチンは根で生合成されたニコチンが葉に移動し、そこで脱メチル化酵素(nicotine N-demethylase)の作用でノルニコチンに変換される。ノルニコチンのタバコに与える影響としては、タバコ黄色種において紅葉(Cherry-red leaf)をもたらす(岡ら、1961, 岡山たばこ試験場報告21,65-71)、あるいは、タバコバーレー種において香りの特性が変わる(生沼ら、1972, 盛岡たばこ試験場報告8,1-10)などが報告されており、いずれもタバコの原料価値を下げる原因となる。また、ノルニコチンはタバコ特異的ニトロソアミン(TSNA)の一種であるN−ニトロソノルニコチン(NNN)の前駆体となるため、タバコ原料中のノルニコチン含量を低減させることが望まれている。
【0003】
本来タバコ栽培種はノルニコチンをほとんど含まないニコチン型タバコである。ニコチン型タバコは2対のノルニコチン劣性遺伝子を持つが、その自殖後代でそのうち1対が優性遺伝子に突然変異し、前述の問題を有するノルニコチン型個体が出現すると考えられている。また、このニコチン型タバコからノルニコチン型タバコへの変異率は0.8%と極めて高いこともわかっている(Wernsman, E. et al.(1970), Tabaccc Science 12, 226-228)。
【0004】
タバコ葉中のノルニコチン含量を低減する試みとして、まずノルニコチン型に変異した個体を可能な限り採種用個体から排除する方法が知られている(非特許文献1)。しかしながら、本方法では多数の個体のノルニコチン解析という煩雑な作業を要する。一方、遺伝子工学的手法を用いてニコチンからノルニコチンへの変換を抑制することも試みられている。例えば、ある種のチトクロームP450遺伝子の発現を抑制することによって、ノルニコチン型タバコのノルニコチン含量を90%以上低減したタバコが作出されている(非特許文献2)。また、quinolate phosphoribosyl transferase (QPTase)遺伝子(NtQPT1)の発現を抑制することによって、非組換え(野生型)タバコと比較してノルニコチン含量を約1/3に低減したという報告がある(非特許文献3)が、この方法ではニコチン含量も1/10以下に低減し、その結果アルカロイド含量が大幅に低減している。アルカロイド含量はタバコの喫味に大きな影響を及ぼすことから、このようなアルカロイド含量の大幅な低減は望ましくない。
【0005】
その他、遺伝子工学的手法を用いてニコチン含量の増減を試みた例もある。例えば、ニコチン含量を増加させる報告としては、Nicotiana sylvestrisにPMTase遺伝子を導入することによってニコチン含量を約40%増加させた例(非特許文献4)、Nicotiana tabacumにVitreoscillaヘモグロンビン(VHb)をコードする遺伝子を導入することによってニコチン含量を34%増加させた例(特許文献1)、チトクロームP450遺伝子の発現を抑制することによってニコチン含量を増加させた例(非特許文献5)、一方、ニコチン含量を低減させる報告としては、ニコチン生合成遺伝子を用いてニコチン含量を低減させる方法(特許文献2)、ADC, ODC, SAMS, PMT, NADHといったニコチン生合成に関与する酵素遺伝子を用いてニコチン含量を低減させる方法(特許文献3、特許文献4)などが挙げられる。
【0006】
【特許文献1】WO 98/12913A
【特許文献2】WO 2002/100199A
【特許文献3】WO 00/67558A
【特許文献4】WO 2005/018307
【非特許文献1】佐藤ら、1982, 磐田たばこ試験場報告14,17-21
【非特許文献2】Siminszky,B. et al. (2005) Proc. Natl. Acad. Sci USA 102, 41,14919-14924
【非特許文献3】Biotechnology:A Tool for Reduced Risk Tobacco Products-The Nicotine Experience from Test Tube to Cigarette Pack, Jiahua Xie et al., Recent Advances in Tobacco Science Vol. 30: Biotechnology: A Tool for Reduced Risk Tobacco Products Symposium Proceedings 58th Meeting Tobacco Science Research Conference, 2004, 17-37
【非特許文献4】Proc. Natl. Acad. Sci. USA. January 2, 2001, Vol.98, No.1, 362-372
【非特許文献5】Gavilano, L. et al. (2006) J.Agric.Food Chem. 54, 9071-9078
【発明の開示】
【発明が解決しようとする課題】
【0007】
従って、本発明の課題は、アルカロイド含量を大きく変化させることなく、ノルニコチン含量を有意に低減させた形質転換タバコ属植物を提供することにある。また本発明のさらなる課題は、アルカロイド含量が調節された形質転換タバコ属植物を提供することにある。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決すべく鋭意研究を重ねた結果、ニコチン型タバコとノルニコチン型タバコの葉において発現量に違いがある遺伝子群の中から、幼苗個体であるか鉢上げ個体であるかを問わず、ノルニコチン型タバコの葉において発現量が高い遺伝子を見出した。また、その遺伝子の発現を抑制するため、RNA干渉 (以下、「RNAi」という)用のベクターを構築してタバコに導入することによって、ノルニコチン含量が低減した形質転換タバコを得ることに成功し、本発明を完成させるに至った。
【0009】
即ち、本発明は以下の発明を包含する。
(1) 以下の(a)〜(d)にポリヌクレオチドからなるNor#35遺伝子。
(a) 配列番号1に示す塩基配列からなるポリヌクレオチド
(b) 配列番号1に示す塩基配列において1若しくは数個の塩基が欠失、置換、若しくは付加された塩基配列からなり、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(c) 配列番号1に示す塩基配列からなるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(d) 配列番号1に示す塩基配列に対して80%以上の相同性を有する塩基配列からなり、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(2) 配列番号3に示す塩基配列を有する(1)に記載のNor#35遺伝子。
(3) (1)または(2)に記載のNor#35遺伝子を含む組換えベクター。
(4) (1)若しくは(2)に記載のNor#35遺伝子または(3)に記載の組換えベクターを導入した形質転換タバコ属植物。
(5) 植物が、植物体、植物器官、植物組織、または植物細胞である、(4)に記載の形質転換タバコ属植物。
(6) 配列番号1に示す塩基配列中の連続した100塩基以上からなる塩基配列をアンチセンス方向に含む、Nor#35遺伝子発現抑制用ベクター。
(7) 配列番号1に示す塩基配列中の連続した21塩基以上の塩基配列と該塩基配列の相補配列とを含む、Nor#35遺伝子発現抑制用ベクター。
(8) 配列番号1に示す塩基配列中の連続した21塩基以上の塩基配列と該塩基配列の相補配列との間にスペーサー配列を挿入した連続配列を含む、Nor#35遺伝子発現抑制用ベクター。
(9) (6)〜(8)のいずれかに記載のNor#35遺伝子発現抑制用ベクターを導入した形質転換タバコ属植物。
(10) 植物が、植物体、植物器官、植物組織、または植物細胞である、(9)に記載の形質転換タバコ属植物。
(11) (1)または(2)に記載のNor#35遺伝子の発現を抑制した植物細胞を作製する工程、および、該植物細胞から植物体を再生させる工程を含む、アルカロイド含量が低減した形質転換タバコ属植物の作製方法。
(12) (1)または(2)に記載のNor#35遺伝子の発現抑制が、RNAi法、アンチセンス法、遺伝子破壊法、および共抑制法から選ばれるいずれかの手法により行なわれることを特徴とする、(11)に記載の方法。
(13) 植物細胞内において(1)または(2)に記載のNor#35遺伝子の発現量を調節することを特徴とする、植物中のアルカロイド量を改変する方法。
【発明の効果】
【0010】
本発明によれば、アルカロイド含量を調節する機能を有する遺伝子が提供される。タバコ属植物において本遺伝子の発現を抑制させることにより、タバコの喫味に影響を与えるアルカロイド含量を大きく変化させることなく、ノルニコチン含量を低減させることが可能となる。従って、本発明は、ノルニコチンに起因するタバコの原料価値を下げる紅葉などの問題が少なく、しかも喫味のよいタバコを製造するのに有用である。
【発明を実施するための最良の形態】
【0011】
1.本発明の遺伝子
本発明の遺伝子であるNor#35遺伝子は、アルカロイド含量を調節する機能を有する遺伝子である。Nor#35遺伝子はノルニコチン型タバコとニコチン型タバコにおいて発現量の異なる遺伝子の1つであり、Amplified fragment length polymorphism Differential Display (以下、「AFLP-DD法」という)によって同定されたものである。
【0012】
Nor#35遺伝子は、配列番号1に示す塩基配列(cDNA配列)を有し、配列番号3に示すゲノムDNA配列を有する。Nor#35遺伝子のオープンリーディングフレーム(ORF)は、配列番号1に示す塩基配列の第107から第1153位までの領域(1047塩基)であり、配列番号2に示すアミノ酸配列(348アミノ酸残基)からなるタンパク質をコードする。従って、本発明の遺伝子には、配列番号1に示す塩基配列の第107から第1153位までの領域を含む限り、いずれの遺伝子も含まれる。
【0013】
本発明の遺伝子は、好ましくは、配列番号1に示す塩基配列からなるポリヌクレオチドからなる遺伝子であるが、当該遺伝子には限定はされず、アルカロイド含量を調節する機能を有する限り、配列番号1に示す塩基配列において1若しくは数個の塩基が欠失、置換、若しくは付加された塩基配列からなるポリヌクレオチド、配列番号1に示す塩基配列からなるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズするポリヌクレオチドであってもよい。
【0014】
ここで、欠失、置換、若しくは付加されてもよい塩基の数としては、部位特異的突然変異誘発法等の公知の変異導入法により欠失、置換、若しくは付加できる程度の数の塩基をいい、好ましくは、1個から数個である。例えば、配列番号1に示す塩基配列の1〜10個、好ましくは1〜5個の塩基が欠失してもよく、配列番号1に示す塩基配列に1〜10個、好ましくは1〜5個の塩基が付加してもよく、あるいは、配列番号1に示す塩基配列の1〜10個、好ましくは1〜5個の塩基が他の塩基に置換してもよい。また、ここにいう「変異」は、主には部位特異的突然変異誘発法等により人為的に導入された変異を意味するが、天然に存在する同様の変異であってもよい。
【0015】
また、ここで、ストリンジェントな条件とは、いわゆる塩基配列に特異的な2本鎖のポリヌクレオチドが形成され、非特異的な2本鎖のポリヌクレオチドが形成されない条件をいう。より具体的には、0.25M Na2HPO4, pH7.2, 7%SDS, 1mM EDTA, 1×デンハルト溶液からなる緩衝液中で温度が60〜68℃、好ましく65℃、さらに好ましくは68℃の条件下で16〜24時間ハイブリダイズさせ、さらに20mM Na2HPO4, pH7.2, 1%SDS, 1mM EDTAからなる緩衝液中で温度が60〜68℃、好ましく65℃、さらに好ましくは68℃の条件下で15分間の洗浄を2回を行う条件をいう。当業者であれば、Molecular Cloning(Sambrook, J. et al., Molecular Cloning :a Laboratory Manual 2nd ed., Cold Spring Harbor Laboratory Press, 10 Skyline Drive Plainview, NY (1989))等を参照することにより、こうしたホモログ遺伝子を容易に取得することができ、また、配列番号1に示す塩基配列との相同性は、FASTA検索やBLAST検索により決定することができる。
【0016】
また、本発明の遺伝子には、配列番号1に示す塩基配列に対して80%以上の相同性を有する塩基配列からなり、かつアルカロイド含量を調節する機能を有するポリヌクレオチドも含まれる。上記80%以上の相同性は、好ましくは85%以上、より好ましくは90%以上、最も好ましくは95%以上の相同性をいう。配列の同一性は、FASTA検索やBLAST検索により決定することができる。
【0017】
ここで、「アルカロイド含量」とは、タバコの主要アルカロイドであるニコチンとノルニコチンの総量をいい、「アルカロイド含量を調節する機能」には、ニコチンまたはノルニコチンのいずれかの含量を調節(増加または低減)する活性、および、ニコチンとノルニコチン両方の含量を調節(増加または低減)する活性を含むものとする。
【0018】
上記のホモログ遺伝子は、配列番号1に示す塩基配列からなるポリヌクレオチドを含む遺伝子を当該技術分野で公知の手法によって改変することによって得ることができる。遺伝子に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法またはこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-K(TAKARA-BIO社)やMutant-G(TAKARA-BIO社))などを用いて、あるいは、TAKARA-BIO社のLA PCR in vitro Mutagenesis シリーズキットを用いて変異が導入される。
【0019】
本発明の遺伝子を取得する方法は特に限定されるものではなく、一般的な方法が採用される。例えば、本発明の遺伝子を、それを有する生物のゲノムDNA、cDNAライブラリーなどから適切な制限酵素で切り出し、精製すればよい。本発明の遺伝子のゲノムDNAは、例えば、植物細胞または組織からゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作製し、このライブラリーから、配列番号1に示す塩基配列を基に設計したプローブあるいはプライマーを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションあるいは3次元PCRを行うことにより取得することができる。
【0020】
また、一端、本発明の遺伝子の塩基配列が確定されると、その後は化学合成によって、または本遺伝子のcDNAないしゲノムDNAを鋳型としたPCRによって、あるいは該塩基配列を有するDNA断片をプローブとしてハイブリダイズさせることにより本発明の遺伝子を得ることができる。
【0021】
2.植物形質転換用組換えベクター
本発明における植物形質転換用組換えベクターには、Nor#35遺伝子を植物細胞内で発現させるためのベクターとNor#35遺伝子の植物細胞内での発現を抑制させるためのベクターが含まれ、植物細胞内のアルカロイド含量を調節(増減)する上で、目的に応じて使用すればよい。
(1) Nor#35遺伝子発現ベクター
植物形質転換に用いる本発明の組換えベクターは、上記の本発明の遺伝子(以下、「目的遺伝子」ともいう)を適当なベクターに導入することにより構築することができる。ここで、ベクターとしては、アグロバクテリウムを介して植物細胞に目的遺伝子を導入することができる、pBI系、pPZP系、pSMA系のベクターなどが好適に用いられる。特に、バイナリーベクター系(pBI121、pBI101、pBI2113、pBI101.2等)、または中間ベクター系(pLGV23Neo、pNCAT等)のプラスミドが好ましい。バイナリーベクターとは大腸菌(Escherichia coli)およびアグロバクテリウムにおいて複製可能なシャトルベクターで、バイナリーベクターを保持するアグロバクテリムを植物細胞に感染させると、ベクター上にあるT-DNA領域と呼ばれているLB配列およびRB配列ではさまれた部分のDNAを植物の核DNAに組み込むことが可能である(EMBO Journal, 10(3), 697-704 (1991))。一方、植物細胞に遺伝子を直接導入も可能であり、例えばpUC系ベクター(pUC18、pUC19、pUC9等)が利用されている。また、カリフラワーモザイクウイルス(CaMV)、インゲンマメモザイクウイルス(BGMV)、タバコモザイクウイルス(TMV)等の植物ウイルスベクターも用いることができる。
【0022】
バイナリーベクター系プラスミドを用いる場合、上記のバイナリーベクターのT-DNA領域内に、目的遺伝子を含む遺伝子カセットを挿入し、この組換えベクターを大腸菌中で増幅する。次いで、増幅した組換えベクターをアグロバクテリウム・ツメファシエンスの菌株C58、LBA4404、EHA101、EHA105等に、凍結融解法、エレクトロポレーション法等用いて導入し、該アグロバクテリウムを植物細胞の形質転換に用いる。
【0023】
また、上記の方法以外にも、三者接合法(Nucleic Acids Research, 12:8711(1984))によって、植物細胞形質転換用アグロバクテリウムを調製することができる。すなわち、目的遺伝子を含むプラスミドを保有する大腸菌、ヘルパープラスミド(例えば、pRK2013等)を保有する大腸菌、およびアグロバクテリウムを混合培養し、リファンピシリンおよびカナマイシンを含む培地上で培養することにより植物細胞形質転換用のアグロバクテリウムを得ることができる。
【0024】
ベクターに目的遺伝子を挿入するには、まず、精製されたDNAを適当な制限酵素で消化し、適当なベクター DNAの制限酵素部位またはマルチクローニングサイトに挿入する方法などが採用される。
【0025】
また、組換えベクターには、目的遺伝子以外にプロモーター配列、エンハンサー配列、ターミネーター配列、ポリA付加シグナル、5'-UTR配列、選抜マーカー遺伝子、レポーター遺伝子、必要に応じて、アグロバクテリウム細菌中で増幅するための複製開始点などを配置することができる。
【0026】
「プロモーター」としては、植物細胞において機能し、植物の特定の組織内あるいは特定の発育段階において発現を導くことのできるDNAであれば、植物由来のものでなくてもよい。具体例としては、カリフラワーモザイクウイルス(CaMV)由来の35SRNAプロモーター、アグロバクテリウム由来のノパリン合成酵素遺伝子のプロモーター(Pnos)、トウモロコシ由来のユビキチン遺伝子プロモーター、イネ由来のアクチン遺伝子プロモーター、タバコ由来のPathogenesis related (PR)タンパク質遺伝子のプロモーター等が挙げられる。
【0027】
エンハンサー配列としては、例えば、目的遺伝子の発現効率を高めるために用いられ、CaMV35Sプロモーター内の上流側の配列を含むエンハンサー領域などが挙げられる。
【0028】
ターミネーター配列としては、プロモーター配列により転写された遺伝子のmRNA合成を終結できる配列であればよく、例えば、ノパリン合成酵素(NOS)遺伝子のターミネーター、オクトピン合成酵素(OCS)遺伝子のターミネーター、CaMV 35S RNA遺伝子のターミネーター等が挙げられる。
【0029】
選抜マーカー遺伝子としては、例えば、アンピシリン耐性遺伝子(Amp)、ネオマイシン耐性遺伝子(NTPII)、ハイグロマイシン耐性遺伝子(htp)、ビアラフォス耐性遺伝子(Bar)遺伝子、クロラムフェニコールアセチルトランスフェラーゼ(CAT)などが挙げられる。これらの選抜マーカー遺伝子を利用して例えば、アンピシリン、ネオマイシン、ハイグロマイシ、ビアラフォス、クロラムフェニコール等の選抜薬剤を含む培地上で目的遺伝子が導入された組換え体を簡単に選抜できるようになる。
【0030】
また、選抜マーカー遺伝子は上記のように目的遺伝子とともに同一の組換えベクター上に配置してもよい。あるいは、選抜マーカー遺伝子を目的遺伝子とそれぞれ別々の組換えベクター上に配置してもよい。別々に配置した場合は、各ベクターを宿主に同時導入(コトランスフェクト)する。
【0031】
(2) Nor#35遺伝子発現抑制用ベクター
本発明にかかる、Nor#35遺伝子発現抑制用ベクターは、Nor#35遺伝子の発現抑制を引き起こすための核酸分子を適当なベクターに挿入したもので、たとえば、アンチセンスベクターまたはRNAiベクターなどをいう。「Nor#35遺伝子の発現抑制」には、Nor#35遺伝子の転写物量の減少およびNor#35遺伝子がコードするタンパク質(翻訳産物)の減少が含まれる。Nor#35遺伝子発現抑制用ベクターは、目的遺伝子がNor#35遺伝子の発現抑制を引き起こす核酸分子である以外、基本的に、プロモーター配列等のその他の要素は前項のNor#35遺伝子発現ベクターと同じである。
【0032】
アンチセンスベクターとは、アンチセンスRNA法にてNor#35遺伝子の発現抑制を引き起こす核酸分子を含むベクターをいう。アンチセンスRNA法とは、目的遺伝子の転写産物と相補的なRNA(アンチセンスRNA)を発現させて、該アンチセンスRNAを目的遺伝子の転写産物と相補的に結合させることによって該遺伝子の発現を抑制する方法をいう。具体的には、植物細胞で機能する前記のプロモーター配列の下流に、Nor#35遺伝子のアンチセンス配列をつないで植物細胞に導入すると、上記mRNAと相補的なアンチセンスRNAを産生することができる。
【0033】
アンチセンスベクターに導入するNor#35遺伝子の発現を抑制する核酸分子は、例えば、Nor#35遺伝子の塩基配列(配列番号1)の全長のアンチセンス核酸分子であってもよいが、当該塩基配列中の連続した100塩基以上、好ましくは500塩基以上、より好ましくは1000塩基以上からなる塩基配列を有するアンチセンス核酸分子でもよい(特開昭60-232062号、特開平2000-23685号)。
【0034】
一方、RNAiベクター(特開2002-51602号、特開2004-52642号)は、RNAiを引き起こす二本鎖RNA(double-strand RNA、dsRNA)を発現するベクターである。前記のRNAiを引き起こすdsRNAは、二本鎖RNA特異的なRNase(Dicer)により21〜25塩基のsiRNAに切断され、その後RNA-induced silencing complex(RISC)と呼ばれる複合体を形成し、最終的にRISCは標的mRNAを塩基配列相同性により認識し分解する。植物のRNAiにはRNAiを引き起こすdsRNAをヘアピン型dsRNAとして発現するベクターが好適に利用される。これは数塩基以上のリンカー(スペーサー)配列の両端にIR(inverted repeat:逆位反復)となるようにdsRNA形成部分に対応したDNA配列を配置し、植物体内で高発現するプロモーターによりヘアピン型dsRNAを転写し、細胞内でsiRNAを産生するシステムである。また、siRNA発現システムには上記のようなヘアピンタイプのほか、タンデムタイプもある。タンデムタイプでは、2つのプロモーターからセンスRNAとアンチセンスRNAが転写され、細胞内でハイブリダイズしてsiRNAを産生する。RNAiを引き起こす二本鎖RNAの配列は、例えば、配列番号1および配列番号3の塩基配列中の連続した21塩基、好ましくは100塩基以上、さらに好ましくは300塩基以上を含む塩基配列およびその相補配列が用いられる。例えばNor#35遺伝子の転写産物の標的となる特定配列は、配列番号4に示す塩基配列に由来するRNAおよびその相補配列からなる。
【0035】
本発明において用いるRNAiベクターとしては、配列番号1に示す塩基配列に相同なDNA配列を、スペーサーをはさんで21塩基以上のIR(inverted repeat:逆位反復)となるように同一のベクター上に含むものが好ましい。
【0036】
このようなRNAiベクターとして、例えば、COMMONWEALTH SCIENTIFIC AND INDUSTRIAL RESEARCH ORGANIZATION 社のベクターが知られている(WO 99/53050、特表2002-511258)。
【0037】
3.形質転換タバコ属植物およびその製造方法
本発明の形質転換タバコ属植物は、上記の組換えベクターを用いて、対象のタバコ属植物を形質転換することによって作出することができる。形質転換タバコ属植物を作出する際には、既に報告され、確立されている種々の方法を適宜利用することができ、その好ましい例として、アグロバクテリウム法、PEG−リン酸カルシウム法、エレクトロポレーション法、リポソーム法、パーティクルガン法、マイクロインジェクション法等が挙げられるが、アグロバクテリウム法が好ましい。アグロバクテリウム法を用いる場合は、プロトプラストを用いる場合、培養細胞を用いる場合、組織を用いる場合がある。
【0038】
プロトプラストを用いる場合は、Tiプラスミドと組換えベクターをもつアグロバクテリウムと共存培養する方法、スフェロプラスト化したアグロバクテリウムと融合する方法(スフェロプラスト法)、培養細胞を用いる場合は、Tiプラスミドと組換えベクターをもつアグロバクテリウムと共存培養する方法、組織を用いる場合は、対象植物の滅菌した葉片に同様にTiプラスミドと組換えベクターをもつアグロバクテリウムを感染させる方法等により行うことができる。
【0039】
植物体が形質転換されたか否かの確認は、PCR法、サザンハイブリダイゼーション法、ノーザンハイブリダイゼーション法、ウェスタンブロッティング法等により行うことができる。例えば、導入した遺伝子配列に特異的なプライマーを設計し、植物体から調製したゲノムDNAを鋳型としてPCRを行う。特異的プライマーとは、導入した遺伝子の全長あるいはその一部を増幅可能なプライマーであって、対照由来のDNA、例えば非組換え体由来のDNAを鋳型としてPCRを行った場合、DNA断片を増幅しないあるいは鎖長の異なるDNA断片を増幅するプライマーである。PCRを行った後は、増幅産物をアガロースゲル電気泳動、ポリアクリルアミドゲル電気泳動またはキャピラリー電気泳動等で分離し、臭化エチジウム、SYBR Green液等により染色して検出することにより、形質転換されたことを確認することができる。また、予め蛍光色素等により標識したプライマーを用いてPCRを行い、増幅産物を検出することもできる。さらに、マイクロプレート等の固相に増幅産物を結合させ、蛍光または酵素反応等により増幅産物を確認する方法でもよい。
【0040】
あるいは、種々のレポーター遺伝子、例えばベータグルクロニダーゼ(GUS)、ルシフェラーゼ(LUC)、緑色蛍光タンパク質(GFP)、ベータガラクトシダーゼ(LacZ)等の遺伝子の発現によっても植物細胞が形質転換されたか否かを確認することができる。該レポーター遺伝子を含む発現カセットを目的遺伝子を含む発現カセットと同一ベクター上に配置したベクターを作製し、該ベクターを導入したアグロバクテリムを用いて上記と同様の方法で植物細胞を形質転換させ、該レポーター遺伝子の機能発現より確認できる。
【0041】
また、レポーター遺伝子を含む発現カセットを目的遺伝子とそれぞれ別々の組換えベクター上に配置してもよい。別々に配置した場合は、各ベクターを宿主に同時導入(コトランスフェクト)する。
【0042】
本発明においてタバコ属植物とは、タバコ(Nicotiana)属植物であれば特に限定はされず、例えば、ニコチアナ・シルベストリス(Nicotiana sylvestris)、ニコチアナ・トメントシフォルミス(Nicotiana tomentosiformis)、ニコチアナ・タバカム(Nicotiana tabacum)、ニコチアナ・プランバギニフォリア(Nicotiana plumbaginifolia)、ハナタバコ(Nicotiana affinis)、マルバタバコ(Nicotiana rustica)などが挙げられる。またニコチアナ・タバカム(Nicotiana tabacum)については、紙巻製品の原料となる黄色種、バーレー種、オリエント種、在来種、葉巻タバコの原料に用いられる葉巻種などを含む。
【0043】
本発明において、形質転換の対象とする植物材料としては、例えば、根、茎、葉、種子、胚、胚珠、子房、茎頂、葯、花粉等の植物組織やその切片、細胞、カルス、それを酵素処理して細胞壁を除いたプロプラスト等の植物細胞が挙げられる。
【0044】
また、本発明において形質転換植物体とは、植物体全体、植物器官(例えば根、茎、葉、花弁、種子、果実等)、植物組織(例えば表皮、篩部、柔組織、木部、維管束等)、植物細胞、カルス等のいずれをも意味するものである。
【0045】
得られた形質転換細胞から形質転換体を再生させるためには既知の組織培養法により器官または個体を再生させればよい。このような操作は、植物細胞から植物体への再生方法として一般的に知られている方法により、当業者であれば容易に行うことができる。植物細胞から植物体への再生については、例えば、以下のように行うことができる。
【0046】
形質転換の対象とする植物材料として植物組織、例えばリーフディスクを用いた場合、アグロバクテリウム感染後、これらを無機塩類、ビタミン類、炭素源(エネルギー源としての糖類など)、植物生長調節物質(オーキシン、サイトカイニン等の植物ホルモン)やカナマイシン等の選抜薬剤等を加えて滅菌した再分化固型培地上で適当な光および温度条件の下、培養することによって茎葉を形成させることができる。次に、上記固型培地より植物生長調節物質を除いた培地(発根培地)上で茎葉を培養することにより不定根を誘導し、完全な植物体へと再生することができる。
【0047】
本発明の形質転換植物体は、形質転換処理を施した再分化当代である「T0世代」やT0世代の植物の自殖種子である「T1世代」などの後代植物や、それらを片親にして交配した雑種植物やその後代植物を含む。
【0048】
本発明において、ノルニコチン含量を低減させた形質転換タパコ属植物を得る場合、上記のような通常の植物形質転換体作製の工程に、Nor#35遺伝子の発現を抑制した植物細胞を作製する工程を含む。Nor#35遺伝子の発現抑制する手段としては、例えば、アンチセンスRNA法、RNAi法、遺伝子破壊法、および共抑制法(Napoli, C. et al.(1990), Plant Cell 2, 279-289、Van Der Krol, A. R. et al (1990), Plant Cell 2, 291-299参照)等の方法を挙げることができる。
【0049】
アンチセンスRNA法、およびRNAi法は、それぞれ前項に記載したアンチセンスベクター、およびRNAiベクターをタバコ属植物細胞に導入することにより行うことができる。タバコ属植物へのベクターの導入は、前述したとおり、アグロバクテリウム法、ポリエチレングリコール法、エレクトロポレーション法等を用いることができ、形質転換されたタバコ属植物は、ベクター内に連結されたレポーター遺伝子や選抜マーカー遺伝子により容易に選抜される。
【0050】
Nor#35遺伝子の発現の抑制する他の手段として、触媒活性を有するRNA分子であるリボザイムを用いてもよい。標的のmRNAを切断できるよう設計されたリボザイムは、植物細胞中で転写されるように前述の組換えベクター内のカリフラワーモザイクウイルス(CaMV)35Sプロモーターなどのプロモーターの下流に連結して用いればよい。
【0051】
4.植物中のアルカロイド量を改変する方法
本発明はまた、植物細胞内においてNor#35遺伝子の発現量を調節することによって植物中のアルカロイド量を改変する方法を提供する。具体的には、Nor#35遺伝子の発現量の調節は、Nor#35遺伝子の遺伝子配列を利用して行ってもよいが、Nor#35遺伝子の転写調節因子の遺伝子配列を利用して行ってもよい。Nor#35遺伝子の発現量を調節する転写調節因子としては、Nor#35遺伝子がMH(マレイン酸ヒドラジド)および重炭酸ナトリウム処理によって発現誘導されることから、本薬剤処理によってNor#35遺伝子の発現に先立って発現誘導される転写調節因子等が挙げられる。
【実施例】
【0052】
以下、実施例によって本発明を更に具体的に説明するが、これらの実施例は本発明を限定するものでない。
(実施例1)ノルニコチン型タバコにおけるノルニコチンの薬剤誘導条件の決定
(1) 薬剤処理によるノルニコチン誘導
準同質遺伝子系統であるニコチン型タバコNS17(黄色種の品種Bright Yellow 103の半数体倍加系統)とノルニコチン型タバコB301(NS17のノルニコチン型に変異した系統)を試験材料として用いた。薬剤誘導処理にはマレイン酸ヒドラジド液剤(商品名OMH-K、大塚化学株式会社:以下、「MH」という)および重炭酸ナトリウムを、それぞれ0.2%(V/V)、0.8%(W/V)含む水溶液を用いた。
【0053】
B301(ノルニコチン型)、NS17(ニコチン型)の播種および植物体の栽培は温室内(昼夜温25℃、自然日長)で行った。播種後3週間目の芽生えは、仮植肥土を入れた25穴連結ビニールポット(信和株式会社、28.5cm×28.5cm)に1個体ずつ植え替え、2週間育苗した後、直径約12.5cmの素焼き鉢(4寸鉢)に各個体を移植した。移植後3週間目にノルニコチンを誘導するために、上記のMHおよび重炭酸ナトリウムを含む水溶液を植物体の地上部全体に均一に噴霧した(以下、本処理を「薬剤誘導処理」という)。薬剤誘導処理後5日間に渡り、毎日処理葉のサンプリングを行い、ガスクロマトグラフィーによるノルニコチンおよびニコチンの定量分析をアルカロイド分析法(Hybrid Method)にて行った(Chen, C. et al.(2005),Beitrage Tabakforschung 21, 369-379)。
【0054】
(2) 薬剤誘導後のノルニコチン蓄積パターンの経時的解析
本試験にはB301(ノルニコチン型)、NS17(ニコチン型)それぞれ21個体を供試し、毎日、各3個体から展開した緑葉をサンプリングした。収穫葉(全葉)は熱風乾燥(70℃、約1日間)した後、直ぐにコーヒーミルで粉砕し、デシケーター内で室温保存した。本粉砕試料をノルニコチン分析に供試した。B301(ノルニコチン型)とNS17(ニコチン型)の薬剤誘導処理後の葉中ノルニコチン含量の経時的変化を図1に示す。ノルニコチン型では薬剤誘導処理後2日後には明らかなノルニコチンの蓄積が観察され始め、以降ノルニコチン含量の増加が確認されたが、ニコチン型では薬剤誘導処理後5日間経過してもノルニコチンの蓄積は認められなかった。本結果から、ノルニコチン蓄積の初期段階にある薬剤誘導処理後2-4日目の植物体から下記AFLP-DD解析に供試するmRNAを調製することとした。
【0055】
(実施例2)AFLP-DD解析
(1) AFLP-DD解析用試料の調製およびmRNAの単離
実施例1に記載した方法に従ってタバコ植物体を栽培し、薬剤誘導処理2-4日目の葉のラミナ部分を収穫し、速やかに液体窒素中で凍結後、マイナス80℃フリーザーで保存した。この凍結試料を全RNAおよびmRNAの単離に供した。全RNAについてはRNA WIZ (Ambion社) とPURE SCRIPT(Gentra社)を、mRNAについてはGene Elute mRNA Miniprep kit(SIGMA社)を用いてキットの指示書に従って単離を行った。その結果B301より45μg、NS17より42.5μgのmRNAを精製した。
【0056】
(2) AFLP-DD解析
Messenger Scape社(東京都渋谷区幡ヶ谷3-20-2 TSビル101)にB301より調製したmRNA18μg、NS17より調製したmRNA 17μgを送付し、AFLP-DD解析を行った。AFLP-DD解析(Fukumura, R. et al. (2003) Nucleic. Acids. Res. 31, 16)の概要を図2に示す。AFLP-DD解析は、1)poly(A)+RNAからcDNAの合成、2)制限酵素Msp Iによる消化、3)Msp Iアダプター配列の付加、4)制限酵素Mse Iによる消化、5)Mse Iアダプター配列の付加、6)蛍光標識したプライマーを用いたPCRによるDNAの増幅、7)ABI PRISM 3100キャピラリー電気泳動装置(アプライドバイオシステムズ社)を用いた電気泳動によるPCR産物の分離およびピークの検出、8)フラグメントアナリシスソフトウェアGeneScan(アプライドバイオシステムズジャパン社)を用いた遺伝子発現プロファイル(横軸にDNA断片の推定鎖長を、縦軸に遺伝子発現量の指標となる各遺伝子断片の蛍光強度をピークとして表示したもの)および各DNA断片の蛍光強度の2サンプル間での比較データの自動作成、といった一連の作業から構成される。
【0057】
(3) ノルニコチン型タバコとニコチン型タバコで発現量の異なる遺伝子の一次選抜
AFLP-DD解析の結果、B301(ノルニコチン型)で発現量が高い275個の遺伝子断片、NS17(ニコチン型)で発現量が高い221個の遺伝子断片を見出した。合計496個の遺伝子断片のうち、蛍光強度比が2倍以上であった19個の遺伝子断片(下記表1記載のA1〜A9、A11〜A20)を選抜し、これらの塩基配列をMessenger Scape社にて決定した。
【0058】
【表1】
【0059】
Messenger Scape社によるフラグメントアナリシスソフトウェアGeneScanによる自動解析結果に加え、葉たばこ研究所においても遺伝子発現プロファイルの目視による解析を行った。何故なら前者の自動解析ではB301(ノルニコチン型)あるいはNS17(ニコチン型)のどちらか片方においてほとんどピークが検出されない場合、そうしたピークは自動選抜されてこないからである。前記の遺伝子発現プロファイルを目視で再解析した結果、ニコチン型でのみピークが検出される遺伝子断片(ニコチン型特異的断片)を15個(下記表2記載のB16〜B30)、ノルニコチン型でのみピークが検出される遺伝子断片(ノルニコチン型特異的断片)を8個(下記表3記載のC31〜C38)の合計23個を見出した。これら遺伝子断片の塩基配列解析をMessenger Scape社にて決定した。
【0060】
【表2】
【0061】
【表3】
【0062】
塩基配列解析はダイレクトシーケンス法にて行った。すなわち、AFLP-DD解析の過程で得られたPCR産物をポリアクリルアミドゲル電気泳動で分離し、選抜した各ピークの鎖長に対応する遺伝子断片を切り取り、ゲル中のDNAを溶出してPCRにて増幅後、シーケンス反応、シーケンス解析をし、塩基配列を決定した。決定した塩基配列をもとに、下記実施例3で用いる定量PCR解析用のプライマーを設計した。
【0063】
(実施例3)定量PCRによる遺伝子発現の再確認
(1) 薬剤誘導処理によりノルニコチン蓄積を誘導したタバコ(幼苗個体および鉢上げ個体)の調製
B301(ノルニコチン型)とNS17(ニコチン型)の1組み合わせ以外に、タバコ品種である、Bright Yellow103 (BY103)、Bright Yellow4 (BY4)、Burley21 (B21) のそれぞれのノルニコチン型とニコチン型を加えた合計4組の遺伝資源を温室内(昼夜温25℃、自然日長)で栽培した。幼苗個体のノルニコチン解析には、播種後3週間目の芽生えを36穴連結ビニールポット(信和株式会社製、28.5cm×28.5cm)に1個体ずつ植え替えた後1週間栽培し、薬剤誘導処理(MHおよび重炭酸ナトリウムを含む水溶液を約200μl/個体ずつ噴霧)後5日目にサンプリングした葉を供試した。鉢上げ個体のノルニコチン解析には、実施例1に記載の方法に従って植物体を4寸鉢で栽培し、薬剤誘導処理(MHおよび重炭酸ナトリウムを含む水溶液を植物体の地上部全体に均一に噴霧した)後5日目にサンプリングした葉を供試した。これらサンプリングした葉は実施例1に記載の方法に従い熱風乾燥および粉砕後、アルカロイド分析法(Hybrid Method)によりノルニコチン含量を解析した。その結果、いずれの遺伝資源においても薬剤誘導処理によってノルニコチン型においてのみノルニコチン蓄積が認められた。よって、本実験において用いた材料および方法(遺伝資源、栽培方法、薬剤誘導処理)を用いて、以下に述べる定量PCR解析用のサンプルを調製した。但し、薬剤誘導処理後5日目の葉はサンプリング後、熱風乾燥は行わず、速やかに液体窒素中で凍結後マイナス80℃のフリーザーで保存した。
【0064】
(2) 定量PCR解析による候補遺伝子の二次選抜
上記4組のノルニコチン型とニコチン型の幼苗個体および鉢上げ個体から調製した薬剤誘導処理後5日目の葉(マイナス80℃のフリーザーで保存した葉)から実施例2に記載の方法に従って全RNAを単離した。これら全RNAを鋳型として、B301(ノルニコチン型)およびNS17(ニコチン型)で発現量の異なった42個の遺伝子断片(表1、表2、表3)の定量PCR解析を行なった。
【0065】
定量PCR解析は、SYBR Green PCR Core Reagents (Applied Biosystems社)を用い、キット添付の指示書に従い、7500リアルタイムPCRシステム(Applied Biosystems社)上で、95℃で10分間処理後、94℃で15秒間および60℃で1分間の処理を40サイクル反応させることによって行った。幼苗個体の解析には、上記4組の遺伝資源に関し、それぞれ1組当りノルニコチン型24個体、ニコチン型24個体を供試し、また鉢上げ個体の解析には、それぞれ1組当りノルニコチン型6個体、ニコチン型6個体を供試した。また幼苗個体の解析では、12個体由来の全ての葉を混合したものを1サンプルとし、薬剤誘導処理直前(0日目)および薬剤誘導処理後5日目の植物体から各1サンプルずつ調製し、全RNA抽出に供試した。また鉢上げ個体の解析では、3個体由来の全ての葉を混合したものを1サンプルとし、薬剤誘導処理直前(0日目)および薬剤誘導処理後5日目の植物体から各1サンプルずつ調製し、全RNA抽出に供試した。一次選抜した42遺伝子断片の定量PCR解析を行った結果、4組全ての遺伝資源において、幼苗個体および鉢上げ個体を問わず、薬剤誘導処理5日目においてニコチン型と比較してノルニコチン型で5倍以上のmRNAの発現量が高い遺伝子断片4個を二次選抜した(下記表4)。これら4個の遺伝子断片のうち、3個は表1記載のピーク番号A2、A3およびA4、残りの1個は表3記載のピーク番号C35に対応する遺伝子断片であった。なお、薬剤誘導処理直前のノルニコチン型およびニコチン型では、これら4個の遺伝子断片の発現量は非常に低く、かつニコチン型とノルニコチン型で発現量の差は認められなかった。
【0066】
【表4】
【0067】
二次選抜した4個の遺伝子断片のうち、ピーク番号C35に対応する遺伝子断片(以下、「C35遺伝子断片」という)について更に詳細な解析を以降の実施例にて行った。C35遺伝子断片の塩基配列を配列番号7に示す。なお、C35遺伝子断片の定量PCR解析には下記の配列番号5のプライマーと配列番号6のプライマーを用いた。
配列番号5:Nor#35_ F1
GGTTCAGTCGGTAAATAGTGAACGGT
配列番号6:Nor#35_ R1
TCAGCGCAAGATTTCCAATTTGTCCGA
【0068】
(実施例4)Nor#35遺伝子のcDNAおよびゲノムDNAクローンの単離および構造解析
(1) Nor#35遺伝子の完全長cDNAの単離と塩基配列の決定
B301(ノルニコチン型)の全RNAより5'/3'RACE法(Loh,Y. et al. (1989) Science 243, 217;Ohara et al. (1989) Proc. Natl. Acad. Sci USA 86, 5673;Frohmann, M. (1994) PCR Methods and Applications, 4, 540-558)を用いて、C35遺伝子断片の塩基配列をその一部に有するcDNAの全長配列(以下、「Nor#35遺伝子」という)を明らかにした。5'/3'RACE法に用いたオリゴプライマーはC35遺伝子断片の塩基配列をもとに化学合成した(配列番号8〜13)。
【0069】
配列番号8:GSP1(3') primer
GGTTCAGTCGGTAAATAGTGAACGGT
配列番号9:GSP1(5') primer
TCAGCGCAAGATTTCCAATTTGTCCGA
配列番号10:GSP2(5') primer
CCGTTCACTATTTACCGACTGAACCCAT
配列番号11:GeneRacerTM 3' Primer(Invitrogen社)
GCTGTCAACGATACGCTACGTAACG
配列番号12:GeneRacerTM 5’Primer(Invitrogen社)
CGACTGGAGCACGAGGACACTGA
配列番号13:GeneRacerTM 5’Nested Primer(Invitrogen社)
GGACACTGACATGGACTGAAGGAGTA
【0070】
5'/3'RACEは、以下の手順で行った。まず、実施例1に記載の方法に従って温室栽培したノルニコチン誘導処理後3,4,5日目のB301(ノルニコチン型)の鉢上げ個体から全RNAを抽出した。次に、各全RNAを5μgずつ混合し、そのうち2μgを下記の1本鎖cDNA合成に用いた。1本鎖cDNA の合成はGeneRacerTMキット(Invitrogen社)を使用し、キットの指示書に従って行った。最終的に20μlの鋳型DNA溶液を得た。続いて、以下の条件で1st-PCRを行った。配列番号8のオリゴプライマー(15pmol)および配列番号11のオリゴプライマー(45pmol)を用いて、上記で得た鋳型DNA溶液1μlからcDNAの3’末端までの領域を含む1st-PCR産物(以下、「3’側1st-PCR産物」という)を増幅した。増幅反応(50μlの反応系)は、PfuUltraTM High-Fidelity DNA Polymerase (Stratagene社)およびGeneAmpTM PCR System 9700(Applied Biosystems社)を用い、以下のプログラムで行った。すなわち、95℃で2分間処理した後、95℃で30秒間および72℃で2分間の処理を5サイクル実施、95℃で30秒間および70℃で2分間の処理を5サイクル実施、95℃で30秒間および68℃で2分間の処理を25サイクル実施、72℃でさらに10分間の処理、からなるプログラムである。同様にして、配列番号9のオリゴプライマー(15pmol)および配列番号12のオリゴプライマー(45pmol)を用いて、cDNAの5’ 末端まで領域を含む1st-PCR産物(以下、「5’側1st-PCR産物」という)を増幅した。さらに,5'RACE では、5’ 側1st-PCR産物1μl、配列番号10のオリゴプライマー(15pmol)および配列番号13のオリゴプライマー(15pmol)を用いて、cDNAの5’側末端まで領域を含むnested-PCR産物(以下「5’側nested-PCR産物」という)を、以下のプログラムで増幅した。すなわち、95℃で2分間処理した後、95℃で30秒間および65℃で2分間の処理を25サイクル実施、72℃でさらに10分間の処理、からなるプログラムである。3’側1st-PCR産物および5’側nested-PCR産物を1.5%アガロースゲルで電気泳動したところ、それぞれ約500bp、約1200bpのバンドを確認した。
【0071】
次に、前述の3’側1st-PCR産物(約500bp)および5’側nested-PCR産物(約1200bp)を、おのおのクローニングベクターpCR 4Blunt-TOPO(Invitrogene社)に挿入した後、大腸菌TOP10株にエレクトロポレーション(Gene Puiser Xcell エレクトロポレーションシステムBIO-RAD社)法を用いて導入した。導入後、大腸菌は50mg/lアンピシリンを含むLuria Bertani (以下、「LB」という)固形培地(寒天1.5%)上で培養し、得られたアンピシリン耐性のコロニー約10個をLB(50mg/lアンピシリンを含む)液体培地中で終夜37℃で振とう培養した。本培養物からQIAprep Spin Miniprep Kit (QIAGEN社)を用いて指示書に従いプラスミドDNAを精製し、以下に述べるシークエンス解析の鋳型DNAとして用いた。BigDyeTMTerminator v3.1 Cycle Sequencing Kit(Applied Biosystems社)および3700 DNA Analyzer(Applied Biosystems社) を用いて、キットおよび装置の指示書に従ってシークエンス解析を実施した結果、3’側1st-PCR産物および5’側nested-PCR産物の塩基配列を決定することができた。また、Nor#35遺伝子の5’側末端までの領域を含む約1200bpのDNAが挿入されたプラスミド(以下、「プラスミド#35_5Race_05」という)は後にRNAiコンストラクトの構築に使用した。
【0072】
(2) Nor#35遺伝子がコードするアミノ酸配列の推定
3’側1st-PCR産物および5’側nested-PCR産物の塩基配列、および配列番号7記載の塩基配列を塩基配列解析ソフトウェアATGC, GENETYX(ゼネティックス社)を用いて結合処理した結果、1552bpからなるNor#35遺伝子の全長cDNA配列(配列番号1、 Nor#35_cDNA)を決定することができた。また、Nor#35遺伝子がコードするアミノ酸配列(配列番号2 、Nor#35_protein)も推定した。Nor#35遺伝子のアミノ酸配列(配列番号2)は、BLASTによる配列比較解析により、Phytophthora infestansの感染により誘導されるSolanum tuberosumのjasmonic acid 2翻訳産物(ACCESSION No.AAU12055)およびjasmonic acidに応答するLycopersicon esculentumのacid 2翻訳産物(ACCESSION No.AAF04915)と約70%の相同性があることが判明した。しかしながら、これら翻訳産物やこれら翻訳産物をコードする遺伝子の機能は解明されていない。
【0073】
(3) Nor#35遺伝子のゲノムDNAクローンの単離と塩基配列の決定
Nor#35遺伝子のゲノムクローン配列を葉たばこ研究所保有のBAC(Bacterial Artificial Chromosome)ライブラリーから単離した。該研究所保有のBACライブラリーは、タバコ品種BY4号のゲノムDNA由来で、平均約150Kbのインサートを有す約10万個のBACプラスミドから構成されている(Noguchi et al. (1999) Mol Gen Genet.262,822-829)。配列番号1記載のNor#35遺伝子配列を基に下記の配列番号14 (#35_F2)および配列番号15 (#35_R1)のプライマーを用いて3次元PCR法により上記BACライブラリーをスクリーニングした。その結果、2個のBACクローン(289F6および902D10)を選抜した。
【0074】
配列番号14:Nor#35_F2
TGGGTTCAGTCGGTAAATAGTGAACGGT
配列番号15:Nor#35_R1
TCAGCGCAAGATTTCCAATTTGTCCGA
【0075】
これら2個のBACクローンからBACプラスミドをQIAGEN Large-Construct Kit(QIAGEN社)を用いて各々精製し、そのうち各1μgのプラスミドをシークエンス解析の鋳型として用いた。BACプラスミドのインサートDNAのシークエンス解析は、BigDyeTMTerminator V3.1 Cycle Sequencing Kit(Applied Biosystems社)、3700 DNA Analyzer(Applied Biosystems社)、および下記に示す9種のプライマー(配列番号16〜24)を用い、キットおよび装置の指示書に従って行った。
【0076】
配列番号16:Nor#35_F6
CTACTTAGGATGACAAACAGCAGGAT
配列番号17:Nor#35_F7
CAAGATTTTGTTCATGTAACTGAGCAC
配列番号18:Nor#35_F8
TGCTGCCAAGAAGAATAATGGGTGTTCA
配列番号19:Nor#35_F9
AGTGCTCTTGTATAGCTACGGGTT
配列番号20:Nor#35_R2
CCGTTCACTATTTACCGACTGAACCCAT
配列番号21:Nor#35_R4
ATCTAGAAAATGACTACACGTTGCCGA
配列番号22:Nor#35_R5
GCCAGCAACTTTGCGACACAAGT
配列番号23:Nor#35_R6
AACCCGTAGCTATACAAGAGCACT
配列番号24:Nor#35_R7
TCCGTCAATTTCATTGTACTCTGCAT
【0077】
シークエンス解析により得た配列データを塩基配列結合ソフトウェアATGC, GENETYX(ゼネティックス社)を用いて結合処理することにより、最終的に3371bpのゲノムクローン配列(配列番号3)を得た。Nor#35遺伝子のゲノムクローン配列(配列番号3)と全長cDNA配列(配列番号1)を塩基配列解析ソフトウェアATGC, GENETYX(ゼネティックス社) (ATGC, GENETYX; ゼネティックス社)を用いて解析した結果、Nor#35遺伝子のゲノムクローン配列は,3つのエキソンと2つのイントロンを含む構造を持つことが分かった(図3)。
【0078】
(実施例5)Nor#35遺伝子の機能解析(遺伝子発現の抑制)
(1) RNAiベクターpSP107_Nor#35の構築
RNAiにおいては植物細胞内で導入遺伝子が転写されdsRNAを形成する必要性がある。dsRNAを形成する配列として配列番号4に示したDNA配列を選択し、本配列の5’および3’末端に異なる制限酵素部位を導入した2種類のDNA断片をPCR法で調製した。プラスミド#35_5Race_05を鋳型DNAとして配列番号25および26に示した#35_F_NotIと#35_R_PstIをプライマーとして、同様にプラスミド#35_5Race_05を鋳型DNAとして配列番号27および28に示した#35_F_BamHIと#35_R_SphIをプライマーとして、図4に示した2種のDNA断片をPCR法で増幅した。
【0079】
配列番号25:Nor#35_F_NotI
ACATAGCGGCCGCGAATGGATCACGACCAAACAGAGTAGCTG
配列番号26:Nor#35_R_PstI
ACTACTGCAGTTACCGACTGAACCCATATTTACTGTTTGGGT
配列番号27:Nor#35_ F_BamHI
CTATGGATCCGAATGGATCACGACCAAACAGAGTAGCTG
配列番号28:Nor#35_ R_SphI
TGTAGCATGCTTACCGACTGAACCCATATTTACTGTTTGGGT
【0080】
増幅した2種のDNA断片は,それぞれ5'/3'末端に、プライマーにより付与されたNotI/PstI、または、BamHI/SphIの制限酵素認識配列を持つ(図4)。増幅した2種のDNA断片は、それぞれこれら制限酵素で二重消化した後MinElute kit(QIAGEN社)で精製し、以下の実験に供試した。
【0081】
逆向反復配列(即ち、特定の配列に関し、正方向配列とその逆方向配列をタンデムに結合)を構築するためのベクターとして、pDNR-1(Clontech社)を以下に示すように改変したpSP148(図5)を利用した。pSP148(図5)の2個のloxP部位の間には、35Sプロモーター,マルチクローニングサイト1(MCS1),スペーサー配列,マルチクローニングサイト2(MCS2),NOSターミネーターがこの順番で存在する。このベクターのMCS1のPstI/NotI制限酵素認識部位およびMCS2のBamHI/SphI制限酵素認識部位に上記の制限酵素で二重消化後精製したDNA断片を挿入することによりpSP148_Nor#35FRを作製した(図6)。
【0082】
次に、pSP148_Nor#35FR上の2個のloxP部位にはさまれた領域を、Cre-loxP組換え(Saue, 1994, Curr. Opin. Biotechnol. 5:521-527;Abremski et al., 1984, J. Biol. Chem. 259:1509-1514)を利用してバイナリーベクターpSP107(図7)上のloxP部位に導入して、RNAiベクターpSP107_Nor#35を作製した(図8)。バイナリーベクターpSP107(図7)はpBI121(Clontech社)のHindIII-Sse8387I 部位にproK-loxP断片(Creator Acceptor Vector Construction Kit, Clontech社) を導入して作製したものである。また、Cre-loxP組換えはCre Recombinase kit(Clontech社)を用い、その指示書に従って行った。本RNAiベクター、pSP107_Nor#35をAgrobacterium tumefaciens LBA4404に導入し、以下の形質転換実験に使用した。
【0083】
(2)形質転換用アグロバクテリウムの調製
RNAiベクターpSP107_Nor#35を、Agrobacterium tumefaciens の菌株LBA4404(Hoekeman,A. et al. (1983) Nature 303)に凍結融解法(Ann,G. et al.(1988)Plant Molecular Biology Manual:1-19)により導入し、カナマイシン耐性であるコロニーを選抜し、本コロニーをタバコ植物の形質転換に用いた。また、形質転換当代(T0世代)植物の解析の際に対照として使用する植物を作出するため、カナマイシン耐性遺伝子およびGUS遺伝子のみからなるバイナリーベクターpBI121を導入したAgrobacterium tumefaciens 菌株LBA4404も調製した。
【0084】
(3)タバコの形質転換
温室内(昼夜温25℃、自然日長)で栽培したBY103(ノルニコチン型)の葉組織を使用して形質転換を行った(Komari,T. et al. (1989) ,Thor. Appl. Genet 77,547-552)。実施例1に記載の方法で栽培した移植後3週間目の鉢上げ植物より上位展開葉を採取し、70%(V/V)エチルアルコ−ルおよび1.0%(V/V)次亜塩素酸ナトリウム溶液で表面殺菌し、続いて十分の滅菌水で2回洗浄した後、剃刀を用いて約1cm角の葉片を調製した。この葉片20枚と上記カナマイシン耐性を示したAgrobacterium tumefaciens約108細胞とをLinsmaier and Skoog(Linsmaier, EM. et al.(1965),Physiol Plant 18, 100-127)の無機塩類と30 g/lのスクロースより成る液体培地中で共存培養(28℃で48時間)を行った。その後、葉片をセフォタキシム250 mg/lを含む滅菌水で1回洗浄し、細菌を洗い落とした後、Linsmaier and Skoog(1965)の無機塩類、インド−ル酢酸0.3 mg/l、6-(γ,γ-Dimethylallylamino)purine 10 mg/l、カナマイシン100 mg/l、セフォタキシム250 mg/lおよび寒天0.9%を含む再分化固型培地に置床し、約1ヶ月後、再分化した茎葉をLinsmaier and Skoog(1965)の無機塩類、30g/lのスクロ−ス、カナマイシン100 mg/l、セフォタキシム250 mg/lおよび寒天0.9%を含む発根培地に置床した。さらに1ヶ月後、発根した植物体(T0世代)を閉鎖系温室内で4寸鉢に移植し、以下の実験に供試した。
【0085】
(4)GUS染色による形質転換体の選抜
レポーター遺伝子であるGUS遺伝子の発現を指標として導入遺伝子が安定して発現していると思われる形質転換体を次のようにして選抜した。閉鎖系温室(昼夜温25℃、自然日長)に移植後3週間栽培した形質転換タバコ植物(T0世代)の展開緑葉から、鋏で約1cm角の葉片を切り出した。これら葉片を1.0%(V/V)Ttiton X-100 を含む50mMリン酸緩衝液(pH7.0)中で37℃で1時間インキュベートした後、50mMリン酸緩衝液(pH7.0)で2回洗浄し、1.0mM 5-bromo-4-chloro-3-indolyl-β-D-glucuronide(X-Gluc)、20%(V/V)メタノールを含む50mMリン酸緩衝液(pH7.0)中で37℃で一晩インキュベートした。葉片は99%(V/V)メタノールで洗浄後、発色の有無を最終確認した。その結果、pSP107_Nor#35のT-DNAを導入した60個体(以下「Nor#35 RNAi個体」という)、pBI121のT-DNAを導入した15個体(以下「対照個体」という)に関しGUS遺伝子の発現を確認した。GUS染色の詳細については樋江井ら(Hiei,Y. et al. (1994),Plant. J 6,271-282)の方法に従った。
【0086】
(5) Nor#35遺伝子の発現抑制の確認(T0世代)
次に、Nor#35 RNAi個体(T0世代)において内生のNor#35遺伝子の発現が低減したか否かを定量PCR法で調査した。Nor#35 RNAi個体から24個体、対照個体から4個体を無作為に選び、その展開緑葉から全RNAを抽出した。まず、実施例1に記載の方法に従って薬剤誘導処理した薬剤誘導処理後7日目の展開緑葉から直径約10mmのリーフディスクを採取した。1個体当り展開緑葉1枚からリーフディスク5枚を金属パンチで打ち抜き、RNAlater (Ambion社)に浸漬および保存したものを全RNA抽出用試料とした。全RNAはRNeasy Plant Mini Kit (QIAGEN社)を用いて抽出し、さらに、Omniscript RT Kit(QIAGEN社)を用いて指示書に従い逆転写反応を行い、その反応液の一部を定量PCR解析に供試した。定量PCR に用いるNor#35遺伝子およびβ-actin遺伝子(ハウスキーピング遺伝子)のTaqManTM(Roche Molecular Systems社)プローブとプライマーは、PrimerExpress (Applied Biosystems社)を用いて設計し(配列番号29〜34)、シグマ アルドリッチ ジャパン 株式会社 ライフサイエンス事業部で合成した。なお、リーフディスクを打ち抜いた葉の上位葉2枚のラミナ部分を、リーフディスクを打ち抜いた時に同時に収穫し、実施例1に記載の方法に従い、熱風乾燥後粉砕した試料をノルニコチン含量およびニコチン含量の分析に供試した。
【0087】
配列番号29:Nor#35_TMF_ver3
TTGGGAAAGGGTAAAATGTGAATG
配列番号30:Nor#35_TMR_ver3
TCCCCAACCAAAGAATATTTGC
配列番号31:Nor#35_TMP_ver3(FAM)
TGTTCGGACAAATTGGAAATCTTGCGC
配列番号32:actb_TMF
TCTGGATTGGAGGYTCTATCTTG
配列番号33:actb_TMR
TCGTCATACTCTGCCTTTGCA
配列番号34:actb_TMP(Cy5)
CCTCAGCACCTTCCAGCAGATGTGG
定量PCR解析は実施例3に記載の方法に従って行い、その結果を図9に示した。Nor#35 RNAi個体3個体 (MH9, MH20, MH21)において、内生のNor#35遺伝子の発現が対照個体(平均値)と比較して16-49%のレベルまで低減していた。
【0088】
(6) 薬剤誘導処理したNor#35遺伝子発現抑制個体(T0世代)のアルカロイド分析
Nor#35遺伝子の発現抑制を確認した3個体 (MH9, MH20, MH21)および対照個体3個体のアルカロイド分析を実施した。
【0089】
前述の熱風乾燥後粉砕した試料を用いてアルカロイド含量(ノルニコチン含量とニコチン含量の総量)、ノルニコチン含量およびノルニコチン指数(ノルニコチン含量/全アルカロイド含量×100)を解析した結果を下記表5に示した。対照個体(平均値)と比較してMH9ではアルカロイド含量は53%、ノルニコチン含量は80%、MH20ではアルカロイド含量は23%、ノルニコチン含量は69%、MH21ではアルカロイド含量は35%、ノルニコチン含量は76%、低減していた。
【0090】
【表5】
【0091】
(7) 黄色乾燥したNor#35遺伝子発現抑制個体(T0世代)のアルカロイド分析
Nor#35 RNAi個体のうち薬剤誘導処理を行わなかった20個体、対照個体のうち薬剤誘導処理を行わなかった6個体、を黄色乾燥実験に供試した。閉鎖系温室内で栽培開始後9週間目に芯止めし、その約1週間後に下位葉および中位葉を収穫し、黄色乾燥後、実施例1に記載のアルカロイド分析法(Hybrid Method)によりノルニコチンおよびニコチン含量を解析した。黄色乾燥は下記の13のステップから構成される。
【0092】
ステップ1:乾燥開始後3時間かけて昇温/加湿し、温度38℃/相対湿度100%に到達後、本条件で8時間処理、
ステップ2:次に1時間かけて昇温/排湿し、温度40℃/相対湿度87%に到達後、本条件で15時間処理、
ステップ3:次に1時間かけて昇温/排湿し、温度43℃/相対湿度63%に到達後、本条件で15時間処理、
ステップ4:次に1時間かけて昇温/排湿し、温度45℃/相対湿度52%に到達後、本条件で15時間処理、
ステップ5:次に1時間かけて昇温/排湿し、温度48℃/相対湿度43%に到達後、本条件で15時間処理、
ステップ6:次に1時間かけて昇温/排湿し、温度50℃/相対湿度37%に到達後、本条件で10時間処理、
ステップ7:次に1時間かけて昇温/排湿し、温度53℃/相対湿度31%に到達後、本条件で8時間処理、
ステップ8:次に1時間かけて昇温/排湿し、温度56℃/相対湿度26%に到達後、本条件で6時間処理、
ステップ9:次に1時間かけて昇温/排湿し、温度60℃/相対湿度20%に到達後、本条件で6時間処理、
ステップ10:次に1時間かけて昇温/排湿し、温度63℃/相対湿度16%に到達後、本条件で6時間処理、
ステップ11:次に2時間かけて昇温し、温度65℃/相対湿度16%に到達後、本条件で2時間処理、
ステップ12:次に1時間で昇温し、温度68℃/相対湿度16%に到達後、本条件で8時間処理、及び
ステップ13:5時間かけて、温度30℃/相対湿度60%まで降温/加湿し乾燥を終了。
【0093】
ノルニコチンおよびニコチン含量の分析結果を下記表6に示した。対照個体(平均値)と比較してYC3ではアルカロイド含量は21%、ノルニコチン含量は45%、YC4ではアルカロイド含量は25%、ノルニコチン含量は52%、YC10ではアルカロイド含量は43%、ノルニコチン含量は60%、低減していた。
【0094】
【表6】
【0095】
(8) 薬剤誘導処理したNor#35遺伝子発現抑制個体(自殖T1世代)のアルカロイド分析
T0世代においてアルカロイド含量およびノルニコチン含量の低減を確認した4個体(MH21、YC3、YC4、YC10)につき自殖T1種子を採種後、閉鎖系温室で栽培し、アルカロイド含量およびノルニコチン含量の解析を再度行った。
【0096】
上記選抜した4個体(MH21、YC3、YC4、YC10)のT1世代を実施例3に記載の方法に従って36穴連結ビニールポット2枚に栽培した。MH21のT1世代を72個体、YC3のT1世代を 72個体、YC4のT1世代を72個体、YC10のT1世代を72個体栽培した。36穴連結ビニールポットに仮植後1週間目に各個体から葉の一部をとって前述の方法に従ってGUS染色を行い、導入されたGUS遺伝子のT1世代における分離比検定を行った。検定の結果、YC10には2遺伝子座以上にGUS遺伝子が導入され、残り3個体(MH21、YC3、YC4)にはGUS遺伝子が1遺伝子座に導入されたものと推定された。以後の解析には、1遺伝子座に遺伝子が導入されたと推定されたMH21、YC3およびYC4のT1世代(3系統群)を供試した。
【0097】
次に、上記3系統群(MH21、YC3およびYC4のT1世代)の各個体は、36穴連結ビニールポットに仮植後3週間目に定量PCR解析用サンプルとして葉片を採取した、また葉片採取後同日内に薬剤誘導処理(MHおよび重炭酸ナトリウムを含む水溶液を約200μlずつ噴霧)を行った。植物体地上部に散布された薬剤が乾燥した後、鋏で各ビニールポットを切り離し、各個体間で葉が重ならないように間隔を空けて栽培した。薬剤誘導処理後7日目に、T1世代の各個体からラミナを採取し(全ての緑葉から採取)、実施例1に記載の方法に従って、熱風乾燥および粉砕後ニコチンおよびノルニコチン含量の解析を行った。なお上記3系統群(T1世代)の中で、GUS染色が認められずかつGUS遺伝子発現も定量PCR(配列番号35および配列番号36に示したプライマー及び配列番号37に示したTaqManプローブを使用)で認められなかった個体を対照個体として用いた。以下、YC3由来の対照個体をYC3-cont、YC4由来の対照個体をYC4-cont、MH21由来の対照個体をMH21-contと称する。一方、GUS染色が認められた個体を#35遺伝子発現抑制個体(T1世代)とした。以下、YC3個体由来の#35遺伝子発現抑制個体(T1世代)をYC3-T1、YC4個体由来の#35遺伝子発現抑制個体(T1世代)をYC4-T1、MH21個体由来の#35遺伝子発現抑制個体(T1世代)をMH21-T1と称する。ニコチンおよびノルニコチン含量の解析には5個体由来のラミナをまとめたものを1サンプルとして、MH21-T1、YC3-T1およびYC4-T1については5サンプル、YC3-contは3サンプル、YC4-contは5サンプル、MH21-contは2サンプルを解析に供試した。
【0098】
配列番号35:GUS_TM_F_ver2
GTTGCAACTGGACAAGGCACTA
配列番号36:GUS_TM_R_ver2
TGTCTGGCTTTTGGCTGTGA
配列番号37:GUS_TM_P_ver2_FAM
CTGGCAACCGGGTGAAGGTTATCTCTATGA
【0099】
薬剤誘導処理後7日目のYC3-T1、YC4-T1、MH21-T1、YC3-cont、YC4-cont、MH21-contのアルカロイド含量、ノルニコチン含量およびノルニコチン指数を下記表7に示した。YC3-T1はその対照である YC3-contに比較してアルカロイド含量は12%、ノルニコチン含量46%、YC4-T1はその対照であるYC4-cont に比較してアルカロイド含量は19%、ノルニコチン含量41%、MH21-T1はその対照であるMH21-cont に比較してアルカロイド含量は27%、ノルニコチン含量41%の低減が認められた。以上のように、Nor#35遺伝子の発現抑制により葉中アルカロイド含量を大きく変化させることなくノルニコチン含量を低減させたタバコ属植物を作出することができた。
【0100】
【表7】
【図面の簡単な説明】
【0101】
【図1】B301(ノルニコチン型)、NS17(ニコチン型)における薬剤処理によるノルニコチン含量の経時変化を示す。
【図2】AFLP-DD法の概要を示す。
【図3】Nor#35遺伝子のゲノム遺伝子の構造を示す。
【図4】Nor#35遺伝子の5’側末端領域を含むプラスミド(Nor #35_5Race_05)を鋳型としてPCR増幅したRNAiのトリガーとなる2種のDNA断片を示す。
【図5】Nor#35遺伝子の逆向反復配列を構築するためのベクターpSP148の構造を示す。
【図6】ヘアピン型dsRNAを発現するカセットを含むpSP148_Nor#35FRの構造を示す。
【図7】バイナリーベクターpSP107の構造を示す。
【図8】dsRNAを発現するカセットを含む断片をCre-lox組換えにより導入したバイナリーベクターpSP107_Nor#35の構造を示す。
【図9】#35組換え体T0世代におけるNor#35遺伝子発現の定量PCRによる解析結果を示す。
【特許請求の範囲】
【請求項1】
以下の(a)〜(d)にポリヌクレオチドからなるNor#35遺伝子。
(a) 配列番号1に示す塩基配列からなるポリヌクレオチド
(b) 配列番号1に示す塩基配列において1若しくは数個の塩基が欠失、置換、若しくは付加された塩基配列からなり、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(c) 配列番号1に示す塩基配列からなるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(d) 配列番号1に示す塩基配列に対して80%以上の相同性を有する塩基配列からなり、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
【請求項2】
配列番号3に示す塩基配列を有する請求項1に記載のNor#35遺伝子。
【請求項3】
請求項1または2に記載のNor#35遺伝子を含む組換えベクター。
【請求項4】
請求項1若しくは2に記載のNor#35遺伝子または請求項3に記載の組換えベクターを導入した形質転換タバコ属植物。
【請求項5】
植物が、植物体、植物器官、植物組織、または植物細胞である、請求項4に記載の形質転換タバコ属植物。
【請求項6】
配列番号1に示す塩基配列中の連続した100塩基以上からなる塩基配列をアンチセンス方向に含む、Nor#35遺伝子発現抑制用ベクター。
【請求項7】
配列番号1に示す塩基配列中の連続した21塩基以上の塩基配列と該塩基配列の相補配列とを含む、Nor#35遺伝子発現抑制用ベクター。
【請求項8】
配列番号1に示す塩基配列中の連続した21塩基以上の塩基配列と該塩基配列の相補配列との間にスペーサー配列を挿入した連続配列を含む、Nor#35遺伝子発現抑制用ベクター。
【請求項9】
請求項6〜8のいずれかに記載のNor#35遺伝子発現抑制用ベクターを導入した形質転換タバコ属植物。
【請求項10】
植物が、植物体、植物器官、植物組織、または植物細胞である、請求項9に記載の形質転換タバコ属植物。
【請求項11】
請求項1または2に記載のNor#35遺伝子の発現を抑制した植物細胞を作製する工程、および、該植物細胞から植物体を再生させる工程を含む、アルカロイド含量が低減した形質転換タバコ属植物の作製方法。
【請求項12】
請求項1または2に記載のNor#35遺伝子の発現抑制が、RNAi法、アンチセンス法、遺伝子破壊法、および共抑制法から選ばれるいずれかの手法により行なわれることを特徴とする、請求項11に記載の方法。
【請求項13】
植物細胞内において請求項1または2に記載のNor#35遺伝子の発現量を調節することを特徴とする、植物中のアルカロイド量を改変する方法。
【請求項1】
以下の(a)〜(d)にポリヌクレオチドからなるNor#35遺伝子。
(a) 配列番号1に示す塩基配列からなるポリヌクレオチド
(b) 配列番号1に示す塩基配列において1若しくは数個の塩基が欠失、置換、若しくは付加された塩基配列からなり、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(c) 配列番号1に示す塩基配列からなるポリヌクレオチドと相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
(d) 配列番号1に示す塩基配列に対して80%以上の相同性を有する塩基配列からなり、かつアルカロイド含量を調節する機能を有するタンパク質をコードするポリヌクレオチド
【請求項2】
配列番号3に示す塩基配列を有する請求項1に記載のNor#35遺伝子。
【請求項3】
請求項1または2に記載のNor#35遺伝子を含む組換えベクター。
【請求項4】
請求項1若しくは2に記載のNor#35遺伝子または請求項3に記載の組換えベクターを導入した形質転換タバコ属植物。
【請求項5】
植物が、植物体、植物器官、植物組織、または植物細胞である、請求項4に記載の形質転換タバコ属植物。
【請求項6】
配列番号1に示す塩基配列中の連続した100塩基以上からなる塩基配列をアンチセンス方向に含む、Nor#35遺伝子発現抑制用ベクター。
【請求項7】
配列番号1に示す塩基配列中の連続した21塩基以上の塩基配列と該塩基配列の相補配列とを含む、Nor#35遺伝子発現抑制用ベクター。
【請求項8】
配列番号1に示す塩基配列中の連続した21塩基以上の塩基配列と該塩基配列の相補配列との間にスペーサー配列を挿入した連続配列を含む、Nor#35遺伝子発現抑制用ベクター。
【請求項9】
請求項6〜8のいずれかに記載のNor#35遺伝子発現抑制用ベクターを導入した形質転換タバコ属植物。
【請求項10】
植物が、植物体、植物器官、植物組織、または植物細胞である、請求項9に記載の形質転換タバコ属植物。
【請求項11】
請求項1または2に記載のNor#35遺伝子の発現を抑制した植物細胞を作製する工程、および、該植物細胞から植物体を再生させる工程を含む、アルカロイド含量が低減した形質転換タバコ属植物の作製方法。
【請求項12】
請求項1または2に記載のNor#35遺伝子の発現抑制が、RNAi法、アンチセンス法、遺伝子破壊法、および共抑制法から選ばれるいずれかの手法により行なわれることを特徴とする、請求項11に記載の方法。
【請求項13】
植物細胞内において請求項1または2に記載のNor#35遺伝子の発現量を調節することを特徴とする、植物中のアルカロイド量を改変する方法。
【図1】

【図2】
【図3】
【図4】
【図5】
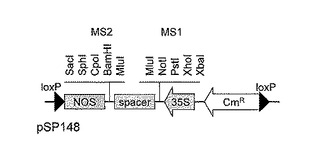
【図6】
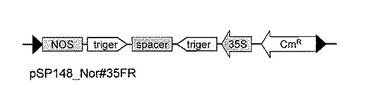
【図7】
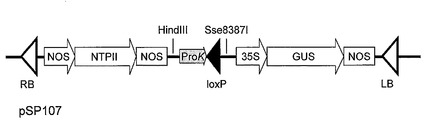
【図8】
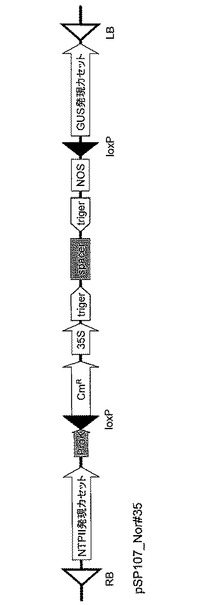
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2008−212065(P2008−212065A)
【公開日】平成20年9月18日(2008.9.18)
【国際特許分類】
【出願番号】特願2007−54591(P2007−54591)
【出願日】平成19年3月5日(2007.3.5)
【出願人】(000004569)日本たばこ産業株式会社 (406)
【Fターム(参考)】
【公開日】平成20年9月18日(2008.9.18)
【国際特許分類】
【出願日】平成19年3月5日(2007.3.5)
【出願人】(000004569)日本たばこ産業株式会社 (406)
【Fターム(参考)】
[ Back to top ]