
アルドラーゼ、それらをコードする核酸ならびにそれらの作製および使用方法
【課題】 HMGおよび/もしくはKHGアルドラーゼ活性などのピルビン酸活性を含むアルドラーゼ活性を有するポリペプチド、これらのポリペプチドをコードするポリヌクレオチドならびにこれらのポリヌクレオチドおよびポリペプチドを作製および使用する方法を提供する。
【解決手段】 アルドラーゼ活性を有するポリペプチド、これらのポリペプチドをコードするポリヌクレオチドを天然の起源から単離する。
【解決手段】 アルドラーゼ活性を有するポリペプチド、これらのポリペプチドをコードするポリヌクレオチドを天然の起源から単離する。
Notice: Undefined index: DEJ in /mnt/www/gzt_disp.php on line 298
【特許請求の範囲】
【請求項1】
(a)少なくとも1個のポリペプチドをコードする核酸であって、該核酸がcDNA、転写物(mRNA)若しくは遺伝子の完全長に渡って、配列番号243の配列に対して、少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上、若しくは100%の配列同一性を有する配列を含み、
該核酸が、アルドラーゼ活性を有する少なくとも1個のポリペプチドをコードするか、又はアルドラーゼ特異的抗体を生成することができるエピトープ若しくは免疫原として作用するポリペプチド若しくはペプチドをコードする、前記核酸;
(b)少なくとも1個のポリペプチドをコードする核酸であって、該核酸が、(a)の配列を含む核酸の相補体に、ストリンジェントな条件下でハイブリダイズする配列を含み、
該核酸が、アルドラーゼ活性を有する少なくとも1個のポリペプチドをコードするか、又はアルドラーゼ特異的抗体を生成することができるエピトープ若しくは免疫原として作用するポリペプチド若しくはペプチドをコードする、前記核酸;
(c)アルドラーゼ活性を有する少なくとも1個のポリペプチドをコードする核酸であって、該ポリペプチドが、配列番号244の配列又はその酵素的に活性な断片を含む、前記核酸;
(d)アルドラーゼ活性を有するが、シグナル配列を欠くポリペプチドをコードする(a)、(b)若しくは(c)の核酸;
(e)アルドラーゼ活性を有するポリペプチドをコードし、異種配列をさらに含む(a)、(b)、(c)若しくは(d)の核酸;又は、
(f)(a)、(b)、(c)、(d)若しくは(e)と相補的な核酸配列、
を含む、単離された、合成又は組換え核酸。
【請求項2】
(a)請求項1に記載の配列を含む核酸を増幅することができるか、又は、
(b)請求項1に記載の配列の最初(5'側)の10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35個以上の残基の配列を有する第1のメンバー、及び該第1のメンバーの相補鎖の最初(5'側)の10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35個以上の残基の配列を有する第2のメンバーを含む、
増幅プライマー対であって、
必要に応じて、(a)の増幅プライマー配列対のメンバーが、該配列の少なくとも10〜50個の連続する塩基、若しくは該配列の約10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35個以上の連続する塩基を含むオリゴヌクレオチドを含む、前記プライマー対。
【請求項3】
請求項1に記載の核酸を含む発現カセット、ベクター又はクローニングビヒクル。
【請求項4】
請求項1に記載の核酸を含むか、又は請求項3に記載の発現カセット、ベクター若しくはクローニングビヒクルを含む形質転換細胞。
【請求項5】
請求項1に記載の核酸を含むか、又は請求項3に記載の発現カセット、ベクター若しくはクローニングビヒクル、請求項4に記載の形質転換細胞を含むトランスジェニック非ヒト動物、植物、植物部分又は種子。
【請求項6】
アルドラーゼ活性を有する単離された、合成又は組換えポリペプチド若しくはペプチドであって、
(a)該ポリペプチド若しくは酵素の完全長に渡って、配列番号244に対して、少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上、若しくは100%の配列同一性を有するアミノ酸配列、及び/又はその酵素的に活性な断片;
(b)ポリペプチドが、(i)アルドラーゼ活性を有するか、若しくは(ii)(a)の配列を有するポリペプチドに特異的に結合する抗体を生成することができる免疫原性活性を有する、請求項1に記載の核酸によりコードされるアミノ酸配列、及び/又はその酵素的に活性な断片;
(c)アルドラーゼ活性を有するが、シグナル配列を欠く(a)又は(b)のポリペプチド;
(d)アルドラーゼ活性を有し、異種配列をさらに含む(a)、(b)又は(c)のポリペプチド;又は、
(e)ポリペプチドが少なくとも1個の糖鎖付加部位を含む、(a)、(b)、(c)若しくは(d)のポリペプチド、
を含む、前記ポリペプチド若しくはペプチド。
【請求項7】
請求項6に記載のポリペプチドに特異的に結合する、単離された、合成又は組換え抗体。
【請求項8】
組換えポリペプチドを製造する方法であって、
(a)請求項1に記載の核酸を提供する工程;及び
(b)ポリペプチドの発現を可能にする条件下で、工程(a)の核酸を発現させることにより、組換えポリペプチドを製造する工程、
を含む、前記方法。
【請求項9】
アルドラーゼ活性を有するポリペプチドをコードする核酸の変異体を作製する方法であって、
(a)請求項1に記載の核酸を含む鋳型核酸を提供する工程;及び
(b)鋳型配列において1個以上のヌクレオチドを改変、欠失若しくは付加するか、又はその組合せを行って、鋳型核酸の変異体を作製する工程、
を含む、前記方法。
【請求項10】
アルドラーゼポリペプチドをコードする核酸中のコドンを改変するための方法である、請求項9に記載の方法であって、下記工程:
(a)請求項1に記載の核酸を提供する工程;及び
(b)工程(a)の核酸中のコドンを同定し、それを、置換されるコドンと同じアミノ酸をコードする異なるコドンと置換し、それによってアルドラーゼをコードする核酸中のコドンを改変する工程、
を含む、前記方法。
【請求項11】
請求項6に記載のアミノ酸配列の、アミノ末端残基1〜14、1〜15、1〜16、1〜17、1〜18、1〜19、1〜20、1〜21、1〜22、1〜23、1〜24、1〜25、1〜26、1〜27、1〜28、1〜29、1〜30、1〜31、1〜32、1〜33、1〜34、1〜35、1〜36、1〜37、1〜38、1〜40、1〜41、1〜42、1〜43若しくは1〜44に記載のアミノ酸配列から成る、単離された、合成若しくは組換えシグナル又はリーダー配列。
【請求項12】
組成物中の炭素-炭素結合を切断する方法であって、下記工程:
(a)請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを提供する工程;
(b)炭素-炭素結合を含む組成物を提供する工程;及び
(c)アルドラーゼが該組成物中の炭素-炭素結合を切断する条件下で、工程(a)のポリペプチドを工程(b)の組成物と接触させる工程、
を含む、前記方法。
【請求項13】
炭素-炭素結合を形成させる方法であって、下記工程:
(a)請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを提供する工程;
(b)供与体及び受容体化合物を提供する工程;及び
(c)アルドラーゼが炭素-炭素結合を形成する条件下で、工程(a)のポリペプチドを工程(b)の化合物と接触させる工程、
を含む、前記方法。
【請求項14】
請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを含む組成物であって、
必要に応じて、アルドラーゼを、錠剤、ゲル、ピル、埋込み物、液体、スプレー、粉末、食品、飼料ペレットとして、若しくはカプセル化製剤として製剤化し、
必要に応じて、該組成物は、食品、飼料、飲料、食品添加物、飼料添加物、飲料添加物、栄養補助食品、パン生地若しくはパン製品であり、並びに
必要に応じて、該組成物は燃料であり、必要に応じて、該燃料はエタノール、メタノール、プロパノール、ブタノール及び/若しくはディーゼルである、
前記組成物。
【請求項15】
請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドと、炭素-炭素結合を含有する化合物を含む組成物とを接触させることを含む、食品、飼料又は飲料を作製する方法。
【請求項16】
4-置換D-グルタミン酸を作製する方法であって、下記工程:
(a)請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを提供する工程;
(b)α-ケト酸受容体及びピルビン酸若しくはピルビン酸供与体を提供する工程;及び
(c)アルドラーゼが、4-置換D-グルタミン酸の合成を触媒する条件下で、工程(a)のポリペプチドを工程(b)の化合物と接触させる工程、
を含み、
必要に応じて、該ポリペプチドが、ピルビン酸アルドラーゼ、HMGアルドラーゼ及び/若しくはKHGアルドラーゼ活性を有し、並びに
必要に応じて、該方法がさらに、D-アミノトランスフェラーゼの使用を含む、
前記方法。
【請求項17】
抗生物質としての請求項16に記載の4-置換D-グルタミン酸の使用。
【請求項18】
3,4-置換2-ケト-グルタル酸を作製する方法であって、下記工程:
(a)請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを提供する工程;
(b)供与体及び受容体化合物を提供する工程;及び
(c)アルドラーゼが、3,4-置換2-ケト-グルタル酸の合成を触媒する条件下で、工程(a)のポリペプチドを工程(b)の化合物と接触させる工程、
を含み、
必要に応じて、該ポリペプチドが、ピルビン酸アルドラーゼ、HMGアルドラーゼ及び/若しくはKHGアルドラーゼ活性を有し、並びに
必要に応じて、供与体及び受容体が、ピルビン酸若しくはピルビン酸供与体並びにα-ケト酸受容体、ケトン及び/若しくはアルデヒドである、
前記方法。
【請求項19】
バイオマス又は任意のリグノセルロース材料を燃料に変換する方法であって、該バイオマス又はリグノセルロース材料を、請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチド又はその酵素的に活性な断片と接触させることを含み、
必要に応じて、該バイオマス又はリグノセルロース材料を予備処理した後、アルドラーゼと接触させ、
必要に応じて、該バイオマス又はリグノセルロース材料を、セルロース及び/若しくはヘミセルロース分解酵素と接触させた後、アルドラーゼと接触させ、並びに
必要に応じて、該燃料が、エタノール、メタノール、プロパノール、ブタノール及び/若しくはディーゼルである、
前記方法。
【請求項1】
(a)少なくとも1個のポリペプチドをコードする核酸であって、該核酸がcDNA、転写物(mRNA)若しくは遺伝子の完全長に渡って、配列番号243の配列に対して、少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上、若しくは100%の配列同一性を有する配列を含み、
該核酸が、アルドラーゼ活性を有する少なくとも1個のポリペプチドをコードするか、又はアルドラーゼ特異的抗体を生成することができるエピトープ若しくは免疫原として作用するポリペプチド若しくはペプチドをコードする、前記核酸;
(b)少なくとも1個のポリペプチドをコードする核酸であって、該核酸が、(a)の配列を含む核酸の相補体に、ストリンジェントな条件下でハイブリダイズする配列を含み、
該核酸が、アルドラーゼ活性を有する少なくとも1個のポリペプチドをコードするか、又はアルドラーゼ特異的抗体を生成することができるエピトープ若しくは免疫原として作用するポリペプチド若しくはペプチドをコードする、前記核酸;
(c)アルドラーゼ活性を有する少なくとも1個のポリペプチドをコードする核酸であって、該ポリペプチドが、配列番号244の配列又はその酵素的に活性な断片を含む、前記核酸;
(d)アルドラーゼ活性を有するが、シグナル配列を欠くポリペプチドをコードする(a)、(b)若しくは(c)の核酸;
(e)アルドラーゼ活性を有するポリペプチドをコードし、異種配列をさらに含む(a)、(b)、(c)若しくは(d)の核酸;又は、
(f)(a)、(b)、(c)、(d)若しくは(e)と相補的な核酸配列、
を含む、単離された、合成又は組換え核酸。
【請求項2】
(a)請求項1に記載の配列を含む核酸を増幅することができるか、又は、
(b)請求項1に記載の配列の最初(5'側)の10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35個以上の残基の配列を有する第1のメンバー、及び該第1のメンバーの相補鎖の最初(5'側)の10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35個以上の残基の配列を有する第2のメンバーを含む、
増幅プライマー対であって、
必要に応じて、(a)の増幅プライマー配列対のメンバーが、該配列の少なくとも10〜50個の連続する塩基、若しくは該配列の約10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35個以上の連続する塩基を含むオリゴヌクレオチドを含む、前記プライマー対。
【請求項3】
請求項1に記載の核酸を含む発現カセット、ベクター又はクローニングビヒクル。
【請求項4】
請求項1に記載の核酸を含むか、又は請求項3に記載の発現カセット、ベクター若しくはクローニングビヒクルを含む形質転換細胞。
【請求項5】
請求項1に記載の核酸を含むか、又は請求項3に記載の発現カセット、ベクター若しくはクローニングビヒクル、請求項4に記載の形質転換細胞を含むトランスジェニック非ヒト動物、植物、植物部分又は種子。
【請求項6】
アルドラーゼ活性を有する単離された、合成又は組換えポリペプチド若しくはペプチドであって、
(a)該ポリペプチド若しくは酵素の完全長に渡って、配列番号244に対して、少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上、若しくは100%の配列同一性を有するアミノ酸配列、及び/又はその酵素的に活性な断片;
(b)ポリペプチドが、(i)アルドラーゼ活性を有するか、若しくは(ii)(a)の配列を有するポリペプチドに特異的に結合する抗体を生成することができる免疫原性活性を有する、請求項1に記載の核酸によりコードされるアミノ酸配列、及び/又はその酵素的に活性な断片;
(c)アルドラーゼ活性を有するが、シグナル配列を欠く(a)又は(b)のポリペプチド;
(d)アルドラーゼ活性を有し、異種配列をさらに含む(a)、(b)又は(c)のポリペプチド;又は、
(e)ポリペプチドが少なくとも1個の糖鎖付加部位を含む、(a)、(b)、(c)若しくは(d)のポリペプチド、
を含む、前記ポリペプチド若しくはペプチド。
【請求項7】
請求項6に記載のポリペプチドに特異的に結合する、単離された、合成又は組換え抗体。
【請求項8】
組換えポリペプチドを製造する方法であって、
(a)請求項1に記載の核酸を提供する工程;及び
(b)ポリペプチドの発現を可能にする条件下で、工程(a)の核酸を発現させることにより、組換えポリペプチドを製造する工程、
を含む、前記方法。
【請求項9】
アルドラーゼ活性を有するポリペプチドをコードする核酸の変異体を作製する方法であって、
(a)請求項1に記載の核酸を含む鋳型核酸を提供する工程;及び
(b)鋳型配列において1個以上のヌクレオチドを改変、欠失若しくは付加するか、又はその組合せを行って、鋳型核酸の変異体を作製する工程、
を含む、前記方法。
【請求項10】
アルドラーゼポリペプチドをコードする核酸中のコドンを改変するための方法である、請求項9に記載の方法であって、下記工程:
(a)請求項1に記載の核酸を提供する工程;及び
(b)工程(a)の核酸中のコドンを同定し、それを、置換されるコドンと同じアミノ酸をコードする異なるコドンと置換し、それによってアルドラーゼをコードする核酸中のコドンを改変する工程、
を含む、前記方法。
【請求項11】
請求項6に記載のアミノ酸配列の、アミノ末端残基1〜14、1〜15、1〜16、1〜17、1〜18、1〜19、1〜20、1〜21、1〜22、1〜23、1〜24、1〜25、1〜26、1〜27、1〜28、1〜29、1〜30、1〜31、1〜32、1〜33、1〜34、1〜35、1〜36、1〜37、1〜38、1〜40、1〜41、1〜42、1〜43若しくは1〜44に記載のアミノ酸配列から成る、単離された、合成若しくは組換えシグナル又はリーダー配列。
【請求項12】
組成物中の炭素-炭素結合を切断する方法であって、下記工程:
(a)請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを提供する工程;
(b)炭素-炭素結合を含む組成物を提供する工程;及び
(c)アルドラーゼが該組成物中の炭素-炭素結合を切断する条件下で、工程(a)のポリペプチドを工程(b)の組成物と接触させる工程、
を含む、前記方法。
【請求項13】
炭素-炭素結合を形成させる方法であって、下記工程:
(a)請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを提供する工程;
(b)供与体及び受容体化合物を提供する工程;及び
(c)アルドラーゼが炭素-炭素結合を形成する条件下で、工程(a)のポリペプチドを工程(b)の化合物と接触させる工程、
を含む、前記方法。
【請求項14】
請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを含む組成物であって、
必要に応じて、アルドラーゼを、錠剤、ゲル、ピル、埋込み物、液体、スプレー、粉末、食品、飼料ペレットとして、若しくはカプセル化製剤として製剤化し、
必要に応じて、該組成物は、食品、飼料、飲料、食品添加物、飼料添加物、飲料添加物、栄養補助食品、パン生地若しくはパン製品であり、並びに
必要に応じて、該組成物は燃料であり、必要に応じて、該燃料はエタノール、メタノール、プロパノール、ブタノール及び/若しくはディーゼルである、
前記組成物。
【請求項15】
請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドと、炭素-炭素結合を含有する化合物を含む組成物とを接触させることを含む、食品、飼料又は飲料を作製する方法。
【請求項16】
4-置換D-グルタミン酸を作製する方法であって、下記工程:
(a)請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを提供する工程;
(b)α-ケト酸受容体及びピルビン酸若しくはピルビン酸供与体を提供する工程;及び
(c)アルドラーゼが、4-置換D-グルタミン酸の合成を触媒する条件下で、工程(a)のポリペプチドを工程(b)の化合物と接触させる工程、
を含み、
必要に応じて、該ポリペプチドが、ピルビン酸アルドラーゼ、HMGアルドラーゼ及び/若しくはKHGアルドラーゼ活性を有し、並びに
必要に応じて、該方法がさらに、D-アミノトランスフェラーゼの使用を含む、
前記方法。
【請求項17】
抗生物質としての請求項16に記載の4-置換D-グルタミン酸の使用。
【請求項18】
3,4-置換2-ケト-グルタル酸を作製する方法であって、下記工程:
(a)請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチドを提供する工程;
(b)供与体及び受容体化合物を提供する工程;及び
(c)アルドラーゼが、3,4-置換2-ケト-グルタル酸の合成を触媒する条件下で、工程(a)のポリペプチドを工程(b)の化合物と接触させる工程、
を含み、
必要に応じて、該ポリペプチドが、ピルビン酸アルドラーゼ、HMGアルドラーゼ及び/若しくはKHGアルドラーゼ活性を有し、並びに
必要に応じて、供与体及び受容体が、ピルビン酸若しくはピルビン酸供与体並びにα-ケト酸受容体、ケトン及び/若しくはアルデヒドである、
前記方法。
【請求項19】
バイオマス又は任意のリグノセルロース材料を燃料に変換する方法であって、該バイオマス又はリグノセルロース材料を、請求項6に記載のポリペプチド若しくは請求項1に記載の核酸によりコードされるポリペプチド又はその酵素的に活性な断片と接触させることを含み、
必要に応じて、該バイオマス又はリグノセルロース材料を予備処理した後、アルドラーゼと接触させ、
必要に応じて、該バイオマス又はリグノセルロース材料を、セルロース及び/若しくはヘミセルロース分解酵素と接触させた後、アルドラーゼと接触させ、並びに
必要に応じて、該燃料が、エタノール、メタノール、プロパノール、ブタノール及び/若しくはディーゼルである、
前記方法。
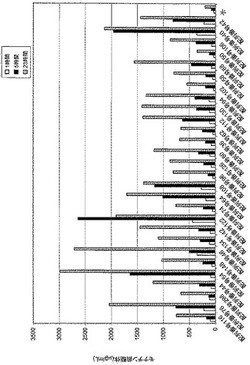
【図1】


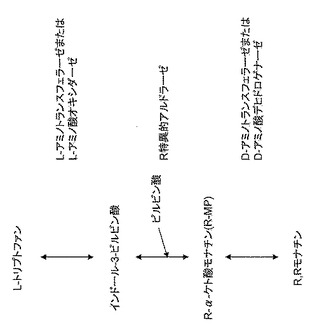
【図2】

【図3】
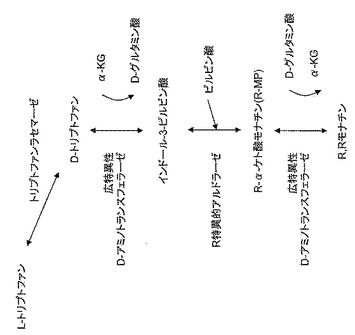
【図4】
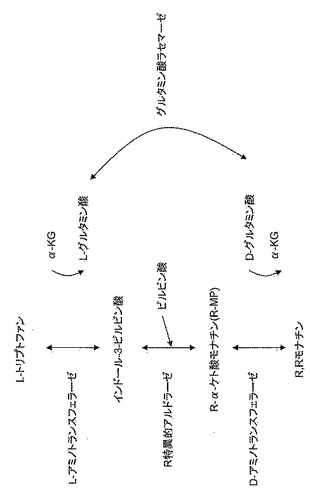
【図5】
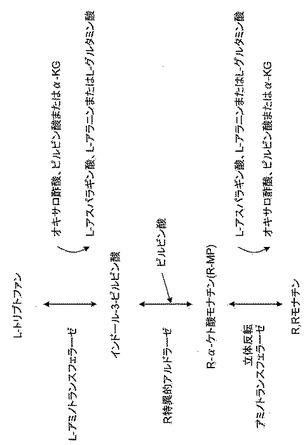
【図6】
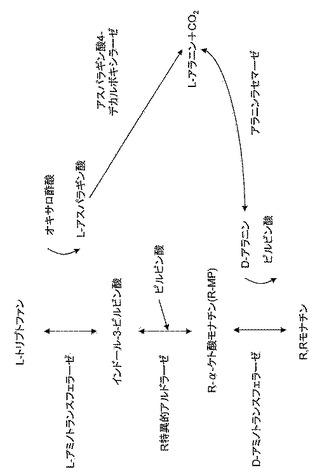
【図7】
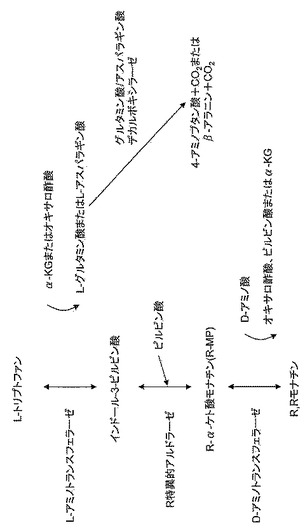
【図8】
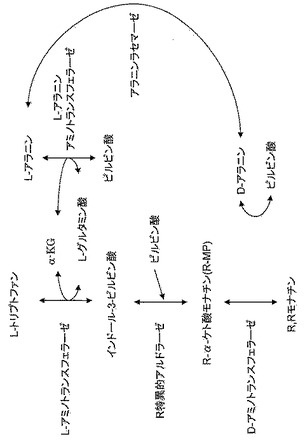
【図9】
【図10】

【図11】
【図12】

【図13】

【図14】
【図15】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2013−66469(P2013−66469A)
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2012−225075(P2012−225075)
【出願日】平成24年10月10日(2012.10.10)
【分割の表示】特願2008−558521(P2008−558521)の分割
【原出願日】平成19年3月7日(2007.3.7)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
【出願人】(508268621)ヴェレニウム コーポレーション (3)
【Fターム(参考)】
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願日】平成24年10月10日(2012.10.10)
【分割の表示】特願2008−558521(P2008−558521)の分割
【原出願日】平成19年3月7日(2007.3.7)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
【出願人】(508268621)ヴェレニウム コーポレーション (3)
【Fターム(参考)】
[ Back to top ]