
アレルギー治療に用いるカベイラクサ(Parietariajudaica)の脂質輸送タンパク質ファミリーに属する低アレルギー誘導性のキメラタンパク質
アレルギーの治療に用いるカベイラクサの脂質輸送ファミリーに属する低アレルギー誘発性のキメラタンパク質。本発明は、アレルギー、特に花粉症の予防及び治療に用いることができるカベイラクサの様々なアレルゲンのキメラポリペプチドをコードする組み換えDNA分子に関する。具体的には、低アレルギー誘発性の特性があるアレルゲンPar j 1及びPar j 2の断片から構成されたキメラポリペプチドが記載される。異種発現系でこれらの組み換えポリペプチドを製造する方法も記載されている。キメラタンパク質を精製するのに有効な方法も記載されている。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アレルギー、特に花粉症、最も詳細には脂質輸送タンパク質ファミリー由来のアレルゲン及びより具体的にはパリエタリア(Parietaria)種由来の花粉で見出されるアレルゲンによって引き起こされるアレルギーの予防及び治療のためのキメラタンパク質産生の分野に関する。
【背景技術】
【0002】
I型アレルギーは、先進国における深刻な健康問題である。この種のアレルギーは、空気によって運ばれる抗原に対するIgE抗体の形成によって引き起こされる。これらのIgE抗体は、肥満細胞及び好塩基球と相互作用し、ヒスタミン等の生物学的メディエータを放出し、集団の約20%でアレルギー性鼻炎、アレルギー性結膜炎及びアレルギー性気管支喘息を引き起こす[(1)Miyamoto, T.(1992)著「日本の花粉症有病率の増加(Increased prevalence of pollen allergy in Japan)」 In Advances in Allergology and Clinical Immunology. P. Godard, J. Bousquet及びF. B. Michel編(Cornforth, UK: The Parthenon Publishing Group), pp. 343-347]。
【0003】
特異的免疫療法(SIT)は、特異的なアレルゲンによって誘発されるアレルギー反応に効果的な治療であり、基本的にはアレルギーを生じるタンパク質(アレルゲン抽出物)を高濃度で定期的に投与することによって患者における免疫反応を調節することから成る。高用量のアレルゲンを注入すると、抗原提示細胞、例えば樹状細胞(好ましくは、協働するバージン細胞(virgin cells)(nTH)であるT細胞がTH1又はTH0に発達することを促進する)によってIL−12の高度合成が誘導される。このことが、TH2細胞に関連するアレルギー応答型の免疫応答から、TH1/TH0型の応答への偏向(deviation)を可能にし、高レベルのIFN−γが産生される[(2)Akdis, CA.及びBlaser, K.(2000)著「アレルゲン特異的免疫療法のメカニズム(Mechanisms of allergen-specific immunotherapy)」 Allergy 55, 522-530]。免疫偏向は、免疫抑制サイトカインIL−10及びTGF−βを産生する制御性T細胞(TR1)の影響下でTH2記憶細胞の寛容(アンエネルギー(anenergy)又はクローン除去)の誘導によって強まる[(3)Akdis, C.A., Joss, A., Akdis, M.及びBlaser, K.(2001)著「アレルギー炎症におけるIL−10誘導性の細胞不活性化及びアレルゲンに対する正常な応答のメカニズム(Mechanism of IL-10 induced cell inactivation in allergic inflammation and normal response to allergens)」 Int. Arch Allergy Immunol. 124; 180-182]。TH2細胞の活性化及び増殖の低減は、IL−4及びB細胞によるIgEの産生の低減によって達成される。TH2細胞の活性、並びに鼻粘液及び気管支粘液への浸潤の低減によって、IL−5合成が低減し、これによって好エオシン性浸潤が低減され、MBPタンパク質及びECPタンパク質等の炎症性メディエータの放出が大幅に低減する。優性表現型アレルゲンTH0のT細胞の新規の特異的なクローンは、B細胞による、大量の特異的なIgGアレルゲン抗体の産生を促進するTH1型サイトカインとTH2型サイトカインとの混合物を産生する。一方で、高レベルのIL−10によって、特異的なIgG4アレルゲン抗体の高度合成が誘導される。これらの2種類の特異的な抗体は、肥満細胞においてこれらの受容体と結合したIgE抗体の交差(intersection)を与え、このようにしてヒスタミンの脱顆粒及び放出を阻害する遮断抗体として作用することができる[(4)Moverate, R.(2003)著「花粉ワクチンによる特異的な免疫療法の免疫機構:診断法に関する示唆及び改善されたワクチン接種戦略の開発(Immunological mechanisms of specific immunotherapy with pollen vaccines: implications for diagnostics and the development of improved vaccination strategies)」 Expert Rev. Vacc. 2, 85-97、(5)Wachholz, P.A., Soni, N. K., Till, S.及びDurham, S. R.(2003)著「イネ科花粉の免疫療法後のIgG抗体によるB細胞とのアレルゲン−IgE結合の阻害(Inhibition of allergen-IgE binding to B cells by IgG antibodies after grass pollen immunotherapy)」 J. Allergy Clin. Immunol. 112; 915-922]。これらは、抗原含有細胞によるIgE媒介性の抗原の回収も阻害し、このことはアレルゲンに対する免疫応答を抑制する。
【0004】
天然源から単離されたアレルゲン抽出物は、タンパク質と他の分子との複雑な混合物である。これらの組成、ひいてはアレルギー誘発性は用いられる物質に依存し、周囲条件、花粉の場合の成熟状態、及び菌類等の場合のダニ等の成長条件によって変わる。幾つかの抽出物は主要なアレルゲンを不十分な濃度でしか含有することができず、また患者がアレルギーではない望ましくない成分で汚染される可能性があるか、又はその両方である。現行の免疫療法は、完全なアレルゲン抽出物を排他的に用い、このことによって、
−ワクチンとエフェクター細胞のIgEとの反応性による深刻な拒絶反応、
−免疫療法後のワクチンに存在する他のアレルゲンに対する新たな感作の発生、
−アレルゲン抽出物の標準化の困難性等の多くの問題が生じる。
【0005】
これは全て、免疫療法が、要望どおりの安全且つ効率的な治療ではないことを意味する。アレルギーの病因及び特異的な免疫療法のメカニズムのより良好な知識によって、上記問題の解決に近づくことができるようになった。特異的なアレルゲン応答性TH2におけるIgE媒介性抗原の影響を知ることで、IgEと結合しないアレルゲンを作製することがますます試みられている。現行の特異的な免疫療法の主な目的は、IgEエピトープを不活性化し、このようにしてIgEとの結合、及びその結果として拒絶反応を低減及びさらには除去するという目的でアレルゲンを修飾することである[(6)Valenta, R.及びLinhart, B.(2005)著「アレルギーワクチンの分子設計(Molecular design of allergy vaccines)」 Curr. Opin. Immunol. 17, 1-10]。このように、修飾アレルゲンは、食作用/飲作用媒介性の抗原回収機構によってT細胞に向けられ、IgEの交差及びIgE依存性抗原の提示を防ぐ。このことは、T細胞によるTH0又はTH1サイトカインの産生と、IgEの産生減少と、B細胞によるIgGの産生増大との平衡化を誘導する。このこと全てによって、アナフィラキシーの危険性なく、TH2型のT細胞寛容が誘導される。アレルゲン及びアレルゲン誘導体を得る組み換え方法の進歩によって、アレルギー治療のために新規のワクチンを開発する能力が大きく増大している。このことは、IgEエピトープの有意なアミノ酸を突然変異又は欠失する可能性、並びに低アレルギー誘発性ワクチンを得るためのこれらの分別及びオリゴマー化により可能になった。IgEと結合する能力が低減したが、T細胞との反応性を維持するこれらの分子は、より大用量で投与することができ、注入回数を少なくして、免疫療法をより迅速に且つ安全にすることができる。さらに、組み換えアレルゲンは、微生物発現系を用いて、発酵タンクで大規模に製造することができ、その精製は、天然の等価物よりも効率的で且つ安価である。免疫療法における低アレルギー誘発性誘導体の使用は、三量体のBet v 1断片[(7)Niederberger, V., Horak, F., Vrtala, S., Spitzauer, S., Krauth, M. T., Valent, P., Reisinger, J., Pelzmann, M., Hayek, B., Kronqvist, M., Gafvelin, G., Groenlund, H., Purohit, A., Suck, R., Fiebig, H., Cromwell, O., Pauli, G., van Hage-Hamsten, M.及びValenta, R.(2004)著「遺伝子操作したアレルゲンによるワクチン接種はアレルギー疾患の進行を防ぐ(Vaccination with genetically engineered allergens prevents progression of allergic disease)」 Proc. Natl. Acad. Sci. U.S.A. 101, 14677-14682]、ハチ毒(bee venon)タンパク質の多アレルギー性ハイブリッド(Api m 1、2、3)[(8)Schmid-Grendelmeier, P., Karamloo, F., Mueller, U., Housley-Marcovic, Z., Soldatova, L., Zumkehr, J., Kemeny, D.M., Kuendig, T., Reimers, A., von Beust, B.R., Salagianni, M., Akdis, M., Kussebi, F., Spangfort, M. D., Blaser, K.及びAkdis, C.A.(2005)著「IgE結合を低減し、T細胞エピトープを保存した、組み換え型の多アレルゲンワクチンによるアレルギーの予防(Prevention of allergy by a recombinant multi-allergen vaccine with reduced IgE binding and preserved T cell epitopes)」 Eur. J. Immunol. 35, 3268-3276]、及びそれらの三次構造を除去するためのPar j 2とPar j 1との突然変異融合物を用いてこれまでに記載されている。
【0006】
何人かの著者は、IgEとの結合が、高親和性及び低親和性の両方のIgEに対する表面受容体を発現する、樹状細胞及び活性化Bリンパ球細胞としてのプロフェッショナル抗原提示細胞によって、アレルゲンの捕捉及び提示を容易にし得るので、アレルギーワクチンは低アレルギー誘発性のアレルゲン(hypoallergens)では構成されていないはずであることを言及してる。このプローチは、免疫療法において舌下経路を用いる場合に特別な意味がある。また、高親和性の受容体の交差によって、アレルゲンに対するT細胞の応答が低減され得る[(9)Allam, J.P., Novak, N., Fuchs, C., Asen, S., Berge, S., Appel, T., et al.(2003)著「ヒトの口腔粘膜由来の樹状細胞の特徴付け:構成的なFcεRIの発現が高い、新規のランゲルハンス細胞型(Characterization of dendritic cells from human oral mucosa: a new Langerhans' cell type with high constitutive FcεRI expression)」 J. Allergy Clin. Immunol. 112, 141-8、(10)von Bubnoff, D., Matz, H., Frahnert, C., Rao, M. L., Hanau, D., de la Salle, H., Bieber, T.(2003)著「FcεRIはT細胞応答の調節に関与するトリプトファン分解経路を誘導する(FcεRI induces the triptophan degradation pathway involved in regulating T cell responses)」 J. Immunol. 169, 1810-1816]。多くの研究者らは未だに、欠失を導入することなくこの種のワクチンを製造している[(11)Batard, T., Didierlaurent, A., Chabre, H., Mothes, N., Bussieres, L., Bohle, B., et al.(2005)著「カバノキ花粉症に対する候補ワクチンとしての野生型の組み換えBet v 1aの特徴付け(Characterization of wild-type recombinant Bet v 1a as a candidate vaccine against birch pollen allergy)」 Int. Arch. Allergy Immunol. 136, 239-249、(12)Jutel, M., Jaeger, L., Suck, R., Meyer, H., Fiebig, H., Cromwell, O.(2005)著「組み換えイネ科花粉アレルゲンによるアレルゲン特異的な免疫療法(Allergen-specific immunotherapy with recombinant grass pollen allergens)」 J. Allergy Clin. Immunol. 116, 608-13、(13)Niederberger, V., Horak, F., Vrtala, S., Spitzauer, S., Krauth, M.T., Valent, P., et al.(2004)著「遺伝子操作したアレルゲンによるワクチン接種はアレルギー疾患の進行を防ぐ(Vaccination with genetically engineered allergens prevents progression of allergic disease)」 Proc. Natl. Acad. Sci. U.S.A. 101, 14677-82、(14)Cromwell, O., Fiebig, H., Suck, R., Kahlert, H., Nandy, A., Kettner, J., et al.(2006)著「組み換えアレルゲンワクチンの戦略及び最初の臨床研究の有益な結果(Strategies for recombinant allergen vaccines and fruitful results from first clinical studies)」 Immunol. Allergy Clin. N. Am. 26, 261-81]。
【0007】
パリエタリアは、草本性双子葉植物綱(genus)イラクサ科イラクサ目である。パリエタリア属の様々な種が地中海沿岸に幅広く豊富に分布している[(15)Colombo, P., Duro, G., Costa, M.A., Izzo, V., Mirisola, M., Locorotondo, G., Cocchiara, R.及びGeraci, D.(1998)著「アレルゲンの最新情報。パリエタリア花粉のアレルゲン(An update on allergens. Parietaria pollen allergens)」 Allergy 53, 917-921]。最も一般的な種は、カベイラクサ及びパリエタリア・オフィキナリス(P. officinalis)であるが、パリエタリア・ルシタニカ(P. lusitanica)、パリエタリア・マウリタニカ(P. mauritanica)、及びパリエタリア・クレティカ(P. cretica)等の他の種が幾つかの地域で幾らか存在し得る。それにもかかわらず、パリエタリア花粉の存在がイングランド南部、オーストリア、中東欧の温帯地方、オーストラリア及びカリフォルニアで示されたので、地中海地域は、パリエタリア花粉が見出された唯一の場所というわけではない[(16)Colombo, P., Bonura, A., Costa, M., Izzo, V., Passantino, R., Licorotondo, G., Amoroso, S.及びGerasi, D.(2003)著「パリエタリアのアレルゲン(The allergens of Parietaria)」 Int. Arch. Allergy Immunol. 130, 173-179、(17)Carreira, J.及びPolo, F.(1995)著「オリーブ及びパリエタリア種のアレルゲン並びに地中海地方におけるこれらの関連性(The allergens of Olea europaea and Parietaria spp. and their relevance in the Mediterranean Area)」 Allergy Clin. Immunol. News 7, 79-84]。パリエタリアの顕著な特徴は、数ヶ月間続く長い受粉期間であり、パリエタリアに対してアレルギーである患者において、軽度の鼻結膜炎から重症喘息までに及ぶ、ほとんど永続的な症状が現れる。顕著な交差反応性が様々な種のパリエタリア間で示されたので、パリエタリアに対する通常の軽い単一特異的な感作は、この属の様々な種に対する感作を伴う。
【0008】
2つの最も一般的な種であるカベイラクサ及びパリエタリア・オフィキナリスのアレルギー誘発性画分の精製及び特徴付けに関して様々な文献が提示されている。これらの画分の分子量は、10kDa〜14kDaの範囲であり、これらの抽出物のアレルギー誘発力全体に実質的に関与している[(16)Colombo, P., Bonura, A., Costa, M., Izzo, V., Passantino, R., Licorotondo, G., Amoroso, S.及びGerasi, D.(2003)著「パリエタリアのアレルゲン(The allergens of Parietaria)」 Int. Arch. Allergy Immunol. 130, 173-179、(18)Ayuso, R., Carreira, J., Lombardero, M., Duffort, O., Peris, A., Basomba, A.及びPolo, F.(1993)著「2つのPar jイソアレルゲンの、mAbに基づいたアフィニティクロマトグラフィによる単離。これらの物理化学、免疫化学及びアレルギー誘発性の特性の比較(Isolation by mAb based affinity chromatography of two Par j isoallergens. Comparison of their physicochemical, immunochemical and allergenic properties)」 Mol. Immunol. 30, 1347-1354、(19)Polo, F., Ayuso, R.及びCarreira, J.(1990)著「カベイラクサ花粉の主要なアレルゲンのHPLC精製(HPLC purification of the main allergen of Parietaria judaica pollen)」 Mol. Immunol. 27, 151-157、(20)Polo, F., Ayuso, R.及びCarreira, J.(1991)著「カベイラクサのアレルゲンIの構造と、IgE結合能との間の関係に対する研究(Studies on the relationship between structure and IgE-binding ability of Parietaria judaica allergen I)」 Mol. Immunol. 28, 169-175]。組み換えDNA技術の発展によって、パリエタリア花粉のアレルゲンの分子の特徴付けを完了させることができた。Par j 1及びPar j 2として知られるカベイラクサ花粉の2つの主要なアレルゲンをクローニング及びシーケンシングした[(21)Duro, G., Colombo, P., Costa, M.A., Izzo, V., Porcasi, R., DiFiore, R., Locorotondo, G., Mirisola, M.G., Cocchiara, R.及びGeraci, D.(1996)著「カベイラクサ花粉の新たな主要なアレルゲンであるPar j 2.0101のcDNAクローニング、配列解析及びアレルギー学的特徴付け(cDNA cloning, sequence analysis and allergological characterization of Par j 2.0101, a new major allergen of the Parietaria judaica pollen)」 FEBS Lett. 399, 295-298、(22)Costa, M.A., Colombo, P., Izzo, V., Kennedy, H., Venturella, S., Cocchiara, R., Mistrello, G., Falagiani, P.及びGeraci, D.(1994)著「カベイラクサ花粉の主要なアレルゲンであるPar j IのcDNAクローニング発現及び一次構造(cDNA cloning expression and primary structure of Par j I, a major allergen of Parietaria judaica pollen)」 FEBS Lett. 341, 182-186、(23)Amoresano, A., Pucci, P., Duro, G., Colombo, P., Costa, M.A., Izzo, V., Lambda, D.及びGeraci, D.(2003)著「カベイラクサ花粉の主要なアレルゲンであるPar j 2.0101のジスルフィド架橋の評価(Assignment of disulphide bridges in Par j 2.0101, a major allergen of Parietaria judaica pollen)」 Biol. Chem. 384, 1165-1172]。両方のアレルゲンが非特異的脂質輸送タンパク質(ns−LTP)のファミリーに属し、これらの末端領域でシグナルペプチドを保有しており、プロセシング後に、分子量がそれぞれ14726Da及び11344Daであり、約45%の同一残基を有するタンパク質を生じる。両方のアレルゲンにおいて構造的関連域に位置する可能性があるIgE結合直線状エピトープが記載されている[(24)Asturias, J.A., Gomez-Bayon, N., Eseverri, J.L.及びMartinez, A.(2003)著「カベイラクサ花粉の主要なアレルゲンであるPar j 1及びPar j 2は、同様の免疫グロブリンEエピトープを有する(Par j 1 and Par j 2, the major allergens from Parietaria judaica pollen, have similar immunoglobulin E epitopes)」 Clinical and Experimental Allergy 33, 518-524]。カベイラクサ花粉に対するアレルギーの治療に最適な低アレルギー誘発性の分子を得ることができるために、これらの領域は作用性の標的である。
【0009】
ns−LTPは、in vitroでの極性脂質の膜間の交換及び/又は移動を完了させることができることが知られている[(25)van Ree, R.(2002)著「食物アレルゲンとしての非特異的な脂質輸送タンパク質の臨床的重要性(Clinical importance of non-specific lipid transfer proteins as food allergens)」 Biochem. Soc. Trans 30, 910-913]。分子質量が約9kDaであるLTP1及び分子質量が約7kDaであるLTP2の2つの主要なファミリーが植物で特徴付けられている。LTPファミリーに属するアレルゲンが、幅広く研究された、食物以外の植物で同定されている。したがって、パラゴムノキ(Hevea brasiliensis)のラテックス由来のHev b 12は、バラ科の果実のアレルギー誘発性LTPと約65%の配列同一性を示す9.3kDaの塩基性タンパク質である[(26)Beezhold, D.H., Hickey, V.L., Kostyal, D.A., and et al.(2003)著「パラゴムノキ由来の脂質輸送タンパク質(Hev b 12)、交差反応性ラテックスタンパク質(Lipid transfer protein from Hevea brasiliensis (Hev b 12), a cross-reactive latex protein)」 Ann Allergy Asthma Immunol 439-445]。さらに、オウショウヨモギ(Artemisia vulgaris)のArt v 3[(27)Diaz-Perales, A., Lombardero, M., Sanchez-Monge, R., and et al.(2000)著「潜在的な植物パンアレルゲンとしての脂質輸送タンパク質:IgE結合能が異なる、ヨモギ属の花粉、クリ属の実、バラ科の果実のタンパク質間の交差反応性(Lipid-transfer proteins as potential plant panallergens: cross-reactivity among proteins of Artemisia pollen, Castanea nut and Rosaceae fruits, with different IgE-binding capacities)」 Clin Exp Allergy 1403-1410]及びオリーブのOle e 7[(28)Tejera, M.L., Villalba, M., Batanero, E.及びRodriguez, R.(1999)著「オリーブの木の花粉の新規のアレルゲンであるOle e 7の同定、単離及び特徴付け(Identification, isolation, and characterization of Ole e 7, a new allergen of olive tree pollen)」 J. Allergy Clin. Immunol. 797-802、(29)Rodriguez, R., Villalba, M., Batanero, E., and et al.(2002)著「オリーブ花粉のアレルギー誘発性の多様性(Allergenic diversity of the olive pollen)」 Allergy 6-16]等の幾つかの花粉アレルゲンがLTPとして記載されており、9kDa〜10kDaであり、食物のアレルギー誘発性LTPと30%〜55%の配列同一性を有する。一方で、互いに約45%の配列同一性を有する、カベイラクサの主要なアレルゲンであるPar j 1及びPar j 2はそれぞれ14.7kDa及び11.3kDaと、LTPファミリーにおける通常のものよりも分子量が高い[(16)Colombo, P., Bonura, A., Costa, M., Izzo, V., Passantino, R., Licorotondo, G., Amoroso, S.及びGerasi, D.(2003)著「パリエタリアのアレルゲン(The allergens of Parietaria)」 Int. Arch. Allergy Immunol. 130, 173-179]。しかし、両方のアレルゲンがLTPと同様の構造を有するが、これらは共通の配列領域で食物LTPとの適度なレベルの同一性を有する(Par j 1とモモのLTPとの間で28%)。
【0010】
国際公開特許第WO2005/085278号は、カベイラクサの2つの主要なアレルゲンを含む融合タンパク質の構築物を開示しており、アレルゲン配列が本質的に同じ長さを維持するように、この融合タンパク質の三次元構造が、それぞれのアレルゲンの一次配列における或る特定のシステイン残基(より具体的には、ジスルフィド架橋の形成を伴うシステイン残基)の置換によって破壊される。後の実験によると、このことが、天然のアレルゲンよりもアレルギー誘発性が1000倍低いタンパク質をもたらすと考えられる。
【0011】
本発明者らは、アレルギー誘発性の驚くほど大きな低減は、アレルゲンの三次元構造を破壊するだけでなく、幾つかのIgE結合部位(Bエピトープとして知られる)の欠失によっても得ることができること、及び最も驚くべきことは、これによって免疫原性が低減されないことを発見している。
【0012】
本発明は、より少数のIgE結合エピトープを含有するカベイラクサの2つのアレルゲン(Par j 1及びPar j 2)の断片を結合させることによって得られた様々なキメラタンパク質、並びにそれらを得るための様々な方法及び中間物を初めて開示する。本発明のキメラタンパク質の三次元構造を破壊するだけでなく、或る特定のBエピトープが欠失されてもいる。本発明によるキメラタンパク質は、i)カベイラクサに対してアレルギーの患者由来の血清混合物を用いるin vitro ELISA、ELISA阻害試験及び免疫検出試験、ii)カベイラクサに対してアレルギーの患者における皮膚反応性のin vivo試験、iii)カベイラクサに対してアレルギーの患者由来の個々の血清によるin vitro EAST阻害試験に基づき、IgE抗体との結合能が低いので、アレルギー誘発性が99.99%低減した低アレルギー誘発性と呼ぶことができる。一方で、カベイラクサに対してアレルギーの13人の患者由来の末梢血単核球(CMSP)のリンパ球増殖試験によって示されるように、本発明によるキメラタンパク質は、免疫原性能を維持する。
【0013】
アレルギー誘発性抽出物は、タンパク質と非タンパク質分子との複雑な混合物である。抽出物成分に対する特定のIgEのレベルを検出する技法の使用を増やすことで、アレルギー患者が通常、様々な成分に対して反応性があることを実証することが可能になっている。単一のアレルゲンにのみ反応するアレルギー患者の症例は珍しい。アレルギー誘発性抽出物には、免疫療法において明らかな問題があるので、単一の分子でできる限り多くの治療的特性をグループ分けすることが1つの解決策である。
【発明の開示】
【発明が解決しようとする課題】
【0014】
本発明者らは、1つの分子中に2つのアレルゲンを混合させることに成功し、このことによって、産業上の製造及び治療の観点から利益を受けるだけではなく、免疫原性を変えない、アレルギー誘発性の有意な低減も示される。
【課題を解決するための手段】
【0015】
これらの理由で、本発明は、アレルゲンPar j 1及びPar j 2の断片から構成されたキメラタンパク質(以下、Q1、Q2、及びQ3と呼ばれる)に関し、これは、幾つかのIgE結合Bエピトープが欠失しているので、免疫原性能を損なうことなく、アレルギー反応性を低減させる。このように、得られたキメラポリペプチドは、2つの個々のタンパク質の合計よりも分子量が小さい。
【0016】
本発明は、上記のキメラポリペプチドをコードするDNAを含むヌクレオチド配列にも関し、発現系は発現及び制御に必要な配列を伴う当該配列を含み、受容体細胞は当該発現系によって形質転換される。
【0017】
本発明は、このキメラポリペプチドの臨床使用に、アレルギーの治療に特異的な免疫療法に、並びにこのキメラポリペプチドが生じる可能性がある組成物及びその様々な投与方法にも関する。
【0018】
免疫療法において用いられることが可能な本発明のキメラタンパク質の低アレルギー特性が広く実証されている。本発明者らによって行われた免疫検査によって、図10及び図11で示されるように、キメラQ2は、カベイラクサに対してアレルギーの患者の血清中でIgEを認識しないことが示される。図12で示されるように、Q2のIgE結合能は、2つの天然タンパク質の混合物のものの10000倍低い。
【0019】
この低いアレルギー誘発性のデータは、皮膚プリックテストでの30人の患者におけるin vivo実験によって制限を受けた。キメラQ1のアレルギー誘発性は、2つの単離された天然タンパク質で得られたものよりも3.5倍低かった(図13)。他方では、記載のBエピトープを全く含有せず、Par j 1及びPar j 2の断片によって形成された、Q2のアレルギー誘発性は、2つの単離された天然タンパク質で得られたものよりも112倍低かった。
【0020】
キメラQ2の低アレルギー誘発性(IgE結合能)は、カベイラクサに対してアレルギーの30人の患者の血清で、この分子の反応性を測定することによって確認された(図14)。驚くべきことに、このアレルギー誘発性の低減は、個々の天然タンパク質を合計したものとは異ならない、キメラQ2の免疫原性能の維持を伴っていた(図16)。
【0021】
免疫原性の維持によって、このキメラを、完全ではあるが、はるかに安全な(アレルギー誘発性が低い)抽出物に対する代替物として用いることが可能になる。
【0022】
株の寄託
本発明に対応する微生物株は、特許手続きのための微生物寄託の国際認識に関するブタペスト条約に従って、バレンシア大学(Universidad de Valencia, Edificio de Investigacion, Campus de Burjasot, 46100 BURJASOT, Valencia)にあるスペインの基準菌株コレクション(Spanish collection of type cultures)(CECT)で2006年3月7日に寄託され、大腸菌株CECT7141の参照番号を有する。
【発明を実施するための最良の形態】
【0023】
本発明の一態様によれば、任意のBエピトープ若しくはIgE結合エピトープ(epitode)の欠失、又は任意のBエピトープ若しくはIgE結合エピトープを用いない合成によって得られた低アレルギー誘発性のキメラタンパク質又はペプチドが提供される。好ましくは、欠失はPar j 1及びPar j 2の28位〜53位でBエピトープに適用される。
【0024】
本明細書で用いられるような「低アレルギー誘発性」という用語及び類似の変化形は相対的な用語であり、アレルギー応答性を刺激する本発明のタンパク質及びペプチドのin vivo及びin vitroの能力が、野生型の免疫原の同じ能力に比べて低減していることに関する。
【0025】
好ましくは、キメラタンパク質は、配列番号2、配列番号4又は配列番号6で示されるアミノ酸配列を含み、最も好ましくは、キメラタンパク質は、配列番号4で示されるアミノ酸配列を含む。
【0026】
代替的に又は付加的に、キメラタンパク質は、配列番号2、配列番号4又は配列番号6で示されるアミノ酸配列と相同な配列を含む。好ましくは、相同配列は、配列番号2、配列番号4又は配列番号6で示されるアミノ酸配列、最も好ましくは配列番号4で示されるアミノ酸配列に対する相同性が少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは100%である。
【0027】
別の好ましい実施形態には、配列番号2、配列番号4又は配列番号6で示されるアミノ酸配列に対する相同性が少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%、最も好ましくは100%であるアミノ酸配列を有するキメラタンパク質又はペプチドが含まれる。
【0028】
キメラタンパク質は、精製を容易にするペプチド配列を含んでいてもよい。このようなペプチドは、当該技術分野で一般的に知られており、例えばポリヒスチジンテイル(polyhistidine tail)である。
【0029】
キメラタンパク質を構成する断片は、ポリメラーゼ連鎖反応(PCR)における増幅による既知のスキームに従って、資格があり訓練を受けた人によって合成することができる。当該断片は、適切な制限酵素で消化した後に、ライゲーションによって発現ベクターに統合することができる。この発現ベクター(市販のベクターpQE32等)は、キメラタンパク質を、精製を助ける配列(例えばアミノ末端に位置する一連のヒスチジン)と融合させる能力を有し得る。キメラタンパク質の構築中に、異なるDNA断片が、異なる制限酵素によって認識配列で形成されたリンカーを介して結合され、したがって天然アレルゲンの元の配列には存在しなかった残基は最終キメラ分子に現れる。タンパク質を正しく読み取るのを妨げなかったこれらの新規の残基は、図2、図5及び図8における配列で標識され、関連の実施例で適切に記載される。同様に、キメラは、元の配列に存在しない14個の残基をアミノ末端域に有し、固体支持体と結合した二価金属との相互作用によって迅速に精製されることができる高ヒスチジン領域に対応する。
【0030】
本発明の第2の態様によれば、本発明のタンパク質又はペプチドをコードするポリヌクレオチドが提供される。
【0031】
好ましくは、ポリヌクレオチドは、配列番号1、配列番号3又は配列番号5で示されるヌクレオチド配列を含み、最も好ましくは、ポリヌクレオチドは、配列番号3で示されるヌクレオチド配列を含む。
【0032】
代替的に又は付加的に、ポリヌクレオチドは、配列番号1、配列番号3又は配列番号5で示されるヌクレオチド配列と相同な配列を含み得る。好ましくは、相同配列は、配列番号1、配列番号3又は配列番号5で示されるヌクレオチド配列に対する相同性が少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%である。
【0033】
別の好ましい実施形態には、配列番号1、配列番号3又は配列番号5で示されるヌクレオチド配列に対する相同性が少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%、最も好ましくは100%であるヌクレオチド配列を有するポリヌクレオチドが含まれる。
【0034】
ポリヌクレオチドは、シグナルペプチドをコードする配列をさらに含み得る。シグナルペプチドは、小胞体の膜を通るタンパク質輸送を開始させるアミノ酸配列である。好適なシグナルペプチドは、本発明の分野における当業者に既知である。
【0035】
本発明には、本発明のポリヌクレオチド配列によってコードされたペプチドも含まれる。
【0036】
本発明の他の態様によれば、本発明のポリヌクレオチド配列を含む発現系、及び本発明のタンパク質又はペプチドを発現することができる当該発現系によって形質転換された宿主細胞が提供される。
【0037】
本発明は、本発明のペプチド又はタンパク質を製造する方法も含み、当該方法は、
(i)宿主細胞において、本発明のタンパク質又はペプチドをコードするヌクレオチド配列を発現することができる複製可能な発現系を調製する工程と、
(ii)宿主細胞を上記発現系で形質転換する工程と、
(iii)上記タンパク質又は上記ペプチドを発現させる条件下で上記形質転換した宿主細胞を培養する工程と、
(iv)任意で、上記タンパク質又は上記ペプチドを回収する工程と、
を含む。
【0038】
本発明は、例えば牛乳又は卵白中に本発明のタンパク質又はペプチドを産生することができるトランスジェニック動物も含む。
【0039】
本発明の別の態様によれば、免疫障害、特にアレルギー等の過敏性障害の治療のための本発明のタンパク質又はペプチドの使用が提供される。
【0040】
本発明のさらなる態様によれば、免疫障害、特にアレルギー等の過敏性障害の治療用の薬物調製のための本発明のタンパク質又はペプチドの使用が提供される。好ましくは薬物はワクチンである。
【0041】
好ましくは、過敏性障害は、パリエタリアの花粉、より好ましくはカベイラクサの花粉に対するアレルギーである。
【0042】
本明細書で用いられるように、治療という用語、及び「治療する(treat)」又は「治療すること(treating)」等の変化形は、ヒト又は非ヒト動物で有益であり得る任意のレジーム(regime)を表す。治療は、現状に対するものであり得るか、又は予防的なものであり得る(予防的治療)。治療には、治癒効果、軽減効果又は予防効果が含まれ得る。好ましくは、治療は予防的治療である。
【0043】
本発明のまたさらなる態様によれば、効果的な量の本発明のタンパク質又はペプチドと、薬学的に許容可能な賦形剤とを含む薬学的組成物が提供される。好ましくは、薬学的組成物はワクチン組成物である。
【0044】
本発明のまたさらなる態様によれば、免疫障害、特にアレルギー等の過敏性障害を治療する方法が提供され、当該方法は、治療が必要な被験者に、効果的な量の本発明のタンパク質又はペプチドを投与する工程を含む。
【0045】
本発明の別の態様によれば、免疫障害、特にアレルギー等の過敏性障害の治療に用いられる本発明のタンパク質又はペプチドが提供される。
【0046】
個々の患者に適切な投与形態及び剤形の選択は当業者には明らかである。
【0047】
本発明は、本発明に従ったキメラ、好ましくはキメラQ2、又は哺乳動物における脱感作治療ためにそれから誘導された合成ペプチドの使用に及ぶ。脱感作方法は、非経口経路(皮下、静脈内、又は筋肉内)、又は経口、経鼻若しくは直腸経路による対象のアレルゲンの反復投与を伴う。これらの(ポリ)ペプチドは、使用に関する現在の(prevailing)法律及びガレノス製剤手順に従って、単独又は他の希釈剤と併用して投与することができる。
【0048】
本発明で記載されるキメラタンパク質Q2及びキメラタンパク質Q3は、図10、図11、図12及び図14で示されるように、完全抽出物又は複合天然タンパク質よりもカベイラクサに対してアレルギーである患者の血清に対する反応性が低く、その上in vivoでの皮膚試験(図13)でもこの低アレルギー誘発性が見られることから、低アレルギー誘発性である。
【0049】
キメラ分子Q1及びキメラ分子Q2は、複合天然タンパク質と同程度の免疫原性能も有している(図16)。両方の特性(低アレルギー誘発性及び免疫原性)のために、キメラQ2は、鼻炎、結膜炎、喘息、蕁麻疹、血管浮腫、湿疹、皮膚炎、又はさらにアナフィラキシーショックとして現れ得るカベイラクサの花粉に対するアレルギーの予防的及び治癒的な治療に対する優れた候補になる。
【0050】
本発明によるキメラタンパク質の免疫学的特徴が以下に記載される。図10は免疫検出試験を示しており、キメラQ2及びキメラQ3(レーン5及びレーン6)が、2つの天然タンパク質(レーン1)、単離した組み換えタンパク質(レーン2及びレーン3)又はキメラ(レーン4)の反応性と比べた場合に、アレルギー患者におけるIgE結合能が大きく低減していることを示し、このことは、完全ではあるが、これらのBエピトープを全て含有する2つのタンパク質の組み換え融合を示す。このことは、29番目の残基と、52番目の残基との間に含まれるBエプトープの欠如(キメラQ2)がアレルギー誘発性の低減に寄与することを示していると考えられる。同様の結果が、キメラタンパク質に対する15個の異なる血清の反応性を研究した際に得られた(図11)。キメラタンパク質Q2及びキメラタンパク質Q3の反応性は、2つの完全なアレルゲンの結合体(Par j 1−Par j 2)であるQ1で見られたものよりもはるかに小さい。このアレルギー誘発性の低減は、カベイラクサに対してアレルギーである患者由来の血清混合物によるELISA阻害によって定量された(図12)。2つの天然タンパク質の混合物(than of)抽出物の60%阻害に達するのに、10000倍以上のタンパク質Q2が必要であった。したがって、アレルギー誘発性が天然タンパク質よりも10000倍低く、アレルギー誘発性がキメラタンパク質Q1よりも約20倍低いことが推測され得た。
【0051】
キメラQ2の低アレルギー誘発性のより直接的な測定は、カベイラクサの花粉に対してアレルギーである30人の患者における皮膚反応性の直接的な測定によって得られた。図13で与えられたデータは、キメラQ2の皮膚反応性が顕著に低減したことを示している。記載されたそれぞれの分布の比較は、キメラQ2の平均パッチサイズが2つの天然タンパク質で見られたものよりも112倍小さいことを示し、このことはアレルギー活性の99%超の低減を示している。
【0052】
また、キメラタンパク質Q2とのIgEの低結合能が、EASTによって測定されたカベイラクサに対してアレルギーであるさらなる30人の患者由来の血清によって実証された(図14)。全ての患者において、IgE結合は、天然タンパク質の混合物に比べて、キメラQ2で大きく低減した。
【0053】
IgE結合能、及びしたがってBエピトープの欠失による拒絶反応の誘発能のこの大きな低減は、免疫原性能の維持を伴った。タンパク質Q2によって、図16で示されるように、天然抽出物及び組み合わされた2つの純粋な天然タンパク質の混合物によって誘導されたものと同程度のリンパ増殖指数が実証された。このことは全て、Par j 1及びPar j 2の断片で構築されたキメラタンパク質Q2が、より少数のIgEエピトープを含有したが、保護的な免疫応答を誘導するのに十分なTエピトープを維持したことを示している。
【0054】
本発明は、本発明の調製及びその質の実証における実験段階に関して、以下の実施例によってより良好に理解される。これらの実施例は単に例示的な実施例であり、本発明を限定しない。
【実施例1】
【0055】
Q1、Q2、及びQ3融合体の構築
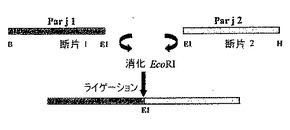
キメラタンパク質は、Gonzalez-Rioja et al.著「組み換えアレルゲンPar j 1及びPar j 2の発現及び精製(Expression and purification of the recombinant allergens Par j 1 and Par j 2)」(XXIII Congreso EAACI, Abstracts Book(2004), 181-182)で記載されたPar j 1.0103及びPar j 2.0101をコードする配列と、それぞれの場合に特異的な誘発因子(triggers)とを含有するプラスミドをマトリクスとして用いるポリメラーゼの連鎖増幅(PCR)によって構築された。この誘発因子は、ハイブリダイゼーション(hybridation)域と、異なる制限エンドヌクレアーゼに対する様々なセクション部位(下線)と、幾つかのアンカーヌクレオチドとから構成される。PCR誘導性の増幅反応は、反応量50μl中に以下の成分を有していた:増幅緩衝液×10、5μl、200μMのdNTP、100pmolの各誘発オリゴヌクレオチド:2.5単位のポリメラーゼTaq(Pfx DNAポリメラーゼ、Invitrogen)、DNAマトリクス1ng、及び50μlまで滅菌蒸留水。増幅反応が、それぞれの場合において記載される特定条件下でRoboCyclerサーモサイクラー(Stratagene)において行われた。反応生成物をアガロースゲル(2%)中で電気泳動にかけ、製造業者によって記載された方法を利用してGeneclean(Bio101)を用いて、対象のバンドをゲルから単離した。単離した断片を適切な制限酵素で消化し、同じ酵素で消化したpBluescriptベクターと結合させた。ライゲーション混合物を用いて、大腸菌DH5α(Invitrogen(Paisley, UK)から入手可能)のコンピテント細胞を形質転換した。プラスミドDNAを単離するために、得られたコロニーを培養させ、これを対象の断片を遊離させるために、適切な酵素で消化した。陽性コロニーがそのシーケンシングで選択された。pBluescriptに挿入されたDNAのシーケンシングは、製造業者の取扱説明書に従って、蛍光ジデオキシヌクレオチドによる使用のために改良されたサンガー法[(30)Hanahan, D.(1983)著「プラスミドによる大腸菌の形質転換に対する研究(Studies on transformation of Escherichia coli with plasmids)」 J. Mol. Biol. 166, 557-580]及びPRISMレディ反応ジデオキシ末端化サイクルシーケンシングキット(Ready Reaction DiDeoxy Termination Cycle Sequencing Kit)(Perkin Elmer)を用いたサーモサイクラーにおける増幅によって行われた。
【0056】
A)キメラタンパク質Q1
この場合、不安定な三次構造を有するが、連続的なIgEエピトープ全てが完全であるタンパク質を得るために、両方のタンパク質(Par j 1及びPar j 2)の完全な配列が融合されると考えられた。Q1として知られている構築物を得るために、ベクターpKS−BluescriptでクローニングしたPar j 1及びPar j 2のcDNAをマトリクスとして用いた。2つの配列を融合するために、Par j 1の配列のC末端と、Par j 2のN末端との両方においてリンカー(この場合EcoRI標的)を有し、その後断片をライゲーションさせる必要があった。当該標的は対応する合成オリゴヌクレオチド(F1R1及びF1F2)に付加され、PCRプロセスにおいて増幅断片に組み込まれた。
【0057】
合成オリゴヌクレオチドには以下のものを用いた:
断片1:F1F1、CG GGATCC TGCAAGAAACCTGCGG(BamHI)及びF1R1、CG GAATTC GGCTTTTTCCGGTGCGG(EcoRI)。条件:94℃、4分(1サイクル)、94℃、30秒 − 53℃、30秒 − 72℃、90秒(5サイクル)、94℃、30秒 − 60℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)。
断片2:F1F2、CG GAATTC GAGGAGGCTTGCGGGA(EcoRI)及びF1R2、CG AAGCTT CTAATAGTAACCTCTGA(HindIII)。条件:94℃、4分(1サイクル)、94℃、30秒 − 51℃、30秒 − 72℃、90秒(5サイクル)、94℃、30秒 − 59℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)。
【0058】
Par j 1及びPar j 2のそれぞれに対応する2つのPCR産物が生成された。これらのうちの第1の断片(Par j 1)に関しては、合成オリゴヌクレオチドF1F1及び合成オリゴヌクレオチドF1R1をそれぞれN末端及びC末端に用いて、約420塩基対(pb)のサイズが得られた。増幅断片がpKS−Bluescriptベクターでクローニングされ、その配列が確認された。Par j 2に対応する第2の断片に関しては、オリゴヌクレオチドF1F2及びオリゴヌクレオチドF1R2をそれぞれN末端及びC末端に用いて、同じ手順を用いた。この際、約300pbの断片が得られ、これをpKS−Bluescriptベクターでクローニングして、その配列を確認した。この第2の断片は、EcoRI標的によるライゲーションによって第1の断片と結合し、Par j 1がN末端で、Par j 2がC末端で維持され、その後得られた断片が市販の発現ベクターpQE−32(Qiagen)でサブクローニングされ、親和性末端(affinity tail)に対応するN末端で13個のアミノ酸の追加配列を含有していた(図2)。当該配列は、256個のアミノ酸を有し、見かけの分子量が28070Daのポリペプチドをコードした。
【0059】
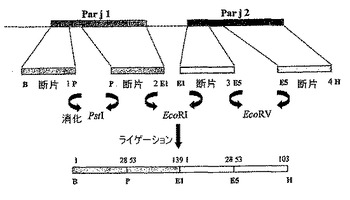
B)キメラタンパク質Q2
このキメラを得る場合、両方のタンパク質の断片は結合されているが、記載の連続的なIgEエピトープを全く含んでいないと考えられた[(18)Asturias, J.A, Gomez-Bayon, N., Eseverri, J. L.及びMartinez, A.(2003)著「カベイラクサ花粉由来の主要なアレルゲンであるPar j 1及びPar j 2は同様の免疫グロブリンEエピトープを有している(Par j 1 and Par j 2, the major allergens from Parietaria judaica pollen, have similar immunoglobulin E epitopes)」 Clinical and Experimental Allergy 33, 518-524]。以下の断片を含んだ幾つかの合成オリゴヌクレオチドを設計した:Par j 1(残基1〜28及び残基53〜139の断片)及びPar j 2(残基1〜28及び残基53〜103の断片)(図3及び図4)。IgEエピトープを含有する配列の一部はこの方法で取り除かれた。
【0060】
4つの断片(それぞれのアレルゲンに対して2つの断片)がこの構築物で増幅され、pKS BluescriptベクターでクローニングしたPar j 1及びPar j 2のcDNAをマトリクスとして用いた。新規のタンパク質の構造機構は、異なる増幅断片の連続結合への制限酵素(この場合、PstI、EcoRi及びEcoRV)の使用である。
【0061】
合成オリゴヌクレオチドには、以下のものを用いた:
断片1:F1F1、CG GGATCC TGCAAGAAACCTGCGG(BamHI)及びF2R1、CG CTGCAG CCCCTTTGACGGCTCTT(PstI)。条件:分(1サイクル)、94℃、30秒 − 54℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)。
断片2:F2F2、CG CTGCAG ATCCAGACCGCCATGAA(PstI)及びF2R2、CG GAATTC GGCTTTTTCCGGTGCGGG(EcoRI)。
断片3:F2F3、CG GAATTC GAGGAGGCTTGCGGGAA(EcoRI)及びF2R3、CG GATATC CTCCTTCGACGGCTCCTT(EcoRV)。
断片4:F2F4、CG GATATC ATAGTGCGCGCCACGAA(EcoRV)及びF1R2、CG AAGCTT CTAATAGTAACCTCTGA(HindIII)。3つの断片の条件は、94℃、4分(1サイクル)、94℃、30秒 − 54℃、30秒 − 72℃、90秒(5サイクル)、94℃、30秒 − 62℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)であった。
【0062】
Par j 1(断片1)に対応する、これらのうちの第1の断片に関しては、合成オリゴヌクレオチドF1F1及び合成オリゴヌクレオチドF2R1をそれぞれN末端及びC末端に用いて、約90塩基対のサイズが得られ、pKS−Bluescriptベクターでクローニングした。Par j 1に対応する第2の断片に関しては、オリゴヌクレオチドF2F2及びオリゴヌクレオチドF2R2をそれぞれN末端及びC末端に用いて、同じ手順を実行した。この際、約260pbの断片が得られ、pKS−Bluescriptベクターでクローニングした。この最後の断片は、PstIによって担持された標的のリンカーによるライゲーションによって第1の断片と結合し、その配列が確認された。
【0063】
Par j 2(断片3及び断片4)の配列から増幅された2つの断片に関しては、断片1及び断片2に記載された手順に従った。N末端及びC末端それぞれに対してオリゴヌクレオチドF2F3/F2R3及びオリゴヌクレオチドF2F4/F1R4をその増幅のために用い、PCR産物のサイズは断片3では約90pb、及び断片4では約150pbであった。断片4は、EcoRVによって担持された標的のリンカーによるライゲーションによって断片3と結合し、その配列が確認された。
【0064】
最終工程において、生成された2つの新規の断片(それぞれのアレルゲンに対して1つの断片)が、EcoRIによって担持された標的のリンカーによるライゲーションによって結合し、Par j 1がN末端で、Par j 2がC末端で維持された。その後、当該断片が市販の発現ベクターpQE−32でサブクローニングされ、親和性末端に対応するN末端で13個のアミノ酸の追加配列を含有していた(図5)。当該配列は、212個のアミノ酸を有し、見かけの分子量が23336Daのポリペプチドをコードした。
【0065】
C)キメラタンパク質Q3
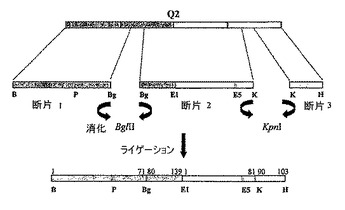
この最後の構築物は、この場合、存在し得る任意のIgEエピトープを取り除こうと試みた以外はQ2と実質的に同一であり、Par j 1(残基71〜80)及びPar j 2(残基72〜81)(図6及び図7)の配列に存在する8つのアミノ酸の別の付加的な配列が取り除かれた。
【0066】
合成オリゴヌクレオチドには以下のものを用いた:
断片1:F1F1、CG GGATCC TGCAAGAAACCTGCGG(BamHI)及びF3R1、CG AGATCT GACCTCGCTGACGAG(BglII)。
断片2:F3F2、CG AGATCT AGCAAGCTCCCGCCC(BglII)及びF3R2、CG GGTACC GGGGACCTCGGCGAC(KpnI)。2つの断片の条件は、94℃、4分(1サイクル)、94℃、30秒 − 54℃、30秒 − 72℃、90秒(5サイクル)、94℃、30秒 − 63℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)であった。
断片3:F3F3、CG GGTACC CTCCCGCCCATCACC(KpnI)及びF3R3、TTTAAAAAGGCCGTAATATCC。条件:94℃、4分(1サイクル)、94℃、30秒 − 56℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)。
【0067】
この場合、構築物Q2−pQE−32をマトリクスとして用い、断片1、断片2及び断片3それぞれに対して、約150pb、約370pb及び約290pbのサイズが得られた。用いられたオリゴヌクレオチドは、断片1に対してF3F1及びF3R1、断片2に対してF3F2及びF3R2、並びに断片3に対してF3F3及びF3R3であった。上記の場合の断片の連続ライゲーションのプロセスを再び繰り返した。用いられる新規の制限酵素はBglII及びKpnIであった。第3のそして最後の断片では、そのサイズが小さかったので(66pb)、その後の精製プロセスにおいてより高い利便性で働くことができるようにより大きなサイズの断片を得るため、ベクターpQE−32−Q2の配列の一部を増幅することになった。それぞれの工程で、ベクターpKS−Bluescriptでクローニングされた、PCRによって得られた断片、及びその配列が確認された。第1のグルタミンアミノ酸(Q)に対応する第1の断片における自然突然変異がプロリン(P)によって生じた。
【0068】
最終的に、得られた断片が発現ベクターpQE−32でサブクローニングされ、親和性末端に対応するN末端で13個のアミノ酸の追加の配列を含有していた(図8)。当該配列は、200個のアミノ酸を有し、見かけの分子量が21938Daのポリペプチドをコードした。
【実施例2】
【0069】
キメラタンパク質Q1、キメラタンパク質Q2、及びキメラタンパク質Q3の発現及び精製
LB(100μg/mlのアンピシリン及び25μg/mlのカナマイシンをそれぞれ補充した)のシートから単離したコロニーから始め、同じ培地50mlの前接種材料(pre-inoculum)を産生し、撹拌しながら(260rpm)、37℃で一晩インキュベートした。光学密度(600nm)を0.2から始めて、同じ培地1Lを当該前接種材料で接種させた。混合物を撹拌しながら37℃で1時間30分インキュベートした後、同じインキュベート条件下で3時間、IPTG(1mMの最終濃度)による誘導を行った。
【0070】
Q2の場合、細胞を10000rpm、4℃で15分間、遠心分離して、溶解緩衝液(20mMのリン酸塩(pH7.4)、50mMのイミダゾール、0.5MのNaCl)50mlに再懸濁させた。撹拌しながら、37℃で30分間、再懸濁質(matter)をリゾチーム(0.1mg/mlの最終濃度)で処理した。それから、超音波処理(5パルス、20秒)を行い、混合物を15000rpm、4℃で15分間、遠心分離した。上清を0.45μmのフィルタ(Millex HV, Millipore)で濾過し、AKTAプライムシステム(Aはウムラウト)(GE-Healthcare)に適合されたHi−Trapキレート化HPカラム(GE-Healthcare)5mlに加えた(2.5ml/分)。このカラムをニッケルでキレート化し、予め10容量の溶解緩衝液で平衡化していた。カラムを15容量の溶解緩衝液で洗浄すると、カラムと結合したタンパク質の溶出を溶出緩衝液(20mMのリン酸塩(pH7.4)、0.5Mのイミダゾール)を用いて100%まで行った。塩を除去し、20mMでpH7のリン酸緩衝液サンプルを交換するために、溶出液をHi−Trap脱塩カラム(GE-Healthcare)5mlに通した。タンパク質は−40℃に維持した。
【0071】
Q1及びQ3の場合、さらなる精製工程が必要であった。その誘導後に、細胞を10000rpm、4℃で15分間、遠心分離し、8Mの尿素を補充した溶解緩衝液50mlで再懸濁して、前の段落で記載された手順を採用した。2番目の精製工程では、予めPBSで平衡化したHi−Load 16/60 Superdex−200カラム(GE-Healthcare)に、得られた溶出液を加えた(1ml/分)。前の段落で記載された緩衝液でタンパク質の溶出を行った。タンパク質は−40℃に維持した。
【0072】
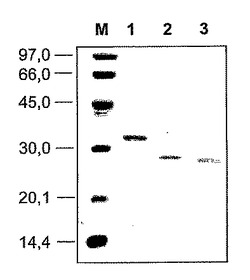
SDSを有するポリアクリルアミドゲル(SDS−PAGE)で、電気泳動を行った。Laemmliによって記載された方法は、MINI−PROTEAN電気泳動装置(Bio-Rad)を用いて、基本的に[(31)Laemmli, U.K.(1970)著「バクテリオファージT4頭部集合化中の構造タンパク質の切断(Cleavage of structural proteins during the assembly of the head of bacteriophage T4)」 Nature 277, 680-685]に従った。10×10cmであり、ポリアクリルアミド濃度が12.5%であるゲルに、トリス−グリシン緩衝液で45分間、200ボルトの電流を流した。参照として用いられるタンパク質は、低分子量に関するBio-Radのキット由来のものであった。分子量の算出及びゲルの濃度測定分析は、画像分析器(Diversity, BioRad)を用いて行った。
【0073】
ハイブリッドタンパク質Q1は、32kDaのHis標識化融合タンパク質として表され(図9)、細菌培養物の最終収率は2mg/Lであった。Q2タンパク質及びQ3タンパク質は、28kDaのHis標識化融合タンパク質として表され、培養物の最終収率はそれぞれ、5mg/L及び7mg/Lであった(図9)。
【実施例3】
【0074】
精製ハイブリッド分子の円偏光二色性分析
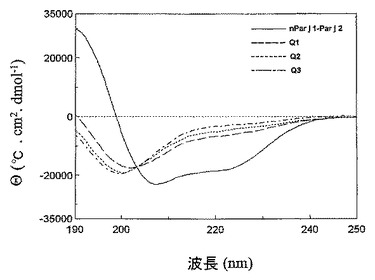
pH7.0、20℃でのFar−UV(190nm〜250nm)のCDスペクトルが、20℃で温度を自動調節したキュベット中に日本分光株式会社のPTC−423S温度制御装置を備えた日本分光株式会社のJ−810分光偏光計によって記録された。タンパク質濃度は、0.2cmキュベットにおいて20mMのリン酸ナトリウム緩衝液中で0.035mg/mlであり、40個の走査が集積した。全てのスペクトルは適切なバックグラウンドを差し引き、平均残余楕円率に変換した。
【0075】
二次構造要素は、天然のパリエタリアのアレルゲン混合物(Par j 1及びPar j 2)、及びハイブリッドタンパク質を適用したCD分光法によって分析された(図10)。ハイブリッドタンパク質のスペクトルはそれらの中でほぼ同一であったが、天然アレルゲンのCDスペクトルとは全く異なっていた。NPAスペクトルは、208nmで最小を示し、222nmで明確に増大し(shoulder)、190nmで最大を示したが、ハイブリッドタンパク質のスペクトルは、ほぼ完全な変性(unfolded)状態になるランダムコイル構造に移行した。
【実施例4】
【0076】
カベイラクサに対してアレルギーである患者の血清混合物に対するキメラタンパク質の低IgE固定の反応性を実証するための免疫試験
A)免疫検出
融合物1、融合物2及び融合物3のIgE結合活性の第1の評価が、カベイラクサに対してアレルギーである患者由来の血清混合物を用いる免疫移動法(immunotransfer method)によって行われた。タンパク質抽出物及び精製タンパク質をポリアクリルアミドゲルに適用したら、Towbin et al[(32)Towbin, H., Staehelin, I.及びGordon, J.(1979)著「ポリアクリルアミドゲルからニトロセルロースシートへのタンパク質の電気泳動転写:手順及び幾つかの用途(Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications)」 Proc. Natl. Acad. Sci. USA 76, 4350-4354]の方法によって電気的移行を行った。この目的のために、SDS−PAGEによって単離したタンパク質をPVDF Hybond−P膜(GE-Healthcare)に電気的に固定した(transfixed)。周囲温度で1時間、膜を遮断したら、4℃で一晩、膜を一次抗体とインキュベートして、同じ洗浄緩衝液で様々な回数(various)洗浄後、周囲温度で1時間、膜を二次ペルオキシダーゼ結合抗体とインキュベートした。製造業者によって記載されたように、膜をフィルム(Hyperfilm.ECL、GE-Healthcare)に曝すことによって、ECL化学発光法(GE-Healthcare)でバンドを検出した。
【0077】
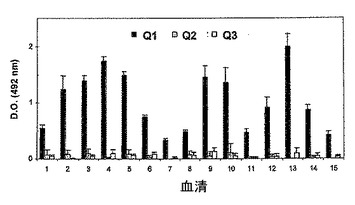
免疫検出試験によって、3つの融合物間でIgE結合能が異なることが実証された。したがって、IgE抗体の認識はQ1の場合でのみ存在した(図11)。Q2の場合では、認識は非常に急激に落ちたが、Q3の場合では、実証された認識は0であった。
【0078】
B)直接ELISA
カベイラクサに対してアレルギーの患者由来の個々の血清を用いたELISA法によって、3つの融合物に対するIgEの反応性を分析した。周囲温度で一晩、ポリスチレンプレート(Greiner)を、PBS緩衝液(10mMのリン酸塩(pH7.2)、137mMのNaCl、2.7mMのKCl)中で、カップ1つ当たり1μgのカベイラクサ抽出物タンパク質又は1μgの純粋なタンパク質とインキュベートした。1%BSA−0.05%Tween20を補充した、200μl/カップのPBSでこれらを遮断し、37℃で1時間維持した。アレルギー患者由来の100μl/カップの血清(10倍希釈) を添加し、37℃で90分間インキュベートした。200μl/カップのPBS−T(PBS+0.05%Tween20)による3回の洗浄の後、ペルオキシダーゼと結合したヒトIgE免疫グロブリン(Dako)に対する100μl/カップの抗血清を添加し(1000倍希釈)、37℃で90分間インキュベートした。PBS−Tで新たに3回洗浄した後、製造業者の取扱説明書に従って調製した、200μl/カップのo−フェニレンジアミン(Sigma−Fastのタブレットセット、Sigma)溶液を添加し、プレートを30分間暗所に維持した。反応を50μl/カップの3MのH2SO4で停止させ、ELISAイージーリーダーEAR−400AT(SLT-Lab Instruments)プレートリーダーにおいて492nmで吸光度を測定した。
【0079】
IgEに対する反応性は、Q1の場合でのみ実証されたが、Q2及びQ3の両方はIge抗体の認識が実質的にないことが実証された(図12)。
【0080】
C)ELISA阻害
ELISA阻害試験に関して、カベイラクサに対してアレルギーの患者由来の血清混合物を、4℃で一晩、所定の濃度の阻害タンパク質(1ng/ml〜1000ng/ml)と予めインキュベートした。後の残りの手順は、ELISA直接試験に記載されたものと同じである。
【0081】
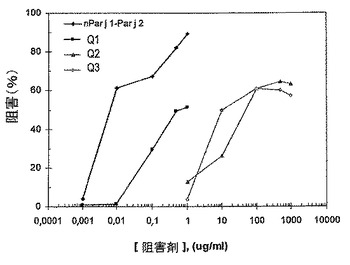
カベイラクサに対してアレルギーの患者由来の血清混合物によるELISA阻害試験において、Q1では、カベイラクサの花粉のIgE結合活性の約50%程度の阻害が実証され、これはnPar j 1−Par j 2のものより幾らか低く(92%)、500倍以上のタンパク質を添加する必要があった(図13)。キメラQ2及びキメラQ3は、最大阻害の約60%程度に達したが、これは同程度の阻害に達するためには、nPar j 1−Par j 2で必要とされるものよりも10000倍高いタンパク質濃度を必要とし、このことは、これらのキメラがアレルギー誘発性を約99.99%低減させたことを示唆していた。
【実施例5】
【0082】
キメラタンパク質Q1、キメラタンパク質Q2、及びキメラタンパク質Q3の低皮膚反応性を実証するためのin vivo実験
in vivo皮膚反応(プリック)試験を行い、カベイラクサ花粉に対するアレルギーに対する満足のいく免疫療法を開発することができるために、候補低アレルギー誘発性の分子を求めるという目的で、キメラ1、2、及び3の低アレルギー誘発性を評価した。
【0083】
カベイラクサ抽出物、nPar j 1−Par j 2、大腸菌で発現したrPar j 1及びrPar j 2、並びにキメラ1、2及び3を用いて、皮膚試験を行った。精製タンパク質は、0.5%フェノール含有及び50%グリセリン含有生理食塩水溶液で希釈した。天然のPar j、Par j 2、Q1の場合に用いる濃度は、0.5μg/ml、5μg/ml、及び50μg/mlであり、Q2又はQ3の場合の濃度は5μg/ml、50μg/ml、及び250μg/mlであった。0.9%NaCl及び塩酸ヒスタミン(10mg/ml)をそれぞれ、陰性対照及び陽性対照に用いた。
【0084】
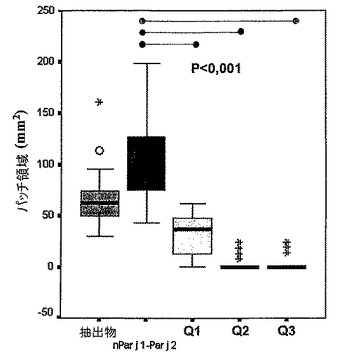
この実験では、試験されるそれぞれのアレルゲンの液滴を、前腕の掌面上に沈着させた後に、ランセットを用いて、液滴を通して穿刺穴を作製した。二重で、増大濃度及び低減濃度の反対の条件(strings)で、それぞれの試験を行った。15分後、パッチの輪郭を先端が細い黒色のマーカーペンで描いた。低アレルギー誘発性の絆創膏の細片をパッチ上に貼り、ゆっくり押し付けて、インクのマーキングを細片に写し、これをパッチ記録シートに転写させた。Summasketchデジタル化タブレット及びコンピュータベースのプログラム(Autocad v.11)を用いてこの記録をスキャンすることによって、パッチ領域を測定した。
【0085】
ブロック図で結果を図示し、関連変数に関してウィルコクスン検定を用いて統計調査を行うことによって、得られた結果を解釈した(図14)。これらの図は、皮膚反応に対応する値の分布(パッチ領域(mm2)で測定された)が、抽出物(平均65.25mm2、信頼範囲(95%):54.98〜75.53)及びnPar j 1−Par j 2(平均106.80mm2、信頼範囲(95%):91.90〜121.70)に対して、3つのキメラの場合で有意に異なることを示している(P<0.001)。キメラ2及びキメラ3の値は実質的に0である:Q2(平均0.95mm2、信頼範囲(95%):0〜2.89)及びQ3(平均0.15mm2、信頼範囲(95%):0〜0.47)。
【実施例6】
【0086】
キメラタンパク質Q1、キメラタンパク質Q2、及びキメラタンパク質Q3の低IgE抗体結合能を実証するための実験
in vivo試験の補足として、EAST直接法を用いて、特異的なIgEを求めることによって、新たにin vitro試験を行った。
【0087】
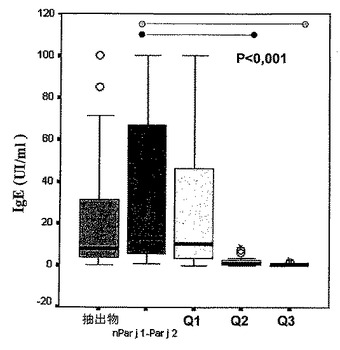
天然タンパク質及び組み換えタンパク質(50μg/ml)、並びにカベイラクサ抽出物(2mg/ml)を、臭化シアンで活性化したディスクにカップリングさせることによって、Ceska et al.[(33)Ceska, M. and Lundkvist, U.(1972)著「IgEの決定のための新規の簡易なラジオイムノアッセイ法(A new and simple radioimmunoassay method for the determination of IgE)」 Immunochemistry 9, 1021-1030]に従って、特異的なIgEを求めた。その後、患者由来の血清50μlを添加し、周囲温度で1時間、インキュベートした。洗浄後、37℃で30分間、ディスクをアルカリホスファターゼと結合したヒト抗IgE抗体50μlとインキュベートして、製造業者(Hycor Biomedical Inc.)によってIgE特異的なハイテックキットEIAで与えられた取扱説明書に従って、定量を行った。
【0088】
得られた結果は、単にin vivoで得られた結果を再確認しているだけである。3つのキメラのうち、Q1はnPar j 1−Par j 2に対して、その免疫原性能を維持したが、Q2及びQ3では当該免疫原性能が非常に顕著に低減した(P<0.001)(図15)。
【実施例7】
【0089】
キメラタンパク質Q1、キメラタンパク質Q2及びキメラタンパク質Q3の免疫原性能を実証する誘導性リンパ増殖実験
免疫療法における低アレルギー誘発性分子の使用に対する本質的な要求は、その抗原性(Tエピトープ)の維持である。したがって、IgE抗体と結合しないことに加えて、本発明のタンパク質が抗原性を維持するか否か確認するために、この実験で用いられる様々なタンパク質によって刺激された末梢血単核球(CMSP)でリンパ増殖試験を行った。比色分析によって測定されるホルマザン化合物を発生させるために、生存細胞のミトコンドリアデヒドロゲナーゼによるテトラゾール塩WST−1の消化に基づく比色法によって、この試験が行われた。
【0090】
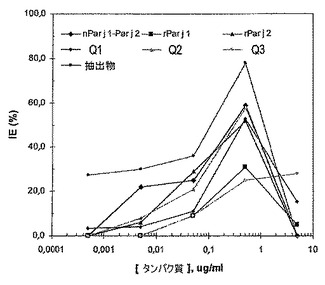
リンパ球(lymphocite)分離溶液(Lymphoprep, Nycomed)を用いる密度勾配における遠心分離によって、カベイラクサに対してアレルギーの13人の患者のCMSPを単離した。それから、培養培地(血清無含有培地AIM V、Gibco)において1×106個の生存細胞/mlでCMSPを再懸濁し、PBS(Sigma Chemical Co.)中の0.25%トリパンブルー(tripan blue)でこれらの生存率を試験した。製造業者によって記載されたように、90%を超える生存率で調製されたCMSPをin vitro増殖試験に直ちに用いた(細胞増殖剤WST−1、Roche Diagnostics)。最終容量が200μlの培地中の2×105個のCMSPで、それらを底が平らなマイクロプレート(Nunclon、NUNC)に沈着させた後に、カベイラクサ抽出物を用いて、5%CO2の加湿雰囲気下、37℃で三重で測定を行い、最終濃度が0.0005μg/ml−0.005μg/ml−0.05μg/ml−0.5μg/ml−5μg/mlになるまで、様々なタンパク質を精製した。三重の対照の刺激を受けない培養物は全ての場合に含まれた。3日後、細胞増殖剤WST−1 20μlを全てのカップに加え、4時間インキュベートした。代謝的に活性のある細胞によって産生されたホルマザンを450nmでの吸光によって定量した。この試験で用いた組み換えタンパク質は、3つのキメラ、並びに大腸菌で発現したrPar j 1及びrPar j 2であった。
【0091】
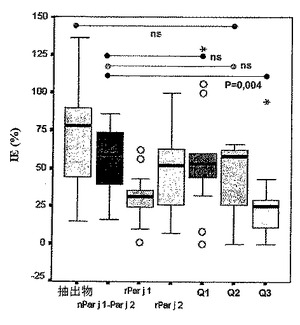
第1の工程において、免疫原性(immunuogenic)タンパク質をスキャンして、その後の試験の進行のために最適濃度を求めた。全ての場合、最大増殖(IE%)を示すタンパク質濃度が0.5μg/mlであることが見出された(図16)。
【0092】
カベイラクサに対してアレルギーの13人の患者による増殖の結果は、ブロック図の統計分析及び2つの対になったサンプルに関するノンパラメトリック試験によって求められた。統計分析によって、対照に用いられたカベイラク抽出物には、nPar j 1−Par j 2とあまり変わらない抗原刺激能があり(P=0.142)、Q1及びQ2は、これらの抽出物(それぞれ、P=0.152及びP=0.294)及びnPar j 1−Par j 2(それぞれ、P=0.484及びP=0.182)と有意差のない値分布を示したことが示された(図17)。一方で、Q3は、この抽出物(P=0.002)及びnPar j 1−Par j 2(P=0.004)に対してほとんど刺激能を有しなかった。またその一方で、この抽出物(それぞれ、P=0.142及びP=0.003)及びnPar j 1−Par j 2(それぞれ、P=0.041及びP=0.002)に対して、rPar j 1へのrPar j 2の抗原力が大きかったことが指摘されるべきである。
【0093】
得られた結果によって、依然として、Q2は上記特性を維持したのに対し、Q3はそれを完全に喪失したことが示される。カベイラクサに対するアレルギーを治療するために様々な低アレルギー誘発性分子を得るプロセスについて記載された全てのものから、Q2がカベイラクサに対するアレルギーの満足のいく免疫療法を開発する候補低アレルギー誘発性分子であることを結論付けることができる。
【0094】
投与方法
本発明は、哺乳動物における減感作治療のための、記載の低アレルギー誘発性キメラ又はこれら由来の合成ペプチドの使用に関する。減感作方法は、非経口経路(皮下、静脈内、又は筋肉内)、経口、舌下、経鼻若しくは直腸経路による対象のアレルゲンの反復投与を伴う。これらの(ポリ)ペプチドは、使用に関する現行の法律及びガレノス製剤手順に従って、単独又は他の薬学的に許容可能な希釈剤及び賦形剤と併用して投与することができる。
【0095】

【図面の簡単な説明】
【0096】
【図1】Q1の構造図である。
【図2】Q1のアミノ酸及びヌクレオチドの配列を示す図である。ベクターpQE−32の親和性末端に対応する残基には下線が引かれている。EcoRI(EF)セクションによって導入された残基は陰影を付けている。システイン残基は二重下線が引かれている。
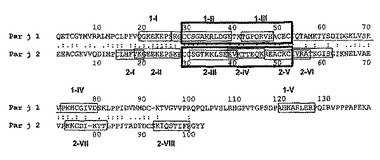
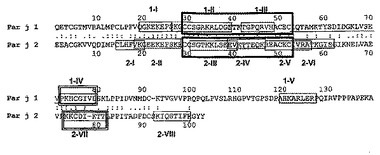
【図3】Par j 1及びPar j 2のIgEエプトープ(囲み線)と、Tエピトープ(下線)との比較を示す図であり、太線の囲み線はQ2欠失エピトープを示す。同一の残基(:)及び同様の残基(.)が標識されている。
【図4】Q2の構造図である。
【図5】Q2のアミノ酸及びヌクレオチドの配列を示す図である。ベクターpQE−32の親和性末端に対応する残基は下線が引かれている。EcoRI(EF)、PstI(LQ)、及びEcorRV(DI)セクションによって導入された残基は陰影を付けている。システイン残基は二重下線が引かれている。
【図6】Par j 1及びPar j 2のIgEエプトープ(囲み線)と、Tエピトープ(下線)との比較を示す図であり、太線の囲み線はQ3欠失エピトープを示す。同一の残基(:)及び同様の残基(.)が標識されている。
【図7】Q3の構造図である。
【図8】Q3のアミノ酸及びヌクレオチドの配列を示す図である。ベクターpQE−32の親和性末端に対応する残基は下線が引かれている。EcoRI(EF)、PstI(LQ)、EcorRV(DI)、Bg1II(RS)及びKpnI(GT)セクションによって導入された残基は陰影を付けている。システイン残基は二重下線が引かれている。
【図9】電気泳動後のポリアクリルアミドゲルのクマシーブルーによる染色を示す図である:Q1(レーン1)、Q2(レーン2)、Q3(レーン3)。
【図10】精製タンパク質nPar j 1−nPar j 2、Q1、Q2、及びQ3の円偏光二色性分析の結果を示す図である。
【図11】カベイラクサに対してアレルギーである患者の血清混合物のIgE抗体による免疫検出を示す図であり、この図は、nPar j 1−nPar j 2(レーン1)、rPar j 1(レーン2)、rPar j 2(レーン3)、Q1(レーン4)、Q2(レーン5)、Q3(レーン6)から成る。
【図12】カベイラクサに対してアレルギーである患者由来の15個の血清(10倍希釈)を用いて、Q1、Q2、及びQ3に対するIgE抗体の結合を示す図である。3つの実験値は偏差で示される。
【図13】固相中のカベイラクサ抽出物、並びにカベイラクサに対してアレルギーである患者の血清混合物を用いて、阻害剤としてnPar j 1−Par j 2、Q1、Q2、及びQ3を用いたELISA阻害試験の結果を示す図である。それぞれの値は、標準偏差が10%未満の3つの実験の平均に対応している。
【図14】カベイラクサ抽出物、nPar j 1−Par j 2及びQ1(50μg/ml)、並びにQ3(250μg/ml)によって行われた皮膚試験の結果を示す図である。示された値は、パッチ面積(mm2)である。
【図15】カベイラクサ抽出物、nPar j 1−Par j 2及びQ1(50μg/ml)、並びにQ2及びQ3(250μg/ml)によって行われた特異的なIgEの測定の結果を示す図である。
【図16】カベイラクサに対してアレルギーである患者におけるTリンパ球の増殖を研究するのに最適な濃度の測定を示す図である。刺激指数(IE)。
【図17】カベイラクサ抽出物及び0.5μg/mlの3つのキメラタンパク質、天然型及び組み換え型のPar j 1及びPar j 2により得られたTリンパ球の増殖を示す図である。示された値は、刺激指数(%)のものである。有意差なし(ns)。
【技術分野】
【0001】
本発明は、アレルギー、特に花粉症、最も詳細には脂質輸送タンパク質ファミリー由来のアレルゲン及びより具体的にはパリエタリア(Parietaria)種由来の花粉で見出されるアレルゲンによって引き起こされるアレルギーの予防及び治療のためのキメラタンパク質産生の分野に関する。
【背景技術】
【0002】
I型アレルギーは、先進国における深刻な健康問題である。この種のアレルギーは、空気によって運ばれる抗原に対するIgE抗体の形成によって引き起こされる。これらのIgE抗体は、肥満細胞及び好塩基球と相互作用し、ヒスタミン等の生物学的メディエータを放出し、集団の約20%でアレルギー性鼻炎、アレルギー性結膜炎及びアレルギー性気管支喘息を引き起こす[(1)Miyamoto, T.(1992)著「日本の花粉症有病率の増加(Increased prevalence of pollen allergy in Japan)」 In Advances in Allergology and Clinical Immunology. P. Godard, J. Bousquet及びF. B. Michel編(Cornforth, UK: The Parthenon Publishing Group), pp. 343-347]。
【0003】
特異的免疫療法(SIT)は、特異的なアレルゲンによって誘発されるアレルギー反応に効果的な治療であり、基本的にはアレルギーを生じるタンパク質(アレルゲン抽出物)を高濃度で定期的に投与することによって患者における免疫反応を調節することから成る。高用量のアレルゲンを注入すると、抗原提示細胞、例えば樹状細胞(好ましくは、協働するバージン細胞(virgin cells)(nTH)であるT細胞がTH1又はTH0に発達することを促進する)によってIL−12の高度合成が誘導される。このことが、TH2細胞に関連するアレルギー応答型の免疫応答から、TH1/TH0型の応答への偏向(deviation)を可能にし、高レベルのIFN−γが産生される[(2)Akdis, CA.及びBlaser, K.(2000)著「アレルゲン特異的免疫療法のメカニズム(Mechanisms of allergen-specific immunotherapy)」 Allergy 55, 522-530]。免疫偏向は、免疫抑制サイトカインIL−10及びTGF−βを産生する制御性T細胞(TR1)の影響下でTH2記憶細胞の寛容(アンエネルギー(anenergy)又はクローン除去)の誘導によって強まる[(3)Akdis, C.A., Joss, A., Akdis, M.及びBlaser, K.(2001)著「アレルギー炎症におけるIL−10誘導性の細胞不活性化及びアレルゲンに対する正常な応答のメカニズム(Mechanism of IL-10 induced cell inactivation in allergic inflammation and normal response to allergens)」 Int. Arch Allergy Immunol. 124; 180-182]。TH2細胞の活性化及び増殖の低減は、IL−4及びB細胞によるIgEの産生の低減によって達成される。TH2細胞の活性、並びに鼻粘液及び気管支粘液への浸潤の低減によって、IL−5合成が低減し、これによって好エオシン性浸潤が低減され、MBPタンパク質及びECPタンパク質等の炎症性メディエータの放出が大幅に低減する。優性表現型アレルゲンTH0のT細胞の新規の特異的なクローンは、B細胞による、大量の特異的なIgGアレルゲン抗体の産生を促進するTH1型サイトカインとTH2型サイトカインとの混合物を産生する。一方で、高レベルのIL−10によって、特異的なIgG4アレルゲン抗体の高度合成が誘導される。これらの2種類の特異的な抗体は、肥満細胞においてこれらの受容体と結合したIgE抗体の交差(intersection)を与え、このようにしてヒスタミンの脱顆粒及び放出を阻害する遮断抗体として作用することができる[(4)Moverate, R.(2003)著「花粉ワクチンによる特異的な免疫療法の免疫機構:診断法に関する示唆及び改善されたワクチン接種戦略の開発(Immunological mechanisms of specific immunotherapy with pollen vaccines: implications for diagnostics and the development of improved vaccination strategies)」 Expert Rev. Vacc. 2, 85-97、(5)Wachholz, P.A., Soni, N. K., Till, S.及びDurham, S. R.(2003)著「イネ科花粉の免疫療法後のIgG抗体によるB細胞とのアレルゲン−IgE結合の阻害(Inhibition of allergen-IgE binding to B cells by IgG antibodies after grass pollen immunotherapy)」 J. Allergy Clin. Immunol. 112; 915-922]。これらは、抗原含有細胞によるIgE媒介性の抗原の回収も阻害し、このことはアレルゲンに対する免疫応答を抑制する。
【0004】
天然源から単離されたアレルゲン抽出物は、タンパク質と他の分子との複雑な混合物である。これらの組成、ひいてはアレルギー誘発性は用いられる物質に依存し、周囲条件、花粉の場合の成熟状態、及び菌類等の場合のダニ等の成長条件によって変わる。幾つかの抽出物は主要なアレルゲンを不十分な濃度でしか含有することができず、また患者がアレルギーではない望ましくない成分で汚染される可能性があるか、又はその両方である。現行の免疫療法は、完全なアレルゲン抽出物を排他的に用い、このことによって、
−ワクチンとエフェクター細胞のIgEとの反応性による深刻な拒絶反応、
−免疫療法後のワクチンに存在する他のアレルゲンに対する新たな感作の発生、
−アレルゲン抽出物の標準化の困難性等の多くの問題が生じる。
【0005】
これは全て、免疫療法が、要望どおりの安全且つ効率的な治療ではないことを意味する。アレルギーの病因及び特異的な免疫療法のメカニズムのより良好な知識によって、上記問題の解決に近づくことができるようになった。特異的なアレルゲン応答性TH2におけるIgE媒介性抗原の影響を知ることで、IgEと結合しないアレルゲンを作製することがますます試みられている。現行の特異的な免疫療法の主な目的は、IgEエピトープを不活性化し、このようにしてIgEとの結合、及びその結果として拒絶反応を低減及びさらには除去するという目的でアレルゲンを修飾することである[(6)Valenta, R.及びLinhart, B.(2005)著「アレルギーワクチンの分子設計(Molecular design of allergy vaccines)」 Curr. Opin. Immunol. 17, 1-10]。このように、修飾アレルゲンは、食作用/飲作用媒介性の抗原回収機構によってT細胞に向けられ、IgEの交差及びIgE依存性抗原の提示を防ぐ。このことは、T細胞によるTH0又はTH1サイトカインの産生と、IgEの産生減少と、B細胞によるIgGの産生増大との平衡化を誘導する。このこと全てによって、アナフィラキシーの危険性なく、TH2型のT細胞寛容が誘導される。アレルゲン及びアレルゲン誘導体を得る組み換え方法の進歩によって、アレルギー治療のために新規のワクチンを開発する能力が大きく増大している。このことは、IgEエピトープの有意なアミノ酸を突然変異又は欠失する可能性、並びに低アレルギー誘発性ワクチンを得るためのこれらの分別及びオリゴマー化により可能になった。IgEと結合する能力が低減したが、T細胞との反応性を維持するこれらの分子は、より大用量で投与することができ、注入回数を少なくして、免疫療法をより迅速に且つ安全にすることができる。さらに、組み換えアレルゲンは、微生物発現系を用いて、発酵タンクで大規模に製造することができ、その精製は、天然の等価物よりも効率的で且つ安価である。免疫療法における低アレルギー誘発性誘導体の使用は、三量体のBet v 1断片[(7)Niederberger, V., Horak, F., Vrtala, S., Spitzauer, S., Krauth, M. T., Valent, P., Reisinger, J., Pelzmann, M., Hayek, B., Kronqvist, M., Gafvelin, G., Groenlund, H., Purohit, A., Suck, R., Fiebig, H., Cromwell, O., Pauli, G., van Hage-Hamsten, M.及びValenta, R.(2004)著「遺伝子操作したアレルゲンによるワクチン接種はアレルギー疾患の進行を防ぐ(Vaccination with genetically engineered allergens prevents progression of allergic disease)」 Proc. Natl. Acad. Sci. U.S.A. 101, 14677-14682]、ハチ毒(bee venon)タンパク質の多アレルギー性ハイブリッド(Api m 1、2、3)[(8)Schmid-Grendelmeier, P., Karamloo, F., Mueller, U., Housley-Marcovic, Z., Soldatova, L., Zumkehr, J., Kemeny, D.M., Kuendig, T., Reimers, A., von Beust, B.R., Salagianni, M., Akdis, M., Kussebi, F., Spangfort, M. D., Blaser, K.及びAkdis, C.A.(2005)著「IgE結合を低減し、T細胞エピトープを保存した、組み換え型の多アレルゲンワクチンによるアレルギーの予防(Prevention of allergy by a recombinant multi-allergen vaccine with reduced IgE binding and preserved T cell epitopes)」 Eur. J. Immunol. 35, 3268-3276]、及びそれらの三次構造を除去するためのPar j 2とPar j 1との突然変異融合物を用いてこれまでに記載されている。
【0006】
何人かの著者は、IgEとの結合が、高親和性及び低親和性の両方のIgEに対する表面受容体を発現する、樹状細胞及び活性化Bリンパ球細胞としてのプロフェッショナル抗原提示細胞によって、アレルゲンの捕捉及び提示を容易にし得るので、アレルギーワクチンは低アレルギー誘発性のアレルゲン(hypoallergens)では構成されていないはずであることを言及してる。このプローチは、免疫療法において舌下経路を用いる場合に特別な意味がある。また、高親和性の受容体の交差によって、アレルゲンに対するT細胞の応答が低減され得る[(9)Allam, J.P., Novak, N., Fuchs, C., Asen, S., Berge, S., Appel, T., et al.(2003)著「ヒトの口腔粘膜由来の樹状細胞の特徴付け:構成的なFcεRIの発現が高い、新規のランゲルハンス細胞型(Characterization of dendritic cells from human oral mucosa: a new Langerhans' cell type with high constitutive FcεRI expression)」 J. Allergy Clin. Immunol. 112, 141-8、(10)von Bubnoff, D., Matz, H., Frahnert, C., Rao, M. L., Hanau, D., de la Salle, H., Bieber, T.(2003)著「FcεRIはT細胞応答の調節に関与するトリプトファン分解経路を誘導する(FcεRI induces the triptophan degradation pathway involved in regulating T cell responses)」 J. Immunol. 169, 1810-1816]。多くの研究者らは未だに、欠失を導入することなくこの種のワクチンを製造している[(11)Batard, T., Didierlaurent, A., Chabre, H., Mothes, N., Bussieres, L., Bohle, B., et al.(2005)著「カバノキ花粉症に対する候補ワクチンとしての野生型の組み換えBet v 1aの特徴付け(Characterization of wild-type recombinant Bet v 1a as a candidate vaccine against birch pollen allergy)」 Int. Arch. Allergy Immunol. 136, 239-249、(12)Jutel, M., Jaeger, L., Suck, R., Meyer, H., Fiebig, H., Cromwell, O.(2005)著「組み換えイネ科花粉アレルゲンによるアレルゲン特異的な免疫療法(Allergen-specific immunotherapy with recombinant grass pollen allergens)」 J. Allergy Clin. Immunol. 116, 608-13、(13)Niederberger, V., Horak, F., Vrtala, S., Spitzauer, S., Krauth, M.T., Valent, P., et al.(2004)著「遺伝子操作したアレルゲンによるワクチン接種はアレルギー疾患の進行を防ぐ(Vaccination with genetically engineered allergens prevents progression of allergic disease)」 Proc. Natl. Acad. Sci. U.S.A. 101, 14677-82、(14)Cromwell, O., Fiebig, H., Suck, R., Kahlert, H., Nandy, A., Kettner, J., et al.(2006)著「組み換えアレルゲンワクチンの戦略及び最初の臨床研究の有益な結果(Strategies for recombinant allergen vaccines and fruitful results from first clinical studies)」 Immunol. Allergy Clin. N. Am. 26, 261-81]。
【0007】
パリエタリアは、草本性双子葉植物綱(genus)イラクサ科イラクサ目である。パリエタリア属の様々な種が地中海沿岸に幅広く豊富に分布している[(15)Colombo, P., Duro, G., Costa, M.A., Izzo, V., Mirisola, M., Locorotondo, G., Cocchiara, R.及びGeraci, D.(1998)著「アレルゲンの最新情報。パリエタリア花粉のアレルゲン(An update on allergens. Parietaria pollen allergens)」 Allergy 53, 917-921]。最も一般的な種は、カベイラクサ及びパリエタリア・オフィキナリス(P. officinalis)であるが、パリエタリア・ルシタニカ(P. lusitanica)、パリエタリア・マウリタニカ(P. mauritanica)、及びパリエタリア・クレティカ(P. cretica)等の他の種が幾つかの地域で幾らか存在し得る。それにもかかわらず、パリエタリア花粉の存在がイングランド南部、オーストリア、中東欧の温帯地方、オーストラリア及びカリフォルニアで示されたので、地中海地域は、パリエタリア花粉が見出された唯一の場所というわけではない[(16)Colombo, P., Bonura, A., Costa, M., Izzo, V., Passantino, R., Licorotondo, G., Amoroso, S.及びGerasi, D.(2003)著「パリエタリアのアレルゲン(The allergens of Parietaria)」 Int. Arch. Allergy Immunol. 130, 173-179、(17)Carreira, J.及びPolo, F.(1995)著「オリーブ及びパリエタリア種のアレルゲン並びに地中海地方におけるこれらの関連性(The allergens of Olea europaea and Parietaria spp. and their relevance in the Mediterranean Area)」 Allergy Clin. Immunol. News 7, 79-84]。パリエタリアの顕著な特徴は、数ヶ月間続く長い受粉期間であり、パリエタリアに対してアレルギーである患者において、軽度の鼻結膜炎から重症喘息までに及ぶ、ほとんど永続的な症状が現れる。顕著な交差反応性が様々な種のパリエタリア間で示されたので、パリエタリアに対する通常の軽い単一特異的な感作は、この属の様々な種に対する感作を伴う。
【0008】
2つの最も一般的な種であるカベイラクサ及びパリエタリア・オフィキナリスのアレルギー誘発性画分の精製及び特徴付けに関して様々な文献が提示されている。これらの画分の分子量は、10kDa〜14kDaの範囲であり、これらの抽出物のアレルギー誘発力全体に実質的に関与している[(16)Colombo, P., Bonura, A., Costa, M., Izzo, V., Passantino, R., Licorotondo, G., Amoroso, S.及びGerasi, D.(2003)著「パリエタリアのアレルゲン(The allergens of Parietaria)」 Int. Arch. Allergy Immunol. 130, 173-179、(18)Ayuso, R., Carreira, J., Lombardero, M., Duffort, O., Peris, A., Basomba, A.及びPolo, F.(1993)著「2つのPar jイソアレルゲンの、mAbに基づいたアフィニティクロマトグラフィによる単離。これらの物理化学、免疫化学及びアレルギー誘発性の特性の比較(Isolation by mAb based affinity chromatography of two Par j isoallergens. Comparison of their physicochemical, immunochemical and allergenic properties)」 Mol. Immunol. 30, 1347-1354、(19)Polo, F., Ayuso, R.及びCarreira, J.(1990)著「カベイラクサ花粉の主要なアレルゲンのHPLC精製(HPLC purification of the main allergen of Parietaria judaica pollen)」 Mol. Immunol. 27, 151-157、(20)Polo, F., Ayuso, R.及びCarreira, J.(1991)著「カベイラクサのアレルゲンIの構造と、IgE結合能との間の関係に対する研究(Studies on the relationship between structure and IgE-binding ability of Parietaria judaica allergen I)」 Mol. Immunol. 28, 169-175]。組み換えDNA技術の発展によって、パリエタリア花粉のアレルゲンの分子の特徴付けを完了させることができた。Par j 1及びPar j 2として知られるカベイラクサ花粉の2つの主要なアレルゲンをクローニング及びシーケンシングした[(21)Duro, G., Colombo, P., Costa, M.A., Izzo, V., Porcasi, R., DiFiore, R., Locorotondo, G., Mirisola, M.G., Cocchiara, R.及びGeraci, D.(1996)著「カベイラクサ花粉の新たな主要なアレルゲンであるPar j 2.0101のcDNAクローニング、配列解析及びアレルギー学的特徴付け(cDNA cloning, sequence analysis and allergological characterization of Par j 2.0101, a new major allergen of the Parietaria judaica pollen)」 FEBS Lett. 399, 295-298、(22)Costa, M.A., Colombo, P., Izzo, V., Kennedy, H., Venturella, S., Cocchiara, R., Mistrello, G., Falagiani, P.及びGeraci, D.(1994)著「カベイラクサ花粉の主要なアレルゲンであるPar j IのcDNAクローニング発現及び一次構造(cDNA cloning expression and primary structure of Par j I, a major allergen of Parietaria judaica pollen)」 FEBS Lett. 341, 182-186、(23)Amoresano, A., Pucci, P., Duro, G., Colombo, P., Costa, M.A., Izzo, V., Lambda, D.及びGeraci, D.(2003)著「カベイラクサ花粉の主要なアレルゲンであるPar j 2.0101のジスルフィド架橋の評価(Assignment of disulphide bridges in Par j 2.0101, a major allergen of Parietaria judaica pollen)」 Biol. Chem. 384, 1165-1172]。両方のアレルゲンが非特異的脂質輸送タンパク質(ns−LTP)のファミリーに属し、これらの末端領域でシグナルペプチドを保有しており、プロセシング後に、分子量がそれぞれ14726Da及び11344Daであり、約45%の同一残基を有するタンパク質を生じる。両方のアレルゲンにおいて構造的関連域に位置する可能性があるIgE結合直線状エピトープが記載されている[(24)Asturias, J.A., Gomez-Bayon, N., Eseverri, J.L.及びMartinez, A.(2003)著「カベイラクサ花粉の主要なアレルゲンであるPar j 1及びPar j 2は、同様の免疫グロブリンEエピトープを有する(Par j 1 and Par j 2, the major allergens from Parietaria judaica pollen, have similar immunoglobulin E epitopes)」 Clinical and Experimental Allergy 33, 518-524]。カベイラクサ花粉に対するアレルギーの治療に最適な低アレルギー誘発性の分子を得ることができるために、これらの領域は作用性の標的である。
【0009】
ns−LTPは、in vitroでの極性脂質の膜間の交換及び/又は移動を完了させることができることが知られている[(25)van Ree, R.(2002)著「食物アレルゲンとしての非特異的な脂質輸送タンパク質の臨床的重要性(Clinical importance of non-specific lipid transfer proteins as food allergens)」 Biochem. Soc. Trans 30, 910-913]。分子質量が約9kDaであるLTP1及び分子質量が約7kDaであるLTP2の2つの主要なファミリーが植物で特徴付けられている。LTPファミリーに属するアレルゲンが、幅広く研究された、食物以外の植物で同定されている。したがって、パラゴムノキ(Hevea brasiliensis)のラテックス由来のHev b 12は、バラ科の果実のアレルギー誘発性LTPと約65%の配列同一性を示す9.3kDaの塩基性タンパク質である[(26)Beezhold, D.H., Hickey, V.L., Kostyal, D.A., and et al.(2003)著「パラゴムノキ由来の脂質輸送タンパク質(Hev b 12)、交差反応性ラテックスタンパク質(Lipid transfer protein from Hevea brasiliensis (Hev b 12), a cross-reactive latex protein)」 Ann Allergy Asthma Immunol 439-445]。さらに、オウショウヨモギ(Artemisia vulgaris)のArt v 3[(27)Diaz-Perales, A., Lombardero, M., Sanchez-Monge, R., and et al.(2000)著「潜在的な植物パンアレルゲンとしての脂質輸送タンパク質:IgE結合能が異なる、ヨモギ属の花粉、クリ属の実、バラ科の果実のタンパク質間の交差反応性(Lipid-transfer proteins as potential plant panallergens: cross-reactivity among proteins of Artemisia pollen, Castanea nut and Rosaceae fruits, with different IgE-binding capacities)」 Clin Exp Allergy 1403-1410]及びオリーブのOle e 7[(28)Tejera, M.L., Villalba, M., Batanero, E.及びRodriguez, R.(1999)著「オリーブの木の花粉の新規のアレルゲンであるOle e 7の同定、単離及び特徴付け(Identification, isolation, and characterization of Ole e 7, a new allergen of olive tree pollen)」 J. Allergy Clin. Immunol. 797-802、(29)Rodriguez, R., Villalba, M., Batanero, E., and et al.(2002)著「オリーブ花粉のアレルギー誘発性の多様性(Allergenic diversity of the olive pollen)」 Allergy 6-16]等の幾つかの花粉アレルゲンがLTPとして記載されており、9kDa〜10kDaであり、食物のアレルギー誘発性LTPと30%〜55%の配列同一性を有する。一方で、互いに約45%の配列同一性を有する、カベイラクサの主要なアレルゲンであるPar j 1及びPar j 2はそれぞれ14.7kDa及び11.3kDaと、LTPファミリーにおける通常のものよりも分子量が高い[(16)Colombo, P., Bonura, A., Costa, M., Izzo, V., Passantino, R., Licorotondo, G., Amoroso, S.及びGerasi, D.(2003)著「パリエタリアのアレルゲン(The allergens of Parietaria)」 Int. Arch. Allergy Immunol. 130, 173-179]。しかし、両方のアレルゲンがLTPと同様の構造を有するが、これらは共通の配列領域で食物LTPとの適度なレベルの同一性を有する(Par j 1とモモのLTPとの間で28%)。
【0010】
国際公開特許第WO2005/085278号は、カベイラクサの2つの主要なアレルゲンを含む融合タンパク質の構築物を開示しており、アレルゲン配列が本質的に同じ長さを維持するように、この融合タンパク質の三次元構造が、それぞれのアレルゲンの一次配列における或る特定のシステイン残基(より具体的には、ジスルフィド架橋の形成を伴うシステイン残基)の置換によって破壊される。後の実験によると、このことが、天然のアレルゲンよりもアレルギー誘発性が1000倍低いタンパク質をもたらすと考えられる。
【0011】
本発明者らは、アレルギー誘発性の驚くほど大きな低減は、アレルゲンの三次元構造を破壊するだけでなく、幾つかのIgE結合部位(Bエピトープとして知られる)の欠失によっても得ることができること、及び最も驚くべきことは、これによって免疫原性が低減されないことを発見している。
【0012】
本発明は、より少数のIgE結合エピトープを含有するカベイラクサの2つのアレルゲン(Par j 1及びPar j 2)の断片を結合させることによって得られた様々なキメラタンパク質、並びにそれらを得るための様々な方法及び中間物を初めて開示する。本発明のキメラタンパク質の三次元構造を破壊するだけでなく、或る特定のBエピトープが欠失されてもいる。本発明によるキメラタンパク質は、i)カベイラクサに対してアレルギーの患者由来の血清混合物を用いるin vitro ELISA、ELISA阻害試験及び免疫検出試験、ii)カベイラクサに対してアレルギーの患者における皮膚反応性のin vivo試験、iii)カベイラクサに対してアレルギーの患者由来の個々の血清によるin vitro EAST阻害試験に基づき、IgE抗体との結合能が低いので、アレルギー誘発性が99.99%低減した低アレルギー誘発性と呼ぶことができる。一方で、カベイラクサに対してアレルギーの13人の患者由来の末梢血単核球(CMSP)のリンパ球増殖試験によって示されるように、本発明によるキメラタンパク質は、免疫原性能を維持する。
【0013】
アレルギー誘発性抽出物は、タンパク質と非タンパク質分子との複雑な混合物である。抽出物成分に対する特定のIgEのレベルを検出する技法の使用を増やすことで、アレルギー患者が通常、様々な成分に対して反応性があることを実証することが可能になっている。単一のアレルゲンにのみ反応するアレルギー患者の症例は珍しい。アレルギー誘発性抽出物には、免疫療法において明らかな問題があるので、単一の分子でできる限り多くの治療的特性をグループ分けすることが1つの解決策である。
【発明の開示】
【発明が解決しようとする課題】
【0014】
本発明者らは、1つの分子中に2つのアレルゲンを混合させることに成功し、このことによって、産業上の製造及び治療の観点から利益を受けるだけではなく、免疫原性を変えない、アレルギー誘発性の有意な低減も示される。
【課題を解決するための手段】
【0015】
これらの理由で、本発明は、アレルゲンPar j 1及びPar j 2の断片から構成されたキメラタンパク質(以下、Q1、Q2、及びQ3と呼ばれる)に関し、これは、幾つかのIgE結合Bエピトープが欠失しているので、免疫原性能を損なうことなく、アレルギー反応性を低減させる。このように、得られたキメラポリペプチドは、2つの個々のタンパク質の合計よりも分子量が小さい。
【0016】
本発明は、上記のキメラポリペプチドをコードするDNAを含むヌクレオチド配列にも関し、発現系は発現及び制御に必要な配列を伴う当該配列を含み、受容体細胞は当該発現系によって形質転換される。
【0017】
本発明は、このキメラポリペプチドの臨床使用に、アレルギーの治療に特異的な免疫療法に、並びにこのキメラポリペプチドが生じる可能性がある組成物及びその様々な投与方法にも関する。
【0018】
免疫療法において用いられることが可能な本発明のキメラタンパク質の低アレルギー特性が広く実証されている。本発明者らによって行われた免疫検査によって、図10及び図11で示されるように、キメラQ2は、カベイラクサに対してアレルギーの患者の血清中でIgEを認識しないことが示される。図12で示されるように、Q2のIgE結合能は、2つの天然タンパク質の混合物のものの10000倍低い。
【0019】
この低いアレルギー誘発性のデータは、皮膚プリックテストでの30人の患者におけるin vivo実験によって制限を受けた。キメラQ1のアレルギー誘発性は、2つの単離された天然タンパク質で得られたものよりも3.5倍低かった(図13)。他方では、記載のBエピトープを全く含有せず、Par j 1及びPar j 2の断片によって形成された、Q2のアレルギー誘発性は、2つの単離された天然タンパク質で得られたものよりも112倍低かった。
【0020】
キメラQ2の低アレルギー誘発性(IgE結合能)は、カベイラクサに対してアレルギーの30人の患者の血清で、この分子の反応性を測定することによって確認された(図14)。驚くべきことに、このアレルギー誘発性の低減は、個々の天然タンパク質を合計したものとは異ならない、キメラQ2の免疫原性能の維持を伴っていた(図16)。
【0021】
免疫原性の維持によって、このキメラを、完全ではあるが、はるかに安全な(アレルギー誘発性が低い)抽出物に対する代替物として用いることが可能になる。
【0022】
株の寄託
本発明に対応する微生物株は、特許手続きのための微生物寄託の国際認識に関するブタペスト条約に従って、バレンシア大学(Universidad de Valencia, Edificio de Investigacion, Campus de Burjasot, 46100 BURJASOT, Valencia)にあるスペインの基準菌株コレクション(Spanish collection of type cultures)(CECT)で2006年3月7日に寄託され、大腸菌株CECT7141の参照番号を有する。
【発明を実施するための最良の形態】
【0023】
本発明の一態様によれば、任意のBエピトープ若しくはIgE結合エピトープ(epitode)の欠失、又は任意のBエピトープ若しくはIgE結合エピトープを用いない合成によって得られた低アレルギー誘発性のキメラタンパク質又はペプチドが提供される。好ましくは、欠失はPar j 1及びPar j 2の28位〜53位でBエピトープに適用される。
【0024】
本明細書で用いられるような「低アレルギー誘発性」という用語及び類似の変化形は相対的な用語であり、アレルギー応答性を刺激する本発明のタンパク質及びペプチドのin vivo及びin vitroの能力が、野生型の免疫原の同じ能力に比べて低減していることに関する。
【0025】
好ましくは、キメラタンパク質は、配列番号2、配列番号4又は配列番号6で示されるアミノ酸配列を含み、最も好ましくは、キメラタンパク質は、配列番号4で示されるアミノ酸配列を含む。
【0026】
代替的に又は付加的に、キメラタンパク質は、配列番号2、配列番号4又は配列番号6で示されるアミノ酸配列と相同な配列を含む。好ましくは、相同配列は、配列番号2、配列番号4又は配列番号6で示されるアミノ酸配列、最も好ましくは配列番号4で示されるアミノ酸配列に対する相同性が少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは100%である。
【0027】
別の好ましい実施形態には、配列番号2、配列番号4又は配列番号6で示されるアミノ酸配列に対する相同性が少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%、最も好ましくは100%であるアミノ酸配列を有するキメラタンパク質又はペプチドが含まれる。
【0028】
キメラタンパク質は、精製を容易にするペプチド配列を含んでいてもよい。このようなペプチドは、当該技術分野で一般的に知られており、例えばポリヒスチジンテイル(polyhistidine tail)である。
【0029】
キメラタンパク質を構成する断片は、ポリメラーゼ連鎖反応(PCR)における増幅による既知のスキームに従って、資格があり訓練を受けた人によって合成することができる。当該断片は、適切な制限酵素で消化した後に、ライゲーションによって発現ベクターに統合することができる。この発現ベクター(市販のベクターpQE32等)は、キメラタンパク質を、精製を助ける配列(例えばアミノ末端に位置する一連のヒスチジン)と融合させる能力を有し得る。キメラタンパク質の構築中に、異なるDNA断片が、異なる制限酵素によって認識配列で形成されたリンカーを介して結合され、したがって天然アレルゲンの元の配列には存在しなかった残基は最終キメラ分子に現れる。タンパク質を正しく読み取るのを妨げなかったこれらの新規の残基は、図2、図5及び図8における配列で標識され、関連の実施例で適切に記載される。同様に、キメラは、元の配列に存在しない14個の残基をアミノ末端域に有し、固体支持体と結合した二価金属との相互作用によって迅速に精製されることができる高ヒスチジン領域に対応する。
【0030】
本発明の第2の態様によれば、本発明のタンパク質又はペプチドをコードするポリヌクレオチドが提供される。
【0031】
好ましくは、ポリヌクレオチドは、配列番号1、配列番号3又は配列番号5で示されるヌクレオチド配列を含み、最も好ましくは、ポリヌクレオチドは、配列番号3で示されるヌクレオチド配列を含む。
【0032】
代替的に又は付加的に、ポリヌクレオチドは、配列番号1、配列番号3又は配列番号5で示されるヌクレオチド配列と相同な配列を含み得る。好ましくは、相同配列は、配列番号1、配列番号3又は配列番号5で示されるヌクレオチド配列に対する相同性が少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%である。
【0033】
別の好ましい実施形態には、配列番号1、配列番号3又は配列番号5で示されるヌクレオチド配列に対する相同性が少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%、最も好ましくは100%であるヌクレオチド配列を有するポリヌクレオチドが含まれる。
【0034】
ポリヌクレオチドは、シグナルペプチドをコードする配列をさらに含み得る。シグナルペプチドは、小胞体の膜を通るタンパク質輸送を開始させるアミノ酸配列である。好適なシグナルペプチドは、本発明の分野における当業者に既知である。
【0035】
本発明には、本発明のポリヌクレオチド配列によってコードされたペプチドも含まれる。
【0036】
本発明の他の態様によれば、本発明のポリヌクレオチド配列を含む発現系、及び本発明のタンパク質又はペプチドを発現することができる当該発現系によって形質転換された宿主細胞が提供される。
【0037】
本発明は、本発明のペプチド又はタンパク質を製造する方法も含み、当該方法は、
(i)宿主細胞において、本発明のタンパク質又はペプチドをコードするヌクレオチド配列を発現することができる複製可能な発現系を調製する工程と、
(ii)宿主細胞を上記発現系で形質転換する工程と、
(iii)上記タンパク質又は上記ペプチドを発現させる条件下で上記形質転換した宿主細胞を培養する工程と、
(iv)任意で、上記タンパク質又は上記ペプチドを回収する工程と、
を含む。
【0038】
本発明は、例えば牛乳又は卵白中に本発明のタンパク質又はペプチドを産生することができるトランスジェニック動物も含む。
【0039】
本発明の別の態様によれば、免疫障害、特にアレルギー等の過敏性障害の治療のための本発明のタンパク質又はペプチドの使用が提供される。
【0040】
本発明のさらなる態様によれば、免疫障害、特にアレルギー等の過敏性障害の治療用の薬物調製のための本発明のタンパク質又はペプチドの使用が提供される。好ましくは薬物はワクチンである。
【0041】
好ましくは、過敏性障害は、パリエタリアの花粉、より好ましくはカベイラクサの花粉に対するアレルギーである。
【0042】
本明細書で用いられるように、治療という用語、及び「治療する(treat)」又は「治療すること(treating)」等の変化形は、ヒト又は非ヒト動物で有益であり得る任意のレジーム(regime)を表す。治療は、現状に対するものであり得るか、又は予防的なものであり得る(予防的治療)。治療には、治癒効果、軽減効果又は予防効果が含まれ得る。好ましくは、治療は予防的治療である。
【0043】
本発明のまたさらなる態様によれば、効果的な量の本発明のタンパク質又はペプチドと、薬学的に許容可能な賦形剤とを含む薬学的組成物が提供される。好ましくは、薬学的組成物はワクチン組成物である。
【0044】
本発明のまたさらなる態様によれば、免疫障害、特にアレルギー等の過敏性障害を治療する方法が提供され、当該方法は、治療が必要な被験者に、効果的な量の本発明のタンパク質又はペプチドを投与する工程を含む。
【0045】
本発明の別の態様によれば、免疫障害、特にアレルギー等の過敏性障害の治療に用いられる本発明のタンパク質又はペプチドが提供される。
【0046】
個々の患者に適切な投与形態及び剤形の選択は当業者には明らかである。
【0047】
本発明は、本発明に従ったキメラ、好ましくはキメラQ2、又は哺乳動物における脱感作治療ためにそれから誘導された合成ペプチドの使用に及ぶ。脱感作方法は、非経口経路(皮下、静脈内、又は筋肉内)、又は経口、経鼻若しくは直腸経路による対象のアレルゲンの反復投与を伴う。これらの(ポリ)ペプチドは、使用に関する現在の(prevailing)法律及びガレノス製剤手順に従って、単独又は他の希釈剤と併用して投与することができる。
【0048】
本発明で記載されるキメラタンパク質Q2及びキメラタンパク質Q3は、図10、図11、図12及び図14で示されるように、完全抽出物又は複合天然タンパク質よりもカベイラクサに対してアレルギーである患者の血清に対する反応性が低く、その上in vivoでの皮膚試験(図13)でもこの低アレルギー誘発性が見られることから、低アレルギー誘発性である。
【0049】
キメラ分子Q1及びキメラ分子Q2は、複合天然タンパク質と同程度の免疫原性能も有している(図16)。両方の特性(低アレルギー誘発性及び免疫原性)のために、キメラQ2は、鼻炎、結膜炎、喘息、蕁麻疹、血管浮腫、湿疹、皮膚炎、又はさらにアナフィラキシーショックとして現れ得るカベイラクサの花粉に対するアレルギーの予防的及び治癒的な治療に対する優れた候補になる。
【0050】
本発明によるキメラタンパク質の免疫学的特徴が以下に記載される。図10は免疫検出試験を示しており、キメラQ2及びキメラQ3(レーン5及びレーン6)が、2つの天然タンパク質(レーン1)、単離した組み換えタンパク質(レーン2及びレーン3)又はキメラ(レーン4)の反応性と比べた場合に、アレルギー患者におけるIgE結合能が大きく低減していることを示し、このことは、完全ではあるが、これらのBエピトープを全て含有する2つのタンパク質の組み換え融合を示す。このことは、29番目の残基と、52番目の残基との間に含まれるBエプトープの欠如(キメラQ2)がアレルギー誘発性の低減に寄与することを示していると考えられる。同様の結果が、キメラタンパク質に対する15個の異なる血清の反応性を研究した際に得られた(図11)。キメラタンパク質Q2及びキメラタンパク質Q3の反応性は、2つの完全なアレルゲンの結合体(Par j 1−Par j 2)であるQ1で見られたものよりもはるかに小さい。このアレルギー誘発性の低減は、カベイラクサに対してアレルギーである患者由来の血清混合物によるELISA阻害によって定量された(図12)。2つの天然タンパク質の混合物(than of)抽出物の60%阻害に達するのに、10000倍以上のタンパク質Q2が必要であった。したがって、アレルギー誘発性が天然タンパク質よりも10000倍低く、アレルギー誘発性がキメラタンパク質Q1よりも約20倍低いことが推測され得た。
【0051】
キメラQ2の低アレルギー誘発性のより直接的な測定は、カベイラクサの花粉に対してアレルギーである30人の患者における皮膚反応性の直接的な測定によって得られた。図13で与えられたデータは、キメラQ2の皮膚反応性が顕著に低減したことを示している。記載されたそれぞれの分布の比較は、キメラQ2の平均パッチサイズが2つの天然タンパク質で見られたものよりも112倍小さいことを示し、このことはアレルギー活性の99%超の低減を示している。
【0052】
また、キメラタンパク質Q2とのIgEの低結合能が、EASTによって測定されたカベイラクサに対してアレルギーであるさらなる30人の患者由来の血清によって実証された(図14)。全ての患者において、IgE結合は、天然タンパク質の混合物に比べて、キメラQ2で大きく低減した。
【0053】
IgE結合能、及びしたがってBエピトープの欠失による拒絶反応の誘発能のこの大きな低減は、免疫原性能の維持を伴った。タンパク質Q2によって、図16で示されるように、天然抽出物及び組み合わされた2つの純粋な天然タンパク質の混合物によって誘導されたものと同程度のリンパ増殖指数が実証された。このことは全て、Par j 1及びPar j 2の断片で構築されたキメラタンパク質Q2が、より少数のIgEエピトープを含有したが、保護的な免疫応答を誘導するのに十分なTエピトープを維持したことを示している。
【0054】
本発明は、本発明の調製及びその質の実証における実験段階に関して、以下の実施例によってより良好に理解される。これらの実施例は単に例示的な実施例であり、本発明を限定しない。
【実施例1】
【0055】
Q1、Q2、及びQ3融合体の構築
キメラタンパク質は、Gonzalez-Rioja et al.著「組み換えアレルゲンPar j 1及びPar j 2の発現及び精製(Expression and purification of the recombinant allergens Par j 1 and Par j 2)」(XXIII Congreso EAACI, Abstracts Book(2004), 181-182)で記載されたPar j 1.0103及びPar j 2.0101をコードする配列と、それぞれの場合に特異的な誘発因子(triggers)とを含有するプラスミドをマトリクスとして用いるポリメラーゼの連鎖増幅(PCR)によって構築された。この誘発因子は、ハイブリダイゼーション(hybridation)域と、異なる制限エンドヌクレアーゼに対する様々なセクション部位(下線)と、幾つかのアンカーヌクレオチドとから構成される。PCR誘導性の増幅反応は、反応量50μl中に以下の成分を有していた:増幅緩衝液×10、5μl、200μMのdNTP、100pmolの各誘発オリゴヌクレオチド:2.5単位のポリメラーゼTaq(Pfx DNAポリメラーゼ、Invitrogen)、DNAマトリクス1ng、及び50μlまで滅菌蒸留水。増幅反応が、それぞれの場合において記載される特定条件下でRoboCyclerサーモサイクラー(Stratagene)において行われた。反応生成物をアガロースゲル(2%)中で電気泳動にかけ、製造業者によって記載された方法を利用してGeneclean(Bio101)を用いて、対象のバンドをゲルから単離した。単離した断片を適切な制限酵素で消化し、同じ酵素で消化したpBluescriptベクターと結合させた。ライゲーション混合物を用いて、大腸菌DH5α(Invitrogen(Paisley, UK)から入手可能)のコンピテント細胞を形質転換した。プラスミドDNAを単離するために、得られたコロニーを培養させ、これを対象の断片を遊離させるために、適切な酵素で消化した。陽性コロニーがそのシーケンシングで選択された。pBluescriptに挿入されたDNAのシーケンシングは、製造業者の取扱説明書に従って、蛍光ジデオキシヌクレオチドによる使用のために改良されたサンガー法[(30)Hanahan, D.(1983)著「プラスミドによる大腸菌の形質転換に対する研究(Studies on transformation of Escherichia coli with plasmids)」 J. Mol. Biol. 166, 557-580]及びPRISMレディ反応ジデオキシ末端化サイクルシーケンシングキット(Ready Reaction DiDeoxy Termination Cycle Sequencing Kit)(Perkin Elmer)を用いたサーモサイクラーにおける増幅によって行われた。
【0056】
A)キメラタンパク質Q1
この場合、不安定な三次構造を有するが、連続的なIgEエピトープ全てが完全であるタンパク質を得るために、両方のタンパク質(Par j 1及びPar j 2)の完全な配列が融合されると考えられた。Q1として知られている構築物を得るために、ベクターpKS−BluescriptでクローニングしたPar j 1及びPar j 2のcDNAをマトリクスとして用いた。2つの配列を融合するために、Par j 1の配列のC末端と、Par j 2のN末端との両方においてリンカー(この場合EcoRI標的)を有し、その後断片をライゲーションさせる必要があった。当該標的は対応する合成オリゴヌクレオチド(F1R1及びF1F2)に付加され、PCRプロセスにおいて増幅断片に組み込まれた。
【0057】
合成オリゴヌクレオチドには以下のものを用いた:
断片1:F1F1、CG GGATCC TGCAAGAAACCTGCGG(BamHI)及びF1R1、CG GAATTC GGCTTTTTCCGGTGCGG(EcoRI)。条件:94℃、4分(1サイクル)、94℃、30秒 − 53℃、30秒 − 72℃、90秒(5サイクル)、94℃、30秒 − 60℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)。
断片2:F1F2、CG GAATTC GAGGAGGCTTGCGGGA(EcoRI)及びF1R2、CG AAGCTT CTAATAGTAACCTCTGA(HindIII)。条件:94℃、4分(1サイクル)、94℃、30秒 − 51℃、30秒 − 72℃、90秒(5サイクル)、94℃、30秒 − 59℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)。
【0058】
Par j 1及びPar j 2のそれぞれに対応する2つのPCR産物が生成された。これらのうちの第1の断片(Par j 1)に関しては、合成オリゴヌクレオチドF1F1及び合成オリゴヌクレオチドF1R1をそれぞれN末端及びC末端に用いて、約420塩基対(pb)のサイズが得られた。増幅断片がpKS−Bluescriptベクターでクローニングされ、その配列が確認された。Par j 2に対応する第2の断片に関しては、オリゴヌクレオチドF1F2及びオリゴヌクレオチドF1R2をそれぞれN末端及びC末端に用いて、同じ手順を用いた。この際、約300pbの断片が得られ、これをpKS−Bluescriptベクターでクローニングして、その配列を確認した。この第2の断片は、EcoRI標的によるライゲーションによって第1の断片と結合し、Par j 1がN末端で、Par j 2がC末端で維持され、その後得られた断片が市販の発現ベクターpQE−32(Qiagen)でサブクローニングされ、親和性末端(affinity tail)に対応するN末端で13個のアミノ酸の追加配列を含有していた(図2)。当該配列は、256個のアミノ酸を有し、見かけの分子量が28070Daのポリペプチドをコードした。
【0059】
B)キメラタンパク質Q2
このキメラを得る場合、両方のタンパク質の断片は結合されているが、記載の連続的なIgEエピトープを全く含んでいないと考えられた[(18)Asturias, J.A, Gomez-Bayon, N., Eseverri, J. L.及びMartinez, A.(2003)著「カベイラクサ花粉由来の主要なアレルゲンであるPar j 1及びPar j 2は同様の免疫グロブリンEエピトープを有している(Par j 1 and Par j 2, the major allergens from Parietaria judaica pollen, have similar immunoglobulin E epitopes)」 Clinical and Experimental Allergy 33, 518-524]。以下の断片を含んだ幾つかの合成オリゴヌクレオチドを設計した:Par j 1(残基1〜28及び残基53〜139の断片)及びPar j 2(残基1〜28及び残基53〜103の断片)(図3及び図4)。IgEエピトープを含有する配列の一部はこの方法で取り除かれた。
【0060】
4つの断片(それぞれのアレルゲンに対して2つの断片)がこの構築物で増幅され、pKS BluescriptベクターでクローニングしたPar j 1及びPar j 2のcDNAをマトリクスとして用いた。新規のタンパク質の構造機構は、異なる増幅断片の連続結合への制限酵素(この場合、PstI、EcoRi及びEcoRV)の使用である。
【0061】
合成オリゴヌクレオチドには、以下のものを用いた:
断片1:F1F1、CG GGATCC TGCAAGAAACCTGCGG(BamHI)及びF2R1、CG CTGCAG CCCCTTTGACGGCTCTT(PstI)。条件:分(1サイクル)、94℃、30秒 − 54℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)。
断片2:F2F2、CG CTGCAG ATCCAGACCGCCATGAA(PstI)及びF2R2、CG GAATTC GGCTTTTTCCGGTGCGGG(EcoRI)。
断片3:F2F3、CG GAATTC GAGGAGGCTTGCGGGAA(EcoRI)及びF2R3、CG GATATC CTCCTTCGACGGCTCCTT(EcoRV)。
断片4:F2F4、CG GATATC ATAGTGCGCGCCACGAA(EcoRV)及びF1R2、CG AAGCTT CTAATAGTAACCTCTGA(HindIII)。3つの断片の条件は、94℃、4分(1サイクル)、94℃、30秒 − 54℃、30秒 − 72℃、90秒(5サイクル)、94℃、30秒 − 62℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)であった。
【0062】
Par j 1(断片1)に対応する、これらのうちの第1の断片に関しては、合成オリゴヌクレオチドF1F1及び合成オリゴヌクレオチドF2R1をそれぞれN末端及びC末端に用いて、約90塩基対のサイズが得られ、pKS−Bluescriptベクターでクローニングした。Par j 1に対応する第2の断片に関しては、オリゴヌクレオチドF2F2及びオリゴヌクレオチドF2R2をそれぞれN末端及びC末端に用いて、同じ手順を実行した。この際、約260pbの断片が得られ、pKS−Bluescriptベクターでクローニングした。この最後の断片は、PstIによって担持された標的のリンカーによるライゲーションによって第1の断片と結合し、その配列が確認された。
【0063】
Par j 2(断片3及び断片4)の配列から増幅された2つの断片に関しては、断片1及び断片2に記載された手順に従った。N末端及びC末端それぞれに対してオリゴヌクレオチドF2F3/F2R3及びオリゴヌクレオチドF2F4/F1R4をその増幅のために用い、PCR産物のサイズは断片3では約90pb、及び断片4では約150pbであった。断片4は、EcoRVによって担持された標的のリンカーによるライゲーションによって断片3と結合し、その配列が確認された。
【0064】
最終工程において、生成された2つの新規の断片(それぞれのアレルゲンに対して1つの断片)が、EcoRIによって担持された標的のリンカーによるライゲーションによって結合し、Par j 1がN末端で、Par j 2がC末端で維持された。その後、当該断片が市販の発現ベクターpQE−32でサブクローニングされ、親和性末端に対応するN末端で13個のアミノ酸の追加配列を含有していた(図5)。当該配列は、212個のアミノ酸を有し、見かけの分子量が23336Daのポリペプチドをコードした。
【0065】
C)キメラタンパク質Q3
この最後の構築物は、この場合、存在し得る任意のIgEエピトープを取り除こうと試みた以外はQ2と実質的に同一であり、Par j 1(残基71〜80)及びPar j 2(残基72〜81)(図6及び図7)の配列に存在する8つのアミノ酸の別の付加的な配列が取り除かれた。
【0066】
合成オリゴヌクレオチドには以下のものを用いた:
断片1:F1F1、CG GGATCC TGCAAGAAACCTGCGG(BamHI)及びF3R1、CG AGATCT GACCTCGCTGACGAG(BglII)。
断片2:F3F2、CG AGATCT AGCAAGCTCCCGCCC(BglII)及びF3R2、CG GGTACC GGGGACCTCGGCGAC(KpnI)。2つの断片の条件は、94℃、4分(1サイクル)、94℃、30秒 − 54℃、30秒 − 72℃、90秒(5サイクル)、94℃、30秒 − 63℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)であった。
断片3:F3F3、CG GGTACC CTCCCGCCCATCACC(KpnI)及びF3R3、TTTAAAAAGGCCGTAATATCC。条件:94℃、4分(1サイクル)、94℃、30秒 − 56℃、30秒 − 72℃、90秒(35サイクル)、72℃、10分(1サイクル)。
【0067】
この場合、構築物Q2−pQE−32をマトリクスとして用い、断片1、断片2及び断片3それぞれに対して、約150pb、約370pb及び約290pbのサイズが得られた。用いられたオリゴヌクレオチドは、断片1に対してF3F1及びF3R1、断片2に対してF3F2及びF3R2、並びに断片3に対してF3F3及びF3R3であった。上記の場合の断片の連続ライゲーションのプロセスを再び繰り返した。用いられる新規の制限酵素はBglII及びKpnIであった。第3のそして最後の断片では、そのサイズが小さかったので(66pb)、その後の精製プロセスにおいてより高い利便性で働くことができるようにより大きなサイズの断片を得るため、ベクターpQE−32−Q2の配列の一部を増幅することになった。それぞれの工程で、ベクターpKS−Bluescriptでクローニングされた、PCRによって得られた断片、及びその配列が確認された。第1のグルタミンアミノ酸(Q)に対応する第1の断片における自然突然変異がプロリン(P)によって生じた。
【0068】
最終的に、得られた断片が発現ベクターpQE−32でサブクローニングされ、親和性末端に対応するN末端で13個のアミノ酸の追加の配列を含有していた(図8)。当該配列は、200個のアミノ酸を有し、見かけの分子量が21938Daのポリペプチドをコードした。
【実施例2】
【0069】
キメラタンパク質Q1、キメラタンパク質Q2、及びキメラタンパク質Q3の発現及び精製
LB(100μg/mlのアンピシリン及び25μg/mlのカナマイシンをそれぞれ補充した)のシートから単離したコロニーから始め、同じ培地50mlの前接種材料(pre-inoculum)を産生し、撹拌しながら(260rpm)、37℃で一晩インキュベートした。光学密度(600nm)を0.2から始めて、同じ培地1Lを当該前接種材料で接種させた。混合物を撹拌しながら37℃で1時間30分インキュベートした後、同じインキュベート条件下で3時間、IPTG(1mMの最終濃度)による誘導を行った。
【0070】
Q2の場合、細胞を10000rpm、4℃で15分間、遠心分離して、溶解緩衝液(20mMのリン酸塩(pH7.4)、50mMのイミダゾール、0.5MのNaCl)50mlに再懸濁させた。撹拌しながら、37℃で30分間、再懸濁質(matter)をリゾチーム(0.1mg/mlの最終濃度)で処理した。それから、超音波処理(5パルス、20秒)を行い、混合物を15000rpm、4℃で15分間、遠心分離した。上清を0.45μmのフィルタ(Millex HV, Millipore)で濾過し、AKTAプライムシステム(Aはウムラウト)(GE-Healthcare)に適合されたHi−Trapキレート化HPカラム(GE-Healthcare)5mlに加えた(2.5ml/分)。このカラムをニッケルでキレート化し、予め10容量の溶解緩衝液で平衡化していた。カラムを15容量の溶解緩衝液で洗浄すると、カラムと結合したタンパク質の溶出を溶出緩衝液(20mMのリン酸塩(pH7.4)、0.5Mのイミダゾール)を用いて100%まで行った。塩を除去し、20mMでpH7のリン酸緩衝液サンプルを交換するために、溶出液をHi−Trap脱塩カラム(GE-Healthcare)5mlに通した。タンパク質は−40℃に維持した。
【0071】
Q1及びQ3の場合、さらなる精製工程が必要であった。その誘導後に、細胞を10000rpm、4℃で15分間、遠心分離し、8Mの尿素を補充した溶解緩衝液50mlで再懸濁して、前の段落で記載された手順を採用した。2番目の精製工程では、予めPBSで平衡化したHi−Load 16/60 Superdex−200カラム(GE-Healthcare)に、得られた溶出液を加えた(1ml/分)。前の段落で記載された緩衝液でタンパク質の溶出を行った。タンパク質は−40℃に維持した。
【0072】
SDSを有するポリアクリルアミドゲル(SDS−PAGE)で、電気泳動を行った。Laemmliによって記載された方法は、MINI−PROTEAN電気泳動装置(Bio-Rad)を用いて、基本的に[(31)Laemmli, U.K.(1970)著「バクテリオファージT4頭部集合化中の構造タンパク質の切断(Cleavage of structural proteins during the assembly of the head of bacteriophage T4)」 Nature 277, 680-685]に従った。10×10cmであり、ポリアクリルアミド濃度が12.5%であるゲルに、トリス−グリシン緩衝液で45分間、200ボルトの電流を流した。参照として用いられるタンパク質は、低分子量に関するBio-Radのキット由来のものであった。分子量の算出及びゲルの濃度測定分析は、画像分析器(Diversity, BioRad)を用いて行った。
【0073】
ハイブリッドタンパク質Q1は、32kDaのHis標識化融合タンパク質として表され(図9)、細菌培養物の最終収率は2mg/Lであった。Q2タンパク質及びQ3タンパク質は、28kDaのHis標識化融合タンパク質として表され、培養物の最終収率はそれぞれ、5mg/L及び7mg/Lであった(図9)。
【実施例3】
【0074】
精製ハイブリッド分子の円偏光二色性分析
pH7.0、20℃でのFar−UV(190nm〜250nm)のCDスペクトルが、20℃で温度を自動調節したキュベット中に日本分光株式会社のPTC−423S温度制御装置を備えた日本分光株式会社のJ−810分光偏光計によって記録された。タンパク質濃度は、0.2cmキュベットにおいて20mMのリン酸ナトリウム緩衝液中で0.035mg/mlであり、40個の走査が集積した。全てのスペクトルは適切なバックグラウンドを差し引き、平均残余楕円率に変換した。
【0075】
二次構造要素は、天然のパリエタリアのアレルゲン混合物(Par j 1及びPar j 2)、及びハイブリッドタンパク質を適用したCD分光法によって分析された(図10)。ハイブリッドタンパク質のスペクトルはそれらの中でほぼ同一であったが、天然アレルゲンのCDスペクトルとは全く異なっていた。NPAスペクトルは、208nmで最小を示し、222nmで明確に増大し(shoulder)、190nmで最大を示したが、ハイブリッドタンパク質のスペクトルは、ほぼ完全な変性(unfolded)状態になるランダムコイル構造に移行した。
【実施例4】
【0076】
カベイラクサに対してアレルギーである患者の血清混合物に対するキメラタンパク質の低IgE固定の反応性を実証するための免疫試験
A)免疫検出
融合物1、融合物2及び融合物3のIgE結合活性の第1の評価が、カベイラクサに対してアレルギーである患者由来の血清混合物を用いる免疫移動法(immunotransfer method)によって行われた。タンパク質抽出物及び精製タンパク質をポリアクリルアミドゲルに適用したら、Towbin et al[(32)Towbin, H., Staehelin, I.及びGordon, J.(1979)著「ポリアクリルアミドゲルからニトロセルロースシートへのタンパク質の電気泳動転写:手順及び幾つかの用途(Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications)」 Proc. Natl. Acad. Sci. USA 76, 4350-4354]の方法によって電気的移行を行った。この目的のために、SDS−PAGEによって単離したタンパク質をPVDF Hybond−P膜(GE-Healthcare)に電気的に固定した(transfixed)。周囲温度で1時間、膜を遮断したら、4℃で一晩、膜を一次抗体とインキュベートして、同じ洗浄緩衝液で様々な回数(various)洗浄後、周囲温度で1時間、膜を二次ペルオキシダーゼ結合抗体とインキュベートした。製造業者によって記載されたように、膜をフィルム(Hyperfilm.ECL、GE-Healthcare)に曝すことによって、ECL化学発光法(GE-Healthcare)でバンドを検出した。
【0077】
免疫検出試験によって、3つの融合物間でIgE結合能が異なることが実証された。したがって、IgE抗体の認識はQ1の場合でのみ存在した(図11)。Q2の場合では、認識は非常に急激に落ちたが、Q3の場合では、実証された認識は0であった。
【0078】
B)直接ELISA
カベイラクサに対してアレルギーの患者由来の個々の血清を用いたELISA法によって、3つの融合物に対するIgEの反応性を分析した。周囲温度で一晩、ポリスチレンプレート(Greiner)を、PBS緩衝液(10mMのリン酸塩(pH7.2)、137mMのNaCl、2.7mMのKCl)中で、カップ1つ当たり1μgのカベイラクサ抽出物タンパク質又は1μgの純粋なタンパク質とインキュベートした。1%BSA−0.05%Tween20を補充した、200μl/カップのPBSでこれらを遮断し、37℃で1時間維持した。アレルギー患者由来の100μl/カップの血清(10倍希釈) を添加し、37℃で90分間インキュベートした。200μl/カップのPBS−T(PBS+0.05%Tween20)による3回の洗浄の後、ペルオキシダーゼと結合したヒトIgE免疫グロブリン(Dako)に対する100μl/カップの抗血清を添加し(1000倍希釈)、37℃で90分間インキュベートした。PBS−Tで新たに3回洗浄した後、製造業者の取扱説明書に従って調製した、200μl/カップのo−フェニレンジアミン(Sigma−Fastのタブレットセット、Sigma)溶液を添加し、プレートを30分間暗所に維持した。反応を50μl/カップの3MのH2SO4で停止させ、ELISAイージーリーダーEAR−400AT(SLT-Lab Instruments)プレートリーダーにおいて492nmで吸光度を測定した。
【0079】
IgEに対する反応性は、Q1の場合でのみ実証されたが、Q2及びQ3の両方はIge抗体の認識が実質的にないことが実証された(図12)。
【0080】
C)ELISA阻害
ELISA阻害試験に関して、カベイラクサに対してアレルギーの患者由来の血清混合物を、4℃で一晩、所定の濃度の阻害タンパク質(1ng/ml〜1000ng/ml)と予めインキュベートした。後の残りの手順は、ELISA直接試験に記載されたものと同じである。
【0081】
カベイラクサに対してアレルギーの患者由来の血清混合物によるELISA阻害試験において、Q1では、カベイラクサの花粉のIgE結合活性の約50%程度の阻害が実証され、これはnPar j 1−Par j 2のものより幾らか低く(92%)、500倍以上のタンパク質を添加する必要があった(図13)。キメラQ2及びキメラQ3は、最大阻害の約60%程度に達したが、これは同程度の阻害に達するためには、nPar j 1−Par j 2で必要とされるものよりも10000倍高いタンパク質濃度を必要とし、このことは、これらのキメラがアレルギー誘発性を約99.99%低減させたことを示唆していた。
【実施例5】
【0082】
キメラタンパク質Q1、キメラタンパク質Q2、及びキメラタンパク質Q3の低皮膚反応性を実証するためのin vivo実験
in vivo皮膚反応(プリック)試験を行い、カベイラクサ花粉に対するアレルギーに対する満足のいく免疫療法を開発することができるために、候補低アレルギー誘発性の分子を求めるという目的で、キメラ1、2、及び3の低アレルギー誘発性を評価した。
【0083】
カベイラクサ抽出物、nPar j 1−Par j 2、大腸菌で発現したrPar j 1及びrPar j 2、並びにキメラ1、2及び3を用いて、皮膚試験を行った。精製タンパク質は、0.5%フェノール含有及び50%グリセリン含有生理食塩水溶液で希釈した。天然のPar j、Par j 2、Q1の場合に用いる濃度は、0.5μg/ml、5μg/ml、及び50μg/mlであり、Q2又はQ3の場合の濃度は5μg/ml、50μg/ml、及び250μg/mlであった。0.9%NaCl及び塩酸ヒスタミン(10mg/ml)をそれぞれ、陰性対照及び陽性対照に用いた。
【0084】
この実験では、試験されるそれぞれのアレルゲンの液滴を、前腕の掌面上に沈着させた後に、ランセットを用いて、液滴を通して穿刺穴を作製した。二重で、増大濃度及び低減濃度の反対の条件(strings)で、それぞれの試験を行った。15分後、パッチの輪郭を先端が細い黒色のマーカーペンで描いた。低アレルギー誘発性の絆創膏の細片をパッチ上に貼り、ゆっくり押し付けて、インクのマーキングを細片に写し、これをパッチ記録シートに転写させた。Summasketchデジタル化タブレット及びコンピュータベースのプログラム(Autocad v.11)を用いてこの記録をスキャンすることによって、パッチ領域を測定した。
【0085】
ブロック図で結果を図示し、関連変数に関してウィルコクスン検定を用いて統計調査を行うことによって、得られた結果を解釈した(図14)。これらの図は、皮膚反応に対応する値の分布(パッチ領域(mm2)で測定された)が、抽出物(平均65.25mm2、信頼範囲(95%):54.98〜75.53)及びnPar j 1−Par j 2(平均106.80mm2、信頼範囲(95%):91.90〜121.70)に対して、3つのキメラの場合で有意に異なることを示している(P<0.001)。キメラ2及びキメラ3の値は実質的に0である:Q2(平均0.95mm2、信頼範囲(95%):0〜2.89)及びQ3(平均0.15mm2、信頼範囲(95%):0〜0.47)。
【実施例6】
【0086】
キメラタンパク質Q1、キメラタンパク質Q2、及びキメラタンパク質Q3の低IgE抗体結合能を実証するための実験
in vivo試験の補足として、EAST直接法を用いて、特異的なIgEを求めることによって、新たにin vitro試験を行った。
【0087】
天然タンパク質及び組み換えタンパク質(50μg/ml)、並びにカベイラクサ抽出物(2mg/ml)を、臭化シアンで活性化したディスクにカップリングさせることによって、Ceska et al.[(33)Ceska, M. and Lundkvist, U.(1972)著「IgEの決定のための新規の簡易なラジオイムノアッセイ法(A new and simple radioimmunoassay method for the determination of IgE)」 Immunochemistry 9, 1021-1030]に従って、特異的なIgEを求めた。その後、患者由来の血清50μlを添加し、周囲温度で1時間、インキュベートした。洗浄後、37℃で30分間、ディスクをアルカリホスファターゼと結合したヒト抗IgE抗体50μlとインキュベートして、製造業者(Hycor Biomedical Inc.)によってIgE特異的なハイテックキットEIAで与えられた取扱説明書に従って、定量を行った。
【0088】
得られた結果は、単にin vivoで得られた結果を再確認しているだけである。3つのキメラのうち、Q1はnPar j 1−Par j 2に対して、その免疫原性能を維持したが、Q2及びQ3では当該免疫原性能が非常に顕著に低減した(P<0.001)(図15)。
【実施例7】
【0089】
キメラタンパク質Q1、キメラタンパク質Q2及びキメラタンパク質Q3の免疫原性能を実証する誘導性リンパ増殖実験
免疫療法における低アレルギー誘発性分子の使用に対する本質的な要求は、その抗原性(Tエピトープ)の維持である。したがって、IgE抗体と結合しないことに加えて、本発明のタンパク質が抗原性を維持するか否か確認するために、この実験で用いられる様々なタンパク質によって刺激された末梢血単核球(CMSP)でリンパ増殖試験を行った。比色分析によって測定されるホルマザン化合物を発生させるために、生存細胞のミトコンドリアデヒドロゲナーゼによるテトラゾール塩WST−1の消化に基づく比色法によって、この試験が行われた。
【0090】
リンパ球(lymphocite)分離溶液(Lymphoprep, Nycomed)を用いる密度勾配における遠心分離によって、カベイラクサに対してアレルギーの13人の患者のCMSPを単離した。それから、培養培地(血清無含有培地AIM V、Gibco)において1×106個の生存細胞/mlでCMSPを再懸濁し、PBS(Sigma Chemical Co.)中の0.25%トリパンブルー(tripan blue)でこれらの生存率を試験した。製造業者によって記載されたように、90%を超える生存率で調製されたCMSPをin vitro増殖試験に直ちに用いた(細胞増殖剤WST−1、Roche Diagnostics)。最終容量が200μlの培地中の2×105個のCMSPで、それらを底が平らなマイクロプレート(Nunclon、NUNC)に沈着させた後に、カベイラクサ抽出物を用いて、5%CO2の加湿雰囲気下、37℃で三重で測定を行い、最終濃度が0.0005μg/ml−0.005μg/ml−0.05μg/ml−0.5μg/ml−5μg/mlになるまで、様々なタンパク質を精製した。三重の対照の刺激を受けない培養物は全ての場合に含まれた。3日後、細胞増殖剤WST−1 20μlを全てのカップに加え、4時間インキュベートした。代謝的に活性のある細胞によって産生されたホルマザンを450nmでの吸光によって定量した。この試験で用いた組み換えタンパク質は、3つのキメラ、並びに大腸菌で発現したrPar j 1及びrPar j 2であった。
【0091】
第1の工程において、免疫原性(immunuogenic)タンパク質をスキャンして、その後の試験の進行のために最適濃度を求めた。全ての場合、最大増殖(IE%)を示すタンパク質濃度が0.5μg/mlであることが見出された(図16)。
【0092】
カベイラクサに対してアレルギーの13人の患者による増殖の結果は、ブロック図の統計分析及び2つの対になったサンプルに関するノンパラメトリック試験によって求められた。統計分析によって、対照に用いられたカベイラク抽出物には、nPar j 1−Par j 2とあまり変わらない抗原刺激能があり(P=0.142)、Q1及びQ2は、これらの抽出物(それぞれ、P=0.152及びP=0.294)及びnPar j 1−Par j 2(それぞれ、P=0.484及びP=0.182)と有意差のない値分布を示したことが示された(図17)。一方で、Q3は、この抽出物(P=0.002)及びnPar j 1−Par j 2(P=0.004)に対してほとんど刺激能を有しなかった。またその一方で、この抽出物(それぞれ、P=0.142及びP=0.003)及びnPar j 1−Par j 2(それぞれ、P=0.041及びP=0.002)に対して、rPar j 1へのrPar j 2の抗原力が大きかったことが指摘されるべきである。
【0093】
得られた結果によって、依然として、Q2は上記特性を維持したのに対し、Q3はそれを完全に喪失したことが示される。カベイラクサに対するアレルギーを治療するために様々な低アレルギー誘発性分子を得るプロセスについて記載された全てのものから、Q2がカベイラクサに対するアレルギーの満足のいく免疫療法を開発する候補低アレルギー誘発性分子であることを結論付けることができる。
【0094】
投与方法
本発明は、哺乳動物における減感作治療のための、記載の低アレルギー誘発性キメラ又はこれら由来の合成ペプチドの使用に関する。減感作方法は、非経口経路(皮下、静脈内、又は筋肉内)、経口、舌下、経鼻若しくは直腸経路による対象のアレルゲンの反復投与を伴う。これらの(ポリ)ペプチドは、使用に関する現行の法律及びガレノス製剤手順に従って、単独又は他の薬学的に許容可能な希釈剤及び賦形剤と併用して投与することができる。
【0095】
【図面の簡単な説明】
【0096】
【図1】Q1の構造図である。
【図2】Q1のアミノ酸及びヌクレオチドの配列を示す図である。ベクターpQE−32の親和性末端に対応する残基には下線が引かれている。EcoRI(EF)セクションによって導入された残基は陰影を付けている。システイン残基は二重下線が引かれている。
【図3】Par j 1及びPar j 2のIgEエプトープ(囲み線)と、Tエピトープ(下線)との比較を示す図であり、太線の囲み線はQ2欠失エピトープを示す。同一の残基(:)及び同様の残基(.)が標識されている。
【図4】Q2の構造図である。
【図5】Q2のアミノ酸及びヌクレオチドの配列を示す図である。ベクターpQE−32の親和性末端に対応する残基は下線が引かれている。EcoRI(EF)、PstI(LQ)、及びEcorRV(DI)セクションによって導入された残基は陰影を付けている。システイン残基は二重下線が引かれている。
【図6】Par j 1及びPar j 2のIgEエプトープ(囲み線)と、Tエピトープ(下線)との比較を示す図であり、太線の囲み線はQ3欠失エピトープを示す。同一の残基(:)及び同様の残基(.)が標識されている。
【図7】Q3の構造図である。
【図8】Q3のアミノ酸及びヌクレオチドの配列を示す図である。ベクターpQE−32の親和性末端に対応する残基は下線が引かれている。EcoRI(EF)、PstI(LQ)、EcorRV(DI)、Bg1II(RS)及びKpnI(GT)セクションによって導入された残基は陰影を付けている。システイン残基は二重下線が引かれている。
【図9】電気泳動後のポリアクリルアミドゲルのクマシーブルーによる染色を示す図である:Q1(レーン1)、Q2(レーン2)、Q3(レーン3)。
【図10】精製タンパク質nPar j 1−nPar j 2、Q1、Q2、及びQ3の円偏光二色性分析の結果を示す図である。
【図11】カベイラクサに対してアレルギーである患者の血清混合物のIgE抗体による免疫検出を示す図であり、この図は、nPar j 1−nPar j 2(レーン1)、rPar j 1(レーン2)、rPar j 2(レーン3)、Q1(レーン4)、Q2(レーン5)、Q3(レーン6)から成る。
【図12】カベイラクサに対してアレルギーである患者由来の15個の血清(10倍希釈)を用いて、Q1、Q2、及びQ3に対するIgE抗体の結合を示す図である。3つの実験値は偏差で示される。
【図13】固相中のカベイラクサ抽出物、並びにカベイラクサに対してアレルギーである患者の血清混合物を用いて、阻害剤としてnPar j 1−Par j 2、Q1、Q2、及びQ3を用いたELISA阻害試験の結果を示す図である。それぞれの値は、標準偏差が10%未満の3つの実験の平均に対応している。
【図14】カベイラクサ抽出物、nPar j 1−Par j 2及びQ1(50μg/ml)、並びにQ3(250μg/ml)によって行われた皮膚試験の結果を示す図である。示された値は、パッチ面積(mm2)である。
【図15】カベイラクサ抽出物、nPar j 1−Par j 2及びQ1(50μg/ml)、並びにQ2及びQ3(250μg/ml)によって行われた特異的なIgEの測定の結果を示す図である。
【図16】カベイラクサに対してアレルギーである患者におけるTリンパ球の増殖を研究するのに最適な濃度の測定を示す図である。刺激指数(IE)。
【図17】カベイラクサ抽出物及び0.5μg/mlの3つのキメラタンパク質、天然型及び組み換え型のPar j 1及びPar j 2により得られたTリンパ球の増殖を示す図である。示された値は、刺激指数(%)のものである。有意差なし(ns)。
【特許請求の範囲】
【請求項1】
キメラタンパク質であって、非特異的なクラス1の脂質輸送タンパク質由来のアミノ酸配列を含み、該キメラタンパク質のアミノ酸配列が、該非特異的なクラス1の脂質輸送タンパク質由来の該配列に対応するIgE抗体と結合する1つ又は複数のエピトープを欠いていることを特徴とする、キメラタンパク質。
【請求項2】
Par j 1及びPar j 2として知られるカベイラクサ(Parietaria judaica)花粉の主要なアレルゲンに対応する配列由来のアミノ酸配列を含むことを特徴とする、請求項1に記載のキメラタンパク質。
【請求項3】
前記キメラタンパク質のアミノ酸配列が、Par j 1又はPar j 2の28位〜53位を含むことを特徴とする、請求項1又は2に記載のキメラタンパク質。
【請求項4】
Par j 1及びPar j 2の28位〜53位のアミノ酸配列を欠いている、Par j 1及びPar j 2由来の配列を含むことを特徴とする、請求項3に記載のキメラタンパク質。
【請求項5】
キメラタンパク質であって、配列番号2、配列番号4、又は配列番号6との相同性が、少なくとも70%であるアミノ酸配列を含むことを特徴とする、キメラタンパク質。
【請求項6】
配列番号4との相同性が少なくとも70%であるアミノ酸配列を含むことを特徴とする、請求項5に記載のキメラタンパク質。
【請求項7】
キメラタンパク質であって、配列番号2、配列番号4、又は配列番号6のアミノ酸配列を含むことを特徴とする、キメラタンパク質。
【請求項8】
配列番号4のアミノ酸配列を含むことを特徴とする、請求項7に記載のキメラタンパク質。
【請求項9】
免疫障害の治療に用いられるキメラタンパク質。
【請求項10】
アレルギーの治療に用いられる、請求項9に記載のキメラタンパク質。
【請求項11】
請求項1から10のいずれか一項に記載のタンパク質をコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項12】
配列番号1、配列番号3、又は配列番号5のヌクレオチド配列に対する相同性が少なくとも70%であるヌクレオチドを含むポリヌクレオチド。
【請求項13】
配列番号3のヌクレオチド配列に対する相同性が少なくとも70%であるヌクレオチドを含むポリヌクレオチド。
【請求項14】
配列番号1、配列番号3、又は配列番号5のヌクレオチド配列を含むポリヌクレオチド。
【請求項15】
配列番号3のヌクレオチド配列を含む、請求項14に記載のポリヌクレオチド。
【請求項16】
形質転換された宿主生物における微生物発現ビヒクルであって、自己複製しており、請求項11から15に記載のポリヌクレオチドを発現するのに用いられる、形質転換された宿主生物における微生物発現ビヒクル。
【請求項17】
請求項16に記載の微生物発現ビヒクルで形質転換された宿主生物。
【請求項18】
原核生物である、請求項17に記載の生物。
【請求項19】
前記原核生物が大腸菌属に属する、請求項18に記載の生物。
【請求項20】
真核生物である、請求項17に記載の生物。
【請求項21】
請求項1〜10のいずれか一項に記載のキメラタンパク質を含有するポリペプチドを製造する方法であって、宿主生物において自己複製しており、且つ請求項1〜10のいずれか一項に記載のキメラタンパク質を発現するのに用いられる微生物発現ビヒクルを含有する該宿主生物の培養を含む、請求項1〜10のいずれか一項に記載のキメラタンパク質を含有するポリペプチドを製造する方法。
【請求項22】
請求項1から10のいずれか一項に記載のキメラタンパク質を精製する方法であって、該キメラタンパク質を培養物、細胞又はその両方から単離することを含む、請求項1〜10のいずれか一項に記載のキメラタンパク質を精製する方法。
【請求項23】
免疫障害の治療のための請求項1から10のいずれか一項に記載のタンパク質の使用。
【請求項24】
免疫障害の治療用の薬物の調製における請求項1から10のいずれか一項に記載のタンパク質の使用。
【請求項25】
前記免疫障害がアレルギーである、請求項23又は24に記載の使用。
【請求項26】
前記アレルギーがカベイラクサ花粉に対するアレルギーである、請求項25に記載の使用。
【請求項27】
効果的な量の請求項1から10のいずれか一項に記載のタンパク質と、薬学的に許容可能な賦形剤とを含む薬学的組成物。
【請求項28】
前記組成物がワクチン組成物である、請求項27に記載の薬学的組成物。
【請求項29】
免疫障害を治療する方法であって、請求項27又は28に記載の薬学的組成物を、それを必要とする被験者に投与する工程を含む、免疫障害を治療する方法
【請求項1】
キメラタンパク質であって、非特異的なクラス1の脂質輸送タンパク質由来のアミノ酸配列を含み、該キメラタンパク質のアミノ酸配列が、該非特異的なクラス1の脂質輸送タンパク質由来の該配列に対応するIgE抗体と結合する1つ又は複数のエピトープを欠いていることを特徴とする、キメラタンパク質。
【請求項2】
Par j 1及びPar j 2として知られるカベイラクサ(Parietaria judaica)花粉の主要なアレルゲンに対応する配列由来のアミノ酸配列を含むことを特徴とする、請求項1に記載のキメラタンパク質。
【請求項3】
前記キメラタンパク質のアミノ酸配列が、Par j 1又はPar j 2の28位〜53位を含むことを特徴とする、請求項1又は2に記載のキメラタンパク質。
【請求項4】
Par j 1及びPar j 2の28位〜53位のアミノ酸配列を欠いている、Par j 1及びPar j 2由来の配列を含むことを特徴とする、請求項3に記載のキメラタンパク質。
【請求項5】
キメラタンパク質であって、配列番号2、配列番号4、又は配列番号6との相同性が、少なくとも70%であるアミノ酸配列を含むことを特徴とする、キメラタンパク質。
【請求項6】
配列番号4との相同性が少なくとも70%であるアミノ酸配列を含むことを特徴とする、請求項5に記載のキメラタンパク質。
【請求項7】
キメラタンパク質であって、配列番号2、配列番号4、又は配列番号6のアミノ酸配列を含むことを特徴とする、キメラタンパク質。
【請求項8】
配列番号4のアミノ酸配列を含むことを特徴とする、請求項7に記載のキメラタンパク質。
【請求項9】
免疫障害の治療に用いられるキメラタンパク質。
【請求項10】
アレルギーの治療に用いられる、請求項9に記載のキメラタンパク質。
【請求項11】
請求項1から10のいずれか一項に記載のタンパク質をコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項12】
配列番号1、配列番号3、又は配列番号5のヌクレオチド配列に対する相同性が少なくとも70%であるヌクレオチドを含むポリヌクレオチド。
【請求項13】
配列番号3のヌクレオチド配列に対する相同性が少なくとも70%であるヌクレオチドを含むポリヌクレオチド。
【請求項14】
配列番号1、配列番号3、又は配列番号5のヌクレオチド配列を含むポリヌクレオチド。
【請求項15】
配列番号3のヌクレオチド配列を含む、請求項14に記載のポリヌクレオチド。
【請求項16】
形質転換された宿主生物における微生物発現ビヒクルであって、自己複製しており、請求項11から15に記載のポリヌクレオチドを発現するのに用いられる、形質転換された宿主生物における微生物発現ビヒクル。
【請求項17】
請求項16に記載の微生物発現ビヒクルで形質転換された宿主生物。
【請求項18】
原核生物である、請求項17に記載の生物。
【請求項19】
前記原核生物が大腸菌属に属する、請求項18に記載の生物。
【請求項20】
真核生物である、請求項17に記載の生物。
【請求項21】
請求項1〜10のいずれか一項に記載のキメラタンパク質を含有するポリペプチドを製造する方法であって、宿主生物において自己複製しており、且つ請求項1〜10のいずれか一項に記載のキメラタンパク質を発現するのに用いられる微生物発現ビヒクルを含有する該宿主生物の培養を含む、請求項1〜10のいずれか一項に記載のキメラタンパク質を含有するポリペプチドを製造する方法。
【請求項22】
請求項1から10のいずれか一項に記載のキメラタンパク質を精製する方法であって、該キメラタンパク質を培養物、細胞又はその両方から単離することを含む、請求項1〜10のいずれか一項に記載のキメラタンパク質を精製する方法。
【請求項23】
免疫障害の治療のための請求項1から10のいずれか一項に記載のタンパク質の使用。
【請求項24】
免疫障害の治療用の薬物の調製における請求項1から10のいずれか一項に記載のタンパク質の使用。
【請求項25】
前記免疫障害がアレルギーである、請求項23又は24に記載の使用。
【請求項26】
前記アレルギーがカベイラクサ花粉に対するアレルギーである、請求項25に記載の使用。
【請求項27】
効果的な量の請求項1から10のいずれか一項に記載のタンパク質と、薬学的に許容可能な賦形剤とを含む薬学的組成物。
【請求項28】
前記組成物がワクチン組成物である、請求項27に記載の薬学的組成物。
【請求項29】
免疫障害を治療する方法であって、請求項27又は28に記載の薬学的組成物を、それを必要とする被験者に投与する工程を含む、免疫障害を治療する方法
【図1】

【図2】

【図3】
【図4】
【図5】

【図6】
【図7】
【図8】

【図9】
【図10】
【図11】

【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2009−533046(P2009−533046A)
【公表日】平成21年9月17日(2009.9.17)
【国際特許分類】
【出願番号】特願2009−504853(P2009−504853)
【出願日】平成19年4月11日(2007.4.11)
【国際出願番号】PCT/IB2007/001025
【国際公開番号】WO2007/116316
【国際公開日】平成19年10月18日(2007.10.18)
【出願人】(508306543)ビアル インダストリアル ファーマキューティカ,エス.エー. (1)
【Fターム(参考)】
【公表日】平成21年9月17日(2009.9.17)
【国際特許分類】
【出願日】平成19年4月11日(2007.4.11)
【国際出願番号】PCT/IB2007/001025
【国際公開番号】WO2007/116316
【国際公開日】平成19年10月18日(2007.10.18)
【出願人】(508306543)ビアル インダストリアル ファーマキューティカ,エス.エー. (1)
【Fターム(参考)】
[ Back to top ]