
アントシアニンとフラボノールを大量に含む果物を生産する手段および方法
図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、栽培トマト植物体または選良種に遺伝子操作的に遺伝子移入されることを特徴とするタンパク質をコードするAFT遺伝子を提供する手段および方法を提供する。このAFT遺伝子は、栽培トマト植物体にAFT遺伝子を移入していない従来の栽培植物体と比較して高い濃度のフラボノイド類をもたらし、AFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体を提供する。トランスジェニック植物は、植物体、植物部位、または種子内にフラボノイド経路の代謝物、特にアントシアニンまたはフラボノール類を発現し、保有する特定のDNA配列が1以上の複数の形質転換、および/または植物の形質転換および/または発現に有益な発現ベクターに組み込まれることが開示される。同様にその生成方法が開示される。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、特にデルフィニジン、ペツニジン、マルビジンといった高濃度のアントシアニン類と特にケルセチン、ケンフェロールといった高フラボノール表現型を有する、トマト果実などの果物を生産する手段および方法に関するものである。
【背景技術】
【0002】
果実や野菜をカロチノイド、フラボノイド化合物、ビタミン類などの機能性代謝物で成分強化することが過去数年間で重要な育成の目標となっている。この傾向の好例としては、果実に高濃度のカロチノイド類、フラボノイド類、ビタミンCおよびEを含めるために商用のトマト品種への高色素(hp)変異体の遺伝子移入が行われることである。
【0003】
S.chilenseに由来する「アントシアニン果実(AFT)」遺伝子型は、表皮および外果皮組織がフラボノイド化合物の分類に属する高濃度のアントシアニン類や代謝物によって紫色を呈する特徴を持っている。アントシアニン濃度の上昇は単一遺伝子によって決定されることが報告されている(ジョーンら著「ジャーナル・ヘレディティー」第94号(2003年)449〜446頁)。フラボノイド類は、多くの植物体で自然に存在するポリフェノール化合物である。フラボノイド類は果物、野菜、植物原料の飲料(茶、赤ワイン)に含まれ、多くの栄養補助食品や植物性生薬に用いられる。その中心的成分であるアグリコンに基づいて、カルコン類、フラバノン類、ジヒドロフラボノール類、フラボノール類、アントシアニン類などの異なった区分に分類できる。これまでに4000以上の異なったフラボノイド類が判明しており、こうした膨大な多様性は、グリコシル化、メチル化、アシル化などのアグリコンの単一変異もしくは組み換え変異に起因する。一分類としてのフラボノイドは、植物体の育成、更には、病原菌抵抗性、色素産生、紫外線保護、花粉成長、種皮発達などの植物開発など多くの側面を担っている(ハルボーン著「フラボノイド」)。「1986年以来の研究の進歩(第一版)」(ボビーら著「植物細胞」第14号(2002年)、2509〜2526頁)。
【0004】
アントシアニン類は、紫、赤、青色の植物色素を持つ最も一般的な種類である。植物体において300を超える異なったアントシアニン化合物が確認されている。これらは、フラボノイド類特有のC6−C3−C6炭素構造を持つ平面分子を持っている。
【0005】
特に、フラボノール類(ケンペロールやケルセチンなど)に属するフラボノイド類は、その高抗酸化能力によって人の食事における潜在的な健康保護機能(ライス・エバンスら著「植物化学の動向」第2号(1997年)152〜159頁、および、プロトジェンテら著「フリーラジカル研究」第36号(2002年)217〜233頁)、更に、ヒトの保護酵素システムを誘発するインビトロでの抗酸化能力(クック&サマン著「栄養生化学ジャーナル」第7号(1996年)66〜76頁)を持っているという証拠が増えてきている。こうした知見から、フラボノイド類によって冠状動脈性心臓病や癌などの大病から防御できることが予測できる(ヘルトーク&ホルマン著「欧州臨床栄養生化学ジャーナル」第50号(1996)63〜71頁)。いくつかの疫学研究によれば、心臓保護とタマネギ、りんご、茶などの食品原料の摂取との間に深い関係があることが分かっている(ヘルトークら著「ランセット342号」(1993)1007〜1011頁)。これに関して、アントシアニン類は、ラジカル吸収能(ORAC)の分析で測定できる強い抗酸化活性酸素によって特に注目されていた。ぶどう(ワンら著「農産食品化学ジャーナル」第44号(1996)701〜705頁)、ブルーベリー、ブラックベリー、ラズベリー、サクランボ(ワンら著「農産食品化学ジャーナル」第45号(1997)304〜309頁)が、他の果実や野菜に比べてアントシアニン類を多量に含み、抗酸化能を持つことが知られている。最も重要な食品のひとつであるトマトが世界中で広く収穫されており、ぶどう、ベリー類、サクランボなどより手軽に広く食せられているので、アントシアニン摂取のための食品として有望であると考えられる。ところが、市販されているトマトには高濃度のフラボノイド類(アントシアニン類を含む)が含まれておらず、高濃度のフラボノイド類を含有する市販トマト植物の実現は重要な目標である。
【0006】
トマト果実のフラボノイド類含有量を増やし、多様化させる施策として以下の事柄に重点的に取り組んでいる。
1.フラボノイド化合物に係わる構造遺伝子の代謝工学
2.代謝経路に影響する転写因子の遺伝子組み換え
3.単一点突然変異(自然発生突然変異または誘発突然変異)および/または植物栄養素量に及ぼす顕著な効果を有する量的形質遺伝子座(レビンら著「イスラエル植物科学ジャーナル」(2006年)近日刊行)
【0007】
35Sプロモーターの制御下におけるP.hybrida由来の「カルコン異性化酵素(CHI)」遺伝子によるトマトの形質転換は、果皮フラボノール濃度を劇的に増加させる。ところが、高フラボノール遺伝子組換え植物体による葉、葉野菜、およびbreaker(未熟果)や熟したトマト果肉中にはフラボノール量の増加は見られなかった(ミューアら著「ネーチャー・バイオテクノロジ」第19号470〜474頁)。
【0008】
トマト果肉中のフラボノール蓄積、および、それによるトマト果実中のフラボノール量の増加は、転写因子LC及びCIをコードするトウモロコシ遺伝子の同時過剰発現によって実現する(ボビーら著「植物細胞」第14号(2002年)、2509〜2526頁)。
【0009】
代替的アプローチでは、フラボノール化を導く、P.hybrida由来の4つの生合成酵素「カルコン合成酵素(CHS)、カルコン合成酵素(CHI)、フラバノン−3−ヒドロキシラーゼ(F3H)、フラボノールシンターゼ(FLS)」をコードする遺伝子を異所的かつ同時にトマト植物体に発現させた。4種のトランス遺伝子を含む約75%の主形質転換体が極めて高濃度のケルセチン配糖体を果皮内や他の部分に蓄積するが、殻軸組織ではケンフェロールやナリンゲニン配糖体が顕著に増加する(ベルフォネンら著「実験的植物ジャーナル」第53号2099〜2106頁)。
【0010】
遺伝子組み換えによるトマト果実の果肉におけるフラボノイドの高濃度化に関しては大成功とはいかなかったことが記されており、加えて、高フラボノイド化植物体を得るにあたって非遺伝子組み換え的アプローチへの要求が高まっている。この要求は、遺伝子組み換え食品(GMO)として知られているような遺伝子組み換え果実や野菜に対する消費者の抵抗感が契機となっている。
【0011】
栽培もののトマト、つまり、S.chilense、S.habrochaites、S.cheesmaniae、S.lycopersicoidesなどのいくつかのトマト種の果実はアントシアニンを著しく多量に含んでいる(リック著「カリフォルニア科学アカデミー・オケージョナル・ペーパー」1964年第44、59号;ジョージフ著「トマト遺伝子協同組合報告書」1972年第22号10頁;リックら著「トマト遺伝子協同組合報告書」1994年第44号29〜30頁)。S.chilense由来のアントシアニン果実(AFT)、S.lycopersicoides由来のナス(ABG)、L.cheesmaniae由来のアトロビオラセウム(atv)はトマト果実においてにアントシアニンの発現をひきおこす。野生種S.pennellii v.puberulumは機能性フラボノイド類でトマト果実を強化する源(ソース)と見られている(ウィリットら著「農業食品化学ジャーナル」第53号1231〜1236頁)が、果皮濃縮は消極的な手法であり、産物は不安定である。
【0012】
果物のフラボノイドを増やす別の試行としては、高色素(hp)変異の遺伝子移入である。トマトhp変異体(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)は、赤熟果物のカロチノイド(リコピンや、カロチン)濃度にプラスに働く効果があることはよく知られている(レビンら著「理論応用遺伝学」第106号(2003年)454〜460頁)。hp−1変異を有する熟した植物果物も、トマト果実果皮のフラボノイド・ケルセチンを同質遺伝子系統の13倍の増加を示すことが分かっている(イェンら著「理論応用遺伝学」第95号(1997年)1069〜1079頁)。トマト変異体hp−2およびhp−2jの果皮中およびトマト変異体hp−2dgの果実中のケルセチンにおける同様の増加も明らかにされている(ビノら著「新植物学」第166号(2005年)427〜438頁、および、レビンら著「イスラエル植物科学ジャーナル」(2006年)近日刊行)。
【0013】
AFT遺伝子型のS.chilense種果実は、果実の表皮および外果皮組織に含まれるアントシアニンによって特徴づけられる。加工トマトとAFT植物体の交配のF2およびBC1群におけるアントシアニン発現の分離比はアントシアニン発現のための単一優性遺伝子の仮説に一致することが分かっている。トマト果実におけるT−DNA賦活標識実験で、アントシアニン生合成のMYB転写制御因子を識別して、ペチュニアAn2と高い相同性を有するANT1と呼ばれている(マシュースら著「植物細胞」第15号(2003年)1689〜1703頁)。
【0014】
ANT1変異トマトは、苗木栽培の芽形成のかなり早期から強烈な紫色素を顕わし、アントシアニン蓄積につながる生合成経路の活性化に反映させた。ANT1植物体の植物性組織は強烈な紫色を呈したが、その果実を66倍率で拡大観察したところ表皮にのみ紫色の斑点が視認できた。こうしたことから、表現型が更に強烈な紫を呈するような表皮および外果皮に更に高濃度のアントシアニンを高アントシアニンのトマトに含ませることが長い間の要望であった。
【0015】
野生型S.chilenseトマト株由来のAFTは高いアントシアニン濃度をもたらすので、市販のhp−1トマトは特定のフラボノイド濃度が若干増大するように見え、多く増加したフラボノイド濃度を持つ安定した品種改良増殖が長年の要望に実用的に応えることになる。
【0016】
AFT−S.chilense遺伝子が、栽培トマト対応物より高いアントシアニン濃度の原因であることが知られている。よって、この遺伝子の特徴、分離、および、フラボノイド濃度を強化したトマトを含む商用植物への形質転換もまた、医薬成分や医薬合成品用のアントシアニンの調製などGMO利用が許容され得る効果が期待できる用途における長年の要求に応えることができる。
【0017】
S.chilenseに加えて、S.habrochaites、S.cheesmaniae、S.lycopersicoides、S.pennelli v.puberelumなどの栽培トマトに関連するいくつかのトマト種果実は、栽培トマトに比較して相当に高いアントシアニン量を含有する。よって、夫々のAFT相同遺伝子の栽培トマトへの遺伝子移入および遺伝子発現が長い間待ち望まれていた新たな進展をもたらす。また、AFT以外の遺伝子または対立遺伝子を強化するフラボノイドを含むトマト増殖へのS.chilenseに由来するAFT遺伝子の遺伝子移入および遺伝子発現が更にフラボノイド含有量を増加させる。
【0018】
上述したように、形質転換遺伝子組み換えによるトマト果実中のフラボノイド含有量のある種の増加は達成されたが、GMO消費に対する消費者の抵抗感は周知のところであり、こうして、非遺伝子品種改良によって実現した高いフラボノイド栽培品種の市場での需要がある。したがって、こうした品種改良を促すためにDNAマーカーのような遺伝子配列を用いるために植物種のAFT突然変異表現型をコード化する遺伝子(または、遺伝子群)の識別が有用、且つ、新規な技術手法であると見られている。
【0019】
アントシアニン及びフラボノールのような食用の抗酸化物質の原料を得る際の関心の高さから見て、アントシアニン、特に、デルフィニジン、ペツニジン、マルビジンなどのアントシアニンと、特に、ケルセチン、ケンフェロールなどのフラボノイド表現型を高濃度に含むトマトを生産する手段および方法の実現が、長い間依然として要求されている。
【発明の概要】
【0020】
本発明の目的は、タンパク質をコードするAFT遺伝子であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、栽培植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするタンパク質をコードするAFT遺伝子を提供することにある。
【0021】
本発明の他の目的は、栽培S.licopersicum種トマト植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培S.licopersicum種トマト植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培S.licopersicum種植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするAFT遺伝子を提供することにある。
【0022】
本発明の他の目的は、S.chilense遺伝子型に由来し、栽培S.licopersicum種トマト植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培S.licopersicum種トマト植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培S.licopersicum種植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするAFT遺伝子を提供することにある。
【0023】
本発明の他の目的は、前記フラボノイド類の少なくとも一部がアントシアニン類および/またはフラボノール類であることを特徴とするAFT遺伝子を提供することにある。
【0024】
本発明の他の目的は、遺伝子がS.peruvianumに由来することを特徴とするAFT遺伝子を提供することにある。
【0025】
本発明の他の目的は、遺伝子が、S.habrochaites、S.cheesmaniae、S.licopersicoides、S.peruvianum、S.pennelli v.puberelumよりなる群から選択されたことを特徴とするAFT遺伝子を提供することにある。
【0026】
本発明の他の目的は、AFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体であって、前記AFT−S.chilense遺伝子型を遺伝子移入していない従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のフラボノイド類によって特徴づけられる遺伝子移入由来トマト植物体を提供することにある。
【0027】
本発明の更に他の目的は、前記AFT−S.chilense遺伝子型を遺伝子移入していない従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のアントシアニン類および/またはフラボノ−ル類によって特徴づけられる遺伝子移入由来トマト植物体を提供することにある。
【0028】
本発明の更に他の目的は、前記AFT−S.chilense遺伝子型がS.peruvianumから遺伝子移入されたことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0029】
本発明の更なる目的は、前記AFT−S.chilense遺伝子型が、S.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群から遺伝子移入されたことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0030】
本発明の更に他の目的は、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のフラボノイド類を有することで特徴づけられる純親系統を得るために、前記F2植物体を自家交配させてF3群および更なる個体群(F4、F5等)を作成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
方法によって遺伝子移入的に得たことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0031】
本発明の更にその他の目的は、前記フラボノイド類が、アントシアニン類およびフラボノール類であるデルフィニジン、ペツニジン、マルビジン、ケルセチン、ケンフェロール、ナリンゲニンを含むことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0032】
本発明の他の目的は、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、hp−1、hp−1w、hp−2、hp−2j、hp−2dgとして定義される1以上の同型対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0033】
本発明の他の目的は、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子の1以上の同型接合対立遺伝子(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0034】
本発明の他の目的は、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子に欠損のあるhp−1、hp−1w、hp−2、hp−2j、hp−2dg変異植物体と等表現型の光形態形成遺伝子の1以上の同型接合対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0035】
本発明の他の目的は、DNAマーカーを用いてS.chilense由来のAFT遺伝子座において同型接合であるF2植物体を選択する付加的手段からなることを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0036】
本発明の他の目的は、AFT遺伝子型がS.peruvianumに由来することを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0037】
本発明の他の目的は、AFT遺伝子型がS.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群から選択されたことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0038】
本発明の更なる目的は、前記DNAマーカーがS.peruvianumに由来することを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0039】
本発明の更なる目的は、前記DNAマーカーが、S.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群に由来することを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0040】
本発明の更なる目的は、フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル含有フラボノイドおよび/またはメトキシ置換、C−メチルフラボノイド、メチレンジオキシ・フラボノイド・カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンよりなる群のうちのいずれかの成分から前記フラボノイドを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0041】
本発明の更なる目的は、デルフィジン、ペツニジン、マルビジンよりなる群から前記アントシアニンを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0042】
本発明の更なる目的は、ケルセチン、ケンフェロールよりなる群から前記フラボノイドを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0043】
本発明の更なる目的は、4,2,4,6−テトラヒドロキシカルコン、ナリンゲニン、ケンフェロール、ジヒドロキシケンフェロール、ミリセチン、ケルセチン、ジヒドロケルセチン、ジヒドロミリセチン、ロイコペラルゴニジン、ロイコシアニジン、ロイコデルフィニジン、ペラルゴニジン−3−グルコシド、シアニジン−3−グルコシド、デルフィニジン−3−グルコシドよりなる群から前記フラボノイドを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0044】
本発明の更なる目的は、(i)2−フェニルクロモン(2−フェニル−1,4−ベンゾピロン)構造由来の二次植物代謝物、(ii)前記代謝物が3−フェニルクロモン(3−フェニル−1,4−ベンゾピロン)構造由来であるイソフラボノイド類、(iii)前記代謝物が4−フェニルクマリン(4−フェニル−1,2−ベンゾピロン)構造由来であるネオフラボノイド類よりなる群から前記フラボノイドを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0045】
本発明の更なる目的は、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、従来の栽培S.licopersicum種トマト植物体と比較してトマト植物体中に高いフラボノイド濃度をもたらすことを特徴とするタンパク質をコードするDNA配列を提供することにある。
【0046】
本発明の更なる目的は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、従来の栽培S.licopersicum種トマト植物体と比較してトマト植物体中に高いフラボノイド濃度をもたらすことを特徴とするDNA配列を提供することにある。
【0047】
本発明の更なる目的は、前記DNA配列は、植物体、植物部位または種子内において、フラボノイド経路の代謝物であるアントシアニンまたはフラボノール類の蓄積又は発現をもたらすことを特徴とするDNA配列を提供することにある。
【0048】
本発明の更なる目的は、前記DNA配列は、S.licopersicumのトマト植物体、トマト植物部位または種子内において、フラボノイド経路の代謝物であるアントシアニンまたはフラボノール類の蓄積又は発現をもたらすことを特徴とするDNA配列を提供することにある。
【0049】
本発明の更なる目的は、前記DNA配列は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、栽培トマト系統におけるAFT遺伝子型の遺伝子移入のための遺伝資源、種子、苗木、発芽、植物体、植物部位のスクリーニングに有用であることを特徴とするDNA配列を提供することにある。
【0050】
本発明の更なる目的は、トランスジェニック植物であって、植物体、植物部位または種子内にフラボノイド経路の代謝物であるアントシアニンまたはフラボノイド類を発現し、前記植物体は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有するDNAからなり、前記DNAは、1以上の複数の形質転換、および/または植物の形質転換および/または発現に有益な発現ベクターに組み込まれていることを特徴とする遺伝子移入植物体を提供することにある。
【0051】
本発明の範疇には、タンパク質をコードするAFT遺伝子を得る方法であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、栽培植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするタンパク質をコードするAFT遺伝子を得る方法を提供することも含まれる。
【0052】
本発明の範疇には、従来の栽培S.licopersicum種トマト植物体と比較して高濃度のアントシアニン類および/またはフラボノイド類を含むことを特徴とするAFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体を作り出す方法であって、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のフラボノイド類を有することで特徴づけられる純親系統を得るために、前記F2植物体を自家交配させてF3群を生成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
ことからなる方法を提供することも含まれる。
【0053】
本発明の範疇には、従来の栽培S.licopersicum種トマト植物体と比較して高濃度のアントシアニン類および/またはフラボノイド類を含むことを特徴とするAFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体を作り出す方法であって、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のアントシアニン類とフラボノール類であるデルフィニジン、ペツニジン、マルビジン、ケルセチン、ケンフェロールを有することで特徴づけられた純親系統を得るために、前記F2植物体を自家交配させてF3群および更なる個体群(F4、F5等)を作成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
ことからなる方法を提供することも含まれる。
【0054】
本発明の範疇には、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、hp−1、hp−1w、hp−2、hp−2j、hp−2dgとして定義される1以上の同型対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする方法を提供することも含まれる。
【0055】
本発明の範疇には、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子における1以上の同型対立遺伝子(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする方法を提供することも含まれる。
【0056】
本発明の範疇には、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子に欠損のあるhp−1、hp−1w、hp−2、hp−2j、hp−2dg変異植物体と等表現型の光形態形成遺伝子の1以上の同型接合対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする方法を提供することも含まれる。
【0057】
更に、本発明の範疇には、HP−1とAFT遺伝子座が同型接合であるF2植物体を付加的にDNAマーカーを用いて選択することを特徴とする方法を提供することも含まれる。
【0058】
更に、本発明の範疇には、デルフィジン、ペツニジン、マルビジンよりなる群から前記アントシアニンを選択することを特徴とする方法を提供することも含まれる。
【0059】
更に、本発明の範疇には、ケルセチン、ケンフェロール、ナリンゲニンよりなる群から前記フラボノイドを選択することを特徴とする方法を提供することも含まれる。
【0060】
更に、本発明の範疇には、4,2,4,6−テトラヒドロキシカルコン、ナリンゲニン、ケンフェロール、ジヒドロキシケンフェロール、ミリセチン、ケルセチン、ジヒドロケルセチン、ジヒドロミリセチン、ロイコペラルゴニジン、ロイコシアニジン、ロイコデルフィニジン、ペラルゴニジン−3−グルコシド、シアニジン−3−グルコシド、デルフィニジン−3−グルコシドよりなる群から前記フラボノイドを選択したことを特徴とする従来の栽培S.licopersicum種トマト植物体と比較して高濃度のフラボノイド類を含むトマト植物体を作り出す方法を提供することも含まれる。
【0061】
更に、本発明の範疇には、(i)2−フェニルクロモン(2−フェニル−1,4−ベンゾピロン)構造由来の二次植物代謝物、(ii)前記代謝物が3−フェニルクロモン(3−フェニル−1,4−ベンゾピロン)構造由来であるイソフラボノイド類、(iii)前記代謝物が4−フェニルクマリン(4−フェニル−1,2−ベンゾピロン)構造由来であるネオフラボノイド類よりなる群から前記フラボノイドを選択したことを特徴とするトマト植物体を作り出す方法を提供することも含まれる。
【0062】
更に、本発明の範疇には、フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル含有フラボノイドおよび/またはメトキシ置換、C−メチルフラボノイド、メチレンジオキシ・フラボノイド・カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンよりなる群のいずれかの成分から前記フラボノイドを選択したことを特徴とするトマト植物体を作り出す方法を提供することも含まれる。
【0063】
更に、本発明の範疇には、タンパク質をコードするDNAを作り出す方法であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、コードされたアミノ酸配列を識別して選択的に検証する処理を含み、前記配列がAFT表現型および増強したフラボノイド濃度に関連するS.chilense中に天然に存在するタンパク質の配列であることを特徴とする方法を提供することも含まれる。
【0064】
更に、本発明の範疇には、核酸を有用に得る方法であって、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、AFT表現型に少なくとも部分的に関連するS.chilense中に天然に存在するタンパク質をコードする前記核酸配列を識別して選択的に検証することからなる方法を提供することも含まれる。
【0065】
更に、本発明の範疇には、従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のフラボノイド類を含むトマト植物体を作り出し、栽培トマト系統におけるAFT−S.chilense遺伝子型の遺伝子移入のために遺伝資源、種子、苗木、植物、発芽、植物体をスクリーニングして容易にした方法であって、
(i)図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有する核酸を作り出し、
(ii)表1に定義されたPCRプライマーを調製し、
(iii)工程(i)のDNAを増幅し、
(iv)それを用いて標的組織を調べることからなる方法を提供することも含まれる。
【0066】
更に、本発明の範疇には、植物体、植物部位、または種子内にフラボノイド経路の代謝物であるアントシアニンまたはフラボノイド類を蓄積または発現させるための遺伝子組み換え方法であって、
(i)図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有するDNAを作り出し、
(ii)前記DNAを1以上の複数の形質転換、および/または植物の形質転換および/または発現に有益な発現ベクターに組み込むことからなる遺伝子組み換え方法を提供することも含まれる。
【0067】
最後に、本発明の範疇には、トマト植物体、トマト部位または種子に有用なことを特徴とする遺伝子組み換え方法を提供することも含まれる。
【発明を実施するための形態】
【0068】
本発明を当分野の技術者が実施可能なように本発明の発明者が考える最良の実施形態に基づいて以下の各章に沿って順に説明するが、トマトや、特に、デルフィニジン、ペツニジン、マルビジンなどの高アントシアニン、および、ケルセチン、ケンフェロールなどの高フラボノイド表現型を含む他の果実を生産するための手段および方法を提供することが本発明の基本概念であり、各種改変が可能なことは当該分野の技術者には自明のことである。
【0069】
用語「hp−1」とは、トマト品種に移入して果物を高濃度のカロチノイド、フラベノイド、ビタミンCおよびEで強化する高色素−1遺伝子変異を意味する。hp−1変異は、トマト紫外線損傷DNA結合タンパク質1(DDB1)および非黄化変異体1(DET1)遺伝子に位置するhp−1w、hp−2、hp−2j、hp−2dgを含む変異表現型のグループに属する。
【0070】
ここでの用語「アントシアニン果実(AFT)」とは、特定単一遺伝子を意味し、S.chilenseから遺伝子移入すると、S.lycopersicumなどの栽培トマトの高濃度のアントシアニンおよび他のフラボノイド代謝産物をもたらす。
【0071】
用語「ANT1−L」とは、アントシアニンおよびフラボノイド蓄積に関連するS.lycopersicumの特定単一遺伝子を意味する。
【0072】
用語「ANT1−C」とは、アントシアニンおよびフラボノイド蓄積に関連するS.chilenseの特定単一遺伝子を意味する。ANT1−Cでコードされているポリペプチドは、8つのアミノ酸が変化している点でANT1−Lとは異なる(図3)。
【0073】
ここでの用語「遺伝子移入」または「遺伝子移入由来」とは、交雑や戻し交雑によって遺伝子がある種から他の種または登録種の遺伝子集団への移動を起こす植物体の育種方法を意味し、所望の遺伝子型や表現型を選択することによって実施できる。DNAマーカーによって、所望の遺伝子型の選択が容易になり、育種を促進する。
【0074】
ここでの「形質転換」とは、異種の植物DNA配列を導入する任意の方法を意味し、異種の植物DNA配列がある種のDNAベクターシステムか構築物に組み込まれ、持続的に標的宿主植物ゲノムまたは細胞質に導入される方法であり、前記植物DNAの構築物の導入は従来から知られている様々な方法で実施できる。
【0075】
ここでの用語「植物体」または「植物部位」とは、限定的ではないが、果実、種子、胚芽、分裂組織、カルス組織、花、枝葉、根、芽、植物配偶体、胞子体花粉、小胞子を含む任意の植物体、植物器官および組織を意味する。植物細胞は、任意の植物器官または組織から取り出し、それらから培養調製ができる。本発明による方法に適用できる植物類は、一般に、形質転換技術を受容できるモノコテリデノス植物体とジコテリデノス植物体の双方を含む高等植物類と同じ程度に広い解釈での植物体である。
【0076】
ここでの用語「フラボノイド」とは、(i)特に、2−フェニルクロモン(2−フェニル−1,4−ベンゾピラン)構造に由来するフラボノイド、(ii)3−フェニルクロモン(3−フェニル−1,4−ベンゾピラン)構造に由来するイソフラボノイド、(iii)4−フェニルクマリン(4−フェニル−1,2−ベンゾピラン)構造に由来するネオフラボノイドなど、IUPAC命名法に従って定めた任意の植物体二次代謝産物を意味する。同様に、この用語には、フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル置換および/またはメトキシ置換フラボノイド、C−メチルフラボノイド、メチレンジオキシフラボノイド、カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリビフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンなどの意味も含まれる。
【0077】
ここでの用語「フラボノール」とは、IUPACが定めた3−ヒドロキシル−2−フェニル−4H−1−ベンゾピラン−4−オン骨格を有する任意のフラボノイドを意味する。これらの多様性は、ケルセチン(3,5,7,3’,4’−ペンタヒドロキシル−2−フェニル−4H−1−ベンゾピラン−4−オン)、ケンペロール(3,5,7,4’−テトラヒドロキシル−2−フェニル−4H−1−ベンゾピラン−4−オン)、ミリセチン(3,5,7,3’,4’,5’−ヘキサヒドロキシル−2−フェニル−4H−1−ベンゾピラン−4−オン)を用いた非限定的方法で典型的に示されるフェノール性(−OH)基の異なった部分に由来する。
【0078】
ここでの用語「アントシアニジン」とは、ベンゼン環に融合した酸素含有ヘテロ環ピランを有する任意のフラベノイドを意味し、ここでのピラン環は2位でフェニル基に結合し、異なる置換基を有することができる。
【0079】
用語「アントシアニン」とは、ある種の糖成分を有するアントシアニジンを意味する。
【0080】
用語「cv.」とは、商用および非商用の品種を意味する。
【0081】
用語「選良種」とは、特にトマトの一般的な商用の交配植物種を意味する。
【0082】
〔植物体材料、交配、生育条件〕
次の植物体材料を準備した: S.chilense−AFTを含むLA1996種、マネーメーカー種(赤色果実・放任受粉型の青果物市場向けトマト)、アイルサ・クレイグ種(赤色果実・放任受粉型トマト・遺伝的にほぼ同質であり、hp−1変異について同型接合)、VF36種(LA0490)(赤色果実・放任受粉型品種)、ラトガーズ種(LA1090)(赤色果実・放任受粉型品種)、LA1589種(赤色果実S.Pimpinellifolium種)、PI128650紫果実S.Peruvianum種。
【0083】
母系として、マネーメーカー種とアイルサ・クレイグ種hp−1/hp−1と、父系LA1996とで異種交配を行った。この異種交配で得られたF1植物体を自家受粉させて、hp−1変異とAFT対立遺伝子が分離するF2群を作成した。ここでのDNAマーカーに基づいてhp−1変異が同型接合でAFT遺伝子座が異型接合する植物体を上記F2群から選別し、hp−1/hp−1の遺伝的背景においてAFTが分離するF3群を生成した。
【0084】
植物体をイスラエル中部の2箇所、ゼライム・ゲデラ・シード社(イスラエル)のバルカニセンターで作付けした。夏季には植物を路地栽培乃至遮光栽培し、冬季には最低摂氏15℃の気温に温度調節したハウス内で栽培した。夏季には、5月の第一週に移植を行い、冬季は11月の第1週に移植を行った。
【0085】
〔ゲノムDNA抽出〕
ゲノムDNAを、フルトンら著「植物分子生物学報告」第13号(1995年)207〜209頁に開示の手法によって個別の植物体から抽出した。
【0086】
〔ポリメラーゼ連鎖反応(PCR)プライマーの設計〕
配列分析および遺伝子座特異プライマーの設計をDNAMAN配列分析ソフトウェアV4.1(リンノン・バイオソフト社(米国ケベック)製)を用いて行った。
【0087】
〔パイロシークエンス遺伝子型決定法〕
hp−1変異の遺伝子型を判定するためにパイロシークエンス方式を用いた。このパイロシークエンス遺伝子型決定方式は、hp−1/hp−1変異植物体とその準遺伝子同質体の相互間のhp−1変異遺伝子型をコードする遺伝子中に見つけられる一塩基多型に基づいている。遺伝子型決定手法として、リーバーマンら著「理論応用遺伝学」第108号(2004年)1574〜1581頁に開示された方法を採用した。
【0088】
〔フラボノイド生合成経路のANT1および他の構造遺伝子の遺伝子型決定法〕
遺伝子型決定は、連鎖解析および/またはPCRの後に制限酵素分解を用いる多型決定を目的として行われた。PCR増幅に用いるプライマーを表1に示す。PCR増幅産物を臭化エチジウムで染色した1.0%アガロースゲル中で電気泳動法によって視覚化した。
【表1】
【0089】
〔リアルタイムPCR分析〕
緑熟トマト果実の果皮からRNAを抽出した。各実験で、各遺伝子型からの3つの1平方ミリ角(LA1996、正常赤色果実遺伝子型マネーメーカー種、VF36、ラトガーズ、LA1996とマネーメーカー種のF1交配種)を分析した。TRIzol試薬システム(インビトロジェン社(米国カリフォルニア州カールズバッド)製)を用いてRNA抽出を行った。混入する可能性のあるゲノムDNAは「TURBO DNA−free」キットを用いて分解させ、「iScript−cDNA」合成キット(バイオ−ラッド研究所(米国カリフォルニア州ハーキュリーズ)製)を使用してcDNA合成のテンプレートとして全RNAを用いた。全RNAの純度および合成cDNAの品質は以下のいずれかのプライマーを用いたPCRによって検証した。
【0090】
「サイバー・グリーンPCR マスター・ミックス」(アプライド・バイオシステム社(米国カリフォルニア州フォスターシティ)製)を用いてリアルタイムPCR分析を行った。PCR反応は初期インキュベーション温度95℃で10分間、続いて、95℃で15秒間の変性、更に61℃で30秒間のアニーリングと、72℃で30秒間の重合化処理を40サイクル行った。
【0091】
この実験調査中、18SリボソームRNAを参照遺伝子として用いて、そのためのプライマーが設計され、同様にその他の遺伝子も分析された。観察対象であるカルコン合成酵素-1(CHS1)、カルコン合成酵素-2(CHS2)、カルコンイソメラーゼ(CHI)、 ジヒドロフラボノール還元酵素(DFR)、フラバノン−3−ヒドロキシラーゼ(FSH)、アントシアニン1(ANT1)を表2に示す。サンプルを、「GeneAmp5700配列検出システム」を用いて通常重複して分析し、「GeneAmp5700SDSソフト(アプライド・バイオシステム社製)」でデータを収集して解析した。検査遺伝子転写物の相対存在量を式[2(CT検査遺伝子-CT参照遺伝子)]で計算した。式において、CTは、蛍光発光が一定の閾値を越える分別サイクル数(通常は0.1に設定)。
【表2】
【0092】
〔アントシアニン、フラボノール抽出、および、定量化〕
新鮮なトマト果皮のサンプル(0.1〜0.3g)を液体窒素中で粉砕し、暗所で冷メタノール、水、酢酸の混合液2ml(11:5:1)を用いて色素を抽出した(マーカム&K.R.オフマン著「植物化学」第34号(1993)679〜685頁)。
【0093】
抽出物20,800g(14,000rpm)で10分間遠心し、上清中にアントシアニン脱離させた。更に、精製物をヘキサン容積2/3とした。サンプルを0.5mlに濃縮し、1時間2N・HCl中で等量のメタノールと共に沸騰させて加水分解し、0.45μm−ポリビニリデンジフロライドフィルター(米国ナルジェ社製)でろ過した。
【0094】
LC−10ATポンプ、SCL-10Aコントローラ、SPD−MlOAVPフォトダイオードアレイ検出器を備えたΗPLC(島津製作所製)を用いてフラボノイドの組成を測定した。抽出物をRP18カラム(Vydac−201TP54)に充填し、次の溶液を用いて27℃で分離した(溶液:(A)H2O、pH2.3、(B)H2O:MeCN:HOAc(107:50:40)、pH2.3)。溶液AとBとの比率が4:1から3:7のリニアに勾配をつくるように45分かけて供給し、流速毎分0.5mlで比率が3:7の状態で更に10分間保持し、保持時間と250〜650nmにおける吸収スペクトルを標準精製フラボノイド(アピン・ケミカルズ、ポリフェノールズ、シグマ)と比較してフラボノイドを識別した。
【0095】
〔トマトゲノムに対するANT1およびAFT遺伝子のマッピング〕
以前発表された資料文献(レビンら著「イスラエル植物科学ジャーナル」第100号(2000年)256〜262頁)のように、ANT1遺伝子と強い関連性を持つこの実験調査で見つかったAFT遺伝子はS.Pennellii遺伝子移入系統を用いてトマトゲノムにマッピングされた(エシェドら著「理論応用遺伝学」第83号(1992年)1027〜1034頁)。
【0096】
元の親種M82とS.Pennelliiを含む遺伝子移入系統の個々の植物体から抽出したDNAをPCR反応の際のテンプレートとして用いた。自動サーモサイクラー(MJリサーチ社(米国マサチューセッツ州ウォルサム)製)で反応を行った。
【0097】
次のDNAプライマーをこれらの反応に用いた(マシュースら著「植物細胞」第15号(2003年)1689〜1703頁)。
F:5’−TCCCCCGGGATGAACAGTACATCTATG−3’
R:5’−GGACTAGTTTAATCAAGTAGATTCCATAAGTCA−3’
【0098】
PCR反応は初期インキュベーションに94℃3分間、引き続いて94℃30秒の変性、60℃30秒間アニーリングし、72℃60秒間の重合化処理を35サイクル行った。72℃の最終伸長反応を7分間行い、上記サイクル処理を完了した。臭化エチジウムで染色した1.0%アガロースゲルを1ml用いた電気泳動法によって得られたPCR産物を視覚化した。親種M−82(LA3475)およびS.Pennellii(LA0716)の間の多型性を得るための制限酵素は必要なかった。
【0099】
〔ANT1遺伝子のクローニング〕
個々のAFT変異(LA1996)植物体および野生遺伝子型(アイルサ・クレイグ)の葉組織100mgから全RNAを抽出した。TRIzol試薬システム(インビトロジェン社(米国カリフォルニア州カールズバッド)製)を用いてRNA抽出を行った。混入の可能性のあるゲノムDNAは「TURBO DNA−free」キットを用いて分解させ、「iScript−cDNA」合成キット(バイオ−ラッド研究所(米国カリフォルニア州ハーキュリーズ)製)を使用したcDNA合成のテンプレートとして全RNAを用いた。ANT1遺伝子配列(GenBank受入番号:AY348870)を変異体種と正常遺伝種の双方で増幅させるためにPCR反応のテンプレートとしてcDNAを用いた。下記遺伝子5’末端および3’末端の特異的プライマーを用いたプルーフリーディングTaqポリメラーゼ(ロッシュ・ダイアゴノスティック社(米国インディアナ州インディアナポリス)製)のPwo-DNAポリメラーゼ)でPCRを実行した。
F: 5’−ATGAACAGTACATCTATGTCTTCATTGG−3’
R: 5’−GGACTAGTTTAATCAAGTAGATTCCATAAGTC−3’
【0100】
TAクローニングの後、結果的に得られた産物をpCRII−TOPOベクター(インビトロジェン社(米国カリフォルニア州カールズバッド)製)に結合し、配列分析で検証した。
【0101】
〔植物形質転換のバイナリーベクター構造〕
カリフラワー・モザイク・ウイルス(CaMV)35S構成プロモーターの制御下に変異および正常ANT1を有する植物形質転換のための構築物を調製した。全ての構築物は、35Sプロモーターの下でNPTII選択マーカー遺伝子を有する。
【0102】
これらの構築物を調製するために、pMON10098プラスミドをEcoRIで分解させた後にエビのアルカリフォスファターゼ(ロッシュ・ダイアゴノスティック社(米国インディアナ州インディアナポリス)製)を用いて処理し、(pCRII−TOPOベクターからの)EcoRI分解ANT1と結合した。pMON−35S−ANT1を並んで含むクローンを単離し、変異および正常ANT1のクローンについて配列を検証した。
【0103】
〔形質転換プロトコル〕
上記構築物の形質転換のためにNicotiana tobacum SRIの葉切除片、および、S.lycopersicum種マネーメーカーの子葉切除片を用いた。
【0104】
培養調整は次の通りである:植物体種子を石鹸(市販洗剤「パルムオリーブ」)と水で洗って水中に置き、その後、流水で1.5時間洗浄した。続いて、種子を96%v/vのエタノール中で1分間、振り混ぜて、3%v/v(+0.01%v/vTween20)の次亜塩素酸ソーダ中に15分間放置し、更に、1.5%v/v(+0.01%v/vツイン20)の次亜塩素酸ソーダ中で30分間激しく撹拌した。最後に種子を滅菌蒸留水で3回洗浄した。処理した葉切除片、および、子葉切除片を上記種子から得られた苗木茎の基部から取り外し、植物成長調整剤成分含有において異なる下記のMS培地に接種した。
【0105】
上記移植片を、本出願人の研究所において標準的な培地でMS無機塩を含む塩基培地(ムラシゲ&スクーグ著「生理学植物」第15(3)号(1962年)473〜497頁)に蔗糖またはグルコース30gと寒天8g/Lを添加した培地に接種した。成長調節剤補充(単位mg/L): タバコ…IAA0.8、キネチン2、ゼアチン1; トマト…IAA0.1、ゼアチン1
【0106】
培地を培養室に23℃で16時間照明(照明状況:50mmol;m−2;s−1の白色蛍光ランプ)の下で保管した。全ての外植片を前記培地に植え、発芽能力を評価した。
【0107】
この実験調査全般で用いた全てのベクターをアグロバクテリウム・ツメフアシェンス菌EHA105に挿入した。葉切除片、および、子葉切除片を滅菌状態でアグロバクテリウム菌と共に、アセトシリンゴン200μMを添加したMS培地で20分間、培養した。細菌ろ紙で組織を吸い取り処理した後、アセトシリンゴン100μMを添加した暗所MS培地において22℃でカルス組織片を共培養した。その後、切除片を洗浄し、選別用にカナマイシン50mg/Lとセフォタキシム200mg/Lを含む固形再生培地に移し、培養容器内で育成した。
【0108】
〔統計分析〕
JMP統計判定ソフト(SASインスティテュート(米国ノースカロライナ州カリー)製)を用いて分散分析(ANOVA)を行った。更に、<http://workbench.sdsc.ede/>に開示された「生物学ワークベンチ」を用いたCLUSRAL W方法(トンプソンら著「核酸研究」第22号(1994)4673〜4680頁)によってヌクレオチドおよびアミノ酸配列を整列させた。
【0109】
図1に、従来技術によるアントシアニンおよびフラボノールの生合成経路の概略図を示している。
【0110】
図2に、アイルサ・クレイグ品種(上図)とLA1996(下図)とのANT1遺伝子のヌクレオチド配列の比較を概略的に示している(開始コドンおよび終止コドンは両配列に下線をつけており、イントロン領域は黄色で彩色している)。
【0111】
図3に、アイルサ・クレイグ品種(上図)とLA1996(下図)とのANT1タンパク質のアミノ酸比較を概略的に示している(2線間で異なるアミノ酸を黄色で彩色している)。
【0112】
図4に、S.lycopersicum(ANTl−L)由来のANT1対立遺伝子とS.chilense(ANTl−C)由来のANT1対立遺伝子間の共優性遺伝子多型の写真画像を示している。
【0113】
図5に、ANT1遺伝子と、ANT1とhp−1に分離するF2群中のアントシアニン蓄積の形質の視覚的表示を示している(各果実は夫々の遺伝子型の個別植物体から得られたものである)。
【0114】
図6に、染色体10のトマトゲノムに対するANT1の制限酵素遺伝子解析の写真および配置図を示している。
【0115】
図7に、カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下のS.chilense(ANTl−C)由来のANT1遺伝子で形質転換されたタバコ再生体およびS.lycopersicum(ANTl−L)由来のANT1遺伝子で形質転換されたタバコ再生体との比較写真を示している。
【0116】
図8に、カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下におけるS.chilense(ANTl−C)由来のANT1遺伝子で形質転換されたトマトおよびS.lycopersicum(ANTl−L)由来のANT1遺伝子で形質転換されたトマト(マネーメーカー品種)再生体の比較写真を示している。
【0117】
図9に、トマト登録種およびコショウからクローンが作られたANT1遺伝子のアミノ酸配列の概略図を示している(果実アントシアニンを蓄積していない登録種:LA1589はS.pimpinellifolium、LA2838AはS.lycopersicumである。果実アントシアニンを蓄積している登録種:PI128650はS.peruvianum、hp−799は未知のS.peruvianumとS.lycopersicumとの交配から生まれた選択系統、LA1996はS.chilense由来のAFT遺伝子型、CAE75745はアントシアニンを蓄積するコショウである)。
【0118】
図10に、hp−1変異およびANT1−C対立遺伝子(ANT1−C/ANT1−C hp−1/hp−1)が同型接合したF3植物体とLA1996植物体(ANT1−C/ANT1−C +/+)から得られたトマト果実を示している。
【0119】
図11に、本発明の別の態様によるS.Peruvianum(ANT1−P/ANT1−P hp−1/hp−1)由来のANT1対立遺伝子とhp−1変異が同型接合した登録品種のトマト植物体および果実を示している。
【実施例1】
【0120】
AFT遺伝子座での同型接合果実は、アントシアニンに加えて高濃度のフラボノール・ケルセチンとケンフェロールを含有している。
【0121】
AFT遺伝子型LA1996種の植物体、赤色果実マネーメーカー種の植物体、および、マネーメーカー種とLA1996種との交配種であるF1植物体を乱塊法計画の開放耕地で育成した。各遺伝子型の5種子を3ブロックに各々に作付けし、赤熟段階で果実をサンプルとして高性能液体クロマトグラフィー分析を行い果皮のフラボノール量とアントシアニン量を測定した。赤熟果実で識別された主要なアントシアニンとその平均濃度を遺伝子型に従って図3に示す。また、同遺伝子型の赤熟果実に含まれる主要なフラボノールとその平均濃度を遺伝子型に従って図4に示す。
【0122】
図3に示す結果から、赤熟果実のマネーメーカー種植物体に較べてAFT/AFT植物体から得られた熟果の果皮に含まれるアントシアニン、デルフィニジン、ペツニジン、マルビジンが統計的に有意な蓄積を見せたことが分かる。これらのことから、同じAFT/AFT植物体と赤熟化トマト植物体UC82Bの果実に含まれるアントシアニン濃度を比較した以前の結果を確認できる(ジョーンら著「ジャーナル・ヘレディティー」第94号(2003年)449〜456頁)。加えて、図4に示す結果によって、AFT/AFT変異植物体の果実が、機能的フラボノール、特に、ケルセチンおよびケンフェロールを統計的に有意に蓄積するという特徴を持っていることが分かる。ケルセチン濃度は、AFT/AFT遺伝型の熟果において、果皮重量(gFW)に基づく赤熟マネーメーカー種植物体の濃度に較べて〜3.6倍高く、果皮部分(cm2)では〜4.3倍高かった。ケンペロール濃度は、AFT/AFT遺伝型の熟果において、果皮重量(gFW)に基づく赤熟マネーメーカー種植物体の濃度に較べて〜2.7倍高く、果皮部分(cm2)では〜3.3倍高かった。
【0123】
【表3】
【0124】
【表4】
【0125】
表3および表4に示す結果から、異型接合のF1植物体の果皮におけるアントシアニンおよびフラボノール濃度は、通常、赤熟遺伝子型のものと較べて高いが、LA1996遺伝子型(AFT/AFT)よりかなり低いことが明らかになった。これらの結果は、AFT遺伝子(または複数AFT遺伝子)の部分的優性効果を示している。
【0126】
しかしながら、こうした統計分析では、対応する赤熟マネーメーカー種植物体と比較した場合において、F1植物体から得られる果実における平均アントシアニン濃度と平均フラボノール濃度との統計的に有意な差を明らかにできなかった。
【実施例2】
【0127】
フラボノイド生合成経路の重要な酵素の転写のアップレギュレーションがAFT植物体の特徴である。
【0128】
上記の予備試験の枠組みの中に作付けしたLA1996種植物体、および、VF36種およびラトガーズ種の2種の赤熟遺伝型から得られた緑熟果からリアルタイムPCRに用いるRNAサンプルを抽出した。cDNA合成およびリアルタイムPCR分析に続いて、フランボノイド生合成経路CHl、CH2、F3H、DFR(プラマーは表3に示す)の4種の構造的な酵素の転写プロフィールに関する3種の遺伝子型を比較した。この結果は、2種の赤熟遺伝型(データは示していない)に比較した時のCHS1、CHS2、DFRの極端な発現上昇、および、LA1996のF3Hの適度な発現減少を示している。特注に値することは、フラボノイド生合成の初期段階で機能する2つのCHS1遺伝子と生合成の後段での酵素活性をコードするDFR遺伝子について極端な発現上昇が見られたことである(図1)。分析は乱塊法計画の開放耕地での実験で得られたサンプルを用いて繰り返し行われた(実施例1参照)。上記3箇所からのLA1996種植物体、赤熟マネーメーカー種植物体、これら2種間の異種交配によるF1植物体からリアルタイムPCRで用いるサンプルを採取した。フラボノイド生合成経路の重要な遺伝子の転写における倍増を示す結果を表5に示す。
【0129】
【表5】
【0130】
これらの結果から、対応する赤熟果実に較べて同型接合および異型接合のAFT植物体の果皮にCHS1、CHS2、DFRをコードする遺伝子が実質的に発現上昇することが確認できた。
【実施例3】
【0131】
CHS1、CHS2、DFRをコードする遺伝子はAFT植物体において多型でない。
【0132】
CHSはフラボノイド生合成経路における第一関与段階で機能する酵素をコードする遺伝子である。上記したような有意の転写発現増加のために、CHS1またはCHS2がAFT表現型をもたらす遺伝子になり得ると仮定できる。この仮説を検証するために、2つの遺伝子(表1)の夫々に遺伝子座特異プライマーを設計し、PCRにVF36種(LA0490)およびラトガーズ種(LA1090)の2つの赤熟品種から対応するゲノム領域を増幅させ、31(CHS1)および35(CHS2)制限酵素でそれらを分解させた。
【0133】
これら2遺伝子について、LA1996種と上記の赤熟品種との間に多型性はなかった。同様に、アントシアニン生合成の後段で働くDFR(制限酵素27)、および、F3H(制限酵素22)にも多型性はなく、前記の実験において転写的に誤った制御であったことが判明した。これらの結果は、制御遺伝子がAFT表現型をコードする遺伝子であるという理論に一致する。
【実施例4】
【0134】
トマトANT1遺伝子は、AFT表現型をコードする有望な遺伝子候補である。
【0135】
トマト果実におけるT−DNA賦活標識実験で、アントシアニン生合成のMYB転写制御因子を識別して、ペチュニアAn2と高い相同性を有することからANT1と呼ばれている(マシュースら著「植物細胞」第15号(2003年)1689〜1703頁)。
【0136】
ant1変異トマト植物体は、栽培の芽形成のかなり早期から強烈な紫色素を顕わし、アントシアニン蓄積につながる生合成経路の活性化に反映させた。ant1植物体の植物性組織は強烈な紫色を呈し、果実は表皮および果皮に紫色の斑点があった。果実の転写結果(実施例2)と同様に、ant1変異実生は、アントシアニン生合成の初期(CHS)および後期(DFR)で機能するタンパク質をコードする遺伝子が発現上昇を示した(マシュースら著「植物細胞」第15号(2003年)1689〜1703頁)。
【0137】
ANT1遺伝子配列は後になってRFLPプローブとして用いられ、295−F2個体を用いて、ANT1遺伝子とコショウA遺伝子、枝葉、花、未熟果実にアントシアニン色素を蓄積する優性遺伝子の間の完全同時分離を示した(ボロフスキーら著「Theor Appl Genet」第109号(2004年)23〜29頁)。A遺伝子はコショウ染色体10にマッピングされ、染色体はLA1996種において初期の段階では多型は認められなかった(ジョーンら著「ジャーナル・ヘレディティー」第94号(2003年)449〜446頁)。それにも拘わらず、LA1996種および赤熟アイルサ・クレイグ種植物体由来のANT1遺伝子を配列で特徴付け、AFT表現型の有望な候補遺伝子としてのANT1遺伝子を強調できる可能性のあるヌクレオチド多型を検出することを決定した。配列分析によって、ANT1遺伝子コード領域と非コード領域における2つの遺伝子型には複数のヌクレオチドに差があることを明らかにした(図2)。特筆すべきは、アイルサ・クレイグ品種の読み取り枠のヌクレオチド配列とミクロトーム種(GenBank受入番号:AY348870<http://www.ncbi.nlm.nih.gov/>で検索))との間で完全な配列同一性が見られたことである。
【0138】
アイルサ・クレイグ品種とLA1996種間のアミノ酸配列を比較すると、2つの遺伝子型間で8つのアミノ酸が異なることが明らかになった(図3)。アイルサ・クレイグ品種のアミノ酸配列と、ミクロトーム種(GenBank受入番号:AAQ55181<http://www.ncbi.nlm.nih.gov/>で検索))からクローニングされた元来のANT1のアミノ酸配列との間に完全な同一性が明確に顕れた。7つのアミノ酸の差異がLA1996種と2つの赤熟遺伝子型(アイルサ・クレイグ種とミクロトーム種)との間の大きな違いであると見なせる(表6)。
【0139】
【表6】
【0140】
ANT1遺伝子のLA1996種と赤熟遺伝子型間のヌクレオチド配列差に基づいて、PCRプライマー(表1)を設計し、PCR増幅反応でうまく利用できた。増幅産物をNcoI制限酵素で分解させ、図4に示すように、S.lycopersicum(ANT1−L)とS.chilense(ANT1−C)由来のANT1対立遺伝子間の共優性多型を示すようにした。
【0141】
上記で詳細に示されたヌクレオチド配列およびタンパク質配列の多型に加えて、ANT1遺伝子は、LA1996から得られたトマトの果皮において、対応する赤熟マネーメーカー種と比較して統計的に有意な4.9倍(S.E.は1.4)の転写のダウンレギュレーションを示した。また、マネーメーカー種とLA1996種との異種交配から得られたF1植物体より採取した果実にも統計的に有意な転写のダウンレギュレーションが見られたが、転写の減少倍率は半分(2.5±0.2)であった。
【実施例5】
【0142】
トマトのANT1遺伝子はトマト果実内のアントシアニン蓄積の形質と綿密に関連する。
【0143】
連鎖解析を行って、ANT1とアントシアニン蓄積量との間の関連性について調べた。そのために、LA1996種とアイルサ・クレイグ品種との異種交配により得られるF2群であって、hp−1変異体のホモ接合体を作成した。hp−1/hp−1変異植物体は、赤熟ホモ接合型hp変異植物体中においても、増強したアントシアニン蓄積がみられることから(イェンら著「理論応用遺伝学」第95号(1997年)1069〜1079頁、ビノら著「新植物学」第166号(2005年)427〜438頁、および、レビンら著「イスラエル植物科学ジャーナル」(2006年)近日刊行)、AFT遺伝子を有するhp−1/hp−1変異植物体中でもアントシアニン蓄積の凝集が見られると仮定した。先に設計されているパイロシークエンス・プライマー(リーバーマンら著「理論応用遺伝学」第108号(2004年)1574〜1581頁)、および、表1に示すPCRプライマーをそれぞれ用いて、合計247のF2植物体のHP−1およびANT1遺伝子についての遺伝子型が特定された。アントシアニン蓄積の形質は緑熟果および赤熟果の目視検査により記録された。
【0144】
【表7】
【0145】
表7および図5に示した結果は、ANT1−Cとアントシアニン蓄積量の形質との間に強い関連性があることを明らかにし、ホモ接合体hp−1/hp−1遺伝子型のホモ接合体との顕著、且つ、完全な関連性を示した。ANT1−CがANT1−Lより優位であると期待した通り、4つの異型接合のANT1−C/ANT1−L植物体は、緑熟果または赤熟果においてアントシアニン蓄積の表現型を示さなかった。遺伝子組み換え体であることを考えると、これらの植物は、ANT1とAFT遺伝子との間隔が〜.8センチモルガンであることを示唆している(F2を基準に算出)。しかしながら、この見識に異論がある。つまり、施行した増殖条件では、LA1996種とアイルサ・クレイグ品種を含む数種の赤熟の開放受粉した栽培品種との交配から得られた異型接合のANT1−C/ANT1−L植物体において如何なる可視表現型も観察できなかったからである。表3および表4に示した本発明に基づく代謝データでは、異型接合植物体が表現型を示す能力がないことも如実に顕しており、ANT1−C/ANT1−LのF1植物体から採取した果実に含まれるアントシアニンおよびフラボノールの平均含有量は、同型接合体ANT1−C/ANT1−Cより同型接合体ANT1−L/ANT1−Lにかなり似ていることを明らかにしている。ANT1とAFTの完全連鎖の可能性に関する見解を更に立証するために、AFT表現型に特徴的な表現型を示さない2つの非hp−1異型接合体ANT1−C/ANT1−LのF2植物体を自家受粉させた。結果的に得られたF3群の夫々の60の植物体を作付けして、熟成果を外観検査したところ、異型接合植物体で推測されたように、AFT形質はこれら2つの群で実際に分離することが明らかとなった。加えて、hp−1変異体が同型接合であり、ANT1遺伝子(hp−1/hp−1、ANT1−C/ANT1−L)が異型接合である植物体を自家受粉させた結果、得られたF3植物体はANT1遺伝子の遺伝子型が特定された。結果的に得られた遺伝子型の夫々を顕す18つの植物体を作付けした。果実成熟した上で、これらの植物体を外観検査したところ、ANT1遺伝子型とAFT表現型との間に完全な関連性が見られ、ここでも、ANT1とAFT遺伝子との間に強い関連性と完全連鎖の可能性が示された。
【実施例6】
【0146】
トマトAFT遺伝子を染色体10にマッピングする。
【0147】
LA1996種から遺伝子移入されたAFT遺伝子とANT1遺伝子配列との強い関連性によって初めてAFT遺伝子の染色体位置がトマトゲノムにおいてマッピングできた。その目的のために、S.pennelliiの遺伝子移入種が用いられた(エシェドら著「理論応用遺伝学」第83号(1992年)1027〜1034頁)。図6で要約した結果では、遺伝子移入種10−3に対してのみANT1がトマト染色体10の長腕にマッピングされている。ここでの実験研究で得られたANT1とAFT形質との強い関連性は、AFT表現型をもたらす遺伝子もまたトマト染色体10の長腕に位置することを表している。
【実施例7】
【0148】
hp−1変異体は、ANT1−C対立遺伝子におけるアントシアニンおよびフラボノール発現を相加的な方法より増強できる。
【0149】
図1に視覚表示したように、hp−1変異体は、ANT1−C対立遺伝子に起因して赤熟果実のアントシアニン発現を増強する。このhp−1の積極的な貢献は、ANT1−C/ANT1−C同型接合植物体およびANT1−C/ANT1−L異型接合植物体で見られた。果実アントシアニン濃度および可能性としてフラボノール濃度にも及ぼすこの相乗作用を定量化するために、赤熟果実を次のF3遺伝子型の夫々の植物体18種から採取した:AFT/AFT hp−1/hp−1、AFT/+ hp−1/hp−1、および、++ hp−1/hp−1。同様に初期親種は、AFT/AFT ++(LA1996種)、および、++ hp−1/hp−1(hp−1変異体に対して同型接合であるアイルサ・クレイグ種)である。
【0150】
【表8】
【0151】
表8に示した結果は、複合遺伝子型AFT/AFT hp−1/hp−1が初期親種に比較して相加的効果以上の有意な効果をアントシアニン、デルフィニジン、ペツニジン、マルビジンに及ぼすことが明らかにした。
【0152】
この遺伝子型は、表9に示すように、フラボノール類のケセルチン、ケンフェロールの濃度を同様に高める傾向を示した。
【0153】
【表9】
【実施例8】
【0154】
タバコおよびトマト植物体の形質転換は、ANT1−Cアントシアニン蓄積の非常に大きな効果を示す。
【0155】
カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下でのS.chilense (ANT1−C)およびS.lycopersicum(ANT1−L)を由来とするANT1遺伝子の形質転換は、トマトおよびタバコの再生体において相当多量かつ早期のアントシアニン産生を示した。これらの結果より、ANT1はおそらく大部分がAFT表現型をコードする遺伝子であり、ANT1−C対立遺伝子は、栽培トマト由来のANT1−L対立遺伝子と比較して、アントシアニン産生に相当大きな効果をもたらすことを明確に示している。
【実施例9】
【0156】
ANT1遺伝子における187位プロリンのグルタミンへの置換 − AFT遺伝子型におけるアントシアニン蓄積の決定因子。
【0157】
コショウと同様に高果実アントシアニンのトマト種からクローニングされたANT1遺伝子のアミノ酸配列を低果実アントシアニンのトマト種およびコショウと比較した。これらの比較の検討のいくつかを図9に示しており、高濃度で果実アントシアニンを蓄積する種と蓄積しない種間での明瞭な差異である、たった一つのアミノ酸としてANT1遺伝子の187位プロリンのグルタミンへの置換を表している。この結果は、この単一アミノ酸置換のみがAFT表現型に見られる果実アントシアニン蓄積の増加の要因になり得ることを明らかにしている。しかし、ANT1遺伝子における他のアミノ酸変化が同程度またはそれ以上の果実アントシアニン蓄積表現型を産生する可能性を有する。
【図面の簡単な説明】
【0158】
本発明を理解し、本発明の実施方法が分かるように、非限定例であるが添付の図面を参照して好ましい実施例をここに開示する。
【0159】
【図1】図1は、アントシアニンおよびフラボノールの生合成経路の概略図である(ホルトンとコーニシュ著「植物細胞」第7号(1995年)1071〜1083頁援用)。
【図2】図2は、アイルサ・クレイグ品種(上図)とLA1996(下図)とのANT1遺伝子のヌクレオチド配列の比較を概略的に示している(開始コドンおよび終止コドンは両配列に下線をつけており、イントロン領域は黄色で彩色している)。
【図3】図3は、アイルサ・クレイグ品種(上図)とLA1996(下図)とのANT1タンパク質のアミノ酸比較を概略的に示している(2線間で異なるアミノ酸を黄色で彩色している)。
【図4】図4は、S.lycopersicum(ANTl−L)由来のANT1対立遺伝子とS.chilense(ANTl−C)由来のANT1対立遺伝子間の共優性遺伝子多型を示す写真画像である。
【図5】図5は、ANT1遺伝子と、ANT1とhp−1に分離するF2群中のアントシアニン蓄積の形質の関連性を視覚表示している(各果実は夫々の遺伝子型の個々の植物体から得られたものである)。
【図6】図6は、トマトゲノムに対するANT1の制限酵素遺伝子解析を表した写真および配置図である(トマト染色体10のマップはネット資料<http://tgrc.ucdavis.edu/pennellii-ILs.pdf>から援用)。
【図7】図7は、カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下において、S.chilense(ANTl−C)由来のANT1遺伝子で形質転換されたタバコ再生体およびS.lycopersicum(ANTl−L)由来のANT1遺伝子で形質転換されたタバコ再生体との比較写真である。
【図8】図8は、カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下におけるS.chilense(ANTl−C)由来のANT1遺伝子で形質転換されたトマトおよびS.lycopersicum(ANTl−L)由来のANT1遺伝子で形質転換されたトマト(マネーメーカー品種)再生体の比較写真である。
【図9】図9は、トマト登録品種およびコショウからクローンが作られたANT1遺伝子のアミノ酸配列の概略図である(果実アントシアニンを蓄積していない登録品種:LA1589はS.Pimpinellifolium、LA2838AはS.lycopersicumである。/果実アントシアニンを蓄積している登録品種:PI128650はS.Peruvianum、hp−799は未知のS.PeruvianumとS.lycopersicumとの交配から生まれた選択系統、LA1996はS.chilense由来のAFT遺伝子型、CAE75745はアントシアニンを蓄積するコショウである)。
【図10】図10は、本発明の一態様によるhp−1変異およびANT1−C対立遺伝子(ANT1−C/ANT1−C hp−1/hp−1)が同型接合したF3植物体とLA1996植物体(ANT1−C/ANT1−C +/+)から得られたトマト果実を示している。
【図11】図11は、本発明の別の態様によるS.Peruvianum(ANT1−P/ANT1−P hp−1/hp−1)由来のANT1対立遺伝子とhp−1変異が同型接合した登録品種のトマト植物および果実を示している。
【技術分野】
【0001】
本発明は、特にデルフィニジン、ペツニジン、マルビジンといった高濃度のアントシアニン類と特にケルセチン、ケンフェロールといった高フラボノール表現型を有する、トマト果実などの果物を生産する手段および方法に関するものである。
【背景技術】
【0002】
果実や野菜をカロチノイド、フラボノイド化合物、ビタミン類などの機能性代謝物で成分強化することが過去数年間で重要な育成の目標となっている。この傾向の好例としては、果実に高濃度のカロチノイド類、フラボノイド類、ビタミンCおよびEを含めるために商用のトマト品種への高色素(hp)変異体の遺伝子移入が行われることである。
【0003】
S.chilenseに由来する「アントシアニン果実(AFT)」遺伝子型は、表皮および外果皮組織がフラボノイド化合物の分類に属する高濃度のアントシアニン類や代謝物によって紫色を呈する特徴を持っている。アントシアニン濃度の上昇は単一遺伝子によって決定されることが報告されている(ジョーンら著「ジャーナル・ヘレディティー」第94号(2003年)449〜446頁)。フラボノイド類は、多くの植物体で自然に存在するポリフェノール化合物である。フラボノイド類は果物、野菜、植物原料の飲料(茶、赤ワイン)に含まれ、多くの栄養補助食品や植物性生薬に用いられる。その中心的成分であるアグリコンに基づいて、カルコン類、フラバノン類、ジヒドロフラボノール類、フラボノール類、アントシアニン類などの異なった区分に分類できる。これまでに4000以上の異なったフラボノイド類が判明しており、こうした膨大な多様性は、グリコシル化、メチル化、アシル化などのアグリコンの単一変異もしくは組み換え変異に起因する。一分類としてのフラボノイドは、植物体の育成、更には、病原菌抵抗性、色素産生、紫外線保護、花粉成長、種皮発達などの植物開発など多くの側面を担っている(ハルボーン著「フラボノイド」)。「1986年以来の研究の進歩(第一版)」(ボビーら著「植物細胞」第14号(2002年)、2509〜2526頁)。
【0004】
アントシアニン類は、紫、赤、青色の植物色素を持つ最も一般的な種類である。植物体において300を超える異なったアントシアニン化合物が確認されている。これらは、フラボノイド類特有のC6−C3−C6炭素構造を持つ平面分子を持っている。
【0005】
特に、フラボノール類(ケンペロールやケルセチンなど)に属するフラボノイド類は、その高抗酸化能力によって人の食事における潜在的な健康保護機能(ライス・エバンスら著「植物化学の動向」第2号(1997年)152〜159頁、および、プロトジェンテら著「フリーラジカル研究」第36号(2002年)217〜233頁)、更に、ヒトの保護酵素システムを誘発するインビトロでの抗酸化能力(クック&サマン著「栄養生化学ジャーナル」第7号(1996年)66〜76頁)を持っているという証拠が増えてきている。こうした知見から、フラボノイド類によって冠状動脈性心臓病や癌などの大病から防御できることが予測できる(ヘルトーク&ホルマン著「欧州臨床栄養生化学ジャーナル」第50号(1996)63〜71頁)。いくつかの疫学研究によれば、心臓保護とタマネギ、りんご、茶などの食品原料の摂取との間に深い関係があることが分かっている(ヘルトークら著「ランセット342号」(1993)1007〜1011頁)。これに関して、アントシアニン類は、ラジカル吸収能(ORAC)の分析で測定できる強い抗酸化活性酸素によって特に注目されていた。ぶどう(ワンら著「農産食品化学ジャーナル」第44号(1996)701〜705頁)、ブルーベリー、ブラックベリー、ラズベリー、サクランボ(ワンら著「農産食品化学ジャーナル」第45号(1997)304〜309頁)が、他の果実や野菜に比べてアントシアニン類を多量に含み、抗酸化能を持つことが知られている。最も重要な食品のひとつであるトマトが世界中で広く収穫されており、ぶどう、ベリー類、サクランボなどより手軽に広く食せられているので、アントシアニン摂取のための食品として有望であると考えられる。ところが、市販されているトマトには高濃度のフラボノイド類(アントシアニン類を含む)が含まれておらず、高濃度のフラボノイド類を含有する市販トマト植物の実現は重要な目標である。
【0006】
トマト果実のフラボノイド類含有量を増やし、多様化させる施策として以下の事柄に重点的に取り組んでいる。
1.フラボノイド化合物に係わる構造遺伝子の代謝工学
2.代謝経路に影響する転写因子の遺伝子組み換え
3.単一点突然変異(自然発生突然変異または誘発突然変異)および/または植物栄養素量に及ぼす顕著な効果を有する量的形質遺伝子座(レビンら著「イスラエル植物科学ジャーナル」(2006年)近日刊行)
【0007】
35Sプロモーターの制御下におけるP.hybrida由来の「カルコン異性化酵素(CHI)」遺伝子によるトマトの形質転換は、果皮フラボノール濃度を劇的に増加させる。ところが、高フラボノール遺伝子組換え植物体による葉、葉野菜、およびbreaker(未熟果)や熟したトマト果肉中にはフラボノール量の増加は見られなかった(ミューアら著「ネーチャー・バイオテクノロジ」第19号470〜474頁)。
【0008】
トマト果肉中のフラボノール蓄積、および、それによるトマト果実中のフラボノール量の増加は、転写因子LC及びCIをコードするトウモロコシ遺伝子の同時過剰発現によって実現する(ボビーら著「植物細胞」第14号(2002年)、2509〜2526頁)。
【0009】
代替的アプローチでは、フラボノール化を導く、P.hybrida由来の4つの生合成酵素「カルコン合成酵素(CHS)、カルコン合成酵素(CHI)、フラバノン−3−ヒドロキシラーゼ(F3H)、フラボノールシンターゼ(FLS)」をコードする遺伝子を異所的かつ同時にトマト植物体に発現させた。4種のトランス遺伝子を含む約75%の主形質転換体が極めて高濃度のケルセチン配糖体を果皮内や他の部分に蓄積するが、殻軸組織ではケンフェロールやナリンゲニン配糖体が顕著に増加する(ベルフォネンら著「実験的植物ジャーナル」第53号2099〜2106頁)。
【0010】
遺伝子組み換えによるトマト果実の果肉におけるフラボノイドの高濃度化に関しては大成功とはいかなかったことが記されており、加えて、高フラボノイド化植物体を得るにあたって非遺伝子組み換え的アプローチへの要求が高まっている。この要求は、遺伝子組み換え食品(GMO)として知られているような遺伝子組み換え果実や野菜に対する消費者の抵抗感が契機となっている。
【0011】
栽培もののトマト、つまり、S.chilense、S.habrochaites、S.cheesmaniae、S.lycopersicoidesなどのいくつかのトマト種の果実はアントシアニンを著しく多量に含んでいる(リック著「カリフォルニア科学アカデミー・オケージョナル・ペーパー」1964年第44、59号;ジョージフ著「トマト遺伝子協同組合報告書」1972年第22号10頁;リックら著「トマト遺伝子協同組合報告書」1994年第44号29〜30頁)。S.chilense由来のアントシアニン果実(AFT)、S.lycopersicoides由来のナス(ABG)、L.cheesmaniae由来のアトロビオラセウム(atv)はトマト果実においてにアントシアニンの発現をひきおこす。野生種S.pennellii v.puberulumは機能性フラボノイド類でトマト果実を強化する源(ソース)と見られている(ウィリットら著「農業食品化学ジャーナル」第53号1231〜1236頁)が、果皮濃縮は消極的な手法であり、産物は不安定である。
【0012】
果物のフラボノイドを増やす別の試行としては、高色素(hp)変異の遺伝子移入である。トマトhp変異体(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)は、赤熟果物のカロチノイド(リコピンや、カロチン)濃度にプラスに働く効果があることはよく知られている(レビンら著「理論応用遺伝学」第106号(2003年)454〜460頁)。hp−1変異を有する熟した植物果物も、トマト果実果皮のフラボノイド・ケルセチンを同質遺伝子系統の13倍の増加を示すことが分かっている(イェンら著「理論応用遺伝学」第95号(1997年)1069〜1079頁)。トマト変異体hp−2およびhp−2jの果皮中およびトマト変異体hp−2dgの果実中のケルセチンにおける同様の増加も明らかにされている(ビノら著「新植物学」第166号(2005年)427〜438頁、および、レビンら著「イスラエル植物科学ジャーナル」(2006年)近日刊行)。
【0013】
AFT遺伝子型のS.chilense種果実は、果実の表皮および外果皮組織に含まれるアントシアニンによって特徴づけられる。加工トマトとAFT植物体の交配のF2およびBC1群におけるアントシアニン発現の分離比はアントシアニン発現のための単一優性遺伝子の仮説に一致することが分かっている。トマト果実におけるT−DNA賦活標識実験で、アントシアニン生合成のMYB転写制御因子を識別して、ペチュニアAn2と高い相同性を有するANT1と呼ばれている(マシュースら著「植物細胞」第15号(2003年)1689〜1703頁)。
【0014】
ANT1変異トマトは、苗木栽培の芽形成のかなり早期から強烈な紫色素を顕わし、アントシアニン蓄積につながる生合成経路の活性化に反映させた。ANT1植物体の植物性組織は強烈な紫色を呈したが、その果実を66倍率で拡大観察したところ表皮にのみ紫色の斑点が視認できた。こうしたことから、表現型が更に強烈な紫を呈するような表皮および外果皮に更に高濃度のアントシアニンを高アントシアニンのトマトに含ませることが長い間の要望であった。
【0015】
野生型S.chilenseトマト株由来のAFTは高いアントシアニン濃度をもたらすので、市販のhp−1トマトは特定のフラボノイド濃度が若干増大するように見え、多く増加したフラボノイド濃度を持つ安定した品種改良増殖が長年の要望に実用的に応えることになる。
【0016】
AFT−S.chilense遺伝子が、栽培トマト対応物より高いアントシアニン濃度の原因であることが知られている。よって、この遺伝子の特徴、分離、および、フラボノイド濃度を強化したトマトを含む商用植物への形質転換もまた、医薬成分や医薬合成品用のアントシアニンの調製などGMO利用が許容され得る効果が期待できる用途における長年の要求に応えることができる。
【0017】
S.chilenseに加えて、S.habrochaites、S.cheesmaniae、S.lycopersicoides、S.pennelli v.puberelumなどの栽培トマトに関連するいくつかのトマト種果実は、栽培トマトに比較して相当に高いアントシアニン量を含有する。よって、夫々のAFT相同遺伝子の栽培トマトへの遺伝子移入および遺伝子発現が長い間待ち望まれていた新たな進展をもたらす。また、AFT以外の遺伝子または対立遺伝子を強化するフラボノイドを含むトマト増殖へのS.chilenseに由来するAFT遺伝子の遺伝子移入および遺伝子発現が更にフラボノイド含有量を増加させる。
【0018】
上述したように、形質転換遺伝子組み換えによるトマト果実中のフラボノイド含有量のある種の増加は達成されたが、GMO消費に対する消費者の抵抗感は周知のところであり、こうして、非遺伝子品種改良によって実現した高いフラボノイド栽培品種の市場での需要がある。したがって、こうした品種改良を促すためにDNAマーカーのような遺伝子配列を用いるために植物種のAFT突然変異表現型をコード化する遺伝子(または、遺伝子群)の識別が有用、且つ、新規な技術手法であると見られている。
【0019】
アントシアニン及びフラボノールのような食用の抗酸化物質の原料を得る際の関心の高さから見て、アントシアニン、特に、デルフィニジン、ペツニジン、マルビジンなどのアントシアニンと、特に、ケルセチン、ケンフェロールなどのフラボノイド表現型を高濃度に含むトマトを生産する手段および方法の実現が、長い間依然として要求されている。
【発明の概要】
【0020】
本発明の目的は、タンパク質をコードするAFT遺伝子であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、栽培植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするタンパク質をコードするAFT遺伝子を提供することにある。
【0021】
本発明の他の目的は、栽培S.licopersicum種トマト植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培S.licopersicum種トマト植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培S.licopersicum種植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするAFT遺伝子を提供することにある。
【0022】
本発明の他の目的は、S.chilense遺伝子型に由来し、栽培S.licopersicum種トマト植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培S.licopersicum種トマト植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培S.licopersicum種植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするAFT遺伝子を提供することにある。
【0023】
本発明の他の目的は、前記フラボノイド類の少なくとも一部がアントシアニン類および/またはフラボノール類であることを特徴とするAFT遺伝子を提供することにある。
【0024】
本発明の他の目的は、遺伝子がS.peruvianumに由来することを特徴とするAFT遺伝子を提供することにある。
【0025】
本発明の他の目的は、遺伝子が、S.habrochaites、S.cheesmaniae、S.licopersicoides、S.peruvianum、S.pennelli v.puberelumよりなる群から選択されたことを特徴とするAFT遺伝子を提供することにある。
【0026】
本発明の他の目的は、AFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体であって、前記AFT−S.chilense遺伝子型を遺伝子移入していない従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のフラボノイド類によって特徴づけられる遺伝子移入由来トマト植物体を提供することにある。
【0027】
本発明の更に他の目的は、前記AFT−S.chilense遺伝子型を遺伝子移入していない従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のアントシアニン類および/またはフラボノ−ル類によって特徴づけられる遺伝子移入由来トマト植物体を提供することにある。
【0028】
本発明の更に他の目的は、前記AFT−S.chilense遺伝子型がS.peruvianumから遺伝子移入されたことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0029】
本発明の更なる目的は、前記AFT−S.chilense遺伝子型が、S.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群から遺伝子移入されたことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0030】
本発明の更に他の目的は、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のフラボノイド類を有することで特徴づけられる純親系統を得るために、前記F2植物体を自家交配させてF3群および更なる個体群(F4、F5等)を作成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
方法によって遺伝子移入的に得たことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0031】
本発明の更にその他の目的は、前記フラボノイド類が、アントシアニン類およびフラボノール類であるデルフィニジン、ペツニジン、マルビジン、ケルセチン、ケンフェロール、ナリンゲニンを含むことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0032】
本発明の他の目的は、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、hp−1、hp−1w、hp−2、hp−2j、hp−2dgとして定義される1以上の同型対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0033】
本発明の他の目的は、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子の1以上の同型接合対立遺伝子(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0034】
本発明の他の目的は、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子に欠損のあるhp−1、hp−1w、hp−2、hp−2j、hp−2dg変異植物体と等表現型の光形態形成遺伝子の1以上の同型接合対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0035】
本発明の他の目的は、DNAマーカーを用いてS.chilense由来のAFT遺伝子座において同型接合であるF2植物体を選択する付加的手段からなることを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0036】
本発明の他の目的は、AFT遺伝子型がS.peruvianumに由来することを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0037】
本発明の他の目的は、AFT遺伝子型がS.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群から選択されたことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0038】
本発明の更なる目的は、前記DNAマーカーがS.peruvianumに由来することを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0039】
本発明の更なる目的は、前記DNAマーカーが、S.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群に由来することを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0040】
本発明の更なる目的は、フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル含有フラボノイドおよび/またはメトキシ置換、C−メチルフラボノイド、メチレンジオキシ・フラボノイド・カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンよりなる群のうちのいずれかの成分から前記フラボノイドを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0041】
本発明の更なる目的は、デルフィジン、ペツニジン、マルビジンよりなる群から前記アントシアニンを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0042】
本発明の更なる目的は、ケルセチン、ケンフェロールよりなる群から前記フラボノイドを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0043】
本発明の更なる目的は、4,2,4,6−テトラヒドロキシカルコン、ナリンゲニン、ケンフェロール、ジヒドロキシケンフェロール、ミリセチン、ケルセチン、ジヒドロケルセチン、ジヒドロミリセチン、ロイコペラルゴニジン、ロイコシアニジン、ロイコデルフィニジン、ペラルゴニジン−3−グルコシド、シアニジン−3−グルコシド、デルフィニジン−3−グルコシドよりなる群から前記フラボノイドを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0044】
本発明の更なる目的は、(i)2−フェニルクロモン(2−フェニル−1,4−ベンゾピロン)構造由来の二次植物代謝物、(ii)前記代謝物が3−フェニルクロモン(3−フェニル−1,4−ベンゾピロン)構造由来であるイソフラボノイド類、(iii)前記代謝物が4−フェニルクマリン(4−フェニル−1,2−ベンゾピロン)構造由来であるネオフラボノイド類よりなる群から前記フラボノイドを選択したことを特徴とする遺伝子移入由来トマト植物体を提供することにある。
【0045】
本発明の更なる目的は、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、従来の栽培S.licopersicum種トマト植物体と比較してトマト植物体中に高いフラボノイド濃度をもたらすことを特徴とするタンパク質をコードするDNA配列を提供することにある。
【0046】
本発明の更なる目的は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、従来の栽培S.licopersicum種トマト植物体と比較してトマト植物体中に高いフラボノイド濃度をもたらすことを特徴とするDNA配列を提供することにある。
【0047】
本発明の更なる目的は、前記DNA配列は、植物体、植物部位または種子内において、フラボノイド経路の代謝物であるアントシアニンまたはフラボノール類の蓄積又は発現をもたらすことを特徴とするDNA配列を提供することにある。
【0048】
本発明の更なる目的は、前記DNA配列は、S.licopersicumのトマト植物体、トマト植物部位または種子内において、フラボノイド経路の代謝物であるアントシアニンまたはフラボノール類の蓄積又は発現をもたらすことを特徴とするDNA配列を提供することにある。
【0049】
本発明の更なる目的は、前記DNA配列は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、栽培トマト系統におけるAFT遺伝子型の遺伝子移入のための遺伝資源、種子、苗木、発芽、植物体、植物部位のスクリーニングに有用であることを特徴とするDNA配列を提供することにある。
【0050】
本発明の更なる目的は、トランスジェニック植物であって、植物体、植物部位または種子内にフラボノイド経路の代謝物であるアントシアニンまたはフラボノイド類を発現し、前記植物体は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有するDNAからなり、前記DNAは、1以上の複数の形質転換、および/または植物の形質転換および/または発現に有益な発現ベクターに組み込まれていることを特徴とする遺伝子移入植物体を提供することにある。
【0051】
本発明の範疇には、タンパク質をコードするAFT遺伝子を得る方法であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、栽培植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするタンパク質をコードするAFT遺伝子を得る方法を提供することも含まれる。
【0052】
本発明の範疇には、従来の栽培S.licopersicum種トマト植物体と比較して高濃度のアントシアニン類および/またはフラボノイド類を含むことを特徴とするAFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体を作り出す方法であって、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のフラボノイド類を有することで特徴づけられる純親系統を得るために、前記F2植物体を自家交配させてF3群を生成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
ことからなる方法を提供することも含まれる。
【0053】
本発明の範疇には、従来の栽培S.licopersicum種トマト植物体と比較して高濃度のアントシアニン類および/またはフラボノイド類を含むことを特徴とするAFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体を作り出す方法であって、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のアントシアニン類とフラボノール類であるデルフィニジン、ペツニジン、マルビジン、ケルセチン、ケンフェロールを有することで特徴づけられた純親系統を得るために、前記F2植物体を自家交配させてF3群および更なる個体群(F4、F5等)を作成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
ことからなる方法を提供することも含まれる。
【0054】
本発明の範疇には、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、hp−1、hp−1w、hp−2、hp−2j、hp−2dgとして定義される1以上の同型対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする方法を提供することも含まれる。
【0055】
本発明の範疇には、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子における1以上の同型対立遺伝子(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする方法を提供することも含まれる。
【0056】
本発明の範疇には、少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子に欠損のあるhp−1、hp−1w、hp−2、hp−2j、hp−2dg変異植物体と等表現型の光形態形成遺伝子の1以上の同型接合対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする方法を提供することも含まれる。
【0057】
更に、本発明の範疇には、HP−1とAFT遺伝子座が同型接合であるF2植物体を付加的にDNAマーカーを用いて選択することを特徴とする方法を提供することも含まれる。
【0058】
更に、本発明の範疇には、デルフィジン、ペツニジン、マルビジンよりなる群から前記アントシアニンを選択することを特徴とする方法を提供することも含まれる。
【0059】
更に、本発明の範疇には、ケルセチン、ケンフェロール、ナリンゲニンよりなる群から前記フラボノイドを選択することを特徴とする方法を提供することも含まれる。
【0060】
更に、本発明の範疇には、4,2,4,6−テトラヒドロキシカルコン、ナリンゲニン、ケンフェロール、ジヒドロキシケンフェロール、ミリセチン、ケルセチン、ジヒドロケルセチン、ジヒドロミリセチン、ロイコペラルゴニジン、ロイコシアニジン、ロイコデルフィニジン、ペラルゴニジン−3−グルコシド、シアニジン−3−グルコシド、デルフィニジン−3−グルコシドよりなる群から前記フラボノイドを選択したことを特徴とする従来の栽培S.licopersicum種トマト植物体と比較して高濃度のフラボノイド類を含むトマト植物体を作り出す方法を提供することも含まれる。
【0061】
更に、本発明の範疇には、(i)2−フェニルクロモン(2−フェニル−1,4−ベンゾピロン)構造由来の二次植物代謝物、(ii)前記代謝物が3−フェニルクロモン(3−フェニル−1,4−ベンゾピロン)構造由来であるイソフラボノイド類、(iii)前記代謝物が4−フェニルクマリン(4−フェニル−1,2−ベンゾピロン)構造由来であるネオフラボノイド類よりなる群から前記フラボノイドを選択したことを特徴とするトマト植物体を作り出す方法を提供することも含まれる。
【0062】
更に、本発明の範疇には、フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル含有フラボノイドおよび/またはメトキシ置換、C−メチルフラボノイド、メチレンジオキシ・フラボノイド・カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンよりなる群のいずれかの成分から前記フラボノイドを選択したことを特徴とするトマト植物体を作り出す方法を提供することも含まれる。
【0063】
更に、本発明の範疇には、タンパク質をコードするDNAを作り出す方法であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、コードされたアミノ酸配列を識別して選択的に検証する処理を含み、前記配列がAFT表現型および増強したフラボノイド濃度に関連するS.chilense中に天然に存在するタンパク質の配列であることを特徴とする方法を提供することも含まれる。
【0064】
更に、本発明の範疇には、核酸を有用に得る方法であって、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、AFT表現型に少なくとも部分的に関連するS.chilense中に天然に存在するタンパク質をコードする前記核酸配列を識別して選択的に検証することからなる方法を提供することも含まれる。
【0065】
更に、本発明の範疇には、従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のフラボノイド類を含むトマト植物体を作り出し、栽培トマト系統におけるAFT−S.chilense遺伝子型の遺伝子移入のために遺伝資源、種子、苗木、植物、発芽、植物体をスクリーニングして容易にした方法であって、
(i)図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有する核酸を作り出し、
(ii)表1に定義されたPCRプライマーを調製し、
(iii)工程(i)のDNAを増幅し、
(iv)それを用いて標的組織を調べることからなる方法を提供することも含まれる。
【0066】
更に、本発明の範疇には、植物体、植物部位、または種子内にフラボノイド経路の代謝物であるアントシアニンまたはフラボノイド類を蓄積または発現させるための遺伝子組み換え方法であって、
(i)図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有するDNAを作り出し、
(ii)前記DNAを1以上の複数の形質転換、および/または植物の形質転換および/または発現に有益な発現ベクターに組み込むことからなる遺伝子組み換え方法を提供することも含まれる。
【0067】
最後に、本発明の範疇には、トマト植物体、トマト部位または種子に有用なことを特徴とする遺伝子組み換え方法を提供することも含まれる。
【発明を実施するための形態】
【0068】
本発明を当分野の技術者が実施可能なように本発明の発明者が考える最良の実施形態に基づいて以下の各章に沿って順に説明するが、トマトや、特に、デルフィニジン、ペツニジン、マルビジンなどの高アントシアニン、および、ケルセチン、ケンフェロールなどの高フラボノイド表現型を含む他の果実を生産するための手段および方法を提供することが本発明の基本概念であり、各種改変が可能なことは当該分野の技術者には自明のことである。
【0069】
用語「hp−1」とは、トマト品種に移入して果物を高濃度のカロチノイド、フラベノイド、ビタミンCおよびEで強化する高色素−1遺伝子変異を意味する。hp−1変異は、トマト紫外線損傷DNA結合タンパク質1(DDB1)および非黄化変異体1(DET1)遺伝子に位置するhp−1w、hp−2、hp−2j、hp−2dgを含む変異表現型のグループに属する。
【0070】
ここでの用語「アントシアニン果実(AFT)」とは、特定単一遺伝子を意味し、S.chilenseから遺伝子移入すると、S.lycopersicumなどの栽培トマトの高濃度のアントシアニンおよび他のフラボノイド代謝産物をもたらす。
【0071】
用語「ANT1−L」とは、アントシアニンおよびフラボノイド蓄積に関連するS.lycopersicumの特定単一遺伝子を意味する。
【0072】
用語「ANT1−C」とは、アントシアニンおよびフラボノイド蓄積に関連するS.chilenseの特定単一遺伝子を意味する。ANT1−Cでコードされているポリペプチドは、8つのアミノ酸が変化している点でANT1−Lとは異なる(図3)。
【0073】
ここでの用語「遺伝子移入」または「遺伝子移入由来」とは、交雑や戻し交雑によって遺伝子がある種から他の種または登録種の遺伝子集団への移動を起こす植物体の育種方法を意味し、所望の遺伝子型や表現型を選択することによって実施できる。DNAマーカーによって、所望の遺伝子型の選択が容易になり、育種を促進する。
【0074】
ここでの「形質転換」とは、異種の植物DNA配列を導入する任意の方法を意味し、異種の植物DNA配列がある種のDNAベクターシステムか構築物に組み込まれ、持続的に標的宿主植物ゲノムまたは細胞質に導入される方法であり、前記植物DNAの構築物の導入は従来から知られている様々な方法で実施できる。
【0075】
ここでの用語「植物体」または「植物部位」とは、限定的ではないが、果実、種子、胚芽、分裂組織、カルス組織、花、枝葉、根、芽、植物配偶体、胞子体花粉、小胞子を含む任意の植物体、植物器官および組織を意味する。植物細胞は、任意の植物器官または組織から取り出し、それらから培養調製ができる。本発明による方法に適用できる植物類は、一般に、形質転換技術を受容できるモノコテリデノス植物体とジコテリデノス植物体の双方を含む高等植物類と同じ程度に広い解釈での植物体である。
【0076】
ここでの用語「フラボノイド」とは、(i)特に、2−フェニルクロモン(2−フェニル−1,4−ベンゾピラン)構造に由来するフラボノイド、(ii)3−フェニルクロモン(3−フェニル−1,4−ベンゾピラン)構造に由来するイソフラボノイド、(iii)4−フェニルクマリン(4−フェニル−1,2−ベンゾピラン)構造に由来するネオフラボノイドなど、IUPAC命名法に従って定めた任意の植物体二次代謝産物を意味する。同様に、この用語には、フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル置換および/またはメトキシ置換フラボノイド、C−メチルフラボノイド、メチレンジオキシフラボノイド、カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリビフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンなどの意味も含まれる。
【0077】
ここでの用語「フラボノール」とは、IUPACが定めた3−ヒドロキシル−2−フェニル−4H−1−ベンゾピラン−4−オン骨格を有する任意のフラボノイドを意味する。これらの多様性は、ケルセチン(3,5,7,3’,4’−ペンタヒドロキシル−2−フェニル−4H−1−ベンゾピラン−4−オン)、ケンペロール(3,5,7,4’−テトラヒドロキシル−2−フェニル−4H−1−ベンゾピラン−4−オン)、ミリセチン(3,5,7,3’,4’,5’−ヘキサヒドロキシル−2−フェニル−4H−1−ベンゾピラン−4−オン)を用いた非限定的方法で典型的に示されるフェノール性(−OH)基の異なった部分に由来する。
【0078】
ここでの用語「アントシアニジン」とは、ベンゼン環に融合した酸素含有ヘテロ環ピランを有する任意のフラベノイドを意味し、ここでのピラン環は2位でフェニル基に結合し、異なる置換基を有することができる。
【0079】
用語「アントシアニン」とは、ある種の糖成分を有するアントシアニジンを意味する。
【0080】
用語「cv.」とは、商用および非商用の品種を意味する。
【0081】
用語「選良種」とは、特にトマトの一般的な商用の交配植物種を意味する。
【0082】
〔植物体材料、交配、生育条件〕
次の植物体材料を準備した: S.chilense−AFTを含むLA1996種、マネーメーカー種(赤色果実・放任受粉型の青果物市場向けトマト)、アイルサ・クレイグ種(赤色果実・放任受粉型トマト・遺伝的にほぼ同質であり、hp−1変異について同型接合)、VF36種(LA0490)(赤色果実・放任受粉型品種)、ラトガーズ種(LA1090)(赤色果実・放任受粉型品種)、LA1589種(赤色果実S.Pimpinellifolium種)、PI128650紫果実S.Peruvianum種。
【0083】
母系として、マネーメーカー種とアイルサ・クレイグ種hp−1/hp−1と、父系LA1996とで異種交配を行った。この異種交配で得られたF1植物体を自家受粉させて、hp−1変異とAFT対立遺伝子が分離するF2群を作成した。ここでのDNAマーカーに基づいてhp−1変異が同型接合でAFT遺伝子座が異型接合する植物体を上記F2群から選別し、hp−1/hp−1の遺伝的背景においてAFTが分離するF3群を生成した。
【0084】
植物体をイスラエル中部の2箇所、ゼライム・ゲデラ・シード社(イスラエル)のバルカニセンターで作付けした。夏季には植物を路地栽培乃至遮光栽培し、冬季には最低摂氏15℃の気温に温度調節したハウス内で栽培した。夏季には、5月の第一週に移植を行い、冬季は11月の第1週に移植を行った。
【0085】
〔ゲノムDNA抽出〕
ゲノムDNAを、フルトンら著「植物分子生物学報告」第13号(1995年)207〜209頁に開示の手法によって個別の植物体から抽出した。
【0086】
〔ポリメラーゼ連鎖反応(PCR)プライマーの設計〕
配列分析および遺伝子座特異プライマーの設計をDNAMAN配列分析ソフトウェアV4.1(リンノン・バイオソフト社(米国ケベック)製)を用いて行った。
【0087】
〔パイロシークエンス遺伝子型決定法〕
hp−1変異の遺伝子型を判定するためにパイロシークエンス方式を用いた。このパイロシークエンス遺伝子型決定方式は、hp−1/hp−1変異植物体とその準遺伝子同質体の相互間のhp−1変異遺伝子型をコードする遺伝子中に見つけられる一塩基多型に基づいている。遺伝子型決定手法として、リーバーマンら著「理論応用遺伝学」第108号(2004年)1574〜1581頁に開示された方法を採用した。
【0088】
〔フラボノイド生合成経路のANT1および他の構造遺伝子の遺伝子型決定法〕
遺伝子型決定は、連鎖解析および/またはPCRの後に制限酵素分解を用いる多型決定を目的として行われた。PCR増幅に用いるプライマーを表1に示す。PCR増幅産物を臭化エチジウムで染色した1.0%アガロースゲル中で電気泳動法によって視覚化した。
【表1】
【0089】
〔リアルタイムPCR分析〕
緑熟トマト果実の果皮からRNAを抽出した。各実験で、各遺伝子型からの3つの1平方ミリ角(LA1996、正常赤色果実遺伝子型マネーメーカー種、VF36、ラトガーズ、LA1996とマネーメーカー種のF1交配種)を分析した。TRIzol試薬システム(インビトロジェン社(米国カリフォルニア州カールズバッド)製)を用いてRNA抽出を行った。混入する可能性のあるゲノムDNAは「TURBO DNA−free」キットを用いて分解させ、「iScript−cDNA」合成キット(バイオ−ラッド研究所(米国カリフォルニア州ハーキュリーズ)製)を使用してcDNA合成のテンプレートとして全RNAを用いた。全RNAの純度および合成cDNAの品質は以下のいずれかのプライマーを用いたPCRによって検証した。
【0090】
「サイバー・グリーンPCR マスター・ミックス」(アプライド・バイオシステム社(米国カリフォルニア州フォスターシティ)製)を用いてリアルタイムPCR分析を行った。PCR反応は初期インキュベーション温度95℃で10分間、続いて、95℃で15秒間の変性、更に61℃で30秒間のアニーリングと、72℃で30秒間の重合化処理を40サイクル行った。
【0091】
この実験調査中、18SリボソームRNAを参照遺伝子として用いて、そのためのプライマーが設計され、同様にその他の遺伝子も分析された。観察対象であるカルコン合成酵素-1(CHS1)、カルコン合成酵素-2(CHS2)、カルコンイソメラーゼ(CHI)、 ジヒドロフラボノール還元酵素(DFR)、フラバノン−3−ヒドロキシラーゼ(FSH)、アントシアニン1(ANT1)を表2に示す。サンプルを、「GeneAmp5700配列検出システム」を用いて通常重複して分析し、「GeneAmp5700SDSソフト(アプライド・バイオシステム社製)」でデータを収集して解析した。検査遺伝子転写物の相対存在量を式[2(CT検査遺伝子-CT参照遺伝子)]で計算した。式において、CTは、蛍光発光が一定の閾値を越える分別サイクル数(通常は0.1に設定)。
【表2】
【0092】
〔アントシアニン、フラボノール抽出、および、定量化〕
新鮮なトマト果皮のサンプル(0.1〜0.3g)を液体窒素中で粉砕し、暗所で冷メタノール、水、酢酸の混合液2ml(11:5:1)を用いて色素を抽出した(マーカム&K.R.オフマン著「植物化学」第34号(1993)679〜685頁)。
【0093】
抽出物20,800g(14,000rpm)で10分間遠心し、上清中にアントシアニン脱離させた。更に、精製物をヘキサン容積2/3とした。サンプルを0.5mlに濃縮し、1時間2N・HCl中で等量のメタノールと共に沸騰させて加水分解し、0.45μm−ポリビニリデンジフロライドフィルター(米国ナルジェ社製)でろ過した。
【0094】
LC−10ATポンプ、SCL-10Aコントローラ、SPD−MlOAVPフォトダイオードアレイ検出器を備えたΗPLC(島津製作所製)を用いてフラボノイドの組成を測定した。抽出物をRP18カラム(Vydac−201TP54)に充填し、次の溶液を用いて27℃で分離した(溶液:(A)H2O、pH2.3、(B)H2O:MeCN:HOAc(107:50:40)、pH2.3)。溶液AとBとの比率が4:1から3:7のリニアに勾配をつくるように45分かけて供給し、流速毎分0.5mlで比率が3:7の状態で更に10分間保持し、保持時間と250〜650nmにおける吸収スペクトルを標準精製フラボノイド(アピン・ケミカルズ、ポリフェノールズ、シグマ)と比較してフラボノイドを識別した。
【0095】
〔トマトゲノムに対するANT1およびAFT遺伝子のマッピング〕
以前発表された資料文献(レビンら著「イスラエル植物科学ジャーナル」第100号(2000年)256〜262頁)のように、ANT1遺伝子と強い関連性を持つこの実験調査で見つかったAFT遺伝子はS.Pennellii遺伝子移入系統を用いてトマトゲノムにマッピングされた(エシェドら著「理論応用遺伝学」第83号(1992年)1027〜1034頁)。
【0096】
元の親種M82とS.Pennelliiを含む遺伝子移入系統の個々の植物体から抽出したDNAをPCR反応の際のテンプレートとして用いた。自動サーモサイクラー(MJリサーチ社(米国マサチューセッツ州ウォルサム)製)で反応を行った。
【0097】
次のDNAプライマーをこれらの反応に用いた(マシュースら著「植物細胞」第15号(2003年)1689〜1703頁)。
F:5’−TCCCCCGGGATGAACAGTACATCTATG−3’
R:5’−GGACTAGTTTAATCAAGTAGATTCCATAAGTCA−3’
【0098】
PCR反応は初期インキュベーションに94℃3分間、引き続いて94℃30秒の変性、60℃30秒間アニーリングし、72℃60秒間の重合化処理を35サイクル行った。72℃の最終伸長反応を7分間行い、上記サイクル処理を完了した。臭化エチジウムで染色した1.0%アガロースゲルを1ml用いた電気泳動法によって得られたPCR産物を視覚化した。親種M−82(LA3475)およびS.Pennellii(LA0716)の間の多型性を得るための制限酵素は必要なかった。
【0099】
〔ANT1遺伝子のクローニング〕
個々のAFT変異(LA1996)植物体および野生遺伝子型(アイルサ・クレイグ)の葉組織100mgから全RNAを抽出した。TRIzol試薬システム(インビトロジェン社(米国カリフォルニア州カールズバッド)製)を用いてRNA抽出を行った。混入の可能性のあるゲノムDNAは「TURBO DNA−free」キットを用いて分解させ、「iScript−cDNA」合成キット(バイオ−ラッド研究所(米国カリフォルニア州ハーキュリーズ)製)を使用したcDNA合成のテンプレートとして全RNAを用いた。ANT1遺伝子配列(GenBank受入番号:AY348870)を変異体種と正常遺伝種の双方で増幅させるためにPCR反応のテンプレートとしてcDNAを用いた。下記遺伝子5’末端および3’末端の特異的プライマーを用いたプルーフリーディングTaqポリメラーゼ(ロッシュ・ダイアゴノスティック社(米国インディアナ州インディアナポリス)製)のPwo-DNAポリメラーゼ)でPCRを実行した。
F: 5’−ATGAACAGTACATCTATGTCTTCATTGG−3’
R: 5’−GGACTAGTTTAATCAAGTAGATTCCATAAGTC−3’
【0100】
TAクローニングの後、結果的に得られた産物をpCRII−TOPOベクター(インビトロジェン社(米国カリフォルニア州カールズバッド)製)に結合し、配列分析で検証した。
【0101】
〔植物形質転換のバイナリーベクター構造〕
カリフラワー・モザイク・ウイルス(CaMV)35S構成プロモーターの制御下に変異および正常ANT1を有する植物形質転換のための構築物を調製した。全ての構築物は、35Sプロモーターの下でNPTII選択マーカー遺伝子を有する。
【0102】
これらの構築物を調製するために、pMON10098プラスミドをEcoRIで分解させた後にエビのアルカリフォスファターゼ(ロッシュ・ダイアゴノスティック社(米国インディアナ州インディアナポリス)製)を用いて処理し、(pCRII−TOPOベクターからの)EcoRI分解ANT1と結合した。pMON−35S−ANT1を並んで含むクローンを単離し、変異および正常ANT1のクローンについて配列を検証した。
【0103】
〔形質転換プロトコル〕
上記構築物の形質転換のためにNicotiana tobacum SRIの葉切除片、および、S.lycopersicum種マネーメーカーの子葉切除片を用いた。
【0104】
培養調整は次の通りである:植物体種子を石鹸(市販洗剤「パルムオリーブ」)と水で洗って水中に置き、その後、流水で1.5時間洗浄した。続いて、種子を96%v/vのエタノール中で1分間、振り混ぜて、3%v/v(+0.01%v/vTween20)の次亜塩素酸ソーダ中に15分間放置し、更に、1.5%v/v(+0.01%v/vツイン20)の次亜塩素酸ソーダ中で30分間激しく撹拌した。最後に種子を滅菌蒸留水で3回洗浄した。処理した葉切除片、および、子葉切除片を上記種子から得られた苗木茎の基部から取り外し、植物成長調整剤成分含有において異なる下記のMS培地に接種した。
【0105】
上記移植片を、本出願人の研究所において標準的な培地でMS無機塩を含む塩基培地(ムラシゲ&スクーグ著「生理学植物」第15(3)号(1962年)473〜497頁)に蔗糖またはグルコース30gと寒天8g/Lを添加した培地に接種した。成長調節剤補充(単位mg/L): タバコ…IAA0.8、キネチン2、ゼアチン1; トマト…IAA0.1、ゼアチン1
【0106】
培地を培養室に23℃で16時間照明(照明状況:50mmol;m−2;s−1の白色蛍光ランプ)の下で保管した。全ての外植片を前記培地に植え、発芽能力を評価した。
【0107】
この実験調査全般で用いた全てのベクターをアグロバクテリウム・ツメフアシェンス菌EHA105に挿入した。葉切除片、および、子葉切除片を滅菌状態でアグロバクテリウム菌と共に、アセトシリンゴン200μMを添加したMS培地で20分間、培養した。細菌ろ紙で組織を吸い取り処理した後、アセトシリンゴン100μMを添加した暗所MS培地において22℃でカルス組織片を共培養した。その後、切除片を洗浄し、選別用にカナマイシン50mg/Lとセフォタキシム200mg/Lを含む固形再生培地に移し、培養容器内で育成した。
【0108】
〔統計分析〕
JMP統計判定ソフト(SASインスティテュート(米国ノースカロライナ州カリー)製)を用いて分散分析(ANOVA)を行った。更に、<http://workbench.sdsc.ede/>に開示された「生物学ワークベンチ」を用いたCLUSRAL W方法(トンプソンら著「核酸研究」第22号(1994)4673〜4680頁)によってヌクレオチドおよびアミノ酸配列を整列させた。
【0109】
図1に、従来技術によるアントシアニンおよびフラボノールの生合成経路の概略図を示している。
【0110】
図2に、アイルサ・クレイグ品種(上図)とLA1996(下図)とのANT1遺伝子のヌクレオチド配列の比較を概略的に示している(開始コドンおよび終止コドンは両配列に下線をつけており、イントロン領域は黄色で彩色している)。
【0111】
図3に、アイルサ・クレイグ品種(上図)とLA1996(下図)とのANT1タンパク質のアミノ酸比較を概略的に示している(2線間で異なるアミノ酸を黄色で彩色している)。
【0112】
図4に、S.lycopersicum(ANTl−L)由来のANT1対立遺伝子とS.chilense(ANTl−C)由来のANT1対立遺伝子間の共優性遺伝子多型の写真画像を示している。
【0113】
図5に、ANT1遺伝子と、ANT1とhp−1に分離するF2群中のアントシアニン蓄積の形質の視覚的表示を示している(各果実は夫々の遺伝子型の個別植物体から得られたものである)。
【0114】
図6に、染色体10のトマトゲノムに対するANT1の制限酵素遺伝子解析の写真および配置図を示している。
【0115】
図7に、カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下のS.chilense(ANTl−C)由来のANT1遺伝子で形質転換されたタバコ再生体およびS.lycopersicum(ANTl−L)由来のANT1遺伝子で形質転換されたタバコ再生体との比較写真を示している。
【0116】
図8に、カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下におけるS.chilense(ANTl−C)由来のANT1遺伝子で形質転換されたトマトおよびS.lycopersicum(ANTl−L)由来のANT1遺伝子で形質転換されたトマト(マネーメーカー品種)再生体の比較写真を示している。
【0117】
図9に、トマト登録種およびコショウからクローンが作られたANT1遺伝子のアミノ酸配列の概略図を示している(果実アントシアニンを蓄積していない登録種:LA1589はS.pimpinellifolium、LA2838AはS.lycopersicumである。果実アントシアニンを蓄積している登録種:PI128650はS.peruvianum、hp−799は未知のS.peruvianumとS.lycopersicumとの交配から生まれた選択系統、LA1996はS.chilense由来のAFT遺伝子型、CAE75745はアントシアニンを蓄積するコショウである)。
【0118】
図10に、hp−1変異およびANT1−C対立遺伝子(ANT1−C/ANT1−C hp−1/hp−1)が同型接合したF3植物体とLA1996植物体(ANT1−C/ANT1−C +/+)から得られたトマト果実を示している。
【0119】
図11に、本発明の別の態様によるS.Peruvianum(ANT1−P/ANT1−P hp−1/hp−1)由来のANT1対立遺伝子とhp−1変異が同型接合した登録品種のトマト植物体および果実を示している。
【実施例1】
【0120】
AFT遺伝子座での同型接合果実は、アントシアニンに加えて高濃度のフラボノール・ケルセチンとケンフェロールを含有している。
【0121】
AFT遺伝子型LA1996種の植物体、赤色果実マネーメーカー種の植物体、および、マネーメーカー種とLA1996種との交配種であるF1植物体を乱塊法計画の開放耕地で育成した。各遺伝子型の5種子を3ブロックに各々に作付けし、赤熟段階で果実をサンプルとして高性能液体クロマトグラフィー分析を行い果皮のフラボノール量とアントシアニン量を測定した。赤熟果実で識別された主要なアントシアニンとその平均濃度を遺伝子型に従って図3に示す。また、同遺伝子型の赤熟果実に含まれる主要なフラボノールとその平均濃度を遺伝子型に従って図4に示す。
【0122】
図3に示す結果から、赤熟果実のマネーメーカー種植物体に較べてAFT/AFT植物体から得られた熟果の果皮に含まれるアントシアニン、デルフィニジン、ペツニジン、マルビジンが統計的に有意な蓄積を見せたことが分かる。これらのことから、同じAFT/AFT植物体と赤熟化トマト植物体UC82Bの果実に含まれるアントシアニン濃度を比較した以前の結果を確認できる(ジョーンら著「ジャーナル・ヘレディティー」第94号(2003年)449〜456頁)。加えて、図4に示す結果によって、AFT/AFT変異植物体の果実が、機能的フラボノール、特に、ケルセチンおよびケンフェロールを統計的に有意に蓄積するという特徴を持っていることが分かる。ケルセチン濃度は、AFT/AFT遺伝型の熟果において、果皮重量(gFW)に基づく赤熟マネーメーカー種植物体の濃度に較べて〜3.6倍高く、果皮部分(cm2)では〜4.3倍高かった。ケンペロール濃度は、AFT/AFT遺伝型の熟果において、果皮重量(gFW)に基づく赤熟マネーメーカー種植物体の濃度に較べて〜2.7倍高く、果皮部分(cm2)では〜3.3倍高かった。
【0123】
【表3】
【0124】
【表4】
【0125】
表3および表4に示す結果から、異型接合のF1植物体の果皮におけるアントシアニンおよびフラボノール濃度は、通常、赤熟遺伝子型のものと較べて高いが、LA1996遺伝子型(AFT/AFT)よりかなり低いことが明らかになった。これらの結果は、AFT遺伝子(または複数AFT遺伝子)の部分的優性効果を示している。
【0126】
しかしながら、こうした統計分析では、対応する赤熟マネーメーカー種植物体と比較した場合において、F1植物体から得られる果実における平均アントシアニン濃度と平均フラボノール濃度との統計的に有意な差を明らかにできなかった。
【実施例2】
【0127】
フラボノイド生合成経路の重要な酵素の転写のアップレギュレーションがAFT植物体の特徴である。
【0128】
上記の予備試験の枠組みの中に作付けしたLA1996種植物体、および、VF36種およびラトガーズ種の2種の赤熟遺伝型から得られた緑熟果からリアルタイムPCRに用いるRNAサンプルを抽出した。cDNA合成およびリアルタイムPCR分析に続いて、フランボノイド生合成経路CHl、CH2、F3H、DFR(プラマーは表3に示す)の4種の構造的な酵素の転写プロフィールに関する3種の遺伝子型を比較した。この結果は、2種の赤熟遺伝型(データは示していない)に比較した時のCHS1、CHS2、DFRの極端な発現上昇、および、LA1996のF3Hの適度な発現減少を示している。特注に値することは、フラボノイド生合成の初期段階で機能する2つのCHS1遺伝子と生合成の後段での酵素活性をコードするDFR遺伝子について極端な発現上昇が見られたことである(図1)。分析は乱塊法計画の開放耕地での実験で得られたサンプルを用いて繰り返し行われた(実施例1参照)。上記3箇所からのLA1996種植物体、赤熟マネーメーカー種植物体、これら2種間の異種交配によるF1植物体からリアルタイムPCRで用いるサンプルを採取した。フラボノイド生合成経路の重要な遺伝子の転写における倍増を示す結果を表5に示す。
【0129】
【表5】
【0130】
これらの結果から、対応する赤熟果実に較べて同型接合および異型接合のAFT植物体の果皮にCHS1、CHS2、DFRをコードする遺伝子が実質的に発現上昇することが確認できた。
【実施例3】
【0131】
CHS1、CHS2、DFRをコードする遺伝子はAFT植物体において多型でない。
【0132】
CHSはフラボノイド生合成経路における第一関与段階で機能する酵素をコードする遺伝子である。上記したような有意の転写発現増加のために、CHS1またはCHS2がAFT表現型をもたらす遺伝子になり得ると仮定できる。この仮説を検証するために、2つの遺伝子(表1)の夫々に遺伝子座特異プライマーを設計し、PCRにVF36種(LA0490)およびラトガーズ種(LA1090)の2つの赤熟品種から対応するゲノム領域を増幅させ、31(CHS1)および35(CHS2)制限酵素でそれらを分解させた。
【0133】
これら2遺伝子について、LA1996種と上記の赤熟品種との間に多型性はなかった。同様に、アントシアニン生合成の後段で働くDFR(制限酵素27)、および、F3H(制限酵素22)にも多型性はなく、前記の実験において転写的に誤った制御であったことが判明した。これらの結果は、制御遺伝子がAFT表現型をコードする遺伝子であるという理論に一致する。
【実施例4】
【0134】
トマトANT1遺伝子は、AFT表現型をコードする有望な遺伝子候補である。
【0135】
トマト果実におけるT−DNA賦活標識実験で、アントシアニン生合成のMYB転写制御因子を識別して、ペチュニアAn2と高い相同性を有することからANT1と呼ばれている(マシュースら著「植物細胞」第15号(2003年)1689〜1703頁)。
【0136】
ant1変異トマト植物体は、栽培の芽形成のかなり早期から強烈な紫色素を顕わし、アントシアニン蓄積につながる生合成経路の活性化に反映させた。ant1植物体の植物性組織は強烈な紫色を呈し、果実は表皮および果皮に紫色の斑点があった。果実の転写結果(実施例2)と同様に、ant1変異実生は、アントシアニン生合成の初期(CHS)および後期(DFR)で機能するタンパク質をコードする遺伝子が発現上昇を示した(マシュースら著「植物細胞」第15号(2003年)1689〜1703頁)。
【0137】
ANT1遺伝子配列は後になってRFLPプローブとして用いられ、295−F2個体を用いて、ANT1遺伝子とコショウA遺伝子、枝葉、花、未熟果実にアントシアニン色素を蓄積する優性遺伝子の間の完全同時分離を示した(ボロフスキーら著「Theor Appl Genet」第109号(2004年)23〜29頁)。A遺伝子はコショウ染色体10にマッピングされ、染色体はLA1996種において初期の段階では多型は認められなかった(ジョーンら著「ジャーナル・ヘレディティー」第94号(2003年)449〜446頁)。それにも拘わらず、LA1996種および赤熟アイルサ・クレイグ種植物体由来のANT1遺伝子を配列で特徴付け、AFT表現型の有望な候補遺伝子としてのANT1遺伝子を強調できる可能性のあるヌクレオチド多型を検出することを決定した。配列分析によって、ANT1遺伝子コード領域と非コード領域における2つの遺伝子型には複数のヌクレオチドに差があることを明らかにした(図2)。特筆すべきは、アイルサ・クレイグ品種の読み取り枠のヌクレオチド配列とミクロトーム種(GenBank受入番号:AY348870<http://www.ncbi.nlm.nih.gov/>で検索))との間で完全な配列同一性が見られたことである。
【0138】
アイルサ・クレイグ品種とLA1996種間のアミノ酸配列を比較すると、2つの遺伝子型間で8つのアミノ酸が異なることが明らかになった(図3)。アイルサ・クレイグ品種のアミノ酸配列と、ミクロトーム種(GenBank受入番号:AAQ55181<http://www.ncbi.nlm.nih.gov/>で検索))からクローニングされた元来のANT1のアミノ酸配列との間に完全な同一性が明確に顕れた。7つのアミノ酸の差異がLA1996種と2つの赤熟遺伝子型(アイルサ・クレイグ種とミクロトーム種)との間の大きな違いであると見なせる(表6)。
【0139】
【表6】
【0140】
ANT1遺伝子のLA1996種と赤熟遺伝子型間のヌクレオチド配列差に基づいて、PCRプライマー(表1)を設計し、PCR増幅反応でうまく利用できた。増幅産物をNcoI制限酵素で分解させ、図4に示すように、S.lycopersicum(ANT1−L)とS.chilense(ANT1−C)由来のANT1対立遺伝子間の共優性多型を示すようにした。
【0141】
上記で詳細に示されたヌクレオチド配列およびタンパク質配列の多型に加えて、ANT1遺伝子は、LA1996から得られたトマトの果皮において、対応する赤熟マネーメーカー種と比較して統計的に有意な4.9倍(S.E.は1.4)の転写のダウンレギュレーションを示した。また、マネーメーカー種とLA1996種との異種交配から得られたF1植物体より採取した果実にも統計的に有意な転写のダウンレギュレーションが見られたが、転写の減少倍率は半分(2.5±0.2)であった。
【実施例5】
【0142】
トマトのANT1遺伝子はトマト果実内のアントシアニン蓄積の形質と綿密に関連する。
【0143】
連鎖解析を行って、ANT1とアントシアニン蓄積量との間の関連性について調べた。そのために、LA1996種とアイルサ・クレイグ品種との異種交配により得られるF2群であって、hp−1変異体のホモ接合体を作成した。hp−1/hp−1変異植物体は、赤熟ホモ接合型hp変異植物体中においても、増強したアントシアニン蓄積がみられることから(イェンら著「理論応用遺伝学」第95号(1997年)1069〜1079頁、ビノら著「新植物学」第166号(2005年)427〜438頁、および、レビンら著「イスラエル植物科学ジャーナル」(2006年)近日刊行)、AFT遺伝子を有するhp−1/hp−1変異植物体中でもアントシアニン蓄積の凝集が見られると仮定した。先に設計されているパイロシークエンス・プライマー(リーバーマンら著「理論応用遺伝学」第108号(2004年)1574〜1581頁)、および、表1に示すPCRプライマーをそれぞれ用いて、合計247のF2植物体のHP−1およびANT1遺伝子についての遺伝子型が特定された。アントシアニン蓄積の形質は緑熟果および赤熟果の目視検査により記録された。
【0144】
【表7】
【0145】
表7および図5に示した結果は、ANT1−Cとアントシアニン蓄積量の形質との間に強い関連性があることを明らかにし、ホモ接合体hp−1/hp−1遺伝子型のホモ接合体との顕著、且つ、完全な関連性を示した。ANT1−CがANT1−Lより優位であると期待した通り、4つの異型接合のANT1−C/ANT1−L植物体は、緑熟果または赤熟果においてアントシアニン蓄積の表現型を示さなかった。遺伝子組み換え体であることを考えると、これらの植物は、ANT1とAFT遺伝子との間隔が〜.8センチモルガンであることを示唆している(F2を基準に算出)。しかしながら、この見識に異論がある。つまり、施行した増殖条件では、LA1996種とアイルサ・クレイグ品種を含む数種の赤熟の開放受粉した栽培品種との交配から得られた異型接合のANT1−C/ANT1−L植物体において如何なる可視表現型も観察できなかったからである。表3および表4に示した本発明に基づく代謝データでは、異型接合植物体が表現型を示す能力がないことも如実に顕しており、ANT1−C/ANT1−LのF1植物体から採取した果実に含まれるアントシアニンおよびフラボノールの平均含有量は、同型接合体ANT1−C/ANT1−Cより同型接合体ANT1−L/ANT1−Lにかなり似ていることを明らかにしている。ANT1とAFTの完全連鎖の可能性に関する見解を更に立証するために、AFT表現型に特徴的な表現型を示さない2つの非hp−1異型接合体ANT1−C/ANT1−LのF2植物体を自家受粉させた。結果的に得られたF3群の夫々の60の植物体を作付けして、熟成果を外観検査したところ、異型接合植物体で推測されたように、AFT形質はこれら2つの群で実際に分離することが明らかとなった。加えて、hp−1変異体が同型接合であり、ANT1遺伝子(hp−1/hp−1、ANT1−C/ANT1−L)が異型接合である植物体を自家受粉させた結果、得られたF3植物体はANT1遺伝子の遺伝子型が特定された。結果的に得られた遺伝子型の夫々を顕す18つの植物体を作付けした。果実成熟した上で、これらの植物体を外観検査したところ、ANT1遺伝子型とAFT表現型との間に完全な関連性が見られ、ここでも、ANT1とAFT遺伝子との間に強い関連性と完全連鎖の可能性が示された。
【実施例6】
【0146】
トマトAFT遺伝子を染色体10にマッピングする。
【0147】
LA1996種から遺伝子移入されたAFT遺伝子とANT1遺伝子配列との強い関連性によって初めてAFT遺伝子の染色体位置がトマトゲノムにおいてマッピングできた。その目的のために、S.pennelliiの遺伝子移入種が用いられた(エシェドら著「理論応用遺伝学」第83号(1992年)1027〜1034頁)。図6で要約した結果では、遺伝子移入種10−3に対してのみANT1がトマト染色体10の長腕にマッピングされている。ここでの実験研究で得られたANT1とAFT形質との強い関連性は、AFT表現型をもたらす遺伝子もまたトマト染色体10の長腕に位置することを表している。
【実施例7】
【0148】
hp−1変異体は、ANT1−C対立遺伝子におけるアントシアニンおよびフラボノール発現を相加的な方法より増強できる。
【0149】
図1に視覚表示したように、hp−1変異体は、ANT1−C対立遺伝子に起因して赤熟果実のアントシアニン発現を増強する。このhp−1の積極的な貢献は、ANT1−C/ANT1−C同型接合植物体およびANT1−C/ANT1−L異型接合植物体で見られた。果実アントシアニン濃度および可能性としてフラボノール濃度にも及ぼすこの相乗作用を定量化するために、赤熟果実を次のF3遺伝子型の夫々の植物体18種から採取した:AFT/AFT hp−1/hp−1、AFT/+ hp−1/hp−1、および、++ hp−1/hp−1。同様に初期親種は、AFT/AFT ++(LA1996種)、および、++ hp−1/hp−1(hp−1変異体に対して同型接合であるアイルサ・クレイグ種)である。
【0150】
【表8】
【0151】
表8に示した結果は、複合遺伝子型AFT/AFT hp−1/hp−1が初期親種に比較して相加的効果以上の有意な効果をアントシアニン、デルフィニジン、ペツニジン、マルビジンに及ぼすことが明らかにした。
【0152】
この遺伝子型は、表9に示すように、フラボノール類のケセルチン、ケンフェロールの濃度を同様に高める傾向を示した。
【0153】
【表9】
【実施例8】
【0154】
タバコおよびトマト植物体の形質転換は、ANT1−Cアントシアニン蓄積の非常に大きな効果を示す。
【0155】
カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下でのS.chilense (ANT1−C)およびS.lycopersicum(ANT1−L)を由来とするANT1遺伝子の形質転換は、トマトおよびタバコの再生体において相当多量かつ早期のアントシアニン産生を示した。これらの結果より、ANT1はおそらく大部分がAFT表現型をコードする遺伝子であり、ANT1−C対立遺伝子は、栽培トマト由来のANT1−L対立遺伝子と比較して、アントシアニン産生に相当大きな効果をもたらすことを明確に示している。
【実施例9】
【0156】
ANT1遺伝子における187位プロリンのグルタミンへの置換 − AFT遺伝子型におけるアントシアニン蓄積の決定因子。
【0157】
コショウと同様に高果実アントシアニンのトマト種からクローニングされたANT1遺伝子のアミノ酸配列を低果実アントシアニンのトマト種およびコショウと比較した。これらの比較の検討のいくつかを図9に示しており、高濃度で果実アントシアニンを蓄積する種と蓄積しない種間での明瞭な差異である、たった一つのアミノ酸としてANT1遺伝子の187位プロリンのグルタミンへの置換を表している。この結果は、この単一アミノ酸置換のみがAFT表現型に見られる果実アントシアニン蓄積の増加の要因になり得ることを明らかにしている。しかし、ANT1遺伝子における他のアミノ酸変化が同程度またはそれ以上の果実アントシアニン蓄積表現型を産生する可能性を有する。
【図面の簡単な説明】
【0158】
本発明を理解し、本発明の実施方法が分かるように、非限定例であるが添付の図面を参照して好ましい実施例をここに開示する。
【0159】
【図1】図1は、アントシアニンおよびフラボノールの生合成経路の概略図である(ホルトンとコーニシュ著「植物細胞」第7号(1995年)1071〜1083頁援用)。
【図2】図2は、アイルサ・クレイグ品種(上図)とLA1996(下図)とのANT1遺伝子のヌクレオチド配列の比較を概略的に示している(開始コドンおよび終止コドンは両配列に下線をつけており、イントロン領域は黄色で彩色している)。
【図3】図3は、アイルサ・クレイグ品種(上図)とLA1996(下図)とのANT1タンパク質のアミノ酸比較を概略的に示している(2線間で異なるアミノ酸を黄色で彩色している)。
【図4】図4は、S.lycopersicum(ANTl−L)由来のANT1対立遺伝子とS.chilense(ANTl−C)由来のANT1対立遺伝子間の共優性遺伝子多型を示す写真画像である。
【図5】図5は、ANT1遺伝子と、ANT1とhp−1に分離するF2群中のアントシアニン蓄積の形質の関連性を視覚表示している(各果実は夫々の遺伝子型の個々の植物体から得られたものである)。
【図6】図6は、トマトゲノムに対するANT1の制限酵素遺伝子解析を表した写真および配置図である(トマト染色体10のマップはネット資料<http://tgrc.ucdavis.edu/pennellii-ILs.pdf>から援用)。
【図7】図7は、カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下において、S.chilense(ANTl−C)由来のANT1遺伝子で形質転換されたタバコ再生体およびS.lycopersicum(ANTl−L)由来のANT1遺伝子で形質転換されたタバコ再生体との比較写真である。
【図8】図8は、カリフラワー・モザイク・ウイルス35S構成プロモーターの制御下におけるS.chilense(ANTl−C)由来のANT1遺伝子で形質転換されたトマトおよびS.lycopersicum(ANTl−L)由来のANT1遺伝子で形質転換されたトマト(マネーメーカー品種)再生体の比較写真である。
【図9】図9は、トマト登録品種およびコショウからクローンが作られたANT1遺伝子のアミノ酸配列の概略図である(果実アントシアニンを蓄積していない登録品種:LA1589はS.Pimpinellifolium、LA2838AはS.lycopersicumである。/果実アントシアニンを蓄積している登録品種:PI128650はS.Peruvianum、hp−799は未知のS.PeruvianumとS.lycopersicumとの交配から生まれた選択系統、LA1996はS.chilense由来のAFT遺伝子型、CAE75745はアントシアニンを蓄積するコショウである)。
【図10】図10は、本発明の一態様によるhp−1変異およびANT1−C対立遺伝子(ANT1−C/ANT1−C hp−1/hp−1)が同型接合したF3植物体とLA1996植物体(ANT1−C/ANT1−C +/+)から得られたトマト果実を示している。
【図11】図11は、本発明の別の態様によるS.Peruvianum(ANT1−P/ANT1−P hp−1/hp−1)由来のANT1対立遺伝子とhp−1変異が同型接合した登録品種のトマト植物および果実を示している。
【特許請求の範囲】
【請求項1】
タンパク質をコードするAFT遺伝子であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、栽培植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするタンパク質をコードするAFT遺伝子。
【請求項2】
栽培S.licopersicum種トマト植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培S.licopersicum種トマト植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培S.licopersicum種植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とする請求項1に記載のAFT遺伝子。
【請求項3】
S.chilense遺伝子型に由来し、栽培S.licopersicum種トマト植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培S.licopersicum種トマト植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培S.licopersicum種植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とする請求項1または請求項2に記載のAFT遺伝子。
【請求項4】
前記フラボノイド類の少なくとも一部がアントシアニン類および/またはフラボノール類であることを特徴とする請求項1に記載のAFT遺伝子。
【請求項5】
前記AFT遺伝子がS.peruvianumに由来することを特徴とする請求項1に記載のAFT遺伝子。
【請求項6】
前記AFT遺伝子が、S.habrochaites、S.cheesmaniae、S.licopersicoides、S.peruvianum、S.pennelli v.puberelumよりなる群から選択されたことを特徴とする請求項1に記載のAFT遺伝子。
【請求項7】
AFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体であって、前記AFT−S.chilense遺伝子型を遺伝子移入していない従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のフラボノイド類によって特徴づけられる遺伝子移入由来トマト植物体。
【請求項8】
前記AFT−S.chilense遺伝子型を遺伝子移入していない従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のアントシアニン類および/またはフラボノ−ル類によって特徴づけられる請求項7に記載の遺伝子移入由来トマト植物体。
【請求項9】
前記AFT−S.chilense遺伝子型がS.peruvianumから遺伝子移入されたことを特徴とする請求項7に記載の遺伝子移入由来トマト植物体。
【請求項10】
前記AFT−S.chilense遺伝子型が、S.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群から遺伝子移入されたことを特徴とする請求項7に記載の遺伝子移入由来トマト植物体。
【請求項11】
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のフラボノイド類を有することで特徴づけられる純親系統を得るために、前記F2植物体を自家交配させてF3群および更なる個体群(F4、F5等)を作成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
方法によって遺伝子移入的に得たことを特徴とする請求項7に記載の遺伝子移入由来トマト植物体。
【請求項12】
前記フラボノイド類が、アントシアニン類およびフラボノール類であるデルフィニジン、ペツニジン、マルビジン、ケルセチン、ケンフェロール、ナリンゲニンを含むことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項13】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、hp−1、hp−1w、hp−2、hp−2j、hp−2dgとして定義される1以上の同型対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項14】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子の1以上の同型接合対立遺伝子(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項15】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子に欠損のあるhp−1、hp−1w、hp−2、hp−2j、hp−2dg変異植物体と等表現型の光形態形成遺伝子の1以上の同型接合対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項16】
DNAマーカーを用いてS.chilense由来のAFT遺伝子座において同型接合であるF2植物体を選択する付加的手段からなることを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項17】
AFT遺伝子型がS.peruvianumに由来することを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項18】
AFT遺伝子型がS.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群から選択されたことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項19】
前記DNAマーカーがS.peruvianumに由来することを特徴とする請求項16に記載の遺伝子移入由来トマト植物体。
【請求項20】
前記DNAマーカーが、S.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群に由来することを特徴とする請求項16に記載の遺伝子移入由来トマト植物体。
【請求項21】
フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル含有フラボノイドおよび/またはメトキシ置換、C−メチルフラボノイド、メチレンジオキシ・フラボノイド・カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンよりなる群のうちのいずれかの成分から前記フラボノイドを選択したことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項22】
デルフィジン、ペツニジン、マルビジンよりなる群から前記アントシアニンを選択したことを特徴とする請求項8に記載の遺伝子移入由来トマト植物体。
【請求項23】
ケルセチン、ケンフェロールよりなる群から前記フラボノイドを選択したことを特徴とする請求項1に記載の遺伝子移入由来トマト植物体。
【請求項24】
4,2,4,6−テトラヒドロキシカルコン、ナリンゲニン、ケンフェロール、ジヒドロキシケンフェロール、ミリセチン、ケルセチン、ジヒドロケルセチン、ジヒドロミリセチン、ロイコペラルゴニジン、ロイコシアニジン、ロイコデルフィニジン、ペラルゴニジン−3−グルコシド、シアニジン−3−グルコシド、デルフィニジン−3−グルコシドよりなる群から前記フラボノイドを選択したことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項25】
(i)2−フェニルクロモン(2−フェニル−1,4−ベンゾピロン)構造由来の二次植物代謝物、(ii)前記代謝物が3−フェニルクロモン(3−フェニル−1,4−ベンゾピロン)構造由来であるイソフラボノイド類、(iii)前記代謝物が4−フェニルクマリン(4−フェニル−1,2−ベンゾピロン)構造由来であるネオフラボノイド類よりなる群から前記フラボノイドを選択したことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項26】
図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、従来の栽培S.licopersicum種トマト植物体と比較してトマト植物体中に高いフラボノイド濃度をもたらすことを特徴とするタンパク質をコードするDNA配列。
【請求項27】
図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、従来の栽培S.licopersicum種トマト植物体と比較してトマト植物体中に高いフラボノイド濃度をもたらすことを特徴とする請求項26に記載のDNA配列。
【請求項28】
前記DNA配列は、植物体、植物部位または種子内において、フラボノイド経路の代謝物であるアントシアニンまたはフラボノール類の蓄積又は発現をもたらすことを特徴とする請求項26または請求項27に記載のDNA配列。
【請求項29】
前記DNA配列は、S.licopersicumのトマト植物体、トマト植物部位または種子内において、フラボノイド経路の代謝物であるアントシアニンまたはフラボノール類の蓄積又は発現をもたらすことを特徴とする請求項26または請求項27に記載のDNA配列。
【請求項30】
前記DNA配列は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、栽培トマト系統におけるAFT遺伝子型の遺伝子移入のための遺伝資源、種子、苗木、発芽、植物体、植物部位のスクリーニングに有用であることを特徴とする請求項26乃至請求項29のいずれかに記載のDNA配列。
【請求項31】
トランスジェニック植物であって、植物体、植物部位または種子内にフラボノイド経路の代謝物であるアントシアニンまたはフラボノイド類を発現し、前記植物体は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有するDNAからなり、前記DNAは、1以上の複数の形質転換、および/または植物の形質転換および/または発現に有益な発現ベクターに組み込まれていることを特徴とする遺伝子移入植物体。
【請求項32】
タンパク質をコードするAFT遺伝子を得る方法であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、栽培植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするタンパク質をコードするAFT遺伝子を得る方法。
【請求項33】
従来の栽培S.licopersicum種トマト植物体と比較して高濃度のアントシアニン類および/またはフラボノイド類を含むことを特徴とするAFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体を作り出す方法であって、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のフラボノイド類を有することで特徴づけられる純親系統を得るために、前記F2植物体を自家交配させてF3群を生成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
ことからなる方法。
【請求項34】
従来の栽培S.licopersicum種トマト植物体と比較して高濃度のアントシアニン類および/またはフラボノイド類を含むことを特徴とするAFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体を作り出す方法であって、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のアントシアニン類とフラボノール類であるデルフィニジン、ペツニジン、マルビジン、ケルセチン、ケンフェロールを有することで特徴づけられた純親系統を得るために、前記F2植物体を自家交配させてF3群および更なる個体群(F4、F5等)を作成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
ことからなる方法。
【請求項35】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、hp−1、hp−1w、hp−2、hp−2j、hp−2dgとして定義される1以上の同型対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする請求項34に記載の方法。
【請求項36】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子における1以上の同型対立遺伝子(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする請求項34に記載の方法。
【請求項37】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子に欠損のあるhp−1、hp−1w、hp−2、hp−2j、hp−2dg変異植物体と等表現型の光形態形成遺伝子の1以上の同型接合対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする請求項34に記載の方法。
【請求項38】
HP−1とAFT遺伝子座が同型接合であるF2植物体を付加的にDNAマーカーを用いて選択することを特徴とする請求項34に記載の方法。
【請求項39】
デルフィジン、ペツニジン、マルビジンよりなる群から前記アントシアニンを選択することを特徴とする請求項34に記載の方法。
【請求項40】
ケルセチン、ケンフェロール、ナリンゲニンよりなる群から前記フラボノイドを選択することを特徴とする請求項34に記載の方法。
【請求項41】
4,2,4,6−テトラヒドロキシカルコン、ナリンゲニン、ケンフェロール、ジヒドロキシケンフェロール、ミリセチン、ケルセチン、ジヒドロケルセチン、ジヒドロミリセチン、ロイコペラルゴニジン、ロイコシアニジン、ロイコデルフィニジン、ペラルゴニジン−3−グルコシド、シアニジン−3−グルコシド、デルフィニジン−3−グルコシドよりなる群から前記フラボノイドを選択したことを特徴とする請求項34に記載の従来の栽培S.licopersicum種トマト植物体と比較して高濃度のフラボノイド類を含むトマト植物体を作り出す方法。
【請求項42】
(i)2−フェニルクロモン(2−フェニル−1,4−ベンゾピロン)構造由来の二次植物代謝物、(ii)前記代謝物が3−フェニルクロモン(3−フェニル−1,4−ベンゾピロン)構造由来であるイソフラボノイド類、(iii)前記代謝物が4−フェニルクマリン(4−フェニル−1,2−ベンゾピロン)構造由来であるネオフラボノイド類よりなる群から前記フラボノイドを選択したことを特徴とする請求項34に記載のトマト植物体を作り出す方法。
【請求項43】
フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル含有フラボノイドおよび/またはメトキシ置換、C−メチルフラボノイド、メチレンジオキシ・フラボノイド・カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンよりなる群のいずれかの成分から前記フラボノイドを選択したことを特徴とする請求項34に記載のトマト植物体を作り出す方法。
【請求項44】
タンパク質をコードするDNAを作り出す方法であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、コードされたアミノ酸配列を識別して選択的に検証する処理を含み、前記配列がAFT表現型および増強したフラボノイド濃度に関連するS.chilense中に天然に存在するタンパク質の配列であることを特徴とする方法。
【請求項45】
核酸を有用に得る方法であって、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、AFT表現型に少なくとも部分的に関連するS.chilense中に天然に存在するタンパク質をコードする前記核酸配列を識別して選択的に検証することからなる方法。
【請求項46】
従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のフラボノイド類を含むトマト植物体を作り出し、栽培トマト系統におけるAFT−S.chilense遺伝子型の遺伝子移入のために遺伝資源、種子、苗木、植物、発芽、植物体をスクリーニングして容易にした方法であって、
(i)図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有する核酸を作り出し、
(ii)表1に定義されたPCRプライマーを調製し、
(iii)工程(i)のDNAを増幅し、
(iv)それを用いて標的組織を調べることからなる請求項34に記載の方法。
【請求項47】
植物体、植物部位、または種子内にフラボノイド経路の代謝物であるアントシアニンまたはフラボノイド類を蓄積または発現させるための遺伝子組み換え方法であって、
(i)図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有するDNAを作り出し、
(ii)前記DNAを1以上の複数の形質転換、および/または植物の形質転換および/または発現に有益な発現ベクターに組み込むことからなる遺伝子組み換え方法。
【請求項48】
トマト植物体、トマト部位または種子に有用なことを特徴とする請求項47に記載の遺伝子組み換え方法。
【請求項1】
タンパク質をコードするAFT遺伝子であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、栽培植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするタンパク質をコードするAFT遺伝子。
【請求項2】
栽培S.licopersicum種トマト植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培S.licopersicum種トマト植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培S.licopersicum種植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とする請求項1に記載のAFT遺伝子。
【請求項3】
S.chilense遺伝子型に由来し、栽培S.licopersicum種トマト植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培S.licopersicum種トマト植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培S.licopersicum種植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とする請求項1または請求項2に記載のAFT遺伝子。
【請求項4】
前記フラボノイド類の少なくとも一部がアントシアニン類および/またはフラボノール類であることを特徴とする請求項1に記載のAFT遺伝子。
【請求項5】
前記AFT遺伝子がS.peruvianumに由来することを特徴とする請求項1に記載のAFT遺伝子。
【請求項6】
前記AFT遺伝子が、S.habrochaites、S.cheesmaniae、S.licopersicoides、S.peruvianum、S.pennelli v.puberelumよりなる群から選択されたことを特徴とする請求項1に記載のAFT遺伝子。
【請求項7】
AFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体であって、前記AFT−S.chilense遺伝子型を遺伝子移入していない従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のフラボノイド類によって特徴づけられる遺伝子移入由来トマト植物体。
【請求項8】
前記AFT−S.chilense遺伝子型を遺伝子移入していない従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のアントシアニン類および/またはフラボノ−ル類によって特徴づけられる請求項7に記載の遺伝子移入由来トマト植物体。
【請求項9】
前記AFT−S.chilense遺伝子型がS.peruvianumから遺伝子移入されたことを特徴とする請求項7に記載の遺伝子移入由来トマト植物体。
【請求項10】
前記AFT−S.chilense遺伝子型が、S.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群から遺伝子移入されたことを特徴とする請求項7に記載の遺伝子移入由来トマト植物体。
【請求項11】
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のフラボノイド類を有することで特徴づけられる純親系統を得るために、前記F2植物体を自家交配させてF3群および更なる個体群(F4、F5等)を作成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
方法によって遺伝子移入的に得たことを特徴とする請求項7に記載の遺伝子移入由来トマト植物体。
【請求項12】
前記フラボノイド類が、アントシアニン類およびフラボノール類であるデルフィニジン、ペツニジン、マルビジン、ケルセチン、ケンフェロール、ナリンゲニンを含むことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項13】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、hp−1、hp−1w、hp−2、hp−2j、hp−2dgとして定義される1以上の同型対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項14】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子の1以上の同型接合対立遺伝子(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項15】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子に欠損のあるhp−1、hp−1w、hp−2、hp−2j、hp−2dg変異植物体と等表現型の光形態形成遺伝子の1以上の同型接合対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配される方法によって遺伝子移入的に得たことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項16】
DNAマーカーを用いてS.chilense由来のAFT遺伝子座において同型接合であるF2植物体を選択する付加的手段からなることを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項17】
AFT遺伝子型がS.peruvianumに由来することを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項18】
AFT遺伝子型がS.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群から選択されたことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項19】
前記DNAマーカーがS.peruvianumに由来することを特徴とする請求項16に記載の遺伝子移入由来トマト植物体。
【請求項20】
前記DNAマーカーが、S.habrochaites、S.cheesmaniae、S.licopersiciodes、S.peruvianum、S.pennelli v.puberelumよりなる群に由来することを特徴とする請求項16に記載の遺伝子移入由来トマト植物体。
【請求項21】
フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル含有フラボノイドおよび/またはメトキシ置換、C−メチルフラボノイド、メチレンジオキシ・フラボノイド・カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンよりなる群のうちのいずれかの成分から前記フラボノイドを選択したことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項22】
デルフィジン、ペツニジン、マルビジンよりなる群から前記アントシアニンを選択したことを特徴とする請求項8に記載の遺伝子移入由来トマト植物体。
【請求項23】
ケルセチン、ケンフェロールよりなる群から前記フラボノイドを選択したことを特徴とする請求項1に記載の遺伝子移入由来トマト植物体。
【請求項24】
4,2,4,6−テトラヒドロキシカルコン、ナリンゲニン、ケンフェロール、ジヒドロキシケンフェロール、ミリセチン、ケルセチン、ジヒドロケルセチン、ジヒドロミリセチン、ロイコペラルゴニジン、ロイコシアニジン、ロイコデルフィニジン、ペラルゴニジン−3−グルコシド、シアニジン−3−グルコシド、デルフィニジン−3−グルコシドよりなる群から前記フラボノイドを選択したことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項25】
(i)2−フェニルクロモン(2−フェニル−1,4−ベンゾピロン)構造由来の二次植物代謝物、(ii)前記代謝物が3−フェニルクロモン(3−フェニル−1,4−ベンゾピロン)構造由来であるイソフラボノイド類、(iii)前記代謝物が4−フェニルクマリン(4−フェニル−1,2−ベンゾピロン)構造由来であるネオフラボノイド類よりなる群から前記フラボノイドを選択したことを特徴とする請求項11に記載の遺伝子移入由来トマト植物体。
【請求項26】
図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、従来の栽培S.licopersicum種トマト植物体と比較してトマト植物体中に高いフラボノイド濃度をもたらすことを特徴とするタンパク質をコードするDNA配列。
【請求項27】
図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、従来の栽培S.licopersicum種トマト植物体と比較してトマト植物体中に高いフラボノイド濃度をもたらすことを特徴とする請求項26に記載のDNA配列。
【請求項28】
前記DNA配列は、植物体、植物部位または種子内において、フラボノイド経路の代謝物であるアントシアニンまたはフラボノール類の蓄積又は発現をもたらすことを特徴とする請求項26または請求項27に記載のDNA配列。
【請求項29】
前記DNA配列は、S.licopersicumのトマト植物体、トマト植物部位または種子内において、フラボノイド経路の代謝物であるアントシアニンまたはフラボノール類の蓄積又は発現をもたらすことを特徴とする請求項26または請求項27に記載のDNA配列。
【請求項30】
前記DNA配列は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、栽培トマト系統におけるAFT遺伝子型の遺伝子移入のための遺伝資源、種子、苗木、発芽、植物体、植物部位のスクリーニングに有用であることを特徴とする請求項26乃至請求項29のいずれかに記載のDNA配列。
【請求項31】
トランスジェニック植物であって、植物体、植物部位または種子内にフラボノイド経路の代謝物であるアントシアニンまたはフラボノイド類を発現し、前記植物体は、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有するDNAからなり、前記DNAは、1以上の複数の形質転換、および/または植物の形質転換および/または発現に有益な発現ベクターに組み込まれていることを特徴とする遺伝子移入植物体。
【請求項32】
タンパク質をコードするAFT遺伝子を得る方法であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、栽培植物体または選良種に遺伝子操作的に遺伝子移入され、前記栽培植物体に前記AFT遺伝子を遺伝子移入していない従来の栽培植物体と比較して高い濃度のフラボノイド類をもたらすことを特徴とするタンパク質をコードするAFT遺伝子を得る方法。
【請求項33】
従来の栽培S.licopersicum種トマト植物体と比較して高濃度のアントシアニン類および/またはフラボノイド類を含むことを特徴とするAFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体を作り出す方法であって、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のフラボノイド類を有することで特徴づけられる純親系統を得るために、前記F2植物体を自家交配させてF3群を生成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
ことからなる方法。
【請求項34】
従来の栽培S.licopersicum種トマト植物体と比較して高濃度のアントシアニン類および/またはフラボノイド類を含むことを特徴とするAFT−S.chilense遺伝子型の遺伝子移入由来トマト植物体を作り出す方法であって、
(i)hp−1/hp−1系統トマトであるS.licopersicum種トマトと、S.chilense由来のAFT遺伝子を含むトマト間で交配し、
(ii)前記交配から得られたF1植物体を自家交配させ、
(iii)hp−1変異とAFT対立遺伝子の双方が分離しているF2群を作成し、
(iv)hp−1変異とS.chilense由来のAFT遺伝子座が同型接合であるF2植物体を選択し、
(v)従来の栽培S.licopersicum種トマト植物体および/または初期親系統と比較して、相加的な手段以上の高濃度のアントシアニン類とフラボノール類であるデルフィニジン、ペツニジン、マルビジン、ケルセチン、ケンフェロールを有することで特徴づけられた純親系統を得るために、前記F2植物体を自家交配させてF3群および更なる個体群(F4、F5等)を作成し、
(vi)前記純親系統を他の同系統または異種の親系統と交配して用いて、商用のF1ハイブリッドを作り出す、
ことからなる方法。
【請求項35】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、hp−1、hp−1w、hp−2、hp−2j、hp−2dgとして定義される1以上の同型対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする請求項34に記載の方法。
【請求項36】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子における1以上の同型対立遺伝子(hp−1、hp−1w、hp−2、hp−2j、hp−2dg)によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする請求項34に記載の方法。
【請求項37】
少なくとも1つの親系統が高色素系統であるS.licopersicumであって、高色素対立遺伝子が、紫外線損傷DNA結合タンパク質1(DDB1)または非黄化変異体1(DET1)遺伝子に欠損のあるhp−1、hp−1w、hp−2、hp−2j、hp−2dg変異植物体と等表現型の光形態形成遺伝子の1以上の同型接合対立遺伝子によって特徴づけられる群から選択され、S.chilense由来のAFT遺伝子を含む植物体と異種交配することによって遺伝子移入植物体を作り出すことを特徴とする請求項34に記載の方法。
【請求項38】
HP−1とAFT遺伝子座が同型接合であるF2植物体を付加的にDNAマーカーを用いて選択することを特徴とする請求項34に記載の方法。
【請求項39】
デルフィジン、ペツニジン、マルビジンよりなる群から前記アントシアニンを選択することを特徴とする請求項34に記載の方法。
【請求項40】
ケルセチン、ケンフェロール、ナリンゲニンよりなる群から前記フラボノイドを選択することを特徴とする請求項34に記載の方法。
【請求項41】
4,2,4,6−テトラヒドロキシカルコン、ナリンゲニン、ケンフェロール、ジヒドロキシケンフェロール、ミリセチン、ケルセチン、ジヒドロケルセチン、ジヒドロミリセチン、ロイコペラルゴニジン、ロイコシアニジン、ロイコデルフィニジン、ペラルゴニジン−3−グルコシド、シアニジン−3−グルコシド、デルフィニジン−3−グルコシドよりなる群から前記フラボノイドを選択したことを特徴とする請求項34に記載の従来の栽培S.licopersicum種トマト植物体と比較して高濃度のフラボノイド類を含むトマト植物体を作り出す方法。
【請求項42】
(i)2−フェニルクロモン(2−フェニル−1,4−ベンゾピロン)構造由来の二次植物代謝物、(ii)前記代謝物が3−フェニルクロモン(3−フェニル−1,4−ベンゾピロン)構造由来であるイソフラボノイド類、(iii)前記代謝物が4−フェニルクマリン(4−フェニル−1,2−ベンゾピロン)構造由来であるネオフラボノイド類よりなる群から前記フラボノイドを選択したことを特徴とする請求項34に記載のトマト植物体を作り出す方法。
【請求項43】
フラボノイドアグリコン、フラボノイド−O−グリコシド、フラボノイド−C−グリコシド、ヒドロキシル含有フラボノイドおよび/またはメトキシ置換、C−メチルフラボノイド、メチレンジオキシ・フラボノイド・カルコン、オーロン、ジヒドロカルコン、フラバノン、ジヒドロフラバノール、アンソクロ、プロアントシアニジン、濃縮プロアントシアニジン、ロイコアントシアニジン、フラバノール−3,4−ols、フラバノール−3−ols、グリコシルフラボノイド、ビフラボノイド、トリフラボノイド、イソフラボノイド、イソフラボン、イソフラバノン、ロテノノイド、プテロカルパン、イソフラバン、キノン誘導体、3−アリール−4−ヒドロキシクマリン、3−アリールクマリン、イソフラボノイ−3−エン、クメスタン、α−メチルデオキシベンゾイン、2−アリールベンゾフラン、イソフラバノール、クマロノクロモンよりなる群のいずれかの成分から前記フラボノイドを選択したことを特徴とする請求項34に記載のトマト植物体を作り出す方法。
【請求項44】
タンパク質をコードするDNAを作り出す方法であって、図9に示すアミノ酸配列(LA1996配列)に対して少なくとも80%の同一性を有し、コードされたアミノ酸配列を識別して選択的に検証する処理を含み、前記配列がAFT表現型および増強したフラボノイド濃度に関連するS.chilense中に天然に存在するタンパク質の配列であることを特徴とする方法。
【請求項45】
核酸を有用に得る方法であって、図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有し、AFT表現型に少なくとも部分的に関連するS.chilense中に天然に存在するタンパク質をコードする前記核酸配列を識別して選択的に検証することからなる方法。
【請求項46】
従来の栽培S.licopersicum種トマト植物体と比較して高い濃度のフラボノイド類を含むトマト植物体を作り出し、栽培トマト系統におけるAFT−S.chilense遺伝子型の遺伝子移入のために遺伝資源、種子、苗木、植物、発芽、植物体をスクリーニングして容易にした方法であって、
(i)図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有する核酸を作り出し、
(ii)表1に定義されたPCRプライマーを調製し、
(iii)工程(i)のDNAを増幅し、
(iv)それを用いて標的組織を調べることからなる請求項34に記載の方法。
【請求項47】
植物体、植物部位、または種子内にフラボノイド経路の代謝物であるアントシアニンまたはフラボノイド類を蓄積または発現させるための遺伝子組み換え方法であって、
(i)図2下列に示す残基1から残基1008の核酸配列(LA1996配列)に対して少なくとも80%の相同性を有するDNAを作り出し、
(ii)前記DNAを1以上の複数の形質転換、および/または植物の形質転換および/または発現に有益な発現ベクターに組み込むことからなる遺伝子組み換え方法。
【請求項48】
トマト植物体、トマト部位または種子に有用なことを特徴とする請求項47に記載の遺伝子組み換え方法。
【図1】

【図2】

【図3】

【図4】
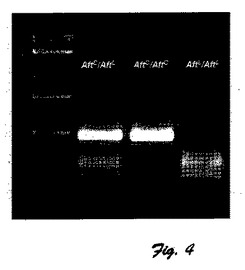
【図5】
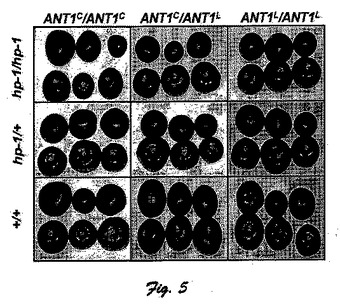
【図6】
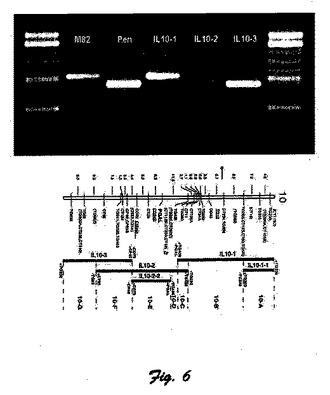
【図7】
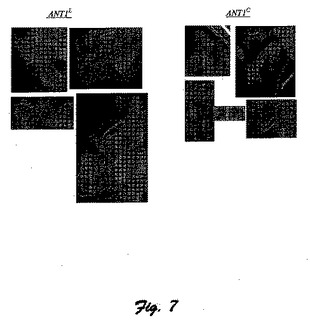
【図8】
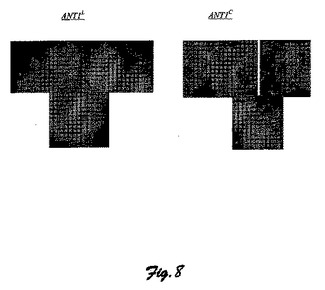
【図9】

【図10】

【図11】
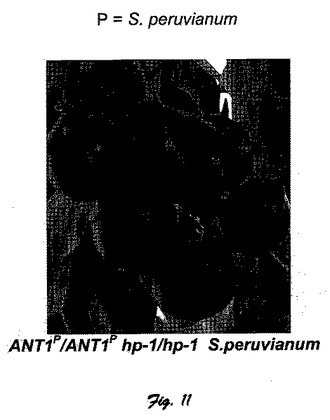
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2010−517526(P2010−517526A)
【公表日】平成22年5月27日(2010.5.27)
【国際特許分類】
【出願番号】特願2009−547812(P2009−547812)
【出願日】平成20年2月6日(2008.2.6)
【国際出願番号】PCT/IL2008/000159
【国際公開番号】WO2008/096354
【国際公開日】平成20年8月14日(2008.8.14)
【出願人】(509219903)
【Fターム(参考)】
【公表日】平成22年5月27日(2010.5.27)
【国際特許分類】
【出願日】平成20年2月6日(2008.2.6)
【国際出願番号】PCT/IL2008/000159
【国際公開番号】WO2008/096354
【国際公開日】平成20年8月14日(2008.8.14)
【出願人】(509219903)
【Fターム(参考)】
[ Back to top ]