
アンドロゲン依存性1−f−アロマターゼ・レポーター遺伝子
本発明は、アンドロゲン依存性1−f−アロマターゼ・レポーター遺伝子、及び1−f−アロマターゼ・レポーター遺伝子の製造方法、及びアンドロゲン受容体のリガンドを同定する方法におけるその使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アンドロゲン依存性1−f−アロマターゼ・レポーター遺伝子、及び1−f−アロマターゼ・レポーター遺伝子の製造方法、及びアンドロゲン受容体のリガンドを同定する方法におけるその使用に関する。
【背景技術】
【0002】
中枢神経系内のアンドロゲンによって誘発されるアロマターゼ発現、ひいてはエストロゲンの形成は、性同一性の刷り込みを含む性特異的な行動学的反応の確立、及び特に雄性における性欲の調節において極めて重要である。加えて、脳内のアンドロゲンによって誘発されるアロマターゼ発現は、学習及び記憶のようなプロセスにおいて重要な役割を果たすと考えられている(Wickelgren, I.; Science, Vol. 276: 675-678, 1997)。神経再生プロセスにおけるエストロゲンから生じる好ましい効果が、記載されている(Abe-Dohmae他, J. Neurochem., Vol. 67: 2087-2095, 1996; I. Azcoitia他, 2001, J. Neurobiol. 47: 318-329)。エストロゲン受容体及び神経エストロゲン産生の誘発又はアップレギュレーションは、種々のニューロン損傷における修復メカニズムの一部であると見られる(Dubal他, 1999, J Neurosci 10:6385-6393; Garcia-Segura他, 1999, Neuroscience 89: 567-578; Peterson他, 2001, J Neuroendocrinol 13: 317-323; Carswell他, 2005, J. Steroid Biochem. Mol. Biol. 96:89-91)。従って、エストロゲンは、神経変性疾患の治療又は予防のための重要な薬理学的選択肢である。しかしこれらの使用は、両方の性において、他の器官に副作用が生じるおそれがあるため、制限されている。
【0003】
目下のところ、アンドロゲンによって誘発されたアロマターゼ発現、ひいては脳特異的なエストロゲン合成を決定するために、そしてアロマターゼ活性に影響を与える物質を同定して特徴付けするために利用可能な試験管内(in vitro)試験システムはない。
【0004】
脳内終点エストロゲン濃度、脳内アロマターゼ発現、脳試料中のアロマターゼ活性、又は両方の性における行動学的試験を用いた試験管内検査は、従来技術から周知である。
【0005】
既に言及されているとおり、アンドロゲンは、アンドロゲン受容体(AR)との結合によって媒介されて、アロマターゼ発現を誘発する。ARは、活性化又は不活化することができる。アンドロゲン欠乏の事例における活性化の例は、例えばサルコペニア、性腺機能低下症、加齢関連性腺機能低下症、及び性欲障害の形態、及び男性の勃起障害において生じる。
【0006】
ARの薬理学的不活化は、前立腺の良性及び悪性疾患において、又はアンドロゲン過剰に関連する疾患、例えば座瘡のある種の形態、脱毛、又は女性における多嚢胞性卵巣及び多毛症において重要である。
【0007】
しかしながら、一般には、アンドロゲン受容体の完全な活性化又は不活化は望ましくなく、むしろ組織選択作用が望ましい。例えば、男性性腺機能低下症の事例では、性欲、気質、及び骨又は筋肉同化作用のAR媒介活性化が望ましいが、しかしこれは活性化というよりも、むしろ前立腺又は皮膚に対するニュートラルな作用又は軽減作用である。ARの不活化が望ましい事例、例えば悪性前立腺癌又は脱毛症の場合、ARの不活化は、性欲、骨代謝などに関して不都合である( S. S. Wolf and M. Obendorf, 2004, “Selective androgen receptor modulators (SARMs)”;E. Nieschlag & H. M. Behre: “Testosterone” 3rd. Edition; Cambridge University Press, ISBN 0521833809; 623-640)。
【0008】
これまで、細胞培養又はin vitroに基づくアッセイ・システムは、これらの組織選択的なアンドロゲン又は抗アンドロゲンを見いだして特徴付けするには不十分であった。このことは、アロマターゼ発現の決定にも当てはまる。
【発明の概要】
【発明が解決しようとする課題】
【0009】
従って、目的は、
−中枢神経系CNS内のアロマテーゼ発現を選択的に増大させ、
且つ/又は
−ARの活性に組織選択的な影響を与える
ことを特徴とする薬理学的活性の物質を同定して特徴付けすることができる方法を提供することである。
【課題を解決するための手段】
【0010】
この目的は、レポーター遺伝子アッセイにおいてアロマターゼ発現を刺激することを介して、脳内のエストロゲン合成を選択的に調節してアンドロゲン受容体に組織選択的な影響を与える物質を同定できる方法によって達成される。1f−アロマターゼ・プロモーターのアンドロゲン依存性調節が存在することが判った。
【0011】
ヒトのアロマターゼは、染色体15上に位置する遺伝子においてコードされている(Chen, S.他, (1988): Molec. Biol., Vol. 7: 27-38)。
【0012】
ヒトP450arom遺伝子(CYP19)は、30kbのコード領域と約93kbの調節領域とを含んでいる(Bulun, S.E.他、Semin Reprod Med. Vol. 22: 5-9, 2004)。転写産物は、10個のエキソンから成っており、エキソンIは非コードである。コード・エキソンII〜Xは、全ての組織内で同一に発現される。今日まで、コード領域内のスプライシング変異体はヒトにおいてはまだ見いだされていない。
【0013】
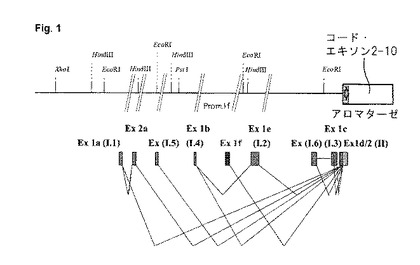
エキソンIに関しては、種々の組織に由来するmRNAの配列決定中にいくつかの変異体が発見された(検討の際には、Harada, N.; Utsumi, T.及びTakagi, Y. Proc. Natl. Acad. Sci. USA, 90: 11312-11316, 1993、及びSimpson, E. R.; Endocrine Reviews, 15: 341-355, 1994参照)。これらの変異体は組織特異的に発現され、そして全ては、エキソンIIの出発点に配置されたのと同じスプライシング部位に連結することができる(図1)。mRNAにおけるエキソン−I配列の組織特異的発生は、選択的プロモーターの使用によって説明することができる。選択的プロモーターは、それぞれのエキソン−I変異体の前の遺伝子内に配置されている。これらの種々のプロモーターの活性化は、まだ完全には解明されていない制御を被る。表1は、Harada他による種々の組織内のヒト・アロマターゼの或る特定の5’−非翻訳エキソンの現在知られている用途の例を挙げる(1993, Proc. Natl. Acad. Sci. USA, Vol. 90: 11312-11316)。
【0014】
特にHarada(Harada, N.他, (1993) Proc. Natl. Acad. Sci. USA, Vol. 90: 11312-11316)及びSimpson(Simpson, E. R.; Endocrine Reviews, Vol. 15: 341-355, 1994)を中心とした2つのチームの今日までの研究が、アロマターゼ−mRNAを配列決定することによって示したところでは、ほとんどの組織内でいくつかのプロモーターを使用することができ、また逆に、異なる組織では1つのプロモーターを使用することもできる(表1)。
【0015】
このことの例外は、プロモーター1fであるように見える。プロモーター1fは知られている限り、ヒトにおいて、専ら神経組織内で使用される。
【0016】
脳内では、アンドロゲンは特に、アロマターゼ発現に対する効果を有している(Abdelgadir, S. E.他, Endocrinol., Vol. 135: 395-401, 1994;Rosselli, C. E.他, (1996) Endocrine, Vol. 5: 59-65;Rosselli, C. E.他, (1997, Brain Research, Vo. 44: 351-357)。この発見にもかかわらず、アンドロゲン受容体(AR)が脳内のアロマターゼ発現を調節する、その正確な分子メカニズムはまだ知られていない。1f−アロマターゼ・プロモーター内の推定上のAR担当要素が文献において述べられているのは事実だが、神経アロマターゼ発現のAR媒介調節における1f−プロモーターの機能関与はまだ示されていない(Honda, S.他、Biochem Biophys Res Commun 198 (1994), 1153-1160)。それどころか、1f−アロマターゼ・プロモーターについて最初に記述したチーム(S. Honda他, Biochem Biophys Res Commun 198 (1994), 1153-1160)は、後の研究で他のレギュレーターを指摘した。これらのレギュレーターの更なる特徴付けはなされていない(Honda, S. I.他, J Steroid Biochem Mol Biol. 79:255-60, 2001; Honda, S.他, Brain Res Mol Brain Res. 66: 122-32, 1999)。
【0017】
本発明の基礎は、1f−アロマターゼ・エキソンの制御が、専ら1f−アロマターゼ・プロモーターの活性によって調節され、そして細胞の脈絡には依存しないという観察である。試験管内試験システムによって、神経組織、神経細胞、動物全体又は動物からの抽出物を使用する必要なしに、神経アロマターゼ発現の調節に対する薬理学的活性物質の影響を示すことが今や可能になった。
【0018】
このような試験システムを用いることによって、中枢神経系内のアロマテーゼ発現を選択的に増大させるか、又はARの活性に組織選択的な影響を与えるのに適した薬理学的活性の物質を発見して特徴付けすることが可能になり、また単純化される。
【0019】
従って本発明の目的は、中枢神経系内のアロマテーゼ発現を選択的に増大させるか、又はARの活性に組織選択的な影響を与えるのに適した物質を同定し特徴付けするための試験管内試験システムであって、前記試験システムが、レポーター遺伝子とアロマターゼ・プロモーターとの融合体を含有する、試験管内試験システムである。
【0020】
アロマターゼ、すなわちエストロゲン合成における鍵酵素の組織特異的な調節は、最終的に、種々の組織内における標的エストロゲン提供に関与する。
【0021】
ホルモン制御されたアロマターゼ発現を特徴付けするためには、確立されたヒト細胞系がこれらの試験管内試験に適しており、組織特異的な生体内(in vivo)発現との相関が、種々のプロモーターを使用することによって保証される。レポーター遺伝子アッセイの開発は、アロマターゼ・プロモーターの場合、個々のプロモーターの活性化による発現に与える特定物質の組織特異的影響を試験するシンプルで洗練された可能性をもたらす。所期の脳特異的発現の場合、これは、本発明による1f−アロマターゼ・プロモーターである。
【0022】
ARによって神経組織内で調節される1f−アロマターゼ・プロモーター(1f−アロ−プロモーター)の活性と、ARによって別の組織内で調節されるプロモーター又は全身的にARによって調節されるプロモーターの活性とを比較することによって、組織選択性に関してARに対して活性の物質を試験管内における特徴付けすることが可能になる。1f−アロマターゼ・プロモーターのプロモーター活性はこの場合、神経組織内のAR媒介型活性のための代理マーカーとして、又はより全身的に、性腺組織以外(例えば男性においては、精巣及び前立腺以外;女性においては卵巣以外)の部位のAR媒介型活性のための代理マーカーとして役立つ。
【0023】
ヒト・アロマターゼの1fプロモーターを用いたレポーター遺伝子アッセイによって、神経アロマターゼ発現の調節を決定することができる。既知の配列(配列番号1)を有する神経1f−アロマターゼ・プロモーターは、レポーター遺伝子の前に置かれている。プロモーター・レポーター遺伝子融合体は、一時的又は安定的に標的系内にトランスフェクトされる。ルシフェラーゼの発現が1f−プロモーターの関数として行われ、そしてアロマターゼ・プロモーターの活性は、レポーター遺伝子によってコードされた酵素の量から決定することができる。本例において、ホタルルシフェラーゼ遺伝子又はウミシイタケ(Renilla)ルシフェラーゼ遺伝子が、レポーター遺伝子の例として選択された。
【図面の簡単な説明】
【0024】
【図1】選択的エキソン−I変異形の周知のスプライシング変異形を有するヒト・アロマターゼ遺伝子のゲノム・プロモーター領域(単純化された表示)。より判りやすくするために、5’領域の部分が切り取られ、省略したものとして示されている(Harada, N.他, Proc. Natl. Acad. Sci. USA, Vol. 90: 11312-11316, 1993、及びSimpson, E. R.; Endocrine Reviews, Vol. 15: 341-355, 1994から修正)
【0025】
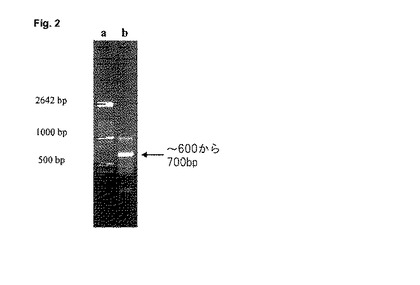
【図2】SHSY5Y細胞のゲノムDNAから増幅された1f−アロ−プロモーターを有するPCR生成物。a:長さ標準、b:1f−アロマターゼ・プロモーターのPCR増幅。
【0026】
【図3】pTAdvベクターのマップ。両端に3’−T突出部を備えたこの線状ベクターを、PCRフラグメントとのライゲートのために使用した。
【0027】
【図4】pTAdvベクター中に増幅1f−アロマターゼ・プロモーターDNAを有する1f−pTadv’’ベクターのベクターマップ。HindIII及びXhoIは、更なるクローニングのために使用された切断部位を意味する。
【0028】
【図5】pGl3−コントロール・ベクターのベクターマップ。HindIII及びXhoIは、更なるクローニングのために使用された切断部位を意味する。
【0029】
【図6】1f−pGl3ベクターのベクターマップ。HindIII及びXhoIは、ベクターの生成のために使用された切断部位を意味する。
【0030】
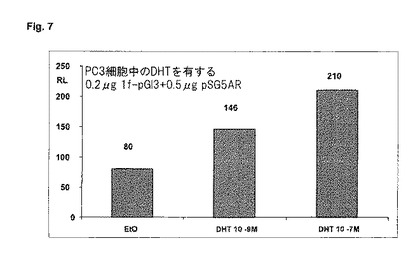
【図7】PC3細胞中のジヒドロテストステロン(DHT)の濃度に対する、1f−プロモーターの活性の依存。0.2μgの1f−pGl3プラスミド及び0.5μgのpSG5−ARプラスミドをトランスフェクトする。DHTと一緒に20時間インキュベートする。二重測定の平均値を、総タンパク質1μg当たりの相対光単位(RLU)で示す。
【0031】
【図8】SH−SY5Y細胞中のジヒドロテストステロン(DHT)の濃度に対する、1f−プロモーターの活性の依存。0.2μgの1f−pGl3プラスミド及び0.5μgのpSG5−ARプラスミドをトランスフェクトする。DHTと一緒に20時間インキュベートする。二重測定の平均値を、総タンパク質1μg当たりの相対光単位(RLU)で示す。
【0032】
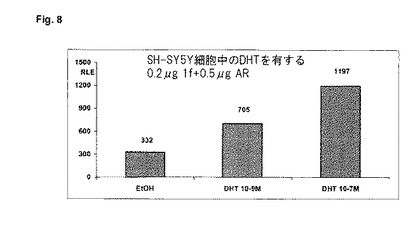
【図9】これは、1f−プロモーターの活性が、PC3細胞中のエストラジオール(E2)の濃度とは無関係であることを示す。0.2μgの1f−pGl3プラスミド及び0.5μgのpSG5−ERプラスミドをトランスフェクトする。E2と一緒に20時間インキュベートする。二重測定の平均値を、総タンパク質1μg当たりの相対光単位(RLU)で示す。
【0033】
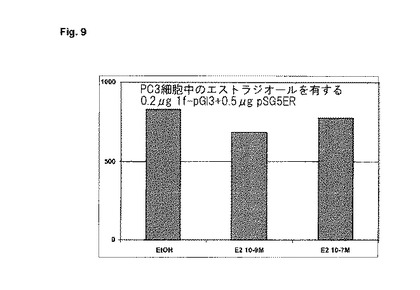
【図10】神経SH−SY5Y細胞内の1f−プロモーターに対する種々のアンドロゲンの影響。0.4μgの1f−pGl3プラスミド及び0.5μgのpSG5−ARプラスミドをトランスフェクトする。a)T(プロピオン酸テストステロン)、b)MENT(7α−メチル−19−ノルテストステロン)、及びc)DHT(ジヒドロテストステロン)と一緒に20時間にわたってインキュベートする。二重測定の平均値を、総タンパク質1μg当たりの相対光単位(RLU)で示す。
【0034】
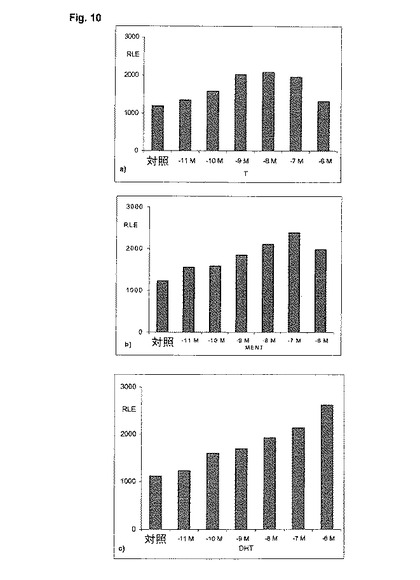
【図11】神経SH−SY5Y細胞内の1f−プロモーター及びMMTV−プロモーターに対する種々のアンドロゲンの影響。1.5μgの1f−pGl3及びMMTV−lucプロモーター及び0.75μgのpSG5−ARプラスミドをトランスフェクトする。オキシメトロン又はDHT(ジヒドロテストステロン)と一緒に20時間にわたってインキュベートする。相対光単位(RLU)の標準化平均値は、標準偏差とともに四重測定からのパーセンテージとして示される。
【0035】
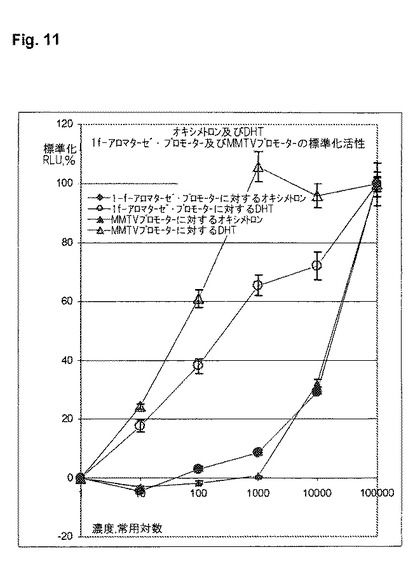
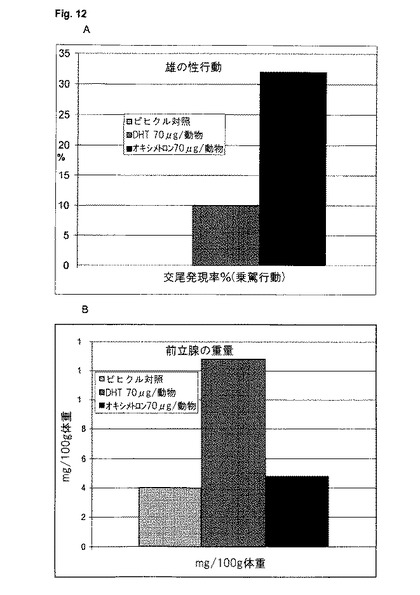
【図12】アンドロゲン作用の神経終点を表す、雄の性行動に対する種々のアンドロゲンのin vivoの影響(A)、及びアンドロゲン作用の性腺終点を表す、前立腺(B)の重量に対する種々のアンドロゲンのin vivoの影響(B)。性的経験のあるインタクトの雄ラットを選び去勢する。7週間後、性的活動がなくなった後、動物たちを1群当たり9〜10匹の3つの群に分け、1050μgのオキシメトロン、DHT又はビヒクルで1週間にわたって処置する。8日目〜120日目まで、動物は毎日、70μgのオキシメトロン(黒)、DHT(暗灰色)又はビヒクル(明灰色)を受ける。各動物の性行動(乗駕行動)を7回試験した。研究開始から4か月後に、器官を取り出し、前立腺の重量及び体重を測定した。
【0036】
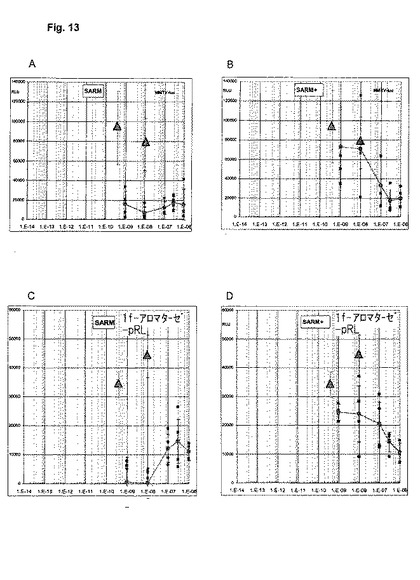
【図13】pC3AR+細胞中の非特異的MMTVプロモーター(A+B)及び神経1f−アロマターゼ・プロモーター(C+D)に対して同時に行われる選択的ARモジュレーター(SARM)のアゴニズム(A+C)及びアンタゴニズム(B+D)の試験。1.5μgの1f−pGl3及びMMTV−lucプロモーター及び0.75μgのpSG5−ARプラスミドをトランスフェクトする。SARMと一緒に20時間にわたってインキュベートする。アンタゴニズム(B+D)の試験では、4 x 10 e-10Mのテストステロンの存在において、SARMを上記濃度でインキュベートする。個々の値を四角で示し、また四重測定からの相対光単位の平均値を、標準偏差とともに円で示す。SARMを添加しないテストステロンの値を三角で示す。非特異的MMTV−プロモーター(A+B)に対して、ここで試験されたSARMは、AR媒介トランス活性化の純粋なアンタゴニストとして挙動するのに対して、神経1f−アロマターゼ・プロモーター(C+D)に対しては部分アゴニスト/部分アンタゴニスト作用が表面化する。
【0037】
本発明は、配列番号1に従う1f−アロマターゼ・プロモーター及びレポーター遺伝子から成るプロモーター・レポーター遺伝子融合体に関し、そして該レポーター遺伝子は、ホタルルシフェラーゼ遺伝子又はウミシイタケ(Renilla)ルシフェラーゼ遺伝子を含む群から選択することができるが、この選択は制限的なものであってはならず、そして融合生成物は、細胞系内に安定的に又は一時的に発現させることができる。
【0038】
さらに、ARが細胞中に発現された場合には、アロマターゼ・プロモーターの活性に、AR活性試験物質の影響がARを介して与えられる。この場合、試験物質の作用は、ルシフェラーゼ酵素活性の変化に基づいて決定することができる。従って一次神経組織又は一次神経細胞を用いて、或いは動物全体又は動物からの抽出物を用いて作業する必要なしに、薬理学的活性物質を模索又は特徴付けすることができる。
【0039】
本発明は、試験管内試験システムにおけるプロモーター・レポーター遺伝子融合体の使用であって、プロモーター・レポーター遺伝子融合体は、配列番号1に従う1f−アロマターゼ・プロモーター及びレポーター遺伝子から成っており、試験物質の影響は、細胞培養におけるハイスループット・レポーター遺伝子アッセイで割り出され、神経組織内のアロマターゼの調節に関する情報を得ることができる、プロモーター・レポーター遺伝子融合体の使用に関する。この方法は、アンドロゲン受容体に結合することにより、アロマターゼ発現に影響を与える物質を同定するために用いられる。
【0040】
本発明はまた、プロモーター・レポーター遺伝子融合体の使用であって、アンドロゲン受容体に結合することによって脳内のアロマターゼ調節に影響を与える物質を同定するために、配列番号1に規定された配列に対して>90%、好ましくは>95%の相同性を有する配列が細胞培養系内で使用される、プロモーター・レポーター遺伝子融合体の使用に関する。
【0041】
さらに、本発明による試験モデルを用いて、ARアゴニスト、アンタゴニスト、及びSARMを含むアンドロゲン性物質の特性を特徴付けすることもできる。ここでは、1f−アロマターゼ・プロモーターの活性が、ARを介して別の組織内で調節される、又は全身的にARによって調節されるプロモーターの活性と比較される。1f−アロマターゼ・プロモーターのプロモーター活性はこの場合、神経組織内のAR媒介型活性のための代理マーカーとして、又はより全身的に、性腺組織以外(例えば男性においては、精巣及び前立腺以外;女性においては卵巣以外)の部位のAR媒介型活性のための代理マーカーとして役立つ。
【0042】
このために、神経1f−アロマターゼ・プロモーターを用いた本発明による試験システムは、ARによって調節される別のプロモーターと組み合わされる。後者のプロモーターは、全身アンドロゲン作用(例えばMMTV−プロモーター)、又は別のアンドロゲン性標的器官内のアンドロゲン作用(例えばアンドロゲン受容体によって前立腺内で活性化されるプロモーター)を反映する。2種の異なるレポーター遺伝子、例えば1f−アロマターゼ・プロモーターに対するウミシイタケ(Renilla)ルシフェラーゼ、及び第2のプロモーターに対するホタルルシフェラーゼを同時に使用することにより、両方のプロモーター活性を同時に測定することが可能になる。これは、ARを介した薬理学的作用による試験物質の組織選択的活性に関する情報を単純かつ洗練された形で提供する。或いは、両プロモーターは、同じレポーター遺伝子の前に連続して挿入することもできる。これもやはり、原則としては動物実験である従来技術と比較してかなり単純なものになる。神経プロモーター活性は、性腺組織以外の部位のプロモーターの代理活性として、より全身的に抽出することができる。
【0043】
本発明はまた、アンドロゲン受容体に結合する物質の組織選択作用を決定する方法であって、以下の段階:
a. アゴニスト活性及びアンタゴニスト活性の両方を同定するために、細胞培養系内のレポーター遺伝子アッセイに該プロモーター・レポーター遺伝子融合体を使用して、アロマターゼ調節のモジュレーションを決定する段階、
b. 別のアンドロゲン依存性プロモーターのモジュレーションを決定する段階、及び
c. 1f−アロマターゼ・プロモーターに対する選択的ARモジュレーションに関連して物質の完全な試験管内における特徴付けを達成するために、段階a及びbを比較するか、あるいは例えば前立腺特異的又は全身的に作用するAR調節型プロモーターと直接的に比較する段階
含んで成る、方法に関する。この方法は、ARに組織選択的な影響を与える物質を同定するために用いることができる。
【0044】
本発明はまた、レポーター遺伝子アッセイにおけるプロモーター・レポーター遺伝子融合体の使用であって、アンドロゲン受容体の活性をモジュレートし、そしてアンドロゲン受容体によって調節される遺伝子の調節の組織選択的モジュレーションを可能にする物質を同定するために、配列番号1に規定された配列に対して>90%、好ましくは>95%の相同性を有する配列が、細胞培養系内のレポーター遺伝子アッセイにおいて使用される、プロモーター・レポーター遺伝子融合体の使用に関する。
【0045】
「アンタゴニスト」は、それらの対応受容体に結合する分子であって、自然発生型リガンドによって、受容体とカップリングされた信号伝達経路が始動するのを阻害する分子として理解されるべきである。通常、アンタゴニストは、受容体との結合を巡って、受容体の自然発生リガンドと競合する。しかしながら、受容体とカップリングされた信号伝達経路が、自然発生型リガンドによって活性化されるのを阻害する分子による受容体の他の変更形も可能である(例えば受容体の非競合的な立体変更形)。
【0046】
受容体アンタゴニストは典型的には、それらの特定の受容体に選択的に結合し、他の受容体には結合しない。通常、受容体アンタゴニストは天然リガンドよりも高い結合親和性を示す。天然リガンドよりも高い、受容体に対する親和性を有するアンタゴニストが好ましいものの、親和性の低いアンタゴニストを使用することもできる。アンタゴニストは、それらの対応受容体に可逆的に結合することが好ましい。
【0047】
「アゴニスト」は、それらの対応受容体に結合し、通常、受容体との結合を巡って、受容体の自然発生リガンドと競合する分子であって、受容体にカップリングされた信号伝達経路の始動を刺激する分子として理解されるべきである。アゴニストは、天然リガンドの結合を支持することもできる。
【0048】
受容体アゴニストは典型的には、それらの特定の受容体に選択的に結合し、他の受容体には結合しない。受容体アゴニストは通常、天然リガンドよりも高い結合親和性を有している。天然リガンドよりも高い、受容体に対する親和性を有するアゴニストが好ましいものの、親和性の低いアゴニストを使用することもできる。アゴニストは、それらの対応受容体に可逆的に結合することが好ましい。
【0049】
アゴニストは、対応受容体の始動及び/又は生理学的作用によって媒介される信号伝達により試験される。
【0050】
「リガンド」は、受容体に結合する化合物又は低分子物質を意味する。これらの結合は通常可逆的である。対応受容体に対するリガンドの結合は、受容体にカップリングされた信号伝達経路を活性化又は不活化する。このようにして、リガンドはその細胞内作用をもたらす。
【0051】
リガンドは、受容体のアゴニスト及びアンタゴニストとして理解されるべきである。
【実施例】
【0052】
生物学的例
1. レポーター遺伝子プラスミド内のヒト1f−アロマターゼ・プロモーターのクローニング
レポーター遺伝子アッセイに際して、ヒト・アロマターゼの1f−プロモーターを、ヒトSH−SY5Y細胞のゲノムDNAから増幅する。QIAamp Blood Kitを、製造業者(Qiagen)によって指定されたプロトコルに従って使用して、ゲノムDNAを単離する。アロマターゼの1f−プロモーターは、刊行物(S. Honda他、Biochem Biophys Res Commun 198 (1994), pp. 1153-1160)、及び遺伝子バンク(CYP19A1)から周知である。特定のプライマー対(表2)を使用して、1f−プロモーターを含むヒト・アロマターゼ・プロモーター領域の部分をPCRによって増幅する。
【0053】
PCRによってDNAフラグメントを増幅するために、各事例において、1μlのゲノムDNA試料を使用する。これを2分間にわたって94℃で変性させ、そして上記DNAプライマーEbrain B4及びEbrain c670を用いて35サイクルで増幅し、ゲル電気泳動法によって分析し(図2)、そしてアガロース・ゲルからのDNA抽出によって副生成物から精製する。
【0054】
1f−プロモーター・フラグメントのPCR生成物は、3’−A突出部を有しており、線状pTAdvクローニング・ベクターとライゲートされる(図3)。pTAdvベクターは、Clontechから得られた。結果として生じる1f−pTAdv2ベクター(図4)の挿入体を配列決定し、そして増幅されたヒト・1f−アロマターゼ・プロモーターの配列を確認する(配列番号1)。これは、以前に発表された配列(S. Honda他, N Biochem Biophys Res Commun 198 (1994), 1153-1160)とは異なるが、しかしより最近の遺伝子バンク登録と符合する。
【0055】
使用されるレポーター遺伝子は例えば、PromegaのpGl3ベクターのルシフェラーゼ遺伝子である。1f−プロモーターを有するプロモーター・レポーター遺伝子構造を生成するために、HindIII及びXhoIを有するpGl3コントロール(図5)から、213bp長のコントロール・プロモーター(SV40)を切り取る。こうして線状化され短縮されたpGl3を、ゲル電気泳動法及びDNA抽出によって精製し、脱リン酸化し、次いで、1f−pTAdv2ベクター(図4)からHindIII及びXhoIを有するように切り取られ精製された1f−プロモーターのフラグメントをスプライシングし、レポーター遺伝子プラスミド1f−pGl3を生成する(図6)。
【0056】
2. 細胞培養内のレポーター遺伝子アッセイにおける1f−プロモーターに対するホルモン活性化合物の影響を決定するためのトランスフェクション試験
PC3(ヒト前立腺癌)細胞(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31870-025、Gibcoから)を含むRPMI 1640培地+200mMのグルタミン及び5mg/lのペニシリン/ストレプトマイシンを含む10%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、PC3細胞を試験培地で2回継代培養する。これはフェノールレッド(Gibco、注文番号:32404-014)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0057】
ルシフェラーゼ活性に基づいて、pGl3融合ベクター1f−pGl3内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、6ウェル・プレート内で、1ウェル当たり200000個の細胞に、0.2μgのこのプラスミドを、0.5μgのpSG5ARプラスミドとともに同時トランスフェクトする。pSG5ARベクターは、pSG5ベクターをベースとしている。このpSG5ベクターはヒト細胞において、根底を成す遺伝子、この事例ではARに対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. and Harris, B.A. 1983, Nucleic Acids Res. 11:7119-36)。レポーター遺伝子プラスミド及びpSG5ARプラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、アンドロゲン・ジヒドロテストステロン(DHT)を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0058】
1f−レポーター遺伝子生成物は、前立腺細胞系PC3中で行われたトランスフェクションにおいて、2.6倍だけDHTによってレポーター遺伝子発現が誘発されたことを示す(図7)。アンドロゲンによる調節は、ここでは試験管内で決定することができる。
【0059】
3. 1f−アロマターゼ・プロモーターのアンドロゲン依存性調節
前立腺PC3に加えて、1f−アロマターゼ・プロモーターに対するアンドロゲン調節型レポーター遺伝子発現を分析するために、神経細胞系を使用する。
【0060】
ヒト神経芽腫細胞系SH−SY5Y(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31885-023、Gibcoから)を含むDulbecco's MOD Eagle Medium+5mg/lのペニシリン/ストレプトマイシンを含む15%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、SH−SY5Y細胞を試験培地で2回継代培養する。これはフェノールレッド(注文番号:11880-028、Gibcoから)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0061】
ルシフェラーゼ活性に基づいて、pGl3融合ベクター1f−pGl3内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、6ウェル・プレート内で、1ウェル当たり300000個の細胞に、0.2μgのこのプラスミドを、0.5μgのpSG5ARプラスミドとともに同時トランスフェクトする。pSG5ARベクターは、pSG5ベクターをベースとしている。このpSG5ベクターはヒト細胞において、根底を成す遺伝子、この事例ではARに対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. & Harris, B.A. 1983; “Plasmids for the cloning and expression of full-length double-stranded cDNAs under control of the SV40 early or late gene promoter”, Nucleic Acids Res. 11:7119-36)。レポーター遺伝子プラスミド及びpSG5ARプラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、アンドロゲン・ジヒドロテストステロン(DHT)を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0062】
1f−レポーター遺伝子生成物は、神経芽腫細胞系SH−SY5Y中で行われたトランスフェクションにおいて、DHTによってレポーター遺伝子発現が濃度に依存して誘発されたことを示している(図8)。
【0063】
使用した両方の細胞系において同様の結果が測定されるように、これらの試験システムにおける1f−プロモーターの活性は、細胞の脈絡とは独立してアンドロゲンによって刺激される。
【0064】
4. 薬理学的活性物質による1f−アロマターゼ・プロモーターのホルモン調節を特徴付けするためのトランスフェクション試験
DHTと同様に、さらなる薬理学的活性物質として1f−ルシフェラーゼ・レポーター遺伝子構造(1f−pGl3)に基づいて、我々は、神経組織内のヒト・アロマターゼの発現に対するエストラジオールの影響を試験した(図9)。PC3(ヒト前立腺癌)細胞(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31870-025、Gibcoから)を含むRPMI 1640培地+200mMのグルタミン及び5mg/lのペニシリン/ストレプトマイシンを含む10%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、PC3細胞を試験培地で2回継代培養する。これはフェノールレッド(Gibco、注文番号:32404-014)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0065】
ルシフェラーゼ活性に基づいて、pGl3融合ベクター1f−pGl3内のクローン型1f−アロマターゼ・プロモーターに対するエストラジオール(E2)の影響を決定するために、6ウェル・プレート内で、1ウェル当たり200000個の細胞に、0.2μgのこのプラスミドを、0.5μgのpSG5−ERプラスミドとともに同時トランスフェクトする。pSG5−ERベクターは、pSG5ベクターをベースとしている。このpSG5ベクターはヒト細胞において、根底を成す遺伝子、この事例ではヒト・エストロゲン受容体アルファ(ERα)に対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. & Harris, B.A. 1983; “Plasmids for the cloning and expression of full-length double-stranded cDNAs under control of the SV40 early or late gene promoter”, Nucleic Acids Res. 11:7119-36)。レポーター遺伝子プラスミド及びpSG5−ERプラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、アンドロゲン・ジヒドロテストステロン(DHT)を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0066】
エストロゲンによる神経アロマターゼ発現又は活性の直接的な影響については、文献に記載されていない。本発明による1fレポーター遺伝子アッセイでも、1f−プロモーター活性に対するエストラジオールの効果は測定することはできない(図9)。この発見は、1f−プロモーターの配列分析とも一致する。この分析では少なくとも、既知のERE配列は含まれていない。脳内のアロマターゼ活性に対するエストロゲン濃度の影響については、文献中で確かに論じられてはいる(Rosselli, C. E.及びResko, J. A. (1993): Aromatase activity in the rat brain: hormonal regulation and sex differences; J. Steroid. Biochem., Vol. 44: 499-508)ものの、これは、この影響がフィードバック・メカニズムによって媒介されるという趣旨のものである。アンドロゲン受容体の存在は、アンドロゲン作用が有効になるために必須であるので、アロマターゼによって生成されたエストロゲンは、アンドロゲン受容体の発現量を低くし、ひいてはアロマターゼの発現量を間接的に低くする。フィードバック調節は、本発明による試験管内レポーター遺伝子アッセイには存在しない。それというのも、pSG5ベクターをベースとする受容体遺伝子プラスミドは、エストロゲン濃度とは独立して、受容体の発現をもたらすからである。複雑なin vivo系内に存在するフィードバック・メカニズムのこのスイッチ・オフは、実施されたトランス活性化アッセイにおいて、プロモーターレベルにおけるアロマターゼ発現の直接的な調節の研究を可能にする。従って、本発明によるレポーター遺伝子アッセイは、神経アロマターゼ発現の直接的な調節に必要な刺激をより詳細に解明するのに最も適している。
【0067】
5. 神経アロマターゼ発現を調節する薬理学的活性物質を同定して特徴付けするためのトランスフェクション試験
DHTと同様に、他の薬理学的活性アンドロゲンを、1f−ルシフェラーゼ・レポーター遺伝子構造(図10)を用いて試験する。ヒト神経芽腫細胞系SH−SY5Y(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31885-023、Gibcoから)を含むDulbecco's MOD Eagle Medium+5mg/lのペニシリン/ストレプトマイシンを含む15%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、SH−SY5Y細胞を試験培地で2回継代培養する。これはフェノールレッド(注文番号:11880-028、Gibcoから)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0068】
ルシフェラーゼ活性に基づいて、pGl3融合ベクター1f−pGl3内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、6ウェル・プレート内で、1ウェル当たり300000個の細胞に、0.4μgのこのプラスミドを、0.5μgのpSG5ARプラスミドとともに同時トランスフェクトする。pSG5ARベクターは、pSG5ベクターをベースとしている。このpSG5ベクターはヒト細胞において、根底を成す遺伝子、この事例ではARに対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. & Harris, B.A. 1983; “Plasmids for the cloning and expression of full-length double-stranded cDNAs under control of the SV40 early or late gene promoter”, Nucleic Acids Res. 11:7119-36)。レポーター遺伝子プラスミド及びpSG5ARプラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、試験物質を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0069】
実施されたトランスフェクションにおいて、1f−レポーター遺伝子プラスミドは、DHTによってレポーター遺伝子発現が濃度に依存して誘発されたことを示している(図8)。DHTと同様に、他のアンドロゲンも、1f−ルシフェラーゼ・レポーター遺伝子構造(図10)で試験する。全てのアンドロゲンは1f−ルシフェラーゼを刺激するが、しかし、その効力はそれぞれの事例において僅かに異なる。MENT及びプロピオン酸テストステロンで活性化を観察することもできる。相対比較におけるプロピオン酸テストステロンによる極めて強力な刺激は、男性の性欲に対するテストステロン誘導体の良好な臨床効果(Gooren, 1987, Arch. Sex. Beh. 16:463-473; Bancroft 1988, In Handbook of Sexology 6, 297-315 Elsevier, Amsterdam, Buna他, 1993, Fertil. Steril. 59: 1118-1123)と符合し、男性の性欲は、霊長類において、主として神経アロマターゼ発現の活性化によってコントロールされる(Zumpe他, 1993, Horm. Behav. 27:200-215)。
【0070】
6. 他の組織と比較して異なった状態で神経組織内のAR調節型遺伝子を調節する組織選択的アンドロゲンを同定して特徴付けするためのトランスフェクション試験
神経1f−アロマターゼ・プロモーターを用いた本発明による試験システムを、全身的なアンドロゲン作用を反映するMMTV−プロモーター(Parker, M. G.他、J Cell Biochem. 1987, 35:285-92)と組み合わせる。ホルモン調節型MMTV−プロモーターを、PromegaのpGL3コントロール・ベクターのホタルルシフェラーゼ遺伝子の前にクローニングすることにより、ベクターMMTV−lucを生成する。
【0071】
ヒト神経芽腫細胞系SH−SY5Y(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31885-023、Gibcoから)を含むDulbecco's MOD Eagle Medium+5mg/lのペニシリン/ストレプトマイシンを含む15%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、SH−SY5Y細胞を試験培地で2回継代培養する。これはフェノールレッド(注文番号:11880-028、Gibcoから)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0072】
MMTV−プロモーターとの比較において、ルシフェラーゼ活性に基づいて、pGl3融合ベクター内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、並行トランスフェクション試験において、6ウェル・プレートの1ウェル当たり300000個のSH−SY5Y細胞中に、1.5μgの1f−pGl3プラスミド又はMMTV−lucプラスミドを、0.75μgのpSG5ARプラスミドと一緒にトランスフェクトする。レポーター遺伝子プラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、試験物質を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、タンパク質1μg当たりの相対光単位を、最大刺激率のパーセンテージとして示す。
【0073】
神経芽腫細胞系SH−SY5Y中で実施されるトランスフェクションにおいて、1f−レポーター遺伝子プラスミドは、被験アンドロゲン、この例ではDHT及びオキシメトロンとともにレポーター遺伝子発現が誘発されることを示す。MMTVレポーター遺伝子プラスミドも、両事例においてレポーター遺伝子発現が誘発されることを示す(図12)。予期されるように、オキシメトロンはDHTよりも弱い。驚くべきことに、また予測できないほどに、これらの物質は2つのプロモーター構造に対する誘発に関して異なっており、時にはかなり異なっている(図11)。オキシメトロンは両被験プロモーターに対して、極めて類似した活性化をもたらすのに対して、DHTは、2つのプロモータ間で顕著な差を示し、そしてMMTVプロモーターと比較して、神経1f−アロマターゼ・プロモーターに対する効力が少ない。これらの差は、in vivoの、ひいてはヒトにおける組織選択的な差の可能性を指摘する。DHTが例えばテストステロンと比較して、雄の性行動に対する作用がかなり小さいのに対して、前立腺に対しては極めて強力なアンドロゲンとして作用することが、動物実験から判っている(Arteaga-Silva, M.他, 2005, Physiol Behav. 85: 571-80)。DHTのこの挙動は、4か月にわたって行われた大規模なin vivo試験においても、オキサンドロロンと比較して確認された(図12)。従って、本発明による試験管内試験システムは、組織選択作用を有する可能性のあるアンドロゲンをシンプルに、且つ洗練された形で、迅速に同定して特徴付けするのを可能にする。従って、これは、ハイスループット試験が可能な、理想的な試験システムである。
【0074】
7. AR調節型遺伝子の組織選択的な示差調節をもたらす組織選択的アンドロゲンを、部分アゴニスト効果及び部分アンタゴニスト効果に関しても、同時に同定して特徴付けするためのトランスフェクション試験
神経1f−アロマターゼ・プロモーターを用いた本発明による試験システムを、全身的なアンドロゲン作用を反映するMMTV−プロモーター(Parker, M. G.他、J Cell Biochem. 1987, 35:285-92)と組み合わせる。両方のアンドロゲン調節プロモーター活性を、トランスフェクション・アッセイ、又は任意には安定的にトランスフェクトされた細胞系内で、2つの異なるレポーター遺伝子構造を使用して、測定することができる。この例において、ホルモン調節型MMTV−プロモーターを、PromegaのpGL3コントロール・ベクターのホタルルシフェラーゼ遺伝子の前にクローニングすることにより、ベクターMMTV−lucを生成するのに対して、1f−アロマターゼ・プロモーターを、PromegaのpRLベクターのウミシイタケ(Renilla)ルシフェラーゼ遺伝子の前にクローニングすることにより、1f−アロマターゼ−pRLを生成する。
【0075】
試験の準備を任意に単純化するために、ARを先ず、細胞系内に安定的にトランスフェクトする。ARのcDNAを発現プラスミドpSG5内にクローニングすることにより、プラスミドpSG5ARを生成する。このベクターは、ヒト細胞において、根底を成す遺伝子、この事例ではARに対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. 及びHarris, B.A. 1983, Nucleic Acids Res. 11:7119-36)。PC3(ヒト前立腺癌)細胞(DSMZ GmbH, Braunschweig)に、pSG5ARベクターを安定的にトランスフェクトし、これとともに、抗生物質G418の抵抗性遺伝子をコードするプラスミドを付加的に使用する。結果としてのPC3−AR+細胞はヒトARを安定的に発現させる。PC3AR+細胞を、フェノールレッド(注文番号:31870-025、Gibcoから)を含むRPMI 1640培地+200mMのグルタミン、5mg/lのペニシリン/ストレプトマイシン、及び200μg/mlのGeniticinを含む10%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、PC3−AR+を試験培地で2回継代培養する。これはGeniticinを含有せず、フェノールレッド(Gibco、注文番号:32404-014)を含有せず、ホルモン残留物を除去するために活性炭で精製されたウシ胎仔血清(DCS)を含有する。
【0076】
MMTV−プロモーターのホタルルシフェラーゼ活性との比較において、ウミシイタケ(Renilla)活性に基づいて、pRL融合ベクター内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、トランスフェクション試験において、175ml瓶1つ当たり300000個のPC3−AR+細胞中に、8μgの1f−アロマターゼ−pRLプラスミド及び20μgのMMTV−lucプラスミドをトランスフェクトする。レポーター遺伝子プラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから6時間後、細胞を洗浄し、そして1ウェル当たりの細胞数が10000となるように、96ウェル・マイクロタイタープレート上に分配する。96ウェル・プレートに移してから16時間後に、アゴニスト活性の測定のために単独で、そして、アンタゴニスト活性の測定のために10 x e-10Mのテストステロンの存在において、選択的アンドロゲン受容体モジュレーター(SARM)を添加する。インキュベータ内でさらに20時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。製造業者の指示書(Promega; Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従って、dual Luciferase Assay Kit(注文番号:E 1960、Promegaから)を用いて相対光単位で、蛍及びウミシイタケ(Renilla)のルシフェラーゼ活性を決定する。光単位で示された測定値を任意に標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0077】
アゴニズムに関する試験において、ここで試験されたSARMは、前の例で既に提示したDHTと同様に、2つのAR依存性プロモーターに対して極めて異なった形で作用する(図13)。しかし、DHTとは異なり、この事例では、全身的なMMTVプロモーターの活性化はないのに対して、性腺以外の部位のAR調節型プロモーターの例として役立つ神経1f−アロマターゼ・プロモーターが活性化される。DHTを用いた前の例と同様に、このこともin vivo及びヒトにおける組織選択性の可能性を指摘する。さらに、アンタゴニズムの並行測定は、AR活性の選択的モジュレーションの可能性を示唆することができる。すなわち、従来技術を凌いで、組織選択作用の可能性に関してSARM活性の示唆を得ることが可能である。
【0078】
本例にも明示されているように、測定はマイクロタイター形式で行うことができる。このことは、本発明による試験システムの基本的なハイスループット能力を強調する。本例におけるように全般的に明らかにアンタゴニスト性を有するが、しかし特異的な組織では或るアゴニスト作用をも示すSARMは、種々様々な指示に関して特に興味深いものである(S.S. Wolf and M. Obendorf, 2004, in E. Nieschlag & H. M. Behre: “Testosterone” 3rd. Edition; Cambridge University Press, ISBN 0521833809; 623-640)。
【技術分野】
【0001】
本発明は、アンドロゲン依存性1−f−アロマターゼ・レポーター遺伝子、及び1−f−アロマターゼ・レポーター遺伝子の製造方法、及びアンドロゲン受容体のリガンドを同定する方法におけるその使用に関する。
【背景技術】
【0002】
中枢神経系内のアンドロゲンによって誘発されるアロマターゼ発現、ひいてはエストロゲンの形成は、性同一性の刷り込みを含む性特異的な行動学的反応の確立、及び特に雄性における性欲の調節において極めて重要である。加えて、脳内のアンドロゲンによって誘発されるアロマターゼ発現は、学習及び記憶のようなプロセスにおいて重要な役割を果たすと考えられている(Wickelgren, I.; Science, Vol. 276: 675-678, 1997)。神経再生プロセスにおけるエストロゲンから生じる好ましい効果が、記載されている(Abe-Dohmae他, J. Neurochem., Vol. 67: 2087-2095, 1996; I. Azcoitia他, 2001, J. Neurobiol. 47: 318-329)。エストロゲン受容体及び神経エストロゲン産生の誘発又はアップレギュレーションは、種々のニューロン損傷における修復メカニズムの一部であると見られる(Dubal他, 1999, J Neurosci 10:6385-6393; Garcia-Segura他, 1999, Neuroscience 89: 567-578; Peterson他, 2001, J Neuroendocrinol 13: 317-323; Carswell他, 2005, J. Steroid Biochem. Mol. Biol. 96:89-91)。従って、エストロゲンは、神経変性疾患の治療又は予防のための重要な薬理学的選択肢である。しかしこれらの使用は、両方の性において、他の器官に副作用が生じるおそれがあるため、制限されている。
【0003】
目下のところ、アンドロゲンによって誘発されたアロマターゼ発現、ひいては脳特異的なエストロゲン合成を決定するために、そしてアロマターゼ活性に影響を与える物質を同定して特徴付けするために利用可能な試験管内(in vitro)試験システムはない。
【0004】
脳内終点エストロゲン濃度、脳内アロマターゼ発現、脳試料中のアロマターゼ活性、又は両方の性における行動学的試験を用いた試験管内検査は、従来技術から周知である。
【0005】
既に言及されているとおり、アンドロゲンは、アンドロゲン受容体(AR)との結合によって媒介されて、アロマターゼ発現を誘発する。ARは、活性化又は不活化することができる。アンドロゲン欠乏の事例における活性化の例は、例えばサルコペニア、性腺機能低下症、加齢関連性腺機能低下症、及び性欲障害の形態、及び男性の勃起障害において生じる。
【0006】
ARの薬理学的不活化は、前立腺の良性及び悪性疾患において、又はアンドロゲン過剰に関連する疾患、例えば座瘡のある種の形態、脱毛、又は女性における多嚢胞性卵巣及び多毛症において重要である。
【0007】
しかしながら、一般には、アンドロゲン受容体の完全な活性化又は不活化は望ましくなく、むしろ組織選択作用が望ましい。例えば、男性性腺機能低下症の事例では、性欲、気質、及び骨又は筋肉同化作用のAR媒介活性化が望ましいが、しかしこれは活性化というよりも、むしろ前立腺又は皮膚に対するニュートラルな作用又は軽減作用である。ARの不活化が望ましい事例、例えば悪性前立腺癌又は脱毛症の場合、ARの不活化は、性欲、骨代謝などに関して不都合である( S. S. Wolf and M. Obendorf, 2004, “Selective androgen receptor modulators (SARMs)”;E. Nieschlag & H. M. Behre: “Testosterone” 3rd. Edition; Cambridge University Press, ISBN 0521833809; 623-640)。
【0008】
これまで、細胞培養又はin vitroに基づくアッセイ・システムは、これらの組織選択的なアンドロゲン又は抗アンドロゲンを見いだして特徴付けするには不十分であった。このことは、アロマターゼ発現の決定にも当てはまる。
【発明の概要】
【発明が解決しようとする課題】
【0009】
従って、目的は、
−中枢神経系CNS内のアロマテーゼ発現を選択的に増大させ、
且つ/又は
−ARの活性に組織選択的な影響を与える
ことを特徴とする薬理学的活性の物質を同定して特徴付けすることができる方法を提供することである。
【課題を解決するための手段】
【0010】
この目的は、レポーター遺伝子アッセイにおいてアロマターゼ発現を刺激することを介して、脳内のエストロゲン合成を選択的に調節してアンドロゲン受容体に組織選択的な影響を与える物質を同定できる方法によって達成される。1f−アロマターゼ・プロモーターのアンドロゲン依存性調節が存在することが判った。
【0011】
ヒトのアロマターゼは、染色体15上に位置する遺伝子においてコードされている(Chen, S.他, (1988): Molec. Biol., Vol. 7: 27-38)。
【0012】
ヒトP450arom遺伝子(CYP19)は、30kbのコード領域と約93kbの調節領域とを含んでいる(Bulun, S.E.他、Semin Reprod Med. Vol. 22: 5-9, 2004)。転写産物は、10個のエキソンから成っており、エキソンIは非コードである。コード・エキソンII〜Xは、全ての組織内で同一に発現される。今日まで、コード領域内のスプライシング変異体はヒトにおいてはまだ見いだされていない。
【0013】
エキソンIに関しては、種々の組織に由来するmRNAの配列決定中にいくつかの変異体が発見された(検討の際には、Harada, N.; Utsumi, T.及びTakagi, Y. Proc. Natl. Acad. Sci. USA, 90: 11312-11316, 1993、及びSimpson, E. R.; Endocrine Reviews, 15: 341-355, 1994参照)。これらの変異体は組織特異的に発現され、そして全ては、エキソンIIの出発点に配置されたのと同じスプライシング部位に連結することができる(図1)。mRNAにおけるエキソン−I配列の組織特異的発生は、選択的プロモーターの使用によって説明することができる。選択的プロモーターは、それぞれのエキソン−I変異体の前の遺伝子内に配置されている。これらの種々のプロモーターの活性化は、まだ完全には解明されていない制御を被る。表1は、Harada他による種々の組織内のヒト・アロマターゼの或る特定の5’−非翻訳エキソンの現在知られている用途の例を挙げる(1993, Proc. Natl. Acad. Sci. USA, Vol. 90: 11312-11316)。
【0014】
特にHarada(Harada, N.他, (1993) Proc. Natl. Acad. Sci. USA, Vol. 90: 11312-11316)及びSimpson(Simpson, E. R.; Endocrine Reviews, Vol. 15: 341-355, 1994)を中心とした2つのチームの今日までの研究が、アロマターゼ−mRNAを配列決定することによって示したところでは、ほとんどの組織内でいくつかのプロモーターを使用することができ、また逆に、異なる組織では1つのプロモーターを使用することもできる(表1)。
【0015】
このことの例外は、プロモーター1fであるように見える。プロモーター1fは知られている限り、ヒトにおいて、専ら神経組織内で使用される。
【0016】
脳内では、アンドロゲンは特に、アロマターゼ発現に対する効果を有している(Abdelgadir, S. E.他, Endocrinol., Vol. 135: 395-401, 1994;Rosselli, C. E.他, (1996) Endocrine, Vol. 5: 59-65;Rosselli, C. E.他, (1997, Brain Research, Vo. 44: 351-357)。この発見にもかかわらず、アンドロゲン受容体(AR)が脳内のアロマターゼ発現を調節する、その正確な分子メカニズムはまだ知られていない。1f−アロマターゼ・プロモーター内の推定上のAR担当要素が文献において述べられているのは事実だが、神経アロマターゼ発現のAR媒介調節における1f−プロモーターの機能関与はまだ示されていない(Honda, S.他、Biochem Biophys Res Commun 198 (1994), 1153-1160)。それどころか、1f−アロマターゼ・プロモーターについて最初に記述したチーム(S. Honda他, Biochem Biophys Res Commun 198 (1994), 1153-1160)は、後の研究で他のレギュレーターを指摘した。これらのレギュレーターの更なる特徴付けはなされていない(Honda, S. I.他, J Steroid Biochem Mol Biol. 79:255-60, 2001; Honda, S.他, Brain Res Mol Brain Res. 66: 122-32, 1999)。
【0017】
本発明の基礎は、1f−アロマターゼ・エキソンの制御が、専ら1f−アロマターゼ・プロモーターの活性によって調節され、そして細胞の脈絡には依存しないという観察である。試験管内試験システムによって、神経組織、神経細胞、動物全体又は動物からの抽出物を使用する必要なしに、神経アロマターゼ発現の調節に対する薬理学的活性物質の影響を示すことが今や可能になった。
【0018】
このような試験システムを用いることによって、中枢神経系内のアロマテーゼ発現を選択的に増大させるか、又はARの活性に組織選択的な影響を与えるのに適した薬理学的活性の物質を発見して特徴付けすることが可能になり、また単純化される。
【0019】
従って本発明の目的は、中枢神経系内のアロマテーゼ発現を選択的に増大させるか、又はARの活性に組織選択的な影響を与えるのに適した物質を同定し特徴付けするための試験管内試験システムであって、前記試験システムが、レポーター遺伝子とアロマターゼ・プロモーターとの融合体を含有する、試験管内試験システムである。
【0020】
アロマターゼ、すなわちエストロゲン合成における鍵酵素の組織特異的な調節は、最終的に、種々の組織内における標的エストロゲン提供に関与する。
【0021】
ホルモン制御されたアロマターゼ発現を特徴付けするためには、確立されたヒト細胞系がこれらの試験管内試験に適しており、組織特異的な生体内(in vivo)発現との相関が、種々のプロモーターを使用することによって保証される。レポーター遺伝子アッセイの開発は、アロマターゼ・プロモーターの場合、個々のプロモーターの活性化による発現に与える特定物質の組織特異的影響を試験するシンプルで洗練された可能性をもたらす。所期の脳特異的発現の場合、これは、本発明による1f−アロマターゼ・プロモーターである。
【0022】
ARによって神経組織内で調節される1f−アロマターゼ・プロモーター(1f−アロ−プロモーター)の活性と、ARによって別の組織内で調節されるプロモーター又は全身的にARによって調節されるプロモーターの活性とを比較することによって、組織選択性に関してARに対して活性の物質を試験管内における特徴付けすることが可能になる。1f−アロマターゼ・プロモーターのプロモーター活性はこの場合、神経組織内のAR媒介型活性のための代理マーカーとして、又はより全身的に、性腺組織以外(例えば男性においては、精巣及び前立腺以外;女性においては卵巣以外)の部位のAR媒介型活性のための代理マーカーとして役立つ。
【0023】
ヒト・アロマターゼの1fプロモーターを用いたレポーター遺伝子アッセイによって、神経アロマターゼ発現の調節を決定することができる。既知の配列(配列番号1)を有する神経1f−アロマターゼ・プロモーターは、レポーター遺伝子の前に置かれている。プロモーター・レポーター遺伝子融合体は、一時的又は安定的に標的系内にトランスフェクトされる。ルシフェラーゼの発現が1f−プロモーターの関数として行われ、そしてアロマターゼ・プロモーターの活性は、レポーター遺伝子によってコードされた酵素の量から決定することができる。本例において、ホタルルシフェラーゼ遺伝子又はウミシイタケ(Renilla)ルシフェラーゼ遺伝子が、レポーター遺伝子の例として選択された。
【図面の簡単な説明】
【0024】
【図1】選択的エキソン−I変異形の周知のスプライシング変異形を有するヒト・アロマターゼ遺伝子のゲノム・プロモーター領域(単純化された表示)。より判りやすくするために、5’領域の部分が切り取られ、省略したものとして示されている(Harada, N.他, Proc. Natl. Acad. Sci. USA, Vol. 90: 11312-11316, 1993、及びSimpson, E. R.; Endocrine Reviews, Vol. 15: 341-355, 1994から修正)
【0025】
【図2】SHSY5Y細胞のゲノムDNAから増幅された1f−アロ−プロモーターを有するPCR生成物。a:長さ標準、b:1f−アロマターゼ・プロモーターのPCR増幅。
【0026】
【図3】pTAdvベクターのマップ。両端に3’−T突出部を備えたこの線状ベクターを、PCRフラグメントとのライゲートのために使用した。
【0027】
【図4】pTAdvベクター中に増幅1f−アロマターゼ・プロモーターDNAを有する1f−pTadv’’ベクターのベクターマップ。HindIII及びXhoIは、更なるクローニングのために使用された切断部位を意味する。
【0028】
【図5】pGl3−コントロール・ベクターのベクターマップ。HindIII及びXhoIは、更なるクローニングのために使用された切断部位を意味する。
【0029】
【図6】1f−pGl3ベクターのベクターマップ。HindIII及びXhoIは、ベクターの生成のために使用された切断部位を意味する。
【0030】
【図7】PC3細胞中のジヒドロテストステロン(DHT)の濃度に対する、1f−プロモーターの活性の依存。0.2μgの1f−pGl3プラスミド及び0.5μgのpSG5−ARプラスミドをトランスフェクトする。DHTと一緒に20時間インキュベートする。二重測定の平均値を、総タンパク質1μg当たりの相対光単位(RLU)で示す。
【0031】
【図8】SH−SY5Y細胞中のジヒドロテストステロン(DHT)の濃度に対する、1f−プロモーターの活性の依存。0.2μgの1f−pGl3プラスミド及び0.5μgのpSG5−ARプラスミドをトランスフェクトする。DHTと一緒に20時間インキュベートする。二重測定の平均値を、総タンパク質1μg当たりの相対光単位(RLU)で示す。
【0032】
【図9】これは、1f−プロモーターの活性が、PC3細胞中のエストラジオール(E2)の濃度とは無関係であることを示す。0.2μgの1f−pGl3プラスミド及び0.5μgのpSG5−ERプラスミドをトランスフェクトする。E2と一緒に20時間インキュベートする。二重測定の平均値を、総タンパク質1μg当たりの相対光単位(RLU)で示す。
【0033】
【図10】神経SH−SY5Y細胞内の1f−プロモーターに対する種々のアンドロゲンの影響。0.4μgの1f−pGl3プラスミド及び0.5μgのpSG5−ARプラスミドをトランスフェクトする。a)T(プロピオン酸テストステロン)、b)MENT(7α−メチル−19−ノルテストステロン)、及びc)DHT(ジヒドロテストステロン)と一緒に20時間にわたってインキュベートする。二重測定の平均値を、総タンパク質1μg当たりの相対光単位(RLU)で示す。
【0034】
【図11】神経SH−SY5Y細胞内の1f−プロモーター及びMMTV−プロモーターに対する種々のアンドロゲンの影響。1.5μgの1f−pGl3及びMMTV−lucプロモーター及び0.75μgのpSG5−ARプラスミドをトランスフェクトする。オキシメトロン又はDHT(ジヒドロテストステロン)と一緒に20時間にわたってインキュベートする。相対光単位(RLU)の標準化平均値は、標準偏差とともに四重測定からのパーセンテージとして示される。
【0035】
【図12】アンドロゲン作用の神経終点を表す、雄の性行動に対する種々のアンドロゲンのin vivoの影響(A)、及びアンドロゲン作用の性腺終点を表す、前立腺(B)の重量に対する種々のアンドロゲンのin vivoの影響(B)。性的経験のあるインタクトの雄ラットを選び去勢する。7週間後、性的活動がなくなった後、動物たちを1群当たり9〜10匹の3つの群に分け、1050μgのオキシメトロン、DHT又はビヒクルで1週間にわたって処置する。8日目〜120日目まで、動物は毎日、70μgのオキシメトロン(黒)、DHT(暗灰色)又はビヒクル(明灰色)を受ける。各動物の性行動(乗駕行動)を7回試験した。研究開始から4か月後に、器官を取り出し、前立腺の重量及び体重を測定した。
【0036】
【図13】pC3AR+細胞中の非特異的MMTVプロモーター(A+B)及び神経1f−アロマターゼ・プロモーター(C+D)に対して同時に行われる選択的ARモジュレーター(SARM)のアゴニズム(A+C)及びアンタゴニズム(B+D)の試験。1.5μgの1f−pGl3及びMMTV−lucプロモーター及び0.75μgのpSG5−ARプラスミドをトランスフェクトする。SARMと一緒に20時間にわたってインキュベートする。アンタゴニズム(B+D)の試験では、4 x 10 e-10Mのテストステロンの存在において、SARMを上記濃度でインキュベートする。個々の値を四角で示し、また四重測定からの相対光単位の平均値を、標準偏差とともに円で示す。SARMを添加しないテストステロンの値を三角で示す。非特異的MMTV−プロモーター(A+B)に対して、ここで試験されたSARMは、AR媒介トランス活性化の純粋なアンタゴニストとして挙動するのに対して、神経1f−アロマターゼ・プロモーター(C+D)に対しては部分アゴニスト/部分アンタゴニスト作用が表面化する。
【0037】
本発明は、配列番号1に従う1f−アロマターゼ・プロモーター及びレポーター遺伝子から成るプロモーター・レポーター遺伝子融合体に関し、そして該レポーター遺伝子は、ホタルルシフェラーゼ遺伝子又はウミシイタケ(Renilla)ルシフェラーゼ遺伝子を含む群から選択することができるが、この選択は制限的なものであってはならず、そして融合生成物は、細胞系内に安定的に又は一時的に発現させることができる。
【0038】
さらに、ARが細胞中に発現された場合には、アロマターゼ・プロモーターの活性に、AR活性試験物質の影響がARを介して与えられる。この場合、試験物質の作用は、ルシフェラーゼ酵素活性の変化に基づいて決定することができる。従って一次神経組織又は一次神経細胞を用いて、或いは動物全体又は動物からの抽出物を用いて作業する必要なしに、薬理学的活性物質を模索又は特徴付けすることができる。
【0039】
本発明は、試験管内試験システムにおけるプロモーター・レポーター遺伝子融合体の使用であって、プロモーター・レポーター遺伝子融合体は、配列番号1に従う1f−アロマターゼ・プロモーター及びレポーター遺伝子から成っており、試験物質の影響は、細胞培養におけるハイスループット・レポーター遺伝子アッセイで割り出され、神経組織内のアロマターゼの調節に関する情報を得ることができる、プロモーター・レポーター遺伝子融合体の使用に関する。この方法は、アンドロゲン受容体に結合することにより、アロマターゼ発現に影響を与える物質を同定するために用いられる。
【0040】
本発明はまた、プロモーター・レポーター遺伝子融合体の使用であって、アンドロゲン受容体に結合することによって脳内のアロマターゼ調節に影響を与える物質を同定するために、配列番号1に規定された配列に対して>90%、好ましくは>95%の相同性を有する配列が細胞培養系内で使用される、プロモーター・レポーター遺伝子融合体の使用に関する。
【0041】
さらに、本発明による試験モデルを用いて、ARアゴニスト、アンタゴニスト、及びSARMを含むアンドロゲン性物質の特性を特徴付けすることもできる。ここでは、1f−アロマターゼ・プロモーターの活性が、ARを介して別の組織内で調節される、又は全身的にARによって調節されるプロモーターの活性と比較される。1f−アロマターゼ・プロモーターのプロモーター活性はこの場合、神経組織内のAR媒介型活性のための代理マーカーとして、又はより全身的に、性腺組織以外(例えば男性においては、精巣及び前立腺以外;女性においては卵巣以外)の部位のAR媒介型活性のための代理マーカーとして役立つ。
【0042】
このために、神経1f−アロマターゼ・プロモーターを用いた本発明による試験システムは、ARによって調節される別のプロモーターと組み合わされる。後者のプロモーターは、全身アンドロゲン作用(例えばMMTV−プロモーター)、又は別のアンドロゲン性標的器官内のアンドロゲン作用(例えばアンドロゲン受容体によって前立腺内で活性化されるプロモーター)を反映する。2種の異なるレポーター遺伝子、例えば1f−アロマターゼ・プロモーターに対するウミシイタケ(Renilla)ルシフェラーゼ、及び第2のプロモーターに対するホタルルシフェラーゼを同時に使用することにより、両方のプロモーター活性を同時に測定することが可能になる。これは、ARを介した薬理学的作用による試験物質の組織選択的活性に関する情報を単純かつ洗練された形で提供する。或いは、両プロモーターは、同じレポーター遺伝子の前に連続して挿入することもできる。これもやはり、原則としては動物実験である従来技術と比較してかなり単純なものになる。神経プロモーター活性は、性腺組織以外の部位のプロモーターの代理活性として、より全身的に抽出することができる。
【0043】
本発明はまた、アンドロゲン受容体に結合する物質の組織選択作用を決定する方法であって、以下の段階:
a. アゴニスト活性及びアンタゴニスト活性の両方を同定するために、細胞培養系内のレポーター遺伝子アッセイに該プロモーター・レポーター遺伝子融合体を使用して、アロマターゼ調節のモジュレーションを決定する段階、
b. 別のアンドロゲン依存性プロモーターのモジュレーションを決定する段階、及び
c. 1f−アロマターゼ・プロモーターに対する選択的ARモジュレーションに関連して物質の完全な試験管内における特徴付けを達成するために、段階a及びbを比較するか、あるいは例えば前立腺特異的又は全身的に作用するAR調節型プロモーターと直接的に比較する段階
含んで成る、方法に関する。この方法は、ARに組織選択的な影響を与える物質を同定するために用いることができる。
【0044】
本発明はまた、レポーター遺伝子アッセイにおけるプロモーター・レポーター遺伝子融合体の使用であって、アンドロゲン受容体の活性をモジュレートし、そしてアンドロゲン受容体によって調節される遺伝子の調節の組織選択的モジュレーションを可能にする物質を同定するために、配列番号1に規定された配列に対して>90%、好ましくは>95%の相同性を有する配列が、細胞培養系内のレポーター遺伝子アッセイにおいて使用される、プロモーター・レポーター遺伝子融合体の使用に関する。
【0045】
「アンタゴニスト」は、それらの対応受容体に結合する分子であって、自然発生型リガンドによって、受容体とカップリングされた信号伝達経路が始動するのを阻害する分子として理解されるべきである。通常、アンタゴニストは、受容体との結合を巡って、受容体の自然発生リガンドと競合する。しかしながら、受容体とカップリングされた信号伝達経路が、自然発生型リガンドによって活性化されるのを阻害する分子による受容体の他の変更形も可能である(例えば受容体の非競合的な立体変更形)。
【0046】
受容体アンタゴニストは典型的には、それらの特定の受容体に選択的に結合し、他の受容体には結合しない。通常、受容体アンタゴニストは天然リガンドよりも高い結合親和性を示す。天然リガンドよりも高い、受容体に対する親和性を有するアンタゴニストが好ましいものの、親和性の低いアンタゴニストを使用することもできる。アンタゴニストは、それらの対応受容体に可逆的に結合することが好ましい。
【0047】
「アゴニスト」は、それらの対応受容体に結合し、通常、受容体との結合を巡って、受容体の自然発生リガンドと競合する分子であって、受容体にカップリングされた信号伝達経路の始動を刺激する分子として理解されるべきである。アゴニストは、天然リガンドの結合を支持することもできる。
【0048】
受容体アゴニストは典型的には、それらの特定の受容体に選択的に結合し、他の受容体には結合しない。受容体アゴニストは通常、天然リガンドよりも高い結合親和性を有している。天然リガンドよりも高い、受容体に対する親和性を有するアゴニストが好ましいものの、親和性の低いアゴニストを使用することもできる。アゴニストは、それらの対応受容体に可逆的に結合することが好ましい。
【0049】
アゴニストは、対応受容体の始動及び/又は生理学的作用によって媒介される信号伝達により試験される。
【0050】
「リガンド」は、受容体に結合する化合物又は低分子物質を意味する。これらの結合は通常可逆的である。対応受容体に対するリガンドの結合は、受容体にカップリングされた信号伝達経路を活性化又は不活化する。このようにして、リガンドはその細胞内作用をもたらす。
【0051】
リガンドは、受容体のアゴニスト及びアンタゴニストとして理解されるべきである。
【実施例】
【0052】
生物学的例
1. レポーター遺伝子プラスミド内のヒト1f−アロマターゼ・プロモーターのクローニング
レポーター遺伝子アッセイに際して、ヒト・アロマターゼの1f−プロモーターを、ヒトSH−SY5Y細胞のゲノムDNAから増幅する。QIAamp Blood Kitを、製造業者(Qiagen)によって指定されたプロトコルに従って使用して、ゲノムDNAを単離する。アロマターゼの1f−プロモーターは、刊行物(S. Honda他、Biochem Biophys Res Commun 198 (1994), pp. 1153-1160)、及び遺伝子バンク(CYP19A1)から周知である。特定のプライマー対(表2)を使用して、1f−プロモーターを含むヒト・アロマターゼ・プロモーター領域の部分をPCRによって増幅する。
【0053】
PCRによってDNAフラグメントを増幅するために、各事例において、1μlのゲノムDNA試料を使用する。これを2分間にわたって94℃で変性させ、そして上記DNAプライマーEbrain B4及びEbrain c670を用いて35サイクルで増幅し、ゲル電気泳動法によって分析し(図2)、そしてアガロース・ゲルからのDNA抽出によって副生成物から精製する。
【0054】
1f−プロモーター・フラグメントのPCR生成物は、3’−A突出部を有しており、線状pTAdvクローニング・ベクターとライゲートされる(図3)。pTAdvベクターは、Clontechから得られた。結果として生じる1f−pTAdv2ベクター(図4)の挿入体を配列決定し、そして増幅されたヒト・1f−アロマターゼ・プロモーターの配列を確認する(配列番号1)。これは、以前に発表された配列(S. Honda他, N Biochem Biophys Res Commun 198 (1994), 1153-1160)とは異なるが、しかしより最近の遺伝子バンク登録と符合する。
【0055】
使用されるレポーター遺伝子は例えば、PromegaのpGl3ベクターのルシフェラーゼ遺伝子である。1f−プロモーターを有するプロモーター・レポーター遺伝子構造を生成するために、HindIII及びXhoIを有するpGl3コントロール(図5)から、213bp長のコントロール・プロモーター(SV40)を切り取る。こうして線状化され短縮されたpGl3を、ゲル電気泳動法及びDNA抽出によって精製し、脱リン酸化し、次いで、1f−pTAdv2ベクター(図4)からHindIII及びXhoIを有するように切り取られ精製された1f−プロモーターのフラグメントをスプライシングし、レポーター遺伝子プラスミド1f−pGl3を生成する(図6)。
【0056】
2. 細胞培養内のレポーター遺伝子アッセイにおける1f−プロモーターに対するホルモン活性化合物の影響を決定するためのトランスフェクション試験
PC3(ヒト前立腺癌)細胞(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31870-025、Gibcoから)を含むRPMI 1640培地+200mMのグルタミン及び5mg/lのペニシリン/ストレプトマイシンを含む10%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、PC3細胞を試験培地で2回継代培養する。これはフェノールレッド(Gibco、注文番号:32404-014)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0057】
ルシフェラーゼ活性に基づいて、pGl3融合ベクター1f−pGl3内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、6ウェル・プレート内で、1ウェル当たり200000個の細胞に、0.2μgのこのプラスミドを、0.5μgのpSG5ARプラスミドとともに同時トランスフェクトする。pSG5ARベクターは、pSG5ベクターをベースとしている。このpSG5ベクターはヒト細胞において、根底を成す遺伝子、この事例ではARに対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. and Harris, B.A. 1983, Nucleic Acids Res. 11:7119-36)。レポーター遺伝子プラスミド及びpSG5ARプラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、アンドロゲン・ジヒドロテストステロン(DHT)を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0058】
1f−レポーター遺伝子生成物は、前立腺細胞系PC3中で行われたトランスフェクションにおいて、2.6倍だけDHTによってレポーター遺伝子発現が誘発されたことを示す(図7)。アンドロゲンによる調節は、ここでは試験管内で決定することができる。
【0059】
3. 1f−アロマターゼ・プロモーターのアンドロゲン依存性調節
前立腺PC3に加えて、1f−アロマターゼ・プロモーターに対するアンドロゲン調節型レポーター遺伝子発現を分析するために、神経細胞系を使用する。
【0060】
ヒト神経芽腫細胞系SH−SY5Y(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31885-023、Gibcoから)を含むDulbecco's MOD Eagle Medium+5mg/lのペニシリン/ストレプトマイシンを含む15%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、SH−SY5Y細胞を試験培地で2回継代培養する。これはフェノールレッド(注文番号:11880-028、Gibcoから)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0061】
ルシフェラーゼ活性に基づいて、pGl3融合ベクター1f−pGl3内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、6ウェル・プレート内で、1ウェル当たり300000個の細胞に、0.2μgのこのプラスミドを、0.5μgのpSG5ARプラスミドとともに同時トランスフェクトする。pSG5ARベクターは、pSG5ベクターをベースとしている。このpSG5ベクターはヒト細胞において、根底を成す遺伝子、この事例ではARに対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. & Harris, B.A. 1983; “Plasmids for the cloning and expression of full-length double-stranded cDNAs under control of the SV40 early or late gene promoter”, Nucleic Acids Res. 11:7119-36)。レポーター遺伝子プラスミド及びpSG5ARプラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、アンドロゲン・ジヒドロテストステロン(DHT)を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0062】
1f−レポーター遺伝子生成物は、神経芽腫細胞系SH−SY5Y中で行われたトランスフェクションにおいて、DHTによってレポーター遺伝子発現が濃度に依存して誘発されたことを示している(図8)。
【0063】
使用した両方の細胞系において同様の結果が測定されるように、これらの試験システムにおける1f−プロモーターの活性は、細胞の脈絡とは独立してアンドロゲンによって刺激される。
【0064】
4. 薬理学的活性物質による1f−アロマターゼ・プロモーターのホルモン調節を特徴付けするためのトランスフェクション試験
DHTと同様に、さらなる薬理学的活性物質として1f−ルシフェラーゼ・レポーター遺伝子構造(1f−pGl3)に基づいて、我々は、神経組織内のヒト・アロマターゼの発現に対するエストラジオールの影響を試験した(図9)。PC3(ヒト前立腺癌)細胞(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31870-025、Gibcoから)を含むRPMI 1640培地+200mMのグルタミン及び5mg/lのペニシリン/ストレプトマイシンを含む10%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、PC3細胞を試験培地で2回継代培養する。これはフェノールレッド(Gibco、注文番号:32404-014)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0065】
ルシフェラーゼ活性に基づいて、pGl3融合ベクター1f−pGl3内のクローン型1f−アロマターゼ・プロモーターに対するエストラジオール(E2)の影響を決定するために、6ウェル・プレート内で、1ウェル当たり200000個の細胞に、0.2μgのこのプラスミドを、0.5μgのpSG5−ERプラスミドとともに同時トランスフェクトする。pSG5−ERベクターは、pSG5ベクターをベースとしている。このpSG5ベクターはヒト細胞において、根底を成す遺伝子、この事例ではヒト・エストロゲン受容体アルファ(ERα)に対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. & Harris, B.A. 1983; “Plasmids for the cloning and expression of full-length double-stranded cDNAs under control of the SV40 early or late gene promoter”, Nucleic Acids Res. 11:7119-36)。レポーター遺伝子プラスミド及びpSG5−ERプラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、アンドロゲン・ジヒドロテストステロン(DHT)を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0066】
エストロゲンによる神経アロマターゼ発現又は活性の直接的な影響については、文献に記載されていない。本発明による1fレポーター遺伝子アッセイでも、1f−プロモーター活性に対するエストラジオールの効果は測定することはできない(図9)。この発見は、1f−プロモーターの配列分析とも一致する。この分析では少なくとも、既知のERE配列は含まれていない。脳内のアロマターゼ活性に対するエストロゲン濃度の影響については、文献中で確かに論じられてはいる(Rosselli, C. E.及びResko, J. A. (1993): Aromatase activity in the rat brain: hormonal regulation and sex differences; J. Steroid. Biochem., Vol. 44: 499-508)ものの、これは、この影響がフィードバック・メカニズムによって媒介されるという趣旨のものである。アンドロゲン受容体の存在は、アンドロゲン作用が有効になるために必須であるので、アロマターゼによって生成されたエストロゲンは、アンドロゲン受容体の発現量を低くし、ひいてはアロマターゼの発現量を間接的に低くする。フィードバック調節は、本発明による試験管内レポーター遺伝子アッセイには存在しない。それというのも、pSG5ベクターをベースとする受容体遺伝子プラスミドは、エストロゲン濃度とは独立して、受容体の発現をもたらすからである。複雑なin vivo系内に存在するフィードバック・メカニズムのこのスイッチ・オフは、実施されたトランス活性化アッセイにおいて、プロモーターレベルにおけるアロマターゼ発現の直接的な調節の研究を可能にする。従って、本発明によるレポーター遺伝子アッセイは、神経アロマターゼ発現の直接的な調節に必要な刺激をより詳細に解明するのに最も適している。
【0067】
5. 神経アロマターゼ発現を調節する薬理学的活性物質を同定して特徴付けするためのトランスフェクション試験
DHTと同様に、他の薬理学的活性アンドロゲンを、1f−ルシフェラーゼ・レポーター遺伝子構造(図10)を用いて試験する。ヒト神経芽腫細胞系SH−SY5Y(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31885-023、Gibcoから)を含むDulbecco's MOD Eagle Medium+5mg/lのペニシリン/ストレプトマイシンを含む15%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、SH−SY5Y細胞を試験培地で2回継代培養する。これはフェノールレッド(注文番号:11880-028、Gibcoから)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0068】
ルシフェラーゼ活性に基づいて、pGl3融合ベクター1f−pGl3内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、6ウェル・プレート内で、1ウェル当たり300000個の細胞に、0.4μgのこのプラスミドを、0.5μgのpSG5ARプラスミドとともに同時トランスフェクトする。pSG5ARベクターは、pSG5ベクターをベースとしている。このpSG5ベクターはヒト細胞において、根底を成す遺伝子、この事例ではARに対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. & Harris, B.A. 1983; “Plasmids for the cloning and expression of full-length double-stranded cDNAs under control of the SV40 early or late gene promoter”, Nucleic Acids Res. 11:7119-36)。レポーター遺伝子プラスミド及びpSG5ARプラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、試験物質を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0069】
実施されたトランスフェクションにおいて、1f−レポーター遺伝子プラスミドは、DHTによってレポーター遺伝子発現が濃度に依存して誘発されたことを示している(図8)。DHTと同様に、他のアンドロゲンも、1f−ルシフェラーゼ・レポーター遺伝子構造(図10)で試験する。全てのアンドロゲンは1f−ルシフェラーゼを刺激するが、しかし、その効力はそれぞれの事例において僅かに異なる。MENT及びプロピオン酸テストステロンで活性化を観察することもできる。相対比較におけるプロピオン酸テストステロンによる極めて強力な刺激は、男性の性欲に対するテストステロン誘導体の良好な臨床効果(Gooren, 1987, Arch. Sex. Beh. 16:463-473; Bancroft 1988, In Handbook of Sexology 6, 297-315 Elsevier, Amsterdam, Buna他, 1993, Fertil. Steril. 59: 1118-1123)と符合し、男性の性欲は、霊長類において、主として神経アロマターゼ発現の活性化によってコントロールされる(Zumpe他, 1993, Horm. Behav. 27:200-215)。
【0070】
6. 他の組織と比較して異なった状態で神経組織内のAR調節型遺伝子を調節する組織選択的アンドロゲンを同定して特徴付けするためのトランスフェクション試験
神経1f−アロマターゼ・プロモーターを用いた本発明による試験システムを、全身的なアンドロゲン作用を反映するMMTV−プロモーター(Parker, M. G.他、J Cell Biochem. 1987, 35:285-92)と組み合わせる。ホルモン調節型MMTV−プロモーターを、PromegaのpGL3コントロール・ベクターのホタルルシフェラーゼ遺伝子の前にクローニングすることにより、ベクターMMTV−lucを生成する。
【0071】
ヒト神経芽腫細胞系SH−SY5Y(DSMZ GmbH, Braunschweig)を、フェノールレッド(注文番号:31885-023、Gibcoから)を含むDulbecco's MOD Eagle Medium+5mg/lのペニシリン/ストレプトマイシンを含む15%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、SH−SY5Y細胞を試験培地で2回継代培養する。これはフェノールレッド(注文番号:11880-028、Gibcoから)を含有せず、ホルモン残留物を除去するために活性炭で予め精製されたウシ胎仔血清(DCS)を含有する。
【0072】
MMTV−プロモーターとの比較において、ルシフェラーゼ活性に基づいて、pGl3融合ベクター内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、並行トランスフェクション試験において、6ウェル・プレートの1ウェル当たり300000個のSH−SY5Y細胞中に、1.5μgの1f−pGl3プラスミド又はMMTV−lucプラスミドを、0.75μgのpSG5ARプラスミドと一緒にトランスフェクトする。レポーター遺伝子プラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから20時間後、試験物質を添加する。インキュベータ内でさらに24時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。Promegaの指示書(Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従ってルシフェラーゼ・アッセイ基質を使用して、ルシフェラーゼ活性を決定する。光単位で示された測定値を標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、タンパク質1μg当たりの相対光単位を、最大刺激率のパーセンテージとして示す。
【0073】
神経芽腫細胞系SH−SY5Y中で実施されるトランスフェクションにおいて、1f−レポーター遺伝子プラスミドは、被験アンドロゲン、この例ではDHT及びオキシメトロンとともにレポーター遺伝子発現が誘発されることを示す。MMTVレポーター遺伝子プラスミドも、両事例においてレポーター遺伝子発現が誘発されることを示す(図12)。予期されるように、オキシメトロンはDHTよりも弱い。驚くべきことに、また予測できないほどに、これらの物質は2つのプロモーター構造に対する誘発に関して異なっており、時にはかなり異なっている(図11)。オキシメトロンは両被験プロモーターに対して、極めて類似した活性化をもたらすのに対して、DHTは、2つのプロモータ間で顕著な差を示し、そしてMMTVプロモーターと比較して、神経1f−アロマターゼ・プロモーターに対する効力が少ない。これらの差は、in vivoの、ひいてはヒトにおける組織選択的な差の可能性を指摘する。DHTが例えばテストステロンと比較して、雄の性行動に対する作用がかなり小さいのに対して、前立腺に対しては極めて強力なアンドロゲンとして作用することが、動物実験から判っている(Arteaga-Silva, M.他, 2005, Physiol Behav. 85: 571-80)。DHTのこの挙動は、4か月にわたって行われた大規模なin vivo試験においても、オキサンドロロンと比較して確認された(図12)。従って、本発明による試験管内試験システムは、組織選択作用を有する可能性のあるアンドロゲンをシンプルに、且つ洗練された形で、迅速に同定して特徴付けするのを可能にする。従って、これは、ハイスループット試験が可能な、理想的な試験システムである。
【0074】
7. AR調節型遺伝子の組織選択的な示差調節をもたらす組織選択的アンドロゲンを、部分アゴニスト効果及び部分アンタゴニスト効果に関しても、同時に同定して特徴付けするためのトランスフェクション試験
神経1f−アロマターゼ・プロモーターを用いた本発明による試験システムを、全身的なアンドロゲン作用を反映するMMTV−プロモーター(Parker, M. G.他、J Cell Biochem. 1987, 35:285-92)と組み合わせる。両方のアンドロゲン調節プロモーター活性を、トランスフェクション・アッセイ、又は任意には安定的にトランスフェクトされた細胞系内で、2つの異なるレポーター遺伝子構造を使用して、測定することができる。この例において、ホルモン調節型MMTV−プロモーターを、PromegaのpGL3コントロール・ベクターのホタルルシフェラーゼ遺伝子の前にクローニングすることにより、ベクターMMTV−lucを生成するのに対して、1f−アロマターゼ・プロモーターを、PromegaのpRLベクターのウミシイタケ(Renilla)ルシフェラーゼ遺伝子の前にクローニングすることにより、1f−アロマターゼ−pRLを生成する。
【0075】
試験の準備を任意に単純化するために、ARを先ず、細胞系内に安定的にトランスフェクトする。ARのcDNAを発現プラスミドpSG5内にクローニングすることにより、プラスミドpSG5ARを生成する。このベクターは、ヒト細胞において、根底を成す遺伝子、この事例ではARに対応する遺伝子の構成的発現をもたらすSV40初期遺伝子プロモーターを有している(Breathnach, R. 及びHarris, B.A. 1983, Nucleic Acids Res. 11:7119-36)。PC3(ヒト前立腺癌)細胞(DSMZ GmbH, Braunschweig)に、pSG5ARベクターを安定的にトランスフェクトし、これとともに、抗生物質G418の抵抗性遺伝子をコードするプラスミドを付加的に使用する。結果としてのPC3−AR+細胞はヒトARを安定的に発現させる。PC3AR+細胞を、フェノールレッド(注文番号:31870-025、Gibcoから)を含むRPMI 1640培地+200mMのグルタミン、5mg/lのペニシリン/ストレプトマイシン、及び200μg/mlのGeniticinを含む10%ウシ胎仔血清(FCS)中で継代培養し、インキュベータ(Heraeus)内で5%のCO2及び100%の相対湿度とともに37℃で培養する。試験前に、PC3−AR+を試験培地で2回継代培養する。これはGeniticinを含有せず、フェノールレッド(Gibco、注文番号:32404-014)を含有せず、ホルモン残留物を除去するために活性炭で精製されたウシ胎仔血清(DCS)を含有する。
【0076】
MMTV−プロモーターのホタルルシフェラーゼ活性との比較において、ウミシイタケ(Renilla)活性に基づいて、pRL融合ベクター内のクローン型1f−アロマターゼ・プロモーターに対するホルモンの影響を決定するために、トランスフェクション試験において、175ml瓶1つ当たり300000個のPC3−AR+細胞中に、8μgの1f−アロマターゼ−pRLプラスミド及び20μgのMMTV−lucプラスミドをトランスフェクトする。レポーター遺伝子プラスミドの一時的トランスフェクションは、GibcoBRLの指示書(Lipofectin-Reagent添付文書、Cat. No.:18292-037)に従って、リポフェクチンを用いて行われる。トランスフェクションから6時間後、細胞を洗浄し、そして1ウェル当たりの細胞数が10000となるように、96ウェル・マイクロタイタープレート上に分配する。96ウェル・プレートに移してから16時間後に、アゴニスト活性の測定のために単独で、そして、アンタゴニスト活性の測定のために10 x e-10Mのテストステロンの存在において、選択的アンドロゲン受容体モジュレーター(SARM)を添加する。インキュベータ内でさらに20時間にわたってインキュベートした後、細胞をPBSで洗浄し、溶解し、そしてレポーター遺伝子活性を決定する。製造業者の指示書(Promega; Luciferase Assay Substrate添付文書、Cat. No.: E151A、Luciferase Assay System with Reporter Lysis Buffer; Part TB161, USA 3/98から)に従って、dual Luciferase Assay Kit(注文番号:E 1960、Promegaから)を用いて相対光単位で、蛍及びウミシイタケ(Renilla)のルシフェラーゼ活性を決定する。光単位で示された測定値を任意に標準化するために、測定値を、用いられるタンパク質の量(μg)で割り算し、相対光単位(RLU)で結果を示す。
【0077】
アゴニズムに関する試験において、ここで試験されたSARMは、前の例で既に提示したDHTと同様に、2つのAR依存性プロモーターに対して極めて異なった形で作用する(図13)。しかし、DHTとは異なり、この事例では、全身的なMMTVプロモーターの活性化はないのに対して、性腺以外の部位のAR調節型プロモーターの例として役立つ神経1f−アロマターゼ・プロモーターが活性化される。DHTを用いた前の例と同様に、このこともin vivo及びヒトにおける組織選択性の可能性を指摘する。さらに、アンタゴニズムの並行測定は、AR活性の選択的モジュレーションの可能性を示唆することができる。すなわち、従来技術を凌いで、組織選択作用の可能性に関してSARM活性の示唆を得ることが可能である。
【0078】
本例にも明示されているように、測定はマイクロタイター形式で行うことができる。このことは、本発明による試験システムの基本的なハイスループット能力を強調する。本例におけるように全般的に明らかにアンタゴニスト性を有するが、しかし特異的な組織では或るアゴニスト作用をも示すSARMは、種々様々な指示に関して特に興味深いものである(S.S. Wolf and M. Obendorf, 2004, in E. Nieschlag & H. M. Behre: “Testosterone” 3rd. Edition; Cambridge University Press, ISBN 0521833809; 623-640)。
【特許請求の範囲】
【請求項1】
配列番号1に従う1f−アロマターゼ・プロモーター及びレポーター遺伝子から成るプロモーター・レポーター遺伝子融合体。
【請求項2】
前記レポーター遺伝子がホタルルシフェラーゼ遺伝子又はウミシイタケルシフェラーゼ遺伝子を含む群から選択することができること、そして前記融合生成物を細胞系内で安定的に又は一時的に発現させることができることを特徴とする、請求項1に記載のプロモーター・レポーター遺伝子融合体。
【請求項3】
細胞培養系内において、アンドロゲン受容体に結合することにより神経組織内のアロマターゼ調節に影響を与える物質を同定することができることを特徴とする、請求項1又は2にプロモーター・レポーター遺伝子融合体の使用。
【請求項4】
前記アンドロゲン受容体に結合することによって脳内のアロマターゼ調節に影響を与える物質を同定するために、配列番号1に規定された配列に対して90%以上、好ましくは95%以上の相同性を有する配列が細胞培養系内で使用されることを特徴とする、請求項3に記載の使用。
【請求項5】
アンドロゲン受容体に結合する物質の組織選択作用の決定方法であって、以下の段階:
a. アゴニスト活性及びアンタゴニスト活性の両方を同定するために、細胞培養系内のレポーター遺伝子アッセイにプロモーター・レポーター遺伝子融合体を使用して、アロマターゼ調節のモジュレーションを決定する段階、
b. 別のアンドロゲン調節プロモーターのモジュレーションを決定する段階、及び
c. 1f−アロマターゼ・プロモーターに対する選択的ARモジュレーションに関連して物質の完全な試験管内における特徴付けを達成するために段階a及びbを比較するか、あるいは他のAR調節型プロモーターと直接的に比較する段階、
を含んで成る、決定方法。
【請求項6】
配列番号1に規定された配列に対して90%以上、好ましくは95%以上の相同性を有する配列が、段階a)で使用されることを特徴とする、請求項5に記載の物質の組織選択作用の決定方法。
【請求項7】
中枢神経系内のアロマテーゼ発現を選択的に増大させ、且つ/又はARの活性に組織選択的な影響を与えるのに適した物質の同定及び特徴付けのための試験管内試験システムであって、該試験システムが、請求項1又は2に記載のレポーター遺伝子とアロマターゼ・プロモーターとの融合体を含有することを特徴とする、試験管内試験システム。
【請求項1】
配列番号1に従う1f−アロマターゼ・プロモーター及びレポーター遺伝子から成るプロモーター・レポーター遺伝子融合体。
【請求項2】
前記レポーター遺伝子がホタルルシフェラーゼ遺伝子又はウミシイタケルシフェラーゼ遺伝子を含む群から選択することができること、そして前記融合生成物を細胞系内で安定的に又は一時的に発現させることができることを特徴とする、請求項1に記載のプロモーター・レポーター遺伝子融合体。
【請求項3】
細胞培養系内において、アンドロゲン受容体に結合することにより神経組織内のアロマターゼ調節に影響を与える物質を同定することができることを特徴とする、請求項1又は2にプロモーター・レポーター遺伝子融合体の使用。
【請求項4】
前記アンドロゲン受容体に結合することによって脳内のアロマターゼ調節に影響を与える物質を同定するために、配列番号1に規定された配列に対して90%以上、好ましくは95%以上の相同性を有する配列が細胞培養系内で使用されることを特徴とする、請求項3に記載の使用。
【請求項5】
アンドロゲン受容体に結合する物質の組織選択作用の決定方法であって、以下の段階:
a. アゴニスト活性及びアンタゴニスト活性の両方を同定するために、細胞培養系内のレポーター遺伝子アッセイにプロモーター・レポーター遺伝子融合体を使用して、アロマターゼ調節のモジュレーションを決定する段階、
b. 別のアンドロゲン調節プロモーターのモジュレーションを決定する段階、及び
c. 1f−アロマターゼ・プロモーターに対する選択的ARモジュレーションに関連して物質の完全な試験管内における特徴付けを達成するために段階a及びbを比較するか、あるいは他のAR調節型プロモーターと直接的に比較する段階、
を含んで成る、決定方法。
【請求項6】
配列番号1に規定された配列に対して90%以上、好ましくは95%以上の相同性を有する配列が、段階a)で使用されることを特徴とする、請求項5に記載の物質の組織選択作用の決定方法。
【請求項7】
中枢神経系内のアロマテーゼ発現を選択的に増大させ、且つ/又はARの活性に組織選択的な影響を与えるのに適した物質の同定及び特徴付けのための試験管内試験システムであって、該試験システムが、請求項1又は2に記載のレポーター遺伝子とアロマターゼ・プロモーターとの融合体を含有することを特徴とする、試験管内試験システム。
【図1】


【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2010−531657(P2010−531657A)
【公表日】平成22年9月30日(2010.9.30)
【国際特許分類】
【出願番号】特願2010−513992(P2010−513992)
【出願日】平成20年7月3日(2008.7.3)
【国際出願番号】PCT/EP2008/058622
【国際公開番号】WO2009/004068
【国際公開日】平成21年1月8日(2009.1.8)
【出願人】(300049958)バイエル・シエーリング・ファーマ アクチエンゲゼルシャフト (357)
【Fターム(参考)】
【公表日】平成22年9月30日(2010.9.30)
【国際特許分類】
【出願日】平成20年7月3日(2008.7.3)
【国際出願番号】PCT/EP2008/058622
【国際公開番号】WO2009/004068
【国際公開日】平成21年1月8日(2009.1.8)
【出願人】(300049958)バイエル・シエーリング・ファーマ アクチエンゲゼルシャフト (357)
【Fターム(参考)】
[ Back to top ]