
イソプレノイドの生成
本発明は、微生物による、イソプレノイド、具体的にはコエンザイムQ−10の生成の方法に関する。より具体的には、本発明は、異なる微生物、好ましくはパラコッカス属、より好ましくはパラコッカス・ゼアキサンチニファシエンス種由来のメバロン酸(mev)オペロンの1つ以上の遺伝子で形質転換され、それによってmevオペロンが変異して向上したコエンザイムQ−10生成につながるロドバクター属、好ましくはロドバクター・スフェロイデス種の微生物による、コエンザイムQ−10の向上した生成の方法に関する。かかる変異を担持する配列ならびにかかる変異mevオペロンを担持する微生物もまた、含まれる。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
本発明は、イソプレノイド、具体的には微生物によるコエンザイムQ−10の生成の方法に関する。より具体的には、本発明は、異なる微生物由来、好ましくはパラコッカス属、より好ましくはパラコッカス・ゼアキサンチニファシエンス種のメバロン酸(mev)オペロンの1つ以上の遺伝子で形質転換された、それによってmevオペロンが変異して、増大したコエンザイムQ−10生成につながるロドバクター属の微生物、好ましくはロドバクター・スフェロイデス(R.sphaeroides)種の微生物による、コエンザイムQ−10の増大した生成の方法に関する。かかる変異を担持する配列ならびにかかる変異mevオペロンを担持する微生物もまた、含まれる。
【0002】
コエンザイムQ−10(2,3−ジメトキシ−ジメチル−6−デカプレニル−1,4−ベンゾキノン)は、ユビキノン10としても公知であり、10個のC−5イソプレノイド単位で構成されるイソプレノイド側鎖を有する脂溶性ベンゾキノンである。コエンザイムQ−10(以下CoQ10と略す)は、微生物および植物において、ならびに動物において見られる。これは、ヒトおよび殆どの哺乳動物における、ユビキノンの最も一般的な形態である。CoQ10がヒトの健康状態および疾患からのその防御における重要な因子であるという、確立され、かつ発展中の証拠がある。CoQ10の医学的および健康上の有益な効果は、その2つの主要な生理学的機能、すなわちミトコンドリアの電子伝達系(アデノシン三リン酸の合成に連結されている)の必須の補因子としての機能および脂溶性抗酸化物質としての作用と関連づけられている。
【0003】
CoQ10の健康上の利益により、この化合物の商業的重要性が高まった。CoQ10は、化学合成によって、または微生物を用いた発酵によって生成され得る。これらの微生物は、遺伝子工学によってCoQ10生成に関して改良された天然のCoQ10生産者であり得るか、またはそれらは天然にCoQ10を生成し得ないが遺伝子工学によってそれを合成できるように操作され得る。
【0004】
細菌においては、ユビキノンのイソプレノイド尾部はC−5化合物イソペンテニルピロリン酸(IPP)に由来するが、ユビキノンのキノイド環は芳香族化合物の生合成における中心的中間体であるコリスミ酸に由来する。キノイド環に付加されるイソプレノイド尾部の長さは、細菌中に存在する特定のプレニルトランスフェラーゼ酵素に依存する。例えば、大腸菌(Escherichia coli)においては、オクタプレニルピロリン酸シンターゼが、ファルネシルピロリン酸(FPP、C−15)および5個のIPP単位からのオクタプレニルピロリン酸(C−40)の形成を触媒する。キノイド環へのこの分子の付加は、結果としてユビキノン−8の形成を生じる。パラコッカスおよびロドバクター種においては、デカプレニルピロリン酸(DPP)シンターゼが、FPP(C−15)および7個のIPP単位からのDPP(C−50)の形成を触媒する。次いで、キノイド環へのDPPの付加が、結果としてユビキノン−10(CoQ10)の形成を生じる。
【0005】
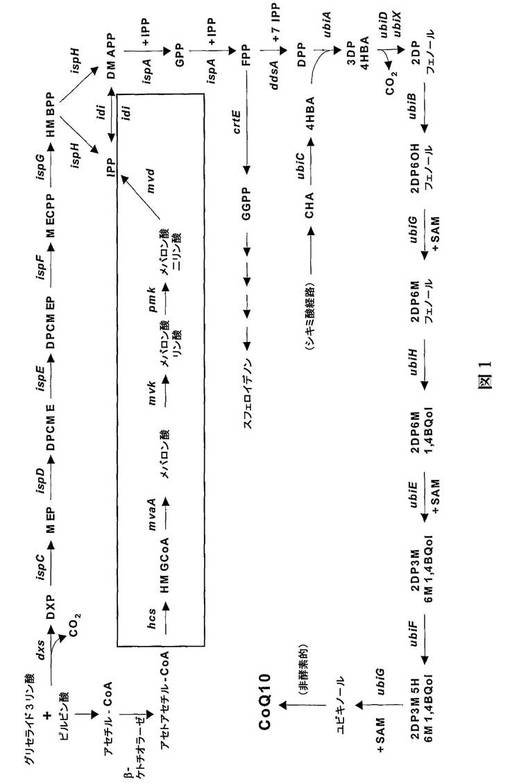
自然界においては、2つの異なる経路がIPPの生合成で知られている(図1)。メバロン酸経路は、その名前が含意するように、中心的中間体としてメバロン酸を利用し、真核生物においてよく研究されている。メバロン酸経路は、長年の間、自然界におけるIPP合成の普遍的な経路と考えられてきた。しかしながら、この十年間に、非メバロン酸経路またはMEP経路(中間体として2C−メチル−D−エリスリトール4−リン酸を有するため)と呼ばれるIPP生合成の第二の経路が発見された。MEP経路は、これまでに、多くの真正細菌中および高等植物のプラスチド区画中に存在することが示されている。
【0006】
ロドバクター・スフェロイデス中の、1−デオキシ−D−キシルロース5−リン酸レダクトイソメラーゼをコードするyaeM遺伝子(現在ispCと呼ばれている)の存在および殆ど完了しているロドバクター・スフェロイデスのゲノム配列の精査に基づくと、この細菌は、IPPの生合成にMEP経路を排他的に使用するようである。ロドバクター・スフェロイデスにおけるMEP経路の排他的使用を支持するさらなる証拠は、密接に関連する種、ロドバクター・カプスラータ(Rhodobacter capsulatus)が、イソプレノイド生合成にMEP経路のみを使用するという発見である。
【0007】
国際公開第02/26933A1号パンフレットは、MEP経路の酵素のいくつかをコードする数個の天然および/または異種の遺伝子を過剰発現することによってロドバクター・スフェロイデスにおけるCoQ10の生成を増大させる方法を開示している。しかしながら、これらの遺伝子の過剰発現は、結果として、CoQ10生成の、非常に穏当な改良を生じた。
【0008】
上述のように、いくつかの細菌は、IPPの生合成に、メバロン酸経路のみを使用する。パラコッカス・ゼアキサンチニファシエンスは、かかる細菌の一例である。パラコッカス・ゼアキサンチニファシエンスにおいて、メバロン酸経路の5つの遺伝子をコードする遺伝子に加えてIPPイソメラーゼをコードする遺伝子(図1参照)が、染色体上の単一の転写単位、すなわち以下メバロン酸(mev)オペロンと呼ばれるオペロンに、共にクラスター化される(フンベリン(Huembelin)ら、ジーン(Gene)297、129−139頁、2002年)。
【0009】
イソプレノイド、具体的にはCoQ10の生成は、mevオペロンの1つ以上の遺伝子を天然に欠損した、すなわち天然にイソプレノイド生成に非メバロン酸経路を使用する微生物への変異mevオペロンの導入によって、相当に増大され得ることがわかっており、ここで、1つ以上の変異を含む、完全なmevオペロンまたは前記mevオペロンの1つ以上の遺伝子のいずれかが導入され得る。1つ以上の変異は、例えば、以下の遺伝子の1つまたは全てにあり得る。ヒドロキシメチルグルタリル−CoAレダクターゼをコードするmvaA、イソペンテニル二リン酸イソメラーゼをコードするidi、ヒドロキシメチルグルタリル−CoAシンターゼをコードするhcs、メバロン酸キナーゼをコードするmvk、ホスホメバロン酸キナーゼをコードするpmk、およびジホスホメバロン酸デカルボキシラーゼをコードするmvd。
【0010】
したがって、本発明は、ヒドロキシメチルグルタリル−CoAレダクターゼ活性、イソペンテニル二リン酸イソメラーゼ活性、ヒドロキシメチルグルタリル−CoAシンターゼ活性、メバロン酸キナーゼ活性、ホスホメバロン酸キナーゼ活性、および/またはジホスホメバロン酸デカルボキシラーゼ活性を有するタンパク質をコードする1つ以上の遺伝子を含むポリヌクレオチド配列を提供し、前記ポリヌクレオチド配列は、微生物中に存在する場合にイソプレノイドの向上した生成につながる1つ以上の変異を担持する。イソプレノイド生成の向上は、例えば、前記ポリヌクレオチド配列を担持しない、すなわちmev経路を使用しない野生型微生物における生成と、本発明におけるポリヌクレオチド配列を担持する微生物との比較によって、測定され得る。
【0011】
ある実施形態において、本発明は、配列番号1または2から入手可能な、すなわち野生型mevオペロンの遺伝子またはその断片を含む、ポリヌクレオチド配列に関し、ここで断片は、mevオペロンの遺伝子、例えばヒドロキシメチルグルタリル−CoAレダクターゼをコードするmvaA、イソペンテニル二リン酸イソメラーゼをコードするidi、ヒドロキシメチルグルタリル−CoAシンターゼをコードするhcs、メバロン酸キナーゼをコードするmvk、ホスホメバロン酸キナーゼをコードするpmk、およびジホスホメバロン酸デカルボキシラーゼをコードするmvdの、少なくとも1つの活性を有する。好ましくは、変異ポリヌクレオチド配列は配列番号3またはその断片によって表され、微生物中に存在する場合に、例えばCoQ10生成の増大につながる。
【0012】
ある態様において、本発明は、変異を担持するmevオペロンを含むDNA配列に関し、前記DNA配列は配列番号3によって表される。
【0013】
用語「向上したイソプレノイド生成」、具体的には向上したCoQ10生成は、本明細書中で使用される場合、例えば、それぞれの野生型ポリヌクレオチドを担持するそれぞれの微生物と比較した場合に、上述のように1つ以上の変異を含むポリヌクレオチドを担持する微生物によって得られる、少なくとも約10%の増大を意味する。イソプレノイドの生成は、標準的な方法、例えばHPLC(実施例2参照)によって測定され、mg/lで、またはmg/l/OD600として表され得る(実施例5参照)。
【0014】
完全なmevオペロンまたは1つ以上の変異を担持するその1つ以上の遺伝子は、例えば、完全に合成されるか、または部分的に単離および合成されるかのいずれかであり得る。単離されたmevオペロンまたはその1つ以上の遺伝子は、メバロン酸経路を使用する任意の微生物から生じ得、すなわち、ここで前記オペロン内に含まれる1つ以上の遺伝子は、天然に生じる。好ましくは、微生物はパラコッカス属に属し、より好ましくは、例えばパラコッカス種R114またはパラコッカス・ゼアキサンチニファシエンスATCC21588等のパラコッカス・ゼアキサンチニファシエンス(Paracoccus zeaxanthinifaciens)に由来する。完全に機能的であるオペロンのDNA配列の対立遺伝子の変異および突然変異もまた、上記の用語に含まれる。パラコッカス・ゼアキサンチニファシエンスATCC21588の完全なmevオペロンは配列番号1に示され、パラコッカス種R114の完全なmevオペロンは配列番号2に示される(国際公開第02/099095号パンフレットの配列番号42も参照)。
【0015】
系統パラコッカス種R114は、パラコッカス・ゼアキサンチニファシエンスATCC21588の派生物であり、ブダペスト条約の合意の下、特許寄託名称PTA−3335で2001年4月24日にATCCと共に寄託されている。本発明に使用され得るパラコッカス種および系統ならびにパラコッカスとしてのフラボバクテリウム種の分類学的再分類に関しては、国際公開第02/099095号パンフレット、47頁が参照される。使用され得るかかる種の例は、パラコッカス・マルクシイ(P.marcusii)、パラコッカス・カロチニファシエンス(P.carotinifaciens)、パラコッカス・ソルベンティボランス(P.solventivorans)、パラコッカス・ゼアキサンチニファシエンスまたはパラコッカス種R114である。
【0016】
本発明のmevオペロンは、オペロン内の任意の位置に位置し得る1つ以上の変異を担持し、前記オペロン内の1つ以上の遺伝子の活性の変化につながり、結果として、かかるポリヌクレオチドを担持する微生物内の、イソプレノイドの向上した生成を生じる。
【0017】
ある実施形態において、mevオペロンは少なくとも1つの変異を担持し、これは好ましくはmevオペロンのhcs遺伝子中、例えばパラコッカス・ゼアキサンチニファシエンスのmevオペロンのhcs遺伝子中、例えばパラコッカス種R114またはパラコッカス・ゼアキサンチニファシエンスATCC21588のmevオペロンのhcs遺伝子中に位置する。より好ましくは、変異mevオペロンは、本発明において使用される場合、配列番号3によって表される。パラコッカス種R114の野生型hcs遺伝子の配列およびそれぞれのタンパク質は、それぞれ配列番号4および5に示される。
【0018】
ある実施形態において、本発明は、hcs遺伝子に1つ以上の変異を含むポリヌクレオチド、好ましくは配列番号7に示されるヒドロキシメチルグルタリル−CoAシンターゼを有するタンパク質をコードする、配列番号6に示されるポリヌクレオチドに関し、ここで配列番号5の90位に存在するグルタミンはリシンに置換される。
【0019】
したがって、mevオペロンの1つ以上の遺伝子を含む上述のようなポリヌクレオチド配列を提供することが本発明の目的であり、前記ポリヌクレオチドは、
(a)配列番号7を一致させるアミノ酸配列を含むポリペプチドをコードするポリヌクレオチド、
(b)配列番号6のヌクレオチド配列を含むポリヌクレオチド、
(c)(a)または(b)のポリヌクレオチドによってコードされるポリペプチドの断片または派生物をコードするヌクレオチド配列を含むポリヌクレオチドであって、ここで前記派生物において、前記ポリペプチドと比較して1つ以上のアミノ酸残基が保存的に置換され、前記断片または派生物がヒドロキシメチルグルタリル−CoAシンターゼ(Hcs)活性を有する、ポリヌクレオチド、
(d)ストリンジェントな条件下でその相補鎖が(a)〜(c)のいずれか1つに定義されるポリペプチドにハイブリダイズし、Hcsタンパク質をコードする、ポリヌクレオチド
からなる群より選択される。
【0020】
本発明のポリペプチドおよびポリヌクレオチドは、好ましくは単離された形態で提供され、好ましくは、均質になるまで精製される。
【0021】
用語「単離された」は、物質がその元々の環境(例えば、それが天然に生じる場合、天然の環境)から除去されることを意味する。例えば、生きている微生物中に存在する天然に生じるポリヌクレオチドまたはポリペプチドは単離されていないが、天然の系に共存する物質の一部または全てから分離された同じポリヌクレオチドまたはポリペプチドは、単離されている。かかるポリヌクレオチドはベクターの一部であり得、かつ/またはかかるポリヌクレオチドもしくはポリペプチドは、組成物の一部であり得、さらに、かかるベクターまたは組成物が天然の環境の一部でないという点で、単離され得る。
【0022】
単離されたポリヌクレオチドまたは核酸は、本明細書中で使用される場合、それが由来する生物の天然に生じるゲノムにおいて直接隣接している(一方は5’末端で、一方は3'末端で)コード配列の両方に直接隣接していないDNAまたはRNAであり得る。したがって、ある実施形態において、核酸は、コード配列に直接隣接する5’非コード(例えばプロモーター)配列の一部または全てを含む。用語「単離されたポリヌクレオチド」は、そのため、例えば、ベクター、自己複製プラスミドもしくはウイルス、または原核生物もしくは真核生物のゲノムDNAに組み込まれたか、または他の配列から独立した分離した分子として存在する(例えば、PCRまたは制限エンドヌクレアーゼ処理によって生じたcDNAまたはゲノムDNA断片)、組み換えDNAを含む。これはまた、実質的に細胞性物質、ウイルス性物質または培地(組み換えDNA技術によって生成された場合)または化学的前駆体もしくは他の化学物質(化学的に合成された場合)を含まないさらなるポリペプチドをコードするハイブリッド遺伝子の一部である組み換えDNAを含む。さらに、「単離された核酸断片」は、断片として天然に生じず、天然の状態で見られない核酸断片である。
【0023】
遺伝子は、コード配列、例えば遺伝子のコード領域の3’および5’末端に位置する非翻訳配列等の非コード配列、ならびに制御配列を含み得る。さらに、遺伝子は、本明細書中に定義される単離された核酸分子をいう。タンパク質のアミノ酸配列の変化につながるDNA配列多型が集団内に存在し得ることが、当業者によってさらに理解される。
【0024】
本発明はまた、ストリンジェントな条件下、好ましくは高度にストリンジェントな条件下で、例えば配列番号6に示されるポリヌクレオチド等の本発明のポリヌクレオチドにハイブリダイズできる、単離されたポリヌクレオチドに関する。
【0025】
本明細書中で使用される場合、用語「ハイブリダイズする」は、互いに少なくとも約50%、少なくとも約60%、少なくとも約70%、より好ましくは少なくとも約80%、さらにより好ましくは少なくとも約85%〜90%、最も好ましくは少なくとも95%相同なヌクレオチド配列が典型的に互いにハイブリダイズしたままである、ハイブリダイゼーションおよび洗浄の条件を説明するよう意図される。
【0026】
かかるハイブリダイゼーション条件の好ましい、非限定的な例は、6×塩化ナトリウム/クエン酸ナトリウム(SSC)中約45℃でのハイブリダイゼーション、それに続く1×SSC、0.1%SDS中50℃、好ましくは55℃、より好ましくは60℃、さらにより好ましくは65℃での1回以上の洗浄である。
【0027】
高度にストリンジェントな条件としては、例えば、100μg/mlサケ精子DNAありもしくはなしのDigイージーハイブ(DigEasyHyb)溶液(ロシュ・ダイアグノスティクス・ゲーエムベーハー(Roche Diagnostics GmbH))、または50%ホルムアミド、5×SSC(150mM NaCl、15mMクエン酸三ナトリウム)、0.02%ドデシル硫酸ナトリウム、0.1%N−ラウロイルサルコシンおよび2%ブロッキング試薬(ロシュ・ダイアグノスティクス・ゲーエムベーハー(Roche Diagnostics GmbH))を含有する溶液等の溶液中のジゴキシゲニン(DIG)標識DNAプローブ(DIG標識システムを用いて調製される、ドイツ連邦共和国マンハイム68298のロシュ・ダイアグノスティクス・ゲーエムベーハー(Roche Diagnostics GmbH、68298Mannheim、Germany))を用いた2時間〜4日間42℃でのインキュベーション、それに続く、5〜15分2×SSCおよび0.1%SDS中室温で2回のフィルターの洗浄ならびにそれに次ぐ15〜30分0.5×SSCおよび0.1%SDSまたは0.1×SSCおよび0.1%SDS中65〜68℃で2回の洗浄が挙げられる。
【0028】
当業者は、ストリンジェントおよび高度にストリンジェントなハイブリダイゼーション条件のいずれの条件を適用すべきかを知るであろう。かかる条件に関するさらなる手引きは、当該分野において、例えばサンブルック(Sambrook)ら、1989年、モレキュラー・クローニング、ラボラトリー・マニュアル、ニューヨーク州のコールドスプリングハーバープレス(Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Press、N.Y.)およびアウスベル(Ausubel)ら(編)、1995年、カレント・プロトコールズ・イン・モレキュラー・バイオロジー(Current Protocols in Molecular Biology)(ニューヨークのジョン・ワイリー・アンド・サンズ(John Wiley & Sons、N.Y.))において容易に入手可能である。勿論、(mRNAの3’末端のポリ(A)域等の)ポリ(A)配列またはT(もしくはU)残基の相補的伸展のみにハイブリダイズするポリヌクレオチドは、かかるポリヌクレオチドはポリ(A)伸展またはその相補物を含む任意の核酸分子(例えば、事実上任意の二本鎖cDNAクローン)にハイブリダイズするので、本発明の核酸の一部に特異的にハイブリダイズするのに用いられる本発明のポリヌクレオチドに含まれない。
【0029】
本発明を、イソプレノイド、具体的にはユビキノン、好ましくはCoQ10の生成から使用し得る。好ましくは、1つ以上の変異を含むポリヌクレオチド、すなわち完全なmevオペロン、またはその断片のみが、mevオペロンの1つ以上の遺伝子が元々欠損している、すなわちイソプレノイドの生成に天然にMEP経路のみを使用している、適した微生物に導入される。
【0030】
変異mevオペロンまたは上述のその1つ以上の遺伝子は、例えば、ロドバクター属の微生物に導入され得る。
【0031】
具体的には、本発明は、(1)本発明におけるポリヌクレオチドを、前記ポリヌクレオチドを元々欠損している微生物に導入する工程、および(2)工程(1)の微生物を、イソプレノイドの生成を可能にする条件下で培養する工程を含む、イソプレノイド、具体的にはユビキノン、好ましくはCoQ10の生成の方法に関する。好ましくは、パラコッカス属に属する微生物の変異mevオペロンまたはその断片が、ロドバクター属に属する微生物に導入される。より好ましくは、ヒドロキシメチルグルタリル−CoAシンターゼをコードする遺伝子に1つ以上の変異を含むポリヌクレオチドが前記微生物に導入され、配列番号3によって表されるポリヌクレオチド配列の導入が最も好ましい。
【0032】
mevオペロンの1つ以上の遺伝子を元々欠損している、すなわちイソプレノイドの生成にMEP経路を使用する任意の微生物が、本発明の目的のために使用され得る。具体的には、ロドバクター属の微生物が、例えばロドバクター・スフェロイデス、ロドバクター・アドリアティクス(R.adriaticus)、ロドバクター・カプスラータ(R.capsulatus)、ロドバクター・スルフィドフィルス(R.sulfidophilus)、またはロドバクター・ベルドカンピイ(R.veldkampii)等の、本発明における変異ポリヌクレオチドの導入に使用され得る。好ましい系統はロドバクター・スフェロイデスであり、ロドバクター・スフェロイデスATCC35053が、さらにより好ましい。
【0033】
本明細書中に挙げられた微生物はまた、国際原核生物命名規約に定義される、同じ物理化学的特性を有するかかる種の異名または基礎異名も含むことが理解される。
【0034】
本発明はさらに、上述のポリヌクレオチドを導入した、すなわち1つ以上の変異を担持するmevオペロンの1つ以上の遺伝子を含む、組み換え微生物を提供し、ここでそれぞれの野生型微生物は、元々mevオペロンの1つ以上の遺伝子を欠損しており、すなわちイソプレノイドの生成にMEP経路を使用しており、それぞれの野生型微生物と比較して、イソプレノイド、具体的にはユビキノン、好ましくはCoQ10の、向上した生成につながる。
【0035】
上述のmevオペロンの1つ以上の遺伝子内の変異の他に、組み換え微生物は、前記微生物内のイソプレノイドの生成の向上につながる限りは、さらなる修飾/変化を含み得る。
【0036】
ある実施形態において、かかるさらなる修飾は、デカプレニル二リン酸シンターゼ活性を有するタンパク質をコードするDNA配列の、好ましくはパラコッカス属の微生物、例えばパラコッカス・ゼアキサンチニファシエンス、具体的にはパラコッカス・ゼアキサンチニファシエンスATCC21588から入手可能な、前記組み換え微生物への導入である。配列番号8におけるポリヌクレオチド配列または配列番号9におけるタンパク質をコードするポリヌクレオチドが最も好ましい。
【0037】
したがって、本発明は、上述のような、デカプレニル二リン酸シンターゼ活性を有するタンパク質をコードするDNA配列が、MEP経路を天然に使用していて上述のように導入された1つ以上の変異を含むmevオペロンの1つ以上の遺伝子を有する微生物、例えばロドバクター属の微生物にさらに導入される、イソプレノイド、好ましくはCoQ10の生成の方法に関する。
【0038】
デカプレニル二リン酸シンターゼ活性を有するタンパク質をコードする任意のDNA配列が、本発明に使用され得る。かかるDNAの例は、パラコッカス属、例えばパラコッカス・ゼアキサンチニファシエンス、より好ましくはパラコッカス・ゼアキサンチニファシエンスATCC21588の微生物のデカプレニル二リン酸シンターゼ活性を有するタンパク質をコードする、ddsA遺伝子またはその部分配列である。デカプレニル二リン酸シンターゼ(配列番号7)をコードする、配列番号6によって表されるパラコッカス・ゼアキサンチニファシエンスATCC21588由来のddsA遺伝子が最も好ましい。
【0039】
したがって、(1)メバロン酸(mev)オペロン、およびパラコッカス属に属する微生物のデカプレニル二リン酸シンターゼ活性を有するタンパク質をコードするDNA配列の両方を、ロドバクター属に属する微生物に導入する工程であって、ここで前記mevオペロンが1つ以上の変異を担持する、工程、ならびに(2)修飾されたロドバクター系統を培養する工程を含む、CoQ10の生成の方法を提供することは、本発明のある態様である。
【0040】
用語「に導入する」は、本明細書および特許請求の範囲において、微生物または宿主生物、例えばロドバクター属の形質転換に関して、遺伝物質、具体的には変異メバロン酸オペロンまたは1つ以上のその遺伝子を宿主生物に効率的にもたらすよう、すなわち生物によって発現されるように使用され得る、当業者に周知の任意の方法を含むよう使用される。導入は、標準的な方法に従って、例えばベクター、好ましくは発現プラスミドによって、または宿主のゲノムへの組み込みによって、実施され得る。好ましい方法は、例えばPcrtEプロモーター制御下の発現プラスミドpBBR−K−PcrtE(このプラスミドの構築は、国際公開第02/099095号パンフレットの91頁12〜27行目、実施例6に詳細に記載されている)にクローニングされる遺伝子等の、プラスミドを介した遺伝子の導入である。例えば発現プラスミド等のDNAを、ロドバクター属の微生物に導入する好ましい方法は、例えばロドバクター系統への大腸菌(E.coli)S17−1由来のプラスミドのコンジュゲーション転移等の、プラスミドのコンジュゲーション転移である(ニシムラ(Nishimura)ら、ニュークレイック・アシッズ・リサーチ(Nucl.Acids Res.)18、6169頁、1990年、サイモン(Simon)ら、バイオ/テクノロジー(Bio/Technology)1983年、784−91頁)。
【0041】
mevオペロンまたはその1つ以上の遺伝子の変異は、例えば、当業者が熟知していて具体的な説明を必要としない方法を用いた、PCR部位特異的変異誘発によって生じ得る。さらなる変異誘発方法は、これも本発明の目的に用いられ得、当業者に公知であるが、例えば、UV照射、転位または化学的変異誘発を含む。増大したイソプレノイド、例えばCoQ10の生産性を示す変異の同定のためのスクリーニング方法は、例えば、UV光、HPLC、NMRまたは薄層クロマトグラフィーによる、イソプレノイド、例えばCoQ10の直接の測定から選択され得る。イソプレノイド、例えばCoQ10の生成はまた、当業者が熟知しているカロテノイド色強度の増大を測定することによって、間接的に測定され得る。
【0042】
本発明における、変異mevオペロンまたはその1つ以上の遺伝子で形質転換された宿主は、公知の方法に従って、すなわち、例えば宿主に同化され得る炭素および窒素源、無機塩等を含有する培地中、効率的な増殖および所望の生成物、具体的にはCoQ10の発現に適した温度、pHおよび通気条件下で、培養され得る。
【0043】
発酵培地および/または形質転換体、すなわち例えばパラコッカス属に属する微生物の変異メバロン酸オペロンが導入された微生物からの単離、ならびに任意に、例えばヒトまたは動物用途のためのかかる生成されたCoQ10の製剤化を含む得られたイソプレノイド、具体的にはCoQ10の、精製およびさらなる処理が、当該分野で周知の方法に従って実施され得る。しかしながら、動物の健康および栄養物摂取における使用のためには、特別の精製が必要でなくてもよい。この場合、CoQ10のような生成されたイソプレノイドは、バイオマスおよび/または発酵培地の他の成分とともに、さらに処理されて商業的に魅力的な製品を生じ得る。
【0044】
本発明の方法は、結果として、高収量の、例えばCoQ10等のイソプレノイドを生じる。増大は、例えばロドバクターの非組み換え系統を用いた、または例えば、例えばパラコッカス系統の野生型mevオペロンを担持するロドバクター等の組み換え系統を用いた方法と比較して、例えば少なくとも約10%であり得る。
【0045】
以下の実施例は、本発明を全く制限することなく、本発明を説明する。
【0046】
実施例1:細菌および培養条件
ロドバクター・スフェロイデス系統ATCC 35053(アメリカ合衆国ヴァージニア州マナッサスのアメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection、Manassas VA、USA)から入手)を、CoQ10の向上した生成を有する組み換え系統の構築の基本宿主として用いた。全てのロドバクター・スフェロイデス系統を、30℃で、培地RS100中で増殖させた。培地RS100の組成および調製を、表1に要約する。50mg/lカナマイシンを、組み換え系統の増殖のために培地に添加した。大腸菌系統を37℃、LB培地(アメリカ合衆国メリーランド州スパークスのベクトン・ディッキンソン(Becton Dickinson、Sparks、MD、USA))中で増殖させた。組み換え大腸菌系統中でのプラスミドの維持のために、アンピシリン(100mg/l)および/またはカナマイシン(25〜50mg/l、プラスミドによる)を、培地に添加した。大腸菌およびロドバクター・スフェロイデスの液体培養物を、通法により、ロータリーシェイカー中、200rpmで好気的に増殖させた。固形培地が必要な場合は、寒天(1.5%終濃度)を添加した。
【0047】
【表1】

【0048】
実施例2:CoQ10の分析的アッセイ
400μlの全培地(実施例5参照)を、使い捨ての15mlポリプロピレン遠心チューブに移した。4mlの滅菌抽出溶液(1:1(v/v)DMSO/THF中の0.5g/l BHT)を添加し、試料をラボラトリーシェイカー(IKA、ドイツ連邦共和国(Germany))中で20分間混合し、抽出を促進した。最後に、試料を遠心分離し、逆相HPLCによる分析のために、上清を琥珀色のガラスバイアルに移した。この方法は、カロテノイドのフィトエン、スフェロイデノン、スフェロイデンおよびニューロスポレンからのCoQ10の明らかな分離を有する、ユビキノンおよびその対応するヒドロキノンの同時測定のために開発された。温度制御自動サンプラーおよびダイオードアレイ検出器を備えたアジレント(Agilent)1100HPLCシステム(アメリカ合衆国のアジレント・テクノロジー(Agilent Technologies、USA))を用いて、クロマトグラフィーを行った。方法パラメータは、以下の通りである。
【0049】
【表2】
【0050】
【表3】
【0051】
実施例3:パラコッカス・ゼアキサンチニファシエンスからの変異mevオペロンのクローニング
プラスチドpBBR−K−mev−op−wtおよびpBBR−K−mev−op−R114の構築
プラスミドpBBR−K−mev−op−up−4(メバロン酸オペロンの最初の4つの遺伝子を含むプラスミド)の構築は、国際公開第02/099095号パンフレットの実施例13(105頁10行目〜106頁8行目)に詳細に記載されている。
【0052】
パラコッカス・ゼアキサンチニファシエンスR114ゲノムDNAを鋳型として使用して、プライマーhcs−5326(配列番号10)およびmvd−9000(配列番号11)を用いてPCRを行う。プライマーmvd−9000は配列番号2のヌクレオチド6977〜6996からの配列の逆の相補物に対応するが、プライマーhcs−5326は、配列番号2のヌクレオチド3321〜3340からのパラコッカス・ゼアキサンチニファシエンスmevオペロンの配列に対応する。3675bpの、得られたPCR産物を、pCR2.1−TOPO(アメリカ合衆国カリフォルニア州カールスバッドのインビトロジェン(Invitrogen、Carlsbad、CA、USA))にクローニングし、結果としてプラスミドTOPO−pCR2.1−mev−op−d−3wtを生じる。このため、このプラスミドは、hcsの3'末端ならびに最後の3つの遺伝子mvk、pmkおよびmvdを含むメバロン酸オペロンの下流半分を含む。
【0053】
プラスミドpBBR−K−mev−op−up−4およびTOPO−pCR2.1−mev−op−d−3wtを、制限エンドヌクレアーゼSacIおよびNdeIで消化し、TOPO−pCR2.1−mev−op−d−3wt由来の得られた3319bp断片を、pBBR−K−mev−op−up−4由来の8027bp断片とライゲートする。得られたプラスミド、pBBR−K−mev−op−R114は、その(推定)プロモーターを含む、パラコッカス・ゼアキサンチニファシエンスR114由来の完全なメバロン酸オペロンを含む。
【0054】
プラスミドpBBR−K−mev−op−wt−PcrtE−crtEおよびpBBR−K−mev−op−R114−PcrtE−crtEの構築
プラスミドpBBR−K−PcrtE−crtEの構築は、国際公開第02/099095号パンフレットの実施例6(92頁10〜17行目)に詳細に記載されている。プラスミドpBBR−K−PcrtE−crtEをNaeIで切断し、1.33kb断片を単離し、pBBR−K−mev−op−up−4のEcl136II部位に挿入した。挿入の方向をチェックし、メバロン酸オペロン遺伝子と同じ方向でcrtE遺伝子を担持するプラスミドを、pBBR−K−mev−op−up−4−PcrtE−crtE−2と呼んだ。
【0055】
プラスミドpBBR−K−mev−op−up−4−PcrtE−crtE−2をSphIおよびSpeIで切断し、crtE遺伝子を含む得られた5566bp断片を単離した。この断片を、同じ酵素を用いたpBBR−K−mev−op−wtまたはpBBR−K−mev−op−R114の制限消化後に得られた7132bpのSphI−SpeI断片とライゲートし、結果としてプラスミドpBBR−K−mev−op−wt−PcrtE−crtEおよびpBBR−K−mev−op−R114−PcrtE−crtEをそれぞれ生じた。
【0056】
プラスミドpBBR−K−mev−op−wt−PcrtE−ddsAwtおよびpBBR−K−mev−op−R114−PcrtE−ddsAwtの構築
プラスミドpBBR−K−PcrtEの構築は、国際公開第02/099095号パンフレットの実施例6(91頁12〜27行目)に詳細に記載されている。
【0057】
パラコッカス・ゼアキサンチニファシエンス系統ATCC21588由来のddsA遺伝子(ddsAwtと呼ぶ)を、プライマーdds−Nde(配列番号12)およびdds−Bam(配列番号13)を用いたPCR(GCリッチPCRシステム、ドイツ連邦共和国マンハイムのロシュ・モレキュラー・バイオケミカルズ(Roche Molecular Biochemicals、Mannheim、Germany))によって増幅し、pCR2.1−TOPO(アメリカ合衆国カリフォルニア州カールスバッドのインビトロジェン(Invitrogen、Carlsbad、CA、USA))にクローニングし、結果としてプラスミドTOPO−ddsAwtを生じた。
【0058】
プラスミドTOPO−ddsAwtをNdeIおよびBamHIで切断し、ddsA遺伝子を含む1005bpの得られた断片を、NdeIおよびBamHIで切断したプラスミドpBBR−K−PcrtEにクローニングし、結果としてプラスミドpBBR−K−PcrtE−ddsAwtを生じた。オリゴヌクレオチドdds−R−1(配列番号14)およびdds−R−2(配列番号15)を用いたクイックチェンジXLサイト・ディレクティド・ミュータジェネシス・キット(QuikChange XL Site−Directed Mutagenesis Kit)(アメリカ合衆国カリフォルニア州ラ・ホーヤのストラタジーン(Stratagene、La Jolla、CA、USA))でサイレント変異を導入することによって、ddsA遺伝子内のエンドヌクレアーゼEcoRIの認識部位を排除した。得られたプラスミドpBBR−K−PcrtE−ddsAwt−Rを、EcoRIおよびMamIで切断し、ddsA遺伝子を含む断片1278bp断片を、EcoRIおよびEcoRVで切断したTOPO−ddsAwtに挿入し、結果としてプラスミドpCR2.1−TOPO−ddsAwt−Rを生じた。このプラスミドをCelIIおよびXbaIで切断し、ddsA遺伝子を含む得られた1211bpの断片を、CelIIおよびBlnIでのpBBR−K−mev−op−wt−PcrtE−crtEおよびpBBR−K−mev−op−R114−PcrtE−crtEそれぞれの消化から得られた11.6kpの制限断片とライゲートした。得られたプラスミドを、それぞれpBBR−K−mev−op−wt−PcrtE−ddsAwtおよびpBBR−K−mev−op−R114−PcrtE−ddsAwtと名づけた。
【0059】
プラスミドpBBR−K−mev−op−4−89−PcrtE−ddsAwtの構築
クイックチェンジXLサイト・ディレクティド・ミュータジェネシス・キット(QuikChange XL Site−Directed Mutagenesis Kit)(アメリカ合衆国カリフォルニア州ラ・ホーヤのストラタジーン(Stratagene、La Jolla、CA、USA))ならびにプライマーmut4−89−1−fw(配列番号16)およびmut4−89−1−rev(配列番号17)を用いた2回のPCR部位特異的変異誘発によって、プラスミドpBBR−K−mev−op−4−89−PcrtE−ddsAwtを得る。両方のプライマーは互いに相補的であり、配列番号4の268位に対応する配列番号3の2949位にCの代わりに所望の変異Aを含む。第一の変異誘発反応を、以下のように製造業者の使用説明書に従って設定する。5μlの10×反応バッファー、10ngのプラスミドDNApBBR−K−mev−op−wt−PcrtE−ddsAwt、125ngのプライマーmut4−89−1−fw、125ngのmut4−89−1−rev、1μlのdNTPミックス、3μlのクイックソリューション(QuikSolution)および2.5UのPfuターボ(PfuTurbo)DNAポリメラーゼを、50μlの最終体積で混合する。以下のパラメータを用いてサイクルを行う。1サイクル:95℃1分、18サイクル:95℃50秒、60℃50秒、68℃30分、1サイクル:68℃7分。反応混合物を37℃まで冷却した後、10Uの制限エンドヌクレアーゼDpnIを添加し、反応物を37℃で2時間インキュベートする。製造業者のプロトコルに従って、大腸菌XL10−ゴールド・ウルトラコンポーネント(Gold Ultracomponent)細胞(アメリカ合衆国カリフォルニア州ラ・ホーヤのストラタジーン(Stratagene、La Jolla、CA、USA))をDpnI処理DNAで形質転換する。
【0060】
得られたプラスミドpBBR−K−mev−op−4−89−PcrtE−ddsAwtを単離し、配列番号3の6948位でTの代わりに所望の変異Cを含むプライマーmut4−89−2−fw(配列番号18)およびmut4−89−2−rev(配列番号19)を用いた二回目のPCR部位特異的変異誘発の鋳型DNAとして使用する。上述のように変異誘発を行う。得られたプラスミドpBBR−K−mev−op−4−89−PcrtE−ddsAwtを単離し、マイクロシンス・ゲーエムベーハー(Microsynth GmbH)(スイス連邦バルガッハ(Balgach、Switzerland))で配列決定した。変異mevオペロンの完全な配列は配列番号3に表され、以下の遺伝子を含む。617〜1639位のmvaA(ヒドロキシメチルグルタリル−CoAレダクターゼをコードする)、1636〜2685位のidi(イソペンテニル二リン酸イソメラーゼをコードする)、2682〜3848位のhcs(ヒドロキシメチルグルタリル−CoAシンターゼをコードする)、3829〜4965位のmvk(メバロン酸キナーゼをコードする)、4965〜5882位のpmk(ホスホメバロン酸キナーゼをコードする)、および5875〜6873位のmvd(ジホスホメバロン酸デカルボキシラーゼをコードする)。
【0061】
実施例4:ロドバクター・スフェロイデスATCC35053への変異mevオペロンの導入
変異mevオペロンを担持するプラスミドでの大腸菌S17−1の形質転換(サイモン(Simon)ら、バイオ/テクノロジー(Bio/Technology)11、784−791頁、1983年)、およびそれに続く、コンジュゲーションによる大腸菌S17−1からロドバクター・スフェロイデスATCC35053へのプラスミドの転移を、標準的な手順を用いて行った(ニシムラ(Nishimura)ら、ニュークレイック・アシッズ・リサーチ(Nucl.Acids Res.)18、6169頁、1990年、サイモン(Simon)ら、バイオ/テクノロジー(Bio/Technology)1983年、784−91頁)。
【0062】
100mg/lリファンピシンを補給したRS100液体培地中で系統ATCC35053を増殖させること、100mg/lリファンピシンを含むRS100プレート上に細胞を平板培養すること、およびシングルコロニーを単離することによって、ロドバクター・スフェロイデスATCC35053の自然発生リファンピシン耐性突然変異をまず単離した。コンジュゲーションのために、100mg/lのリファンピシンおよびドナー細胞(50mg/lカナマイシンを含有するLB培地中で増殖させた、転移されるプラスミドを担持する大腸菌S17−1)を含有するRS100培地中で増殖させたレシピエント細胞(リファンピシン耐性ロドバクター・スフェロイデスATCC35053)の培養物の1ミリリットルのアリコートを、遠心分離によってペレット化した。上清を廃棄し、新鮮なRS100培地で細胞を2回洗浄して抗生物質を除去した。次いで、1mlの新鮮なRS100培地に各ペレットを再懸濁した。50μlのドナー細胞および0.45mlのレシピエント細胞を混合し、遠心分離によってペレット化し、0.03mlの新鮮なRS100培地に再懸濁し、RS100プレート上に点在させた。30℃で一晩のインキュベーションの後、接種ループで細胞を採取し、0.3mlのRS100培地に再懸濁した。この懸濁液の希釈溶液を、100mg/lリファンピシンおよび50mg/lカナマイシンを含むRS100プレート上に広げ、30℃でインキュベーションした。コロニー(ロドバクター・スフェロイデスATCC35053の、推定の形質転換細胞)をプレートから選び取り、50mg/lカナマイシンを含有する液体RS100培地中で増殖させ、56℃のアニーリング温度および以下の2つの異なるプライマー対を用いた1分、15秒の伸長時間でPCR反応においてプラスミドの存在を試験した。
pBBR−K−up(配列番号20)/PcrtE−2442(配列番号21)
Kan3out(配列番号22)/mvaA3256(配列番号23)
【0063】
50mg/lカナマイシンを含むRS100プレート上に、陽性のクローンで筋をつけ、シングルコロニーを得た。各クローン由来の1つのシングルコロニーを、50mg/lカナマイシンを含有する液体RS100培地中で再度増殖させ、上述のように、期待されるプラスミドの存在をPCRによって確認した。得られた組み換え系統を、ATCC35053/pBBR−K−mev−op−4−89−PcrtE−ddsAwtと呼んだ。
【0064】
実施例5:ロドバクター・スフェロイデスATCC35053の形質転換系統におけるCoQ10の生成
ロドバクター・スフェロイデス系統ATCC35053、ATCC35053/pBBR−K−mev−op−R114−PcrtE−ddsAwtおよびATCC35053/pBBR−K−mev−op−4−89−PcrtE−ddsAwtを、RS100培地中の振とうフラスコ培養において増殖させた。組み換えロドバクター・スフェロイデスを含む培養物は、50mg/lカナマイシンを含んでいた。25mlの培養物を30℃で250mlのバッフル三角フラスコ中、200rpmで振とうしながら増殖させた。CoQ10生成を試験するために、ロドバクター・スフェロイデス系統の凍結したグリセロール化ストック培養物を用い、25mlの種培養物を接種した。24〜28時間の種培養物の増殖の後、適した体積の培養物を用いて、660ナノメートルでの初期光学密度(OD660)が0.16であるように、実験フラスコを接種した。2mlの試料を、24時間間隔で無菌的に採取した。分析は、増殖(OD660として測定される)、pH、培養物上清中のグルコースならびに実施例2に記載されるCoQ10およびカロテノイド(HPLCによって測定される)を含んだ。結果を表3に要約する。これらの結果は、パラコッカス・ゼアキサンチニファシエンス由来のクローン化変異メバロン酸オペロンの発現がロドバクター・スフェロイデスにおけるCoQ10生成を有意に向上させたことを明らかに示す。
【0065】
【表4】
【図面の簡単な説明】
【0066】
【図1】図1は、ロドバクター・スフェロイデスにおけるCoQ10生合成の経路を表し、これはIPP形成にMEP経路を使用する。囲まれた領域は、(IPPの形成につながる)メバロン酸経路に加えてIPPイソメラーゼ段階を含む、反応順序を示す。メバロン酸経路は、ロドバクター・スフェロイデスにおいては天然に生じない。パラコッカス・ゼアキサンチニファシエンスにおいては、メバロン酸経路の5つの酵素に加えてIPPイソメラーゼをコードする遺伝子が、以下メバロン酸オペロンと呼ばれる、オペロンを形成する。
【発明の詳細な説明】
【0001】
本発明は、イソプレノイド、具体的には微生物によるコエンザイムQ−10の生成の方法に関する。より具体的には、本発明は、異なる微生物由来、好ましくはパラコッカス属、より好ましくはパラコッカス・ゼアキサンチニファシエンス種のメバロン酸(mev)オペロンの1つ以上の遺伝子で形質転換された、それによってmevオペロンが変異して、増大したコエンザイムQ−10生成につながるロドバクター属の微生物、好ましくはロドバクター・スフェロイデス(R.sphaeroides)種の微生物による、コエンザイムQ−10の増大した生成の方法に関する。かかる変異を担持する配列ならびにかかる変異mevオペロンを担持する微生物もまた、含まれる。
【0002】
コエンザイムQ−10(2,3−ジメトキシ−ジメチル−6−デカプレニル−1,4−ベンゾキノン)は、ユビキノン10としても公知であり、10個のC−5イソプレノイド単位で構成されるイソプレノイド側鎖を有する脂溶性ベンゾキノンである。コエンザイムQ−10(以下CoQ10と略す)は、微生物および植物において、ならびに動物において見られる。これは、ヒトおよび殆どの哺乳動物における、ユビキノンの最も一般的な形態である。CoQ10がヒトの健康状態および疾患からのその防御における重要な因子であるという、確立され、かつ発展中の証拠がある。CoQ10の医学的および健康上の有益な効果は、その2つの主要な生理学的機能、すなわちミトコンドリアの電子伝達系(アデノシン三リン酸の合成に連結されている)の必須の補因子としての機能および脂溶性抗酸化物質としての作用と関連づけられている。
【0003】
CoQ10の健康上の利益により、この化合物の商業的重要性が高まった。CoQ10は、化学合成によって、または微生物を用いた発酵によって生成され得る。これらの微生物は、遺伝子工学によってCoQ10生成に関して改良された天然のCoQ10生産者であり得るか、またはそれらは天然にCoQ10を生成し得ないが遺伝子工学によってそれを合成できるように操作され得る。
【0004】
細菌においては、ユビキノンのイソプレノイド尾部はC−5化合物イソペンテニルピロリン酸(IPP)に由来するが、ユビキノンのキノイド環は芳香族化合物の生合成における中心的中間体であるコリスミ酸に由来する。キノイド環に付加されるイソプレノイド尾部の長さは、細菌中に存在する特定のプレニルトランスフェラーゼ酵素に依存する。例えば、大腸菌(Escherichia coli)においては、オクタプレニルピロリン酸シンターゼが、ファルネシルピロリン酸(FPP、C−15)および5個のIPP単位からのオクタプレニルピロリン酸(C−40)の形成を触媒する。キノイド環へのこの分子の付加は、結果としてユビキノン−8の形成を生じる。パラコッカスおよびロドバクター種においては、デカプレニルピロリン酸(DPP)シンターゼが、FPP(C−15)および7個のIPP単位からのDPP(C−50)の形成を触媒する。次いで、キノイド環へのDPPの付加が、結果としてユビキノン−10(CoQ10)の形成を生じる。
【0005】
自然界においては、2つの異なる経路がIPPの生合成で知られている(図1)。メバロン酸経路は、その名前が含意するように、中心的中間体としてメバロン酸を利用し、真核生物においてよく研究されている。メバロン酸経路は、長年の間、自然界におけるIPP合成の普遍的な経路と考えられてきた。しかしながら、この十年間に、非メバロン酸経路またはMEP経路(中間体として2C−メチル−D−エリスリトール4−リン酸を有するため)と呼ばれるIPP生合成の第二の経路が発見された。MEP経路は、これまでに、多くの真正細菌中および高等植物のプラスチド区画中に存在することが示されている。
【0006】
ロドバクター・スフェロイデス中の、1−デオキシ−D−キシルロース5−リン酸レダクトイソメラーゼをコードするyaeM遺伝子(現在ispCと呼ばれている)の存在および殆ど完了しているロドバクター・スフェロイデスのゲノム配列の精査に基づくと、この細菌は、IPPの生合成にMEP経路を排他的に使用するようである。ロドバクター・スフェロイデスにおけるMEP経路の排他的使用を支持するさらなる証拠は、密接に関連する種、ロドバクター・カプスラータ(Rhodobacter capsulatus)が、イソプレノイド生合成にMEP経路のみを使用するという発見である。
【0007】
国際公開第02/26933A1号パンフレットは、MEP経路の酵素のいくつかをコードする数個の天然および/または異種の遺伝子を過剰発現することによってロドバクター・スフェロイデスにおけるCoQ10の生成を増大させる方法を開示している。しかしながら、これらの遺伝子の過剰発現は、結果として、CoQ10生成の、非常に穏当な改良を生じた。
【0008】
上述のように、いくつかの細菌は、IPPの生合成に、メバロン酸経路のみを使用する。パラコッカス・ゼアキサンチニファシエンスは、かかる細菌の一例である。パラコッカス・ゼアキサンチニファシエンスにおいて、メバロン酸経路の5つの遺伝子をコードする遺伝子に加えてIPPイソメラーゼをコードする遺伝子(図1参照)が、染色体上の単一の転写単位、すなわち以下メバロン酸(mev)オペロンと呼ばれるオペロンに、共にクラスター化される(フンベリン(Huembelin)ら、ジーン(Gene)297、129−139頁、2002年)。
【0009】
イソプレノイド、具体的にはCoQ10の生成は、mevオペロンの1つ以上の遺伝子を天然に欠損した、すなわち天然にイソプレノイド生成に非メバロン酸経路を使用する微生物への変異mevオペロンの導入によって、相当に増大され得ることがわかっており、ここで、1つ以上の変異を含む、完全なmevオペロンまたは前記mevオペロンの1つ以上の遺伝子のいずれかが導入され得る。1つ以上の変異は、例えば、以下の遺伝子の1つまたは全てにあり得る。ヒドロキシメチルグルタリル−CoAレダクターゼをコードするmvaA、イソペンテニル二リン酸イソメラーゼをコードするidi、ヒドロキシメチルグルタリル−CoAシンターゼをコードするhcs、メバロン酸キナーゼをコードするmvk、ホスホメバロン酸キナーゼをコードするpmk、およびジホスホメバロン酸デカルボキシラーゼをコードするmvd。
【0010】
したがって、本発明は、ヒドロキシメチルグルタリル−CoAレダクターゼ活性、イソペンテニル二リン酸イソメラーゼ活性、ヒドロキシメチルグルタリル−CoAシンターゼ活性、メバロン酸キナーゼ活性、ホスホメバロン酸キナーゼ活性、および/またはジホスホメバロン酸デカルボキシラーゼ活性を有するタンパク質をコードする1つ以上の遺伝子を含むポリヌクレオチド配列を提供し、前記ポリヌクレオチド配列は、微生物中に存在する場合にイソプレノイドの向上した生成につながる1つ以上の変異を担持する。イソプレノイド生成の向上は、例えば、前記ポリヌクレオチド配列を担持しない、すなわちmev経路を使用しない野生型微生物における生成と、本発明におけるポリヌクレオチド配列を担持する微生物との比較によって、測定され得る。
【0011】
ある実施形態において、本発明は、配列番号1または2から入手可能な、すなわち野生型mevオペロンの遺伝子またはその断片を含む、ポリヌクレオチド配列に関し、ここで断片は、mevオペロンの遺伝子、例えばヒドロキシメチルグルタリル−CoAレダクターゼをコードするmvaA、イソペンテニル二リン酸イソメラーゼをコードするidi、ヒドロキシメチルグルタリル−CoAシンターゼをコードするhcs、メバロン酸キナーゼをコードするmvk、ホスホメバロン酸キナーゼをコードするpmk、およびジホスホメバロン酸デカルボキシラーゼをコードするmvdの、少なくとも1つの活性を有する。好ましくは、変異ポリヌクレオチド配列は配列番号3またはその断片によって表され、微生物中に存在する場合に、例えばCoQ10生成の増大につながる。
【0012】
ある態様において、本発明は、変異を担持するmevオペロンを含むDNA配列に関し、前記DNA配列は配列番号3によって表される。
【0013】
用語「向上したイソプレノイド生成」、具体的には向上したCoQ10生成は、本明細書中で使用される場合、例えば、それぞれの野生型ポリヌクレオチドを担持するそれぞれの微生物と比較した場合に、上述のように1つ以上の変異を含むポリヌクレオチドを担持する微生物によって得られる、少なくとも約10%の増大を意味する。イソプレノイドの生成は、標準的な方法、例えばHPLC(実施例2参照)によって測定され、mg/lで、またはmg/l/OD600として表され得る(実施例5参照)。
【0014】
完全なmevオペロンまたは1つ以上の変異を担持するその1つ以上の遺伝子は、例えば、完全に合成されるか、または部分的に単離および合成されるかのいずれかであり得る。単離されたmevオペロンまたはその1つ以上の遺伝子は、メバロン酸経路を使用する任意の微生物から生じ得、すなわち、ここで前記オペロン内に含まれる1つ以上の遺伝子は、天然に生じる。好ましくは、微生物はパラコッカス属に属し、より好ましくは、例えばパラコッカス種R114またはパラコッカス・ゼアキサンチニファシエンスATCC21588等のパラコッカス・ゼアキサンチニファシエンス(Paracoccus zeaxanthinifaciens)に由来する。完全に機能的であるオペロンのDNA配列の対立遺伝子の変異および突然変異もまた、上記の用語に含まれる。パラコッカス・ゼアキサンチニファシエンスATCC21588の完全なmevオペロンは配列番号1に示され、パラコッカス種R114の完全なmevオペロンは配列番号2に示される(国際公開第02/099095号パンフレットの配列番号42も参照)。
【0015】
系統パラコッカス種R114は、パラコッカス・ゼアキサンチニファシエンスATCC21588の派生物であり、ブダペスト条約の合意の下、特許寄託名称PTA−3335で2001年4月24日にATCCと共に寄託されている。本発明に使用され得るパラコッカス種および系統ならびにパラコッカスとしてのフラボバクテリウム種の分類学的再分類に関しては、国際公開第02/099095号パンフレット、47頁が参照される。使用され得るかかる種の例は、パラコッカス・マルクシイ(P.marcusii)、パラコッカス・カロチニファシエンス(P.carotinifaciens)、パラコッカス・ソルベンティボランス(P.solventivorans)、パラコッカス・ゼアキサンチニファシエンスまたはパラコッカス種R114である。
【0016】
本発明のmevオペロンは、オペロン内の任意の位置に位置し得る1つ以上の変異を担持し、前記オペロン内の1つ以上の遺伝子の活性の変化につながり、結果として、かかるポリヌクレオチドを担持する微生物内の、イソプレノイドの向上した生成を生じる。
【0017】
ある実施形態において、mevオペロンは少なくとも1つの変異を担持し、これは好ましくはmevオペロンのhcs遺伝子中、例えばパラコッカス・ゼアキサンチニファシエンスのmevオペロンのhcs遺伝子中、例えばパラコッカス種R114またはパラコッカス・ゼアキサンチニファシエンスATCC21588のmevオペロンのhcs遺伝子中に位置する。より好ましくは、変異mevオペロンは、本発明において使用される場合、配列番号3によって表される。パラコッカス種R114の野生型hcs遺伝子の配列およびそれぞれのタンパク質は、それぞれ配列番号4および5に示される。
【0018】
ある実施形態において、本発明は、hcs遺伝子に1つ以上の変異を含むポリヌクレオチド、好ましくは配列番号7に示されるヒドロキシメチルグルタリル−CoAシンターゼを有するタンパク質をコードする、配列番号6に示されるポリヌクレオチドに関し、ここで配列番号5の90位に存在するグルタミンはリシンに置換される。
【0019】
したがって、mevオペロンの1つ以上の遺伝子を含む上述のようなポリヌクレオチド配列を提供することが本発明の目的であり、前記ポリヌクレオチドは、
(a)配列番号7を一致させるアミノ酸配列を含むポリペプチドをコードするポリヌクレオチド、
(b)配列番号6のヌクレオチド配列を含むポリヌクレオチド、
(c)(a)または(b)のポリヌクレオチドによってコードされるポリペプチドの断片または派生物をコードするヌクレオチド配列を含むポリヌクレオチドであって、ここで前記派生物において、前記ポリペプチドと比較して1つ以上のアミノ酸残基が保存的に置換され、前記断片または派生物がヒドロキシメチルグルタリル−CoAシンターゼ(Hcs)活性を有する、ポリヌクレオチド、
(d)ストリンジェントな条件下でその相補鎖が(a)〜(c)のいずれか1つに定義されるポリペプチドにハイブリダイズし、Hcsタンパク質をコードする、ポリヌクレオチド
からなる群より選択される。
【0020】
本発明のポリペプチドおよびポリヌクレオチドは、好ましくは単離された形態で提供され、好ましくは、均質になるまで精製される。
【0021】
用語「単離された」は、物質がその元々の環境(例えば、それが天然に生じる場合、天然の環境)から除去されることを意味する。例えば、生きている微生物中に存在する天然に生じるポリヌクレオチドまたはポリペプチドは単離されていないが、天然の系に共存する物質の一部または全てから分離された同じポリヌクレオチドまたはポリペプチドは、単離されている。かかるポリヌクレオチドはベクターの一部であり得、かつ/またはかかるポリヌクレオチドもしくはポリペプチドは、組成物の一部であり得、さらに、かかるベクターまたは組成物が天然の環境の一部でないという点で、単離され得る。
【0022】
単離されたポリヌクレオチドまたは核酸は、本明細書中で使用される場合、それが由来する生物の天然に生じるゲノムにおいて直接隣接している(一方は5’末端で、一方は3'末端で)コード配列の両方に直接隣接していないDNAまたはRNAであり得る。したがって、ある実施形態において、核酸は、コード配列に直接隣接する5’非コード(例えばプロモーター)配列の一部または全てを含む。用語「単離されたポリヌクレオチド」は、そのため、例えば、ベクター、自己複製プラスミドもしくはウイルス、または原核生物もしくは真核生物のゲノムDNAに組み込まれたか、または他の配列から独立した分離した分子として存在する(例えば、PCRまたは制限エンドヌクレアーゼ処理によって生じたcDNAまたはゲノムDNA断片)、組み換えDNAを含む。これはまた、実質的に細胞性物質、ウイルス性物質または培地(組み換えDNA技術によって生成された場合)または化学的前駆体もしくは他の化学物質(化学的に合成された場合)を含まないさらなるポリペプチドをコードするハイブリッド遺伝子の一部である組み換えDNAを含む。さらに、「単離された核酸断片」は、断片として天然に生じず、天然の状態で見られない核酸断片である。
【0023】
遺伝子は、コード配列、例えば遺伝子のコード領域の3’および5’末端に位置する非翻訳配列等の非コード配列、ならびに制御配列を含み得る。さらに、遺伝子は、本明細書中に定義される単離された核酸分子をいう。タンパク質のアミノ酸配列の変化につながるDNA配列多型が集団内に存在し得ることが、当業者によってさらに理解される。
【0024】
本発明はまた、ストリンジェントな条件下、好ましくは高度にストリンジェントな条件下で、例えば配列番号6に示されるポリヌクレオチド等の本発明のポリヌクレオチドにハイブリダイズできる、単離されたポリヌクレオチドに関する。
【0025】
本明細書中で使用される場合、用語「ハイブリダイズする」は、互いに少なくとも約50%、少なくとも約60%、少なくとも約70%、より好ましくは少なくとも約80%、さらにより好ましくは少なくとも約85%〜90%、最も好ましくは少なくとも95%相同なヌクレオチド配列が典型的に互いにハイブリダイズしたままである、ハイブリダイゼーションおよび洗浄の条件を説明するよう意図される。
【0026】
かかるハイブリダイゼーション条件の好ましい、非限定的な例は、6×塩化ナトリウム/クエン酸ナトリウム(SSC)中約45℃でのハイブリダイゼーション、それに続く1×SSC、0.1%SDS中50℃、好ましくは55℃、より好ましくは60℃、さらにより好ましくは65℃での1回以上の洗浄である。
【0027】
高度にストリンジェントな条件としては、例えば、100μg/mlサケ精子DNAありもしくはなしのDigイージーハイブ(DigEasyHyb)溶液(ロシュ・ダイアグノスティクス・ゲーエムベーハー(Roche Diagnostics GmbH))、または50%ホルムアミド、5×SSC(150mM NaCl、15mMクエン酸三ナトリウム)、0.02%ドデシル硫酸ナトリウム、0.1%N−ラウロイルサルコシンおよび2%ブロッキング試薬(ロシュ・ダイアグノスティクス・ゲーエムベーハー(Roche Diagnostics GmbH))を含有する溶液等の溶液中のジゴキシゲニン(DIG)標識DNAプローブ(DIG標識システムを用いて調製される、ドイツ連邦共和国マンハイム68298のロシュ・ダイアグノスティクス・ゲーエムベーハー(Roche Diagnostics GmbH、68298Mannheim、Germany))を用いた2時間〜4日間42℃でのインキュベーション、それに続く、5〜15分2×SSCおよび0.1%SDS中室温で2回のフィルターの洗浄ならびにそれに次ぐ15〜30分0.5×SSCおよび0.1%SDSまたは0.1×SSCおよび0.1%SDS中65〜68℃で2回の洗浄が挙げられる。
【0028】
当業者は、ストリンジェントおよび高度にストリンジェントなハイブリダイゼーション条件のいずれの条件を適用すべきかを知るであろう。かかる条件に関するさらなる手引きは、当該分野において、例えばサンブルック(Sambrook)ら、1989年、モレキュラー・クローニング、ラボラトリー・マニュアル、ニューヨーク州のコールドスプリングハーバープレス(Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Press、N.Y.)およびアウスベル(Ausubel)ら(編)、1995年、カレント・プロトコールズ・イン・モレキュラー・バイオロジー(Current Protocols in Molecular Biology)(ニューヨークのジョン・ワイリー・アンド・サンズ(John Wiley & Sons、N.Y.))において容易に入手可能である。勿論、(mRNAの3’末端のポリ(A)域等の)ポリ(A)配列またはT(もしくはU)残基の相補的伸展のみにハイブリダイズするポリヌクレオチドは、かかるポリヌクレオチドはポリ(A)伸展またはその相補物を含む任意の核酸分子(例えば、事実上任意の二本鎖cDNAクローン)にハイブリダイズするので、本発明の核酸の一部に特異的にハイブリダイズするのに用いられる本発明のポリヌクレオチドに含まれない。
【0029】
本発明を、イソプレノイド、具体的にはユビキノン、好ましくはCoQ10の生成から使用し得る。好ましくは、1つ以上の変異を含むポリヌクレオチド、すなわち完全なmevオペロン、またはその断片のみが、mevオペロンの1つ以上の遺伝子が元々欠損している、すなわちイソプレノイドの生成に天然にMEP経路のみを使用している、適した微生物に導入される。
【0030】
変異mevオペロンまたは上述のその1つ以上の遺伝子は、例えば、ロドバクター属の微生物に導入され得る。
【0031】
具体的には、本発明は、(1)本発明におけるポリヌクレオチドを、前記ポリヌクレオチドを元々欠損している微生物に導入する工程、および(2)工程(1)の微生物を、イソプレノイドの生成を可能にする条件下で培養する工程を含む、イソプレノイド、具体的にはユビキノン、好ましくはCoQ10の生成の方法に関する。好ましくは、パラコッカス属に属する微生物の変異mevオペロンまたはその断片が、ロドバクター属に属する微生物に導入される。より好ましくは、ヒドロキシメチルグルタリル−CoAシンターゼをコードする遺伝子に1つ以上の変異を含むポリヌクレオチドが前記微生物に導入され、配列番号3によって表されるポリヌクレオチド配列の導入が最も好ましい。
【0032】
mevオペロンの1つ以上の遺伝子を元々欠損している、すなわちイソプレノイドの生成にMEP経路を使用する任意の微生物が、本発明の目的のために使用され得る。具体的には、ロドバクター属の微生物が、例えばロドバクター・スフェロイデス、ロドバクター・アドリアティクス(R.adriaticus)、ロドバクター・カプスラータ(R.capsulatus)、ロドバクター・スルフィドフィルス(R.sulfidophilus)、またはロドバクター・ベルドカンピイ(R.veldkampii)等の、本発明における変異ポリヌクレオチドの導入に使用され得る。好ましい系統はロドバクター・スフェロイデスであり、ロドバクター・スフェロイデスATCC35053が、さらにより好ましい。
【0033】
本明細書中に挙げられた微生物はまた、国際原核生物命名規約に定義される、同じ物理化学的特性を有するかかる種の異名または基礎異名も含むことが理解される。
【0034】
本発明はさらに、上述のポリヌクレオチドを導入した、すなわち1つ以上の変異を担持するmevオペロンの1つ以上の遺伝子を含む、組み換え微生物を提供し、ここでそれぞれの野生型微生物は、元々mevオペロンの1つ以上の遺伝子を欠損しており、すなわちイソプレノイドの生成にMEP経路を使用しており、それぞれの野生型微生物と比較して、イソプレノイド、具体的にはユビキノン、好ましくはCoQ10の、向上した生成につながる。
【0035】
上述のmevオペロンの1つ以上の遺伝子内の変異の他に、組み換え微生物は、前記微生物内のイソプレノイドの生成の向上につながる限りは、さらなる修飾/変化を含み得る。
【0036】
ある実施形態において、かかるさらなる修飾は、デカプレニル二リン酸シンターゼ活性を有するタンパク質をコードするDNA配列の、好ましくはパラコッカス属の微生物、例えばパラコッカス・ゼアキサンチニファシエンス、具体的にはパラコッカス・ゼアキサンチニファシエンスATCC21588から入手可能な、前記組み換え微生物への導入である。配列番号8におけるポリヌクレオチド配列または配列番号9におけるタンパク質をコードするポリヌクレオチドが最も好ましい。
【0037】
したがって、本発明は、上述のような、デカプレニル二リン酸シンターゼ活性を有するタンパク質をコードするDNA配列が、MEP経路を天然に使用していて上述のように導入された1つ以上の変異を含むmevオペロンの1つ以上の遺伝子を有する微生物、例えばロドバクター属の微生物にさらに導入される、イソプレノイド、好ましくはCoQ10の生成の方法に関する。
【0038】
デカプレニル二リン酸シンターゼ活性を有するタンパク質をコードする任意のDNA配列が、本発明に使用され得る。かかるDNAの例は、パラコッカス属、例えばパラコッカス・ゼアキサンチニファシエンス、より好ましくはパラコッカス・ゼアキサンチニファシエンスATCC21588の微生物のデカプレニル二リン酸シンターゼ活性を有するタンパク質をコードする、ddsA遺伝子またはその部分配列である。デカプレニル二リン酸シンターゼ(配列番号7)をコードする、配列番号6によって表されるパラコッカス・ゼアキサンチニファシエンスATCC21588由来のddsA遺伝子が最も好ましい。
【0039】
したがって、(1)メバロン酸(mev)オペロン、およびパラコッカス属に属する微生物のデカプレニル二リン酸シンターゼ活性を有するタンパク質をコードするDNA配列の両方を、ロドバクター属に属する微生物に導入する工程であって、ここで前記mevオペロンが1つ以上の変異を担持する、工程、ならびに(2)修飾されたロドバクター系統を培養する工程を含む、CoQ10の生成の方法を提供することは、本発明のある態様である。
【0040】
用語「に導入する」は、本明細書および特許請求の範囲において、微生物または宿主生物、例えばロドバクター属の形質転換に関して、遺伝物質、具体的には変異メバロン酸オペロンまたは1つ以上のその遺伝子を宿主生物に効率的にもたらすよう、すなわち生物によって発現されるように使用され得る、当業者に周知の任意の方法を含むよう使用される。導入は、標準的な方法に従って、例えばベクター、好ましくは発現プラスミドによって、または宿主のゲノムへの組み込みによって、実施され得る。好ましい方法は、例えばPcrtEプロモーター制御下の発現プラスミドpBBR−K−PcrtE(このプラスミドの構築は、国際公開第02/099095号パンフレットの91頁12〜27行目、実施例6に詳細に記載されている)にクローニングされる遺伝子等の、プラスミドを介した遺伝子の導入である。例えば発現プラスミド等のDNAを、ロドバクター属の微生物に導入する好ましい方法は、例えばロドバクター系統への大腸菌(E.coli)S17−1由来のプラスミドのコンジュゲーション転移等の、プラスミドのコンジュゲーション転移である(ニシムラ(Nishimura)ら、ニュークレイック・アシッズ・リサーチ(Nucl.Acids Res.)18、6169頁、1990年、サイモン(Simon)ら、バイオ/テクノロジー(Bio/Technology)1983年、784−91頁)。
【0041】
mevオペロンまたはその1つ以上の遺伝子の変異は、例えば、当業者が熟知していて具体的な説明を必要としない方法を用いた、PCR部位特異的変異誘発によって生じ得る。さらなる変異誘発方法は、これも本発明の目的に用いられ得、当業者に公知であるが、例えば、UV照射、転位または化学的変異誘発を含む。増大したイソプレノイド、例えばCoQ10の生産性を示す変異の同定のためのスクリーニング方法は、例えば、UV光、HPLC、NMRまたは薄層クロマトグラフィーによる、イソプレノイド、例えばCoQ10の直接の測定から選択され得る。イソプレノイド、例えばCoQ10の生成はまた、当業者が熟知しているカロテノイド色強度の増大を測定することによって、間接的に測定され得る。
【0042】
本発明における、変異mevオペロンまたはその1つ以上の遺伝子で形質転換された宿主は、公知の方法に従って、すなわち、例えば宿主に同化され得る炭素および窒素源、無機塩等を含有する培地中、効率的な増殖および所望の生成物、具体的にはCoQ10の発現に適した温度、pHおよび通気条件下で、培養され得る。
【0043】
発酵培地および/または形質転換体、すなわち例えばパラコッカス属に属する微生物の変異メバロン酸オペロンが導入された微生物からの単離、ならびに任意に、例えばヒトまたは動物用途のためのかかる生成されたCoQ10の製剤化を含む得られたイソプレノイド、具体的にはCoQ10の、精製およびさらなる処理が、当該分野で周知の方法に従って実施され得る。しかしながら、動物の健康および栄養物摂取における使用のためには、特別の精製が必要でなくてもよい。この場合、CoQ10のような生成されたイソプレノイドは、バイオマスおよび/または発酵培地の他の成分とともに、さらに処理されて商業的に魅力的な製品を生じ得る。
【0044】
本発明の方法は、結果として、高収量の、例えばCoQ10等のイソプレノイドを生じる。増大は、例えばロドバクターの非組み換え系統を用いた、または例えば、例えばパラコッカス系統の野生型mevオペロンを担持するロドバクター等の組み換え系統を用いた方法と比較して、例えば少なくとも約10%であり得る。
【0045】
以下の実施例は、本発明を全く制限することなく、本発明を説明する。
【0046】
実施例1:細菌および培養条件
ロドバクター・スフェロイデス系統ATCC 35053(アメリカ合衆国ヴァージニア州マナッサスのアメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection、Manassas VA、USA)から入手)を、CoQ10の向上した生成を有する組み換え系統の構築の基本宿主として用いた。全てのロドバクター・スフェロイデス系統を、30℃で、培地RS100中で増殖させた。培地RS100の組成および調製を、表1に要約する。50mg/lカナマイシンを、組み換え系統の増殖のために培地に添加した。大腸菌系統を37℃、LB培地(アメリカ合衆国メリーランド州スパークスのベクトン・ディッキンソン(Becton Dickinson、Sparks、MD、USA))中で増殖させた。組み換え大腸菌系統中でのプラスミドの維持のために、アンピシリン(100mg/l)および/またはカナマイシン(25〜50mg/l、プラスミドによる)を、培地に添加した。大腸菌およびロドバクター・スフェロイデスの液体培養物を、通法により、ロータリーシェイカー中、200rpmで好気的に増殖させた。固形培地が必要な場合は、寒天(1.5%終濃度)を添加した。
【0047】
【表1】
【0048】
実施例2:CoQ10の分析的アッセイ
400μlの全培地(実施例5参照)を、使い捨ての15mlポリプロピレン遠心チューブに移した。4mlの滅菌抽出溶液(1:1(v/v)DMSO/THF中の0.5g/l BHT)を添加し、試料をラボラトリーシェイカー(IKA、ドイツ連邦共和国(Germany))中で20分間混合し、抽出を促進した。最後に、試料を遠心分離し、逆相HPLCによる分析のために、上清を琥珀色のガラスバイアルに移した。この方法は、カロテノイドのフィトエン、スフェロイデノン、スフェロイデンおよびニューロスポレンからのCoQ10の明らかな分離を有する、ユビキノンおよびその対応するヒドロキノンの同時測定のために開発された。温度制御自動サンプラーおよびダイオードアレイ検出器を備えたアジレント(Agilent)1100HPLCシステム(アメリカ合衆国のアジレント・テクノロジー(Agilent Technologies、USA))を用いて、クロマトグラフィーを行った。方法パラメータは、以下の通りである。
【0049】
【表2】
【0050】
【表3】
【0051】
実施例3:パラコッカス・ゼアキサンチニファシエンスからの変異mevオペロンのクローニング
プラスチドpBBR−K−mev−op−wtおよびpBBR−K−mev−op−R114の構築
プラスミドpBBR−K−mev−op−up−4(メバロン酸オペロンの最初の4つの遺伝子を含むプラスミド)の構築は、国際公開第02/099095号パンフレットの実施例13(105頁10行目〜106頁8行目)に詳細に記載されている。
【0052】
パラコッカス・ゼアキサンチニファシエンスR114ゲノムDNAを鋳型として使用して、プライマーhcs−5326(配列番号10)およびmvd−9000(配列番号11)を用いてPCRを行う。プライマーmvd−9000は配列番号2のヌクレオチド6977〜6996からの配列の逆の相補物に対応するが、プライマーhcs−5326は、配列番号2のヌクレオチド3321〜3340からのパラコッカス・ゼアキサンチニファシエンスmevオペロンの配列に対応する。3675bpの、得られたPCR産物を、pCR2.1−TOPO(アメリカ合衆国カリフォルニア州カールスバッドのインビトロジェン(Invitrogen、Carlsbad、CA、USA))にクローニングし、結果としてプラスミドTOPO−pCR2.1−mev−op−d−3wtを生じる。このため、このプラスミドは、hcsの3'末端ならびに最後の3つの遺伝子mvk、pmkおよびmvdを含むメバロン酸オペロンの下流半分を含む。
【0053】
プラスミドpBBR−K−mev−op−up−4およびTOPO−pCR2.1−mev−op−d−3wtを、制限エンドヌクレアーゼSacIおよびNdeIで消化し、TOPO−pCR2.1−mev−op−d−3wt由来の得られた3319bp断片を、pBBR−K−mev−op−up−4由来の8027bp断片とライゲートする。得られたプラスミド、pBBR−K−mev−op−R114は、その(推定)プロモーターを含む、パラコッカス・ゼアキサンチニファシエンスR114由来の完全なメバロン酸オペロンを含む。
【0054】
プラスミドpBBR−K−mev−op−wt−PcrtE−crtEおよびpBBR−K−mev−op−R114−PcrtE−crtEの構築
プラスミドpBBR−K−PcrtE−crtEの構築は、国際公開第02/099095号パンフレットの実施例6(92頁10〜17行目)に詳細に記載されている。プラスミドpBBR−K−PcrtE−crtEをNaeIで切断し、1.33kb断片を単離し、pBBR−K−mev−op−up−4のEcl136II部位に挿入した。挿入の方向をチェックし、メバロン酸オペロン遺伝子と同じ方向でcrtE遺伝子を担持するプラスミドを、pBBR−K−mev−op−up−4−PcrtE−crtE−2と呼んだ。
【0055】
プラスミドpBBR−K−mev−op−up−4−PcrtE−crtE−2をSphIおよびSpeIで切断し、crtE遺伝子を含む得られた5566bp断片を単離した。この断片を、同じ酵素を用いたpBBR−K−mev−op−wtまたはpBBR−K−mev−op−R114の制限消化後に得られた7132bpのSphI−SpeI断片とライゲートし、結果としてプラスミドpBBR−K−mev−op−wt−PcrtE−crtEおよびpBBR−K−mev−op−R114−PcrtE−crtEをそれぞれ生じた。
【0056】
プラスミドpBBR−K−mev−op−wt−PcrtE−ddsAwtおよびpBBR−K−mev−op−R114−PcrtE−ddsAwtの構築
プラスミドpBBR−K−PcrtEの構築は、国際公開第02/099095号パンフレットの実施例6(91頁12〜27行目)に詳細に記載されている。
【0057】
パラコッカス・ゼアキサンチニファシエンス系統ATCC21588由来のddsA遺伝子(ddsAwtと呼ぶ)を、プライマーdds−Nde(配列番号12)およびdds−Bam(配列番号13)を用いたPCR(GCリッチPCRシステム、ドイツ連邦共和国マンハイムのロシュ・モレキュラー・バイオケミカルズ(Roche Molecular Biochemicals、Mannheim、Germany))によって増幅し、pCR2.1−TOPO(アメリカ合衆国カリフォルニア州カールスバッドのインビトロジェン(Invitrogen、Carlsbad、CA、USA))にクローニングし、結果としてプラスミドTOPO−ddsAwtを生じた。
【0058】
プラスミドTOPO−ddsAwtをNdeIおよびBamHIで切断し、ddsA遺伝子を含む1005bpの得られた断片を、NdeIおよびBamHIで切断したプラスミドpBBR−K−PcrtEにクローニングし、結果としてプラスミドpBBR−K−PcrtE−ddsAwtを生じた。オリゴヌクレオチドdds−R−1(配列番号14)およびdds−R−2(配列番号15)を用いたクイックチェンジXLサイト・ディレクティド・ミュータジェネシス・キット(QuikChange XL Site−Directed Mutagenesis Kit)(アメリカ合衆国カリフォルニア州ラ・ホーヤのストラタジーン(Stratagene、La Jolla、CA、USA))でサイレント変異を導入することによって、ddsA遺伝子内のエンドヌクレアーゼEcoRIの認識部位を排除した。得られたプラスミドpBBR−K−PcrtE−ddsAwt−Rを、EcoRIおよびMamIで切断し、ddsA遺伝子を含む断片1278bp断片を、EcoRIおよびEcoRVで切断したTOPO−ddsAwtに挿入し、結果としてプラスミドpCR2.1−TOPO−ddsAwt−Rを生じた。このプラスミドをCelIIおよびXbaIで切断し、ddsA遺伝子を含む得られた1211bpの断片を、CelIIおよびBlnIでのpBBR−K−mev−op−wt−PcrtE−crtEおよびpBBR−K−mev−op−R114−PcrtE−crtEそれぞれの消化から得られた11.6kpの制限断片とライゲートした。得られたプラスミドを、それぞれpBBR−K−mev−op−wt−PcrtE−ddsAwtおよびpBBR−K−mev−op−R114−PcrtE−ddsAwtと名づけた。
【0059】
プラスミドpBBR−K−mev−op−4−89−PcrtE−ddsAwtの構築
クイックチェンジXLサイト・ディレクティド・ミュータジェネシス・キット(QuikChange XL Site−Directed Mutagenesis Kit)(アメリカ合衆国カリフォルニア州ラ・ホーヤのストラタジーン(Stratagene、La Jolla、CA、USA))ならびにプライマーmut4−89−1−fw(配列番号16)およびmut4−89−1−rev(配列番号17)を用いた2回のPCR部位特異的変異誘発によって、プラスミドpBBR−K−mev−op−4−89−PcrtE−ddsAwtを得る。両方のプライマーは互いに相補的であり、配列番号4の268位に対応する配列番号3の2949位にCの代わりに所望の変異Aを含む。第一の変異誘発反応を、以下のように製造業者の使用説明書に従って設定する。5μlの10×反応バッファー、10ngのプラスミドDNApBBR−K−mev−op−wt−PcrtE−ddsAwt、125ngのプライマーmut4−89−1−fw、125ngのmut4−89−1−rev、1μlのdNTPミックス、3μlのクイックソリューション(QuikSolution)および2.5UのPfuターボ(PfuTurbo)DNAポリメラーゼを、50μlの最終体積で混合する。以下のパラメータを用いてサイクルを行う。1サイクル:95℃1分、18サイクル:95℃50秒、60℃50秒、68℃30分、1サイクル:68℃7分。反応混合物を37℃まで冷却した後、10Uの制限エンドヌクレアーゼDpnIを添加し、反応物を37℃で2時間インキュベートする。製造業者のプロトコルに従って、大腸菌XL10−ゴールド・ウルトラコンポーネント(Gold Ultracomponent)細胞(アメリカ合衆国カリフォルニア州ラ・ホーヤのストラタジーン(Stratagene、La Jolla、CA、USA))をDpnI処理DNAで形質転換する。
【0060】
得られたプラスミドpBBR−K−mev−op−4−89−PcrtE−ddsAwtを単離し、配列番号3の6948位でTの代わりに所望の変異Cを含むプライマーmut4−89−2−fw(配列番号18)およびmut4−89−2−rev(配列番号19)を用いた二回目のPCR部位特異的変異誘発の鋳型DNAとして使用する。上述のように変異誘発を行う。得られたプラスミドpBBR−K−mev−op−4−89−PcrtE−ddsAwtを単離し、マイクロシンス・ゲーエムベーハー(Microsynth GmbH)(スイス連邦バルガッハ(Balgach、Switzerland))で配列決定した。変異mevオペロンの完全な配列は配列番号3に表され、以下の遺伝子を含む。617〜1639位のmvaA(ヒドロキシメチルグルタリル−CoAレダクターゼをコードする)、1636〜2685位のidi(イソペンテニル二リン酸イソメラーゼをコードする)、2682〜3848位のhcs(ヒドロキシメチルグルタリル−CoAシンターゼをコードする)、3829〜4965位のmvk(メバロン酸キナーゼをコードする)、4965〜5882位のpmk(ホスホメバロン酸キナーゼをコードする)、および5875〜6873位のmvd(ジホスホメバロン酸デカルボキシラーゼをコードする)。
【0061】
実施例4:ロドバクター・スフェロイデスATCC35053への変異mevオペロンの導入
変異mevオペロンを担持するプラスミドでの大腸菌S17−1の形質転換(サイモン(Simon)ら、バイオ/テクノロジー(Bio/Technology)11、784−791頁、1983年)、およびそれに続く、コンジュゲーションによる大腸菌S17−1からロドバクター・スフェロイデスATCC35053へのプラスミドの転移を、標準的な手順を用いて行った(ニシムラ(Nishimura)ら、ニュークレイック・アシッズ・リサーチ(Nucl.Acids Res.)18、6169頁、1990年、サイモン(Simon)ら、バイオ/テクノロジー(Bio/Technology)1983年、784−91頁)。
【0062】
100mg/lリファンピシンを補給したRS100液体培地中で系統ATCC35053を増殖させること、100mg/lリファンピシンを含むRS100プレート上に細胞を平板培養すること、およびシングルコロニーを単離することによって、ロドバクター・スフェロイデスATCC35053の自然発生リファンピシン耐性突然変異をまず単離した。コンジュゲーションのために、100mg/lのリファンピシンおよびドナー細胞(50mg/lカナマイシンを含有するLB培地中で増殖させた、転移されるプラスミドを担持する大腸菌S17−1)を含有するRS100培地中で増殖させたレシピエント細胞(リファンピシン耐性ロドバクター・スフェロイデスATCC35053)の培養物の1ミリリットルのアリコートを、遠心分離によってペレット化した。上清を廃棄し、新鮮なRS100培地で細胞を2回洗浄して抗生物質を除去した。次いで、1mlの新鮮なRS100培地に各ペレットを再懸濁した。50μlのドナー細胞および0.45mlのレシピエント細胞を混合し、遠心分離によってペレット化し、0.03mlの新鮮なRS100培地に再懸濁し、RS100プレート上に点在させた。30℃で一晩のインキュベーションの後、接種ループで細胞を採取し、0.3mlのRS100培地に再懸濁した。この懸濁液の希釈溶液を、100mg/lリファンピシンおよび50mg/lカナマイシンを含むRS100プレート上に広げ、30℃でインキュベーションした。コロニー(ロドバクター・スフェロイデスATCC35053の、推定の形質転換細胞)をプレートから選び取り、50mg/lカナマイシンを含有する液体RS100培地中で増殖させ、56℃のアニーリング温度および以下の2つの異なるプライマー対を用いた1分、15秒の伸長時間でPCR反応においてプラスミドの存在を試験した。
pBBR−K−up(配列番号20)/PcrtE−2442(配列番号21)
Kan3out(配列番号22)/mvaA3256(配列番号23)
【0063】
50mg/lカナマイシンを含むRS100プレート上に、陽性のクローンで筋をつけ、シングルコロニーを得た。各クローン由来の1つのシングルコロニーを、50mg/lカナマイシンを含有する液体RS100培地中で再度増殖させ、上述のように、期待されるプラスミドの存在をPCRによって確認した。得られた組み換え系統を、ATCC35053/pBBR−K−mev−op−4−89−PcrtE−ddsAwtと呼んだ。
【0064】
実施例5:ロドバクター・スフェロイデスATCC35053の形質転換系統におけるCoQ10の生成
ロドバクター・スフェロイデス系統ATCC35053、ATCC35053/pBBR−K−mev−op−R114−PcrtE−ddsAwtおよびATCC35053/pBBR−K−mev−op−4−89−PcrtE−ddsAwtを、RS100培地中の振とうフラスコ培養において増殖させた。組み換えロドバクター・スフェロイデスを含む培養物は、50mg/lカナマイシンを含んでいた。25mlの培養物を30℃で250mlのバッフル三角フラスコ中、200rpmで振とうしながら増殖させた。CoQ10生成を試験するために、ロドバクター・スフェロイデス系統の凍結したグリセロール化ストック培養物を用い、25mlの種培養物を接種した。24〜28時間の種培養物の増殖の後、適した体積の培養物を用いて、660ナノメートルでの初期光学密度(OD660)が0.16であるように、実験フラスコを接種した。2mlの試料を、24時間間隔で無菌的に採取した。分析は、増殖(OD660として測定される)、pH、培養物上清中のグルコースならびに実施例2に記載されるCoQ10およびカロテノイド(HPLCによって測定される)を含んだ。結果を表3に要約する。これらの結果は、パラコッカス・ゼアキサンチニファシエンス由来のクローン化変異メバロン酸オペロンの発現がロドバクター・スフェロイデスにおけるCoQ10生成を有意に向上させたことを明らかに示す。
【0065】
【表4】
【図面の簡単な説明】
【0066】
【図1】図1は、ロドバクター・スフェロイデスにおけるCoQ10生合成の経路を表し、これはIPP形成にMEP経路を使用する。囲まれた領域は、(IPPの形成につながる)メバロン酸経路に加えてIPPイソメラーゼ段階を含む、反応順序を示す。メバロン酸経路は、ロドバクター・スフェロイデスにおいては天然に生じない。パラコッカス・ゼアキサンチニファシエンスにおいては、メバロン酸経路の5つの酵素に加えてIPPイソメラーゼをコードする遺伝子が、以下メバロン酸オペロンと呼ばれる、オペロンを形成する。
【特許請求の範囲】
【請求項1】
ヒドロキシメチルグルタリル−CoAレダクターゼ活性、イソペンテニル二リン酸イソメラーゼ活性、ヒドロキシメチルグルタリル−CoAシンターゼ活性、メバロン酸キナーゼ活性、ホスホメバロン酸キナーゼ活性、および/またはジホスホメバロン酸デカルボキシラーゼ活性を有するタンパク質をコードする1つ以上の遺伝子を含むポリヌクレオチド配列であって、前記ポリヌクレオチド配列が、微生物中に存在する場合にイソプレノイドの向上した生成につながる1つ以上の変異を担持する、ポリヌクレオチド配列。
【請求項2】
配列番号1もしくは2またはその断片から入手可能であり、かつ1つ以上の変異を担持する、請求項1に記載のポリヌクレオチド配列。
【請求項3】
ヒドロキシメチルグルタリル−CoAシンターゼ活性を有するタンパク質をコードするhcs遺伝子に1つ以上の変異を含む、請求項1または2に記載のポリヌクレオチド配列。
【請求項4】
配列番号3またはその断片によって表される、請求項1〜3のいずれか一項に記載のポリヌクレオチド配列。
【請求項5】
(a)配列番号7に記載のアミノ酸配列を含むポリペプチドをコードするポリヌクレオチド、
(b)配列番号6に記載のヌクレオチド配列を含むポリヌクレオチド、
(c)(a)または(b)のポリヌクレオチドによってコードされるポリペプチドの断片または誘導体をコードするヌクレオチド配列を含むポリヌクレオチドであって、ここで前記誘導体において、1つ以上のアミノ酸残基が前記ポリペプチドと比較して保存的に置換され、前記断片または誘導体がヒドロキシメチルグルタリル−CoAシンターゼ活性を有する、ポリヌクレオチド、
(d)ストリンジェントな条件下でその相補鎖が(a)〜(c)のいずれか1つに定義されるポリペプチドにハイブリダイズし、Hcsタンパク質をコードする、ポリヌクレオチド
からなる群より選択される、請求項1〜4のいずれか一項に記載のポリヌクレオチド配列。
【請求項6】
イソプレノイド、好ましくはコエンザイムQ10の生成のための、請求項1〜5のいずれか一項に記載のポリヌクレオチドの使用。
【請求項7】
(a)請求項1〜5のいずれか一項に記載のポリヌクレオチド配列を、元々前記ポリヌクレオチドを欠損した微生物に導入する工程、および
(b)工程(a)の微生物を、イソプレノイドの生成を可能にする条件下で培養する工程
を含む、イソプレノイドの生成の方法。
【請求項8】
ポリヌクレオチドが導入される微生物が、ロドバクター(Rhodobacter)属、好ましくはロドバクター・スフェロイデス(Rhodobacter sphaeroides)から選択される、請求項7に記載の方法。
【請求項9】
導入されたポリヌクレオチドが、パラコッカス(Paracoccus)属、好ましくはパラコッカス・ゼアキサンチニファシエンス(P.zeaxanthinifaciens)から選択される微生物から入手可能である、請求項7または8に記載の方法。
【請求項10】
請求項1〜5のいずれか一項に記載のポリヌクレオチド配列を含む微生物。
【請求項11】
ロドバクター属、好ましくはロドバクター・スフェロイデスから選択される、請求項10に記載の微生物。
【請求項12】
パラコッカス属から選択される微生物から入手可能である請求項1〜5のいずれか一項に記載のポリヌクレオチドが導入される、請求項10または11に記載の微生物。
【請求項13】
イソプレノイドの生成のための、請求項10〜12のいずれか一項に記載の微生物の使用。
【請求項1】
ヒドロキシメチルグルタリル−CoAレダクターゼ活性、イソペンテニル二リン酸イソメラーゼ活性、ヒドロキシメチルグルタリル−CoAシンターゼ活性、メバロン酸キナーゼ活性、ホスホメバロン酸キナーゼ活性、および/またはジホスホメバロン酸デカルボキシラーゼ活性を有するタンパク質をコードする1つ以上の遺伝子を含むポリヌクレオチド配列であって、前記ポリヌクレオチド配列が、微生物中に存在する場合にイソプレノイドの向上した生成につながる1つ以上の変異を担持する、ポリヌクレオチド配列。
【請求項2】
配列番号1もしくは2またはその断片から入手可能であり、かつ1つ以上の変異を担持する、請求項1に記載のポリヌクレオチド配列。
【請求項3】
ヒドロキシメチルグルタリル−CoAシンターゼ活性を有するタンパク質をコードするhcs遺伝子に1つ以上の変異を含む、請求項1または2に記載のポリヌクレオチド配列。
【請求項4】
配列番号3またはその断片によって表される、請求項1〜3のいずれか一項に記載のポリヌクレオチド配列。
【請求項5】
(a)配列番号7に記載のアミノ酸配列を含むポリペプチドをコードするポリヌクレオチド、
(b)配列番号6に記載のヌクレオチド配列を含むポリヌクレオチド、
(c)(a)または(b)のポリヌクレオチドによってコードされるポリペプチドの断片または誘導体をコードするヌクレオチド配列を含むポリヌクレオチドであって、ここで前記誘導体において、1つ以上のアミノ酸残基が前記ポリペプチドと比較して保存的に置換され、前記断片または誘導体がヒドロキシメチルグルタリル−CoAシンターゼ活性を有する、ポリヌクレオチド、
(d)ストリンジェントな条件下でその相補鎖が(a)〜(c)のいずれか1つに定義されるポリペプチドにハイブリダイズし、Hcsタンパク質をコードする、ポリヌクレオチド
からなる群より選択される、請求項1〜4のいずれか一項に記載のポリヌクレオチド配列。
【請求項6】
イソプレノイド、好ましくはコエンザイムQ10の生成のための、請求項1〜5のいずれか一項に記載のポリヌクレオチドの使用。
【請求項7】
(a)請求項1〜5のいずれか一項に記載のポリヌクレオチド配列を、元々前記ポリヌクレオチドを欠損した微生物に導入する工程、および
(b)工程(a)の微生物を、イソプレノイドの生成を可能にする条件下で培養する工程
を含む、イソプレノイドの生成の方法。
【請求項8】
ポリヌクレオチドが導入される微生物が、ロドバクター(Rhodobacter)属、好ましくはロドバクター・スフェロイデス(Rhodobacter sphaeroides)から選択される、請求項7に記載の方法。
【請求項9】
導入されたポリヌクレオチドが、パラコッカス(Paracoccus)属、好ましくはパラコッカス・ゼアキサンチニファシエンス(P.zeaxanthinifaciens)から選択される微生物から入手可能である、請求項7または8に記載の方法。
【請求項10】
請求項1〜5のいずれか一項に記載のポリヌクレオチド配列を含む微生物。
【請求項11】
ロドバクター属、好ましくはロドバクター・スフェロイデスから選択される、請求項10に記載の微生物。
【請求項12】
パラコッカス属から選択される微生物から入手可能である請求項1〜5のいずれか一項に記載のポリヌクレオチドが導入される、請求項10または11に記載の微生物。
【請求項13】
イソプレノイドの生成のための、請求項10〜12のいずれか一項に記載の微生物の使用。
【図1】

【公表番号】特表2008−509689(P2008−509689A)
【公表日】平成20年4月3日(2008.4.3)
【国際特許分類】
【出願番号】特願2007−526356(P2007−526356)
【出願日】平成17年8月11日(2005.8.11)
【国際出願番号】PCT/EP2005/008702
【国際公開番号】WO2006/018211
【国際公開日】平成18年2月23日(2006.2.23)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成20年4月3日(2008.4.3)
【国際特許分類】
【出願日】平成17年8月11日(2005.8.11)
【国際出願番号】PCT/EP2005/008702
【国際公開番号】WO2006/018211
【国際公開日】平成18年2月23日(2006.2.23)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]