
イソマルターゼ
【課題】イソマルトースに作用し、マルトースには作用しないイソマルターゼを提供する。
【解決手段】活性中心にGFRMDVINFのアミノ酸配列を有し、糸状菌に由来する、マルトースに作用しないイソマルターゼ。
【解決手段】活性中心にGFRMDVINFのアミノ酸配列を有し、糸状菌に由来する、マルトースに作用しないイソマルターゼ。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、デンプンからのグルコース生産に有用なイソマルターゼに関する。
【背景技術】
【0002】
デンプンからの工業的グルコース生産において、α−アミラーゼによる液化、グルコアミラーゼによる糖化、イソアミラーゼやプルラナーゼなどによる枝切り処理などに、様々な酵素が利用されている。このグルコース生産の一連の糖化工程において、糖化酵素であるグルコアミラーゼが有する糖転移、縮合反応等の副反応によって、副産物としてイソマルトースが生じ、デンプンをすべてグルコースに変換することができず、デンプンからのグルコース生産性が低下するという問題点がある。
【0003】
イソマルトースは、グルコースがα−1,6グルコシド結合したものであるが、これをグルコースに分解することができれば、グルコースの収率上昇が期待できると考えられている。イソマルトースをグルコースに分解するために、α−1,6グルコシド結合を分解する酵素、イソマルターゼ(イソマルトシダーゼ、オリゴ−1,6−グルコシダーゼ)の働きが期待されている。
しかしながら、これまでに報告のあるイソマルターゼはイソマルトースを分解できるものの、糖転移反応により様々なオリゴ糖が生じ、効果的にグルコースを生産するために前述の目的のためには不向きであった。またこれらのイソマルターゼはマルトースにも作用し、グルコースを経て結果的にイソマルトースを合成してしまう。そこで、イソマルトースには作用するがマルトースには作用せず、また、グルコースからイソマルトースは合成しないという性質を有する新規なイソマルターゼが望まれていた。
【0004】
デンプンの枝きり酵素といわれるイソアミラーゼやプルラナーゼは、α−1,6グルコシド結合を分解する酵素であるが、低分子のイソマルトースには作用しない。
糸状菌の一種である麹菌Aspergillus oryzaeは清酒、味噌、醤油などの製造に古来より利用されている。これまでに全ゲノム配列が解析され(非特許文献1、2)、有用タンパク質や抗生物質の生産などの産業利用に向けて現在も研究が進められている(非特許文献3)。A.oryzaeは、アミラーゼなどのデンプン分解酵素を多く生産する微生物として知られ(非特許文献4)、ゲノム上にはいくつかのα−アミラーゼ様遺伝子の存在が推定されているが、それらのすべてがα−アミラーゼとして機能しているかは不明である。
一般的にα−アミラーゼやイソマルターゼは、糖質加水分解酵素ファミリー13(GH Family 13)に属しており、GH Family 13は、ほとんどがα−1,4、α−1,6グルコシド結合に作用するとされている。すなわち、マルトースとイソマルトース両方に作用する。GH Family 13に属する酵素は、3つの触媒残基を含む4つの高度に保存されたアミノ酸領域を持つとされている(非特許文献5、6)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Machida,M.他「Genome sequencing and analysis of Aspergillus oryzae.」Nature,438,1157−61(2005).
【非特許文献2】Galagan,JE.他「Sequencing of Aspergillus nidulans and comparative analysis with A.fumigatus and A.oryzae.」Nature,438,1105−15(2005).
【非特許文献3】Abe,K.他「Impact of Aspergillus oryzae genomics on industrial production of metabolites.」Mycopathologia,162,143−53(2006).
【非特許文献4】Machida,M.他「Genomics of Aspergillus oryzae:learning from the history of Koji mold and exploration of its future.」DNA Res 15,173−83(2008).
【非特許文献5】MacGregor EA他「Relationship of sequence and structure to specificity in the alpha−amylase family of enzymes.」Biochim Biophys Acta 1546 1−20(2001).
【非特許文献6】Nakajima,T.他「Comparison of amino acid sequences of eleven different alpha−amylases.」Appl Microbiol Biotechnol 23 355−360(1986).
【非特許文献7】Keizo Y他「Val216 decides the substrate specificity of α−glucosidase in Saccharomyces cerevisiae.」Eur.J.Biochem.271,3414−3420(2004).
【非特許文献8】Garcia,Bianca他「Molecular cloning of an α−glucosidase−like gene from Penicillium minioluteum and structure prediction of its gene product」Biochemical and Biophysical Research Communications(2001),281(1),151−158.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の課題は、イソマルトースには作用するがマルトースには作用せず、また、グルコースからイソマルトースは合成しないという性質を有する新規なイソマルターゼ、及びデンプンからのグルコースの効率的な生産方法を提供することにある。
【課題を解決するための手段】
【0007】
そこで本発明者は、前記糸状菌に着目し、イソマルトースには作用するがマルトースには作用しないイソマルターゼを見出すべく種々検討を行った。まず糸状菌とは全く相違する酵母に着目し、酵母の一つであるSaccharomyces cerevisiae由来のイソマルターゼについて検討し、保存領域中の216番目のバリンが基質特異性を決定することが報告されている(非特許文献7)ことから、このイソマルターゼと同じ位置にバリンを持つ配列を有する糸状菌を麹菌(Aspergillus oryzae)ゲノムデータベース(DOGAN)で探索した。その結果、全く意外にも、イソマルターゼを産生することが知られていない糸状菌である麹菌のゲノム上に、バリンを持つ配列が存在することを見出した。そして当該配列を有する遺伝子を大腸菌で発現させたところ、イソマルトースを分解する作用を有し、かつマルトースには作用しないイソマルターゼが得られることを見出し、本発明を完成した。なお、イソマルトースを分解できる酵素(オリゴ−1,6−グリコシダーゼ、デキストラングルコシダーゼ、ネオプルラナーゼ、DexCタンパク等)は、細菌、酵母、糸状菌等から既に多数報告されている(非特許文献8)が、前述の構造を有していないものは、イソマルトースとマルトース両方に作用すると考えられる。
【0008】
すなわち、本発明は、活性中心にGFRMDVINF(配列番号1)のアミノ酸配列を有し、糸状菌に由来する、マルトースに作用しないイソマルターゼを提供するものである。
【0009】
また、本発明は、次のアミノ酸配列(a)又は(b)を有し、マルトースに作用しないイソマルターゼを提供するものである。
(a)配列番号3又は配列番号5
(b)配列番号3又は配列番号5のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列。
【0010】
また、本発明は、上記のイソマルターゼをコードする遺伝子を提供するものである。
【0011】
さらに本発明は、デンプンに、α−アミラーゼ、グルコアミラーゼ、イソアミラーゼ及びプルラナーゼから選ばれる酵素と、上記のイソマルターゼを作用させることを特徴とするグルコースの製造法を提供するものである。
【発明の効果】
【0012】
本発明のイソマルターゼは、イソマルトースには作用するがマルトースには作用せず、また、グルコースからイソマルトースは合成しないという性質を有する。従って、このイソマルターゼを用いれば、デンプンから高収率でグルコースを生産することができる。
【図面の簡単な説明】
【0013】
【図1】市販グルコアミラーゼ1/100水溶液の基質特異性を示す。
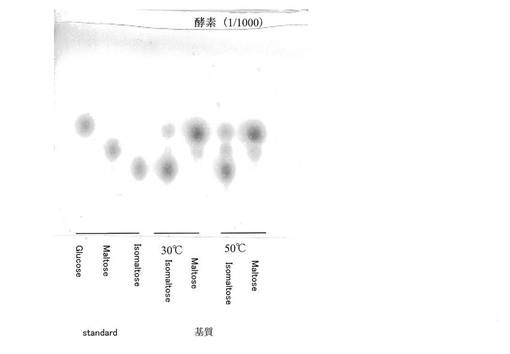
【図2】市販グルコアミラーゼ1/1000水溶液の基質特異性を示す。
【図3】形質転換体(+agl1)とネガティブコントロール株(−agl1)の細胞抽出液(可溶性画分)のSDS−PAGE結果を示す。(ゲル濃度:12%、AはCBB染色、Bでは一次抗体にHis−Tag Monoclonal Antibody、二次抗体にGoat Anti−mouse HRP conjugateを使用)
【図4】形質転換体(+agl1)とネガティブコントロール株(−agl1)の細胞抽出液(可溶性画分)のウエスタンブロッティング結果を示す。(ゲル濃度:12%、AはCBB染色、Bでは一次抗体にHis−Tag Monoclonal Antibody、二次抗体にGoat Anti−mouse HRP conjugateを使用)
【図5】本発明Aspergillus oryzaeイソマルターゼ(AGL1)の反応温度範囲を示す。
【図6】本発明Aspergillus oryzaeイソマルターゼ(AGL1)の反応pH範囲を示す。
【図7】本発明Aspergillus oryzaeイソマルターゼ(AGL1)の基質特異性を示す。
【図8】本発明Fusarium oxysporumイソマルターゼ(FOAGL1)の反応温度範囲を示す。
【図9】本発明Fusarium oxysporumイソマルターゼ(FOAGL1)の反応pH範囲を示す。
【図10】本発明Fusarium oxysporumイソマルターゼ(FOAGL1)の基質特異性を示す。
【図11】本発明Aspergillus oryzaeイソマルターゼ(AGL1)と市販糖化酵素を併用した場合の糖化結果を示す。
【図12】本発明Fusarium oxysporumイソマルターゼ(FOAGL1)と市販糖化酵素を併用した場合の糖化結果を示す。
【発明を実施するための形態】
【0014】
本発明のイソマルターゼは、活性中心にGFRMDVINF(配列番号1)のアミノ酸配列を有する。このアミノ酸配列は、本発明のイソマルターゼが、イソマルトースに作用してグルコースに分解するが、マルトースには作用しないという特徴を有する活性中心であると考えられる。また、本発明のイソマルターゼは、グルコースからイソマルトース等のオリゴ糖を合成しない。従って、本発明のイソマルターゼは、イソマルトースからグルコースを生成するという基質特異性の極めて高い酵素であるという特徴を有する。
【0015】
本発明のイソマルターゼは、25〜40℃で50%以上の相対活性を示し、最適温度は約30℃である。また、本発明イソマルターゼは、pH5〜8で50%以上の相対活性を示し、最適pHは約7.0である。本発明のイソマルターゼの基質特異性は明確であり、イソマルトースに作用するが、マルトース、ニゲロース、トレハロース、デキストラン及びスターチに作用しない。なお、コウジビオースはイソマルトースの20%程度作用する。またグルコースにも作用しないし、オリゴ糖を生成しない。
【0016】
本発明のイソマルターゼは、活性中心に前記のようなアミノ酸配列を有することを特徴とするが、その具体例としては、次のアミノ酸配列(a)又は(b)を有し、マルトースに作用しないものが挙げられる。
(a)配列番号3又は配列番号5
(b)配列番号3又は配列番号5のアミノ酸配列において1若しくは数個のアミノ酸配列が欠失、置換若しくは付加されたアミノ酸配列。
【0017】
ここで配列番号3のアミノ酸配列を有するイソマルターゼは、Aspergillus oryzae由来のイソマルターゼであり、配列番号5のアミノ酸配列を有するイソマルターゼはFusarium oxysporum由来のイソマルターゼである。アミノ酸配列(b)とアミノ酸配列(a)との相同性(同一性)は、配列番号3及び5とそれらの欠失、置換又は付加体との間で、それぞれ、70%以上であればよいが、80%以上が好ましく、90%以上がより好ましく、95%以上がさらに好ましい。
【0018】
本発明の遺伝子は、前記のイソマルターゼをコードする遺伝子である。当該遺伝子は、前記活性中心のアミノ酸配列をコードするものであればよいが、前記アミノ酸配列(a)又は(b)をコードするものが好ましい。さらに好ましい遺伝子は、次の塩基配列(c)又は(d)を有するものである。
(c)配列番号2又は配列番号4
(d)配列番号2又は配列番号4からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAの塩基配列。
【0019】
ここでストリンジェントな条件とは、50%ホルムアミドの存在下で700mMのNaCl中42℃、又はこれと同等の条件をいう。また、塩基配列(c)と塩基配列(d)との相同性(同一性)は、配列番号2又は4の塩基配列と、それぞれ、70%以上であればよいが、80%以上が好ましく、90%以上がより好ましく、95%以上がさらに好ましい。
【0020】
本発明のイソマルターゼは、例えば糸状菌由来の本発明イソマルターゼ遺伝子をクローニングし、当該遺伝子を発現ベクターに組み込んで組み換えプラスミドを調製し、得られた組み換えプラスミドを大腸菌その他の宿主に形質転換し、該形質転換体を培養することにより製造できる。
【0021】
本発明イソマルターゼ遺伝子は、例えば糸状菌由来のcDNAライブラリーからPCR法、または糸状菌ゲノムよりPCR後、イントロンを除去する方法等によってクローニングすることができる。その一例としては、Saccharomyces cerevisiae由来のイソマルターゼのアミノ酸配列に基づき、保存領域中の216番目のバリン残基を有する領域と同様な領域を、糸状菌のデータベースから探索した。その結果、Aspergillus oryzaeのゲノムデータベースからGFRMDVINF(配列番号1)の配列を見出した。この配列がイソマルターゼの活性中心と推定した。そして、この活性中心を有する遺伝子が増幅されるようにプライマーを設計し、Aspergillus oryzaeのcDNAライブラリーを鋳型としてPCR法により目的遺伝子を得た。
【0022】
イソマルターゼ遺伝子を組み込むための発現ベクターとしては、プラスミドベクターが挙げられ、例えば大腸菌用としてはpET−14b、pBR322等が挙げられる。枯草菌用としては、pUB110等が挙げられる。糸状菌用としては、pPTRI等が挙げられる。また、酵母用としては、pRS403等が挙げられる。
【0023】
得られた組み換えプラスミドは、大腸菌、枯草菌、糸状菌、酵母等に形質転換し、当該形質転換体を培養すれば、本発明イソマルターゼが得られる。
【0024】
本発明のイソマルターゼは、前記のようにイソマルトースをグルコースに分解し、マルトースに作用せず、グルコースからオリゴ糖を合成しない。従って、デンプンに、α−アミラーゼ、グルコアミラーゼ、イソアミラーゼ及びプルラナーゼから選ばれる酵素と、本発明のイソマルターゼを作用させれば、グルコースが選択的に製造できる。ここで、α−アミラーゼとしては、例えばクライスターゼT10(大和化成株式会社)を用いることができる。グルコアミラーゼとしては、例えばGODO−ANGH(合同酒精株式会社)を用いることができる。イソアミラーゼとしては、例えばGODO−FIA(合同酒精株式会社)を用いることができる。プルラナーゼとしては、例えばプルラナーゼ「アマノ」3(天野エンザイム株式会社)を用いることができる。
【0025】
反応は、例えばデンプンおよびアミラーゼ等のデンプン糖化酵素に前記酵素を添加し、当該酵素が作用するpH、温度条件にて混合撹拌することにより行なわれる。本発明方法によれば、イソマルトースが副生しないため、高収率でグルコースを製造することができる。あるいは、一般的な糖化反応後に、イソマルトースとグルコースを分離し、これに前記酵素を添加し、当該酵素が作用するpH、温度条件にて混合攪拌する。本発明方法によれば、副生したイソマルトースをグルコースに変換することが可能になり、高収率でグルコースを製造することができる。
【実施例】
【0026】
次に実施例を挙げて本発明をより詳細に説明するが、本発明は何らこれに限定されるものではない。
【0027】
実施例1(グルコアミラーゼによる分解)
市販グルコアミラーゼ(合同酒精株式会社:GODO−ANGH)の1/100あるいは1/1000水溶液を15μL及び、0.1M酢酸緩衝液pH6の5μLを、2%マルトースもしくは2%イソマルトース水溶液に混合し、30℃で18時間作用させた。5分間沸騰水中で反応を止め、反応液を真空濃縮後、シリカゲルTLCに供した。展開溶媒はイソプロパノール/n−ブタノール/水(10/5/4)で、展開後、風乾し、アンスロン硫酸液を噴霧し、105℃で加熱して発色させた。
【0028】
図1(市販グルコアミラーゼ1/100水溶液の結果)及び図2(市販グルコアミラーゼ1/1000水溶液の結果)に示したように、市販グルコアミラーゼは、マルトースによく作用したが、イソマルトースも分解することができた。また、マルトースやイソマルトースを分解するとともに、マルトースからは、イソマルトースが、イソマルトースからはマルトースが、転移あるいはグルコースの再結合により生じることがわかった。
【0029】
実施例2(データベースからのagl1遺伝子発見)
Saccharomyces cerevisiaeのイソマルターゼと相同位置にバリンを持ち、他種のイソマルターゼと高い相同性を示した推定α−アミラーゼ様遺伝子を麹菌(Aspergillus oryzae)ゲノムデータベース(DOGAN)上で見出し、agl1と名づけた(DOGAN ID:Ao090020000176)。agl1はα−アミラーゼではなく、イソマルターゼをコードしていると推測した。今日まで糸状菌からのイソマルターゼの報告は知られていない。
また、同様の検索によりFusarium oxysporumのagl1遺伝子(foagl1)を見出した。
【0030】
【表1】

【0031】
実施例3(Aspergillus oryzaeイソマルターゼ遺伝子による大腸菌の形質転換)
大腸菌発現用ベクターとして、pET−14b (Novagen)を用いた。
agl1遺伝子増幅のためのPCRに使用したプライマーの配列は以下の通りである。
【0032】
glupha1.6F
5’−acgggatcccatggccacacc−3’(配列番号10)
glupha1.6R
5’−ctggggatcctaatcaggcctc−3’(配列番号11)
【0033】
麹菌より調製したcDNAライブラリーを鋳型としてプライマーglupha1.6F及びプライマーglupha1.6Rを用いてPCRを行い、agl1遺伝子を含む断片を得た。取得したDNA断片はエタノール沈殿等を行い、回収した。agl1遺伝子を含むDNA断片をBamH I処理し、エタノール沈殿等にてDNAを回収した。BamH I処理及び脱リン酸化処理を行ったpET−14b とライゲーション反応させ、大腸菌の形質転換を行った。得られたコロニーからプラスミドを調製し、agl1遺伝子を含むプラスミドpET−14b−agl1を得た。agl1遺伝子の塩基配列を配列番号2に、また該遺伝子がコードするイソマルターゼのアミノ酸配列を配列番号3に示す。
【0034】
実施例4(大腸菌でのAspergillus oryzaeイソマルターゼの発現)
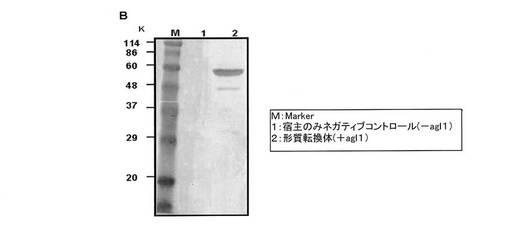
E.coli BL21(DE3)、E.coli AD494(DE3)、E.coli Origami(DE3)pLysS及びE.coli Rosetta−gami(DE3)をプラスミドpET−14b−agl1により形質転換し、形質転換株を取得した。また、ネガティブコントロールとしてagl1を持たないpET−14bベクターを用いて形質転換株を取得した。各形質転換株のコロニーをLB培地に植菌し、前培養を行った。40mLのLB培地に前培養液を接種し、20℃で9時間培養した。その後、最終濃度0.1mMとなるようにIPTGを添加し、20℃で12時間誘導培養を行った。発現誘導後、菌体を7,000rpm、20分間の遠心分離で回収し、1mLの10mMリン酸緩衝液(pH7.0)に縣濁した。菌体は、超音波破砕機にて破砕した。破砕した菌体溶液を12,000rpmで20分間遠心分離し、上清を粗酵素溶液として回収した。各菌株由来の粗酵素について、アクリルアミドゲル濃度12%にてSDS−PAGE及びウエスタンブロッティングを行い、発現したAGL1の評価を行った。検出はHis−Tag Monoclonal Antibody及びGoat Anti−Mouse HRP conjugateにより行った。各形質転換株で発現されたAGL1の活性についてTLCを用いて評価したところ、イソマルトース分解活性はE.coli Origami(DE3)pLysS及びE.coli Rosetta−gami(DE3)を宿主とした形質転換体で強く見られたが、マルトース分解活性がすべての形質転換株で見られた。このことから、観察されたマルトース分解活性は宿主である大腸菌由来のものである、もしくはAGL1のマルトース分解活性に大腸菌由来のマルトース分解活性が加算されたものではないかと推測した。大腸菌由来のマルターゼについては報告されている。よって、最適宿主であるE.coli Origami(DE3)pLysSを宿主とした形質転換株(+agl1)とE.coli Origami(DE3)pLysS(−agl1,ネガティブコントロール)との比較実験を行った。AGL1タンパク質の発現状況について、SDS−PAGE及びWestern blottingにて形質転換株とネガティブコントロール株とを比較した(図3、図4)。形質転換株ではAGL1と推定できるバンドを確認できたが、ネガティブコントロールでは確認できなかった。即ち宿主大腸菌はAGL1を産生せず、形質転換によってAGL1を産生するようになった事を確認した。
【0035】
実施例5(大腸菌で発現したAspergillus oryzaeイソマルターゼの性質:AGL1の精製)
TOYOPEARL AF−Chelate−650Mカラム(φ10×45mm)に4mLの200mM NiSO4を注入した後、40mLの蒸留水及び40mLの10mMリン酸緩衝液(pH7.0)で平衡化を行った。4mLのAGL1粗酵素溶液をニッケルキレートTOYOPEARL AF−Chelate−650Mカラム(φ10×45mm)にアプライし、0.3M NaClを含む10mMリン酸緩衝液(pH7.0)で非吸着画分を溶出し、0.3M NaCl、20mM イミダゾールを含む10mMリン酸緩衝液(pH7.0)で吸着画分を溶出した。SDS−PAGEにて単一バンドを確認できた画分を回収し、10 mM酢酸緩衝液(pH6.0)に対して透析を行った。
タンパク質の定量にはブラッドフォード法を用いた。
【0036】
調製したE.coli Origami(DE3)pLysSを宿主とした形質転換体由来の粗酵素4mLをニッケルキレート TOYOPEARL AF−Chalate−650Mカラム(φ10×45mm)に供し、20mMイミダゾールで吸着タンパク質の溶出を行った。非吸着画分と吸着画分についてSDS−PAGEを行ったところ、Fraction No.11にAGL1と思われるタンパク質が確認された。20mMリン酸緩衝液(pH7.0)に対して、最終濃度が0.041%GOD、0.01%POD、0.5%O−ジアニシジンとなるように添加・混合し、GOD試薬とした。20mg/mLイソマルトース10μl、200mM リン酸緩衝液(pH7.0)5μlの混合液に、精製酵素溶液5μlを加え反応液とした(全量20μl)。また、精製酵素溶液を蒸留水にした溶液をコントロールとした。酵素反応液を30℃で3時間インキュベートした。酵素反応液を100℃で3分間処理した後、GOD試薬200μl加えて30℃で10分インキュベートした。4N HClを4μl加えた後、400nmにおける吸光度を測定した。GOD法の原理については以下の通りである。
【0037】
【化1】
【0038】
α−グルコシダーゼ活性の1Uは、本条件において、1分間に1μmolのグルコースを生成させる活性と定義し、以下の式より酵素活性を算出した。
【0039】
【数1】
【0040】
AGL1はNiキレートカラムクロマトグラフィーにより27.6倍に精製され、収率は76.6%となった。精製酵素はSDS−PAGEにて単一なバンドを示した。
【0041】
実施例6(大腸菌で発現したAspergillus oryzaeイソマルターゼの性質:最適温度)
20mg/mLイソマルトース10μl、200mM リン酸緩衝液(pH7.0)5μlの混合液に、精製酵素溶液5μlを加え反応液とした(全量20μl)。反応液を20〜60℃の温度で3時間インキュベートし、GOD法によって活性を測定した。
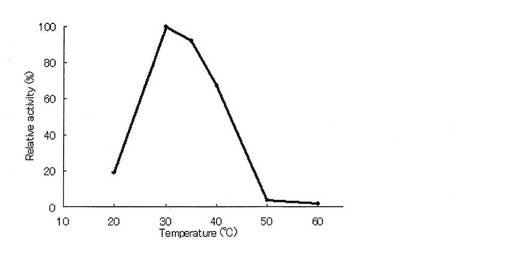
精製酵素を用いて、GOD法により最適温度について評価した(図5)。最適温度は30℃であった。
【0042】
実施例7(大腸菌で発現したAspergillus oryzaeイソマルターゼの性質:至適pH)
20mg/mLのイソマルトース10μl、200mMの各種緩衝液の混合液に、精製酵素溶液5μlを加え反応液とした(全量20μl)。反応液を30℃で3時間インキュベートし、GOD法によって活性を測定した。
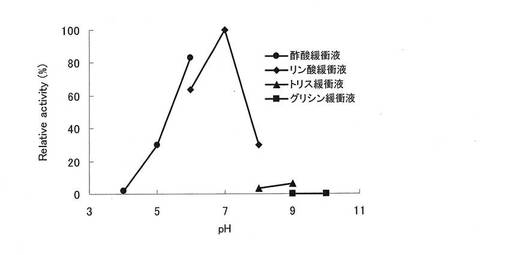
精製酵素を用いて、GOD法により最適pHについて評価した(図6)。最適pHは7.0であった。
【0043】
実施例8(大腸菌で発現したAspergillus oryzaeイソマルターゼの性質:特異性)
5mg/mLの各基質10μl、200mMリン酸緩衝液(pH7.0)5μlの混合液に、精製酵素溶液5μlを加え反応液とした(全量20μl)。酵素反応液を30℃で3時間インキュベートし、GOD法によって活性を測定した。
精製酵素を用いて、GOD法により基質特異性について評価した(表2)。AGL1はイソマルトースに特異的に作用したが、コウジビオースに対してもわずかに活性を示した。
【0044】
【表2】
【0045】
実施例9(糸状菌イソマルターゼによる分解)
実施例5で得られた糸状菌由来イソマルターゼ粗酵素16μLに、10mg/mLとなるように、グルコース、マルトース、イソマルトースの0.1M酢酸緩衝液pH6溶液を4μLをそれぞれ加えた。30℃にて、8、24時間反応し、反応液を実施例1と同様にTLC分析に供した。図7に示したように、グルコースやマルトースを基質とした場合に、分解や転移物は検出されなかった。一方、イソマルトースは分解し、グルコースのスポットが増加した。また、イソマルトースから他の転移物は生じなかった。
【0046】
実施例10(Fusarium oxysporumイソマルターゼ遺伝子のクローニング)
実施例3と同様にしてFusarium oxysporum由来のイソマルターゼ遺伝子をクローニングした。その遺伝子の塩基配列を配列番号4に、また該遺伝子がコードするイソマルターゼのアミノ酸配列を配列番号5に示す。
Fusarium oxysporumより調製したcDNAライブラリーを鋳型としてPCRを行い、foagl1配列を含むDNA断片を得た。得られた断片を大腸菌の発現用プラスミドpET-14bへ組み込み、発現プラスミドpET−14b−foagl1を調製した。
【0047】
実施例11(Fusarium oxysporumイソマルターゼ遺伝子から取得した酵素の性質:最適温度、至適pH、基質特異性)
実施例4と同様にして、発現プラスミドpET−14b−foagl1により形質転換された大腸菌を培養後、菌体を回収して、超音波破砕を行った結果、イソマルトース分解活性を確認した。この菌体破砕抽出物を実施例5の精製方法により精製して得られた、Fusarium oxysporumイソマルターゼ酵素(FOAGL1)を以下の試験に供した。
得られたFusarium oxysporumイソマルターゼについて、実施例6と同様の方法で最適温度、実施例7と同様の方法で至適pHを調べた結果、Fusarium oxysporumイソマルターゼの最適温度は30℃(図8)、最適pHはpH6.5(図9)であった。
また、実施例8と同様の方法であるGOD法により、基質特異性について評価した(表3)。FOAGL1はイソマルトースに特異的に作用した。マルトースやトレハロースには作用せず、コウジビオースやパノースに対してわずかに活性を示した。
【0048】
【表3】
【0049】
実施例12 (Fusarium oxysporumイソマルターゼの分解特性)
実施例11で得られたFusarium oxysporumイソマルターゼ酵素(FOAGL1)のグルコース、マルトース、イソマルトースの分解について調べた。
Fusarium oxysporum由来イソマルターゼ(FOAGL1)粗酵素16μLに、10mg/mLとなるように、グルコース、マルトース、イソマルトースの0.1M酢酸緩衝液pH6溶液を4μLをそれぞれ加えた。30℃にて、8、24時間反応し、反応液を実施例1と同様にTLC分析に供した。図10に示したように、グルコースやマルトースを基質とした場合に、分解や転移物は検出されなかった。一方、イソマルトースは分解し、グルコースのスポットが増加した。また、イソマルトースから他の転移物は生じなかった。
【0050】
実施例13(本発明酵素と市販糖化用酵素との併用)
溶性デンプンを濃度30%(w/w)となるように、0.1M 酢酸緩衝液(pH4.2)に加熱溶解した。溶性デンプン液50mLに対して、市販糖化用酵素OPTIMAX 4060(グルコアミラーゼ・プルラナーゼ混合酵素)を40μL加え、60℃にて16時間酵素反応を行った。反応後、試料を100℃加熱し、酵素反応を停止させた(デンプン分解物)。
デンプン分解物を0.1Mリン酸緩衝液(pH6.8)にて10倍希釈した後、実施例4または実施例5により得られたAspergillus oryzaeイソマルターゼイソマルターゼ(AGL1)、あるいは実施例10によりFusarium oxysporum由来イソマルターゼ(FOAGL1)を加え、30℃で5時間反応させた。反応後、5分間沸騰水中で反応を止め、シリカゲルTLCに供した。展開溶媒はイソプロパノール/n−ブタノール/水(10/5/4)で、展開後、風乾し、アンスロン硫酸液を噴霧し、105℃で加熱して発色させた。
【0051】
OPTIMAX4060のみを作用させた図11と図12の各第2レーンでは、イソマルトースのスポットが確認された。それに対して、イソマルターゼを作用させた図11と図12の各第3レーンでは、そのスポットが薄くなり、イソマルトースが分解されていることがわかった。
この結果から、本酵素を使用することにより糖化工程におけるグルコースの収率を向上させることができることがわかった。
【産業上の利用可能性】
【0052】
本発明のイソマルターゼにより、デンプンからの工業的グルコース生産工程で、グルコースの収率を飛躍的に向上させることが可能になる。
【技術分野】
【0001】
本発明は、デンプンからのグルコース生産に有用なイソマルターゼに関する。
【背景技術】
【0002】
デンプンからの工業的グルコース生産において、α−アミラーゼによる液化、グルコアミラーゼによる糖化、イソアミラーゼやプルラナーゼなどによる枝切り処理などに、様々な酵素が利用されている。このグルコース生産の一連の糖化工程において、糖化酵素であるグルコアミラーゼが有する糖転移、縮合反応等の副反応によって、副産物としてイソマルトースが生じ、デンプンをすべてグルコースに変換することができず、デンプンからのグルコース生産性が低下するという問題点がある。
【0003】
イソマルトースは、グルコースがα−1,6グルコシド結合したものであるが、これをグルコースに分解することができれば、グルコースの収率上昇が期待できると考えられている。イソマルトースをグルコースに分解するために、α−1,6グルコシド結合を分解する酵素、イソマルターゼ(イソマルトシダーゼ、オリゴ−1,6−グルコシダーゼ)の働きが期待されている。
しかしながら、これまでに報告のあるイソマルターゼはイソマルトースを分解できるものの、糖転移反応により様々なオリゴ糖が生じ、効果的にグルコースを生産するために前述の目的のためには不向きであった。またこれらのイソマルターゼはマルトースにも作用し、グルコースを経て結果的にイソマルトースを合成してしまう。そこで、イソマルトースには作用するがマルトースには作用せず、また、グルコースからイソマルトースは合成しないという性質を有する新規なイソマルターゼが望まれていた。
【0004】
デンプンの枝きり酵素といわれるイソアミラーゼやプルラナーゼは、α−1,6グルコシド結合を分解する酵素であるが、低分子のイソマルトースには作用しない。
糸状菌の一種である麹菌Aspergillus oryzaeは清酒、味噌、醤油などの製造に古来より利用されている。これまでに全ゲノム配列が解析され(非特許文献1、2)、有用タンパク質や抗生物質の生産などの産業利用に向けて現在も研究が進められている(非特許文献3)。A.oryzaeは、アミラーゼなどのデンプン分解酵素を多く生産する微生物として知られ(非特許文献4)、ゲノム上にはいくつかのα−アミラーゼ様遺伝子の存在が推定されているが、それらのすべてがα−アミラーゼとして機能しているかは不明である。
一般的にα−アミラーゼやイソマルターゼは、糖質加水分解酵素ファミリー13(GH Family 13)に属しており、GH Family 13は、ほとんどがα−1,4、α−1,6グルコシド結合に作用するとされている。すなわち、マルトースとイソマルトース両方に作用する。GH Family 13に属する酵素は、3つの触媒残基を含む4つの高度に保存されたアミノ酸領域を持つとされている(非特許文献5、6)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Machida,M.他「Genome sequencing and analysis of Aspergillus oryzae.」Nature,438,1157−61(2005).
【非特許文献2】Galagan,JE.他「Sequencing of Aspergillus nidulans and comparative analysis with A.fumigatus and A.oryzae.」Nature,438,1105−15(2005).
【非特許文献3】Abe,K.他「Impact of Aspergillus oryzae genomics on industrial production of metabolites.」Mycopathologia,162,143−53(2006).
【非特許文献4】Machida,M.他「Genomics of Aspergillus oryzae:learning from the history of Koji mold and exploration of its future.」DNA Res 15,173−83(2008).
【非特許文献5】MacGregor EA他「Relationship of sequence and structure to specificity in the alpha−amylase family of enzymes.」Biochim Biophys Acta 1546 1−20(2001).
【非特許文献6】Nakajima,T.他「Comparison of amino acid sequences of eleven different alpha−amylases.」Appl Microbiol Biotechnol 23 355−360(1986).
【非特許文献7】Keizo Y他「Val216 decides the substrate specificity of α−glucosidase in Saccharomyces cerevisiae.」Eur.J.Biochem.271,3414−3420(2004).
【非特許文献8】Garcia,Bianca他「Molecular cloning of an α−glucosidase−like gene from Penicillium minioluteum and structure prediction of its gene product」Biochemical and Biophysical Research Communications(2001),281(1),151−158.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の課題は、イソマルトースには作用するがマルトースには作用せず、また、グルコースからイソマルトースは合成しないという性質を有する新規なイソマルターゼ、及びデンプンからのグルコースの効率的な生産方法を提供することにある。
【課題を解決するための手段】
【0007】
そこで本発明者は、前記糸状菌に着目し、イソマルトースには作用するがマルトースには作用しないイソマルターゼを見出すべく種々検討を行った。まず糸状菌とは全く相違する酵母に着目し、酵母の一つであるSaccharomyces cerevisiae由来のイソマルターゼについて検討し、保存領域中の216番目のバリンが基質特異性を決定することが報告されている(非特許文献7)ことから、このイソマルターゼと同じ位置にバリンを持つ配列を有する糸状菌を麹菌(Aspergillus oryzae)ゲノムデータベース(DOGAN)で探索した。その結果、全く意外にも、イソマルターゼを産生することが知られていない糸状菌である麹菌のゲノム上に、バリンを持つ配列が存在することを見出した。そして当該配列を有する遺伝子を大腸菌で発現させたところ、イソマルトースを分解する作用を有し、かつマルトースには作用しないイソマルターゼが得られることを見出し、本発明を完成した。なお、イソマルトースを分解できる酵素(オリゴ−1,6−グリコシダーゼ、デキストラングルコシダーゼ、ネオプルラナーゼ、DexCタンパク等)は、細菌、酵母、糸状菌等から既に多数報告されている(非特許文献8)が、前述の構造を有していないものは、イソマルトースとマルトース両方に作用すると考えられる。
【0008】
すなわち、本発明は、活性中心にGFRMDVINF(配列番号1)のアミノ酸配列を有し、糸状菌に由来する、マルトースに作用しないイソマルターゼを提供するものである。
【0009】
また、本発明は、次のアミノ酸配列(a)又は(b)を有し、マルトースに作用しないイソマルターゼを提供するものである。
(a)配列番号3又は配列番号5
(b)配列番号3又は配列番号5のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列。
【0010】
また、本発明は、上記のイソマルターゼをコードする遺伝子を提供するものである。
【0011】
さらに本発明は、デンプンに、α−アミラーゼ、グルコアミラーゼ、イソアミラーゼ及びプルラナーゼから選ばれる酵素と、上記のイソマルターゼを作用させることを特徴とするグルコースの製造法を提供するものである。
【発明の効果】
【0012】
本発明のイソマルターゼは、イソマルトースには作用するがマルトースには作用せず、また、グルコースからイソマルトースは合成しないという性質を有する。従って、このイソマルターゼを用いれば、デンプンから高収率でグルコースを生産することができる。
【図面の簡単な説明】
【0013】
【図1】市販グルコアミラーゼ1/100水溶液の基質特異性を示す。
【図2】市販グルコアミラーゼ1/1000水溶液の基質特異性を示す。
【図3】形質転換体(+agl1)とネガティブコントロール株(−agl1)の細胞抽出液(可溶性画分)のSDS−PAGE結果を示す。(ゲル濃度:12%、AはCBB染色、Bでは一次抗体にHis−Tag Monoclonal Antibody、二次抗体にGoat Anti−mouse HRP conjugateを使用)
【図4】形質転換体(+agl1)とネガティブコントロール株(−agl1)の細胞抽出液(可溶性画分)のウエスタンブロッティング結果を示す。(ゲル濃度:12%、AはCBB染色、Bでは一次抗体にHis−Tag Monoclonal Antibody、二次抗体にGoat Anti−mouse HRP conjugateを使用)
【図5】本発明Aspergillus oryzaeイソマルターゼ(AGL1)の反応温度範囲を示す。
【図6】本発明Aspergillus oryzaeイソマルターゼ(AGL1)の反応pH範囲を示す。
【図7】本発明Aspergillus oryzaeイソマルターゼ(AGL1)の基質特異性を示す。
【図8】本発明Fusarium oxysporumイソマルターゼ(FOAGL1)の反応温度範囲を示す。
【図9】本発明Fusarium oxysporumイソマルターゼ(FOAGL1)の反応pH範囲を示す。
【図10】本発明Fusarium oxysporumイソマルターゼ(FOAGL1)の基質特異性を示す。
【図11】本発明Aspergillus oryzaeイソマルターゼ(AGL1)と市販糖化酵素を併用した場合の糖化結果を示す。
【図12】本発明Fusarium oxysporumイソマルターゼ(FOAGL1)と市販糖化酵素を併用した場合の糖化結果を示す。
【発明を実施するための形態】
【0014】
本発明のイソマルターゼは、活性中心にGFRMDVINF(配列番号1)のアミノ酸配列を有する。このアミノ酸配列は、本発明のイソマルターゼが、イソマルトースに作用してグルコースに分解するが、マルトースには作用しないという特徴を有する活性中心であると考えられる。また、本発明のイソマルターゼは、グルコースからイソマルトース等のオリゴ糖を合成しない。従って、本発明のイソマルターゼは、イソマルトースからグルコースを生成するという基質特異性の極めて高い酵素であるという特徴を有する。
【0015】
本発明のイソマルターゼは、25〜40℃で50%以上の相対活性を示し、最適温度は約30℃である。また、本発明イソマルターゼは、pH5〜8で50%以上の相対活性を示し、最適pHは約7.0である。本発明のイソマルターゼの基質特異性は明確であり、イソマルトースに作用するが、マルトース、ニゲロース、トレハロース、デキストラン及びスターチに作用しない。なお、コウジビオースはイソマルトースの20%程度作用する。またグルコースにも作用しないし、オリゴ糖を生成しない。
【0016】
本発明のイソマルターゼは、活性中心に前記のようなアミノ酸配列を有することを特徴とするが、その具体例としては、次のアミノ酸配列(a)又は(b)を有し、マルトースに作用しないものが挙げられる。
(a)配列番号3又は配列番号5
(b)配列番号3又は配列番号5のアミノ酸配列において1若しくは数個のアミノ酸配列が欠失、置換若しくは付加されたアミノ酸配列。
【0017】
ここで配列番号3のアミノ酸配列を有するイソマルターゼは、Aspergillus oryzae由来のイソマルターゼであり、配列番号5のアミノ酸配列を有するイソマルターゼはFusarium oxysporum由来のイソマルターゼである。アミノ酸配列(b)とアミノ酸配列(a)との相同性(同一性)は、配列番号3及び5とそれらの欠失、置換又は付加体との間で、それぞれ、70%以上であればよいが、80%以上が好ましく、90%以上がより好ましく、95%以上がさらに好ましい。
【0018】
本発明の遺伝子は、前記のイソマルターゼをコードする遺伝子である。当該遺伝子は、前記活性中心のアミノ酸配列をコードするものであればよいが、前記アミノ酸配列(a)又は(b)をコードするものが好ましい。さらに好ましい遺伝子は、次の塩基配列(c)又は(d)を有するものである。
(c)配列番号2又は配列番号4
(d)配列番号2又は配列番号4からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAの塩基配列。
【0019】
ここでストリンジェントな条件とは、50%ホルムアミドの存在下で700mMのNaCl中42℃、又はこれと同等の条件をいう。また、塩基配列(c)と塩基配列(d)との相同性(同一性)は、配列番号2又は4の塩基配列と、それぞれ、70%以上であればよいが、80%以上が好ましく、90%以上がより好ましく、95%以上がさらに好ましい。
【0020】
本発明のイソマルターゼは、例えば糸状菌由来の本発明イソマルターゼ遺伝子をクローニングし、当該遺伝子を発現ベクターに組み込んで組み換えプラスミドを調製し、得られた組み換えプラスミドを大腸菌その他の宿主に形質転換し、該形質転換体を培養することにより製造できる。
【0021】
本発明イソマルターゼ遺伝子は、例えば糸状菌由来のcDNAライブラリーからPCR法、または糸状菌ゲノムよりPCR後、イントロンを除去する方法等によってクローニングすることができる。その一例としては、Saccharomyces cerevisiae由来のイソマルターゼのアミノ酸配列に基づき、保存領域中の216番目のバリン残基を有する領域と同様な領域を、糸状菌のデータベースから探索した。その結果、Aspergillus oryzaeのゲノムデータベースからGFRMDVINF(配列番号1)の配列を見出した。この配列がイソマルターゼの活性中心と推定した。そして、この活性中心を有する遺伝子が増幅されるようにプライマーを設計し、Aspergillus oryzaeのcDNAライブラリーを鋳型としてPCR法により目的遺伝子を得た。
【0022】
イソマルターゼ遺伝子を組み込むための発現ベクターとしては、プラスミドベクターが挙げられ、例えば大腸菌用としてはpET−14b、pBR322等が挙げられる。枯草菌用としては、pUB110等が挙げられる。糸状菌用としては、pPTRI等が挙げられる。また、酵母用としては、pRS403等が挙げられる。
【0023】
得られた組み換えプラスミドは、大腸菌、枯草菌、糸状菌、酵母等に形質転換し、当該形質転換体を培養すれば、本発明イソマルターゼが得られる。
【0024】
本発明のイソマルターゼは、前記のようにイソマルトースをグルコースに分解し、マルトースに作用せず、グルコースからオリゴ糖を合成しない。従って、デンプンに、α−アミラーゼ、グルコアミラーゼ、イソアミラーゼ及びプルラナーゼから選ばれる酵素と、本発明のイソマルターゼを作用させれば、グルコースが選択的に製造できる。ここで、α−アミラーゼとしては、例えばクライスターゼT10(大和化成株式会社)を用いることができる。グルコアミラーゼとしては、例えばGODO−ANGH(合同酒精株式会社)を用いることができる。イソアミラーゼとしては、例えばGODO−FIA(合同酒精株式会社)を用いることができる。プルラナーゼとしては、例えばプルラナーゼ「アマノ」3(天野エンザイム株式会社)を用いることができる。
【0025】
反応は、例えばデンプンおよびアミラーゼ等のデンプン糖化酵素に前記酵素を添加し、当該酵素が作用するpH、温度条件にて混合撹拌することにより行なわれる。本発明方法によれば、イソマルトースが副生しないため、高収率でグルコースを製造することができる。あるいは、一般的な糖化反応後に、イソマルトースとグルコースを分離し、これに前記酵素を添加し、当該酵素が作用するpH、温度条件にて混合攪拌する。本発明方法によれば、副生したイソマルトースをグルコースに変換することが可能になり、高収率でグルコースを製造することができる。
【実施例】
【0026】
次に実施例を挙げて本発明をより詳細に説明するが、本発明は何らこれに限定されるものではない。
【0027】
実施例1(グルコアミラーゼによる分解)
市販グルコアミラーゼ(合同酒精株式会社:GODO−ANGH)の1/100あるいは1/1000水溶液を15μL及び、0.1M酢酸緩衝液pH6の5μLを、2%マルトースもしくは2%イソマルトース水溶液に混合し、30℃で18時間作用させた。5分間沸騰水中で反応を止め、反応液を真空濃縮後、シリカゲルTLCに供した。展開溶媒はイソプロパノール/n−ブタノール/水(10/5/4)で、展開後、風乾し、アンスロン硫酸液を噴霧し、105℃で加熱して発色させた。
【0028】
図1(市販グルコアミラーゼ1/100水溶液の結果)及び図2(市販グルコアミラーゼ1/1000水溶液の結果)に示したように、市販グルコアミラーゼは、マルトースによく作用したが、イソマルトースも分解することができた。また、マルトースやイソマルトースを分解するとともに、マルトースからは、イソマルトースが、イソマルトースからはマルトースが、転移あるいはグルコースの再結合により生じることがわかった。
【0029】
実施例2(データベースからのagl1遺伝子発見)
Saccharomyces cerevisiaeのイソマルターゼと相同位置にバリンを持ち、他種のイソマルターゼと高い相同性を示した推定α−アミラーゼ様遺伝子を麹菌(Aspergillus oryzae)ゲノムデータベース(DOGAN)上で見出し、agl1と名づけた(DOGAN ID:Ao090020000176)。agl1はα−アミラーゼではなく、イソマルターゼをコードしていると推測した。今日まで糸状菌からのイソマルターゼの報告は知られていない。
また、同様の検索によりFusarium oxysporumのagl1遺伝子(foagl1)を見出した。
【0030】
【表1】
【0031】
実施例3(Aspergillus oryzaeイソマルターゼ遺伝子による大腸菌の形質転換)
大腸菌発現用ベクターとして、pET−14b (Novagen)を用いた。
agl1遺伝子増幅のためのPCRに使用したプライマーの配列は以下の通りである。
【0032】
glupha1.6F
5’−acgggatcccatggccacacc−3’(配列番号10)
glupha1.6R
5’−ctggggatcctaatcaggcctc−3’(配列番号11)
【0033】
麹菌より調製したcDNAライブラリーを鋳型としてプライマーglupha1.6F及びプライマーglupha1.6Rを用いてPCRを行い、agl1遺伝子を含む断片を得た。取得したDNA断片はエタノール沈殿等を行い、回収した。agl1遺伝子を含むDNA断片をBamH I処理し、エタノール沈殿等にてDNAを回収した。BamH I処理及び脱リン酸化処理を行ったpET−14b とライゲーション反応させ、大腸菌の形質転換を行った。得られたコロニーからプラスミドを調製し、agl1遺伝子を含むプラスミドpET−14b−agl1を得た。agl1遺伝子の塩基配列を配列番号2に、また該遺伝子がコードするイソマルターゼのアミノ酸配列を配列番号3に示す。
【0034】
実施例4(大腸菌でのAspergillus oryzaeイソマルターゼの発現)
E.coli BL21(DE3)、E.coli AD494(DE3)、E.coli Origami(DE3)pLysS及びE.coli Rosetta−gami(DE3)をプラスミドpET−14b−agl1により形質転換し、形質転換株を取得した。また、ネガティブコントロールとしてagl1を持たないpET−14bベクターを用いて形質転換株を取得した。各形質転換株のコロニーをLB培地に植菌し、前培養を行った。40mLのLB培地に前培養液を接種し、20℃で9時間培養した。その後、最終濃度0.1mMとなるようにIPTGを添加し、20℃で12時間誘導培養を行った。発現誘導後、菌体を7,000rpm、20分間の遠心分離で回収し、1mLの10mMリン酸緩衝液(pH7.0)に縣濁した。菌体は、超音波破砕機にて破砕した。破砕した菌体溶液を12,000rpmで20分間遠心分離し、上清を粗酵素溶液として回収した。各菌株由来の粗酵素について、アクリルアミドゲル濃度12%にてSDS−PAGE及びウエスタンブロッティングを行い、発現したAGL1の評価を行った。検出はHis−Tag Monoclonal Antibody及びGoat Anti−Mouse HRP conjugateにより行った。各形質転換株で発現されたAGL1の活性についてTLCを用いて評価したところ、イソマルトース分解活性はE.coli Origami(DE3)pLysS及びE.coli Rosetta−gami(DE3)を宿主とした形質転換体で強く見られたが、マルトース分解活性がすべての形質転換株で見られた。このことから、観察されたマルトース分解活性は宿主である大腸菌由来のものである、もしくはAGL1のマルトース分解活性に大腸菌由来のマルトース分解活性が加算されたものではないかと推測した。大腸菌由来のマルターゼについては報告されている。よって、最適宿主であるE.coli Origami(DE3)pLysSを宿主とした形質転換株(+agl1)とE.coli Origami(DE3)pLysS(−agl1,ネガティブコントロール)との比較実験を行った。AGL1タンパク質の発現状況について、SDS−PAGE及びWestern blottingにて形質転換株とネガティブコントロール株とを比較した(図3、図4)。形質転換株ではAGL1と推定できるバンドを確認できたが、ネガティブコントロールでは確認できなかった。即ち宿主大腸菌はAGL1を産生せず、形質転換によってAGL1を産生するようになった事を確認した。
【0035】
実施例5(大腸菌で発現したAspergillus oryzaeイソマルターゼの性質:AGL1の精製)
TOYOPEARL AF−Chelate−650Mカラム(φ10×45mm)に4mLの200mM NiSO4を注入した後、40mLの蒸留水及び40mLの10mMリン酸緩衝液(pH7.0)で平衡化を行った。4mLのAGL1粗酵素溶液をニッケルキレートTOYOPEARL AF−Chelate−650Mカラム(φ10×45mm)にアプライし、0.3M NaClを含む10mMリン酸緩衝液(pH7.0)で非吸着画分を溶出し、0.3M NaCl、20mM イミダゾールを含む10mMリン酸緩衝液(pH7.0)で吸着画分を溶出した。SDS−PAGEにて単一バンドを確認できた画分を回収し、10 mM酢酸緩衝液(pH6.0)に対して透析を行った。
タンパク質の定量にはブラッドフォード法を用いた。
【0036】
調製したE.coli Origami(DE3)pLysSを宿主とした形質転換体由来の粗酵素4mLをニッケルキレート TOYOPEARL AF−Chalate−650Mカラム(φ10×45mm)に供し、20mMイミダゾールで吸着タンパク質の溶出を行った。非吸着画分と吸着画分についてSDS−PAGEを行ったところ、Fraction No.11にAGL1と思われるタンパク質が確認された。20mMリン酸緩衝液(pH7.0)に対して、最終濃度が0.041%GOD、0.01%POD、0.5%O−ジアニシジンとなるように添加・混合し、GOD試薬とした。20mg/mLイソマルトース10μl、200mM リン酸緩衝液(pH7.0)5μlの混合液に、精製酵素溶液5μlを加え反応液とした(全量20μl)。また、精製酵素溶液を蒸留水にした溶液をコントロールとした。酵素反応液を30℃で3時間インキュベートした。酵素反応液を100℃で3分間処理した後、GOD試薬200μl加えて30℃で10分インキュベートした。4N HClを4μl加えた後、400nmにおける吸光度を測定した。GOD法の原理については以下の通りである。
【0037】
【化1】
【0038】
α−グルコシダーゼ活性の1Uは、本条件において、1分間に1μmolのグルコースを生成させる活性と定義し、以下の式より酵素活性を算出した。
【0039】
【数1】
【0040】
AGL1はNiキレートカラムクロマトグラフィーにより27.6倍に精製され、収率は76.6%となった。精製酵素はSDS−PAGEにて単一なバンドを示した。
【0041】
実施例6(大腸菌で発現したAspergillus oryzaeイソマルターゼの性質:最適温度)
20mg/mLイソマルトース10μl、200mM リン酸緩衝液(pH7.0)5μlの混合液に、精製酵素溶液5μlを加え反応液とした(全量20μl)。反応液を20〜60℃の温度で3時間インキュベートし、GOD法によって活性を測定した。
精製酵素を用いて、GOD法により最適温度について評価した(図5)。最適温度は30℃であった。
【0042】
実施例7(大腸菌で発現したAspergillus oryzaeイソマルターゼの性質:至適pH)
20mg/mLのイソマルトース10μl、200mMの各種緩衝液の混合液に、精製酵素溶液5μlを加え反応液とした(全量20μl)。反応液を30℃で3時間インキュベートし、GOD法によって活性を測定した。
精製酵素を用いて、GOD法により最適pHについて評価した(図6)。最適pHは7.0であった。
【0043】
実施例8(大腸菌で発現したAspergillus oryzaeイソマルターゼの性質:特異性)
5mg/mLの各基質10μl、200mMリン酸緩衝液(pH7.0)5μlの混合液に、精製酵素溶液5μlを加え反応液とした(全量20μl)。酵素反応液を30℃で3時間インキュベートし、GOD法によって活性を測定した。
精製酵素を用いて、GOD法により基質特異性について評価した(表2)。AGL1はイソマルトースに特異的に作用したが、コウジビオースに対してもわずかに活性を示した。
【0044】
【表2】
【0045】
実施例9(糸状菌イソマルターゼによる分解)
実施例5で得られた糸状菌由来イソマルターゼ粗酵素16μLに、10mg/mLとなるように、グルコース、マルトース、イソマルトースの0.1M酢酸緩衝液pH6溶液を4μLをそれぞれ加えた。30℃にて、8、24時間反応し、反応液を実施例1と同様にTLC分析に供した。図7に示したように、グルコースやマルトースを基質とした場合に、分解や転移物は検出されなかった。一方、イソマルトースは分解し、グルコースのスポットが増加した。また、イソマルトースから他の転移物は生じなかった。
【0046】
実施例10(Fusarium oxysporumイソマルターゼ遺伝子のクローニング)
実施例3と同様にしてFusarium oxysporum由来のイソマルターゼ遺伝子をクローニングした。その遺伝子の塩基配列を配列番号4に、また該遺伝子がコードするイソマルターゼのアミノ酸配列を配列番号5に示す。
Fusarium oxysporumより調製したcDNAライブラリーを鋳型としてPCRを行い、foagl1配列を含むDNA断片を得た。得られた断片を大腸菌の発現用プラスミドpET-14bへ組み込み、発現プラスミドpET−14b−foagl1を調製した。
【0047】
実施例11(Fusarium oxysporumイソマルターゼ遺伝子から取得した酵素の性質:最適温度、至適pH、基質特異性)
実施例4と同様にして、発現プラスミドpET−14b−foagl1により形質転換された大腸菌を培養後、菌体を回収して、超音波破砕を行った結果、イソマルトース分解活性を確認した。この菌体破砕抽出物を実施例5の精製方法により精製して得られた、Fusarium oxysporumイソマルターゼ酵素(FOAGL1)を以下の試験に供した。
得られたFusarium oxysporumイソマルターゼについて、実施例6と同様の方法で最適温度、実施例7と同様の方法で至適pHを調べた結果、Fusarium oxysporumイソマルターゼの最適温度は30℃(図8)、最適pHはpH6.5(図9)であった。
また、実施例8と同様の方法であるGOD法により、基質特異性について評価した(表3)。FOAGL1はイソマルトースに特異的に作用した。マルトースやトレハロースには作用せず、コウジビオースやパノースに対してわずかに活性を示した。
【0048】
【表3】
【0049】
実施例12 (Fusarium oxysporumイソマルターゼの分解特性)
実施例11で得られたFusarium oxysporumイソマルターゼ酵素(FOAGL1)のグルコース、マルトース、イソマルトースの分解について調べた。
Fusarium oxysporum由来イソマルターゼ(FOAGL1)粗酵素16μLに、10mg/mLとなるように、グルコース、マルトース、イソマルトースの0.1M酢酸緩衝液pH6溶液を4μLをそれぞれ加えた。30℃にて、8、24時間反応し、反応液を実施例1と同様にTLC分析に供した。図10に示したように、グルコースやマルトースを基質とした場合に、分解や転移物は検出されなかった。一方、イソマルトースは分解し、グルコースのスポットが増加した。また、イソマルトースから他の転移物は生じなかった。
【0050】
実施例13(本発明酵素と市販糖化用酵素との併用)
溶性デンプンを濃度30%(w/w)となるように、0.1M 酢酸緩衝液(pH4.2)に加熱溶解した。溶性デンプン液50mLに対して、市販糖化用酵素OPTIMAX 4060(グルコアミラーゼ・プルラナーゼ混合酵素)を40μL加え、60℃にて16時間酵素反応を行った。反応後、試料を100℃加熱し、酵素反応を停止させた(デンプン分解物)。
デンプン分解物を0.1Mリン酸緩衝液(pH6.8)にて10倍希釈した後、実施例4または実施例5により得られたAspergillus oryzaeイソマルターゼイソマルターゼ(AGL1)、あるいは実施例10によりFusarium oxysporum由来イソマルターゼ(FOAGL1)を加え、30℃で5時間反応させた。反応後、5分間沸騰水中で反応を止め、シリカゲルTLCに供した。展開溶媒はイソプロパノール/n−ブタノール/水(10/5/4)で、展開後、風乾し、アンスロン硫酸液を噴霧し、105℃で加熱して発色させた。
【0051】
OPTIMAX4060のみを作用させた図11と図12の各第2レーンでは、イソマルトースのスポットが確認された。それに対して、イソマルターゼを作用させた図11と図12の各第3レーンでは、そのスポットが薄くなり、イソマルトースが分解されていることがわかった。
この結果から、本酵素を使用することにより糖化工程におけるグルコースの収率を向上させることができることがわかった。
【産業上の利用可能性】
【0052】
本発明のイソマルターゼにより、デンプンからの工業的グルコース生産工程で、グルコースの収率を飛躍的に向上させることが可能になる。
【特許請求の範囲】
【請求項1】
活性中心にGFRMDVINFのアミノ酸配列を有し、糸状菌に由来する、マルトースに作用しないイソマルターゼ。
【請求項2】
グルコースからオリゴ糖を合成しない請求項1記載のイソマルターゼ。
【請求項3】
糸状菌が、Aspergillus oryzae又はFusarium oxysporumである請求項1又は2記載のイソマルターゼ。
【請求項4】
次のアミノ酸配列(a)又は(b)を有し、マルトースに作用しないイソマルターゼ。
(a)配列番号3又は配列番号5
(b)配列番号3又は配列番号5のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列。
【請求項5】
糸状菌に由来するものである請求項4記載のイソマルターゼ。
【請求項6】
糸状菌が、Aspergillus oryzae又はFusarium oxysporumである請求項5記載のイソマルターゼ。
【請求項7】
グルコースからオリゴ糖を合成しない請求項4〜6のいずれか1項記載のイソマルターゼ。
【請求項8】
請求項1〜7のいずれか1項記載のイソマルターゼをコードする遺伝子。
【請求項9】
次の塩基配列(c)又は(d)を有するものである請求項8記載の遺伝子。
(c)配列番号2又は配列番号4
(d)配列番号2又は配列番号4からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAの塩基配列。
【請求項10】
デンプンに、α−アミラーゼ、グルコアミラーゼ、イソアミラーゼ及びプルラナーゼから選ばれる酵素と、請求項1〜7のいずれか1項記載のイソマルターゼとを作用させることを特徴とするグルコースの製造法。
【請求項1】
活性中心にGFRMDVINFのアミノ酸配列を有し、糸状菌に由来する、マルトースに作用しないイソマルターゼ。
【請求項2】
グルコースからオリゴ糖を合成しない請求項1記載のイソマルターゼ。
【請求項3】
糸状菌が、Aspergillus oryzae又はFusarium oxysporumである請求項1又は2記載のイソマルターゼ。
【請求項4】
次のアミノ酸配列(a)又は(b)を有し、マルトースに作用しないイソマルターゼ。
(a)配列番号3又は配列番号5
(b)配列番号3又は配列番号5のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列。
【請求項5】
糸状菌に由来するものである請求項4記載のイソマルターゼ。
【請求項6】
糸状菌が、Aspergillus oryzae又はFusarium oxysporumである請求項5記載のイソマルターゼ。
【請求項7】
グルコースからオリゴ糖を合成しない請求項4〜6のいずれか1項記載のイソマルターゼ。
【請求項8】
請求項1〜7のいずれか1項記載のイソマルターゼをコードする遺伝子。
【請求項9】
次の塩基配列(c)又は(d)を有するものである請求項8記載の遺伝子。
(c)配列番号2又は配列番号4
(d)配列番号2又は配列番号4からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAの塩基配列。
【請求項10】
デンプンに、α−アミラーゼ、グルコアミラーゼ、イソアミラーゼ及びプルラナーゼから選ばれる酵素と、請求項1〜7のいずれか1項記載のイソマルターゼとを作用させることを特徴とするグルコースの製造法。
【図1】

【図2】

【図3】

【図4】
【図5】
【図6】
【図7】
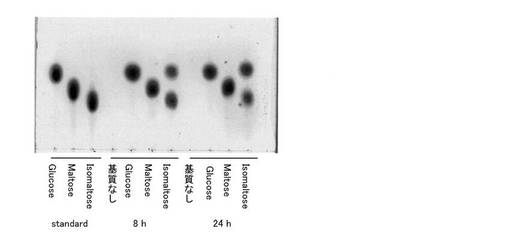
【図8】
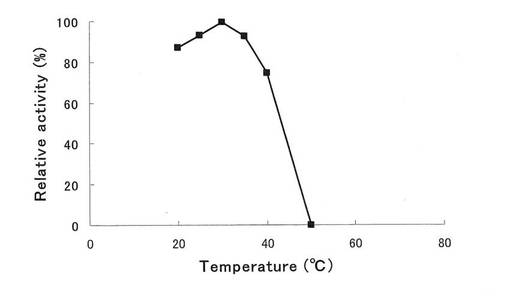
【図9】

【図10】
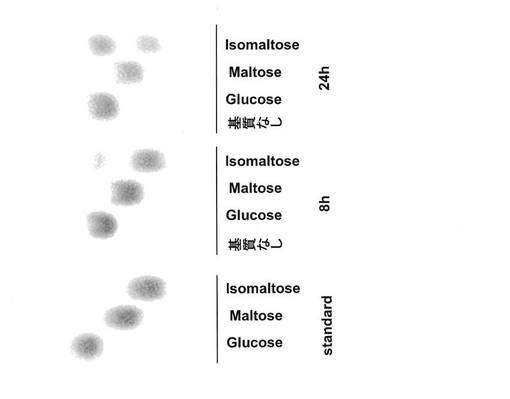
【図11】

【図12】
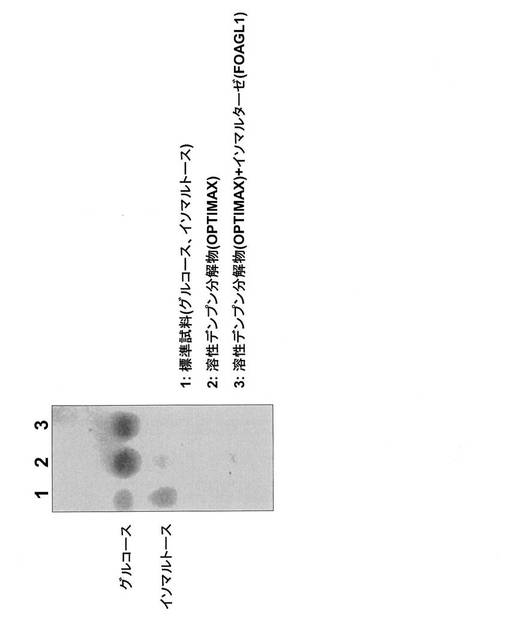
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公開番号】特開2012−223189(P2012−223189A)
【公開日】平成24年11月15日(2012.11.15)
【国際特許分類】
【出願番号】特願2012−78986(P2012−78986)
【出願日】平成24年3月30日(2012.3.30)
【出願人】(504229284)国立大学法人弘前大学 (162)
【出願人】(303036670)合同酒精株式会社 (10)
【Fターム(参考)】
【公開日】平成24年11月15日(2012.11.15)
【国際特許分類】
【出願日】平成24年3月30日(2012.3.30)
【出願人】(504229284)国立大学法人弘前大学 (162)
【出願人】(303036670)合同酒精株式会社 (10)
【Fターム(参考)】
[ Back to top ]