
イネいもち病罹病性遺伝子Pi21および抵抗性遺伝子pi21ならびにそれらの利用
【課題】新規な植物の病斑進展制御遺伝子の提供ならびに該遺伝子を利用した植物の耐病性の改変方法を提供する。
【解決手段】連鎖解析によりイネの圃場抵抗性遺伝子pi21を単離することに成功し、該遺伝子の導入や発現制御により植物のいもち病圃場抵抗性を改変し得る可能性を見出した。これにより、植物に圃場抵抗性を効率的に付与することが可能となった。また、いもち病圃場抵抗性であるイネを早期に選抜することが可能となった。さらには圃場抵抗性に関与する遺伝子の組織発現特異性や発現レベルを変更することにより、抵抗性と実用性の高い特性を兼ね備えた品種を育成することからなる。
【解決手段】連鎖解析によりイネの圃場抵抗性遺伝子pi21を単離することに成功し、該遺伝子の導入や発現制御により植物のいもち病圃場抵抗性を改変し得る可能性を見出した。これにより、植物に圃場抵抗性を効率的に付与することが可能となった。また、いもち病圃場抵抗性であるイネを早期に選抜することが可能となった。さらには圃場抵抗性に関与する遺伝子の組織発現特異性や発現レベルを変更することにより、抵抗性と実用性の高い特性を兼ね備えた品種を育成することからなる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、イネいもち病圃場抵抗性遺伝子pi21、ならびに該遺伝子を利用した植物のいもち病圃場抵抗性を改変する方法に関する。
【背景技術】
【0002】
イネのいもち病菌に対する抵抗性は真性抵抗性と圃場抵抗性の2種類に分類される(非特許文献1)。前者は過敏感反応に基づく抵抗性であり、効果が大きくレースに対する特異性が高い質的な抵抗性である。1個の抵抗性遺伝子を導入した品種は、その遺伝子に親和性の菌が出現することによって、数年でその効果を失うことが経験的に知られる。一方圃場抵抗性は、真性抵抗性が機能しない条件下で観察される抵抗性の品種間差と定義される。真性抵抗性に比べて効果が小さいものの、レースに対する特異性が低いことから、品種に持続性のある抵抗性を付与できる点で実用性が高い。
【0003】
真性抵抗性に関与する遺伝子は30種類以上が知られ、そのうち、Pib、Pita遺伝子が単離されている(非特許文献2)。これらの遺伝子は、既報の植物の病害抵抗性遺伝子と類似した構造である nucleotide binding site (NBS)や leucine-rich repeats (LRRs)をもつNBS-LRR クラス遺伝子であることが明らかとなっている。他の病害抵抗性遺伝子と同様、植物の抵抗性遺伝子の産物は、それに対応する病原体の非病原性遺伝子の産物を直接的あるいは間接的に認識する受容体としての機能を果たすものと考えられている。実際、Pitaは非病原性遺伝子産物と物理的に直接結合することが明らかにされている。
【0004】
圃場抵抗性については、日本陸稲品種が優れた特性を有することが知られ、関与する複数の遺伝子座に関して、染色体上の位置が明らかにされている(非特許文献3)。しかしながら、遺伝子の構造や発現機構は明らかにされておらず、真性抵抗性に比べてこれを効率的に育種選抜に利用できる状況にはない。また、圃場抵抗性と類似した概念である不完全抵抗性(incomplete resistance)に関しては、西アフリカの陸稲品種Moroberekanの複数の染色体領域の関与が報告されている(非特許文献4)ものの、遺伝子は特定されていない。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特願平10-271807
【特許文献2】特願平11-153146
【特許文献3】特許第3376453号(P3376453)
【特許文献4】特開2003-88379(P2003-88379A)
【特許文献5】特開2003-199577(P2003-199577A)
【特許文献6】特開2003-199448(P2003-199448A)
【特許文献7】特開2004-329215(P2004-329215A)
【非特許文献】
【0006】
【非特許文献1】イネのいもち病と抵抗性育種 p175-186 高坂・山崎編 1980博友社
【非特許文献2】Wang et al. Plant J 19:55-64, 1999,Bryan et al. Plant Cell. 12:2033-46, 2000
【非特許文献3】Fukuoka and Okuno 2001 Theor Appl Genet. 03:185-190
【非特許文献4】Wang et al. Genet 136:1421-1434, 1994
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明はこのような状況を鑑みてなされたものであり、本発明が解決しようとする課題は、いもち病圃場抵抗性に関わる遺伝子をマップベースクローニング法により単離・同定すること、ならびに該遺伝子を利用した植物のいもち病圃場抵抗性の改変方法を提供することである。
【課題を解決するための手段】
【0008】
本発明は、植物のいもち病に対する抵抗性を制御する遺伝子に関する。Pi21遺伝子の対立遺伝子pi21は、いもち病圃場抵抗性という形質をイネ(Oryza sativa L)に付与する遺伝子として、イネ第4染色体という広大な領域のいずれかの場所に存在することが知られている。本発明者らは、その存在領域を解明し、単一の遺伝子として単離することを試みた。
【0009】
本発明者らはまず、pi21領域の遺伝地図を作成するために、マップベースクローニングに不可欠な大規模分離集団によるpi21領域の詳細な連鎖解析を行った。病斑進展を抑制する抵抗性対立遺伝子pi21を持つ日本陸稲品種オワリハタモチに、病斑進展を抑制しない罹病性対立遺伝子Pi21を持つ水稲品種日本晴あるいは愛知旭を連続戻し交雑して得た集団に対してRFLPマーカーによる連鎖解析を試みたところ、pi21遺伝子座がRFLPマーカーG271とG317の間に存在することが確認された。
【0010】
次に本発明者らは、より精度の高いpi21領域の遺伝地図を作成するために、pi21の両側に存在するRFLPマーカーRA3591と13S1を利用して、pi21座近傍の染色体組み換え型個体を選抜した。同時に、日本水稲品種の遺伝的背景にインド型水稲品種Kasalathの罹病性対立遺伝子を持つ系統とオワリハタモチ由来の抵抗性対立遺伝子を持つ系統を交雑して得たF2集団を用いて検索を行い、pi21座近傍の染色体組み換え型個体の選抜も行った。これらの個体および作出されたDNAマーカーを用いて詳細な連鎖地図を作成した結果、pi21遺伝子座がSSCPマーカーPa102484およびSNPマーカーP702D3_#12に挟まれる約25kbのゲノム領域に存在することがわかった。さらに、pi21遺伝子を含むと考えられるPACクローンP702D03の塩基配列を決定し、また、25kbのゲノム候補領域の抵抗性品種オワリハタモチと罹病性品種愛知旭およびKasalathの塩基配列の解析も行うことにより、pi21遺伝子が、SNPマーカーP702D03_#38とP702D03_#80に挟み込まれる約1.8kbのゲノム領域に存在することが明らかとなった。
【0011】
そこで本発明者らは、既に得られている日本晴の塩基配列情報を利用して該当部分を増幅できるプライマーを設計し、ゲノムPCRおよび RT-PCR産物を罹病性品種日本晴および愛知旭と抵抗性品種オワリハタモチの間で塩基配列の比較を行った。その結果、遺伝子のエクソン領域において、抵抗性品種と罹病性品種との間に2ヶ所のDNA変異があることが明らかとなった。罹病性の品種に対して抵抗性の品種では、7アミノ酸および16アミノ酸の欠失が起こっており、これらの変異が感染したいもち病の病斑進展に関係することが証明された。すなわち、本発明は、植物のいもち病に対する抵抗性を制御するpi21遺伝子に関し、具体的には以下の発明を提供するものである。
〔1〕以下(a)〜(h)のいずれかに記載のDNA;
(a)配列番号:3または22に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:1、2、20または21に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:3または22に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(d)配列番号:1、2、20または21に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(e)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(f)配列番号:4または5に記載の塩基配列のコード領域を含むDNA、
(g)配列番号:6に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(h)配列番号:4または5に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA。
〔2〕植物においていもち病圃場抵抗性能を有する、以下(i)〜(iv)のいずれかに記載のDNA;
(i)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物と相補的なRNAをコードするDNA、
(ii)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA、
(iii)〔1〕の(a)〜(d)のいずれかに記載のDNAの発現を共抑制効果により阻害するRNAをコードするDNA、
(iv)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物を特異的に切断するRNAi活性を有するRNAをコードするDNA。
〔3〕植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである、〔2〕記載のDNA。
〔4〕〔1〕〜〔3〕のいずれかに記載のDNAを含むベクター。
〔5〕〔1〕〜〔3〕のいずれかに記載のDNAを発現可能に保持する形質転換細胞。
〔6〕〔1〕の(a)〜(d)のいずれかに記載のDNAが導入された形質転換植物細胞。
〔7〕〔2〕もしくは〔3〕に記載のDNAが導入された形質転換植物細胞。
〔8〕植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである、〔6〕または〔7〕に記載の形質転換植物細胞。
〔9〕〔6〕〜〔8〕に記載の形質転換細胞を含む形質転換植物体。
〔10〕〔8〕に記載の形質転換植物体の子孫またはクローンである、形質転換植物体。
〔11〕〔9〕または〔10〕のいずれかに記載の形質転換植物体の繁殖材料。
〔12〕〔9〕または〔10〕のいずれかに記載の形質転換植物体の製造方法であって、〔1〕の(a)〜(d)、〔2〕または〔3〕に記載のDNAを植物細胞に導入し、該植物細胞から植物体を再生させる工程を含む方法。
〔13〕〔2〕または〔3〕に記載のDNAを植物体の細胞内で発現させる工程を含む、植物にいもち病圃場抵抗性を付与する方法。
〔14〕植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである〔13〕に記載の方法。
〔15〕〔1〕の(a)〜(d)のいずれかに記載のDNAによりコードされるタンパク質。
〔16〕〔1〕の(a)〜(d)のいずれかに記載のDNAを含むベクターを含む形質転換細胞を培養し、該細胞またはその培養上清から組換えタンパク質を回収する工程を含む、〔15〕に記載のタンパク質の製造方法。
〔17〕〔15〕に記載のタンパク質に結合する抗体。
〔18〕〔1〕に記載のDNAまたはその相補配列に相補的な少なくとも15の連続する塩基を含むDNA。
〔19〕〔2〕もしくは〔3〕に記載のDNA、または該DNAを含むベクターのいずれか1つを含む、植物のいもち病圃場抵抗性を増加させる薬剤。
〔20〕配列番号:1、4または20に記載の塩基配列の全部または一部を増幅するプライマーセット。
〔21〕(a)〜(c)のいずれかに示す、少なくとも1つのプライマーセット
(a)配列番号:8に記載の塩基配列からなるDNAおよび配列番号:9に記載の塩基配列からなるDNA、
(b)配列番号:16に記載の塩基配列からなるDNAおよび配列番号:17に記載の塩基配列からなるDNA、
(c)配列番号:26に記載の塩基配列からなるDNAおよび配列番号:27に記載の塩基配列からなるDNA。
〔22〕配列番号:7、10、18、19、23または25に記載の塩基配列からなるDNA。
〔23〕以下の(a)〜(c)の工程を含む方法であって、分子量または塩基配列が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から〔1〕に記載のDNA領域を増幅する工程、
(c)増幅したDNA断片の分子量または塩基配列を、〔1〕の(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
〔24〕以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から〔1〕に記載のDNA領域を増幅する工程、
(c)増幅した二本鎖のDNAを非変性ゲル上で分離する工程、
(d)分離した二本鎖DNAのゲル上での移動度を、〔1〕の(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
〔25〕以下の(a)〜(e)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から〔1〕に記載のDNA領域を増幅する工程、
(c)増幅したDNAを一本鎖DNAに解離させる工程、
(d)解離させた一本鎖DNAを非変性ゲル上で分離する工程、
(e)分離した一本鎖DNAのゲル上での移動度を、〔1〕の(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
〔26〕以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から〔1〕に記載のDNA領域を増幅する工程、
(c)増幅したDNAを、DNA変性剤の濃度が次第に高まるゲル上で分離する工程、
(d)分離したDNAのゲル上での移動度と、〔1〕の(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
〔27〕以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性である植物を選抜する方法;
(a)いもち病圃場抵抗性能である植物と任意の機能を有する植物とが交配された品種を作製する工程、
(b)〔23〕〜〔26〕のいずれかに記載の方法により、工程(a)で作製された植物がいもち病圃場抵抗性であるか否かを判定する工程。
〔28〕〔1〕に記載のDNAと連鎖する分子マーカーであって、該分子マーカーがいもち病圃場抵抗性の形質を有するイネと同様の遺伝子型を示す場合に、被検イネがいもち病圃場抵抗性であると判定する方法。
〔29〕分子マーカーが配列番号:10に記載のDNAからなる分子マーカーである、〔28〕に記載の方法。
〔30〕以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性であるイネを選抜する方法;
(a)いもち病圃場抵抗性能であるイネと任意の機能を有するイネとが交配された品種を作製する工程、
(b)〔28〕または〔29〕のいずれかに記載の方法により、工程(a)で作製されたイネがいもち病圃場抵抗性であるか否かを判定する工程。
〔31〕以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物に被検化合物を接触させる工程、
(b)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物と被検化合物の結合を検出する工程、
(c)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物と結合する被検化合物を選択する工程。
〔32〕以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)植物から採取した細胞に、被検化合物を接触させる工程、
(b)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物の発現レベルを測定する工程、
(c)被検化合物を接触させていない場合と比較して、上記転写産物の発現レベルを減少させた被検化合物を選択する工程。
〔33〕以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)〔1〕の(a)〜(d)のいずれかに記載のDNAのプロモーター領域の下流にレポーター遺伝子が機能的に結合したDNAを有する細胞または細胞抽出液を提供する工程、
(b)該細胞または該細胞抽出液に被検化合物を接触させる工程、
(c)該細胞または該細胞抽出液における該レポーター遺伝子の発現レベルを測定する工程、
(d)被検化合物を接触させていない場合と比較して、該レポーター遺伝子の発現レベルを減少させた被検化合物を選択する工程。
〔34〕以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)〔6〕に記載の形質転換植物細胞より形質転換植物体を再生させる工程、
(b)該形質転換植物体に、いもち病菌および被検化合物を接触させる工程、
(c)被検化合物を接触させていない場合と比較して、該形質転換植物体のいもち病を抑制する被検化合物を選択する工程。
〔35〕〔31〕〜〔34〕のいずれかに記載のスクリーニング方法に用いるためのキット。
【発明の効果】
【0012】
Pi21遺伝子の特性は、植物のいもち病圃場抵抗性品種の作出に特に適している。これまでは、いもち病圃場抵抗性を付与するためには、圃場抵抗性を本来持つが劣悪な特性をもちあわせた品種とこれを持たないが多くの優れた特性を持つ品種を交雑し、その子孫より圃場抵抗性の優れた特性と他の優れた特性を併せ持った個体を選び出す作業が必要であった。しかしながら、圃場抵抗性を的確に評価するためには多大な労力を必要とし、かつ、これを付与する遺伝子の正確な染色体上の位置が明らかではない場合、これを効率的かつ的確に選抜し、実用性の高い品種に導入することは困難であった。そして実際、これまで成功してこなかった。
【0013】
本発明によって、圃場抵抗性に関与する遺伝子の染色体上の位置および遺伝子の構造が提供された。これにより、植物に圃場抵抗性を効率的に付与することが可能となった。また、圃場抵抗性に関与する遺伝子の組織発現特異性や発現レベルを変更することにより、抵抗性と実用性の高い特性を兼ね備えた品種を育成することも可能となった。このように本発明の遺伝子は、実用性が高く、安全性の高い農業を実現する上で有用である。また、本発明の方法で作製された植物は、例えば有用農作物では高い収量が安定的に得られるだけではなく、鑑賞用植物では新たな美的価値が付加されることも大いに期待される。
【図面の簡単な説明】
【0014】
【図1】愛知旭の遺伝的背景にpi21遺伝子を持つ系統AA-pi21(図左)および愛知旭(図右)におけるいもち病の病斑を示す写真である。
【図2】pi21遺伝子領域の詳細な連鎖地図およびゲノムクローンの整列地図を示す図である。AおよびBは、72個体および1014個体の分離集団により作成した遺伝地図を示す。Cは、日本晴のPACクローンによる整列地図を示す。Dは、pi21遺伝子領域の詳細な遺伝地図と候補ゲノム領域を示す。
【図3】pi21候補遺伝子の構造と日本晴・愛知旭とオワリハタモチのゲノム塩基配列の比較を示す図である。
【図4】抵抗性系統AApi21に日本晴由来のPi21遺伝子を導入した形質転換体に生じた病斑を示す写真である。A:ベクターのみを導入。B:Pi21遺伝子1コピー導入。C: Pi21遺伝子3コピー以上導入。
【発明を実施するための形態】
【0015】
イネにおいていもち病の進展を抑制しない罹病性遺伝子Pi21の対立遺伝子pi21遺伝子は、いもち病圃場抵抗性という形質をイネに付与する遺伝子として、これまでイネ第4染色体という広大な領域にいずれかの場所に存在することが知られていた。本発明者らは、マップベースクローニングの手法を利用することによりイネ第4染色体におけるpi21遺伝子の存在領域の絞り込みを行い、遂に単一の遺伝子として同定することに成功した。またpi21遺伝子の対立遺伝子Pi21遺伝子を単離することにも成功した。
【0016】
本発明において「いもち病」とは、植物がいもち病菌に感染し、その作用によってその一部あるいは全体が変色・壊死すること、あるいはそれによって認識できる病徴(病斑)をいう。いもち病の病斑は植物の各部位に発生し、発生の部位によりそれぞれ、苗いもち、葉いもち、穂いもち、籾いもち、節いもち、葉節(葉舌)いもちなどと呼ばれる。本発明における「いもち病」とは、これら全ての部位に発生するいもち病を含む。またイネにいもち病を引き起こす「イネいもち病菌」は、現在のところ統一された学名はないものの、Magnaporthe griseaあるいはMagnaporthe oryzaeなどと呼ばれるものがある。またいもち病菌には完全世代名と不完全世代名があり、完全世代名Magnaporthe oryzaeに対する不完全世代名としてPyricularia oryzaeが与えられ、使い分けられている。本発明におけるいもち病菌には、名称のいかんを問わずこれら全てのいもち病菌が含まれる。
【0017】
また本明細書において「いもち病罹病性」とは、植物がいもち病に感染する性質(まれにその病徴が著しいこと)を意味する。さらに「いもち病圃場抵抗性」とは、植物がいもち病菌に感染したときの(同一植物種内の)品種や系統間での病斑の数や大きさの差として認識される病徴の違いまたは病斑の数や大きさを抑制する性質を意味する。また「真性抵抗性」とは、植物がいもち病菌の侵入した細胞において過敏感反応による細胞死を起こし,感染を防ぐ性質をいう。
【0018】
本発明は、植物のいもち病に関与するいもち病罹病性遺伝子Pi21、およびいもち病圃場抵抗性遺伝子pi21を提供する。
【0019】
本発明のPi21遺伝子には、より具体的には、
(a)配列番号:3または22に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:1、2、20または21に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:3または22に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(d)配列番号:1、2、20または21に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
が含まれる。
【0020】
また本発明のpi21遺伝子には、より具体的には、
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4または5に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(d)配列番号:4または5に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
が含まれる。
【0021】
本発明のPi21遺伝子またはpi21遺伝子を利用することにより、例えば、組み換えタンパク質の調製やいもち病圃場抵抗性が改変された形質転換植物体を作出することなどが可能となる。
【0022】
本発明において、本発明の遺伝子が由来する植物としては、特に制限はないが、例えばイネ、トウモロコシ、コムギ、オオムギ、エンバク、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビ、パールミレット等の単子葉植物や、ナタネ、ダイズ、ワタ、トマト、ジャガイモ等の双子葉植物が挙げられる。また例えば花卉植物としては、キク、バラ、カーネーション、シクラメン等が挙げられるが、これらに限定されない。
【0023】
本発明の「Pi21遺伝子」、「pi21遺伝子」はそれぞれ、「Pi21タンパク質」、「pi21タンパク質」をコードしうるものであれば、その形態に特に制限はなく、「Pi21遺伝子」、「pi21遺伝子」にはそれぞれ、cDNAの他、ゲノムDNA、化学合成DNAなども含まれる。また、Pi21遺伝子はPi21タンパク質を、pi21遺伝子はpi21タンパク質をコードするものであれば、遺伝暗号の縮重に基づく任意の塩基配列を有するDNAが含まれる。
【0024】
ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、植物からゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作成し、これを展開して、Pi21遺伝子またはpi21遺伝子(例えば、配列番号:1、2、4、5、20または21のいずれかに記載のDNA)を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより調製することが可能である。また、Pi21遺伝子またはpi21遺伝子に特異的なプライマーを作成し、これを利用したPCRをおこなうことによって調製することも可能である。また、cDNAは、例えば、植物から抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。
【0025】
さらに、Pi21遺伝子またはpi21遺伝子は広く植物界に存在すると考えられるため、Pi21遺伝子またはpi21遺伝子には、イネのみならず、種々の植物に存在する相同遺伝子も含まれる。ここで「相同遺伝子」とは、種々の植物において、イネにおけるPi21遺伝子またはpi21遺伝子産物と同様の生理機能(例えばいもち病罹病性またはいもち病圃場抵抗性)を有するタンパク質をコードする遺伝子をさす。
【0026】
相同遺伝子を単離するための当業者によく知られた方法としては、ハイブリダイゼーション技術(Southern, E. M., Journal of Molecular Biology, Vol. 98, 503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R. K., et al. Science, vol. 230, 1350-1354, 1985, Saiki, R. K. et al. Science, vol.239, 487-491,1988)が挙げられる。即ち、当業者にとっては、イネPi21遺伝子またはpi21遺伝子の塩基配列(例えば、配列番号:1、2、4、5、20または21のいずれかに記載の配列)もしくはその一部をプローブとして、またPi21遺伝子またはpi21遺伝子に特異的にハイブリダイズするオリゴヌクレオチドをプライマーとして、種々の植物からPi21遺伝子またはpi21遺伝子の相同遺伝子を単離することは通常行いうることである。
【0027】
このような相同遺伝子をコードするDNAを単離するためには、通常ストリンジェントな条件下でハイブリダイゼーション反応を行なう。ストリンジェントなハイブリダイゼーション条件としては、6M 尿素、0.4%SDS、0.5 x SSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を例示できる。よりストリンジェンシーの高い条件、例えば、6M 尿素、0.4%SDS、0.1 x SSCの条件を用いれば、より相同性の高いDNAの単離を期待することができる。単離したDNAの配列の決定は、公知の方法で行うことができる。単離されたDNAの相同性は、アミノ酸配列全体で、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の相同性は、BLASTN(核酸レベル)やBLASTX(アミノ酸レベル)のプログラム(Altschul et al. J. Mol. Biol., 215: 403-410, 1990)を利用して決定することができる。該プログラムは、Karlin及びAltschulによるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA, 87:2264-2268, 1990, Proc. Natl. Acad. Sci. USA, 90: 5873-5877, 1993) に基づいている。BLASTNによって塩基配列を解析する場合には、パラメーターは例えばscore = 100、wordlength =12とする。また、BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えばscore = 50、wordlength = 3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res. 25: 3389-3402, 1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0028】
また本発明は、植物の内因性のPi21遺伝子の発現を抑制するために用いるDNAであって、
(a)Pi21遺伝子の転写産物と相補的なRNAをコードするDNA、
(b)Pi21遺伝子の転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA、
(c)Pi21遺伝子の発現を共抑制効果により阻害するRNAをコードするDNA、
(d)Pi21遺伝子の転写産物を特異的に切断するRNAi活性を有するRNAをコードするDNA、
を提供する。
これらのDNAにより、植物のいもち病による病斑進展を抑制することが可能である。
【0029】
本発明において、Pi21遺伝子の発現を抑制する植物には特に制限はなく、いもち病圃場抵抗性を付与させたい所望の植物を用いることができるが、産業的な観点からは農作物や鑑賞用植物が好適である。有用農作物としては、特に制限はないが、例えばイネ、トウモロコシ、コムギ、オオムギ、エンバク、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビ、パールミレット等の単子葉植物や、ナタネ、ダイズ、ワタ、トマト、ジャガイモ等の双子葉植物が挙げられる。また、観賞用植物としては、例えばキク、バラ、カーネーション、シクラメン等の花卉植物が挙げられるが、これらに限定されない。またイネいもち病菌に感受性を有する植物として、オオムギ、イタリアンライグ.ラス、メドーフェスクなどの牧草、トウモロコシがあり、それ以外にも、イネより分離したいもち病菌が寄生性を有する植物としてエゾノサヤヌカグサ、マコモなどのイネ族、ウシノケグサ族、オオムギ族、エンバクなどのカラスムギ族、シナダレスズメガヤなどのヒゲシバ族、アワ、メヒシバなどのキビ族など多くの植物が報告されおり、これらの植物もまたいもち病圃場抵抗性を付与させる植物に含まれる。
【0030】
本明細書における「Pi21遺伝子の発現を抑制」には、遺伝子の転写の抑制およびタンパク質への翻訳の抑制が含まれる。また、DNAの発現の完全な停止のみならず発現の減少も含まれる。
【0031】
「Pi21遺伝子の発現を抑制するために用いるDNA」の一つの態様は、Pi21遺伝子と相補的なアンチセンスRNAをコードするDNAである。植物細胞におけるアンチセンス効果は、一時的遺伝子発現法を用いて、電気穿孔法で導入したアンチセンスRNAが植物においてアンチセンス効果を発揮することにより、初めて実証された(Ecker and Davis, Proc. Natl. Acad. USA, 83: 5372, 1986)。その後、タバコやペチュニアにおいても、アンチセンスRNAの発現によって標的遺伝子の発現を低下させる例が報告されており(Krol et. al., Nature 333: 866, 1988)、現在では植物における遺伝子発現を抑制させる手段として確立している。
【0032】
アンチセンス核酸が標的遺伝子の発現を抑制する作用としては、以下のような複数の要因が存在する。すなわち、三重鎖形成による転写開始阻害、RNAポリメラーゼによって局部的に開状ループ構造がつくられた部位とのハイブリッド形成による転写抑制、合成の進みつつあるRNAとのハイブリッド形成による転写阻害、イントロンとエキソンとの接合点でのハイブリッド形成によるスプライシング抑制、スプライソソーム形成部位とのハイブリッド形成によるスプライシング抑制、mRNAとのハイブリッド形成による核から細胞質への移行抑制、キャッピング部位やポリ(A)付加部位とのハイブリッド形成によるスプライシング抑制、翻訳開始因子結合部位とのハイブリッド形成による翻訳開始抑制、開始コドン近傍のリボソーム結合部位とのハイブリッド形成による翻訳抑制、mRNAの翻訳領域やポリソーム結合部位とのハイブリッド形成によるペプチド鎖の伸長阻止、および核酸とタンパク質との相互作用部位とのハイブリッド形成による遺伝子発現抑制などである。これらは、転写、スプライシング、または翻訳の過程を阻害して、標的遺伝子の発現を抑制する(平島および井上「新生化学実験講座2 核酸IV 遺伝子の複製と発現」,日本生化学会編,東京化学同人, pp.319-347, 1993)。
【0033】
本発明で用いられるアンチセンス配列は、上記のいずれの作用で標的遺伝子の発現を抑制してもよい。一つの態様としては、遺伝子のmRNAの5'端近傍の非翻訳領域に相補的なアンチセンス配列を設計すれば、遺伝子の翻訳阻害に効果的であろう。しかし、コード領域もしくは3'側の非翻訳領域に相補的な配列も使用し得る。このように、遺伝子の翻訳領域だけでなく非翻訳領域の配列のアンチセンス配列を含むDNAも、本発明で利用されるアンチセンスDNAに含まれる。使用されるアンチセンスDNAは、適当なプロモーターの下流に連結され、好ましくは3'側に転写終結シグナルを含む配列が連結される。
【0034】
アンチセンスDNAは、例えば、配列番号:1、2、20または21に記載のDNAの配列情報を基にホスホロチオネート法(Stein, Nucleic Acids Res., 16: 3209-3221, 1988)などにより調製することが可能である。調製されたDNAは、公知の方法で、所望の植物へ形質転換できる。アンチセンスDNAの配列は、形質転換する植物が持つ内因性遺伝子の転写産物と相補的な配列であることが好ましいが、遺伝子の発現を有効に阻害できる限り、完全に相補的でなくてもよい。転写されたRNAは、標的とする遺伝子の転写産物に対して好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の相補性を有する。アンチセンス配列を用いて、効果的に標的遺伝子の発現を阻害するには、アンチセンスDNAの長さは、少なくとも15塩基以上であり、好ましくは100塩基以上であり、さらに好ましくは500塩基以上である。通常、用いられるアンチセンスDNAの長さは5kbよりも短く、好ましくは2.5kbよりも短い。
【0035】
内因性のPi21遺伝子の発現の抑制は、リボザイムをコードするDNAを利用して行うことも可能である。リボザイムとは触媒活性を有するRNA分子のことをいう。リボザイムには種々の活性を有するものがあるが、中でもRNAを切断する酵素としてのリボザイムの研究により、RNAの部位特異的な切断を目的とするリボザイムの設計が可能となった。リボザイムには、グループIイントロン型や、RNasePに含まれるM1RNAのように400ヌクレオチド以上の大きさのものもあるが、ハンマーヘッド型やヘアピン型と呼ばれる40ヌクレオチド程度の活性ドメインを有するものもある(小泉誠および大塚栄子、蛋白質核酸酵素, 35: 2191, 1990)。
【0036】
例えば、ハンマーヘッド型リボザイムの自己切断ドメインは、G13U14C15のC15の3'側を切断するが、活性にはU14が9位のAと塩基対を形成することが重要とされ、15位の塩基はCの他にAまたはUでも切断されることが示されている(Koizumi et. al., FEBS Lett. 228: 225, 1988)。リボザイムの基質結合部を標的部位近傍のRNA配列と相補的になるように設計すれば、標的RNA中のUC、UUまたはUAという配列を認識する制限酵素的なRNA切断リボザイムを作出することが可能である(Koizumi et. al., FEBS Lett. 239: 285,1988、小泉誠および大塚栄子, 蛋白質核酸酵素,35: 2191, 1990、Koizumi et. al., Nucleic. Acids. Res. 17: 7059, 1989)。
【0037】
また、ヘアピン型リボザイムも、本発明の目的のために有用である。ヘアピン型リボザイムは、例えばタバコリングスポットウイルスのサテライトRNAのマイナス鎖に見出される(Buzayan, Nature 323: 349,1986)。このリボザイムも、標的特異的なRNA切断を起こすように設計できることが示されている(Kikuchi and Sasaki, Nucleic Acids Res. 19: 6751, 1992, 及び菊池洋,化学と生物 30: 112, 1992)。
【0038】
標的を切断できるよう設計されたリボザイムは、植物細胞中で転写されるようにカリフラワーモザイクウイルスの35Sプロモーターなどのプロモーターおよび転写終結配列に連結される。しかし、その際、転写されたRNAの5'末端や3'末端に余分な配列が付加されていると、リボザイムの活性が失われてしまうことがある。このようなとき、転写されたリボザイムを含むRNAからリボザイム部分だけを正確に切り出すために、リボザイム部分の5'側や3'側に、トリミングを行うためのシスに働く別のトリミングリボザイムを配置させることも可能である (Taira et. al., Protein Eng. 3: 733, 1990、Dzianott and Bujarski, Proc. Natl. Acad. Sci. USA, 86: 4823, 1989、Grosshans and Cech, Nucleic Acids Res. 19: 3875, 1991、Taira et. al., Nucleic Acids Res. 19: 5125, 1991)。
【0039】
また、このような構成単位をタンデムに並べ、標的遺伝子内の複数の部位を切断できるようにして、より効果を高めることもできる(Yuyama et al., Biochem. Biophys. Res. Commun. 186: 1271, 1992)。このようなリボザイムを用いて本発明で標的となる遺伝子の転写産物を特異的に切断し、該遺伝子の発現を抑制することができる。
【0040】
内在性遺伝子の発現の抑制はさらに、標的遺伝子配列と同一もしくは類似した配列を有するDNAの形質転換によってもたらされる「共抑制」によっても達成されうる。「共抑制」とは、植物に標的内在性遺伝子と同一若しくは類似した配列を有する遺伝子を形質転換により導入すると、導入する外来遺伝子および標的内在性遺伝子の両方の発現が抑制される現象のことをいう。共抑制の機構の詳細は明らかではないが、植物においてはしばしば観察される(Curr. Biol. 7: R793, 1997、Curr. Biol. 6: 810, 1996)。
【0041】
例えば、Pi21遺伝子が共抑制された植物体を得るためには、Pi21遺伝子若しくはこれと類似した配列を有するDNAを発現できるように作製したベクターDNAを目的の植物へ形質転換し、得られた植物体からPi21変異体の形質を有する植物、即ちいもち病圃場抵抗性である植物を選択すればよい。共抑制に用いる遺伝子は、標的遺伝子と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。
【0042】
さらに、本発明における内在性遺伝子の発現の抑制は、標的遺伝子のドミナントネガティブの形質を有する遺伝子を植物へ形質転換することによっても達成することができる。ドミナントネガティブの形質を有する遺伝子とは、該遺伝子を発現させることによって、植物体が本来持つ内在性の野生型遺伝子の活性を消失もしくは低下させる機能を有する遺伝子のことをいう。
【0043】
「Pi21遺伝子の発現を抑制するために用いるDNA」の他の一つの態様は、内因性のPi21遺伝子の転写産物と相補的なdsRNA(二重鎖RNA)をコードするDNAである。標的遺伝子配列と同一もしくは類似した配列を有するdsRNAを細胞内に導入することにより、導入した外来遺伝子および標的内因性遺伝子の発現がいずれも抑制される、RNAi(RNA干渉、RNA interference)と呼ばれる現象を引き起こすことができる。細胞に約40〜数百塩基対のdsRNAが導入されると、ヘリカーゼドメインを持つダイサー(Dicer)と呼ばれるRNaseIII様のヌクレアーゼがATP存在下で、dsRNAを3'末端から約21〜23塩基対ずつ切り出し、siRNA(short interference RNA)を生じる。このsiRNAに特異的なタンパク質が結合して、ヌクレアーゼ複合体(RISC: RNA-induced silencing complex)が形成される。この複合体はsiRNAと同じ配列を認識して結合し、RNaseIII様の酵素活性によってsiRNAの中央部で標的遺伝子の転写産物(mRNA)を切断する。また、この経路とは別にsiRNAのアンチセンス鎖がmRNAに結合してRNA依存性RNAポリメラーゼ(RsRP)のプライマーとして作用し、dsRNAが合成される。このdsRNAが再びダイサーの基質となって、新たなsiRNAを生じて作用を増幅する経路も考えられている。
【0044】
RNAiは当初、線虫において発見されたが(Fire, A. et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391, 806-811, 1998)、現在では、線虫のみならず、植物、線形動物、ショウジョウバエ、原生動物などの種々の生物において観察されている(Fire, A. RNA-triggered gene silencing. Trends Genet. 15, 358-363 (1999)、Sharp, P. A. RNA interference 2001. Genes Dev. 15, 485-490 (2001)、Hammond, S. M., Caudy, A. A. & Hannon, G. J. Post-transcriptional gene silencing by double-stranded RNA. Nature Rev. Genet. 2, 110-119 (2001)、Zamore, P. D. RNA interference: listening to the sound of silence. Nat Struct Biol. 8, 746-750 (2001))。これら生物では、実際に外来よりdsRNAを導入することにより標的遺伝子の発現が抑制されることが確認され、さらにはノックアウト個体を創生する方法としても利用されつつある。
【0045】
RNAiの登場当初は、dsRNAはある程度の長さ(40塩基)以上でなければ効果がないと考えられていたが、米ロックフェラー大のTuschlらは21塩基対前後の単鎖dsRNA(siRNA)を細胞に導入すれば、哺乳動物細胞においてもPKRによる抗ウイルス反応を起こさず、RNAiの効果があることを報告し(Tuschl, Nature, 411, 494-498 (2001))、RNAiは分化したヒトなどの哺乳動物細胞に応用可能な技術として俄然注目を集めることになった。
【0046】
本発明のDNAは、標的遺伝子の転写産物(mRNA)のいずれかの領域に対するアンチセンスRNAをコードしたアンチセンスコードDNAと、前記mRNAのいずれかの領域のセンスRNAをコードしたセンスコードDNAを含み、前記アンチセンスコードDNAおよび前記センスコードDNAより前記アンチセンスRNAおよび前記センスRNAを発現させることができる。また、これらのアンチセンスRNAおよびセンスRNAよりdsRNAを作成することもできる。本発明における標的配列は、標的配列と同一もしくは類似した配列を有するdsRNAを細胞内に導入した結果Pi21遺伝子の発現が抑制されるものであれば特に制限されない。標的配列の一例としては、Pi21遺伝子の3'非翻訳領域配列が挙げられる。Pi21遺伝子の3'非翻訳領域配列を配列番号:11および24に示す。
【0047】
本発明のdsRNAの発現システムをベクター等に保持させる場合の構成としては、同一のベクターからアンチセンスRNA、センスRNAを発現させる場合と、異なるベクターからそれぞれアンチセンスRNA、センスRNAを発現させる場合がある。例えば、同一のベクターからアンチセンスRNA、センスRNAを発現させる構成としては、アンチセンスコードDNAおよびセンスコードDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモータを連結させたアンチセンスRNA発現カセット、センスRNA発現カセットをそれぞれ構築し、これらカセットを同方向にあるいは逆方向にベクターに挿入することにより構成することができる。
【0048】
また、異なる鎖上に対向するように、アンチセンスコードDNAとセンスコードDNAとを逆向きに配置した発現システムを構成することもできる。この構成では、アンチセンスRNAコード鎖とセンスRNAコード鎖とが対となった一つの二本鎖DNA(siRNAコードDNA)が備えられ、その両側にそれぞれの鎖からアンチセンスRNA、センスRNAとを発現し得るようにプロモータを対向して備えられる。この場合には、センスRNA、アンチセンスRNAの下流に余分な配列が付加されることを避けるために、それぞれの鎖(アンチセンスRNAコード鎖、センスRNAコード鎖)の3'末端にターミネーターをそれぞれ備えることが好ましい。このターミネーターは、A(アデニン)塩基を4つ以上連続させた配列などを用いることができる。また、このパリンドロームスタイルの発現システムでは、二つのプロモータの種類は異なっていることが好ましい。
【0049】
また、異なるベクターからアンチセンスRNA、センスRNAを発現させる構成としては、例えば、アンチセンスコードDNAおよびセンスコードDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモータを連結させたアンチセンスRNA発現カセット、センスRNA発現カセットをそれぞれ構築し、これらカセットを異なるベクターに保持させることにより構成することができる。
【0050】
RNAiにおいては、dsRNAとしてsiRNAが使用されたものであってもよい。「siRNA」は、細胞内で毒性を示さない範囲の短鎖からなる二重鎖RNAを意味し、Tuschlら(前掲)により報告された全長21〜23塩基対に限定されるものではなく、毒性を示さない範囲の長さであれば特に限定はなく、例えば、15〜49塩基対と、好適には15〜35塩基対と、さらに好適には21〜30塩基対とすることができる。あるいは、発現されるsiRNAが転写され最終的な二重鎖RNA部分の長さが、例えば、15〜49塩基対、好適には15〜35塩基対、さらに好適には21〜30塩基対とすることができる。
【0051】
本発明のDNAとしては、標的配列のインバーテッドリピートの間に適当な配列(イントロン配列が望ましい)を挿入し、ヘアピン構造を持つダブルストランドRNA(self-complementary 'hairpin' RNA(hpRNA))を作るようなコンストラクト(Smith, N.A., et al. Nature, 407: 319, 2000、Wesley, S. V. et al. Plant J. 27: 581, 2001、Piccin, A. et al. Nucleic Acids Res. 29:E55, 2001)を用いることもできる。
【0052】
RNAiに用いるDNAは、標的遺伝子と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。また、配列の同一性は上述した手法により決定できる。
【0053】
dsRNAにおけるRNA同士が対合した二重鎖RNAの部分は、完全に対合しているものに限らず、ミスマッチ(対応する塩基が相補的でない)、バルジ(一方の鎖に対応する塩基がない)などにより不対合部分が含まれていてもよい。本発明においては、dsRNAにおけるRNA同士が対合する二重鎖RNA領域中に、バルジおよびミスマッチの両方が含まれていてもよい。
【0054】
また本発明は、Pi21遺伝子、pi21遺伝子、またはPi21遺伝子の発現を抑制するDNAのいずれか1つを含むベクターならびに形質転換細胞を提供する。
上記ベクターとしては、例えば、大腸菌を宿主とする場合には、ベクターを大腸菌(例えば、JM109、DH5α、HB101、XL1Blue)等で大量に増幅させ大量調製するために、大腸菌で増幅されるための「ori」をもち、さらに形質転換された大腸菌の選抜遺伝子(例えば、なんらかの薬剤(アンピシリンやテトラサイクリン、カナマイシン、クロラムフェニコールにより判別できるような薬剤耐性遺伝子)を有すれば特に制限はない。ベクターの例としては、M13系ベクター、pUC系ベクター、pBR322、pBluescript、pCR-Script等が挙げられる。また、cDNAのサブクローニング、切り出しを目的とした場合、上記ベクターの他に、例えば、pGEM-T、pDIRECT、pT7等が挙げられる。Pi21遺伝子、pi21遺伝子、またはPi21遺伝子の発現を抑制するDNAを生産する目的においてベクターを使用する場合には、特に、発現ベクターが有用である。発現ベクターとしては、例えば、大腸菌での発現を目的とした場合は、ベクターが大腸菌で増幅されるような上記特徴を持つほかに、宿主をJM109、DH5α、HB101、XL1-Blue等の大腸菌とした場合においては、大腸菌で効率よく発現できるようなプロモーター、例えば、lacZプロモーター(Wardら, Nature (1989) 341, 544-546;FASEB J. (1992) 6, 2422-2427)、araBプロモーター(Betterら, Science (1988) 240, 1041-1043 )、またはT7プロモーター等を持っていることが不可欠である。このようなベクターとしては、上記ベクターの他にpGEX-5X-1(ファルマシア社製)、「QIAexpress system」(キアゲン社製)、pEGFP、またはpET等が挙げられる。
【0055】
また、ベクターには、ポリペプチド分泌のためのシグナル配列が含まれていてもよい。ポリペプチド分泌のためのシグナル配列としては、大腸菌のペリプラズムに産生させる場合、pelBシグナル配列(Lei, S. P. et al J. Bacteriol. (1987) 169, 4379)を使用すればよい。宿主細胞へのベクターの導入は、例えば塩化カルシウム法、エレクトロポレーション法を用いて行うことができる。
【0056】
大腸菌以外にも、例えば、Pi21遺伝子、pi21遺伝子、またはPi21遺伝子の発現を抑制するDNAを製造するためのベクターとしては、哺乳動物由来の発現ベクター(例えば、pcDNA3 (インビトロゲン社製)や、pEGF-BOS (Nucleic Acids. Res.1990, 18(17),p5322)、pEF 、pCDM8 )、昆虫細胞由来の発現ベクター(例えば「Bac-to-BAC baculovairus expression system」(ギブコBRL社製)、pBacPAK8)、植物由来の発現ベクター(例えばpMH1、pMH2)、動物ウィルス由来の発現ベクター(例えば、pHSV、pMV、pAdexLcw )、レトロウィルス由来の発現ベクター(例えば、pZIPneo)、酵母由来の発現ベクター(例えば、「Pichia Expression Kit」(インビトロゲン社製)、pNV11、SP-Q01)、枯草菌由来の発現ベクター(例えば、pPL608、pKTH50)等が挙げられる。
【0057】
CHO細胞、COS細胞、NIH3T3細胞等の動物細胞での発現を目的とした場合には、細胞内で発現させるために必要なプロモーター、例えばSV40プロモーター(Mulliganら, Nature (1979) 277, 108)、MMLV-LTRプロモーター、EF1αプロモーター(Mizushimaら, Nucleic Acids Res. (1990) 18, 5322)、CMVプロモーター等を持っていることが不可欠であり、細胞への形質転換を選抜するための遺伝子(例えば、薬剤(ネオマイシン、G418等)により判別できるような薬剤耐性遺伝子)を有すればさらに好ましい。このような特性を有するベクターとしては、例えば、pMAM、pDR2、pBK-RSV、pBK-CMV、pOPRSV、pOP13等が挙げられる。
【0058】
本発明のDNAの細胞への導入は、当業者においては、公知の方法、例えば電気穿孔法(エレクトロポーレーション法)などにより実施することができる。
【0059】
さらに本発明は、Pi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが導入された形質転換植物細胞、これらの形質転換植物体、該形質転換植物体の子孫またはクローンである形質転換植物体、該形質転換植物体の繁殖材料を提供する。また、上記形質転換体、繁殖材料の製造方法を提供する。
【0060】
Pi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAの植物細胞への導入は、上記の方法によって行うことが可能である。
【0061】
また植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である(Toki et. al., Plant Physiol. 100: 1503-1507, 1995)。例えば、イネにおいては、形質転換植物体を作出する手法については、ポリエチレングリコールによりプロトプラストへ遺伝子導入し、植物体(インド型イネ品種が適している)を再生させる方法(Datta et. al., In Gene Transfer To Plants (Potrykus I and Spangenberg Eds.) pp66-74, 1995)、電気パルスによりプロトプラストへ遺伝子導入し、植物体(日本型イネ品種が適している)を再生させる方法(Toki et. al., Plant Physiol. 100: 1503-1507, 1992)、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法(Christou et. al., Bio/technology, 9: 957-962, 1991)およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法(Hiei et. al., Plant J. 6: 271-282, 1994)など、いくつかの技術が既に確立し、本願発明の技術分野において広く用いられている。本発明においては、これらの方法を好適に用いることができる。
【0062】
上記アグロバクテリウム法を用いる場合、例えばNagelらの方法(Microbiol. Lett. 67: 325, 1990)が用いられる。この方法によれば、組み換えベクターをアグロバクテリウム細菌中に形質転換して、次いで形質転換されたアグロバクテリウムを、リーフディスク法等の公知の方法により細胞に導入する。上記ベクターは、例えば植物体に導入した後、本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが植物体中で発現するように、発現プロモーターを含む。一般に、該プロモーターの下流には本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが位置し、さらに該DNAの下流にはターミネーターが位置する。この目的に用いられる組み換えベクターは、植物への導入方法、または植物の種類に応じて、当業者によって適宜選択される。上記プロモーターとして、例えばカリフラワーモザイクウイルス由来のCaMV35S、トウモロコシのユビキチンプロモーター(特開平2-79983号公報)等を挙げることができる。
【0063】
また、上記ターミネーターは、カリフラワーモザイクウイルス由来のターミネーター、あるいはノパリン合成酵素遺伝子由来のターミネーター等を例示することができるが、植物体中で機能するプロモーターやターミネーターであれば、これらに限定されない。
【0064】
また、本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAを導入する植物は、外植片であってもよく、これらの植物から培養細胞を調製し、得られた培養細胞に導入してもよい。本発明の「植物細胞」は、例えば葉、根、茎、花および種子中の胚盤等の植物の細胞、カルス、懸濁培養細胞等が挙げられる。
【0065】
また、本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAの導入により形質転換した細胞を効率的に選択するために、上記組み換えベクターは、適当な選抜マーカー遺伝子、もしくは選抜マーカー遺伝子を含むプラスミドベクターと共に植物細胞へ導入するのが好ましい。この目的に使用する選抜マーカー遺伝子は、例えば、抗生物質ハイグロマイシンに耐性であるハイグロマイシンホスホトランスフェラーゼ遺伝子、カナマイシンまたはゲンタマイシンに耐性であるネオマイシンホスホトランスフェラーゼ、および除草剤ホスフィノスリシンに耐性であるアセチルトランスフェラーゼ遺伝子等が挙げられる。
【0066】
組み換えベクターを導入した細胞は、導入された選抜マーカー遺伝子の種類に従って適当な選抜用薬剤を含む公知の選抜用培地に置床し培養する。これにより形質転換された植物培養細胞を得ることができる。
【0067】
形質転換細胞から再生させた植物体は、次いで順化用培地で培養する。その後、順化した再生植物体を、通常の栽培条件で栽培すると、いもち病圃場抵抗性である植物体が得られ、成熟して結実して種子を得ることができる。
【0068】
なお、このように再生され、かつ栽培した形質転換植物体中の導入された外来DNAの存在は、公知のPCR法やサザンハイブリダイゼーション法によって、または植物体中のDNAの塩基配列を解析することによって確認することができる。
この場合、形質転換植物体からのDNAの抽出は、公知のJ.Sambrookらの方法(Molecular Cloning、第2版、Cold Spring Harbor Laboratory Press, 1989)に準じて実施することができる。
【0069】
再生させた植物体中に存在する本発明のDNAよりなる外来遺伝子を、PCR法を用いて解析する場合には、上記のように再生植物体から抽出したDNAを鋳型として増幅反応を行う。また、本発明のDNA、あるいは本発明により改変されたDNAの塩基配列に従って適当に選択された塩基配列をもつ合成したオリゴヌクレオチドをプライマーとして用い、これらを混合させた反応液中において増幅反応を行うこともできる。増幅反応においては、DNAの変性、アニーリング、伸張反応を数十回繰り返すと、本発明のDNA配列を含むDNA断片の増幅生成物を得ることができる。増幅生成物を含む反応液を例えばアガロース電気泳動にかけると、増幅された各種のDNA断片が分画されて、そのDNA断片が本発明のDNAに対応することを確認することが可能である。
【0070】
一旦、染色体内に本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫を得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラスト等を得て、それらを基に該植物体を量産することも可能である。本発明には、Pi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが導入された植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、並びに該植物体、その子孫およびクローンの繁殖材料が含まれる。これらの植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、並びに該植物体、その子孫およびクローンの繁殖材料は、植物にいもち病圃場抵抗性を付与する方法に使用することが可能である。
【0071】
本発明はさらに、Pi21遺伝子の発現を抑制するDNAを植物体の細胞内で発現させる工程を含む、植物にいもち病圃場抵抗性を付与する方法を提供する。植物にいもち病圃場抵抗性を付与することは、上述の方法を用いてPi21遺伝子の発現を抑制するDNAを発現可能に含むベクターを植物細胞に導入し、これを再生することにより可能である。
【0072】
本明細書においていもち病圃場抵抗性を「付与する」とは、いもち病圃場抵抗性能を全く有していない植物にいもち病圃場抵抗性能を持たせることのみならず、すでにいもち病圃場抵抗性能を有している植物のいもち病抵抗性能を、さらに増大させることも意味する。
本発明において、Pi21遺伝子の発現を抑制していもち病圃場抵抗性を付与させる植物には特に制限はなく、例えば上述の植物を挙げることが出来る。
【0073】
本発明はまた、本発明のPi21遺伝子によりコードされるタンパク質、該タンパク質の製造方法、該タンパク質に結合する抗体を提供する。
組み換えタンパク質を調製する場合には、通常、本発明のタンパク質をコードするDNAを適当な発現ベクターに挿入し、該ベクターを適当な細胞に導入し、形質転換細胞を培養して発現させたタンパク質を精製する。組み換えタンパク質は、精製を容易にするなどの目的で、他のタンパク質との融合タンパク質として発現させることも可能である。例えば、大腸菌を宿主としてマルトース結合タンパク質との融合タンパク質として調製する方法(米国New England BioLabs社発売のベクターpMALシリーズ)、グルタチオン-S-トランスフェラーゼ(GST)との融合タンパク質として調製する方法(Amersham Pharmacia Biotech社発売のベクターpGEXシリーズ)、ヒスチジンタグを付加して調製する方法(Novagen社のpETシリーズ)などを利用することが可能である。宿主細胞としては、組み換えタンパク質の発現に適した細胞であれば特に制限はなく、上記の大腸菌の他、例えば、酵母、種々の動植物細胞、昆虫細胞などを用いることが可能である。宿主細胞へのベクターの導入には、当業者に公知の種々の方法を用いることが可能である。例えば、大腸菌への導入には、カルシウムイオンを利用した導入方法(Mandel, M. & Higa, A. Journal of Molecular Biology, Vol.53, 158-162,(1970)、Hanahan, D. Journal of Molecular Biology, Vol.166, 557-580,(1983))を用いることができる。宿主細胞内で発現させた組み換えタンパク質は、該宿主細胞またはその培養上清から、当業者に公知の方法により精製し、回収することが可能である。組み換えタンパク質を上述したマルトース結合タンパク質などとの融合タンパク質として発現させた場合には、容易にアフィニティー精製を行うことが可能である。
【0074】
得られた組換えタンパク質を用いれば、これに結合する抗体を調製することができる。ポリクローナル抗体であれば、例えば、次のようにして取得することができる。Pi21タンパク質、あるいはGSTとの融合タンパク質として大腸菌等の微生物において発現させたリコンビナントタンパク質、またはその部分ペプチドをウサギ等の小動物に免疫し血清を得る。これを、例えば、硫安沈殿、プロテインA、プロテインGカラム、DEAEイオン交換クロマトグラフィー、Pi21タンパク質や合成ペプチドをカップリングしたアフィニティーカラム等により精製することにより調製する。また、モノクローナル抗体であれば、例えば、Pi21タンパク質またはその部分ペプチドをマウス等の小動物に免疫を行い、同マウスより脾臓を摘出し、これをすりつぶして細胞を分離し、該細胞とマウスミエローマ細胞とをポリエチレングリコール等の試薬を用いて融合させ、これによりできた融合細胞(ハイブリドーマ)の中から、Pi21タンパク質に結合する抗体を産生するクローンを選択する。次いで、得られたハイブリドーマをマウス腹腔内に移植し、同マウスより腹水を回収し、得られたモノクローナル抗体を、例えば、硫安沈殿、プロテインA、プロテインGカラム、DEAEイオン交換クロマトグラフィー、Pi21タンパク質や合成ペプチドをカップリングしたアフィニティーカラム等により精製することで、調製することが可能である。これにより得られた抗体は、本発明のタンパク質の精製や検出などに利用することが可能である。本発明には、本発明のタンパク質に結合する抗体が含まれる。
【0075】
本発明はまた、Pi21遺伝子またはpi21遺伝子からなるDNAまたはその相補鎖に相補的な少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを提供する。
ここで「相補鎖」とは、A:T(ただしRNAの場合はU)、G:Cの塩基対からなる2本鎖核酸の一方の鎖に対する他方の鎖を指す。また、「相補的」とは、少なくとも15個の連続したヌクレオチド領域で完全に相補配列である場合に限られず、少なくとも70%、好ましくは少なくとも80%、より好ましくは90%、さらに好ましくは95%以上の塩基配列上の相同性を有すればよい。相同性を決定するためのアルゴリズムは当業者に周知のものを使用すればよい。
【0076】
本発明のオリゴヌクレオチドは、配列番号:1、2、4、5、20または21に記載の塩基配列からなるDNAの検出や増幅に用いるプローブやプライマーとして使用することができる。また、本発明のオリゴヌクレオチドは、DNAアレイの基板の形態で使用することができる。
【0077】
該オリゴヌクレオチドをプライマーとして用いる場合、その長さは、通常15bp〜100bpであり、好ましくは17bp〜30bpである。プライマーは、本発明のDNAまたはその相補鎖の少なくとも一部を増幅しうるものであれば、特に制限されない。また、プライマーとして用いる場合、3'側の領域は相補的とし、5'側には制限酵素認識配列やタグなどを付加することができる。
【0078】
また、上記オリゴヌクレオチドをプローブとして使用する場合、該プローブは、配列番号:1、2、4、5、20または21に記載の塩基配列からなるDNAまたはその相補鎖の少なくとも一部に特異的にハイブリダイズするものであれば、特に制限されない。該プローブは、合成オリゴヌクレオチドであってもよく、通常少なくとも15bp以上の鎖長を有する。
【0079】
本発明のオリゴヌクレオチドをプローブとして用いる場合は、適宜標識して用いることが好ましい。標識する方法としては、T4ポリヌクレオチドキナーゼを用いて、オリゴヌクレオチドの5'端を32Pでリン酸化することにより標識する方法、およびクレノウ酵素等のDNAポリメラーゼを用い、ランダムヘキサマーオリゴヌクレオチド等をプライマーとして32P等のアイソトープ、蛍光色素、またはビオチン等によって標識された基質塩基を取り込ませる方法(ランダムプライム法等)を例示することができる。
【0080】
本発明のオリゴヌクレオチドは、例えば市販のオリゴヌクレオチド合成機により作製することができる。プローブは、制限酵素処理等によって取得される二本鎖DNA断片として作製することもできる。
【0081】
さらに本発明は、Pi21遺伝子の発現を抑制するDNA、該DNAを含むベクターの用途を提供するものである。すなわち本発明は、Pi21遺伝子の発現を抑制するDNA、該DNAを含むベクターのいずれか1つを有効成分として含む、植物のいもち病圃場抵抗性を増加させるための薬剤に関する。また本発明は、植物のいもち病圃場抵抗性を増加させるための薬剤の製造における、Pi21遺伝子の発現を抑制するDNA、該DNAを含むベクターの使用に関する。
【0082】
本発明の植物にいもち病圃場抵抗性を増加させるための薬剤においては、有効成分であるオリゴヌクレオチド以外に、例えば、滅菌水、生理食塩水、植物油、界面活性剤、脂質、溶解補助剤、緩衝剤、保存剤等が必要に応じて混合されていてもよい。
【0083】
本発明はまた、罹病性遺伝子Pi21または抵抗性遺伝子pi21に連鎖する分子マーカーを提供する。
本発明における「分子マーカー」とは、Pi21遺伝子またはpi21遺伝子と遺伝的に連鎖するDNA領域であって、他のDNA領域と識別可能なDNA領域をいう。
【0084】
一般に分子マーカーとは、単位cMで表す地図距離が短いほどその遺伝子の近傍に位置し、その遺伝子と同時に遺伝するため、有用性が高い。pi21はマーカー「Pa102484」からマーカー「P702D3_#12」の間に存在することが示された(図2c)。したがって本発明の方法においては、図2cに記載の分子マーカーの中でも、上記2つのマーカー間および該2つのマーカーの間に存在するマーカー(「P702D03_#38」、「P702D03_#79」、「P702D03_#80」)は好ましいマーカーである。その中でも「P702D03_#79」は特に好ましいマーカーであり、配列番号:7または23(罹病性遺伝子Pi21と連鎖)または配列番号:10(抵抗性遺伝子pi21と連鎖)に記載のDNAとして例示できる。
【0085】
本発明の分子マーカーとしては、STS(Sequence Tagged Site)マーカーを挙げることができる。STSマーカーとは、DNA上の配列タグ部位(STS)の多型の有無の判定に利用できるDNA領域をいい、STSとは、DNA上の特定位置に特異的な配列部位をいう。STSの多型は、該特定配列部位を含むDNA領域をPCR法等の核酸増幅法により増幅し、該増幅産物をアガロースまたはポリアクリルアミドゲル電気泳動にかけることにより、バンドの有無や位置の違いとして検出することができる。
【0086】
本発明の分子マーカーとしてSTSマーカーを用いる場合は、本発明の識別方法を、例えば以下のようにして行うことができる。まず、被検イネおよびいもち病圃場抵抗性の形質を有するイネからDNA試料を周知方法によって調製する。次に、調製したDNAを鋳型として、プライマーDNAを用いて核酸増幅反応(例えば、PCR法)を行う。増幅したDNA断片の大きさを、電気泳動等により被検イネといもち病圃場抵抗性遺伝子と連鎖するマーカー(例えば配列番号:10に記載のマーカー)との間で比較し、同様の遺伝子型を示す場合には、該被検植物は、いもち病圃場抵抗性の形質を有すると判定される。
【0087】
本発明の識別方法に使用するプライマーDNAは、当業者においては、各種分子マーカーについての配列情報を考慮して、最適なプライマーを適宜設計することが可能である。通常、上記プライマーとは、イネに特異的に存在し、Pi21遺伝子またはpi21遺伝子と連鎖する塩基配列を挟み込むように設計された、該塩基配列を増幅するための一対のプライマーセットである。
【0088】
具体的には、STSマーカーのプライマーセットとしては、
(a)プライマー5’-AGA AGG TGG AGT ACG ACG TGA AGA-3’(配列番号:8)および5’-AGT TTA GTG AGC CTC TCC ACG ATT A-3’(配列番号:9)からなるプライマーセット、
(b)プライマー5’- GTA CGA CGT GAA GAA CAA CAG G -3’(配列番号:16)および5’- GCT TGG GCT TGC AGT CC-3’(配列番号:17)からなるプライマーセット、
(c)プライマー5’-GAT CCT CAT CGT CGA CGT CTG GC-3’(配列番号:26)および5’-AGG GTA CGG CAC CAG CTT G-3’(配列番号:27)からなるプライマーセット
を例示することが出来る。これらのプライマーセットによって増幅されるDNA配列によって特徴付けられる情報を、本発明の分子マーカーと比較することによって、被検イネがいもち病に対する圃場抵抗性の有無を判定することが可能である。
【0089】
また上記のプライマー以外にも、当業者であれば、配列番号:1、4もしくは20に記載の塩基配列を利用して、同様の機能を有するプライマーセットを作成することが可能である。本発明のプライマーには、このようなプライマーもまた含まれる。
【0090】
本発明のPCRプライマーは、当業者においては、例えば、自動オリゴヌクレオチド合成機等を利用して作製することができる。また、当業者においては周知の多型検出方法、例えば、上記PCRプライマーを用いた後述のPCR-SSCP法等によっても本発明の方法を実施することが可能である。
【0091】
また、本発明の分子マーカーがゲノムDNAのエクソン中に存在する場合には、mRNAを鋳型としたRT-PCRを利用することも可能である。また、Taqman(量的PCR検出)システム(Roche社)を利用すれば、蛍光により増幅産物の有無を検出することが可能である。このシステムによれば、電気泳動の手間も省けるため短時間で本発明の識別方法を行うことが可能である。
【0092】
さらに本発明は、以下の(a)〜(c)の工程を含む方法であって、分子量または塩基配列が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)増幅したDNA断片の分子量または塩基配列を、pi21遺伝子のそれと比較する工程
【0093】
本発明の上記DNA試料の調製(抽出)方法としては、当業者においては、公知の方法によって行うことができる。好ましい調製方法として、例えば、CTAB法を用いてDNAを抽出する方法を挙げることができる。
また、本発明の識別方法に供される、DNA試料は、特に制限されるものではないが、通常、被検植物であるイネから抽出するゲノムDNAを用いる。また、ゲノムDNAの採取源としては特に限定されるものではなく、イネのいずれの組織からも抽出できる。例えば、穂、葉、根、茎、種子、胚乳部、フスマ、胚等から抽出することができる。
【0094】
本発明の植物のいもち病圃場抵抗性を識別する方法では、次に、調製したDNAを鋳型として、プライマーDNAを用いて核酸増幅反応(例えば、PCR法)を行う。増幅したDNA断片を制限酵素で切断し、切断されたDNA断片の大きさを、電気泳動等により被検植物といもち病圃場抵抗性の植物との間で比較し、分子量または塩基配列が一致する場合に、該被検植物は、いもち病圃場抵抗性の形質を有すると判定される。「いもち病圃場抵抗性の植物」としては、実施例に記載のオワリハタモチが挙げられるが、これに限定されない。
【0095】
本発明における、被検植物がいもち病圃場抵抗性であると判定する方法において、「一致する」とは、対立遺伝子の両方の遺伝子の分子量または塩基配列がいもち病圃場抵抗性植物のそれと一致するときあるいはアミノ酸配列において一致することを意味する。したがって、対立遺伝子の一方の遺伝子の分子量、塩基配列またはアミノ酸配列がいもち病圃場抵抗性植物のそれと異なるが、もう一方がいもち病圃場抵抗性植物のそれと同じである場合には、「一致する」に含まれない。
上記電気泳動分析は、常法にしたがって行えばよい。例えば、アガロースまたはポリアクリルアミドのゲル中で電圧をかけて電気泳動し、分離したDNAパターンを分析する。
【0096】
また本発明は、以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)増幅した二本鎖のDNAを非変性ゲル上で分離する工程
(d)分離した二本鎖DNAのゲル上での移動度を、pi21遺伝子のそれと比較する工程
【0097】
さらに本発明は、以下の(a)〜(e)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)増幅したDNAを一本鎖DNAに解離させる工程
(d)解離させた一本鎖DNAを非変性ゲル上で分離する工程
(e)分離した一本鎖DNAのゲル上での移動度を、pi21遺伝子のそれと比較する工程
【0098】
このような方法として、PCR-SSCP(single-strand conformation polymorphism、一本鎖高次構造多型)法(Cloning and polymerase chain reaction-single-strand conformation polymorphism analysis of anonymous Alu repeats on chromosome 11. Genomics. 1992 Jan 1; 12(1): 139-146.、Detection of p53 gene mutations in human brain tumors by single-strand conformation polymorphism analysis of polymerase chain reaction products. Oncogene. 1991 Aug 1; 6(8): 1313-1318.、Multiple fluorescence-based PCR-SSCP analysis with postlabeling.、PCR Methods Appl. 1995 Apr 1; 4(5): 275-282.)が挙げられる。この方法は操作が比較的簡便であり、また試料の量も少なくてすむなどの利点を有するため、特に多数のDNAサンプルをスクリーニングするのに好適である。その原理は以下の如くである。二本鎖DNA断片を一本鎖に解離すると、各鎖はその塩基配列に依存した独自の高次構造を形成する。この解離したDNA鎖を変性剤を含まないポリアクリルアミドゲル中で電気泳動すると、それぞれの高次構造の差に応じて、相補的な同じ鎖長の一本鎖DNAが異なる位置に移動する。一塩基の置換によってもこの一本鎖DNAの高次構造は変化し、ポリアクリルアミドゲル電気泳動において異なる移動度を示す。従って、この移動度の変化を検出することによりDNA断片に点突然変異や欠失、あるいは挿入などによる変異が存在することを検出することができる。
【0099】
具体的には、まず、Pi21遺伝子またはpi21遺伝子の標的部位を含む領域をPCR法などによって増幅する。増幅される範囲としては、100〜600bp程度の長さが好ましい。PCRによる遺伝子断片増幅の際、32Pなどのアイソトープ、あるいは蛍光色素やビオチンなどによって標識したプライマーを用いるか、あるいはPCR反応液に32Pなどのアイソトープ、あるいは蛍光色素やビオチンなどによって標識した基質塩基を加えてPCRを行うことによって合成されるDNA断片を標識する。あるいはPCR反応後にクレノウ酵素などを用いて32Pなどのアイソトープ、あるいは蛍光色素やビオチンなどによって標識した基質塩基を合成されたDNA断片に付加することによっても標識を行うことができる。このようにして得られたDNA断片を2本鎖のまま、尿素などの変性剤を含まないポリアクリルアミドゲルによって電気泳動を行う。もしくは、このようなDNA断片に熱を加えることなどにより変性させてから、尿素などの変性剤を含まないポリアクリルアミドゲルによって電気泳動を行ってもよい。この際、ポリアクリルアミドゲルに適量(5から10%程度)のグリセロールを添加することにより、DNA断片の分離の条件を改善することができる。また、泳動条件は各DNA断片の性質により変動するが、通常、室温(20から25℃)で行い、好ましい分離が得られないときには4から30℃までの温度で最適の移動度を与える温度の検討を行う。電気泳動後、DNA断片の移動度を、X線フィルムを用いたオートラジオグラフィーや、蛍光を検出するスキャナー等で検出し、解析する。移動度に差があるバンドが検出された場合、このバンドを直接ゲルから切り出し、PCRによって再度増幅し、それを直接シークエンシングすることにより、変異の存在を確認することができる。また、標識したDNAを使わない場合においても、電気泳動後のゲルをエチジウムブロマイドや銀染色法などによって染色することによって、バンドを検出することができる。
【0100】
さらに本発明は、以下の(a)〜(e)の工程を含む方法であって、検出されたDNA断片の大きさが一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)調製したDNA試料を制限酵素により切断する工程
(d)DNA断片をその大きさに応じて分離する工程
(e)検出されたDNA断片の大きさを、pi21遺伝子のそれと比較する工程
【0101】
このような方法としては、制限酵素断片長多型(Restriction Fragment Length Polymorphism/RFLP)を利用したRFLP法やPCR−RFLP法などが挙げられる。DNAを切断する酵素としては、通常、制限酵素を用いる。具体的には、制限酵素の認識部位に塩基の付加、欠失が存在する場合、あるいは制限酵素処理によって生じるDNA断片内に塩基挿入、または欠失がある場合、制限酵素処理後に生じる断片の大きさが、いもち病罹病性の植物といもち病圃場抵抗性の植物との間で変化する。この変異部位を含む部分をPCR法によって増幅し、それぞれの制限酵素で処理することによって、これらの多型部位を電気泳動後のバンドの移動度の差として検出することができる。あるいは、染色体DNAをこれらの制限酵素によって処理し、電気泳動した後、プローブDNAを用いてサザンブロッティングを行うことにより、多型部位を検出することができる。用いられる制限酵素は、それぞれの変異部位に応じて適宜選択することができる。この方法では、ゲノムDNA以外にも被験者から調製したRNAを逆転写酵素でcDNAにし、これをそのまま制限酵素で切断した後サザンブロッティングを行うこともできる。また、このcDNAを鋳型としてPCRでPi21遺伝子またはpi21遺伝子の一部、あるいは全部を増幅し、それを制限酵素で切断した後、移動度の差を調べることもできる。
【0102】
さらに本発明は、以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)増幅したDNAを、DNA変性剤の濃度が次第に高まるゲル上で分離する工程
(d)分離したDNAのゲル上での移動度と、pi21遺伝子のそれと比較する工程
【0103】
このような方法としては、変性剤濃度勾配ゲル電気泳動法(denaturant gradient gel electrophoresis:DGGE)が挙げられる。Pi21遺伝子またはpi21遺伝子の標的部位を含む領域を本発明のプライマーなどを用いたPCR法などによって増幅し、これを尿素などの変性剤の濃度が移動するに従って徐々に高くなっているポリアクリルアミドゲル中で電気泳動し、健常者と比較する。変異が存在するDNA断片の場合、より低い変性剤濃度位置でDNA断片が一本鎖になり、極端に移動速度が遅くなるため、この移動度の差を検出することにより多型の存在を検出することができる。
【0104】
これら方法以外にも、アレル特異的オリゴヌクレオチド(Allele Specific Oligonucleotide/ASO)ハイブリダイゼーション法が利用できる。多型が存在すると考えられる塩基配列を含むオリゴヌクレオチドを作製し、これと試料DNAでハイブリダイゼーションを行わせると、該オリゴヌクレオチドと異なる多型塩基が、ハイブリダイズする試料DNAに存在する場合、ハイブリッド形成の効率が低下する。それをサザンブロット法や、特殊な蛍光試薬がハイブリッドのギャップにインターカレーションすることにより消光する性質を利用した方法などにより検出できる。
【0105】
また、リボヌクレアーゼAミスマッチ切断法による検出も可能である。具体的には、Pi21遺伝子あるいはpi21遺伝子の標的部位を含む領域をPCR法などによって増幅し、これに対し、プラスミドベクター等に組み込んだ健常者型のcDNA等から調製した標識RNAとハイブリダイゼーションを行わしめる。健常者型と異なる塩基が存在する部分においてはハイブリッドが一本鎖構造となるので、この部分をリボヌクレアーゼAによって切断し、これをオートラジオグラフィーなどで検出することによって多型の存在を検出することができる。
【0106】
本発明において「被検植物」とは、特に限定されるものではなく、いもち病菌に感染しうる全ての植物が含まれる。好ましい一例としては、イネが挙げられる。イネであれば特に制限されず、インディカ種であっても、ジャポニカ種であってもよく、または、インディカ種とジャポニカ種との交配品種・系統や野生イネ、あるいは栽培品種と野生イネの交配・交雑品種であってもよい。
【0107】
さらに本発明は、pi21遺伝子と連鎖する分子マーカーであって、少なくとも配列番号:7、10、または23に記載のDNAからなる分子マーカーを指標として、イネのいもち病圃場抵抗性を判定する方法を提供する。本発明の好ましい分子マーカーとしては、上述のように、「P702D03_#38」、「P702D03_#79」、「P702D03_#80」が挙げられる。その中でも「P702D03_#79」は特に好ましいマーカーであり、配列番号:7、23(罹病性遺伝子Pi21と連鎖)、または10(抵抗性遺伝子pi21と連鎖)に記載のDNAとして例示できる。本発明の識別方法においては、これらの分子マーカーのうち、少なくとも「P702D03_#79」を指標とした識別方法である。従って本発明の識別方法においては、「P702D03_#79」を単独で用いてもよく、「P702D03_#79」と他のマーカーを組み合わせて用いてもよい。「P702D03_#79」と他のマーカーを組み合わせとしては、「P702D03_#38」の組み合わせ、「P702D03_#79」と「P702D03_#80」の組み合わせ、「P702D03_#79」とその他任意のマーカーの組み合わせが挙げられる。
【0108】
本発明の識別方法においては、pi21遺伝子と連鎖する分子マーカーを有するか否かを調べることにより、被検イネのいもち病圃場抵抗性を特異的かつ効率的に判定できる。本発明の判定方法においては、いもち病圃場抵抗性の有無を判定したい所望のイネにおいて、配列番号:10に記載の塩基配列を有する場合に、被検イネはいもち病圃場抵抗性であるイネであるものと判定され、配列番号:10に記載の塩基配列を有さない場合(配列番号:7または23に記載の塩基配列を有する場合)に、被検イネはいもち病罹病性であるイネと判定される。
【0109】
被検イネにおける分子マーカーと本発明の分子マーカーとの比較は、分子マーカーのDNA配列の比較だけではなく、該DNA配列によって特徴付けられる情報の比較によっても実施することができる。分子マーカーのDNA配列によって特徴づけられる情報としては、分子マーカーの分子量についての情報、分子マーカーに含まれる変異部位や多型部位の存在の有無についての情報が挙げられる。当業者であれば、周知の方法によって、配列番号:10と、配列番号:7または配列番号:23に記載の塩基配列とを比較することにより、多型部位(欠失部位および1塩基置換部位)を同定することができる。
本発明の判定方法は、このような、分子マーカーに含まれる変異部位や多型部位の存在の有無についての情報を検出することによっても行うことが出来る。
【0110】
上記のような変異部位や多型部位の存在の有無についての情報の検出は、配列を直接シーケンシングすることはもちろん、変異部位や多型部位を含む領域を増幅しうるプライマーを用いることによって、または変異部位や多型部位にハイブリダイズしうるプローブ(例えば、配列番号:18、19または25に記載の塩基配列の全部または一部を含むDNA)を用いることによって、検出することが可能である。
【0111】
また、本発明の判定方法を利用すれば、いもち病圃場抵抗性であると識別される植物(例えばイネ)を早期に選抜することが可能となる。
すなわち本発明は、以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性である植物を選抜する方法を提供する。
(a)いもち病圃場抵抗性である植物(例えばイネ)と任意の機能を有する植物(例えばイネ)とが交配された品種を作製する工程
(b)本明細書に記載の、被検植物がいもち病圃場抵抗性であるか否かを判定する方法により、工程(a)で得られた植物がいもち病圃場抵抗性であるか否かを判定する工程
本発明において植物とは、特に限定されるものではないが、好ましくはイネである。イネの具体的な態様は上述の通りである。
【0112】
本発明の選抜方法を利用すれば、いもち病圃場抵抗性と識別される植物(例えばイネ)を早期に選抜することが可能となる。本発明はこのようないもち病圃場抵抗性と識別される植物を早期に選抜する方法も提供する。ここでいう「早期」とは、例えばイネの出穂より前の状態を指し、好ましくは発芽直後の状態を指す。本発明の選抜方法を利用すれば、いもち病圃場抵抗性の形質を有する品種の育成を従来よりも短期間で成し遂げることが可能となる。
【0113】
本発明は、植物のいもち病を予防または改善するための薬剤のスクリーニング方法に関する。
本発明のスクリーニング方法の第一の態様としては、以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法が挙げられる。
(a)Pi21遺伝子の転写産物に被検化合物を接触させる工程
(b)Pi21遺伝子の転写産物と被検化合物の結合を検出する工程
(c)Pi21遺伝子の転写産物と結合する被検化合物を選択する工程
【0114】
第一の態様では、まず、Pi21遺伝子の転写産物に被検化合物を接触させる。本発明のスクリーニング方法における「Pi21遺伝子の転写産物」としては、Pi21遺伝子の転写産物に加えて該転写産物より翻訳された翻訳産物も含まれる。
【0115】
本発明の方法における「被検化合物」としては、特に制限はなく、例えば、天然化合物、有機化合物、無機化合物、タンパク質、ペプチド等の単一化合物、並びに、化合物ライブラリー、遺伝子ライブラリーの発現産物、細胞抽出物、細胞培養上清、発酵微生物産生物、海洋生物抽出物、植物抽出物、原核細胞抽出物、真核単細胞抽出物もしくは動物細胞抽出物等を挙げることができる。上記被検化合物は必要に応じて適宜標識して用いることができる。標識としては、例えば、放射標識、蛍光標識等を挙げることができる。
【0116】
本発明において「接触」は、以下のようにして行う。例えば、Pi21遺伝子の転写産物が精製された状態であれば、精製標品に被検化合物を添加することにより行うことができる。また、細胞内に発現した状態または細胞抽出液内に発現した状態であれば、それぞれ、細胞の培養液または該細胞抽出液に被検化合物を添加することにより行うことができる。本発明における細胞としては、特に制限されないが、イネを含む植物由来の細胞が好ましい。被検化合物がタンパク質の場合には、例えば、該タンパク質をコードするDNAを含むベクターを、Pi21遺伝子が発現している細胞へ導入する、または該ベクターをPi21遺伝子が発現している細胞抽出液に添加することで行うことも可能である。また、例えば、酵母または動物細胞等を用いた 2ハイブリッド法を利用することも可能である。
【0117】
第一の態様では、次いで、上記Pi21遺伝子の転写産物と被検化合物の結合を検出する。タンパク質間の結合を検出または測定する手段は、例えばタンパク質に付した標識を利用することにより行うことができる。標識の種類は、例えば、蛍光標識、放射標識等が挙げられる。また、酵母ツーハイブリット法や、BIACOREを用いた測定方法等、公知の方法によって測定することもできる。本方法においては、ついで、上記生合成酵素と結合した被検化合物を選択する。選択された被検化合物の中には、植物のいもち病を予防または改善するための薬剤が含まれる。また、選択された被検化合物を、以下のスクリーニングの被検化合物として用いてもよい。
【0118】
また、本発明のスクリーニング方法の第二の態様として、以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法を提供する。
(a)植物から採取した細胞に、被検化合物を接触させる工程
(b)Pi21遺伝子の転写産物の発現レベルを測定する工程
(c)被検化合物を接触させていない場合と比較して、上記転写産物の発現レベルを減少させた被検化合物を選択する工程
【0119】
第二の態様では、まず、植物から採取した細胞に被検化合物を接触させる。ここで「植物から採取した細胞」は、いもち病罹病性遺伝子を有することが明らかな任意の植物であってよい。「被検化合物」、「接触」の記載に関しては上記の説明の通りである。
【0120】
第二の態様では、次いで、「Pi21タンパク質」の発現レベルを測定する。Pi21タンパク質の発現レベルの測定は、当業者に公知の方法によって行うことができる。例えば、Pi21タンパク質をコードするmRNAを定法に従って抽出し、このmRNAを鋳型としたノーザンハイブリダイゼーション法、またはRT-PCR法を実施することによって該遺伝子の転写レベルの測定を行うことができる。さらに、DNAアレイ技術を用いて、Pi21タンパク質の発現レベルを測定することも可能である。
【0121】
また、Pi21タンパク質を含む画分を定法に従って回収し、Pi21タンパク質の発現をSDS-PAGE等の電気泳動法で検出することにより、遺伝子の翻訳レベルの測定を行うこともできる。また、Pi21タンパク質に対する抗体を用いて、ウェスタンブロッティング法を実施し、Pi21タンパク質の発現を検出することにより、遺伝子の翻訳レベルの測定を行うことも可能である。
【0122】
Pi21タンパク質の検出に用いる抗体としては、検出可能な抗体であれば、特に制限はないが、例えばモノクローナル抗体、またはポリクローナル抗体の両方を利用することができる。該抗体は、上述のように、当業者に公知の方法により調製することが可能である。
【0123】
第二の態様においては、ついで、Pi21タンパク質の発現レベルが、被検化合物を接触させないときに比べ減少する場合に、被検化合物を、植物のいもち病を予防または改善するための薬剤として選択する。
【0124】
本発明のスクリーニング方法の第三の態様は、以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法である。
(a)Pi21遺伝子のプロモーター領域の下流にレポーター遺伝子が機能的に結合したDNAを有する細胞または細胞抽出液を提供する工程
(b)該細胞または該細胞抽出液に被検化合物を接触させる工程
(c)該細胞または該細胞抽出液における該レポーター遺伝子の発現レベルを測定する工程
(d)被検化合物を接触させていない場合と比較して、該レポーター遺伝子の発現レベルを減少させた被検化合物を選択する工程
【0125】
第三の態様では、まず、Pi21遺伝子のプロモータ―領域の下流にレポーター遺伝子が機能的に結合したDNAを有する細胞または細胞抽出液を提供する。
第三の態様において、「機能的に結合した」とは、Pi21遺伝子のプロモータ―領域に転写因子が結合することにより、レポーター遺伝子の発現が誘導されるように、Pi21遺伝子のプロモータ―領域とレポーター遺伝子とが結合していることをいう。従って、レポーター遺伝子が他の遺伝子と結合しており、他の遺伝子産物との融合タンパク質を形成する場合であっても、Pi21遺伝子のプロモータ―領域に転写因子が結合することによって、該融合タンパク質の発現が誘導されるものであれば、上記「機能的に結合した」の 意に含まれる。
【0126】
上記レポーター遺伝子としては、その発現が検出可能なものであれば特に制限されず、例えば、当業者において一般的に使用されるCAT遺伝子、lacZ遺伝子、ルシフェラーゼ遺伝子、β-グルクロニダーゼ遺伝子(GUS)およびGFP遺伝子等を挙げることができる。
【0127】
第三の態様では、次いで、上記細胞または上記細胞抽出液に被検化合物を接触させる。次いで、該細胞または該細胞抽出液における上記レポーター遺伝子の発現レベルを測定する。「被検化合物」および「接触」の記載に関しては上記の説明の通りである。
【0128】
レポーター遺伝子の発現レベルは、使用するレポーター遺伝子の種類に応じて、当業者に公知の方法により測定することができる。例えば、レポーター遺伝子がCAT遺伝子である場合には、該遺伝子産物によるクロラムフェニコールのアセチル化を検出することによって、レポーター遺伝子の発現レベルを測定することができる。レポーター遺伝子がlacZ遺伝子である場合には、該遺伝子発現産物の触媒作用による色素化合物の発色を検出することにより、また、ルシフェラーゼ遺伝子である場合には、該遺伝子発現産物の触媒作用による蛍光化合物の蛍光を検出することにより、また、β-グルクロニダーゼ遺伝子(GUS)である場合には、該遺伝子発現産物の触媒作用によるGlucuron(ICN社)の発光や5-ブロモ-4-クロロ-3-インドリル-β-グルクロニド(X-Gluc)の発色を検出することにより、さらに、GFP遺伝子である場合には、GFPタンパク質による蛍光を検出することにより、レポーター遺伝子の発現レベルを測定することができる。
【0129】
第三の態様においては、ついで、上記レポーター遺伝子の発現レベルが、被検化合物を接触させないときに比べ減少する場合に、被検化合物を、植物のいもち病を予防または改善するための薬剤として選択する。
【0130】
本発明のスクリーニング方法の第四の態様は、以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法である。
(a)Pi21遺伝子が導入された形質転換植物細胞より形質転換植物体を再生させる工程
(b)該形質転換植物体に、いもち病菌および被検化合物を接触させる工程
(c)被検化合物を接触させていない場合と比較して、該形質転換植物体のいもち病を抑制する被検化合物を選択する工程
【0131】
第四の態様では、まず、Pi21遺伝子を含む形質転換植物細胞より形質転換植物体を再生させる。該形質転換植物体の再生は、上述したように当業者に公知の方法によって行うことが出来る。
第四の態様では次に、工程(a)で再生させた該形質転換植物体に、いもち病菌および被検化合物を接触させる。「いもち病菌」および「被検化合物」は上記の説明の通りである。「接触」の一例としては、霧吹きを用いて被検化合物を植物に直接噴霧する方法が挙げられる。しかし第四の態様における「接触」はこれに限定されず、植物と被検化合物が物理的に触れることが出来る限り、あらゆる方法が含まれる。本発明の接触は、いもち病菌に感染した形質転換植物体に被検化合物を接触させてもよいし、被検化合物が接触された形質転換植物体にいもち病菌を感染させてもよい。
【0132】
第四の態様においては、ついで、被検化合物を接触させていない場合と比較して該形質転換植物体のいもち病を抑制する被検化合物を選択する。いもち病が抑制されたか否かは、該形質転換植物体の表現型を指標として判断することが出来る。該形質転換植物体の表現型としては、特に制限はないが、植物の各部位においてその一部あるいは全体が変色・壊死することが例示できる。また、該形質転換植物体のいもち病の抑制には、完全な抑制だけでなく、部分的な抑制も含まれる。
【0133】
本発明はまた、上記に記載のスクリーニング方法に用いるためのキットに関する。このようなキットには、上記に記載のスクリーニング方法の検出工程や測定工程に使用されるものを含みうる。例えば、Pi21遺伝子の発現レベルの測定に必要とされるプローブ、プライマー、抗体、染色液等を挙げることができる。その他、蒸留水、塩、緩衝液、タンパク質安定剤、保存剤等が含まれていてもよい。
【実施例】
【0134】
以下、本発明を実施例によりさらに具体的に説明するが本発明はこれら実施例に制限されるものではない。
【0135】
〔実施例1〕遺伝地図作成
マップベースクローニングに不可欠な大規模分離集団によるpi21領域の詳細な連鎖解析を行った。連鎖解析用の集団では病斑進展を抑制する抵抗性対立遺伝子pi21を持つ日本陸稲品種オワリハタモチに、病斑進展を抑制しない罹病性対立遺伝子Pi21を持つ水稲品種日本晴あるいは愛知旭(図1)を連続戻し交雑して得たBC1F2集団72個体を用いた。RFLPマーカーによる連鎖解析の結果、pi21遺伝子座はRFLPマーカーG271およびG317の間に存在することが明らかとなった(図2A)。
【0136】
精度の高いpi21領域の遺伝地図を作成するために、上述の交雑組合せ229個体と後代BC1F4集団643個体を加え、合計1014個体を用いて、pi21の両側に存在するRFLPマーカーRA3591と13S1を利用して、pi21座近傍の染色体組み換え型27個体を選抜した(図2B)。さらに、日本水稲品種の遺伝的背景にインド型水稲品種Kasalathの罹病性対立遺伝子を持つ系統とオワリハタモチ由来の抵抗性対立遺伝子を持つ系統を交雑して得たF2集団2703個体を用いた検索を行った。pi21座の両側に存在するPCRマーカー14T1と4S1を利用して、pi21座近傍の染色体組み換え型24個体が選抜できた。さらにそれらの個体を用いて、以下の取り組みにおいて作出されたDNAマーカーによる詳細な連鎖地図作成を行った。
【0137】
〔実施例2〕P1由来人工染色体(PAC)クローンによるpi21遺伝子領域の整列化
イネゲノム解析研究において作出された日本晴PACクローンの整列化地図を利用して、pi21遺伝子座近傍に存在するDNAマーカーRA3591およびC975塩基配列を有するPACクローンを特定した(図2C)。さらに特定されたPACクローンP479G02、P415D09、P473G08、P703E11、P434F09、P702D03、P419B08、P472G09およびP502G01の末端断片をカセット法により単離し、特定できたPACクローンを整列化した。その結果、PACクローンP032D02、P678A02、P405D12、P689F04、P479G02、P415D09、P473G08、P703E11、P434F09、およびP702D03はpi21遺伝子領域を含むことが明かとなった(図2C)。
【0138】
〔実施例3〕pi21遺伝子領域の絞り込み
pi21領域に整列化されたPACクローンの末端断片をクローン化し、それらを新たなRFLPマーカーあるいはCAPSマーカーとして、詳細な遺伝地図を作成したところ、pi21遺伝子座はSSCPマーカーPa102484およびSNPマーカーP702D3_#12に挟み込まれるゲノム領域に存在することが明らかとなった。この結果、pi21遺伝子座は2つのマーカーに挟まれる約25kbのゲノム領域に存在することが明らかとなった(図2D)。
【0139】
〔実施例4〕塩基配列解析による候補遺伝子領域の特定
pi21遺伝子を含むと考えられるPACクローンP702D03の塩基配列を決定し、25kbのゲノム候補領域の抵抗性品種オワリハタモチと罹病性品種愛知旭およびKasalathの塩基配列解析を行なった。塩基配列の解析は日本晴の候補領域の配列を利用して設計したプライマーを用いて上述3品種より増幅したDNA断片を使用し、dye-terminater法により行った。連鎖解析により特定された候補遺伝子領域内の塩基多型情報を利用して、候補領域のさらなる絞り込みを行った。その結果、pi21遺伝子はSTSマーカーP702D03_#79(プライマー5’-AGA AGG TGG AGT ACG ACG TGA AGA-3’(配列番号:8)およびAGT TTA GTG AGC CTC TCC ACG ATT A-3’(配列番号:9))と共分離し、SNPマーカーP702D03_#38(プライマーTTT TCC TGA GAA ATT TGT AAA GA-3’ (配列番号:12)およびCGT CGA CGA TGA GGA TCT-3’ (配列番号:13))と、P702D03_#80(プライマー5’-CTC CCA ATG TGT TTA GCA TC-3’ (配列番号:14)および5’-CAA CCA TAT GTC CCT AAG GAT-3’(配列番号:15))との間にそれぞれ1個の組み換え個体を検出した。以上の結果から、pi21遺伝子はSNPマーカーP702D03_#38およびP702D03_#80に挟み込まれる約1.8kbのゲノム領域に存在することが明らかとなった(図2D)。
【0140】
単離されたイネ(Oryza sativa L、愛知旭および日本晴)由来Pi21遺伝子の遺伝子の塩基配列を配列番号:1に、cDNAの塩基配列を配列番号:2に、該cDNAがコードするタンパク質(「Pi21タンパク質」)のアミノ酸配列を配列番号:3に示した。なお、愛知旭および日本晴のPi21遺伝子に対応する、Kasalath由来のPi21遺伝子の遺伝子の塩基配列を配列番号:20に、cDNAの塩基配列を配列番号:21に、該cDNAがコードするタンパク質(「Pi21タンパク質」)のアミノ酸配列を配列番号:22に示した。
【0141】
〔実施例5〕pi21候補遺伝子の塩基配列解析
1.8kb候補ゲノム領域の品種日本晴の配列について遺伝子予測ならびに類似性検索を行ったところ、日本晴の完全長cDNAクローン(AK106153,AK070581,AK072320)が見出されたが、アラビドプシスなどで類似の遺伝子はなく相同性から遺伝子の機能を予測することはできなかった。しかしながら、この遺伝子は予想翻訳開始点から約10アミノ酸の部位にある金属結合部位は、アラビドプシスで報告されているエチレンシグナル伝達系における金属を運搬するシャペロンの機能を持つ遺伝子(Hirayama et al 1999 Cell)を連想させ、実際に準同質遺伝子系統AA-pi21では愛知旭に比べてエチレンに対する感受性が変化していることから類似の機能を持つ可能性がある。既に得られている日本晴の塩基配列情報から、該当部分を増幅できるプライマーを設計し、ゲノムPCRおよび RT-PCR産物を罹病性品種日本晴および愛知旭と抵抗性品種オワリハタモチの間で塩基配列を比較したところ、遺伝子のエクソン領域に抵抗性品種と罹病性品種との間に2ヶ所のDNA変異があった。罹病性の品種に対して抵抗性の品種では、7アミノ酸および16アミノ酸の欠失が起こっており、これらの変異が感染したいもち病の病斑進展に関係するものと考えられた(図3)。
【0142】
単離されたpi21遺伝子の遺伝子の塩基配列を配列番号:4に、cDNAの塩基配列を配列番号:5に、cDNAがコードするタンパク質(「pi21タンパク質」)のアミノ酸配列を配列番号:6に示す。
【0143】
〔実施例6〕形質転換による候補遺伝子の機能の同定
(1)罹病性遺伝子のAA-pi21への導入
pi21遺伝子の候補として特定された5’上流予測プロモーター領域を含む罹病性品種日本晴のゲノム領域XbaI 4.7kb断片を、アグロバクテリウムを介して形質転換可能なベクターpPZP2H-lacに組み込んだ。この断片を導入したベクターならびにベクターのみを用い、土岐の方法(文献 Plant Mol. Biol. Rep. 15:16-21, 1997)により形質転換を行った。形質転換する系統にはpi21準同質遺伝子系統AA-pi21を用いた。XbaI 4.7kb断片が導入されたベクターからは36個体、ベクターのみからは12個体のハイグロマイシン耐性個体を得た。候補遺伝子に特異的なプライマー(センス鎖5'- GTA CGA CGT GAA GAA CAA CAG G-3'(配列番号:16))および (アンチセンス鎖5'- GCT TGG GCT TGC AGT CC 3'(配列番号:17))を用いて、導入した領域が組み込まれたか否かをPCR法により調査した。その結果、すべての形質転換体について、候補遺伝子が組み込まれていることが判明した。これらの個体を、隔離温室で栽培して、自殖後代系統においていもち病菌(レース007)を接種した。その結果、ベクターのみを導入した個体および分離によって導入遺伝子が伝達されなかったT1個体では、準同質遺伝子系統AA-pi21と同様に、病斑の進展が罹病性品種愛知旭に比べていもち病の病斑の進展が抑制されたのに対して、候補遺伝子が導入されているT1個体では、より病斑が進展した(図4)。特に、導入遺伝子のコピー数の多い系統では感受性の度合いが増していた。
【0144】
(2)抵抗性遺伝子の罹病性品種への導入
一方、抵抗性品種オワリハタモチのXbaI 4.7kb断片についても同様にベクターに組み込み、罹病性品種愛知旭を用いて形質転換を行った。XbaI 4.7kb断片が導入されたベクターからは56個体、ベクターのみからは24個体のハイグロマイシン耐性個体を得た。(1)と同様、導入した領域が組み込まれたか否かをPCR法により調査した結果、すべての形質転換体について、候補遺伝子が組み込まれていることが判明した。これらの個体を、(1)と同様の方法で隔離温室で栽培して、自殖後代系統においていもち病菌(レース007)を接種した。その結果、ベクターのみを導入した個体、導入遺伝子が伝達されなかったT1個体および、候補遺伝子が導入されているT1個体の全てにおいて罹病性品種愛知旭と同様のいもち病の進展が見られた。
【0145】
(3)候補遺伝子の機能の同定
以上の結果から、日本晴由来の候補遺伝子領域(XbaI 4.7kb)には、準同質遺伝子系統AA-pi21の病斑形成を促進する機能を有することが判明し、候補遺伝子がPi21遺伝子であると判断した。
【0146】
〔実施例7〕イネにおける候補遺伝子の変異
世界のイネ品種79品種を用いて候補遺伝子の変異を検索したところ、日本晴と愛知旭およびオワリハタモチに見出されたタイプ以外に、エクソン領域に挿入欠失のある10種類の変異があった。これらは主として日本晴・愛知旭とオワリハタモチの間で見出した2ヶ所の欠失部位について、その挿入/欠失の有無とその大きさによって規定される。シロイヌナズナのエチレンシグナル伝達の系で提唱される金属分子のシャペロンとの類似から、既知の遺伝子とは相同性がないこの領域は他の分子と結合するものと予想され、この部位の変異がシグナル伝達の効率を微妙に制御し、機能の変異をもたらす可能性がある。
【0147】
以上の結果から、マップベースクローニング法により候補として絞り込まれた遺伝子がイネいもち病の病斑進展を制御する遺伝子pi21であることが判明した。本成果は植物の量的抵抗性の生物的機能を証明した初めてのケースである。RT-PCR分析によってpi21あるいはPi21遺伝子の発現を調査したところ、地上部の全ての組織で恒常的に発現していることから、この遺伝子が植物の生育の基本的な役割を果たしているもの予想される。コピー数の変化が表現型の変化をもたらしたことから、プロモーターによる発現量、発現組織を変化させることが機能改変に重要な要素となる。すなわち、単離したpi21遺伝子あるいは種内に認められる別の対立遺伝子を利用して、本来植物がもつ耐病性を効率的に高めることが可能であると考えられる。
【技術分野】
【0001】
本発明は、イネいもち病圃場抵抗性遺伝子pi21、ならびに該遺伝子を利用した植物のいもち病圃場抵抗性を改変する方法に関する。
【背景技術】
【0002】
イネのいもち病菌に対する抵抗性は真性抵抗性と圃場抵抗性の2種類に分類される(非特許文献1)。前者は過敏感反応に基づく抵抗性であり、効果が大きくレースに対する特異性が高い質的な抵抗性である。1個の抵抗性遺伝子を導入した品種は、その遺伝子に親和性の菌が出現することによって、数年でその効果を失うことが経験的に知られる。一方圃場抵抗性は、真性抵抗性が機能しない条件下で観察される抵抗性の品種間差と定義される。真性抵抗性に比べて効果が小さいものの、レースに対する特異性が低いことから、品種に持続性のある抵抗性を付与できる点で実用性が高い。
【0003】
真性抵抗性に関与する遺伝子は30種類以上が知られ、そのうち、Pib、Pita遺伝子が単離されている(非特許文献2)。これらの遺伝子は、既報の植物の病害抵抗性遺伝子と類似した構造である nucleotide binding site (NBS)や leucine-rich repeats (LRRs)をもつNBS-LRR クラス遺伝子であることが明らかとなっている。他の病害抵抗性遺伝子と同様、植物の抵抗性遺伝子の産物は、それに対応する病原体の非病原性遺伝子の産物を直接的あるいは間接的に認識する受容体としての機能を果たすものと考えられている。実際、Pitaは非病原性遺伝子産物と物理的に直接結合することが明らかにされている。
【0004】
圃場抵抗性については、日本陸稲品種が優れた特性を有することが知られ、関与する複数の遺伝子座に関して、染色体上の位置が明らかにされている(非特許文献3)。しかしながら、遺伝子の構造や発現機構は明らかにされておらず、真性抵抗性に比べてこれを効率的に育種選抜に利用できる状況にはない。また、圃場抵抗性と類似した概念である不完全抵抗性(incomplete resistance)に関しては、西アフリカの陸稲品種Moroberekanの複数の染色体領域の関与が報告されている(非特許文献4)ものの、遺伝子は特定されていない。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特願平10-271807
【特許文献2】特願平11-153146
【特許文献3】特許第3376453号(P3376453)
【特許文献4】特開2003-88379(P2003-88379A)
【特許文献5】特開2003-199577(P2003-199577A)
【特許文献6】特開2003-199448(P2003-199448A)
【特許文献7】特開2004-329215(P2004-329215A)
【非特許文献】
【0006】
【非特許文献1】イネのいもち病と抵抗性育種 p175-186 高坂・山崎編 1980博友社
【非特許文献2】Wang et al. Plant J 19:55-64, 1999,Bryan et al. Plant Cell. 12:2033-46, 2000
【非特許文献3】Fukuoka and Okuno 2001 Theor Appl Genet. 03:185-190
【非特許文献4】Wang et al. Genet 136:1421-1434, 1994
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明はこのような状況を鑑みてなされたものであり、本発明が解決しようとする課題は、いもち病圃場抵抗性に関わる遺伝子をマップベースクローニング法により単離・同定すること、ならびに該遺伝子を利用した植物のいもち病圃場抵抗性の改変方法を提供することである。
【課題を解決するための手段】
【0008】
本発明は、植物のいもち病に対する抵抗性を制御する遺伝子に関する。Pi21遺伝子の対立遺伝子pi21は、いもち病圃場抵抗性という形質をイネ(Oryza sativa L)に付与する遺伝子として、イネ第4染色体という広大な領域のいずれかの場所に存在することが知られている。本発明者らは、その存在領域を解明し、単一の遺伝子として単離することを試みた。
【0009】
本発明者らはまず、pi21領域の遺伝地図を作成するために、マップベースクローニングに不可欠な大規模分離集団によるpi21領域の詳細な連鎖解析を行った。病斑進展を抑制する抵抗性対立遺伝子pi21を持つ日本陸稲品種オワリハタモチに、病斑進展を抑制しない罹病性対立遺伝子Pi21を持つ水稲品種日本晴あるいは愛知旭を連続戻し交雑して得た集団に対してRFLPマーカーによる連鎖解析を試みたところ、pi21遺伝子座がRFLPマーカーG271とG317の間に存在することが確認された。
【0010】
次に本発明者らは、より精度の高いpi21領域の遺伝地図を作成するために、pi21の両側に存在するRFLPマーカーRA3591と13S1を利用して、pi21座近傍の染色体組み換え型個体を選抜した。同時に、日本水稲品種の遺伝的背景にインド型水稲品種Kasalathの罹病性対立遺伝子を持つ系統とオワリハタモチ由来の抵抗性対立遺伝子を持つ系統を交雑して得たF2集団を用いて検索を行い、pi21座近傍の染色体組み換え型個体の選抜も行った。これらの個体および作出されたDNAマーカーを用いて詳細な連鎖地図を作成した結果、pi21遺伝子座がSSCPマーカーPa102484およびSNPマーカーP702D3_#12に挟まれる約25kbのゲノム領域に存在することがわかった。さらに、pi21遺伝子を含むと考えられるPACクローンP702D03の塩基配列を決定し、また、25kbのゲノム候補領域の抵抗性品種オワリハタモチと罹病性品種愛知旭およびKasalathの塩基配列の解析も行うことにより、pi21遺伝子が、SNPマーカーP702D03_#38とP702D03_#80に挟み込まれる約1.8kbのゲノム領域に存在することが明らかとなった。
【0011】
そこで本発明者らは、既に得られている日本晴の塩基配列情報を利用して該当部分を増幅できるプライマーを設計し、ゲノムPCRおよび RT-PCR産物を罹病性品種日本晴および愛知旭と抵抗性品種オワリハタモチの間で塩基配列の比較を行った。その結果、遺伝子のエクソン領域において、抵抗性品種と罹病性品種との間に2ヶ所のDNA変異があることが明らかとなった。罹病性の品種に対して抵抗性の品種では、7アミノ酸および16アミノ酸の欠失が起こっており、これらの変異が感染したいもち病の病斑進展に関係することが証明された。すなわち、本発明は、植物のいもち病に対する抵抗性を制御するpi21遺伝子に関し、具体的には以下の発明を提供するものである。
〔1〕以下(a)〜(h)のいずれかに記載のDNA;
(a)配列番号:3または22に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:1、2、20または21に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:3または22に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(d)配列番号:1、2、20または21に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(e)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(f)配列番号:4または5に記載の塩基配列のコード領域を含むDNA、
(g)配列番号:6に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(h)配列番号:4または5に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA。
〔2〕植物においていもち病圃場抵抗性能を有する、以下(i)〜(iv)のいずれかに記載のDNA;
(i)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物と相補的なRNAをコードするDNA、
(ii)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA、
(iii)〔1〕の(a)〜(d)のいずれかに記載のDNAの発現を共抑制効果により阻害するRNAをコードするDNA、
(iv)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物を特異的に切断するRNAi活性を有するRNAをコードするDNA。
〔3〕植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである、〔2〕記載のDNA。
〔4〕〔1〕〜〔3〕のいずれかに記載のDNAを含むベクター。
〔5〕〔1〕〜〔3〕のいずれかに記載のDNAを発現可能に保持する形質転換細胞。
〔6〕〔1〕の(a)〜(d)のいずれかに記載のDNAが導入された形質転換植物細胞。
〔7〕〔2〕もしくは〔3〕に記載のDNAが導入された形質転換植物細胞。
〔8〕植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである、〔6〕または〔7〕に記載の形質転換植物細胞。
〔9〕〔6〕〜〔8〕に記載の形質転換細胞を含む形質転換植物体。
〔10〕〔8〕に記載の形質転換植物体の子孫またはクローンである、形質転換植物体。
〔11〕〔9〕または〔10〕のいずれかに記載の形質転換植物体の繁殖材料。
〔12〕〔9〕または〔10〕のいずれかに記載の形質転換植物体の製造方法であって、〔1〕の(a)〜(d)、〔2〕または〔3〕に記載のDNAを植物細胞に導入し、該植物細胞から植物体を再生させる工程を含む方法。
〔13〕〔2〕または〔3〕に記載のDNAを植物体の細胞内で発現させる工程を含む、植物にいもち病圃場抵抗性を付与する方法。
〔14〕植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである〔13〕に記載の方法。
〔15〕〔1〕の(a)〜(d)のいずれかに記載のDNAによりコードされるタンパク質。
〔16〕〔1〕の(a)〜(d)のいずれかに記載のDNAを含むベクターを含む形質転換細胞を培養し、該細胞またはその培養上清から組換えタンパク質を回収する工程を含む、〔15〕に記載のタンパク質の製造方法。
〔17〕〔15〕に記載のタンパク質に結合する抗体。
〔18〕〔1〕に記載のDNAまたはその相補配列に相補的な少なくとも15の連続する塩基を含むDNA。
〔19〕〔2〕もしくは〔3〕に記載のDNA、または該DNAを含むベクターのいずれか1つを含む、植物のいもち病圃場抵抗性を増加させる薬剤。
〔20〕配列番号:1、4または20に記載の塩基配列の全部または一部を増幅するプライマーセット。
〔21〕(a)〜(c)のいずれかに示す、少なくとも1つのプライマーセット
(a)配列番号:8に記載の塩基配列からなるDNAおよび配列番号:9に記載の塩基配列からなるDNA、
(b)配列番号:16に記載の塩基配列からなるDNAおよび配列番号:17に記載の塩基配列からなるDNA、
(c)配列番号:26に記載の塩基配列からなるDNAおよび配列番号:27に記載の塩基配列からなるDNA。
〔22〕配列番号:7、10、18、19、23または25に記載の塩基配列からなるDNA。
〔23〕以下の(a)〜(c)の工程を含む方法であって、分子量または塩基配列が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から〔1〕に記載のDNA領域を増幅する工程、
(c)増幅したDNA断片の分子量または塩基配列を、〔1〕の(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
〔24〕以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から〔1〕に記載のDNA領域を増幅する工程、
(c)増幅した二本鎖のDNAを非変性ゲル上で分離する工程、
(d)分離した二本鎖DNAのゲル上での移動度を、〔1〕の(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
〔25〕以下の(a)〜(e)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から〔1〕に記載のDNA領域を増幅する工程、
(c)増幅したDNAを一本鎖DNAに解離させる工程、
(d)解離させた一本鎖DNAを非変性ゲル上で分離する工程、
(e)分離した一本鎖DNAのゲル上での移動度を、〔1〕の(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
〔26〕以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から〔1〕に記載のDNA領域を増幅する工程、
(c)増幅したDNAを、DNA変性剤の濃度が次第に高まるゲル上で分離する工程、
(d)分離したDNAのゲル上での移動度と、〔1〕の(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
〔27〕以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性である植物を選抜する方法;
(a)いもち病圃場抵抗性能である植物と任意の機能を有する植物とが交配された品種を作製する工程、
(b)〔23〕〜〔26〕のいずれかに記載の方法により、工程(a)で作製された植物がいもち病圃場抵抗性であるか否かを判定する工程。
〔28〕〔1〕に記載のDNAと連鎖する分子マーカーであって、該分子マーカーがいもち病圃場抵抗性の形質を有するイネと同様の遺伝子型を示す場合に、被検イネがいもち病圃場抵抗性であると判定する方法。
〔29〕分子マーカーが配列番号:10に記載のDNAからなる分子マーカーである、〔28〕に記載の方法。
〔30〕以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性であるイネを選抜する方法;
(a)いもち病圃場抵抗性能であるイネと任意の機能を有するイネとが交配された品種を作製する工程、
(b)〔28〕または〔29〕のいずれかに記載の方法により、工程(a)で作製されたイネがいもち病圃場抵抗性であるか否かを判定する工程。
〔31〕以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物に被検化合物を接触させる工程、
(b)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物と被検化合物の結合を検出する工程、
(c)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物と結合する被検化合物を選択する工程。
〔32〕以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)植物から採取した細胞に、被検化合物を接触させる工程、
(b)〔1〕の(a)〜(d)のいずれかに記載のDNAの転写産物の発現レベルを測定する工程、
(c)被検化合物を接触させていない場合と比較して、上記転写産物の発現レベルを減少させた被検化合物を選択する工程。
〔33〕以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)〔1〕の(a)〜(d)のいずれかに記載のDNAのプロモーター領域の下流にレポーター遺伝子が機能的に結合したDNAを有する細胞または細胞抽出液を提供する工程、
(b)該細胞または該細胞抽出液に被検化合物を接触させる工程、
(c)該細胞または該細胞抽出液における該レポーター遺伝子の発現レベルを測定する工程、
(d)被検化合物を接触させていない場合と比較して、該レポーター遺伝子の発現レベルを減少させた被検化合物を選択する工程。
〔34〕以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)〔6〕に記載の形質転換植物細胞より形質転換植物体を再生させる工程、
(b)該形質転換植物体に、いもち病菌および被検化合物を接触させる工程、
(c)被検化合物を接触させていない場合と比較して、該形質転換植物体のいもち病を抑制する被検化合物を選択する工程。
〔35〕〔31〕〜〔34〕のいずれかに記載のスクリーニング方法に用いるためのキット。
【発明の効果】
【0012】
Pi21遺伝子の特性は、植物のいもち病圃場抵抗性品種の作出に特に適している。これまでは、いもち病圃場抵抗性を付与するためには、圃場抵抗性を本来持つが劣悪な特性をもちあわせた品種とこれを持たないが多くの優れた特性を持つ品種を交雑し、その子孫より圃場抵抗性の優れた特性と他の優れた特性を併せ持った個体を選び出す作業が必要であった。しかしながら、圃場抵抗性を的確に評価するためには多大な労力を必要とし、かつ、これを付与する遺伝子の正確な染色体上の位置が明らかではない場合、これを効率的かつ的確に選抜し、実用性の高い品種に導入することは困難であった。そして実際、これまで成功してこなかった。
【0013】
本発明によって、圃場抵抗性に関与する遺伝子の染色体上の位置および遺伝子の構造が提供された。これにより、植物に圃場抵抗性を効率的に付与することが可能となった。また、圃場抵抗性に関与する遺伝子の組織発現特異性や発現レベルを変更することにより、抵抗性と実用性の高い特性を兼ね備えた品種を育成することも可能となった。このように本発明の遺伝子は、実用性が高く、安全性の高い農業を実現する上で有用である。また、本発明の方法で作製された植物は、例えば有用農作物では高い収量が安定的に得られるだけではなく、鑑賞用植物では新たな美的価値が付加されることも大いに期待される。
【図面の簡単な説明】
【0014】
【図1】愛知旭の遺伝的背景にpi21遺伝子を持つ系統AA-pi21(図左)および愛知旭(図右)におけるいもち病の病斑を示す写真である。
【図2】pi21遺伝子領域の詳細な連鎖地図およびゲノムクローンの整列地図を示す図である。AおよびBは、72個体および1014個体の分離集団により作成した遺伝地図を示す。Cは、日本晴のPACクローンによる整列地図を示す。Dは、pi21遺伝子領域の詳細な遺伝地図と候補ゲノム領域を示す。
【図3】pi21候補遺伝子の構造と日本晴・愛知旭とオワリハタモチのゲノム塩基配列の比較を示す図である。
【図4】抵抗性系統AApi21に日本晴由来のPi21遺伝子を導入した形質転換体に生じた病斑を示す写真である。A:ベクターのみを導入。B:Pi21遺伝子1コピー導入。C: Pi21遺伝子3コピー以上導入。
【発明を実施するための形態】
【0015】
イネにおいていもち病の進展を抑制しない罹病性遺伝子Pi21の対立遺伝子pi21遺伝子は、いもち病圃場抵抗性という形質をイネに付与する遺伝子として、これまでイネ第4染色体という広大な領域にいずれかの場所に存在することが知られていた。本発明者らは、マップベースクローニングの手法を利用することによりイネ第4染色体におけるpi21遺伝子の存在領域の絞り込みを行い、遂に単一の遺伝子として同定することに成功した。またpi21遺伝子の対立遺伝子Pi21遺伝子を単離することにも成功した。
【0016】
本発明において「いもち病」とは、植物がいもち病菌に感染し、その作用によってその一部あるいは全体が変色・壊死すること、あるいはそれによって認識できる病徴(病斑)をいう。いもち病の病斑は植物の各部位に発生し、発生の部位によりそれぞれ、苗いもち、葉いもち、穂いもち、籾いもち、節いもち、葉節(葉舌)いもちなどと呼ばれる。本発明における「いもち病」とは、これら全ての部位に発生するいもち病を含む。またイネにいもち病を引き起こす「イネいもち病菌」は、現在のところ統一された学名はないものの、Magnaporthe griseaあるいはMagnaporthe oryzaeなどと呼ばれるものがある。またいもち病菌には完全世代名と不完全世代名があり、完全世代名Magnaporthe oryzaeに対する不完全世代名としてPyricularia oryzaeが与えられ、使い分けられている。本発明におけるいもち病菌には、名称のいかんを問わずこれら全てのいもち病菌が含まれる。
【0017】
また本明細書において「いもち病罹病性」とは、植物がいもち病に感染する性質(まれにその病徴が著しいこと)を意味する。さらに「いもち病圃場抵抗性」とは、植物がいもち病菌に感染したときの(同一植物種内の)品種や系統間での病斑の数や大きさの差として認識される病徴の違いまたは病斑の数や大きさを抑制する性質を意味する。また「真性抵抗性」とは、植物がいもち病菌の侵入した細胞において過敏感反応による細胞死を起こし,感染を防ぐ性質をいう。
【0018】
本発明は、植物のいもち病に関与するいもち病罹病性遺伝子Pi21、およびいもち病圃場抵抗性遺伝子pi21を提供する。
【0019】
本発明のPi21遺伝子には、より具体的には、
(a)配列番号:3または22に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:1、2、20または21に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:3または22に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(d)配列番号:1、2、20または21に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
が含まれる。
【0020】
また本発明のpi21遺伝子には、より具体的には、
(a)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:4または5に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:6に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(d)配列番号:4または5に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
が含まれる。
【0021】
本発明のPi21遺伝子またはpi21遺伝子を利用することにより、例えば、組み換えタンパク質の調製やいもち病圃場抵抗性が改変された形質転換植物体を作出することなどが可能となる。
【0022】
本発明において、本発明の遺伝子が由来する植物としては、特に制限はないが、例えばイネ、トウモロコシ、コムギ、オオムギ、エンバク、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビ、パールミレット等の単子葉植物や、ナタネ、ダイズ、ワタ、トマト、ジャガイモ等の双子葉植物が挙げられる。また例えば花卉植物としては、キク、バラ、カーネーション、シクラメン等が挙げられるが、これらに限定されない。
【0023】
本発明の「Pi21遺伝子」、「pi21遺伝子」はそれぞれ、「Pi21タンパク質」、「pi21タンパク質」をコードしうるものであれば、その形態に特に制限はなく、「Pi21遺伝子」、「pi21遺伝子」にはそれぞれ、cDNAの他、ゲノムDNA、化学合成DNAなども含まれる。また、Pi21遺伝子はPi21タンパク質を、pi21遺伝子はpi21タンパク質をコードするものであれば、遺伝暗号の縮重に基づく任意の塩基配列を有するDNAが含まれる。
【0024】
ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、植物からゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作成し、これを展開して、Pi21遺伝子またはpi21遺伝子(例えば、配列番号:1、2、4、5、20または21のいずれかに記載のDNA)を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより調製することが可能である。また、Pi21遺伝子またはpi21遺伝子に特異的なプライマーを作成し、これを利用したPCRをおこなうことによって調製することも可能である。また、cDNAは、例えば、植物から抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。
【0025】
さらに、Pi21遺伝子またはpi21遺伝子は広く植物界に存在すると考えられるため、Pi21遺伝子またはpi21遺伝子には、イネのみならず、種々の植物に存在する相同遺伝子も含まれる。ここで「相同遺伝子」とは、種々の植物において、イネにおけるPi21遺伝子またはpi21遺伝子産物と同様の生理機能(例えばいもち病罹病性またはいもち病圃場抵抗性)を有するタンパク質をコードする遺伝子をさす。
【0026】
相同遺伝子を単離するための当業者によく知られた方法としては、ハイブリダイゼーション技術(Southern, E. M., Journal of Molecular Biology, Vol. 98, 503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R. K., et al. Science, vol. 230, 1350-1354, 1985, Saiki, R. K. et al. Science, vol.239, 487-491,1988)が挙げられる。即ち、当業者にとっては、イネPi21遺伝子またはpi21遺伝子の塩基配列(例えば、配列番号:1、2、4、5、20または21のいずれかに記載の配列)もしくはその一部をプローブとして、またPi21遺伝子またはpi21遺伝子に特異的にハイブリダイズするオリゴヌクレオチドをプライマーとして、種々の植物からPi21遺伝子またはpi21遺伝子の相同遺伝子を単離することは通常行いうることである。
【0027】
このような相同遺伝子をコードするDNAを単離するためには、通常ストリンジェントな条件下でハイブリダイゼーション反応を行なう。ストリンジェントなハイブリダイゼーション条件としては、6M 尿素、0.4%SDS、0.5 x SSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を例示できる。よりストリンジェンシーの高い条件、例えば、6M 尿素、0.4%SDS、0.1 x SSCの条件を用いれば、より相同性の高いDNAの単離を期待することができる。単離したDNAの配列の決定は、公知の方法で行うことができる。単離されたDNAの相同性は、アミノ酸配列全体で、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の相同性は、BLASTN(核酸レベル)やBLASTX(アミノ酸レベル)のプログラム(Altschul et al. J. Mol. Biol., 215: 403-410, 1990)を利用して決定することができる。該プログラムは、Karlin及びAltschulによるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA, 87:2264-2268, 1990, Proc. Natl. Acad. Sci. USA, 90: 5873-5877, 1993) に基づいている。BLASTNによって塩基配列を解析する場合には、パラメーターは例えばscore = 100、wordlength =12とする。また、BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えばscore = 50、wordlength = 3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res. 25: 3389-3402, 1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0028】
また本発明は、植物の内因性のPi21遺伝子の発現を抑制するために用いるDNAであって、
(a)Pi21遺伝子の転写産物と相補的なRNAをコードするDNA、
(b)Pi21遺伝子の転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA、
(c)Pi21遺伝子の発現を共抑制効果により阻害するRNAをコードするDNA、
(d)Pi21遺伝子の転写産物を特異的に切断するRNAi活性を有するRNAをコードするDNA、
を提供する。
これらのDNAにより、植物のいもち病による病斑進展を抑制することが可能である。
【0029】
本発明において、Pi21遺伝子の発現を抑制する植物には特に制限はなく、いもち病圃場抵抗性を付与させたい所望の植物を用いることができるが、産業的な観点からは農作物や鑑賞用植物が好適である。有用農作物としては、特に制限はないが、例えばイネ、トウモロコシ、コムギ、オオムギ、エンバク、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビ、パールミレット等の単子葉植物や、ナタネ、ダイズ、ワタ、トマト、ジャガイモ等の双子葉植物が挙げられる。また、観賞用植物としては、例えばキク、バラ、カーネーション、シクラメン等の花卉植物が挙げられるが、これらに限定されない。またイネいもち病菌に感受性を有する植物として、オオムギ、イタリアンライグ.ラス、メドーフェスクなどの牧草、トウモロコシがあり、それ以外にも、イネより分離したいもち病菌が寄生性を有する植物としてエゾノサヤヌカグサ、マコモなどのイネ族、ウシノケグサ族、オオムギ族、エンバクなどのカラスムギ族、シナダレスズメガヤなどのヒゲシバ族、アワ、メヒシバなどのキビ族など多くの植物が報告されおり、これらの植物もまたいもち病圃場抵抗性を付与させる植物に含まれる。
【0030】
本明細書における「Pi21遺伝子の発現を抑制」には、遺伝子の転写の抑制およびタンパク質への翻訳の抑制が含まれる。また、DNAの発現の完全な停止のみならず発現の減少も含まれる。
【0031】
「Pi21遺伝子の発現を抑制するために用いるDNA」の一つの態様は、Pi21遺伝子と相補的なアンチセンスRNAをコードするDNAである。植物細胞におけるアンチセンス効果は、一時的遺伝子発現法を用いて、電気穿孔法で導入したアンチセンスRNAが植物においてアンチセンス効果を発揮することにより、初めて実証された(Ecker and Davis, Proc. Natl. Acad. USA, 83: 5372, 1986)。その後、タバコやペチュニアにおいても、アンチセンスRNAの発現によって標的遺伝子の発現を低下させる例が報告されており(Krol et. al., Nature 333: 866, 1988)、現在では植物における遺伝子発現を抑制させる手段として確立している。
【0032】
アンチセンス核酸が標的遺伝子の発現を抑制する作用としては、以下のような複数の要因が存在する。すなわち、三重鎖形成による転写開始阻害、RNAポリメラーゼによって局部的に開状ループ構造がつくられた部位とのハイブリッド形成による転写抑制、合成の進みつつあるRNAとのハイブリッド形成による転写阻害、イントロンとエキソンとの接合点でのハイブリッド形成によるスプライシング抑制、スプライソソーム形成部位とのハイブリッド形成によるスプライシング抑制、mRNAとのハイブリッド形成による核から細胞質への移行抑制、キャッピング部位やポリ(A)付加部位とのハイブリッド形成によるスプライシング抑制、翻訳開始因子結合部位とのハイブリッド形成による翻訳開始抑制、開始コドン近傍のリボソーム結合部位とのハイブリッド形成による翻訳抑制、mRNAの翻訳領域やポリソーム結合部位とのハイブリッド形成によるペプチド鎖の伸長阻止、および核酸とタンパク質との相互作用部位とのハイブリッド形成による遺伝子発現抑制などである。これらは、転写、スプライシング、または翻訳の過程を阻害して、標的遺伝子の発現を抑制する(平島および井上「新生化学実験講座2 核酸IV 遺伝子の複製と発現」,日本生化学会編,東京化学同人, pp.319-347, 1993)。
【0033】
本発明で用いられるアンチセンス配列は、上記のいずれの作用で標的遺伝子の発現を抑制してもよい。一つの態様としては、遺伝子のmRNAの5'端近傍の非翻訳領域に相補的なアンチセンス配列を設計すれば、遺伝子の翻訳阻害に効果的であろう。しかし、コード領域もしくは3'側の非翻訳領域に相補的な配列も使用し得る。このように、遺伝子の翻訳領域だけでなく非翻訳領域の配列のアンチセンス配列を含むDNAも、本発明で利用されるアンチセンスDNAに含まれる。使用されるアンチセンスDNAは、適当なプロモーターの下流に連結され、好ましくは3'側に転写終結シグナルを含む配列が連結される。
【0034】
アンチセンスDNAは、例えば、配列番号:1、2、20または21に記載のDNAの配列情報を基にホスホロチオネート法(Stein, Nucleic Acids Res., 16: 3209-3221, 1988)などにより調製することが可能である。調製されたDNAは、公知の方法で、所望の植物へ形質転換できる。アンチセンスDNAの配列は、形質転換する植物が持つ内因性遺伝子の転写産物と相補的な配列であることが好ましいが、遺伝子の発現を有効に阻害できる限り、完全に相補的でなくてもよい。転写されたRNAは、標的とする遺伝子の転写産物に対して好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の相補性を有する。アンチセンス配列を用いて、効果的に標的遺伝子の発現を阻害するには、アンチセンスDNAの長さは、少なくとも15塩基以上であり、好ましくは100塩基以上であり、さらに好ましくは500塩基以上である。通常、用いられるアンチセンスDNAの長さは5kbよりも短く、好ましくは2.5kbよりも短い。
【0035】
内因性のPi21遺伝子の発現の抑制は、リボザイムをコードするDNAを利用して行うことも可能である。リボザイムとは触媒活性を有するRNA分子のことをいう。リボザイムには種々の活性を有するものがあるが、中でもRNAを切断する酵素としてのリボザイムの研究により、RNAの部位特異的な切断を目的とするリボザイムの設計が可能となった。リボザイムには、グループIイントロン型や、RNasePに含まれるM1RNAのように400ヌクレオチド以上の大きさのものもあるが、ハンマーヘッド型やヘアピン型と呼ばれる40ヌクレオチド程度の活性ドメインを有するものもある(小泉誠および大塚栄子、蛋白質核酸酵素, 35: 2191, 1990)。
【0036】
例えば、ハンマーヘッド型リボザイムの自己切断ドメインは、G13U14C15のC15の3'側を切断するが、活性にはU14が9位のAと塩基対を形成することが重要とされ、15位の塩基はCの他にAまたはUでも切断されることが示されている(Koizumi et. al., FEBS Lett. 228: 225, 1988)。リボザイムの基質結合部を標的部位近傍のRNA配列と相補的になるように設計すれば、標的RNA中のUC、UUまたはUAという配列を認識する制限酵素的なRNA切断リボザイムを作出することが可能である(Koizumi et. al., FEBS Lett. 239: 285,1988、小泉誠および大塚栄子, 蛋白質核酸酵素,35: 2191, 1990、Koizumi et. al., Nucleic. Acids. Res. 17: 7059, 1989)。
【0037】
また、ヘアピン型リボザイムも、本発明の目的のために有用である。ヘアピン型リボザイムは、例えばタバコリングスポットウイルスのサテライトRNAのマイナス鎖に見出される(Buzayan, Nature 323: 349,1986)。このリボザイムも、標的特異的なRNA切断を起こすように設計できることが示されている(Kikuchi and Sasaki, Nucleic Acids Res. 19: 6751, 1992, 及び菊池洋,化学と生物 30: 112, 1992)。
【0038】
標的を切断できるよう設計されたリボザイムは、植物細胞中で転写されるようにカリフラワーモザイクウイルスの35Sプロモーターなどのプロモーターおよび転写終結配列に連結される。しかし、その際、転写されたRNAの5'末端や3'末端に余分な配列が付加されていると、リボザイムの活性が失われてしまうことがある。このようなとき、転写されたリボザイムを含むRNAからリボザイム部分だけを正確に切り出すために、リボザイム部分の5'側や3'側に、トリミングを行うためのシスに働く別のトリミングリボザイムを配置させることも可能である (Taira et. al., Protein Eng. 3: 733, 1990、Dzianott and Bujarski, Proc. Natl. Acad. Sci. USA, 86: 4823, 1989、Grosshans and Cech, Nucleic Acids Res. 19: 3875, 1991、Taira et. al., Nucleic Acids Res. 19: 5125, 1991)。
【0039】
また、このような構成単位をタンデムに並べ、標的遺伝子内の複数の部位を切断できるようにして、より効果を高めることもできる(Yuyama et al., Biochem. Biophys. Res. Commun. 186: 1271, 1992)。このようなリボザイムを用いて本発明で標的となる遺伝子の転写産物を特異的に切断し、該遺伝子の発現を抑制することができる。
【0040】
内在性遺伝子の発現の抑制はさらに、標的遺伝子配列と同一もしくは類似した配列を有するDNAの形質転換によってもたらされる「共抑制」によっても達成されうる。「共抑制」とは、植物に標的内在性遺伝子と同一若しくは類似した配列を有する遺伝子を形質転換により導入すると、導入する外来遺伝子および標的内在性遺伝子の両方の発現が抑制される現象のことをいう。共抑制の機構の詳細は明らかではないが、植物においてはしばしば観察される(Curr. Biol. 7: R793, 1997、Curr. Biol. 6: 810, 1996)。
【0041】
例えば、Pi21遺伝子が共抑制された植物体を得るためには、Pi21遺伝子若しくはこれと類似した配列を有するDNAを発現できるように作製したベクターDNAを目的の植物へ形質転換し、得られた植物体からPi21変異体の形質を有する植物、即ちいもち病圃場抵抗性である植物を選択すればよい。共抑制に用いる遺伝子は、標的遺伝子と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。
【0042】
さらに、本発明における内在性遺伝子の発現の抑制は、標的遺伝子のドミナントネガティブの形質を有する遺伝子を植物へ形質転換することによっても達成することができる。ドミナントネガティブの形質を有する遺伝子とは、該遺伝子を発現させることによって、植物体が本来持つ内在性の野生型遺伝子の活性を消失もしくは低下させる機能を有する遺伝子のことをいう。
【0043】
「Pi21遺伝子の発現を抑制するために用いるDNA」の他の一つの態様は、内因性のPi21遺伝子の転写産物と相補的なdsRNA(二重鎖RNA)をコードするDNAである。標的遺伝子配列と同一もしくは類似した配列を有するdsRNAを細胞内に導入することにより、導入した外来遺伝子および標的内因性遺伝子の発現がいずれも抑制される、RNAi(RNA干渉、RNA interference)と呼ばれる現象を引き起こすことができる。細胞に約40〜数百塩基対のdsRNAが導入されると、ヘリカーゼドメインを持つダイサー(Dicer)と呼ばれるRNaseIII様のヌクレアーゼがATP存在下で、dsRNAを3'末端から約21〜23塩基対ずつ切り出し、siRNA(short interference RNA)を生じる。このsiRNAに特異的なタンパク質が結合して、ヌクレアーゼ複合体(RISC: RNA-induced silencing complex)が形成される。この複合体はsiRNAと同じ配列を認識して結合し、RNaseIII様の酵素活性によってsiRNAの中央部で標的遺伝子の転写産物(mRNA)を切断する。また、この経路とは別にsiRNAのアンチセンス鎖がmRNAに結合してRNA依存性RNAポリメラーゼ(RsRP)のプライマーとして作用し、dsRNAが合成される。このdsRNAが再びダイサーの基質となって、新たなsiRNAを生じて作用を増幅する経路も考えられている。
【0044】
RNAiは当初、線虫において発見されたが(Fire, A. et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391, 806-811, 1998)、現在では、線虫のみならず、植物、線形動物、ショウジョウバエ、原生動物などの種々の生物において観察されている(Fire, A. RNA-triggered gene silencing. Trends Genet. 15, 358-363 (1999)、Sharp, P. A. RNA interference 2001. Genes Dev. 15, 485-490 (2001)、Hammond, S. M., Caudy, A. A. & Hannon, G. J. Post-transcriptional gene silencing by double-stranded RNA. Nature Rev. Genet. 2, 110-119 (2001)、Zamore, P. D. RNA interference: listening to the sound of silence. Nat Struct Biol. 8, 746-750 (2001))。これら生物では、実際に外来よりdsRNAを導入することにより標的遺伝子の発現が抑制されることが確認され、さらにはノックアウト個体を創生する方法としても利用されつつある。
【0045】
RNAiの登場当初は、dsRNAはある程度の長さ(40塩基)以上でなければ効果がないと考えられていたが、米ロックフェラー大のTuschlらは21塩基対前後の単鎖dsRNA(siRNA)を細胞に導入すれば、哺乳動物細胞においてもPKRによる抗ウイルス反応を起こさず、RNAiの効果があることを報告し(Tuschl, Nature, 411, 494-498 (2001))、RNAiは分化したヒトなどの哺乳動物細胞に応用可能な技術として俄然注目を集めることになった。
【0046】
本発明のDNAは、標的遺伝子の転写産物(mRNA)のいずれかの領域に対するアンチセンスRNAをコードしたアンチセンスコードDNAと、前記mRNAのいずれかの領域のセンスRNAをコードしたセンスコードDNAを含み、前記アンチセンスコードDNAおよび前記センスコードDNAより前記アンチセンスRNAおよび前記センスRNAを発現させることができる。また、これらのアンチセンスRNAおよびセンスRNAよりdsRNAを作成することもできる。本発明における標的配列は、標的配列と同一もしくは類似した配列を有するdsRNAを細胞内に導入した結果Pi21遺伝子の発現が抑制されるものであれば特に制限されない。標的配列の一例としては、Pi21遺伝子の3'非翻訳領域配列が挙げられる。Pi21遺伝子の3'非翻訳領域配列を配列番号:11および24に示す。
【0047】
本発明のdsRNAの発現システムをベクター等に保持させる場合の構成としては、同一のベクターからアンチセンスRNA、センスRNAを発現させる場合と、異なるベクターからそれぞれアンチセンスRNA、センスRNAを発現させる場合がある。例えば、同一のベクターからアンチセンスRNA、センスRNAを発現させる構成としては、アンチセンスコードDNAおよびセンスコードDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモータを連結させたアンチセンスRNA発現カセット、センスRNA発現カセットをそれぞれ構築し、これらカセットを同方向にあるいは逆方向にベクターに挿入することにより構成することができる。
【0048】
また、異なる鎖上に対向するように、アンチセンスコードDNAとセンスコードDNAとを逆向きに配置した発現システムを構成することもできる。この構成では、アンチセンスRNAコード鎖とセンスRNAコード鎖とが対となった一つの二本鎖DNA(siRNAコードDNA)が備えられ、その両側にそれぞれの鎖からアンチセンスRNA、センスRNAとを発現し得るようにプロモータを対向して備えられる。この場合には、センスRNA、アンチセンスRNAの下流に余分な配列が付加されることを避けるために、それぞれの鎖(アンチセンスRNAコード鎖、センスRNAコード鎖)の3'末端にターミネーターをそれぞれ備えることが好ましい。このターミネーターは、A(アデニン)塩基を4つ以上連続させた配列などを用いることができる。また、このパリンドロームスタイルの発現システムでは、二つのプロモータの種類は異なっていることが好ましい。
【0049】
また、異なるベクターからアンチセンスRNA、センスRNAを発現させる構成としては、例えば、アンチセンスコードDNAおよびセンスコードDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモータを連結させたアンチセンスRNA発現カセット、センスRNA発現カセットをそれぞれ構築し、これらカセットを異なるベクターに保持させることにより構成することができる。
【0050】
RNAiにおいては、dsRNAとしてsiRNAが使用されたものであってもよい。「siRNA」は、細胞内で毒性を示さない範囲の短鎖からなる二重鎖RNAを意味し、Tuschlら(前掲)により報告された全長21〜23塩基対に限定されるものではなく、毒性を示さない範囲の長さであれば特に限定はなく、例えば、15〜49塩基対と、好適には15〜35塩基対と、さらに好適には21〜30塩基対とすることができる。あるいは、発現されるsiRNAが転写され最終的な二重鎖RNA部分の長さが、例えば、15〜49塩基対、好適には15〜35塩基対、さらに好適には21〜30塩基対とすることができる。
【0051】
本発明のDNAとしては、標的配列のインバーテッドリピートの間に適当な配列(イントロン配列が望ましい)を挿入し、ヘアピン構造を持つダブルストランドRNA(self-complementary 'hairpin' RNA(hpRNA))を作るようなコンストラクト(Smith, N.A., et al. Nature, 407: 319, 2000、Wesley, S. V. et al. Plant J. 27: 581, 2001、Piccin, A. et al. Nucleic Acids Res. 29:E55, 2001)を用いることもできる。
【0052】
RNAiに用いるDNAは、標的遺伝子と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。また、配列の同一性は上述した手法により決定できる。
【0053】
dsRNAにおけるRNA同士が対合した二重鎖RNAの部分は、完全に対合しているものに限らず、ミスマッチ(対応する塩基が相補的でない)、バルジ(一方の鎖に対応する塩基がない)などにより不対合部分が含まれていてもよい。本発明においては、dsRNAにおけるRNA同士が対合する二重鎖RNA領域中に、バルジおよびミスマッチの両方が含まれていてもよい。
【0054】
また本発明は、Pi21遺伝子、pi21遺伝子、またはPi21遺伝子の発現を抑制するDNAのいずれか1つを含むベクターならびに形質転換細胞を提供する。
上記ベクターとしては、例えば、大腸菌を宿主とする場合には、ベクターを大腸菌(例えば、JM109、DH5α、HB101、XL1Blue)等で大量に増幅させ大量調製するために、大腸菌で増幅されるための「ori」をもち、さらに形質転換された大腸菌の選抜遺伝子(例えば、なんらかの薬剤(アンピシリンやテトラサイクリン、カナマイシン、クロラムフェニコールにより判別できるような薬剤耐性遺伝子)を有すれば特に制限はない。ベクターの例としては、M13系ベクター、pUC系ベクター、pBR322、pBluescript、pCR-Script等が挙げられる。また、cDNAのサブクローニング、切り出しを目的とした場合、上記ベクターの他に、例えば、pGEM-T、pDIRECT、pT7等が挙げられる。Pi21遺伝子、pi21遺伝子、またはPi21遺伝子の発現を抑制するDNAを生産する目的においてベクターを使用する場合には、特に、発現ベクターが有用である。発現ベクターとしては、例えば、大腸菌での発現を目的とした場合は、ベクターが大腸菌で増幅されるような上記特徴を持つほかに、宿主をJM109、DH5α、HB101、XL1-Blue等の大腸菌とした場合においては、大腸菌で効率よく発現できるようなプロモーター、例えば、lacZプロモーター(Wardら, Nature (1989) 341, 544-546;FASEB J. (1992) 6, 2422-2427)、araBプロモーター(Betterら, Science (1988) 240, 1041-1043 )、またはT7プロモーター等を持っていることが不可欠である。このようなベクターとしては、上記ベクターの他にpGEX-5X-1(ファルマシア社製)、「QIAexpress system」(キアゲン社製)、pEGFP、またはpET等が挙げられる。
【0055】
また、ベクターには、ポリペプチド分泌のためのシグナル配列が含まれていてもよい。ポリペプチド分泌のためのシグナル配列としては、大腸菌のペリプラズムに産生させる場合、pelBシグナル配列(Lei, S. P. et al J. Bacteriol. (1987) 169, 4379)を使用すればよい。宿主細胞へのベクターの導入は、例えば塩化カルシウム法、エレクトロポレーション法を用いて行うことができる。
【0056】
大腸菌以外にも、例えば、Pi21遺伝子、pi21遺伝子、またはPi21遺伝子の発現を抑制するDNAを製造するためのベクターとしては、哺乳動物由来の発現ベクター(例えば、pcDNA3 (インビトロゲン社製)や、pEGF-BOS (Nucleic Acids. Res.1990, 18(17),p5322)、pEF 、pCDM8 )、昆虫細胞由来の発現ベクター(例えば「Bac-to-BAC baculovairus expression system」(ギブコBRL社製)、pBacPAK8)、植物由来の発現ベクター(例えばpMH1、pMH2)、動物ウィルス由来の発現ベクター(例えば、pHSV、pMV、pAdexLcw )、レトロウィルス由来の発現ベクター(例えば、pZIPneo)、酵母由来の発現ベクター(例えば、「Pichia Expression Kit」(インビトロゲン社製)、pNV11、SP-Q01)、枯草菌由来の発現ベクター(例えば、pPL608、pKTH50)等が挙げられる。
【0057】
CHO細胞、COS細胞、NIH3T3細胞等の動物細胞での発現を目的とした場合には、細胞内で発現させるために必要なプロモーター、例えばSV40プロモーター(Mulliganら, Nature (1979) 277, 108)、MMLV-LTRプロモーター、EF1αプロモーター(Mizushimaら, Nucleic Acids Res. (1990) 18, 5322)、CMVプロモーター等を持っていることが不可欠であり、細胞への形質転換を選抜するための遺伝子(例えば、薬剤(ネオマイシン、G418等)により判別できるような薬剤耐性遺伝子)を有すればさらに好ましい。このような特性を有するベクターとしては、例えば、pMAM、pDR2、pBK-RSV、pBK-CMV、pOPRSV、pOP13等が挙げられる。
【0058】
本発明のDNAの細胞への導入は、当業者においては、公知の方法、例えば電気穿孔法(エレクトロポーレーション法)などにより実施することができる。
【0059】
さらに本発明は、Pi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが導入された形質転換植物細胞、これらの形質転換植物体、該形質転換植物体の子孫またはクローンである形質転換植物体、該形質転換植物体の繁殖材料を提供する。また、上記形質転換体、繁殖材料の製造方法を提供する。
【0060】
Pi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAの植物細胞への導入は、上記の方法によって行うことが可能である。
【0061】
また植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である(Toki et. al., Plant Physiol. 100: 1503-1507, 1995)。例えば、イネにおいては、形質転換植物体を作出する手法については、ポリエチレングリコールによりプロトプラストへ遺伝子導入し、植物体(インド型イネ品種が適している)を再生させる方法(Datta et. al., In Gene Transfer To Plants (Potrykus I and Spangenberg Eds.) pp66-74, 1995)、電気パルスによりプロトプラストへ遺伝子導入し、植物体(日本型イネ品種が適している)を再生させる方法(Toki et. al., Plant Physiol. 100: 1503-1507, 1992)、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法(Christou et. al., Bio/technology, 9: 957-962, 1991)およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法(Hiei et. al., Plant J. 6: 271-282, 1994)など、いくつかの技術が既に確立し、本願発明の技術分野において広く用いられている。本発明においては、これらの方法を好適に用いることができる。
【0062】
上記アグロバクテリウム法を用いる場合、例えばNagelらの方法(Microbiol. Lett. 67: 325, 1990)が用いられる。この方法によれば、組み換えベクターをアグロバクテリウム細菌中に形質転換して、次いで形質転換されたアグロバクテリウムを、リーフディスク法等の公知の方法により細胞に導入する。上記ベクターは、例えば植物体に導入した後、本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが植物体中で発現するように、発現プロモーターを含む。一般に、該プロモーターの下流には本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが位置し、さらに該DNAの下流にはターミネーターが位置する。この目的に用いられる組み換えベクターは、植物への導入方法、または植物の種類に応じて、当業者によって適宜選択される。上記プロモーターとして、例えばカリフラワーモザイクウイルス由来のCaMV35S、トウモロコシのユビキチンプロモーター(特開平2-79983号公報)等を挙げることができる。
【0063】
また、上記ターミネーターは、カリフラワーモザイクウイルス由来のターミネーター、あるいはノパリン合成酵素遺伝子由来のターミネーター等を例示することができるが、植物体中で機能するプロモーターやターミネーターであれば、これらに限定されない。
【0064】
また、本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAを導入する植物は、外植片であってもよく、これらの植物から培養細胞を調製し、得られた培養細胞に導入してもよい。本発明の「植物細胞」は、例えば葉、根、茎、花および種子中の胚盤等の植物の細胞、カルス、懸濁培養細胞等が挙げられる。
【0065】
また、本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAの導入により形質転換した細胞を効率的に選択するために、上記組み換えベクターは、適当な選抜マーカー遺伝子、もしくは選抜マーカー遺伝子を含むプラスミドベクターと共に植物細胞へ導入するのが好ましい。この目的に使用する選抜マーカー遺伝子は、例えば、抗生物質ハイグロマイシンに耐性であるハイグロマイシンホスホトランスフェラーゼ遺伝子、カナマイシンまたはゲンタマイシンに耐性であるネオマイシンホスホトランスフェラーゼ、および除草剤ホスフィノスリシンに耐性であるアセチルトランスフェラーゼ遺伝子等が挙げられる。
【0066】
組み換えベクターを導入した細胞は、導入された選抜マーカー遺伝子の種類に従って適当な選抜用薬剤を含む公知の選抜用培地に置床し培養する。これにより形質転換された植物培養細胞を得ることができる。
【0067】
形質転換細胞から再生させた植物体は、次いで順化用培地で培養する。その後、順化した再生植物体を、通常の栽培条件で栽培すると、いもち病圃場抵抗性である植物体が得られ、成熟して結実して種子を得ることができる。
【0068】
なお、このように再生され、かつ栽培した形質転換植物体中の導入された外来DNAの存在は、公知のPCR法やサザンハイブリダイゼーション法によって、または植物体中のDNAの塩基配列を解析することによって確認することができる。
この場合、形質転換植物体からのDNAの抽出は、公知のJ.Sambrookらの方法(Molecular Cloning、第2版、Cold Spring Harbor Laboratory Press, 1989)に準じて実施することができる。
【0069】
再生させた植物体中に存在する本発明のDNAよりなる外来遺伝子を、PCR法を用いて解析する場合には、上記のように再生植物体から抽出したDNAを鋳型として増幅反応を行う。また、本発明のDNA、あるいは本発明により改変されたDNAの塩基配列に従って適当に選択された塩基配列をもつ合成したオリゴヌクレオチドをプライマーとして用い、これらを混合させた反応液中において増幅反応を行うこともできる。増幅反応においては、DNAの変性、アニーリング、伸張反応を数十回繰り返すと、本発明のDNA配列を含むDNA断片の増幅生成物を得ることができる。増幅生成物を含む反応液を例えばアガロース電気泳動にかけると、増幅された各種のDNA断片が分画されて、そのDNA断片が本発明のDNAに対応することを確認することが可能である。
【0070】
一旦、染色体内に本発明のPi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫を得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラスト等を得て、それらを基に該植物体を量産することも可能である。本発明には、Pi21タンパク質をコードするDNAまたはPi21遺伝子の発現を抑制するDNAが導入された植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、並びに該植物体、その子孫およびクローンの繁殖材料が含まれる。これらの植物細胞、該細胞を含む植物体、該植物体の子孫およびクローン、並びに該植物体、その子孫およびクローンの繁殖材料は、植物にいもち病圃場抵抗性を付与する方法に使用することが可能である。
【0071】
本発明はさらに、Pi21遺伝子の発現を抑制するDNAを植物体の細胞内で発現させる工程を含む、植物にいもち病圃場抵抗性を付与する方法を提供する。植物にいもち病圃場抵抗性を付与することは、上述の方法を用いてPi21遺伝子の発現を抑制するDNAを発現可能に含むベクターを植物細胞に導入し、これを再生することにより可能である。
【0072】
本明細書においていもち病圃場抵抗性を「付与する」とは、いもち病圃場抵抗性能を全く有していない植物にいもち病圃場抵抗性能を持たせることのみならず、すでにいもち病圃場抵抗性能を有している植物のいもち病抵抗性能を、さらに増大させることも意味する。
本発明において、Pi21遺伝子の発現を抑制していもち病圃場抵抗性を付与させる植物には特に制限はなく、例えば上述の植物を挙げることが出来る。
【0073】
本発明はまた、本発明のPi21遺伝子によりコードされるタンパク質、該タンパク質の製造方法、該タンパク質に結合する抗体を提供する。
組み換えタンパク質を調製する場合には、通常、本発明のタンパク質をコードするDNAを適当な発現ベクターに挿入し、該ベクターを適当な細胞に導入し、形質転換細胞を培養して発現させたタンパク質を精製する。組み換えタンパク質は、精製を容易にするなどの目的で、他のタンパク質との融合タンパク質として発現させることも可能である。例えば、大腸菌を宿主としてマルトース結合タンパク質との融合タンパク質として調製する方法(米国New England BioLabs社発売のベクターpMALシリーズ)、グルタチオン-S-トランスフェラーゼ(GST)との融合タンパク質として調製する方法(Amersham Pharmacia Biotech社発売のベクターpGEXシリーズ)、ヒスチジンタグを付加して調製する方法(Novagen社のpETシリーズ)などを利用することが可能である。宿主細胞としては、組み換えタンパク質の発現に適した細胞であれば特に制限はなく、上記の大腸菌の他、例えば、酵母、種々の動植物細胞、昆虫細胞などを用いることが可能である。宿主細胞へのベクターの導入には、当業者に公知の種々の方法を用いることが可能である。例えば、大腸菌への導入には、カルシウムイオンを利用した導入方法(Mandel, M. & Higa, A. Journal of Molecular Biology, Vol.53, 158-162,(1970)、Hanahan, D. Journal of Molecular Biology, Vol.166, 557-580,(1983))を用いることができる。宿主細胞内で発現させた組み換えタンパク質は、該宿主細胞またはその培養上清から、当業者に公知の方法により精製し、回収することが可能である。組み換えタンパク質を上述したマルトース結合タンパク質などとの融合タンパク質として発現させた場合には、容易にアフィニティー精製を行うことが可能である。
【0074】
得られた組換えタンパク質を用いれば、これに結合する抗体を調製することができる。ポリクローナル抗体であれば、例えば、次のようにして取得することができる。Pi21タンパク質、あるいはGSTとの融合タンパク質として大腸菌等の微生物において発現させたリコンビナントタンパク質、またはその部分ペプチドをウサギ等の小動物に免疫し血清を得る。これを、例えば、硫安沈殿、プロテインA、プロテインGカラム、DEAEイオン交換クロマトグラフィー、Pi21タンパク質や合成ペプチドをカップリングしたアフィニティーカラム等により精製することにより調製する。また、モノクローナル抗体であれば、例えば、Pi21タンパク質またはその部分ペプチドをマウス等の小動物に免疫を行い、同マウスより脾臓を摘出し、これをすりつぶして細胞を分離し、該細胞とマウスミエローマ細胞とをポリエチレングリコール等の試薬を用いて融合させ、これによりできた融合細胞(ハイブリドーマ)の中から、Pi21タンパク質に結合する抗体を産生するクローンを選択する。次いで、得られたハイブリドーマをマウス腹腔内に移植し、同マウスより腹水を回収し、得られたモノクローナル抗体を、例えば、硫安沈殿、プロテインA、プロテインGカラム、DEAEイオン交換クロマトグラフィー、Pi21タンパク質や合成ペプチドをカップリングしたアフィニティーカラム等により精製することで、調製することが可能である。これにより得られた抗体は、本発明のタンパク質の精製や検出などに利用することが可能である。本発明には、本発明のタンパク質に結合する抗体が含まれる。
【0075】
本発明はまた、Pi21遺伝子またはpi21遺伝子からなるDNAまたはその相補鎖に相補的な少なくとも15ヌクレオチドの鎖長を有するオリゴヌクレオチドを提供する。
ここで「相補鎖」とは、A:T(ただしRNAの場合はU)、G:Cの塩基対からなる2本鎖核酸の一方の鎖に対する他方の鎖を指す。また、「相補的」とは、少なくとも15個の連続したヌクレオチド領域で完全に相補配列である場合に限られず、少なくとも70%、好ましくは少なくとも80%、より好ましくは90%、さらに好ましくは95%以上の塩基配列上の相同性を有すればよい。相同性を決定するためのアルゴリズムは当業者に周知のものを使用すればよい。
【0076】
本発明のオリゴヌクレオチドは、配列番号:1、2、4、5、20または21に記載の塩基配列からなるDNAの検出や増幅に用いるプローブやプライマーとして使用することができる。また、本発明のオリゴヌクレオチドは、DNAアレイの基板の形態で使用することができる。
【0077】
該オリゴヌクレオチドをプライマーとして用いる場合、その長さは、通常15bp〜100bpであり、好ましくは17bp〜30bpである。プライマーは、本発明のDNAまたはその相補鎖の少なくとも一部を増幅しうるものであれば、特に制限されない。また、プライマーとして用いる場合、3'側の領域は相補的とし、5'側には制限酵素認識配列やタグなどを付加することができる。
【0078】
また、上記オリゴヌクレオチドをプローブとして使用する場合、該プローブは、配列番号:1、2、4、5、20または21に記載の塩基配列からなるDNAまたはその相補鎖の少なくとも一部に特異的にハイブリダイズするものであれば、特に制限されない。該プローブは、合成オリゴヌクレオチドであってもよく、通常少なくとも15bp以上の鎖長を有する。
【0079】
本発明のオリゴヌクレオチドをプローブとして用いる場合は、適宜標識して用いることが好ましい。標識する方法としては、T4ポリヌクレオチドキナーゼを用いて、オリゴヌクレオチドの5'端を32Pでリン酸化することにより標識する方法、およびクレノウ酵素等のDNAポリメラーゼを用い、ランダムヘキサマーオリゴヌクレオチド等をプライマーとして32P等のアイソトープ、蛍光色素、またはビオチン等によって標識された基質塩基を取り込ませる方法(ランダムプライム法等)を例示することができる。
【0080】
本発明のオリゴヌクレオチドは、例えば市販のオリゴヌクレオチド合成機により作製することができる。プローブは、制限酵素処理等によって取得される二本鎖DNA断片として作製することもできる。
【0081】
さらに本発明は、Pi21遺伝子の発現を抑制するDNA、該DNAを含むベクターの用途を提供するものである。すなわち本発明は、Pi21遺伝子の発現を抑制するDNA、該DNAを含むベクターのいずれか1つを有効成分として含む、植物のいもち病圃場抵抗性を増加させるための薬剤に関する。また本発明は、植物のいもち病圃場抵抗性を増加させるための薬剤の製造における、Pi21遺伝子の発現を抑制するDNA、該DNAを含むベクターの使用に関する。
【0082】
本発明の植物にいもち病圃場抵抗性を増加させるための薬剤においては、有効成分であるオリゴヌクレオチド以外に、例えば、滅菌水、生理食塩水、植物油、界面活性剤、脂質、溶解補助剤、緩衝剤、保存剤等が必要に応じて混合されていてもよい。
【0083】
本発明はまた、罹病性遺伝子Pi21または抵抗性遺伝子pi21に連鎖する分子マーカーを提供する。
本発明における「分子マーカー」とは、Pi21遺伝子またはpi21遺伝子と遺伝的に連鎖するDNA領域であって、他のDNA領域と識別可能なDNA領域をいう。
【0084】
一般に分子マーカーとは、単位cMで表す地図距離が短いほどその遺伝子の近傍に位置し、その遺伝子と同時に遺伝するため、有用性が高い。pi21はマーカー「Pa102484」からマーカー「P702D3_#12」の間に存在することが示された(図2c)。したがって本発明の方法においては、図2cに記載の分子マーカーの中でも、上記2つのマーカー間および該2つのマーカーの間に存在するマーカー(「P702D03_#38」、「P702D03_#79」、「P702D03_#80」)は好ましいマーカーである。その中でも「P702D03_#79」は特に好ましいマーカーであり、配列番号:7または23(罹病性遺伝子Pi21と連鎖)または配列番号:10(抵抗性遺伝子pi21と連鎖)に記載のDNAとして例示できる。
【0085】
本発明の分子マーカーとしては、STS(Sequence Tagged Site)マーカーを挙げることができる。STSマーカーとは、DNA上の配列タグ部位(STS)の多型の有無の判定に利用できるDNA領域をいい、STSとは、DNA上の特定位置に特異的な配列部位をいう。STSの多型は、該特定配列部位を含むDNA領域をPCR法等の核酸増幅法により増幅し、該増幅産物をアガロースまたはポリアクリルアミドゲル電気泳動にかけることにより、バンドの有無や位置の違いとして検出することができる。
【0086】
本発明の分子マーカーとしてSTSマーカーを用いる場合は、本発明の識別方法を、例えば以下のようにして行うことができる。まず、被検イネおよびいもち病圃場抵抗性の形質を有するイネからDNA試料を周知方法によって調製する。次に、調製したDNAを鋳型として、プライマーDNAを用いて核酸増幅反応(例えば、PCR法)を行う。増幅したDNA断片の大きさを、電気泳動等により被検イネといもち病圃場抵抗性遺伝子と連鎖するマーカー(例えば配列番号:10に記載のマーカー)との間で比較し、同様の遺伝子型を示す場合には、該被検植物は、いもち病圃場抵抗性の形質を有すると判定される。
【0087】
本発明の識別方法に使用するプライマーDNAは、当業者においては、各種分子マーカーについての配列情報を考慮して、最適なプライマーを適宜設計することが可能である。通常、上記プライマーとは、イネに特異的に存在し、Pi21遺伝子またはpi21遺伝子と連鎖する塩基配列を挟み込むように設計された、該塩基配列を増幅するための一対のプライマーセットである。
【0088】
具体的には、STSマーカーのプライマーセットとしては、
(a)プライマー5’-AGA AGG TGG AGT ACG ACG TGA AGA-3’(配列番号:8)および5’-AGT TTA GTG AGC CTC TCC ACG ATT A-3’(配列番号:9)からなるプライマーセット、
(b)プライマー5’- GTA CGA CGT GAA GAA CAA CAG G -3’(配列番号:16)および5’- GCT TGG GCT TGC AGT CC-3’(配列番号:17)からなるプライマーセット、
(c)プライマー5’-GAT CCT CAT CGT CGA CGT CTG GC-3’(配列番号:26)および5’-AGG GTA CGG CAC CAG CTT G-3’(配列番号:27)からなるプライマーセット
を例示することが出来る。これらのプライマーセットによって増幅されるDNA配列によって特徴付けられる情報を、本発明の分子マーカーと比較することによって、被検イネがいもち病に対する圃場抵抗性の有無を判定することが可能である。
【0089】
また上記のプライマー以外にも、当業者であれば、配列番号:1、4もしくは20に記載の塩基配列を利用して、同様の機能を有するプライマーセットを作成することが可能である。本発明のプライマーには、このようなプライマーもまた含まれる。
【0090】
本発明のPCRプライマーは、当業者においては、例えば、自動オリゴヌクレオチド合成機等を利用して作製することができる。また、当業者においては周知の多型検出方法、例えば、上記PCRプライマーを用いた後述のPCR-SSCP法等によっても本発明の方法を実施することが可能である。
【0091】
また、本発明の分子マーカーがゲノムDNAのエクソン中に存在する場合には、mRNAを鋳型としたRT-PCRを利用することも可能である。また、Taqman(量的PCR検出)システム(Roche社)を利用すれば、蛍光により増幅産物の有無を検出することが可能である。このシステムによれば、電気泳動の手間も省けるため短時間で本発明の識別方法を行うことが可能である。
【0092】
さらに本発明は、以下の(a)〜(c)の工程を含む方法であって、分子量または塩基配列が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)増幅したDNA断片の分子量または塩基配列を、pi21遺伝子のそれと比較する工程
【0093】
本発明の上記DNA試料の調製(抽出)方法としては、当業者においては、公知の方法によって行うことができる。好ましい調製方法として、例えば、CTAB法を用いてDNAを抽出する方法を挙げることができる。
また、本発明の識別方法に供される、DNA試料は、特に制限されるものではないが、通常、被検植物であるイネから抽出するゲノムDNAを用いる。また、ゲノムDNAの採取源としては特に限定されるものではなく、イネのいずれの組織からも抽出できる。例えば、穂、葉、根、茎、種子、胚乳部、フスマ、胚等から抽出することができる。
【0094】
本発明の植物のいもち病圃場抵抗性を識別する方法では、次に、調製したDNAを鋳型として、プライマーDNAを用いて核酸増幅反応(例えば、PCR法)を行う。増幅したDNA断片を制限酵素で切断し、切断されたDNA断片の大きさを、電気泳動等により被検植物といもち病圃場抵抗性の植物との間で比較し、分子量または塩基配列が一致する場合に、該被検植物は、いもち病圃場抵抗性の形質を有すると判定される。「いもち病圃場抵抗性の植物」としては、実施例に記載のオワリハタモチが挙げられるが、これに限定されない。
【0095】
本発明における、被検植物がいもち病圃場抵抗性であると判定する方法において、「一致する」とは、対立遺伝子の両方の遺伝子の分子量または塩基配列がいもち病圃場抵抗性植物のそれと一致するときあるいはアミノ酸配列において一致することを意味する。したがって、対立遺伝子の一方の遺伝子の分子量、塩基配列またはアミノ酸配列がいもち病圃場抵抗性植物のそれと異なるが、もう一方がいもち病圃場抵抗性植物のそれと同じである場合には、「一致する」に含まれない。
上記電気泳動分析は、常法にしたがって行えばよい。例えば、アガロースまたはポリアクリルアミドのゲル中で電圧をかけて電気泳動し、分離したDNAパターンを分析する。
【0096】
また本発明は、以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)増幅した二本鎖のDNAを非変性ゲル上で分離する工程
(d)分離した二本鎖DNAのゲル上での移動度を、pi21遺伝子のそれと比較する工程
【0097】
さらに本発明は、以下の(a)〜(e)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)増幅したDNAを一本鎖DNAに解離させる工程
(d)解離させた一本鎖DNAを非変性ゲル上で分離する工程
(e)分離した一本鎖DNAのゲル上での移動度を、pi21遺伝子のそれと比較する工程
【0098】
このような方法として、PCR-SSCP(single-strand conformation polymorphism、一本鎖高次構造多型)法(Cloning and polymerase chain reaction-single-strand conformation polymorphism analysis of anonymous Alu repeats on chromosome 11. Genomics. 1992 Jan 1; 12(1): 139-146.、Detection of p53 gene mutations in human brain tumors by single-strand conformation polymorphism analysis of polymerase chain reaction products. Oncogene. 1991 Aug 1; 6(8): 1313-1318.、Multiple fluorescence-based PCR-SSCP analysis with postlabeling.、PCR Methods Appl. 1995 Apr 1; 4(5): 275-282.)が挙げられる。この方法は操作が比較的簡便であり、また試料の量も少なくてすむなどの利点を有するため、特に多数のDNAサンプルをスクリーニングするのに好適である。その原理は以下の如くである。二本鎖DNA断片を一本鎖に解離すると、各鎖はその塩基配列に依存した独自の高次構造を形成する。この解離したDNA鎖を変性剤を含まないポリアクリルアミドゲル中で電気泳動すると、それぞれの高次構造の差に応じて、相補的な同じ鎖長の一本鎖DNAが異なる位置に移動する。一塩基の置換によってもこの一本鎖DNAの高次構造は変化し、ポリアクリルアミドゲル電気泳動において異なる移動度を示す。従って、この移動度の変化を検出することによりDNA断片に点突然変異や欠失、あるいは挿入などによる変異が存在することを検出することができる。
【0099】
具体的には、まず、Pi21遺伝子またはpi21遺伝子の標的部位を含む領域をPCR法などによって増幅する。増幅される範囲としては、100〜600bp程度の長さが好ましい。PCRによる遺伝子断片増幅の際、32Pなどのアイソトープ、あるいは蛍光色素やビオチンなどによって標識したプライマーを用いるか、あるいはPCR反応液に32Pなどのアイソトープ、あるいは蛍光色素やビオチンなどによって標識した基質塩基を加えてPCRを行うことによって合成されるDNA断片を標識する。あるいはPCR反応後にクレノウ酵素などを用いて32Pなどのアイソトープ、あるいは蛍光色素やビオチンなどによって標識した基質塩基を合成されたDNA断片に付加することによっても標識を行うことができる。このようにして得られたDNA断片を2本鎖のまま、尿素などの変性剤を含まないポリアクリルアミドゲルによって電気泳動を行う。もしくは、このようなDNA断片に熱を加えることなどにより変性させてから、尿素などの変性剤を含まないポリアクリルアミドゲルによって電気泳動を行ってもよい。この際、ポリアクリルアミドゲルに適量(5から10%程度)のグリセロールを添加することにより、DNA断片の分離の条件を改善することができる。また、泳動条件は各DNA断片の性質により変動するが、通常、室温(20から25℃)で行い、好ましい分離が得られないときには4から30℃までの温度で最適の移動度を与える温度の検討を行う。電気泳動後、DNA断片の移動度を、X線フィルムを用いたオートラジオグラフィーや、蛍光を検出するスキャナー等で検出し、解析する。移動度に差があるバンドが検出された場合、このバンドを直接ゲルから切り出し、PCRによって再度増幅し、それを直接シークエンシングすることにより、変異の存在を確認することができる。また、標識したDNAを使わない場合においても、電気泳動後のゲルをエチジウムブロマイドや銀染色法などによって染色することによって、バンドを検出することができる。
【0100】
さらに本発明は、以下の(a)〜(e)の工程を含む方法であって、検出されたDNA断片の大きさが一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)調製したDNA試料を制限酵素により切断する工程
(d)DNA断片をその大きさに応じて分離する工程
(e)検出されたDNA断片の大きさを、pi21遺伝子のそれと比較する工程
【0101】
このような方法としては、制限酵素断片長多型(Restriction Fragment Length Polymorphism/RFLP)を利用したRFLP法やPCR−RFLP法などが挙げられる。DNAを切断する酵素としては、通常、制限酵素を用いる。具体的には、制限酵素の認識部位に塩基の付加、欠失が存在する場合、あるいは制限酵素処理によって生じるDNA断片内に塩基挿入、または欠失がある場合、制限酵素処理後に生じる断片の大きさが、いもち病罹病性の植物といもち病圃場抵抗性の植物との間で変化する。この変異部位を含む部分をPCR法によって増幅し、それぞれの制限酵素で処理することによって、これらの多型部位を電気泳動後のバンドの移動度の差として検出することができる。あるいは、染色体DNAをこれらの制限酵素によって処理し、電気泳動した後、プローブDNAを用いてサザンブロッティングを行うことにより、多型部位を検出することができる。用いられる制限酵素は、それぞれの変異部位に応じて適宜選択することができる。この方法では、ゲノムDNA以外にも被験者から調製したRNAを逆転写酵素でcDNAにし、これをそのまま制限酵素で切断した後サザンブロッティングを行うこともできる。また、このcDNAを鋳型としてPCRでPi21遺伝子またはpi21遺伝子の一部、あるいは全部を増幅し、それを制限酵素で切断した後、移動度の差を調べることもできる。
【0102】
さらに本発明は、以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法を提供する。
(a)被検植物からDNA試料を調製する工程
(b)該DNA試料からPi21遺伝子またはpi21遺伝子の領域を増幅する工程
(c)増幅したDNAを、DNA変性剤の濃度が次第に高まるゲル上で分離する工程
(d)分離したDNAのゲル上での移動度と、pi21遺伝子のそれと比較する工程
【0103】
このような方法としては、変性剤濃度勾配ゲル電気泳動法(denaturant gradient gel electrophoresis:DGGE)が挙げられる。Pi21遺伝子またはpi21遺伝子の標的部位を含む領域を本発明のプライマーなどを用いたPCR法などによって増幅し、これを尿素などの変性剤の濃度が移動するに従って徐々に高くなっているポリアクリルアミドゲル中で電気泳動し、健常者と比較する。変異が存在するDNA断片の場合、より低い変性剤濃度位置でDNA断片が一本鎖になり、極端に移動速度が遅くなるため、この移動度の差を検出することにより多型の存在を検出することができる。
【0104】
これら方法以外にも、アレル特異的オリゴヌクレオチド(Allele Specific Oligonucleotide/ASO)ハイブリダイゼーション法が利用できる。多型が存在すると考えられる塩基配列を含むオリゴヌクレオチドを作製し、これと試料DNAでハイブリダイゼーションを行わせると、該オリゴヌクレオチドと異なる多型塩基が、ハイブリダイズする試料DNAに存在する場合、ハイブリッド形成の効率が低下する。それをサザンブロット法や、特殊な蛍光試薬がハイブリッドのギャップにインターカレーションすることにより消光する性質を利用した方法などにより検出できる。
【0105】
また、リボヌクレアーゼAミスマッチ切断法による検出も可能である。具体的には、Pi21遺伝子あるいはpi21遺伝子の標的部位を含む領域をPCR法などによって増幅し、これに対し、プラスミドベクター等に組み込んだ健常者型のcDNA等から調製した標識RNAとハイブリダイゼーションを行わしめる。健常者型と異なる塩基が存在する部分においてはハイブリッドが一本鎖構造となるので、この部分をリボヌクレアーゼAによって切断し、これをオートラジオグラフィーなどで検出することによって多型の存在を検出することができる。
【0106】
本発明において「被検植物」とは、特に限定されるものではなく、いもち病菌に感染しうる全ての植物が含まれる。好ましい一例としては、イネが挙げられる。イネであれば特に制限されず、インディカ種であっても、ジャポニカ種であってもよく、または、インディカ種とジャポニカ種との交配品種・系統や野生イネ、あるいは栽培品種と野生イネの交配・交雑品種であってもよい。
【0107】
さらに本発明は、pi21遺伝子と連鎖する分子マーカーであって、少なくとも配列番号:7、10、または23に記載のDNAからなる分子マーカーを指標として、イネのいもち病圃場抵抗性を判定する方法を提供する。本発明の好ましい分子マーカーとしては、上述のように、「P702D03_#38」、「P702D03_#79」、「P702D03_#80」が挙げられる。その中でも「P702D03_#79」は特に好ましいマーカーであり、配列番号:7、23(罹病性遺伝子Pi21と連鎖)、または10(抵抗性遺伝子pi21と連鎖)に記載のDNAとして例示できる。本発明の識別方法においては、これらの分子マーカーのうち、少なくとも「P702D03_#79」を指標とした識別方法である。従って本発明の識別方法においては、「P702D03_#79」を単独で用いてもよく、「P702D03_#79」と他のマーカーを組み合わせて用いてもよい。「P702D03_#79」と他のマーカーを組み合わせとしては、「P702D03_#38」の組み合わせ、「P702D03_#79」と「P702D03_#80」の組み合わせ、「P702D03_#79」とその他任意のマーカーの組み合わせが挙げられる。
【0108】
本発明の識別方法においては、pi21遺伝子と連鎖する分子マーカーを有するか否かを調べることにより、被検イネのいもち病圃場抵抗性を特異的かつ効率的に判定できる。本発明の判定方法においては、いもち病圃場抵抗性の有無を判定したい所望のイネにおいて、配列番号:10に記載の塩基配列を有する場合に、被検イネはいもち病圃場抵抗性であるイネであるものと判定され、配列番号:10に記載の塩基配列を有さない場合(配列番号:7または23に記載の塩基配列を有する場合)に、被検イネはいもち病罹病性であるイネと判定される。
【0109】
被検イネにおける分子マーカーと本発明の分子マーカーとの比較は、分子マーカーのDNA配列の比較だけではなく、該DNA配列によって特徴付けられる情報の比較によっても実施することができる。分子マーカーのDNA配列によって特徴づけられる情報としては、分子マーカーの分子量についての情報、分子マーカーに含まれる変異部位や多型部位の存在の有無についての情報が挙げられる。当業者であれば、周知の方法によって、配列番号:10と、配列番号:7または配列番号:23に記載の塩基配列とを比較することにより、多型部位(欠失部位および1塩基置換部位)を同定することができる。
本発明の判定方法は、このような、分子マーカーに含まれる変異部位や多型部位の存在の有無についての情報を検出することによっても行うことが出来る。
【0110】
上記のような変異部位や多型部位の存在の有無についての情報の検出は、配列を直接シーケンシングすることはもちろん、変異部位や多型部位を含む領域を増幅しうるプライマーを用いることによって、または変異部位や多型部位にハイブリダイズしうるプローブ(例えば、配列番号:18、19または25に記載の塩基配列の全部または一部を含むDNA)を用いることによって、検出することが可能である。
【0111】
また、本発明の判定方法を利用すれば、いもち病圃場抵抗性であると識別される植物(例えばイネ)を早期に選抜することが可能となる。
すなわち本発明は、以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性である植物を選抜する方法を提供する。
(a)いもち病圃場抵抗性である植物(例えばイネ)と任意の機能を有する植物(例えばイネ)とが交配された品種を作製する工程
(b)本明細書に記載の、被検植物がいもち病圃場抵抗性であるか否かを判定する方法により、工程(a)で得られた植物がいもち病圃場抵抗性であるか否かを判定する工程
本発明において植物とは、特に限定されるものではないが、好ましくはイネである。イネの具体的な態様は上述の通りである。
【0112】
本発明の選抜方法を利用すれば、いもち病圃場抵抗性と識別される植物(例えばイネ)を早期に選抜することが可能となる。本発明はこのようないもち病圃場抵抗性と識別される植物を早期に選抜する方法も提供する。ここでいう「早期」とは、例えばイネの出穂より前の状態を指し、好ましくは発芽直後の状態を指す。本発明の選抜方法を利用すれば、いもち病圃場抵抗性の形質を有する品種の育成を従来よりも短期間で成し遂げることが可能となる。
【0113】
本発明は、植物のいもち病を予防または改善するための薬剤のスクリーニング方法に関する。
本発明のスクリーニング方法の第一の態様としては、以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法が挙げられる。
(a)Pi21遺伝子の転写産物に被検化合物を接触させる工程
(b)Pi21遺伝子の転写産物と被検化合物の結合を検出する工程
(c)Pi21遺伝子の転写産物と結合する被検化合物を選択する工程
【0114】
第一の態様では、まず、Pi21遺伝子の転写産物に被検化合物を接触させる。本発明のスクリーニング方法における「Pi21遺伝子の転写産物」としては、Pi21遺伝子の転写産物に加えて該転写産物より翻訳された翻訳産物も含まれる。
【0115】
本発明の方法における「被検化合物」としては、特に制限はなく、例えば、天然化合物、有機化合物、無機化合物、タンパク質、ペプチド等の単一化合物、並びに、化合物ライブラリー、遺伝子ライブラリーの発現産物、細胞抽出物、細胞培養上清、発酵微生物産生物、海洋生物抽出物、植物抽出物、原核細胞抽出物、真核単細胞抽出物もしくは動物細胞抽出物等を挙げることができる。上記被検化合物は必要に応じて適宜標識して用いることができる。標識としては、例えば、放射標識、蛍光標識等を挙げることができる。
【0116】
本発明において「接触」は、以下のようにして行う。例えば、Pi21遺伝子の転写産物が精製された状態であれば、精製標品に被検化合物を添加することにより行うことができる。また、細胞内に発現した状態または細胞抽出液内に発現した状態であれば、それぞれ、細胞の培養液または該細胞抽出液に被検化合物を添加することにより行うことができる。本発明における細胞としては、特に制限されないが、イネを含む植物由来の細胞が好ましい。被検化合物がタンパク質の場合には、例えば、該タンパク質をコードするDNAを含むベクターを、Pi21遺伝子が発現している細胞へ導入する、または該ベクターをPi21遺伝子が発現している細胞抽出液に添加することで行うことも可能である。また、例えば、酵母または動物細胞等を用いた 2ハイブリッド法を利用することも可能である。
【0117】
第一の態様では、次いで、上記Pi21遺伝子の転写産物と被検化合物の結合を検出する。タンパク質間の結合を検出または測定する手段は、例えばタンパク質に付した標識を利用することにより行うことができる。標識の種類は、例えば、蛍光標識、放射標識等が挙げられる。また、酵母ツーハイブリット法や、BIACOREを用いた測定方法等、公知の方法によって測定することもできる。本方法においては、ついで、上記生合成酵素と結合した被検化合物を選択する。選択された被検化合物の中には、植物のいもち病を予防または改善するための薬剤が含まれる。また、選択された被検化合物を、以下のスクリーニングの被検化合物として用いてもよい。
【0118】
また、本発明のスクリーニング方法の第二の態様として、以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法を提供する。
(a)植物から採取した細胞に、被検化合物を接触させる工程
(b)Pi21遺伝子の転写産物の発現レベルを測定する工程
(c)被検化合物を接触させていない場合と比較して、上記転写産物の発現レベルを減少させた被検化合物を選択する工程
【0119】
第二の態様では、まず、植物から採取した細胞に被検化合物を接触させる。ここで「植物から採取した細胞」は、いもち病罹病性遺伝子を有することが明らかな任意の植物であってよい。「被検化合物」、「接触」の記載に関しては上記の説明の通りである。
【0120】
第二の態様では、次いで、「Pi21タンパク質」の発現レベルを測定する。Pi21タンパク質の発現レベルの測定は、当業者に公知の方法によって行うことができる。例えば、Pi21タンパク質をコードするmRNAを定法に従って抽出し、このmRNAを鋳型としたノーザンハイブリダイゼーション法、またはRT-PCR法を実施することによって該遺伝子の転写レベルの測定を行うことができる。さらに、DNAアレイ技術を用いて、Pi21タンパク質の発現レベルを測定することも可能である。
【0121】
また、Pi21タンパク質を含む画分を定法に従って回収し、Pi21タンパク質の発現をSDS-PAGE等の電気泳動法で検出することにより、遺伝子の翻訳レベルの測定を行うこともできる。また、Pi21タンパク質に対する抗体を用いて、ウェスタンブロッティング法を実施し、Pi21タンパク質の発現を検出することにより、遺伝子の翻訳レベルの測定を行うことも可能である。
【0122】
Pi21タンパク質の検出に用いる抗体としては、検出可能な抗体であれば、特に制限はないが、例えばモノクローナル抗体、またはポリクローナル抗体の両方を利用することができる。該抗体は、上述のように、当業者に公知の方法により調製することが可能である。
【0123】
第二の態様においては、ついで、Pi21タンパク質の発現レベルが、被検化合物を接触させないときに比べ減少する場合に、被検化合物を、植物のいもち病を予防または改善するための薬剤として選択する。
【0124】
本発明のスクリーニング方法の第三の態様は、以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法である。
(a)Pi21遺伝子のプロモーター領域の下流にレポーター遺伝子が機能的に結合したDNAを有する細胞または細胞抽出液を提供する工程
(b)該細胞または該細胞抽出液に被検化合物を接触させる工程
(c)該細胞または該細胞抽出液における該レポーター遺伝子の発現レベルを測定する工程
(d)被検化合物を接触させていない場合と比較して、該レポーター遺伝子の発現レベルを減少させた被検化合物を選択する工程
【0125】
第三の態様では、まず、Pi21遺伝子のプロモータ―領域の下流にレポーター遺伝子が機能的に結合したDNAを有する細胞または細胞抽出液を提供する。
第三の態様において、「機能的に結合した」とは、Pi21遺伝子のプロモータ―領域に転写因子が結合することにより、レポーター遺伝子の発現が誘導されるように、Pi21遺伝子のプロモータ―領域とレポーター遺伝子とが結合していることをいう。従って、レポーター遺伝子が他の遺伝子と結合しており、他の遺伝子産物との融合タンパク質を形成する場合であっても、Pi21遺伝子のプロモータ―領域に転写因子が結合することによって、該融合タンパク質の発現が誘導されるものであれば、上記「機能的に結合した」の 意に含まれる。
【0126】
上記レポーター遺伝子としては、その発現が検出可能なものであれば特に制限されず、例えば、当業者において一般的に使用されるCAT遺伝子、lacZ遺伝子、ルシフェラーゼ遺伝子、β-グルクロニダーゼ遺伝子(GUS)およびGFP遺伝子等を挙げることができる。
【0127】
第三の態様では、次いで、上記細胞または上記細胞抽出液に被検化合物を接触させる。次いで、該細胞または該細胞抽出液における上記レポーター遺伝子の発現レベルを測定する。「被検化合物」および「接触」の記載に関しては上記の説明の通りである。
【0128】
レポーター遺伝子の発現レベルは、使用するレポーター遺伝子の種類に応じて、当業者に公知の方法により測定することができる。例えば、レポーター遺伝子がCAT遺伝子である場合には、該遺伝子産物によるクロラムフェニコールのアセチル化を検出することによって、レポーター遺伝子の発現レベルを測定することができる。レポーター遺伝子がlacZ遺伝子である場合には、該遺伝子発現産物の触媒作用による色素化合物の発色を検出することにより、また、ルシフェラーゼ遺伝子である場合には、該遺伝子発現産物の触媒作用による蛍光化合物の蛍光を検出することにより、また、β-グルクロニダーゼ遺伝子(GUS)である場合には、該遺伝子発現産物の触媒作用によるGlucuron(ICN社)の発光や5-ブロモ-4-クロロ-3-インドリル-β-グルクロニド(X-Gluc)の発色を検出することにより、さらに、GFP遺伝子である場合には、GFPタンパク質による蛍光を検出することにより、レポーター遺伝子の発現レベルを測定することができる。
【0129】
第三の態様においては、ついで、上記レポーター遺伝子の発現レベルが、被検化合物を接触させないときに比べ減少する場合に、被検化合物を、植物のいもち病を予防または改善するための薬剤として選択する。
【0130】
本発明のスクリーニング方法の第四の態様は、以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法である。
(a)Pi21遺伝子が導入された形質転換植物細胞より形質転換植物体を再生させる工程
(b)該形質転換植物体に、いもち病菌および被検化合物を接触させる工程
(c)被検化合物を接触させていない場合と比較して、該形質転換植物体のいもち病を抑制する被検化合物を選択する工程
【0131】
第四の態様では、まず、Pi21遺伝子を含む形質転換植物細胞より形質転換植物体を再生させる。該形質転換植物体の再生は、上述したように当業者に公知の方法によって行うことが出来る。
第四の態様では次に、工程(a)で再生させた該形質転換植物体に、いもち病菌および被検化合物を接触させる。「いもち病菌」および「被検化合物」は上記の説明の通りである。「接触」の一例としては、霧吹きを用いて被検化合物を植物に直接噴霧する方法が挙げられる。しかし第四の態様における「接触」はこれに限定されず、植物と被検化合物が物理的に触れることが出来る限り、あらゆる方法が含まれる。本発明の接触は、いもち病菌に感染した形質転換植物体に被検化合物を接触させてもよいし、被検化合物が接触された形質転換植物体にいもち病菌を感染させてもよい。
【0132】
第四の態様においては、ついで、被検化合物を接触させていない場合と比較して該形質転換植物体のいもち病を抑制する被検化合物を選択する。いもち病が抑制されたか否かは、該形質転換植物体の表現型を指標として判断することが出来る。該形質転換植物体の表現型としては、特に制限はないが、植物の各部位においてその一部あるいは全体が変色・壊死することが例示できる。また、該形質転換植物体のいもち病の抑制には、完全な抑制だけでなく、部分的な抑制も含まれる。
【0133】
本発明はまた、上記に記載のスクリーニング方法に用いるためのキットに関する。このようなキットには、上記に記載のスクリーニング方法の検出工程や測定工程に使用されるものを含みうる。例えば、Pi21遺伝子の発現レベルの測定に必要とされるプローブ、プライマー、抗体、染色液等を挙げることができる。その他、蒸留水、塩、緩衝液、タンパク質安定剤、保存剤等が含まれていてもよい。
【実施例】
【0134】
以下、本発明を実施例によりさらに具体的に説明するが本発明はこれら実施例に制限されるものではない。
【0135】
〔実施例1〕遺伝地図作成
マップベースクローニングに不可欠な大規模分離集団によるpi21領域の詳細な連鎖解析を行った。連鎖解析用の集団では病斑進展を抑制する抵抗性対立遺伝子pi21を持つ日本陸稲品種オワリハタモチに、病斑進展を抑制しない罹病性対立遺伝子Pi21を持つ水稲品種日本晴あるいは愛知旭(図1)を連続戻し交雑して得たBC1F2集団72個体を用いた。RFLPマーカーによる連鎖解析の結果、pi21遺伝子座はRFLPマーカーG271およびG317の間に存在することが明らかとなった(図2A)。
【0136】
精度の高いpi21領域の遺伝地図を作成するために、上述の交雑組合せ229個体と後代BC1F4集団643個体を加え、合計1014個体を用いて、pi21の両側に存在するRFLPマーカーRA3591と13S1を利用して、pi21座近傍の染色体組み換え型27個体を選抜した(図2B)。さらに、日本水稲品種の遺伝的背景にインド型水稲品種Kasalathの罹病性対立遺伝子を持つ系統とオワリハタモチ由来の抵抗性対立遺伝子を持つ系統を交雑して得たF2集団2703個体を用いた検索を行った。pi21座の両側に存在するPCRマーカー14T1と4S1を利用して、pi21座近傍の染色体組み換え型24個体が選抜できた。さらにそれらの個体を用いて、以下の取り組みにおいて作出されたDNAマーカーによる詳細な連鎖地図作成を行った。
【0137】
〔実施例2〕P1由来人工染色体(PAC)クローンによるpi21遺伝子領域の整列化
イネゲノム解析研究において作出された日本晴PACクローンの整列化地図を利用して、pi21遺伝子座近傍に存在するDNAマーカーRA3591およびC975塩基配列を有するPACクローンを特定した(図2C)。さらに特定されたPACクローンP479G02、P415D09、P473G08、P703E11、P434F09、P702D03、P419B08、P472G09およびP502G01の末端断片をカセット法により単離し、特定できたPACクローンを整列化した。その結果、PACクローンP032D02、P678A02、P405D12、P689F04、P479G02、P415D09、P473G08、P703E11、P434F09、およびP702D03はpi21遺伝子領域を含むことが明かとなった(図2C)。
【0138】
〔実施例3〕pi21遺伝子領域の絞り込み
pi21領域に整列化されたPACクローンの末端断片をクローン化し、それらを新たなRFLPマーカーあるいはCAPSマーカーとして、詳細な遺伝地図を作成したところ、pi21遺伝子座はSSCPマーカーPa102484およびSNPマーカーP702D3_#12に挟み込まれるゲノム領域に存在することが明らかとなった。この結果、pi21遺伝子座は2つのマーカーに挟まれる約25kbのゲノム領域に存在することが明らかとなった(図2D)。
【0139】
〔実施例4〕塩基配列解析による候補遺伝子領域の特定
pi21遺伝子を含むと考えられるPACクローンP702D03の塩基配列を決定し、25kbのゲノム候補領域の抵抗性品種オワリハタモチと罹病性品種愛知旭およびKasalathの塩基配列解析を行なった。塩基配列の解析は日本晴の候補領域の配列を利用して設計したプライマーを用いて上述3品種より増幅したDNA断片を使用し、dye-terminater法により行った。連鎖解析により特定された候補遺伝子領域内の塩基多型情報を利用して、候補領域のさらなる絞り込みを行った。その結果、pi21遺伝子はSTSマーカーP702D03_#79(プライマー5’-AGA AGG TGG AGT ACG ACG TGA AGA-3’(配列番号:8)およびAGT TTA GTG AGC CTC TCC ACG ATT A-3’(配列番号:9))と共分離し、SNPマーカーP702D03_#38(プライマーTTT TCC TGA GAA ATT TGT AAA GA-3’ (配列番号:12)およびCGT CGA CGA TGA GGA TCT-3’ (配列番号:13))と、P702D03_#80(プライマー5’-CTC CCA ATG TGT TTA GCA TC-3’ (配列番号:14)および5’-CAA CCA TAT GTC CCT AAG GAT-3’(配列番号:15))との間にそれぞれ1個の組み換え個体を検出した。以上の結果から、pi21遺伝子はSNPマーカーP702D03_#38およびP702D03_#80に挟み込まれる約1.8kbのゲノム領域に存在することが明らかとなった(図2D)。
【0140】
単離されたイネ(Oryza sativa L、愛知旭および日本晴)由来Pi21遺伝子の遺伝子の塩基配列を配列番号:1に、cDNAの塩基配列を配列番号:2に、該cDNAがコードするタンパク質(「Pi21タンパク質」)のアミノ酸配列を配列番号:3に示した。なお、愛知旭および日本晴のPi21遺伝子に対応する、Kasalath由来のPi21遺伝子の遺伝子の塩基配列を配列番号:20に、cDNAの塩基配列を配列番号:21に、該cDNAがコードするタンパク質(「Pi21タンパク質」)のアミノ酸配列を配列番号:22に示した。
【0141】
〔実施例5〕pi21候補遺伝子の塩基配列解析
1.8kb候補ゲノム領域の品種日本晴の配列について遺伝子予測ならびに類似性検索を行ったところ、日本晴の完全長cDNAクローン(AK106153,AK070581,AK072320)が見出されたが、アラビドプシスなどで類似の遺伝子はなく相同性から遺伝子の機能を予測することはできなかった。しかしながら、この遺伝子は予想翻訳開始点から約10アミノ酸の部位にある金属結合部位は、アラビドプシスで報告されているエチレンシグナル伝達系における金属を運搬するシャペロンの機能を持つ遺伝子(Hirayama et al 1999 Cell)を連想させ、実際に準同質遺伝子系統AA-pi21では愛知旭に比べてエチレンに対する感受性が変化していることから類似の機能を持つ可能性がある。既に得られている日本晴の塩基配列情報から、該当部分を増幅できるプライマーを設計し、ゲノムPCRおよび RT-PCR産物を罹病性品種日本晴および愛知旭と抵抗性品種オワリハタモチの間で塩基配列を比較したところ、遺伝子のエクソン領域に抵抗性品種と罹病性品種との間に2ヶ所のDNA変異があった。罹病性の品種に対して抵抗性の品種では、7アミノ酸および16アミノ酸の欠失が起こっており、これらの変異が感染したいもち病の病斑進展に関係するものと考えられた(図3)。
【0142】
単離されたpi21遺伝子の遺伝子の塩基配列を配列番号:4に、cDNAの塩基配列を配列番号:5に、cDNAがコードするタンパク質(「pi21タンパク質」)のアミノ酸配列を配列番号:6に示す。
【0143】
〔実施例6〕形質転換による候補遺伝子の機能の同定
(1)罹病性遺伝子のAA-pi21への導入
pi21遺伝子の候補として特定された5’上流予測プロモーター領域を含む罹病性品種日本晴のゲノム領域XbaI 4.7kb断片を、アグロバクテリウムを介して形質転換可能なベクターpPZP2H-lacに組み込んだ。この断片を導入したベクターならびにベクターのみを用い、土岐の方法(文献 Plant Mol. Biol. Rep. 15:16-21, 1997)により形質転換を行った。形質転換する系統にはpi21準同質遺伝子系統AA-pi21を用いた。XbaI 4.7kb断片が導入されたベクターからは36個体、ベクターのみからは12個体のハイグロマイシン耐性個体を得た。候補遺伝子に特異的なプライマー(センス鎖5'- GTA CGA CGT GAA GAA CAA CAG G-3'(配列番号:16))および (アンチセンス鎖5'- GCT TGG GCT TGC AGT CC 3'(配列番号:17))を用いて、導入した領域が組み込まれたか否かをPCR法により調査した。その結果、すべての形質転換体について、候補遺伝子が組み込まれていることが判明した。これらの個体を、隔離温室で栽培して、自殖後代系統においていもち病菌(レース007)を接種した。その結果、ベクターのみを導入した個体および分離によって導入遺伝子が伝達されなかったT1個体では、準同質遺伝子系統AA-pi21と同様に、病斑の進展が罹病性品種愛知旭に比べていもち病の病斑の進展が抑制されたのに対して、候補遺伝子が導入されているT1個体では、より病斑が進展した(図4)。特に、導入遺伝子のコピー数の多い系統では感受性の度合いが増していた。
【0144】
(2)抵抗性遺伝子の罹病性品種への導入
一方、抵抗性品種オワリハタモチのXbaI 4.7kb断片についても同様にベクターに組み込み、罹病性品種愛知旭を用いて形質転換を行った。XbaI 4.7kb断片が導入されたベクターからは56個体、ベクターのみからは24個体のハイグロマイシン耐性個体を得た。(1)と同様、導入した領域が組み込まれたか否かをPCR法により調査した結果、すべての形質転換体について、候補遺伝子が組み込まれていることが判明した。これらの個体を、(1)と同様の方法で隔離温室で栽培して、自殖後代系統においていもち病菌(レース007)を接種した。その結果、ベクターのみを導入した個体、導入遺伝子が伝達されなかったT1個体および、候補遺伝子が導入されているT1個体の全てにおいて罹病性品種愛知旭と同様のいもち病の進展が見られた。
【0145】
(3)候補遺伝子の機能の同定
以上の結果から、日本晴由来の候補遺伝子領域(XbaI 4.7kb)には、準同質遺伝子系統AA-pi21の病斑形成を促進する機能を有することが判明し、候補遺伝子がPi21遺伝子であると判断した。
【0146】
〔実施例7〕イネにおける候補遺伝子の変異
世界のイネ品種79品種を用いて候補遺伝子の変異を検索したところ、日本晴と愛知旭およびオワリハタモチに見出されたタイプ以外に、エクソン領域に挿入欠失のある10種類の変異があった。これらは主として日本晴・愛知旭とオワリハタモチの間で見出した2ヶ所の欠失部位について、その挿入/欠失の有無とその大きさによって規定される。シロイヌナズナのエチレンシグナル伝達の系で提唱される金属分子のシャペロンとの類似から、既知の遺伝子とは相同性がないこの領域は他の分子と結合するものと予想され、この部位の変異がシグナル伝達の効率を微妙に制御し、機能の変異をもたらす可能性がある。
【0147】
以上の結果から、マップベースクローニング法により候補として絞り込まれた遺伝子がイネいもち病の病斑進展を制御する遺伝子pi21であることが判明した。本成果は植物の量的抵抗性の生物的機能を証明した初めてのケースである。RT-PCR分析によってpi21あるいはPi21遺伝子の発現を調査したところ、地上部の全ての組織で恒常的に発現していることから、この遺伝子が植物の生育の基本的な役割を果たしているもの予想される。コピー数の変化が表現型の変化をもたらしたことから、プロモーターによる発現量、発現組織を変化させることが機能改変に重要な要素となる。すなわち、単離したpi21遺伝子あるいは種内に認められる別の対立遺伝子を利用して、本来植物がもつ耐病性を効率的に高めることが可能であると考えられる。
【特許請求の範囲】
【請求項1】
以下(a)〜(h)のいずれかに記載のDNA;
(a)配列番号:3または22に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:1、2、20または21に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:3または22に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(d)配列番号:1、2、20または21に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(e)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(f)配列番号:4または5に記載の塩基配列のコード領域を含むDNA、
(g)配列番号:6に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(h)配列番号:4または5に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA。
【請求項2】
植物においていもち病圃場抵抗性能を有する、以下(i)〜(iv)のいずれかに記載のDNA;
(i)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物と相補的なRNAをコードするDNA、
(ii)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA、
(iii)請求項1(a)〜(d)のいずれかに記載のDNAの発現を共抑制効果により阻害するRNAをコードするDNA、
(iv)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物を特異的に切断するRNAi活性を有するRNAをコードするDNA。
【請求項3】
植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである、請求項2に記載のDNA。
【請求項4】
請求項1〜3のいずれかに記載のDNAを含むベクター。
【請求項5】
請求項1〜3のいずれかに記載のDNAを発現可能に保持する形質転換細胞。
【請求項6】
請求項1(a)〜(d)のいずれかに記載のDNAが導入された形質転換植物細胞。
【請求項7】
請求項2もしくは3に記載のDNAが導入された形質転換植物細胞。
【請求項8】
植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである、請求項6または7に記載の形質転換植物細胞。
【請求項9】
請求項6〜8に記載の形質転換細胞を含む形質転換植物体。
【請求項10】
請求項8に記載の形質転換植物体の子孫またはクローンである、形質転換植物体。
【請求項11】
請求項9または10のいずれかに記載の形質転換植物体の繁殖材料。
【請求項12】
請求項9または10のいずれかに記載の形質転換植物体の製造方法であって、請求項1(a)〜(d)、請求項2または3に記載のDNAを植物細胞に導入し、該植物細胞から植物体を再生させる工程を含む方法。
【請求項13】
請求項2または3に記載のDNAを植物体の細胞内で発現させる工程を含む、植物にいもち病圃場抵抗性を付与する方法。
【請求項14】
植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである請求項13に記載の方法。
【請求項15】
請求項1(a)〜(d)のいずれかに記載のDNAによりコードされるタンパク質。
【請求項16】
請求項1(a)〜(d)のいずれかに記載のDNAを含むベクターを含む形質転換細胞を培養し、該細胞またはその培養上清から組換えタンパク質を回収する工程を含む、請求項15に記載のタンパク質の製造方法。
【請求項17】
請求項15に記載のタンパク質に結合する抗体。
【請求項18】
請求項1に記載のDNAまたはその相補配列に相補的な少なくとも15の連続する塩基を含むDNA。
【請求項19】
請求項2もしくは3に記載のDNA、または該DNAを含むベクターのいずれか1つを含む、植物のいもち病圃場抵抗性を増加させる薬剤。
【請求項20】
配列番号:1、4または20に記載の塩基配列の全部または一部を増幅するプライマーセット。
【請求項21】
(a)〜(c)のいずれかに示す、少なくとも1つのプライマーセット
(a)配列番号:8に記載の塩基配列からなるDNAおよび配列番号:9に記載の塩基配列からなるDNA、
(b)配列番号:16に記載の塩基配列からなるDNAおよび配列番号:17に記載の塩基配列からなるDNA、
(c)配列番号:26に記載の塩基配列からなるDNAおよび配列番号:27に記載の塩基配列からなるDNA。
【請求項22】
配列番号:7、10、18、19、23または25に記載の塩基配列からなるDNA。
【請求項23】
以下の(a)〜(c)の工程を含む方法であって、分子量または塩基配列が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から請求項1に記載のDNA領域を増幅する工程、
(c)増幅したDNA断片の分子量または塩基配列を、請求項1(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
【請求項24】
以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から請求項1に記載のDNA領域を増幅する工程、
(c)増幅した二本鎖のDNAを非変性ゲル上で分離する工程、
(d)分離した二本鎖DNAのゲル上での移動度を、請求項1(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
【請求項25】
以下の(a)〜(e)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から請求項1に記載のDNA領域を増幅する工程、
(c)増幅したDNAを一本鎖DNAに解離させる工程、
(d)解離させた一本鎖DNAを非変性ゲル上で分離する工程、
(e)分離した一本鎖DNAのゲル上での移動度を、請求項1(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
【請求項26】
以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から請求項1に記載のDNA領域を増幅する工程、
(c)増幅したDNAを、DNA変性剤の濃度が次第に高まるゲル上で分離する工程、
(d)分離したDNAのゲル上での移動度と、請求項1(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
【請求項27】
以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性である植物を選抜する方法;
(a)いもち病圃場抵抗性能である植物と任意の機能を有する植物とが交配された品種を作製する工程、
(b)請求項23〜26のいずれかに記載の方法により、工程(a)で作製された植物がいもち病圃場抵抗性であるか否かを判定する工程。
【請求項28】
請求項1に記載のDNAと連鎖する分子マーカーであって、該分子マーカーがいもち病圃場抵抗性の形質を有するイネと同様の遺伝子型を示す場合に、被検イネがいもち病圃場抵抗性であると判定する方法。
【請求項29】
分子マーカーが配列番号:10に記載のDNAからなる分子マーカーである、請求項28に記載の方法。
【請求項30】
以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性であるイネを選抜する方法;
(a)いもち病圃場抵抗性能であるイネと任意の機能を有するイネとが交配された品種を作製する工程、
(b)請求項28または29のいずれかに記載の方法により、工程(a)で作製されたイネがいもち病圃場抵抗性であるか否かを判定する工程。
【請求項31】
以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物に被検化合物を接触させる工程、
(b)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物と被検化合物の結合を検出する工程、
(c)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物と結合する被検化合物を選択する工程。
【請求項32】
以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)植物から採取した細胞に、被検化合物を接触させる工程、
(b)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物の発現レベルを測定する工程、
(c)被検化合物を接触させていない場合と比較して、上記転写産物の発現レベルを減少させた被検化合物を選択する工程。
【請求項33】
以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)請求項1(a)〜(d)のいずれかに記載のDNAのプロモーター領域の下流にレポーター遺伝子が機能的に結合したDNAを有する細胞または細胞抽出液を提供する工程、
(b)該細胞または該細胞抽出液に被検化合物を接触させる工程、
(c)該細胞または該細胞抽出液における該レポーター遺伝子の発現レベルを測定する工程、
(d)被検化合物を接触させていない場合と比較して、該レポーター遺伝子の発現レベルを減少させた被検化合物を選択する工程。
【請求項34】
以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)請求項6に記載の形質転換植物細胞より形質転換植物体を再生させる工程、
(b)該形質転換植物体に、いもち病菌および被検化合物を接触させる工程、
(c)被検化合物を接触させていない場合と比較して、該形質転換植物体のいもち病を抑制する被検化合物を選択する工程。
【請求項35】
請求項31〜34のいずれかに記載のスクリーニング方法に用いるためのキット。
【請求項1】
以下(a)〜(h)のいずれかに記載のDNA;
(a)配列番号:3または22に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号:1、2、20または21に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:3または22に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(d)配列番号:1、2、20または21に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:3または22に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(e)配列番号:6に記載のアミノ酸配列からなるタンパク質をコードするDNA、
(f)配列番号:4または5に記載の塩基配列のコード領域を含むDNA、
(g)配列番号:6に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質であって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA、
(h)配列番号:4または5に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAであって、配列番号:6に記載のアミノ酸配列からなるタンパク質と同等の機能を有するタンパク質をコードするDNA。
【請求項2】
植物においていもち病圃場抵抗性能を有する、以下(i)〜(iv)のいずれかに記載のDNA;
(i)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物と相補的なRNAをコードするDNA、
(ii)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA、
(iii)請求項1(a)〜(d)のいずれかに記載のDNAの発現を共抑制効果により阻害するRNAをコードするDNA、
(iv)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物を特異的に切断するRNAi活性を有するRNAをコードするDNA。
【請求項3】
植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである、請求項2に記載のDNA。
【請求項4】
請求項1〜3のいずれかに記載のDNAを含むベクター。
【請求項5】
請求項1〜3のいずれかに記載のDNAを発現可能に保持する形質転換細胞。
【請求項6】
請求項1(a)〜(d)のいずれかに記載のDNAが導入された形質転換植物細胞。
【請求項7】
請求項2もしくは3に記載のDNAが導入された形質転換植物細胞。
【請求項8】
植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである、請求項6または7に記載の形質転換植物細胞。
【請求項9】
請求項6〜8に記載の形質転換細胞を含む形質転換植物体。
【請求項10】
請求項8に記載の形質転換植物体の子孫またはクローンである、形質転換植物体。
【請求項11】
請求項9または10のいずれかに記載の形質転換植物体の繁殖材料。
【請求項12】
請求項9または10のいずれかに記載の形質転換植物体の製造方法であって、請求項1(a)〜(d)、請求項2または3に記載のDNAを植物細胞に導入し、該植物細胞から植物体を再生させる工程を含む方法。
【請求項13】
請求項2または3に記載のDNAを植物体の細胞内で発現させる工程を含む、植物にいもち病圃場抵抗性を付与する方法。
【請求項14】
植物がイネ、コムギ、オオムギ、エンバク、トウモロコシ、ハトムギ、イタリアンライグラス、ペレニアルライグラス、チモシー、メドーフェスク、キビ、アワ、サトウキビである請求項13に記載の方法。
【請求項15】
請求項1(a)〜(d)のいずれかに記載のDNAによりコードされるタンパク質。
【請求項16】
請求項1(a)〜(d)のいずれかに記載のDNAを含むベクターを含む形質転換細胞を培養し、該細胞またはその培養上清から組換えタンパク質を回収する工程を含む、請求項15に記載のタンパク質の製造方法。
【請求項17】
請求項15に記載のタンパク質に結合する抗体。
【請求項18】
請求項1に記載のDNAまたはその相補配列に相補的な少なくとも15の連続する塩基を含むDNA。
【請求項19】
請求項2もしくは3に記載のDNA、または該DNAを含むベクターのいずれか1つを含む、植物のいもち病圃場抵抗性を増加させる薬剤。
【請求項20】
配列番号:1、4または20に記載の塩基配列の全部または一部を増幅するプライマーセット。
【請求項21】
(a)〜(c)のいずれかに示す、少なくとも1つのプライマーセット
(a)配列番号:8に記載の塩基配列からなるDNAおよび配列番号:9に記載の塩基配列からなるDNA、
(b)配列番号:16に記載の塩基配列からなるDNAおよび配列番号:17に記載の塩基配列からなるDNA、
(c)配列番号:26に記載の塩基配列からなるDNAおよび配列番号:27に記載の塩基配列からなるDNA。
【請求項22】
配列番号:7、10、18、19、23または25に記載の塩基配列からなるDNA。
【請求項23】
以下の(a)〜(c)の工程を含む方法であって、分子量または塩基配列が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から請求項1に記載のDNA領域を増幅する工程、
(c)増幅したDNA断片の分子量または塩基配列を、請求項1(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
【請求項24】
以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から請求項1に記載のDNA領域を増幅する工程、
(c)増幅した二本鎖のDNAを非変性ゲル上で分離する工程、
(d)分離した二本鎖DNAのゲル上での移動度を、請求項1(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
【請求項25】
以下の(a)〜(e)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から請求項1に記載のDNA領域を増幅する工程、
(c)増幅したDNAを一本鎖DNAに解離させる工程、
(d)解離させた一本鎖DNAを非変性ゲル上で分離する工程、
(e)分離した一本鎖DNAのゲル上での移動度を、請求項1(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
【請求項26】
以下の(a)〜(d)の工程を含む方法であって、ゲル上での移動度が一致するときに、被検植物がいもち病圃場抵抗性であると判定する方法;
(a)被検植物からDNA試料を調製する工程、
(b)該DNA試料から請求項1に記載のDNA領域を増幅する工程、
(c)増幅したDNAを、DNA変性剤の濃度が次第に高まるゲル上で分離する工程、
(d)分離したDNAのゲル上での移動度と、請求項1(e)または(f)のいずれかに記載のDNAのそれと比較する工程。
【請求項27】
以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性である植物を選抜する方法;
(a)いもち病圃場抵抗性能である植物と任意の機能を有する植物とが交配された品種を作製する工程、
(b)請求項23〜26のいずれかに記載の方法により、工程(a)で作製された植物がいもち病圃場抵抗性であるか否かを判定する工程。
【請求項28】
請求項1に記載のDNAと連鎖する分子マーカーであって、該分子マーカーがいもち病圃場抵抗性の形質を有するイネと同様の遺伝子型を示す場合に、被検イネがいもち病圃場抵抗性であると判定する方法。
【請求項29】
分子マーカーが配列番号:10に記載のDNAからなる分子マーカーである、請求項28に記載の方法。
【請求項30】
以下の(a)および(b)に記載の工程を含む、いもち病圃場抵抗性であるイネを選抜する方法;
(a)いもち病圃場抵抗性能であるイネと任意の機能を有するイネとが交配された品種を作製する工程、
(b)請求項28または29のいずれかに記載の方法により、工程(a)で作製されたイネがいもち病圃場抵抗性であるか否かを判定する工程。
【請求項31】
以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物に被検化合物を接触させる工程、
(b)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物と被検化合物の結合を検出する工程、
(c)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物と結合する被検化合物を選択する工程。
【請求項32】
以下の(a)〜(c)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)植物から採取した細胞に、被検化合物を接触させる工程、
(b)請求項1(a)〜(d)のいずれかに記載のDNAの転写産物の発現レベルを測定する工程、
(c)被検化合物を接触させていない場合と比較して、上記転写産物の発現レベルを減少させた被検化合物を選択する工程。
【請求項33】
以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)請求項1(a)〜(d)のいずれかに記載のDNAのプロモーター領域の下流にレポーター遺伝子が機能的に結合したDNAを有する細胞または細胞抽出液を提供する工程、
(b)該細胞または該細胞抽出液に被検化合物を接触させる工程、
(c)該細胞または該細胞抽出液における該レポーター遺伝子の発現レベルを測定する工程、
(d)被検化合物を接触させていない場合と比較して、該レポーター遺伝子の発現レベルを減少させた被検化合物を選択する工程。
【請求項34】
以下の(a)〜(d)の工程を含む、植物のいもち病を予防または改善するための薬剤のスクリーニング方法;
(a)請求項6に記載の形質転換植物細胞より形質転換植物体を再生させる工程、
(b)該形質転換植物体に、いもち病菌および被検化合物を接触させる工程、
(c)被検化合物を接触させていない場合と比較して、該形質転換植物体のいもち病を抑制する被検化合物を選択する工程。
【請求項35】
請求項31〜34のいずれかに記載のスクリーニング方法に用いるためのキット。
【図2】


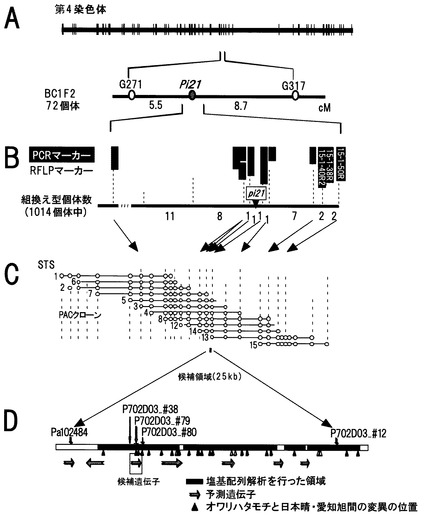
【図3】
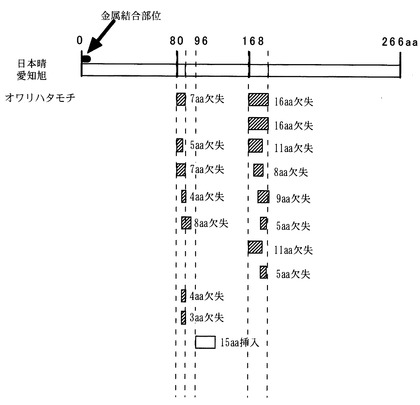
【図1】

【図4】
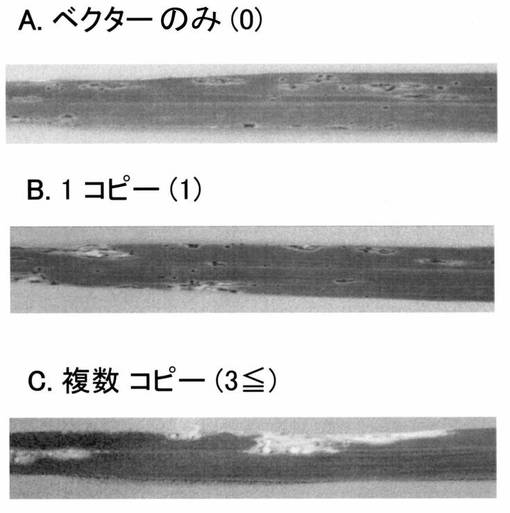
【図3】
【図1】
【図4】
【公開番号】特開2011−160806(P2011−160806A)
【公開日】平成23年8月25日(2011.8.25)
【国際特許分類】
【出願番号】特願2011−58457(P2011−58457)
【出願日】平成23年3月16日(2011.3.16)
【分割の表示】特願2005−187867(P2005−187867)の分割
【原出願日】平成17年6月28日(2005.6.28)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
【公開日】平成23年8月25日(2011.8.25)
【国際特許分類】
【出願日】平成23年3月16日(2011.3.16)
【分割の表示】特願2005−187867(P2005−187867)の分割
【原出願日】平成17年6月28日(2005.6.28)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
[ Back to top ]