
イネ品種コシヒカリの染色体の一部が、オリザ・グラベリマの染色体断片で置換された染色体部分置換系統群
【課題】イネ品種コシヒカリの染色体の一部が、オリザ・グラベリマの染色体断片で置換された染色体部分置換系統群からなるゲノムライブラリーの提供。
【解決手段】34種類の、イネ品種コシヒカリの染色体の一部がオリザ・グラベリマの染色体断片で置換された染色体部分置換系統を有することを特徴とするゲノムライブラリー、及び、オリザ・グラベリマの染色体中、前記34種のいずれかの染色体部分置換系統の染色体中に含有されている領域が、全染色体の90%以上であることを特徴とする前記記載のゲノムライブラリー。
【解決手段】34種類の、イネ品種コシヒカリの染色体の一部がオリザ・グラベリマの染色体断片で置換された染色体部分置換系統を有することを特徴とするゲノムライブラリー、及び、オリザ・グラベリマの染色体中、前記34種のいずれかの染色体部分置換系統の染色体中に含有されている領域が、全染色体の90%以上であることを特徴とする前記記載のゲノムライブラリー。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、イネ品種コシヒカリの染色体の一部が、アフリカ栽培品種オリザ・グラベリマの染色体断片で置換された染色体部分置換系統群からなるゲノムライブラリーに関する。
【背景技術】
【0002】
オリザ・グラベリマ(Oryza glaberrima Steud)は野生種のオリザ・バータイ(Oryza barthii)から栽培化された西アフリカに固有のイネの栽培品種である。より詳細には、オリザ・グラベリマは約3,500年前に、西アフリカの中でも主にナイジェリアのリバーデルタの湿潤地帯で栽培化された後、紀元前1,000年頃にギニアの海岸近くで第2の栽培化が起こったと考えられている。アジアのイネの栽培品種であるオリザ・サティバ(Oryza sativa L.)と比較すると、オリザ・グラベリマは、遺伝的に多様性が少なく(非特許文献7、37、及び31参照。)、栽培分布も狭く、地域起源を越えては広がっていない(非特許文献30及び21参照。)。これにもかかわらず、オリザ・グラベリマは、人の介入なしに過酷な西アフリカの環境で成長させる多くの新規遺伝子を潜在的に持っている(非特許文献20参照。)。雑草競争、耐干性、低い成長投入量に対応する能力、不利な生態系への広い適合性はアフリカ型イネ固有の特徴である(非特許文献29、6、25、13、19、20、及び8参照。)。このイネの品種はモザイク病(非特許文献28、1、及び5参照。)、枯れ病(非特許文献32、30、及び12参照。)、もん枯れ病(非特許文献12及び36参照。)、アフリカ小昆虫(非特許文献36及び20参照。)を含むいくつかの生物的ストレスにも耐性を持っている。
【0003】
現在までに、アフリカ型イネでの重要な特徴に対するいくつかのQTL(量的形質座)が既に同定されている(非特許文献24、2、及び23参照。)。一般的にF1若しくはF2/F3系統、組換え近交系、倍加半数系、戻し交配が、これらの特徴の検出に使われている。しかしながら、F1系統の使用により成功する場合の多くは、主要な効果を持つQTLを検出する場合のみである。表現型の効果が弱いQTLと他の遺伝子との相互関係をもつエピスタシスは、これらの系統では容易に検出されない。興味深いQTLの検出するのを成功させるためには、準同質系統をQTLクローニングとマッピングのために開発する必要がある(非特許文献33、11、39、及び40参照。)。QTL検出とマッピングは、多大労力と多くの時間を要するため、遺伝子クローニングのスタンダードなストラテジーとしてマップベースクローニングへの使用に限られている。
【0004】
これらの課題に取り組むため、イネにおいても、染色体部分置換系統群の形で遺伝子資源が開発された(非特許文献3、11、17、及び10参照。)。CSSLsは、ドナー系統を受容系統と交雑させ、F1及びBCnF1を反復親に戻し交配することによって作出される。一般的には、野生イネ又は一般的な栽培品種がドナー系統となり、通常、優秀な系統が受容系統となる。これらの繰返し戻し交配及びその後の3〜4後代世代の育成によって得られた後代個体は、反復親の遺伝的背景中に、1又は2のホモ接合性のドナー系統の遺伝子型の染色体断片が含まれている。このように、ドナー系統の全ゲノムは、互いに連続的又はオーバーラップする(重なり部分がある)小さな染色体断片とし、これらが導入されている選択された系統群であらわされることができる(非特許文献14及び39参照。)。
【0005】
遺伝的に多様なイネ品種と野生型イネ品種とからなる染色体部分置換系統(以下、CSSLと略記することがある。)は、重要な農業形質を制御する新規な遺伝子の同定に有用な遺伝資源である。特に、CSSLは、複合的、量的形質を解析のターゲットとする場合に有効である。CSSLは、表現型と導入された染色体断片とが強い相関を有するため、個々の座の本当の表現型の影響をわかりにくくする表現型のノイズとして知られているQTL間の干渉の影響が、取り除かれるためである。ドナーゲノムの非常に短い、連続した又はオーバーラップする染色体断片を含む一連のCSSLからなる染色体置換系統群(以下、CSSLsと略記することがある。)セットは、均一の遺伝的背景を有するため、CSSLsセットを用いることにより、単純な統計分析を通じてQTLを同定することができる(非特許文献27及び14参照。)。
【0006】
一方で近年、ゲノム解析技術は格段の進歩を遂げ、作物の改良を、高精度かつ容易に行えるようになった。とりわけ、DNAマーカー技術の進捗は著しく、同技術によって、有用な形質を備える新品種を作製することができるようになった。また、DNAマーカーを利用して、これまでに同定された重要遺伝子の有用アレルを含む染色体領域を置換することにより、他の多くの形質に対してほとんど影響を及ぼさずに、目的形質を特異的に改良することが出来るようになった(例えば、特許文献1参照。)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許第4409610号公報
【非特許文献】
【0008】
【非特許文献1】Abo, E. M., et al. (1998) J. Sustain. Agric. Vol.11: pp.85-111.
【非特許文献2】Aluko, G., C. et al. (2004) Theor. Appl. Genet. Vol.109: pp.630-639.
【非特許文献3】Ando, T., et al. (2008) Theor. Appl. Genet. Vol.116: pp.881-890.
【非特許文献4】Ashikari, M., et al. (2005) Science. Vol.309: pp.741-745.
【非特許文献5】Attere, C. and C. A. Fatokun (1983) Plant Divers. Vol.67: pp.420-421.
【非特許文献6】Baggie, I., et al. (2002) Int. Rice Res. Notes. Vol.27: pp.42.
【非特許文献7】Barry, M. B., et al. (2007) Euphytica Vol.154: pp.127-137.
【非特許文献8】Dingkuhn, M., et al. (1998) Field Crops Res. Vol.57: pp.57-69.
【非特許文献9】Doi, K., et al. (1998) Breed Science Vol.49: pp.395-399.
【非特許文献10】Doi, K., et al. (1997) Rice Genet. Newsl. Vol.14: pp.39-41.
【非特許文献11】Ebitani, T., et al. (2005). Breeding Science Vol.55: pp.65-73.
【非特許文献12】Eizenga, G. C., et al. (2006) Crop Sci. Vol.46: pp.1870-1878.
【非特許文献13】Fofana, B. and R. Rauber (2000) Weed Res. Vol.40: pp.271-280
【非特許文献14】Gutierrez, A. G., et al. (2010) BMC Plant Biol. Vol.10: pp.6.
【非特許文献15】Hu, F., et al. (2006) Euphytica Vol.151: pp.273-278.
【非特許文献16】Ikeda, K., M. et al. (2007) Plant Journal Vol.51(6): pp.1030-1040.
【非特許文献17】Kubo, T., et al. (2002) Breed. Sci. Vol.52: pp.319-325.
【非特許文献18】Jiao, Y., et al. (2010) Nat. Genet. Vol.42: pp.541-544.
【非特許文献19】Johnson, D. E., et al. (1998) Weed Res. Vol.38: pp.207-216.
【非特許文献20】Jones, M. P., et al. (1997) Breeding Science Vol.47: pp.395-398.
【非特許文献21】Khush, G.S. (1997) Plant Mol. Biol. Vol.35: pp.25-34.
【非特許文献22】Li, J., et al. (2008) Euphytica Vol.164: pp.699-708.
【非特許文献23】Li, J., et al. (2004) Genome Vol.47: pp.697-704.
【非特許文献24】Lorieux, M., et al. (2000) Theor. Appl. Genet. Vol.100: pp.593-601.
【非特許文献25】Maji, T., et al. (2001) Oryza Vol.38: pp.17-23.
【非特許文献26】Miura, K., et al. (2010) Nat. Genet. Vol.42: pp.545-549.
【非特許文献27】Nadeau, J. H., et al. (2000) Nat. Genet. Vol.24: pp.221-225.
【非特許文献28】Ndjiondjop, M. N., et al. (2001) Mol. Plant. Pathol. Vol.59: pp.309-316.
【非特許文献29】Sahrawat, K. L. and M. Sitka (2002) Res. Notes Vol.27: pp.30-31.
【非特許文献30】Sarla, N. and B. P. M Swamy (2005) Curr. Sci. Vol.89(6): pp.955-963.
【非特許文献31】Second, G. (1986) The International Rice Research Institute, Los Banos, Laguna, Philippines. pp.27-39.
【非特許文献32】Silue, D. and J. Notteghem (1991) Int. Rice Res. Notes Vol.16: pp.13-14.
【非特許文献33】Takai, T., et al. (2007) Breed. Sci. Vol.57: pp.257-261.
【非特許文献34】Taguchi, K. K. D. and A. Yoshimura (1999) Rice Genet. News. Vol.16: pp.70-71
【非特許文献35】Terao, T., et al. (2010) Theor. Appl. Genet. 120(5): pp.875-893.
【非特許文献36】Ukwungwu, M. N., et al. (1999) Journ. Food. Tech. Afr. Vol.4(3): pp.108-110.
【非特許文献37】Wang, Z., et al. (1992) Theor. Appl. Genet. Vol.83: pp.565-581.
【非特許文献38】Wasano, K. and Y. Hirota (1986) Bull. Fac. Agr. Saga Univ. Vol.60: pp.49-59.
【非特許文献39】Yano, M. (2001) Curr. Opin. Plant Biol. Vol.4: pp.130-135.
【非特許文献40】Yano, M. and T. Sasaki (1997) Plant Mol. Biol. Vol.35: pp.145-153.
【発明の概要】
【発明が解決しようとする課題】
【0009】
オリザ・グラベリマをはじめとする外国稲は、遺伝資源としての潜在能力に期待はあるものの、劣悪形質が多いため、これまでの育種事業では病害抵抗性のような主導遺伝子の利用にとどまっている。
【0010】
本発明は、オリザ・サティバ育種のための遺伝資源として、オリザ・サティバの優秀な栽培種であるジャポニカ栽培種コシヒカリ(以下、イネ品種コシヒカリ)の遺伝的背景に、多くの新規遺伝子を潜在的に持つ可能性のあるオリザ・グラベリマの染色体断片を置換したイネ品種コシヒカリの染色体部分置換系統群を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者は、上記課題を解決すべく鋭意研究した結果、イネ品種コシヒカリとオリザ・グラベリマとを識別するための新規なSNPからなるDNAマーカーを開発し、これらのDNAマーカーを利用して、イネ品種コシヒカリの遺伝的背景に、オリザ・グラベリマの染色体断片が置換された34系統の染色体部分置換系統群を作出し、本発明を完成させた。
【0012】
すなわち、本発明は、下記(1)〜(34)の、イネ品種コシヒカリの染色体の一部がオリザ・グラベリマの染色体断片で置換された染色体部分置換系統を有することを特徴とするゲノムライブラリーを提供するものである。
(1)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(2)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の11,026,011番目の塩基から20,366,961番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の8,896,816番目の塩基から11,026,011番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の20,366,961番目の塩基から24,412,980番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(3)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の17,803,741番目の塩基から31,371,175番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の13,813,982番目の塩基から17,803,741番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の31,371,175番目の塩基から35,598,471番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(4)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の25,979,477番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の24,412,982番目の塩基から25,979,477番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の35,598,472番目の塩基から39,677,013番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(5)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の35,598,472番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の31,371,176番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(6)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の上流末端から3,922,571番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第2染色体の3,922,571番目の塩基から7,030,896番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(7)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の3,922,571番目の塩基から14,655,638番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の2,010,335番目の塩基から3,922,571番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の14,655,638番目の塩基から18,765,983番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(8)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の11,165,933番目の塩基から22,086,348番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の9,256,585番目の塩基から11,165,933番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の22,086,348番目の塩基から25,263,252番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(9)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の18,765,984番目の塩基から25,263,253番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の14,655,639番目の塩基から18,765,984番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の25,263,253番目の塩基から27,919,902番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(10)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の30,860,986番目の塩基から33,189,264番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の29,345,358番目の塩基から30,860,986番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の33,189,264番目の塩基から35,267,218番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(11)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の上流末端から11,073,623番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第3染色体の11,073,623番目の塩基から14,311,069番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(12)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の11,073,623番目の塩基から33,640,115番目の塩基までを 含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第3染色体の8,197,229番目の塩基から11,073,623番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第3染色体の33,640,115番目の塩基から35,945,010番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(13)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の上流末端から22,125,293番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第4染色体の22,125,293番目の塩基から25,877,140番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(14)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の19,650,008番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第4染色体の14,903,263番目の塩基から19,650,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(15)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の上流末端から5,346,476番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第5染色体の5,346,476番目の塩基から7,305,955番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(16)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の3,173,133番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の1,047,931番目の塩基から3,173,133番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第5染色体の21,023,551番目の塩基から24,520,268番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(17)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の21,023,551番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の18,878,115番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(18)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の上流末端から2,186,456番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第6染色体の2,186,456番目の塩基から5,904,223番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(19)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の2,186,456番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の372,259番目の塩基から2,186,456番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第6染色体の22,907,151番目の塩基から25,711,514番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(20)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の22,907,151番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の20,235,923番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(21)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の上流末端から6,843,696番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第7染色体の6,843,696番目の塩基から10,130,031番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(22)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の1,745,458番目の塩基から18,571,959番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第7染色体の260,865番目の塩基から1,745,458番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第7染色体の18,571,959番目の塩基から21,942,075番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(23)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の21,942,076番目の塩基から24,516,006番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第1の染色体断片にホモ置換されており、
当該第7染色体の28,847,226番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第2の染色体断片にホモ置換されており、
かつ前記第1の染色体断片の上流端が、イネ品種日本晴の第7染色体の18,571,960番目の塩基から21,942,076番目の塩基までを含む領域に相当する領域に存在し、前記第1の染色体断片の下流端が、イネ品種日本晴の第7染色体の24,516,006番目の塩基から25,490,052番目の塩基までを含む領域に相当する領域に存在し、
前記第2の染色体断片の上流端が、イネ品種日本晴の第7染色体の25,490,054番目の塩基から28,847,226番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(24)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の上流末端から8,094,582番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第8染色体の8,094,582番目の塩基から10,572,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(25)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の8,094,582番目の塩基から19,903,905番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の5,278,856番目の塩基から8,094,582番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第8染色体の19,903,905番目の塩基から23,047,466番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(26)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の18,114,324番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の15,484,341番目の塩基から18,114,324番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(27)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の上流末端から13,254,003番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第9染色体の13,254,003番目の塩基から15,063,824番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(28)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の13,254,003番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第9染色体の10,483,156番目の塩基から13,254,003番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(29)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の上流末端から14,142,299番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第10染色体の14,142,299番目の塩基から17,555,158番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(30)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の14,142,299番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第10染色体の11,761,139番目の塩基から14,142,299番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(31)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の上流末端から21,896,773番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第11染色体の21,896,773番目の塩基から25,149,243番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(32)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の21,896,773番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第11染色体の19,050,306番目の塩基から21,896,773番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(33)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の上流末端から14,689,883番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にヘテロ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第12染色体の14,689,883番目の塩基から17,499,934番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(34)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の21,257,971番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第12染色体の17,499,936番目の塩基から21,257,971番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統。
【発明の効果】
【0013】
本発明のゲノムライブラリー、及びこれを構成する各染色体部分置換系統は、イネ品種コシヒカリの遺伝的背景に、従来、イネ品種コシヒカリの形質改善には利用されてこなかったオリザ・グラベリマの染色体断片が組み込まれている。このため、本発明のゲノムライブラリー、及びこれを構成する各染色体部分置換系統を用いることにより、オリザ・グラベリマが有する様々な遺伝子や形質を、遺伝的背景を揃えた条件で精度よく解析することができる。さらに、本発明のゲノムライブラリーは、オリザ・グラベリマが有している好ましい農業形質を制御するQTLの検出や、当該形質の原因となる新規遺伝子の同定、新品種育成等に好適に利用することができる。
【図面の簡単な説明】
【0014】
【図1】本発明のゲノムライブラリーを構成する34系統(GLSL−1〜GLSL−34)のゲノムを模式的に示した図である。
【図2】本発明のゲノムライブラリーを構成する34系統(GLSL−1〜GLSL−34)の作出工程を模式的に示した図である。
【発明を実施するための形態】
【0015】
本発明及び本願明細書において「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域」は、米国ゲノム研究所(TIGR)によるRice Genome Anootation Databaseにおいて公開されているイネ品種日本晴のゲノムDNAの塩基配列(バージョン2)に基づいて決定される領域である。
【0016】
また、本発明及び本願明細書において、「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域に相当する領域」とは、イネ個体の染色体中のイネ品種日本晴の染色体中の当該領域と相同性の高い領域であり、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントすることにより決定することができる。また、イネ品種日本晴以外のイネ個体中の「イネ品種日本晴のSNP(Single Nucleotide Polymorphism、一遺伝子多型)に相当するSNP」は、当該SNPを含む領域において、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントした場合に、当該SNPに対応する位置にある塩基を意味する。
【0017】
本発明において品種とは、同一種の植物であって、遺伝的構成が異なるために、ある形質において同種内の他品種から明確に識別し得る集団を意味する。また、イネ品種日本晴及びイネ品種コシヒカリの遺伝子情報等は、例えば、国際的な塩基配列データベースであるNCBI(National center for Biotechnology Information)やDDBJ(DNA Data Bank of Japan)等において入手することができる。特にイネの各品種の遺伝子情報は、KOME(Knowledge−based Oryza Molecular biological Encyclopedia、http://cdna01.dna.affrc.go.jp/cDNA/)等において入手することができる。
【0018】
本発明及び本願明細書において、DNAマーカーとは、種や品種が異なる生物由来の染色体断片同士を識別し得る染色体上のDNA配列の差異を検出し得るものである。具体的には、SNPである。
【0019】
DNAマーカーによる、ある品種由来アレルとその他の種又は品種由来アレルとの識別は、常法により行うことができる。例えば、各個体から抽出したDNAを鋳型とし、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等を用いてPCRを行い、電気泳動法等を用いてPCR産物の有無を検出し、各多型を識別することができる。また、各個体から抽出したDNAを制限酵素処理した後、電気泳動法等を用いてDNA断片のパターンを検出し、各多型を識別することができる。なお、特定のSNPと特異的にハイブリダイズし得るプライマー等は、該SNPの塩基配列に応じて、汎用されているプライマー設計ツール等を用いて常法により設計することができる。また、設計されたプライマー等は、当該技術分野においてよく知られている方法のいずれを用いても合成することができる。
なお、これらのDNAマーカーによる、ある品種由来アレルとその他の種又は品種由来アレルとの識別は、常法により行うことができる。例えば、各個体から抽出したDNAを鋳型とし、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等を用いてPCRを行い、電気泳動法等を用いてPCR産物の有無を検出し、各多型を識別することができる。また、各個体から抽出したDNAを制限酵素処理した後、電気泳動法等を用いてDNA断片のパターンを検出し、各多型を識別することができる。なお、特定のSNPと特異的にハイブリダイズし得るプライマー等は、該SNPの塩基配列に応じて、汎用されているプライマー設計ツール等を用いて常法により設計することができる。また、設計されたプライマー等は、当該技術分野においてよく知られている方法のいずれを用いても合成することができる。
【0020】
<DNAマーカー>
本発明者らは、まず、オリザ・グラベリマ由来の染色体断片とイネ品種コシヒカリ由来の染色体断片とを識別するために、イネ品種日本晴のゲノムの塩基配列に基づいて、第1〜第12染色体の全染色体領域において、イネ品種コシヒカリとオリザ・グラベリマの間で多型を示すSNPからなる142のDNAマーカーを作製した。DNAマーカー間の距離は、平均2.75Mbであった。各DNAマーカーの位置を、表1〜4に示す。各表の染色体中の位置情報は、イネ品種日本晴のゲノム中の相当するSNPの位置である。なお、オリザ・グラベリマの遺伝子解析はほとんど行われておらず、これらの142のDNAマーカーは、本発明者らによって初めて見出されたものである。
【0021】
【表1】

【0022】
【表2】
【0023】
【表3】
【0024】
【表4】
【0025】
表1〜4に記載のDNAマーカーは、染色体中にイネ品種コシヒカリ由来の染色体断片とオリザ・グラベリマ由来の染色体断片の両方が含まれているイネ個体のジェノタイピング(遺伝子型の同定)に特に好適に用いられる。イネ個体のジェノタイピングにおいて、表1〜4に記載のDNAマーカーの遺伝子型をタイピングすることによって、得られたタイピング結果から、当該イネ個体の染色体中に、イネ品種コシヒカリ由来の染色体断片又はオリザ・グラベリマ由来の染色体断片の有無や、当該断片が存在する染色体中の位置等を調べることができる。
【0026】
<染色体部分置換系統(CSSL)及びゲノムライブラリー>
本発明のゲノムライブラリーは、イネ品種コシヒカリの染色体が、部分的にオリザ・グラベリマの染色体断片で置換された34系統(GLSL−1〜GLSL−34)のCSSLsを有する。本発明のゲノムライブラリー中のCSSLがそれぞれ含有するオリザ・グラベリマ由来の染色体断片は、互いに連続的又はオーバーラップしており、34系統全てを合わせるとオリザ・グラベリマの全ゲノムの90%以上が保持されている。
【0027】
本発明のゲノムライブラリーを構成するCSSLは、1セットとして使用することができ、それぞれ単独で使用してもよい。これらのCSSLは、イネの粒特性や他の有用な農業的特徴を調節するQTLの検出に特に有用である。また、検出されたQTLを利用することにより、育種の幅が広がることも期待できる。例えば、検出されたQTLのピラミティングにより、新規な品種を容易に育種することが可能になる。さらに、本発明のゲノムライブラリーを構成する各CSSLは、染色体中のオリザ・グラベリマ由来の染色体断片の位置が、前記の142のDNAマーカーによって明らかであるため、育種の材料としても好適である。
【0028】
図1に、本発明のゲノムライブラリーを構成する34系統(GLSL−1〜GLSL−34)のゲノムを模式的に示す。図1の各染色体中、白抜き部分はイネ品種コシヒカリのホモ接合型の染色体領域を、黒塗部分はオリザ・グラベリマのホモ接合型の染色体領域を、ハッチング部分はイネ品種コシヒカリとオリザ・グラベリマのヘテロ接合型の染色体領域を、それぞれ表す。
【0029】
オリザ・グラベリマの第1染色体及び第2染色体の染色体断片は、それぞれ5のCSSLに置き換えられており、第3染色体〜第12染色体の染色体断片は2又は3のCSSLに置き換えられていた。ドナーゲノム(オリザ・グラベリマのゲノム)のうちのおよそ6%は、34系統(GLSL−1〜GLSL−34)(以下、GLSLセット、ということがある。)のいずれにおいても、イネ品種コシヒカリの相当する領域に置換されなかった。この置換されなかった領域は、図1各染色体において、GLSL−1〜GLSL−34のいずれにおいても黒塗部分が存在していない、小さなギャップとして示される。これらのギャップは、具体的には、以下の6部分領域である;第1染色体中のSP−185を含み、かつSP−2077及びSP−192を含まない部分領域;第2染色体中のSP−1444及びSP−1450を含み、かつSP−2941及びSP−1459を含まない部分領域;第2染色体中のSP−1471よりも下流の、SP−3571を含む末端部分領域;第3染色体中のSP−348よりも下流の、SP−362を含む末端部分領域;第7染色体中のSP−741を含み、かつSP−2545及びSP−4152を含まない部分領域;第12染色体中のSP−3747を含み、かつSP−2687及びSP−2697を含まない部分領域。
【0030】
また、GLSLセットのうちの33系統は、オリザ・グラベリマ由来の染色体断片がホモ置換されている(ホモ接合型で置換されている)が、第12染色体に置換が生じているGLSL−33のみは、オリザ・グラベリマ由来の染色体断片がヘテロ(ヘテロ接合型で)置換されている。GLSL−33中のオリザ・グラベリマ由来の染色体断片の長さは約17.5Mbであり、当該染色体断片の両脇にはDNAマーカーSP−5151及びSP−2687が存在する。
【0031】
GLSL−1は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基(SP−2077)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基(SP−185の−1塩基(1塩基上流の塩基))までを含む領域に相当する領域に存在しているCSSLである。
GLSL−2は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の11,026,011番目の塩基(SP−192)から20,366,961番目の塩基(SP−2016)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の8,896,816番目の塩基(SP−185の+1塩基(1塩基下流の塩基))から11,026,011番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の20,366,961番目の塩基から24,412,980番目の塩基(SP−1217の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−3は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の17,803,741番目の塩基(SP−1208)から31,371,175番目の塩基(SP−262)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の13,813,982番目の塩基(SP−2430の+1塩基)から17,803,741番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の31,371,175番目の塩基から35,598,471番目の塩基(SP−1244の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−4は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の25,979,477番目の塩基(SP−2478)から35,598,472番目の塩基(SP−1244)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の24,412,982番目の塩基(SP−1217の+1塩基)から25,979,477番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の35,598,472番目の塩基から39,677,013番目の塩基(SP−2056の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−5は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の35,598,472番目の塩基(SP−1244)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の31,371,176番目の塩基(SP−262の+1塩基)から35,598,472番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0032】
GLSL−6は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の上流末端から3,922,571番目の塩基(SP−1387)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第2染色体の3,922,571番目の塩基から7,030,896番目の塩基(SP−1398の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−7は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の3,922,571番目の塩基(SP−1387)から14,655,638番目の塩基(SP−1413)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の2,010,335番目の塩基(SP−6の+1塩基)から3,922,571番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の14,655,638番目の塩基から18,765,983番目の塩基(SP−2495の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−8は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の11,165,933番目の塩基(SP−1406)から22,086,348番目の塩基(SP−954)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の9,256,585番目の塩基(SP−29の+1塩基)から11,165,933番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の22,086,348番目の塩基から25,263,252番目の塩基(SP−2941の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−9は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の18,765,984番目の塩基(SP−2495)から25,263,253番目の塩基(SP−2941)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の14,655,639番目の塩基(SP−1413の+1塩基)から18,765,984番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の25,263,253番目の塩基から27,919,902番目の塩基(SP−1444の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−10は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の30,860,986番目の塩基(SP−1459)から33,189,264番目の塩基(SP−1471)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の29,345,358番目の塩基(SP−1450の+1塩基)から30,860,986番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の33,189,264番目の塩基から35,267,218番目の塩基(SP−3571の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
【0033】
GLSL−11は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の上流末端から11,073,623番目の塩基(SP−1323)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第3染色体の11,073,623番目の塩基から14,311,069番目の塩基(SP−1334の−1塩基)までを含む領域に相当する領域に存在している染色体部分置換系統CSSLである。
GLSL−12は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の11,073,623番目の塩基(SP−1323)から33,640,115番目の塩基(SP−348)までを 含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第3染色体の8,197,229番目の塩基(SP−1308の+1塩基)から11,073,623番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第3染色体の33,640,115番目の塩基から35,945,010番目の塩基(SP−362の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−13は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の上流末端から22,125,293番目の塩基(SP−392)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第4染色体の22,125,293番目の塩基から25,877,140番目の塩基(SP−402の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−14は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の19,650,008番目の塩基(SP−386)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第4染色体の14,903,263番目の塩基(SP−3250の+1塩基)から19,650,008番目の塩基までを含む領域に相当する領域に存在している
【0034】
GLSL−15は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の上流末端から5,346,476番目の塩基(SP−517)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第5染色体の5,346,476番目の塩基から7,305,955番目の塩基(SP−1526の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−16は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の3,173,133番目の塩基(SP−505)から21,023,551番目の塩基(SP−547)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の1,047,931番目の塩基(SP−495の+1塩基)から3,173,133番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第5染色体の21,023,551番目の塩基から24,520,268番目の塩基(SP−1561の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−17は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の21,023,551番目の塩基(SP−547)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の18,878,115番目の塩基(SP−1546の+1塩基)から21,023,551番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
GLSL−18は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の上流末端から2,186,456番目の塩基(SP−2503)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第6染色体の2,186,456番目の塩基から5,904,223番目の塩基(SP−1596の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−19は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の2,186,456番目の塩基(SP−2503)から22,907,151番目の塩基(SP−1613)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の372,259番目の塩基(SP−590の+1塩基)から2,186,456番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第6染色体の22,907,151番目の塩基から25,711,514番目の塩基(SP−3026の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−20は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の22,907,151番目の塩基(SP−1613)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の20,235,923番目の塩基(SP−3022の+1塩基)から22,907,151番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0035】
GLSL−21は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の上流末端から6,843,696番目の塩基(SP−1648)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第7染色体の6,843,696番目の塩基から10,130,031番目の塩基(SP−3066の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−22は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の1,745,458番目の塩基(SP−2315)から18,571,959番目の塩基(SP−1662)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第7染色体の260,865番目の塩基(SP−676の+1塩基)から1,745,458番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第7染色体の18,571,959番目の塩基から21,942,075番目の塩基(SP−3095の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−23は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の21,942,076番目の塩基(SP−3095)から24,516,006番目の塩基(SP−2545)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第1の染色体断片にホモ置換されており、当該第7染色体の28,847,226番目の塩基(SP−4152)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第2の染色体断片にホモ置換されており、かつ前記第1の染色体断片の上流端が、イネ品種日本晴の第7染色体の18,571,960番目の塩基(SP−1662の+1塩基)から21,942,076番目の塩基までを含む領域に相当する領域に存在し、前記第1の染色体断片の下流端が、イネ品種日本晴の第7染色体の24,516,006番目の塩基から25,490,052番目の塩基(SP−741の−1塩基)までを含む領域に相当する領域に存在し、前記第2の染色体断片の上流端が、イネ品種日本晴の第7染色体の25,490,054番目の塩基(SP−741の+1塩基)から28,847,226番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
GLSL−24は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の上流末端から8,094,582番目の塩基(SP−795)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第8染色体の8,094,582番目の塩基から10,572,008番目の塩基(SP−1707の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−25は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の8,094,582番目の塩基(SP−795)から19,903,905番目の塩基(SP−3199)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の5,278,856番目の塩基(SP−3598の+1塩基)から8,094,582番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第8染色体の19,903,905番目の塩基から23,047,466番目の塩基(SP−2407の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−26は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の18,114,324番目の塩基(SP−809)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の15,484,341番目の塩基(SP−806の+1塩基)から18,114,324番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0036】
GLSL−27は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の上流末端から13,254,003番目の塩基(SP−2180)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第9染色体の13,254,003番目の塩基から15,063,824番目の塩基(SP−1747の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−28は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の13,254,003番目の塩基(SP−2180)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第9染色体の10,483,156番目の塩基(SP−875の+1塩基)から13,254,003番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
GLSL−29は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の上流末端から14,142,299番目の塩基(SP−966)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第10染色体の14,142,299番目の塩基から17,555,158番目の塩基(SP−981の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−30は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の14,142,299番目の塩基(SP−966)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第10染色体の11,761,139番目の塩基(SP−3357の+1塩基)から14,142,299番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0037】
GLSL−31は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の上流末端から21,896,773番目の塩基(SP−1073)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第11染色体の21,896,773番目の塩基から25,149,243番目の塩基(SP−2678の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−32は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の21,896,773番目の塩基(SP−1073)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第11染色体の19,050,306番目の塩基(SP−1062の+1塩基)から21,896,773番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
GLSL−33は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の上流末端から14,689,883番目の塩基(SP−2687)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にヘテロ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第12染色体の14,689,883番目の塩基から17,499,934番目の塩基(SP−3747の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−34は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の21,257,971番目の塩基(SP−2697)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第12染色体の17,499,936番目の塩基(SP−3747の+1塩基)から21,257,971番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0038】
GLSL−1〜GLSL−34は、後記実施例に記載の方法で作出することができるが、その他、公知のいずれの手法により作成したものであってもよい。例えば、イネ品種コシヒカリとオリザ・グラベリマを交配し、得られた後代個体に対して自家交配又は戻し交配を繰返し、得られた後代個体をジェノタイピングし、その結果に基づいて、染色体中の所望の領域のみがオリザ・グラベリマ由来の染色体領域であり、その他がイネ品種コシヒカリ由来の染色体で構成されているイネ個体を選抜することにより、GLSL−1〜GLSL−34を作出することができる。選抜に用いるDNAマーカーは、表1〜4に記載されているものを用いてもよく、その他のDNAマーカーを用いてもよい。
【0039】
例えば、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基までを含む領域に相当する領域(当該領域の下流端は、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基までを含む領域に相当する領域に存在している。)のみがオリザ・グラベリマ由来の染色体断片にホモ置換されているイネ個体GLSL−1は、図1中のGLSL−1の染色体中で黒塗部分に存在するSNP(SP−1177、SP−155、SP−2077)が両アレルともにオリザ・グラベリマタイプであり、白抜部分に存在するその他のSNPが両アレルともイネ品種コシヒカリタイプであるイネ個体を選抜することにより得られる。
【0040】
同様に、GLSL−2〜GLSL−32及びGLSL−34も、図1中の各染色体中で黒塗部分に存在するSNPが両アレルともにオリザ・グラベリマタイプであり、白抜部分に存在するその他のSNPが両アレルともにイネ品種コシヒカリタイプであるイネ個体を選抜することにより得られる。また、GLSL−33は、図1中の染色体中でハッチング部分に存在するSNPが、一方のアレルがオリザ・グラベリマタイプ、他方のアレルがイネ品種コシヒカリタイプであり、白抜部分に存在するその他のSNPが両アレルともにイネ品種コシヒカリタイプであるイネ個体を選抜することにより得られる。各染色体中の黒塗部分(又はハッチング部分)に存在するSNPを、表5に示す。
【0041】
【表5】
【0042】
GLSL−1〜GLSL−34は、イネ品種コシヒカリと同様の手法により、栽培し、自家交配や人工交配により米を収穫することができる。また、GLSL−1〜GLSL−34及びその後代個体は、イネ品種コシヒカリと同様に、新品種育成の親個体とすることができる。例えば、GLSL−1〜GLSL−34中の互いに異なるCSSL同士を交配したり、GLSL−1〜GLSL−34の各個体と別の品種の個体とを交配することによって、後代個体を得ることができる。さらに、得られた後代個体を、自殖したり、GLSL−1〜GLSL−34の個体と戻し交配することにより、新品種の育種を試みることもできる。
【0043】
また、GLSL−1〜GLSL−34は、公知の染色体部分置換系統と同様に、新品種育成のための材料として用いることができる。GLSL−1〜GLSL−34を材料とした場合の新品種の育種方法としては、例えば、特許文献1等に記載の新品種の製造方法等が挙げられる。
【0044】
鑑別対象であるイネ個体に対して、表1〜4に記載のDNAマーカーの1又は2以上についてジェノタイピングし、得られたタイピング結果に基づき、当該イネ個体が、GLSL−1〜GLSL−34のいずれかであるかを鑑別することができる。表1〜4に記載の142種類全てのDNAマーカーを用いることによって、確実に鑑別することができる。また、142種類全てのDNAマーカーを用いずに鑑別してもよい。全てのDNAマーカーを用いない場合には、鑑別に用いるDNAマーカーは、表5に記載のDNAマーカーを用いることが好ましく、図1の黒塗部分(又はハッチング部分)と白抜き部分との境界付近のものであることがより好ましい。
【0045】
例えば、あるイネ個体がGLSL−2であるか否かは、黒塗部分中の白抜き部分との境界に近いSP−192及びSP−2016、並びに白抜き部分中の黒塗部分との境界に近いSP−185及びSP−1217について遺伝子型をタイピングする。得られたタイピング結果が、SP−192及びSP−2016が両アレルともにオリザ・グラベリマタイプであり、SP−185及びSP−1217が両アレルともにイネ品種コシヒカリタイプであった場合には、当該イネ個体はGLSL−2であり、それ以外のタイピング結果の場合には、当該イネ個体はGLSL−2ではない、と鑑別できる。その他のCSSLについても同様にして鑑別することができる。
【実施例】
【0046】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0047】
[実施例1]
<ゲノムライブラリーの製造>
優秀なジャポニカイネ栽培種であるイネ品種コシヒカリを反復親(種子親)とし、アフリカ型品種オリザ・グラベリマをドナー親(花粉親)として、交配することによって、GLSL−1〜GLSL−34を作出した。また、以下の実施例において、GLSL−1〜GLSL−34を纏めて「GLSLs」といい、個々のCSSLをGLSLということがある。
【0048】
具体的には、まず、イネ品種コシヒカリとオリザ・グラベリマを交配し、得られたF1交雑種を反復親イネ品種コシヒカリと戻し交配してBC1F1世代(54個体)を得、続く反復親との連続戻し交配により、BC2F1世代(54系統の個体)、BC3F1世代(155系統の個体)、BC4F1世代(155系統の個体)、BC5F1世代(112系統の個体)を得た。
次いで、13系統のBC3F1世代、155系統のBC4F1世代、及び112系統のBC5F1世代から、それぞれ10個体ずつ育成し、表1〜4に記載の142種のSNPを、AcycloPrime−FD検出システムと蛍光ポラライゼーションアナライザー(Perkin Elmer Life Sciences社製)によってジェノタイピングした。タイピング結果に基づいて、オリザ・グラベリマ由来の染色体断片を2以上含むGLSLの候補個体として、BC3F1世代から2個体、BC4F1世代から17個体、BC5F1世代から5個体を選抜した。さらにこれらの選抜した個体を戻し交配し、それぞれBC4F1世代、BC5F1世代、又はBC6F1世代を得た。
これらの集団のうち、未だオリザ・グラベリマ由来の染色体断片を2以上含んでいたGLSLの候補個体に対して、反復親との戻し交配を行い、それぞれBC5F1世代、BC6F1世代、又はBC7F1世代を得た。戻し交配により得られたGLSLの候補個体を自殖し、ドナーゲノム(オリザ・グラベリマ)由来の染色体部分置換を固定した。
以上の工程により、本発明のゲノムライブラリーを構成するGLSL−1〜GLSL−34が作出された。
このGLSLの作出工程の概要を、図2に模式的に示した。図2中、括弧内の数値は、各戻し交配世代によって得られた系統数を示し、円で囲まれた数値は、オリザ・グラベリマ由来の染色体断片を2以上含んでいるGLSLの候補個体の数を示す。四角で囲まれた数値は、1又は2のオリザ・グラベリマ由来の染色体断片を含んでいるGLSLの候補個体の数を示し、角括弧内の数値は、先行する戻し交配世代から得られたGLSLの総数を示す。
【0049】
[実施例2]
実施例1で作出されたGLSLsについて、収量性、草型、成熟性、籾外観に関する10形質の有用農業形質の評価を行い、QTLを検出した。GLSLsは、コシヒカリの遺伝的背景をもち、染色体の一部のみがアフリカ型品種オリザ・グラベリマ由来の染色体断片に置換されているため、これらを用いることにより、遺伝的背景を揃えた条件でのQTLの検出を試みた。
【0050】
<GLSLsの形質測定>
まず、GLSLsの様々な農業形質の表現型の観察を、2007年夏、千葉県木更津市研究ほ場にて行った。各GLSLから24の苗を、対照とするイネ品種コシヒカリの苗と共に、ほ場の栽培区画に2条植えし、成熟するまで育てた。苗は、最初温室で栽培し、播種から30日後に、栽培区画に移植した。定植幅は株間20cm、条間30cmで行った。
成熟後、各GLSL及びイネ品種コシヒカリの、それぞれの栽培区画から予め定められた5個体(第1列の3〜7番目の株)を個別に収穫し、籾長(Grain length)、籾幅(Grain width)、籾厚(Grain thickness)、一穂当たりの籾数(Grain number/panicle)、籾の100粒重(100−grain weight)を含む、籾外観形質を調べた。これらの個体について、さらに、稈長(Culm length)、穂長(Panicle length)、一穂当たりの枝こう数(No. of primary branches/panicle)、及び種子稔性(Fertility)といったその他の農業形質についても調べた。各GLSLの出穂日数(Heading date)は、種子の播種から各系統の50%の個体が実際に出穂した日までの日数とした。籾の100粒重は、少なくとも2ヶ月間温室内で風乾させた種子を用いて決定した。
GLSLsについての籾外観形質の測定結果を、その他の重要な農業形質の結果と共に表6及び7に示した。
【0051】
【表6】
【0052】
【表7】
【0053】
<GLSLsの形質評価及びQTL検出>
一元配置分散分析、及び95%信頼区画(P<0.05)によるダネットの多重比較テストを使う比較手法により、GLSLsにおける推定QTLの同定を行った。ある形質の
算出された平均値が、いずれかのGLSLにおいて、イネ品種コシヒカリよりも明らかに相違した場合には、当該GLSLが含有しているオリザ・グラベリマ由来の染色体断片にQTLが存在していると判断された。
【0054】
(籾外観形質)
籾長に対する推定QTLは、オリザ・グラベリマの第1、3、4、及び8染色体由来の置換染色体断片を有する8つのGLSLに検出された。これらのGLSLのうち7系統(GLSL−3、11、12、13、24、25、26)は、籾長が4〜14%ほど、イネ品種コシヒカリよりも有意に長くなっていたが、残る1系統(GLSL−2)は、当該形質に対して負の効果を示すQTLを有していた。つまり、籾長に対し正の効果を示す推定QTLと、籾長に対し負の効果を示す推定QTLの両方が、第1染色体に存在していた。
【0055】
20のGLSL(GLSL−1、4、5、11、12、13、14、20、21、22、23、24、25、26、27、28、30、31、32、33)が、イネ品種コシヒカリ(平均籾幅:3.21mm)と比べてより幅の大きい籾を産生した。これらの系統は、オリザ・グラベリマの第1、3、4、5、6、7、8、9、10、11、及び12染色体由来の断片で置換されており、イネ品種コシヒカリよりも4〜10%も籾幅が有意に増大していた。これに対して、オリザ・グラベリマの第5及び6染色体由来の断片で置換された2系統(GLSL−17、18)では、イネ品種コシヒカリよりも7〜8%も籾幅が有意に減少していた。
【0056】
籾厚は、オリザ・グラベリマの第1、7、及び8染色体由来の断片で置換された4つのGLSL(GLSL−4、22、25、26)では3〜4%も有意に増大していたが、オリザ・グラベリマの第1、5、6、及び7染色体由来の断片で置換された5つのGLSL(GLSL−2、16、17、18,21)では3〜5%も有意に減少していた。
【0057】
一穂当たりの籾粒数に対する推定QTLは、イネ品種コシヒカリに比べて平均値が有意に高い8のGLSLにおいて検出された。一穂当たりの籾粒数における26〜80%の増大が、これらのオリザ・グラベリマの第1、2、及び7染色体由来の断片で置換された系統(GLSL−1、2、3、6、7、8、21、22)において記録された。一方で、当該形質に対して負の効果を示す2つの推定QTLが、オリザ・グラベリマの第5及び8染色体由来の断片で置換された系統(GLSL−16、25)において観察された。
【0058】
3つの系統(GLSL−4、25、31)が、イネ品種コシヒカリ(平均100粒重:2.23g)と比べてより籾の100粒重が有意に高い値を示した。これらの系統は、オリザ・グラベリマの第1、8、及び11染色体由来の断片で置換されており、イネ品種コシヒカリよりも11〜15%も100粒重が有意に増大していた。しかしながら、オリザ・グラベリマの第1、5及び10染色体由来の断片で置換された4系統(GLSL−2、16、17、29)では、当該形質に対して負の効果が観察され、イネ品種コシヒカリよりも7〜12%程度も減少していた。
【0059】
(その他の農業形質)
稈長が有意に低くなることに対する推定QTLは、オリザ・グラベリマの第1、2、4、5、6、8、9、10、11、及び12染色体由来の断片で置換された20のGLSL(GLSL−1、6、7、8、9、14、15、16、17、18、19、20、24、25、27、28、29、30、31、33)において同定された。これらのGLSLの稈長は、イネ品種コシヒカリ(平均稈長:95.72cm)に比べて10〜27%短かった。
【0060】
イネ品種コシヒカリの平均出穂期は、播種から93.20日であったが、9つのGLSL(GLSL−3、11、12、13、21、23、24、25、31)は、イネ品種コシヒカリに比べて有意に出穂期が遅かった。オリザ・グラベリマの第1、3、4、5、及び8染色体から挿入された断片中にある推定QTLが、イネ品種コシヒカリの出穂期を4.6〜18.2日遅らせていた。
【0061】
穂長に対する推定QTLが、イネ品種コシヒカリと比べて穂長の平均値が有意に長い8つのGLSL(GLSL−2、3、4、6、7、8、11、22)において検出された。これらの系統は、オリザ・グラベリマの第1、2、3、及び7染色体由来の断片が挿入されており、穂長が15〜24%増加することが記録された。しかしながら、オリザ・グラベリマの第5染色体由来の断片が挿入された1系統(GLSL−16)は、イネ品種コシヒカリよりも穂長が有意に減少(13%)したことが記録された。
【0062】
穂における枝こう数については、推定QTLが、オリザ・グラベリマの第1、2、3、及び7染色体由来の断片で置換された5つのGLSL(GLSL−2、7、11、12、22)において検出された。これらのGLSLは、イネ品種コシヒカリ(一穂当たりの平均枝こう数:10.40)に比べて、一穂当たりの枝こう数が25〜36%増大していた。
【0063】
種子稔性については、23〜84%の範囲内の有意な減少が観察されたのは6系統(GLSL−12、17、18、21、29、31)であったが、2系統(GLSL−2、19)を除く全てのGLSLが、イネ品種コシヒカリに比べてより低い種子稔性を示した。
【0064】
(GLSLsにおいて観察されたその他の形質)
幾つかのGLSLにおいて明らかに現れていた他の形質についても、同定した。これらの形質は、表現型上測定してQTL解析に影響するものではないが、明らかに主要な遺伝子、又はGLSLsに挿入されたオリザ・グラベリマ由来の染色体断片に存在するQTLと関連している可能性がある。例えば、オリザ・グラベリマの第5染色体のおよそ3.15Mbを含むGLSL−15では、無毛の葉が観察された。無毛の葉を制御する遺伝子は、イネ品種コシヒカリの準同質遺伝子系統を用いることによって、当該染色体断片からより狭い領域にマッピングされ、続いてクローニングされる。その他の同定された形質としては、GLSL−13及び15において長い芒が観察された。GLSL−13及び15は、それぞれ、オリザ・グラベリマの第4及び8染色体由来の断片を含んでいた。
【0065】
農業的特長に対する推定QTLの染色体上の位置を、表8に示す。表8中、「GL」は籾長を、「GW」は籾幅を、「GT」は籾厚を、「GN」は一穂当たりの籾数を、「GWe」は籾の100粒重を、「CL」は稈長を、「PL」は穂長を、「PB」は一穂当たりの枝こう数を、「F」は種子稔性を、「HD」は出穂日数を、それぞれ意味する。
【0066】
【表8】
【0067】
(総括)
GLSLsから、12本の染色体中に分散している全105の推定QTLが同定された。調査された全ての形質は、これらのQTLのうちの5又はそれ以上によって調節されていることがわかった。1の形質の調節に関わる複数のQTLは、同一の染色体上の2〜3部位にある場合もあり、複数の染色体中に分散された位置にある場合もあった。これらのQTLは、染色体の他の部位のみならず、環境とも影響しあう。イネの多くの重要な農業形質は、2〜3のQTLによって調節されているという報告がある。
同定されたQTLは、観察した農業形質に対して正又は負の影響を示した。オリザ・グラベリマの保持する105個のQTLのうち、84個のQTLはイネ品種コシヒカリに対し正の効果を示し、21個のQTLはイネ品種コシヒカリに対し負の効果を示した。正の効果を示した84個のQTLのうち、55個は籾収量に関係する形質に影響しており、20個は植物高さに、9個は成熟期に、それぞれ影響していた。これらの結果から、オリザ・グラベリマ由来の染色体が、現存するイネ品種の収量や草型に関する形質を改良するために有用であることが示唆された。
【0068】
籾長、籾幅、籾厚、出穂期、穂における枝こう数に対するQTLは、オリザ・グラベリマの第3染色体をカバーする2GLSL(GLSL−11、12)において検出された。これらの2系統において導入された染色体断片は、およそ11と28Mbであり、7Mbオーバーラップしていた。
イネの育種に対する有用性の点からは、第1染色体に位置しており、2つのGLSLにおいて同定された2個のQTLが、前途有望と考えられる。両QTLともに一穂当たりの籾粒数を強くコントロールすることが示され、穂長と籾幅に対して多面的な効果も示していた。これとは別に、この2つのQTLは、2つのGLSLにおいて、イネ品種コシヒカリと比べて70%以上も一穂当たりの籾粒数を増大させた。もしも、特に広い生態学的条件下でこれらの機能と発現が確認されたら、GN1A(非特許文献4参照。)、APO1(非特許文献16及び35参照。)、及び最近報告されたWFP及びIPA1(非特許文献26及び18参照。)と同様に、これらのQTLは、クローニングされ、より収量が改善されたイネ品種の育種に使用することができるだろう。
既に同定され報告されている幾つかのオリザ・グラベリマ由来のQTLも、本実施例のGLSLsにおいて同様に検出された。これは、新しく作出された本実施例のGLSLsが、オリザ・グラベリマの全ゲノムを正確に含んでいることを示す。既に報告され、本実施例で確認されたQTLには、不稔性に影響する4つの座(第3、6、7、及び10染色体に位置する。)(非特許文献10、34、15及び22参照。)、及び第1染色体上にあり、出穂期に関係する1のQTL(非特許文献9参照。)が含まれていた。
【0069】
本実施例で作出された34のGLSLは、オリザ・グラベリマが有する大量の多様な遺伝子に対するアクセスを可能にし得る、価値のある遺伝資源であることについては疑いはない。これらGLSLsからは、本実施例において同定された籾収量や草高、成熟性についてのQTLの他にも、より多くの重要な農業形質を調節する遺伝子が発見されるであろう。アフリカ型イネは、最も厳しい環境下でも良好に成育することが知られており、生物的及び非生物的ストレスに対する有用な新規遺伝子を含んでいる可能性は高い。害虫及び病気に対する抵抗性についてのGLSLsの評価については、問題のある土壌や極限環境条件が、これらの有用な遺伝子の同定を容易にするだろう。
【産業上の利用可能性】
【0070】
本発明のゲノムライブラリー、及びこれを構成する各CSSLは、有用農業形質を調節するQTLの検出に特に好適に利用することができ、さらに検出されたQTLのピラミティングが、シンクサイズ等の形質が改善された新品種育成にも利用することができるため、特に農業の分野において利用が可能である。
【技術分野】
【0001】
本発明は、イネ品種コシヒカリの染色体の一部が、アフリカ栽培品種オリザ・グラベリマの染色体断片で置換された染色体部分置換系統群からなるゲノムライブラリーに関する。
【背景技術】
【0002】
オリザ・グラベリマ(Oryza glaberrima Steud)は野生種のオリザ・バータイ(Oryza barthii)から栽培化された西アフリカに固有のイネの栽培品種である。より詳細には、オリザ・グラベリマは約3,500年前に、西アフリカの中でも主にナイジェリアのリバーデルタの湿潤地帯で栽培化された後、紀元前1,000年頃にギニアの海岸近くで第2の栽培化が起こったと考えられている。アジアのイネの栽培品種であるオリザ・サティバ(Oryza sativa L.)と比較すると、オリザ・グラベリマは、遺伝的に多様性が少なく(非特許文献7、37、及び31参照。)、栽培分布も狭く、地域起源を越えては広がっていない(非特許文献30及び21参照。)。これにもかかわらず、オリザ・グラベリマは、人の介入なしに過酷な西アフリカの環境で成長させる多くの新規遺伝子を潜在的に持っている(非特許文献20参照。)。雑草競争、耐干性、低い成長投入量に対応する能力、不利な生態系への広い適合性はアフリカ型イネ固有の特徴である(非特許文献29、6、25、13、19、20、及び8参照。)。このイネの品種はモザイク病(非特許文献28、1、及び5参照。)、枯れ病(非特許文献32、30、及び12参照。)、もん枯れ病(非特許文献12及び36参照。)、アフリカ小昆虫(非特許文献36及び20参照。)を含むいくつかの生物的ストレスにも耐性を持っている。
【0003】
現在までに、アフリカ型イネでの重要な特徴に対するいくつかのQTL(量的形質座)が既に同定されている(非特許文献24、2、及び23参照。)。一般的にF1若しくはF2/F3系統、組換え近交系、倍加半数系、戻し交配が、これらの特徴の検出に使われている。しかしながら、F1系統の使用により成功する場合の多くは、主要な効果を持つQTLを検出する場合のみである。表現型の効果が弱いQTLと他の遺伝子との相互関係をもつエピスタシスは、これらの系統では容易に検出されない。興味深いQTLの検出するのを成功させるためには、準同質系統をQTLクローニングとマッピングのために開発する必要がある(非特許文献33、11、39、及び40参照。)。QTL検出とマッピングは、多大労力と多くの時間を要するため、遺伝子クローニングのスタンダードなストラテジーとしてマップベースクローニングへの使用に限られている。
【0004】
これらの課題に取り組むため、イネにおいても、染色体部分置換系統群の形で遺伝子資源が開発された(非特許文献3、11、17、及び10参照。)。CSSLsは、ドナー系統を受容系統と交雑させ、F1及びBCnF1を反復親に戻し交配することによって作出される。一般的には、野生イネ又は一般的な栽培品種がドナー系統となり、通常、優秀な系統が受容系統となる。これらの繰返し戻し交配及びその後の3〜4後代世代の育成によって得られた後代個体は、反復親の遺伝的背景中に、1又は2のホモ接合性のドナー系統の遺伝子型の染色体断片が含まれている。このように、ドナー系統の全ゲノムは、互いに連続的又はオーバーラップする(重なり部分がある)小さな染色体断片とし、これらが導入されている選択された系統群であらわされることができる(非特許文献14及び39参照。)。
【0005】
遺伝的に多様なイネ品種と野生型イネ品種とからなる染色体部分置換系統(以下、CSSLと略記することがある。)は、重要な農業形質を制御する新規な遺伝子の同定に有用な遺伝資源である。特に、CSSLは、複合的、量的形質を解析のターゲットとする場合に有効である。CSSLは、表現型と導入された染色体断片とが強い相関を有するため、個々の座の本当の表現型の影響をわかりにくくする表現型のノイズとして知られているQTL間の干渉の影響が、取り除かれるためである。ドナーゲノムの非常に短い、連続した又はオーバーラップする染色体断片を含む一連のCSSLからなる染色体置換系統群(以下、CSSLsと略記することがある。)セットは、均一の遺伝的背景を有するため、CSSLsセットを用いることにより、単純な統計分析を通じてQTLを同定することができる(非特許文献27及び14参照。)。
【0006】
一方で近年、ゲノム解析技術は格段の進歩を遂げ、作物の改良を、高精度かつ容易に行えるようになった。とりわけ、DNAマーカー技術の進捗は著しく、同技術によって、有用な形質を備える新品種を作製することができるようになった。また、DNAマーカーを利用して、これまでに同定された重要遺伝子の有用アレルを含む染色体領域を置換することにより、他の多くの形質に対してほとんど影響を及ぼさずに、目的形質を特異的に改良することが出来るようになった(例えば、特許文献1参照。)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許第4409610号公報
【非特許文献】
【0008】
【非特許文献1】Abo, E. M., et al. (1998) J. Sustain. Agric. Vol.11: pp.85-111.
【非特許文献2】Aluko, G., C. et al. (2004) Theor. Appl. Genet. Vol.109: pp.630-639.
【非特許文献3】Ando, T., et al. (2008) Theor. Appl. Genet. Vol.116: pp.881-890.
【非特許文献4】Ashikari, M., et al. (2005) Science. Vol.309: pp.741-745.
【非特許文献5】Attere, C. and C. A. Fatokun (1983) Plant Divers. Vol.67: pp.420-421.
【非特許文献6】Baggie, I., et al. (2002) Int. Rice Res. Notes. Vol.27: pp.42.
【非特許文献7】Barry, M. B., et al. (2007) Euphytica Vol.154: pp.127-137.
【非特許文献8】Dingkuhn, M., et al. (1998) Field Crops Res. Vol.57: pp.57-69.
【非特許文献9】Doi, K., et al. (1998) Breed Science Vol.49: pp.395-399.
【非特許文献10】Doi, K., et al. (1997) Rice Genet. Newsl. Vol.14: pp.39-41.
【非特許文献11】Ebitani, T., et al. (2005). Breeding Science Vol.55: pp.65-73.
【非特許文献12】Eizenga, G. C., et al. (2006) Crop Sci. Vol.46: pp.1870-1878.
【非特許文献13】Fofana, B. and R. Rauber (2000) Weed Res. Vol.40: pp.271-280
【非特許文献14】Gutierrez, A. G., et al. (2010) BMC Plant Biol. Vol.10: pp.6.
【非特許文献15】Hu, F., et al. (2006) Euphytica Vol.151: pp.273-278.
【非特許文献16】Ikeda, K., M. et al. (2007) Plant Journal Vol.51(6): pp.1030-1040.
【非特許文献17】Kubo, T., et al. (2002) Breed. Sci. Vol.52: pp.319-325.
【非特許文献18】Jiao, Y., et al. (2010) Nat. Genet. Vol.42: pp.541-544.
【非特許文献19】Johnson, D. E., et al. (1998) Weed Res. Vol.38: pp.207-216.
【非特許文献20】Jones, M. P., et al. (1997) Breeding Science Vol.47: pp.395-398.
【非特許文献21】Khush, G.S. (1997) Plant Mol. Biol. Vol.35: pp.25-34.
【非特許文献22】Li, J., et al. (2008) Euphytica Vol.164: pp.699-708.
【非特許文献23】Li, J., et al. (2004) Genome Vol.47: pp.697-704.
【非特許文献24】Lorieux, M., et al. (2000) Theor. Appl. Genet. Vol.100: pp.593-601.
【非特許文献25】Maji, T., et al. (2001) Oryza Vol.38: pp.17-23.
【非特許文献26】Miura, K., et al. (2010) Nat. Genet. Vol.42: pp.545-549.
【非特許文献27】Nadeau, J. H., et al. (2000) Nat. Genet. Vol.24: pp.221-225.
【非特許文献28】Ndjiondjop, M. N., et al. (2001) Mol. Plant. Pathol. Vol.59: pp.309-316.
【非特許文献29】Sahrawat, K. L. and M. Sitka (2002) Res. Notes Vol.27: pp.30-31.
【非特許文献30】Sarla, N. and B. P. M Swamy (2005) Curr. Sci. Vol.89(6): pp.955-963.
【非特許文献31】Second, G. (1986) The International Rice Research Institute, Los Banos, Laguna, Philippines. pp.27-39.
【非特許文献32】Silue, D. and J. Notteghem (1991) Int. Rice Res. Notes Vol.16: pp.13-14.
【非特許文献33】Takai, T., et al. (2007) Breed. Sci. Vol.57: pp.257-261.
【非特許文献34】Taguchi, K. K. D. and A. Yoshimura (1999) Rice Genet. News. Vol.16: pp.70-71
【非特許文献35】Terao, T., et al. (2010) Theor. Appl. Genet. 120(5): pp.875-893.
【非特許文献36】Ukwungwu, M. N., et al. (1999) Journ. Food. Tech. Afr. Vol.4(3): pp.108-110.
【非特許文献37】Wang, Z., et al. (1992) Theor. Appl. Genet. Vol.83: pp.565-581.
【非特許文献38】Wasano, K. and Y. Hirota (1986) Bull. Fac. Agr. Saga Univ. Vol.60: pp.49-59.
【非特許文献39】Yano, M. (2001) Curr. Opin. Plant Biol. Vol.4: pp.130-135.
【非特許文献40】Yano, M. and T. Sasaki (1997) Plant Mol. Biol. Vol.35: pp.145-153.
【発明の概要】
【発明が解決しようとする課題】
【0009】
オリザ・グラベリマをはじめとする外国稲は、遺伝資源としての潜在能力に期待はあるものの、劣悪形質が多いため、これまでの育種事業では病害抵抗性のような主導遺伝子の利用にとどまっている。
【0010】
本発明は、オリザ・サティバ育種のための遺伝資源として、オリザ・サティバの優秀な栽培種であるジャポニカ栽培種コシヒカリ(以下、イネ品種コシヒカリ)の遺伝的背景に、多くの新規遺伝子を潜在的に持つ可能性のあるオリザ・グラベリマの染色体断片を置換したイネ品種コシヒカリの染色体部分置換系統群を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者は、上記課題を解決すべく鋭意研究した結果、イネ品種コシヒカリとオリザ・グラベリマとを識別するための新規なSNPからなるDNAマーカーを開発し、これらのDNAマーカーを利用して、イネ品種コシヒカリの遺伝的背景に、オリザ・グラベリマの染色体断片が置換された34系統の染色体部分置換系統群を作出し、本発明を完成させた。
【0012】
すなわち、本発明は、下記(1)〜(34)の、イネ品種コシヒカリの染色体の一部がオリザ・グラベリマの染色体断片で置換された染色体部分置換系統を有することを特徴とするゲノムライブラリーを提供するものである。
(1)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(2)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の11,026,011番目の塩基から20,366,961番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の8,896,816番目の塩基から11,026,011番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の20,366,961番目の塩基から24,412,980番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(3)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の17,803,741番目の塩基から31,371,175番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の13,813,982番目の塩基から17,803,741番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の31,371,175番目の塩基から35,598,471番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(4)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の25,979,477番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の24,412,982番目の塩基から25,979,477番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の35,598,472番目の塩基から39,677,013番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(5)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の35,598,472番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の31,371,176番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(6)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の上流末端から3,922,571番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第2染色体の3,922,571番目の塩基から7,030,896番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(7)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の3,922,571番目の塩基から14,655,638番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の2,010,335番目の塩基から3,922,571番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の14,655,638番目の塩基から18,765,983番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(8)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の11,165,933番目の塩基から22,086,348番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の9,256,585番目の塩基から11,165,933番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の22,086,348番目の塩基から25,263,252番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(9)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の18,765,984番目の塩基から25,263,253番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の14,655,639番目の塩基から18,765,984番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の25,263,253番目の塩基から27,919,902番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(10)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の30,860,986番目の塩基から33,189,264番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の29,345,358番目の塩基から30,860,986番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の33,189,264番目の塩基から35,267,218番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(11)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の上流末端から11,073,623番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第3染色体の11,073,623番目の塩基から14,311,069番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(12)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の11,073,623番目の塩基から33,640,115番目の塩基までを 含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第3染色体の8,197,229番目の塩基から11,073,623番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第3染色体の33,640,115番目の塩基から35,945,010番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(13)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の上流末端から22,125,293番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第4染色体の22,125,293番目の塩基から25,877,140番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(14)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の19,650,008番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第4染色体の14,903,263番目の塩基から19,650,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(15)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の上流末端から5,346,476番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第5染色体の5,346,476番目の塩基から7,305,955番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(16)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の3,173,133番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の1,047,931番目の塩基から3,173,133番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第5染色体の21,023,551番目の塩基から24,520,268番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(17)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の21,023,551番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の18,878,115番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(18)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の上流末端から2,186,456番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第6染色体の2,186,456番目の塩基から5,904,223番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(19)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の2,186,456番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の372,259番目の塩基から2,186,456番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第6染色体の22,907,151番目の塩基から25,711,514番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(20)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の22,907,151番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の20,235,923番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(21)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の上流末端から6,843,696番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第7染色体の6,843,696番目の塩基から10,130,031番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(22)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の1,745,458番目の塩基から18,571,959番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第7染色体の260,865番目の塩基から1,745,458番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第7染色体の18,571,959番目の塩基から21,942,075番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(23)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の21,942,076番目の塩基から24,516,006番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第1の染色体断片にホモ置換されており、
当該第7染色体の28,847,226番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第2の染色体断片にホモ置換されており、
かつ前記第1の染色体断片の上流端が、イネ品種日本晴の第7染色体の18,571,960番目の塩基から21,942,076番目の塩基までを含む領域に相当する領域に存在し、前記第1の染色体断片の下流端が、イネ品種日本晴の第7染色体の24,516,006番目の塩基から25,490,052番目の塩基までを含む領域に相当する領域に存在し、
前記第2の染色体断片の上流端が、イネ品種日本晴の第7染色体の25,490,054番目の塩基から28,847,226番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(24)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の上流末端から8,094,582番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第8染色体の8,094,582番目の塩基から10,572,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(25)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の8,094,582番目の塩基から19,903,905番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の5,278,856番目の塩基から8,094,582番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第8染色体の19,903,905番目の塩基から23,047,466番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(26)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の18,114,324番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の15,484,341番目の塩基から18,114,324番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(27)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の上流末端から13,254,003番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第9染色体の13,254,003番目の塩基から15,063,824番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(28)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の13,254,003番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第9染色体の10,483,156番目の塩基から13,254,003番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(29)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の上流末端から14,142,299番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第10染色体の14,142,299番目の塩基から17,555,158番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(30)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の14,142,299番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第10染色体の11,761,139番目の塩基から14,142,299番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(31)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の上流末端から21,896,773番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第11染色体の21,896,773番目の塩基から25,149,243番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(32)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の21,896,773番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第11染色体の19,050,306番目の塩基から21,896,773番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(33)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の上流末端から14,689,883番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にヘテロ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第12染色体の14,689,883番目の塩基から17,499,934番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(34)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の21,257,971番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第12染色体の17,499,936番目の塩基から21,257,971番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統。
【発明の効果】
【0013】
本発明のゲノムライブラリー、及びこれを構成する各染色体部分置換系統は、イネ品種コシヒカリの遺伝的背景に、従来、イネ品種コシヒカリの形質改善には利用されてこなかったオリザ・グラベリマの染色体断片が組み込まれている。このため、本発明のゲノムライブラリー、及びこれを構成する各染色体部分置換系統を用いることにより、オリザ・グラベリマが有する様々な遺伝子や形質を、遺伝的背景を揃えた条件で精度よく解析することができる。さらに、本発明のゲノムライブラリーは、オリザ・グラベリマが有している好ましい農業形質を制御するQTLの検出や、当該形質の原因となる新規遺伝子の同定、新品種育成等に好適に利用することができる。
【図面の簡単な説明】
【0014】
【図1】本発明のゲノムライブラリーを構成する34系統(GLSL−1〜GLSL−34)のゲノムを模式的に示した図である。
【図2】本発明のゲノムライブラリーを構成する34系統(GLSL−1〜GLSL−34)の作出工程を模式的に示した図である。
【発明を実施するための形態】
【0015】
本発明及び本願明細書において「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域」は、米国ゲノム研究所(TIGR)によるRice Genome Anootation Databaseにおいて公開されているイネ品種日本晴のゲノムDNAの塩基配列(バージョン2)に基づいて決定される領域である。
【0016】
また、本発明及び本願明細書において、「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域に相当する領域」とは、イネ個体の染色体中のイネ品種日本晴の染色体中の当該領域と相同性の高い領域であり、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントすることにより決定することができる。また、イネ品種日本晴以外のイネ個体中の「イネ品種日本晴のSNP(Single Nucleotide Polymorphism、一遺伝子多型)に相当するSNP」は、当該SNPを含む領域において、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントした場合に、当該SNPに対応する位置にある塩基を意味する。
【0017】
本発明において品種とは、同一種の植物であって、遺伝的構成が異なるために、ある形質において同種内の他品種から明確に識別し得る集団を意味する。また、イネ品種日本晴及びイネ品種コシヒカリの遺伝子情報等は、例えば、国際的な塩基配列データベースであるNCBI(National center for Biotechnology Information)やDDBJ(DNA Data Bank of Japan)等において入手することができる。特にイネの各品種の遺伝子情報は、KOME(Knowledge−based Oryza Molecular biological Encyclopedia、http://cdna01.dna.affrc.go.jp/cDNA/)等において入手することができる。
【0018】
本発明及び本願明細書において、DNAマーカーとは、種や品種が異なる生物由来の染色体断片同士を識別し得る染色体上のDNA配列の差異を検出し得るものである。具体的には、SNPである。
【0019】
DNAマーカーによる、ある品種由来アレルとその他の種又は品種由来アレルとの識別は、常法により行うことができる。例えば、各個体から抽出したDNAを鋳型とし、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等を用いてPCRを行い、電気泳動法等を用いてPCR産物の有無を検出し、各多型を識別することができる。また、各個体から抽出したDNAを制限酵素処理した後、電気泳動法等を用いてDNA断片のパターンを検出し、各多型を識別することができる。なお、特定のSNPと特異的にハイブリダイズし得るプライマー等は、該SNPの塩基配列に応じて、汎用されているプライマー設計ツール等を用いて常法により設計することができる。また、設計されたプライマー等は、当該技術分野においてよく知られている方法のいずれを用いても合成することができる。
なお、これらのDNAマーカーによる、ある品種由来アレルとその他の種又は品種由来アレルとの識別は、常法により行うことができる。例えば、各個体から抽出したDNAを鋳型とし、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等を用いてPCRを行い、電気泳動法等を用いてPCR産物の有無を検出し、各多型を識別することができる。また、各個体から抽出したDNAを制限酵素処理した後、電気泳動法等を用いてDNA断片のパターンを検出し、各多型を識別することができる。なお、特定のSNPと特異的にハイブリダイズし得るプライマー等は、該SNPの塩基配列に応じて、汎用されているプライマー設計ツール等を用いて常法により設計することができる。また、設計されたプライマー等は、当該技術分野においてよく知られている方法のいずれを用いても合成することができる。
【0020】
<DNAマーカー>
本発明者らは、まず、オリザ・グラベリマ由来の染色体断片とイネ品種コシヒカリ由来の染色体断片とを識別するために、イネ品種日本晴のゲノムの塩基配列に基づいて、第1〜第12染色体の全染色体領域において、イネ品種コシヒカリとオリザ・グラベリマの間で多型を示すSNPからなる142のDNAマーカーを作製した。DNAマーカー間の距離は、平均2.75Mbであった。各DNAマーカーの位置を、表1〜4に示す。各表の染色体中の位置情報は、イネ品種日本晴のゲノム中の相当するSNPの位置である。なお、オリザ・グラベリマの遺伝子解析はほとんど行われておらず、これらの142のDNAマーカーは、本発明者らによって初めて見出されたものである。
【0021】
【表1】
【0022】
【表2】
【0023】
【表3】
【0024】
【表4】
【0025】
表1〜4に記載のDNAマーカーは、染色体中にイネ品種コシヒカリ由来の染色体断片とオリザ・グラベリマ由来の染色体断片の両方が含まれているイネ個体のジェノタイピング(遺伝子型の同定)に特に好適に用いられる。イネ個体のジェノタイピングにおいて、表1〜4に記載のDNAマーカーの遺伝子型をタイピングすることによって、得られたタイピング結果から、当該イネ個体の染色体中に、イネ品種コシヒカリ由来の染色体断片又はオリザ・グラベリマ由来の染色体断片の有無や、当該断片が存在する染色体中の位置等を調べることができる。
【0026】
<染色体部分置換系統(CSSL)及びゲノムライブラリー>
本発明のゲノムライブラリーは、イネ品種コシヒカリの染色体が、部分的にオリザ・グラベリマの染色体断片で置換された34系統(GLSL−1〜GLSL−34)のCSSLsを有する。本発明のゲノムライブラリー中のCSSLがそれぞれ含有するオリザ・グラベリマ由来の染色体断片は、互いに連続的又はオーバーラップしており、34系統全てを合わせるとオリザ・グラベリマの全ゲノムの90%以上が保持されている。
【0027】
本発明のゲノムライブラリーを構成するCSSLは、1セットとして使用することができ、それぞれ単独で使用してもよい。これらのCSSLは、イネの粒特性や他の有用な農業的特徴を調節するQTLの検出に特に有用である。また、検出されたQTLを利用することにより、育種の幅が広がることも期待できる。例えば、検出されたQTLのピラミティングにより、新規な品種を容易に育種することが可能になる。さらに、本発明のゲノムライブラリーを構成する各CSSLは、染色体中のオリザ・グラベリマ由来の染色体断片の位置が、前記の142のDNAマーカーによって明らかであるため、育種の材料としても好適である。
【0028】
図1に、本発明のゲノムライブラリーを構成する34系統(GLSL−1〜GLSL−34)のゲノムを模式的に示す。図1の各染色体中、白抜き部分はイネ品種コシヒカリのホモ接合型の染色体領域を、黒塗部分はオリザ・グラベリマのホモ接合型の染色体領域を、ハッチング部分はイネ品種コシヒカリとオリザ・グラベリマのヘテロ接合型の染色体領域を、それぞれ表す。
【0029】
オリザ・グラベリマの第1染色体及び第2染色体の染色体断片は、それぞれ5のCSSLに置き換えられており、第3染色体〜第12染色体の染色体断片は2又は3のCSSLに置き換えられていた。ドナーゲノム(オリザ・グラベリマのゲノム)のうちのおよそ6%は、34系統(GLSL−1〜GLSL−34)(以下、GLSLセット、ということがある。)のいずれにおいても、イネ品種コシヒカリの相当する領域に置換されなかった。この置換されなかった領域は、図1各染色体において、GLSL−1〜GLSL−34のいずれにおいても黒塗部分が存在していない、小さなギャップとして示される。これらのギャップは、具体的には、以下の6部分領域である;第1染色体中のSP−185を含み、かつSP−2077及びSP−192を含まない部分領域;第2染色体中のSP−1444及びSP−1450を含み、かつSP−2941及びSP−1459を含まない部分領域;第2染色体中のSP−1471よりも下流の、SP−3571を含む末端部分領域;第3染色体中のSP−348よりも下流の、SP−362を含む末端部分領域;第7染色体中のSP−741を含み、かつSP−2545及びSP−4152を含まない部分領域;第12染色体中のSP−3747を含み、かつSP−2687及びSP−2697を含まない部分領域。
【0030】
また、GLSLセットのうちの33系統は、オリザ・グラベリマ由来の染色体断片がホモ置換されている(ホモ接合型で置換されている)が、第12染色体に置換が生じているGLSL−33のみは、オリザ・グラベリマ由来の染色体断片がヘテロ(ヘテロ接合型で)置換されている。GLSL−33中のオリザ・グラベリマ由来の染色体断片の長さは約17.5Mbであり、当該染色体断片の両脇にはDNAマーカーSP−5151及びSP−2687が存在する。
【0031】
GLSL−1は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基(SP−2077)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基(SP−185の−1塩基(1塩基上流の塩基))までを含む領域に相当する領域に存在しているCSSLである。
GLSL−2は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の11,026,011番目の塩基(SP−192)から20,366,961番目の塩基(SP−2016)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の8,896,816番目の塩基(SP−185の+1塩基(1塩基下流の塩基))から11,026,011番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の20,366,961番目の塩基から24,412,980番目の塩基(SP−1217の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−3は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の17,803,741番目の塩基(SP−1208)から31,371,175番目の塩基(SP−262)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の13,813,982番目の塩基(SP−2430の+1塩基)から17,803,741番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の31,371,175番目の塩基から35,598,471番目の塩基(SP−1244の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−4は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の25,979,477番目の塩基(SP−2478)から35,598,472番目の塩基(SP−1244)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の24,412,982番目の塩基(SP−1217の+1塩基)から25,979,477番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の35,598,472番目の塩基から39,677,013番目の塩基(SP−2056の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−5は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の35,598,472番目の塩基(SP−1244)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の31,371,176番目の塩基(SP−262の+1塩基)から35,598,472番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0032】
GLSL−6は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の上流末端から3,922,571番目の塩基(SP−1387)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第2染色体の3,922,571番目の塩基から7,030,896番目の塩基(SP−1398の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−7は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の3,922,571番目の塩基(SP−1387)から14,655,638番目の塩基(SP−1413)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の2,010,335番目の塩基(SP−6の+1塩基)から3,922,571番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の14,655,638番目の塩基から18,765,983番目の塩基(SP−2495の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−8は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の11,165,933番目の塩基(SP−1406)から22,086,348番目の塩基(SP−954)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の9,256,585番目の塩基(SP−29の+1塩基)から11,165,933番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の22,086,348番目の塩基から25,263,252番目の塩基(SP−2941の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−9は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の18,765,984番目の塩基(SP−2495)から25,263,253番目の塩基(SP−2941)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の14,655,639番目の塩基(SP−1413の+1塩基)から18,765,984番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の25,263,253番目の塩基から27,919,902番目の塩基(SP−1444の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−10は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の30,860,986番目の塩基(SP−1459)から33,189,264番目の塩基(SP−1471)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の29,345,358番目の塩基(SP−1450の+1塩基)から30,860,986番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の33,189,264番目の塩基から35,267,218番目の塩基(SP−3571の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
【0033】
GLSL−11は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の上流末端から11,073,623番目の塩基(SP−1323)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第3染色体の11,073,623番目の塩基から14,311,069番目の塩基(SP−1334の−1塩基)までを含む領域に相当する領域に存在している染色体部分置換系統CSSLである。
GLSL−12は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の11,073,623番目の塩基(SP−1323)から33,640,115番目の塩基(SP−348)までを 含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第3染色体の8,197,229番目の塩基(SP−1308の+1塩基)から11,073,623番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第3染色体の33,640,115番目の塩基から35,945,010番目の塩基(SP−362の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−13は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の上流末端から22,125,293番目の塩基(SP−392)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第4染色体の22,125,293番目の塩基から25,877,140番目の塩基(SP−402の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−14は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の19,650,008番目の塩基(SP−386)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第4染色体の14,903,263番目の塩基(SP−3250の+1塩基)から19,650,008番目の塩基までを含む領域に相当する領域に存在している
【0034】
GLSL−15は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の上流末端から5,346,476番目の塩基(SP−517)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第5染色体の5,346,476番目の塩基から7,305,955番目の塩基(SP−1526の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−16は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の3,173,133番目の塩基(SP−505)から21,023,551番目の塩基(SP−547)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の1,047,931番目の塩基(SP−495の+1塩基)から3,173,133番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第5染色体の21,023,551番目の塩基から24,520,268番目の塩基(SP−1561の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−17は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の21,023,551番目の塩基(SP−547)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の18,878,115番目の塩基(SP−1546の+1塩基)から21,023,551番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
GLSL−18は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の上流末端から2,186,456番目の塩基(SP−2503)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第6染色体の2,186,456番目の塩基から5,904,223番目の塩基(SP−1596の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−19は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の2,186,456番目の塩基(SP−2503)から22,907,151番目の塩基(SP−1613)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の372,259番目の塩基(SP−590の+1塩基)から2,186,456番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第6染色体の22,907,151番目の塩基から25,711,514番目の塩基(SP−3026の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−20は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の22,907,151番目の塩基(SP−1613)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の20,235,923番目の塩基(SP−3022の+1塩基)から22,907,151番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0035】
GLSL−21は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の上流末端から6,843,696番目の塩基(SP−1648)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第7染色体の6,843,696番目の塩基から10,130,031番目の塩基(SP−3066の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−22は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の1,745,458番目の塩基(SP−2315)から18,571,959番目の塩基(SP−1662)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第7染色体の260,865番目の塩基(SP−676の+1塩基)から1,745,458番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第7染色体の18,571,959番目の塩基から21,942,075番目の塩基(SP−3095の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−23は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の21,942,076番目の塩基(SP−3095)から24,516,006番目の塩基(SP−2545)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第1の染色体断片にホモ置換されており、当該第7染色体の28,847,226番目の塩基(SP−4152)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第2の染色体断片にホモ置換されており、かつ前記第1の染色体断片の上流端が、イネ品種日本晴の第7染色体の18,571,960番目の塩基(SP−1662の+1塩基)から21,942,076番目の塩基までを含む領域に相当する領域に存在し、前記第1の染色体断片の下流端が、イネ品種日本晴の第7染色体の24,516,006番目の塩基から25,490,052番目の塩基(SP−741の−1塩基)までを含む領域に相当する領域に存在し、前記第2の染色体断片の上流端が、イネ品種日本晴の第7染色体の25,490,054番目の塩基(SP−741の+1塩基)から28,847,226番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
GLSL−24は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の上流末端から8,094,582番目の塩基(SP−795)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第8染色体の8,094,582番目の塩基から10,572,008番目の塩基(SP−1707の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−25は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の8,094,582番目の塩基(SP−795)から19,903,905番目の塩基(SP−3199)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の5,278,856番目の塩基(SP−3598の+1塩基)から8,094,582番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第8染色体の19,903,905番目の塩基から23,047,466番目の塩基(SP−2407の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−26は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の18,114,324番目の塩基(SP−809)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の15,484,341番目の塩基(SP−806の+1塩基)から18,114,324番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0036】
GLSL−27は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の上流末端から13,254,003番目の塩基(SP−2180)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第9染色体の13,254,003番目の塩基から15,063,824番目の塩基(SP−1747の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−28は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の13,254,003番目の塩基(SP−2180)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第9染色体の10,483,156番目の塩基(SP−875の+1塩基)から13,254,003番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
GLSL−29は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の上流末端から14,142,299番目の塩基(SP−966)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の下流端が、イネ品種日本晴の第10染色体の14,142,299番目の塩基から17,555,158番目の塩基(SP−981の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−30は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の14,142,299番目の塩基(SP−966)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第10染色体の11,761,139番目の塩基(SP−3357の+1塩基)から14,142,299番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0037】
GLSL−31は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の上流末端から21,896,773番目の塩基(SP−1073)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第11染色体の21,896,773番目の塩基から25,149,243番目の塩基(SP−2678の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−32は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の21,896,773番目の塩基(SP−1073)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、かつ前記染色体断片の上流端が、イネ品種日本晴の第11染色体の19,050,306番目の塩基(SP−1062の+1塩基)から21,896,773番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
GLSL−33は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の上流末端から14,689,883番目の塩基(SP−2687)までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にヘテロ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第12染色体の14,689,883番目の塩基から17,499,934番目の塩基(SP−3747の−1塩基)までを含む領域に相当する領域に存在しているCSSLである。
GLSL−34は、イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の21,257,971番目の塩基(SP−2697)から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第12染色体の17,499,936番目の塩基(SP−3747の+1塩基)から21,257,971番目の塩基までを含む領域に相当する領域に存在しているCSSLである。
【0038】
GLSL−1〜GLSL−34は、後記実施例に記載の方法で作出することができるが、その他、公知のいずれの手法により作成したものであってもよい。例えば、イネ品種コシヒカリとオリザ・グラベリマを交配し、得られた後代個体に対して自家交配又は戻し交配を繰返し、得られた後代個体をジェノタイピングし、その結果に基づいて、染色体中の所望の領域のみがオリザ・グラベリマ由来の染色体領域であり、その他がイネ品種コシヒカリ由来の染色体で構成されているイネ個体を選抜することにより、GLSL−1〜GLSL−34を作出することができる。選抜に用いるDNAマーカーは、表1〜4に記載されているものを用いてもよく、その他のDNAマーカーを用いてもよい。
【0039】
例えば、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基までを含む領域に相当する領域(当該領域の下流端は、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基までを含む領域に相当する領域に存在している。)のみがオリザ・グラベリマ由来の染色体断片にホモ置換されているイネ個体GLSL−1は、図1中のGLSL−1の染色体中で黒塗部分に存在するSNP(SP−1177、SP−155、SP−2077)が両アレルともにオリザ・グラベリマタイプであり、白抜部分に存在するその他のSNPが両アレルともイネ品種コシヒカリタイプであるイネ個体を選抜することにより得られる。
【0040】
同様に、GLSL−2〜GLSL−32及びGLSL−34も、図1中の各染色体中で黒塗部分に存在するSNPが両アレルともにオリザ・グラベリマタイプであり、白抜部分に存在するその他のSNPが両アレルともにイネ品種コシヒカリタイプであるイネ個体を選抜することにより得られる。また、GLSL−33は、図1中の染色体中でハッチング部分に存在するSNPが、一方のアレルがオリザ・グラベリマタイプ、他方のアレルがイネ品種コシヒカリタイプであり、白抜部分に存在するその他のSNPが両アレルともにイネ品種コシヒカリタイプであるイネ個体を選抜することにより得られる。各染色体中の黒塗部分(又はハッチング部分)に存在するSNPを、表5に示す。
【0041】
【表5】
【0042】
GLSL−1〜GLSL−34は、イネ品種コシヒカリと同様の手法により、栽培し、自家交配や人工交配により米を収穫することができる。また、GLSL−1〜GLSL−34及びその後代個体は、イネ品種コシヒカリと同様に、新品種育成の親個体とすることができる。例えば、GLSL−1〜GLSL−34中の互いに異なるCSSL同士を交配したり、GLSL−1〜GLSL−34の各個体と別の品種の個体とを交配することによって、後代個体を得ることができる。さらに、得られた後代個体を、自殖したり、GLSL−1〜GLSL−34の個体と戻し交配することにより、新品種の育種を試みることもできる。
【0043】
また、GLSL−1〜GLSL−34は、公知の染色体部分置換系統と同様に、新品種育成のための材料として用いることができる。GLSL−1〜GLSL−34を材料とした場合の新品種の育種方法としては、例えば、特許文献1等に記載の新品種の製造方法等が挙げられる。
【0044】
鑑別対象であるイネ個体に対して、表1〜4に記載のDNAマーカーの1又は2以上についてジェノタイピングし、得られたタイピング結果に基づき、当該イネ個体が、GLSL−1〜GLSL−34のいずれかであるかを鑑別することができる。表1〜4に記載の142種類全てのDNAマーカーを用いることによって、確実に鑑別することができる。また、142種類全てのDNAマーカーを用いずに鑑別してもよい。全てのDNAマーカーを用いない場合には、鑑別に用いるDNAマーカーは、表5に記載のDNAマーカーを用いることが好ましく、図1の黒塗部分(又はハッチング部分)と白抜き部分との境界付近のものであることがより好ましい。
【0045】
例えば、あるイネ個体がGLSL−2であるか否かは、黒塗部分中の白抜き部分との境界に近いSP−192及びSP−2016、並びに白抜き部分中の黒塗部分との境界に近いSP−185及びSP−1217について遺伝子型をタイピングする。得られたタイピング結果が、SP−192及びSP−2016が両アレルともにオリザ・グラベリマタイプであり、SP−185及びSP−1217が両アレルともにイネ品種コシヒカリタイプであった場合には、当該イネ個体はGLSL−2であり、それ以外のタイピング結果の場合には、当該イネ個体はGLSL−2ではない、と鑑別できる。その他のCSSLについても同様にして鑑別することができる。
【実施例】
【0046】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0047】
[実施例1]
<ゲノムライブラリーの製造>
優秀なジャポニカイネ栽培種であるイネ品種コシヒカリを反復親(種子親)とし、アフリカ型品種オリザ・グラベリマをドナー親(花粉親)として、交配することによって、GLSL−1〜GLSL−34を作出した。また、以下の実施例において、GLSL−1〜GLSL−34を纏めて「GLSLs」といい、個々のCSSLをGLSLということがある。
【0048】
具体的には、まず、イネ品種コシヒカリとオリザ・グラベリマを交配し、得られたF1交雑種を反復親イネ品種コシヒカリと戻し交配してBC1F1世代(54個体)を得、続く反復親との連続戻し交配により、BC2F1世代(54系統の個体)、BC3F1世代(155系統の個体)、BC4F1世代(155系統の個体)、BC5F1世代(112系統の個体)を得た。
次いで、13系統のBC3F1世代、155系統のBC4F1世代、及び112系統のBC5F1世代から、それぞれ10個体ずつ育成し、表1〜4に記載の142種のSNPを、AcycloPrime−FD検出システムと蛍光ポラライゼーションアナライザー(Perkin Elmer Life Sciences社製)によってジェノタイピングした。タイピング結果に基づいて、オリザ・グラベリマ由来の染色体断片を2以上含むGLSLの候補個体として、BC3F1世代から2個体、BC4F1世代から17個体、BC5F1世代から5個体を選抜した。さらにこれらの選抜した個体を戻し交配し、それぞれBC4F1世代、BC5F1世代、又はBC6F1世代を得た。
これらの集団のうち、未だオリザ・グラベリマ由来の染色体断片を2以上含んでいたGLSLの候補個体に対して、反復親との戻し交配を行い、それぞれBC5F1世代、BC6F1世代、又はBC7F1世代を得た。戻し交配により得られたGLSLの候補個体を自殖し、ドナーゲノム(オリザ・グラベリマ)由来の染色体部分置換を固定した。
以上の工程により、本発明のゲノムライブラリーを構成するGLSL−1〜GLSL−34が作出された。
このGLSLの作出工程の概要を、図2に模式的に示した。図2中、括弧内の数値は、各戻し交配世代によって得られた系統数を示し、円で囲まれた数値は、オリザ・グラベリマ由来の染色体断片を2以上含んでいるGLSLの候補個体の数を示す。四角で囲まれた数値は、1又は2のオリザ・グラベリマ由来の染色体断片を含んでいるGLSLの候補個体の数を示し、角括弧内の数値は、先行する戻し交配世代から得られたGLSLの総数を示す。
【0049】
[実施例2]
実施例1で作出されたGLSLsについて、収量性、草型、成熟性、籾外観に関する10形質の有用農業形質の評価を行い、QTLを検出した。GLSLsは、コシヒカリの遺伝的背景をもち、染色体の一部のみがアフリカ型品種オリザ・グラベリマ由来の染色体断片に置換されているため、これらを用いることにより、遺伝的背景を揃えた条件でのQTLの検出を試みた。
【0050】
<GLSLsの形質測定>
まず、GLSLsの様々な農業形質の表現型の観察を、2007年夏、千葉県木更津市研究ほ場にて行った。各GLSLから24の苗を、対照とするイネ品種コシヒカリの苗と共に、ほ場の栽培区画に2条植えし、成熟するまで育てた。苗は、最初温室で栽培し、播種から30日後に、栽培区画に移植した。定植幅は株間20cm、条間30cmで行った。
成熟後、各GLSL及びイネ品種コシヒカリの、それぞれの栽培区画から予め定められた5個体(第1列の3〜7番目の株)を個別に収穫し、籾長(Grain length)、籾幅(Grain width)、籾厚(Grain thickness)、一穂当たりの籾数(Grain number/panicle)、籾の100粒重(100−grain weight)を含む、籾外観形質を調べた。これらの個体について、さらに、稈長(Culm length)、穂長(Panicle length)、一穂当たりの枝こう数(No. of primary branches/panicle)、及び種子稔性(Fertility)といったその他の農業形質についても調べた。各GLSLの出穂日数(Heading date)は、種子の播種から各系統の50%の個体が実際に出穂した日までの日数とした。籾の100粒重は、少なくとも2ヶ月間温室内で風乾させた種子を用いて決定した。
GLSLsについての籾外観形質の測定結果を、その他の重要な農業形質の結果と共に表6及び7に示した。
【0051】
【表6】
【0052】
【表7】
【0053】
<GLSLsの形質評価及びQTL検出>
一元配置分散分析、及び95%信頼区画(P<0.05)によるダネットの多重比較テストを使う比較手法により、GLSLsにおける推定QTLの同定を行った。ある形質の
算出された平均値が、いずれかのGLSLにおいて、イネ品種コシヒカリよりも明らかに相違した場合には、当該GLSLが含有しているオリザ・グラベリマ由来の染色体断片にQTLが存在していると判断された。
【0054】
(籾外観形質)
籾長に対する推定QTLは、オリザ・グラベリマの第1、3、4、及び8染色体由来の置換染色体断片を有する8つのGLSLに検出された。これらのGLSLのうち7系統(GLSL−3、11、12、13、24、25、26)は、籾長が4〜14%ほど、イネ品種コシヒカリよりも有意に長くなっていたが、残る1系統(GLSL−2)は、当該形質に対して負の効果を示すQTLを有していた。つまり、籾長に対し正の効果を示す推定QTLと、籾長に対し負の効果を示す推定QTLの両方が、第1染色体に存在していた。
【0055】
20のGLSL(GLSL−1、4、5、11、12、13、14、20、21、22、23、24、25、26、27、28、30、31、32、33)が、イネ品種コシヒカリ(平均籾幅:3.21mm)と比べてより幅の大きい籾を産生した。これらの系統は、オリザ・グラベリマの第1、3、4、5、6、7、8、9、10、11、及び12染色体由来の断片で置換されており、イネ品種コシヒカリよりも4〜10%も籾幅が有意に増大していた。これに対して、オリザ・グラベリマの第5及び6染色体由来の断片で置換された2系統(GLSL−17、18)では、イネ品種コシヒカリよりも7〜8%も籾幅が有意に減少していた。
【0056】
籾厚は、オリザ・グラベリマの第1、7、及び8染色体由来の断片で置換された4つのGLSL(GLSL−4、22、25、26)では3〜4%も有意に増大していたが、オリザ・グラベリマの第1、5、6、及び7染色体由来の断片で置換された5つのGLSL(GLSL−2、16、17、18,21)では3〜5%も有意に減少していた。
【0057】
一穂当たりの籾粒数に対する推定QTLは、イネ品種コシヒカリに比べて平均値が有意に高い8のGLSLにおいて検出された。一穂当たりの籾粒数における26〜80%の増大が、これらのオリザ・グラベリマの第1、2、及び7染色体由来の断片で置換された系統(GLSL−1、2、3、6、7、8、21、22)において記録された。一方で、当該形質に対して負の効果を示す2つの推定QTLが、オリザ・グラベリマの第5及び8染色体由来の断片で置換された系統(GLSL−16、25)において観察された。
【0058】
3つの系統(GLSL−4、25、31)が、イネ品種コシヒカリ(平均100粒重:2.23g)と比べてより籾の100粒重が有意に高い値を示した。これらの系統は、オリザ・グラベリマの第1、8、及び11染色体由来の断片で置換されており、イネ品種コシヒカリよりも11〜15%も100粒重が有意に増大していた。しかしながら、オリザ・グラベリマの第1、5及び10染色体由来の断片で置換された4系統(GLSL−2、16、17、29)では、当該形質に対して負の効果が観察され、イネ品種コシヒカリよりも7〜12%程度も減少していた。
【0059】
(その他の農業形質)
稈長が有意に低くなることに対する推定QTLは、オリザ・グラベリマの第1、2、4、5、6、8、9、10、11、及び12染色体由来の断片で置換された20のGLSL(GLSL−1、6、7、8、9、14、15、16、17、18、19、20、24、25、27、28、29、30、31、33)において同定された。これらのGLSLの稈長は、イネ品種コシヒカリ(平均稈長:95.72cm)に比べて10〜27%短かった。
【0060】
イネ品種コシヒカリの平均出穂期は、播種から93.20日であったが、9つのGLSL(GLSL−3、11、12、13、21、23、24、25、31)は、イネ品種コシヒカリに比べて有意に出穂期が遅かった。オリザ・グラベリマの第1、3、4、5、及び8染色体から挿入された断片中にある推定QTLが、イネ品種コシヒカリの出穂期を4.6〜18.2日遅らせていた。
【0061】
穂長に対する推定QTLが、イネ品種コシヒカリと比べて穂長の平均値が有意に長い8つのGLSL(GLSL−2、3、4、6、7、8、11、22)において検出された。これらの系統は、オリザ・グラベリマの第1、2、3、及び7染色体由来の断片が挿入されており、穂長が15〜24%増加することが記録された。しかしながら、オリザ・グラベリマの第5染色体由来の断片が挿入された1系統(GLSL−16)は、イネ品種コシヒカリよりも穂長が有意に減少(13%)したことが記録された。
【0062】
穂における枝こう数については、推定QTLが、オリザ・グラベリマの第1、2、3、及び7染色体由来の断片で置換された5つのGLSL(GLSL−2、7、11、12、22)において検出された。これらのGLSLは、イネ品種コシヒカリ(一穂当たりの平均枝こう数:10.40)に比べて、一穂当たりの枝こう数が25〜36%増大していた。
【0063】
種子稔性については、23〜84%の範囲内の有意な減少が観察されたのは6系統(GLSL−12、17、18、21、29、31)であったが、2系統(GLSL−2、19)を除く全てのGLSLが、イネ品種コシヒカリに比べてより低い種子稔性を示した。
【0064】
(GLSLsにおいて観察されたその他の形質)
幾つかのGLSLにおいて明らかに現れていた他の形質についても、同定した。これらの形質は、表現型上測定してQTL解析に影響するものではないが、明らかに主要な遺伝子、又はGLSLsに挿入されたオリザ・グラベリマ由来の染色体断片に存在するQTLと関連している可能性がある。例えば、オリザ・グラベリマの第5染色体のおよそ3.15Mbを含むGLSL−15では、無毛の葉が観察された。無毛の葉を制御する遺伝子は、イネ品種コシヒカリの準同質遺伝子系統を用いることによって、当該染色体断片からより狭い領域にマッピングされ、続いてクローニングされる。その他の同定された形質としては、GLSL−13及び15において長い芒が観察された。GLSL−13及び15は、それぞれ、オリザ・グラベリマの第4及び8染色体由来の断片を含んでいた。
【0065】
農業的特長に対する推定QTLの染色体上の位置を、表8に示す。表8中、「GL」は籾長を、「GW」は籾幅を、「GT」は籾厚を、「GN」は一穂当たりの籾数を、「GWe」は籾の100粒重を、「CL」は稈長を、「PL」は穂長を、「PB」は一穂当たりの枝こう数を、「F」は種子稔性を、「HD」は出穂日数を、それぞれ意味する。
【0066】
【表8】
【0067】
(総括)
GLSLsから、12本の染色体中に分散している全105の推定QTLが同定された。調査された全ての形質は、これらのQTLのうちの5又はそれ以上によって調節されていることがわかった。1の形質の調節に関わる複数のQTLは、同一の染色体上の2〜3部位にある場合もあり、複数の染色体中に分散された位置にある場合もあった。これらのQTLは、染色体の他の部位のみならず、環境とも影響しあう。イネの多くの重要な農業形質は、2〜3のQTLによって調節されているという報告がある。
同定されたQTLは、観察した農業形質に対して正又は負の影響を示した。オリザ・グラベリマの保持する105個のQTLのうち、84個のQTLはイネ品種コシヒカリに対し正の効果を示し、21個のQTLはイネ品種コシヒカリに対し負の効果を示した。正の効果を示した84個のQTLのうち、55個は籾収量に関係する形質に影響しており、20個は植物高さに、9個は成熟期に、それぞれ影響していた。これらの結果から、オリザ・グラベリマ由来の染色体が、現存するイネ品種の収量や草型に関する形質を改良するために有用であることが示唆された。
【0068】
籾長、籾幅、籾厚、出穂期、穂における枝こう数に対するQTLは、オリザ・グラベリマの第3染色体をカバーする2GLSL(GLSL−11、12)において検出された。これらの2系統において導入された染色体断片は、およそ11と28Mbであり、7Mbオーバーラップしていた。
イネの育種に対する有用性の点からは、第1染色体に位置しており、2つのGLSLにおいて同定された2個のQTLが、前途有望と考えられる。両QTLともに一穂当たりの籾粒数を強くコントロールすることが示され、穂長と籾幅に対して多面的な効果も示していた。これとは別に、この2つのQTLは、2つのGLSLにおいて、イネ品種コシヒカリと比べて70%以上も一穂当たりの籾粒数を増大させた。もしも、特に広い生態学的条件下でこれらの機能と発現が確認されたら、GN1A(非特許文献4参照。)、APO1(非特許文献16及び35参照。)、及び最近報告されたWFP及びIPA1(非特許文献26及び18参照。)と同様に、これらのQTLは、クローニングされ、より収量が改善されたイネ品種の育種に使用することができるだろう。
既に同定され報告されている幾つかのオリザ・グラベリマ由来のQTLも、本実施例のGLSLsにおいて同様に検出された。これは、新しく作出された本実施例のGLSLsが、オリザ・グラベリマの全ゲノムを正確に含んでいることを示す。既に報告され、本実施例で確認されたQTLには、不稔性に影響する4つの座(第3、6、7、及び10染色体に位置する。)(非特許文献10、34、15及び22参照。)、及び第1染色体上にあり、出穂期に関係する1のQTL(非特許文献9参照。)が含まれていた。
【0069】
本実施例で作出された34のGLSLは、オリザ・グラベリマが有する大量の多様な遺伝子に対するアクセスを可能にし得る、価値のある遺伝資源であることについては疑いはない。これらGLSLsからは、本実施例において同定された籾収量や草高、成熟性についてのQTLの他にも、より多くの重要な農業形質を調節する遺伝子が発見されるであろう。アフリカ型イネは、最も厳しい環境下でも良好に成育することが知られており、生物的及び非生物的ストレスに対する有用な新規遺伝子を含んでいる可能性は高い。害虫及び病気に対する抵抗性についてのGLSLsの評価については、問題のある土壌や極限環境条件が、これらの有用な遺伝子の同定を容易にするだろう。
【産業上の利用可能性】
【0070】
本発明のゲノムライブラリー、及びこれを構成する各CSSLは、有用農業形質を調節するQTLの検出に特に好適に利用することができ、さらに検出されたQTLのピラミティングが、シンクサイズ等の形質が改善された新品種育成にも利用することができるため、特に農業の分野において利用が可能である。
【特許請求の範囲】
【請求項1】
下記(1)〜(34)の、イネ品種コシヒカリの染色体の一部がオリザ・グラベリマの染色体断片で置換された染色体部分置換系統を有することを特徴とするゲノムライブラリー;
(1)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(2)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の11,026,011番目の塩基から20,366,961番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の8,896,816番目の塩基から11,026,011番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の20,366,961番目の塩基から24,412,980番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(3)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の17,803,741番目の塩基から31,371,175番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の13,813,982番目の塩基から17,803,741番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の31,371,175番目の塩基から35,598,471番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(4)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の25,979,477番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の24,412,982番目の塩基から25,979,477番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の35,598,472番目の塩基から39,677,013番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(5)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の35,598,472番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の31,371,176番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(6)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の上流末端から3,922,571番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第2染色体の3,922,571番目の塩基から7,030,896番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(7)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の3,922,571番目の塩基から14,655,638番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の2,010,335番目の塩基から3,922,571番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の14,655,638番目の塩基から18,765,983番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(8)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の11,165,933番目の塩基から22,086,348番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の9,256,585番目の塩基から11,165,933番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の22,086,348番目の塩基から25,263,252番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(9)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の18,765,984番目の塩基から25,263,253番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の14,655,639番目の塩基から18,765,984番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の25,263,253番目の塩基から27,919,902番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(10)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の30,860,986番目の塩基から33,189,264番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の29,345,358番目の塩基から30,860,986番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の33,189,264番目の塩基から35,267,218番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(11)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の上流末端から11,073,623番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第3染色体の11,073,623番目の塩基から14,311,069番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(12)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の11,073,623番目の塩基から33,640,115番目の塩基までを 含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第3染色体の8,197,229番目の塩基から11,073,623番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第3染色体の33,640,115番目の塩基から35,945,010番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(13)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の上流末端から22,125,293番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第4染色体の22,125,293番目の塩基から25,877,140番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(14)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の19,650,008番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第4染色体の14,903,263番目の塩基から19,650,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(15)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の上流末端から5,346,476番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第5染色体の5,346,476番目の塩基から7,305,955番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(16)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の3,173,133番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の1,047,931番目の塩基から3,173,133番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第5染色体の21,023,551番目の塩基から24,520,268番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(17)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の21,023,551番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の18,878,115番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(18)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の上流末端から2,186,456番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第6染色体の2,186,456番目の塩基から5,904,223番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(19)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の2,186,456番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の372,259番目の塩基から2,186,456番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第6染色体の22,907,151番目の塩基から25,711,514番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(20)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の22,907,151番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の20,235,923番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(21)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の上流末端から6,843,696番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第7染色体の6,843,696番目の塩基から10,130,031番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(22)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の1,745,458番目の塩基から18,571,959番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第7染色体の260,865番目の塩基から1,745,458番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第7染色体の18,571,959番目の塩基から21,942,075番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(23)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の21,942,076番目の塩基から24,516,006番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第1の染色体断片にホモ置換されており、
当該第7染色体の28,847,226番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第2の染色体断片にホモ置換されており、
かつ前記第1の染色体断片の上流端が、イネ品種日本晴の第7染色体の18,571,960番目の塩基から21,942,076番目の塩基までを含む領域に相当する領域に存在し、前記第1の染色体断片の下流端が、イネ品種日本晴の第7染色体の24,516,006番目の塩基から25,490,052番目の塩基までを含む領域に相当する領域に存在し、
前記第2の染色体断片の上流端が、イネ品種日本晴の第7染色体の25,490,054番目の塩基から28,847,226番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(24)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の上流末端から8,094,582番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第8染色体の8,094,582番目の塩基から10,572,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(25)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の8,094,582番目の塩基から19,903,905番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の5,278,856番目の塩基から8,094,582番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第8染色体の19,903,905番目の塩基から23,047,466番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(26)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の18,114,324番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の15,484,341番目の塩基から18,114,324番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(27)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の上流末端から13,254,003番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第9染色体の13,254,003番目の塩基から15,063,824番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(28)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の13,254,003番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第9染色体の10,483,156番目の塩基から13,254,003番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(29)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の上流末端から14,142,299番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第10染色体の14,142,299番目の塩基から17,555,158番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(30)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の14,142,299番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第10染色体の11,761,139番目の塩基から14,142,299番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(31)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の上流末端から21,896,773番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第11染色体の21,896,773番目の塩基から25,149,243番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(32)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の21,896,773番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第11染色体の19,050,306番目の塩基から21,896,773番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(33)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の上流末端から14,689,883番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にヘテロ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第12染色体の14,689,883番目の塩基から17,499,934番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(34)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の21,257,971番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第12染色体の17,499,936番目の塩基から21,257,971番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統。
【請求項2】
オリザ・グラベリマの染色体中、前記(1)〜(34)のいずれかの染色体部分置換系統の染色体中に含有されている領域が、全染色体の90%以上であることを特徴とする請求項1に記載のゲノムライブラリー。
【請求項3】
(1)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(2)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の11,026,011番目の塩基から20,366,961番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の8,896,816番目の塩基から11,026,011番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の20,366,961番目の塩基から24,412,980番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(3)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の17,803,741番目の塩基から31,371,175番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の13,813,982番目の塩基から17,803,741番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の31,371,175番目の塩基から35,598,471番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(4)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の25,979,477番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の24,412,982番目の塩基から25,979,477番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の35,598,472番目の塩基から39,677,013番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(5)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の35,598,472番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の31,371,176番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(6)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の上流末端から3,922,571番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第2染色体の3,922,571番目の塩基から7,030,896番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(7)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の3,922,571番目の塩基から14,655,638番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の2,010,335番目の塩基から3,922,571番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の14,655,638番目の塩基から18,765,983番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(8)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の11,165,933番目の塩基から22,086,348番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の9,256,585番目の塩基から11,165,933番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の22,086,348番目の塩基から25,263,252番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(9)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の18,765,984番目の塩基から25,263,253番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の14,655,639番目の塩基から18,765,984番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の25,263,253番目の塩基から27,919,902番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(10)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の30,860,986番目の塩基から33,189,264番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の29,345,358番目の塩基から30,860,986番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の33,189,264番目の塩基から35,267,218番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(11)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の上流末端から11,073,623番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第3染色体の11,073,623番目の塩基から14,311,069番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(12)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の11,073,623番目の塩基から33,640,115番目の塩基までを 含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第3染色体の8,197,229番目の塩基から11,073,623番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第3染色体の33,640,115番目の塩基から35,945,010番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(13)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の上流末端から22,125,293番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第4染色体の22,125,293番目の塩基から25,877,140番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(14)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の19,650,008番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第4染色体の14,903,263番目の塩基から19,650,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(15)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の上流末端から5,346,476番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第5染色体の5,346,476番目の塩基から7,305,955番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(16)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の3,173,133番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の1,047,931番目の塩基から3,173,133番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第5染色体の21,023,551番目の塩基から24,520,268番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(17)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の21,023,551番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の18,878,115番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(18)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の上流末端から2,186,456番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第6染色体の2,186,456番目の塩基から5,904,223番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(19)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の2,186,456番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の372,259番目の塩基から2,186,456番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第6染色体の22,907,151番目の塩基から25,711,514番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(20)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の22,907,151番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の20,235,923番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(21)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の上流末端から6,843,696番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第7染色体の6,843,696番目の塩基から10,130,031番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(22)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の1,745,458番目の塩基から18,571,959番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第7染色体の260,865番目の塩基から1,745,458番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第7染色体の18,571,959番目の塩基から21,942,075番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(23)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の21,942,076番目の塩基から24,516,006番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第1の染色体断片にホモ置換されており、
当該第7染色体の28,847,226番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第2の染色体断片にホモ置換されており、
かつ前記第1の染色体断片の上流端が、イネ品種日本晴の第7染色体の18,571,960番目の塩基から21,942,076番目の塩基までを含む領域に相当する領域に存在し、前記第1の染色体断片の下流端が、イネ品種日本晴の第7染色体の24,516,006番目の塩基から25,490,052番目の塩基までを含む領域に相当する領域に存在し、
前記第2の染色体断片の上流端が、イネ品種日本晴の第7染色体の25,490,054番目の塩基から28,847,226番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(24)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の上流末端から8,094,582番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第8染色体の8,094,582番目の塩基から10,572,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(25)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の8,094,582番目の塩基から19,903,905番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の5,278,856番目の塩基から8,094,582番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第8染色体の19,903,905番目の塩基から23,047,466番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(26)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の18,114,324番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の15,484,341番目の塩基から18,114,324番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(27)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の上流末端から13,254,003番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第9染色体の13,254,003番目の塩基から15,063,824番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(28)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の13,254,003番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第9染色体の10,483,156番目の塩基から13,254,003番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(29)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の上流末端から14,142,299番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第10染色体の14,142,299番目の塩基から17,555,158番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(30)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の14,142,299番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第10染色体の11,761,139番目の塩基から14,142,299番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(31)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の上流末端から21,896,773番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第11染色体の21,896,773番目の塩基から25,149,243番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(32)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の21,896,773番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第11染色体の19,050,306番目の塩基から21,896,773番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(33)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の上流末端から14,689,883番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にヘテロ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第12染色体の14,689,883番目の塩基から17,499,934番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、又は
(34)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の21,257,971番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第12染色体の17,499,936番目の塩基から21,257,971番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統
であることを特徴とするイネ。
【請求項4】
請求項3記載のイネの個体と、請求項3記載のイネの個体又は請求項3記載のイネ以外のイネの個体とを交配して得られる後代個体。
【請求項5】
下記(1−1)〜(12−9)のSNPからなる群より選択される1以上のDNAマーカーをタイピングすることを特徴とする、イネのジェノタイピング方法;
(1−1)イネ品種日本晴の第1染色体の590,339番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(1−2)イネ品種日本晴の第1染色体の1,946,270番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(1−3)イネ品種日本晴の第1染色体の6,522,242番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(1−4)イネ品種日本晴の第1染色体の8,896,815番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(1−5)イネ品種日本晴の第1染色体の11,026,011番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(1−6)イネ品種日本晴の第1染色体の13,813,981番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(1−7)イネ品種日本晴の第1染色体の17,803,741番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(1−8)イネ品種日本晴の第1染色体の20,366,961番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(1−9)イネ品種日本晴の第1染色体の24,412,981番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(1−10)イネ品種日本晴の第1染色体の25,979,477番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(1−11)イネ品種日本晴の第1染色体の27,840,561番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(1−12)イネ品種日本晴の第1染色体の31,371,175番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(1−13)イネ品種日本晴の第1染色体の35,598,472番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(1−14)イネ品種日本晴の第1染色体の39,677,014番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(1−15)イネ品種日本晴の第1染色体の42,000,585番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(2−1)イネ品種日本晴の第2染色体の151,976番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(2−2)イネ品種日本晴の第2染色体の2,010,334番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(2−3)イネ品種日本晴の第2染色体の3,922,571番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(2−4)イネ品種日本晴の第2染色体の7,030,897番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(2−5)イネ品種日本晴の第2染色体の9,256,584番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではC)、
(2−6)イネ品種日本晴の第2染色体の11,165,933番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(2−7)イネ品種日本晴の第2染色体の14,655,638番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(2−8)イネ品種日本晴の第2染色体の18,765,984番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(2−9)イネ品種日本晴の第2染色体の22,086,348番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(2−10)イネ品種日本晴の第2染色体の25,263,253番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(2−11)イネ品種日本晴の第2染色体の27,919,903番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(2−12)イネ品種日本晴の第2染色体の29,345,357番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(2−13)イネ品種日本晴の第2染色体の30,860,986番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではC)、
(2−14)イネ品種日本晴の第2染色体の33,189,264番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(2−15)イネ品種日本晴の第2染色体の35,267,219番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(3−1)イネ品種日本晴の第3染色体の577,726番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(3−2)イネ品種日本晴の第3染色体の3,979,349番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(3−3)イネ品種日本晴の第3染色体の6,252,925番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(3−4)イネ品種日本晴の第3染色体の8,197,228番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(3−5)イネ品種日本晴の第3染色体の11,073,623番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(3−6)イネ品種日本晴の第3染色体の14,311,070番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(3−7)イネ品種日本晴の第3染色体の19,846,318番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(3−8)イネ品種日本晴の第3染色体の22,287,129番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(3−9)イネ品種日本晴の第3染色体の24,639,787番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(3−10)イネ品種日本晴の第3染色体の26,784,345番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(3−11)イネ品種日本晴の第3染色体の30,021,442番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(3−12)イネ品種日本晴の第3染色体の31,580,078番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(3−13)イネ品種日本晴の第3染色体の33,640,115番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(3−14)イネ品種日本晴の第3染色体の35,945,011番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(4−1)イネ品種日本晴の第4染色体の218,809番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(4−2)イネ品種日本晴の第4染色体の2,471,151番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(4−3)イネ品種日本晴の第4染色体の7,120,309番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(4−4)イネ品種日本晴の第4染色体の12,350,862番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(4−5)イネ品種日本晴の第4染色体の14,903,262番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(4−6)イネ品種日本晴の第4染色体の19,650,008番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(4−7)イネ品種日本晴の第4染色体の22,125,293番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(4−8)イネ品種日本晴の第4染色体の25,877,141番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(4−9)イネ品種日本晴の第4染色体の28,542,313番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(4−10)イネ品種日本晴の第4染色体の32,633,649番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(4−11)イネ品種日本晴の第4染色体の34,498,729番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(5−1)イネ品種日本晴の第5染色体の40,031番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(5−2)イネ品種日本晴の第5染色体の1,047,930番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(5−3)イネ品種日本晴の第5染色体の3,173,133番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(5−4)イネ品種日本晴の第5染色体の5,346,476番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(5−5)イネ品種日本晴の第5染色体の7,305,956番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(5−6)イネ品種日本晴の第5染色体の12,908,872番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではC)、
(5−7)イネ品種日本晴の第5染色体の15,459,785番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではC)、
(5−8)イネ品種日本晴の第5染色体の18,878,114番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(5−9)イネ品種日本晴の第5染色体の21,023,551番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(5−10)イネ品種日本晴の第5染色体の24,520,269番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(5−11)イネ品種日本晴の第5染色体の26,499,154番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(5−12)イネ品種日本晴の第5染色体の28,699,770番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(6−1)イネ品種日本晴の第6染色体の372,258番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(6−2)イネ品種日本晴の第6染色体の2,186,456番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(6−3)イネ品種日本晴の第6染色体の5,904,224番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(6−4)イネ品種日本晴の第6染色体の7,974,364番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(6−5)イネ品種日本晴の第6染色体の11,653,694番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(6−6)イネ品種日本晴の第6染色体の13,689,109番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(6−7)イネ品種日本晴の第6染色体の16,525,923番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(6−8)イネ品種日本晴の第6染色体の20,235,922番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(6−9)イネ品種日本晴の第6染色体の22,907,151番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(6−10)イネ品種日本晴の第6染色体の25,711,515番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(6−11)イネ品種日本晴の第6染色体の28,329,768番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではC)、
(6−12)イネ品種日本晴の第6染色体の29,901,305番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(7−1)イネ品種日本晴の第7染色体の260,864番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(7−2)イネ品種日本晴の第7染色体の1,745,458番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(7−3)イネ品種日本晴の第7染色体の3,898,132番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(7−4)イネ品種日本晴の第7染色体の6,843,696番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(7−5)イネ品種日本晴の第7染色体の10,130,032番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(7−6)イネ品種日本晴の第7染色体の13,302,019番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(7−7)イネ品種日本晴の第7染色体の17,327,528番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(7−8)イネ品種日本晴の第7染色体の18,571,959番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(7−9)イネ品種日本晴の第7染色体の21,942,076番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(7−10)イネ品種日本晴の第7染色体の24,516,006番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(7−11)イネ品種日本晴の第7染色体の25,490,053番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(7−12)イネ品種日本晴の第7染色体の28,847,226番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(8−1)イネ品種日本晴の第8染色体の454,665番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(8−2)イネ品種日本晴の第8染色体の1,669,653番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(8−3)イネ品種日本晴の第8染色体の5,278,855番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(8−4)イネ品種日本晴の第8染色体の8,094,582番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(8−5)イネ品種日本晴の第8染色体の10,572,009番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(8−6)イネ品種日本晴の第8染色体の12,249,318番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(8−7)イネ品種日本晴の第8染色体の15,484,340番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(8−8)イネ品種日本晴の第8染色体の18,114,324番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(8−9)イネ品種日本晴の第8染色体の19,903,905番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(8−10)イネ品種日本晴の第8染色体の23,047,467番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(8−11)イネ品種日本晴の第8染色体の25,921,712番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではC)、
(8−12)イネ品種日本晴の第8染色体の27,758,654番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−1)イネ品種日本晴の第9染色体の29,179番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(9−2)イネ品種日本晴の第9染色体の676,400番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(9−3)イネ品種日本晴の第9染色体の3,066,192番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(9−4)イネ品種日本晴の第9染色体の5,167,966番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−5)イネ品種日本晴の第9染色体の6,999,573番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(9−6)イネ品種日本晴の第9染色体の10,483,155番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−7)イネ品種日本晴の第9染色体の13,254,003番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−8)イネ品種日本晴の第9染色体の15,063,825番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(9−9)イネ品種日本晴の第9染色体の17,880,593番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−10)イネ品種日本晴の第9染色体の20,652,100番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−1)イネ品種日本晴の第10染色体の94,506番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではC)、
(10−2)イネ品種日本晴の第10染色体の3,643,898番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(10−3)イネ品種日本晴の第10染色体の7,069,227番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(10−4)イネ品種日本晴の第10染色体の9,791,595番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−5)イネ品種日本晴の第10染色体の11,761,138番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−6)イネ品種日本晴の第10染色体の14,142,299番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−7)イネ品種日本晴の第10染色体の17,555,159番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−8)イネ品種日本晴の第10染色体の19,468,326番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではC)、
(10−9)イネ品種日本晴の第10染色体の21,612,698番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(10−10)イネ品種日本晴の第10染色体の22,686,557番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(11−1)イネ品種日本晴の第11染色体の268,939番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(11−2)イネ品種日本晴の第11染色体の2,489,967番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(11−3)イネ品種日本晴の第11染色体の3,979,301番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(11−4)イネ品種日本晴の第11染色体の7,481,468番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(11−5)イネ品種日本晴の第11染色体の9,892,301番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(11−6)イネ品種日本晴の第11染色体の16,132,370番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(11−7)イネ品種日本晴の第11染色体の19,050,305番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(11−8)イネ品種日本晴の第11染色体の21,896,773番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(11−9)イネ品種日本晴の第11染色体の25,149,244番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(11−10)イネ品種日本晴の第11染色体の27,560,928番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(12−1)イネ品種日本晴の第12染色体の240,790番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(12−2)イネ品種日本晴の第12染色体の2,930,909番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(12−3)イネ品種日本晴の第12染色体の7,634,664番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(12−4)イネ品種日本晴の第12染色体の10,417,940番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(12−5)イネ品種日本晴の第12染色体の14,689,883番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(12−6)イネ品種日本晴の第12染色体の17,499,935番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(12−7)イネ品種日本晴の第12染色体の21,257,971番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(12−8)イネ品種日本晴の第12染色体の23,828,918番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(12−9)イネ品種日本晴の第12染色体の27,011,209番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)。
【請求項6】
あるイネ個体が、請求項3に記載のイネ品種コシヒカリの染色体部分置換系統であるか否かを鑑別する方法であって、
鑑別対象であるイネ個体に対して、請求項5に記載のイネ個体のジェノタイピング方法を行い、得られたタイピング結果に基づき、当該イネ個体が請求項3に記載のイネ品種コシヒカリの染色体部分置換系統であるか否かを鑑別することを特徴とする、イネの鑑別方法。
【請求項1】
下記(1)〜(34)の、イネ品種コシヒカリの染色体の一部がオリザ・グラベリマの染色体断片で置換された染色体部分置換系統を有することを特徴とするゲノムライブラリー;
(1)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(2)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の11,026,011番目の塩基から20,366,961番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の8,896,816番目の塩基から11,026,011番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の20,366,961番目の塩基から24,412,980番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(3)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の17,803,741番目の塩基から31,371,175番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の13,813,982番目の塩基から17,803,741番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の31,371,175番目の塩基から35,598,471番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(4)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の25,979,477番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の24,412,982番目の塩基から25,979,477番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の35,598,472番目の塩基から39,677,013番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(5)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の35,598,472番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の31,371,176番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(6)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の上流末端から3,922,571番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第2染色体の3,922,571番目の塩基から7,030,896番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(7)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の3,922,571番目の塩基から14,655,638番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の2,010,335番目の塩基から3,922,571番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の14,655,638番目の塩基から18,765,983番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(8)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の11,165,933番目の塩基から22,086,348番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の9,256,585番目の塩基から11,165,933番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の22,086,348番目の塩基から25,263,252番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(9)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の18,765,984番目の塩基から25,263,253番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の14,655,639番目の塩基から18,765,984番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の25,263,253番目の塩基から27,919,902番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(10)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の30,860,986番目の塩基から33,189,264番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の29,345,358番目の塩基から30,860,986番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の33,189,264番目の塩基から35,267,218番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(11)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の上流末端から11,073,623番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第3染色体の11,073,623番目の塩基から14,311,069番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(12)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の11,073,623番目の塩基から33,640,115番目の塩基までを 含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第3染色体の8,197,229番目の塩基から11,073,623番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第3染色体の33,640,115番目の塩基から35,945,010番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(13)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の上流末端から22,125,293番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第4染色体の22,125,293番目の塩基から25,877,140番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(14)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の19,650,008番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第4染色体の14,903,263番目の塩基から19,650,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(15)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の上流末端から5,346,476番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第5染色体の5,346,476番目の塩基から7,305,955番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(16)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の3,173,133番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の1,047,931番目の塩基から3,173,133番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第5染色体の21,023,551番目の塩基から24,520,268番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(17)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の21,023,551番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の18,878,115番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(18)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の上流末端から2,186,456番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第6染色体の2,186,456番目の塩基から5,904,223番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(19)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の2,186,456番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の372,259番目の塩基から2,186,456番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第6染色体の22,907,151番目の塩基から25,711,514番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(20)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の22,907,151番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の20,235,923番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(21)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の上流末端から6,843,696番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第7染色体の6,843,696番目の塩基から10,130,031番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(22)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の1,745,458番目の塩基から18,571,959番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第7染色体の260,865番目の塩基から1,745,458番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第7染色体の18,571,959番目の塩基から21,942,075番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(23)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の21,942,076番目の塩基から24,516,006番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第1の染色体断片にホモ置換されており、
当該第7染色体の28,847,226番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第2の染色体断片にホモ置換されており、
かつ前記第1の染色体断片の上流端が、イネ品種日本晴の第7染色体の18,571,960番目の塩基から21,942,076番目の塩基までを含む領域に相当する領域に存在し、前記第1の染色体断片の下流端が、イネ品種日本晴の第7染色体の24,516,006番目の塩基から25,490,052番目の塩基までを含む領域に相当する領域に存在し、
前記第2の染色体断片の上流端が、イネ品種日本晴の第7染色体の25,490,054番目の塩基から28,847,226番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(24)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の上流末端から8,094,582番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第8染色体の8,094,582番目の塩基から10,572,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(25)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の8,094,582番目の塩基から19,903,905番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の5,278,856番目の塩基から8,094,582番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第8染色体の19,903,905番目の塩基から23,047,466番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(26)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の18,114,324番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の15,484,341番目の塩基から18,114,324番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(27)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の上流末端から13,254,003番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第9染色体の13,254,003番目の塩基から15,063,824番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(28)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の13,254,003番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第9染色体の10,483,156番目の塩基から13,254,003番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(29)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の上流末端から14,142,299番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第10染色体の14,142,299番目の塩基から17,555,158番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(30)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の14,142,299番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第10染色体の11,761,139番目の塩基から14,142,299番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(31)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の上流末端から21,896,773番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第11染色体の21,896,773番目の塩基から25,149,243番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(32)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の21,896,773番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第11染色体の19,050,306番目の塩基から21,896,773番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(33)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の上流末端から14,689,883番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にヘテロ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第12染色体の14,689,883番目の塩基から17,499,934番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(34)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の21,257,971番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第12染色体の17,499,936番目の塩基から21,257,971番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統。
【請求項2】
オリザ・グラベリマの染色体中、前記(1)〜(34)のいずれかの染色体部分置換系統の染色体中に含有されている領域が、全染色体の90%以上であることを特徴とする請求項1に記載のゲノムライブラリー。
【請求項3】
(1)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の上流末端から6,522,242番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第1染色体の6,522,242番目の塩基から8,896,814番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(2)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の11,026,011番目の塩基から20,366,961番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の8,896,816番目の塩基から11,026,011番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の20,366,961番目の塩基から24,412,980番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(3)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の17,803,741番目の塩基から31,371,175番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の13,813,982番目の塩基から17,803,741番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の31,371,175番目の塩基から35,598,471番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(4)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の25,979,477番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の24,412,982番目の塩基から25,979,477番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第1染色体の35,598,472番目の塩基から39,677,013番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(5)イネ品種コシヒカリの染色体中、イネ品種日本晴の第1染色体の35,598,472番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第1染色体の31,371,176番目の塩基から35,598,472番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(6)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の上流末端から3,922,571番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第2染色体の3,922,571番目の塩基から7,030,896番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(7)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の3,922,571番目の塩基から14,655,638番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の2,010,335番目の塩基から3,922,571番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の14,655,638番目の塩基から18,765,983番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(8)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の11,165,933番目の塩基から22,086,348番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の9,256,585番目の塩基から11,165,933番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の22,086,348番目の塩基から25,263,252番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(9)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の18,765,984番目の塩基から25,263,253番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の14,655,639番目の塩基から18,765,984番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の25,263,253番目の塩基から27,919,902番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(10)イネ品種コシヒカリの染色体中、イネ品種日本晴の第2染色体の30,860,986番目の塩基から33,189,264番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第2染色体の29,345,358番目の塩基から30,860,986番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第2染色体の33,189,264番目の塩基から35,267,218番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(11)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の上流末端から11,073,623番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第3染色体の11,073,623番目の塩基から14,311,069番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(12)イネ品種コシヒカリの染色体中、イネ品種日本晴の第3染色体の11,073,623番目の塩基から33,640,115番目の塩基までを 含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第3染色体の8,197,229番目の塩基から11,073,623番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第3染色体の33,640,115番目の塩基から35,945,010番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(13)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の上流末端から22,125,293番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第4染色体の22,125,293番目の塩基から25,877,140番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(14)イネ品種コシヒカリの染色体中、イネ品種日本晴の第4染色体の19,650,008番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第4染色体の14,903,263番目の塩基から19,650,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(15)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の上流末端から5,346,476番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第5染色体の5,346,476番目の塩基から7,305,955番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(16)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の3,173,133番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の1,047,931番目の塩基から3,173,133番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第5染色体の21,023,551番目の塩基から24,520,268番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(17)イネ品種コシヒカリの染色体中、イネ品種日本晴の第5染色体の21,023,551番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第5染色体の18,878,115番目の塩基から21,023,551番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(18)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の上流末端から2,186,456番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第6染色体の2,186,456番目の塩基から5,904,223番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(19)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の2,186,456番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の372,259番目の塩基から2,186,456番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第6染色体の22,907,151番目の塩基から25,711,514番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(20)イネ品種コシヒカリの染色体中、イネ品種日本晴の第6染色体の22,907,151番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第6染色体の20,235,923番目の塩基から22,907,151番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(21)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の上流末端から6,843,696番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第7染色体の6,843,696番目の塩基から10,130,031番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(22)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の1,745,458番目の塩基から18,571,959番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第7染色体の260,865番目の塩基から1,745,458番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第7染色体の18,571,959番目の塩基から21,942,075番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(23)イネ品種コシヒカリの染色体中、イネ品種日本晴の第7染色体の21,942,076番目の塩基から24,516,006番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第1の染色体断片にホモ置換されており、
当該第7染色体の28,847,226番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる第2の染色体断片にホモ置換されており、
かつ前記第1の染色体断片の上流端が、イネ品種日本晴の第7染色体の18,571,960番目の塩基から21,942,076番目の塩基までを含む領域に相当する領域に存在し、前記第1の染色体断片の下流端が、イネ品種日本晴の第7染色体の24,516,006番目の塩基から25,490,052番目の塩基までを含む領域に相当する領域に存在し、
前記第2の染色体断片の上流端が、イネ品種日本晴の第7染色体の25,490,054番目の塩基から28,847,226番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(24)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の上流末端から8,094,582番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第8染色体の8,094,582番目の塩基から10,572,008番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(25)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の8,094,582番目の塩基から19,903,905番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の5,278,856番目の塩基から8,094,582番目の塩基までを含む領域に相当する領域に存在し、前記染色体断片の下流端が、イネ品種日本晴の第8染色体の19,903,905番目の塩基から23,047,466番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(26)イネ品種コシヒカリの染色体中、イネ品種日本晴の第8染色体の18,114,324番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第8染色体の15,484,341番目の塩基から18,114,324番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(27)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の上流末端から13,254,003番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第9染色体の13,254,003番目の塩基から15,063,824番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(28)イネ品種コシヒカリの染色体中、イネ品種日本晴の第9染色体の13,254,003番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第9染色体の10,483,156番目の塩基から13,254,003番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(29)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の上流末端から14,142,299番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第10染色体の14,142,299番目の塩基から17,555,158番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(30)イネ品種コシヒカリの染色体中、イネ品種日本晴の第10染色体の14,142,299番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第10染色体の11,761,139番目の塩基から14,142,299番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(31)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の上流末端から21,896,773番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第11染色体の21,896,773番目の塩基から25,149,243番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(32)イネ品種コシヒカリの染色体中、イネ品種日本晴の第11染色体の21,896,773番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第11染色体の19,050,306番目の塩基から21,896,773番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、
(33)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の上流末端から14,689,883番目の塩基までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にヘテロ置換されており、
かつ前記染色体断片の下流端が、イネ品種日本晴の第12染色体の14,689,883番目の塩基から17,499,934番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統、又は
(34)イネ品種コシヒカリの染色体中、イネ品種日本晴の第12染色体の21,257,971番目の塩基から下流末端までを含む領域に相当する領域が、オリザ・グラベリマの当該領域からなる染色体断片にホモ置換されており、
かつ前記染色体断片の上流端が、イネ品種日本晴の第12染色体の17,499,936番目の塩基から21,257,971番目の塩基までを含む領域に相当する領域に存在している染色体部分置換系統
であることを特徴とするイネ。
【請求項4】
請求項3記載のイネの個体と、請求項3記載のイネの個体又は請求項3記載のイネ以外のイネの個体とを交配して得られる後代個体。
【請求項5】
下記(1−1)〜(12−9)のSNPからなる群より選択される1以上のDNAマーカーをタイピングすることを特徴とする、イネのジェノタイピング方法;
(1−1)イネ品種日本晴の第1染色体の590,339番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(1−2)イネ品種日本晴の第1染色体の1,946,270番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(1−3)イネ品種日本晴の第1染色体の6,522,242番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(1−4)イネ品種日本晴の第1染色体の8,896,815番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(1−5)イネ品種日本晴の第1染色体の11,026,011番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(1−6)イネ品種日本晴の第1染色体の13,813,981番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(1−7)イネ品種日本晴の第1染色体の17,803,741番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(1−8)イネ品種日本晴の第1染色体の20,366,961番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(1−9)イネ品種日本晴の第1染色体の24,412,981番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(1−10)イネ品種日本晴の第1染色体の25,979,477番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(1−11)イネ品種日本晴の第1染色体の27,840,561番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(1−12)イネ品種日本晴の第1染色体の31,371,175番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(1−13)イネ品種日本晴の第1染色体の35,598,472番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(1−14)イネ品種日本晴の第1染色体の39,677,014番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(1−15)イネ品種日本晴の第1染色体の42,000,585番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(2−1)イネ品種日本晴の第2染色体の151,976番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(2−2)イネ品種日本晴の第2染色体の2,010,334番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(2−3)イネ品種日本晴の第2染色体の3,922,571番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(2−4)イネ品種日本晴の第2染色体の7,030,897番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(2−5)イネ品種日本晴の第2染色体の9,256,584番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではC)、
(2−6)イネ品種日本晴の第2染色体の11,165,933番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(2−7)イネ品種日本晴の第2染色体の14,655,638番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(2−8)イネ品種日本晴の第2染色体の18,765,984番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(2−9)イネ品種日本晴の第2染色体の22,086,348番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(2−10)イネ品種日本晴の第2染色体の25,263,253番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(2−11)イネ品種日本晴の第2染色体の27,919,903番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(2−12)イネ品種日本晴の第2染色体の29,345,357番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(2−13)イネ品種日本晴の第2染色体の30,860,986番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではC)、
(2−14)イネ品種日本晴の第2染色体の33,189,264番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(2−15)イネ品種日本晴の第2染色体の35,267,219番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(3−1)イネ品種日本晴の第3染色体の577,726番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(3−2)イネ品種日本晴の第3染色体の3,979,349番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(3−3)イネ品種日本晴の第3染色体の6,252,925番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(3−4)イネ品種日本晴の第3染色体の8,197,228番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(3−5)イネ品種日本晴の第3染色体の11,073,623番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(3−6)イネ品種日本晴の第3染色体の14,311,070番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(3−7)イネ品種日本晴の第3染色体の19,846,318番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(3−8)イネ品種日本晴の第3染色体の22,287,129番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(3−9)イネ品種日本晴の第3染色体の24,639,787番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(3−10)イネ品種日本晴の第3染色体の26,784,345番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(3−11)イネ品種日本晴の第3染色体の30,021,442番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(3−12)イネ品種日本晴の第3染色体の31,580,078番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(3−13)イネ品種日本晴の第3染色体の33,640,115番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(3−14)イネ品種日本晴の第3染色体の35,945,011番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(4−1)イネ品種日本晴の第4染色体の218,809番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(4−2)イネ品種日本晴の第4染色体の2,471,151番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(4−3)イネ品種日本晴の第4染色体の7,120,309番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(4−4)イネ品種日本晴の第4染色体の12,350,862番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(4−5)イネ品種日本晴の第4染色体の14,903,262番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(4−6)イネ品種日本晴の第4染色体の19,650,008番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(4−7)イネ品種日本晴の第4染色体の22,125,293番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(4−8)イネ品種日本晴の第4染色体の25,877,141番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(4−9)イネ品種日本晴の第4染色体の28,542,313番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(4−10)イネ品種日本晴の第4染色体の32,633,649番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(4−11)イネ品種日本晴の第4染色体の34,498,729番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(5−1)イネ品種日本晴の第5染色体の40,031番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(5−2)イネ品種日本晴の第5染色体の1,047,930番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(5−3)イネ品種日本晴の第5染色体の3,173,133番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(5−4)イネ品種日本晴の第5染色体の5,346,476番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(5−5)イネ品種日本晴の第5染色体の7,305,956番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(5−6)イネ品種日本晴の第5染色体の12,908,872番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではC)、
(5−7)イネ品種日本晴の第5染色体の15,459,785番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではC)、
(5−8)イネ品種日本晴の第5染色体の18,878,114番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(5−9)イネ品種日本晴の第5染色体の21,023,551番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(5−10)イネ品種日本晴の第5染色体の24,520,269番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(5−11)イネ品種日本晴の第5染色体の26,499,154番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(5−12)イネ品種日本晴の第5染色体の28,699,770番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(6−1)イネ品種日本晴の第6染色体の372,258番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(6−2)イネ品種日本晴の第6染色体の2,186,456番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(6−3)イネ品種日本晴の第6染色体の5,904,224番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(6−4)イネ品種日本晴の第6染色体の7,974,364番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(6−5)イネ品種日本晴の第6染色体の11,653,694番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(6−6)イネ品種日本晴の第6染色体の13,689,109番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(6−7)イネ品種日本晴の第6染色体の16,525,923番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(6−8)イネ品種日本晴の第6染色体の20,235,922番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(6−9)イネ品種日本晴の第6染色体の22,907,151番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(6−10)イネ品種日本晴の第6染色体の25,711,515番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(6−11)イネ品種日本晴の第6染色体の28,329,768番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではC)、
(6−12)イネ品種日本晴の第6染色体の29,901,305番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(7−1)イネ品種日本晴の第7染色体の260,864番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(7−2)イネ品種日本晴の第7染色体の1,745,458番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(7−3)イネ品種日本晴の第7染色体の3,898,132番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(7−4)イネ品種日本晴の第7染色体の6,843,696番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(7−5)イネ品種日本晴の第7染色体の10,130,032番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(7−6)イネ品種日本晴の第7染色体の13,302,019番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(7−7)イネ品種日本晴の第7染色体の17,327,528番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(7−8)イネ品種日本晴の第7染色体の18,571,959番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(7−9)イネ品種日本晴の第7染色体の21,942,076番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(7−10)イネ品種日本晴の第7染色体の24,516,006番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(7−11)イネ品種日本晴の第7染色体の25,490,053番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(7−12)イネ品種日本晴の第7染色体の28,847,226番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(8−1)イネ品種日本晴の第8染色体の454,665番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(8−2)イネ品種日本晴の第8染色体の1,669,653番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(8−3)イネ品種日本晴の第8染色体の5,278,855番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(8−4)イネ品種日本晴の第8染色体の8,094,582番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(8−5)イネ品種日本晴の第8染色体の10,572,009番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(8−6)イネ品種日本晴の第8染色体の12,249,318番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(8−7)イネ品種日本晴の第8染色体の15,484,340番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(8−8)イネ品種日本晴の第8染色体の18,114,324番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(8−9)イネ品種日本晴の第8染色体の19,903,905番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(8−10)イネ品種日本晴の第8染色体の23,047,467番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(8−11)イネ品種日本晴の第8染色体の25,921,712番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではC)、
(8−12)イネ品種日本晴の第8染色体の27,758,654番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−1)イネ品種日本晴の第9染色体の29,179番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(9−2)イネ品種日本晴の第9染色体の676,400番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(9−3)イネ品種日本晴の第9染色体の3,066,192番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(9−4)イネ品種日本晴の第9染色体の5,167,966番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−5)イネ品種日本晴の第9染色体の6,999,573番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(9−6)イネ品種日本晴の第9染色体の10,483,155番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−7)イネ品種日本晴の第9染色体の13,254,003番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−8)イネ品種日本晴の第9染色体の15,063,825番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(9−9)イネ品種日本晴の第9染色体の17,880,593番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(9−10)イネ品種日本晴の第9染色体の20,652,100番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−1)イネ品種日本晴の第10染色体の94,506番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではC)、
(10−2)イネ品種日本晴の第10染色体の3,643,898番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではA)、
(10−3)イネ品種日本晴の第10染色体の7,069,227番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(10−4)イネ品種日本晴の第10染色体の9,791,595番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−5)イネ品種日本晴の第10染色体の11,761,138番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−6)イネ品種日本晴の第10染色体の14,142,299番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−7)イネ品種日本晴の第10染色体の17,555,159番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(10−8)イネ品種日本晴の第10染色体の19,468,326番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではC)、
(10−9)イネ品種日本晴の第10染色体の21,612,698番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではT)、
(10−10)イネ品種日本晴の第10染色体の22,686,557番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(11−1)イネ品種日本晴の第11染色体の268,939番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(11−2)イネ品種日本晴の第11染色体の2,489,967番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(11−3)イネ品種日本晴の第11染色体の3,979,301番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(11−4)イネ品種日本晴の第11染色体の7,481,468番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(11−5)イネ品種日本晴の第11染色体の9,892,301番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(11−6)イネ品種日本晴の第11染色体の16,132,370番目のSNPに相当するSNP(イネ品種コシヒカリではG、オリザ・グラベリマではA)、
(11−7)イネ品種日本晴の第11染色体の19,050,305番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(11−8)イネ品種日本晴の第11染色体の21,896,773番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(11−9)イネ品種日本晴の第11染色体の25,149,244番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(11−10)イネ品種日本晴の第11染色体の27,560,928番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではG)、
(12−1)イネ品種日本晴の第12染色体の240,790番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(12−2)イネ品種日本晴の第12染色体の2,930,909番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(12−3)イネ品種日本晴の第12染色体の7,634,664番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(12−4)イネ品種日本晴の第12染色体の10,417,940番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)、
(12−5)イネ品種日本晴の第12染色体の14,689,883番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではT)、
(12−6)イネ品種日本晴の第12染色体の17,499,935番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではG)、
(12−7)イネ品種日本晴の第12染色体の21,257,971番目のSNPに相当するSNP(イネ品種コシヒカリではC、オリザ・グラベリマではT)、
(12−8)イネ品種日本晴の第12染色体の23,828,918番目のSNPに相当するSNP(イネ品種コシヒカリではT、オリザ・グラベリマではC)、
(12−9)イネ品種日本晴の第12染色体の27,011,209番目のSNPに相当するSNP(イネ品種コシヒカリではA、オリザ・グラベリマではG)。
【請求項6】
あるイネ個体が、請求項3に記載のイネ品種コシヒカリの染色体部分置換系統であるか否かを鑑別する方法であって、
鑑別対象であるイネ個体に対して、請求項5に記載のイネ個体のジェノタイピング方法を行い、得られたタイピング結果に基づき、当該イネ個体が請求項3に記載のイネ品種コシヒカリの染色体部分置換系統であるか否かを鑑別することを特徴とする、イネの鑑別方法。
【図1】

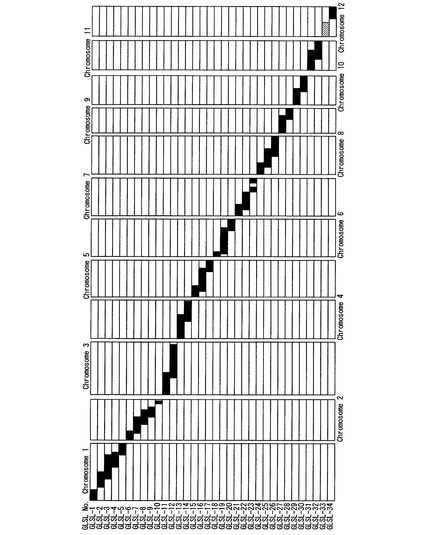
【図2】
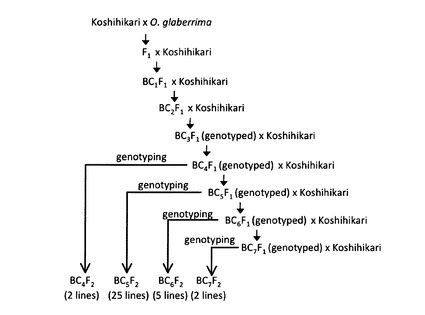
【図2】
【公開番号】特開2012−107012(P2012−107012A)
【公開日】平成24年6月7日(2012.6.7)
【国際特許分類】
【出願番号】特願2011−250745(P2011−250745)
【出願日】平成23年11月16日(2011.11.16)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【Fターム(参考)】
【公開日】平成24年6月7日(2012.6.7)
【国際特許分類】
【出願日】平成23年11月16日(2011.11.16)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【Fターム(参考)】
[ Back to top ]