
イネ科植物の高温障害を低減させる方法、高温耐性イネ科植物作出用ベクター及びイネ科植物
【課題】イネ科植物の高温障害を低減させる新たな技術を提供する。
【解決手段】イネ科植物の高温障害を低減させる方法であって、イネ科植物において、下記(A)〜(C)のいずれかの遺伝子の発現を抑制する:(A)特定のアミノ酸配列からなるタンパク質をコードする遺伝子;(B)特定のアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;(C)特定のアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【解決手段】イネ科植物の高温障害を低減させる方法であって、イネ科植物において、下記(A)〜(C)のいずれかの遺伝子の発現を抑制する:(A)特定のアミノ酸配列からなるタンパク質をコードする遺伝子;(B)特定のアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;(C)特定のアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、イネ科植物の高温障害を低減させる方法、高温耐性イネ科植物作出用ベクター、及び高温障害が低減されているイネ科植物に関する。
【背景技術】
【0002】
水稲は、登熟期に高温に曝されると白未熟粒を形成するという品質障害(以下、「高温障害」ともいう。)を起こすことが知られている。このような品質障害が起こると、米の一等米比率が著しく低下してしまうという問題がある。そのため、これまでに、高温障害が起こりにくい品種の選抜、栽培法の研究等が行なわれている(非特許文献1)。また、イネについて、高温下での遺伝子発現解析が試みられている(非特許文献2)。
【0003】
また、特許文献1には、ホスホリパーゼD欠失性であるイネ系統について記載されており、特許文献2には、植物由来のホスホリパーゼD遺伝子について記載されている。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開2008−245638号公報(2008年10月16日公開)
【特許文献2】国際公開第95/09234号パンフレット(1995年4月6日公開)
【非特許文献】
【0005】
【非特許文献1】千葉ら,2009,日作紀 78:4555−464
【非特許文献2】Yamakawa H., Kuroda M., Hirose T., Yamaguchi T. 2007. Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiol. 144:258-277.
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかしながら、従来における高温障害を低減させるための技術は、品種改良等に頼ったものが多く、高温障害を大幅に低減させることができるような顕著な成果を得られる技術ではない。特に、遺伝子を改変したり、ある一つの遺伝子の発現を抑制したりすることによって、イネの高温障害を低減させるような技術は、これまで全く報告されていない。そのため、確実に高温障害を低減させるためのさらなる技術の開発が望まれている。
【0007】
また、上述した特許文献1及び2には、イネのホスホリパーゼDと高温障害との関連性については一切記載も示唆もされていない。
【0008】
本発明は、上述した問題に鑑みてなされたものであり、その目的は、イネ科植物の高温障害を低減させることができる新たな技術を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、上述した課題を解決するために鋭意検討を行なった結果、イネ科植物の登熟過程の種子組織においてホスホリパーゼ(PLD)が活性化されること、及び高温条件下において通常の5倍以上のPLD活性が検出されることを見出した。そこで、イネ科植物のホスホリパーゼDについてさらに解析を進めたところ、複数のホスホリパーゼDのうちの1つの遺伝子の発現を抑制させた場合に、高温障害が大幅に低減されることを見出し、本発明を完成させた。
【0010】
本発明に係るイネ科植物の高温障害を低減させる方法は、イネ科植物において、下記(A)〜(C)のいずれかの遺伝子の発現を抑制することを特徴とする:
(A)配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子;
(B)配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;
(C)配列番号1に示すアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【0011】
本発明に係るイネ科植物の高温障害を低減させる方法は、イネ科植物において、下記(A)〜(C)のいずれかのDNAが含む遺伝子の発現を抑制することを特徴とする:
(A)配列番号2〜4のいずれかに示す塩基配列からなるDNA;
(B)配列番号2〜4のいずれかに示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA;
(C)配列番号2〜4のいずれかに示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA。
【0012】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることで、当該遺伝子の発現を抑制することが好ましい。
【0013】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記挿入は、T−DNAの挿入であることが好ましい。
【0014】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記遺伝子の発現をRNAi法により抑制することが好ましい。
【0015】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記イネ科植物は、イネであることが好ましい。
【0016】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記高温障害は、登熟期に高温に曝された場合における白未熟粒の形成であることが好ましい。
【0017】
本発明に係る高温耐性イネ科植物作出用ベクターは、下記(A)〜(C)のいずれかの遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制することを特徴とする:
(A)配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子;
(B)配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;
(C)配列番号1に示すアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【0018】
本発明に係る高温耐性イネ科植物作出用ベクターは、下記(A)〜(C)のいずれかのDNAが含む遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制することを特徴とする:
(A)配列番号2に示す塩基配列からなるDNA;
(B)配列番号2に示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA;
(C)配列番号2に示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA。
【0019】
本発明に係る高温耐性イネ科植物作出用ベクターは、配列番号4に示す塩基配列における少なくとも一部のセンス鎖配列と、当該センス鎖配列に相補的なアンチセンス鎖配列とを含み、当該塩基配列が含む遺伝子の発現をRNAi法により抑制することを特徴とする。
【0020】
本発明に係るイネ科植物は、上述したいずれかの方法により高温障害が低減されていることを特徴とする。
【0021】
本発明に係るイネは、上述したいずれかの方法により高温障害が低減されていることを特徴とする。
【0022】
また、本発明に係るイネでは、上記高温障害は、登熟期に高温に曝された場合における白未熟粒の形成であることが好ましい。
【発明の効果】
【0023】
本発明によれば、イネ科植物の高温障害を低減させることができる新たな方法、高温耐性イネ科植物作出用ベクター、及び高温障害が低減されているイネ科植物を提供することができる。
【図面の簡単な説明】
【0024】
【図1】イネのゲノムDNAにおけるOsPLDbeta2遺伝子領域の構成を示す図である。
【図2】OsPLDbeta2遺伝子のmRNAの構成を示す図である。
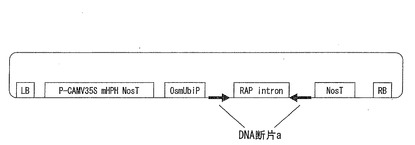
【図3】RNAiによるOsPLDbeta2遺伝子の発現抑制用ベクターのコンストラクトを示す図である。
【図4】OsPLDbeta2の定量PCRにおいて増幅した領域の位置を示す図である。
【図5】OsPLDbeta2−RNAi系統とベクターコントロールとのそれぞれにおける、OsPLDbeta2の発現量(mRNA量)を示すグラフである。
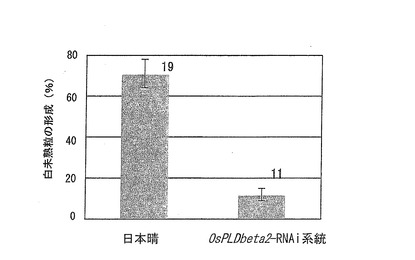
【図6】OsPLDbeta2−RNAi系統における高温処理による白未熟粒の形成割合(%)を示すグラフである。
【図7】OsPLDbeta2−Tos17系統におけるOsPLDbeta2遺伝子領域を示す図である。
【図8】OsPLDbeta2−Tos17系統を閉鎖系温室において栽培した場合の、高温処理による白未熟粒の形成割合(%)を示すグラフである。
【図9】OsPLDbeta2−Tos17系統を圃場栽培した場合の、高温処理による白未熟粒の形成割合(%)を示すグラフである。
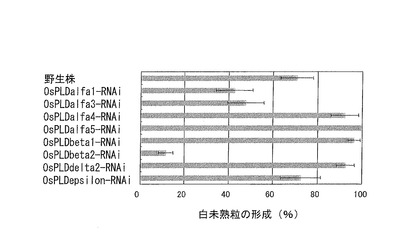
【図10】イネのホスホリパーゼD遺伝子の各抑制系統における高温処理による白未熟粒の形成割合(%)を示すグラフである。
【発明を実施するための形態】
【0025】
〔高温障害を低減させる方法〕
本発明に係る方法は、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子の発現を抑制することにより、イネ科植物の高温障害を低減させる方法である。イネ科植物には、イネ、コムギ、トウモロコシ、オオムギ等が含まれる。
【0026】
配列番号1に示すアミノ酸配列からなるタンパク質は、イネのホスホリパーゼDの1つであり、OsPLDbeta2とも称される。
【0027】
配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子としては、例えば、配列番号2〜4のいずれかに示す塩基配列、又は配列番号2〜4のいずれかに示す塩基配列の1個もしくは数個の塩基が欠失、置換もしくは付加された塩基配列からなるDNAが含む遺伝子であってもよい。
【0028】
配列番号2に示す塩基配列は、OsPLDbeta2遺伝子(RAP locus ID:Os03g0119100)を含む配列(Accession number:AF411221)であり、この遺伝子のエキソン配列とイントロン配列とを含む。
【0029】
図1は、イネのゲノムDNAにおけるOsPLDbeta2遺伝子領域の構成を示す図である。図1には、OsPLDbeta2遺伝子のエキソン領域を黒く塗りつぶした四角で示し、イントロン領域を細線で示し、5’側と3’側とにある非翻訳領域(UTR)を太線で示す。
【0030】
配列番号2に示す塩基配列からなるDNAは、全長4572bpであり、図1に示すように、10個のエキソンと、このエキソン間にある9個のイントロンとを含んでいる。なお、配列番号2に示す塩基配列のうち、213−1337、1557−1673、1832−2026、2126−2374、2459−2572、2615−2762、2989−3137、3269−3439、3541−3738、3816−4109bpがエキソンである。
【0031】
配列番号3に示す塩基配列は、OsPLDbeta2遺伝子の翻訳領域(2715bp)のみを含む塩基配列である。また、配列番号4に示す塩基配列は、OsPLDbeta2遺伝子の転写産物であるmRNAを逆転写して得られるcDNAの塩基配列である。
【0032】
図2は、OsPLDbeta2遺伝子のmRNAの構成を示す図である。図2に示すように、OsPLDbeta2遺伝子のmRNAは、翻訳領域と、その翻訳領域の5’側と3’側とにある非翻訳領域(UTR)とを含む配列からなる。
【0033】
配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子の相同遺伝子としては、当該遺伝子の塩基配列と相同の塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子であればよい。
【0034】
相同遺伝子としては、例えば配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であってもよい。
【0035】
アミノ酸配列中のいくつかのアミノ酸が、このアミノ酸配列からなるタンパク質の構造又は機能に有意に影響することなく容易に改変され得ることは、当該分野において周知である。さらに、人為的に改変させるだけではなく、天然のタンパク質において、当該タンパク質の構造又は機能を有意に変化させない変異体が存在することもまた周知である。
【0036】
また、相同遺伝子は、例えば配列番号1に示すアミノ酸配列と70%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましくは95%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であってもよい。
【0037】
また、相同遺伝子は、配列番号2〜4のいずれかに示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなるDNAが含む遺伝子であっていてもよい。
【0038】
また、相同遺伝子は、配列番号2〜4のいずれかに示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNAが含む遺伝子であってもよい。
【0039】
ハイブリダイゼーションは、Sambrookら、Molecular Cloning,A Laboratory Manual,2d Ed.,Cold Spring
Harbor Laboratory(1989)に記載されている方法のような周知の方法で行うことができる。通常、温度が高いほど、塩濃度が低いほどストリンジェンシーは高くなり(ハイブリダイズし難くなる)、より相同なポリヌクレオチドを取得することができる。適切なハイブリダイゼーション温度は、塩基配列やその塩基配列の長さによって異なり、例えば、アミノ酸6個をコードする18塩基からなるDNAフラグメントをプローブとして用いる場合、50℃以下の温度が好ましい。
【0040】
本明細書中で使用される場合、用語「ストリンジェントな条件」は、ハイブリダイゼーション溶液(50%ホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20μg/mlの変性剪断サケ精子DNAを含む)中にて42℃で一晩インキュベーションした後、約65℃にて0.1×SSC中でフィルターを洗浄する条件が意図される。
【0041】
また、相同遺伝子は、配列番号2〜4のいずれかに示す塩基配列と70%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましくは95%以上の相同性を有する塩基配列からなるDNAが含む遺伝子であってもよい。
【0042】
本明細書中、「DNA」とは、cDNA又はゲノムDNAの形態を含む。また、DNAは、二本鎖又は一本鎖であってもよい。一本鎖DNAは、センス鎖であってもよく、アンチセンス鎖であってもよい。また、DNAは、非翻訳領域(UTR)の配列、ベクター配列(発現用ベクター配列を含む)などを含むものであってもよい。
【0043】
DNAを取得するための供給源としては、特に限定されないが、植物であることが好ましく、より好ましくはイネ科植物であり、さらに好ましくはイネである。
【0044】
「イネ科植物において、遺伝子の発現を抑制する」とは、DNAからmRNAへの転写、又はmRNAからタンパク質への翻訳の少なくとも一方のステップを人為的に抑制することをさす。
【0045】
遺伝子の発現を抑制する方法としては、(a)抑制したい遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせる方法、(b)RNAiにより抑制する方法、などの公知の方法を用いることができる。
【0046】
抑制したい遺伝子に対し、塩基の欠失、置換又は挿入を生じさせる場合、欠失、置換又は挿入は、遺伝子のコード領域(特にエキソン)内でなされることにより、この遺伝子を機能破壊するものであってもよい。また、欠失、置換又は挿入は、遺伝子の転写調節領域内でなされることにより、この遺伝子の転写を抑制するものであってもよい。
【0047】
塩基の欠失又は置換を生じさせる場合には、例えば従来公知の突然変異体誘導方法を用いてもよい。また、塩基の挿入を生じさせる場合には、例えば従来公知のT−DNAを挿入する方法を用いてもよい。目的の遺伝子に対して塩基の欠失、置換又は挿入がなされたかどうかは、サザンハイブリダイゼーション、PCR法、DNAシークエンス解析等の従来の方法により調べることができる。
【0048】
RNAi法により抑制する場合には、従来公知の方法を用いることができる。例えば、抑制したい遺伝子のmRNAに相当する塩基配列における少なくとも一部のセンス鎖配列と、当該センス鎖配列に相補的なアンチセンス鎖配列とが、逆方向反復塩基配列構造となっているベクターを用いてもよい。このベクターを公知の方法によりイネ科植物に導入することによって、OsPLDbeta2遺伝子の発現をRNAi法により抑制することができる。
【0049】
ベクターをイネ科植物に導入する方法としては、公知の方法を用いることができる。例えば、マイクロインジェクション法、エレクトロポレーション法、パーティクルガン法、アグロバクテリウム法などを用いることができる。
【0050】
「高温障害」とは、イネ科植物が高温に曝されることにより発生する品質障害をさす。例えばイネであれば、高温障害とは、登熟期に高温に曝された場合における白未熟粒の形成をさす。「高温に曝される」とは、気温の平均が約28℃以上の環境に置かれることをいう。また、登熟期とは、出穂後約40日の間をいう。
【0051】
「高温障害を低減させる」とは、イネ科植物が高温に曝されることにより発生する品質障害を低減させることをいう。例えばイネであれば、上述した遺伝子の発現を抑制していない植物と比較して、登熟期に高温に曝された場合における白未熟粒の形成割合を低減させることをいう。
【0052】
白未熟粒の形成割合とは、採取した米の全数に対する白未熟粒の割合(%)である。白未熟粒か否かの判断は、玄米の外観を目視することにより、あるいは穀粒判別機を用いて、行なうことができる。「白未熟粒の形成割合を低減させる」とは、少なくとも親植物における白未熟粒の形成割合よりも低減させることをいう。
【0053】
なお、本発明に係る方法は、下記の本発明に係る高温耐性イネ科植物作出用ベクターを用いて実現することが可能である。
【0054】
〔高温耐性イネ科植物作出用ベクター〕
本発明は、本発明に係る方法に用いるための高温耐性イネ科植物作出用ベクターを提供する。本発明に係る高温耐性イネ科植物作出用ベクターは、イネ科植物において、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子の発現を抑制させるためのベクターである。
【0055】
本発明に係る高温耐性イネ科植物作出用ベクターを用いれば、イネ科植物の高温障害を低減させることができるので、高温耐性のイネ科植物を作出することが可能である。すなわち、後述する本発明に係るイネ科植物を作出することができる。
【0056】
高温耐性イネ科植物作出用ベクターに用いるベクターとしては、公知の中間ベクター、バイナリーベクター等を用いることができる。
【0057】
本発明に係る高温耐性イネ科植物作出用ベクターは、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制するものであってもよい。
【0058】
また、本発明に係る高温耐性イネ科植物作出用ベクターは、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子の発現を、RNAi法により抑制するものであってもよい。
【0059】
このような高温耐性イネ科植物作出用ベクターの例として、例えばプロモーター配列の下流に、抑制したい遺伝子の転写領域の少なくとも一部のセンス鎖配列とアンチセンス鎖配列とが逆方向反復塩基配列構造をとっているベクターが挙げられる。この高温耐性イネ科植物作出用ベクターをイネ科植物に導入することによって、OsPLDbeta2遺伝子の発現をRNAi法により抑制することができる。
【0060】
プロモーター配列としては、植物細胞内で転写可能なプロモーターであればよく、例えばカリフラワーモザイクウイルス35Sプロモーター等を用いることができる。
【0061】
センス鎖配列及びアンチセンス鎖配列としては、例えば配列番号4に示す塩基配列における少なくとも一部のセンス鎖配列及びアンチセンス鎖配列であってもよい。センス鎖配列としては、配列番号4に示す塩基配列における任意の部分の配列であってよく、またセンス鎖配列の長さについても特に限定されない。好ましくは、センス鎖配列は、配列番号4における、転写開始点を1としたときの2900bp〜3240bpに相当する341bpの配列である。
【0062】
アンチセンス鎖配列は、組み合わせられるセンス鎖配列に相補的な配列であればよい。
【0063】
センス鎖配列とアンチセンス鎖配列との間のスペーサー配列は、特に限定されないが、例えばイントロン配列等であることが好ましい。
【0064】
〔イネ科植物〕
本発明は、本発明に係る方法により高温障害が低減されているイネ科植物を提供する。特に、本発明は、高温障害が低減されているイネを提供する。
【0065】
なお、本明細書中「イネ科植物」又は「イネ」とは、種子又は植物体そのものだけでなく、植物体をなす細胞、組織、器官、ならびに誘導カルス、プロトプラスト、スフェロプラストなどをも含む概念とする。
【0066】
本発明に係るイネ科植物は、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子の発現が抑制されていることにより、高温障害が低減されている。
【0067】
「高温障害が低減されている」とは、イネ科植物において、高温に曝されることにより発生する品質障害が低減されていることをいう。例えばイネであれば、上述した遺伝子の発現が抑制されていない植物と比較して、登熟期に高温に曝された場合における白未熟粒の形成割合が低減されていることをいい、好ましくは、白未熟粒の形成割合が40%以下に低減されていることをいう。
【0068】
以下に実施例を示し、本発明の実施の形態についてさらに詳しく説明する。もちろん、本発明は以下の実施例に限定されるものではなく、細部については様々な態様が可能であることはいうまでもない。
【実施例】
【0069】
〔実施例1〕
本実施例では、イネのホスホリパーゼD遺伝子OsPLDbeta2の発現がRNAiにより抑制されている系統(OsPLDbeta2−RNAi系統)を作製した。
【0070】
(RNAiによる発現抑制用ベクターの構築)
まず、以下の方法により、OsPLDbeta2遺伝子のRNA干渉(RNAi)による発現抑制用のベクターを構築した。
【0071】
なお、本実施例では、ベクターとしてpZH2Bikベクター(Kuroda M, Kimizu M, Mikami C. 2010. A Simple Set of Plasmids for the Production of Transgenic Plants. Biosci. Biotech. Biochem. 74: 2348-2351)を用いた。このベクターは、アグロバクテリウムによる形質転換に用いることができるバイナリーベクターである。
【0072】
まず、OsPLDbeta2遺伝子のmRNAの一部の配列からなるDNA断片aを調製した。調製したDNA断片aは、OsPLDbeta2遺伝子のcDNA(配列番号4)における、転写開始点を1としたときの2900bp〜3240bpに相当する配列からなる、341bpのDNA断片aである。
【0073】
SDS−Phenol法を用いて、イネ(日本晴)の葉身より全RNAを抽出した。この全RNAから、SuperScript II Reverse Transcriptase(Invitrogen社)を用いて一本鎖DNAを調製した。この一本鎖DNAをテンプレートとして、プライマー1F(配列番号5:GCCCATCCAAGAAAACCTCAC)及びプライマー1R(配列番号6:CGGAGAACCCTAATCCCTCA)を用いてPCRによりDNA断片aを調製した。
【0074】
調製したDNA断片aを、pZH2Bikベクター上の2つの領域に、互いに逆方向となるように挿入し、RNAiによる発現抑制用ベクターを構築した。図3は、RNAiによるOsPLDbeta2遺伝子の発現抑制用ベクターのコンストラクトを示す図である。
【0075】
なお、図3中、LB及びRBは、T−DNA左右境界領域(約0.7kbp)を示す。P−CAMV35Sは、カリフラワーウイルス35Sプロモーター(約0.8kbp)を示す。HPHは、ハイグロマイシンホスホトランスフェラーゼ遺伝子(約1.0kbp)を示す。NosTは、Nosターミネーター(約0.3kbp)を示す。OsmUbiPは、改変イネポリユビキチンプロモーター(約1.8kbp)を示す。RAP intronは、イネアスパラギン酸プロテアーゼ(Oryzasin1)遺伝子(D32165)のイントロン(約1.0kbp)を示す。
【0076】
(OsPLDbeta2−RNAi系統の作製)
次に、作製した発現抑制用ベクターを用いて、OsPLDbeta2−RNAi系統を作製した。
【0077】
発現抑制用ベクターを、アグロバクテリウム法(国際公開第01/06844号パンフレットを参照)により日本晴カルスに導入し、再分化させた植物体からT0世代の種子を取得した。世代促進により導入された遺伝子を固定し、OsPLDbeta2−RNAi系統を確立した。
【0078】
(定量PCR)
作製したOsPLDbeta2−RNAi系統におけるOsPLDbeta2の発現を定量PCRにより解析した。また、コントロールとして、ベクターコントロール(日本晴にpZH2Bikベクターのみを導入したイネ)を用いた。
【0079】
OsPLDbeta2−RNAi系統とベクターコントロールとの種子をMS培地に無菌播種し、15日後のそれぞれの苗(Seedling)から全RNAを抽出した。この全RNAから、1本鎖DNAを作製し、定量PCRを行なった。定量PCRでは、プライマー2F(配列番号7:TTCCAATGTGGCCTGAAGGT)及びプライマー2R(配列番号8:CTAGGACTGCCGCCTGCTTC)を用いた。図4は、OsPLDbeta2の定量PCRにおいて増幅した領域の位置を示す図である。
【0080】
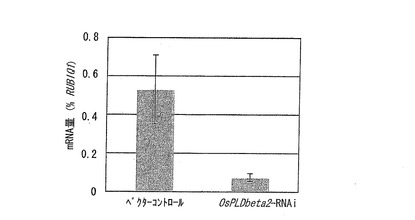
定量PCRの結果を図5に示す。図5は、OsPLDbeta2−RNAi系統とベクターコントロールとのそれぞれにおける、OsPLDbeta2の発現量(mRNA量)を示すグラフである。なお、mRNA量は、イネのポリユビキチン遺伝子(RUBIQ1)(Wang et al. 2000 Plant Science 156:201-211)の発現量を100(%)として示した。
【0081】
定量PCRの結果、OsPLDbeta2−RNAi系統では、ベクターコントロールと比較してOsPLDbeta2の発現が大幅に低下していることが示された(図5)。
【0082】
(高温処理による白未熟粒の形成)
次に、作製したOsPLDbeta2−RNAi系統について、高温処理による白未熟粒の形成割合(%)を調べた。コントロールとして、野生株の日本晴を用いた。
【0083】
OsPLDbeta2−RNAi系統と野生株の日本晴とを、閉鎖系温室(隔離温室)において平温(27℃/25℃)で栽培し、出穂後、高温(33℃/28℃)で栽培した。出穂後40日後に種子を採取し、米の白未熟粒の形成割合を計測した。なお、白未熟粒か否かは、玄米の外観を目視することにより、あるいは穀粒判別機(静岡精機)を用いて自動的に計測することにより、判断した。目視する方法を用いた場合には、少しでも乳白が存在する場合に白未熟粒と認定した。また、白未熟粒の形成割合は、採取した米の全数に対する白未熟粒の割合(%)として算出した。
【0084】
この結果を図6に示す。図6は、OsPLDbeta2−RNAi系統における高温処理による白未熟粒の形成割合(%)を示すグラフである。また、グラフ上に示した数字は、栽培試験回数を表す。
【0085】
OsPLDbeta2−RNAi系統における高温処理による白未熟粒の形成割合は、野生株の日本晴に比べて大幅に低減していた(図6)。このことから、OsPLDbeta2の発現抑制によって、高温による品質障害を低減させることができることが示された。
【0086】
〔実施例2〕
本実施例では、OsPLDbeta2遺伝子にレトロトランスポゾンTos17が挿入されたOsPLDbeta2欠損系統(OsPLDbeta2−Tos17系統)を作製した。
【0087】
(OsPLDbeta2−Tos17系統の作製)
日本晴において、レトロトランスポゾンTos17を増幅させた突然変異集団(ミュータントパネル)の中から、OsPLDbeta2遺伝子にTos17が挿入された系統(NE3817)について、独立行政法人農業生物資源研究所ゲノムリソースセンターより譲渡を受けた。この系統の世代促進による遺伝子固定系統(Mutant homo)を、OsPLDbeta2−Tos17系統とした。
【0088】
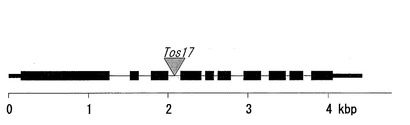
図7は、OsPLDbeta2−Tos17系統におけるOsPLDbeta2遺伝子領域を示す図である。OsPLDbeta2−Tos17系統では、配列番号2に示す塩基配列の2075bpの位置にTos17が挿入されている。
【0089】
(高温処理による白未熟粒の形成−閉鎖系温室)
このOsPLDbeta2−Tos17系統を用いて、実施例1と同様の方法により、閉鎖系温室において栽培した後高温処理を行ない、白未熟粒の形成割合を計測した。
【0090】
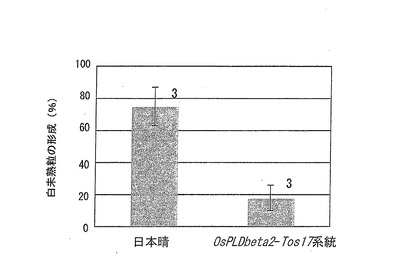
この結果を図8に示す。図8は、OsPLDbeta2−Tos17系統を閉鎖系温室において栽培した場合の、高温処理による白未熟粒の形成割合(%)を示すグラフである。また、グラフ上に示した数字は、栽培試験回数を表す。
【0091】
OsPLDbeta2−Tos17系統を閉鎖系温室において栽培した場合の、高温処理による白未熟粒の形成割合は、野生株の日本晴に比べて大幅に低減していた(図8)。このことから、OsPLDbeta2を欠損させることによって、高温による品質障害を低減させることができることが示された。
【0092】
(高温処理による白未熟粒の形成−圃場)
次に、OsPLDbeta2−Tos17系統について、圃場栽培した場合の高温処理による白未熟粒の形成割合(%)を調べた。コントロールとして、野生株の日本晴を用いた。
【0093】
OsPLDbeta2−Tos17系統と日本晴とを圃場栽培した。その際、出穂直前(約10日前)に高さ1.8メートルのビニールで栽培区を囲むことにより、高温処理を行なった(播種:2010年4月10日、圃場移植:2010年5月18日、高温処理開始:2010年8月2日、出穂日:日本晴 8月9〜11日;OsPLDbeta2−Tos17系統 8月12〜13日)。出穂後40日後に種子を採取し、実施例1と同様の方法により米の白未熟粒の形成割合を計測した。
【0094】
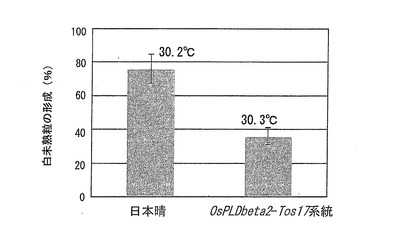
この結果を図9に示す。図9は、OsPLDbeta2−Tos17系統を圃場栽培した場合の、高温処理による白未熟粒の形成割合(%)を示すグラフである。また、グラフ上に示した数字は、出穂後20日間の平均気温を表す。
【0095】
OsPLDbeta2−Tos17系統を圃場栽培した場合の、高温処理による白未熟粒の形成割合は、野生株の日本晴に比べて大幅に低減していた(図9)。このことから、OsPLDbeta2を欠損させることによって、圃場栽培においても高温による品質障害を低減させることができることが示された。
【0096】
したがって、本発明は、農業の実用形質としても有望であり、高温耐性素材の開発に有用であることが示唆された。
【0097】
〔比較例〕
イネにおけるOsPLDbeta2以外の以下の7個のホスホリパーゼD遺伝子について、それぞれ発現がRNAiにより抑制されている系統を作製した。なお、それぞれの遺伝子のRAP locus IDを括弧内に示す。
OsPLDalfa1(Os01g0172400)
OsPLDalfa3(Os06g0604400)
OsPLDalfa4(Os06g0604200)
OsPLDalfa5(Os06g0604300)
OsPLDbeta1(Os10g0524400)
OsPLDdelta2(Os03g0840800)
OsPLDepsilon(Os09g0421300)
これらの抑制系統の作製については、実施例1と同様の方法を用いた。
【0098】
作製した各抑制系統について、実施例1と同様の方法により、高温処理による白未熟粒の形成割合(%)を計測した。コントロールとしては、野生株の日本晴を用いた。この結果を図10に示す。図10は、イネのホスホリパーゼD遺伝子の各抑制系統における高温処理による白未熟粒の形成割合(%)を示すグラフである。
【0099】
図10に示すように、今回調べたホスホリパーゼD遺伝子のうち、OsPLDbeta2以外の遺伝子については、発現を抑制しても高温障害を低減させることができないか、その効果が低いことが示された。
【産業上の利用可能性】
【0100】
本発明は、イネの高温障害を効率的に低減させることができるため、実用的な農業技術の開発に有用である。
【技術分野】
【0001】
本発明は、イネ科植物の高温障害を低減させる方法、高温耐性イネ科植物作出用ベクター、及び高温障害が低減されているイネ科植物に関する。
【背景技術】
【0002】
水稲は、登熟期に高温に曝されると白未熟粒を形成するという品質障害(以下、「高温障害」ともいう。)を起こすことが知られている。このような品質障害が起こると、米の一等米比率が著しく低下してしまうという問題がある。そのため、これまでに、高温障害が起こりにくい品種の選抜、栽培法の研究等が行なわれている(非特許文献1)。また、イネについて、高温下での遺伝子発現解析が試みられている(非特許文献2)。
【0003】
また、特許文献1には、ホスホリパーゼD欠失性であるイネ系統について記載されており、特許文献2には、植物由来のホスホリパーゼD遺伝子について記載されている。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開2008−245638号公報(2008年10月16日公開)
【特許文献2】国際公開第95/09234号パンフレット(1995年4月6日公開)
【非特許文献】
【0005】
【非特許文献1】千葉ら,2009,日作紀 78:4555−464
【非特許文献2】Yamakawa H., Kuroda M., Hirose T., Yamaguchi T. 2007. Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiol. 144:258-277.
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかしながら、従来における高温障害を低減させるための技術は、品種改良等に頼ったものが多く、高温障害を大幅に低減させることができるような顕著な成果を得られる技術ではない。特に、遺伝子を改変したり、ある一つの遺伝子の発現を抑制したりすることによって、イネの高温障害を低減させるような技術は、これまで全く報告されていない。そのため、確実に高温障害を低減させるためのさらなる技術の開発が望まれている。
【0007】
また、上述した特許文献1及び2には、イネのホスホリパーゼDと高温障害との関連性については一切記載も示唆もされていない。
【0008】
本発明は、上述した問題に鑑みてなされたものであり、その目的は、イネ科植物の高温障害を低減させることができる新たな技術を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、上述した課題を解決するために鋭意検討を行なった結果、イネ科植物の登熟過程の種子組織においてホスホリパーゼ(PLD)が活性化されること、及び高温条件下において通常の5倍以上のPLD活性が検出されることを見出した。そこで、イネ科植物のホスホリパーゼDについてさらに解析を進めたところ、複数のホスホリパーゼDのうちの1つの遺伝子の発現を抑制させた場合に、高温障害が大幅に低減されることを見出し、本発明を完成させた。
【0010】
本発明に係るイネ科植物の高温障害を低減させる方法は、イネ科植物において、下記(A)〜(C)のいずれかの遺伝子の発現を抑制することを特徴とする:
(A)配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子;
(B)配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;
(C)配列番号1に示すアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【0011】
本発明に係るイネ科植物の高温障害を低減させる方法は、イネ科植物において、下記(A)〜(C)のいずれかのDNAが含む遺伝子の発現を抑制することを特徴とする:
(A)配列番号2〜4のいずれかに示す塩基配列からなるDNA;
(B)配列番号2〜4のいずれかに示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA;
(C)配列番号2〜4のいずれかに示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA。
【0012】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることで、当該遺伝子の発現を抑制することが好ましい。
【0013】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記挿入は、T−DNAの挿入であることが好ましい。
【0014】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記遺伝子の発現をRNAi法により抑制することが好ましい。
【0015】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記イネ科植物は、イネであることが好ましい。
【0016】
また、本発明に係るイネ科植物の高温障害を低減させる方法では、上記高温障害は、登熟期に高温に曝された場合における白未熟粒の形成であることが好ましい。
【0017】
本発明に係る高温耐性イネ科植物作出用ベクターは、下記(A)〜(C)のいずれかの遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制することを特徴とする:
(A)配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子;
(B)配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;
(C)配列番号1に示すアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【0018】
本発明に係る高温耐性イネ科植物作出用ベクターは、下記(A)〜(C)のいずれかのDNAが含む遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制することを特徴とする:
(A)配列番号2に示す塩基配列からなるDNA;
(B)配列番号2に示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA;
(C)配列番号2に示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA。
【0019】
本発明に係る高温耐性イネ科植物作出用ベクターは、配列番号4に示す塩基配列における少なくとも一部のセンス鎖配列と、当該センス鎖配列に相補的なアンチセンス鎖配列とを含み、当該塩基配列が含む遺伝子の発現をRNAi法により抑制することを特徴とする。
【0020】
本発明に係るイネ科植物は、上述したいずれかの方法により高温障害が低減されていることを特徴とする。
【0021】
本発明に係るイネは、上述したいずれかの方法により高温障害が低減されていることを特徴とする。
【0022】
また、本発明に係るイネでは、上記高温障害は、登熟期に高温に曝された場合における白未熟粒の形成であることが好ましい。
【発明の効果】
【0023】
本発明によれば、イネ科植物の高温障害を低減させることができる新たな方法、高温耐性イネ科植物作出用ベクター、及び高温障害が低減されているイネ科植物を提供することができる。
【図面の簡単な説明】
【0024】
【図1】イネのゲノムDNAにおけるOsPLDbeta2遺伝子領域の構成を示す図である。
【図2】OsPLDbeta2遺伝子のmRNAの構成を示す図である。
【図3】RNAiによるOsPLDbeta2遺伝子の発現抑制用ベクターのコンストラクトを示す図である。
【図4】OsPLDbeta2の定量PCRにおいて増幅した領域の位置を示す図である。
【図5】OsPLDbeta2−RNAi系統とベクターコントロールとのそれぞれにおける、OsPLDbeta2の発現量(mRNA量)を示すグラフである。
【図6】OsPLDbeta2−RNAi系統における高温処理による白未熟粒の形成割合(%)を示すグラフである。
【図7】OsPLDbeta2−Tos17系統におけるOsPLDbeta2遺伝子領域を示す図である。
【図8】OsPLDbeta2−Tos17系統を閉鎖系温室において栽培した場合の、高温処理による白未熟粒の形成割合(%)を示すグラフである。
【図9】OsPLDbeta2−Tos17系統を圃場栽培した場合の、高温処理による白未熟粒の形成割合(%)を示すグラフである。
【図10】イネのホスホリパーゼD遺伝子の各抑制系統における高温処理による白未熟粒の形成割合(%)を示すグラフである。
【発明を実施するための形態】
【0025】
〔高温障害を低減させる方法〕
本発明に係る方法は、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子の発現を抑制することにより、イネ科植物の高温障害を低減させる方法である。イネ科植物には、イネ、コムギ、トウモロコシ、オオムギ等が含まれる。
【0026】
配列番号1に示すアミノ酸配列からなるタンパク質は、イネのホスホリパーゼDの1つであり、OsPLDbeta2とも称される。
【0027】
配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子としては、例えば、配列番号2〜4のいずれかに示す塩基配列、又は配列番号2〜4のいずれかに示す塩基配列の1個もしくは数個の塩基が欠失、置換もしくは付加された塩基配列からなるDNAが含む遺伝子であってもよい。
【0028】
配列番号2に示す塩基配列は、OsPLDbeta2遺伝子(RAP locus ID:Os03g0119100)を含む配列(Accession number:AF411221)であり、この遺伝子のエキソン配列とイントロン配列とを含む。
【0029】
図1は、イネのゲノムDNAにおけるOsPLDbeta2遺伝子領域の構成を示す図である。図1には、OsPLDbeta2遺伝子のエキソン領域を黒く塗りつぶした四角で示し、イントロン領域を細線で示し、5’側と3’側とにある非翻訳領域(UTR)を太線で示す。
【0030】
配列番号2に示す塩基配列からなるDNAは、全長4572bpであり、図1に示すように、10個のエキソンと、このエキソン間にある9個のイントロンとを含んでいる。なお、配列番号2に示す塩基配列のうち、213−1337、1557−1673、1832−2026、2126−2374、2459−2572、2615−2762、2989−3137、3269−3439、3541−3738、3816−4109bpがエキソンである。
【0031】
配列番号3に示す塩基配列は、OsPLDbeta2遺伝子の翻訳領域(2715bp)のみを含む塩基配列である。また、配列番号4に示す塩基配列は、OsPLDbeta2遺伝子の転写産物であるmRNAを逆転写して得られるcDNAの塩基配列である。
【0032】
図2は、OsPLDbeta2遺伝子のmRNAの構成を示す図である。図2に示すように、OsPLDbeta2遺伝子のmRNAは、翻訳領域と、その翻訳領域の5’側と3’側とにある非翻訳領域(UTR)とを含む配列からなる。
【0033】
配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子の相同遺伝子としては、当該遺伝子の塩基配列と相同の塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子であればよい。
【0034】
相同遺伝子としては、例えば配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であってもよい。
【0035】
アミノ酸配列中のいくつかのアミノ酸が、このアミノ酸配列からなるタンパク質の構造又は機能に有意に影響することなく容易に改変され得ることは、当該分野において周知である。さらに、人為的に改変させるだけではなく、天然のタンパク質において、当該タンパク質の構造又は機能を有意に変化させない変異体が存在することもまた周知である。
【0036】
また、相同遺伝子は、例えば配列番号1に示すアミノ酸配列と70%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましくは95%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であってもよい。
【0037】
また、相同遺伝子は、配列番号2〜4のいずれかに示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなるDNAが含む遺伝子であっていてもよい。
【0038】
また、相同遺伝子は、配列番号2〜4のいずれかに示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNAが含む遺伝子であってもよい。
【0039】
ハイブリダイゼーションは、Sambrookら、Molecular Cloning,A Laboratory Manual,2d Ed.,Cold Spring
Harbor Laboratory(1989)に記載されている方法のような周知の方法で行うことができる。通常、温度が高いほど、塩濃度が低いほどストリンジェンシーは高くなり(ハイブリダイズし難くなる)、より相同なポリヌクレオチドを取得することができる。適切なハイブリダイゼーション温度は、塩基配列やその塩基配列の長さによって異なり、例えば、アミノ酸6個をコードする18塩基からなるDNAフラグメントをプローブとして用いる場合、50℃以下の温度が好ましい。
【0040】
本明細書中で使用される場合、用語「ストリンジェントな条件」は、ハイブリダイゼーション溶液(50%ホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20μg/mlの変性剪断サケ精子DNAを含む)中にて42℃で一晩インキュベーションした後、約65℃にて0.1×SSC中でフィルターを洗浄する条件が意図される。
【0041】
また、相同遺伝子は、配列番号2〜4のいずれかに示す塩基配列と70%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましくは95%以上の相同性を有する塩基配列からなるDNAが含む遺伝子であってもよい。
【0042】
本明細書中、「DNA」とは、cDNA又はゲノムDNAの形態を含む。また、DNAは、二本鎖又は一本鎖であってもよい。一本鎖DNAは、センス鎖であってもよく、アンチセンス鎖であってもよい。また、DNAは、非翻訳領域(UTR)の配列、ベクター配列(発現用ベクター配列を含む)などを含むものであってもよい。
【0043】
DNAを取得するための供給源としては、特に限定されないが、植物であることが好ましく、より好ましくはイネ科植物であり、さらに好ましくはイネである。
【0044】
「イネ科植物において、遺伝子の発現を抑制する」とは、DNAからmRNAへの転写、又はmRNAからタンパク質への翻訳の少なくとも一方のステップを人為的に抑制することをさす。
【0045】
遺伝子の発現を抑制する方法としては、(a)抑制したい遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせる方法、(b)RNAiにより抑制する方法、などの公知の方法を用いることができる。
【0046】
抑制したい遺伝子に対し、塩基の欠失、置換又は挿入を生じさせる場合、欠失、置換又は挿入は、遺伝子のコード領域(特にエキソン)内でなされることにより、この遺伝子を機能破壊するものであってもよい。また、欠失、置換又は挿入は、遺伝子の転写調節領域内でなされることにより、この遺伝子の転写を抑制するものであってもよい。
【0047】
塩基の欠失又は置換を生じさせる場合には、例えば従来公知の突然変異体誘導方法を用いてもよい。また、塩基の挿入を生じさせる場合には、例えば従来公知のT−DNAを挿入する方法を用いてもよい。目的の遺伝子に対して塩基の欠失、置換又は挿入がなされたかどうかは、サザンハイブリダイゼーション、PCR法、DNAシークエンス解析等の従来の方法により調べることができる。
【0048】
RNAi法により抑制する場合には、従来公知の方法を用いることができる。例えば、抑制したい遺伝子のmRNAに相当する塩基配列における少なくとも一部のセンス鎖配列と、当該センス鎖配列に相補的なアンチセンス鎖配列とが、逆方向反復塩基配列構造となっているベクターを用いてもよい。このベクターを公知の方法によりイネ科植物に導入することによって、OsPLDbeta2遺伝子の発現をRNAi法により抑制することができる。
【0049】
ベクターをイネ科植物に導入する方法としては、公知の方法を用いることができる。例えば、マイクロインジェクション法、エレクトロポレーション法、パーティクルガン法、アグロバクテリウム法などを用いることができる。
【0050】
「高温障害」とは、イネ科植物が高温に曝されることにより発生する品質障害をさす。例えばイネであれば、高温障害とは、登熟期に高温に曝された場合における白未熟粒の形成をさす。「高温に曝される」とは、気温の平均が約28℃以上の環境に置かれることをいう。また、登熟期とは、出穂後約40日の間をいう。
【0051】
「高温障害を低減させる」とは、イネ科植物が高温に曝されることにより発生する品質障害を低減させることをいう。例えばイネであれば、上述した遺伝子の発現を抑制していない植物と比較して、登熟期に高温に曝された場合における白未熟粒の形成割合を低減させることをいう。
【0052】
白未熟粒の形成割合とは、採取した米の全数に対する白未熟粒の割合(%)である。白未熟粒か否かの判断は、玄米の外観を目視することにより、あるいは穀粒判別機を用いて、行なうことができる。「白未熟粒の形成割合を低減させる」とは、少なくとも親植物における白未熟粒の形成割合よりも低減させることをいう。
【0053】
なお、本発明に係る方法は、下記の本発明に係る高温耐性イネ科植物作出用ベクターを用いて実現することが可能である。
【0054】
〔高温耐性イネ科植物作出用ベクター〕
本発明は、本発明に係る方法に用いるための高温耐性イネ科植物作出用ベクターを提供する。本発明に係る高温耐性イネ科植物作出用ベクターは、イネ科植物において、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子の発現を抑制させるためのベクターである。
【0055】
本発明に係る高温耐性イネ科植物作出用ベクターを用いれば、イネ科植物の高温障害を低減させることができるので、高温耐性のイネ科植物を作出することが可能である。すなわち、後述する本発明に係るイネ科植物を作出することができる。
【0056】
高温耐性イネ科植物作出用ベクターに用いるベクターとしては、公知の中間ベクター、バイナリーベクター等を用いることができる。
【0057】
本発明に係る高温耐性イネ科植物作出用ベクターは、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制するものであってもよい。
【0058】
また、本発明に係る高温耐性イネ科植物作出用ベクターは、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子の発現を、RNAi法により抑制するものであってもよい。
【0059】
このような高温耐性イネ科植物作出用ベクターの例として、例えばプロモーター配列の下流に、抑制したい遺伝子の転写領域の少なくとも一部のセンス鎖配列とアンチセンス鎖配列とが逆方向反復塩基配列構造をとっているベクターが挙げられる。この高温耐性イネ科植物作出用ベクターをイネ科植物に導入することによって、OsPLDbeta2遺伝子の発現をRNAi法により抑制することができる。
【0060】
プロモーター配列としては、植物細胞内で転写可能なプロモーターであればよく、例えばカリフラワーモザイクウイルス35Sプロモーター等を用いることができる。
【0061】
センス鎖配列及びアンチセンス鎖配列としては、例えば配列番号4に示す塩基配列における少なくとも一部のセンス鎖配列及びアンチセンス鎖配列であってもよい。センス鎖配列としては、配列番号4に示す塩基配列における任意の部分の配列であってよく、またセンス鎖配列の長さについても特に限定されない。好ましくは、センス鎖配列は、配列番号4における、転写開始点を1としたときの2900bp〜3240bpに相当する341bpの配列である。
【0062】
アンチセンス鎖配列は、組み合わせられるセンス鎖配列に相補的な配列であればよい。
【0063】
センス鎖配列とアンチセンス鎖配列との間のスペーサー配列は、特に限定されないが、例えばイントロン配列等であることが好ましい。
【0064】
〔イネ科植物〕
本発明は、本発明に係る方法により高温障害が低減されているイネ科植物を提供する。特に、本発明は、高温障害が低減されているイネを提供する。
【0065】
なお、本明細書中「イネ科植物」又は「イネ」とは、種子又は植物体そのものだけでなく、植物体をなす細胞、組織、器官、ならびに誘導カルス、プロトプラスト、スフェロプラストなどをも含む概念とする。
【0066】
本発明に係るイネ科植物は、配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子又はその相同遺伝子の発現が抑制されていることにより、高温障害が低減されている。
【0067】
「高温障害が低減されている」とは、イネ科植物において、高温に曝されることにより発生する品質障害が低減されていることをいう。例えばイネであれば、上述した遺伝子の発現が抑制されていない植物と比較して、登熟期に高温に曝された場合における白未熟粒の形成割合が低減されていることをいい、好ましくは、白未熟粒の形成割合が40%以下に低減されていることをいう。
【0068】
以下に実施例を示し、本発明の実施の形態についてさらに詳しく説明する。もちろん、本発明は以下の実施例に限定されるものではなく、細部については様々な態様が可能であることはいうまでもない。
【実施例】
【0069】
〔実施例1〕
本実施例では、イネのホスホリパーゼD遺伝子OsPLDbeta2の発現がRNAiにより抑制されている系統(OsPLDbeta2−RNAi系統)を作製した。
【0070】
(RNAiによる発現抑制用ベクターの構築)
まず、以下の方法により、OsPLDbeta2遺伝子のRNA干渉(RNAi)による発現抑制用のベクターを構築した。
【0071】
なお、本実施例では、ベクターとしてpZH2Bikベクター(Kuroda M, Kimizu M, Mikami C. 2010. A Simple Set of Plasmids for the Production of Transgenic Plants. Biosci. Biotech. Biochem. 74: 2348-2351)を用いた。このベクターは、アグロバクテリウムによる形質転換に用いることができるバイナリーベクターである。
【0072】
まず、OsPLDbeta2遺伝子のmRNAの一部の配列からなるDNA断片aを調製した。調製したDNA断片aは、OsPLDbeta2遺伝子のcDNA(配列番号4)における、転写開始点を1としたときの2900bp〜3240bpに相当する配列からなる、341bpのDNA断片aである。
【0073】
SDS−Phenol法を用いて、イネ(日本晴)の葉身より全RNAを抽出した。この全RNAから、SuperScript II Reverse Transcriptase(Invitrogen社)を用いて一本鎖DNAを調製した。この一本鎖DNAをテンプレートとして、プライマー1F(配列番号5:GCCCATCCAAGAAAACCTCAC)及びプライマー1R(配列番号6:CGGAGAACCCTAATCCCTCA)を用いてPCRによりDNA断片aを調製した。
【0074】
調製したDNA断片aを、pZH2Bikベクター上の2つの領域に、互いに逆方向となるように挿入し、RNAiによる発現抑制用ベクターを構築した。図3は、RNAiによるOsPLDbeta2遺伝子の発現抑制用ベクターのコンストラクトを示す図である。
【0075】
なお、図3中、LB及びRBは、T−DNA左右境界領域(約0.7kbp)を示す。P−CAMV35Sは、カリフラワーウイルス35Sプロモーター(約0.8kbp)を示す。HPHは、ハイグロマイシンホスホトランスフェラーゼ遺伝子(約1.0kbp)を示す。NosTは、Nosターミネーター(約0.3kbp)を示す。OsmUbiPは、改変イネポリユビキチンプロモーター(約1.8kbp)を示す。RAP intronは、イネアスパラギン酸プロテアーゼ(Oryzasin1)遺伝子(D32165)のイントロン(約1.0kbp)を示す。
【0076】
(OsPLDbeta2−RNAi系統の作製)
次に、作製した発現抑制用ベクターを用いて、OsPLDbeta2−RNAi系統を作製した。
【0077】
発現抑制用ベクターを、アグロバクテリウム法(国際公開第01/06844号パンフレットを参照)により日本晴カルスに導入し、再分化させた植物体からT0世代の種子を取得した。世代促進により導入された遺伝子を固定し、OsPLDbeta2−RNAi系統を確立した。
【0078】
(定量PCR)
作製したOsPLDbeta2−RNAi系統におけるOsPLDbeta2の発現を定量PCRにより解析した。また、コントロールとして、ベクターコントロール(日本晴にpZH2Bikベクターのみを導入したイネ)を用いた。
【0079】
OsPLDbeta2−RNAi系統とベクターコントロールとの種子をMS培地に無菌播種し、15日後のそれぞれの苗(Seedling)から全RNAを抽出した。この全RNAから、1本鎖DNAを作製し、定量PCRを行なった。定量PCRでは、プライマー2F(配列番号7:TTCCAATGTGGCCTGAAGGT)及びプライマー2R(配列番号8:CTAGGACTGCCGCCTGCTTC)を用いた。図4は、OsPLDbeta2の定量PCRにおいて増幅した領域の位置を示す図である。
【0080】
定量PCRの結果を図5に示す。図5は、OsPLDbeta2−RNAi系統とベクターコントロールとのそれぞれにおける、OsPLDbeta2の発現量(mRNA量)を示すグラフである。なお、mRNA量は、イネのポリユビキチン遺伝子(RUBIQ1)(Wang et al. 2000 Plant Science 156:201-211)の発現量を100(%)として示した。
【0081】
定量PCRの結果、OsPLDbeta2−RNAi系統では、ベクターコントロールと比較してOsPLDbeta2の発現が大幅に低下していることが示された(図5)。
【0082】
(高温処理による白未熟粒の形成)
次に、作製したOsPLDbeta2−RNAi系統について、高温処理による白未熟粒の形成割合(%)を調べた。コントロールとして、野生株の日本晴を用いた。
【0083】
OsPLDbeta2−RNAi系統と野生株の日本晴とを、閉鎖系温室(隔離温室)において平温(27℃/25℃)で栽培し、出穂後、高温(33℃/28℃)で栽培した。出穂後40日後に種子を採取し、米の白未熟粒の形成割合を計測した。なお、白未熟粒か否かは、玄米の外観を目視することにより、あるいは穀粒判別機(静岡精機)を用いて自動的に計測することにより、判断した。目視する方法を用いた場合には、少しでも乳白が存在する場合に白未熟粒と認定した。また、白未熟粒の形成割合は、採取した米の全数に対する白未熟粒の割合(%)として算出した。
【0084】
この結果を図6に示す。図6は、OsPLDbeta2−RNAi系統における高温処理による白未熟粒の形成割合(%)を示すグラフである。また、グラフ上に示した数字は、栽培試験回数を表す。
【0085】
OsPLDbeta2−RNAi系統における高温処理による白未熟粒の形成割合は、野生株の日本晴に比べて大幅に低減していた(図6)。このことから、OsPLDbeta2の発現抑制によって、高温による品質障害を低減させることができることが示された。
【0086】
〔実施例2〕
本実施例では、OsPLDbeta2遺伝子にレトロトランスポゾンTos17が挿入されたOsPLDbeta2欠損系統(OsPLDbeta2−Tos17系統)を作製した。
【0087】
(OsPLDbeta2−Tos17系統の作製)
日本晴において、レトロトランスポゾンTos17を増幅させた突然変異集団(ミュータントパネル)の中から、OsPLDbeta2遺伝子にTos17が挿入された系統(NE3817)について、独立行政法人農業生物資源研究所ゲノムリソースセンターより譲渡を受けた。この系統の世代促進による遺伝子固定系統(Mutant homo)を、OsPLDbeta2−Tos17系統とした。
【0088】
図7は、OsPLDbeta2−Tos17系統におけるOsPLDbeta2遺伝子領域を示す図である。OsPLDbeta2−Tos17系統では、配列番号2に示す塩基配列の2075bpの位置にTos17が挿入されている。
【0089】
(高温処理による白未熟粒の形成−閉鎖系温室)
このOsPLDbeta2−Tos17系統を用いて、実施例1と同様の方法により、閉鎖系温室において栽培した後高温処理を行ない、白未熟粒の形成割合を計測した。
【0090】
この結果を図8に示す。図8は、OsPLDbeta2−Tos17系統を閉鎖系温室において栽培した場合の、高温処理による白未熟粒の形成割合(%)を示すグラフである。また、グラフ上に示した数字は、栽培試験回数を表す。
【0091】
OsPLDbeta2−Tos17系統を閉鎖系温室において栽培した場合の、高温処理による白未熟粒の形成割合は、野生株の日本晴に比べて大幅に低減していた(図8)。このことから、OsPLDbeta2を欠損させることによって、高温による品質障害を低減させることができることが示された。
【0092】
(高温処理による白未熟粒の形成−圃場)
次に、OsPLDbeta2−Tos17系統について、圃場栽培した場合の高温処理による白未熟粒の形成割合(%)を調べた。コントロールとして、野生株の日本晴を用いた。
【0093】
OsPLDbeta2−Tos17系統と日本晴とを圃場栽培した。その際、出穂直前(約10日前)に高さ1.8メートルのビニールで栽培区を囲むことにより、高温処理を行なった(播種:2010年4月10日、圃場移植:2010年5月18日、高温処理開始:2010年8月2日、出穂日:日本晴 8月9〜11日;OsPLDbeta2−Tos17系統 8月12〜13日)。出穂後40日後に種子を採取し、実施例1と同様の方法により米の白未熟粒の形成割合を計測した。
【0094】
この結果を図9に示す。図9は、OsPLDbeta2−Tos17系統を圃場栽培した場合の、高温処理による白未熟粒の形成割合(%)を示すグラフである。また、グラフ上に示した数字は、出穂後20日間の平均気温を表す。
【0095】
OsPLDbeta2−Tos17系統を圃場栽培した場合の、高温処理による白未熟粒の形成割合は、野生株の日本晴に比べて大幅に低減していた(図9)。このことから、OsPLDbeta2を欠損させることによって、圃場栽培においても高温による品質障害を低減させることができることが示された。
【0096】
したがって、本発明は、農業の実用形質としても有望であり、高温耐性素材の開発に有用であることが示唆された。
【0097】
〔比較例〕
イネにおけるOsPLDbeta2以外の以下の7個のホスホリパーゼD遺伝子について、それぞれ発現がRNAiにより抑制されている系統を作製した。なお、それぞれの遺伝子のRAP locus IDを括弧内に示す。
OsPLDalfa1(Os01g0172400)
OsPLDalfa3(Os06g0604400)
OsPLDalfa4(Os06g0604200)
OsPLDalfa5(Os06g0604300)
OsPLDbeta1(Os10g0524400)
OsPLDdelta2(Os03g0840800)
OsPLDepsilon(Os09g0421300)
これらの抑制系統の作製については、実施例1と同様の方法を用いた。
【0098】
作製した各抑制系統について、実施例1と同様の方法により、高温処理による白未熟粒の形成割合(%)を計測した。コントロールとしては、野生株の日本晴を用いた。この結果を図10に示す。図10は、イネのホスホリパーゼD遺伝子の各抑制系統における高温処理による白未熟粒の形成割合(%)を示すグラフである。
【0099】
図10に示すように、今回調べたホスホリパーゼD遺伝子のうち、OsPLDbeta2以外の遺伝子については、発現を抑制しても高温障害を低減させることができないか、その効果が低いことが示された。
【産業上の利用可能性】
【0100】
本発明は、イネの高温障害を効率的に低減させることができるため、実用的な農業技術の開発に有用である。
【特許請求の範囲】
【請求項1】
イネ科植物において、下記(A)〜(C)のいずれかの遺伝子の発現を抑制することを特徴とする、イネ科植物の高温障害を低減させる方法:
(A)配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子;
(B)配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;
(C)配列番号1に示すアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【請求項2】
イネ科植物において、下記(A)〜(C)のいずれかのDNAが含む遺伝子の発現を抑制することを特徴とする、イネ科植物の高温障害を低減させる方法:
(A)配列番号2〜4のいずれかに示す塩基配列からなるDNA;
(B)配列番号2〜4のいずれかに示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA;
(C)配列番号2〜4のいずれかに示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA。
【請求項3】
上記遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることで、当該遺伝子の発現を抑制することを特徴とする請求項1又は2に記載の方法。
【請求項4】
上記挿入は、T−DNAの挿入であることを特徴とする請求項3に記載の方法。
【請求項5】
上記遺伝子の発現をRNAi法により抑制することを特徴とする請求項1又は2に記載の方法。
【請求項6】
上記イネ科植物は、イネであることを特徴とする請求項1〜5のいずれか1項に記載の方法。
【請求項7】
上記高温障害は、登熟期に高温に曝された場合における白未熟粒の形成であることを特徴とする請求項6に記載の方法。
【請求項8】
下記(A)〜(C)のいずれかの遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制することを特徴とする高温耐性イネ科植物作出用ベクター:
(A)配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子;
(B)配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;
(C)配列番号1に示すアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【請求項9】
下記(A)〜(C)のいずれかのDNAが含む遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制することを特徴とする高温耐性イネ科植物作出用ベクター:
(A)配列番号2に示す塩基配列からなるDNA;
(B)配列番号2に示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA;
(C)配列番号2に示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA。
【請求項10】
配列番号4に示す塩基配列における少なくとも一部のセンス鎖配列と、当該センス鎖配列に相補的なアンチセンス鎖配列とを含み、当該塩基配列が含む遺伝子の発現をRNAi法により抑制することを特徴とする高温耐性イネ科植物作出用ベクター。
【請求項11】
請求項1〜7のいずれか1項に記載の方法により高温障害が低減されていることを特徴とするイネ科植物。
【請求項12】
請求項1〜7のいずれか1項に記載の方法により高温障害が低減されていることを特徴とするイネ。
【請求項13】
上記高温障害は、登熟期に高温に曝された場合における白未熟粒の形成であることを特徴とする請求項12に記載のイネ。
【請求項1】
イネ科植物において、下記(A)〜(C)のいずれかの遺伝子の発現を抑制することを特徴とする、イネ科植物の高温障害を低減させる方法:
(A)配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子;
(B)配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;
(C)配列番号1に示すアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【請求項2】
イネ科植物において、下記(A)〜(C)のいずれかのDNAが含む遺伝子の発現を抑制することを特徴とする、イネ科植物の高温障害を低減させる方法:
(A)配列番号2〜4のいずれかに示す塩基配列からなるDNA;
(B)配列番号2〜4のいずれかに示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA;
(C)配列番号2〜4のいずれかに示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA。
【請求項3】
上記遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることで、当該遺伝子の発現を抑制することを特徴とする請求項1又は2に記載の方法。
【請求項4】
上記挿入は、T−DNAの挿入であることを特徴とする請求項3に記載の方法。
【請求項5】
上記遺伝子の発現をRNAi法により抑制することを特徴とする請求項1又は2に記載の方法。
【請求項6】
上記イネ科植物は、イネであることを特徴とする請求項1〜5のいずれか1項に記載の方法。
【請求項7】
上記高温障害は、登熟期に高温に曝された場合における白未熟粒の形成であることを特徴とする請求項6に記載の方法。
【請求項8】
下記(A)〜(C)のいずれかの遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制することを特徴とする高温耐性イネ科植物作出用ベクター:
(A)配列番号1に示すアミノ酸配列からなるタンパク質をコードする遺伝子;
(B)配列番号1に示すアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子;
(C)配列番号1に示すアミノ酸配列と70%以上の相同性を有するアミノ酸配列からなるタンパク質をコードする遺伝子であって、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子。
【請求項9】
下記(A)〜(C)のいずれかのDNAが含む遺伝子に対し、塩基の欠失、置換及び挿入から選択される少なくとも一つを生じさせることにより、当該遺伝子の発現を抑制することを特徴とする高温耐性イネ科植物作出用ベクター:
(A)配列番号2に示す塩基配列からなるDNA;
(B)配列番号2に示す塩基配列の1個又は数個の塩基が欠失、置換又は付加された塩基配列からなり、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA;
(C)配列番号2に示す塩基配列と相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、イネ科植物において発現が抑制された場合に高温障害を低減させる遺伝子を含むDNA。
【請求項10】
配列番号4に示す塩基配列における少なくとも一部のセンス鎖配列と、当該センス鎖配列に相補的なアンチセンス鎖配列とを含み、当該塩基配列が含む遺伝子の発現をRNAi法により抑制することを特徴とする高温耐性イネ科植物作出用ベクター。
【請求項11】
請求項1〜7のいずれか1項に記載の方法により高温障害が低減されていることを特徴とするイネ科植物。
【請求項12】
請求項1〜7のいずれか1項に記載の方法により高温障害が低減されていることを特徴とするイネ。
【請求項13】
上記高温障害は、登熟期に高温に曝された場合における白未熟粒の形成であることを特徴とする請求項12に記載のイネ。
【図1】



【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2012−187048(P2012−187048A)
【公開日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願番号】特願2011−53487(P2011−53487)
【出願日】平成23年3月10日(2011.3.10)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
【公開日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願日】平成23年3月10日(2011.3.10)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
[ Back to top ]