
イネF1種子の生産方法、イネF1種子、及びイネ雄性不稔系統
【課題】F1雑種育種法において好適なコシヒカリの雄性不稔系統、及び、当該イネ雄性不稔系統を用いたイネF1種子の生産方法の提供。
【解決手段】イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統、又は半糯性を示すイネ雄性不稔系統を母本とし、イネ稔性回復系統を花粉親として交配し、交配後の母本から雑種第1代種子(F1種子)を採取することを特徴とするイネF1種子の生産方法、前記記載のイネF1種子の生産方法により得られたことを特徴とするイネF1種子、及びイネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有することを特徴とするイネ雄性不稔系統。
【解決手段】イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統、又は半糯性を示すイネ雄性不稔系統を母本とし、イネ稔性回復系統を花粉親として交配し、交配後の母本から雑種第1代種子(F1種子)を採取することを特徴とするイネF1種子の生産方法、前記記載のイネF1種子の生産方法により得られたことを特徴とするイネF1種子、及びイネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有することを特徴とするイネ雄性不稔系統。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、良好な形質を有するイネ雄性不稔系統、当該イネ雄性不稔系統を用いたイネF1種子の生産方法、及び当該方法により得られたイネF1種子に関する。
【背景技術】
【0002】
近年、ゲノム解析技術は格段の進歩を遂げ、作物の改良を、高精度かつ容易に行えるようになった。とりわけ、DNAマーカー技術の進捗は著しく、同技術によって、有用な形質を備える新品種を作製することができるようになった。例えば、これまでに、DNAマーカーを用いて、灰色カビ病耐性を備えるトマトや(例えば、特許文献1参照。)、耐倒伏性と玄米粒の大きさが改善されたイネ(Oryza sativa)(例えば、特許文献2参照。)が作出されている。
【0003】
また、DNAマーカーを利用して、これまでに同定された重要遺伝子の有用アレルを含む染色体領域を置換することにより、他の多くの形質に対してほとんど影響を及ぼさずに、目的形質を特異的に改良することが出来るようになった(例えば、特許文献3参照。)。例えばイネにおいては、稈長(sd1遺伝子近傍染色体領域)、到穂日数(hd1遺伝子近傍染色体領域)、1穂粒数(Gn1遺伝子近傍染色体領域)などが改良されたイネが作出されている(例えば、特許文献4参照。)。イネ品種コシヒカリの染色体中のsd1遺伝子が、ハバタキ由来のsd1遺伝子に置換されることにより、イネ品種コシヒカリよりも稈長が有意に短くなり、耐倒伏性が改善される。また、イネ品種コシヒカリの染色体中のGn1遺伝子が、ハバタキ由来のGn1遺伝子に置換されることにより、イネ品種コシヒカリよりも着粒密度が高くなる。イネ品種コシヒカリの染色体中のhd1遺伝子が、ハバタキ由来のhd1遺伝子に置換されることにより、イネ品種コシヒカリよりも早生化される。
【0004】
より優れた形質を備える作物を作出する方法として、雄性不稔細胞質等を利用して母本の花粉合成能を失わせることで遠縁系統間の交配を実現し、その雑種種子を品種として利用するF1雑種育種法がある。例えば、レタスにおいては、F1雑種育種法において母本として使用可能なレタス雄性不稔系統が作出されている(例えば、特許文献5参照。)。
【0005】
F1雑種育種法は、雑種強勢を利用して収量性を容易にかつ非常に高いレベルで改良できる技術として利用されている。わが国のイネ育種においても、1970年に実用的な雄性不稔細胞質が発見されて以来、利用が試みられてきた。但し、当時育成された系統の食味品質が十分に高くなかったこと、及び、その後のコメ余りの時代の中で多収性に関する必要性が低下したことなどにより、次第に利用されなくなってきた歴史がある。
【0006】
しかしながら、近年では、作物の収量ポテンシャルを向上させることは、食糧の増産や、栽培コスト、及び栽培時投入エネルギーについての効率化を図る上で、再び重要になってきており、今後ますます重要な育種目標となる。また、生産能の増強によって植物体そのものを大きくすることは、第2世代のバイオエタノール原材料として目される作物残渣の生産性を高め、作物栽培行程にて排出されるGHG量を相対的に削減することを通して、エネルギー問題・環境問題の解決にも貢献できる。
【0007】
収量ポテンシャルの向上が重要視されるようになってきた現在、改めてF1雑種育種技術を見直す機運が高まりつつある。F1雑種育種法では、組み合わせ検定における選抜候補系統として非常に多くの系統間でF1雑種を作る必要があるため、高い選抜効率を維持するためには、母本となる雄性不稔系統の選定が非常に重要となる。
【0008】
我が国のリーディングバラエティであるイネ品種コシヒカリは、食味品質に対する評価が高く、これを育成母本とした系統には、多くの良食味系統が存在する。また、食味品質以外にも、我が国で最も広範に栽培されていることが示すような広域栽培適応性を備えており、また、強い難穂発芽性などの優れた形質を多数有している。さらに、多くの試験において研究対象に用いられることによって学術的知識が蓄積されており、改良の糸口が見つけやすい利点もある。以上のことを考慮すると、コシヒカリは、F1雑種の母本を育成するに当たり最も有望な系統の1つであると言える。
【0009】
また、これまで特異的に改良することが困難であった食味品質についても、半糯性突然変異形質を利用することで、アミロース含量を低下させ、コメの粘りを強くすることを通して品質向上が図れるようになった。半糯性のコメの多くは胚乳が白濁するため、通常のコメと容易に区別できる。過去に報告されている半糯性突然変異には、アミロース含量を最も大きく左右するとされるwx遺伝子の変異の他、7つの異なるdu座が関係したものがある(例えば、非特許文献1参照。)
【先行技術文献】
【特許文献】
【0010】
【特許文献1】特許第4248881号公報
【特許文献2】特許第4368391号公報
【特許文献3】特許第4409610号公報
【特許文献4】特許第4352102号公報
【特許文献5】特許第3949637号公報
【非特許文献】
【0011】
【非特許文献1】鈴木保宏、“米のアミロース含量の変動−気候と調節−”、「農業及び園芸」、2006年、第81巻、第183〜190ページ。
【発明の概要】
【発明が解決しようとする課題】
【0012】
しかしながら、コシヒカリを片親にしてF1雑種を育成しようとする場合には、しばしば問題が発生する。例えば、インディカ種を含むコシヒカリとは遠縁となる育成系統との組み合わせ検定においては、晩生化や長稈化してしまう結果、実用系統としての適性を外す系統が頻繁に出現し、選抜の効率を大きく低下させる結果となる。
また、F1雑種では雑種強勢により収量性が向上するが、一般の多収系統と比較してコシヒカリ自身の収量ポテンシャルが十分に高くないために、選抜に値するような多収系統が得られる比率は高くない。これについて、例えば、コシヒカリの1穂に着生する粒数を増やすことなどにより、そのポテンシャルをさらに向上させことが望まれる。
さらに、F1雑種を育成する場合には、通常、より大きな雑種強勢を得るために、コシヒカリより食味品質が劣る遠縁系統を片親にする必要がある。遠縁系統を片親とすると、多くの場合、当該遠縁系統が有する食味に関する劣悪形質が遺伝し、選抜の効率を大きく落とす結果となる。
【0013】
本発明は、F1雑種育種法において好適なコシヒカリの雄性不稔系統、及び、当該イネ雄性不稔系統を用いたイネF1種子の生産方法を提供することを目的とする。
【課題を解決するための手段】
【0014】
本発明者らは、上記課題を解決すべく鋭意研究した結果、母本として、染色体の一部を外来品種由来の染色体断片で置換することや突然変異によって、特定の形質を改善させた雄性不稔系統を用いることにより、優れたF1雑種をより効率よく作出可能であることを見出し、本発明を完成させた。
【0015】
すなわち、本発明は、
(1) イネ(Oryza sativa)品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統、又は半糯性を示すイネ雄性不稔系統を母本とし、イネ稔性回復系統を花粉親として交配し、交配後の母本から雑種第1代種子(F1種子)を採取することを特徴とするイネF1種子の生産方法、
(2) 前記イネ雄性不稔系統が、受託番号がFERM P−22217であるイネ細胞質雄性不稔系統CMS−コシヒカリえいち2号(Oryza sativa L.cultivar Koshihikari−Eichi−2go)、イネ細胞質雄性不稔系統CMS−コシヒカリえいち3号、イネ細胞質雄性不稔系統CMS−コシヒカリえいち4号、イネ細胞質雄性不稔系統CMS−コシヒカリかずさ1号、イネ細胞質雄性不稔系統CMS−コシヒカリかずさ2号、及び受託番号がFERM P−22218であるイネ細胞質雄性不稔系統CMS−コシヒカリかずさ3号からなる群より選択される細胞質雄性不稔系統であることを特徴とする前記(1)に記載のイネF1種子の生産方法、
(3) 前記(1)又は(2)に記載のイネF1種子の生産方法により得られたことを特徴とするイネF1種子、
(4) イネ(Oryza sativa)品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有することを特徴とするイネ雄性不稔系統、
(5) 受託番号がFERM P−22217であるイネ細胞質雄性不稔系統CMS−コシヒカリえいち2号(Oryza sativa L.cultivar Koshihikari−Eichi−2go)、
(6) イネ細胞質雄性不稔系統CMS−コシヒカリえいち3号(Oryza sativa L.cultivar Koshihikari−Eichi−3go)、
(7) イネ細胞質雄性不稔系統CMS−コシヒカリえいち4号(Oryza sativa L.cultivar Koshihikari−Eichi−4go)、
(8) イネ細胞質雄性不稔系統CMS−コシヒカリかずさ1号(Oryza sativa L.cultivar Koshihikari−Kazusa−1go)、
(9) イネ細胞質雄性不稔系統CMS−コシヒカリかずさ2号(Oryza sativa L.cultivar Koshihikari−Kazusa−2go)、
(10) 受託番号がFERM P−22218であるイネ細胞質雄性不稔系統CMS−コシヒカリかずさ3号(Oryza sativa L.cultivar Koshihikari−Kazusa−3go)、
を、提供するものである。
【発明の効果】
【0016】
本発明のイネF1種子の生産方法は、特定の形質が改善されたイネ雄性不稔系統を母本とすることで、当該形質を備えるF1雑種の種子を生産することができる。特に、本発明において、イネ品種ハバタキ由来のsd1遺伝子を含有するイネ雄性不稔系統を母本として用いた場合には、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、稈長が有意に短く、耐倒伏性が改善されたF1雑種を作出することができる。また、イネ品種ハバタキ由来のGn1遺伝子を含有するイネ雄性不稔系統を母本として用いた場合には、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、着粒密度の高いF1雑種を作出することができる。また、イネ品種ハバタキ由来のhd1遺伝子を含有するイネ雄性不稔系統を母本として用いた場合には、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、早生化されたF1雑種を作出することができる。さらに、半糯性を有するイネ雄性不稔系統を母本として用いた場合には、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、食味に優れたF1雑種を作出することができる。
【図面の簡単な説明】
【0017】
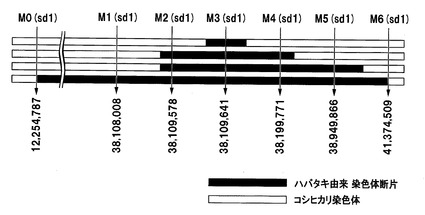
【図1】イネの第1染色体中のsd1遺伝子がコードされている付近のDNAマーカー(SNP)を示した図である。
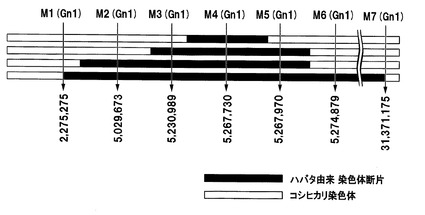
【図2】イネの第1染色体中のGn1遺伝子がコードされている付近のDNAマーカー(SNP)を示した図である。
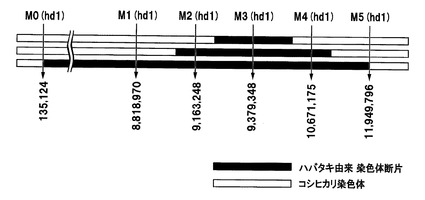
【図3】イネの第6染色体中のhd1遺伝子がコードされている付近のDNAマーカー(SNP)を示した図である。
【図4】実施例1において用いられたイネ品種コシヒカリえいち2号のゲノムを模式的に表した図である。
【図5】実施例1において用いられたイネ品種コシヒカリえいち2号_長領域のゲノムを模式的に表した図である。
【図6】実施例1において用いられたイネ品種コシヒカリえいち3号のゲノムを模式的に表した図である。
【図7】実施例1において用いられたイネ品種コシヒカリえいち4号のゲノムを模式的に表した図である。
【図8】実施例1において用いられたイネ品種コシヒカリえいち4号_長領域のゲノムを模式的に表した図である。
【図9】実施例1において用いられたイネ品種コシヒカリかずさ1号のゲノムを模式的に表した図である。
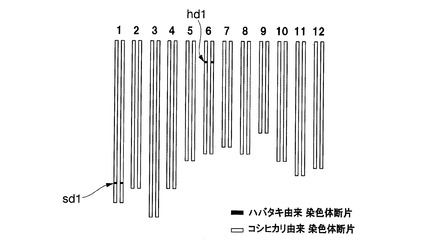
【図10】実施例1において用いられたイネ品種コシヒカリかずさ2号のゲノムを模式的に表した図である。
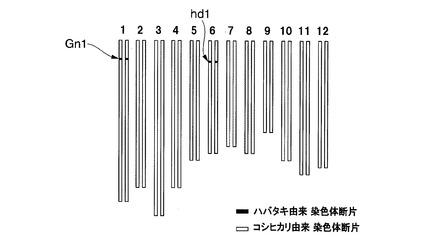
【図11】実施例1において用いられたイネ品種コシヒカリかずさ3号のゲノムを模式的に表した図である。
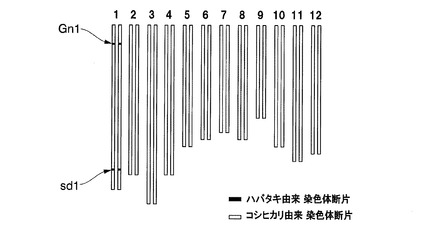
【図12】実施例1において、CMS−コシヒカリえいち4号を母本として得られたF1雑種、CMS−コシヒカリを母本として得られたF1雑種、及びイネ品種コシヒカリの稈長の測定結果を示した図である。
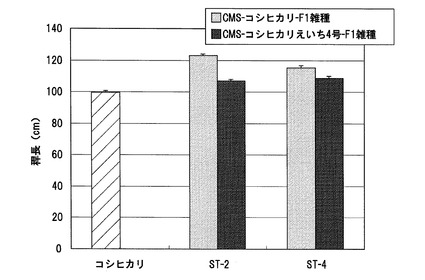
【図13】実施例1において、CMS−コシヒカリとST−1から得られたF1雑種、CMS−コシヒカリえいち4号とST−1から得られたF1雑種、CMS−コシヒカリえいち4号_長領域とST−1から得られたF1雑種、及びイネ品種コシヒカリの稈長の測定結果を示した図である。
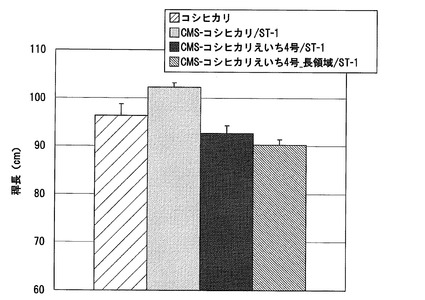
【図14】実施例1において、CMS−コシヒカリえいち3号又はCMS−コシヒカリを母本とし、ST−2又はST−4を花粉親として得られたF1雑種の到穂日数の測定結果を示した図である。
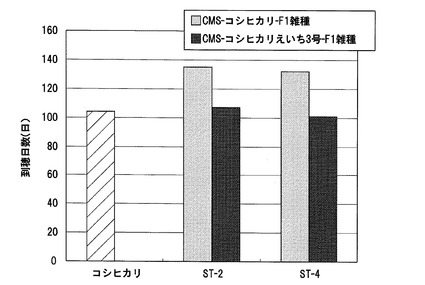
【図15】実施例1において、CMS−コシヒカリえいち2号又はCMS−コシヒカリを母本とし、ST−2又はST−4を花粉親として得られたF1雑種の1穂当たりの着粒数の測定結果を示した図である。
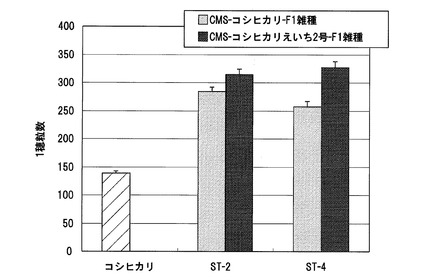
【図16】実施例1において、CMS−コシヒカリえいち2号、CMS−コシヒカリえいち2号_長領域、又はCMS−コシヒカリを母本とし、ST−2を花粉親として得られたF1雑種の1穂当たりの着粒数の測定結果を示した図である。
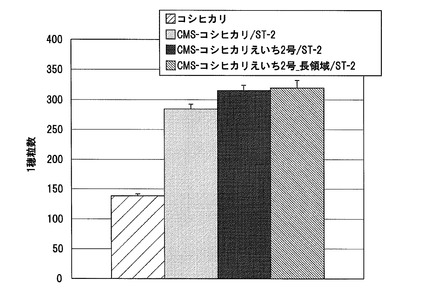
【図17】実施例3において、各F1雑種及びその花粉親の到穂日数の測定結果を、花粉親ごとに示した図である。
【発明を実施するための形態】
【0018】
本発明において準同質遺伝子系統とは、元品種の染色体の一部のみが外来品種由来の染色体断片に置換されている系統を意味する。ここで、外来品種は、元品種以外の品種であれば特に限定されるものではなく、元品種と同一種の植物の品種であってもよく、元品種と異なる種の植物の品種であってもよく、動物等の植物以外の品種であってもよい。なお、本発明において品種とは、同一種の植物であって、遺伝的構成が異なるために、ある形質において同種内の他品種から明確に識別し得る集団を意味する。
【0019】
本発明においてDNAマーカーは、元品種由来の染色体と外来品種由来の染色体を識別し得る染色体上のDNA配列の差異を検出し得るものであれば、特に限定されるものではなく、遺伝子解析分野で通常用いられているDNAマーカーを用いることができる。該DNAマーカーとして、例えば、SNP(Single Nucleotide Polymorphism、一遺伝子多型)やSSR(Simple Sequence Repeats、単純反覆配列)の繰り返し数の違い等の遺伝子多型を検出し得るマーカーであってもよく、RFLP(Restriction Fragment Length Polymorphism、制限酵素断片長多型)マーカーであってもよい。なお、これらのDNAマーカーによる、元品種由来アレルと外来品種由来アレルとの識別は、常法により行うことができる。例えば、各個体から抽出したDNAを鋳型とし、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等を用いてPCRを行い、電気泳動法等を用いてPCR産物の有無を検出し、各多型を識別することができる。また、各個体から抽出したDNAを制限酵素処理した後、電気泳動法等を用いてDNA断片のパターンを検出し、各多型を識別することができる。なお、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等は、該SNPやSSRの塩基配列に応じて、汎用されているプライマー設計ツール等を用いて常法により設計することができる。また、設計されたプライマー等は、当該技術分野においてよく知られている方法のいずれを用いても合成することができる。
【0020】
これらのDNAマーカーは、公知のDNAマーカーを適宜用いることができる。また、新規に作製したDNAマーカーであってもよい。公知のDNAマーカーとして、例えば、イネにおいては、国際公開第2003/070934号パンフレット等において開示されているSNPマーカーや、Rice Genome Research Program(RGB:http://rgp.dna.affrc.go.jp/publicdata.html)において公開されているDNAマーカーを用いることができる。
【0021】
なお、各品種の遺伝子情報等は、例えば、国際的な塩基配列データベースであるNCBI(National center for Biotechnology Information)やDDBJ(DNA Data Bank of Japan)等において入手することができる。特にイネの各品種の遺伝子情報は、KOME(Knowledge−based Oryza Molecular biological Encyclopedia、http://cdna01.dna.affrc.go.jp/cDNA/)等において入手することができる。
【0022】
本発明及び本願明細書において「イネ品種日本晴の染色体のX番目の塩基」は、TIGR(The Institute for Genomic Research;http://www.tigr.org/tdb/e2k1/osa1/blastsearch.shtml)において公開されているイネ品種日本晴のゲノムDNAの塩基配列(バージョン2)に基づいて決定される領域である。
【0023】
また、本発明及び本願明細書において、「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域に相当する領域」とは、イネ個体の染色体中のイネ品種日本晴の染色体中の当該領域と相同性の高い領域であり、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントすることにより決定することができる。また、イネ品種日本晴以外のイネ個体中の「イネ品種日本晴のSNPに相当するSNP」は、当該SNPを含む領域において、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントした場合に、当該SNPに対応する位置にある塩基を意味する。
【0024】
本発明のイネF1種子の生産方法は、特定の形質が改善されたイネ品種コシヒカリの雄性不稔系統を母本とし、イネ稔性回復系統を花粉親として交配し、交配後の母本から雑種第1代種子(F1種子)を採取することを特徴とする。
【0025】
まず、本発明において用いられるイネ雄性不稔系統について説明する。本発明において用いられるイネ雄性不稔系統は、イネ品種コシヒカリの染色体の一部が外来品種由来の染色体断片に置換されることによって、又は突然変異によって、特定の形質が改善されている準同質遺伝子系統の雄性不稔系統である。
【0026】
準同質遺伝子系統の雄性不稔系統は、常法により作出することができる。例えば、雄性不稔細胞質であること以外はイネ品種コシヒカリと同じ形質を備えるコシヒカリ細胞質雄性不稔系統と、所望の領域が外来品種由来の染色体断片で置換された、若しくは所望の突然変異がなされたイネ品種コシヒカリの準同質遺伝子系統とを交配し、得られたF1雑種に対して、当該イネ品種コシヒカリの準同質遺伝子系統を花粉親として用いた連続戻し交配を行うことにより、雄性不稔細胞質であること以外は当該イネ品種コシヒカリの準同質遺伝子系統と同じ形質を備えるイネ細胞質雄性不稔系統を得ることができる。なお、コシヒカリ細胞質雄性不稔系統は、例えば、イネ品種コシヒカリとイネ細胞質雄性不稔系統とを交配し、得られたF1雑種に対して、イネ品種コシヒカリを花粉親として戻し交配を繰り返すことにより、作出することができる。イネ細胞質雄性不稔系統としては、雄性不稔細胞質性を示すイネ科の品種であれば特に限定されるものではなく、例えば、BT型の雄性不稔細胞質であるイネ品種CHINSURAH BORO 2、WA型の雄性不稔細胞質であるイネ品種Male sterile wild rice、GA型の雄性不稔細胞質であるイネ品種Gambiaca、Di型の雄性不稔細胞質であるイネ品種Dissi等を挙げることができる。
【0027】
その他、準同質遺伝子系統の雄性不稔系統としては、特定の環境条件下で不稔を誘発する突然変異遺伝子による環境条件依存型雄性不稔系統であってもよい。環境条件依存型雄性不稔系統としては、長日条件で雄性不稔を引き起こすPMS1遺伝子やPMS2遺伝子を利用した日長感応性雄性不稔(PGMS)系統や、高温条件で雄性不稔を引き起こすTMS1遺伝子やTMS2遺伝子を利用した温度感応性雄性不稔(TGMS)系統等がある。これらの変異遺伝子を有する環境条件依存型雄性不稔系統に、イネ品種コシヒカリの準同質遺伝子系統を交配し、得られたF1雑種に対して、当該イネ品種コシヒカリの準同質遺伝子系統を花粉親として用いた連続戻し交配を行うことにより、当該変異遺伝子による環境条件依存型雄性不稔性を示す以外は、当該イネ品種コシヒカリの準同質遺伝子系統と同じ形質を備えるイネ雄性不稔系統を得ることができる。
【0028】
まず、母本として、イネ品種コシヒカリの染色体の一部が外来品種由来の染色体断片に置換されたイネ品種コシヒカリの準同質遺伝子系統について説明する。
イネ品種コシヒカリの準同質遺伝子系統において挿入されている外来品種由来の染色体断片は、当該染色体断片が挿入されることによって特定の形質をイネ品種コシヒカリよりも改善されるものであれば、特に限定されるものではない。例えば、挿入される外来品種由来の染色体断片は、所望の形質改善に直接寄与する遺伝子(原因遺伝子)をコードする領域を含んでいればよく、当該原因遺伝子のみを含む領域であってもよく、当該原因遺伝子とその他の遺伝子を含む領域(例えば、14.6Mbp〜29.2Mbpの長さからなる領域)であってもよい。
【0029】
本発明及び本願明細書において、「イネ品種“X”由来の“Y”遺伝子」には、イネ品種“X”自体に由来する“Y”遺伝子(すなわち、イネ品種“X”の染色体中に存在している“Y”遺伝子)に加えて、イネ品種“X”と実質的に同等な“Y”遺伝子を有するイネ品種由来の“Y”遺伝子も含まれる。イネ品種“X”由来の“Y”遺伝子に代えて、イネ品種“X”由来の“Y”遺伝子と実質的に同等であるイネ品種“X”以外のイネ品種由来の“Y”遺伝子を染色体に組み込んだ場合であっても、本発明にと同じ効果が奏されるためである。ここで、イネ品種“X”由来の“Y”遺伝子と実質的に同等な“Y”遺伝子とは、イネ品種“X”以外のイネ品種由来の“Y”遺伝子であって、イネ品種“X”由来の“Y”遺伝子と同程度の機能を有する遺伝子である。具体的には、イネ品種“X”の後代品種であって、イネ品種“X”から“Y”遺伝子が存在する領域のアレルを受け継いだイネ品種や、イネ品種“X”の先祖に当たるイネ品種であって、イネ品種“X”へ“Y”遺伝子が存在する領域のアレルを共通して有するイネ品種、これらのイネ品種“X”と実質的に同等な“Y”遺伝子を有するイネ品種が有する“Y”遺伝子が存在する領域の染色体断片が組み込まれたイネ品種等が挙げられる。
【0030】
すなわち、本発明及び本願明細書においては、特に記載が無い限り、「イネ品種ハバタキ由来のsd1遺伝子」には、イネ品種ハバタキ自体に由来するsd1遺伝子のみならず、該遺伝子と実質的に同等なsd1遺伝子、例えば、イネ品種低脚烏尖、イネ品種IR8、イネ品種キヌヒカリ、イネ品種ゆめひたち、イネ品種コシヒカリえいち4号、イネ品種コシヒカリかずさ2号、イネ品種コシヒカリかずさ3号、又はイネ品種コシヒカリかずさ4号等のイネ品種由来のsd1遺伝子が含まれる。
【0031】
同様に、本発明及び本願明細書においては、特に記載が無い限り、「イネ品種ハバタキ由来のGn1遺伝子」には、イネ品種ハバタキ自体に由来するGn1遺伝子のみならず、該遺伝子と実質的に同等なGn1遺伝子、例えば、イネ品種コシヒカリえいち2号、イネ品種コシヒカリかずさ2号、イネ品種コシヒカリかずさ3号、又はイネ品種コシヒカリかずさ4号等のイネ品種由来のGn1遺伝子が含まれる。
【0032】
同様に、本発明及び本願明細書においては、特に記載が無い限り、「イネ品種ハバタキ由来のhd1遺伝子」には、イネ品種ハバタキ自体に由来するhd1遺伝子のみならず、該遺伝子と実質的に同等なhd1遺伝子、例えば、イネ品種コシヒカリえいち3号、イネ品種コシヒカリかずさ1号、イネ品種コシヒカリかずさ2号、又はイネ品種コシヒカリかずさ4号等のイネ品種由来のhd1遺伝子が含まれる。
【0033】
本発明においては、イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統を母本として用いる。これらの3種の遺伝子のうち、2種類の遺伝子を適宜組み合わせて含有させた雄性不稔系統であってもよく、3種類全ての遺伝子を含む雄性不稔系統であってもよい。
【0034】
イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統は、イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統と、コシヒカリ雄性不稔系統とから前述の方法により作出することができる。イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統は、例えば、適当なDNAマーカーを用いて、特許文献3及び4に記載の方法等によって作出することができる。なお、本発明において母本として用いるイネ品種コシヒカリの準同質遺伝子系統は、新たに作出されたものであってもよく、既存の系統を用いてもよい。
【0035】
イネ品種ハバタキ由来のsd1遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来sd1含有準同質遺伝子系統)は、イネ品種コシヒカリの染色体中のsd1遺伝子がコードされている領域が、イネ品種ハバタキ由来のsd1遺伝子をコードする領域を含む染色体断片に置換されている。ハバタキ由来sd1含有準同質遺伝子系統に含まれるハバタキ由来染色体断片は、sd1遺伝子がコードされている領域を含んでいれば特に限定されるものではなく、sd1遺伝子がコードされている領域のみを含んでいてもよく、sd1遺伝子の近傍に存在している遺伝子もsd1遺伝子と共にイネ品種コシヒカリに挿入されてもよい。図1に、イネの第1染色体中のsd1遺伝子がコードされている38.11Mbp付近のDNAマーカー(SNP)を示す。当該ハバタキ由来染色体断片の長さは、DNAマーカーを用いて決定することできる。例えば、図1に示すように、ハバタキ由来sd1含有準同質遺伝子系統において、挿入されたハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の38,109,578番目の塩基配列に依存する多型(PCRにより、イネ品種コシヒカリではPCR産物が得られ、イネ品種ハバタキではPCR産物が得られない)(以下、G2003)とイネ品種日本晴の第1染色体の38,109,641番目の塩基配列に依存する多型(PCRにより、イネ品種コシヒカリではPCR産物が得られ、イネ品種ハバタキではPCR産物が得られない)(以下、G2002)との間にあり、当該ハバタキ由来染色体断片の下流端が、G2003とイネ品種日本晴の第1染色体の38,199,771番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではT)(以下、SP−462)との間にあってもよい(図1の1段目)。また、当該ハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の38,108,008番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)(以下、SP−4009)とG2003との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−462とイネ品種日本晴の第1染色体の38,949,866番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではC)(以下、SP−1259)との間にあってもよい(図1の2段目)。当該ハバタキ由来染色体断片の上流端が、SP−4009とG2003との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−1259とイネ品種日本晴の第1染色体の41,374,509番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではG)(以下、SP−477)との間にあってもよい(図1の3段目)。さらに、イネ品種ハバタキ由来のsd1遺伝子をコードする領域を含む、よりさらに長い領域が、ハバタキ由来染色体断片により置換されていてもよい。例えば、イネ品種日本晴の第1染色体の12,254,787番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)(以下、SP−2058)からSP−477までの約29.1Mbpの領域を含む領域が、ハバタキ由来染色体断片により置換されていてもよい(図1の4段目)。各DNAマーカー及び識別に使用可能なプライマーの塩基配列を表1に示す。
【0036】
【表1】

【0037】
ハバタキ由来sd1遺伝子を含有するイネ雄性不稔系統を母本とすることにより得られたF1雑種は、ハバタキ由来のsd1遺伝子を含有しているため、コシヒカリ雄性不稔系統を母本として得られたF1雑種よりも、稈長が有意に低く、耐倒伏性が大幅に改善されている。このため、本発明のイネF1種子の生産方法において、ハバタキ由来のsd1遺伝子を含有するイネ雄性不稔系統を用いることにより、耐倒伏性が改善されたF1雑種の種子を効率よく生産することができ、F1雑種育成のための組み合わせ検定の効率を向上させることができる。
【0038】
イネ品種ハバタキ由来のGn1遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来Gn1含有準同質遺伝子系統)は、イネ品種コシヒカリの染色体中のGn1遺伝子がコードされている領域が、イネ品種ハバタキ由来のGn1遺伝子をコードする領域を含む染色体断片に置換されている。ハバタキ由来Gn1含有準同質遺伝子系統に含まれるハバタキ由来染色体断片は、Gn1遺伝子がコードされている領域を含んでいれば特に限定されるものではなく、Gn1遺伝子がコードされている領域のみを含んでいてもよく、Gn1遺伝子の近傍に存在している遺伝子もGn1遺伝子と共にイネ品種コシヒカリに挿入されてもよい。図2に、イネの第1染色体中のGn1遺伝子がコードされている5.267Mbp付近のDNAマーカー(SNP)を示す。当該ハバタキ由来染色体断片の長さは、DNAマーカーを用いて決定することできる。例えば、図2に示すように、ハバタキ由来Gn1含有準同質遺伝子系統において、挿入されたハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の5,230,989番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではA)(以下、SP−170)とイネ品種日本晴の第1染色体の5,267,730番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではC)(以下、SP−4028)との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−4028とイネ品種日本晴の第1染色体の5,267,970番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)(以下、SP−4038)との間にあってもよい(図2の1段目)。また、当該ハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の5,029,673番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではG)(以下、SP−2032)とSP−170との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−4038とイネ品種日本晴の第1染色体の5,274,879番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではT)(以下、SP−4030)との間にあってもよい(図2の2段目)。当該ハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の2,275,275番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)(以下、SP−158)とSP−2032との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−4038とSP−4030との間にあってもよい(図2の3段目)。さらに、イネ品種ハバタキ由来のGn1遺伝子をコードする領域を含む、よりさらに長い領域が、ハバタキ由来染色体断片により置換されていてもよい。例えば、SP−158からイネ品種日本晴の第1染色体の31,371,175番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではA)(以下、SP−262)までの約29.1Mbpの領域を含む領域が、ハバタキ由来染色体断片により置換されていてもよい(図2の4段目)。各DNAマーカー及び識別に使用可能なプライマーの塩基配列を表2に示す。
【0039】
【表2】
【0040】
ハバタキ由来Gn1遺伝子を含有するイネ雄性不稔系統を母本とすることにより得られたF1雑種は、ハバタキ由来のGn1遺伝子を含有しているため、コシヒカリ雄性不稔系統を母本として得られたF1雑種よりも着粒密度が向上している。このため、本発明のイネF1種子の生産方法において、ハバタキ由来のGn1遺伝子を含有するイネ雄性不稔系統を用いることにより、着粒密度が改善されたF1雑種の種子を効率よく生産することができ、F1雑種育成のための組み合わせ検定の効率を向上させることができる。
【0041】
イネ品種ハバタキ由来のhd1遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来hd1含有準同質遺伝子系統)は、イネ品種コシヒカリの染色体中のhd1遺伝子がコードされている領域が、イネ品種ハバタキ由来のhd1遺伝子をコードする領域を含む染色体断片に置換されている。ハバタキ由来hd1含有準同質遺伝子系統に含まれるハバタキ由来染色体断片はhd1遺伝子がコードされている領域を含んでいれば特に限定されるものではなく、hd1遺伝子がコードされている領域のみを含んでいてもよく、hd1遺伝子の近傍に存在している遺伝子もhd1遺伝子と共にイネ品種コシヒカリに挿入されてもよい。図3に、イネの第1染色体中のhd1遺伝子がコードされている9.38Mbp付近のDNAマーカー(SNP)を示す。当該ハバタキ由来染色体断片の長さは、DNAマーカーを用いて決定することできる。例えば、図3に示すように、ハバタキ由来hd1含有準同質遺伝子系統において、挿入されたハバタキ由来染色体断片の上流端が、イネ品種日本晴の第6染色体の9,163,248番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではA)(以下、SP−586)とイネ品種日本晴の第6染色体の9,379,348番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではG)(以下、SP−2254)との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−2254とイネ品種日本晴の第6染色体の10,671,175番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではC)(以下、SP−1603)との間にあってもよい(図3の上段)。また、当該ハバタキ由来染色体断片の上流端が、イネ品種日本晴の第6染色体の8,818,970番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではT)(以下、SP−2513)とSP−586との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−1603とイネ品種日本晴の第6染色体の11,949,796番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではC)(以下、SP−604)との間にあってもよい(図3の中段)。さらに、イネ品種ハバタキ由来のhd1遺伝子をコードする領域を含む、よりさらに長い領域が、ハバタキ由来染色体断片により置換されていてもよい。例えば、イネ品種日本晴の第6染色体の135,124番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではG)(以下、SP−2229)からイネ品種日本晴の第6染色体の29,016,207番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではT)(以下、SP−1635)までの約28.9Mbpの領域を含む領域が、ハバタキ由来染色体断片により置換されていてもよい(図3の下段)。各DNAマーカー及び識別に使用可能なプライマーの塩基配列を表3に示す。
【0042】
【表3】
【0043】
ハバタキ由来hd1遺伝子を含有するイネ雄性不稔系統を母本とすることにより得られたF1雑種は、ハバタキ由来のhd1遺伝子を含有しているため、コシヒカリ雄性不稔系統を母本として得られたF1雑種よりも早生となる。このため、本発明のイネF1種子の生産方法において、ハバタキ由来のhd1遺伝子を含有するイネ雄性不稔系統を用いることにより、出穂期が早められたF1雑種の種子を効率よく生産することができ、F1雑種育成のための組み合わせ検定の効率を向上させることができる。
【0044】
sd1遺伝子、Gn1遺伝子、及びhd1遺伝子のうちの2以上の遺伝子がハバタキ由来の遺伝子に置換されたイネ品種コシヒカリの準同質遺伝子系統は、異なる種類の遺伝子がハバタキ由来の遺伝子に置換されている準同質遺伝子系統同士を交配し、得られたF1雑種を自家交配することによって得られたF2雑種の中から、DNAマーカーを用いて、イネ品種コシヒカリの染色体中に導入された外来遺伝子由来の遺伝子が、両方の相同染色体に導入されているホモ個体を選抜することによって得られる。例えば、sd1遺伝子とGn1遺伝子がハバタキ由来の遺伝子に置換されたイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来sd1/ハバタキ由来Gn1含有準同質遺伝子系統)は、ハバタキ由来sd1含有準同質遺伝子系統とハバタキ由来Gn1含有準同質遺伝子系統を交配し、得られたF1雑種を自家交配させて得られた雑種第2代(F2雑種)の中から、DNAマーカーを指標として、両方の相同染色体において、sd1遺伝子がコードされている領域及びGn1遺伝子がコードされている領域が、いずれもハバタキ由来の領域である個体を選抜することにより、作出することができる。同様に、sd1遺伝子、Gn1遺伝子、及びhd1遺伝子がハバタキ由来の遺伝子に置換されたイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来sd1/ハバタキ由来Gn1/ハバタキ由来hd1含有準同質遺伝子系統)は、ハバタキ由来sd1/ハバタキ由来Gn1含有準同質遺伝子系統とハバタキ由来hd1含有準同質遺伝子系統を交配し、得られたF1雑種を自家交配させて得られた雑種第2代(F2雑種)の中から、DNAマーカーを指標として、両方の相同染色体において、sd1遺伝子がコードされている領域、Gn1遺伝子がコードされている領域、及びhd1遺伝子がコードされている領域が、いずれもハバタキ由来の領域である個体を選抜することにより、作出することができる。
【0045】
本発明において用いられるイネ雄性不稔系統としては、半糯性を示すものであってもよい。半糯性を示すイネ雄性不稔系統は、半糯性を示すイネ品種コシヒカリの準同質遺伝子系統を、雄性不稔系統と2回戻し交配し、得られたF3雑種の中から全ての種子において半糯性を示すイネ個体を選抜することによって得られる。
【0046】
半糯性遺伝子を有する染色体断片を導入することにより、玄米のアミロース含量を低下させることを通して、食味品質を向上させることができる。特に半糯性変異遺伝子を片親にのみ持たせたF1雑種系統においては、半糯性を示す玄米が通常形質を有する玄米と1:3の比率で出現するため、穏やかな低アミロース化を実現することができ、半糯性が全くないイコシヒカリよりも食味が非常に良好となる。このため、本発明のイネF1種子の生産方法において、半糯性を示すイネ品種コシヒカリの準同質遺伝子系統を用いることにより、食味品質が向上されたF1雑種の種子を効率よく生産することができ、F1雑種育成のための組み合わせ検定の効率を向上させることができる。
【0047】
半糯性を示すイネ品種コシヒカリの準同質遺伝子系統は、例えば、イネ品種コシヒカリの突然変異群から、半糯性を示す表現形質や、DNAマーカーを利用した半糯性に寄与する遺伝子(半糯性遺伝子)のタイプによって選抜することにより得ることができる。半糯性遺伝子としては、例えば、イネ品種の第6染色体に存在するwaxy−mq遺伝子等が挙げられる。その他、半糯性を示すイネ品種コシヒカリの準同質遺伝子系統としては、waxy−mq遺伝子の突然変異株であるイネ品種ミルキークイーン等の公知のイネ品種コシヒカリの突然変異種であってもよく、イネ品種コシヒカリ以外の半糯性を示すイネ品種(例えば、他品種由来の突然変異体)に対して、イネ品種コシヒカリを連続戻し交配することにより準同質遺伝子系統化したものであってもよい。
【0048】
本発明において用いられるイネ雄性不稔系統としては、イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有しており、かつ半糯性を有するものであってもよい。このような半糯性を示すイネ雄性不稔系統は、具体的には、以下のようにして作出できる。まず、sd1遺伝子、Gn1遺伝子、及びhd1遺伝子のうちの少なくとも1以上の遺伝子がハバタキ由来の遺伝子に置換されたイネ品種コシヒカリの準同質遺伝子系統と、半糯性を示すイネ品種コシヒカリの準同質遺伝子系統とを交配し、得られたF1雑種を自家交配することによって得られたF2雑種の中から、sd1遺伝子、Gn1遺伝子、及びhd1遺伝子のうちの少なくとも1以上の遺伝子がハバタキ由来の遺伝子に置換されており、かつ半糯性を示すイネ個体を選抜する。得られたイネ個体とコシヒカリ雄性不稔系統と交配し、得られたF1雑種に対して連続戻し交配を行うことにより、sd1遺伝子、Gn1遺伝子、及びhd1遺伝子のうちの少なくとも1以上の遺伝子がハバタキ由来の遺伝子に置換されており、かつ半糯性を示すイネ雄性不稔系統を得ることができる。
【0049】
本発明のイネF1種子の生産方法において花粉親として用いられるイネ稔性回復系統は、母本として用いるイネ雄性不稔系統の稔性を回復し得るイネ系統であれば特に限定されるものではない。例えば、母本がBT型の雄性不稔細胞質である場合には、イネ品種JFR-4、イネ品種ST−1、イネ品種ST−2、イネ品種ST−4、イネ品種タカナリ、イネ品種桂朝2号、イネ品種水原258号、及びイネ品種ハバタキ等がイネ稔性回復系統として挙げられる。なお、あるイネ品種があるコシヒカリ雄性不稔系統に対するイネ稔性回復系統か否かは、当該イネ品種と、前記コシヒカリ雄性不稔系統とを交配させ、得られたF1雑種の雄性稔性に基づいて調べることができる。当該F1雑種において雄性不稔性が回復されている場合には、当該イネ品種は当該コシヒカリ雄性不稔系統に対するイネ稔性回復系統であると分かる。また、母本が環境条件依存型雄性不稔である場合には、母本が備える雄性不稔性の原因となる変異遺伝子を有していないイネ系統であれば、イネ稔性回復系統として用いることができる。当該変異遺伝子はF1(ヘテロの状態)の場合には変異形質が現れないためである。
【0050】
本発明のイネF1種子の生産方法において、イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統と、イネ稔性回復系統を花粉親との交配は、自然交配であってもよく、人工的に交配させてもよい。
【実施例】
【0051】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0052】
[実施例1]
<イネ品種コシヒカリの準同質遺伝子系統>
母本として用いるイネ雄性不稔系統を作出するためのイネ品種コシヒカリの準同質遺伝子系統として、イネ品種コシヒカリえいち2号、イネ品種コシヒカリえいち2号_長領域、イネ品種コシヒカリえいち3号、イネ品種コシヒカリえいち4号、イネ品種コシヒカリえいち4号_長領域、イネ品種コシヒカリかずさ1号、イネ品種コシヒカリかずさ2号、イネ品種コシヒカリかずさ3号、及びイネ品種コシヒカリかずさ4号を得た。
【0053】
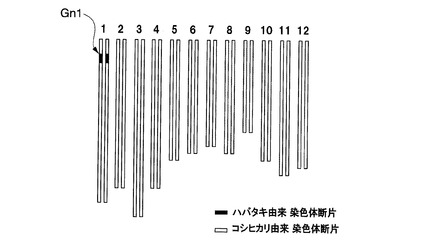
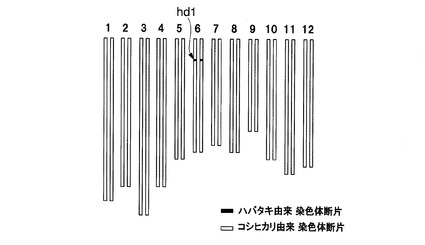
これらの準同質遺伝子系統のうち、イネ品種コシヒカリえいち2号、イネ品種コシヒカリえいち3号、イネ品種コシヒカリえいち4号、及びイネ品種コシヒカリかずさ4号(寄託番号:FERM P−21596)は、特許文献3及び4に記載されているものを用いた。これらの系統は、ハバタキにコシヒカリを複数回戻し交配した後に、DNAマーカー選抜によって育成された。イネ品種コシヒカリえいち4号の染色体中のハバタキ由来染色体断片に置換されている領域と表1に記載のDNAマーカーとの位置関係は図1の2段目の通りであり、イネ品種コシヒカリえいち2号の染色体中のハバタキ由来染色体断片に置換されている領域と表2に記載のDNAマーカーとの位置関係は図2の2段目の通りであり、イネ品種コシヒカリえいち3号の染色体中のハバタキ由来染色体断片に置換されている領域と表3に記載のDNAマーカーとの位置関係は図3の中段の通りである。
【0054】
また、特許文献3に記載の方法に準拠し、イネ品種コシヒカリえいち4号よりも長い領域がハバタキ由来染色体断片により置換されたイネ品種コシヒカリえいち4号_長領域を作出した。より詳細には、表1に記載のDNAマーカーのうち、SP−4009、G2003、G2002、SP−1259、及びSP−477を用いて、目的のゲノムを有す個体を選抜した。
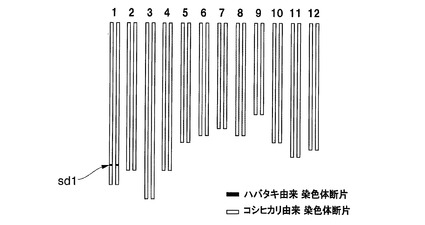
具体的には、イネ品種ハバタキに対してイネ品種コシヒカリを複数回戻し交配させた後、得られた雑種の種子をさらに栽培し、圃場に移植できる程度に成育させた後、各栽培個体の葉からDNAを回収し、SP−4009及びSP−477がコシヒカリ由来アレルのホモ染色体領域であり、G2003、G2002、及びSP−1259がハバタキ由来アレルのホモ染色体領域である栽培個体1個を選抜した。この選抜された栽培個体が、sd1遺伝子を含有する領域をハバタキ由来染色体断片に置換した新品種であり、本発明者はこの新品種を「イネ品種コシヒカリえいち4号_長領域」と命名した。イネ品種コシヒカリえいち4号_長領域の染色体中のハバタキ由来染色体断片に置換されている領域と表1に記載のDNAマーカーとの位置関係は、図1の3段目の通りである。
【0055】
同様に、特許文献3に記載の方法に準拠し、イネ品種コシヒカリえいち2号よりも長い領域がハバタキ由来染色体断片により置換されたイネ品種コシヒカリえいち2号_長領域を作出した。より詳細には、表2に記載のDNAマーカーのうち、SP−156、SP−2032、SP−4028、SP−4038、及びSP−4030を用いて、目的のゲノムを有す個体を選抜した。
具体的には、イネ品種ハバタキに対してイネ品種コシヒカリを複数回戻し交配させた後、得られた雑種の種子をさらに栽培し、圃場に移植できる程度に成育させた後、各栽培個体の葉からDNAを回収し、SP−156及びSP−4030がコシヒカリ由来アレルのホモ染色体領域であり、SP−2032、SP−4028、及びSP−4038がハバタキ由来アレルのホモ染色体領域である栽培個体1個を選抜した。この選抜された栽培個体が、Gn1遺伝子を含有する領域をハバタキ由来染色体断片に置換した新品種であり、本発明者はこの新品種を「イネ品種コシヒカリえいち2号_長領域」と命名した。イネ品種コシヒカリえいち2号_長領域の染色体中のハバタキ由来染色体断片に置換されている領域と表2に記載のDNAマーカーとの位置関係は、図2の3段目の通りである。
【0056】
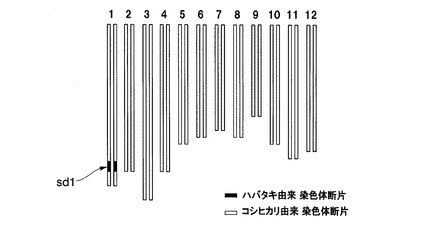
さらに、sd1遺伝子とhd1遺伝子がハバタキ由来の遺伝子である準同質遺伝子系統を作出した。具体的には、イネ品種コシヒカリえいち4号とイネ品種コシヒカリえいち3号を交配し、得られた後代個体(種子)のうち2個を栽培し、自殖(自家交配)させ、さらに後代個体である種子を100個得た。この100個の種子を全て栽培し、各後代個体のDNAマーカーを調べ、SP−462〔M4(sd1)〕がハバタキ由来アレルのホモ染色体領域であり、かつSP−2254〔M3(hd1)〕がハバタキ由来アレルのホモ染色体領域である栽培個体を1個体選抜した。この選抜された栽培個体が、sd1遺伝子を含有する領域及びhd1遺伝子を含有する領域がいずれも、ハバタキ由来染色体断片(ホモ)に置換された新品種であり、本発明者はこの新品種を「イネ品種コシヒカリかずさ1号」と命名した。
【0057】
さらに、Gn1遺伝子とhd1遺伝子がハバタキ由来の遺伝子である準同質遺伝子系統を作出した。具体的には、イネ品種コシヒカリえいち2号とイネ品種コシヒカリえいち3号を交配し、得られた後代個体(種子)のうち2個を栽培し、自殖(自家交配)させ、さらに後代個体である種子を100個得た。この100個の種子を全て栽培し、各後代個体のDNAマーカーを調べ、SP−4028〔M4(Gn1)〕がハバタキ由来アレルのホモ染色体領域であり、かつSP−2254〔M3(hd1)〕がハバタキ由来アレルのホモ染色体領域である栽培個体を1個体選抜した。この選抜された栽培個体が、Gn1遺伝子を含有する領域及びhd1遺伝子を含有する領域がいずれも、ハバタキ由来染色体断片(ホモ)に置換された新品種であり、本発明者はこの新品種を「イネ品種コシヒカリかずさ2号」と命名した。
【0058】
さらに、sd1遺伝子とGn1遺伝子がハバタキ由来の遺伝子である準同質遺伝子系統を作出した。具体的には、イネ品種コシヒカリえいち4号とイネ品種コシヒカリえいち2号を交配し、得られた後代個体(種子)のうち2個を栽培し、自殖(自家交配)させ、さらに後代個体である種子を100個得た。この100個の種子を全て栽培し、各後代個体のDNAマーカーを調べ、SP−462〔M4(sd1)〕がハバタキ由来アレルのホモ染色体領域であり、かつSP−4028〔M4(Gn1)〕がハバタキ由来アレルのホモ染色体領域である栽培個体を1個体選抜した。この選抜された栽培個体が、sd1遺伝子を含有する領域及びGn1遺伝子を含有する領域がいずれも、ハバタキ由来染色体断片(ホモ)に置換された新品種であり、本発明者はこの新品種を「イネ品種コシヒカリかずさ3号」と命名した。
【0059】
図4にイネ品種コシヒカリえいち2号のゲノムを、図5にイネ品種コシヒカリえいち2号_長領域のゲノムを、図6にイネ品種コシヒカリえいち3号のゲノムを、図7にイネ品種コシヒカリえいち4号のゲノムを、図8にイネ品種コシヒカリえいち4号_長領域のゲノムを、図9にイネ品種コシヒカリかずさ1号のゲノムを、図10にイネ品種コシヒカリかずさ2号のゲノムを、図11にイネ品種コシヒカリかずさ3号のゲノムを、それぞれ模式的に示す。
【0060】
<イネ品種コシヒカリの細胞質雄性不稔系統(CMS−コシヒカリ)>
イネ品種CHINSURAH BORO 2にイネ品種コシヒカリを6回戻し交配し、圃場における生育特性が、雄性不稔であることを除きコシヒカリと同等の形質を示すCMS−コシヒカリを育成した。
【0061】
<イネ品種ミルキークイーンの細胞質雄性不稔系統(CMS−ミルキークイーン)>
CMS−コシヒカリを母本として、コシヒカリの半糯性突然変異であるミルキークイーンを2回戻し交配を行い、玄米形質が粳型と半糯型に分離する種子を得た。その後、この種子から半糯性を示す玄米を選抜した。この選抜された玄米が、半糯性を有するイネ品種コシヒカリの細胞質雄性不稔系統であり、該系統をCMS−ミルキークイーンとして育成した。
【0062】
<イネ細胞質雄性不稔系統(CMS系統)>
CMS−コシヒカリを母本とし、イネ品種コシヒカリえいち2号、イネ品種コシヒカリえいち2号_長領域、イネ品種コシヒカリえいち3号、イネ品種コシヒカリえいち4号、イネ品種コシヒカリえいち4号_長領域、イネ品種コシヒカリかずさ2号、イネ品種コシヒカリかずさ3号、又はイネ品種コシヒカリかずさ4号を花粉親として連続戻し交配を行った。得られた後代個体の中から、雄性不稔性を示す栽培個体を選抜した。これらの選抜された各栽培個体の葉からDNAを回収し、DNAマーカーを調べ、花粉親と同じ領域が、外来品種由来アレルのホモ染色体領域である栽培個体をそれぞれ1個体ずつ選抜した。これらの選抜された栽培個体は、雄性不稔である以外は、基本的に花粉親と同じ形質を備える新品種である。本発明者は、イネ品種コシヒカリえいち2号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち2号」、イネ品種コシヒカリえいち2号_長領域を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち2号_長領域」、イネ品種コシヒカリえいち3号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち3号」、イネ品種コシヒカリえいち4号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち4号」、イネ品種コシヒカリえいち4号_長領域を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち4号_長領域」、イネ品種コシヒカリかずさ1号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリかずさ1号」、イネ品種コシヒカリかずさ2号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリかずさ2号」、及びイネ品種コシヒカリかずさ3号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリかずさ3号」と命名した。
【0063】
なお、出願人は、実施例1において得られた新品種のうち、CMS−イネ品種コシヒカリえいち2号、及びCMS−イネ品種コシヒカリかずさ3号を、独立行政法人産業技術総合研究所特許生物寄託センター(茨城県つくば市東1−1−1 つくばセンター中央第6)に新規植物として寄託した。イネ細胞質雄性不稔系統CMS−イネ品種コシヒカリえいち2号の受託番号はFERM P−22217であり、イネ細胞質雄性不稔系統CMS−イネ品種コシヒカリかずさ3号の受託番号はFERM P−22218である。
【0064】
<F1種子の生産>
上記で得られたイネ細胞質雄性不稔系統(CMS系統)を母本とし、花粉親として独自に育成した回復系中間母本系統ST−1、ST−2、又はST−4をそれぞれ交配してF1雑種の種子を得た。対照として、CMS−コシヒカリを母本とし、ST−1、ST−2、又はST−4を花粉親として交配してF1雑種の種子を得た。得られたF1種子を栽培し、2006年度、2007年度、及び2008年度の愛知県における圃場試験において形質の検討を行った。検討は、種苗法(平成10年法律第83号)第5条第1項に基づく品種登録出願のための特性審査に準拠して行った。
【0065】
<F1雑種の稈長>
CMS−コシヒカリえいち4号を母本として得られたF1雑種と、CMS−コシヒカリを母本として得られたF1雑種と、イネ品種コシヒカリの稈長を比較した。各系統の稈長の測定結果を図12に示す。この結果、対照品種であるコシヒカリの稈長は99.6cmであったのに対して、CMS−コシヒカリとST−2又はST−4を交配させて得られたF1雑種の稈長は、それぞれ123.0cm、115.3cmであった。一方、CMS−コシヒカリえいち4号と、ST−2又はST−4とを交配させて得られたF1雑種の稈長は、それぞれ106.9cm、108.8cmであり、CMS−コシヒカリを母本としたF1雑種よりも6.5〜16.1cm短くなっており、同時に収穫時における倒伏程度が軽減されていた。また、コシヒカリえいち4号を母本として得られたF1雑種系統は、稈長が短くなる以外は、コシヒカリを母本として得られたF1雑種系統と同じ形質を示した。
【0066】
さらに別の年に、CMS−コシヒカリとST−1から得られたF1雑種、CMS−コシヒカリえいち4号とST−1から得られたF1雑種、CMS−コシヒカリえいち4号_長領域とST−1から得られたF1雑種、及びイネ品種コシヒカリの稈長を測定し、比較した。各系統の稈長の測定結果を図13に示す。この結果、CMS−コシヒカリを母本として得られたF1雑種は、対照品種であるコシヒカリよりも稈長が長くなっていたのに対して、CMS−コシヒカリえいち4号又はCMS−コシヒカリえいち4号_長領域を母本として得られたF1雑種は、コシヒカリよりも稈長が短くなっていた。また、CMS−コシヒカリえいち4号から得られたF1雑種と、CMS−コシヒカリえいち4号_長領域から得られたF1雑種を比較したところ、両者に有意な差異は見られなかった。
これらの結果から、sd1遺伝子がコードされる領域がハバタキ由来の染色体断片で置換されているイネ雄性不稔系統を用いることにより、ハバタキ由来のsd1遺伝子を有さないイネ雄性不稔系統を用いた場合よりも、稈長が短く、耐倒伏性が改善されたF1雑種が得られることが明らかである。
【0067】
<F1雑種の到穂日数>
CMS−コシヒカリえいち3号又はCMS−コシヒカリを母本とし、ST−2又はST−4を花粉親としてF1雑種を得、これらのF1雑種の到穂日数を調べた。各系統の到穂日数の測定結果を図14に示す。4月16日に播種した本試験における対照品種コシヒカリの到穂日数は104日であったのに対して、CMS−コシヒカリとST−2又はST−4から得られたF1雑種の到穂日数は、135日又は132日であった。一方、CMS−コシヒカリえいち3号とST−2又はST−4から得られたF1雑種の到穂日数は、107日又は101日であり、CMS−コシヒカリを母本としたF1雑種よりも10〜28日程度短く、早生化されていた。このように、CMS−コシヒカリえいち3号を母本としたF1雑種は、コシヒカリとほぼ同程度の到穂日数であり、愛知県における栽培に適応可能な熟期を示した。また、CMS−コシヒカリえいち3号を母本としたF1雑種系統の形質は、到穂日数が小さくなること、及び、早生になることにより植物体が小さくなる傾向を示したが、それ以外の形質については、基本的にCMS−コシヒカリを母本としたF1雑種と同じであった。
これらの結果から、hd1遺伝子がコードされる領域がハバタキ由来の染色体断片で置換されているイネ細胞質雄性不稔系統を用いることにより、ハバタキ由来のhd1遺伝子を有さないイネ細胞質雄性不稔系統を用いた場合よりも、早生化されたF1雑種が得られることが明らかである。
【0068】
<F1雑種の着粒数>
CMS−コシヒカリえいち2号又はCMS−コシヒカリを母本とし、ST−2又はST−4を花粉親としてF1雑種得、これらのF1雑種の着粒数を調べた。各系統の着粒数の測定結果を図15に示す。この結果、対照品種であるコシヒカリの主茎における1穂粒数は138.5粒であったのに対して、CMS−コシヒカリとST−2又はST−4から得られたF1雑種の主茎における1穂粒数は、それぞれ284.0粒、257.4粒であった。一方、CMS−コシヒカリえいち2号とST−2又はST−4から得られたF1雑種の1穂粒数は、それぞれ314.8粒、327.0粒であり、CMS−コシヒカリを母本としたF1雑種よりも30〜70粒多くなっており、1穂重、及び収量性が向上していた。また、CMS−コシヒカリえいち2号を母本としたF1雑種系統の形質は、着粒数が多くなった以外は、基本的にCMS−コシヒカリを母本としたF1雑種と同じであった。
【0069】
また、CMS−コシヒカリえいち2号、CMS−コシヒカリえいち2号_長領域、又はCMS−コシヒカリを母本とし、ST−2を花粉親として得られたF1雑種の着粒数を測定し、比較した。各系統の着粒数の測定結果を図16に示す。この結果、CMS−コシヒカリえいち2号から得られたF1雑種と、CMS−コシヒカリえいち2号_長領域から得られたF1雑種との1穂粒数を比較したところ、両者に有意な差異は見られなかった。
【0070】
<複数の遺伝子を置換したイネ細胞質雄性不稔系統から得られたF1雑種系統の形質>
CMS−コシヒカリ、CMS−コシヒカリえいち2号、CMS−コシヒカリえいち3号、CMS−コシヒカリえいち4号、CMS−コシヒカリかずさ1号、CMS−コシヒカリかずさ2号、又はCMS−コシヒカリかずさ3号を母本とし、ST−2を花粉親として得られたF1雑種の、稈長、1穂粒数、及び到穂日数を表4に示す。対照として、コシヒカリの結果も合わせて示した。この結果、ハバタキ由来のsd1遺伝子を含むCMS−コシヒカリかずさ1号及びCMS−コシヒカリかずさ3号から得られたF1雑種系統は、CMS−コシヒカリえいち4号から得られたF1雑種系統と同様に、CMS−コシヒカリから得られたF1雑種系統よりも稈長が有意に短くなっていた。また、ハバタキ由来のhd1遺伝子を含むCMS−コシヒカリかずさ1号及びCMS−コシヒカリかずさ2号から得られたF1雑種系統は、CMS−コシヒカリえいち3号から得られたF1雑種系統と同様に、CMS−コシヒカリから得られたF1雑種系統よりも到穂日数が短くなっていた。また、ハバタキ由来のGn1遺伝子を含むCMS−コシヒカリかずさ3号から得られたF1雑種系統は、CMS−コシヒカリえいち2号から得られたF1雑種系統と同様に、CMS−コシヒカリから得られたF1雑種系統よりも1穂粒数が多くなっていた。ハバタキ由来のGn1遺伝子とハバタキ由来のhd1遺伝子を含むCMS−コシヒカリかずさ2号から得られたF1雑種系統では、元品種であるコシヒカリよりも若干晩生であるものの、CMS−コシヒカリから得られたF1雑種系統よりも顕著に早生であることに加えて、1穂粒数がコシヒカリよりも非常に多くなっていた。
これらの結果から、母本において、それぞれの領域が単独に置換された場合と同じように、複数の領域が置換された場合でも、それぞれの領域の効果は、F1雑種においても得られることが明らかである。
【0071】
【表4】
【0072】
[実施例2]
<食味の評価>
CMS−コシヒカリ又はCMS−ミルキークイーンを母本とし、ST−1、ST−2、又はST−4を花粉親として交配させてF1雑種の種子を得た。得られたF1雑種を栽培して得られた玄米を、食味官能試験へ供試した。食味官能試験においては、対照として、複数産地で栽培されたコシヒカリの玄米をブレンドしたものを用いた。試験結果を表5に示す。この結果、CMS−コシヒカリから得られたF1雑種の玄米の食味総合値は、それぞれ、コシヒカリと同等、または、やや劣るという結果であった。これに対して、CMS−ミルキークイーンから得られたF1雑種の玄米の食味総合値は、いずれも、CMS−コシヒカリから得られたF1雑種よりも良好であった。粘りの項目が強くなる傾向が観察されたことから、食味総合値の向上をもたらしたのは、半糯性遺伝子の導入であることが推測された。なお、CMS系統の違いによる、玄米形質以外における有意な形質の差異は観察されなかった。
これらの結果から、半糯性を示すイネ細胞質雄性不稔系統を母本として用いることにより、半糯性を示さないイネ細胞質雄性不稔系統を母本として用いた場合よりも、F1雑種の食味を改善し得ることが明らかである。
【0073】
【表5】
【0074】
[実施例3]
<F1雑種の到穂日数>
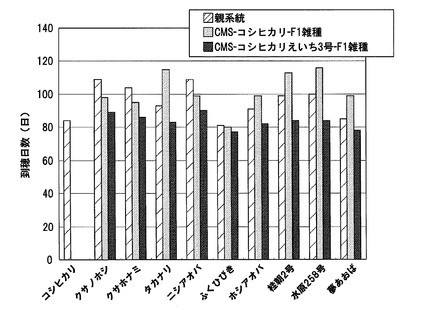
CMS−コシヒカリ又はCMS−コシヒカリえいち3号を母本とし、イネ品種クサノホシ、イネ品種クサホナミ、イネ品種タカナリ、イネ品種ニシアオバ、イネ品種ふくひびき、イネ品種ホシアオバ、イネ品種桂朝2号、イネ品種水原258号、又はイネ品種夢あおば花粉親として交配させてF1雑種の種子を得た。得られたF1雑種を栽培し、到穂日数を調べた。対照として、コシヒカリと各花粉親も栽培し、それぞれの到穂日数を調べた。各系統の到穂日数の測定結果を図17に示す。5月13日に播種した本試験における対照品種コシヒカリの到穂日数は84日であったのに対して、供試したCMS−コシヒカリを母本として得られたF1雑種系統の多くは、倒穂日数が長くなり、晩生化し、95日以内を示したのはわずか1系統のみであった。これに対して、CMS−コシヒカリえいち3号を母本として得られたF1雑種系統では、いずれも花粉親やCMS−コシヒカリを母本として得られたF1雑種系統よりも倒穂日数が短くなっており、早生化していた。特に、CMS−コシヒカリえいち3号を母本として得られたF1雑種系統は、全て倒穂日数が90日以内に抑えられていた。
これらの結果から、ハバタキ由来のhd1遺伝子を含むイネ雄性不稔系統を母本として用いることにより、CMS−コシヒカリを母本とした場合よりも、F1雑種を早生化し得ること、よって、コシヒカリに近い熟期の系統を育成する上で、選抜効率を大きく向上させる効果があることが明らかとなった。
【産業上の利用可能性】
【0075】
本発明のイネF1種子の生産方法は、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、イネF1雑種の種子をより高い選抜効率で生産することができるため、当該方法は、特に植物の育種の分野において利用が可能である。
【受託番号】
【0076】
FERM P−22217
FERM P−22218
【技術分野】
【0001】
本発明は、良好な形質を有するイネ雄性不稔系統、当該イネ雄性不稔系統を用いたイネF1種子の生産方法、及び当該方法により得られたイネF1種子に関する。
【背景技術】
【0002】
近年、ゲノム解析技術は格段の進歩を遂げ、作物の改良を、高精度かつ容易に行えるようになった。とりわけ、DNAマーカー技術の進捗は著しく、同技術によって、有用な形質を備える新品種を作製することができるようになった。例えば、これまでに、DNAマーカーを用いて、灰色カビ病耐性を備えるトマトや(例えば、特許文献1参照。)、耐倒伏性と玄米粒の大きさが改善されたイネ(Oryza sativa)(例えば、特許文献2参照。)が作出されている。
【0003】
また、DNAマーカーを利用して、これまでに同定された重要遺伝子の有用アレルを含む染色体領域を置換することにより、他の多くの形質に対してほとんど影響を及ぼさずに、目的形質を特異的に改良することが出来るようになった(例えば、特許文献3参照。)。例えばイネにおいては、稈長(sd1遺伝子近傍染色体領域)、到穂日数(hd1遺伝子近傍染色体領域)、1穂粒数(Gn1遺伝子近傍染色体領域)などが改良されたイネが作出されている(例えば、特許文献4参照。)。イネ品種コシヒカリの染色体中のsd1遺伝子が、ハバタキ由来のsd1遺伝子に置換されることにより、イネ品種コシヒカリよりも稈長が有意に短くなり、耐倒伏性が改善される。また、イネ品種コシヒカリの染色体中のGn1遺伝子が、ハバタキ由来のGn1遺伝子に置換されることにより、イネ品種コシヒカリよりも着粒密度が高くなる。イネ品種コシヒカリの染色体中のhd1遺伝子が、ハバタキ由来のhd1遺伝子に置換されることにより、イネ品種コシヒカリよりも早生化される。
【0004】
より優れた形質を備える作物を作出する方法として、雄性不稔細胞質等を利用して母本の花粉合成能を失わせることで遠縁系統間の交配を実現し、その雑種種子を品種として利用するF1雑種育種法がある。例えば、レタスにおいては、F1雑種育種法において母本として使用可能なレタス雄性不稔系統が作出されている(例えば、特許文献5参照。)。
【0005】
F1雑種育種法は、雑種強勢を利用して収量性を容易にかつ非常に高いレベルで改良できる技術として利用されている。わが国のイネ育種においても、1970年に実用的な雄性不稔細胞質が発見されて以来、利用が試みられてきた。但し、当時育成された系統の食味品質が十分に高くなかったこと、及び、その後のコメ余りの時代の中で多収性に関する必要性が低下したことなどにより、次第に利用されなくなってきた歴史がある。
【0006】
しかしながら、近年では、作物の収量ポテンシャルを向上させることは、食糧の増産や、栽培コスト、及び栽培時投入エネルギーについての効率化を図る上で、再び重要になってきており、今後ますます重要な育種目標となる。また、生産能の増強によって植物体そのものを大きくすることは、第2世代のバイオエタノール原材料として目される作物残渣の生産性を高め、作物栽培行程にて排出されるGHG量を相対的に削減することを通して、エネルギー問題・環境問題の解決にも貢献できる。
【0007】
収量ポテンシャルの向上が重要視されるようになってきた現在、改めてF1雑種育種技術を見直す機運が高まりつつある。F1雑種育種法では、組み合わせ検定における選抜候補系統として非常に多くの系統間でF1雑種を作る必要があるため、高い選抜効率を維持するためには、母本となる雄性不稔系統の選定が非常に重要となる。
【0008】
我が国のリーディングバラエティであるイネ品種コシヒカリは、食味品質に対する評価が高く、これを育成母本とした系統には、多くの良食味系統が存在する。また、食味品質以外にも、我が国で最も広範に栽培されていることが示すような広域栽培適応性を備えており、また、強い難穂発芽性などの優れた形質を多数有している。さらに、多くの試験において研究対象に用いられることによって学術的知識が蓄積されており、改良の糸口が見つけやすい利点もある。以上のことを考慮すると、コシヒカリは、F1雑種の母本を育成するに当たり最も有望な系統の1つであると言える。
【0009】
また、これまで特異的に改良することが困難であった食味品質についても、半糯性突然変異形質を利用することで、アミロース含量を低下させ、コメの粘りを強くすることを通して品質向上が図れるようになった。半糯性のコメの多くは胚乳が白濁するため、通常のコメと容易に区別できる。過去に報告されている半糯性突然変異には、アミロース含量を最も大きく左右するとされるwx遺伝子の変異の他、7つの異なるdu座が関係したものがある(例えば、非特許文献1参照。)
【先行技術文献】
【特許文献】
【0010】
【特許文献1】特許第4248881号公報
【特許文献2】特許第4368391号公報
【特許文献3】特許第4409610号公報
【特許文献4】特許第4352102号公報
【特許文献5】特許第3949637号公報
【非特許文献】
【0011】
【非特許文献1】鈴木保宏、“米のアミロース含量の変動−気候と調節−”、「農業及び園芸」、2006年、第81巻、第183〜190ページ。
【発明の概要】
【発明が解決しようとする課題】
【0012】
しかしながら、コシヒカリを片親にしてF1雑種を育成しようとする場合には、しばしば問題が発生する。例えば、インディカ種を含むコシヒカリとは遠縁となる育成系統との組み合わせ検定においては、晩生化や長稈化してしまう結果、実用系統としての適性を外す系統が頻繁に出現し、選抜の効率を大きく低下させる結果となる。
また、F1雑種では雑種強勢により収量性が向上するが、一般の多収系統と比較してコシヒカリ自身の収量ポテンシャルが十分に高くないために、選抜に値するような多収系統が得られる比率は高くない。これについて、例えば、コシヒカリの1穂に着生する粒数を増やすことなどにより、そのポテンシャルをさらに向上させことが望まれる。
さらに、F1雑種を育成する場合には、通常、より大きな雑種強勢を得るために、コシヒカリより食味品質が劣る遠縁系統を片親にする必要がある。遠縁系統を片親とすると、多くの場合、当該遠縁系統が有する食味に関する劣悪形質が遺伝し、選抜の効率を大きく落とす結果となる。
【0013】
本発明は、F1雑種育種法において好適なコシヒカリの雄性不稔系統、及び、当該イネ雄性不稔系統を用いたイネF1種子の生産方法を提供することを目的とする。
【課題を解決するための手段】
【0014】
本発明者らは、上記課題を解決すべく鋭意研究した結果、母本として、染色体の一部を外来品種由来の染色体断片で置換することや突然変異によって、特定の形質を改善させた雄性不稔系統を用いることにより、優れたF1雑種をより効率よく作出可能であることを見出し、本発明を完成させた。
【0015】
すなわち、本発明は、
(1) イネ(Oryza sativa)品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統、又は半糯性を示すイネ雄性不稔系統を母本とし、イネ稔性回復系統を花粉親として交配し、交配後の母本から雑種第1代種子(F1種子)を採取することを特徴とするイネF1種子の生産方法、
(2) 前記イネ雄性不稔系統が、受託番号がFERM P−22217であるイネ細胞質雄性不稔系統CMS−コシヒカリえいち2号(Oryza sativa L.cultivar Koshihikari−Eichi−2go)、イネ細胞質雄性不稔系統CMS−コシヒカリえいち3号、イネ細胞質雄性不稔系統CMS−コシヒカリえいち4号、イネ細胞質雄性不稔系統CMS−コシヒカリかずさ1号、イネ細胞質雄性不稔系統CMS−コシヒカリかずさ2号、及び受託番号がFERM P−22218であるイネ細胞質雄性不稔系統CMS−コシヒカリかずさ3号からなる群より選択される細胞質雄性不稔系統であることを特徴とする前記(1)に記載のイネF1種子の生産方法、
(3) 前記(1)又は(2)に記載のイネF1種子の生産方法により得られたことを特徴とするイネF1種子、
(4) イネ(Oryza sativa)品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有することを特徴とするイネ雄性不稔系統、
(5) 受託番号がFERM P−22217であるイネ細胞質雄性不稔系統CMS−コシヒカリえいち2号(Oryza sativa L.cultivar Koshihikari−Eichi−2go)、
(6) イネ細胞質雄性不稔系統CMS−コシヒカリえいち3号(Oryza sativa L.cultivar Koshihikari−Eichi−3go)、
(7) イネ細胞質雄性不稔系統CMS−コシヒカリえいち4号(Oryza sativa L.cultivar Koshihikari−Eichi−4go)、
(8) イネ細胞質雄性不稔系統CMS−コシヒカリかずさ1号(Oryza sativa L.cultivar Koshihikari−Kazusa−1go)、
(9) イネ細胞質雄性不稔系統CMS−コシヒカリかずさ2号(Oryza sativa L.cultivar Koshihikari−Kazusa−2go)、
(10) 受託番号がFERM P−22218であるイネ細胞質雄性不稔系統CMS−コシヒカリかずさ3号(Oryza sativa L.cultivar Koshihikari−Kazusa−3go)、
を、提供するものである。
【発明の効果】
【0016】
本発明のイネF1種子の生産方法は、特定の形質が改善されたイネ雄性不稔系統を母本とすることで、当該形質を備えるF1雑種の種子を生産することができる。特に、本発明において、イネ品種ハバタキ由来のsd1遺伝子を含有するイネ雄性不稔系統を母本として用いた場合には、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、稈長が有意に短く、耐倒伏性が改善されたF1雑種を作出することができる。また、イネ品種ハバタキ由来のGn1遺伝子を含有するイネ雄性不稔系統を母本として用いた場合には、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、着粒密度の高いF1雑種を作出することができる。また、イネ品種ハバタキ由来のhd1遺伝子を含有するイネ雄性不稔系統を母本として用いた場合には、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、早生化されたF1雑種を作出することができる。さらに、半糯性を有するイネ雄性不稔系統を母本として用いた場合には、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、食味に優れたF1雑種を作出することができる。
【図面の簡単な説明】
【0017】
【図1】イネの第1染色体中のsd1遺伝子がコードされている付近のDNAマーカー(SNP)を示した図である。
【図2】イネの第1染色体中のGn1遺伝子がコードされている付近のDNAマーカー(SNP)を示した図である。
【図3】イネの第6染色体中のhd1遺伝子がコードされている付近のDNAマーカー(SNP)を示した図である。
【図4】実施例1において用いられたイネ品種コシヒカリえいち2号のゲノムを模式的に表した図である。
【図5】実施例1において用いられたイネ品種コシヒカリえいち2号_長領域のゲノムを模式的に表した図である。
【図6】実施例1において用いられたイネ品種コシヒカリえいち3号のゲノムを模式的に表した図である。
【図7】実施例1において用いられたイネ品種コシヒカリえいち4号のゲノムを模式的に表した図である。
【図8】実施例1において用いられたイネ品種コシヒカリえいち4号_長領域のゲノムを模式的に表した図である。
【図9】実施例1において用いられたイネ品種コシヒカリかずさ1号のゲノムを模式的に表した図である。
【図10】実施例1において用いられたイネ品種コシヒカリかずさ2号のゲノムを模式的に表した図である。
【図11】実施例1において用いられたイネ品種コシヒカリかずさ3号のゲノムを模式的に表した図である。
【図12】実施例1において、CMS−コシヒカリえいち4号を母本として得られたF1雑種、CMS−コシヒカリを母本として得られたF1雑種、及びイネ品種コシヒカリの稈長の測定結果を示した図である。
【図13】実施例1において、CMS−コシヒカリとST−1から得られたF1雑種、CMS−コシヒカリえいち4号とST−1から得られたF1雑種、CMS−コシヒカリえいち4号_長領域とST−1から得られたF1雑種、及びイネ品種コシヒカリの稈長の測定結果を示した図である。
【図14】実施例1において、CMS−コシヒカリえいち3号又はCMS−コシヒカリを母本とし、ST−2又はST−4を花粉親として得られたF1雑種の到穂日数の測定結果を示した図である。
【図15】実施例1において、CMS−コシヒカリえいち2号又はCMS−コシヒカリを母本とし、ST−2又はST−4を花粉親として得られたF1雑種の1穂当たりの着粒数の測定結果を示した図である。
【図16】実施例1において、CMS−コシヒカリえいち2号、CMS−コシヒカリえいち2号_長領域、又はCMS−コシヒカリを母本とし、ST−2を花粉親として得られたF1雑種の1穂当たりの着粒数の測定結果を示した図である。
【図17】実施例3において、各F1雑種及びその花粉親の到穂日数の測定結果を、花粉親ごとに示した図である。
【発明を実施するための形態】
【0018】
本発明において準同質遺伝子系統とは、元品種の染色体の一部のみが外来品種由来の染色体断片に置換されている系統を意味する。ここで、外来品種は、元品種以外の品種であれば特に限定されるものではなく、元品種と同一種の植物の品種であってもよく、元品種と異なる種の植物の品種であってもよく、動物等の植物以外の品種であってもよい。なお、本発明において品種とは、同一種の植物であって、遺伝的構成が異なるために、ある形質において同種内の他品種から明確に識別し得る集団を意味する。
【0019】
本発明においてDNAマーカーは、元品種由来の染色体と外来品種由来の染色体を識別し得る染色体上のDNA配列の差異を検出し得るものであれば、特に限定されるものではなく、遺伝子解析分野で通常用いられているDNAマーカーを用いることができる。該DNAマーカーとして、例えば、SNP(Single Nucleotide Polymorphism、一遺伝子多型)やSSR(Simple Sequence Repeats、単純反覆配列)の繰り返し数の違い等の遺伝子多型を検出し得るマーカーであってもよく、RFLP(Restriction Fragment Length Polymorphism、制限酵素断片長多型)マーカーであってもよい。なお、これらのDNAマーカーによる、元品種由来アレルと外来品種由来アレルとの識別は、常法により行うことができる。例えば、各個体から抽出したDNAを鋳型とし、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等を用いてPCRを行い、電気泳動法等を用いてPCR産物の有無を検出し、各多型を識別することができる。また、各個体から抽出したDNAを制限酵素処理した後、電気泳動法等を用いてDNA断片のパターンを検出し、各多型を識別することができる。なお、特定のSNPやSSRと特異的にハイブリダイズし得るプライマー等は、該SNPやSSRの塩基配列に応じて、汎用されているプライマー設計ツール等を用いて常法により設計することができる。また、設計されたプライマー等は、当該技術分野においてよく知られている方法のいずれを用いても合成することができる。
【0020】
これらのDNAマーカーは、公知のDNAマーカーを適宜用いることができる。また、新規に作製したDNAマーカーであってもよい。公知のDNAマーカーとして、例えば、イネにおいては、国際公開第2003/070934号パンフレット等において開示されているSNPマーカーや、Rice Genome Research Program(RGB:http://rgp.dna.affrc.go.jp/publicdata.html)において公開されているDNAマーカーを用いることができる。
【0021】
なお、各品種の遺伝子情報等は、例えば、国際的な塩基配列データベースであるNCBI(National center for Biotechnology Information)やDDBJ(DNA Data Bank of Japan)等において入手することができる。特にイネの各品種の遺伝子情報は、KOME(Knowledge−based Oryza Molecular biological Encyclopedia、http://cdna01.dna.affrc.go.jp/cDNA/)等において入手することができる。
【0022】
本発明及び本願明細書において「イネ品種日本晴の染色体のX番目の塩基」は、TIGR(The Institute for Genomic Research;http://www.tigr.org/tdb/e2k1/osa1/blastsearch.shtml)において公開されているイネ品種日本晴のゲノムDNAの塩基配列(バージョン2)に基づいて決定される領域である。
【0023】
また、本発明及び本願明細書において、「イネ品種日本晴の染色体のX番目の塩基からY番目の塩基までの領域に相当する領域」とは、イネ個体の染色体中のイネ品種日本晴の染色体中の当該領域と相同性の高い領域であり、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントすることにより決定することができる。また、イネ品種日本晴以外のイネ個体中の「イネ品種日本晴のSNPに相当するSNP」は、当該SNPを含む領域において、イネ品種日本晴の公知のゲノムDNAと当該イネ個体のゲノムDNAの塩基配列を、最もホモロジーが高くなるようにアラインメントした場合に、当該SNPに対応する位置にある塩基を意味する。
【0024】
本発明のイネF1種子の生産方法は、特定の形質が改善されたイネ品種コシヒカリの雄性不稔系統を母本とし、イネ稔性回復系統を花粉親として交配し、交配後の母本から雑種第1代種子(F1種子)を採取することを特徴とする。
【0025】
まず、本発明において用いられるイネ雄性不稔系統について説明する。本発明において用いられるイネ雄性不稔系統は、イネ品種コシヒカリの染色体の一部が外来品種由来の染色体断片に置換されることによって、又は突然変異によって、特定の形質が改善されている準同質遺伝子系統の雄性不稔系統である。
【0026】
準同質遺伝子系統の雄性不稔系統は、常法により作出することができる。例えば、雄性不稔細胞質であること以外はイネ品種コシヒカリと同じ形質を備えるコシヒカリ細胞質雄性不稔系統と、所望の領域が外来品種由来の染色体断片で置換された、若しくは所望の突然変異がなされたイネ品種コシヒカリの準同質遺伝子系統とを交配し、得られたF1雑種に対して、当該イネ品種コシヒカリの準同質遺伝子系統を花粉親として用いた連続戻し交配を行うことにより、雄性不稔細胞質であること以外は当該イネ品種コシヒカリの準同質遺伝子系統と同じ形質を備えるイネ細胞質雄性不稔系統を得ることができる。なお、コシヒカリ細胞質雄性不稔系統は、例えば、イネ品種コシヒカリとイネ細胞質雄性不稔系統とを交配し、得られたF1雑種に対して、イネ品種コシヒカリを花粉親として戻し交配を繰り返すことにより、作出することができる。イネ細胞質雄性不稔系統としては、雄性不稔細胞質性を示すイネ科の品種であれば特に限定されるものではなく、例えば、BT型の雄性不稔細胞質であるイネ品種CHINSURAH BORO 2、WA型の雄性不稔細胞質であるイネ品種Male sterile wild rice、GA型の雄性不稔細胞質であるイネ品種Gambiaca、Di型の雄性不稔細胞質であるイネ品種Dissi等を挙げることができる。
【0027】
その他、準同質遺伝子系統の雄性不稔系統としては、特定の環境条件下で不稔を誘発する突然変異遺伝子による環境条件依存型雄性不稔系統であってもよい。環境条件依存型雄性不稔系統としては、長日条件で雄性不稔を引き起こすPMS1遺伝子やPMS2遺伝子を利用した日長感応性雄性不稔(PGMS)系統や、高温条件で雄性不稔を引き起こすTMS1遺伝子やTMS2遺伝子を利用した温度感応性雄性不稔(TGMS)系統等がある。これらの変異遺伝子を有する環境条件依存型雄性不稔系統に、イネ品種コシヒカリの準同質遺伝子系統を交配し、得られたF1雑種に対して、当該イネ品種コシヒカリの準同質遺伝子系統を花粉親として用いた連続戻し交配を行うことにより、当該変異遺伝子による環境条件依存型雄性不稔性を示す以外は、当該イネ品種コシヒカリの準同質遺伝子系統と同じ形質を備えるイネ雄性不稔系統を得ることができる。
【0028】
まず、母本として、イネ品種コシヒカリの染色体の一部が外来品種由来の染色体断片に置換されたイネ品種コシヒカリの準同質遺伝子系統について説明する。
イネ品種コシヒカリの準同質遺伝子系統において挿入されている外来品種由来の染色体断片は、当該染色体断片が挿入されることによって特定の形質をイネ品種コシヒカリよりも改善されるものであれば、特に限定されるものではない。例えば、挿入される外来品種由来の染色体断片は、所望の形質改善に直接寄与する遺伝子(原因遺伝子)をコードする領域を含んでいればよく、当該原因遺伝子のみを含む領域であってもよく、当該原因遺伝子とその他の遺伝子を含む領域(例えば、14.6Mbp〜29.2Mbpの長さからなる領域)であってもよい。
【0029】
本発明及び本願明細書において、「イネ品種“X”由来の“Y”遺伝子」には、イネ品種“X”自体に由来する“Y”遺伝子(すなわち、イネ品種“X”の染色体中に存在している“Y”遺伝子)に加えて、イネ品種“X”と実質的に同等な“Y”遺伝子を有するイネ品種由来の“Y”遺伝子も含まれる。イネ品種“X”由来の“Y”遺伝子に代えて、イネ品種“X”由来の“Y”遺伝子と実質的に同等であるイネ品種“X”以外のイネ品種由来の“Y”遺伝子を染色体に組み込んだ場合であっても、本発明にと同じ効果が奏されるためである。ここで、イネ品種“X”由来の“Y”遺伝子と実質的に同等な“Y”遺伝子とは、イネ品種“X”以外のイネ品種由来の“Y”遺伝子であって、イネ品種“X”由来の“Y”遺伝子と同程度の機能を有する遺伝子である。具体的には、イネ品種“X”の後代品種であって、イネ品種“X”から“Y”遺伝子が存在する領域のアレルを受け継いだイネ品種や、イネ品種“X”の先祖に当たるイネ品種であって、イネ品種“X”へ“Y”遺伝子が存在する領域のアレルを共通して有するイネ品種、これらのイネ品種“X”と実質的に同等な“Y”遺伝子を有するイネ品種が有する“Y”遺伝子が存在する領域の染色体断片が組み込まれたイネ品種等が挙げられる。
【0030】
すなわち、本発明及び本願明細書においては、特に記載が無い限り、「イネ品種ハバタキ由来のsd1遺伝子」には、イネ品種ハバタキ自体に由来するsd1遺伝子のみならず、該遺伝子と実質的に同等なsd1遺伝子、例えば、イネ品種低脚烏尖、イネ品種IR8、イネ品種キヌヒカリ、イネ品種ゆめひたち、イネ品種コシヒカリえいち4号、イネ品種コシヒカリかずさ2号、イネ品種コシヒカリかずさ3号、又はイネ品種コシヒカリかずさ4号等のイネ品種由来のsd1遺伝子が含まれる。
【0031】
同様に、本発明及び本願明細書においては、特に記載が無い限り、「イネ品種ハバタキ由来のGn1遺伝子」には、イネ品種ハバタキ自体に由来するGn1遺伝子のみならず、該遺伝子と実質的に同等なGn1遺伝子、例えば、イネ品種コシヒカリえいち2号、イネ品種コシヒカリかずさ2号、イネ品種コシヒカリかずさ3号、又はイネ品種コシヒカリかずさ4号等のイネ品種由来のGn1遺伝子が含まれる。
【0032】
同様に、本発明及び本願明細書においては、特に記載が無い限り、「イネ品種ハバタキ由来のhd1遺伝子」には、イネ品種ハバタキ自体に由来するhd1遺伝子のみならず、該遺伝子と実質的に同等なhd1遺伝子、例えば、イネ品種コシヒカリえいち3号、イネ品種コシヒカリかずさ1号、イネ品種コシヒカリかずさ2号、又はイネ品種コシヒカリかずさ4号等のイネ品種由来のhd1遺伝子が含まれる。
【0033】
本発明においては、イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統を母本として用いる。これらの3種の遺伝子のうち、2種類の遺伝子を適宜組み合わせて含有させた雄性不稔系統であってもよく、3種類全ての遺伝子を含む雄性不稔系統であってもよい。
【0034】
イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統は、イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統と、コシヒカリ雄性不稔系統とから前述の方法により作出することができる。イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統は、例えば、適当なDNAマーカーを用いて、特許文献3及び4に記載の方法等によって作出することができる。なお、本発明において母本として用いるイネ品種コシヒカリの準同質遺伝子系統は、新たに作出されたものであってもよく、既存の系統を用いてもよい。
【0035】
イネ品種ハバタキ由来のsd1遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来sd1含有準同質遺伝子系統)は、イネ品種コシヒカリの染色体中のsd1遺伝子がコードされている領域が、イネ品種ハバタキ由来のsd1遺伝子をコードする領域を含む染色体断片に置換されている。ハバタキ由来sd1含有準同質遺伝子系統に含まれるハバタキ由来染色体断片は、sd1遺伝子がコードされている領域を含んでいれば特に限定されるものではなく、sd1遺伝子がコードされている領域のみを含んでいてもよく、sd1遺伝子の近傍に存在している遺伝子もsd1遺伝子と共にイネ品種コシヒカリに挿入されてもよい。図1に、イネの第1染色体中のsd1遺伝子がコードされている38.11Mbp付近のDNAマーカー(SNP)を示す。当該ハバタキ由来染色体断片の長さは、DNAマーカーを用いて決定することできる。例えば、図1に示すように、ハバタキ由来sd1含有準同質遺伝子系統において、挿入されたハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の38,109,578番目の塩基配列に依存する多型(PCRにより、イネ品種コシヒカリではPCR産物が得られ、イネ品種ハバタキではPCR産物が得られない)(以下、G2003)とイネ品種日本晴の第1染色体の38,109,641番目の塩基配列に依存する多型(PCRにより、イネ品種コシヒカリではPCR産物が得られ、イネ品種ハバタキではPCR産物が得られない)(以下、G2002)との間にあり、当該ハバタキ由来染色体断片の下流端が、G2003とイネ品種日本晴の第1染色体の38,199,771番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではT)(以下、SP−462)との間にあってもよい(図1の1段目)。また、当該ハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の38,108,008番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)(以下、SP−4009)とG2003との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−462とイネ品種日本晴の第1染色体の38,949,866番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではC)(以下、SP−1259)との間にあってもよい(図1の2段目)。当該ハバタキ由来染色体断片の上流端が、SP−4009とG2003との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−1259とイネ品種日本晴の第1染色体の41,374,509番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではG)(以下、SP−477)との間にあってもよい(図1の3段目)。さらに、イネ品種ハバタキ由来のsd1遺伝子をコードする領域を含む、よりさらに長い領域が、ハバタキ由来染色体断片により置換されていてもよい。例えば、イネ品種日本晴の第1染色体の12,254,787番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)(以下、SP−2058)からSP−477までの約29.1Mbpの領域を含む領域が、ハバタキ由来染色体断片により置換されていてもよい(図1の4段目)。各DNAマーカー及び識別に使用可能なプライマーの塩基配列を表1に示す。
【0036】
【表1】
【0037】
ハバタキ由来sd1遺伝子を含有するイネ雄性不稔系統を母本とすることにより得られたF1雑種は、ハバタキ由来のsd1遺伝子を含有しているため、コシヒカリ雄性不稔系統を母本として得られたF1雑種よりも、稈長が有意に低く、耐倒伏性が大幅に改善されている。このため、本発明のイネF1種子の生産方法において、ハバタキ由来のsd1遺伝子を含有するイネ雄性不稔系統を用いることにより、耐倒伏性が改善されたF1雑種の種子を効率よく生産することができ、F1雑種育成のための組み合わせ検定の効率を向上させることができる。
【0038】
イネ品種ハバタキ由来のGn1遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来Gn1含有準同質遺伝子系統)は、イネ品種コシヒカリの染色体中のGn1遺伝子がコードされている領域が、イネ品種ハバタキ由来のGn1遺伝子をコードする領域を含む染色体断片に置換されている。ハバタキ由来Gn1含有準同質遺伝子系統に含まれるハバタキ由来染色体断片は、Gn1遺伝子がコードされている領域を含んでいれば特に限定されるものではなく、Gn1遺伝子がコードされている領域のみを含んでいてもよく、Gn1遺伝子の近傍に存在している遺伝子もGn1遺伝子と共にイネ品種コシヒカリに挿入されてもよい。図2に、イネの第1染色体中のGn1遺伝子がコードされている5.267Mbp付近のDNAマーカー(SNP)を示す。当該ハバタキ由来染色体断片の長さは、DNAマーカーを用いて決定することできる。例えば、図2に示すように、ハバタキ由来Gn1含有準同質遺伝子系統において、挿入されたハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の5,230,989番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではA)(以下、SP−170)とイネ品種日本晴の第1染色体の5,267,730番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではC)(以下、SP−4028)との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−4028とイネ品種日本晴の第1染色体の5,267,970番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)(以下、SP−4038)との間にあってもよい(図2の1段目)。また、当該ハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の5,029,673番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではG)(以下、SP−2032)とSP−170との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−4038とイネ品種日本晴の第1染色体の5,274,879番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではT)(以下、SP−4030)との間にあってもよい(図2の2段目)。当該ハバタキ由来染色体断片の上流端が、イネ品種日本晴の第1染色体の2,275,275番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではC)(以下、SP−158)とSP−2032との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−4038とSP−4030との間にあってもよい(図2の3段目)。さらに、イネ品種ハバタキ由来のGn1遺伝子をコードする領域を含む、よりさらに長い領域が、ハバタキ由来染色体断片により置換されていてもよい。例えば、SP−158からイネ品種日本晴の第1染色体の31,371,175番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではA)(以下、SP−262)までの約29.1Mbpの領域を含む領域が、ハバタキ由来染色体断片により置換されていてもよい(図2の4段目)。各DNAマーカー及び識別に使用可能なプライマーの塩基配列を表2に示す。
【0039】
【表2】
【0040】
ハバタキ由来Gn1遺伝子を含有するイネ雄性不稔系統を母本とすることにより得られたF1雑種は、ハバタキ由来のGn1遺伝子を含有しているため、コシヒカリ雄性不稔系統を母本として得られたF1雑種よりも着粒密度が向上している。このため、本発明のイネF1種子の生産方法において、ハバタキ由来のGn1遺伝子を含有するイネ雄性不稔系統を用いることにより、着粒密度が改善されたF1雑種の種子を効率よく生産することができ、F1雑種育成のための組み合わせ検定の効率を向上させることができる。
【0041】
イネ品種ハバタキ由来のhd1遺伝子を含有するイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来hd1含有準同質遺伝子系統)は、イネ品種コシヒカリの染色体中のhd1遺伝子がコードされている領域が、イネ品種ハバタキ由来のhd1遺伝子をコードする領域を含む染色体断片に置換されている。ハバタキ由来hd1含有準同質遺伝子系統に含まれるハバタキ由来染色体断片はhd1遺伝子がコードされている領域を含んでいれば特に限定されるものではなく、hd1遺伝子がコードされている領域のみを含んでいてもよく、hd1遺伝子の近傍に存在している遺伝子もhd1遺伝子と共にイネ品種コシヒカリに挿入されてもよい。図3に、イネの第1染色体中のhd1遺伝子がコードされている9.38Mbp付近のDNAマーカー(SNP)を示す。当該ハバタキ由来染色体断片の長さは、DNAマーカーを用いて決定することできる。例えば、図3に示すように、ハバタキ由来hd1含有準同質遺伝子系統において、挿入されたハバタキ由来染色体断片の上流端が、イネ品種日本晴の第6染色体の9,163,248番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではA)(以下、SP−586)とイネ品種日本晴の第6染色体の9,379,348番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではG)(以下、SP−2254)との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−2254とイネ品種日本晴の第6染色体の10,671,175番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではC)(以下、SP−1603)との間にあってもよい(図3の上段)。また、当該ハバタキ由来染色体断片の上流端が、イネ品種日本晴の第6染色体の8,818,970番目のSNPに相当するSNP(イネ品種コシヒカリではC、イネ品種ハバタキではT)(以下、SP−2513)とSP−586との間にあり、当該ハバタキ由来染色体断片の下流端が、SP−1603とイネ品種日本晴の第6染色体の11,949,796番目のSNPに相当するSNP(イネ品種コシヒカリではT、イネ品種ハバタキではC)(以下、SP−604)との間にあってもよい(図3の中段)。さらに、イネ品種ハバタキ由来のhd1遺伝子をコードする領域を含む、よりさらに長い領域が、ハバタキ由来染色体断片により置換されていてもよい。例えば、イネ品種日本晴の第6染色体の135,124番目のSNPに相当するSNP(イネ品種コシヒカリではA、イネ品種ハバタキではG)(以下、SP−2229)からイネ品種日本晴の第6染色体の29,016,207番目のSNPに相当するSNP(イネ品種コシヒカリではG、イネ品種ハバタキではT)(以下、SP−1635)までの約28.9Mbpの領域を含む領域が、ハバタキ由来染色体断片により置換されていてもよい(図3の下段)。各DNAマーカー及び識別に使用可能なプライマーの塩基配列を表3に示す。
【0042】
【表3】
【0043】
ハバタキ由来hd1遺伝子を含有するイネ雄性不稔系統を母本とすることにより得られたF1雑種は、ハバタキ由来のhd1遺伝子を含有しているため、コシヒカリ雄性不稔系統を母本として得られたF1雑種よりも早生となる。このため、本発明のイネF1種子の生産方法において、ハバタキ由来のhd1遺伝子を含有するイネ雄性不稔系統を用いることにより、出穂期が早められたF1雑種の種子を効率よく生産することができ、F1雑種育成のための組み合わせ検定の効率を向上させることができる。
【0044】
sd1遺伝子、Gn1遺伝子、及びhd1遺伝子のうちの2以上の遺伝子がハバタキ由来の遺伝子に置換されたイネ品種コシヒカリの準同質遺伝子系統は、異なる種類の遺伝子がハバタキ由来の遺伝子に置換されている準同質遺伝子系統同士を交配し、得られたF1雑種を自家交配することによって得られたF2雑種の中から、DNAマーカーを用いて、イネ品種コシヒカリの染色体中に導入された外来遺伝子由来の遺伝子が、両方の相同染色体に導入されているホモ個体を選抜することによって得られる。例えば、sd1遺伝子とGn1遺伝子がハバタキ由来の遺伝子に置換されたイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来sd1/ハバタキ由来Gn1含有準同質遺伝子系統)は、ハバタキ由来sd1含有準同質遺伝子系統とハバタキ由来Gn1含有準同質遺伝子系統を交配し、得られたF1雑種を自家交配させて得られた雑種第2代(F2雑種)の中から、DNAマーカーを指標として、両方の相同染色体において、sd1遺伝子がコードされている領域及びGn1遺伝子がコードされている領域が、いずれもハバタキ由来の領域である個体を選抜することにより、作出することができる。同様に、sd1遺伝子、Gn1遺伝子、及びhd1遺伝子がハバタキ由来の遺伝子に置換されたイネ品種コシヒカリの準同質遺伝子系統(ハバタキ由来sd1/ハバタキ由来Gn1/ハバタキ由来hd1含有準同質遺伝子系統)は、ハバタキ由来sd1/ハバタキ由来Gn1含有準同質遺伝子系統とハバタキ由来hd1含有準同質遺伝子系統を交配し、得られたF1雑種を自家交配させて得られた雑種第2代(F2雑種)の中から、DNAマーカーを指標として、両方の相同染色体において、sd1遺伝子がコードされている領域、Gn1遺伝子がコードされている領域、及びhd1遺伝子がコードされている領域が、いずれもハバタキ由来の領域である個体を選抜することにより、作出することができる。
【0045】
本発明において用いられるイネ雄性不稔系統としては、半糯性を示すものであってもよい。半糯性を示すイネ雄性不稔系統は、半糯性を示すイネ品種コシヒカリの準同質遺伝子系統を、雄性不稔系統と2回戻し交配し、得られたF3雑種の中から全ての種子において半糯性を示すイネ個体を選抜することによって得られる。
【0046】
半糯性遺伝子を有する染色体断片を導入することにより、玄米のアミロース含量を低下させることを通して、食味品質を向上させることができる。特に半糯性変異遺伝子を片親にのみ持たせたF1雑種系統においては、半糯性を示す玄米が通常形質を有する玄米と1:3の比率で出現するため、穏やかな低アミロース化を実現することができ、半糯性が全くないイコシヒカリよりも食味が非常に良好となる。このため、本発明のイネF1種子の生産方法において、半糯性を示すイネ品種コシヒカリの準同質遺伝子系統を用いることにより、食味品質が向上されたF1雑種の種子を効率よく生産することができ、F1雑種育成のための組み合わせ検定の効率を向上させることができる。
【0047】
半糯性を示すイネ品種コシヒカリの準同質遺伝子系統は、例えば、イネ品種コシヒカリの突然変異群から、半糯性を示す表現形質や、DNAマーカーを利用した半糯性に寄与する遺伝子(半糯性遺伝子)のタイプによって選抜することにより得ることができる。半糯性遺伝子としては、例えば、イネ品種の第6染色体に存在するwaxy−mq遺伝子等が挙げられる。その他、半糯性を示すイネ品種コシヒカリの準同質遺伝子系統としては、waxy−mq遺伝子の突然変異株であるイネ品種ミルキークイーン等の公知のイネ品種コシヒカリの突然変異種であってもよく、イネ品種コシヒカリ以外の半糯性を示すイネ品種(例えば、他品種由来の突然変異体)に対して、イネ品種コシヒカリを連続戻し交配することにより準同質遺伝子系統化したものであってもよい。
【0048】
本発明において用いられるイネ雄性不稔系統としては、イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有しており、かつ半糯性を有するものであってもよい。このような半糯性を示すイネ雄性不稔系統は、具体的には、以下のようにして作出できる。まず、sd1遺伝子、Gn1遺伝子、及びhd1遺伝子のうちの少なくとも1以上の遺伝子がハバタキ由来の遺伝子に置換されたイネ品種コシヒカリの準同質遺伝子系統と、半糯性を示すイネ品種コシヒカリの準同質遺伝子系統とを交配し、得られたF1雑種を自家交配することによって得られたF2雑種の中から、sd1遺伝子、Gn1遺伝子、及びhd1遺伝子のうちの少なくとも1以上の遺伝子がハバタキ由来の遺伝子に置換されており、かつ半糯性を示すイネ個体を選抜する。得られたイネ個体とコシヒカリ雄性不稔系統と交配し、得られたF1雑種に対して連続戻し交配を行うことにより、sd1遺伝子、Gn1遺伝子、及びhd1遺伝子のうちの少なくとも1以上の遺伝子がハバタキ由来の遺伝子に置換されており、かつ半糯性を示すイネ雄性不稔系統を得ることができる。
【0049】
本発明のイネF1種子の生産方法において花粉親として用いられるイネ稔性回復系統は、母本として用いるイネ雄性不稔系統の稔性を回復し得るイネ系統であれば特に限定されるものではない。例えば、母本がBT型の雄性不稔細胞質である場合には、イネ品種JFR-4、イネ品種ST−1、イネ品種ST−2、イネ品種ST−4、イネ品種タカナリ、イネ品種桂朝2号、イネ品種水原258号、及びイネ品種ハバタキ等がイネ稔性回復系統として挙げられる。なお、あるイネ品種があるコシヒカリ雄性不稔系統に対するイネ稔性回復系統か否かは、当該イネ品種と、前記コシヒカリ雄性不稔系統とを交配させ、得られたF1雑種の雄性稔性に基づいて調べることができる。当該F1雑種において雄性不稔性が回復されている場合には、当該イネ品種は当該コシヒカリ雄性不稔系統に対するイネ稔性回復系統であると分かる。また、母本が環境条件依存型雄性不稔である場合には、母本が備える雄性不稔性の原因となる変異遺伝子を有していないイネ系統であれば、イネ稔性回復系統として用いることができる。当該変異遺伝子はF1(ヘテロの状態)の場合には変異形質が現れないためである。
【0050】
本発明のイネF1種子の生産方法において、イネ品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統と、イネ稔性回復系統を花粉親との交配は、自然交配であってもよく、人工的に交配させてもよい。
【実施例】
【0051】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0052】
[実施例1]
<イネ品種コシヒカリの準同質遺伝子系統>
母本として用いるイネ雄性不稔系統を作出するためのイネ品種コシヒカリの準同質遺伝子系統として、イネ品種コシヒカリえいち2号、イネ品種コシヒカリえいち2号_長領域、イネ品種コシヒカリえいち3号、イネ品種コシヒカリえいち4号、イネ品種コシヒカリえいち4号_長領域、イネ品種コシヒカリかずさ1号、イネ品種コシヒカリかずさ2号、イネ品種コシヒカリかずさ3号、及びイネ品種コシヒカリかずさ4号を得た。
【0053】
これらの準同質遺伝子系統のうち、イネ品種コシヒカリえいち2号、イネ品種コシヒカリえいち3号、イネ品種コシヒカリえいち4号、及びイネ品種コシヒカリかずさ4号(寄託番号:FERM P−21596)は、特許文献3及び4に記載されているものを用いた。これらの系統は、ハバタキにコシヒカリを複数回戻し交配した後に、DNAマーカー選抜によって育成された。イネ品種コシヒカリえいち4号の染色体中のハバタキ由来染色体断片に置換されている領域と表1に記載のDNAマーカーとの位置関係は図1の2段目の通りであり、イネ品種コシヒカリえいち2号の染色体中のハバタキ由来染色体断片に置換されている領域と表2に記載のDNAマーカーとの位置関係は図2の2段目の通りであり、イネ品種コシヒカリえいち3号の染色体中のハバタキ由来染色体断片に置換されている領域と表3に記載のDNAマーカーとの位置関係は図3の中段の通りである。
【0054】
また、特許文献3に記載の方法に準拠し、イネ品種コシヒカリえいち4号よりも長い領域がハバタキ由来染色体断片により置換されたイネ品種コシヒカリえいち4号_長領域を作出した。より詳細には、表1に記載のDNAマーカーのうち、SP−4009、G2003、G2002、SP−1259、及びSP−477を用いて、目的のゲノムを有す個体を選抜した。
具体的には、イネ品種ハバタキに対してイネ品種コシヒカリを複数回戻し交配させた後、得られた雑種の種子をさらに栽培し、圃場に移植できる程度に成育させた後、各栽培個体の葉からDNAを回収し、SP−4009及びSP−477がコシヒカリ由来アレルのホモ染色体領域であり、G2003、G2002、及びSP−1259がハバタキ由来アレルのホモ染色体領域である栽培個体1個を選抜した。この選抜された栽培個体が、sd1遺伝子を含有する領域をハバタキ由来染色体断片に置換した新品種であり、本発明者はこの新品種を「イネ品種コシヒカリえいち4号_長領域」と命名した。イネ品種コシヒカリえいち4号_長領域の染色体中のハバタキ由来染色体断片に置換されている領域と表1に記載のDNAマーカーとの位置関係は、図1の3段目の通りである。
【0055】
同様に、特許文献3に記載の方法に準拠し、イネ品種コシヒカリえいち2号よりも長い領域がハバタキ由来染色体断片により置換されたイネ品種コシヒカリえいち2号_長領域を作出した。より詳細には、表2に記載のDNAマーカーのうち、SP−156、SP−2032、SP−4028、SP−4038、及びSP−4030を用いて、目的のゲノムを有す個体を選抜した。
具体的には、イネ品種ハバタキに対してイネ品種コシヒカリを複数回戻し交配させた後、得られた雑種の種子をさらに栽培し、圃場に移植できる程度に成育させた後、各栽培個体の葉からDNAを回収し、SP−156及びSP−4030がコシヒカリ由来アレルのホモ染色体領域であり、SP−2032、SP−4028、及びSP−4038がハバタキ由来アレルのホモ染色体領域である栽培個体1個を選抜した。この選抜された栽培個体が、Gn1遺伝子を含有する領域をハバタキ由来染色体断片に置換した新品種であり、本発明者はこの新品種を「イネ品種コシヒカリえいち2号_長領域」と命名した。イネ品種コシヒカリえいち2号_長領域の染色体中のハバタキ由来染色体断片に置換されている領域と表2に記載のDNAマーカーとの位置関係は、図2の3段目の通りである。
【0056】
さらに、sd1遺伝子とhd1遺伝子がハバタキ由来の遺伝子である準同質遺伝子系統を作出した。具体的には、イネ品種コシヒカリえいち4号とイネ品種コシヒカリえいち3号を交配し、得られた後代個体(種子)のうち2個を栽培し、自殖(自家交配)させ、さらに後代個体である種子を100個得た。この100個の種子を全て栽培し、各後代個体のDNAマーカーを調べ、SP−462〔M4(sd1)〕がハバタキ由来アレルのホモ染色体領域であり、かつSP−2254〔M3(hd1)〕がハバタキ由来アレルのホモ染色体領域である栽培個体を1個体選抜した。この選抜された栽培個体が、sd1遺伝子を含有する領域及びhd1遺伝子を含有する領域がいずれも、ハバタキ由来染色体断片(ホモ)に置換された新品種であり、本発明者はこの新品種を「イネ品種コシヒカリかずさ1号」と命名した。
【0057】
さらに、Gn1遺伝子とhd1遺伝子がハバタキ由来の遺伝子である準同質遺伝子系統を作出した。具体的には、イネ品種コシヒカリえいち2号とイネ品種コシヒカリえいち3号を交配し、得られた後代個体(種子)のうち2個を栽培し、自殖(自家交配)させ、さらに後代個体である種子を100個得た。この100個の種子を全て栽培し、各後代個体のDNAマーカーを調べ、SP−4028〔M4(Gn1)〕がハバタキ由来アレルのホモ染色体領域であり、かつSP−2254〔M3(hd1)〕がハバタキ由来アレルのホモ染色体領域である栽培個体を1個体選抜した。この選抜された栽培個体が、Gn1遺伝子を含有する領域及びhd1遺伝子を含有する領域がいずれも、ハバタキ由来染色体断片(ホモ)に置換された新品種であり、本発明者はこの新品種を「イネ品種コシヒカリかずさ2号」と命名した。
【0058】
さらに、sd1遺伝子とGn1遺伝子がハバタキ由来の遺伝子である準同質遺伝子系統を作出した。具体的には、イネ品種コシヒカリえいち4号とイネ品種コシヒカリえいち2号を交配し、得られた後代個体(種子)のうち2個を栽培し、自殖(自家交配)させ、さらに後代個体である種子を100個得た。この100個の種子を全て栽培し、各後代個体のDNAマーカーを調べ、SP−462〔M4(sd1)〕がハバタキ由来アレルのホモ染色体領域であり、かつSP−4028〔M4(Gn1)〕がハバタキ由来アレルのホモ染色体領域である栽培個体を1個体選抜した。この選抜された栽培個体が、sd1遺伝子を含有する領域及びGn1遺伝子を含有する領域がいずれも、ハバタキ由来染色体断片(ホモ)に置換された新品種であり、本発明者はこの新品種を「イネ品種コシヒカリかずさ3号」と命名した。
【0059】
図4にイネ品種コシヒカリえいち2号のゲノムを、図5にイネ品種コシヒカリえいち2号_長領域のゲノムを、図6にイネ品種コシヒカリえいち3号のゲノムを、図7にイネ品種コシヒカリえいち4号のゲノムを、図8にイネ品種コシヒカリえいち4号_長領域のゲノムを、図9にイネ品種コシヒカリかずさ1号のゲノムを、図10にイネ品種コシヒカリかずさ2号のゲノムを、図11にイネ品種コシヒカリかずさ3号のゲノムを、それぞれ模式的に示す。
【0060】
<イネ品種コシヒカリの細胞質雄性不稔系統(CMS−コシヒカリ)>
イネ品種CHINSURAH BORO 2にイネ品種コシヒカリを6回戻し交配し、圃場における生育特性が、雄性不稔であることを除きコシヒカリと同等の形質を示すCMS−コシヒカリを育成した。
【0061】
<イネ品種ミルキークイーンの細胞質雄性不稔系統(CMS−ミルキークイーン)>
CMS−コシヒカリを母本として、コシヒカリの半糯性突然変異であるミルキークイーンを2回戻し交配を行い、玄米形質が粳型と半糯型に分離する種子を得た。その後、この種子から半糯性を示す玄米を選抜した。この選抜された玄米が、半糯性を有するイネ品種コシヒカリの細胞質雄性不稔系統であり、該系統をCMS−ミルキークイーンとして育成した。
【0062】
<イネ細胞質雄性不稔系統(CMS系統)>
CMS−コシヒカリを母本とし、イネ品種コシヒカリえいち2号、イネ品種コシヒカリえいち2号_長領域、イネ品種コシヒカリえいち3号、イネ品種コシヒカリえいち4号、イネ品種コシヒカリえいち4号_長領域、イネ品種コシヒカリかずさ2号、イネ品種コシヒカリかずさ3号、又はイネ品種コシヒカリかずさ4号を花粉親として連続戻し交配を行った。得られた後代個体の中から、雄性不稔性を示す栽培個体を選抜した。これらの選抜された各栽培個体の葉からDNAを回収し、DNAマーカーを調べ、花粉親と同じ領域が、外来品種由来アレルのホモ染色体領域である栽培個体をそれぞれ1個体ずつ選抜した。これらの選抜された栽培個体は、雄性不稔である以外は、基本的に花粉親と同じ形質を備える新品種である。本発明者は、イネ品種コシヒカリえいち2号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち2号」、イネ品種コシヒカリえいち2号_長領域を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち2号_長領域」、イネ品種コシヒカリえいち3号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち3号」、イネ品種コシヒカリえいち4号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち4号」、イネ品種コシヒカリえいち4号_長領域を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリえいち4号_長領域」、イネ品種コシヒカリかずさ1号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリかずさ1号」、イネ品種コシヒカリかずさ2号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリかずさ2号」、及びイネ品種コシヒカリかずさ3号を花粉親として得られたイネ細胞質雄性不稔系統を「CMS−イネ品種コシヒカリかずさ3号」と命名した。
【0063】
なお、出願人は、実施例1において得られた新品種のうち、CMS−イネ品種コシヒカリえいち2号、及びCMS−イネ品種コシヒカリかずさ3号を、独立行政法人産業技術総合研究所特許生物寄託センター(茨城県つくば市東1−1−1 つくばセンター中央第6)に新規植物として寄託した。イネ細胞質雄性不稔系統CMS−イネ品種コシヒカリえいち2号の受託番号はFERM P−22217であり、イネ細胞質雄性不稔系統CMS−イネ品種コシヒカリかずさ3号の受託番号はFERM P−22218である。
【0064】
<F1種子の生産>
上記で得られたイネ細胞質雄性不稔系統(CMS系統)を母本とし、花粉親として独自に育成した回復系中間母本系統ST−1、ST−2、又はST−4をそれぞれ交配してF1雑種の種子を得た。対照として、CMS−コシヒカリを母本とし、ST−1、ST−2、又はST−4を花粉親として交配してF1雑種の種子を得た。得られたF1種子を栽培し、2006年度、2007年度、及び2008年度の愛知県における圃場試験において形質の検討を行った。検討は、種苗法(平成10年法律第83号)第5条第1項に基づく品種登録出願のための特性審査に準拠して行った。
【0065】
<F1雑種の稈長>
CMS−コシヒカリえいち4号を母本として得られたF1雑種と、CMS−コシヒカリを母本として得られたF1雑種と、イネ品種コシヒカリの稈長を比較した。各系統の稈長の測定結果を図12に示す。この結果、対照品種であるコシヒカリの稈長は99.6cmであったのに対して、CMS−コシヒカリとST−2又はST−4を交配させて得られたF1雑種の稈長は、それぞれ123.0cm、115.3cmであった。一方、CMS−コシヒカリえいち4号と、ST−2又はST−4とを交配させて得られたF1雑種の稈長は、それぞれ106.9cm、108.8cmであり、CMS−コシヒカリを母本としたF1雑種よりも6.5〜16.1cm短くなっており、同時に収穫時における倒伏程度が軽減されていた。また、コシヒカリえいち4号を母本として得られたF1雑種系統は、稈長が短くなる以外は、コシヒカリを母本として得られたF1雑種系統と同じ形質を示した。
【0066】
さらに別の年に、CMS−コシヒカリとST−1から得られたF1雑種、CMS−コシヒカリえいち4号とST−1から得られたF1雑種、CMS−コシヒカリえいち4号_長領域とST−1から得られたF1雑種、及びイネ品種コシヒカリの稈長を測定し、比較した。各系統の稈長の測定結果を図13に示す。この結果、CMS−コシヒカリを母本として得られたF1雑種は、対照品種であるコシヒカリよりも稈長が長くなっていたのに対して、CMS−コシヒカリえいち4号又はCMS−コシヒカリえいち4号_長領域を母本として得られたF1雑種は、コシヒカリよりも稈長が短くなっていた。また、CMS−コシヒカリえいち4号から得られたF1雑種と、CMS−コシヒカリえいち4号_長領域から得られたF1雑種を比較したところ、両者に有意な差異は見られなかった。
これらの結果から、sd1遺伝子がコードされる領域がハバタキ由来の染色体断片で置換されているイネ雄性不稔系統を用いることにより、ハバタキ由来のsd1遺伝子を有さないイネ雄性不稔系統を用いた場合よりも、稈長が短く、耐倒伏性が改善されたF1雑種が得られることが明らかである。
【0067】
<F1雑種の到穂日数>
CMS−コシヒカリえいち3号又はCMS−コシヒカリを母本とし、ST−2又はST−4を花粉親としてF1雑種を得、これらのF1雑種の到穂日数を調べた。各系統の到穂日数の測定結果を図14に示す。4月16日に播種した本試験における対照品種コシヒカリの到穂日数は104日であったのに対して、CMS−コシヒカリとST−2又はST−4から得られたF1雑種の到穂日数は、135日又は132日であった。一方、CMS−コシヒカリえいち3号とST−2又はST−4から得られたF1雑種の到穂日数は、107日又は101日であり、CMS−コシヒカリを母本としたF1雑種よりも10〜28日程度短く、早生化されていた。このように、CMS−コシヒカリえいち3号を母本としたF1雑種は、コシヒカリとほぼ同程度の到穂日数であり、愛知県における栽培に適応可能な熟期を示した。また、CMS−コシヒカリえいち3号を母本としたF1雑種系統の形質は、到穂日数が小さくなること、及び、早生になることにより植物体が小さくなる傾向を示したが、それ以外の形質については、基本的にCMS−コシヒカリを母本としたF1雑種と同じであった。
これらの結果から、hd1遺伝子がコードされる領域がハバタキ由来の染色体断片で置換されているイネ細胞質雄性不稔系統を用いることにより、ハバタキ由来のhd1遺伝子を有さないイネ細胞質雄性不稔系統を用いた場合よりも、早生化されたF1雑種が得られることが明らかである。
【0068】
<F1雑種の着粒数>
CMS−コシヒカリえいち2号又はCMS−コシヒカリを母本とし、ST−2又はST−4を花粉親としてF1雑種得、これらのF1雑種の着粒数を調べた。各系統の着粒数の測定結果を図15に示す。この結果、対照品種であるコシヒカリの主茎における1穂粒数は138.5粒であったのに対して、CMS−コシヒカリとST−2又はST−4から得られたF1雑種の主茎における1穂粒数は、それぞれ284.0粒、257.4粒であった。一方、CMS−コシヒカリえいち2号とST−2又はST−4から得られたF1雑種の1穂粒数は、それぞれ314.8粒、327.0粒であり、CMS−コシヒカリを母本としたF1雑種よりも30〜70粒多くなっており、1穂重、及び収量性が向上していた。また、CMS−コシヒカリえいち2号を母本としたF1雑種系統の形質は、着粒数が多くなった以外は、基本的にCMS−コシヒカリを母本としたF1雑種と同じであった。
【0069】
また、CMS−コシヒカリえいち2号、CMS−コシヒカリえいち2号_長領域、又はCMS−コシヒカリを母本とし、ST−2を花粉親として得られたF1雑種の着粒数を測定し、比較した。各系統の着粒数の測定結果を図16に示す。この結果、CMS−コシヒカリえいち2号から得られたF1雑種と、CMS−コシヒカリえいち2号_長領域から得られたF1雑種との1穂粒数を比較したところ、両者に有意な差異は見られなかった。
【0070】
<複数の遺伝子を置換したイネ細胞質雄性不稔系統から得られたF1雑種系統の形質>
CMS−コシヒカリ、CMS−コシヒカリえいち2号、CMS−コシヒカリえいち3号、CMS−コシヒカリえいち4号、CMS−コシヒカリかずさ1号、CMS−コシヒカリかずさ2号、又はCMS−コシヒカリかずさ3号を母本とし、ST−2を花粉親として得られたF1雑種の、稈長、1穂粒数、及び到穂日数を表4に示す。対照として、コシヒカリの結果も合わせて示した。この結果、ハバタキ由来のsd1遺伝子を含むCMS−コシヒカリかずさ1号及びCMS−コシヒカリかずさ3号から得られたF1雑種系統は、CMS−コシヒカリえいち4号から得られたF1雑種系統と同様に、CMS−コシヒカリから得られたF1雑種系統よりも稈長が有意に短くなっていた。また、ハバタキ由来のhd1遺伝子を含むCMS−コシヒカリかずさ1号及びCMS−コシヒカリかずさ2号から得られたF1雑種系統は、CMS−コシヒカリえいち3号から得られたF1雑種系統と同様に、CMS−コシヒカリから得られたF1雑種系統よりも到穂日数が短くなっていた。また、ハバタキ由来のGn1遺伝子を含むCMS−コシヒカリかずさ3号から得られたF1雑種系統は、CMS−コシヒカリえいち2号から得られたF1雑種系統と同様に、CMS−コシヒカリから得られたF1雑種系統よりも1穂粒数が多くなっていた。ハバタキ由来のGn1遺伝子とハバタキ由来のhd1遺伝子を含むCMS−コシヒカリかずさ2号から得られたF1雑種系統では、元品種であるコシヒカリよりも若干晩生であるものの、CMS−コシヒカリから得られたF1雑種系統よりも顕著に早生であることに加えて、1穂粒数がコシヒカリよりも非常に多くなっていた。
これらの結果から、母本において、それぞれの領域が単独に置換された場合と同じように、複数の領域が置換された場合でも、それぞれの領域の効果は、F1雑種においても得られることが明らかである。
【0071】
【表4】
【0072】
[実施例2]
<食味の評価>
CMS−コシヒカリ又はCMS−ミルキークイーンを母本とし、ST−1、ST−2、又はST−4を花粉親として交配させてF1雑種の種子を得た。得られたF1雑種を栽培して得られた玄米を、食味官能試験へ供試した。食味官能試験においては、対照として、複数産地で栽培されたコシヒカリの玄米をブレンドしたものを用いた。試験結果を表5に示す。この結果、CMS−コシヒカリから得られたF1雑種の玄米の食味総合値は、それぞれ、コシヒカリと同等、または、やや劣るという結果であった。これに対して、CMS−ミルキークイーンから得られたF1雑種の玄米の食味総合値は、いずれも、CMS−コシヒカリから得られたF1雑種よりも良好であった。粘りの項目が強くなる傾向が観察されたことから、食味総合値の向上をもたらしたのは、半糯性遺伝子の導入であることが推測された。なお、CMS系統の違いによる、玄米形質以外における有意な形質の差異は観察されなかった。
これらの結果から、半糯性を示すイネ細胞質雄性不稔系統を母本として用いることにより、半糯性を示さないイネ細胞質雄性不稔系統を母本として用いた場合よりも、F1雑種の食味を改善し得ることが明らかである。
【0073】
【表5】
【0074】
[実施例3]
<F1雑種の到穂日数>
CMS−コシヒカリ又はCMS−コシヒカリえいち3号を母本とし、イネ品種クサノホシ、イネ品種クサホナミ、イネ品種タカナリ、イネ品種ニシアオバ、イネ品種ふくひびき、イネ品種ホシアオバ、イネ品種桂朝2号、イネ品種水原258号、又はイネ品種夢あおば花粉親として交配させてF1雑種の種子を得た。得られたF1雑種を栽培し、到穂日数を調べた。対照として、コシヒカリと各花粉親も栽培し、それぞれの到穂日数を調べた。各系統の到穂日数の測定結果を図17に示す。5月13日に播種した本試験における対照品種コシヒカリの到穂日数は84日であったのに対して、供試したCMS−コシヒカリを母本として得られたF1雑種系統の多くは、倒穂日数が長くなり、晩生化し、95日以内を示したのはわずか1系統のみであった。これに対して、CMS−コシヒカリえいち3号を母本として得られたF1雑種系統では、いずれも花粉親やCMS−コシヒカリを母本として得られたF1雑種系統よりも倒穂日数が短くなっており、早生化していた。特に、CMS−コシヒカリえいち3号を母本として得られたF1雑種系統は、全て倒穂日数が90日以内に抑えられていた。
これらの結果から、ハバタキ由来のhd1遺伝子を含むイネ雄性不稔系統を母本として用いることにより、CMS−コシヒカリを母本とした場合よりも、F1雑種を早生化し得ること、よって、コシヒカリに近い熟期の系統を育成する上で、選抜効率を大きく向上させる効果があることが明らかとなった。
【産業上の利用可能性】
【0075】
本発明のイネF1種子の生産方法は、イネ品種コシヒカリの雄性不稔系統を母本として用いた場合よりも、イネF1雑種の種子をより高い選抜効率で生産することができるため、当該方法は、特に植物の育種の分野において利用が可能である。
【受託番号】
【0076】
FERM P−22217
FERM P−22218
【特許請求の範囲】
【請求項1】
イネ(Oryza sativa)品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統、又は半糯性を示すイネ雄性不稔系統を母本とし、イネ稔性回復系統を花粉親として交配し、交配後の母本から雑種第1代種子(F1種子)を採取することを特徴とするイネF1種子の生産方法。
【請求項2】
前記イネ雄性不稔系統が、受託番号がFERM P−22217であるイネ細胞質雄性不稔系統CMS−コシヒカリえいち2号(Oryza sativa L.cultivar Koshihikari−Eichi−2go)、イネ細胞質雄性不稔系統CMS−コシヒカリえいち3号、イネ細胞質雄性不稔系統CMS−コシヒカリえいち4号、イネ細胞質雄性不稔系統CMS−コシヒカリかずさ1号、イネ細胞質雄性不稔系統CMS−コシヒカリかずさ2号、及び受託番号がFERM P−22218であるイネ細胞質雄性不稔系統CMS−コシヒカリかずさ3号からなる群より選択される細胞質雄性不稔系統であることを特徴とする請求項1に記載のイネF1種子の生産方法。
【請求項3】
請求項1又は2に記載のイネF1種子の生産方法により得られたことを特徴とするイネF1種子。
【請求項4】
イネ(Oryza sativa)品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有することを特徴とするイネ雄性不稔系統。
【請求項5】
受託番号がFERM P−22217であるイネ細胞質雄性不稔系統CMS−コシヒカリえいち2号(Oryza sativa L.cultivar Koshihikari−Eichi−2go)。
【請求項6】
イネ細胞質雄性不稔系統CMS−コシヒカリえいち3号(Oryza sativa L.cultivar Koshihikari−Eichi−3go)。
【請求項7】
イネ細胞質雄性不稔系統CMS−コシヒカリえいち4号(Oryza sativa L.cultivar Koshihikari−Eichi−4go)。
【請求項8】
イネ細胞質雄性不稔系統CMS−コシヒカリかずさ1号(Oryza sativa L.cultivar Koshihikari−Kazusa−1go)。
【請求項9】
イネ細胞質雄性不稔系統CMS−コシヒカリかずさ2号(Oryza sativa L.cultivar Koshihikari−Kazusa−2go)。
【請求項10】
受託番号がFERM P−22218であるイネ細胞質雄性不稔系統CMS−コシヒカリかずさ3号(Oryza sativa L.cultivar Koshihikari−Kazusa−3go)。
【請求項1】
イネ(Oryza sativa)品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有するイネ雄性不稔系統、又は半糯性を示すイネ雄性不稔系統を母本とし、イネ稔性回復系統を花粉親として交配し、交配後の母本から雑種第1代種子(F1種子)を採取することを特徴とするイネF1種子の生産方法。
【請求項2】
前記イネ雄性不稔系統が、受託番号がFERM P−22217であるイネ細胞質雄性不稔系統CMS−コシヒカリえいち2号(Oryza sativa L.cultivar Koshihikari−Eichi−2go)、イネ細胞質雄性不稔系統CMS−コシヒカリえいち3号、イネ細胞質雄性不稔系統CMS−コシヒカリえいち4号、イネ細胞質雄性不稔系統CMS−コシヒカリかずさ1号、イネ細胞質雄性不稔系統CMS−コシヒカリかずさ2号、及び受託番号がFERM P−22218であるイネ細胞質雄性不稔系統CMS−コシヒカリかずさ3号からなる群より選択される細胞質雄性不稔系統であることを特徴とする請求項1に記載のイネF1種子の生産方法。
【請求項3】
請求項1又は2に記載のイネF1種子の生産方法により得られたことを特徴とするイネF1種子。
【請求項4】
イネ(Oryza sativa)品種ハバタキ由来のsd1遺伝子、イネ品種ハバタキ由来のGn1遺伝子、及びイネ品種ハバタキ由来のhd1遺伝子からなる群より選択される1以上の遺伝子を含有することを特徴とするイネ雄性不稔系統。
【請求項5】
受託番号がFERM P−22217であるイネ細胞質雄性不稔系統CMS−コシヒカリえいち2号(Oryza sativa L.cultivar Koshihikari−Eichi−2go)。
【請求項6】
イネ細胞質雄性不稔系統CMS−コシヒカリえいち3号(Oryza sativa L.cultivar Koshihikari−Eichi−3go)。
【請求項7】
イネ細胞質雄性不稔系統CMS−コシヒカリえいち4号(Oryza sativa L.cultivar Koshihikari−Eichi−4go)。
【請求項8】
イネ細胞質雄性不稔系統CMS−コシヒカリかずさ1号(Oryza sativa L.cultivar Koshihikari−Kazusa−1go)。
【請求項9】
イネ細胞質雄性不稔系統CMS−コシヒカリかずさ2号(Oryza sativa L.cultivar Koshihikari−Kazusa−2go)。
【請求項10】
受託番号がFERM P−22218であるイネ細胞質雄性不稔系統CMS−コシヒカリかずさ3号(Oryza sativa L.cultivar Koshihikari−Kazusa−3go)。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公開番号】特開2012−210206(P2012−210206A)
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願番号】特願2012−59240(P2012−59240)
【出願日】平成24年3月15日(2012.3.15)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願日】平成24年3月15日(2012.3.15)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
[ Back to top ]