
インターフェロンα産生阻害剤
【課題】本発明は、形質細胞様樹状細胞からのインターフェロンα産生阻害剤や、形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤や、全身性エリテマトーデスの予防・治療剤を提供することを目的とする。
【解決手段】Hsp90阻害物質を用いることを特徴とする。Hsp90阻害物質としては、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されるものを好適に例示することができる。
【解決手段】Hsp90阻害物質を用いることを特徴とする。Hsp90阻害物質としては、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されるものを好適に例示することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、形質細胞様樹状細胞からのインターフェロンα産生阻害剤や、形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤や、全身性エリテマトーデスの予防・治療剤に関する。
【背景技術】
【0002】
樹状細胞(Dendritic cell)(以下、単に「DC」とも表示する。)は、抗原の取り込み、抗原の提示、サイトカインの産生など、自然免疫において重要な役割を担う細胞である。DCは、従来型樹状細胞(conventional DC)(以下、単に「cDC」とも表示する。)と、形質細胞様樹状細胞(plasmacytoid DC)(以下、単に「pDC」とも表示する。)に大別される。
【0003】
pDCは、B細胞と似た特徴を持つ樹状細胞で、典型的な樹状突起を持たず、生体内を循環している。また、pDCのエンドソームやリソソームには、核酸を認識するToll-like receptor(以下、単に「TLR」とも表示する。)のうち、TLR7及びTLR9が選択的に発現している。TLR7はウイルスの一本鎖RNAをリガンドとしており、TLR9は非メチル化CpGをリガンドとしている。pDCは、TLR7やTLR9を介するこれらのリガンド刺激によって、体細胞の100〜1000倍もの大量のインターフェロン(以下、単に「IFN」とも表示する。)を生産することができるため、近年注目を集めている。かかるIFNの中でも、インターフェロンα(IFNα)は、DC自身の活性化、細胞傷害性T細胞(cytotoxic T lymphocyte)(以下、単に「CTL」とも表示する。)の誘導、及び、B細胞からの抗体産生を促進する。
【0004】
一方、全身性エリテマトーデス(systemic lupus erythematosus)(以下、単に「SLE」とも表示する。)では、血清中のインターフェロンα高値が病態増悪の原因の一つと考えられている。SLE患者の血清では、抗DNA抗体が存在し、自己ゲノムDNA−抗DNA抗体からなる抗原抗体複合体が形成される。この複合体が、Fc受容体を介してB細胞やpDCに取り込まれてそれらの細胞のエンドソームへ効率よく輸送され、その結果、TLR9依存性に樹状細胞を強く活性化し、インターフェロンαを産生することがSLEの病態増悪の大きな原因であることが知られている(非特許文献1及び2)。
【0005】
また、ヒートショックプロテイン(HSP)は、熱ショックにより発現が増加する細胞内タンパク質で、熱以外の放射線・低栄養などのストレスにも反応し増加することが知られている。ヒートショックプロテインの機能としては、細胞内でさまざまなタンパク質と複合体を形成し、そのタンパク質を安定化させ正常に機能にさせる分子シャペロンとしての機能があり、特にヒートショックプロテイン90(Hsp90)のクライアントタンパク質にはプロテインキナーゼやステロイドホルモン受容体などの細胞増殖や分化に重要な役割を果たすシグナル伝達分子が多い。また、ヒートショックプロテインは、ペプチドと複合体を形成し、ストレスに曝露されると腫瘍細胞や感染細胞から細胞外に放出されて、免疫応答に寄与していることも知られている。
【0006】
Hsp90阻害剤は様々な用途に用いられており、例えば特許文献1には、Hsp90阻害剤を抗癌剤として用いることが記載されている。また、非特許文献3には、Hsp90がインターフェロンγやインターフェロンαを活性化することや、Hsp90の阻害物質であるゲルダナマイシンにより、インターフェロンγやインターフェロンαの抗ウイルス効果が抑制されることが記載されている。また、特許文献2には、Hsp90が腫瘍壊死因子α(Tumor Necrosis Factor α:TNFα)とインターロイキン6(IL−6)を活性化することが記載されており、そのクレーム21には、Hsp90阻害物質を全身性エリテマトーデス(SLE)等の患者に投与することにより、TNFα及びIL−6を低下させる方法が記載されている。
【0007】
しかし、Hsp90阻害物質が、pDCからのインターフェロンαの産生自体を阻害することや、インビボにおいて、全身性エリテマトーデスに対して予防・治療効果を実際に発揮することはこれまで知られていなかった。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特表2004−529188号公報
【特許文献2】国際公開第2007/077454号パンフレット
【非特許文献】
【0009】
【非特許文献1】J. Clin.Invest. 2005 Feb;115(2):407-17
【非特許文献2】Ann. N. Y. Acad. Sci. 2005 Dec;1062:242-51
【非特許文献3】J. Biol. Chem. 2006;281:1876-1884
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明の課題は、形質細胞様樹状細胞からのインターフェロンα産生阻害剤や、形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤や、全身性エリテマトーデスの予防・治療剤を提供することにある。
【課題を解決するための手段】
【0011】
本発明者らは、前述の背景技術に記載したような状況下、鋭意研究を行った結果、(a)Hsp90阻害物質が、全身性エリテマトーデス(SLE)のモデルマウスの病態を改善し得ること、(b)Hsp90阻害物質が、マウスやヒト由来の形質細胞様樹状細胞からのインターフェロンα産生を阻害し得ること、(c)かかるインターフェロンα産生の阻害が、Aktタンパク質やmTORタンパク質のリン酸化が抑制されることによるものであること、及び、(d)かかるインターフェロンα産生の阻害が、TLR9の小胞体から初期エンドソームへの移行が阻害されることによるものであることを見いだし、本発明を完成するに至った。
【0012】
すなわち、本発明は、(1)Hsp90阻害物質を含有することを特徴とする、形質細胞様樹状細胞からのインターフェロンα産生阻害剤や、(2)Hsp90阻害物質が、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されることを特徴とする上記(1)に記載のインターフェロンα産生阻害剤に関する。
【0013】
また、本発明は、(3)Hsp90阻害物質を含有することを特徴とする、形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤や、(4)Hsp90阻害物質が、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されることを特徴とする上記(3)に記載の移行阻害剤に関する。
【0014】
さらに、本発明は、(5)上記(1)若しくは(2)に記載のインターフェロンα産生阻害剤、又は、上記(3)若しくは(4)に記載の移行阻害剤を含有する全身性エリテマトーデスの予防・治療剤に関する。
【発明の効果】
【0015】
本発明によれば、形質細胞様樹状細胞からのインターフェロンα産生を阻害することや、形質細胞様樹状細胞におけるTLR9の小胞体から初期エンドソームへの移行を阻害することができる。また、本発明によれば、全身性エリテマトーデスを予防・治療することができる。
【図面の簡単な説明】
【0016】
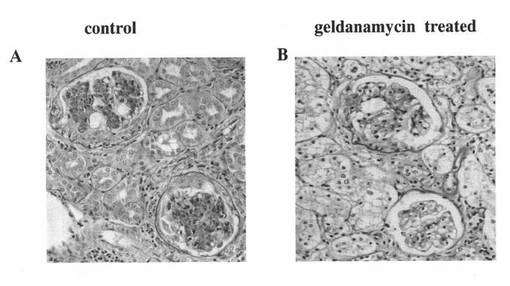
【図1】SLE発症モデルマウスの腎臓組織切片をヘマトキシリン・エオシン染色し、光学顕微鏡で観察した結果を示す図である。図1A:コントロール群のマウスの結果を表す。図1B:17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン塩酸塩(17−DMAG)を投与したゲルダナマイシン処理群のマウスの結果を表す。
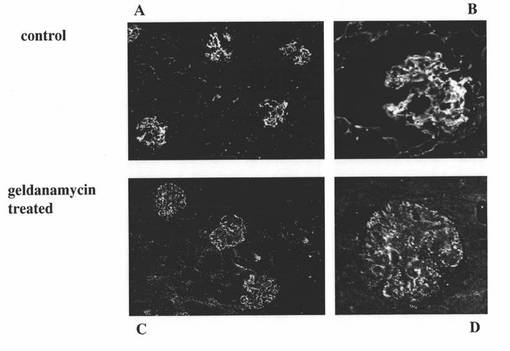
【図2】SLE発症モデルマウスの腎組織切片をFITC標識ヤギ抗マウスIgG抗体で染色し、蛍光顕微鏡で観察した結果を示す図である。図2A:コントロール群のマウスの結果を表す。図2B:図2Aの一部を拡大した結果を表す。図2C:17−DMAGを投与したゲルダナマイシン処理群のマウスの結果を表す。図2D:図2Cの一部を拡大した結果を表す。
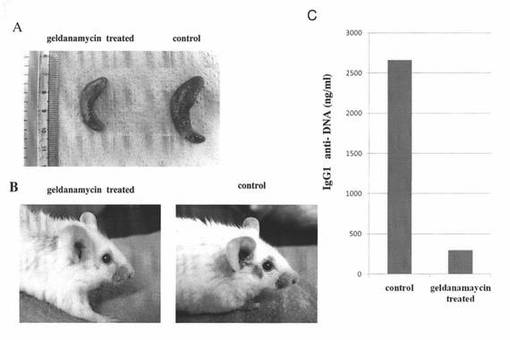
【図3】SLE発症モデルマウスの脾臓及び皮膚の観察結果、並びに、血清中の抗DNA抗体濃度の測定結果を示す図である。図3A:左は17−DMAGを投与したゲルダナマイシン処理群のマウスの脾臓を表し、右はコントロール群のマウスの脾臓を表す。図3B:左は17−DMAGを投与したゲルダナマイシン処理群のマウスの頭部の様子を表し、右はコントロール群のマウスの頭部の様子を表す。図3C:“control”はコントロール群のマウスの血清中の抗DNA抗体の濃度を表し、“geldanamycin treated”は17−DMAGを投与したゲルダナマイシン処理群のマウスの血清中の抗DNA抗体の濃度を表す。
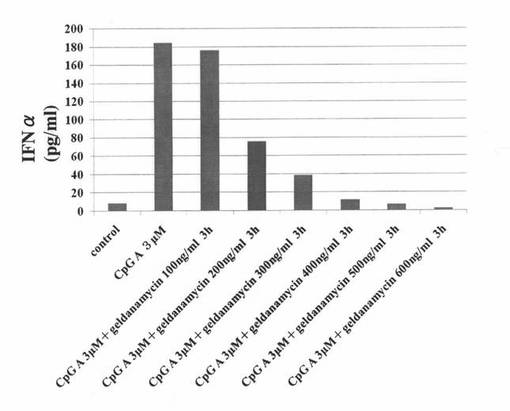
【図4】マウスpDCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養した培養液中に含まれるインターフェロンα濃度を測定した結果を示す図である。“control”はCpG−Aもゲルダナマイシンも添加せずに培養したマウスpDCの結果を表し、“CpG A 3μM”はゲルダナマイシンを添加せず、CpG−Aを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 100ng/ml 3h”はCpG−Aと、100ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 200ng/ml 3h”はCpG−Aと、200ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 300ng/ml 3h”はCpG−Aと、300ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 400ng/ml 3h”はCpG−Aと、400ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 500ng/ml 3h”はCpG−Aと、500ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 600ng/ml 3h”はCpG−Aと、600ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表す。
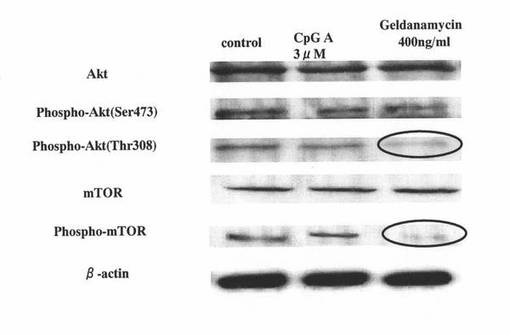
【図5】マウスpDCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養したマウスpDCについて、リン酸化Aktタンパク質及びリン酸化mTORタンパク質等に対するウエスタンブロッティング解析を行った結果を示す図である。左のレーンは、CpG−Aもゲルダナマイシンも添加せずに培養したマウスpDCの結果を表し、真ん中のレーンは、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したマウスpDCの結果を表し、右のレーンは、CpG−Aと、400ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表す。
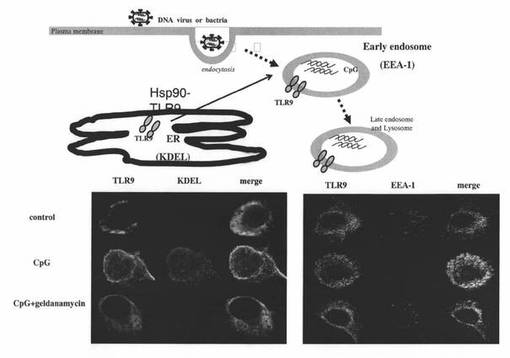
【図6】マウスpDCにおけるTLR9、KDEL(小胞体のマーカー)、EEA−1(初期エンドソームのマーカー)を免疫蛍光染色し、蛍光顕微鏡で観察した結果を示す図などである。上段は、ウイルスや細菌が細胞に感染したときの、TLR9の局在の変化を示した模式図である。下段左パネルの左列はTLR9の緑色蛍光を表し、中央列はKDEL(小胞体のマーカー)の赤色蛍光を表し、右列は対応する左列と中央列の蛍光を重ね合わせた蛍光を表し、上段列は“control”の場合の蛍光を表し、中段列は“CpG”の場合の蛍光を表し、下段列は“CpG+geldanamycin”の場合の蛍光を表す。下段右パネルの左列はTLR9の緑色蛍光を表し、中央列はEEA−1(初期エンドソームのマーカー)の赤色蛍光を表し、右列は対応する左列と中央列の蛍光を重ね合わせた蛍光を表し、上段列は“control”の場合の蛍光を表し、中段列は“CpG”の場合の蛍光を表し、下段列は“CpG+geldanamycin”の場合の蛍光を表す。
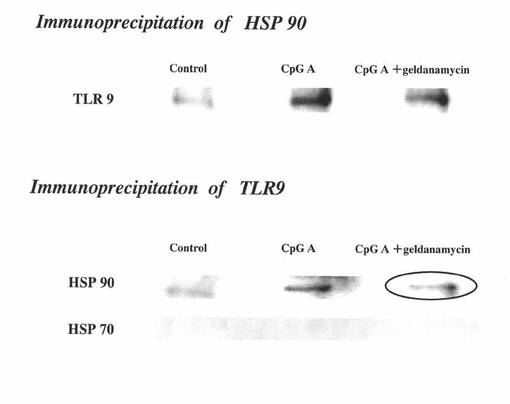
【図7】マウスpDCのライセートを抗TLR9抗体、又は、抗Hsp90抗体を用いて免疫沈降した物を電気泳動した後にウエスタンブロッティングを行った結果を示す図である。上段パネルは抗Hsp90抗体を用いた場合の結果を表し、中段パネル及び下段パネルは抗TLR9抗体を用いた場合の結果を表す。また、各パネルの左列は“control”の場合の結果を表し、中央列は“CpGA”の場合の結果を表し、右列は“CpGA+geldanamycin”の場合の結果を表す。
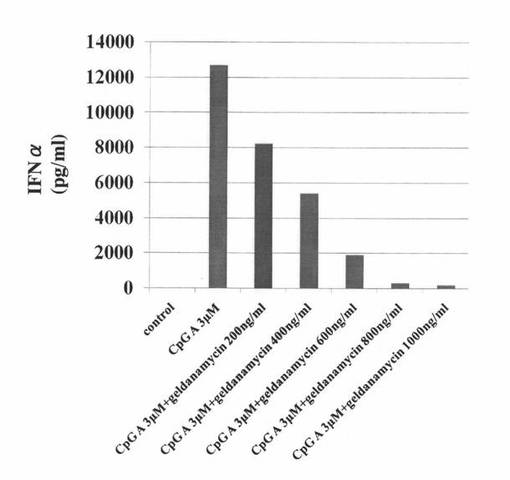
【図8】ヒトpDCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養した培養液中に含まれるインターフェロンα濃度を測定した結果を示す図である。“control”はCpG−Aもゲルダナマイシンも添加せずに培養したヒトpDCの結果を表し、“CpG A 3μM”はゲルダナマイシンを添加せず、CpG−Aを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 200ng/ml”はCpG−Aと、200ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 400ng/ml”はCpG−Aと、400ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 600ng/ml”はCpG−Aと、600ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 800ng/ml”はCpG−Aと、800ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 1000ng/ml”はCpG−Aと、1000ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表す。
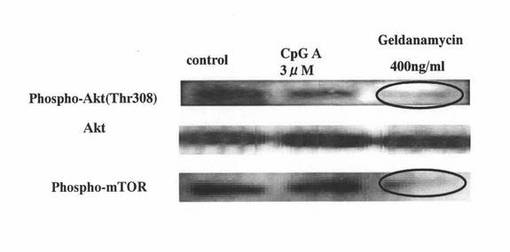
【図9】ヒトpDCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養したヒトpDCについて、リン酸化Aktタンパク質及びリン酸化mTORタンパク質に対するウエスタンブロッティング解析を行った結果を示す図である。左のレーンは、CpG−Aもゲルダナマイシンも添加せずに培養したヒトpDCの結果を表し、真ん中のレーンは、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したヒトpDCの結果を表し、右のレーンは、CpG−Aと、400ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表す。
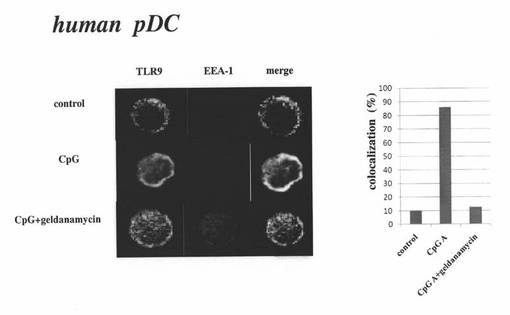
【図10】ヒトpDCにおけるTLR9、EEA−1(初期エンドソームのマーカー)を免疫蛍光染色し、蛍光顕微鏡で観察した結果を示す図などである。左パネルの左列はTLR9の緑色蛍光を表し、中央列はEEA−1の赤色蛍光を表し、右列は対応する左列と中央列の蛍光を重ね合わせた蛍光を表し、上段列は“control”の場合の蛍光を表し、中段列は“CpG”の場合の蛍光を表し、下段列は“CpG+geldanamycin”の場合の蛍光を表す。右パネルは、蛍光画像中のpDCをランダムに100個選択し、そのうち、TLR9(緑色蛍光)と、EEA−1(赤色蛍光)とが共局在しているpDCを計測して、それらが共局在しているpDCの割合(%)を示した図である。
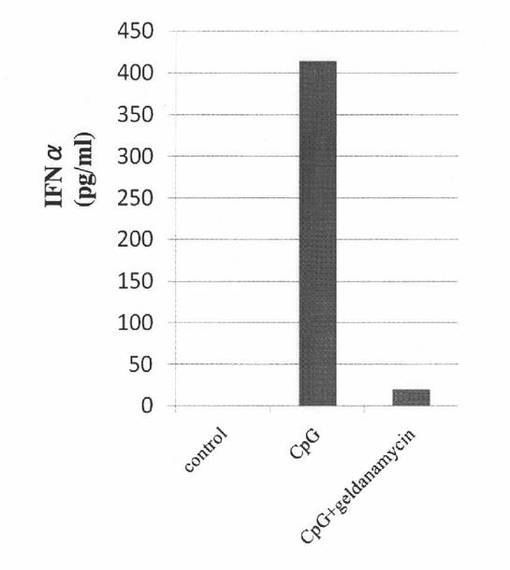
【図11】ヒトPBMCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養した培養液中に含まれるインターフェロンα濃度を測定した結果を示す図である。“control”はCpG−Aもゲルダナマイシンも添加せずに培養したヒトPBMCの結果を表し、“CpG”はゲルダナマイシンを添加せず、CpG−Aを添加して培養したヒトPBMCの結果を表し、“CpG + geldanamycin”はCpG−Aと、ゲルダナマイシンを添加して培養したヒトPBMCの結果を表す。
【発明を実施するための形態】
【0017】
本発明の「形質細胞様樹状細胞からのインターフェロンα産生阻害剤」(以下、単に「本発明のインターフェロンα産生阻害剤」とも表示する。)や、本発明の「形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤」(以下、単に「本発明の移行阻害剤」とも表示する。)としては、Hsp90阻害物質を含有している限り特に制限されない。また、かかる本発明のインターフェロンα産生阻害剤や、本発明の移行阻害剤は、全身性エリテマトーデス(SLE)の予防・治療剤(以下、「本発明のSLE予防・治療剤」とも表示する。)としても用いることができる。これらの剤の作用機序の詳細は明らかではないが、Hsp90阻害物質が、形質細胞様樹状細胞におけるTLR9の、小胞体から初期エンドソームへの移行を抑制し、かかるTLR9が自己DNAを認識することを抑制し、インターフェロンα産生に重要なAktタンパク質やmTORタンパク質を介するシグナル伝達を抑制することによって、インターフェロンα産生を阻害し、その結果、SLEに対する予防及び/又は治療効果が発揮されるものと考えられる。なお、本明細書において、「阻害」なる語と、「抑制」なる語は特に区別なく用いられる。
【0018】
上記のHsp90阻害物質とは、いずれかの哺乳動物のHsp90の機能を阻害する物質を意味するが、かかるHsp90の機能としては、形質細胞様樹状細胞におけるTLR9の、小胞体から初期エンドソームへの移行を介助する機能や、形質細胞様樹状細胞におけるAktタンパク質及び/又はmTORタンパク質をリン酸化する機能を好適に例示することができる。上記のHsp90阻害物質としては、Hsp90阻害物質である限り特に制限されないが、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体、及びエーテル体(以下、まとめて「ゲルダナマイシン等」とも表示する。)を好適に例示することができ、中でも、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、及び、それらの塩をより好適に例示することができる。また、Hsp90阻害物質を有効成分とする抗がん剤として、現在、KW−2478(American Association for Cancer Research, Published OnlineFirst on April 20, 2010, 10.1158/1078-0432.CCR-09-3112)の臨床試験が行われている。KW−2478では、Hsp90阻害物質として、2-{2-エチル-3,5-ジヒドロキシ-6-[3-メトキシ-4-(2-モルホリン-4-イルエトキシ)ベンゾイル]フェニル}-N,N-ビス(2-メトキシエチル)アセトアミド モノヒドロクロライドが用いられている。かかる物質は、医薬としての実用化の可能性が高いため、本発明に用いるHsp90阻害物質として特に好適に例示することができる。本発明のインターフェロンα産生阻害剤や移行阻害剤やSLE予防・治療剤(以下、これらを3つまとめて「本発明の剤」とも表示する。)には、本発明に用いるHsp90阻害物質を2種類以上併用してもよい。また、本発明におけるゲルダナマイシン等は市販のものを使用することができ、また、適宜エステル体やエーテル体とすることができる。
【0019】
上記の哺乳動物としては、特に制限されないが、ヒト、サル、マウス、ラット、ハムスター、モルモット、ウシ、ブタ、ウマ、ウサギ、ヒツジ、ヤギ、ネコ、イヌ等を好適に例示することができ、中でもヒトをより好適に例示することができる。
【0020】
ある物質がHsp90阻害物質であるかどうかは、例えば、
哺乳動物(好ましくはヒト)の形質細胞様樹状細胞を、被検物質存在下で培養する工程A;
前述の形質細胞様樹状細胞を刺激するCpG−A存在下で培養する工程B;
前述の形質細胞様樹状細胞についてウエスタンブロッティングを行い、リン酸化Aktタンパク質及び/又はリン酸化mTORタンパク質のシグナル強度を検出する工程C;
工程Cにおいて検出したシグナル強度を、被検物質の非存在下において検出したシグナル強度と比較する工程D;
工程Cにおいて検出したシグナル強度が、被検物質の非存在下において検出したシグナル強度よりも低い場合に、その被検物質をHsp90阻害物質と評価する工程E;
を有する方法Xや、
哺乳動物(好ましくはヒト)の形質細胞様樹状細胞を、被検物質存在下で培養する工程A;
前述の形質細胞様樹状細胞を刺激するCpG−A存在下で培養する工程B;
培養液中のインターフェロンα濃度を測定する工程C’;
工程C’で測定したインターフェロンα濃度の測定値を、被検物質非存在下で培養した場合のインターフェロンα濃度の測定値と比較する工程D’;
工程C’で測定したインターフェロンα濃度の測定値が、被検物質非存在下で培養した場合の測定値よりも低い場合に、その被検物質をHsp90阻害物質と評価する工程E’;
を有する方法Yや、
SLEモデル哺乳動物に被検物質を投与する工程F;
SLEモデル哺乳動物から血清を採取し、血清中の抗DNA抗体濃度を測定する工程G;
工程Gで測定した抗DNA抗体濃度の測定値を、被検物質非投与の場合の抗DNA抗体濃度の測定値と比較する工程H;
工程Gで測定した抗DNA抗体濃度の測定値が、被検物質非投与の場合の抗DNA抗体濃度の測定値よりも低い場合に、その被検物質をHsp90阻害物質と評価する工程I;
を有する方法Zや、
哺乳動物(好ましくはヒト)の形質細胞様樹状細胞を、被検物質存在下で培養する工程A;
前述の形質細胞様樹状細胞を刺激するCpG−A存在下で培養する工程B;
前述の形質細胞様樹状細胞において、TLR9が初期エンドソームに局在する割合を測定する工程C”;
工程C”において測定した割合を、被検物質の非存在下において測定した割合と比較する工程D”;
工程C”において測定した割合が、被検物質の非存在下において測定した割合よりも低い場合に、その被検物質をHsp90阻害物質と評価する工程E”;
を有する方法V等により、容易に確認することができる。
【0021】
前述の哺乳動物の形質細胞様樹状細胞は、例えば、かかる形質細胞様樹状細胞特異的な抗体等を用いて、哺乳動物の末梢血から分離、精製することができる。また、前述のリン酸化Aktタンパク質や、リン酸化mTORタンパク質の検出は、リン酸化Aktタンパク質に対する特異的抗体や、リン酸化mTORタンパク質に対する特異的抗体等を利用して行うことができる。また、前述のインターフェロンα濃度の測定は、そのインターフェロンαに特異的な抗体を含む市販のキット等を用いて、ELISA法により行うことができる。また、SLEモデル哺乳動物は、公知のものや、市販のものを利用することができる。さらに、抗DNA抗体濃度の測定は、ELISA法により行うことができる。また、形質細胞様樹状細胞において、TLR9が初期エンドソームに局在する割合の測定は、形質細胞様樹状細胞を抗TLR9抗体又は抗EEA−1抗体で蛍光染色し、抗TLR9抗体の蛍光と抗EEA−1抗体の蛍光とが共局在している形質細胞様樹状細胞の割合を測定すること等により行うことができる。
【0022】
本発明の剤は、所望の「インターフェロンα抑制効果」や、所望の「形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行抑制効果(以下、単に「移行抑制効果」とも表示する。)や、所望の「SLEの予防・治療効果」が得られる限り、前述のHsp90阻害物質の他に、他のインターフェロンα産生阻害剤や、他の移行阻害剤や、他のSLEの予防・治療剤等の任意成分を含んでいてもよい。
【0023】
Hsp90阻害物質のインターフェロンα産生阻害効果は、例えば、前述の方法Yにおいて、工程C’で測定したインターフェロンα濃度の測定値が、被検物質非存在下で培養した場合の測定値と比較して低下した程度を調べることによって算出することができる。また、Hsp90阻害物質の移行抑制効果は、例えば、前述の方法Vにおいて、工程C”において測定した割合が、被検物質の非存在下において測定した割合と比較して低下した程度を調べることによって算出することができる。さらに、Hsp90阻害物質のSLEの予防・治療効果は、例えば、前述の方法Zにおいて、工程Gで測定した抗DNA抗体濃度の測定値が、被検物質非投与の場合の抗DNA抗体濃度の測定値と比較して低下した程度を調べることによって算出することができる。
【0024】
本発明におけるHsp90阻害物質のインターフェロンα産生阻害効果の好適な程度としては、Hsp90阻害物質を600ng/ml含む培養液にて、ヒトの形質細胞様樹状細胞を3時間培養し、次いで、CpG−A(3μM)を培養液に添加し、添加してから24時間後に測定した培養液中のインターフェロンα濃度が、該Hsp90阻害物質を添加しなかった場合のインターフェロンα濃度に対して、割合として好ましくは30%以上、より好ましくは50%以上、さらに好ましくは70%以上、より好ましくは80%以上低下することを例示することができる。
【0025】
本発明におけるHsp90阻害物質の移行抑制効果の好適な程度としては、Hsp90阻害物質を400ng/ml含む培養液にて、ヒトの形質細胞様樹状細胞を3時間培養し、次いで、CpG−A(3μM)を培養液に添加し、添加してから24時間後に測定した培養液から分離した形質細胞様樹状細胞のうち、抗TLR9抗体の蛍光と抗EEA−1抗体の蛍光とが共局在している形質細胞様樹状細胞の割合(%)が、該Hsp90阻害物質を添加しなかった場合のその割合(%)に対して、割合として好ましくは30%以上、より好ましくは50%以上、さらに好ましくは70%以上、より好ましくは80%以上低下することを例示することができる。
【0026】
また、本発明におけるHsp90阻害物質のSLEの予防・治療効果の好適な程度としては、21週齢のMRL/lprマウス(Jackson Laboratory社製)に、0.125mg/dayのHsp90阻害物質を50日間連続して経口投与した後に採取した血清中の抗DNA抗体の濃度が、Hsp90阻害物質非投与の場合の抗DNA抗体の濃度に対して、割合として好ましくは30%以上、より好ましくは50%以上、さらに好ましくは70%以上、より好ましくは80%以上、さらに好ましくは90%以上低下することを例示することができる。
【0027】
本発明の剤に含有されるHsp90阻害物質は、常法によって適宜の製剤とすることができる。製剤の剤型としては散剤、顆粒剤などの固形製剤であってもよいが、優れたインターフェロンα産生効果や、移行抑制効果や、SLEの予防・治療効果を得る観点からは、溶液剤、乳剤、懸濁剤などの液剤とすることが好ましい。前述の液剤の製造方法としては、例えばHsp90阻害物質を溶剤と混合する方法や、さらに懸濁化剤や乳化剤を混合する方法を好適に例示することができる。以上のように、本発明におけるHsp90阻害物質を製剤とする場合には、製剤上の必要に応じて、適宜の薬学的に許容される担体、例えば、賦形剤、結合剤、溶剤、溶解補助剤、懸濁化剤、乳化剤、等張化剤、緩衝剤、安定化剤、無痛化剤、防腐剤、抗酸化剤、着色剤、滑沢剤、崩壊剤、湿潤剤、吸着剤、甘味剤、希釈剤などの任意成分を配合することができる。
【0028】
本発明の剤の投与方法としては、所望のインターフェロンα産生阻害効果や、所望の移行抑制効果や、所望のSLEの予防・治療効果が得られる限り特に制限されず、静脈内投与、経口投与、筋肉内投与、皮下投与、経皮投与、経鼻投与、経肺投与等を例示することができる。また本発明の剤の投与量や投与回数や投与濃度は、投与対象の体重等に応じて、適宜調節することができる。
【0029】
本発明のSLE予防・治療剤が対象とする疾患としては全身性エリテマトーデスである限り特に制限されない。また、本発明の剤の投与対象となる哺乳動物としては、特に制限されないが、ヒト、サル、マウス、ラット、ハムスター、モルモット、ウシ、ブタ、ウマ、ウサギ、ヒツジ、ヤギ、ネコ、イヌ等を好適に例示することができ、中でもヒトをより好適に例示することができる。また、本発明の剤に含有されるHsp90阻害物質が阻害作用を発揮するHsp90の由来である哺乳動物の種類は、本発明の剤の投与対象となる哺乳動物の種類と一致していることが、より安定して優れたインターフェロンα産生阻害効果や、移行抑制効果や、SLEの予防・治療効果を得る観点から、好ましい。
【0030】
なお、本発明の他の態様として、本発明のインターフェロンα産生阻害剤、移行阻害剤、又は、SLE予防・治療剤の製造における、本発明におけるHsp90阻害物質の使用;や、本発明におけるHsp90阻害物質を、本発明のインターフェロンα産生阻害剤、移行阻害剤、又は、SLE予防・治療剤に使用する方法;や、SLEの予防・治療における、本発明におけるHsp90阻害物質の使用;や、本発明のインターフェロンα産生阻害剤を対象に投与することにより、対象の形質細胞様樹状細胞からのインターフェロンα産生を阻害する方法;や、本発明の移行阻害剤を対象に投与することにより、対象の形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行を抑制する方法;や、本発明のSLE予防・治療剤を対象に投与することにより、対象のSLEを予防・治療する方法;も例示することができる。これらの使用や方法における文言の内容やその好ましい態様は、前述したとおりである。また、本発明の移行阻害剤は、形質細胞様樹状細胞におけるHsp90とTLR9の分子会合阻害剤としても利用することができる。
【0031】
以下に実施例により本発明を詳細に説明するが、本発明はこれらの実施例に限定される
ものではない。
【実施例1】
【0032】
[SLE発症モデルマウスに対するHsp90阻害物質の投与アッセイ]
Hsp90阻害物質が、SLEに対してどのような効果を発揮するかを調べるために、以下のようなアッセイを行った。
【0033】
まず、SLE発症モデルマウスであるMRL/lprマウス(Jackson Laboratory社製)を用意し、21週齢になるまで生育させた。これらのマウスを2つのグループに分けて、一方のグループをゲルダナマイシン処理群とし、他方をコントロール群とした。そして、ゲルダナマイシン処理群のマウスには、0.125mg/dayの17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン塩酸塩(17-dimethyl aminoethylamino-17-demethoxygeldanamycin hydrochloride:17−DMAG)を、50日間連続して経口投与し、コントロール群のマウスにはPBSを同様に投与した。投与終了後、両群のマウスから腎臓を取り出し、腎臓組織の凍結切片を作製した。以下、作製したこれらの凍結切片を凍結切片Xとも表示する。
【0034】
これらの凍結切片Xをヘマトキシリン・エオシン染色した後、光学顕微鏡で観察して得られた腎臓組織所見を図1に示す。図1Bは、ゲルダナマイシン処理群(geldanamycin treated)のマウスの腎臓組織所見を示し、図1Aは、コントロール群(control)のマウスの腎臓組織所見を示す。コントロール群のマウスの腎臓組織(図1A)では、糸球体基底膜は明らかに肥厚し、半月体形成を示す糸球体が目立ったのに対し、ゲルダナマイシン処理群のマウスの腎臓組織(図1B)では、糸球体基底膜肥厚の改善、半月体形成の抑制が明らかに認められ、著明な組織学的改善を認めた。
【0035】
また、前述の投与終了後の両群のマウスにおけるIgGの腎糸球体基底膜への沈着の程度を調べるために、前述の凍結切片Xを、FITC標識ヤギ抗マウスIgG抗体で染色した後、蛍光顕微鏡で観察した。その結果を図2に示す。図2Cは、ゲルダナマイシン処理群(geldanamycin treated)のマウスの腎臓組織切片の蛍光画像を表し、図2Dは、図2Cの一部を拡大した蛍光画像を表す。また、図2Aは、コントロール群(control)のマウスの腎臓組織切片の蛍光画像を表し、図2Bは、図2Aの一部を拡大した蛍光画像を表す。コントロール群のマウスの腎臓組織(図2A及びB)では、腎糸球体基底膜への厚いIgG沈着が見られたのに対し、ゲルダナマイシン処理群のマウスの腎臓組織(図2C及びD)では、明らかにIgG沈着が軽度で、腎糸球体基底膜の肥厚も軽度であった。
【0036】
さらに、前述の投与終了後の両群のマウスにおける脾腫の程度を調べるために、前述の投与終了後の両群のマウスから脾臓を取り出した。両群のマウスの脾臓の所見を図3Aに示し、皮膚の所見を図3Bに示す。ゲルダナマイシン処理群(geldanamycin treated)のマウスの脾臓(図3A左)は、コントロール群(control)のそれ(図3A右)と比較してサイズが小さく、脾腫は明らかに改善していた。また、皮膚に関して言えば、コントロール群のマウス(図3B右)では、眼瞼周囲の紅斑が明らかに認められたのに対し、ゲルダナマイシン処理群のマウス(図3B左)ではその紅斑は非常に軽度であった。
【0037】
また、前述の投与終了後の両群のマウスの血清中の抗DNA抗体濃度を調べるために、前述の投与終了後の両群のマウスから血清を採取し、血清中の抗DNA抗体の濃度をELISA(Mouse Anti ds ELISA KIT(AKRDD-061,Shibayagi(Gunma,Japan))で測定した。その結果を図3Cに示す。SLEでは血清中の抗DNA抗体が高濃度になることが知られているが、ゲルダナマイシン処理群(geldanamycin treated)のマウスの血清中の抗DNA抗体の濃度は、コントロール群(control)のそれと比較して明らかに低下していた(図3C)。
【0038】
図1〜図3の結果から、Hsp90阻害物質である17−DMAGの経口投与によって、明らかにSLEによる腎病変、皮膚病変の改善が認められた。また、SLEの活動度の指標となる抗DNA抗体についても、明らかに低値を示した。これらの結果は、SLEの治療薬としてHsp90阻害物質が有効であることを示すものである。
【実施例2】
【0039】
[マウスの形質細胞様樹状細胞を用いたHsp90阻害アッセイ]
Hsp90阻害物質による効果とその作用機序を検討するために、以下のようなアッセイを行った。
【0040】
(1)インターフェロンα測定アッセイ
C57/BL6マウスの骨髄細胞を、顆粒球単球コロニー刺激因子(GM−CSF;5μg/ml)を用いて培養し、培養から5日目に、pDC特異的な単クローン抗体mPDCA1を用いて、形質細胞様樹状細胞(pDC)を分離、精製し、精製pDCを得た。Hsp90阻害物質であるゲルダナマイシンを各種濃度(100ng/ml、200ng/ml、300ng/ml、400ng/ml、500ng/ml、又は、600ng/ml)で含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加し、培養を継続した。24時間後に、培養液を採取し、これらの培養液中に含まれるインターフェロンα濃度をELISA法にて測定した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したもの(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したもの(CpG-A 3μM)についても、同様に、インターフェロンα濃度を測定した。これらの結果を図4に示す。
【0041】
図4の結果から分かるように、CpG−Aの添加によって誘導されたインターフェロンα産生は、Hsp90阻害物質濃度依存性に抑制された。なお、Hsp90阻害物質として、ゲルダナマイシンに代えて、17−アリルアミノ−デメトキシゲルダナマイシン(17-allylamino-17-demethoxygeldanamycin:17−AAG)、又は、17−DMAGを用いて同様のアッセイを行ったところ、17−AAGや17−DMAGを用いた場合も、ゲルダナマイシンを用いた場合と同様に、Hsp90阻害物質濃度依存性に、インターフェロンα産生が抑制されることが示された。
【0042】
(2)シグナル伝達分子に関するウエスタンブロッティング解析
CpG−AによるpDCからのインターフェロンα産生には、シグナル伝達分子であるAktタンパク質及びmTORタンパク質のリン酸化が重要であることが知られている。そこで、pDCにおけるこれらの分子のリン酸化をウエスタンブロッティング法(イムノブロット法)により解析することとした。
【0043】
前述の実施例2の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDCを分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpG-A 3μM)についても、同様に分離した。分離したこれらの各pDC(それぞれ約1×106cellずつ)からライセートを調製し、SDS−PAGEゲルにて電気泳動を行った。このSDS−PAGEゲル中のタンパク質をメンブレンに転写し、次いで、かかるメンブレンをブロッキングバッファーにてブロッキングした。その後、メンブレンに各種の1次抗体を反応させてから、メンブレンを洗浄した。1次抗体としては、Aktタンパク質に対する抗体、473番目のセリンがリン酸化されたAktタンパク質に対する抗体、308番目のスレオニンがリン酸化されたAktタンパク質に対する抗体、mTORタンパク質に対する抗体、リン酸化されたmTORタンパク質に対する抗体、又は、β−アクチンに対する抗体をそれぞれ用いた。洗浄したメンブレンに、1次抗体を認識する標識2次抗体を反応させて、各種の1次抗体のシグナルを検出した。その結果を図5に示す。
【0044】
図5の結果から分かるように、左のレーン(control)や真ん中のレーン(CpG-A 3μM)などのコントロール群のpDCでは、308位におけるリン酸化Aktタンパク質に対するシグナル(Phospho-Akt(Thr308))や、リン酸化mTORタンパク質に対するシグナル(Phospho-mTOR)が認められたのに対して、ゲルダナマイシン処理したpDC(右のレーン:Geldanamycin 400ng/ml)では、308位におけるリン酸化Aktタンパク質に対するシグナル(Phospho-Akt(Thr308))や、リン酸化mTORタンパク質に対するシグナル(Phospho-mTOR)の低下が認められ、それらの分子のリン酸化が抑制されたことが示された。
【0045】
(3)pDCにおけるTLR9等の蛍光アッセイ
Hsp90阻害物質が、pDCにおけるTLR9の局在に影響を与えるかどうかを調べるために、以下のような蛍光アッセイを行うこととした。
【0046】
前述の実施例2の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDC(CpG+geldanamycin)を分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpG)についても、同様に分離した。分離したこれらの各pDCにおいて蛍光色素ラベルしたTLR9、KDEL(小胞体のマーカー)、及び、EEA−1(初期エンドソームのマーカー)を蛍光顕微鏡で観察した結果を図6の下段に示す。なお、TLR9(緑色蛍光)と、KDEL又はEEA−1(赤色蛍光)とが共局在すると、黄色の蛍光が確認される。
【0047】
コントロールでは、TLR9はほとんどが小胞体(Endoplasmic Reticulum: ER)に存在し、初期エンドソーム(Early endosome)では見られなかった(図6下段の左パネル及び右パネルの上段列(control))が、CpG−Aを添加培養したpDCでは、TLR9が小胞体から初期エンドソームに移動したことが示された(図6下段の左パネル及び右パネルの中段列(CpG))。これに対し、あらかじめゲルダナマイシン処理をしたpDCでは、CpG−Aを添加培養しても、TLR9は小胞体にとどまっており、初期エンドソームへの移行が抑制された(図6下段の左パネル及び右パネルの下段列(CpG+geldanamycin))。これらの結果から、Hsp90は、TLR9の小胞体から初期エンドソームへの移行の際の、分子シャペロンとして働いていると考えられた。
【0048】
(4)TLR9とHsp90の免疫沈降
前述の実施例2の(3)の結果を踏まえて、TLR9とHsp90の分子会合の有無を調べるために、以下のような免疫沈降を行うこととした。
【0049】
前述の実施例2の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDC(CpGA+geldanamycin)を分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpGA)についても、同様に分離した。分離したこれらの各pDCからそれぞれライセートを調製し、抗TLR9抗体、又は、抗Hsp90抗体を用いて免疫沈降を行った。免疫沈降物をSDS−PAGEにより電気泳動した後にウエスタンブロッティングを行った結果を図7に示す。
【0050】
図7の結果から分かるように、Hsp90とTLR9は分子会合しており(上段パネル及び中段パネルの中央列(CpGA))、その分子会合はゲルダナマイシンで抑制されることが示された(上段パネル及び中段パネルの右列(CpGA+geldanamycin))。
【実施例3】
【0051】
[ヒトの形質細胞様樹状細胞を用いたHsp90阻害アッセイ]
前述の実施例2では、マウスの形質細胞様樹状細胞を用いたアッセイを行った。そこで、ヒトの形質細胞様樹状細胞を用いた場合についても、Hsp90阻害物質による効果とその作用機序を検討するために、以下のようなアッセイを行った。
【0052】
(1)インターフェロンα測定アッセイ
まず、ヒト末梢血を用意した。このヒト末梢血から、pDC特異的な単クローン抗体BDCA−4を用いて、形質細胞様樹状細胞(pDC)を分離、精製し、精製pDCを得た。Hsp90阻害物質であるゲルダナマイシンを各種濃度(200ng/ml、400ng/ml、600ng/ml、800ng/ml、1000ng/ml)で含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加し、培養を継続した。24時間後に、培養液を採取し、これらの培養液中に含まれるインターフェロンα濃度をELISA法にて測定した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したもの(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したもの(CpG-A 3μM)についても、同様に、インターフェロンα濃度を測定した。これらの結果を図8に示す。
【0053】
図8の結果から分かるように、CpG−Aの添加によって誘導されたインターフェロンα産生は、ヒトpDCにおいても、Hsp90阻害物質濃度依存性に抑制された。なお、Hsp90阻害物質として、ゲルダナマイシンに代えて、17−AAG、又は、17−DMAGを用いて同様のアッセイを行ったところ、17−AAGや17−DMAGを用いた場合も、ゲルダナマイシンを用いた場合と同様に、Hsp90阻害物質濃度依存性に、インターフェロンα産生が抑制されることが示された。
【0054】
(2)シグナル伝達分子に関するウエスタンブロッティング解析
前述の実施例3の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDCを分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpG-A 3μM)についても、同様に分離した。分離したこれらの各pDC(それぞれ約1×106cellずつ)からライセートを調製し、SDS−PAGEゲルにて電気泳動を行った。このSDS−PAGEゲル中のタンパク質をメンブレンに転写し、次いで、かかるメンブレンをブロッキングバッファーにてブロッキングした。その後、メンブレンに各種の1次抗体を反応させてから、メンブレンを洗浄した。1次抗体としては、308番目のスレオニンがリン酸化されたAktタンパク質に対する抗体、Aktタンパク質に対する抗体、又は、リン酸化されたmTORタンパク質に対する抗体をそれぞれ用いた。洗浄したメンブレンに、1次抗体を認識する標識2次抗体を反応させて、各種の1次抗体のシグナルを検出した。その結果を図9に示す。
【0055】
図9の結果から分かるように、左のレーン(control)や真ん中のレーン(CpG-A 3μM)などのコントロール群のpDCでは、308位におけるリン酸化Aktタンパク質に対するシグナル(Phospho-Akt(Thr308))や、リン酸化mTORタンパク質に対するシグナル(Phospho-mTOR)が認められたのに対して、ゲルダナマイシン処理したpDC(右のレーン:Geldanamycin 400ng/ml)では、308位におけるリン酸化Aktタンパク質に対するシグナル(Phospho-Akt(Thr308))や、リン酸化mTORタンパク質に対するシグナル(Phospho-mTOR)の低下が認められ、それらの分子のリン酸化が抑制されたことが示された。この結果は、マウスpDCを用いた前述の実施例2(2)のウエスタンブロッティング解析の結果と同様であった。
【0056】
(3)pDCにおけるTLR9等の蛍光アッセイ
前述の実施例2の(3)において、あらかじめゲルダナマイシン処理をしたマウスpDCでは、CpG−Aを添加培養しても、TLR9は小胞体にとどまっており、初期エンドソームへの移行が抑制することが示された。この現象が、ヒトpDCでも生じるかを確認するために、以下のような蛍光アッセイを行うこととした。
【0057】
前述の実施例3の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDCを分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpG-A 3μM)についても、同様に分離した。分離したこれらの各pDCにおいて蛍光色素ラベルしたTLR9、EEA−1(初期エンドソームのマーカー)を蛍光顕微鏡で観察した結果を図10の左パネルに示す。なお、TLR9(緑色蛍光)と、EEA−1(赤色蛍光)とが共局在すると、黄色の蛍光が確認される。また、蛍光画像中のpDCをランダムに100個選択し、そのうち、TLR9(緑色蛍光)と、EEA−1(赤色蛍光)とが共局在しているpDCを計測して、それらが共局在しているpDCの割合(%)を図10の右パネルに示す。
【0058】
図10の左パネル及び右パネルから分かるように、ヒトpDCでも、CpG−Aの添加培養により、TLR9の初期エンドソームへの移行が生じ(図10左パネルの中段列(CpG))、その移行はゲルダナマイシンにより抑制されることが示された。これらの結果から、ヒトpDCにおいても、Hsp90は、TLR9の小胞体から初期エンドソームへの移行の際の、分子シャペロンとして働いていると考えられた。
【0059】
[実施例1〜3の結果のまとめ]
前述の実施例2や3の結果から分かるように、Hsp90阻害物質の投与により、マウスpDCやヒトpDCからのインターフェロンα産生が抑制された。そして、これは、pDCにおけるTLR9の、小胞体から初期エンドソームへの移行が抑制され、かかるTLR9が自己DNAを認識することが抑制されることや、インターフェロンα産生に重要なAktタンパク質やmTORタンパク質を介するシグナル伝達が抑制されることによることが示された。また、前述の実施例1におけるSLEマウスモデルの結果と総合すると、Hsp90阻害物質によってSLEの病態が改善した機序の1つは、Hsp90阻害物質によりpDCからのインターフェロンα産生が抑制されたことによるものと考えられた。
【実施例4】
【0060】
[ヒトの末梢血単核球を用いたHsp90阻害アッセイ]
前述の実施例3では、ヒトの形質細胞様樹状細胞を用いたアッセイを行った。そこで、ヒトの未成熟形質細胞様樹状細胞を含む末梢血単核球(PBMC)についても、インターフェロンα測定アッセイを行った。
【0061】
(1)インターフェロンα測定アッセイ
まず、ヒト末梢血を用意した。このヒト末梢血から、pDC特異的な単クローン抗体BDCA−4を用いて、形質細胞様樹状細胞(pDC)を分離、精製し、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加し、培養を継続した。24時間後に、培養液を採取し、これらの培養液中に含まれるインターフェロンα濃度をELISA法にて測定した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したもの(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したもの(CpG-A)についても、同様に、インターフェロンα濃度を測定した。これらの結果を図11に示す。
【0062】
図11の結果から分かるように、CpG−Aの添加によって誘導されたインターフェロンα産生は、ヒトpDCを含むヒトPBMCにおいても、Hsp90阻害物質により抑制された。このように、Hsp90阻害物質はpDCを含む末梢血に作用して、CpG−Aによるインターフェロンα産生を抑制する効果を示すことが明らかとなった。
【産業上の利用可能性】
【0063】
本発明は、形質細胞様樹状細胞からのインターフェロンα産生阻害剤の分野や、形質細胞様樹状細胞におけるAktタンパク質又はmTORタンパク質のリン酸化抑制剤の分野や、全身性エリテマトーデスの予防・治療の分野に好適に例示することができる。
【技術分野】
【0001】
本発明は、形質細胞様樹状細胞からのインターフェロンα産生阻害剤や、形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤や、全身性エリテマトーデスの予防・治療剤に関する。
【背景技術】
【0002】
樹状細胞(Dendritic cell)(以下、単に「DC」とも表示する。)は、抗原の取り込み、抗原の提示、サイトカインの産生など、自然免疫において重要な役割を担う細胞である。DCは、従来型樹状細胞(conventional DC)(以下、単に「cDC」とも表示する。)と、形質細胞様樹状細胞(plasmacytoid DC)(以下、単に「pDC」とも表示する。)に大別される。
【0003】
pDCは、B細胞と似た特徴を持つ樹状細胞で、典型的な樹状突起を持たず、生体内を循環している。また、pDCのエンドソームやリソソームには、核酸を認識するToll-like receptor(以下、単に「TLR」とも表示する。)のうち、TLR7及びTLR9が選択的に発現している。TLR7はウイルスの一本鎖RNAをリガンドとしており、TLR9は非メチル化CpGをリガンドとしている。pDCは、TLR7やTLR9を介するこれらのリガンド刺激によって、体細胞の100〜1000倍もの大量のインターフェロン(以下、単に「IFN」とも表示する。)を生産することができるため、近年注目を集めている。かかるIFNの中でも、インターフェロンα(IFNα)は、DC自身の活性化、細胞傷害性T細胞(cytotoxic T lymphocyte)(以下、単に「CTL」とも表示する。)の誘導、及び、B細胞からの抗体産生を促進する。
【0004】
一方、全身性エリテマトーデス(systemic lupus erythematosus)(以下、単に「SLE」とも表示する。)では、血清中のインターフェロンα高値が病態増悪の原因の一つと考えられている。SLE患者の血清では、抗DNA抗体が存在し、自己ゲノムDNA−抗DNA抗体からなる抗原抗体複合体が形成される。この複合体が、Fc受容体を介してB細胞やpDCに取り込まれてそれらの細胞のエンドソームへ効率よく輸送され、その結果、TLR9依存性に樹状細胞を強く活性化し、インターフェロンαを産生することがSLEの病態増悪の大きな原因であることが知られている(非特許文献1及び2)。
【0005】
また、ヒートショックプロテイン(HSP)は、熱ショックにより発現が増加する細胞内タンパク質で、熱以外の放射線・低栄養などのストレスにも反応し増加することが知られている。ヒートショックプロテインの機能としては、細胞内でさまざまなタンパク質と複合体を形成し、そのタンパク質を安定化させ正常に機能にさせる分子シャペロンとしての機能があり、特にヒートショックプロテイン90(Hsp90)のクライアントタンパク質にはプロテインキナーゼやステロイドホルモン受容体などの細胞増殖や分化に重要な役割を果たすシグナル伝達分子が多い。また、ヒートショックプロテインは、ペプチドと複合体を形成し、ストレスに曝露されると腫瘍細胞や感染細胞から細胞外に放出されて、免疫応答に寄与していることも知られている。
【0006】
Hsp90阻害剤は様々な用途に用いられており、例えば特許文献1には、Hsp90阻害剤を抗癌剤として用いることが記載されている。また、非特許文献3には、Hsp90がインターフェロンγやインターフェロンαを活性化することや、Hsp90の阻害物質であるゲルダナマイシンにより、インターフェロンγやインターフェロンαの抗ウイルス効果が抑制されることが記載されている。また、特許文献2には、Hsp90が腫瘍壊死因子α(Tumor Necrosis Factor α:TNFα)とインターロイキン6(IL−6)を活性化することが記載されており、そのクレーム21には、Hsp90阻害物質を全身性エリテマトーデス(SLE)等の患者に投与することにより、TNFα及びIL−6を低下させる方法が記載されている。
【0007】
しかし、Hsp90阻害物質が、pDCからのインターフェロンαの産生自体を阻害することや、インビボにおいて、全身性エリテマトーデスに対して予防・治療効果を実際に発揮することはこれまで知られていなかった。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特表2004−529188号公報
【特許文献2】国際公開第2007/077454号パンフレット
【非特許文献】
【0009】
【非特許文献1】J. Clin.Invest. 2005 Feb;115(2):407-17
【非特許文献2】Ann. N. Y. Acad. Sci. 2005 Dec;1062:242-51
【非特許文献3】J. Biol. Chem. 2006;281:1876-1884
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明の課題は、形質細胞様樹状細胞からのインターフェロンα産生阻害剤や、形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤や、全身性エリテマトーデスの予防・治療剤を提供することにある。
【課題を解決するための手段】
【0011】
本発明者らは、前述の背景技術に記載したような状況下、鋭意研究を行った結果、(a)Hsp90阻害物質が、全身性エリテマトーデス(SLE)のモデルマウスの病態を改善し得ること、(b)Hsp90阻害物質が、マウスやヒト由来の形質細胞様樹状細胞からのインターフェロンα産生を阻害し得ること、(c)かかるインターフェロンα産生の阻害が、Aktタンパク質やmTORタンパク質のリン酸化が抑制されることによるものであること、及び、(d)かかるインターフェロンα産生の阻害が、TLR9の小胞体から初期エンドソームへの移行が阻害されることによるものであることを見いだし、本発明を完成するに至った。
【0012】
すなわち、本発明は、(1)Hsp90阻害物質を含有することを特徴とする、形質細胞様樹状細胞からのインターフェロンα産生阻害剤や、(2)Hsp90阻害物質が、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されることを特徴とする上記(1)に記載のインターフェロンα産生阻害剤に関する。
【0013】
また、本発明は、(3)Hsp90阻害物質を含有することを特徴とする、形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤や、(4)Hsp90阻害物質が、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されることを特徴とする上記(3)に記載の移行阻害剤に関する。
【0014】
さらに、本発明は、(5)上記(1)若しくは(2)に記載のインターフェロンα産生阻害剤、又は、上記(3)若しくは(4)に記載の移行阻害剤を含有する全身性エリテマトーデスの予防・治療剤に関する。
【発明の効果】
【0015】
本発明によれば、形質細胞様樹状細胞からのインターフェロンα産生を阻害することや、形質細胞様樹状細胞におけるTLR9の小胞体から初期エンドソームへの移行を阻害することができる。また、本発明によれば、全身性エリテマトーデスを予防・治療することができる。
【図面の簡単な説明】
【0016】
【図1】SLE発症モデルマウスの腎臓組織切片をヘマトキシリン・エオシン染色し、光学顕微鏡で観察した結果を示す図である。図1A:コントロール群のマウスの結果を表す。図1B:17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン塩酸塩(17−DMAG)を投与したゲルダナマイシン処理群のマウスの結果を表す。
【図2】SLE発症モデルマウスの腎組織切片をFITC標識ヤギ抗マウスIgG抗体で染色し、蛍光顕微鏡で観察した結果を示す図である。図2A:コントロール群のマウスの結果を表す。図2B:図2Aの一部を拡大した結果を表す。図2C:17−DMAGを投与したゲルダナマイシン処理群のマウスの結果を表す。図2D:図2Cの一部を拡大した結果を表す。
【図3】SLE発症モデルマウスの脾臓及び皮膚の観察結果、並びに、血清中の抗DNA抗体濃度の測定結果を示す図である。図3A:左は17−DMAGを投与したゲルダナマイシン処理群のマウスの脾臓を表し、右はコントロール群のマウスの脾臓を表す。図3B:左は17−DMAGを投与したゲルダナマイシン処理群のマウスの頭部の様子を表し、右はコントロール群のマウスの頭部の様子を表す。図3C:“control”はコントロール群のマウスの血清中の抗DNA抗体の濃度を表し、“geldanamycin treated”は17−DMAGを投与したゲルダナマイシン処理群のマウスの血清中の抗DNA抗体の濃度を表す。
【図4】マウスpDCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養した培養液中に含まれるインターフェロンα濃度を測定した結果を示す図である。“control”はCpG−Aもゲルダナマイシンも添加せずに培養したマウスpDCの結果を表し、“CpG A 3μM”はゲルダナマイシンを添加せず、CpG−Aを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 100ng/ml 3h”はCpG−Aと、100ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 200ng/ml 3h”はCpG−Aと、200ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 300ng/ml 3h”はCpG−Aと、300ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 400ng/ml 3h”はCpG−Aと、400ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 500ng/ml 3h”はCpG−Aと、500ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表し、“CpG A 3μM + geldanamycin 600ng/ml 3h”はCpG−Aと、600ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表す。
【図5】マウスpDCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養したマウスpDCについて、リン酸化Aktタンパク質及びリン酸化mTORタンパク質等に対するウエスタンブロッティング解析を行った結果を示す図である。左のレーンは、CpG−Aもゲルダナマイシンも添加せずに培養したマウスpDCの結果を表し、真ん中のレーンは、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したマウスpDCの結果を表し、右のレーンは、CpG−Aと、400ng/mlのゲルダナマイシンを添加して培養したマウスpDCの結果を表す。
【図6】マウスpDCにおけるTLR9、KDEL(小胞体のマーカー)、EEA−1(初期エンドソームのマーカー)を免疫蛍光染色し、蛍光顕微鏡で観察した結果を示す図などである。上段は、ウイルスや細菌が細胞に感染したときの、TLR9の局在の変化を示した模式図である。下段左パネルの左列はTLR9の緑色蛍光を表し、中央列はKDEL(小胞体のマーカー)の赤色蛍光を表し、右列は対応する左列と中央列の蛍光を重ね合わせた蛍光を表し、上段列は“control”の場合の蛍光を表し、中段列は“CpG”の場合の蛍光を表し、下段列は“CpG+geldanamycin”の場合の蛍光を表す。下段右パネルの左列はTLR9の緑色蛍光を表し、中央列はEEA−1(初期エンドソームのマーカー)の赤色蛍光を表し、右列は対応する左列と中央列の蛍光を重ね合わせた蛍光を表し、上段列は“control”の場合の蛍光を表し、中段列は“CpG”の場合の蛍光を表し、下段列は“CpG+geldanamycin”の場合の蛍光を表す。
【図7】マウスpDCのライセートを抗TLR9抗体、又は、抗Hsp90抗体を用いて免疫沈降した物を電気泳動した後にウエスタンブロッティングを行った結果を示す図である。上段パネルは抗Hsp90抗体を用いた場合の結果を表し、中段パネル及び下段パネルは抗TLR9抗体を用いた場合の結果を表す。また、各パネルの左列は“control”の場合の結果を表し、中央列は“CpGA”の場合の結果を表し、右列は“CpGA+geldanamycin”の場合の結果を表す。
【図8】ヒトpDCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養した培養液中に含まれるインターフェロンα濃度を測定した結果を示す図である。“control”はCpG−Aもゲルダナマイシンも添加せずに培養したヒトpDCの結果を表し、“CpG A 3μM”はゲルダナマイシンを添加せず、CpG−Aを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 200ng/ml”はCpG−Aと、200ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 400ng/ml”はCpG−Aと、400ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 600ng/ml”はCpG−Aと、600ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 800ng/ml”はCpG−Aと、800ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表し、“CpG A 3μM + geldanamycin 1000ng/ml”はCpG−Aと、1000ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表す。
【図9】ヒトpDCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養したヒトpDCについて、リン酸化Aktタンパク質及びリン酸化mTORタンパク質に対するウエスタンブロッティング解析を行った結果を示す図である。左のレーンは、CpG−Aもゲルダナマイシンも添加せずに培養したヒトpDCの結果を表し、真ん中のレーンは、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したヒトpDCの結果を表し、右のレーンは、CpG−Aと、400ng/mlのゲルダナマイシンを添加して培養したヒトpDCの結果を表す。
【図10】ヒトpDCにおけるTLR9、EEA−1(初期エンドソームのマーカー)を免疫蛍光染色し、蛍光顕微鏡で観察した結果を示す図などである。左パネルの左列はTLR9の緑色蛍光を表し、中央列はEEA−1の赤色蛍光を表し、右列は対応する左列と中央列の蛍光を重ね合わせた蛍光を表し、上段列は“control”の場合の蛍光を表し、中段列は“CpG”の場合の蛍光を表し、下段列は“CpG+geldanamycin”の場合の蛍光を表す。右パネルは、蛍光画像中のpDCをランダムに100個選択し、そのうち、TLR9(緑色蛍光)と、EEA−1(赤色蛍光)とが共局在しているpDCを計測して、それらが共局在しているpDCの割合(%)を示した図である。
【図11】ヒトPBMCにCpG−Aを添加して培養した後、さらにゲルダナマイシンを添加して培養した培養液中に含まれるインターフェロンα濃度を測定した結果を示す図である。“control”はCpG−Aもゲルダナマイシンも添加せずに培養したヒトPBMCの結果を表し、“CpG”はゲルダナマイシンを添加せず、CpG−Aを添加して培養したヒトPBMCの結果を表し、“CpG + geldanamycin”はCpG−Aと、ゲルダナマイシンを添加して培養したヒトPBMCの結果を表す。
【発明を実施するための形態】
【0017】
本発明の「形質細胞様樹状細胞からのインターフェロンα産生阻害剤」(以下、単に「本発明のインターフェロンα産生阻害剤」とも表示する。)や、本発明の「形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤」(以下、単に「本発明の移行阻害剤」とも表示する。)としては、Hsp90阻害物質を含有している限り特に制限されない。また、かかる本発明のインターフェロンα産生阻害剤や、本発明の移行阻害剤は、全身性エリテマトーデス(SLE)の予防・治療剤(以下、「本発明のSLE予防・治療剤」とも表示する。)としても用いることができる。これらの剤の作用機序の詳細は明らかではないが、Hsp90阻害物質が、形質細胞様樹状細胞におけるTLR9の、小胞体から初期エンドソームへの移行を抑制し、かかるTLR9が自己DNAを認識することを抑制し、インターフェロンα産生に重要なAktタンパク質やmTORタンパク質を介するシグナル伝達を抑制することによって、インターフェロンα産生を阻害し、その結果、SLEに対する予防及び/又は治療効果が発揮されるものと考えられる。なお、本明細書において、「阻害」なる語と、「抑制」なる語は特に区別なく用いられる。
【0018】
上記のHsp90阻害物質とは、いずれかの哺乳動物のHsp90の機能を阻害する物質を意味するが、かかるHsp90の機能としては、形質細胞様樹状細胞におけるTLR9の、小胞体から初期エンドソームへの移行を介助する機能や、形質細胞様樹状細胞におけるAktタンパク質及び/又はmTORタンパク質をリン酸化する機能を好適に例示することができる。上記のHsp90阻害物質としては、Hsp90阻害物質である限り特に制限されないが、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体、及びエーテル体(以下、まとめて「ゲルダナマイシン等」とも表示する。)を好適に例示することができ、中でも、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、及び、それらの塩をより好適に例示することができる。また、Hsp90阻害物質を有効成分とする抗がん剤として、現在、KW−2478(American Association for Cancer Research, Published OnlineFirst on April 20, 2010, 10.1158/1078-0432.CCR-09-3112)の臨床試験が行われている。KW−2478では、Hsp90阻害物質として、2-{2-エチル-3,5-ジヒドロキシ-6-[3-メトキシ-4-(2-モルホリン-4-イルエトキシ)ベンゾイル]フェニル}-N,N-ビス(2-メトキシエチル)アセトアミド モノヒドロクロライドが用いられている。かかる物質は、医薬としての実用化の可能性が高いため、本発明に用いるHsp90阻害物質として特に好適に例示することができる。本発明のインターフェロンα産生阻害剤や移行阻害剤やSLE予防・治療剤(以下、これらを3つまとめて「本発明の剤」とも表示する。)には、本発明に用いるHsp90阻害物質を2種類以上併用してもよい。また、本発明におけるゲルダナマイシン等は市販のものを使用することができ、また、適宜エステル体やエーテル体とすることができる。
【0019】
上記の哺乳動物としては、特に制限されないが、ヒト、サル、マウス、ラット、ハムスター、モルモット、ウシ、ブタ、ウマ、ウサギ、ヒツジ、ヤギ、ネコ、イヌ等を好適に例示することができ、中でもヒトをより好適に例示することができる。
【0020】
ある物質がHsp90阻害物質であるかどうかは、例えば、
哺乳動物(好ましくはヒト)の形質細胞様樹状細胞を、被検物質存在下で培養する工程A;
前述の形質細胞様樹状細胞を刺激するCpG−A存在下で培養する工程B;
前述の形質細胞様樹状細胞についてウエスタンブロッティングを行い、リン酸化Aktタンパク質及び/又はリン酸化mTORタンパク質のシグナル強度を検出する工程C;
工程Cにおいて検出したシグナル強度を、被検物質の非存在下において検出したシグナル強度と比較する工程D;
工程Cにおいて検出したシグナル強度が、被検物質の非存在下において検出したシグナル強度よりも低い場合に、その被検物質をHsp90阻害物質と評価する工程E;
を有する方法Xや、
哺乳動物(好ましくはヒト)の形質細胞様樹状細胞を、被検物質存在下で培養する工程A;
前述の形質細胞様樹状細胞を刺激するCpG−A存在下で培養する工程B;
培養液中のインターフェロンα濃度を測定する工程C’;
工程C’で測定したインターフェロンα濃度の測定値を、被検物質非存在下で培養した場合のインターフェロンα濃度の測定値と比較する工程D’;
工程C’で測定したインターフェロンα濃度の測定値が、被検物質非存在下で培養した場合の測定値よりも低い場合に、その被検物質をHsp90阻害物質と評価する工程E’;
を有する方法Yや、
SLEモデル哺乳動物に被検物質を投与する工程F;
SLEモデル哺乳動物から血清を採取し、血清中の抗DNA抗体濃度を測定する工程G;
工程Gで測定した抗DNA抗体濃度の測定値を、被検物質非投与の場合の抗DNA抗体濃度の測定値と比較する工程H;
工程Gで測定した抗DNA抗体濃度の測定値が、被検物質非投与の場合の抗DNA抗体濃度の測定値よりも低い場合に、その被検物質をHsp90阻害物質と評価する工程I;
を有する方法Zや、
哺乳動物(好ましくはヒト)の形質細胞様樹状細胞を、被検物質存在下で培養する工程A;
前述の形質細胞様樹状細胞を刺激するCpG−A存在下で培養する工程B;
前述の形質細胞様樹状細胞において、TLR9が初期エンドソームに局在する割合を測定する工程C”;
工程C”において測定した割合を、被検物質の非存在下において測定した割合と比較する工程D”;
工程C”において測定した割合が、被検物質の非存在下において測定した割合よりも低い場合に、その被検物質をHsp90阻害物質と評価する工程E”;
を有する方法V等により、容易に確認することができる。
【0021】
前述の哺乳動物の形質細胞様樹状細胞は、例えば、かかる形質細胞様樹状細胞特異的な抗体等を用いて、哺乳動物の末梢血から分離、精製することができる。また、前述のリン酸化Aktタンパク質や、リン酸化mTORタンパク質の検出は、リン酸化Aktタンパク質に対する特異的抗体や、リン酸化mTORタンパク質に対する特異的抗体等を利用して行うことができる。また、前述のインターフェロンα濃度の測定は、そのインターフェロンαに特異的な抗体を含む市販のキット等を用いて、ELISA法により行うことができる。また、SLEモデル哺乳動物は、公知のものや、市販のものを利用することができる。さらに、抗DNA抗体濃度の測定は、ELISA法により行うことができる。また、形質細胞様樹状細胞において、TLR9が初期エンドソームに局在する割合の測定は、形質細胞様樹状細胞を抗TLR9抗体又は抗EEA−1抗体で蛍光染色し、抗TLR9抗体の蛍光と抗EEA−1抗体の蛍光とが共局在している形質細胞様樹状細胞の割合を測定すること等により行うことができる。
【0022】
本発明の剤は、所望の「インターフェロンα抑制効果」や、所望の「形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行抑制効果(以下、単に「移行抑制効果」とも表示する。)や、所望の「SLEの予防・治療効果」が得られる限り、前述のHsp90阻害物質の他に、他のインターフェロンα産生阻害剤や、他の移行阻害剤や、他のSLEの予防・治療剤等の任意成分を含んでいてもよい。
【0023】
Hsp90阻害物質のインターフェロンα産生阻害効果は、例えば、前述の方法Yにおいて、工程C’で測定したインターフェロンα濃度の測定値が、被検物質非存在下で培養した場合の測定値と比較して低下した程度を調べることによって算出することができる。また、Hsp90阻害物質の移行抑制効果は、例えば、前述の方法Vにおいて、工程C”において測定した割合が、被検物質の非存在下において測定した割合と比較して低下した程度を調べることによって算出することができる。さらに、Hsp90阻害物質のSLEの予防・治療効果は、例えば、前述の方法Zにおいて、工程Gで測定した抗DNA抗体濃度の測定値が、被検物質非投与の場合の抗DNA抗体濃度の測定値と比較して低下した程度を調べることによって算出することができる。
【0024】
本発明におけるHsp90阻害物質のインターフェロンα産生阻害効果の好適な程度としては、Hsp90阻害物質を600ng/ml含む培養液にて、ヒトの形質細胞様樹状細胞を3時間培養し、次いで、CpG−A(3μM)を培養液に添加し、添加してから24時間後に測定した培養液中のインターフェロンα濃度が、該Hsp90阻害物質を添加しなかった場合のインターフェロンα濃度に対して、割合として好ましくは30%以上、より好ましくは50%以上、さらに好ましくは70%以上、より好ましくは80%以上低下することを例示することができる。
【0025】
本発明におけるHsp90阻害物質の移行抑制効果の好適な程度としては、Hsp90阻害物質を400ng/ml含む培養液にて、ヒトの形質細胞様樹状細胞を3時間培養し、次いで、CpG−A(3μM)を培養液に添加し、添加してから24時間後に測定した培養液から分離した形質細胞様樹状細胞のうち、抗TLR9抗体の蛍光と抗EEA−1抗体の蛍光とが共局在している形質細胞様樹状細胞の割合(%)が、該Hsp90阻害物質を添加しなかった場合のその割合(%)に対して、割合として好ましくは30%以上、より好ましくは50%以上、さらに好ましくは70%以上、より好ましくは80%以上低下することを例示することができる。
【0026】
また、本発明におけるHsp90阻害物質のSLEの予防・治療効果の好適な程度としては、21週齢のMRL/lprマウス(Jackson Laboratory社製)に、0.125mg/dayのHsp90阻害物質を50日間連続して経口投与した後に採取した血清中の抗DNA抗体の濃度が、Hsp90阻害物質非投与の場合の抗DNA抗体の濃度に対して、割合として好ましくは30%以上、より好ましくは50%以上、さらに好ましくは70%以上、より好ましくは80%以上、さらに好ましくは90%以上低下することを例示することができる。
【0027】
本発明の剤に含有されるHsp90阻害物質は、常法によって適宜の製剤とすることができる。製剤の剤型としては散剤、顆粒剤などの固形製剤であってもよいが、優れたインターフェロンα産生効果や、移行抑制効果や、SLEの予防・治療効果を得る観点からは、溶液剤、乳剤、懸濁剤などの液剤とすることが好ましい。前述の液剤の製造方法としては、例えばHsp90阻害物質を溶剤と混合する方法や、さらに懸濁化剤や乳化剤を混合する方法を好適に例示することができる。以上のように、本発明におけるHsp90阻害物質を製剤とする場合には、製剤上の必要に応じて、適宜の薬学的に許容される担体、例えば、賦形剤、結合剤、溶剤、溶解補助剤、懸濁化剤、乳化剤、等張化剤、緩衝剤、安定化剤、無痛化剤、防腐剤、抗酸化剤、着色剤、滑沢剤、崩壊剤、湿潤剤、吸着剤、甘味剤、希釈剤などの任意成分を配合することができる。
【0028】
本発明の剤の投与方法としては、所望のインターフェロンα産生阻害効果や、所望の移行抑制効果や、所望のSLEの予防・治療効果が得られる限り特に制限されず、静脈内投与、経口投与、筋肉内投与、皮下投与、経皮投与、経鼻投与、経肺投与等を例示することができる。また本発明の剤の投与量や投与回数や投与濃度は、投与対象の体重等に応じて、適宜調節することができる。
【0029】
本発明のSLE予防・治療剤が対象とする疾患としては全身性エリテマトーデスである限り特に制限されない。また、本発明の剤の投与対象となる哺乳動物としては、特に制限されないが、ヒト、サル、マウス、ラット、ハムスター、モルモット、ウシ、ブタ、ウマ、ウサギ、ヒツジ、ヤギ、ネコ、イヌ等を好適に例示することができ、中でもヒトをより好適に例示することができる。また、本発明の剤に含有されるHsp90阻害物質が阻害作用を発揮するHsp90の由来である哺乳動物の種類は、本発明の剤の投与対象となる哺乳動物の種類と一致していることが、より安定して優れたインターフェロンα産生阻害効果や、移行抑制効果や、SLEの予防・治療効果を得る観点から、好ましい。
【0030】
なお、本発明の他の態様として、本発明のインターフェロンα産生阻害剤、移行阻害剤、又は、SLE予防・治療剤の製造における、本発明におけるHsp90阻害物質の使用;や、本発明におけるHsp90阻害物質を、本発明のインターフェロンα産生阻害剤、移行阻害剤、又は、SLE予防・治療剤に使用する方法;や、SLEの予防・治療における、本発明におけるHsp90阻害物質の使用;や、本発明のインターフェロンα産生阻害剤を対象に投与することにより、対象の形質細胞様樹状細胞からのインターフェロンα産生を阻害する方法;や、本発明の移行阻害剤を対象に投与することにより、対象の形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行を抑制する方法;や、本発明のSLE予防・治療剤を対象に投与することにより、対象のSLEを予防・治療する方法;も例示することができる。これらの使用や方法における文言の内容やその好ましい態様は、前述したとおりである。また、本発明の移行阻害剤は、形質細胞様樹状細胞におけるHsp90とTLR9の分子会合阻害剤としても利用することができる。
【0031】
以下に実施例により本発明を詳細に説明するが、本発明はこれらの実施例に限定される
ものではない。
【実施例1】
【0032】
[SLE発症モデルマウスに対するHsp90阻害物質の投与アッセイ]
Hsp90阻害物質が、SLEに対してどのような効果を発揮するかを調べるために、以下のようなアッセイを行った。
【0033】
まず、SLE発症モデルマウスであるMRL/lprマウス(Jackson Laboratory社製)を用意し、21週齢になるまで生育させた。これらのマウスを2つのグループに分けて、一方のグループをゲルダナマイシン処理群とし、他方をコントロール群とした。そして、ゲルダナマイシン処理群のマウスには、0.125mg/dayの17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン塩酸塩(17-dimethyl aminoethylamino-17-demethoxygeldanamycin hydrochloride:17−DMAG)を、50日間連続して経口投与し、コントロール群のマウスにはPBSを同様に投与した。投与終了後、両群のマウスから腎臓を取り出し、腎臓組織の凍結切片を作製した。以下、作製したこれらの凍結切片を凍結切片Xとも表示する。
【0034】
これらの凍結切片Xをヘマトキシリン・エオシン染色した後、光学顕微鏡で観察して得られた腎臓組織所見を図1に示す。図1Bは、ゲルダナマイシン処理群(geldanamycin treated)のマウスの腎臓組織所見を示し、図1Aは、コントロール群(control)のマウスの腎臓組織所見を示す。コントロール群のマウスの腎臓組織(図1A)では、糸球体基底膜は明らかに肥厚し、半月体形成を示す糸球体が目立ったのに対し、ゲルダナマイシン処理群のマウスの腎臓組織(図1B)では、糸球体基底膜肥厚の改善、半月体形成の抑制が明らかに認められ、著明な組織学的改善を認めた。
【0035】
また、前述の投与終了後の両群のマウスにおけるIgGの腎糸球体基底膜への沈着の程度を調べるために、前述の凍結切片Xを、FITC標識ヤギ抗マウスIgG抗体で染色した後、蛍光顕微鏡で観察した。その結果を図2に示す。図2Cは、ゲルダナマイシン処理群(geldanamycin treated)のマウスの腎臓組織切片の蛍光画像を表し、図2Dは、図2Cの一部を拡大した蛍光画像を表す。また、図2Aは、コントロール群(control)のマウスの腎臓組織切片の蛍光画像を表し、図2Bは、図2Aの一部を拡大した蛍光画像を表す。コントロール群のマウスの腎臓組織(図2A及びB)では、腎糸球体基底膜への厚いIgG沈着が見られたのに対し、ゲルダナマイシン処理群のマウスの腎臓組織(図2C及びD)では、明らかにIgG沈着が軽度で、腎糸球体基底膜の肥厚も軽度であった。
【0036】
さらに、前述の投与終了後の両群のマウスにおける脾腫の程度を調べるために、前述の投与終了後の両群のマウスから脾臓を取り出した。両群のマウスの脾臓の所見を図3Aに示し、皮膚の所見を図3Bに示す。ゲルダナマイシン処理群(geldanamycin treated)のマウスの脾臓(図3A左)は、コントロール群(control)のそれ(図3A右)と比較してサイズが小さく、脾腫は明らかに改善していた。また、皮膚に関して言えば、コントロール群のマウス(図3B右)では、眼瞼周囲の紅斑が明らかに認められたのに対し、ゲルダナマイシン処理群のマウス(図3B左)ではその紅斑は非常に軽度であった。
【0037】
また、前述の投与終了後の両群のマウスの血清中の抗DNA抗体濃度を調べるために、前述の投与終了後の両群のマウスから血清を採取し、血清中の抗DNA抗体の濃度をELISA(Mouse Anti ds ELISA KIT(AKRDD-061,Shibayagi(Gunma,Japan))で測定した。その結果を図3Cに示す。SLEでは血清中の抗DNA抗体が高濃度になることが知られているが、ゲルダナマイシン処理群(geldanamycin treated)のマウスの血清中の抗DNA抗体の濃度は、コントロール群(control)のそれと比較して明らかに低下していた(図3C)。
【0038】
図1〜図3の結果から、Hsp90阻害物質である17−DMAGの経口投与によって、明らかにSLEによる腎病変、皮膚病変の改善が認められた。また、SLEの活動度の指標となる抗DNA抗体についても、明らかに低値を示した。これらの結果は、SLEの治療薬としてHsp90阻害物質が有効であることを示すものである。
【実施例2】
【0039】
[マウスの形質細胞様樹状細胞を用いたHsp90阻害アッセイ]
Hsp90阻害物質による効果とその作用機序を検討するために、以下のようなアッセイを行った。
【0040】
(1)インターフェロンα測定アッセイ
C57/BL6マウスの骨髄細胞を、顆粒球単球コロニー刺激因子(GM−CSF;5μg/ml)を用いて培養し、培養から5日目に、pDC特異的な単クローン抗体mPDCA1を用いて、形質細胞様樹状細胞(pDC)を分離、精製し、精製pDCを得た。Hsp90阻害物質であるゲルダナマイシンを各種濃度(100ng/ml、200ng/ml、300ng/ml、400ng/ml、500ng/ml、又は、600ng/ml)で含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加し、培養を継続した。24時間後に、培養液を採取し、これらの培養液中に含まれるインターフェロンα濃度をELISA法にて測定した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したもの(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したもの(CpG-A 3μM)についても、同様に、インターフェロンα濃度を測定した。これらの結果を図4に示す。
【0041】
図4の結果から分かるように、CpG−Aの添加によって誘導されたインターフェロンα産生は、Hsp90阻害物質濃度依存性に抑制された。なお、Hsp90阻害物質として、ゲルダナマイシンに代えて、17−アリルアミノ−デメトキシゲルダナマイシン(17-allylamino-17-demethoxygeldanamycin:17−AAG)、又は、17−DMAGを用いて同様のアッセイを行ったところ、17−AAGや17−DMAGを用いた場合も、ゲルダナマイシンを用いた場合と同様に、Hsp90阻害物質濃度依存性に、インターフェロンα産生が抑制されることが示された。
【0042】
(2)シグナル伝達分子に関するウエスタンブロッティング解析
CpG−AによるpDCからのインターフェロンα産生には、シグナル伝達分子であるAktタンパク質及びmTORタンパク質のリン酸化が重要であることが知られている。そこで、pDCにおけるこれらの分子のリン酸化をウエスタンブロッティング法(イムノブロット法)により解析することとした。
【0043】
前述の実施例2の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDCを分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpG-A 3μM)についても、同様に分離した。分離したこれらの各pDC(それぞれ約1×106cellずつ)からライセートを調製し、SDS−PAGEゲルにて電気泳動を行った。このSDS−PAGEゲル中のタンパク質をメンブレンに転写し、次いで、かかるメンブレンをブロッキングバッファーにてブロッキングした。その後、メンブレンに各種の1次抗体を反応させてから、メンブレンを洗浄した。1次抗体としては、Aktタンパク質に対する抗体、473番目のセリンがリン酸化されたAktタンパク質に対する抗体、308番目のスレオニンがリン酸化されたAktタンパク質に対する抗体、mTORタンパク質に対する抗体、リン酸化されたmTORタンパク質に対する抗体、又は、β−アクチンに対する抗体をそれぞれ用いた。洗浄したメンブレンに、1次抗体を認識する標識2次抗体を反応させて、各種の1次抗体のシグナルを検出した。その結果を図5に示す。
【0044】
図5の結果から分かるように、左のレーン(control)や真ん中のレーン(CpG-A 3μM)などのコントロール群のpDCでは、308位におけるリン酸化Aktタンパク質に対するシグナル(Phospho-Akt(Thr308))や、リン酸化mTORタンパク質に対するシグナル(Phospho-mTOR)が認められたのに対して、ゲルダナマイシン処理したpDC(右のレーン:Geldanamycin 400ng/ml)では、308位におけるリン酸化Aktタンパク質に対するシグナル(Phospho-Akt(Thr308))や、リン酸化mTORタンパク質に対するシグナル(Phospho-mTOR)の低下が認められ、それらの分子のリン酸化が抑制されたことが示された。
【0045】
(3)pDCにおけるTLR9等の蛍光アッセイ
Hsp90阻害物質が、pDCにおけるTLR9の局在に影響を与えるかどうかを調べるために、以下のような蛍光アッセイを行うこととした。
【0046】
前述の実施例2の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDC(CpG+geldanamycin)を分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpG)についても、同様に分離した。分離したこれらの各pDCにおいて蛍光色素ラベルしたTLR9、KDEL(小胞体のマーカー)、及び、EEA−1(初期エンドソームのマーカー)を蛍光顕微鏡で観察した結果を図6の下段に示す。なお、TLR9(緑色蛍光)と、KDEL又はEEA−1(赤色蛍光)とが共局在すると、黄色の蛍光が確認される。
【0047】
コントロールでは、TLR9はほとんどが小胞体(Endoplasmic Reticulum: ER)に存在し、初期エンドソーム(Early endosome)では見られなかった(図6下段の左パネル及び右パネルの上段列(control))が、CpG−Aを添加培養したpDCでは、TLR9が小胞体から初期エンドソームに移動したことが示された(図6下段の左パネル及び右パネルの中段列(CpG))。これに対し、あらかじめゲルダナマイシン処理をしたpDCでは、CpG−Aを添加培養しても、TLR9は小胞体にとどまっており、初期エンドソームへの移行が抑制された(図6下段の左パネル及び右パネルの下段列(CpG+geldanamycin))。これらの結果から、Hsp90は、TLR9の小胞体から初期エンドソームへの移行の際の、分子シャペロンとして働いていると考えられた。
【0048】
(4)TLR9とHsp90の免疫沈降
前述の実施例2の(3)の結果を踏まえて、TLR9とHsp90の分子会合の有無を調べるために、以下のような免疫沈降を行うこととした。
【0049】
前述の実施例2の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDC(CpGA+geldanamycin)を分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpGA)についても、同様に分離した。分離したこれらの各pDCからそれぞれライセートを調製し、抗TLR9抗体、又は、抗Hsp90抗体を用いて免疫沈降を行った。免疫沈降物をSDS−PAGEにより電気泳動した後にウエスタンブロッティングを行った結果を図7に示す。
【0050】
図7の結果から分かるように、Hsp90とTLR9は分子会合しており(上段パネル及び中段パネルの中央列(CpGA))、その分子会合はゲルダナマイシンで抑制されることが示された(上段パネル及び中段パネルの右列(CpGA+geldanamycin))。
【実施例3】
【0051】
[ヒトの形質細胞様樹状細胞を用いたHsp90阻害アッセイ]
前述の実施例2では、マウスの形質細胞様樹状細胞を用いたアッセイを行った。そこで、ヒトの形質細胞様樹状細胞を用いた場合についても、Hsp90阻害物質による効果とその作用機序を検討するために、以下のようなアッセイを行った。
【0052】
(1)インターフェロンα測定アッセイ
まず、ヒト末梢血を用意した。このヒト末梢血から、pDC特異的な単クローン抗体BDCA−4を用いて、形質細胞様樹状細胞(pDC)を分離、精製し、精製pDCを得た。Hsp90阻害物質であるゲルダナマイシンを各種濃度(200ng/ml、400ng/ml、600ng/ml、800ng/ml、1000ng/ml)で含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加し、培養を継続した。24時間後に、培養液を採取し、これらの培養液中に含まれるインターフェロンα濃度をELISA法にて測定した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したもの(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したもの(CpG-A 3μM)についても、同様に、インターフェロンα濃度を測定した。これらの結果を図8に示す。
【0053】
図8の結果から分かるように、CpG−Aの添加によって誘導されたインターフェロンα産生は、ヒトpDCにおいても、Hsp90阻害物質濃度依存性に抑制された。なお、Hsp90阻害物質として、ゲルダナマイシンに代えて、17−AAG、又は、17−DMAGを用いて同様のアッセイを行ったところ、17−AAGや17−DMAGを用いた場合も、ゲルダナマイシンを用いた場合と同様に、Hsp90阻害物質濃度依存性に、インターフェロンα産生が抑制されることが示された。
【0054】
(2)シグナル伝達分子に関するウエスタンブロッティング解析
前述の実施例3の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDCを分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpG-A 3μM)についても、同様に分離した。分離したこれらの各pDC(それぞれ約1×106cellずつ)からライセートを調製し、SDS−PAGEゲルにて電気泳動を行った。このSDS−PAGEゲル中のタンパク質をメンブレンに転写し、次いで、かかるメンブレンをブロッキングバッファーにてブロッキングした。その後、メンブレンに各種の1次抗体を反応させてから、メンブレンを洗浄した。1次抗体としては、308番目のスレオニンがリン酸化されたAktタンパク質に対する抗体、Aktタンパク質に対する抗体、又は、リン酸化されたmTORタンパク質に対する抗体をそれぞれ用いた。洗浄したメンブレンに、1次抗体を認識する標識2次抗体を反応させて、各種の1次抗体のシグナルを検出した。その結果を図9に示す。
【0055】
図9の結果から分かるように、左のレーン(control)や真ん中のレーン(CpG-A 3μM)などのコントロール群のpDCでは、308位におけるリン酸化Aktタンパク質に対するシグナル(Phospho-Akt(Thr308))や、リン酸化mTORタンパク質に対するシグナル(Phospho-mTOR)が認められたのに対して、ゲルダナマイシン処理したpDC(右のレーン:Geldanamycin 400ng/ml)では、308位におけるリン酸化Aktタンパク質に対するシグナル(Phospho-Akt(Thr308))や、リン酸化mTORタンパク質に対するシグナル(Phospho-mTOR)の低下が認められ、それらの分子のリン酸化が抑制されたことが示された。この結果は、マウスpDCを用いた前述の実施例2(2)のウエスタンブロッティング解析の結果と同様であった。
【0056】
(3)pDCにおけるTLR9等の蛍光アッセイ
前述の実施例2の(3)において、あらかじめゲルダナマイシン処理をしたマウスpDCでは、CpG−Aを添加培養しても、TLR9は小胞体にとどまっており、初期エンドソームへの移行が抑制することが示された。この現象が、ヒトpDCでも生じるかを確認するために、以下のような蛍光アッセイを行うこととした。
【0057】
前述の実施例3の(1)と同じ方法で、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加して15分間培養した後、培養液からpDCを分離した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したpDC(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したpDC(CpG-A 3μM)についても、同様に分離した。分離したこれらの各pDCにおいて蛍光色素ラベルしたTLR9、EEA−1(初期エンドソームのマーカー)を蛍光顕微鏡で観察した結果を図10の左パネルに示す。なお、TLR9(緑色蛍光)と、EEA−1(赤色蛍光)とが共局在すると、黄色の蛍光が確認される。また、蛍光画像中のpDCをランダムに100個選択し、そのうち、TLR9(緑色蛍光)と、EEA−1(赤色蛍光)とが共局在しているpDCを計測して、それらが共局在しているpDCの割合(%)を図10の右パネルに示す。
【0058】
図10の左パネル及び右パネルから分かるように、ヒトpDCでも、CpG−Aの添加培養により、TLR9の初期エンドソームへの移行が生じ(図10左パネルの中段列(CpG))、その移行はゲルダナマイシンにより抑制されることが示された。これらの結果から、ヒトpDCにおいても、Hsp90は、TLR9の小胞体から初期エンドソームへの移行の際の、分子シャペロンとして働いていると考えられた。
【0059】
[実施例1〜3の結果のまとめ]
前述の実施例2や3の結果から分かるように、Hsp90阻害物質の投与により、マウスpDCやヒトpDCからのインターフェロンα産生が抑制された。そして、これは、pDCにおけるTLR9の、小胞体から初期エンドソームへの移行が抑制され、かかるTLR9が自己DNAを認識することが抑制されることや、インターフェロンα産生に重要なAktタンパク質やmTORタンパク質を介するシグナル伝達が抑制されることによることが示された。また、前述の実施例1におけるSLEマウスモデルの結果と総合すると、Hsp90阻害物質によってSLEの病態が改善した機序の1つは、Hsp90阻害物質によりpDCからのインターフェロンα産生が抑制されたことによるものと考えられた。
【実施例4】
【0060】
[ヒトの末梢血単核球を用いたHsp90阻害アッセイ]
前述の実施例3では、ヒトの形質細胞様樹状細胞を用いたアッセイを行った。そこで、ヒトの未成熟形質細胞様樹状細胞を含む末梢血単核球(PBMC)についても、インターフェロンα測定アッセイを行った。
【0061】
(1)インターフェロンα測定アッセイ
まず、ヒト末梢血を用意した。このヒト末梢血から、pDC特異的な単クローン抗体BDCA−4を用いて、形質細胞様樹状細胞(pDC)を分離、精製し、精製pDCを得た。400ng/mlのゲルダナマイシンを含む培養液にて、あらかじめ、前述の精製pDCを37℃で3時間培養した。次いで、pDCを刺激するCpG−A(3μM)を培養液に添加し、培養を継続した。24時間後に、培養液を採取し、これらの培養液中に含まれるインターフェロンα濃度をELISA法にて測定した。また、コントロールとして、CpG−Aもゲルダナマイシンも添加せずに培養したもの(control)や、ゲルダナマイシンを添加せず、CpG−Aを添加して培養したもの(CpG-A)についても、同様に、インターフェロンα濃度を測定した。これらの結果を図11に示す。
【0062】
図11の結果から分かるように、CpG−Aの添加によって誘導されたインターフェロンα産生は、ヒトpDCを含むヒトPBMCにおいても、Hsp90阻害物質により抑制された。このように、Hsp90阻害物質はpDCを含む末梢血に作用して、CpG−Aによるインターフェロンα産生を抑制する効果を示すことが明らかとなった。
【産業上の利用可能性】
【0063】
本発明は、形質細胞様樹状細胞からのインターフェロンα産生阻害剤の分野や、形質細胞様樹状細胞におけるAktタンパク質又はmTORタンパク質のリン酸化抑制剤の分野や、全身性エリテマトーデスの予防・治療の分野に好適に例示することができる。
【特許請求の範囲】
【請求項1】
Hsp90阻害物質を含有することを特徴とする、形質細胞様樹状細胞からのインターフェロンα産生阻害剤。
【請求項2】
Hsp90阻害物質が、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されることを特徴とする請求項1に記載のインターフェロンα産生阻害剤。
【請求項3】
Hsp90阻害物質を含有することを特徴とする、形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤。
【請求項4】
Hsp90阻害物質が、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されることを特徴とする請求項3に記載の移行阻害剤。
【請求項5】
請求項1若しくは2に記載のインターフェロンα産生阻害剤、又は、請求項3若しくは4に記載の移行阻害剤を含有する全身性エリテマトーデスの予防・治療剤。
【請求項1】
Hsp90阻害物質を含有することを特徴とする、形質細胞様樹状細胞からのインターフェロンα産生阻害剤。
【請求項2】
Hsp90阻害物質が、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されることを特徴とする請求項1に記載のインターフェロンα産生阻害剤。
【請求項3】
Hsp90阻害物質を含有することを特徴とする、形質細胞様樹状細胞におけるTLR9の初期エンドソームへの移行阻害剤。
【請求項4】
Hsp90阻害物質が、ゲルダナマイシン、17−ジメチルアミノエチルアミノ−17−デメトキシゲルダナマイシン、17−アリルアミノ−デメトキシゲルダナマイシン、並びに、それらの塩、エステル体及びエーテル体からなる群から選択されることを特徴とする請求項3に記載の移行阻害剤。
【請求項5】
請求項1若しくは2に記載のインターフェロンα産生阻害剤、又は、請求項3若しくは4に記載の移行阻害剤を含有する全身性エリテマトーデスの予防・治療剤。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2012−72097(P2012−72097A)
【公開日】平成24年4月12日(2012.4.12)
【国際特許分類】
【出願番号】特願2010−219422(P2010−219422)
【出願日】平成22年9月29日(2010.9.29)
【出願人】(307014555)北海道公立大学法人 札幌医科大学 (31)
【Fターム(参考)】
【公開日】平成24年4月12日(2012.4.12)
【国際特許分類】
【出願日】平成22年9月29日(2010.9.29)
【出願人】(307014555)北海道公立大学法人 札幌医科大学 (31)
【Fターム(参考)】
[ Back to top ]