
インターフェロン−α発現調節剤
【課題】内在のI型インターフェロン(IFN)の産生を増強させ得る物質を同定し、該物質を利用したウイルス感染症や癌などの疾患の新規予防・治療手段を提供すること。内在のI型IFNの産生を増強させ得る物質を探索するためのスクリーニング系を提供すること。
【解決手段】IFN-α mRNAに対する内在性アンチセンスRNA(AS RNA)に相補的な配列、特にIFN-α mRNAのSLIまたはSLII領域内の塩基配列に相同な配列を含み、IFN-αの発現を増強させ得るセンスオリゴヌクレオチド。該センスオリゴヌクレオチドを含有してなるIFN-α発現増強剤、ウイルス感染症または癌の予防または治療剤。被験物質の存在下および非存在下で、IFN-α mRNAとそれに対するAS RNAとのハイブリダイゼーションを検出・比較することを特徴とする、IFN-α発現調節物質のスクリーニング方法。
【解決手段】IFN-α mRNAに対する内在性アンチセンスRNA(AS RNA)に相補的な配列、特にIFN-α mRNAのSLIまたはSLII領域内の塩基配列に相同な配列を含み、IFN-αの発現を増強させ得るセンスオリゴヌクレオチド。該センスオリゴヌクレオチドを含有してなるIFN-α発現増強剤、ウイルス感染症または癌の予防または治療剤。被験物質の存在下および非存在下で、IFN-α mRNAとそれに対するAS RNAとのハイブリダイゼーションを検出・比較することを特徴とする、IFN-α発現調節物質のスクリーニング方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、インターフェロン−αの内在性アンチセンスRNAに特異的なセンスオリゴヌクレオチド、並びにそのインターフェロン−α発現増強作用の利用に関する。
【背景技術】
【0002】
ウイルス感染に対する生体防御において、自然免疫は感染初期の速やかな防御機構として重要な役割を果たしている。I型インターフェロン(IFN)は自然免疫における抗ウイルス活性の中心的な役割を担うサイトカインであり、ウイルス感染によって一過的に分泌され、周囲の細胞に働きかけて抗ウイルス活性、細胞増殖抑制活性などの多様な生理活性を発揮する。I型IFNはIFN-α、-β、-ω、-ε、-κに分けられ、このうちIFN-αには20数種類のサブタイプが存在する。IFN-αおよび-βは、これらの生理活性に基づき、血漿由来の製剤および遺伝子工学的に製造された組換えタンパク製剤が、B型肝炎、C型肝炎等のウイルス感染症、あるいは腫瘍などの種々の疾患に対する治療(もしくは補助療法)に利用されている。
【0003】
しかし、IFNの長期投与は、インフルエンザ様症状(頭痛、発熱、関節痛、筋肉痛、食欲不振、全身倦怠感、悪心・嘔吐等)、自己免疫疾患、うつ症状(不眠や焦燥感等)、脱毛、甲状腺機能異常、痴呆等、多様な副作用を生じる場合がある。そのため、IFNを投与することなく、生体内のIFN産生を増強させることによって、ウイルス感染の防御や、癌等の改善・治療を図るための研究も進められている。しかし、IFN産生を増強する物質の多くはウイルスや病原菌などで、安全性に問題のあるものが多い。
【0004】
ウイルス感染に対するIFN-α、-βなどのI型IFNの発現応答を早期に高めることができれば、未知のウイルスを含めた種々のウイルス感染症の予防・治療に有効であると考えられる。しかしながら、ウイルス応答における内在I型IFNの発現の立ち上がりを早める作用を有する物質は、これまで報告されていない。最近、Toll様受容体(TLR)を介したウイルス感染の認識からキナーゼカスケードを介してI型IFNの発現誘導に至るシグナル伝達経路が解明されてきているが、IFN遺伝子の転写後調節については、未だ十分に理解されていない。
【0005】
アンチセンスRNAとは、mRNAに対して、相補的な塩基配列を持ったRNAであり、具体的には、センス遺伝子をコードするDNA鎖(すなわちmRNAの非鋳型鎖)を鋳型として合成されたRNAである。センス-アンチセンスRNA間には2本鎖形成能があり、例えば、2本鎖RNAはRNA干渉が働く際に必要であること、及びマイクロRNAと呼ばれる小さなRNAによるタンパク質の翻訳制御に2本鎖RNAが関与することなどが知られている。最近の網羅的なcDNA解析により、かなり多くのアンチセンスRNAが転写されていることが明らかとなった(非特許文献1、2)。例えば、マウスでは約2,500対(非特許文献3)、ヒトでは約2,600対(非特許文献4)のセンス-アンチセンスRNAペアが存在することが示唆されており、それらの中にはタンパク質をコードしない非翻訳性のアンチセンスRNAが多く含まれている。
【0006】
アンチセンスRNAの生理機能についてはほとんど解明されていないが、内皮型一酸化窒素合成酵素(eNOS)mRNAに相補的なアンチセンスRNAは、ヒストン脱アセチル化酵素阻害剤の存在下で増加してeNOS mRNA量を減少させることが報告されている(非特許文献5)。また、酵母においても、PHO84遺伝子のアンチセンスRNAはヒストン脱アセチル化を介してmRNAの発現を抑制することが知られている(非特許文献6)。
一方、本発明者らは、誘導型一酸化窒素合成酵素(iNOS)mRNAの3’-非翻訳領域(UTR)に相補的な内在性アンチセンスRNA(「AS RNA」ともいう)が存在し、このアンチセンスRNAがiNOS mRNAとハイブリダイズすることによって該mRNAが安定化されiNOSの産生量およびNOの合成量が増大すること、該AS RNAに相補的なセンスオリゴヌクレオチドは、AS RNAのiNOS mRNAへの結合を阻害することにより該mRNAを不安定化し、iNOSの産生およびiNOSによるNO合成を抑制し得ることを報告した(特許文献1および2、非特許文献7)。
【0007】
しかしながら、IFN-α遺伝子については、内在性AS RNAが存在するかどうかすら、これまで報告されていなかった。
【特許文献1】WO 2007/142303号パンフレット
【特許文献2】WO 2007/142304号パンフレット
【非特許文献1】Cheng et al., Science, 308: 1149-1154 (2005)
【非特許文献2】Katayama et al., Science, 309: 1564-1566 (2005)
【非特許文献3】Kiyosawa et al., Genome Res., 13: 1324-1334 (2003)
【非特許文献4】Yelin et al., Nat. Biotechnol., 21: 379-386 (2003)
【非特許文献5】Robb et al., J. Biol. Chem., 279: 37982-37996 (2004)
【非特許文献6】Camblong et al., Cell, 131: 706-717 (2007)
【非特許文献7】Matsui et al., Hepatology, 47: 686-697 (2008)
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明の目的は、内在のI型IFNの産生を増強させ得る物質を同定し、該物質を利用したウイルス感染症や癌、自己免疫疾患などの疾患の新規予防・治療手段を提供することである。特に、ウイルス感染に対するI型IFN応答の立ち上がりを早めることによる、ウイルス感染症の有効な予防・治療手段を提供することである。加えて、膠原病やクローン病のように、免疫システムのバランスの破綻から引き起こされるI型IFN産生調節異常が、疾患発症の引き金の一つになると考えられる自己免疫疾患についても治療手段を提供することである。また、本発明の別の目的は、内在のI型IFNの産生を増強させ得る物質を探索するためのスクリーニング系を提供することである。
【課題を解決するための手段】
【0009】
本発明者らは、IFN-α mRNAの3’-UTRに、iNOS mRNAと同様、mRNAの早い代謝回転に関与すると考えられているAU-rich element(ARE)モチーフが存在することに着目し、IFN-αの発現調節にも内在性AS RNAが関与しているのではないかと発想してRT-PCR解析を行った結果、IFN-α AS RNAの存在が確認された。次に、ウイルス感染後のIFN-α mRNAおよびAS RNAの発現変動を解析したところ、IFN-α AS RNAは構成的に発現しているが、ウイルス感染後速やかに発現が低下し、それに伴ってIFN-α mRNAの発現レベルが一過的に上昇することが明らかとなった。これらの結果はIFN-α AS RNAがmRNAの発現を抑制していることを示唆するものであり、iNOSにおけるAS RNAによる発現調節とは異なっていた。
【0010】
本発明者らは、IFN-α mRNAの核外輸送に重要な、2つのステムループ(SLIおよびSLII(SLIIは内部にBulged SLを含む))を有する二次構造を形成する領域(図4参照)を同定していたことから、IFN-α AS RNAは、3’-UTRのAREモチーフではなく、この二次構造を形成する領域をターゲットとしてIFN-α mRNAにハイブリダイズし、mRNAの核外輸送を阻害することにより、IFN-αタンパク質への翻訳を抑制しているのではないかと考えた。そこで、SLIIおよびBulged SL領域のループ部分の配列を有するセンスオリゴヌクレオチドを作製し、IFN-α AS RNAの発現阻害効果およびIFN-αの発現とウイルス増殖に及ぼす効果を調べたところ、これらのセンスオリゴヌクレオチドはIFN-α AS RNAの発現を効率よく阻害し、IFN-αの産生量を増大させ、ウイルス増殖を抑制した。
本発明者らは、これらの知見に基づいてさらに研究を重ねた結果、本発明を完成するに至った。
【0011】
即ち、本発明は以下の通りである。
〔1〕IFN-α mRNAに対する内在性アンチセンスRNAに相補的な配列を含み、IFN-αの発現を増強させ得るセンスオリゴヌクレオチド。
〔2〕該配列がIFN-α mRNAのSLIまたはSLII領域内の塩基配列に相同な配列である、上記〔1〕記載のセンスオリゴヌクレオチド。
〔3〕該配列がBulged SL領域内の塩基配列に相同な配列である、上記〔2〕記載のセンスオリゴヌクレオチド。
〔4〕上記〔1〕〜〔3〕のいずれかに記載のセンスオリゴヌクレオチドを含有してなるIFN-α発現増強剤。
〔5〕ウイルス感染症または癌の予防または治療用である、上記〔4〕記載の剤。
〔6〕被験物質の存在下および非存在下で、IFN-α mRNAとそれに対する内在性アンチセンスRNAとのハイブリダイゼーションを検出・比較することを特徴とする、IFN-α発現調節物質のスクリーニング方法。
〔7〕IFN-α mRNAとそれに対する内在性アンチセンスRNAとを発現する細胞に被験物質を接触させ、該細胞における該mRNA量および/またはIFN-αタンパク質量の変化を測定することを特徴とする、上記〔6〕記載の方法。
【発明の効果】
【0012】
本発明のセンスオリゴヌクレオチドは、IFN-α mRNAに対する内在性AS RNAとハイブリダイズしてそのmRNAに対する作用を阻害することにより、IFN-αの産生を増強することができる。特に、ウイルス感染初期における迅速なIFN-αの産生誘導を可能にする。さらに、IFN-α製剤と異なり、吸入等による患者自身による投与が可能なため、患者負担を軽減することもできる。
【発明を実施するための最良の形態】
【0013】
本発明は、IFN-αの発現増強活性を有するIFN-αのセンスオリゴヌクレオチドを提供する。本発明のセンスオリゴヌクレオチドは、IFN-α mRNAに対する内在性AS RNAに相補的な配列を含み、かつIFN-αの発現を増強させ得るオリゴヌクレオチドであれば、いかなるものであってもよい。ここで「相補的」な配列とは、AS RNAに対して完全相補的な配列のみならず、細胞の生理的な条件下でAS RNAとハイブリダイズしてmRNAへのAS RNAの作用を阻害し得る限り、1ないし数(2, 3, 4 もしくは 5)塩基のミスマッチを含んでもよい。好ましくは、IFN-α AS RNAに相補的な配列とは、ストリンジェントな条件、例えば、Current Protocols in Molecular Biology, John Wiley & Sons,6.3.1-6.3.6, 1999に記載される条件(例えば、6×SSC(sodium chloride/sodium citrate)/45℃でのハイブリダイゼーション、次いで0.2×SSC/0.1% SDS/50〜65℃での一回以上の洗浄等が挙げられる)下で、該AS RNAとハイブリダイズし得る配列である。
具体的には、IFN-α mRNAに対する内在性AS RNAは、該mRNAの熱力学的に安定でない部分と相互作用して該mRNAを不安定化するので、本発明のセンスオリゴヌクレオチドは、IFN-α mRNA中の熱力学的に安定でない部分と相同な配列を含むことが好ましい。熱力学的に安定でない部分としては、mRNAが二次構造をとった際に一本鎖の状態にある(例えば、ステムループ構造のループ部分にあたる)領域が挙げられる。IFN-α mRNAの二次構造は、該mRNAの塩基配列情報をもとに、mfold(GCG Software; Proc. Natl. Acad. Sci. USA, 86: 7706-10 (1989) 参照)に代表されるような既存のRNA二次構造予測プログラムを用いて予測することができる。IFN-α mRNAには20数種類のサブタイプが存在するが(例えば、Science, 209: 1343-1347 (1980)、Gene, 11: 181-186 (1980)、Nature, 290: 20-26 (1981)、Nature, 313: 698-700 (1985)、J. Invest. Dermatol., 83: 128s-136s (1984)、J. Interferon Res., 2: 575-585 (1982)、J. Interferon Res., 13: 227-231 (1993)、J. Biol. Chem., 268: 12565-12569 (1993)、Acta Virol., 38: 101-104 (1994)、Biochim. Biophys. Acta, 1264: 363-368 (1995) 参照)、それらの塩基配列情報はいずれも容易に入手可能である。例えば、ヒトIFN-α1 mRNA(cDNA)の塩基配列は、DDBJ/EMBL/GenBank ヌクレオチドデータベースにAccession No. AB445100として登録されている。当該塩基配列を配列番号1に、当該塩基配列より推定されるヒトIFN-α1のアミノ酸配列を配列番号2に、それぞれ示す。
【0014】
さらに、本発明のセンスオリゴヌクレオチドは、配列非特異的な反応を起こす配列(例えば、5’-CG-3’、5’-GGGG-3’、5’-GGGGG-3’等)を含まないものから選択することが好ましく、また、IFN-α mRNA以外のRNA中に類似の配列が存在しないものから選択することが好ましい。他のRNA中に類似の配列が存在しないことは、センスオリゴヌクレオチドの候補配列をクエリーとして、対象とする哺乳動物のゲノム配列に対して相同性検索をかけることにより確認することができる。ここで相同性検索手段としては、公知の核酸の相同性検索ソフトウェア(例えば、NCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)NBLASTおよびXBLASTプログラム(version 2.0)、GCGソフトウェアパッケージ中のFASTAプログラム等)を用いることができる。また、ゲノムDNAデータセットとしては、例えば、Celera社が提供する全ヒトゲノムデータを用いることができる。
【0015】
好ましくは、本発明のセンスオリゴヌクレオチドは、IFN-α mRNAのSLI(配列番号1に示されるヒトIFN-α1 mRNA塩基配列にあっては、塩基番号229〜305で示される塩基配列)またはSLII領域(配列番号1に示されるヒトIFN-α1 mRNA塩基配列にあっては、塩基番号308〜434で示される塩基配列)内の塩基配列に相同な配列を含む。ヒトIFN-α1 mRNAの種々の欠失変異体を用いた実験から、該mRNAの核外輸送には、配列番号1に示される塩基配列中塩基番号208〜452で示される領域が形成する2つのステムループ(SLIおよびSLII)からなる二次構造を、核外輸送因子が認識することが必要であることが明らかとなった。IFN-α mRNAに対する内在性AS RNAは、この核外輸送責任領域(CSSともいう)をターゲットとして該mRNAと相互作用することにより、該mRNAの核外への輸送およびその後の翻訳を阻害するとともに、RNaseによるmRNAの分解を促進すると考えられるので、CSS中の配列のうちで上記の条件に適合する領域、即ち、CSSが二次構造をとった際にSLIまたはSLII領域内でループ構造を形成する部分の塩基配列に相同な配列を含むオリゴヌクレオチド(図4および5参照)は、IFN-α AS RNAにハイブリダイズすることで該AS RNAのIFN-α mRNAへの作用を阻害し、該mRNAレベルを増大させ、該mRNAの核外輸送の抑制を解除することにより、IFN-αタンパク質の産生を増強することができる。特に、SLII領域内のBulged SL(配列番号1に示されるヒトIFN-α1 mRNA塩基配列にあっては、塩基番号322〜352で示される塩基配列)がIFN-α mRNAの発現を左右することから、Bulged SL領域内の塩基配列に相同な配列を含むセンスオリゴヌクレオチドがより好ましい。
尚、「相同な」配列とは、IFN-α mRNAの特定の部分塩基配列と完全に同一な配列のみならず、該mRNAに対する内在性AS RNAと細胞の生理的な条件下でハイブリダイズしてmRNAに対するAS RNAの作用を阻害し得る限り、1ないし数(2, 3, 4もしくは 5)個の塩基が異なっていてもよい。
【0016】
より具体的なセンスオリゴヌクレオチドの例として、例えば、以下のものが挙げられる(括弧内の数字は配列番号1に示される塩基配列中の対応する領域を示す)。
配列番号3:5’-CCAGCAGATCTTCAACCTCT-3’(塩基番号319〜338)
配列番号4:5’-ATCTTCAACCTCTTTACCAC-3’(塩基番号326〜345)
配列番号5:5’-GATGAGGACCTCCTAGACAA-3’(塩基番号368〜387)
配列番号6:5’-GACCTCCTAGACAAATTCTG-3’(塩基番号374〜393)
【0017】
本発明のセンスオリゴヌクレオチドの長さに特に制限はないが、配列特異性の面から、標的配列に相補的な部分を少なくとも10塩基以上、好ましくは約12塩基以上、より好ましくは約15塩基以上含むものである。また、合成の容易さ、製造コスト、投与のし易さ等の面から、50塩基以下、好ましくは40塩基以下、より好ましくは30塩基以下の塩基長を有するものが挙げられる。
【0018】
本発明のセンスオリゴヌクレオチドは、一本鎖DNA、一本鎖RNA、DNA/RNAキメラのいずれであってもよく、さらに公知の修飾の付加されたものであってもよい。ここで「ヌクレオチド」とは、プリンおよびピリミジン塩基を含有するのみでなく、修飾されたその他の複素環型塩基をもつようなものを含んでいてもよい。センスオリゴヌクレオチドがDNA(ODN)の場合、標的AS RNAとセンスODNとによって形成されるRNA:DNAハイブリッドは、内在性RNase Hに認識されて標的AS RNAの選択的な分解を引き起こすことができる。
【0019】
センスオリゴヌクレオチドを構成するヌクレオチド分子は、天然型のDNAもしくはRNAでもよいが、安定性(化学的および/または対酵素)や比活性(RNAとの親和性)を向上させるために、種々の化学修飾を含むことができる。例えば、ヌクレアーゼなどによる分解を防ぐために、センスオリゴヌクレオチドを構成する各ヌクレオチドのリン酸残基(ホスフェート)を、例えば、ホスホロチオエート(PS)、メチルホスホネート、ホスホロジチオネートなどの化学修飾リン酸残基に置換することができる。また、各ヌクレオチドの糖(リボース)の2'位の水酸基を、-OR(Rは、例えばCH3(2'-O-Me)、CH2CH2OCH3(2'-O-MOE)、CH2CH2NHC(NH)NH2、CH2CONHCH3、CH2CH2CN等を示す)に置換してもよい。さらに、塩基部分(ピリミジン、プリン)に化学修飾を施してもよく、例えば、ピリミジン塩基の5位へのメチル基やカチオン性官能基の導入、あるいは2位のカルボニル基のチオカルボニルへの置換などが挙げられる。
RNAの糖部のコンフォーメーションはC2'-endo(S型)とC3'-endo(N型)の2つが支配的であり、一本鎖RNAではこの両者の平衡として存在するが、二本鎖を形成するとN型に固定される。したがって、標的AS RNAに対して強い結合能を付与するために、2'酸素と4’炭素を架橋することにより、糖部のコンフォーメーションをN型に固定したRNA誘導体であるBNA (LNA)(Imanishi, T. et al., Chem. Commun., 1653-9, 2002; Jepsen, J.S. et al., Oligonucleotides, 14, 130-46, 2004)やENA(Morita, K. et al., Nucleosides Nucleotides Nucleic Acids, 22, 1619-21, 2003)もまた、好ましく用いられ得る。
【0020】
本発明のセンスオリゴヌクレオチドは、IFN-αのmRNA(cDNA)配列に基づいて、市販のDNA/RNA自動合成機(アプライド・バイオシステムズ社、ベックマン社等)を用いて、これに相同な配列を合成することにより調製することができる。
【0021】
本発明のセンスオリゴヌクレオチドは、リポソーム、ミクロスフェアのような特殊な形態で供与されたり、ポリリジンのようなポリカチオン体、脂質(例、ホスホリピド、コレステロールなど)などの疎水性物質が付加された形態で提供され得る。あるいはまた、本発明のセンスオリゴヌクレオチドを、膜透過機能を有するペプチド(例えば、ショウジョウバエ由来のAntennapediaホメオドメイン(AntP)、ヒト免疫不全ウイルス(HIV)由来のTAT、単純ヘルペスウイルス(HSV)由来のVP22等の細胞通過ドメイン)などで修飾することにより、該オリゴヌクレオチドの細胞への取り込みを促進することができる。
【0022】
本発明のセンスオリゴヌクレオチドは、IFN-α mRNAに対する内在性AS RNAの作用を阻害することにより、IFN-αmRNAおよびIFN-αタンパク質の発現を増強することができる。IFN-αは、抗ウイルス作用、細胞増殖抑制作用、ナチュラルキラー細胞の活性化作用などの多様な生理活性を有するので、本発明のセンスオリゴヌクレオチドを含有する医薬は、IFN-αの発現増強剤として、種々の疾患の予防および/または治療に利用することができる。そのような疾患としては、例えば、B型肝炎、C型肝炎(活動性、非活動性)、ヘルペス感染症(性器ヘルペス、角膜ヘルペス炎、口腔ヘルペスなど)、尖圭コンジローマ、AIDS等のウイルス感染症、腎臓癌、腎細胞癌、乳癌、膀胱癌、基底細胞癌、頭頸部癌、頸管異形成、皮膚悪性腫瘍、カポジ肉腫、悪性黒色腫、非ホジキン型リンパ腫、幼児血管腫、慢性肉芽腫症、慢性骨髄性白血病(CML)、成人T細胞白血病、ヘアリー細胞白血病、毛様細胞白血病、T細胞白血病ウイルス(HTLV−1)骨髄症、多発性骨髄腫、リンパ腫などの癌、亜急性硬化性全脳炎(SSPE)、クローン病、関節リウマチ、シェーグレン症候群、多発性硬化症(MS)、口内炎、性器疣贅、膣内疣贅、赤血球増加症、血小板増加症、乾癬、菌状息肉症、突発性難聴、老年性円盤状黄斑変性症、ページェット病などが挙げられるが、これらに限定されない。
IFN-α AS RNAは通常、白血球あるいはリンパ芽球様細胞などのIFN-α産生細胞において構成的に発現し、IFN-αの発現を抑制しているが、ウイルス感染に応答してその発現が抑制されることによってIFN-αの発現抑制が解除されてIFN-αが産生・分泌され、自然免疫が誘導される。したがって、ウイルス感染初期に際して、本発明のセンスオリゴヌクレオチドを含有する医薬は、IFN-α AS RNAの発現を速やかに抑制することにより、IFN-α産生の立ち上がりを早め、より迅速に自然免疫系を誘導することができる。
【0023】
本発明のセンスオリゴヌクレオチドを含有する医薬は低毒性であり、そのまま液剤として、または適当な剤型の医薬組成物として、ヒトまたは非ヒト哺乳動物(例、マウス、ラット、ウサギ、ヒツジ、ブタ、ウシ、ネコ、イヌ、サルなど)に対して経口的または非経口的(例、吸入投与、血管内投与、皮下投与、経粘膜投与など)に投与することができる。
これらの核酸を上記のウイルス感染症や癌などの予防・治療剤として使用する場合、自体公知の方法に従って製剤化し、投与することができる。即ち、本発明のセンスオリゴヌクレオチドを、単独で、あるいはレトロウイルスベクター、レンチウイルスベクター、アデノウイルスベクター、アデノ随伴ウイルスベクターなどの適当な哺乳動物細胞用の発現ベクターに機能可能な態様で挿入した後、常套手段に従って製剤化することができる。該オリゴヌクレオチドは、そのままで、あるいは摂取促進のための補助剤とともに、遺伝子銃やハイドロゲルカテーテルのようなカテーテルによって投与することもできる。あるいは、エアロゾル化して吸入剤として気管内に局所投与することもできる。
さらに、体内動態の改良、半減期の長期化、細胞内取り込み効率の改善を目的に、前記オリゴヌクレオチドを単独またはリポソームなどの担体とともに製剤(注射剤)化し、静脈、皮下等に投与してもよい。
【0024】
本発明のセンスオリゴヌクレオチドは、それ自体を投与してもよいし、または適当な医薬組成物として投与してもよい。投与に用いられる医薬組成物としては、本発明のセンスオリゴヌクレオチドと薬理学的に許容され得る担体、希釈剤もしくは賦形剤とを含むものであってよい。このような医薬組成物は、経口または非経口投与に適する剤形として提供される。
【0025】
非経口投与のための組成物としては、例えば、注射剤、エアロゾル剤、坐剤等が用いられ、注射剤は静脈注射剤、皮下注射剤、皮内注射剤、筋肉注射剤、点滴注射剤等の剤形を包含してもよい。このような注射剤は、公知の方法に従って調製できる。エアロゾル製剤はジクロロジフロロメタン、プロパン、窒素などのような圧縮された許容しうる抛射薬内に入れることができる。あるいはネブライザーやアトマイザーのような非圧縮性製剤用医薬品として製剤化してもよい。直腸投与に用いられる坐剤は、上記核酸を通常の坐薬用基剤に混合することによって調製されてもよい。
【0026】
経口投与のための組成物としては、固体または液体の剤形、具体的には錠剤(糖衣錠、フィルムコーティング錠を含む)、丸剤、顆粒剤、散剤、カプセル剤(ソフトカプセル剤を含む)、シロップ剤、乳剤、懸濁剤等が挙げられる。このような組成物は公知の方法によって製造され、製剤分野において通常用いられる担体、希釈剤もしくは賦形剤を含有していても良い。錠剤用の担体、賦形剤としては、例えば、乳糖、でんぷん、蔗糖、ステアリン酸マグネシウムが用いられる。
【0027】
上記の非経口用または経口用医薬組成物は、活性成分の投与量に適合するような投薬単位の剤形に調製されることが好都合である。このような投薬単位の剤形としては、例えば、錠剤、丸剤、カプセル剤、注射剤(アンプル)、エアロゾル剤、坐剤が挙げられる。本発明のセンスオリゴヌクレオチドは、例えば、投薬単位剤形当たり通常5〜500mg、とりわけ注射剤では5〜100mg、その他の剤形では10〜250mg含有されていることが好ましい。
【0028】
本発明のセンスオリゴヌクレオチドを含有する上記医薬の投与量は、投与対象、対象疾患、症状、投与ルートなどによっても異なるが、例えば、成人の急性C型肝炎の予防および/または治療に使用する場合には、本発明のセンスオリゴヌクレオチドを1回量として、通常0.01〜20mg/kg体重程度、好ましくは0.1〜10mg/kg体重程度、さらに好ましくは0.1〜5mg/kg体重程度を、1日1回〜数回程度、静脈注射や吸入等により投与するのが好都合である。他の非経口投与および経口投与の場合もこれに準ずる量を投与することができる。症状が特に重い場合には、その症状に応じて増量してもよい。
【0029】
なお前記した医薬組成物は、本発明のセンスオリゴヌクレオチドとの配合により好ましくない相互作用を生じない限り、他の薬剤を含有してもよい。他の薬剤としては、例えば、抗ウイルス薬、抗腫瘍薬、抗菌薬、抗真菌薬、抗原虫薬、抗生物質、抗セプシス薬、抗セプティックショック薬、エンドトキシン拮抗薬、免疫調節薬、非ステロイド性抗炎症薬、ステロイド薬、炎症性メディエーター作用抑制薬、炎症性メディエーター産生抑制薬、抗炎症性メディエーター作用抑制薬、抗炎症性メディエーター産生抑制薬などが挙げられる。
【0030】
本発明はまた、IFN-α mRNAに対する内在性AS RNAの抑制作用を調節することによりIFN-αの発現を調節する物質をスクリーニングする方法を提供する。本発明のスクリーニング方法は、被験物質の存在下および非存在下で、IFN-α mRNAとそれに対する内在性AS RNAとのハイブリダイゼーションを検出し、その程度を比較することを特徴とする。
例えば、IFN-α mRNAとIFN-α AS RNAとを常法により単離し、いずれか一方を固相化し、他方を適当な標識剤で標識して、RNAが生理的な二次構造を形成し得る条件下で、被験物質の存在下および非存在下に両者をハイブリダイズさせ、固相に結合した標識量を両条件下で比較する方法が挙げられる。ここでmRNAおよびAS RNAとしては、それぞれその全長を用いてもよいし、あるいはmRNAの核外輸送責任領域(CSS;配列番号1に示されるヒトIFN-α1 mRNAの塩基配列にあっては、塩基番号208〜452で示される領域)、SLII領域(配列番号1に示されるヒトIFN-α1 mRNAの塩基配列にあっては、塩基番号308〜434で示される領域)あるいはSLI領域(配列番号1に示されるヒトIFN-α1 mRNAの塩基配列にあっては、塩基番号229〜305で示される領域)、並びにAS RNAのそれらの領域に相補的な配列を含むそれらのフラグメントを用いてもよい。
固相の材料としては、シリコンなどの半導体、ガラス、ダイアモンドなどの無機物、ポリエチレンテレフタレート、ポリプロピレン等の高分子物質を主成分とするフィルムなどが挙げられ、また固相の形状としては、スライドガラス、マイクロウェルプレート、マイクロビーズ、繊維型などが挙げられるが、それらに制限されない。固相上にmRNAもしくはAS RNAを固定化する方法としては、予め該RNAにアミノ基、アルデヒド基、SH基、ビオチンなどの官能基を導入しておき、一方、固相上にも該RNAと反応し得る官能基(例:アルデヒド基、アミノ基、SH基、ストレプトアビジンなど)を導入し、両官能基間の共有結合で固相とRNAを架橋したり、ポリアニオン性のRNAに対して、固相をポリカチオンコーティングして静電結合を利用してRNAを固定化するなどの方法が挙げられるが、これらに限定されない。固相化RNAの調製法としては、フォトリソグラフィー法を用いてRNAを基板(ガラス、シリコンなど)上で1ヌクレオチドずつ合成するAffymetrix方式と、マイクロスポッティング法、インクジェット法、バブルジェット(登録商標)法などを用いて、予め調製されたRNAを基板上にスポッティングするStanford方式とが挙げられるが、使用するRNAの塩基長を考慮すれば、Stanford方式あるいは両者を組み合わせた手法を用いるのが好ましい。
標識剤としては、例えば、放射性同位元素、酵素、蛍光物質、発光物質などが用いられる。放射性同位元素としては、例えば、〔32P〕、〔3H〕、〔14C〕などが用いられる。酵素としては、安定で比活性の大きなものが好ましく、例えば、β−ガラクトシダーゼ、β-グルコシダーゼ、アルカリフォスファターゼ、パーオキシダーゼ、リンゴ酸脱水素酵素などが用いられる。蛍光物質としては、例えば、フルオレスカミン、フルオレッセンイソチオシアネート、Cy3、Cy5などが用いられる。発光物質としては、例えば、ルミノール、ルミノール誘導体、ルシフェリン、ルシゲニンなどが用いられる。さらに、プローブと標識剤との結合にビオチン-(ストレプト)アビジンを用いることもできる。
被験物質としては、いかなる公知物質および新規物質であってもよく、例えば、核酸、糖質、脂質、蛋白質、ペプチド、有機低分子化合物、コンビナトリアルケミストリー技術を用いて作製された化合物ライブラリー、固相合成やファージディスプレイ法により作製されたランダムペプチドライブラリー、あるいは微生物、動植物、海洋生物等由来の天然成分などがあげられる。添加される被験物質の濃度は化合物の種類(溶解度、毒性等)により異なるが、例えば、約0.1nM〜約100nMの範囲で適宜選択される。インキュベート時間としては、例えば、約1〜約24時間が挙げられる。
固相上のRNAと、標識したRNA(および被験物質)とを接触させて、インキュベートした後、固相に結合しなかったRNAを洗い流し、固相に結合したRNAの標識量を検出する。被験物質の存在下で、非存在下に比べて固相に結合した標識量が有意に減少した場合、該被験物質をIFN-α発現増強物質の候補として選択することができる。
【0031】
好ましい実施態様においては、IFN-α mRNAとIFN-α AS RNAとを発現する細胞に被験物質を接触させ、該細胞における該mRNA量および/またはIFN-αタンパク質量の変化を測定することにより、より直接的にIFN-α発現調節物質を選択することができる。
IFN-α mRNAとIFN-α AS RNAとを発現する細胞は、両RNAを生来発現し得る細胞(例えば、白血球、リンパ芽球細胞等)であってもよいし、それらのいずれか一方もしくは両方を発現するDNAを導入した組換え細胞、あるいはウイルスを感染させた細胞であってもよい。組換え細胞の場合、宿主細胞として、例えば、H4IIE-C3細胞、HepG2細胞、293T細胞、HEK293細胞、COS7細胞、2B4T細胞、CHO細胞、MCF-7細胞、H295R細胞などの動物細胞をあげることができる。IFN-α mRNAおよびIFN-α AS RNAをコードするDNAは、両RNAを常法により単離し、逆転写反応等によって二本鎖DNAに変換した後、宿主細胞内で機能しうるプロモーターを有する発現ベクターに挿入して、例えば、リン酸カルシウム共沈殿法、PEG法、エレクトロポレーション法、マイクロインジェクション法、リポフェクション法などにより、このベクターを宿主細胞に導入することにより作製することができる。
【0032】
被験物質としては、前記したとおりのものが用いられる。被験物質と上記細胞との接触は、例えば、該細胞の培養に適した培地(例えば、約5〜20%の胎仔ウシ血清を含む最小必須培地(MEM)、ダルベッコ改変イーグル培地(DMEM)、RPMI1640培地、199培地、F12培地など)や各種緩衝液(例えば、HEPES緩衝液、リン酸緩衝液、リン酸緩衝生理食塩水、トリス塩酸緩衝液、ホウ酸緩衝液、酢酸緩衝液など)の中に被験物質を添加して、細胞を一定時間インキュベートすることにより実施することができる。添加される被験物質の濃度は化合物の種類(溶解度、毒性等)により異なるが、例えば、約0.1nM〜約100nMの範囲で適宜選択される。インキュベート時間としては、例えば、約1〜約24時間が挙げられる。
【0033】
インキュベーション終了後(あるいは経時的に)、細胞からRNAを抽出してRT-PCR、リアルタイムPCRやノーザンブロット解析によりIFN-α mRNA量を測定するか、あるいは培養上清を回収して、自体公知の各種イムノアッセイやウェスタンブロッティング等によりIFN-αタンパク質量を測定する。被験物質の添加により細胞におけるIFN-α mRNA量および/またはIFN-αタンパク質量が有意に増加した場合、該被験物質をIFN-α発現増強物質の候補として選択することができる。
【0034】
あるいはまた、ウイルス感染によりIFN-α mRNAとIFN-α AS RNAとを発現する細胞において、被験物質の存在下および非存在下で、該細胞におけるIFN-αの産生および/またはウイルス力価の変動を指標として、IFN-α発現調節物質をスクリーニングすることもできる。被験物質の存在下で、非存在下に比べてIFN-αの産生量が有意に増加した場合および/またはウイルス増殖が有意に抑制された場合、該被験物質をIFN-α発現増強物質の候補として選択することができる。
【0035】
上記のスクリーニング法により選択されたIFN-α発現増強物質は、本発明のセンスオリゴヌクレオチドと同様に、IFN-α mRNAに対する内在性AS RNAの作用を阻害することにより、IFN-α mRNAおよびIFN-αタンパク質の発現を増強することができる。したがって、上記IFN-α発現増強物質を含有する医薬は、例えば、B型肝炎、C型肝炎(活動性、非活動性)、ヘルペス感染症(性器ヘルペス、角膜ヘルペス炎、口腔ヘルペスなど)、尖圭コンジローマ、AIDS等のウイルス感染症、腎臓癌、腎細胞癌、乳癌、膀胱癌、基底細胞癌、頭頸部癌、頸管異形成、皮膚悪性腫瘍、カポジ肉腫、悪性黒色腫、非ホジキン型リンパ腫、幼児血管腫、慢性肉芽腫症、慢性骨髄性白血病(CML)、成人T細胞白血病、ヘアリー細胞白血病、毛様細胞白血病、T細胞白血病ウイルス(HTLV−1)骨髄症、多発性骨髄腫、リンパ腫などの癌、亜急性硬化性全脳炎(SSPE)、クローン病、関節リウマチ、シェーグレン症候群、多発性硬化症(MS)、口内炎、性器疣贅、膣内疣贅、赤血球増加症、血小板増加症、乾癬、菌状息肉症、突発性難聴、老年性円盤状黄斑変性症、ページェット病などの予防および/または治療に有用である。
【0036】
上記のスクリーニング法により選択された物質を含有する医薬は低毒性であり、そのまま液剤として、または適当な剤型の医薬組成物として、ヒトまたは非ヒト哺乳動物(例、マウス、ラット、ウサギ、ヒツジ、ブタ、ウシ、ネコ、イヌ、サルなど)に対して経口的または非経口的(例、吸入投与、血管内投与、皮下投与など)に投与することができる。投与に用いられる医薬組成物としては、選択された物質と薬理学的に許容され得る担体、希釈剤もしくは賦形剤とを含むものであってよい。
【0037】
非経口投与のための組成物としては、例えば、注射剤、エアロゾル剤、坐剤等が用いられ、注射剤は静脈注射剤、皮下注射剤、皮内注射剤、筋肉注射剤、点滴注射剤等の剤形を包含してもよい。このような注射剤は、公知の方法に従って調製できる。注射剤の調製方法としては、例えば、選択されたIFN-α発現増強物質を通常注射剤に用いられる無菌の水性液、または油性液に溶解、懸濁または乳化することによって調製できる。注射用の水性液としては、例えば、生理食塩水、ブドウ糖やその他の補助薬を含む等張液等が用いられ、適当な溶解補助剤、例えば、アルコール(例、エタノール)、ポリアルコール(例、プロピレングリコール、ポリエチレングリコール)、非イオン界面活性剤〔例、ポリソルベート80、HCO−50(polyoxyethylene(50mol)adduct of hydrogenated castor oil)〕等と併用してもよい。油性液としては、例えば、ゴマ油、大豆油等が用いられ、溶解補助剤として安息香酸ベンジル、ベンジルアルコール等を併用してもよい。調製された注射液は、適当なアンプルに充填されることが好ましい。エアロゾル製剤はジクロロジフロロメタン、プロパン、窒素などのような圧縮された許容しうる抛射薬内に入れることができる。あるいはネブライザーやアトマイザーのような非圧縮性製剤用医薬品して製剤化してもよい。直腸投与に用いられる坐剤は、上記IFN-α発現増強物質を通常の坐薬用基剤に混合することによって調製されても良い。
【0038】
経口投与のための組成物としては、固体または液体の剤形、具体的には錠剤(糖衣錠、フィルムコーティング錠を含む)、丸剤、顆粒剤、散剤、カプセル剤(ソフトカプセル剤を含む)、シロップ剤、乳剤、懸濁剤等が挙げられる。このような組成物は公知の方法によって製造され、製剤分野において通常用いられる担体、希釈剤もしくは賦形剤を含有していても良い。錠剤用の担体、賦形剤としては、例えば、乳糖、でんぷん、蔗糖、ステアリン酸マグネシウムが用いられる。
【0039】
上記の非経口用または経口用医薬組成物は、活性成分の投与量に適合するような投薬単位の剤形に調製されることが好都合である。このような投薬単位の剤形としては、例えば、錠剤、丸剤、カプセル剤、注射剤(アンプル)、エアロゾル剤、坐剤が挙げられる。IFN-α発現増強物質は、投薬単位剤形当たり通常5〜500mg、とりわけ注射剤では5〜100mg、その他の剤形では10〜250mg含有されていることが好ましい。
【0040】
上記のIFN-α発現増強物質を含有する上記医薬の投与量は、投与対象、対象疾患、症状、投与ルートなどによっても異なるが、例えば、成人の急性C型肝炎の予防および/または治療に使用する場合には、IFN-α発現増強物質を1回量として、通常0.01〜20mg/kg体重程度、好ましくは0.1〜10mg/kg体重程度、さらに好ましくは0.1〜5mg/kg体重程度を、1日1回〜数回程度、静脈注射により投与するのが好都合である。他の非経口投与および経口投与の場合もこれに準ずる量を投与することができる。症状が特に重い場合には、その症状に応じて増量してもよい。
【0041】
以下に実施例を挙げて本発明をさらに具体的に説明するが、これらは単なる例示であって本発明の範囲を何ら限定するものではない。
【実施例】
【0042】
材料と方法
1. 細胞培養
ヒトBリンパ芽球細胞株のNamalwa細胞 (ATCC CRL-1432)を、ウシ胎児血清 (FCS; GIBCO)を10%含むRPMI1640培地 (Wako) (R10)を用い、37℃, 5% CO2条件下で培養した。
【0043】
2. 細胞感染に用いるウイルスの用意と力価の測定
孵化鶏卵(10-12日齢)の漿尿膜腔に10,000倍希釈したSendai virus (SenV) (原液力価 5,120 HA unit/ ml)を200μl接種し、35℃で72時間感染後、4℃に一夜冷蔵した。この感染孵化鶏卵から回収した漿尿液中のSenVの力価を、初日ヒナ赤血球を用いた赤血球凝集試験により測定した。
【0044】
3. DNAトランスフェクション/ AS RNA knock-down (KD)とKD細胞
実験前日に培養Namalwa細胞を、6-ウェルプレート(Nunc)に3 x 106 cells/ 2 ml/ wellとなるように播種し、37℃, 5% CO2条件下で24時間培養した。その後、室温で150 x g, 4分間遠心沈澱させ、7ウェル分相当のNamalwa細胞を回収した。この細胞沈渣を7mlのR10に浮遊後、210μlのMATra-S Immobilizer (IBA; 7-2002-100)を添加し、5分毎に撹拌しながら、室温で15分間反応させた。その後、6-ウェルプレートを2枚使い、R10培地を予め1 ml宛分注した7ウェルに対しImmobilizerを吸着させたNamalwa細胞浮遊液各1mlを分注し、専用のマグネットプレート (IBA 7-2004-000)上に室温で、20分間留置した。この間に、RPMI1640 200μlあたり、センスオリゴヌクレオチド(hIFN-α/Se1-4)または陰性対照センスオリゴヌクレオチド (hIFN-α/NSe0-3, hIFN-α/ScrblSe1-2) 3μgとMATra-A Reagent (IBA 7-2001-100) 3μlを加え、混和後、室温で15分間留置し、トランスフェクション ミックスを作製した。所定の時間が経過したマグネットプレート上のImmobilizer吸着のNamalwa細胞に対し、このトランスフェクション ミックスを加え、10分間静置したのち、CO2インキュベーターに移し、37℃で6時間培養した。
センスオリゴヌクレオチド(*はS化したヌクレオチドを示す)
hIFN-α/Se1: 5’-C*C*A*GCAGATCTTCAACC*T*C*T-3’(配列番号3)
hIFN-α/Se2: 5’-A*T*C*TTCAACCTCTTTAC*C*A*C-3’(配列番号4)
hIFN-α/Se3: 5’-G*A*T*GAGGACCTCCTAGA*C*A*A-3’(配列番号5)
hIFN-α/Se4: 5’-G*A*C*CTCCTAGACAAATT*C*T*G-3’(配列番号6)
陰性対照センスオリゴヌクレオチド
hIFN-α/NSe0: 5’-G*A*C*CTTGATGCTCCTGG*C*A*C-3’(配列番号7)
hIFN-α/NSe1: 5’-A*A*T*CTGTCCTTCCTCCT*G*T*C-3’(配列番号8)
hIFN-α/NSe2: 5’-C*C*A*GGAGGAGTTTGATG*G*C*A-3’(配列番号9)
hIFN-α/NSe3: 5’-G*C*A*GAAATCATGAGATC*C*C*T-3’(配列番号10)
hIFN-α/ScrblSe1: 5’-G*C*T*CCCTAGCCCAACTA*T*T*A-3’(配列番号11)
hIFN-α/ScrblSe2: 5’-G*T*C*CCTACAATACCCAC*T*G*T-3’(配列番号12)
【0045】
4. Namalwa細胞へのSenV感染
上述のセンスオリゴヌクレオチドのトランスフェクションによりAS RNA発現をノックダウンしたNamalwa細胞 (AS RNA-minus) (6-ウェルプレートで、1ウェルあたり3x106 cells)あるいは、同一細胞数でセンスオリゴヌクレオチドをトランスフェクションしていないNamalwa細胞 (AS RNA-plus)をそれぞれ、室温で150 x g, 4分間、2回遠心・洗浄し、最終沈渣を2mlのRPMI1640に再浮遊した。その後SenVを50 HA unit/106 cellsの割合で加え、室温で1時間感染させた(それぞれAS RNA-minus/Inf細胞並びにAS RNA-plus/Inf細胞とする)。終了後、再度室温で150 x g, 4分間、2回遠心・洗浄し、3 x 106 cells/2 ml/wellとなるようにR10培地に浮遊後、6-ウェルプレートに再度播種した。各感染細胞は37℃で静置培養し、0, 0.5, 1, 3, 6, 12並びに24時間経過後に回収し、以下のRNA抽出、精製実験に供した。
【0046】
5. 細胞総RNA分画の抽出並びに各種酵素を用いたRNAの精製
5-a. 細胞総RNA分画の抽出
上記所定の時間経過後、AS RNA-minus/Inf細胞並びにAS RNA-plus/Inf細胞を回収し、4℃で150 x g, 4分間遠心後、各細胞沈渣に対しTRIzol (Invitrogen; 15596-018) 1mlを加えた。TRIzol中でピペッティングにより細胞を溶解後、室温で5分間留置した。その後、クロロフォルム (Wako; 試薬特級、 038-02606)0.2mlを加え、15秒間用手にて震盪混和後室温に再度2-3分間静置した。続いて、4℃にて12,000 x g, 15分間遠心分離した。この操作により試料は三層に分離するので、最上層の無色の水層(約600μl)から400μlを新しい滅菌済み1.5 ml微量遠心チューブに分取した。このチューブに、使用したTRIzol 1mlあたり0.5mlのイソプロピルアルコール(Wako; 試薬特級)を加え、室温にて混和、静置10分後、再度4℃にて12,000 x g, 10分間遠心した。上清を吸引、除去し、RNA沈渣に1mlの75%エチルアルコールを加えた。4℃にて7,500 x g, 5分間遠心し、上清を吸引、除去後、1mlの99.5%エチルアルコール(Wako; 試薬特級)を加え、再度4℃にて7,500 x g, 5分間遠心した。上清を吸引、除去後、沈渣を風乾し、滅菌再蒸留水に溶解した。このRNA溶液に対し、滅菌再蒸留水を最終210μlになるように添加後、20mg/mlのグリコーゲン (Roche Applied Science; 901 393)を1μl, 7.5Mの酢酸アンモニウム (Wako; 分子生物学用 01820485)を1/2容加え、撹拌後2.5容の99.5%エチルアルコールを添加し、4℃にて10分間留置した。その後、4℃, 13,000 rpmにて30分間遠心し、得られたRNA沈渣を上述のように75%, 99.5%のエチルアルコールで洗浄、風乾後、50μlのTE8.0(10 mM Tris・HCl, pH 8.0/ 0.1 mM EDTA)に溶解し、以下の酵素反応に供する迄-20℃に保存した。
【0047】
5-b. プロテアーゼ消化
上述の粗抽出した細胞総RNAにProteinase K (PCR grade; Roche Applied Science, 03115 887 001) 20μgを加え、Tris/SDS buffer (100mM Tris-HCl, pH7.4/ 50 mM EDTA, 2.5% SDS) 100μl中で、37℃, 15分間混入するNamalwa細胞由来の蛋白質成分を消化した。その後RNase-freeのTE8.0飽和フェノール(日本ジーン, 319 90093):クロロフォルム:イソアミルアルコール (Wako; 試薬特級; 135-12015)=25:24:1混合液 (PCIAA)を1容加え、4℃で15,300 x g, 2分間遠心した。分取した水層のRNA溶液に対し、イソアミルアルコール=24:1混合液 (CIAA)をさらに1容加え同様に遠心後、5-aで行ったと同様に水層中の細胞総RNAを塩析し、RNA沈渣を50μlのTE8.0に溶解した。
【0048】
5-c. DNase I 消化
この細胞総RNA溶液に対し、TURBO DNA-freeTM Kit (Applied Biosystems, AM1907) 添付のTURBO DNase2単位を加え、添付のバッファー100μl中で、37℃, 30分間混入するゲノムDNAを消化した。その後、さらに酵素を2単位追加し、計60分経過後に添付のDNase Inactivation Reagentを10μl加え、十分に撹拌後、室温に2分間放置し、酵素反応を停止させた。その後、4℃にて、9,100 x gで2分間遠心し、上清中の細胞総RNAを5-aで行ったと同様に塩析させた。AS-plus/Inf並びにAS-minus/Inf由来のRNA沈渣を各々60μl並びに30μlのTE8.0に溶解し、260, 280, 320nmの吸光度の測定から、RNA濃度並びにフェノールと蛋白質成分の混入度を測定した。精製したRNA標品のA260/A280比は常に1.95-2.05であり、A320値は0.006以下であった。
【0049】
6. 逆転写反応並びにPCR
6-a. 逆転写反応
前項で精製したAS-minus/Inf並びにAS-plus/Infの全細胞RNAの各2μgに対し最終14μlとなるように滅菌再蒸留水を加えた。続いて、mRNAの逆転写用に5μMのoligo(dT)12-18 primer (invitrogen; 18418-012)を、AS RNAの逆転写用に2μMのhIFN-α/F1 primerを各1μl加え、70℃で10分間保温後、4℃に留置した。これらのRNA溶液に対し、5倍濃度の逆転写酵素バッファー(東洋紡、TRT-1B)を5μl、10mM dNTPsを25μl、滅菌再蒸留水1.2μl、RNase阻害剤(RNase OUT; invitrogen、10777-019)0.3μl、逆転写酵素 (ReverTra Ace; 東洋紡、TRT-101) 1μlを各々加え、47℃で30分間逆転写反応を進めた。その後、70℃に15分間保温し酵素を失活させた後に4℃に留置した。次いで、上述の逆転写酵素バッファーで5単位/μlとなるように希釈したRNaseH (TakaraBio, 2150A)を1μl添加し、37℃、20分間鋳型RNAを消化後、前記のように作製したcDNAを塩析した。得られたcDNA沈渣は、mRNA由来を40μl、AS RNA由来は20μlのTE8.0に溶解した。
【0050】
6-b. PCRと Agarose gel 電気泳動
6-aで得たcDNA溶液の各2μlに対し、滅菌再蒸留水29.6μl, 10倍濃度のGene taqバッファー(日本ジーン、314-02873)を4μl、2.5mMのdNTPsを3.2μl、100μMのForward primerとReverse primer を各0.4μl加えた。次に、0.2μlのanti-Taq high (東洋紡、TCP-101)と0.2μlのtaq polymerase (Gene taq; 日本ジーン、314-02873)を混和後、室温で5分間保温し、この混合液0.4μlを各サンプルに添加後、MJ Mini Gradient Thermal Cycler (Bio-Rad)を用いて、PCRを以下のように行った。
1サイクル: 94℃, 1分
10サイクル: 94℃, 1分/ 72℃, 1分30秒
15サイクル: 94℃, 1分/ 65℃, 1分30秒/ 72℃, 10秒
5+αサイクル: 94℃, 1分/ 60℃, 1分30秒/ 72℃, 10秒
使用プライマー
IFN-α1 mRNA由来のcDNA用:
IFN-α/ F1:5’-TGGTGCTCAGCTGCAAGTCAAGC-3’ (配列番号13)
IFN-α/ R1:5’-AGAGATGGCTGGAGCCTTCTG-3’ (配列番号14)
AS RNA由来のcDNA用:
IFN-α/ F1B:5’-CTGGGCTGTGATCTCCCTGAGAC-3’ (配列番号15)
IFN-α/ R1:同上
IFN-β mRNA由来のcDNA用:
IFN-β/F1:5’-GATTCATCTAGCACTGGCTGG-3’ (配列番号16)
IFN-β/R1:5’-CCTTCAGGTAATGCAGAATCC-3’ (配列番号17)
EF mRNA由来のcDNA用:
EF/F1:5’-TCTGGTTGGAATGGTGACAACATGC-3’ (配列番号18)
EF/R1:5’-CCAGGAAGAGCTTCACTCAAAGCTT-3’ (配列番号19)
各精製RNA試料濃度を検定するための内部標準として増幅したEF mRNA由来のcDNAは30サイクル、IFN-α1 mRNA由来のcDNAは35+αサイクル、AS RNA由来のcDNA並びにIFN-β mRNA由来のcDNAは38+αサイクル増幅し、PCR産物は3%アガロース(アガロースS;日本ジーン、312-01193)による電気泳動で解析した。
尚、mRNA画分叉はAS RNA画分に混入するゲノムDNA由来の増幅シグナルを検出する目的で、逆転写反応に用いたAS-minus/Inf並びにAS-plus/Infの全細胞RNAを各々100ngまたは200ng使用して同様にPCRを行った。
【0051】
7. ELISAによるhIFN-αの定量
AS RNAノックダウンがIFN-α蛋白質発現に与える影響を検定する目的で、4のSenV感染Namalawa細胞培養上清を、感染後0, 0.5, 1, 3, 6, 12並びに24時間に回収し、上清中に分泌されたIFN-αをヒトインターフェロン−α ELISAキット(PBL Biomedical Laboratories, #41100-4)を用いて定量した。具体的には、添付のヒト精製IFN-αを適宜に希釈して作製した0-500pg/ml溶液と各時間毎に回収した培養上清の希釈液を、キット添付の抗IFN-α特異抗体を吸着させた96ウェルプレートに一定量添加後添付試薬を用いて呈色反応させ、450nmの吸光度を各々測定した。既知濃度のIFN-αの吸光度の測定結果から作製した検量線を用い、各時間毎の回収サンプルの吸光度から培養上清中に分泌されたIFN-α量を算出した。
【0052】
結果
1. RT-PCR法によるアンチセンスRNAの増幅、同定を最適化するプライマーの決定
図1にインターフェロン−α1 (IFNA1) 遺伝子及び実験に用いたプライマーの配列と、プライマー部位を遺伝子上に図示した模式図を示す (SLI, SLII並びにBulged SLは、図4のmRNA核外輸送責任領域が形成する二次構造を参照のこと)。
これらのプライマーを組み合わせ、RT-PCR法によりNamalwa細胞で構成的に発現しているIFNA1遺伝子由来のアンチセンス (AS) RNAを同定した結果を表1に要約した。
【0053】
【表1】

【0054】
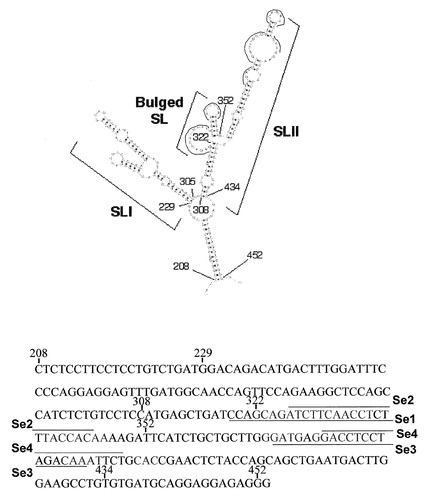
最適な組合せは、RT反応プライマーにはF1を、PCRプライマーにF1B/R1を組み合わせた場合(1)であり、AS cDNAは35サイクルから検出可能となり、40サイクルでは明瞭に観察できた。この組み合わせは、増幅量は約1/10程度に留まるものの、やはり40サイクルでAS cDNAが観察できたF1逆転写プライマー、F2B/R2 PCRプライマーペアの組合せ(2)を除き、検出効率においてその他の組合せより10サイクル分、即ち210倍高かった。図2に(1)並びに(2)の組み合わせを用い、40サイクル増幅したAS RNA由来のcDNA (AS cDNA)を示す。RT (minus)は、逆転写を行っていないRNAサンプルを用いて実施したPCRの結果を示し、RNAサンプルに混入するゲノムDNA由来のシグナルの混入は無視できる程度であった。
【0055】
2. センダイウイルス(SenV) に伴うNamalwa細胞におけるAS RNAとIFN-α1 mRNAの発現動態の変動(図3)
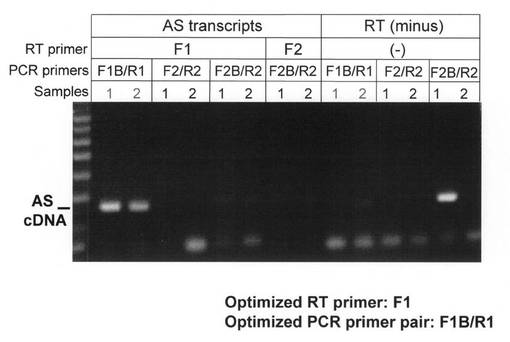
Namalwa細胞において構成的に発現するAS RNA (IFN-α1 AS RNA; 0 hr) は、SenV感染により発現が急速に減衰 (IFN-α1 AS RNA; 1 hr) し、感染後3時間には消失した (IFN-α1 AS RNA; 3 hr)が、12時間後より再発現を認め (IFN-α1 AS RNA; 12 hr)、24時間経過後には感染前の発現レベルに復した (IFN-α1 AS RNA; 24 hr)。
これに対し、IFN-α1 mRNAの発現はAS RNAの発現動態と逆相関し、感染後1時間経過時点 (IFN-α1 mRNA; 1 hr)で第1のピークに到達後減衰し、6時間経過時点(IFN-α1 mRNA; 6 hr)では認められなくなった。興味深いことに、この時点でIFN-β mRNAの発現が一時的に認められ (IFN-β mRNA; 6 hr)、その後感染12時間の時点でIFN-α1 mRNAの発現を再度認めた (IFN-α1 mRNA; 12 hr)。この時点が同mRNA発現の第2で最大のピークであった。その後、その発現は感染24時間 (IFN-α1 mRNA: 24 hr)に向けて減衰し、この減衰と期を一にしてAS RNAの再発現が認められた (IFN-α1 AS RNA: 12, 24 hr)。
以上の結果から、AS RNA発現とSenV感染に伴うIFN-α1 mRNAの発現の間に相補関係が示唆されたため、これを検証する目的で以下に述べるセンスオリゴヌクレオチドによるAS RNA発現のノックダウンを行い、IFN-α蛋白質発現並びに感染ウイルス増殖に及ぼす影響を検証した。
【0056】
3. IFN-α1 mRNAの核外輸送責任領域が形成するRNA二次構造とセンスオリゴヌクレオチドによるAS RNA発現の阻害
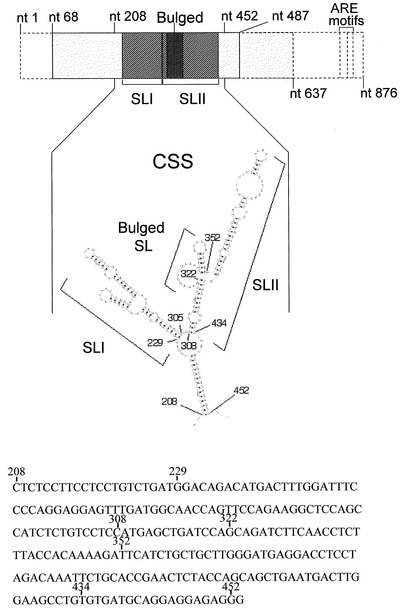
これまでの研究から、IFN-α1 mRNAが、核内で転写、プロセッシングを受けた後核外輸送されるにあたっては、同mRNA上の輸送責任領域(CSS: IFNA1遺伝子 nt208-452)が形成する二つのステムループ (SLI, SLII)からなるRNA二次構造(図4)の核外輸送因子による認識が必要な事を明らかにした(Kimura et al., 投稿準備中)。その過程で、SLII構造中のBulged SL(図4)の有無がIFN-α1 mRNAの発現を左右する事が証明されたため(Kimura et al., 投稿準備中)、AS RNAとIFN-α1 mRNA間の相補的発現動態を考えあわせ、Bulged SL構造を形成する遺伝子配列に相同なセンスオリゴヌクレオチドを作製し、AS RNA発現ヘの影響を検討した。用いたセンスオリゴヌクレオチドSe1〜Se4のシークエンス(配列番号3〜6)とSLII構造中の位置を図5に示す。Se1(配列番号3)、Se2(配列番号4)は相互に重複しつつBulged SL構造を形成する配列と相補であり、Se3(配列番号5)とSe4(配列番号6)はこの領域を除く残りのSLII構造部分中Bulge/Bubble或いはループを形成する配列に相補となるように設計した。
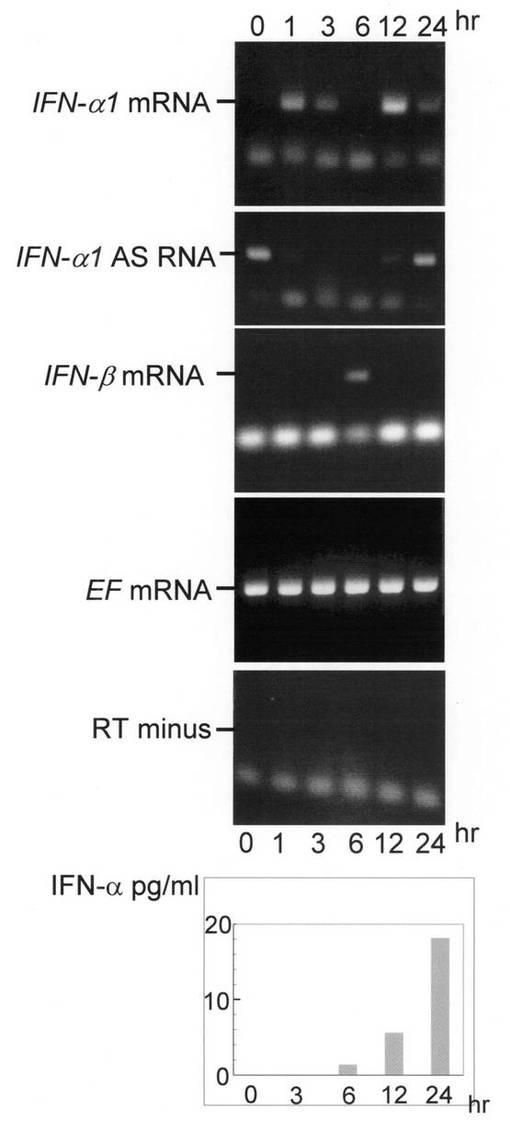
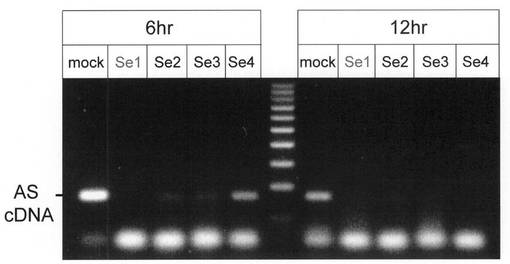
これらを用いたAS RNA阻害実験結果を図6に示す。導入したセンスオリゴヌクレオチド中AS RNA発現の阻害効果はSe1が最大であり、その効果は導入12時間後も確認できた。
【0057】
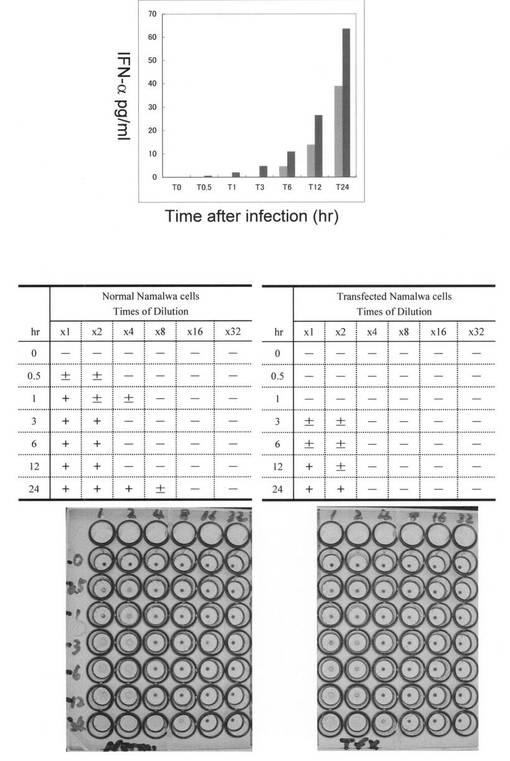
4. AS RNA発現阻害(ノックダウン、KD)がIFN-αの発現とSenV増殖に及ぼす効果
Se1とSe3を等量ずつ導入したSenV感染Namalwa細胞から産生されたIFN-α量を、感染後経時的に測定した結果を図7に示す。IFN-αは未導入細胞においては感染6時間を経過して初めて検出可能であったが、センスオリゴヌクレオチドの導入により産生が早期に繰り上がった結果感染後30分の時点で既に検出できるようになり、感染24時間後に産生されたIFN-α量は未導入細胞に比較し70%増大していた。続いて、各経過時間に培養液中に放出されたSenVのウイルス力価を赤血球凝集試験で測定した。対照感染細胞では、感染後24時間の培養液中のSenV力価が8単位/mlであるのに対し、Se1とSe3を導入したNamalwa細胞においては2-4単位/mlに半減するのが観察され、感染細胞中に発現するIFN-α量の増加との逆相関が証明できた。
以上の結果から、Namalwa細胞中に構成的に発現し、SenV感染に伴うIFN-α1 mRNAの発現に逆相関的に異動する同mRNAに対するAS RNAの発現を、センスオリゴヌクレオチドを用いノックダウンしたところ、IFN-α蛋白質の発現の増大を通じ、感染ウイルスの増殖が抑制されることが明らかにできた。
【産業上の利用可能性】
【0058】
本発明のセンスオリゴヌクレオチドおよび本発明のスクリーニング方法により選択されるIFN-α発現増強物質は、ウイルス感染症や癌の新規治療薬として有用である。
【図面の簡単な説明】
【0059】
【図1】IFN-α AS RNAの増幅効率の至適化に用いたプライマーとIFNA1遺伝子上の位置を示す模式図である。
【図2】IFN-α AS RNA増幅プライマー至適化実験で決定した最良プライマーを用いたAS RNA増幅効果を示す図である。
【図3】SenV感染によるNamalwa細胞におけるIFN-α1 mRNAとAS RNAの発現動態の変動を示す図である。下のグラフはIFN-αタンパク質量の変動を示す。
【図4】IFN-α1 mRNAの核外輸送責任領域(CSS)の位置、該領域が形成するRNA二次構造並びに該領域の塩基配列を示す図である。
【図5】IFN-α AS RNAのノックダウン実験に用いたセンスオリゴヌクレオチドのCSSに対する位置とその配列を示す図である。
【図6】センスオリゴヌクレオチドによるIFN-α AS RNA発現のノックダウン効果を示す図である。
【図7】IFN-α AS RNAのノックダウンに伴うIFN-α1の早期出現とその増大、並びにウイルス増殖の遅延・抑制効果を示す図である。
【技術分野】
【0001】
本発明は、インターフェロン−αの内在性アンチセンスRNAに特異的なセンスオリゴヌクレオチド、並びにそのインターフェロン−α発現増強作用の利用に関する。
【背景技術】
【0002】
ウイルス感染に対する生体防御において、自然免疫は感染初期の速やかな防御機構として重要な役割を果たしている。I型インターフェロン(IFN)は自然免疫における抗ウイルス活性の中心的な役割を担うサイトカインであり、ウイルス感染によって一過的に分泌され、周囲の細胞に働きかけて抗ウイルス活性、細胞増殖抑制活性などの多様な生理活性を発揮する。I型IFNはIFN-α、-β、-ω、-ε、-κに分けられ、このうちIFN-αには20数種類のサブタイプが存在する。IFN-αおよび-βは、これらの生理活性に基づき、血漿由来の製剤および遺伝子工学的に製造された組換えタンパク製剤が、B型肝炎、C型肝炎等のウイルス感染症、あるいは腫瘍などの種々の疾患に対する治療(もしくは補助療法)に利用されている。
【0003】
しかし、IFNの長期投与は、インフルエンザ様症状(頭痛、発熱、関節痛、筋肉痛、食欲不振、全身倦怠感、悪心・嘔吐等)、自己免疫疾患、うつ症状(不眠や焦燥感等)、脱毛、甲状腺機能異常、痴呆等、多様な副作用を生じる場合がある。そのため、IFNを投与することなく、生体内のIFN産生を増強させることによって、ウイルス感染の防御や、癌等の改善・治療を図るための研究も進められている。しかし、IFN産生を増強する物質の多くはウイルスや病原菌などで、安全性に問題のあるものが多い。
【0004】
ウイルス感染に対するIFN-α、-βなどのI型IFNの発現応答を早期に高めることができれば、未知のウイルスを含めた種々のウイルス感染症の予防・治療に有効であると考えられる。しかしながら、ウイルス応答における内在I型IFNの発現の立ち上がりを早める作用を有する物質は、これまで報告されていない。最近、Toll様受容体(TLR)を介したウイルス感染の認識からキナーゼカスケードを介してI型IFNの発現誘導に至るシグナル伝達経路が解明されてきているが、IFN遺伝子の転写後調節については、未だ十分に理解されていない。
【0005】
アンチセンスRNAとは、mRNAに対して、相補的な塩基配列を持ったRNAであり、具体的には、センス遺伝子をコードするDNA鎖(すなわちmRNAの非鋳型鎖)を鋳型として合成されたRNAである。センス-アンチセンスRNA間には2本鎖形成能があり、例えば、2本鎖RNAはRNA干渉が働く際に必要であること、及びマイクロRNAと呼ばれる小さなRNAによるタンパク質の翻訳制御に2本鎖RNAが関与することなどが知られている。最近の網羅的なcDNA解析により、かなり多くのアンチセンスRNAが転写されていることが明らかとなった(非特許文献1、2)。例えば、マウスでは約2,500対(非特許文献3)、ヒトでは約2,600対(非特許文献4)のセンス-アンチセンスRNAペアが存在することが示唆されており、それらの中にはタンパク質をコードしない非翻訳性のアンチセンスRNAが多く含まれている。
【0006】
アンチセンスRNAの生理機能についてはほとんど解明されていないが、内皮型一酸化窒素合成酵素(eNOS)mRNAに相補的なアンチセンスRNAは、ヒストン脱アセチル化酵素阻害剤の存在下で増加してeNOS mRNA量を減少させることが報告されている(非特許文献5)。また、酵母においても、PHO84遺伝子のアンチセンスRNAはヒストン脱アセチル化を介してmRNAの発現を抑制することが知られている(非特許文献6)。
一方、本発明者らは、誘導型一酸化窒素合成酵素(iNOS)mRNAの3’-非翻訳領域(UTR)に相補的な内在性アンチセンスRNA(「AS RNA」ともいう)が存在し、このアンチセンスRNAがiNOS mRNAとハイブリダイズすることによって該mRNAが安定化されiNOSの産生量およびNOの合成量が増大すること、該AS RNAに相補的なセンスオリゴヌクレオチドは、AS RNAのiNOS mRNAへの結合を阻害することにより該mRNAを不安定化し、iNOSの産生およびiNOSによるNO合成を抑制し得ることを報告した(特許文献1および2、非特許文献7)。
【0007】
しかしながら、IFN-α遺伝子については、内在性AS RNAが存在するかどうかすら、これまで報告されていなかった。
【特許文献1】WO 2007/142303号パンフレット
【特許文献2】WO 2007/142304号パンフレット
【非特許文献1】Cheng et al., Science, 308: 1149-1154 (2005)
【非特許文献2】Katayama et al., Science, 309: 1564-1566 (2005)
【非特許文献3】Kiyosawa et al., Genome Res., 13: 1324-1334 (2003)
【非特許文献4】Yelin et al., Nat. Biotechnol., 21: 379-386 (2003)
【非特許文献5】Robb et al., J. Biol. Chem., 279: 37982-37996 (2004)
【非特許文献6】Camblong et al., Cell, 131: 706-717 (2007)
【非特許文献7】Matsui et al., Hepatology, 47: 686-697 (2008)
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明の目的は、内在のI型IFNの産生を増強させ得る物質を同定し、該物質を利用したウイルス感染症や癌、自己免疫疾患などの疾患の新規予防・治療手段を提供することである。特に、ウイルス感染に対するI型IFN応答の立ち上がりを早めることによる、ウイルス感染症の有効な予防・治療手段を提供することである。加えて、膠原病やクローン病のように、免疫システムのバランスの破綻から引き起こされるI型IFN産生調節異常が、疾患発症の引き金の一つになると考えられる自己免疫疾患についても治療手段を提供することである。また、本発明の別の目的は、内在のI型IFNの産生を増強させ得る物質を探索するためのスクリーニング系を提供することである。
【課題を解決するための手段】
【0009】
本発明者らは、IFN-α mRNAの3’-UTRに、iNOS mRNAと同様、mRNAの早い代謝回転に関与すると考えられているAU-rich element(ARE)モチーフが存在することに着目し、IFN-αの発現調節にも内在性AS RNAが関与しているのではないかと発想してRT-PCR解析を行った結果、IFN-α AS RNAの存在が確認された。次に、ウイルス感染後のIFN-α mRNAおよびAS RNAの発現変動を解析したところ、IFN-α AS RNAは構成的に発現しているが、ウイルス感染後速やかに発現が低下し、それに伴ってIFN-α mRNAの発現レベルが一過的に上昇することが明らかとなった。これらの結果はIFN-α AS RNAがmRNAの発現を抑制していることを示唆するものであり、iNOSにおけるAS RNAによる発現調節とは異なっていた。
【0010】
本発明者らは、IFN-α mRNAの核外輸送に重要な、2つのステムループ(SLIおよびSLII(SLIIは内部にBulged SLを含む))を有する二次構造を形成する領域(図4参照)を同定していたことから、IFN-α AS RNAは、3’-UTRのAREモチーフではなく、この二次構造を形成する領域をターゲットとしてIFN-α mRNAにハイブリダイズし、mRNAの核外輸送を阻害することにより、IFN-αタンパク質への翻訳を抑制しているのではないかと考えた。そこで、SLIIおよびBulged SL領域のループ部分の配列を有するセンスオリゴヌクレオチドを作製し、IFN-α AS RNAの発現阻害効果およびIFN-αの発現とウイルス増殖に及ぼす効果を調べたところ、これらのセンスオリゴヌクレオチドはIFN-α AS RNAの発現を効率よく阻害し、IFN-αの産生量を増大させ、ウイルス増殖を抑制した。
本発明者らは、これらの知見に基づいてさらに研究を重ねた結果、本発明を完成するに至った。
【0011】
即ち、本発明は以下の通りである。
〔1〕IFN-α mRNAに対する内在性アンチセンスRNAに相補的な配列を含み、IFN-αの発現を増強させ得るセンスオリゴヌクレオチド。
〔2〕該配列がIFN-α mRNAのSLIまたはSLII領域内の塩基配列に相同な配列である、上記〔1〕記載のセンスオリゴヌクレオチド。
〔3〕該配列がBulged SL領域内の塩基配列に相同な配列である、上記〔2〕記載のセンスオリゴヌクレオチド。
〔4〕上記〔1〕〜〔3〕のいずれかに記載のセンスオリゴヌクレオチドを含有してなるIFN-α発現増強剤。
〔5〕ウイルス感染症または癌の予防または治療用である、上記〔4〕記載の剤。
〔6〕被験物質の存在下および非存在下で、IFN-α mRNAとそれに対する内在性アンチセンスRNAとのハイブリダイゼーションを検出・比較することを特徴とする、IFN-α発現調節物質のスクリーニング方法。
〔7〕IFN-α mRNAとそれに対する内在性アンチセンスRNAとを発現する細胞に被験物質を接触させ、該細胞における該mRNA量および/またはIFN-αタンパク質量の変化を測定することを特徴とする、上記〔6〕記載の方法。
【発明の効果】
【0012】
本発明のセンスオリゴヌクレオチドは、IFN-α mRNAに対する内在性AS RNAとハイブリダイズしてそのmRNAに対する作用を阻害することにより、IFN-αの産生を増強することができる。特に、ウイルス感染初期における迅速なIFN-αの産生誘導を可能にする。さらに、IFN-α製剤と異なり、吸入等による患者自身による投与が可能なため、患者負担を軽減することもできる。
【発明を実施するための最良の形態】
【0013】
本発明は、IFN-αの発現増強活性を有するIFN-αのセンスオリゴヌクレオチドを提供する。本発明のセンスオリゴヌクレオチドは、IFN-α mRNAに対する内在性AS RNAに相補的な配列を含み、かつIFN-αの発現を増強させ得るオリゴヌクレオチドであれば、いかなるものであってもよい。ここで「相補的」な配列とは、AS RNAに対して完全相補的な配列のみならず、細胞の生理的な条件下でAS RNAとハイブリダイズしてmRNAへのAS RNAの作用を阻害し得る限り、1ないし数(2, 3, 4 もしくは 5)塩基のミスマッチを含んでもよい。好ましくは、IFN-α AS RNAに相補的な配列とは、ストリンジェントな条件、例えば、Current Protocols in Molecular Biology, John Wiley & Sons,6.3.1-6.3.6, 1999に記載される条件(例えば、6×SSC(sodium chloride/sodium citrate)/45℃でのハイブリダイゼーション、次いで0.2×SSC/0.1% SDS/50〜65℃での一回以上の洗浄等が挙げられる)下で、該AS RNAとハイブリダイズし得る配列である。
具体的には、IFN-α mRNAに対する内在性AS RNAは、該mRNAの熱力学的に安定でない部分と相互作用して該mRNAを不安定化するので、本発明のセンスオリゴヌクレオチドは、IFN-α mRNA中の熱力学的に安定でない部分と相同な配列を含むことが好ましい。熱力学的に安定でない部分としては、mRNAが二次構造をとった際に一本鎖の状態にある(例えば、ステムループ構造のループ部分にあたる)領域が挙げられる。IFN-α mRNAの二次構造は、該mRNAの塩基配列情報をもとに、mfold(GCG Software; Proc. Natl. Acad. Sci. USA, 86: 7706-10 (1989) 参照)に代表されるような既存のRNA二次構造予測プログラムを用いて予測することができる。IFN-α mRNAには20数種類のサブタイプが存在するが(例えば、Science, 209: 1343-1347 (1980)、Gene, 11: 181-186 (1980)、Nature, 290: 20-26 (1981)、Nature, 313: 698-700 (1985)、J. Invest. Dermatol., 83: 128s-136s (1984)、J. Interferon Res., 2: 575-585 (1982)、J. Interferon Res., 13: 227-231 (1993)、J. Biol. Chem., 268: 12565-12569 (1993)、Acta Virol., 38: 101-104 (1994)、Biochim. Biophys. Acta, 1264: 363-368 (1995) 参照)、それらの塩基配列情報はいずれも容易に入手可能である。例えば、ヒトIFN-α1 mRNA(cDNA)の塩基配列は、DDBJ/EMBL/GenBank ヌクレオチドデータベースにAccession No. AB445100として登録されている。当該塩基配列を配列番号1に、当該塩基配列より推定されるヒトIFN-α1のアミノ酸配列を配列番号2に、それぞれ示す。
【0014】
さらに、本発明のセンスオリゴヌクレオチドは、配列非特異的な反応を起こす配列(例えば、5’-CG-3’、5’-GGGG-3’、5’-GGGGG-3’等)を含まないものから選択することが好ましく、また、IFN-α mRNA以外のRNA中に類似の配列が存在しないものから選択することが好ましい。他のRNA中に類似の配列が存在しないことは、センスオリゴヌクレオチドの候補配列をクエリーとして、対象とする哺乳動物のゲノム配列に対して相同性検索をかけることにより確認することができる。ここで相同性検索手段としては、公知の核酸の相同性検索ソフトウェア(例えば、NCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)NBLASTおよびXBLASTプログラム(version 2.0)、GCGソフトウェアパッケージ中のFASTAプログラム等)を用いることができる。また、ゲノムDNAデータセットとしては、例えば、Celera社が提供する全ヒトゲノムデータを用いることができる。
【0015】
好ましくは、本発明のセンスオリゴヌクレオチドは、IFN-α mRNAのSLI(配列番号1に示されるヒトIFN-α1 mRNA塩基配列にあっては、塩基番号229〜305で示される塩基配列)またはSLII領域(配列番号1に示されるヒトIFN-α1 mRNA塩基配列にあっては、塩基番号308〜434で示される塩基配列)内の塩基配列に相同な配列を含む。ヒトIFN-α1 mRNAの種々の欠失変異体を用いた実験から、該mRNAの核外輸送には、配列番号1に示される塩基配列中塩基番号208〜452で示される領域が形成する2つのステムループ(SLIおよびSLII)からなる二次構造を、核外輸送因子が認識することが必要であることが明らかとなった。IFN-α mRNAに対する内在性AS RNAは、この核外輸送責任領域(CSSともいう)をターゲットとして該mRNAと相互作用することにより、該mRNAの核外への輸送およびその後の翻訳を阻害するとともに、RNaseによるmRNAの分解を促進すると考えられるので、CSS中の配列のうちで上記の条件に適合する領域、即ち、CSSが二次構造をとった際にSLIまたはSLII領域内でループ構造を形成する部分の塩基配列に相同な配列を含むオリゴヌクレオチド(図4および5参照)は、IFN-α AS RNAにハイブリダイズすることで該AS RNAのIFN-α mRNAへの作用を阻害し、該mRNAレベルを増大させ、該mRNAの核外輸送の抑制を解除することにより、IFN-αタンパク質の産生を増強することができる。特に、SLII領域内のBulged SL(配列番号1に示されるヒトIFN-α1 mRNA塩基配列にあっては、塩基番号322〜352で示される塩基配列)がIFN-α mRNAの発現を左右することから、Bulged SL領域内の塩基配列に相同な配列を含むセンスオリゴヌクレオチドがより好ましい。
尚、「相同な」配列とは、IFN-α mRNAの特定の部分塩基配列と完全に同一な配列のみならず、該mRNAに対する内在性AS RNAと細胞の生理的な条件下でハイブリダイズしてmRNAに対するAS RNAの作用を阻害し得る限り、1ないし数(2, 3, 4もしくは 5)個の塩基が異なっていてもよい。
【0016】
より具体的なセンスオリゴヌクレオチドの例として、例えば、以下のものが挙げられる(括弧内の数字は配列番号1に示される塩基配列中の対応する領域を示す)。
配列番号3:5’-CCAGCAGATCTTCAACCTCT-3’(塩基番号319〜338)
配列番号4:5’-ATCTTCAACCTCTTTACCAC-3’(塩基番号326〜345)
配列番号5:5’-GATGAGGACCTCCTAGACAA-3’(塩基番号368〜387)
配列番号6:5’-GACCTCCTAGACAAATTCTG-3’(塩基番号374〜393)
【0017】
本発明のセンスオリゴヌクレオチドの長さに特に制限はないが、配列特異性の面から、標的配列に相補的な部分を少なくとも10塩基以上、好ましくは約12塩基以上、より好ましくは約15塩基以上含むものである。また、合成の容易さ、製造コスト、投与のし易さ等の面から、50塩基以下、好ましくは40塩基以下、より好ましくは30塩基以下の塩基長を有するものが挙げられる。
【0018】
本発明のセンスオリゴヌクレオチドは、一本鎖DNA、一本鎖RNA、DNA/RNAキメラのいずれであってもよく、さらに公知の修飾の付加されたものであってもよい。ここで「ヌクレオチド」とは、プリンおよびピリミジン塩基を含有するのみでなく、修飾されたその他の複素環型塩基をもつようなものを含んでいてもよい。センスオリゴヌクレオチドがDNA(ODN)の場合、標的AS RNAとセンスODNとによって形成されるRNA:DNAハイブリッドは、内在性RNase Hに認識されて標的AS RNAの選択的な分解を引き起こすことができる。
【0019】
センスオリゴヌクレオチドを構成するヌクレオチド分子は、天然型のDNAもしくはRNAでもよいが、安定性(化学的および/または対酵素)や比活性(RNAとの親和性)を向上させるために、種々の化学修飾を含むことができる。例えば、ヌクレアーゼなどによる分解を防ぐために、センスオリゴヌクレオチドを構成する各ヌクレオチドのリン酸残基(ホスフェート)を、例えば、ホスホロチオエート(PS)、メチルホスホネート、ホスホロジチオネートなどの化学修飾リン酸残基に置換することができる。また、各ヌクレオチドの糖(リボース)の2'位の水酸基を、-OR(Rは、例えばCH3(2'-O-Me)、CH2CH2OCH3(2'-O-MOE)、CH2CH2NHC(NH)NH2、CH2CONHCH3、CH2CH2CN等を示す)に置換してもよい。さらに、塩基部分(ピリミジン、プリン)に化学修飾を施してもよく、例えば、ピリミジン塩基の5位へのメチル基やカチオン性官能基の導入、あるいは2位のカルボニル基のチオカルボニルへの置換などが挙げられる。
RNAの糖部のコンフォーメーションはC2'-endo(S型)とC3'-endo(N型)の2つが支配的であり、一本鎖RNAではこの両者の平衡として存在するが、二本鎖を形成するとN型に固定される。したがって、標的AS RNAに対して強い結合能を付与するために、2'酸素と4’炭素を架橋することにより、糖部のコンフォーメーションをN型に固定したRNA誘導体であるBNA (LNA)(Imanishi, T. et al., Chem. Commun., 1653-9, 2002; Jepsen, J.S. et al., Oligonucleotides, 14, 130-46, 2004)やENA(Morita, K. et al., Nucleosides Nucleotides Nucleic Acids, 22, 1619-21, 2003)もまた、好ましく用いられ得る。
【0020】
本発明のセンスオリゴヌクレオチドは、IFN-αのmRNA(cDNA)配列に基づいて、市販のDNA/RNA自動合成機(アプライド・バイオシステムズ社、ベックマン社等)を用いて、これに相同な配列を合成することにより調製することができる。
【0021】
本発明のセンスオリゴヌクレオチドは、リポソーム、ミクロスフェアのような特殊な形態で供与されたり、ポリリジンのようなポリカチオン体、脂質(例、ホスホリピド、コレステロールなど)などの疎水性物質が付加された形態で提供され得る。あるいはまた、本発明のセンスオリゴヌクレオチドを、膜透過機能を有するペプチド(例えば、ショウジョウバエ由来のAntennapediaホメオドメイン(AntP)、ヒト免疫不全ウイルス(HIV)由来のTAT、単純ヘルペスウイルス(HSV)由来のVP22等の細胞通過ドメイン)などで修飾することにより、該オリゴヌクレオチドの細胞への取り込みを促進することができる。
【0022】
本発明のセンスオリゴヌクレオチドは、IFN-α mRNAに対する内在性AS RNAの作用を阻害することにより、IFN-αmRNAおよびIFN-αタンパク質の発現を増強することができる。IFN-αは、抗ウイルス作用、細胞増殖抑制作用、ナチュラルキラー細胞の活性化作用などの多様な生理活性を有するので、本発明のセンスオリゴヌクレオチドを含有する医薬は、IFN-αの発現増強剤として、種々の疾患の予防および/または治療に利用することができる。そのような疾患としては、例えば、B型肝炎、C型肝炎(活動性、非活動性)、ヘルペス感染症(性器ヘルペス、角膜ヘルペス炎、口腔ヘルペスなど)、尖圭コンジローマ、AIDS等のウイルス感染症、腎臓癌、腎細胞癌、乳癌、膀胱癌、基底細胞癌、頭頸部癌、頸管異形成、皮膚悪性腫瘍、カポジ肉腫、悪性黒色腫、非ホジキン型リンパ腫、幼児血管腫、慢性肉芽腫症、慢性骨髄性白血病(CML)、成人T細胞白血病、ヘアリー細胞白血病、毛様細胞白血病、T細胞白血病ウイルス(HTLV−1)骨髄症、多発性骨髄腫、リンパ腫などの癌、亜急性硬化性全脳炎(SSPE)、クローン病、関節リウマチ、シェーグレン症候群、多発性硬化症(MS)、口内炎、性器疣贅、膣内疣贅、赤血球増加症、血小板増加症、乾癬、菌状息肉症、突発性難聴、老年性円盤状黄斑変性症、ページェット病などが挙げられるが、これらに限定されない。
IFN-α AS RNAは通常、白血球あるいはリンパ芽球様細胞などのIFN-α産生細胞において構成的に発現し、IFN-αの発現を抑制しているが、ウイルス感染に応答してその発現が抑制されることによってIFN-αの発現抑制が解除されてIFN-αが産生・分泌され、自然免疫が誘導される。したがって、ウイルス感染初期に際して、本発明のセンスオリゴヌクレオチドを含有する医薬は、IFN-α AS RNAの発現を速やかに抑制することにより、IFN-α産生の立ち上がりを早め、より迅速に自然免疫系を誘導することができる。
【0023】
本発明のセンスオリゴヌクレオチドを含有する医薬は低毒性であり、そのまま液剤として、または適当な剤型の医薬組成物として、ヒトまたは非ヒト哺乳動物(例、マウス、ラット、ウサギ、ヒツジ、ブタ、ウシ、ネコ、イヌ、サルなど)に対して経口的または非経口的(例、吸入投与、血管内投与、皮下投与、経粘膜投与など)に投与することができる。
これらの核酸を上記のウイルス感染症や癌などの予防・治療剤として使用する場合、自体公知の方法に従って製剤化し、投与することができる。即ち、本発明のセンスオリゴヌクレオチドを、単独で、あるいはレトロウイルスベクター、レンチウイルスベクター、アデノウイルスベクター、アデノ随伴ウイルスベクターなどの適当な哺乳動物細胞用の発現ベクターに機能可能な態様で挿入した後、常套手段に従って製剤化することができる。該オリゴヌクレオチドは、そのままで、あるいは摂取促進のための補助剤とともに、遺伝子銃やハイドロゲルカテーテルのようなカテーテルによって投与することもできる。あるいは、エアロゾル化して吸入剤として気管内に局所投与することもできる。
さらに、体内動態の改良、半減期の長期化、細胞内取り込み効率の改善を目的に、前記オリゴヌクレオチドを単独またはリポソームなどの担体とともに製剤(注射剤)化し、静脈、皮下等に投与してもよい。
【0024】
本発明のセンスオリゴヌクレオチドは、それ自体を投与してもよいし、または適当な医薬組成物として投与してもよい。投与に用いられる医薬組成物としては、本発明のセンスオリゴヌクレオチドと薬理学的に許容され得る担体、希釈剤もしくは賦形剤とを含むものであってよい。このような医薬組成物は、経口または非経口投与に適する剤形として提供される。
【0025】
非経口投与のための組成物としては、例えば、注射剤、エアロゾル剤、坐剤等が用いられ、注射剤は静脈注射剤、皮下注射剤、皮内注射剤、筋肉注射剤、点滴注射剤等の剤形を包含してもよい。このような注射剤は、公知の方法に従って調製できる。エアロゾル製剤はジクロロジフロロメタン、プロパン、窒素などのような圧縮された許容しうる抛射薬内に入れることができる。あるいはネブライザーやアトマイザーのような非圧縮性製剤用医薬品として製剤化してもよい。直腸投与に用いられる坐剤は、上記核酸を通常の坐薬用基剤に混合することによって調製されてもよい。
【0026】
経口投与のための組成物としては、固体または液体の剤形、具体的には錠剤(糖衣錠、フィルムコーティング錠を含む)、丸剤、顆粒剤、散剤、カプセル剤(ソフトカプセル剤を含む)、シロップ剤、乳剤、懸濁剤等が挙げられる。このような組成物は公知の方法によって製造され、製剤分野において通常用いられる担体、希釈剤もしくは賦形剤を含有していても良い。錠剤用の担体、賦形剤としては、例えば、乳糖、でんぷん、蔗糖、ステアリン酸マグネシウムが用いられる。
【0027】
上記の非経口用または経口用医薬組成物は、活性成分の投与量に適合するような投薬単位の剤形に調製されることが好都合である。このような投薬単位の剤形としては、例えば、錠剤、丸剤、カプセル剤、注射剤(アンプル)、エアロゾル剤、坐剤が挙げられる。本発明のセンスオリゴヌクレオチドは、例えば、投薬単位剤形当たり通常5〜500mg、とりわけ注射剤では5〜100mg、その他の剤形では10〜250mg含有されていることが好ましい。
【0028】
本発明のセンスオリゴヌクレオチドを含有する上記医薬の投与量は、投与対象、対象疾患、症状、投与ルートなどによっても異なるが、例えば、成人の急性C型肝炎の予防および/または治療に使用する場合には、本発明のセンスオリゴヌクレオチドを1回量として、通常0.01〜20mg/kg体重程度、好ましくは0.1〜10mg/kg体重程度、さらに好ましくは0.1〜5mg/kg体重程度を、1日1回〜数回程度、静脈注射や吸入等により投与するのが好都合である。他の非経口投与および経口投与の場合もこれに準ずる量を投与することができる。症状が特に重い場合には、その症状に応じて増量してもよい。
【0029】
なお前記した医薬組成物は、本発明のセンスオリゴヌクレオチドとの配合により好ましくない相互作用を生じない限り、他の薬剤を含有してもよい。他の薬剤としては、例えば、抗ウイルス薬、抗腫瘍薬、抗菌薬、抗真菌薬、抗原虫薬、抗生物質、抗セプシス薬、抗セプティックショック薬、エンドトキシン拮抗薬、免疫調節薬、非ステロイド性抗炎症薬、ステロイド薬、炎症性メディエーター作用抑制薬、炎症性メディエーター産生抑制薬、抗炎症性メディエーター作用抑制薬、抗炎症性メディエーター産生抑制薬などが挙げられる。
【0030】
本発明はまた、IFN-α mRNAに対する内在性AS RNAの抑制作用を調節することによりIFN-αの発現を調節する物質をスクリーニングする方法を提供する。本発明のスクリーニング方法は、被験物質の存在下および非存在下で、IFN-α mRNAとそれに対する内在性AS RNAとのハイブリダイゼーションを検出し、その程度を比較することを特徴とする。
例えば、IFN-α mRNAとIFN-α AS RNAとを常法により単離し、いずれか一方を固相化し、他方を適当な標識剤で標識して、RNAが生理的な二次構造を形成し得る条件下で、被験物質の存在下および非存在下に両者をハイブリダイズさせ、固相に結合した標識量を両条件下で比較する方法が挙げられる。ここでmRNAおよびAS RNAとしては、それぞれその全長を用いてもよいし、あるいはmRNAの核外輸送責任領域(CSS;配列番号1に示されるヒトIFN-α1 mRNAの塩基配列にあっては、塩基番号208〜452で示される領域)、SLII領域(配列番号1に示されるヒトIFN-α1 mRNAの塩基配列にあっては、塩基番号308〜434で示される領域)あるいはSLI領域(配列番号1に示されるヒトIFN-α1 mRNAの塩基配列にあっては、塩基番号229〜305で示される領域)、並びにAS RNAのそれらの領域に相補的な配列を含むそれらのフラグメントを用いてもよい。
固相の材料としては、シリコンなどの半導体、ガラス、ダイアモンドなどの無機物、ポリエチレンテレフタレート、ポリプロピレン等の高分子物質を主成分とするフィルムなどが挙げられ、また固相の形状としては、スライドガラス、マイクロウェルプレート、マイクロビーズ、繊維型などが挙げられるが、それらに制限されない。固相上にmRNAもしくはAS RNAを固定化する方法としては、予め該RNAにアミノ基、アルデヒド基、SH基、ビオチンなどの官能基を導入しておき、一方、固相上にも該RNAと反応し得る官能基(例:アルデヒド基、アミノ基、SH基、ストレプトアビジンなど)を導入し、両官能基間の共有結合で固相とRNAを架橋したり、ポリアニオン性のRNAに対して、固相をポリカチオンコーティングして静電結合を利用してRNAを固定化するなどの方法が挙げられるが、これらに限定されない。固相化RNAの調製法としては、フォトリソグラフィー法を用いてRNAを基板(ガラス、シリコンなど)上で1ヌクレオチドずつ合成するAffymetrix方式と、マイクロスポッティング法、インクジェット法、バブルジェット(登録商標)法などを用いて、予め調製されたRNAを基板上にスポッティングするStanford方式とが挙げられるが、使用するRNAの塩基長を考慮すれば、Stanford方式あるいは両者を組み合わせた手法を用いるのが好ましい。
標識剤としては、例えば、放射性同位元素、酵素、蛍光物質、発光物質などが用いられる。放射性同位元素としては、例えば、〔32P〕、〔3H〕、〔14C〕などが用いられる。酵素としては、安定で比活性の大きなものが好ましく、例えば、β−ガラクトシダーゼ、β-グルコシダーゼ、アルカリフォスファターゼ、パーオキシダーゼ、リンゴ酸脱水素酵素などが用いられる。蛍光物質としては、例えば、フルオレスカミン、フルオレッセンイソチオシアネート、Cy3、Cy5などが用いられる。発光物質としては、例えば、ルミノール、ルミノール誘導体、ルシフェリン、ルシゲニンなどが用いられる。さらに、プローブと標識剤との結合にビオチン-(ストレプト)アビジンを用いることもできる。
被験物質としては、いかなる公知物質および新規物質であってもよく、例えば、核酸、糖質、脂質、蛋白質、ペプチド、有機低分子化合物、コンビナトリアルケミストリー技術を用いて作製された化合物ライブラリー、固相合成やファージディスプレイ法により作製されたランダムペプチドライブラリー、あるいは微生物、動植物、海洋生物等由来の天然成分などがあげられる。添加される被験物質の濃度は化合物の種類(溶解度、毒性等)により異なるが、例えば、約0.1nM〜約100nMの範囲で適宜選択される。インキュベート時間としては、例えば、約1〜約24時間が挙げられる。
固相上のRNAと、標識したRNA(および被験物質)とを接触させて、インキュベートした後、固相に結合しなかったRNAを洗い流し、固相に結合したRNAの標識量を検出する。被験物質の存在下で、非存在下に比べて固相に結合した標識量が有意に減少した場合、該被験物質をIFN-α発現増強物質の候補として選択することができる。
【0031】
好ましい実施態様においては、IFN-α mRNAとIFN-α AS RNAとを発現する細胞に被験物質を接触させ、該細胞における該mRNA量および/またはIFN-αタンパク質量の変化を測定することにより、より直接的にIFN-α発現調節物質を選択することができる。
IFN-α mRNAとIFN-α AS RNAとを発現する細胞は、両RNAを生来発現し得る細胞(例えば、白血球、リンパ芽球細胞等)であってもよいし、それらのいずれか一方もしくは両方を発現するDNAを導入した組換え細胞、あるいはウイルスを感染させた細胞であってもよい。組換え細胞の場合、宿主細胞として、例えば、H4IIE-C3細胞、HepG2細胞、293T細胞、HEK293細胞、COS7細胞、2B4T細胞、CHO細胞、MCF-7細胞、H295R細胞などの動物細胞をあげることができる。IFN-α mRNAおよびIFN-α AS RNAをコードするDNAは、両RNAを常法により単離し、逆転写反応等によって二本鎖DNAに変換した後、宿主細胞内で機能しうるプロモーターを有する発現ベクターに挿入して、例えば、リン酸カルシウム共沈殿法、PEG法、エレクトロポレーション法、マイクロインジェクション法、リポフェクション法などにより、このベクターを宿主細胞に導入することにより作製することができる。
【0032】
被験物質としては、前記したとおりのものが用いられる。被験物質と上記細胞との接触は、例えば、該細胞の培養に適した培地(例えば、約5〜20%の胎仔ウシ血清を含む最小必須培地(MEM)、ダルベッコ改変イーグル培地(DMEM)、RPMI1640培地、199培地、F12培地など)や各種緩衝液(例えば、HEPES緩衝液、リン酸緩衝液、リン酸緩衝生理食塩水、トリス塩酸緩衝液、ホウ酸緩衝液、酢酸緩衝液など)の中に被験物質を添加して、細胞を一定時間インキュベートすることにより実施することができる。添加される被験物質の濃度は化合物の種類(溶解度、毒性等)により異なるが、例えば、約0.1nM〜約100nMの範囲で適宜選択される。インキュベート時間としては、例えば、約1〜約24時間が挙げられる。
【0033】
インキュベーション終了後(あるいは経時的に)、細胞からRNAを抽出してRT-PCR、リアルタイムPCRやノーザンブロット解析によりIFN-α mRNA量を測定するか、あるいは培養上清を回収して、自体公知の各種イムノアッセイやウェスタンブロッティング等によりIFN-αタンパク質量を測定する。被験物質の添加により細胞におけるIFN-α mRNA量および/またはIFN-αタンパク質量が有意に増加した場合、該被験物質をIFN-α発現増強物質の候補として選択することができる。
【0034】
あるいはまた、ウイルス感染によりIFN-α mRNAとIFN-α AS RNAとを発現する細胞において、被験物質の存在下および非存在下で、該細胞におけるIFN-αの産生および/またはウイルス力価の変動を指標として、IFN-α発現調節物質をスクリーニングすることもできる。被験物質の存在下で、非存在下に比べてIFN-αの産生量が有意に増加した場合および/またはウイルス増殖が有意に抑制された場合、該被験物質をIFN-α発現増強物質の候補として選択することができる。
【0035】
上記のスクリーニング法により選択されたIFN-α発現増強物質は、本発明のセンスオリゴヌクレオチドと同様に、IFN-α mRNAに対する内在性AS RNAの作用を阻害することにより、IFN-α mRNAおよびIFN-αタンパク質の発現を増強することができる。したがって、上記IFN-α発現増強物質を含有する医薬は、例えば、B型肝炎、C型肝炎(活動性、非活動性)、ヘルペス感染症(性器ヘルペス、角膜ヘルペス炎、口腔ヘルペスなど)、尖圭コンジローマ、AIDS等のウイルス感染症、腎臓癌、腎細胞癌、乳癌、膀胱癌、基底細胞癌、頭頸部癌、頸管異形成、皮膚悪性腫瘍、カポジ肉腫、悪性黒色腫、非ホジキン型リンパ腫、幼児血管腫、慢性肉芽腫症、慢性骨髄性白血病(CML)、成人T細胞白血病、ヘアリー細胞白血病、毛様細胞白血病、T細胞白血病ウイルス(HTLV−1)骨髄症、多発性骨髄腫、リンパ腫などの癌、亜急性硬化性全脳炎(SSPE)、クローン病、関節リウマチ、シェーグレン症候群、多発性硬化症(MS)、口内炎、性器疣贅、膣内疣贅、赤血球増加症、血小板増加症、乾癬、菌状息肉症、突発性難聴、老年性円盤状黄斑変性症、ページェット病などの予防および/または治療に有用である。
【0036】
上記のスクリーニング法により選択された物質を含有する医薬は低毒性であり、そのまま液剤として、または適当な剤型の医薬組成物として、ヒトまたは非ヒト哺乳動物(例、マウス、ラット、ウサギ、ヒツジ、ブタ、ウシ、ネコ、イヌ、サルなど)に対して経口的または非経口的(例、吸入投与、血管内投与、皮下投与など)に投与することができる。投与に用いられる医薬組成物としては、選択された物質と薬理学的に許容され得る担体、希釈剤もしくは賦形剤とを含むものであってよい。
【0037】
非経口投与のための組成物としては、例えば、注射剤、エアロゾル剤、坐剤等が用いられ、注射剤は静脈注射剤、皮下注射剤、皮内注射剤、筋肉注射剤、点滴注射剤等の剤形を包含してもよい。このような注射剤は、公知の方法に従って調製できる。注射剤の調製方法としては、例えば、選択されたIFN-α発現増強物質を通常注射剤に用いられる無菌の水性液、または油性液に溶解、懸濁または乳化することによって調製できる。注射用の水性液としては、例えば、生理食塩水、ブドウ糖やその他の補助薬を含む等張液等が用いられ、適当な溶解補助剤、例えば、アルコール(例、エタノール)、ポリアルコール(例、プロピレングリコール、ポリエチレングリコール)、非イオン界面活性剤〔例、ポリソルベート80、HCO−50(polyoxyethylene(50mol)adduct of hydrogenated castor oil)〕等と併用してもよい。油性液としては、例えば、ゴマ油、大豆油等が用いられ、溶解補助剤として安息香酸ベンジル、ベンジルアルコール等を併用してもよい。調製された注射液は、適当なアンプルに充填されることが好ましい。エアロゾル製剤はジクロロジフロロメタン、プロパン、窒素などのような圧縮された許容しうる抛射薬内に入れることができる。あるいはネブライザーやアトマイザーのような非圧縮性製剤用医薬品して製剤化してもよい。直腸投与に用いられる坐剤は、上記IFN-α発現増強物質を通常の坐薬用基剤に混合することによって調製されても良い。
【0038】
経口投与のための組成物としては、固体または液体の剤形、具体的には錠剤(糖衣錠、フィルムコーティング錠を含む)、丸剤、顆粒剤、散剤、カプセル剤(ソフトカプセル剤を含む)、シロップ剤、乳剤、懸濁剤等が挙げられる。このような組成物は公知の方法によって製造され、製剤分野において通常用いられる担体、希釈剤もしくは賦形剤を含有していても良い。錠剤用の担体、賦形剤としては、例えば、乳糖、でんぷん、蔗糖、ステアリン酸マグネシウムが用いられる。
【0039】
上記の非経口用または経口用医薬組成物は、活性成分の投与量に適合するような投薬単位の剤形に調製されることが好都合である。このような投薬単位の剤形としては、例えば、錠剤、丸剤、カプセル剤、注射剤(アンプル)、エアロゾル剤、坐剤が挙げられる。IFN-α発現増強物質は、投薬単位剤形当たり通常5〜500mg、とりわけ注射剤では5〜100mg、その他の剤形では10〜250mg含有されていることが好ましい。
【0040】
上記のIFN-α発現増強物質を含有する上記医薬の投与量は、投与対象、対象疾患、症状、投与ルートなどによっても異なるが、例えば、成人の急性C型肝炎の予防および/または治療に使用する場合には、IFN-α発現増強物質を1回量として、通常0.01〜20mg/kg体重程度、好ましくは0.1〜10mg/kg体重程度、さらに好ましくは0.1〜5mg/kg体重程度を、1日1回〜数回程度、静脈注射により投与するのが好都合である。他の非経口投与および経口投与の場合もこれに準ずる量を投与することができる。症状が特に重い場合には、その症状に応じて増量してもよい。
【0041】
以下に実施例を挙げて本発明をさらに具体的に説明するが、これらは単なる例示であって本発明の範囲を何ら限定するものではない。
【実施例】
【0042】
材料と方法
1. 細胞培養
ヒトBリンパ芽球細胞株のNamalwa細胞 (ATCC CRL-1432)を、ウシ胎児血清 (FCS; GIBCO)を10%含むRPMI1640培地 (Wako) (R10)を用い、37℃, 5% CO2条件下で培養した。
【0043】
2. 細胞感染に用いるウイルスの用意と力価の測定
孵化鶏卵(10-12日齢)の漿尿膜腔に10,000倍希釈したSendai virus (SenV) (原液力価 5,120 HA unit/ ml)を200μl接種し、35℃で72時間感染後、4℃に一夜冷蔵した。この感染孵化鶏卵から回収した漿尿液中のSenVの力価を、初日ヒナ赤血球を用いた赤血球凝集試験により測定した。
【0044】
3. DNAトランスフェクション/ AS RNA knock-down (KD)とKD細胞
実験前日に培養Namalwa細胞を、6-ウェルプレート(Nunc)に3 x 106 cells/ 2 ml/ wellとなるように播種し、37℃, 5% CO2条件下で24時間培養した。その後、室温で150 x g, 4分間遠心沈澱させ、7ウェル分相当のNamalwa細胞を回収した。この細胞沈渣を7mlのR10に浮遊後、210μlのMATra-S Immobilizer (IBA; 7-2002-100)を添加し、5分毎に撹拌しながら、室温で15分間反応させた。その後、6-ウェルプレートを2枚使い、R10培地を予め1 ml宛分注した7ウェルに対しImmobilizerを吸着させたNamalwa細胞浮遊液各1mlを分注し、専用のマグネットプレート (IBA 7-2004-000)上に室温で、20分間留置した。この間に、RPMI1640 200μlあたり、センスオリゴヌクレオチド(hIFN-α/Se1-4)または陰性対照センスオリゴヌクレオチド (hIFN-α/NSe0-3, hIFN-α/ScrblSe1-2) 3μgとMATra-A Reagent (IBA 7-2001-100) 3μlを加え、混和後、室温で15分間留置し、トランスフェクション ミックスを作製した。所定の時間が経過したマグネットプレート上のImmobilizer吸着のNamalwa細胞に対し、このトランスフェクション ミックスを加え、10分間静置したのち、CO2インキュベーターに移し、37℃で6時間培養した。
センスオリゴヌクレオチド(*はS化したヌクレオチドを示す)
hIFN-α/Se1: 5’-C*C*A*GCAGATCTTCAACC*T*C*T-3’(配列番号3)
hIFN-α/Se2: 5’-A*T*C*TTCAACCTCTTTAC*C*A*C-3’(配列番号4)
hIFN-α/Se3: 5’-G*A*T*GAGGACCTCCTAGA*C*A*A-3’(配列番号5)
hIFN-α/Se4: 5’-G*A*C*CTCCTAGACAAATT*C*T*G-3’(配列番号6)
陰性対照センスオリゴヌクレオチド
hIFN-α/NSe0: 5’-G*A*C*CTTGATGCTCCTGG*C*A*C-3’(配列番号7)
hIFN-α/NSe1: 5’-A*A*T*CTGTCCTTCCTCCT*G*T*C-3’(配列番号8)
hIFN-α/NSe2: 5’-C*C*A*GGAGGAGTTTGATG*G*C*A-3’(配列番号9)
hIFN-α/NSe3: 5’-G*C*A*GAAATCATGAGATC*C*C*T-3’(配列番号10)
hIFN-α/ScrblSe1: 5’-G*C*T*CCCTAGCCCAACTA*T*T*A-3’(配列番号11)
hIFN-α/ScrblSe2: 5’-G*T*C*CCTACAATACCCAC*T*G*T-3’(配列番号12)
【0045】
4. Namalwa細胞へのSenV感染
上述のセンスオリゴヌクレオチドのトランスフェクションによりAS RNA発現をノックダウンしたNamalwa細胞 (AS RNA-minus) (6-ウェルプレートで、1ウェルあたり3x106 cells)あるいは、同一細胞数でセンスオリゴヌクレオチドをトランスフェクションしていないNamalwa細胞 (AS RNA-plus)をそれぞれ、室温で150 x g, 4分間、2回遠心・洗浄し、最終沈渣を2mlのRPMI1640に再浮遊した。その後SenVを50 HA unit/106 cellsの割合で加え、室温で1時間感染させた(それぞれAS RNA-minus/Inf細胞並びにAS RNA-plus/Inf細胞とする)。終了後、再度室温で150 x g, 4分間、2回遠心・洗浄し、3 x 106 cells/2 ml/wellとなるようにR10培地に浮遊後、6-ウェルプレートに再度播種した。各感染細胞は37℃で静置培養し、0, 0.5, 1, 3, 6, 12並びに24時間経過後に回収し、以下のRNA抽出、精製実験に供した。
【0046】
5. 細胞総RNA分画の抽出並びに各種酵素を用いたRNAの精製
5-a. 細胞総RNA分画の抽出
上記所定の時間経過後、AS RNA-minus/Inf細胞並びにAS RNA-plus/Inf細胞を回収し、4℃で150 x g, 4分間遠心後、各細胞沈渣に対しTRIzol (Invitrogen; 15596-018) 1mlを加えた。TRIzol中でピペッティングにより細胞を溶解後、室温で5分間留置した。その後、クロロフォルム (Wako; 試薬特級、 038-02606)0.2mlを加え、15秒間用手にて震盪混和後室温に再度2-3分間静置した。続いて、4℃にて12,000 x g, 15分間遠心分離した。この操作により試料は三層に分離するので、最上層の無色の水層(約600μl)から400μlを新しい滅菌済み1.5 ml微量遠心チューブに分取した。このチューブに、使用したTRIzol 1mlあたり0.5mlのイソプロピルアルコール(Wako; 試薬特級)を加え、室温にて混和、静置10分後、再度4℃にて12,000 x g, 10分間遠心した。上清を吸引、除去し、RNA沈渣に1mlの75%エチルアルコールを加えた。4℃にて7,500 x g, 5分間遠心し、上清を吸引、除去後、1mlの99.5%エチルアルコール(Wako; 試薬特級)を加え、再度4℃にて7,500 x g, 5分間遠心した。上清を吸引、除去後、沈渣を風乾し、滅菌再蒸留水に溶解した。このRNA溶液に対し、滅菌再蒸留水を最終210μlになるように添加後、20mg/mlのグリコーゲン (Roche Applied Science; 901 393)を1μl, 7.5Mの酢酸アンモニウム (Wako; 分子生物学用 01820485)を1/2容加え、撹拌後2.5容の99.5%エチルアルコールを添加し、4℃にて10分間留置した。その後、4℃, 13,000 rpmにて30分間遠心し、得られたRNA沈渣を上述のように75%, 99.5%のエチルアルコールで洗浄、風乾後、50μlのTE8.0(10 mM Tris・HCl, pH 8.0/ 0.1 mM EDTA)に溶解し、以下の酵素反応に供する迄-20℃に保存した。
【0047】
5-b. プロテアーゼ消化
上述の粗抽出した細胞総RNAにProteinase K (PCR grade; Roche Applied Science, 03115 887 001) 20μgを加え、Tris/SDS buffer (100mM Tris-HCl, pH7.4/ 50 mM EDTA, 2.5% SDS) 100μl中で、37℃, 15分間混入するNamalwa細胞由来の蛋白質成分を消化した。その後RNase-freeのTE8.0飽和フェノール(日本ジーン, 319 90093):クロロフォルム:イソアミルアルコール (Wako; 試薬特級; 135-12015)=25:24:1混合液 (PCIAA)を1容加え、4℃で15,300 x g, 2分間遠心した。分取した水層のRNA溶液に対し、イソアミルアルコール=24:1混合液 (CIAA)をさらに1容加え同様に遠心後、5-aで行ったと同様に水層中の細胞総RNAを塩析し、RNA沈渣を50μlのTE8.0に溶解した。
【0048】
5-c. DNase I 消化
この細胞総RNA溶液に対し、TURBO DNA-freeTM Kit (Applied Biosystems, AM1907) 添付のTURBO DNase2単位を加え、添付のバッファー100μl中で、37℃, 30分間混入するゲノムDNAを消化した。その後、さらに酵素を2単位追加し、計60分経過後に添付のDNase Inactivation Reagentを10μl加え、十分に撹拌後、室温に2分間放置し、酵素反応を停止させた。その後、4℃にて、9,100 x gで2分間遠心し、上清中の細胞総RNAを5-aで行ったと同様に塩析させた。AS-plus/Inf並びにAS-minus/Inf由来のRNA沈渣を各々60μl並びに30μlのTE8.0に溶解し、260, 280, 320nmの吸光度の測定から、RNA濃度並びにフェノールと蛋白質成分の混入度を測定した。精製したRNA標品のA260/A280比は常に1.95-2.05であり、A320値は0.006以下であった。
【0049】
6. 逆転写反応並びにPCR
6-a. 逆転写反応
前項で精製したAS-minus/Inf並びにAS-plus/Infの全細胞RNAの各2μgに対し最終14μlとなるように滅菌再蒸留水を加えた。続いて、mRNAの逆転写用に5μMのoligo(dT)12-18 primer (invitrogen; 18418-012)を、AS RNAの逆転写用に2μMのhIFN-α/F1 primerを各1μl加え、70℃で10分間保温後、4℃に留置した。これらのRNA溶液に対し、5倍濃度の逆転写酵素バッファー(東洋紡、TRT-1B)を5μl、10mM dNTPsを25μl、滅菌再蒸留水1.2μl、RNase阻害剤(RNase OUT; invitrogen、10777-019)0.3μl、逆転写酵素 (ReverTra Ace; 東洋紡、TRT-101) 1μlを各々加え、47℃で30分間逆転写反応を進めた。その後、70℃に15分間保温し酵素を失活させた後に4℃に留置した。次いで、上述の逆転写酵素バッファーで5単位/μlとなるように希釈したRNaseH (TakaraBio, 2150A)を1μl添加し、37℃、20分間鋳型RNAを消化後、前記のように作製したcDNAを塩析した。得られたcDNA沈渣は、mRNA由来を40μl、AS RNA由来は20μlのTE8.0に溶解した。
【0050】
6-b. PCRと Agarose gel 電気泳動
6-aで得たcDNA溶液の各2μlに対し、滅菌再蒸留水29.6μl, 10倍濃度のGene taqバッファー(日本ジーン、314-02873)を4μl、2.5mMのdNTPsを3.2μl、100μMのForward primerとReverse primer を各0.4μl加えた。次に、0.2μlのanti-Taq high (東洋紡、TCP-101)と0.2μlのtaq polymerase (Gene taq; 日本ジーン、314-02873)を混和後、室温で5分間保温し、この混合液0.4μlを各サンプルに添加後、MJ Mini Gradient Thermal Cycler (Bio-Rad)を用いて、PCRを以下のように行った。
1サイクル: 94℃, 1分
10サイクル: 94℃, 1分/ 72℃, 1分30秒
15サイクル: 94℃, 1分/ 65℃, 1分30秒/ 72℃, 10秒
5+αサイクル: 94℃, 1分/ 60℃, 1分30秒/ 72℃, 10秒
使用プライマー
IFN-α1 mRNA由来のcDNA用:
IFN-α/ F1:5’-TGGTGCTCAGCTGCAAGTCAAGC-3’ (配列番号13)
IFN-α/ R1:5’-AGAGATGGCTGGAGCCTTCTG-3’ (配列番号14)
AS RNA由来のcDNA用:
IFN-α/ F1B:5’-CTGGGCTGTGATCTCCCTGAGAC-3’ (配列番号15)
IFN-α/ R1:同上
IFN-β mRNA由来のcDNA用:
IFN-β/F1:5’-GATTCATCTAGCACTGGCTGG-3’ (配列番号16)
IFN-β/R1:5’-CCTTCAGGTAATGCAGAATCC-3’ (配列番号17)
EF mRNA由来のcDNA用:
EF/F1:5’-TCTGGTTGGAATGGTGACAACATGC-3’ (配列番号18)
EF/R1:5’-CCAGGAAGAGCTTCACTCAAAGCTT-3’ (配列番号19)
各精製RNA試料濃度を検定するための内部標準として増幅したEF mRNA由来のcDNAは30サイクル、IFN-α1 mRNA由来のcDNAは35+αサイクル、AS RNA由来のcDNA並びにIFN-β mRNA由来のcDNAは38+αサイクル増幅し、PCR産物は3%アガロース(アガロースS;日本ジーン、312-01193)による電気泳動で解析した。
尚、mRNA画分叉はAS RNA画分に混入するゲノムDNA由来の増幅シグナルを検出する目的で、逆転写反応に用いたAS-minus/Inf並びにAS-plus/Infの全細胞RNAを各々100ngまたは200ng使用して同様にPCRを行った。
【0051】
7. ELISAによるhIFN-αの定量
AS RNAノックダウンがIFN-α蛋白質発現に与える影響を検定する目的で、4のSenV感染Namalawa細胞培養上清を、感染後0, 0.5, 1, 3, 6, 12並びに24時間に回収し、上清中に分泌されたIFN-αをヒトインターフェロン−α ELISAキット(PBL Biomedical Laboratories, #41100-4)を用いて定量した。具体的には、添付のヒト精製IFN-αを適宜に希釈して作製した0-500pg/ml溶液と各時間毎に回収した培養上清の希釈液を、キット添付の抗IFN-α特異抗体を吸着させた96ウェルプレートに一定量添加後添付試薬を用いて呈色反応させ、450nmの吸光度を各々測定した。既知濃度のIFN-αの吸光度の測定結果から作製した検量線を用い、各時間毎の回収サンプルの吸光度から培養上清中に分泌されたIFN-α量を算出した。
【0052】
結果
1. RT-PCR法によるアンチセンスRNAの増幅、同定を最適化するプライマーの決定
図1にインターフェロン−α1 (IFNA1) 遺伝子及び実験に用いたプライマーの配列と、プライマー部位を遺伝子上に図示した模式図を示す (SLI, SLII並びにBulged SLは、図4のmRNA核外輸送責任領域が形成する二次構造を参照のこと)。
これらのプライマーを組み合わせ、RT-PCR法によりNamalwa細胞で構成的に発現しているIFNA1遺伝子由来のアンチセンス (AS) RNAを同定した結果を表1に要約した。
【0053】
【表1】
【0054】
最適な組合せは、RT反応プライマーにはF1を、PCRプライマーにF1B/R1を組み合わせた場合(1)であり、AS cDNAは35サイクルから検出可能となり、40サイクルでは明瞭に観察できた。この組み合わせは、増幅量は約1/10程度に留まるものの、やはり40サイクルでAS cDNAが観察できたF1逆転写プライマー、F2B/R2 PCRプライマーペアの組合せ(2)を除き、検出効率においてその他の組合せより10サイクル分、即ち210倍高かった。図2に(1)並びに(2)の組み合わせを用い、40サイクル増幅したAS RNA由来のcDNA (AS cDNA)を示す。RT (minus)は、逆転写を行っていないRNAサンプルを用いて実施したPCRの結果を示し、RNAサンプルに混入するゲノムDNA由来のシグナルの混入は無視できる程度であった。
【0055】
2. センダイウイルス(SenV) に伴うNamalwa細胞におけるAS RNAとIFN-α1 mRNAの発現動態の変動(図3)
Namalwa細胞において構成的に発現するAS RNA (IFN-α1 AS RNA; 0 hr) は、SenV感染により発現が急速に減衰 (IFN-α1 AS RNA; 1 hr) し、感染後3時間には消失した (IFN-α1 AS RNA; 3 hr)が、12時間後より再発現を認め (IFN-α1 AS RNA; 12 hr)、24時間経過後には感染前の発現レベルに復した (IFN-α1 AS RNA; 24 hr)。
これに対し、IFN-α1 mRNAの発現はAS RNAの発現動態と逆相関し、感染後1時間経過時点 (IFN-α1 mRNA; 1 hr)で第1のピークに到達後減衰し、6時間経過時点(IFN-α1 mRNA; 6 hr)では認められなくなった。興味深いことに、この時点でIFN-β mRNAの発現が一時的に認められ (IFN-β mRNA; 6 hr)、その後感染12時間の時点でIFN-α1 mRNAの発現を再度認めた (IFN-α1 mRNA; 12 hr)。この時点が同mRNA発現の第2で最大のピークであった。その後、その発現は感染24時間 (IFN-α1 mRNA: 24 hr)に向けて減衰し、この減衰と期を一にしてAS RNAの再発現が認められた (IFN-α1 AS RNA: 12, 24 hr)。
以上の結果から、AS RNA発現とSenV感染に伴うIFN-α1 mRNAの発現の間に相補関係が示唆されたため、これを検証する目的で以下に述べるセンスオリゴヌクレオチドによるAS RNA発現のノックダウンを行い、IFN-α蛋白質発現並びに感染ウイルス増殖に及ぼす影響を検証した。
【0056】
3. IFN-α1 mRNAの核外輸送責任領域が形成するRNA二次構造とセンスオリゴヌクレオチドによるAS RNA発現の阻害
これまでの研究から、IFN-α1 mRNAが、核内で転写、プロセッシングを受けた後核外輸送されるにあたっては、同mRNA上の輸送責任領域(CSS: IFNA1遺伝子 nt208-452)が形成する二つのステムループ (SLI, SLII)からなるRNA二次構造(図4)の核外輸送因子による認識が必要な事を明らかにした(Kimura et al., 投稿準備中)。その過程で、SLII構造中のBulged SL(図4)の有無がIFN-α1 mRNAの発現を左右する事が証明されたため(Kimura et al., 投稿準備中)、AS RNAとIFN-α1 mRNA間の相補的発現動態を考えあわせ、Bulged SL構造を形成する遺伝子配列に相同なセンスオリゴヌクレオチドを作製し、AS RNA発現ヘの影響を検討した。用いたセンスオリゴヌクレオチドSe1〜Se4のシークエンス(配列番号3〜6)とSLII構造中の位置を図5に示す。Se1(配列番号3)、Se2(配列番号4)は相互に重複しつつBulged SL構造を形成する配列と相補であり、Se3(配列番号5)とSe4(配列番号6)はこの領域を除く残りのSLII構造部分中Bulge/Bubble或いはループを形成する配列に相補となるように設計した。
これらを用いたAS RNA阻害実験結果を図6に示す。導入したセンスオリゴヌクレオチド中AS RNA発現の阻害効果はSe1が最大であり、その効果は導入12時間後も確認できた。
【0057】
4. AS RNA発現阻害(ノックダウン、KD)がIFN-αの発現とSenV増殖に及ぼす効果
Se1とSe3を等量ずつ導入したSenV感染Namalwa細胞から産生されたIFN-α量を、感染後経時的に測定した結果を図7に示す。IFN-αは未導入細胞においては感染6時間を経過して初めて検出可能であったが、センスオリゴヌクレオチドの導入により産生が早期に繰り上がった結果感染後30分の時点で既に検出できるようになり、感染24時間後に産生されたIFN-α量は未導入細胞に比較し70%増大していた。続いて、各経過時間に培養液中に放出されたSenVのウイルス力価を赤血球凝集試験で測定した。対照感染細胞では、感染後24時間の培養液中のSenV力価が8単位/mlであるのに対し、Se1とSe3を導入したNamalwa細胞においては2-4単位/mlに半減するのが観察され、感染細胞中に発現するIFN-α量の増加との逆相関が証明できた。
以上の結果から、Namalwa細胞中に構成的に発現し、SenV感染に伴うIFN-α1 mRNAの発現に逆相関的に異動する同mRNAに対するAS RNAの発現を、センスオリゴヌクレオチドを用いノックダウンしたところ、IFN-α蛋白質の発現の増大を通じ、感染ウイルスの増殖が抑制されることが明らかにできた。
【産業上の利用可能性】
【0058】
本発明のセンスオリゴヌクレオチドおよび本発明のスクリーニング方法により選択されるIFN-α発現増強物質は、ウイルス感染症や癌の新規治療薬として有用である。
【図面の簡単な説明】
【0059】
【図1】IFN-α AS RNAの増幅効率の至適化に用いたプライマーとIFNA1遺伝子上の位置を示す模式図である。
【図2】IFN-α AS RNA増幅プライマー至適化実験で決定した最良プライマーを用いたAS RNA増幅効果を示す図である。
【図3】SenV感染によるNamalwa細胞におけるIFN-α1 mRNAとAS RNAの発現動態の変動を示す図である。下のグラフはIFN-αタンパク質量の変動を示す。
【図4】IFN-α1 mRNAの核外輸送責任領域(CSS)の位置、該領域が形成するRNA二次構造並びに該領域の塩基配列を示す図である。
【図5】IFN-α AS RNAのノックダウン実験に用いたセンスオリゴヌクレオチドのCSSに対する位置とその配列を示す図である。
【図6】センスオリゴヌクレオチドによるIFN-α AS RNA発現のノックダウン効果を示す図である。
【図7】IFN-α AS RNAのノックダウンに伴うIFN-α1の早期出現とその増大、並びにウイルス増殖の遅延・抑制効果を示す図である。
【特許請求の範囲】
【請求項1】
インターフェロン−α mRNAに対する内在性アンチセンスRNAに相補的な配列を含み、インターフェロン−αの発現を増強させ得るセンスオリゴヌクレオチド。
【請求項2】
該配列がインターフェロン−α mRNAのSLIまたはSLII領域内の塩基配列に相同な配列である、請求項1記載のセンスオリゴヌクレオチド。
【請求項3】
該配列がBulged SL領域内の塩基配列に相同な配列である、請求項2記載のセンスオリゴヌクレオチド。
【請求項4】
請求項1〜3のいずれか1項に記載のセンスオリゴヌクレオチドを含有してなるインターフェロン−α発現増強剤。
【請求項5】
ウイルス感染症または癌の予防または治療用である、請求項4記載の剤。
【請求項6】
被験物質の存在下および非存在下で、インターフェロン−α mRNAとそれに対する内在性アンチセンスRNAとのハイブリダイゼーションを検出・比較することを特徴とする、インターフェロン−α発現調節物質のスクリーニング方法。
【請求項7】
インターフェロン−α mRNAとそれに対する内在性アンチセンスRNAとを発現する細胞に被験物質を接触させ、該細胞における該mRNA量および/またはインターフェロン−αタンパク質量の変化を測定することを特徴とする、請求項6記載の方法。
【請求項1】
インターフェロン−α mRNAに対する内在性アンチセンスRNAに相補的な配列を含み、インターフェロン−αの発現を増強させ得るセンスオリゴヌクレオチド。
【請求項2】
該配列がインターフェロン−α mRNAのSLIまたはSLII領域内の塩基配列に相同な配列である、請求項1記載のセンスオリゴヌクレオチド。
【請求項3】
該配列がBulged SL領域内の塩基配列に相同な配列である、請求項2記載のセンスオリゴヌクレオチド。
【請求項4】
請求項1〜3のいずれか1項に記載のセンスオリゴヌクレオチドを含有してなるインターフェロン−α発現増強剤。
【請求項5】
ウイルス感染症または癌の予防または治療用である、請求項4記載の剤。
【請求項6】
被験物質の存在下および非存在下で、インターフェロン−α mRNAとそれに対する内在性アンチセンスRNAとのハイブリダイゼーションを検出・比較することを特徴とする、インターフェロン−α発現調節物質のスクリーニング方法。
【請求項7】
インターフェロン−α mRNAとそれに対する内在性アンチセンスRNAとを発現する細胞に被験物質を接触させ、該細胞における該mRNA量および/またはインターフェロン−αタンパク質量の変化を測定することを特徴とする、請求項6記載の方法。
【図1】


【図4】
【図5】
【図2】
【図3】
【図6】
【図7】
【図4】
【図5】
【図2】
【図3】
【図6】
【図7】
【公開番号】特開2010−104280(P2010−104280A)
【公開日】平成22年5月13日(2010.5.13)
【国際特許分類】
【出願番号】特願2008−278954(P2008−278954)
【出願日】平成20年10月29日(2008.10.29)
【出願人】(593006630)学校法人立命館 (359)
【出願人】(500409219)学校法人関西医科大学 (36)
【出願人】(592196156)株式会社アミノアップ化学 (7)
【Fターム(参考)】
【公開日】平成22年5月13日(2010.5.13)
【国際特許分類】
【出願日】平成20年10月29日(2008.10.29)
【出願人】(593006630)学校法人立命館 (359)
【出願人】(500409219)学校法人関西医科大学 (36)
【出願人】(592196156)株式会社アミノアップ化学 (7)
【Fターム(参考)】
[ Back to top ]