
インフルエンザウイルスに対する中和分子
本発明は、インフルエンザA型ウイルスに対する中和抗体を同定、産生、および操作するための方法および手段、ならびに産生された中和抗体に関する。具体的には、本発明は、種々のインフルエンザA型ウイルス亜型に対する中和抗体、ならびにかかる抗体を作製する方法および手段に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、インフルエンザA型ウイルスに対する中和分子を同定、産生、および設計するための方法および手段、ならびに産生された中和分子に関する。本発明は、さらに、標的インフルエンザA型ウイルス上の本発明の中和分子の結合部位を使用したワクチンの設計および産生を含む、産生された分子の種々の使用に関する。
【背景技術】
【0002】
流感は、インフルエンザウイルスにより引き起こされる感染性呼吸器疾患である。インフルエンザは軽度から重度の疾患を引き起こし、時には死に至り得る。米国では年間、人口の5〜20%がインフルエンザに罹患し、約200,000人が入院し、約36,000の死者をもたらす。
【0003】
インフルエンザウイルスは咳およびくしゃみによって生じた呼吸器飛沫で広がり、それが通常人から人へ伝播する。インフルエンザ表面抗原、特に、血球凝集素に対する免疫は、感染の可能性、および感染した場合の疾患の重症度を軽減する。インフルエンザワクチンを利用可能ではあるが、1つのインフルエンザウイルスの型または亜型に対するワクチンは別のインフルエンザの型または亜型に対する限定的な防御を付与するか、または全く防御しないため、毎年インフルエンザワクチンの中に1つまたは複数の新しい株を組みこむ必要がある。
【0004】
インフルエンザウイルスは分節負鎖RNAウイルスであり、オルトミクスウイルス科に属する。インフルエンザA型ウイルスは、9種類の構造タンパク質からなり、調節機能を有する1つの非構造NS1タンパク質をさらにコードする。非構造NS1タンパク質は、再生サイクル中に大量に合成され、感染細胞のサイトゾルおよび細胞核内に局在する。ウイルスゲノムの分節性により、異なるウイルス株との細胞の混合感染中に遺伝子再集合(ゲノムセグメントの交換)機能が生じる。インフルエンザA型ウイルスは、その表面上に提示される異なる血球凝集素(HA)およびノイラミニダーゼ(NA)ウイルスタンパク質に応じて種々の亜型にさらに分類することができる。インフルエンザA型ウイルス亜型は、2つのウイルス表面糖タンパク質である、血球凝集素(HAまたはH)およびノイラミニダーゼ(NAまたはN)によって同定される。各インフルエンザウイルス亜型は、HおよびNタンパク質のその組み合わせによって同定される。16種のHA亜型および9種のNA亜型が公知である。インフルエンザA型ウイルスは、ヒト、トリ、ブタ、ウマ、および他の動物に感染し得るが、野鳥がこれらのウイルスの天然の宿主である。いくつかのインフルエンザA型亜型(すなわち、H1N1、H1N2、およびH3N2)のみが、現在ヒト間で流布しているが、16種のH亜型と9種のNA亜型とのすべての組み合わせが鳥類、特に、野生の水鳥および浜に生息する鳥で同定されている。さらに、H5およびH7インフルエンザウイルスもヒト疾病を引き起こし得るという証拠が増えつつある。
【0005】
インフルエンザA型ウイルスのHAは、2つの構造的に異なる領域、すなわち、球状頭部領域(globular head region)およびステム領域(stem region)を含む。球状頭部領域は、標的細胞へのウイルス付着を担い、かつHAの血球凝集活性に関与する受容体結合部位を含む。ステム領域は、ウイルスエンベロープと細胞のエンドソーム膜との間の細胞融合に必要であり、したがって融合活性に関連する融合ペプチドを含む(Wiley et al,Ann.Rev.Biochem.,56:365−394(1987))。
【0006】
パンデミック(世界的流行病)は、世界的な疾患の発生である。インフルエンザパンデミックは、新規のインフルエンザA型ウイルスが、(1)ヒト集団においてほとんどまたは全く免疫が存在しないものとして出現した場合、(2)重篤な疾病を引き起こし始め、次いで、(3)世界規模で人から人へ容易に伝播する場合に起こる。20世紀には、かかるインフルエンザパンデミックが3回起こっている。第1に、1918年に、「スペイン風邪」インフルエンザパンデミックにより、米国で少なくとも500,000人が死亡し、世界で4000万人までが死亡した。このパンデミックは、インフルエンザA型H1N1亜型に起因していた。第2に、1957年に、インフルエンザA型H2N2亜型に起因する「アジア風邪」インフルエンザパンデミックにより、米国で少なくとも70,000人が死亡し、世界で100〜200万人が死亡する結果となった。最も最近では、1968年に、インフルエンザA型H3N2亜型に起因する「香港風邪」インフルエンザパンデミックにより、米国で約34,000人が死亡し、世界で700,000人が死亡する結果となった。
【0007】
1997年に、最初のインフルエンザA型H5N1の症例が香港で報告された。これは、このトリウイルス型のヒトへの最初の直接感染であったが、ヒトからヒトへの感染が認められなかったので、パンデミックという結果にならなかった。
【0008】
Lu et al,Resp.Res.7:43(2006)(doi:10.1186/1465−992−7−43)は、不活化H5N1ウイルスを接種したウマ由来の抗H5N1 IgG、およびH5N1特異的F(ab’)2フラグメントの調製を報告しており、これらは、H5N1ウイルスを感染させたBALB/cマウスを防御すると説明されていた。
【0009】
Hanson et al.,Resp.Res.7:126(doi:10.1186/1465−9921−7−126)は、マウスの受動免疫化のためのインフルエンザA型ウイルスH5血球凝集素に特異的なキメラモノクローナル抗体の使用を記載している。
【0010】
インフルエンザウイルスに対する中和抗体は、2008年1月17日に公開された米国特許出願公開第20080014205号に開示されている。
【0011】
あるインフルエンザA型ウイルスに起因する呼吸器疾患の重症度および潜在的パンデミックの脅威を考慮して、有効な防止方法および治療方法が非常に必要とされている。本発明は、インフルエンザA型ウイルスのH1亜型、H3亜型、およびH5亜型が含まれるが、これらに限定されない、ウイルスの種々のH亜型に対するインフルエンザA型中和分子を提供することによって、この必要性に取り組んでいる。本発明は、さらに、種々のヒトおよび非ヒト種から得た分離株ならびに種々のインフルエンザのエピデミックおよび/またはパンデミックの犠牲者および/または生存者由来の分離株が含まれるが、これらに限定されない、インフルエンザA型ウイルスの所与の亜型の1つを超える分離株(株)、好ましくはすべての分離株(株)を中和することができる分子を提供する。
【0012】
かかる交差反応性中和分子は、感染集団またはエピデミックもしくはパンデミックの場合のリスク集団の受動免疫化を含む、インフルエンザウイルス感染の防止および/または治療のために使用することができる。さらに、交差反応性抗体は、将来のワクチン発見のための設計ガイド、および現在のワクチン臨床開発のための評価ツールとして使用することができる。
【発明の概要】
【0013】
1つの態様では、本発明は、重鎖および軽鎖ポリペプチド由来の超可変領域を含む結合分子を提供する。1つの実施形態では、結合分子は、配列番号7、配列番号8、および配列番号9からなる群より選択される重鎖からの1つ、2つ、もしくは3つの超可変領域配列、またはその機能的に活性なフラグメントを含む。別の実施形態では、結合分子は、配列番号7、配列番号8、および配列番号9のすべての超可変領域配列を含む。1つの他の実施形態では、結合分子は、軽鎖と会合したときに、標的に結合することが可能である、結合分子である。1つの実施形態では、軽鎖または結合分子は、配列番号13、配列番号14、および配列番号15のポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む。別の実施形態では、軽鎖または結合分子は、配列番号16、配列番号17、および配列番号18のポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む。1つの他の実施形態では、軽鎖または結合分子は、配列番号19、配列番号20、および配列番号21のポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む。別の実施形態では、軽鎖または結合分子は、配列番号13、配列番号14、および配列番号15のすべての超可変配列を含む。別の実施形態では、軽鎖または結合分子は、配列番号16、配列番号17、および配列番号18のすべての超可変配列を含む。1つの他の実施形態では、軽鎖または結合分子は、配列番号19、配列番号20、および配列番号21のすべての超可変配列を含む。
【0014】
別の実施形態では、結合分子は、配列番号10、配列番号11、および配列番号12からなる群より選択される重鎖からの1つ、2つ、もしくは3つの超可変配列、またはその機能的に活性なフラグメントを含む。1つの他の実施形態では、結合分子は、配列番号10、配列番号11、および配列番号12のすべての超可変領域配列を含む。1つの他の実施形態では、結合分子は、軽鎖と会合したときに、標的に結合することが可能である結合分子である。1つの他の実施形態では、軽鎖または結合分子は、配列番号22、配列番号23、および配列番号24のポリペプチド配列の1つ、2つ、もしくは3つの超可変配列を含む。1つの他の実施形態では、軽鎖または結合分子は、配列番号22、配列番号23、および配列番号24のすべての超可変領域配列を含む。
【0015】
1つの実施形態では、結合分子は抗体である。別の実施形態では、結合分子はサロボディである。
【0016】
別の態様では、本発明は、VpreB配列および/またはλ5配列を含む結合分子を提供する。1つの実施形態では、結合分子は、VpreB配列および/またはλ5配列を含むポリペプチドを含む。別の実施形態では、結合分子は、λ5配列に融合したVpreB配列を含むポリペプチドをさらに含む。1つの他の実施形態では、結合分子は、Vκ様配列および/またはJCκ配列を含むκ様代替軽鎖(SLC)構築物をさらに含む。
【0017】
すべての実施形態では、結合分子は、(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する。別の実施形態では、結合分子は、H1およびH3インフルエンザA型ウイルス亜型の少なくとも1つを中和する。1つの実施形態では、結合分子は、H1およびH3インフルエンザA型ウイルス亜型を中和する。別の実施形態では、結合分子は、インフルエンザA型ウイルスの球状頭部が細胞表面に結合するのを阻止する。1つの他の実施形態では、結合分子は、インフルエンザA型ウイルスが感染される細胞に付着するのを阻止する。別の実施形態では、結合分子は、H1HA抗原に結合する。1つの実施形態では、結合分子はまた、少なくとも1つのさらなるHA抗原に結合する。別の実施形態では、さらなるHA抗原は、H3である。別の実施形態では、結合分子は、H2HA抗原に結合する。
【0018】
1つの実施形態では、本発明は、重鎖を含む抗体を提供し、重鎖は、配列番号1として示したアミノ酸配列を含む。別の実施形態では、抗体は、配列番号3、配列番号4、および配列番号5からなる群より選択されるアミノ酸配列を含む軽鎖をさらに含む。1つの他の実施形態では、抗体は、(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する、抗体である。別の実施形態では、抗体は、H1およびH3インフルエンザA型ウイルス亜型の少なくとも1つを中和する抗体である。1つの実施形態では、抗体は、H1およびH3インフルエンザA型ウイルス亜型を中和する抗体である。別の実施形態では、抗体は、インフルエンザA型ウイルスの球状頭部が細胞表面に結合するのを阻止する抗体である。1つの他の実施形態では、抗体は、インフルエンザA型ウイルスが感染される細胞に付着するのを阻止する抗体である。別の実施形態では、抗体は、H1HA抗原に結合する抗体である。1つの実施形態では、抗体は、少なくとも1つのさらなるHA抗原に結合する抗体である。別の実施形態では、さらなるHA抗原は、H3である。
【0019】
別の実施形態では、抗体は、重鎖を含み、重鎖は、配列番号2として示したアミノ酸配列を含む。1つの他の実施形態では、抗体は、配列番号6として示したアミノ酸配列を含む軽鎖をさらに含む。1つの他の実施形態では、抗体は、(i)インフルエンザA型ウイルスの少なくとも1つの亜型および/または少なくとも1つの分離株を中和し、(ii)ウイルスの血球凝集素(HA)抗原に結合する、抗体である。1つの実施形態では、抗体は、HA抗原のH5亜型のエピトープに結合する抗体である。別の実施形態では、抗体は、H1HA抗原を中和する、H5HA抗原を中和する、またはH1およびH5インフルエンザA型ウイルス亜型を中和する、抗体である。
【0020】
別の実施形態では、抗体または結合分子は、インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株に結合する、および/またはそれに対して反応性である、および/またはそれを中和する。1つの実施形態では、ウイルスは、ヒトに感染する能力を有するウイルスである。別の実施形態では、分離株は、ヒト被験体から得られた分離株である。1つの他の実施形態では、分離株は、非ヒト動物から得られた分離株である。別の実施形態では、非ヒト動物は、トリである。1つの実施形態では、前記トリは、野鳥またはニワトリである。1つの他の実施形態では、非ヒト動物は、ブタである。
【0021】
別の実施形態では、抗体または結合分子は、HA抗原のH1亜型のエピトープに結合する、抗体または結合分子である。1つの他の実施形態では、抗体または結合分子は、HA抗原のH3亜型のエピトープに結合する、抗体または結合分子である。別の実施形態では、抗体または結合分子は、HA抗原のH1亜型のエピトープ、およびHA抗原のH3亜型のエピトープに結合する、抗体または結合分子である。1つの実施形態では、抗体または結合分子は、HA抗原のH9亜型のエピトープに結合する、抗体または結合分子である。別の実施形態では、抗体または結合分子は、HA抗原のH5亜型のエピトープに結合する、抗体または結合分子である。別の実施形態では、抗体または結合分子は、HA抗原のH2亜型のエピトープに結合する、抗体または結合分子である。
【0022】
すべての実施形態では、抗体または結合分子は、インフルエンザA型ウイルスの表面上に提示されるエピトープに結合する。
【0023】
すべての実施形態では、H1亜型、もしくは前記HAは、H1ウイルスのニューカレドニア/20/99分離株に由来するか、H1亜型、もしくは前記HAは、H1ウイルスのソロモン諸島/3/06分離株、またはH1ウイルスのメンフィス/3/2008分離株に由来する。
【0024】
すべての実施形態では、H3亜型、もしくは前記HAは、H3ウイルスのウィスコンシン/67/05分離株に由来するか、またはH3亜型、もしくは前記HAは、H3ウイルスの香港/68分離株に由来する。
【0025】
すべての実施形態では、H9亜型、もしくは前記HAは、H9ウイルスの香港/1073/99分離株に由来する。
【0026】
すべての実施形態では、H5亜型、もしくは前記HAは、H5ウイルスのベトナム/1203/04分離株に由来する。
【0027】
すべての実施形態では、H2亜型、もしくは前記HAは、H2ウイルスのアダチ/1/1957分離株に由来する。
【0028】
すべての実施形態では、抗体または結合分子は、H1HA抗原およびH3抗原と交差反応性である、抗体または結合分子である。
【0029】
1つの他の実施形態では、本発明は、配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する抗体または結合分子を提供する。別の実施形態では、抗体または結合分子は、配列番号3、配列番号4、および配列番号5からなる群より選択されるアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む軽鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する。1つの他の実施形態では、抗体または結合分子は、(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する、抗体または結合分子である。
【0030】
1つの実施形態では、本発明は、配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子を提供する。別の実施形態では、抗体または結合分子は、配列番号6として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む軽鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する。1つの他の実施形態では、抗体または結合分子は、(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)ウイルスの血球凝集素(HA)に結合する、抗体または結合分子である。
【0031】
別の実施形態では、本発明は、本明細書中に記載の結合分子または抗体を含む組成物を提供する。
【0032】
別の態様では、本発明は、インフルエンザA型ウイルス感染の治療および/または防止を必要とする被験体における、インフルエンザA型ウイルス感染の治療および/または防止のための方法を提供する。1つの実施形態では、前記方法は、有効量の本明細書中に記載の組成物を前記被験体に投与することを含む。別の実施形態では、前記方法は、有効量の本明細書中に記載の中和抗体または結合分子を前記被験体に投与することを含む。1つの他の実施形態では、前記被験体はヒト患者である。
【0033】
別の態様では、本発明は、インフルエンザA型ウイルス感染に有効なワクチンを提供する。1つの実施形態では、ワクチンは、本明細書中に記載の抗体または結合分子に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む。別の実施形態では、ワクチンは合成ワクチンである。1つの他の実施形態では、ワクチンは弱毒化インフルエンザA型ウイルスまたはその一部を含む。別の実施形態では、ワクチンは死滅させたインフルエンザA型ウイルスまたはその一部を含む。1つの他の実施形態では、抗体または結合分子は、(a)配列番号1として示したアミノ酸配列、または該アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子、(b)配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む重鎖ポリペプチドを含む抗体、(c)配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子、および(d)配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体、からなる群より選択される。
【0034】
すべての実施形態では、抗体はHA抗原に結合する。HA抗原は、H3亜型、H1亜型、H2亜型、H1亜型およびH3亜型、H5亜型、H9亜型、ならびにそれらの任意の組み合わせからなる群より選択され得る。1つの実施形態では、抗原は、インフルエンザA型ウイルスの表面上に提示される。別の実施形態では、本明細書中に記載の抗体または結合分子に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドは、中和抗体を惹起する抗原決定基を含む。別の実施形態では、ワクチンは、経口投与、経皮投与、または非経口投与に適切である。別の実施形態では、ワクチンは、経粘膜送達に好適である。1つの他の実施形態では、経粘膜送達は、鼻腔内投与である。別の実施形態では、ワクチンは、小児期免疫化用ワクチンである。
【0035】
1つの態様では、本発明は、長さが修正された重鎖ループを有するインフルエンザ中和抗体または結合分子を提供する。長さが修正された鎖は、延長重鎖ループまたは短縮重鎖ループであり得る。1つの実施形態では、抗体または結合分子は、延長重鎖ループを有するインフルエンザA型ウイルスの少なくとも1つの分離株を中和する、ヒトに感染する能力を有するインフルエンザA型ウイルスの血球凝集素へ結合する、中和抗体または結合分子である。1つの他の実施形態では、長さが修正された重鎖ループは、CDR3ループまたはCDR1ループである。別の実施形態では、CDR3ループは、配列番号9のアミノ酸配列を含む。さらに別の抗体では、CDR1ループは、配列番号7のアミノ酸配列を含む。
【0036】
別の態様では、本発明は、低下した酸化能を有する設計抗体または結合分子を提供する。1つの実施形態では、低下した酸化能を有する設計抗体または結合分子は、重鎖可変ドメイン中の1つまたは複数のメチオニン置換を有するインフルエンザA型ウイルスの少なくとも1つの分離株を中和する、ヒトに感染する能力を有するインフルエンザA型ウイルスの血球凝集素へ結合する抗体である。別の実施形態では、重鎖可変ドメインは、CDR3領域である。1つの他の実施形態では、メチオニン置換は、カバットナンバリングシステムによる96位および/または98位である。さらに別の実施形態では、メチオニンは、ロイシンで置換される。1つの実施形態では、重鎖可変ドメインは、配列番号29のアミノ酸配列を含む。
【図面の簡単な説明】
【0037】
【図1】交差反応性を生じる/増加させるための並行した発見プールの組換えによる、2つの異なる標的(標的AおよびB)に対する反応強度を増加させるための戦略を示す。組換え抗体ライブラリーの各選択ラウンドにより、両標的に対する得られたプールの反応強度を増加させる。
【図2】標的Bに対する交差反応性を増加させる一方で、標的Aに対する反応性を維持する戦略を示す。第1に、標的Aに反応性を示すクローンを選択し、次いで、標的Aに反応性を示すクローンの変異誘発ライブラリーを調製し、示すように選択が行われ、標的Aおよび標的Bの両方との強い反応性を示す1つまたは複数の抗体を得る。
【図3】「目的変異誘発」法によって多様な多機能抗体集団を生成するための代表的な変異誘発法を示す。
【図4】H1、H3、およびH9から血球凝集素抗原への1286−C5抗体の結合能力を示す。
【図5】H1、H3、H5、およびH9から血球凝集素抗原への1286−A11抗体の結合能力を示す。
【図6】合体された抗体ライブラリーを生成するための代表的な方法を示す。
【図7A】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Aは、高力価の致死的H3N2ウイルス攻撃に対するC05抗体による予防効果を示す。
【図7B】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Bは、高力価の致死的H3N2ウイルス攻撃に対するC05抗体による予防効果を示す。
【図7C】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Cは、致死的H3N2ウイルス攻撃に対するC05抗体による治療効果を示す。
【図7D】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Dは、致死的H3N2ウイルス攻撃に対するC05抗体による治療効果を示す。
【図7E】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Eは、高力価の季節性H1N1ウイルス攻撃に対するC05抗体による予防効果を示す。
【図8】C05変異体がH1およびH3HAタンパク質の認識を維持することを示す。
【図9】C05変異体がH1およびH3HAタンパク質の認識を維持することを示す。
【発明を実施するための形態】
【0038】
A.定義
他で定義されない限り、本明細書中で使用される技術用語および科学用語は、本発明の属する分野の当業者によって一般に理解される意味と同様の意味を有する。Singleton et al.,Dictionary of Microbiology and Molecular Biology 2nd ed.,J.Wiley & Sons(New York,NY 1994)は、本出願で使用した多数の用語に対する一般的な指針を当業者に提供している。
【0039】
当業者は、本発明の実施の際に使用することができる本明細書中に記載のものと類似または等価の多数の方法および材料を認識するであろう。実際、本発明は、記載の方法および材料に決して制限されない。本発明の目的のために、以下の用語を下記のように定義する。
【0040】
「インフルエンザA型亜型」または「インフルエンザA型ウイルス亜型」という用語は、交換可能に使用され、これらは、血球凝集素(H)ウイルス表面タンパク質によって特徴づけられ、それにより、H番号、例えば、H1、H3、H5、およびH9等によって分類されるインフルエンザA型ウイルス変異体を表す。加えて、亜型を、N番号、例えば、N1およびN2等によって示したノイラミニダーゼ(N)ウイルス表面タンパク質によってさらに特徴づけることができる。したがって、亜型を、H番号およびN番号の両方、例えば、H1N1、H2N2、H3N2、H5N1、H5N2、およびH9N2等によって表すことができる。本用語には、具体的には、通常は変異に起因し、異なる病理学的プロファイルを示す各亜型内のすべての株(絶滅した株が含まれる)が含まれる。かかる株はまた、過去、現在、および未来の分離株のすべてを含む、ウイルス亜型の種々の「分離株」と表されるであろう。したがって、本文脈中で、「株」および「分離株」という用語は、交換可能に使用される。亜型は、インフルエンザA型ウイルスに基づいた抗原を含む。抗原は、血球凝集素ウイルス表面タンパク質に基づくことができ、この抗原を「HA抗原」と指定することができる。いくつかの例では、かかる抗原は、特定の亜型、例えば、H1亜型およびH3亜型等のタンパク質に基づき、それぞれ、H1抗原およびH3抗原と指定することができる。
【0041】
「インフルエンザ」という用語は、インフルエンザウイルスに起因する接触伝染病を表すために使用される。
【0042】
本発明の文脈において、「結合分子」という用語は、最も広い意味で使用され、標的に特異的に結合するポリペプチド配列を含む任意の分子を含む。定義には、ホモ二量体またはヘテロ二量体形態等の単量体または多量体であれ、抗体および抗体フラグメント、抗体様分子およびそのフラグメントが含まれるが、これらに限定されない。多量体結合分子は、共有結合性および/または非共有結合性相互作用を介してそれらの配座を保持することができ、かつそれらが標的(例えば、抗体の場合では、抗原)に結合する必要特性を保持する限り、互いに、および/または分子もしくは部分に抱合し得る。
特定のポリペプチドまたは特定のポリペプチド上のエピトープ「に特異的に結合する」、または「それに特異的」である結合分子は、任意の他のポリペプチドまたはポリペプチドエピトープに実質的に結合することなく、その特定のポリペプチドまたは特定のポリペプチド上のエピトープに結合するものである。
【0043】
「抱合体」、「抱合した」、および「抱合」という用語は、任意のおよびすべての共有結合または非共有結合の形態を表し、直接的な遺伝子融合または化学的融合、リンカーまたは架橋剤によるカップリング、および、例えば、ファンデルワールス力またはロイシンジッパーの使用による非共有結合が含まれるが、これらに限定されない。
【0044】
「抗体」(Ab)という用語は、最も広い意味で使用され、特定の抗原、ならびに免疫グロブリンおよび抗原特異性が欠乏する他の抗体様分子に対して結合特異性を示すポリペプチドが含まれる。後者の種類のポリペプチドは、例えば、リンパ系によって低レベルで、かつ骨髄腫により増大したレベルで産生される。本用途では、「抗体」という用語は、単クローン抗体、多クローン抗体、および抗体フラグメントを具体的に含むが、これらに限定されない。
【0045】
「低下した酸化能」または「減少した不均質性酸化能」という用語は、酸化的アミノ酸から非酸化的アミノ酸への少なくとも1つのアミノ酸置換を有するポリペプチドを含有する抗体を表す。アミノ酸配列は、メチオニン置換であり得る。抗体ポリペプチドは、メチオニンアミノ酸残基を非酸化的アミノ酸残基で置換するために、選択的に操作され得、それによって、低下した酸化能を有する抗体を提供する。例えば、メチオニンは、ロイシン、セリン、またはアラニンで置換され得る。
【0046】
「未変性の抗体」は、通常、2つの同一の軽(L)鎖および2つの同一の重(H)鎖から構成される約150,000ダルトンのヘテロ四量体糖タンパク質である。各軽鎖は、共有結合性のジスルフィド結合によって重鎖に連結されている一方で、ジスルフィド結合数は異なる免疫グロブリンアイソタイプの重鎖間で異なる。各重鎖および軽鎖はまた、規則的間隔の鎖間ジスルフィド架橋を有する。各重鎖は、一方の末端に可変ドメイン(VH)を有し、その後に多数の定常ドメインを有する。各軽鎖は、一方の末端に可変ドメイン(VL)を有し、その他方の末端に定常ドメインを有し、軽鎖の定常ドメインは、重鎖の第1の定常ドメインと整列し、軽鎖可変ドメインは、重鎖の可変ドメインと整列している。特定のアミノ酸残基は、軽鎖と重鎖の可変ドメイン間の界面を形成すると考えられている(Chothia et al,J.Mol.Biol.186:651(1985)、Novotny and Haber,Proc.Natl.Acad.Sci.U.S.A.82:4592(1985))。
【0047】
抗体鎖に関する「可変」という用語は、抗体間の配列が非常に異なり、かつ特定の各抗体のその特定の抗原に対する結合および特異性が関連する抗体鎖部分を表すために使用される。かかる可変性は、軽鎖および重鎖の可変ドメインの両方において、超可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存されている部分は、フレームワーク領域(FR)と呼ばれる。未変性の重鎖および軽鎖の可変ドメインはそれぞれ、4つのFR(それぞれ、FR1、FR2、FR3、およびFR4)を含み、FRはそのほとんどが3つの超可変領域と連結したβシート配置の形態をとり、それにより、超可変領域はβシート構造に連結してループを形成し、βシート構造の一部を形成することもある。各鎖の超可変領域は、FRによって他の鎖由来の超可変領域とともに極近傍に保持され、これが抗体の抗原結合部位の形成に寄与している(Kabat et al,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.(1991),pages647−669を参照)。定常ドメインは、抗体の抗原への結合に直接は関与しないが、抗体依存性細胞毒性における抗体の関与等の種々のエフェクター機能を示す。
【0048】
「超可変領域」という用語は、本明細書中で抗原結合を担う抗体のアミノ酸残基を表す。超可変領域は、「相補性決定領域」または「CDR」由来のアミノ酸残基(すなわち、軽鎖可変ドメイン中の残基30〜36(L1)、46〜55(L2)、および86〜96(L3)、ならびに重鎖可変ドメイン中の30〜35(H1)、47〜58(H2)、および93〜101(H3))を含む(MacCallum et al, J Mol Biol.1996)。「フレームワーク」または「FR」残基は、本明細書中に定義の超可変領域残基以外の可変ドメイン残基である。
【0049】
それらの重鎖の定常ドメインのアミノ酸配列に応じて、抗体を異なるクラスに割り当てることができる。5つの主な抗体クラスのIgA、IgD、IgE、IgG、およびIgMがあり、これらのうちのいくつかは、さらに、サブクラス(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgA、およびIgA2に分けられ得る。
【0050】
異なる免疫グロブリンクラスに対応する重鎖定常ドメインはそれぞれ、α、δ、ε、γ、およびμと呼ばれる。
【0051】
任意の脊椎動物種由来の抗体の「軽鎖」を、それらの定常ドメインのアミノ酸配列に基づいて、カッパ(κ)およびラムダ(λ)と呼ばれる2つの明確に異なる型の1つに割り当てることができる。
【0052】
「抗体フラグメント」という用語は、全長抗体の一部、一般に、その抗原結合ドメインまたは可変ドメインである。抗体フラグメントの例には、Fab、Fab’、F(ab’)2、およびFvフラグメント、線状抗体、一本鎖抗体分子、ディアボディ、ならびに抗体フラグメントから形成された多重特異性抗体が含まれるが、これらに限定されない。抗体フラグメントのさらなる例には、scFv、(scFv)2、dAb(単一ドメイン抗体)、および相補性決定領域(CDR)フラグメント、ならびにその2つのループが組み合わせ変異誘発の影響を受けやすい最小化可変ドメインであるミニボディが含まれるが、これらに限定されない。
【0053】
「モノクローナル抗体」という用語は、B細胞の単一クローンによって合成される抗体分子を表すために使用される。修飾語「モノクローナル」は、実質的に均一な抗体集団から得られた抗体の特徴を示し、任意の特定の方法によって抗体産生が必要であると解釈されるべきではない。したがって、モノクローナル抗体を、Kohler and Milstein,Nature 256:495(1975)、Eur.J.Immunol.6:511(1976)に最初に記載されたハイブリドーマ法、組換えDNA技術によって作製することができるか、あるいはファージ抗体ライブラリーから単離することもできる。
【0054】
「ポリクローナル抗体」という用語は、B細胞集団によって合成された抗体分子集団を表すために使用される。
【0055】
「単鎖Fv」または「sFv」抗体フラグメントは、抗体のVHドメインおよびVLドメインを含み、これらのドメインは、単一のポリペプチド鎖中に存在する。一般に、Fvポリペプチドは、さらに、VHドメインとVLドメインとの間にポリペプチドリンカーを含み、それにより、sFvが抗原結合に所望の構造を形成することができる。sFvの概説については、Pluckthun in The Pharmacology of Monoclonal Antibodies,vol.113,RosenburgおよびMoore eds.Springer−Verlag,New York,pp.269−315(1994)を参照。単鎖抗体は、例えば、WO88/06630号およびWO92/01047号に開示されている。
【0056】
「ディアボディ」という用語は、2つの抗原結合部位を有する小さな抗体フラグメントを表し、このフラグメントは、同一ポリペプチド鎖中に軽鎖可変ドメイン(VL)に連結した重鎖可変ドメイン(VH)を含む(VH−VL)。短すぎて同一鎖上の2ドメイン間で対合が不可能なリンカーの使用により、ドメインは別の鎖の相補ドメインと強制的に対合して2つの抗原結合部位が作製される。ディアボディは、例えば、欧州特許第404,097号、WO93/11161号、およびHollinger et al,Proc.Natl.Acad.Sci.USA 90:6444−6448(1993)において、より完全に記載されている。
【0057】
「ミニボディ」という用語は、自己アセンブリして80kDaの2価の二量体(scFv−CH3)2を形成するscFv−CH3融合タンパク質を表すために使用される。
【0058】
「アプタマー」という用語は、高い特異性および親和性を有するタンパク質標的に結合する合成核酸リガンドを表すために本明細書中で使用される。アプタマーは、タンパク質機能の有力な阻害剤として公知である。
【0059】
dAbフラグメント(Ward et al.,Nature 341:544 546(1989))は、VHドメインまたはVLドメインからなる。
【0060】
本明細書中で使用する場合、「抗体結合領域」という用語は、抗原に結合することが可能な免疫グロブリンまたは抗体可変領域の1つまたは複数の部分を表す。典型的には、抗体結合領域は、例えば、抗体軽鎖(VL)(またはその可変領域)、抗体重鎖(VH)(またはその可変領域)、重鎖Fd領域、Fab、F(ab’)2等の抗体軽鎖と重鎖(またはその可変領域)との組み合わせ、単一ドメイン、または一本鎖抗体(scFv)、または全長抗体、例えば、IgG(例えば、IgG1、IgG2、IgG3、またはIgG4亜型)、IgA1、IgA2、IgD、IgE、またはIgM抗体等である。
【0061】
「二重特異性抗体」という用語は、2つの異なる抗原型に特異性を示す抗体を表す。この用語には、本明細書中で具体的に使用する場合、標的抗原、および特定の組織への送達を容易にする別の標的に対して結合特異性を示す抗体が含まれるが、これらに限定されない。同様に、多重特異性抗体は、2つ以上の結合特異性を有する。
【0062】
「線状抗体」という表現は、抗原結合領域対を形成するタンデムFdセグメント対(VH−CH1−VH−CH1)を含むことを表すために使用される。線状抗体は、二重特異性または単一特性であり得、例えば、Zapata et al,Protein Eng.8(10):1057−1062(1995)に記載されている。
【0063】
本発明の目的のために、「抗体様分子」という用語には、ウイルス抗原に結合してそれを中和することができる上記定義の抗体フラグメント以外の任意の分子が含まれる。本用語には、具体的には、例えば、2008年10月2日に公開されたPCT公開第WO2008/118970号、およびXu et al.,Proc.Natl.Acad.Sci.USA,105(31):10756−61(2008)に記載されるような、代替軽鎖(SLC)エレメントを含む「サロボディ」と呼ばれるプレB細胞受容体(pre−BCR)様構造が含まれるが、これらに限定されない。SLCは、非共有結合したVpre−Bおよびλ5タンパク質から構成される非分散型のヘテロ二量体である。それぞれ、VpreB鎖はVλIgドメインに相同であり、λ5鎖は基準抗体のCλドメインに相同である。ヘテロ二量体SLCは、pre−BCR HCのCλドメインと第1の定常ドメインとの間のジスルフィド結合によってpre−BCR複合体中の重鎖と共有結合している。SLCの固有の特徴は、VpreB1およびλ5ドメインの非基準ペプチドがそれぞれ伸長していることである。VpreB1はそのC末端にさらなる21残基を有し、λ5はそのN末端上に50アミノ酸長のテールを有する(例えば、Vettermann et al.,Semin.Immunol.18:44−55(2006)を参照)。サロボディ構造には、具体的には、pre−BCRの未変性三量体pre−BCR様機能単位、VpreB1とλ5との融合物、およびλ5のN末端50アミノ酸、VpreB1のC末端21アミノ酸、または両方のペプチド伸長物のいずれかが排除された三量体が含まれるが、これらに限定されない。加えて、古典的抗体軽鎖の定常成分を使用したキメラ構築物は、サロボディの定義内に明確に含まれる。
【0064】
他の代表的な「抗体様分子」は、本明細書中に定義されるように、抗体代替κ軽鎖配列を含む類似の構造であり、このκ軽鎖配列は、任意に、別のポリペプチド、例えば、抗体重鎖および/または軽鎖ドメイン配列等とパートナーを組む。κ様B細胞受容体(κ様BCR)は、κ様代替軽鎖(κ様SLC)を使用して同定されている(Frances et al.,EMBO J 13:5937−43(1994)、Thompson et al.,Immunogenetics 48:305−11(1998)、Rangel et al.,J Biol Chem 280:17807−14(2005))。Rangelら、J Biol Chem 280(18):17807−17814(2005)は、非再配列Vκ遺伝子の産物であるVκ様タンパク質の同定および分子の特徴づけを報告しており、これは、Thompsonら、Immunogenetics 48:305−311(1998)によって以前に報告されたcDNA配列と同一であることが判明した。それに対して、Francesら、EMBO J 13:5937−43(1994)は、B細胞前駆体表面でμ重鎖と会合する能力を有し、それにより、B細胞成長のためのλ5経路の代替物が得られる再配列生殖系列JCkの同定および特徴づけを報告した。κ様およびλ様pre−BCRが協力して軽鎖の再配列を促進するように作用し、B細胞前駆体の成熟を確実にすることが示唆されている。概説については、McKeller and Martinez−Valdez Seminars in Immunology 18:4043(2006)を参照。
【0065】
「λ5」という用語は、本明細書中で最も広い意味で使用され、任意の未変性配列または変異体λ5ポリペプチドを表し、具体的には、未変性配列のヒトおよび他の哺乳動物のλ5ポリペプチド、翻訳後修飾によって形成された変異体、ならびにかかる未変性配列のポリペプチドの変異体が含まれるが、これらに限定されない。
【0066】
「変異体VpreBポリペプチド」および「VpreBポリペプチドの変異体」という用語は、交換可能に使用され、アミノ酸修飾の結果として、1つまたは複数のアミノ酸位置における未変性配列のVpreBポリペプチドと異なるポリペプチドであると本明細書中で定義する。「変異体VpreBポリペプチド」は、本明細書中に定義されるように、未変性の抗体λまたはK軽鎖配列またはそのフラグメントと異なるであろう。「変異体VpreBポリペプチド」は、好ましくは、未変性配列のVpreBポリペプチドと少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約98%の配列同一性を保持するであろう。別の好ましい実施形態では、「変異体VpreBポリペプチド」は、未変性抗体のλまたはK軽鎖配列とそのアミノ酸配列が95%未満、90%未満、85%未満、80%未満、75%未満、70%未満、65%未満、または60%未満同一であろう。変異体VpreBポリペプチドには、具体的には、VpreB配列のC末端の非Ig様の固有のテールが部分的または完全に除去されたVpreBポリペプチドが含まれるが、これらに限定されない。「変異体λ5ポリペプチド」および「λ5ポリペプチドの変異体」という用語は、交換可能に使用され、アミノ酸修飾の結果として、1つまたは複数のアミノ酸位置における未変性配列のλ5ポリペプチドと異なるポリペプチドであると本明細書中で定義する。「変異体λ5ポリペプチド」は、本明細書中に定義されるように、未変性抗体のλもしくはκ軽鎖配列、またはそのフラグメントと異なるであろう。「変異体λ5ポリペプチド」は、好ましくは、未変性配列のλ5ポリペプチドと少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約98%の配列同一性を保持するであろう。別の好ましい実施形態では、「変異体λ5ポリペプチド」は、未変性の抗体のλまたはκ軽鎖配列とそのアミノ酸配列が95%未満、90%未満、85%未満、80%未満、75%未満、70%未満、65%未満、または60%未満同一であろう。変異体λ5ポリペプチドは、具体的には、λ5配列のN末端の固有のテールが部分的または完全に除去されたλ5ポリペプチドが含まれるが、これらに限定されない。
【0067】
「VpreB配列」という用語は、上記定義の「VpreB」の配列またはそのフラグメントを表すために本明細書中で使用される。
【0068】
「λ5配列」という用語は、上記定義の「λ5」の配列またはそのフラグメントを表すために本明細書中で使用される。
【0069】
「代替軽鎖配列」という用語は、本明細書中に定義されるように、上記定義の「VpreB配列」および/または「λ5配列」を含む任意のポリペプチド配列を意味する。
【0070】
「κ様代替軽鎖可変ドメイン」、「Vκ様SLC」、および「Vκ様」という用語は、交換可能に使用され、これらは、非再配列Vκ遺伝子およびその変異体の産物である任意の未変性配列のポリペプチドを表す。1つの実施形態では、未変性配列のVκ様ポリペプチドの変異体は、抗体κ軽鎖配列に関連するC末端伸長(テール)を含む。特定の実施形態では、未変性配列のVκ様ポリペプチドの変異体は、Vκ様ポリペプチドと対応する抗体κ軽鎖とを区別する固有のC末端伸長(テール)の少なくとも一部、好ましくはすべてを保持する。別の実施形態では、変異体Vκ様ポリペプチドのC末端テールは、残りの配列と天然には関連しない配列である。後者の実施形態では、未変性Vκ様配列中に天然に存在するC末端テールと変異体配列との相違は、1つまたは複数のアミノ酸の改変(置換、挿入、欠失、および/または付加)に起因し得るか、またはC末端テールが異なるVκ様タンパク質中に天然に存在するテールと同一であり得る。Vκ様ポリペプチドは、1つまたは複数の抗体κ軽鎖CDR1、CDR2、およびCDR3配列に対応する領域中にアミノ酸の改変を含み得る。すべての例では、変異体は、未変性の抗体のκ軽鎖可変領域配列と比較して少なくとも4個、少なくとも5個、少なくとも6個、少なくとも7個、少なくとも8個、少なくとも9個、または少なくとも10個のアミノ酸、好ましくは4〜100個、4〜90個、4〜80個、4〜70個、4〜60個、4〜50個、4〜45個、4〜40個、4〜35個、4〜30個、4〜25個、4〜20個、4〜15、または4〜10個のアミノ酸残基のC末端伸長を含むことができるか、好ましくは含む。本明細書中に定義されるように、Vκ様ポリペプチド変異体は、未変性の抗体のκもしくはλ軽鎖配列またはそのフラグメントと異なり、好ましくは、未変性配列のVκポリペプチドと少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約98%の配列同一性を保持するであろう。別の好ましい実施形態では、Vκ様ポリペプチド変異体は、未変性の抗体のλまたはκ軽鎖配列とそのアミノ酸配列が95%未満、90%未満、85%未満、80%未満、75%未満、70%未満、65%未満、60%未満、55%未満、50%未満、45%未満、または40%未満同一であろう。他の実施形態では、配列同一性は、約40%と約95%との間、約45%と約90%との間、約50%と約85%との間、約55%と約80%との間、約60%と約75%との間、約60%と約80%との間、約65%と約85%との間、約65%と約90%との間、または約65%と約95%との間である。すべての実施形態では、好ましくは、Vκ様ポリペプチドは標的に結合することができる。
【0071】
「JCκ」および「JCκ様」という用語は、交換可能に使用され、これらは、未変性配列のκJ定常(C)領域セグメントおよび固有のN末端伸長(テール)、ならびにその変異体と同一の部分を含む未変性配列のポリペプチドを表す。1つの実施形態では、未変性配列のJCκ様ポリペプチドの変異体は、抗体JCセグメントと区別されるN末端伸長(テール)を含む。特定の実施形態では、未変性配列のJCκ様ポリペプチドの変異体は、JCκ様ポリペプチドを対応する抗体κ軽鎖JCセグメントと区別する固有のN末端伸長(テール)の少なくとも一部、好ましくはすべてを保持する。別の実施形態では、変異体JCκ様ポリペプチドのN末端テールは、残りの配列と天然には関連しない配列である。後者の実施形態では、未変性JCκ様配列中に天然に存在するN末端テールと変異体配列との相違は、1つまたは複数のアミノ酸の改変(置換、挿入、欠失、および/または付加)に起因し得るか、またはN末端テールは異なるJCκ様タンパク質中に天然に存在するテールと同一であり得る。未変性配列のJCκ様ポリペプチドの変異体は、未変性の抗体のκ可変ドメインJC配列と同一の配列の一部に1つまたは複数のアミノ酸の改変を含み得る。すべての場合、変異体は、未変性の抗体のκ軽鎖JC配列と比較して少なくとも4個、少なくとも5個、少なくとも6個、少なくとも7個、少なくとも8個、少なくとも9個、または少なくとも10個のアミノ酸、好ましくは4〜100個、または4〜90個、または4〜80個、または4〜70個、または4〜60個、または4〜50個、または4〜45個、または4〜40個、または4〜35個、または4〜30個、または4〜25個、または4〜20個、または4〜15個、または4〜10個のアミノ酸残基のN末端伸長(固有のN末端)を含むことができるか、好ましくは含む。JCκ様ポリペプチド変異体は、本明細書中に定義されるように、未変性の抗体のλもしくはκ軽鎖JC配列またはそのフラグメントと異なり、好ましくは、未変性配列のJCポリペプチドと少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約98%の配列同一性を保持するであろう。別の好ましい実施形態では、JCκ様ポリペプチド変異体は、未変性の抗体のλもしくはκ軽鎖JC配列とそのアミノ酸配列が95%未満、90%未満、85%未満、80%未満、75%未満、70%未満、65%未満、または60%未満同一であろう。他の実施形態では、配列同一性は、約40%と約95%との間、約45%と約90%との間、約50%と約85%との間、約55%と約80%との間、約60%と約75%との間、約60%と約80%との間、約65%と約85%との間、約65%と約90%との間、または約65%と〜約95%との間である。
【0072】
アミノ酸配列同一率を、配列比較プログラムNCBI−BLAST2を用いて決定することができる(Altschul et al.,Nucleic Acids Res.25:3389−3402(1997))。NCBI−BLAST2配列比較プログラムを、http://www.ncbi.nlm.nih.govからダウンロードすることができるか、またはそうでなければ、National Institute of Health,Bethesda,MDから入手することができる。NCBI−BLAST2は、いくつかの検索パラメーターを使用し、これらの検索パラメーターのすべてを、例えば、アンマスク=イエス、鎖=すべて、予想される発生数=10、最小低複雑性長=15/5、マルチパスe値=0.01、マルチパス定数=25、最終ギャップアラインメントのドロップオフ=25、およびスコアリング行列=BLOSUM62を含む、デフォルト値に設定する。
【0073】
「融合」という用語は、そのヌクレオチドコード配列のインフレームでの組み合わせによる1つのポリペプチド鎖中の異なる起源のアミノ酸の組み合わせを表すために本明細書中で使用される。「融合」という用語は、その末端の1つへの融合に加えて、内部融合、すなわち、ポリペプチド鎖内の異なる起源の配列の挿入を明確に含む。
【0074】
本明細書中で使用する場合、「ペプチド」、「ポリペプチド」、および「タンパク質」という用語は、すべて、共有結合性の「ペプチド結合」によって連結されたアミノ酸の一次配列を表す。一般に、ペプチドは、少数のアミノ酸、典型的には、約2〜約50個のアミノ酸からなり、タンパク質より短い。「ポリペプチド」という用語は、本明細書中に定義されるように、ペプチドおよびタンパク質を含む。
【0075】
「アミノ酸」または「アミノ酸残基」という用語は、典型的に、その当該分野で認識された定義を有するアミノ酸、例えば、アラニン(Ala)、アルギニン(Arg)、アスパラギン(Asn)、アスパラギン酸(Asp)、システイン(Cys)、グルタミン(Gln)、グルタミン酸(Glu)、グリシン(Gly)、ヒスチジン(His)、イソロイシン(Ile)、ロイシン(Leu)、リシン(Lys)、メチオニン(Met)、フェニルアラニン(Phe)、プロリン(Pro)、セリン(Ser)、トレオニン(Thr)、トリプトファン(Trp)、チロシン(Tyr)、およびバリン(Val)からなる群より選択されるアミノ酸等を表すが、必要に応じて、修飾アミノ酸、合成アミノ酸、または希アミノ酸を使用することができる。したがって、連邦規則集第37巻1.822(b)(4)に列挙した修飾アミノ酸および通常でないアミノ酸は、本定義内に具体的に含まれ、参照により本明細書に明確に組み込まれる。アミノ酸を、種々の亜群に分類することができる。したがって、アミノ酸を、非極性側鎖(例えば、Ala、Cys、Ile、Leu、Met、Phe、Pro、Val)、負電荷の側鎖(例えば、Asp、Glu)、正電荷の側鎖(例えば、Arg、His、Lys)、または無電荷極性側鎖(例えば、Asn、Cys、Gln、Gly、His、Met、Phe、Ser、Thr、Trp、およびTyr)を有するアミノ酸に分類することができる。アミノ酸を、小アミノ酸(Gly、Ala)、求核性アミノ酸(Ser、His、Thr、Cys)、疎水性アミノ酸(Val、Leu、Ile、Met、Pro)、芳香族アミノ酸(Phe、Tyr、Trp、Asp、Glu)、アミド(Asp、Glu)、および塩基性アミノ酸(Lys、Arg)に分類することもできる。
【0076】
「ポリヌクレオチド」という用語は、DNA分子およびRNA分子等の核酸、ならびにその類似体(例えば、ヌクレオチド類似体または核酸化学を用いて生成したDNAまたはRNA)を表す。必要に応じて、ポリヌクレオチドを、例えば、当該分野で認識された核酸化学を用いて合成的に作製するか、または例えば、ポリメラーゼを用いて酵素的に作製するか、必要に応じて、修飾することができる。典型的な修飾には、メチル化、ビオチン化、および他の当該分野で公知の修飾が含まれる。加えて、核酸分子は、必要に応じて、検出可能な部分に連結した一本鎖または二本鎖であり得る。
【0077】
基準ポリペプチドに関する「変異体」という用語は、未変性ポリペプチドと比較して、少なくとも1つのアミノ酸の変異または修飾(すなわち、改変)を保有するポリペプチドを表す。「アミノ酸修飾」によって生成された変異体を、例えば、未変性アミノ酸配列中の少なくとも1つのアミノ酸の置換、欠失、挿入、および/または化学修飾によって産生することができる。
【0078】
「アミノ酸修飾」は、所定のアミノ酸配列のアミノ酸配列の変化を表す。例示的な修飾には、アミノ酸の置換、挿入、および/または欠失が含まれる。
【0079】
「特定の位置でのアミノ酸修飾」は、特定の残基の置換もしくは欠失、または特定の残基に隣接する少なくとも1つのアミノ酸残基の挿入を表す。「隣接する」特定の残基の挿入は、その1〜2個の残基内の挿入を意味する。挿入は、特定の残基のN末端またはC末端であり得る。
【0080】
「アミノ酸置換」は、所定のアミノ酸配列中の少なくとも1つの既存のアミノ酸残基の別の異なる「置換」アミノ酸残基との置換を表す。置換残基または残基は、「天然に存在するアミノ酸残基」(すなわち、遺伝子コードによってコードされる)であり得、アラニン(Ala)、アルギニン(Arg)、アスパラギン(Asn)、アスパラギン酸(Asp)、システイン(Cys)、グルタミン(Gln)、グルタミン酸(Glu)、グリシン(Gly)、ヒスチジン(His)、イソロイシン(Ile)、ロイシン(Leu)、リシン(Lys)、メチオニン(Met)、フェニルアラニン(Phe)、プロリン(Pro)、セリン(Ser)、トレオニン(Thr)、トリプトファン(Trp)、チロシン(Tyr)、およびバリン(Val)からなる群より選択され得る。1つまたは複数の天然に存在しないアミノ酸残基との置換はまた、本明細書中のアミノ酸置換の定義に含まれる。
【0081】
「天然に存在しないアミノ酸残基」は、ポリペプチド鎖中の隣接アミノ酸残基に共有結合することができる上記列挙の天然に存在するアミノ酸残基以外の残基を表す。天然に存在しないアミノ酸残基の例には、ノルロイシン、オルニチン、ノルバリン、ホモセリン、およびEllmanらのMeth.Enzvm.202:301 336(1991)に記載されるもの等の他のアミノ酸残基類似体が含まれる。かかる天然に存在しないアミノ酸残基を生成するために、Norenら Science 244:182(1989)およびEllmanら(前出)の手順を使用することができる。簡潔に述べれば、これらの手順は、天然に存在しないアミノ酸残基での抑制性tRNAの化学的活性化およびその後のRNAのインビトロ転写および翻訳を含む。
【0082】
「アミノ酸挿入」は、所定のアミノ酸配列への少なくとも1つのアミノ酸の組み込みを表す。挿入は、通常、1つまたは2つのアミノ酸残基の挿入からなる一方で、本出願は、より巨大な「ペプチド挿入」、例えば、約3〜約5個、またはさらに約10個までのアミノ酸残基の挿入を企図する。挿入される残基は、上記開示の天然に存在する残基または天然に存在しない残基であり得る。
【0083】
「アミノ酸欠失」は、所定のアミノ酸配列からの少なくとも1つのアミノ酸残基の除去を表す。
【0084】
「変異誘発」という用語は、他で特定しない限り、任意の当該分野で認識されたポリヌクレオチド配列またはポリペプチド配列を改変させる技術を表す。好ましい変異誘発型には、エラー多発性PCR変異誘発、飽和変異誘発、または他の部位特異的変異誘発が含まれる。
【0085】
「部位特異的変異誘発」は、当該分野で標準的な技術であり、少数のミスマッチを除いて所望の変異を示す変異誘発すべき一本鎖ファージDNAに相補的な合成オリゴヌクレオチドプライマーを用いて行う。簡潔に述べれば、合成オリゴヌクレオチドを、一本鎖ファージDNAに相補的な鎖の合成を指示するためのプライマーとして使用し、得られた二本鎖DNAを、ファージ支持宿主細菌に形質転換する。形質転換された細菌の培養物を上層寒天にプレートし、ファージを保有する単一の細胞からプラークを形成させる。理論的には、新規のプラークの50%が一本鎖として変異形態を有するファージを含み、50%が元の配列を有するであろう。目的のプラークを、正確に適合するハイブリッド形成が可能であるが、元の鎖とのミスマッチがハイブリッド形成を阻止するのに十分である温度でのキナーゼ化合成プライマーとのハイブリッド形成によって選択する。次いで、プローブとハイブリッド形成するプラークを選択し、配列決定し、培養し、DNAを回収する。
【0086】
「中和分子」という用語は、本明細書中でその最も広い意味で使用され、これは、中和を達成する機構と無関係にウイルスが標的細胞に複製的に感染するのを阻害する任意の分子を表す。中和分子は、好ましくは、上記定義の抗体または抗体様分子もしくは結合分子である。中和は、例えば、細胞表面へのウイルスの付着または接着を阻害することによって、例えば、ウイルスの付着または接着を担う部位に直接結合するか、またはその近傍で結合する抗体または抗体様分子もしくは結合分子等の分子を操作することによって、達成することができる。中和はまた、ビリオン表面に指向してビリオンの凝縮をもたらす抗体または抗体様分子等の分子によって達成することができる。中和は、標的細胞へのウイルスの付着後のウイルス膜と細胞膜との融合の阻害、エンドサイトーシスの阻害、および感染細胞由来の子孫ウイルスの阻害等によってさらに生じ得る。本発明の、抗体または抗体様分子もしくは結合分子等の中和分子は、中和を達成する機構によって制限されない。
【0087】
「抗体レパートリー」という用語は、本明細書中で最も広い意味で使用され、これは、結合能力、結合特異性、胃腸管輸送能力、安定性、および親和性等の特定の性質をスクリーニングするために使用することができる抗体または抗体フラグメントの集団を表す。本用語には、具体的には、例えば、抗体ファージ提示ライブラリー等の組み合わせライブラリーのすべての型を含む、抗体ライブラリーが含まれ、これには、ナイーブライブラリー、合成ライブラリー、および半合成ライブラリーを含む、任意の供給源由来の単鎖Fv(scFv)およびFab抗体ファージ提示ライブラリーが含まれるが、これらに限定されない。
【0088】
同様に、(上記定義のように)「抗体様分子のレパートリー」は、結合能力、結合特異性、胃腸管輸送能力、安定性、および親和性等の特定の性質をスクリーニングするために使用することができる、かかる分子の集団を表す。本用語には、具体的には、サロボディライブラリーおよびκ様軽鎖構築物のライブラリー(上記定義)が含まれ、これらには、例えば、抗体ファージ提示ライブラリー等の組み合わせライブラリーのすべての型が含まれる。組み合わせサロボディライブラリーは、例えば、Xu et al.,(2008)、(前出)に開示されている。
【0089】
「抗体レパートリー」という用語は、本明細書中で最も広い意味で使用され、これは、結合能力、結合特異性、胃腸管輸送能力、安定性、および親和性等の特定の性質をスクリーニングするために使用することができる抗体または抗体フラグメントの集団を表す。本用語には、具体的には、例えば、抗体ファージ提示ライブラリー等の組み合わせライブラリーのすべての型を含む、抗体ライブラリーが含まれ、これには、ナイーブライブラリー、合成ライブラリー、および半合成ライブラリーを含む、任意の供給源由来の単鎖Fv(scFv)およびFab抗体ファージ提示ライブラリーが含まれるが、これらに限定されない。
【0090】
「ファージ提示ライブラリー」は、ファージコートタンパク質との融合物としてのクローン化タンパク質配列集団を発現するタンパク質発現ライブラリーである。したがって、「ファージ提示ライブラリー」という語句は、本明細書中で、ファージ(例えば、繊維状ファージ)の集団を表し、このファージは外部(典型的には、異種)タンパク質を発現する。外部タンパク質は、ファージが接触する他の部分と自由に相互作用する(または結合する)。外部タンパク質を提示する各ファージは、ファージ提示ライブラリーの「メンバー」である。
【0091】
「抗体ファージ提示ライブラリー」は、抗体または抗体フラグメントを提示するファージ提示ライブラリーを表す。抗体ライブラリーには、ファージ集団、かかるファージ集団をコードするベクター集団、またはかかるファージ集団もしくはベクター集団を保有する細胞が含まれる。ライブラリーは、ファージ粒子あたり平均して1つの一本鎖抗体または抗体フラグメントを提示する1価であり得るか、ウイルス粒子あたり平均して2つ以上の抗体または抗体フラグメントを提示する2価であり得る。「抗体フラグメント」という用語には、単鎖Fv(scFv)フラグメントおよびFabフラグメントが含まれるが、これらに限定されない。好ましい抗体ライブラリーは、平均して106個超、107個超、108個超、または109個超の異なるメンバーを含む。
【0092】
「繊維状ファージ」という用語は、その表面上に異種ポリペプチドを提示することができるウイルス粒子を表し、繊維状ファージには、f1、fd、Pf1、およびM13が含まれるが、これらに限定されない。繊維状ファージは、テトラサイクリン(例えば、「fd−tet」)等の選択マーカーを含むことができる。種々の繊維状ファージ提示系が当業者に周知である(例えば、Zacher et al,Gene 9:127−140(1980)、Smith et al,Science 228:1315−1317(1985)、およびParmley and Smith,Gene 73:305−318(1988)を参照)。
【0093】
「パニング」という用語は、標的に対して高い親和性および特異性を有する抗体等の化合物を保有するファージの同定および単離における複数ラウンドのスクリーニング過程を表すために使用される。
【0094】
本明細書中で使用する、「非ヒト動物」という用語は、哺乳動物、例えば、非ヒト霊長類、げっ歯類(例えば、マウスおよびラット)、および例えば、ウサギ、ブタ、ヒツジ、ヤギ、ウシ、ブタ、ウマ、およびロバ等の非げっ歯類動物を含むが、これらに限定されない。鳥類(例えば、ニワトリ、シチメンチョウ、アヒル、およびガチョウ等)も含まれる。本明細書中で使用する、「非霊長類動物」という用語は、特に上記列挙の哺乳動物を含むが、これらに限定されず、霊長類以外の哺乳動物を表す。
【0095】
「機能的に異なる抗体」という語句、およびその文法上の変異体は、結合特異性、結合親和性、および例えば、標的を中和する能力、もしくは生物学的活性の範囲または質等の任意の免疫学的機能または生物学的機能を含むが、これらに限定されず、少なくとも1つの性質が互い異なる抗体を表すために使用される。
【0096】
「保存アミノ酸残基」という語句は、互いに整列している2つ以上のアミノ酸配列間で同一のアミノ酸残基を表すために使用される。
【0097】
本明細書中で使用する場合、「エピトープ」という用語は、少なくとも約3〜5個、好ましくは少なくとも約5〜10個、または少なくとも約5〜15個のアミノ酸、典型的には、せいぜい500個または約1,000個のアミノ酸の配列を表し、それ自体またはより大きな配列の一部として、かかる配列に応答して生成される抗体に結合する配列を定義する。エピトープは、それが由来する親タンパク質の一部と同一の配列を有するポリペプチドに制限されない。実際、ウイルスゲノムは、その状態が絶えず変化し、分離株間で比較的高い変異性を示す。したがって、「エピトープ」という用語は、未変性配列と同一の配列、および未変性配列の欠失、置換および/または挿入等の修飾物を含む。一般に、かかる修飾物は事実上保存されるが、非保存的修飾も企図される。この用語には、具体的には、「ミモトープ」、すなわち、連続的な線状の未変性配列を同定しないか、あるいは必ずしも未変性タンパク質中に生じないが、未変性タンパク質上のエピトープを機能的に模倣する配列が含まれる。「エピトープ」という用語には、具体的には、線状および立体構造エピトープが含まれる。
【0098】
B.一般的技術
本発明の方法を実施するための技術当該分野で周知であり、標準的な実験テキスト(例えば、Ausubel et al.,Current Protocols of Molecular Biology,John Wiley and Sons(1997)、Molecular Cloning:A Laboratory Manual,Third Edition,J.Sambrook and D.W.Russelleds.,Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001、Antibody Phage Display:Methods and Protocols,P.M.O‘Brian and R.Aitken eds.,Human Press,In:Methods in Molecular Biology,Vol.178、Phage Display:A Laboratory Manual,C.F.Barbas III et al.eds.,Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001、およびAntibodies,G.Subramanian,ed.,Kluwer Academic,2004に記載されている。変異誘発を、例えば、部位特異的変異誘発(Kunkel et al,Proc.Natl.Acad.Sci USA 82:488−492(1985))を用いて行うことができる。
【0099】
1つの態様では、本発明のウイルス抗原中和分子は、典型的には、抗体または多様なポリペプチドライブラリーを用いて選択される抗体である。以下の記載では、本発明は、ある抗体ライブラリー型を参照して例示しているが、本発明は、任意の特定の抗体または多様なポリペプチドライブラリー型の使用に制限されない。組換えモノクローナル抗体ライブラリーは、免疫フラグメントまたはナイーブフラグメントに基づき得る。免疫抗体ライブラリー由来の抗体は、典型的には、無作為な組み合わせライブラリーを産生するために、元のB細胞から発現に適切なベクターにクローン化されるVHおよびVL遺伝子プールを使用して構築し、その後に抗体を選択および/またはスクリーニングすることができる。他のライブラリー型は、抗原に結合するクローンに明確に偏っていない遺伝子供給源由来の抗体フラグメントから構成され得る。したがって、ナイーブ抗体ライブラリーは、天然で免疫化されていない再配列V遺伝子に由来する。合成抗体ライブラリーを、1つまたは複数のV遺伝子のCDRへの完全または適合した縮重領域の導入するインビトロ法によって完全に構築する。半合成ライブラリーは、天然の多様性と合成の多様性が組み合わされており、しばしば、天然の多様性が増加する一方で、所望のレベルの機能的多様性が保持されるように作製される。したがって、かかるライブラリーは、例えば、天然CDR領域のシャフリング(Soderlind et al,Nat.Biotechnol.18:852−856(2000))またはヒトB細胞由来の天然に再配列されたCDR配列と合成CDR1およびCDR2多様性との組み合わせ(Hoet et al,Nat.Biotechnol.23:455−38(2005))によって作製することができる。本発明は、ナイーブ抗体ライブラリー、合成抗体ライブラリー、および半合成抗体ライブラリー、またはその任意の組み合わせの使用を含む。
【0100】
同様に、本発明の方法は、抗体の提示のために使用されるいかなる特定の技術にも制限されない。本発明はファージ提示を参照して例示されているが、本発明の抗体を、他の提示および富化技術によって同定することもできる。抗体フラグメントは、抗体遺伝子をコードする繊維状ファージの表面上に提示されている(Hoogenboom and Winter J.Mol.Biol,222:381 388(1992)、McCafferty et al,Nature 348(6301):552 554(1990)、Griffiths et al EMBO J.,13(14):3245−3260(1994))。抗体ライブラリーの選択およびスクリーニング技術の概説については、例えば、Hoogenboom,Nature Biotechnol.23(9):1105−1116(2005)を参照。加えて、Escherichia coli(Agterberg et al,Gene 88:37−45(1990)、Charbit et al,Gene 70:181−189(1988)、Francisco et al,Proc.Natl.Acad.Sci.USA 89:2713−2717(1992))およびSaccharomyces cerevisiae等の酵母(Boder and Wittrup,Nat.Biotechnol.15:553−557(1997)、Kieke et al,Protein Eng.10:1303−1310(1997))の表面上の異種タンパク質およびそのフラグメントの提示について当該分野で公知の系が存在する。他の公知の提示技術には、リボゾームまたはmRNA提示(Mattheakis et al,Proc.Natl,Acad.Sci,USA 91:9022−9026(1994)、Hanes and Pluckthun,Proc.Natl,Acad.Sci,USA 94:4937−4942(1997))、DNA提示(Yonezawa et al,Nucl.Acid Res.31(19):e118(2003))、細菌提示等の微生物細胞提示(Georgiou et al,Nature Biotech.15:29−34(1997))、哺乳動物細胞上の提示、胞子提示(Isticato et al,J.Bacteriol.183:6294−6301(2001)、Cheng et al,Appl.Environ,Microbiol 71:3337−3341(2005)、および2006年11月13日に出願された同時係属の米国仮出願第60/865,574号))、レトロウイルス提示等のウイルス提示(Urban et al,Nucleic Acids Res.33:e35(2005)、タンパク質−DNA結合に基づいた提示(Odegrip et al,Proc.Acad.Natl、Sci.USA 101:2806−2810(2004)、Reiersen et al,Nucleic Acids Res.33:e10(2005)、ならびにマイクロビーズ提示(Sepp et al,FEBS Lett.532:455−458(2002))が含まれる。
【0101】
C.好ましい実施形態の詳細な説明
1つの態様では、本発明は、インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、ウイルスの血球凝集素(HA)抗原に結合するが、血球凝集を阻害しないモノクローナル抗体および抗体様分子の選択、産生、および使用に関する。
【0102】
インフルエンザA型ウイルスのビリオンは、8セグメントの線状の負のセンス一本鎖RNAを含む。総ゲノム長は13600ヌクレオチドであり、この8セグメントは、2350ヌクレオチド長、2350ヌクレオチド長、2250ヌクレオチド長、1780ヌクレオチド長、1575ヌクレオチド長、1420ヌクレオチド長、1050ヌクレオチド長、および900ヌクレオチド長である。インフルエンザA型ウイルスの宿主特異性および弱毒化は、ウイルス血球凝集素(H、HA)、核タンパク質(NP)、マトリックス(M)、および非構造(NS)遺伝子にそれぞれまたはウイルス遺伝子の組み合わせで寄与している(例えば、Rogers et al,Virology 127:361−373(1983)、Scholtissek et al,Virology 147:287−294(1985)、Snyder et al,J.Clin.Microbiol.24:467−469(1986)、Tian et al,J.Virol.53:771−775(1985)、Treanor et al,Virology 171:1−9(1989)を参照)。
【0103】
血球凝集素およびノイラミニダーゼタンパク質を含む、インフルエンザA型ウイルスおよびその表面タンパク質のヌクレオチド配列およびアミノ酸配列は、例えば、Theoretical Biology and Biophysics Group of Los Alamos National Laboratoryが保持しているインフルエンザ配列データベース等のGenBankおよび他の配列データベースから利用可能である。インフルエンザA型ウイルス血球凝集素の15種の公知のH亜型(H1〜H15)のアミノ酸配列は、2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)に示されている。さらなるインフルエンザA型ウイルス血球凝集素亜型(H16)が、最近スウェーデンにてユリカモメから単離され、Fouchier et al.,J.Virol.79(5):2814−22(2005)によって報告された。各H亜型の非常に多様な株も公知である。例えば、H5 A/香港/156/97と指定されたHAタンパク質配列は、1997年5月に香港でヒトから単離されたインフルエンザA型H5N1ウイルスから決定され、Suarez et al.,J.Virol.72:6678−6688(1998)に他の関連H5N1分離株から得たいくつかのさらなる株の配列と比較して示されている。
【0104】
インフルエンザウイルスノイラミニダーゼの触媒部位および抗原部位の構造はColman et al.,Nature 303:41−4(1983)によって公開されており、ノイラミニダーゼ配列はGenBankおよび他の配列データベースから利用可能である。
【0105】
感染個体の免疫応答に起因するウイルス特異的抗体が、典型的にはウイルス血球凝集素との相互作用を介してウイルスを中和することが公知である(Ada et al.,Curr.Top.Microbiol.Immunol.128:1−54(1986)、Couch et al.,Annu.Rev.Micobiol.37:529−549(1983))。インフルエンザウイルス血球凝集素の三次元構造およびインフルエンザウイルス血球凝集素と中和抗体との複合体の結晶構造も決定され、公開されている(例えば、Wilson et al.,Nature 289:366−73(1981)、Ruigrok et al.,J.Gen.Virol.69(Pt 11):2785−95(1988)、Wrigley et al.,Virology 131(2):308−14(1983)、Daniels et al.,EMBO J.6:1459−1465(1987)、およびBizebard et al.,Nature 376:92−94(2002)を参照)。
【0106】
本発明によれば、所望の性質を有する抗体を1つまたは複数の抗体ライブラリーから特定し、この抗体は、種々の供給源に由来し、異なる型であり得る。
【0107】
包括的ヒトインフルエンザ抗体ライブラリー
包括的ヒトインフルエンザ抗体ライブラリーは、種々の以前のインフルエンザ、季節性の大流行、エピデミック、ならびに1968年の香港風邪(H3N2)、1957年のアジア風邪(H2N2)、1918年のスペイン風邪(H1N1)、および2004/2005年のトリインフルエンザ(H5N1)を含む、パンデミックの回復期患者から得た抗体から作製することができる。例えば、2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照。かかるライブラリーを調製するために、血液または骨髄のサンプルを、インフルエンザウイルスに感染していることが知られているか疑いのある個体から採取する。特に地理的に離れた供給源由来の末梢血サンプルを、輸送および使用前に安定化することが必要であり得る。この目的のためのキットは、周知かつ市販されており、例えば、BD Vacutainer(登録商標)CPT(商標)細胞調製管等をリンパ球の遠心精製のために使用することができ、グアニジウム、トリゾール、またはRNAlaterをサンプル安定化のために使用した。安定化リンパ球または全骨髄の受け取りの際、当該分野で公知の免疫グロブリンオリゴプライマーを使用してRT−RCPを行い、重鎖および軽鎖レパートリーをレスキューする。当該分野で公知の手順に従って、PCRレパートリー産物をリンカーオリゴと組み合わせてscFvライブラリーを生成し、m13pIIIタンパク質を使用してインフレームで直接クローン化する。
【0108】
典型的なプロトコルでは、ヒト血清中の抗体を、例えば、周知の血球凝集素阻害(HAI)アッセイ(Kendal,A.P.、M.S.Pereira and J.J.Skehel.1982.Concepts and procedures for laboratory−based influenza surveillance.U.S.Department of Health and Human Services,Public Health Service,Centers for Disease Control,Atlanta,Georgia)またはマイクロ中和アッセイ(Harmon et al.,J.Clin.Microbiol.26:333−337(1988))を含む、周知の血清学的アッセイによって検出することができる。血清サンプルがインフルエンザ中和抗体を含むと既に確認されている場合、この検出工程は必要ないかもしれない。全血由来のリンパ球または骨髄中に存在するリンパ球を、次に、当該分野で公知の方法によって処理する。新鮮な組織またはRNAlaterで安定化した組織由来の全RNAを、TriBD試薬(Sigma)によって抽出する。その後、単離したドナー総RNAを、Oligotex精製(Qiagen)を用いて、mRNAにさらに精製する。次に、AccuScript逆転写酵素(Stratagene)のプロトコルに従ったランダム九量体オリゴヌクレオチドおよびまたはオリゴ(dT)18プライマーの使用によって第1のcDNA鎖を合成する。簡潔に述べれば、100ngのmRNA、0.5mMのdNTP、および300ngのランダム九量体およびまたは500ngのオリゴ(dT)18プライマーを含むAccuscriptRT緩衝液(Stratagene)を65℃で5分間インキュベートし、その後に4℃に急冷する。次いで、100mMのDTT、Accuscript RT、およびRNAse Blockを各反応物に添加し、42℃で1時間インキュベートし、逆転写酵素を70℃で15分間の加熱によって不活性化する。得られたcDNAを抗体重鎖および軽鎖V遺伝子のRT−PCR増幅のテンプレートとして使用することができ、次いで、ベクターにクローン化することができるか、またはファージ提示ライブラリーを意図する場合、ファージベクターにクローン化することができる。この手順により、抗体重鎖および軽鎖可変領域クローンのレパートリー(VHおよびVLライブラリー)が得られ、これを分離したままにするか、スクリーニングのために組み合わせることができる。
【0109】
1918年のスペイン風邪等の先のエピデミックおよびパンデミックの生存者の末梢リンパ球由来の免疫グロブリンレパートリーを、上記に類似の様式で回収し、安定化し、レスキューすることができる。さらなるH1およびH3ライブラリーレパートリーを、適切にタイミングを合わせてワクチン接種を行った局所起源のドナーから回収することができる。さらなる選択肢として、市販の骨髄総RNAまたはmRNAを商業的供給源から購入し、H2抗体スクリーニングにも適切なドナーの背景に応じてH1およびH3に適切なライブラリーを産生することができる。
【0110】
合成ヒト様レパートリー
本発明の方法では、合成ヒト抗体レパートリーを、合成抗体ライブラリーによって示すことができ、このレパートリーを、当該分野で公知の方法によって作製することができるか、または商業的供給源から入手することができる。したがって、例えば、完全に合成のヒトレパートリーは、2009年3月26日に出願されたHorowitzらの米国特許出願公開第20090082213号(参照によりその全体が本明細書に組み込まれる)に記載されている。簡潔に述べれば、この特許出願は、所定のアミノ酸が、目的の免疫グロブリンの1つまたは複数の相補性決定領域に組み合わせて導入された免疫グロブリンのライブラリーを記載している。さらに、例えば、かかるライブラリーのサブセットを含むユニバーサル免疫グロブリンライブラリーは、2003年12月11日に公開された米国特許出願公開第20030228302号(参照によりその全体が本明細書に組み込まれる)に記載されている。
【0111】
種々の変異を有する抗体重鎖および軽鎖の特定のサブライブラリーを組み合わせて、本発明の抗体のフレームワーク構築物を得て、その後に重鎖および軽鎖の両方のCDRに多様性を導入することができる。この多様性を、例えば、Kunkel変異誘発等の当該分野で公知の方法によって達成することができ、これを数回繰り返して多様性をさらに増大させることができる。したがって、例えば、複数のKunkel変異誘発ラウンドによって重鎖および軽鎖CDR1およびCD2領域に多様性を個別または同時に導入することができる。必要に応じて、種々のKunkelクローンを、CDRの長さによって分離することができ、そして/またはターゲティングしたCDR(例えば、CDR1またはCDR3)中に多様性を欠くクローンを、例えば、テンプレート特異的制限酵素での消化によって除去することができる。これらの工程の完了の際、ライブラリーのサイズは、約109メンバーを超えるべきであるが、より少数のメンバーを有するライブラリーも有用である。
【0112】
特定の実施形態では、免疫化抗体ライブラリーおよび合成抗体ライブラリーの両方を、本発明の中和抗体の同定のために使用する。2つのライブラリー型は基本的に異なる。合成抗体ライブラリーは、抗原に結合する能力が予想されるヒト抗体集団を合成する一方で、免疫化レパートリーはトリH5血球凝集素、および/またはH1、H2、またはH3血球凝集素(場合による)を特異的に認識するための配列を含むであろう。したがって、免疫化レパートリーは、理論的には、ターゲティングしたインフルエンザ亜型の重要成分を認識するように最適化される。これらの相違の結果として、2つの方法によって異なる抗体組が産生され、したがって、所望の中和抗体を同定するためのより有効なアプローチが得られる。
【0113】
高免疫化非ヒト霊長類抗体ライブラリー
本方法では、抗体ライブラリーを、例えば、マカクまたはヒヒ等の高免疫化非ヒト霊長類から救済する。具体的には、非ヒト霊長類を、インフルエンザA型ウイルスの種々の亜型または種々の血球凝集素(H)タンパク質で免疫化する。動物が免疫化されるインフルエンザA型ウイルス亜型または血球凝集素を認識する抗体の力価を生じる動物を屠殺し、その脾臓を採取する。上の包括的インフルエンザ抗体ライブラリーについて記載されるように、免疫化した動物の血液または骨髄を採取し、産生された抗体を回収し、増幅する。
【0114】
本発明の中和抗体の単離のための戦略
使用した抗体ライブラリーの型と無関係に、例えば、2つの異なるインフルエンザA型亜型および/または同一亜型の2つの株(分離株)、ならびに/またはヒトおよび非ヒト分離株との反応を示す等の二重特異性を有する抗体を、調節された交差反応選択および/または定方向組み合わせおよび/または変異誘発操作によって発見および最適化することができる。
【0115】
図1に例示する典型的な富化スキームでは、標的AおよびBと指定した2つの標的に交差反応性を示す抗体を含むライブラリーを、複数の富化ラウンドに供する(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照)。富化が標的Aとの反応性に基づく場合、各富化ラウンドにより、標的Aに対してプールの反応強度が増加するであろう。同様に、富化が標的Bとの反応性に基づく場合、各富化ラウンドにより、標的Bに対してプールの反応強度が増加するであろう。このアプローチが、ファージ提示ライブラリーをスクリーニングする場合に使用される選択方法であるパニング(以下を参照)を表すにもかかわらず、アプローチは、上記で考察した任意のライブラリー型、当該分野で公知の他の型、および任意の提示技術型に等しく適用可能である。標的AおよびBには、インフルエンザウイルスの種々の分離株、型、および亜型が含まれるが、これらに限定されない、抗体が結合する任意の標的が含まれる。
【0116】
本発明の目的が複数の特異性を有する中和抗体を同定することであるので、交差反応発見選択スキームが開発された。簡潔にするために、このスキームを、二重特異性を有する抗体の選択を示す図2に例示する。この場合、2つの標的である標的AおよびBとの反応性を示す抗体を含む抗体ライブラリーを、一方の標的(例えば、標的A)との反応性について最初に選択し、その後に他方の標的(例えば、標的B)との反応性について選択する。それぞれの連続的な選択ラウンドにより、両標的に対する得られたプールの反応強度を増加させる(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)も参照されたい)。したがって、本方法は、二重特異性を有する抗体の同定に特に有用である。もちろん、本方法を、さらなる標的に対するさらなる富化ラウンドを含めることによって、さらなる標的に対する反応性を示す抗体の同定に拡大することができる。また、スクリーニングされるライブラリーがファージ提示ライブラリーである場合、交差反応パニングによって選択を行うが、他のライブラリーおよび他の選択方法を使用することもできる。
【0117】
上記で考察した2つの方法の組み合わせは、上記のように、それぞれ標的Aおよび標的Bに対する反応性についての2つの個別の富化ラウンド、得られた2つのプールの組換え、およびその後の交差反応選択ラウンドを含む(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照)。このアプローチを、図1に例示する。純粋な交差反応と同様に、組換えられたライブラリーの各選択ラウンドにより、両標的に対する得られたプールの反応強度が増加する。
【0118】
図2に例示するさらなる実施形態では、第1に、標的Aと強い反応性を示し、かつ標的Bと検出可能な交差反応性を有するクローンを同定する。このクローンに基づいて、変異誘発ライブラリーを調製し、次いで、交互のラウンドにおいてそれぞれ標的Bおよび標的Aとの反応性について選択する。このスキームにより、標的Aとの強い反応性を保持し、かつ標的Bとの反応性が増加した抗体が得られる(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照)。これまでと同様に、スクリーニングされるライブラリーがファージ提示ライブラリーである場合、パニングによって選択を行うが、同一の戦略に従って、他のライブラリー、他の提示技術、および他の選択方法を使用することもできる。
【0119】
上記で考察するように、標的AおよびBは、例えば、インフルエンザA型ウイルスの2つの異なる亜型、同一のインフルエンザA型ウイルスの2つの異なる株(分離株)、2つの異なる種(一方の種は好ましくはヒトである)由来の亜型または分離株であり得る。したがって、例えば、標的Aは、H1N1ウイルスのニューカレドニア分離株の分離株であり得、標的Bは、H3N2ウイルスのウィスコンシン分離株であり得る。これらの例は例示のみを目的とし、任意の2つまたは複数の標的に対する二重および複数の特異性を有する抗体を類似の様式で同定、選択、および最適化することができることを強調しておく。
【0120】
代替として、個別のフレームワークおよびCDRの長さを分離することが可能なUAL等の抗体ライブラリーを使用して、標的Aに対する抗体を見出す場合、抗原Bをスクリーニングし、ライブラリーを類似のパラメーターの多様な集団に制限することができる。一旦抗原Bに対する抗体が見出された場合、各A抗体およびB抗体に基づいたキメラ抗体または変異誘発抗体を使用して二重特異性集団を操作することができる。
【0121】
ファージ提示
特定の実施形態では、本発明は、複数の(二重を含む)特異性を有するモノクローナル中和抗体を機能的に発見するためにファージ提示抗体ライブラリーを使用する。かかる抗体は、例えば、H1、H3、および/またはH9亜型、H1およびH3、H5およびH1等のH5、H7、および/またはH9亜型;H5およびH2;H5およびH3;H5、H1、およびH2;H5、H1、およびH3;H5、H2、およびH3;H1、H2およびH3等の亜型を含む、1つを超えるインフルエンザA型ウイルス亜型、ならびに/または同一亜型の1つを超える株(分離株)を中和することができるモノクローナル抗体であり得る。
【0122】
ファージ抗体ライブラリーを生成するために、上記考察のライブラリーを含む、任意の供給源から得たcDNAライブラリーをファージベクターにクローン化する。
【0123】
したがって、例えば、上記のようにRT−PCRによってリンパ球または骨髄からレスキューした抗体重鎖および軽鎖レパートリー集団を、m13 pIIIタンパク質に融合したscFvライブラリーとして再アセンブリする。組み合わせライブラリーは、約106超、107超、108超、または109超の異なるメンバーを含み、107超もしくはそれ以上の異なるメンバーが好ましい。品質管理のために、ランダムクローンを配列決定して、全体的なレパートリーの複雑さを評価する。
【0124】
同様に、ナイーブ、免疫化ヒト、または高免疫化非ヒト霊長類の抗体ライブラリー由来の重鎖および軽鎖可変領域の最初のPCR救済後、PCR産物をリンカーオリゴと組み合わせて、M13 pIIIコートタンパク質を使用してインフレームで直接クローン化するためのscFvライブラリーを生成する。このライブラリーは、約106超、107超、108超、または109超の異なるメンバーを含み、107超もしくはそれ以上の異なるメンバーが好ましい。品質管理のために、ランダムクローンを配列決定して、全体的なレパートリーのサイズおよび複雑さを評価する。
【0125】
抗体ファージ提示ライブラリーは、単鎖Fv(scFv)形式またはFab形式等の種々の形式の抗体を含むことができる。概説については、例えば、Hoogenboom,Methods Mol.Biol.178:1−37(2002)を参照。
【0126】
スクリーニング
所望の中和特性を有する抗体を同定するためのスクリーニング方法を、上記している。所望の血球凝集素タンパク質への直接結合に基づいて反応性を評価することができる。
【0127】
血球凝集素(HA)タンパク質の産生
血球凝集素(HA)タンパク質を、組換えDNAテクノロジーによって産生することができる。この方法では、HA遺伝子を、適切なベクター、好ましくは、Spodoptera frugiperda(Sf9)細胞等のバキュロウイルス感染昆虫細胞における発現のためのバキュロウイルス発現ベクターにクローン化する。
【0128】
HAタンパク質をコードする核酸を、ポリ−his(ヘキサヒスチジンタグ)等のC末端エピトープタグを使用するか使用しないで、Bac−to−Bac(Invitrogen)等のバキュロウイルス発現ベクターに挿入する。ポリ−hisタグは、ニッケルキレートクロマトグラフィーによる容易な精製を提供する。
【0129】
一般に、クローニングは、アセンブリPCRによって個別に合成されたオリゴから基準cDNAを作製する工程を含む。対応する分離株の変異体HAタンパク質を、さらなるアセンブリPCRへの適切な変異オリゴの置換、あるいはKunkel変異誘発等の変異誘発技術のいずれかによって作製する。
【0130】
組換えバキュロウイルスを、リポフェクチン(Gibco−BRLから市販されている)を使用した上記バクミドのSf9細胞(ATCC CRL 1711)へのトランスフェクションによって生成する。28℃での4〜5日間のインキュベーション後、放出されたウイルスを採取し、さらなる増幅のために使用する。ウイルス感染およびタンパク質発現を、O’Reilley et al.,Baculovirus Expression Vectors:A Laboratory Manual(Oxford:Oxford University Press,1994)に記載のように行う。
【0131】
次いで、発現したポリ−Hisタグ化HAポリペプチドを、例えば、以下のように、Ni2+−キレートアフィニティクロマトグラフィーによって精製することができる。Rupert et al.,Nature 362:175−179(1993)に記載のように、組換えウイルス感染Sf9細胞から上清を回収する。総容積5mLを用いて、Ni2+−NTAアガロースカラム(Qiagenから市販されている)を調製し、25mLの水で洗浄し、25mLのローディング緩衝液で平衡化する。濾過した細胞抽出物を、0.5mL/分でカラムに充填する。カラムをローディング緩衝液でベースラインA280まで洗浄し、その時点で画分回収を開始する。次に、カラムを二次洗浄緩衝液(50mMリン酸塩;300mM NaCl、10%グリセロール(pH6.0))で洗浄し、非特異的結合タンパク質を溶離する。A280ベースラインに再度到達した後、カラムを、二次洗浄緩衝液を含む0〜500mMイミダゾール勾配を使用して展開する。1mLの画分を回収し、SDS−PAGEおよび銀染色またはアルカリホスファターゼ抱合Ni2+−NTA(Qiagen)を使用したウェスタンブロットによって分析する。溶離したHis10タグ化HAポリペプチドを含む画分をプールし、ローディング緩衝液に対して透析する。
【0132】
代替として、IgGタグ化(またはFcタグ化)HAポリペプチドの精製を、例えば、プロテインAまたはプロテインGカラムクロマトグラフィーを含む公知のクロマトグラフィー技術を用いて行うことができる。
【0133】
Sf9細胞の代替物として、HAタンパク質を、他の組換え宿主細胞、原核生物、酵母、または高等真核細胞中で産生することもできる。適切な原核生物には、グラム陰性生物またはグラム陽性生物等の真正細菌、例えば、Enterobacteriaceae(Escherichia(例えば、E.coli)、Enterobacter、Erwinia、Klebsiella、Proteus、Salmonella(例えば、Salmonella typhimurium)、Serratia(例えば、Serratia marcescans)、およびShigella等)、ならびにBacilli(B.subtilisおよびB.licheniformis(例えば、1989年4月12日に公開された第DD266,710号に開示のB.licheniformis 41P)等)、P.aeruginosa等のPseudomonas、ならびにStreptomycesが含まれるが、これらに限定されない。E.coli K12 MM294株(ATCC 31,446)、E.coli X1776(ATCC 31,537)、E.coli W3110株(ATCC 27,325)、およびK5 772(ATCC 53,635)等の種々のE.coli株は、公的に利用可能である。
【0134】
原核生物に加えて、糸状菌または酵母等の真核微生物は、HAポリペプチドをコードする核酸を含むベクターの適切なクローニング宿主または発現宿主である。Saccharomyces cerevisiaeは、一般的に使用されている下等真核宿主微生物である。しかし、多数の他の属、種、および株、例えば、Schizosaccharomyces pombe(Beach and Nurse,Nature 290:140(1981)、1985年5月2日に公開された欧州特許第139,383号);Kluyveromyces宿主(米国特許第4,943,529号;Fleer et al.,Bio/Technology 9:968−975(1991))、例えば、K.Lactis(MW98−8C、CBS683、CBS4574、Louvencourt et al.,J.Bacteriol 737(1983))、K.fragilis(ATCC 12,424)、K.Bulgaricus(ATCC 16,045)、K.wickeramii(ATCC 24,178)、K.waltii(ATCC 56,500)、K.drosophilarum(ATCC 36,906)、Van den Berg et al.,Bio/Technology 8:135(1990))、K.thermotolerans、およびK.marxianus等;yarrowia(欧州特許第402,226号);Pichia pastoris(欧州特許第183,070号、Screekrishna et al.,J.Basic Microbiol.28:265−278(1988));Candida;Trichoderma reesia(欧州特許第244,234号);Neurospora crassa(Case et al.,Proc.Natl.Acad.Sci.USA 76:5259−5263(1979));Schwanniomyces occidentalis(1990年10月31日に公開された欧州特許第394,538号)等のSchwanniomyces;ならびに例えば、Neurospora、Penicillium、Tolypocladium(1991年1月10日に公開されたWO91/00357号等の糸状菌、およびA.nidulans(Ballance et al.,Biochem.Biophys.Res.Commun.112:284−289(1983)、Tilburn et al.,Gene 26:205−221(1983)、Yelton et al.,Proc.Natl.Acad.Sci.USA 81:1470−1474(1984)等のAspergillus宿主、およびA.niger(KellyおよびHynes、EMBO J.4:475−479(1985)等)を、本明細書中で一般的に利用可能であり、かつ有用である。メチロトロープ酵母が本明細書中で適切であり、HAnsenula、Candida、Kloeckera、Pichia、Saccharomyces、Torulopsis、およびRhodotorulaからなる属から選択される、メタノール中で成長することができる酵母が含まれるが、これらに限定されない。この酵母クラスの例である特定の種のリストを、C.Anthony、The Biochemistry of Methylotrophs 269(1982)中に見出すことができる。
【0135】
HAタンパク質発現に適切な宿主細胞には、多細胞生物細胞が含まれる。無脊椎動物細胞の例には、Drosophila S2およびSpodoptera Sf9等の上記昆虫細胞、ならびに植物細胞が含まれる。有用な哺乳動物宿主細胞株の例には、チャイニーズハムスター卵巣(CHO)細胞およびCOS細胞が含まれる。より具体的な例には、SV40によって形質転換されたサル腎臓CV1株(COS−7、ATCC CRL 1651)、ヒト胚腎臓株(HEK293細胞または懸濁培養での成長についてサブクローン化されたHEK293細胞(Graham et al,J.Gen Virol.36:59(1977)))、チャイニーズハムスター卵巣細胞/−DHFR(CHO、Urlaub and Chasin,Proc.Natl.Acad.Sci.USA 77:4216(1980))、マウスセルトリ細胞(TM4、Mather,Biol.Reprod.23:243−251(1980))、ヒト肺細胞(W138、ATCC CCL75)、ヒト肝臓細胞(Hep G2、HB 8065)、およびマウス乳腺腫瘍(MMT 060562、ATCC CCL51)が含まれる。適切な宿主細胞の選択は、当業者の技術範囲内と見なされる。
【0136】
血球凝集素(HA)タンパク質パニング
HAタンパク質を、マイクロタイターウェルまたは磁性ビーズの表面上に固定して、上記ライブラリーをパニングする。特定の実施形態では、各ライブラリーを、1つまたは複数のHタンパク質に4℃で2時間結合させ、次いで、冷PBSで広範囲に洗浄し、その後に0.2Mグリシン−HCl緩衝液(pH2.5)を使用してHA特異的結合クローンを溶離する。回収したファージを中性pHにし、感受性宿主E.coliの感染によって増幅させる。その後、陽性クローン富化を繰り返すためにファージミド産生を誘導し、その後に選別のためにクローンを単離することができる。十分な富化の際に、全プールを、感染によって、HB2151等の非アンバー抑制性E.coli株に導入して可溶性scFvタンパク質を発現させる。代替として、下記のように、プールを、pBAD等の単量体scFv発現ベクターにサブクローン化することができ、組換え可溶性scFvタンパク質をインビトロでの分析および特徴づけのために発現させる。
【0137】
特徴づけ
クローンを、上記にように、1つまたは複数のHタンパク質への結合親和性について試験する。例えば、H1タンパク質(Refseq ABQ10137、分離株ニューカレドニア/20/99(H1N1)、および/またはRefseq ABU99069、分離株ソロモン諸島/3/06(H1N1))への結合を試験し、かつH3タンパク質(Refseq ACC67032、分離株ウィスコンシン/67/05(H3N2)、および/またはRefseq CAA24269、香港/68(H3N2))への結合を平行して試験するが、他の分離株も単独または任意の組み合わせで使用することができる。証明された結合に基づき得た陽性クローンは、中和能力について試験することができる。中和についての典型的な機能試験は、赤血球への全ウイルス結合を使用した血球凝集阻害アッセイを含む。代替として、組替え血球凝集素タンパク質および赤血球を使用した血球凝集アッセイが可能である。全血の必要性を排除するために、気道上皮細胞に対して血球凝集素結合阻害アッセイを行うことができる。結合アッセイは、任意のフローサイトメトリーまたは細胞ELISA(cELISA)ベースのアッセイが含まれるが、これらに限定されない、任意の形態で行うことができる。高価なフローサイトメトリー装置を使用する必要がなく、クローン評価がより自動化され、より多数のデータを収集することができるという点で、cELISAの使用は有利である。他方では、フローサイトメトリーは、より優れた感度、一貫性、および速度が可能である。
【0138】
1つの態様では、本発明の抗体は、インフルエンザウイルスを含有するH1HA、および/またはインフルエンザウイルスを含有するH3HAに対する結合親和性を有する。本発明の抗体の結合親和性を、例えば、Munson et al.,Anal.Biochem.,107:220(1980)のスキャッチャード分析等の当業者に公知の方法によって決定することができる。1つの実施形態では、結合親和性は、<100pMである。1つの実施形態では、抗体の結合親和性は、約1×10−7〜約1×10−13M、約1×10−8〜約1×10−12M、または約1×10−9〜約1×10−11Mである。他の実施形態では、抗体の結合親和性は、約1×10−7M、約1×10−8M、約1×10−9M、約1×10−1×10M、約1×10−11M、約1×10−12M、または約1×10−13Mである。
【0139】
最適化:変異誘発ライブラリー
インフルエンザエピデミックおよび非ヒト動物ウイルスに起因するヒト感染に関連する潜在的パンデミックを含むパンデミックの有効な管理のために、Hタンパク質の現在の分離株および未来の変異体を有効に中和する抗体が必要である。この目的を達成するために、ターゲティングした血球凝集素亜型のすべての公知の分離株に結合する多様なH(例えば、H1、H3、H5等)中和クローンを単離する必要がある。
【0140】
いくつかの例では、交差反応性抗体は、単一の抗原に対して対象としたスクリーニングを介して出現する。分離交差反応性クローンの可能性を増大するために、複数の抗原に対して前進的にスクリーニングすることによって複数の選択的圧力を印加するであろう。いずれかの事象では、交差反応性は、当該分野において公知の抗体最適化法によってさらに改良することができる。例えば、本明細書中に記載する免疫グロブリン鎖の可変領域のある領域は、選択したCDRおよび/またはフレームワーク領域の軽鎖シャッフリング、目的変異誘発、CDR合体、および直接変異誘発を含む、1つまたは複数の最適化戦略を対象とし得る。
【0141】
1つを超えるインフルエンザA型亜型および/または同一亜型の1つを超える分離株との本明細書中の抗体の交差反応性を意図的に導入するために設計された1つの変異誘発法を、本明細書中で「目的」変異誘発と表す。目的変異誘発を使用して、好ましくは反応性の異なる1つまたは複数の抗体クローンに基づいて抗体の集団を合理的に操作することができる。本発明の文脈では、目的変異誘発を使用して、抗体の個別のCDR中の配列等の類似の配列上の類似の位置によって特定された単一または複数の残基をコードする。この場合、これらの集団を、類似の位置に見出される残基の範囲を捕捉するためのオリゴ縮重を使用して生成する。この集団内で、親クローンの特異性の間またはそれを超えて一連の特異性が存在すると予想される。目的変異誘発の目的は、2つ以上の個別の物質または集団の間に多様な多機能抗体集団またはライブラリーを生成することである。インフルエンザの場合、本方法を、2つの異なるエピトープ、分離株、または亜型を認識する2つの抗体を使用し、両機能的質を単一の抗体に変えるために利用することができる。例として、第1のインフルエンザA型抗体は、H1亜型の分離株に特異性を示し得、第2の抗体は、インフルエンザA型ウイルスのH3亜型の分離株に特異性を示す。目的変異誘発ライブラリーを作製するために、両抗体についてのCDR配列を最初に獲得し、アラインメントする。次に、同一性が保存されたすべての位置を、適合した残基に単一コドンを使用して固定する。非保存位置で、両残基をコードするために縮重コドンを組み込む。いくつかの例では、縮重コドンは、この位置で2つの親残基のみをコードするであろう。しかし、いくつかの例では、さらなる副産物が産生される。副産物の産生レベルを、副産物産生を強制するように調節するか、サイズの限度または目的に応じてこの産生を排除することができる。
【0142】
したがって、例えば、2つの抗体の第1の位置がそれぞれ、トレオニンおよびアラニンである場合、第1の2つの位置中にA/G−C−を有する縮重コドンは、第3の位置中の塩基と無関係にトレオニンまたはアラニンのみをコードするであろう。例えば、次の位置の残基がリジンおよびアルギニンである場合、縮重コドンA−A/G−A/Gは、リジンまたはアルギニンのみをコードするであろう。しかし、縮重コドンA/C−A/G−A/G/C/Tを使用した場合、アスパラギン、ヒスチジン、グルタミン、およびセリン副産物も生成されるであろう。
【0143】
便宜上、適合したCDR長を有する抗体のみを使用することがより簡潔である。これを強制するための1つの方法は、最初に発見された抗体によって与えられたCERの長さおよび潜在的なさらなるフレームワーク制限に基づいて、第2の抗原についてサイズが制限されたライブラリーをスクリーングすることである。しかし、同じ長さのCDRの使用は単に便利であるが、必要条件ではないことを付け加えておく。この方法がインフルエンザA型ウイルスの中和抗体の機能的に多様な巨大ライブラリーを作製するのに有用である一方で、その適用性は遥かにより広いことが明白である。この変異誘発技術を使用して、任意の抗体の機能的に多様なライブラリーまたは集団を産生することができる(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照)。したがって、図3は、変異誘発された親配列としてTNF−α抗体およびCD11a抗体のCDRを使用した目的変異誘発法の使用を例示するために本明細書中に含まれる。
【0144】
交差反応性が自然に一般に選択されないため、典型的な変異戦略を実行することは、強力な交差反応性を可能にし得ない可能性が高い。目的変異は、交差反応性タンパク質を有する抗体の範囲を生成するための対象方法として考案された。代替として、CDR合体は、交差反応性抗体の生成および/または最適化のための別の迅速かつ強力な戦略を提供し得る。抗原結合および特異性が、いずれかまたは両方の鎖から選択されたCDRの組み合わせが異なることによって大きく影響されることが確立されている。既存の抗体に含有されるCDRが、既に、標的に対して最適化され得るため、図6に示されるように、不均質性標的に対する有効性を証明し得る、単一の抗体または複数の抗体に由来するCDR合体の集団を作製することができる。
【0145】
図6では、CDRが合体された抗体は、単一のフレームワーク時にCERの組み合わせ重ね合わせとして示すが、それらは、複数の関連フレームワーク、および非関連フレームワークの際に、またはそのキメラフレームワークの際でさえ、CDRが重ね合わされない可能性があり、得られる合体された抗体または複数の抗体の全体の多様性および生産性を増大する。合体ライブラリーは、既存の抗体に見られる生産の多様性、および合体集合体あたりの多くの新規抗体を同定する能力を活用する利点を有する。付加的な重鎖フレームワークのこのより広域の使用は、CDRの文脈における結合のサンプリング、およびより高い交差反応性および潜在性が誘導体化される、従来のB細胞成熟において達成可能ではない組み合わせを可能にするフレームワーク変異体を可能にする。CDRが標的に係合するための相互作用ループとして機能するため、それぞれの本来の配置に関し、かつそれに関わらず、両方の重鎖と軽鎖との間でCDRを混合することによって、遥かに広範囲の組み合わせを作成することができる。
【0146】
より従来の最適化が活性の効力または範囲を増加させるのに十分である場合、次いで、標的としたランダム変異誘発、飽和変異誘発、またはさらには、エラー多発性PCRを利用することができる。
【0147】
曖昧に合成されたオリゴヌクレオチドを使用した標的化ランダム変異誘発(Matteuchi and Heyneker,Nucleic Acids Research 11:3113−3121(1983))は、意図するコドンおよびすべての可能なコドンを、特定の比で、互いに、指定の位置に生成する技術である。曖昧に合成されたオリゴヌクレオチドにより、指定の場所への非「野生型」塩基、または、コドンの特異的付加によるヌクレオチド付加の正確さが低下する。これを、典型的には、オリゴヌクレオチド合成機における野生型塩基と非野生型塩基との比の固定および合成時の2つの試薬の混合物の指定によって行う。
【0148】
飽和変異誘発(Hayashi et al.,Biotechniques 17:310−315(1994))は、タンパク質中の特定の位置の20種すべてのアミノ酸を置換し、各変異体に対応するクローンを特定の表現型についてアッセイする技術である(米国特許第6,171,820号、同第6,358,709号、および同第6,361,974号も参照されたい)。
【0149】
エラー多発性PCR(Leung et al.,Technique 1:11−15(1989)、Cadwell and Joyce,PCR Method Applic.2:28−33(1992))は、クローン化した遺伝子にランダム点変異を導入する修正ポリメラーゼ連鎖反応(PCR)技術である。得られたPCR産物をクローン化してランダム変異ライブラリーを産生することができるか、またはT7プロモーターが適切なPCRプライマー内に組み込まれる場合に直接転写することができる。
【0150】
他の変異誘発技術も当該分野で周知であり、例えば、In Vitro Mutagenesis Protocols,J.Braman,Ed.,Humana Press,2001に記載されている。
【0151】
最適化:変異誘発ライブラリースクリーニングのための選択的考察
この場合、主な目的の1つは、現在のH1および/もしくはH3(またはH5もしくはH9)分離株ならびに将来の変異体を有効に処置するために、集団からの1つの抗体(または複数の抗体)を操作し、単離することである。新規の分離株H1/H3中の変異を認識することができる耐性を有する抗体を操作するために、種々のH1/H3分離株に結合する中和クローンを同定すべきである。第1のH1/H3分離株に対してクローンを選択する場合、より低いレベルで、第2のH1/H3分離株に結合/中和すると予想される。この場合、目的は、第1のH1/H3分離株結合の改善(または少なくとも維持)という状況において、第2のH1/H3分離株の認識を劇的に改善することである。したがって、第2のH1/H3分離株の改善のために最初に選択を行い、その後に第1のH1/H3タンパク質を選択する。それにより、新規の株に対してより高い選択圧が得られる一方で、第2のパラメーターに対する選択圧が維持される。
【0152】
他のH中和抗体を類似の様式で最適化することができる。この場合、出発点または目的のいずれかとして他の単離株(例えば、H5、H9等)由来の任意の基準タンパク質配列を用いて、選択および最適化することができる。
【0153】
加えて、中和抗体クローンを使用して、型間認識を試験する。型間認識の例は、非H1起源のクローンまたは最適化クローン由来の同時または操作されたH1結合である。
【0154】
概して、複数の変異誘発集合およびスクリーンは、C5およびA11抗体、C5様およびA11様抗体、C5およびA11抗体様分子、C5およびC5様サロボディ、ならびにA11およびA11様サロボディに基づくことができる。それによって、先述の分子のそれぞれは、広い範囲の反応性および高い効力を有する適切な反応性分子を単離するために、上記に記載した任意の、かつすべての選択を対象とすることができる。
【0155】
中和抗体のエピトープマッピング
一旦所望の性質を有する中和抗体が同定されると、かかる抗体の大部分によって認識される優性のエピトープを同定することが望ましいかもしれない。エピトープマッピング法は、当該分野で周知であり、例えば、Morris,Glenn E.,Epitope Mapping Protocols,Totowa,N.J.ed.,Humana Press,1996、およびEpitope Mapping:A Practical Approach,Westwood and Hay,eds.,Oxford University Press,2001に開示されている。
【0156】
エピトープマッピングは、抗体が結合するエピトープの同定に関する。抗体−抗原複合体の結晶学的分析、競合アッセイ、遺伝子フラグメント発現アッセイ、および合成ペプチドベースのアッセイを含む、タンパク質上のエピトープの位置を決定するための当業者に公知の方法が多数存在する(例えば、HarlowおよびLaneの11章中の、Using Antibodies,a Laboratory Manual,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.,1999、米国特許第7,332,579号(それぞれ、参照によりその全体が本明細書に組み込まれる)を参照)。2つの抗体が同一のエピトープまたは立体的に重複するエピトープを認識する場合に、抗体は基準抗体と「本質的に同一のエピトープ」に結合する。同一または立体的に重複するエピトープに2つの抗体が結合するかどうかを決定するための一般的に使用されている方法は、競合アッセイであり、このアッセイを、標識した抗原あるいは標識した抗体のいずれかを用いて、すべての異なる形式で形成することができる。通常、抗原を96ウェルプレート上に固定し、非標識抗体が標識抗体の結合を遮断する能力を、放射性標識または酵素標識を使用して測定する。
【0157】
中和抗体の産生
一旦所望の中和特性を有する抗体が同定されると、抗体フラグメントを含むかかる抗体を、例えば、ハイブリドーマ技術または組換えDNA技術を含む当該分野で周知の方法によって産生することができる。
【0158】
ハイブリドーマ法では、マウスまたはハムスター等の他の適切な宿主動物を免疫化して、免疫化のために使用されるタンパク質に特異的に結合する抗体を産生するか産生することができるリンパ球を惹起する。代替として、リンパ球をインビトロで免疫化することができる。次いで、ポリエチレングリコール等の適切な融合剤を使用してリンパ球を骨髄腫細胞と融合し、ハイブリドーマ細胞を形成する(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。
【0159】
このようにして調製したハイブリドーマ細胞を、適切な培養培地中に播種し、成長させ、これには、非融合の親骨髄腫細胞の成長または生存を阻害する1つまたは複数の物質を含むことが好ましい。例えば、親骨髄腫細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠く場合、ハイブリドーマ用の培養培地は、典型的には、ヒポキサンチン、アミノプテリン、およびチミジン(HAT培地)を含み、これらの物質はHGPRT欠損細胞の成長を阻害する。
【0160】
好ましい骨髄腫細胞は、効率的に融合し、選択した抗体産生細胞による抗体の安定な高レベル産生を支持し、HAT培地等の培地に感受性を示す骨髄腫細胞である。これらの細胞株のうち、好ましい骨髄腫細胞株は、Salk Institute Cell Distribution Center,San Diego,California USAから利用可能なMOPC−21およびMPC−11マウス腫瘍由来の細胞株、およびAmerican Type Culture Collection,Rockville,Maryland USAから利用可能なSP−2細胞またはX63−Ag8−653細胞等のマウス骨髄腫株である。ヒトモノクローナル抗体の産生のためのヒト骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞株も記載されている(Kozbor,J.Immunol.133:3001(19.84)、およびBrodeur et al,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0161】
ハイブリドーマ細胞が成長する培養培地を、抗原に対するモノクローナル抗体の産生について評価分析する。好ましくは、ハイブリドーマ細胞によって産生されたモノクローナル抗体の結合特異性を、放射免疫アッセイ(RIA)または酵素結合免疫吸着アッセイ(ELISA)等の免疫沈降またはインビトロ結合アッセイによって決定する。
【0162】
組換えモノクローナル抗体を、例えば、必要な抗体鎖をコードするDNAの単離、周知の組換え発現ベクターを用いた組換え宿主細胞と同時発現用のコード配列との共トランスフェクションによって産生することができる。組換え宿主細胞は、上記の細胞等の原核細胞および真核細胞であり得る。
【0163】
ヒト化抗体の作製で使用すべきヒト可変ドメイン(軽鎖および重鎖の両方)の選択は、抗原性の減少に非常に重要である。いわゆる「ベストフィット」法によれば、げっ歯類抗体の可変ドメイン配列を、公知のヒト可変ドメイン配列の全ライブラリーに対してスクリーニングする。次いで、げっ歯類配列に最も近いヒト配列を、ヒト化抗体のヒトフレームワーク領域(FR)として許容する(Sims et al.,J.Immunol.151:2296(1993)、Chothia et al.,J.Mol.Biol.196:901(1987))。抗体を抗原に対する高親和性および他の好ましい生物学的性質を保持してヒト化することが重要である。この目的を達成するために、好ましい方法に従って、ヒト化抗体を、親配列およびヒト化配列の三次元モデルを使用した親配列および種々の概念ヒト化産物の分析過程によって調製する。
【0164】
加えて、当該分野で公知の方法に従って、ヒト抗体を生成することができる。例えば、免疫化の際に内因性免疫グロブリン産生の非存在下でヒト抗体の全レパートリーを産生することができるトランスジェニック動物(例えば、マウス)を作製することができる。例えば、Jakobovits et al.,Proc.Natl.Acad.Sci.USA 90:2551(1993)、Jakobovits et al.,Nature 362:255−258(1993)、Bruggermann et al.,Year in Immuno.7:33(1993)、および米国特許第5,591,669号、同第5,589,369号、および同第5,545,807号を参照。
【0165】
中和抗体
以下の実施例に記載されるものを含む、本明細書中に記載の技術の使用によって多数の中和抗体が同定されている。1つの態様では、本発明は、血球凝集素タンパク質エピトープに結合する中和抗体を提供する。1つの実施形態では、中和抗体は、血球凝集素タンパク質のHA1サブユニット上の少なくとも1つのエピトープに結合する。別の実施形態では、中和抗体は、血球凝集素タンパク質のHA1サブユニット上の少なくとも2つ、少なくとも3つ、少なくとも4つ、少なくとも5つ、または少なくとも6つのエピトープに結合する。
【0166】
いくつかの実施形態では、本発明の抗体は、H3および/またはH1を含むウイルスを中和する。他の実施形態では、抗体は、H3およびH1の両方を中和する。1つの実施形態では、本発明の抗体は、血球凝集を阻止する。他の実施形態では、抗体は、感染すべき標的細胞へのインフルエンザA型ウイルスの結合を阻止する。別の実施形態では、抗血球凝集素抗体は、インフルエンザA型ウイルスのHAの球状頭部上の受容体結合部位が標的細胞に付着してHAの血球凝集素活性が生じるのを阻害する。
【0167】
以下の実施例に記載される実験に基づいて、多数の抗血球凝集素抗体の重鎖/軽鎖対合を同定した。別の実施形態では、本発明の抗体は、2つまたは複数のインフルエンザA型ウイルス亜型に対して交差反応性である。1つの実施形態では、抗体は、配列番号1もしくは配列番号2として示したアミノ酸配列を含有する重鎖ポリペプチド、および配列番号3、配列番号4、配列番号5、もしくは配列番号6として示したアミノ酸配列を含有する軽鎖ポリペプチドを含有する。好ましい実施形態では、本発明の中和抗体は、(i)配列番号2として示した重鎖アミノ酸配列、および配列番号6として示した軽鎖アミノ酸配列を含む抗体(下記の実施例において、および表1に示すとおり、クローン1286−A11)、(ii)配列番号1として示した重鎖アミノ酸配列、および配列番号3として示した軽鎖アミノ酸配列を含む抗体(下記の実施例において、および表1に示すとおり、クローン1286−C5)、(iii)配列番号1として示した重鎖アミノ酸配列、および配列番号4として示した軽鎖アミノ酸配列を含む抗体(下記の実施例において、および表1に示すとおり、クローン1286−C5)、または(iv)配列番号1として示した重鎖アミノ酸配列、および配列番号5として示した軽鎖アミノ酸配列を含む抗体(下記の実施例において、かつ表1に示すとおり、クローン1286−C5)のエピトープと実質的に同一であるエピトープに結合する。

【0168】
典型的な重鎖ループよりも長い重鎖ループを有する抗体は、ある特性を示すとして報告されている。例えば、典型的な重鎖CDR3ループよりも長いループは、多反応性(Schettino et al.,J.Immunol.1997;158;2477−2489)に関係付けられており、ごく近年では、それらは、多くの抗H1V抗体(Saphire et al.,Science 2001;293;1151−1159、Kunert et al.,AIDS Res.Hum.Retroviruses;1998;14(13);1115−1128、Barbas et al.,J.Mol Biol 1993;230(3):812−823)、および抗肺炎球菌抗体(Baxendale et al.,2008;Clin.Exper.Immunol.2007;151;51−60)と関連している。延長ループは、より深いプロービング、および病原性抗原との相互作用を促進し得る。19アミノ酸長の重鎖CDR3を含有するH1V抗体の場合では、標的は、gp120上の感受性凹型領域と接触する、指状突起物の形成を介して非常に特異的に係合する。実施例7に記載されるように、C05抗体重鎖配列は、11アミノ酸の極めて非定型の長さの重鎖CDR1ループ、および25アミノ酸のCDR3ループを有する。
【0169】
1つの態様では、本発明は、長さが修正された重鎖CDRループを有する、インフルエンザ中和抗体または結合分子に関する。通常、抗原に接触するヒト重鎖CDR1ループの長さは、典型的には、約6または8アミノ酸のいずれかであり、抗原に接触するヒト重鎖CDR3ループの典型的な長さは、約13アミノ酸である(Kabat et al.,Sequences of Proteins of Immunological Interest,National Institutes of Health,Bethesda,MD,ed.5,(1991))、MacCallum et al., 1996,J.Mol.Biol.262,732−745)。1つの実施形態では、長さが修正された重鎖は、抗体の典型的な重鎖CDRループよりも長いCDRループを指す、「延長」重鎖CDRループである。
【0170】
1つの実施形態では、抗体または結合分子は、延長重鎖CDR3ループ、および/または延長重鎖CDR1ループを有する。1つの実施形態では、延長重鎖CDR3ループは、典型的な重鎖CDR3ループよりも長い、約1アミノ酸、約2アミノ酸、約3アミノ酸、約4アミノ酸、約5アミノ酸、約6アミノ酸、約7アミノ酸、約8アミノ酸、約9アミノ酸、約10アミノ酸、約11アミノ酸、約12アミノ酸、約13アミノ酸、約14アミノ酸、約15アミノ酸、約16アミノ酸、約17アミノ酸、約18アミノ酸、約19アミノ酸、または約20アミノ酸である。別の実施形態では、延長重鎖CDR3ループは、典型的な重鎖CDR3ループよりも長い、約1〜約20アミノ酸、約1〜約19アミノ酸、約1〜約18アミノ酸、約1〜約17アミノ酸、約1〜約16アミノ酸、約1〜約15アミノ酸、約1〜約14アミノ酸、約1〜約13アミノ酸、約1〜約12アミノ酸、約1〜約11アミノ酸、約1〜約10アミノ酸、約1〜約9アミノ酸、約1〜約8アミノ酸、約1〜約7アミノ酸v1〜約6アミノ酸、約1〜約5アミノ酸、約1〜約4アミノ酸、約1〜約3アミノ酸、または約1〜約2アミノ酸である。さらに別の実施形態では、延長重鎖CDR3ループは、約14、約15、約16、約17、約18、約19、約20、約21、約22、約23、約24、約25、約26、約27、約28、約29、約30、約31、約32、約33、約34、または約35アミノ酸の長さである。
【0171】
好ましい実施形態では、延長重鎖CDR3ループは、約25アミノ酸の長さである。別の好ましい実施形態では、延長重鎖CDR3ループは、アミノ酸配列AKHMSMQQVVSAGWERADLVGDAFD(配列番号9)を含む。記載される抗体が血球凝集を阻害することが可能であるため、この長さの重鎖CDR3ループもまた、球状頭部内の深部にプローブする突出物を形成することが可能である。球状頭部のかかる深部プローブは、血球凝集素の球状頭部ドメインの凹部内でシアル酸に介入する新規手段を提供し得る。これは、本明細書中に記載の延長CDR3ループを有する抗体に認められた活性の顕著な全幅に寄与し得る。
【0172】
1つの実施形態では、抗体または結合分子は、延長重鎖CDR1ループを有する。1つの実施形態では、延長重鎖CDR1ループは、典型的な重鎖CDR1ループよりも長い、約1アミノ酸、約2アミノ酸、約3アミノ酸、約4アミノ酸、約5アミノ酸、約6アミノ酸、約7アミノ酸、約8アミノ酸、約9アミノ酸、約10アミノ酸、約11アミノ酸、約12アミノ酸、約13アミノ酸、約14アミノ酸、または約15アミノ酸である。別の実施形態では、延長 重鎖CDR1ループは、典型的な重鎖CDR1ループよりも長い、約1〜約15アミノ酸、約1〜約14アミノ酸、約1〜約13アミノ酸、約1〜約12アミノ酸、約1〜約11アミノ酸、約1〜約10アミノ酸、約1〜約9アミノ酸、約1〜約8アミノ酸、約1〜約7アミノ酸、約1〜約6アミノ酸、約1〜約5アミノ酸、約1〜約4アミノ酸、約1〜約3アミノ酸、または約1〜約2アミノ酸である。さらに別の実施形態では、延長重鎖CDR3ループは、約3、約4、約5、約6、約7、約8、約9、約10、約11、約12、約13、約14、または約15アミノ酸の長さである。
【0173】
好ましい実施形態では、延長重鎖CDR1ループは、約11アミノ酸の長さである。別の好ましい実施形態では、延長重鎖CDR1ループは、アミノ酸配列GESTLSYYAVS(配列番号7)を含む。重鎖CDR1領域の配置が、典型的に、CDR3の近位であり、結合表面の縁に向かうため、付加的な長さ(例えば、3〜5アミノ酸)は、赤血球凝集素への抗体のより優れた安定性を作成する、付加的な物理的ドッキング表面積を与え得ることが可能である。興味深いことに、VH遺伝子の分析は、かかる延長された長さのCDR1の使用を明らかにしない。同様に、BLAST検索は、任意の以前に報告されたかかるCDR1の長さを有する抗体の同定において失敗に終わった。ポジティブ選択が、かかる新規CDR1長および組成物を強化する役割を担ったという考察を生じる。
【0174】
1つの他の態様では、長さが修正された重鎖ループは、短縮された重鎖ループである。短縮重鎖ループは、欠失を含み得る。1つの実施形態では、CDR1ループおよび/またはCDR3ループは、欠失を含む。別の実施形態では、配列番号7および/または配列番号9は、欠失を含む。1つの実施形態では、重鎖CDR1またはCDR3ループ内の欠失は、約1アミノ酸、約2アミノ酸、約3アミノ酸、約4アミノ酸、約5アミノ酸、約6アミノ酸、約7アミノ酸、約8アミノ酸、約9アミノ酸、約10アミノ酸、約11アミノ酸、約12アミノ酸、約13アミノ酸、約14アミノ酸、または約15アミノ酸、または約16アミノ酸、または約16アミノ酸、または約17アミノ酸、または約18アミノ酸、または約19アミノ酸、または約20アミノ酸、または約21アミノ酸、または約22アミノ酸、または約23アミノ酸、または約24アミノ酸である。
【0175】
1つの態様では、本発明は、低下した酸化能または減少した不均質性酸化能を有するインフルエンザ中和抗体または結合分子に関する。ポリペプチドの水性製剤中の過酸化物によるメチオニンの酸化は、タンパク質ベースの治療薬の開発に有害であると見なされる。この懸念は、少なくとも2倍である。第1に、メチオニンが必須である場合、次いで、酸化は、抗体が活性を維持することができるように制御されなければならない。しかし、それらが活性に必須ではない場合、次いで、非酸化的置換は、減少した不均質性を有する抗体または結合分子を産生するために好ましい。1つの実施形態では、低下した酸化能を有する抗体または結合分子は、変異体重鎖アミノ酸配列を有する。別の実施形態では、変異体アミノ酸配列は、CDR3ループ内である。1つの実施形態では、変異体アミノ酸配列は、カバットナンバリング変換による96位および/または98位におけるアミノ酸に対応する、少なくとも1つの置換を含む。好ましい実施形態では、96位および/または98位におけるメチオニンは、置換アミノ酸である。さらに別の実施形態では、変異体重鎖アミノ酸配列は、メチオニン残基の少なくとも1つの置換を含有する。1つの実施形態では、変異体重鎖アミノ酸配列は、メチオニン残基の少なくとも2つの置換を含有する。好ましい実施形態では、変異体配列は、ロイシン置換に対して少なくとも1つのメチオニン、より好ましくは、ロイシン置換に対して少なくとも2つのメチオニンを含有する。別の実施形態では、変異体配列は、以下のうちの1つを含む:
AKHMSLQQVVSAGWERADLVGDAFD(配列番号25)、
AKHLSMQQVVSAGWERADLVGDAFD(配列番号26)、
AKHASLQQVVSAGWERADLVGDAFD(配列番号27)、および
AKHSSLQQVVSAGWERADLVGDAFD(配列番号28)、好ましくは
AKHLSLQQVVSAGWERADLVGDAFD(配列番号29)。
【0176】
1つの実施形態では、減少した不均質性酸化能を有する抗体または結合分子は、以下の特性((i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和する、(ii)ウイルスの血球凝集素(HA)抗原に結合する、(iii)血球凝集を阻害する、またはそれらの任意の組み合わせ)のうちの1つまたは複数を保持する。
【0177】
1つの態様では、本発明は、配列番号1〜29として示した1つまたは複数のアミノ酸配列を含むか、本質的にこれからなるか、またはこれからなる、ポリペプチドを有する中和抗体または結合分子に関する。
【0178】
別の態様では、本発明は、本明細書中に記載の重鎖または軽鎖アミノ酸配列の組み合わせを有する中和抗体または結合分子に関する。例えば、1286−C05および1286−A11抗体の重鎖および軽鎖アミノ酸配列が特徴づけられており(実施例2)、本発明は、それらの種々の重鎖および軽鎖、それらのCDR領域、および/またはそれらの機能的に活性なフラグメントの間の組み合わせを検討する。本明細書中に記載の抗体または結合分子は、任意の組み合わせにおいて、重鎖および軽鎖、および/または重鎖および軽鎖超可変CDR領域(配列番号1〜29)からなり得る。1つの実施形態では、抗体または結合分子は、重鎖(配列番号1もしくは2)、および軽鎖(配列番号6)、または重鎖(配列番号1もしくは2)、および1つまたは複数の軽鎖(配列番号3、4、もしくは5)を含む。別の実施形態では、抗体または結合分子は、(i)重鎖(配列番号1)、および1つまたは複数の重鎖超可変CDR領域(配列番号10〜12)、(ii)重鎖(配列番号2)、および1つまたは複数の重鎖超可変CDR領域(配列番号7〜9)、(iii)軽鎖(配列番号13〜21)のうちの1つ、および1つまたは複数の重鎖超可変CDR領域(配列番号10〜12)、および(iv)軽鎖(配列番号22〜24)のうちの1つ、および1つまたは複数の重鎖超可変CDR領域(配列番号7〜9)を含む。他の実施形態では、抗体または結合分子は、(i)1つまたは複数の重鎖超可変CDR領域(配列番号7、8、もしくは9)、および1つまたは複数の重鎖超可変CDR領域(配列番号10、11、または12)、(ii)1つまたは複数の重鎖超可変CDR領域(配列番号7、8、もしくは9)、および1つまたは複数の軽鎖超可変CDR領域(配列番号22、23、もしくは24)、および(iii)1つまたは複数の重鎖超可変CDR領域(配列番号10、11、もしくは12)、および1つまたは複数の軽鎖超可変CDR領域(配列番号13〜21)を含む。当該分野の当業者は、配列番号1〜29、またはそれらの機能的に活性なフラグメントの他の組み合わせも理解するであろう。
【0179】
中和抗体の使用
本発明のインフルエンザ中和抗体を、インフルエンザA型感染の防止および/または治療のために、ならびに適切な交差中和エピトープを提示するワクチンの開発のために使用することができる。治療への応用ために、抗体または抗体ベースの輸送配列を使用することによってその送達が容易になった抗体または他の分子を、通常、薬学的組成物の形態で使用する。技術および製剤を、一般に、Remington’s_ PHArmaceutical Sciences,18th Edition,Mack Publishing Co.(Easton,Pa.1990)に見出すことができる。Wang and Hanson “Parenteral Formulations of Proteins and Peptides:Stability and Stabilizers,” Journal of Parenteral Science and Technology,Technical Report No.10,Supp.42−2S(1988)も参照。
【0180】
1つの態様では、本発明は、治療または防止を必要とする被験者におけるインフルエンザの治療または防止する方法に関する。1つの実施形態では、方法は、本明細書に記載のインフルエンザ中和抗体を必要とする被験者にインフルエンザ中和抗体を投与するステップを含む。抗体は、C05抗体であり得る。実施例6に記載するように、C05抗体は、インフルエンザ感染、例えば、H3N2感染に対する治療的または予防的効果を提供し得る。
【0181】
抗体は、典型的には、凍結乾燥製剤または水溶液の形態で製剤化される。許容可能な担体、賦形剤、または安定剤は、使用される投薬量および濃度で受容者に無毒であり、これらには、リン酸、クエン酸、および他の有機酸の緩衝液等の緩衝液;アスコルビン酸およびメチオニンを含む抗酸化剤;オクタデシルジメチルベンジルアンモニウムクロリド、塩化ヘキサメトニウム、塩化ベンザルコニウム、塩化ベンゼトニウム、フェノール、ブチルアルコールまたはベンジルアルコール、メチルパラベンまたはプロピルパラベン等のアルキルパラベン、カテコール、レゾルシノール、シクロヘキサノール、3−ペンタノール、およびm−クレゾール等の防腐剤;低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチン、または免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、またはリジン等のアミノ酸;モノサッカリド、ジサッカリド、およびグルコース、マンノース、またはデキストリンを含む他の炭水化物;EDTA等のキレート剤;スクロース、マンニトール、トレハロース、またはソルビトール等の糖;ナトリウム等の塩形成対イオン;金属錯体(例えば、Zn−タンパク質錯体);および/またはTWEEN(商標)、PLURONICS(商標)、またはポリエチレングリコール(PEG)等の非イオン性界面活性剤が含まれる。
【0182】
抗体を、例えば、コアセルベーション技術または界面重合によって調製されたマイクロカプセル(例えば、それぞれ、ヒドロキシメチルセルロースまたはゼラチン−マイクロカプセルおよびポリ−(メチルメタクリラート)マイクロカプセル)、コロイド状薬物送達系(例えば、リポソーム、アルブミンミクロスフィア、マイクロエマルジョン、ナノ粒子、およびナノカプセル)、またはマクロエマルジョン中に捕捉することもできる。かかる技術は、Remington’s Pharmaceutical Sciences(前出)に開示されている。
【0183】
本明細書中に開示の中和抗体を、免疫リポソームとして製剤化することもできる。抗体を含むリポソームを、当該分野で公知の方法(Epstein et al.,Proc.Natl.Acad.Sci.USA 82:3688(1985)、Hwang et al.,Proc.Natl Acad.Sci.USA 77:4030(1980)、米国特許第4,485,045号および同第4,544,545号、および1997年10月23日に公開されたWO97/38731号に記載の方法等)によって調製する。循環期間が増加したリポソームは、米国特許第5,013,556号に開示されている。
【0184】
特に有用なリポソームを、ホスファチジルコリン、コレステロール、およびPEG由来ホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物を使用した逆相蒸発法によって生成することができる。リポソームを、規定の孔サイズのフィルターを介して押出して所望の直径を有するリポソームを得る。本発明の抗体のFab’フラグメントを、ジスルフィド鎖間反応によって、Martin et al.,J.Biol.Chem.257:286−288(1982)に記載のリポソームに抱合することができる。化学療法薬を任意にリポソーム内に含める。Gabizon et al.,J.National Cancer Inst.81(19)1484(1989)を参照。
【0185】
疾患の防止または治療のために、抗体の適切な投薬量は、治療すべき感染の型、疾患の重症度および経過、ならびに抗体投与が防止または治療のいずれを目的とするかに依存するであろう。抗体を、1回または一連の治療にわたって患者に適切に投与する。疾患の型および重症度に応じて、約1μg/kg〜約15mg/kgの抗体は、例えば、1回または複数回の個別の投与または連続注入のいずれかでの抗体への投与のための典型的な最初の候補投薬量である。
【0186】
本発明の中和抗体を、インフルエンザA型ウイルスの抗原決定基のエピトープマッピングのためのツールとしてさらに使用することができ、これはワクチン開発に有用である。実際、以下の実施例に示すように、本明細書の発明者らは、ワクチン設計のガイドとして使用することができるいくつかの広範に反応性を示す中和抗体を同定した。
【0187】
したがって、本発明の中和抗体を使用して、抗体が結合する中和エピトープを機能的に模倣するペプチドまたはポリペプチドを選択することができ、それにより、インフルエンザA型ウイルス感染のワクチンを開発することができる。1つの実施形態では、本発明は、本明細書中に記載の抗体に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む、インフルエンザA型ウイルスに有効なワクチンを提供する。1つの実施形態では、ワクチンは、血球凝集素(HA)抗原に結合する抗体に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む。別の実施形態では、ワクチンは合成ワクチンであり得る。他の実施形態では、ワクチンは、(i)弱毒化インフルエンザA型ウイルスまたはその一部、または(ii)死滅させたインフルエンザA型ウイルスまたはその一部を含むことができる。1つの他の実施形態では、ワクチンは、血球凝集素(HA)抗原に結合する抗体に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む。HA抗原は、H3亜型またはH1亜型であり得る。別の実施形態では、HA抗原は、インフルエンザA型ウイルスの表面上に提示される。
【0188】
別の実施形態では、ワクチンのペプチドまたはポリペプチドは、インフルエンザA型ウイルス中和抗体を惹起する抗原決定基を含む。
【0189】
1つの態様では、本発明は、疾患の防止または治療を必要とする被験者の疾患の防止または治療に有用な、またはそのための薬剤または薬学的組成物の調製のための本明細書に記載する抗体の使用を提供する。別の実施形態では、本発明は、疾患の防止または治療を必要とする被験者の疾患を治療または防止するための薬学的組成物を提供し、前記組成物は、本明細書に記載のインフルエンザ中和抗体を含む。
【0190】
中和特性を有する非抗体分子
前述で本発明は抗体ライブラリーを参照して例示されているが、サロボディ等の他の非抗体分子のライブラリーを、類似の様式で調製し、使用し、最適化することができる。したがって、プレB細胞受容体(pre−BCR)に基づいた固有の組み合わせタンパク質ライブラリー(「サロボディライブラリー」)の構築は、Xu et al.,2008(前出)に記載されている。前に考察のように、pre−BCRは、抗体レパートリーの通常の作製中に産生されるタンパク質である。基準抗体と異なり、pre−BCRサブユニットは、2つの代替軽鎖(SLC)成分と対合した抗体重鎖から構成される三量体である。多様な重鎖が固定SLCと対合するこれらのpre−BCRタンパク質に基づいた組み合わせライブラリーを、哺乳動物系、Escherichia coli系、およびファージミド系で発現させた。これらのライブラリーは、標的抗原に対してナノモルの親和性を有するメンバーを含む。ライブラリーの構築、選択的富化、およびライブラリーメンバーの生物物理学的特徴づけの説明は、Xu et al.,(2008)(前出)の材料と方法の項に詳述されている。本明細書に記載の抗体配列のいずれかを使用して、かかる結合、または非抗体分子(例えば、サロボディ等)を構築し得る。
【0191】
本発明のさらなる詳細を、以下の非限定的な実施例によって例示する。
【実施例】
【0192】
実施例1−インフルエンザドナー由来の抗体ライブラリー
包括的インフルエンザライブラリーに含有するために選択されたドナーは、以前にインフルエンザ感染しており、1957年のH2N2、または1968年のH3N2インフルエンザパンデミックの時点で約5歳であり、現在、健康状態が良好であることが確認された。H1N1 A/ニューカレドニア/20/99、H3N2 A/パナマ/2007/99、およびH5N1 A/ベトナム/1203/2004ウイルスのパネルまたは血球凝集素タンパク質上で血清学を実施して、血球凝集素タンパク質に対する抗体の存在を確認した。
【0193】
第1に、5〜20mLの骨髄を、選択条件を満たしている各ドナーから採取し、製造者の指示毎のRNAlater(Applied Biosystems)で混合して、細胞RNAの完全性を保持した。RNAを、TRI−BD試薬プロトコール(Sigma−Aldrich)を使用して単離した。
【0194】
重鎖および軽鎖レパートリーを、重鎖のためのランダム感作されたcDNAテンプレート、軽鎖のためのオリゴdT感作されたcDNAテンプレート、および遺伝子特異的可変ドメインプライマーを使用するRT−PCRによって、各ドナー由来のRNAから回収させた。
【0195】
次に、ドナーあたり1μgの各プールされたκ軽鎖、およびプールされたλ軽鎖をNotlおよびBamH1、ならびにQiagenゲル抽出キットを使用して精製されたゲルで消化した。κおよびλ軽鎖クローニングでは、5μgの各ベクターを、NotlおよびBamH1で消化し、Qiagenゲル抽出キットを使用してゲル精製した。200ngのゲル精製κまたはλインサートおよび1μgのゲル精製ベクターを使用して、軽鎖ライブラリーライゲーションを行う。インキュベーションは、14℃で一晩である。クローニング効果を決定するために、対照連結反応を、軽鎖インサートの追加なく、1つのエレクトロポレーションの量(200〜300ngのベクターDNA)と同等に設定する。転換前に、Edge Biosystem Perfromaスピンカラムを使用して、ライゲーション物を脱塩する。それぞれのライブラリーを、80μLのDh5αエレクトロコンピテント細胞アリコートを使用して、3〜5回のエレクトロポレーションで変換し、それぞれ1mLのSOC中に回収し、プールし、37℃で1時間増殖させた。各ライブラリーのサンプルを、選択的媒体上でプレーティングし、クローニングの効果および形質転換体の総数を決定するために使用した。残部を200mLの2YT+100μg/mLアンピシリン+2%グルコースに移し、37℃で一晩成長させた。良好なライブラリーは、10%未満のバックグラウンド、および1×106メンバーを超える総数の形質転換体を有する。翌日、Qiagen High Speed Maxiprepキットを使用して軽鎖ライブラリープラスミドを単離した。
【0196】
重鎖集合をクローニングするために、1.5〜2μgの各5ドナー特異的重鎖可変遺伝子(VH1/7、VH2、5、6、VH3、およびVH4)を、SfiIおよびXhoIを40単位/μgのDNAで消化し、Qiagenゲル抽出キットを使用してゲル精製する。受容プラスミドを調製するために、15μgの各軽鎖ライブラリーベクターを、SfiIおよびXhoIを40単位余剰/μgのDNAで消化し、Qiagenゲル抽出キットを使用してゲル精製する。ライブラリーライゲーション物を、消化した1.2μgのSfiI/XhoIを組み合わせることによって達成し、ドナーあたりのゲル精製された重鎖DNAを、5ngの各軽鎖ライブラリー、κ、およびλをそれぞれ有する、300ngの各5重鎖改変遺伝子ファミリーを含有するようにプールした。対照ライゲーション反応は、重鎖インサートの添加なく、1つのエレクトロポレーションの量(300〜600ngのベクターDNA)と同等に設定する。ライゲーション物を、14℃で一晩インキュベートし、次いで、Edge Bio System Pefromaスピンカラムで脱塩した。ライゲーション物を、80μLのTG−1細胞を使用して行う、ライブラリーあたり8〜12回のエレクトロポレーションに移し、それぞれ1mLのSOC中に回収し、プールし、37℃で1時間増殖させた。それぞれのサンプルを使用して、クローニングの有効性および形質転換体の総数を決定した。形質転換体の標的数/ライブラリーは、10%未満のバックグラウンドを有する、少なくとも1×107でなければならない。残部を、300mLの2YT+100μg/mLアンピシリン+2%グルコースに移し、OD600が約0.3になるまで成長させた。次に、m13K07ヘルパーファージを、5:1の感染(MOI)の多重度で添加し、振動せずに、37℃で1時間インキュベートした。ヘルパー感染後、遠心分離によって細胞を採取し、300mLの2YT+100μg/mLのアンピリシン+2%のグルコース+70μg/mLのカナマイシン中で再懸濁し、ストックファージ産生用に振動して一晩37℃で増殖を続けた。
【0197】
培養上清を含有する得られたファージを、4℃で10分間の間、6000RPMでの遠心分離によって採取する。次に、ファージを、各上清への0.2容量の20%のPEG/2.5MのNaCl溶液の添加、および1時間の間、氷上でのインキュベーションによって沈殿させる。次いで、ファージを、4℃で15分間の間、7900RPMでの遠心分離によって採取する。上清を除去し、ファージパレットを、30mLの無菌1XPBS中で再懸濁する。長期の−20℃の保管では、PBSを、50%のグリセロールで補充する。
【0198】
実施例2−中和抗体の調製
前述のように、ヒト骨髄ファージ提示抗体ライブラリー由来の抗体(実施例1を参照)を、変換し、哺乳動物発現免疫グロブリンとして試験した(Kashyap AK et al.,Proc Natl Acad Sci USA.2008 Apr 22;105(16):5986−91を参照)。重鎖を2つの配列クラスに分類した:
QVQLQESGGGLVQPGESRLSCVGSGSSFGESTLSYYAVSWVRQAPGKGLEWLSIINAGGGDIDYADSVEGRFTISRDNSKETLYLQMTNLRVEDTGVYYCAKHMSMQQVVSAGWERADLVGDAFDVWGQGTMVTVSS(配列番号1)
QVOLQQSGPRLVKPSQTLSLTCAISGDSVSGDSGTWNWROSPSRGLEWLGRTYYRSKWYNDYAESVKSRIVIKADTSKNEFSLQLNSVTPEDTAIYYCARAGVKIFGLIVGALDNWGRGTLVTVSS(配列番号2)。
下線の超可変CDR領域を、以下の重鎖に示す。
GESTLSYYAVS(配列番号7)
WLSIINAGGGDID(配列番号8)
AKHMSMQQVVSAGWERADLVGDAFD(配列番号9)
SGDSGTWN(配列番号10)
WLGRTYYRSKWYND(配列番号11)
ARAGVKIFGLIVGALD(配列番号12)
【0199】
配列番号1によって記載される重鎖は、1つのλ軽鎖(配列番号3)、およびクローン1286−C5によって例示される2つのκ軽鎖(配列番号4〜5)と対を成すことが見出された。
QSVLTQPPSVSGAPGQRVTISCTGSSSNIGAGYDVHWYQQLPGTAPKLLIYDNNNRPSGVPDRFSGSKSGASASLAITGLQAEDEAHYYCQSYDNSLSGSVFGGGTQTLTVLS(配列番号3)
DIQLTOSPSSLSASVGDRVTLTCQASODIRKFLNWYQQKPGKGPKLLIYDASNLQRGVPSRFSGGGSGTDFTLIISSLQPEDVGTYYCQQYDGLPFTFGGGTKLEIK(配列番号4)
DIQLTQSPSSLSASIGDRVTITCQASQDIRNSLNWYEHKPGKAPKLLIHDASNLETGVPSRFSGGGSGTDFTLTISSLQPEDFATYYCQQANSFPLTFGGGTKVEIK(配列番号5)
下線の超可変CDR領域を、以下の軽鎖に示す。
IGAGYDVHWY(配列番号13)
LLIYDNNNRP(配列番号14)
QSYDNSLSGS(配列番号15)
IRKFLNWY(配列番号16)
LLIYDASNLQ(配列番号17)
QQYDGLPF(配列番号18)
IRNSLNWY(配列番号19)
LLIHDASNLE(配列番号20)
QQANSFPL(配列番号21)
【0200】
配列番号2によって記載され、単一のκ軽鎖(配列番号6)とのクローン1286−A11対によって例示される重鎖。
EIVMTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASTRATGVPDRFHGGGSGTDFTLTISRLEPEDFAVYYCQQYDTSSGLTFGGGTKVEIK(配列番号6)
下線の超可変CDR領域を、以下の軽鎖に示す。
SSYLAWY(配列番号22)
LLIYGASTRA(配列番号23)
QQYDTSSGL(配列番号24)
【0201】
図4〜5は、クローン1286−C5抗体およびクローン1286−A11の結合能力を示す。先述のように、これらの抗体の機能を特徴づけるために、対応するIgGsを、異なる抗原のパネル上で結合する能力に対して試験した(図4および5)(Kashyap AK et al.,2008(前出)も参照されたい)。クローン1286−A11および1286−C5抗体の結合能力。両1286−A11および1286−C5は、H1N1、H3N2、およびH9N2血球凝集素に結合することができた。加えて、1286−A11は、H5N1血球凝集素に結合することが可能であった。種々のインフルエンザ亜型ウイルスのマイクロ中和もまた、先述のように実施した(同文献も参照されたい)。
【0202】
下記の表2に示すように、各抗体は、MDCK細胞中でインビトロH1ウイルス複製を中和することが可能であった。
【0203】
1286−C5もまた、H3ウイルス複製を中和する顕著な能力を示した。1286−A11は、H3亜型ウイルスを測定可能に中和しなかったが、H5亜型中和を提示した。1286−C5抗体の作用の機構は、血球凝集阻害を介するものであるが、1286−A11の作用の機構は、決定されなかった。
これらの抗体のそれぞれは、飽和変異誘発および誤りがちのPCR等の標準的指向およびランダム化抗体最適化技術のそれぞれを介して、増加した活性の効力および範囲に最適化されるであろう。おそらく、これらの抗体は、種々のフラグメント、ならびに単一特異性および多特異性サロボディに変換された場合に有用であろう。
【0204】
実施例3−ユニバーサルインフルエンザワクチンの生成
異種病原体のワクチン設計の目的は、有効かつ広範な防御抗原を同定および設計することである。インフルエンザの場合、相当な歴史的努力によって保存された線状配列および領域の経験的試験が行われてきたが、ほとんど成功していない。これらの失敗についてのもっともらしい理由は、抗原性被験物質に対する注目される応答が実際の感染条件での病原体上の抗原の中和のための実際の真の産生部位であるという知識不足である。インフルエンザのために、インフルエンザ感染の生存者のレパートリー内にこれらの真の解決策が見出されると期待されるであろう。本発明者らの場合、多数の抗体の集団の中の一定の抗体は、インフルエンザの複数の亜型を中和することができることが証明された。これらの抗体のうちのいくつかは、血球凝集の古典的阻害を介して、インフルエンザを中和する。まとめると、真の生存者由来のかかる交差中和抗体を使用したワクチンの設計および評価が交差反応性または「ユニバーサル」インフルエンザワクチンの設計および妥当性に役に立つと期待される。
【0205】
具体的には、交差中和モノクローナル抗体を、現在および将来的に製造されるワクチン中の交差中和エピトープの質および抗原性を維持または増強するワクチン産生過程の設計および確証で使用することができる。ワクチンへの抗体結合が構造的完全性および抗原潜在性を反映すると仮定すると、かかるワクチン過程の誘導体への交差中和抗体の結合を評価して、その交差中和潜在性を定量的に評価するであろう。
【0206】
これらのユニバーサルエピトープに対する応答を最大にするために、胞子殻または胞子外膜等の抗原性補強剤を通してこれらのユニバーサルエピトープに対する免疫原性を増加させるための誘導体を作製するであろう。代替として、予測モデルおよび支持試験を介して、それらの免疫原性を増大するために、これらの交差中和エピトープを操作および最適化することが出来る。どんな場合でも、得られた抗原を再度試験して、標的への結合有効性を維持するだけでなく、定方向の免疫原性も達成されることを保証するであろう。これは、おそらく競合ELISAによって免疫化した個体または試験動物由来の血清中に含まれる特異的なユニバーサル中和力価の決定を含むであろう。インビトロ代替物として、免疫原性のために抗原−抗体結合データをインビトロまたはコンピュータを用いて予測モデルの抗体結合データと組み合わせるであろう。ユニバーサルエピトープに対する応答をさらに指示するために、公知の非中和および非交差反応性血球凝集素エピトープを脱免疫化し得る。
血球凝集素のキメラ、フラグメント、および立体構造模倣物の設計および確証のために本抗体を拡大することが妥当である。例えば、インフルエンザが非中和応答を生じる多数の免疫原性エピトープを含むことが十分に確立されている。交差防御抗体を使用して、これらの免疫優性非中和エピトープが部分的または完全に脱免疫化されたワクチンの抗原変異体が維持されるか、ユニバーサル保護エピトープの認識が強化されるのかどうかを評価することが可能である。
【0207】
また、これらのワクチン設計の結果として、抗原エピトープを最小にし、さらに、血球凝集素の構成から除去して、立体構造交差特異性抗原を作製することができる。
【0208】
この戦略は、中和エピトープまたはエレメントを最小にするために応答を案内するために上に詳細の方法を概説した。立体構造的に依存している可能性が高く、不連続な配列空間内に存在するかかる最小化したエレメントの知識から、前に記載のように(Horowitz et al.,Combinatorial Libraries of Conformationally Constrained Polypeptide Sequences、PCT公開第WO/2008/089073号を参照)、より小さなポリペプチド内に組み合わせ様式で立体構造中和エピトープを作製する可能性があり、不連続なエピトープのみの近位の配置、または設計した構造支持体の文脈では、立体構造エピトープの必須の性質を再生することができる。
【0209】
かかる設計では、立体構造エピトープの形態をとり、これらを血球凝集素関連および非関連構造足場上で発現させるか、立体構造依存性抗体によって選択することができるライブラリー内の立体構造エピトープ集団としての形態もとるであろう。
【0210】
不連続エピトープの立体構造エピトープへの減少により、従来のタンパク質操作を使用して可能なペプチド免疫原よりもさらにより小さなサイズのペプチド免疫原が得られるであろう。さらに、これらの構造エピトープを、非ペプチド模倣物の使用によって、さらに増強、サイズの減少、または置換することができる。いずれにしても、任意のこれらの立体構造誘導体または模倣物を、配列番号1〜6のうちの1つまたは複数、配列番号1〜6のうちの少なくとも1つを含む関連抗体、または選択するインフルエンザウイルスに対応する抗体によって確証するであろう。
【0211】
方法と材料。インフルエンザ融合エピトープ胞子ワクチン標的。
1.分泌タンパク質または哺乳動物細胞としての標的の哺乳動物発現
a.単一分離株由来の球状HA1変異体
b.関連分離株由来の球状HA1キメラ
c.非関連分離株由来の球状HA1キメラ
d.関連分離株および非関連分離株由来の球状HA1キメラ
2.分泌タンパク質または哺乳動物細胞の配列番号1〜6の抗体または関連抗体での立体構造エピトープの検出。
3.首尾の良い立体構造抗原の胞子発現への導入。
4.配列番号1〜6の抗体または関連抗体での胞子結合の試験。
5.インビボ交差反応性免疫原性の評価。
【0212】
実施例4−交差亜型中和抗体の効力および範囲の増加
実施例1に記載される抗体の重鎖および軽鎖の配列情報に基づき、変異誘発方法を使用して、個別またはライブラリー中の集団の中で試験するための改良された変異体を作製する。有益な変異を導入するために一般に使用される方法は、結合を担うことが公知の領域にわたって結合またはエラー多発性PCR変異誘発を担う部位での飽和変異誘発であり得る。
【0213】
従来の抗体最適化戦略の固有の限界のため、交差反応性および効力が不十分である場合、目的変異誘発、合体抗体ライブラリー、または互いに、もしくは以前に記載した従来の最適化戦略を用いるこれらの方法のうちのいずれかもしくは両方の組み合わせを考慮するであろう。
【0214】
実施例5−防御の効力および範囲を増加させるためのワクチンおよび抗体の同時投与
抗体と抗原との複合体は、動物における多数の微生物タンパク質および他のタンパク質に対する応答を潜在的に誘導することが公知である。1つの可能な説明は、ワクチン抗体複合体の強制的な取り込みは抗原提示細胞上のFc受容体によって生じるということである。交差反応性抗体と季節性ワクチンとの複合体により、効力を年々増大させることが可能であり、新規のウイルス分離株を各季節のインフルエンザワクチンのために選択する場合、交差反応性抗体が多数の血球凝集素抗原を認識するため、新規の抗体を再度作製する必要がなくなるからである。さらに、これらの抗体が保存された中和領域に指向するので、抗原と複合体を形成した場合、抗体はこれらの非常に保存された感受性領域へのより有効な防御応答を実際に指示することができる。前述のように、ワクチンは、従来の生ウイルスまたは死滅ウイルス、組換えタンパク質またはタンパク質フラグメント、またはさらに最小化されたペプチド、または抗体、抗体フラグメント、または誘導体と複合体化した非ペプチドの立体構造エピトープ、またはサロボディであり得る。
【0215】
実施例6−インフルエンザ攻撃に対する保護効果
メスの生後6〜8週のDBA/2(Charles River)マウスをABSL3+コンテインメント内にケージあたり5〜6匹で収容した。餌および水を不断で提供した。マウス(グループあたり5〜6匹)は、腹腔内(IP)注射によって、約200〜300μLの無菌リン酸緩衝食塩水(PBS)中の体重kgあたり1、2.5、10、または25mgの抗体C05(C05−1286)を受容した。対照グループは、IP注射によって、200〜300μLのPBS中のPBS25mgの非免疫性ヒトIgGまたはPBSのみのいずれかが注入された。抗体および対照は、ウイルス攻撃の24時間前にX−31(H3N2)またはA/メンフィス/3/2008(H1N1)ウイルスが投与された。致死的ウイルス攻撃では、マウスは、30〜50μLのPBS中の33MLD50(50%のマウス致死量)インフルエンザウイルスでの鼻腔内投与によって接種された。H3N2およびH1N1ウイルスの両方は、DBA/2マウスにおいて高病原性である。死亡前に起こる症状は、30%超の体重減少および一般的な不活発である。体重、罹患率、および死亡率を、14日間の間毎日監視した。
【0216】
図7A〜Bは、高力価の致死的H3N2ウイルス攻撃に対するC05抗体の予防効果を例証する。高力価攻撃(33MLD50)では、以下の結果が得られた。PBSでの処置:0%の生存率。2.5mg/kgでの処置:80%の生存率。10mg/kgでの処置:100%の生存率。25mg/kgでの処置:100%の生存率。25mg/kgのIgGアイソタイプでの処置:0%の生存率。図7Eは、H1N1メンフィス/3/2008ウイルス攻撃に対するC05抗体の予防効果を例証する。
【0217】
図7C〜Dは、致死的H3N2ウイルス攻撃に対するC05抗体の処置効果を示す。DBA/2マウスは、まず、30〜50μLのPBS中の33MLD50(50%のマウス致死量)のX−31(H3N2)インフルエンザウイルスでの鼻腔内投与によって接種された。マウス(グループあたり5〜6匹)は、腹腔内(IP)注射によって、約200〜300μLの無菌リン酸緩衝食塩水(PBS)中の体重kgあたり15mgの抗体C05(C05−1286)を受容した。対照グループは、IP注射によって、200〜300μLのPBS中のPBS25mgの非免疫性ヒトIgGまたはPBSのみのいずれかが注入された。C05抗体は、感染後、24時間、48時間、72時間、または96時間で投与された。対照物の投与は、感染後24時間で生じた。体重、罹患率、死亡率を、14日間毎日監視した。
【0218】
実施例7−異なる長さの重鎖ループを有する変異体の生成
下記に示すように、C05抗体重鎖配列(配列番号1)は、アミノ酸の非常に非定型の長さのCDR1(配列番号7)ループを有する。C05は、VH3−23生殖系列と比較して、CDR1においてさらに5個のアミノ酸を有する。
【0219】
重鎖CDR1領域の配置が、典型的には、CDR3に近位であり、結合表面の縁に向かうため、付加的な長さの3〜5アミノ酸が、血球凝集素に抗体に対してより高い安定性を作製する付加的な物理的ドッキング表面積を与え得ることが可能である。VH遺伝レパートリーの分析は、かかる延長した長さのCDR1の使用を明らかにしない。さらに、BLAST検索は、かかるCDR1の長さを有する任意の抗体を同定しない。したがって、ポジティブ選択が、かかる新規のCDR1の長さおよびその組成物の存在および強化において役割を担い得るという考察を生じる。重鎖は、CDR1およびCDR3の両方において、かかる独自のループの長さを特長とするため、これらのループの組成物を変化させるだけでなく、これらのループの長さを変化させることによって、効力および全幅の増加を考慮するであろう。改良を行うことに関して、まず、上述(パネルの左下)のように、親CDR1ループ中のアミノ酸延長の欠失、および対応するより短い生殖系列CDR1ペプチド配列での置換によって、および/または生成中間の長さのループ、またはその多種多様な集団を介して開始することができるであろう。依然として延長ループが有益であるが、完全に最適ではない場合、段階的または組み合わせ手段で、CDR1内、FR1接合、および/またはCDR1のFR2接合における(1〜20)ランダムまたは選択したアミノ酸の挿入によるループの延長を考慮するであろう。延長ループを有する、かかる抗体の組み合わせライブラリーを作製することによって、両感受性および非感受性インフルエンザ分離株、株、および型へのより優れた、かつ広域なバインダーに対して区別的にスクリーニングすることができる。
【0220】
加えて、C05抗体は、25アミノ酸の典型的な重鎖CDR3(配列番号9)ループよりも長いループを特徴とする。上述のCDR1挿入および欠失の例と同様に、CDR3領域は、同様に修正することができる。下記(右下のパネル)に示すように、CDR3配列は、段階的組み合わせ手段(C05のゲノム配列が示される)で、ループ内またはCDR3のFR3およびFR4接合における1〜20アミノ酸によって、同様に、縮小または拡張することができるであろう。
【0221】
他の相互利益の変異誘発への類似により、異なるループの長さのCDR1およびCDR3ライブラリーを組み合わせて、現在の感受性および非感受性インフルエンザ分離株、株、および型に対する、新規、かつより広域な効力の抗インフルエンザ抗体をインテロゲートすることができるであろう。重要なことには、これらの戦略もまた、親C05CDRだけでなく、対象の抗体に交差されたVh3−23生殖系列または他のVh生殖系列のCDR1ループに適用することができるであろう。
【0222】
実施例8−減少した不均質性酸化能を有する変異体
抗体1286−C05は、カバット残基96および98において、重鎖CDR3ループ内に2つのメチオニンを含有する。定義により、このループは、表面曝露され、したがって、酸化を受けやすい。カバット残基96において、アラニン、ロイシン、またはセリンをメチオニンに、カバット残基98において、ロイシンをメチオニンに置換する二重点突然変異を生成することによって、メチオニンのいずれかまたは両方が必須であったかどうかを試験した。対応するタンパク質を、一過性哺乳動物系を産生し、先述のように精製した(Kashyap AK et al.(前出)2008)。得られたタンパク質を、H1(ニューカレドニア/20/99)血球凝集素への結合に対して試験し、親タンパク質のフォールド内で結合することを見出した。次に、これらのタンパク質を、それらのH3(ウィスコンシン/67/2005)血球凝集素へ結合する能力に対して試験し、それは、ロイシン96/ロイシン98変異体が、実質的に、アラニン96/ロイシン98およびセリン96/ロイシン98変異体よりもよく結合したことを示した。図8は、C05非酸化的「XL」変異体が、H1およびH3HAタンパク質の認識を維持することを例証する。
【0223】
図9は、H1およびH3への非変異体C05の結合と比較して、H1(ニューカレドニア/20/99)およびH3(ウィスコンシン/67/2005)血球凝集素へのロイシン96/ロイシン98C05変異体の結合を示す。C05非酸化的「LL」は、H1およびH3HAタンパク質の認識を維持する。
【0224】
表3は、血球凝集阻害(HAI)アッセイによって試験されたように、C05−LL変異体が活性の同様の効力および全幅を提示したことを例証し、そこでは、血球凝集アッセイおよび血球凝集阻害アッセイは、本質的に、EdwardsおよびDimmock(Journal of Virology,v75pp、10208−18、2001)に説明するとおりであり、そこでは、組換えウイルスは、Kashyapら(2008)(前出)によって説明するとおりに生成された。値は、0.5%のcRBCの血球凝集を阻害する抗体の最小濃度を表す。
【0225】
実施例8−活性の増大した親和性および全幅を有する変異体の作製
変異誘発の種々の方法を使用して、個別またはライブラリー中の集団を試験するための改良された変異体を作製する。直接的構造分析を介して特異的に発見されたCDRまたはこれらの接触残基等の結合を担う部位において有益な変異を導入するために使用される方法は、飽和変異誘発、ルックスルー変異誘発、または倹約変異誘発があるが、これらに限定されない。
【0226】
対象ステップとして、上述の方法によって、CDR3の変異誘発に基づき変異体の集団を作製するであろう。他のヒト抗体を使用する先行研究は、軽鎖、他の重鎖CDR、または同時に一度にこれらの領域のうちのいくつかまたはすべてへの点変異体を調査し、かつ生成して、合成改良を産生することによって、多大な利益を証明した。通常、CDRの組成物のみを変化させながら、CDRの親フレームワークおよび長さを維持することによって、これを達成することができる。従来の誤りがちなPCR変異誘発および他の確率プロセスを、同様の領域にわたって、かつ重鎖可変ドメインの他の領域においても使用して、変異体の集団を生成することができるであろう。任意の事象では、得られる集団またはクローンは、活性の増大した親和性、中和、および/または全幅に対して選択することができるであろう。
【0227】
上述のように、かかる最適化集団を生成するが、重鎖が、CDR1およびCDR3の両方において、かかる独自のループの長さを特徴とするため、これらのループの組成物を変化させるだけでなく、これらのループの長さも変化させることによって、活性の効力および全幅を試験することがさらに重要であることができるであろう。例えば、対応するより短い生殖系列CDR1ペプチド配列、および/または変異集団での置換によって、延長CDR1ループの効果を試験するであろう。加えて、ループが適合し、さらには既存の延長ループの長さを超えるまで、段階的ライブラリーの方法で、CDR1内、またはCDR1のFR1およびFR接合におけるランダムアミノ酸の挿入を行い、より優れたさらに広域のバインダーに対して区別的にスクリーニングすることができるであろう。同様に、CDR3配列は、段階的ライブラリーの方法で、ループ内またはCDR3のFR3およびFR4接合において、収縮または拡張することができるであろう。他の相互に有益な変異誘発への類似により、異なったループの長さのCDR1およびCFR3ライブラリーを組み合わせて、新規かつより広域な効力の抗インフルエンザ抗体を調べることができるであろう。
【0228】
上記の説明中で本発明は、ある実施形態を参照して例示しているが、本発明はこれらに限定されない。実際に、本明細書中に示し、かつ記載した実施形態に加えて、本発明の種々の変更が上記の説明から当業者に明らかとなり、これらの変更は添付の特許請求の範囲の権利範囲内である。
【0229】
本明細書を通して引用されたすべての参照は、参照によりその全体が本明細書に組み込まれる。
【技術分野】
【0001】
本発明は、インフルエンザA型ウイルスに対する中和分子を同定、産生、および設計するための方法および手段、ならびに産生された中和分子に関する。本発明は、さらに、標的インフルエンザA型ウイルス上の本発明の中和分子の結合部位を使用したワクチンの設計および産生を含む、産生された分子の種々の使用に関する。
【背景技術】
【0002】
流感は、インフルエンザウイルスにより引き起こされる感染性呼吸器疾患である。インフルエンザは軽度から重度の疾患を引き起こし、時には死に至り得る。米国では年間、人口の5〜20%がインフルエンザに罹患し、約200,000人が入院し、約36,000の死者をもたらす。
【0003】
インフルエンザウイルスは咳およびくしゃみによって生じた呼吸器飛沫で広がり、それが通常人から人へ伝播する。インフルエンザ表面抗原、特に、血球凝集素に対する免疫は、感染の可能性、および感染した場合の疾患の重症度を軽減する。インフルエンザワクチンを利用可能ではあるが、1つのインフルエンザウイルスの型または亜型に対するワクチンは別のインフルエンザの型または亜型に対する限定的な防御を付与するか、または全く防御しないため、毎年インフルエンザワクチンの中に1つまたは複数の新しい株を組みこむ必要がある。
【0004】
インフルエンザウイルスは分節負鎖RNAウイルスであり、オルトミクスウイルス科に属する。インフルエンザA型ウイルスは、9種類の構造タンパク質からなり、調節機能を有する1つの非構造NS1タンパク質をさらにコードする。非構造NS1タンパク質は、再生サイクル中に大量に合成され、感染細胞のサイトゾルおよび細胞核内に局在する。ウイルスゲノムの分節性により、異なるウイルス株との細胞の混合感染中に遺伝子再集合(ゲノムセグメントの交換)機能が生じる。インフルエンザA型ウイルスは、その表面上に提示される異なる血球凝集素(HA)およびノイラミニダーゼ(NA)ウイルスタンパク質に応じて種々の亜型にさらに分類することができる。インフルエンザA型ウイルス亜型は、2つのウイルス表面糖タンパク質である、血球凝集素(HAまたはH)およびノイラミニダーゼ(NAまたはN)によって同定される。各インフルエンザウイルス亜型は、HおよびNタンパク質のその組み合わせによって同定される。16種のHA亜型および9種のNA亜型が公知である。インフルエンザA型ウイルスは、ヒト、トリ、ブタ、ウマ、および他の動物に感染し得るが、野鳥がこれらのウイルスの天然の宿主である。いくつかのインフルエンザA型亜型(すなわち、H1N1、H1N2、およびH3N2)のみが、現在ヒト間で流布しているが、16種のH亜型と9種のNA亜型とのすべての組み合わせが鳥類、特に、野生の水鳥および浜に生息する鳥で同定されている。さらに、H5およびH7インフルエンザウイルスもヒト疾病を引き起こし得るという証拠が増えつつある。
【0005】
インフルエンザA型ウイルスのHAは、2つの構造的に異なる領域、すなわち、球状頭部領域(globular head region)およびステム領域(stem region)を含む。球状頭部領域は、標的細胞へのウイルス付着を担い、かつHAの血球凝集活性に関与する受容体結合部位を含む。ステム領域は、ウイルスエンベロープと細胞のエンドソーム膜との間の細胞融合に必要であり、したがって融合活性に関連する融合ペプチドを含む(Wiley et al,Ann.Rev.Biochem.,56:365−394(1987))。
【0006】
パンデミック(世界的流行病)は、世界的な疾患の発生である。インフルエンザパンデミックは、新規のインフルエンザA型ウイルスが、(1)ヒト集団においてほとんどまたは全く免疫が存在しないものとして出現した場合、(2)重篤な疾病を引き起こし始め、次いで、(3)世界規模で人から人へ容易に伝播する場合に起こる。20世紀には、かかるインフルエンザパンデミックが3回起こっている。第1に、1918年に、「スペイン風邪」インフルエンザパンデミックにより、米国で少なくとも500,000人が死亡し、世界で4000万人までが死亡した。このパンデミックは、インフルエンザA型H1N1亜型に起因していた。第2に、1957年に、インフルエンザA型H2N2亜型に起因する「アジア風邪」インフルエンザパンデミックにより、米国で少なくとも70,000人が死亡し、世界で100〜200万人が死亡する結果となった。最も最近では、1968年に、インフルエンザA型H3N2亜型に起因する「香港風邪」インフルエンザパンデミックにより、米国で約34,000人が死亡し、世界で700,000人が死亡する結果となった。
【0007】
1997年に、最初のインフルエンザA型H5N1の症例が香港で報告された。これは、このトリウイルス型のヒトへの最初の直接感染であったが、ヒトからヒトへの感染が認められなかったので、パンデミックという結果にならなかった。
【0008】
Lu et al,Resp.Res.7:43(2006)(doi:10.1186/1465−992−7−43)は、不活化H5N1ウイルスを接種したウマ由来の抗H5N1 IgG、およびH5N1特異的F(ab’)2フラグメントの調製を報告しており、これらは、H5N1ウイルスを感染させたBALB/cマウスを防御すると説明されていた。
【0009】
Hanson et al.,Resp.Res.7:126(doi:10.1186/1465−9921−7−126)は、マウスの受動免疫化のためのインフルエンザA型ウイルスH5血球凝集素に特異的なキメラモノクローナル抗体の使用を記載している。
【0010】
インフルエンザウイルスに対する中和抗体は、2008年1月17日に公開された米国特許出願公開第20080014205号に開示されている。
【0011】
あるインフルエンザA型ウイルスに起因する呼吸器疾患の重症度および潜在的パンデミックの脅威を考慮して、有効な防止方法および治療方法が非常に必要とされている。本発明は、インフルエンザA型ウイルスのH1亜型、H3亜型、およびH5亜型が含まれるが、これらに限定されない、ウイルスの種々のH亜型に対するインフルエンザA型中和分子を提供することによって、この必要性に取り組んでいる。本発明は、さらに、種々のヒトおよび非ヒト種から得た分離株ならびに種々のインフルエンザのエピデミックおよび/またはパンデミックの犠牲者および/または生存者由来の分離株が含まれるが、これらに限定されない、インフルエンザA型ウイルスの所与の亜型の1つを超える分離株(株)、好ましくはすべての分離株(株)を中和することができる分子を提供する。
【0012】
かかる交差反応性中和分子は、感染集団またはエピデミックもしくはパンデミックの場合のリスク集団の受動免疫化を含む、インフルエンザウイルス感染の防止および/または治療のために使用することができる。さらに、交差反応性抗体は、将来のワクチン発見のための設計ガイド、および現在のワクチン臨床開発のための評価ツールとして使用することができる。
【発明の概要】
【0013】
1つの態様では、本発明は、重鎖および軽鎖ポリペプチド由来の超可変領域を含む結合分子を提供する。1つの実施形態では、結合分子は、配列番号7、配列番号8、および配列番号9からなる群より選択される重鎖からの1つ、2つ、もしくは3つの超可変領域配列、またはその機能的に活性なフラグメントを含む。別の実施形態では、結合分子は、配列番号7、配列番号8、および配列番号9のすべての超可変領域配列を含む。1つの他の実施形態では、結合分子は、軽鎖と会合したときに、標的に結合することが可能である、結合分子である。1つの実施形態では、軽鎖または結合分子は、配列番号13、配列番号14、および配列番号15のポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む。別の実施形態では、軽鎖または結合分子は、配列番号16、配列番号17、および配列番号18のポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む。1つの他の実施形態では、軽鎖または結合分子は、配列番号19、配列番号20、および配列番号21のポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む。別の実施形態では、軽鎖または結合分子は、配列番号13、配列番号14、および配列番号15のすべての超可変配列を含む。別の実施形態では、軽鎖または結合分子は、配列番号16、配列番号17、および配列番号18のすべての超可変配列を含む。1つの他の実施形態では、軽鎖または結合分子は、配列番号19、配列番号20、および配列番号21のすべての超可変配列を含む。
【0014】
別の実施形態では、結合分子は、配列番号10、配列番号11、および配列番号12からなる群より選択される重鎖からの1つ、2つ、もしくは3つの超可変配列、またはその機能的に活性なフラグメントを含む。1つの他の実施形態では、結合分子は、配列番号10、配列番号11、および配列番号12のすべての超可変領域配列を含む。1つの他の実施形態では、結合分子は、軽鎖と会合したときに、標的に結合することが可能である結合分子である。1つの他の実施形態では、軽鎖または結合分子は、配列番号22、配列番号23、および配列番号24のポリペプチド配列の1つ、2つ、もしくは3つの超可変配列を含む。1つの他の実施形態では、軽鎖または結合分子は、配列番号22、配列番号23、および配列番号24のすべての超可変領域配列を含む。
【0015】
1つの実施形態では、結合分子は抗体である。別の実施形態では、結合分子はサロボディである。
【0016】
別の態様では、本発明は、VpreB配列および/またはλ5配列を含む結合分子を提供する。1つの実施形態では、結合分子は、VpreB配列および/またはλ5配列を含むポリペプチドを含む。別の実施形態では、結合分子は、λ5配列に融合したVpreB配列を含むポリペプチドをさらに含む。1つの他の実施形態では、結合分子は、Vκ様配列および/またはJCκ配列を含むκ様代替軽鎖(SLC)構築物をさらに含む。
【0017】
すべての実施形態では、結合分子は、(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する。別の実施形態では、結合分子は、H1およびH3インフルエンザA型ウイルス亜型の少なくとも1つを中和する。1つの実施形態では、結合分子は、H1およびH3インフルエンザA型ウイルス亜型を中和する。別の実施形態では、結合分子は、インフルエンザA型ウイルスの球状頭部が細胞表面に結合するのを阻止する。1つの他の実施形態では、結合分子は、インフルエンザA型ウイルスが感染される細胞に付着するのを阻止する。別の実施形態では、結合分子は、H1HA抗原に結合する。1つの実施形態では、結合分子はまた、少なくとも1つのさらなるHA抗原に結合する。別の実施形態では、さらなるHA抗原は、H3である。別の実施形態では、結合分子は、H2HA抗原に結合する。
【0018】
1つの実施形態では、本発明は、重鎖を含む抗体を提供し、重鎖は、配列番号1として示したアミノ酸配列を含む。別の実施形態では、抗体は、配列番号3、配列番号4、および配列番号5からなる群より選択されるアミノ酸配列を含む軽鎖をさらに含む。1つの他の実施形態では、抗体は、(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する、抗体である。別の実施形態では、抗体は、H1およびH3インフルエンザA型ウイルス亜型の少なくとも1つを中和する抗体である。1つの実施形態では、抗体は、H1およびH3インフルエンザA型ウイルス亜型を中和する抗体である。別の実施形態では、抗体は、インフルエンザA型ウイルスの球状頭部が細胞表面に結合するのを阻止する抗体である。1つの他の実施形態では、抗体は、インフルエンザA型ウイルスが感染される細胞に付着するのを阻止する抗体である。別の実施形態では、抗体は、H1HA抗原に結合する抗体である。1つの実施形態では、抗体は、少なくとも1つのさらなるHA抗原に結合する抗体である。別の実施形態では、さらなるHA抗原は、H3である。
【0019】
別の実施形態では、抗体は、重鎖を含み、重鎖は、配列番号2として示したアミノ酸配列を含む。1つの他の実施形態では、抗体は、配列番号6として示したアミノ酸配列を含む軽鎖をさらに含む。1つの他の実施形態では、抗体は、(i)インフルエンザA型ウイルスの少なくとも1つの亜型および/または少なくとも1つの分離株を中和し、(ii)ウイルスの血球凝集素(HA)抗原に結合する、抗体である。1つの実施形態では、抗体は、HA抗原のH5亜型のエピトープに結合する抗体である。別の実施形態では、抗体は、H1HA抗原を中和する、H5HA抗原を中和する、またはH1およびH5インフルエンザA型ウイルス亜型を中和する、抗体である。
【0020】
別の実施形態では、抗体または結合分子は、インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株に結合する、および/またはそれに対して反応性である、および/またはそれを中和する。1つの実施形態では、ウイルスは、ヒトに感染する能力を有するウイルスである。別の実施形態では、分離株は、ヒト被験体から得られた分離株である。1つの他の実施形態では、分離株は、非ヒト動物から得られた分離株である。別の実施形態では、非ヒト動物は、トリである。1つの実施形態では、前記トリは、野鳥またはニワトリである。1つの他の実施形態では、非ヒト動物は、ブタである。
【0021】
別の実施形態では、抗体または結合分子は、HA抗原のH1亜型のエピトープに結合する、抗体または結合分子である。1つの他の実施形態では、抗体または結合分子は、HA抗原のH3亜型のエピトープに結合する、抗体または結合分子である。別の実施形態では、抗体または結合分子は、HA抗原のH1亜型のエピトープ、およびHA抗原のH3亜型のエピトープに結合する、抗体または結合分子である。1つの実施形態では、抗体または結合分子は、HA抗原のH9亜型のエピトープに結合する、抗体または結合分子である。別の実施形態では、抗体または結合分子は、HA抗原のH5亜型のエピトープに結合する、抗体または結合分子である。別の実施形態では、抗体または結合分子は、HA抗原のH2亜型のエピトープに結合する、抗体または結合分子である。
【0022】
すべての実施形態では、抗体または結合分子は、インフルエンザA型ウイルスの表面上に提示されるエピトープに結合する。
【0023】
すべての実施形態では、H1亜型、もしくは前記HAは、H1ウイルスのニューカレドニア/20/99分離株に由来するか、H1亜型、もしくは前記HAは、H1ウイルスのソロモン諸島/3/06分離株、またはH1ウイルスのメンフィス/3/2008分離株に由来する。
【0024】
すべての実施形態では、H3亜型、もしくは前記HAは、H3ウイルスのウィスコンシン/67/05分離株に由来するか、またはH3亜型、もしくは前記HAは、H3ウイルスの香港/68分離株に由来する。
【0025】
すべての実施形態では、H9亜型、もしくは前記HAは、H9ウイルスの香港/1073/99分離株に由来する。
【0026】
すべての実施形態では、H5亜型、もしくは前記HAは、H5ウイルスのベトナム/1203/04分離株に由来する。
【0027】
すべての実施形態では、H2亜型、もしくは前記HAは、H2ウイルスのアダチ/1/1957分離株に由来する。
【0028】
すべての実施形態では、抗体または結合分子は、H1HA抗原およびH3抗原と交差反応性である、抗体または結合分子である。
【0029】
1つの他の実施形態では、本発明は、配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する抗体または結合分子を提供する。別の実施形態では、抗体または結合分子は、配列番号3、配列番号4、および配列番号5からなる群より選択されるアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む軽鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する。1つの他の実施形態では、抗体または結合分子は、(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する、抗体または結合分子である。
【0030】
1つの実施形態では、本発明は、配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子を提供する。別の実施形態では、抗体または結合分子は、配列番号6として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む軽鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する。1つの他の実施形態では、抗体または結合分子は、(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)ウイルスの血球凝集素(HA)に結合する、抗体または結合分子である。
【0031】
別の実施形態では、本発明は、本明細書中に記載の結合分子または抗体を含む組成物を提供する。
【0032】
別の態様では、本発明は、インフルエンザA型ウイルス感染の治療および/または防止を必要とする被験体における、インフルエンザA型ウイルス感染の治療および/または防止のための方法を提供する。1つの実施形態では、前記方法は、有効量の本明細書中に記載の組成物を前記被験体に投与することを含む。別の実施形態では、前記方法は、有効量の本明細書中に記載の中和抗体または結合分子を前記被験体に投与することを含む。1つの他の実施形態では、前記被験体はヒト患者である。
【0033】
別の態様では、本発明は、インフルエンザA型ウイルス感染に有効なワクチンを提供する。1つの実施形態では、ワクチンは、本明細書中に記載の抗体または結合分子に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む。別の実施形態では、ワクチンは合成ワクチンである。1つの他の実施形態では、ワクチンは弱毒化インフルエンザA型ウイルスまたはその一部を含む。別の実施形態では、ワクチンは死滅させたインフルエンザA型ウイルスまたはその一部を含む。1つの他の実施形態では、抗体または結合分子は、(a)配列番号1として示したアミノ酸配列、または該アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子、(b)配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む重鎖ポリペプチドを含む抗体、(c)配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子、および(d)配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体、からなる群より選択される。
【0034】
すべての実施形態では、抗体はHA抗原に結合する。HA抗原は、H3亜型、H1亜型、H2亜型、H1亜型およびH3亜型、H5亜型、H9亜型、ならびにそれらの任意の組み合わせからなる群より選択され得る。1つの実施形態では、抗原は、インフルエンザA型ウイルスの表面上に提示される。別の実施形態では、本明細書中に記載の抗体または結合分子に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドは、中和抗体を惹起する抗原決定基を含む。別の実施形態では、ワクチンは、経口投与、経皮投与、または非経口投与に適切である。別の実施形態では、ワクチンは、経粘膜送達に好適である。1つの他の実施形態では、経粘膜送達は、鼻腔内投与である。別の実施形態では、ワクチンは、小児期免疫化用ワクチンである。
【0035】
1つの態様では、本発明は、長さが修正された重鎖ループを有するインフルエンザ中和抗体または結合分子を提供する。長さが修正された鎖は、延長重鎖ループまたは短縮重鎖ループであり得る。1つの実施形態では、抗体または結合分子は、延長重鎖ループを有するインフルエンザA型ウイルスの少なくとも1つの分離株を中和する、ヒトに感染する能力を有するインフルエンザA型ウイルスの血球凝集素へ結合する、中和抗体または結合分子である。1つの他の実施形態では、長さが修正された重鎖ループは、CDR3ループまたはCDR1ループである。別の実施形態では、CDR3ループは、配列番号9のアミノ酸配列を含む。さらに別の抗体では、CDR1ループは、配列番号7のアミノ酸配列を含む。
【0036】
別の態様では、本発明は、低下した酸化能を有する設計抗体または結合分子を提供する。1つの実施形態では、低下した酸化能を有する設計抗体または結合分子は、重鎖可変ドメイン中の1つまたは複数のメチオニン置換を有するインフルエンザA型ウイルスの少なくとも1つの分離株を中和する、ヒトに感染する能力を有するインフルエンザA型ウイルスの血球凝集素へ結合する抗体である。別の実施形態では、重鎖可変ドメインは、CDR3領域である。1つの他の実施形態では、メチオニン置換は、カバットナンバリングシステムによる96位および/または98位である。さらに別の実施形態では、メチオニンは、ロイシンで置換される。1つの実施形態では、重鎖可変ドメインは、配列番号29のアミノ酸配列を含む。
【図面の簡単な説明】
【0037】
【図1】交差反応性を生じる/増加させるための並行した発見プールの組換えによる、2つの異なる標的(標的AおよびB)に対する反応強度を増加させるための戦略を示す。組換え抗体ライブラリーの各選択ラウンドにより、両標的に対する得られたプールの反応強度を増加させる。
【図2】標的Bに対する交差反応性を増加させる一方で、標的Aに対する反応性を維持する戦略を示す。第1に、標的Aに反応性を示すクローンを選択し、次いで、標的Aに反応性を示すクローンの変異誘発ライブラリーを調製し、示すように選択が行われ、標的Aおよび標的Bの両方との強い反応性を示す1つまたは複数の抗体を得る。
【図3】「目的変異誘発」法によって多様な多機能抗体集団を生成するための代表的な変異誘発法を示す。
【図4】H1、H3、およびH9から血球凝集素抗原への1286−C5抗体の結合能力を示す。
【図5】H1、H3、H5、およびH9から血球凝集素抗原への1286−A11抗体の結合能力を示す。
【図6】合体された抗体ライブラリーを生成するための代表的な方法を示す。
【図7A】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Aは、高力価の致死的H3N2ウイルス攻撃に対するC05抗体による予防効果を示す。
【図7B】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Bは、高力価の致死的H3N2ウイルス攻撃に対するC05抗体による予防効果を示す。
【図7C】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Cは、致死的H3N2ウイルス攻撃に対するC05抗体による治療効果を示す。
【図7D】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Dは、致死的H3N2ウイルス攻撃に対するC05抗体による治療効果を示す。
【図7E】インフルエンザ中和抗体の治療効果および保護効果を示す。図7Eは、高力価の季節性H1N1ウイルス攻撃に対するC05抗体による予防効果を示す。
【図8】C05変異体がH1およびH3HAタンパク質の認識を維持することを示す。
【図9】C05変異体がH1およびH3HAタンパク質の認識を維持することを示す。
【発明を実施するための形態】
【0038】
A.定義
他で定義されない限り、本明細書中で使用される技術用語および科学用語は、本発明の属する分野の当業者によって一般に理解される意味と同様の意味を有する。Singleton et al.,Dictionary of Microbiology and Molecular Biology 2nd ed.,J.Wiley & Sons(New York,NY 1994)は、本出願で使用した多数の用語に対する一般的な指針を当業者に提供している。
【0039】
当業者は、本発明の実施の際に使用することができる本明細書中に記載のものと類似または等価の多数の方法および材料を認識するであろう。実際、本発明は、記載の方法および材料に決して制限されない。本発明の目的のために、以下の用語を下記のように定義する。
【0040】
「インフルエンザA型亜型」または「インフルエンザA型ウイルス亜型」という用語は、交換可能に使用され、これらは、血球凝集素(H)ウイルス表面タンパク質によって特徴づけられ、それにより、H番号、例えば、H1、H3、H5、およびH9等によって分類されるインフルエンザA型ウイルス変異体を表す。加えて、亜型を、N番号、例えば、N1およびN2等によって示したノイラミニダーゼ(N)ウイルス表面タンパク質によってさらに特徴づけることができる。したがって、亜型を、H番号およびN番号の両方、例えば、H1N1、H2N2、H3N2、H5N1、H5N2、およびH9N2等によって表すことができる。本用語には、具体的には、通常は変異に起因し、異なる病理学的プロファイルを示す各亜型内のすべての株(絶滅した株が含まれる)が含まれる。かかる株はまた、過去、現在、および未来の分離株のすべてを含む、ウイルス亜型の種々の「分離株」と表されるであろう。したがって、本文脈中で、「株」および「分離株」という用語は、交換可能に使用される。亜型は、インフルエンザA型ウイルスに基づいた抗原を含む。抗原は、血球凝集素ウイルス表面タンパク質に基づくことができ、この抗原を「HA抗原」と指定することができる。いくつかの例では、かかる抗原は、特定の亜型、例えば、H1亜型およびH3亜型等のタンパク質に基づき、それぞれ、H1抗原およびH3抗原と指定することができる。
【0041】
「インフルエンザ」という用語は、インフルエンザウイルスに起因する接触伝染病を表すために使用される。
【0042】
本発明の文脈において、「結合分子」という用語は、最も広い意味で使用され、標的に特異的に結合するポリペプチド配列を含む任意の分子を含む。定義には、ホモ二量体またはヘテロ二量体形態等の単量体または多量体であれ、抗体および抗体フラグメント、抗体様分子およびそのフラグメントが含まれるが、これらに限定されない。多量体結合分子は、共有結合性および/または非共有結合性相互作用を介してそれらの配座を保持することができ、かつそれらが標的(例えば、抗体の場合では、抗原)に結合する必要特性を保持する限り、互いに、および/または分子もしくは部分に抱合し得る。
特定のポリペプチドまたは特定のポリペプチド上のエピトープ「に特異的に結合する」、または「それに特異的」である結合分子は、任意の他のポリペプチドまたはポリペプチドエピトープに実質的に結合することなく、その特定のポリペプチドまたは特定のポリペプチド上のエピトープに結合するものである。
【0043】
「抱合体」、「抱合した」、および「抱合」という用語は、任意のおよびすべての共有結合または非共有結合の形態を表し、直接的な遺伝子融合または化学的融合、リンカーまたは架橋剤によるカップリング、および、例えば、ファンデルワールス力またはロイシンジッパーの使用による非共有結合が含まれるが、これらに限定されない。
【0044】
「抗体」(Ab)という用語は、最も広い意味で使用され、特定の抗原、ならびに免疫グロブリンおよび抗原特異性が欠乏する他の抗体様分子に対して結合特異性を示すポリペプチドが含まれる。後者の種類のポリペプチドは、例えば、リンパ系によって低レベルで、かつ骨髄腫により増大したレベルで産生される。本用途では、「抗体」という用語は、単クローン抗体、多クローン抗体、および抗体フラグメントを具体的に含むが、これらに限定されない。
【0045】
「低下した酸化能」または「減少した不均質性酸化能」という用語は、酸化的アミノ酸から非酸化的アミノ酸への少なくとも1つのアミノ酸置換を有するポリペプチドを含有する抗体を表す。アミノ酸配列は、メチオニン置換であり得る。抗体ポリペプチドは、メチオニンアミノ酸残基を非酸化的アミノ酸残基で置換するために、選択的に操作され得、それによって、低下した酸化能を有する抗体を提供する。例えば、メチオニンは、ロイシン、セリン、またはアラニンで置換され得る。
【0046】
「未変性の抗体」は、通常、2つの同一の軽(L)鎖および2つの同一の重(H)鎖から構成される約150,000ダルトンのヘテロ四量体糖タンパク質である。各軽鎖は、共有結合性のジスルフィド結合によって重鎖に連結されている一方で、ジスルフィド結合数は異なる免疫グロブリンアイソタイプの重鎖間で異なる。各重鎖および軽鎖はまた、規則的間隔の鎖間ジスルフィド架橋を有する。各重鎖は、一方の末端に可変ドメイン(VH)を有し、その後に多数の定常ドメインを有する。各軽鎖は、一方の末端に可変ドメイン(VL)を有し、その他方の末端に定常ドメインを有し、軽鎖の定常ドメインは、重鎖の第1の定常ドメインと整列し、軽鎖可変ドメインは、重鎖の可変ドメインと整列している。特定のアミノ酸残基は、軽鎖と重鎖の可変ドメイン間の界面を形成すると考えられている(Chothia et al,J.Mol.Biol.186:651(1985)、Novotny and Haber,Proc.Natl.Acad.Sci.U.S.A.82:4592(1985))。
【0047】
抗体鎖に関する「可変」という用語は、抗体間の配列が非常に異なり、かつ特定の各抗体のその特定の抗原に対する結合および特異性が関連する抗体鎖部分を表すために使用される。かかる可変性は、軽鎖および重鎖の可変ドメインの両方において、超可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存されている部分は、フレームワーク領域(FR)と呼ばれる。未変性の重鎖および軽鎖の可変ドメインはそれぞれ、4つのFR(それぞれ、FR1、FR2、FR3、およびFR4)を含み、FRはそのほとんどが3つの超可変領域と連結したβシート配置の形態をとり、それにより、超可変領域はβシート構造に連結してループを形成し、βシート構造の一部を形成することもある。各鎖の超可変領域は、FRによって他の鎖由来の超可変領域とともに極近傍に保持され、これが抗体の抗原結合部位の形成に寄与している(Kabat et al,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.(1991),pages647−669を参照)。定常ドメインは、抗体の抗原への結合に直接は関与しないが、抗体依存性細胞毒性における抗体の関与等の種々のエフェクター機能を示す。
【0048】
「超可変領域」という用語は、本明細書中で抗原結合を担う抗体のアミノ酸残基を表す。超可変領域は、「相補性決定領域」または「CDR」由来のアミノ酸残基(すなわち、軽鎖可変ドメイン中の残基30〜36(L1)、46〜55(L2)、および86〜96(L3)、ならびに重鎖可変ドメイン中の30〜35(H1)、47〜58(H2)、および93〜101(H3))を含む(MacCallum et al, J Mol Biol.1996)。「フレームワーク」または「FR」残基は、本明細書中に定義の超可変領域残基以外の可変ドメイン残基である。
【0049】
それらの重鎖の定常ドメインのアミノ酸配列に応じて、抗体を異なるクラスに割り当てることができる。5つの主な抗体クラスのIgA、IgD、IgE、IgG、およびIgMがあり、これらのうちのいくつかは、さらに、サブクラス(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgA、およびIgA2に分けられ得る。
【0050】
異なる免疫グロブリンクラスに対応する重鎖定常ドメインはそれぞれ、α、δ、ε、γ、およびμと呼ばれる。
【0051】
任意の脊椎動物種由来の抗体の「軽鎖」を、それらの定常ドメインのアミノ酸配列に基づいて、カッパ(κ)およびラムダ(λ)と呼ばれる2つの明確に異なる型の1つに割り当てることができる。
【0052】
「抗体フラグメント」という用語は、全長抗体の一部、一般に、その抗原結合ドメインまたは可変ドメインである。抗体フラグメントの例には、Fab、Fab’、F(ab’)2、およびFvフラグメント、線状抗体、一本鎖抗体分子、ディアボディ、ならびに抗体フラグメントから形成された多重特異性抗体が含まれるが、これらに限定されない。抗体フラグメントのさらなる例には、scFv、(scFv)2、dAb(単一ドメイン抗体)、および相補性決定領域(CDR)フラグメント、ならびにその2つのループが組み合わせ変異誘発の影響を受けやすい最小化可変ドメインであるミニボディが含まれるが、これらに限定されない。
【0053】
「モノクローナル抗体」という用語は、B細胞の単一クローンによって合成される抗体分子を表すために使用される。修飾語「モノクローナル」は、実質的に均一な抗体集団から得られた抗体の特徴を示し、任意の特定の方法によって抗体産生が必要であると解釈されるべきではない。したがって、モノクローナル抗体を、Kohler and Milstein,Nature 256:495(1975)、Eur.J.Immunol.6:511(1976)に最初に記載されたハイブリドーマ法、組換えDNA技術によって作製することができるか、あるいはファージ抗体ライブラリーから単離することもできる。
【0054】
「ポリクローナル抗体」という用語は、B細胞集団によって合成された抗体分子集団を表すために使用される。
【0055】
「単鎖Fv」または「sFv」抗体フラグメントは、抗体のVHドメインおよびVLドメインを含み、これらのドメインは、単一のポリペプチド鎖中に存在する。一般に、Fvポリペプチドは、さらに、VHドメインとVLドメインとの間にポリペプチドリンカーを含み、それにより、sFvが抗原結合に所望の構造を形成することができる。sFvの概説については、Pluckthun in The Pharmacology of Monoclonal Antibodies,vol.113,RosenburgおよびMoore eds.Springer−Verlag,New York,pp.269−315(1994)を参照。単鎖抗体は、例えば、WO88/06630号およびWO92/01047号に開示されている。
【0056】
「ディアボディ」という用語は、2つの抗原結合部位を有する小さな抗体フラグメントを表し、このフラグメントは、同一ポリペプチド鎖中に軽鎖可変ドメイン(VL)に連結した重鎖可変ドメイン(VH)を含む(VH−VL)。短すぎて同一鎖上の2ドメイン間で対合が不可能なリンカーの使用により、ドメインは別の鎖の相補ドメインと強制的に対合して2つの抗原結合部位が作製される。ディアボディは、例えば、欧州特許第404,097号、WO93/11161号、およびHollinger et al,Proc.Natl.Acad.Sci.USA 90:6444−6448(1993)において、より完全に記載されている。
【0057】
「ミニボディ」という用語は、自己アセンブリして80kDaの2価の二量体(scFv−CH3)2を形成するscFv−CH3融合タンパク質を表すために使用される。
【0058】
「アプタマー」という用語は、高い特異性および親和性を有するタンパク質標的に結合する合成核酸リガンドを表すために本明細書中で使用される。アプタマーは、タンパク質機能の有力な阻害剤として公知である。
【0059】
dAbフラグメント(Ward et al.,Nature 341:544 546(1989))は、VHドメインまたはVLドメインからなる。
【0060】
本明細書中で使用する場合、「抗体結合領域」という用語は、抗原に結合することが可能な免疫グロブリンまたは抗体可変領域の1つまたは複数の部分を表す。典型的には、抗体結合領域は、例えば、抗体軽鎖(VL)(またはその可変領域)、抗体重鎖(VH)(またはその可変領域)、重鎖Fd領域、Fab、F(ab’)2等の抗体軽鎖と重鎖(またはその可変領域)との組み合わせ、単一ドメイン、または一本鎖抗体(scFv)、または全長抗体、例えば、IgG(例えば、IgG1、IgG2、IgG3、またはIgG4亜型)、IgA1、IgA2、IgD、IgE、またはIgM抗体等である。
【0061】
「二重特異性抗体」という用語は、2つの異なる抗原型に特異性を示す抗体を表す。この用語には、本明細書中で具体的に使用する場合、標的抗原、および特定の組織への送達を容易にする別の標的に対して結合特異性を示す抗体が含まれるが、これらに限定されない。同様に、多重特異性抗体は、2つ以上の結合特異性を有する。
【0062】
「線状抗体」という表現は、抗原結合領域対を形成するタンデムFdセグメント対(VH−CH1−VH−CH1)を含むことを表すために使用される。線状抗体は、二重特異性または単一特性であり得、例えば、Zapata et al,Protein Eng.8(10):1057−1062(1995)に記載されている。
【0063】
本発明の目的のために、「抗体様分子」という用語には、ウイルス抗原に結合してそれを中和することができる上記定義の抗体フラグメント以外の任意の分子が含まれる。本用語には、具体的には、例えば、2008年10月2日に公開されたPCT公開第WO2008/118970号、およびXu et al.,Proc.Natl.Acad.Sci.USA,105(31):10756−61(2008)に記載されるような、代替軽鎖(SLC)エレメントを含む「サロボディ」と呼ばれるプレB細胞受容体(pre−BCR)様構造が含まれるが、これらに限定されない。SLCは、非共有結合したVpre−Bおよびλ5タンパク質から構成される非分散型のヘテロ二量体である。それぞれ、VpreB鎖はVλIgドメインに相同であり、λ5鎖は基準抗体のCλドメインに相同である。ヘテロ二量体SLCは、pre−BCR HCのCλドメインと第1の定常ドメインとの間のジスルフィド結合によってpre−BCR複合体中の重鎖と共有結合している。SLCの固有の特徴は、VpreB1およびλ5ドメインの非基準ペプチドがそれぞれ伸長していることである。VpreB1はそのC末端にさらなる21残基を有し、λ5はそのN末端上に50アミノ酸長のテールを有する(例えば、Vettermann et al.,Semin.Immunol.18:44−55(2006)を参照)。サロボディ構造には、具体的には、pre−BCRの未変性三量体pre−BCR様機能単位、VpreB1とλ5との融合物、およびλ5のN末端50アミノ酸、VpreB1のC末端21アミノ酸、または両方のペプチド伸長物のいずれかが排除された三量体が含まれるが、これらに限定されない。加えて、古典的抗体軽鎖の定常成分を使用したキメラ構築物は、サロボディの定義内に明確に含まれる。
【0064】
他の代表的な「抗体様分子」は、本明細書中に定義されるように、抗体代替κ軽鎖配列を含む類似の構造であり、このκ軽鎖配列は、任意に、別のポリペプチド、例えば、抗体重鎖および/または軽鎖ドメイン配列等とパートナーを組む。κ様B細胞受容体(κ様BCR)は、κ様代替軽鎖(κ様SLC)を使用して同定されている(Frances et al.,EMBO J 13:5937−43(1994)、Thompson et al.,Immunogenetics 48:305−11(1998)、Rangel et al.,J Biol Chem 280:17807−14(2005))。Rangelら、J Biol Chem 280(18):17807−17814(2005)は、非再配列Vκ遺伝子の産物であるVκ様タンパク質の同定および分子の特徴づけを報告しており、これは、Thompsonら、Immunogenetics 48:305−311(1998)によって以前に報告されたcDNA配列と同一であることが判明した。それに対して、Francesら、EMBO J 13:5937−43(1994)は、B細胞前駆体表面でμ重鎖と会合する能力を有し、それにより、B細胞成長のためのλ5経路の代替物が得られる再配列生殖系列JCkの同定および特徴づけを報告した。κ様およびλ様pre−BCRが協力して軽鎖の再配列を促進するように作用し、B細胞前駆体の成熟を確実にすることが示唆されている。概説については、McKeller and Martinez−Valdez Seminars in Immunology 18:4043(2006)を参照。
【0065】
「λ5」という用語は、本明細書中で最も広い意味で使用され、任意の未変性配列または変異体λ5ポリペプチドを表し、具体的には、未変性配列のヒトおよび他の哺乳動物のλ5ポリペプチド、翻訳後修飾によって形成された変異体、ならびにかかる未変性配列のポリペプチドの変異体が含まれるが、これらに限定されない。
【0066】
「変異体VpreBポリペプチド」および「VpreBポリペプチドの変異体」という用語は、交換可能に使用され、アミノ酸修飾の結果として、1つまたは複数のアミノ酸位置における未変性配列のVpreBポリペプチドと異なるポリペプチドであると本明細書中で定義する。「変異体VpreBポリペプチド」は、本明細書中に定義されるように、未変性の抗体λまたはK軽鎖配列またはそのフラグメントと異なるであろう。「変異体VpreBポリペプチド」は、好ましくは、未変性配列のVpreBポリペプチドと少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約98%の配列同一性を保持するであろう。別の好ましい実施形態では、「変異体VpreBポリペプチド」は、未変性抗体のλまたはK軽鎖配列とそのアミノ酸配列が95%未満、90%未満、85%未満、80%未満、75%未満、70%未満、65%未満、または60%未満同一であろう。変異体VpreBポリペプチドには、具体的には、VpreB配列のC末端の非Ig様の固有のテールが部分的または完全に除去されたVpreBポリペプチドが含まれるが、これらに限定されない。「変異体λ5ポリペプチド」および「λ5ポリペプチドの変異体」という用語は、交換可能に使用され、アミノ酸修飾の結果として、1つまたは複数のアミノ酸位置における未変性配列のλ5ポリペプチドと異なるポリペプチドであると本明細書中で定義する。「変異体λ5ポリペプチド」は、本明細書中に定義されるように、未変性抗体のλもしくはκ軽鎖配列、またはそのフラグメントと異なるであろう。「変異体λ5ポリペプチド」は、好ましくは、未変性配列のλ5ポリペプチドと少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約98%の配列同一性を保持するであろう。別の好ましい実施形態では、「変異体λ5ポリペプチド」は、未変性の抗体のλまたはκ軽鎖配列とそのアミノ酸配列が95%未満、90%未満、85%未満、80%未満、75%未満、70%未満、65%未満、または60%未満同一であろう。変異体λ5ポリペプチドは、具体的には、λ5配列のN末端の固有のテールが部分的または完全に除去されたλ5ポリペプチドが含まれるが、これらに限定されない。
【0067】
「VpreB配列」という用語は、上記定義の「VpreB」の配列またはそのフラグメントを表すために本明細書中で使用される。
【0068】
「λ5配列」という用語は、上記定義の「λ5」の配列またはそのフラグメントを表すために本明細書中で使用される。
【0069】
「代替軽鎖配列」という用語は、本明細書中に定義されるように、上記定義の「VpreB配列」および/または「λ5配列」を含む任意のポリペプチド配列を意味する。
【0070】
「κ様代替軽鎖可変ドメイン」、「Vκ様SLC」、および「Vκ様」という用語は、交換可能に使用され、これらは、非再配列Vκ遺伝子およびその変異体の産物である任意の未変性配列のポリペプチドを表す。1つの実施形態では、未変性配列のVκ様ポリペプチドの変異体は、抗体κ軽鎖配列に関連するC末端伸長(テール)を含む。特定の実施形態では、未変性配列のVκ様ポリペプチドの変異体は、Vκ様ポリペプチドと対応する抗体κ軽鎖とを区別する固有のC末端伸長(テール)の少なくとも一部、好ましくはすべてを保持する。別の実施形態では、変異体Vκ様ポリペプチドのC末端テールは、残りの配列と天然には関連しない配列である。後者の実施形態では、未変性Vκ様配列中に天然に存在するC末端テールと変異体配列との相違は、1つまたは複数のアミノ酸の改変(置換、挿入、欠失、および/または付加)に起因し得るか、またはC末端テールが異なるVκ様タンパク質中に天然に存在するテールと同一であり得る。Vκ様ポリペプチドは、1つまたは複数の抗体κ軽鎖CDR1、CDR2、およびCDR3配列に対応する領域中にアミノ酸の改変を含み得る。すべての例では、変異体は、未変性の抗体のκ軽鎖可変領域配列と比較して少なくとも4個、少なくとも5個、少なくとも6個、少なくとも7個、少なくとも8個、少なくとも9個、または少なくとも10個のアミノ酸、好ましくは4〜100個、4〜90個、4〜80個、4〜70個、4〜60個、4〜50個、4〜45個、4〜40個、4〜35個、4〜30個、4〜25個、4〜20個、4〜15、または4〜10個のアミノ酸残基のC末端伸長を含むことができるか、好ましくは含む。本明細書中に定義されるように、Vκ様ポリペプチド変異体は、未変性の抗体のκもしくはλ軽鎖配列またはそのフラグメントと異なり、好ましくは、未変性配列のVκポリペプチドと少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約98%の配列同一性を保持するであろう。別の好ましい実施形態では、Vκ様ポリペプチド変異体は、未変性の抗体のλまたはκ軽鎖配列とそのアミノ酸配列が95%未満、90%未満、85%未満、80%未満、75%未満、70%未満、65%未満、60%未満、55%未満、50%未満、45%未満、または40%未満同一であろう。他の実施形態では、配列同一性は、約40%と約95%との間、約45%と約90%との間、約50%と約85%との間、約55%と約80%との間、約60%と約75%との間、約60%と約80%との間、約65%と約85%との間、約65%と約90%との間、または約65%と約95%との間である。すべての実施形態では、好ましくは、Vκ様ポリペプチドは標的に結合することができる。
【0071】
「JCκ」および「JCκ様」という用語は、交換可能に使用され、これらは、未変性配列のκJ定常(C)領域セグメントおよび固有のN末端伸長(テール)、ならびにその変異体と同一の部分を含む未変性配列のポリペプチドを表す。1つの実施形態では、未変性配列のJCκ様ポリペプチドの変異体は、抗体JCセグメントと区別されるN末端伸長(テール)を含む。特定の実施形態では、未変性配列のJCκ様ポリペプチドの変異体は、JCκ様ポリペプチドを対応する抗体κ軽鎖JCセグメントと区別する固有のN末端伸長(テール)の少なくとも一部、好ましくはすべてを保持する。別の実施形態では、変異体JCκ様ポリペプチドのN末端テールは、残りの配列と天然には関連しない配列である。後者の実施形態では、未変性JCκ様配列中に天然に存在するN末端テールと変異体配列との相違は、1つまたは複数のアミノ酸の改変(置換、挿入、欠失、および/または付加)に起因し得るか、またはN末端テールは異なるJCκ様タンパク質中に天然に存在するテールと同一であり得る。未変性配列のJCκ様ポリペプチドの変異体は、未変性の抗体のκ可変ドメインJC配列と同一の配列の一部に1つまたは複数のアミノ酸の改変を含み得る。すべての場合、変異体は、未変性の抗体のκ軽鎖JC配列と比較して少なくとも4個、少なくとも5個、少なくとも6個、少なくとも7個、少なくとも8個、少なくとも9個、または少なくとも10個のアミノ酸、好ましくは4〜100個、または4〜90個、または4〜80個、または4〜70個、または4〜60個、または4〜50個、または4〜45個、または4〜40個、または4〜35個、または4〜30個、または4〜25個、または4〜20個、または4〜15個、または4〜10個のアミノ酸残基のN末端伸長(固有のN末端)を含むことができるか、好ましくは含む。JCκ様ポリペプチド変異体は、本明細書中に定義されるように、未変性の抗体のλもしくはκ軽鎖JC配列またはそのフラグメントと異なり、好ましくは、未変性配列のJCポリペプチドと少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、少なくとも約95%、または少なくとも約98%の配列同一性を保持するであろう。別の好ましい実施形態では、JCκ様ポリペプチド変異体は、未変性の抗体のλもしくはκ軽鎖JC配列とそのアミノ酸配列が95%未満、90%未満、85%未満、80%未満、75%未満、70%未満、65%未満、または60%未満同一であろう。他の実施形態では、配列同一性は、約40%と約95%との間、約45%と約90%との間、約50%と約85%との間、約55%と約80%との間、約60%と約75%との間、約60%と約80%との間、約65%と約85%との間、約65%と約90%との間、または約65%と〜約95%との間である。
【0072】
アミノ酸配列同一率を、配列比較プログラムNCBI−BLAST2を用いて決定することができる(Altschul et al.,Nucleic Acids Res.25:3389−3402(1997))。NCBI−BLAST2配列比較プログラムを、http://www.ncbi.nlm.nih.govからダウンロードすることができるか、またはそうでなければ、National Institute of Health,Bethesda,MDから入手することができる。NCBI−BLAST2は、いくつかの検索パラメーターを使用し、これらの検索パラメーターのすべてを、例えば、アンマスク=イエス、鎖=すべて、予想される発生数=10、最小低複雑性長=15/5、マルチパスe値=0.01、マルチパス定数=25、最終ギャップアラインメントのドロップオフ=25、およびスコアリング行列=BLOSUM62を含む、デフォルト値に設定する。
【0073】
「融合」という用語は、そのヌクレオチドコード配列のインフレームでの組み合わせによる1つのポリペプチド鎖中の異なる起源のアミノ酸の組み合わせを表すために本明細書中で使用される。「融合」という用語は、その末端の1つへの融合に加えて、内部融合、すなわち、ポリペプチド鎖内の異なる起源の配列の挿入を明確に含む。
【0074】
本明細書中で使用する場合、「ペプチド」、「ポリペプチド」、および「タンパク質」という用語は、すべて、共有結合性の「ペプチド結合」によって連結されたアミノ酸の一次配列を表す。一般に、ペプチドは、少数のアミノ酸、典型的には、約2〜約50個のアミノ酸からなり、タンパク質より短い。「ポリペプチド」という用語は、本明細書中に定義されるように、ペプチドおよびタンパク質を含む。
【0075】
「アミノ酸」または「アミノ酸残基」という用語は、典型的に、その当該分野で認識された定義を有するアミノ酸、例えば、アラニン(Ala)、アルギニン(Arg)、アスパラギン(Asn)、アスパラギン酸(Asp)、システイン(Cys)、グルタミン(Gln)、グルタミン酸(Glu)、グリシン(Gly)、ヒスチジン(His)、イソロイシン(Ile)、ロイシン(Leu)、リシン(Lys)、メチオニン(Met)、フェニルアラニン(Phe)、プロリン(Pro)、セリン(Ser)、トレオニン(Thr)、トリプトファン(Trp)、チロシン(Tyr)、およびバリン(Val)からなる群より選択されるアミノ酸等を表すが、必要に応じて、修飾アミノ酸、合成アミノ酸、または希アミノ酸を使用することができる。したがって、連邦規則集第37巻1.822(b)(4)に列挙した修飾アミノ酸および通常でないアミノ酸は、本定義内に具体的に含まれ、参照により本明細書に明確に組み込まれる。アミノ酸を、種々の亜群に分類することができる。したがって、アミノ酸を、非極性側鎖(例えば、Ala、Cys、Ile、Leu、Met、Phe、Pro、Val)、負電荷の側鎖(例えば、Asp、Glu)、正電荷の側鎖(例えば、Arg、His、Lys)、または無電荷極性側鎖(例えば、Asn、Cys、Gln、Gly、His、Met、Phe、Ser、Thr、Trp、およびTyr)を有するアミノ酸に分類することができる。アミノ酸を、小アミノ酸(Gly、Ala)、求核性アミノ酸(Ser、His、Thr、Cys)、疎水性アミノ酸(Val、Leu、Ile、Met、Pro)、芳香族アミノ酸(Phe、Tyr、Trp、Asp、Glu)、アミド(Asp、Glu)、および塩基性アミノ酸(Lys、Arg)に分類することもできる。
【0076】
「ポリヌクレオチド」という用語は、DNA分子およびRNA分子等の核酸、ならびにその類似体(例えば、ヌクレオチド類似体または核酸化学を用いて生成したDNAまたはRNA)を表す。必要に応じて、ポリヌクレオチドを、例えば、当該分野で認識された核酸化学を用いて合成的に作製するか、または例えば、ポリメラーゼを用いて酵素的に作製するか、必要に応じて、修飾することができる。典型的な修飾には、メチル化、ビオチン化、および他の当該分野で公知の修飾が含まれる。加えて、核酸分子は、必要に応じて、検出可能な部分に連結した一本鎖または二本鎖であり得る。
【0077】
基準ポリペプチドに関する「変異体」という用語は、未変性ポリペプチドと比較して、少なくとも1つのアミノ酸の変異または修飾(すなわち、改変)を保有するポリペプチドを表す。「アミノ酸修飾」によって生成された変異体を、例えば、未変性アミノ酸配列中の少なくとも1つのアミノ酸の置換、欠失、挿入、および/または化学修飾によって産生することができる。
【0078】
「アミノ酸修飾」は、所定のアミノ酸配列のアミノ酸配列の変化を表す。例示的な修飾には、アミノ酸の置換、挿入、および/または欠失が含まれる。
【0079】
「特定の位置でのアミノ酸修飾」は、特定の残基の置換もしくは欠失、または特定の残基に隣接する少なくとも1つのアミノ酸残基の挿入を表す。「隣接する」特定の残基の挿入は、その1〜2個の残基内の挿入を意味する。挿入は、特定の残基のN末端またはC末端であり得る。
【0080】
「アミノ酸置換」は、所定のアミノ酸配列中の少なくとも1つの既存のアミノ酸残基の別の異なる「置換」アミノ酸残基との置換を表す。置換残基または残基は、「天然に存在するアミノ酸残基」(すなわち、遺伝子コードによってコードされる)であり得、アラニン(Ala)、アルギニン(Arg)、アスパラギン(Asn)、アスパラギン酸(Asp)、システイン(Cys)、グルタミン(Gln)、グルタミン酸(Glu)、グリシン(Gly)、ヒスチジン(His)、イソロイシン(Ile)、ロイシン(Leu)、リシン(Lys)、メチオニン(Met)、フェニルアラニン(Phe)、プロリン(Pro)、セリン(Ser)、トレオニン(Thr)、トリプトファン(Trp)、チロシン(Tyr)、およびバリン(Val)からなる群より選択され得る。1つまたは複数の天然に存在しないアミノ酸残基との置換はまた、本明細書中のアミノ酸置換の定義に含まれる。
【0081】
「天然に存在しないアミノ酸残基」は、ポリペプチド鎖中の隣接アミノ酸残基に共有結合することができる上記列挙の天然に存在するアミノ酸残基以外の残基を表す。天然に存在しないアミノ酸残基の例には、ノルロイシン、オルニチン、ノルバリン、ホモセリン、およびEllmanらのMeth.Enzvm.202:301 336(1991)に記載されるもの等の他のアミノ酸残基類似体が含まれる。かかる天然に存在しないアミノ酸残基を生成するために、Norenら Science 244:182(1989)およびEllmanら(前出)の手順を使用することができる。簡潔に述べれば、これらの手順は、天然に存在しないアミノ酸残基での抑制性tRNAの化学的活性化およびその後のRNAのインビトロ転写および翻訳を含む。
【0082】
「アミノ酸挿入」は、所定のアミノ酸配列への少なくとも1つのアミノ酸の組み込みを表す。挿入は、通常、1つまたは2つのアミノ酸残基の挿入からなる一方で、本出願は、より巨大な「ペプチド挿入」、例えば、約3〜約5個、またはさらに約10個までのアミノ酸残基の挿入を企図する。挿入される残基は、上記開示の天然に存在する残基または天然に存在しない残基であり得る。
【0083】
「アミノ酸欠失」は、所定のアミノ酸配列からの少なくとも1つのアミノ酸残基の除去を表す。
【0084】
「変異誘発」という用語は、他で特定しない限り、任意の当該分野で認識されたポリヌクレオチド配列またはポリペプチド配列を改変させる技術を表す。好ましい変異誘発型には、エラー多発性PCR変異誘発、飽和変異誘発、または他の部位特異的変異誘発が含まれる。
【0085】
「部位特異的変異誘発」は、当該分野で標準的な技術であり、少数のミスマッチを除いて所望の変異を示す変異誘発すべき一本鎖ファージDNAに相補的な合成オリゴヌクレオチドプライマーを用いて行う。簡潔に述べれば、合成オリゴヌクレオチドを、一本鎖ファージDNAに相補的な鎖の合成を指示するためのプライマーとして使用し、得られた二本鎖DNAを、ファージ支持宿主細菌に形質転換する。形質転換された細菌の培養物を上層寒天にプレートし、ファージを保有する単一の細胞からプラークを形成させる。理論的には、新規のプラークの50%が一本鎖として変異形態を有するファージを含み、50%が元の配列を有するであろう。目的のプラークを、正確に適合するハイブリッド形成が可能であるが、元の鎖とのミスマッチがハイブリッド形成を阻止するのに十分である温度でのキナーゼ化合成プライマーとのハイブリッド形成によって選択する。次いで、プローブとハイブリッド形成するプラークを選択し、配列決定し、培養し、DNAを回収する。
【0086】
「中和分子」という用語は、本明細書中でその最も広い意味で使用され、これは、中和を達成する機構と無関係にウイルスが標的細胞に複製的に感染するのを阻害する任意の分子を表す。中和分子は、好ましくは、上記定義の抗体または抗体様分子もしくは結合分子である。中和は、例えば、細胞表面へのウイルスの付着または接着を阻害することによって、例えば、ウイルスの付着または接着を担う部位に直接結合するか、またはその近傍で結合する抗体または抗体様分子もしくは結合分子等の分子を操作することによって、達成することができる。中和はまた、ビリオン表面に指向してビリオンの凝縮をもたらす抗体または抗体様分子等の分子によって達成することができる。中和は、標的細胞へのウイルスの付着後のウイルス膜と細胞膜との融合の阻害、エンドサイトーシスの阻害、および感染細胞由来の子孫ウイルスの阻害等によってさらに生じ得る。本発明の、抗体または抗体様分子もしくは結合分子等の中和分子は、中和を達成する機構によって制限されない。
【0087】
「抗体レパートリー」という用語は、本明細書中で最も広い意味で使用され、これは、結合能力、結合特異性、胃腸管輸送能力、安定性、および親和性等の特定の性質をスクリーニングするために使用することができる抗体または抗体フラグメントの集団を表す。本用語には、具体的には、例えば、抗体ファージ提示ライブラリー等の組み合わせライブラリーのすべての型を含む、抗体ライブラリーが含まれ、これには、ナイーブライブラリー、合成ライブラリー、および半合成ライブラリーを含む、任意の供給源由来の単鎖Fv(scFv)およびFab抗体ファージ提示ライブラリーが含まれるが、これらに限定されない。
【0088】
同様に、(上記定義のように)「抗体様分子のレパートリー」は、結合能力、結合特異性、胃腸管輸送能力、安定性、および親和性等の特定の性質をスクリーニングするために使用することができる、かかる分子の集団を表す。本用語には、具体的には、サロボディライブラリーおよびκ様軽鎖構築物のライブラリー(上記定義)が含まれ、これらには、例えば、抗体ファージ提示ライブラリー等の組み合わせライブラリーのすべての型が含まれる。組み合わせサロボディライブラリーは、例えば、Xu et al.,(2008)、(前出)に開示されている。
【0089】
「抗体レパートリー」という用語は、本明細書中で最も広い意味で使用され、これは、結合能力、結合特異性、胃腸管輸送能力、安定性、および親和性等の特定の性質をスクリーニングするために使用することができる抗体または抗体フラグメントの集団を表す。本用語には、具体的には、例えば、抗体ファージ提示ライブラリー等の組み合わせライブラリーのすべての型を含む、抗体ライブラリーが含まれ、これには、ナイーブライブラリー、合成ライブラリー、および半合成ライブラリーを含む、任意の供給源由来の単鎖Fv(scFv)およびFab抗体ファージ提示ライブラリーが含まれるが、これらに限定されない。
【0090】
「ファージ提示ライブラリー」は、ファージコートタンパク質との融合物としてのクローン化タンパク質配列集団を発現するタンパク質発現ライブラリーである。したがって、「ファージ提示ライブラリー」という語句は、本明細書中で、ファージ(例えば、繊維状ファージ)の集団を表し、このファージは外部(典型的には、異種)タンパク質を発現する。外部タンパク質は、ファージが接触する他の部分と自由に相互作用する(または結合する)。外部タンパク質を提示する各ファージは、ファージ提示ライブラリーの「メンバー」である。
【0091】
「抗体ファージ提示ライブラリー」は、抗体または抗体フラグメントを提示するファージ提示ライブラリーを表す。抗体ライブラリーには、ファージ集団、かかるファージ集団をコードするベクター集団、またはかかるファージ集団もしくはベクター集団を保有する細胞が含まれる。ライブラリーは、ファージ粒子あたり平均して1つの一本鎖抗体または抗体フラグメントを提示する1価であり得るか、ウイルス粒子あたり平均して2つ以上の抗体または抗体フラグメントを提示する2価であり得る。「抗体フラグメント」という用語には、単鎖Fv(scFv)フラグメントおよびFabフラグメントが含まれるが、これらに限定されない。好ましい抗体ライブラリーは、平均して106個超、107個超、108個超、または109個超の異なるメンバーを含む。
【0092】
「繊維状ファージ」という用語は、その表面上に異種ポリペプチドを提示することができるウイルス粒子を表し、繊維状ファージには、f1、fd、Pf1、およびM13が含まれるが、これらに限定されない。繊維状ファージは、テトラサイクリン(例えば、「fd−tet」)等の選択マーカーを含むことができる。種々の繊維状ファージ提示系が当業者に周知である(例えば、Zacher et al,Gene 9:127−140(1980)、Smith et al,Science 228:1315−1317(1985)、およびParmley and Smith,Gene 73:305−318(1988)を参照)。
【0093】
「パニング」という用語は、標的に対して高い親和性および特異性を有する抗体等の化合物を保有するファージの同定および単離における複数ラウンドのスクリーニング過程を表すために使用される。
【0094】
本明細書中で使用する、「非ヒト動物」という用語は、哺乳動物、例えば、非ヒト霊長類、げっ歯類(例えば、マウスおよびラット)、および例えば、ウサギ、ブタ、ヒツジ、ヤギ、ウシ、ブタ、ウマ、およびロバ等の非げっ歯類動物を含むが、これらに限定されない。鳥類(例えば、ニワトリ、シチメンチョウ、アヒル、およびガチョウ等)も含まれる。本明細書中で使用する、「非霊長類動物」という用語は、特に上記列挙の哺乳動物を含むが、これらに限定されず、霊長類以外の哺乳動物を表す。
【0095】
「機能的に異なる抗体」という語句、およびその文法上の変異体は、結合特異性、結合親和性、および例えば、標的を中和する能力、もしくは生物学的活性の範囲または質等の任意の免疫学的機能または生物学的機能を含むが、これらに限定されず、少なくとも1つの性質が互い異なる抗体を表すために使用される。
【0096】
「保存アミノ酸残基」という語句は、互いに整列している2つ以上のアミノ酸配列間で同一のアミノ酸残基を表すために使用される。
【0097】
本明細書中で使用する場合、「エピトープ」という用語は、少なくとも約3〜5個、好ましくは少なくとも約5〜10個、または少なくとも約5〜15個のアミノ酸、典型的には、せいぜい500個または約1,000個のアミノ酸の配列を表し、それ自体またはより大きな配列の一部として、かかる配列に応答して生成される抗体に結合する配列を定義する。エピトープは、それが由来する親タンパク質の一部と同一の配列を有するポリペプチドに制限されない。実際、ウイルスゲノムは、その状態が絶えず変化し、分離株間で比較的高い変異性を示す。したがって、「エピトープ」という用語は、未変性配列と同一の配列、および未変性配列の欠失、置換および/または挿入等の修飾物を含む。一般に、かかる修飾物は事実上保存されるが、非保存的修飾も企図される。この用語には、具体的には、「ミモトープ」、すなわち、連続的な線状の未変性配列を同定しないか、あるいは必ずしも未変性タンパク質中に生じないが、未変性タンパク質上のエピトープを機能的に模倣する配列が含まれる。「エピトープ」という用語には、具体的には、線状および立体構造エピトープが含まれる。
【0098】
B.一般的技術
本発明の方法を実施するための技術当該分野で周知であり、標準的な実験テキスト(例えば、Ausubel et al.,Current Protocols of Molecular Biology,John Wiley and Sons(1997)、Molecular Cloning:A Laboratory Manual,Third Edition,J.Sambrook and D.W.Russelleds.,Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001、Antibody Phage Display:Methods and Protocols,P.M.O‘Brian and R.Aitken eds.,Human Press,In:Methods in Molecular Biology,Vol.178、Phage Display:A Laboratory Manual,C.F.Barbas III et al.eds.,Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001、およびAntibodies,G.Subramanian,ed.,Kluwer Academic,2004に記載されている。変異誘発を、例えば、部位特異的変異誘発(Kunkel et al,Proc.Natl.Acad.Sci USA 82:488−492(1985))を用いて行うことができる。
【0099】
1つの態様では、本発明のウイルス抗原中和分子は、典型的には、抗体または多様なポリペプチドライブラリーを用いて選択される抗体である。以下の記載では、本発明は、ある抗体ライブラリー型を参照して例示しているが、本発明は、任意の特定の抗体または多様なポリペプチドライブラリー型の使用に制限されない。組換えモノクローナル抗体ライブラリーは、免疫フラグメントまたはナイーブフラグメントに基づき得る。免疫抗体ライブラリー由来の抗体は、典型的には、無作為な組み合わせライブラリーを産生するために、元のB細胞から発現に適切なベクターにクローン化されるVHおよびVL遺伝子プールを使用して構築し、その後に抗体を選択および/またはスクリーニングすることができる。他のライブラリー型は、抗原に結合するクローンに明確に偏っていない遺伝子供給源由来の抗体フラグメントから構成され得る。したがって、ナイーブ抗体ライブラリーは、天然で免疫化されていない再配列V遺伝子に由来する。合成抗体ライブラリーを、1つまたは複数のV遺伝子のCDRへの完全または適合した縮重領域の導入するインビトロ法によって完全に構築する。半合成ライブラリーは、天然の多様性と合成の多様性が組み合わされており、しばしば、天然の多様性が増加する一方で、所望のレベルの機能的多様性が保持されるように作製される。したがって、かかるライブラリーは、例えば、天然CDR領域のシャフリング(Soderlind et al,Nat.Biotechnol.18:852−856(2000))またはヒトB細胞由来の天然に再配列されたCDR配列と合成CDR1およびCDR2多様性との組み合わせ(Hoet et al,Nat.Biotechnol.23:455−38(2005))によって作製することができる。本発明は、ナイーブ抗体ライブラリー、合成抗体ライブラリー、および半合成抗体ライブラリー、またはその任意の組み合わせの使用を含む。
【0100】
同様に、本発明の方法は、抗体の提示のために使用されるいかなる特定の技術にも制限されない。本発明はファージ提示を参照して例示されているが、本発明の抗体を、他の提示および富化技術によって同定することもできる。抗体フラグメントは、抗体遺伝子をコードする繊維状ファージの表面上に提示されている(Hoogenboom and Winter J.Mol.Biol,222:381 388(1992)、McCafferty et al,Nature 348(6301):552 554(1990)、Griffiths et al EMBO J.,13(14):3245−3260(1994))。抗体ライブラリーの選択およびスクリーニング技術の概説については、例えば、Hoogenboom,Nature Biotechnol.23(9):1105−1116(2005)を参照。加えて、Escherichia coli(Agterberg et al,Gene 88:37−45(1990)、Charbit et al,Gene 70:181−189(1988)、Francisco et al,Proc.Natl.Acad.Sci.USA 89:2713−2717(1992))およびSaccharomyces cerevisiae等の酵母(Boder and Wittrup,Nat.Biotechnol.15:553−557(1997)、Kieke et al,Protein Eng.10:1303−1310(1997))の表面上の異種タンパク質およびそのフラグメントの提示について当該分野で公知の系が存在する。他の公知の提示技術には、リボゾームまたはmRNA提示(Mattheakis et al,Proc.Natl,Acad.Sci,USA 91:9022−9026(1994)、Hanes and Pluckthun,Proc.Natl,Acad.Sci,USA 94:4937−4942(1997))、DNA提示(Yonezawa et al,Nucl.Acid Res.31(19):e118(2003))、細菌提示等の微生物細胞提示(Georgiou et al,Nature Biotech.15:29−34(1997))、哺乳動物細胞上の提示、胞子提示(Isticato et al,J.Bacteriol.183:6294−6301(2001)、Cheng et al,Appl.Environ,Microbiol 71:3337−3341(2005)、および2006年11月13日に出願された同時係属の米国仮出願第60/865,574号))、レトロウイルス提示等のウイルス提示(Urban et al,Nucleic Acids Res.33:e35(2005)、タンパク質−DNA結合に基づいた提示(Odegrip et al,Proc.Acad.Natl、Sci.USA 101:2806−2810(2004)、Reiersen et al,Nucleic Acids Res.33:e10(2005)、ならびにマイクロビーズ提示(Sepp et al,FEBS Lett.532:455−458(2002))が含まれる。
【0101】
C.好ましい実施形態の詳細な説明
1つの態様では、本発明は、インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、ウイルスの血球凝集素(HA)抗原に結合するが、血球凝集を阻害しないモノクローナル抗体および抗体様分子の選択、産生、および使用に関する。
【0102】
インフルエンザA型ウイルスのビリオンは、8セグメントの線状の負のセンス一本鎖RNAを含む。総ゲノム長は13600ヌクレオチドであり、この8セグメントは、2350ヌクレオチド長、2350ヌクレオチド長、2250ヌクレオチド長、1780ヌクレオチド長、1575ヌクレオチド長、1420ヌクレオチド長、1050ヌクレオチド長、および900ヌクレオチド長である。インフルエンザA型ウイルスの宿主特異性および弱毒化は、ウイルス血球凝集素(H、HA)、核タンパク質(NP)、マトリックス(M)、および非構造(NS)遺伝子にそれぞれまたはウイルス遺伝子の組み合わせで寄与している(例えば、Rogers et al,Virology 127:361−373(1983)、Scholtissek et al,Virology 147:287−294(1985)、Snyder et al,J.Clin.Microbiol.24:467−469(1986)、Tian et al,J.Virol.53:771−775(1985)、Treanor et al,Virology 171:1−9(1989)を参照)。
【0103】
血球凝集素およびノイラミニダーゼタンパク質を含む、インフルエンザA型ウイルスおよびその表面タンパク質のヌクレオチド配列およびアミノ酸配列は、例えば、Theoretical Biology and Biophysics Group of Los Alamos National Laboratoryが保持しているインフルエンザ配列データベース等のGenBankおよび他の配列データベースから利用可能である。インフルエンザA型ウイルス血球凝集素の15種の公知のH亜型(H1〜H15)のアミノ酸配列は、2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)に示されている。さらなるインフルエンザA型ウイルス血球凝集素亜型(H16)が、最近スウェーデンにてユリカモメから単離され、Fouchier et al.,J.Virol.79(5):2814−22(2005)によって報告された。各H亜型の非常に多様な株も公知である。例えば、H5 A/香港/156/97と指定されたHAタンパク質配列は、1997年5月に香港でヒトから単離されたインフルエンザA型H5N1ウイルスから決定され、Suarez et al.,J.Virol.72:6678−6688(1998)に他の関連H5N1分離株から得たいくつかのさらなる株の配列と比較して示されている。
【0104】
インフルエンザウイルスノイラミニダーゼの触媒部位および抗原部位の構造はColman et al.,Nature 303:41−4(1983)によって公開されており、ノイラミニダーゼ配列はGenBankおよび他の配列データベースから利用可能である。
【0105】
感染個体の免疫応答に起因するウイルス特異的抗体が、典型的にはウイルス血球凝集素との相互作用を介してウイルスを中和することが公知である(Ada et al.,Curr.Top.Microbiol.Immunol.128:1−54(1986)、Couch et al.,Annu.Rev.Micobiol.37:529−549(1983))。インフルエンザウイルス血球凝集素の三次元構造およびインフルエンザウイルス血球凝集素と中和抗体との複合体の結晶構造も決定され、公開されている(例えば、Wilson et al.,Nature 289:366−73(1981)、Ruigrok et al.,J.Gen.Virol.69(Pt 11):2785−95(1988)、Wrigley et al.,Virology 131(2):308−14(1983)、Daniels et al.,EMBO J.6:1459−1465(1987)、およびBizebard et al.,Nature 376:92−94(2002)を参照)。
【0106】
本発明によれば、所望の性質を有する抗体を1つまたは複数の抗体ライブラリーから特定し、この抗体は、種々の供給源に由来し、異なる型であり得る。
【0107】
包括的ヒトインフルエンザ抗体ライブラリー
包括的ヒトインフルエンザ抗体ライブラリーは、種々の以前のインフルエンザ、季節性の大流行、エピデミック、ならびに1968年の香港風邪(H3N2)、1957年のアジア風邪(H2N2)、1918年のスペイン風邪(H1N1)、および2004/2005年のトリインフルエンザ(H5N1)を含む、パンデミックの回復期患者から得た抗体から作製することができる。例えば、2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照。かかるライブラリーを調製するために、血液または骨髄のサンプルを、インフルエンザウイルスに感染していることが知られているか疑いのある個体から採取する。特に地理的に離れた供給源由来の末梢血サンプルを、輸送および使用前に安定化することが必要であり得る。この目的のためのキットは、周知かつ市販されており、例えば、BD Vacutainer(登録商標)CPT(商標)細胞調製管等をリンパ球の遠心精製のために使用することができ、グアニジウム、トリゾール、またはRNAlaterをサンプル安定化のために使用した。安定化リンパ球または全骨髄の受け取りの際、当該分野で公知の免疫グロブリンオリゴプライマーを使用してRT−RCPを行い、重鎖および軽鎖レパートリーをレスキューする。当該分野で公知の手順に従って、PCRレパートリー産物をリンカーオリゴと組み合わせてscFvライブラリーを生成し、m13pIIIタンパク質を使用してインフレームで直接クローン化する。
【0108】
典型的なプロトコルでは、ヒト血清中の抗体を、例えば、周知の血球凝集素阻害(HAI)アッセイ(Kendal,A.P.、M.S.Pereira and J.J.Skehel.1982.Concepts and procedures for laboratory−based influenza surveillance.U.S.Department of Health and Human Services,Public Health Service,Centers for Disease Control,Atlanta,Georgia)またはマイクロ中和アッセイ(Harmon et al.,J.Clin.Microbiol.26:333−337(1988))を含む、周知の血清学的アッセイによって検出することができる。血清サンプルがインフルエンザ中和抗体を含むと既に確認されている場合、この検出工程は必要ないかもしれない。全血由来のリンパ球または骨髄中に存在するリンパ球を、次に、当該分野で公知の方法によって処理する。新鮮な組織またはRNAlaterで安定化した組織由来の全RNAを、TriBD試薬(Sigma)によって抽出する。その後、単離したドナー総RNAを、Oligotex精製(Qiagen)を用いて、mRNAにさらに精製する。次に、AccuScript逆転写酵素(Stratagene)のプロトコルに従ったランダム九量体オリゴヌクレオチドおよびまたはオリゴ(dT)18プライマーの使用によって第1のcDNA鎖を合成する。簡潔に述べれば、100ngのmRNA、0.5mMのdNTP、および300ngのランダム九量体およびまたは500ngのオリゴ(dT)18プライマーを含むAccuscriptRT緩衝液(Stratagene)を65℃で5分間インキュベートし、その後に4℃に急冷する。次いで、100mMのDTT、Accuscript RT、およびRNAse Blockを各反応物に添加し、42℃で1時間インキュベートし、逆転写酵素を70℃で15分間の加熱によって不活性化する。得られたcDNAを抗体重鎖および軽鎖V遺伝子のRT−PCR増幅のテンプレートとして使用することができ、次いで、ベクターにクローン化することができるか、またはファージ提示ライブラリーを意図する場合、ファージベクターにクローン化することができる。この手順により、抗体重鎖および軽鎖可変領域クローンのレパートリー(VHおよびVLライブラリー)が得られ、これを分離したままにするか、スクリーニングのために組み合わせることができる。
【0109】
1918年のスペイン風邪等の先のエピデミックおよびパンデミックの生存者の末梢リンパ球由来の免疫グロブリンレパートリーを、上記に類似の様式で回収し、安定化し、レスキューすることができる。さらなるH1およびH3ライブラリーレパートリーを、適切にタイミングを合わせてワクチン接種を行った局所起源のドナーから回収することができる。さらなる選択肢として、市販の骨髄総RNAまたはmRNAを商業的供給源から購入し、H2抗体スクリーニングにも適切なドナーの背景に応じてH1およびH3に適切なライブラリーを産生することができる。
【0110】
合成ヒト様レパートリー
本発明の方法では、合成ヒト抗体レパートリーを、合成抗体ライブラリーによって示すことができ、このレパートリーを、当該分野で公知の方法によって作製することができるか、または商業的供給源から入手することができる。したがって、例えば、完全に合成のヒトレパートリーは、2009年3月26日に出願されたHorowitzらの米国特許出願公開第20090082213号(参照によりその全体が本明細書に組み込まれる)に記載されている。簡潔に述べれば、この特許出願は、所定のアミノ酸が、目的の免疫グロブリンの1つまたは複数の相補性決定領域に組み合わせて導入された免疫グロブリンのライブラリーを記載している。さらに、例えば、かかるライブラリーのサブセットを含むユニバーサル免疫グロブリンライブラリーは、2003年12月11日に公開された米国特許出願公開第20030228302号(参照によりその全体が本明細書に組み込まれる)に記載されている。
【0111】
種々の変異を有する抗体重鎖および軽鎖の特定のサブライブラリーを組み合わせて、本発明の抗体のフレームワーク構築物を得て、その後に重鎖および軽鎖の両方のCDRに多様性を導入することができる。この多様性を、例えば、Kunkel変異誘発等の当該分野で公知の方法によって達成することができ、これを数回繰り返して多様性をさらに増大させることができる。したがって、例えば、複数のKunkel変異誘発ラウンドによって重鎖および軽鎖CDR1およびCD2領域に多様性を個別または同時に導入することができる。必要に応じて、種々のKunkelクローンを、CDRの長さによって分離することができ、そして/またはターゲティングしたCDR(例えば、CDR1またはCDR3)中に多様性を欠くクローンを、例えば、テンプレート特異的制限酵素での消化によって除去することができる。これらの工程の完了の際、ライブラリーのサイズは、約109メンバーを超えるべきであるが、より少数のメンバーを有するライブラリーも有用である。
【0112】
特定の実施形態では、免疫化抗体ライブラリーおよび合成抗体ライブラリーの両方を、本発明の中和抗体の同定のために使用する。2つのライブラリー型は基本的に異なる。合成抗体ライブラリーは、抗原に結合する能力が予想されるヒト抗体集団を合成する一方で、免疫化レパートリーはトリH5血球凝集素、および/またはH1、H2、またはH3血球凝集素(場合による)を特異的に認識するための配列を含むであろう。したがって、免疫化レパートリーは、理論的には、ターゲティングしたインフルエンザ亜型の重要成分を認識するように最適化される。これらの相違の結果として、2つの方法によって異なる抗体組が産生され、したがって、所望の中和抗体を同定するためのより有効なアプローチが得られる。
【0113】
高免疫化非ヒト霊長類抗体ライブラリー
本方法では、抗体ライブラリーを、例えば、マカクまたはヒヒ等の高免疫化非ヒト霊長類から救済する。具体的には、非ヒト霊長類を、インフルエンザA型ウイルスの種々の亜型または種々の血球凝集素(H)タンパク質で免疫化する。動物が免疫化されるインフルエンザA型ウイルス亜型または血球凝集素を認識する抗体の力価を生じる動物を屠殺し、その脾臓を採取する。上の包括的インフルエンザ抗体ライブラリーについて記載されるように、免疫化した動物の血液または骨髄を採取し、産生された抗体を回収し、増幅する。
【0114】
本発明の中和抗体の単離のための戦略
使用した抗体ライブラリーの型と無関係に、例えば、2つの異なるインフルエンザA型亜型および/または同一亜型の2つの株(分離株)、ならびに/またはヒトおよび非ヒト分離株との反応を示す等の二重特異性を有する抗体を、調節された交差反応選択および/または定方向組み合わせおよび/または変異誘発操作によって発見および最適化することができる。
【0115】
図1に例示する典型的な富化スキームでは、標的AおよびBと指定した2つの標的に交差反応性を示す抗体を含むライブラリーを、複数の富化ラウンドに供する(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照)。富化が標的Aとの反応性に基づく場合、各富化ラウンドにより、標的Aに対してプールの反応強度が増加するであろう。同様に、富化が標的Bとの反応性に基づく場合、各富化ラウンドにより、標的Bに対してプールの反応強度が増加するであろう。このアプローチが、ファージ提示ライブラリーをスクリーニングする場合に使用される選択方法であるパニング(以下を参照)を表すにもかかわらず、アプローチは、上記で考察した任意のライブラリー型、当該分野で公知の他の型、および任意の提示技術型に等しく適用可能である。標的AおよびBには、インフルエンザウイルスの種々の分離株、型、および亜型が含まれるが、これらに限定されない、抗体が結合する任意の標的が含まれる。
【0116】
本発明の目的が複数の特異性を有する中和抗体を同定することであるので、交差反応発見選択スキームが開発された。簡潔にするために、このスキームを、二重特異性を有する抗体の選択を示す図2に例示する。この場合、2つの標的である標的AおよびBとの反応性を示す抗体を含む抗体ライブラリーを、一方の標的(例えば、標的A)との反応性について最初に選択し、その後に他方の標的(例えば、標的B)との反応性について選択する。それぞれの連続的な選択ラウンドにより、両標的に対する得られたプールの反応強度を増加させる(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)も参照されたい)。したがって、本方法は、二重特異性を有する抗体の同定に特に有用である。もちろん、本方法を、さらなる標的に対するさらなる富化ラウンドを含めることによって、さらなる標的に対する反応性を示す抗体の同定に拡大することができる。また、スクリーニングされるライブラリーがファージ提示ライブラリーである場合、交差反応パニングによって選択を行うが、他のライブラリーおよび他の選択方法を使用することもできる。
【0117】
上記で考察した2つの方法の組み合わせは、上記のように、それぞれ標的Aおよび標的Bに対する反応性についての2つの個別の富化ラウンド、得られた2つのプールの組換え、およびその後の交差反応選択ラウンドを含む(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照)。このアプローチを、図1に例示する。純粋な交差反応と同様に、組換えられたライブラリーの各選択ラウンドにより、両標的に対する得られたプールの反応強度が増加する。
【0118】
図2に例示するさらなる実施形態では、第1に、標的Aと強い反応性を示し、かつ標的Bと検出可能な交差反応性を有するクローンを同定する。このクローンに基づいて、変異誘発ライブラリーを調製し、次いで、交互のラウンドにおいてそれぞれ標的Bおよび標的Aとの反応性について選択する。このスキームにより、標的Aとの強い反応性を保持し、かつ標的Bとの反応性が増加した抗体が得られる(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照)。これまでと同様に、スクリーニングされるライブラリーがファージ提示ライブラリーである場合、パニングによって選択を行うが、同一の戦略に従って、他のライブラリー、他の提示技術、および他の選択方法を使用することもできる。
【0119】
上記で考察するように、標的AおよびBは、例えば、インフルエンザA型ウイルスの2つの異なる亜型、同一のインフルエンザA型ウイルスの2つの異なる株(分離株)、2つの異なる種(一方の種は好ましくはヒトである)由来の亜型または分離株であり得る。したがって、例えば、標的Aは、H1N1ウイルスのニューカレドニア分離株の分離株であり得、標的Bは、H3N2ウイルスのウィスコンシン分離株であり得る。これらの例は例示のみを目的とし、任意の2つまたは複数の標的に対する二重および複数の特異性を有する抗体を類似の様式で同定、選択、および最適化することができることを強調しておく。
【0120】
代替として、個別のフレームワークおよびCDRの長さを分離することが可能なUAL等の抗体ライブラリーを使用して、標的Aに対する抗体を見出す場合、抗原Bをスクリーニングし、ライブラリーを類似のパラメーターの多様な集団に制限することができる。一旦抗原Bに対する抗体が見出された場合、各A抗体およびB抗体に基づいたキメラ抗体または変異誘発抗体を使用して二重特異性集団を操作することができる。
【0121】
ファージ提示
特定の実施形態では、本発明は、複数の(二重を含む)特異性を有するモノクローナル中和抗体を機能的に発見するためにファージ提示抗体ライブラリーを使用する。かかる抗体は、例えば、H1、H3、および/またはH9亜型、H1およびH3、H5およびH1等のH5、H7、および/またはH9亜型;H5およびH2;H5およびH3;H5、H1、およびH2;H5、H1、およびH3;H5、H2、およびH3;H1、H2およびH3等の亜型を含む、1つを超えるインフルエンザA型ウイルス亜型、ならびに/または同一亜型の1つを超える株(分離株)を中和することができるモノクローナル抗体であり得る。
【0122】
ファージ抗体ライブラリーを生成するために、上記考察のライブラリーを含む、任意の供給源から得たcDNAライブラリーをファージベクターにクローン化する。
【0123】
したがって、例えば、上記のようにRT−PCRによってリンパ球または骨髄からレスキューした抗体重鎖および軽鎖レパートリー集団を、m13 pIIIタンパク質に融合したscFvライブラリーとして再アセンブリする。組み合わせライブラリーは、約106超、107超、108超、または109超の異なるメンバーを含み、107超もしくはそれ以上の異なるメンバーが好ましい。品質管理のために、ランダムクローンを配列決定して、全体的なレパートリーの複雑さを評価する。
【0124】
同様に、ナイーブ、免疫化ヒト、または高免疫化非ヒト霊長類の抗体ライブラリー由来の重鎖および軽鎖可変領域の最初のPCR救済後、PCR産物をリンカーオリゴと組み合わせて、M13 pIIIコートタンパク質を使用してインフレームで直接クローン化するためのscFvライブラリーを生成する。このライブラリーは、約106超、107超、108超、または109超の異なるメンバーを含み、107超もしくはそれ以上の異なるメンバーが好ましい。品質管理のために、ランダムクローンを配列決定して、全体的なレパートリーのサイズおよび複雑さを評価する。
【0125】
抗体ファージ提示ライブラリーは、単鎖Fv(scFv)形式またはFab形式等の種々の形式の抗体を含むことができる。概説については、例えば、Hoogenboom,Methods Mol.Biol.178:1−37(2002)を参照。
【0126】
スクリーニング
所望の中和特性を有する抗体を同定するためのスクリーニング方法を、上記している。所望の血球凝集素タンパク質への直接結合に基づいて反応性を評価することができる。
【0127】
血球凝集素(HA)タンパク質の産生
血球凝集素(HA)タンパク質を、組換えDNAテクノロジーによって産生することができる。この方法では、HA遺伝子を、適切なベクター、好ましくは、Spodoptera frugiperda(Sf9)細胞等のバキュロウイルス感染昆虫細胞における発現のためのバキュロウイルス発現ベクターにクローン化する。
【0128】
HAタンパク質をコードする核酸を、ポリ−his(ヘキサヒスチジンタグ)等のC末端エピトープタグを使用するか使用しないで、Bac−to−Bac(Invitrogen)等のバキュロウイルス発現ベクターに挿入する。ポリ−hisタグは、ニッケルキレートクロマトグラフィーによる容易な精製を提供する。
【0129】
一般に、クローニングは、アセンブリPCRによって個別に合成されたオリゴから基準cDNAを作製する工程を含む。対応する分離株の変異体HAタンパク質を、さらなるアセンブリPCRへの適切な変異オリゴの置換、あるいはKunkel変異誘発等の変異誘発技術のいずれかによって作製する。
【0130】
組換えバキュロウイルスを、リポフェクチン(Gibco−BRLから市販されている)を使用した上記バクミドのSf9細胞(ATCC CRL 1711)へのトランスフェクションによって生成する。28℃での4〜5日間のインキュベーション後、放出されたウイルスを採取し、さらなる増幅のために使用する。ウイルス感染およびタンパク質発現を、O’Reilley et al.,Baculovirus Expression Vectors:A Laboratory Manual(Oxford:Oxford University Press,1994)に記載のように行う。
【0131】
次いで、発現したポリ−Hisタグ化HAポリペプチドを、例えば、以下のように、Ni2+−キレートアフィニティクロマトグラフィーによって精製することができる。Rupert et al.,Nature 362:175−179(1993)に記載のように、組換えウイルス感染Sf9細胞から上清を回収する。総容積5mLを用いて、Ni2+−NTAアガロースカラム(Qiagenから市販されている)を調製し、25mLの水で洗浄し、25mLのローディング緩衝液で平衡化する。濾過した細胞抽出物を、0.5mL/分でカラムに充填する。カラムをローディング緩衝液でベースラインA280まで洗浄し、その時点で画分回収を開始する。次に、カラムを二次洗浄緩衝液(50mMリン酸塩;300mM NaCl、10%グリセロール(pH6.0))で洗浄し、非特異的結合タンパク質を溶離する。A280ベースラインに再度到達した後、カラムを、二次洗浄緩衝液を含む0〜500mMイミダゾール勾配を使用して展開する。1mLの画分を回収し、SDS−PAGEおよび銀染色またはアルカリホスファターゼ抱合Ni2+−NTA(Qiagen)を使用したウェスタンブロットによって分析する。溶離したHis10タグ化HAポリペプチドを含む画分をプールし、ローディング緩衝液に対して透析する。
【0132】
代替として、IgGタグ化(またはFcタグ化)HAポリペプチドの精製を、例えば、プロテインAまたはプロテインGカラムクロマトグラフィーを含む公知のクロマトグラフィー技術を用いて行うことができる。
【0133】
Sf9細胞の代替物として、HAタンパク質を、他の組換え宿主細胞、原核生物、酵母、または高等真核細胞中で産生することもできる。適切な原核生物には、グラム陰性生物またはグラム陽性生物等の真正細菌、例えば、Enterobacteriaceae(Escherichia(例えば、E.coli)、Enterobacter、Erwinia、Klebsiella、Proteus、Salmonella(例えば、Salmonella typhimurium)、Serratia(例えば、Serratia marcescans)、およびShigella等)、ならびにBacilli(B.subtilisおよびB.licheniformis(例えば、1989年4月12日に公開された第DD266,710号に開示のB.licheniformis 41P)等)、P.aeruginosa等のPseudomonas、ならびにStreptomycesが含まれるが、これらに限定されない。E.coli K12 MM294株(ATCC 31,446)、E.coli X1776(ATCC 31,537)、E.coli W3110株(ATCC 27,325)、およびK5 772(ATCC 53,635)等の種々のE.coli株は、公的に利用可能である。
【0134】
原核生物に加えて、糸状菌または酵母等の真核微生物は、HAポリペプチドをコードする核酸を含むベクターの適切なクローニング宿主または発現宿主である。Saccharomyces cerevisiaeは、一般的に使用されている下等真核宿主微生物である。しかし、多数の他の属、種、および株、例えば、Schizosaccharomyces pombe(Beach and Nurse,Nature 290:140(1981)、1985年5月2日に公開された欧州特許第139,383号);Kluyveromyces宿主(米国特許第4,943,529号;Fleer et al.,Bio/Technology 9:968−975(1991))、例えば、K.Lactis(MW98−8C、CBS683、CBS4574、Louvencourt et al.,J.Bacteriol 737(1983))、K.fragilis(ATCC 12,424)、K.Bulgaricus(ATCC 16,045)、K.wickeramii(ATCC 24,178)、K.waltii(ATCC 56,500)、K.drosophilarum(ATCC 36,906)、Van den Berg et al.,Bio/Technology 8:135(1990))、K.thermotolerans、およびK.marxianus等;yarrowia(欧州特許第402,226号);Pichia pastoris(欧州特許第183,070号、Screekrishna et al.,J.Basic Microbiol.28:265−278(1988));Candida;Trichoderma reesia(欧州特許第244,234号);Neurospora crassa(Case et al.,Proc.Natl.Acad.Sci.USA 76:5259−5263(1979));Schwanniomyces occidentalis(1990年10月31日に公開された欧州特許第394,538号)等のSchwanniomyces;ならびに例えば、Neurospora、Penicillium、Tolypocladium(1991年1月10日に公開されたWO91/00357号等の糸状菌、およびA.nidulans(Ballance et al.,Biochem.Biophys.Res.Commun.112:284−289(1983)、Tilburn et al.,Gene 26:205−221(1983)、Yelton et al.,Proc.Natl.Acad.Sci.USA 81:1470−1474(1984)等のAspergillus宿主、およびA.niger(KellyおよびHynes、EMBO J.4:475−479(1985)等)を、本明細書中で一般的に利用可能であり、かつ有用である。メチロトロープ酵母が本明細書中で適切であり、HAnsenula、Candida、Kloeckera、Pichia、Saccharomyces、Torulopsis、およびRhodotorulaからなる属から選択される、メタノール中で成長することができる酵母が含まれるが、これらに限定されない。この酵母クラスの例である特定の種のリストを、C.Anthony、The Biochemistry of Methylotrophs 269(1982)中に見出すことができる。
【0135】
HAタンパク質発現に適切な宿主細胞には、多細胞生物細胞が含まれる。無脊椎動物細胞の例には、Drosophila S2およびSpodoptera Sf9等の上記昆虫細胞、ならびに植物細胞が含まれる。有用な哺乳動物宿主細胞株の例には、チャイニーズハムスター卵巣(CHO)細胞およびCOS細胞が含まれる。より具体的な例には、SV40によって形質転換されたサル腎臓CV1株(COS−7、ATCC CRL 1651)、ヒト胚腎臓株(HEK293細胞または懸濁培養での成長についてサブクローン化されたHEK293細胞(Graham et al,J.Gen Virol.36:59(1977)))、チャイニーズハムスター卵巣細胞/−DHFR(CHO、Urlaub and Chasin,Proc.Natl.Acad.Sci.USA 77:4216(1980))、マウスセルトリ細胞(TM4、Mather,Biol.Reprod.23:243−251(1980))、ヒト肺細胞(W138、ATCC CCL75)、ヒト肝臓細胞(Hep G2、HB 8065)、およびマウス乳腺腫瘍(MMT 060562、ATCC CCL51)が含まれる。適切な宿主細胞の選択は、当業者の技術範囲内と見なされる。
【0136】
血球凝集素(HA)タンパク質パニング
HAタンパク質を、マイクロタイターウェルまたは磁性ビーズの表面上に固定して、上記ライブラリーをパニングする。特定の実施形態では、各ライブラリーを、1つまたは複数のHタンパク質に4℃で2時間結合させ、次いで、冷PBSで広範囲に洗浄し、その後に0.2Mグリシン−HCl緩衝液(pH2.5)を使用してHA特異的結合クローンを溶離する。回収したファージを中性pHにし、感受性宿主E.coliの感染によって増幅させる。その後、陽性クローン富化を繰り返すためにファージミド産生を誘導し、その後に選別のためにクローンを単離することができる。十分な富化の際に、全プールを、感染によって、HB2151等の非アンバー抑制性E.coli株に導入して可溶性scFvタンパク質を発現させる。代替として、下記のように、プールを、pBAD等の単量体scFv発現ベクターにサブクローン化することができ、組換え可溶性scFvタンパク質をインビトロでの分析および特徴づけのために発現させる。
【0137】
特徴づけ
クローンを、上記にように、1つまたは複数のHタンパク質への結合親和性について試験する。例えば、H1タンパク質(Refseq ABQ10137、分離株ニューカレドニア/20/99(H1N1)、および/またはRefseq ABU99069、分離株ソロモン諸島/3/06(H1N1))への結合を試験し、かつH3タンパク質(Refseq ACC67032、分離株ウィスコンシン/67/05(H3N2)、および/またはRefseq CAA24269、香港/68(H3N2))への結合を平行して試験するが、他の分離株も単独または任意の組み合わせで使用することができる。証明された結合に基づき得た陽性クローンは、中和能力について試験することができる。中和についての典型的な機能試験は、赤血球への全ウイルス結合を使用した血球凝集阻害アッセイを含む。代替として、組替え血球凝集素タンパク質および赤血球を使用した血球凝集アッセイが可能である。全血の必要性を排除するために、気道上皮細胞に対して血球凝集素結合阻害アッセイを行うことができる。結合アッセイは、任意のフローサイトメトリーまたは細胞ELISA(cELISA)ベースのアッセイが含まれるが、これらに限定されない、任意の形態で行うことができる。高価なフローサイトメトリー装置を使用する必要がなく、クローン評価がより自動化され、より多数のデータを収集することができるという点で、cELISAの使用は有利である。他方では、フローサイトメトリーは、より優れた感度、一貫性、および速度が可能である。
【0138】
1つの態様では、本発明の抗体は、インフルエンザウイルスを含有するH1HA、および/またはインフルエンザウイルスを含有するH3HAに対する結合親和性を有する。本発明の抗体の結合親和性を、例えば、Munson et al.,Anal.Biochem.,107:220(1980)のスキャッチャード分析等の当業者に公知の方法によって決定することができる。1つの実施形態では、結合親和性は、<100pMである。1つの実施形態では、抗体の結合親和性は、約1×10−7〜約1×10−13M、約1×10−8〜約1×10−12M、または約1×10−9〜約1×10−11Mである。他の実施形態では、抗体の結合親和性は、約1×10−7M、約1×10−8M、約1×10−9M、約1×10−1×10M、約1×10−11M、約1×10−12M、または約1×10−13Mである。
【0139】
最適化:変異誘発ライブラリー
インフルエンザエピデミックおよび非ヒト動物ウイルスに起因するヒト感染に関連する潜在的パンデミックを含むパンデミックの有効な管理のために、Hタンパク質の現在の分離株および未来の変異体を有効に中和する抗体が必要である。この目的を達成するために、ターゲティングした血球凝集素亜型のすべての公知の分離株に結合する多様なH(例えば、H1、H3、H5等)中和クローンを単離する必要がある。
【0140】
いくつかの例では、交差反応性抗体は、単一の抗原に対して対象としたスクリーニングを介して出現する。分離交差反応性クローンの可能性を増大するために、複数の抗原に対して前進的にスクリーニングすることによって複数の選択的圧力を印加するであろう。いずれかの事象では、交差反応性は、当該分野において公知の抗体最適化法によってさらに改良することができる。例えば、本明細書中に記載する免疫グロブリン鎖の可変領域のある領域は、選択したCDRおよび/またはフレームワーク領域の軽鎖シャッフリング、目的変異誘発、CDR合体、および直接変異誘発を含む、1つまたは複数の最適化戦略を対象とし得る。
【0141】
1つを超えるインフルエンザA型亜型および/または同一亜型の1つを超える分離株との本明細書中の抗体の交差反応性を意図的に導入するために設計された1つの変異誘発法を、本明細書中で「目的」変異誘発と表す。目的変異誘発を使用して、好ましくは反応性の異なる1つまたは複数の抗体クローンに基づいて抗体の集団を合理的に操作することができる。本発明の文脈では、目的変異誘発を使用して、抗体の個別のCDR中の配列等の類似の配列上の類似の位置によって特定された単一または複数の残基をコードする。この場合、これらの集団を、類似の位置に見出される残基の範囲を捕捉するためのオリゴ縮重を使用して生成する。この集団内で、親クローンの特異性の間またはそれを超えて一連の特異性が存在すると予想される。目的変異誘発の目的は、2つ以上の個別の物質または集団の間に多様な多機能抗体集団またはライブラリーを生成することである。インフルエンザの場合、本方法を、2つの異なるエピトープ、分離株、または亜型を認識する2つの抗体を使用し、両機能的質を単一の抗体に変えるために利用することができる。例として、第1のインフルエンザA型抗体は、H1亜型の分離株に特異性を示し得、第2の抗体は、インフルエンザA型ウイルスのH3亜型の分離株に特異性を示す。目的変異誘発ライブラリーを作製するために、両抗体についてのCDR配列を最初に獲得し、アラインメントする。次に、同一性が保存されたすべての位置を、適合した残基に単一コドンを使用して固定する。非保存位置で、両残基をコードするために縮重コドンを組み込む。いくつかの例では、縮重コドンは、この位置で2つの親残基のみをコードするであろう。しかし、いくつかの例では、さらなる副産物が産生される。副産物の産生レベルを、副産物産生を強制するように調節するか、サイズの限度または目的に応じてこの産生を排除することができる。
【0142】
したがって、例えば、2つの抗体の第1の位置がそれぞれ、トレオニンおよびアラニンである場合、第1の2つの位置中にA/G−C−を有する縮重コドンは、第3の位置中の塩基と無関係にトレオニンまたはアラニンのみをコードするであろう。例えば、次の位置の残基がリジンおよびアルギニンである場合、縮重コドンA−A/G−A/Gは、リジンまたはアルギニンのみをコードするであろう。しかし、縮重コドンA/C−A/G−A/G/C/Tを使用した場合、アスパラギン、ヒスチジン、グルタミン、およびセリン副産物も生成されるであろう。
【0143】
便宜上、適合したCDR長を有する抗体のみを使用することがより簡潔である。これを強制するための1つの方法は、最初に発見された抗体によって与えられたCERの長さおよび潜在的なさらなるフレームワーク制限に基づいて、第2の抗原についてサイズが制限されたライブラリーをスクリーングすることである。しかし、同じ長さのCDRの使用は単に便利であるが、必要条件ではないことを付け加えておく。この方法がインフルエンザA型ウイルスの中和抗体の機能的に多様な巨大ライブラリーを作製するのに有用である一方で、その適用性は遥かにより広いことが明白である。この変異誘発技術を使用して、任意の抗体の機能的に多様なライブラリーまたは集団を産生することができる(2008年1月17日に公開された米国特許出願公開第20080014205号(参照によりその全体が本明細書に組み込まれる)を参照)。したがって、図3は、変異誘発された親配列としてTNF−α抗体およびCD11a抗体のCDRを使用した目的変異誘発法の使用を例示するために本明細書中に含まれる。
【0144】
交差反応性が自然に一般に選択されないため、典型的な変異戦略を実行することは、強力な交差反応性を可能にし得ない可能性が高い。目的変異は、交差反応性タンパク質を有する抗体の範囲を生成するための対象方法として考案された。代替として、CDR合体は、交差反応性抗体の生成および/または最適化のための別の迅速かつ強力な戦略を提供し得る。抗原結合および特異性が、いずれかまたは両方の鎖から選択されたCDRの組み合わせが異なることによって大きく影響されることが確立されている。既存の抗体に含有されるCDRが、既に、標的に対して最適化され得るため、図6に示されるように、不均質性標的に対する有効性を証明し得る、単一の抗体または複数の抗体に由来するCDR合体の集団を作製することができる。
【0145】
図6では、CDRが合体された抗体は、単一のフレームワーク時にCERの組み合わせ重ね合わせとして示すが、それらは、複数の関連フレームワーク、および非関連フレームワークの際に、またはそのキメラフレームワークの際でさえ、CDRが重ね合わされない可能性があり、得られる合体された抗体または複数の抗体の全体の多様性および生産性を増大する。合体ライブラリーは、既存の抗体に見られる生産の多様性、および合体集合体あたりの多くの新規抗体を同定する能力を活用する利点を有する。付加的な重鎖フレームワークのこのより広域の使用は、CDRの文脈における結合のサンプリング、およびより高い交差反応性および潜在性が誘導体化される、従来のB細胞成熟において達成可能ではない組み合わせを可能にするフレームワーク変異体を可能にする。CDRが標的に係合するための相互作用ループとして機能するため、それぞれの本来の配置に関し、かつそれに関わらず、両方の重鎖と軽鎖との間でCDRを混合することによって、遥かに広範囲の組み合わせを作成することができる。
【0146】
より従来の最適化が活性の効力または範囲を増加させるのに十分である場合、次いで、標的としたランダム変異誘発、飽和変異誘発、またはさらには、エラー多発性PCRを利用することができる。
【0147】
曖昧に合成されたオリゴヌクレオチドを使用した標的化ランダム変異誘発(Matteuchi and Heyneker,Nucleic Acids Research 11:3113−3121(1983))は、意図するコドンおよびすべての可能なコドンを、特定の比で、互いに、指定の位置に生成する技術である。曖昧に合成されたオリゴヌクレオチドにより、指定の場所への非「野生型」塩基、または、コドンの特異的付加によるヌクレオチド付加の正確さが低下する。これを、典型的には、オリゴヌクレオチド合成機における野生型塩基と非野生型塩基との比の固定および合成時の2つの試薬の混合物の指定によって行う。
【0148】
飽和変異誘発(Hayashi et al.,Biotechniques 17:310−315(1994))は、タンパク質中の特定の位置の20種すべてのアミノ酸を置換し、各変異体に対応するクローンを特定の表現型についてアッセイする技術である(米国特許第6,171,820号、同第6,358,709号、および同第6,361,974号も参照されたい)。
【0149】
エラー多発性PCR(Leung et al.,Technique 1:11−15(1989)、Cadwell and Joyce,PCR Method Applic.2:28−33(1992))は、クローン化した遺伝子にランダム点変異を導入する修正ポリメラーゼ連鎖反応(PCR)技術である。得られたPCR産物をクローン化してランダム変異ライブラリーを産生することができるか、またはT7プロモーターが適切なPCRプライマー内に組み込まれる場合に直接転写することができる。
【0150】
他の変異誘発技術も当該分野で周知であり、例えば、In Vitro Mutagenesis Protocols,J.Braman,Ed.,Humana Press,2001に記載されている。
【0151】
最適化:変異誘発ライブラリースクリーニングのための選択的考察
この場合、主な目的の1つは、現在のH1および/もしくはH3(またはH5もしくはH9)分離株ならびに将来の変異体を有効に処置するために、集団からの1つの抗体(または複数の抗体)を操作し、単離することである。新規の分離株H1/H3中の変異を認識することができる耐性を有する抗体を操作するために、種々のH1/H3分離株に結合する中和クローンを同定すべきである。第1のH1/H3分離株に対してクローンを選択する場合、より低いレベルで、第2のH1/H3分離株に結合/中和すると予想される。この場合、目的は、第1のH1/H3分離株結合の改善(または少なくとも維持)という状況において、第2のH1/H3分離株の認識を劇的に改善することである。したがって、第2のH1/H3分離株の改善のために最初に選択を行い、その後に第1のH1/H3タンパク質を選択する。それにより、新規の株に対してより高い選択圧が得られる一方で、第2のパラメーターに対する選択圧が維持される。
【0152】
他のH中和抗体を類似の様式で最適化することができる。この場合、出発点または目的のいずれかとして他の単離株(例えば、H5、H9等)由来の任意の基準タンパク質配列を用いて、選択および最適化することができる。
【0153】
加えて、中和抗体クローンを使用して、型間認識を試験する。型間認識の例は、非H1起源のクローンまたは最適化クローン由来の同時または操作されたH1結合である。
【0154】
概して、複数の変異誘発集合およびスクリーンは、C5およびA11抗体、C5様およびA11様抗体、C5およびA11抗体様分子、C5およびC5様サロボディ、ならびにA11およびA11様サロボディに基づくことができる。それによって、先述の分子のそれぞれは、広い範囲の反応性および高い効力を有する適切な反応性分子を単離するために、上記に記載した任意の、かつすべての選択を対象とすることができる。
【0155】
中和抗体のエピトープマッピング
一旦所望の性質を有する中和抗体が同定されると、かかる抗体の大部分によって認識される優性のエピトープを同定することが望ましいかもしれない。エピトープマッピング法は、当該分野で周知であり、例えば、Morris,Glenn E.,Epitope Mapping Protocols,Totowa,N.J.ed.,Humana Press,1996、およびEpitope Mapping:A Practical Approach,Westwood and Hay,eds.,Oxford University Press,2001に開示されている。
【0156】
エピトープマッピングは、抗体が結合するエピトープの同定に関する。抗体−抗原複合体の結晶学的分析、競合アッセイ、遺伝子フラグメント発現アッセイ、および合成ペプチドベースのアッセイを含む、タンパク質上のエピトープの位置を決定するための当業者に公知の方法が多数存在する(例えば、HarlowおよびLaneの11章中の、Using Antibodies,a Laboratory Manual,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.,1999、米国特許第7,332,579号(それぞれ、参照によりその全体が本明細書に組み込まれる)を参照)。2つの抗体が同一のエピトープまたは立体的に重複するエピトープを認識する場合に、抗体は基準抗体と「本質的に同一のエピトープ」に結合する。同一または立体的に重複するエピトープに2つの抗体が結合するかどうかを決定するための一般的に使用されている方法は、競合アッセイであり、このアッセイを、標識した抗原あるいは標識した抗体のいずれかを用いて、すべての異なる形式で形成することができる。通常、抗原を96ウェルプレート上に固定し、非標識抗体が標識抗体の結合を遮断する能力を、放射性標識または酵素標識を使用して測定する。
【0157】
中和抗体の産生
一旦所望の中和特性を有する抗体が同定されると、抗体フラグメントを含むかかる抗体を、例えば、ハイブリドーマ技術または組換えDNA技術を含む当該分野で周知の方法によって産生することができる。
【0158】
ハイブリドーマ法では、マウスまたはハムスター等の他の適切な宿主動物を免疫化して、免疫化のために使用されるタンパク質に特異的に結合する抗体を産生するか産生することができるリンパ球を惹起する。代替として、リンパ球をインビトロで免疫化することができる。次いで、ポリエチレングリコール等の適切な融合剤を使用してリンパ球を骨髄腫細胞と融合し、ハイブリドーマ細胞を形成する(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。
【0159】
このようにして調製したハイブリドーマ細胞を、適切な培養培地中に播種し、成長させ、これには、非融合の親骨髄腫細胞の成長または生存を阻害する1つまたは複数の物質を含むことが好ましい。例えば、親骨髄腫細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠く場合、ハイブリドーマ用の培養培地は、典型的には、ヒポキサンチン、アミノプテリン、およびチミジン(HAT培地)を含み、これらの物質はHGPRT欠損細胞の成長を阻害する。
【0160】
好ましい骨髄腫細胞は、効率的に融合し、選択した抗体産生細胞による抗体の安定な高レベル産生を支持し、HAT培地等の培地に感受性を示す骨髄腫細胞である。これらの細胞株のうち、好ましい骨髄腫細胞株は、Salk Institute Cell Distribution Center,San Diego,California USAから利用可能なMOPC−21およびMPC−11マウス腫瘍由来の細胞株、およびAmerican Type Culture Collection,Rockville,Maryland USAから利用可能なSP−2細胞またはX63−Ag8−653細胞等のマウス骨髄腫株である。ヒトモノクローナル抗体の産生のためのヒト骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞株も記載されている(Kozbor,J.Immunol.133:3001(19.84)、およびBrodeur et al,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0161】
ハイブリドーマ細胞が成長する培養培地を、抗原に対するモノクローナル抗体の産生について評価分析する。好ましくは、ハイブリドーマ細胞によって産生されたモノクローナル抗体の結合特異性を、放射免疫アッセイ(RIA)または酵素結合免疫吸着アッセイ(ELISA)等の免疫沈降またはインビトロ結合アッセイによって決定する。
【0162】
組換えモノクローナル抗体を、例えば、必要な抗体鎖をコードするDNAの単離、周知の組換え発現ベクターを用いた組換え宿主細胞と同時発現用のコード配列との共トランスフェクションによって産生することができる。組換え宿主細胞は、上記の細胞等の原核細胞および真核細胞であり得る。
【0163】
ヒト化抗体の作製で使用すべきヒト可変ドメイン(軽鎖および重鎖の両方)の選択は、抗原性の減少に非常に重要である。いわゆる「ベストフィット」法によれば、げっ歯類抗体の可変ドメイン配列を、公知のヒト可変ドメイン配列の全ライブラリーに対してスクリーニングする。次いで、げっ歯類配列に最も近いヒト配列を、ヒト化抗体のヒトフレームワーク領域(FR)として許容する(Sims et al.,J.Immunol.151:2296(1993)、Chothia et al.,J.Mol.Biol.196:901(1987))。抗体を抗原に対する高親和性および他の好ましい生物学的性質を保持してヒト化することが重要である。この目的を達成するために、好ましい方法に従って、ヒト化抗体を、親配列およびヒト化配列の三次元モデルを使用した親配列および種々の概念ヒト化産物の分析過程によって調製する。
【0164】
加えて、当該分野で公知の方法に従って、ヒト抗体を生成することができる。例えば、免疫化の際に内因性免疫グロブリン産生の非存在下でヒト抗体の全レパートリーを産生することができるトランスジェニック動物(例えば、マウス)を作製することができる。例えば、Jakobovits et al.,Proc.Natl.Acad.Sci.USA 90:2551(1993)、Jakobovits et al.,Nature 362:255−258(1993)、Bruggermann et al.,Year in Immuno.7:33(1993)、および米国特許第5,591,669号、同第5,589,369号、および同第5,545,807号を参照。
【0165】
中和抗体
以下の実施例に記載されるものを含む、本明細書中に記載の技術の使用によって多数の中和抗体が同定されている。1つの態様では、本発明は、血球凝集素タンパク質エピトープに結合する中和抗体を提供する。1つの実施形態では、中和抗体は、血球凝集素タンパク質のHA1サブユニット上の少なくとも1つのエピトープに結合する。別の実施形態では、中和抗体は、血球凝集素タンパク質のHA1サブユニット上の少なくとも2つ、少なくとも3つ、少なくとも4つ、少なくとも5つ、または少なくとも6つのエピトープに結合する。
【0166】
いくつかの実施形態では、本発明の抗体は、H3および/またはH1を含むウイルスを中和する。他の実施形態では、抗体は、H3およびH1の両方を中和する。1つの実施形態では、本発明の抗体は、血球凝集を阻止する。他の実施形態では、抗体は、感染すべき標的細胞へのインフルエンザA型ウイルスの結合を阻止する。別の実施形態では、抗血球凝集素抗体は、インフルエンザA型ウイルスのHAの球状頭部上の受容体結合部位が標的細胞に付着してHAの血球凝集素活性が生じるのを阻害する。
【0167】
以下の実施例に記載される実験に基づいて、多数の抗血球凝集素抗体の重鎖/軽鎖対合を同定した。別の実施形態では、本発明の抗体は、2つまたは複数のインフルエンザA型ウイルス亜型に対して交差反応性である。1つの実施形態では、抗体は、配列番号1もしくは配列番号2として示したアミノ酸配列を含有する重鎖ポリペプチド、および配列番号3、配列番号4、配列番号5、もしくは配列番号6として示したアミノ酸配列を含有する軽鎖ポリペプチドを含有する。好ましい実施形態では、本発明の中和抗体は、(i)配列番号2として示した重鎖アミノ酸配列、および配列番号6として示した軽鎖アミノ酸配列を含む抗体(下記の実施例において、および表1に示すとおり、クローン1286−A11)、(ii)配列番号1として示した重鎖アミノ酸配列、および配列番号3として示した軽鎖アミノ酸配列を含む抗体(下記の実施例において、および表1に示すとおり、クローン1286−C5)、(iii)配列番号1として示した重鎖アミノ酸配列、および配列番号4として示した軽鎖アミノ酸配列を含む抗体(下記の実施例において、および表1に示すとおり、クローン1286−C5)、または(iv)配列番号1として示した重鎖アミノ酸配列、および配列番号5として示した軽鎖アミノ酸配列を含む抗体(下記の実施例において、かつ表1に示すとおり、クローン1286−C5)のエピトープと実質的に同一であるエピトープに結合する。
【0168】
典型的な重鎖ループよりも長い重鎖ループを有する抗体は、ある特性を示すとして報告されている。例えば、典型的な重鎖CDR3ループよりも長いループは、多反応性(Schettino et al.,J.Immunol.1997;158;2477−2489)に関係付けられており、ごく近年では、それらは、多くの抗H1V抗体(Saphire et al.,Science 2001;293;1151−1159、Kunert et al.,AIDS Res.Hum.Retroviruses;1998;14(13);1115−1128、Barbas et al.,J.Mol Biol 1993;230(3):812−823)、および抗肺炎球菌抗体(Baxendale et al.,2008;Clin.Exper.Immunol.2007;151;51−60)と関連している。延長ループは、より深いプロービング、および病原性抗原との相互作用を促進し得る。19アミノ酸長の重鎖CDR3を含有するH1V抗体の場合では、標的は、gp120上の感受性凹型領域と接触する、指状突起物の形成を介して非常に特異的に係合する。実施例7に記載されるように、C05抗体重鎖配列は、11アミノ酸の極めて非定型の長さの重鎖CDR1ループ、および25アミノ酸のCDR3ループを有する。
【0169】
1つの態様では、本発明は、長さが修正された重鎖CDRループを有する、インフルエンザ中和抗体または結合分子に関する。通常、抗原に接触するヒト重鎖CDR1ループの長さは、典型的には、約6または8アミノ酸のいずれかであり、抗原に接触するヒト重鎖CDR3ループの典型的な長さは、約13アミノ酸である(Kabat et al.,Sequences of Proteins of Immunological Interest,National Institutes of Health,Bethesda,MD,ed.5,(1991))、MacCallum et al., 1996,J.Mol.Biol.262,732−745)。1つの実施形態では、長さが修正された重鎖は、抗体の典型的な重鎖CDRループよりも長いCDRループを指す、「延長」重鎖CDRループである。
【0170】
1つの実施形態では、抗体または結合分子は、延長重鎖CDR3ループ、および/または延長重鎖CDR1ループを有する。1つの実施形態では、延長重鎖CDR3ループは、典型的な重鎖CDR3ループよりも長い、約1アミノ酸、約2アミノ酸、約3アミノ酸、約4アミノ酸、約5アミノ酸、約6アミノ酸、約7アミノ酸、約8アミノ酸、約9アミノ酸、約10アミノ酸、約11アミノ酸、約12アミノ酸、約13アミノ酸、約14アミノ酸、約15アミノ酸、約16アミノ酸、約17アミノ酸、約18アミノ酸、約19アミノ酸、または約20アミノ酸である。別の実施形態では、延長重鎖CDR3ループは、典型的な重鎖CDR3ループよりも長い、約1〜約20アミノ酸、約1〜約19アミノ酸、約1〜約18アミノ酸、約1〜約17アミノ酸、約1〜約16アミノ酸、約1〜約15アミノ酸、約1〜約14アミノ酸、約1〜約13アミノ酸、約1〜約12アミノ酸、約1〜約11アミノ酸、約1〜約10アミノ酸、約1〜約9アミノ酸、約1〜約8アミノ酸、約1〜約7アミノ酸v1〜約6アミノ酸、約1〜約5アミノ酸、約1〜約4アミノ酸、約1〜約3アミノ酸、または約1〜約2アミノ酸である。さらに別の実施形態では、延長重鎖CDR3ループは、約14、約15、約16、約17、約18、約19、約20、約21、約22、約23、約24、約25、約26、約27、約28、約29、約30、約31、約32、約33、約34、または約35アミノ酸の長さである。
【0171】
好ましい実施形態では、延長重鎖CDR3ループは、約25アミノ酸の長さである。別の好ましい実施形態では、延長重鎖CDR3ループは、アミノ酸配列AKHMSMQQVVSAGWERADLVGDAFD(配列番号9)を含む。記載される抗体が血球凝集を阻害することが可能であるため、この長さの重鎖CDR3ループもまた、球状頭部内の深部にプローブする突出物を形成することが可能である。球状頭部のかかる深部プローブは、血球凝集素の球状頭部ドメインの凹部内でシアル酸に介入する新規手段を提供し得る。これは、本明細書中に記載の延長CDR3ループを有する抗体に認められた活性の顕著な全幅に寄与し得る。
【0172】
1つの実施形態では、抗体または結合分子は、延長重鎖CDR1ループを有する。1つの実施形態では、延長重鎖CDR1ループは、典型的な重鎖CDR1ループよりも長い、約1アミノ酸、約2アミノ酸、約3アミノ酸、約4アミノ酸、約5アミノ酸、約6アミノ酸、約7アミノ酸、約8アミノ酸、約9アミノ酸、約10アミノ酸、約11アミノ酸、約12アミノ酸、約13アミノ酸、約14アミノ酸、または約15アミノ酸である。別の実施形態では、延長 重鎖CDR1ループは、典型的な重鎖CDR1ループよりも長い、約1〜約15アミノ酸、約1〜約14アミノ酸、約1〜約13アミノ酸、約1〜約12アミノ酸、約1〜約11アミノ酸、約1〜約10アミノ酸、約1〜約9アミノ酸、約1〜約8アミノ酸、約1〜約7アミノ酸、約1〜約6アミノ酸、約1〜約5アミノ酸、約1〜約4アミノ酸、約1〜約3アミノ酸、または約1〜約2アミノ酸である。さらに別の実施形態では、延長重鎖CDR3ループは、約3、約4、約5、約6、約7、約8、約9、約10、約11、約12、約13、約14、または約15アミノ酸の長さである。
【0173】
好ましい実施形態では、延長重鎖CDR1ループは、約11アミノ酸の長さである。別の好ましい実施形態では、延長重鎖CDR1ループは、アミノ酸配列GESTLSYYAVS(配列番号7)を含む。重鎖CDR1領域の配置が、典型的に、CDR3の近位であり、結合表面の縁に向かうため、付加的な長さ(例えば、3〜5アミノ酸)は、赤血球凝集素への抗体のより優れた安定性を作成する、付加的な物理的ドッキング表面積を与え得ることが可能である。興味深いことに、VH遺伝子の分析は、かかる延長された長さのCDR1の使用を明らかにしない。同様に、BLAST検索は、任意の以前に報告されたかかるCDR1の長さを有する抗体の同定において失敗に終わった。ポジティブ選択が、かかる新規CDR1長および組成物を強化する役割を担ったという考察を生じる。
【0174】
1つの他の態様では、長さが修正された重鎖ループは、短縮された重鎖ループである。短縮重鎖ループは、欠失を含み得る。1つの実施形態では、CDR1ループおよび/またはCDR3ループは、欠失を含む。別の実施形態では、配列番号7および/または配列番号9は、欠失を含む。1つの実施形態では、重鎖CDR1またはCDR3ループ内の欠失は、約1アミノ酸、約2アミノ酸、約3アミノ酸、約4アミノ酸、約5アミノ酸、約6アミノ酸、約7アミノ酸、約8アミノ酸、約9アミノ酸、約10アミノ酸、約11アミノ酸、約12アミノ酸、約13アミノ酸、約14アミノ酸、または約15アミノ酸、または約16アミノ酸、または約16アミノ酸、または約17アミノ酸、または約18アミノ酸、または約19アミノ酸、または約20アミノ酸、または約21アミノ酸、または約22アミノ酸、または約23アミノ酸、または約24アミノ酸である。
【0175】
1つの態様では、本発明は、低下した酸化能または減少した不均質性酸化能を有するインフルエンザ中和抗体または結合分子に関する。ポリペプチドの水性製剤中の過酸化物によるメチオニンの酸化は、タンパク質ベースの治療薬の開発に有害であると見なされる。この懸念は、少なくとも2倍である。第1に、メチオニンが必須である場合、次いで、酸化は、抗体が活性を維持することができるように制御されなければならない。しかし、それらが活性に必須ではない場合、次いで、非酸化的置換は、減少した不均質性を有する抗体または結合分子を産生するために好ましい。1つの実施形態では、低下した酸化能を有する抗体または結合分子は、変異体重鎖アミノ酸配列を有する。別の実施形態では、変異体アミノ酸配列は、CDR3ループ内である。1つの実施形態では、変異体アミノ酸配列は、カバットナンバリング変換による96位および/または98位におけるアミノ酸に対応する、少なくとも1つの置換を含む。好ましい実施形態では、96位および/または98位におけるメチオニンは、置換アミノ酸である。さらに別の実施形態では、変異体重鎖アミノ酸配列は、メチオニン残基の少なくとも1つの置換を含有する。1つの実施形態では、変異体重鎖アミノ酸配列は、メチオニン残基の少なくとも2つの置換を含有する。好ましい実施形態では、変異体配列は、ロイシン置換に対して少なくとも1つのメチオニン、より好ましくは、ロイシン置換に対して少なくとも2つのメチオニンを含有する。別の実施形態では、変異体配列は、以下のうちの1つを含む:
AKHMSLQQVVSAGWERADLVGDAFD(配列番号25)、
AKHLSMQQVVSAGWERADLVGDAFD(配列番号26)、
AKHASLQQVVSAGWERADLVGDAFD(配列番号27)、および
AKHSSLQQVVSAGWERADLVGDAFD(配列番号28)、好ましくは
AKHLSLQQVVSAGWERADLVGDAFD(配列番号29)。
【0176】
1つの実施形態では、減少した不均質性酸化能を有する抗体または結合分子は、以下の特性((i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和する、(ii)ウイルスの血球凝集素(HA)抗原に結合する、(iii)血球凝集を阻害する、またはそれらの任意の組み合わせ)のうちの1つまたは複数を保持する。
【0177】
1つの態様では、本発明は、配列番号1〜29として示した1つまたは複数のアミノ酸配列を含むか、本質的にこれからなるか、またはこれからなる、ポリペプチドを有する中和抗体または結合分子に関する。
【0178】
別の態様では、本発明は、本明細書中に記載の重鎖または軽鎖アミノ酸配列の組み合わせを有する中和抗体または結合分子に関する。例えば、1286−C05および1286−A11抗体の重鎖および軽鎖アミノ酸配列が特徴づけられており(実施例2)、本発明は、それらの種々の重鎖および軽鎖、それらのCDR領域、および/またはそれらの機能的に活性なフラグメントの間の組み合わせを検討する。本明細書中に記載の抗体または結合分子は、任意の組み合わせにおいて、重鎖および軽鎖、および/または重鎖および軽鎖超可変CDR領域(配列番号1〜29)からなり得る。1つの実施形態では、抗体または結合分子は、重鎖(配列番号1もしくは2)、および軽鎖(配列番号6)、または重鎖(配列番号1もしくは2)、および1つまたは複数の軽鎖(配列番号3、4、もしくは5)を含む。別の実施形態では、抗体または結合分子は、(i)重鎖(配列番号1)、および1つまたは複数の重鎖超可変CDR領域(配列番号10〜12)、(ii)重鎖(配列番号2)、および1つまたは複数の重鎖超可変CDR領域(配列番号7〜9)、(iii)軽鎖(配列番号13〜21)のうちの1つ、および1つまたは複数の重鎖超可変CDR領域(配列番号10〜12)、および(iv)軽鎖(配列番号22〜24)のうちの1つ、および1つまたは複数の重鎖超可変CDR領域(配列番号7〜9)を含む。他の実施形態では、抗体または結合分子は、(i)1つまたは複数の重鎖超可変CDR領域(配列番号7、8、もしくは9)、および1つまたは複数の重鎖超可変CDR領域(配列番号10、11、または12)、(ii)1つまたは複数の重鎖超可変CDR領域(配列番号7、8、もしくは9)、および1つまたは複数の軽鎖超可変CDR領域(配列番号22、23、もしくは24)、および(iii)1つまたは複数の重鎖超可変CDR領域(配列番号10、11、もしくは12)、および1つまたは複数の軽鎖超可変CDR領域(配列番号13〜21)を含む。当該分野の当業者は、配列番号1〜29、またはそれらの機能的に活性なフラグメントの他の組み合わせも理解するであろう。
【0179】
中和抗体の使用
本発明のインフルエンザ中和抗体を、インフルエンザA型感染の防止および/または治療のために、ならびに適切な交差中和エピトープを提示するワクチンの開発のために使用することができる。治療への応用ために、抗体または抗体ベースの輸送配列を使用することによってその送達が容易になった抗体または他の分子を、通常、薬学的組成物の形態で使用する。技術および製剤を、一般に、Remington’s_ PHArmaceutical Sciences,18th Edition,Mack Publishing Co.(Easton,Pa.1990)に見出すことができる。Wang and Hanson “Parenteral Formulations of Proteins and Peptides:Stability and Stabilizers,” Journal of Parenteral Science and Technology,Technical Report No.10,Supp.42−2S(1988)も参照。
【0180】
1つの態様では、本発明は、治療または防止を必要とする被験者におけるインフルエンザの治療または防止する方法に関する。1つの実施形態では、方法は、本明細書に記載のインフルエンザ中和抗体を必要とする被験者にインフルエンザ中和抗体を投与するステップを含む。抗体は、C05抗体であり得る。実施例6に記載するように、C05抗体は、インフルエンザ感染、例えば、H3N2感染に対する治療的または予防的効果を提供し得る。
【0181】
抗体は、典型的には、凍結乾燥製剤または水溶液の形態で製剤化される。許容可能な担体、賦形剤、または安定剤は、使用される投薬量および濃度で受容者に無毒であり、これらには、リン酸、クエン酸、および他の有機酸の緩衝液等の緩衝液;アスコルビン酸およびメチオニンを含む抗酸化剤;オクタデシルジメチルベンジルアンモニウムクロリド、塩化ヘキサメトニウム、塩化ベンザルコニウム、塩化ベンゼトニウム、フェノール、ブチルアルコールまたはベンジルアルコール、メチルパラベンまたはプロピルパラベン等のアルキルパラベン、カテコール、レゾルシノール、シクロヘキサノール、3−ペンタノール、およびm−クレゾール等の防腐剤;低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチン、または免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、またはリジン等のアミノ酸;モノサッカリド、ジサッカリド、およびグルコース、マンノース、またはデキストリンを含む他の炭水化物;EDTA等のキレート剤;スクロース、マンニトール、トレハロース、またはソルビトール等の糖;ナトリウム等の塩形成対イオン;金属錯体(例えば、Zn−タンパク質錯体);および/またはTWEEN(商標)、PLURONICS(商標)、またはポリエチレングリコール(PEG)等の非イオン性界面活性剤が含まれる。
【0182】
抗体を、例えば、コアセルベーション技術または界面重合によって調製されたマイクロカプセル(例えば、それぞれ、ヒドロキシメチルセルロースまたはゼラチン−マイクロカプセルおよびポリ−(メチルメタクリラート)マイクロカプセル)、コロイド状薬物送達系(例えば、リポソーム、アルブミンミクロスフィア、マイクロエマルジョン、ナノ粒子、およびナノカプセル)、またはマクロエマルジョン中に捕捉することもできる。かかる技術は、Remington’s Pharmaceutical Sciences(前出)に開示されている。
【0183】
本明細書中に開示の中和抗体を、免疫リポソームとして製剤化することもできる。抗体を含むリポソームを、当該分野で公知の方法(Epstein et al.,Proc.Natl.Acad.Sci.USA 82:3688(1985)、Hwang et al.,Proc.Natl Acad.Sci.USA 77:4030(1980)、米国特許第4,485,045号および同第4,544,545号、および1997年10月23日に公開されたWO97/38731号に記載の方法等)によって調製する。循環期間が増加したリポソームは、米国特許第5,013,556号に開示されている。
【0184】
特に有用なリポソームを、ホスファチジルコリン、コレステロール、およびPEG由来ホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物を使用した逆相蒸発法によって生成することができる。リポソームを、規定の孔サイズのフィルターを介して押出して所望の直径を有するリポソームを得る。本発明の抗体のFab’フラグメントを、ジスルフィド鎖間反応によって、Martin et al.,J.Biol.Chem.257:286−288(1982)に記載のリポソームに抱合することができる。化学療法薬を任意にリポソーム内に含める。Gabizon et al.,J.National Cancer Inst.81(19)1484(1989)を参照。
【0185】
疾患の防止または治療のために、抗体の適切な投薬量は、治療すべき感染の型、疾患の重症度および経過、ならびに抗体投与が防止または治療のいずれを目的とするかに依存するであろう。抗体を、1回または一連の治療にわたって患者に適切に投与する。疾患の型および重症度に応じて、約1μg/kg〜約15mg/kgの抗体は、例えば、1回または複数回の個別の投与または連続注入のいずれかでの抗体への投与のための典型的な最初の候補投薬量である。
【0186】
本発明の中和抗体を、インフルエンザA型ウイルスの抗原決定基のエピトープマッピングのためのツールとしてさらに使用することができ、これはワクチン開発に有用である。実際、以下の実施例に示すように、本明細書の発明者らは、ワクチン設計のガイドとして使用することができるいくつかの広範に反応性を示す中和抗体を同定した。
【0187】
したがって、本発明の中和抗体を使用して、抗体が結合する中和エピトープを機能的に模倣するペプチドまたはポリペプチドを選択することができ、それにより、インフルエンザA型ウイルス感染のワクチンを開発することができる。1つの実施形態では、本発明は、本明細書中に記載の抗体に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む、インフルエンザA型ウイルスに有効なワクチンを提供する。1つの実施形態では、ワクチンは、血球凝集素(HA)抗原に結合する抗体に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む。別の実施形態では、ワクチンは合成ワクチンであり得る。他の実施形態では、ワクチンは、(i)弱毒化インフルエンザA型ウイルスまたはその一部、または(ii)死滅させたインフルエンザA型ウイルスまたはその一部を含むことができる。1つの他の実施形態では、ワクチンは、血球凝集素(HA)抗原に結合する抗体に結合した中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む。HA抗原は、H3亜型またはH1亜型であり得る。別の実施形態では、HA抗原は、インフルエンザA型ウイルスの表面上に提示される。
【0188】
別の実施形態では、ワクチンのペプチドまたはポリペプチドは、インフルエンザA型ウイルス中和抗体を惹起する抗原決定基を含む。
【0189】
1つの態様では、本発明は、疾患の防止または治療を必要とする被験者の疾患の防止または治療に有用な、またはそのための薬剤または薬学的組成物の調製のための本明細書に記載する抗体の使用を提供する。別の実施形態では、本発明は、疾患の防止または治療を必要とする被験者の疾患を治療または防止するための薬学的組成物を提供し、前記組成物は、本明細書に記載のインフルエンザ中和抗体を含む。
【0190】
中和特性を有する非抗体分子
前述で本発明は抗体ライブラリーを参照して例示されているが、サロボディ等の他の非抗体分子のライブラリーを、類似の様式で調製し、使用し、最適化することができる。したがって、プレB細胞受容体(pre−BCR)に基づいた固有の組み合わせタンパク質ライブラリー(「サロボディライブラリー」)の構築は、Xu et al.,2008(前出)に記載されている。前に考察のように、pre−BCRは、抗体レパートリーの通常の作製中に産生されるタンパク質である。基準抗体と異なり、pre−BCRサブユニットは、2つの代替軽鎖(SLC)成分と対合した抗体重鎖から構成される三量体である。多様な重鎖が固定SLCと対合するこれらのpre−BCRタンパク質に基づいた組み合わせライブラリーを、哺乳動物系、Escherichia coli系、およびファージミド系で発現させた。これらのライブラリーは、標的抗原に対してナノモルの親和性を有するメンバーを含む。ライブラリーの構築、選択的富化、およびライブラリーメンバーの生物物理学的特徴づけの説明は、Xu et al.,(2008)(前出)の材料と方法の項に詳述されている。本明細書に記載の抗体配列のいずれかを使用して、かかる結合、または非抗体分子(例えば、サロボディ等)を構築し得る。
【0191】
本発明のさらなる詳細を、以下の非限定的な実施例によって例示する。
【実施例】
【0192】
実施例1−インフルエンザドナー由来の抗体ライブラリー
包括的インフルエンザライブラリーに含有するために選択されたドナーは、以前にインフルエンザ感染しており、1957年のH2N2、または1968年のH3N2インフルエンザパンデミックの時点で約5歳であり、現在、健康状態が良好であることが確認された。H1N1 A/ニューカレドニア/20/99、H3N2 A/パナマ/2007/99、およびH5N1 A/ベトナム/1203/2004ウイルスのパネルまたは血球凝集素タンパク質上で血清学を実施して、血球凝集素タンパク質に対する抗体の存在を確認した。
【0193】
第1に、5〜20mLの骨髄を、選択条件を満たしている各ドナーから採取し、製造者の指示毎のRNAlater(Applied Biosystems)で混合して、細胞RNAの完全性を保持した。RNAを、TRI−BD試薬プロトコール(Sigma−Aldrich)を使用して単離した。
【0194】
重鎖および軽鎖レパートリーを、重鎖のためのランダム感作されたcDNAテンプレート、軽鎖のためのオリゴdT感作されたcDNAテンプレート、および遺伝子特異的可変ドメインプライマーを使用するRT−PCRによって、各ドナー由来のRNAから回収させた。
【0195】
次に、ドナーあたり1μgの各プールされたκ軽鎖、およびプールされたλ軽鎖をNotlおよびBamH1、ならびにQiagenゲル抽出キットを使用して精製されたゲルで消化した。κおよびλ軽鎖クローニングでは、5μgの各ベクターを、NotlおよびBamH1で消化し、Qiagenゲル抽出キットを使用してゲル精製した。200ngのゲル精製κまたはλインサートおよび1μgのゲル精製ベクターを使用して、軽鎖ライブラリーライゲーションを行う。インキュベーションは、14℃で一晩である。クローニング効果を決定するために、対照連結反応を、軽鎖インサートの追加なく、1つのエレクトロポレーションの量(200〜300ngのベクターDNA)と同等に設定する。転換前に、Edge Biosystem Perfromaスピンカラムを使用して、ライゲーション物を脱塩する。それぞれのライブラリーを、80μLのDh5αエレクトロコンピテント細胞アリコートを使用して、3〜5回のエレクトロポレーションで変換し、それぞれ1mLのSOC中に回収し、プールし、37℃で1時間増殖させた。各ライブラリーのサンプルを、選択的媒体上でプレーティングし、クローニングの効果および形質転換体の総数を決定するために使用した。残部を200mLの2YT+100μg/mLアンピシリン+2%グルコースに移し、37℃で一晩成長させた。良好なライブラリーは、10%未満のバックグラウンド、および1×106メンバーを超える総数の形質転換体を有する。翌日、Qiagen High Speed Maxiprepキットを使用して軽鎖ライブラリープラスミドを単離した。
【0196】
重鎖集合をクローニングするために、1.5〜2μgの各5ドナー特異的重鎖可変遺伝子(VH1/7、VH2、5、6、VH3、およびVH4)を、SfiIおよびXhoIを40単位/μgのDNAで消化し、Qiagenゲル抽出キットを使用してゲル精製する。受容プラスミドを調製するために、15μgの各軽鎖ライブラリーベクターを、SfiIおよびXhoIを40単位余剰/μgのDNAで消化し、Qiagenゲル抽出キットを使用してゲル精製する。ライブラリーライゲーション物を、消化した1.2μgのSfiI/XhoIを組み合わせることによって達成し、ドナーあたりのゲル精製された重鎖DNAを、5ngの各軽鎖ライブラリー、κ、およびλをそれぞれ有する、300ngの各5重鎖改変遺伝子ファミリーを含有するようにプールした。対照ライゲーション反応は、重鎖インサートの添加なく、1つのエレクトロポレーションの量(300〜600ngのベクターDNA)と同等に設定する。ライゲーション物を、14℃で一晩インキュベートし、次いで、Edge Bio System Pefromaスピンカラムで脱塩した。ライゲーション物を、80μLのTG−1細胞を使用して行う、ライブラリーあたり8〜12回のエレクトロポレーションに移し、それぞれ1mLのSOC中に回収し、プールし、37℃で1時間増殖させた。それぞれのサンプルを使用して、クローニングの有効性および形質転換体の総数を決定した。形質転換体の標的数/ライブラリーは、10%未満のバックグラウンドを有する、少なくとも1×107でなければならない。残部を、300mLの2YT+100μg/mLアンピシリン+2%グルコースに移し、OD600が約0.3になるまで成長させた。次に、m13K07ヘルパーファージを、5:1の感染(MOI)の多重度で添加し、振動せずに、37℃で1時間インキュベートした。ヘルパー感染後、遠心分離によって細胞を採取し、300mLの2YT+100μg/mLのアンピリシン+2%のグルコース+70μg/mLのカナマイシン中で再懸濁し、ストックファージ産生用に振動して一晩37℃で増殖を続けた。
【0197】
培養上清を含有する得られたファージを、4℃で10分間の間、6000RPMでの遠心分離によって採取する。次に、ファージを、各上清への0.2容量の20%のPEG/2.5MのNaCl溶液の添加、および1時間の間、氷上でのインキュベーションによって沈殿させる。次いで、ファージを、4℃で15分間の間、7900RPMでの遠心分離によって採取する。上清を除去し、ファージパレットを、30mLの無菌1XPBS中で再懸濁する。長期の−20℃の保管では、PBSを、50%のグリセロールで補充する。
【0198】
実施例2−中和抗体の調製
前述のように、ヒト骨髄ファージ提示抗体ライブラリー由来の抗体(実施例1を参照)を、変換し、哺乳動物発現免疫グロブリンとして試験した(Kashyap AK et al.,Proc Natl Acad Sci USA.2008 Apr 22;105(16):5986−91を参照)。重鎖を2つの配列クラスに分類した:
QVQLQESGGGLVQPGESRLSCVGSGSSFGESTLSYYAVSWVRQAPGKGLEWLSIINAGGGDIDYADSVEGRFTISRDNSKETLYLQMTNLRVEDTGVYYCAKHMSMQQVVSAGWERADLVGDAFDVWGQGTMVTVSS(配列番号1)
QVOLQQSGPRLVKPSQTLSLTCAISGDSVSGDSGTWNWROSPSRGLEWLGRTYYRSKWYNDYAESVKSRIVIKADTSKNEFSLQLNSVTPEDTAIYYCARAGVKIFGLIVGALDNWGRGTLVTVSS(配列番号2)。
下線の超可変CDR領域を、以下の重鎖に示す。
GESTLSYYAVS(配列番号7)
WLSIINAGGGDID(配列番号8)
AKHMSMQQVVSAGWERADLVGDAFD(配列番号9)
SGDSGTWN(配列番号10)
WLGRTYYRSKWYND(配列番号11)
ARAGVKIFGLIVGALD(配列番号12)
【0199】
配列番号1によって記載される重鎖は、1つのλ軽鎖(配列番号3)、およびクローン1286−C5によって例示される2つのκ軽鎖(配列番号4〜5)と対を成すことが見出された。
QSVLTQPPSVSGAPGQRVTISCTGSSSNIGAGYDVHWYQQLPGTAPKLLIYDNNNRPSGVPDRFSGSKSGASASLAITGLQAEDEAHYYCQSYDNSLSGSVFGGGTQTLTVLS(配列番号3)
DIQLTOSPSSLSASVGDRVTLTCQASODIRKFLNWYQQKPGKGPKLLIYDASNLQRGVPSRFSGGGSGTDFTLIISSLQPEDVGTYYCQQYDGLPFTFGGGTKLEIK(配列番号4)
DIQLTQSPSSLSASIGDRVTITCQASQDIRNSLNWYEHKPGKAPKLLIHDASNLETGVPSRFSGGGSGTDFTLTISSLQPEDFATYYCQQANSFPLTFGGGTKVEIK(配列番号5)
下線の超可変CDR領域を、以下の軽鎖に示す。
IGAGYDVHWY(配列番号13)
LLIYDNNNRP(配列番号14)
QSYDNSLSGS(配列番号15)
IRKFLNWY(配列番号16)
LLIYDASNLQ(配列番号17)
QQYDGLPF(配列番号18)
IRNSLNWY(配列番号19)
LLIHDASNLE(配列番号20)
QQANSFPL(配列番号21)
【0200】
配列番号2によって記載され、単一のκ軽鎖(配列番号6)とのクローン1286−A11対によって例示される重鎖。
EIVMTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASTRATGVPDRFHGGGSGTDFTLTISRLEPEDFAVYYCQQYDTSSGLTFGGGTKVEIK(配列番号6)
下線の超可変CDR領域を、以下の軽鎖に示す。
SSYLAWY(配列番号22)
LLIYGASTRA(配列番号23)
QQYDTSSGL(配列番号24)
【0201】
図4〜5は、クローン1286−C5抗体およびクローン1286−A11の結合能力を示す。先述のように、これらの抗体の機能を特徴づけるために、対応するIgGsを、異なる抗原のパネル上で結合する能力に対して試験した(図4および5)(Kashyap AK et al.,2008(前出)も参照されたい)。クローン1286−A11および1286−C5抗体の結合能力。両1286−A11および1286−C5は、H1N1、H3N2、およびH9N2血球凝集素に結合することができた。加えて、1286−A11は、H5N1血球凝集素に結合することが可能であった。種々のインフルエンザ亜型ウイルスのマイクロ中和もまた、先述のように実施した(同文献も参照されたい)。
【0202】
下記の表2に示すように、各抗体は、MDCK細胞中でインビトロH1ウイルス複製を中和することが可能であった。
【0203】
1286−C5もまた、H3ウイルス複製を中和する顕著な能力を示した。1286−A11は、H3亜型ウイルスを測定可能に中和しなかったが、H5亜型中和を提示した。1286−C5抗体の作用の機構は、血球凝集阻害を介するものであるが、1286−A11の作用の機構は、決定されなかった。
これらの抗体のそれぞれは、飽和変異誘発および誤りがちのPCR等の標準的指向およびランダム化抗体最適化技術のそれぞれを介して、増加した活性の効力および範囲に最適化されるであろう。おそらく、これらの抗体は、種々のフラグメント、ならびに単一特異性および多特異性サロボディに変換された場合に有用であろう。
【0204】
実施例3−ユニバーサルインフルエンザワクチンの生成
異種病原体のワクチン設計の目的は、有効かつ広範な防御抗原を同定および設計することである。インフルエンザの場合、相当な歴史的努力によって保存された線状配列および領域の経験的試験が行われてきたが、ほとんど成功していない。これらの失敗についてのもっともらしい理由は、抗原性被験物質に対する注目される応答が実際の感染条件での病原体上の抗原の中和のための実際の真の産生部位であるという知識不足である。インフルエンザのために、インフルエンザ感染の生存者のレパートリー内にこれらの真の解決策が見出されると期待されるであろう。本発明者らの場合、多数の抗体の集団の中の一定の抗体は、インフルエンザの複数の亜型を中和することができることが証明された。これらの抗体のうちのいくつかは、血球凝集の古典的阻害を介して、インフルエンザを中和する。まとめると、真の生存者由来のかかる交差中和抗体を使用したワクチンの設計および評価が交差反応性または「ユニバーサル」インフルエンザワクチンの設計および妥当性に役に立つと期待される。
【0205】
具体的には、交差中和モノクローナル抗体を、現在および将来的に製造されるワクチン中の交差中和エピトープの質および抗原性を維持または増強するワクチン産生過程の設計および確証で使用することができる。ワクチンへの抗体結合が構造的完全性および抗原潜在性を反映すると仮定すると、かかるワクチン過程の誘導体への交差中和抗体の結合を評価して、その交差中和潜在性を定量的に評価するであろう。
【0206】
これらのユニバーサルエピトープに対する応答を最大にするために、胞子殻または胞子外膜等の抗原性補強剤を通してこれらのユニバーサルエピトープに対する免疫原性を増加させるための誘導体を作製するであろう。代替として、予測モデルおよび支持試験を介して、それらの免疫原性を増大するために、これらの交差中和エピトープを操作および最適化することが出来る。どんな場合でも、得られた抗原を再度試験して、標的への結合有効性を維持するだけでなく、定方向の免疫原性も達成されることを保証するであろう。これは、おそらく競合ELISAによって免疫化した個体または試験動物由来の血清中に含まれる特異的なユニバーサル中和力価の決定を含むであろう。インビトロ代替物として、免疫原性のために抗原−抗体結合データをインビトロまたはコンピュータを用いて予測モデルの抗体結合データと組み合わせるであろう。ユニバーサルエピトープに対する応答をさらに指示するために、公知の非中和および非交差反応性血球凝集素エピトープを脱免疫化し得る。
血球凝集素のキメラ、フラグメント、および立体構造模倣物の設計および確証のために本抗体を拡大することが妥当である。例えば、インフルエンザが非中和応答を生じる多数の免疫原性エピトープを含むことが十分に確立されている。交差防御抗体を使用して、これらの免疫優性非中和エピトープが部分的または完全に脱免疫化されたワクチンの抗原変異体が維持されるか、ユニバーサル保護エピトープの認識が強化されるのかどうかを評価することが可能である。
【0207】
また、これらのワクチン設計の結果として、抗原エピトープを最小にし、さらに、血球凝集素の構成から除去して、立体構造交差特異性抗原を作製することができる。
【0208】
この戦略は、中和エピトープまたはエレメントを最小にするために応答を案内するために上に詳細の方法を概説した。立体構造的に依存している可能性が高く、不連続な配列空間内に存在するかかる最小化したエレメントの知識から、前に記載のように(Horowitz et al.,Combinatorial Libraries of Conformationally Constrained Polypeptide Sequences、PCT公開第WO/2008/089073号を参照)、より小さなポリペプチド内に組み合わせ様式で立体構造中和エピトープを作製する可能性があり、不連続なエピトープのみの近位の配置、または設計した構造支持体の文脈では、立体構造エピトープの必須の性質を再生することができる。
【0209】
かかる設計では、立体構造エピトープの形態をとり、これらを血球凝集素関連および非関連構造足場上で発現させるか、立体構造依存性抗体によって選択することができるライブラリー内の立体構造エピトープ集団としての形態もとるであろう。
【0210】
不連続エピトープの立体構造エピトープへの減少により、従来のタンパク質操作を使用して可能なペプチド免疫原よりもさらにより小さなサイズのペプチド免疫原が得られるであろう。さらに、これらの構造エピトープを、非ペプチド模倣物の使用によって、さらに増強、サイズの減少、または置換することができる。いずれにしても、任意のこれらの立体構造誘導体または模倣物を、配列番号1〜6のうちの1つまたは複数、配列番号1〜6のうちの少なくとも1つを含む関連抗体、または選択するインフルエンザウイルスに対応する抗体によって確証するであろう。
【0211】
方法と材料。インフルエンザ融合エピトープ胞子ワクチン標的。
1.分泌タンパク質または哺乳動物細胞としての標的の哺乳動物発現
a.単一分離株由来の球状HA1変異体
b.関連分離株由来の球状HA1キメラ
c.非関連分離株由来の球状HA1キメラ
d.関連分離株および非関連分離株由来の球状HA1キメラ
2.分泌タンパク質または哺乳動物細胞の配列番号1〜6の抗体または関連抗体での立体構造エピトープの検出。
3.首尾の良い立体構造抗原の胞子発現への導入。
4.配列番号1〜6の抗体または関連抗体での胞子結合の試験。
5.インビボ交差反応性免疫原性の評価。
【0212】
実施例4−交差亜型中和抗体の効力および範囲の増加
実施例1に記載される抗体の重鎖および軽鎖の配列情報に基づき、変異誘発方法を使用して、個別またはライブラリー中の集団の中で試験するための改良された変異体を作製する。有益な変異を導入するために一般に使用される方法は、結合を担うことが公知の領域にわたって結合またはエラー多発性PCR変異誘発を担う部位での飽和変異誘発であり得る。
【0213】
従来の抗体最適化戦略の固有の限界のため、交差反応性および効力が不十分である場合、目的変異誘発、合体抗体ライブラリー、または互いに、もしくは以前に記載した従来の最適化戦略を用いるこれらの方法のうちのいずれかもしくは両方の組み合わせを考慮するであろう。
【0214】
実施例5−防御の効力および範囲を増加させるためのワクチンおよび抗体の同時投与
抗体と抗原との複合体は、動物における多数の微生物タンパク質および他のタンパク質に対する応答を潜在的に誘導することが公知である。1つの可能な説明は、ワクチン抗体複合体の強制的な取り込みは抗原提示細胞上のFc受容体によって生じるということである。交差反応性抗体と季節性ワクチンとの複合体により、効力を年々増大させることが可能であり、新規のウイルス分離株を各季節のインフルエンザワクチンのために選択する場合、交差反応性抗体が多数の血球凝集素抗原を認識するため、新規の抗体を再度作製する必要がなくなるからである。さらに、これらの抗体が保存された中和領域に指向するので、抗原と複合体を形成した場合、抗体はこれらの非常に保存された感受性領域へのより有効な防御応答を実際に指示することができる。前述のように、ワクチンは、従来の生ウイルスまたは死滅ウイルス、組換えタンパク質またはタンパク質フラグメント、またはさらに最小化されたペプチド、または抗体、抗体フラグメント、または誘導体と複合体化した非ペプチドの立体構造エピトープ、またはサロボディであり得る。
【0215】
実施例6−インフルエンザ攻撃に対する保護効果
メスの生後6〜8週のDBA/2(Charles River)マウスをABSL3+コンテインメント内にケージあたり5〜6匹で収容した。餌および水を不断で提供した。マウス(グループあたり5〜6匹)は、腹腔内(IP)注射によって、約200〜300μLの無菌リン酸緩衝食塩水(PBS)中の体重kgあたり1、2.5、10、または25mgの抗体C05(C05−1286)を受容した。対照グループは、IP注射によって、200〜300μLのPBS中のPBS25mgの非免疫性ヒトIgGまたはPBSのみのいずれかが注入された。抗体および対照は、ウイルス攻撃の24時間前にX−31(H3N2)またはA/メンフィス/3/2008(H1N1)ウイルスが投与された。致死的ウイルス攻撃では、マウスは、30〜50μLのPBS中の33MLD50(50%のマウス致死量)インフルエンザウイルスでの鼻腔内投与によって接種された。H3N2およびH1N1ウイルスの両方は、DBA/2マウスにおいて高病原性である。死亡前に起こる症状は、30%超の体重減少および一般的な不活発である。体重、罹患率、および死亡率を、14日間の間毎日監視した。
【0216】
図7A〜Bは、高力価の致死的H3N2ウイルス攻撃に対するC05抗体の予防効果を例証する。高力価攻撃(33MLD50)では、以下の結果が得られた。PBSでの処置:0%の生存率。2.5mg/kgでの処置:80%の生存率。10mg/kgでの処置:100%の生存率。25mg/kgでの処置:100%の生存率。25mg/kgのIgGアイソタイプでの処置:0%の生存率。図7Eは、H1N1メンフィス/3/2008ウイルス攻撃に対するC05抗体の予防効果を例証する。
【0217】
図7C〜Dは、致死的H3N2ウイルス攻撃に対するC05抗体の処置効果を示す。DBA/2マウスは、まず、30〜50μLのPBS中の33MLD50(50%のマウス致死量)のX−31(H3N2)インフルエンザウイルスでの鼻腔内投与によって接種された。マウス(グループあたり5〜6匹)は、腹腔内(IP)注射によって、約200〜300μLの無菌リン酸緩衝食塩水(PBS)中の体重kgあたり15mgの抗体C05(C05−1286)を受容した。対照グループは、IP注射によって、200〜300μLのPBS中のPBS25mgの非免疫性ヒトIgGまたはPBSのみのいずれかが注入された。C05抗体は、感染後、24時間、48時間、72時間、または96時間で投与された。対照物の投与は、感染後24時間で生じた。体重、罹患率、死亡率を、14日間毎日監視した。
【0218】
実施例7−異なる長さの重鎖ループを有する変異体の生成
下記に示すように、C05抗体重鎖配列(配列番号1)は、アミノ酸の非常に非定型の長さのCDR1(配列番号7)ループを有する。C05は、VH3−23生殖系列と比較して、CDR1においてさらに5個のアミノ酸を有する。
【0219】
重鎖CDR1領域の配置が、典型的には、CDR3に近位であり、結合表面の縁に向かうため、付加的な長さの3〜5アミノ酸が、血球凝集素に抗体に対してより高い安定性を作製する付加的な物理的ドッキング表面積を与え得ることが可能である。VH遺伝レパートリーの分析は、かかる延長した長さのCDR1の使用を明らかにしない。さらに、BLAST検索は、かかるCDR1の長さを有する任意の抗体を同定しない。したがって、ポジティブ選択が、かかる新規のCDR1の長さおよびその組成物の存在および強化において役割を担い得るという考察を生じる。重鎖は、CDR1およびCDR3の両方において、かかる独自のループの長さを特長とするため、これらのループの組成物を変化させるだけでなく、これらのループの長さを変化させることによって、効力および全幅の増加を考慮するであろう。改良を行うことに関して、まず、上述(パネルの左下)のように、親CDR1ループ中のアミノ酸延長の欠失、および対応するより短い生殖系列CDR1ペプチド配列での置換によって、および/または生成中間の長さのループ、またはその多種多様な集団を介して開始することができるであろう。依然として延長ループが有益であるが、完全に最適ではない場合、段階的または組み合わせ手段で、CDR1内、FR1接合、および/またはCDR1のFR2接合における(1〜20)ランダムまたは選択したアミノ酸の挿入によるループの延長を考慮するであろう。延長ループを有する、かかる抗体の組み合わせライブラリーを作製することによって、両感受性および非感受性インフルエンザ分離株、株、および型へのより優れた、かつ広域なバインダーに対して区別的にスクリーニングすることができる。
【0220】
加えて、C05抗体は、25アミノ酸の典型的な重鎖CDR3(配列番号9)ループよりも長いループを特徴とする。上述のCDR1挿入および欠失の例と同様に、CDR3領域は、同様に修正することができる。下記(右下のパネル)に示すように、CDR3配列は、段階的組み合わせ手段(C05のゲノム配列が示される)で、ループ内またはCDR3のFR3およびFR4接合における1〜20アミノ酸によって、同様に、縮小または拡張することができるであろう。
【0221】
他の相互利益の変異誘発への類似により、異なるループの長さのCDR1およびCDR3ライブラリーを組み合わせて、現在の感受性および非感受性インフルエンザ分離株、株、および型に対する、新規、かつより広域な効力の抗インフルエンザ抗体をインテロゲートすることができるであろう。重要なことには、これらの戦略もまた、親C05CDRだけでなく、対象の抗体に交差されたVh3−23生殖系列または他のVh生殖系列のCDR1ループに適用することができるであろう。
【0222】
実施例8−減少した不均質性酸化能を有する変異体
抗体1286−C05は、カバット残基96および98において、重鎖CDR3ループ内に2つのメチオニンを含有する。定義により、このループは、表面曝露され、したがって、酸化を受けやすい。カバット残基96において、アラニン、ロイシン、またはセリンをメチオニンに、カバット残基98において、ロイシンをメチオニンに置換する二重点突然変異を生成することによって、メチオニンのいずれかまたは両方が必須であったかどうかを試験した。対応するタンパク質を、一過性哺乳動物系を産生し、先述のように精製した(Kashyap AK et al.(前出)2008)。得られたタンパク質を、H1(ニューカレドニア/20/99)血球凝集素への結合に対して試験し、親タンパク質のフォールド内で結合することを見出した。次に、これらのタンパク質を、それらのH3(ウィスコンシン/67/2005)血球凝集素へ結合する能力に対して試験し、それは、ロイシン96/ロイシン98変異体が、実質的に、アラニン96/ロイシン98およびセリン96/ロイシン98変異体よりもよく結合したことを示した。図8は、C05非酸化的「XL」変異体が、H1およびH3HAタンパク質の認識を維持することを例証する。
【0223】
図9は、H1およびH3への非変異体C05の結合と比較して、H1(ニューカレドニア/20/99)およびH3(ウィスコンシン/67/2005)血球凝集素へのロイシン96/ロイシン98C05変異体の結合を示す。C05非酸化的「LL」は、H1およびH3HAタンパク質の認識を維持する。
【0224】
表3は、血球凝集阻害(HAI)アッセイによって試験されたように、C05−LL変異体が活性の同様の効力および全幅を提示したことを例証し、そこでは、血球凝集アッセイおよび血球凝集阻害アッセイは、本質的に、EdwardsおよびDimmock(Journal of Virology,v75pp、10208−18、2001)に説明するとおりであり、そこでは、組換えウイルスは、Kashyapら(2008)(前出)によって説明するとおりに生成された。値は、0.5%のcRBCの血球凝集を阻害する抗体の最小濃度を表す。
【0225】
実施例8−活性の増大した親和性および全幅を有する変異体の作製
変異誘発の種々の方法を使用して、個別またはライブラリー中の集団を試験するための改良された変異体を作製する。直接的構造分析を介して特異的に発見されたCDRまたはこれらの接触残基等の結合を担う部位において有益な変異を導入するために使用される方法は、飽和変異誘発、ルックスルー変異誘発、または倹約変異誘発があるが、これらに限定されない。
【0226】
対象ステップとして、上述の方法によって、CDR3の変異誘発に基づき変異体の集団を作製するであろう。他のヒト抗体を使用する先行研究は、軽鎖、他の重鎖CDR、または同時に一度にこれらの領域のうちのいくつかまたはすべてへの点変異体を調査し、かつ生成して、合成改良を産生することによって、多大な利益を証明した。通常、CDRの組成物のみを変化させながら、CDRの親フレームワークおよび長さを維持することによって、これを達成することができる。従来の誤りがちなPCR変異誘発および他の確率プロセスを、同様の領域にわたって、かつ重鎖可変ドメインの他の領域においても使用して、変異体の集団を生成することができるであろう。任意の事象では、得られる集団またはクローンは、活性の増大した親和性、中和、および/または全幅に対して選択することができるであろう。
【0227】
上述のように、かかる最適化集団を生成するが、重鎖が、CDR1およびCDR3の両方において、かかる独自のループの長さを特徴とするため、これらのループの組成物を変化させるだけでなく、これらのループの長さも変化させることによって、活性の効力および全幅を試験することがさらに重要であることができるであろう。例えば、対応するより短い生殖系列CDR1ペプチド配列、および/または変異集団での置換によって、延長CDR1ループの効果を試験するであろう。加えて、ループが適合し、さらには既存の延長ループの長さを超えるまで、段階的ライブラリーの方法で、CDR1内、またはCDR1のFR1およびFR接合におけるランダムアミノ酸の挿入を行い、より優れたさらに広域のバインダーに対して区別的にスクリーニングすることができるであろう。同様に、CDR3配列は、段階的ライブラリーの方法で、ループ内またはCDR3のFR3およびFR4接合において、収縮または拡張することができるであろう。他の相互に有益な変異誘発への類似により、異なったループの長さのCDR1およびCFR3ライブラリーを組み合わせて、新規かつより広域な効力の抗インフルエンザ抗体を調べることができるであろう。
【0228】
上記の説明中で本発明は、ある実施形態を参照して例示しているが、本発明はこれらに限定されない。実際に、本明細書中に示し、かつ記載した実施形態に加えて、本発明の種々の変更が上記の説明から当業者に明らかとなり、これらの変更は添付の特許請求の範囲の権利範囲内である。
【0229】
本明細書を通して引用されたすべての参照は、参照によりその全体が本明細書に組み込まれる。
【特許請求の範囲】
【請求項1】
配列番号7、配列番号8、および配列番号9からなる群より選択される重鎖からの1つ、2つ、もしくは3つの超可変領域配列を含む、結合分子、またはその機能的に活性なフラグメント。
【請求項2】
配列番号10、配列番号11、および配列番号12からなる群より選択される重鎖からの1つ、2つ、もしくは3つの超可変領域配列を含む、結合分子、またはその機能的に活性なフラグメント。
【請求項3】
配列番号7、配列番号8、もしくは配列番号9からのすべての超可変領域配列を含む、請求項1に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項4】
配列番号10、配列番号11、もしくは配列番号12からのすべての超可変領域配列を含む、請求項2に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項5】
軽鎖と会合するときに、標的に結合することが可能である、請求項1に記載の結合分子。
【請求項6】
軽鎖と会合するときに、標的に結合することが可能である、請求項2に記載の結合分子。
【請求項7】
前記軽鎖は、配列番号13、配列番号14、もしくは配列番号15の前記ポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む、請求項5に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項8】
前記軽鎖は、配列番号16、配列番号17、もしくは配列番号18の前記ポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む、請求項5に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項9】
前記軽鎖は、配列番号19、配列番号20、もしくは配列番号21の前記ポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む、請求項5に記載の結合分子。
【請求項10】
前記軽鎖は、配列番号22、配列番号23、もしくは配列番号24の前記ポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む、請求項6に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項11】
配列番号13、配列番号14、もしくは配列番号15のすべての超可変領域配列を含む、請求項7に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項12】
配列番号16、配列番号17、もしくは配列番号18のすべての超可変領域配列を含む、請求項7に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項13】
配列番号19、配列番号20、もしくは配列番号21のすべての超可変領域配列を含む、請求項7に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項14】
配列番号22、配列番号23、もしくは配列番号24のすべての超可変領域配列を含む、請求項8に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項15】
VpreB配列および/またはλ5配列を含むポリペプチドをさらに含む、請求項1もしくは2に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項16】
λ5配列に融合されたVpreB配列を含むポリペプチドをさらに含む、請求項1もしくは2に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項17】
Vic様配列および/またはJCκ配列を含むκ様代替軽鎖(SLC)構築物をさらに含む、請求項1もしくは2に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項18】
抗体、またはその機能的に活性なフラグメントである、請求項1もしくは2に記載の結合分子。
【請求項19】
重鎖を含む、結合分子であって、前記重鎖は、配列番号1として示した前記アミノ酸配列を含む、結合分子、またはその機能的に活性なフラグメント。
【請求項20】
重鎖を含む、結合分子であって、前記重鎖は、配列番号2として示した前記アミノ酸配列を含む、結合分子、またはその機能的に活性なフラグメント。
【請求項21】
配列番号3、配列番号4、および配列番号5からなる群より選択されるアミノ酸配列を含む軽鎖をさらに含む、請求項19に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項22】
配列番号6として示した前記アミノ酸配列を含む軽鎖をさらに含む、請求項20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項23】
(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)前記ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する、請求項19に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項24】
(i)インフルエンザA型ウイルスの少なくとも1つの亜型および/または少なくとも1つの分離株を中和し、(ii)前記ウイルスの血球凝集素(HA)抗原に結合する、請求項20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項25】
前記HA抗原のH1亜型のエピトープに結合する、請求項19または20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項26】
前記HA抗原のH3亜型のエピトープに結合する、請求項19もしくは20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項27】
前記HA抗原のH1亜型のエピトープに、および前記HA抗原のH3亜型のエピトープに結合する、請求項19もしくは20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項28】
前記HA抗原のH9亜型のエピトープに結合する、請求項19もしくは20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項29】
前記HA抗原のH5亜型のエピトープに結合する、請求項20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項30】
前記エピトープは、インフルエンザA型ウイルス上に提示される、請求項25から29のいずれか1項に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項31】
H1HA抗原およびH3抗原と交差反応性である、請求項19もしくは20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項32】
前記H1およびH3インフルエンザA型ウイルス亜型の少なくとも1つを中和する、請求項19に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項33】
前記H1およびH3インフルエンザA型ウイルス亜型を中和する、請求項19に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項34】
前記H1HA抗原を中和する、請求項20に記載の結合分子、または機能的に活性なフラグメント。
【請求項35】
前記H5HA抗原を中和する、請求項20に記載の結合分子、または機能的に活性なフラグメント。
【請求項36】
前記H1およびH5HAインフルエンザA型ウイルス亜型を中和する、請求項20に記載の結合分子、または機能的に活性なフラグメント。
【請求項37】
前記H1亜型、または前記HAは、前記H1ウイルスのニューカレドニア/20/99分離株に由来する、請求項25に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項38】
前記H1亜型、または前記HAは、前記H1ウイルスのソロモン諸島/3/06分離株に由来する、請求項25に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項39】
前記H3亜型、または前記HAは、前記H3ウイルスのウィスコンシン/67/05分離株に由来する、請求項26に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項40】
前記H3亜型、または前記HAは、前記H3ウイルスの香港/68分離株に由来する、請求項26に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項41】
前記H9亜型、または前記HAは、前記H9ウイルスの香港/1073/99分離株に由来する、請求項28に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項42】
前記H5亜型、または前記HAは、前記H5ウイルスのベトナム/1203/04分離株に由来する、請求項29に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項43】
前記インフルエンザA型ウイルスの球状頭部が細胞表面に結合するのを阻止する、請求項23に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項44】
前記インフルエンザA型ウイルスが感染される細胞に付着するのを阻止する、請求項23に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項45】
前記ウイルスの少なくとも1つがヒトに感染する能力を有する、請求項23もしくは24に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項46】
前記分離株の少なくとも1つがヒト被験体から得たものである、請求項23もしくは24に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項47】
前記分離株の少なくとも1つが非ヒト動物から得たものである、請求項23もしくは24に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項48】
前記非ヒト動物がトリである、請求項47に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項49】
前記トリが野鳥またはニワトリである、請求項48に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項50】
前記非ヒト動物は、ブタである、請求項47に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項51】
H1HA抗原に結合する、請求項19に記載の結合分子、または機能的に活性なフラグメント。
【請求項52】
少なくとも1つのさらなるHA抗原に結合する、請求項51に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項53】
前記さらなるHA抗原は、H3である、請求項52に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項54】
配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体。
【請求項55】
配列番号3、配列番号4、および配列番号5からなる群より選択されるアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む軽鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、請求項54に記載の抗体。
【請求項56】
配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体。
【請求項57】
配列番号6として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む軽鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、請求項56に記載の抗体。
【請求項58】
(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)前記ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する、請求項54もしくは55に記載の抗体。
【請求項59】
(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)前記ウイルスの血球凝集素(HA)抗原に結合する、請求項56もしくは57に記載の抗体。
【請求項60】
請求項1から59のいずれか1項に記載の結合分子または抗体を含む、組成物。
【請求項61】
被験体におけるインフルエンザA型ウイルス感染の防止および/または治療のための方法であって、請求項60に記載の有効量の組成物を前記被験体に投与することを含む、方法。
【請求項62】
被験体におけるインフルエンザA型ウイルス感染を治療するための方法であって、請求項1から59のうちのいずれか1つの中和抗体または結合分子の有効量を前記被験体に投与することを含む、方法。
【請求項63】
前記被験体は、ヒト患者である、請求項61もしくは62に記載の方法。
【請求項64】
請求項1から59のいずれか1項に記載の抗体または結合分子によって結合された中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む、インフルエンザA型ウイルスに有効なワクチン。
【請求項65】
合成ワクチンである、請求項64に記載のワクチン。
【請求項66】
弱毒化インフルエンザA型ウイルスまたはその一部を含む、請求項64に記載のワクチン。
【請求項67】
死滅させたインフルエンザA型ウイルスまたはその一部を含む、請求項64に記載のワクチン。
【請求項68】
前記抗体または結合分子は、
(a)配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子、
(b)配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む重鎖ポリペプチドを含む、抗体、
(c)配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子、またそのフラグメント、および
(d)配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む、抗体、またはそのフラグメント、からなる群より選択される、請求項64に記載のワクチン。
【請求項69】
前記抗体または結合分子は、HA抗原に結合する、請求項64に記載のワクチン。
【請求項70】
前記HA抗原は、H3亜型である、請求項69に記載のワクチン。
【請求項71】
前記HA抗原は、H1亜型である、請求項69に記載のワクチン。
【請求項72】
前記HA抗原は、H1亜型およびH3亜型である、請求項69に記載のワクチン。
【請求項73】
前記HA抗原は、H5亜型である、請求項69に記載のワクチン。
【請求項74】
前記HA抗原は、H9亜型である、請求項69に記載のワクチン。
【請求項75】
前記抗原は、インフルエンザA型ウイルスの表面上に提示される、請求項69に記載のワクチン。
【請求項76】
前記ペプチドまたはポリペプチドは、中和抗体を惹起する抗原決定基を含む、請求項64から75のいずれか1項に記載のワクチン。
【請求項77】
経口投与に好適な、請求項64に記載のワクチン。
【請求項78】
経粘膜送達に好適な、請求項64に記載のワクチン。
【請求項79】
前記経粘膜送達は、鼻腔内投与である、請求項78に記載のワクチン。
【請求項80】
非経口投与に好適な、請求項64に記載のワクチン。
【請求項81】
経皮投与に好適な、請求項64に記載のワクチン。
【請求項82】
前記ワクチンは、小児期免疫化用である、請求項64に記載のワクチン。
【請求項83】
長さが修正された重鎖ループを含むインフルエンザA型ウイルスの少なくとも1つの分離株を中和する、ヒトに感染する能力を有するインフルエンザA型ウイルスの血球凝集素へ結合する、中和抗体または結合分子。
【請求項84】
前記延長重鎖ループは、CDR3ループである、請求項83に記載の抗体または結合分子。
【請求項85】
前記CDR3ループは、配列番号9のアミノ酸配列を含む、請求項84に記載の抗体または結合分子。
【請求項86】
前記延長重鎖ループは、CDR1ループである、請求項85に記載の抗体または結合分子。
【請求項87】
前記CDR1ループは、配列番号7のアミノ酸配列を含む、請求項86に記載の抗体または結合分子。
【請求項88】
前記長さの修正は、延長重鎖ループを含む、請求項83に記載の抗体または結合分子。
【請求項89】
前記長さの修正は、前記重鎖ループにおける欠失を含む、請求項83に記載の抗体または結合分子。
【請求項90】
重鎖可変ドメイン中の1つまたは複数のメチオニン置換を含むインフルエンザA型ウイルスの少なくとも1つの分離株を中和する、ヒトに感染する能力を有するインフルエンザA型ウイルスの血球凝集素に結合する、低下した酸化能を有する、設計抗体または結合分子。
【請求項91】
前記重鎖可変ドメインは、CDR3領域である、請求項90に記載の設計抗体または結合分子。
【請求項92】
前記メチオニン置換は、カバットナンバリングシステムによる96位においてである、請求項90に記載の設計抗体または結合分子。
【請求項93】
前記メチオニン置換は、カバットナンバリングシステムによる98位である、請求項90に記載の設計抗体または結合分子。
【請求項94】
前記メチオニン置換は、カバットナンバリングシステムによる96位および98位である、請求項90に記載の設計抗体または結合分子。
【請求項95】
前記メチオニンは、ロイシンで置換される、請求項90から94のいずれか1項に記載の設計抗体または結合分子。
【請求項96】
前記重鎖可変ドメインは、配列番号29のアミノ酸配列を含む、請求項90に記載の設計抗体または結合分子。
【請求項1】
配列番号7、配列番号8、および配列番号9からなる群より選択される重鎖からの1つ、2つ、もしくは3つの超可変領域配列を含む、結合分子、またはその機能的に活性なフラグメント。
【請求項2】
配列番号10、配列番号11、および配列番号12からなる群より選択される重鎖からの1つ、2つ、もしくは3つの超可変領域配列を含む、結合分子、またはその機能的に活性なフラグメント。
【請求項3】
配列番号7、配列番号8、もしくは配列番号9からのすべての超可変領域配列を含む、請求項1に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項4】
配列番号10、配列番号11、もしくは配列番号12からのすべての超可変領域配列を含む、請求項2に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項5】
軽鎖と会合するときに、標的に結合することが可能である、請求項1に記載の結合分子。
【請求項6】
軽鎖と会合するときに、標的に結合することが可能である、請求項2に記載の結合分子。
【請求項7】
前記軽鎖は、配列番号13、配列番号14、もしくは配列番号15の前記ポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む、請求項5に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項8】
前記軽鎖は、配列番号16、配列番号17、もしくは配列番号18の前記ポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む、請求項5に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項9】
前記軽鎖は、配列番号19、配列番号20、もしくは配列番号21の前記ポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む、請求項5に記載の結合分子。
【請求項10】
前記軽鎖は、配列番号22、配列番号23、もしくは配列番号24の前記ポリペプチド配列のうちの1つ、2つ、もしくは3つの超可変配列を含む、請求項6に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項11】
配列番号13、配列番号14、もしくは配列番号15のすべての超可変領域配列を含む、請求項7に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項12】
配列番号16、配列番号17、もしくは配列番号18のすべての超可変領域配列を含む、請求項7に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項13】
配列番号19、配列番号20、もしくは配列番号21のすべての超可変領域配列を含む、請求項7に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項14】
配列番号22、配列番号23、もしくは配列番号24のすべての超可変領域配列を含む、請求項8に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項15】
VpreB配列および/またはλ5配列を含むポリペプチドをさらに含む、請求項1もしくは2に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項16】
λ5配列に融合されたVpreB配列を含むポリペプチドをさらに含む、請求項1もしくは2に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項17】
Vic様配列および/またはJCκ配列を含むκ様代替軽鎖(SLC)構築物をさらに含む、請求項1もしくは2に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項18】
抗体、またはその機能的に活性なフラグメントである、請求項1もしくは2に記載の結合分子。
【請求項19】
重鎖を含む、結合分子であって、前記重鎖は、配列番号1として示した前記アミノ酸配列を含む、結合分子、またはその機能的に活性なフラグメント。
【請求項20】
重鎖を含む、結合分子であって、前記重鎖は、配列番号2として示した前記アミノ酸配列を含む、結合分子、またはその機能的に活性なフラグメント。
【請求項21】
配列番号3、配列番号4、および配列番号5からなる群より選択されるアミノ酸配列を含む軽鎖をさらに含む、請求項19に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項22】
配列番号6として示した前記アミノ酸配列を含む軽鎖をさらに含む、請求項20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項23】
(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)前記ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する、請求項19に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項24】
(i)インフルエンザA型ウイルスの少なくとも1つの亜型および/または少なくとも1つの分離株を中和し、(ii)前記ウイルスの血球凝集素(HA)抗原に結合する、請求項20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項25】
前記HA抗原のH1亜型のエピトープに結合する、請求項19または20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項26】
前記HA抗原のH3亜型のエピトープに結合する、請求項19もしくは20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項27】
前記HA抗原のH1亜型のエピトープに、および前記HA抗原のH3亜型のエピトープに結合する、請求項19もしくは20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項28】
前記HA抗原のH9亜型のエピトープに結合する、請求項19もしくは20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項29】
前記HA抗原のH5亜型のエピトープに結合する、請求項20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項30】
前記エピトープは、インフルエンザA型ウイルス上に提示される、請求項25から29のいずれか1項に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項31】
H1HA抗原およびH3抗原と交差反応性である、請求項19もしくは20に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項32】
前記H1およびH3インフルエンザA型ウイルス亜型の少なくとも1つを中和する、請求項19に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項33】
前記H1およびH3インフルエンザA型ウイルス亜型を中和する、請求項19に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項34】
前記H1HA抗原を中和する、請求項20に記載の結合分子、または機能的に活性なフラグメント。
【請求項35】
前記H5HA抗原を中和する、請求項20に記載の結合分子、または機能的に活性なフラグメント。
【請求項36】
前記H1およびH5HAインフルエンザA型ウイルス亜型を中和する、請求項20に記載の結合分子、または機能的に活性なフラグメント。
【請求項37】
前記H1亜型、または前記HAは、前記H1ウイルスのニューカレドニア/20/99分離株に由来する、請求項25に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項38】
前記H1亜型、または前記HAは、前記H1ウイルスのソロモン諸島/3/06分離株に由来する、請求項25に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項39】
前記H3亜型、または前記HAは、前記H3ウイルスのウィスコンシン/67/05分離株に由来する、請求項26に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項40】
前記H3亜型、または前記HAは、前記H3ウイルスの香港/68分離株に由来する、請求項26に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項41】
前記H9亜型、または前記HAは、前記H9ウイルスの香港/1073/99分離株に由来する、請求項28に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項42】
前記H5亜型、または前記HAは、前記H5ウイルスのベトナム/1203/04分離株に由来する、請求項29に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項43】
前記インフルエンザA型ウイルスの球状頭部が細胞表面に結合するのを阻止する、請求項23に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項44】
前記インフルエンザA型ウイルスが感染される細胞に付着するのを阻止する、請求項23に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項45】
前記ウイルスの少なくとも1つがヒトに感染する能力を有する、請求項23もしくは24に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項46】
前記分離株の少なくとも1つがヒト被験体から得たものである、請求項23もしくは24に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項47】
前記分離株の少なくとも1つが非ヒト動物から得たものである、請求項23もしくは24に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項48】
前記非ヒト動物がトリである、請求項47に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項49】
前記トリが野鳥またはニワトリである、請求項48に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項50】
前記非ヒト動物は、ブタである、請求項47に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項51】
H1HA抗原に結合する、請求項19に記載の結合分子、または機能的に活性なフラグメント。
【請求項52】
少なくとも1つのさらなるHA抗原に結合する、請求項51に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項53】
前記さらなるHA抗原は、H3である、請求項52に記載の結合分子、またはその機能的に活性なフラグメント。
【請求項54】
配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体。
【請求項55】
配列番号3、配列番号4、および配列番号5からなる群より選択されるアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む軽鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、請求項54に記載の抗体。
【請求項56】
配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体。
【請求項57】
配列番号6として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む軽鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、請求項56に記載の抗体。
【請求項58】
(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)前記ウイルスの血球凝集素(HA)抗原に結合し、かつ(iii)血球凝集を阻害する、請求項54もしくは55に記載の抗体。
【請求項59】
(i)インフルエンザA型ウイルスの1つを超える亜型および/または1つを超える分離株を中和し、(ii)前記ウイルスの血球凝集素(HA)抗原に結合する、請求項56もしくは57に記載の抗体。
【請求項60】
請求項1から59のいずれか1項に記載の結合分子または抗体を含む、組成物。
【請求項61】
被験体におけるインフルエンザA型ウイルス感染の防止および/または治療のための方法であって、請求項60に記載の有効量の組成物を前記被験体に投与することを含む、方法。
【請求項62】
被験体におけるインフルエンザA型ウイルス感染を治療するための方法であって、請求項1から59のうちのいずれか1つの中和抗体または結合分子の有効量を前記被験体に投与することを含む、方法。
【請求項63】
前記被験体は、ヒト患者である、請求項61もしくは62に記載の方法。
【請求項64】
請求項1から59のいずれか1項に記載の抗体または結合分子によって結合された中和エピトープを機能的に模倣するペプチドまたはポリペプチドを含む、インフルエンザA型ウイルスに有効なワクチン。
【請求項65】
合成ワクチンである、請求項64に記載のワクチン。
【請求項66】
弱毒化インフルエンザA型ウイルスまたはその一部を含む、請求項64に記載のワクチン。
【請求項67】
死滅させたインフルエンザA型ウイルスまたはその一部を含む、請求項64に記載のワクチン。
【請求項68】
前記抗体または結合分子は、
(a)配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子、
(b)配列番号1として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む重鎖ポリペプチドを含む、抗体、
(c)配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む抗体のエピトープと本質的に同一のエピトープに結合する、抗体または結合分子、またそのフラグメント、および
(d)配列番号2として示したアミノ酸配列、または前記アミノ酸配列に基づいたコンセンサス配列もしくは変異体配列を含む重鎖ポリペプチドを含む、抗体、またはそのフラグメント、からなる群より選択される、請求項64に記載のワクチン。
【請求項69】
前記抗体または結合分子は、HA抗原に結合する、請求項64に記載のワクチン。
【請求項70】
前記HA抗原は、H3亜型である、請求項69に記載のワクチン。
【請求項71】
前記HA抗原は、H1亜型である、請求項69に記載のワクチン。
【請求項72】
前記HA抗原は、H1亜型およびH3亜型である、請求項69に記載のワクチン。
【請求項73】
前記HA抗原は、H5亜型である、請求項69に記載のワクチン。
【請求項74】
前記HA抗原は、H9亜型である、請求項69に記載のワクチン。
【請求項75】
前記抗原は、インフルエンザA型ウイルスの表面上に提示される、請求項69に記載のワクチン。
【請求項76】
前記ペプチドまたはポリペプチドは、中和抗体を惹起する抗原決定基を含む、請求項64から75のいずれか1項に記載のワクチン。
【請求項77】
経口投与に好適な、請求項64に記載のワクチン。
【請求項78】
経粘膜送達に好適な、請求項64に記載のワクチン。
【請求項79】
前記経粘膜送達は、鼻腔内投与である、請求項78に記載のワクチン。
【請求項80】
非経口投与に好適な、請求項64に記載のワクチン。
【請求項81】
経皮投与に好適な、請求項64に記載のワクチン。
【請求項82】
前記ワクチンは、小児期免疫化用である、請求項64に記載のワクチン。
【請求項83】
長さが修正された重鎖ループを含むインフルエンザA型ウイルスの少なくとも1つの分離株を中和する、ヒトに感染する能力を有するインフルエンザA型ウイルスの血球凝集素へ結合する、中和抗体または結合分子。
【請求項84】
前記延長重鎖ループは、CDR3ループである、請求項83に記載の抗体または結合分子。
【請求項85】
前記CDR3ループは、配列番号9のアミノ酸配列を含む、請求項84に記載の抗体または結合分子。
【請求項86】
前記延長重鎖ループは、CDR1ループである、請求項85に記載の抗体または結合分子。
【請求項87】
前記CDR1ループは、配列番号7のアミノ酸配列を含む、請求項86に記載の抗体または結合分子。
【請求項88】
前記長さの修正は、延長重鎖ループを含む、請求項83に記載の抗体または結合分子。
【請求項89】
前記長さの修正は、前記重鎖ループにおける欠失を含む、請求項83に記載の抗体または結合分子。
【請求項90】
重鎖可変ドメイン中の1つまたは複数のメチオニン置換を含むインフルエンザA型ウイルスの少なくとも1つの分離株を中和する、ヒトに感染する能力を有するインフルエンザA型ウイルスの血球凝集素に結合する、低下した酸化能を有する、設計抗体または結合分子。
【請求項91】
前記重鎖可変ドメインは、CDR3領域である、請求項90に記載の設計抗体または結合分子。
【請求項92】
前記メチオニン置換は、カバットナンバリングシステムによる96位においてである、請求項90に記載の設計抗体または結合分子。
【請求項93】
前記メチオニン置換は、カバットナンバリングシステムによる98位である、請求項90に記載の設計抗体または結合分子。
【請求項94】
前記メチオニン置換は、カバットナンバリングシステムによる96位および98位である、請求項90に記載の設計抗体または結合分子。
【請求項95】
前記メチオニンは、ロイシンで置換される、請求項90から94のいずれか1項に記載の設計抗体または結合分子。
【請求項96】
前記重鎖可変ドメインは、配列番号29のアミノ酸配列を含む、請求項90に記載の設計抗体または結合分子。
【図1】

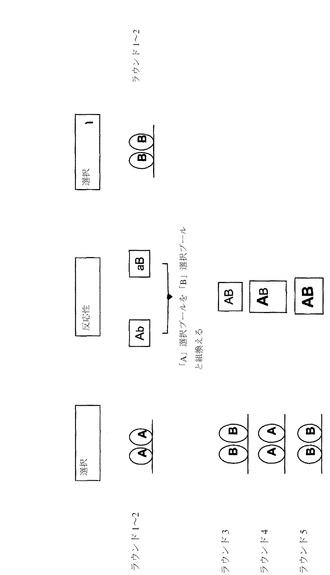
【図2】
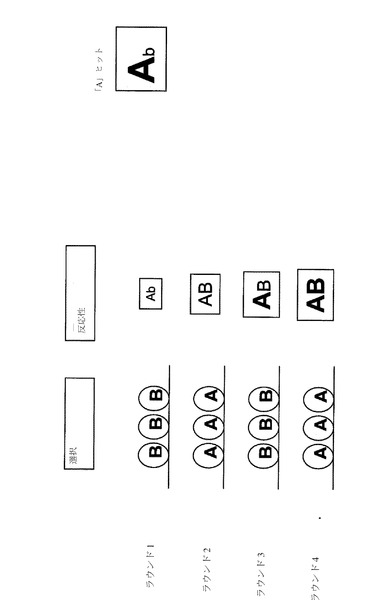
【図3】
【図4】
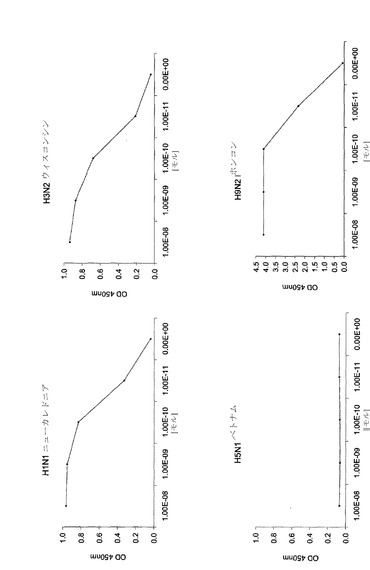
【図5】
【図6】
【図7A】
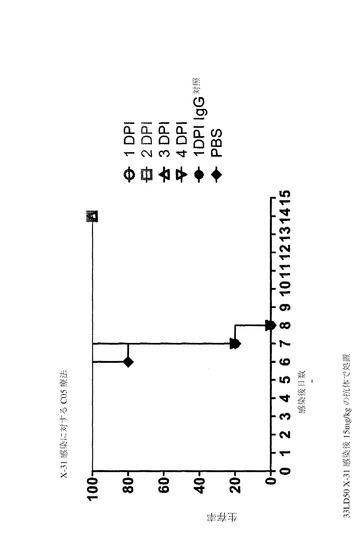
【図7B−1】
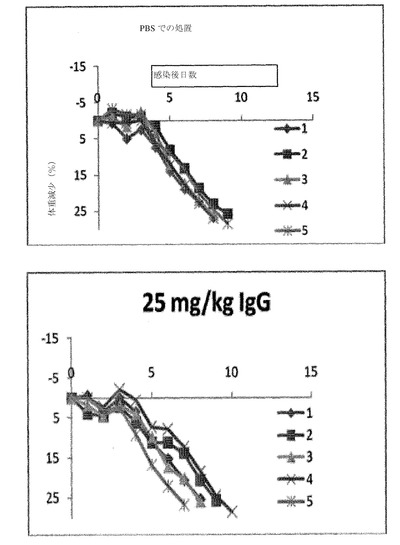
【図7B−2】
【図7B−3】
【図7C】
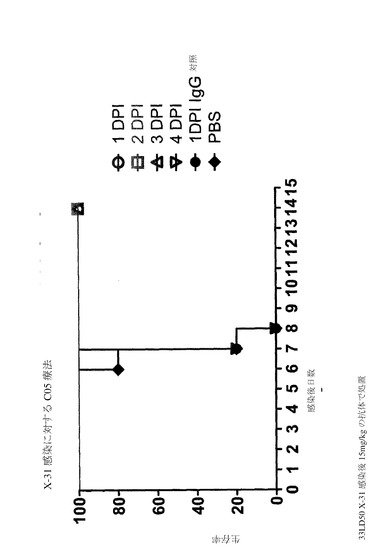
【図7D−1】
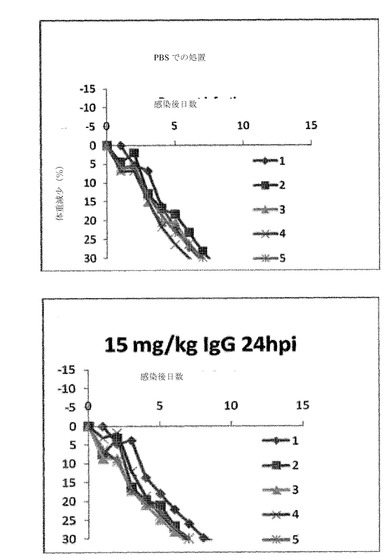
【図7D−2】
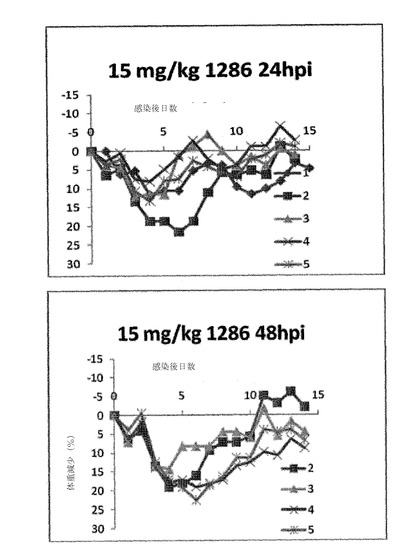
【図7D−3】
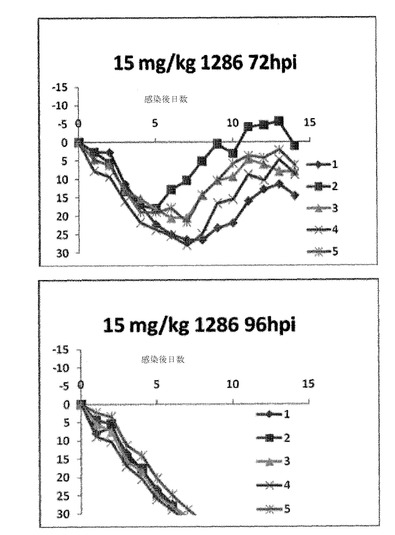
【図7E】
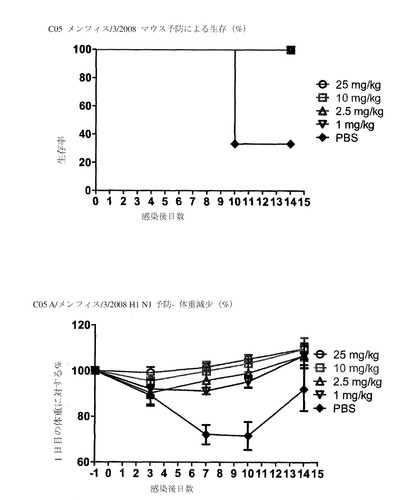
【図8】

【図9】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7A】
【図7B−1】
【図7B−2】
【図7B−3】
【図7C】
【図7D−1】
【図7D−2】
【図7D−3】
【図7E】
【図8】
【図9】
【公表番号】特表2012−526839(P2012−526839A)
【公表日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願番号】特願2012−510989(P2012−510989)
【出願日】平成22年5月12日(2010.5.12)
【国際出願番号】PCT/US2010/034604
【国際公開番号】WO2010/132604
【国際公開日】平成22年11月18日(2010.11.18)
【出願人】(508339161)シー レーン バイオテクノロジーズ, エルエルシー (10)
【Fターム(参考)】
【公表日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願日】平成22年5月12日(2010.5.12)
【国際出願番号】PCT/US2010/034604
【国際公開番号】WO2010/132604
【国際公開日】平成22年11月18日(2010.11.18)
【出願人】(508339161)シー レーン バイオテクノロジーズ, エルエルシー (10)
【Fターム(参考)】
[ Back to top ]