
インフルエンザウイルスに対する中和抗体
【課題】インフルエンザA型ウイルスに対する中和抗体を同定し、産生し、さらに、遺伝子操作する方法および手段、ならびに産生された中和抗体を提供すること。
【解決手段】本発明は、インフルエンザA型ウイルスに対する中和抗体を同定し、産生し、さらに、遺伝子操作する方法および手段、ならびに産生された中和抗体に関する。特に、本発明は、例えば、H1、H2、H3、およびH5サブタイプのすべてなど2つ以上のH1、H2、H3、H5、H7およびH9に対する中和抗体を含む、種々のインフルエンザA型ウイルスサブタイプに対する中和抗体およびこの種の抗体を作る方法および手段に関する。さらに具体的に述べると、本発明は、インフルエンザA型ウイルスサブタイプの分離株を1つより多く、好ましくはすべてを中和し得る抗体に関する。
【解決手段】本発明は、インフルエンザA型ウイルスに対する中和抗体を同定し、産生し、さらに、遺伝子操作する方法および手段、ならびに産生された中和抗体に関する。特に、本発明は、例えば、H1、H2、H3、およびH5サブタイプのすべてなど2つ以上のH1、H2、H3、H5、H7およびH9に対する中和抗体を含む、種々のインフルエンザA型ウイルスサブタイプに対する中和抗体およびこの種の抗体を作る方法および手段に関する。さらに具体的に述べると、本発明は、インフルエンザA型ウイルスサブタイプの分離株を1つより多く、好ましくはすべてを中和し得る抗体に関する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、インフルエンザA型ウイルスに対する中和抗体を同定し、産生し、さらに、遺伝子操作する方法および手段、ならびに産生された中和抗体に関する。特に、本発明は、例えば、H1、H2、H3、およびH5サブタイプのすべてなど2つ以上のH1、H2、H3、H5、H7およびH9に対する中和抗体を含む、種々のインフルエンザA型ウイルスサブタイプに対する中和抗体およびこの種の抗体を作る方法および手段に関する。さらに具体的に述べると、本発明は、インフルエンザA型ウイルスサブタイプの分離株を1つより多く、好ましくはすべてを中和し得る抗体に関する。
【背景技術】
【0002】
発明の背景
インフルエンザは、インフルエンザウイルスにより引き起こされる接触伝染性の呼吸器疾患である。該ウイルスは軽症から重篤までの疾患を引き起こし、時には死に至ることもある。毎年米国では、国民の5〜20%がインフルエンザにかかり、約200,000人が入院させられ、約36,000人が死亡する。
【0003】
インフルエンザウイルスは、せきやくしゃみをした時に生じる呼吸器系のしぶきで広がり、該ウイルスは、通常、ヒトからヒトへ伝染する。インフルエンザ表面抗原に対する免疫、特にヘマグルチニンは、感染が起きた場合、感染の可能性および感染しても疾患の重篤性を低減する。インフルエンザワクチンは利用できるが、1つの型またはサブタイプのインフルエンザウイルスに対する抗体は、別の型またはサブタイプのインフルエンザに対する保護は限定的または全く保護できず、毎年インフルエンザワクチンに1つより多くの株を組み込む必要がある。
【0004】
インフルエンザウイルスは、分節マイナス鎖(negative−strand)RNAウイルスであり、オルトミクス・ウイルス(Orthomyxoviridae)科に属する。インフルエンザA型ウイルスは9つの構造タンパク質からなり、さらに、制御的機能を有する1つの非構造NS1タンパク質をコードする。該非構造NS1タンパク質は、再現サイクルの間に大量に合成され、感染細胞の細胞質ゾルおよび核の中に局在化される。ウイルスゲノムの分節性により、種々のウイルス株を有する細胞の混合感染の間に起きる遺伝子的再集合(ゲノム分節の交換)のメカニズムが可能になる。インフルエンザA型ウイルスは、さらに、ウイルスタンパク質表面上に表示された種々のヘマグルチニン(HA:hemagglutinin)およびノイラミニダーゼ(NA:neuraminidase)ウイルスタンパク質により種々のサブタイプに分類される。
【0005】
インフルエンザA型ウイルスサブタイプは、2つのウイルス表面糖タンパク質である、ヘマグルチニン(HAまたはH)およびノイラミニダーゼ(NAまたはN)により同定される。各々のインフルエンザウイルスのサブタイプは、HおよびNのタンパク質のその組み合わせにより同定される。HAサブタイプが16、NAサブタイプが9種知られている。インフルエンザA型ウイルスは、ヒト、トリ、ブタ、ウマ、および他の動物に感染させることができるが、野鳥は、これらのウイルスの自然の宿主である。一部のインフルエンザAサブタイプのみ(すなわち、H1N1、H1N2、およびH3N2)が、現在ヒトの間に循環しているが、16のHおよび9のNAサブタイプのすべての組み合わせは、鳥類、特に原野にいる水鳥および海岸で生息しているトリにおいて同定されている。さらに、H5およびH7インフルエンザウイルスがヒトを発病させうることを示す証拠も蓄積されつつある。
【0006】
インフルエンザA型ウイルスのHAは、2つの構造的に異なる領域、すなわち、球状頭部領域およびステム領域を含んでいる。球状頭部領域は、標的細胞へのウイルスの取り付けに関与する受容体結合部位を含み、HAのヘマグルチニン化活性に関与している。ステム領域は融合ペプチドを含み、該ペプチドは、細胞のウイルス外被とエンドソーム膜との間の膜の融合に必要であり、したがって、融合活性に関係する(Wiley et al.,Ann.Rev.Biochem.,56:365−394(1987))。
【0007】
パンデミックは、世界的な疾患の大流行である。インフルエンザのパンデミックは、新しいインフルエンザA型ウイルスが、(1)人間集団において該ウイルスに対して免疫がほとんどないか、全くないウイルスが出現し、(2)重病を引き起こし始め、(3)ヒトとヒトとの間で世界的に容易に広がる時に起きる。20世紀の間に、この種のインフルエンザのパンデミックが3度発生した。最初は、1918年の「スペイン風邪」で、このインフルエンザパンデミックにより米国で少なくとも500,000人死亡し、世界の死者は4000万人に上った。このパンデミックは、インフルエンザA H1N1サブタイプにより発生した。2回目のインフルエンザパンデミックは、1957年の「アジア風邪」で、インフルエンザA H2N2サブタイプにより発生し、死者は、米国で少なくとも70,000人、世界では100〜200万人に上った。最も最近のインフルエンザパンデミックは、1968年の「香港風邪」で、インフルエンザA H3N2サブタイプにより発生し、死者は、米国で約34,000人、世界では70万人であった。
【0008】
1997年に、インフルエンザA H5N1の最初の症例が、香港で報告された。この鳥類型ウイルスが直接ヒトに感染したのは、これが初めてであったが、ヒトからヒトへの感染が認められなかったので、パンデミックは起きなかった。
【0009】
非特許文献1は、不活性化したH5N1ウイルスを予防接種したウマからの抗H51 IgGsおよびH5N1に感染したBALB/cマウスを保護すると説明されたH5N1−特異性F(ab’)2断片の調製について報告している。
【0010】
非特許文献2は、マウスの受動免疫法のインフルエンザA H5 ウイルスヘマグルチニンに対して特異的なキメラモノクローナル抗体の使用について説明している。
【0011】
特定のインフルエンザA型ウイルスにより生じた呼吸器疾患の重篤性、および潜在的なパンデミックの脅威に鑑みて、効率的な予防および処置方法を求める大きなニーズがある。本発明は、インフルエンザA型ウイルスのH1、およびH3サブタイプ、およびH5サブタイプを含むがこれらに限定されない、該ウイルスの種々のHサブタイプに対するインフルエンザA中和抗体を提供することによりこのニーズに取り組んでいる。本発明は、さらに、種々のヒトおよびヒト以外の種から得られた分離株および種々のインフルエンザの大流行および/またはパンデミックの犠牲者および/または生存者からの分離株を含むがこれらに限定されないインフルエンザA型ウイルスの任意のサブタイプの分離株の1つより多い、好ましくはすべてを中和し得る抗体を提供する。
【0012】
この種の中和抗体は、大流行またはパンデミックの場合、感染した人々または感染のリスクがある人々の受動免疫法を含むインフルエンザウイルス感染の予防および/または処置に利用することができる。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】Lu et al.,Resp.Res.7:43(2006)(doi:10.1186/1465−992−7−43)
【非特許文献2】Hanson et al.,Resp.Res.7:126(doi:10.1186/1465−9921−7−126)
【発明の概要】
【課題を解決するための手段】
【0014】
1つの態様では、本発明は、インフルエンザA型ウイルスサブタイプの1つより多くの分離株またはインフルエンザA型ウイルスの1つより多くのサブタイプを中和する中和抗体に関する。
【0015】
1つの実施形態では、該抗体は、H5、H7およびH9サブタイプの1つ以上などインフルエンザA型ウイルスサブタイプの実質的にすべての分離株を中和する。
【0016】
別の実施形態では、該抗体は、H5、H7およびH9サブタイプの1つ以上など特定のインフルエンザA型ウイルスサブタイプの1つより多くの分離株を中和する。
【0017】
さらに別の実施形態では、該抗体は、インフルエンザA型ウイルスの1つより多くのサブタイプ、および少なくとも1つのサブタイプの1つより多くの分離株を中和する。
【0018】
さらなる実施形態では、本明細書の該抗体により中和された少なくとも1つのサブタイプおよび/または分離株が、ヒトに感染する能力を有する。
【0019】
別の実施形態では、少なくとも1つの分離株は、例えば、野鳥およびニワトリを含む鳥類に由来するものである。
【0020】
特定の実施形態では、本明細書の抗体は、インフルエンザA型ウイルスのH5N1サブタイプを中和する。好ましくは、該抗体は、このインフルエンザA型ウイルスサブタイプの1つより多くの分離株、より好ましくは実質的にすべての分離株を中和する。
【0021】
別の実施形態では、本明細書の該抗体は、H5N1サブタイプ、ならびにH1N1、H1N2、およびH3N2サブタイプからなる群から選択される少なくとも1つの追加されたサブタイプを中和する。
【0022】
追加の実施形態では、本明細書の抗体は、追加されたサブタイプ(単数および複数)の1つより多くの分離株、好ましくは実質的にすべての分離株を中和する。
【0023】
別の実施形態では、本発明の中和抗体は、H5タンパク質に結合する。好ましくは、該抗体は、H5タンパク質の1つより多くの改変体、より一層好ましくは、H5タンパク質の実質的にすべての分離株を中和する。
【0024】
他の実施形態では、本明細書の抗体は、H5タンパク質およびH1、H2および/またはH3のタンパク質など少なくとも1つの追加されたHタンパク質に結合する。
【0025】
異なる態様では、本発明は、本明細書で説明した該中和抗体を含む組成物に関する。
【0026】
さらなる態様では、本発明は、単一のインフルエンザA型ウイルスサブタイプまたは複数のインフルエンザA型ウイルスサブタイプの1つより多くの分離株を中和し得る抗体を同定する方法に関する。この方法は、インフルエンザA型ウイルスサブタイプの第1および第2の分離株の両方、または該インフルエンザA型ウイルスの第1および第2のサブタイプと反応する抗体ライブラリーにおける抗体の同定、および第1および第2の分離株、または第1および第2のサブタイプ、それぞれ、に結合する該抗体の能力に基づいて、同定した該抗体を選択の連続的交互ラウンドを受けることを含む。
【0027】
ある実施形態では、第1および第2のインフルエンザA型ウイルスサブタイプ分離株の両方と反応する抗体は、第1および第2の分離株と、それぞれ、反応し、同定された抗体を組み換える抗体の少なくとも2ラウンドの別々の濃縮により同定された。
【0028】
別の実施形態では、第1および第2のインフルエンザA型ウイルスサブタイプ分離株の両方と反応することができる抗体は、第1および第2の分離株に、それぞれ、結合する該抗体の能力に基づいて、選択の連続的交互ラウンドを受ける前に突然変異誘発を受ける。望ましいならば、第1および第2の分離株に結合し得る該抗体は、さらに、1つより多くのインフルエンザA型サブタイプに結合する該抗体の能力に基づいて選択される。
【0029】
この種の濃縮技法の適用は、一般に、抗体が結合する標的とは関係なく、同様に抗体に適用することができる。この種の一般的な濃縮/選択の方法は、本発明の一部として明らかに含まれている。
【0030】
さらなる態様では、本発明は、本発明の中和抗体が共有する配列コレクションに関する。
【0031】
さらにさらなる態様では、本発明は、本明細書の有効量の中和抗体または抗体組成物を対象に投与することを含むA型インフルエンザに感染している対象を処置する方法に関する。
【0032】
別の態様では、本発明は、A型インフルエンザに感染するリスクのある対象に本発明の中和抗体の有効量を投与することを含む、A型インフルエンザの感染を予防する方法に関する。
【0033】
異なる態様では、本発明は、(a)少なくとも2つの機能が異なる抗体のCDR配列を整列させ、(b)整列させられたCDR配列間に保存されたアミノ酸残基を同定し、(c)整列させられたCDR配列の複数の改変体を産生するために突然変異を誘発された非保存位置における機能が異なる抗体に存在する、少なくともアミノ酸残基をコードする縮重オリゴヌクレオチドプローブを用いて、整列させられたCDR配列の少なくとも1つの配列において複数の非保存アミノ酸残基の突然変異誘発を行い、ならびに、望ましいならば、該抗体コレクションが望ましい程度の多様性および/またはサイズに達するまで改変体の1つ以上と共にステップ(b)および(c)を反復することを含む、様々な多機能性抗体コレクションを作成する方法に関する。
【0034】
特定の実施形態では、整列させられたCDR配列は同じ長さを有する。
【0035】
別の実施形態では、保存された該アミノ酸残基類は、整列させられた少なくとも2つのCDR配列に保持される。
【0036】
さらなる態様では、本発明は、少なくとも1つの特性が互いに異なる、複数の中和抗体を含む抗体コレクションに関する。
【0037】
本発明は、さらに、この種のコレクションに存在する核酸の配列に連結されたか、または該配列に組み込まれる独特のバーコードで標識した核酸を含むコレクション中の核酸を独自に同定する方法に関する。
本発明はまた、以下の項目を提供する。
(項目1)
インフルエンザA型ウイルスサブタイプの1つより多くの分離株および/または上記インフルエンザA型ウイルスの1つより多くのサブタイプを中和する中和抗体。
(項目2)
インフルエンザA型ウイルスH1サブタイプの1つより多くの分離株を中和する、項目1に記載の中和抗体。
(項目3)
インフルエンザA型ウイルスH3サブタイプの1つより多くの分離株を中和する、項目1に記載の中和抗体。
(項目4)
インフルエンザA型ウイルスH1およびH3サブタイプを中和する、項目1に記載の中和抗体。
(項目5)
インフルエンザA型ウイルスH1および/またはH3サブタイプの1つより多くの分離株を中和する、項目4に記載の中和抗体。
(項目6)
インフルエンザA型ウイルスサブタイプの実質的にすべての分離株を中和する、項目1に記載の中和抗体。
(項目7)
上記サブタイプが、H5、H7およびH9サブタイプからなる群から選択される、項目1または項目6に記載の中和抗体。
(項目8)
上記サブタイプが、上記H5サブタイプである、項目7に記載の中和抗体。
(項目9)
上記抗体が、インフルエンザA型ウイルスH5サブタイプの実質的にすべての分離株を中和する、項目8に記載の中和抗体。
(項目10)
上記サブタイプが、上記H7サブタイプである、項目7に記載の中和抗体。
(項目11)
上記抗体が、上記インフルエンザA型ウイルスH7サブタイプの実質的にすべての分離株を中和する、項目10に記載の中和抗体。
(項目12)
上記サブタイプが、上記H9サブタイプである、項目7に記載の中和抗体。
(項目13)
上記抗体が、上記インフルエンザA型ウイルスH9サブタイプの実質的にすべての分離株を中和する、項目12に記載の中和抗体。
(項目14)
上記抗体が、さらに、インフルエンザA型ウイルスの追加のHサブタイプを少なくとも1つ中和する、項目7に記載の中和抗体。
(項目15)
上記追加Hサブタイプが、H1、H2およびH3サブタイプからなる群から選択される、項目14に記載の中和抗体。
(項目16)
上記インフルエンザA型ウイルスの追加Hサブタイプの1つより多くの分離株を中和する、項目15に記載の中和抗体。
(項目17)
上記インフルエンザA型ウイルスのH5N1サブタイプを中和する、項目1に記載の中和抗体。
(項目18)
上記インフルエンザA型ウイルスのH5N1サブタイプの1つより多くの分離株を中和する、項目17に記載の中和抗体。
(項目19)
上記分離株の少なくとも1つがヒトに感染する能力を有する、項目18に記載の中和抗体。
(項目20)
上記分離株の少なくとも1つがヒト対象から得られた、項目19に記載の中和抗体。
(項目21)
上記ヒト対象が病気に罹患している、項目20に記載の中和抗体。
(項目22)
上記ヒト対象が上記インフルエンザA型ウイルスのH5N1サブタイプ感染から回復した、項目20に記載の中和抗体。
(項目23)
上記分離株の少なくとも1つがヒト以外の動物から得られた、項目19に記載の中和抗体。
(項目24)
上記ヒト以外の動物が鳥類である、項目23に記載の中和抗体。
(項目25)
上記ヒト以外の動物が野鳥である、項目24に記載の中和抗体。
(項目26)
上記ヒト以外の動物がニワトリである、項目24に記載の中和抗体。
(項目27)
上記インフルエンザA型ウイルスのH5N1サブタイプの実質的にすべての分離株を中和する、項目18に記載の中和抗体。
(項目28)
上記H5N1サブタイプおよびH1N1、H2N2、およびH3N2サブタイプからなる群から選択された少なくとも1つの追加サブタイプを中和する、項目17に記載の中和抗体。
(項目29)
上記インフルエンザA型ウイルスのH5N1サブタイプのより多くの分離株を中和する、項目28に記載の中和抗体。
(項目30)
上記インフルエンザA型ウイルスのH5N1サブタイプの実質的にすべての分離株を中和する、項目29に記載の中和抗体。
(項目31)
上記追加サブタイプの1つより多くの分離株を中和する、項目30に記載の中和抗体。(項目32)
上記追加サブタイプの実質的にすべての分離株を中和する、項目31に記載の中和抗体。
(項目33)
上記抗体がH5タンパク質に結合する、項目1に記載の中和抗体。
(項目34)
上記抗体が上記H5タンパク質の1つより多くの改変体に結合する、項目33に記載の中和抗体。
(項目35)
上記抗体が上記H5タンパク質のすべての改変体に結合する、項目34に記載の中和抗体。
(項目36)
上記抗体が少なくとも1つの追加のHタンパク質に結合する、項目35に記載の中和抗体。
(項目37)
上記追加のHタンパク質が、H1、H2およびH3タンパク質からなる群から選択される、項目36に記載の中和抗体。
(項目38)
上記抗体が上記追加のHタンパク質の1つより多くの改変体に結合する、項目37に記載の中和抗体。
(項目39)
上記抗体が上記追加のHタンパク質の実質的にすべての改変体に結合する、項目38に記載の中和抗体。
(項目40)
項目1から39までのいずれか1項に記載の中和抗体を含む組成物。
(項目41)
インフルエンザA型ウイルスサブタイプの1つより多くの分離株またはインフルエンザA型ウイルスの1つより多くのサブタイプを中和し得る抗体を同定する方法であって、
上記インフルエンザA型ウイルスサブタイプの第1および第2の分離株の両方、または上記インフルエンザA型ウイルスの第1および第2のサブタイプと反応する抗体を、抗体ライブラリーにおいて同定すること、および上記第1および第2の分離株、または上記第1および第2のサブタイプそれぞれに結合する抗体の能力に基づいて、同定した上記抗体に選択の連続的交互ラウンドを受けさせることを含む、方法。
(項目42)
少なくとも2ラウンドの選択を含む、項目41に記載の方法。
(項目43)
上記第1および第2の分離株が、上記インフルエンザA型ウイルスのH5N1サブタイプの異なる分離株である、項目41に記載の方法。
(項目44)
第1および第2のインフルエンザA型ウイルスサブタイプ分離株の両方と反応する上記抗体が、それぞれ、上記第1分離株および上記第2分離株と反応する抗体の少なくとも2ラウンドの別々の濃縮により同定され、同定された上記抗体を組み換える、項目41に記載の方法。
(項目45)
上記第1および上記第2のインフルエンザA型サブタイプ分離株の両方と反応し得る上記抗体が、上記第1および第2の分離株それぞれに結合する抗体の能力に基づいて、選択の上記連続交互ラウンドを受ける前に突然変異誘発を受ける、項目41に記載の方法。
(項目46)
上記抗体ライブラリーがファージディスプレイ・ライブラリーである、項目41に記載の方法。
(項目47)
選択がバイオパニングによってなされる、項目46に記載の方法。
(項目48)
上記インフルエンザA型ウイルスサブタイプがH5N1サブタイプである、項目41に記載の方法。
(項目49)
上記第1分離株が上記H5N1ウイルスの2006年のトルコ分離株である、項目48に記載の方法。
(項目50)
上記第1分離株が上記H5N1ウイルスの2003/2004年のベトナム分離株である、項目48に記載の方法。
(項目51)
上記第2分離株が上記H5N1ウイルスの2003/2004年のベトナム分離株である、項目48に記載の方法。
(項目52)
上記第2分離株が上記H5N1ウイルスの1997年の香港分離株である、項目50に記載の方法。
(項目53)
上記第1および第2の分離株が異なる種に由来する、項目48に記載の方法。
(項目54)
上記種の少なくとも1つがヒトである、項目53に記載の方法。
(項目55)
上記種の少なくとも1つが鳥類である、項目53に記載の方法。
(項目56)
上記第1および第2の分離株に結合し得る上記抗体が、さらに、1つより多くのインフルエンザA型サブタイプに結合する上記抗体の能力に基づいて選択される、項目41に記載の方法。
(項目57)
項目41〜56のいずれか1項に記載の方法により同定された上記中和抗体により共有される配列コレクション。
(項目58)
配列コレクションであって、図11、12、13、および14A−Dに示された独特の1つ以上の重鎖および/または軽鎖の配列、または上記配列に基づくコンセンサス配列または改変体配列を含む、コレクション。
(項目59)
項目41〜56のいずれか1項に記載の方法により同定可能な中和抗体またはその断片。
(項目60)
図11、12、13、および14A−Dに示された独特の配列から選択された重鎖および/または軽鎖の配列、または上記配列に基づくコンセンサス配列または改変体配列またはこれらの断片を含む、項目59に記載の中和抗体。
(項目61)
インフルエンザA型ウイルス感染に対して鳥類または哺乳類の対象に受動免疫を与えることができる、項目59または項目60に記載の中和抗体または抗体断片。
(項目62)
上記哺乳類の対象がヒトである、項目61に記載の中和抗体または抗体断片。
(項目63)
上記インフルエンザA型ウイルス感染が、H5N1、H1N1、H2N2、およびH3N2サブタイプからなる群から選択されるウイルスにより引き起こされる、項目62に記載の中和抗体または抗体断片。
(項目64)
対象においてA型インフルエンザ感染を予防および/または処置する方法であって、上記対象に項目40に記載の組成物を有効量投与することを含む、方法。
(項目65)
対象のA型インフルエンザ感染を処置する方法であって、上記対象に項目59に記載の中和抗体を有効量投与することを含む、方法。
(項目66)
上記対象がヒトの患者である、項目64または項目65に記載の方法。
(項目67)
A型インフルエンザ感染を予防する方法であって、A型インフルエンザ感染を発症するリスクのある対象に項目40に記載の組成物を有効量投与することを含む、方法。
(項目68)
A型インフルエンザ感染を予防する方法であって、A型インフルエンザ感染を発症するリスクのある対象に項目59に記載の中和抗体を有効量投与することを含む、方法。
(項目69)
上記対象がヒトの患者である、項目67または項目68に記載の方法。
(項目70)
多様な多機能性抗体コレクションを作る方法であって、
(a)少なくとも2つの機能が異なる抗体のCDR配列を整列させ、(b)上記整列させられたCDR配列の間に保存されたアミノ酸残基を同定し、(c)上記整列させられたCDR配列の複数の改変体を産生するために、突然変異誘発された非保存位置における機能が異なる抗体に存在する少なくとも上記アミノ酸残基をコードする縮重オリゴヌクレオチドプローブを用いて、上記整列させられたCDR配列の少なくとも1つの配列において複数の非保存アミノ酸残基の突然変異誘発を行い、そして、望ましいならば、上記抗体コレクションが望ましい程度の多様性またはサイズに達するまで上記改変体の1つ以上を用いてステップ(b)および(c)を反復することを含む、方法。
(項目71)
上記整列させられたCDR配列が同じ長さを有する、項目70に記載の方法。
(項目72)
ステップ(c)において産生された上記突然変異誘発された改変体が、上記整列させられたCDR配列の少なくとも2つに存在する保存されたすべての残基を保持する、項目70に記載の方法。
(項目73)
ステップ(c)において産生された上記突然変異誘発された改変体が、上記整列させられたCDR配列のすべてに存在する保存されたすべての残基を保持する、項目70に記載の方法。
(項目74)
上記機能的に異なる抗体が標的抗原上で異なるエピトープに結合する、項目70に記載の方法。
(項目75)
上記機能的に異なる抗体が異なる標的抗原に結合する、項目70に記載の方法。
(項目76)
上記異なる標的抗原が上記同じ抗原の改変体である、項目75に記載の方法。
(項目77)
上記機能的に異なる抗体が異なる結合親和性を有する、項目70に記載の方法。
(項目78)
上記機能的に異なる抗体が異なる生物学的特性を有する、項目70に記載の方法。
(項目79)
上記機能的に異なる抗体がインフルエンザA型ウイルスに結合する、項目70に記載の方法。
(項目80)
上記機能的に異なる抗体の少なくとも2つが同じインフルエンザA型ウイルスについて異なるエピトープに結合する、項目79に記載の方法。
(項目81)
上記機能的に異なる抗体が異なるインフルエンザA型ウイルスサブタイプに結合する、項目79に記載の方法。
(項目82)
上記機能的に異なる抗体の少なくとも2つが同じインフルエンザA型ウイルスサブタイプの異なる分離株に結合する、項目79に記載の方法。
(項目83)
上記機能的に異なる抗体の少なくとも2つが同じインフルエンザA型ウイルスサブタイプの異なる分離株および異なるインフルエンザA型ウイルスサブタイプに結合する、項目79に記載の方法。
(項目84)
上記機能的に異なる抗体の少なくとも2つが異なる親和性を有する、項目70〜83のいずれか1項に記載の方法。
(項目85)
上記機能的に異なる抗体の少なくとも2つは、これらが結合するインフルエンザA型ウイルスを中和する能力に違いがある、項目70〜83のいずれか1項に記載の方法。
(項目86)
少なくとも1つの特性が互いに異なる複数の中和抗体を含む、抗体コレクション。
(項目87)
少なくとも約100の中和抗体を含む、項目86に記載の抗体コレクション。
(項目88)
項目70〜83のいずれか1項に記載の方法により調製された項目87に記載の抗体コレクション。
(項目89)
コレクション中の核酸を独自に同定する方法であって、上記コレクション中に存在する核酸配列に連結または上記配列に組み込まれる独特のバーコードで上記核酸を標識することを含む、方法。
(項目90)
上記バーコードが、1〜約24個のヌクレオチド長の非コードヌクレオチド配列である、項目89に記載の方法。
(項目91)
上記非コードヌクレオチド配列が、標識された上記核酸配列の3’非コード領域に連結される、項目90に記載の方法。
(項目92)
上記バーコードが、標識された上記核酸配列内に組み込まれる1つ以上のサイレント突然変異のコード配列である、項目89に記載の方法。
(項目93)
上記バーコードが、ペプチドまたはポリペプチドの配列である、項目89に記載の方法。
【図面の簡単な説明】
【0038】
【図1A】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1B】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1C】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1D】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1E】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1F】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1G】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1H】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1I】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1J】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図2】2種類の標的、AおよびBに対する反応性強度を高めるための典型的なパニング濃縮スキームを示している。各濃縮ラウンドは、個々の標的(単数および複数)に対するプールの反応性強度を高める。
【図3】標的AおよびBと交差反応するクローンを選択する方法を示し、この方法では、各連続ラウンドは両標的に対して得られるプールの反応性強度を高める。
【図4】交差反応性を発生および/または高めるために並行発見プールを組み換えることにより2種類の標的(標的AおよびB)に対する反応性強度を高める方法を示している。組み換え抗体ライブラリーの各選択ラウンドは、両標的に対して得られるプールの反応性強度を高める。
【図5】標的Aに対する反応性を維持しながら標的Bに対する交差反応性を高める方法を示している。第1に、標的Aと反応するクローンが選択され、次いで、標的Aと反応するクローンの突然変異誘発ライブラリーが調製され、選択が示されたようになされ、標的Aおよび標的Bの両方と強い反応性を示す1つ以上の抗体クローンを生じる。
【図6】「目標突然変異誘発」法により多様な多機能性抗体コレクションを作る代表的な突然変異誘発法を示している。
【図7】トルコのH5N1鳥インフルエンザ大流行の6名の生存者から得られた血液試料についてH5ヘマグルチニン(HA)の血清学的研究結果を示している。これらのデータは、HA抗原に対する抗体の存在を実証している。
【図8】H5抗原(A/Vietnam/1203/2004)およびH1N1(A/New Caledonia/20/99)およびH3N2(A/Panama/2007/99)ウイルスについてテストした、12名のローカルドナーの血清試料を用いて得られた血清学的研究結果を示している。
【図9】抗体ファージライブラリーの構築において使われた独特のバーコード法を示している。
【図10】H5タンパク質およびH5N1ウイルスについてトルコの鳥インフルエンザ生存者のプールされたライブラリーから得られた5種類のクローンのscFvELISAテストの結果を示している。
【図11】報告されたトルコの分離株からH5ヘマグルチニンタンパク質の配列およびLos Alamos National Laboratoryの配列データベースからダウンロードした1つのベトナムの分離株を比較している配列アラインメントを示している。
【図12】対応する軽鎖および生殖系列起源配列と共に、トルコドナーのプールされた抗体ライブラリーにおいて同定された独特のクローンの重鎖可変領域配列を示している。この図に示した配列(3〜23重鎖クローン)は、3ラウンドのパニング後の、すべてのトルコドナーのすべての重鎖および軽鎖のプールされたライブラリーに由来する。
【図13】対応する軽鎖および生殖系列起源配列と共に、トルコドナーのプールされた抗体ライブラリーにおいて同定された独特のクローンの重鎖可変領域配列を示している。この図に示した配列(3〜30重鎖クローン)は、2ラウンドのパニング後の、すべてのトルコドナーのすべての重鎖および軽鎖のプールされたライブラリーに由来する。
【図14A】4ラウンドのパニング後、個々のトルコドナーの抗体ライブラリーから同定された、追加された独特のH5N1に特異な抗体重鎖可変領域配列を示している。
【図14B】4ラウンドのパニング後、個々のトルコドナーの抗体ライブラリーから同定された、追加された独特のH5N1に特異な抗体重鎖可変領域配列を示している。
【図14C】4ラウンドのパニング後、個々のトルコドナーの抗体ライブラリーから同定された、追加された独特のH5N1に特異な抗体重鎖可変領域配列を示している。
【図14D】4ラウンドのパニング後、個々のトルコドナーの抗体ライブラリーから同定された、追加された独特のH5N1に特異な抗体重鎖可変領域配列を示している。
【図15】トルコの鳥インフルエンザ生存者の血清および骨髄の分析により同定された抗体重鎖配列を用いて多様な抗体重鎖および軽鎖ライブラリーを作り出すために目標の突然変異誘発の使用を例示している。
【図16】トルコの鳥インフルエンザ生存者の血清および骨髄の分析により同定された抗体軽鎖配列を用いて多様な抗体重鎖および軽鎖ライブラリーを作り出すために目標の突然変異誘発の使用を例示している。
【図17】HAタンパク質のトルコおよびインドネシアの改変体と共にH5N1ベトナムウイルスscFv抗体から得られた特定のFab断片の交差反応性を確認しているELISAの結果を示している。
【図18】HAタンパク質のトルコおよびインドネシアの改変体と共にH5N1ベトナムウイルスscFv抗体から得られた特定のFab断片の交差反応性を確認しているELISAの結果を示している。
【発明を実施するための形態】
【0039】
A.定義
特に明記しない限り、本明細書で使われる技術・科学用語は、本発明が属する技術分野の当業者が普通に理解している意味と同じ意味を有する。Singleton et al.,Dictionary of Microbiology and Molecular Biology 2nd ed.,J.Wiley & Sons(New York,NY 1994)は、本出願において使われる多くの用語の手引書を当業者に提供する。
【0040】
当業者は、本発明を実行する場合に使われる可能性がある、本明細書で説明した方法および物質と類似または同等な方法および物質が多数あることを認めるであろう。実際に、本発明は、説明した方法および物質に決して限定しない。本発明では、次のように用語を定義する。
【0041】
用語「インフルエンザA型サブタイプ」または「インフルエンザA型ウイルスサブタイプ」は、相互互換的に使われ、ヘマグルチニン(H)およびノイラミニダーゼ(N)のウイルス表面タンパク質の種々の組み合わせにより特徴づけられるインフルエンザA型ウイルス改変体を意味し、したがって、例えば、H1N1およびH3N2などのH数とN数の組み合わせにより呼ばれる。これらの用語は、通常突然変異から生じ、異なる病原性特性を示す、各サブタイプ内のすべての菌株(絶滅菌株を含む)を明らかに含んでいる。この種の菌株は、過去、現在および未来の分離株をすべて含む、ウイルスのサブタイプの種々の「分離株」も意味する。したがって、この文脈では、用語「菌株」と「分離株」は相互互換的に使われる。
【0042】
用語「インフルエンザ」は、インフルエンザウイルスによって引き起こされた伝染性疾患を意味するのに使われる。
【0043】
本発明の文脈では、用語「抗体」(Ab:antibody)は、最も広い意味で使われ、特定の抗原ならびに免疫グロブリンおよび抗原特異性に欠ける他の抗体状分子に対して結合選択性を示すポリペプチド類を含む。後の種類のポリペプチド類は、例えば、低レベルではリンパ系により、高レベルでは骨髄腫により産生される。本出願では、「抗体」は、限定なしに、モノクローナル抗体、ポリクローナル抗体、および抗体断片をカバーしている。
【0044】
「未変性抗体」は、通常、2つの同一軽(L:light)鎖と2つの同一重(H:heavy)鎖とからなる、約150,000ドールトンのヘテロテトラマー性糖タンパク質である。各軽鎖は、共有ジサルファイド結合(単数および複数)により重鎖に連結し、ジサルファイド結合の数は、種々の免疫グロブリンのアイソタイプの重鎖間で変わる。各重鎖および軽鎖は、規則的に間隔をあけた鎖内ジサルファイド橋も有する。各重鎖は、1つの終端において可変ドメイン(VH)とそれに続く多数の定常ドメインを有する。各軽鎖は、1つの終端において可変ドメイン(VL)およびその他の終端において定常ドメインを有し;軽鎖の定常ドメインは重鎖の第1定常ドメインを用いて整列させられ、軽鎖可変ドメインは、重鎖の可変ドメインを用いて整列させられている。特定のアミノ酸残基は、軽鎖および重鎖の可変ドメインの間で界面を形成すると考えられている。Chothia et al.,J.Mol.Biol.186:651(1985);Novotny and Haber,Proc.Natl.Acad.Sci.U.S.A.82:4592(1985).
抗体鎖に関連した用語「可変」は、抗体内の配列で広範に違いがあり、その特定の抗原に対する特定の各抗体の結合および選択性に関与する該抗体の部分を意味するのに使われる。この種の可変性は、軽鎖および重鎖の両方の可変ドメインにおいて超可変領域と呼ばれる3つのセグメントに濃縮されている。可変ドメイン類のより高度に保存された部分は、フレームワーク領域(FR:framework region)と呼ばれる。未変性重鎖および軽鎖の可変ドメイン類は、各々が、4つのFR(FR1、FR2、FR3およびFR4、それぞれ)を含み、大抵が3つの超可変領域により接続されたβ−シート配置を採用し、該超可変領域は、ループ接続、および一部の場合にはβ−シート構造の一部を形成している。各鎖の超可変領域は、FR類により、および他の鎖からの超可変領域を用いて近接して一緒に保持され、抗体の抗原結合部位の形成に寄与する(Kabat et
al.,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.(1991),pages 647−669を参照のこと)。定常ドメイン類は、抗体の抗原への結合には直接には関与しないが、抗体依存性細胞毒性における抗体の関与など種々のエフェクター機能を示す。
【0045】
用語「超可変領域」は、本明細書で使われる場合、抗原結合に関与している抗体のアミノ酸残基類を意味している。超可変領域は、「相補性決定領域」または「CDR」からのアミノ酸残基類を含む(すなわち、軽鎖可変ドメインにおける残基30−36(L1)、46−55(L2)および86−96(L3)、ならびに重鎖可変ドメインにおける30−35(H1)、47−58(H2)および93−101(H3);MacCallum
et al.,J Mol Biol.1996。「フレームワーク」または「FR」残基類は、本明細書で定義された超可変領域残基類以外の可変ドメイン残基類である。
【0046】
これらの重鎖の定常ドメインのアミノ酸配列により、抗体は種々のクラスに帰属させることができる。抗体のクラスには、主なものが5つ、すなわち、IgA、IgD、IgE、IgG、およびIgMがあり、これらのいくつかは、さらに、サブクラス(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgA、およびIgA2に分類することができる。
【0047】
種々のクラスの免疫グロブリンに対応する重鎖定常ドメイン類は、それぞれ、α、δ、ε、γ、およびμと呼ばれる。
【0048】
脊椎動物に由来する抗体の「軽鎖」は、これらの定常ドメインのアミノ酸配列に基づいて、カッパ(κ)およびラムダ(λ)と呼ばれる2つの明確に異なる型の1つに帰属させることができる。
【0049】
「抗体断片」は、全長抗体の一部、一般に、抗体の抗原結合または可変ドメインを含む。抗体断片の例には、Fab、Fab’、F(ab’)2、およびFv断片、線状抗体、単一鎖抗体分子、二重特異性抗体、および抗体断片から形成された多選択性抗体があるが、これらに限定されない。
【0050】
用語「モノクローナル抗体」は、B細胞の単一クローンにより合成された抗体分子を意味するのに使われる。修飾語句「モノクローナル」は、ほぼ均一な抗体集団からえられている抗体のキャラクターを示しており、特定の方法により抗体の産生を必要とするものと解釈すべきではない。したがって、モノクローナル抗体は、Kohler and Milstein,Nature 256:495(1975);Eur.J.Immunol.6:511(1976)により最初に報告されたハイブリドーマ法、組換えDNA法により作るか、またはファージ抗体ライブラリーから分離してもよい。
【0051】
用語「ポリクローナル抗体」は、B細胞の集団により合成された抗体分子の集団を呼ぶのに使われる。
【0052】
「単一鎖Fv」または「sFv」抗体断片は、抗体のVHおよびVL両ドメインを含み、これらのドメインは、単一ポリペプチド鎖中に存在する。一般に、Fvポリペプチドは、さらに、VHおよびVL両ドメインの間にポリペプチドリンカーを含み、該リンカーは、sFvが、抗原結合について望ましい構造を形成することを可能にする。sFvの総説については、The Pharmacology of Monoclonal Antibodies,vol.113,Rosenburg and Moore eds.Springer−Verlag,New York,pp.269−315(1994)のPlueckthunを参照のこと。単一鎖抗体は、例えば、国際公開第88/06630号および国際公開第92/01047号で開示されている。
【0053】
用語「二重特異性抗体」は、2つの抗原結合部位を有する小さな抗体断片を意味し、該断片は、同じポリペプチド鎖(VH−VL)において、軽鎖可変ドメイン(VL)に接続した重鎖可変ドメイン(VH)を含む。同じ鎖上で2つのドメイン間にペアを作るには短かすぎるリンカーを用いることにより、これらのドメインは別の鎖の相補性ドメインとペアを作ることを強いられる。二重特異性抗体は、例えば、欧州特許第404,097号;国際公開第93/11161号;およびHollinger et al.,Proc.Natl.Acad.Sci.USA 90:6444−6448(1993)により詳細に説明されている。
【0054】
用語「二重特異性抗体」は、2つの異なる型の抗原に対して特異性を示す抗体を意味している。本明細書で使われる用語は、標的抗原および特定の組織への送達を容易にする別の標的に対して結合特異性を示す抗体を、無制限に特異的に含んでいる。同様に、多選択性抗体は、2つ以上の結合特異性を有する。
【0055】
表現「線状抗体」は、抗原結合領域のペアを形成する一対の直列型のFdセグメント(VH−CH1−VH−CH1)を含むことを意味するのに使われる。線状抗体は、二重特異性または単一特異性であり、例えば、Zapata et al.,Protein Eng.8(10):1057−1062(1995)により説明されている。
【0056】
用語「中和抗体」は、本明細書では最も広い意味で使われ、インフルエンザウイルスは、中和が行われるメカニズムとは関係なく、標的細胞を複製により感染することを抑制するあらゆる抗体を意味している。したがって、例えば、ウイルスの連結または接着に関与する部位に、直接またはすぐそばに結合する抗体を遺伝子操作することにより、該ウイルスの細胞表面への連結または接着を抑制することにより、中和を実現することができる。ウイルス表面に向けられた抗体によりウイルスを凝集させることにより、中和を実現することもできる。中和は、さらに、標的細胞にウイルスを連結後ウイルスおよび細胞膜の融合の抑制、エンドサイトーシスの抑制、感染細胞から子孫ウイルスの抑制などにより、中和をさらに行うことができる。本発明の中和抗体は、中和が行われるメカニズムにより限定されない。
【0057】
用語「抗体レパートリー」は、本明細書では最も広い意味で使われ、結合能力、結合特異性、胃腸内を輸送する能力、安定性、親和性などの特性を選別するのに使われる抗体または抗体断片のコレクションを意味している。この用語は、具体的には、例えば、制約なしに、単一鎖Fv(scFv)を含む、抗体ファージディスプレイ・ライブラリー、およびナイーブな合成、半合成ライブラリーを含むあらゆる供給源に由来するFab抗体ファージディスプレイ・ライブラリーなどすべての形のコンビナトリアルなライブラリーを含む抗体ライブラリーを含んでいる。
【0058】
「ファージディスプレイ・ライブラリー」は、ファージ被覆タンパク質との融合物としてクローンタンパク質配列コレクションを発現するタンパク質発現ライブラリーである。したがって、フレーズ「ファージディスプレイ・ライブラリー」は、本明細書では、ファージ(例えば、糸状のファージ)のコレクションを意味し、該ファージは外部(通常異種)タンパク質を発現している。この外部タンパク質は、該ファージが接触している他の部分と自由に相互作用(該部分に結合)している。外部タンパク質を示している各ファージは、該ファージディスプレイ・ライブラリーの「メンバー」である。
【0059】
「抗体ファージディスプレイ・ライブラリー」は、抗体または抗体断片を示すファージディスプレイ・ライブラリーを意味している。この抗体ライブラリーは、ファージの集団またはこの種のファージ集団をコードしているベクターのコレクション、またはこの種のファージまたはベクターのコレクションを宿す細胞を含んでいる。このライブラリーは、ファージ粒子につき平均して1つの単一鎖抗体または抗体断片を示す一価、あるいはファージ粒子につき平均して2個以上の抗体または抗体断片を示す多価でよい。用語「抗体断片」は、単一鎖Fv(scFv)断片およびFab断片を無制限に含んでいる。好ましい抗体ライブラリーは、平均して106より多い、または107より多い、または108より多い、または109より多い種類のメンバーを含む。
【0060】
用語「糸状ファージ」は、その表面に不均一なポリペプチドを示し得るウイルス粒子を意味し、f1、fd、Pf1、およびM13を無制限に含んでいる。この糸状ファージは、テトラサイクリン(例えば、「fd−tet」)などの選択可能なマーカーを含んでよい。種々の糸状ファージディスプレイシステムが、当業者には周知である(例えば、Zacher et al.,Gene 9:127−140(1980),Smith et al.,Science 228:1315−1317(1985);and Parmley and Smith,Gene 73:305−318(1988)参照のこと)。
【0061】
用語「パニング」は、標的に対して高い親和性および特異性を有する抗体などのファージ搬送化合物の同定および分離における複数ラウンドの選別プロセスを意味している。
【0062】
本明細書で使われる用語「ヒト以外の動物」には、例えば、ヒト以外の霊長類、げっ歯類(例えば、マウスおよびラット)、ならびに、ウサギ、ブタ、ヒツジ、ヤギ、ウシ、ブタ、ウマおよびロバなどのげっ歯類以外の動物があるが、これらに限定されない。「ヒト以外の動物」には、鳥類(例えば、ニワトリ、七面鳥、アヒル、ガチョウ、など)もある。本明細書で使われる用語「霊長類以外の動物」は、上で具体的に並べた哺乳類を含むがこれらに限定されない霊長類以外の哺乳類を意味している。
【0063】
フレーズ「機能的に異なる抗体」およびこれらの抗体の文法的な改変体は、例えば、標的を中和する能力、生物活性の範囲または特質などの、結合特異性、結合親和性、および何らかの免疫学的または生物学的機能を無制限に含む少なくとも1つの特性において互いに異なる抗体を意味するのに使われる。
【0064】
フレーズ「保存されたアミノ酸残基類」は、互いに整列させられた2つ以上のアミノ酸配列の間で同一であるアミノ酸残基類を意味するのに使われる。
B.一般的技法
本発明の方法を行う技法は、当該技術において周知であり、例えば、Ausubel et al.,Current Protcols of Molecular Biology,John Wiley and Sons(1997);Molecular
Cloning:A Laboratory Manual,Third Edition,J.Sambrook and D.W.Russel,eds.Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001;Antibody Phage
Display:Methods and Protocols,P.M.O’Brian and R.Aitken,eds.,Humana Press,In:Methods in Molecular Biology,Vol.178;Phage Display:A Laboratory Manual,C.F.Barbas III et al.,eds.Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001;and Antibodies,G.Subramanian,ed.,Kluwer Academic,2004;を含む研究室用標準教科書に記載されている。突然変異誘発は、例えば、部位特異的な突然変異誘発を用いて行うことができる(Kunkcl et al.,Proc.Natl.Acad.Sci.USA 82:488−492(1985))。
【0065】
次の説明では、本発明は、特定の型の抗体ライブラリーを参照しながら説明しているが、本発明は、特定の型の抗体ライブラリーの使用に限定されない。組換えモノクローナル抗体ライブラリーは、免疫断片またはナイーブな断片をベースにすることができる。免疫抗体ライブラリーに由来する抗体は、通常、VHおよびVL遺伝子プールを用いて構築され、該プールは、供給源のB細胞から、ランダムにコンビナトリアルなライブラリーを産生するために発現する適切なベクター内にクローン化され、該ライブラリーは、次いで、選択および/または選別される。他の型のライブラリーには、抗原に結合するクローンに対して明らかに偏っていない遺伝子の供給源に由来する抗原断片が含まれている。したがって、ナイーブな抗体ライブラリーは、天然の、免疫されていない、再整理されたV遺伝子から導かれる。合成抗体ライブラリーは、完全なまたは目的に合わせられた縮重のエリアを1つ以上のV遺伝子のCDR内に導入して、インビトロ法により完全に構築される。半合成ライブラリーは、天然と合成の多様性を併用し、所望のレベルの機能的多様性を維持しながら、天然の多様性を増すためにしばしば作り出される。したがって、この種のライブラリーは、例えば、天然のCDR領域を混ぜ合わせる(Soderlind et al.,Nat.Biotechnol.18:852−856(2000))か、または合成CDR1およびCDR2の多様性(Hoet et al.,Nat.Biotechnol.23:455−38(2005))を用いてヒトのB細胞に由来する自然に再整理されたCDR配列を併用することにより作り出すことができる。本発明は、ナイーブ、合成および半合成のライブラリーまたはこれらの併用使用を包含する。
【0066】
同様に、本発明の方法は、抗体のディスプレイに使われる特定の技術により限定されない。本発明は、ファージディスプレイに関連して説明しているが、本発明の抗体は、リボソームまたはmRNAディスプレイ(Mattheakis et al.,Proc.Natl.Acad.Sci.USA 91:9022−9026(1994);Hanes and Pluckthun,Proc.Natl.Acad.Sci.USA 94:4937−4942(1997))、細菌ディスプレイなどの微生物細胞ディスプレイ(Georgiou et al.,Nature Biotech.15:29−34(1997)),または酵母細胞ディスプレイ(Kieke et al.,Protein Eng.10:1303−1310(1997)),レトロウイルスディスプレイなどの哺乳類細胞上のディスプレイ、胞子ディスプレイ、ウイルスディスプレイ(Urban et al.,Nucleic Acids Res.33:e35(2005),タンパク質−DNA結合をベースにしたディスプレイ(Odegrip et al.,Proc.Natl.Acad.Sci.USA 101:2806−2810(2004);Reiersen et al.,Nucleic Acids Res.33:e10(2005)),およびマイクロビーズディスプレイ(Sepp et al.,FEBS Lett.532;455−458(2002))など他のディスプレイおよび濃縮技術によっても同定することができる。
【0067】
リボソームディスプレイでは、該抗体およびコードmRNAはリボソームにより連結され、ディスプレイは、mRNAの翻訳の終端においてポリペプチドを放出することなく停止される。選択は、全体として3元錯体に基づいている。
【0068】
mRNAディスプレイライブラリーでは、抗体と該コードmRNAとの間の共有結合は、アダプター分子として使われる、プロマイシンにより作られる(Wilson et al.,Proc.Natl.Acad.Sci.USA 98:3750−3755(2001))。ディスプレイ抗体にこの技法を使用する場合は、例えば、Lipovsek and Pluckthun,J.Immunol.Methods,290:51−67(2004)を参照のこと。
【0069】
微生物細胞ディスプレイ技法には、サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)などの酵母上の表面ディスプレイがある(Boder and Wittrup,Nat.Biotechnol.15:553−557(1997))。したがって、例えば、抗体は、α−アグルチニン酵母接着受容体に融合することによりサッカロマイセス・セレヴィシエの表面にディスプレイすることができ、該受容体は酵母の細胞壁上に配置される。この方法は、フローサイトメトリーによりレパートリーを選択する可能性を提供する。蛍光標識付きの抗原および抗エピトープ標識試薬を用いて細胞を染色することにより、酵母細胞は、細胞表面上の抗原結合および抗体発現のレベルにより分類することができる。酵母ディスプレイプラットフォームは、ファージと併用することもできる(例えば、Van den Beucken et al.,FEBS Lett.546:288−294(2003))。
【0070】
抗体ライブラリーを選択し、選別する技法の総説については、例えば、Hoogenboom,Nature Biotechnol.23(9):1105−1116(2005)を参照のこと。
C.好ましい実施形態に関する詳細な説明
本発明は、絶滅株の分離株を含むインフルエンザA型サブタイプの1つより多くの株(分離株)を中和するモノクローナル抗体、ならびにH5ヘマグルチニンの存在を特徴とするサブタイプを含む、1つより多くのインフルエンザA型サブタイプに対する中和抗体の選択、産生および使用に関する。特定の実施形態では、本発明は、1つより多くのインフルエンザA型のサブタイプおよび/または1つより多くの分離株、または2つより多い分離株、または3つより多い分離株、または4つより多い分離株、または5つより多い分離株、など、最も好ましくは1つ以上のサブタイプのすべての分離株を中和するモノクローナル抗体の選択、産生および使用に関する。
【0071】
インフルエンザA型ウイルスのビリオンは、線状マイナス一本鎖RNAの8個のセグメントを含んでいる。ゲノム全長は、13600ヌクレオチドであり、該8個のセグメントの長さは、それぞれ、2350ヌクレオチド、2350ヌクレオチド、2250ヌクレオチド、1780ヌクレオチド、1575ヌクレオチド、1420ヌクレオチド、1050ヌクレオチド、900ヌクレオチドである。インフルエンザA型ウイルスの宿主特異性および減弱は、個別にまたはウイルス遺伝子の併用において、ウイルスヘマグルチニン(H,HA)、核タンパク質(NP)、マトリックス(M)、および非構造(NS)遺伝子に起因するとされた(例えば、Rogers et al.,Virology 127:361−373(1983);Scholtissek et al.,Virology 147:287−294(1985);Snyder et al.,J.Clin.Microbiol.24:467−469(1986);Tian et al.,J.Virol.53:771−775(1985);Treanor et al.,Virology 171:1−9(1989)を参照のこと。
【0072】
ヘマグルチニンおよびノイラミニダーゼのタンパク質を含む、インフルエンザA型ウイルスおよびこれらの表面タンパク質のヌクレオチドおよびアミノ酸配列は、GenBankおよび、例えば、the Theoretical Biology and Biophysics Group of Los Alamos National Laboratoryにより管理された、配列データベースthe Influenza Sequence Databaseなど他の配列データベースから入手できる。インフルエンザA型ウイルス・ヘマグルチニン(H1−H15)の15の既知Hサブタイプのアミノ酸配列は、図1(配列番号:1−15)に示している。追加のインフルエンザA型ウイルス・ヘマグルチニンサブタイプ(H16)が、最近、スエーデンでユリカモメから単離され、Fouchier et al.,J.Virol.79(5):2814−22(2005)により報告された。各Hサブタイプには、非常に多様な株があることも知られている。例えば、図1においてH5 A/Hong Kong/156/97と指定されたHAタンパク質の配列は、1997年5月に香港においてヒトから単離されたインフルエンザA H5N1ウイルスから決定され、Suarez et al.,J.Virol.72:6678−6688(1998)において他の関連H5N1分離株から得られたいくつかの追加株の配列と比較して示されている。
【0073】
インフルエンザウイルスであるノイラミニダーゼの触媒および抗原の部位の構造は、Colman et al.,Nature 303:41−4(1983)により公表され、ノイラミニダーゼの配列は、GenBankおよび他の配列データベースから入手できる。
【0074】
感染個体の免疫反応から得られるウイルス特異性抗体は、通常、ウイルスのヘマグルチニンとの相互作用により該ウイルスを中和することが知られている(Ada et al.,Curr.Top.Microbiol.Immunol.128:1−54(1986):Couch et al.,Annu.Rev.Micobiol.37:529−549(1983))。インフルエンザウイルス・ヘマグルチニンの3次元構造およびインフルエンザウイルス・ヘマグルチニンと中和抗体との間の錯体の結晶構造も、測定され、公表された、例えば、Wilson et al.,Nature 289:366−73(1981);Ruigrok et al.,J.Gen.Virol.69(Pt11):2785−95(1988);Wrigley et al.,Virology 131(2):308−14(1983);Daniels et al.,EMBO J.6:1459−1465(1987);and Bizebard et
al.,Nature 376:92−94(2002)を参照のこと。
【0075】
本発明によると、所望の特性を有する抗体は、1つ以上の抗体ライブラリーから同定され、該ライブラリーは、種々の供給源に由来し、種々の型をとることができる。
【0076】
包括的なヒトインフルエンザ抗体ライブラリー
包括的なヒトインフルエンザ抗体ライブラリーは、以前のいろいろなインフルエンザ、季節的突発的流行、および1968年の香港風邪(H3N2)、1957年のアジア風邪(H2N2)、1918年のスペイン風邪(H1N1)、および2004/2005年の鳥インフルエンザ(H5N1)を含むパンデミックの回復期の患者から得られた抗体から作り出すことができる。この種のライブラリーを作るために、インフルエンザウイルスに感染したことが分かっているか、またはその疑いがある個体から血液または骨髄の試料を集める。周辺の血液試料、特に地理学的に離れた供給源からの試料は、輸送、および使用の前に安定化させる必要がある。このためのキットが周知であり、リンパ球の遠心精製の場合、BD Vacutainer(登録商標)CPT(商標)細胞調製チューブが商業的に入手でき、グアニジウム、Trizol、またはRNAlaterを用いて該試料を安定化する。安定化されたリンパ球または全骨髄を受領すると、当該技術分野において周知の免疫グロブリンオリゴプライマーを用いて、重鎖および軽鎖のレパートリーを救出するためにRT−PCRが行われる。PCRレパートリー製品は、当該技術分野で周知の手順に続いて、m13pIIIを用いてフレーム内で直接クローン化するために、リンカーオリゴと併用してscFvライブラリーを作成する。
【0077】
典型的プロトコルでは、ヒト血清内の抗体は、例えば、周知のヘマグルチニン抑制(HAI:hemagglutinin inhibition)アッセイを含む周知の血清学的アッセイにより検出することができる(Kendal,A.P.,M.S.Pereira,and J.J.Skehel.)1982.Concepts and procedures for laboratory−based influenza surveillance.U.S.Department of Health and Human Services,Public Health Service,Centers for Disease Control,Atlanta,Georgia),or the microneutralization assay(Harmon et al.,J.Clin.Microbiol.26:333−337(1988))。この検出ステップは、該血清試料がインフルエンザ中和抗体を含むことがすでに確認されているならば、必要なさそうである。全血からのリンパ球または骨髄に存在するリンパ球は、次に、当該技術分野で周知の方法により処理される。全RNAは、フレッシュな組織またはRNAlater安定化組織からTri BD試薬(Sigma)により抽出する。次いで、単離ドナー全RNAは、Oligotex精製(Quiagen)を用いてさらにmRNAに精製する。次に、第1ストランドcDNA合成は、AccuScript逆転写酵素(Stratagene)のプロトコルに従いランダム九量体オリゴヌクレオチドおよび/またはオリゴ(dT)18プライマーを用いて行う。要するに、AccuscriptのRTバッファー(Stratagene)中で、100ngのmRNA、0.5mMのdNTPおよび300ngのランダム九量体および/または500ngのオリゴ(dT)18プライマーを65℃で5分間インキュベートし、次いで、4℃へ急冷する。次いで、100mMのDTT、Accuscript RT、およびRNAse Blockを、各反応物に添加し、42℃で1時間インキュベートし、70℃で15分間加熱して逆転写酵素を不活性化する。得られたcDNAは、該抗体重鎖および軽鎖のV遺伝子のRT−PCR増幅の鋳型として用いることができ、これらの遺伝子は、次いで、ベクター、またはファージディスプレイ・ライブラリーが意図されるならば、ファージミドベクターにクローン化することができる。この手順は、抗体重鎖および軽鎖の可変領域クローンのレパートリー(VHおよびVLのライブラリー)を作り、このレパートリーは選別するために個別にまたは組み合わせて保持することができる。
【0078】
早期の流行および1918年のスペイン風邪などのパンデミックの生存者の周辺リンパ球からの免疫グロブリンレパートリーは、上で説明した方法と類似した方法で回収され、安定化され、救出することができる。追加のH1およびH3のライブラリーでは、レパートリーは、適切な時間にワクチンを注射した局所調達ドナーから回収することができる。追加の選択肢として、商業的に利用できる骨髄総RNAまたはmRNAは、H1およびH3に適したライブラリー、およびドナーのバックグラウンドによっては、H2抗体選別にも適したライブラリー、を作るために販売元から購入することができる。
【0079】
普遍的抗体ライブラリー(UAL:Universal Antibody Library)−合成ヒト状レパートリー
本発明の方法では、合成ヒト抗体レパートリーは、普遍的抗体ライブラリーにより表すことができ、このライブラリーは、当該技術分野で周知または販売元から得た方法により作ることができる。したがって、この種のライブラリーのサブセットを含む普遍的免疫グロブリンライブラリーは、2003年12月11日に発行された米国特許出願公開第2003−0228302号で説明され、その全体の開示は、引用により明らかに本明細書に組み込まれている。要するに、この特許発行物は、興味深い原型免疫グロブリンのライブラリーについて説明しており、該免疫グロブリンでは、所定の単一アミノ酸が、興味深い免疫グロブリンの1つ以上の相補性決定領域における1つ以上の位置において置換されている。この種のライブラリーのサブセットには、突然変異した免疫グロブリンがあり、該免疫グロブリンでは、所定のアミノ酸が、可能なすべての組み合わせにおいて免疫グロブリンの6つの相補性決定領域における1つ以上の位置において置換されている。この種の突然変異は、例えば、米国特許第5,798,208、5,830,650、6,649,340号、および米国特許出願公開第2003−0194807号に記載されたウォークスルー突然変異誘発により作ることができ、これらの開示内容の全体が引用により本明細書に明らかに組み込まれている。ウォークスルー突然変異誘発では、免疫グロブリンのライブラリーは、該ライブラリーにおいて所定の単一アミノ酸が、該免疫グロブリンの1つ以上の相補性決定領域(CDR:complementarity determining region)またはフレームワーク(FR)領域などの該免疫グロブリンにおいて興味深い、規定された領域、またはいくつかの規定された領域の各位置に少なくとも一度組み込まれる。得られる突然変異免疫グロブリンは、原型免疫グロブリンにおける同じ位置(単数および複数)に存在した「未変性」または「野生型」アミノ酸の代わりに、該免疫グロブリンの1つ以上の領域(例えば、CDRまたはFR領域)内で、これらの免疫グロブリンが1つ以上の位置に組み込まれた所定の単一アミノ酸を有することに、原型免疫グロブリンとの違いがある。突然変異された免疫グロブリンのセットには、興味深い規定された領域の各位置に対して個別に突然変異された免疫グロブリンがあり、したがって、興味深い規定された領域の各位置に対して(例えば、CDRまたはFR)、突然変異された各免疫グロブリンは、原型免疫グロブリンに存在するアミノ酸か、または所定のアミノ酸および可能なすべての改変体を含む突然変異されたすべての免疫グロブリンの混合物を有する。
【0080】
種々の突然変異を有する抗体重鎖および軽鎖の特定のサブライブラリーは、併用すると本発明の抗体のフレームワーク・コンストラクトを得ることができ、このコンストラクトは、次いで、重鎖および軽鎖両方のCDRにおいて多様性を導入する。この多様性は、例えば、Kunkelによる突然変異誘発などの当該技術分野において周知の方法により実現することができ、多様性をさらに増すために数回繰り返すことができる。したがって、重鎖および軽鎖のCDR1およびCD2領域内の多様性は、個別または同時に、複数ラウンドのKunkel突然変異誘発により導入することができる。必要ならば、種々のKunkelクローンは、CDR長さにより分離することができ、および/または標的CDR(例えば、CDR1またはCDR3)において多様性に欠けているクローンは、例えば、鋳型特異性制限酵素を用いた消化により除去することができる。これらのステップが完了すると、ライブラリーのサイズは、約109メンバーを超えるはずであるが、メンバーが比較的少ないライブラリーも有効である。
【0081】
特定の実施形態では、免疫抗体ライブラリーおよび汎用抗体ライブラリーの両方が、本発明の中和抗体の同定に使われる。これら2つの型のライブラリーは、基本的には異なっている。汎用抗体ライブラリーは、タンパク質とペプチドとを結合する予想した能力を有するヒト様抗体の遡及的に合成したコレクションであり、一方、免疫レパートリーは、鳥類H5ヘマグルチニン、および/または場合によっては、H1、H2、またはH3ヘマグルチニンを特異的に認識する配列を含むであろう。したがって、免疫レパートリーは、標的インフルエンザサブタイプ(単数および複数)の必須成分を承認するために理論的に最適化される。これらの差の結果として、これら2つの方法は、抗体の種々のセットを産生し、このようにして、所望の中和抗体を同定するより効率的な方法を提供する。
【0082】
過剰免疫を受けたヒト以外の霊長類の抗体ライブラリー
この方法では、抗体ライブラリーは、マカクまたはヒヒなどの過剰免疫を受けたヒト以外の霊長類から救出される。具体的には、ヒト以外の霊長類は、インフルエンザA型ウイルスの種々のサブタイプまたは種々のヘマグルチニン(H)タンパク質の免疫がある。免疫があるインフルエンザA型ウイルスまたはヘマグルチニンを認識している抗体のタイターを発現している動物は、犠牲にされ、該動物の脾臓を収集した。該免疫動物の血液または骨髄を集め、包括的なインフルエンザ抗体ライブラリーのために、上で説明したように産生された抗体を集め、増幅した。
【0083】
本発明の中和抗体を単離する方法
用いた単数または複数の抗体ライブラリーの型とは無関係に、2種類のインフルエンザAサブタイプおよび/または同じサブタイプの2つの株(分離株)、および/またはヒトおよびヒト以外の分離株と反応性を示すなどの2元特異性を有する抗体は、発見し、制御された交差反応性選択および/または定方向のコンビナトリアルな遺伝子操作および/または突然変異誘発遺伝子操作によって最適化することができる。
【0084】
図2に例示された、典型的な濃縮スキームでは、標的AおよびBとして指定された2つの標的に対して交差反応性を示す抗体を含むライブラリーは、複数ラウンドの濃縮を受ける。濃縮が標的Aとの反応性に基づいているならば、各ラウンドの濃縮は、標的Aに対するプールの反応性強度を高めるであろう。同様に、濃縮が標的Bとの反応性に基づいているならば、各ラウンドの濃縮は、標的Bに対するプールの反応性強度を高めるであろう。図2は、ファージディスプレイ・ライブラリーを選別する場合(下を参照のこと)、使われる選択方法であるパニングを指しているが、この方法は、当該技術分野において周知のもの以外の、上で説明したあらゆる型のライブラリー、およびあらゆる型のディスプレイ技法に同等に適用できる。標的AおよびBは、インフルエンザウイルスの種々の分離株、型およびサブタイプを含むがこれらに限定されない、抗体が結合するあらゆる標的を含んでいる。
【0085】
本発明の目的は、複数の特異性を有する中和抗体を同定することなので、交差反応発見選択スキームが開発された。単純にするために、このスキームは、二重特異性を有する抗体の選択を示す図3で例示する。この場合、2つの標的、すなわち、標的AおよびBとの反応性を示している抗体を含む抗体ライブラリーは、まず、標的の1つ、例えば、標的Aとの反応性について選択され、次いで、他の標的、例えば、標的Bとの反応性について選択される。各連続選択ラウンドが、両方の標的に対して得られるプールの反応性強度を高める。したがって、この方法は、二重特異性を有する抗体を同定する場合に特に有効である。もちろん、この方法は、追加の標的(単数および複数)に対する追加の濃縮ラウンドを含むことにより、さらなる標的に対して反応性を示す抗体の同定に拡大することができる。やはり、選別されたライブラリーがファージディスプレイ・ライブラリーであるならば、選択は、交差反応パニングによりなされるが、他のライブラリーおよび他の選択方法も使用することができる。
【0086】
上で説明した2つの方法の併用には、標的Aおよび標的B、それぞれ、に対する反応性について、2つの別々の濃縮ラウンドがある。すなわち、これら2つのラウンドは、上で説明したように、得られた2つのプールを組み換えるラウンドと、次の交差反応選択ラウンドである。この方法は、図4に例示されている。純粋な交差反応選択におけるように、組換えライブラリーの選択の各ラウンドは、両方の標的に対して得られるプールの反応性強度を高める。
【0087】
図5で例示されたさらなる実施形態では、まず、標的Aと強い反応性を示し、標的Bと検出可能な交差反応性を有するクローンが同定される。このクローンに基づいて、突然変異誘発ライブラリーが作られ、次いで、該ライブラリーは、標的Bおよび標的Aとの反応性に対し、それぞれ、交互ラウンドにおいて選択される。このスキームは、標的Aと強い反応性を維持しながら、標的Bとの反応性を高めた抗体を生じる結果になる。直前に記載されているように、選択はパニングによりなされるが、選別されたライブラリーがファージディスプレイ・ライブラリーであるとしても、他のライブラリー、他のディスプレイ技法、および選択方法も、同じ方法に続いて使用することもできる。
【0088】
上で説明したように、標的AおよびBは、例えば、インフルエンザA型ウイルスの2種類のサブタイプ、同じインフルエンザA型ウイルスの2種類の株(分離株)、2種類の種に由来するサブタイプまたは分離株でよいが、1つの種はヒトであることが好ましい。したがって、例えば、標的Aは、H5N1ウイルスの2004年ベトナム分離株の分離株でよく、標的Bは、H5N1ウイルスの1997年香港の分離株でよい。これらの実施例は、単なる例示であり、2個または3個以上の標的に対して二重および多重特異性を有する抗体は、類似の方法で同定、選択および最適化されうることが強調されている。
【0089】
あるいは、個々のフレームワークおよびCDR長さの分離を可能にするUALなどの抗体ライブラリーが、標的Aに対する抗体の発見に使われるならば、抗原Bは選別され、ライブラリーは類似のパラメータの多様なコレクションに限定されることもありうる。一旦、抗原Bに対する抗体が発見され、次いで、それぞれのAおよびB抗体に基づいてキメラ抗体または突然変異抗体は二重特異性コレクションの遺伝子操作に使われる。
【0090】
ファージディスプレイ
特定の実施形態では、本発明は、多重(二重を含む)特異性を有する中和モノクローナル抗体を機能的に発見するためにファージディスプレイ抗体ライブラリーを利用する。この種の抗体は、H5およびH1,H5およびH2,H5およびH3,H5、H1およびH2,H5、H1およびH3,H5、H2およびH3,H1、H2およびH3などのサブタイプなどの、H5、H7および/またはH9サブタイプを含む、例えば、1つより多くのインフルエンザA型ウイルスサブタイプ、および/または同じサブタイプの1つより多くの株(分離株)を中和し得るモノクローナル抗体でよい。
【0091】
ファージ抗体ライブラリーを作るために、上で説明したライブラリーを含むあらゆる供給源から得られたcDNAライブラリーを、ファージミドベクター内にクローン化する。
【0092】
したがって、例えば、上で説明したように、RT−PCRによりリンパ球または骨髄から救出した抗体重鎖および軽鎖レパートリーのコレクションは、m13 pIIIタンパク質に融合したscFvライブラリーとして再構築する。コンビナトリアルなライブラリーは、約106より多い、または107より多い、または108より多い、または109より多い種々のメンバーを含み、107より多い種々のメンバーまたはそれより上が好ましい。品質管理のために、ランダムクローンを配列し、レパートリー全体の複雑さを評価する。
【0093】
同様に、ナイーブまたは免疫のあるヒト、あるいは過剰免疫のあるヒト以外の霊長類抗体ライブラリーから、重鎖および軽鎖可変領域の早期PCR救出後、PCR産生物をリンカーオリゴと結合し、scFvライブラリーを作りM13 pIII被覆タンパク質を有するフレーム内で直接クローン化する。このライブラリーは、約106より多い、または107より多い、または108より多い、または109より多い種々のメンバーを含み、107より多い種々のメンバーまたはそれより上が好ましい。品質管理のステップとして、レパートリー全体のサイズおよび複雑さを評価するために、ランダムクローンの配列を決める。
【0094】
抗体ファージディスプレイ・ライブラリーは、単一鎖Fv(scFv)またはFabフォーマットなどの種々のフォーマットにおいて抗体を含むことがある。総説については、例えば、Hoogenboom,Methods Mol.Biol.178:1−37(2002)を参照のこと。
【0095】
選別
所望の中和特性を有する抗体を選別する方法は、上で説明した。反応性は、所望のヘマグルチニンタンパク質への直接結合に基づいて評価することができる。
【0096】
ヘマグルチニン(HA)タンパク質の産生
ヘマグルチニン(HA)タンパク質は、組換えDNA技術により産生することができる。この方法では、HA遺伝子は、適切なベクター、ヨトウガ(Sf9)細胞などのバキュロウイルス感染昆虫細胞において発現するために、バキュロウイルス発現ベクターにクローン化されるのが好ましい。
【0097】
HAタンパク質をコードする核酸は、ポリ−his(ヘキサヒスチジン標識)などの
C−末端エピトープ標識がある場合とない場合がある、Bac−to−Bac(Invitrogen)などのバキュロウイルス発現ベクター内に挿入される。ポリ−his標識は、ニッケルキレート・クロマトグラフィによる簡易精製を提供する。
【0098】
一般に、クローン化は、個々に合成されたオリゴからアセンブリーPCRによる基準cDNAの作製を含む。分離株に対応する改変体HAタンパク質は、適切な突然変異体オリゴを追加のアセンブリーPCR内に置換するか、またはKunkel突然変異誘発による突然変異誘発技法により作られる。HAタンパク質配列の2つのクラスターは、H5については、1997年および2004年のサブタイプ分離株が存在する。したがって、単一基準タンパク質は、各クラスターについて作られる。同様に、基準タンパク質は、1918年のスペイン風邪(H1)、1958年のアジア風邪(H2)、1968年の香港風邪(H3)、および現在のH1、H2、H3分離株について作られる。
【0099】
組換えバキュロウイルスは、リポフェクチン(Gibco−BRLから市販されている)を用いて上のBacmidをSf9細胞(ATCC CRL 1711)に形質移入することにより作られる。28℃で4〜5日インキュベート後、放出されたウイルスを取得し、さらなる増幅に使われる。ウイルス感染およびタンパク質発現は、O’Reilley et al.,Baculovirus Expression Vectors:A Laboratory Manual(Oxford:Oxford University Press,1994)により説明された通りになされる。
【0100】
次いで、発現されたポリ−His−標識付きHAポリペプチドは、例えば、次に説明するように、Ni2+キレート親和性クロマトグラフィにより精製することができる。上澄みは、Rupert et al.Nature 362:175−179(1993)が説明した、組換えウイルス感染Sf9細胞から集める。Ni2+−NTAアガロースカラム(Quiagenから市販されている)は、ベッド容積5mLにて用意し、25mLの水で洗浄し、25mLのローディングバッファーを用いて平衡化する。ろ過した細胞抽出物は、0.5mL/分にてカラムに入れる。カラムは、二次洗浄バッファーを用いてベースラインA280まで洗浄し、その時点でフラクションの収集を開始する。次に、このカラムを二次洗浄バッファー(50mMホスフェート、300mMのNaCl、10%グリセロール、pH6.0)を用いて洗浄し、非特異的に結合したタンパク質を溶離する。A280ベースラインに再度到達後、このカラムは、二次洗浄バッファー中の0〜500mMのイミダゾールグラジエントを用いて展開する。1mLの各フラクションを集め、SDS−PAGEおよび銀染色またはアルカリ性ホスファターゼに結合したNi2+−NTA(Qiagen)を用いたウエスタン・ブロットにより分析した。溶離したHis10−標識付きHAポリペプチドを含むフラクションは、プールし、ローディングバッファーに対して透析する。
【0101】
あるいは、IgG標識付き(またはFc−標識付き)HAポリペプチドの精製は、例えば、Protein Aまたはprotein Gカラムクロマトグラフィを含む既知のクロマトグラフィ技法を用いて行うことができる。
【0102】
Sf9細胞HAタンパク質を用いる代替として、他の組換え宿主細胞中に、原核生物、酵母、または比較的高い真核生物細胞を産生することもできる。適切な原核生物には、グラム陰性菌またはグラム陽性菌などの真正細菌(eubacteria)、例えば、エシェリキア属(Escherichia)、例えば、大腸菌(E.coli)などの腸内細菌科(Enterobacteriaceae)、エンテロバクター属(Enterobacter)、エルビニア菌属(Erwinia)、クレブシエラ属(Klebsiella)、プロテウス属(Proteus)、サルモネラ属(Salmonella)、例えば、ネズミチフス菌(Salmonella typhimurium)、セラチア属(Serratia)、例えば、セラチア・マルセッセンス(Serratia marcescans)、および赤痢菌(Shigella)、ならびに枯草菌(B.subtilis)およびリケニホルミス菌(B.licheniformis)(例えば、1989年4月12日に発行されたDD266,710で開示されたリケニホルミス菌41P)などのバシラス属(Bacilli)、緑膿菌(P.aeruginosa)などのシュードモナス属(Pseudomonas)およびストレプトマイセス属(Streptmyces)があるが、これらに限定されない。大腸菌K12株MM294(ATCC31,446)、大腸菌X1776(ATCC31,537)、大腸菌株W3110(ATCC27,325)、およびK5772(ATCC53,635)などの種々の大腸菌株が、公然と利用できる。
【0103】
原核生物に加えて、糸状菌または酵母菌などの真核微生物が、HAポリペプチドをコードする核酸を含むベクターの適切なクローニングまたは発現宿主である。サッカロマイセス・セレヴィシエは、普通に使われる比較的低級の真核宿主微生物である。しかし、多数の他の属、種、および株が、通常、利用でき、本明細書では、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)(Beach and Nurse,Nature 290:140(1981);1985年5月2日に発行された欧州特許第139,383号);例えば、K.ラクティス(Kluyveromyces lactis)(MW98−8C,CBS683,CBS4574;Louvencourt et al.,J.Bacteriol.737(1983)),K.フラジリス(fragilis)(ATCC12,424),K.ブルガリカス(bulgaricus)(ATCC16,045),K.ウイッケラミイ(wickeramii)(ATCC24,178),K.ワルティー(waltii)(ATCC56,500),K.ドロソフィララム(drosophilaraum)(ATCC36,906;Van den Berg et al.,Bio/Technology8:135(1990))、K.サーモトレランス(thermotolerans)、およびK.マルキシアヌス(marxianus)などのクリベロマイセス宿主(Kluyveromyces host)(米国特許第4,943,529号;Fleer et al.,Bio/Technology9:968−975);ヤロウイア属(yarrowia)(欧州特許第402,226号);ピキア・パストリス(Pichia pastoris)(欧州特許第183,070号;Sreekrishna et al.,J.Basic Microbiol.28:265−278(1988));カンジダ属(Candida);トリコデルマ・レージア(Trichoderma reesia(欧州特許第244,234号);アスバンカビ(Neurospora crassa)(Case et al.,Proc.Natl.Acad.Sci.USA 76:5259−5263(1979));シュワニオマイセス・オクシデンタルス(Schwanniomyces occidentalis)(1990年10月31日に発行された欧州特許第394,538号)などのシュワニオマイセス(Schwanniomyces);およびアカバンカビ(Neurospora)、ペニシリウム属(Penicillium)、トリポクラジウム(Tolypocladium)(1991年1月10日に発行された国際公開第91/00357号)などの糸状菌、および偽巣コウジ菌(A.nidulans(Ballance et al.,Biochem.Biophys.Res.Commun.112:284−289(1983);Tilburn et al.,Gene26:205−221(1983);Yelton et al.,Proc.Natl.Acad.Sci.USA 81:1470−1474(1984))およびA.ニガー(niger)Kelly and Hynes,EMBO J.4:475−479(1985)などのアスペルギルス属宿主(Aspergillus)なども本明細書で有効である。メチルトローフ酵母は、本明細書に適しており、ハンゼヌラ(Hansenula)、カンジダ、クロエケラ属(kloeckera)、ピキア属、酵母菌属(Saccharomyces)、トルロプシス属(Torulopsis)、およびロドトルラ属(Rhodotorula)からなる属から選択されたメタノール上で増殖し得る酵母を含むが、これらに限定されない。このクラスの酵母を代表する特異的な種のリストは、C.Anthony,The Biochemistry of Methylotrophs 269(1982)に存在しているであろう。
【0104】
HAタンパク質を発現させる適切な宿主細胞には、多細胞微生物の細胞がある。無脊椎動物の例には、上述のショウジョウバエ(Drosophila)S2およびプテラ属(Spodoptera)Sf9、ならびに植物細胞がある。有効な哺乳類の宿主細胞株の例には、中国ハムスターの卵巣(CHO:Chinese hamster ovary)およびCOS細胞がある。より特異的な例には、SV40(COS−7,ATCC CRL1651)により形質転換されたサルの腎臓CV1株;ヒトの胚腎臓株(HEK293または懸濁培養において増殖するためにサブクローン化されたHEK293細胞(Graham et al.,J.Gen.Virol.36:59(1977));中国ハムスターの卵巣細胞/−DHFR(CHO,Urlaub and Chasin,Proc.Natl.Acad.Sci.USA 77:4216(1980));マウスのセルトリ細胞(TM4,Mather,Biol.Reprod.23:243−251(1980));ヒトの肺細胞(W138,ATCC CCL75);ヒトの肝細胞(Hep G2,HB8065);およびマウス乳腺腫瘍(MMT060562,ATCC CCL51)がある。適切な宿主細胞の選択は、当該技術分野のスキル内にあると考えられている。
【0105】
ヘマグルチニン(HA)タンパク質パニング
HAタンパク質は、上で説明したライブラリーを選別するために、マイクロタイターウエルまたは磁気ビードの表面に固定する。特定の実施形態では、各ライブラリーは、4度で2時間H5タンパク質を結合することができ、次いで、冷PBSを用いて十分洗浄し、0.2Mグリシン−HClバッファー(pH2.5)を用いてHA特異結合クローンを溶離する。回収されたファージは、pHは中性で、感受性宿主大腸菌を感染させることにより増幅される。次いで、陽性クローンの濃縮および死傷者分類のためのその後のクローン単離を繰り返すために、ファージミド産生を誘発することができる。十分濃縮されると、プール全体は、可溶性scFvタンパク質を発現するためにHB2151などの非アンバー抑制大腸菌株内に感染により移される。一方、プール(単数および複数)は、pBADなどのモノマー性scFv発現ベクター内にサブクローン化され、組換え可溶性scFvタンパク質は、下で説明するように、インビトロ分析およびキャラクテリゼーションのために発現される。
【0106】
キャラクテリゼーション
H5クローンは、まず、上で説明した、産生されたH5タンパク質への結合親和性についてテストする。特定の実施例では、2004H5タンパク質(Refseq AAS65618,Isolate;A/Thailand/2(SP−33)/2004(H5N1))への結合がテストされ、および1997H5タンパク質(Refseq AAF74331,Isolate;A/Hong Kong/486/97(H5N1))に対する並行テストがなされ、他の分離株も単独または何らかの組み合わせで使用することができる。2004および1997H5タンパク質を用いて得られた陽性クローンは、2つの広義のカテゴリー;2004selectiveおよび2004/1997nonselectiveに入る。中和に関する典型的な機能テストは、赤血球に対する全ウイルス結合を用いたヘマグルチニン抑制アッセイを含む。安全面の懸念により、組換えタンパク質および赤血球を用いた、代わりのヘマグルチニンアッセイが好ましい。全血の必要性を排除するために、ヘマグルチニン結合抑制アッセイは気道上皮細胞について行うことができる。この結合アッセイは、フローサイトメトリーまたは細胞ELISA(cELISA)をベースにしたアッセイを含む、無制限に、何らかの形態で行うことができる。cELISAを用いると、高価なフローサイトメトリー装置を使用しなくてすみ、より高度に自動化されたクローン・アセスメントおよびより大きなデータコレクションが得られる利点がある。一方、フローサイトメトリーには、感度が高く、一貫性に優れ、高速である利点もある。
【0107】
HIクローンは、現在の2004H1への結合を含む、あらゆるH1タンパク質への結合、および並行して、1918年および1976年のタンパク質への結合についてテストすることができる。陽性クローンは、2つの広義のカテゴリー;2004selectiveおよび2004nonselectiveに入るであろう。やはり、上で説明した方法と類似な方法を用いて中和についてテストすることは必須である。
【0108】
H2およびH3などの他のHAタンパク質は、類似の方法で特性を明らかにすることができる。
【0109】
最適化
鳥類ウイルス(H5)により引き起こされたヒトの感染と関連した潜在的パンデミックを含むインフルエンザの流行およびパンデミックに効率的に対応するために、H5タンパク質、ならびに将来の突然変異など、Hタンパク質の現在の分離株を効率的に中和する抗体が必要である。この目標を達成するために、標的となるヘマグルチニンサブタイプ(単数および複数)の既知のすべての分離株に結合する、様々なH(例えば、H5)中和クローンを同定する必要がある。
【0110】
望ましいならば、例えば、2005年6月23日に公開された、米国特許出願公開第2005−0136428号において説明された、ルックスルー・ムタジェネシス(LTM:Look Through Mutagenesis)など、当該技術分野において周知の方法により、交差反応性をさらに改良することができる。なお、この特許発行物は、引用により明らかに本明細書に組み込まれている。
【0111】
ルックスルー・ムタジェネシス(LTM)は、選択されたアミノ酸のコンビナトリアルな突然変異を同時に評価および最適化する多次元突然変異法である。このプロセスは、1つ以上の相補性決定領域(CDR)ドメイン内の正確な分布に焦点を当て、アミノ酸側鎖に関する化学の相乗的寄与を探っている。LTMは、CDR内の単一変異の位置シリーズを作り、そこでは各野生型残基が、選択された多数のアミノ酸の1つにより系統的に置換される。突然変異CDRは併用され、すべての改変体の定量的ディスプレイに対して禁止的になることなく複雑さおよびサイズを増しながらコンビナトリアルな単一鎖可変断片(scFv)ライブラリーを作る。正の選択後、改善された特性を有するクローンの配列が決定され、これらの有益な突然変異が位置づけられる。改善されたHA結合特性の相乗的突然変異を同定するために、すべての有益な順列を発現しているコンビナトリアルなライブラリー(コンビナトリアルで有益な突然変異、CBM:combinatorial beneficial mutation)は、最適化されたscFv候補のパネルを同定するために正に選択され、分析された混合DNAプローブにより産生することができる。この操作は、Fvおよび他の抗体ライブラリーを用いて類似の方法で行うことができる。
【0112】
突然変異誘発は、上で説明したウォークスルー・ムタジェネシス(WTM:walk through mutagenesis)により行うこともできる。
【0113】
1つより多くのA型インフルエンザサブタイプおよび/または1つより多くの同じサブタイプの分離株を用いて、本明細書の抗体の交差反応性を意図的にデザインするための別の有効な突然変異誘発法は、本明細書では目標突然変異誘発と呼ばれる。目標突然変異誘発は、1つ以上の好ましくは反応性の異なる抗体クローンに基づく抗体のコレクションを合理的に遺伝子操作するために使うことができる。本発明の文脈において、目標突然変異誘発は、抗体の個々のCDRにおける配列など同様の配列上の類似の位置により規定された単一または複数の残基をコードするのに使われる。この場合、これらのコレクションは、同等の位置に存在した残基の範囲を捕獲するためにオリゴ縮重を用いて作られる。このコレクション内には特異性の連続体が、親クローンのコレクションの間またはコレクションを超えて存在するであろうと期待される。目標突然変異誘発の目的は、2つ以上の分離実体またはコレクションの間に多様な多機能性抗体コレクションまたはライブラリーを作ることである。インフルエンザの場合、この方法は、2つの異なるエピトープ、分離株、またはサブタイプを認識する2つの抗体を使用し、両方の機能性品質を単一抗体に変えるために利用することができる。1例として、第1のインフルエンザA型抗体は、H5サブタイプのベトナム分離株に対して特異的であり、第2の抗体は、インフルエンザA型ウイルスのH5サブタイプのタイランドまたはトルコ分離株に対して特異的である。目標突然変異誘発ライブラリーを作成するために、両方の抗体のCDR配列がまず達成され、整列させられる。次に、保存されたアイデンティティのすべての位置は、適合残基に対する単一コドンを用いて固定される。非保存位置では、縮重コドンは、両方の残基をコードするために組み込まれる。一部の例では、縮重コドンは、この位置において2つの親残基をコードするのみである。しかし、一部の例では、追加の副産物が産生される。副産物産生のレベルは、副産物の産生を強制するか、またはサイズの限界または目標に依存するこの産生を排除するためにダイヤルを回すことができる。
【0114】
したがって、例えば、これら2つの抗体の最初の位置が、それぞれ、スレオニンおよびアラニンであるならば、最初の2つの位置においてA/G−G−を有する縮重コドンは、第3の位置の塩基とは関係なく、スレオニンまたはアラニンをコードするだけである。例えば、もし次の位置の残基がリジンおよびアルギニンであるならば、縮重コドンA−A/G−A/Gは、リジンまたはアルギニンをコードするだけであろう。しかし、縮重コドンA/C−A/G−A/G/C/Tが使われたならば、その場合は、アスパラギン、ヒスチジン、グルタミン、およびその上、セリン副産物が作られるであろう。
【0115】
便宜上、適合したCDR長さを有する抗体のみを用いると、より単純である。これを強制する1つの方法は、CDR長さおよび最初に発見された抗体により与えられた潜在的に等しいフレームワーク制限に基づいて第2抗原のサイズ制限ライブラリーを選別することである。しかし、等しい長さのCDRを用いることは、単なる便宜であり必要条件ではないことを指摘する必要がある。この方法は、インフルエンザA型ウイルス中和抗体の機能的に多様な大きなライブラリーを作成するためには有効であるが、その適用性が非常に広いことは、簡単に分かる。この突然変異誘発技法は、何らかの抗体の機能的に多様なライブラリーまたはコレクションを作るために使用することができる。したがって、図6は、突然変異誘発された親配列としてTNF−α抗体およびCD11a抗体のCDRを用いた目標変異誘発方法の使用を例示するために本明細書に含まれている。
【0116】
他の代表的な突然変異誘発法には、飽和突然変異誘発および誤りがちなPCRがある。
【0117】
飽和突然変異誘発(Hayashi et al.,Biotechniques 17:310−315(1994))は、全20のアミノ酸類が、タンパク質内の特定の位置で置換され、各改変体に対応するクローンが特定の表現についてアッセイされる技法である。(米国特許第6,171,820;6,358,709および6,361,974号も参照のこと。)
誤りがちなPCR(Leung et al.,Technique 1:11−15(1989);Cadwell and Joyce,PCR Method Applic.2:28−33(1992))は、クローン化遺伝子内にランダムポイント突然変異を導入する改変ポリメラーゼ連鎖反応(PCR:polymerase chain reaction)技法である。得られるPCR産生物は、クローン化してランダム突然変異体ライブラリーを作るか、またはT7プロモーターが適切なPCRプライマー内に組み込まれるならば、直接転写することができる。
【0118】
他の突然変異誘発技法も周知の技法であり、例えば、In Vitro Mutagenesis Protocols,J.Braman,Ed.,Humana Press,2001で説明されている。
【0119】
この場合、主な目標の1つは、現在のH5(またはH7またはH9)分離株ならびに将来の突然変異を効率的に処置するために抗体(単数および複数)を遺伝子操作することである。例えば、最近の2004年分離株および以前の1997年分離株の両方を含む、種々のH5分離株に結合するクローンを中和する新しい分離株H5における突然変異を承認し得る耐性を有する抗体を遺伝子操作することは、確認すべきである。クローンが2004年分離株について選択されるならば、そのクローンは、それほどの重要性はないが、1997年分離株を結合および/または中和するであろうことが期待される。この場合、目標は、2004年分離株結合を改善する(または少なくとも維持する)文脈内で1997年承認を劇的に改善することである。したがって、選択はまず1997年基準タンパク質について改善がなされ、次いで、2004年タンパク質に関する選択がこれに続く。そのようにすると、第2パラメータに対する圧力を維持しながら、新しい株に対する選択圧力をより大きなものにする。
【0120】
上で説明したライブラリーのいずれか、または当該技術分野で周知の何か他の型のライブラリーを、単独またはいずれかの組み合わせに基づいて最適化することができる。特定の実施形態において、最適化は、3つの型のLTMライブラリー、すなわち、3種類の突然変異誘発軽鎖ライブラリー、3種類の突然変異誘発重鎖ライブラリー、および6種類の突然変異誘発(軽鎖+重鎖)ライブラリー、を選別することにより開始することができる。H5は、小さな改変は望ましいかもしれないが、上で説明したように、本質的には、選別される。例えば、グリシン−HCl溶離の前に、次の方法:室温または37度における十分な洗浄、または過剰の可溶性親scFvの存在下における長時間のインキュベーション、のいずれかまたは両方によりラウンドごとに洗浄の厳しさを高めることにより改善された結合を選択することができる。これらの選択改変は、得られるクローンにおけるオフ率の動態を改善するはずである。3〜4ラウンドの選択後、我々はランダムクローンの配列を決め、ELISAにより結合についてテストすることになる。改善されたクローンの配列分析後、許容できる改善された突然変異のすべては、H5分離株の両方の結合を相乗的に改善する場合に選択するコンビナトリアルな有益突然変異誘発(CBM:combinatorial beneficial mutagenesis)ライブラリーに一体化される。CBMライブラリーは、すべての位置において改善された、オリジナルな親の残基をすべて表すために縮重オリゴヌクレオチド類を合成することにより作られる。得られるライブラリーは、LTM選別と同様に、厳しさを高めながら選択される。十分な選択後、上で説明した結合および中和のアッセイの大腸菌からのモノマー性scFvタンパク質を発現し、精製するために、プールはpBAD発現ベクター内にサブクローン化される。
【0121】
H1中和抗体は、類似な方法で最適化することができる。この場合、1918年、1976年、および出発点または目標としての現在からの何らかの基準タンパク質配列を用いて選択し、最適化することができる。
【0122】
さらに、型間の承認は、中和抗体クローンを用いてテストする。型間の承認の1例は、H5から供給または最適化されたクローンからの同時発生的または遺伝子操作したH1結合である。
【0123】
所望の特性を有する中和抗体が、一旦、同定されると、この種の抗体の大部分により承認された優勢なエピトープ(単数および複数)を同定することが望ましい。エピトープをマッピングする方法は、当該技術分野で周知であり、例えば、Morris,Glenn E.,Epitope Mapping Protocols,Totowa,N.J.ed.,Humana Press,1996;and Epitope Mapping:A Practical Approach,Westwood and Hay,eds.,Oxford University Press,2001において開示されている。
【0124】
種々のドナーからのライブラリーなどの、またはインフルエンザウイルスを含むが、これらに限定されない抗体ライブラリーのハンドリングは、種々の抗体コレクションを区別している独特のバーコードを適用することにより大いに促進される。これらのバーコードは、これらが標識付きのクローン(単数および複数)と同調して伝播し得るように選択されるのが好ましい。
【0125】
したがって、これらのバーコードは、配列決定または特定のPCRプライマーによりデコンボリュートすることができる長さが約1〜24個の非コードヌクレオチド類に等しい非コード配列でよい。このように、抗体レパートリーなどの核酸のコレクションは、クローン化ステップにおいて連結することができる。
【0126】
別の実施例では、これらのバーコードは、サイレント突然変異のコード配列である。これらのライブラリーが、中断回分配列(例えば、Sfi GGCCNNNNNGGCC)を認識する制限酵素を利用するならば、抗体ライブラリーなどのクローンの種々のコレクションを区別するために「N」の代わりに別のヌクレオチド類を組み込むことができる。このバーコード法は、このレパートリーが増幅ステップにおいて連結される利点がある。
【0127】
別の例では、これらのバーコードは、免疫学的に異なるペプチドまたはファージ粒子に融合したタンパク質配列コードするコード配列である。実施例には、例えば、pIII、pVIII、pVII、またはpIXファージへのエピトープ(例えば、Myc、HA、FLAG)融合物がある。これらのエピトープは、単独、または種々の組み合わせで使用することができる、シス(ライブラリーコードプラスミドで)配置として、またはトランス(特異的に改変されたヘルパーファージ)配置として提供することができる。
【0128】
可能なバーコード類の他の実施例には、無制限に、ハプテン類または蛍光染色体を有する化学的および酵素ファージ改変がある。この種の標識は、選択の単一ラウンドの場合好ましい。
【0129】
本明細書では、バーコードは、抗体ライブラリーを区別するものとして例示されるが、当業者は、説明された方法が、独自に標識する場合、および一般に、核酸分子および核酸のコレクションを区別する場合に広く適用できることを十分理解するであろう。
【0130】
中和抗体の産生
所望の中和特性を有する抗体が、一旦、同定されると、抗体断片を含むこの種の抗体は、例えば、ハイブリドーマ技法または組換えDNA技術を含む当該技術分野で周知の方法により産生することができる。
【0131】
ハイブリドーマ法では、マウスまたはハムスターなど他の適切な宿主動物が、免疫付与に使われるタンパク質に特異的に結合する抗体を産生または産生し得るリンパ球を誘発するために免疫される。一方、リンパ球は、インビトロで免疫にされる。次いで、リンパ球は、ポリエチレングリコールなどの適切な融合剤を用いて骨髄腫細胞と融合し、ハイブリドーマ細胞を形成する(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。
【0132】
したがって、このようにして調製されたハイブリドーマ細胞は、適切な培養媒体に播種され、増殖され、該媒体は、未融合の親の骨髄腫細胞の増殖または生存を抑制する1つ以上の物質を含むのが好ましい。例えば、親の骨髄腫細胞に構造ヒボキサンチングアニンホスホリボシル転換酵素(HGPRT:hypoxathine guanine phosphoribosyl transferaseまたはHPRT)が欠けているならば、該ハイブリドーマの培養媒体は、通常、ヒボキサンチン、アミノプテリン、およびチミジン(HAT:hypoxanthine,aminoprotein,thymidine媒体)を含み、これらの物質は、HGPRTに欠けている細胞の増殖を防止する。
【0133】
好ましい骨髄腫細胞は、効率的に融合し、選択された抗体産生細胞により抗体の安定した高レベル産生を支え、HAT媒体などの媒体に敏感な骨髄腫細胞である。これらの中で、好ましい骨髄腫細胞株は、Salk Institute Cell Distribution Center,San Diego,California USAから入手できるMOPC−21およびMPC−11マウス腫瘍から導かれた細胞株などのマウス骨髄腫株およびAmerican Type Culture Collection,Rockville,Maryland USAから入手できるSP−2またはX−63−Ag8−653細胞などのマウス骨髄腫株である。ヒトの骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞株も、ヒトのモノクローナル抗体の産生に対して説明されている(Kozbor,J.Immunol.133:3001(1984);and Brodeur
et al.,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0134】
その中でハイブリドーマ細胞を増殖する培養媒体は、抗原に対抗するモノクローナル抗体の産生についてアッセイされる。ハイブリドーマ細胞により産生されたモノクローナル抗体の結合特異性は、免疫沈降または放射免疫アッセイ(RIA:radioimmunoassay)または酵素免疫測定吸着法(ELISA:enzyme−linked immunoabsorbent assay)などのインビトロ結合アッセイにより測定されるのが好ましい。
【0135】
組換えモノクローナル抗体は、例えば、必要な抗体鎖をコードするDNAを単離し、周知の組換え発現ベクターを用いた同時発現のコード発現を用いた組換え宿主細胞を同時導入することにより産生することができる。組換え宿主細胞は、上で説明した原核細胞および真核細胞などでよい。
【0136】
ヒト化抗体を作る場合に使われるヒト可変ドメインの選択は、軽重両方ともに、抗原性を下げることが非常に重要である。いわゆる「最適」方法により、げっ歯類抗体の可変ドメインの配列は、既知ヒト可変ドメイン配列の全ライブラリーに対して選別される。次いで、げっ歯類の配列に最も近いヒト配列が、ヒト化抗体のヒトフレームワーク領域(FR)として受け入れられる(Sims et al.,J.Immunol.151:2296(1993);Chopita et al.,J.Mol.Biol.196:901(1987))。抗体が、抗原および他の好ましい生物学的特性に対して高い親和性を保持したまま、ヒト化されることが重要である。好ましい方法によりこの目標を達成するために、ヒト化された抗体が、親の配列およびヒト化された配列の3次元モデルを用いて種々の概念的なヒト化された産生物および親の配列を分析するプロセスにより作られる。
【0137】
さらに、ヒトの抗体は、当該技術分野で周知の次のような方法で作ることができる。例えば、内在性免疫グロブリンの産生がない場合にヒトの抗体のレパートリーの全体を、免疫付与の際に、産生し得る遺伝子導入動物(例えば、マウス)を作ることができる。例えば、Jakobovits et al.,Proc.Natl.Acad.Sci.USA 90:2551(1993);Jakobovits et al.,Nature 362:255−258(1993);Bruggermann et al.,Year in Immuno,7:33(1993);および米国特許第5,591,669、5,589,369および5,545,807号を参照のこと。
【0138】
中和抗体の使用
本発明のインフルエンザ中和抗体は、A型インフルエンザ感染の予防および/または処置に使用することができる。治療に使う場合は、抗体または分子類は、それらの送達が、抗体(単数および複数)をベースにする輸送配列を用いることにより容易になり、通常、医薬組成物の形で使われる。技法および処方は、一般に、Remington’s Pharmaceutical Sciences,18th Edition, Mack
Publishing Co.(Easton,Pa.1990)に記載されている。また、Wang and Hanson”Parenteral Formulations of Proteins and Peptides;Stability and Stabilizers,”Journal of Parenteral Science and Technology,Technical Report No.10,Supp.42−2S(1988)も参照のこと。
【0139】
抗体は、通常、凍結乾燥された製剤または水溶液の形で処方される。許容できる担体、賦形剤、または安定剤は、用いる用量および濃度において受容者に対して無毒であり、燐酸塩、クエン酸塩、および他の有機酸などのバッファー、アスコルビン酸およびメチオニンを含む抗酸化剤、保存剤(オクタデシルジメチルベンジルアンモニウムクロライド、ヘキサメトニウムクロライド、ベンザルコニウムクロライド、ベンゼトニウムクロライド、フェノール、ブチルアルコールまたはベンジルアルコール、メチルパラベンまたはプロピルパラベンなどのパラベン類、カテコール、レゾルシノール、シクロヘキサノール、3−ペンタノール、およびm−クレゾールなど)、低分子量(残基約10個未満)ポリペプチド類;血清アルブミン、ゼラチン、または免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、またはリジンなどのアミノ酸類;単糖類、二糖類、およびグルコース、マノース、またはデキストリン類を含む他の炭水化物;EDTAなどのキレート化剤;スクロース、マニトール、トレハロースまたはソルビトールなどの砂糖類;ナトリウムなどの塩形成対イオン類;金属錯体類(Zn−タンパク質錯体);および/またはTWEEN(商標)、PLURONICS(商標)またはポリエチレングリコール(PEG)を含む。
【0140】
これらの抗体は、例えば、コロイド状薬剤デリバリーシステム(例えば、リポソーム、アルブミンマイクロスフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)、またはマクロエマルジョンにおいて、液滴形成技法または界面重合(例えば、ヒドロキシメチルセルロースまたはゼラチンマイクロカプセルおよびポリ−(メチルメタクリレート)マイクロカプセル、それぞれ)により調製されたマイクロカプセルに捕捉されることもありうる。この種の技法は、上記、Remington’s Pharmaceutical Sciencesで開示されている。
【0141】
本明細書で開示された中和抗体は、免疫リポソーム類として処方することもできる。抗体を含むリポソーム類は、Epstein et al.,Proc.Natl.Acad.Sci.USA 82:3688(1985);Hwang et al.,Proc.Natl.Acad.Sci.USA 77:4030(1980);米国特許第4,485,045および4,544,545号;および1997年10月23日に発行された国際公開第97/38731号において説明されているような当該技術分野で周知の方法により調製される。循環時間の長いリポソーム類は、米国特許第5,013,556号で開示されている。
【0142】
特に有効なリポソーム類は、ホスファチジルコリン、コレステロールおよびPEG−誘導体化されたホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物を用いて逆相蒸発法により作ることができる。リポソーム類は、規定されたポアサイズのフィルターを通して押し出され所望の直径を有するリポソーム類が得られる。本発明の抗体のFab’断片は、ジサルファイド交換反応によりMartin et al.,J.Biol.Chem.257:286−288(1982)で説明されたようにリポソーム類に接合することができる。化学療法剤は、任意に、リポソーム内に含まれる。Gabizon et al.,J.National Cancer Inst.81(19)1484(1989)を参照のこと。
【0143】
疾患の予防または処置のために、抗体の適切な用量は、処置されるべき感染の型、疾患の重篤さおよびコース、および抗体を投与する目的が予防であるか、治療であるか、により変わる。抗体は、一度または処置シリーズにわたり、患者に適切に投与される。疾患の型および重篤さにより、抗体の患者への初期用量は、約1μg/kgから約15mg/kgまでの範囲にあり、1回以上個別に投与する場合と、連続注入する場合とがある。
【0144】
本発明のより詳細な説明は、次の無制限にある実施例によりなされる。
【実施例】
【0145】
以前の鳥類インフルエンザ大流行の生存者からの抗体ライブラリーおよび中和抗体の調製
物質および方法
骨髄プロトコルおよび血清製剤
血液は、標準静脈穿刺により採血し、凝血させ、処理して血清を回収した。血清は、ドライアイスにのせて出荷するまで、3〜4日間−20℃で貯蔵した。ドナーは、局所麻酔薬を注入して麻酔にかけ、5mlの骨髄を各H5N1生存者の骨盤から採取した。次に、5mlの骨髄を45mlのRNAlater(Ambion)を含む無菌の50mlのチューブに入れた。この混合物を、目視できる凝集塊がなくなるまで、約8〜20回丁寧に反転させ、骨髄とRNAlaterとをよく混合させた。次に、この試料を2〜10℃で一晩冷蔵した。一晩冷蔵後、試料は、ドライアイスにのせて出荷するまで、3〜4日間−20℃で貯蔵した。受け取り次第直ぐに、RNAlater/髄および血清含有チューブは、処理するまで−80℃で貯蔵した。
【0146】
血清学:HA ELISA
ELISAプレート(Thermo,Immulon 4HBX 96W)は、1X ELISAプレートコーティング溶液(BioFX)中で100μlの100ng/mLのH5ヘマグルチニン(Protein Sciences,A/Vietnam/1203/2004)で被覆し、室温で一晩インキュベーションした。次の日複数のプレートを300μlのPBS/0.05%Tween−20(PBST)で3度洗浄した。洗浄後、300μlのブロッキング溶液(PBS/0.05%Tween−20中4%無脂肪ドライミルク)を添加し、室温で1時間インキュベートした。ブロッキングステップの後、これらのプレートは、300μlのPBS/0.05%Tween−20(PBST)で3度洗浄した。次に、PBS/0.05%Tween中で1:20,000に希釈した100μlの血清試料を、室温で1〜2時間インキュベートし、次いで、300μlのPBS/0.05%Tween−20で3度洗浄した。PBS/0.05%Tween中で1:5,000に希釈した抗ヒトFc−HRP接合体の100μlを室温で1〜2時間インキュベートし、次いで、300μlのPBS/0.05%Tween−20で3度洗浄した。この最終洗浄の後、100μlの発色基質溶液(TMB1基質、BioFX)を添加し、十分な時間が経過した後、100μlのSTOP溶液(BioFX)を添加して終結させた。450nmにおける吸光度をプレートリーダー(Softmax Proソフトウエアを備えたMolecular Devices Thermomaxマイクロプレートリーダー)で読み取り、データを記録し、次いで、エクセル(マイクロソフト)を用いてプロットした。
【0147】
骨髄:RNA抽出およびmRNAの精製
予め−80℃で貯蔵した骨髄(20mlのRNAlater中約2.5ml)を、RNAlaterを除去するために遠心分離により回収し、次いで、300μlの酢酸を含む11.25mlのTRIBD試薬(Sigma)に再懸濁した。次いで、このペレットは、激しく渦流状にした。次に、1.5mlのBCP(1−ブロモ−3−クロロプロパン,Sigma)を添加し、渦流にすることにより混合し、室温で5分間インキュベートし、次いで、4℃で15分間12000×gで遠心分離した。界面を乱さないように、水相を注意して除去した。次に、水相からの全RNA25mlのイソプロパノールを添加して沈殿させ、室温で10分間インキュベートし、4℃で10分間12000×gで遠心分離した。イソプロパノールを添加後、残留RNAlaterにより2相が形成され、界面に沈殿RNAが沈降しているのが認められた。残留RNAlaterを排除し、RNAを最大限に回収するために、H2O中50%のイソプロパノールの5mlアリコートを添加し、相分離が認められなくなるまで混合し、その点で4℃で10分間12000×gで遠心分離によりRNAをペレット化した。RNAペレットを75%EtOHで洗浄し、RNAseフリーの1.6mlマイクロ遠心チューブに移し、遠心分離により再び回収した。最後に、RNAペレットを100μlの1mM燐酸ナトリウム、pH8.2に再懸濁させ、RNA純度を決めるためにA260およびA280を読みとった。
【0148】
逆転写の前に、mRNAは、Qiagen Oligotex mRNA精製キットにより、全RNAから精製した。要するに、50〜200μgの骨髄RNAをRNaseフリーの水を用いて250μlにし、250μlのOBBバッファーおよびOligotex懸濁液と混合し、次いで、70℃で3分間インキュベーションした。Oligotex粒子のオリゴdT30とmRNAポリ−A−テールとの間のハイブリッド形成は、室温で10分間行った。次いで、ハイブリッド形成された懸濁液は、スピンカラムに移し、1分間遠心分離した。スピンカラムは、400μlのバッファーOW2で2度洗浄した。次いで、精製したmRNAは、20μlのホットな(70℃)バッファーOEBを用いて遠心分離により2度溶離した。典型的な収量は、全RNAとして500ng〜1.5μgであった。
【0149】
骨髄mRNAについてN9およびオリゴdTを用いた逆転写
逆転写(RT:reverse transcription)反応は、75〜100ngのmRNAを、2μlの10X Accuscript RT Buffer(Stratagene)、0.8μlの100mM dNTP、およびN9(300ng)とオリゴdTプライマー(100ng)とのいずれかとを一緒に混合し、次いで、水で最後に、容積を17μlにした。これらの混合物を65℃で5分間加熱し、次いで、室温まで冷却した。次に、2μlのDTT、0.5μlのRNase Block(Stratagene)、0.5μlのAccuscript RT(Stratagene)を、各反応物に添加した。次に、N9で予め刺激を受けた反応物を室温で10分間インキュベートし、オリゴ−dTで予め刺激を受けた反応物を氷上で10分間インキュベートした。最後に、両方の反応物を42℃で60分間、次いで、70℃で15分間インキュベートし、酵素を撲滅した。
【0150】
骨髄誘導cDNAからのPCR
抗体の重鎖および軽鎖のレパートリーは、ヒトの生殖細胞VおよびJ領域に基づいて先に説明した方法および縮重プライマーを用いて本質的に骨髄cDNAから増幅された(O’Brien,P.M.,Aitken R.Standard protocols for the construction of scFv Libraries.Antibody Phage Display−Methods and Protocols,vol 178,59−71,2001,Humana Press)。
【0151】
要するに、ラムダ軽鎖に対して先に刺激されたオリゴdT cDNA(75ngのmRNAから)および先に刺激されたN9 cDNA(カッパ軽鎖の75ngのmRNAから、重鎖の100mgのmRNAから)を用いたPCR反応は、5μlの10X増幅バッファー(Invitrogen)、1.5μlのdNTP(10mM)、1μlのMgSO4(50mM)、2.5μlのVregionプライマー(10μM)および2.5μlプライマー(10μM)−VHの場合10μM、0.5μlの白金Pfxポリメラーゼ(Invitrogen)、および最終容積を50μlにする無菌dH2Oと一緒に混合した。PCRパラメータは、ステップ1−95℃5分、ステップ2−95℃30秒、ステップ3−58℃30秒、ステップ4−68℃1分、ステップ5−サイクルステップ2−4を40回、ステップ6−68℃5分であった。軽鎖PCR産生物は、Qiagen PCR
Cleanupキットを用いてクリーンアップした。重鎖PCR産生物は、Qiagen Gel Extraction Kitを用いて1.5%アガロースゲルから精製し、次いで、再増幅したゲルであつた。重鎖再増幅は、次のようにして行った。すなわち、10μlの10X増幅バッファー(Invitrogen)、3μlのdNTP(10mM)、2μlのMgSO4(50mM)、5μlのVHプライマー(10μM)および5μlのJHプライマー(10μM)、5μlの重鎖Primary PCR産生物、1μlの白金Pfxを混合し、水で容積を100μlに調節した。サイクリングパラメータは、ステップ1−95℃5分、ステップ2−95℃30秒、ステップ3−58℃30秒、ステップ4−68℃1分、ステップ5−サイクルステップ2−4を20回、ステップ6−68℃5分であった。再増幅した重鎖PCR産生物は、Qiagen Extraction Kitを用いて1.5%アガロースTAEゲルからクリーンアップした。
【0152】
抗体ファージライブラリーの構築
個々の鳥類インフルエンザの生存者に関する個別の抗体ライブラリーは、末端のpIIIストップコドンに続いて未翻訳領域に挿入した3−ヌクレオチドを同定する独特のバーコードを用いて構築した。
【0153】
軽鎖クローン化
プールされたカッパ軽鎖およびラムダ軽鎖をドナーにつき各々1μgは、Qiagen
Gel Extraction Kitを用いて1.5%アガロースTAEゲルから精製したゲル、ならびにNotIおよびBamHIを用いて消化した。5μgの各ベクターは、Qiagen Gel Extraction Kitを用いて1%アガロースTAEゲルから精製したゲル、ならびにNotIおよびBamHIを用いて消化した。ライブラリーの連結は、200mgのゲル精製カッパまたはラムダの挿入物および1μgのゲル精製ベクターを用いて、60μl中RTで1時間または14℃で一晩行った。連結は、Edge BioSystem Perfromaスピンカラムを用いて脱塩した。このライブラリーは、80μlのTG−1またはXL−1ブルーのアリコートにおいて5つの電気穿孔中で形質転換し、各々1mlのSOCとして回収し、プールし、37℃で1時間増殖させた。形質転換体の総数は、形質転換の各々からのアリコートを蒔くことによりこの増殖後に測定した。残りの電気穿孔は、200mlの2YT+50μg/mlのAmpicillin+2%グルコース中37℃で一晩増殖することにより増幅させた。次の軽鎖ライブラリーは、Qiagen高速Maxiprepキットを用いてこれらの一晩培養物からプラスミド精製により回収した。
【0154】
重鎖クローン化
各々が1.5〜2μgのドナー特異性重鎖類(VH1、VH2、5、6プール、VH3、およびVH4)は、Qiagen Gel Extraction Kitを用いて1.5%アガロースTAEゲルから精製したゲル、ならびにSfiIおよびXhoIを有する
40ユニット過剰/μgのDNAを用いて消化した。15μgの各軽鎖ライブラリーベクターは、Qiagen Gel Extraction Kitを用いて1.5%アガロースTAEゲルから精製したゲル、ならびにSfiIおよびXhoIを有する40ユニット/μgのDNAを用いて消化した。ライブラリー連結は、消化された1.2μgのSfiI/XhoI、ゲル精製重鎖ドナーコレクションおよび5μgの14℃で一晩置かれた各軽鎖ライブラリー(カッパおよびラムダ)を併用して作成した。次いで、ライブラリー連結物は、Edge BioSystem Pefromaスピンカラムを用いて脱塩し、80μlのTG−1アリコート中でライブラリーあたり20個の電気穿孔を介して形質転換され、各々は1mlのSOC中に回収され、プールされ、37℃で1時間増殖された。再び、この増殖の後、アリコートの各々を用いて、形質転換体の総数を測定し、残りは、1Lの2YT+50μg/mlのAmpicillin+2%グルコースに移され、OD600が約0.3になるまで激しく曝気処理しながら、37℃で増殖させた。次のM13K07ヘルパーファージは、5:1の感染多重度(MOI:multiplicity
of infection)で添加し、撹拌せずに37℃で1時間インキュベートした。次に、遠心分離により細胞を取得し、1Lの2YT+50μg/mlのAmpicillin、70μg/mlのKanamycin中に再懸濁させ、scFvファージミドを産生させるために激しく曝気しながら37℃で一晩増殖させた。次の朝、遠心分離により細胞を集め、ファージミドを含む上澄みを集めた。ファージミドは、0.2容量の20%PEG/5MのNaCl溶液を添加し、氷上で1時間インキュベーションして上澄みから沈殿させた。次いで、ファージミド・ライブラリーストックは、遠心分離により取得し、20mlの無菌PBSに再懸濁させた。残りの細菌は、追加の遠心分離により除去し、最終ファージミド・ライブラリーは、PBS+50%グリセロール中−20℃で貯蔵した。
【0155】
ファージミドパニングおよび増幅
ELISAプレート(Immulon 4HBX平底、Nunc)を、ELISAプレートコーティング溶液(BioFX)中室温で一晩インキュベーションして100μlの100ng/mLのH5ヘマグルチニンタンパク質(Protein Sciences,A/Vietnam/1203/2004)で被覆した。次の日プレート類は、300μlのPBSTで3度洗浄した。洗浄後、300μlのブロッキング溶液(PBS/0.05%Tween−20中4%無脂肪ドライミルク)を添加し、氷上で30分間インキュベートした。ブロッキングステップの後、これらのプレートは300μlのPBSTで3度洗浄した。ファージパニングの直前に、Millipore Amicon Ultraカラムを用いて、凍結ファージミドストックからグリセロールを除去し、次いで、4%無脂肪ドライミルク中に15分間ブロックした。次に、ファージミドの100μlのアリコートを8つのウエル中に分配し、(全ファージ約1×1012CFU)4℃で2時間インキュベートし、次いで、300μlのPBSTで8度洗浄した。ファージミドは、室温で10分後に100μl/ウエルの溶離バッファー(0.2Mグリシン−HCl、pH2.2、1mg/mlBSA)に集めた。次いで、溶離液を、溶離液mlあたり56.25μlの2Mトリス塩基を添加して中和した。中和後、5mlのTG1細胞(OD600約0.3)を振とうなしで2−YT中37℃で30分間0.5mlの中和ファージで感染させた。このステップの後、一部の細胞をLB AMPグルコースプレート上に蒔き、ファージミドの総回収率を測定した。残りの接種液は、10mlのYATG(最終濃度2%グルコースおよび50μg/mlアンピシリン)に入れ、OD600が約0.3になるまで激しく曝気しながら37℃で増殖させた。次に、培養物を、5:1のMOIにてM13K07ヘルパーファージに感染させ、撹拌せずに37℃で30〜60分間インキュベートした。これらの細胞は、遠心分離により集め、25mlの2−YTAK(Ampicilin 50μg/ml,Kanamycin 70μg/ml)に再懸濁し、フレッシュな培養フラスコに移し、振とうしながら37℃で成長させた。その後のラウンドは、同様に回収し、増幅した。
【0156】
scFvELISA
バイオパンされたファージからの大腸菌のHB2151形質転換細胞の個々のコロニーを、1mlの2YT+100μg/mlのAMP中37℃で一晩増殖させた。次の朝、これらの細胞は遠心分離により取得し、1.5mlの周辺溶解バッファー(1mlのBBS(Teknova)+0.5mlの10mg/mlのリゾチーム+最終濃度が10mMになるEDTA)に再懸濁させた。これらの細胞は、遠心分離により再びペレット化し、周辺可溶化液を含むscFvを集めた。scFv可溶化液は、希釈バッファー(PBS/0.05%BSA)と1:1で混合し、先に抗原が希釈バッファーで被覆され、ブロックされたウエルに100μlを添加した。これらの試料を、室温で2時間インキュベートし、次いで、PBS/0.05%Tweenで3度洗浄した。次に、希釈バッファー中の1:5000に希釈したビオチン抗ヒスチジンマウス(Serotec)を100μl各ウエルに添加し、室温で1時間インキュベートした。このインキュベーションの後、これらのウエルはPBS/0.05%Tweenで3度洗浄し、次いで、各ウエルに100μlの1:2500のストレプトアビジン:HRP(Serotec)を添加し、室温で2時間インキュベートし、次いで、PBS/0.05%Tweenで3度洗浄した。この最終洗浄の後、100μlの発色基質溶液(TMB1基質、BioFX)を添加し、十分な時間が経過した後、100μlのSTOP溶液(BioFX)を添加して終結させた。450nmにおける吸光度は、プレートリーダー(Softmax Proソフトウエアを備えたMolecular Devices Thermomaxマイクロプレートリーダー)で読み取り、データを記録し、次いで、エクセル(マイクロソフト)を用いてプロットした。
【0157】
塩基配列の決定
重鎖および軽鎖の配列を推定するために、個々のクローンを増殖させ、プラスミドDNAを抽出した(Qiagen)。プラスミドDNAは、標準DNA塩基配列決定法により配列を決めた。
【0158】
ヘマグルチニン抑制(HAI)アッセイ
ヘマグルチニン抑制は、本質的には、Rogers et al.,Virology131:394−408(1983)の方法により、ウイルスまたはタンパク質/ウエルの4HAU(ヘマグルチネート化ユニット)を用いて丸底マイクロタイタープレート(Corning)において行った。HAI測定では、精製単一鎖可変断片(scFv)の25μlの試料が、各マイクロタイターウエルにおいてテストウイルスの4HAUを含むがこれらに限定されない25μlのPBSと混合した。室温における15分の予備インキュベーションの後、25μlの0.75%ヒト赤血球を添加し、混合した。HAI抗体活性は、室温で60分インキュベーション後目視検査により測定した。
【0159】
結果
骨髄および血液の試料は、2006年1月トルコで起きたH5N1鳥類インフルエンザの大流行の6名の生存者から、大流行の約4ヶ月後に採取した。6名の生存者全員について、トルコの保健省により承認された、理学的検査、臨床試験、および分子診断測定に続いて鳥インフルエンザの初期診断がなされた。これらの生存者のうち4名は、さらに、世界保健機関(WHO)により確認された。血清試料は、上で説明した血清学プロトコルを用いてH5ヘマグルチニン(A/Vietnam/1203/2004)に対する抗体の存在を確認するために分析した。図7に示したように、6名の患者全員の血液試料(それぞれ、SLB H1−H6と指定した)は、H5抗原に対する抗体の存在を示した。このことを確認後、これらの個体の骨髄試料からRNAを抽出し、上で説明したプロトコルを用いて骨髄mRNAを精製し、逆転写した。次いで、抗体の重鎖および軽鎖のレパートリーを、上で説明した骨髄cDNAから増幅し、個別の抗体の重鎖および軽鎖のファージミドライブラリーを、個々のライブラリーを区別するために、上で説明した3ヌクレオチドバーコードを用いて各生存者について別々にクローン化した。
【0160】
骨髄および血液試料は、2006年にインフルエンザ症状で処置を受けた12名のローカルドナーからも採取した。H1、H3およびH5ヘマグルチニンに対する抗体の存在を確認するために上で説明した血清学的研究が、それぞれ、なされた。図8に示したように、テストしたすべての血清試料は、H1および/またはH3のヘマグルチニンの抗体について陽性であり、どのサブタイプが優勢であるかは、個々のドナーが自身の生活時間を通して最も多くさらされたインフルエンザA型ウイルスのサブタイプに左右された。その上、興味深いことに、血清にH5ヘマグルチニンの抗体を、なりのレベル含む患者がいた(図8のドナーSLB1およびSLB5)。この確認の後、ドナーの骨髄試料からRNAを抽出し、上で説明したプロトコルを用いて骨髄mRNAを精製し、逆転写した。次いで、抗体の重鎖および軽鎖のレパートリーを、上で説明した骨髄cDNAから増幅し、個別の抗体の重鎖および軽鎖のライブラリーを、個々のライブラリーを区別するために、上で説明した3ヌクレオチドバーコードを用いて各生存者について別々にクローン化した。
【0161】
図9に例示したように、利用できる4つのヌクレオチド類の3個を用いると、64の独特のバーコードを作ることができる。
【0162】
トルコ鳥インフルエンザの生存者の骨髄試料から調製し、プールされた抗体ライブラリーの3ラウンドのパニングの後、得られた48のランダムクローンから、40をH5ヘマグルチニンタンパク質(Protein Sciences,A/Vietnam/1203/2004)への結合についてELISAによりテストし、ベトナムH5N1ウイルス(CBER,A/Vietnam/1203/2004)を不活性化した。これらのクローンの配列を決めた。40のクローンの内、5つは異なっていることが判明した。図10に示したように、全部で5つの異なるクローン(クローンF5およびG1は同じ配列を有する)は、H5タンパク質およびベトナムのH5N1ウイルスの両方に結合していた。図11は、上の実験において使われたベトナムの分離株のH5ヘマグルチニン配列に対してトルコのドナーからのH5ヘマグルチニンタンパク質の配列を比較している配列アラインメントを示している。これらの実験の結果は、配列の違いにも関わらず、テストした抗体は、トルコおよびベトナムの両方のH5タンパク質およびウイルスに結合し、したがって、1つより多くのH5N1ウイルスの分離株と交差反応性を示した。
【0163】
追加された4つの独特のクローンは、第2ラウンドのパニングにより産生された12のクローンの中から同定された。
【0164】
トルコドナーのプールされた抗体ライブラリーにおいて同定された独特のクローンの重鎖可変領域配列は、対応する軽鎖および生殖系列起源の配列と共に、図12および13に示している。特に、図12に示した配列(3〜23重鎖クローン)は、3ラウンドのパニング後の、すべてのトルコドナーのすべての重鎖および軽鎖のプールされたライブラリーに由来する。図13に示した配列(3〜30重鎖クローン)は、2ラウンドのパニング後の、すべてのトルコドナーのすべての重鎖および軽鎖のプールされたライブラリーに由来する。
【0165】
追加された独特のH5N1に特異な抗体重鎖可変領域配列は、4ラウンドのパニング後、上で説明したELISAプロトコルを用いて個々のトルコドナーの抗体ライブラリーから同定した。これらのH5N1のELISA陽性クローンは、図14A−Dに示している。
【0166】
図15および16は、上で説明したトルコの鳥インフルエンザ生存者の血清および骨髄の分析により同定した抗体重鎖(図15)および軽鎖(図16)配列を用いて多様な抗体重鎖および軽鎖ライブラリーを作り出すために目標の突然変異誘発の使用を例示している。
【0167】
図17および18は、HAタンパク質のトルコおよびインドネシアの改変体と共にH5N1ベトナムウイルスscFv抗体から得られた特定のFab断片交差反応性を確認しているELISAの結果を示している。
【0168】
前述の説明では本発明は、特定の実施形態を参照しながら説明しているが、本発明はそのように限定されない。実際に、本発明で示しおよび説明した発明に加えて、前述の説明から当業者に明らかになり、添付した特許請求の範囲内に入る、本発明の種々の改変がある。
【0169】
本明細書を通じて引用された引用文献は、すべて、引用により本明細書に明らかに組み込まれている。
【技術分野】
【0001】
発明の分野
本発明は、インフルエンザA型ウイルスに対する中和抗体を同定し、産生し、さらに、遺伝子操作する方法および手段、ならびに産生された中和抗体に関する。特に、本発明は、例えば、H1、H2、H3、およびH5サブタイプのすべてなど2つ以上のH1、H2、H3、H5、H7およびH9に対する中和抗体を含む、種々のインフルエンザA型ウイルスサブタイプに対する中和抗体およびこの種の抗体を作る方法および手段に関する。さらに具体的に述べると、本発明は、インフルエンザA型ウイルスサブタイプの分離株を1つより多く、好ましくはすべてを中和し得る抗体に関する。
【背景技術】
【0002】
発明の背景
インフルエンザは、インフルエンザウイルスにより引き起こされる接触伝染性の呼吸器疾患である。該ウイルスは軽症から重篤までの疾患を引き起こし、時には死に至ることもある。毎年米国では、国民の5〜20%がインフルエンザにかかり、約200,000人が入院させられ、約36,000人が死亡する。
【0003】
インフルエンザウイルスは、せきやくしゃみをした時に生じる呼吸器系のしぶきで広がり、該ウイルスは、通常、ヒトからヒトへ伝染する。インフルエンザ表面抗原に対する免疫、特にヘマグルチニンは、感染が起きた場合、感染の可能性および感染しても疾患の重篤性を低減する。インフルエンザワクチンは利用できるが、1つの型またはサブタイプのインフルエンザウイルスに対する抗体は、別の型またはサブタイプのインフルエンザに対する保護は限定的または全く保護できず、毎年インフルエンザワクチンに1つより多くの株を組み込む必要がある。
【0004】
インフルエンザウイルスは、分節マイナス鎖(negative−strand)RNAウイルスであり、オルトミクス・ウイルス(Orthomyxoviridae)科に属する。インフルエンザA型ウイルスは9つの構造タンパク質からなり、さらに、制御的機能を有する1つの非構造NS1タンパク質をコードする。該非構造NS1タンパク質は、再現サイクルの間に大量に合成され、感染細胞の細胞質ゾルおよび核の中に局在化される。ウイルスゲノムの分節性により、種々のウイルス株を有する細胞の混合感染の間に起きる遺伝子的再集合(ゲノム分節の交換)のメカニズムが可能になる。インフルエンザA型ウイルスは、さらに、ウイルスタンパク質表面上に表示された種々のヘマグルチニン(HA:hemagglutinin)およびノイラミニダーゼ(NA:neuraminidase)ウイルスタンパク質により種々のサブタイプに分類される。
【0005】
インフルエンザA型ウイルスサブタイプは、2つのウイルス表面糖タンパク質である、ヘマグルチニン(HAまたはH)およびノイラミニダーゼ(NAまたはN)により同定される。各々のインフルエンザウイルスのサブタイプは、HおよびNのタンパク質のその組み合わせにより同定される。HAサブタイプが16、NAサブタイプが9種知られている。インフルエンザA型ウイルスは、ヒト、トリ、ブタ、ウマ、および他の動物に感染させることができるが、野鳥は、これらのウイルスの自然の宿主である。一部のインフルエンザAサブタイプのみ(すなわち、H1N1、H1N2、およびH3N2)が、現在ヒトの間に循環しているが、16のHおよび9のNAサブタイプのすべての組み合わせは、鳥類、特に原野にいる水鳥および海岸で生息しているトリにおいて同定されている。さらに、H5およびH7インフルエンザウイルスがヒトを発病させうることを示す証拠も蓄積されつつある。
【0006】
インフルエンザA型ウイルスのHAは、2つの構造的に異なる領域、すなわち、球状頭部領域およびステム領域を含んでいる。球状頭部領域は、標的細胞へのウイルスの取り付けに関与する受容体結合部位を含み、HAのヘマグルチニン化活性に関与している。ステム領域は融合ペプチドを含み、該ペプチドは、細胞のウイルス外被とエンドソーム膜との間の膜の融合に必要であり、したがって、融合活性に関係する(Wiley et al.,Ann.Rev.Biochem.,56:365−394(1987))。
【0007】
パンデミックは、世界的な疾患の大流行である。インフルエンザのパンデミックは、新しいインフルエンザA型ウイルスが、(1)人間集団において該ウイルスに対して免疫がほとんどないか、全くないウイルスが出現し、(2)重病を引き起こし始め、(3)ヒトとヒトとの間で世界的に容易に広がる時に起きる。20世紀の間に、この種のインフルエンザのパンデミックが3度発生した。最初は、1918年の「スペイン風邪」で、このインフルエンザパンデミックにより米国で少なくとも500,000人死亡し、世界の死者は4000万人に上った。このパンデミックは、インフルエンザA H1N1サブタイプにより発生した。2回目のインフルエンザパンデミックは、1957年の「アジア風邪」で、インフルエンザA H2N2サブタイプにより発生し、死者は、米国で少なくとも70,000人、世界では100〜200万人に上った。最も最近のインフルエンザパンデミックは、1968年の「香港風邪」で、インフルエンザA H3N2サブタイプにより発生し、死者は、米国で約34,000人、世界では70万人であった。
【0008】
1997年に、インフルエンザA H5N1の最初の症例が、香港で報告された。この鳥類型ウイルスが直接ヒトに感染したのは、これが初めてであったが、ヒトからヒトへの感染が認められなかったので、パンデミックは起きなかった。
【0009】
非特許文献1は、不活性化したH5N1ウイルスを予防接種したウマからの抗H51 IgGsおよびH5N1に感染したBALB/cマウスを保護すると説明されたH5N1−特異性F(ab’)2断片の調製について報告している。
【0010】
非特許文献2は、マウスの受動免疫法のインフルエンザA H5 ウイルスヘマグルチニンに対して特異的なキメラモノクローナル抗体の使用について説明している。
【0011】
特定のインフルエンザA型ウイルスにより生じた呼吸器疾患の重篤性、および潜在的なパンデミックの脅威に鑑みて、効率的な予防および処置方法を求める大きなニーズがある。本発明は、インフルエンザA型ウイルスのH1、およびH3サブタイプ、およびH5サブタイプを含むがこれらに限定されない、該ウイルスの種々のHサブタイプに対するインフルエンザA中和抗体を提供することによりこのニーズに取り組んでいる。本発明は、さらに、種々のヒトおよびヒト以外の種から得られた分離株および種々のインフルエンザの大流行および/またはパンデミックの犠牲者および/または生存者からの分離株を含むがこれらに限定されないインフルエンザA型ウイルスの任意のサブタイプの分離株の1つより多い、好ましくはすべてを中和し得る抗体を提供する。
【0012】
この種の中和抗体は、大流行またはパンデミックの場合、感染した人々または感染のリスクがある人々の受動免疫法を含むインフルエンザウイルス感染の予防および/または処置に利用することができる。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】Lu et al.,Resp.Res.7:43(2006)(doi:10.1186/1465−992−7−43)
【非特許文献2】Hanson et al.,Resp.Res.7:126(doi:10.1186/1465−9921−7−126)
【発明の概要】
【課題を解決するための手段】
【0014】
1つの態様では、本発明は、インフルエンザA型ウイルスサブタイプの1つより多くの分離株またはインフルエンザA型ウイルスの1つより多くのサブタイプを中和する中和抗体に関する。
【0015】
1つの実施形態では、該抗体は、H5、H7およびH9サブタイプの1つ以上などインフルエンザA型ウイルスサブタイプの実質的にすべての分離株を中和する。
【0016】
別の実施形態では、該抗体は、H5、H7およびH9サブタイプの1つ以上など特定のインフルエンザA型ウイルスサブタイプの1つより多くの分離株を中和する。
【0017】
さらに別の実施形態では、該抗体は、インフルエンザA型ウイルスの1つより多くのサブタイプ、および少なくとも1つのサブタイプの1つより多くの分離株を中和する。
【0018】
さらなる実施形態では、本明細書の該抗体により中和された少なくとも1つのサブタイプおよび/または分離株が、ヒトに感染する能力を有する。
【0019】
別の実施形態では、少なくとも1つの分離株は、例えば、野鳥およびニワトリを含む鳥類に由来するものである。
【0020】
特定の実施形態では、本明細書の抗体は、インフルエンザA型ウイルスのH5N1サブタイプを中和する。好ましくは、該抗体は、このインフルエンザA型ウイルスサブタイプの1つより多くの分離株、より好ましくは実質的にすべての分離株を中和する。
【0021】
別の実施形態では、本明細書の該抗体は、H5N1サブタイプ、ならびにH1N1、H1N2、およびH3N2サブタイプからなる群から選択される少なくとも1つの追加されたサブタイプを中和する。
【0022】
追加の実施形態では、本明細書の抗体は、追加されたサブタイプ(単数および複数)の1つより多くの分離株、好ましくは実質的にすべての分離株を中和する。
【0023】
別の実施形態では、本発明の中和抗体は、H5タンパク質に結合する。好ましくは、該抗体は、H5タンパク質の1つより多くの改変体、より一層好ましくは、H5タンパク質の実質的にすべての分離株を中和する。
【0024】
他の実施形態では、本明細書の抗体は、H5タンパク質およびH1、H2および/またはH3のタンパク質など少なくとも1つの追加されたHタンパク質に結合する。
【0025】
異なる態様では、本発明は、本明細書で説明した該中和抗体を含む組成物に関する。
【0026】
さらなる態様では、本発明は、単一のインフルエンザA型ウイルスサブタイプまたは複数のインフルエンザA型ウイルスサブタイプの1つより多くの分離株を中和し得る抗体を同定する方法に関する。この方法は、インフルエンザA型ウイルスサブタイプの第1および第2の分離株の両方、または該インフルエンザA型ウイルスの第1および第2のサブタイプと反応する抗体ライブラリーにおける抗体の同定、および第1および第2の分離株、または第1および第2のサブタイプ、それぞれ、に結合する該抗体の能力に基づいて、同定した該抗体を選択の連続的交互ラウンドを受けることを含む。
【0027】
ある実施形態では、第1および第2のインフルエンザA型ウイルスサブタイプ分離株の両方と反応する抗体は、第1および第2の分離株と、それぞれ、反応し、同定された抗体を組み換える抗体の少なくとも2ラウンドの別々の濃縮により同定された。
【0028】
別の実施形態では、第1および第2のインフルエンザA型ウイルスサブタイプ分離株の両方と反応することができる抗体は、第1および第2の分離株に、それぞれ、結合する該抗体の能力に基づいて、選択の連続的交互ラウンドを受ける前に突然変異誘発を受ける。望ましいならば、第1および第2の分離株に結合し得る該抗体は、さらに、1つより多くのインフルエンザA型サブタイプに結合する該抗体の能力に基づいて選択される。
【0029】
この種の濃縮技法の適用は、一般に、抗体が結合する標的とは関係なく、同様に抗体に適用することができる。この種の一般的な濃縮/選択の方法は、本発明の一部として明らかに含まれている。
【0030】
さらなる態様では、本発明は、本発明の中和抗体が共有する配列コレクションに関する。
【0031】
さらにさらなる態様では、本発明は、本明細書の有効量の中和抗体または抗体組成物を対象に投与することを含むA型インフルエンザに感染している対象を処置する方法に関する。
【0032】
別の態様では、本発明は、A型インフルエンザに感染するリスクのある対象に本発明の中和抗体の有効量を投与することを含む、A型インフルエンザの感染を予防する方法に関する。
【0033】
異なる態様では、本発明は、(a)少なくとも2つの機能が異なる抗体のCDR配列を整列させ、(b)整列させられたCDR配列間に保存されたアミノ酸残基を同定し、(c)整列させられたCDR配列の複数の改変体を産生するために突然変異を誘発された非保存位置における機能が異なる抗体に存在する、少なくともアミノ酸残基をコードする縮重オリゴヌクレオチドプローブを用いて、整列させられたCDR配列の少なくとも1つの配列において複数の非保存アミノ酸残基の突然変異誘発を行い、ならびに、望ましいならば、該抗体コレクションが望ましい程度の多様性および/またはサイズに達するまで改変体の1つ以上と共にステップ(b)および(c)を反復することを含む、様々な多機能性抗体コレクションを作成する方法に関する。
【0034】
特定の実施形態では、整列させられたCDR配列は同じ長さを有する。
【0035】
別の実施形態では、保存された該アミノ酸残基類は、整列させられた少なくとも2つのCDR配列に保持される。
【0036】
さらなる態様では、本発明は、少なくとも1つの特性が互いに異なる、複数の中和抗体を含む抗体コレクションに関する。
【0037】
本発明は、さらに、この種のコレクションに存在する核酸の配列に連結されたか、または該配列に組み込まれる独特のバーコードで標識した核酸を含むコレクション中の核酸を独自に同定する方法に関する。
本発明はまた、以下の項目を提供する。
(項目1)
インフルエンザA型ウイルスサブタイプの1つより多くの分離株および/または上記インフルエンザA型ウイルスの1つより多くのサブタイプを中和する中和抗体。
(項目2)
インフルエンザA型ウイルスH1サブタイプの1つより多くの分離株を中和する、項目1に記載の中和抗体。
(項目3)
インフルエンザA型ウイルスH3サブタイプの1つより多くの分離株を中和する、項目1に記載の中和抗体。
(項目4)
インフルエンザA型ウイルスH1およびH3サブタイプを中和する、項目1に記載の中和抗体。
(項目5)
インフルエンザA型ウイルスH1および/またはH3サブタイプの1つより多くの分離株を中和する、項目4に記載の中和抗体。
(項目6)
インフルエンザA型ウイルスサブタイプの実質的にすべての分離株を中和する、項目1に記載の中和抗体。
(項目7)
上記サブタイプが、H5、H7およびH9サブタイプからなる群から選択される、項目1または項目6に記載の中和抗体。
(項目8)
上記サブタイプが、上記H5サブタイプである、項目7に記載の中和抗体。
(項目9)
上記抗体が、インフルエンザA型ウイルスH5サブタイプの実質的にすべての分離株を中和する、項目8に記載の中和抗体。
(項目10)
上記サブタイプが、上記H7サブタイプである、項目7に記載の中和抗体。
(項目11)
上記抗体が、上記インフルエンザA型ウイルスH7サブタイプの実質的にすべての分離株を中和する、項目10に記載の中和抗体。
(項目12)
上記サブタイプが、上記H9サブタイプである、項目7に記載の中和抗体。
(項目13)
上記抗体が、上記インフルエンザA型ウイルスH9サブタイプの実質的にすべての分離株を中和する、項目12に記載の中和抗体。
(項目14)
上記抗体が、さらに、インフルエンザA型ウイルスの追加のHサブタイプを少なくとも1つ中和する、項目7に記載の中和抗体。
(項目15)
上記追加Hサブタイプが、H1、H2およびH3サブタイプからなる群から選択される、項目14に記載の中和抗体。
(項目16)
上記インフルエンザA型ウイルスの追加Hサブタイプの1つより多くの分離株を中和する、項目15に記載の中和抗体。
(項目17)
上記インフルエンザA型ウイルスのH5N1サブタイプを中和する、項目1に記載の中和抗体。
(項目18)
上記インフルエンザA型ウイルスのH5N1サブタイプの1つより多くの分離株を中和する、項目17に記載の中和抗体。
(項目19)
上記分離株の少なくとも1つがヒトに感染する能力を有する、項目18に記載の中和抗体。
(項目20)
上記分離株の少なくとも1つがヒト対象から得られた、項目19に記載の中和抗体。
(項目21)
上記ヒト対象が病気に罹患している、項目20に記載の中和抗体。
(項目22)
上記ヒト対象が上記インフルエンザA型ウイルスのH5N1サブタイプ感染から回復した、項目20に記載の中和抗体。
(項目23)
上記分離株の少なくとも1つがヒト以外の動物から得られた、項目19に記載の中和抗体。
(項目24)
上記ヒト以外の動物が鳥類である、項目23に記載の中和抗体。
(項目25)
上記ヒト以外の動物が野鳥である、項目24に記載の中和抗体。
(項目26)
上記ヒト以外の動物がニワトリである、項目24に記載の中和抗体。
(項目27)
上記インフルエンザA型ウイルスのH5N1サブタイプの実質的にすべての分離株を中和する、項目18に記載の中和抗体。
(項目28)
上記H5N1サブタイプおよびH1N1、H2N2、およびH3N2サブタイプからなる群から選択された少なくとも1つの追加サブタイプを中和する、項目17に記載の中和抗体。
(項目29)
上記インフルエンザA型ウイルスのH5N1サブタイプのより多くの分離株を中和する、項目28に記載の中和抗体。
(項目30)
上記インフルエンザA型ウイルスのH5N1サブタイプの実質的にすべての分離株を中和する、項目29に記載の中和抗体。
(項目31)
上記追加サブタイプの1つより多くの分離株を中和する、項目30に記載の中和抗体。(項目32)
上記追加サブタイプの実質的にすべての分離株を中和する、項目31に記載の中和抗体。
(項目33)
上記抗体がH5タンパク質に結合する、項目1に記載の中和抗体。
(項目34)
上記抗体が上記H5タンパク質の1つより多くの改変体に結合する、項目33に記載の中和抗体。
(項目35)
上記抗体が上記H5タンパク質のすべての改変体に結合する、項目34に記載の中和抗体。
(項目36)
上記抗体が少なくとも1つの追加のHタンパク質に結合する、項目35に記載の中和抗体。
(項目37)
上記追加のHタンパク質が、H1、H2およびH3タンパク質からなる群から選択される、項目36に記載の中和抗体。
(項目38)
上記抗体が上記追加のHタンパク質の1つより多くの改変体に結合する、項目37に記載の中和抗体。
(項目39)
上記抗体が上記追加のHタンパク質の実質的にすべての改変体に結合する、項目38に記載の中和抗体。
(項目40)
項目1から39までのいずれか1項に記載の中和抗体を含む組成物。
(項目41)
インフルエンザA型ウイルスサブタイプの1つより多くの分離株またはインフルエンザA型ウイルスの1つより多くのサブタイプを中和し得る抗体を同定する方法であって、
上記インフルエンザA型ウイルスサブタイプの第1および第2の分離株の両方、または上記インフルエンザA型ウイルスの第1および第2のサブタイプと反応する抗体を、抗体ライブラリーにおいて同定すること、および上記第1および第2の分離株、または上記第1および第2のサブタイプそれぞれに結合する抗体の能力に基づいて、同定した上記抗体に選択の連続的交互ラウンドを受けさせることを含む、方法。
(項目42)
少なくとも2ラウンドの選択を含む、項目41に記載の方法。
(項目43)
上記第1および第2の分離株が、上記インフルエンザA型ウイルスのH5N1サブタイプの異なる分離株である、項目41に記載の方法。
(項目44)
第1および第2のインフルエンザA型ウイルスサブタイプ分離株の両方と反応する上記抗体が、それぞれ、上記第1分離株および上記第2分離株と反応する抗体の少なくとも2ラウンドの別々の濃縮により同定され、同定された上記抗体を組み換える、項目41に記載の方法。
(項目45)
上記第1および上記第2のインフルエンザA型サブタイプ分離株の両方と反応し得る上記抗体が、上記第1および第2の分離株それぞれに結合する抗体の能力に基づいて、選択の上記連続交互ラウンドを受ける前に突然変異誘発を受ける、項目41に記載の方法。
(項目46)
上記抗体ライブラリーがファージディスプレイ・ライブラリーである、項目41に記載の方法。
(項目47)
選択がバイオパニングによってなされる、項目46に記載の方法。
(項目48)
上記インフルエンザA型ウイルスサブタイプがH5N1サブタイプである、項目41に記載の方法。
(項目49)
上記第1分離株が上記H5N1ウイルスの2006年のトルコ分離株である、項目48に記載の方法。
(項目50)
上記第1分離株が上記H5N1ウイルスの2003/2004年のベトナム分離株である、項目48に記載の方法。
(項目51)
上記第2分離株が上記H5N1ウイルスの2003/2004年のベトナム分離株である、項目48に記載の方法。
(項目52)
上記第2分離株が上記H5N1ウイルスの1997年の香港分離株である、項目50に記載の方法。
(項目53)
上記第1および第2の分離株が異なる種に由来する、項目48に記載の方法。
(項目54)
上記種の少なくとも1つがヒトである、項目53に記載の方法。
(項目55)
上記種の少なくとも1つが鳥類である、項目53に記載の方法。
(項目56)
上記第1および第2の分離株に結合し得る上記抗体が、さらに、1つより多くのインフルエンザA型サブタイプに結合する上記抗体の能力に基づいて選択される、項目41に記載の方法。
(項目57)
項目41〜56のいずれか1項に記載の方法により同定された上記中和抗体により共有される配列コレクション。
(項目58)
配列コレクションであって、図11、12、13、および14A−Dに示された独特の1つ以上の重鎖および/または軽鎖の配列、または上記配列に基づくコンセンサス配列または改変体配列を含む、コレクション。
(項目59)
項目41〜56のいずれか1項に記載の方法により同定可能な中和抗体またはその断片。
(項目60)
図11、12、13、および14A−Dに示された独特の配列から選択された重鎖および/または軽鎖の配列、または上記配列に基づくコンセンサス配列または改変体配列またはこれらの断片を含む、項目59に記載の中和抗体。
(項目61)
インフルエンザA型ウイルス感染に対して鳥類または哺乳類の対象に受動免疫を与えることができる、項目59または項目60に記載の中和抗体または抗体断片。
(項目62)
上記哺乳類の対象がヒトである、項目61に記載の中和抗体または抗体断片。
(項目63)
上記インフルエンザA型ウイルス感染が、H5N1、H1N1、H2N2、およびH3N2サブタイプからなる群から選択されるウイルスにより引き起こされる、項目62に記載の中和抗体または抗体断片。
(項目64)
対象においてA型インフルエンザ感染を予防および/または処置する方法であって、上記対象に項目40に記載の組成物を有効量投与することを含む、方法。
(項目65)
対象のA型インフルエンザ感染を処置する方法であって、上記対象に項目59に記載の中和抗体を有効量投与することを含む、方法。
(項目66)
上記対象がヒトの患者である、項目64または項目65に記載の方法。
(項目67)
A型インフルエンザ感染を予防する方法であって、A型インフルエンザ感染を発症するリスクのある対象に項目40に記載の組成物を有効量投与することを含む、方法。
(項目68)
A型インフルエンザ感染を予防する方法であって、A型インフルエンザ感染を発症するリスクのある対象に項目59に記載の中和抗体を有効量投与することを含む、方法。
(項目69)
上記対象がヒトの患者である、項目67または項目68に記載の方法。
(項目70)
多様な多機能性抗体コレクションを作る方法であって、
(a)少なくとも2つの機能が異なる抗体のCDR配列を整列させ、(b)上記整列させられたCDR配列の間に保存されたアミノ酸残基を同定し、(c)上記整列させられたCDR配列の複数の改変体を産生するために、突然変異誘発された非保存位置における機能が異なる抗体に存在する少なくとも上記アミノ酸残基をコードする縮重オリゴヌクレオチドプローブを用いて、上記整列させられたCDR配列の少なくとも1つの配列において複数の非保存アミノ酸残基の突然変異誘発を行い、そして、望ましいならば、上記抗体コレクションが望ましい程度の多様性またはサイズに達するまで上記改変体の1つ以上を用いてステップ(b)および(c)を反復することを含む、方法。
(項目71)
上記整列させられたCDR配列が同じ長さを有する、項目70に記載の方法。
(項目72)
ステップ(c)において産生された上記突然変異誘発された改変体が、上記整列させられたCDR配列の少なくとも2つに存在する保存されたすべての残基を保持する、項目70に記載の方法。
(項目73)
ステップ(c)において産生された上記突然変異誘発された改変体が、上記整列させられたCDR配列のすべてに存在する保存されたすべての残基を保持する、項目70に記載の方法。
(項目74)
上記機能的に異なる抗体が標的抗原上で異なるエピトープに結合する、項目70に記載の方法。
(項目75)
上記機能的に異なる抗体が異なる標的抗原に結合する、項目70に記載の方法。
(項目76)
上記異なる標的抗原が上記同じ抗原の改変体である、項目75に記載の方法。
(項目77)
上記機能的に異なる抗体が異なる結合親和性を有する、項目70に記載の方法。
(項目78)
上記機能的に異なる抗体が異なる生物学的特性を有する、項目70に記載の方法。
(項目79)
上記機能的に異なる抗体がインフルエンザA型ウイルスに結合する、項目70に記載の方法。
(項目80)
上記機能的に異なる抗体の少なくとも2つが同じインフルエンザA型ウイルスについて異なるエピトープに結合する、項目79に記載の方法。
(項目81)
上記機能的に異なる抗体が異なるインフルエンザA型ウイルスサブタイプに結合する、項目79に記載の方法。
(項目82)
上記機能的に異なる抗体の少なくとも2つが同じインフルエンザA型ウイルスサブタイプの異なる分離株に結合する、項目79に記載の方法。
(項目83)
上記機能的に異なる抗体の少なくとも2つが同じインフルエンザA型ウイルスサブタイプの異なる分離株および異なるインフルエンザA型ウイルスサブタイプに結合する、項目79に記載の方法。
(項目84)
上記機能的に異なる抗体の少なくとも2つが異なる親和性を有する、項目70〜83のいずれか1項に記載の方法。
(項目85)
上記機能的に異なる抗体の少なくとも2つは、これらが結合するインフルエンザA型ウイルスを中和する能力に違いがある、項目70〜83のいずれか1項に記載の方法。
(項目86)
少なくとも1つの特性が互いに異なる複数の中和抗体を含む、抗体コレクション。
(項目87)
少なくとも約100の中和抗体を含む、項目86に記載の抗体コレクション。
(項目88)
項目70〜83のいずれか1項に記載の方法により調製された項目87に記載の抗体コレクション。
(項目89)
コレクション中の核酸を独自に同定する方法であって、上記コレクション中に存在する核酸配列に連結または上記配列に組み込まれる独特のバーコードで上記核酸を標識することを含む、方法。
(項目90)
上記バーコードが、1〜約24個のヌクレオチド長の非コードヌクレオチド配列である、項目89に記載の方法。
(項目91)
上記非コードヌクレオチド配列が、標識された上記核酸配列の3’非コード領域に連結される、項目90に記載の方法。
(項目92)
上記バーコードが、標識された上記核酸配列内に組み込まれる1つ以上のサイレント突然変異のコード配列である、項目89に記載の方法。
(項目93)
上記バーコードが、ペプチドまたはポリペプチドの配列である、項目89に記載の方法。
【図面の簡単な説明】
【0038】
【図1A】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1B】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1C】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1D】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1E】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1F】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1G】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1H】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1I】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図1J】15の既知のヘマグルチニン(H)タンパク質サブタイプのアミノ酸配列を示している。
【図2】2種類の標的、AおよびBに対する反応性強度を高めるための典型的なパニング濃縮スキームを示している。各濃縮ラウンドは、個々の標的(単数および複数)に対するプールの反応性強度を高める。
【図3】標的AおよびBと交差反応するクローンを選択する方法を示し、この方法では、各連続ラウンドは両標的に対して得られるプールの反応性強度を高める。
【図4】交差反応性を発生および/または高めるために並行発見プールを組み換えることにより2種類の標的(標的AおよびB)に対する反応性強度を高める方法を示している。組み換え抗体ライブラリーの各選択ラウンドは、両標的に対して得られるプールの反応性強度を高める。
【図5】標的Aに対する反応性を維持しながら標的Bに対する交差反応性を高める方法を示している。第1に、標的Aと反応するクローンが選択され、次いで、標的Aと反応するクローンの突然変異誘発ライブラリーが調製され、選択が示されたようになされ、標的Aおよび標的Bの両方と強い反応性を示す1つ以上の抗体クローンを生じる。
【図6】「目標突然変異誘発」法により多様な多機能性抗体コレクションを作る代表的な突然変異誘発法を示している。
【図7】トルコのH5N1鳥インフルエンザ大流行の6名の生存者から得られた血液試料についてH5ヘマグルチニン(HA)の血清学的研究結果を示している。これらのデータは、HA抗原に対する抗体の存在を実証している。
【図8】H5抗原(A/Vietnam/1203/2004)およびH1N1(A/New Caledonia/20/99)およびH3N2(A/Panama/2007/99)ウイルスについてテストした、12名のローカルドナーの血清試料を用いて得られた血清学的研究結果を示している。
【図9】抗体ファージライブラリーの構築において使われた独特のバーコード法を示している。
【図10】H5タンパク質およびH5N1ウイルスについてトルコの鳥インフルエンザ生存者のプールされたライブラリーから得られた5種類のクローンのscFvELISAテストの結果を示している。
【図11】報告されたトルコの分離株からH5ヘマグルチニンタンパク質の配列およびLos Alamos National Laboratoryの配列データベースからダウンロードした1つのベトナムの分離株を比較している配列アラインメントを示している。
【図12】対応する軽鎖および生殖系列起源配列と共に、トルコドナーのプールされた抗体ライブラリーにおいて同定された独特のクローンの重鎖可変領域配列を示している。この図に示した配列(3〜23重鎖クローン)は、3ラウンドのパニング後の、すべてのトルコドナーのすべての重鎖および軽鎖のプールされたライブラリーに由来する。
【図13】対応する軽鎖および生殖系列起源配列と共に、トルコドナーのプールされた抗体ライブラリーにおいて同定された独特のクローンの重鎖可変領域配列を示している。この図に示した配列(3〜30重鎖クローン)は、2ラウンドのパニング後の、すべてのトルコドナーのすべての重鎖および軽鎖のプールされたライブラリーに由来する。
【図14A】4ラウンドのパニング後、個々のトルコドナーの抗体ライブラリーから同定された、追加された独特のH5N1に特異な抗体重鎖可変領域配列を示している。
【図14B】4ラウンドのパニング後、個々のトルコドナーの抗体ライブラリーから同定された、追加された独特のH5N1に特異な抗体重鎖可変領域配列を示している。
【図14C】4ラウンドのパニング後、個々のトルコドナーの抗体ライブラリーから同定された、追加された独特のH5N1に特異な抗体重鎖可変領域配列を示している。
【図14D】4ラウンドのパニング後、個々のトルコドナーの抗体ライブラリーから同定された、追加された独特のH5N1に特異な抗体重鎖可変領域配列を示している。
【図15】トルコの鳥インフルエンザ生存者の血清および骨髄の分析により同定された抗体重鎖配列を用いて多様な抗体重鎖および軽鎖ライブラリーを作り出すために目標の突然変異誘発の使用を例示している。
【図16】トルコの鳥インフルエンザ生存者の血清および骨髄の分析により同定された抗体軽鎖配列を用いて多様な抗体重鎖および軽鎖ライブラリーを作り出すために目標の突然変異誘発の使用を例示している。
【図17】HAタンパク質のトルコおよびインドネシアの改変体と共にH5N1ベトナムウイルスscFv抗体から得られた特定のFab断片の交差反応性を確認しているELISAの結果を示している。
【図18】HAタンパク質のトルコおよびインドネシアの改変体と共にH5N1ベトナムウイルスscFv抗体から得られた特定のFab断片の交差反応性を確認しているELISAの結果を示している。
【発明を実施するための形態】
【0039】
A.定義
特に明記しない限り、本明細書で使われる技術・科学用語は、本発明が属する技術分野の当業者が普通に理解している意味と同じ意味を有する。Singleton et al.,Dictionary of Microbiology and Molecular Biology 2nd ed.,J.Wiley & Sons(New York,NY 1994)は、本出願において使われる多くの用語の手引書を当業者に提供する。
【0040】
当業者は、本発明を実行する場合に使われる可能性がある、本明細書で説明した方法および物質と類似または同等な方法および物質が多数あることを認めるであろう。実際に、本発明は、説明した方法および物質に決して限定しない。本発明では、次のように用語を定義する。
【0041】
用語「インフルエンザA型サブタイプ」または「インフルエンザA型ウイルスサブタイプ」は、相互互換的に使われ、ヘマグルチニン(H)およびノイラミニダーゼ(N)のウイルス表面タンパク質の種々の組み合わせにより特徴づけられるインフルエンザA型ウイルス改変体を意味し、したがって、例えば、H1N1およびH3N2などのH数とN数の組み合わせにより呼ばれる。これらの用語は、通常突然変異から生じ、異なる病原性特性を示す、各サブタイプ内のすべての菌株(絶滅菌株を含む)を明らかに含んでいる。この種の菌株は、過去、現在および未来の分離株をすべて含む、ウイルスのサブタイプの種々の「分離株」も意味する。したがって、この文脈では、用語「菌株」と「分離株」は相互互換的に使われる。
【0042】
用語「インフルエンザ」は、インフルエンザウイルスによって引き起こされた伝染性疾患を意味するのに使われる。
【0043】
本発明の文脈では、用語「抗体」(Ab:antibody)は、最も広い意味で使われ、特定の抗原ならびに免疫グロブリンおよび抗原特異性に欠ける他の抗体状分子に対して結合選択性を示すポリペプチド類を含む。後の種類のポリペプチド類は、例えば、低レベルではリンパ系により、高レベルでは骨髄腫により産生される。本出願では、「抗体」は、限定なしに、モノクローナル抗体、ポリクローナル抗体、および抗体断片をカバーしている。
【0044】
「未変性抗体」は、通常、2つの同一軽(L:light)鎖と2つの同一重(H:heavy)鎖とからなる、約150,000ドールトンのヘテロテトラマー性糖タンパク質である。各軽鎖は、共有ジサルファイド結合(単数および複数)により重鎖に連結し、ジサルファイド結合の数は、種々の免疫グロブリンのアイソタイプの重鎖間で変わる。各重鎖および軽鎖は、規則的に間隔をあけた鎖内ジサルファイド橋も有する。各重鎖は、1つの終端において可変ドメイン(VH)とそれに続く多数の定常ドメインを有する。各軽鎖は、1つの終端において可変ドメイン(VL)およびその他の終端において定常ドメインを有し;軽鎖の定常ドメインは重鎖の第1定常ドメインを用いて整列させられ、軽鎖可変ドメインは、重鎖の可変ドメインを用いて整列させられている。特定のアミノ酸残基は、軽鎖および重鎖の可変ドメインの間で界面を形成すると考えられている。Chothia et al.,J.Mol.Biol.186:651(1985);Novotny and Haber,Proc.Natl.Acad.Sci.U.S.A.82:4592(1985).
抗体鎖に関連した用語「可変」は、抗体内の配列で広範に違いがあり、その特定の抗原に対する特定の各抗体の結合および選択性に関与する該抗体の部分を意味するのに使われる。この種の可変性は、軽鎖および重鎖の両方の可変ドメインにおいて超可変領域と呼ばれる3つのセグメントに濃縮されている。可変ドメイン類のより高度に保存された部分は、フレームワーク領域(FR:framework region)と呼ばれる。未変性重鎖および軽鎖の可変ドメイン類は、各々が、4つのFR(FR1、FR2、FR3およびFR4、それぞれ)を含み、大抵が3つの超可変領域により接続されたβ−シート配置を採用し、該超可変領域は、ループ接続、および一部の場合にはβ−シート構造の一部を形成している。各鎖の超可変領域は、FR類により、および他の鎖からの超可変領域を用いて近接して一緒に保持され、抗体の抗原結合部位の形成に寄与する(Kabat et
al.,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.(1991),pages 647−669を参照のこと)。定常ドメイン類は、抗体の抗原への結合には直接には関与しないが、抗体依存性細胞毒性における抗体の関与など種々のエフェクター機能を示す。
【0045】
用語「超可変領域」は、本明細書で使われる場合、抗原結合に関与している抗体のアミノ酸残基類を意味している。超可変領域は、「相補性決定領域」または「CDR」からのアミノ酸残基類を含む(すなわち、軽鎖可変ドメインにおける残基30−36(L1)、46−55(L2)および86−96(L3)、ならびに重鎖可変ドメインにおける30−35(H1)、47−58(H2)および93−101(H3);MacCallum
et al.,J Mol Biol.1996。「フレームワーク」または「FR」残基類は、本明細書で定義された超可変領域残基類以外の可変ドメイン残基類である。
【0046】
これらの重鎖の定常ドメインのアミノ酸配列により、抗体は種々のクラスに帰属させることができる。抗体のクラスには、主なものが5つ、すなわち、IgA、IgD、IgE、IgG、およびIgMがあり、これらのいくつかは、さらに、サブクラス(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgA、およびIgA2に分類することができる。
【0047】
種々のクラスの免疫グロブリンに対応する重鎖定常ドメイン類は、それぞれ、α、δ、ε、γ、およびμと呼ばれる。
【0048】
脊椎動物に由来する抗体の「軽鎖」は、これらの定常ドメインのアミノ酸配列に基づいて、カッパ(κ)およびラムダ(λ)と呼ばれる2つの明確に異なる型の1つに帰属させることができる。
【0049】
「抗体断片」は、全長抗体の一部、一般に、抗体の抗原結合または可変ドメインを含む。抗体断片の例には、Fab、Fab’、F(ab’)2、およびFv断片、線状抗体、単一鎖抗体分子、二重特異性抗体、および抗体断片から形成された多選択性抗体があるが、これらに限定されない。
【0050】
用語「モノクローナル抗体」は、B細胞の単一クローンにより合成された抗体分子を意味するのに使われる。修飾語句「モノクローナル」は、ほぼ均一な抗体集団からえられている抗体のキャラクターを示しており、特定の方法により抗体の産生を必要とするものと解釈すべきではない。したがって、モノクローナル抗体は、Kohler and Milstein,Nature 256:495(1975);Eur.J.Immunol.6:511(1976)により最初に報告されたハイブリドーマ法、組換えDNA法により作るか、またはファージ抗体ライブラリーから分離してもよい。
【0051】
用語「ポリクローナル抗体」は、B細胞の集団により合成された抗体分子の集団を呼ぶのに使われる。
【0052】
「単一鎖Fv」または「sFv」抗体断片は、抗体のVHおよびVL両ドメインを含み、これらのドメインは、単一ポリペプチド鎖中に存在する。一般に、Fvポリペプチドは、さらに、VHおよびVL両ドメインの間にポリペプチドリンカーを含み、該リンカーは、sFvが、抗原結合について望ましい構造を形成することを可能にする。sFvの総説については、The Pharmacology of Monoclonal Antibodies,vol.113,Rosenburg and Moore eds.Springer−Verlag,New York,pp.269−315(1994)のPlueckthunを参照のこと。単一鎖抗体は、例えば、国際公開第88/06630号および国際公開第92/01047号で開示されている。
【0053】
用語「二重特異性抗体」は、2つの抗原結合部位を有する小さな抗体断片を意味し、該断片は、同じポリペプチド鎖(VH−VL)において、軽鎖可変ドメイン(VL)に接続した重鎖可変ドメイン(VH)を含む。同じ鎖上で2つのドメイン間にペアを作るには短かすぎるリンカーを用いることにより、これらのドメインは別の鎖の相補性ドメインとペアを作ることを強いられる。二重特異性抗体は、例えば、欧州特許第404,097号;国際公開第93/11161号;およびHollinger et al.,Proc.Natl.Acad.Sci.USA 90:6444−6448(1993)により詳細に説明されている。
【0054】
用語「二重特異性抗体」は、2つの異なる型の抗原に対して特異性を示す抗体を意味している。本明細書で使われる用語は、標的抗原および特定の組織への送達を容易にする別の標的に対して結合特異性を示す抗体を、無制限に特異的に含んでいる。同様に、多選択性抗体は、2つ以上の結合特異性を有する。
【0055】
表現「線状抗体」は、抗原結合領域のペアを形成する一対の直列型のFdセグメント(VH−CH1−VH−CH1)を含むことを意味するのに使われる。線状抗体は、二重特異性または単一特異性であり、例えば、Zapata et al.,Protein Eng.8(10):1057−1062(1995)により説明されている。
【0056】
用語「中和抗体」は、本明細書では最も広い意味で使われ、インフルエンザウイルスは、中和が行われるメカニズムとは関係なく、標的細胞を複製により感染することを抑制するあらゆる抗体を意味している。したがって、例えば、ウイルスの連結または接着に関与する部位に、直接またはすぐそばに結合する抗体を遺伝子操作することにより、該ウイルスの細胞表面への連結または接着を抑制することにより、中和を実現することができる。ウイルス表面に向けられた抗体によりウイルスを凝集させることにより、中和を実現することもできる。中和は、さらに、標的細胞にウイルスを連結後ウイルスおよび細胞膜の融合の抑制、エンドサイトーシスの抑制、感染細胞から子孫ウイルスの抑制などにより、中和をさらに行うことができる。本発明の中和抗体は、中和が行われるメカニズムにより限定されない。
【0057】
用語「抗体レパートリー」は、本明細書では最も広い意味で使われ、結合能力、結合特異性、胃腸内を輸送する能力、安定性、親和性などの特性を選別するのに使われる抗体または抗体断片のコレクションを意味している。この用語は、具体的には、例えば、制約なしに、単一鎖Fv(scFv)を含む、抗体ファージディスプレイ・ライブラリー、およびナイーブな合成、半合成ライブラリーを含むあらゆる供給源に由来するFab抗体ファージディスプレイ・ライブラリーなどすべての形のコンビナトリアルなライブラリーを含む抗体ライブラリーを含んでいる。
【0058】
「ファージディスプレイ・ライブラリー」は、ファージ被覆タンパク質との融合物としてクローンタンパク質配列コレクションを発現するタンパク質発現ライブラリーである。したがって、フレーズ「ファージディスプレイ・ライブラリー」は、本明細書では、ファージ(例えば、糸状のファージ)のコレクションを意味し、該ファージは外部(通常異種)タンパク質を発現している。この外部タンパク質は、該ファージが接触している他の部分と自由に相互作用(該部分に結合)している。外部タンパク質を示している各ファージは、該ファージディスプレイ・ライブラリーの「メンバー」である。
【0059】
「抗体ファージディスプレイ・ライブラリー」は、抗体または抗体断片を示すファージディスプレイ・ライブラリーを意味している。この抗体ライブラリーは、ファージの集団またはこの種のファージ集団をコードしているベクターのコレクション、またはこの種のファージまたはベクターのコレクションを宿す細胞を含んでいる。このライブラリーは、ファージ粒子につき平均して1つの単一鎖抗体または抗体断片を示す一価、あるいはファージ粒子につき平均して2個以上の抗体または抗体断片を示す多価でよい。用語「抗体断片」は、単一鎖Fv(scFv)断片およびFab断片を無制限に含んでいる。好ましい抗体ライブラリーは、平均して106より多い、または107より多い、または108より多い、または109より多い種類のメンバーを含む。
【0060】
用語「糸状ファージ」は、その表面に不均一なポリペプチドを示し得るウイルス粒子を意味し、f1、fd、Pf1、およびM13を無制限に含んでいる。この糸状ファージは、テトラサイクリン(例えば、「fd−tet」)などの選択可能なマーカーを含んでよい。種々の糸状ファージディスプレイシステムが、当業者には周知である(例えば、Zacher et al.,Gene 9:127−140(1980),Smith et al.,Science 228:1315−1317(1985);and Parmley and Smith,Gene 73:305−318(1988)参照のこと)。
【0061】
用語「パニング」は、標的に対して高い親和性および特異性を有する抗体などのファージ搬送化合物の同定および分離における複数ラウンドの選別プロセスを意味している。
【0062】
本明細書で使われる用語「ヒト以外の動物」には、例えば、ヒト以外の霊長類、げっ歯類(例えば、マウスおよびラット)、ならびに、ウサギ、ブタ、ヒツジ、ヤギ、ウシ、ブタ、ウマおよびロバなどのげっ歯類以外の動物があるが、これらに限定されない。「ヒト以外の動物」には、鳥類(例えば、ニワトリ、七面鳥、アヒル、ガチョウ、など)もある。本明細書で使われる用語「霊長類以外の動物」は、上で具体的に並べた哺乳類を含むがこれらに限定されない霊長類以外の哺乳類を意味している。
【0063】
フレーズ「機能的に異なる抗体」およびこれらの抗体の文法的な改変体は、例えば、標的を中和する能力、生物活性の範囲または特質などの、結合特異性、結合親和性、および何らかの免疫学的または生物学的機能を無制限に含む少なくとも1つの特性において互いに異なる抗体を意味するのに使われる。
【0064】
フレーズ「保存されたアミノ酸残基類」は、互いに整列させられた2つ以上のアミノ酸配列の間で同一であるアミノ酸残基類を意味するのに使われる。
B.一般的技法
本発明の方法を行う技法は、当該技術において周知であり、例えば、Ausubel et al.,Current Protcols of Molecular Biology,John Wiley and Sons(1997);Molecular
Cloning:A Laboratory Manual,Third Edition,J.Sambrook and D.W.Russel,eds.Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001;Antibody Phage
Display:Methods and Protocols,P.M.O’Brian and R.Aitken,eds.,Humana Press,In:Methods in Molecular Biology,Vol.178;Phage Display:A Laboratory Manual,C.F.Barbas III et al.,eds.Cold Spring Harbor,New York,USA,Cold Spring Harbor Laboratory Press,2001;and Antibodies,G.Subramanian,ed.,Kluwer Academic,2004;を含む研究室用標準教科書に記載されている。突然変異誘発は、例えば、部位特異的な突然変異誘発を用いて行うことができる(Kunkcl et al.,Proc.Natl.Acad.Sci.USA 82:488−492(1985))。
【0065】
次の説明では、本発明は、特定の型の抗体ライブラリーを参照しながら説明しているが、本発明は、特定の型の抗体ライブラリーの使用に限定されない。組換えモノクローナル抗体ライブラリーは、免疫断片またはナイーブな断片をベースにすることができる。免疫抗体ライブラリーに由来する抗体は、通常、VHおよびVL遺伝子プールを用いて構築され、該プールは、供給源のB細胞から、ランダムにコンビナトリアルなライブラリーを産生するために発現する適切なベクター内にクローン化され、該ライブラリーは、次いで、選択および/または選別される。他の型のライブラリーには、抗原に結合するクローンに対して明らかに偏っていない遺伝子の供給源に由来する抗原断片が含まれている。したがって、ナイーブな抗体ライブラリーは、天然の、免疫されていない、再整理されたV遺伝子から導かれる。合成抗体ライブラリーは、完全なまたは目的に合わせられた縮重のエリアを1つ以上のV遺伝子のCDR内に導入して、インビトロ法により完全に構築される。半合成ライブラリーは、天然と合成の多様性を併用し、所望のレベルの機能的多様性を維持しながら、天然の多様性を増すためにしばしば作り出される。したがって、この種のライブラリーは、例えば、天然のCDR領域を混ぜ合わせる(Soderlind et al.,Nat.Biotechnol.18:852−856(2000))か、または合成CDR1およびCDR2の多様性(Hoet et al.,Nat.Biotechnol.23:455−38(2005))を用いてヒトのB細胞に由来する自然に再整理されたCDR配列を併用することにより作り出すことができる。本発明は、ナイーブ、合成および半合成のライブラリーまたはこれらの併用使用を包含する。
【0066】
同様に、本発明の方法は、抗体のディスプレイに使われる特定の技術により限定されない。本発明は、ファージディスプレイに関連して説明しているが、本発明の抗体は、リボソームまたはmRNAディスプレイ(Mattheakis et al.,Proc.Natl.Acad.Sci.USA 91:9022−9026(1994);Hanes and Pluckthun,Proc.Natl.Acad.Sci.USA 94:4937−4942(1997))、細菌ディスプレイなどの微生物細胞ディスプレイ(Georgiou et al.,Nature Biotech.15:29−34(1997)),または酵母細胞ディスプレイ(Kieke et al.,Protein Eng.10:1303−1310(1997)),レトロウイルスディスプレイなどの哺乳類細胞上のディスプレイ、胞子ディスプレイ、ウイルスディスプレイ(Urban et al.,Nucleic Acids Res.33:e35(2005),タンパク質−DNA結合をベースにしたディスプレイ(Odegrip et al.,Proc.Natl.Acad.Sci.USA 101:2806−2810(2004);Reiersen et al.,Nucleic Acids Res.33:e10(2005)),およびマイクロビーズディスプレイ(Sepp et al.,FEBS Lett.532;455−458(2002))など他のディスプレイおよび濃縮技術によっても同定することができる。
【0067】
リボソームディスプレイでは、該抗体およびコードmRNAはリボソームにより連結され、ディスプレイは、mRNAの翻訳の終端においてポリペプチドを放出することなく停止される。選択は、全体として3元錯体に基づいている。
【0068】
mRNAディスプレイライブラリーでは、抗体と該コードmRNAとの間の共有結合は、アダプター分子として使われる、プロマイシンにより作られる(Wilson et al.,Proc.Natl.Acad.Sci.USA 98:3750−3755(2001))。ディスプレイ抗体にこの技法を使用する場合は、例えば、Lipovsek and Pluckthun,J.Immunol.Methods,290:51−67(2004)を参照のこと。
【0069】
微生物細胞ディスプレイ技法には、サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)などの酵母上の表面ディスプレイがある(Boder and Wittrup,Nat.Biotechnol.15:553−557(1997))。したがって、例えば、抗体は、α−アグルチニン酵母接着受容体に融合することによりサッカロマイセス・セレヴィシエの表面にディスプレイすることができ、該受容体は酵母の細胞壁上に配置される。この方法は、フローサイトメトリーによりレパートリーを選択する可能性を提供する。蛍光標識付きの抗原および抗エピトープ標識試薬を用いて細胞を染色することにより、酵母細胞は、細胞表面上の抗原結合および抗体発現のレベルにより分類することができる。酵母ディスプレイプラットフォームは、ファージと併用することもできる(例えば、Van den Beucken et al.,FEBS Lett.546:288−294(2003))。
【0070】
抗体ライブラリーを選択し、選別する技法の総説については、例えば、Hoogenboom,Nature Biotechnol.23(9):1105−1116(2005)を参照のこと。
C.好ましい実施形態に関する詳細な説明
本発明は、絶滅株の分離株を含むインフルエンザA型サブタイプの1つより多くの株(分離株)を中和するモノクローナル抗体、ならびにH5ヘマグルチニンの存在を特徴とするサブタイプを含む、1つより多くのインフルエンザA型サブタイプに対する中和抗体の選択、産生および使用に関する。特定の実施形態では、本発明は、1つより多くのインフルエンザA型のサブタイプおよび/または1つより多くの分離株、または2つより多い分離株、または3つより多い分離株、または4つより多い分離株、または5つより多い分離株、など、最も好ましくは1つ以上のサブタイプのすべての分離株を中和するモノクローナル抗体の選択、産生および使用に関する。
【0071】
インフルエンザA型ウイルスのビリオンは、線状マイナス一本鎖RNAの8個のセグメントを含んでいる。ゲノム全長は、13600ヌクレオチドであり、該8個のセグメントの長さは、それぞれ、2350ヌクレオチド、2350ヌクレオチド、2250ヌクレオチド、1780ヌクレオチド、1575ヌクレオチド、1420ヌクレオチド、1050ヌクレオチド、900ヌクレオチドである。インフルエンザA型ウイルスの宿主特異性および減弱は、個別にまたはウイルス遺伝子の併用において、ウイルスヘマグルチニン(H,HA)、核タンパク質(NP)、マトリックス(M)、および非構造(NS)遺伝子に起因するとされた(例えば、Rogers et al.,Virology 127:361−373(1983);Scholtissek et al.,Virology 147:287−294(1985);Snyder et al.,J.Clin.Microbiol.24:467−469(1986);Tian et al.,J.Virol.53:771−775(1985);Treanor et al.,Virology 171:1−9(1989)を参照のこと。
【0072】
ヘマグルチニンおよびノイラミニダーゼのタンパク質を含む、インフルエンザA型ウイルスおよびこれらの表面タンパク質のヌクレオチドおよびアミノ酸配列は、GenBankおよび、例えば、the Theoretical Biology and Biophysics Group of Los Alamos National Laboratoryにより管理された、配列データベースthe Influenza Sequence Databaseなど他の配列データベースから入手できる。インフルエンザA型ウイルス・ヘマグルチニン(H1−H15)の15の既知Hサブタイプのアミノ酸配列は、図1(配列番号:1−15)に示している。追加のインフルエンザA型ウイルス・ヘマグルチニンサブタイプ(H16)が、最近、スエーデンでユリカモメから単離され、Fouchier et al.,J.Virol.79(5):2814−22(2005)により報告された。各Hサブタイプには、非常に多様な株があることも知られている。例えば、図1においてH5 A/Hong Kong/156/97と指定されたHAタンパク質の配列は、1997年5月に香港においてヒトから単離されたインフルエンザA H5N1ウイルスから決定され、Suarez et al.,J.Virol.72:6678−6688(1998)において他の関連H5N1分離株から得られたいくつかの追加株の配列と比較して示されている。
【0073】
インフルエンザウイルスであるノイラミニダーゼの触媒および抗原の部位の構造は、Colman et al.,Nature 303:41−4(1983)により公表され、ノイラミニダーゼの配列は、GenBankおよび他の配列データベースから入手できる。
【0074】
感染個体の免疫反応から得られるウイルス特異性抗体は、通常、ウイルスのヘマグルチニンとの相互作用により該ウイルスを中和することが知られている(Ada et al.,Curr.Top.Microbiol.Immunol.128:1−54(1986):Couch et al.,Annu.Rev.Micobiol.37:529−549(1983))。インフルエンザウイルス・ヘマグルチニンの3次元構造およびインフルエンザウイルス・ヘマグルチニンと中和抗体との間の錯体の結晶構造も、測定され、公表された、例えば、Wilson et al.,Nature 289:366−73(1981);Ruigrok et al.,J.Gen.Virol.69(Pt11):2785−95(1988);Wrigley et al.,Virology 131(2):308−14(1983);Daniels et al.,EMBO J.6:1459−1465(1987);and Bizebard et
al.,Nature 376:92−94(2002)を参照のこと。
【0075】
本発明によると、所望の特性を有する抗体は、1つ以上の抗体ライブラリーから同定され、該ライブラリーは、種々の供給源に由来し、種々の型をとることができる。
【0076】
包括的なヒトインフルエンザ抗体ライブラリー
包括的なヒトインフルエンザ抗体ライブラリーは、以前のいろいろなインフルエンザ、季節的突発的流行、および1968年の香港風邪(H3N2)、1957年のアジア風邪(H2N2)、1918年のスペイン風邪(H1N1)、および2004/2005年の鳥インフルエンザ(H5N1)を含むパンデミックの回復期の患者から得られた抗体から作り出すことができる。この種のライブラリーを作るために、インフルエンザウイルスに感染したことが分かっているか、またはその疑いがある個体から血液または骨髄の試料を集める。周辺の血液試料、特に地理学的に離れた供給源からの試料は、輸送、および使用の前に安定化させる必要がある。このためのキットが周知であり、リンパ球の遠心精製の場合、BD Vacutainer(登録商標)CPT(商標)細胞調製チューブが商業的に入手でき、グアニジウム、Trizol、またはRNAlaterを用いて該試料を安定化する。安定化されたリンパ球または全骨髄を受領すると、当該技術分野において周知の免疫グロブリンオリゴプライマーを用いて、重鎖および軽鎖のレパートリーを救出するためにRT−PCRが行われる。PCRレパートリー製品は、当該技術分野で周知の手順に続いて、m13pIIIを用いてフレーム内で直接クローン化するために、リンカーオリゴと併用してscFvライブラリーを作成する。
【0077】
典型的プロトコルでは、ヒト血清内の抗体は、例えば、周知のヘマグルチニン抑制(HAI:hemagglutinin inhibition)アッセイを含む周知の血清学的アッセイにより検出することができる(Kendal,A.P.,M.S.Pereira,and J.J.Skehel.)1982.Concepts and procedures for laboratory−based influenza surveillance.U.S.Department of Health and Human Services,Public Health Service,Centers for Disease Control,Atlanta,Georgia),or the microneutralization assay(Harmon et al.,J.Clin.Microbiol.26:333−337(1988))。この検出ステップは、該血清試料がインフルエンザ中和抗体を含むことがすでに確認されているならば、必要なさそうである。全血からのリンパ球または骨髄に存在するリンパ球は、次に、当該技術分野で周知の方法により処理される。全RNAは、フレッシュな組織またはRNAlater安定化組織からTri BD試薬(Sigma)により抽出する。次いで、単離ドナー全RNAは、Oligotex精製(Quiagen)を用いてさらにmRNAに精製する。次に、第1ストランドcDNA合成は、AccuScript逆転写酵素(Stratagene)のプロトコルに従いランダム九量体オリゴヌクレオチドおよび/またはオリゴ(dT)18プライマーを用いて行う。要するに、AccuscriptのRTバッファー(Stratagene)中で、100ngのmRNA、0.5mMのdNTPおよび300ngのランダム九量体および/または500ngのオリゴ(dT)18プライマーを65℃で5分間インキュベートし、次いで、4℃へ急冷する。次いで、100mMのDTT、Accuscript RT、およびRNAse Blockを、各反応物に添加し、42℃で1時間インキュベートし、70℃で15分間加熱して逆転写酵素を不活性化する。得られたcDNAは、該抗体重鎖および軽鎖のV遺伝子のRT−PCR増幅の鋳型として用いることができ、これらの遺伝子は、次いで、ベクター、またはファージディスプレイ・ライブラリーが意図されるならば、ファージミドベクターにクローン化することができる。この手順は、抗体重鎖および軽鎖の可変領域クローンのレパートリー(VHおよびVLのライブラリー)を作り、このレパートリーは選別するために個別にまたは組み合わせて保持することができる。
【0078】
早期の流行および1918年のスペイン風邪などのパンデミックの生存者の周辺リンパ球からの免疫グロブリンレパートリーは、上で説明した方法と類似した方法で回収され、安定化され、救出することができる。追加のH1およびH3のライブラリーでは、レパートリーは、適切な時間にワクチンを注射した局所調達ドナーから回収することができる。追加の選択肢として、商業的に利用できる骨髄総RNAまたはmRNAは、H1およびH3に適したライブラリー、およびドナーのバックグラウンドによっては、H2抗体選別にも適したライブラリー、を作るために販売元から購入することができる。
【0079】
普遍的抗体ライブラリー(UAL:Universal Antibody Library)−合成ヒト状レパートリー
本発明の方法では、合成ヒト抗体レパートリーは、普遍的抗体ライブラリーにより表すことができ、このライブラリーは、当該技術分野で周知または販売元から得た方法により作ることができる。したがって、この種のライブラリーのサブセットを含む普遍的免疫グロブリンライブラリーは、2003年12月11日に発行された米国特許出願公開第2003−0228302号で説明され、その全体の開示は、引用により明らかに本明細書に組み込まれている。要するに、この特許発行物は、興味深い原型免疫グロブリンのライブラリーについて説明しており、該免疫グロブリンでは、所定の単一アミノ酸が、興味深い免疫グロブリンの1つ以上の相補性決定領域における1つ以上の位置において置換されている。この種のライブラリーのサブセットには、突然変異した免疫グロブリンがあり、該免疫グロブリンでは、所定のアミノ酸が、可能なすべての組み合わせにおいて免疫グロブリンの6つの相補性決定領域における1つ以上の位置において置換されている。この種の突然変異は、例えば、米国特許第5,798,208、5,830,650、6,649,340号、および米国特許出願公開第2003−0194807号に記載されたウォークスルー突然変異誘発により作ることができ、これらの開示内容の全体が引用により本明細書に明らかに組み込まれている。ウォークスルー突然変異誘発では、免疫グロブリンのライブラリーは、該ライブラリーにおいて所定の単一アミノ酸が、該免疫グロブリンの1つ以上の相補性決定領域(CDR:complementarity determining region)またはフレームワーク(FR)領域などの該免疫グロブリンにおいて興味深い、規定された領域、またはいくつかの規定された領域の各位置に少なくとも一度組み込まれる。得られる突然変異免疫グロブリンは、原型免疫グロブリンにおける同じ位置(単数および複数)に存在した「未変性」または「野生型」アミノ酸の代わりに、該免疫グロブリンの1つ以上の領域(例えば、CDRまたはFR領域)内で、これらの免疫グロブリンが1つ以上の位置に組み込まれた所定の単一アミノ酸を有することに、原型免疫グロブリンとの違いがある。突然変異された免疫グロブリンのセットには、興味深い規定された領域の各位置に対して個別に突然変異された免疫グロブリンがあり、したがって、興味深い規定された領域の各位置に対して(例えば、CDRまたはFR)、突然変異された各免疫グロブリンは、原型免疫グロブリンに存在するアミノ酸か、または所定のアミノ酸および可能なすべての改変体を含む突然変異されたすべての免疫グロブリンの混合物を有する。
【0080】
種々の突然変異を有する抗体重鎖および軽鎖の特定のサブライブラリーは、併用すると本発明の抗体のフレームワーク・コンストラクトを得ることができ、このコンストラクトは、次いで、重鎖および軽鎖両方のCDRにおいて多様性を導入する。この多様性は、例えば、Kunkelによる突然変異誘発などの当該技術分野において周知の方法により実現することができ、多様性をさらに増すために数回繰り返すことができる。したがって、重鎖および軽鎖のCDR1およびCD2領域内の多様性は、個別または同時に、複数ラウンドのKunkel突然変異誘発により導入することができる。必要ならば、種々のKunkelクローンは、CDR長さにより分離することができ、および/または標的CDR(例えば、CDR1またはCDR3)において多様性に欠けているクローンは、例えば、鋳型特異性制限酵素を用いた消化により除去することができる。これらのステップが完了すると、ライブラリーのサイズは、約109メンバーを超えるはずであるが、メンバーが比較的少ないライブラリーも有効である。
【0081】
特定の実施形態では、免疫抗体ライブラリーおよび汎用抗体ライブラリーの両方が、本発明の中和抗体の同定に使われる。これら2つの型のライブラリーは、基本的には異なっている。汎用抗体ライブラリーは、タンパク質とペプチドとを結合する予想した能力を有するヒト様抗体の遡及的に合成したコレクションであり、一方、免疫レパートリーは、鳥類H5ヘマグルチニン、および/または場合によっては、H1、H2、またはH3ヘマグルチニンを特異的に認識する配列を含むであろう。したがって、免疫レパートリーは、標的インフルエンザサブタイプ(単数および複数)の必須成分を承認するために理論的に最適化される。これらの差の結果として、これら2つの方法は、抗体の種々のセットを産生し、このようにして、所望の中和抗体を同定するより効率的な方法を提供する。
【0082】
過剰免疫を受けたヒト以外の霊長類の抗体ライブラリー
この方法では、抗体ライブラリーは、マカクまたはヒヒなどの過剰免疫を受けたヒト以外の霊長類から救出される。具体的には、ヒト以外の霊長類は、インフルエンザA型ウイルスの種々のサブタイプまたは種々のヘマグルチニン(H)タンパク質の免疫がある。免疫があるインフルエンザA型ウイルスまたはヘマグルチニンを認識している抗体のタイターを発現している動物は、犠牲にされ、該動物の脾臓を収集した。該免疫動物の血液または骨髄を集め、包括的なインフルエンザ抗体ライブラリーのために、上で説明したように産生された抗体を集め、増幅した。
【0083】
本発明の中和抗体を単離する方法
用いた単数または複数の抗体ライブラリーの型とは無関係に、2種類のインフルエンザAサブタイプおよび/または同じサブタイプの2つの株(分離株)、および/またはヒトおよびヒト以外の分離株と反応性を示すなどの2元特異性を有する抗体は、発見し、制御された交差反応性選択および/または定方向のコンビナトリアルな遺伝子操作および/または突然変異誘発遺伝子操作によって最適化することができる。
【0084】
図2に例示された、典型的な濃縮スキームでは、標的AおよびBとして指定された2つの標的に対して交差反応性を示す抗体を含むライブラリーは、複数ラウンドの濃縮を受ける。濃縮が標的Aとの反応性に基づいているならば、各ラウンドの濃縮は、標的Aに対するプールの反応性強度を高めるであろう。同様に、濃縮が標的Bとの反応性に基づいているならば、各ラウンドの濃縮は、標的Bに対するプールの反応性強度を高めるであろう。図2は、ファージディスプレイ・ライブラリーを選別する場合(下を参照のこと)、使われる選択方法であるパニングを指しているが、この方法は、当該技術分野において周知のもの以外の、上で説明したあらゆる型のライブラリー、およびあらゆる型のディスプレイ技法に同等に適用できる。標的AおよびBは、インフルエンザウイルスの種々の分離株、型およびサブタイプを含むがこれらに限定されない、抗体が結合するあらゆる標的を含んでいる。
【0085】
本発明の目的は、複数の特異性を有する中和抗体を同定することなので、交差反応発見選択スキームが開発された。単純にするために、このスキームは、二重特異性を有する抗体の選択を示す図3で例示する。この場合、2つの標的、すなわち、標的AおよびBとの反応性を示している抗体を含む抗体ライブラリーは、まず、標的の1つ、例えば、標的Aとの反応性について選択され、次いで、他の標的、例えば、標的Bとの反応性について選択される。各連続選択ラウンドが、両方の標的に対して得られるプールの反応性強度を高める。したがって、この方法は、二重特異性を有する抗体を同定する場合に特に有効である。もちろん、この方法は、追加の標的(単数および複数)に対する追加の濃縮ラウンドを含むことにより、さらなる標的に対して反応性を示す抗体の同定に拡大することができる。やはり、選別されたライブラリーがファージディスプレイ・ライブラリーであるならば、選択は、交差反応パニングによりなされるが、他のライブラリーおよび他の選択方法も使用することができる。
【0086】
上で説明した2つの方法の併用には、標的Aおよび標的B、それぞれ、に対する反応性について、2つの別々の濃縮ラウンドがある。すなわち、これら2つのラウンドは、上で説明したように、得られた2つのプールを組み換えるラウンドと、次の交差反応選択ラウンドである。この方法は、図4に例示されている。純粋な交差反応選択におけるように、組換えライブラリーの選択の各ラウンドは、両方の標的に対して得られるプールの反応性強度を高める。
【0087】
図5で例示されたさらなる実施形態では、まず、標的Aと強い反応性を示し、標的Bと検出可能な交差反応性を有するクローンが同定される。このクローンに基づいて、突然変異誘発ライブラリーが作られ、次いで、該ライブラリーは、標的Bおよび標的Aとの反応性に対し、それぞれ、交互ラウンドにおいて選択される。このスキームは、標的Aと強い反応性を維持しながら、標的Bとの反応性を高めた抗体を生じる結果になる。直前に記載されているように、選択はパニングによりなされるが、選別されたライブラリーがファージディスプレイ・ライブラリーであるとしても、他のライブラリー、他のディスプレイ技法、および選択方法も、同じ方法に続いて使用することもできる。
【0088】
上で説明したように、標的AおよびBは、例えば、インフルエンザA型ウイルスの2種類のサブタイプ、同じインフルエンザA型ウイルスの2種類の株(分離株)、2種類の種に由来するサブタイプまたは分離株でよいが、1つの種はヒトであることが好ましい。したがって、例えば、標的Aは、H5N1ウイルスの2004年ベトナム分離株の分離株でよく、標的Bは、H5N1ウイルスの1997年香港の分離株でよい。これらの実施例は、単なる例示であり、2個または3個以上の標的に対して二重および多重特異性を有する抗体は、類似の方法で同定、選択および最適化されうることが強調されている。
【0089】
あるいは、個々のフレームワークおよびCDR長さの分離を可能にするUALなどの抗体ライブラリーが、標的Aに対する抗体の発見に使われるならば、抗原Bは選別され、ライブラリーは類似のパラメータの多様なコレクションに限定されることもありうる。一旦、抗原Bに対する抗体が発見され、次いで、それぞれのAおよびB抗体に基づいてキメラ抗体または突然変異抗体は二重特異性コレクションの遺伝子操作に使われる。
【0090】
ファージディスプレイ
特定の実施形態では、本発明は、多重(二重を含む)特異性を有する中和モノクローナル抗体を機能的に発見するためにファージディスプレイ抗体ライブラリーを利用する。この種の抗体は、H5およびH1,H5およびH2,H5およびH3,H5、H1およびH2,H5、H1およびH3,H5、H2およびH3,H1、H2およびH3などのサブタイプなどの、H5、H7および/またはH9サブタイプを含む、例えば、1つより多くのインフルエンザA型ウイルスサブタイプ、および/または同じサブタイプの1つより多くの株(分離株)を中和し得るモノクローナル抗体でよい。
【0091】
ファージ抗体ライブラリーを作るために、上で説明したライブラリーを含むあらゆる供給源から得られたcDNAライブラリーを、ファージミドベクター内にクローン化する。
【0092】
したがって、例えば、上で説明したように、RT−PCRによりリンパ球または骨髄から救出した抗体重鎖および軽鎖レパートリーのコレクションは、m13 pIIIタンパク質に融合したscFvライブラリーとして再構築する。コンビナトリアルなライブラリーは、約106より多い、または107より多い、または108より多い、または109より多い種々のメンバーを含み、107より多い種々のメンバーまたはそれより上が好ましい。品質管理のために、ランダムクローンを配列し、レパートリー全体の複雑さを評価する。
【0093】
同様に、ナイーブまたは免疫のあるヒト、あるいは過剰免疫のあるヒト以外の霊長類抗体ライブラリーから、重鎖および軽鎖可変領域の早期PCR救出後、PCR産生物をリンカーオリゴと結合し、scFvライブラリーを作りM13 pIII被覆タンパク質を有するフレーム内で直接クローン化する。このライブラリーは、約106より多い、または107より多い、または108より多い、または109より多い種々のメンバーを含み、107より多い種々のメンバーまたはそれより上が好ましい。品質管理のステップとして、レパートリー全体のサイズおよび複雑さを評価するために、ランダムクローンの配列を決める。
【0094】
抗体ファージディスプレイ・ライブラリーは、単一鎖Fv(scFv)またはFabフォーマットなどの種々のフォーマットにおいて抗体を含むことがある。総説については、例えば、Hoogenboom,Methods Mol.Biol.178:1−37(2002)を参照のこと。
【0095】
選別
所望の中和特性を有する抗体を選別する方法は、上で説明した。反応性は、所望のヘマグルチニンタンパク質への直接結合に基づいて評価することができる。
【0096】
ヘマグルチニン(HA)タンパク質の産生
ヘマグルチニン(HA)タンパク質は、組換えDNA技術により産生することができる。この方法では、HA遺伝子は、適切なベクター、ヨトウガ(Sf9)細胞などのバキュロウイルス感染昆虫細胞において発現するために、バキュロウイルス発現ベクターにクローン化されるのが好ましい。
【0097】
HAタンパク質をコードする核酸は、ポリ−his(ヘキサヒスチジン標識)などの
C−末端エピトープ標識がある場合とない場合がある、Bac−to−Bac(Invitrogen)などのバキュロウイルス発現ベクター内に挿入される。ポリ−his標識は、ニッケルキレート・クロマトグラフィによる簡易精製を提供する。
【0098】
一般に、クローン化は、個々に合成されたオリゴからアセンブリーPCRによる基準cDNAの作製を含む。分離株に対応する改変体HAタンパク質は、適切な突然変異体オリゴを追加のアセンブリーPCR内に置換するか、またはKunkel突然変異誘発による突然変異誘発技法により作られる。HAタンパク質配列の2つのクラスターは、H5については、1997年および2004年のサブタイプ分離株が存在する。したがって、単一基準タンパク質は、各クラスターについて作られる。同様に、基準タンパク質は、1918年のスペイン風邪(H1)、1958年のアジア風邪(H2)、1968年の香港風邪(H3)、および現在のH1、H2、H3分離株について作られる。
【0099】
組換えバキュロウイルスは、リポフェクチン(Gibco−BRLから市販されている)を用いて上のBacmidをSf9細胞(ATCC CRL 1711)に形質移入することにより作られる。28℃で4〜5日インキュベート後、放出されたウイルスを取得し、さらなる増幅に使われる。ウイルス感染およびタンパク質発現は、O’Reilley et al.,Baculovirus Expression Vectors:A Laboratory Manual(Oxford:Oxford University Press,1994)により説明された通りになされる。
【0100】
次いで、発現されたポリ−His−標識付きHAポリペプチドは、例えば、次に説明するように、Ni2+キレート親和性クロマトグラフィにより精製することができる。上澄みは、Rupert et al.Nature 362:175−179(1993)が説明した、組換えウイルス感染Sf9細胞から集める。Ni2+−NTAアガロースカラム(Quiagenから市販されている)は、ベッド容積5mLにて用意し、25mLの水で洗浄し、25mLのローディングバッファーを用いて平衡化する。ろ過した細胞抽出物は、0.5mL/分にてカラムに入れる。カラムは、二次洗浄バッファーを用いてベースラインA280まで洗浄し、その時点でフラクションの収集を開始する。次に、このカラムを二次洗浄バッファー(50mMホスフェート、300mMのNaCl、10%グリセロール、pH6.0)を用いて洗浄し、非特異的に結合したタンパク質を溶離する。A280ベースラインに再度到達後、このカラムは、二次洗浄バッファー中の0〜500mMのイミダゾールグラジエントを用いて展開する。1mLの各フラクションを集め、SDS−PAGEおよび銀染色またはアルカリ性ホスファターゼに結合したNi2+−NTA(Qiagen)を用いたウエスタン・ブロットにより分析した。溶離したHis10−標識付きHAポリペプチドを含むフラクションは、プールし、ローディングバッファーに対して透析する。
【0101】
あるいは、IgG標識付き(またはFc−標識付き)HAポリペプチドの精製は、例えば、Protein Aまたはprotein Gカラムクロマトグラフィを含む既知のクロマトグラフィ技法を用いて行うことができる。
【0102】
Sf9細胞HAタンパク質を用いる代替として、他の組換え宿主細胞中に、原核生物、酵母、または比較的高い真核生物細胞を産生することもできる。適切な原核生物には、グラム陰性菌またはグラム陽性菌などの真正細菌(eubacteria)、例えば、エシェリキア属(Escherichia)、例えば、大腸菌(E.coli)などの腸内細菌科(Enterobacteriaceae)、エンテロバクター属(Enterobacter)、エルビニア菌属(Erwinia)、クレブシエラ属(Klebsiella)、プロテウス属(Proteus)、サルモネラ属(Salmonella)、例えば、ネズミチフス菌(Salmonella typhimurium)、セラチア属(Serratia)、例えば、セラチア・マルセッセンス(Serratia marcescans)、および赤痢菌(Shigella)、ならびに枯草菌(B.subtilis)およびリケニホルミス菌(B.licheniformis)(例えば、1989年4月12日に発行されたDD266,710で開示されたリケニホルミス菌41P)などのバシラス属(Bacilli)、緑膿菌(P.aeruginosa)などのシュードモナス属(Pseudomonas)およびストレプトマイセス属(Streptmyces)があるが、これらに限定されない。大腸菌K12株MM294(ATCC31,446)、大腸菌X1776(ATCC31,537)、大腸菌株W3110(ATCC27,325)、およびK5772(ATCC53,635)などの種々の大腸菌株が、公然と利用できる。
【0103】
原核生物に加えて、糸状菌または酵母菌などの真核微生物が、HAポリペプチドをコードする核酸を含むベクターの適切なクローニングまたは発現宿主である。サッカロマイセス・セレヴィシエは、普通に使われる比較的低級の真核宿主微生物である。しかし、多数の他の属、種、および株が、通常、利用でき、本明細書では、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)(Beach and Nurse,Nature 290:140(1981);1985年5月2日に発行された欧州特許第139,383号);例えば、K.ラクティス(Kluyveromyces lactis)(MW98−8C,CBS683,CBS4574;Louvencourt et al.,J.Bacteriol.737(1983)),K.フラジリス(fragilis)(ATCC12,424),K.ブルガリカス(bulgaricus)(ATCC16,045),K.ウイッケラミイ(wickeramii)(ATCC24,178),K.ワルティー(waltii)(ATCC56,500),K.ドロソフィララム(drosophilaraum)(ATCC36,906;Van den Berg et al.,Bio/Technology8:135(1990))、K.サーモトレランス(thermotolerans)、およびK.マルキシアヌス(marxianus)などのクリベロマイセス宿主(Kluyveromyces host)(米国特許第4,943,529号;Fleer et al.,Bio/Technology9:968−975);ヤロウイア属(yarrowia)(欧州特許第402,226号);ピキア・パストリス(Pichia pastoris)(欧州特許第183,070号;Sreekrishna et al.,J.Basic Microbiol.28:265−278(1988));カンジダ属(Candida);トリコデルマ・レージア(Trichoderma reesia(欧州特許第244,234号);アスバンカビ(Neurospora crassa)(Case et al.,Proc.Natl.Acad.Sci.USA 76:5259−5263(1979));シュワニオマイセス・オクシデンタルス(Schwanniomyces occidentalis)(1990年10月31日に発行された欧州特許第394,538号)などのシュワニオマイセス(Schwanniomyces);およびアカバンカビ(Neurospora)、ペニシリウム属(Penicillium)、トリポクラジウム(Tolypocladium)(1991年1月10日に発行された国際公開第91/00357号)などの糸状菌、および偽巣コウジ菌(A.nidulans(Ballance et al.,Biochem.Biophys.Res.Commun.112:284−289(1983);Tilburn et al.,Gene26:205−221(1983);Yelton et al.,Proc.Natl.Acad.Sci.USA 81:1470−1474(1984))およびA.ニガー(niger)Kelly and Hynes,EMBO J.4:475−479(1985)などのアスペルギルス属宿主(Aspergillus)なども本明細書で有効である。メチルトローフ酵母は、本明細書に適しており、ハンゼヌラ(Hansenula)、カンジダ、クロエケラ属(kloeckera)、ピキア属、酵母菌属(Saccharomyces)、トルロプシス属(Torulopsis)、およびロドトルラ属(Rhodotorula)からなる属から選択されたメタノール上で増殖し得る酵母を含むが、これらに限定されない。このクラスの酵母を代表する特異的な種のリストは、C.Anthony,The Biochemistry of Methylotrophs 269(1982)に存在しているであろう。
【0104】
HAタンパク質を発現させる適切な宿主細胞には、多細胞微生物の細胞がある。無脊椎動物の例には、上述のショウジョウバエ(Drosophila)S2およびプテラ属(Spodoptera)Sf9、ならびに植物細胞がある。有効な哺乳類の宿主細胞株の例には、中国ハムスターの卵巣(CHO:Chinese hamster ovary)およびCOS細胞がある。より特異的な例には、SV40(COS−7,ATCC CRL1651)により形質転換されたサルの腎臓CV1株;ヒトの胚腎臓株(HEK293または懸濁培養において増殖するためにサブクローン化されたHEK293細胞(Graham et al.,J.Gen.Virol.36:59(1977));中国ハムスターの卵巣細胞/−DHFR(CHO,Urlaub and Chasin,Proc.Natl.Acad.Sci.USA 77:4216(1980));マウスのセルトリ細胞(TM4,Mather,Biol.Reprod.23:243−251(1980));ヒトの肺細胞(W138,ATCC CCL75);ヒトの肝細胞(Hep G2,HB8065);およびマウス乳腺腫瘍(MMT060562,ATCC CCL51)がある。適切な宿主細胞の選択は、当該技術分野のスキル内にあると考えられている。
【0105】
ヘマグルチニン(HA)タンパク質パニング
HAタンパク質は、上で説明したライブラリーを選別するために、マイクロタイターウエルまたは磁気ビードの表面に固定する。特定の実施形態では、各ライブラリーは、4度で2時間H5タンパク質を結合することができ、次いで、冷PBSを用いて十分洗浄し、0.2Mグリシン−HClバッファー(pH2.5)を用いてHA特異結合クローンを溶離する。回収されたファージは、pHは中性で、感受性宿主大腸菌を感染させることにより増幅される。次いで、陽性クローンの濃縮および死傷者分類のためのその後のクローン単離を繰り返すために、ファージミド産生を誘発することができる。十分濃縮されると、プール全体は、可溶性scFvタンパク質を発現するためにHB2151などの非アンバー抑制大腸菌株内に感染により移される。一方、プール(単数および複数)は、pBADなどのモノマー性scFv発現ベクター内にサブクローン化され、組換え可溶性scFvタンパク質は、下で説明するように、インビトロ分析およびキャラクテリゼーションのために発現される。
【0106】
キャラクテリゼーション
H5クローンは、まず、上で説明した、産生されたH5タンパク質への結合親和性についてテストする。特定の実施例では、2004H5タンパク質(Refseq AAS65618,Isolate;A/Thailand/2(SP−33)/2004(H5N1))への結合がテストされ、および1997H5タンパク質(Refseq AAF74331,Isolate;A/Hong Kong/486/97(H5N1))に対する並行テストがなされ、他の分離株も単独または何らかの組み合わせで使用することができる。2004および1997H5タンパク質を用いて得られた陽性クローンは、2つの広義のカテゴリー;2004selectiveおよび2004/1997nonselectiveに入る。中和に関する典型的な機能テストは、赤血球に対する全ウイルス結合を用いたヘマグルチニン抑制アッセイを含む。安全面の懸念により、組換えタンパク質および赤血球を用いた、代わりのヘマグルチニンアッセイが好ましい。全血の必要性を排除するために、ヘマグルチニン結合抑制アッセイは気道上皮細胞について行うことができる。この結合アッセイは、フローサイトメトリーまたは細胞ELISA(cELISA)をベースにしたアッセイを含む、無制限に、何らかの形態で行うことができる。cELISAを用いると、高価なフローサイトメトリー装置を使用しなくてすみ、より高度に自動化されたクローン・アセスメントおよびより大きなデータコレクションが得られる利点がある。一方、フローサイトメトリーには、感度が高く、一貫性に優れ、高速である利点もある。
【0107】
HIクローンは、現在の2004H1への結合を含む、あらゆるH1タンパク質への結合、および並行して、1918年および1976年のタンパク質への結合についてテストすることができる。陽性クローンは、2つの広義のカテゴリー;2004selectiveおよび2004nonselectiveに入るであろう。やはり、上で説明した方法と類似な方法を用いて中和についてテストすることは必須である。
【0108】
H2およびH3などの他のHAタンパク質は、類似の方法で特性を明らかにすることができる。
【0109】
最適化
鳥類ウイルス(H5)により引き起こされたヒトの感染と関連した潜在的パンデミックを含むインフルエンザの流行およびパンデミックに効率的に対応するために、H5タンパク質、ならびに将来の突然変異など、Hタンパク質の現在の分離株を効率的に中和する抗体が必要である。この目標を達成するために、標的となるヘマグルチニンサブタイプ(単数および複数)の既知のすべての分離株に結合する、様々なH(例えば、H5)中和クローンを同定する必要がある。
【0110】
望ましいならば、例えば、2005年6月23日に公開された、米国特許出願公開第2005−0136428号において説明された、ルックスルー・ムタジェネシス(LTM:Look Through Mutagenesis)など、当該技術分野において周知の方法により、交差反応性をさらに改良することができる。なお、この特許発行物は、引用により明らかに本明細書に組み込まれている。
【0111】
ルックスルー・ムタジェネシス(LTM)は、選択されたアミノ酸のコンビナトリアルな突然変異を同時に評価および最適化する多次元突然変異法である。このプロセスは、1つ以上の相補性決定領域(CDR)ドメイン内の正確な分布に焦点を当て、アミノ酸側鎖に関する化学の相乗的寄与を探っている。LTMは、CDR内の単一変異の位置シリーズを作り、そこでは各野生型残基が、選択された多数のアミノ酸の1つにより系統的に置換される。突然変異CDRは併用され、すべての改変体の定量的ディスプレイに対して禁止的になることなく複雑さおよびサイズを増しながらコンビナトリアルな単一鎖可変断片(scFv)ライブラリーを作る。正の選択後、改善された特性を有するクローンの配列が決定され、これらの有益な突然変異が位置づけられる。改善されたHA結合特性の相乗的突然変異を同定するために、すべての有益な順列を発現しているコンビナトリアルなライブラリー(コンビナトリアルで有益な突然変異、CBM:combinatorial beneficial mutation)は、最適化されたscFv候補のパネルを同定するために正に選択され、分析された混合DNAプローブにより産生することができる。この操作は、Fvおよび他の抗体ライブラリーを用いて類似の方法で行うことができる。
【0112】
突然変異誘発は、上で説明したウォークスルー・ムタジェネシス(WTM:walk through mutagenesis)により行うこともできる。
【0113】
1つより多くのA型インフルエンザサブタイプおよび/または1つより多くの同じサブタイプの分離株を用いて、本明細書の抗体の交差反応性を意図的にデザインするための別の有効な突然変異誘発法は、本明細書では目標突然変異誘発と呼ばれる。目標突然変異誘発は、1つ以上の好ましくは反応性の異なる抗体クローンに基づく抗体のコレクションを合理的に遺伝子操作するために使うことができる。本発明の文脈において、目標突然変異誘発は、抗体の個々のCDRにおける配列など同様の配列上の類似の位置により規定された単一または複数の残基をコードするのに使われる。この場合、これらのコレクションは、同等の位置に存在した残基の範囲を捕獲するためにオリゴ縮重を用いて作られる。このコレクション内には特異性の連続体が、親クローンのコレクションの間またはコレクションを超えて存在するであろうと期待される。目標突然変異誘発の目的は、2つ以上の分離実体またはコレクションの間に多様な多機能性抗体コレクションまたはライブラリーを作ることである。インフルエンザの場合、この方法は、2つの異なるエピトープ、分離株、またはサブタイプを認識する2つの抗体を使用し、両方の機能性品質を単一抗体に変えるために利用することができる。1例として、第1のインフルエンザA型抗体は、H5サブタイプのベトナム分離株に対して特異的であり、第2の抗体は、インフルエンザA型ウイルスのH5サブタイプのタイランドまたはトルコ分離株に対して特異的である。目標突然変異誘発ライブラリーを作成するために、両方の抗体のCDR配列がまず達成され、整列させられる。次に、保存されたアイデンティティのすべての位置は、適合残基に対する単一コドンを用いて固定される。非保存位置では、縮重コドンは、両方の残基をコードするために組み込まれる。一部の例では、縮重コドンは、この位置において2つの親残基をコードするのみである。しかし、一部の例では、追加の副産物が産生される。副産物産生のレベルは、副産物の産生を強制するか、またはサイズの限界または目標に依存するこの産生を排除するためにダイヤルを回すことができる。
【0114】
したがって、例えば、これら2つの抗体の最初の位置が、それぞれ、スレオニンおよびアラニンであるならば、最初の2つの位置においてA/G−G−を有する縮重コドンは、第3の位置の塩基とは関係なく、スレオニンまたはアラニンをコードするだけである。例えば、もし次の位置の残基がリジンおよびアルギニンであるならば、縮重コドンA−A/G−A/Gは、リジンまたはアルギニンをコードするだけであろう。しかし、縮重コドンA/C−A/G−A/G/C/Tが使われたならば、その場合は、アスパラギン、ヒスチジン、グルタミン、およびその上、セリン副産物が作られるであろう。
【0115】
便宜上、適合したCDR長さを有する抗体のみを用いると、より単純である。これを強制する1つの方法は、CDR長さおよび最初に発見された抗体により与えられた潜在的に等しいフレームワーク制限に基づいて第2抗原のサイズ制限ライブラリーを選別することである。しかし、等しい長さのCDRを用いることは、単なる便宜であり必要条件ではないことを指摘する必要がある。この方法は、インフルエンザA型ウイルス中和抗体の機能的に多様な大きなライブラリーを作成するためには有効であるが、その適用性が非常に広いことは、簡単に分かる。この突然変異誘発技法は、何らかの抗体の機能的に多様なライブラリーまたはコレクションを作るために使用することができる。したがって、図6は、突然変異誘発された親配列としてTNF−α抗体およびCD11a抗体のCDRを用いた目標変異誘発方法の使用を例示するために本明細書に含まれている。
【0116】
他の代表的な突然変異誘発法には、飽和突然変異誘発および誤りがちなPCRがある。
【0117】
飽和突然変異誘発(Hayashi et al.,Biotechniques 17:310−315(1994))は、全20のアミノ酸類が、タンパク質内の特定の位置で置換され、各改変体に対応するクローンが特定の表現についてアッセイされる技法である。(米国特許第6,171,820;6,358,709および6,361,974号も参照のこと。)
誤りがちなPCR(Leung et al.,Technique 1:11−15(1989);Cadwell and Joyce,PCR Method Applic.2:28−33(1992))は、クローン化遺伝子内にランダムポイント突然変異を導入する改変ポリメラーゼ連鎖反応(PCR:polymerase chain reaction)技法である。得られるPCR産生物は、クローン化してランダム突然変異体ライブラリーを作るか、またはT7プロモーターが適切なPCRプライマー内に組み込まれるならば、直接転写することができる。
【0118】
他の突然変異誘発技法も周知の技法であり、例えば、In Vitro Mutagenesis Protocols,J.Braman,Ed.,Humana Press,2001で説明されている。
【0119】
この場合、主な目標の1つは、現在のH5(またはH7またはH9)分離株ならびに将来の突然変異を効率的に処置するために抗体(単数および複数)を遺伝子操作することである。例えば、最近の2004年分離株および以前の1997年分離株の両方を含む、種々のH5分離株に結合するクローンを中和する新しい分離株H5における突然変異を承認し得る耐性を有する抗体を遺伝子操作することは、確認すべきである。クローンが2004年分離株について選択されるならば、そのクローンは、それほどの重要性はないが、1997年分離株を結合および/または中和するであろうことが期待される。この場合、目標は、2004年分離株結合を改善する(または少なくとも維持する)文脈内で1997年承認を劇的に改善することである。したがって、選択はまず1997年基準タンパク質について改善がなされ、次いで、2004年タンパク質に関する選択がこれに続く。そのようにすると、第2パラメータに対する圧力を維持しながら、新しい株に対する選択圧力をより大きなものにする。
【0120】
上で説明したライブラリーのいずれか、または当該技術分野で周知の何か他の型のライブラリーを、単独またはいずれかの組み合わせに基づいて最適化することができる。特定の実施形態において、最適化は、3つの型のLTMライブラリー、すなわち、3種類の突然変異誘発軽鎖ライブラリー、3種類の突然変異誘発重鎖ライブラリー、および6種類の突然変異誘発(軽鎖+重鎖)ライブラリー、を選別することにより開始することができる。H5は、小さな改変は望ましいかもしれないが、上で説明したように、本質的には、選別される。例えば、グリシン−HCl溶離の前に、次の方法:室温または37度における十分な洗浄、または過剰の可溶性親scFvの存在下における長時間のインキュベーション、のいずれかまたは両方によりラウンドごとに洗浄の厳しさを高めることにより改善された結合を選択することができる。これらの選択改変は、得られるクローンにおけるオフ率の動態を改善するはずである。3〜4ラウンドの選択後、我々はランダムクローンの配列を決め、ELISAにより結合についてテストすることになる。改善されたクローンの配列分析後、許容できる改善された突然変異のすべては、H5分離株の両方の結合を相乗的に改善する場合に選択するコンビナトリアルな有益突然変異誘発(CBM:combinatorial beneficial mutagenesis)ライブラリーに一体化される。CBMライブラリーは、すべての位置において改善された、オリジナルな親の残基をすべて表すために縮重オリゴヌクレオチド類を合成することにより作られる。得られるライブラリーは、LTM選別と同様に、厳しさを高めながら選択される。十分な選択後、上で説明した結合および中和のアッセイの大腸菌からのモノマー性scFvタンパク質を発現し、精製するために、プールはpBAD発現ベクター内にサブクローン化される。
【0121】
H1中和抗体は、類似な方法で最適化することができる。この場合、1918年、1976年、および出発点または目標としての現在からの何らかの基準タンパク質配列を用いて選択し、最適化することができる。
【0122】
さらに、型間の承認は、中和抗体クローンを用いてテストする。型間の承認の1例は、H5から供給または最適化されたクローンからの同時発生的または遺伝子操作したH1結合である。
【0123】
所望の特性を有する中和抗体が、一旦、同定されると、この種の抗体の大部分により承認された優勢なエピトープ(単数および複数)を同定することが望ましい。エピトープをマッピングする方法は、当該技術分野で周知であり、例えば、Morris,Glenn E.,Epitope Mapping Protocols,Totowa,N.J.ed.,Humana Press,1996;and Epitope Mapping:A Practical Approach,Westwood and Hay,eds.,Oxford University Press,2001において開示されている。
【0124】
種々のドナーからのライブラリーなどの、またはインフルエンザウイルスを含むが、これらに限定されない抗体ライブラリーのハンドリングは、種々の抗体コレクションを区別している独特のバーコードを適用することにより大いに促進される。これらのバーコードは、これらが標識付きのクローン(単数および複数)と同調して伝播し得るように選択されるのが好ましい。
【0125】
したがって、これらのバーコードは、配列決定または特定のPCRプライマーによりデコンボリュートすることができる長さが約1〜24個の非コードヌクレオチド類に等しい非コード配列でよい。このように、抗体レパートリーなどの核酸のコレクションは、クローン化ステップにおいて連結することができる。
【0126】
別の実施例では、これらのバーコードは、サイレント突然変異のコード配列である。これらのライブラリーが、中断回分配列(例えば、Sfi GGCCNNNNNGGCC)を認識する制限酵素を利用するならば、抗体ライブラリーなどのクローンの種々のコレクションを区別するために「N」の代わりに別のヌクレオチド類を組み込むことができる。このバーコード法は、このレパートリーが増幅ステップにおいて連結される利点がある。
【0127】
別の例では、これらのバーコードは、免疫学的に異なるペプチドまたはファージ粒子に融合したタンパク質配列コードするコード配列である。実施例には、例えば、pIII、pVIII、pVII、またはpIXファージへのエピトープ(例えば、Myc、HA、FLAG)融合物がある。これらのエピトープは、単独、または種々の組み合わせで使用することができる、シス(ライブラリーコードプラスミドで)配置として、またはトランス(特異的に改変されたヘルパーファージ)配置として提供することができる。
【0128】
可能なバーコード類の他の実施例には、無制限に、ハプテン類または蛍光染色体を有する化学的および酵素ファージ改変がある。この種の標識は、選択の単一ラウンドの場合好ましい。
【0129】
本明細書では、バーコードは、抗体ライブラリーを区別するものとして例示されるが、当業者は、説明された方法が、独自に標識する場合、および一般に、核酸分子および核酸のコレクションを区別する場合に広く適用できることを十分理解するであろう。
【0130】
中和抗体の産生
所望の中和特性を有する抗体が、一旦、同定されると、抗体断片を含むこの種の抗体は、例えば、ハイブリドーマ技法または組換えDNA技術を含む当該技術分野で周知の方法により産生することができる。
【0131】
ハイブリドーマ法では、マウスまたはハムスターなど他の適切な宿主動物が、免疫付与に使われるタンパク質に特異的に結合する抗体を産生または産生し得るリンパ球を誘発するために免疫される。一方、リンパ球は、インビトロで免疫にされる。次いで、リンパ球は、ポリエチレングリコールなどの適切な融合剤を用いて骨髄腫細胞と融合し、ハイブリドーマ細胞を形成する(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。
【0132】
したがって、このようにして調製されたハイブリドーマ細胞は、適切な培養媒体に播種され、増殖され、該媒体は、未融合の親の骨髄腫細胞の増殖または生存を抑制する1つ以上の物質を含むのが好ましい。例えば、親の骨髄腫細胞に構造ヒボキサンチングアニンホスホリボシル転換酵素(HGPRT:hypoxathine guanine phosphoribosyl transferaseまたはHPRT)が欠けているならば、該ハイブリドーマの培養媒体は、通常、ヒボキサンチン、アミノプテリン、およびチミジン(HAT:hypoxanthine,aminoprotein,thymidine媒体)を含み、これらの物質は、HGPRTに欠けている細胞の増殖を防止する。
【0133】
好ましい骨髄腫細胞は、効率的に融合し、選択された抗体産生細胞により抗体の安定した高レベル産生を支え、HAT媒体などの媒体に敏感な骨髄腫細胞である。これらの中で、好ましい骨髄腫細胞株は、Salk Institute Cell Distribution Center,San Diego,California USAから入手できるMOPC−21およびMPC−11マウス腫瘍から導かれた細胞株などのマウス骨髄腫株およびAmerican Type Culture Collection,Rockville,Maryland USAから入手できるSP−2またはX−63−Ag8−653細胞などのマウス骨髄腫株である。ヒトの骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞株も、ヒトのモノクローナル抗体の産生に対して説明されている(Kozbor,J.Immunol.133:3001(1984);and Brodeur
et al.,Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0134】
その中でハイブリドーマ細胞を増殖する培養媒体は、抗原に対抗するモノクローナル抗体の産生についてアッセイされる。ハイブリドーマ細胞により産生されたモノクローナル抗体の結合特異性は、免疫沈降または放射免疫アッセイ(RIA:radioimmunoassay)または酵素免疫測定吸着法(ELISA:enzyme−linked immunoabsorbent assay)などのインビトロ結合アッセイにより測定されるのが好ましい。
【0135】
組換えモノクローナル抗体は、例えば、必要な抗体鎖をコードするDNAを単離し、周知の組換え発現ベクターを用いた同時発現のコード発現を用いた組換え宿主細胞を同時導入することにより産生することができる。組換え宿主細胞は、上で説明した原核細胞および真核細胞などでよい。
【0136】
ヒト化抗体を作る場合に使われるヒト可変ドメインの選択は、軽重両方ともに、抗原性を下げることが非常に重要である。いわゆる「最適」方法により、げっ歯類抗体の可変ドメインの配列は、既知ヒト可変ドメイン配列の全ライブラリーに対して選別される。次いで、げっ歯類の配列に最も近いヒト配列が、ヒト化抗体のヒトフレームワーク領域(FR)として受け入れられる(Sims et al.,J.Immunol.151:2296(1993);Chopita et al.,J.Mol.Biol.196:901(1987))。抗体が、抗原および他の好ましい生物学的特性に対して高い親和性を保持したまま、ヒト化されることが重要である。好ましい方法によりこの目標を達成するために、ヒト化された抗体が、親の配列およびヒト化された配列の3次元モデルを用いて種々の概念的なヒト化された産生物および親の配列を分析するプロセスにより作られる。
【0137】
さらに、ヒトの抗体は、当該技術分野で周知の次のような方法で作ることができる。例えば、内在性免疫グロブリンの産生がない場合にヒトの抗体のレパートリーの全体を、免疫付与の際に、産生し得る遺伝子導入動物(例えば、マウス)を作ることができる。例えば、Jakobovits et al.,Proc.Natl.Acad.Sci.USA 90:2551(1993);Jakobovits et al.,Nature 362:255−258(1993);Bruggermann et al.,Year in Immuno,7:33(1993);および米国特許第5,591,669、5,589,369および5,545,807号を参照のこと。
【0138】
中和抗体の使用
本発明のインフルエンザ中和抗体は、A型インフルエンザ感染の予防および/または処置に使用することができる。治療に使う場合は、抗体または分子類は、それらの送達が、抗体(単数および複数)をベースにする輸送配列を用いることにより容易になり、通常、医薬組成物の形で使われる。技法および処方は、一般に、Remington’s Pharmaceutical Sciences,18th Edition, Mack
Publishing Co.(Easton,Pa.1990)に記載されている。また、Wang and Hanson”Parenteral Formulations of Proteins and Peptides;Stability and Stabilizers,”Journal of Parenteral Science and Technology,Technical Report No.10,Supp.42−2S(1988)も参照のこと。
【0139】
抗体は、通常、凍結乾燥された製剤または水溶液の形で処方される。許容できる担体、賦形剤、または安定剤は、用いる用量および濃度において受容者に対して無毒であり、燐酸塩、クエン酸塩、および他の有機酸などのバッファー、アスコルビン酸およびメチオニンを含む抗酸化剤、保存剤(オクタデシルジメチルベンジルアンモニウムクロライド、ヘキサメトニウムクロライド、ベンザルコニウムクロライド、ベンゼトニウムクロライド、フェノール、ブチルアルコールまたはベンジルアルコール、メチルパラベンまたはプロピルパラベンなどのパラベン類、カテコール、レゾルシノール、シクロヘキサノール、3−ペンタノール、およびm−クレゾールなど)、低分子量(残基約10個未満)ポリペプチド類;血清アルブミン、ゼラチン、または免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、またはリジンなどのアミノ酸類;単糖類、二糖類、およびグルコース、マノース、またはデキストリン類を含む他の炭水化物;EDTAなどのキレート化剤;スクロース、マニトール、トレハロースまたはソルビトールなどの砂糖類;ナトリウムなどの塩形成対イオン類;金属錯体類(Zn−タンパク質錯体);および/またはTWEEN(商標)、PLURONICS(商標)またはポリエチレングリコール(PEG)を含む。
【0140】
これらの抗体は、例えば、コロイド状薬剤デリバリーシステム(例えば、リポソーム、アルブミンマイクロスフェア、マイクロエマルジョン、ナノ粒子およびナノカプセル)、またはマクロエマルジョンにおいて、液滴形成技法または界面重合(例えば、ヒドロキシメチルセルロースまたはゼラチンマイクロカプセルおよびポリ−(メチルメタクリレート)マイクロカプセル、それぞれ)により調製されたマイクロカプセルに捕捉されることもありうる。この種の技法は、上記、Remington’s Pharmaceutical Sciencesで開示されている。
【0141】
本明細書で開示された中和抗体は、免疫リポソーム類として処方することもできる。抗体を含むリポソーム類は、Epstein et al.,Proc.Natl.Acad.Sci.USA 82:3688(1985);Hwang et al.,Proc.Natl.Acad.Sci.USA 77:4030(1980);米国特許第4,485,045および4,544,545号;および1997年10月23日に発行された国際公開第97/38731号において説明されているような当該技術分野で周知の方法により調製される。循環時間の長いリポソーム類は、米国特許第5,013,556号で開示されている。
【0142】
特に有効なリポソーム類は、ホスファチジルコリン、コレステロールおよびPEG−誘導体化されたホスファチジルエタノールアミン(PEG−PE)を含む脂質組成物を用いて逆相蒸発法により作ることができる。リポソーム類は、規定されたポアサイズのフィルターを通して押し出され所望の直径を有するリポソーム類が得られる。本発明の抗体のFab’断片は、ジサルファイド交換反応によりMartin et al.,J.Biol.Chem.257:286−288(1982)で説明されたようにリポソーム類に接合することができる。化学療法剤は、任意に、リポソーム内に含まれる。Gabizon et al.,J.National Cancer Inst.81(19)1484(1989)を参照のこと。
【0143】
疾患の予防または処置のために、抗体の適切な用量は、処置されるべき感染の型、疾患の重篤さおよびコース、および抗体を投与する目的が予防であるか、治療であるか、により変わる。抗体は、一度または処置シリーズにわたり、患者に適切に投与される。疾患の型および重篤さにより、抗体の患者への初期用量は、約1μg/kgから約15mg/kgまでの範囲にあり、1回以上個別に投与する場合と、連続注入する場合とがある。
【0144】
本発明のより詳細な説明は、次の無制限にある実施例によりなされる。
【実施例】
【0145】
以前の鳥類インフルエンザ大流行の生存者からの抗体ライブラリーおよび中和抗体の調製
物質および方法
骨髄プロトコルおよび血清製剤
血液は、標準静脈穿刺により採血し、凝血させ、処理して血清を回収した。血清は、ドライアイスにのせて出荷するまで、3〜4日間−20℃で貯蔵した。ドナーは、局所麻酔薬を注入して麻酔にかけ、5mlの骨髄を各H5N1生存者の骨盤から採取した。次に、5mlの骨髄を45mlのRNAlater(Ambion)を含む無菌の50mlのチューブに入れた。この混合物を、目視できる凝集塊がなくなるまで、約8〜20回丁寧に反転させ、骨髄とRNAlaterとをよく混合させた。次に、この試料を2〜10℃で一晩冷蔵した。一晩冷蔵後、試料は、ドライアイスにのせて出荷するまで、3〜4日間−20℃で貯蔵した。受け取り次第直ぐに、RNAlater/髄および血清含有チューブは、処理するまで−80℃で貯蔵した。
【0146】
血清学:HA ELISA
ELISAプレート(Thermo,Immulon 4HBX 96W)は、1X ELISAプレートコーティング溶液(BioFX)中で100μlの100ng/mLのH5ヘマグルチニン(Protein Sciences,A/Vietnam/1203/2004)で被覆し、室温で一晩インキュベーションした。次の日複数のプレートを300μlのPBS/0.05%Tween−20(PBST)で3度洗浄した。洗浄後、300μlのブロッキング溶液(PBS/0.05%Tween−20中4%無脂肪ドライミルク)を添加し、室温で1時間インキュベートした。ブロッキングステップの後、これらのプレートは、300μlのPBS/0.05%Tween−20(PBST)で3度洗浄した。次に、PBS/0.05%Tween中で1:20,000に希釈した100μlの血清試料を、室温で1〜2時間インキュベートし、次いで、300μlのPBS/0.05%Tween−20で3度洗浄した。PBS/0.05%Tween中で1:5,000に希釈した抗ヒトFc−HRP接合体の100μlを室温で1〜2時間インキュベートし、次いで、300μlのPBS/0.05%Tween−20で3度洗浄した。この最終洗浄の後、100μlの発色基質溶液(TMB1基質、BioFX)を添加し、十分な時間が経過した後、100μlのSTOP溶液(BioFX)を添加して終結させた。450nmにおける吸光度をプレートリーダー(Softmax Proソフトウエアを備えたMolecular Devices Thermomaxマイクロプレートリーダー)で読み取り、データを記録し、次いで、エクセル(マイクロソフト)を用いてプロットした。
【0147】
骨髄:RNA抽出およびmRNAの精製
予め−80℃で貯蔵した骨髄(20mlのRNAlater中約2.5ml)を、RNAlaterを除去するために遠心分離により回収し、次いで、300μlの酢酸を含む11.25mlのTRIBD試薬(Sigma)に再懸濁した。次いで、このペレットは、激しく渦流状にした。次に、1.5mlのBCP(1−ブロモ−3−クロロプロパン,Sigma)を添加し、渦流にすることにより混合し、室温で5分間インキュベートし、次いで、4℃で15分間12000×gで遠心分離した。界面を乱さないように、水相を注意して除去した。次に、水相からの全RNA25mlのイソプロパノールを添加して沈殿させ、室温で10分間インキュベートし、4℃で10分間12000×gで遠心分離した。イソプロパノールを添加後、残留RNAlaterにより2相が形成され、界面に沈殿RNAが沈降しているのが認められた。残留RNAlaterを排除し、RNAを最大限に回収するために、H2O中50%のイソプロパノールの5mlアリコートを添加し、相分離が認められなくなるまで混合し、その点で4℃で10分間12000×gで遠心分離によりRNAをペレット化した。RNAペレットを75%EtOHで洗浄し、RNAseフリーの1.6mlマイクロ遠心チューブに移し、遠心分離により再び回収した。最後に、RNAペレットを100μlの1mM燐酸ナトリウム、pH8.2に再懸濁させ、RNA純度を決めるためにA260およびA280を読みとった。
【0148】
逆転写の前に、mRNAは、Qiagen Oligotex mRNA精製キットにより、全RNAから精製した。要するに、50〜200μgの骨髄RNAをRNaseフリーの水を用いて250μlにし、250μlのOBBバッファーおよびOligotex懸濁液と混合し、次いで、70℃で3分間インキュベーションした。Oligotex粒子のオリゴdT30とmRNAポリ−A−テールとの間のハイブリッド形成は、室温で10分間行った。次いで、ハイブリッド形成された懸濁液は、スピンカラムに移し、1分間遠心分離した。スピンカラムは、400μlのバッファーOW2で2度洗浄した。次いで、精製したmRNAは、20μlのホットな(70℃)バッファーOEBを用いて遠心分離により2度溶離した。典型的な収量は、全RNAとして500ng〜1.5μgであった。
【0149】
骨髄mRNAについてN9およびオリゴdTを用いた逆転写
逆転写(RT:reverse transcription)反応は、75〜100ngのmRNAを、2μlの10X Accuscript RT Buffer(Stratagene)、0.8μlの100mM dNTP、およびN9(300ng)とオリゴdTプライマー(100ng)とのいずれかとを一緒に混合し、次いで、水で最後に、容積を17μlにした。これらの混合物を65℃で5分間加熱し、次いで、室温まで冷却した。次に、2μlのDTT、0.5μlのRNase Block(Stratagene)、0.5μlのAccuscript RT(Stratagene)を、各反応物に添加した。次に、N9で予め刺激を受けた反応物を室温で10分間インキュベートし、オリゴ−dTで予め刺激を受けた反応物を氷上で10分間インキュベートした。最後に、両方の反応物を42℃で60分間、次いで、70℃で15分間インキュベートし、酵素を撲滅した。
【0150】
骨髄誘導cDNAからのPCR
抗体の重鎖および軽鎖のレパートリーは、ヒトの生殖細胞VおよびJ領域に基づいて先に説明した方法および縮重プライマーを用いて本質的に骨髄cDNAから増幅された(O’Brien,P.M.,Aitken R.Standard protocols for the construction of scFv Libraries.Antibody Phage Display−Methods and Protocols,vol 178,59−71,2001,Humana Press)。
【0151】
要するに、ラムダ軽鎖に対して先に刺激されたオリゴdT cDNA(75ngのmRNAから)および先に刺激されたN9 cDNA(カッパ軽鎖の75ngのmRNAから、重鎖の100mgのmRNAから)を用いたPCR反応は、5μlの10X増幅バッファー(Invitrogen)、1.5μlのdNTP(10mM)、1μlのMgSO4(50mM)、2.5μlのVregionプライマー(10μM)および2.5μlプライマー(10μM)−VHの場合10μM、0.5μlの白金Pfxポリメラーゼ(Invitrogen)、および最終容積を50μlにする無菌dH2Oと一緒に混合した。PCRパラメータは、ステップ1−95℃5分、ステップ2−95℃30秒、ステップ3−58℃30秒、ステップ4−68℃1分、ステップ5−サイクルステップ2−4を40回、ステップ6−68℃5分であった。軽鎖PCR産生物は、Qiagen PCR
Cleanupキットを用いてクリーンアップした。重鎖PCR産生物は、Qiagen Gel Extraction Kitを用いて1.5%アガロースゲルから精製し、次いで、再増幅したゲルであつた。重鎖再増幅は、次のようにして行った。すなわち、10μlの10X増幅バッファー(Invitrogen)、3μlのdNTP(10mM)、2μlのMgSO4(50mM)、5μlのVHプライマー(10μM)および5μlのJHプライマー(10μM)、5μlの重鎖Primary PCR産生物、1μlの白金Pfxを混合し、水で容積を100μlに調節した。サイクリングパラメータは、ステップ1−95℃5分、ステップ2−95℃30秒、ステップ3−58℃30秒、ステップ4−68℃1分、ステップ5−サイクルステップ2−4を20回、ステップ6−68℃5分であった。再増幅した重鎖PCR産生物は、Qiagen Extraction Kitを用いて1.5%アガロースTAEゲルからクリーンアップした。
【0152】
抗体ファージライブラリーの構築
個々の鳥類インフルエンザの生存者に関する個別の抗体ライブラリーは、末端のpIIIストップコドンに続いて未翻訳領域に挿入した3−ヌクレオチドを同定する独特のバーコードを用いて構築した。
【0153】
軽鎖クローン化
プールされたカッパ軽鎖およびラムダ軽鎖をドナーにつき各々1μgは、Qiagen
Gel Extraction Kitを用いて1.5%アガロースTAEゲルから精製したゲル、ならびにNotIおよびBamHIを用いて消化した。5μgの各ベクターは、Qiagen Gel Extraction Kitを用いて1%アガロースTAEゲルから精製したゲル、ならびにNotIおよびBamHIを用いて消化した。ライブラリーの連結は、200mgのゲル精製カッパまたはラムダの挿入物および1μgのゲル精製ベクターを用いて、60μl中RTで1時間または14℃で一晩行った。連結は、Edge BioSystem Perfromaスピンカラムを用いて脱塩した。このライブラリーは、80μlのTG−1またはXL−1ブルーのアリコートにおいて5つの電気穿孔中で形質転換し、各々1mlのSOCとして回収し、プールし、37℃で1時間増殖させた。形質転換体の総数は、形質転換の各々からのアリコートを蒔くことによりこの増殖後に測定した。残りの電気穿孔は、200mlの2YT+50μg/mlのAmpicillin+2%グルコース中37℃で一晩増殖することにより増幅させた。次の軽鎖ライブラリーは、Qiagen高速Maxiprepキットを用いてこれらの一晩培養物からプラスミド精製により回収した。
【0154】
重鎖クローン化
各々が1.5〜2μgのドナー特異性重鎖類(VH1、VH2、5、6プール、VH3、およびVH4)は、Qiagen Gel Extraction Kitを用いて1.5%アガロースTAEゲルから精製したゲル、ならびにSfiIおよびXhoIを有する
40ユニット過剰/μgのDNAを用いて消化した。15μgの各軽鎖ライブラリーベクターは、Qiagen Gel Extraction Kitを用いて1.5%アガロースTAEゲルから精製したゲル、ならびにSfiIおよびXhoIを有する40ユニット/μgのDNAを用いて消化した。ライブラリー連結は、消化された1.2μgのSfiI/XhoI、ゲル精製重鎖ドナーコレクションおよび5μgの14℃で一晩置かれた各軽鎖ライブラリー(カッパおよびラムダ)を併用して作成した。次いで、ライブラリー連結物は、Edge BioSystem Pefromaスピンカラムを用いて脱塩し、80μlのTG−1アリコート中でライブラリーあたり20個の電気穿孔を介して形質転換され、各々は1mlのSOC中に回収され、プールされ、37℃で1時間増殖された。再び、この増殖の後、アリコートの各々を用いて、形質転換体の総数を測定し、残りは、1Lの2YT+50μg/mlのAmpicillin+2%グルコースに移され、OD600が約0.3になるまで激しく曝気処理しながら、37℃で増殖させた。次のM13K07ヘルパーファージは、5:1の感染多重度(MOI:multiplicity
of infection)で添加し、撹拌せずに37℃で1時間インキュベートした。次に、遠心分離により細胞を取得し、1Lの2YT+50μg/mlのAmpicillin、70μg/mlのKanamycin中に再懸濁させ、scFvファージミドを産生させるために激しく曝気しながら37℃で一晩増殖させた。次の朝、遠心分離により細胞を集め、ファージミドを含む上澄みを集めた。ファージミドは、0.2容量の20%PEG/5MのNaCl溶液を添加し、氷上で1時間インキュベーションして上澄みから沈殿させた。次いで、ファージミド・ライブラリーストックは、遠心分離により取得し、20mlの無菌PBSに再懸濁させた。残りの細菌は、追加の遠心分離により除去し、最終ファージミド・ライブラリーは、PBS+50%グリセロール中−20℃で貯蔵した。
【0155】
ファージミドパニングおよび増幅
ELISAプレート(Immulon 4HBX平底、Nunc)を、ELISAプレートコーティング溶液(BioFX)中室温で一晩インキュベーションして100μlの100ng/mLのH5ヘマグルチニンタンパク質(Protein Sciences,A/Vietnam/1203/2004)で被覆した。次の日プレート類は、300μlのPBSTで3度洗浄した。洗浄後、300μlのブロッキング溶液(PBS/0.05%Tween−20中4%無脂肪ドライミルク)を添加し、氷上で30分間インキュベートした。ブロッキングステップの後、これらのプレートは300μlのPBSTで3度洗浄した。ファージパニングの直前に、Millipore Amicon Ultraカラムを用いて、凍結ファージミドストックからグリセロールを除去し、次いで、4%無脂肪ドライミルク中に15分間ブロックした。次に、ファージミドの100μlのアリコートを8つのウエル中に分配し、(全ファージ約1×1012CFU)4℃で2時間インキュベートし、次いで、300μlのPBSTで8度洗浄した。ファージミドは、室温で10分後に100μl/ウエルの溶離バッファー(0.2Mグリシン−HCl、pH2.2、1mg/mlBSA)に集めた。次いで、溶離液を、溶離液mlあたり56.25μlの2Mトリス塩基を添加して中和した。中和後、5mlのTG1細胞(OD600約0.3)を振とうなしで2−YT中37℃で30分間0.5mlの中和ファージで感染させた。このステップの後、一部の細胞をLB AMPグルコースプレート上に蒔き、ファージミドの総回収率を測定した。残りの接種液は、10mlのYATG(最終濃度2%グルコースおよび50μg/mlアンピシリン)に入れ、OD600が約0.3になるまで激しく曝気しながら37℃で増殖させた。次に、培養物を、5:1のMOIにてM13K07ヘルパーファージに感染させ、撹拌せずに37℃で30〜60分間インキュベートした。これらの細胞は、遠心分離により集め、25mlの2−YTAK(Ampicilin 50μg/ml,Kanamycin 70μg/ml)に再懸濁し、フレッシュな培養フラスコに移し、振とうしながら37℃で成長させた。その後のラウンドは、同様に回収し、増幅した。
【0156】
scFvELISA
バイオパンされたファージからの大腸菌のHB2151形質転換細胞の個々のコロニーを、1mlの2YT+100μg/mlのAMP中37℃で一晩増殖させた。次の朝、これらの細胞は遠心分離により取得し、1.5mlの周辺溶解バッファー(1mlのBBS(Teknova)+0.5mlの10mg/mlのリゾチーム+最終濃度が10mMになるEDTA)に再懸濁させた。これらの細胞は、遠心分離により再びペレット化し、周辺可溶化液を含むscFvを集めた。scFv可溶化液は、希釈バッファー(PBS/0.05%BSA)と1:1で混合し、先に抗原が希釈バッファーで被覆され、ブロックされたウエルに100μlを添加した。これらの試料を、室温で2時間インキュベートし、次いで、PBS/0.05%Tweenで3度洗浄した。次に、希釈バッファー中の1:5000に希釈したビオチン抗ヒスチジンマウス(Serotec)を100μl各ウエルに添加し、室温で1時間インキュベートした。このインキュベーションの後、これらのウエルはPBS/0.05%Tweenで3度洗浄し、次いで、各ウエルに100μlの1:2500のストレプトアビジン:HRP(Serotec)を添加し、室温で2時間インキュベートし、次いで、PBS/0.05%Tweenで3度洗浄した。この最終洗浄の後、100μlの発色基質溶液(TMB1基質、BioFX)を添加し、十分な時間が経過した後、100μlのSTOP溶液(BioFX)を添加して終結させた。450nmにおける吸光度は、プレートリーダー(Softmax Proソフトウエアを備えたMolecular Devices Thermomaxマイクロプレートリーダー)で読み取り、データを記録し、次いで、エクセル(マイクロソフト)を用いてプロットした。
【0157】
塩基配列の決定
重鎖および軽鎖の配列を推定するために、個々のクローンを増殖させ、プラスミドDNAを抽出した(Qiagen)。プラスミドDNAは、標準DNA塩基配列決定法により配列を決めた。
【0158】
ヘマグルチニン抑制(HAI)アッセイ
ヘマグルチニン抑制は、本質的には、Rogers et al.,Virology131:394−408(1983)の方法により、ウイルスまたはタンパク質/ウエルの4HAU(ヘマグルチネート化ユニット)を用いて丸底マイクロタイタープレート(Corning)において行った。HAI測定では、精製単一鎖可変断片(scFv)の25μlの試料が、各マイクロタイターウエルにおいてテストウイルスの4HAUを含むがこれらに限定されない25μlのPBSと混合した。室温における15分の予備インキュベーションの後、25μlの0.75%ヒト赤血球を添加し、混合した。HAI抗体活性は、室温で60分インキュベーション後目視検査により測定した。
【0159】
結果
骨髄および血液の試料は、2006年1月トルコで起きたH5N1鳥類インフルエンザの大流行の6名の生存者から、大流行の約4ヶ月後に採取した。6名の生存者全員について、トルコの保健省により承認された、理学的検査、臨床試験、および分子診断測定に続いて鳥インフルエンザの初期診断がなされた。これらの生存者のうち4名は、さらに、世界保健機関(WHO)により確認された。血清試料は、上で説明した血清学プロトコルを用いてH5ヘマグルチニン(A/Vietnam/1203/2004)に対する抗体の存在を確認するために分析した。図7に示したように、6名の患者全員の血液試料(それぞれ、SLB H1−H6と指定した)は、H5抗原に対する抗体の存在を示した。このことを確認後、これらの個体の骨髄試料からRNAを抽出し、上で説明したプロトコルを用いて骨髄mRNAを精製し、逆転写した。次いで、抗体の重鎖および軽鎖のレパートリーを、上で説明した骨髄cDNAから増幅し、個別の抗体の重鎖および軽鎖のファージミドライブラリーを、個々のライブラリーを区別するために、上で説明した3ヌクレオチドバーコードを用いて各生存者について別々にクローン化した。
【0160】
骨髄および血液試料は、2006年にインフルエンザ症状で処置を受けた12名のローカルドナーからも採取した。H1、H3およびH5ヘマグルチニンに対する抗体の存在を確認するために上で説明した血清学的研究が、それぞれ、なされた。図8に示したように、テストしたすべての血清試料は、H1および/またはH3のヘマグルチニンの抗体について陽性であり、どのサブタイプが優勢であるかは、個々のドナーが自身の生活時間を通して最も多くさらされたインフルエンザA型ウイルスのサブタイプに左右された。その上、興味深いことに、血清にH5ヘマグルチニンの抗体を、なりのレベル含む患者がいた(図8のドナーSLB1およびSLB5)。この確認の後、ドナーの骨髄試料からRNAを抽出し、上で説明したプロトコルを用いて骨髄mRNAを精製し、逆転写した。次いで、抗体の重鎖および軽鎖のレパートリーを、上で説明した骨髄cDNAから増幅し、個別の抗体の重鎖および軽鎖のライブラリーを、個々のライブラリーを区別するために、上で説明した3ヌクレオチドバーコードを用いて各生存者について別々にクローン化した。
【0161】
図9に例示したように、利用できる4つのヌクレオチド類の3個を用いると、64の独特のバーコードを作ることができる。
【0162】
トルコ鳥インフルエンザの生存者の骨髄試料から調製し、プールされた抗体ライブラリーの3ラウンドのパニングの後、得られた48のランダムクローンから、40をH5ヘマグルチニンタンパク質(Protein Sciences,A/Vietnam/1203/2004)への結合についてELISAによりテストし、ベトナムH5N1ウイルス(CBER,A/Vietnam/1203/2004)を不活性化した。これらのクローンの配列を決めた。40のクローンの内、5つは異なっていることが判明した。図10に示したように、全部で5つの異なるクローン(クローンF5およびG1は同じ配列を有する)は、H5タンパク質およびベトナムのH5N1ウイルスの両方に結合していた。図11は、上の実験において使われたベトナムの分離株のH5ヘマグルチニン配列に対してトルコのドナーからのH5ヘマグルチニンタンパク質の配列を比較している配列アラインメントを示している。これらの実験の結果は、配列の違いにも関わらず、テストした抗体は、トルコおよびベトナムの両方のH5タンパク質およびウイルスに結合し、したがって、1つより多くのH5N1ウイルスの分離株と交差反応性を示した。
【0163】
追加された4つの独特のクローンは、第2ラウンドのパニングにより産生された12のクローンの中から同定された。
【0164】
トルコドナーのプールされた抗体ライブラリーにおいて同定された独特のクローンの重鎖可変領域配列は、対応する軽鎖および生殖系列起源の配列と共に、図12および13に示している。特に、図12に示した配列(3〜23重鎖クローン)は、3ラウンドのパニング後の、すべてのトルコドナーのすべての重鎖および軽鎖のプールされたライブラリーに由来する。図13に示した配列(3〜30重鎖クローン)は、2ラウンドのパニング後の、すべてのトルコドナーのすべての重鎖および軽鎖のプールされたライブラリーに由来する。
【0165】
追加された独特のH5N1に特異な抗体重鎖可変領域配列は、4ラウンドのパニング後、上で説明したELISAプロトコルを用いて個々のトルコドナーの抗体ライブラリーから同定した。これらのH5N1のELISA陽性クローンは、図14A−Dに示している。
【0166】
図15および16は、上で説明したトルコの鳥インフルエンザ生存者の血清および骨髄の分析により同定した抗体重鎖(図15)および軽鎖(図16)配列を用いて多様な抗体重鎖および軽鎖ライブラリーを作り出すために目標の突然変異誘発の使用を例示している。
【0167】
図17および18は、HAタンパク質のトルコおよびインドネシアの改変体と共にH5N1ベトナムウイルスscFv抗体から得られた特定のFab断片交差反応性を確認しているELISAの結果を示している。
【0168】
前述の説明では本発明は、特定の実施形態を参照しながら説明しているが、本発明はそのように限定されない。実際に、本発明で示しおよび説明した発明に加えて、前述の説明から当業者に明らかになり、添付した特許請求の範囲内に入る、本発明の種々の改変がある。
【0169】
本明細書を通じて引用された引用文献は、すべて、引用により本明細書に明らかに組み込まれている。
【特許請求の範囲】
【請求項1】
本明細書に記載の発明。
【請求項1】
本明細書に記載の発明。
【図1A】



【図1B】

【図1C】

【図1D】

【図1E】

【図1F】

【図1G】

【図1H】

【図1I】

【図1J】

【図2】
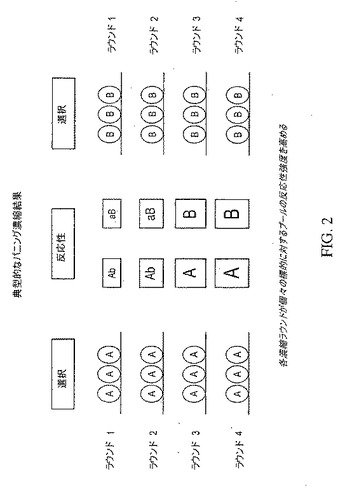
【図3】
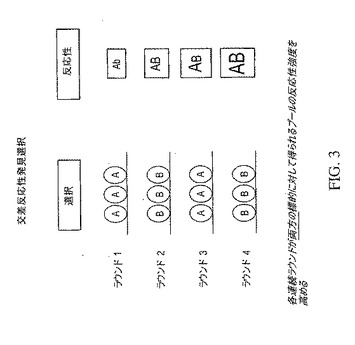
【図4】
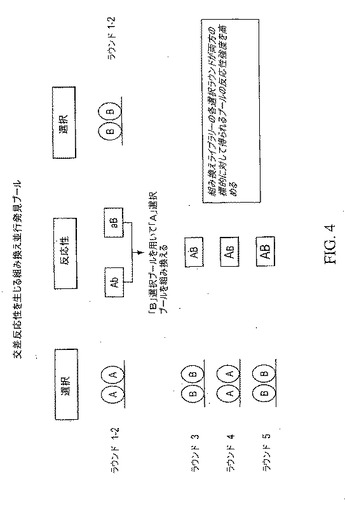
【図5】
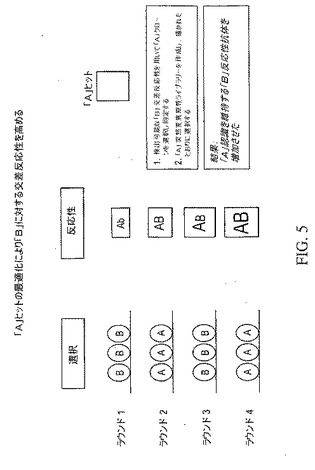
【図6】
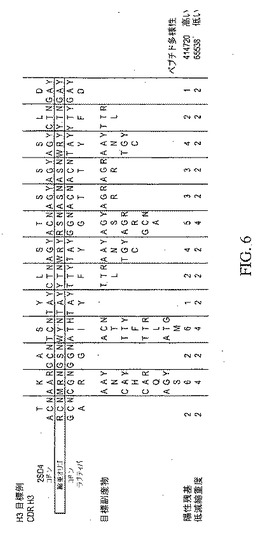
【図7】
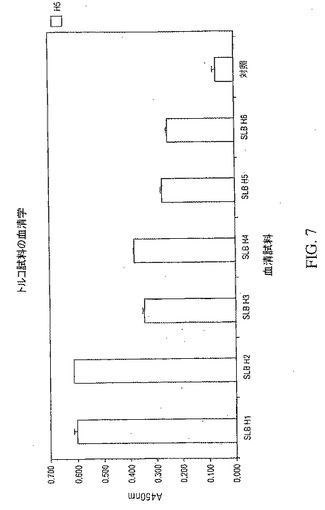
【図8】
【図9】
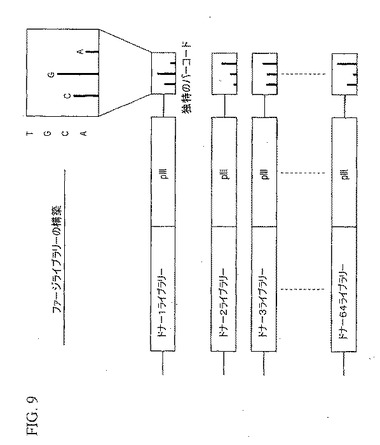
【図10】
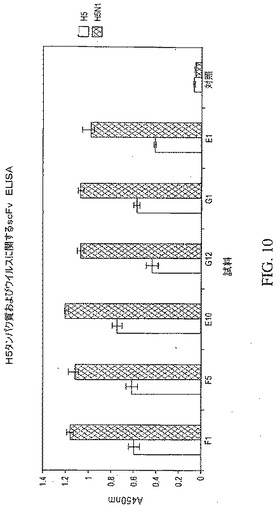
【図11】

【図12】
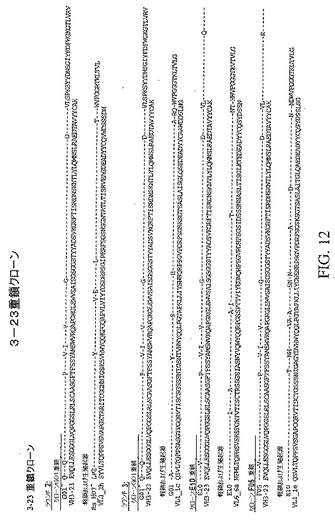
【図13】
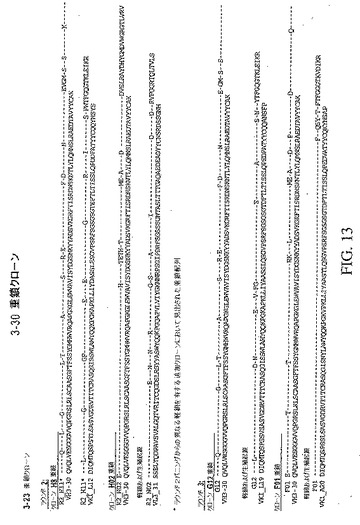
【図14A】
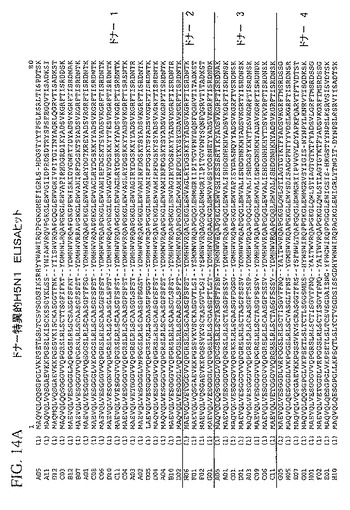
【図14B】
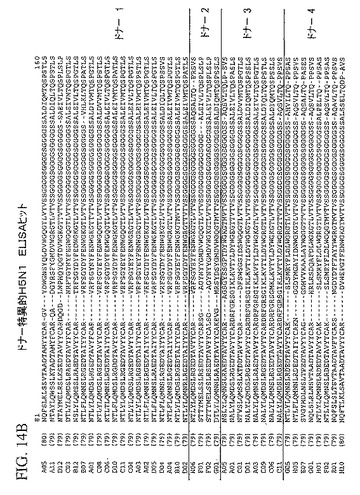
【図14C】
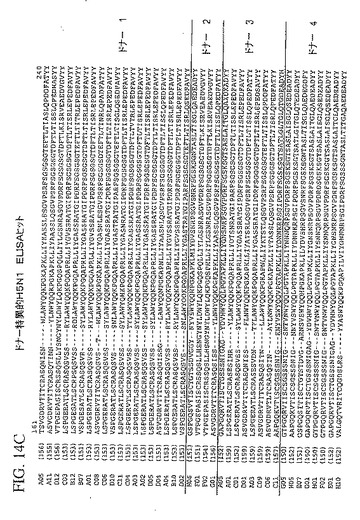
【図14D】
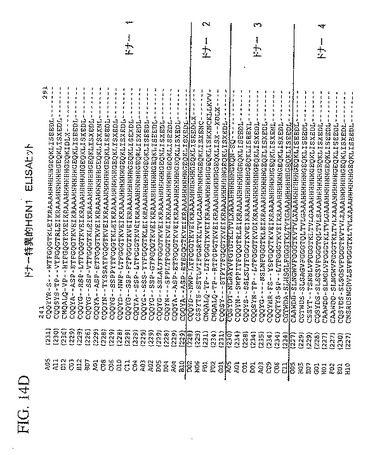
【図15】
【図16】
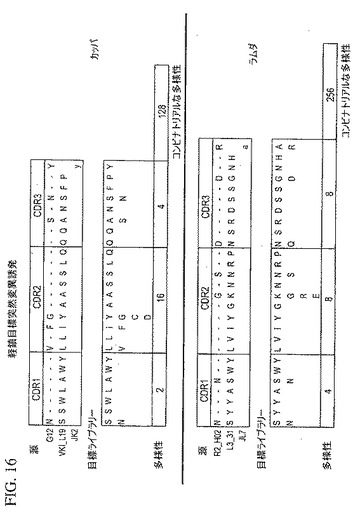
【図17】
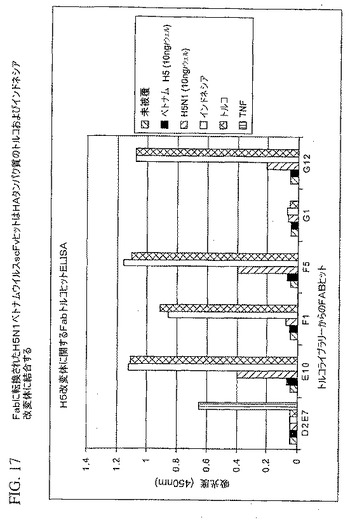
【図18】
【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図1H】
【図1I】
【図1J】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14A】
【図14B】
【図14C】
【図14D】
【図15】
【図16】
【図17】
【図18】
【公開番号】特開2013−67660(P2013−67660A)
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【外国語出願】
【出願番号】特願2013−2320(P2013−2320)
【出願日】平成25年1月10日(2013.1.10)
【分割の表示】特願2009−511210(P2009−511210)の分割
【原出願日】平成19年5月15日(2007.5.15)
【出願人】(508339161)シー レーン バイオテクノロジーズ, エルエルシー (10)
【Fターム(参考)】
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2013−2320(P2013−2320)
【出願日】平成25年1月10日(2013.1.10)
【分割の表示】特願2009−511210(P2009−511210)の分割
【原出願日】平成19年5月15日(2007.5.15)
【出願人】(508339161)シー レーン バイオテクノロジーズ, エルエルシー (10)
【Fターム(参考)】
[ Back to top ]