
インフルエンザウイルスの調製方法
本発明は、インフルエンザウイルス感染の予防用または/および治療用の医薬組成物の調製方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、インフルエンザウイルス感染の予防用または/および治療用の医薬組成物の作製方法に関する。
【0002】
さらに、本発明は、インフルエンザウイルス感染の予防用または/および治療用の医薬組成物に関する。
【背景技術】
【0003】
脅迫的なインフルエンザのパンデミックから見て、利用可能な持続的に有効な薬剤を開発および製造する緊急の必要性がある。ドイツ単独で、インフルエンザの年間の発症により、1年に5,000〜20,000例の死亡が引き起こされている(情報源:Robert-Koch研究所)。繰り返されている大きなインフルエンザのパンデミックは、特に恐れられている。第1の大きなパンデミック、いわゆる「スペインかぜ」では、1918〜1919年に、高い割合の健康な中年のヒトを含む約4千万の命が犠牲になった。H5N1インフルエンザウイルスは、現在は鳥類において主に複製されるが、後天的な変異によってこのウイルスがヒトからヒトへと伝染できるようになった場合、これによって同様なパンデミックが引き起こされる恐れがある(2、3)。より最近では、新規のインフルエンザウイルス異型が出現し、すなわち、A型インフルエンザ(H1N1)「豚インフルエンザ」株(4)が、予測不可能なパンデミックの脅威をもたらしている。ヒトにおけるパンデミックの可能性は、近年、鳥インフルエンザ(H5N1)の世界的な広がりおよび家畜の感染と共に、より急激に増大している。病原性の高いヒトインフルエンザ組換え体が現れるのは、単に時間の問題である。インフルエンザ感染の予防または療法のための瞬間に利用可能な方法、例えばウイルス表面タンパク質を用いた予防接種または抗ウイルス薬(ノイラミニダーゼ阻害剤またはイオンチャネル遮断薬)の使用などは、様々な欠点を有する。既にこの早期の段階で、我々の最も有効な製剤のうちの1種(タミフル)に対する耐性が出現しており、その結果、汎流行を封じ込めるのに不適当になることもある。インフルエンザに対するワクチンおよび薬剤の使用における中心的な課題は、病原体の多様性である。これまで、有効なワクチンの開発には、病原体異型の正確な予測が必要とされてきた。ウイルス構成要素に対する薬剤は、病原体の変異により、急激にその有効性を失うことがある。
【0004】
これまでほとんど注意されてこなかった研究の領域は、宿主細胞における重大な意味を持つ標的構造の同定である。ウイルスは、固有の細胞内タンパク質に依存して宿主内で複製することができるかが決まる。こうした細胞内因子の知識は、ウイルス複製に不可欠であるが、新規薬剤の開発を導きうるヒトには(少なくとも一時的に)不要である。概算では、ヒトゲノムにおいてウイルス増殖に必須な約500遺伝子が予測されている。これらのうち、少なくとも10%は、ヒトの生体にとって一時的にまたは恒久的にも不必要であろう。これらの遺伝子およびこれらの産物の阻害は、ウイルス標的とは対照的に、これらの構造において一定であり、最短の時間での抗ウイルス薬の新しい作製の開発を可能にするであろう。こうした遺伝子産物の阻害は、抗ウイルス薬に対する感受性がより短いウイルスエスケープ変異体の発生を克服しうる。他の遺伝子ファミリーキナーゼの中で細胞内で重要な調節タンパク質は、ウイルスによって乗っ取られて宿主細胞の組成が操作されることが多い。
【0005】
A型インフルエンザは、哺乳動物宿主細胞内での生存の成功を容易にする一連の戦略を示すマイナス鎖RNAウイルスである(5)。感染すると、先天性免疫性レセプター、例えば細胞内タンパク質レチノイン酸誘導性遺伝子I(RIG-I)などのそれらの同族リガンドへの結合が何十もの免疫および炎症に関連した遺伝子の一過的発現を誘発する(6、7)。特に、その後のI型インターフェロンの誘導は、ミクソウイルス耐性(Mx)タンパク質などの、内因性抗ウイルス活性を有するGTPアーゼの上方制御を刺激する。オルソミクソウイルスファミリーのメンバーに対するMxタンパク質の抗ウイルス活性は、マウスにおいて最初に観察された(8)。核に位置するMx1タンパク質は、それがない場合には、パンデミック1918の株およびきわめて致死的なH5N1インフルエンザウイルスを含むインフルエンザウイルスによる致死感染に対して保護を与える(9、10)。ヒトオーソログ、MxAは、細胞質に局在し、かつ入ってくるウイルスヌクレオカプシドに結合して不活性化することによって作用すると考えられている(11)。報告によると、興味深いことに、ヒトMxAは、遺伝子導入マウスにおいて多様なRNAウイルスに対する保護機能を示す(12)。これらの先天性応答戦略に対抗するために、インフルエンザウイルスは、例えば、インターフェロン-β(IFN-β)の産生を低減することによってまたは抗ウイルス性タンパク質2'-5'オリゴアデニレートシンセターゼ(OAS)およびプロテインキナーゼR(PKR)の発現を阻害することによって、そのNS1タンパク質を用いる。(13)。しかし、in vitroおよびin vivoでのA型インフルエンザ感染中に既に観察されているMxAの能動的抑制は、現在完全には理解されていない。
【0006】
ウイルス剤が、その感染能力を調節するために用いる興味深い戦略は、遺伝子発現の重要な転写後制御因子として知られる(14)、タンパク質をコードしない干渉する約22nt長の短いRNA分子の一クラスである、マイクロRNA(miRNA)の使用である。大きなゲノムを有するウイルスは、宿主の生理機能を変えて複製を増強するためのそれらの自身のmiRNAをコードすることができる(15)。逆に、小さなRNAゲノムのC型肝炎ウイルスは、その複製を促進するために、宿主細胞miR-122の発現を操作することができる(16)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO 02/44321
【非特許文献】
【0008】
【非特許文献1】Bartel D、Cell 136:215〜233、2009
【非特許文献2】Kohlerら、1975、Nature 256:495〜497
【非特許文献3】Kozborら、1985、J. Immunol. Methods 81、31〜42
【非特許文献4】Coteら、PNAS、80:2026〜2030
【非特許文献5】Coleら、1984、Mol. Cell Biol. 62:109〜120
【非特許文献6】Morrisonら、1984、PNAS、81:6851〜6855
【非特許文献7】Neubergerら、1984、312:604〜608
【非特許文献8】Takedaら、1985、Nature 314:452〜454
【非特許文献9】Orlandiら、1989、PNAS 86:3833〜3837
【非特許文献10】Winterら、1991、Nature 349:293〜299
【非特許文献11】Burton、1991、PNAS、88:11120〜11123
【非特許文献12】Huseら、1989、Science 254:1275〜1281
【発明の概要】
【発明が解決しようとする課題】
【0009】
インフルエンザウイルスワクチンの作製のための一般的な戦略は、有胚(embryonated)鶏卵または細胞培養におけるインフルエンザウイルス複製に基づいている。細胞培養または有胚卵におけるウイルス複製は、時間のかかる高価な方法である。したがって、インフルエンザワクチンの作製方法を改良することが本発明の課題である。
【課題を解決するための手段】
【0010】
本発明の一目的は、以下のステップ:
(a)改変のない場合のインフルエンザウイルス複製と比較してインフルエンザウイルス複製の能力が増大された、インフルエンザウイルスを複製することのできる改変細胞、改変有胚卵または/および改変非ヒト生物を提供するステップ、
(b)(a)の該細胞、該有胚卵または/および該生物をインフルエンザウイルスと接触させるステップ、
(c)該インフルエンザウイルスの複製を可能にする条件下で、該細胞、該有胚卵または/および該非ヒト生物を培養するステップ、ならびに
(d)ステップ(c)において生成された該インフルエンザウイルスまたは/およびその少なくとも1種の構成要素を分離するステップ
を含む、インフルエンザウイルスの調製方法である。
【0011】
ステップ(d)のインフルエンザウイルスから、インフルエンザウイルス感染の予防用または/および治療用の医薬組成物を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、調製することもできる。
【0012】
本発明のもう1つの目的は、インフルエンザウイルス感染の予防用または/および治療用の医薬組成物の調製方法であって、以下のステップ:
(a)改変のない場合のインフルエンザウイルス複製と比較してインフルエンザウイルス複製の能力が増大された、インフルエンザウイルスを複製することのできる改変細胞、改変有胚卵または/および改変非ヒト生物を提供するステップ、
(b)(a)の該細胞、該有胚卵または/および該生物をインフルエンザウイルスと接触させるステップ、
(c)該インフルエンザウイルスの複製を可能にする条件下で、該細胞、該有胚卵または/および該非ヒト生物を培養するステップ、
(d)ステップ(c)において産生された該インフルエンザウイルスまたは/およびその少なくとも1種の構成要素を分離するステップ、ならびに、
(e)ステップ(d)において分離された該インフルエンザウイルスまたは/およびその構成要素から、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、該医薬組成物を調製するステップ
を含む方法である。
【0013】
本明細書における「方法」または「本発明の方法」に対する参照とは、インフルエンザウイルスの調製方法ならびにインフルエンザウイルス感染の予防用または/および治療用の医薬組成物の調製方法に対する参照である。
【0014】
ステップ(a)において用いられる細胞は、インフルエンザウイルスに感染させることができるいかなる細胞であってもよい。インフルエンザウイルスの産生に好適な細胞系は知られている。好ましくは、該細胞は、哺乳動物細胞または鳥類細胞である。ヒトの細胞も好ましい。上皮細胞、例えば肺上皮細胞なども好ましい。該細胞は、細胞系であってもよい。好適な肺上皮細胞系は、A594である。もう1つの好適な細胞は、ヒト胚性腎細胞系293Tである。本発明の一実施形態において、本発明の方法は、本明細書に記載の細胞を用いる。
【0015】
ステップ(a)において用いられる非ヒト生物は、インフルエンザウイルスに感染させることができるいかなる生物であってもよい。好ましくは、該生物は、インフルエンザワクチンの作製に用いられる生物である。より好ましくは、該生物は、例えば有胚鶏卵のような、有胚卵である。当業者は、こうした生物を得る方法を知っている。受精によって有胚卵を得る方法は知られている。インフルエンザウイルスの接種によるインフルエンザウイルス複製の誘導は知られている。本発明の一実施形態において、本発明の方法は、本明細書に記載のように、非ヒト生物または/および有胚卵を用いる。
【0016】
本発明のステップ(a)は、本明細書に記載のように改変された細胞、有胚卵もしくは/および非ヒト生物の提供を含んでいてもよく、または改変のステップを含んでいてもよい。
【0017】
改変細胞または/および改変有胚卵は、本明細書に記載のように、ステップ(a)において提供され、ステップ(b)、(c)および(d)において、またはステップ(b)、(c)(d)および(e)において用いられることが好ましい。
【0018】
本明細書中で使用される場合、「該細胞、該有胚卵または/および非ヒト生物の改変」としては、該細胞、該卵または/および該生物における少なくとも1種の遺伝子または/および遺伝子産物の発現または/および活性の下方制御または/および上方制御が挙げられる。
【0019】
本明細書中で使用される場合、「該細胞、該有胚卵または/および該非ヒト生物の改変」は、該細胞、該有胚卵または/および該非ヒト生物を、モジュレーターのない場合のインフルエンザウイルス複製と比較して、該細胞または/および該生物におけるインフルエンザウイルス複製を増大させることができる少なくとも1種のモジュレーターと接触させることを含んでいてもよく、このとき、接触は、ステップ(b)の前もしくは後に、またはステップ(b)と同時に行われてもよい。
【0020】
本明細書中で使用される場合、「該細胞、該有胚卵または/および非ヒト生物の改変」は、組換え細胞、組換え有胚卵または/および組換え非ヒト生物の作製または/および提供を含んでいてもよく、このとき、少なくとも1種の遺伝子または/および遺伝子産物の発現または/および活性が、改変のない場合のインフルエンザウイルス複製と比較して該細胞、該有胚卵または/および該非ヒト生物のインフルエンザウイルス複製の能力が増大されるように改変される。
【0021】
組換え細胞、組換え有胚卵または/および組換え非ヒト生物の調製は、該細胞内、該有胚卵内もしくは/および該非ヒト生物内への核酸分子の導入、または/ならびに該細胞内、該卵内もしくは/および該生物内の核酸配列の欠失を含んでいてもよい。該核酸分子は、該細胞の、該有胚卵のまたは/および該非ヒト生物のゲノム内に組み込まれてもよい。それによって、該細胞、該卵または/および該生物の配列が、改変、置換または/および欠失されうる。該核酸分子は、該細胞または/および該生物と異種の配列を含有していてもよい。核酸分子の組み込みは、恒久的にまたは一過的に行われてもよい。組換え有胚卵または/および組換え非ヒト生物は、生殖細胞系列の操作によって調製されてもよい。本発明の文脈において、「有胚卵」とは、特にその胚を指す。例えば、「有胚卵の改変」は、特にその胚の改変である。
【0022】
当業者は、細胞内、有胚卵内もしくは/および生物内に核酸分子を導入する方法、または/ならびに該細胞内、該卵内もしくは/および該生物内の核酸配列を欠失させる方法(本明細書中で用いているような、「組換え技術」)を知っている。これらの方法は、好適なベクター、例えばプラスミドなどを用いる、トランスフェクションを含んでいてもよい。これらの方法は、該細胞または/および該生物のゲノム内の核酸分子の相同組換えも含んでいてもよい。また、該核酸分子は、該細胞、該有胚卵または/および該生物のゲノム内に無作為に挿入されてもよい。
【0023】
表1a、1b、4および5に、インフルエンザウイルス複製の調整のための標的を記載しており、ここで、該標的は、本明細書に記載のように、モジュレーターとの接触、または組換え技術のいずれかによる、該細胞、該有胚卵または/および非ヒト生物の改変に好適でありうる。
【0024】
本発明の文脈における「調整」とは、「活性化」または「阻害」であってもよい。
【0025】
下方制御によりインフルエンザウイルス複製を増大させる遺伝子の例を、表1aおよび5に記載する。したがって、これらの遺伝子または/およびその遺伝子産物の発現または/および活性を増大させることによって、インフルエンザウイルス複製を低減させることができる。これらの遺伝子または/および遺伝子産物の低減された発現または/および活性は、ウイルス作製の改良によって本発明の方法において利用することができる。
【0026】
したがって、ステップ(a)において提供される該細胞、該有胚卵または/および非ヒト生物は、組換え細胞、組換え有胚卵または/および組換え非ヒト生物であってもよく、このとき、表1aおよび5から選択される遺伝子の遺伝子発現または/および活性は下方制御される。
【0027】
下方制御によりインフルエンザウイルス複製を低減させる遺伝子の例を、表1bおよび4に記載する。したがって、これらの遺伝子または/および遺伝子産物の発現または/および活性を低減させることによって、インフルエンザウイルス複製を低減させることができる。これらの遺伝子または/および遺伝子産物の増大された発現または/および活性は、ウイルス作製の改良によって本発明の方法において利用することができる。
【0028】
したがって、ステップ(a)において提供される該細胞、有胚卵または/および非ヒト生物は、組換え細胞、組換え有胚卵または/および組換え非ヒト生物であってもよく、このとき、表1bおよび表4から選択される遺伝子の遺伝子発現または/および活性は上方制御される。特に、表1bおよび表4から選択される遺伝子の上方制御は、前記遺伝子の過剰発現である。
【0029】
本発明の文脈において、「標的」としては、
(a)遺伝子または/およびゲノム内、特に、ヒト遺伝子または/およびヒトゲノム内のヌクレオチド配列、
(b)核酸、または/および(a)のヌクレオチド配列によってコードされるポリペプチド
などが挙げられる。
(a)または/および(b)の配列は、宿主細胞におけるインフルエンザウイルス複製の調節に関与していてもよい。該標的は、直接的または間接的に、インフルエンザウイルス複製の調節に関与していてもよい。特に、標的は、その標的の活性化またはその標的の阻害のいずれかによって、インフルエンザウイルス複製を増大させるのに好適である。
【0030】
標的の例は、遺伝子および遺伝子の部分的な配列、例えば調節配列などである。本発明による標的としては、RNA、特にmRNA、tRNA、rRNA、miRNA、piRNAなどの遺伝子産物も挙げられる。標的は、該標的遺伝子によってコードされるポリペプチドまたは/およびタンパク質も含みうる。標的遺伝子の好ましい遺伝子産物は、その標的遺伝子によってコードされるmRNA、miRNA、ポリペプチドまたは/およびタンパク質から選択される。最も好ましい遺伝子産物は、標的遺伝子によってコードされるポリペプチドまたはタンパク質である。標的タンパク質または標的ポリペプチドは、翻訳後に修飾されてもよく、されなくてもよい。
【0031】
本明細書中で使用される場合、「遺伝子産物」は、RNA、特にmRNA、tRNA、rRNA、miRNA、およびpiRNAから選択されてもよい。また、「遺伝子産物」は、前記遺伝子によってコードされるポリペプチドまたは/およびタンパク質であってもよい。
【0032】
本発明の文脈において、該遺伝子または/および遺伝子産物の「活性」としては、転写、翻訳、翻訳後修飾、転写後調節、遺伝子または/および遺伝子産物の活性の調整などが挙げられる。該活性は、リガンド結合によって調節されてもよく、そのリガンドは、活性化剤または阻害剤であってもよい。また、該活性は、本明細書に記載のように、miRNA分子、shRNA分子、siRNA分子、アンチセンス核酸、デコイ核酸または/および他の任意の核酸によっても調節されうる。該遺伝子の活性は、本明細書に記載のように、組換え技術によっても調節されうる。調節は、小分子、抗体、アプタマー、または/およびスピーゲルマー(spiegelmer)(鏡像アプタマー)によっても行われうる。
【0033】
本発明の方法は、あらゆるインフルエンザウイルスでの感染の予防用または/および治療用の医薬組成物の作製に好適でありうる。
【0034】
該インフルエンザウイルスは、ワクチン作製に好適ないずれのインフルエンザウイルスであってもよい。該インフルエンザウイルスは、A型インフルエンザウイルスであってもよい。該A型インフルエンザウイルスは、鳥類および哺乳類の生物からこれまでに単離されたA型インフルエンザウイルスから選択されてもよい。詳細には、該A型インフルエンザウイルスは、H1N1、H1N2、H1N3、H1N4、H1N5、H1N6、H1N7、H1N9、H2N1、H2N2、H2N3、H2N4、H2N5、H2N7、H2N8、H2N9、H3N1、H3N2、H3N3、H3N4、H3N5、H3N6、H3N8、H4N1、H4N2、H4N3、H4N4、H4N5、H4N6、H4N8、H4N9、H5N1、H5N2、H5N3、H5N6、H5N7、H5N8、H5N9、H6N1、H6N2、H6N3、H6N4、H6N5、H6N6、H6N7、H6N8、H6N9、H7N1、H7N2、H7N3、H7N4、H7N5、H7N7、H7N8、H7N9、H9N1、H9N2、H9N3、H9N5、H9N6、H9N7、H9N8、H10N1、H10N3、H10N4、H10N6、H10N7、H10N8、H10N9、H11N2、H11N3、H11N6、H11N9、H12N1、H12N4、H12N5、H12N9、H13N2、H13N6、H13N8、H13N9、H14N5、H15N2、H15N8、H15N9およびH16N3から選択されてもよい。より詳細には、該A型インフルエンザウイルスは、H1N1、H3N2、H7N7、H5N1から選択される。さらにより詳細には、該A型インフルエンザウイルスは、株Puerto Rico/8/34、鳥インフルエンザウイルス分離株H5N1、鳥インフルエンザ株A/FPV/Bratislava/79(H7N7)、株A/WSN/33(H1N1)、株A/Panama/99(H3N2)、または豚インフルエンザ株H1N1である。
【0035】
該インフルエンザウイルスは、B型インフルエンザウイルスであってもよい。詳細には、該B型インフルエンザウイルスは、ビクトリア系統の代表種および山形系統の代表種から選択されてもよい。
【0036】
本発明の方法において、インフルエンザウイルス複製を増大させるためのステップ(a)に従った該細胞または/および生物の改変としては、表1A、表1B、表4および表5から選択される遺伝子または/およびそれらの遺伝子産物の発現を調節することが挙げられる。特に、該細胞または/および生物の改変は、表1Bおよび表4から選択される遺伝子または/およびそれらの遺伝子産物の発現を活性化してもよく、あるいは該細胞または/および生物の改変は、表1Aおよび5から選択される遺伝子または/およびそれらの遺伝子産物の発現を阻害してもよい。この発現の調節は、該細胞、該有胚卵もしくは/および該生物を、本明細書に記載のモジュレーターと接触させることによって行われてもよく、または組換え細胞、組換え有胚卵もしくは/および組換え生物において行われてもよく、これらの作製については本明細書に記載している。
【0037】
本発明の方法において、該細胞、該有胚卵または/および該非ヒト生物の改変は、MxA遺伝子の発現または/および遺伝子産物活性の阻害を含んでいてもよい。インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、MxA遺伝子の発現または/および遺伝子産物活性を阻害する能力があってもよい。本明細書に記載のように、該細胞、該有胚卵または/および該非ヒト生物は、MxA遺伝子の発現または/および遺伝子産物活性が、miR-141、miR-141*、miR-200c、miR-200c*、これらの前駆体、これらの誘導体、アンチセンス核酸、siRNA、shRNAおよびインフルエンザウイルス配列から選択される少なくとも1種のモジュレーターによって阻害されるように、組換えによって改変されてもよい。特に、本発明の方法において、MxA遺伝子は、miR-141、miR-141*、miR-200c、miR-200c*、これらの前駆体、これらの誘導体、アンチセンス核酸、siRNA、shRNAおよびインフルエンザウイルス配列から選択される少なくとも1種のモジュレーターによって翻訳後に阻害される。
【0038】
本発明の方法において、miR-141、miR-141*、miR-200c、miR-200c*、または/およびこれらの前駆体は、該細胞内、該有胚卵内または/および該非ヒト生物内で過剰発現されてもよい。過剰発現は、一過的または恒久的であってもよい。
【0039】
RNAレベルにおいて、阻害は、アンチセンス核酸、siRNA、shRNA、デコイ核酸または/およびこれらの誘導体によって行われてもよい。MxAポリペプチドのレベルにおいて、阻害は、小分子、抗体、アプタマー、スピーゲルマー(鏡像アプタマー)によって行われてもよい。
【0040】
miR-141、miR-141*、miR-200c、およびmiR-200c*の配列、これらの前駆体ならびに該前駆体のヘアピン構造については、図18に記載している。miR-141およびmiR-141*は、単一の前駆体によって細胞内で共発現させることもできる。miR-141およびmiR-141*は、該前駆体のヘアピン構造を形成しうる相補配列を含有する。miR-200cおよびmiR-200c*は、単一の前駆体によって細胞内で共発現させることもできる。miR-200cおよびmiR-200c*は、該前駆体のヘアピン構造を形成しうる相補配列を含有する。
【0041】
本発明の文脈において、A型インフルエンザの配列は、MxA遺伝子と高度な同一性を有することが確認された。したがって、本発明の方法において、MxA遺伝子が阻害されてもよく、または/かつA型インフルエンザウイルス配列もしくは/およびA型インフルエンザウイルス配列に由来する配列を含有する少なくとも1種のマイクロRNAもしくは/および少なくとも1種のアンチセンスRNAによってウイルス複製が活性化されてもよい。
【0042】
該細胞の、該有胚卵のまたは/および該非ヒト生物の改変は、遺伝子の発現または/および遺伝子産物活性の阻害を含んでいてもよく、このとき、該遺伝子は、
(a)表1Aおよび5の配列から選択されるヌクレオチド配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含有する。
【0043】
該細胞、該有胚卵または/および該非ヒト生物の改変は、遺伝子の発現または/および遺伝子産物活性の活性化を含んでいてもよく、このとき、該遺伝子は、
(i)表1Bおよび表4の配列から選択されるヌクレオチド配列、
(ii)(i)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の配列の断片、
(iii)(i)もしくは/および(ii)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含有する。
【0044】
インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、遺伝子の発現または/および遺伝子産物活性を阻害する能力があってもよく、このとき、該遺伝子は、
(a)表1Aおよび5の配列から選択されるヌクレオチド配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含有する。
【0045】
インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、遺伝子の発現または/および遺伝子産物活性を活性化する能力があってもよく、このとき、該遺伝子は、
(i)表1Bおよび表4の配列から選択されるヌクレオチド配列、
(ii)(i)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の配列の断片、
(iii)(i)もしくは/および(ii)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含有する。
【0046】
該細胞、該有胚卵または/および非ヒト生物は、本明細書に記載のように、遺伝子の発現または/および遺伝子産物活性が阻害されるように、組換えによって改変されてもよく、このとき、該遺伝子は、
(a)表1Aおよび5の配列から選択されるヌクレオチド配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含有する。
【0047】
該細胞、該有胚卵または/および非ヒト生物は、本明細書に記載のように、遺伝子の発現または/および遺伝子産物活性が活性化されるように、組換えによって改変されてもよく、このとき、該遺伝子は、
(i)表1Bおよび表4の配列から選択されるヌクレオチド配列、
(ii)(i)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の配列の断片、
(iii)(i)もしくは/および(ii)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含有する。
【0048】
本明細書中で使用される場合、本発明の1つまたは複数の表に開示しているヌクレオチド配列または/および遺伝子に対する参照は、前記表に開示している特定の配列に対する参照、および前記表に開示している受託番号、遺伝子名、Locus Link、記号、GeneID、GeneSymbol、または/およびGenbankIDによって特徴付けられる配列に対する参照であると理解される。受託番号、遺伝子名、Locus Link、記号、GeneID、GeneSymbol、または/およびGenbankIDに対する参照によって、当業者は、対応するヌクレオチド配列または/およびアミノ酸配列を特定することができる。特定の配列は、表に示しているような、受託番号、遺伝子名、Locus Link、記号、GeneID、GeneSymbol、およびGenbankIDのうちの1つまたは複数によって特徴付けることができる。本発明の1つまたは複数の表に開示している遺伝子に対する参照は、特に、配列、例えば、前記表に開示している受託番号、遺伝子名、Locus Link、記号、GeneID、GeneSymbol、または/およびGenbankIDによって特徴付けられる遺伝子配列などに対する参照であると理解される。
【0049】
改変(調整および組換え改変を含む)は、キナーゼの改変または/およびキナーゼ結合ポリペプチドのモジュレーターの改変であってもよく、このとき、少なくとも1種のキナーゼまたは/およびキナーゼ結合ポリペプチドは、表1Aおよび表1Bから選択される核酸または/および遺伝子によってコードされる。
【0050】
本発明の方法において、インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、
(i)表1Bおよび表4から選択されるヌクレオチド配列、
(ii)(i)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の配列の断片、
(iii)(i)もしくは/および(ii)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含有する活性化剤であってもよい。
【0051】
少なくとも1種の該活性化剤は、配列(i)、(ii)(iii)または/および(iv)を含有する遺伝子の発現または/および遺伝子産物活性を活性化する能力があってもよい。
【0052】
本発明の方法において、インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、
(a)表1Aおよび5から選択されるヌクレオチド配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含有する阻害剤であってもよい。
【0053】
少なくとも1種の該阻害剤は、配列(a)、(b) (c)または/および(d)を含有する遺伝子の発現または/および遺伝子産物活性を阻害する能力があってもよい。
【0054】
本発明の方法において用いられるインフルエンザウイルス複製の少なくともモジュレーターは、核酸、例えばリボザイム、ペプチド、ポリペプチド、抗体、アプタマー、スピーゲルマー、小分子のような核酸類似体、およびデコイ核酸からなる群から選択されてもよい。
【0055】
インフルエンザウイルス複製の該モジュレーターは、1000ダルトンより小さいまたは500ダルトンより小さい分子量を有する化合物であってもよい。本発明の文脈において、「小分子」とは、1000ダルトンより小さいまたは500ダルトンより小さい分子量を有する化合物を指す。本発明の方法において、該小分子は、
(a)表1A、表1B、表4、および表5から選択される核酸または/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、または/ならびに
(c)(a)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対して指向されてもよい。
【0056】
本発明のモジュレーターは、核酸を含有することが好ましく、このとき、該核酸は、表2および表4の配列ならびにこれらの断片から選択されるヌクレオチド配列を含有する。
【0057】
好ましくは、該核酸は、
(a)RNA、その類似体および誘導体、
(b)DNA、その類似体および誘導体、ならびに
(c)(a)と(b)との組合せ
から選択される。
【0058】
好適な阻害剤は、RNA干渉の能力のあるRNA分子である。本発明のモジュレーター、特に、本発明の阻害剤は、
(i)RNA干渉の能力のあるRNA分子、例えば、siRNAもしくは/およびshRNAなど、
(ii)miRNA、
(iii)該RNA分子(i)もしくは/および(ii)の前駆体、
(iv)該RNA分子(i)、(ii)もしくは/および(iii)の断片、
(v)(i)、(ii)(iii)または/および(iv)の該RNA分子の誘導体、または/ならびに
(vi)(i)、(ii)(iii)または/および(iv)の該RNA分子をコードするDNA分子
を含有していてもよい。
【0059】
好ましいモジュレーターは、
(i)miRNA、
(ii)該RNA分子(i)の前駆体、または/ならびに
(iii)該RNA分子(i)もしくは/および該前駆体(ii)をコードするDNA分子
である。
【0060】
さらにもう1つの好ましいモジュレーターは、
(i)RNA干渉の能力のあるRNA分子、例えば、siRNAもしくは/およびshRNAなど、
(ii)該RNA分子(i)の前駆体、または/ならびに
(iii)該RNA分子(i)もしくは/および該前駆体(ii)をコードするDNA分子
である。
【0061】
RNA干渉の能力のあるRNA分子は、WO 02/44321に記載されており、その開示は、参照により本明細書に含まれている。マイクロRNAは、Bartel D(Cell 136:215〜233、2009)に記載されており、その開示は、参照により本明細書に含まれている。
【0062】
本発明のRNA分子は、二本鎖RNA分子、好ましくは、一方の末端に単独でまたは双方の末端に一本鎖オーバーハングのあるまたはない二本鎖siRNA分子であってもよい。該siRNA分子は、少なくとも1つのヌクレオチド類似体または/およびデオキシリボヌクレオチドを含有していてもよい。
【0063】
本発明のRNA分子は、shRNA分子であってもよい。該shRNA分子は、少なくとも1つのヌクレオチド類似体または/およびデオキシリボヌクレオチドを含有していてもよい。
【0064】
本発明において用いられるDNA分子は、ベクターであってもよい。
【0065】
本発明において用いられる核酸は、アンチセンス核酸または該アンチセンス核酸をコードするDNAであってもよい。
【0066】
本発明において用いられる核酸または/および核酸断片は、少なくとも15、好ましくは少なくとも17、より好ましくは少なくとも19、最も好ましくは少なくとも21ヌクレオチドの長さを有していてもよい。該核酸または/および該核酸断片は、最大で29、好ましくは最大で27、より好ましくは最大で25、特により好ましくは最大で23、最も好ましくは最大で22ヌクレオチドの長さを有していてもよい。
【0067】
本発明において用いられる核酸は、マイクロRNA(miRNA)、その前駆体、断片、または誘導体であってもよい。該miRNAは、本明細書に記載の核酸の長さを有していてもよい。該miRNAは、特に、約22ヌクレオチド、より好ましくは22ヌクレオチドの長さを有していてもよい。
【0068】
本発明のモジュレーターは、抗体を含有していてもよく、このとき、該抗体は、キナーゼまたは/およびキナーゼ結合ポリペプチドに対して指向されてもよい。
【0069】
該抗体は、
(a)表1A、および表1Bから選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、または/ならびに
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含有するキナーゼまたは/およびキナーゼ結合ポリペプチドに対するものであることが好ましい。
【0070】
他の好ましい一実施形態では、該抗体は、
(a)表4から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、または/ならびに
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対するものである。
【0071】
さらに他の好ましい一実施形態では、該抗体は、
(a)表5から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、または/ならびに
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対するものである。
【0072】
本発明の抗体は、モノクローナル抗体もしくはポリクローナル抗体、キメラ抗体、キメラ単鎖抗体、FabフラグメントまたはFab発現ライブラリーによって産生される断片であってもよい。
【0073】
本発明の抗体を作製する技術は、当業者に知られている。モノクローナル抗体は、ヒトB細胞ハイブリドーマ技術またはEBV-ハイブリドーマ技術によって作製することができる(Kohlerら、1975、Nature 256:495〜497、Kozborら、1985、J. Immunol. Methods 81、31〜42、Coteら、PNAS、80:2026〜2030、Coleら、1984、Mol. Cell Biol. 62:109〜120)。キメラ抗体(マウス/ヒト)は、Morrisonら(1984、PNAS、81:6851〜6855)、Neubergerら(1984、312:604〜608)およびTakedaら(1985、Nature 314:452〜454)の方法を行うことによって作製することができる。単鎖抗体は、当業者に知られている技術によって作製することができる。
【0074】
組換え免疫グロブリンライブラリー(Orlandiら、1989、PNAS 86:3833〜3837、Winterら、1991、Nature 349:293〜299)をスクリーニングして本発明の抗体を得ることもできる。ランダム結合性免疫グロブリンライブラリー(Burton、1991、PNAS、88:11120〜11123)を使用して、異なるイディオタイプの組成を有する、関連した特異性を有する抗体を作製することもできる。
【0075】
抗体作製のためのもう1つの戦略は、in vivoでのリンパ球集団の刺激である。
【0076】
さらに、本発明の(F(ab')2フラグメントを含有する)抗体フラグメントは、抗体のプロテアーゼ消化によって、例えばペプシンによって、作製することができる。こうしたF(ab')2フラグメントのジスルフィド結合を減少させることにより、結果としてFabフラグメントが得られる。もう1つのアプローチでは、該Fabフラグメントは、Fab発現ライブラリーから直接得ることもできる(Huseら、1989、Science 254:1275〜1281)。
【0077】
本発明のポリクローナル抗体は、表1A、表1B、表4および表5から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列またはこれらの免疫原性断片を抗原として用いて、標準的な免疫プロトコルが当業者に知られている、例えば、ウマ、ヤギ、ウサギ、ヒトなどの、宿主の標準的な免疫プロトコルによって調製することができる。
【0078】
該抗体は、標的遺伝子の遺伝子産物に特異的な抗体、特に、標的遺伝子によってコードされるポリペプチドまたはタンパク質に特異的な抗体であってもよい。
【0079】
アプタマーおよびスピーゲルマーは、抗体と結合特性を共有する。アプタマーおよびスピーゲルマーは、標的分子を特異的に結合するように設計される。
【0080】
本発明の核酸は、(a)アプタマー、(b)アプタマーをコードするDNA分子、および(c)スピーゲルマーから選択されてもよい。
【0081】
当業者は、アプタマーについて知っている。本発明において、「アプタマー」は、標的分子に結合することができる核酸であってもよい。アプタマーは、(例えば、>1015種の異なる核酸配列を含有する)結合核酸ライブラリーにおいて、固定化された標的分子への結合およびその後のその核酸配列の同定によって、特定することができる。この選択方法は、特異性を高めるために一回または複数回繰り返されてもよい。当業者は、所定の分子を特異的に結合するアプタマーを作製するための好適な方法を知っている。該アプタマーは、本明細書に記載のような核酸の長さを有しうる。該アプタマーは、多くとも300、多くとも200、多くとも100、または多くとも50ヌクレオチドまでの長さを有しうる。該アプタマーは、少なくとも10、少なくとも15、または少なくとも20ヌクレオチドの長さを有していてもよい。該アプタマーは、DNA分子によってコードされていてもよい。該アプタマーは、本明細書に記載のように、少なくとも1つのヌクレオチド類似体または/および少なくとも1つのヌクレオチド誘導体を含有していてもよい。
【0082】
当業者は、スピーゲルマーについて知っている。本発明において、「スピーゲルマー」は、標的分子に結合することができる核酸であってもよい。当業者は、所定の分子を特異的に結合するスピーゲルマーを作製するための好適な方法を知っている。該スピーゲルマーは、ヌクレアーゼ耐性の結合を形成することができるヌクレオチドを含有する。好ましくは、該スピーゲルマーは、Lヌクレオチドを含有する。より好ましくは、該スピーゲルマーは、L-オリゴヌクレオチドである。該スピーゲルマーは、本明細書に記載のような核酸の長さを有しうる。該スピーゲルマーは、多くとも300、多くとも200、多くとも100、または多くとも50ヌクレオチドまでの長さを有しうる。該スピーゲルマーは、少なくとも10、少なくとも15、または少なくとも20ヌクレオチドの長さを有しうる。該スピーゲルマーは、本明細書に記載のように、少なくとも1つのヌクレオチド類似体または/および少なくとも1つのヌクレオチド誘導体を含有していてもよい。
【0083】
当業者は、デコイ核酸について知っている。本発明において、「デコイ」または「デコイ核酸」は、核酸結合タンパク質、例えばDNA結合タンパク質などを特異的に結合する能力のある核酸であってもよい。該デコイ核酸は、DNA分子、好ましくは、二本鎖DNA分子であってもよい。該デコイ核酸は、核酸結合タンパク質によって認識されうる「認識配列」と呼ばれる配列を含有する。該認識配列は、少なくとも3、少なくとも5、または少なくとも10ヌクレオチドの長さを有していることが好ましい。該認識配列は、多くとも15、多くとも20、または多くとも25ヌクレオチドまでの長さを有していることが好ましい。核酸結合タンパク質の例は、転写因子であり、これは、二本鎖DNA分子を結合することが好ましい。デコイ核酸を用いた、本明細書に記載のような、細胞、有胚卵、または/および非ヒト動物のトランスフェクションは、デコイ核酸が結合する核酸結合タンパク質の活性を低下させる結果になってもよい。本明細書に記載のデコイ核酸は、本明細書に記載のような核酸分子の長さを有しうる。該デコイ核酸分子は、多くとも300、多くとも200、多くとも100、多くとも50、多くとも40、または多くとも30ヌクレオチドまでの長さを有しうる。該デコイ核酸は、少なくとも3、少なくとも5、少なくとも10、少なくとも15、または少なくとも20ヌクレオチドの長さを有していてもよい。該デコイ核酸は、DNA分子によってコードされていてもよい。該デコイ核酸は、本明細書に記載のように、少なくとも1つのヌクレオチド類似体または/および少なくとも1つのヌクレオチド誘導体を含有していてもよい。
【0084】
本明細書に記載のRNA分子または/およびDNA分子は、少なくとも1つのヌクレオチド類似体を含有していてもよい。本明細書中で使用される場合、「ヌクレオチド類似体」とは、その核酸内の主鎖、少なくとも1つのリボース、少なくとも1つの塩基、3'末端または/および5'末端における修飾に好適な構成単位を指しうる。主鎖修飾としては、ホスホロチオエート結合(PT);ペプチド核酸(PNA);モルホリノ核酸;ホスホロアミデート結合DNA(PA)などが挙げられ、これは、主鎖窒素を含む。リボース修飾としては、例えば、リボースの2'酸素を4'炭素と結合させているメチレン架橋での、ロックド核酸(LNA);2'-デオキシ-2'-フルオロウリジン;2'-フルオロ(2'-F);2'-O-アルキル-RNA(2-O-RNA)、例えば、2'-O-メチル(2'-O-Me)、2'-O-メトキシエチル(2'-O-MOE)などが挙げられる。修飾塩基は、2'-フルオロピリミジンであってもよい。5'修飾としては、5'-TAMRA-ヘキシルリンカー、5'-ホスフェート、5'-アミノ、5'-アミノ-C6リンカー、5'-ビオチン、5'-フルオレセイン、5'-テトラクロロ-フルオレセイン、5'-ピレン、5'-チオール、5'-アミノ、(12炭素)リンカー、5'-ダブシル、5'-コレステロール、5'-DY547(Cy3(商標)代替物)などが挙げられる。3'末端修飾としては、3'反転デオキシチミジン、3'-ピューロマイシン、3'-ジデオキシシチジン、3'-コレステロール、3'-アミノモディファイヤー(6原子)、3'-DY547(Cy3(商標)代替物)などが挙げられる。
【0085】
特に、本明細書に記載のヌクレオチド類似体は、siRNA、アンチセンスRNA、およびアプタマー中の好適な構成単位である。
【0086】
本明細書中で使用される場合、「核酸類似体」とは、本明細書に記載の少なくとも1つのヌクレオチド類似体を含有する核酸を指す。さらに、本明細書に記載の核酸分子は、少なくとも1つのデオキシリボヌクレオチドおよび少なくとも1つのリボヌクレオチドを含有していてもよい。
【0087】
本発明のRNA分子は、少なくとも1つのデオキシリボヌクレオチドまたは/および少なくとも1つのヌクレオチド類似体を含有していてもよい。本発明のDNA分子は、少なくとも1つのリボヌクレオチドまたは/および少なくとも1つのヌクレオチド類似体を含有していてもよい。
【0088】
本明細書に記載の誘導体とは、化学的に修飾された化合物を指す。本明細書に記載の核酸分子の誘導体とは、化学的に修飾された核酸分子を指す。修飾は、核酸分子中に、または/および該核酸の作製において用いられる少なくとも1個の核酸構成単位中に導入されてもよい。
【0089】
本発明において、「断片」という用語は、核酸、ポリペプチドおよびタンパク質の断片を指す。また、「断片」は、核酸、ポリペプチドおよびタンパク質の部分的な配列も指す。
【0090】
本発明において用いられるポリペプチドまたは/およびペプチドの断片、特に、表1A、表1B、表4および表5から選択される核酸または/および遺伝子によってコードされるアミノ酸配列の断片は、少なくとも5アミノ酸残基、少なくとも10、または少なくとも20アミノ酸残基の長さを有していてもよい。前記断片の長さは、最大で200アミノ酸残基、最大で100アミノ酸残基、最大で60アミノ酸残基、または最大で40アミノ酸残基であってもよい。
【0091】
本明細書に記載のアミノ酸配列の断片は、該配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有していてもよい。
【0092】
本明細書に記載のヌクレオチド配列の断片は、該配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有していてもよい。
【0093】
表1A、1B、4および5に示している核酸分子の断片は、多くとも1000、多くとも2000、または多くとも3000ヌクレオチドまでの長さを有しうる。核酸断片は、本明細書に記載のようなsiRNA分子、miRNA分子、アプタマー、スピーゲルマー、または/およびデコイの長さを有しうる。また、核酸断片は、多くとも300、多くとも200、多くとも100、または多くとも50ヌクレオチドまでの長さも有しうる。また、核酸断片は、少なくとも3、少なくとも5、少なくとも10、少なくとも15、または少なくとも20ヌクレオチドの長さも有しうる。
【0094】
本発明の方法において、遺伝子の発現の調節は、特に転写または/および翻訳の、下方制御または上方制御であってもよい。
【0095】
当業者は、遺伝子が上方制御されるかどうかまたは下方制御されるかどうかを容易に決定することができる。本発明の文脈において、遺伝子発現の上方制御(活性化)は、少なくとも2倍、好ましくは少なくとも4倍までの上方制御であってもよい。本発明の文脈における下方制御(阻害)は、少なくとも2分の1、好ましくは少なくとも4分の1までの、遺伝子発現の低減であってもよい。最も好ましいものは、例えばRNA干渉による、遺伝子発現のほぼ完全な阻害である。
【0096】
遺伝子の活性の調整は、活性の低減または増大であってもよい。「活性の阻害」は、少なくとも2分の1、好ましくは少なくとも4分の1までの、遺伝子または遺伝子産物の活性の低減であってもよい。「活性の阻害」は、活性のほぼ完全な阻害を含む。「活性の活性化」は、少なくとも2倍、好ましくは少なくとも4倍までの、遺伝子または遺伝子産物の活性の増大であってもよい。
【0097】
本発明において、本明細書に記載の方法、細胞、生物、および医薬組成物の特定の実施形態は、本出願に記載のあらゆる個々の遺伝子、核酸配列または/および遺伝子産物に関する。特定の一実施形態において、個々の遺伝子は、表1、4、および5に記載の遺伝子から選択される。他の特定の実施形態は、表1、4、および5に記載の個々の遺伝子に関する。他の特定の一実施形態において、個々の遺伝子産物は、表1、4、および5に記載の遺伝子によって産生される遺伝子産物から選択される。他の特定の実施形態は、表1、4、および5に記載の遺伝子によって産生される個々の遺伝子産物に関する。さらに他の特定の一実施形態において、個々の核酸配列または核酸分子は、表1、2、4および5に記載の核酸分子または核酸配列から選択される。他の特定の実施形態は、表1、2、4、および5に記載の個々の核酸分子または核酸配列に関する。さらなる特定の実施形態は、表1、2、3、4、および5に記載の遺伝子、遺伝子産物および核酸分子のあらゆる組合せに関する。組合せは、2、3、4、5、6、7、8、9、10またはより多くの異なる種を含んでいてもよい。表3は、核酸分子の特定の組合せに関する。
【0098】
本発明のさらなる特定の実施形態は、表5に開示の配列に関する。本発明の特定の実施形態は、表5に記載のあらゆる個々の遺伝子、核酸分子または/および遺伝子産物に関する。特定の一実施形態において、個々の遺伝子は、表5に記載の遺伝子から選択される。他の特定の実施形態は、表5に記載の個々の遺伝子に関する。他の特定の一実施形態において、個々の遺伝子産物は、表5に記載の遺伝子によって産生される遺伝子産物から選択される。他の特定の実施形態は、表5に記載の遺伝子によって産生される個々の遺伝子産物に関する。さらに他の特定の一実施形態において、個々の核酸分子または核酸配列は、表5に記載の核酸分子または核酸配列から選択される。他の特定の実施形態は、表5に記載の個々の核酸分子または核酸配列に関する。さらなる特定の実施形態は、表5に記載の遺伝子、遺伝子産物および核酸分子のあらゆる組合せに関する。組合せは、2、3、4、5、6、7、8、9、10またはより多くの異なる種を含んでいてもよい。
【0099】
本発明の特定の実施形態は、MxA遺伝子、MxAポリペプチド、およびこれらの断片に関する。さらに特定の実施形態は、miR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体、ならびにmiR-141、miR-141*、miR-200c、miR-200c*または/およびこれらの前駆体をコードするDNA分子に関する。
【0100】
修飾は、単一の核酸種によって行われてもよく、または、表1a、1b、2、4もしくは/および5ならびにその断片から選択されうる、2、3、4、5、6もしくはより多くの異なる核酸種を含有する核酸の組合せによって行われてもよい。好ましい組合せは、表3に記載している(本明細書では「プール」とも呼ぶ)。表3は、少なくとも2種のキナーゼ遺伝子または/およびキナーゼ結合ポリペプチド遺伝子の組合せを含む。この組合せが、例えば表1a、b、4および5から選択される、単一の遺伝子の発現を変更することも好ましい。2つの核酸種の組合せが好ましい。より好ましいのは、表2から選択される2種の核酸の組合せである。さらにより好ましいのは、表2に開示の特定の組合せから選択される2種の核酸の組合せであり、このとき、この2種の核酸は、単一の遺伝子の発現を変更する。
【0101】
改変、特に調整は、RNA干渉によって行われるノックダウンであってもよい。該核酸または核酸種の該組合せは、表2、表4および表5の配列ならびにそれらの断片から選択される配列を含有していてもよい、siRNAであってもよい。該組合せは、例えば表1bおよび表4から選択される、単一の遺伝子をノックダウンすることが好ましい。2つのsiRNA種の組合せが好ましく、これは、表1bの遺伝子に由来する、表2の配列、ならびに表4および表5の配列、から選択されてもよく、このとき、該組合せは、単一の遺伝子をノックダウンすることが好ましい。
【0102】
本発明のステップ(a)において用いることができる、組換え技術による「遺伝子または/および遺伝子産物の活性化」または「遺伝子または/および遺伝子産物の阻害」は、当業者が知っているいかなる好適な方法を含んでいてもよい。
【0103】
目的の遺伝子または/およびその遺伝子産物の活性化の好ましい方法は、
-活性化する遺伝子の少なくとも1個のさらなるコピーを、恒久的または一過的のいずれかで、該細胞または/および生物内に導入すること、
-転写を増強すること、
-過剰発現、
-遺伝子内に強力なプロモーター、例えばCMVプロモーターを導入すること、
-好適なエンハンサーを導入すること、
-転写的に活性なマイクロRNAの阻害、このとき、該マイクロRNAは、活性化する遺伝子の活性を阻害し、このとき、阻害は、好適な核酸分子によって行うことができる、
-miRNA結合部位の欠失、
-例えば、例えばヘパドナウイルスからの、転写後調節エレメントを3'末端に導入することによる、または、例えばD型レトロウイルスからの、1つもしくは複数の構成的な輸送エレメントを3'末端に導入することによる、または/およびスプライシングを受けうるイントロンを用いることによる、核からの輸出を含むRNAプロセシングの改良、
-例えば、潜在性スプライシング部位の欠失、GC含量の最適化、キラーモチーフおよびリピートの欠失、構造の最適化による、リボソーム結合の改良ならびにコード配列または/および3'UTRの最適化による、翻訳の改良、
から選択されてもよい。
【0104】
目的の遺伝子または/およびその遺伝子産物の阻害の好ましい方法は、
-阻害する遺伝子の少なくとも1個のさらなるコピーを該細胞または/および生物において欠失させること、このとき、該遺伝子は、完全にまたは部分的に欠失される。例えば、調節配列または/およびコード配列が、完全にまたは部分的に、欠失される、
-転写を低減させること、
-存在する場合には、エンハンサーを欠失させること、
-転写的に活性なマイクロRNAの導入または/および活性化、このとき、該マイクロRNAは、阻害する遺伝子の活性を阻害し、このとき、活性化は、内因性マイクロRNAコード配列の活性化であってもよく、導入は、外因性マイクロRNA分子の導入であってもよい、
-miRNA結合部位の導入、
-存在する場合には、3'末端に導入している転写後調節エレメントまたは3'末端に導入している1つもしくは複数の構成的な輸送エレメントの欠失もしくは/および修飾による、あるいはイントロン-エキソン構造を変更することによる、核からの輸出を含むRNAプロセシングの低減、
-例えば、潜在性スプライシング部位の導入、GC含量の変更、キラーモチーフおよびリピートの導入による、リボソーム結合ならびにコード配列または/および3'UTRの改変による、翻訳の低減、
から選択されてもよい。
【0105】
本発明の多様な実施形態において用いられる遺伝子は、表1A、1B、2、4および5、またはこれらの任意の組合せのいずれから選択されてもよい。
【0106】
ステップ(b)に従って該細胞または/および該生物をインフルエンザウイルスと接触させることは知られている。該非ヒト生物が有胚卵である場合、当業者は、例えば受精後の定められた間隔で、インフルエンザウイルスを該卵に接種する好適な方法を知っている。既知の接種技術も、有胚卵へのモジュレーターの投与のためにまたは/および有胚卵の組換え改変のために適用することができる。
【0107】
当業者は、インフルエンザウイルスの複製を可能にする条件下で該細胞、該有胚卵または/および該非ヒト生物を培養するステップ(c)に従った方法を知っている。好適な細胞培養法を適用することができる。該非ヒト生物が有胚卵である場合、当業者は、高温でのインキュベートなどの、インフルエンザウイルス複製を可能にする好適な方法を知っている。
【0108】
インフルエンザウイルスまたは/およびその構成要素をステップ(d)に従って単離することは、ウイルスまたは/およびその構成要素の、当業者に知られるあらゆる単離方法に関する。「単離」は、ウイルスまたは/およびその構成要素のほぼ純性または未精製の調製物または製剤の作製を含む。ウイルスの構成要素としては、ウイルスのタンパク質、ポリペプチド、ならびにウイルスのタンパク質または/およびポリペプチドをコードする核酸などが挙げられる。また、生ウイルスを単離することもできる。
【0109】
当業者は、ステップ(e)に従って、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、医薬組成物を調製する方法を知っている。本発明の方法によって作製される医薬組成物は、免疫原性の組成物であってもよい。また、本発明の方法によって作製される医薬組成物は、ワクチンであってもよい。
【0110】
本発明のさらにもう1つの主題は、MxA遺伝子の発現または/および遺伝子産物活性の阻害剤を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含有する医薬組成物である。該医薬組成物は、miR-141、miR-141*、miR-200c、miR-200c*、これらの前駆体および誘導体から選択される少なくとも1種の阻害剤を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含有することが好ましい。また、該阻害剤は、アンチセンス核酸、siRNA、shRNA、および小分子から選択されてもよい。
【0111】
本発明のさらにもう1つの主題は、MxA遺伝子の発現または/および遺伝子産物活性の活性化剤を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含有する医薬組成物である。該医薬組成物は、miR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害する能力のある少なくとも1種の阻害剤を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含有することが好ましい。
【0112】
miR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる該阻害剤は、アプタマー、スピーゲルマーおよびデコイ核酸から選択されることが好ましい。該医薬組成物における好ましい阻害剤であるアプタマーは、miR-141、miR-141*、miR-200cまたは/およびmiR-200c*ならびにこれらの前駆体の活性を阻害することができる。該医薬組成物における好ましい阻害剤であるスピーゲルマーは、miR-141、miR-141*、miR-200cまたは/およびmiR-200c*ならびにこれらの前駆体の活性を阻害することができる。該医薬組成物中の好ましい阻害剤であるデコイ核酸分子は、miR-141、miR-141*、miR-200cまたは/およびmiR-200c*ならびにこれらの前駆体の転写に関わる転写因子を結合することができる配列を含有する。
【0113】
本明細書に記載の医薬組成物(本発明の方法によって作製されたか、またはMxA遺伝子の発現もしくは/および遺伝子産物活性の活性化剤を含有する該医薬組成物のいずれか)は、好ましくは、ヒトにおいて使用されるものであるかまたは獣医学的医薬品である。該医薬組成物は、好ましくは、インフルエンザウイルス感染の予防、軽減または/および治療のために使用されるものである。
【0114】
該医薬組成物中の担体は、送達系を含有していてもよい。当業者は、本発明の医薬組成物に好適な送達系を知っている。該医薬組成物は、ネイキッド核酸の形態で、ウイルスベクター、リポソームを含む非ウイルスベクター、ナノ粒子または/およびポリマーとの組合せで、送達されてもよい。該医薬組成物または/および該核酸は、エレクトロポレーションによって送達されてもよい。
【0115】
ネイキッド核酸としては、RNA、改変RNA、DNA、改変DNA、RNA-DNA-ハイブリッド、アプタマー融合体、プラスミドDNA、ミニサークル、トランスポゾンなどが挙げられる。
【0116】
ウイルスベクターとしては、ポックスウイルス、アデノウイルス、アデノ随伴ウイルス、水疱性口内炎ウイルス、アルファウイルス、はしかウイルス、ポリオウイルス、B型肝炎ウイルス、レトロウイルス、およびレンチウイルスなどが挙げられる。
【0117】
リポソームとしては、安定な核酸-脂質粒子(SNALP)、カチオン性リポソーム、カチオン性カルジオリピン類似体ベースのリポソーム、中性リポソーム、リポソーム-ポリカチオン-DNA、カチオン性イムノリポソーム、イムノリポソーム、コレステロールの疎水性誘導体、ラウリン酸およびリトコール酸を含有するリポソームなどが挙げられる。リポソーム形成に好適な化合物の例は、1,2-ジラウロイル-sn-グリセロ-3-ホスホエタノールアミン(DLPE);1,2-ジオレオイル-sn-グリセロ-3-ホスホ-L-セリン(DOPS);コレステロール(CHOL);1,2-ジオレイル-sn-グリセロ-3-ホスホコリン(DOPC)である。
【0118】
ナノ粒子としては、CaCO3ナノ粒子、キトサン-コーティングされたナノ粒子、葉酸化された脂質ナノ粒子、ナノサイズの核酸担体などが挙げられる。
【0119】
ポリマーとしては、ポリエチレンイミン(PEI)、ポリエステルアミン(PEA)、ポリエチレングリコール(PEG)-オリゴ結合体、PEGリポソーム、ポリマーナノ球体などが挙げられる。
【0120】
該医薬組成物は、アテロコラーゲン、カーボンナノチューブ、シクロデキストリン含有ポリカチオン、融合タンパク質(例えば、プロタミン-抗体結合体)と組み合わせて送達されてもよい。
【0121】
MxA遺伝子の発現もしくは/および遺伝子産物活性の活性化剤、または/ならびにmiR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる阻害剤は、インフルエンザウイルス感染の予防用、軽減用または/および治療用の医薬組成物の製造に使用されてもよい。本明細書に記載の送達系および送達法を使用してもよい。
【0122】
本発明のもう1つの主題は、MxA遺伝子の発現もしくは/および遺伝子産物活性の活性化剤、または/ならびにmiR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる阻害剤の、インフルエンザウイルス感染の予防用、軽減用または/および治療用の使用である。本明細書に記載の送達系および送達法を使用してもよい。
【0123】
本発明のもう1つの主題は、治療有効量の、MxA遺伝子の発現もしくは/および遺伝子産物活性の活性化剤、または/ならびにmiR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる阻害剤を、これを必要としている対象に投与することを含む、インフルエンザウイルス感染の予防、軽減または/および治療の方法である。インフルエンザウイルス感染の予防、軽減または/および治療の該方法において、本明細書に記載の送達系および送達法を使用してもよい。
【0124】
本発明のさらにもう1つの主題は、本明細書に記載のように、本発明の方法のステップ(a)に従って作製される組換え細胞である。
【0125】
本発明のさらにもう1つの主題は、本明細書に記載のように、本発明の方法のステップ(a)に従って作製される組換え非ヒト生物である。
【0126】
本発明のさらにもう1つの主題は、本明細書に記載のように、本発明の方法のステップ(a)に従って作製される組換え有胚卵である。該組換え有胚卵は、好ましくは、組換え有胚鶏卵である。
【0127】
以下の図面、表および実施例によって本発明をさらに説明する。
【図面の簡単な説明】
【0128】
【図1】実施例のsiRNAキナーゼスクリーニングの試験的設定を示した図である。
【図2】発光データに関する、トランスフェクトした(対照)-siRNAの効果を示した図である。このダイアグラムは、1枚の96ウェルプレートからの典型的なスクリーニング結果を示している。全ての試験に、いくつかの対照を、例えば、未感染のもの、ルシフェラーゼに対するsiRNAでトランスフェクトしたもの、モック処理したものおよびA型インフルエンザウイルスからのウイルス核タンパク質遺伝子(NP)に対するsiRNAのように三重試験で含めた。ルシフェラーゼsiRNAおよび抗NP siRNAで処理した細胞間の発光の相違を、定義による100%阻害に設定した。
【図3】実施例において試験した全てのsiRNAについてのインフルエンザウイルス複製の阻害を示した図である。
【図4】解析した全てのsiRNAからの値「%阻害」を使用してz-スコアを算出した図である。きわめて効率的なsiRNAを桃色にラベルしており、ルシフェラーゼsiRNAをトランスフェクトした対照細胞と比較して50%を超える阻害を示している。
【図5】ゲノム全体のsiRNAスクリーニングの試験的設定を示した図である(実施例4を参照されたい)。
【図6】miR-141の発現は、ヒト肺上皮細胞においてA型インフルエンザでの感染に応じて増強され、ウイルス複製を増強することを示した図である。(A)A549ヒト肺上皮細胞を、ヒトインフルエンザ株であるA/WSN/33(H1N1)、A/Puerto Rico/8/34(H1N1)、A/Panama/99(H3N2)および鳥インフルエンザ株であるA/FPV/Bratislava/79(H7N7)で感染させることにより、感染24時間後のmiR-141の上方制御がqRT-PCRによって検出される。miR-141の発現を、非感染(NI)細胞に対して標準化している。(B)miR-141特異的な前駆体もしくは阻害剤、または非特異的なRNA阻害剤(NS)でのトランスフェクション後のA549細胞におけるウイルス増殖(A/WSN/33)(MOI 0.05)。トランスフェクトして感染させたA549細胞からのウイルス含有上清を、インフルエンザ依存的ルシフェラーゼアッセイを使用して定量した。(C、D)安定に形質導入された異なるmiRNAを過剰発現しているA549細胞におけるウイルス複製。初代細胞培養物における感染細胞の百分率(C)および結果として生じるウイルス後代(D)を、自動顕微鏡法およびルシフェラーゼアッセイをそれぞれ使用して定量した。データは、3つの重複した試料(A、C)または3つの独立した試験(B、D)の平均値+SDである。*t-検定(P<0.05)。NT、トランスフェクトなし。
【図7】A型インフルエンザ感染に応じた、MxAおよびmiR-141の遺伝子発現を示した図である。(A〜C)miR-141とMxA発現との間の時間依存的関連性を解析するために、A549細胞をインフルエンザA/WSN/33(MOI 1)に感染させた。表示の各時点で、MxA mRNA(A)、MxAタンパク質(B)およびmiR-141(C)のレベルをqRT-PCR(RNA)およびイムノブロット解析(タンパク質)によって決定した。β-アクチンは、イムノブロットのローディング対照としての役割を果たす。(D〜F)A549細胞を、表示のMOIでインフルエンザA/WSN/33に感染させて、感染24時間後に細胞を溶解した。MxA mRNA(D)およびMxAタンパク質(E)のレベルを、それぞれqRT-PCRおよびイムノブロットによって決定した。β-アクチンは、イムノブロットのローディング対照としての役割を果たす。(F)免疫蛍光法による、表示のMOIでのMxA(緑色)およびインフルエンザA/WSN/33の核タンパク質(NP)(赤色)の検出(感染24時間後)。スケールバー、10μm。A、CおよびDにおいて、MxA発現は、非感染(NI)対照に相対的なものである。データは、3つの重複した試料の平均値+SDである。
【図8A】miR-141機能の阻害は、MxA遺伝子発現を回復させることを示した図である。(A)A549細胞におけるMxA(緑色シグナル)および核タンパク質(NP、赤色シグナル)の検出(感染24時間後)。細胞を、miR-141阻害剤および対照阻害剤(miR-198;非特異的、NS)でそれぞれトランスフェクトして、8時間後にインフルエンザA/WSN/33またはA/FPV/Bratislava/79(MOI 1)に感染させた。スケールバー、10μm。矢印は、MxA-陽性のインフルエンザ感染細胞を示す。MxA-陽性のインフルエンザ感染細胞の定量については、図12に示している。
【図8BC】miR-141機能の阻害は、MxA遺伝子発現を回復させることを示した図である。 (B)qRT-PCRによって定量される、異なるMOIでのA型インフルエンザ感染に応じたmiR-141のレベル。上方制御は、非感染細胞(NI)に相対的なものである。データは、3つの重複した試料の平均値+SDである。(C)MxA遺伝子発現のイムノブロット解析。A549細胞からの細胞溶解液を、特定量のmiR-141および非特異的阻害剤でトランスフェクトして、表示のMOIでインフルエンザA/WSN/33に感染させた。β-アクチンは、ローディング対照としての役割を果たす。ブロットは、3つの独立した試験の代表である。
【図9AB】MxAの下方制御は、miR-141によって直接仲介されることを示した図である。(A)内因性MxAタンパク質および外因性eGFP-MxA融合タンパク質のイムノブロット解析(感染24時間後)。A549細胞を、表示のMOIでインフルエンザA/WSN/33に感染させて、1時間後にpEGFP-MxAでトランスフェクトした。β-アクチンは、ローディング対照としての役割を果たす。示しているブロットは、3つの独立した試験の代表である。MxAおよびeGFP-MxAのバンド強度をブロットの下に示している。(B)ウイルスのNP染色(青色)、mCherry発現(赤色)およびeGFP-MxA発現(緑色)を用いた、感染させトランスフェクトしたA549細胞の感染24時間後の代表的な共焦点画像。スケールバー、10μm。eGFP-MxA陽性細胞の定量については、図15に示している。
【図9C】MxAの下方制御は、miR-141によって直接仲介されることを示した図である。(C)トランスフェクトした細胞またはインフルエンザA/WSN/33を感染させた細胞におけるMxAおよびeGFP-MxA融合タンパク質の発現。A549細胞を、表示のMOIで感染させて、未処理のままにしておくか、または異なるmiRNAの過剰発現プラスミド(空、miR-198およびmiR-144)で6時間トランスフェクトし、その後、pEGFP-MxAでのトランスフェクションとIFN-β(500U/ml)での同時の処理とを行った。β-アクチンは、ローディング対照としての役割を果たす。示しているブロットは、3つの独立した試験の代表である。MxAおよびeGFP-MxAのバンド強度をブロットの下に示している。
【図9D】MxAの下方制御は、miR-141によって直接仲介されることを示した図である。 (D)異なるmiRNAを過剰発現している細胞または対照細胞におけるmCherry(赤色)、外因性eGFP-MxA(緑色)および内因性MxA(青色)のレベルを示している代表的な共焦点顕微鏡写真。矢印は、内因性MxAの下方制御と、eGFP-MxAの発現がないまたは弱いこととの双方を示す、トランスフェクトされた細胞(mCherry陽性)を示している。スケールバー、10μm。eGFP-MxA陽性細胞の定量については、図16に示している。
【図10】MxAを欠損した、インフルエンザA/WSN/33感染A549細胞(MOI 0.05)における一次感染のレベルを示した図である。インフルエンザNP特異抗体で細胞を染色することによって、感染24時間後に一次感染を決定し、その後、核を染色するためにヘキスト色素で処理した。自動顕微鏡法を使用して感染細胞および非感染細胞の数を定量して、感染細胞の割合を算出した(図10)。エラーバーは、三重試験の標準偏差(SD)を示す。NT、トランスフェクトなし。*/t-検定/(P<0.05)。
【図11】MxAを欠損した、インフルエンザA/WSN/33感染A549細胞(MOI 0.05)におけるウイルス後代のレベルを示した図である。インフルエンザ依存的ルシフェラーゼアッセイを使用して、細胞上清中のウイルス後代を検出した(図11)。エラーバーは、三重試験の標準偏差(SD)を示す。NT、トランスフェクトなし。*/t-検定/(P<0.05)。
【図12】インフルエンザ感染細胞における内因性MxAの定量を示した図である。A549細胞を、miR-141および対照阻害剤(非特異的(NS);miR-181)でそれぞれトランスフェクトして、トランスフェクション後8時間でインフルエンザA/WSN/33またはA/FPV/Bratislava/79に感染させた(MOI 1)。感染24時間後に、免疫蛍光共焦点顕微鏡法を使用して、インフルエンザ感染細胞(処理ごとに細胞約100個)でMxAの発現を解析した。MxA陽性細胞の数は、総細胞数の平均百分率+SDとして示している。miR-141阻害剤でトランスフェクトした細胞は、1のMOIでの感染後に、MxA陽性細胞の数が対照阻害剤と比較して有意に増加した。*/t-検定/(/P/<0.05)。
【図13】安定にmiRNAを過剰発現しているA549細胞におけるMxAの発現を示した図である。表示のmiRNAをコードするレトロウイルスベクターまたは空の対照ベクターで予め形質導入されている細胞を、未処理のままにしておくか、またはIFN-β(500U/ml)で24時間処理した。3つの独立した試験からの代表的なブロットを示している。MxAのバンド強度は、Aidaイメージアナライザープログラム(V.4.03)を使用して決定した。
【図14】ウェブツールRNAhybrid(/9/)によって最も小さい最小自由エネルギー(mfe)で予測されるhas-miR-141(UAACACUGUCUGGUAAAGAUGG、miRBase配列データベース(/8/)とMxAの完全なmRNA配列(NM_002462)との間の相互作用を示した図である。
【図15】eGFP-MxA陽性細胞の定量を示した図である。A549細胞を、表示のMOIでインフルエンザA/WSN/33に感染させた。1時間後、細胞を、外因性MxAを検出するためのeGFP-MxA融合プラスミドと、トランスフェクション効率を標準化するための構成的に発現されるmCherry構築物(モル比6:1)とでコトランスフェクトした。感染24時間後に、免疫蛍光共焦点顕微鏡法を使用して処理ごとに100細胞でeGFP-MxAの発現を解析した。eGFP-MxAおよびmCherry陽性細胞の数は、総細胞数の平均百分率+SDとして示している。1のMOIにおいて、eGFP-MxA陽性細胞の百分率は、非感染(NI)細胞培養物および部分感染細胞培養物(MOI 0.05)と比較して有意に低下する。*/t-検定/(/P/<0.05)。
【図16】感染のない場合のeGFP-MxA陽性細胞の定量を示した図である。A549細胞を、異なる過剰発現プラスミド(空、miR-198およびmiR-141)で6時間トランスフェクトし、その後、(外因性MxAを検出するための)eGFP-MxA融合プラスミドでのトランスフェクションとIFN-β(500U/ml)での同時の処理とを行った。免疫蛍光共焦点顕微鏡法を使用して処理ごとに約100個のmCherry陽性細胞でeGFP-MxAの発現を解析した。eGFP-MxA陽性細胞の数は、総細胞数の平均百分率+SDとして示している。miR-141を過剰発現するプラスミドでトランスフェクトした細胞は、対照(トランスフェクトなし(NT)、空ベクターおよびmiR-198)と比較して、eGFP-MxA陽性細胞の有意な減少を示す。*/t-検定/(/P/<0.05)。
【図17】IFN-β刺激(1000U/ml)に応じた、安定に形質導入されたmiRNA過剰発現細胞におけるデュアルルシフェラーゼアッセイによって検出されるISREプロモーター活性を示した図である。これらのA549細胞を、最小プロモーター内にISREモチーフを含むプロモーターか、または対照として構成的なSV40プロモーター(pGL3-対照)のいずれかによって働くホタルルシフェラーゼ遺伝子をコードするプラスミドでトランスフェクトした。さらに、細胞を、トランスフェクション対照として/レニラ/ルシフェラーゼプラスミドでトランスフェクトし、直後に、IFN-βで刺激した。/レニラ/発光シグナルに対するホタルの比を示している。エラーバーは、四重試験の標準偏差(SD)を示す。
【図18】miR-141/141*およびmiR-200c/200c*、ならびにこれらの前駆体の配列を示した図である。さらに、これらの前駆体のヘアピン構造を記載している。
【発明を実施するための形態】
【0129】
表1:siRNAキナーゼスクリーニングの結果を示した表である:a:細胞数に対して標準化し、4つの独立の試験を使用して標準偏差を算出した、%でのウイルス複製の活性化(「負の」阻害)、b:細胞数に対して標準化し、4つの独立の試験を使用して標準偏差を算出した、%でのウイルス複製の阻害。プールXは、表3に記載の組合せを指し、ここで、Xはプールの番号を意味する。
表2:実施例1のsiRNAキナーゼスクリーニングにおいて用いたオリゴヌクレオチド配列を示した表である。(a)前記遺伝子に特異的な2種のオリゴヌクレオチド配列(「標的1」および「標的2」)の組合せによって、または(b)別の遺伝子に特異的なプールされたオリゴヌクレオチド(「プールX」、ここで、Xは、表3に記載のプールの番号を意味する)によって、特定の遺伝子のノックダウンを行った。
表3:実施例のsiRNAキナーゼスクリーニングにおいて用いたオリゴヌクレオチドプールを示した表である。
表4:実施例4のsiRNAスクリーニングにおいて用いたオリゴヌクレオチド配列を示した表である。遺伝子に特異的な4種までのオリゴヌクレオチド配列(「標的配列1」、「標的配列2」、「標的配列3」、および「標的配列4」)を(別々の試験においてそれぞれ)用いた。
表5:実施例4のsiRNAスクリーニングにおいて用いたオリゴヌクレオチド配列を示した表である。遺伝子に特異的な4種までのオリゴヌクレオチド配列(「標的配列1」、「標的配列2」、「標的配列3」、および「標的配列4」)を(別々の試験においてそれぞれ)用いた。この表に記載している遺伝子のノックダウンは、結果としてウイルス複製の増大になった。
【0130】
[実施例1]
キナーゼは、ウイルス後代に影響を及ぼしうる最も有望な候補のうちの1つであるので、本発明者らは、この群の遺伝子に対するsiRNAを使用して、インフルエンザウイルスの変更された複製に関して、それぞれのキナーゼまたはキナーゼ結合ポリペプチドの個々の役割を特定した。全てのsiRNAを、4つの独立の試験において試験した。キナーゼに対するsiRNAは、細胞の複製に影響を及ぼしうるまたはさらに細胞毒性であるので、自動顕微鏡を使用して、細胞数に関する個々のsiRNAトランスフェクションの効果を解析した。インフルエンザA/WSN/33核タンパク質(NP)セグメントの非翻訳領域に隣接するホタルルシフェラーゼをコードするRNA転写産物を発現させるためのRNAポリメラーゼIプロモーター/ターミネーターカセットを使用して構築されたインフルエンザレポータープラスミドを用いて、複製コンピテントインフルエンザウイルスの量を定量した。インフルエンザ感染の1日前に、ヒト胚性腎細胞(293T)をこの指標プラスミドでトランスフェクトした。この細胞を選択したのは、それがA型インフルエンザウイルス感染後にルシフェラーゼ発現のきわめて強力な増幅を示すからである。この細胞ベースのアッセイは、以下のステップを含んでいた(図1にも、siRNAキナーゼスクリーニングの試験的設定を説明している):
1日目: 96ウェルプレート中でのA549細胞(肺上皮細胞)の播種
2日目: キナーゼまたはキナーゼ結合タンパク質に対するものであるsiRNAでのトランスフェクション
3日目: インフルエンザA/WSN/33での感染+インフルエンザ指標プラスミドでの293T細胞のトランスフェクション
4日目: A549細胞の上清での293T細胞の感染+自動顕微鏡による細胞数の決定
5日目: 指標細胞の溶解、およびウイルス複製を定量化するためのルシフェラーゼアッセイの実施
【0131】
インフルエンザに関連するキナーゼの同定のために、発光値を、(siRNAトランスフェクションおよびウイルス感染の後に測定した)細胞数に対して標準化した。このことにより、より少ない(またはより多い)細胞数に起因する非特異的な効果を最小限に抑えることができる。
【0132】
全てのスクリーニング法において正確なアッセイを実証できるようにするために、いくつかの対照を含めた(図2)。ウイルス核タンパク質に対する対照siRNAは、ほぼ非感染細胞のレベルまで複製を低減させることができた。
【0133】
百分率の阻害の実例は、いくつかのsiRNAがインフルエンザウイルス複製を増強することができるのに対して、別のsiRNAはインフルエンザNP遺伝子に対する抗ウイルス性対照siRNAよりも強力に(>113%)複製を阻害することができることを示す(図3)。これに関して、47種のsiRNAは、50%を超えて複製を低減させ、9種のsiRNAは、80%を超える阻害を示した。細胞数に対して標準化し、4つの独立の試験を使用して標準偏差を算出した、%でのウイルス複製の活性化(表1a、「負の」阻害)および阻害(表1b)を示している、結果の表を表1aおよび1bに示している。
【0134】
z-スコアの算出を用いて、同様な結果が得られた。z-スコアは、生のスコアと標準偏差の単位における母平均との間の距離を表す。z-スコアは、以下の式を用いて算出した:
【0135】
【数1】

【0136】
式中、Xは、標準化する生のスコアであり、σは、母集団の標準偏差であり、かつμは、母集団の平均である。
【0137】
[実施例2]
今後の試験において、この抗ウイルス作用は、プールされたsiRNAの代わりに個々のsiRNAまたはshRNAを使用することによってより詳細に検証されるであろう。実施例1の試験的設定を使用して、さらに新しいsiRNA(特定された遺伝子につき少なくとも2種のさらなるsiRNA)およびshRNAを試験することになる。インフルエンザウイルスの複製に重要であると思われるこれらの確認された遺伝子を、次いで、マウスにおいて、鼻腔内に投与するsiRNAまたはshRNAを使用してノックダウンすることになる。この抗ウイルス療法の評価には、肺上皮組織への化合物の輸送の効率を決定することが最も重要である。療法の成功は、効率の高いキナーゼ阻害剤と適切な輸送システムとの組合せに依存している。適合する可能性があって費用効率のよい作用剤は、本発明者らがin vivoでの試験においてsiRNAまたはshRNAの送達に成功して適用しているキトサンである。本発明者らは、該化合物を鼻腔内に使用するかまたはこれらを直接肺内に投与するであろう。
【0138】
効率的なsiRNAまたはshRNAは、肺組織内のウイルス力価の低下を導くはずであり、このために、動物は、それがない場合には致死のインフルエンザウイルス感染に対して保護されるはずである。
【0139】
キナーゼ阻害剤の生物学的作用を試験するために、本発明者らは、試験を4つの部分に分けることになる:
1.化合物/キトサンナノ粒子の鼻腔内適用後の呼吸器におけるキナーゼ阻害剤の分布の解析。最良の有効性のための化合物/キトサン濃度の最適化。成功した場合のみに、さらなる試験が行われることになる。
2.LD50試験において、ウイルス分離株インフルエンザA/Puerto Rico/8/34および鳥インフルエンザ分離株(試験4のため)の絶対的な病原性を評価することになる。
3.肺組織におけるウイルス力価または(特定の場合において)生存率を解析することによる、鼻腔内適用およびインフルエンザA/Puerto Rico/8/34での感染後の、選択したsiRNAまたはshRNAの抗ウイルス作用の試験。
4.肺組織におけるウイルス力価または(特定の場合において)生存率を解析することによる、鼻腔内適用および(H5N1のような)病原性の高い鳥インフルエンザウイルス分離株での感染後の、選択したsiRNAまたはshRNAの抗ウイルス作用の試験。
【0140】
使用されるウイルス分離株は、鳥インフルエンザの現在の発生および広がりに依存する。本発明者らは、現在流行している株の複製をin vivoで効率よく阻害することを目的とする。
【0141】
確認された遺伝子に対するキナーゼ阻害剤も、損なわれたウイルス複製についてマウスで試験されることになる。
【0142】
Max-Planck感染生物学研究所、Berlin、Germanyは、好適な細胞培養物(A549細胞)における個々の全てのヒト遺伝子の遮断を、原則として、可能にする、ゲノム全体のRNAiライブラリーを有する。したがって、次の段階では、このスクリーニングは、ゲノム全体の規模まで拡張されるであろう。ウイルスの吸着、複製および出芽に関与する多くのさらなる細胞内因子は、依然として未知であるからである。
【0143】
[実施例3]
(キナーゼまたはキナーゼ結合タンパク質に対するsiRNAだけでなく)さらなるsiRNAおよびshRNAも、A型インフルエンザウイルスの複製の低減について検証されるであろう。これらのsiRNAおよびshRNAの評価には、(自動顕微鏡を使用する代わりに)市販の細胞生存率アッセイを使用して細胞数を間接的に定量すること、ならびにこれらのsiRNAおよびshRNAをリバーストランスフェクトする、すなわち、384ウェルプレート中に既に調製されているトランスフェクション混合物に細胞を添加することになることを除き、実施例1に記載したのと同じ試験的設定を用いることになる。
【0144】
[実施例4]
ヒトゲノムの中で何百もの遺伝子がインフルエンザウイルスの複製におそらく関連している。そのため、約59886種のsiRNAを使用することにより、(実施例1に記載の)キナーゼおよびキナーゼ結合因子のスクリーニング方法を、全ての既知のヒト遺伝子を解析するゲノム全体の規模に拡張した。
【0145】
以下を除き、実施例1に記載したのと同様の方法で、試験的設定を行った:
●59886種のsiRNAを使用して、スクリーニングをゲノム全体のレベルに拡張した
●細胞を384ウェルプレート中に播種した。
●トランスフェクトした細胞の数が膨大なため、必ずしも全ての細胞数を自動顕微鏡法によって解析できたわけではない。
●トランスフェクション試薬HiperFect(Qiagen、Hilden、Germany)を使用して、siRNAを、新たに播種したA549細胞においてリバーストランスフェクトした。
●特定の遺伝子のノックダウンを、特定の遺伝子に特異的な4種までのsiRNA(表4にある「標的配列1」、「標的配列2」、「標的配列3」、および「標的配列4」)によって独立に行った。
●さらなる対照:陰性対照として、「AllStars陰性対照siRNA」(Qiagen、Hilden、Germany、注文番号1027280)、陽性対照(インフルエンザ複製に対する細胞内因子)として、PKMYT(GenelD:9088、GenBank受託番号:NM_182687、標的配列:CTGGGAGGAACTTACCGTCTA)に対して指向されたsiRNA、トランスフェクション対照(トランスフェクション後の細胞毒性)として、PLK(GenelD:5347、GenBank受託番号:BC014135、標的配列:CCGGATCAAGAAGAATGAATA)に対して指向されたsiRNAを含めた。
●感染過程における早期または後期の事象に対する阻害作用を精査することができるように、選択したウェル中のトランスフェクトしたA549細胞の感染率を自動顕微鏡法によって測定する。
●(「Allstars陰性対照siRNA」でトランスフェクトした対照ウェルを基準に)B-スコア標準化方法を使用して、Michael Butros、Ligia BrasおよびWolfgang Huberによって開発された、統計的R-パッケージ「cellHTS」ソフトウェアによって結果を解析した。
●読み出される情報は、ウイルス複製の阻害である。
【0146】
強力な抗ウイルス活性(z-スコア<-2.0)を示したsiRNAおよび対応する遺伝子の一覧を表4に記載している。
【0147】
この細胞ベースのアッセイは、以下のステップを含んでいた(ゲノム全体のsiRNAスクリーニングの試験的設定を説明している図5も参照されたい):
1日目: A549細胞(肺上皮細胞)の播種+siRNAのリバーストランスフェクション
3日目: インフルエンザA/WSN/33での感染+指標プラスミドでの293T細胞zqのトランスフェクション
4日目: A549細胞の上清での293T細胞の感染+ホルムアルデヒドでのA549細胞の固定
5日目: 293T細胞におけるウイルス複製を定量化するためのルシフェラーゼアッセイ
x日目: 自動顕微鏡による感染率の決定。
【0148】
[実施例5]
A型インフルエンザウイルスは、内因性のmiR-141を誘導して、MxAの抗ウイルス活性を次第に低下させる
哺乳動物細胞におけるグローバルな制御因子として知られる、マイクロRNA(miRNA)も、ウイルスを感染させて宿主細胞の機能を奪うための魅力的なツールである。宿主のmiRNA機能に対するインフルエンザウイルスの影響力を評価するために、本発明者らは、A549ヒト肺上皮細胞におけるmiRNAの発現解析を行った。1種の内因性miRNA(miR-141)は、感染細胞において強く誘導され、過剰発現されると、ウイルス複製を増強したのに対して、miR-141の阻害は複製を低減させた。本発明者らは、miR-141の重要な標的として、抗ウイルス性MxA宿主遺伝子を同定した。したがって、インフルエンザウイルス感染は、MxAの転写後サイレンシングを引き起こしたが、隣接した非感染細胞におけるインターフェロンで誘導されるMxA合成は可能であった。このように、本発明者らは、宿主細胞を介した抗ウイルス応答からA型インフルエンザウイルスを保護する、miRNAに基づく重要な調節機構を発見し、これは、抗ウイルス介入の新規選択肢を構成するものである。
【0149】
本発明者らは、A型インフルエンザウイルスも細胞内miRNAの使用によって利点を得ているかどうかを知ることに興味を持った。始めに、2種の異なるマイクロアレイプラットフォームを使用して、ヒトmiRNAの発現パターンにおける変化をモニタリングした。双方のアレイ型(AmbionおよびExiqon)により、感染されたA549ヒト肺上皮細胞において、単一のmiRNA、miR-141が、非感染細胞と比較して有意に上方制御されたことが明らかにされた。マイクロアレイ結果をリアルタイム逆転写-PCR(qRT-PCR)によって確認し、A549細胞をヒトおよび鳥類の一区画のA型インフルエンザウイルス株に感染させた後のmiR-141の劇的な増加(5〜15倍)が示された(図6A)。
【0150】
増加した細胞内miR-141がインフルエンザ複製に対する影響力を有するかどうかを調べるために、本発明者らは、細胞を、miR-141-特異的な化学的に修飾された核酸阻害剤またはインフルエンザウイルスA/WSN/33で感染後のmiR-141前駆体のいずれかでトランスフェクトした。感染効率を示す、ウイルス依存的ルシフェラーゼアッセイ(17)を使用して、感染力をもつ粒子の数を定量した。miR-141前駆体でのA549細胞の処理は、感染力をもつ粒子の形成を有意に増加させたのに対して、miR-141阻害剤は、逆の作用を導いた(図6B)。この知見により、細胞内miR-141はインフルエンザウイルス複製の効率の調節における役割を果たしているという最初の証拠が提供された。
【0151】
高レベルのmiR-141の存在下でウイルス複製が増強されることを確認するために、本発明者らは、ヒトmiR-141または異なる対照(miR-198または空ベクター)のいずれかを安定に過剰発現する細胞系を作製した。次いで、細胞を、低い感染多重度(MOI 0.05)でインフルエンザA/WSN/33に感染させた。免疫蛍光顕微鏡法により、感染24時間後(h p.i.:hours post infection)における感染細胞の数は、miR-141陽性細胞において、多様な対照と比較して有意に多かったことが明らかになった(図6C)。これらの細胞上清中の感染力をもつ粒子の定量(図6D)により、A型インフルエンザウイルスは、miR-141のレベルが増大されているA549細胞においてより効率よく複製することができることがさらに確認された。合わせると、これらのデータにより、インフルエンザウイルス複製におけるmiR-141発現の刺激性の効果が確証される。
【0152】
主に腫瘍の発生に関連する(18)、miR-141の細胞内標的遺伝子は、以前に報告されているが、これらの遺伝子のうち、ウイルス複製におけるこのmiRNAの影響について真実味のある説明を提供したものはない。miRNAは、翻訳抑制だけではなく、それらのmRNA標的の安定性にも影響しうるという仮定に基づき(19)、本発明者らは、市販の全ヒトゲノムマイクロアレイを使用して、miR-141を過剰発現しているA549細胞におけるmRNA発現プロファイルを対照細胞と比較した。miR-141を過剰発現している細胞において3倍を超える影響を受ける標的mRNAの中で(データは示していない)、本発明者らは、抗ウイルス因子MxAを同定した(12)。MxAの抗ウイルス作用を確認するために、本発明者らは、shRNAを介したMxAのノックダウンを示す(タンパク質レベルにおけるノックダウンそれぞれ約85%)2種の安定なA549細胞系を作製した。予想通り、MxAのサイレンシングは、一次ウイルス複製(図10)および上清に放出される感染力をもつウイルスの力価(図11)を有意に増大させた。これらの知見により、本発明者らは、MxAが、miR-141機能とウイルス複製との間の重要な接点を構成しているのではないかと考えた。
【0153】
MxAに関連してmiR-141に増強されるウイルス複製の分子基盤を調べるために、本発明者らは、感染中のA549細胞におけるMxA mRNA、MxAタンパク質およびmiR-141のレベルを、それぞれqRT-PCRおよびイムノブロットを使用して解析した(図7A〜C)。MxA遺伝子発現はI型インターフェロンによる刺激を必要とするので(20)、感染していないA549細胞では、MxA mRNAもMxAタンパク質もいずれも検出されなかった。1のMOIで感染させた細胞において、有意なレベルのMxAのmRNAが感染8時間後という早い時期に転写され、感染24時間後にピークに達した(図7A)。驚くべきことに、MxAタンパク質は、このMOIを使用して感染の24時間後でさえも検出されなかった(図7B)。同じ感染時間にわたり、miR-141のレベルは着実に上昇して、感染していないA549細胞と比較して、20倍の上方制御を導いた(図7C)。これらの知見により、インフルエンザウイルスでの細胞の感染に応じて、miR-141の発現の増加に付随して、転写後の機構により、MxAタンパク質合成が妨げられるが、mRNA合成は妨げられないことが示された。しかし、興味深い追加の現象が明らかとなった-低いウイルス力価(MOI 0.1)において、本発明者らは、感染24時間後に強いMxAタンパク質シグナルを認めた(図7B)。MOIを高めることに応じた感染24時間後のMxAタンパク質合成の漸増により、0.1のMOIまでのMxAタンパク質レベルの用量依存的上昇が明らかとなった。MxAタンパク質レベルの上昇は、低いMOIでのMxA転写と関連していた;これに対して、0.1を超えるMOIでは、持続性のmRNA転写にもかかわらず、MxAタンパク質レベルが急激に低下した(図7Dおよび7E)。したがって、MxAタンパク質の転写後抑制は、高いMOIでは明白であったが、低いMOIではそうではなかった。
【0154】
この現象を説明するために、本発明者らは、免疫蛍光顕微鏡法を使用して、単細胞レベルで、ウイルスに感染してMxAを発現している細胞を可視化した。感染していない(NI)細胞および完全に感染した培養物(MOI 1)において、MxAタンパク質は検出されなかった(図7F)。しかし、MxAタンパク質は、部分的に感染した培養物(MOI 0.1)の非感染細胞において検出された(図7F)。これらの非感染細胞におけるMxAの合成は、非感染細胞においてMxA合成を刺激することが知られる(21)、I型インターフェロンに関連しているパラクリン調節ループに起因すると考えることができる。合わせると、これらのデータより、MxAタンパク質の効率的な転写後下方制御は全てのウイルス感染細胞において生じること;および、最も可能性の高いのはIFN-βの産生を介して、感染細胞が非感染細胞におけるMxAの上方制御を誘発することが明らかに実証される。
【0155】
ウイルス感染細胞において観察されたMxAタンパク質の下方制御が、miR-141による転写後サイレンシングに依存的であるかどうかを試験するために、細胞を特定のmiRNA阻害剤で処理した(図8A)。著しいことに、MxA陽性細胞は、miR-141阻害剤での処理後に、ウイルスに感染した、すなわち、ウイルス核タンパク質(NP)を発現している細胞のみにおいて認められた(図8A、矢印を参照されたい)。阻害剤のない場合、または、関連のないmiR-198阻害剤でのトランスフェクションによる場合には、NPおよびMxAタンパク質の発現は、相互排他的であった。H7N7鳥インフルエンザ株A/FPV/Bratislava/79でも同様の結果が得られた(図8A)。MxA陽性細胞の定量により、単細胞での知見が確証された(図12)。miR-141がMxAの転写サイレンシングを誘導するという見解と一致して、本発明者らは、細胞培養物中のmiR-141レベルが、MOIが高まるとともに着実に上昇して、約1のMOI、すなわち、感染24時間後に全ての細胞が感染しているところでピークに達することを観察した(図8B)。MxAの制御におけるmiR-141の重要な役割は、イムノブロット解析によってさらに支持されており、miR-141特異的阻害剤のトランスフェクションは、全ての細胞が感染したときでさえも、MxA発現を促進することが明らかになった(図8C)。これらの知見により、miR-141は、ウイルスに誘導される、抗ウイルス性MxAタンパク質の下方制御に重要であることが明らかに示される。
【0156】
感染のない場合のmiR-141の機能を実証するために、本発明者らは、IFN-βを使用して、異なるmiRNAを過剰発現している多様なA549細胞系においてMxA発現を刺激した。より高レベルのmiR-141を発現している細胞は、miR-198または空ベクターでトランスフェクトした対照細胞と比較して、よりわずかな程度のMxA誘導(約30%)を示し、IFN-β依存的な、しかしウイルス非依存的な、MxAサイレンシングにおけるmiR-141の作用が示唆された。
【0157】
miR-141を介したMxAサイレンシングの分子基盤を研究するために、本発明者らは、MxA mRNA配列を解析した。典型的には、miRNAは、その標的mRNAの3'非翻訳領域(UTR)と相互作用することによって機能して、遺伝子発現におけるその調節作用を発揮する(22)。しかし、MxA 3'-UTRに連結したルシフェラーゼレポーター構築物で一過的にトランスフェクトしたA549細胞は、miR-141過剰発現に応答したルシフェラーゼ活性の低下を示さなかった(データは示していない)。さらに、miR-141とMxA mRNAとの配列アライメントにより、MxAの3'-UTR領域ではなく、コード配列(CDS)中に相補的な領域が明らかにされた(図14)。これらの知見により、miR-141を介したMxAの抑制にはCDSが関与することが示唆される。MxA下方制御におけるmiR-141の間接的な調節機能の可能性を除外するために、本発明者らは、構成的なサイトメガロウイルス(CMV)プロモーターの制御下に置かれる、増強された緑色蛍光タンパク質(eGFP)とMxAとの遺伝子融合体を作製した。eGFP-MxA融合体により、遺伝子導入したMxAタンパク質と内因性のMxAタンパク質との、これらの異なる分子サイズによる、並行したモニタリングが可能となった。このアプローチにより、本発明者らは、(例えば、miR-141に調節される推定上の転写因子を介して)天然のMxAプロモーターからの転写に影響を及ぼす、miR-141の間接的な調節機能と、MxAの転写産物、その3'-UTRを含む、に影響を及ぼす、直接的な転写後遺伝子サイレンシング機構とを区別できるようになった。eGFP-MxA融合体は、A549細胞のトランスフェクション後すぐに発現された;しかし、感染に応じたその発現は、インフルエンザA/WSN/33のMOIが高まるとともに着実に低下した(図9A)。同様に、内因性MxAは、約0.1のMOIで最も高いタンパク質レベルを示し(図6Eと比較されたい)、MOIが高まるとともに低下した(図9A)。ウエスタンブロットデータの定量により、1のMOIでのウイルス感染に応じた、トランスフェクトした組換えMxAタンパク質および内因性MxAタンパク質の約10分の1の減少が示された(図9A;ブロットの下のバンド強度の値を参照されたい)。したがって、インフルエンザウイルスに誘発された、遺伝子導入したeGFP-MxAと内因性のMxAとの減少は、eGFP-MxAの転写を導く構成的なCMVプロモーターにもかかわらず、ほぼ同じであった。
【0158】
これらの知見により、転写後の直接的なMxA合成の干渉が、共焦点顕微鏡法を使用した単細胞解析によってさらに支持されたことが示される:構成的に発現されるmCherry構築物と、eGFP-MxA融合タンパク質(モル比1:6)との同時発現は、好結果のトランスフェクション戦略を示した(図9B)。しかし、MOIを高めてこれらの細胞をインフルエンザウイルスに感染させることにより、eGFP-MxA融合体タンパク質の選択的減少が導かれたが、mCherryタンパク質では導かれなかった(図9B)。eGFP-MxA陽性細胞の定量により、これらの知見が確証された(図15)。この試験により、インフルエンザウイルス感染に応じた、MxAの特異的な転写後サイレンシングが明らかに実証される。
【0159】
MxA発現の転写後サイレンシングにおけるmiR-141の直接的な機能を支持するさらなる証拠を集めるために、本発明者らは、ウイルス感染のない場合のMxA発現に対するmiRNAの作用を調べた。ヒトMxAプロモーターはIFNに刺激される2つの機能的な応答エレメント(ISRE)(23)を含有しているので、本発明者らは、MxAの発現を誘導するためにIFN-βを使用した。レトロウイルスのmiR-141のトランスフェクションにより、コトランスフェクトしたeGFP-MxA融合タンパク質とIFN-βに誘導される内因性MxAとの発現における並行した低下が引き起こされたが(図9C)、miR-198または空ベクターでは引き起こされなかった。MxAタンパク質発現の定量により、miR-141の過剰発現に応じて、対照プラスミドを有する細胞と比較して、eGFP-MxAの約3分の1の減少および内因性MxAの約10分の1の減少が示された(図9C;ブロットの下のバンド強度の値を参照されたい)。ここでも、単細胞レベルでのこれらの交差した結果を、共焦点顕微鏡法によって解析および確認し、IFN-βで刺激してmiR-141でトランスフェクトした細胞において外因性MxAおよび内因性MxAの双方の量が減少していることが示された(図9D)。図9Bにおける試験的設定と同一である、mCherryプラスミドのコトランスフェクションにより、eGFP-MxAをトランスフェクトした細胞の追跡が可能となった。eGFP-MxA陽性細胞の定量により、miR-141をコードするプラスミドで細胞をトランスフェクトした後の有意に低減されたMxAのレベルが明らかにされた(図16)。最も注目すべきことに、レトロウイルスのmiR-141構築物でトランスフェクトされた個々の細胞および内因性MxAの下方制御を示した個々の細胞によっても、eGFP-MxAの発現がないまたは弱いことが示された(図9D中の矢印を参照されたい)。miR-141の直接的な機能は、miR-141、または関連のないmiR-198を過剰発現している、IFN-βに刺激されたA549細胞におけるISREプロモーター活性の解析によっても実証され、MxAの上流のシグナル伝達はmiR-141によって影響されないことが示された(図17)。考え合わせると、これらのデータにより、RNA干渉に基づく機構を介して転写後レベルで作用するMxA合成におけるmiR-141の直接的な作用の概念が強く支持される。
【0160】
ウイルスによるMxAの能動的な抑制(1)の根底にある分子機構は、このように、全く謎のままだった。ここで、本発明者らは、ヒト細胞において抗ウイルス性MxAタンパク質の下方制御に関与する重要な分子として、A型インフルエンザウイルスに誘導されるmiR-141を同定した。本研究において行った広範囲な単細胞解析により、感染細胞におけるMxAの能動的なmiR-141-依存的阻害が示唆される;しかし、感染細胞は、β-インターフェロンに関連しているパラクリンループを介して、隣接するまだ感染していない細胞における抗ウイルス性のMxA合成を依然として誘導する。それにもかかわらず、かつ最もよく知られているmiRNA翻訳制御機構に反して(24、25)、miR-141による干渉はきわめて強力であり、miR-141は、インフルエンザ感染を治療するための優れた標的を構成しうることが示唆される。さらに、ヒトおよび鳥類の一区画のA型インフルエンザ分離株に応答したmiR-141上方制御についての本発明者らの知見は、広範囲に効能がある治療選択肢のために有望である。
【0161】
(参考文献)
【0162】
【表1a−1】
【表1a−2】
【表1a−3】
【表1a−4】
【表1a−5】
【表1a−6】
【0163】
【表1b−1】
【表1b−2】
【表1b−3】
【表1b−4】
【表1b−5】
【表1b−6】
【表1b−7】
【表1b−8】
【表1b−9】
【表1b−10】
【0164】
【表2−1】
【表2−2】
【表2−3】
【表2−4】
【表2−5】
【表2−6】
【表2−7】
【表2−8】
【表2−9】
【表2−10】
【表2−11】
【表2−12】
【表2−13】
【表2−14】
【表2−15】
【表2−16】
【0165】
【表3−1】
【表3−2】
【表3−3】
【表3−4】
【表3−5】
【表3−6】
【表3−7】
【表3−8】
【表3−9】
【表3−10】
【表3−11】
【0166】
【表4−1】
【表4−2】
【表4−3】
【表4−4】
【表4−5】
【表4−6】
【表4−7】
【表4−8】
【表4−9】
【表4−10】
【表4−11】
【表4−12】
【表4−13】
【表4−14】
【表4−15】
【表4−16】
【表4−17】
【表4−18】
【表4−19】
【表4−20】
【表4−21】
【表4−22】
【表4−23】
【表4−24】
【表4−25】
【表4−26】
【表4−27】
【表4−28】
【表4−29】
【表4−30】
【表4−31】
【表4−32】
【表4−33】
【表4−34】
【表4−35】
【表4−36】
【表4−37】
【表4−38】
【表4−39】
【表4−40】
【表4−41】
【表4−42】
【表4−43】
【表4−44】
【表4−45】
【表4−46】
【表4−47】
【表4−48】
【表4−49】
【表4−50】
【表4−51】
【表4−52】
【表4−53】
【表4−54】
【表4−55】
【表4−56】
【表4−57】
【表4−58】
【表4−59】
【表4−60】
【表4−61】
【表4−62】
【表4−63】
【表4−64】
【表4−65】
【表4−66】
【表4−67】
【表4−68】
【表4−69】
【表4−70】
【表4−71】
【表4−72】
【表4−73】
【表4−74】
【表4−75】
【表4−76】
【表4−77】
【表4−78】
【表4−79】
【表4−80】
【表4−81】
【表4−82】
【表4−83】
【表4−84】
【表4−85】
【表4−86】
【表4−87】
【表4−88】
【表4−89】
【表4−90】
【表4−91】
【表4−92】
【表4−93】
【表4−94】
【表4−95】
【表4−96】
【表4−97】
【表4−98】
【表4−99】
【表4−100】
【表4−101】
【表4−102】
【表4−103】
【表4−104】
【表4−105】
【表4−106】
【表4−107】
【表4−108】
【表4−109】
【表4−110】
【表4−111】
【表4−112】
【表4−113】
【表4−114】
【表4−115】
【表4−116】
【表4−117】
【表4−118】
【表4−119】
【表4−120】
【表4−121】
【表4−122】
【表4−123】
【表4−124】
【表4−125】
【表4−126】
【表4−127】
【表4−128】
【表4−129】
【表4−130】
【表4−131】
【表4−132】
【表4−133】
【表4−134】
【表4−135】
【表4−136】
【表4−137】
【表4−138】
【表4−139】
【表4−140】
【表4−141】
【表4−142】
【表4−143】
【表4−144】
【表4−145】
【表4−146】
【表4−147】
【表4−148】
【表4−149】
【表4−150】
【表4−151】
【表4−152】
【表4−153】
【表4−154】
【技術分野】
【0001】
本発明は、インフルエンザウイルス感染の予防用または/および治療用の医薬組成物の作製方法に関する。
【0002】
さらに、本発明は、インフルエンザウイルス感染の予防用または/および治療用の医薬組成物に関する。
【背景技術】
【0003】
脅迫的なインフルエンザのパンデミックから見て、利用可能な持続的に有効な薬剤を開発および製造する緊急の必要性がある。ドイツ単独で、インフルエンザの年間の発症により、1年に5,000〜20,000例の死亡が引き起こされている(情報源:Robert-Koch研究所)。繰り返されている大きなインフルエンザのパンデミックは、特に恐れられている。第1の大きなパンデミック、いわゆる「スペインかぜ」では、1918〜1919年に、高い割合の健康な中年のヒトを含む約4千万の命が犠牲になった。H5N1インフルエンザウイルスは、現在は鳥類において主に複製されるが、後天的な変異によってこのウイルスがヒトからヒトへと伝染できるようになった場合、これによって同様なパンデミックが引き起こされる恐れがある(2、3)。より最近では、新規のインフルエンザウイルス異型が出現し、すなわち、A型インフルエンザ(H1N1)「豚インフルエンザ」株(4)が、予測不可能なパンデミックの脅威をもたらしている。ヒトにおけるパンデミックの可能性は、近年、鳥インフルエンザ(H5N1)の世界的な広がりおよび家畜の感染と共に、より急激に増大している。病原性の高いヒトインフルエンザ組換え体が現れるのは、単に時間の問題である。インフルエンザ感染の予防または療法のための瞬間に利用可能な方法、例えばウイルス表面タンパク質を用いた予防接種または抗ウイルス薬(ノイラミニダーゼ阻害剤またはイオンチャネル遮断薬)の使用などは、様々な欠点を有する。既にこの早期の段階で、我々の最も有効な製剤のうちの1種(タミフル)に対する耐性が出現しており、その結果、汎流行を封じ込めるのに不適当になることもある。インフルエンザに対するワクチンおよび薬剤の使用における中心的な課題は、病原体の多様性である。これまで、有効なワクチンの開発には、病原体異型の正確な予測が必要とされてきた。ウイルス構成要素に対する薬剤は、病原体の変異により、急激にその有効性を失うことがある。
【0004】
これまでほとんど注意されてこなかった研究の領域は、宿主細胞における重大な意味を持つ標的構造の同定である。ウイルスは、固有の細胞内タンパク質に依存して宿主内で複製することができるかが決まる。こうした細胞内因子の知識は、ウイルス複製に不可欠であるが、新規薬剤の開発を導きうるヒトには(少なくとも一時的に)不要である。概算では、ヒトゲノムにおいてウイルス増殖に必須な約500遺伝子が予測されている。これらのうち、少なくとも10%は、ヒトの生体にとって一時的にまたは恒久的にも不必要であろう。これらの遺伝子およびこれらの産物の阻害は、ウイルス標的とは対照的に、これらの構造において一定であり、最短の時間での抗ウイルス薬の新しい作製の開発を可能にするであろう。こうした遺伝子産物の阻害は、抗ウイルス薬に対する感受性がより短いウイルスエスケープ変異体の発生を克服しうる。他の遺伝子ファミリーキナーゼの中で細胞内で重要な調節タンパク質は、ウイルスによって乗っ取られて宿主細胞の組成が操作されることが多い。
【0005】
A型インフルエンザは、哺乳動物宿主細胞内での生存の成功を容易にする一連の戦略を示すマイナス鎖RNAウイルスである(5)。感染すると、先天性免疫性レセプター、例えば細胞内タンパク質レチノイン酸誘導性遺伝子I(RIG-I)などのそれらの同族リガンドへの結合が何十もの免疫および炎症に関連した遺伝子の一過的発現を誘発する(6、7)。特に、その後のI型インターフェロンの誘導は、ミクソウイルス耐性(Mx)タンパク質などの、内因性抗ウイルス活性を有するGTPアーゼの上方制御を刺激する。オルソミクソウイルスファミリーのメンバーに対するMxタンパク質の抗ウイルス活性は、マウスにおいて最初に観察された(8)。核に位置するMx1タンパク質は、それがない場合には、パンデミック1918の株およびきわめて致死的なH5N1インフルエンザウイルスを含むインフルエンザウイルスによる致死感染に対して保護を与える(9、10)。ヒトオーソログ、MxAは、細胞質に局在し、かつ入ってくるウイルスヌクレオカプシドに結合して不活性化することによって作用すると考えられている(11)。報告によると、興味深いことに、ヒトMxAは、遺伝子導入マウスにおいて多様なRNAウイルスに対する保護機能を示す(12)。これらの先天性応答戦略に対抗するために、インフルエンザウイルスは、例えば、インターフェロン-β(IFN-β)の産生を低減することによってまたは抗ウイルス性タンパク質2'-5'オリゴアデニレートシンセターゼ(OAS)およびプロテインキナーゼR(PKR)の発現を阻害することによって、そのNS1タンパク質を用いる。(13)。しかし、in vitroおよびin vivoでのA型インフルエンザ感染中に既に観察されているMxAの能動的抑制は、現在完全には理解されていない。
【0006】
ウイルス剤が、その感染能力を調節するために用いる興味深い戦略は、遺伝子発現の重要な転写後制御因子として知られる(14)、タンパク質をコードしない干渉する約22nt長の短いRNA分子の一クラスである、マイクロRNA(miRNA)の使用である。大きなゲノムを有するウイルスは、宿主の生理機能を変えて複製を増強するためのそれらの自身のmiRNAをコードすることができる(15)。逆に、小さなRNAゲノムのC型肝炎ウイルスは、その複製を促進するために、宿主細胞miR-122の発現を操作することができる(16)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO 02/44321
【非特許文献】
【0008】
【非特許文献1】Bartel D、Cell 136:215〜233、2009
【非特許文献2】Kohlerら、1975、Nature 256:495〜497
【非特許文献3】Kozborら、1985、J. Immunol. Methods 81、31〜42
【非特許文献4】Coteら、PNAS、80:2026〜2030
【非特許文献5】Coleら、1984、Mol. Cell Biol. 62:109〜120
【非特許文献6】Morrisonら、1984、PNAS、81:6851〜6855
【非特許文献7】Neubergerら、1984、312:604〜608
【非特許文献8】Takedaら、1985、Nature 314:452〜454
【非特許文献9】Orlandiら、1989、PNAS 86:3833〜3837
【非特許文献10】Winterら、1991、Nature 349:293〜299
【非特許文献11】Burton、1991、PNAS、88:11120〜11123
【非特許文献12】Huseら、1989、Science 254:1275〜1281
【発明の概要】
【発明が解決しようとする課題】
【0009】
インフルエンザウイルスワクチンの作製のための一般的な戦略は、有胚(embryonated)鶏卵または細胞培養におけるインフルエンザウイルス複製に基づいている。細胞培養または有胚卵におけるウイルス複製は、時間のかかる高価な方法である。したがって、インフルエンザワクチンの作製方法を改良することが本発明の課題である。
【課題を解決するための手段】
【0010】
本発明の一目的は、以下のステップ:
(a)改変のない場合のインフルエンザウイルス複製と比較してインフルエンザウイルス複製の能力が増大された、インフルエンザウイルスを複製することのできる改変細胞、改変有胚卵または/および改変非ヒト生物を提供するステップ、
(b)(a)の該細胞、該有胚卵または/および該生物をインフルエンザウイルスと接触させるステップ、
(c)該インフルエンザウイルスの複製を可能にする条件下で、該細胞、該有胚卵または/および該非ヒト生物を培養するステップ、ならびに
(d)ステップ(c)において生成された該インフルエンザウイルスまたは/およびその少なくとも1種の構成要素を分離するステップ
を含む、インフルエンザウイルスの調製方法である。
【0011】
ステップ(d)のインフルエンザウイルスから、インフルエンザウイルス感染の予防用または/および治療用の医薬組成物を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、調製することもできる。
【0012】
本発明のもう1つの目的は、インフルエンザウイルス感染の予防用または/および治療用の医薬組成物の調製方法であって、以下のステップ:
(a)改変のない場合のインフルエンザウイルス複製と比較してインフルエンザウイルス複製の能力が増大された、インフルエンザウイルスを複製することのできる改変細胞、改変有胚卵または/および改変非ヒト生物を提供するステップ、
(b)(a)の該細胞、該有胚卵または/および該生物をインフルエンザウイルスと接触させるステップ、
(c)該インフルエンザウイルスの複製を可能にする条件下で、該細胞、該有胚卵または/および該非ヒト生物を培養するステップ、
(d)ステップ(c)において産生された該インフルエンザウイルスまたは/およびその少なくとも1種の構成要素を分離するステップ、ならびに、
(e)ステップ(d)において分離された該インフルエンザウイルスまたは/およびその構成要素から、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、該医薬組成物を調製するステップ
を含む方法である。
【0013】
本明細書における「方法」または「本発明の方法」に対する参照とは、インフルエンザウイルスの調製方法ならびにインフルエンザウイルス感染の予防用または/および治療用の医薬組成物の調製方法に対する参照である。
【0014】
ステップ(a)において用いられる細胞は、インフルエンザウイルスに感染させることができるいかなる細胞であってもよい。インフルエンザウイルスの産生に好適な細胞系は知られている。好ましくは、該細胞は、哺乳動物細胞または鳥類細胞である。ヒトの細胞も好ましい。上皮細胞、例えば肺上皮細胞なども好ましい。該細胞は、細胞系であってもよい。好適な肺上皮細胞系は、A594である。もう1つの好適な細胞は、ヒト胚性腎細胞系293Tである。本発明の一実施形態において、本発明の方法は、本明細書に記載の細胞を用いる。
【0015】
ステップ(a)において用いられる非ヒト生物は、インフルエンザウイルスに感染させることができるいかなる生物であってもよい。好ましくは、該生物は、インフルエンザワクチンの作製に用いられる生物である。より好ましくは、該生物は、例えば有胚鶏卵のような、有胚卵である。当業者は、こうした生物を得る方法を知っている。受精によって有胚卵を得る方法は知られている。インフルエンザウイルスの接種によるインフルエンザウイルス複製の誘導は知られている。本発明の一実施形態において、本発明の方法は、本明細書に記載のように、非ヒト生物または/および有胚卵を用いる。
【0016】
本発明のステップ(a)は、本明細書に記載のように改変された細胞、有胚卵もしくは/および非ヒト生物の提供を含んでいてもよく、または改変のステップを含んでいてもよい。
【0017】
改変細胞または/および改変有胚卵は、本明細書に記載のように、ステップ(a)において提供され、ステップ(b)、(c)および(d)において、またはステップ(b)、(c)(d)および(e)において用いられることが好ましい。
【0018】
本明細書中で使用される場合、「該細胞、該有胚卵または/および非ヒト生物の改変」としては、該細胞、該卵または/および該生物における少なくとも1種の遺伝子または/および遺伝子産物の発現または/および活性の下方制御または/および上方制御が挙げられる。
【0019】
本明細書中で使用される場合、「該細胞、該有胚卵または/および該非ヒト生物の改変」は、該細胞、該有胚卵または/および該非ヒト生物を、モジュレーターのない場合のインフルエンザウイルス複製と比較して、該細胞または/および該生物におけるインフルエンザウイルス複製を増大させることができる少なくとも1種のモジュレーターと接触させることを含んでいてもよく、このとき、接触は、ステップ(b)の前もしくは後に、またはステップ(b)と同時に行われてもよい。
【0020】
本明細書中で使用される場合、「該細胞、該有胚卵または/および非ヒト生物の改変」は、組換え細胞、組換え有胚卵または/および組換え非ヒト生物の作製または/および提供を含んでいてもよく、このとき、少なくとも1種の遺伝子または/および遺伝子産物の発現または/および活性が、改変のない場合のインフルエンザウイルス複製と比較して該細胞、該有胚卵または/および該非ヒト生物のインフルエンザウイルス複製の能力が増大されるように改変される。
【0021】
組換え細胞、組換え有胚卵または/および組換え非ヒト生物の調製は、該細胞内、該有胚卵内もしくは/および該非ヒト生物内への核酸分子の導入、または/ならびに該細胞内、該卵内もしくは/および該生物内の核酸配列の欠失を含んでいてもよい。該核酸分子は、該細胞の、該有胚卵のまたは/および該非ヒト生物のゲノム内に組み込まれてもよい。それによって、該細胞、該卵または/および該生物の配列が、改変、置換または/および欠失されうる。該核酸分子は、該細胞または/および該生物と異種の配列を含有していてもよい。核酸分子の組み込みは、恒久的にまたは一過的に行われてもよい。組換え有胚卵または/および組換え非ヒト生物は、生殖細胞系列の操作によって調製されてもよい。本発明の文脈において、「有胚卵」とは、特にその胚を指す。例えば、「有胚卵の改変」は、特にその胚の改変である。
【0022】
当業者は、細胞内、有胚卵内もしくは/および生物内に核酸分子を導入する方法、または/ならびに該細胞内、該卵内もしくは/および該生物内の核酸配列を欠失させる方法(本明細書中で用いているような、「組換え技術」)を知っている。これらの方法は、好適なベクター、例えばプラスミドなどを用いる、トランスフェクションを含んでいてもよい。これらの方法は、該細胞または/および該生物のゲノム内の核酸分子の相同組換えも含んでいてもよい。また、該核酸分子は、該細胞、該有胚卵または/および該生物のゲノム内に無作為に挿入されてもよい。
【0023】
表1a、1b、4および5に、インフルエンザウイルス複製の調整のための標的を記載しており、ここで、該標的は、本明細書に記載のように、モジュレーターとの接触、または組換え技術のいずれかによる、該細胞、該有胚卵または/および非ヒト生物の改変に好適でありうる。
【0024】
本発明の文脈における「調整」とは、「活性化」または「阻害」であってもよい。
【0025】
下方制御によりインフルエンザウイルス複製を増大させる遺伝子の例を、表1aおよび5に記載する。したがって、これらの遺伝子または/およびその遺伝子産物の発現または/および活性を増大させることによって、インフルエンザウイルス複製を低減させることができる。これらの遺伝子または/および遺伝子産物の低減された発現または/および活性は、ウイルス作製の改良によって本発明の方法において利用することができる。
【0026】
したがって、ステップ(a)において提供される該細胞、該有胚卵または/および非ヒト生物は、組換え細胞、組換え有胚卵または/および組換え非ヒト生物であってもよく、このとき、表1aおよび5から選択される遺伝子の遺伝子発現または/および活性は下方制御される。
【0027】
下方制御によりインフルエンザウイルス複製を低減させる遺伝子の例を、表1bおよび4に記載する。したがって、これらの遺伝子または/および遺伝子産物の発現または/および活性を低減させることによって、インフルエンザウイルス複製を低減させることができる。これらの遺伝子または/および遺伝子産物の増大された発現または/および活性は、ウイルス作製の改良によって本発明の方法において利用することができる。
【0028】
したがって、ステップ(a)において提供される該細胞、有胚卵または/および非ヒト生物は、組換え細胞、組換え有胚卵または/および組換え非ヒト生物であってもよく、このとき、表1bおよび表4から選択される遺伝子の遺伝子発現または/および活性は上方制御される。特に、表1bおよび表4から選択される遺伝子の上方制御は、前記遺伝子の過剰発現である。
【0029】
本発明の文脈において、「標的」としては、
(a)遺伝子または/およびゲノム内、特に、ヒト遺伝子または/およびヒトゲノム内のヌクレオチド配列、
(b)核酸、または/および(a)のヌクレオチド配列によってコードされるポリペプチド
などが挙げられる。
(a)または/および(b)の配列は、宿主細胞におけるインフルエンザウイルス複製の調節に関与していてもよい。該標的は、直接的または間接的に、インフルエンザウイルス複製の調節に関与していてもよい。特に、標的は、その標的の活性化またはその標的の阻害のいずれかによって、インフルエンザウイルス複製を増大させるのに好適である。
【0030】
標的の例は、遺伝子および遺伝子の部分的な配列、例えば調節配列などである。本発明による標的としては、RNA、特にmRNA、tRNA、rRNA、miRNA、piRNAなどの遺伝子産物も挙げられる。標的は、該標的遺伝子によってコードされるポリペプチドまたは/およびタンパク質も含みうる。標的遺伝子の好ましい遺伝子産物は、その標的遺伝子によってコードされるmRNA、miRNA、ポリペプチドまたは/およびタンパク質から選択される。最も好ましい遺伝子産物は、標的遺伝子によってコードされるポリペプチドまたはタンパク質である。標的タンパク質または標的ポリペプチドは、翻訳後に修飾されてもよく、されなくてもよい。
【0031】
本明細書中で使用される場合、「遺伝子産物」は、RNA、特にmRNA、tRNA、rRNA、miRNA、およびpiRNAから選択されてもよい。また、「遺伝子産物」は、前記遺伝子によってコードされるポリペプチドまたは/およびタンパク質であってもよい。
【0032】
本発明の文脈において、該遺伝子または/および遺伝子産物の「活性」としては、転写、翻訳、翻訳後修飾、転写後調節、遺伝子または/および遺伝子産物の活性の調整などが挙げられる。該活性は、リガンド結合によって調節されてもよく、そのリガンドは、活性化剤または阻害剤であってもよい。また、該活性は、本明細書に記載のように、miRNA分子、shRNA分子、siRNA分子、アンチセンス核酸、デコイ核酸または/および他の任意の核酸によっても調節されうる。該遺伝子の活性は、本明細書に記載のように、組換え技術によっても調節されうる。調節は、小分子、抗体、アプタマー、または/およびスピーゲルマー(spiegelmer)(鏡像アプタマー)によっても行われうる。
【0033】
本発明の方法は、あらゆるインフルエンザウイルスでの感染の予防用または/および治療用の医薬組成物の作製に好適でありうる。
【0034】
該インフルエンザウイルスは、ワクチン作製に好適ないずれのインフルエンザウイルスであってもよい。該インフルエンザウイルスは、A型インフルエンザウイルスであってもよい。該A型インフルエンザウイルスは、鳥類および哺乳類の生物からこれまでに単離されたA型インフルエンザウイルスから選択されてもよい。詳細には、該A型インフルエンザウイルスは、H1N1、H1N2、H1N3、H1N4、H1N5、H1N6、H1N7、H1N9、H2N1、H2N2、H2N3、H2N4、H2N5、H2N7、H2N8、H2N9、H3N1、H3N2、H3N3、H3N4、H3N5、H3N6、H3N8、H4N1、H4N2、H4N3、H4N4、H4N5、H4N6、H4N8、H4N9、H5N1、H5N2、H5N3、H5N6、H5N7、H5N8、H5N9、H6N1、H6N2、H6N3、H6N4、H6N5、H6N6、H6N7、H6N8、H6N9、H7N1、H7N2、H7N3、H7N4、H7N5、H7N7、H7N8、H7N9、H9N1、H9N2、H9N3、H9N5、H9N6、H9N7、H9N8、H10N1、H10N3、H10N4、H10N6、H10N7、H10N8、H10N9、H11N2、H11N3、H11N6、H11N9、H12N1、H12N4、H12N5、H12N9、H13N2、H13N6、H13N8、H13N9、H14N5、H15N2、H15N8、H15N9およびH16N3から選択されてもよい。より詳細には、該A型インフルエンザウイルスは、H1N1、H3N2、H7N7、H5N1から選択される。さらにより詳細には、該A型インフルエンザウイルスは、株Puerto Rico/8/34、鳥インフルエンザウイルス分離株H5N1、鳥インフルエンザ株A/FPV/Bratislava/79(H7N7)、株A/WSN/33(H1N1)、株A/Panama/99(H3N2)、または豚インフルエンザ株H1N1である。
【0035】
該インフルエンザウイルスは、B型インフルエンザウイルスであってもよい。詳細には、該B型インフルエンザウイルスは、ビクトリア系統の代表種および山形系統の代表種から選択されてもよい。
【0036】
本発明の方法において、インフルエンザウイルス複製を増大させるためのステップ(a)に従った該細胞または/および生物の改変としては、表1A、表1B、表4および表5から選択される遺伝子または/およびそれらの遺伝子産物の発現を調節することが挙げられる。特に、該細胞または/および生物の改変は、表1Bおよび表4から選択される遺伝子または/およびそれらの遺伝子産物の発現を活性化してもよく、あるいは該細胞または/および生物の改変は、表1Aおよび5から選択される遺伝子または/およびそれらの遺伝子産物の発現を阻害してもよい。この発現の調節は、該細胞、該有胚卵もしくは/および該生物を、本明細書に記載のモジュレーターと接触させることによって行われてもよく、または組換え細胞、組換え有胚卵もしくは/および組換え生物において行われてもよく、これらの作製については本明細書に記載している。
【0037】
本発明の方法において、該細胞、該有胚卵または/および該非ヒト生物の改変は、MxA遺伝子の発現または/および遺伝子産物活性の阻害を含んでいてもよい。インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、MxA遺伝子の発現または/および遺伝子産物活性を阻害する能力があってもよい。本明細書に記載のように、該細胞、該有胚卵または/および該非ヒト生物は、MxA遺伝子の発現または/および遺伝子産物活性が、miR-141、miR-141*、miR-200c、miR-200c*、これらの前駆体、これらの誘導体、アンチセンス核酸、siRNA、shRNAおよびインフルエンザウイルス配列から選択される少なくとも1種のモジュレーターによって阻害されるように、組換えによって改変されてもよい。特に、本発明の方法において、MxA遺伝子は、miR-141、miR-141*、miR-200c、miR-200c*、これらの前駆体、これらの誘導体、アンチセンス核酸、siRNA、shRNAおよびインフルエンザウイルス配列から選択される少なくとも1種のモジュレーターによって翻訳後に阻害される。
【0038】
本発明の方法において、miR-141、miR-141*、miR-200c、miR-200c*、または/およびこれらの前駆体は、該細胞内、該有胚卵内または/および該非ヒト生物内で過剰発現されてもよい。過剰発現は、一過的または恒久的であってもよい。
【0039】
RNAレベルにおいて、阻害は、アンチセンス核酸、siRNA、shRNA、デコイ核酸または/およびこれらの誘導体によって行われてもよい。MxAポリペプチドのレベルにおいて、阻害は、小分子、抗体、アプタマー、スピーゲルマー(鏡像アプタマー)によって行われてもよい。
【0040】
miR-141、miR-141*、miR-200c、およびmiR-200c*の配列、これらの前駆体ならびに該前駆体のヘアピン構造については、図18に記載している。miR-141およびmiR-141*は、単一の前駆体によって細胞内で共発現させることもできる。miR-141およびmiR-141*は、該前駆体のヘアピン構造を形成しうる相補配列を含有する。miR-200cおよびmiR-200c*は、単一の前駆体によって細胞内で共発現させることもできる。miR-200cおよびmiR-200c*は、該前駆体のヘアピン構造を形成しうる相補配列を含有する。
【0041】
本発明の文脈において、A型インフルエンザの配列は、MxA遺伝子と高度な同一性を有することが確認された。したがって、本発明の方法において、MxA遺伝子が阻害されてもよく、または/かつA型インフルエンザウイルス配列もしくは/およびA型インフルエンザウイルス配列に由来する配列を含有する少なくとも1種のマイクロRNAもしくは/および少なくとも1種のアンチセンスRNAによってウイルス複製が活性化されてもよい。
【0042】
該細胞の、該有胚卵のまたは/および該非ヒト生物の改変は、遺伝子の発現または/および遺伝子産物活性の阻害を含んでいてもよく、このとき、該遺伝子は、
(a)表1Aおよび5の配列から選択されるヌクレオチド配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含有する。
【0043】
該細胞、該有胚卵または/および該非ヒト生物の改変は、遺伝子の発現または/および遺伝子産物活性の活性化を含んでいてもよく、このとき、該遺伝子は、
(i)表1Bおよび表4の配列から選択されるヌクレオチド配列、
(ii)(i)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の配列の断片、
(iii)(i)もしくは/および(ii)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含有する。
【0044】
インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、遺伝子の発現または/および遺伝子産物活性を阻害する能力があってもよく、このとき、該遺伝子は、
(a)表1Aおよび5の配列から選択されるヌクレオチド配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含有する。
【0045】
インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、遺伝子の発現または/および遺伝子産物活性を活性化する能力があってもよく、このとき、該遺伝子は、
(i)表1Bおよび表4の配列から選択されるヌクレオチド配列、
(ii)(i)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の配列の断片、
(iii)(i)もしくは/および(ii)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含有する。
【0046】
該細胞、該有胚卵または/および非ヒト生物は、本明細書に記載のように、遺伝子の発現または/および遺伝子産物活性が阻害されるように、組換えによって改変されてもよく、このとき、該遺伝子は、
(a)表1Aおよび5の配列から選択されるヌクレオチド配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含有する。
【0047】
該細胞、該有胚卵または/および非ヒト生物は、本明細書に記載のように、遺伝子の発現または/および遺伝子産物活性が活性化されるように、組換えによって改変されてもよく、このとき、該遺伝子は、
(i)表1Bおよび表4の配列から選択されるヌクレオチド配列、
(ii)(i)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の配列の断片、
(iii)(i)もしくは/および(ii)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含有する。
【0048】
本明細書中で使用される場合、本発明の1つまたは複数の表に開示しているヌクレオチド配列または/および遺伝子に対する参照は、前記表に開示している特定の配列に対する参照、および前記表に開示している受託番号、遺伝子名、Locus Link、記号、GeneID、GeneSymbol、または/およびGenbankIDによって特徴付けられる配列に対する参照であると理解される。受託番号、遺伝子名、Locus Link、記号、GeneID、GeneSymbol、または/およびGenbankIDに対する参照によって、当業者は、対応するヌクレオチド配列または/およびアミノ酸配列を特定することができる。特定の配列は、表に示しているような、受託番号、遺伝子名、Locus Link、記号、GeneID、GeneSymbol、およびGenbankIDのうちの1つまたは複数によって特徴付けることができる。本発明の1つまたは複数の表に開示している遺伝子に対する参照は、特に、配列、例えば、前記表に開示している受託番号、遺伝子名、Locus Link、記号、GeneID、GeneSymbol、または/およびGenbankIDによって特徴付けられる遺伝子配列などに対する参照であると理解される。
【0049】
改変(調整および組換え改変を含む)は、キナーゼの改変または/およびキナーゼ結合ポリペプチドのモジュレーターの改変であってもよく、このとき、少なくとも1種のキナーゼまたは/およびキナーゼ結合ポリペプチドは、表1Aおよび表1Bから選択される核酸または/および遺伝子によってコードされる。
【0050】
本発明の方法において、インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、
(i)表1Bおよび表4から選択されるヌクレオチド配列、
(ii)(i)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の配列の断片、
(iii)(i)もしくは/および(ii)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含有する活性化剤であってもよい。
【0051】
少なくとも1種の該活性化剤は、配列(i)、(ii)(iii)または/および(iv)を含有する遺伝子の発現または/および遺伝子産物活性を活性化する能力があってもよい。
【0052】
本発明の方法において、インフルエンザウイルス複製を増大させることができる少なくとも1種の該モジュレーターは、
(a)表1Aおよび5から選択されるヌクレオチド配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含有する阻害剤であってもよい。
【0053】
少なくとも1種の該阻害剤は、配列(a)、(b) (c)または/および(d)を含有する遺伝子の発現または/および遺伝子産物活性を阻害する能力があってもよい。
【0054】
本発明の方法において用いられるインフルエンザウイルス複製の少なくともモジュレーターは、核酸、例えばリボザイム、ペプチド、ポリペプチド、抗体、アプタマー、スピーゲルマー、小分子のような核酸類似体、およびデコイ核酸からなる群から選択されてもよい。
【0055】
インフルエンザウイルス複製の該モジュレーターは、1000ダルトンより小さいまたは500ダルトンより小さい分子量を有する化合物であってもよい。本発明の文脈において、「小分子」とは、1000ダルトンより小さいまたは500ダルトンより小さい分子量を有する化合物を指す。本発明の方法において、該小分子は、
(a)表1A、表1B、表4、および表5から選択される核酸または/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、または/ならびに
(c)(a)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対して指向されてもよい。
【0056】
本発明のモジュレーターは、核酸を含有することが好ましく、このとき、該核酸は、表2および表4の配列ならびにこれらの断片から選択されるヌクレオチド配列を含有する。
【0057】
好ましくは、該核酸は、
(a)RNA、その類似体および誘導体、
(b)DNA、その類似体および誘導体、ならびに
(c)(a)と(b)との組合せ
から選択される。
【0058】
好適な阻害剤は、RNA干渉の能力のあるRNA分子である。本発明のモジュレーター、特に、本発明の阻害剤は、
(i)RNA干渉の能力のあるRNA分子、例えば、siRNAもしくは/およびshRNAなど、
(ii)miRNA、
(iii)該RNA分子(i)もしくは/および(ii)の前駆体、
(iv)該RNA分子(i)、(ii)もしくは/および(iii)の断片、
(v)(i)、(ii)(iii)または/および(iv)の該RNA分子の誘導体、または/ならびに
(vi)(i)、(ii)(iii)または/および(iv)の該RNA分子をコードするDNA分子
を含有していてもよい。
【0059】
好ましいモジュレーターは、
(i)miRNA、
(ii)該RNA分子(i)の前駆体、または/ならびに
(iii)該RNA分子(i)もしくは/および該前駆体(ii)をコードするDNA分子
である。
【0060】
さらにもう1つの好ましいモジュレーターは、
(i)RNA干渉の能力のあるRNA分子、例えば、siRNAもしくは/およびshRNAなど、
(ii)該RNA分子(i)の前駆体、または/ならびに
(iii)該RNA分子(i)もしくは/および該前駆体(ii)をコードするDNA分子
である。
【0061】
RNA干渉の能力のあるRNA分子は、WO 02/44321に記載されており、その開示は、参照により本明細書に含まれている。マイクロRNAは、Bartel D(Cell 136:215〜233、2009)に記載されており、その開示は、参照により本明細書に含まれている。
【0062】
本発明のRNA分子は、二本鎖RNA分子、好ましくは、一方の末端に単独でまたは双方の末端に一本鎖オーバーハングのあるまたはない二本鎖siRNA分子であってもよい。該siRNA分子は、少なくとも1つのヌクレオチド類似体または/およびデオキシリボヌクレオチドを含有していてもよい。
【0063】
本発明のRNA分子は、shRNA分子であってもよい。該shRNA分子は、少なくとも1つのヌクレオチド類似体または/およびデオキシリボヌクレオチドを含有していてもよい。
【0064】
本発明において用いられるDNA分子は、ベクターであってもよい。
【0065】
本発明において用いられる核酸は、アンチセンス核酸または該アンチセンス核酸をコードするDNAであってもよい。
【0066】
本発明において用いられる核酸または/および核酸断片は、少なくとも15、好ましくは少なくとも17、より好ましくは少なくとも19、最も好ましくは少なくとも21ヌクレオチドの長さを有していてもよい。該核酸または/および該核酸断片は、最大で29、好ましくは最大で27、より好ましくは最大で25、特により好ましくは最大で23、最も好ましくは最大で22ヌクレオチドの長さを有していてもよい。
【0067】
本発明において用いられる核酸は、マイクロRNA(miRNA)、その前駆体、断片、または誘導体であってもよい。該miRNAは、本明細書に記載の核酸の長さを有していてもよい。該miRNAは、特に、約22ヌクレオチド、より好ましくは22ヌクレオチドの長さを有していてもよい。
【0068】
本発明のモジュレーターは、抗体を含有していてもよく、このとき、該抗体は、キナーゼまたは/およびキナーゼ結合ポリペプチドに対して指向されてもよい。
【0069】
該抗体は、
(a)表1A、および表1Bから選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、または/ならびに
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含有するキナーゼまたは/およびキナーゼ結合ポリペプチドに対するものであることが好ましい。
【0070】
他の好ましい一実施形態では、該抗体は、
(a)表4から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、または/ならびに
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対するものである。
【0071】
さらに他の好ましい一実施形態では、該抗体は、
(a)表5から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の配列の断片、または/ならびに
(c)(a)もしくは/および(b)の配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対するものである。
【0072】
本発明の抗体は、モノクローナル抗体もしくはポリクローナル抗体、キメラ抗体、キメラ単鎖抗体、FabフラグメントまたはFab発現ライブラリーによって産生される断片であってもよい。
【0073】
本発明の抗体を作製する技術は、当業者に知られている。モノクローナル抗体は、ヒトB細胞ハイブリドーマ技術またはEBV-ハイブリドーマ技術によって作製することができる(Kohlerら、1975、Nature 256:495〜497、Kozborら、1985、J. Immunol. Methods 81、31〜42、Coteら、PNAS、80:2026〜2030、Coleら、1984、Mol. Cell Biol. 62:109〜120)。キメラ抗体(マウス/ヒト)は、Morrisonら(1984、PNAS、81:6851〜6855)、Neubergerら(1984、312:604〜608)およびTakedaら(1985、Nature 314:452〜454)の方法を行うことによって作製することができる。単鎖抗体は、当業者に知られている技術によって作製することができる。
【0074】
組換え免疫グロブリンライブラリー(Orlandiら、1989、PNAS 86:3833〜3837、Winterら、1991、Nature 349:293〜299)をスクリーニングして本発明の抗体を得ることもできる。ランダム結合性免疫グロブリンライブラリー(Burton、1991、PNAS、88:11120〜11123)を使用して、異なるイディオタイプの組成を有する、関連した特異性を有する抗体を作製することもできる。
【0075】
抗体作製のためのもう1つの戦略は、in vivoでのリンパ球集団の刺激である。
【0076】
さらに、本発明の(F(ab')2フラグメントを含有する)抗体フラグメントは、抗体のプロテアーゼ消化によって、例えばペプシンによって、作製することができる。こうしたF(ab')2フラグメントのジスルフィド結合を減少させることにより、結果としてFabフラグメントが得られる。もう1つのアプローチでは、該Fabフラグメントは、Fab発現ライブラリーから直接得ることもできる(Huseら、1989、Science 254:1275〜1281)。
【0077】
本発明のポリクローナル抗体は、表1A、表1B、表4および表5から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列またはこれらの免疫原性断片を抗原として用いて、標準的な免疫プロトコルが当業者に知られている、例えば、ウマ、ヤギ、ウサギ、ヒトなどの、宿主の標準的な免疫プロトコルによって調製することができる。
【0078】
該抗体は、標的遺伝子の遺伝子産物に特異的な抗体、特に、標的遺伝子によってコードされるポリペプチドまたはタンパク質に特異的な抗体であってもよい。
【0079】
アプタマーおよびスピーゲルマーは、抗体と結合特性を共有する。アプタマーおよびスピーゲルマーは、標的分子を特異的に結合するように設計される。
【0080】
本発明の核酸は、(a)アプタマー、(b)アプタマーをコードするDNA分子、および(c)スピーゲルマーから選択されてもよい。
【0081】
当業者は、アプタマーについて知っている。本発明において、「アプタマー」は、標的分子に結合することができる核酸であってもよい。アプタマーは、(例えば、>1015種の異なる核酸配列を含有する)結合核酸ライブラリーにおいて、固定化された標的分子への結合およびその後のその核酸配列の同定によって、特定することができる。この選択方法は、特異性を高めるために一回または複数回繰り返されてもよい。当業者は、所定の分子を特異的に結合するアプタマーを作製するための好適な方法を知っている。該アプタマーは、本明細書に記載のような核酸の長さを有しうる。該アプタマーは、多くとも300、多くとも200、多くとも100、または多くとも50ヌクレオチドまでの長さを有しうる。該アプタマーは、少なくとも10、少なくとも15、または少なくとも20ヌクレオチドの長さを有していてもよい。該アプタマーは、DNA分子によってコードされていてもよい。該アプタマーは、本明細書に記載のように、少なくとも1つのヌクレオチド類似体または/および少なくとも1つのヌクレオチド誘導体を含有していてもよい。
【0082】
当業者は、スピーゲルマーについて知っている。本発明において、「スピーゲルマー」は、標的分子に結合することができる核酸であってもよい。当業者は、所定の分子を特異的に結合するスピーゲルマーを作製するための好適な方法を知っている。該スピーゲルマーは、ヌクレアーゼ耐性の結合を形成することができるヌクレオチドを含有する。好ましくは、該スピーゲルマーは、Lヌクレオチドを含有する。より好ましくは、該スピーゲルマーは、L-オリゴヌクレオチドである。該スピーゲルマーは、本明細書に記載のような核酸の長さを有しうる。該スピーゲルマーは、多くとも300、多くとも200、多くとも100、または多くとも50ヌクレオチドまでの長さを有しうる。該スピーゲルマーは、少なくとも10、少なくとも15、または少なくとも20ヌクレオチドの長さを有しうる。該スピーゲルマーは、本明細書に記載のように、少なくとも1つのヌクレオチド類似体または/および少なくとも1つのヌクレオチド誘導体を含有していてもよい。
【0083】
当業者は、デコイ核酸について知っている。本発明において、「デコイ」または「デコイ核酸」は、核酸結合タンパク質、例えばDNA結合タンパク質などを特異的に結合する能力のある核酸であってもよい。該デコイ核酸は、DNA分子、好ましくは、二本鎖DNA分子であってもよい。該デコイ核酸は、核酸結合タンパク質によって認識されうる「認識配列」と呼ばれる配列を含有する。該認識配列は、少なくとも3、少なくとも5、または少なくとも10ヌクレオチドの長さを有していることが好ましい。該認識配列は、多くとも15、多くとも20、または多くとも25ヌクレオチドまでの長さを有していることが好ましい。核酸結合タンパク質の例は、転写因子であり、これは、二本鎖DNA分子を結合することが好ましい。デコイ核酸を用いた、本明細書に記載のような、細胞、有胚卵、または/および非ヒト動物のトランスフェクションは、デコイ核酸が結合する核酸結合タンパク質の活性を低下させる結果になってもよい。本明細書に記載のデコイ核酸は、本明細書に記載のような核酸分子の長さを有しうる。該デコイ核酸分子は、多くとも300、多くとも200、多くとも100、多くとも50、多くとも40、または多くとも30ヌクレオチドまでの長さを有しうる。該デコイ核酸は、少なくとも3、少なくとも5、少なくとも10、少なくとも15、または少なくとも20ヌクレオチドの長さを有していてもよい。該デコイ核酸は、DNA分子によってコードされていてもよい。該デコイ核酸は、本明細書に記載のように、少なくとも1つのヌクレオチド類似体または/および少なくとも1つのヌクレオチド誘導体を含有していてもよい。
【0084】
本明細書に記載のRNA分子または/およびDNA分子は、少なくとも1つのヌクレオチド類似体を含有していてもよい。本明細書中で使用される場合、「ヌクレオチド類似体」とは、その核酸内の主鎖、少なくとも1つのリボース、少なくとも1つの塩基、3'末端または/および5'末端における修飾に好適な構成単位を指しうる。主鎖修飾としては、ホスホロチオエート結合(PT);ペプチド核酸(PNA);モルホリノ核酸;ホスホロアミデート結合DNA(PA)などが挙げられ、これは、主鎖窒素を含む。リボース修飾としては、例えば、リボースの2'酸素を4'炭素と結合させているメチレン架橋での、ロックド核酸(LNA);2'-デオキシ-2'-フルオロウリジン;2'-フルオロ(2'-F);2'-O-アルキル-RNA(2-O-RNA)、例えば、2'-O-メチル(2'-O-Me)、2'-O-メトキシエチル(2'-O-MOE)などが挙げられる。修飾塩基は、2'-フルオロピリミジンであってもよい。5'修飾としては、5'-TAMRA-ヘキシルリンカー、5'-ホスフェート、5'-アミノ、5'-アミノ-C6リンカー、5'-ビオチン、5'-フルオレセイン、5'-テトラクロロ-フルオレセイン、5'-ピレン、5'-チオール、5'-アミノ、(12炭素)リンカー、5'-ダブシル、5'-コレステロール、5'-DY547(Cy3(商標)代替物)などが挙げられる。3'末端修飾としては、3'反転デオキシチミジン、3'-ピューロマイシン、3'-ジデオキシシチジン、3'-コレステロール、3'-アミノモディファイヤー(6原子)、3'-DY547(Cy3(商標)代替物)などが挙げられる。
【0085】
特に、本明細書に記載のヌクレオチド類似体は、siRNA、アンチセンスRNA、およびアプタマー中の好適な構成単位である。
【0086】
本明細書中で使用される場合、「核酸類似体」とは、本明細書に記載の少なくとも1つのヌクレオチド類似体を含有する核酸を指す。さらに、本明細書に記載の核酸分子は、少なくとも1つのデオキシリボヌクレオチドおよび少なくとも1つのリボヌクレオチドを含有していてもよい。
【0087】
本発明のRNA分子は、少なくとも1つのデオキシリボヌクレオチドまたは/および少なくとも1つのヌクレオチド類似体を含有していてもよい。本発明のDNA分子は、少なくとも1つのリボヌクレオチドまたは/および少なくとも1つのヌクレオチド類似体を含有していてもよい。
【0088】
本明細書に記載の誘導体とは、化学的に修飾された化合物を指す。本明細書に記載の核酸分子の誘導体とは、化学的に修飾された核酸分子を指す。修飾は、核酸分子中に、または/および該核酸の作製において用いられる少なくとも1個の核酸構成単位中に導入されてもよい。
【0089】
本発明において、「断片」という用語は、核酸、ポリペプチドおよびタンパク質の断片を指す。また、「断片」は、核酸、ポリペプチドおよびタンパク質の部分的な配列も指す。
【0090】
本発明において用いられるポリペプチドまたは/およびペプチドの断片、特に、表1A、表1B、表4および表5から選択される核酸または/および遺伝子によってコードされるアミノ酸配列の断片は、少なくとも5アミノ酸残基、少なくとも10、または少なくとも20アミノ酸残基の長さを有していてもよい。前記断片の長さは、最大で200アミノ酸残基、最大で100アミノ酸残基、最大で60アミノ酸残基、または最大で40アミノ酸残基であってもよい。
【0091】
本明細書に記載のアミノ酸配列の断片は、該配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有していてもよい。
【0092】
本明細書に記載のヌクレオチド配列の断片は、該配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有していてもよい。
【0093】
表1A、1B、4および5に示している核酸分子の断片は、多くとも1000、多くとも2000、または多くとも3000ヌクレオチドまでの長さを有しうる。核酸断片は、本明細書に記載のようなsiRNA分子、miRNA分子、アプタマー、スピーゲルマー、または/およびデコイの長さを有しうる。また、核酸断片は、多くとも300、多くとも200、多くとも100、または多くとも50ヌクレオチドまでの長さも有しうる。また、核酸断片は、少なくとも3、少なくとも5、少なくとも10、少なくとも15、または少なくとも20ヌクレオチドの長さも有しうる。
【0094】
本発明の方法において、遺伝子の発現の調節は、特に転写または/および翻訳の、下方制御または上方制御であってもよい。
【0095】
当業者は、遺伝子が上方制御されるかどうかまたは下方制御されるかどうかを容易に決定することができる。本発明の文脈において、遺伝子発現の上方制御(活性化)は、少なくとも2倍、好ましくは少なくとも4倍までの上方制御であってもよい。本発明の文脈における下方制御(阻害)は、少なくとも2分の1、好ましくは少なくとも4分の1までの、遺伝子発現の低減であってもよい。最も好ましいものは、例えばRNA干渉による、遺伝子発現のほぼ完全な阻害である。
【0096】
遺伝子の活性の調整は、活性の低減または増大であってもよい。「活性の阻害」は、少なくとも2分の1、好ましくは少なくとも4分の1までの、遺伝子または遺伝子産物の活性の低減であってもよい。「活性の阻害」は、活性のほぼ完全な阻害を含む。「活性の活性化」は、少なくとも2倍、好ましくは少なくとも4倍までの、遺伝子または遺伝子産物の活性の増大であってもよい。
【0097】
本発明において、本明細書に記載の方法、細胞、生物、および医薬組成物の特定の実施形態は、本出願に記載のあらゆる個々の遺伝子、核酸配列または/および遺伝子産物に関する。特定の一実施形態において、個々の遺伝子は、表1、4、および5に記載の遺伝子から選択される。他の特定の実施形態は、表1、4、および5に記載の個々の遺伝子に関する。他の特定の一実施形態において、個々の遺伝子産物は、表1、4、および5に記載の遺伝子によって産生される遺伝子産物から選択される。他の特定の実施形態は、表1、4、および5に記載の遺伝子によって産生される個々の遺伝子産物に関する。さらに他の特定の一実施形態において、個々の核酸配列または核酸分子は、表1、2、4および5に記載の核酸分子または核酸配列から選択される。他の特定の実施形態は、表1、2、4、および5に記載の個々の核酸分子または核酸配列に関する。さらなる特定の実施形態は、表1、2、3、4、および5に記載の遺伝子、遺伝子産物および核酸分子のあらゆる組合せに関する。組合せは、2、3、4、5、6、7、8、9、10またはより多くの異なる種を含んでいてもよい。表3は、核酸分子の特定の組合せに関する。
【0098】
本発明のさらなる特定の実施形態は、表5に開示の配列に関する。本発明の特定の実施形態は、表5に記載のあらゆる個々の遺伝子、核酸分子または/および遺伝子産物に関する。特定の一実施形態において、個々の遺伝子は、表5に記載の遺伝子から選択される。他の特定の実施形態は、表5に記載の個々の遺伝子に関する。他の特定の一実施形態において、個々の遺伝子産物は、表5に記載の遺伝子によって産生される遺伝子産物から選択される。他の特定の実施形態は、表5に記載の遺伝子によって産生される個々の遺伝子産物に関する。さらに他の特定の一実施形態において、個々の核酸分子または核酸配列は、表5に記載の核酸分子または核酸配列から選択される。他の特定の実施形態は、表5に記載の個々の核酸分子または核酸配列に関する。さらなる特定の実施形態は、表5に記載の遺伝子、遺伝子産物および核酸分子のあらゆる組合せに関する。組合せは、2、3、4、5、6、7、8、9、10またはより多くの異なる種を含んでいてもよい。
【0099】
本発明の特定の実施形態は、MxA遺伝子、MxAポリペプチド、およびこれらの断片に関する。さらに特定の実施形態は、miR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体、ならびにmiR-141、miR-141*、miR-200c、miR-200c*または/およびこれらの前駆体をコードするDNA分子に関する。
【0100】
修飾は、単一の核酸種によって行われてもよく、または、表1a、1b、2、4もしくは/および5ならびにその断片から選択されうる、2、3、4、5、6もしくはより多くの異なる核酸種を含有する核酸の組合せによって行われてもよい。好ましい組合せは、表3に記載している(本明細書では「プール」とも呼ぶ)。表3は、少なくとも2種のキナーゼ遺伝子または/およびキナーゼ結合ポリペプチド遺伝子の組合せを含む。この組合せが、例えば表1a、b、4および5から選択される、単一の遺伝子の発現を変更することも好ましい。2つの核酸種の組合せが好ましい。より好ましいのは、表2から選択される2種の核酸の組合せである。さらにより好ましいのは、表2に開示の特定の組合せから選択される2種の核酸の組合せであり、このとき、この2種の核酸は、単一の遺伝子の発現を変更する。
【0101】
改変、特に調整は、RNA干渉によって行われるノックダウンであってもよい。該核酸または核酸種の該組合せは、表2、表4および表5の配列ならびにそれらの断片から選択される配列を含有していてもよい、siRNAであってもよい。該組合せは、例えば表1bおよび表4から選択される、単一の遺伝子をノックダウンすることが好ましい。2つのsiRNA種の組合せが好ましく、これは、表1bの遺伝子に由来する、表2の配列、ならびに表4および表5の配列、から選択されてもよく、このとき、該組合せは、単一の遺伝子をノックダウンすることが好ましい。
【0102】
本発明のステップ(a)において用いることができる、組換え技術による「遺伝子または/および遺伝子産物の活性化」または「遺伝子または/および遺伝子産物の阻害」は、当業者が知っているいかなる好適な方法を含んでいてもよい。
【0103】
目的の遺伝子または/およびその遺伝子産物の活性化の好ましい方法は、
-活性化する遺伝子の少なくとも1個のさらなるコピーを、恒久的または一過的のいずれかで、該細胞または/および生物内に導入すること、
-転写を増強すること、
-過剰発現、
-遺伝子内に強力なプロモーター、例えばCMVプロモーターを導入すること、
-好適なエンハンサーを導入すること、
-転写的に活性なマイクロRNAの阻害、このとき、該マイクロRNAは、活性化する遺伝子の活性を阻害し、このとき、阻害は、好適な核酸分子によって行うことができる、
-miRNA結合部位の欠失、
-例えば、例えばヘパドナウイルスからの、転写後調節エレメントを3'末端に導入することによる、または、例えばD型レトロウイルスからの、1つもしくは複数の構成的な輸送エレメントを3'末端に導入することによる、または/およびスプライシングを受けうるイントロンを用いることによる、核からの輸出を含むRNAプロセシングの改良、
-例えば、潜在性スプライシング部位の欠失、GC含量の最適化、キラーモチーフおよびリピートの欠失、構造の最適化による、リボソーム結合の改良ならびにコード配列または/および3'UTRの最適化による、翻訳の改良、
から選択されてもよい。
【0104】
目的の遺伝子または/およびその遺伝子産物の阻害の好ましい方法は、
-阻害する遺伝子の少なくとも1個のさらなるコピーを該細胞または/および生物において欠失させること、このとき、該遺伝子は、完全にまたは部分的に欠失される。例えば、調節配列または/およびコード配列が、完全にまたは部分的に、欠失される、
-転写を低減させること、
-存在する場合には、エンハンサーを欠失させること、
-転写的に活性なマイクロRNAの導入または/および活性化、このとき、該マイクロRNAは、阻害する遺伝子の活性を阻害し、このとき、活性化は、内因性マイクロRNAコード配列の活性化であってもよく、導入は、外因性マイクロRNA分子の導入であってもよい、
-miRNA結合部位の導入、
-存在する場合には、3'末端に導入している転写後調節エレメントまたは3'末端に導入している1つもしくは複数の構成的な輸送エレメントの欠失もしくは/および修飾による、あるいはイントロン-エキソン構造を変更することによる、核からの輸出を含むRNAプロセシングの低減、
-例えば、潜在性スプライシング部位の導入、GC含量の変更、キラーモチーフおよびリピートの導入による、リボソーム結合ならびにコード配列または/および3'UTRの改変による、翻訳の低減、
から選択されてもよい。
【0105】
本発明の多様な実施形態において用いられる遺伝子は、表1A、1B、2、4および5、またはこれらの任意の組合せのいずれから選択されてもよい。
【0106】
ステップ(b)に従って該細胞または/および該生物をインフルエンザウイルスと接触させることは知られている。該非ヒト生物が有胚卵である場合、当業者は、例えば受精後の定められた間隔で、インフルエンザウイルスを該卵に接種する好適な方法を知っている。既知の接種技術も、有胚卵へのモジュレーターの投与のためにまたは/および有胚卵の組換え改変のために適用することができる。
【0107】
当業者は、インフルエンザウイルスの複製を可能にする条件下で該細胞、該有胚卵または/および該非ヒト生物を培養するステップ(c)に従った方法を知っている。好適な細胞培養法を適用することができる。該非ヒト生物が有胚卵である場合、当業者は、高温でのインキュベートなどの、インフルエンザウイルス複製を可能にする好適な方法を知っている。
【0108】
インフルエンザウイルスまたは/およびその構成要素をステップ(d)に従って単離することは、ウイルスまたは/およびその構成要素の、当業者に知られるあらゆる単離方法に関する。「単離」は、ウイルスまたは/およびその構成要素のほぼ純性または未精製の調製物または製剤の作製を含む。ウイルスの構成要素としては、ウイルスのタンパク質、ポリペプチド、ならびにウイルスのタンパク質または/およびポリペプチドをコードする核酸などが挙げられる。また、生ウイルスを単離することもできる。
【0109】
当業者は、ステップ(e)に従って、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、医薬組成物を調製する方法を知っている。本発明の方法によって作製される医薬組成物は、免疫原性の組成物であってもよい。また、本発明の方法によって作製される医薬組成物は、ワクチンであってもよい。
【0110】
本発明のさらにもう1つの主題は、MxA遺伝子の発現または/および遺伝子産物活性の阻害剤を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含有する医薬組成物である。該医薬組成物は、miR-141、miR-141*、miR-200c、miR-200c*、これらの前駆体および誘導体から選択される少なくとも1種の阻害剤を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含有することが好ましい。また、該阻害剤は、アンチセンス核酸、siRNA、shRNA、および小分子から選択されてもよい。
【0111】
本発明のさらにもう1つの主題は、MxA遺伝子の発現または/および遺伝子産物活性の活性化剤を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含有する医薬組成物である。該医薬組成物は、miR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害する能力のある少なくとも1種の阻害剤を、場合によっては、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含有することが好ましい。
【0112】
miR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる該阻害剤は、アプタマー、スピーゲルマーおよびデコイ核酸から選択されることが好ましい。該医薬組成物における好ましい阻害剤であるアプタマーは、miR-141、miR-141*、miR-200cまたは/およびmiR-200c*ならびにこれらの前駆体の活性を阻害することができる。該医薬組成物における好ましい阻害剤であるスピーゲルマーは、miR-141、miR-141*、miR-200cまたは/およびmiR-200c*ならびにこれらの前駆体の活性を阻害することができる。該医薬組成物中の好ましい阻害剤であるデコイ核酸分子は、miR-141、miR-141*、miR-200cまたは/およびmiR-200c*ならびにこれらの前駆体の転写に関わる転写因子を結合することができる配列を含有する。
【0113】
本明細書に記載の医薬組成物(本発明の方法によって作製されたか、またはMxA遺伝子の発現もしくは/および遺伝子産物活性の活性化剤を含有する該医薬組成物のいずれか)は、好ましくは、ヒトにおいて使用されるものであるかまたは獣医学的医薬品である。該医薬組成物は、好ましくは、インフルエンザウイルス感染の予防、軽減または/および治療のために使用されるものである。
【0114】
該医薬組成物中の担体は、送達系を含有していてもよい。当業者は、本発明の医薬組成物に好適な送達系を知っている。該医薬組成物は、ネイキッド核酸の形態で、ウイルスベクター、リポソームを含む非ウイルスベクター、ナノ粒子または/およびポリマーとの組合せで、送達されてもよい。該医薬組成物または/および該核酸は、エレクトロポレーションによって送達されてもよい。
【0115】
ネイキッド核酸としては、RNA、改変RNA、DNA、改変DNA、RNA-DNA-ハイブリッド、アプタマー融合体、プラスミドDNA、ミニサークル、トランスポゾンなどが挙げられる。
【0116】
ウイルスベクターとしては、ポックスウイルス、アデノウイルス、アデノ随伴ウイルス、水疱性口内炎ウイルス、アルファウイルス、はしかウイルス、ポリオウイルス、B型肝炎ウイルス、レトロウイルス、およびレンチウイルスなどが挙げられる。
【0117】
リポソームとしては、安定な核酸-脂質粒子(SNALP)、カチオン性リポソーム、カチオン性カルジオリピン類似体ベースのリポソーム、中性リポソーム、リポソーム-ポリカチオン-DNA、カチオン性イムノリポソーム、イムノリポソーム、コレステロールの疎水性誘導体、ラウリン酸およびリトコール酸を含有するリポソームなどが挙げられる。リポソーム形成に好適な化合物の例は、1,2-ジラウロイル-sn-グリセロ-3-ホスホエタノールアミン(DLPE);1,2-ジオレオイル-sn-グリセロ-3-ホスホ-L-セリン(DOPS);コレステロール(CHOL);1,2-ジオレイル-sn-グリセロ-3-ホスホコリン(DOPC)である。
【0118】
ナノ粒子としては、CaCO3ナノ粒子、キトサン-コーティングされたナノ粒子、葉酸化された脂質ナノ粒子、ナノサイズの核酸担体などが挙げられる。
【0119】
ポリマーとしては、ポリエチレンイミン(PEI)、ポリエステルアミン(PEA)、ポリエチレングリコール(PEG)-オリゴ結合体、PEGリポソーム、ポリマーナノ球体などが挙げられる。
【0120】
該医薬組成物は、アテロコラーゲン、カーボンナノチューブ、シクロデキストリン含有ポリカチオン、融合タンパク質(例えば、プロタミン-抗体結合体)と組み合わせて送達されてもよい。
【0121】
MxA遺伝子の発現もしくは/および遺伝子産物活性の活性化剤、または/ならびにmiR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる阻害剤は、インフルエンザウイルス感染の予防用、軽減用または/および治療用の医薬組成物の製造に使用されてもよい。本明細書に記載の送達系および送達法を使用してもよい。
【0122】
本発明のもう1つの主題は、MxA遺伝子の発現もしくは/および遺伝子産物活性の活性化剤、または/ならびにmiR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる阻害剤の、インフルエンザウイルス感染の予防用、軽減用または/および治療用の使用である。本明細書に記載の送達系および送達法を使用してもよい。
【0123】
本発明のもう1つの主題は、治療有効量の、MxA遺伝子の発現もしくは/および遺伝子産物活性の活性化剤、または/ならびにmiR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる阻害剤を、これを必要としている対象に投与することを含む、インフルエンザウイルス感染の予防、軽減または/および治療の方法である。インフルエンザウイルス感染の予防、軽減または/および治療の該方法において、本明細書に記載の送達系および送達法を使用してもよい。
【0124】
本発明のさらにもう1つの主題は、本明細書に記載のように、本発明の方法のステップ(a)に従って作製される組換え細胞である。
【0125】
本発明のさらにもう1つの主題は、本明細書に記載のように、本発明の方法のステップ(a)に従って作製される組換え非ヒト生物である。
【0126】
本発明のさらにもう1つの主題は、本明細書に記載のように、本発明の方法のステップ(a)に従って作製される組換え有胚卵である。該組換え有胚卵は、好ましくは、組換え有胚鶏卵である。
【0127】
以下の図面、表および実施例によって本発明をさらに説明する。
【図面の簡単な説明】
【0128】
【図1】実施例のsiRNAキナーゼスクリーニングの試験的設定を示した図である。
【図2】発光データに関する、トランスフェクトした(対照)-siRNAの効果を示した図である。このダイアグラムは、1枚の96ウェルプレートからの典型的なスクリーニング結果を示している。全ての試験に、いくつかの対照を、例えば、未感染のもの、ルシフェラーゼに対するsiRNAでトランスフェクトしたもの、モック処理したものおよびA型インフルエンザウイルスからのウイルス核タンパク質遺伝子(NP)に対するsiRNAのように三重試験で含めた。ルシフェラーゼsiRNAおよび抗NP siRNAで処理した細胞間の発光の相違を、定義による100%阻害に設定した。
【図3】実施例において試験した全てのsiRNAについてのインフルエンザウイルス複製の阻害を示した図である。
【図4】解析した全てのsiRNAからの値「%阻害」を使用してz-スコアを算出した図である。きわめて効率的なsiRNAを桃色にラベルしており、ルシフェラーゼsiRNAをトランスフェクトした対照細胞と比較して50%を超える阻害を示している。
【図5】ゲノム全体のsiRNAスクリーニングの試験的設定を示した図である(実施例4を参照されたい)。
【図6】miR-141の発現は、ヒト肺上皮細胞においてA型インフルエンザでの感染に応じて増強され、ウイルス複製を増強することを示した図である。(A)A549ヒト肺上皮細胞を、ヒトインフルエンザ株であるA/WSN/33(H1N1)、A/Puerto Rico/8/34(H1N1)、A/Panama/99(H3N2)および鳥インフルエンザ株であるA/FPV/Bratislava/79(H7N7)で感染させることにより、感染24時間後のmiR-141の上方制御がqRT-PCRによって検出される。miR-141の発現を、非感染(NI)細胞に対して標準化している。(B)miR-141特異的な前駆体もしくは阻害剤、または非特異的なRNA阻害剤(NS)でのトランスフェクション後のA549細胞におけるウイルス増殖(A/WSN/33)(MOI 0.05)。トランスフェクトして感染させたA549細胞からのウイルス含有上清を、インフルエンザ依存的ルシフェラーゼアッセイを使用して定量した。(C、D)安定に形質導入された異なるmiRNAを過剰発現しているA549細胞におけるウイルス複製。初代細胞培養物における感染細胞の百分率(C)および結果として生じるウイルス後代(D)を、自動顕微鏡法およびルシフェラーゼアッセイをそれぞれ使用して定量した。データは、3つの重複した試料(A、C)または3つの独立した試験(B、D)の平均値+SDである。*t-検定(P<0.05)。NT、トランスフェクトなし。
【図7】A型インフルエンザ感染に応じた、MxAおよびmiR-141の遺伝子発現を示した図である。(A〜C)miR-141とMxA発現との間の時間依存的関連性を解析するために、A549細胞をインフルエンザA/WSN/33(MOI 1)に感染させた。表示の各時点で、MxA mRNA(A)、MxAタンパク質(B)およびmiR-141(C)のレベルをqRT-PCR(RNA)およびイムノブロット解析(タンパク質)によって決定した。β-アクチンは、イムノブロットのローディング対照としての役割を果たす。(D〜F)A549細胞を、表示のMOIでインフルエンザA/WSN/33に感染させて、感染24時間後に細胞を溶解した。MxA mRNA(D)およびMxAタンパク質(E)のレベルを、それぞれqRT-PCRおよびイムノブロットによって決定した。β-アクチンは、イムノブロットのローディング対照としての役割を果たす。(F)免疫蛍光法による、表示のMOIでのMxA(緑色)およびインフルエンザA/WSN/33の核タンパク質(NP)(赤色)の検出(感染24時間後)。スケールバー、10μm。A、CおよびDにおいて、MxA発現は、非感染(NI)対照に相対的なものである。データは、3つの重複した試料の平均値+SDである。
【図8A】miR-141機能の阻害は、MxA遺伝子発現を回復させることを示した図である。(A)A549細胞におけるMxA(緑色シグナル)および核タンパク質(NP、赤色シグナル)の検出(感染24時間後)。細胞を、miR-141阻害剤および対照阻害剤(miR-198;非特異的、NS)でそれぞれトランスフェクトして、8時間後にインフルエンザA/WSN/33またはA/FPV/Bratislava/79(MOI 1)に感染させた。スケールバー、10μm。矢印は、MxA-陽性のインフルエンザ感染細胞を示す。MxA-陽性のインフルエンザ感染細胞の定量については、図12に示している。
【図8BC】miR-141機能の阻害は、MxA遺伝子発現を回復させることを示した図である。 (B)qRT-PCRによって定量される、異なるMOIでのA型インフルエンザ感染に応じたmiR-141のレベル。上方制御は、非感染細胞(NI)に相対的なものである。データは、3つの重複した試料の平均値+SDである。(C)MxA遺伝子発現のイムノブロット解析。A549細胞からの細胞溶解液を、特定量のmiR-141および非特異的阻害剤でトランスフェクトして、表示のMOIでインフルエンザA/WSN/33に感染させた。β-アクチンは、ローディング対照としての役割を果たす。ブロットは、3つの独立した試験の代表である。
【図9AB】MxAの下方制御は、miR-141によって直接仲介されることを示した図である。(A)内因性MxAタンパク質および外因性eGFP-MxA融合タンパク質のイムノブロット解析(感染24時間後)。A549細胞を、表示のMOIでインフルエンザA/WSN/33に感染させて、1時間後にpEGFP-MxAでトランスフェクトした。β-アクチンは、ローディング対照としての役割を果たす。示しているブロットは、3つの独立した試験の代表である。MxAおよびeGFP-MxAのバンド強度をブロットの下に示している。(B)ウイルスのNP染色(青色)、mCherry発現(赤色)およびeGFP-MxA発現(緑色)を用いた、感染させトランスフェクトしたA549細胞の感染24時間後の代表的な共焦点画像。スケールバー、10μm。eGFP-MxA陽性細胞の定量については、図15に示している。
【図9C】MxAの下方制御は、miR-141によって直接仲介されることを示した図である。(C)トランスフェクトした細胞またはインフルエンザA/WSN/33を感染させた細胞におけるMxAおよびeGFP-MxA融合タンパク質の発現。A549細胞を、表示のMOIで感染させて、未処理のままにしておくか、または異なるmiRNAの過剰発現プラスミド(空、miR-198およびmiR-144)で6時間トランスフェクトし、その後、pEGFP-MxAでのトランスフェクションとIFN-β(500U/ml)での同時の処理とを行った。β-アクチンは、ローディング対照としての役割を果たす。示しているブロットは、3つの独立した試験の代表である。MxAおよびeGFP-MxAのバンド強度をブロットの下に示している。
【図9D】MxAの下方制御は、miR-141によって直接仲介されることを示した図である。 (D)異なるmiRNAを過剰発現している細胞または対照細胞におけるmCherry(赤色)、外因性eGFP-MxA(緑色)および内因性MxA(青色)のレベルを示している代表的な共焦点顕微鏡写真。矢印は、内因性MxAの下方制御と、eGFP-MxAの発現がないまたは弱いこととの双方を示す、トランスフェクトされた細胞(mCherry陽性)を示している。スケールバー、10μm。eGFP-MxA陽性細胞の定量については、図16に示している。
【図10】MxAを欠損した、インフルエンザA/WSN/33感染A549細胞(MOI 0.05)における一次感染のレベルを示した図である。インフルエンザNP特異抗体で細胞を染色することによって、感染24時間後に一次感染を決定し、その後、核を染色するためにヘキスト色素で処理した。自動顕微鏡法を使用して感染細胞および非感染細胞の数を定量して、感染細胞の割合を算出した(図10)。エラーバーは、三重試験の標準偏差(SD)を示す。NT、トランスフェクトなし。*/t-検定/(P<0.05)。
【図11】MxAを欠損した、インフルエンザA/WSN/33感染A549細胞(MOI 0.05)におけるウイルス後代のレベルを示した図である。インフルエンザ依存的ルシフェラーゼアッセイを使用して、細胞上清中のウイルス後代を検出した(図11)。エラーバーは、三重試験の標準偏差(SD)を示す。NT、トランスフェクトなし。*/t-検定/(P<0.05)。
【図12】インフルエンザ感染細胞における内因性MxAの定量を示した図である。A549細胞を、miR-141および対照阻害剤(非特異的(NS);miR-181)でそれぞれトランスフェクトして、トランスフェクション後8時間でインフルエンザA/WSN/33またはA/FPV/Bratislava/79に感染させた(MOI 1)。感染24時間後に、免疫蛍光共焦点顕微鏡法を使用して、インフルエンザ感染細胞(処理ごとに細胞約100個)でMxAの発現を解析した。MxA陽性細胞の数は、総細胞数の平均百分率+SDとして示している。miR-141阻害剤でトランスフェクトした細胞は、1のMOIでの感染後に、MxA陽性細胞の数が対照阻害剤と比較して有意に増加した。*/t-検定/(/P/<0.05)。
【図13】安定にmiRNAを過剰発現しているA549細胞におけるMxAの発現を示した図である。表示のmiRNAをコードするレトロウイルスベクターまたは空の対照ベクターで予め形質導入されている細胞を、未処理のままにしておくか、またはIFN-β(500U/ml)で24時間処理した。3つの独立した試験からの代表的なブロットを示している。MxAのバンド強度は、Aidaイメージアナライザープログラム(V.4.03)を使用して決定した。
【図14】ウェブツールRNAhybrid(/9/)によって最も小さい最小自由エネルギー(mfe)で予測されるhas-miR-141(UAACACUGUCUGGUAAAGAUGG、miRBase配列データベース(/8/)とMxAの完全なmRNA配列(NM_002462)との間の相互作用を示した図である。
【図15】eGFP-MxA陽性細胞の定量を示した図である。A549細胞を、表示のMOIでインフルエンザA/WSN/33に感染させた。1時間後、細胞を、外因性MxAを検出するためのeGFP-MxA融合プラスミドと、トランスフェクション効率を標準化するための構成的に発現されるmCherry構築物(モル比6:1)とでコトランスフェクトした。感染24時間後に、免疫蛍光共焦点顕微鏡法を使用して処理ごとに100細胞でeGFP-MxAの発現を解析した。eGFP-MxAおよびmCherry陽性細胞の数は、総細胞数の平均百分率+SDとして示している。1のMOIにおいて、eGFP-MxA陽性細胞の百分率は、非感染(NI)細胞培養物および部分感染細胞培養物(MOI 0.05)と比較して有意に低下する。*/t-検定/(/P/<0.05)。
【図16】感染のない場合のeGFP-MxA陽性細胞の定量を示した図である。A549細胞を、異なる過剰発現プラスミド(空、miR-198およびmiR-141)で6時間トランスフェクトし、その後、(外因性MxAを検出するための)eGFP-MxA融合プラスミドでのトランスフェクションとIFN-β(500U/ml)での同時の処理とを行った。免疫蛍光共焦点顕微鏡法を使用して処理ごとに約100個のmCherry陽性細胞でeGFP-MxAの発現を解析した。eGFP-MxA陽性細胞の数は、総細胞数の平均百分率+SDとして示している。miR-141を過剰発現するプラスミドでトランスフェクトした細胞は、対照(トランスフェクトなし(NT)、空ベクターおよびmiR-198)と比較して、eGFP-MxA陽性細胞の有意な減少を示す。*/t-検定/(/P/<0.05)。
【図17】IFN-β刺激(1000U/ml)に応じた、安定に形質導入されたmiRNA過剰発現細胞におけるデュアルルシフェラーゼアッセイによって検出されるISREプロモーター活性を示した図である。これらのA549細胞を、最小プロモーター内にISREモチーフを含むプロモーターか、または対照として構成的なSV40プロモーター(pGL3-対照)のいずれかによって働くホタルルシフェラーゼ遺伝子をコードするプラスミドでトランスフェクトした。さらに、細胞を、トランスフェクション対照として/レニラ/ルシフェラーゼプラスミドでトランスフェクトし、直後に、IFN-βで刺激した。/レニラ/発光シグナルに対するホタルの比を示している。エラーバーは、四重試験の標準偏差(SD)を示す。
【図18】miR-141/141*およびmiR-200c/200c*、ならびにこれらの前駆体の配列を示した図である。さらに、これらの前駆体のヘアピン構造を記載している。
【発明を実施するための形態】
【0129】
表1:siRNAキナーゼスクリーニングの結果を示した表である:a:細胞数に対して標準化し、4つの独立の試験を使用して標準偏差を算出した、%でのウイルス複製の活性化(「負の」阻害)、b:細胞数に対して標準化し、4つの独立の試験を使用して標準偏差を算出した、%でのウイルス複製の阻害。プールXは、表3に記載の組合せを指し、ここで、Xはプールの番号を意味する。
表2:実施例1のsiRNAキナーゼスクリーニングにおいて用いたオリゴヌクレオチド配列を示した表である。(a)前記遺伝子に特異的な2種のオリゴヌクレオチド配列(「標的1」および「標的2」)の組合せによって、または(b)別の遺伝子に特異的なプールされたオリゴヌクレオチド(「プールX」、ここで、Xは、表3に記載のプールの番号を意味する)によって、特定の遺伝子のノックダウンを行った。
表3:実施例のsiRNAキナーゼスクリーニングにおいて用いたオリゴヌクレオチドプールを示した表である。
表4:実施例4のsiRNAスクリーニングにおいて用いたオリゴヌクレオチド配列を示した表である。遺伝子に特異的な4種までのオリゴヌクレオチド配列(「標的配列1」、「標的配列2」、「標的配列3」、および「標的配列4」)を(別々の試験においてそれぞれ)用いた。
表5:実施例4のsiRNAスクリーニングにおいて用いたオリゴヌクレオチド配列を示した表である。遺伝子に特異的な4種までのオリゴヌクレオチド配列(「標的配列1」、「標的配列2」、「標的配列3」、および「標的配列4」)を(別々の試験においてそれぞれ)用いた。この表に記載している遺伝子のノックダウンは、結果としてウイルス複製の増大になった。
【0130】
[実施例1]
キナーゼは、ウイルス後代に影響を及ぼしうる最も有望な候補のうちの1つであるので、本発明者らは、この群の遺伝子に対するsiRNAを使用して、インフルエンザウイルスの変更された複製に関して、それぞれのキナーゼまたはキナーゼ結合ポリペプチドの個々の役割を特定した。全てのsiRNAを、4つの独立の試験において試験した。キナーゼに対するsiRNAは、細胞の複製に影響を及ぼしうるまたはさらに細胞毒性であるので、自動顕微鏡を使用して、細胞数に関する個々のsiRNAトランスフェクションの効果を解析した。インフルエンザA/WSN/33核タンパク質(NP)セグメントの非翻訳領域に隣接するホタルルシフェラーゼをコードするRNA転写産物を発現させるためのRNAポリメラーゼIプロモーター/ターミネーターカセットを使用して構築されたインフルエンザレポータープラスミドを用いて、複製コンピテントインフルエンザウイルスの量を定量した。インフルエンザ感染の1日前に、ヒト胚性腎細胞(293T)をこの指標プラスミドでトランスフェクトした。この細胞を選択したのは、それがA型インフルエンザウイルス感染後にルシフェラーゼ発現のきわめて強力な増幅を示すからである。この細胞ベースのアッセイは、以下のステップを含んでいた(図1にも、siRNAキナーゼスクリーニングの試験的設定を説明している):
1日目: 96ウェルプレート中でのA549細胞(肺上皮細胞)の播種
2日目: キナーゼまたはキナーゼ結合タンパク質に対するものであるsiRNAでのトランスフェクション
3日目: インフルエンザA/WSN/33での感染+インフルエンザ指標プラスミドでの293T細胞のトランスフェクション
4日目: A549細胞の上清での293T細胞の感染+自動顕微鏡による細胞数の決定
5日目: 指標細胞の溶解、およびウイルス複製を定量化するためのルシフェラーゼアッセイの実施
【0131】
インフルエンザに関連するキナーゼの同定のために、発光値を、(siRNAトランスフェクションおよびウイルス感染の後に測定した)細胞数に対して標準化した。このことにより、より少ない(またはより多い)細胞数に起因する非特異的な効果を最小限に抑えることができる。
【0132】
全てのスクリーニング法において正確なアッセイを実証できるようにするために、いくつかの対照を含めた(図2)。ウイルス核タンパク質に対する対照siRNAは、ほぼ非感染細胞のレベルまで複製を低減させることができた。
【0133】
百分率の阻害の実例は、いくつかのsiRNAがインフルエンザウイルス複製を増強することができるのに対して、別のsiRNAはインフルエンザNP遺伝子に対する抗ウイルス性対照siRNAよりも強力に(>113%)複製を阻害することができることを示す(図3)。これに関して、47種のsiRNAは、50%を超えて複製を低減させ、9種のsiRNAは、80%を超える阻害を示した。細胞数に対して標準化し、4つの独立の試験を使用して標準偏差を算出した、%でのウイルス複製の活性化(表1a、「負の」阻害)および阻害(表1b)を示している、結果の表を表1aおよび1bに示している。
【0134】
z-スコアの算出を用いて、同様な結果が得られた。z-スコアは、生のスコアと標準偏差の単位における母平均との間の距離を表す。z-スコアは、以下の式を用いて算出した:
【0135】
【数1】
【0136】
式中、Xは、標準化する生のスコアであり、σは、母集団の標準偏差であり、かつμは、母集団の平均である。
【0137】
[実施例2]
今後の試験において、この抗ウイルス作用は、プールされたsiRNAの代わりに個々のsiRNAまたはshRNAを使用することによってより詳細に検証されるであろう。実施例1の試験的設定を使用して、さらに新しいsiRNA(特定された遺伝子につき少なくとも2種のさらなるsiRNA)およびshRNAを試験することになる。インフルエンザウイルスの複製に重要であると思われるこれらの確認された遺伝子を、次いで、マウスにおいて、鼻腔内に投与するsiRNAまたはshRNAを使用してノックダウンすることになる。この抗ウイルス療法の評価には、肺上皮組織への化合物の輸送の効率を決定することが最も重要である。療法の成功は、効率の高いキナーゼ阻害剤と適切な輸送システムとの組合せに依存している。適合する可能性があって費用効率のよい作用剤は、本発明者らがin vivoでの試験においてsiRNAまたはshRNAの送達に成功して適用しているキトサンである。本発明者らは、該化合物を鼻腔内に使用するかまたはこれらを直接肺内に投与するであろう。
【0138】
効率的なsiRNAまたはshRNAは、肺組織内のウイルス力価の低下を導くはずであり、このために、動物は、それがない場合には致死のインフルエンザウイルス感染に対して保護されるはずである。
【0139】
キナーゼ阻害剤の生物学的作用を試験するために、本発明者らは、試験を4つの部分に分けることになる:
1.化合物/キトサンナノ粒子の鼻腔内適用後の呼吸器におけるキナーゼ阻害剤の分布の解析。最良の有効性のための化合物/キトサン濃度の最適化。成功した場合のみに、さらなる試験が行われることになる。
2.LD50試験において、ウイルス分離株インフルエンザA/Puerto Rico/8/34および鳥インフルエンザ分離株(試験4のため)の絶対的な病原性を評価することになる。
3.肺組織におけるウイルス力価または(特定の場合において)生存率を解析することによる、鼻腔内適用およびインフルエンザA/Puerto Rico/8/34での感染後の、選択したsiRNAまたはshRNAの抗ウイルス作用の試験。
4.肺組織におけるウイルス力価または(特定の場合において)生存率を解析することによる、鼻腔内適用および(H5N1のような)病原性の高い鳥インフルエンザウイルス分離株での感染後の、選択したsiRNAまたはshRNAの抗ウイルス作用の試験。
【0140】
使用されるウイルス分離株は、鳥インフルエンザの現在の発生および広がりに依存する。本発明者らは、現在流行している株の複製をin vivoで効率よく阻害することを目的とする。
【0141】
確認された遺伝子に対するキナーゼ阻害剤も、損なわれたウイルス複製についてマウスで試験されることになる。
【0142】
Max-Planck感染生物学研究所、Berlin、Germanyは、好適な細胞培養物(A549細胞)における個々の全てのヒト遺伝子の遮断を、原則として、可能にする、ゲノム全体のRNAiライブラリーを有する。したがって、次の段階では、このスクリーニングは、ゲノム全体の規模まで拡張されるであろう。ウイルスの吸着、複製および出芽に関与する多くのさらなる細胞内因子は、依然として未知であるからである。
【0143】
[実施例3]
(キナーゼまたはキナーゼ結合タンパク質に対するsiRNAだけでなく)さらなるsiRNAおよびshRNAも、A型インフルエンザウイルスの複製の低減について検証されるであろう。これらのsiRNAおよびshRNAの評価には、(自動顕微鏡を使用する代わりに)市販の細胞生存率アッセイを使用して細胞数を間接的に定量すること、ならびにこれらのsiRNAおよびshRNAをリバーストランスフェクトする、すなわち、384ウェルプレート中に既に調製されているトランスフェクション混合物に細胞を添加することになることを除き、実施例1に記載したのと同じ試験的設定を用いることになる。
【0144】
[実施例4]
ヒトゲノムの中で何百もの遺伝子がインフルエンザウイルスの複製におそらく関連している。そのため、約59886種のsiRNAを使用することにより、(実施例1に記載の)キナーゼおよびキナーゼ結合因子のスクリーニング方法を、全ての既知のヒト遺伝子を解析するゲノム全体の規模に拡張した。
【0145】
以下を除き、実施例1に記載したのと同様の方法で、試験的設定を行った:
●59886種のsiRNAを使用して、スクリーニングをゲノム全体のレベルに拡張した
●細胞を384ウェルプレート中に播種した。
●トランスフェクトした細胞の数が膨大なため、必ずしも全ての細胞数を自動顕微鏡法によって解析できたわけではない。
●トランスフェクション試薬HiperFect(Qiagen、Hilden、Germany)を使用して、siRNAを、新たに播種したA549細胞においてリバーストランスフェクトした。
●特定の遺伝子のノックダウンを、特定の遺伝子に特異的な4種までのsiRNA(表4にある「標的配列1」、「標的配列2」、「標的配列3」、および「標的配列4」)によって独立に行った。
●さらなる対照:陰性対照として、「AllStars陰性対照siRNA」(Qiagen、Hilden、Germany、注文番号1027280)、陽性対照(インフルエンザ複製に対する細胞内因子)として、PKMYT(GenelD:9088、GenBank受託番号:NM_182687、標的配列:CTGGGAGGAACTTACCGTCTA)に対して指向されたsiRNA、トランスフェクション対照(トランスフェクション後の細胞毒性)として、PLK(GenelD:5347、GenBank受託番号:BC014135、標的配列:CCGGATCAAGAAGAATGAATA)に対して指向されたsiRNAを含めた。
●感染過程における早期または後期の事象に対する阻害作用を精査することができるように、選択したウェル中のトランスフェクトしたA549細胞の感染率を自動顕微鏡法によって測定する。
●(「Allstars陰性対照siRNA」でトランスフェクトした対照ウェルを基準に)B-スコア標準化方法を使用して、Michael Butros、Ligia BrasおよびWolfgang Huberによって開発された、統計的R-パッケージ「cellHTS」ソフトウェアによって結果を解析した。
●読み出される情報は、ウイルス複製の阻害である。
【0146】
強力な抗ウイルス活性(z-スコア<-2.0)を示したsiRNAおよび対応する遺伝子の一覧を表4に記載している。
【0147】
この細胞ベースのアッセイは、以下のステップを含んでいた(ゲノム全体のsiRNAスクリーニングの試験的設定を説明している図5も参照されたい):
1日目: A549細胞(肺上皮細胞)の播種+siRNAのリバーストランスフェクション
3日目: インフルエンザA/WSN/33での感染+指標プラスミドでの293T細胞zqのトランスフェクション
4日目: A549細胞の上清での293T細胞の感染+ホルムアルデヒドでのA549細胞の固定
5日目: 293T細胞におけるウイルス複製を定量化するためのルシフェラーゼアッセイ
x日目: 自動顕微鏡による感染率の決定。
【0148】
[実施例5]
A型インフルエンザウイルスは、内因性のmiR-141を誘導して、MxAの抗ウイルス活性を次第に低下させる
哺乳動物細胞におけるグローバルな制御因子として知られる、マイクロRNA(miRNA)も、ウイルスを感染させて宿主細胞の機能を奪うための魅力的なツールである。宿主のmiRNA機能に対するインフルエンザウイルスの影響力を評価するために、本発明者らは、A549ヒト肺上皮細胞におけるmiRNAの発現解析を行った。1種の内因性miRNA(miR-141)は、感染細胞において強く誘導され、過剰発現されると、ウイルス複製を増強したのに対して、miR-141の阻害は複製を低減させた。本発明者らは、miR-141の重要な標的として、抗ウイルス性MxA宿主遺伝子を同定した。したがって、インフルエンザウイルス感染は、MxAの転写後サイレンシングを引き起こしたが、隣接した非感染細胞におけるインターフェロンで誘導されるMxA合成は可能であった。このように、本発明者らは、宿主細胞を介した抗ウイルス応答からA型インフルエンザウイルスを保護する、miRNAに基づく重要な調節機構を発見し、これは、抗ウイルス介入の新規選択肢を構成するものである。
【0149】
本発明者らは、A型インフルエンザウイルスも細胞内miRNAの使用によって利点を得ているかどうかを知ることに興味を持った。始めに、2種の異なるマイクロアレイプラットフォームを使用して、ヒトmiRNAの発現パターンにおける変化をモニタリングした。双方のアレイ型(AmbionおよびExiqon)により、感染されたA549ヒト肺上皮細胞において、単一のmiRNA、miR-141が、非感染細胞と比較して有意に上方制御されたことが明らかにされた。マイクロアレイ結果をリアルタイム逆転写-PCR(qRT-PCR)によって確認し、A549細胞をヒトおよび鳥類の一区画のA型インフルエンザウイルス株に感染させた後のmiR-141の劇的な増加(5〜15倍)が示された(図6A)。
【0150】
増加した細胞内miR-141がインフルエンザ複製に対する影響力を有するかどうかを調べるために、本発明者らは、細胞を、miR-141-特異的な化学的に修飾された核酸阻害剤またはインフルエンザウイルスA/WSN/33で感染後のmiR-141前駆体のいずれかでトランスフェクトした。感染効率を示す、ウイルス依存的ルシフェラーゼアッセイ(17)を使用して、感染力をもつ粒子の数を定量した。miR-141前駆体でのA549細胞の処理は、感染力をもつ粒子の形成を有意に増加させたのに対して、miR-141阻害剤は、逆の作用を導いた(図6B)。この知見により、細胞内miR-141はインフルエンザウイルス複製の効率の調節における役割を果たしているという最初の証拠が提供された。
【0151】
高レベルのmiR-141の存在下でウイルス複製が増強されることを確認するために、本発明者らは、ヒトmiR-141または異なる対照(miR-198または空ベクター)のいずれかを安定に過剰発現する細胞系を作製した。次いで、細胞を、低い感染多重度(MOI 0.05)でインフルエンザA/WSN/33に感染させた。免疫蛍光顕微鏡法により、感染24時間後(h p.i.:hours post infection)における感染細胞の数は、miR-141陽性細胞において、多様な対照と比較して有意に多かったことが明らかになった(図6C)。これらの細胞上清中の感染力をもつ粒子の定量(図6D)により、A型インフルエンザウイルスは、miR-141のレベルが増大されているA549細胞においてより効率よく複製することができることがさらに確認された。合わせると、これらのデータにより、インフルエンザウイルス複製におけるmiR-141発現の刺激性の効果が確証される。
【0152】
主に腫瘍の発生に関連する(18)、miR-141の細胞内標的遺伝子は、以前に報告されているが、これらの遺伝子のうち、ウイルス複製におけるこのmiRNAの影響について真実味のある説明を提供したものはない。miRNAは、翻訳抑制だけではなく、それらのmRNA標的の安定性にも影響しうるという仮定に基づき(19)、本発明者らは、市販の全ヒトゲノムマイクロアレイを使用して、miR-141を過剰発現しているA549細胞におけるmRNA発現プロファイルを対照細胞と比較した。miR-141を過剰発現している細胞において3倍を超える影響を受ける標的mRNAの中で(データは示していない)、本発明者らは、抗ウイルス因子MxAを同定した(12)。MxAの抗ウイルス作用を確認するために、本発明者らは、shRNAを介したMxAのノックダウンを示す(タンパク質レベルにおけるノックダウンそれぞれ約85%)2種の安定なA549細胞系を作製した。予想通り、MxAのサイレンシングは、一次ウイルス複製(図10)および上清に放出される感染力をもつウイルスの力価(図11)を有意に増大させた。これらの知見により、本発明者らは、MxAが、miR-141機能とウイルス複製との間の重要な接点を構成しているのではないかと考えた。
【0153】
MxAに関連してmiR-141に増強されるウイルス複製の分子基盤を調べるために、本発明者らは、感染中のA549細胞におけるMxA mRNA、MxAタンパク質およびmiR-141のレベルを、それぞれqRT-PCRおよびイムノブロットを使用して解析した(図7A〜C)。MxA遺伝子発現はI型インターフェロンによる刺激を必要とするので(20)、感染していないA549細胞では、MxA mRNAもMxAタンパク質もいずれも検出されなかった。1のMOIで感染させた細胞において、有意なレベルのMxAのmRNAが感染8時間後という早い時期に転写され、感染24時間後にピークに達した(図7A)。驚くべきことに、MxAタンパク質は、このMOIを使用して感染の24時間後でさえも検出されなかった(図7B)。同じ感染時間にわたり、miR-141のレベルは着実に上昇して、感染していないA549細胞と比較して、20倍の上方制御を導いた(図7C)。これらの知見により、インフルエンザウイルスでの細胞の感染に応じて、miR-141の発現の増加に付随して、転写後の機構により、MxAタンパク質合成が妨げられるが、mRNA合成は妨げられないことが示された。しかし、興味深い追加の現象が明らかとなった-低いウイルス力価(MOI 0.1)において、本発明者らは、感染24時間後に強いMxAタンパク質シグナルを認めた(図7B)。MOIを高めることに応じた感染24時間後のMxAタンパク質合成の漸増により、0.1のMOIまでのMxAタンパク質レベルの用量依存的上昇が明らかとなった。MxAタンパク質レベルの上昇は、低いMOIでのMxA転写と関連していた;これに対して、0.1を超えるMOIでは、持続性のmRNA転写にもかかわらず、MxAタンパク質レベルが急激に低下した(図7Dおよび7E)。したがって、MxAタンパク質の転写後抑制は、高いMOIでは明白であったが、低いMOIではそうではなかった。
【0154】
この現象を説明するために、本発明者らは、免疫蛍光顕微鏡法を使用して、単細胞レベルで、ウイルスに感染してMxAを発現している細胞を可視化した。感染していない(NI)細胞および完全に感染した培養物(MOI 1)において、MxAタンパク質は検出されなかった(図7F)。しかし、MxAタンパク質は、部分的に感染した培養物(MOI 0.1)の非感染細胞において検出された(図7F)。これらの非感染細胞におけるMxAの合成は、非感染細胞においてMxA合成を刺激することが知られる(21)、I型インターフェロンに関連しているパラクリン調節ループに起因すると考えることができる。合わせると、これらのデータより、MxAタンパク質の効率的な転写後下方制御は全てのウイルス感染細胞において生じること;および、最も可能性の高いのはIFN-βの産生を介して、感染細胞が非感染細胞におけるMxAの上方制御を誘発することが明らかに実証される。
【0155】
ウイルス感染細胞において観察されたMxAタンパク質の下方制御が、miR-141による転写後サイレンシングに依存的であるかどうかを試験するために、細胞を特定のmiRNA阻害剤で処理した(図8A)。著しいことに、MxA陽性細胞は、miR-141阻害剤での処理後に、ウイルスに感染した、すなわち、ウイルス核タンパク質(NP)を発現している細胞のみにおいて認められた(図8A、矢印を参照されたい)。阻害剤のない場合、または、関連のないmiR-198阻害剤でのトランスフェクションによる場合には、NPおよびMxAタンパク質の発現は、相互排他的であった。H7N7鳥インフルエンザ株A/FPV/Bratislava/79でも同様の結果が得られた(図8A)。MxA陽性細胞の定量により、単細胞での知見が確証された(図12)。miR-141がMxAの転写サイレンシングを誘導するという見解と一致して、本発明者らは、細胞培養物中のmiR-141レベルが、MOIが高まるとともに着実に上昇して、約1のMOI、すなわち、感染24時間後に全ての細胞が感染しているところでピークに達することを観察した(図8B)。MxAの制御におけるmiR-141の重要な役割は、イムノブロット解析によってさらに支持されており、miR-141特異的阻害剤のトランスフェクションは、全ての細胞が感染したときでさえも、MxA発現を促進することが明らかになった(図8C)。これらの知見により、miR-141は、ウイルスに誘導される、抗ウイルス性MxAタンパク質の下方制御に重要であることが明らかに示される。
【0156】
感染のない場合のmiR-141の機能を実証するために、本発明者らは、IFN-βを使用して、異なるmiRNAを過剰発現している多様なA549細胞系においてMxA発現を刺激した。より高レベルのmiR-141を発現している細胞は、miR-198または空ベクターでトランスフェクトした対照細胞と比較して、よりわずかな程度のMxA誘導(約30%)を示し、IFN-β依存的な、しかしウイルス非依存的な、MxAサイレンシングにおけるmiR-141の作用が示唆された。
【0157】
miR-141を介したMxAサイレンシングの分子基盤を研究するために、本発明者らは、MxA mRNA配列を解析した。典型的には、miRNAは、その標的mRNAの3'非翻訳領域(UTR)と相互作用することによって機能して、遺伝子発現におけるその調節作用を発揮する(22)。しかし、MxA 3'-UTRに連結したルシフェラーゼレポーター構築物で一過的にトランスフェクトしたA549細胞は、miR-141過剰発現に応答したルシフェラーゼ活性の低下を示さなかった(データは示していない)。さらに、miR-141とMxA mRNAとの配列アライメントにより、MxAの3'-UTR領域ではなく、コード配列(CDS)中に相補的な領域が明らかにされた(図14)。これらの知見により、miR-141を介したMxAの抑制にはCDSが関与することが示唆される。MxA下方制御におけるmiR-141の間接的な調節機能の可能性を除外するために、本発明者らは、構成的なサイトメガロウイルス(CMV)プロモーターの制御下に置かれる、増強された緑色蛍光タンパク質(eGFP)とMxAとの遺伝子融合体を作製した。eGFP-MxA融合体により、遺伝子導入したMxAタンパク質と内因性のMxAタンパク質との、これらの異なる分子サイズによる、並行したモニタリングが可能となった。このアプローチにより、本発明者らは、(例えば、miR-141に調節される推定上の転写因子を介して)天然のMxAプロモーターからの転写に影響を及ぼす、miR-141の間接的な調節機能と、MxAの転写産物、その3'-UTRを含む、に影響を及ぼす、直接的な転写後遺伝子サイレンシング機構とを区別できるようになった。eGFP-MxA融合体は、A549細胞のトランスフェクション後すぐに発現された;しかし、感染に応じたその発現は、インフルエンザA/WSN/33のMOIが高まるとともに着実に低下した(図9A)。同様に、内因性MxAは、約0.1のMOIで最も高いタンパク質レベルを示し(図6Eと比較されたい)、MOIが高まるとともに低下した(図9A)。ウエスタンブロットデータの定量により、1のMOIでのウイルス感染に応じた、トランスフェクトした組換えMxAタンパク質および内因性MxAタンパク質の約10分の1の減少が示された(図9A;ブロットの下のバンド強度の値を参照されたい)。したがって、インフルエンザウイルスに誘発された、遺伝子導入したeGFP-MxAと内因性のMxAとの減少は、eGFP-MxAの転写を導く構成的なCMVプロモーターにもかかわらず、ほぼ同じであった。
【0158】
これらの知見により、転写後の直接的なMxA合成の干渉が、共焦点顕微鏡法を使用した単細胞解析によってさらに支持されたことが示される:構成的に発現されるmCherry構築物と、eGFP-MxA融合タンパク質(モル比1:6)との同時発現は、好結果のトランスフェクション戦略を示した(図9B)。しかし、MOIを高めてこれらの細胞をインフルエンザウイルスに感染させることにより、eGFP-MxA融合体タンパク質の選択的減少が導かれたが、mCherryタンパク質では導かれなかった(図9B)。eGFP-MxA陽性細胞の定量により、これらの知見が確証された(図15)。この試験により、インフルエンザウイルス感染に応じた、MxAの特異的な転写後サイレンシングが明らかに実証される。
【0159】
MxA発現の転写後サイレンシングにおけるmiR-141の直接的な機能を支持するさらなる証拠を集めるために、本発明者らは、ウイルス感染のない場合のMxA発現に対するmiRNAの作用を調べた。ヒトMxAプロモーターはIFNに刺激される2つの機能的な応答エレメント(ISRE)(23)を含有しているので、本発明者らは、MxAの発現を誘導するためにIFN-βを使用した。レトロウイルスのmiR-141のトランスフェクションにより、コトランスフェクトしたeGFP-MxA融合タンパク質とIFN-βに誘導される内因性MxAとの発現における並行した低下が引き起こされたが(図9C)、miR-198または空ベクターでは引き起こされなかった。MxAタンパク質発現の定量により、miR-141の過剰発現に応じて、対照プラスミドを有する細胞と比較して、eGFP-MxAの約3分の1の減少および内因性MxAの約10分の1の減少が示された(図9C;ブロットの下のバンド強度の値を参照されたい)。ここでも、単細胞レベルでのこれらの交差した結果を、共焦点顕微鏡法によって解析および確認し、IFN-βで刺激してmiR-141でトランスフェクトした細胞において外因性MxAおよび内因性MxAの双方の量が減少していることが示された(図9D)。図9Bにおける試験的設定と同一である、mCherryプラスミドのコトランスフェクションにより、eGFP-MxAをトランスフェクトした細胞の追跡が可能となった。eGFP-MxA陽性細胞の定量により、miR-141をコードするプラスミドで細胞をトランスフェクトした後の有意に低減されたMxAのレベルが明らかにされた(図16)。最も注目すべきことに、レトロウイルスのmiR-141構築物でトランスフェクトされた個々の細胞および内因性MxAの下方制御を示した個々の細胞によっても、eGFP-MxAの発現がないまたは弱いことが示された(図9D中の矢印を参照されたい)。miR-141の直接的な機能は、miR-141、または関連のないmiR-198を過剰発現している、IFN-βに刺激されたA549細胞におけるISREプロモーター活性の解析によっても実証され、MxAの上流のシグナル伝達はmiR-141によって影響されないことが示された(図17)。考え合わせると、これらのデータにより、RNA干渉に基づく機構を介して転写後レベルで作用するMxA合成におけるmiR-141の直接的な作用の概念が強く支持される。
【0160】
ウイルスによるMxAの能動的な抑制(1)の根底にある分子機構は、このように、全く謎のままだった。ここで、本発明者らは、ヒト細胞において抗ウイルス性MxAタンパク質の下方制御に関与する重要な分子として、A型インフルエンザウイルスに誘導されるmiR-141を同定した。本研究において行った広範囲な単細胞解析により、感染細胞におけるMxAの能動的なmiR-141-依存的阻害が示唆される;しかし、感染細胞は、β-インターフェロンに関連しているパラクリンループを介して、隣接するまだ感染していない細胞における抗ウイルス性のMxA合成を依然として誘導する。それにもかかわらず、かつ最もよく知られているmiRNA翻訳制御機構に反して(24、25)、miR-141による干渉はきわめて強力であり、miR-141は、インフルエンザ感染を治療するための優れた標的を構成しうることが示唆される。さらに、ヒトおよび鳥類の一区画のA型インフルエンザ分離株に応答したmiR-141上方制御についての本発明者らの知見は、広範囲に効能がある治療選択肢のために有望である。
【0161】
(参考文献)
【0162】
【表1a−1】
【表1a−2】
【表1a−3】
【表1a−4】
【表1a−5】
【表1a−6】
【0163】
【表1b−1】
【表1b−2】
【表1b−3】
【表1b−4】
【表1b−5】
【表1b−6】
【表1b−7】
【表1b−8】
【表1b−9】
【表1b−10】
【0164】
【表2−1】
【表2−2】
【表2−3】
【表2−4】
【表2−5】
【表2−6】
【表2−7】
【表2−8】
【表2−9】
【表2−10】
【表2−11】
【表2−12】
【表2−13】
【表2−14】
【表2−15】
【表2−16】
【0165】
【表3−1】
【表3−2】
【表3−3】
【表3−4】
【表3−5】
【表3−6】
【表3−7】
【表3−8】
【表3−9】
【表3−10】
【表3−11】
【0166】
【表4−1】
【表4−2】
【表4−3】
【表4−4】
【表4−5】
【表4−6】
【表4−7】
【表4−8】
【表4−9】
【表4−10】
【表4−11】
【表4−12】
【表4−13】
【表4−14】
【表4−15】
【表4−16】
【表4−17】
【表4−18】
【表4−19】
【表4−20】
【表4−21】
【表4−22】
【表4−23】
【表4−24】
【表4−25】
【表4−26】
【表4−27】
【表4−28】
【表4−29】
【表4−30】
【表4−31】
【表4−32】
【表4−33】
【表4−34】
【表4−35】
【表4−36】
【表4−37】
【表4−38】
【表4−39】
【表4−40】
【表4−41】
【表4−42】
【表4−43】
【表4−44】
【表4−45】
【表4−46】
【表4−47】
【表4−48】
【表4−49】
【表4−50】
【表4−51】
【表4−52】
【表4−53】
【表4−54】
【表4−55】
【表4−56】
【表4−57】
【表4−58】
【表4−59】
【表4−60】
【表4−61】
【表4−62】
【表4−63】
【表4−64】
【表4−65】
【表4−66】
【表4−67】
【表4−68】
【表4−69】
【表4−70】
【表4−71】
【表4−72】
【表4−73】
【表4−74】
【表4−75】
【表4−76】
【表4−77】
【表4−78】
【表4−79】
【表4−80】
【表4−81】
【表4−82】
【表4−83】
【表4−84】
【表4−85】
【表4−86】
【表4−87】
【表4−88】
【表4−89】
【表4−90】
【表4−91】
【表4−92】
【表4−93】
【表4−94】
【表4−95】
【表4−96】
【表4−97】
【表4−98】
【表4−99】
【表4−100】
【表4−101】
【表4−102】
【表4−103】
【表4−104】
【表4−105】
【表4−106】
【表4−107】
【表4−108】
【表4−109】
【表4−110】
【表4−111】
【表4−112】
【表4−113】
【表4−114】
【表4−115】
【表4−116】
【表4−117】
【表4−118】
【表4−119】
【表4−120】
【表4−121】
【表4−122】
【表4−123】
【表4−124】
【表4−125】
【表4−126】
【表4−127】
【表4−128】
【表4−129】
【表4−130】
【表4−131】
【表4−132】
【表4−133】
【表4−134】
【表4−135】
【表4−136】
【表4−137】
【表4−138】
【表4−139】
【表4−140】
【表4−141】
【表4−142】
【表4−143】
【表4−144】
【表4−145】
【表4−146】
【表4−147】
【表4−148】
【表4−149】
【表4−150】
【表4−151】
【表4−152】
【表4−153】
【表4−154】
【特許請求の範囲】
【請求項1】
以下のステップ:
(a)改変のない場合のインフルエンザウイルス複製と比較してインフルエンザウイルス複製の能力が増大された、インフルエンザウイルスを複製することのできる改変細胞、改変有胚卵または/および改変非ヒト生物を提供するステップ、
(b)(a)の前記細胞、前記有胚卵または/および前記生物をインフルエンザウイルスと接触させるステップ、
(c)前記インフルエンザウイルスの複製を可能にする条件下で、前記細胞、前記有胚卵または/および前記非ヒト生物を培養するステップ、ならびに
(d)ステップ(c)において生成された前記インフルエンザウイルスまたは/およびその少なくとも1種の構成要素を分離するステップ
を含む、インフルエンザウイルスの調製方法。
【請求項2】
ステップ(a)が、モジュレーターのない場合のインフルエンザウイルス複製と比較して、前記細胞または/および前記生物におけるインフルエンザウイルス複製を増大させることができる少なくとも1種のモジュレーターと、前記細胞、前記有胚卵または/および前記非ヒト生物を接触させることを含む、請求項1に記載の方法。
【請求項3】
ステップ(a)が、組換え細胞、組換え有胚卵または/および組換え非ヒト生物の作製または/および提供を含み、改変のない場合のインフルエンザウイルス複製と比較して、前記細胞、前記有胚卵または/および前記非ヒト生物のインフルエンザウイルス複製の能力が増大するように、少なくとも1種の遺伝子または/および遺伝子産物の発現または/および活性を改変する、請求項1または2に記載の方法。
【請求項4】
前記インフルエンザウイルスが、A型インフルエンザウイルスまたは/およびB型インフルエンザウイルス、好ましくは、H1N1、H3N2、H7N7、H5N1から選択される株である、請求項1から3のいずれかに記載の方法。
【請求項5】
前記細胞、前記有胚卵または/および非ヒト生物の改変が、MxA遺伝子の発現または/および遺伝子産物活性の阻害を含む、請求項1から4のいずれかに記載の方法。
【請求項6】
MxA遺伝子の発現または/および遺伝子産物活性が、miR-141、miR-141*、miR-200c、miR-200c*、これらの前駆体、これらの誘導体、アンチセンス核酸、siRNA、shRNAおよびインフルエンザウイルス配列から選択される少なくとも1種のモジュレーターによって阻害される、請求項5に記載の方法。
【請求項7】
miR-141、miR-141*、miR-200c、miR-200c*およびこれらの前駆体のうちの少なくとも1種が、前記細胞内、前記有胚卵内または/および前記非ヒト生物内で過剰発現される、請求項5または6に記載の方法。
【請求項8】
前記細胞、前記有胚卵または/および前記非ヒト生物の改変が、遺伝子の発現または/および遺伝子産物活性の阻害を含み、前記遺伝子が、
(a)表1Aおよび表5の配列から選択されるヌクレオチド配列、
(b)(a)の前記配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の前記配列の断片、
(c)(a)もしくは/および(b)の前記配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含む、請求項1から4のいずれかに記載の方法。
【請求項9】
前記細胞の、前記有胚卵のまたは/および前記非ヒト生物の改変が、遺伝子の発現または/および遺伝子産物活性の活性化を含み、前記遺伝子が、
(i)表1Bおよび表4の配列から選択されるヌクレオチド配列、
(ii)(i)の前記配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の前記配列の断片、
(iii)(i)もしくは/および(ii)の前記配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含む、請求項1から4のいずれかに記載の方法。
【請求項10】
前記少なくとも1種のモジュレーターが、核酸、核酸類似体、ペプチド、ポリペプチド、抗体、アプタマー、スピーゲルマー、小分子およびデコイ核酸からなる群から選択される、請求項2に記載の方法。
【請求項11】
前記核酸が、
(a)RNA、その類似体および誘導体、
(b)DNA、その類似体および誘導体、ならびに
(c)(a)と(b)との組合せ
から選択される、請求項10に記載の方法。
【請求項12】
前記核酸が、
(i)RNA干渉の能力のあるRNA分子、例えば、siRNAもしくは/およびshRNAなど、
(ii)miRNA、
(iii)前記RNA分子(i)もしくは/および(ii)の前駆体、
(iv)前記RNA分子(i)、(ii)もしくは/および(iii)の断片、
(v)(i)、(ii)(iii)もしくは/および(iv)の前記RNA分子の誘導体、または/ならびに
(vi)(i)、(ii)(iii)もしくは/および(iv)の前記RNA分子をコードするDNA分子
である、請求項10から11のいずれかに記載の方法。
【請求項13】
前記RNA分子が、二本鎖RNA分子、好ましくは、一方の末端に単独でまたは双方の末端に一本鎖オーバーハングのあるまたはない二本鎖siRNA分子である、請求項10から12のいずれかに記載の方法。
【請求項14】
前記RNA分子が、少なくとも1つのヌクレオチド類似体または/およびデオキシリボヌクレオチドを含有する、請求項10から13のいずれかに記載の方法。
【請求項15】
前記核酸が、(a)アプタマー、(b)アプタマーをコードするDNA分子、および(c)スピーゲルマーから選択される、請求項10から14のいずれかに記載の方法。
【請求項16】
前記核酸が、アンチセンス核酸または/および前記アンチセンス核酸をコードするDNAである、請求項10から14のいずれかに記載の方法。
【請求項17】
前記核酸が、少なくとも15、好ましくは少なくとも17、より好ましくは少なくとも19、最も好ましくは少なくとも21ヌクレオチドの長さを有する、請求項10から13のいずれかに記載の方法。
【請求項18】
前記核酸が、最大で29、好ましくは最大で27、より好ましくは最大で25、特により好ましくは最大で23、最も好ましくは最大で22ヌクレオチドの長さを有する、請求項10から14のいずれかに記載の方法。
【請求項19】
前記抗体が、
(a)表1A、表1B、表4、および表5から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の前記配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の前記配列の断片、または/ならびに
(c)(a)の前記配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対するものである、請求項10に記載の方法。
【請求項20】
前記小分子が、
(a)表1A、表1B、表4、および表5から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の前記配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の前記配列の断片、または/ならびに
(c)(a)の前記配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対するものである、請求項10に記載の方法。
【請求項21】
請求項3に記載の方法で作製された組換え細胞。
【請求項22】
請求項3に記載の方法で作製された組換え有胚卵。
【請求項23】
請求項3に記載の方法で作製された組換え非ヒト生物。
【請求項24】
MxA遺伝子の発現または/および遺伝子産物活性の活性化剤を、場合によって、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含む医薬組成物。
【請求項25】
前記発現または/および遺伝子産物活性の前記活性化剤が、miR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる少なくとも1種の阻害剤である、請求項21に記載の医薬組成物。
【請求項1】
以下のステップ:
(a)改変のない場合のインフルエンザウイルス複製と比較してインフルエンザウイルス複製の能力が増大された、インフルエンザウイルスを複製することのできる改変細胞、改変有胚卵または/および改変非ヒト生物を提供するステップ、
(b)(a)の前記細胞、前記有胚卵または/および前記生物をインフルエンザウイルスと接触させるステップ、
(c)前記インフルエンザウイルスの複製を可能にする条件下で、前記細胞、前記有胚卵または/および前記非ヒト生物を培養するステップ、ならびに
(d)ステップ(c)において生成された前記インフルエンザウイルスまたは/およびその少なくとも1種の構成要素を分離するステップ
を含む、インフルエンザウイルスの調製方法。
【請求項2】
ステップ(a)が、モジュレーターのない場合のインフルエンザウイルス複製と比較して、前記細胞または/および前記生物におけるインフルエンザウイルス複製を増大させることができる少なくとも1種のモジュレーターと、前記細胞、前記有胚卵または/および前記非ヒト生物を接触させることを含む、請求項1に記載の方法。
【請求項3】
ステップ(a)が、組換え細胞、組換え有胚卵または/および組換え非ヒト生物の作製または/および提供を含み、改変のない場合のインフルエンザウイルス複製と比較して、前記細胞、前記有胚卵または/および前記非ヒト生物のインフルエンザウイルス複製の能力が増大するように、少なくとも1種の遺伝子または/および遺伝子産物の発現または/および活性を改変する、請求項1または2に記載の方法。
【請求項4】
前記インフルエンザウイルスが、A型インフルエンザウイルスまたは/およびB型インフルエンザウイルス、好ましくは、H1N1、H3N2、H7N7、H5N1から選択される株である、請求項1から3のいずれかに記載の方法。
【請求項5】
前記細胞、前記有胚卵または/および非ヒト生物の改変が、MxA遺伝子の発現または/および遺伝子産物活性の阻害を含む、請求項1から4のいずれかに記載の方法。
【請求項6】
MxA遺伝子の発現または/および遺伝子産物活性が、miR-141、miR-141*、miR-200c、miR-200c*、これらの前駆体、これらの誘導体、アンチセンス核酸、siRNA、shRNAおよびインフルエンザウイルス配列から選択される少なくとも1種のモジュレーターによって阻害される、請求項5に記載の方法。
【請求項7】
miR-141、miR-141*、miR-200c、miR-200c*およびこれらの前駆体のうちの少なくとも1種が、前記細胞内、前記有胚卵内または/および前記非ヒト生物内で過剰発現される、請求項5または6に記載の方法。
【請求項8】
前記細胞、前記有胚卵または/および前記非ヒト生物の改変が、遺伝子の発現または/および遺伝子産物活性の阻害を含み、前記遺伝子が、
(a)表1Aおよび表5の配列から選択されるヌクレオチド配列、
(b)(a)の前記配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の前記配列の断片、
(c)(a)もしくは/および(b)の前記配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(d)(a)、(b)もしくは/および(c)の配列と相補的な配列
を含む、請求項1から4のいずれかに記載の方法。
【請求項9】
前記細胞の、前記有胚卵のまたは/および前記非ヒト生物の改変が、遺伝子の発現または/および遺伝子産物活性の活性化を含み、前記遺伝子が、
(i)表1Bおよび表4の配列から選択されるヌクレオチド配列、
(ii)(i)の前記配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(i)の前記配列の断片、
(iii)(i)もしくは/および(ii)の前記配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一である配列、または/ならびに
(iv)(i)、(ii)もしくは/および(iii)の配列と相補的な配列
を含む、請求項1から4のいずれかに記載の方法。
【請求項10】
前記少なくとも1種のモジュレーターが、核酸、核酸類似体、ペプチド、ポリペプチド、抗体、アプタマー、スピーゲルマー、小分子およびデコイ核酸からなる群から選択される、請求項2に記載の方法。
【請求項11】
前記核酸が、
(a)RNA、その類似体および誘導体、
(b)DNA、その類似体および誘導体、ならびに
(c)(a)と(b)との組合せ
から選択される、請求項10に記載の方法。
【請求項12】
前記核酸が、
(i)RNA干渉の能力のあるRNA分子、例えば、siRNAもしくは/およびshRNAなど、
(ii)miRNA、
(iii)前記RNA分子(i)もしくは/および(ii)の前駆体、
(iv)前記RNA分子(i)、(ii)もしくは/および(iii)の断片、
(v)(i)、(ii)(iii)もしくは/および(iv)の前記RNA分子の誘導体、または/ならびに
(vi)(i)、(ii)(iii)もしくは/および(iv)の前記RNA分子をコードするDNA分子
である、請求項10から11のいずれかに記載の方法。
【請求項13】
前記RNA分子が、二本鎖RNA分子、好ましくは、一方の末端に単独でまたは双方の末端に一本鎖オーバーハングのあるまたはない二本鎖siRNA分子である、請求項10から12のいずれかに記載の方法。
【請求項14】
前記RNA分子が、少なくとも1つのヌクレオチド類似体または/およびデオキシリボヌクレオチドを含有する、請求項10から13のいずれかに記載の方法。
【請求項15】
前記核酸が、(a)アプタマー、(b)アプタマーをコードするDNA分子、および(c)スピーゲルマーから選択される、請求項10から14のいずれかに記載の方法。
【請求項16】
前記核酸が、アンチセンス核酸または/および前記アンチセンス核酸をコードするDNAである、請求項10から14のいずれかに記載の方法。
【請求項17】
前記核酸が、少なくとも15、好ましくは少なくとも17、より好ましくは少なくとも19、最も好ましくは少なくとも21ヌクレオチドの長さを有する、請求項10から13のいずれかに記載の方法。
【請求項18】
前記核酸が、最大で29、好ましくは最大で27、より好ましくは最大で25、特により好ましくは最大で23、最も好ましくは最大で22ヌクレオチドの長さを有する、請求項10から14のいずれかに記載の方法。
【請求項19】
前記抗体が、
(a)表1A、表1B、表4、および表5から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の前記配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の前記配列の断片、または/ならびに
(c)(a)の前記配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対するものである、請求項10に記載の方法。
【請求項20】
前記小分子が、
(a)表1A、表1B、表4、および表5から選択される核酸もしくは/および遺伝子によってコードされるアミノ酸配列、
(b)(a)の前記配列の少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%の長さを有する、(a)の前記配列の断片、または/ならびに
(c)(a)の前記配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%同一であるアミノ酸配列
を含むポリペプチドに対するものである、請求項10に記載の方法。
【請求項21】
請求項3に記載の方法で作製された組換え細胞。
【請求項22】
請求項3に記載の方法で作製された組換え有胚卵。
【請求項23】
請求項3に記載の方法で作製された組換え非ヒト生物。
【請求項24】
MxA遺伝子の発現または/および遺伝子産物活性の活性化剤を、場合によって、薬学的に許容される担体、アジュバント、賦形剤または/および添加物と共に、含む医薬組成物。
【請求項25】
前記発現または/および遺伝子産物活性の前記活性化剤が、miR-141、miR-141*、miR-200c、miR-200c*、およびこれらの前駆体から選択されるmiRNAの活性を阻害することができる少なくとも1種の阻害剤である、請求項21に記載の医薬組成物。
【図14】

【図18】

【図1】

【図2】

【図3】
【図4】

【図5】

【図6】

【図7】
【図8A】
【図8BC】

【図9AB】
【図9C】

【図9D】
【図10】

【図11】
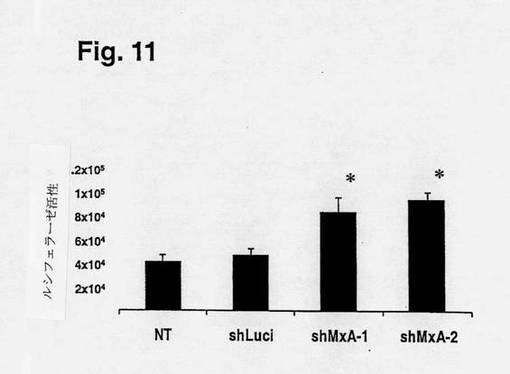
【図12】
【図13】

【図15】
【図16】
【図17】
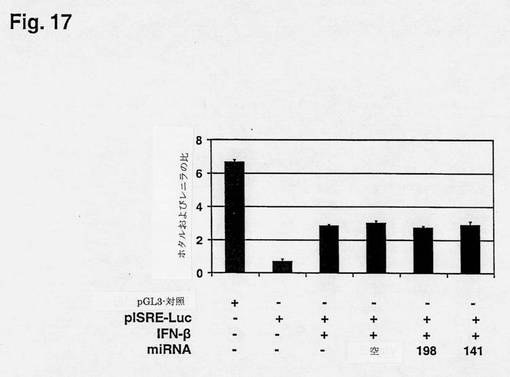
【図18】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8A】
【図8BC】
【図9AB】
【図9C】
【図9D】
【図10】
【図11】
【図12】
【図13】
【図15】
【図16】
【図17】
【公表番号】特表2013−504311(P2013−504311A)
【公表日】平成25年2月7日(2013.2.7)
【国際特許分類】
【出願番号】特願2012−528375(P2012−528375)
【出願日】平成22年9月10日(2010.9.10)
【国際出願番号】PCT/EP2010/063333
【国際公開番号】WO2011/029914
【国際公開日】平成23年3月17日(2011.3.17)
【出願人】(509289113)
【Fターム(参考)】
【公表日】平成25年2月7日(2013.2.7)
【国際特許分類】
【出願日】平成22年9月10日(2010.9.10)
【国際出願番号】PCT/EP2010/063333
【国際公開番号】WO2011/029914
【国際公開日】平成23年3月17日(2011.3.17)
【出願人】(509289113)
【Fターム(参考)】
[ Back to top ]