
ウイルス性疾患に対して抵抗の鳥および他の動物の作製
【課題】ウイルス配列標的とするsiRNAをコードする生殖細胞およびこの生殖細胞を得る方法の提供。
【解決手段】本発明はウイルス感染に対して抵抗性である遺伝子改変動物に関する。また提供されるのは、ウイルス感染に対して抵抗性である動物を作製する方法である。本発明により、ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含む構築物を含んでいる、非ヒト脊椎動物生殖細胞であって、該配列がプロモーターに作動可能に連結されている、生殖細胞が提供される。本発明によってまた、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物であって、ここで、該脊椎動物中の大多数の細胞は、該疾患の原因となるウイルスのゲノムの保存された領域に対するsiRNAをコードしている配列を含み、該配列はプロモーターに作動可能に連結されている、非ヒト脊椎動物もまた提供される。
【解決手段】本発明はウイルス感染に対して抵抗性である遺伝子改変動物に関する。また提供されるのは、ウイルス感染に対して抵抗性である動物を作製する方法である。本発明により、ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含む構築物を含んでいる、非ヒト脊椎動物生殖細胞であって、該配列がプロモーターに作動可能に連結されている、生殖細胞が提供される。本発明によってまた、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物であって、ここで、該脊椎動物中の大多数の細胞は、該疾患の原因となるウイルスのゲノムの保存された領域に対するsiRNAをコードしている配列を含み、該配列はプロモーターに作動可能に連結されている、非ヒト脊椎動物もまた提供される。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の引用)
この引用は、2005年3月31日に出願した米国仮出願第60/666,636号に対する優先権を主張する。米国仮出願第60/666,636号は、すべての目的のために、本明細書中に参考として援用される。
【0002】
(発明の分野)
本発明は、ウイルス感染に対して抵抗性の動物を作製するためのRNA干渉技術の使用に関する。
【背景技術】
【0003】
(発明の背景)
疾病管理センター(Centers for Disease Control;CDC)および世界保健機構は、致死率の高いトリインフルエンザウイルスH5N1株が、少なくとも3つの大陸にまたがって広がって、47カ国を超える国々で報告されたと報告した。非特許文献1を参照のこと。トリインフルエンザは、インフルエンザ(フルー)ウイルスによって引き起こされる感染症である。これらのインフルエンザウイルスは、鳥の中で自然に発生する。野鳥は、それらの腸野中で世界中にウイルスを運び、そして通常は無症候性である。しかしながら、トリインフルエンザは、鳥の間では非常に接触感染性であり、一部の家畜化された鳥(ニワトリ、アヒルおよびシチメンチョウを含む)を非常に病んだ状態にし得、そして死を引き起こし得る。
【0004】
感染した鳥は、それらの唾液、鼻の分泌物および糞中にインフルエンザウイルスを脱落する。感染しやすい鳥が、汚染された分泌物もしくは排出物と、または感染した鳥からの分泌物もしくは排出物によって汚染された表面と接触した場合、この感染しやすい鳥は感染されることになる。家畜化された鳥は、感染した水鳥もしくは他の感染した家禽との直接接触によって、またはトリインフルエンザウイルスによって汚染された表面(例えばほこりまたはケージ)もしくは材料(例えば、水または餌)との接触によって、トリインフルエンザウイルスに感染されることになり得る。
【0005】
家禽におけるトリインフルエンザウイルスによる感染によって、極端に低い毒性および極端に高い毒性によって区別される2つの主要な型の疾患が引き起こされる。「病原性が低い」形態は非検出となり得、そして通常、軽度の症状(例えば、乱れた羽毛および産卵の低下)のみを引き起こす。しかしながら、非常に病原性が高い形態は、家禽の群れによって、より急速に広がる。この形態は、複数の内臓に影響を及ぼして、48時間以内にしばしば90〜100%に達し得る死亡率を有する疾患を引き起こし得る。従って、トリインフルエンザに抵抗性を有する家禽を作製する方法が存在すれば、家禽飼育場での巨大な経済的損失の可能性が激減し得る。これらのウイルス抵抗性家禽は、代々引き続いてそのウイルス抵抗性を世代間で受け渡す新種の家禽を作製するために交配され得る。
【0006】
家禽の世界市場は、1年あたり500億羽を超える家禽を生産すると推定される。それゆえ、さらにより重要なことには、ウイルス抵抗性動物(例えば、家禽)を作製する能力は、恐れられる「トリインフルエンザ」ウイルスのヒトへの広がりを防止し得、それゆえ、ヒトにおけるトリインフルエンザの汎発流行を回避し得る。
【0007】
家禽産業にとってまた問題がある、他のタイプのウイルス性疾患が存在する。例えば、マレク病は、しばしば若いめん鳥の群れの重篤な死亡損失を引き起こし、ブロイラ加工工場の不良認定の主な原因であった。マレク病は、ヘルペスウイルスファミリーに属するMDウイルス(MDV)によって引き起こされるリンパ増殖性疾患であり、そして過去50年間にわたって家禽産業にとって大きな課題である。3つの血清型のMDV株が認識されている。腫瘍性株はすべて、血清型1(MDV−I)と分類され、一方、元々非腫瘍性のニワトリ株および七面鳥のヘルペスウイルス(HVT)はそれぞれ、血清型2(MDV−2)および血清型3に属する。世界全体でのマレク病に起因する毎年の経済的損失は、大きなワクチン接種キャンペーンにもかかわらず、毎年10億アメリカドルの範囲であると推定される。ワクチン接種キャンペーンは世界全体での罹患した群れの数を減らしたにもかかわらず、多数の疾患発生が世界の大部分の部分に存在し、このことは、この疾患を制御する、より良好な方法を必要とする。
【0008】
成功裏に処置され得るウイルス性疾患は、限られた数しか存在しない。現在、ワクチン接種は、動物におけるウイルス性疾患と戦うために用いられる唯一の方法である。多くの場合、特に家畜のウイルス性疾患においては、このアプローチは、効力の高いワクチンを生産することができないことに起因して適用できない。いくつかのワクチンが無効である別の理由は、抗原連続変異またはワクチン効果の喪失をもたらす、高い変異率である。一旦ワクチンがその効果を失うと、この疾患が出現し得、そして家禽および家畜のウイルス性疾患の場合、大きな経済的損失を負わせ得る。
【0009】
2つの古典的形態のワクチンが存在する;1つ目のものは、弱毒生ワクチンであり、こ
れは、免疫系の体液性免疫および細胞性免疫の両方を誘導するので、これは最も強力なものである。この種のワクチンの欠点は、非常に毒性が高い逸脱変異の再出現の危険性である。この理由のために、多くの国は、この種のワクチンの使用を承認するのを渋る。第2のタイプは、不活化ウイルスワクチンであり、これは体液性免疫系しか誘発しないので、これは多くの場合、それほど有効ではない。過去数年にわたって、サブユニットワクチンおよびDNAワクチンのようないくつかタイプの組換えワクチンが開発された。残念なことに、それらのほとんどは、幅広くは使用されていない。マレク病の場合に示されるように、ワクチン接種の欠点のうちの1つは、高い年間の経済的な支出である。World Organization for Animal Health(OIE)によると、2002年にマレク病に対するワクチン接種を受けた動物の総数は、24億5700万であった。例えば、非特許文献2を参照のこと。
【0010】
ワクチン接種をするには、2つの公知の方法が存在する。第1の方法は、約$3/1000ユニットの費用がかかる、無細胞(凍結乾燥)形態である。第2の形態は、約$8/1000ユニットの費用がかかる、細胞関連の(「湿った」)形態である。2002年のマレク病のためのワクチン接種について見積もられた世界支出は、約74億ドル〜197億ドルの間にあった。
【0011】
1970年代初頭から、大部分のニワトリは、弱毒化血清型1株またはHVTを使用して、MDに対してワクチン接種を受けた。1983年に始まった、二価の組合せおよび多価の組合せが、より毒性の実地での株から保護するために用いられている。ワクチン接種はニワトリを保護する際に著しく有効であるにもかかわらず、MDは疾患の優先順位のリストの中で順位が高いままである。優先順位が高い主要な理由は、ワクチンの破壊を引き起こす毒性の増加に向かってMDV株が継続して進化していることである。
【0012】
したがって、例えば、動物におけるウイルス性疾患(例えば、家禽におけるトリインフルエンザおよびマレク病)の伝染を防止するかまたは低下させる方法の必要が存在する。トリインフルエンザウイルスの現在の発生を考慮すると、ウイルス感染(例えば、トリインフルエンザ)に対して抵抗性である、遺伝子が改変された動物(例えば、家禽)を作製し得る方法についての強い必要性が存在する。
【0013】
過去10年において、「RNA干渉」(RNAi)と称される、遺伝子サイレンシングを経る遺伝子不活化の新規なアプローチが開示された。例えば、非特許文献3および特許文献1を参照のこと。RNA干渉とは、RNAポリヌクレオチドが内因性細胞プロセスを通して作用する場合に起こる、RNAの配列に対応する配列の遺伝子発現を特異的に抑制する事象をいう。標的遺伝子のサイレンシングは、時々はRNAse IIIエンドヌクレアーゼ消化による、宿主動物による二本鎖(ds)RNAによるmRNAの分解の際に起こる。この消化は、分子のサイズは30塩基程度の大きさであり得るにもかかわらず、長さ(またはサイズ)が約21〜23ヌクレオチド(または塩基)である分子をもたらす。
【0014】
これらの短いRNA種は、おそらく、RNA誘導性サイレンシング複合体(RISC)と呼ばれるRNAiヌクレアーゼ複合体を経て、対応するRNAメッセージおよび転写産物の分解を媒介する。RISCは、小さいdsRNAが塩基対合相互作用によって相補的なmRNAを認識するのを助ける。その基質とのsiRNA相互作用後に、おそらくRISC中に存在する酵素によって、mRNAは、分解についての標的とされる。このタイプの機構は、ウイルス感染、トランスポゾンジャンピングおよび類似の現象を抑制する際に生物体にとって、ならび内因性遺伝子の発現を制御するために、有用なようである。RNAi活性はこれまでのところ、他の生物体の中でも、植物、昆虫、ネマトーダおよび脊椎動物において実証されている。一般的な背景情報については、非特許文献4;非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;非特許文献10;非特許文献11;非特許文献12;非特許文献13;非特許文献14;および非特許文献15を参照のこと。
【先行技術文献】
【特許文献】
【0015】
【特許文献1】米国特許第6,506,559号明細書
【非特許文献】
【0016】
【非特許文献1】インターネット<URL:http://www.cdc.gov/flu/avian/outbreaks/current.htm>
【非特許文献2】インターネット<URL:www.oie.com>
【非特許文献3】Fireら,Nature 391:806−811(1998)
【非特許文献4】Schutzら,Virology 344(1):151−7(2006)
【非特許文献5】Leonardら,Gene Ther.13(6):532−40(2006)
【非特許文献6】Colbere−Garapinら,Microbes Infect.7(4):767−75(2005)
【非特許文献7】Wall,Theriogenology 57(1):189−201(2002)
【非特許文献8】El−Bashirら,Nature 411:494−498(2001)
【非特許文献9】Fire,A.ら,Science 391:806−811(1998)
【非特許文献10】Gitlinら,Nature 418:430−434(2002)
【非特許文献11】Gitlinら,J.Virol.79:1027−1035(2005)
【非特許文献12】Kahanaら,J.Gen.Virol.85,3213−3217(2004)
【非特許文献13】Kronkeら,J.Virol.78:3436−3446(2004)
【非特許文献14】Leonardら,J.Virol.79:1645−1654(2005)
【非特許文献15】Yokotaら,EMBO Rep.4:602−608(2003)
【発明の概要】
【発明が解決しようとする課題】
【0017】
動物(特に家禽)における様々なウイルス性疾患の広範囲にわたる課題を考慮すると、家禽および家畜のウイルス性疾患を効果的に制御するための様々なアプローチは、この課題を解決するために非常に有益である。そのため、本願明細書において開示される本発明は、病原性ウイルスのウイルス複製を著しく阻害する、重要なウイルス機能をブロックすることを標的とする分子を保有および発現するトランスジェニック非ヒト脊椎動物(例えば、家禽および家畜)を作製するための方法、ならびに遺伝子改変された家禽および家畜自体を提供することによって、この課題に対する解決策を提供する。
【課題を解決するための手段】
【0018】
(発明の簡単な説明)
本発明は、ウイルス配列標的とするsiRNAをコードする生殖細胞およびこの生殖細胞を得る方法を提供する。本発明はまた、ウイルス感染に対して抵抗性である非ヒト脊椎動物およびこの動物を得る方法を提供する。
【0019】
一つの態様では、本発明は、ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含む構築物を含んでいる、非ヒト脊椎動物の生殖細胞であり、ここで、この配列はプロモーターに作動可能に連結されている。一つの実施形態では、この構築物は、このウイルスゲノムに対する複数のsiRNAをコードしている配列を含む。別の実施形態では、この細胞は、ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含み、ここで、この配列は、プロモーターに作動可能に連結されている。別の実施形態では、この脊椎動物は、非ヒト哺乳類である。別の実施形態では、このウイルスは、口蹄疫ウイルス(FMDV)である。別の実施形態では、保存された配列は、配列番号1、配列番号2および配列番号3からなる群から選択される。別の実施形態では、この脊椎動物は、鳥類種のものである。別の実施形態では、このウイルスは、トリインフルエンザウイルスである。別の実施形態では、保存された配列は、図1〜図16中の配列からなる群から選択される。他の実施形態では、このウイルスは、マレク病ウイルス(MDV)である。別の実施形態では、この生殖細胞は、精子である。
【0020】
別の態様においては、本発明は、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物を提供し、ここで、この脊椎動物中の大多数の細胞は、この疾患の原因となるウイルスのゲノムの保存された領域に対するsiRNAをコードしている配列を含み、ここで、この配列はプロモーターに作動可能に連結されている。一つの実施形態では、この構築物は、このウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む。別の実施形態では、この細胞は、このウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含む。別の実施形態では、この脊椎動物は、非ヒト哺乳類である。別の実施形態では、このウイルス性疾患は、FMDVである。別の実施形態では、保存された配列は、配列番号1、配列番号2および配列番号3からなる群から選択される。別の実施形態では、この脊椎動物は、鳥類種のものである。別の実施形態では、このウイルスは、トリインフルエンザウイルスである。別の実施形態では、保存された配列は、図1〜図16中の配列からなる群から選択される。別の実施形態では、このウイルスは、MDVである。
【0021】
別の態様においては、本発明は、非ヒト脊椎動物の生殖細胞を作製する方法を提供し、この方法は、保存された領域またはウイルスゲノムに対するsiRNAをコードしている配列を含む構築物とこの生殖細胞とを、この構築物をこの細胞に取り込ませる条件下でインキュベートする工程を包含し、ここでこの配列は、プロモーターに作動可能に連結されている。一つの実施形態では、この構築物は、宿主細胞ゲノムに組み込まれる。
【0022】
別の態様においては、本発明は、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物を作製するための方法を提供し、この方法は、(a)保存された領域またはウイルスゲノムに対するsiRNAをコードしている配列を含む構築物と生殖細胞とを、この生殖細胞が二倍体細胞を形成する条件下でインキュベートする工程であって、ここでこの配列は、プロモーターに作動可能に連結されている、工程;および(b)(a)に記載の二倍体細胞を、この二倍体細胞が非ヒト脊椎動物を形成する条件下でインキュベートする工程を包含する。
例えば、本願発明は以下の項目を提供する。
(項目1)
ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含む構築物を含んでいる、非ヒト脊椎動物生殖細胞であって、該配列がプロモーターに作動可能に連結されている、生殖細胞。
(項目2)
前記構築物が、前記ウイルスゲノムに対する複数のsiRNAをコードしている配列を含む、項目1に記載の生殖細胞。
(項目3)
前記細胞が、前記ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含んでおり、ここで、該配列はプロモーターに作動可能に連結されている、項目1に記載の生殖細胞。
(項目4)
前記脊椎動物が非ヒト哺乳類である、項目1、または項目2、または項目3に記載の生殖細胞。
(項目5)
前記ウイルスが口蹄疫ウイルス(FMDV)である、項目4に記載の生殖細胞。
(項目6)
保存された配列が配列番号1、配列番号2および配列番号3からなる群より選択される、項目5に記載の生殖細胞。
(項目7)
前記脊椎動物が鳥類種のものである、項目1または項目2または項目3に記載の生殖細胞。
(項目8)
前記ウイルスがトリインフルエンザウイルスである、項目7に記載の生殖細胞。
(項目9)
保存された配列が図1〜図16中の配列からなる群より選択される、項目8に記載の生殖細胞。
(項目10)
前記ウイルスがマレク病ウイルス(MDV)である、項目7に記載の生殖細胞。
(項目11)
前記生殖細胞が精子である、項目1または項目2または項目3または項目4または項目5または項目6または項目7または項目8または項目9または項目10に記載の生殖細胞。
(項目12)
ウイルス性疾患に対して抵抗性である非ヒト脊椎動物であって、ここで、該脊椎動物中の大多数の細胞は、該疾患の原因となるウイルスのゲノムの保存された領域に対するsiRNAをコードしている配列を含み、該配列はプロモーターに作動可能に連結されている、非ヒト脊椎動物。
(項目13)
前記構築物が、前記ウイルスのゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む、項目12に記載の非ヒト脊椎動物。
(項目14)
前記細胞が、前記ウイルスのゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含む、項目12に記載の非ヒト脊椎動物。
(項目15)
前記脊椎動物が非ヒト哺乳類である、項目12または項目13または項目14に記載の非ヒト脊椎動物。
(項目16)
前記ウイルス性疾患がFMDVである、項目15に記載の非ヒト脊椎動物。
(項目17)
保存された配列が配列番号1、配列番号2および配列番号3からなる群より選択される、項目16に記載の非ヒト脊椎動物。
(項目18)
前記脊椎動物が鳥類種のものである、項目12または項目13または項目14に記載の非ヒト脊椎動物。
(項目19)
前記ウイルスがトリインフルエンザウイルスである、項目18に記載の非ヒト脊椎動物。
(項目20)
保存された配列が図1〜図16中の配列からなる群より選択される、項目19に記載の非ヒト脊椎動物。
(項目21)
前記ウイルスがMDVである、項目12に記載の非ヒト脊椎動物。
(項目22)
非ヒト脊椎動物の生殖細胞を作製する方法であって、該生殖細胞が項目1に記載の生殖細胞であり、該方法は、保存された領域またはウイルスゲノムに対するsiRNAをコードしている配列を含む構築物と該生殖細胞とを、該構築物を該細胞に取り込ませる条件下でインキュベートする工程を包含し、ここで該配列は、プロモーターに作動可能に連結されている、方法。
(項目23)
前記構築物が宿主細胞ゲノム中に組み込まれる、項目22に記載の方法。
(項目24)
項目12に記載の非ヒト脊椎動物を作製するための方法であって、
(a)項目1または項目2または項目3または項目4または項目5または項目6または項目7または項目8または項目9または項目10または項目11に記載の生殖細胞を、該生殖細胞が二倍体細胞を形成する条件下でインキュベートする工程;および
(b)(a)に記載の二倍体細胞を、該二倍体細胞が非ヒト脊椎動物を形成する条件下でインキュベートする工程
を包含する、方法。
【図面の簡単な説明】
【0023】
【図1−1】図1−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−2】図1−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−3】図1−3は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−4】図1−4は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−5】図1−5は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−6】図1−6は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図2−1】図2−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図2−2】図2−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図3−1】図3−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図3−2】図3−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図4−1】図4−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図4−2】図4−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図4−3】図4−3は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図5−1】図5−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図5−2】図5−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図6−1】図6−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図6−2】図6−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図7】図7は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図8−1】図8−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図8−2】図8−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図8−3】図8−3は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図8−4】図8−4は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図9−1】図9−1は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図9−2】図9−2は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図10】図10は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図11】図11は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図12】図12は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図13−1】図13−1は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図13−2】図13−2は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図14】図14は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図15】図15は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図16−1】図16−1は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図16−2】図16−2は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図16−3】図16−3は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【発明を実施するための形態】
【0024】
(発明の詳細な説明)
本発明は、ウイルス感染に対して抵抗性である動物およびこのようなタイプのトランスジェニック動物を作製する方法を提供する。標的ウイルス遺伝子は、サイレンシング分子(例えば、本願明細書において開示されるsiRNA分子)の存在によってサイレンシングされるので、ウイルスの生活環においてそれらの役割を果たすことができない。このプロセスの結果は、これらの動物において羅病率または/および死亡率を誘導する感染をすることができないこととなる、ウイルス複製の有意なブロックである。それゆえ、本発明は、肉および他の畜産物の絶え間ない供給を確実にし、全体として、農民にとって、および社会にとって、相当な経済効果を有する。
【0025】
本発明の実施は、特に明記しない限り、分子生物学(組換え技術を含む)、微生物学、細胞生物学、生化学および免疫学の従来の技術を使用する。これらは当業者の技術範囲内にある。このような技術は、例えば、Molecular Cloning:A Laboratory Manual,第2版(Sambrookら,1989)Cold Spring Harbor Press;Oligonucleotide Synthesis(MJ.Gait編,1984);Animal Cell Culture(R.I.Freshney編,1987);Methods in Enzymology(Academic Press,Inc.);Handbook of Experimental Immunology(D.M.Weir & C.C.Blackwell編);Gene Transfer Vectors for Mammalian Cells(J.M.Miller & M.P.Calos編,1987);Current Protocols in Molecular Biology(F.M.Ausubelら編,1987);PCR:The Polymerase Chain Reaction,(Mullisら編,1994);Current Protocols in Immunology(J.E.Coliganら編,1991)およびShort Protocols in Molecular Biology(Wiley and Sons,1999)のような文献において充分に説明されている。
【0026】
この特許出願において引用されるすべての参考文献、特許および特許出願は、それぞれがすべての目的のためにその全体が本明細書中に参考として援用される。
【0027】
(定義)
本出願において使用されるすべての科学用語および専門用語は、他に特定されない限り、当該分野において一般に使用される意味を有する。本出願において用いられる場合、以下の語または語句は、指定した意味を有する。
【0028】
交換可能に本明細書中で使用される用語「ポリヌクレオチド」および「核酸」とは、リボヌクレオチドまたはデオキシリボヌクレオチドのいずれであってもよい、任意の長さのポリマー形態のヌクレオチドをいう。これらの用語は、一本鎖、二本鎖または三本鎖の、DNA、ゲノムDNA、cDNA、RNA、DNA−RNAハイブリッド、またはプリン塩基およびピリミジン塩基を含むポリマー、または天然のものであるか、化学的に改変されているか、生化学的に改変されているか、非天然のものであるか、もしくは誘導体化されているヌクレオチド塩基を包含する。
【0029】
「プロモーター」は、転写開始および転写速度が制御されるポリヌクレオチド配列領域である制御配列である。プロモーターは、調節タンパク質および調節分子(例えば、RNAポリメラーゼおよび他の転写因子)が結合し得る遺伝要素を含み得る。プロモーターは例えば、構成性であってもよく、誘導性であってもよく、抑制性であってもよく、または組織特異的であってもよい。語句「作動可能に連結された」、「作動するように配置された」、「作動するように連結された」、「制御下にある」および「転写制御下にある」とは、プロモーターがその配列の転写開始および/または発現を制御するために核酸配列に関して適切な機能的な位置および/または方向にあることを意味する。プロモーターは、「エンハンサー」とともに用いられてもよく、そうでなくてもよい。エンハンサーとは、核酸配列の転写活性化に関与するシス作用性調節配列をいう。1つのプロモーターは、一つ以上の遺伝子の発現を調節し得る。
【0030】
(RNAi標的)
本発明は、ウイルス抵抗性動物が作製され得るように遺伝子をサイレンシングするための方法を提供する。一つの実施形態では、本発明は、免疫トランスジェニックの家禽および家畜を内部的に作製するリポフェクションに関連した遺伝子サイレンシング技術(例えば、RNAi)を使用する。好ましい実施形態では、本発明は、ウイルスに抵抗性の動物を作製するために、例えば、WO 99/42569において開示されるとおりの制限酵素媒介組込み(「REMI」)を使用する。これらの動物は、ウイルス遺伝子(例えば、複製に欠くことのできないウイルス遺伝子)を標的とする1つまたは複数のサイレンシング分子を保有および発現する。
【0031】
本発明によって提供される短い干渉RNA(「siRNA」)は、標的遺伝子が細胞内に存在して発現し得る場合に、標的遺伝子の発現の調節および/または減衰を可能にする。発現調節は、遺伝子機能の部分的阻害であってもよく、より好ましくは、遺伝子機能の完全阻害であるか、または他の第2の標的遺伝子のアップレギュレーションもしくは一次標的遺伝子の阻害に応答するこのような遺伝子の発現の強化ですらあり得る。
【0032】
遺伝子発現の減衰は、遺伝子機能、転写産物のプロセシングまたは転写産物の翻訳の部分的または完全な抑制または阻害を含み得る。RNA干渉の情況では、遺伝子発現の調節は、タンパク質とRNA(特に、「ガイド」RNAとして作用し得る小さいdsRNA)との複合体を通して進行すると考えられている。それゆえ、siRNAは、そのヌクレオチド配列が標的遺伝子のヌクレオチド配列の少なくとも一部分と十分に一致する場合、効果的であると考えられる。本発明はこのメカニズムの仮説によって束縛されないが、siRNAにおけるヌクレオチド配列が、標的遺伝子配列の少なくとも一部分と実質的に同じであることが非常に好ましい。一つの実施形態では、標的ウイルスヌクレオチドとsiRNAとの間の配列同一性は、100%(すなわち、正確に相同)である。別の実施形態では、1塩基対(bp)の相違が存在する。別の実施形態では、2塩基対の相違が存在する。別の実施形態では、3塩基対の相違が存在する。
【0033】
「標的遺伝子」または「標的配列」は一般に、遺伝子産物(例えば、ポリペチド)をコードする領域を含むポリヌクレオチド、または複製、転写もしくは翻訳、もしくはポリペチドの発現にとって重要な他のプロセスを調節するポリヌクレオチド領域、または、ポリペチドをコードする領域およびそれに対して作動可能に連結された発現調節領域を含むポリヌクレオチドを意味する。標的配列は、遺伝子産物全体をコードする必要が必ずしもあるわけではない。本発明の情況では、「作動可能に連結される」とは、その配列の開始および/または発現を制御するためにポリヌクレオチド配列に関して適切な機能的位置および/または方向にあるプロモーターをいう。
【0034】
一つの態様では、siRNAについての標的遺伝子は、ウイルス遺伝子である。一つの実施形態では、ウイルス遺伝子は、鳥類のウイルス性疾患(例えば、トリインフルエンザおよびマレク病)の伝播に関与する遺伝子である。本発明の範囲内に含まれる遺伝子配列としては、図1〜図16に示される配列が挙げられる。
【0035】
siRNAは、ウイルス複製を有意にブロックするが、細胞崩壊を引き起こさないのに十分な長さのものであり、そして通常、約19塩基対〜約27塩基対の間の範囲にある。一つの実施形態では、siRNAは、長さが21bpである。別の実施形態では、siRNAは、長さが22bpである。なお他の実施形態では、siRNAは、長さが約19bp、約20bp、約23bp、約24bp、約25bp、約26bpまたは約27bpである。なお別の実施形態では、siRNAは、長さが約28bp、約29bpまたは約30bpである。siRNAが存在する細胞にとっての一種の自己破壊機構を誘発するので、siRNAの長さを長くしすぎることはできないことが、当業者にとって明らかである。例えば、Elbashirらは、長さ21bpという長さが、細胞の抗ウイルス機構(例えば、インターフェロン応答)を回避するために効果的であることを教示する(Elbashirら,Nature 411:494−498(2001)を参照のこと)。
【0036】
図1〜図16に示すように、種々の長さが、干渉RNAとして使われ得る。1つの好ましい実施形態は、長さが21bpである。当業者は、図1〜図16において示される配列を使用し得、そして異なる21bpのsiRNAに達するために、一度に1塩基対ずつ、段階的に移動し得る。順列のうちのいくつかは、これらの図に示される。しかしながら、図1〜図16において示されるように、いったん一般的な配列が示されると、配列を段階的に移動することは当業者にとっては慣用的であるので、配列が示される可能な21bpのsiRNAのすべてが示されたわけではない。さらに、21bp以外の長さもまた、本発明に包含される。上記で議論したように、一つの実施形態では、siRNAは、長さが約21bp〜約27bpである。当業者は、図1〜図16において開示される配列を使用し、そして所望の長さを使用し、そして一度に1bpずつ配列を移動して、異なるフレームを生成する。例えば、23bpのsiRNAが作製されるべきである場合、当業者は、配列の一端において出発し、siRNAの1つの順列として最初の23bpを使用する。次いで、当業者は、配列を1bpを移動し、そして次の23bpが次の順列である、などである。
【0037】
配列データは、実験的に決定されるのであろうとまたは公共データバンクから入手可能であるのであろうと、生命情報科学ツールによってスクリーニングされ得る。相同性分析は、公に入手可能であるプログラムまたは市販のプログラム(例えば、GCGパッケージ中のPILEUPプログラムおよびPRETTYプログラム)を用いて達成される。相同性検索は、PILEUPプログラムおよびPRETTYプログラムまたはFASTAプログラム(Pearsonら,PNAS 85:2444−2448(1988))を使用して、公に入手可能な配列(例えば、Genbank)について必要に応じて実施される。
【0038】
ウイルス遺伝子配列(特に、動物の健康および安全性に影響を及ぼすもの)が本発明の標的遺伝子と意図される。標的として役立つ理想的なウイルス配列は、非常に保存された配列である。保存程度が高いということは一般に、保存された配列から離れて変異しないという選択圧の指標である。多くのウイルスについて、非構造遺伝子は、そのさまざまな血清型を通して非常に保存される傾向があり、従って、RNAi技術の良好な標的である。良好な標的でもある他のウイルス遺伝子は、ウイルスの複製およびその後の増殖にとって重要であるかまたは必須の遺伝子である。一つの態様では、アカバネウイルスの構造(S)遺伝子は、siRNAを作製するために用いられる。
【0039】
他の実施態様では、標的配列は、トリインフルエンザの異なった株を通して保存される短いストレッチの配列である。一つの実施形態では、siRNAのために使用される配列は、トリインフルエンザウイルスのH5N1株のゲノム1に由来する。この非限定的な例は、図9〜図12に示される。太字のヌクレオチドは、相同性分析が行わったときに塩基対の相違が検出された場所を示す。別の実施形態では、siRNAのために使用される標的配列は、トリインフルエンザウイルスのH5N1株のゲノム5由来である。この非限定的な例は、図13〜図16に示される。別の実施形態では、siRNAのために使用される標的配列は、トリインフルエンザウイルスのH5N1株のゲノム7由来である。この非限定的な例は、図1〜図8に示される。他の実施態様において、標的配列は、H5N1の他のゲノム型に由来する。さらに、トリインフルエンザの他の株(例えば、H5N2、H5N3など)に立ち向かうsiRNAもまた、本発明の範囲内に包含され、そして本明細書中の教示に従って、当業者によって作製され得る。
【0040】
別の態様においては、口蹄疫ウイルス(FMDV)の保存されたストレッチが、siRNAを構築するための標的配列として用いられる。なお他の態様では、以下の疾患を引き起こすウイルスの非限定的なリストが、siRNA構築のための標的配列として使用される:ニューカッスル病、西ナイル熱、鶏痘、トリ感染性気管支炎、トリ脳脊髄炎、トリ白血病、アヒルウイルス性肝炎、アヒルウイルス腸炎、ランピースキン病、ウシの感染性鼻気管炎、ウシウイルス下痢、ウシ白血病、リフトバレー熱、牛疫、ブルータング、小反芻獣疫(PPR)、羊痘およびヤギ痘、伝染性膿疱性皮膚炎(膿瘡)、ボーダー疾患、マエディ−ビスナ、伝染性胃腸炎(TGE)、ウマインフルエンザ、アフリカ馬疫、ベネズエラウマ脳脊髄炎およびコイ春ウイルス血症。
【0041】
(siRNAをコードする構築物)
siRNAをコードしている構築物は、プロモーターおよびsiRNAを細胞内の適切な標的へと「導く」配列を含むがこれらに限定されない、いくつかの構成要素を含む。用いられ得るプロモーターとしては、RNAポリメラーゼIIプロモーターおよびRNAポリメラーゼIIIプロモーターが挙げられるがこれに限定されない。RNA pol IIIプロモーターは、siRNAの構築のためのプロモーターとして特によく働く。用いられ得るプロモーター(pol IIおよびpol IIIの両方)の非限定的な例としては、以下が挙げられる:5Sリボソーム(r)RNA、マウスU6、ヒトU6、マウスHI、ヒトHI、サイトメガロウイルスプロモーター、ニワトリユビキチンC、ヒト7SKプロモーター、ウシU6プロモーター、ニワトリU6プロモーター、マレク病ウイルス38kdリン酸化タンパク質(pp38)遺伝子、1.8kbのmRNAのニワトリβ−アクチン、ヒトβ−アクチン、ウシβ−アクチン、ニワトリPRLプロモーター、ニワトリSPATA4遺伝子プロモーター、ニワトリPolIプロモーター、マレク病ウイルスのUS1遺伝子のプロモーター、リボヌクレオチドレダクターゼ遺伝子のマレク病ウイルス小サブユニッ、トリ白血病ウイルスおよび肉腫ウイルスのLTRト、RSV−LTR、合成ポックスウイルスプロモーター、ワクシニアウイルスP1lプロモーター、ワクシニアウイルスP174およびP190、鶏痘初期/後期プロモーターP.E/L、鶏痘ウイルスチミジンキナーゼプロモーター、ワクシニアp7.5プロモーターおよび鶏痘−PFLlプロモーター。
【0042】
標準的な分子生物学の技術を使用して、これらのプロモーターは、ウイルス遺伝子を標的として、それらをサイレンシングする配列に作動可能に連結される。1つのプロモーターは、一つ以上の標的配列の発現を調節し得る。プロモーターおよび標的配列は、標準的なクローニングベクターまたは発現ベクター中にクローニングされる。本明細書中で使用する場合、「ベクター」とは、他の核酸配列を細胞へと送達し得る核酸分子をいう。ベクターは、プラスミド、バクテリオファージ、植物または他の動物ウイルスに由来し得る。
【0043】
siRNA構築物は、siRNAを発現し得るように作製される。当業者は、特定のタイプのプロモーターが、脊椎動物の生理に依存して、一部の脊椎動物において他の脊椎動物においてよりも良好なsiRNAの発現を調節し、そしてその脊椎動物にとって最適であるプロモーターを使用する手段を講じるべきであることを認識している。プロモーターはまた、一つ以上のsiRNAの発現を調節し得る。ベクターは、一つ以上のsiRNAをコードする配列を含み得る。エンハンサー、選択マーカーおよび他の標準的な分子ツールもまた、より簡単な分子操作のために用いられ得る。REMI技術を使用する場合、ベクターは、REMI技術の使用を容易にするように構築されるべきである。その技術についての適切なガイダンスは、WO 99/42569において見出される。
【0044】
本発明の一態様において、ウイルスゲノム上の異なる標的に対応するいくつかのsiRNA遺伝子を有するベクターが用いられることができる。このタイプのベクター構造は、さまざまな部位での同時切断に起因して、有意な阻害からほぼ完全な阻害を確実にする。さらに、たとえウイルスの標的配列のうちの一つが変異または変化した場合であっても、これはまだ活性である。二本鎖siRNAに関しては、標準的な2bp突出を有すること、ならびに1bp突出を有すること、または突出が全く無いことが可能である。
【0045】
(生殖細胞の構築)
本明細書中で使用される場合、「生殖細胞」は、精子および卵細胞ならびにそれらの前駆体と定義される。生殖細胞は半数体であり、1セットの染色体しか有さないが、他の非生殖細胞は、2セットの染色体を有する。本発明は、ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含む構築物を含んでいる非ヒト脊椎動物の生殖細胞の構築を提供し、ここで、この配列はプロモーターに作動可能に連結されている。生殖細胞は、ウイルスゲノムに対する複数のsiRNAをコードしている配列を含む構築物を含み得る。用いられ得るウイルス配列の例は、上記に記載されている。一つの実施形態では、このウイルス配列は、口蹄疫ウイルス(FDMV)である。別の実施形態では、このウイルス配列は、トリインフルエンザウイルスである。
【0046】
本発明は、非ヒト脊椎動物の生殖細胞を作製する方法を提供し、ここで、この生殖細胞は、ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含んでいる構築物を含む。本発明の方法を実施するために、当業者は、保存された領域またはウイルスゲノムに対するsiRNAをコードしている配列を含む構築物とともに生殖細胞を、この構築物が細胞中に取り込まれる条件下でインキュベートし、ここで、この配列は、プロモーターに作動可能に連結されている。一つの実施形態では、この構築物は、REMI技術を使用して宿主細胞ゲノムに組み込まれる。
【0047】
一つの態様では、生殖細胞は、ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含み、ここで、この配列は、プロモーターに作動可能に連結されている。別の実施形態では、非ヒト脊椎動物の生殖細胞は、非ヒト哺乳類である。非ヒト脊椎動物の非限定的な例としては、ニワトリ、アヒル、ガチョウ、家禽、畜牛、ウシ、ブタ、魚、ヒツジ、ヤギおよびウマが挙げられる。別の実施形態では、脊椎動物は、鳥類種のものである。別の態様においては、生殖細胞は、異なるプロモーターの制御下のいくつかの遺伝子を含んでいる1つのベクターを含む。
【0048】
一つの実施形態では、生殖細胞は、口蹄疫ウイルス(FMDV)を標的とするsiRNAをコードしている配列を含む構築物を含む。このような標的配列の例は、以下の通りである:i)3B領域に位置する、nt4900〜4922の5’−CCTGTCGCTTTGAAAGTGAAAGC−3’(配列番号1);(ii)3D領域に位置する、nt6934〜6992の5’−GAGATTCCAAGCTACAGATCACTTTACCTGCGTTGGGTGAACGCCGTGTGCGGTGACGC−3’(配列番号2);および(iii)3D領域に位置する、nt6892〜6917の5’−GACGAGTACCGGCGTCTCTTTGAGCC−3’(配列番号3)。全ての位置を、FMDV血清型O1(G)配列(GenBank登録番号AF189157)についていう。
【0049】
一つの態様では、生殖細胞は、トリインフルエンザを標的とするsiRNAをコードしている配列を含む構築物を含む。この態様の実施形態において、生殖細胞は、保存された配列が図1〜図16に示される配列のうちのいずれかから選択される構築物を含む。
【0050】
別の態様では、生殖細胞は、マレク病ウイルス(MDV)を標的とするsiRNAをコードしている配列を含む構築物を含む。上記のいずれかにおいて、生殖細胞は、精子である。
【0051】
(トランスジェニック動物の構築)
次いで、上記の通りの構築物は、多数の標準技術(例えば、リポフェクション)を用いることにより、動物細胞に導入される。しかしながら、以前の技術(例えば、リポフェクチン)のうちのいくつかは、極めて低い成功率を生じる。発明者らは、WO 99/42569において教示されるとおり、これらのsiRNAコード構築物の成功裏の導入を生じる優れた方法が、制限酵素媒介組込み(「REMI」)を使用することであることを見出した。したがって、一つの実施形態では、siRNAをコードしているポリヌクレオチドは、REMIを使用して動物の生殖細胞(例えば、精子)に導入することである。これらの生殖細胞の使用から得られる動物は、これらの疾患に対して抵抗性にされる。これらのトランスジェニック動物は、ウイルス抵抗性動物のストック全体を作製するために交配され得る。家禽の場合には、siRNAが標的とするウイルスとしては、マレク病、ガンボロおよびトリインフルエンザが挙げられるが、これらに限定されない。
【0052】
動物の細胞(例えば、生殖細胞)に導入されるsiRNAの数は、変化し得る。一つの実施形態では、1つのsiRNA構築物が導入される。他の実施形態では、1つ以上のsiRNAが導入される。当業者は、siRNAの異なる組合せが、標的とされるウイルス性疾患に基づいて導入され得ることを認識している。
【0053】
本発明は、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物の作製を提供し、ここで、この脊椎動物中の大多数の細胞は、この疾患の原因となるウイルスのゲノムの保存された領域に対するsiRNAをコードしている配列を含み、ここで、この配列は、プロモーターに作動可能に連結されている。ウイルス配列の例は、上記で考察されている。一つの実施形態では、この構築物は、ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む。別の実施形態では、これらの細胞は、ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含む。別の実施形態では、この非ヒト脊椎動物は、非ヒト哺乳類である。
【0054】
一つの実施形態では、非ヒト脊椎動物中の大多数の細胞は、口蹄疫ウイルス(FMDV)を標的とするsiRNAをコードしている配列を含む構築物を含む。このような標的配列の例は、以下の通りである:i)3B領域に位置する、nt4900〜4922の5’−CCTGTCGCTTTGAAAGTGAAAGC−3’(配列番号1);(ii)3D領域に位置する、nt6934〜6992の5’−GAGATTCCAAGCTACAGATCACTTTACCTGCGTTGGGTGAACGCCGTGTGCGGTGACGC−3’(配列番号2);および(iii)3D領域に位置する、nt6892〜6917の5’−GACGAGTACCGGCGTCTCTTTGAGCC−3’(配列番号3)。全ての位置を、FMDV血清型O1(G)配列(GenBank登録番号AF189157)についていう。
【0055】
非ヒト脊椎動物の非限定的な例としては、ニワトリ、アヒル、ガチョウ、家禽、畜牛、ウシ、ブタ、魚、ヒツジ、ヤギおよびウマが挙げられる。別の実施形態では、脊椎動物は、鳥類種のものである。さらに別の実施形態では、非ヒト脊椎動物は、保存された配列が、図1〜図16において示される配列のうちの1つから選択されるsiRNAをコードしている構築物を含んでいる細胞を有する。別の実施形態では、非ヒト脊椎動物は、MDVに対して抵抗性である。
【0056】
本発明は、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物を作製するための方法を提供し、この方法は、以下の工程を包含する:(a)上記の生殖細胞を、この生殖細胞が二倍体細胞を形成する条件下でインキュベートする工程;および(b)(a)に記載の二倍体細胞を、この二倍体細胞が非ヒト脊椎動物を形成する条件下でインキュベートする工程。
【0057】
本発明はまた、クローン化による、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物の作製を意図する。そのような場合、一つ以上のsiRNAは、クローン化されるべき細胞のゲノムに挿入される。この技術のさまざまな方法は、市販される(例えば、Advanced Cell Technologyの核移植プロトコル)。
【0058】
(適用)
本発明は、ウイルス性疾患に対して抵抗性である家禽を作製するための方法を本願明細書において提供する。これらの疾患としては、ニューカッスル病、西ナイル熱、鶏痘、鳥伝染性気管支炎、トリ脳脊髄炎、トリ白血病、アヒルウイルス性肝炎およびアヒルウイルス腸炎が挙げられるが、これらに限定されない。
【0059】
本発明は、ウイルス性疾患に対して抵抗性であるウシを作製するための方法を本願明細書において提供する。これらの疾患としては、ランピースキン病、ウシの感染性鼻気管炎、ウシウイルス下痢、ウシ白血病、リフトバレー熱、牛疫およびブルータングが挙げられるが、これらに限定されない。
【0060】
本発明は、ウイルス性疾患に対して抵抗性であるヒツジおよびヤギを作製するための方法を本願明細書において提供する。これらの疾患としては、小反芻獣疫(PPR)、羊痘およびヤギ痘、伝染性膿疱性皮膚炎(膿瘡)、ボーダー病、ブルータング、マエディ−ビスナおよびリフトバレー熱に接しているが挙げられるが、これらに限定されない。
【0061】
本発明は、ウイルス性疾患に対して抵抗性であるウマを作製するための方法を本願明細書において提供する。これらの疾患としては、ウマインフルエンザ、アフリカ馬疫およびベネズエラウマ脳脊髄炎が挙げられるが、これらに限定されない。
【0062】
本発明は、ウイルス性疾患に対して抵抗性である魚を作製するための方法を本願明細書において提供する。これらの疾患としては、コイ春ウイルス血症が挙げられるが、これに限定されない。
【0063】
本発明は、古典的ブタコレラ病、アフリカブタコレラ病、伝染性胃腸炎(TGE)および口蹄病に対する抵抗性を生じるために、ブタに適用され得る。一般に、本願明細書における教示は、ウイルス性疾患に感染しやすい任意の動物に対して適用され得る。
【0064】
病気による大きな経済的損失を受けることに耐えている大規模養鶏産業の場合、実用的な解決案は、トリインフルエンザおよびマレク病、伝染性ファブリキウス嚢病および他のトリウイルスに対して抵抗性である若鶏および産卵鶏の種畜を作製するために、siRNA技術の実施にある。本願明細書において開示される本発明の使用は、安全な家禽供給源を消費者に提供し、そして、鳥から人間へのトリウイルスの広がりとしての一般集団が止まる。
【0065】
以下は、本発明を例示するために提供され、本発明を限定するために提供されるわけではない。
【実施例】
【0066】
(実施例1 口蹄疫に対して抵抗性の動物の作製)
口蹄病に感染しやすいトランスジェニック動物を、口蹄疫ウイルス(FMDV)のウイルス配列から構築されるsiRNAを動物の生殖細胞に挿入することによって作製する。一般に、FMDVに感染しやすい動物は、偶蹄動物である。
【0067】
(口蹄疫siRNAの構築)
siRNAを構築するために、標的ウイルス配列を最初に同定する。これが達成され得る一つの方法は、異なる株または血清型からのFMDV配列について局所的相同分析を実行し、そして配列相同性の短いストレッチを同定することによる。一つの実施形態では、この相同性は、100%(すなわち、すべての血清型にわたって正確に同じ配列)である。他の実施形態では、1塩基対または2塩基対の相違がみられる。相同分析は、公に入手可能なプログラムまたは市販のプログラム(例えば、GCGパッケージ中のPILEUPプログラムおよびPRETTYプログラム)を用いて達成される。相同性検索は、PILEUPプログラムおよびPRETTYプログラムまたはFASTAプログラム(Pearsonら,PNAS 85:2444−2448(1988))を使用して、公に入手可能な配列(例えば、Genbank)について必要に応じて実施される。
【0068】
一旦短い相同性ストレッチが同定されると、siRNAが、商業的に利用可能なサービスを利用して合成される。siRNAは、種々の長さで合成される。いくつかのsiRNAは、長さが19bpであり、一方で、他のものは、長さが20bp、21bp、22bp、23bp、24bp、25bp、26bp、または27bpである。これらのsiRNAの各々は、血清型をとおして100%保存されるか、または各siRNA中に1bp〜2bpの相違を有する。これらの保存された配列は、生殖細胞への送達に適しているベクター中の少なくとも一つのプロモーターに作動可能に連結されている。別の代替法は、すでに合成されたsiRNAを使用することである。例えば、Kahanaら,J.Gen.Virol.85:3213−3217(2004)において開示されるsiRNAを参照のこと。
【0069】
次いで、これらのsiRNA構築物を、WO 99/42569において開示されるREMI技術を使用して偶蹄動物の生殖細胞(例えば、精子または卵子)に挿入し、安定して組み込まれたsiRNAを有する生殖細胞が得られる。次いで、生殖細胞を用いて、当該分野で公知の方法(例えば、体外受精、人工授精)によってトランスジェニック動物が得られ、口蹄疫に対して抵抗性である動物が得られる。
【0070】
(実施例2 アカバネウイルスについてのsiRNAを含む精子の作製)
非ヒト脊椎動物からの精子系統は、アカバネウイルスの保存された領域に対するsiRNAをコードしている配列を含む構築物とともに精子を、この構築物を細胞に取り込ませる条件下でインキュベートすることによって作製される(ここで、この配列は、プロモーターに作動可能に連結されている)。非限定的な例としては、リポフェクチンまたは類似の機構、塩化カルシウムまたはプロタミンを使用することが挙げられる。代替法では、この構築物は、REMI技術を使用して、宿主細胞ゲノムに安定して組み込まれる。siRNAは、2つの基準に従って設計される:異なる単離株間で高い相同性を共有している領域、およびsiRNA標的発見プログラムによって決定した場合のサイレンシング活性の高い類似。保存された配列を標的とするsiRNA分子の使用は、これらの分子が、それらの起源に関係なく、同じ株のすべてのウイルスでなかったとしてもほとんどのウイルスを切断する能力を確実にしなければならない。第2の、そしてより重要な理由は、配列の保存が、変化に対して強い選択圧を示すということである。選択圧は、これらの標的領域を不変に保つと期待されるが、それらが変化する場合、それらは、siRNA分子の抗ウイルス活性を抑制し得る。
【0071】
(実施例3 口蹄疫ウイルスについてのsiRNAを含む精子の作製)
非ヒト脊椎動物からの精子系統は、口蹄疫ウイルスの保存された領域に対するsiRNAをコードしている配列を含む構築物とともに精子を、この構築物を細胞に取り込ませる条件下でインキュベートすることによって作製される(ここで、この配列は、プロモーターに作動可能に連結されている)。非限定的な例としては、リポフェクチンまたは類似の機構、塩化カルシウムまたはプロタミンを使用することが挙げられる。代替法では、この構築物は、REMI技術を使用して、宿主細胞ゲノムに安定して組み込まれる。
【0072】
(実施例4 トリインフルエンザウイルスについてのsiRNAを含む精子の作製)
非ヒト脊椎動物からの精子系統は、トリインフルエンザウイルスの保存された領域に対するsiRNAをコードしている配列を含む構築物とともに精子を、この構築物を細胞に取り込ませる条件下でインキュベートすることによって作製される(ここで、この配列は、プロモーターに作動可能に連結されている)。非限定的な例としては、リポフェクチンまたは類似の機構、塩化カルシウムまたはプロタミンを使用することが挙げられる。代替法では、この構築物は、REMI技術を使用して、宿主細胞ゲノムに安定して組み込まれる。
【技術分野】
【0001】
(関連出願の引用)
この引用は、2005年3月31日に出願した米国仮出願第60/666,636号に対する優先権を主張する。米国仮出願第60/666,636号は、すべての目的のために、本明細書中に参考として援用される。
【0002】
(発明の分野)
本発明は、ウイルス感染に対して抵抗性の動物を作製するためのRNA干渉技術の使用に関する。
【背景技術】
【0003】
(発明の背景)
疾病管理センター(Centers for Disease Control;CDC)および世界保健機構は、致死率の高いトリインフルエンザウイルスH5N1株が、少なくとも3つの大陸にまたがって広がって、47カ国を超える国々で報告されたと報告した。非特許文献1を参照のこと。トリインフルエンザは、インフルエンザ(フルー)ウイルスによって引き起こされる感染症である。これらのインフルエンザウイルスは、鳥の中で自然に発生する。野鳥は、それらの腸野中で世界中にウイルスを運び、そして通常は無症候性である。しかしながら、トリインフルエンザは、鳥の間では非常に接触感染性であり、一部の家畜化された鳥(ニワトリ、アヒルおよびシチメンチョウを含む)を非常に病んだ状態にし得、そして死を引き起こし得る。
【0004】
感染した鳥は、それらの唾液、鼻の分泌物および糞中にインフルエンザウイルスを脱落する。感染しやすい鳥が、汚染された分泌物もしくは排出物と、または感染した鳥からの分泌物もしくは排出物によって汚染された表面と接触した場合、この感染しやすい鳥は感染されることになる。家畜化された鳥は、感染した水鳥もしくは他の感染した家禽との直接接触によって、またはトリインフルエンザウイルスによって汚染された表面(例えばほこりまたはケージ)もしくは材料(例えば、水または餌)との接触によって、トリインフルエンザウイルスに感染されることになり得る。
【0005】
家禽におけるトリインフルエンザウイルスによる感染によって、極端に低い毒性および極端に高い毒性によって区別される2つの主要な型の疾患が引き起こされる。「病原性が低い」形態は非検出となり得、そして通常、軽度の症状(例えば、乱れた羽毛および産卵の低下)のみを引き起こす。しかしながら、非常に病原性が高い形態は、家禽の群れによって、より急速に広がる。この形態は、複数の内臓に影響を及ぼして、48時間以内にしばしば90〜100%に達し得る死亡率を有する疾患を引き起こし得る。従って、トリインフルエンザに抵抗性を有する家禽を作製する方法が存在すれば、家禽飼育場での巨大な経済的損失の可能性が激減し得る。これらのウイルス抵抗性家禽は、代々引き続いてそのウイルス抵抗性を世代間で受け渡す新種の家禽を作製するために交配され得る。
【0006】
家禽の世界市場は、1年あたり500億羽を超える家禽を生産すると推定される。それゆえ、さらにより重要なことには、ウイルス抵抗性動物(例えば、家禽)を作製する能力は、恐れられる「トリインフルエンザ」ウイルスのヒトへの広がりを防止し得、それゆえ、ヒトにおけるトリインフルエンザの汎発流行を回避し得る。
【0007】
家禽産業にとってまた問題がある、他のタイプのウイルス性疾患が存在する。例えば、マレク病は、しばしば若いめん鳥の群れの重篤な死亡損失を引き起こし、ブロイラ加工工場の不良認定の主な原因であった。マレク病は、ヘルペスウイルスファミリーに属するMDウイルス(MDV)によって引き起こされるリンパ増殖性疾患であり、そして過去50年間にわたって家禽産業にとって大きな課題である。3つの血清型のMDV株が認識されている。腫瘍性株はすべて、血清型1(MDV−I)と分類され、一方、元々非腫瘍性のニワトリ株および七面鳥のヘルペスウイルス(HVT)はそれぞれ、血清型2(MDV−2)および血清型3に属する。世界全体でのマレク病に起因する毎年の経済的損失は、大きなワクチン接種キャンペーンにもかかわらず、毎年10億アメリカドルの範囲であると推定される。ワクチン接種キャンペーンは世界全体での罹患した群れの数を減らしたにもかかわらず、多数の疾患発生が世界の大部分の部分に存在し、このことは、この疾患を制御する、より良好な方法を必要とする。
【0008】
成功裏に処置され得るウイルス性疾患は、限られた数しか存在しない。現在、ワクチン接種は、動物におけるウイルス性疾患と戦うために用いられる唯一の方法である。多くの場合、特に家畜のウイルス性疾患においては、このアプローチは、効力の高いワクチンを生産することができないことに起因して適用できない。いくつかのワクチンが無効である別の理由は、抗原連続変異またはワクチン効果の喪失をもたらす、高い変異率である。一旦ワクチンがその効果を失うと、この疾患が出現し得、そして家禽および家畜のウイルス性疾患の場合、大きな経済的損失を負わせ得る。
【0009】
2つの古典的形態のワクチンが存在する;1つ目のものは、弱毒生ワクチンであり、こ
れは、免疫系の体液性免疫および細胞性免疫の両方を誘導するので、これは最も強力なものである。この種のワクチンの欠点は、非常に毒性が高い逸脱変異の再出現の危険性である。この理由のために、多くの国は、この種のワクチンの使用を承認するのを渋る。第2のタイプは、不活化ウイルスワクチンであり、これは体液性免疫系しか誘発しないので、これは多くの場合、それほど有効ではない。過去数年にわたって、サブユニットワクチンおよびDNAワクチンのようないくつかタイプの組換えワクチンが開発された。残念なことに、それらのほとんどは、幅広くは使用されていない。マレク病の場合に示されるように、ワクチン接種の欠点のうちの1つは、高い年間の経済的な支出である。World Organization for Animal Health(OIE)によると、2002年にマレク病に対するワクチン接種を受けた動物の総数は、24億5700万であった。例えば、非特許文献2を参照のこと。
【0010】
ワクチン接種をするには、2つの公知の方法が存在する。第1の方法は、約$3/1000ユニットの費用がかかる、無細胞(凍結乾燥)形態である。第2の形態は、約$8/1000ユニットの費用がかかる、細胞関連の(「湿った」)形態である。2002年のマレク病のためのワクチン接種について見積もられた世界支出は、約74億ドル〜197億ドルの間にあった。
【0011】
1970年代初頭から、大部分のニワトリは、弱毒化血清型1株またはHVTを使用して、MDに対してワクチン接種を受けた。1983年に始まった、二価の組合せおよび多価の組合せが、より毒性の実地での株から保護するために用いられている。ワクチン接種はニワトリを保護する際に著しく有効であるにもかかわらず、MDは疾患の優先順位のリストの中で順位が高いままである。優先順位が高い主要な理由は、ワクチンの破壊を引き起こす毒性の増加に向かってMDV株が継続して進化していることである。
【0012】
したがって、例えば、動物におけるウイルス性疾患(例えば、家禽におけるトリインフルエンザおよびマレク病)の伝染を防止するかまたは低下させる方法の必要が存在する。トリインフルエンザウイルスの現在の発生を考慮すると、ウイルス感染(例えば、トリインフルエンザ)に対して抵抗性である、遺伝子が改変された動物(例えば、家禽)を作製し得る方法についての強い必要性が存在する。
【0013】
過去10年において、「RNA干渉」(RNAi)と称される、遺伝子サイレンシングを経る遺伝子不活化の新規なアプローチが開示された。例えば、非特許文献3および特許文献1を参照のこと。RNA干渉とは、RNAポリヌクレオチドが内因性細胞プロセスを通して作用する場合に起こる、RNAの配列に対応する配列の遺伝子発現を特異的に抑制する事象をいう。標的遺伝子のサイレンシングは、時々はRNAse IIIエンドヌクレアーゼ消化による、宿主動物による二本鎖(ds)RNAによるmRNAの分解の際に起こる。この消化は、分子のサイズは30塩基程度の大きさであり得るにもかかわらず、長さ(またはサイズ)が約21〜23ヌクレオチド(または塩基)である分子をもたらす。
【0014】
これらの短いRNA種は、おそらく、RNA誘導性サイレンシング複合体(RISC)と呼ばれるRNAiヌクレアーゼ複合体を経て、対応するRNAメッセージおよび転写産物の分解を媒介する。RISCは、小さいdsRNAが塩基対合相互作用によって相補的なmRNAを認識するのを助ける。その基質とのsiRNA相互作用後に、おそらくRISC中に存在する酵素によって、mRNAは、分解についての標的とされる。このタイプの機構は、ウイルス感染、トランスポゾンジャンピングおよび類似の現象を抑制する際に生物体にとって、ならび内因性遺伝子の発現を制御するために、有用なようである。RNAi活性はこれまでのところ、他の生物体の中でも、植物、昆虫、ネマトーダおよび脊椎動物において実証されている。一般的な背景情報については、非特許文献4;非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;非特許文献10;非特許文献11;非特許文献12;非特許文献13;非特許文献14;および非特許文献15を参照のこと。
【先行技術文献】
【特許文献】
【0015】
【特許文献1】米国特許第6,506,559号明細書
【非特許文献】
【0016】
【非特許文献1】インターネット<URL:http://www.cdc.gov/flu/avian/outbreaks/current.htm>
【非特許文献2】インターネット<URL:www.oie.com>
【非特許文献3】Fireら,Nature 391:806−811(1998)
【非特許文献4】Schutzら,Virology 344(1):151−7(2006)
【非特許文献5】Leonardら,Gene Ther.13(6):532−40(2006)
【非特許文献6】Colbere−Garapinら,Microbes Infect.7(4):767−75(2005)
【非特許文献7】Wall,Theriogenology 57(1):189−201(2002)
【非特許文献8】El−Bashirら,Nature 411:494−498(2001)
【非特許文献9】Fire,A.ら,Science 391:806−811(1998)
【非特許文献10】Gitlinら,Nature 418:430−434(2002)
【非特許文献11】Gitlinら,J.Virol.79:1027−1035(2005)
【非特許文献12】Kahanaら,J.Gen.Virol.85,3213−3217(2004)
【非特許文献13】Kronkeら,J.Virol.78:3436−3446(2004)
【非特許文献14】Leonardら,J.Virol.79:1645−1654(2005)
【非特許文献15】Yokotaら,EMBO Rep.4:602−608(2003)
【発明の概要】
【発明が解決しようとする課題】
【0017】
動物(特に家禽)における様々なウイルス性疾患の広範囲にわたる課題を考慮すると、家禽および家畜のウイルス性疾患を効果的に制御するための様々なアプローチは、この課題を解決するために非常に有益である。そのため、本願明細書において開示される本発明は、病原性ウイルスのウイルス複製を著しく阻害する、重要なウイルス機能をブロックすることを標的とする分子を保有および発現するトランスジェニック非ヒト脊椎動物(例えば、家禽および家畜)を作製するための方法、ならびに遺伝子改変された家禽および家畜自体を提供することによって、この課題に対する解決策を提供する。
【課題を解決するための手段】
【0018】
(発明の簡単な説明)
本発明は、ウイルス配列標的とするsiRNAをコードする生殖細胞およびこの生殖細胞を得る方法を提供する。本発明はまた、ウイルス感染に対して抵抗性である非ヒト脊椎動物およびこの動物を得る方法を提供する。
【0019】
一つの態様では、本発明は、ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含む構築物を含んでいる、非ヒト脊椎動物の生殖細胞であり、ここで、この配列はプロモーターに作動可能に連結されている。一つの実施形態では、この構築物は、このウイルスゲノムに対する複数のsiRNAをコードしている配列を含む。別の実施形態では、この細胞は、ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含み、ここで、この配列は、プロモーターに作動可能に連結されている。別の実施形態では、この脊椎動物は、非ヒト哺乳類である。別の実施形態では、このウイルスは、口蹄疫ウイルス(FMDV)である。別の実施形態では、保存された配列は、配列番号1、配列番号2および配列番号3からなる群から選択される。別の実施形態では、この脊椎動物は、鳥類種のものである。別の実施形態では、このウイルスは、トリインフルエンザウイルスである。別の実施形態では、保存された配列は、図1〜図16中の配列からなる群から選択される。他の実施形態では、このウイルスは、マレク病ウイルス(MDV)である。別の実施形態では、この生殖細胞は、精子である。
【0020】
別の態様においては、本発明は、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物を提供し、ここで、この脊椎動物中の大多数の細胞は、この疾患の原因となるウイルスのゲノムの保存された領域に対するsiRNAをコードしている配列を含み、ここで、この配列はプロモーターに作動可能に連結されている。一つの実施形態では、この構築物は、このウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む。別の実施形態では、この細胞は、このウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含む。別の実施形態では、この脊椎動物は、非ヒト哺乳類である。別の実施形態では、このウイルス性疾患は、FMDVである。別の実施形態では、保存された配列は、配列番号1、配列番号2および配列番号3からなる群から選択される。別の実施形態では、この脊椎動物は、鳥類種のものである。別の実施形態では、このウイルスは、トリインフルエンザウイルスである。別の実施形態では、保存された配列は、図1〜図16中の配列からなる群から選択される。別の実施形態では、このウイルスは、MDVである。
【0021】
別の態様においては、本発明は、非ヒト脊椎動物の生殖細胞を作製する方法を提供し、この方法は、保存された領域またはウイルスゲノムに対するsiRNAをコードしている配列を含む構築物とこの生殖細胞とを、この構築物をこの細胞に取り込ませる条件下でインキュベートする工程を包含し、ここでこの配列は、プロモーターに作動可能に連結されている。一つの実施形態では、この構築物は、宿主細胞ゲノムに組み込まれる。
【0022】
別の態様においては、本発明は、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物を作製するための方法を提供し、この方法は、(a)保存された領域またはウイルスゲノムに対するsiRNAをコードしている配列を含む構築物と生殖細胞とを、この生殖細胞が二倍体細胞を形成する条件下でインキュベートする工程であって、ここでこの配列は、プロモーターに作動可能に連結されている、工程;および(b)(a)に記載の二倍体細胞を、この二倍体細胞が非ヒト脊椎動物を形成する条件下でインキュベートする工程を包含する。
例えば、本願発明は以下の項目を提供する。
(項目1)
ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含む構築物を含んでいる、非ヒト脊椎動物生殖細胞であって、該配列がプロモーターに作動可能に連結されている、生殖細胞。
(項目2)
前記構築物が、前記ウイルスゲノムに対する複数のsiRNAをコードしている配列を含む、項目1に記載の生殖細胞。
(項目3)
前記細胞が、前記ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含んでおり、ここで、該配列はプロモーターに作動可能に連結されている、項目1に記載の生殖細胞。
(項目4)
前記脊椎動物が非ヒト哺乳類である、項目1、または項目2、または項目3に記載の生殖細胞。
(項目5)
前記ウイルスが口蹄疫ウイルス(FMDV)である、項目4に記載の生殖細胞。
(項目6)
保存された配列が配列番号1、配列番号2および配列番号3からなる群より選択される、項目5に記載の生殖細胞。
(項目7)
前記脊椎動物が鳥類種のものである、項目1または項目2または項目3に記載の生殖細胞。
(項目8)
前記ウイルスがトリインフルエンザウイルスである、項目7に記載の生殖細胞。
(項目9)
保存された配列が図1〜図16中の配列からなる群より選択される、項目8に記載の生殖細胞。
(項目10)
前記ウイルスがマレク病ウイルス(MDV)である、項目7に記載の生殖細胞。
(項目11)
前記生殖細胞が精子である、項目1または項目2または項目3または項目4または項目5または項目6または項目7または項目8または項目9または項目10に記載の生殖細胞。
(項目12)
ウイルス性疾患に対して抵抗性である非ヒト脊椎動物であって、ここで、該脊椎動物中の大多数の細胞は、該疾患の原因となるウイルスのゲノムの保存された領域に対するsiRNAをコードしている配列を含み、該配列はプロモーターに作動可能に連結されている、非ヒト脊椎動物。
(項目13)
前記構築物が、前記ウイルスのゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む、項目12に記載の非ヒト脊椎動物。
(項目14)
前記細胞が、前記ウイルスのゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含む、項目12に記載の非ヒト脊椎動物。
(項目15)
前記脊椎動物が非ヒト哺乳類である、項目12または項目13または項目14に記載の非ヒト脊椎動物。
(項目16)
前記ウイルス性疾患がFMDVである、項目15に記載の非ヒト脊椎動物。
(項目17)
保存された配列が配列番号1、配列番号2および配列番号3からなる群より選択される、項目16に記載の非ヒト脊椎動物。
(項目18)
前記脊椎動物が鳥類種のものである、項目12または項目13または項目14に記載の非ヒト脊椎動物。
(項目19)
前記ウイルスがトリインフルエンザウイルスである、項目18に記載の非ヒト脊椎動物。
(項目20)
保存された配列が図1〜図16中の配列からなる群より選択される、項目19に記載の非ヒト脊椎動物。
(項目21)
前記ウイルスがMDVである、項目12に記載の非ヒト脊椎動物。
(項目22)
非ヒト脊椎動物の生殖細胞を作製する方法であって、該生殖細胞が項目1に記載の生殖細胞であり、該方法は、保存された領域またはウイルスゲノムに対するsiRNAをコードしている配列を含む構築物と該生殖細胞とを、該構築物を該細胞に取り込ませる条件下でインキュベートする工程を包含し、ここで該配列は、プロモーターに作動可能に連結されている、方法。
(項目23)
前記構築物が宿主細胞ゲノム中に組み込まれる、項目22に記載の方法。
(項目24)
項目12に記載の非ヒト脊椎動物を作製するための方法であって、
(a)項目1または項目2または項目3または項目4または項目5または項目6または項目7または項目8または項目9または項目10または項目11に記載の生殖細胞を、該生殖細胞が二倍体細胞を形成する条件下でインキュベートする工程;および
(b)(a)に記載の二倍体細胞を、該二倍体細胞が非ヒト脊椎動物を形成する条件下でインキュベートする工程
を包含する、方法。
【図面の簡単な説明】
【0023】
【図1−1】図1−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−2】図1−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−3】図1−3は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−4】図1−4は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−5】図1−5は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図1−6】図1−6は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図2−1】図2−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図2−2】図2−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図3−1】図3−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図3−2】図3−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図4−1】図4−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図4−2】図4−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図4−3】図4−3は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図5−1】図5−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図5−2】図5−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図6−1】図6−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図6−2】図6−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図7】図7は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図8−1】図8−1は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図8−2】図8−2は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図8−3】図8−3は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図8−4】図8−4は、トリインフルエンザH5N1株のゲノム7について用いられ得るトリインフルエンザH5N1配列を示す。
【図9−1】図9−1は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図9−2】図9−2は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図10】図10は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図11】図11は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図12】図12は、トリインフルエンザH5N1株のゲノム1について用いられ得るトリインフルエンザH5N1配列を示す。
【図13−1】図13−1は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図13−2】図13−2は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図14】図14は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図15】図15は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図16−1】図16−1は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図16−2】図16−2は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【図16−3】図16−3は、トリインフルエンザH5N1株のゲノム5について用いられ得るトリインフルエンザH5N1配列を示す。
【発明を実施するための形態】
【0024】
(発明の詳細な説明)
本発明は、ウイルス感染に対して抵抗性である動物およびこのようなタイプのトランスジェニック動物を作製する方法を提供する。標的ウイルス遺伝子は、サイレンシング分子(例えば、本願明細書において開示されるsiRNA分子)の存在によってサイレンシングされるので、ウイルスの生活環においてそれらの役割を果たすことができない。このプロセスの結果は、これらの動物において羅病率または/および死亡率を誘導する感染をすることができないこととなる、ウイルス複製の有意なブロックである。それゆえ、本発明は、肉および他の畜産物の絶え間ない供給を確実にし、全体として、農民にとって、および社会にとって、相当な経済効果を有する。
【0025】
本発明の実施は、特に明記しない限り、分子生物学(組換え技術を含む)、微生物学、細胞生物学、生化学および免疫学の従来の技術を使用する。これらは当業者の技術範囲内にある。このような技術は、例えば、Molecular Cloning:A Laboratory Manual,第2版(Sambrookら,1989)Cold Spring Harbor Press;Oligonucleotide Synthesis(MJ.Gait編,1984);Animal Cell Culture(R.I.Freshney編,1987);Methods in Enzymology(Academic Press,Inc.);Handbook of Experimental Immunology(D.M.Weir & C.C.Blackwell編);Gene Transfer Vectors for Mammalian Cells(J.M.Miller & M.P.Calos編,1987);Current Protocols in Molecular Biology(F.M.Ausubelら編,1987);PCR:The Polymerase Chain Reaction,(Mullisら編,1994);Current Protocols in Immunology(J.E.Coliganら編,1991)およびShort Protocols in Molecular Biology(Wiley and Sons,1999)のような文献において充分に説明されている。
【0026】
この特許出願において引用されるすべての参考文献、特許および特許出願は、それぞれがすべての目的のためにその全体が本明細書中に参考として援用される。
【0027】
(定義)
本出願において使用されるすべての科学用語および専門用語は、他に特定されない限り、当該分野において一般に使用される意味を有する。本出願において用いられる場合、以下の語または語句は、指定した意味を有する。
【0028】
交換可能に本明細書中で使用される用語「ポリヌクレオチド」および「核酸」とは、リボヌクレオチドまたはデオキシリボヌクレオチドのいずれであってもよい、任意の長さのポリマー形態のヌクレオチドをいう。これらの用語は、一本鎖、二本鎖または三本鎖の、DNA、ゲノムDNA、cDNA、RNA、DNA−RNAハイブリッド、またはプリン塩基およびピリミジン塩基を含むポリマー、または天然のものであるか、化学的に改変されているか、生化学的に改変されているか、非天然のものであるか、もしくは誘導体化されているヌクレオチド塩基を包含する。
【0029】
「プロモーター」は、転写開始および転写速度が制御されるポリヌクレオチド配列領域である制御配列である。プロモーターは、調節タンパク質および調節分子(例えば、RNAポリメラーゼおよび他の転写因子)が結合し得る遺伝要素を含み得る。プロモーターは例えば、構成性であってもよく、誘導性であってもよく、抑制性であってもよく、または組織特異的であってもよい。語句「作動可能に連結された」、「作動するように配置された」、「作動するように連結された」、「制御下にある」および「転写制御下にある」とは、プロモーターがその配列の転写開始および/または発現を制御するために核酸配列に関して適切な機能的な位置および/または方向にあることを意味する。プロモーターは、「エンハンサー」とともに用いられてもよく、そうでなくてもよい。エンハンサーとは、核酸配列の転写活性化に関与するシス作用性調節配列をいう。1つのプロモーターは、一つ以上の遺伝子の発現を調節し得る。
【0030】
(RNAi標的)
本発明は、ウイルス抵抗性動物が作製され得るように遺伝子をサイレンシングするための方法を提供する。一つの実施形態では、本発明は、免疫トランスジェニックの家禽および家畜を内部的に作製するリポフェクションに関連した遺伝子サイレンシング技術(例えば、RNAi)を使用する。好ましい実施形態では、本発明は、ウイルスに抵抗性の動物を作製するために、例えば、WO 99/42569において開示されるとおりの制限酵素媒介組込み(「REMI」)を使用する。これらの動物は、ウイルス遺伝子(例えば、複製に欠くことのできないウイルス遺伝子)を標的とする1つまたは複数のサイレンシング分子を保有および発現する。
【0031】
本発明によって提供される短い干渉RNA(「siRNA」)は、標的遺伝子が細胞内に存在して発現し得る場合に、標的遺伝子の発現の調節および/または減衰を可能にする。発現調節は、遺伝子機能の部分的阻害であってもよく、より好ましくは、遺伝子機能の完全阻害であるか、または他の第2の標的遺伝子のアップレギュレーションもしくは一次標的遺伝子の阻害に応答するこのような遺伝子の発現の強化ですらあり得る。
【0032】
遺伝子発現の減衰は、遺伝子機能、転写産物のプロセシングまたは転写産物の翻訳の部分的または完全な抑制または阻害を含み得る。RNA干渉の情況では、遺伝子発現の調節は、タンパク質とRNA(特に、「ガイド」RNAとして作用し得る小さいdsRNA)との複合体を通して進行すると考えられている。それゆえ、siRNAは、そのヌクレオチド配列が標的遺伝子のヌクレオチド配列の少なくとも一部分と十分に一致する場合、効果的であると考えられる。本発明はこのメカニズムの仮説によって束縛されないが、siRNAにおけるヌクレオチド配列が、標的遺伝子配列の少なくとも一部分と実質的に同じであることが非常に好ましい。一つの実施形態では、標的ウイルスヌクレオチドとsiRNAとの間の配列同一性は、100%(すなわち、正確に相同)である。別の実施形態では、1塩基対(bp)の相違が存在する。別の実施形態では、2塩基対の相違が存在する。別の実施形態では、3塩基対の相違が存在する。
【0033】
「標的遺伝子」または「標的配列」は一般に、遺伝子産物(例えば、ポリペチド)をコードする領域を含むポリヌクレオチド、または複製、転写もしくは翻訳、もしくはポリペチドの発現にとって重要な他のプロセスを調節するポリヌクレオチド領域、または、ポリペチドをコードする領域およびそれに対して作動可能に連結された発現調節領域を含むポリヌクレオチドを意味する。標的配列は、遺伝子産物全体をコードする必要が必ずしもあるわけではない。本発明の情況では、「作動可能に連結される」とは、その配列の開始および/または発現を制御するためにポリヌクレオチド配列に関して適切な機能的位置および/または方向にあるプロモーターをいう。
【0034】
一つの態様では、siRNAについての標的遺伝子は、ウイルス遺伝子である。一つの実施形態では、ウイルス遺伝子は、鳥類のウイルス性疾患(例えば、トリインフルエンザおよびマレク病)の伝播に関与する遺伝子である。本発明の範囲内に含まれる遺伝子配列としては、図1〜図16に示される配列が挙げられる。
【0035】
siRNAは、ウイルス複製を有意にブロックするが、細胞崩壊を引き起こさないのに十分な長さのものであり、そして通常、約19塩基対〜約27塩基対の間の範囲にある。一つの実施形態では、siRNAは、長さが21bpである。別の実施形態では、siRNAは、長さが22bpである。なお他の実施形態では、siRNAは、長さが約19bp、約20bp、約23bp、約24bp、約25bp、約26bpまたは約27bpである。なお別の実施形態では、siRNAは、長さが約28bp、約29bpまたは約30bpである。siRNAが存在する細胞にとっての一種の自己破壊機構を誘発するので、siRNAの長さを長くしすぎることはできないことが、当業者にとって明らかである。例えば、Elbashirらは、長さ21bpという長さが、細胞の抗ウイルス機構(例えば、インターフェロン応答)を回避するために効果的であることを教示する(Elbashirら,Nature 411:494−498(2001)を参照のこと)。
【0036】
図1〜図16に示すように、種々の長さが、干渉RNAとして使われ得る。1つの好ましい実施形態は、長さが21bpである。当業者は、図1〜図16において示される配列を使用し得、そして異なる21bpのsiRNAに達するために、一度に1塩基対ずつ、段階的に移動し得る。順列のうちのいくつかは、これらの図に示される。しかしながら、図1〜図16において示されるように、いったん一般的な配列が示されると、配列を段階的に移動することは当業者にとっては慣用的であるので、配列が示される可能な21bpのsiRNAのすべてが示されたわけではない。さらに、21bp以外の長さもまた、本発明に包含される。上記で議論したように、一つの実施形態では、siRNAは、長さが約21bp〜約27bpである。当業者は、図1〜図16において開示される配列を使用し、そして所望の長さを使用し、そして一度に1bpずつ配列を移動して、異なるフレームを生成する。例えば、23bpのsiRNAが作製されるべきである場合、当業者は、配列の一端において出発し、siRNAの1つの順列として最初の23bpを使用する。次いで、当業者は、配列を1bpを移動し、そして次の23bpが次の順列である、などである。
【0037】
配列データは、実験的に決定されるのであろうとまたは公共データバンクから入手可能であるのであろうと、生命情報科学ツールによってスクリーニングされ得る。相同性分析は、公に入手可能であるプログラムまたは市販のプログラム(例えば、GCGパッケージ中のPILEUPプログラムおよびPRETTYプログラム)を用いて達成される。相同性検索は、PILEUPプログラムおよびPRETTYプログラムまたはFASTAプログラム(Pearsonら,PNAS 85:2444−2448(1988))を使用して、公に入手可能な配列(例えば、Genbank)について必要に応じて実施される。
【0038】
ウイルス遺伝子配列(特に、動物の健康および安全性に影響を及ぼすもの)が本発明の標的遺伝子と意図される。標的として役立つ理想的なウイルス配列は、非常に保存された配列である。保存程度が高いということは一般に、保存された配列から離れて変異しないという選択圧の指標である。多くのウイルスについて、非構造遺伝子は、そのさまざまな血清型を通して非常に保存される傾向があり、従って、RNAi技術の良好な標的である。良好な標的でもある他のウイルス遺伝子は、ウイルスの複製およびその後の増殖にとって重要であるかまたは必須の遺伝子である。一つの態様では、アカバネウイルスの構造(S)遺伝子は、siRNAを作製するために用いられる。
【0039】
他の実施態様では、標的配列は、トリインフルエンザの異なった株を通して保存される短いストレッチの配列である。一つの実施形態では、siRNAのために使用される配列は、トリインフルエンザウイルスのH5N1株のゲノム1に由来する。この非限定的な例は、図9〜図12に示される。太字のヌクレオチドは、相同性分析が行わったときに塩基対の相違が検出された場所を示す。別の実施形態では、siRNAのために使用される標的配列は、トリインフルエンザウイルスのH5N1株のゲノム5由来である。この非限定的な例は、図13〜図16に示される。別の実施形態では、siRNAのために使用される標的配列は、トリインフルエンザウイルスのH5N1株のゲノム7由来である。この非限定的な例は、図1〜図8に示される。他の実施態様において、標的配列は、H5N1の他のゲノム型に由来する。さらに、トリインフルエンザの他の株(例えば、H5N2、H5N3など)に立ち向かうsiRNAもまた、本発明の範囲内に包含され、そして本明細書中の教示に従って、当業者によって作製され得る。
【0040】
別の態様においては、口蹄疫ウイルス(FMDV)の保存されたストレッチが、siRNAを構築するための標的配列として用いられる。なお他の態様では、以下の疾患を引き起こすウイルスの非限定的なリストが、siRNA構築のための標的配列として使用される:ニューカッスル病、西ナイル熱、鶏痘、トリ感染性気管支炎、トリ脳脊髄炎、トリ白血病、アヒルウイルス性肝炎、アヒルウイルス腸炎、ランピースキン病、ウシの感染性鼻気管炎、ウシウイルス下痢、ウシ白血病、リフトバレー熱、牛疫、ブルータング、小反芻獣疫(PPR)、羊痘およびヤギ痘、伝染性膿疱性皮膚炎(膿瘡)、ボーダー疾患、マエディ−ビスナ、伝染性胃腸炎(TGE)、ウマインフルエンザ、アフリカ馬疫、ベネズエラウマ脳脊髄炎およびコイ春ウイルス血症。
【0041】
(siRNAをコードする構築物)
siRNAをコードしている構築物は、プロモーターおよびsiRNAを細胞内の適切な標的へと「導く」配列を含むがこれらに限定されない、いくつかの構成要素を含む。用いられ得るプロモーターとしては、RNAポリメラーゼIIプロモーターおよびRNAポリメラーゼIIIプロモーターが挙げられるがこれに限定されない。RNA pol IIIプロモーターは、siRNAの構築のためのプロモーターとして特によく働く。用いられ得るプロモーター(pol IIおよびpol IIIの両方)の非限定的な例としては、以下が挙げられる:5Sリボソーム(r)RNA、マウスU6、ヒトU6、マウスHI、ヒトHI、サイトメガロウイルスプロモーター、ニワトリユビキチンC、ヒト7SKプロモーター、ウシU6プロモーター、ニワトリU6プロモーター、マレク病ウイルス38kdリン酸化タンパク質(pp38)遺伝子、1.8kbのmRNAのニワトリβ−アクチン、ヒトβ−アクチン、ウシβ−アクチン、ニワトリPRLプロモーター、ニワトリSPATA4遺伝子プロモーター、ニワトリPolIプロモーター、マレク病ウイルスのUS1遺伝子のプロモーター、リボヌクレオチドレダクターゼ遺伝子のマレク病ウイルス小サブユニッ、トリ白血病ウイルスおよび肉腫ウイルスのLTRト、RSV−LTR、合成ポックスウイルスプロモーター、ワクシニアウイルスP1lプロモーター、ワクシニアウイルスP174およびP190、鶏痘初期/後期プロモーターP.E/L、鶏痘ウイルスチミジンキナーゼプロモーター、ワクシニアp7.5プロモーターおよび鶏痘−PFLlプロモーター。
【0042】
標準的な分子生物学の技術を使用して、これらのプロモーターは、ウイルス遺伝子を標的として、それらをサイレンシングする配列に作動可能に連結される。1つのプロモーターは、一つ以上の標的配列の発現を調節し得る。プロモーターおよび標的配列は、標準的なクローニングベクターまたは発現ベクター中にクローニングされる。本明細書中で使用する場合、「ベクター」とは、他の核酸配列を細胞へと送達し得る核酸分子をいう。ベクターは、プラスミド、バクテリオファージ、植物または他の動物ウイルスに由来し得る。
【0043】
siRNA構築物は、siRNAを発現し得るように作製される。当業者は、特定のタイプのプロモーターが、脊椎動物の生理に依存して、一部の脊椎動物において他の脊椎動物においてよりも良好なsiRNAの発現を調節し、そしてその脊椎動物にとって最適であるプロモーターを使用する手段を講じるべきであることを認識している。プロモーターはまた、一つ以上のsiRNAの発現を調節し得る。ベクターは、一つ以上のsiRNAをコードする配列を含み得る。エンハンサー、選択マーカーおよび他の標準的な分子ツールもまた、より簡単な分子操作のために用いられ得る。REMI技術を使用する場合、ベクターは、REMI技術の使用を容易にするように構築されるべきである。その技術についての適切なガイダンスは、WO 99/42569において見出される。
【0044】
本発明の一態様において、ウイルスゲノム上の異なる標的に対応するいくつかのsiRNA遺伝子を有するベクターが用いられることができる。このタイプのベクター構造は、さまざまな部位での同時切断に起因して、有意な阻害からほぼ完全な阻害を確実にする。さらに、たとえウイルスの標的配列のうちの一つが変異または変化した場合であっても、これはまだ活性である。二本鎖siRNAに関しては、標準的な2bp突出を有すること、ならびに1bp突出を有すること、または突出が全く無いことが可能である。
【0045】
(生殖細胞の構築)
本明細書中で使用される場合、「生殖細胞」は、精子および卵細胞ならびにそれらの前駆体と定義される。生殖細胞は半数体であり、1セットの染色体しか有さないが、他の非生殖細胞は、2セットの染色体を有する。本発明は、ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含む構築物を含んでいる非ヒト脊椎動物の生殖細胞の構築を提供し、ここで、この配列はプロモーターに作動可能に連結されている。生殖細胞は、ウイルスゲノムに対する複数のsiRNAをコードしている配列を含む構築物を含み得る。用いられ得るウイルス配列の例は、上記に記載されている。一つの実施形態では、このウイルス配列は、口蹄疫ウイルス(FDMV)である。別の実施形態では、このウイルス配列は、トリインフルエンザウイルスである。
【0046】
本発明は、非ヒト脊椎動物の生殖細胞を作製する方法を提供し、ここで、この生殖細胞は、ウイルスゲノムの保存された領域に対するsiRNAをコードしている配列を含んでいる構築物を含む。本発明の方法を実施するために、当業者は、保存された領域またはウイルスゲノムに対するsiRNAをコードしている配列を含む構築物とともに生殖細胞を、この構築物が細胞中に取り込まれる条件下でインキュベートし、ここで、この配列は、プロモーターに作動可能に連結されている。一つの実施形態では、この構築物は、REMI技術を使用して宿主細胞ゲノムに組み込まれる。
【0047】
一つの態様では、生殖細胞は、ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含み、ここで、この配列は、プロモーターに作動可能に連結されている。別の実施形態では、非ヒト脊椎動物の生殖細胞は、非ヒト哺乳類である。非ヒト脊椎動物の非限定的な例としては、ニワトリ、アヒル、ガチョウ、家禽、畜牛、ウシ、ブタ、魚、ヒツジ、ヤギおよびウマが挙げられる。別の実施形態では、脊椎動物は、鳥類種のものである。別の態様においては、生殖細胞は、異なるプロモーターの制御下のいくつかの遺伝子を含んでいる1つのベクターを含む。
【0048】
一つの実施形態では、生殖細胞は、口蹄疫ウイルス(FMDV)を標的とするsiRNAをコードしている配列を含む構築物を含む。このような標的配列の例は、以下の通りである:i)3B領域に位置する、nt4900〜4922の5’−CCTGTCGCTTTGAAAGTGAAAGC−3’(配列番号1);(ii)3D領域に位置する、nt6934〜6992の5’−GAGATTCCAAGCTACAGATCACTTTACCTGCGTTGGGTGAACGCCGTGTGCGGTGACGC−3’(配列番号2);および(iii)3D領域に位置する、nt6892〜6917の5’−GACGAGTACCGGCGTCTCTTTGAGCC−3’(配列番号3)。全ての位置を、FMDV血清型O1(G)配列(GenBank登録番号AF189157)についていう。
【0049】
一つの態様では、生殖細胞は、トリインフルエンザを標的とするsiRNAをコードしている配列を含む構築物を含む。この態様の実施形態において、生殖細胞は、保存された配列が図1〜図16に示される配列のうちのいずれかから選択される構築物を含む。
【0050】
別の態様では、生殖細胞は、マレク病ウイルス(MDV)を標的とするsiRNAをコードしている配列を含む構築物を含む。上記のいずれかにおいて、生殖細胞は、精子である。
【0051】
(トランスジェニック動物の構築)
次いで、上記の通りの構築物は、多数の標準技術(例えば、リポフェクション)を用いることにより、動物細胞に導入される。しかしながら、以前の技術(例えば、リポフェクチン)のうちのいくつかは、極めて低い成功率を生じる。発明者らは、WO 99/42569において教示されるとおり、これらのsiRNAコード構築物の成功裏の導入を生じる優れた方法が、制限酵素媒介組込み(「REMI」)を使用することであることを見出した。したがって、一つの実施形態では、siRNAをコードしているポリヌクレオチドは、REMIを使用して動物の生殖細胞(例えば、精子)に導入することである。これらの生殖細胞の使用から得られる動物は、これらの疾患に対して抵抗性にされる。これらのトランスジェニック動物は、ウイルス抵抗性動物のストック全体を作製するために交配され得る。家禽の場合には、siRNAが標的とするウイルスとしては、マレク病、ガンボロおよびトリインフルエンザが挙げられるが、これらに限定されない。
【0052】
動物の細胞(例えば、生殖細胞)に導入されるsiRNAの数は、変化し得る。一つの実施形態では、1つのsiRNA構築物が導入される。他の実施形態では、1つ以上のsiRNAが導入される。当業者は、siRNAの異なる組合せが、標的とされるウイルス性疾患に基づいて導入され得ることを認識している。
【0053】
本発明は、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物の作製を提供し、ここで、この脊椎動物中の大多数の細胞は、この疾患の原因となるウイルスのゲノムの保存された領域に対するsiRNAをコードしている配列を含み、ここで、この配列は、プロモーターに作動可能に連結されている。ウイルス配列の例は、上記で考察されている。一つの実施形態では、この構築物は、ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む。別の実施形態では、これらの細胞は、ウイルスゲノムの保存された領域に対する複数のsiRNAをコードしている配列を含む複数の構築物を含む。別の実施形態では、この非ヒト脊椎動物は、非ヒト哺乳類である。
【0054】
一つの実施形態では、非ヒト脊椎動物中の大多数の細胞は、口蹄疫ウイルス(FMDV)を標的とするsiRNAをコードしている配列を含む構築物を含む。このような標的配列の例は、以下の通りである:i)3B領域に位置する、nt4900〜4922の5’−CCTGTCGCTTTGAAAGTGAAAGC−3’(配列番号1);(ii)3D領域に位置する、nt6934〜6992の5’−GAGATTCCAAGCTACAGATCACTTTACCTGCGTTGGGTGAACGCCGTGTGCGGTGACGC−3’(配列番号2);および(iii)3D領域に位置する、nt6892〜6917の5’−GACGAGTACCGGCGTCTCTTTGAGCC−3’(配列番号3)。全ての位置を、FMDV血清型O1(G)配列(GenBank登録番号AF189157)についていう。
【0055】
非ヒト脊椎動物の非限定的な例としては、ニワトリ、アヒル、ガチョウ、家禽、畜牛、ウシ、ブタ、魚、ヒツジ、ヤギおよびウマが挙げられる。別の実施形態では、脊椎動物は、鳥類種のものである。さらに別の実施形態では、非ヒト脊椎動物は、保存された配列が、図1〜図16において示される配列のうちの1つから選択されるsiRNAをコードしている構築物を含んでいる細胞を有する。別の実施形態では、非ヒト脊椎動物は、MDVに対して抵抗性である。
【0056】
本発明は、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物を作製するための方法を提供し、この方法は、以下の工程を包含する:(a)上記の生殖細胞を、この生殖細胞が二倍体細胞を形成する条件下でインキュベートする工程;および(b)(a)に記載の二倍体細胞を、この二倍体細胞が非ヒト脊椎動物を形成する条件下でインキュベートする工程。
【0057】
本発明はまた、クローン化による、ウイルス性疾患に対して抵抗性である非ヒト脊椎動物の作製を意図する。そのような場合、一つ以上のsiRNAは、クローン化されるべき細胞のゲノムに挿入される。この技術のさまざまな方法は、市販される(例えば、Advanced Cell Technologyの核移植プロトコル)。
【0058】
(適用)
本発明は、ウイルス性疾患に対して抵抗性である家禽を作製するための方法を本願明細書において提供する。これらの疾患としては、ニューカッスル病、西ナイル熱、鶏痘、鳥伝染性気管支炎、トリ脳脊髄炎、トリ白血病、アヒルウイルス性肝炎およびアヒルウイルス腸炎が挙げられるが、これらに限定されない。
【0059】
本発明は、ウイルス性疾患に対して抵抗性であるウシを作製するための方法を本願明細書において提供する。これらの疾患としては、ランピースキン病、ウシの感染性鼻気管炎、ウシウイルス下痢、ウシ白血病、リフトバレー熱、牛疫およびブルータングが挙げられるが、これらに限定されない。
【0060】
本発明は、ウイルス性疾患に対して抵抗性であるヒツジおよびヤギを作製するための方法を本願明細書において提供する。これらの疾患としては、小反芻獣疫(PPR)、羊痘およびヤギ痘、伝染性膿疱性皮膚炎(膿瘡)、ボーダー病、ブルータング、マエディ−ビスナおよびリフトバレー熱に接しているが挙げられるが、これらに限定されない。
【0061】
本発明は、ウイルス性疾患に対して抵抗性であるウマを作製するための方法を本願明細書において提供する。これらの疾患としては、ウマインフルエンザ、アフリカ馬疫およびベネズエラウマ脳脊髄炎が挙げられるが、これらに限定されない。
【0062】
本発明は、ウイルス性疾患に対して抵抗性である魚を作製するための方法を本願明細書において提供する。これらの疾患としては、コイ春ウイルス血症が挙げられるが、これに限定されない。
【0063】
本発明は、古典的ブタコレラ病、アフリカブタコレラ病、伝染性胃腸炎(TGE)および口蹄病に対する抵抗性を生じるために、ブタに適用され得る。一般に、本願明細書における教示は、ウイルス性疾患に感染しやすい任意の動物に対して適用され得る。
【0064】
病気による大きな経済的損失を受けることに耐えている大規模養鶏産業の場合、実用的な解決案は、トリインフルエンザおよびマレク病、伝染性ファブリキウス嚢病および他のトリウイルスに対して抵抗性である若鶏および産卵鶏の種畜を作製するために、siRNA技術の実施にある。本願明細書において開示される本発明の使用は、安全な家禽供給源を消費者に提供し、そして、鳥から人間へのトリウイルスの広がりとしての一般集団が止まる。
【0065】
以下は、本発明を例示するために提供され、本発明を限定するために提供されるわけではない。
【実施例】
【0066】
(実施例1 口蹄疫に対して抵抗性の動物の作製)
口蹄病に感染しやすいトランスジェニック動物を、口蹄疫ウイルス(FMDV)のウイルス配列から構築されるsiRNAを動物の生殖細胞に挿入することによって作製する。一般に、FMDVに感染しやすい動物は、偶蹄動物である。
【0067】
(口蹄疫siRNAの構築)
siRNAを構築するために、標的ウイルス配列を最初に同定する。これが達成され得る一つの方法は、異なる株または血清型からのFMDV配列について局所的相同分析を実行し、そして配列相同性の短いストレッチを同定することによる。一つの実施形態では、この相同性は、100%(すなわち、すべての血清型にわたって正確に同じ配列)である。他の実施形態では、1塩基対または2塩基対の相違がみられる。相同分析は、公に入手可能なプログラムまたは市販のプログラム(例えば、GCGパッケージ中のPILEUPプログラムおよびPRETTYプログラム)を用いて達成される。相同性検索は、PILEUPプログラムおよびPRETTYプログラムまたはFASTAプログラム(Pearsonら,PNAS 85:2444−2448(1988))を使用して、公に入手可能な配列(例えば、Genbank)について必要に応じて実施される。
【0068】
一旦短い相同性ストレッチが同定されると、siRNAが、商業的に利用可能なサービスを利用して合成される。siRNAは、種々の長さで合成される。いくつかのsiRNAは、長さが19bpであり、一方で、他のものは、長さが20bp、21bp、22bp、23bp、24bp、25bp、26bp、または27bpである。これらのsiRNAの各々は、血清型をとおして100%保存されるか、または各siRNA中に1bp〜2bpの相違を有する。これらの保存された配列は、生殖細胞への送達に適しているベクター中の少なくとも一つのプロモーターに作動可能に連結されている。別の代替法は、すでに合成されたsiRNAを使用することである。例えば、Kahanaら,J.Gen.Virol.85:3213−3217(2004)において開示されるsiRNAを参照のこと。
【0069】
次いで、これらのsiRNA構築物を、WO 99/42569において開示されるREMI技術を使用して偶蹄動物の生殖細胞(例えば、精子または卵子)に挿入し、安定して組み込まれたsiRNAを有する生殖細胞が得られる。次いで、生殖細胞を用いて、当該分野で公知の方法(例えば、体外受精、人工授精)によってトランスジェニック動物が得られ、口蹄疫に対して抵抗性である動物が得られる。
【0070】
(実施例2 アカバネウイルスについてのsiRNAを含む精子の作製)
非ヒト脊椎動物からの精子系統は、アカバネウイルスの保存された領域に対するsiRNAをコードしている配列を含む構築物とともに精子を、この構築物を細胞に取り込ませる条件下でインキュベートすることによって作製される(ここで、この配列は、プロモーターに作動可能に連結されている)。非限定的な例としては、リポフェクチンまたは類似の機構、塩化カルシウムまたはプロタミンを使用することが挙げられる。代替法では、この構築物は、REMI技術を使用して、宿主細胞ゲノムに安定して組み込まれる。siRNAは、2つの基準に従って設計される:異なる単離株間で高い相同性を共有している領域、およびsiRNA標的発見プログラムによって決定した場合のサイレンシング活性の高い類似。保存された配列を標的とするsiRNA分子の使用は、これらの分子が、それらの起源に関係なく、同じ株のすべてのウイルスでなかったとしてもほとんどのウイルスを切断する能力を確実にしなければならない。第2の、そしてより重要な理由は、配列の保存が、変化に対して強い選択圧を示すということである。選択圧は、これらの標的領域を不変に保つと期待されるが、それらが変化する場合、それらは、siRNA分子の抗ウイルス活性を抑制し得る。
【0071】
(実施例3 口蹄疫ウイルスについてのsiRNAを含む精子の作製)
非ヒト脊椎動物からの精子系統は、口蹄疫ウイルスの保存された領域に対するsiRNAをコードしている配列を含む構築物とともに精子を、この構築物を細胞に取り込ませる条件下でインキュベートすることによって作製される(ここで、この配列は、プロモーターに作動可能に連結されている)。非限定的な例としては、リポフェクチンまたは類似の機構、塩化カルシウムまたはプロタミンを使用することが挙げられる。代替法では、この構築物は、REMI技術を使用して、宿主細胞ゲノムに安定して組み込まれる。
【0072】
(実施例4 トリインフルエンザウイルスについてのsiRNAを含む精子の作製)
非ヒト脊椎動物からの精子系統は、トリインフルエンザウイルスの保存された領域に対するsiRNAをコードしている配列を含む構築物とともに精子を、この構築物を細胞に取り込ませる条件下でインキュベートすることによって作製される(ここで、この配列は、プロモーターに作動可能に連結されている)。非限定的な例としては、リポフェクチンまたは類似の機構、塩化カルシウムまたはプロタミンを使用することが挙げられる。代替法では、この構築物は、REMI技術を使用して、宿主細胞ゲノムに安定して組み込まれる。
【特許請求の範囲】
【請求項1】
本願明細書に記載された発明。
【請求項1】
本願明細書に記載された発明。
【図1−1】



【図1−2】

【図1−3】

【図1−4】

【図1−5】

【図1−6】

【図2−1】

【図2−2】

【図3−1】

【図3−2】

【図4−1】

【図4−2】

【図4−3】

【図5−1】

【図5−2】

【図6−1】

【図6−2】

【図7】

【図8−1】

【図8−2】

【図8−3】

【図8−4】

【図9−1】

【図9−2】

【図10】

【図11】
【図12】

【図13−1】

【図13−2】

【図14】

【図15】
【図16−1】

【図16−2】

【図16−3】
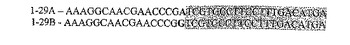
【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図1−6】
【図2−1】
【図2−2】
【図3−1】
【図3−2】
【図4−1】
【図4−2】
【図4−3】
【図5−1】
【図5−2】
【図6−1】
【図6−2】
【図7】
【図8−1】
【図8−2】
【図8−3】
【図8−4】
【図9−1】
【図9−2】
【図10】
【図11】
【図12】
【図13−1】
【図13−2】
【図14】
【図15】
【図16−1】
【図16−2】
【図16−3】
【公開番号】特開2012−125256(P2012−125256A)
【公開日】平成24年7月5日(2012.7.5)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−77496(P2012−77496)
【出願日】平成24年3月29日(2012.3.29)
【分割の表示】特願2008−503622(P2008−503622)の分割
【原出願日】平成18年3月31日(2006.3.31)
【出願人】(507324692)
【Fターム(参考)】
【公開日】平成24年7月5日(2012.7.5)
【国際特許分類】
【出願番号】特願2012−77496(P2012−77496)
【出願日】平成24年3月29日(2012.3.29)
【分割の表示】特願2008−503622(P2008−503622)の分割
【原出願日】平成18年3月31日(2006.3.31)
【出願人】(507324692)
【Fターム(参考)】
[ Back to top ]