
ウイルス粒子様ナノカプセル
【課題】特定の粒子構造を有するナノカプセルを、生物宿主を用いて高効率、且つ簡便に製造する。
【解決手段】下記の(1)〜(4)のいずれかに示すアミノ酸配列からなるタンパク質を含むナノカプセル:(1)特定のアミノ酸配列;(2)上記(1)に示されるアミノ酸配列に、更に生体分子を認識するアミノ酸配列が付加されたアミノ酸配列;(3)(1)と異なる特定のアミノ酸配列;(4)上記(1)〜(3)のいずれかに示されるアミノ酸配列において、1個又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
【解決手段】下記の(1)〜(4)のいずれかに示すアミノ酸配列からなるタンパク質を含むナノカプセル:(1)特定のアミノ酸配列;(2)上記(1)に示されるアミノ酸配列に、更に生体分子を認識するアミノ酸配列が付加されたアミノ酸配列;(3)(1)と異なる特定のアミノ酸配列;(4)上記(1)〜(3)のいずれかに示されるアミノ酸配列において、1個又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ウイルス粒子様の形状を有するナノカプセルに関するものである。
【背景技術】
【0002】
ヒトB型肝炎ウイルス等のようなウイルスエンベロープタンパク質を宿主細胞にて生合成させると、当該タンパク質が宿主細胞から脂質二重膜といった脂質を取り込みつつ、自己凝集してウイルス粒子様のナノカプセルを形成することが既に知られている。
【0003】
例えば、特許文献1、2に示すように、ヒトB型肝炎ウイルス表面抗原(HBsAg)タンパク質を生合成させることで、ウイルス粒子様のナノカプセルが作製できることが知られている。しかしながら、当該ナノカプセルの作製のために用いられる宿主細胞としては、酵母細胞、動物細胞、昆虫細胞等といった真核細胞に限られており、原核細胞を用いてウイルス様の粒子を形成した報告は未だ存在しない。
【0004】
一方、特許文献3ではHBsAgタンパク質のトランケートミュータントを、大腸菌を宿主細胞として用いて生合成させる技術が開示されているが、抗原としてのみ用いられる技術しか開示されておらず、ウイルス粒子様のナノカプセルが製造できるかどうかについては何ら明らかにされていない。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特許第4085231号
【特許文献2】特許第4212921号
【特許文献3】特開2002−112797号公報
【発明の概要】
【発明が解決しようとする課題】
【0006】
生物宿主を用いて有用物質等を製造する場合、宿主として真核生物由来の細胞を用いるよりも原核生物由来の細胞を用いる方が、遥かに効率が良く、且つ簡便である。特に、タンパク質を主成分とするような有用物質を製造する際には、タンパク質を構成するアミノ酸に種々の変異を施して、機能を改良することが一般的な開発手段であるために、数多くの変異体を作製してその機能を評価するためには、効率が良く、簡便な製造手法が求められる。以上の点から、動物細胞、昆虫細胞、酵母細胞等の真核生物由来の細胞を宿主として有用物質を製造することは、あまり好ましくない。
【0007】
一方で、大腸菌などに代表される原核生物は、生産効率やコストの面において、有用物質を生産する宿主生物として非常に優れており、更に簡便な方法が提供できる。しかし複雑な立体構造等を有する有用物質の生産には向いておらず、立体構造を補正するシャペロン等といった特殊なタンパク質の存在下にて生産する方法や、化学的な試薬を用いて正常な立体構造に戻す操作が必要となり、結局のところ原核生物を用いる際に得られる効率の良さを相殺してしまうか、若しくはその効率を余計に悪くしてしまう虞もある。また、原核生物では生産量そのものが極めて低い有用物質も多い。
【0008】
さらに、結局、有用物質そのものでは機能を十分に発揮できないものが製造されてしまい、特定の機能ドメインのみに限定された有用物質の一部分のみの生産しか実現しない場合もある。
【0009】
本発明の課題は、特定の粒子構造を有するナノカプセルを、生物宿主を用いて高効率、且つ簡便に製造することである。
【課題を解決するための手段】
【0010】
上記の課題を解決すべく、出願人は鋭意研究を重ねた結果、特定のアミノ酸配列からなるタンパク質を含むナノカプセルが、従来のナノカプセルと同等か、又はそれ以上の効果を有しながらも、従来の製造方法よりも容易で、且つ簡便な方法によって製造できることを見出した。
【0011】
本発明は、係る知見に基づいて完成されたものであり、以下に示す態様の発明を広く包含するものである。
【0012】
項1 下記の(1)〜(4)のいずれかに示すアミノ酸配列からなるタンパク質を含むナノカプセル:
(1)配列番号1に示されるアミノ酸配列;
(2)上記(1)に示されるアミノ酸配列に、更に生体分子を認識するアミノ酸配列が付加されたアミノ酸配列;
(3)配列番号2に示されるアミノ酸配列;
(4)上記(1)〜(3)のいずれかに示されるアミノ酸配列において、1個又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
【0013】
項2 下記の(5)〜(10)のいずれかに示す塩基配列からなるポリヌクレオチド:
(5)配列番号1に示されるアミノ酸配列をコードする塩基配列;
(6)配列番号3に示される塩基配列;
(7)上記(5)または(6)に示される塩基配列に、更に生体分子を認識するアミノ酸配列をコードする塩基配列が付加された塩基配列;
(8)配列番号2に示されるアミノ酸配列をコードする塩基配列;
(9)配列番号4に示される塩基配列;
(10)上記(5)〜(9)のいずれかに示される塩基配列において、1個又は数個の塩基が置換、欠失、挿入、及び/又は付加された塩基配列。
【0014】
項3 上記項2に記載のポリヌクレオチドを含む組換えベクター。
【0015】
項4 上記項3に記載の組換えベクターを含む形質転換体。
【0016】
項5 上記項1に記載のナノカプセルを製造する方法。
【発明の効果】
【0017】
本発明のナノカプセルは、従前の大量生産に用いられていた酵母細胞ではなく、大腸菌を宿主細胞として用いて製造することが可能であるために、簡便、迅速、且つ低コストにナノカプセルを製造することが可能である。
【0018】
従って、本発明のナノカプセルに新たな機能を付与するために、ナノカプセルの構成要素であるタンパク質に変異を施して、有用な機能を有する様々なナノカプセルの開発にかかる時間は、従来に比べて大きく短縮させることが可能となる。
【0019】
また、本発明のナノカプセルは、従来のウイルス粒子様のナノカプセルとほぼ同等の粒径を有する。そして、本発明のナノカプセルには、従来のウイルス粒子様のナノカプセルと同様に、生体分子を認識するアミノ酸配列からなるペプチドをカプセルの外側に提示することが可能であり、ワクチンとしての利用のみならず、DDS、診断薬、イメージング試薬、免疫測定試薬、バイオセンサー試薬等の分野に有用である。
【0020】
そして、本発明のナノカプセルは、従来のナノカプセルと比較して短いアミノ酸配列からなるタンパク質を構成成分とするために、タンパク質の重量が比較的少ないナノカプセルが提供されるので、従来のナノカプセルと比較して、堅牢な構造を取り難くなる。従って、本発明のナノカプセルは、特に内部に物質を包含させる点において、従来のウイルス粒子様のナノカプセルよりも優れた効果を示す傾向となり、DDS等の用途に特に有用である。
【0021】
さらに、本発明のナノカプセルは、従来のウイルス粒子様のナノカプセルと比較して、抗原性が低いことから人体に投与した際の安全性が高いことが明らかであるため、特にDDSの分野において有用である。
【図面の簡単な説明】
【0022】
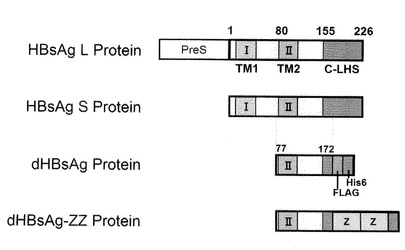
【図1】本発明の各種ナノカプセルが有するタンパク質のドメイン等を説明する模式図。
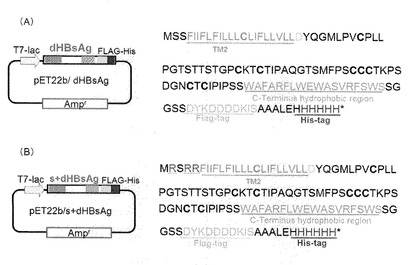
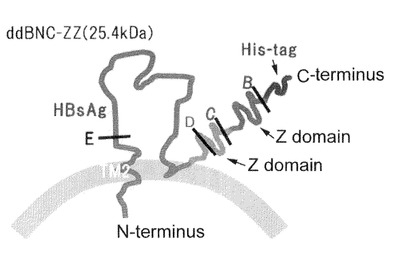
【図2】本発明の各種ナノカプセルが有するタンパク質のアミノ酸配列と、それをコードするポリヌクレオチドを含む発現ベクター等を説明する模式図。(A)はdHBsAgを示し、(B)はs+dHBsAgを示す。
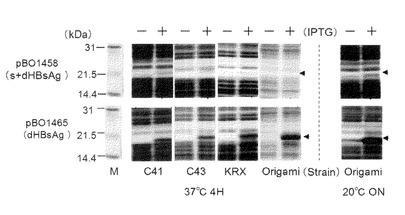
【図3】本発明のナノカプセルが有するタンパク質を、各種大腸菌株を用いた合成、及び培養条件等を説明する図。
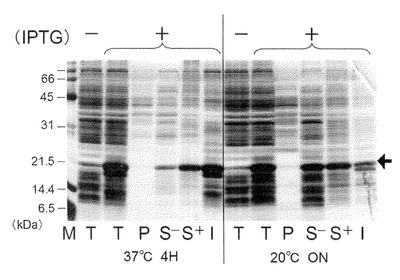
【図4】本発明のナノカプセルが有するタンパク質を、大腸菌を用いて合成させた際の菌体内局在等を説明する図。Mはマーカー、Tは全菌、Pはペリプラズム画分、S−は超音波破砕による可溶性画分、S+はTritonX−100存在下での超音波破砕による可溶性画分、I不溶性画分を示す。
【図5】本発明のナノカプセルが有するタンパク質を、大腸菌を用いて合成させた際の菌体内局在等を説明するグラフ。Cは培養上清、Pはペリプラズム画分、S−は超音波破砕による可溶性画分、S+はTritonX−100存在下での超音波破砕による可溶性画分を示す。
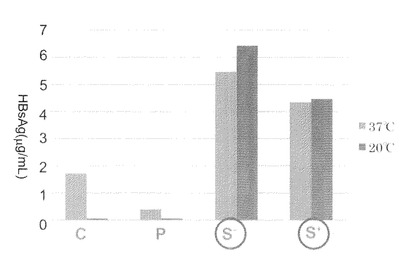
【図6−1】本発明のナノカプセルの精製時における、抗FLAG抗体カラムクロマトグラフィー溶出画分のSDS−PAGE解析を示す。Mはマーカー、S−は超音波破砕による可溶性画分、Fは素通り画分、IWはカラムの初回洗浄後画分、LWはカラムの最終洗浄画分を示す。
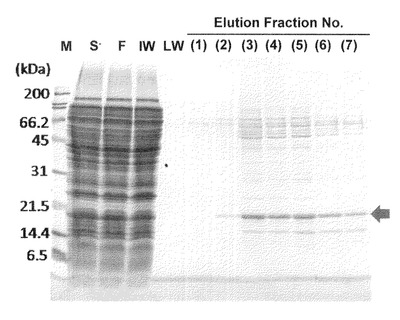
【図6−2】本発明のナノカプセル精製時の、ゲル濾過カラムクロマトグラムチャートを示す。
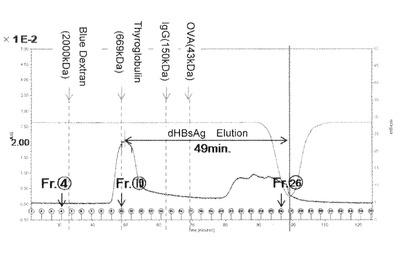
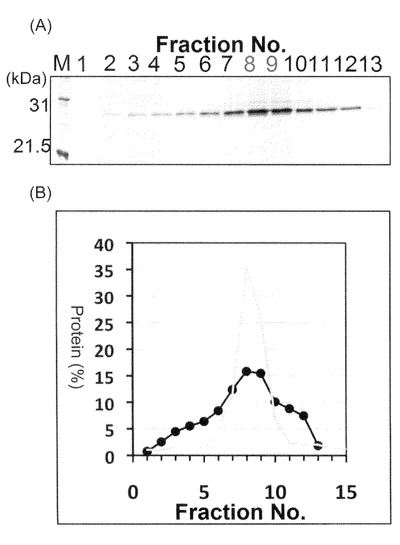
【図6−3】本発明のナノカプセル精製時の、ゲル濾過ピーク画分(Fr.10)のSDS−PAGE(CBB染色)、及び各種抗体を用いたウエスタンブロッティング解析を示す。
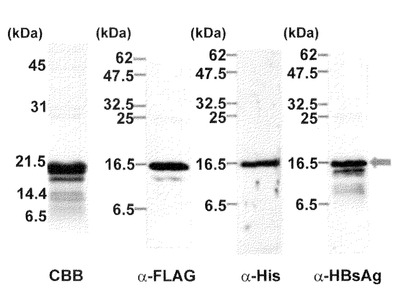
【図7】本発明のナノカプセルをショ糖密度勾配遠心解析した結果を示す図。黒丸は、dHBsAgの結果を表し、灰色の領域は従来法で作製したHBsAgを表す。グラフの左軸は、全タンパク質中のdHBsAg又はHBsAgタンパク質の重量割合を示す。
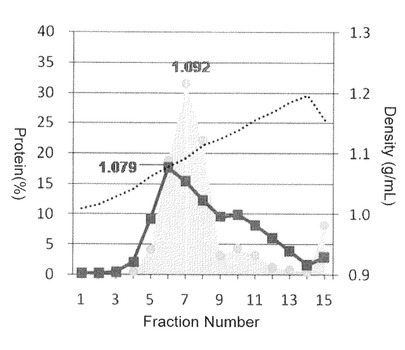
【図8】本発明のナノカプセルの電子顕微鏡写真像。(A)は市販の精製HBsAg粒子(ビークル社製)を示し、(B)はdHBsAg粒子を示す。
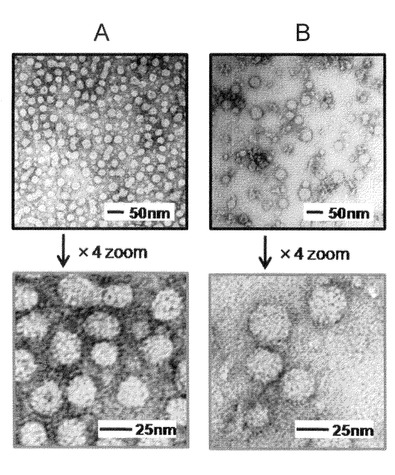
【図9】本発明のナノカプセルが有するタンパク質を、大腸菌を用いて合成させた際の培養条件、菌体内局在、及び精製条件等を説明する図。(A)培養条件及び菌体内局在を示す。Mはマーカー、S−は超音波破砕による可溶性画分、S+はTriton X−100存在下での超音波破砕による可溶性画分、Iは不溶性画分を示し、IPTGは、全菌試料を表す。(B)精製条件を表す。Mはマーカー、S−は超音波破砕による可溶性画分、EはNi−カラム溶出画分を示す。
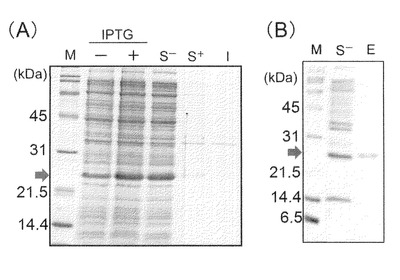
【図10】本発明のナノカプセルと抗体の結合能等を説明する図。△はdBNCと抗体との結合を示し、○はdBNCーZZと抗体との結合を示す。
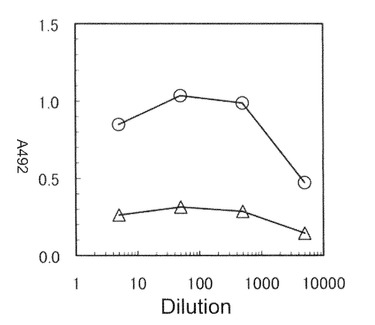
【図11】本発明のナノカプセルをショ糖密度勾配遠心解析した結果を示す図。(A)SDS−PAGE解析結果であり、Mはマーカーを示す。(B)SDS−PAGE解析における28kDaのバンドを数値化したもの(黒丸)。灰色は従来法で作製したHBsAgーZZを表す。グラフの左軸は、全タンパク質中の本発明のナノカプセル又はHBsAgタンパク質量の割合を示す。
【図12】本発明のナノカプセルの電子顕微鏡写真像。(A)市販の精製HBsAg粒子、(B)dHBsAg−ZZ粒子。
【図13−1】本発明のナノカプセルのトリプシン消化実験を説明する模式図。
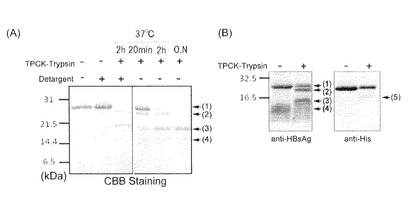
【図13−2】本発明のナノカプセルのトリプシン消化実験の結果を示す図。(A)SDS−PAGE解析結果、(B)各種抗体を用いたウエスタンブロッティング解析。
【図14】本発明のナノカプセルのAFM写真像。(A)AFM写真像、(B)縦断面解析結果。
【発明を実施するための形態】
【0023】
本発明を実施するために使用する様々な技術は、特にその出典を明示した技術を除いては、公知の文献等に基づいて当業者であれば容易かつ確実に実施可能である。例えば、遺伝子工学及び分子生物学的技術であれば、Sambrook and Russell,「Molecular Cloning A LABORATORY MANUAL」, Cold Spring Harbor Laboratory Press, New York, 2001; Ausubel, F. M. et al. 「Current Protocols in Molecular Biology」, John Wiley & Sons, New York, N.Y等の文献を参照すればよい。
【0024】
本発明のナノカプセル
本発明のナノカプセルは、下記の(1)〜(4)のいずれかに示すアミノ酸配列からなるタンパク質を含む。
【0025】
(1)配列番号1に示されるアミノ酸配列;
(2)上記(1)に示されるアミノ酸配列に、更に生体分子を認識するアミノ酸配列が付加されたアミノ酸配列;
(3)配列番号2に示されるアミノ酸配列;
(4)上記(1)〜(3)のいずれかに示されるアミノ酸配列において、1個又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
【0026】
本発明のナノ粒子には、(1)〜(4)のいずれかに記載のアミノ酸配列からなるタンパク質のうち、一種のみのタンパク質が含まれていても良く、二種以上のタンパク質が含まれていても良い。例えば、(1)に示すアミノ酸配列からなるタンパク質と、(2)に示すアミノ酸配列からなるタンパク質を一定の比率で含むナノ粒子は、生体認識分子の提示能に優れることから、特に免疫測定試薬等の分野にて優れた効果を示す傾向にある。
【0027】
上記(1)〜(4)にて示すアミノ酸配列からなるタンパク質は、自己凝集することによってウイルス粒子様の形状を形成する性質を有する。このような性質を本明細書では粒子形成能と定義する。
【0028】
本発明のナノカプセルは、ウイルスエンベロープ様の粒子形状を有する。ウイルス粒子様の形状とは、脂質二重膜によって構成される殻の構造に、上記のタンパク質が包埋される形状である。本発明のナノカプセルは、リポソームといった脂質のみから構成される粒子形状の構造物と比較して、タンパク質が構成成分に含まれるため、リポソームよりも構造安定性が高いという性質を有する。このような形状は、単にウイルス様粒子とも称され、具体的には球状、楕円球状、ラグビーボール状、略球状等とも理解される形状である。
【0029】
本発明のナノカプセルには、上述のタンパク質以外に、製造する際の使用する宿主細胞に存在する、タンパク質、脂質、核酸等といった成分が含まれる。なお、これらの成分は上述のタンパク質に結合した状態でナノカプセルに含まれていても良い。
【0030】
本発明のナノカプセルにおける上記タンパク質の含有量は、特に限定はされないが、ナノカプセル100重量部に対して通常は75〜重量部90重量部程度である。
【0031】
本発明のナノカプセルに含まれる脂質の含有量は、特に限定されないが、ナノカプセル100重量部に対して通常は10〜重量部25重量部程度である。
【0032】
本発明のナノカプセルの内部は、中空であってもよく、水分、脂質成分、イオン成分等が含まれていてもよい。
【0033】
本発明のナノカプセルの粒子径は、通常は10〜200nm程度であり、より好ましくは13〜100nm程度、より好ましくは16〜50nm程度であり、最も好ましくは20〜30nm程度である。ここで、粒子径の測定方法は、電子顕微鏡像から、無作為に抽出したナノカプセルの直径を測定し、それらの平均値によって算出したものである。なお、本発明のナノカプセルは、エクストルーダーを用いて、孔径の小さいフィルターを通過させることによって適宜調節することが可能である。
【0034】
本発明のナノカプセルは、(a)及び(c)に上述したように従来のHBsAgタンパク質を含むナノカプセルと比較して、アミノ酸配列を大幅に欠失しながらも、粒子形成能を保持している。このようなアミノ酸からなるタンパク質を含む本発明のナノカプセルは、内部に蛍光物質、放射性物質、磁性物質等の標識物質;DNA、RNA、PNA等の核酸;薬剤といったDDS、生体内でのキャリアー等として送達又は移送させるべき分子を、より高効率に内包させることが可能である。
【0035】
なお、ここでいう内包とは、ナノカプセルの内部に当該分子の全てが完全に包み込まれている必要はなく、当該分子の一部がナノカプセルの外部に突出していてもよい。
【0036】
本発明のナノカプセルに、上述した分子を内包させる方法は、特に限定はされないが、脂質融合を利用した方法、エレクトロポーレーション法等が挙げられる。
【0037】
その他に、本発明のナノカプセルを構成する粒子形成能を有するタンパク質を、界面活性剤やその他手段を用いて粒子構造を破壊し、次いで所望の内包分子と混和した後に、再度、粒子形成させて得られるナノカプセル中に、分子を内包する方法が挙げられる。
【0038】
上記(1)にて定義する配列番号1に示されるアミノ酸配列は、NCBIのAccession No.Q9W966に示されるB型肝炎ウイルス表面抗原(以後、HBsAgと称することがある。)のSタンパク質のアミノ酸配列(アミノ酸残基数226)に対して、以下の(a)〜(c)に示す変異を施したアミノ酸配列である。
【0039】
(a)2〜77番目のアミノ酸残基の欠失。
(b)78番目及び79番目のアルギニン残基の、それぞれセリン残基への置換。
(c)173−226番目のアミノ酸残基を欠失。
【0040】
特に、上記の(b)に示すような変異を導入することにより、本発明のタンパク質が宿主細胞内にて効率的に作製され易くなり、ひいては当該タンパク質を含むナノカプセルの形成に有利となる。
【0041】
上記(2)にて定義する生体分子は特に限定されないが、具体的には表面抗原、レセプター、抗体、サイトカイン、転写因子等の糖タンパク質若しくはタンパク質、又はペプチド若しくは糖ペプチド;ヘパリン、シアルルイスX等の糖若しくは糖鎖等が挙げられる。
【0042】
上記(2)にて定義する生体分子を認識するアミノ酸配列は、特に限定はされないが、例えば生体分子がレセプターであれば、当該レセプターに対して相互作用する各種成長因子等のアミノ酸配列とすればよい。
【0043】
例えば、生体分子がビオチンであれば、アビジン、ストレプトアビジン、ニュートラアビジン等に代表されるアビジン系タンパク質のアミノ酸配列とすればよい。
【0044】
例えば、生体分子が糖鎖である場合には、アグルチニン、セレクチン等のレクチンのアミノ酸配列とすればよい。
【0045】
例えば、生体分子が抗体である場合には、配列番号5に示すStaphyrococcus aureusu由来のProtein AにおけるイムノグロブリンGのFcドメインに結合する領域に存在するアミノ酸配列であるZタグのアミノ酸配列とすればよい。なお、ここで言う抗体とは、上述のFcドメインを有さないFab、F(ab′)2等の構造を有する抗体は含まれない。
【0046】
例えば、標的生体分子を認識するペプチドあるいはアフィボディーを、ライブラリーから選んで、そのアミノ酸配列を用いてもよい。
【0047】
上記(2)にて定義する生体分子を認識するアミノ酸配列は、上記(1)に示されるアミノ酸配列の内部、N末端側、又はC末端側のいずれの位置に付加されていてもよいが、当該アミノ酸配列からなるタンパク質を含むナノカプセルの構造の安定性、又は生体分子を認識するアミノ酸配列からなるタンパク質若しくはペプチドをナノカプセルの外側に提示させることに鑑みて、C末端側に付加されることが好ましい。
【0048】
ここで、「C末端側」とは、C末端から1〜5番目程度のアミノ酸残基の位置、又はC末端側に0〜15個程度のアミノ酸配列(いわゆるスペーサー配列)を付加したさらに後ろ側の位置を意味する。なお、「N末端側」も同様に説明される。
【0049】
このように、生体分子を認識するアミノ酸配列が、ナノカプセルの外側に提示された上記(2)にて定義するアミノ酸配列としては、配列番号2に示すアミノ酸配列又に示すアミノ酸配列などが挙げられる。
【0050】
なお、生体分子を認識するアミノ酸配列は一種でも二種以上を組み合わせて付加されていてもよい。
【0051】
上記(4)にて定義する置換、欠失、挿入、及び/又は付加されるアミノ酸の数は、(4)に定義するアミノ酸配列からなるタンパク質が、粒子形成能を失わない範囲とすればよく、特には限定されないが、通常は20個程度以下とすればよく、好ましくは15個程度以下、より好ましくは10個程度以下、更に好ましくは5個程度以下であり、最も好ましくは1個である。
【0052】
なお、置換に関しては、上記(4)にて定義するアミノ酸配列からなるタンパク質が粒子形成能を失わないように、置換対象となるアミノ酸そのものの性質を変化させないような類似アミノ酸の範疇において適宜選択すればよい。ここで、類似アミノ酸とは、Phe、Trp、Tyr等といった芳香族アミノ酸;Ala、Leu、Ile、Val等といった脂肪族アミノ酸;Gln、Asn等といった極性アミノ酸;Lys、Arg、His等といった塩基性アミノ酸;Glu、Asp等といった酸性アミノ酸;Ser、Thr等といった水酸基を有するアミノ酸;Gly、Ala、Ser、Thr、Met等といった側鎖の小さいアミノ酸等に挙げられる各グループ内のアミノ酸である。例えばLeuは、同じ脂肪族アミノ酸の範疇に含まれる類似アミノ酸であるIleに置換すればよい。
【0053】
なお、上述の(1)〜(4)のいずれかに示すアミノ酸配列からなる本発明のタンパク質には、当該タンパク質を精製すること、当該タンパク質を検出すること等を目的として、各種タグポリペプチドが付加されていてもよい。具体的なタグポリペプチドとして、Hisタグ、FLAGタグ、HAタグ、mycタグ、MBPタグ、GSTタグ、GFPタグ、Zタグ、ストレプタグ等が挙げられる。 これらのタグポリペプチドは、単独で付加されていても、二種以上を組み合わせて付加されていてもよい。なお、上記のタグポリペプチドは、上述した(1)〜(4)のいずれかに示すアミノ酸配列内部に挿入される形で付加されていても、当該アミノ酸配列のC末端側に付加されていても、N末端側に付加されていてもよい。本発明のナノカプセルの構造の安定性、又は当該タグポリペプチドをナノカプセルの外側に提示することに鑑みて、C末端側に付加されることが好ましい。なお、「N末端側」、及び「C末端側」なる用語は、既に定義したとおりである。
【0054】
本発明のポリヌクレオチド
本発明のポリヌクレオチドは下記の(5)〜(10)のいずれかに示す塩基配列からなる。
【0055】
(5)配列番号1に示されるアミノ酸配列をコードする塩基配列。
(6)配列番号3に示される塩基配列。
(7)上記(5)または(6)に示される塩基配列に、更に生体分子を認識するアミノ酸配列をコードする塩基配列が付加された塩基配列。
(8)配列番号2に示されるアミノ酸配列をコードする塩基配列。
(9)配列番号4に示される塩基配列。
(10)上記(5)〜(9)のいずれかに示される塩基配列において、1個又は数個の塩基が置換、欠失、挿入、及び/又は付加された塩基配列。
【0056】
上記(5)にて規定する塩基配列は、配列番号1に示されるアミノ酸配列を基に、in silico技術を採用することによって塩基配列を決定することができる。in silico技術を用いれば、1つのアミノ酸配列から、そのアミノ酸配列をコードする多数の塩基配列が決定されることになるが、当該アミノ酸配列をコードする遺伝子を発現させる際に用いる宿主細胞に対して適当なコドン頻度を考慮すれば、配列番号1に示すアミノ酸配列をコードする、より好ましい態様の塩基配列を決定することができる。
【0057】
上記(6)にて規定する配列番号3に示される塩基配列は、上記(5)に示す配列番号1に示されるアミノ酸配列をコードする塩基配列の一態様である。
【0058】
上記(7)にて規定する生体分子、及び生体分子を認識するアミノ酸配列は、上述した本発明のナノカプセルの(2)に定義したものと同様にすればよく、例えば上述したZタグをコードする塩基配列は、配列番号6にて示される。
【0059】
なお、当該アミノ酸配列をコードする塩基配列も、上述の(5)にて説明したin silico技術を採用することによって決定できる。そして、当該塩基配列の好ましい態様も、上述のようにコドン頻度を考慮して決定できる。
【0060】
上記(8)にて規定する塩基配列も上記(5)と同様に、上述の配列番号2に示すアミノ酸配列を基に、in silico技術並びにコドン頻度を考慮して決定することができる。
【0061】
上記(9)にて規定する配列番号4に示される塩基配列は、上記(8)に示す配列番号2を基に決定した塩基配列の一態様である。
【0062】
上記(10)にて定義する置換、欠失、挿入、及び/又は付加される塩基の数は、(10)に定義する塩基配列がコードするアミノ酸配列からなるタンパク質が、粒子形成能を失わない範囲とすればよく、特には限定されないが、通常は30個以下とすればよく、好ましくは20個以下、より好ましくは10個以下、更に好ましくは5個以下であり、最も好ましくは3個以下である。
【0063】
なお、置換に関しては、上記(10)にて定義する塩基配列がコードするアミノ酸配列において、上記(4)にて定義した他の類似アミノ酸となるように塩基配列を置換すればよい。例えば、塩基配列中において、脂肪族アミノ酸の範疇に含まれるLeuを、同じく脂肪族アミノ酸の範疇に含まれる類似アミノ酸であるIleに置換するように、Leuをコードする「CTT」なる塩基配列を、「ATT」と置換すればよい。
【0064】
本発明の組換えベクター
本発明の本発明の組換えベクターは、上述した本発明のポリヌクレオチドを含むものである。組換えベクターとは生物宿主に導入することによって、特定の遺伝子を発現するものであり、本発明においては、上述した本発明のポリヌクレオチドを発現するものである。
【0065】
上記の生物宿主としては、特に限定はされないが、原核細胞であることが好ましい。具体的には、大腸菌、枯草菌、乳酸菌等が挙げられるが上述した本発明のポリヌクレオチドを効率的に発現させることを勘案すれば、大腸菌であることが好ましい。即ち、本発明の組換えベクターは、上述した本発明のポリヌクレオチドを含む大腸菌発現用ベクターが好ましい。
【0066】
具体的な大腸菌発現ベクターは、公知のものを採用すればよく、特に限定はされないが、IPTG、ラムノース等の存在下において、上述した本発明のポリヌクレオチドの発現を誘導することができるベクターを用いることが好ましい。例えば、pETベクターシリーズや、pGEXベクターシリーズ等を挙げることができる。
【0067】
本発明の形質転換体
本発明の形質転換体は、上述した本発明の組換えベクターを含むものであり、上述した組換えベクターによって上述の生物宿主が形質転換されたものを意味する。
【0068】
形質転換する方法は特には限定されず、塩化カルシウム法や塩化ルビジウム法で作成したコンピテントセルを用いる方法、エレクトロポーレーション法、プロトプラスト法等が挙げられる。
【0069】
なお、上述の組換えベクターが、IPTGによって特定の遺伝子の発現を誘導するものであり、用いる宿主細胞が大腸菌である場合には、T7RNAポリメラーゼ遺伝子がゲノム中に配置されたDE3株の大腸菌であることが好ましい。
【0070】
本発明のナノカプセルを製造する方法
本発明のナノカプセルを製造する方法は、以下の3つの工程を含むものである。
(i)上述した本発明の形質転換体を培養する工程、
(ii)当該形質転換体の培養物の可溶化液を回収する工程、及び
(iii)当該可溶化液からナノカプセルを精製する工程。
【0071】
上記(i)における、上述した形質転換体の培養方法は、それぞれの形質転換体に適合した公知の培養方法を採用すればよい。例えば、上述のように形質転換体が大腸菌であり、IPTG、ラムノース等による発現誘導を可能とする組換えベクターを保持している場合には、20℃程度の低温条件下で長時間の培養を行うことが好ましい。
【0072】
上記(ii)における、形質転換体の培養物からの可溶化液の回収方法は特に限定されず、Triton−Xシリーズ、Tweenシリーズ、NP40等といった公知の界面活性剤を用いる方法;超音波処理に供する方法;液体窒素等の冷媒を用い、特定の機器を用いて細胞膜、細胞壁等を破砕する凍結融解法;ガラスビーズ法;ザイモリエースなどのような細胞壁を消化する酵素を用いたスフェロプラスト法;高圧化にて処理を行うフレンチプレス法等の公知の可溶化方法を培養物に対して採用した後に、遠心分離処理に供して得られる液相分画を回収すればよい。なお、これらの可溶化方法は、単独であっても、2種類以上を組み合わせて採用しても良い。
【0073】
上記(iii)における、可溶化液から本発明のナノカプセルを精製する方法は、特に限定されることはなく、公知のタンパク質の精製法を採用すればよい。
【0074】
例えば、上述の可溶化液を塩化セシウム、スクロース、グリセロール、OptiPrep、Percol等の成分を各種濃度又は線形グラジエント濃度勾配にて含有する緩衝液と、超遠心分離処理を採用した密度勾配遠心分離法に供して、本発明のナノカプセル以外の成分を除去する方法;上述の可溶化液に熱処理を与えて、主に夾雑タンパク質等を変性させて本発明のナノカプセル以外の成分を除去する方法;上述の可溶化液に対して硫酸アンモニウム、エタノール酢酸、アセトン等を作用させて、主に夾雑タンパク質を変性させることにより、本発明のナノカプセル以外の成分を除去する方法;上述の可溶化液に対して硫酸ナトリウム、硫酸アンモニウム、ポリエチレングリコールを加えて沈殿させる事で、本発明のナノカプセル以外の成分を除去する方法;上述の可溶化液に対してクロマトグラフィー用いて、本発明のナノカプセル以外の成分を除去する方法等が挙げられ、これらの方法は単独で採用しても、二種類以上を組み合わせて採用しても良い。
【0075】
上述したクロマトグラフィーの具体的な種類は、特に限定されないが、陽イオン交換クロマトグラフィー、陰イオン交換クロマトグラフィー、アフィニティクロマトグラフィー、サイズ排除ゲル濾過クロマトグラフィー、逆相クロマトグラフィー、疎水クロマトグラフィー等を挙げることができ、これらのクロマトグラフィーは単独で用いても、二種以上を組み合わせて用いてもよい。
【0076】
本発明の組換えベクターが、上述したタグポリペプチドをコードするポリヌクレオチド配列が含む場合には、当該タグポリペプチドとの相互作用に基づいたアフィニティカラムを用いることが好ましい。
【0077】
例えば、FLAGタグ、HAタグ、mycタグ等のタグポリペプチドをコードするポリヌクレオチド配列を含む組換えベクターを含む形質転換体から得られた可溶化液より精製する場合、それぞれ抗FLAG抗体、抗HA抗体、抗myc抗体等といった、タグポリペプチドに対して特異的に結合する抗体を担持するポリマーを担持したアフィニティクロマトグラフィーを用いることが好ましい。このようなアフィニティクロマトグラフィーを用いる場合の溶出液としては、当該ペプチドを高濃度で含有する緩衝液、又はGlysin−HCl緩衝液のような、pHが1〜5程度の酸性緩衝液を用いることが好ましい。
【0078】
また、本発明のナノカプセルがMBPタグを有する場合には、マルトース等といった糖類を担持するポリマーを採用したアフィニティクロマトグラフィーを用いることが好ましい。このようなアフィニティクロマトグラフィーを用いる場合の溶出液としては、糖類を高濃度で含む緩衝液を用いることが好ましい。
【0079】
さらに、本発明のナノカプセルがHisタグを有する場合には、Ni−NTAといったヒスチジンとキレート結合が可能となる金属化合物を担持するポリマーを採用したアフィニティクロマトグラフィーを用いることが好ましい。このようなアフィニティクロマトグラフィーを用いる場合の溶出液としては、イミダゾールを高濃度で含有する緩衝液を用いることが好ましい。
【0080】
このようなタグポリペプチドは、精製後適当なプロテアーゼで本発明のナノカプセルから切断除去してもよい。
【0081】
なお、本発明のナノカプセルが、上述した生体分子を認識するアミノ酸配列が付加されたアミノ酸配列からなるタンパク質を含む場合、当該生体分子を担持するポリマーを採用したアフィニティクロマトグラフィーを用いることが有効である。このようなアフィニティクロマトグラフィーを用いる場合の溶出液としては、当該生体分子を高濃度で含有する緩衝液を用いることが好ましい。
【0082】
例えば、生体分子がストレプトアビジンのようなアビジン系タンパク質であり、生体分子を認識するアミノ酸がビオチンである場合には、アビジン系のタンパク質を担持するポリマーを採用したアフィニティクロマトグラフィーを用いることが有効であり、溶出液としてはビオチンを高濃度で含有する緩衝液を用いる方法が挙げられる。
【0083】
なお、本発明のナノカプセルを製造する過程では、当該ナノカプセルを構成するタンパク質の分解を抑えるために、低温条件下での操作とすることが好ましく、必要に応じてEDTA、PMSF等のタンパク質分解酵素阻害剤を1種又は2種以上で組み合わせて用いてもよい。
【0084】
上述のように、本発明のナノカプセルは、上述の本発明の粒子形成能を有するタンパク質が自己凝集することによってウイルス様粒子となり形成されるが、ウイルスエンベロープ様粒子を形成する場は、宿主細胞内、試験管内、培養容器内等といった当該タンパク質の合成の場か、タンパク質合成の終了後の精製、回収、保存等の場であり、特に限定はされない。
【実施例】
【0085】
1)発現ベクターの構築
C末端54残基を欠失させFLAG−tag配列で置換したHBsAg抗原タンパク質をコードするプラスミドDNAである、ベクターpBO749(国際公開番号WO2005/049824)を鋳型とし、primer#1575(配列番号7)と#1555(配列番号8)とを用いたPCRにより、N末端76残基とC末端54残基とを同時に欠失した、配列番号9に示す塩基配列を含む欠失型HBsAg遺伝子(以後、dHBsAgと称することがある。)断片を増幅した。
【0086】
増幅された遺伝子断片をNdeI及びNotIで制限酵素消化処理した後、アガロースゲル電気泳動でdHBsAg遺伝子断片を単離及び精製した。大腸菌発現用ベクターpET22b(Novagen社)をNdeI及びNotIで制限酵素消化処理した後、dHBsAg遺伝子断片のNdeI及びNotIによる制限酵素消化処理後物と連結し、dHBsAg大腸菌発現用ベクターpBO1465を得た。
【0087】
構築されたdHBsAg遺伝子には、C末端側にFLAG−tagとHis6−tag配列が融合されている(図1及び図2)。
【0088】
同様に、ベクターpBO749を鋳型とし、primer#1554(配列番号10)と#1555(配列番号8)とを用いたPCRにより、N末端76残基とC末端54残基とを同時に欠失した、配列番号11に示す塩基配列からなる欠失型HBsAg遺伝子(以後s+dHBsAgと称することがある。)断片を増幅した。
【0089】
増幅された遺伝子断片をNdeI及びNotIで制限酵素処理した後、アガロースゲル電気泳動でdHBsAg遺伝子断片を単離及び精製した。大腸菌発現用ベクターpET22bも同様にNdeI及びNotIで制限酵素処理した後、制限酵素処理後のdHBsAg遺伝子断片と連結し、s+dHBsAg大腸菌発現用ベクターpBO1458を得た。
【0090】
なお図2にて示すように、構築されたs+dHBsAg遺伝子は、上述のdHBsAg遺伝子と比較すれば、アミノ酸に翻訳された際にN末端側のアミノ酸配列のみが異なっている。
【0091】
2)大腸菌によるdHBsAgの発現
上記のdHBsAg大腸菌発現用ベクターpBO1465及びs+dHBsAg大腸菌発現ベクターpBO1458を遺伝子発現用大腸菌Origami2(DE3)株、C41(DE3)株、C43(DE3)株、並びにKRXに形質転換し、翌日出現したコロニー数十個を、0.5%glucoseと終濃度で200μg/mlのアンピシリンを含む4mL若しくは30mLのLB培地で、37℃で2〜5時間振盪培養した。
【0092】
1.5mL若しくは12mLの菌懸濁液を、それぞれ終濃度で500μg/mlアンピシリンをする含有50mL若しくは400mLのLB培地に植菌し、さらに37℃、2〜5時間振盪培養した。OD600が0.8〜1.0になったところでIPTGを終濃度が1mMとなるように加えた。なお、KRX株の場合は、IPTGに換えて0.1%のラムノースを添加した。低温培養検討(20℃で4時間〜一晩培養)時には、IPTG添加前に培養温度を20℃に下げた。
【0093】
IPTGの添加後、37℃で4時間若しくは20℃で一晩(低温培養時)培養した後に菌体を回収した。IPTG添加前と培養終了時における1mlの培養液分の菌体を超純水100μLに再懸濁した後、当容量の2×sample buffer[20%のglycerol、4%SDS、0.02%BPB、及び10%の2−MEを含む50mMのTris−HCl(pH6.8)]を加えて溶解し、95℃で5分間加熱処理した後の20μL(全菌試料:Tとする)をSDS−PAGE処理に供し、続いてクマシーブリリアントブルー染色(CBB染色)を行った。結果を図3に示す。
【0094】
s+dHBsAgは、いずれの大腸菌株においてもIPTG誘導に基づくバンドの出現は観察されなかったが、dHBsAgでは図中の矢印の位置に示されるように、全ての大腸菌株においてIPTG誘導に基づくバンドの出現は観察された。以上のことから、大腸菌を用いてナノカプセルを構成するタンパク質を生合成するには、そのN末端側のアミノ酸配列が重要であることが明らかとなった。
【0095】
3)dHBsAg発現における大腸菌体内の局在
dHBsAgを発現する大腸菌Origami2(DE3)株を上記の各種培養条件で培養し、菌体回収時の遠心後の上清を培養上清(C)とした。1mLの培地による培養にて得られた菌体を、20%のsucroseを含む30mMのTris−HCl(pH8)を75mL用いて再懸濁した。ここに、0.5MのEDTA(pH8)を1.5μL加え、室温条件下で10分間Vortex処理を行った。
【0096】
次いで、10000rpm、4℃の条件で10分間遠心し、上清を取り除いた。得られた沈殿画分を、氷冷した5mMのMgSO4に再懸濁し、4℃で10分間vortex処理を行った。次いで、10000rpm、4℃の条件にて、10分間遠心し、得られた上清をペリプラズム画分(P)とした。一方で、得られた沈殿画分を、0.75mLの氷冷Lysis buffer[5%のglycerol、及び150mMのNaClを含む50mMのTris−HCl(pH7.5)]に再懸濁し、終濃度1mMのEDTA、及び1mMのPMSFを加えた。
【0097】
このサンプルを、2回凍結融解処理に供した後、氷上で30秒ずつ2回の超音波破砕処理を行った。次いで、14000rpm、4℃の条件にて、10分間遠心し、得られた上清画分を可溶性画分(S−)とした。
【0098】
一方で、得られた沈殿画分を0.75mLの氷冷Lysis bufferに懸濁し、終濃度1mMのEDTA、及び1mMのPMSFを加えた。さらに終濃度が0.5%となるようにTriotnX−100を加え、30秒間の超音波破砕処理を1回行った。次いで、14000rpm、4℃の条件にて10分間遠心し、得られた上清画分を可溶性画分(S+)とした。
【0099】
一方で、得られた沈殿画分を0.75mLの氷冷Lysis bufferに再懸濁したものを不溶性画分(I)とした。以上の各画分をSDS−PAGE処理に供し、次いでCBB染色を行った。結果を図4に示す。
【0100】
図中の矢印の位置に示されるように、dHBsAgタンパク質はペリプラズム画分(P)には認められず、TritonX−100非存在下並びに存在下の可溶性画分(S−並びにS+)、及び不溶性画分(I)に分散して認められた。また、37℃における培養では不溶性画分(I)へ、20℃における培養では、可溶性画分(S−並びにS+)により多く局在することが明らかとなった。
【0101】
また、培養時に得られた菌体回収時の上清画分(C)、ペリプラズム画分(P)、及びTritonX−100存在下、並びに非存在下の可溶性画分(S−並びにS+)を、抗HBsAg抗体を用いた酵素免疫測定法(IMx−HBsAg;ダイナボッド社)にて、HBsAgタンパク質を測定した。結果を図5に示す。
【0102】
図4にて示される結果と同様に、HBsAgタンパク質はペリプラズム画分(P)には殆どは局在が認められず、TritonX−100などのような界面活性剤の存在下並びに非存在下の可溶性画分(S−並びにS+)として抽出できることが明らかとなった。
【0103】
4)dHBsAgタンパク質を含むナノカプセルの精製
引き続いて、dHBsAgタンパク質を含むナノカプセル(以後、dBNCと呼ぶことがある。)の精製を行った。
【0104】
IPTGによる誘導下(培養温度は37℃)にてdHBsAgを発現した菌体をLysis Buffer[150mMのNaClを含む50mMのTris−HCl(pH8.0)]に再懸濁し、最終濃度が1mMになるようにPMSFを加えた。次いで、1分ずつ5回氷上で冷却しながら超音波破砕した。その後、8000rpm、4℃の条件下で15分間遠心し上清を可溶性画分(S−)とした。ここで得られた沈殿を0.1%のTween80を含むLysis Bufferに再懸濁し、最終濃度が1mMとなるようにPMSFを加えて、1分ずつ5回氷上で冷却しながら超音波破砕した。超音波破砕後の溶液を同様の条件で遠心して、上清を可溶性画分(S+)とした。残った沈澱を超純水に懸濁して不溶性画分(I)とした。これらの試料(100μLの培養液分に相当)に当容量の2×sample bufferを加えて溶解し、95℃で5分間加熱処理した後SDS−PAGEのサンプルとした。
【0105】
TBS[150mMのNaClを含む50mMのTris−HCl(pH7.4)]で平衡化した抗FLAG M2 Agarose Affinity Gel (Sigma社)カラム(bed量:1mL)に、上記の可溶性画分(S−)をインジェクションし、4℃で2時間rotate後、ゆっくり流し素通り液を回収した。回収した画分を素通り画分(F)とした。
【0106】
次に、0.75mLのTBS500[NaCl濃度を500mMとした上記TBS]でカラムを数回洗浄後(初回の洗浄後画分を(IW)とし、最終洗浄画分を(LW)とする。)、0.4mLのElution Buffer[1mMのEDTAを含む0.1MのGlycine−HCl(pH3.5)]で8回に分けて溶出した。得られた溶出画分をそれぞれ溶出画分(1)〜(8)とした。
【0107】
溶出画分には直ちに1MのTris−HCl(pH8.0)を14μL混合して中和し、その後、各画分をSDS−PAGEに供し、次いでCBB染色を行った。結果を図6−1に示す。
【0108】
図中の矢印に示すように、抗FLAGカラム溶出画分の(2)〜(7)において、dHBsAgタンパク質のバンドが検出された。これらの溶出液のそれぞれ2mLを、0.15MのNaClを含む50mMのTris−HCl(pH7.4)で平衡化したSephacryl S−300HRゲル濾過カラム(GE Healthcare;直径1.6cm×高さ40cm;bed量24mL)にインジェクションし、0.75mLを1画分として回収した。
【0109】
図6−2に示すように、HBsAgタンパク質は、10番目の画分にて最も多く溶出されることが明らかとなった。当該画分は669kDaの分子量を有するサイログロブリンと同様の位置で流出することが明らかとなった。そして、図6−3に示すように、10番目の当該溶出ピーク画分はSDS−PAGE解析により80%以上の純度でdHBsAgタンパク質を含有することを示した。
【0110】
また図6−3に示すようにHRP標識化抗FLAG抗体(抗FLAG M2 PEROXIDASE CONJUGATE、SIGMA)、ヤギ由来のビオチン標識化抗HBs抗体とウサギ由来のアルカリフォスファターゼ標識化抗ビオチン抗体(IMx HBsAg試薬;ダイナボット)、あるいはHisProbeTM−HRP結合体(Thermo社)を各々用いたウエスタンブロッティングでも検出された事から、目的のdHBsAgバンドである事が確認された。
【0111】
以上の結果より、dHBsAgタンパク質は自己凝集して700kDa程度の複合体を形成していることが確認された。
【0112】
5)dBNCの性状解析
<Sucrose密度勾配遠心解析>
16mLの遠心管(16PA TUBE ASSY;HITACHI社)に底からスクロース濃度を50、40、30、20、及び10重量%含むBufferA[68.4mMのNa2HPO4、31.6mMのNaH2PO4、15mMの EDTA・2Na、及び0.85%のNaCl]を2.5mLずつ順に重層し、その上に上記4)にて示す可溶性画分(S−)を2.5mL重層した後、超遠心機(himac CP70MXX;HITACHI社)を用いて4℃、24000rpmの条件で14時間遠心した。
【0113】
陽性対照として、Cos7細胞によって一過的に製造されたHBsAg粒子(培養上清から回収したもの)を用いた。また、陰性対照として、2mgのウサギ血清由来のIgG(SIGMA、分子量150kDa)をBufferAで2.5mLに希釈したものを用いた。
【0114】
遠心処理の後、あらかじめ重量を測定しておいた1.5mLチューブに1mLずつ回収し、重量を測定して密度を求めた。各画分のHBsAg量をIMx HBsAg System(ダイナボット社)を用いて測定した。ウサギ血清由来IgGの濃度はUV法により測定した。その結果を図6に示す。
【0115】
dHBsAgタンパク質はCos7細胞を用いて作製したHBsAg粒子と同様に6−8番目の画分に沈降ピークを示した。ウサギ由来IgGの沈降ピークは2番目画分であった。以上の結果から、dHBsAgタンパク質は自己凝集し、従来のHBsAg粒子と同様に粒子形成していることが示唆された。
【0116】
<電子顕微鏡観察>
抗FLAG抗体溶出画分について、ネガティブ染色法を用いて透過型電子顕微鏡(ELECTRON MICROSCOPE H−7100S;HITACHI)観察した。比較対象として野生型HBsAg粒子(ビークル社製)を用いた。結果を図5に示す。
【0117】
dHBsAgタンパク質は、比較対象に用いた野生型HBsAg粒子と同様に、ウイルスエンベロープ様ナノ粒子(dBNC)が形成する事が明らかとなった。また、dBNCの粒子径は25±5nmと、比較対象に用いた野生型HBsAg粒子(21±2nm)とほぼ同等の粒子径を有することも明らかとなった。この数値は、電子顕微鏡像から50個の粒子を選択し、その直径を測定した平均値である。
【0118】
<dBNCの抗原性>
2種の市販のHBV体外診断用酵素免疫測定法キット(IMx HBsAg System;Abbott社、及びEnzygnost HBsAg 5.0 kit;SIMENS社)を用いて、精製dBNCの抗原性を市販のHBsAgS粒子(HBsAg−adr recombinant protein;GenWay Biotech社、10−663−45362)を標準試料として測定した。表1に結果を示す。
【0119】
【表1】

【0120】
dBNCの抗原性は、野生型S粒子と比較してIMxHBsAgキットによる測定では1/7、Enzygnostキットによる測定では約1/200に低下していた。
【0121】
従って、本発明のdBNCは、特にDDSとして体内に投与した場合、より安全に用いることができることが明らかとなった。
【0122】
6)ZZタグ融合HBsAg欠失体(dHBsAg−ZZ)発現ベクターの構築
まず、dHBsAg遺伝子発現ベクターpBO1465を鋳型とし、primer#361(配列番号12)と、#1633(配列番号13)とを用いたPCRを行いて遺伝子断片を増幅し、その反応液をフェノール抽出後エタノール沈殿した後、NdeI及びNotIで制限酵素消化処理を行うことにより、C末端にFLAGタグを含まない約0.3kbのdHBsAgの遺伝子断片を調製した。
【0123】
この遺伝子断片と、NdeI及びNotIで制限酵素消化処理を施したpBO1465ベクターの断片とをT4−ligaseで連結することにより、C末端にHis6−tagのみが付いたdHBsAg遺伝子を発現するベクターpBO1501を構築した。
【0124】
一方、10ngの酵母発現用ベクターpGLD−ZZ−d50N(Specific protein delivery to target cells by antibody−displaying bionanocapsules. Kurata N, Shishido T, Muraoka M, Tanaka T, Ogino C, Fukuda H, Kondo A. J Biochem. 2008 Dec;144(6):701−7.)を鋳型とし、primer #1732(配列番号14)と、#1733(配列番号15)とをそれぞれ10pmol用い、さらにTaq DNA polymarase(NEB社;1units/μL)を用いPCRにより、ZZtag遺伝子を増幅した後、72℃で14分間アデニン付加反応を行った。
【0125】
この増幅DNA反応液をフェノール抽出後、エタノール沈澱し、30μLの滅菌超純水に再溶解した。これを1.2%アガロースゲル電気泳動で電気泳動後、目的のZZ−tag遺伝子断片を含むゲル片を切り出した。具体的には、QIAEXII Gel− Extractionキット(QIAGEN社)を用いて、アガロースゲルから目的DNA断片を抽出精製した。このZZtag遺伝子断片とpTAC−1 vector(BioDynamics Labs)をT4−ligaseで連結することにより、ZZtag遺伝子をpTAC−1ベクターにクローニングして、pBO1594ベクターを作製した。
【0126】
得られたpBO1594ベクターをNotIで制限酵素消化処理し、1.5%アガロースゲル電気泳動した後、ZZtag断片を含むゲル片を切り出した。QIAEXII Gel Extractionキットで、目的となるZZtagDNA断片を抽出精製し、insert DNAとした。
【0127】
一方、pBO1501ベクターをNotIで制限酵素消化処理した後、65℃で20分間酵素失活反応させた。このNotIによって制限酵素消化処理したpBO1501の断片と、NotIによって制限酵素消化処理したZZtag断片とを、Ligation High ver.2(TOYOBO社)を用いて連結することにより、配列番号16に示す塩基配列を含むdHBsAg−ZZ遺伝子発現用ベクターpBO1595を構築した。
【0128】
このようにして作製したpBO1595ベクターには、dHBsAg−ZZのC末端に検出精製用のHis6−tag配列が融合されたタンパク質をコードする遺伝子が含まれている。
【0129】
7)dHBsAg−ZZの大腸菌による発現と精製
上述のdHBsAg−ZZ発現ベクターpBO1595を、大腸菌株Origami2(DE3)に形質転換した。翌日出現したコロニー数十個について、実施例2と同様の方法で遺伝子の発現及び培養を行い、得られた遺伝子発現菌体について、Tween80を含まない緩衝液にて超音波破砕処理によって可溶化して得られる可溶性画分Tween−(S−)、Tween80を含む緩衝液にて超音波破砕処理によって可溶化して得られる可溶性画分(S+)、及び不溶性画分(I)を調製した。これらの試料(100μLの培養液分)に当容量の2×sample bufferを加えて溶解し、95℃で5分間加熱処理した後、SDS−PAGE処理に供し、次いでCBB染色を行った。結果を図9(A)に示す。
【0130】
図中の矢印にて示されるように、IPTG誘導下による培養を行った場合に、明瞭なdHBsAg−ZZの発現バンドが見られ、主に可溶性画分(S−)に局在することが明らかとなった。またこの発現バンドは、ヤギ由来ビオチン標識化抗HBsAg抗体と、ウサギ由来アルカリフォスファターゼ標識化抗ビオチン抗体、あるいはHisProbeTM−HRP結合体を各々用いたウエスタンブロッティングでも検出された事から、目的のdHBsAg−ZZバンドである事が確認された。
【0131】
この可溶性画分(S−)を、平衡化緩衝液[0.5MのNaCl、及び30mMのイミダゾールを含む50mMリン酸緩衝液(pH7.0)]にて平衡化したNi−Sepharose 6 Fast flowカラムに添加し、室温で5分間rotate後、ゆっくり流し素通り液を回収した(素通り画分)。次に約1mlの上記平衡化緩衝液で7回、カラムを洗浄した後、イミダゾールの濃度を500mMに変えた上記平衡化緩衝液を用いて0.1mLずつ10回溶出させた(Ni−溶出画分:E)。各画分をSDS−PAGE処理に供し、次いでCBB染色を行った結果を図9(B)に示す。
【0132】
図中の矢印に示すように、Ni−カラム溶出画分(E)において、約70%を超える高純度のdHBsAg−ZZタンパク質が回収されることが明らかとなった。また、回収されたdHBsAg−ZZタンパク質は、dHBsAgタンパク質と同様に従来のHBsAgタンパク質と比較して、N末端側を欠失する変異が施されていることから、自己凝集することによってウイルス様の粒子を形成しているものと考えられる。以後、この粒子をdBNC−ZZと呼ぶことがある。
【0133】
また、発現用プラスミドによる大腸菌の形質転換から、高純度dBNC−ZZの取得までにかかった時間は約4日以内であり、迅速にB型肝炎ウイルス粒子様のナノカプセルを生産できることが明らかとなった。
【0134】
7) dBNC−ZZ粒子の性状解析
<抗体認識能>
ウサギ血清由来のIgGを固相化しスキムミルクでブロッキングしたマイクロウェルプレートに、TBSで希釈したdHBsAg−ZZ遺伝子の発現菌体の可溶性画分、及びdHBsAg遺伝子の発現菌体の可溶性画分を添加し、室温で2時間反応させた。各ウェルを3回洗浄後、標識抗体としてTBSで希釈したウサギ由来HRP標識化IgGを添加して室温で2時間反応させた。各ウェルを6回洗浄後、基質溶液[6mgのo−Phenylenediamine、12mLの0.1Mクエン酸緩衝液(pH5.0)、及び12μLの30%H2O2を混合して要事調製したもの]を加えて30分間反応させ、492nmの吸光度を測定した。その結果を図10に示す。
【0135】
dHBsAg試料では吸光度の上昇が全く見られなかったが、dHBsAg−ZZ試料では顕著な吸光度の上昇が観察された事から、dBNC−ZZ粒子では抗体結合能が維持されたZZドメインが多価でその粒子表面に提示されている事が示された。
【0136】
<Sucrose密度勾配遠心解析>
精製したdBNC−ZZ粒子を、上記5)のdHBsAgナノカプセルの性状解析にて示したsucrose密度勾配遠心処理と同様の手法を用いて解析した結果を図11に示す。比較対象として、酵母細胞を用いて生産した野生型HBsAg−ZZ粒子(ビークル社製)を用いた。
【0137】
dHBsAg−ZZ粒子は、比較対象の野生型BNC−ZZ粒子と同じ8番目の画分に沈降ピークを示すことが明らかとなった。この結果から、dHBsAg−ZZ粒子も従来のHBsAg粒子や、dHBsAgと同様に、dHBsAg−ZZタンパク質が自己凝集することによってウイルス粒子様の形状を有していることが示唆された。
【0138】
<電子顕微鏡観察>
精製したdBNC−ZZを電子顕微鏡観察した。観察結果を図12に示す。
【0139】
dBNC−ZZは、従来の野生型HBsAg粒子と同様の表面形状を示す事から、dHBsAgタンパク質が自己凝集することによって、エンベロープ様ナノ粒子構造形成する事が確認された。また、粒子径は22±4nmと、比較対象に用いた野生型HBsAg粒子(21±2nm)とほぼ同等の粒子径を有することも明らかとなった。この数値は、電子顕微鏡像から50個の粒子を選択し、その直径を測定した平均値である。
【0140】
<トリプシン切断実験>
4.1μg/10μLの濃度の精製したdBNC−ZZ粒子に、終濃度が12.5ng/μLとなるようにTPCK-トリプシン(Worthington Biochemial Corporation)を加えた後、37℃で、20分、2時間若しくは16時間消化反応を行った。
【0141】
反応後の液に5μLの4×Sample Buffer(5%の2−メルカプトエタノールを含有)を加えて、95℃で5分処理した後、15%アクリルアミドゲルを用いてSDS−PAGEを行い、CBB染色で全断片を、そしてHRP標識化抗HisProbe抗体を用いたウエスタンブロッティングによりC末端His-tagを含む切断断片を、HRP標識化抗HBsAg抗体を用いたウエスタンブロッティングによりN末端側HBsAg配列を含む切断断片を、各々検出した。結果を図13−2に示す。
【0142】
(A)及び(B)において、(1)に示すバンドから、dBNC−ZZタンパク質の全長の存在が示唆される。(2)に示すバンドから、図13−1に示すBの位置でトリプシン消化され、C末端側のHis−tagが欠失したタンパク質の存在が示唆される。(3)のバンドから、図13−1のCの位置でトリプシン消化され、ZZドメインのうち、C末端側のZドメインを欠失下タンパク質の存在が示唆される。(4)のバンドから、図13−1に示すDの位置でトリプシン消化され2つのZドメインが欠失したタンパク質の存在が示唆される。
【0143】
これらのバンドは、抗HBsAg抗体を用いたウエスタンブロッティングによっても確認されるバンドであることが(B)から明らかであり、本発明のdBNC−ZZにおいて、C末端側のZZドメイン及びHis−tagは、図13−1に示すようにウイルス様粒子表面に露出している構造を有しており、即ちC末端側のアミノ酸配列がウイルス粒子様構造の外側に提示されていることが明らかとなった。なお、図13−2の(B)において(5)にて示されるバンドは、抗His−tag抗体によって確認されるバンドであることから、図13−1のEの位置でトリプシン消化されたものと考えられる。
【0144】
<AFM観察>
精製したdBNC−ZZ粒子を金属板に固定したマイカ(雲母)に4μl添加した後に15分間乾燥させ、大塚蒸留水(大塚製薬,徳島)により3回洗浄した。再び乾燥させ、この試料を走査型プローブ顕微鏡(SPA400−DFM、SII NanoTechnology Inc.;千葉)により観察した。比較対象として、酵母にて作製した野生型BNC−ZZ粒子を用いた。結果を図14に示す。
【0145】
精製したdBNC−ZZ粒子は、高さが6.7−15.5nm程度で、直径は28.7−57.4nm程度であり、酵母で作製した野生型BNCーZZ粒子と類似した大きさであった。
【0146】
通常、脂質二重膜の厚さは5nm程度と考えられることから、10nm以上の厚さを示すのは、脂質二重膜が2層以上重なっていると考えられ、dHBsAgーZZ粒子は、従来のBNC−ZZ粒子と同様に、中空球状物の構造を有することが考えられる。
【技術分野】
【0001】
本発明は、ウイルス粒子様の形状を有するナノカプセルに関するものである。
【背景技術】
【0002】
ヒトB型肝炎ウイルス等のようなウイルスエンベロープタンパク質を宿主細胞にて生合成させると、当該タンパク質が宿主細胞から脂質二重膜といった脂質を取り込みつつ、自己凝集してウイルス粒子様のナノカプセルを形成することが既に知られている。
【0003】
例えば、特許文献1、2に示すように、ヒトB型肝炎ウイルス表面抗原(HBsAg)タンパク質を生合成させることで、ウイルス粒子様のナノカプセルが作製できることが知られている。しかしながら、当該ナノカプセルの作製のために用いられる宿主細胞としては、酵母細胞、動物細胞、昆虫細胞等といった真核細胞に限られており、原核細胞を用いてウイルス様の粒子を形成した報告は未だ存在しない。
【0004】
一方、特許文献3ではHBsAgタンパク質のトランケートミュータントを、大腸菌を宿主細胞として用いて生合成させる技術が開示されているが、抗原としてのみ用いられる技術しか開示されておらず、ウイルス粒子様のナノカプセルが製造できるかどうかについては何ら明らかにされていない。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特許第4085231号
【特許文献2】特許第4212921号
【特許文献3】特開2002−112797号公報
【発明の概要】
【発明が解決しようとする課題】
【0006】
生物宿主を用いて有用物質等を製造する場合、宿主として真核生物由来の細胞を用いるよりも原核生物由来の細胞を用いる方が、遥かに効率が良く、且つ簡便である。特に、タンパク質を主成分とするような有用物質を製造する際には、タンパク質を構成するアミノ酸に種々の変異を施して、機能を改良することが一般的な開発手段であるために、数多くの変異体を作製してその機能を評価するためには、効率が良く、簡便な製造手法が求められる。以上の点から、動物細胞、昆虫細胞、酵母細胞等の真核生物由来の細胞を宿主として有用物質を製造することは、あまり好ましくない。
【0007】
一方で、大腸菌などに代表される原核生物は、生産効率やコストの面において、有用物質を生産する宿主生物として非常に優れており、更に簡便な方法が提供できる。しかし複雑な立体構造等を有する有用物質の生産には向いておらず、立体構造を補正するシャペロン等といった特殊なタンパク質の存在下にて生産する方法や、化学的な試薬を用いて正常な立体構造に戻す操作が必要となり、結局のところ原核生物を用いる際に得られる効率の良さを相殺してしまうか、若しくはその効率を余計に悪くしてしまう虞もある。また、原核生物では生産量そのものが極めて低い有用物質も多い。
【0008】
さらに、結局、有用物質そのものでは機能を十分に発揮できないものが製造されてしまい、特定の機能ドメインのみに限定された有用物質の一部分のみの生産しか実現しない場合もある。
【0009】
本発明の課題は、特定の粒子構造を有するナノカプセルを、生物宿主を用いて高効率、且つ簡便に製造することである。
【課題を解決するための手段】
【0010】
上記の課題を解決すべく、出願人は鋭意研究を重ねた結果、特定のアミノ酸配列からなるタンパク質を含むナノカプセルが、従来のナノカプセルと同等か、又はそれ以上の効果を有しながらも、従来の製造方法よりも容易で、且つ簡便な方法によって製造できることを見出した。
【0011】
本発明は、係る知見に基づいて完成されたものであり、以下に示す態様の発明を広く包含するものである。
【0012】
項1 下記の(1)〜(4)のいずれかに示すアミノ酸配列からなるタンパク質を含むナノカプセル:
(1)配列番号1に示されるアミノ酸配列;
(2)上記(1)に示されるアミノ酸配列に、更に生体分子を認識するアミノ酸配列が付加されたアミノ酸配列;
(3)配列番号2に示されるアミノ酸配列;
(4)上記(1)〜(3)のいずれかに示されるアミノ酸配列において、1個又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
【0013】
項2 下記の(5)〜(10)のいずれかに示す塩基配列からなるポリヌクレオチド:
(5)配列番号1に示されるアミノ酸配列をコードする塩基配列;
(6)配列番号3に示される塩基配列;
(7)上記(5)または(6)に示される塩基配列に、更に生体分子を認識するアミノ酸配列をコードする塩基配列が付加された塩基配列;
(8)配列番号2に示されるアミノ酸配列をコードする塩基配列;
(9)配列番号4に示される塩基配列;
(10)上記(5)〜(9)のいずれかに示される塩基配列において、1個又は数個の塩基が置換、欠失、挿入、及び/又は付加された塩基配列。
【0014】
項3 上記項2に記載のポリヌクレオチドを含む組換えベクター。
【0015】
項4 上記項3に記載の組換えベクターを含む形質転換体。
【0016】
項5 上記項1に記載のナノカプセルを製造する方法。
【発明の効果】
【0017】
本発明のナノカプセルは、従前の大量生産に用いられていた酵母細胞ではなく、大腸菌を宿主細胞として用いて製造することが可能であるために、簡便、迅速、且つ低コストにナノカプセルを製造することが可能である。
【0018】
従って、本発明のナノカプセルに新たな機能を付与するために、ナノカプセルの構成要素であるタンパク質に変異を施して、有用な機能を有する様々なナノカプセルの開発にかかる時間は、従来に比べて大きく短縮させることが可能となる。
【0019】
また、本発明のナノカプセルは、従来のウイルス粒子様のナノカプセルとほぼ同等の粒径を有する。そして、本発明のナノカプセルには、従来のウイルス粒子様のナノカプセルと同様に、生体分子を認識するアミノ酸配列からなるペプチドをカプセルの外側に提示することが可能であり、ワクチンとしての利用のみならず、DDS、診断薬、イメージング試薬、免疫測定試薬、バイオセンサー試薬等の分野に有用である。
【0020】
そして、本発明のナノカプセルは、従来のナノカプセルと比較して短いアミノ酸配列からなるタンパク質を構成成分とするために、タンパク質の重量が比較的少ないナノカプセルが提供されるので、従来のナノカプセルと比較して、堅牢な構造を取り難くなる。従って、本発明のナノカプセルは、特に内部に物質を包含させる点において、従来のウイルス粒子様のナノカプセルよりも優れた効果を示す傾向となり、DDS等の用途に特に有用である。
【0021】
さらに、本発明のナノカプセルは、従来のウイルス粒子様のナノカプセルと比較して、抗原性が低いことから人体に投与した際の安全性が高いことが明らかであるため、特にDDSの分野において有用である。
【図面の簡単な説明】
【0022】
【図1】本発明の各種ナノカプセルが有するタンパク質のドメイン等を説明する模式図。
【図2】本発明の各種ナノカプセルが有するタンパク質のアミノ酸配列と、それをコードするポリヌクレオチドを含む発現ベクター等を説明する模式図。(A)はdHBsAgを示し、(B)はs+dHBsAgを示す。
【図3】本発明のナノカプセルが有するタンパク質を、各種大腸菌株を用いた合成、及び培養条件等を説明する図。
【図4】本発明のナノカプセルが有するタンパク質を、大腸菌を用いて合成させた際の菌体内局在等を説明する図。Mはマーカー、Tは全菌、Pはペリプラズム画分、S−は超音波破砕による可溶性画分、S+はTritonX−100存在下での超音波破砕による可溶性画分、I不溶性画分を示す。
【図5】本発明のナノカプセルが有するタンパク質を、大腸菌を用いて合成させた際の菌体内局在等を説明するグラフ。Cは培養上清、Pはペリプラズム画分、S−は超音波破砕による可溶性画分、S+はTritonX−100存在下での超音波破砕による可溶性画分を示す。
【図6−1】本発明のナノカプセルの精製時における、抗FLAG抗体カラムクロマトグラフィー溶出画分のSDS−PAGE解析を示す。Mはマーカー、S−は超音波破砕による可溶性画分、Fは素通り画分、IWはカラムの初回洗浄後画分、LWはカラムの最終洗浄画分を示す。
【図6−2】本発明のナノカプセル精製時の、ゲル濾過カラムクロマトグラムチャートを示す。
【図6−3】本発明のナノカプセル精製時の、ゲル濾過ピーク画分(Fr.10)のSDS−PAGE(CBB染色)、及び各種抗体を用いたウエスタンブロッティング解析を示す。
【図7】本発明のナノカプセルをショ糖密度勾配遠心解析した結果を示す図。黒丸は、dHBsAgの結果を表し、灰色の領域は従来法で作製したHBsAgを表す。グラフの左軸は、全タンパク質中のdHBsAg又はHBsAgタンパク質の重量割合を示す。
【図8】本発明のナノカプセルの電子顕微鏡写真像。(A)は市販の精製HBsAg粒子(ビークル社製)を示し、(B)はdHBsAg粒子を示す。
【図9】本発明のナノカプセルが有するタンパク質を、大腸菌を用いて合成させた際の培養条件、菌体内局在、及び精製条件等を説明する図。(A)培養条件及び菌体内局在を示す。Mはマーカー、S−は超音波破砕による可溶性画分、S+はTriton X−100存在下での超音波破砕による可溶性画分、Iは不溶性画分を示し、IPTGは、全菌試料を表す。(B)精製条件を表す。Mはマーカー、S−は超音波破砕による可溶性画分、EはNi−カラム溶出画分を示す。
【図10】本発明のナノカプセルと抗体の結合能等を説明する図。△はdBNCと抗体との結合を示し、○はdBNCーZZと抗体との結合を示す。
【図11】本発明のナノカプセルをショ糖密度勾配遠心解析した結果を示す図。(A)SDS−PAGE解析結果であり、Mはマーカーを示す。(B)SDS−PAGE解析における28kDaのバンドを数値化したもの(黒丸)。灰色は従来法で作製したHBsAgーZZを表す。グラフの左軸は、全タンパク質中の本発明のナノカプセル又はHBsAgタンパク質量の割合を示す。
【図12】本発明のナノカプセルの電子顕微鏡写真像。(A)市販の精製HBsAg粒子、(B)dHBsAg−ZZ粒子。
【図13−1】本発明のナノカプセルのトリプシン消化実験を説明する模式図。
【図13−2】本発明のナノカプセルのトリプシン消化実験の結果を示す図。(A)SDS−PAGE解析結果、(B)各種抗体を用いたウエスタンブロッティング解析。
【図14】本発明のナノカプセルのAFM写真像。(A)AFM写真像、(B)縦断面解析結果。
【発明を実施するための形態】
【0023】
本発明を実施するために使用する様々な技術は、特にその出典を明示した技術を除いては、公知の文献等に基づいて当業者であれば容易かつ確実に実施可能である。例えば、遺伝子工学及び分子生物学的技術であれば、Sambrook and Russell,「Molecular Cloning A LABORATORY MANUAL」, Cold Spring Harbor Laboratory Press, New York, 2001; Ausubel, F. M. et al. 「Current Protocols in Molecular Biology」, John Wiley & Sons, New York, N.Y等の文献を参照すればよい。
【0024】
本発明のナノカプセル
本発明のナノカプセルは、下記の(1)〜(4)のいずれかに示すアミノ酸配列からなるタンパク質を含む。
【0025】
(1)配列番号1に示されるアミノ酸配列;
(2)上記(1)に示されるアミノ酸配列に、更に生体分子を認識するアミノ酸配列が付加されたアミノ酸配列;
(3)配列番号2に示されるアミノ酸配列;
(4)上記(1)〜(3)のいずれかに示されるアミノ酸配列において、1個又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
【0026】
本発明のナノ粒子には、(1)〜(4)のいずれかに記載のアミノ酸配列からなるタンパク質のうち、一種のみのタンパク質が含まれていても良く、二種以上のタンパク質が含まれていても良い。例えば、(1)に示すアミノ酸配列からなるタンパク質と、(2)に示すアミノ酸配列からなるタンパク質を一定の比率で含むナノ粒子は、生体認識分子の提示能に優れることから、特に免疫測定試薬等の分野にて優れた効果を示す傾向にある。
【0027】
上記(1)〜(4)にて示すアミノ酸配列からなるタンパク質は、自己凝集することによってウイルス粒子様の形状を形成する性質を有する。このような性質を本明細書では粒子形成能と定義する。
【0028】
本発明のナノカプセルは、ウイルスエンベロープ様の粒子形状を有する。ウイルス粒子様の形状とは、脂質二重膜によって構成される殻の構造に、上記のタンパク質が包埋される形状である。本発明のナノカプセルは、リポソームといった脂質のみから構成される粒子形状の構造物と比較して、タンパク質が構成成分に含まれるため、リポソームよりも構造安定性が高いという性質を有する。このような形状は、単にウイルス様粒子とも称され、具体的には球状、楕円球状、ラグビーボール状、略球状等とも理解される形状である。
【0029】
本発明のナノカプセルには、上述のタンパク質以外に、製造する際の使用する宿主細胞に存在する、タンパク質、脂質、核酸等といった成分が含まれる。なお、これらの成分は上述のタンパク質に結合した状態でナノカプセルに含まれていても良い。
【0030】
本発明のナノカプセルにおける上記タンパク質の含有量は、特に限定はされないが、ナノカプセル100重量部に対して通常は75〜重量部90重量部程度である。
【0031】
本発明のナノカプセルに含まれる脂質の含有量は、特に限定されないが、ナノカプセル100重量部に対して通常は10〜重量部25重量部程度である。
【0032】
本発明のナノカプセルの内部は、中空であってもよく、水分、脂質成分、イオン成分等が含まれていてもよい。
【0033】
本発明のナノカプセルの粒子径は、通常は10〜200nm程度であり、より好ましくは13〜100nm程度、より好ましくは16〜50nm程度であり、最も好ましくは20〜30nm程度である。ここで、粒子径の測定方法は、電子顕微鏡像から、無作為に抽出したナノカプセルの直径を測定し、それらの平均値によって算出したものである。なお、本発明のナノカプセルは、エクストルーダーを用いて、孔径の小さいフィルターを通過させることによって適宜調節することが可能である。
【0034】
本発明のナノカプセルは、(a)及び(c)に上述したように従来のHBsAgタンパク質を含むナノカプセルと比較して、アミノ酸配列を大幅に欠失しながらも、粒子形成能を保持している。このようなアミノ酸からなるタンパク質を含む本発明のナノカプセルは、内部に蛍光物質、放射性物質、磁性物質等の標識物質;DNA、RNA、PNA等の核酸;薬剤といったDDS、生体内でのキャリアー等として送達又は移送させるべき分子を、より高効率に内包させることが可能である。
【0035】
なお、ここでいう内包とは、ナノカプセルの内部に当該分子の全てが完全に包み込まれている必要はなく、当該分子の一部がナノカプセルの外部に突出していてもよい。
【0036】
本発明のナノカプセルに、上述した分子を内包させる方法は、特に限定はされないが、脂質融合を利用した方法、エレクトロポーレーション法等が挙げられる。
【0037】
その他に、本発明のナノカプセルを構成する粒子形成能を有するタンパク質を、界面活性剤やその他手段を用いて粒子構造を破壊し、次いで所望の内包分子と混和した後に、再度、粒子形成させて得られるナノカプセル中に、分子を内包する方法が挙げられる。
【0038】
上記(1)にて定義する配列番号1に示されるアミノ酸配列は、NCBIのAccession No.Q9W966に示されるB型肝炎ウイルス表面抗原(以後、HBsAgと称することがある。)のSタンパク質のアミノ酸配列(アミノ酸残基数226)に対して、以下の(a)〜(c)に示す変異を施したアミノ酸配列である。
【0039】
(a)2〜77番目のアミノ酸残基の欠失。
(b)78番目及び79番目のアルギニン残基の、それぞれセリン残基への置換。
(c)173−226番目のアミノ酸残基を欠失。
【0040】
特に、上記の(b)に示すような変異を導入することにより、本発明のタンパク質が宿主細胞内にて効率的に作製され易くなり、ひいては当該タンパク質を含むナノカプセルの形成に有利となる。
【0041】
上記(2)にて定義する生体分子は特に限定されないが、具体的には表面抗原、レセプター、抗体、サイトカイン、転写因子等の糖タンパク質若しくはタンパク質、又はペプチド若しくは糖ペプチド;ヘパリン、シアルルイスX等の糖若しくは糖鎖等が挙げられる。
【0042】
上記(2)にて定義する生体分子を認識するアミノ酸配列は、特に限定はされないが、例えば生体分子がレセプターであれば、当該レセプターに対して相互作用する各種成長因子等のアミノ酸配列とすればよい。
【0043】
例えば、生体分子がビオチンであれば、アビジン、ストレプトアビジン、ニュートラアビジン等に代表されるアビジン系タンパク質のアミノ酸配列とすればよい。
【0044】
例えば、生体分子が糖鎖である場合には、アグルチニン、セレクチン等のレクチンのアミノ酸配列とすればよい。
【0045】
例えば、生体分子が抗体である場合には、配列番号5に示すStaphyrococcus aureusu由来のProtein AにおけるイムノグロブリンGのFcドメインに結合する領域に存在するアミノ酸配列であるZタグのアミノ酸配列とすればよい。なお、ここで言う抗体とは、上述のFcドメインを有さないFab、F(ab′)2等の構造を有する抗体は含まれない。
【0046】
例えば、標的生体分子を認識するペプチドあるいはアフィボディーを、ライブラリーから選んで、そのアミノ酸配列を用いてもよい。
【0047】
上記(2)にて定義する生体分子を認識するアミノ酸配列は、上記(1)に示されるアミノ酸配列の内部、N末端側、又はC末端側のいずれの位置に付加されていてもよいが、当該アミノ酸配列からなるタンパク質を含むナノカプセルの構造の安定性、又は生体分子を認識するアミノ酸配列からなるタンパク質若しくはペプチドをナノカプセルの外側に提示させることに鑑みて、C末端側に付加されることが好ましい。
【0048】
ここで、「C末端側」とは、C末端から1〜5番目程度のアミノ酸残基の位置、又はC末端側に0〜15個程度のアミノ酸配列(いわゆるスペーサー配列)を付加したさらに後ろ側の位置を意味する。なお、「N末端側」も同様に説明される。
【0049】
このように、生体分子を認識するアミノ酸配列が、ナノカプセルの外側に提示された上記(2)にて定義するアミノ酸配列としては、配列番号2に示すアミノ酸配列又に示すアミノ酸配列などが挙げられる。
【0050】
なお、生体分子を認識するアミノ酸配列は一種でも二種以上を組み合わせて付加されていてもよい。
【0051】
上記(4)にて定義する置換、欠失、挿入、及び/又は付加されるアミノ酸の数は、(4)に定義するアミノ酸配列からなるタンパク質が、粒子形成能を失わない範囲とすればよく、特には限定されないが、通常は20個程度以下とすればよく、好ましくは15個程度以下、より好ましくは10個程度以下、更に好ましくは5個程度以下であり、最も好ましくは1個である。
【0052】
なお、置換に関しては、上記(4)にて定義するアミノ酸配列からなるタンパク質が粒子形成能を失わないように、置換対象となるアミノ酸そのものの性質を変化させないような類似アミノ酸の範疇において適宜選択すればよい。ここで、類似アミノ酸とは、Phe、Trp、Tyr等といった芳香族アミノ酸;Ala、Leu、Ile、Val等といった脂肪族アミノ酸;Gln、Asn等といった極性アミノ酸;Lys、Arg、His等といった塩基性アミノ酸;Glu、Asp等といった酸性アミノ酸;Ser、Thr等といった水酸基を有するアミノ酸;Gly、Ala、Ser、Thr、Met等といった側鎖の小さいアミノ酸等に挙げられる各グループ内のアミノ酸である。例えばLeuは、同じ脂肪族アミノ酸の範疇に含まれる類似アミノ酸であるIleに置換すればよい。
【0053】
なお、上述の(1)〜(4)のいずれかに示すアミノ酸配列からなる本発明のタンパク質には、当該タンパク質を精製すること、当該タンパク質を検出すること等を目的として、各種タグポリペプチドが付加されていてもよい。具体的なタグポリペプチドとして、Hisタグ、FLAGタグ、HAタグ、mycタグ、MBPタグ、GSTタグ、GFPタグ、Zタグ、ストレプタグ等が挙げられる。 これらのタグポリペプチドは、単独で付加されていても、二種以上を組み合わせて付加されていてもよい。なお、上記のタグポリペプチドは、上述した(1)〜(4)のいずれかに示すアミノ酸配列内部に挿入される形で付加されていても、当該アミノ酸配列のC末端側に付加されていても、N末端側に付加されていてもよい。本発明のナノカプセルの構造の安定性、又は当該タグポリペプチドをナノカプセルの外側に提示することに鑑みて、C末端側に付加されることが好ましい。なお、「N末端側」、及び「C末端側」なる用語は、既に定義したとおりである。
【0054】
本発明のポリヌクレオチド
本発明のポリヌクレオチドは下記の(5)〜(10)のいずれかに示す塩基配列からなる。
【0055】
(5)配列番号1に示されるアミノ酸配列をコードする塩基配列。
(6)配列番号3に示される塩基配列。
(7)上記(5)または(6)に示される塩基配列に、更に生体分子を認識するアミノ酸配列をコードする塩基配列が付加された塩基配列。
(8)配列番号2に示されるアミノ酸配列をコードする塩基配列。
(9)配列番号4に示される塩基配列。
(10)上記(5)〜(9)のいずれかに示される塩基配列において、1個又は数個の塩基が置換、欠失、挿入、及び/又は付加された塩基配列。
【0056】
上記(5)にて規定する塩基配列は、配列番号1に示されるアミノ酸配列を基に、in silico技術を採用することによって塩基配列を決定することができる。in silico技術を用いれば、1つのアミノ酸配列から、そのアミノ酸配列をコードする多数の塩基配列が決定されることになるが、当該アミノ酸配列をコードする遺伝子を発現させる際に用いる宿主細胞に対して適当なコドン頻度を考慮すれば、配列番号1に示すアミノ酸配列をコードする、より好ましい態様の塩基配列を決定することができる。
【0057】
上記(6)にて規定する配列番号3に示される塩基配列は、上記(5)に示す配列番号1に示されるアミノ酸配列をコードする塩基配列の一態様である。
【0058】
上記(7)にて規定する生体分子、及び生体分子を認識するアミノ酸配列は、上述した本発明のナノカプセルの(2)に定義したものと同様にすればよく、例えば上述したZタグをコードする塩基配列は、配列番号6にて示される。
【0059】
なお、当該アミノ酸配列をコードする塩基配列も、上述の(5)にて説明したin silico技術を採用することによって決定できる。そして、当該塩基配列の好ましい態様も、上述のようにコドン頻度を考慮して決定できる。
【0060】
上記(8)にて規定する塩基配列も上記(5)と同様に、上述の配列番号2に示すアミノ酸配列を基に、in silico技術並びにコドン頻度を考慮して決定することができる。
【0061】
上記(9)にて規定する配列番号4に示される塩基配列は、上記(8)に示す配列番号2を基に決定した塩基配列の一態様である。
【0062】
上記(10)にて定義する置換、欠失、挿入、及び/又は付加される塩基の数は、(10)に定義する塩基配列がコードするアミノ酸配列からなるタンパク質が、粒子形成能を失わない範囲とすればよく、特には限定されないが、通常は30個以下とすればよく、好ましくは20個以下、より好ましくは10個以下、更に好ましくは5個以下であり、最も好ましくは3個以下である。
【0063】
なお、置換に関しては、上記(10)にて定義する塩基配列がコードするアミノ酸配列において、上記(4)にて定義した他の類似アミノ酸となるように塩基配列を置換すればよい。例えば、塩基配列中において、脂肪族アミノ酸の範疇に含まれるLeuを、同じく脂肪族アミノ酸の範疇に含まれる類似アミノ酸であるIleに置換するように、Leuをコードする「CTT」なる塩基配列を、「ATT」と置換すればよい。
【0064】
本発明の組換えベクター
本発明の本発明の組換えベクターは、上述した本発明のポリヌクレオチドを含むものである。組換えベクターとは生物宿主に導入することによって、特定の遺伝子を発現するものであり、本発明においては、上述した本発明のポリヌクレオチドを発現するものである。
【0065】
上記の生物宿主としては、特に限定はされないが、原核細胞であることが好ましい。具体的には、大腸菌、枯草菌、乳酸菌等が挙げられるが上述した本発明のポリヌクレオチドを効率的に発現させることを勘案すれば、大腸菌であることが好ましい。即ち、本発明の組換えベクターは、上述した本発明のポリヌクレオチドを含む大腸菌発現用ベクターが好ましい。
【0066】
具体的な大腸菌発現ベクターは、公知のものを採用すればよく、特に限定はされないが、IPTG、ラムノース等の存在下において、上述した本発明のポリヌクレオチドの発現を誘導することができるベクターを用いることが好ましい。例えば、pETベクターシリーズや、pGEXベクターシリーズ等を挙げることができる。
【0067】
本発明の形質転換体
本発明の形質転換体は、上述した本発明の組換えベクターを含むものであり、上述した組換えベクターによって上述の生物宿主が形質転換されたものを意味する。
【0068】
形質転換する方法は特には限定されず、塩化カルシウム法や塩化ルビジウム法で作成したコンピテントセルを用いる方法、エレクトロポーレーション法、プロトプラスト法等が挙げられる。
【0069】
なお、上述の組換えベクターが、IPTGによって特定の遺伝子の発現を誘導するものであり、用いる宿主細胞が大腸菌である場合には、T7RNAポリメラーゼ遺伝子がゲノム中に配置されたDE3株の大腸菌であることが好ましい。
【0070】
本発明のナノカプセルを製造する方法
本発明のナノカプセルを製造する方法は、以下の3つの工程を含むものである。
(i)上述した本発明の形質転換体を培養する工程、
(ii)当該形質転換体の培養物の可溶化液を回収する工程、及び
(iii)当該可溶化液からナノカプセルを精製する工程。
【0071】
上記(i)における、上述した形質転換体の培養方法は、それぞれの形質転換体に適合した公知の培養方法を採用すればよい。例えば、上述のように形質転換体が大腸菌であり、IPTG、ラムノース等による発現誘導を可能とする組換えベクターを保持している場合には、20℃程度の低温条件下で長時間の培養を行うことが好ましい。
【0072】
上記(ii)における、形質転換体の培養物からの可溶化液の回収方法は特に限定されず、Triton−Xシリーズ、Tweenシリーズ、NP40等といった公知の界面活性剤を用いる方法;超音波処理に供する方法;液体窒素等の冷媒を用い、特定の機器を用いて細胞膜、細胞壁等を破砕する凍結融解法;ガラスビーズ法;ザイモリエースなどのような細胞壁を消化する酵素を用いたスフェロプラスト法;高圧化にて処理を行うフレンチプレス法等の公知の可溶化方法を培養物に対して採用した後に、遠心分離処理に供して得られる液相分画を回収すればよい。なお、これらの可溶化方法は、単独であっても、2種類以上を組み合わせて採用しても良い。
【0073】
上記(iii)における、可溶化液から本発明のナノカプセルを精製する方法は、特に限定されることはなく、公知のタンパク質の精製法を採用すればよい。
【0074】
例えば、上述の可溶化液を塩化セシウム、スクロース、グリセロール、OptiPrep、Percol等の成分を各種濃度又は線形グラジエント濃度勾配にて含有する緩衝液と、超遠心分離処理を採用した密度勾配遠心分離法に供して、本発明のナノカプセル以外の成分を除去する方法;上述の可溶化液に熱処理を与えて、主に夾雑タンパク質等を変性させて本発明のナノカプセル以外の成分を除去する方法;上述の可溶化液に対して硫酸アンモニウム、エタノール酢酸、アセトン等を作用させて、主に夾雑タンパク質を変性させることにより、本発明のナノカプセル以外の成分を除去する方法;上述の可溶化液に対して硫酸ナトリウム、硫酸アンモニウム、ポリエチレングリコールを加えて沈殿させる事で、本発明のナノカプセル以外の成分を除去する方法;上述の可溶化液に対してクロマトグラフィー用いて、本発明のナノカプセル以外の成分を除去する方法等が挙げられ、これらの方法は単独で採用しても、二種類以上を組み合わせて採用しても良い。
【0075】
上述したクロマトグラフィーの具体的な種類は、特に限定されないが、陽イオン交換クロマトグラフィー、陰イオン交換クロマトグラフィー、アフィニティクロマトグラフィー、サイズ排除ゲル濾過クロマトグラフィー、逆相クロマトグラフィー、疎水クロマトグラフィー等を挙げることができ、これらのクロマトグラフィーは単独で用いても、二種以上を組み合わせて用いてもよい。
【0076】
本発明の組換えベクターが、上述したタグポリペプチドをコードするポリヌクレオチド配列が含む場合には、当該タグポリペプチドとの相互作用に基づいたアフィニティカラムを用いることが好ましい。
【0077】
例えば、FLAGタグ、HAタグ、mycタグ等のタグポリペプチドをコードするポリヌクレオチド配列を含む組換えベクターを含む形質転換体から得られた可溶化液より精製する場合、それぞれ抗FLAG抗体、抗HA抗体、抗myc抗体等といった、タグポリペプチドに対して特異的に結合する抗体を担持するポリマーを担持したアフィニティクロマトグラフィーを用いることが好ましい。このようなアフィニティクロマトグラフィーを用いる場合の溶出液としては、当該ペプチドを高濃度で含有する緩衝液、又はGlysin−HCl緩衝液のような、pHが1〜5程度の酸性緩衝液を用いることが好ましい。
【0078】
また、本発明のナノカプセルがMBPタグを有する場合には、マルトース等といった糖類を担持するポリマーを採用したアフィニティクロマトグラフィーを用いることが好ましい。このようなアフィニティクロマトグラフィーを用いる場合の溶出液としては、糖類を高濃度で含む緩衝液を用いることが好ましい。
【0079】
さらに、本発明のナノカプセルがHisタグを有する場合には、Ni−NTAといったヒスチジンとキレート結合が可能となる金属化合物を担持するポリマーを採用したアフィニティクロマトグラフィーを用いることが好ましい。このようなアフィニティクロマトグラフィーを用いる場合の溶出液としては、イミダゾールを高濃度で含有する緩衝液を用いることが好ましい。
【0080】
このようなタグポリペプチドは、精製後適当なプロテアーゼで本発明のナノカプセルから切断除去してもよい。
【0081】
なお、本発明のナノカプセルが、上述した生体分子を認識するアミノ酸配列が付加されたアミノ酸配列からなるタンパク質を含む場合、当該生体分子を担持するポリマーを採用したアフィニティクロマトグラフィーを用いることが有効である。このようなアフィニティクロマトグラフィーを用いる場合の溶出液としては、当該生体分子を高濃度で含有する緩衝液を用いることが好ましい。
【0082】
例えば、生体分子がストレプトアビジンのようなアビジン系タンパク質であり、生体分子を認識するアミノ酸がビオチンである場合には、アビジン系のタンパク質を担持するポリマーを採用したアフィニティクロマトグラフィーを用いることが有効であり、溶出液としてはビオチンを高濃度で含有する緩衝液を用いる方法が挙げられる。
【0083】
なお、本発明のナノカプセルを製造する過程では、当該ナノカプセルを構成するタンパク質の分解を抑えるために、低温条件下での操作とすることが好ましく、必要に応じてEDTA、PMSF等のタンパク質分解酵素阻害剤を1種又は2種以上で組み合わせて用いてもよい。
【0084】
上述のように、本発明のナノカプセルは、上述の本発明の粒子形成能を有するタンパク質が自己凝集することによってウイルス様粒子となり形成されるが、ウイルスエンベロープ様粒子を形成する場は、宿主細胞内、試験管内、培養容器内等といった当該タンパク質の合成の場か、タンパク質合成の終了後の精製、回収、保存等の場であり、特に限定はされない。
【実施例】
【0085】
1)発現ベクターの構築
C末端54残基を欠失させFLAG−tag配列で置換したHBsAg抗原タンパク質をコードするプラスミドDNAである、ベクターpBO749(国際公開番号WO2005/049824)を鋳型とし、primer#1575(配列番号7)と#1555(配列番号8)とを用いたPCRにより、N末端76残基とC末端54残基とを同時に欠失した、配列番号9に示す塩基配列を含む欠失型HBsAg遺伝子(以後、dHBsAgと称することがある。)断片を増幅した。
【0086】
増幅された遺伝子断片をNdeI及びNotIで制限酵素消化処理した後、アガロースゲル電気泳動でdHBsAg遺伝子断片を単離及び精製した。大腸菌発現用ベクターpET22b(Novagen社)をNdeI及びNotIで制限酵素消化処理した後、dHBsAg遺伝子断片のNdeI及びNotIによる制限酵素消化処理後物と連結し、dHBsAg大腸菌発現用ベクターpBO1465を得た。
【0087】
構築されたdHBsAg遺伝子には、C末端側にFLAG−tagとHis6−tag配列が融合されている(図1及び図2)。
【0088】
同様に、ベクターpBO749を鋳型とし、primer#1554(配列番号10)と#1555(配列番号8)とを用いたPCRにより、N末端76残基とC末端54残基とを同時に欠失した、配列番号11に示す塩基配列からなる欠失型HBsAg遺伝子(以後s+dHBsAgと称することがある。)断片を増幅した。
【0089】
増幅された遺伝子断片をNdeI及びNotIで制限酵素処理した後、アガロースゲル電気泳動でdHBsAg遺伝子断片を単離及び精製した。大腸菌発現用ベクターpET22bも同様にNdeI及びNotIで制限酵素処理した後、制限酵素処理後のdHBsAg遺伝子断片と連結し、s+dHBsAg大腸菌発現用ベクターpBO1458を得た。
【0090】
なお図2にて示すように、構築されたs+dHBsAg遺伝子は、上述のdHBsAg遺伝子と比較すれば、アミノ酸に翻訳された際にN末端側のアミノ酸配列のみが異なっている。
【0091】
2)大腸菌によるdHBsAgの発現
上記のdHBsAg大腸菌発現用ベクターpBO1465及びs+dHBsAg大腸菌発現ベクターpBO1458を遺伝子発現用大腸菌Origami2(DE3)株、C41(DE3)株、C43(DE3)株、並びにKRXに形質転換し、翌日出現したコロニー数十個を、0.5%glucoseと終濃度で200μg/mlのアンピシリンを含む4mL若しくは30mLのLB培地で、37℃で2〜5時間振盪培養した。
【0092】
1.5mL若しくは12mLの菌懸濁液を、それぞれ終濃度で500μg/mlアンピシリンをする含有50mL若しくは400mLのLB培地に植菌し、さらに37℃、2〜5時間振盪培養した。OD600が0.8〜1.0になったところでIPTGを終濃度が1mMとなるように加えた。なお、KRX株の場合は、IPTGに換えて0.1%のラムノースを添加した。低温培養検討(20℃で4時間〜一晩培養)時には、IPTG添加前に培養温度を20℃に下げた。
【0093】
IPTGの添加後、37℃で4時間若しくは20℃で一晩(低温培養時)培養した後に菌体を回収した。IPTG添加前と培養終了時における1mlの培養液分の菌体を超純水100μLに再懸濁した後、当容量の2×sample buffer[20%のglycerol、4%SDS、0.02%BPB、及び10%の2−MEを含む50mMのTris−HCl(pH6.8)]を加えて溶解し、95℃で5分間加熱処理した後の20μL(全菌試料:Tとする)をSDS−PAGE処理に供し、続いてクマシーブリリアントブルー染色(CBB染色)を行った。結果を図3に示す。
【0094】
s+dHBsAgは、いずれの大腸菌株においてもIPTG誘導に基づくバンドの出現は観察されなかったが、dHBsAgでは図中の矢印の位置に示されるように、全ての大腸菌株においてIPTG誘導に基づくバンドの出現は観察された。以上のことから、大腸菌を用いてナノカプセルを構成するタンパク質を生合成するには、そのN末端側のアミノ酸配列が重要であることが明らかとなった。
【0095】
3)dHBsAg発現における大腸菌体内の局在
dHBsAgを発現する大腸菌Origami2(DE3)株を上記の各種培養条件で培養し、菌体回収時の遠心後の上清を培養上清(C)とした。1mLの培地による培養にて得られた菌体を、20%のsucroseを含む30mMのTris−HCl(pH8)を75mL用いて再懸濁した。ここに、0.5MのEDTA(pH8)を1.5μL加え、室温条件下で10分間Vortex処理を行った。
【0096】
次いで、10000rpm、4℃の条件で10分間遠心し、上清を取り除いた。得られた沈殿画分を、氷冷した5mMのMgSO4に再懸濁し、4℃で10分間vortex処理を行った。次いで、10000rpm、4℃の条件にて、10分間遠心し、得られた上清をペリプラズム画分(P)とした。一方で、得られた沈殿画分を、0.75mLの氷冷Lysis buffer[5%のglycerol、及び150mMのNaClを含む50mMのTris−HCl(pH7.5)]に再懸濁し、終濃度1mMのEDTA、及び1mMのPMSFを加えた。
【0097】
このサンプルを、2回凍結融解処理に供した後、氷上で30秒ずつ2回の超音波破砕処理を行った。次いで、14000rpm、4℃の条件にて、10分間遠心し、得られた上清画分を可溶性画分(S−)とした。
【0098】
一方で、得られた沈殿画分を0.75mLの氷冷Lysis bufferに懸濁し、終濃度1mMのEDTA、及び1mMのPMSFを加えた。さらに終濃度が0.5%となるようにTriotnX−100を加え、30秒間の超音波破砕処理を1回行った。次いで、14000rpm、4℃の条件にて10分間遠心し、得られた上清画分を可溶性画分(S+)とした。
【0099】
一方で、得られた沈殿画分を0.75mLの氷冷Lysis bufferに再懸濁したものを不溶性画分(I)とした。以上の各画分をSDS−PAGE処理に供し、次いでCBB染色を行った。結果を図4に示す。
【0100】
図中の矢印の位置に示されるように、dHBsAgタンパク質はペリプラズム画分(P)には認められず、TritonX−100非存在下並びに存在下の可溶性画分(S−並びにS+)、及び不溶性画分(I)に分散して認められた。また、37℃における培養では不溶性画分(I)へ、20℃における培養では、可溶性画分(S−並びにS+)により多く局在することが明らかとなった。
【0101】
また、培養時に得られた菌体回収時の上清画分(C)、ペリプラズム画分(P)、及びTritonX−100存在下、並びに非存在下の可溶性画分(S−並びにS+)を、抗HBsAg抗体を用いた酵素免疫測定法(IMx−HBsAg;ダイナボッド社)にて、HBsAgタンパク質を測定した。結果を図5に示す。
【0102】
図4にて示される結果と同様に、HBsAgタンパク質はペリプラズム画分(P)には殆どは局在が認められず、TritonX−100などのような界面活性剤の存在下並びに非存在下の可溶性画分(S−並びにS+)として抽出できることが明らかとなった。
【0103】
4)dHBsAgタンパク質を含むナノカプセルの精製
引き続いて、dHBsAgタンパク質を含むナノカプセル(以後、dBNCと呼ぶことがある。)の精製を行った。
【0104】
IPTGによる誘導下(培養温度は37℃)にてdHBsAgを発現した菌体をLysis Buffer[150mMのNaClを含む50mMのTris−HCl(pH8.0)]に再懸濁し、最終濃度が1mMになるようにPMSFを加えた。次いで、1分ずつ5回氷上で冷却しながら超音波破砕した。その後、8000rpm、4℃の条件下で15分間遠心し上清を可溶性画分(S−)とした。ここで得られた沈殿を0.1%のTween80を含むLysis Bufferに再懸濁し、最終濃度が1mMとなるようにPMSFを加えて、1分ずつ5回氷上で冷却しながら超音波破砕した。超音波破砕後の溶液を同様の条件で遠心して、上清を可溶性画分(S+)とした。残った沈澱を超純水に懸濁して不溶性画分(I)とした。これらの試料(100μLの培養液分に相当)に当容量の2×sample bufferを加えて溶解し、95℃で5分間加熱処理した後SDS−PAGEのサンプルとした。
【0105】
TBS[150mMのNaClを含む50mMのTris−HCl(pH7.4)]で平衡化した抗FLAG M2 Agarose Affinity Gel (Sigma社)カラム(bed量:1mL)に、上記の可溶性画分(S−)をインジェクションし、4℃で2時間rotate後、ゆっくり流し素通り液を回収した。回収した画分を素通り画分(F)とした。
【0106】
次に、0.75mLのTBS500[NaCl濃度を500mMとした上記TBS]でカラムを数回洗浄後(初回の洗浄後画分を(IW)とし、最終洗浄画分を(LW)とする。)、0.4mLのElution Buffer[1mMのEDTAを含む0.1MのGlycine−HCl(pH3.5)]で8回に分けて溶出した。得られた溶出画分をそれぞれ溶出画分(1)〜(8)とした。
【0107】
溶出画分には直ちに1MのTris−HCl(pH8.0)を14μL混合して中和し、その後、各画分をSDS−PAGEに供し、次いでCBB染色を行った。結果を図6−1に示す。
【0108】
図中の矢印に示すように、抗FLAGカラム溶出画分の(2)〜(7)において、dHBsAgタンパク質のバンドが検出された。これらの溶出液のそれぞれ2mLを、0.15MのNaClを含む50mMのTris−HCl(pH7.4)で平衡化したSephacryl S−300HRゲル濾過カラム(GE Healthcare;直径1.6cm×高さ40cm;bed量24mL)にインジェクションし、0.75mLを1画分として回収した。
【0109】
図6−2に示すように、HBsAgタンパク質は、10番目の画分にて最も多く溶出されることが明らかとなった。当該画分は669kDaの分子量を有するサイログロブリンと同様の位置で流出することが明らかとなった。そして、図6−3に示すように、10番目の当該溶出ピーク画分はSDS−PAGE解析により80%以上の純度でdHBsAgタンパク質を含有することを示した。
【0110】
また図6−3に示すようにHRP標識化抗FLAG抗体(抗FLAG M2 PEROXIDASE CONJUGATE、SIGMA)、ヤギ由来のビオチン標識化抗HBs抗体とウサギ由来のアルカリフォスファターゼ標識化抗ビオチン抗体(IMx HBsAg試薬;ダイナボット)、あるいはHisProbeTM−HRP結合体(Thermo社)を各々用いたウエスタンブロッティングでも検出された事から、目的のdHBsAgバンドである事が確認された。
【0111】
以上の結果より、dHBsAgタンパク質は自己凝集して700kDa程度の複合体を形成していることが確認された。
【0112】
5)dBNCの性状解析
<Sucrose密度勾配遠心解析>
16mLの遠心管(16PA TUBE ASSY;HITACHI社)に底からスクロース濃度を50、40、30、20、及び10重量%含むBufferA[68.4mMのNa2HPO4、31.6mMのNaH2PO4、15mMの EDTA・2Na、及び0.85%のNaCl]を2.5mLずつ順に重層し、その上に上記4)にて示す可溶性画分(S−)を2.5mL重層した後、超遠心機(himac CP70MXX;HITACHI社)を用いて4℃、24000rpmの条件で14時間遠心した。
【0113】
陽性対照として、Cos7細胞によって一過的に製造されたHBsAg粒子(培養上清から回収したもの)を用いた。また、陰性対照として、2mgのウサギ血清由来のIgG(SIGMA、分子量150kDa)をBufferAで2.5mLに希釈したものを用いた。
【0114】
遠心処理の後、あらかじめ重量を測定しておいた1.5mLチューブに1mLずつ回収し、重量を測定して密度を求めた。各画分のHBsAg量をIMx HBsAg System(ダイナボット社)を用いて測定した。ウサギ血清由来IgGの濃度はUV法により測定した。その結果を図6に示す。
【0115】
dHBsAgタンパク質はCos7細胞を用いて作製したHBsAg粒子と同様に6−8番目の画分に沈降ピークを示した。ウサギ由来IgGの沈降ピークは2番目画分であった。以上の結果から、dHBsAgタンパク質は自己凝集し、従来のHBsAg粒子と同様に粒子形成していることが示唆された。
【0116】
<電子顕微鏡観察>
抗FLAG抗体溶出画分について、ネガティブ染色法を用いて透過型電子顕微鏡(ELECTRON MICROSCOPE H−7100S;HITACHI)観察した。比較対象として野生型HBsAg粒子(ビークル社製)を用いた。結果を図5に示す。
【0117】
dHBsAgタンパク質は、比較対象に用いた野生型HBsAg粒子と同様に、ウイルスエンベロープ様ナノ粒子(dBNC)が形成する事が明らかとなった。また、dBNCの粒子径は25±5nmと、比較対象に用いた野生型HBsAg粒子(21±2nm)とほぼ同等の粒子径を有することも明らかとなった。この数値は、電子顕微鏡像から50個の粒子を選択し、その直径を測定した平均値である。
【0118】
<dBNCの抗原性>
2種の市販のHBV体外診断用酵素免疫測定法キット(IMx HBsAg System;Abbott社、及びEnzygnost HBsAg 5.0 kit;SIMENS社)を用いて、精製dBNCの抗原性を市販のHBsAgS粒子(HBsAg−adr recombinant protein;GenWay Biotech社、10−663−45362)を標準試料として測定した。表1に結果を示す。
【0119】
【表1】
【0120】
dBNCの抗原性は、野生型S粒子と比較してIMxHBsAgキットによる測定では1/7、Enzygnostキットによる測定では約1/200に低下していた。
【0121】
従って、本発明のdBNCは、特にDDSとして体内に投与した場合、より安全に用いることができることが明らかとなった。
【0122】
6)ZZタグ融合HBsAg欠失体(dHBsAg−ZZ)発現ベクターの構築
まず、dHBsAg遺伝子発現ベクターpBO1465を鋳型とし、primer#361(配列番号12)と、#1633(配列番号13)とを用いたPCRを行いて遺伝子断片を増幅し、その反応液をフェノール抽出後エタノール沈殿した後、NdeI及びNotIで制限酵素消化処理を行うことにより、C末端にFLAGタグを含まない約0.3kbのdHBsAgの遺伝子断片を調製した。
【0123】
この遺伝子断片と、NdeI及びNotIで制限酵素消化処理を施したpBO1465ベクターの断片とをT4−ligaseで連結することにより、C末端にHis6−tagのみが付いたdHBsAg遺伝子を発現するベクターpBO1501を構築した。
【0124】
一方、10ngの酵母発現用ベクターpGLD−ZZ−d50N(Specific protein delivery to target cells by antibody−displaying bionanocapsules. Kurata N, Shishido T, Muraoka M, Tanaka T, Ogino C, Fukuda H, Kondo A. J Biochem. 2008 Dec;144(6):701−7.)を鋳型とし、primer #1732(配列番号14)と、#1733(配列番号15)とをそれぞれ10pmol用い、さらにTaq DNA polymarase(NEB社;1units/μL)を用いPCRにより、ZZtag遺伝子を増幅した後、72℃で14分間アデニン付加反応を行った。
【0125】
この増幅DNA反応液をフェノール抽出後、エタノール沈澱し、30μLの滅菌超純水に再溶解した。これを1.2%アガロースゲル電気泳動で電気泳動後、目的のZZ−tag遺伝子断片を含むゲル片を切り出した。具体的には、QIAEXII Gel− Extractionキット(QIAGEN社)を用いて、アガロースゲルから目的DNA断片を抽出精製した。このZZtag遺伝子断片とpTAC−1 vector(BioDynamics Labs)をT4−ligaseで連結することにより、ZZtag遺伝子をpTAC−1ベクターにクローニングして、pBO1594ベクターを作製した。
【0126】
得られたpBO1594ベクターをNotIで制限酵素消化処理し、1.5%アガロースゲル電気泳動した後、ZZtag断片を含むゲル片を切り出した。QIAEXII Gel Extractionキットで、目的となるZZtagDNA断片を抽出精製し、insert DNAとした。
【0127】
一方、pBO1501ベクターをNotIで制限酵素消化処理した後、65℃で20分間酵素失活反応させた。このNotIによって制限酵素消化処理したpBO1501の断片と、NotIによって制限酵素消化処理したZZtag断片とを、Ligation High ver.2(TOYOBO社)を用いて連結することにより、配列番号16に示す塩基配列を含むdHBsAg−ZZ遺伝子発現用ベクターpBO1595を構築した。
【0128】
このようにして作製したpBO1595ベクターには、dHBsAg−ZZのC末端に検出精製用のHis6−tag配列が融合されたタンパク質をコードする遺伝子が含まれている。
【0129】
7)dHBsAg−ZZの大腸菌による発現と精製
上述のdHBsAg−ZZ発現ベクターpBO1595を、大腸菌株Origami2(DE3)に形質転換した。翌日出現したコロニー数十個について、実施例2と同様の方法で遺伝子の発現及び培養を行い、得られた遺伝子発現菌体について、Tween80を含まない緩衝液にて超音波破砕処理によって可溶化して得られる可溶性画分Tween−(S−)、Tween80を含む緩衝液にて超音波破砕処理によって可溶化して得られる可溶性画分(S+)、及び不溶性画分(I)を調製した。これらの試料(100μLの培養液分)に当容量の2×sample bufferを加えて溶解し、95℃で5分間加熱処理した後、SDS−PAGE処理に供し、次いでCBB染色を行った。結果を図9(A)に示す。
【0130】
図中の矢印にて示されるように、IPTG誘導下による培養を行った場合に、明瞭なdHBsAg−ZZの発現バンドが見られ、主に可溶性画分(S−)に局在することが明らかとなった。またこの発現バンドは、ヤギ由来ビオチン標識化抗HBsAg抗体と、ウサギ由来アルカリフォスファターゼ標識化抗ビオチン抗体、あるいはHisProbeTM−HRP結合体を各々用いたウエスタンブロッティングでも検出された事から、目的のdHBsAg−ZZバンドである事が確認された。
【0131】
この可溶性画分(S−)を、平衡化緩衝液[0.5MのNaCl、及び30mMのイミダゾールを含む50mMリン酸緩衝液(pH7.0)]にて平衡化したNi−Sepharose 6 Fast flowカラムに添加し、室温で5分間rotate後、ゆっくり流し素通り液を回収した(素通り画分)。次に約1mlの上記平衡化緩衝液で7回、カラムを洗浄した後、イミダゾールの濃度を500mMに変えた上記平衡化緩衝液を用いて0.1mLずつ10回溶出させた(Ni−溶出画分:E)。各画分をSDS−PAGE処理に供し、次いでCBB染色を行った結果を図9(B)に示す。
【0132】
図中の矢印に示すように、Ni−カラム溶出画分(E)において、約70%を超える高純度のdHBsAg−ZZタンパク質が回収されることが明らかとなった。また、回収されたdHBsAg−ZZタンパク質は、dHBsAgタンパク質と同様に従来のHBsAgタンパク質と比較して、N末端側を欠失する変異が施されていることから、自己凝集することによってウイルス様の粒子を形成しているものと考えられる。以後、この粒子をdBNC−ZZと呼ぶことがある。
【0133】
また、発現用プラスミドによる大腸菌の形質転換から、高純度dBNC−ZZの取得までにかかった時間は約4日以内であり、迅速にB型肝炎ウイルス粒子様のナノカプセルを生産できることが明らかとなった。
【0134】
7) dBNC−ZZ粒子の性状解析
<抗体認識能>
ウサギ血清由来のIgGを固相化しスキムミルクでブロッキングしたマイクロウェルプレートに、TBSで希釈したdHBsAg−ZZ遺伝子の発現菌体の可溶性画分、及びdHBsAg遺伝子の発現菌体の可溶性画分を添加し、室温で2時間反応させた。各ウェルを3回洗浄後、標識抗体としてTBSで希釈したウサギ由来HRP標識化IgGを添加して室温で2時間反応させた。各ウェルを6回洗浄後、基質溶液[6mgのo−Phenylenediamine、12mLの0.1Mクエン酸緩衝液(pH5.0)、及び12μLの30%H2O2を混合して要事調製したもの]を加えて30分間反応させ、492nmの吸光度を測定した。その結果を図10に示す。
【0135】
dHBsAg試料では吸光度の上昇が全く見られなかったが、dHBsAg−ZZ試料では顕著な吸光度の上昇が観察された事から、dBNC−ZZ粒子では抗体結合能が維持されたZZドメインが多価でその粒子表面に提示されている事が示された。
【0136】
<Sucrose密度勾配遠心解析>
精製したdBNC−ZZ粒子を、上記5)のdHBsAgナノカプセルの性状解析にて示したsucrose密度勾配遠心処理と同様の手法を用いて解析した結果を図11に示す。比較対象として、酵母細胞を用いて生産した野生型HBsAg−ZZ粒子(ビークル社製)を用いた。
【0137】
dHBsAg−ZZ粒子は、比較対象の野生型BNC−ZZ粒子と同じ8番目の画分に沈降ピークを示すことが明らかとなった。この結果から、dHBsAg−ZZ粒子も従来のHBsAg粒子や、dHBsAgと同様に、dHBsAg−ZZタンパク質が自己凝集することによってウイルス粒子様の形状を有していることが示唆された。
【0138】
<電子顕微鏡観察>
精製したdBNC−ZZを電子顕微鏡観察した。観察結果を図12に示す。
【0139】
dBNC−ZZは、従来の野生型HBsAg粒子と同様の表面形状を示す事から、dHBsAgタンパク質が自己凝集することによって、エンベロープ様ナノ粒子構造形成する事が確認された。また、粒子径は22±4nmと、比較対象に用いた野生型HBsAg粒子(21±2nm)とほぼ同等の粒子径を有することも明らかとなった。この数値は、電子顕微鏡像から50個の粒子を選択し、その直径を測定した平均値である。
【0140】
<トリプシン切断実験>
4.1μg/10μLの濃度の精製したdBNC−ZZ粒子に、終濃度が12.5ng/μLとなるようにTPCK-トリプシン(Worthington Biochemial Corporation)を加えた後、37℃で、20分、2時間若しくは16時間消化反応を行った。
【0141】
反応後の液に5μLの4×Sample Buffer(5%の2−メルカプトエタノールを含有)を加えて、95℃で5分処理した後、15%アクリルアミドゲルを用いてSDS−PAGEを行い、CBB染色で全断片を、そしてHRP標識化抗HisProbe抗体を用いたウエスタンブロッティングによりC末端His-tagを含む切断断片を、HRP標識化抗HBsAg抗体を用いたウエスタンブロッティングによりN末端側HBsAg配列を含む切断断片を、各々検出した。結果を図13−2に示す。
【0142】
(A)及び(B)において、(1)に示すバンドから、dBNC−ZZタンパク質の全長の存在が示唆される。(2)に示すバンドから、図13−1に示すBの位置でトリプシン消化され、C末端側のHis−tagが欠失したタンパク質の存在が示唆される。(3)のバンドから、図13−1のCの位置でトリプシン消化され、ZZドメインのうち、C末端側のZドメインを欠失下タンパク質の存在が示唆される。(4)のバンドから、図13−1に示すDの位置でトリプシン消化され2つのZドメインが欠失したタンパク質の存在が示唆される。
【0143】
これらのバンドは、抗HBsAg抗体を用いたウエスタンブロッティングによっても確認されるバンドであることが(B)から明らかであり、本発明のdBNC−ZZにおいて、C末端側のZZドメイン及びHis−tagは、図13−1に示すようにウイルス様粒子表面に露出している構造を有しており、即ちC末端側のアミノ酸配列がウイルス粒子様構造の外側に提示されていることが明らかとなった。なお、図13−2の(B)において(5)にて示されるバンドは、抗His−tag抗体によって確認されるバンドであることから、図13−1のEの位置でトリプシン消化されたものと考えられる。
【0144】
<AFM観察>
精製したdBNC−ZZ粒子を金属板に固定したマイカ(雲母)に4μl添加した後に15分間乾燥させ、大塚蒸留水(大塚製薬,徳島)により3回洗浄した。再び乾燥させ、この試料を走査型プローブ顕微鏡(SPA400−DFM、SII NanoTechnology Inc.;千葉)により観察した。比較対象として、酵母にて作製した野生型BNC−ZZ粒子を用いた。結果を図14に示す。
【0145】
精製したdBNC−ZZ粒子は、高さが6.7−15.5nm程度で、直径は28.7−57.4nm程度であり、酵母で作製した野生型BNCーZZ粒子と類似した大きさであった。
【0146】
通常、脂質二重膜の厚さは5nm程度と考えられることから、10nm以上の厚さを示すのは、脂質二重膜が2層以上重なっていると考えられ、dHBsAgーZZ粒子は、従来のBNC−ZZ粒子と同様に、中空球状物の構造を有することが考えられる。
【特許請求の範囲】
【請求項1】
下記の(1)〜(4)のいずれかに示すアミノ酸配列からなるタンパク質を含むナノカプセル:
(1)配列番号1に示されるアミノ酸配列;
(2)上記(1)に示されるアミノ酸配列に、更に生体分子を認識するアミノ酸配列が付加されたアミノ酸配列;
(3)配列番号2に示されるアミノ酸配列;
(4)上記(1)〜(3)のいずれかに示されるアミノ酸配列において、1個又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
【請求項2】
下記の(5)〜(10)のいずれかに示す塩基配列からなるポリヌクレオチド:
(5)配列番号1に示されるアミノ酸配列をコードする塩基配列;
(6)配列番号3に示される塩基配列;
(7)上記(5)または(6)に示される塩基配列に、更に生体分子を認識するアミノ酸配列をコードする塩基配列が付加された塩基配列;
(8)配列番号2に示されるアミノ酸配列をコードする塩基配列;
(9)配列番号4に示される塩基配列;
(10)上記(5)〜(9)のいずれかに示される塩基配列において、1個又は数個の塩基が置換、欠失、挿入、及び/又は付加された塩基配列。
【請求項3】
請求項2に記載のポリヌクレオチドを含む組換えベクター。
【請求項4】
請求項3に記載の組換えベクターを含む形質転換体。
【請求項5】
請求項1に記載のナノカプセルを製造する方法。
【請求項1】
下記の(1)〜(4)のいずれかに示すアミノ酸配列からなるタンパク質を含むナノカプセル:
(1)配列番号1に示されるアミノ酸配列;
(2)上記(1)に示されるアミノ酸配列に、更に生体分子を認識するアミノ酸配列が付加されたアミノ酸配列;
(3)配列番号2に示されるアミノ酸配列;
(4)上記(1)〜(3)のいずれかに示されるアミノ酸配列において、1個又は数個のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列。
【請求項2】
下記の(5)〜(10)のいずれかに示す塩基配列からなるポリヌクレオチド:
(5)配列番号1に示されるアミノ酸配列をコードする塩基配列;
(6)配列番号3に示される塩基配列;
(7)上記(5)または(6)に示される塩基配列に、更に生体分子を認識するアミノ酸配列をコードする塩基配列が付加された塩基配列;
(8)配列番号2に示されるアミノ酸配列をコードする塩基配列;
(9)配列番号4に示される塩基配列;
(10)上記(5)〜(9)のいずれかに示される塩基配列において、1個又は数個の塩基が置換、欠失、挿入、及び/又は付加された塩基配列。
【請求項3】
請求項2に記載のポリヌクレオチドを含む組換えベクター。
【請求項4】
請求項3に記載の組換えベクターを含む形質転換体。
【請求項5】
請求項1に記載のナノカプセルを製造する方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6−1】
【図6−2】
【図6−3】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】

【図13−1】
【図13−2】
【図14】

【図2】
【図3】
【図4】
【図5】
【図6−1】
【図6−2】
【図6−3】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13−1】
【図13−2】
【図14】
【公開番号】特開2013−21999(P2013−21999A)
【公開日】平成25年2月4日(2013.2.4)
【国際特許分類】
【出願番号】特願2011−162596(P2011−162596)
【出願日】平成23年7月25日(2011.7.25)
【出願人】(504147243)国立大学法人 岡山大学 (444)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
【公開日】平成25年2月4日(2013.2.4)
【国際特許分類】
【出願日】平成23年7月25日(2011.7.25)
【出願人】(504147243)国立大学法人 岡山大学 (444)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
[ Back to top ]