
ウシアデノウイルスタイプ3ゲノム
【課題】新規ウシアデノウイルス発現ベクター系を提供すること。
【解決手段】1つの実施態様において、本発明は、ウシアデノウイルスゲノムの以下の領域を利用する新規ウシアデノウイルス発現ベクター系に関する:ヌクレオチド4,092〜5,234、ヌクレオチド5,892〜17,735、ヌクレオチド21,198〜26,033およびヌクレオチド31,133〜34,445。これらの領域は、とりわけ、外来配列の挿入のため、転写調節配列および翻訳調節配列を含むDNA制御配列の提供のため、または被験体または生物学的サンプルにおいて、これらの領域によってコードされるウイルス核酸またはタンパク質の存在を検出する診断目的のために使用され得る。別の実施態様において、本発明は、E3領域において欠失(および必要に応じて異種配列の挿入を)含む組換えウシアデノウイルス(BAV)の構築、単離、および増殖のための新規の方法に関する。病原体の防御決定因子をコードする遺伝子の挿入物を含む組換えBAVは、全身および粘膜の免疫応答の増強を刺激し、そして病原体によるチャレンジから宿主動物を防御する。
【解決手段】1つの実施態様において、本発明は、ウシアデノウイルスゲノムの以下の領域を利用する新規ウシアデノウイルス発現ベクター系に関する:ヌクレオチド4,092〜5,234、ヌクレオチド5,892〜17,735、ヌクレオチド21,198〜26,033およびヌクレオチド31,133〜34,445。これらの領域は、とりわけ、外来配列の挿入のため、転写調節配列および翻訳調節配列を含むDNA制御配列の提供のため、または被験体または生物学的サンプルにおいて、これらの領域によってコードされるウイルス核酸またはタンパク質の存在を検出する診断目的のために使用され得る。別の実施態様において、本発明は、E3領域において欠失(および必要に応じて異種配列の挿入を)含む組換えウシアデノウイルス(BAV)の構築、単離、および増殖のための新規の方法に関する。病原体の防御決定因子をコードする遺伝子の挿入物を含む組換えBAVは、全身および粘膜の免疫応答の増強を刺激し、そして病原体によるチャレンジから宿主動物を防御する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願への相互参照
本願は、1997年6月23日に出願された米国特許出願番号08/880,234に対して優先権を主張する。
【0002】
技術分野
本発明は、初期領域1(E1)および初期領域3(E3)領域遺伝子欠失の一方または両方が外来遺伝子によって置換されており、そして新規組換え哺乳動物細胞株がBAV E1配列を用いて安定に形質転換されており、それゆえE1遺伝子産物を発現して外来遺伝子でE1遺伝子欠失が置換されたウシアデノウイルスがそこで複製可能になる、新規のウシアデノウイルス(BAV)発現ベクター系に関する。これらの物質は、生の組換えウイルスまたはサブユニットワクチンの目的のためまたは他の治療目的のための、組換えBAV発現異種(抗原性)ポリペプチドまたはフラグメントの産生において使用される。
【0003】
本発明はまた、本明細書において開示されるBAVゲノム配列を含む新規のウシアデノウイルス(BAV)発現ベクター系に関する。BAVゲノム配列は、生の組換えウイルスまたはサブユニットワクチンを生産する目的のためまたは他の治療目的のための、組換えBAV発現異種(抗原性)ポリペプチドまたはフラグメントを生成するために1以上の外来遺伝子によって置換され得る。さらに、種々のBAV転写調節シグナルおよび翻訳調節シグナルを用いて、本発明のベクター系へと挿入された外来遺伝子の発現を調節し得る。さらに、本発明の新規の配列は、診断目的のために、被験体または生物学的サンプルにおいてBAVの存在を決定するために使用され得る。
【背景技術】
【0004】
発明の背景
アデノウイルスは、ヒトならびに家畜および実験動物において腸感染または呼吸器感染を引き起こす。
【0005】
ウシアデノウイルス(BAV)は、2つのサブグループに分割される、少なくとも9つの血清型を含む。これらのサブグループは、酵素結合免疫吸着アッセイ(ELISA)、免疫蛍光アッセイを用いた血清学的研究、ウイルス中和試験、免疫電子顕微鏡法、それらの宿主特異性によって、および臨床症候群に基づいて特徴付けられている。サブグループIウイルスは、BAV1、2、3および9を含み、そして、BAV4、5、6、7および8を含むサブグループ2と比較して、樹立されたウシ細胞において比較的良好に増殖する。
【0006】
BAV3は、1965年に初めて単離された。そして、BAV遺伝子型が最も良く特徴付けられており、これは、約35kbのゲノムを含む(Kurokawaら、(1978)J.Virol.28:212-218)。BAV3は、BAVのサブグループ1の代表例であり(Bartha(1969)Acta Vet.Acad.Sci.Hung.19:319−321)、通常無症状の感染をもたらす、ウシに共通する病原体である(Darbyshireら(1965)、J.Comp.Pathol.75:327−330)。しかし、これは時折、より重篤な呼吸管感染を伴う(Darbyshireら、1966 Res.Vet.Sci.7:81-93;Mattsonら、1988 J.Vet Res 49:67−69)。他のアデノウイルスと同様に、BAV3は、直鎖の二本鎖DNA分子を含む直径75nmの非莢膜性の正二十面体粒子である(Niiyamaら、(1975)J.Virol.16:621−633)。BAV3は、ハムスターに注射される場合に、腫瘍を形成し得(Darbyshireら、1966 Nature 211:102)、そしてウイルスDNAは、培養中のマウス、ハムスターまたはラットの細胞の形態学的な形質転換を効率的にもたらし得る(TsukamotoおよびSugino、1972、J.Virol.9:465−473);Motoiら、1972 Gann 63:415−418;M.Hitt、私信)。交叉ハイブリダイゼーションは、BAV3とヒトアデノウイルスタイプ2(HAd2)との間において、ゲノムの左端に近いいくつかの領域を含むゲノムのほとんどの領域で観察されたがゲノムの左端では観察されなかった(Huら、1984 J.Virol.49:604-608)。
【0007】
ヒトアデノウイルス(HAd)ゲノムにおいて、2つの重要な領域であるE1およびE3が存在する。E1およびE3では、外来遺伝子が組換えアデノウイルスを生成するために挿入され得る(BerknerおよびSharp(1984)Nuc.Acid Res.12:1925−1941ならびにHaj-AhmadおよびGraham(1986)J.Virol.57:267-274)。E1タンパク質は、組織培養においてウイルス複製に必須であるが、外来DNAをE1領域に含む条件付(conditional)ヘルパーアデノウイルス組換え体は、E1を構成的に発現する細胞株において生成され得る(Grahamら、(1977)J.Gen.Virol.36:59-72)。対照的に、HAd2および五Ad5のE3遺伝子産物は、インビトロでもインビボでも感染性ビリオン産生には必要ではないが、ウイルス感染に対する宿主免疫応答において重要な役割を果たしている(Anderssonら、(1985)Cell 43;215−222;Burgertら、(1987)EMBO J.6:2019−2026;Carlinら、(1989)Cell 57:135−144;Ginsbergら、(1989)PNAS、USA 86:3823−3827;Goodingら(1988)Cell 53:341−346;Tollefsonら、(1991)J.Virol.65:3095−3105;WoldおよびGooding(1989)Mol.Biol.Med.6:433−452ならびにWoldおよびGooding(1991)Virology 184:1−8)。ヒトアデノウイルスタイプ2(HAd2)のE3−19キロダルトン(kDa)糖タンパク質(gp19)は、多数のクラスI主要組織適合性複合体(MHC)抗原の重鎖と、小胞体において結合し、従って、これらの原形質膜への輸送を阻害する(Anderssonら(1985)Cell 43:215−222;BurgertおよびKvist、(1985)Cell 41:987−997;BurgertおよびKvist、(1987)EMBO J.6:2019−2026)。HAd2またはHAd5のE3-14.7kDaタンパク質は、ウイルス感染したマウス細胞の腫瘍壊死因子(TNF)による溶解を予防する(Goodingら(1988)Cell 53:341−346)。さらに、E3-10.4kDaおよびE3-14.5kDaタンパク質は、ウイルス感染した細胞において、エンドソーム媒介性インターナリゼーションおよび上皮増殖因子レセプター(EGF−R)の分解を誘発する複合体を形成する(Carlinら、Cell 57:135−144;Tollefsonら、(1991)J.Virol.65:3095−3105)。E3領域に外来遺伝子を有するヘルパー非依存性組換えアデノウイルスは、すべての許容性細胞株において非常に良好に複製しそして発現する(Chandaら(1990)Virology 175:535−547;Dewarら(1989)J.Virol.63:129−136;Johnsonら(1988)Virology 164:1−14;Lubeckら(1989)PNAS USA 86:6763−6767;McDermottら(1989)Virology 169:244−247;Mittalら(1993)Virus Res.28:67−90;Morinら(1987)PNAS.USA 84:4626−4630;Prevecら(1990)J.Inf.Dis.161:27−30;Prevecら(1989)J.Gen.Virol.70:429−434;Schneiderら(1989)J.Gen.Virol.70:417−427ならびにYuasaら(1991)J.Gen.Virol.72:1927−1934)。上記の研究およびアデノウイルスが野生型(wt)アデノウイルスゲノムの約105%をパッケージングし得るという示唆(Bettら(1993)J.Virol.67:5911−5921およびGhosh-Choudhuryら(1987)EMBO J.6:1733-1739)に基づけば、1.8kbまでの外来DNAの挿入物が欠失をなんら補償することなく外来タンパク質について発現ベクターとしての使用のためにアデノウイルス粒子中にパッケージングされ得る。
【0008】
グループCヒトアデノウイルスのE1A遺伝子産物は、非常に広範囲に研究されており、そしてウイルス遺伝子および細胞性遺伝子の両方のトランス活性化を媒介して(Berkら1979 Cell 17:935-944;JonesおよびShenk、1979 Cell 16:683-689;Nevins、1981 Cell 26:213-220;Nevins、1982 Cell 29:913-919、Berk、1986 Ann.Res.Genet.20:45-79に概説されている)、培養中の細胞の形質転換をもたらすこと(Graham、F.L.(1984)「Transformation by and oncogenicity of human adenoviruses」the Adenoviruses、H.S.Ginsberg編、Plenum Press、New York;Brantonら、1985 Biochim.Biophys.Acta 780:67-94に概説されている)、ならびに細胞のDNA合成および有糸分裂を誘発すること(Zerlerら、1987 Mol.Cell.Biol.7.821-929;Belletら、1989 J.Virol.63:303-310;Howeら、1990 PNAS.USA 87:5883-5887;HoweおよびBayley、1992 Virology 186:15-24)が示されている。E1A転写ユニットは、プロセシングを受けたすべてのE1A転写物からは欠失されるイントロン領域によって分離された2つのコード配列を含む。E1A転写ユニットから産生された2つの最も大きなmRNA種において、第一のコード領域は、さらに12sおよび13sのmRNA種の両方において見いだされれる配列であるエキソン1と、13s mRNA種においてのみ見出される特有の領域とにさらに細分割される。ヒトアデノウイルスのE1Aタンパク質とシミアンアデノウイルスのE1Aタンパク質との間の比較によって、3つの領域のいくらか保存されたタンパク質配列(CR)が規定された(Kimelmanら、1985 J.Virol.53:399-409)。CR1およびCR2は、エキソン1にコードされるが、CR3は、特有の配列およびエキソン2の小部分にコードされる。網状赤血球タンパク質Rb、サイクリンAおよび付随タンパク質キナーゼp33cdk2および未だに割り当てられていないタンパク質を含む多数の細胞タンパク質についての結合部位が、E1Aタンパク質のエキソン1がコードする領域において規定されている(YeeおよびBranton、1985 Virology 147:142-153;Harlowら、1986 Mol.Cell.Biol.6:1579-1589;Barbeauら、1992 Biochem.Cell Biol.70:1123-1134)。E1Aとこれらの細胞タンパク質との相互作用は、E1Aが不死化およびガンへの転換に関与する機構として関連付けられている(Eganら、1989 Oncogene 4:383-388;Whyteら、1988、Nature 334:124-129;Whyteら、1988 J.Virol.62:257-265)。E1A単独は、培養中の細胞を形質転換または不死化し得るが、E1Aと、E1B-19kタンパク質またはE1B-55kタンパク質のいずれかとを両方別個または一緒に共に発現することは、通常、培養中の齧歯類細胞の高度に頻繁な形質転換を必要とする(Graham、1984、前出、Brantonら、1985前出;McLorieら、1991 J.Gen Virol.72:1467-1471に概説される)。
【0009】
ヒト細胞の許容性感染における他のウイルス初期遺伝子のトランス活性化は、主に、E1AのCR3領域においてコードされるアミノ酸配列によって媒介される(Lillieら、1986 Cell 46:1043-1051)。特有な領域におけるCysX2CysX13CysX2Cys配列モチーフにおいて保存されたシステイン残基は、金属イオン結合活性と関連している(Berg 1986、上記)。そして、トランス活性化に必須である(Jelsmaら、1988 Virology 163:494-502;Culpら、1988 PNAS USA 85:6450−6454)。金属結合ドメインのアミノ(N)末端の直近に存在するCR3におけるアミノ酸もまた、転写活性に重要であることが示されているが、金属結合ドメインのカルボキシ(C)末端の直近の配列は、プロモーター領域と会合を形成するのに重要であることが示されている(LillieおよびGreen、1989 Nature 338:39-44;図3を参照のこと)。
【0010】
遺伝子操作の適用は、ワクチンを得るためのアデノウイルス発現系を調製することのいくつかの試みをもたらした。このような研究の例は、酵母宿主における発現のためのアデノウイルス主要後期プロモーターについての米国特許第4,510,245号;欠失された初期領域3に位置するB型肝炎表面抗原をコードする遺伝子を有する生の組換えアデノウイルスタイプ7についての米国特許第4,920,209号;HCMV主要莢膜糖タンパク質についてのヒト細胞における非欠損性ヒトアデノウイルス5組換え発現系についての欧州特許第389 286;E1Aタンパク質を発現する細胞における非病原性免疫原性の生の生存イヌアデノウイルスについてのWO91/11525;ならびにアデノウイルス2のE3領域由来のリーダーおよび/またはプロモーターを含むベクターについての仏国特許第2 642 767号の開示を含む。
【0011】
常在性のアデノウイルスベクターが、ヒト起源のアデノウイルスに比較して、非ヒト動物種における生存組換えウイルスワクチンとしての使用により良好に適すると推定されている。これは、異種配列の挿入に適する領域が常在性アデノウイルスベクターに同定されること、および異種配列の挿入、組換え体の単離および組換え体の増殖のための組成物および方法が開発されることを必要とする。挿入に適する領域は、適切なヘルパー機能が提供される場合、ウイルスゲノム非必須領域または必須領域を含み得る。例えば、HAdとの類似性によって、他のアデノウイルスにおけるE3領域が培養細胞におけるウイルス複製に必須でない場合、外来遺伝子挿入物をE3領域に含むアデノウイルス組換え体が生成され得る。
【0012】
外来遺伝子発現のためのベクターとして作用する適切なウイルスの選択、遺伝子挿入のための部位として適切な領域の同定、ならびに組換えウイルスの構築、単離および増殖は、組換えウイルスワクチンベクターの開発にとって意義深い挑戦を提起する。詳細には、好ましい挿入部位は、ウイルスの生存複製および組織培養およびインビボでもその有効な操作のためには必須ではない。さらに、挿入部位は、ウイルスが複製を継続することを保証しつつ、新たな遺伝的物質を受け入れることができなければならない。ウイルスゲノムの必須領域もまた、組換えウイルスが、トランスでその特定の必須領域の機能を補完する細胞株において増殖する場合は、外来遺伝子の挿入のために利用され得る。
【0013】
ウイルスゲノムにおける適切な挿入部位を決定するための効率よい方法は、そのゲノムの完全なヌクレオチド配列を入手することである。これは、種々のコード領域が規定されることを可能にし、挿入部位としてそれらの可能な使用を容易にする。必須でない非コード領域の規定はまた、配列分析によって示され、そしてこれらはまた、可能な挿入部位として使用され得る。BAV-3ゲノムの特定の領域のヌクレオチド配列は決定されている。ゲノムの究極の左端の配列(これは、逆末端反復(ITR)、パッケージングシグナル、E1およびpIXを含む)は、いくつかのグループによって決定されている:Shinagawaら、1987 Gene55:85−93によるヌクレオチド1〜195(ITR);Zhengら、1994、Virus Research 31:163−186によるヌクレオチド1〜4060(ITR、パッケージングシグナル、E1およびpIX);Elgadiら、1993、Intervirology 36:113−120によるヌクレオチド1〜4091(ITR、パッケージングシグナル、E1およびpIX)。(ヌクレオチド1は、線状の34.4kb BAV−3ゲノムの最も左のヌクレオチドを示す。)決定されているBAV−3ゲノムのさらなる配列は、以下を含む:ヌクレオチド5,235−5,891(主要後期プロモーター、Songら、1996、Virology 220;390−401);ヌクレオチド17,736〜20,584(ヘキソン遺伝子。Huら、1984、J.Virology 49:604−608);ヌクレオチド20,408〜21,197(プロテイナーゼ遺伝子、Caiら、1990、Nucleic Acids Res.18:5568);ならびにヌクレオチド26,034〜31,132(E3領域、pVIIIおよびファイバー遺伝子、Mittalら、1992、J.Gen.Virol.73:3295−3300)。
【0014】
組換えウイルスおよびウイルスゲノムが適用され得る多くの使用の一つは、それらが利用可能である場合、組換えサブユニットワクチンの開発においてである。ワクチン接種は、生の減弱化されたウイルスワクチンが使用される場合は特に、呼吸器系ウイルス疾患および腸ウイルス疾患の制御のための最も有効な手段であることが証明されている。これらのワクチンは、経口または鼻腔内に投与される場合、強力な粘膜免疫を誘導し、これは、これらのウイルスによって引き起こされる初期感染をブロックし、それらによって引き起こされる疾患の進行を減少させるために必要とされる。このアプローチは、遺伝子操作された(毒性遺伝子欠失)ウイルスゲノムをベクターとして使用して他の病原体の遺伝子をインビボで送達および発現することによって拡張されている。Ertlら(1996)J.Immunol.156:3579−3582。
【0015】
ヒトアデノウイルス(HAV)に基づく組換えウイルスベクター系が近年開発されている。Grahamら(1992)、「Vaccines:New approaches to immunological problems」(R.W.Ellis編)、Butterworth-Heineman、Stoneham、363-390頁。複製欠損HAVベクターおよび複製適合HAVベクターの両方が、種々の外来抗原を発現するように操作されている。概説については、Grunhausら(1992)Seminar in Virol.3:237-252;Imler(1995)Vaccine 13:1143-1151を参照のこと.安定な外来遺伝子発現を提供することに加えて、操作されたアデノウイルスは、体液性、細胞性および粘膜性の免疫応答を誘発することが示されている。Bugeら(1997)J.Virol.71:8531-8541。
【0016】
ヒトアデノウイルスの遺伝子治療のためのベクターとしての使用は、宿主において、すでに存在するHAVに対する中和抗体(これは、組換えウイルスの侵入および複製と干渉し得る)の存在のため、および組換えウイルスと既に宿主に存在する野生型HAVとの間の組換えおよび/または補完の可能性のために、妨害されている。従って、高度の種特異的であるHAV以外の動物アデノウイルスが、遺伝子治療および組換えワクチンのためのベクターとして考慮されつつある。
【0017】
ウシアデノウイルス-3(BAV-3)の分子的特徴付けは、ヒトおよび他の哺乳動物種におけるワクチンおよび遺伝子治療のための、ウシアデノウイルスの生のウイルスベクターとしての開発を補佐する。近年、BAV3ゲノムの完全なDNA配列および転写マップが報告されている。この配列は、1997年6月23日に出願された親の米国特許出願U.S.S.N.08/880,234において、およびいくつかの刊行物において開示された。Baxiら、(1998)Virus Genes 16:1-4;Leeら、(1998)、Virus Genes 17:99-100;およびReddyら(1998)J.Virol.72:1394-1402。
【発明の概要】
【課題を解決するための手段】
【0018】
発明の開示
本発明者らは、今や、34,446ヌクレオチドを含む全長BAV−3ゲノムの配列を完成し、それによって外来遺伝子の挿入のため、および診断プローブとしての使用のための両方に適切な領域を同定した。本発明者らはまた、これら領域中に外来遺伝子を挿入してBAV組換え体を生成し、そしてこの組換え体を増殖させた。例えば、このような組換え体は、種々の病原体に対する組換えサブユニットワクチンとして、組換えポリペプチドの過剰発現のために、および遺伝子治療目的のために有用である。
【0019】
1つの実施態様では、本発明は、新規ウシアデノウイルス発現ベクターシステムに関し、ここでE1およびE3遺伝子領域の1つまたは両方が、一部または全てを欠失している。本発明はまた、好ましくはBAVのE1配列であるE1配列で形質転換されたウシ起源の組換え哺乳動物細胞株に関し、これは、1つまたはそれ以上のE1遺伝子産物を構成的に発現し、外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列によって置換されたE1遺伝子領域の一部分またはすべての欠失を有するウシアデノウイルスをその中で複製させる。本発明は、さらに、異種(抗原性)ポリペプチドまたはそのフラグメントの産生におけるこれら材料の使用に関する。
【0020】
別の実施態様では、本発明は、ウシアデノウイルスゲノムまたはそのフラグメントの以下の領域を利用する新規なウシアデノウイルス発現ベクター系に関する:ヌクレオチド4,092-5,234;ヌクレオチド5,892-17,735;ヌクレオチド21,198-26,033およびヌクレオチド31,133-34,445。これらの領域(およびそのフラグメント)は、とりわけ、外来配列の挿入のために、転写および翻訳調節配列を含むDNA制御配列の準備のために、または、被験体または生物学的試料中のウイルス核酸またはこれらの領域によりコードされるタンパク質の存在を検出する診断目的のために用いられ得る。
【0021】
本発明はまた、ウシ、ヒトおよびその他の哺乳動物を含む、哺乳動物において、感染性生物に対する抗体、細胞媒介性および/または粘膜免疫を産生するための生存組換えウイルス、またはサブユニットワクチンの調製方法に関する。この方法は、ウシアデノウイルスゲノム中に、上記抗体に対応するか、または有効なプロモーターとともにまたはなしで上記細胞媒介性および/または粘膜免疫を誘導する抗原をコードする遺伝子または遺伝子フラグメントを挿入し、BAV組換え体を産生する工程を包含する。
【0022】
別の局面では、本発明は、組換えウイルス、および細胞系における目的のDNA配列またはアミノ酸配列の発現のための組換えウイルスベクターの使用を含む。
【0023】
一般に、外来遺伝子構築物は、1つまたはそれ以上の適切な欠失を有する全長ウイルスゲノムの一部分のみを示すヌクレオチド配列中にクローン化される。このキメラDNA配列は、通常、成功するクローニングを可能にし、この配列の多くのコピーを産生するプラスミド中に存在する。次いで、クローン化された外来遺伝子構築物は、例えば、インビボ組換え、次いでDNA媒介同時トランスフェクション技法により完全ウイルスゲノムに含められ得る。このクローン化された外来遺伝子構築物のウイルスゲノム中への取り込みは、外来遺伝子を、複製およびパッケージングシグナルを含むDNA分子中に配置し、感染性ウイルス粒子にパッケージングされ得る組換えアデノウイルスゲノムの多コピーの生成を可能にする。コード配列または1つ以上のコード配列の多コピーは、組換えベクターが1つ以上外来タンパク質を発現し得るように挿入され得る。外来遺伝子は、付加、欠失または置換を有し、発現タンパク質の発現および/または免疫学的効果を増大し得る。
【0024】
本発明はまた、ウシアデノウイルス発現ベクターを含む発現系を含み、ここで任意の外因性調節要素を伴うかまたは伴わない異種ヌクレオチド配列が、E1遺伝子領域および/またはE3遺伝子領域の一部分もしくはすべてを置き換える。
【0025】
別の実施態様では、本発明は、BAVゲノムの1つまたはそれ以上の領域が異種配列により置き換えられるか、または異種ヌクレオチド配列が任意のBAV配列を取り除くことなくBAVゲノム中に導入される発現系を含む。DNA調節配列を含むBAVゲノムの遺伝子内領域は、本発明の実施において、同種および異種(即ち外来)遺伝子の発現に有用である。
【0026】
本発明はまた、(A)全長BAV DNA、および適切な細胞株の同時トランスフェクション後インビボ相同組換えにより組換えウイルスを生成し得るプラスミド(単数)または2つのプラスミドを含む組換えベクター系であって、全長の野生型BAVゲノムを示すBAV DNA、およびE1またはE3遺伝子領域を含むウシアデノウイルス左または右末端配列を含むプラスミドまたはウシアデノウイルスE2、E4、L1、L2、L3、L4、L5、L6もしくはL7配列を、E1、E2、E3、E4、L1、L2、L3、L4、L5、L6もしくはL7遺伝子領域の一部または全てで置換された外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列(即ち、挿入カセット)とともに含む、組換えベクター系;(B)(a)E1遺伝子領域の一部分またはすべてが、外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列により置き換えられている系;(b)E3遺伝子領域の一部分またはすべてが、外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列により置き換えられている系;および(c)E1遺伝子領域の一部分またはすべておよびE3遺伝子領域の一部分またはすべてが欠失し、かつ外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列が少なくとも1つの欠失中に挿入されている系からなる群から選択される生存組換えウシアデノウイルスベクター(BAV)系;(C)E1遺伝子領域の一部分またはすべての欠失、E3遺伝子領域の一部分またはすべての欠失、または両方の欠失を含み、かつ少なくとも1つの欠失中に、疾患を起こす生物の抗原決定基をコードする異種ヌクレオチド配列が挿入される組換えウシアデノウイルス(BAV);(D)E1の一部分またはすべての欠失、E3の一部分またはすべての欠失、または両方の欠失を含み、かつ少なくとも1つの欠失中に、発現プロモーターの制御下にある外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列が挿入される組換えウシアデノウイルス発現系;または(E)(1)所望の免疫応答を得るために必要な抗原決定基をコードする異種ヌクレオチド配列を、(2)生存組換えウイルスまたは組換えタンパク質またはサブユニットワクチンとしての使用のために免疫原性量の上記抗原決定基の発現を提供する有効プロモーターをともにまたはなしで含むBAV組換え体を含む、哺乳動物宿主において免疫応答を産生するための組換え体ウシアデノウイルス(BAV);(F)E1の一部分またはすべての欠失、および/またはE3の一部分またはすべての欠失、および/またはBAVゲノムの以下の領域の少なくとも1つの一部分またはすべての欠失:E2、E4、L1、L2、L3、L4、L5、L6またはL7:を含む変異体ウシアデノウイルス(BAV)を含む。
【0027】
E1およびE3領域に加えて、BAVゲノム内のその他の部位もまた、外来ヌクレオチド配列の挿入に有用である。これらは、限定されないが、E2領域、E4領域、E4プロモーターとゲノムの右末端との間の領域、後期領域(L1−L7)、33kD、52kD、100kD、DBP、pol、pTPおよびペントン遺伝子、ならびに遺伝子IIIA、pV、pVI、pVII、pVIII、およびpXを含む。
【0028】
本発明はまた、ゲノムのE3領域に欠失を含むウイルス、およびゲノムの欠失したE3領域中に異種配列の挿入を含むウイルスを、高効率で得るための方法および組成物を提供する。1つの実施態様では、E3が欠失したウイルスゲノム(異種配列の挿入をともなうまたはなし)が、適切な細胞株、例えば、アデノウイルスE1機能を発現するMDBK細胞または相当する細胞中にトランスフェクトされ、そして組換えウイルスがトランスフェクトされた細胞から回収される。別の実施態様では、欠失したE3領域を(異種配列の挿入をともなうまたはなしで)含むBAVゲノムのセグメントが、原核生物細胞中で、BAVゲノムでの組換えを受け、組換えBAVゲノムを生成することが可能である。本発明の目的には、BAVゲノムは完全長BAVゲノムであり得るか、またはそれは、1つもしくはそれ以上のBAV複製および/またはパッケージング配列を含むことを条件に、1つもしくはそれ以上の欠失を含み得る。欠失したE3領域を含むBAVゲノムセグメントはまた、1つまたはそれ以上のBAV複製および/またはパッケージング配列を含み得る。そうでなければ、組換えBAVゲノムは、特定のゲノム領域における1つまたはそれ以上の欠失、ならびに欠失または欠失のない、異種配列が挿入されたBAVゲノムとともに全長のBAVゲノムを含む。次いで、組換えBAVゲノムを、例えば、ウシ仔胎網膜初代(PFBR)細胞のような適切な細胞株中にトランスフェクトし、そしてトランスフェクトされた細胞から組換えウイルスを回収する。
【0029】
別の局面では、本発明は、BAV E1遺伝子領域配列で安定に形質転換された組換え哺乳動物細胞株を提供し、この組換え細胞株は、これによって、その中で、外来遺伝子またはそのフラグメントをコードする異種または同種ヌクレオチド配列によって置き換えられたE1またはE3遺伝子領域の一部分またはすべての欠失を含むウシアデノウイルスの複製を可能にし得る。本発明はまた、BAVE1遺伝子産物を発現する組換えウシ細胞株からの、成長因子、レセプターおよびその他の細胞タンパク質のようなポリペプチドまたはそのフラグメントの産生、単離および精製を含む。
【0030】
本発明はまた、遺伝子治療が必要な哺乳動物に遺伝子治療を提供する方法を包含する。例えば、遺伝子治療は、遺伝子欠陥を制御するために、宿主細胞中に治療遺伝子を導入するために、変異遺伝子の配列を変え野生型機能を回復させるために、遺伝子の配列を変えその機能を不活性化するために、外因性遺伝子機能を提供するためなどに用いられ得る。例えば、遺伝子治療は、癌、AIDS、その他のウイルスで誘導される病因、感染性疾患、遺伝病などの処置において用いられる。遺伝子治療のプロセスは、上記哺乳動物に異種ヌクレオチド配列を含む生存組換えウシアデノウイルスを、組換えウイルスベクターゲノムが上記哺乳動物ゲノム中に取り込まれるか、または独立にかつ染色体外で維持される条件下で投与し、標的器官または組織中で異種配列の発現を提供する工程を包含する。
【0031】
本発明の別の局面は薬学的組成物を提供し、この組成物は、治療的に有効な量の組換えウイルス、組換えウイルスベクターまたは組換えタンパク質を、薬学的に受容可能なキャリアとともにまたはなしで含む。このような薬学的組成物の1つの例は、組換えウイルスワクチンである。組換えウイルスワクチンは、経口用量(例えば、溶腸被覆錠剤)によるか、注射によるか、またはその他の様式による投与のために処方され得る。より詳細には、これらは、感染に対して哺乳動物宿主を保護するためのワクチンを含み、生存組換えアデノウイルス、または外来遺伝子またはフラグメントが抗原をコードし、そして薬学的に受容可能なキャリアとともにまたはなしで処方される本発明の組換えアデノウイルスにより産生される組換えタンパク質を含む。これらの組成物は、抗原性ポリペプチドまたは保護的抗原を発現し得、それによって目的のポリペプチドまたは抗原に応答する免疫を惹起し、そして感染からの保護を提供する。本発明の実施に有用な薬学的組成物はまた、目的のポリペプチドまたは抗原を発現する組換えBAVベクターを保持する細胞、またはBAVポリペプチドまたは抗原を発現するベクターを保持する細胞を含み得る。
【0032】
本発明はまた、哺乳動物において、抗体、細胞媒介性および/または粘膜免疫を生成する方法を含み、(1)哺乳動物宿主において感染に対する免疫応答を惹起する方法であって:本発明の生存BAV組換え体であって、外来遺伝子またはフラグメントが抗原をコードするBAV組換え体を含むワクチンを、薬学的に受容可能なキャリアとともにまたはなしで投与する工程を包含する方法、および(2)哺乳動物宿主において感染に対する免疫応答を惹起する方法であって:外来遺伝子またはフラグメントが所望の抗原をコードするBAV組換え体を培養することによって調製された組換え抗原を含むワクチンを、薬学的に受容可能なキャリアとともにまたはなしで投与する工程を包含する方法を含む。
【0033】
本発明はさらに、感染細胞または哺乳動物被験体、またはこれら供給源からの核酸調製物のような生物学的試料中の、BAV DNAおよび/またはBAVがコードするタンパク質および抗原の存在を検出するための診断手順の実施において有用な組成物および方法を提供する。これらは、制限されずに、BAV遺伝子およびコード配列およびそれらのフラグメント、ならびにBAVゲノムおよびそれらのフラグメントによりコードされるアミノ酸配列を含む。
【0034】
以下の開示は、本発明のこれらおよびその他の実施態様を、当業者にとって容易に明白にする。しばしば、本開示は、ウシアデノウイルスタイプ3(BAV3)に言及するが、これは、例示の目的のためであって、しかも、他の型のウシアデノウイルス、例えば、1、2、4、5、6、7、8および9に同じ特徴が適用され;しかも本明細書に記載されそして請求の範囲に記載された発明が、これらのウシアデノウイルス型のすべてをカバーすることが意図されることを理解すべきである。
【図面の簡単な説明】
【0035】
【図1】図1.ウシアデノウイルスタイプ3(BAV3)ゲノムの完全なヌクレオチド配列。
【図2】図2.BAV3の転写マップ。ゲノムは0から100まで(マップ単位)番号付られた実線で表される。転写物は、転写の長さおよび方向に関して矢印により表される。E1A、E1B、E2A、E2B、E3、E4、L1、L2、L3、L4、L5、L6およびL7領域の位置が示される。パッケージングおよび複製配列は、195bpITR配列中および左ITRとE1領域との間の領域に位置する。
【図3】図3.E3欠失BAV3ゲノムDNAを含むプラスミドの構築。プラスミドpBAV302を上記の実施例3で記載のように異なるゲノムクローンから構築した。DNA配列の起源は以下の通りである:プラスミドDNA、細線;BAV3ゲノムDNA配列、太線。中空の矢印は以下のPCRプライマーを表す:a:5'−ACGCGTCGACTCCTCCTCA(配列番号2);b:5'−TTGACAGCTAGCTTGTTC(配列番号3);c:5'−CCAAGCTTGCATGCCTG(配列番号4);およびd:5'−GGCGATATCTCAGCTATAACCGCTC(配列番号5)。
【0036】
【図4】図4.組換えBAV3ゲノムの制限酵素分析。DNAは、BAV3(レーンb)、BAV3.E3d(レーンaおよびe)、BAV3.E3gD(レーンcおよびf)またはBAV3.gDt(レーンdおよびg)で感染させたMDBK細胞から得た。 DNAは、Hirtの方法(Hirt(1967)J.Mol.Biol.26:365−369)により抽出し、そしてBamHI(レーンaおよびb)、NheI(レーンcおよびd)またはNdeI(レーンe、fおよびg)で消化した。1kb+DNAラダー(Gibco/BRL)を、ウイルスDNAフラグメントをサイズ分けするために用いた。
【図5】図5.組換えBAV3ウイルスで感染させたMDBK細胞におけるgDタンパク質の発現。(A)BHV−I(レーンa)、BAV3.E3d(レーンe、fおよびg)、またはBAV3.E3gD(レーンb、cおよびd)で感染させたか、または非感染(レーンh)の放射標識されたMDBK細胞の溶解液からのタンパク質を、gD特異的MAbのプールで免疫沈降し、そして還元条件下でSDS−PAGEにより分析した。タンパク質は、感染後6〜16時間(レーンaおよびh)、36〜48時間(レーンbおよびe)、48〜50時間(レーンcおよびf)、または60〜62時間(レーンdおよびg)標識した。(B)BHV−I(レーンa)、BAV3.E3d(レーンe、fおよびg)、またはBAV3.E3gDt(レーンb、cおよびd)で感染させた放射標識されたMDBK細胞の培養培地からのタンパク質を、gD特異的MAbのプールで免疫沈降し、そして還元条件下でSDS−PAGEにより分析した。タンパク質を、感染後、6〜16時間(レーンa)、12〜14時間(レーンbおよびe)、16〜18時間(レーンcおよびf)、または22〜26時間(レーンdおよびg)標識した。分子サイズマーカー(MW)はkDaである。
【図6】図6.組換えgDおよびgDtタンパク質の抗原性分析。BAV3.E3gD(レーンa−e)またはBAV3.E3gDt(レーンf−j)組換えウイルスで感染させた放射標識MDBK細胞の溶解液(レーンa−e)および培養培地(レーンf−j)からのタンパク質を、MAb136(レーンaおよびf)、MAb3E7(レーンbおよびg)、MAb4C1(レーンcおよびh)、MAb2C8(レーンdおよびi)、またはMAb3C1(レーンeおよびj)で免疫沈降し、そして還元条件下でSDS−PAGEにより分析した。分子サイズマーカー(MW)はkDaである。
【図7】図7.MDBK細胞におけるgD発現に対するAraCの影響。100μg/ml AraCの存在下(レーンa、b)または非存在下(レーンc、d)でBAV3.E3gD(レーンa−d)で感染させ、そして感染後22時間(レーンaおよびc)または34時間(レーンbおよびd)に2時間放射標識したMDBK細胞の溶解液からのタンパク質を、gD特異的MAbのプールで免疫沈降し、そしてSDS−PAGEにより分析した。分子サイズマーカー(MW)はkDaである。
【図8】図8.コットンラットにおける抗体応答。組換えBAVを用いた二次免疫の12日後の血清、肺洗浄液(l.w)および鼻洗浄液(n.w)中の糖タンパク質gD(A、B)またはBAV3特異的(C、D)IgA(A、C)またはIgG(B、D)のELISA力価。白棒は、BAV3.E3gDを示し、黒棒はBAV3.E3gDtを示し、そして斜線の棒はBAV3.E3dを示す。誤差の棒は、1群あたり4匹の動物の平均の標準誤差を示す。
【図9】図9.異なるBAV組換え体を用いた免疫後の異なる時点における子ウシ血清中の抗-gD IgG力価(gD特異的ELISAにより測定)。斜線の棒:BAV3.E3dで免疫化した動物;白棒:BAV3.E3gDで免疫化した動物;黒棒:BAV3.E3gDtでワクチン接種した動物。
【図10】図10.異なるBAV組換え体を用いた免疫後の異なる時点における子ウシ鼻スワブにおける抗−gD IgA力価(gD特異的ELISAにより測定)。斜線の棒:BAV3.E3dで免疫化した動物;白棒:BAV3.E3gDで免疫化した動物;黒棒:BAV3.E3gDtでワクチン接種した動物。
【図11】図11.BHV−1抗原投与後に観察されたワクチン接種子ウシの温度。黒四角:BAV3.E3dで免疫化した動物;白三角:BAV3.E3gDで免疫化した動物;白丸:BAV3.E3gDtでワクチン接種した動物。
【図12】図12.ワクチン接種子ウシにおけるBHV−1抗原投与後の鼻損傷の外観および程度の観察。黒四角:BAV3.E3dで免疫化した動物;白三角:BAV3.E3gDで免疫化した動物;白丸:BAV3.E3gDtでワクチン接種した動物。
【図13】図13.ワクチン接種子ウシの試験およびコントロール群で観察された低下の程度。黒四角:BAV3.E3dで免疫化した動物;白三角:BAV3.E3gDで免疫化した動物。
【図14】図14.ワクチン接種子ウシの試験およびコントロール群のBHV−1抗原投与後のBHV−1の単離。黒四角:BAV3.E3dで免疫化した動物;白三角:BAV3.E3gDで免疫化した動物;白丸:BAV3.E3gDtでワクチン接種した動物。
【発明を実施するための形態】
【0037】
発明を実施する様式
本発明の実施は、その他に示されなければ、当該分野内の技術である、従来の微生物学、免疫学、ウイルス学、分子生物学、および組換えDNA技法を利用する。これらの技術は、文献に十分説明されている。例えば、Maniatisら、Molecular Cloning:ALaboratory Manual(1982);DNA Cloning:APractical Approach、I巻およびII巻(D.Glover編);Oligonucleotide Synthesis(N.Gait編(1984));Nucleic Acid Hybridization(B.HamesおよびS.Higgins編(1985)):Transcription and Translation(B.HamesおよびS.Higgins編(1984));Animal Cell Culture(R.Freshney編(1986));Peroal、A Practical Guide to Molecular Cloning(1984).Sambrookら、Molecular Cloning:A Laboratory Manual(第2版);第I、IIおよびIII巻(1989)を参照のこと。
【0038】
A.定義
本発明を記載することにおいて、次の用語が、以下に規定されるように用いられる。
【0039】
「レプリコン」は、インビボでDNA複製の自律単位として機能する、即ち、それ自身の制御の下で複製し得る任意の遺伝要素(例えば、プラスミド、染色体、ウイルス)である。
【0040】
「ベクター」は、プラスミド、ファージ、コスミドまたはウイルスのような、それに別のDNAセグメントが、連結したセグメントの複製を引き起こすように連結され得るレプリコンである。
【0041】
「生存ウイルス」は、「死滅した」ウイルスとは対照差別して、組織培養および接種動物において同一の子孫を産生し得るウイルスを意味する。
【0042】
「ヘルパーフリーウイルスベクター」は、ベクターに欠損するあるものを供給する第2のウイルスまたは細胞株を必要としないベクターである。
【0043】
「2本鎖DNA分子」は、その通常の2本鎖ヘリックス中のデオキシリボヌクレオチド(アデニン、グアニン、チミン、またはシトシン)のポリマー形態をいう。この用語は、分子の1次および2次構造のみについて言及し、そしてそれを任意の特定の三次形態に限定しない。従って、この用語は、とりわけ、直線状DNA分子で見出される2本鎖DNAを含む(例えば、ウイルス、プラスミド、および染色体からのDNAの制限フラグメント)。特定の2本鎖DNA分子の構造を論議することにおいて、本明細書において、配列は、DNAの非転写鎖(即ち、mRNAに相同な配列を有する鎖)に沿って5'〜3'方向にある配列のみを与える通常の慣習に従って記載され得る。
【0044】
DNA「コード配列」は、適切な調節配列の制御下に置かれたとき、インビボで転写されかつポリペプチドに翻訳されるDNA配列である。コード配列の境界は、5'(アミノ)末端の開始コドンおよび3'(カルボキシ)末端の翻訳終結コドンにより決定される。コード配列は、原核生物配列、真核生物mRNAからのcDNA、真核生物(例えば哺乳動物)DNAからのゲノムDNA配列、ウイルスDNA、および合成DNA配列さえ含み得るが、これらに制限されない。ポリアデニル化シグナルおよび転写終結配列は、通常、コード配列の3'側にある。
【0045】
「転写プロモーター配列」は、細胞において、RNAポリメラーゼに結合し、かつ下流(3'方向)コード配列の転写を開始し得るDNA調節領域である。本発明を規定する目的のために、プロモーター配列は、コード配列の翻訳開始コドン(ATG)が3'末端に結合し、そして上流(5'方向)に伸び、バックグラウンドを超える検出可能なレベルで転写を開始するために必要な最小数の塩基または要素を含む。プロモーター配列内には、転写開始部位(ヌクレアーゼS1を用いたマッピングにより簡単に規定される)、およびRNAポリメラーゼの結合を担うタンパク質結合ドメイン(コンセンサス配列)が見出される。真核生物プロモーターは、しばしば、常にではないが、「TATA」ボックスおよび「CAAT」ボックスを含む。原核生物プロモーターは、−10および−35コンセンサス配列に加えて、Shine−Dalgarno配列を含む。
【0046】
DNA「制御配列」は、集合的に、プロモーター配列、リボソーム結合部位、スプライシングシグナル、ポリアデニル化シグナル、転写終結配列、上流調節ドメイン、エンハンサー、翻訳終結配列などをいい、これは、集合的に、宿主細胞中のコード配列の転写および翻訳を提供する。
【0047】
RNAポリメラーゼがプロモーター配列に結合し、そしてコード配列をmRNAに転写し、次いで、コード配列によりコードされたポリペプチドに翻訳される場合、コード配列または配列コードは、細胞において、制御配列に「作動可能に連結」またはその「制御下」にある。
【0048】
「宿主細胞」は、外因性DNA配列により、形質転換されたか、または形質転換し得る細胞である。
【0049】
外因性DNAが細胞膜内側に導入されたとき、細胞はこのような外因性DNAにより「形質転換」されている。外因性DNAは、細胞のゲノムを構成する染色体DNAに組み込まれてもよい(共有結合する)し、または組み込まれなくてもよい。例えば、原核生物および酵母では、外因性DNAは、プラスミドのようなエピソーム要素上で維持され得る。安定に形質転換された細胞は、外因性DNAが染色体中に組み込まれ、その結果それは染色体複製を通じて娘細胞により受け継がれる細胞である。哺乳動物細胞については、この安定性は、外因性DNAを含む娘細胞の集団からなる細胞株またはクローンを確立する細胞の能力により実証される。
【0050】
「クローン」は、単一細胞または共通の祖先に由来する娘細胞の集団である。
【0051】
「細胞株」は、多世代の間インビトロで安定に増殖し得る初代細胞のクローンである。
【0052】
2つのポリペプチド配列は、アミノ酸の少なくとも約80%(好ましくは少なくとも約90%、および最も好ましくは少なくとも約95%)が規定された長さの分子に対して一致する場合「実質的に相同」である。
【0053】
2つのDNA配列は、それらが、ヌクレオチドと同一であるか、またはヌクレオチドの40%より多くは異ならず、好ましくはヌクレオチドの約30%より多くは異ならず(即ち、少なくとも約70%相同)、より好ましくはヌクレオチドの約20%よりは異ならず、そして最も好ましくはヌクレオチドの約10%より多くは異ならない場合に、「実質的に相同」である。
【0054】
実質的に相同であるDNA配列は、例えば、特定の系について規定されるストリンジェントな条件下のサザンハイブリダイゼーション実験において同定され得る。適切なハイブリダイゼーション条件を規定することは、当該分野の技術である。例えば、Maniatisら、前述;DNA Cloning、第IおよびII巻、前述;Nucleic Acid Hybridization、前述、を参照のこと。
【0055】
DNA構築物の「異質」領域は、天然に他の分子と結合して見出されない別のDNA分子の中にあるか、またはそれに結合したDNAの識別可能なセグメントである。従って、異質領域がウイルス遺伝子をコードする場合、この遺伝子は、通常、供給源ウイルスまたはウイルス感染細胞のゲノム中のウイルスゲノムに隣接しないDNAにより隣接される。異種コード配列の別の例は、コード配列自身が天然に見出されない構築物である(例えば、ネイティブ遺伝子とは異なるコドンをもつ合成配列)。本明細書で用いるとき、対立遺伝子異形または自発変異事象はDNAの異種領域を生じない。
【0056】
「ウシ宿主」は、任意の系統、成体または子ウシをいう。
【0057】
用語「タンパク質」は、本明細書において、その他に注記されなければ、ポリペプチドまたはグリコシル化ポリペプチドをそれぞれ示すために用いられる。用語「ポリペプチド」は、その最も広い意味で用いられる(即ち、ペプチド結合を通じて連結される任意のアミノ酸ポリマー(ジペプチドまたはそれより大きい))。従って、用語「ポリペプチド」は、タンパク質、オリゴペプチド、タンパク質フラグメント、アナログ、ムテイン、融合タンパク質などを含む。
【0058】
「融合タンパク質」は、通常、リーダー配列または安定化ポリペプチドをコードする第1の領域、および異種タンパク質をコードする第2の領域を含む遺伝子の発現産物として規定される。それは、抗原性タンパク質フラグメントまたは完全長BAVタンパク質配列および異種配列(代表的には、細胞内で発現されたポリペプチドについて組換え宿主において分泌のために機能的なリーダー配列、またはSODのような宿主細胞プロテアーゼからタンパク質を保護するN末端配列)を含むポリペプチドを含む。抗原性タンパク質フラグメントは、通常、長さが約5−7アミノ酸である。
【0059】
「ネイティブ」タンパク質またはポリペプチドは、BAVまたはBAV感染細胞から回収されるタンパク質またはポリペプチドをいう。従って、用語「ネイティブBAVポリペプチド」は、天然に存在するBAVタンパク質およびそのフラグメントを含む。「非ネイティブ」ポリペプチドは、組換えDNA法によるかまたは直接合成により産生されたポリペプチドをいう。「組換え」ポリペプチドは、組換えDNA技法により産生された、即ち、所望のポリペプチドをコードする外因性DNA構築物により形質転換された細胞から産生されるポリペプチドをいう。
【0060】
「実質的に純粋」なタンパク質は、他のタンパク質がなく、好ましくは少なくとも10%均一、より好ましくは60%均一、そして最も好ましくは95%均一である。
【0061】
「抗原」は、宿主の免疫系を刺激し体液性および/または細胞性抗原特異的応答を刺激する1つまたはそれ以上のエピトープを含む分子をいう。この用語はまた、「免疫原」と交換可能に用いられる。
【0062】
「ハプテン」は、キャリアに連結されなければ体液性または細胞性応答をつくる宿主の免疫系を刺激しない1つまたはそれ以上のエピトープを含む分子である。
【0063】
用語「エピトープ」は、特異的抗体分子が結合するか、またはT細胞により認識される抗原またはハプテン上の部位をいう。この用語はまた、「抗原性決定基」または「抗原性決定部位」と交換可能に用いられる。
【0064】
組成物またはワクチンに対する「免疫学的応答」は、宿主における、目的の組成物またはワクチンに対する細胞性および/または抗体媒介免疫応答の発生である。通常、このような応答は、目的の組成物またはワクチンに含められる抗原に特異的に惹起された、応答を生成する抗体、B細胞、ヘルパーT細胞、サプレッサーT細胞、および/または細胞傷害性T細胞から構成される。
【0065】
用語「免疫原性ポリペプチド」および「免疫原性アミノ酸配列」は、ポリペプチドまたはアミノ酸のそれぞれをいい、これは、ウイルス感染性を中和する抗体を惹起し、および/または免疫化宿主の保護を提供するために抗体-補体または抗体-依存性細胞傷害性を媒介する。本明細書で用いられる「免疫原性ポリペプチド」は、所望のタンパク質の完全長(またはほぼ完全長)配列またはその免疫原性フラグメントを含む。
【0066】
「免疫原性フラグメント」は、1つまたはそれ以上のエピトープを含み、そしてそれ故ウイルス感染性を中和する抗体を惹起するか、および/または、抗体-補体または抗体−依存性細胞傷害性を媒介し、免疫化宿主の保護を提供するポリペプチドのフクラグメントを意味する。このようなフラグメントは、通常、長さが少なくとも約5アミノ酸、そして好ましくは長さが少なくとも約10〜15アミノ酸である。フラグメントの長さには臨界の上限はなく、それはタンパク質配列のほぼ完全長、または2つまたはそれ以上の抗原のフラグメントを含む融合タンパク質さえ含み得る。本明細書で用いる用語「処置」は、ウシまたはヒトまたはその他の哺乳動物のような哺乳動物の処置をいい、(i)感染または再感染の防止(予防)、または(ii)感染の症状の軽減または除去のいずれかをいう。ワクチンは、組換えBAVにより産生される組換えBAV自身または組換え抗原を含む。
【0067】
「感染性」は、ウイルスゲノムを細胞中に送達する能力を有することを意味する。
【0068】
B.一般的方法
本発明は、BAV3ゲノムの完全ヌクレオチド配列を開示する。図1を参照のこと。mRNAの転写マッピングおよびcDNAクローンの配列決定に由来する、BAV3ゲノムの転写マップを図2に示す。BAV3ゲノムのサイズ(34,446bp)および全体構成は、HAVのそれと類似しているようであるが、ある種の差異はある。Reddyら(1998)前述。BAV3ゲノムの特質を示す特徴の1つは、E3コード領域の比較的小さなサイズ(1517bp)である。Mittalら(1992)J.Gen.Virol.73:3295−3300;Mittalら(1993)J.Gen.Virol.74:2825;およびReddyら(1998)前述。BAV3 E3領域およびそのRNA転写物の配列の分析は、BAV3 E3が少なくとも4つのタンパク質をコードし得、その1つ(121R)は、HAV5の14.7kDaタンパク質と限られた相同性を示すことを示唆する。Idamakanti(1988)「ウシアデノウイルス3のE3領域の分子特徴付け」M.Sc.学位論文、University of Saskatchewan、Saskatoon、Saskatchewan。
【0069】
1つの実施態様では、本発明は、ウシアデノウイルスE1および/またはE3領域のヌクレオチド配列の一部分またはすべてを欠失する手段を同定および提供し、外来遺伝子またはそのフラグメントをコードする異種または同種ヌクレオチド配列を挿入し、ウシアデノウイルス組換え体を生成し得る部位を提供する。ヌクレオチド配列の「一部分を欠失すること」は、E1および/またはE3領域の一部分のヌクレオチド配列を欠失するために従来の遺伝子工学技法を用いることを意味する。
【0070】
別の実施態様では、本発明は、E3欠失組換え体BAV3(異種配列の挿入ありまたはなし)を、高い効率で、構築、単離、および増殖するための組成物および方法を提供する。これらは、適切な細胞株(例えば、アデノウイルスE1機能を発現するMDBK細胞または等価な細胞株)における組換え体ウイルスの単離、ならびに組換え体BAVゲノムが原核生物細胞における相同組換えを介して構築され、それにより得られる組換え体ゲノムが適切な細胞株(例えば、初代ウシ胎仔血清網膜細胞またはその等価物)にトランスフェクトされ、そして組換え体ウイルスがトランスフェクト細胞から単離される方法を含む。
【0071】
さらに、異なる形態のBHV-1糖タンパク質gDを発現する組換え体BAV3の構築が提供され、そしてgD発現組換え体BAVウイルスでのコトンラットの鼻腔内免疫がgD特異的な粘膜および全身の免疫応答の誘導をもたらすことが示される。実施例3を参照のこと。BHV-1 gD遺伝子を含むBAV組換え体でのウシ宿主の鼻腔内免疫は、腓におけるBHV-1チャレンジ(抗原投与)に対する防御を提供し、臨床的徴候の出現を減少させ、ウイルスのより迅速なクリアランスを促進し、そしてIgGおよびIgAの両方の力価の増加を提供する。実施例4を参照のこと。類似の様式で、哺乳動物病原体の防御決定因子をコードする任意の遺伝子は、E3欠失BAVに挿入され得、そして得られる組換え体BAVは、ワクチンとして使用され得る。
【0072】
本発明の1つの実施態様では、組換え体BAV発現カセットは、野生型BAVゲノムを1つ以上の適切な制限酵素で切断して、それぞれ、E1またはE3領域の配列を含むBAV制限フラグメントを生じることにより得られ得る。BAV制限フラグメントは、クローニングビヒクル(例えば、プラスミド)に挿入され得、そしてその後少なくとも1つの異種配列(これは、外来タンパク質をコードしてもよいしコードしなくともよい)は、作動可能に連結された真核生物転写調節配列を有するかまたは有さないE1またはE3領域に挿入され得る。組換え体発現カセットは、BAVゲノムと接触され得、そして相同組換えまたはタンパク質の従来の遺伝子操作方法を介して、所望の組換え体が得られる。発現カセットがE1領域または何らかの他の必須領域を含む場合、発現カセットとBAVゲノムとの間の組換えは、適切なヘルパー細胞株(例えば、E1形質転換細胞株)内で生じ得る。E1またはE3領域を含む制限フラグメント以外のBAVゲノムの制限フラグメントはまた、本発明の実施において有用であり、そして異種配列がBAV配列中に挿入され得るように、クローニングビヒクル中に挿入され得る。次いで、これらのDNA構築物は、インビトロまたはインビボで、適切な宿主細胞の形質転換またはトランスフェクションの前または後でBAVゲノムとの組換えを被り得る。
【0073】
適切な宿主細胞は、BAVゲノムと、BAV配列を含むプラスミドとの組換え、またはそれぞれがBAV配列を含む2つ以上のプラスミドの間の組換えを支持する任意の細胞を含む。組換えは一般に、原核生物細胞(例えば、E.coliなど)において行なわれるが、ウイルス粒子を生じる、ウイルスゲノムを含むプラスミドのトランスフェクションは、真核生物細胞、好ましくは哺乳動物細胞、より好ましくはウシ細胞培養物、最も好ましくはMDBKまたはPFBR細胞、およびそれらの等価物において実施される。細菌細胞培養物の増殖、ならびに真核生物細胞および哺乳動物細胞株の培養および維持は、当業者に周知である手順である。
【0074】
1つ以上の異種配列は、BAVゲノムの1つ以上の領域に挿入されて組換えBAVベクターを生じ得る。これは、BAVゲノムの挿入能力および組換えBAVベクターが挿入された異種配列を発現する能力にのみ制限される。一般に、アデノウイルスゲノムは、ゲノム長の約5%の挿入物を受け入れ得、そしてウイルス粒子にパッケージングされ得るまmであり得る。挿入能力は、非必須領域の欠失および/または機能がヘルパー細胞株により提供される必須領域の欠失により増加し得る。
【0075】
1つの実施態様では、挿入は、BAVゲノムのうちの挿入が所望される領域を含むプラスミドを構築することにより達成され得る。次いで、プラスミドが、プラスミドのBAVR部分における認識配列を有する制限酵素を用いて消化され、そして異種配列は制限消化の部位に挿入される。挿入された異種配列とともにBAVゲノムの一部を含むプラスミドは、BAVゲノムまたはBAVゲノムを含む線状化プラスミドとともに細菌細胞(例えば、E.coliなど)に同時形質転換される。ここで、BAVゲノムは、全長ゲノムであり得るか、または1つ以上の欠失を含み得る。プラスミド間の相同組換えは、挿入された異種配列を含む組換え体BAVゲノムを生じる。
【0076】
異種配列の挿入部位を提供するため、または異なる部位での挿入のためのさらなる能力を提供するためのBAV配列の欠失は、当業者に周知の方法により達成され得る。例えば、プラスミド中にクローニングされたBAV配列については、1つ以上の制限酵素(BAV挿入物中に少なくとも1つの認識配列を有する)での消化、およびその後の連結は、いくつかの場合、制限酵素認識部位の間の配列の欠失をもたらす。あるいは、BAV挿入物中の単一制限酵素認識部位での消化、続いてエキソヌクレアーゼ処置、続いて連結は、制限部位に近接したBAV配列の欠失をもたらす。上記の通りに構築した、1つ以上の欠失を有するBAVゲノムの1つ以上の部分を含むプラスミドは、BAVゲノム(全長もしくは欠失型)、または全長もしくは欠失型のいずれかのBAVゲノムを含むプラスミドとともに細菌細胞中に同時トランスフェクトされて、相同組換えにより、1つ以上の特異的部位に欠失を有する組換えBAVゲノムを含むプラスミドを生じ得る。次いで、欠失を含むBAVビリオンは、組換え体BAVゲノムを含むプラスミドを用いる哺乳動物細胞(MDBK細胞またはPFBR細胞およびそれらの等価物を含むがこれらに限定されない)のトランスフェクションにより得られ得る。
【0077】
本発明の1つの実施態様では、挿入部位は、BAVプロモーターに近接し、そして(転写の意味で)その下流にある。BAVプロモーターの位置、およびBAVプロモーターの下流の制限酵素認識配列(挿入部位として使用するため)は、当業者によって、本明細書中に提供されるBAVヌクレオチド配列から容易に決定され得る。
【0078】
あるいは、種々のインビトロ技術は、特定の部位での制限酵素認識配列の挿入のために、または制限酵素認識配列を含まない部位での異種配列の挿入のために用いられ得る。このような方法は、1つ以上の制限酵素認識配列の挿入のためのオリゴヌクレオチド媒介ヘテロ二重鎖形成(例えば、Zollerら(1982)Nucleic Acids Res.10:6487-6500;Brennanら(1990)Roux's Arch.Dev.Biol.199:89-96;およびKunkelら(1987)Meth.Enzymology 154:367-382を参照のこと)、およびより長い配列の挿入のためのPCR媒介方法を含むがこれらに限定されない。例えば、Zhengら(1994)Virus Research 31:163-186を参照のこと。
【0079】
異種配列が真核生物細胞中に活性である転写調節配列をさらに含むならば、BAVプロモーターの下流ではない部位で挿入された異種配列の発現を得ることもまた可能である。このような転写調節配列は、細胞プロモーター(例えば、ウシhsp70プロモーターなど)およびウイルス性プロモーター(例えば、ヘルペスウイルスプロモーター、アデノウイルスプロモーター、およびパポーバウイルスプロモーターなど)、ならびにレトロウイルス長末端反復(LTR)配列のDNAコピーを含み得る。
【0080】
別の実施態様では、原核生物細胞における相同組換えは、クローニングされたBAVゲノムを生じるために用いられ得;そしてクローニングされたBAVゲノムは、プラスミドとして増殖され得る。感染性ウイルスは、プラスミド含有細胞からレスキューされた、クローニングされたBAVゲノムでの哺乳動物細胞のトランスフェクションにより入手され得る。
【0081】
本発明はまた、異種遺伝子の発現を調節するために使用され得るBAV調節配列を提供する。調節配列は、例えば、転写調節配列、プロモーター、エンハンサー、上流調節ドメイン、スプライシングシグナル、ポリアデニル化シグナル、転写終結配列、翻訳調節配列、リボソーム結合部位、および翻訳終結配列であり得る。
【0082】
別の実施態様では、本発明は、外来遺伝子またはそのフラグメントをコードする異種または同種のヌクレオチド配列を挿入してBAV組換え体を生じるのに適切なBAVゲノム(およびそのフラグメント)のさらなる領域を同定および提供する。これらの領域は、ヌクレオチド4,092〜5,234;ヌクレオチド5,892〜17,735;ヌクレオチド21,198〜26,033;ならびにヌクレオチド31,133からBAVゲノムの右端にまたがり、そしてE2領域、E4領域、後期領域、33kD、52kD、100kD、DBP、pol、pTP、およびペントン遺伝子、ならびに遺伝子IIIA、pV、pVI、pVII、pVIII、およびpXを含む領域を含む。BAVゲノムのこれらの領域は、とりわけ、外来配列の挿入、DNA調節配列(転写調節配列および翻訳調節配列を含む)の提供、または生物学的サンプル中のウイルス核酸またはこれらの領域によりコードされるタンパク質の存在を検出する診断目的に用いられ得る。
【0083】
別の実施態様では、クローニングされるBAV-3ゲノムは、プラスミドとして増殖され得、そして感染性ウイルスは、プラスミド含有細胞からレスキューされ得る。
【0084】
ウイルス核酸の存在は、ハイブリダイゼーションアッセイ、ポリメラーゼ連鎖反応、および他の型の増幅反応を含むがこれらに限定されない、当業者に公知の技術により検出され得る。同様に、タンパク質の検出方法は、当業者に周知であり、そして種々の型の免疫圧制、ELISA、ウェスタンブロッティング、酵素アッセイ、免疫組織化学などを含むがこれらに限定されない。本発明のヌクレオチド配列を含む診断キットはまた、細胞破壊および核酸精製のための試薬、ならびにハイブリッドの形成、選択、および検出のための緩衝液および溶媒を含む。本発明のポリペプチドまたはアミノ酸配列を含む診断キットはまた、タンパク質単離ならびに免疫複合体の形成、単離、精製、および/または検出のための試薬を含み得る。
【0085】
種々の外来遺伝子またはヌクレオチド配列またはコード配列(原核生物および真核生物)は、上記のアデノウイルスヌクレオチド配列(例えば、DNA)中に、本発明に従って、広範囲な疾患に対する防御を提供するために挿入され得、そしてこのような多くの遺伝子は既に当該分野において公知である。これまでの問題は、遺伝子または配列についての安全な、便利な、および有効なワクチンベクター、ならびに種々の遺伝子治療適用において用いられる遺伝子移入のために有効な手段を提供することであった。
【0086】
外因性(すなわち、外来)ヌクレオチド配列は、1つ以上の目的の遺伝子から、そして好ましくは1つ以上の目的の治療遺伝子からなり得る。本発明の状況では、目的の遺伝子は、アンチセンスRNA、リボザイム、または目的のタンパク質に翻訳されるmRNAをコードし得る。目的の遺伝子は、ゲノム方の、相補的なDNA(cDNA)型の、または混合型のもの(少なくとも1つのイントロンが欠失されたミニ遺伝子)であり得る。これは、成熟タンパク質、成熟タンパク質の前駆体(特に分泌されることが意図される、従ってシグナルペプチドを含む前駆体)、多様な起源の配列の融合物に由来するキメラタンパク質、または改善もしくは修飾された生物学的特性を示す、天然タンパク質の変異体をコードし得る。このような変異体は、天然タンパク質をコードする遺伝子の1つ以上のヌクレオチドの欠失、置換、および/もしくは付加、または天然タンパク質をコードする配列の他の任意の型の配列変化(例えば、トランジションもしくはインバージョン)により得られ得る。
【0087】
目的の遺伝子は、宿主細胞におけるその発現に適切なエレメント(DNA調節配列)の制御下に置かれ得る。適切なDNA調節配列は、RNAへの遺伝子の転写(アンチセンスRNAもしくはmRNA)またはタンパク質へのmRNAの翻訳に必要なエレメントのセットを意味すると理解される。転写に必要とされるエレメントの中では、当然、プロモーターは特別に重要であると考えられる。プロモーターは、構成性プロモーターまたは調節性プロモーターであり得、そして真核生物、原核生物、またはウイルス起源、そしてさらにアデノウイルス起源の任意の遺伝子から単離され得る。あるいは、プロモーターは、目的の遺伝子の天然のプロモーターであり得る。概して、本発明で用いられるプロモーターは、調節配列を含むように改変され得る。例えば、本発明において使用される目的の遺伝子は、宿主のリンパ球細胞への遺伝子の移入を標的化することが所望される場合、免疫グロブリン遺伝子のプロモーターの制御下に置かれる。例えば、多数の細胞型における発現を可能にする、HSV-1 TK(ヘルペスウイルスタイプ1チミジンキナーゼ)遺伝子プロモーター、 (特に、ヒトアデノウイルスタイプ2の)アデノウイルスMLP(主要後期プロモーター)、RSV(ラウス肉腫ウイルス)LTR(長末端反復)、CMV(サイトメガロウイルス)初期プロモーター、およびPGK(ホスホグリセリン酸キナーゼ)遺伝子プロモーターもまた挙げられ得る。
【0088】
あるいは、特定の細胞型への組換え体BAVベクターの標的化は、組換え体ヘキソンおよび/または繊維遺伝子を構築することにより達成され得る。これらの遺伝子のタンパク質産物は、宿主細胞の認識に関与する;それゆえ、遺伝子は、ウイルスが別の宿主細胞を認識するのを可能にするペプチド配列を含むように改変され得る。
【0089】
本発明の状況で使用可能である目的の遺伝子の中では、以下のものが挙げられる:
【0090】
−サイトカイン(例えば、インターフェロンおよびインターロイキン)をコードする遺伝子;
【0091】
−リンホカインをコードする遺伝子;
【0092】
−病原性生物(例えば、ウイルス、細菌、または寄生生物)、好ましくはHIVウイルス(ヒト免疫不全ウイルス)により認識されるレセプターのような膜レセプターをコードする遺伝子;
【0093】
−凝固因子(例えば、第VIII因子および第IX遺伝子)をコードする遺伝子;
【0094】
−ジストロフィンをコードする遺伝子;
【0095】
−インスリンをコードする遺伝子;
【0096】
−細胞イオンチャネルに直接的または間接的に関与するタンパク質(例えば、CFTR(嚢胞性線維症膜貫通調節タンパク質)タンパク質)をコードする遺伝子;
【0097】
−アンチセンスRNA、病原性生物のゲノムに存在する病原性遺伝子により生成されるタンパク質の活性を阻害し得るタンパク質、または発現が統制解除された細胞遺伝子(例えば、オンコジーン)の活性を阻害し得るタンパク質(もしくはそれらをコードする遺伝子)をコードする遺伝子;
【0098】
−例えば、酵素活性を阻害するタンパク質(例えば、α1-アンチトリプシンまたはウイルスプロテアーゼインヒビター)をコードする遺伝子;
【0099】
−病原性タンパク質の生物学的機能を損なうように変異された病原性タンパク質の改変体(例えば、標的配列への結合について天然タンパク質と競合し得、それによりHIVの活性化を防止し得る、HIVウイルスのtatタンパク質のトランスドミナント改変体など)をコードする遺伝子;
【0100】
−宿主細胞の免疫を増加させるための抗原性エピトープをコードする遺伝子;
【0101】
−主要組織適合遺伝子複合体クラスIおよびIIタンパク質をコードする遺伝子、ならびにこれらの遺伝子のインデューサーであるタンパク質をコードする遺伝子;
【0102】
−抗体をコードする遺伝子;
【0103】
−イムノトキシンをコードする遺伝子;
【0104】
−トキシンをコードする遺伝子;
【0105】
−増殖因子または増殖ホルモンをコードする遺伝子;
【0106】
−細胞レセプターおよびそれらのリガンドをコードする遺伝子;
【0107】
−腫瘍サプレッサーをコードする遺伝子;
【0108】
−オンコジーンを含むがこれらに限定されない、心臓血管病に関与する遺伝子;線維芽細胞増殖因子(FGF)、血管内皮増殖因子(VEGF)、および神経成長因子(NGF)を含むがこれらに限定されない、増殖因子をコードする遺伝子;Rb(網膜芽細胞種)遺伝子を含むがこれらに限定されない、e-nos腫瘍サプレッサー遺伝子;リポタンパク質リパーゼ;スーパーオキシドジスムターゼ(SOD);カタラーゼ;酸素および遊離基スカベンジャー;アポリポタンパタ質;ならびにpai-1(プラスミノゲンアクチベーターインヒビター1);
【0109】
−細胞酵素または病原性生物により生成される細胞酵素をコードする遺伝子;
ならびに
【0110】
−自殺遺伝子。HSV-1 TKは自殺遺伝子が、例として挙げられ得る。
このウイルスTK酵素は、特定のヌクレオシドアナログ(例えば、アシクロビルまたはガンシクロビル)に対して、細胞性TK酵素と比較して著しく高い親和性を示す。このウイルスTK酵素は、これらを一リン酸化分子に変換し、これはそれ自体が細胞酵素によりヌクレオチド前駆体に変換され得る。これらのヌクレオチドアナログは、複製中のDNA分子に取りこまれ得、それゆえ、取りこみは主に分裂細胞のDNAに生じる。この取りこみは、分裂細胞(例えば、ガン細胞)の特異的破壊をもたらし得る。
【0111】
このリストは限定的ではなく、他の目的の遺伝子が本発明の状況において用いられ得る。
【0112】
野生型生物に見出されるとおりの完全な配列ではなく、遺伝子のヌクレオチド配列のフラグメントのみが用いられ得る(ここで、これらは、防御免疫応答または特異的生物学的効果を生じるのに十分である)こともまた可能である。利用可能な場合には、合成遺伝子またはそのフラグメントもまた用いられ得る。しかし、本発明は、広範な種々の遺伝子、フラグメントなどに用いられ得、そして上記に記載したものに限定されない。
【0113】
いくつかの場合には、特定の抗原の遺伝子は、多数のイントロンを含み得るか、またはRNAウイルス由来であり得、これらの場合、相補的なDNAコピー(cDNA)が用いられ得る。
【0114】
遺伝子の首尾良い発現を生じるために、遺伝子は、エンハンサ一エレメントおよびポリアデニル化配列を含む適切なプロモーターとともに発現ベクターに挿入され得る。哺乳動物細胞における外来遺伝子の首尾良い発現を提供する多数の真核生物性のプロモーターおよびポリアデニル化配列、ならびに発現カセットを構築する方法が、当該分野で(例えば、米国特許第5,151,267号(この開示は本明細書中で参考として援用される)において)公知である。プロモーターは、体液性、細胞媒介性、および粘膜性の免疫応答を満足にもたらす免疫原性タンパク質の最適な発現を与えるように既知の基準に従って選択される。
【0115】
組換え体ウイルス感染細胞におけるインビボでの発現により生成される外来タンパク質は、それ自体が免疫原性であり得る。1つより多くの外来遺伝子が、ウイルスゲノムに挿入されて、1つより多くの有効なタンパク質の首尾良い生成が得られ得る。
【0116】
従って、本発明の組換え体ウイルスを用いて、ウシ、ヒト、および他の哺乳動物に罹患する広範な種々の疾患に対する防御を提供することが可能である。本発明の任意の組換え体抗原性決定基または組換え体生ウイルスは、抗原性決定基ワクチンまたは生ワクチンベクターについて記載されたのと実質的に同じ様式で処方および使用され得る。
【0117】
本発明はまた、本発明の方法に従って調製された、治療有効量の組換え体ベクター、組換え体ウイルス、または組換え体タンパク質を、薬学的に受容可能なビヒクルおよび/またはアジュバントと組合せて含む薬学的組成物を含む。このような薬学的組成物は、当該分野で周知である技術に従って調製され得、そして投薬量は当該分野で周知である技術に従って決定され得る。本発明の薬学的組成物は、全身的(例えば、静脈内、気管内、脈管内、肺内、腹腔内、鼻腔内、非経口、腸、筋肉内、皮下、肺瘍内、もしくは頭蓋内)経路を含むがこれらに限定されない、公知の任意の投与経路により、またはエーロゾル適用もしくは肺内点滴注入により投与され得る。投与は、単回用量で、または特定の時間間隔の後に1回以上反復される用量で起こり得る。適切な投与経路および投薬量は、状況(例えば、処置される個体、処置される障害、または目的の遺伝子もしくはポリペプチド)によって変化するが、当業者により決定され得る。
【0118】
本発明はまた、本発明の治療有効量のBAVベクター、組換え体BAV、または宿主細胞が、処置を必要とする哺乳動物非験体に投与される処置方法を包含する。
【0119】
本発明で用いられる抗原は、ネイティブまたは組換え体のいずれかの抗原性ポリペプチドまたはフラグメントであり得る。これらは、部分配列、全長配列、または融合物(例えば、組換え体宿主に適切なリーダー配列を有するか、または別の病原体についてのさらなる抗原配列を有する)でさえあり得る。本発明のウイルス系により発現される好ましい抗原性ポリペプチドは、抗原をコードする全長(またはほぼ全長)の配列を含む。あるいは、抗原性である(すなわち、1つ以上のエピトープをコードする)、より短い配列が用いられ得る。より短い配列は、インビトロアッセイにおいてウイルス感染性を中和するエピトープと定義される「中和エピトープ」をコードし得る。好ましくは、ペプチドは、宿主において「防御免疫応答」(すなわち、免疫された宿主を感染から防御する、抗体媒介性および/または細胞媒介性の免疫応答)を生じ得る、「防御エピトープ」をコードすべきである。
【0120】
本発明で用いられる抗原(特に、短いオリゴペプチドから構成される場合)は、ワクチンキャリアに結合体化され得る。ワクチンキャリアは、当該分野で周知である;例えば、ウシ血清アルブミン(BSA)、ヒト血清アルブミン(HSA)、およびキーホールリンペットヘモシアニン(KLH)。好ましいキャリアタンパク質であるロタウイルスVP6は、EPO公報第0259149号(この開示は本明細書中に参考として援用される)に開示される。
【0121】
挿入され得る所望の抗原の遺伝子またはそのコード配列は、哺乳動物において疾患を生じる生物(特に、ウシロタウイルス、ウシコロナウイルス、ウシヘルペスウイルスタイプ1、ウシRSウイルス、ウシパラインフルエンザウイルスタイプ3(BPI-3)、ウシ下痢ウイルス、Pasteurella haemolytica、Haemophilus somnusなどのようなウシ病原体)のものを含む。外来遺伝子またはフラグメントを有する本発明のワクチンはまた、例えば、腸溶性投薬形態のような適切な経口キャリアにおいて経口投与され得る。経口処方物は、薬学的グレードのマンニトール、ラクトース、スターチ、ステアリン酸マグネシウム、サッカリンナトリウムセルロース、炭酸マグネシウムなどのような通常用いられる賦形剤を含む。経口ワクチン組成物は、約10%〜約95%、好ましくは約25%〜約70%の活性成分を含む、溶液、懸濁液、錠剤、丸剤、カプセル剤、持続放出処方物、または散剤の形態を取り得る。経口および/または鼻腔内ワクチン接種は、全身免疫との組合せで、粘膜免疫(これは、気道および胃腸管に感染する病原体に対する防御において重要な役割を果たす)を惹起するために好ましくあり得る。
【0122】
さらに、ワクチンは、坐剤中に処方され得る。坐剤について、ワクチン組成物は、伝統的なバインダーおよびキャリア(例えば、ポリアルカリグリコールまたはトリグリセリド)を含む。このような坐剤は、約0.5%(w/w)〜約10%(w/w)、好ましくは約1%(w/w)〜約2%(w/w)の範囲で活性成分を含有する混合物から形成され得る。
【0123】
動物に本発明のワクチン組成物を投与するためのプロトコルは、本開示を考慮すれば当該分野の技術範囲内である。当業者は、抗原性フラグメントに対する抗体および/またはT細胞媒介性免疫応答を惹起するのに有効な用量のワクチン組成物の濃度を選択する。広範な制限内では、投薬量は重要であるとは考えられない。代表的には、ワクチン組成物は、約1マイクログラム〜約1,000マイクログラムのサブユニット抗原を便利な容量(例えば、約1〜10cc)のビヒクル中で送達する様式で投与される。好ましくは、単回免疫における投薬量は、約1マイクログラム〜約500マイクログラム、より好ましくは約5〜10マイクログラムから約100〜200マイクログラム(例えば、5〜200マイクログラム)のサブユニット抗原を送達する。
【0124】
投与時期はまた、重要であり得る。例えば、一次接種は、好ましくは、必要に応じて、その後のブースター接種が続き得る。必要に応じてであるが、2回目であるブースター免疫を、初期免疫の数週間後から数ヶ月後に動物に投与することが好ましくあり得る。疾患に対する持続した高レベルの防御を保証するために、ブースター免疫を動物に対して定期的な間隔(例えば、数年毎に一度)で、再投与することが有益であり得る。あるいは、最初の用量は、経口的に、続いて後の接種により、またはその逆で投与され得る。好ましいワクチン接種プロトコルは、日常的なワクチン接種プロトコル実験を介して達成され得る。
【0125】
インビボでの組換え体ウイルスワクチンの全ての投与経路についての投薬量は、患者の大きさ、防御が必要とされる感染の性質、キャリアなどを含む種々の因子に依存し、そして当業者により容易に決定され得る。非限定的な例により、103pfuおよび1015pfu、好ましくは105pfu〜1013pfu、より好ましくは106pfu〜1011pfuなどの投薬量が用いられ得る。インビトロでのサブユニットワクチンについてのように、さらなる投薬量は、関与する臨床的因子により決定された通りに与えられ得る。
【0126】
本発明の1つの実施態様では、多数の組換え体細胞株が、BAV E1領域を含む発現カセットを構築し、そしてこれにより宿主細胞を形質転換して、E1タンパク質を発現する補完細胞株または培養物を提供することにより、本発明に従って生成される。これらの組換え体補完細胞株は、E1配列が欠失した組換え体欠損BAVが複製し、そして必要に応じて組換え体BAV内にコードされる所望の外来遺伝子またはそのフラグメントを発現するのを可能にし得る。これらの細胞株はまた、外来遺伝子またはフラグメントをコードする異種ヌクレオチド配列により置換されたE3遺伝子欠失を有する組換え体BAVを、DNA媒介コトランスフェクション後のインビボ組換えにより生成するのに非常に有用である。より一般的には、BAVゲノムによりコードされる1つ以上の必須機能を欠く組換え体欠損BAVベクターは、適切な補完細胞株中で増殖され得、ここで、特定の補完細胞株は、特定の組換え体欠損BAVベクターにおいて欠けている機能を提供する。補完細胞株は、例えば、ヘルパーウイルスとともに同時感染することにより、または特定のウイルス機能をコードするウイルスゲノムの安定な形態のフラグメントを組込むかさもなければ維持することにより、ウイルス機能を提供し得る。
【0127】
本発明の1つの実施態様では、組換え体発現カセットは、BAVゲノムを適切な制限酵素で切断して、それぞれ、E1もしくはE3遺伝子領域配列を含む、ゲノムの左側末端または右側末端を提示するDNAフラグメントを生成し、そして左または右の末端フラグメントをクローニングビヒクル(例えば、プラスミド)に挿入し、その後少なくとも1つの異種DNA配列を外因性プロモーターの制御ありまたはなしでE1またはE3の欠失に挿入することにより得られ得る。組換え発現カセットは、適切な細胞内でBAVゲノムと接触され、そして相同組換えまたはタンパク質の従来の遺伝子操作法を介して組換え体BAVゲノムが得られる。適切な細胞は、原核生物細胞(例えば、E.coliなど)および真核生物細胞の両方を含む。適切な真核生物細胞の例は、MDBK細胞、アデノウイルスE1機能を発現するMDBK細胞、初代ウシ胎仔網膜細胞、および先に記載した細胞の機能と等価である機能を発現する細胞を含むがこれらに限定されない。E1またはE3領域を含む制限フラグメント以外のBAVゲノムの制限フラグメントもまた、本発明の実施において有用であり、そして異種配列が非E1および非E3 BAV配列に挿入され得るようにクローニングビヒクルに挿入され得る。次いで、これらのDNA構築物は、インビトロまたはインビボで、BAVゲノムとの組換えを、上記の適切な宿主細胞の形質転換またはトランスフェクションの前または後に受け得る。本発明の目的のために、得られる組換え体BAVゲノムが、複製およびパッケージングに要求されるBAV配列を含む限りは、BAVゲノムは、全長ゲノム、または組換わるフラグメントで欠失している領域以外の領域に欠失を含むゲノムのいずれかであり得る。トランスフェクションの方法、細胞培養、ならびに原核生物および真核生物の細胞における組換え(例えば、上記に記載されるもの)は、当業者に周知である。
【0128】
別の実施態様では、E1機能(または任意の特定のウイルスベクターにおいて変異もしくは欠失され得る任意の他のウイルス領域の機能)は、ベクターが欠く機能を発現するウイルスでの細胞の同時感染により供給(補完細胞株を提供)され得る。
【0129】
本発明はまた、ウシアデノウイルス発現系を含む発現系を含み、個々で異種ヌクレオチド配列(例えば、DNA)は、E3領域の一部もしくは全部、E1領域の一部もしくは全部、E2領域の一部もしくは全部、E4領域の一部もしくは全部、E4とゲノムの右末端との間の領域の一部もしくは全部、後期領域(L1〜L7)の一部もしくは全部、および/または33kD、52kD、100kD、DBP、pol、pTP、およびペントン遺伝子、ならびに遺伝子IIIA、pV、pVI、pVII、pVIII、およびpXにより占められる領域の一部もしくは全部を置きかえる。発現系が用いられ得、ここで外来ヌクレオチド配列(例えば、DNA)は、他の任意の異種プロモーターのコントロールを有するかまたは有さない。BAV発現ベクターはまた、逆方向末端反復(ITR)配列およびパッケージング配列を含み得る。
【0130】
BAVの33kD、52kD、100kD、DBP、pTP、ペントン(III)、pIIIA、pIVa2、pV、pVI、pVII、pVIII、およびpX遺伝子は、ウイルス複製に必須である。それゆえ、これらの任意の遺伝子に欠失を含むが、またはこれらの任意の遺伝子によりコードされる機能を欠くBAVベクターは、適切な補完細胞株(すなわち、ヘルパー細胞株)中で増殖させねばならない。ヒトアデノウイルスでは、E4領域における特定のオープンリーディングフレーム(ORF3およびORF6/7)はウイルス複製に必須である。BAV-3のE4領域における類似のオープンリーディングフレームにおける欠失は、ウイルスベクターの増殖のためにヘルパー細胞株の使用を必要とし得る。
【0131】
本発明のアデノウイルスのBAV E1遺伝子産物は、大部分の細胞遺伝子をトランス活性化し、それゆえ、E1タンパク質を構成的に発現する細胞株は、通常の細胞株よりも高いレベルで細胞ポリペプチドを発現し得る。本発明の組換え体哺乳動物(特にウシ)細胞株は、ポリペプチドを調製および単離するために用いられ得る。ポリペプチドは、以下の通りのポリペプチドを含む:(a)アデノウイルスEIAタンパク質に関連するタンパク質(例えば、p300、網膜芽細胞(Rb)タンパク質、サイクリン、キナーゼなど);(b)アデノウイルスE1Bタンパク質に関連するタンパク質(例えば、p53など);(c)増殖因子(例えば、上皮増殖因子(EGF)、トランスフォーミング増殖因子(TGF)など);(d)レセプター(例えば、上皮増殖因子レセプター(EGF-R)、線維芽細胞増殖因子レセプター(FGF-R)、腫瘍壊死因子レセプター(TNF-R)、インスリン様増殖因子レセプター(IGF-R)、主要組織適合遺伝子複合体クラスIレセプターなど);(e)プロトオンコジーンによりコードされるタンパク質(例えば、プロテインキナーゼ(チロシン特異的タンパク質キナーゼおよびセリンもしくはトレオニンに特異的なタンパク質キナーゼ)、p21タンパク質(GTPase活性を有するグアニンヌクレオチド結合タンパク質)など);(f)他の細胞タンパク質(例えば、アクチン、コラーゲン、フィブロネクチン、インテグリン、ホスホプロテイン、プロテオグリカン、ヒストンなど);ならびに(g)転写調節に関与するタンパタ質(例えば、TATAボックス結合タンパク質(TBP)、TBP関連因子(TAF)、Sp1結合タンパク質など)。
【0132】
本発明はまた、遺伝子治療を哺乳動物(例えば、ウシもしくはヒトまたはその必要性がある哺乳動物)に提供して、遺伝子欠損を制御し、治療遺伝子もしくはヌクレオチド配列を提供し、そして/または遺伝子変異を誘導もしくは訂正するための方法を含む。この方法は、例えば、遺伝性疾患、感染性疾患、心臓血管疾患、およびウイルス感染を含むがこれらに限定されない状態の処置において用いられ得る。この方法は、上記哺乳動物に、非欠損形態の上記遺伝子を、組換え体ウイルスベクターのゲノムが上記哺乳動物ゲノムへと取り込まれるか、または独立してかつ染色体外で維持される条件下で外来ヌクレオチド配列を含む生組換え体ウシアデノウイルスを投与して、標的器官または組織において要求される遺伝子の発現を提供する工程を含む。これらの種類の技術は、現在、種々の疾患状態の処置(この非限定的な例は上記に提供される)のために当業者によって用いられている。従来の遺伝子治療における使用のために取りこまれ得る外来遺伝子、ヌクレオチド配列、またはその部分の例として、嚢胞性線維症膜貫通調節タンパク質遺伝子、ヒトミニジストロフィン遺伝子、α-1アンチトリプシン遺伝子、心臓血管疾患において関与し得る。
【0133】
特に、遺伝性疾患(例えば、血友病、サラセミア、気腫、ゴシェ病、嚢胞性線維症、デュシェーヌ筋ジストロフィー、デュシェーヌまたはベッカーのミオパシーなど)、ガン、ウイルス性疾患(例えば、AIDS、ヘルペスウイルス感染、サイトメガロウイルス感染、およびパピローマウイルス感染)、心臓血管病などを含むがこれらに限定されない、ヒトにおける遺伝子治療に関する本発明の実施は、疾患の防御または処置が意図される。本発明の目的のためには、本発明の方法により調製されたベクター、細胞、およびウイルス粒子は、被験体にエクスビボ(すなわち、細胞中または患者から取り出した細胞)で、または処置される身体にインビボで直接のいずれかで、導入され得る。好ましくは、宿主細胞はヒト細胞であり、そして最も好ましくは、肺、線維芽細胞、筋肉、肝臓、またはリンパ球または造血直系の細胞である。
【実施例】
【0134】
実施例
本発明の実施例を以下記載する。これらの実施例は、例示の目的にのみ提供され、そしていかなる方法においても本発明の範囲を限定することを意図しない。本開示に照らして、請求の範囲内の多数の実施態様は、当業者に明白である。発明の詳細な説明において引用された参考文献の内容は、本明細書において参考として援用される。さらなる方法および技術は、共有に係る出願PCT/CA94/00678(WO95/16048として公開された)に見い出され得る。
【0135】
実施例1:ヌクレオチド4,092〜5,234、ヌクレオチド5,892〜17,735、ヌクレオチド21,198〜26,033、およびヌクレオチド31,133〜34,445のヌクレオチド配列の決定。
ウシアデノウイルスタイプ3ゲノムのヌクレオチド配列を完了するために、以下のBAV-3制限フラグメントを、細菌プラスミドにクローニングし、そしてそれらのヌクレオチド配列を当業者に公知の方法によって決定した。
【0136】

【0137】
実施例2:ヌクレオチド4,092〜5,234、ヌクレオチド5,892〜17,735、ヌクレオチド21,198〜26,033、およびヌクレオチド31,133〜34,445によって規定されるBAV-3ゲノムの領域における挿入。
挿入を、当該分野で認識される技術によって行う。これは、制限消化、ヌクレアーゼ消化、連結、キナーゼおよびホスファターゼ処理、DNAポリメラーゼ処理、逆転写酵素処理、およびオリゴヌクレオチド化学合成を含むがこれらに限定されない。目的の外来核酸配列を、その外来配列が、挿入が配向されるBAVゲノムの領域と実質的な相同性を有する配列と隣接するようにプラスミドベクターにクローニングする。次いで、これらの構築物を、BAV-3と同時感染されている宿主細胞へと導入する。感染の間、これらの構築物とBAVゲノムとの間の相同組換えが起こって組換えBAVベクターが生成される。挿入がBAVゲノムの必須の領域において生じる場合、組換えBAVベクターを、ヘルパー細胞株において増殖させる。これは、挿入に起因して消失したウイルス機能を供給する。ウイルスにとっては必須ではないE4領域における挿入については、ヘルパー細胞株におけるBAVベクターの増殖は必要ではない。
【0138】
実施例3:全長および短縮型形態のウシヘルペスウイルス1糖タンパク質gDを発現するE3欠失BAV3の構築および特徴付け
本実施例は、E.coliの相同組換え機構を用いた、BAV3のE3領域における1,245kbの欠失の構築を示す。第一に、1,245kb欠失を、プラスミドにクローニングされたウシアデノウイルス-3(BAV3)ゲノムDNAのE3領域に導入した。線状の制限酵素で切除したE3欠失BAV3ゲノムNDAのウシ仔胎網膜初代細胞へのトランスフェクションは、感染性ウイルス(BAV3.E3d)を産生した。これは、すべてのBAV E3特異的なオープンリーディングフレームがインビトロにおけるウイルス複製について必須ではないことを示唆する。同様のアプローチを用いて、ウシヘルペスウイルス-1(BHV-1)の全長(BAV3.E3gD)または短縮型(BAV3.E3gDt)の糖タンパク質Dを発現する複製適合BAV-3組換え体を構築した。BAV3.E3gDおよびBAV3.E3gDtによって発現された組換えgDおよびgDtタンパク質は、コンホメーション的エピトープに対して惹起されたgD特異的モノクローナル抗体によって認識された。このことは、組換えgDおよびgDtの抗原性が、BHV-1に感染した細胞において発現されたネイティブなgDの抗原性に類似していたことを示唆する。コットンラット(cotton rat)の鼻腔内免疫は、強力なgDおよびBAV3特異的IgAおよびIgG免疫応答を誘発した。これらの結果は、動物の粘膜表面に対するワクチン抗原の送達のための、複製適合ウシアデノウイルス-3ベースのベクターの使用を例示する。
【0139】
材料および方法
細胞およびウイルス。Madin Darbyウシ腎臓(MDBK)細胞およびウシ仔胎網膜初代(PFBR)細胞を、5%ウシ仔胎血清(FBS)を補充したEagle最小必須培地(MEM)において増殖させた。野生型(WBR-1株)および組換えBAV3ウイルスを、MDBK細胞において、以前に記載したとおりに増殖させた。Mittalら(1995)J.Gen.Virol.76:93-102.BHV-1のP8-2株を、増殖させ、そして記載のとおりに定量した。Rouseら(1974)J Immunol.113:1391-1398。
【0140】
動物。Veterinary Infectious Disease Organization(Saskatoon)において維持されたコットンラット(Sigmodon hispidus)の近交コロニーを本研究の動物の供給源として用いた。
【0141】
組換えプラスミドの構築
a)プラスミドpBAV302の構築。図3を参照のこと。T4ポリメラーゼ処理した587bpのフラグメント(これは、オリゴヌクレオチドE3C5'ACGCGTCGACTCCTCCTCA(配列番号2)およびE3C3'TTGACAGCTAGCTTGTTG(配列番号3)ならびにテンプレートとしてプラスミドpSM14(Mittalら(1995)前出)を用いるPCR増幅によって単離された)を、SacIIで消化し、そしてEco47III-SacII消化したプラスミドpSL301に連結して、プラスミドpE3Aを作製した。164bpのフラグメント(これは、オリゴヌクレオチドE3N5'CCAAGCTTGCATGCCTG(配列番号4)およびE3N3'GGCGATATCTCAGCTATAACCGCTC(配列番号5)ならびにプラスミドpSM14(Mittalら(1995)前出)を用いるPCR増幅によって単離された)を、SphIおよびEcoRVで消化し、次いでpE3Aの531bp EcoRV-HindIIIフラグメントおよびSphI-HindIII消化したプラスミドpSL301に連結して、プラスミドpE3Bを作製した。614bp SphI-SacIIフラグメントを、プラスミドpE3Bから単離し、そしてSphI-SacII消化pSM14に連結して、プラスミドpE3Cを作製した。プラスミドpE3Cを、EcoRVで消化し、そしてSrflリンカー(TTGCCCGGGCTT、配列番号6)に連結して、プラスミドpE3CIを作製した。pE3CIの1.755kbのBamHIフラグメントを単離し、そしてBamHI消化pSM17(Mittalら、(1995)前出)に連結して、プラスミドpE3Dを作製した。最後に、プラスミドpE3Dの8783bpのKpnI-XbaIフラグメントを単離し、そしてKpnI/XbaI消化プラスミドpTG5453(これは、全長BAV3ゲノムDNAを含む)に連結して、プラスミドpE3Eを作製した。プラスミドpE3Eは、BAV3ゲノムの末端フラグメント(0-19.7m.u.および76.6-100m.u.)を、E3領域に1,245kbの欠失およびベクター配列内に位置する特有のKpnI部位とともに含む。
【0142】
E3領域に1,245kbの欠失を有するBAV3ゲノムを含むプラスミド(pBAV302)を、E.coli BJ5183において、KpnI消化pE3Eと、脱タンパク質化したBAV3ゲノムDNAとの間での相同DNA組換えによって生成した。
【0143】
b)プラスミドpBAV302gDおよびpBAV302gDtの構築。E3領域における外来遺伝子を発現する組換えBAV3の生成のためのトランスファープラスミドを、pBAV302の8783bp KpnI-XbaIフラグメントを、KpnI/XbaI消化プラスミドpGEM3zf(-)に連結することによって構築して、プラスミドpBAV300を作製した。プラスミドpRSV1.3(Tikooら、(1993)J.Virol.67:2103-2109)の1.3kb BgIIIフラグメント(これは、全長BHV-1gD遺伝子を含む)を、T4 DNAポリメラーゼで処理し、そしてSrfl消化pBAV300に連結して、プラスミドpBAV300.gDを作製した。同様に、プラスミドpRSV1.3XN(Tikooら、前出)の1.3kb BgIIIフラグメント(これは、短縮型BHV-1gD遺伝子を含む)を、T4 DNAポリメラーゼで処理し、そしてSrfI消化pBAV300に連結して、プラスミドpBAV300.gDtを作製した。
【0144】
全長または短縮型gDタンパク質のいずれかをコードする遺伝子を含む組換えBAVゲノムを、E.coli BJ5183において、SrfI線状化pBAV302とpBAV300.gDの10kb KpnI/XbaIフラグメントとの間(これは、プラスミドpBAV302.gDを作製する)、またはSrfI線状化pBAV302とpBAV300.gDtの10kb KpnI/XbaIフラグメントとの間(これは、プラスミドpBAV302.gDtを作製する)の、相同DNA組換えによって生成した。
【0145】
組換えBAV3の構築。60mmディッシュ中のPFBR細胞単層を、10μgのPacI消化pBAV302、pBAV302.gDまたはpBAV302.gDtの組換えプラスミドDNAを用いて、リン酸カルシウム方法を用いて、トランスフェクトした。Grahamら(1973)Virology 52:456-467。37℃でのインキュベーションの15〜20日後、50%の細胞変性効果を示すトランスフェクトされた細胞を採取し、凍結融解を2回行い、そして組換えウイルスを、MDBK細胞においてプラーク精製した。Mittalら(1995)、前出。
【0146】
タンパク質の放射標識および免疫沈降。28cm2ウエル中の約70〜80%のコンフルエントのMDBK細胞単層に、1細胞当たり10PFUの組換えまたは野生型BAV3を感染させた。ウイルスを60分間吸収させた後、細胞を、5%FBSを含むMEM中でインキュベートした。感染後の種々の時点で、細胞を、メチオニンおよびシステインを含まないDulbeccoMEM中で60分間インキュベートし、その後、[35S]メチオニン−システイン(1ウェル当たり100μCi)で標識した。2〜12時間の標識の後、細胞または培地を採集した。タンパク質を、培地から免疫沈降し、そして細胞を、改変RIPA緩衝液で溶解し、その後記載のようにSDS-PAGEによってタンパク質を分析した。Tikoooら、(1993)前出。
【0147】
動物接種。合計25匹の4〜6週齢のコットンラット(性別はいずれか)を3つの群に分けた。ハロタンでの麻酔後、動物に、1日目と21日目の2回、107PFUの個々の組換えウイルスを含む種菌100μlを鼻腔内経路により接種した。血液サンプルを、初回の接種の0、21、および28日後に採取して、BHV-1 gDおよびBAV3に対する抗体応答の発達を、酵素結合免疫吸着アッセイ(ELISA)およびウイルス中和(VN)アッセイによって試験した。各群の4匹の動物を、初回接種21日目および28日目にハロタンの過剰投与により安楽死させた。肺および鼻の切片を別個に採取して、BHV-1 gD特異的粘膜IgGおよびIgA抗体応答およびBAV3特異的粘膜IgGおよびIgA抗体応答の発達をELISAでモニターした。Pappら(1997)J.Gen.Virol.78:2933-2943。さらに、肺を採取し、そしてBHV-1 gD特異的IgA抗体分泌細胞およびBAV3特異的IgA抗体分泌細胞の頻度を、酵素結合免疫スポット(ELISPOT)により決定した。Pappら、前出。
【0148】
肺リンパ球の調製。無菌的に取り出した肺組織を、小片に切断し、そして完全培地(10%FBS、2mM L-アルギニン、1mMピルビン酸ナトリウム、100μMの非必須アミノ酸、10mM HEPES緩衝剤、50μM 2-メルカプトエタノール、100U/mlペニシリンG、100μg/mlストレプトマイシン溶液、150U/mlコラゲナーゼAおよび50U/mlDNaseIを補充したMEM)中で1時間インキュベートした。次いで、組織を、プラスチックメッシュを通して押した。肺細胞懸濁物を、不連続なPercoll勾配を通して遠心分離し、そしてMEMで洗浄した。細胞を、完全培地に再懸濁し、そしてフラスコ中で1時間インキュベートして、接着細胞を付着させた。次いで、非接着性の細胞集団を、再懸濁し、そして以前に記載のように抗原特異的ELISPOTアッセイにおいて使用した。Pappら、前出。
【0149】
ELISA。血清、肺分泌物、および鼻分泌物中のBHV-1 gDおよびBAV3に対して特異的な抗体を、以前に記載のようにELISAによって決定した。Pappら、前出。手短には、96ウェルImmunol-2マイクロタイタープレートを、精製した短縮型gD(0.01μg/ウェル)またはBAV3(0.5μg/ウェル)のいずれかでコートし、そして異なる希釈の各サンプルとインキュベートした。抗原特異的なIgGを、ビオチン化ウサギ抗ラットIgGを用いて検出した。抗原特異的IgAを、ウサギ抗ラットIgAおよび西洋ワサビペルオキシダーゼ結合体化したヤギ抗ウサギIgGを用いて測定した。
【0150】
ウイルス中和。2倍連続希釈の熱非働化血清サンプルを、100PFUのBHV-1と、37℃で1時間インキュベートした。次いで、ウイルスサンプル混合物を、12ウェル組織培養プレート中のコンフルエントなMDBK細胞に配置し、そして2日間インキュベートした。力価を、コントロールと比較したプラーク数の50%減少を引き起こした最高の抗体希釈の逆数として表した。
【0151】
結果
E3欠失組換えBAV3の構築。当初、ウイルス複製におけるBAV3 E3の役割は、HAVのE3領域の役割と類似していると推定された。Woldら(1991)Virology 184:1-8。従って、BAV E3ベースのベクターを、E3配列の欠失を行うことによって構築した。しかし、種々のウシ細胞株(MDBKを含む)における、E3欠失したBAV3組換え体の単離の試みは成功しなかった。ルシフェラーゼ遺伝子を発現する、部分的に欠失したBAV3組換え体(BAV3-Luc)は、BAV3 E1形質転換したMDBK細胞を、トランスフェクションについて使用した場合にのみ単離され得た。Mittalら(1995)前出。しかし、一旦単離されると、BAV3-Lucは、正常ウシ細胞上で増殖され得た。このことは、BAV3のE3領域がインビトロでのウイルス複製については必須ではないことを示唆する。Mittalら(1995)前出。BAV3組換え体の単離の効率を上昇させるために、PBFR細胞を、BAV3組換え体を産生するための新規の手順と共に使用した。Degryse(1996)Gene 170:45-50。この方法を用いて、標的化された改変を、E.coliの高度に効率よい相同組換え機構を用いて、プラスミドが保有するウイルス配列へと導入し、そして感染性のビリオンを、プラスミドベクターから切り出された改変されたゲノムを適切な宿主細胞へとトランスフェクションした後に単離した。Chartierら(1996)J.Virol.70:4805-4610。
【0152】
E.coliの相同組換え機構を利用して、プラスミド(pBAV302)を構築した。これは、1,245kb欠失(ヌクレオチド26456〜27701)およびSrfI制限酵素部位を含んだ(図21)。PacI消化pBAV302 DNAは、PFBR細胞にトランスフェクトされる場合に、14日後に細胞変性効果をもたらした。ウイルスを、プラーク精製し、そしてMDBK細胞において増殖させ、そしてBAV3.E3dと命名した。ウイルスDNAを抽出し、そしてBamHI制限酵素で消化した後にアガロースゲル電気泳動によって分析した。野生型(図4、レーンb)と比較すると、組換えBAV3.E3dゲノムの3,019kb BamHI「D」フラグメントは、1,245kb小さかった(図4、レーンa)。このことは、E3が欠失された組換えBAV3が単離されたことを確認する。この組換えウイルスの増殖特徴と、野生型BAV3の増殖特徴との比較によっては、プラークの大きさまたは複製における有意な差異されなかった。なぜなら、E3欠失組換え体は、野生型BAV3と類似する反応速度で複製されたからである。
【0153】
BHV-1糖タンパク質Dを発現する組換えBAV3の構築。E3欠失の複製適合BAV3組換え体の、生の組換えワクチン抗原についての送達ビヒクルとしての有用性を決定するために、種々の形態のBHV-1糖タンパク質gDを発現する組換えBAV3ウイルスを(Tikooら、(1993)前出)を構築した。全長形態および短縮型形熊のgD遺伝子(外因性プロモーターを何ら含まない)を、BAV3.E3dゲノムのE3領域に、E.coliの相同組換え機構を用いて、平行方向に個々に挿入した。Degryseら(1996)前出。PacI消化pBAV302.gDまたはpBAV302.gDtプラスミドDNAは、ウシ網膜初代細胞にトランスフェクトされた場合に、14日後に細胞変性効果をもたらした。50%の細胞変性効果を示す感染した細胞単層を採取し、凍結融解し、そして組換えウイルスをプラーク精製し、そしてMDBK細胞において増殖させた。組換えBAV3を、BAV3.E3gD(E3領域における全長gD遺伝子の挿入)およびBAV3.E3gDt(E3領域における短縮型gD遺伝子の挿入)と名づけた。ウイルスDNAを抽出し、そして種々の制限酵素での消化後にアガロースゲル電気泳動によって分析した。gD遺伝子がNdeI部位を有するので、組換えウイルスDNAをNdeIで切断した。図4に見られるように、BAV3.E3d(図4、レーンe)と比較して、BAV3.E3gD(図4、レーンf)、およびBAV3.E3gDt(図4、レーンg)ゲノムは、4.6kbのさらなる予測されたバンドを含み、これは、組換えBAV3.E3gDおよびBAV3.gDtが、E3領域にgDまたはgDt遺伝子を含んでいたことを示唆する。gD遺伝子とgDt遺伝子との間を識別するために、組換えウイルスDNAをNheIで消化した、なぜなら、gDt遺伝子は、特有のNheI制限酵素部位を有するがgD遺伝子は有さないからである。Tikooら(1993)前出。予測されるように、5.4kb BAV3.E3gD DNAフラグメント(図4、レーンc)は、BAV3.E3gDtにおける5.0kbのフラグメントと置換されていた(図4、レーンd)。このことは、組換えBAV3.E3gDおよびBAV3.E3gDtが、それぞれgDおよびgDtを含んでいたことを示唆する。これらの組換え体と野生型またはE3欠失BAV3との増殖特徴の比較によっては、産生されたウイルスの複製の反応速度または力価のいずれにおいても有意な差異は示されなかった。
【0154】
BAV3.E3gDおよびBAV3.E3gDtによるgDの発現の分析。BHV-1 gDまたはgDt遺伝子を含む組換えBAV3ウイルスによって発現させる産物を試験するために、MDBK細胞に、組換えBAV3.E3gD、BAV3.E3gDtまたはBAV3.E3dを感染させ、そして異なる時間の間[35S]メチオニン−システインで代謝標識した。抗原性gDとの比較のために、MDBK細胞に、BHV-1を感染させ、そして[35S]メチオニン−システインで同様に標識した。放射標識したタンパク質を、gD特異的なモノクローナル抗体(MAb;Hughesら、(1988)Arch.Virol.103:47−60)で免疫沈降し、そして還元条件下のSDS-PAGEによって分析した。組換えBAV3.E3gD感染した細胞の免疫沈降によって、約71kDaの主要なバンドが示された(図5A、レーンb−d)。これは、BHV-1感染した細胞において産生されたgDタンパク質と共移動した(図5A、レーンa)。このことは、組換えgDが、真正なgDと類似する翻訳後修飾を含んでいたことを示唆する。感染していない細胞(図5A、レーンh)にも組換えBAV3.E3dに感染させた細胞(図5A、レーンe-g)にも類似のバンドは観察されなかった。組換えBAV3.E3gDt感染させた細胞上清の放射免疫沈降は、61kDaの主要バンドを示した(図5B、レーンb-d)。両方の組換えタンパク質は、MDBK細胞の感染の間中発現されていた(図5)。
【0155】
組換えgDタンパク質の抗原性を試験するために、放射標識したタンパク質を、組換えBAV3感染した細胞溶解物(BAV3.E3gD)または上清(BAV3.E3gDt)から、gD特異的なモノクローナル抗体(Highesら(1988)前出)を用いて免疫沈降し、そして還元条件下のSDS-PAGEにより分析した。図6に示すように、gDタンパク質およびgDtタンパク質の両方は、不連続のエピトープIb(MAb 136;レーンaおよびf)、II(MAb 3E7;レーンbおよびg)、IIIb(MAb 4C1;レーンcおよびh)、IIIC(MAb 2C8;レーンdおよびi)およびIIId(MAb 3C1;レーンeおよびj)に対して惹起されたMAbによって認識された。これらの結果は、組換えタンパク質gDおよびgDtの抗原性構造が、MDBK細胞において産生された真正のgD産物の構造と類似することを示唆する。Tikooら(1993)、前出。
【0156】
DNA合成の非存在下でgD発現が生じるか否かを決定するために、BAV3.E3gD感染したMDBK細胞において産生されたgDの量を、DNA合成のインヒビターである1-β-D-アラビノフラノシルシトシン(AraC)の存在下(図7、レーンaおよびb)および非存在下(図7、レーンcおよびd)で比較した。この結果は、gD発現が、AraCの存在下で減少することを示唆する。
【0157】
動物における抗体応答。BAV3組換え体がgD特異的な免疫応答を誘発する能力を決定するために、コットンラットに、2回3週間間隔をおいて、鼻腔内に、107PFUのBAV3.E3gD、BAV3.E3gDtまたはBAV3.E3d組換え体を接種した。血清、肺洗浄物および鼻洗浄物を、IgGおよびIgA抗体の分析のために採取し、その一方肺を、IgA抗体分泌細胞(ASC)の数を分析するために採取した。BAV3.E3gDおよびBAV3.E3gDtの両方は、gD特異的なIgG抗体応答を血清および肺洗浄物において誘発した(図8B)。これは、BAV3.E3d免疫した動物(コントロール)において誘発された応答よりも有意に高かった(P<0.05)。しかし、BAV3.E3gDtによって誘発されたgD特異的なIgG応答は、BAV3.E3gDによって誘発された応答よりも高かった(P<0.05)。
【0158】
BAV3.E3gDおよびBAV3.E3gDtでの免疫は、BAV3.E3dでの免疫よりも、血清および肺洗浄物においてgDに対してより高い(p<0.05)IgA抗体応答を誘発した(図8A)。しかし、BAV3.E3gD免疫群とBAV3.E3gDt免疫群との間のIgA抗体応答においては、有意な差異は存在しなかった。これらの組換え体はまた、血清および肺洗浄物においてBAV3特異的なIgG抗体応答(図8D)、および血清、肺洗浄物および鼻洗浄物においてIgA抗体応答(図8C)を誘発した。これは、群間では有意な差異は存在しなかった。
【0159】
興味深いことに、鼻洗浄物は、gD(図26Aおよび図26B)またはBAV3(図26Cおよび26D)に対して特異的なIgA抗体のみを含んだ。さらに、gDおよびBAV3の両方に特異的なIgA抗体分泌細胞が免疫された動物の肺において検出され得、この細胞の数は、追加免疫後に有意に増加した。
【0160】
gD特異的な血清抗体の生物学的な活性を測定するために、抗BHV-1力価を決定した。BAV3.E3gDtでの免疫は、4.3±0.5のBHV-1 log2力価を誘発し、これは、BAV3.E3gDおよびBAV3.E3dによって誘発された、それぞれ3.0±0.6および0.8±0.3の力価よりも有意に高かった(p<0.05)。
【0161】
考察
家畜でのヒトアデノウイルスのワクチン送達系としての使用は、限定されている。非ヒトアデノウイルスは種特異的であるので、動物特異的なアデノウイルスの、ワクチン送達系としての開発が、論理的な選択肢である。本明細書において、動物の粘膜表面へのワクチン抗原の送達における使用のための複製適合(E3欠失)組換えBAV3の開発が記載される。さらに、BHV-1 gDまたはgDt糖タンパク質を発現する複製適合BAV3を構築し、そしてそれらがコットンラットにおける粘膜免疫応答および全身免疫応答を誘発する能力について試験した。
【0162】
種々のウシ細胞株におけるE3欠失BAV3組換え体を単離する最初の試み(これは、BAV3 DNA媒介トランスフェクション後の感染性子孫の形成を支持する)は、成功しなかった。しかし、ルシフェラーゼ遺伝子を発現する部分的にE3を欠失したBAV3組換え体が、BAV3 E1で形質転換したMDBK細胞株を、トランスフェクションについて使用した場合に単離された。Mittalら(1995)前出。このことは、E1配列の存在に関連するいくつかの機能が、E3欠失BAV3の単離に必要であり得ることを示唆した。しかし、これらの単離の後、それらのE3領域にルシフェラーゼ遺伝子を発現する組換えBAV3ウイルスは、正常のMDBK細胞において効率よく複製した。Mittalら(1995)前出。従って、正常なMDBK細胞がBAV組換え体の単離を可能にするには低すぎるトランスフェクション効率および/または増殖速度を有し得るという別の可能性が存在する。
【0163】
BAV3 E1形質転換したMDBK細胞を用いずに複製適合BAV3組換え体を単離する、信頼がありそして効率的な方法を開発するために、E.coliの相同組換え機構(Chartierら(1996)前出;およびDegryse(1996)前出)をまず使用して、プラスミドにおいてクローニングされた全長BAV3ゲノムの1,245kb欠失をE3領域に導入した。第二に、組換えウイルスを単離するために、MDBK細胞の代わりにPFBR細胞を、プラスミドから切り出された改変されたBAV3ゲノムのトランスフェクションに用いた。
【0164】
野生型ゲノムの105%までがHAV5ビリオンに、次の回の複製において外来遺伝子の再構成も欠失も何ら生じさせずにパッケージングされ得ることが報告されている。Bettら(1993)、J.Virol.67:5911−5921.近年、ヒツジアデノウイルス(OAV)ベクターの挿入容量が野生型ゲノムの114%であることが報告された。Xuら(1997)Virology 230:62−71。BAV3ゲノムのサイズは、HAV5ゲノムのサイズと類似しているので、BAV3ゲノムの挿入能力は、HAV5の能力と類似し得る。従って、本明細書において記載されるBAV3.E3dベクターが1,245kbのE3欠失を有するので、その挿入能力は、少なくとも3.0kbである。
【0165】
以前に、E3領域に0,696kbの欠失を含む組換えBAV3.lucが細胞培養物において野生型BAV3よりも効率が悪く複製されたことが報告された。Mittalら(1995)前出。対照的に、E3領域に1,245kb欠失を含む組換えBAV3.E3dは、細胞培養物中で野生型BAV3と同程度に効率よく複製した。この差異は、ルシフェラーゼ遺伝子の組換えBAV3.lucにおけるルシフェラーゼ遺伝子の挿入に起因し得る。この挿入は、E3領域の下流の遺伝子の発現に影響を与え得る。Mittalら(1995)前出。しかし、組換えBAV3.E3gDまたはBAv3.E3gDtもまた、野生型BAV3と同様に効率よく複製された。このことは、異なる外来遺伝子産物が、細胞培養物において組換えBAV3ウイルスの複製に異なって影響し得ることを示唆する。
【0166】
可能なワクチン抗原の発現のためのBAVベクター系の有用性を確認するために、組換えBAV3.E3gDおよびBAV3.E3gDtを構築した。これらはそれぞれ、BHV-1gDおよびgDt糖タンパク質を発現する。予想されたように、BAV3.E3gDによって発現されたgDは、予測された分子量を有し、そしてBHV-12感染した細胞によって発現された真正のgDと共移動した。同様に、BAV3.E3gDtによって発現されたgDtは、推定の分子量を有していた。Tikooら(1993)前出。さらにBAV3組換え体によって発現されたgDおよびgDtは、コンホメーション的なエピトープに対して特に惹起されたMAbによって認識された。Hughesら(1988)前出。これらの結果は、組換えgDおよびgDtが、ウイルス感染後に合成された真正のgDのものと、識別不可能である翻訳後修飾および抗原性プロフィールを有していたことを示唆する。Hughesら(1988)前出。
【0167】
ヒトアデノウイルスのE3領域において左から右の方向に挿入された、隣接する調節配列を何ら伴わない外来遺伝子は、ウイルスゲノムの上流MLPから、またはE3プロモーターからのいずれかで効率よく発現される。Droninら(1993)Gene 126 247-250;Morinら(1987)Proc.Natl.Acad.Sci.USA 84:4626-4630およびXuら(1995)J.Gen.Virol.76:1971-1980。本発明の実験において、糖タンパク質gD発現は、複製インヒビターであるAraCの存在下で部分的に減少した。このことは、外来遺伝子発現が、MLP(複製依存性プロモーター)およびE3プロモーター(初期複製非依存性プロモーター)の両方によって駆動されたことを示唆している。類似の結果が以前に報告されている。Mittalら(1995)前出。
【0168】
コットンラットのBAV3.E3gDまたはBAV3.E3gDtでの鼻腔内免疫は、gD特異的な粘膜または全身性免疫応答を誘発した。gDtは、gDよりも強力なIgG抗体応答を誘発した。対照的に、コットンラットの組換えHAV5での皮内免疫は、gDに対するよりもgDtに対してより低い免疫応答を誘発した。Mittalら(1996)Virology、222:299-309。さらに、アジュバントとともに組み込まれた組換えgDtの精製した調製物は、マウスに注射された場合、真正のgDの応答に類似する二次免疫応答をもたらした。Baca-Estradaら(1996)Viral Immunol.9:11-22。これらの結果は、免疫およびワクチン処方物の経路が、免疫原が膜にアンカーされる(gD)か、または可溶性タンパク質として分泌される(gDt)か否かよりも高い程度で、免疫原が有効な免疫応答を誘発する能力に影響を与え得ることを強力に示唆する。
【0169】
驚くべきことに、gDおよびgDtは、類似のIgAを誘発したが、有意に異なるIgG応答を、鼻通路および血清において誘発した。粘膜免疫応答および全身免疫応答は、かなりの程度の非依存性を伴って誘発されそして調節されることが示された。Alleyら(1988)The mucosal immune system、「B lymphocytes in human disease」(G.BirdおよびJ.E.Calvert編)、Oxford University Press、222〜254頁;およびConleyら(1987)Ann.Intern.Med.106:892-899。これらのその1つにおける免疫応答の誘導は、必ずしも他における応答を誘導しない。従って、2つの異なるgDの形熊(全長および短縮型)が粘膜区画および全身区画において異なって認識されている可能性がある。
【0170】
粘膜免疫の誘導は、呼吸器感染または腸感染から宿主を防御する際に重要であると考えられる。さらに、分泌IgAの存在は、通常、このような感染に対する耐性に相関していることが見出されている。Murphy(1994)Mucosal immnuity toviruses、「Handbook of mucosal immunity」(P.L.Orga、J.Mestecky、M.E.Lamm、W.strober、J.R.McGhec、およびJ.Bienenstock編)。Academic Press、San Diego、333〜339頁。興味深いことに、鼻腔内免疫は、gD特異的なIgA抗体応答を、コットンラットの血清および肺のみならず、鼻洗浄物においても誘発した。肺における高レベルのIgAおよびASCの存在は、抗体が局所的に産生されることを示唆する。
【0171】
結論として、本発明は、BAV3組換え体の産生のための効率よくかつ信頼性の高い系を記載する。さらに、複製適合組換えBAV3を有するコットンラットの鼻腔内免疫は、ワクチン抗原特異的な、粘膜免疫応答および全身免疫応答を誘発する。
【0172】
実施例4:BHV-1糖タンパク質Dを発現する組換えBAV3でのウシの免疫
本実施例において、BHV-1 gD(全長および短縮型)を発現する複製適合組換えBAV3ウイルスを、それらがBHV-1チャレンジから防御する能力について評価した。3つの3〜4月齢のウシの群を、以下のプロトコルに従って鼻腔内に免疫した。群1を、全長BHV-1 gDを発現するウイルスであるBAV3.E3gDで免疫した。実施例3を参照のこと。群2を、短縮型BHV-1gDを発現するウイルスであるBAV3.E3gDtで免疫した。実施例3およびTikooら(1993)前出を参照のこと。群3を、欠失E3領域を有するが、挿入された異種配列は有さないウイルスであるBAV3.E3dで免疫した。実施例3を参照のこと。
【0173】
1日目および28日目に、動物にワクチン接種し、そして42日目にエアロゾル化したBHV-1でチャレンジした。1、28、40および52日目に血液を血清学のために採取し、そして28日目の血液サンプルにおいてリンパ球増殖を測定した。臨床的症状(温度、鼻の病変および抑鬱を含む)を、チャレンジ後10日間(42日目〜52日目)毎日評価した。1日目と15日目との間に、ウイルス単離のために鼻のスワブを3日毎に採取した。チャレンジ後、ウイルス力価を、2日毎に10日間、鼻のスワブおよび鼻のタンポンによって測定した。22および40日目、および42〜52日目に、抗体力価を、鼻のタンポンを用いて決定した。
【0174】
結果は、gD発現BAV組換え体またはgDt発現BAV組換え体で免疫した動物において、IgG力価が、最初のワクチン接種および追加のワクチン接種の両方の後に増加し、そしてチャレンジ後にさらに増加したことを示す(図9)。比較すると、挿入されたgD遺伝子を欠く、E3欠失したBAV3でワクチン接種した動物はチャレンジ後にのみIgG力価の増加を示し、そしてワクチン接種した動物において得られたものよりも少なくとも1対数分低かった。IgA力価を測定し、そして結果を図10に示す。見られるように、第一の免疫後にはIgAにおいて検出可能な増加は存在しなかった。しかし、gD特異的IgA力価は、第二の免疫後およびBHV-1チャレンジ後に増加した。再度、挿入されたgD遺伝子を欠く、E3欠失したBAV3でワクチン接種した動物は、チャレンジ後にのみIgA力価の増加を、そしてgD発現BAVまたはgDt発現BAVでワクチン処理した動物よりも低い程度で示した。
【0175】
臨床的症状は、E3欠失BAVでワクチン接種した動物と比較して、gD発現BAVおよびgDt発現BAVでワクチン接種した動物において減少した。これらは、発熱(図11)、鼻病変の出現および程度(図12)および抑鬱(図13)を含む。最後に、鼻液におけるウイルス力価は、コントロールに比較して、BAV3.E3gDワクチン接種した動物およびBAV3.E3gDtワクチン接種した動物におけるチャレンジ後により迅速に減少した(図14)。
【0176】
これらの結果は、BHV-1糖タンパク質を発現する組換えBAVウイルスは、ウシにおけるBHV-1チャレンジからの防御を提供し、臨床徴候の出現を減少させ、より迅速にウイルスを排除することを容易にし、そしてIgGおよびIgAの両方の力価の増加を提供した。このことは、ウシ宿主における体液性および粘膜性の免疫の両方の誘導を示唆する。
【0177】
生物学的な材料の寄託
以下の材料は、Veterinary Infectious Disease Organization(VIDO)、120Veterinary Road、Saskatoon、Saskatchewan、S7N 5E3、Canadaに寄託されそして維持されている。
【0178】
寄託された材料のヌクレオチド配列およびそれによってコードされるポリペプチドの配列は、本明細書において参考として援用される。本明細書において明示的に開示された配列と、寄託された配列との間に何らかの差異が存在する場合、寄託された配列が支配する。
【0179】
物質 内部受託番号 寄託日
組換えプラスミド
pSM51 pSM51 1993年12月6日
pSM71 pSM71 1993年12月6日
組換え細胞株
BAV3 E1配列で形質転換したMDBK細胞
(MDBK-BAVE1) 1993年12月6日
BAV E1配列で形質転換したウシ仔胎腎臓細胞
(FBK-BAV-E1) 1993年12月6日
【0180】
本発明を、上記に特定の具体的な実施態様によって例示したが、これらの特定の実施例は、添付の請求項において記載されるような本発明の範囲を制限することを意図するものではない。
【0181】
本明細書は、以下の発明を開示する。
【0182】
1.ウシアデノウイルスタイプ3(BAV-3)のゲノムまたはそのフラグメントに実質的に相同である、ヌクレオチド配列。
【0183】
2.ウシアデノウイルスタイプ3(BAV-3)のゲノムの前記フラグメントに実質的に相同である、前記1に記載のヌクレオチド配列であって、 該フラグメントが、BAV-3ゲノムのヌクレオチド4,092〜5,234、ヌクレオチド5,892〜17,735、ヌクレオチド21,198〜26,033、およびヌクレオチド31,133〜34,445からなる群より選択されるか、またはそのフラグメントである、配列。
【0184】
3.前記1または2に記載のヌクレオチド配列であって、前記そのフラグメントと実質的に相同であり、E2領域、E4領域、後期領域、33kD、52kD、100kD、DBP、pol、pTPおよびペントン遺伝子、ならびに遺伝子11IA、pV、pVI、pVII、pVIII、およびpXからなる群より選択される、配列。
【0185】
4.前記3に記載のヌクレオチド配列であって、E2領域、E4領域、後期領域、33kD、52kD、100kD、DBP、pol、pTPおよびペントン遺伝子、ならびに遺伝子IIIA、pV、pVI、pVII、pVIII、およびpX、ならびにそれらのフラグメントからなる群より選択されるBAV-3ゲノムの領域由来のBAV-3コード配列を含む、配列。
【0186】
5.BAV-3ゲノムの遺伝子間領域を含むヌクレオチド配列と実質的に相同である、ヌクレオチド配列。
【0187】
6.前記1〜5のいずれかに記載のヌクレオチド配列であって、BAV-3 DNA制御配列を含む、配列。
【0188】
7.前記DNA制御配列が、転写調節配列、プロモーター、エンハンサー、上流調節ドメイン、スプラシシングシグナル、ポリアデニル化シグナル、転写終結配列、翻訳調節配列、リボソーム結合部位、および翻訳終結配列からなる群より選択される、前記6に記載のヌクレオチド配列。
【0189】
8.前記1〜7のいずれかに記載のヌクレオチド配列を含む、ベクター。
【0190】
9.ITR配列、BAVパッケージング配列、および少なくとも1つの外来ヌクレオチド配列を含む欠損組換えBAVベクターであって、ここで、該BAVベクターがE1機能を欠き、そしてE2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択される領域によってコードされる1以上の機能をさらに欠いている、欠損組換えBAVベクター。
【0191】
10.前記9に記載の欠損組換えBAVベクターであって、E1領域のすべてまたは一部分が欠失しており、そしてE2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択される領域のすべてまたは一部分がさらに欠失している、欠損組換えBAVベクター。
【0192】
11.前記10に記載の欠損組換えBAVベクターであって、以下:
a)E1領域のすべてまたは一部分が欠失し、そしてE2領域のすべてまたは一部分が欠失しているBAVベクター;
b)E1領域のすべてまたは一部分が欠失し、そしてE4領域のすべてまたは一部分が欠失しているBAVベクター;
c)E1領域のすべてまたは一部分が欠失し、そしてE2領域のすべてまたは一部分が欠失し、そしてE4領域のすべてまたは一部分が欠失しているBAVベクター、からなる群より選択される、欠損組換えBAVベクター。
【0193】
12.さらに、E3領域のすべてまたは一部分が欠失している、前記11に記載の欠損組換えBAVベクター。
【0194】
13.前記外来ヌクレオチド配列が、治療目的のポリペプチドをコードする、前記9に記載の欠損組換えBAVベクター。
【0195】
14.前記治療目的のポリペプチドが、凝固因子、成長ホルモン、サイトカイン、リンホカイン、オンコジーン産物、腫瘍サプレッサー、細胞レセプター、細胞レセプターに対するリガンド、プロテアーゼインヒビター、抗体、毒素、免疫毒素、ジストロフィン、嚢胞性線維症膜貫通調節タンパク質(CFTR)、免疫原性ポリペプチド、および心臓血管疾患に関与するポリペプチドからなる群より選択される、前記13に記載の欠損組換えBAVベクター。
【0196】
15.前記心臓血管疾患に関与するポリペプチドが、線維芽細胞増殖因子(FGF)、血管内皮増殖因子(VEGF)、神経成長因子(NGF)、e-nos産物、pRb、リポタンパク質リパーゼ、スーパーオキサイドジスムターゼ、カタラーゼ、酸素スカベンジャー、フリーラジカルスカベンジャー、アポリポタンパク質およびプラスミノーゲン活性化因子インヒビター1からなる群より選択される、前記14に記載の欠損組換えBAVベクター。
【0197】
16.挿入部位に挿入された外来遺伝子を含む前記9に記載の欠損組換えBAVベクターを構築する方法であって、以下の工程:
a)BAVゲノムと実質的に相同である配列を提供する工程;
b)外来ヌクレオチド配列を提供する工程;
c)該挿入部位の周囲のBAV配列と実質的に相同である配列に該外来ヌクレオチド配列を連結して挿入カセットを形成する工程;
d)該挿入カセットおよび該BAVゲノムと実質的に相同である配列を細胞に導入する工程;
e)該挿入カセットと該BAVゲノムと実質的に相同である配列との間に相同組換えを生じさせて欠損組換えBAVベクターを生成する工程;
f)該欠損組換えBAVベクターが複製される条件下で該細胞を培養する工程;ならびに
g)該欠損組換えBAVベクターを該細胞または該培養培地から収集する工程、を包含する、方法。
【0198】
17.前記挿入部位が、E2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択されるBAVゲノムの領域に位置する、前記16に記載の欠損組換えBAVベクターを構築する方法。
【0199】
18.前記挿入部位が、E1領域において欠失を含む、前記16に記載の欠損組換えBAVベクターを構築する方法。
【0200】
19.前記9に記載のBAVベクターを、適切な補完細胞株に導入する工程、および該感染細胞からウイルスを回収する工程を包含する、組換えBAVを産生する方法。
【0201】
20.前記補完細胞株が、ヘルパーウイルスに感染している、前記19に記載の方法。
【0202】
21.前記補完細胞株が、E1領域、E2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択されるBAVゲノムの領域の1つ以上によってコードされる機能を提供する、前記19に記載の方法。
【0203】
22.前記8または9に記載のベクターを含む、宿主細胞。
【0204】
23.ウシアデノウイルスE1遺伝子領域を含む組換え哺乳動物細胞株であって、BAV E1機能を提供し得、そしてさらに、E2、E4、L1、L2、L3、L4、L5、L6およびL7からなる群より選択されるBAVゲノムの領域に由来する遺伝子によってコードされる機能を提供し得る、組換え哺乳動物細胞株。
【0205】
24.BAV E1およびE2機能を提供し得る、前記23に記載の組換え哺乳動物細胞株。
【0206】
25.BAV E1およびE4機能を提供し得る、前記23に記載の組換え哺乳動物細胞株。
【0207】
26.組換えBAVを産生する方法であって、以下の工程:
前記23に記載の細胞株に、ITR配列、BAVパッケージング配列、および少なくとも1つの外来遺伝子を含むBAVベクターを導入する工程であって、ここで、該ベクターがE1領域において欠失されており、そして該ベクターがE2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択される領域においてさらに欠失されている、工程;ならびに
該感染した細胞からウイルスを回収する工程、を包含する、方法。
【0208】
27.組換えポリペプチドを産生する方法であって、以下の工程:
(a)前記22に記載の宿主細胞集団を提供する工程;および
(b)ポリペプチドが発現される条件下で該細胞集団を増殖させる工程、を包含する、方法。
【0209】
28.前記2〜4のいずれかに記載のヌクレオチド配列によってコードされる、BAVポリペプチド。
【0210】
29.前記27に記載の方法によって産生される、組換えポリペプチド。
【0211】
30.生物学的サンプル中のBAVヌクレオチド配列の存在を検出する方法であって、以下の工程:
(a)生物学的サンプルを提供する工程;および
(b)前記1〜3のいずれかに記載の配列の少なくとも10の連続するヌクレオチドを含む標識されたプローブを使用して、該サンプルをハイブリダイゼーションに供する工程、を包含する、方法。
【0212】
31.生物学的サンプル中のBAVヌクレオチド配列の存在を検出する方法であって、以下の工程:
(a)生物学的サンプルを提供する工程;および
(b)該サンプルを、前記1〜3のいずれかに記載の配列の少なくとも10の連続するヌクレオチドを含むプライマーを用いて、ポリメラーゼ連鎖反応に供する工程、を包含する、方法。
【0213】
32.1以上のプローブを含む、生物学的サンプル中のBAVヌクレオチド配列の存在を検出するためのキットであって、該プローブが前記1〜3のいずれかに記載の配列の少なくとも10の連続するヌクレオチドを含む、キット。
【0214】
33.生物学的サンプル中のBAV抗原の存在を検出する方法であって、以下の工程:
(a)生物学的サンプルを提供する工程;および
(b)前記29に記載のポリペプチドに対して惹起された抗体を用いて、該サンプルを免疫アッセイに供する工程、を包含する、方法。
【0215】
34.生物学的サンプル中のBAV抗原の存在を検出するためのキットであって、前記29に記載のポリペプチドに対して惹起された抗体を1以上含む、キット。
【0216】
35.哺乳動物被験体において免疫応答を誘発し得る薬学的組成物であって、前記2または3に記載のヌクレオチド配列または前記8に記載のベクターを含む、薬学的組成物。
【0217】
36.哺乳動物被験体において免疫応答を誘発し得る薬学的組成物であって、前記28または29に記載のポリペプチドを含む、薬学的組成物。
【0218】
37.哺乳動物被験体において免疫応答を誘発し得る薬学的組成物であって、免疫原性ポリペプチドを発現する前記9に記載の欠損組換えBAVベクターを含む、薬学的組成物。
【0219】
38.前記8〜15のいずれかに記載のベクターを含む、薬学的組成物。
【0220】
39.前記22の宿主細胞を含む、薬学的組成物。
【0221】
40.哺乳動物宿主において免疫応答を誘発して感染から防御する方法であって、防御抗原を発現し得る組換えBAVベクターを含む薬学的組成物を投与する工程を包含する、方法。
【0222】
41.哺乳動物宿主における遺伝子治療の方法であって、前記8に記載のベクターを該宿主に投与する工程を包含する、方法。
【0223】
42.哺乳動物宿主における遺伝子治療の方法であって、前記9に記載の欠損組換えBAVベクターを該宿主に投与する工程を包含する、方法。
【0224】
43.哺乳動物宿主における遺伝子治療の方法であって、前記22に記載の宿主細胞を該宿主に投与する工程を包含する、方法。
【0225】
44.前記28または29に記載のポリペプチドに特異的に結合する、抗体。
【0226】
45.E3特異的配列が欠失している、組換えウシアデノウイルス(BAV)ベクター。
【0227】
46.挿入された異種配列を含む、前記45に記載のベクター。
【0228】
47.前記異種配列が、前記欠失された配列によって以前に占有されていた部位で該E3領域に挿入されている、前記46に記載のベクター。
【0229】
48.前記45に記載のBAVベクターを含む、宿主細胞。
【0230】
49.挿入部位に挿入された異種配列を含む欠損組換えBAVベクターを構築する方法であって、以下の工程:
(a)該挿入部位の周囲のBAV配列と実質的に相同である配列に、該異種配列を連結させて、挿入カセットを形成する工程;
(b)該挿入カセットを、BAVゲノムと実質的に相同である配列を含むポリヌクレオチドとともに、細胞に導入する工程;および
(c)該挿入カセットと、該ポリヌクレオチドとの間に相同組換えを生じさせて、該欠損組換えBAVベクターを生成する工程、を包含する、方法。
【0231】
50.E3特異的配列が、少なくとも1つのセグメントにおいて欠失している、前記49に記載の方法。
【0232】
51.前記細胞が原核生物細胞である、前記49に記載の方法。
【0233】
52.前記挿入部位の周囲のBAV配列と実質的に相同である前記配列が、プラスミドに存在し、そして前記連結する工程が前記異種配列を含む制限フラグメントを該プラスミドに挿入することによって達成される、前記51に記載の方法。
【0234】
53.前記細胞がE.coliである、前記51に記載の方法。
【0235】
54.前記49に記載の方法に従って得られた、組換えBAVゲノム。
【0236】
55.組換えBAVウイルスを得る方法であって、以下の工程:
(a)前記54に記載の組換えBAVゲノムを得る工程;
(b)工程(a)の該組換えBAVゲノムを、適切な哺乳動物宿主細胞に導入する工程;
(c)ウイルス複製を生じさせる工程;および
(d)該宿主細胞から組換えBAVウイルスを精製する工程、を包含する、方法。
【0237】
56.さらに、以下の工程:
前記工程(a)の後に制限酵素で前記組換えBAVを消化する工程であって、該制限酵素消化が前記ポリヌクレオチドに存在する他の配列からBAV配列を分離する、工程、
を包含する、前記55に記載の方法。
【0238】
57.前記宿主細胞がMDBK細胞である、前記55に記載の方法。
【0239】
58.前記MDBK細胞がアデノウイルスE1機能を発現する、前記57に記載の方法。
【0240】
59.前記宿主細胞が、ウシ仔胎網膜初代(PBFR)細胞である、前記55に記載の方法。
【0241】
60.前記セグメントがBAVゲノムを含む、前記55に記載の方法。
【0242】
61.前記E3特異的な配列が、前記組換えBAVゲノムにおいて欠失している、前記60に記載の方法。
【0243】
62.前記異種配列が前記欠損された配列によって以前に占有されていた前記部位で前記E3領域に挿入される、前記61に記載の方法。
【0244】
63.前記62に記載の方法により得られる、組換えBAV。
【0245】
64.前記63に記載の組換えBAVを含む、免疫原性組成物。
【0246】
65.疾患の症状を予防または改善させる方法であって、前記64に記載の免疫原性組成物を、哺乳動物被験体に導入する工程を包含する、方法。
【0247】
66.BAVゲノムおよび少なくとも1つの異種ヌクレオチド配列を含む組換えBAVベクターであって、該異種配列が、E3領域に挿入されている、BAVベクター。
【0248】
67.前記BAVゲノムがE3特異的な配列について欠失されている、前記66に記載のBAVベクター。
【0249】
68.前記異種配列が、哺乳動物病原体の防御決定因子をコードする、前記67に記載のBAVベクター。
【0250】
69.哺乳動物被験体における免疫応答を誘発し得る薬学的組成物であって、前記68に記載の組換えBAVベクターを含む、組成物。
【0251】
70.哺乳動物宿主において免疫応答を誘発して感染から防御する方法であって、前記69に記載の薬学的組成物を投与する工程を包含する、方法。
【0252】
71.哺乳動物宿主における遺伝子治療の方法であって、前記66に記載のベクターを該宿主に投与する工程を包含する、方法。
【0253】
72.前記異種配列が、線維芽細胞増殖因子(FGF)、血管内皮増殖因子(VEGF)、神経成長因子(NGF)、e-nos産物、pRb、リポタンパク質リパーゼ、スーパーオキサイドジスムターゼ、カタラーゼ、酸素スカベンジャー、フリーラジカルスカベンジャー、アポリポタンパク質およびプラスミノーゲン活性化因子インヒビター1からなる群より選択されるポリペプチドをコードする、前記71に記載の方法。
【0254】
【0255】
【0256】
【0257】
【0258】
【0259】
【0260】
【0261】
【0262】
【0263】
【0264】
【0265】
【0266】
【0267】
【0268】
【0269】
【0270】
【0271】
【0272】
【0273】
【0274】
【0275】
【図1−1】
【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図1−6】
【図1−7】
【図1−8】
【図1−9】
【図1−10】
【図1−11】
【図1−12】
【図1−13】
【図1−14】
【図1−15】
【図1−16】
【図1−17】
【図1−18】
【図1−19】
【図1−20】
【図1−21】
【図1−22】
【図1−23】
【図1−24】
【図1−25】
【図1−26】
【図1−27】
【図1−28】
【技術分野】
【0001】
関連出願への相互参照
本願は、1997年6月23日に出願された米国特許出願番号08/880,234に対して優先権を主張する。
【0002】
技術分野
本発明は、初期領域1(E1)および初期領域3(E3)領域遺伝子欠失の一方または両方が外来遺伝子によって置換されており、そして新規組換え哺乳動物細胞株がBAV E1配列を用いて安定に形質転換されており、それゆえE1遺伝子産物を発現して外来遺伝子でE1遺伝子欠失が置換されたウシアデノウイルスがそこで複製可能になる、新規のウシアデノウイルス(BAV)発現ベクター系に関する。これらの物質は、生の組換えウイルスまたはサブユニットワクチンの目的のためまたは他の治療目的のための、組換えBAV発現異種(抗原性)ポリペプチドまたはフラグメントの産生において使用される。
【0003】
本発明はまた、本明細書において開示されるBAVゲノム配列を含む新規のウシアデノウイルス(BAV)発現ベクター系に関する。BAVゲノム配列は、生の組換えウイルスまたはサブユニットワクチンを生産する目的のためまたは他の治療目的のための、組換えBAV発現異種(抗原性)ポリペプチドまたはフラグメントを生成するために1以上の外来遺伝子によって置換され得る。さらに、種々のBAV転写調節シグナルおよび翻訳調節シグナルを用いて、本発明のベクター系へと挿入された外来遺伝子の発現を調節し得る。さらに、本発明の新規の配列は、診断目的のために、被験体または生物学的サンプルにおいてBAVの存在を決定するために使用され得る。
【背景技術】
【0004】
発明の背景
アデノウイルスは、ヒトならびに家畜および実験動物において腸感染または呼吸器感染を引き起こす。
【0005】
ウシアデノウイルス(BAV)は、2つのサブグループに分割される、少なくとも9つの血清型を含む。これらのサブグループは、酵素結合免疫吸着アッセイ(ELISA)、免疫蛍光アッセイを用いた血清学的研究、ウイルス中和試験、免疫電子顕微鏡法、それらの宿主特異性によって、および臨床症候群に基づいて特徴付けられている。サブグループIウイルスは、BAV1、2、3および9を含み、そして、BAV4、5、6、7および8を含むサブグループ2と比較して、樹立されたウシ細胞において比較的良好に増殖する。
【0006】
BAV3は、1965年に初めて単離された。そして、BAV遺伝子型が最も良く特徴付けられており、これは、約35kbのゲノムを含む(Kurokawaら、(1978)J.Virol.28:212-218)。BAV3は、BAVのサブグループ1の代表例であり(Bartha(1969)Acta Vet.Acad.Sci.Hung.19:319−321)、通常無症状の感染をもたらす、ウシに共通する病原体である(Darbyshireら(1965)、J.Comp.Pathol.75:327−330)。しかし、これは時折、より重篤な呼吸管感染を伴う(Darbyshireら、1966 Res.Vet.Sci.7:81-93;Mattsonら、1988 J.Vet Res 49:67−69)。他のアデノウイルスと同様に、BAV3は、直鎖の二本鎖DNA分子を含む直径75nmの非莢膜性の正二十面体粒子である(Niiyamaら、(1975)J.Virol.16:621−633)。BAV3は、ハムスターに注射される場合に、腫瘍を形成し得(Darbyshireら、1966 Nature 211:102)、そしてウイルスDNAは、培養中のマウス、ハムスターまたはラットの細胞の形態学的な形質転換を効率的にもたらし得る(TsukamotoおよびSugino、1972、J.Virol.9:465−473);Motoiら、1972 Gann 63:415−418;M.Hitt、私信)。交叉ハイブリダイゼーションは、BAV3とヒトアデノウイルスタイプ2(HAd2)との間において、ゲノムの左端に近いいくつかの領域を含むゲノムのほとんどの領域で観察されたがゲノムの左端では観察されなかった(Huら、1984 J.Virol.49:604-608)。
【0007】
ヒトアデノウイルス(HAd)ゲノムにおいて、2つの重要な領域であるE1およびE3が存在する。E1およびE3では、外来遺伝子が組換えアデノウイルスを生成するために挿入され得る(BerknerおよびSharp(1984)Nuc.Acid Res.12:1925−1941ならびにHaj-AhmadおよびGraham(1986)J.Virol.57:267-274)。E1タンパク質は、組織培養においてウイルス複製に必須であるが、外来DNAをE1領域に含む条件付(conditional)ヘルパーアデノウイルス組換え体は、E1を構成的に発現する細胞株において生成され得る(Grahamら、(1977)J.Gen.Virol.36:59-72)。対照的に、HAd2および五Ad5のE3遺伝子産物は、インビトロでもインビボでも感染性ビリオン産生には必要ではないが、ウイルス感染に対する宿主免疫応答において重要な役割を果たしている(Anderssonら、(1985)Cell 43;215−222;Burgertら、(1987)EMBO J.6:2019−2026;Carlinら、(1989)Cell 57:135−144;Ginsbergら、(1989)PNAS、USA 86:3823−3827;Goodingら(1988)Cell 53:341−346;Tollefsonら、(1991)J.Virol.65:3095−3105;WoldおよびGooding(1989)Mol.Biol.Med.6:433−452ならびにWoldおよびGooding(1991)Virology 184:1−8)。ヒトアデノウイルスタイプ2(HAd2)のE3−19キロダルトン(kDa)糖タンパク質(gp19)は、多数のクラスI主要組織適合性複合体(MHC)抗原の重鎖と、小胞体において結合し、従って、これらの原形質膜への輸送を阻害する(Anderssonら(1985)Cell 43:215−222;BurgertおよびKvist、(1985)Cell 41:987−997;BurgertおよびKvist、(1987)EMBO J.6:2019−2026)。HAd2またはHAd5のE3-14.7kDaタンパク質は、ウイルス感染したマウス細胞の腫瘍壊死因子(TNF)による溶解を予防する(Goodingら(1988)Cell 53:341−346)。さらに、E3-10.4kDaおよびE3-14.5kDaタンパク質は、ウイルス感染した細胞において、エンドソーム媒介性インターナリゼーションおよび上皮増殖因子レセプター(EGF−R)の分解を誘発する複合体を形成する(Carlinら、Cell 57:135−144;Tollefsonら、(1991)J.Virol.65:3095−3105)。E3領域に外来遺伝子を有するヘルパー非依存性組換えアデノウイルスは、すべての許容性細胞株において非常に良好に複製しそして発現する(Chandaら(1990)Virology 175:535−547;Dewarら(1989)J.Virol.63:129−136;Johnsonら(1988)Virology 164:1−14;Lubeckら(1989)PNAS USA 86:6763−6767;McDermottら(1989)Virology 169:244−247;Mittalら(1993)Virus Res.28:67−90;Morinら(1987)PNAS.USA 84:4626−4630;Prevecら(1990)J.Inf.Dis.161:27−30;Prevecら(1989)J.Gen.Virol.70:429−434;Schneiderら(1989)J.Gen.Virol.70:417−427ならびにYuasaら(1991)J.Gen.Virol.72:1927−1934)。上記の研究およびアデノウイルスが野生型(wt)アデノウイルスゲノムの約105%をパッケージングし得るという示唆(Bettら(1993)J.Virol.67:5911−5921およびGhosh-Choudhuryら(1987)EMBO J.6:1733-1739)に基づけば、1.8kbまでの外来DNAの挿入物が欠失をなんら補償することなく外来タンパク質について発現ベクターとしての使用のためにアデノウイルス粒子中にパッケージングされ得る。
【0008】
グループCヒトアデノウイルスのE1A遺伝子産物は、非常に広範囲に研究されており、そしてウイルス遺伝子および細胞性遺伝子の両方のトランス活性化を媒介して(Berkら1979 Cell 17:935-944;JonesおよびShenk、1979 Cell 16:683-689;Nevins、1981 Cell 26:213-220;Nevins、1982 Cell 29:913-919、Berk、1986 Ann.Res.Genet.20:45-79に概説されている)、培養中の細胞の形質転換をもたらすこと(Graham、F.L.(1984)「Transformation by and oncogenicity of human adenoviruses」the Adenoviruses、H.S.Ginsberg編、Plenum Press、New York;Brantonら、1985 Biochim.Biophys.Acta 780:67-94に概説されている)、ならびに細胞のDNA合成および有糸分裂を誘発すること(Zerlerら、1987 Mol.Cell.Biol.7.821-929;Belletら、1989 J.Virol.63:303-310;Howeら、1990 PNAS.USA 87:5883-5887;HoweおよびBayley、1992 Virology 186:15-24)が示されている。E1A転写ユニットは、プロセシングを受けたすべてのE1A転写物からは欠失されるイントロン領域によって分離された2つのコード配列を含む。E1A転写ユニットから産生された2つの最も大きなmRNA種において、第一のコード領域は、さらに12sおよび13sのmRNA種の両方において見いだされれる配列であるエキソン1と、13s mRNA種においてのみ見出される特有の領域とにさらに細分割される。ヒトアデノウイルスのE1Aタンパク質とシミアンアデノウイルスのE1Aタンパク質との間の比較によって、3つの領域のいくらか保存されたタンパク質配列(CR)が規定された(Kimelmanら、1985 J.Virol.53:399-409)。CR1およびCR2は、エキソン1にコードされるが、CR3は、特有の配列およびエキソン2の小部分にコードされる。網状赤血球タンパク質Rb、サイクリンAおよび付随タンパク質キナーゼp33cdk2および未だに割り当てられていないタンパク質を含む多数の細胞タンパク質についての結合部位が、E1Aタンパク質のエキソン1がコードする領域において規定されている(YeeおよびBranton、1985 Virology 147:142-153;Harlowら、1986 Mol.Cell.Biol.6:1579-1589;Barbeauら、1992 Biochem.Cell Biol.70:1123-1134)。E1Aとこれらの細胞タンパク質との相互作用は、E1Aが不死化およびガンへの転換に関与する機構として関連付けられている(Eganら、1989 Oncogene 4:383-388;Whyteら、1988、Nature 334:124-129;Whyteら、1988 J.Virol.62:257-265)。E1A単独は、培養中の細胞を形質転換または不死化し得るが、E1Aと、E1B-19kタンパク質またはE1B-55kタンパク質のいずれかとを両方別個または一緒に共に発現することは、通常、培養中の齧歯類細胞の高度に頻繁な形質転換を必要とする(Graham、1984、前出、Brantonら、1985前出;McLorieら、1991 J.Gen Virol.72:1467-1471に概説される)。
【0009】
ヒト細胞の許容性感染における他のウイルス初期遺伝子のトランス活性化は、主に、E1AのCR3領域においてコードされるアミノ酸配列によって媒介される(Lillieら、1986 Cell 46:1043-1051)。特有な領域におけるCysX2CysX13CysX2Cys配列モチーフにおいて保存されたシステイン残基は、金属イオン結合活性と関連している(Berg 1986、上記)。そして、トランス活性化に必須である(Jelsmaら、1988 Virology 163:494-502;Culpら、1988 PNAS USA 85:6450−6454)。金属結合ドメインのアミノ(N)末端の直近に存在するCR3におけるアミノ酸もまた、転写活性に重要であることが示されているが、金属結合ドメインのカルボキシ(C)末端の直近の配列は、プロモーター領域と会合を形成するのに重要であることが示されている(LillieおよびGreen、1989 Nature 338:39-44;図3を参照のこと)。
【0010】
遺伝子操作の適用は、ワクチンを得るためのアデノウイルス発現系を調製することのいくつかの試みをもたらした。このような研究の例は、酵母宿主における発現のためのアデノウイルス主要後期プロモーターについての米国特許第4,510,245号;欠失された初期領域3に位置するB型肝炎表面抗原をコードする遺伝子を有する生の組換えアデノウイルスタイプ7についての米国特許第4,920,209号;HCMV主要莢膜糖タンパク質についてのヒト細胞における非欠損性ヒトアデノウイルス5組換え発現系についての欧州特許第389 286;E1Aタンパク質を発現する細胞における非病原性免疫原性の生の生存イヌアデノウイルスについてのWO91/11525;ならびにアデノウイルス2のE3領域由来のリーダーおよび/またはプロモーターを含むベクターについての仏国特許第2 642 767号の開示を含む。
【0011】
常在性のアデノウイルスベクターが、ヒト起源のアデノウイルスに比較して、非ヒト動物種における生存組換えウイルスワクチンとしての使用により良好に適すると推定されている。これは、異種配列の挿入に適する領域が常在性アデノウイルスベクターに同定されること、および異種配列の挿入、組換え体の単離および組換え体の増殖のための組成物および方法が開発されることを必要とする。挿入に適する領域は、適切なヘルパー機能が提供される場合、ウイルスゲノム非必須領域または必須領域を含み得る。例えば、HAdとの類似性によって、他のアデノウイルスにおけるE3領域が培養細胞におけるウイルス複製に必須でない場合、外来遺伝子挿入物をE3領域に含むアデノウイルス組換え体が生成され得る。
【0012】
外来遺伝子発現のためのベクターとして作用する適切なウイルスの選択、遺伝子挿入のための部位として適切な領域の同定、ならびに組換えウイルスの構築、単離および増殖は、組換えウイルスワクチンベクターの開発にとって意義深い挑戦を提起する。詳細には、好ましい挿入部位は、ウイルスの生存複製および組織培養およびインビボでもその有効な操作のためには必須ではない。さらに、挿入部位は、ウイルスが複製を継続することを保証しつつ、新たな遺伝的物質を受け入れることができなければならない。ウイルスゲノムの必須領域もまた、組換えウイルスが、トランスでその特定の必須領域の機能を補完する細胞株において増殖する場合は、外来遺伝子の挿入のために利用され得る。
【0013】
ウイルスゲノムにおける適切な挿入部位を決定するための効率よい方法は、そのゲノムの完全なヌクレオチド配列を入手することである。これは、種々のコード領域が規定されることを可能にし、挿入部位としてそれらの可能な使用を容易にする。必須でない非コード領域の規定はまた、配列分析によって示され、そしてこれらはまた、可能な挿入部位として使用され得る。BAV-3ゲノムの特定の領域のヌクレオチド配列は決定されている。ゲノムの究極の左端の配列(これは、逆末端反復(ITR)、パッケージングシグナル、E1およびpIXを含む)は、いくつかのグループによって決定されている:Shinagawaら、1987 Gene55:85−93によるヌクレオチド1〜195(ITR);Zhengら、1994、Virus Research 31:163−186によるヌクレオチド1〜4060(ITR、パッケージングシグナル、E1およびpIX);Elgadiら、1993、Intervirology 36:113−120によるヌクレオチド1〜4091(ITR、パッケージングシグナル、E1およびpIX)。(ヌクレオチド1は、線状の34.4kb BAV−3ゲノムの最も左のヌクレオチドを示す。)決定されているBAV−3ゲノムのさらなる配列は、以下を含む:ヌクレオチド5,235−5,891(主要後期プロモーター、Songら、1996、Virology 220;390−401);ヌクレオチド17,736〜20,584(ヘキソン遺伝子。Huら、1984、J.Virology 49:604−608);ヌクレオチド20,408〜21,197(プロテイナーゼ遺伝子、Caiら、1990、Nucleic Acids Res.18:5568);ならびにヌクレオチド26,034〜31,132(E3領域、pVIIIおよびファイバー遺伝子、Mittalら、1992、J.Gen.Virol.73:3295−3300)。
【0014】
組換えウイルスおよびウイルスゲノムが適用され得る多くの使用の一つは、それらが利用可能である場合、組換えサブユニットワクチンの開発においてである。ワクチン接種は、生の減弱化されたウイルスワクチンが使用される場合は特に、呼吸器系ウイルス疾患および腸ウイルス疾患の制御のための最も有効な手段であることが証明されている。これらのワクチンは、経口または鼻腔内に投与される場合、強力な粘膜免疫を誘導し、これは、これらのウイルスによって引き起こされる初期感染をブロックし、それらによって引き起こされる疾患の進行を減少させるために必要とされる。このアプローチは、遺伝子操作された(毒性遺伝子欠失)ウイルスゲノムをベクターとして使用して他の病原体の遺伝子をインビボで送達および発現することによって拡張されている。Ertlら(1996)J.Immunol.156:3579−3582。
【0015】
ヒトアデノウイルス(HAV)に基づく組換えウイルスベクター系が近年開発されている。Grahamら(1992)、「Vaccines:New approaches to immunological problems」(R.W.Ellis編)、Butterworth-Heineman、Stoneham、363-390頁。複製欠損HAVベクターおよび複製適合HAVベクターの両方が、種々の外来抗原を発現するように操作されている。概説については、Grunhausら(1992)Seminar in Virol.3:237-252;Imler(1995)Vaccine 13:1143-1151を参照のこと.安定な外来遺伝子発現を提供することに加えて、操作されたアデノウイルスは、体液性、細胞性および粘膜性の免疫応答を誘発することが示されている。Bugeら(1997)J.Virol.71:8531-8541。
【0016】
ヒトアデノウイルスの遺伝子治療のためのベクターとしての使用は、宿主において、すでに存在するHAVに対する中和抗体(これは、組換えウイルスの侵入および複製と干渉し得る)の存在のため、および組換えウイルスと既に宿主に存在する野生型HAVとの間の組換えおよび/または補完の可能性のために、妨害されている。従って、高度の種特異的であるHAV以外の動物アデノウイルスが、遺伝子治療および組換えワクチンのためのベクターとして考慮されつつある。
【0017】
ウシアデノウイルス-3(BAV-3)の分子的特徴付けは、ヒトおよび他の哺乳動物種におけるワクチンおよび遺伝子治療のための、ウシアデノウイルスの生のウイルスベクターとしての開発を補佐する。近年、BAV3ゲノムの完全なDNA配列および転写マップが報告されている。この配列は、1997年6月23日に出願された親の米国特許出願U.S.S.N.08/880,234において、およびいくつかの刊行物において開示された。Baxiら、(1998)Virus Genes 16:1-4;Leeら、(1998)、Virus Genes 17:99-100;およびReddyら(1998)J.Virol.72:1394-1402。
【発明の概要】
【課題を解決するための手段】
【0018】
発明の開示
本発明者らは、今や、34,446ヌクレオチドを含む全長BAV−3ゲノムの配列を完成し、それによって外来遺伝子の挿入のため、および診断プローブとしての使用のための両方に適切な領域を同定した。本発明者らはまた、これら領域中に外来遺伝子を挿入してBAV組換え体を生成し、そしてこの組換え体を増殖させた。例えば、このような組換え体は、種々の病原体に対する組換えサブユニットワクチンとして、組換えポリペプチドの過剰発現のために、および遺伝子治療目的のために有用である。
【0019】
1つの実施態様では、本発明は、新規ウシアデノウイルス発現ベクターシステムに関し、ここでE1およびE3遺伝子領域の1つまたは両方が、一部または全てを欠失している。本発明はまた、好ましくはBAVのE1配列であるE1配列で形質転換されたウシ起源の組換え哺乳動物細胞株に関し、これは、1つまたはそれ以上のE1遺伝子産物を構成的に発現し、外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列によって置換されたE1遺伝子領域の一部分またはすべての欠失を有するウシアデノウイルスをその中で複製させる。本発明は、さらに、異種(抗原性)ポリペプチドまたはそのフラグメントの産生におけるこれら材料の使用に関する。
【0020】
別の実施態様では、本発明は、ウシアデノウイルスゲノムまたはそのフラグメントの以下の領域を利用する新規なウシアデノウイルス発現ベクター系に関する:ヌクレオチド4,092-5,234;ヌクレオチド5,892-17,735;ヌクレオチド21,198-26,033およびヌクレオチド31,133-34,445。これらの領域(およびそのフラグメント)は、とりわけ、外来配列の挿入のために、転写および翻訳調節配列を含むDNA制御配列の準備のために、または、被験体または生物学的試料中のウイルス核酸またはこれらの領域によりコードされるタンパク質の存在を検出する診断目的のために用いられ得る。
【0021】
本発明はまた、ウシ、ヒトおよびその他の哺乳動物を含む、哺乳動物において、感染性生物に対する抗体、細胞媒介性および/または粘膜免疫を産生するための生存組換えウイルス、またはサブユニットワクチンの調製方法に関する。この方法は、ウシアデノウイルスゲノム中に、上記抗体に対応するか、または有効なプロモーターとともにまたはなしで上記細胞媒介性および/または粘膜免疫を誘導する抗原をコードする遺伝子または遺伝子フラグメントを挿入し、BAV組換え体を産生する工程を包含する。
【0022】
別の局面では、本発明は、組換えウイルス、および細胞系における目的のDNA配列またはアミノ酸配列の発現のための組換えウイルスベクターの使用を含む。
【0023】
一般に、外来遺伝子構築物は、1つまたはそれ以上の適切な欠失を有する全長ウイルスゲノムの一部分のみを示すヌクレオチド配列中にクローン化される。このキメラDNA配列は、通常、成功するクローニングを可能にし、この配列の多くのコピーを産生するプラスミド中に存在する。次いで、クローン化された外来遺伝子構築物は、例えば、インビボ組換え、次いでDNA媒介同時トランスフェクション技法により完全ウイルスゲノムに含められ得る。このクローン化された外来遺伝子構築物のウイルスゲノム中への取り込みは、外来遺伝子を、複製およびパッケージングシグナルを含むDNA分子中に配置し、感染性ウイルス粒子にパッケージングされ得る組換えアデノウイルスゲノムの多コピーの生成を可能にする。コード配列または1つ以上のコード配列の多コピーは、組換えベクターが1つ以上外来タンパク質を発現し得るように挿入され得る。外来遺伝子は、付加、欠失または置換を有し、発現タンパク質の発現および/または免疫学的効果を増大し得る。
【0024】
本発明はまた、ウシアデノウイルス発現ベクターを含む発現系を含み、ここで任意の外因性調節要素を伴うかまたは伴わない異種ヌクレオチド配列が、E1遺伝子領域および/またはE3遺伝子領域の一部分もしくはすべてを置き換える。
【0025】
別の実施態様では、本発明は、BAVゲノムの1つまたはそれ以上の領域が異種配列により置き換えられるか、または異種ヌクレオチド配列が任意のBAV配列を取り除くことなくBAVゲノム中に導入される発現系を含む。DNA調節配列を含むBAVゲノムの遺伝子内領域は、本発明の実施において、同種および異種(即ち外来)遺伝子の発現に有用である。
【0026】
本発明はまた、(A)全長BAV DNA、および適切な細胞株の同時トランスフェクション後インビボ相同組換えにより組換えウイルスを生成し得るプラスミド(単数)または2つのプラスミドを含む組換えベクター系であって、全長の野生型BAVゲノムを示すBAV DNA、およびE1またはE3遺伝子領域を含むウシアデノウイルス左または右末端配列を含むプラスミドまたはウシアデノウイルスE2、E4、L1、L2、L3、L4、L5、L6もしくはL7配列を、E1、E2、E3、E4、L1、L2、L3、L4、L5、L6もしくはL7遺伝子領域の一部または全てで置換された外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列(即ち、挿入カセット)とともに含む、組換えベクター系;(B)(a)E1遺伝子領域の一部分またはすべてが、外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列により置き換えられている系;(b)E3遺伝子領域の一部分またはすべてが、外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列により置き換えられている系;および(c)E1遺伝子領域の一部分またはすべておよびE3遺伝子領域の一部分またはすべてが欠失し、かつ外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列が少なくとも1つの欠失中に挿入されている系からなる群から選択される生存組換えウシアデノウイルスベクター(BAV)系;(C)E1遺伝子領域の一部分またはすべての欠失、E3遺伝子領域の一部分またはすべての欠失、または両方の欠失を含み、かつ少なくとも1つの欠失中に、疾患を起こす生物の抗原決定基をコードする異種ヌクレオチド配列が挿入される組換えウシアデノウイルス(BAV);(D)E1の一部分またはすべての欠失、E3の一部分またはすべての欠失、または両方の欠失を含み、かつ少なくとも1つの欠失中に、発現プロモーターの制御下にある外来遺伝子またはそのフラグメントをコードする異種ヌクレオチド配列が挿入される組換えウシアデノウイルス発現系;または(E)(1)所望の免疫応答を得るために必要な抗原決定基をコードする異種ヌクレオチド配列を、(2)生存組換えウイルスまたは組換えタンパク質またはサブユニットワクチンとしての使用のために免疫原性量の上記抗原決定基の発現を提供する有効プロモーターをともにまたはなしで含むBAV組換え体を含む、哺乳動物宿主において免疫応答を産生するための組換え体ウシアデノウイルス(BAV);(F)E1の一部分またはすべての欠失、および/またはE3の一部分またはすべての欠失、および/またはBAVゲノムの以下の領域の少なくとも1つの一部分またはすべての欠失:E2、E4、L1、L2、L3、L4、L5、L6またはL7:を含む変異体ウシアデノウイルス(BAV)を含む。
【0027】
E1およびE3領域に加えて、BAVゲノム内のその他の部位もまた、外来ヌクレオチド配列の挿入に有用である。これらは、限定されないが、E2領域、E4領域、E4プロモーターとゲノムの右末端との間の領域、後期領域(L1−L7)、33kD、52kD、100kD、DBP、pol、pTPおよびペントン遺伝子、ならびに遺伝子IIIA、pV、pVI、pVII、pVIII、およびpXを含む。
【0028】
本発明はまた、ゲノムのE3領域に欠失を含むウイルス、およびゲノムの欠失したE3領域中に異種配列の挿入を含むウイルスを、高効率で得るための方法および組成物を提供する。1つの実施態様では、E3が欠失したウイルスゲノム(異種配列の挿入をともなうまたはなし)が、適切な細胞株、例えば、アデノウイルスE1機能を発現するMDBK細胞または相当する細胞中にトランスフェクトされ、そして組換えウイルスがトランスフェクトされた細胞から回収される。別の実施態様では、欠失したE3領域を(異種配列の挿入をともなうまたはなしで)含むBAVゲノムのセグメントが、原核生物細胞中で、BAVゲノムでの組換えを受け、組換えBAVゲノムを生成することが可能である。本発明の目的には、BAVゲノムは完全長BAVゲノムであり得るか、またはそれは、1つもしくはそれ以上のBAV複製および/またはパッケージング配列を含むことを条件に、1つもしくはそれ以上の欠失を含み得る。欠失したE3領域を含むBAVゲノムセグメントはまた、1つまたはそれ以上のBAV複製および/またはパッケージング配列を含み得る。そうでなければ、組換えBAVゲノムは、特定のゲノム領域における1つまたはそれ以上の欠失、ならびに欠失または欠失のない、異種配列が挿入されたBAVゲノムとともに全長のBAVゲノムを含む。次いで、組換えBAVゲノムを、例えば、ウシ仔胎網膜初代(PFBR)細胞のような適切な細胞株中にトランスフェクトし、そしてトランスフェクトされた細胞から組換えウイルスを回収する。
【0029】
別の局面では、本発明は、BAV E1遺伝子領域配列で安定に形質転換された組換え哺乳動物細胞株を提供し、この組換え細胞株は、これによって、その中で、外来遺伝子またはそのフラグメントをコードする異種または同種ヌクレオチド配列によって置き換えられたE1またはE3遺伝子領域の一部分またはすべての欠失を含むウシアデノウイルスの複製を可能にし得る。本発明はまた、BAVE1遺伝子産物を発現する組換えウシ細胞株からの、成長因子、レセプターおよびその他の細胞タンパク質のようなポリペプチドまたはそのフラグメントの産生、単離および精製を含む。
【0030】
本発明はまた、遺伝子治療が必要な哺乳動物に遺伝子治療を提供する方法を包含する。例えば、遺伝子治療は、遺伝子欠陥を制御するために、宿主細胞中に治療遺伝子を導入するために、変異遺伝子の配列を変え野生型機能を回復させるために、遺伝子の配列を変えその機能を不活性化するために、外因性遺伝子機能を提供するためなどに用いられ得る。例えば、遺伝子治療は、癌、AIDS、その他のウイルスで誘導される病因、感染性疾患、遺伝病などの処置において用いられる。遺伝子治療のプロセスは、上記哺乳動物に異種ヌクレオチド配列を含む生存組換えウシアデノウイルスを、組換えウイルスベクターゲノムが上記哺乳動物ゲノム中に取り込まれるか、または独立にかつ染色体外で維持される条件下で投与し、標的器官または組織中で異種配列の発現を提供する工程を包含する。
【0031】
本発明の別の局面は薬学的組成物を提供し、この組成物は、治療的に有効な量の組換えウイルス、組換えウイルスベクターまたは組換えタンパク質を、薬学的に受容可能なキャリアとともにまたはなしで含む。このような薬学的組成物の1つの例は、組換えウイルスワクチンである。組換えウイルスワクチンは、経口用量(例えば、溶腸被覆錠剤)によるか、注射によるか、またはその他の様式による投与のために処方され得る。より詳細には、これらは、感染に対して哺乳動物宿主を保護するためのワクチンを含み、生存組換えアデノウイルス、または外来遺伝子またはフラグメントが抗原をコードし、そして薬学的に受容可能なキャリアとともにまたはなしで処方される本発明の組換えアデノウイルスにより産生される組換えタンパク質を含む。これらの組成物は、抗原性ポリペプチドまたは保護的抗原を発現し得、それによって目的のポリペプチドまたは抗原に応答する免疫を惹起し、そして感染からの保護を提供する。本発明の実施に有用な薬学的組成物はまた、目的のポリペプチドまたは抗原を発現する組換えBAVベクターを保持する細胞、またはBAVポリペプチドまたは抗原を発現するベクターを保持する細胞を含み得る。
【0032】
本発明はまた、哺乳動物において、抗体、細胞媒介性および/または粘膜免疫を生成する方法を含み、(1)哺乳動物宿主において感染に対する免疫応答を惹起する方法であって:本発明の生存BAV組換え体であって、外来遺伝子またはフラグメントが抗原をコードするBAV組換え体を含むワクチンを、薬学的に受容可能なキャリアとともにまたはなしで投与する工程を包含する方法、および(2)哺乳動物宿主において感染に対する免疫応答を惹起する方法であって:外来遺伝子またはフラグメントが所望の抗原をコードするBAV組換え体を培養することによって調製された組換え抗原を含むワクチンを、薬学的に受容可能なキャリアとともにまたはなしで投与する工程を包含する方法を含む。
【0033】
本発明はさらに、感染細胞または哺乳動物被験体、またはこれら供給源からの核酸調製物のような生物学的試料中の、BAV DNAおよび/またはBAVがコードするタンパク質および抗原の存在を検出するための診断手順の実施において有用な組成物および方法を提供する。これらは、制限されずに、BAV遺伝子およびコード配列およびそれらのフラグメント、ならびにBAVゲノムおよびそれらのフラグメントによりコードされるアミノ酸配列を含む。
【0034】
以下の開示は、本発明のこれらおよびその他の実施態様を、当業者にとって容易に明白にする。しばしば、本開示は、ウシアデノウイルスタイプ3(BAV3)に言及するが、これは、例示の目的のためであって、しかも、他の型のウシアデノウイルス、例えば、1、2、4、5、6、7、8および9に同じ特徴が適用され;しかも本明細書に記載されそして請求の範囲に記載された発明が、これらのウシアデノウイルス型のすべてをカバーすることが意図されることを理解すべきである。
【図面の簡単な説明】
【0035】
【図1】図1.ウシアデノウイルスタイプ3(BAV3)ゲノムの完全なヌクレオチド配列。
【図2】図2.BAV3の転写マップ。ゲノムは0から100まで(マップ単位)番号付られた実線で表される。転写物は、転写の長さおよび方向に関して矢印により表される。E1A、E1B、E2A、E2B、E3、E4、L1、L2、L3、L4、L5、L6およびL7領域の位置が示される。パッケージングおよび複製配列は、195bpITR配列中および左ITRとE1領域との間の領域に位置する。
【図3】図3.E3欠失BAV3ゲノムDNAを含むプラスミドの構築。プラスミドpBAV302を上記の実施例3で記載のように異なるゲノムクローンから構築した。DNA配列の起源は以下の通りである:プラスミドDNA、細線;BAV3ゲノムDNA配列、太線。中空の矢印は以下のPCRプライマーを表す:a:5'−ACGCGTCGACTCCTCCTCA(配列番号2);b:5'−TTGACAGCTAGCTTGTTC(配列番号3);c:5'−CCAAGCTTGCATGCCTG(配列番号4);およびd:5'−GGCGATATCTCAGCTATAACCGCTC(配列番号5)。
【0036】
【図4】図4.組換えBAV3ゲノムの制限酵素分析。DNAは、BAV3(レーンb)、BAV3.E3d(レーンaおよびe)、BAV3.E3gD(レーンcおよびf)またはBAV3.gDt(レーンdおよびg)で感染させたMDBK細胞から得た。 DNAは、Hirtの方法(Hirt(1967)J.Mol.Biol.26:365−369)により抽出し、そしてBamHI(レーンaおよびb)、NheI(レーンcおよびd)またはNdeI(レーンe、fおよびg)で消化した。1kb+DNAラダー(Gibco/BRL)を、ウイルスDNAフラグメントをサイズ分けするために用いた。
【図5】図5.組換えBAV3ウイルスで感染させたMDBK細胞におけるgDタンパク質の発現。(A)BHV−I(レーンa)、BAV3.E3d(レーンe、fおよびg)、またはBAV3.E3gD(レーンb、cおよびd)で感染させたか、または非感染(レーンh)の放射標識されたMDBK細胞の溶解液からのタンパク質を、gD特異的MAbのプールで免疫沈降し、そして還元条件下でSDS−PAGEにより分析した。タンパク質は、感染後6〜16時間(レーンaおよびh)、36〜48時間(レーンbおよびe)、48〜50時間(レーンcおよびf)、または60〜62時間(レーンdおよびg)標識した。(B)BHV−I(レーンa)、BAV3.E3d(レーンe、fおよびg)、またはBAV3.E3gDt(レーンb、cおよびd)で感染させた放射標識されたMDBK細胞の培養培地からのタンパク質を、gD特異的MAbのプールで免疫沈降し、そして還元条件下でSDS−PAGEにより分析した。タンパク質を、感染後、6〜16時間(レーンa)、12〜14時間(レーンbおよびe)、16〜18時間(レーンcおよびf)、または22〜26時間(レーンdおよびg)標識した。分子サイズマーカー(MW)はkDaである。
【図6】図6.組換えgDおよびgDtタンパク質の抗原性分析。BAV3.E3gD(レーンa−e)またはBAV3.E3gDt(レーンf−j)組換えウイルスで感染させた放射標識MDBK細胞の溶解液(レーンa−e)および培養培地(レーンf−j)からのタンパク質を、MAb136(レーンaおよびf)、MAb3E7(レーンbおよびg)、MAb4C1(レーンcおよびh)、MAb2C8(レーンdおよびi)、またはMAb3C1(レーンeおよびj)で免疫沈降し、そして還元条件下でSDS−PAGEにより分析した。分子サイズマーカー(MW)はkDaである。
【図7】図7.MDBK細胞におけるgD発現に対するAraCの影響。100μg/ml AraCの存在下(レーンa、b)または非存在下(レーンc、d)でBAV3.E3gD(レーンa−d)で感染させ、そして感染後22時間(レーンaおよびc)または34時間(レーンbおよびd)に2時間放射標識したMDBK細胞の溶解液からのタンパク質を、gD特異的MAbのプールで免疫沈降し、そしてSDS−PAGEにより分析した。分子サイズマーカー(MW)はkDaである。
【図8】図8.コットンラットにおける抗体応答。組換えBAVを用いた二次免疫の12日後の血清、肺洗浄液(l.w)および鼻洗浄液(n.w)中の糖タンパク質gD(A、B)またはBAV3特異的(C、D)IgA(A、C)またはIgG(B、D)のELISA力価。白棒は、BAV3.E3gDを示し、黒棒はBAV3.E3gDtを示し、そして斜線の棒はBAV3.E3dを示す。誤差の棒は、1群あたり4匹の動物の平均の標準誤差を示す。
【図9】図9.異なるBAV組換え体を用いた免疫後の異なる時点における子ウシ血清中の抗-gD IgG力価(gD特異的ELISAにより測定)。斜線の棒:BAV3.E3dで免疫化した動物;白棒:BAV3.E3gDで免疫化した動物;黒棒:BAV3.E3gDtでワクチン接種した動物。
【図10】図10.異なるBAV組換え体を用いた免疫後の異なる時点における子ウシ鼻スワブにおける抗−gD IgA力価(gD特異的ELISAにより測定)。斜線の棒:BAV3.E3dで免疫化した動物;白棒:BAV3.E3gDで免疫化した動物;黒棒:BAV3.E3gDtでワクチン接種した動物。
【図11】図11.BHV−1抗原投与後に観察されたワクチン接種子ウシの温度。黒四角:BAV3.E3dで免疫化した動物;白三角:BAV3.E3gDで免疫化した動物;白丸:BAV3.E3gDtでワクチン接種した動物。
【図12】図12.ワクチン接種子ウシにおけるBHV−1抗原投与後の鼻損傷の外観および程度の観察。黒四角:BAV3.E3dで免疫化した動物;白三角:BAV3.E3gDで免疫化した動物;白丸:BAV3.E3gDtでワクチン接種した動物。
【図13】図13.ワクチン接種子ウシの試験およびコントロール群で観察された低下の程度。黒四角:BAV3.E3dで免疫化した動物;白三角:BAV3.E3gDで免疫化した動物。
【図14】図14.ワクチン接種子ウシの試験およびコントロール群のBHV−1抗原投与後のBHV−1の単離。黒四角:BAV3.E3dで免疫化した動物;白三角:BAV3.E3gDで免疫化した動物;白丸:BAV3.E3gDtでワクチン接種した動物。
【発明を実施するための形態】
【0037】
発明を実施する様式
本発明の実施は、その他に示されなければ、当該分野内の技術である、従来の微生物学、免疫学、ウイルス学、分子生物学、および組換えDNA技法を利用する。これらの技術は、文献に十分説明されている。例えば、Maniatisら、Molecular Cloning:ALaboratory Manual(1982);DNA Cloning:APractical Approach、I巻およびII巻(D.Glover編);Oligonucleotide Synthesis(N.Gait編(1984));Nucleic Acid Hybridization(B.HamesおよびS.Higgins編(1985)):Transcription and Translation(B.HamesおよびS.Higgins編(1984));Animal Cell Culture(R.Freshney編(1986));Peroal、A Practical Guide to Molecular Cloning(1984).Sambrookら、Molecular Cloning:A Laboratory Manual(第2版);第I、IIおよびIII巻(1989)を参照のこと。
【0038】
A.定義
本発明を記載することにおいて、次の用語が、以下に規定されるように用いられる。
【0039】
「レプリコン」は、インビボでDNA複製の自律単位として機能する、即ち、それ自身の制御の下で複製し得る任意の遺伝要素(例えば、プラスミド、染色体、ウイルス)である。
【0040】
「ベクター」は、プラスミド、ファージ、コスミドまたはウイルスのような、それに別のDNAセグメントが、連結したセグメントの複製を引き起こすように連結され得るレプリコンである。
【0041】
「生存ウイルス」は、「死滅した」ウイルスとは対照差別して、組織培養および接種動物において同一の子孫を産生し得るウイルスを意味する。
【0042】
「ヘルパーフリーウイルスベクター」は、ベクターに欠損するあるものを供給する第2のウイルスまたは細胞株を必要としないベクターである。
【0043】
「2本鎖DNA分子」は、その通常の2本鎖ヘリックス中のデオキシリボヌクレオチド(アデニン、グアニン、チミン、またはシトシン)のポリマー形態をいう。この用語は、分子の1次および2次構造のみについて言及し、そしてそれを任意の特定の三次形態に限定しない。従って、この用語は、とりわけ、直線状DNA分子で見出される2本鎖DNAを含む(例えば、ウイルス、プラスミド、および染色体からのDNAの制限フラグメント)。特定の2本鎖DNA分子の構造を論議することにおいて、本明細書において、配列は、DNAの非転写鎖(即ち、mRNAに相同な配列を有する鎖)に沿って5'〜3'方向にある配列のみを与える通常の慣習に従って記載され得る。
【0044】
DNA「コード配列」は、適切な調節配列の制御下に置かれたとき、インビボで転写されかつポリペプチドに翻訳されるDNA配列である。コード配列の境界は、5'(アミノ)末端の開始コドンおよび3'(カルボキシ)末端の翻訳終結コドンにより決定される。コード配列は、原核生物配列、真核生物mRNAからのcDNA、真核生物(例えば哺乳動物)DNAからのゲノムDNA配列、ウイルスDNA、および合成DNA配列さえ含み得るが、これらに制限されない。ポリアデニル化シグナルおよび転写終結配列は、通常、コード配列の3'側にある。
【0045】
「転写プロモーター配列」は、細胞において、RNAポリメラーゼに結合し、かつ下流(3'方向)コード配列の転写を開始し得るDNA調節領域である。本発明を規定する目的のために、プロモーター配列は、コード配列の翻訳開始コドン(ATG)が3'末端に結合し、そして上流(5'方向)に伸び、バックグラウンドを超える検出可能なレベルで転写を開始するために必要な最小数の塩基または要素を含む。プロモーター配列内には、転写開始部位(ヌクレアーゼS1を用いたマッピングにより簡単に規定される)、およびRNAポリメラーゼの結合を担うタンパク質結合ドメイン(コンセンサス配列)が見出される。真核生物プロモーターは、しばしば、常にではないが、「TATA」ボックスおよび「CAAT」ボックスを含む。原核生物プロモーターは、−10および−35コンセンサス配列に加えて、Shine−Dalgarno配列を含む。
【0046】
DNA「制御配列」は、集合的に、プロモーター配列、リボソーム結合部位、スプライシングシグナル、ポリアデニル化シグナル、転写終結配列、上流調節ドメイン、エンハンサー、翻訳終結配列などをいい、これは、集合的に、宿主細胞中のコード配列の転写および翻訳を提供する。
【0047】
RNAポリメラーゼがプロモーター配列に結合し、そしてコード配列をmRNAに転写し、次いで、コード配列によりコードされたポリペプチドに翻訳される場合、コード配列または配列コードは、細胞において、制御配列に「作動可能に連結」またはその「制御下」にある。
【0048】
「宿主細胞」は、外因性DNA配列により、形質転換されたか、または形質転換し得る細胞である。
【0049】
外因性DNAが細胞膜内側に導入されたとき、細胞はこのような外因性DNAにより「形質転換」されている。外因性DNAは、細胞のゲノムを構成する染色体DNAに組み込まれてもよい(共有結合する)し、または組み込まれなくてもよい。例えば、原核生物および酵母では、外因性DNAは、プラスミドのようなエピソーム要素上で維持され得る。安定に形質転換された細胞は、外因性DNAが染色体中に組み込まれ、その結果それは染色体複製を通じて娘細胞により受け継がれる細胞である。哺乳動物細胞については、この安定性は、外因性DNAを含む娘細胞の集団からなる細胞株またはクローンを確立する細胞の能力により実証される。
【0050】
「クローン」は、単一細胞または共通の祖先に由来する娘細胞の集団である。
【0051】
「細胞株」は、多世代の間インビトロで安定に増殖し得る初代細胞のクローンである。
【0052】
2つのポリペプチド配列は、アミノ酸の少なくとも約80%(好ましくは少なくとも約90%、および最も好ましくは少なくとも約95%)が規定された長さの分子に対して一致する場合「実質的に相同」である。
【0053】
2つのDNA配列は、それらが、ヌクレオチドと同一であるか、またはヌクレオチドの40%より多くは異ならず、好ましくはヌクレオチドの約30%より多くは異ならず(即ち、少なくとも約70%相同)、より好ましくはヌクレオチドの約20%よりは異ならず、そして最も好ましくはヌクレオチドの約10%より多くは異ならない場合に、「実質的に相同」である。
【0054】
実質的に相同であるDNA配列は、例えば、特定の系について規定されるストリンジェントな条件下のサザンハイブリダイゼーション実験において同定され得る。適切なハイブリダイゼーション条件を規定することは、当該分野の技術である。例えば、Maniatisら、前述;DNA Cloning、第IおよびII巻、前述;Nucleic Acid Hybridization、前述、を参照のこと。
【0055】
DNA構築物の「異質」領域は、天然に他の分子と結合して見出されない別のDNA分子の中にあるか、またはそれに結合したDNAの識別可能なセグメントである。従って、異質領域がウイルス遺伝子をコードする場合、この遺伝子は、通常、供給源ウイルスまたはウイルス感染細胞のゲノム中のウイルスゲノムに隣接しないDNAにより隣接される。異種コード配列の別の例は、コード配列自身が天然に見出されない構築物である(例えば、ネイティブ遺伝子とは異なるコドンをもつ合成配列)。本明細書で用いるとき、対立遺伝子異形または自発変異事象はDNAの異種領域を生じない。
【0056】
「ウシ宿主」は、任意の系統、成体または子ウシをいう。
【0057】
用語「タンパク質」は、本明細書において、その他に注記されなければ、ポリペプチドまたはグリコシル化ポリペプチドをそれぞれ示すために用いられる。用語「ポリペプチド」は、その最も広い意味で用いられる(即ち、ペプチド結合を通じて連結される任意のアミノ酸ポリマー(ジペプチドまたはそれより大きい))。従って、用語「ポリペプチド」は、タンパク質、オリゴペプチド、タンパク質フラグメント、アナログ、ムテイン、融合タンパク質などを含む。
【0058】
「融合タンパク質」は、通常、リーダー配列または安定化ポリペプチドをコードする第1の領域、および異種タンパク質をコードする第2の領域を含む遺伝子の発現産物として規定される。それは、抗原性タンパク質フラグメントまたは完全長BAVタンパク質配列および異種配列(代表的には、細胞内で発現されたポリペプチドについて組換え宿主において分泌のために機能的なリーダー配列、またはSODのような宿主細胞プロテアーゼからタンパク質を保護するN末端配列)を含むポリペプチドを含む。抗原性タンパク質フラグメントは、通常、長さが約5−7アミノ酸である。
【0059】
「ネイティブ」タンパク質またはポリペプチドは、BAVまたはBAV感染細胞から回収されるタンパク質またはポリペプチドをいう。従って、用語「ネイティブBAVポリペプチド」は、天然に存在するBAVタンパク質およびそのフラグメントを含む。「非ネイティブ」ポリペプチドは、組換えDNA法によるかまたは直接合成により産生されたポリペプチドをいう。「組換え」ポリペプチドは、組換えDNA技法により産生された、即ち、所望のポリペプチドをコードする外因性DNA構築物により形質転換された細胞から産生されるポリペプチドをいう。
【0060】
「実質的に純粋」なタンパク質は、他のタンパク質がなく、好ましくは少なくとも10%均一、より好ましくは60%均一、そして最も好ましくは95%均一である。
【0061】
「抗原」は、宿主の免疫系を刺激し体液性および/または細胞性抗原特異的応答を刺激する1つまたはそれ以上のエピトープを含む分子をいう。この用語はまた、「免疫原」と交換可能に用いられる。
【0062】
「ハプテン」は、キャリアに連結されなければ体液性または細胞性応答をつくる宿主の免疫系を刺激しない1つまたはそれ以上のエピトープを含む分子である。
【0063】
用語「エピトープ」は、特異的抗体分子が結合するか、またはT細胞により認識される抗原またはハプテン上の部位をいう。この用語はまた、「抗原性決定基」または「抗原性決定部位」と交換可能に用いられる。
【0064】
組成物またはワクチンに対する「免疫学的応答」は、宿主における、目的の組成物またはワクチンに対する細胞性および/または抗体媒介免疫応答の発生である。通常、このような応答は、目的の組成物またはワクチンに含められる抗原に特異的に惹起された、応答を生成する抗体、B細胞、ヘルパーT細胞、サプレッサーT細胞、および/または細胞傷害性T細胞から構成される。
【0065】
用語「免疫原性ポリペプチド」および「免疫原性アミノ酸配列」は、ポリペプチドまたはアミノ酸のそれぞれをいい、これは、ウイルス感染性を中和する抗体を惹起し、および/または免疫化宿主の保護を提供するために抗体-補体または抗体-依存性細胞傷害性を媒介する。本明細書で用いられる「免疫原性ポリペプチド」は、所望のタンパク質の完全長(またはほぼ完全長)配列またはその免疫原性フラグメントを含む。
【0066】
「免疫原性フラグメント」は、1つまたはそれ以上のエピトープを含み、そしてそれ故ウイルス感染性を中和する抗体を惹起するか、および/または、抗体-補体または抗体−依存性細胞傷害性を媒介し、免疫化宿主の保護を提供するポリペプチドのフクラグメントを意味する。このようなフラグメントは、通常、長さが少なくとも約5アミノ酸、そして好ましくは長さが少なくとも約10〜15アミノ酸である。フラグメントの長さには臨界の上限はなく、それはタンパク質配列のほぼ完全長、または2つまたはそれ以上の抗原のフラグメントを含む融合タンパク質さえ含み得る。本明細書で用いる用語「処置」は、ウシまたはヒトまたはその他の哺乳動物のような哺乳動物の処置をいい、(i)感染または再感染の防止(予防)、または(ii)感染の症状の軽減または除去のいずれかをいう。ワクチンは、組換えBAVにより産生される組換えBAV自身または組換え抗原を含む。
【0067】
「感染性」は、ウイルスゲノムを細胞中に送達する能力を有することを意味する。
【0068】
B.一般的方法
本発明は、BAV3ゲノムの完全ヌクレオチド配列を開示する。図1を参照のこと。mRNAの転写マッピングおよびcDNAクローンの配列決定に由来する、BAV3ゲノムの転写マップを図2に示す。BAV3ゲノムのサイズ(34,446bp)および全体構成は、HAVのそれと類似しているようであるが、ある種の差異はある。Reddyら(1998)前述。BAV3ゲノムの特質を示す特徴の1つは、E3コード領域の比較的小さなサイズ(1517bp)である。Mittalら(1992)J.Gen.Virol.73:3295−3300;Mittalら(1993)J.Gen.Virol.74:2825;およびReddyら(1998)前述。BAV3 E3領域およびそのRNA転写物の配列の分析は、BAV3 E3が少なくとも4つのタンパク質をコードし得、その1つ(121R)は、HAV5の14.7kDaタンパク質と限られた相同性を示すことを示唆する。Idamakanti(1988)「ウシアデノウイルス3のE3領域の分子特徴付け」M.Sc.学位論文、University of Saskatchewan、Saskatoon、Saskatchewan。
【0069】
1つの実施態様では、本発明は、ウシアデノウイルスE1および/またはE3領域のヌクレオチド配列の一部分またはすべてを欠失する手段を同定および提供し、外来遺伝子またはそのフラグメントをコードする異種または同種ヌクレオチド配列を挿入し、ウシアデノウイルス組換え体を生成し得る部位を提供する。ヌクレオチド配列の「一部分を欠失すること」は、E1および/またはE3領域の一部分のヌクレオチド配列を欠失するために従来の遺伝子工学技法を用いることを意味する。
【0070】
別の実施態様では、本発明は、E3欠失組換え体BAV3(異種配列の挿入ありまたはなし)を、高い効率で、構築、単離、および増殖するための組成物および方法を提供する。これらは、適切な細胞株(例えば、アデノウイルスE1機能を発現するMDBK細胞または等価な細胞株)における組換え体ウイルスの単離、ならびに組換え体BAVゲノムが原核生物細胞における相同組換えを介して構築され、それにより得られる組換え体ゲノムが適切な細胞株(例えば、初代ウシ胎仔血清網膜細胞またはその等価物)にトランスフェクトされ、そして組換え体ウイルスがトランスフェクト細胞から単離される方法を含む。
【0071】
さらに、異なる形態のBHV-1糖タンパク質gDを発現する組換え体BAV3の構築が提供され、そしてgD発現組換え体BAVウイルスでのコトンラットの鼻腔内免疫がgD特異的な粘膜および全身の免疫応答の誘導をもたらすことが示される。実施例3を参照のこと。BHV-1 gD遺伝子を含むBAV組換え体でのウシ宿主の鼻腔内免疫は、腓におけるBHV-1チャレンジ(抗原投与)に対する防御を提供し、臨床的徴候の出現を減少させ、ウイルスのより迅速なクリアランスを促進し、そしてIgGおよびIgAの両方の力価の増加を提供する。実施例4を参照のこと。類似の様式で、哺乳動物病原体の防御決定因子をコードする任意の遺伝子は、E3欠失BAVに挿入され得、そして得られる組換え体BAVは、ワクチンとして使用され得る。
【0072】
本発明の1つの実施態様では、組換え体BAV発現カセットは、野生型BAVゲノムを1つ以上の適切な制限酵素で切断して、それぞれ、E1またはE3領域の配列を含むBAV制限フラグメントを生じることにより得られ得る。BAV制限フラグメントは、クローニングビヒクル(例えば、プラスミド)に挿入され得、そしてその後少なくとも1つの異種配列(これは、外来タンパク質をコードしてもよいしコードしなくともよい)は、作動可能に連結された真核生物転写調節配列を有するかまたは有さないE1またはE3領域に挿入され得る。組換え体発現カセットは、BAVゲノムと接触され得、そして相同組換えまたはタンパク質の従来の遺伝子操作方法を介して、所望の組換え体が得られる。発現カセットがE1領域または何らかの他の必須領域を含む場合、発現カセットとBAVゲノムとの間の組換えは、適切なヘルパー細胞株(例えば、E1形質転換細胞株)内で生じ得る。E1またはE3領域を含む制限フラグメント以外のBAVゲノムの制限フラグメントはまた、本発明の実施において有用であり、そして異種配列がBAV配列中に挿入され得るように、クローニングビヒクル中に挿入され得る。次いで、これらのDNA構築物は、インビトロまたはインビボで、適切な宿主細胞の形質転換またはトランスフェクションの前または後でBAVゲノムとの組換えを被り得る。
【0073】
適切な宿主細胞は、BAVゲノムと、BAV配列を含むプラスミドとの組換え、またはそれぞれがBAV配列を含む2つ以上のプラスミドの間の組換えを支持する任意の細胞を含む。組換えは一般に、原核生物細胞(例えば、E.coliなど)において行なわれるが、ウイルス粒子を生じる、ウイルスゲノムを含むプラスミドのトランスフェクションは、真核生物細胞、好ましくは哺乳動物細胞、より好ましくはウシ細胞培養物、最も好ましくはMDBKまたはPFBR細胞、およびそれらの等価物において実施される。細菌細胞培養物の増殖、ならびに真核生物細胞および哺乳動物細胞株の培養および維持は、当業者に周知である手順である。
【0074】
1つ以上の異種配列は、BAVゲノムの1つ以上の領域に挿入されて組換えBAVベクターを生じ得る。これは、BAVゲノムの挿入能力および組換えBAVベクターが挿入された異種配列を発現する能力にのみ制限される。一般に、アデノウイルスゲノムは、ゲノム長の約5%の挿入物を受け入れ得、そしてウイルス粒子にパッケージングされ得るまmであり得る。挿入能力は、非必須領域の欠失および/または機能がヘルパー細胞株により提供される必須領域の欠失により増加し得る。
【0075】
1つの実施態様では、挿入は、BAVゲノムのうちの挿入が所望される領域を含むプラスミドを構築することにより達成され得る。次いで、プラスミドが、プラスミドのBAVR部分における認識配列を有する制限酵素を用いて消化され、そして異種配列は制限消化の部位に挿入される。挿入された異種配列とともにBAVゲノムの一部を含むプラスミドは、BAVゲノムまたはBAVゲノムを含む線状化プラスミドとともに細菌細胞(例えば、E.coliなど)に同時形質転換される。ここで、BAVゲノムは、全長ゲノムであり得るか、または1つ以上の欠失を含み得る。プラスミド間の相同組換えは、挿入された異種配列を含む組換え体BAVゲノムを生じる。
【0076】
異種配列の挿入部位を提供するため、または異なる部位での挿入のためのさらなる能力を提供するためのBAV配列の欠失は、当業者に周知の方法により達成され得る。例えば、プラスミド中にクローニングされたBAV配列については、1つ以上の制限酵素(BAV挿入物中に少なくとも1つの認識配列を有する)での消化、およびその後の連結は、いくつかの場合、制限酵素認識部位の間の配列の欠失をもたらす。あるいは、BAV挿入物中の単一制限酵素認識部位での消化、続いてエキソヌクレアーゼ処置、続いて連結は、制限部位に近接したBAV配列の欠失をもたらす。上記の通りに構築した、1つ以上の欠失を有するBAVゲノムの1つ以上の部分を含むプラスミドは、BAVゲノム(全長もしくは欠失型)、または全長もしくは欠失型のいずれかのBAVゲノムを含むプラスミドとともに細菌細胞中に同時トランスフェクトされて、相同組換えにより、1つ以上の特異的部位に欠失を有する組換えBAVゲノムを含むプラスミドを生じ得る。次いで、欠失を含むBAVビリオンは、組換え体BAVゲノムを含むプラスミドを用いる哺乳動物細胞(MDBK細胞またはPFBR細胞およびそれらの等価物を含むがこれらに限定されない)のトランスフェクションにより得られ得る。
【0077】
本発明の1つの実施態様では、挿入部位は、BAVプロモーターに近接し、そして(転写の意味で)その下流にある。BAVプロモーターの位置、およびBAVプロモーターの下流の制限酵素認識配列(挿入部位として使用するため)は、当業者によって、本明細書中に提供されるBAVヌクレオチド配列から容易に決定され得る。
【0078】
あるいは、種々のインビトロ技術は、特定の部位での制限酵素認識配列の挿入のために、または制限酵素認識配列を含まない部位での異種配列の挿入のために用いられ得る。このような方法は、1つ以上の制限酵素認識配列の挿入のためのオリゴヌクレオチド媒介ヘテロ二重鎖形成(例えば、Zollerら(1982)Nucleic Acids Res.10:6487-6500;Brennanら(1990)Roux's Arch.Dev.Biol.199:89-96;およびKunkelら(1987)Meth.Enzymology 154:367-382を参照のこと)、およびより長い配列の挿入のためのPCR媒介方法を含むがこれらに限定されない。例えば、Zhengら(1994)Virus Research 31:163-186を参照のこと。
【0079】
異種配列が真核生物細胞中に活性である転写調節配列をさらに含むならば、BAVプロモーターの下流ではない部位で挿入された異種配列の発現を得ることもまた可能である。このような転写調節配列は、細胞プロモーター(例えば、ウシhsp70プロモーターなど)およびウイルス性プロモーター(例えば、ヘルペスウイルスプロモーター、アデノウイルスプロモーター、およびパポーバウイルスプロモーターなど)、ならびにレトロウイルス長末端反復(LTR)配列のDNAコピーを含み得る。
【0080】
別の実施態様では、原核生物細胞における相同組換えは、クローニングされたBAVゲノムを生じるために用いられ得;そしてクローニングされたBAVゲノムは、プラスミドとして増殖され得る。感染性ウイルスは、プラスミド含有細胞からレスキューされた、クローニングされたBAVゲノムでの哺乳動物細胞のトランスフェクションにより入手され得る。
【0081】
本発明はまた、異種遺伝子の発現を調節するために使用され得るBAV調節配列を提供する。調節配列は、例えば、転写調節配列、プロモーター、エンハンサー、上流調節ドメイン、スプライシングシグナル、ポリアデニル化シグナル、転写終結配列、翻訳調節配列、リボソーム結合部位、および翻訳終結配列であり得る。
【0082】
別の実施態様では、本発明は、外来遺伝子またはそのフラグメントをコードする異種または同種のヌクレオチド配列を挿入してBAV組換え体を生じるのに適切なBAVゲノム(およびそのフラグメント)のさらなる領域を同定および提供する。これらの領域は、ヌクレオチド4,092〜5,234;ヌクレオチド5,892〜17,735;ヌクレオチド21,198〜26,033;ならびにヌクレオチド31,133からBAVゲノムの右端にまたがり、そしてE2領域、E4領域、後期領域、33kD、52kD、100kD、DBP、pol、pTP、およびペントン遺伝子、ならびに遺伝子IIIA、pV、pVI、pVII、pVIII、およびpXを含む領域を含む。BAVゲノムのこれらの領域は、とりわけ、外来配列の挿入、DNA調節配列(転写調節配列および翻訳調節配列を含む)の提供、または生物学的サンプル中のウイルス核酸またはこれらの領域によりコードされるタンパク質の存在を検出する診断目的に用いられ得る。
【0083】
別の実施態様では、クローニングされるBAV-3ゲノムは、プラスミドとして増殖され得、そして感染性ウイルスは、プラスミド含有細胞からレスキューされ得る。
【0084】
ウイルス核酸の存在は、ハイブリダイゼーションアッセイ、ポリメラーゼ連鎖反応、および他の型の増幅反応を含むがこれらに限定されない、当業者に公知の技術により検出され得る。同様に、タンパク質の検出方法は、当業者に周知であり、そして種々の型の免疫圧制、ELISA、ウェスタンブロッティング、酵素アッセイ、免疫組織化学などを含むがこれらに限定されない。本発明のヌクレオチド配列を含む診断キットはまた、細胞破壊および核酸精製のための試薬、ならびにハイブリッドの形成、選択、および検出のための緩衝液および溶媒を含む。本発明のポリペプチドまたはアミノ酸配列を含む診断キットはまた、タンパク質単離ならびに免疫複合体の形成、単離、精製、および/または検出のための試薬を含み得る。
【0085】
種々の外来遺伝子またはヌクレオチド配列またはコード配列(原核生物および真核生物)は、上記のアデノウイルスヌクレオチド配列(例えば、DNA)中に、本発明に従って、広範囲な疾患に対する防御を提供するために挿入され得、そしてこのような多くの遺伝子は既に当該分野において公知である。これまでの問題は、遺伝子または配列についての安全な、便利な、および有効なワクチンベクター、ならびに種々の遺伝子治療適用において用いられる遺伝子移入のために有効な手段を提供することであった。
【0086】
外因性(すなわち、外来)ヌクレオチド配列は、1つ以上の目的の遺伝子から、そして好ましくは1つ以上の目的の治療遺伝子からなり得る。本発明の状況では、目的の遺伝子は、アンチセンスRNA、リボザイム、または目的のタンパク質に翻訳されるmRNAをコードし得る。目的の遺伝子は、ゲノム方の、相補的なDNA(cDNA)型の、または混合型のもの(少なくとも1つのイントロンが欠失されたミニ遺伝子)であり得る。これは、成熟タンパク質、成熟タンパク質の前駆体(特に分泌されることが意図される、従ってシグナルペプチドを含む前駆体)、多様な起源の配列の融合物に由来するキメラタンパク質、または改善もしくは修飾された生物学的特性を示す、天然タンパク質の変異体をコードし得る。このような変異体は、天然タンパク質をコードする遺伝子の1つ以上のヌクレオチドの欠失、置換、および/もしくは付加、または天然タンパク質をコードする配列の他の任意の型の配列変化(例えば、トランジションもしくはインバージョン)により得られ得る。
【0087】
目的の遺伝子は、宿主細胞におけるその発現に適切なエレメント(DNA調節配列)の制御下に置かれ得る。適切なDNA調節配列は、RNAへの遺伝子の転写(アンチセンスRNAもしくはmRNA)またはタンパク質へのmRNAの翻訳に必要なエレメントのセットを意味すると理解される。転写に必要とされるエレメントの中では、当然、プロモーターは特別に重要であると考えられる。プロモーターは、構成性プロモーターまたは調節性プロモーターであり得、そして真核生物、原核生物、またはウイルス起源、そしてさらにアデノウイルス起源の任意の遺伝子から単離され得る。あるいは、プロモーターは、目的の遺伝子の天然のプロモーターであり得る。概して、本発明で用いられるプロモーターは、調節配列を含むように改変され得る。例えば、本発明において使用される目的の遺伝子は、宿主のリンパ球細胞への遺伝子の移入を標的化することが所望される場合、免疫グロブリン遺伝子のプロモーターの制御下に置かれる。例えば、多数の細胞型における発現を可能にする、HSV-1 TK(ヘルペスウイルスタイプ1チミジンキナーゼ)遺伝子プロモーター、 (特に、ヒトアデノウイルスタイプ2の)アデノウイルスMLP(主要後期プロモーター)、RSV(ラウス肉腫ウイルス)LTR(長末端反復)、CMV(サイトメガロウイルス)初期プロモーター、およびPGK(ホスホグリセリン酸キナーゼ)遺伝子プロモーターもまた挙げられ得る。
【0088】
あるいは、特定の細胞型への組換え体BAVベクターの標的化は、組換え体ヘキソンおよび/または繊維遺伝子を構築することにより達成され得る。これらの遺伝子のタンパク質産物は、宿主細胞の認識に関与する;それゆえ、遺伝子は、ウイルスが別の宿主細胞を認識するのを可能にするペプチド配列を含むように改変され得る。
【0089】
本発明の状況で使用可能である目的の遺伝子の中では、以下のものが挙げられる:
【0090】
−サイトカイン(例えば、インターフェロンおよびインターロイキン)をコードする遺伝子;
【0091】
−リンホカインをコードする遺伝子;
【0092】
−病原性生物(例えば、ウイルス、細菌、または寄生生物)、好ましくはHIVウイルス(ヒト免疫不全ウイルス)により認識されるレセプターのような膜レセプターをコードする遺伝子;
【0093】
−凝固因子(例えば、第VIII因子および第IX遺伝子)をコードする遺伝子;
【0094】
−ジストロフィンをコードする遺伝子;
【0095】
−インスリンをコードする遺伝子;
【0096】
−細胞イオンチャネルに直接的または間接的に関与するタンパク質(例えば、CFTR(嚢胞性線維症膜貫通調節タンパク質)タンパク質)をコードする遺伝子;
【0097】
−アンチセンスRNA、病原性生物のゲノムに存在する病原性遺伝子により生成されるタンパク質の活性を阻害し得るタンパク質、または発現が統制解除された細胞遺伝子(例えば、オンコジーン)の活性を阻害し得るタンパク質(もしくはそれらをコードする遺伝子)をコードする遺伝子;
【0098】
−例えば、酵素活性を阻害するタンパク質(例えば、α1-アンチトリプシンまたはウイルスプロテアーゼインヒビター)をコードする遺伝子;
【0099】
−病原性タンパク質の生物学的機能を損なうように変異された病原性タンパク質の改変体(例えば、標的配列への結合について天然タンパク質と競合し得、それによりHIVの活性化を防止し得る、HIVウイルスのtatタンパク質のトランスドミナント改変体など)をコードする遺伝子;
【0100】
−宿主細胞の免疫を増加させるための抗原性エピトープをコードする遺伝子;
【0101】
−主要組織適合遺伝子複合体クラスIおよびIIタンパク質をコードする遺伝子、ならびにこれらの遺伝子のインデューサーであるタンパク質をコードする遺伝子;
【0102】
−抗体をコードする遺伝子;
【0103】
−イムノトキシンをコードする遺伝子;
【0104】
−トキシンをコードする遺伝子;
【0105】
−増殖因子または増殖ホルモンをコードする遺伝子;
【0106】
−細胞レセプターおよびそれらのリガンドをコードする遺伝子;
【0107】
−腫瘍サプレッサーをコードする遺伝子;
【0108】
−オンコジーンを含むがこれらに限定されない、心臓血管病に関与する遺伝子;線維芽細胞増殖因子(FGF)、血管内皮増殖因子(VEGF)、および神経成長因子(NGF)を含むがこれらに限定されない、増殖因子をコードする遺伝子;Rb(網膜芽細胞種)遺伝子を含むがこれらに限定されない、e-nos腫瘍サプレッサー遺伝子;リポタンパク質リパーゼ;スーパーオキシドジスムターゼ(SOD);カタラーゼ;酸素および遊離基スカベンジャー;アポリポタンパタ質;ならびにpai-1(プラスミノゲンアクチベーターインヒビター1);
【0109】
−細胞酵素または病原性生物により生成される細胞酵素をコードする遺伝子;
ならびに
【0110】
−自殺遺伝子。HSV-1 TKは自殺遺伝子が、例として挙げられ得る。
このウイルスTK酵素は、特定のヌクレオシドアナログ(例えば、アシクロビルまたはガンシクロビル)に対して、細胞性TK酵素と比較して著しく高い親和性を示す。このウイルスTK酵素は、これらを一リン酸化分子に変換し、これはそれ自体が細胞酵素によりヌクレオチド前駆体に変換され得る。これらのヌクレオチドアナログは、複製中のDNA分子に取りこまれ得、それゆえ、取りこみは主に分裂細胞のDNAに生じる。この取りこみは、分裂細胞(例えば、ガン細胞)の特異的破壊をもたらし得る。
【0111】
このリストは限定的ではなく、他の目的の遺伝子が本発明の状況において用いられ得る。
【0112】
野生型生物に見出されるとおりの完全な配列ではなく、遺伝子のヌクレオチド配列のフラグメントのみが用いられ得る(ここで、これらは、防御免疫応答または特異的生物学的効果を生じるのに十分である)こともまた可能である。利用可能な場合には、合成遺伝子またはそのフラグメントもまた用いられ得る。しかし、本発明は、広範な種々の遺伝子、フラグメントなどに用いられ得、そして上記に記載したものに限定されない。
【0113】
いくつかの場合には、特定の抗原の遺伝子は、多数のイントロンを含み得るか、またはRNAウイルス由来であり得、これらの場合、相補的なDNAコピー(cDNA)が用いられ得る。
【0114】
遺伝子の首尾良い発現を生じるために、遺伝子は、エンハンサ一エレメントおよびポリアデニル化配列を含む適切なプロモーターとともに発現ベクターに挿入され得る。哺乳動物細胞における外来遺伝子の首尾良い発現を提供する多数の真核生物性のプロモーターおよびポリアデニル化配列、ならびに発現カセットを構築する方法が、当該分野で(例えば、米国特許第5,151,267号(この開示は本明細書中で参考として援用される)において)公知である。プロモーターは、体液性、細胞媒介性、および粘膜性の免疫応答を満足にもたらす免疫原性タンパク質の最適な発現を与えるように既知の基準に従って選択される。
【0115】
組換え体ウイルス感染細胞におけるインビボでの発現により生成される外来タンパク質は、それ自体が免疫原性であり得る。1つより多くの外来遺伝子が、ウイルスゲノムに挿入されて、1つより多くの有効なタンパク質の首尾良い生成が得られ得る。
【0116】
従って、本発明の組換え体ウイルスを用いて、ウシ、ヒト、および他の哺乳動物に罹患する広範な種々の疾患に対する防御を提供することが可能である。本発明の任意の組換え体抗原性決定基または組換え体生ウイルスは、抗原性決定基ワクチンまたは生ワクチンベクターについて記載されたのと実質的に同じ様式で処方および使用され得る。
【0117】
本発明はまた、本発明の方法に従って調製された、治療有効量の組換え体ベクター、組換え体ウイルス、または組換え体タンパク質を、薬学的に受容可能なビヒクルおよび/またはアジュバントと組合せて含む薬学的組成物を含む。このような薬学的組成物は、当該分野で周知である技術に従って調製され得、そして投薬量は当該分野で周知である技術に従って決定され得る。本発明の薬学的組成物は、全身的(例えば、静脈内、気管内、脈管内、肺内、腹腔内、鼻腔内、非経口、腸、筋肉内、皮下、肺瘍内、もしくは頭蓋内)経路を含むがこれらに限定されない、公知の任意の投与経路により、またはエーロゾル適用もしくは肺内点滴注入により投与され得る。投与は、単回用量で、または特定の時間間隔の後に1回以上反復される用量で起こり得る。適切な投与経路および投薬量は、状況(例えば、処置される個体、処置される障害、または目的の遺伝子もしくはポリペプチド)によって変化するが、当業者により決定され得る。
【0118】
本発明はまた、本発明の治療有効量のBAVベクター、組換え体BAV、または宿主細胞が、処置を必要とする哺乳動物非験体に投与される処置方法を包含する。
【0119】
本発明で用いられる抗原は、ネイティブまたは組換え体のいずれかの抗原性ポリペプチドまたはフラグメントであり得る。これらは、部分配列、全長配列、または融合物(例えば、組換え体宿主に適切なリーダー配列を有するか、または別の病原体についてのさらなる抗原配列を有する)でさえあり得る。本発明のウイルス系により発現される好ましい抗原性ポリペプチドは、抗原をコードする全長(またはほぼ全長)の配列を含む。あるいは、抗原性である(すなわち、1つ以上のエピトープをコードする)、より短い配列が用いられ得る。より短い配列は、インビトロアッセイにおいてウイルス感染性を中和するエピトープと定義される「中和エピトープ」をコードし得る。好ましくは、ペプチドは、宿主において「防御免疫応答」(すなわち、免疫された宿主を感染から防御する、抗体媒介性および/または細胞媒介性の免疫応答)を生じ得る、「防御エピトープ」をコードすべきである。
【0120】
本発明で用いられる抗原(特に、短いオリゴペプチドから構成される場合)は、ワクチンキャリアに結合体化され得る。ワクチンキャリアは、当該分野で周知である;例えば、ウシ血清アルブミン(BSA)、ヒト血清アルブミン(HSA)、およびキーホールリンペットヘモシアニン(KLH)。好ましいキャリアタンパク質であるロタウイルスVP6は、EPO公報第0259149号(この開示は本明細書中に参考として援用される)に開示される。
【0121】
挿入され得る所望の抗原の遺伝子またはそのコード配列は、哺乳動物において疾患を生じる生物(特に、ウシロタウイルス、ウシコロナウイルス、ウシヘルペスウイルスタイプ1、ウシRSウイルス、ウシパラインフルエンザウイルスタイプ3(BPI-3)、ウシ下痢ウイルス、Pasteurella haemolytica、Haemophilus somnusなどのようなウシ病原体)のものを含む。外来遺伝子またはフラグメントを有する本発明のワクチンはまた、例えば、腸溶性投薬形態のような適切な経口キャリアにおいて経口投与され得る。経口処方物は、薬学的グレードのマンニトール、ラクトース、スターチ、ステアリン酸マグネシウム、サッカリンナトリウムセルロース、炭酸マグネシウムなどのような通常用いられる賦形剤を含む。経口ワクチン組成物は、約10%〜約95%、好ましくは約25%〜約70%の活性成分を含む、溶液、懸濁液、錠剤、丸剤、カプセル剤、持続放出処方物、または散剤の形態を取り得る。経口および/または鼻腔内ワクチン接種は、全身免疫との組合せで、粘膜免疫(これは、気道および胃腸管に感染する病原体に対する防御において重要な役割を果たす)を惹起するために好ましくあり得る。
【0122】
さらに、ワクチンは、坐剤中に処方され得る。坐剤について、ワクチン組成物は、伝統的なバインダーおよびキャリア(例えば、ポリアルカリグリコールまたはトリグリセリド)を含む。このような坐剤は、約0.5%(w/w)〜約10%(w/w)、好ましくは約1%(w/w)〜約2%(w/w)の範囲で活性成分を含有する混合物から形成され得る。
【0123】
動物に本発明のワクチン組成物を投与するためのプロトコルは、本開示を考慮すれば当該分野の技術範囲内である。当業者は、抗原性フラグメントに対する抗体および/またはT細胞媒介性免疫応答を惹起するのに有効な用量のワクチン組成物の濃度を選択する。広範な制限内では、投薬量は重要であるとは考えられない。代表的には、ワクチン組成物は、約1マイクログラム〜約1,000マイクログラムのサブユニット抗原を便利な容量(例えば、約1〜10cc)のビヒクル中で送達する様式で投与される。好ましくは、単回免疫における投薬量は、約1マイクログラム〜約500マイクログラム、より好ましくは約5〜10マイクログラムから約100〜200マイクログラム(例えば、5〜200マイクログラム)のサブユニット抗原を送達する。
【0124】
投与時期はまた、重要であり得る。例えば、一次接種は、好ましくは、必要に応じて、その後のブースター接種が続き得る。必要に応じてであるが、2回目であるブースター免疫を、初期免疫の数週間後から数ヶ月後に動物に投与することが好ましくあり得る。疾患に対する持続した高レベルの防御を保証するために、ブースター免疫を動物に対して定期的な間隔(例えば、数年毎に一度)で、再投与することが有益であり得る。あるいは、最初の用量は、経口的に、続いて後の接種により、またはその逆で投与され得る。好ましいワクチン接種プロトコルは、日常的なワクチン接種プロトコル実験を介して達成され得る。
【0125】
インビボでの組換え体ウイルスワクチンの全ての投与経路についての投薬量は、患者の大きさ、防御が必要とされる感染の性質、キャリアなどを含む種々の因子に依存し、そして当業者により容易に決定され得る。非限定的な例により、103pfuおよび1015pfu、好ましくは105pfu〜1013pfu、より好ましくは106pfu〜1011pfuなどの投薬量が用いられ得る。インビトロでのサブユニットワクチンについてのように、さらなる投薬量は、関与する臨床的因子により決定された通りに与えられ得る。
【0126】
本発明の1つの実施態様では、多数の組換え体細胞株が、BAV E1領域を含む発現カセットを構築し、そしてこれにより宿主細胞を形質転換して、E1タンパク質を発現する補完細胞株または培養物を提供することにより、本発明に従って生成される。これらの組換え体補完細胞株は、E1配列が欠失した組換え体欠損BAVが複製し、そして必要に応じて組換え体BAV内にコードされる所望の外来遺伝子またはそのフラグメントを発現するのを可能にし得る。これらの細胞株はまた、外来遺伝子またはフラグメントをコードする異種ヌクレオチド配列により置換されたE3遺伝子欠失を有する組換え体BAVを、DNA媒介コトランスフェクション後のインビボ組換えにより生成するのに非常に有用である。より一般的には、BAVゲノムによりコードされる1つ以上の必須機能を欠く組換え体欠損BAVベクターは、適切な補完細胞株中で増殖され得、ここで、特定の補完細胞株は、特定の組換え体欠損BAVベクターにおいて欠けている機能を提供する。補完細胞株は、例えば、ヘルパーウイルスとともに同時感染することにより、または特定のウイルス機能をコードするウイルスゲノムの安定な形態のフラグメントを組込むかさもなければ維持することにより、ウイルス機能を提供し得る。
【0127】
本発明の1つの実施態様では、組換え体発現カセットは、BAVゲノムを適切な制限酵素で切断して、それぞれ、E1もしくはE3遺伝子領域配列を含む、ゲノムの左側末端または右側末端を提示するDNAフラグメントを生成し、そして左または右の末端フラグメントをクローニングビヒクル(例えば、プラスミド)に挿入し、その後少なくとも1つの異種DNA配列を外因性プロモーターの制御ありまたはなしでE1またはE3の欠失に挿入することにより得られ得る。組換え発現カセットは、適切な細胞内でBAVゲノムと接触され、そして相同組換えまたはタンパク質の従来の遺伝子操作法を介して組換え体BAVゲノムが得られる。適切な細胞は、原核生物細胞(例えば、E.coliなど)および真核生物細胞の両方を含む。適切な真核生物細胞の例は、MDBK細胞、アデノウイルスE1機能を発現するMDBK細胞、初代ウシ胎仔網膜細胞、および先に記載した細胞の機能と等価である機能を発現する細胞を含むがこれらに限定されない。E1またはE3領域を含む制限フラグメント以外のBAVゲノムの制限フラグメントもまた、本発明の実施において有用であり、そして異種配列が非E1および非E3 BAV配列に挿入され得るようにクローニングビヒクルに挿入され得る。次いで、これらのDNA構築物は、インビトロまたはインビボで、BAVゲノムとの組換えを、上記の適切な宿主細胞の形質転換またはトランスフェクションの前または後に受け得る。本発明の目的のために、得られる組換え体BAVゲノムが、複製およびパッケージングに要求されるBAV配列を含む限りは、BAVゲノムは、全長ゲノム、または組換わるフラグメントで欠失している領域以外の領域に欠失を含むゲノムのいずれかであり得る。トランスフェクションの方法、細胞培養、ならびに原核生物および真核生物の細胞における組換え(例えば、上記に記載されるもの)は、当業者に周知である。
【0128】
別の実施態様では、E1機能(または任意の特定のウイルスベクターにおいて変異もしくは欠失され得る任意の他のウイルス領域の機能)は、ベクターが欠く機能を発現するウイルスでの細胞の同時感染により供給(補完細胞株を提供)され得る。
【0129】
本発明はまた、ウシアデノウイルス発現系を含む発現系を含み、個々で異種ヌクレオチド配列(例えば、DNA)は、E3領域の一部もしくは全部、E1領域の一部もしくは全部、E2領域の一部もしくは全部、E4領域の一部もしくは全部、E4とゲノムの右末端との間の領域の一部もしくは全部、後期領域(L1〜L7)の一部もしくは全部、および/または33kD、52kD、100kD、DBP、pol、pTP、およびペントン遺伝子、ならびに遺伝子IIIA、pV、pVI、pVII、pVIII、およびpXにより占められる領域の一部もしくは全部を置きかえる。発現系が用いられ得、ここで外来ヌクレオチド配列(例えば、DNA)は、他の任意の異種プロモーターのコントロールを有するかまたは有さない。BAV発現ベクターはまた、逆方向末端反復(ITR)配列およびパッケージング配列を含み得る。
【0130】
BAVの33kD、52kD、100kD、DBP、pTP、ペントン(III)、pIIIA、pIVa2、pV、pVI、pVII、pVIII、およびpX遺伝子は、ウイルス複製に必須である。それゆえ、これらの任意の遺伝子に欠失を含むが、またはこれらの任意の遺伝子によりコードされる機能を欠くBAVベクターは、適切な補完細胞株(すなわち、ヘルパー細胞株)中で増殖させねばならない。ヒトアデノウイルスでは、E4領域における特定のオープンリーディングフレーム(ORF3およびORF6/7)はウイルス複製に必須である。BAV-3のE4領域における類似のオープンリーディングフレームにおける欠失は、ウイルスベクターの増殖のためにヘルパー細胞株の使用を必要とし得る。
【0131】
本発明のアデノウイルスのBAV E1遺伝子産物は、大部分の細胞遺伝子をトランス活性化し、それゆえ、E1タンパク質を構成的に発現する細胞株は、通常の細胞株よりも高いレベルで細胞ポリペプチドを発現し得る。本発明の組換え体哺乳動物(特にウシ)細胞株は、ポリペプチドを調製および単離するために用いられ得る。ポリペプチドは、以下の通りのポリペプチドを含む:(a)アデノウイルスEIAタンパク質に関連するタンパク質(例えば、p300、網膜芽細胞(Rb)タンパク質、サイクリン、キナーゼなど);(b)アデノウイルスE1Bタンパク質に関連するタンパク質(例えば、p53など);(c)増殖因子(例えば、上皮増殖因子(EGF)、トランスフォーミング増殖因子(TGF)など);(d)レセプター(例えば、上皮増殖因子レセプター(EGF-R)、線維芽細胞増殖因子レセプター(FGF-R)、腫瘍壊死因子レセプター(TNF-R)、インスリン様増殖因子レセプター(IGF-R)、主要組織適合遺伝子複合体クラスIレセプターなど);(e)プロトオンコジーンによりコードされるタンパク質(例えば、プロテインキナーゼ(チロシン特異的タンパク質キナーゼおよびセリンもしくはトレオニンに特異的なタンパク質キナーゼ)、p21タンパク質(GTPase活性を有するグアニンヌクレオチド結合タンパク質)など);(f)他の細胞タンパク質(例えば、アクチン、コラーゲン、フィブロネクチン、インテグリン、ホスホプロテイン、プロテオグリカン、ヒストンなど);ならびに(g)転写調節に関与するタンパタ質(例えば、TATAボックス結合タンパク質(TBP)、TBP関連因子(TAF)、Sp1結合タンパク質など)。
【0132】
本発明はまた、遺伝子治療を哺乳動物(例えば、ウシもしくはヒトまたはその必要性がある哺乳動物)に提供して、遺伝子欠損を制御し、治療遺伝子もしくはヌクレオチド配列を提供し、そして/または遺伝子変異を誘導もしくは訂正するための方法を含む。この方法は、例えば、遺伝性疾患、感染性疾患、心臓血管疾患、およびウイルス感染を含むがこれらに限定されない状態の処置において用いられ得る。この方法は、上記哺乳動物に、非欠損形態の上記遺伝子を、組換え体ウイルスベクターのゲノムが上記哺乳動物ゲノムへと取り込まれるか、または独立してかつ染色体外で維持される条件下で外来ヌクレオチド配列を含む生組換え体ウシアデノウイルスを投与して、標的器官または組織において要求される遺伝子の発現を提供する工程を含む。これらの種類の技術は、現在、種々の疾患状態の処置(この非限定的な例は上記に提供される)のために当業者によって用いられている。従来の遺伝子治療における使用のために取りこまれ得る外来遺伝子、ヌクレオチド配列、またはその部分の例として、嚢胞性線維症膜貫通調節タンパク質遺伝子、ヒトミニジストロフィン遺伝子、α-1アンチトリプシン遺伝子、心臓血管疾患において関与し得る。
【0133】
特に、遺伝性疾患(例えば、血友病、サラセミア、気腫、ゴシェ病、嚢胞性線維症、デュシェーヌ筋ジストロフィー、デュシェーヌまたはベッカーのミオパシーなど)、ガン、ウイルス性疾患(例えば、AIDS、ヘルペスウイルス感染、サイトメガロウイルス感染、およびパピローマウイルス感染)、心臓血管病などを含むがこれらに限定されない、ヒトにおける遺伝子治療に関する本発明の実施は、疾患の防御または処置が意図される。本発明の目的のためには、本発明の方法により調製されたベクター、細胞、およびウイルス粒子は、被験体にエクスビボ(すなわち、細胞中または患者から取り出した細胞)で、または処置される身体にインビボで直接のいずれかで、導入され得る。好ましくは、宿主細胞はヒト細胞であり、そして最も好ましくは、肺、線維芽細胞、筋肉、肝臓、またはリンパ球または造血直系の細胞である。
【実施例】
【0134】
実施例
本発明の実施例を以下記載する。これらの実施例は、例示の目的にのみ提供され、そしていかなる方法においても本発明の範囲を限定することを意図しない。本開示に照らして、請求の範囲内の多数の実施態様は、当業者に明白である。発明の詳細な説明において引用された参考文献の内容は、本明細書において参考として援用される。さらなる方法および技術は、共有に係る出願PCT/CA94/00678(WO95/16048として公開された)に見い出され得る。
【0135】
実施例1:ヌクレオチド4,092〜5,234、ヌクレオチド5,892〜17,735、ヌクレオチド21,198〜26,033、およびヌクレオチド31,133〜34,445のヌクレオチド配列の決定。
ウシアデノウイルスタイプ3ゲノムのヌクレオチド配列を完了するために、以下のBAV-3制限フラグメントを、細菌プラスミドにクローニングし、そしてそれらのヌクレオチド配列を当業者に公知の方法によって決定した。
【0136】
【0137】
実施例2:ヌクレオチド4,092〜5,234、ヌクレオチド5,892〜17,735、ヌクレオチド21,198〜26,033、およびヌクレオチド31,133〜34,445によって規定されるBAV-3ゲノムの領域における挿入。
挿入を、当該分野で認識される技術によって行う。これは、制限消化、ヌクレアーゼ消化、連結、キナーゼおよびホスファターゼ処理、DNAポリメラーゼ処理、逆転写酵素処理、およびオリゴヌクレオチド化学合成を含むがこれらに限定されない。目的の外来核酸配列を、その外来配列が、挿入が配向されるBAVゲノムの領域と実質的な相同性を有する配列と隣接するようにプラスミドベクターにクローニングする。次いで、これらの構築物を、BAV-3と同時感染されている宿主細胞へと導入する。感染の間、これらの構築物とBAVゲノムとの間の相同組換えが起こって組換えBAVベクターが生成される。挿入がBAVゲノムの必須の領域において生じる場合、組換えBAVベクターを、ヘルパー細胞株において増殖させる。これは、挿入に起因して消失したウイルス機能を供給する。ウイルスにとっては必須ではないE4領域における挿入については、ヘルパー細胞株におけるBAVベクターの増殖は必要ではない。
【0138】
実施例3:全長および短縮型形態のウシヘルペスウイルス1糖タンパク質gDを発現するE3欠失BAV3の構築および特徴付け
本実施例は、E.coliの相同組換え機構を用いた、BAV3のE3領域における1,245kbの欠失の構築を示す。第一に、1,245kb欠失を、プラスミドにクローニングされたウシアデノウイルス-3(BAV3)ゲノムDNAのE3領域に導入した。線状の制限酵素で切除したE3欠失BAV3ゲノムNDAのウシ仔胎網膜初代細胞へのトランスフェクションは、感染性ウイルス(BAV3.E3d)を産生した。これは、すべてのBAV E3特異的なオープンリーディングフレームがインビトロにおけるウイルス複製について必須ではないことを示唆する。同様のアプローチを用いて、ウシヘルペスウイルス-1(BHV-1)の全長(BAV3.E3gD)または短縮型(BAV3.E3gDt)の糖タンパク質Dを発現する複製適合BAV-3組換え体を構築した。BAV3.E3gDおよびBAV3.E3gDtによって発現された組換えgDおよびgDtタンパク質は、コンホメーション的エピトープに対して惹起されたgD特異的モノクローナル抗体によって認識された。このことは、組換えgDおよびgDtの抗原性が、BHV-1に感染した細胞において発現されたネイティブなgDの抗原性に類似していたことを示唆する。コットンラット(cotton rat)の鼻腔内免疫は、強力なgDおよびBAV3特異的IgAおよびIgG免疫応答を誘発した。これらの結果は、動物の粘膜表面に対するワクチン抗原の送達のための、複製適合ウシアデノウイルス-3ベースのベクターの使用を例示する。
【0139】
材料および方法
細胞およびウイルス。Madin Darbyウシ腎臓(MDBK)細胞およびウシ仔胎網膜初代(PFBR)細胞を、5%ウシ仔胎血清(FBS)を補充したEagle最小必須培地(MEM)において増殖させた。野生型(WBR-1株)および組換えBAV3ウイルスを、MDBK細胞において、以前に記載したとおりに増殖させた。Mittalら(1995)J.Gen.Virol.76:93-102.BHV-1のP8-2株を、増殖させ、そして記載のとおりに定量した。Rouseら(1974)J Immunol.113:1391-1398。
【0140】
動物。Veterinary Infectious Disease Organization(Saskatoon)において維持されたコットンラット(Sigmodon hispidus)の近交コロニーを本研究の動物の供給源として用いた。
【0141】
組換えプラスミドの構築
a)プラスミドpBAV302の構築。図3を参照のこと。T4ポリメラーゼ処理した587bpのフラグメント(これは、オリゴヌクレオチドE3C5'ACGCGTCGACTCCTCCTCA(配列番号2)およびE3C3'TTGACAGCTAGCTTGTTG(配列番号3)ならびにテンプレートとしてプラスミドpSM14(Mittalら(1995)前出)を用いるPCR増幅によって単離された)を、SacIIで消化し、そしてEco47III-SacII消化したプラスミドpSL301に連結して、プラスミドpE3Aを作製した。164bpのフラグメント(これは、オリゴヌクレオチドE3N5'CCAAGCTTGCATGCCTG(配列番号4)およびE3N3'GGCGATATCTCAGCTATAACCGCTC(配列番号5)ならびにプラスミドpSM14(Mittalら(1995)前出)を用いるPCR増幅によって単離された)を、SphIおよびEcoRVで消化し、次いでpE3Aの531bp EcoRV-HindIIIフラグメントおよびSphI-HindIII消化したプラスミドpSL301に連結して、プラスミドpE3Bを作製した。614bp SphI-SacIIフラグメントを、プラスミドpE3Bから単離し、そしてSphI-SacII消化pSM14に連結して、プラスミドpE3Cを作製した。プラスミドpE3Cを、EcoRVで消化し、そしてSrflリンカー(TTGCCCGGGCTT、配列番号6)に連結して、プラスミドpE3CIを作製した。pE3CIの1.755kbのBamHIフラグメントを単離し、そしてBamHI消化pSM17(Mittalら、(1995)前出)に連結して、プラスミドpE3Dを作製した。最後に、プラスミドpE3Dの8783bpのKpnI-XbaIフラグメントを単離し、そしてKpnI/XbaI消化プラスミドpTG5453(これは、全長BAV3ゲノムDNAを含む)に連結して、プラスミドpE3Eを作製した。プラスミドpE3Eは、BAV3ゲノムの末端フラグメント(0-19.7m.u.および76.6-100m.u.)を、E3領域に1,245kbの欠失およびベクター配列内に位置する特有のKpnI部位とともに含む。
【0142】
E3領域に1,245kbの欠失を有するBAV3ゲノムを含むプラスミド(pBAV302)を、E.coli BJ5183において、KpnI消化pE3Eと、脱タンパク質化したBAV3ゲノムDNAとの間での相同DNA組換えによって生成した。
【0143】
b)プラスミドpBAV302gDおよびpBAV302gDtの構築。E3領域における外来遺伝子を発現する組換えBAV3の生成のためのトランスファープラスミドを、pBAV302の8783bp KpnI-XbaIフラグメントを、KpnI/XbaI消化プラスミドpGEM3zf(-)に連結することによって構築して、プラスミドpBAV300を作製した。プラスミドpRSV1.3(Tikooら、(1993)J.Virol.67:2103-2109)の1.3kb BgIIIフラグメント(これは、全長BHV-1gD遺伝子を含む)を、T4 DNAポリメラーゼで処理し、そしてSrfl消化pBAV300に連結して、プラスミドpBAV300.gDを作製した。同様に、プラスミドpRSV1.3XN(Tikooら、前出)の1.3kb BgIIIフラグメント(これは、短縮型BHV-1gD遺伝子を含む)を、T4 DNAポリメラーゼで処理し、そしてSrfI消化pBAV300に連結して、プラスミドpBAV300.gDtを作製した。
【0144】
全長または短縮型gDタンパク質のいずれかをコードする遺伝子を含む組換えBAVゲノムを、E.coli BJ5183において、SrfI線状化pBAV302とpBAV300.gDの10kb KpnI/XbaIフラグメントとの間(これは、プラスミドpBAV302.gDを作製する)、またはSrfI線状化pBAV302とpBAV300.gDtの10kb KpnI/XbaIフラグメントとの間(これは、プラスミドpBAV302.gDtを作製する)の、相同DNA組換えによって生成した。
【0145】
組換えBAV3の構築。60mmディッシュ中のPFBR細胞単層を、10μgのPacI消化pBAV302、pBAV302.gDまたはpBAV302.gDtの組換えプラスミドDNAを用いて、リン酸カルシウム方法を用いて、トランスフェクトした。Grahamら(1973)Virology 52:456-467。37℃でのインキュベーションの15〜20日後、50%の細胞変性効果を示すトランスフェクトされた細胞を採取し、凍結融解を2回行い、そして組換えウイルスを、MDBK細胞においてプラーク精製した。Mittalら(1995)、前出。
【0146】
タンパク質の放射標識および免疫沈降。28cm2ウエル中の約70〜80%のコンフルエントのMDBK細胞単層に、1細胞当たり10PFUの組換えまたは野生型BAV3を感染させた。ウイルスを60分間吸収させた後、細胞を、5%FBSを含むMEM中でインキュベートした。感染後の種々の時点で、細胞を、メチオニンおよびシステインを含まないDulbeccoMEM中で60分間インキュベートし、その後、[35S]メチオニン−システイン(1ウェル当たり100μCi)で標識した。2〜12時間の標識の後、細胞または培地を採集した。タンパク質を、培地から免疫沈降し、そして細胞を、改変RIPA緩衝液で溶解し、その後記載のようにSDS-PAGEによってタンパク質を分析した。Tikoooら、(1993)前出。
【0147】
動物接種。合計25匹の4〜6週齢のコットンラット(性別はいずれか)を3つの群に分けた。ハロタンでの麻酔後、動物に、1日目と21日目の2回、107PFUの個々の組換えウイルスを含む種菌100μlを鼻腔内経路により接種した。血液サンプルを、初回の接種の0、21、および28日後に採取して、BHV-1 gDおよびBAV3に対する抗体応答の発達を、酵素結合免疫吸着アッセイ(ELISA)およびウイルス中和(VN)アッセイによって試験した。各群の4匹の動物を、初回接種21日目および28日目にハロタンの過剰投与により安楽死させた。肺および鼻の切片を別個に採取して、BHV-1 gD特異的粘膜IgGおよびIgA抗体応答およびBAV3特異的粘膜IgGおよびIgA抗体応答の発達をELISAでモニターした。Pappら(1997)J.Gen.Virol.78:2933-2943。さらに、肺を採取し、そしてBHV-1 gD特異的IgA抗体分泌細胞およびBAV3特異的IgA抗体分泌細胞の頻度を、酵素結合免疫スポット(ELISPOT)により決定した。Pappら、前出。
【0148】
肺リンパ球の調製。無菌的に取り出した肺組織を、小片に切断し、そして完全培地(10%FBS、2mM L-アルギニン、1mMピルビン酸ナトリウム、100μMの非必須アミノ酸、10mM HEPES緩衝剤、50μM 2-メルカプトエタノール、100U/mlペニシリンG、100μg/mlストレプトマイシン溶液、150U/mlコラゲナーゼAおよび50U/mlDNaseIを補充したMEM)中で1時間インキュベートした。次いで、組織を、プラスチックメッシュを通して押した。肺細胞懸濁物を、不連続なPercoll勾配を通して遠心分離し、そしてMEMで洗浄した。細胞を、完全培地に再懸濁し、そしてフラスコ中で1時間インキュベートして、接着細胞を付着させた。次いで、非接着性の細胞集団を、再懸濁し、そして以前に記載のように抗原特異的ELISPOTアッセイにおいて使用した。Pappら、前出。
【0149】
ELISA。血清、肺分泌物、および鼻分泌物中のBHV-1 gDおよびBAV3に対して特異的な抗体を、以前に記載のようにELISAによって決定した。Pappら、前出。手短には、96ウェルImmunol-2マイクロタイタープレートを、精製した短縮型gD(0.01μg/ウェル)またはBAV3(0.5μg/ウェル)のいずれかでコートし、そして異なる希釈の各サンプルとインキュベートした。抗原特異的なIgGを、ビオチン化ウサギ抗ラットIgGを用いて検出した。抗原特異的IgAを、ウサギ抗ラットIgAおよび西洋ワサビペルオキシダーゼ結合体化したヤギ抗ウサギIgGを用いて測定した。
【0150】
ウイルス中和。2倍連続希釈の熱非働化血清サンプルを、100PFUのBHV-1と、37℃で1時間インキュベートした。次いで、ウイルスサンプル混合物を、12ウェル組織培養プレート中のコンフルエントなMDBK細胞に配置し、そして2日間インキュベートした。力価を、コントロールと比較したプラーク数の50%減少を引き起こした最高の抗体希釈の逆数として表した。
【0151】
結果
E3欠失組換えBAV3の構築。当初、ウイルス複製におけるBAV3 E3の役割は、HAVのE3領域の役割と類似していると推定された。Woldら(1991)Virology 184:1-8。従って、BAV E3ベースのベクターを、E3配列の欠失を行うことによって構築した。しかし、種々のウシ細胞株(MDBKを含む)における、E3欠失したBAV3組換え体の単離の試みは成功しなかった。ルシフェラーゼ遺伝子を発現する、部分的に欠失したBAV3組換え体(BAV3-Luc)は、BAV3 E1形質転換したMDBK細胞を、トランスフェクションについて使用した場合にのみ単離され得た。Mittalら(1995)前出。しかし、一旦単離されると、BAV3-Lucは、正常ウシ細胞上で増殖され得た。このことは、BAV3のE3領域がインビトロでのウイルス複製については必須ではないことを示唆する。Mittalら(1995)前出。BAV3組換え体の単離の効率を上昇させるために、PBFR細胞を、BAV3組換え体を産生するための新規の手順と共に使用した。Degryse(1996)Gene 170:45-50。この方法を用いて、標的化された改変を、E.coliの高度に効率よい相同組換え機構を用いて、プラスミドが保有するウイルス配列へと導入し、そして感染性のビリオンを、プラスミドベクターから切り出された改変されたゲノムを適切な宿主細胞へとトランスフェクションした後に単離した。Chartierら(1996)J.Virol.70:4805-4610。
【0152】
E.coliの相同組換え機構を利用して、プラスミド(pBAV302)を構築した。これは、1,245kb欠失(ヌクレオチド26456〜27701)およびSrfI制限酵素部位を含んだ(図21)。PacI消化pBAV302 DNAは、PFBR細胞にトランスフェクトされる場合に、14日後に細胞変性効果をもたらした。ウイルスを、プラーク精製し、そしてMDBK細胞において増殖させ、そしてBAV3.E3dと命名した。ウイルスDNAを抽出し、そしてBamHI制限酵素で消化した後にアガロースゲル電気泳動によって分析した。野生型(図4、レーンb)と比較すると、組換えBAV3.E3dゲノムの3,019kb BamHI「D」フラグメントは、1,245kb小さかった(図4、レーンa)。このことは、E3が欠失された組換えBAV3が単離されたことを確認する。この組換えウイルスの増殖特徴と、野生型BAV3の増殖特徴との比較によっては、プラークの大きさまたは複製における有意な差異されなかった。なぜなら、E3欠失組換え体は、野生型BAV3と類似する反応速度で複製されたからである。
【0153】
BHV-1糖タンパク質Dを発現する組換えBAV3の構築。E3欠失の複製適合BAV3組換え体の、生の組換えワクチン抗原についての送達ビヒクルとしての有用性を決定するために、種々の形態のBHV-1糖タンパク質gDを発現する組換えBAV3ウイルスを(Tikooら、(1993)前出)を構築した。全長形態および短縮型形熊のgD遺伝子(外因性プロモーターを何ら含まない)を、BAV3.E3dゲノムのE3領域に、E.coliの相同組換え機構を用いて、平行方向に個々に挿入した。Degryseら(1996)前出。PacI消化pBAV302.gDまたはpBAV302.gDtプラスミドDNAは、ウシ網膜初代細胞にトランスフェクトされた場合に、14日後に細胞変性効果をもたらした。50%の細胞変性効果を示す感染した細胞単層を採取し、凍結融解し、そして組換えウイルスをプラーク精製し、そしてMDBK細胞において増殖させた。組換えBAV3を、BAV3.E3gD(E3領域における全長gD遺伝子の挿入)およびBAV3.E3gDt(E3領域における短縮型gD遺伝子の挿入)と名づけた。ウイルスDNAを抽出し、そして種々の制限酵素での消化後にアガロースゲル電気泳動によって分析した。gD遺伝子がNdeI部位を有するので、組換えウイルスDNAをNdeIで切断した。図4に見られるように、BAV3.E3d(図4、レーンe)と比較して、BAV3.E3gD(図4、レーンf)、およびBAV3.E3gDt(図4、レーンg)ゲノムは、4.6kbのさらなる予測されたバンドを含み、これは、組換えBAV3.E3gDおよびBAV3.gDtが、E3領域にgDまたはgDt遺伝子を含んでいたことを示唆する。gD遺伝子とgDt遺伝子との間を識別するために、組換えウイルスDNAをNheIで消化した、なぜなら、gDt遺伝子は、特有のNheI制限酵素部位を有するがgD遺伝子は有さないからである。Tikooら(1993)前出。予測されるように、5.4kb BAV3.E3gD DNAフラグメント(図4、レーンc)は、BAV3.E3gDtにおける5.0kbのフラグメントと置換されていた(図4、レーンd)。このことは、組換えBAV3.E3gDおよびBAV3.E3gDtが、それぞれgDおよびgDtを含んでいたことを示唆する。これらの組換え体と野生型またはE3欠失BAV3との増殖特徴の比較によっては、産生されたウイルスの複製の反応速度または力価のいずれにおいても有意な差異は示されなかった。
【0154】
BAV3.E3gDおよびBAV3.E3gDtによるgDの発現の分析。BHV-1 gDまたはgDt遺伝子を含む組換えBAV3ウイルスによって発現させる産物を試験するために、MDBK細胞に、組換えBAV3.E3gD、BAV3.E3gDtまたはBAV3.E3dを感染させ、そして異なる時間の間[35S]メチオニン−システインで代謝標識した。抗原性gDとの比較のために、MDBK細胞に、BHV-1を感染させ、そして[35S]メチオニン−システインで同様に標識した。放射標識したタンパク質を、gD特異的なモノクローナル抗体(MAb;Hughesら、(1988)Arch.Virol.103:47−60)で免疫沈降し、そして還元条件下のSDS-PAGEによって分析した。組換えBAV3.E3gD感染した細胞の免疫沈降によって、約71kDaの主要なバンドが示された(図5A、レーンb−d)。これは、BHV-1感染した細胞において産生されたgDタンパク質と共移動した(図5A、レーンa)。このことは、組換えgDが、真正なgDと類似する翻訳後修飾を含んでいたことを示唆する。感染していない細胞(図5A、レーンh)にも組換えBAV3.E3dに感染させた細胞(図5A、レーンe-g)にも類似のバンドは観察されなかった。組換えBAV3.E3gDt感染させた細胞上清の放射免疫沈降は、61kDaの主要バンドを示した(図5B、レーンb-d)。両方の組換えタンパク質は、MDBK細胞の感染の間中発現されていた(図5)。
【0155】
組換えgDタンパク質の抗原性を試験するために、放射標識したタンパク質を、組換えBAV3感染した細胞溶解物(BAV3.E3gD)または上清(BAV3.E3gDt)から、gD特異的なモノクローナル抗体(Highesら(1988)前出)を用いて免疫沈降し、そして還元条件下のSDS-PAGEにより分析した。図6に示すように、gDタンパク質およびgDtタンパク質の両方は、不連続のエピトープIb(MAb 136;レーンaおよびf)、II(MAb 3E7;レーンbおよびg)、IIIb(MAb 4C1;レーンcおよびh)、IIIC(MAb 2C8;レーンdおよびi)およびIIId(MAb 3C1;レーンeおよびj)に対して惹起されたMAbによって認識された。これらの結果は、組換えタンパク質gDおよびgDtの抗原性構造が、MDBK細胞において産生された真正のgD産物の構造と類似することを示唆する。Tikooら(1993)、前出。
【0156】
DNA合成の非存在下でgD発現が生じるか否かを決定するために、BAV3.E3gD感染したMDBK細胞において産生されたgDの量を、DNA合成のインヒビターである1-β-D-アラビノフラノシルシトシン(AraC)の存在下(図7、レーンaおよびb)および非存在下(図7、レーンcおよびd)で比較した。この結果は、gD発現が、AraCの存在下で減少することを示唆する。
【0157】
動物における抗体応答。BAV3組換え体がgD特異的な免疫応答を誘発する能力を決定するために、コットンラットに、2回3週間間隔をおいて、鼻腔内に、107PFUのBAV3.E3gD、BAV3.E3gDtまたはBAV3.E3d組換え体を接種した。血清、肺洗浄物および鼻洗浄物を、IgGおよびIgA抗体の分析のために採取し、その一方肺を、IgA抗体分泌細胞(ASC)の数を分析するために採取した。BAV3.E3gDおよびBAV3.E3gDtの両方は、gD特異的なIgG抗体応答を血清および肺洗浄物において誘発した(図8B)。これは、BAV3.E3d免疫した動物(コントロール)において誘発された応答よりも有意に高かった(P<0.05)。しかし、BAV3.E3gDtによって誘発されたgD特異的なIgG応答は、BAV3.E3gDによって誘発された応答よりも高かった(P<0.05)。
【0158】
BAV3.E3gDおよびBAV3.E3gDtでの免疫は、BAV3.E3dでの免疫よりも、血清および肺洗浄物においてgDに対してより高い(p<0.05)IgA抗体応答を誘発した(図8A)。しかし、BAV3.E3gD免疫群とBAV3.E3gDt免疫群との間のIgA抗体応答においては、有意な差異は存在しなかった。これらの組換え体はまた、血清および肺洗浄物においてBAV3特異的なIgG抗体応答(図8D)、および血清、肺洗浄物および鼻洗浄物においてIgA抗体応答(図8C)を誘発した。これは、群間では有意な差異は存在しなかった。
【0159】
興味深いことに、鼻洗浄物は、gD(図26Aおよび図26B)またはBAV3(図26Cおよび26D)に対して特異的なIgA抗体のみを含んだ。さらに、gDおよびBAV3の両方に特異的なIgA抗体分泌細胞が免疫された動物の肺において検出され得、この細胞の数は、追加免疫後に有意に増加した。
【0160】
gD特異的な血清抗体の生物学的な活性を測定するために、抗BHV-1力価を決定した。BAV3.E3gDtでの免疫は、4.3±0.5のBHV-1 log2力価を誘発し、これは、BAV3.E3gDおよびBAV3.E3dによって誘発された、それぞれ3.0±0.6および0.8±0.3の力価よりも有意に高かった(p<0.05)。
【0161】
考察
家畜でのヒトアデノウイルスのワクチン送達系としての使用は、限定されている。非ヒトアデノウイルスは種特異的であるので、動物特異的なアデノウイルスの、ワクチン送達系としての開発が、論理的な選択肢である。本明細書において、動物の粘膜表面へのワクチン抗原の送達における使用のための複製適合(E3欠失)組換えBAV3の開発が記載される。さらに、BHV-1 gDまたはgDt糖タンパク質を発現する複製適合BAV3を構築し、そしてそれらがコットンラットにおける粘膜免疫応答および全身免疫応答を誘発する能力について試験した。
【0162】
種々のウシ細胞株におけるE3欠失BAV3組換え体を単離する最初の試み(これは、BAV3 DNA媒介トランスフェクション後の感染性子孫の形成を支持する)は、成功しなかった。しかし、ルシフェラーゼ遺伝子を発現する部分的にE3を欠失したBAV3組換え体が、BAV3 E1で形質転換したMDBK細胞株を、トランスフェクションについて使用した場合に単離された。Mittalら(1995)前出。このことは、E1配列の存在に関連するいくつかの機能が、E3欠失BAV3の単離に必要であり得ることを示唆した。しかし、これらの単離の後、それらのE3領域にルシフェラーゼ遺伝子を発現する組換えBAV3ウイルスは、正常のMDBK細胞において効率よく複製した。Mittalら(1995)前出。従って、正常なMDBK細胞がBAV組換え体の単離を可能にするには低すぎるトランスフェクション効率および/または増殖速度を有し得るという別の可能性が存在する。
【0163】
BAV3 E1形質転換したMDBK細胞を用いずに複製適合BAV3組換え体を単離する、信頼がありそして効率的な方法を開発するために、E.coliの相同組換え機構(Chartierら(1996)前出;およびDegryse(1996)前出)をまず使用して、プラスミドにおいてクローニングされた全長BAV3ゲノムの1,245kb欠失をE3領域に導入した。第二に、組換えウイルスを単離するために、MDBK細胞の代わりにPFBR細胞を、プラスミドから切り出された改変されたBAV3ゲノムのトランスフェクションに用いた。
【0164】
野生型ゲノムの105%までがHAV5ビリオンに、次の回の複製において外来遺伝子の再構成も欠失も何ら生じさせずにパッケージングされ得ることが報告されている。Bettら(1993)、J.Virol.67:5911−5921.近年、ヒツジアデノウイルス(OAV)ベクターの挿入容量が野生型ゲノムの114%であることが報告された。Xuら(1997)Virology 230:62−71。BAV3ゲノムのサイズは、HAV5ゲノムのサイズと類似しているので、BAV3ゲノムの挿入能力は、HAV5の能力と類似し得る。従って、本明細書において記載されるBAV3.E3dベクターが1,245kbのE3欠失を有するので、その挿入能力は、少なくとも3.0kbである。
【0165】
以前に、E3領域に0,696kbの欠失を含む組換えBAV3.lucが細胞培養物において野生型BAV3よりも効率が悪く複製されたことが報告された。Mittalら(1995)前出。対照的に、E3領域に1,245kb欠失を含む組換えBAV3.E3dは、細胞培養物中で野生型BAV3と同程度に効率よく複製した。この差異は、ルシフェラーゼ遺伝子の組換えBAV3.lucにおけるルシフェラーゼ遺伝子の挿入に起因し得る。この挿入は、E3領域の下流の遺伝子の発現に影響を与え得る。Mittalら(1995)前出。しかし、組換えBAV3.E3gDまたはBAv3.E3gDtもまた、野生型BAV3と同様に効率よく複製された。このことは、異なる外来遺伝子産物が、細胞培養物において組換えBAV3ウイルスの複製に異なって影響し得ることを示唆する。
【0166】
可能なワクチン抗原の発現のためのBAVベクター系の有用性を確認するために、組換えBAV3.E3gDおよびBAV3.E3gDtを構築した。これらはそれぞれ、BHV-1gDおよびgDt糖タンパク質を発現する。予想されたように、BAV3.E3gDによって発現されたgDは、予測された分子量を有し、そしてBHV-12感染した細胞によって発現された真正のgDと共移動した。同様に、BAV3.E3gDtによって発現されたgDtは、推定の分子量を有していた。Tikooら(1993)前出。さらにBAV3組換え体によって発現されたgDおよびgDtは、コンホメーション的なエピトープに対して特に惹起されたMAbによって認識された。Hughesら(1988)前出。これらの結果は、組換えgDおよびgDtが、ウイルス感染後に合成された真正のgDのものと、識別不可能である翻訳後修飾および抗原性プロフィールを有していたことを示唆する。Hughesら(1988)前出。
【0167】
ヒトアデノウイルスのE3領域において左から右の方向に挿入された、隣接する調節配列を何ら伴わない外来遺伝子は、ウイルスゲノムの上流MLPから、またはE3プロモーターからのいずれかで効率よく発現される。Droninら(1993)Gene 126 247-250;Morinら(1987)Proc.Natl.Acad.Sci.USA 84:4626-4630およびXuら(1995)J.Gen.Virol.76:1971-1980。本発明の実験において、糖タンパク質gD発現は、複製インヒビターであるAraCの存在下で部分的に減少した。このことは、外来遺伝子発現が、MLP(複製依存性プロモーター)およびE3プロモーター(初期複製非依存性プロモーター)の両方によって駆動されたことを示唆している。類似の結果が以前に報告されている。Mittalら(1995)前出。
【0168】
コットンラットのBAV3.E3gDまたはBAV3.E3gDtでの鼻腔内免疫は、gD特異的な粘膜または全身性免疫応答を誘発した。gDtは、gDよりも強力なIgG抗体応答を誘発した。対照的に、コットンラットの組換えHAV5での皮内免疫は、gDに対するよりもgDtに対してより低い免疫応答を誘発した。Mittalら(1996)Virology、222:299-309。さらに、アジュバントとともに組み込まれた組換えgDtの精製した調製物は、マウスに注射された場合、真正のgDの応答に類似する二次免疫応答をもたらした。Baca-Estradaら(1996)Viral Immunol.9:11-22。これらの結果は、免疫およびワクチン処方物の経路が、免疫原が膜にアンカーされる(gD)か、または可溶性タンパク質として分泌される(gDt)か否かよりも高い程度で、免疫原が有効な免疫応答を誘発する能力に影響を与え得ることを強力に示唆する。
【0169】
驚くべきことに、gDおよびgDtは、類似のIgAを誘発したが、有意に異なるIgG応答を、鼻通路および血清において誘発した。粘膜免疫応答および全身免疫応答は、かなりの程度の非依存性を伴って誘発されそして調節されることが示された。Alleyら(1988)The mucosal immune system、「B lymphocytes in human disease」(G.BirdおよびJ.E.Calvert編)、Oxford University Press、222〜254頁;およびConleyら(1987)Ann.Intern.Med.106:892-899。これらのその1つにおける免疫応答の誘導は、必ずしも他における応答を誘導しない。従って、2つの異なるgDの形熊(全長および短縮型)が粘膜区画および全身区画において異なって認識されている可能性がある。
【0170】
粘膜免疫の誘導は、呼吸器感染または腸感染から宿主を防御する際に重要であると考えられる。さらに、分泌IgAの存在は、通常、このような感染に対する耐性に相関していることが見出されている。Murphy(1994)Mucosal immnuity toviruses、「Handbook of mucosal immunity」(P.L.Orga、J.Mestecky、M.E.Lamm、W.strober、J.R.McGhec、およびJ.Bienenstock編)。Academic Press、San Diego、333〜339頁。興味深いことに、鼻腔内免疫は、gD特異的なIgA抗体応答を、コットンラットの血清および肺のみならず、鼻洗浄物においても誘発した。肺における高レベルのIgAおよびASCの存在は、抗体が局所的に産生されることを示唆する。
【0171】
結論として、本発明は、BAV3組換え体の産生のための効率よくかつ信頼性の高い系を記載する。さらに、複製適合組換えBAV3を有するコットンラットの鼻腔内免疫は、ワクチン抗原特異的な、粘膜免疫応答および全身免疫応答を誘発する。
【0172】
実施例4:BHV-1糖タンパク質Dを発現する組換えBAV3でのウシの免疫
本実施例において、BHV-1 gD(全長および短縮型)を発現する複製適合組換えBAV3ウイルスを、それらがBHV-1チャレンジから防御する能力について評価した。3つの3〜4月齢のウシの群を、以下のプロトコルに従って鼻腔内に免疫した。群1を、全長BHV-1 gDを発現するウイルスであるBAV3.E3gDで免疫した。実施例3を参照のこと。群2を、短縮型BHV-1gDを発現するウイルスであるBAV3.E3gDtで免疫した。実施例3およびTikooら(1993)前出を参照のこと。群3を、欠失E3領域を有するが、挿入された異種配列は有さないウイルスであるBAV3.E3dで免疫した。実施例3を参照のこと。
【0173】
1日目および28日目に、動物にワクチン接種し、そして42日目にエアロゾル化したBHV-1でチャレンジした。1、28、40および52日目に血液を血清学のために採取し、そして28日目の血液サンプルにおいてリンパ球増殖を測定した。臨床的症状(温度、鼻の病変および抑鬱を含む)を、チャレンジ後10日間(42日目〜52日目)毎日評価した。1日目と15日目との間に、ウイルス単離のために鼻のスワブを3日毎に採取した。チャレンジ後、ウイルス力価を、2日毎に10日間、鼻のスワブおよび鼻のタンポンによって測定した。22および40日目、および42〜52日目に、抗体力価を、鼻のタンポンを用いて決定した。
【0174】
結果は、gD発現BAV組換え体またはgDt発現BAV組換え体で免疫した動物において、IgG力価が、最初のワクチン接種および追加のワクチン接種の両方の後に増加し、そしてチャレンジ後にさらに増加したことを示す(図9)。比較すると、挿入されたgD遺伝子を欠く、E3欠失したBAV3でワクチン接種した動物はチャレンジ後にのみIgG力価の増加を示し、そしてワクチン接種した動物において得られたものよりも少なくとも1対数分低かった。IgA力価を測定し、そして結果を図10に示す。見られるように、第一の免疫後にはIgAにおいて検出可能な増加は存在しなかった。しかし、gD特異的IgA力価は、第二の免疫後およびBHV-1チャレンジ後に増加した。再度、挿入されたgD遺伝子を欠く、E3欠失したBAV3でワクチン接種した動物は、チャレンジ後にのみIgA力価の増加を、そしてgD発現BAVまたはgDt発現BAVでワクチン処理した動物よりも低い程度で示した。
【0175】
臨床的症状は、E3欠失BAVでワクチン接種した動物と比較して、gD発現BAVおよびgDt発現BAVでワクチン接種した動物において減少した。これらは、発熱(図11)、鼻病変の出現および程度(図12)および抑鬱(図13)を含む。最後に、鼻液におけるウイルス力価は、コントロールに比較して、BAV3.E3gDワクチン接種した動物およびBAV3.E3gDtワクチン接種した動物におけるチャレンジ後により迅速に減少した(図14)。
【0176】
これらの結果は、BHV-1糖タンパク質を発現する組換えBAVウイルスは、ウシにおけるBHV-1チャレンジからの防御を提供し、臨床徴候の出現を減少させ、より迅速にウイルスを排除することを容易にし、そしてIgGおよびIgAの両方の力価の増加を提供した。このことは、ウシ宿主における体液性および粘膜性の免疫の両方の誘導を示唆する。
【0177】
生物学的な材料の寄託
以下の材料は、Veterinary Infectious Disease Organization(VIDO)、120Veterinary Road、Saskatoon、Saskatchewan、S7N 5E3、Canadaに寄託されそして維持されている。
【0178】
寄託された材料のヌクレオチド配列およびそれによってコードされるポリペプチドの配列は、本明細書において参考として援用される。本明細書において明示的に開示された配列と、寄託された配列との間に何らかの差異が存在する場合、寄託された配列が支配する。
【0179】
物質 内部受託番号 寄託日
組換えプラスミド
pSM51 pSM51 1993年12月6日
pSM71 pSM71 1993年12月6日
組換え細胞株
BAV3 E1配列で形質転換したMDBK細胞
(MDBK-BAVE1) 1993年12月6日
BAV E1配列で形質転換したウシ仔胎腎臓細胞
(FBK-BAV-E1) 1993年12月6日
【0180】
本発明を、上記に特定の具体的な実施態様によって例示したが、これらの特定の実施例は、添付の請求項において記載されるような本発明の範囲を制限することを意図するものではない。
【0181】
本明細書は、以下の発明を開示する。
【0182】
1.ウシアデノウイルスタイプ3(BAV-3)のゲノムまたはそのフラグメントに実質的に相同である、ヌクレオチド配列。
【0183】
2.ウシアデノウイルスタイプ3(BAV-3)のゲノムの前記フラグメントに実質的に相同である、前記1に記載のヌクレオチド配列であって、 該フラグメントが、BAV-3ゲノムのヌクレオチド4,092〜5,234、ヌクレオチド5,892〜17,735、ヌクレオチド21,198〜26,033、およびヌクレオチド31,133〜34,445からなる群より選択されるか、またはそのフラグメントである、配列。
【0184】
3.前記1または2に記載のヌクレオチド配列であって、前記そのフラグメントと実質的に相同であり、E2領域、E4領域、後期領域、33kD、52kD、100kD、DBP、pol、pTPおよびペントン遺伝子、ならびに遺伝子11IA、pV、pVI、pVII、pVIII、およびpXからなる群より選択される、配列。
【0185】
4.前記3に記載のヌクレオチド配列であって、E2領域、E4領域、後期領域、33kD、52kD、100kD、DBP、pol、pTPおよびペントン遺伝子、ならびに遺伝子IIIA、pV、pVI、pVII、pVIII、およびpX、ならびにそれらのフラグメントからなる群より選択されるBAV-3ゲノムの領域由来のBAV-3コード配列を含む、配列。
【0186】
5.BAV-3ゲノムの遺伝子間領域を含むヌクレオチド配列と実質的に相同である、ヌクレオチド配列。
【0187】
6.前記1〜5のいずれかに記載のヌクレオチド配列であって、BAV-3 DNA制御配列を含む、配列。
【0188】
7.前記DNA制御配列が、転写調節配列、プロモーター、エンハンサー、上流調節ドメイン、スプラシシングシグナル、ポリアデニル化シグナル、転写終結配列、翻訳調節配列、リボソーム結合部位、および翻訳終結配列からなる群より選択される、前記6に記載のヌクレオチド配列。
【0189】
8.前記1〜7のいずれかに記載のヌクレオチド配列を含む、ベクター。
【0190】
9.ITR配列、BAVパッケージング配列、および少なくとも1つの外来ヌクレオチド配列を含む欠損組換えBAVベクターであって、ここで、該BAVベクターがE1機能を欠き、そしてE2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択される領域によってコードされる1以上の機能をさらに欠いている、欠損組換えBAVベクター。
【0191】
10.前記9に記載の欠損組換えBAVベクターであって、E1領域のすべてまたは一部分が欠失しており、そしてE2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択される領域のすべてまたは一部分がさらに欠失している、欠損組換えBAVベクター。
【0192】
11.前記10に記載の欠損組換えBAVベクターであって、以下:
a)E1領域のすべてまたは一部分が欠失し、そしてE2領域のすべてまたは一部分が欠失しているBAVベクター;
b)E1領域のすべてまたは一部分が欠失し、そしてE4領域のすべてまたは一部分が欠失しているBAVベクター;
c)E1領域のすべてまたは一部分が欠失し、そしてE2領域のすべてまたは一部分が欠失し、そしてE4領域のすべてまたは一部分が欠失しているBAVベクター、からなる群より選択される、欠損組換えBAVベクター。
【0193】
12.さらに、E3領域のすべてまたは一部分が欠失している、前記11に記載の欠損組換えBAVベクター。
【0194】
13.前記外来ヌクレオチド配列が、治療目的のポリペプチドをコードする、前記9に記載の欠損組換えBAVベクター。
【0195】
14.前記治療目的のポリペプチドが、凝固因子、成長ホルモン、サイトカイン、リンホカイン、オンコジーン産物、腫瘍サプレッサー、細胞レセプター、細胞レセプターに対するリガンド、プロテアーゼインヒビター、抗体、毒素、免疫毒素、ジストロフィン、嚢胞性線維症膜貫通調節タンパク質(CFTR)、免疫原性ポリペプチド、および心臓血管疾患に関与するポリペプチドからなる群より選択される、前記13に記載の欠損組換えBAVベクター。
【0196】
15.前記心臓血管疾患に関与するポリペプチドが、線維芽細胞増殖因子(FGF)、血管内皮増殖因子(VEGF)、神経成長因子(NGF)、e-nos産物、pRb、リポタンパク質リパーゼ、スーパーオキサイドジスムターゼ、カタラーゼ、酸素スカベンジャー、フリーラジカルスカベンジャー、アポリポタンパク質およびプラスミノーゲン活性化因子インヒビター1からなる群より選択される、前記14に記載の欠損組換えBAVベクター。
【0197】
16.挿入部位に挿入された外来遺伝子を含む前記9に記載の欠損組換えBAVベクターを構築する方法であって、以下の工程:
a)BAVゲノムと実質的に相同である配列を提供する工程;
b)外来ヌクレオチド配列を提供する工程;
c)該挿入部位の周囲のBAV配列と実質的に相同である配列に該外来ヌクレオチド配列を連結して挿入カセットを形成する工程;
d)該挿入カセットおよび該BAVゲノムと実質的に相同である配列を細胞に導入する工程;
e)該挿入カセットと該BAVゲノムと実質的に相同である配列との間に相同組換えを生じさせて欠損組換えBAVベクターを生成する工程;
f)該欠損組換えBAVベクターが複製される条件下で該細胞を培養する工程;ならびに
g)該欠損組換えBAVベクターを該細胞または該培養培地から収集する工程、を包含する、方法。
【0198】
17.前記挿入部位が、E2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択されるBAVゲノムの領域に位置する、前記16に記載の欠損組換えBAVベクターを構築する方法。
【0199】
18.前記挿入部位が、E1領域において欠失を含む、前記16に記載の欠損組換えBAVベクターを構築する方法。
【0200】
19.前記9に記載のBAVベクターを、適切な補完細胞株に導入する工程、および該感染細胞からウイルスを回収する工程を包含する、組換えBAVを産生する方法。
【0201】
20.前記補完細胞株が、ヘルパーウイルスに感染している、前記19に記載の方法。
【0202】
21.前記補完細胞株が、E1領域、E2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択されるBAVゲノムの領域の1つ以上によってコードされる機能を提供する、前記19に記載の方法。
【0203】
22.前記8または9に記載のベクターを含む、宿主細胞。
【0204】
23.ウシアデノウイルスE1遺伝子領域を含む組換え哺乳動物細胞株であって、BAV E1機能を提供し得、そしてさらに、E2、E4、L1、L2、L3、L4、L5、L6およびL7からなる群より選択されるBAVゲノムの領域に由来する遺伝子によってコードされる機能を提供し得る、組換え哺乳動物細胞株。
【0205】
24.BAV E1およびE2機能を提供し得る、前記23に記載の組換え哺乳動物細胞株。
【0206】
25.BAV E1およびE4機能を提供し得る、前記23に記載の組換え哺乳動物細胞株。
【0207】
26.組換えBAVを産生する方法であって、以下の工程:
前記23に記載の細胞株に、ITR配列、BAVパッケージング配列、および少なくとも1つの外来遺伝子を含むBAVベクターを導入する工程であって、ここで、該ベクターがE1領域において欠失されており、そして該ベクターがE2領域、E4領域、L1領域、L2領域、L3領域、L4領域、L5領域、L6領域、およびL7領域からなる群より選択される領域においてさらに欠失されている、工程;ならびに
該感染した細胞からウイルスを回収する工程、を包含する、方法。
【0208】
27.組換えポリペプチドを産生する方法であって、以下の工程:
(a)前記22に記載の宿主細胞集団を提供する工程;および
(b)ポリペプチドが発現される条件下で該細胞集団を増殖させる工程、を包含する、方法。
【0209】
28.前記2〜4のいずれかに記載のヌクレオチド配列によってコードされる、BAVポリペプチド。
【0210】
29.前記27に記載の方法によって産生される、組換えポリペプチド。
【0211】
30.生物学的サンプル中のBAVヌクレオチド配列の存在を検出する方法であって、以下の工程:
(a)生物学的サンプルを提供する工程;および
(b)前記1〜3のいずれかに記載の配列の少なくとも10の連続するヌクレオチドを含む標識されたプローブを使用して、該サンプルをハイブリダイゼーションに供する工程、を包含する、方法。
【0212】
31.生物学的サンプル中のBAVヌクレオチド配列の存在を検出する方法であって、以下の工程:
(a)生物学的サンプルを提供する工程;および
(b)該サンプルを、前記1〜3のいずれかに記載の配列の少なくとも10の連続するヌクレオチドを含むプライマーを用いて、ポリメラーゼ連鎖反応に供する工程、を包含する、方法。
【0213】
32.1以上のプローブを含む、生物学的サンプル中のBAVヌクレオチド配列の存在を検出するためのキットであって、該プローブが前記1〜3のいずれかに記載の配列の少なくとも10の連続するヌクレオチドを含む、キット。
【0214】
33.生物学的サンプル中のBAV抗原の存在を検出する方法であって、以下の工程:
(a)生物学的サンプルを提供する工程;および
(b)前記29に記載のポリペプチドに対して惹起された抗体を用いて、該サンプルを免疫アッセイに供する工程、を包含する、方法。
【0215】
34.生物学的サンプル中のBAV抗原の存在を検出するためのキットであって、前記29に記載のポリペプチドに対して惹起された抗体を1以上含む、キット。
【0216】
35.哺乳動物被験体において免疫応答を誘発し得る薬学的組成物であって、前記2または3に記載のヌクレオチド配列または前記8に記載のベクターを含む、薬学的組成物。
【0217】
36.哺乳動物被験体において免疫応答を誘発し得る薬学的組成物であって、前記28または29に記載のポリペプチドを含む、薬学的組成物。
【0218】
37.哺乳動物被験体において免疫応答を誘発し得る薬学的組成物であって、免疫原性ポリペプチドを発現する前記9に記載の欠損組換えBAVベクターを含む、薬学的組成物。
【0219】
38.前記8〜15のいずれかに記載のベクターを含む、薬学的組成物。
【0220】
39.前記22の宿主細胞を含む、薬学的組成物。
【0221】
40.哺乳動物宿主において免疫応答を誘発して感染から防御する方法であって、防御抗原を発現し得る組換えBAVベクターを含む薬学的組成物を投与する工程を包含する、方法。
【0222】
41.哺乳動物宿主における遺伝子治療の方法であって、前記8に記載のベクターを該宿主に投与する工程を包含する、方法。
【0223】
42.哺乳動物宿主における遺伝子治療の方法であって、前記9に記載の欠損組換えBAVベクターを該宿主に投与する工程を包含する、方法。
【0224】
43.哺乳動物宿主における遺伝子治療の方法であって、前記22に記載の宿主細胞を該宿主に投与する工程を包含する、方法。
【0225】
44.前記28または29に記載のポリペプチドに特異的に結合する、抗体。
【0226】
45.E3特異的配列が欠失している、組換えウシアデノウイルス(BAV)ベクター。
【0227】
46.挿入された異種配列を含む、前記45に記載のベクター。
【0228】
47.前記異種配列が、前記欠失された配列によって以前に占有されていた部位で該E3領域に挿入されている、前記46に記載のベクター。
【0229】
48.前記45に記載のBAVベクターを含む、宿主細胞。
【0230】
49.挿入部位に挿入された異種配列を含む欠損組換えBAVベクターを構築する方法であって、以下の工程:
(a)該挿入部位の周囲のBAV配列と実質的に相同である配列に、該異種配列を連結させて、挿入カセットを形成する工程;
(b)該挿入カセットを、BAVゲノムと実質的に相同である配列を含むポリヌクレオチドとともに、細胞に導入する工程;および
(c)該挿入カセットと、該ポリヌクレオチドとの間に相同組換えを生じさせて、該欠損組換えBAVベクターを生成する工程、を包含する、方法。
【0231】
50.E3特異的配列が、少なくとも1つのセグメントにおいて欠失している、前記49に記載の方法。
【0232】
51.前記細胞が原核生物細胞である、前記49に記載の方法。
【0233】
52.前記挿入部位の周囲のBAV配列と実質的に相同である前記配列が、プラスミドに存在し、そして前記連結する工程が前記異種配列を含む制限フラグメントを該プラスミドに挿入することによって達成される、前記51に記載の方法。
【0234】
53.前記細胞がE.coliである、前記51に記載の方法。
【0235】
54.前記49に記載の方法に従って得られた、組換えBAVゲノム。
【0236】
55.組換えBAVウイルスを得る方法であって、以下の工程:
(a)前記54に記載の組換えBAVゲノムを得る工程;
(b)工程(a)の該組換えBAVゲノムを、適切な哺乳動物宿主細胞に導入する工程;
(c)ウイルス複製を生じさせる工程;および
(d)該宿主細胞から組換えBAVウイルスを精製する工程、を包含する、方法。
【0237】
56.さらに、以下の工程:
前記工程(a)の後に制限酵素で前記組換えBAVを消化する工程であって、該制限酵素消化が前記ポリヌクレオチドに存在する他の配列からBAV配列を分離する、工程、
を包含する、前記55に記載の方法。
【0238】
57.前記宿主細胞がMDBK細胞である、前記55に記載の方法。
【0239】
58.前記MDBK細胞がアデノウイルスE1機能を発現する、前記57に記載の方法。
【0240】
59.前記宿主細胞が、ウシ仔胎網膜初代(PBFR)細胞である、前記55に記載の方法。
【0241】
60.前記セグメントがBAVゲノムを含む、前記55に記載の方法。
【0242】
61.前記E3特異的な配列が、前記組換えBAVゲノムにおいて欠失している、前記60に記載の方法。
【0243】
62.前記異種配列が前記欠損された配列によって以前に占有されていた前記部位で前記E3領域に挿入される、前記61に記載の方法。
【0244】
63.前記62に記載の方法により得られる、組換えBAV。
【0245】
64.前記63に記載の組換えBAVを含む、免疫原性組成物。
【0246】
65.疾患の症状を予防または改善させる方法であって、前記64に記載の免疫原性組成物を、哺乳動物被験体に導入する工程を包含する、方法。
【0247】
66.BAVゲノムおよび少なくとも1つの異種ヌクレオチド配列を含む組換えBAVベクターであって、該異種配列が、E3領域に挿入されている、BAVベクター。
【0248】
67.前記BAVゲノムがE3特異的な配列について欠失されている、前記66に記載のBAVベクター。
【0249】
68.前記異種配列が、哺乳動物病原体の防御決定因子をコードする、前記67に記載のBAVベクター。
【0250】
69.哺乳動物被験体における免疫応答を誘発し得る薬学的組成物であって、前記68に記載の組換えBAVベクターを含む、組成物。
【0251】
70.哺乳動物宿主において免疫応答を誘発して感染から防御する方法であって、前記69に記載の薬学的組成物を投与する工程を包含する、方法。
【0252】
71.哺乳動物宿主における遺伝子治療の方法であって、前記66に記載のベクターを該宿主に投与する工程を包含する、方法。
【0253】
72.前記異種配列が、線維芽細胞増殖因子(FGF)、血管内皮増殖因子(VEGF)、神経成長因子(NGF)、e-nos産物、pRb、リポタンパク質リパーゼ、スーパーオキサイドジスムターゼ、カタラーゼ、酸素スカベンジャー、フリーラジカルスカベンジャー、アポリポタンパク質およびプラスミノーゲン活性化因子インヒビター1からなる群より選択されるポリペプチドをコードする、前記71に記載の方法。
【0254】
【0255】
【0256】
【0257】
【0258】
【0259】
【0260】
【0261】
【0262】
【0263】
【0264】
【0265】
【0266】
【0267】
【0268】
【0269】
【0270】
【0271】
【0272】
【0273】
【0274】
【0275】
【図1−1】
【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図1−6】
【図1−7】
【図1−8】
【図1−9】
【図1−10】
【図1−11】
【図1−12】
【図1−13】
【図1−14】
【図1−15】
【図1−16】
【図1−17】
【図1−18】
【図1−19】
【図1−20】
【図1−21】
【図1−22】
【図1−23】
【図1−24】
【図1−25】
【図1−26】
【図1−27】
【図1−28】
【特許請求の範囲】
【請求項1】
配列番号1に示されるBAV−3ゲノムのヌクレオチド31,133〜34,445である必須でない領域中の挿入部位に挿入された異種ヌクレオチド配列を含む組換えBAV−3ベクターを構築するための方法であって、以下の工程:
a)該異種ヌクレオチド配列を、第1のポリヌクレオチドに連結させて挿入カセットを生成する工程であって、
該第1のポリヌクレオチドのヌクレオチド配列は、挿入部位の周りで、配列番号1に示されるBAV−3ゲノムのヌクレオチド配列に対して少なくとも90%同一であり、
該第1のポリヌクレオチドは、配列番号1に示すウイルスゲノム全体のうちの一部のみを表し、
該挿入部位は、該BAV−3ゲノムのヌクレオチド31,133〜34,445の中にある
工程;
b)該挿入カセットを、配列番号1に示すヌクレオチド配列からなるBAV−3ゲノムと少なくとも90%同一である第二のポリヌクレオチドとともに細胞に導入する工程であって、
該BAV−3ゲノムのヌクレオチド31,133〜34,445の全部または一部が欠失している、工程;および
c)該挿入カセットと該第二のポリヌクレオチドとの間に相同組み換えを起こさせて該組み換えBAV−3ベクターを生成する工程、
を包含する、方法。
【請求項2】
前記細胞が原核生物細胞である、請求項1に記載の方法。
【請求項3】
前記挿入部位の周囲の前記第1のポリヌクレオチドが、プラスミド中に存在し、そして前記連結する工程が前記異種ヌクレオチド配列を含む制限フラグメントを該プラスミド中に挿入することによって達成される、請求項2に記載の方法。
【請求項4】
組換えBAV−3ウイルスを得る方法であって、以下の工程:
(a)請求項1〜3のいずれか1項に記載の方法により組換えBAV−3ベクターを構築する工程;
(b)工程(a)の該組換えBAV−3ベクターを、適切な宿主細胞に導入する工程;および
(c)ウイルス複製を生じさせる工程、
を包含する、方法。
【請求項5】
更に、
(d)該宿主細胞から組換えBAV−3ウイルスを精製する工程、
を含む、請求項4に記載の方法。
【請求項6】
前記宿主細胞がMDBK細胞であるか、または、アデノウイルスE1機能を発現するMDBK細胞であるか、またはウシ仔胎網膜初代(PBFR)細胞である請求項4に記載の方法。
【請求項7】
請求項1〜6のいずれか1項に記載の方法により得られる、組換えBAV−3またはBAV−3ベクター。
【請求項8】
請求項7に記載の組み換えBAV−3ベクターであって、前記異種ヌクレオチド配列が治療目的のポリペプチドをコードし、該治療目的のポリペプチドが、凝固因子、成長ホルモン、サイトカイン、リンホカイン、オンコジーン産物、腫瘍サプレッサー、細胞レセプター、細胞レセプターに対するリガンド、プロテアーゼインヒビター、抗体、毒素、免疫毒素、ジストロフィン、嚢胞性線維症膜貫通調節タンパク質(CFTR)、免疫原性ポリペプチド、および心臓血管疾患に関与するポリペプチドからなる群より選択される、組み換えBAV−3ベクター。
【請求項9】
前記心臓血管疾患に関与するポリペプチドが、線維芽細胞増殖因子(FGF)、e−nos産物、pRb、リポタンパク質リパーゼ、スーパーオキサイドジスムターゼ、カタラーゼ、酸素スカベンジャー、フリーラジカルスカベンジャー、アポリポタンパク質およびプラスミノーゲン活性化因子インヒビター1からなる群より選択される、請求項8に記載の組み換えBAV−3ベクター。
【請求項10】
請求項7に記載のベクターを含む、宿主細胞。
【請求項11】
組み換えポリペプチドを生成する方法であって、該方法は:
請求項10に記載の宿主細胞の集団を該ポリペプチドが発現される条件下において増殖させる工程、
を包含する、方法。
【請求項12】
組み換えBAV−3ベクターであって、配列番号1に示すヌクレオチド配列に対して少なくとも90%の同一性を有するポリヌクレオチドを含み、ここで、該ポリヌクレオチドのヌクレオチド31,133〜34,445である必須でない領域に異種ヌクレオチド配列が挿入された、BAV−3ベクター。
【請求項13】
該BAV−3ゲノムのヌクレオチド31,133〜34,445の全部または一部が欠失している、請求項12に記載の組み換えBAV−3ベクター。
【請求項14】
前記異種ヌクレオチド配列が、治療目的のポリペプチドをコードし、該治療目的のポリペプチドが、凝固因子、成長ホルモン、サイトカイン、リンホカイン、オンコジーン産物、腫瘍サプレッサー、細胞レセプター、細胞レセプターに対するリガンド、プロテアーゼインヒビター、抗体、毒素、免疫毒素、ジストロフィン、嚢胞性線維症膜貫通調節タンパク質(CFTR)、免疫原性ポリペプチド、および心臓血管疾患に関与するポリペプチドからなる群より選択される、請求項12又は13に記載の組み換えBAV−3ベクター。
【請求項15】
前記心臓血管疾患に関与するポリペプチドが、線維芽細胞増殖因子(FGF)、e−nos産物、pRb、リポタンパク質リパーゼ、スーパーオキサイドジスムターゼ、カタラーゼ、酸素スカベンジャー、フリーラジカルスカベンジャー、アポリポタンパク質およびプラスミノーゲン活性化因子インヒビター1からなる群より選択される、請求項14に記載の組み換えBAV−3ベクター。
【請求項1】
配列番号1に示されるBAV−3ゲノムのヌクレオチド31,133〜34,445である必須でない領域中の挿入部位に挿入された異種ヌクレオチド配列を含む組換えBAV−3ベクターを構築するための方法であって、以下の工程:
a)該異種ヌクレオチド配列を、第1のポリヌクレオチドに連結させて挿入カセットを生成する工程であって、
該第1のポリヌクレオチドのヌクレオチド配列は、挿入部位の周りで、配列番号1に示されるBAV−3ゲノムのヌクレオチド配列に対して少なくとも90%同一であり、
該第1のポリヌクレオチドは、配列番号1に示すウイルスゲノム全体のうちの一部のみを表し、
該挿入部位は、該BAV−3ゲノムのヌクレオチド31,133〜34,445の中にある
工程;
b)該挿入カセットを、配列番号1に示すヌクレオチド配列からなるBAV−3ゲノムと少なくとも90%同一である第二のポリヌクレオチドとともに細胞に導入する工程であって、
該BAV−3ゲノムのヌクレオチド31,133〜34,445の全部または一部が欠失している、工程;および
c)該挿入カセットと該第二のポリヌクレオチドとの間に相同組み換えを起こさせて該組み換えBAV−3ベクターを生成する工程、
を包含する、方法。
【請求項2】
前記細胞が原核生物細胞である、請求項1に記載の方法。
【請求項3】
前記挿入部位の周囲の前記第1のポリヌクレオチドが、プラスミド中に存在し、そして前記連結する工程が前記異種ヌクレオチド配列を含む制限フラグメントを該プラスミド中に挿入することによって達成される、請求項2に記載の方法。
【請求項4】
組換えBAV−3ウイルスを得る方法であって、以下の工程:
(a)請求項1〜3のいずれか1項に記載の方法により組換えBAV−3ベクターを構築する工程;
(b)工程(a)の該組換えBAV−3ベクターを、適切な宿主細胞に導入する工程;および
(c)ウイルス複製を生じさせる工程、
を包含する、方法。
【請求項5】
更に、
(d)該宿主細胞から組換えBAV−3ウイルスを精製する工程、
を含む、請求項4に記載の方法。
【請求項6】
前記宿主細胞がMDBK細胞であるか、または、アデノウイルスE1機能を発現するMDBK細胞であるか、またはウシ仔胎網膜初代(PBFR)細胞である請求項4に記載の方法。
【請求項7】
請求項1〜6のいずれか1項に記載の方法により得られる、組換えBAV−3またはBAV−3ベクター。
【請求項8】
請求項7に記載の組み換えBAV−3ベクターであって、前記異種ヌクレオチド配列が治療目的のポリペプチドをコードし、該治療目的のポリペプチドが、凝固因子、成長ホルモン、サイトカイン、リンホカイン、オンコジーン産物、腫瘍サプレッサー、細胞レセプター、細胞レセプターに対するリガンド、プロテアーゼインヒビター、抗体、毒素、免疫毒素、ジストロフィン、嚢胞性線維症膜貫通調節タンパク質(CFTR)、免疫原性ポリペプチド、および心臓血管疾患に関与するポリペプチドからなる群より選択される、組み換えBAV−3ベクター。
【請求項9】
前記心臓血管疾患に関与するポリペプチドが、線維芽細胞増殖因子(FGF)、e−nos産物、pRb、リポタンパク質リパーゼ、スーパーオキサイドジスムターゼ、カタラーゼ、酸素スカベンジャー、フリーラジカルスカベンジャー、アポリポタンパク質およびプラスミノーゲン活性化因子インヒビター1からなる群より選択される、請求項8に記載の組み換えBAV−3ベクター。
【請求項10】
請求項7に記載のベクターを含む、宿主細胞。
【請求項11】
組み換えポリペプチドを生成する方法であって、該方法は:
請求項10に記載の宿主細胞の集団を該ポリペプチドが発現される条件下において増殖させる工程、
を包含する、方法。
【請求項12】
組み換えBAV−3ベクターであって、配列番号1に示すヌクレオチド配列に対して少なくとも90%の同一性を有するポリヌクレオチドを含み、ここで、該ポリヌクレオチドのヌクレオチド31,133〜34,445である必須でない領域に異種ヌクレオチド配列が挿入された、BAV−3ベクター。
【請求項13】
該BAV−3ゲノムのヌクレオチド31,133〜34,445の全部または一部が欠失している、請求項12に記載の組み換えBAV−3ベクター。
【請求項14】
前記異種ヌクレオチド配列が、治療目的のポリペプチドをコードし、該治療目的のポリペプチドが、凝固因子、成長ホルモン、サイトカイン、リンホカイン、オンコジーン産物、腫瘍サプレッサー、細胞レセプター、細胞レセプターに対するリガンド、プロテアーゼインヒビター、抗体、毒素、免疫毒素、ジストロフィン、嚢胞性線維症膜貫通調節タンパク質(CFTR)、免疫原性ポリペプチド、および心臓血管疾患に関与するポリペプチドからなる群より選択される、請求項12又は13に記載の組み換えBAV−3ベクター。
【請求項15】
前記心臓血管疾患に関与するポリペプチドが、線維芽細胞増殖因子(FGF)、e−nos産物、pRb、リポタンパク質リパーゼ、スーパーオキサイドジスムターゼ、カタラーゼ、酸素スカベンジャー、フリーラジカルスカベンジャー、アポリポタンパク質およびプラスミノーゲン活性化因子インヒビター1からなる群より選択される、請求項14に記載の組み換えBAV−3ベクター。
【図2】

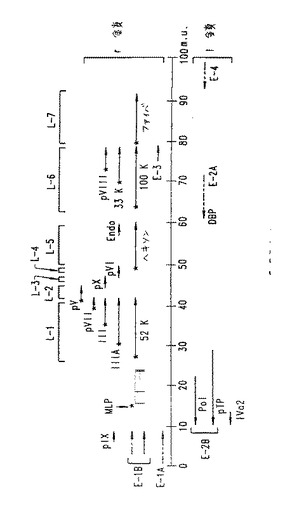
【図3】
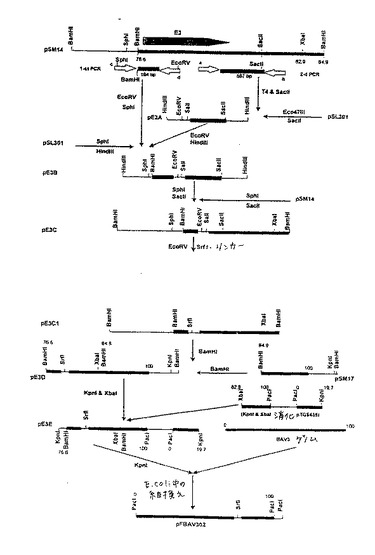
【図4】
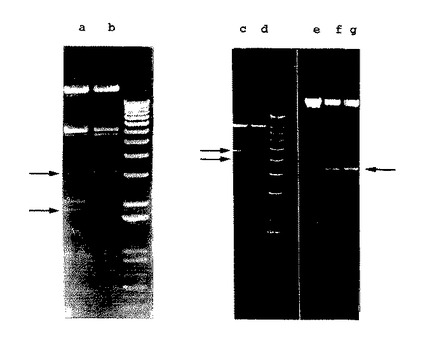
【図5】
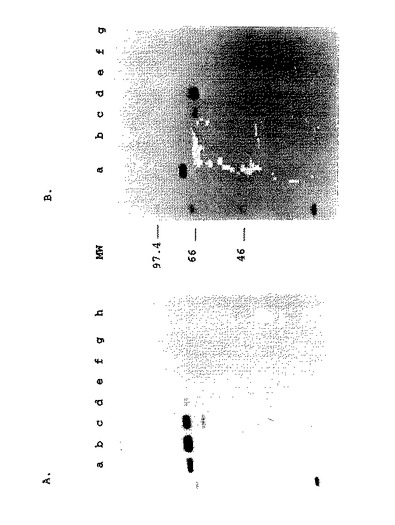
【図6】
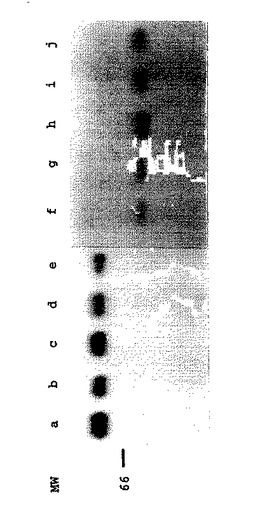
【図7】
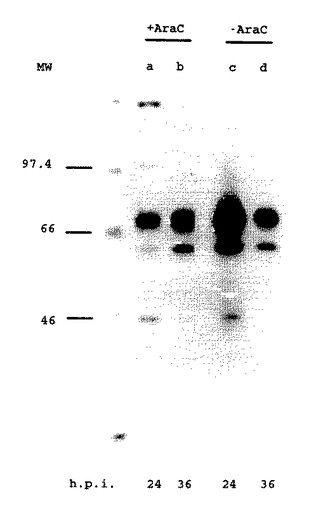
【図8】
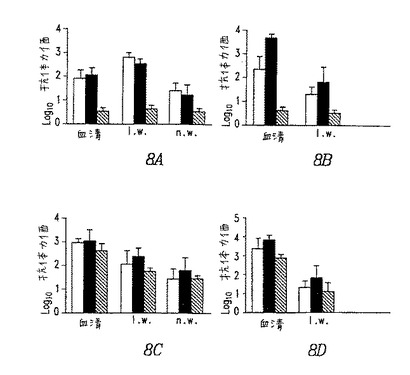
【図9】
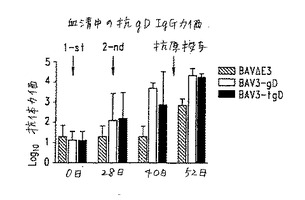
【図10】
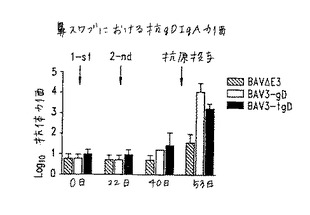
【図11】
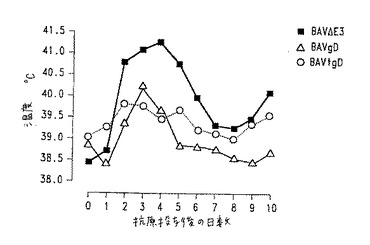
【図12】
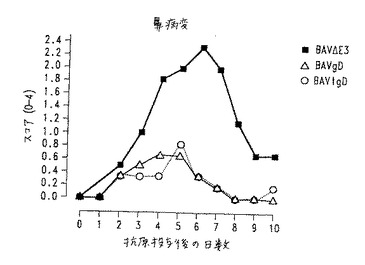
【図13】
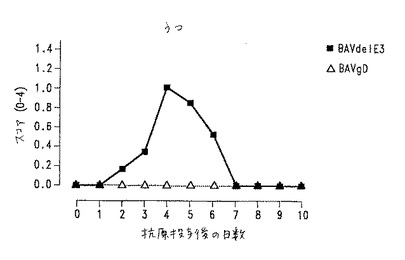
【図14】
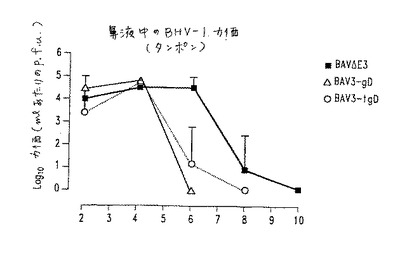
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公開番号】特開2011−167204(P2011−167204A)
【公開日】平成23年9月1日(2011.9.1)
【国際特許分類】
【出願番号】特願2011−115171(P2011−115171)
【出願日】平成23年5月23日(2011.5.23)
【分割の表示】特願平11−503471の分割
【原出願日】平成10年6月23日(1998.6.23)
【出願人】(501176255)ユニバーシティ オブ サスカチュワン (2)
【Fターム(参考)】
【公開日】平成23年9月1日(2011.9.1)
【国際特許分類】
【出願日】平成23年5月23日(2011.5.23)
【分割の表示】特願平11−503471の分割
【原出願日】平成10年6月23日(1998.6.23)
【出願人】(501176255)ユニバーシティ オブ サスカチュワン (2)
【Fターム(参考)】
[ Back to top ]