
ウロキナーゼ型プラスミノーゲン活性化因子(uPA)活性の阻害
本発明はウロキナーゼ型プラスミノーゲン活性化因子(uPA)のその受容体uPARへの結合を阻害するための方法、および/またはuPAの生物学的活性を阻害するための方法に関する。本発明はさらに、腫瘍の形成または転移、血管形成、例えば腫瘍血管形成を阻害するための方法、およびCYTL1アゴニストを同定するためのスクリーニングアッセイに関する。例えば、ウロキナーゼ型プラスミノーゲン活性化因子(uPA)とウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)との相互作用を阻害する方法であって、サイトカイン様1(CYTL1)ポリペプチドまたはそのアゴニストにuPAおよびuPARを含む混合物を接触させることを含む、方法が提供される。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明はウロキナーゼ型プラスミノーゲン活性化因子(uPA)のその受容体uPARへの結合を阻害するための方法、および/またはuPAの生物学的活性を阻害するための方法に関する。本発明はさらに、腫瘍の形成または転移、血管形成、例えば腫瘍血管形成を阻害するための方法、およびCYTL1アゴニストを同定するためのスクリーニングアッセイに関する。
【背景技術】
【0002】
(発明の背景)
ウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)は如何なる既知の造血系サイトカイン受容体にも構造的に類似していない。これは広範な種類の遊走性の細胞型により発現されるグリコシルホスファチジルイノシトール(GPI)アンカリング細胞表面タンパク質である(非特許文献1;非特許文献2;非特許文献3)。uPARは2つの異なる作用を有しており、即ち、第1にはそれが不活性な前ウロキナーゼ型プラスミノーゲン活性化因子(プロ−uPA)を運搬して細胞表面プロテアーゼに接近させ、それにより分解されてウロキナーゼ型プラスミノーゲン活性化因子(uPA)となり、これは細胞表面に係留されて残存し(非特許文献4)、そこでそれはセリンプロテアーゼカスケードを開始し、細胞周囲のタンパク質分解をもたらすことになり(非特許文献5;非特許文献6);そして第2にそれは細胞外マトリックス成分ビトロネクチンに結合し(非特許文献7)、これが次に細胞表面インテグリンに結合する(非特許文献8)。細胞外マトリックスへの接着とその分解の両方に影響することにより、uPAのuPARへの結合は、uPAR欠損マウスにおける好中球リクルートメントの不全により顕在化される通り、細胞の遊走において重要な役割を果たしている(非特許文献9;非特許文献10)。
【0003】
uPA−uPAR相互作用の競合的阻害剤の生物学的効果は多くの系において検討されている。uPAのタンパク質分解的に不活性な突然変異体でトランスフェクトされた腫瘍細胞は腫瘍転移の能力の低減を示しており(非特許文献11)、uPA−ATFは微小血管内皮細胞による管形成(非特許文献12)および網膜傷害後の血管形成(非特許文献13)を阻害する。
【0004】
CYTL1(サイトカイン様1)は新規の分泌タンパク質を同定して分析するための大規模な研究の一部としてクローニングされており(非特許文献14)、そして米国特許出願公開20050037465において「PRO4425」として開示されている。これは造血幹先祖細胞の希少なCD34+サブセットにおいて高度に発現される転写物の産物であるC17と同一であることがわかっている(非特許文献15)。CYTL1特異的cDNAの原料組織の分析、プロテオーム研究(非特許文献16)、および大規模な遺伝子発現分析(非特許文献17;非特許文献18)によれば軟骨および骨における豊富なCYTL1発現が示されている。それと対応して、ヒトにおいてCYTL1をコードする遺伝子(NM_018659)は骨と軟骨の発達に関与を示唆されている遺伝子がリッチ化されている領域(4p16−15)である染色体4:5,067,217−5,072,098にマッピングされている(非特許文献18;非特許文献19;非特許文献20;非特許文献21)。両親媒性の分析によれば4つのα−ヘリックスの存在が示唆されている(非特許文献15)。CYTL1のサイトカイン様性質は造血系の4つのα−ヘリックス束のサイトカインを連想させるパターンにおけるCYTL1鎖中の両親媒性のα−ヘリックスの予測に基づいている(非特許文献22)。本ファミリーのメンバーは有意な配列の類似性の非存在下に十分に保存されたコア折畳み部を有しており(非特許文献23)、そしてこのタンパク質の構造は、特殊化された膜貫通受容体のある集団とのそれらの相互作用を指向する傾向がある(非特許文献24)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Pepperら(1993)J Cell Biol 122(3),673−684
【非特許文献2】Danoら(1999)Apmis 107(1),120−127
【非特許文献3】Gyetkoら、(1994)J Clin Invest 93(4),1380−1387
【非特許文献4】Cubellisら(1986)J Biol Chem 261(34),15819−15822
【非特許文献5】Danoら、(2005)Thrombosis and Haemostasis 93(4),676−681
【非特許文献6】Ploug,M.(2003)Current Pharmaceutical Design 9(19),1499−1528
【非特許文献7】Weiら(1994)J Biol Chem 269(51),32380−32388
【非特許文献8】Madsenら(2007)J Cell Biol 177(5),927−939
【非特許文献9】Rijneveldら(2002)J Immunol 168(7),3507−3511
【非特許文献10】Gyetkoら(2000)J Immunol 165(3),1513−1519
【非特許文献11】Crowlyら(1993)Proc Natl Acad Sci USA 90(11),5021−5025
【非特許文献12】Kroonら(1999)Am J Pathol 154(6),1731−1742
【非特許文献13】L Gatら(2003)Gene Ther 10(25),2098−2103
【非特許文献14】Clarkら(2003)Genome research 13(10),2265−2270
【非特許文献15】Liuら(2000)Genomics 65(3),283−292(2)
【非特許文献16】Hermanssonら、(2004)J Biol Chem 279(42),43514−43521
【非特許文献17】Kumarら(2001)Osteoarthritis and cartilage/OARS,Osteoarthritis Research Society 9(7),641−653
【非特許文献18】Yagerら(2004)Genomics 84(3),524−535
【非特許文献19】Mangionら(1999)American Journal of Human Genetics 65(1),151−157
【非特許文献20】Polymeropoulosら(1996)Genomics 35(1),1−5
【非特許文献21】Shiangら(1994)Cell 78(2),335−342
【非特許文献22】Bazan,J.F.(1990)Immunology today 11(10),350−354
【非特許文献23】Hillら、(2002)J Mol Biol 322(1),205−233
【非特許文献24】Sprang and Bazan,(1993)Current Opinion in Structural Biology 3(6),815−827
【発明の概要】
【課題を解決するための手段】
【0006】
上記において考察したとおり、uPAとその受容体uPARとの間の相互作用は、セリンプロテアーゼカスケードの開始を細胞膜に局在化させること、そして細胞表面受容体と細胞外マトリックスとの間の会合をモジュレートすることの両方により、種々の細胞型の遊走において重要な役割を果たしている。本発明は少なくとも部分的には、uPAR結合に関してuPAと競合するuPARに対する追加的なリガンドとしての分泌サイトカイン様タンパク質CYTL1/C17の同定に基づいている。
【0007】
1つの態様において、本発明はサイトカイン様1(CYTL1)ポリペプチドまたはそのアゴニストにウロキナーゼ型プラスミノーゲン活性化因子(uPA)およびウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)を含む混合物を接触させることを含むuPAとuPARとの相互作用を阻害する方法に関する。
【0008】
別の態様において、本発明は有効量のCYTL1またはそのアゴニストにインビボでウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)およびウロキナーゼ型プラスミノーゲン活性化因子(uPA)を発現する細胞を接触させることを含むuPAの生物学的活性を阻害する方法に関する。
【0009】
さらに別の態様において、本発明は、哺乳類被験体における腫瘍形成または腫瘍転移を阻害するための方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む方法に関する。
【0010】
さらなる態様において、本発明は、哺乳類被験体における血管形成を阻害するための方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む方法に関する。
【0011】
またさらに別の態様において、本発明は(a)ウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)およびCYT1またはそのアゴニストを含有する混合物を候補拮抗剤とともにインキュベートすること、および(b)uPARへのuPAまたはそのアゴニストの結合を競合的に阻害する候補拮抗剤の能力を計測すること、を含むウロキナーゼ型プラスミノーゲン活性化因子(uPA)の拮抗剤のスクリーニングの方法に関する。
【0012】
全ての態様において、アゴニストは例えば、ポリペプチド、ペプチド、非ペプチド性低分子、またはアゴニストCYTL1抗体またはそのフラグメントであってよい。
【0013】
全ての態様において、ポリペプチドは例えばCYTL1変異体(variant)、例えば配列番号2のCYTL1のようなネイティブ配列CYTL1分子の配列と少なくとも約60%、または65%、または70%、または75%、または80%、または85%、または90%、または95%、または98%、または99%のアミノ酸配列同一性を有するCYTL1変異体であってよい。
【0014】
全ての態様において、アゴニストCYTL1抗体または抗体フラグメントは好ましくはモノクローナルであり、そしてキメラ、ヒト化、またはヒト型であることができる。
【0015】
全ての態様において、抗体フラグメントは例えばFab、Fab’、F(ab’)2、およびFvフラグメント;ダイアボディ;直鎖状抗体;単鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体を包含する。
【0016】
全ての態様において、哺乳類被験体は好ましくはヒト患者である。
【0017】
全ての態様において、腫瘍または癌は例えば、癌腫、リンパ腫、芽腫、肉腫、および白血病、例えば、限定しないが、乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択されてよい。
【図面の簡単な説明】
【0018】
【図1】CYTL1のインビボ発現。CYTL1 mRNA発現は(a〜d)ホールマウント標本および(e〜p)薄片に対して実施したインサイチュのハイブリダイゼーションにより分析した。全ての組織は特段の記載が無い限りマウスおよび成人より入手した。(e〜j)においては、上パネルは暗視野像であり、下パネルは相当する明視野像である。a)第12日胚の脊椎骨、b)第13日胚の手掌、c)第12日胚の気管および肺、d)第12日胚の肋骨、e)腱鞘(腱(T)、中足骨(M))、f)気管、g)肺動脈、h)第18日胚中足骨、i)表面軟骨層におけるCYTL1発現を示す(h)における関節表面のより高い倍率、およびj)ヒト気管。白色のバーは100□mを示す。
【図2】軟骨細胞分化の間のCYTL1の発現。ATDC5前軟骨細胞をコンフルエントとなるまで生育させ、次にインスリンおよびアスコルビン酸で処置して分化を促進した。記載した時点において、RNAを採取し、そしてTaqManにより遺伝子発現をアッセイした。記載した結果はGAPDHに対して標準化(normalized)されており、そして観察された最大値に尺度を合わせてある。丸印−コラーゲンII、三角印−アグレカン、四角印−CYTL1。結果は3つの独立した実験の代表である。
【図3】CIAにおけるCYTL1のダウンレギュレーション。(A)IL−1βおよびCYTL1の発現レベルはCIAプロトコル開始してから種々の日数の後にマウスの関節から抽出したRNAのマイクロアレイ分析により計測した。(B)RNAは健常マウスの手掌およびCIAを有するマウスの炎症手掌から抽出した。CYTL1発現はqRT−PCRでアッセイし、そしてβ−アクチンに対して標準化した。平均およびSEM(N=5対照/3CIA)。(C)軟骨細胞の分化は11日間ATDC5細胞において誘導した。次に細胞を0.1ng/mlのIL−1β(黒色バー)または培地単独(白色バー)で24時間処置した。RNAを採取しqRT−PCRでアッセイし、そしてβ−アクチンで標準化した。4つの独立した実験に由来する標準化したデータの平均およびSEM。「相対的発現」とは対照と比較した場合の発現である。
【図4】uPARはCYTL1に対する受容体である。(A)擬似トランスフェクトされたCOS細胞およびuPAR発現コンストラクトでトランスフェクトされたCOS細胞をCYTL1−APまたは対照であるTACI−APとともにインキュベートし、そして結合したAP活性を染料付着により検出した。(B)ヒトおよびマウスCYTL1−APを未トランスフェクトのCOS細胞(白色バー)、またはネズミ(斜線バー)またはヒト(黒色バー)のuPARでトランスフェクトしたCOS細胞とともにインキュベートした。結合したPA活性を比色酵素アッセイにより計測した。(C)uPARトランスフェクトした細胞をCYTL1−APおよび記載濃度の精製CYTL1とともにインキュベートした。CYTL1−APの結合は比色酵素アッセイにより計測し、そしてバックグラウンド(未トランスフェクト細胞)を差し引いた後の精製されたCYTL1非存在下に観察された結合のパーセンテージとして表示した。
【図5】CYTL1−uPAR相互作用の表面プラズモン共鳴分析。記載した濃度における精製されたCYTL1をA)固定化uPAR、B)対照のフローセル上に注入した。C)平衡結合分析−ピークuPAR依存性応答を注入したCYTL1の濃度に対してプロットする。曲線は読み取り値の3つの独立したセットに対するラングミュア方程式の最良フィットを示す。D)以下に示すもの、即ちi)CYTL1(1μm)、ii)CYTL1(1μm)+ヘパリン(160μg/ml)、iii)緩衝液単独、およびiv)ヘパリン(160μg/ml)を固定化したuPAR上に注入した。各条件の3つの独立した調製物の重複応答曲線(対照フローセル差し引き後)を、記載した平衡時の平均の応答とともに示す。
【図6】CYTL1およびuPAは競合してuPARに結合する。CYTL1−APを記載濃度のA)uPA−ATF、B)プロ−uPAまたはC)DFP不活性化uPAの存在下にuPARトランスフェクト細胞と共にインキュベートした。細胞表面結合CYTL1−APを酵素アッセイにより計測し、そして競合物質非存在下の結合パーセントとして表示した。示された曲線によれば、単一の結合部位に対する可逆的競合が推定される。R2値は0.998、0.987および0.880である。D)組み換えヒトuPAR(4776RU)をBIAcoreセンサーチップの1つのフローセル中に固定化した。CYTL1(1μm)の3つの同一注入を80μg/mlプロ−uPAの注入の前後、およびプロ−uPAの解離(洗浄)後に実施した。対照フローセル(BSA固定化、5157RU)の差し引き後の平衡応答を各CYTL1注入に関して記載する。
【図7A】図7Aおよび図7B:ネイティブ配列のヒトPRO4425(CYTL1)cDNAのヌクレオチド配列(配列番号1)およびネイティブヒトPRO4425(CYTL1)ポリペプチドの推定アミノ酸配列(配列番号2)。
【図7B】図7Aおよび図7B:ネイティブ配列のヒトPRO4425(CYTL1)cDNAのヌクレオチド配列(配列番号1)およびネイティブヒトPRO4425(CYTL1)ポリペプチドの推定アミノ酸配列(配列番号2)。
【図8】CYTL1−APを用いてプローブしたアビジンコーティングプレート上に固定化した酵素的にビオチン化されたuPARを示し、結合はAP基質を用いて検出した。CYTL1−APは他のタンパク質の非存在下でuPARに直接結合する。
【図9】CYTL1−APを用いてプローブしたアビジンコーティングプレート上に固定化した酵素的にビオチン化されたuPARを示し、結合はAP基質を用いて検出した。uPARへのCYTL1−APの結合は抗iPARポリクローナルにより阻害される。
【図10】CYTL1−AP融合タンパク質を用いたBIAcore試験。
【図11】CYTL1はPC−3細胞によるマトリゲル侵襲を阻害する。
【図12】CYTL1はuPA依存性細胞増殖を阻害する。
【図13】CYTL1の存在下で成育させた軟骨細胞において低減したコラーゲンII発現が観察される。
【図14】CYTL1KOマウスは肉視的には正常な軟骨および骨を発達させている。
【図15】CYTL1KOマウスは正常な骨鉱物質密度を生じている。
【図16】CYTL1KOマウスは重症度の低い関節炎を有するように観察される。
【図17】組み換えヒトCYTL1に関する精製スキーム。
【図18】CYTL1の安定性を最適化するためのスクリーニング緩衝液。
【図19】CYTL1の安定性を最適化するためのスクリーニング緩衝液。
【図20】CYTL1の安定性を最適化するためのスクリーニング緩衝液。
【図21】CYTL1の安定性を最適化するためのスクリーニング緩衝液。
【図22】マウスCYTL1を精製するための精製スキーム。
【図23】CYTL1結晶回折の結果。
【図24】補足図1。CYTL1の二次構造の推定およびゲノム構造。CYTL1種の相同体のアライメントを示しており、推定二次構造は下部に示す通りであり、H:ヘリックス、E:延長コンホメーション(β−鎖)を示す。「Jnet Rel」とは各残基における推定に関する信頼性を示す(9=最良)。3つのヘリックスが容易に推定され、それに伴い、短いα−ヘリックス(僅か4残基のみ信頼性をもって推定)およびβ−鎖がABループ中に、そして短いβ−鎖がC−Dループ中に在り、これは多くの短鎖サイトカインに特徴的である。注目すべきはC末端領域におけるプロリン残基であり、これはα−ヘリックスの形成とは相反する可能性がある。ヒトCYTL1のエキソン境界に相当する位置は矢印で示す通りであり、矢印上方の数字はエキソン境界相およびイントロンの長さを示す。4ヘリックスサイトカインファミリーにおいて、エキソン境界は特徴的にはヘリックスAの後、ヘリックスBの前、そしてヘリックスCの後に観察され、全てがゼロ相であり、これはCYTL1に関して観察される通りである。
【図25】補足図2。CYTL1のATDC5細胞への結合。A)材料および方法においてトランスフェクトされたCOS細胞に関して記載した通り種々の時点においてATDC5細胞をプローブするためにマウスCYTL1−APを用いた。成熟軟骨細胞の形態を有する細胞への融合タンパク質の結合は第4日より観察され、より後期の時点においてはさらに広範化した。B)ATDC5成熟化の間の種々の時点においてqRT−PCRにより遺伝子発現をアッセイした。記載した結果はGAPDHに対して標準化されており、そして観察された最大値に尺度を合わせてある。丸印−コラーゲンII、三角印−uPAR、四角印−CYTL1。
【図26】表1。qRT−PCRに関するプライマーおよびプローブ。全プローブはFAMレポーター染料およびTAMRAクエンチャーで標識した。
【発明を実施するための形態】
【0019】
I.定義
特段の記載が無い限り、本明細書で使用する専門技術用語は本発明が属する技術における通常の技術者が一般的に理解するものと同様の意味を有する。例えばSingletonら、Dictionary of Microbiology and Molecular Biology第2版、J.Wiley&Sons(New York,NY 1994);Sambrookら、Molecular Cloning,A Laboratory Manual,Cold Springs Harbor Press(Cold Springs Harbor,NY 1989)を参照。本発明の目的のためには以下の用語を下記記載の通り定義する。
【0020】
「ウロキナーゼ型プラスミノーゲン活性化因子」即ち「u−PA」という用語は本明細書においては、431アミノ酸のヒト前プロu−PA(NP_002649;Moroi and Aoki,J.Biol.Chem.251(19),5956−5965(1976))および相当する313アミノ酸の成熟ヒトポリペプチドを21アミノ酸シグナル配列を伴うか伴うことなく(Roldanら、EMBO J.1990;9:467-474)包含するネイティブ配列のu−PAポリペプチド、および、オルタナティブスプライシングされた、そして、対立遺伝子の変異体およびアイソフォーム、並びにその可溶性形態のような全ての天然に存在する変異体を包含する非ヒト哺乳類のそのネイティブ配列相同体を指す。
【0021】
「CYTL1」、「サイトカイン様1」および「PRO4425」という用語は本明細書においては互換的に使用され、そして種々の原料から、例えばヒト組織型から、または他の原料から単離してよく、または組み換えまたは合成方法により製造してよい。「CYTL1ポリペプチド」または「PRO4425ポリペプチド」に言及する本明細書における全ての開示内容はポリペプチドの各々を個別に、または総括して指すものとする。例えば調製、精製、誘導体化、対応または対抗する抗体の形成、投与、含有組成物、関連疾患の処置等の記載は、個別に本発明の各ポリペプチドに関する。「CYTL1ポリペプチド」または「PRO4425ポリペプチド」という用語はまた、本明細書に開示したCYTL1/PRO4425ポリペプチドの変異体を包含する。
【0022】
「ネイティブ配列CYTL1/PRO4425ポリペプチド」は天然から誘導された相当するCYTL1ポリペプチドと同じアミノ酸配列を有するポリペプチドを含む。そのようなネイティブ配列CYTL1ポリペプチドは天然より単離することができ、或いは組み換えまたは合成手段により製造できる。「ネイティブ配列CYTL1ポリペプチド」という用語は特記すれば特定のCYTL1ポリペプチドの天然に存在するトランケーションされた、または分泌された形態(例えば細胞外ドメイン配列)、ポリペプチドの天然に存在する変異体型(例えばオルタナティブスプライシング型)および天然に存在する対立遺伝子変異体を包含する。本発明の種々の実施形態において、本明細書に開示したネイティブ配列CYTL1ポリペプチドは添付図面に示す完全長アミノ酸配列を含む成熟または完全長のネイティブ配列ポリペプチドである。開始および終止コドンは図中太字および下線で示す。しかしながら、添付の図7に開示するCYTL1ポリペプチドは1により示されるメチオニンで開始するように記載されているが、図7中のアミノ酸の1位より上流または下流の何れかに位置する他のメチオニン残基をCYTL1ポリペプチドに関する開始アミノ酸残基として使用してもよいことが考えられ、かつ可能である。
【0023】
本明細書に開示したCYTL1ポリペプチドの「シグナルペプチド」の概ねの位置は図7に示す通りである。しかしながら、留意すべきは、シグナルペプチドのC末端境界は、シグナルペプチドのC末端境界がその型のアミノ酸配列エレメントを同定するために当該分野で定型的に使用されている基準に従って同定してよい、本発明において当初同定されたシグナルペプチドC末端境界の何れかの側において、殆どは約5アミノ酸を超えないが、変動する場合がある(例えばNielsenら、Prot.Eng.10:1−6(1997)およびvon Heinjeら、Nucl.Acids.Res.14:4683−4690(1986))。さらにまた、同様に認識されていることとして、一部の場合においては、分泌されたポリペプチドからのシグナル配列の切断は完全に均一ではなく、1つより多い分泌物質種をもたらす。シグナルペプチドが本発明において同定されたシグナルペプチドのC末端境界の何れかの側において約5アミノ酸を超えない範囲で切断されるこの成熟ポリペプチド、およびそれらをコードするポリヌクレオチドは本発明の意図するものである。
【0024】
「CYTL1変異体」は、本明細書に開示した完全長ネイティブ配列CYTL1ポリペプチド配列と少なくとも約80%アミノ酸配列同一性を有する上述および後述の通り定義される活性なCYTL1ポリペプチド、本明細書に開示したシグナルペプチドを欠いたCYTL1ポリペプチド配列、本明細書に開示したシグナルペプチドを伴うか伴わないCYTL1ポリペプチドの細胞外ドメイン、または本明細書に開示した完全長CYTL1ポリペプチドの何れかの他のフラグメントを意味する。そのようなCYTL1ポリペプチド変異体は例えば、完全長ネイティブアミノ酸配列のNまたはC末端においてアミノ酸残基1つ以上が付加または欠失しているCYTL1ポリペプチドを包含する。通常は、CYTL1ポリペプチド変異体は、少なくとも約80%アミノ酸配列同一性、或いは少なくとも約81%アミノ酸配列同一性、或いは少なくとも約82%アミノ酸配列同一性、或いは少なくとも約83%アミノ酸配列同一性、或いは少なくとも約84%アミノ酸配列同一性、或いは少なくとも約85%アミノ酸配列同一性、或いは少なくとも約86%アミノ酸配列同一性、或いは少なくとも約87%アミノ酸配列同一性、或いは少なくとも約88%アミノ酸配列同一性、或いは少なくとも約89%アミノ酸配列同一性、或いは少なくとも約90%アミノ酸配列同一性、或いは少なくとも約91%アミノ酸配列同一性、或いは少なくとも約92%アミノ酸配列同一性、或いは少なくとも約93%アミノ酸配列同一性、或いは少なくとも約94%アミノ酸配列同一性、或いは少なくとも約95%アミノ酸配列同一性、或いは少なくとも約96%アミノ酸配列同一性、或いは少なくとも約97%アミノ酸配列同一性、或いは少なくとも約98%アミノ酸配列同一性および或いは少なくとも約99%アミノ酸配列同一性を、本明細書に開示した完全長ネイティブ配列CYTL1ポリペプチド配列、本明細書に開示したシグナルペプチドを欠いたCYTL1ポリペプチド配列、本明細書に開示したシグナルペプチドを伴うか伴わないCYTL1ポリペプチドの細胞外ドメイン、または本明細書に開示した完全長CYTL1ポリペプチド配列の何れかの他の特に定義されたフラグメントに対して有することになる。通常は、CYTL1変異体ポリペプチドは少なくとも約10アミノ酸長、或いは少なくとも約20アミノ酸長、或いは少なくとも約30アミノ酸長、或いは少なくとも約40アミノ酸長、或いは少なくとも約50アミノ酸長、或いは少なくとも約60アミノ酸長、或いは少なくとも約70アミノ酸長、或いは少なくとも約80アミノ酸長、或いは少なくとも約90アミノ酸長、或いは少なくとも約100アミノ酸長、或いは少なくとも約150アミノ酸長、或いは少なくとも約200アミノ酸長、或いは少なくとも約300アミノ酸長、またはそれより長いものである。
【0025】
本発明において同定されたCYTL1ポリペプチド配列に関する「パーセント(%)アミノ酸配列同一性」は、配列をアラインし、そして、最大パーセント配列同一性を達成するために必要に応じてギャップを導入した後、そして、如何なる保存的置換も配列同一性の部分として考慮しない場合に、特定のCYTL1ポリペプチド配列におけるアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセンテージとして定義される。パーセントアミノ酸配列同一性を決定する目的のためのアライメントは、当業者の能力の範囲内の種々の方法において、例えば公に入手できるコンピュータソフトウエア、例えばBLAST、BLAST−2、ALIGNまたはMegalign(DNASTAR)ソフトウエアを用いて達成できる。当業者であれば比較すべき配列の完全長に渡って最大アライメントを達成するために必要な何れかのアルゴリズムを包含するアライメントを計測するための適切なパラメータを決定できる。しかしながら本発明の目的のためには、%アミノ酸配列同一性の値は配列比較コンピュータプログラムALIGN−2を用いて得られ、その場合、ALIGN−2プログラムに関する完全なソースコードは後に示す表Iに記載する通りである。ALIGN−2配列比較コンピュータプログラムはGenentech,Inc.の著作であり、そして後に示す表Iに記載するソースコードは米国著作権局(Washington D.C.,20559)においてユーザマニュアルと共にファイルされており、そこで米国著作権登録番号TXU510087の下に登録されている。ALIGN−2プログラムはGenentech,Inc.,South San Francisco,Calif.から公的に入手するか、または後に示す表Iに記載するソースコードからコンパイルしてよい。ALIGN−2プログラムはUNIX(登録商標)のオペレーティングシステム、好ましくはデジタルUNIX(登録商標)V4.0D上の使用のためにコンパイルしなければならない。全ての配列比較パラメータはALIGN−2プログラムにより設定され、変動しない。
【0026】
アミノ酸配列比較のためにALIGN−2を使用する状況において、所定のアミノ酸配列Aの、所定のアミノ酸配列Bへの、Bとの、またはBに対抗した%アミノ酸配列同一性(これは言い換えれば、所定のアミノ酸配列Bへの、Bとの、またはBに対抗した特定の%アミノ酸配列同一性を有するか、または含んでいる所定のアミノ酸配列Aと表現できる)は以下の通り計算される。
【0027】
100×(分数X/Y)
式中、Xは配列アライメントプログラムALIGN−2によりそのプログラムのAおよびBのアライメントにおいて同一マッチとして採点されたアミノ酸残基の数であり、そして式中、YはBにおけるアミノ酸残基の総数である。当然ながら、アミノ酸配列Aの長さがアミノ酸配列Bの長さと等しくない場合、AのBへの%アミノ酸配列同一性はBのAへの%アミノ酸配列同一性と等しくならない。この方法を用いた%アミノ酸配列同一性の計算の例として、表2および3は「CYTL1」と表記されたアミノ酸配列への「比較タンパク質」と表記されたアミノ酸配列の%アミノ酸配列同一性をどのようにして計算するかを示しており、ここで、「CYTL1」は目的の仮説的CYTL1ポリペプチドのアミノ酸配列を示し、「比較タンパク質」は目的の「CYTL1」ポリペプチドを比較すべき相手であるポリペプチドのアミノ酸配列を示し、そして「X」、「Y」および「Z」は各々異なる仮説的アミノ酸残基を示す。
【0028】
特段の記載が無い限り、本明細書において使用する全ての%アミノ酸配列同一性の値はALIGN−2コンピュータプログラムを用いてこの直前に記載した段落において説明した通りに得られる。しかしながら、%アミノ酸配列同一性の値はまた、WU−BLAST−2コンピュータプログラムを用いて後述する通りに得てもよい(Altschulら、Methods in Enzymology 266:460−480(1996))。WU−BLAST−2検索パラメータの大部分はデフォルト値に設定される。デフォルト値に設定されないもの、即ち調節可能なパラメータは、以下の値、即ち:オーバーラップスパン=1、オーバーラップ割合=0.125、ワード閾値(T)=11、および採点マトリックス=BLOSUM62で設定される。WU−BLAST−2を使用する場合、%アミノ酸配列同一性の値は(a)WU−BLAST−2により決定されたものとしての、ネイティブCYTL1ポリペプチドから誘導された配列を有する目的のCYTL1ポリペプチドのアミノ酸配列と目的の比較アミノ酸配列(即ちCYTL1変異体ポリペプチドであってよい目的のCYTL1ポリペプチドが比較される相手となる配列)との間のマッチする同一アミノ酸残基の数を(b)目的のCYTL1ポリペプチドのアミノ酸残基の総数で割ることにより求められる。例えば、「アミノ酸配列Bへの少なくとも80%のアミノ酸配列同一性を有するアミノ酸配列Aを含むポリペプチド」という文言において、アミノ酸配列Aは目的の比較アミノ酸配列であり、そしてアミノ酸配列Bは目的のCYTL1ポリペプチドのアミノ酸配列である。
【0029】
パーセントアミノ酸配列同一性はまた、配列比較プログラムNCBI−BLAST2(Altschulら、Nucleic Acids Res.25:3389−3402(1997))を用いて決定してもよい。NCBI−BLAST2配列比較プログラムはhttp://www.ncbi.nlm.nih.govからダウンロードしてよく、或いは別様にNational Institute of Health,Bethesda,Mdから得てよい。NCBI−BLAST2はいくつかの検索パラメータを使用しており、この場合、これらの検索パラメータの全てはデフォルト値、例えばアンマスク=有り、鎖=全て、予測発生度=10、最小の低複雑度長=15/5、多重通過e値=0.01、多重通過の定数=25、最終ギャップ付きアライメントに関するドロップオフ=25および採点マトリックス=BLOSUM62に設定されている。
【0030】
アミノ酸配列比較のためにNCBI−BLAST2を使用する状況において、所定のアミノ酸配列Aの、所定のアミノ酸配列Bへの、Bとの、またはBに対抗した%アミノ酸配列同一性(これは言い換えれば、所定のアミノ酸配列Bへの、Bとの、またはBに対抗した特定の%アミノ酸配列同一性を有するか、または含んでいる所定のアミノ酸配列Aと表現できる)は以下の通り計算される。
【0031】
100×(分数X/Y)
式中、Xは配列アライメントプログラムNCBI−BLAST2によりそのプログラムのAおよびBのアライメントにおいて同一マッチとして採点されたアミノ酸残基の数であり、そして式中、YはBにおけるアミノ酸残基の総数である。当然ながら、アミノ酸配列Aの長さがアミノ酸配列Bの長さと等しくない場合、AのBへの%アミノ酸配列同一性はBのAへの%アミノ酸配列同一性と等しくならない。
【0032】
「単離された」とは、本明細書に開示した種々のポリペプチドを説明するために用いられる場合、自身の天然の環境の成分から同定および分離および/または回収されたポリペプチドを意味する。自身の天然の環境の夾雑成分はポリペプチドに関する診断または治療用途を典型的には妨害する物質であり、そして酵素、ホルモン、および他のタンパク質性または非タンパク質性の溶質を包含してよい。好ましい実施形態においては、ポリペプチドは(1)スピニングカップシークエネーターの使用によりN末端または内部のアミノ酸配列の少なくとも15残基を得るために十分な程度にまで、または(2)クーマシーブルーまたは、好ましくは銀染色を用いた非還元または還元条件下のSDS−PAGEによる均一性にまで、精製されることになる。単離されたポリペプチドは、CYTL1ポリペプチドの少なくとも1つの天然の環境の成分が存在しないことから、組み換え細胞内部のインサイチュのポリペプチドを包含する。しかしながら通常は単離されたポリペプチドは少なくとも1つの精製工程により調製されることになる。
【0033】
「単離された」CYTL1ポリペプチドをコードする核酸または他のポリペプチドをコードする核酸はポリペプチドをコードする核酸の天然の原料において自身が通常会合している少なくとも1つの夾雑核酸分子から同定され、そして分離される核酸分子である。単離されたポリペプチドをコードする核酸分子は自身が天然に存在する形態またはセッティングとは異なる。従って単離されたポリペプチドをコードする核酸分子は、自身が天然の細胞中に存在することから特定のポリペプチドをコードする核酸分子からは区別される。しかしながら、単離されたポリペプチドをコードする核酸分子は例えば天然の細胞の場合とは異なる染色体位置に核酸分子があるポリペプチドを通常発現する細胞中に含有されたポリペプチドをコードする核酸分子を包含する。
【0034】
「対照配列」という用語は特定の宿主生物中の作動可能に連結したコード配列の発現のために必要なDNA配列を指す。原核生物に適する対照配列は例えばプロモーター、場合によりオペレーター配列、およびリボソーム結合部位を包含する。真核生物の細胞はプロモーター、ポリアデニル化シグナル、およびエンハンサーを利用することが知られている。
【0035】
核酸はそれが別の核酸配列と機能的関係を有するように位置づけられれば「作動可能に連結」されている。例えば、プレ配列または分泌リーダーに関するDNAは、それがポリペプチドの分泌に関与するプレタンパク質として発現されればポリペプチドに関するDNAに作動可能に連結しており、プロモーターまたはエンハンサーはそれが配列の転写に影響すればコード配列に作動可能に連結しており、或いはリボソーム結合部位はそれが翻訳を促進するように位置づけられていればコード配列に作動可能に連結している。一般的に「作動可能に連結」とは連結されるべきDNA配列が近接しており、そして分泌リーダーの場合は、近接し、そして読み相にあることを意味している。しかしながら、エンハンサーは近接している必要はない。連結は好都合な制限部位におけるライゲーションにより達成される。そのような部位が存在しない場合は、合成のオリゴヌクレオチドアダプターまたはリンカーを従来の慣行に従って使用する。
【0036】
ハイブリダイゼーション反応の「ストリンジェンシー」は当業者により容易に決定され得、そして一般的にはプローブ長、洗浄温度、および塩濃度に依存した実験的計算である。一般的に、より長いプローブは適切なアニーリングのためにより高温を必要とするのに対し、より短いプローブはより低い温度を必要とする。ハイブリダイゼーションは一般的に、相補鎖がそれらの融点より低値の環境に存在する場合に再アニーリングする変性DNAの能力に依存している。プローブとハイブリダイズ可能な配列との間の所望の相同性の程度が高値であるほど、使用できる相対的な温度は高値となる。その結果、より高値の相対的温度は反応条件をよりストリンジェントとする傾向があるのに対し、より低値の温度のそのような傾向は低い。ハイブリダイゼーション反応のストリンジェンシーのさらに詳細な説明および解説に関しては、Ausubelら、Current Protocols in Molecular Biology,Wiley Interscience Publishers,(1995)を参照。
【0037】
「ストリンジェントな条件」または「高ストリンジェンシー条件」とは、本明細書において定義される場合は、以下のもの、即ち:(1)洗浄のために低イオン強度および高温を使用、例えば、0.015M塩化ナトリウム/0.0015Mクエン酸ナトリウム/0.1%ドデシル硫酸ナトリウムを50℃で使用;(2)ハイブリダイゼーション中に変性剤、例えばホルムアミド、例えば50%(v/v)ホルムアミド+0.1%ウシ血清アルブミン/0.1%Ficoll/0.1%ポリビニルピロリドン/50mMリン酸ナトリウム緩衝液pH6.5+750mM塩化ナトリウム、75mMクエン酸ナトリウムを42℃で使用;または(3)50%ホルムアミド、5xSSC(0.75MNaCl、0.075Mクエン酸ナトリウム)、50mMリン酸ナトリウム(pH6.8)、0.1%ピロリン酸ナトリウム、5xDenhardt溶液、超音波処理サケ精子DNA(50μg/ml)、0.1%SDS、および10%デキストランスルフェートを42℃で使用し、そして洗浄は42℃において0.2xSSC(塩化ナトリウム/クエン酸ナトリウム)および50%ホルムアミド中55℃、ついで高ストリンジェンシー洗浄を0.1xSSC+EDTAを用いて55℃で実施、により同定してよい。
【0038】
「中ストリンジェント条件」はSambrookら、Molecular Cloning:A Laboratory Manual,New York:Cold Spring Harbor Press,1989に記載の通り同定してよく、そして、上記したものよりも低ストリンジェントな洗浄溶液およびハイブリダイゼーションの条件(例えば温度、イオン強度および%SDS)の使用を包含する。中ストリンジェント条件の例は20%ホルムアミド、5xSSC(150mMNaCl、15mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5xDenhardt溶液、10%デキストランスルフェート、および20mg/ml変性粉砕サケ精子DNAを含む溶液中37℃で一夜インキュベーション、ついで約37〜50℃において1xSSC中フィルター洗浄である。プローブ長等のような要素に適合するために必要に応じて温度、イオン強度などを調節する方法は当業者の知る通りである。
【0039】
「タグ付けされたエピトープ」という用語は本明細書において使用する場合、「タグポリペプチド」に融合したCYTL1ポリペプチドを含むキメラポリペプチドを指す。タグポリペプチドはそれに対抗して抗体が形成できるエピトープをもたらすために十分な残基を有するが、なお、自身が融合する相手であるポリペプチドの活性を妨害しない程度に十分短い。タグポリペプチドはさらに抗体が他のエピトープと実質的に交差反応しないように十分独特である。適当なタグポリペプチドは一般的に少なくとも6つのアミノ酸残基、そして通常は約8〜50アミノ酸残基(好ましくは約10〜20アミノ酸残基)を有する。
【0040】
本発明の目的のための「活性な」または「活性」とは、ネイティブまたは天然に存在するCYTL1の生物学的および/または免疫学的な活性を保持しているCYTL1ポリペプチドの型を指し、その場合、「生物学的」活性はネイティブまたは天然に存在するCYTL1により保有される抗原性のエピトープに対抗する抗体の産生を誘導する能力以外のネイティブまたは天然に存在するCYTL1により誘発される生物学的機能(阻害性または促進性の何れか)を指し、そして「免疫学的」活性はネイティブまたは天然に存在するCYTL1により保有される抗原性のエピトープに対抗する抗体の産生を誘導する能力を指す。本発明の目的のためには、好ましい生物学的活性はウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)に結合し、uPARへのuPAの結合を競合的に阻害し、および/またはuPAの生物学的活性を阻害する能力を包含する。
【0041】
「uPAの生物学的活性」という用語は最も広範な意味において使用され、そして限定しないが、uPARに結合する能力、細胞遊走のモジュレーション、細胞膜へのセリンプロテアーゼカスケードの開始の局在化、細胞表面受容体と細胞外マトリックスとの間の会合のモジュレーション、腫瘍転移の媒介への関与、および血管形成活性を包含する。
【0042】
「アゴニスト」という用語は本明細書においては元も広範な意味において使用される。CYTL1アゴニストは伏在する機序に関わらずネイティブ配列のCYTL1により媒介される生物学的活性を模倣する何れかの分子である。本発明の目的のためには生物学的活性は好ましくは上記定義したuPA生物学的活性を阻害する能力である。CYTL1アゴニストの例は限定しないが、アゴニスト抗CYTL1抗体、ペプチドおよび非ペプチド性有機低分子を包含する。
【0043】
「抗体」という用語は本明細書においては、最も広範な意味において使用され、そして特に、所望の生物学的活性を呈する限りにおいて、インタクトな(intact)抗体、モノクローナル抗体、ポリクローナル抗体、少なくとも2つのインタクトな抗体から形成された多重特異性抗体(例えば二重特異性抗体)、および抗体フラグメントを網羅している。
【0044】
「モノクローナル抗体」という用語は本明細書においては、実質的に均質な抗体の集団から得られた抗体を指し、即ち、集団を構成する個々の抗体は、少量で存在する場合がある可能な天然に存在する突然変異を除き、同一である。モノクローナル抗体は高度に特異的であり、単一の抗原部位に対するものである。さらにまた、異なる決定基(エピトープ)に対する異なる抗体を包含するポリクローナル抗体調製物とは対照的に、各モノクローナル抗体は抗原上の単一の決定基に対するものである。それらの特異性に加えて、モノクローナル抗体は、それらが他の抗体による夾雑を被らずに合成される点において好都合である。「モノクローナル」という修飾語は抗体の実質的に均質な集団から得られるものとしての抗体の特徴を示しており、そして何れかの特定の方法による抗体の産生を要求するものとみなしてはならない。例えば本発明に従って使用するべきモノクローナル抗体はKohlerら、Nature,256:495(1975)により最初に報告されたハイブリドーマ法により作成してよく、或いは組み換えDNA法により作成してよい(例えば米国特許4,816,567参照)。「モノクローナル抗体」はまた例えばClacksonら、Nature,352:624−628(1991)およびMarksら、J.Mol.Biol.,222:581−597(1991)に記載の手法を用いてファージ抗体ライブラリから単離してよい。
【0045】
抗体は特に、所望の生物学的活性を呈する限り、重鎖および/または軽鎖の一部分が特定の種から誘導されるか、または特定の抗体クラスまたはサブクラスに属する抗体の相当する配列と同一または相同であるが、鎖の残余は別の種から誘導されるか、または別の抗体クラスまたはサブクラスに属する抗体の相当する配列と同一または相同である、「キメラ」抗体、並びにそのような抗体のフラグメントを包含する(米国特許4,816,567;およびMorrisonら、Proc.Natl.Acad.Sci.USA,81:6851−6855(1984))。本明細書において目的とされるキメラ抗体は非ヒト霊長類(例えば旧世界猿、類人猿等)およびヒトの定常領域配列から誘導した可変ドメイン抗原結合配列を含む霊長類化抗体を包含する。
【0046】
「抗体フラグメント」はインタクトな抗体の一部分、好ましくはその抗原結合または可変領域を含むものを含む。抗体フラグメントの例はFab、Fab’、F(ab’)2、およびFvフラグメント;ダイアボディ;直鎖状抗体;単鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体を包含する。
【0047】
「インタクトな」抗体は抗原結合可変領域並びに軽鎖定常ドメイン(CL)および重鎖定常ドメインCH1、CH2およびCH3を含むものである。定常ドメインはネイティブ配列定常ドメイン(例えばヒトネイティブ配列定常ドメイン)またはそのアミノ酸配列変異体であってよい。好ましくは、インタクトな抗体は1つ以上のエフェクター機能を有する。
【0048】
非ヒト(例えばげっ歯類)抗体の「ヒト化」型は非ヒト免疫グロブリンから誘導した最小配列を含有するキメラ抗体である。大部分において、ヒト化抗体は、レシピエントの超可変領域に由来する残基が所望の特異性、親和性、および能力を有するマウス、ラット、ウサギまたは非ヒト霊長類のような非ヒト種(ドナー抗体)の超可変領域に由来する残基により置き換えられているヒト免疫グロブリン(レシピエント抗体)である。一部の場合においては、ヒト免疫グロブリンのフレームワーク領域(FR)残基は相当する非ヒト残基により置き換えられている。さらにまた、ヒト化抗体はレシピエント抗体中、またはドナー抗体中には存在しない残基を含んでよい。そのような修飾は抗体の性能をさらに精鋭化するために行われる。一般的に、ヒト化抗体は、少なくとも1つ、そして典型的には2つの可変ドメイン(Fab、Fab’、F(ab’)2、Fabc、Fv)の実質的に全てを含むことになり、その場合、超可変ループの全てまたは実質的に全てが非ヒト免疫グロブリンのものに相当し、そしてFRの全てまたは実質的に全てがヒト免疫グロブリン配列のものである。ヒト化抗体は場合によりやはり免疫グロブリン定常領域の少なくとも一部分(Fc)、典型的にはヒト免疫グロブリンのものを含むことになる。さらに詳細な説明はJonesら、Nature 321:522−525(1986);Riechmannら、Nature 332:323−329(1988);およびPresta,Curr.Op.Struct.Biol.2:593−596(1992)を参照。
【0049】
「超可変領域」という用語は本明細書において使用する場合、配列において超可変であり、および/または構造的に定義されたループを形成する抗体可変ドメインの領域を指す。超可変領域は「相補性決定領域」即ち「CDR」に由来するアミノ酸残基(即ち軽鎖可変ドメインの残基24〜34、50〜56、および89〜97、および重鎖可変ドメインの残基31〜35、50〜65、および95〜102;Kabatら、Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,MD.(1991))および/または「超可変ループ」に由来する残基(即ち軽鎖可変ドメインの残基26〜32、50〜52、および91〜96および重鎖可変ドメインの残基26〜32、53〜55、および96〜101;Chothia and Lesk J.Mol.Biol.196:901−917(1987))を含む。両方の場合において、可変ドメイン残基は後述においてさらに詳細に考察する通り、上記したKabatらに従ってナンバリングされる。「フレームワーク」即ち「FR」残基は本明細書において定義する通り超可変領域における残基以外の可変ドメイン残基である。
【0050】
「親抗体」または「野生型」抗体は本明細書に開示した抗体変異体と比較した場合に1つ以上のアミノ酸配列改変を欠いているアミノ酸配列を含む抗体である。即ち、親抗体は一般的に本明細書に開示した抗体変異体の相当する超可変領域のアミノ酸配列とはアミノ酸配列が異なる少なくとも1つの超可変領域を有する。親ポリペプチドはネイティブ配列(即ち天然に存在する)抗体(天然に存在する対立遺伝子変異体を包含する)、または天然に存在する配列の既存のアミノ酸配列修飾(例えば挿入、欠失および/または他の改変)を有する抗体を含んでよい。本開示全体に渡って、「野生型」、「WT」、「wt」および「親」または「親の」抗体は、互換的に使用される。
【0051】
本明細書において使用する場合、「抗体変異体」または「変異体抗体」とは、親抗体のアミノ酸配列とは異なるアミノ酸配列を有する抗体を指す。好ましくは抗体変異体は天然には存在しないアミノ酸配列を有する重鎖可変ドメインまたは軽鎖可変ドメインを含む。そのような変異体は必然的に親抗体と100%未満の配列の同一性または類似性を有する。好ましい実施形態においては、抗体変異体は親抗体の重鎖または軽鎖の何れかの可変ドメインのアミノ酸配列と約75%〜100%未満、より好ましくは約80%〜100%未満、より好ましくは約85%〜100%未満、より好ましくは約90%〜100%未満、そして最も好ましくは約95%〜100%未満のアミノ酸配列の同一性または類似性のアミノ酸配列を有することになる。抗体変異体は一般的に、1つ以上の自身の超可変領域の中、またはそれに隣接して1つ以上のアミノ酸改変を含むものである。
【0052】
「アミノ酸改変」とは所定のアミノ酸配列のアミノ酸配列における変化を指す。例示される改変は挿入、置換および欠失を包含する。「アミノ酸置換」は所定のアミノ酸配列における別の異なるアミノ酸残基による既存のアミノ酸残基の置き換えを指す。
【0053】
「置き換え」アミノ酸残基はアミノ酸配列において別のアミノ酸残基を置き換えるか置換するアミノ酸残基を指す。置き換え残基は天然に存在する、または天然に存在しないアミノ酸残基であってよい。
【0054】
「アミノ酸挿入」は所定のアミノ酸配列内部への1つ以上のアミノ酸残基の導入を指す。アミノ酸挿入は「ペプチド挿入」を包含し得、この場合は、ペプチド結合で連結された2つ以上のアミノ酸残基を含むペプチドが所定のアミノ酸配列内部に導入される。アミノ酸挿入にペプチドの挿入が関与する場合、挿入されるペプチドは、天然には存在しないアミノ酸配列を有するようにランダム突然変異誘発により形成してよい。「超可変領域に隣接する」アミノ酸改変とは、超可変領域のN末端および/またはC末端における1つ以上のアミノ酸残基の導入または置換であって、挿入または置き換えのアミノ酸残基の少なくとも1つが当該超可変領域のN末端またはC末端のアミノ酸残基とペプチド結合を形成するようなものを指す。
【0055】
「天然に存在するアミノ酸残基」とはアラニン(Ala);アルギニン(Arg);アスパラギン(Asn);アスパラギン酸(Asp);システイン(Cys);グルタミン(Gln);グルタミン酸(Glu);グリシン(Gly);ヒスチジン(His);イソロイシン(Ile):ロイシン(Leu);リジン(Lys);メチオニン(Met);フェニルアラニン(Phe);プロリン(Pro);セリン(Ser);スレオニン(Thr);トリプトファン(Trp);チロシン(Tyr);およびバリン(Val)よりなる群から一般的に選択される遺伝子コードによりコードされるものである。
【0056】
「天然に存在しないアミノ酸残基」とは本明細書においては、ポリペプチド鎖中で隣接するアミノ酸残基に共有結合することができる上記列挙した天然に存在するアミノ酸残基以外のアミノ酸残基である。天然に存在しないアミノ酸残基の例はノルロイシン、オルニチン、ノルバリン、ホモセリンおよび他のアミノ酸残基類縁体、例えばEllmanら、Meth.Enzym.202:301−336(1991)に記載されているものを包含する。そのような天然に存在しないアミノ酸残基を形成するためには、Norenら、Science 244:182(1989)および上記したEllmanらの操作法を使用できる。慨すれば、これらの操作法では天然に存在しないアミノ酸残基を用いてサプレッサーtRNAを化学的に活性化し、その後、RNAのインビトロの転写および翻訳を行う。
【0057】
本開示全体に渡り、Kabat,E.A.ら、Sequences of Proteins of Immunological Interest(National Institutes of Health,Bethesda,Md.(1987)および(1991)のナンバリングシステムを参照することとする。これらの解説書において、Kabatは各サブクラスに関する抗体の多くのアミノ酸配列を列挙し、そしてそのサブクラスにおける各残基位置に関して最も一般的に生じているアミノ酸を列挙している。Kabatは列挙された配列における各アミノ酸に残基番号を割り付けるための方法を使用しており、そして残基番号を割り付けるためのこの方法は本分野において標準的なものとなっている。Kabatナンバリングスキームは本明細書において後述する通りである。本発明の目的のために、Kabatの解説書に包含されない候補抗体アミノ酸配列に残基番号を割り付けるためには、以下の工程に従う。一般的に、候補配列はKabatの何れかの免疫グロブリン配列または何れかのコンセンサス配列とアラインされる。アライメントは手作業によるか、または一般的に受け入れられているコンピュータプログラムを用いてコンピュータにより実施してよく、そのようなプログラムの例はAlign2プログラムである。アライメントは大部分のFab配列にとって共通である一部のアミノ酸残基を用いることにより容易にしてよい。例えば軽鎖および重鎖は各々典型的には、同じ残基番号を有する2つのシステインを有し;VLドメインにおいては2つのシステインは典型的には残基番号23および88にあり、そしてVHドメインにおいては2つのシステイン残基は典型的には22および92と番号付けされている。フレームワーク残基は常時ではないが一般的には概ね同じ数の残基を有するが、CDRの大きさは変動することになる。例えば自身がアラインされるKabatの配列におけるCDRよりも長い候補配列に由来するCDRの場合、典型的には接尾辞を残基番号に付加することにより追加の残基の挿入を示す(例えば図1Bの残基100abc参照)。例えば残基34および36に関してKabat配列とアラインするが、それらの間に残基35とアラインすべき残基を有しない候補配列の場合、番号35は単に残基に割り付けられない。
【0058】
本明細書において使用する場合、「高親和性」を有する抗体はナノモル(nM)範囲かそれより良好なKD、即ち解離定数を有する抗体である。「ナノモル範囲かそれより良好」なKDはXnMと表記してよく、ここでXは約10未満の数である。
【0059】
「フィラメント状ファージ」という用語は自身の表面上に非相同ポリペプチドをディスプレイできるウィルス粒子を指し、そして限定しないがf1、fd、Pf1、およびM13を包含する。フィラメント状ファージはテトラサイクリンのような選択可能なマーカーを含有してよい(例えば「fd−tet」)。種々のフィラメント状ファージディスプレイ系が当業者に良く知られている(例えばZacherら、Gene9:127−140(1980),Smithら、Science 228:1315−1317(1985);およびParmley and Smith,Gene 73:305−318(1988)参照)。
【0060】
「パニング」という用語は標的に対する高い親和性および特異性を有する抗体のようなファージ担持化合物の同定および単離におけるスクリーニングプロセスの多数回実施を指す。
【0061】
「処置する」、「処置」および「療法」という用語は本明細書において使用する場合、治癒的療法、予防的療法、および予防療法を指す。連続処置または投与は1日以上の処置における中断を伴わない少なくとも毎日を基準とした処置を指す。間歇処置または投与、または間歇的様式における処置または投与は連続的ではなく、むしろ周期的な性質にある処置を指す。
【0062】
「哺乳類」という用語は本明細書において使用する場合、哺乳類に分類される何れかの哺乳類、例えばヒト、高等な非ヒト霊長類、げっ歯類、家畜および牧場動物、例えばウシ、ウマ、イヌおよびネコを指す。本発明の好ましい実施形態においては哺乳類はヒトである。
【0063】
1つ以上の別の治療薬「と組み合わせた」投与は、同時(併用)および何れかの順序の連続投与を包含する。
【0064】
「有効量」とは有益な、または所望の治療(予防を包含)結果を起こすために十分な量である。有効量は1回以上の投与において投与できる。
【0065】
本明細書において使用する場合、「細胞」、「細胞系統」および「細胞培養物」という表現は互換的に使用され、そして全てのそのような表記は子孫を包含する。即ち、「形質転換体」および「形質転換された細胞」という単語は初期の対象細胞および形質転換の回数に関わらずそれより誘導された培養物を包含する。さらにまた、全ての子孫は意図的または偶発的な突然変異に起因してDNA含有量において厳密には同一とならない場合がある。「子孫」という用語は元来形質転換された細胞または細胞系統の後の各世代の全ての新生仔を指す。初回形質転換された細胞においてスクリーニングの対象となるものと同じ機能または生物学的活性を有する突然変異体子孫が包含される。異なる表記が意図される場合は、それは文脈から明らかになるはずである。
【0066】
「癌」および「癌性の」という用語は典型的には調節されない細胞の成育を特徴とする哺乳類における生理学的状態を指す、または説明するものである。癌の例は癌腫、リンパ腫、芽腫、肉腫、および白血病を包含するがこれらに限定されない。そのような癌のより特定的な例は、限定しないが、乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌を包含する。
【0067】
「制御配列」という用語は特定の宿主生物において作動可能に連結したコード配列の発現のために必要なDNA配列を指す。原核生物に適する制御配列は例えばプロモーター、場合によりオペレーター配列、およびリボソーム結合部位を包含する。真核生物の細胞はプロモーター、ポリアデニル化シグナルおよびエンハンサーを利用することが知られている。
【0068】
核酸はそれが別の核酸配列と機能的関連性に有るようにおかれた場合に「作動可能に連結」している。例えば、プレ配列または分泌リーダーに関するDNAは、それがポリペプチドの分泌に関与するプレタンパク質として発現される場合には、そのポリペプチドに関するDNAに作動可能に連結しており;プロモーターまたはエンハンサーはそれが配列の転写に影響する場合にコード配列に作動可能に連結しており;或いは、リボソーム結合部位はそれが翻訳を促進するように位置づけられていればコード配列に作動可能に連結している。一般的に「作動可能に連結」とは連結されるべきDNA配列が近接しており、そして分泌リーダーの場合は、近接し、そして読み相にあることを意味している。しかしながら、エンハンサーは近接している必要はない。連結は好都合な制限部位におけるライゲーションにより達成される。そのような部位が存在しない場合は、合成のオリゴヌクレオチドアダプターまたはリンカーを従来の慣行に従って使用する。
【0069】
「低分子」とは約1000ダルトン未満、好ましくは約500ダルトン未満の分子量を有するものとして本明細書においては定義される。
【0070】
「抗血管形成剤」とは血管の発生をブロック、またはある程度まで妨害する化合物を指す。抗血管形成因子は例えば血管形成を促進することに関与する成長因子または成長因子受容体に結合する低分子または抗体であってよい。本発明において好ましい抗血管形成因子はベバシズマブ(AVASTIN(登録商標))のような血管内皮成長因子(VEGF)に結合する抗体である。
【0071】
「抗新生物性組成物」という用語は少なくとも1つの活性な治療薬、例えば「抗癌剤」を含む癌を処置する場合に有用な組成物を指す。治療薬(抗癌剤)の例は、限定しないが例えば、化学療法剤、成長阻害剤、細胞毒性剤、放射線療法において使用される薬剤、抗血管形成剤、アポトーシス剤、抗チューブリン剤、および癌を処置するための他の薬剤、例えば抗HER−2抗体、抗CD20抗体、表皮成長因子受容体(EGFR)拮抗剤(例えばチロシンキナーゼ阻害剤)、HER1/EGFR阻害剤(例えばエルロチニブ(TarcevaTM)、血小板誘導成長因子阻害剤(例えばGleevecTM(イマチニブメシレート))、COX−2阻害剤(例えばセレコキシブ)、インターフェロン、サイトカイン、以下の標的ErbB2、ErbB3、ErbB4、PDGFR−β、BlyS、APRIL、BCMAまたはVEGF受容体の1つ以上に結合する拮抗剤(例えば中和抗体)、TRAIL/Apo2、および他の生物活性および有機化学の薬剤等を包含する。これらの組み合わせもまた本発明に包含される。
【0072】
本明細書において使用する「細胞毒性剤」という用語は、細胞の機能を阻害または防止する、および/または、細胞の破壊を誘発する物質を指す。該用語は放射性同位体(例えばAt211、I131、I125、Y90、Re186、Re188、Sm153、Bi212、P32およびLuの放射性同位体)、化学療法剤、および毒素、例えば低分子毒素または細菌、カビ、植物または動物起源の酵素的に活性な毒素、およびそのフラグメントおよび/または変異体を包含することを意図している。
【0073】
「化学療法剤」は癌の処置において有用な化合物である。(登録商標)化学療法剤の例はアルキル化剤、例えばチオテパおよびCYTOXANシクロホスファミド;アルキルスルホネート、例えばブスルファン、イムプロスルファンおよびピポスルファン;アジリジン、例えばベンゾドパ、カルボクオン、メツレドパ、およびウレドパ;エチレンイミンおよびメチラメラミン、例えばアルトレタミン、トリエチレンメラミン、トリエチレンホスホルアミド、トリエチレンチオホスホルアミドおよびトリメチローロメラミン;TLK 286(TELCYTATM);アセトゲニン(特にブラタシンおよびブラタシノン);デルタ−9−テトラヒドロカンナビノール(ドロナビノール、MARINOL(登録商標));β−ラパコン;ラパコール;コルヒチン;ベツリン酸;カンプトテシン(例えば合成の類縁体トポテカン(HYCAMTIN(登録商標))、CPT−11(イリノテカン、CAMPTOSAR(登録商標))、アセチルカンプトテシン、スコポレクチン、および9−アミノカンプトテシン);ブリオスタチン;カリスタチン;CC−1065(例えばそのアドゼレシン、カルゼレシンおよびビゼレシン合成類縁体);ポドフィロトキシン;ポドフィリン酸;テニポシド;クリプトフィシン(特にクリプトフィシン1およびクリプトフィシン8);ドラスタチン;デュオカルマイシン(例えば合成類縁体、KW−2189およびCB1−TM1);エレウセロビン;パンクラチスタチン;サルコジクチン;スポンジスタチン;窒素マスタード、例えばクロラムブシル、クロルナファジン、コロホスファミド、エストラムスチン、イフォスファミド、メクロレタミン、塩酸メクロレタミンオキシド、メルファラン、ノベムビシン、フェネステリン、プレドニムスチン、トロフォスファミド、ウラシルマスタード;ニトロソ尿素、例えばカルムスチン、クロロゾトシン、フォテムスチン、ロムスチン、ニムスチン、およびラニムヌスチン;ビスホスホネート、例えばクロドロネート;抗生物質、例えばエネジン抗生物質(例えばカリケアマイシン、特にカリケアマイシンγ1IおよびカリケアマイシンオメガI1(例えばAgnew、Chem Intl.編Engl.、33:183−186(1994)参照)およびアントラサイクリン、例えばアナマイシン、AD32、アルカルビシン、ダウノルビシン、デキスラゾキサン、DX−52−1、エピルビシン、GPX−100、イダルビシン、KRN5500、メノガリル、ジネミシン、例えばジネミシンA、エスペラミシン、ネオカルジノスタチン発色団および関連の発色タンパク質エネジン抗生物質(antiobiotic)発色団、アクラシノマイシン、アクチノマイシン、オースラマイシン、アザセリン、ブレオマイシン、カクチノマイシン、カラビシン、カルミノマイシン、カルジノフィリン、クロモマイシニス、ダクチノマイシン、デトルビシン、6−ジアゾ−5−オキソ−L−ノルロイシン、ADRIAMYCIN(登録商標)ドキソルビシン(例えばモルホリノ−ドキソルビシン、シアノモルホリノ−ドキソルビシン、2−ピロリノ−ドキソルビシン、リポソームドキソルビシン、およびデオキシドキソルビシン)、エソルビシン、マルセロマイシン、マイトマイシン、例えばマイトマイシンC、ミコフェノール酸、ノガラマイシン、オリボマイシン、ペプロマイシン、ポトフィロマイシン、ピューロマイシン、ケラマイシン、ロドルビシン、ストレプトニグリン、ストレプトゾシン、ツベルシジン、ウベニメックス、ジノスタチン、およびゾルビシン;葉酸類縁体、例えばデノプテリン、プテロプテリン、およびトリメトレキセート;プリン類縁体、例えばフルダラビン、6−メルカプトプリン、チアミプリン、およびチオグアニン;ピリミジン類縁体、例えばアンシタビン、アザシチジン、6−アザウリジン、カルモフル、シタラビン、ジデオキシウリジン、ドキシフルリジン、エノシタビン、およびフロクスウリジン;アンドロゲン、例えばカルステロン、ドロモスタノロンプロピオネート、エピチオスタノール、メピチオスタン、およびテストラクトン;抗アドレナール、例えばアミノグルテチミド、ミトタン、およびトリロスタン;葉酸補給剤、例えばフォリン酸(ロイコボリン);アセグラトン;抗フォレート抗新生物性剤、例えばALIMTA7、LY231514ペメトレクスド、ジヒドロフォレートレダクターゼ阻害剤、例えばメトトレキセート、代謝拮抗剤、例えば5−フルオロウラシル(5−FU)およびそのプロドラッグ、例えばUFT、S−1およびカペシタビン、およびチミジレートシンターゼ阻害剤およびグリシンアミドリボヌクレオチドホルミルトランスフェラーゼ阻害剤、例えばラルチトレキセド(TOMUDEXTM、TDX);ジヒドロピリミジンデヒドロゲナーゼの阻害剤、例えばエニルウラシル;アルドホスファミドグリコシド;アミノレブリン酸;アムサクリン;ベストラブシル;ビサントレン;エダトラキセート;デフォファミン;デメコルチン;ジアジコン;エルホルニチン;酢酸エリプチニウム;エポチロン;エトグルシド;硝酸ガリウム;ヒドロキシ尿素;レンチナン;ロニダイニン;マイタンシノイド、例えばマイタンシンおよびアンサミトシン;ミトグアゾン;ミトキサントロン;モピダンモール;ニトラエリン;ペントスタチン;フェナメット;ピラルビシン;ロソキサントロン;2−エチルヒドラジド;プロカルバジン;PSK7多糖類複合体(JHS Natural Products,Eugene,OR);ラゾキサン;リゾキシン;シゾフィラン;スピロゲルマニウム;テヌアゾン酸;トリアジコン;2,2’,2’’−トリクロロトリエチルアミン;トリコテセン(特にT−2毒素、ベラキュリンA、ロリジンAおよびアングイジン);ウレタン;ビンデシン(ELDISINE(登録商標)、FILDESIN(登録商標));ダカルバジン;マンノムスチン;ミトブロニトール;ミトラクトール;ピポブロマン;ガシトシン;アラビノシド(「Ara−C」);シクロホスファミド;チオテパ;タキソイドおよびタキサン、例えばTAXOL(登録商標)パクリタキセル(Bristol−Myers Squibb Oncology,Princeton,N.J.)、ABRAXANETMCremophor非含有、パクリタキセルのアルブミン操作ナノ粒子製剤(American Pharmaceutical Partners,Schaumberg,Illinois)、およびTAXOTERE(登録商標)ドセタキセル(Rho^ne−Poulenc Rorer,Antony,France);クロラムブシル;ゲムシタビン(GEMZAR(登録商標));6−チオグアニン;メルカプトプリン;白金;白金類縁体または白金系類縁体、例えばシスプラチン、オキサリプラチンおよびカルボプラチン;ビンブラスチン(VELBAN(登録商標));エトポシド(VP−16);イフォスファミド;ミトキサントロン;ビンクリスチン(ONCOVIN(登録商標));ビンカアルカロイド;ビノレルビン(NAVELBINE(登録商標));ノバントロン;エダトレキセート;ダウノマイシン;アミノプテリン;キセロダ;イバンドロネート;トポイソメラーゼ阻害剤RFS2000;ジフルオロメチルオルニチン(DMFO);レチノイド、例えばレチノイン酸;上記何れかの製薬上許容しうる塩、酸または誘導体;並びに上記2つ以上の組み合わせ、例えばCHOP、即ちシクロホスファミド、ドキソルビシン、ビンクリスチン、およびプレドニゾロンの複合療法の略語、およびFOLFOX、即ち5−FUおよびロイコボリンと組み合わせたオキサリプラチン(ELOXATINTM)を用いた処置計画の略語を包含する。
【0074】
この定義に同様に包含されるものは、腫瘍に対するホルモンの作用を調節または阻害するように作用する抗ホルモン剤、例えば抗エストロゲンおよび選択的エストロゲン受容体モジュレーター(SERM)、例えばタモキシフェン(例えばNOLVADEX(登録商標)タモキシフェン)、ラロキシフェン、ドロロキシフェン、4−ヒドロキシタモキシフェン、トリオキシフェン、ケオキシフェン、LY117018、オナプリストン、およびFARESTON(登録商標)トレミフェン;副腎におけるエストロゲン産生を調節する酵素アロマターゼを阻害するアロマターゼ阻害剤、例えば4(5)−イミダゾール、アミノグルテチミド、MEGASE(登録商標)酢酸メゲストロール、AROMASIN(登録商標)エキセメスタン、ホルメスタニー、ファドロゾール、RIVISOR(登録商標)ボロゾール、FEMARA(登録商標)レトロゾール、およびARIMIDEX(登録商標)アナストロゾール;および抗アンドロゲン、例えばフルタミド、ニルタミド、ビカルタミド、ロイプロリド、およびゴセレリン;並びにトロキサシタビン(1,3−ジオキソランヌクレオシドシトシン類縁体);アンチセンスオリゴヌクレオチド、特に異常な細胞増殖への関与を示唆されているシグナリング経路における遺伝子の発現を阻害するもの、例えばPKC−α、Raf、H−Ras、および表皮成長因子受容体(EGF−R);ワクチン、例えば遺伝子療法ワクチン、例えばALLOVECTIN(登録商標)ワクチン、LEUVECTIN(登録商標)ワクチン、およびVAXID(登録商標)ワクチン;PROLEUKIN(登録商標)rIL−2;LURTOTECAN(登録商標)トポイソメラーゼ1阻害剤;ABARELIX(登録商標)rmRH;および上記何れかの製薬上許容しうる塩、酸または誘導体を包含する。
【0075】
「代謝拮抗物質化学療法剤」は代謝産物に構造的には同様であるが生産的な様式では身体により使用され得ない薬剤である。多くの代謝拮抗物質化学療法剤は核酸、RNAおよびDNAの産生を妨害する。代謝拮抗物質化学療法剤の例は、ゲムシタビン(GEMZAR(登録商標))、5−フルオロウラシル(5−FU)、カペシタビン(XELODATM)、6−メルカプトプリン、メトトレキセート、6−チオグアニン、ペメトレクスド、ラルチトレクスド、アラビノシルシトシンARA−Cシタラビン(CYTOSAR−U(登録商標))、ダカルバジン(DTIC−DOME(登録商標))、アゾシトシン、デオキシシトシン、ピリミジン、フルダラビン(FLUDARA(登録商標))、クラドラビン、2−デオキシ−D−グルコース等を包含する。好ましい代謝拮抗物質化学療法剤はゲムシタビンである。
【0076】
「ゲムシタビン」即ち「2’−デオキシ−2’,2’−ジフルオロシチジン一塩酸塩(b異性体)」は抗腫瘍活性を示すヌクレオシド類縁体である。ゲムシタビンHClに関する実験式はC9H11F2N3O4 A HClである。ゲムシタビンHClは商標GEMZAR(登録商標)の下にEli Lillyから販売されている。
【0077】
「白金系化学療法剤」は分子の一体的部分として白金を含有する有機化合物を含む。白金系化学療法剤の例はカルボプラチン、シスプラチン、およびオキサリプラチンを包含する。
【0078】
「白金系化学療法」とは、場合により1つ以上の他の化学療法剤と組み合わせた1つ以上の白金系化学療法剤を用いた療法を意図する。
【0079】
II.発明を実施するための形態
本発明の実施は特段の記載が無い限り当業者の技量の範囲内の分子生物学(組み換え手法を含む)、微生物学、細胞生物学、生物化学および免疫学の従来の手法を使用することになる。そのような手法は文献、例えば「Molecular Cloning:A Laboratory Manual」,第2版(Sambrookら、1989);「Oligonucleotide Synthesis」(M.J.Gait編、1984);「Animal Cell Culture」(R.I.Freshney編、1987);「Methods in Enzymology」(Academic Press,Inc.);「Handbook of Experimental Immunology」,第4版(D.M.Weir&C.C.Blackwell編、Blackwell Science Inc.,1987);「GeneTransfer Vectors for Mammalian Cells」(J.M.Miller&M.P.Calos編、1987);「Current Protocols in Molecular Biology」(F.M.Ausubelら編、1987);「PCR:The Polymerase Chain Reaction」(Mullisら編、1994);および「Current Protocols in Immunology」(J.E.Coliganら編、1991)において詳細に説明されている。
【0080】
1.アゴニスト抗CYTL1抗体の調製
抗CYTL1抗体は組み換えDNA技術の手法を包含する当該分野で知られた方法により製造できる。
【0081】
i)抗原の調製
場合により他の分子にコンジュゲートされた可溶性抗原またはそのフラグメントは抗体を形成するための免疫原として使用できる。膜貫通分子、例えば受容体に関しては、これらのフラグメント(例えば受容体の細胞外ドメイン)を免疫原として使用できる。或いは、膜貫通分子を発現する細胞を免疫原として使用できる。そのような細胞は天然原料(例えば癌細胞系統)から誘導することができ、或いは膜貫通分子を発現するように組み換え手法により形質転換されている細胞であってよい。抗体を調製するために有用な他の抗原およびその形態は当業者の知る通りである。
【0082】
(ii)ポリクローナル抗体
ポリクローナル抗体は好ましくは該当する抗原およびアジュバントの多数回の皮下(sc)または腹腔内(ip)注射により動物中で育成される。2官能性または誘導体化剤、例えばマレイミドベンゾイルスルホスクシンイミドエステル(システイン残基を介したコンジュゲーション)、N−ヒドロキシスクシンイミド(リジン残基を介する)、グルタルアルデヒド、無水コハク酸、SOCl2またはR1N=C=NR、ただし式中RおよびR1は異なるアルキル基であるもの、を用いて、免疫すべき種において免疫原性であるタンパク質、例えばキーホールリンペットヘモシアニン、血清アルブミン、ウシサイログロブリン、または大豆トリプシン阻害剤に該当する抗原をコンジュゲートすることが有用である場合がある。
【0083】
動物は100μgまたは5μgのタンパク質またはコンジュゲート(それぞれウサギまたはマウスに関する)を3容量のフロイント完全アジュバントと混合し、溶液を多数部位において皮内注射することにより抗原、免疫原性コンジュゲート、または誘導体に対抗して免疫される。1ヵ月後、多数部位における皮下注射によりフロイントの完全アジュバント中のペプチドまたはコンジュゲートの元の量の1/5〜1/10を用いて動物をブーストする。7〜14日後、動物から出血させ、そして血清の抗体力価をアッセイする。動物は力価が平衡に達するまでブーストする。好ましくは、同じ抗原であるが、異なるタンパク質に、および/または異なる交差結合試薬を介してコンジュゲートされているコンジュゲートで動物をブーストする。コンジュゲートはまた、タンパク質融合物として組み換え細胞培養物中に作成できる。さらにまた、ミョウバンのような凝集剤を適宜使用して免疫応答を増強する。
【0084】
(iii)モノクローナル抗体
モノクローナル抗体はKohlerら、Nature,256:495(1975)により最初に記載されたハイブリドーマ法を用いて作成してよく、或いは、組み換えDNA法(米国特許4,816,567)により作成してよい。ハイブリドーマ法においては、マウスまたは他の適切な宿主動物、例えばハムスターまたはマカクサルを上記した通り免疫することにより、免疫のために使用されるタンパク質に特異的に結合することになる抗体を産生するか産生生産できるリンパ球を生じさせる。或いは、リンパ球はインビトロで免疫してよい。次にリンパ球を適切な融合剤、例えばポリエチレングリコールを用いて骨髄腫細胞と融合させることによりハイブリドーマ細胞を形成する(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。
【0085】
このようにして調製されたハイブリドーマ細胞は、未融合の親骨髄腫細胞の成育または生存を阻害する1つ以上の物質を好ましくは含有する適当な培養培地中に播種して生育させる。例えば、親骨髄腫細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠いている場合、ハイブリドーマのための培養培地は典型的にはヒポキサンチン、アミノプテリン、およびチミジンを包含することになり(HAT培地)、これらの物質はHGPRT欠損細胞の成育を防止する。
【0086】
好ましい骨髄腫細胞は、効率的に融合し、選択された抗体産生細胞による抗体の安定な高レベル産生を支援し、そしてHAT培地のような培地に対して感受性であるものである。それらのうち、好ましい骨髄腫細胞系統はネズミ骨髄腫系統、例えばSalk Institute Cell Distribution Center,San Diego,Calif.USAから入手可能なMOPC−21およびMPC−11マウス腫瘍、および、American Type Culture Collection,Rockville,Md.USAから入手可能なSP−2またはX63−Ag8−653細胞から誘導したものである。ヒト骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞系統もまた、ヒトモノクローナル抗体の産生に関して記載されている(Kozbor,J.Immunol.,133:3001(1984);Brodeurら、Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0087】
ハイブリドーマ細胞を成育させている培養培地は抗原に対するモノクローナル抗体の産生に関してアッセイされる。好ましくは、ハイブリドーマ細胞により産生されたモノクローナル抗体の結合特異性は、免疫沈降により、またはインビトロの結合アッセイ、例えばラジオイムノアッセイ(RIA)または酵素結合免疫吸着アッセイ(ELISA)により、測定される。
【0088】
所望の特異性、親和性、および/または活性の抗体を産生するハイブリドーマ細胞が同定された後、クローンは限界希釈操作法によりサブクローニングし、そして標準的な方法により生育させてよい(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。この目的のための適当な培養培地は例えばD−MEMまたはRPMI−1640培地を包含する。さらにまたハイブリドーマ細胞は動物中の腹水腫瘍としてインビボで成育させてよい。
【0089】
サブクローンにより分泌されたモノクローナル抗体は例えばプロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、またはアフィニティークロマトグラフィーのような従来の免疫グロブリン精製操作法により培養培地、腹水、または血清から適当に分離される。
【0090】
モノクローナル抗体をコードするDNAは従来の操作法を用いて容易に単離して配列決定される(例えばモノクローナル抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合できるオリゴヌクレオチドプローブを使用することによる)。ハイブリドーマ細胞はそのようなDNAの好ましい原料として供される。単離された後、DNAは発現ベクター内に入れてよく、そしてこれを次に宿主細胞、例えばE.coli細胞、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、または別様には免疫グロブリンタンパク質を産生しない骨髄腫細胞内にトランスフェクトすることにより、組み換え宿主細胞内のモノクローナル抗体の合成を達成できる。抗体の組み換え産生は後述においてより詳細に説明する。
【0091】
別の実施形態においては、抗体または抗体フラグメントはMcCaffertyら、Nature,348:552−554(1990)に記載の手法を用いて作成された抗体ファージライブラリから単離できる。
【0092】
Clacksonら、Nature,352:624−628(1991)およびMarksら、J.Mol.Biol.,222:581−597(1991)はファージライブラリを用いたそれぞれネズミおよびヒトの抗体の単離を記載している。その後の出版物は鎖シャフリングによる高親和性(nM範囲)のヒト抗体の産生(Marksら、Bio/Technology,10:779−783(1992))並びに極めて大型のファージライブラリを構築するための方策としてのコンビナトリアルな感染およびインビボの組み換え(Waterhouseら、Nuc.Acids.Res.,21:2265−2266(1993))を記載している。即ち、これらの手法はモノクローナル抗体の単離のための伝統的なモノクローナル抗体ハイブリドーマ手法の実行可能な代替法である。
【0093】
DNAはまた、相同ネズミ配列の代わりにヒト重鎖および軽鎖定常ドメインに関するコード配列を代替とすること(米国特許4,816,567;Morrisonら、Proc.Natl Acad.Sci.USA,81:6851(1984))によるか、または、非免疫グロブリンポリペプチドに関するコード配列の全てまたは部分を免疫グロブリンコード配列に共有結合することにより、修飾してよい。
【0094】
典型的には、そのような非免疫グロブリンポリペプチドは抗体の定常ドメインの代替とされるか、またはそれらは、抗体の1つの抗原複合化部位の可変ドメインの代替とされ、これにより抗原に対する特異性を有する1つの抗原複合化部位および異なる抗原に対する特異性を有する別の抗原複合化部位を含むキメラ2価抗体を作製する。
【0095】
(iv)ヒト化およびヒト抗体
ヒト化抗体は非ヒトである原料から自身内に導入された1つ以上のアミノ酸残基を有する。これらの非ヒトアミノ酸残基はしばしば「インポート」残基と称され、これは典型的には「インポート」可変ドメインから取り出される。ヒト化は本質的にはWinterらの方法(Jonesら、Nature,321:522−525(1986);Riechmannら、Nature,332:323−327(1988);Verhoeyenら、Science,239:1534−1536(1988))に従って、ヒト抗体の相当する配列に対してげっ歯類のCDRまたはCDR配列を置換することにより実施することができる。従って、そのような「ヒト化」抗体は実質的にはインタクトなヒト可変ドメインに満たない部分が非ヒト種由来の相当する配列により置換されているキメラ抗体(米国特許4,816,567)である。実際には、ヒト化抗体は典型的には一部のCDR残基および可能性として一部のFR残基がげっ歯類抗体中の類似の部位に由来する残基により置換されているヒト抗体である。
【0096】
ヒト化抗体を作成する場合に使用するべき軽鎖および重鎖の両方のヒト可変ドメインの選択は抗原性を低減するためには極めて重要である。所謂「ベストフィット」法に従って、げっ歯類抗体の可変ドメインの配列を既知ヒト可変ドメイン配列の完全なライブラリに対してスクリーニングする。次にげっ歯類のものと最も近似しているヒト配列をヒト化抗体のためのヒトフレームワーク(FR)として許容する(Simsら、J.Immunol.,151:2296(1993);Chothiaら、J.Mol.Biol.,196:901(1987))。別の方法は軽鎖または重鎖の特定のサブグループの全ヒト抗体のコンセンサス配列から誘導された特定のフレームワークを使用する。同じフレームワークをいくつかの異なるヒト化抗体に対して使用してよい(Carterら、Proc.Natl.Acad Sci.USA,89:4285(1992);Prestaら、J.Immnol.,151:2623(1993))。
【0097】
さらにまた、抗原に対する高親和性および他の望ましい生物学的特性を保持して抗体をヒト化することが重要である。この目標を達成するためには、好ましい方法に従って、親およびヒト化配列の3次元モデルを用いて親配列および種々の概念的なヒト化産物の分析のプロセスによりヒト化抗体を調製する。3次元免疫グロブリンモデルは公的に入手可能であり、そして当業者に良く知られている。選択された候補免疫グロブリン配列の推定の3次元コンホメーション構造を図示して表示できるコンピュータプログラムが使用可能である。これらの表示物の検討により候補免疫グロブリン配列の機能における残基の考えられる役割の分析、即ち、候補免疫グロブリンがその抗原に結合する能力に影響する残基の分析が可能となる。このような方法により、FR残基を選択し、そして、所望の抗体特性、例えば標的抗原に対する増大した親和性を達成できるようにレシピエントおよびインポート配列と組み合わせることができる。一般的に、CDR残基は、抗原結合に対して影響を与えることにおいては、直接、そして最も大きく関与している。
【0098】
或いは、内因性の免疫グロブリンの産生の非存在下においてヒト抗体の完全なレパートリーの産生が免疫により可能となるトランスジェニック動物(例えばマウス)を作成することが、現在可能である。例えば、キメラおよび生殖細胞系統の突然変異体マウスにおける抗体重鎖連結領域(JH)遺伝子のホモ接合欠失は内因性抗体産生の完全な阻害をもたらすことが記載されている。そのような生殖細胞系統突然変異体マウスにおけるヒト生殖細胞系統免疫グロブリン遺伝子アレイの転移は、抗原攻撃時のヒト抗体の産生をもたらすことになる。例えばJakobovitsら、Proc.Natl.Acad.Sci.USA,90:2551(1993);Jakobovitsら、Nature,362:255−258(1993);Bruggermannら、Year in Immuno.,7:33(1993);およびDuchosalら、Nature,355:258(1992)を参照。ヒト抗体はまたファージディスプレイライブラリから誘導できる(Hoogenboomら、J.Mol.Biol.,227:381(1991);Marksら、J.MoL Biol.,222:581−597(1991);Vaughanら、Nature Biotech.,14:309(1996))。抗体ファージディスプレイライブラリからのヒト抗体の形成を以下にさらに説明する。
【0099】
(v)抗体フラグメント
種々の手法が抗体フラグメントの産生のために開発されている。伝統的にはこれらのフラグメントはインタクトな抗体のタンパク質分解性消化を介して誘導されている(例えばMorimotoら、Journal of Biochemical and Biophysical Methods,24:107−117(1992)およびBrennanら、Science,229:81(1985)参照)。しかしながらこれらのフラグメントは現在では組み換え宿主細胞により直接産生できる。例えば抗体フラグメントは上記考察した抗体ファージライブラリから単離できる。或いはFab’−SHフラグメントをE.coliから直接回収し、そして化学的にカップリングさせることによりF(ab’)2フラグメントを形成できる(Carterら、Bio/Technology,10:163−167(1992))。後述する実施例において説明する通り別の実施形態においては、F(ab’)2はF(ab’)2分子の組み立てを促進するためにロイシンジッパーGCN4を用いて形成される。別の手順によれば、F(ab’)2フラグメントは組み換え宿主細胞培養物から直接単離できる。抗体フラグメントの産生のための他の手法は当業者に良く知られている。他の実施形態においては選択される抗体は単鎖Fvフラグメント(scFv)である。WO93/16185参照。
【0100】
(vi)多重特異性抗体
多重特異性抗体はエピトープが通常は異なる抗原に由来する少なくとも2つの異なるエピトープに対する結合特異性を有する。そのような分子は通常は2つの異なるエピトープに結合するのみとなる(即ち二重特異性抗体、BsAb)が、三重特異性抗体のような追加的特異性を有する抗体も本明細書において使用する場合にこの表現に包含される。
【0101】
二重特異性抗体を作成するための方法は当該分野において知られている。完全長二重特異性抗体の伝統的な産生は2つの鎖が異なる特異性を有する2つの免疫グロブリン重鎖−軽鎖の対の同時発現に基づいている(Millsteinら、Nature,305:537−539(1983))。免疫グロブリン重鎖および軽鎖のランダムな組み合わせのために、これらのハイブリドーマ(クアドローマ)は10種の異なる抗体分子の潜在的混合物を産生し、そのうち僅か1つのみが正しい二重特異性の構造を有する。通常はアフィニティークロマトグラフィー工程により行われる正しい分子の精製はむしろ負担が大きく、そして生成物の収率は低値である。同様の操作法がWO93/08829、およびTrauneckerら、EMBO J.,10:3655−3659(1991)に開示されている。異なる手順によれば、所望の結合特異性を有する抗体可変ドメイン(抗体−抗原複合化部位)を免疫グロブリン定常ドメイン配列に融合させる。融合は好ましくはヒンジ、CH2、およびCH3領域の少なくとも一部分を含む免疫グロブリン重鎖定常ドメインと行う。軽鎖結合のために必要な部位を含有する第1の重鎖定常領域(CH1)が融合物の少なくとも1つにおいて存在することが好ましい。免疫グロブリン重鎖融合物、および所望により免疫グロブリン軽鎖をコードするDNAを別個の発現ベクター内に挿入し、そして適当な宿主生物内に同時トランスフェクトする。これにより、構築において使用される3つのポリペプチド鎖の等しくない比率が最適な収率をもたらす場合の実施形態において3つのポリペプチドフラグメントの相互の割合を調節する場合に、大きい柔軟性がもたらされる。しかしながら、等しい比率における少なくとも2つのポリペプチド鎖の発現が高い収率をもたらす場合、または比率が特に重要ではない場合に、1つの発現ベクター中に2つまたは3つ全てのポリペプチド鎖に関するコード配列を挿入することが可能である。
【0102】
この手順の好ましい実施形態において、二重特異性抗体は、1つのアームにおいて第1の結合特異性を有するハイブリッド免疫グロブリン重鎖、および別のアームにおいてハイブリッド免疫グロブリン重鎖−軽鎖対(第2の結合特異性を与える)を有している。この非対称の構造は、二重特異性分子の半分のみにおける免疫グロブリン軽鎖の存在が容易な分離方法を可能とすることから、望ましくない免疫グロブリン鎖の組み合わせからの所望の二重特異性の化合物の分離を容易にする。この手順はWO94/04690に開示されている。二重特異性抗体の形成はさらに詳細には例えばSureshら、Methods in Enzymology,121:210(1986)を参照。
【0103】
WO96/27011に記載されている別の手順に従えば、抗体分子の対の間の界面は、組み換え細胞培養物から回収されるヘテロ二量体のパーセントを最大限とするように操作できる。好ましい界面は抗体定常ドメインのCH3ドメインの少なくとも一部分を含む。この方法においては、第1の抗体分子の界面に由来する1つ以上の小型アミノ酸側鎖をより大型の側鎖(例えばチロシンまたはトリプトファン)と置き換える。大型側鎖と同一または同様のサイズの代償分の「空洞」は、大型アミノ酸側鎖をより小さいもの(例えばアラニンまたはスレオニン)で置き換えることにより第2の抗体分子の界面上に形成される。これによりホモ二量体のような他の望ましくない最終生成物を超えてヘテロ二量体の収率を増大させるための機序が与えられる。
【0104】
二重特異性抗体は交差結合または「ヘテロコンジュゲート」抗体を包含する。例えばヘテロコンジュゲートにおける抗体の1つをアビジンに、もう1つをビオチンにカップリングできる。そのような抗体は例えば、望ましくない細胞に免疫系細胞をターゲティングするため(米国特許4,676,980)およびHIV感染症の処置のため(WO91/00360、WO92/200373)に提案されている。ヘテロコンジュゲート抗体は何れかの好都合な交差結合法を用いて作製してよい。適当な交差結合剤は当該分野で良く知られており、そして多くの交差結合手法と共に米国特許4,676,980に開示されている。
【0105】
抗体フラグメントから二重特異性抗体を形成するための手法もまた、文献に記載されている。例えば二重特異性抗体は化学結合を用いて調製できる。Brennanら、Science,229:81(1985)はインタクトな抗体をタンパク質分解的に切断することによりF(ab’)2フラグメントを形成する操作法を記載している。これらのフラグメントをジチオール錯化剤(dithiol complexing agent)である亜ヒ酸ナトリウムの存在下に還元することにより近接したジチオールを安定化させ、そして分子内のジスルフィド形成を防止する。次に形成されたFab’フラグメントをチオニトロベンゾエート(TNB)誘導体に変換する。次にFab’−TNB誘導体の一方をメルカプトエチルアミンを用いた還元によりFab’−チオールに再変換し、そして等モル量の他のFab’−TNB誘導体と混合することにより二重特異性抗体を形成する。生成した二重特異性抗体は酵素の選択的固定化のための薬剤として使用できる。
【0106】
Fab’−SHフラグメントはまたE.coliから直接回収することができ、そして化学的にカップリングすることにより二重特異性抗体を形成できる。Shalabyら、J.Exp.Med.,175:217−225(1992)は完全にヒト化された二重特異性抗体F(ab’)2分子の産生を記載している。各Fab’フラグメントをE.coliから別個に分泌させ、そしてインビトロの指向的化学カップリングに付すことにより二重特異性抗体を形成している。
【0107】
組み換え細胞培養物から直接二重特異性抗体フラグメントを作成して単離するための種々の手法もまた記載されている。例えば、二重特異性抗体はロイシンジッパーを用いて産生されている。Kostelnyら、J.Immunol.,148(5):1547−1553(1992)。FosおよびJunタンパク質に由来するロイシンジッパーペプチド(pepbdes)を遺伝子融合により2つの異なる抗体のFab’部分に連結している。抗体ホモ二量体をヒンジ領域で還元することにより単量体を形成し、そして次に再酸化して抗体ヘテロ二量体を形成している。この方法はまた、抗体ホモ二量体の産生のためにも利用できる。Hollingerら、Proc.Nati.Acad.Sci.USA,90:6444−6448(1993)の記載している「ダイアボディ」技術は二重特異性抗体フラグメントを作成するための代替的な機序を与えている。フラグメントは同じ鎖上の2ドメインの間の対形成を可能にするには短すぎるリンカーにより軽鎖可変ドメイン(VL)に結合した重鎖可変ドメイン(VH)を含む。従って、1つのフラグメントのVHおよびVLドメインは別のフラグメントの相補なVLおよびVHドメインと強制的に対形成させられ、これにより2つの抗原結合部位が形成される。単鎖Fv(sFv)二量体の使用により二重特異性抗体フラグメントを形成するための別の方策もまた報告されている。Gruberら、J.Immunol.,152:5368(1994)を参照。
【0108】
2より多い結合価を有する抗体もまた意図される。例えば三重特異性抗体を調製できる。Tuftら、J.Immunol.,147:60(1991)。
【0109】
(vii)エフェクター機能の操作
抗体の有効性を増強するように、エフェクター機能に関して本発明の抗体を修飾することが望ましい場合がある。例えば、システイン残基をFc領域に導入することによりこの領域における鎖間ジスルフィド結合の形成を可能にしてよい。このようにして形成されたホモ二量体抗体は向上した内在化能力および/または増大した補体媒介細胞殺傷および抗体依存性細胞毒性(ADCC)を有する場合がある。Caronら、J.Exp Med.,176:1191−1195(1992)およびShopes,B.J.Immunol.,148:2918−2922(1992)を参照。増強された抗腫瘍活性を有するホモ二量体抗体はまた、Wolffら、Cancer Research,53:2560−2565(1993)において記載される通りヘテロ2官能性交差結合剤を用いて調製してよい。或いは、抗体は二重のFc領域を有するように操作することができ、そしてこれにより、増強された補体溶解およびADCC能力を有する場合がある。Stevensonら、Anti−Cancer Drug Design 3:219−230(1989)を参照。
【0110】
(viii)抗体サルベージ受容体結合エピトープ融合物
本発明の特定の実施形態においては、例えば腫瘍浸透性を増大させるためにインタクトな抗体よりもむしろ抗体フラグメントを使用することが望ましい場合がある。そのような場合、抗体フラグメントはその血清中半減期を増大させるために修飾することが望ましい場合がある。これは例えば抗体フラグメント内へのサルベージ受容体結合エピトープの取り込みにより達成してよい(例えば抗体フラグメント内の適切な領域の突然変異により、或いは、エピトープをペプチドタグ内に取り込ませ、次にこれを、例えばDNAまたはペプチド合成により、抗体フラグメントに何れかの末端または中央部において融合する)。
【0111】
サルベージ受容体結合エピトープは好ましくは、Fcドメインの1つまたは2つのループに由来するアミノ酸残基の何れか1つ以上が抗体フラグメントの類似の位置に転移する領域を構築する。さらに好ましくは、Fcドメインの1つまたは2つのループに由来する残基3つ以上が転移する。なおさらに好ましくは、エピトープはFc領域のCH2ドメイン(例えばIgGのもの)から取り出し、そして、抗体のCH1、CH3、またはVH領域、または1つより多くのそのような領域に転移させる。或いは、エピトープをFc領域のCH2ドメインから取り出し、そして抗体フラグメントのCL領域またはVL領域、または両方に転移させる。
【0112】
(ix)抗体の他の共有結合的修飾
抗体の共有結合的修飾は本発明の範囲内に包含される。それらは適宜、化学合成によるか、または酵素的または化学的な抗体切断により行ってよい。抗体の共有結合的修飾の他の型は、選択された側鎖またはNまたはC末端残基と反応できる有機の誘導体化剤に抗体のターゲティングされたアミノ酸残基を反応させることにより分子内に導入される。共有結合的修飾の例は参照により本明細書に組み込まれる米国特許5,534,615に記載されている。抗体の共有結合的修飾の好ましい型は米国特許4,640,835;4,496,689;4,301,144;4,670,417;4,791,192または4,179,337に記載されている様式において、種々の非タンパク質性重合体、例えばポリエチレングリコール、ポリプロピレングリコール、またはポリオキシアルキレンの1つに抗体を連結させることを含む。
【0113】
(x)合成抗体ファージライブラリからの抗体の形成
好ましい実施形態においては、本発明はユニークなファージディスプレイの手順を用いて新規な抗体を形成および選択するための方法を提供する。該手順においては、単一のフレームワーク鋳型に基づく合成抗体ファージライブラリの形成、可変ドメイン内部の十分な多様性の設計、多様化された可変ドメインを有するポリペプチドのディスプレイ、抗原をターゲティングするための高親和性を有する候補抗体の選択、および選択された抗体の単離を行う。
【0114】
ファージディスプレイの方法の詳細は例えば参照により全体が本明細書に組み込まれる2003年12月11日に公開されたWO03/102157に記載されている。
【0115】
1つの態様において、本発明において使用される抗体ライブラリは抗体可変ドメインの少なくとも1つのCDRにおいて溶媒接触性および/または高多様性の位置を突然変異させることにより形成できる。CDRの一部または全てを本明細書に記載した方法を用いて突然変異させることができる。一部の実施形態においては、多様な抗体ライブラリは、単一のライブラリを形成するためにCDRH1、CDRH2およびCDRH3における位置を突然変異させることによるか、または単一のライブラリを形成するためにCDRL3およびCDRH3における位置を突然変異させることによるか、または単一のライブラリを形成するためにCDRL3およびCDRH1、CDRH2およびCDRH3における位置を突然変異させることにより形成することが好ましい場合がある。
【0116】
抗体可変ドメインのライブラリは例えばCDRH1、CDRH2およびCDRH3の溶媒接触性および/または高多様性の位置において突然変異を有するように形成できる。別のライブラリはCDRL1、CDRL2およびCDRL3において突然変異を有するように形成できる。これらのライブラリはまた、所望の親和性を有するバインダーを形成するために相互に組み合わせて使用できる。例えば標的抗原への結合のための重鎖ライブラリの選択を1回以上行った後、軽鎖ライブラリを重鎖バインダーの集団内に入れ換え、選択回数をさらに重ねることによりバインダーの親和性を増大させることができる。
【0117】
好ましくは、重鎖配列の可変領域のCDRH3領域において変異体アミノ酸により元のアミノ酸を置換することによりライブラリを形成する。得られたライブラリは複数の抗体配列を含有することができ、そこにおいては配列の多様性は重鎖配列のCDRH3領域に主に存在する。
【0118】
1つの態様において、ライブラリはヒト化抗体4D5配列、またはヒト化抗体4D5配列のフレームワークアミノ酸の配列との関連で形成される。好ましくはライブラリはDVKコドンセットによりコードされるアミノ酸による重鎖の少なくとも残基95〜100aの置換により形成され、その場合、DVKコドンセットはこれらの位置の各々に対して変異体アミノ酸のセットをコードするために使用される。これらの置換を形成するために有用なオリゴヌクレオチドのセットの例は配列(DVK)7を含む。一部の実施形態においては、ライブラリはDVKおよびNNKコドンセットの両方でコードされたアミノ酸による残基95〜100aの置換により形成する。これらの置換を形成するために有用なオリゴヌクレオチドのセットの例は配列(DVK)6(NNK)を含む。別の実施形態においては、ライブラリはDVKおよびNNKコドンセットの両方でコードされたアミノ酸による少なくとも残基95〜100aの置換により形成する。これらの置換を形成するために有用なオリゴヌクレオチドのセットの例は配列(DVK)5(NNK)を含む。これらの置換を形成するために有用なオリゴヌクレオチドセットの別の例は配列(NNK)6を含む。適当なオリゴヌクレオチド配列の他の例は本明細書に記載する基準に従って当業者が決定できる。
【0119】
別の実施形態においては、異なるCDRH3設計を利用することにより高親和性バインダーを単離し、そして種々のエピトープに対するバインダーを単離する。このライブラリにおいて形成されるCDRH3の長さの範囲は11〜13アミノ酸であるが、これとは異なる長さも形成することができる。H3の多様性はNNK、DVKおよびNVKコドンセット、並びにNおよび/またはC末端におけるより限定された多様性を使用することにより拡張できる。
【0120】
多様性はまた、CDRH1およびCDRH2において形成することもできる。CDR−H1およびH2の多様性の設計は以前の設計よりも天然の多様性により緊密にマッチしている多様性に着目した変更を伴って記載した天然の抗体レパートリーを模倣するためのターゲティングの方策に従う。
【0121】
CDRH3における多様性に関しては、異なる長さのH3を有するように多数のライブラリを別個に構築し、そして次に、抗原をターゲティングするためのバインダーを求める選別のために組み合わせることができる。多数のライブラリをプールし、そして明細書中前述および後述する固体支持体選択および溶液分類方法を用いて分類することができる。マルチソーティングの方策を使用してよい。例えば1つの変法では固体に結合した標的に対する分類を行い、その後、融合ポリペプチド上に存在する場合があるタグ(例えば抗gDタグ)に対する分類を行い、そしてその後、固体に結合した標的に対する別の分類を行う。或いは、ライブラリを固体表面に結合した標的に対して先ず分類することができ、次に溶出したバインダーを標的抗原の漸減濃度における溶液相結合を用いて分類する。異なる分類方法の組み合わせを利用することは高度に発現される配列のみの選択の最小限化を可能にし、そして、多くの異なる高親和性クローンの選択を可能にする。
【0122】
標的抗原に対する高親和性バインダーをライブラリから単離できる。H1/H2領域における限定された多様性は縮重性を約104〜105倍低下させ、そしてより高値のH3多様性を可能にすれば、より高親和性のバインダーが得られる。CDRH3における異なる型の多様性を有するライブラリを利用することにより(例えばDVKまたはNVTを利用することにより)、標的抗原の異なるエピトープに結合する場合があるバインダーの単離が可能となる。
【0123】
上記した通りプールされたライブラリから単離されたバインダーのうち、親和性は軽鎖において限定された多様性をもたらすことによりさらに向上する場合があることが発見された。本実施形態において軽鎖の多様性は以下の通り、即ちCDRL1において:アミノ酸位置28はRDTによりコードされ;アミノ酸位置29はRKTによりコードされ;アミノ酸位置30はRVWによりコードされ;アミノ酸位置31はANWによりコードされ;アミノ酸位置32はTHTによりコードされ;場合により、アミノ酸位置33はCTGによりコードされ;CDRL2において:アミノ酸位置50はKBGによりコードされ;アミノ酸位置53はAVCによりコードされ;そして場合により、アミノ酸位置55はGMAによりコードされ;CDRL3において:アミノ酸位置91はTMTまたはSRTまたは両方によりコードされ;アミノ酸位置92はDMCによりコードされ;アミノ酸位置93はRVTによりコードされ;アミノ酸位置94はNHTによりコードされ;そしてアミノ酸位置96はTWTまたはYKGまたは両方によりコードされるように、形成される。
【0124】
別の実施形態においては、CDRH1、CDRH2およびCDRH3領域における多様性を有するライブラリを形成する。本実施形態においてCDRH3における多様性は種々の長さのH3領域を用いて、そして主にコドンセットXYZおよびNNKまたはNNSを用いて形成する。ライブラリは個々のオリゴヌクレオチドを用いて形成してプールすることができ、或いはオリゴヌクレオチドをプールしてライブラリのサブセットを形成することができる。本実施形態のライブラリは固体に結合した標的に対抗して分類できる。多数の分類から単離されたクローンはELISAアッセイを用いて特異性および親和性に関してスクリーニングできる。特異性に関しては、クローンは所望の標的抗原並びに他の非標的抗原に対抗してスクリーニングできる。次に標的抗原に対するこのようなバインダーを溶液結合競合ELISAアッセイまたはスポット競合アッセイにおいて親和性に関してスクリーニングできる。高親和性のバインダーは上記した通り調製したXYZコドンセットを利用してライブラリから単離できる。これらのバインダーは細胞培養物中において高収率で抗体または抗原結合フラグメントとして容易に製造できる。
【0125】
一部の実施形態においては、CDRH3領域における長さにおいてより大きい多様性を有するライブラリを形成することが望ましい場合がある。例えば約7〜19アミノ酸の範囲のCDRH3領域を有するライブラリを形成することが望ましい場合がある。
【0126】
これらの実施形態のライブラリから単離された高親和性バインダーは高収率で細菌および真核生物細胞の培養物中において容易に製造される。ベクターはgDタグ、ウィルス被膜タンパク質成分配列のような配列を容易に除去するように、および/または、高収率で完全長抗体または抗原結合フラグメントの産生を可能にする定常領域配列において付加するために設計できる。
【0127】
CDRH3において突然変異を有するライブラリは他のCDR、例えばCDRL1、CDRL2、CDRL3、CDRH1および/またはCDRH2の変異体型を含有するライブラリと組み合わせることができる。即ち、例えば、1つの実施形態において、CDRH3ライブラリを、所定のコドンセットを用いて28、29、30、31、および/または32位において変異体アミノ酸を有するヒト化4D5抗体配列との関連で形成されたCDRL3ライブラリと組み合わせる。別の実施形態においては、CDRH3への突然変異を有するライブラリは、変異体CDRH1および/またはCDRH2重鎖可変ドメインを含むライブラリと組み合わせることができる。1つの実施形態において、CDRH1ライブラリは28、30、31、32および33位において変異体アミノ酸を有するヒト化抗体4D5配列を有するように形成される。CDRH2ライブラリは所定のコドンセットを用いて50、52、53、54、56および58位において変異体アミノ酸を有するヒト化抗体4D5配列を有するように形成される。
【0128】
(xi)抗体突然変異体
抗体突然変異を形成するためには、1つ以上のアミノ酸改変(例えば置換)を親抗体の1つ以上の超可変領域に導入する。或いは、または追加的に、フレームワーク領域残基の1つ以上の改変(例えば置換)を親抗体に導入してよく、その場合、これらは第2の哺乳類種に由来する抗原に対する抗体突然変異体の結合親和性の向上をもたらす。修飾すべきフレームワーク領域の残基の例は、直接抗原に非共有結合的に結合し(Amitら、(1986)Science 233:747−753);CDRのコンホメーションに相互作用/影響し(Chothiaら、(1987)J.Mol.Biol.196:901−917);および/またはVL−VH界面に関与する(EP239 400B1)ものを包含する。特定の実施形態においては、1つ以上のそのようなフレームワーク領域残基の修飾は第2の哺乳類種に由来する抗原に対する抗体の結合親和性の増強をもたらす。例えば、約1〜約5フレームワーク残基を本発明の本実施形態において改変してよい。場合により、これは、超可変領域の残基の何れも改変されていない場合であっても、前臨床治験における使用のために適する抗体突然変異体をもたらすためには十分である場合がある。しかしながら通常は、抗体突然変異体は追加的な超可変領域の改変を含むことになる。
【0129】
改変される超可変領域残基は、特に親抗体の元の結合親和性がランダムに産生された抗体突然変異体が容易にスクリーニングされ得るようなものである場合には、ランダムに変更してよい。
【0130】
そのような抗体突然変異体を形成するための1つの有用な操作法は「アラニンスキャニング突然変異誘発」と称されている(Cunningham and Wells(1989)Science 244:1081−1085)。ここでは1つ以上の超可変領域残基をアラニンまたはポリアラニン残基により置き換えることにより第2の哺乳類種に由来する抗原とのアミノ酸の相互作用に影響を及ぼす。次に置換に対して機能的感受性を示している超可変領域残基を、置換部位において、またはそれに対して追加の、または他の突然変異を導入することにより純化する。即ち、アミノ酸配列変異を導入するための部位は予め決定されているが、突然変異の性質自体は予め決定されている必要はない。この方法で産生したala突然変異体を本明細書に記載する通りそれらの生物学的活性に関してスクリーニングする。
【0131】
通常は「好ましい置換」の見出しの下に以下に示すもののような保存的置換により開始する。そのような置換が生物学的活性(例えば結合親和性)の変化をもたらす場合、次に、以下の表において「例示される置換」と命名されている、またはアミノ酸クラスを参照して後述においてさらに説明する通り、より実質的な変化を導入し、そして生成物をスクリーニングする。
【0132】
【表2】

【0133】
抗体の生物学的特性におけるより実質的な修飾は以下、即ち:(a)例えばシートまたはヘリックス型のコンホメーションとしての置換の領域におけるポリペプチド骨格の構造、(b)標的部位における分子の電荷または疎水性、または(c)側鎖の嵩、を維持することに対する自身の作用において有意に異なっている置換を選択することにより達成される。天然に存在する残基は共通の側鎖の特性、即ち:
(1)疎水性:ノルロイシン、met、ala、val、leu、ile;
(2)中性親水性:cys、ser、thr、asn、gln;
(3)酸性:asp、glu;
(4)塩基性:his、lys、arg;
(5)鎖の配向に影響を与える残基:gly、pro;および、
(6)芳香族:trp、tyr、phe
に基づいて群に分割される。
【0134】
非保存的置換はこれらのクラスの1つのメンバーを別のクラスに交換することを包含することになる。
【0135】
別の実施形態においては、修飾のために選択される部位はファージディスプレイを用いてアフィニティー成熟化される(上記参照)。
【0136】
アミノ酸配列突然変異体をコードする核酸分子は当該分野で知られる種々の方法により調製される。これらの方法は限定しないがオリゴヌクレオチド媒介(または部位指向性)突然変異誘発、PCR突然変異誘発、およびより早期に調製された突然変異体または親抗体の非突然変異体型のカセット突然変異誘発を包含する。突然変異体を作成するための好ましい方法は部位指向性突然変異誘発である(例えばKunkel(1985)Proc.Natl.Acad.Sci.USA 82:488参照)。
【0137】
特定の実施形態においては、抗体突然変異体は単一の超可変領域残基が置換されているのみとなる。他の実施形態においては、親抗体の1つ以上の超可変領域残基が置換されることになり、例えば約2〜約10超可変領域置換となる。
【0138】
通常は、向上した生物学的特性を有する抗体突然変異体は親抗体の重鎖または軽鎖可変ドメインの何れかのアミノ酸配列と少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、そして最も好ましくは少なくとも95%のアミノ酸配列同一性または類似性を有するアミノ酸配列を有することになる。この配列に関する同一性または類似性は、最大パーセント配列同一性を達成するために、配列をアラインし、そして必要に応じてギャップを導入した後に、親抗体残基と同一(即ち同じ残基)または類似(即ち共通の側鎖特性に基づいた同じ群に由来するアミノ酸残基、上記参照)である候補配列中のアミノ酸残基のパーセントとして本明細書においては定義される。可変ドメインの外部の抗体配列内へのN末端、C末端、または内部の伸長、欠失、または挿入の何れも、配列の同一性または類似性に影響するものとみなされない。
【0139】
抗体突然変異体の産生の後、親抗体と比較した場合のその分子の生物学的活性を測定する。上記した通り、ここでは抗体の結合親和性および/または他の生物学的活性の測定を行ってよい。本発明の好ましい実施形態においては、抗体突然変異体のパネルを調製し、そして抗原またはそのフラグメントに対する結合親和性に関してスクリーニングする。この初回スクリーニングから選択された1つ以上の抗体突然変異体を、場合により1つ以上の別の生物学的活性アッセイに付すことにより、増強された結合親和性を有する抗体突然変異体が実際に、例えば前臨床試験のために有用であることを確認する。
【0140】
このようにして選択された抗体突然変異体は、多くの場合抗体の意図する用途に応じてさらに修飾に付してよい。そのような修飾では、アミノ酸配列をさらに改変し、非相同ポリペプチドに融合し、および/または後述の通り操作する共有結合的修飾を行ってよい。アミノ酸配列改変に関しては、例示される修飾は上記の通り操作される。例えば抗体突然変異体の適切なコンホメーションの維持に関与する何れかのシステイン残基もまた、一般的にはセリンで置換することにより、分子の酸化安定性を向上させ、そして異常な交差結合を防止してよい。逆に、システインの結合を抗体に付加することによりその安定性を向上させてよい(特に抗体がFvフラグメントのような抗体フラグメントである場合)。別の型のアミノ酸突然変異体は改変されたグリコシル化パターンを有する。これは、抗体中に観察される1つ以上の炭水化物部分を欠失させることにより、および/または、抗体中に存在しない1つ以上のグリコシル化部位を付加することにより、達成してよい。抗体のグリコシル化は典型的にはN連結またはO連結の何れかである。N連結はアスパラギン残基の側鎖への炭水化物部分の連結を指す。Xがプロリン以外の何れかのアミノ酸であるトリペプチド配列であるアスパラギン−X−セリンおよびアスパラギン−X−スレオニンは、アスパラギン側鎖への炭水化物部分の酵素的連結のための認識配列である。即ち、ポリペプチド中のこれらのトリペプチド配列の何れかの存在は、潜在的なグリコシル部位を生じさせる。O連結グリコシル化は糖類であるN−アセチルガラクトサミン、ガラクトース、またはキシロースのうちの1つの、最も一般的にはセリンまたはスレオニンであるが5−ヒドロキシプロリンまたは5−ヒドロキシリジンでもよいヒドロキシアミノ酸への連結を指す。抗体へのグリコシル化部位の付加は、アミノ酸配列をそれが上記トリペプチド配列のうち1つ以上を含有するように改変することにより、好都合に達成される(N連結グリコシル化部位の場合)。改変はまた、元の抗体の配列に対する1つ以上のセリンまたはスレオニン残基の付加または置換により行ってもよい(O連結グリコシル化部位の場合)。
【0141】
同じ型の操作法および改変を用いることによりネイティブ配列CYTL1ポリペプチドのアゴニストとして作用するCYTL1変異体を作成できる。
【0142】
(xii)抗体および他のポリペプチドの組み換え産生
抗体の組み換え産生のためには、それをコードする核酸を単離し、そして後のクローニング(DNAの増幅)または発現のために複製可能なベクター内に挿入する。モノクローナル抗体をコードするDNAは従来の操作法を用いて(例えば抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合できるオリゴヌクレオチドプローブを使用することにより)容易に単離および配列決定される。多くのベクターが入手可能である。ベクター成分は一般的に、限定しないが、以下のもの、即ちシグナル配列、複製起点、1つ以上のマーカー遺伝子、エンハンサーエレメント、プロモーターおよび転写終止配列(例えば参照により本明細書に組み込まれる米国特許5,534,615記載)のうちの1つ以上を包含する。
【0143】
本発明におけるベクター中でのDNAのクローニングまたは発現のための適当な宿主細胞は、上記した原核生物、酵母、または他の高等な真核生物の細胞である。この目的のための適当な原核生物は、真正細菌、例えばグラム陰性またはグラム陽性の生物、例えば腸内細菌科、例えばエシェリシア、例えばE.coli、エンテロバクター、エルウィニア、クレブシエラ、プロテウス、サルモネラ、例えばサルモネラ・チフィムリウム、セラチア、例えばセラチア・マルセスカンス、およびシゲラ、並びにバチルス、例えばB.サブチルスおよびB.リチェニホルミス(例えば1989年4月12日公開のDD266,710に開示されているB.リチェニホルミス41P)、シュードモナス、例えばP.アエルギノーサ、およびストレプトマイセスを包含する。1つの好ましいE.coliクローニング宿主は、E.coli 294(ATCC31,446)であるが、他の菌株、例えばE.coli B、E.coli X1776(ATCC31,537)、およびE.coli W3110(ATCC27,325)も適している。これらの例は限定的なものではなく、例示に過ぎない。
【0144】
原核生物に加えて、真核生物の微生物、例えば糸状カビまたは酵母も抗体をコードするベクターのための適当なクローニングまたは発現の宿主である。サッカロマイセス・セレビシアエ、即ち一般的にはパン酵母が、下等な真核生物宿主微生物の中で最も一般的に使用されている。しかしながら多くの他の属、種、および系統が本発明において一般的に入手可能であり、そして有用であり、例えばスキゾサッカロマイセス・ポンベ;クルイベロマイセス宿主、例えばK.ラクチス、K.フラジリス(ATCC12,424)、K.ブルガリクス(ATCC16,045)、K.ウイケラミ(ATCC24,178)、K.ワルチ(ATCC56,500)、K.ドロソフィラルム(ATCC36,906)、K.サーモトレランス、およびK.マルキシアヌス;ヤロウイニア(EP402,226);ピチア・パストリス(EP183,070);カンジダ;トリコデルマ・リーシア(EP244,234);ニューロスポラ・クラッサ;シュワニオマイセス、例えばシュワニオマイセス・オクシデンタリス;および糸状カビ、例えばニューロスポラ、ペニシリウム、トリポクラジウム、およびアスペルギルス宿主、例えばA.ニヅランスおよびA.ナイジャーが挙げられる。
【0145】
グリコシル化抗体の発現のための適当な宿主細胞は多細胞生物から誘導される。無脊椎動物細胞の例は植物および昆虫の細胞を包含する。多くのバキュロウィルス菌株および変異体およびスポドプテラ・フルギペルダ(イモムシ)、アエデス・アエギプチ(カ)、アエデス・アルボピクツス(カ)、ドロソフィリア・メラノガスター(ミバエ)、およびボンビックス・モリのような宿主に由来する相当する許容性昆虫宿主細胞が同定されている。トランスフェクションのための種々のウィルス菌株、例えばオートグラファ・カリフォルニカNPVのL−1変異体およびボンビックス・モリNPVのBm−5菌株が公的に入手可能であり、そしてそのようなウィルスは、特にスポドプテラ・フルギペルダ細胞のトランスフェクションのために、本発明に従ってここでウィルスとして使用してよい。綿、トウモロコシ、バレイショ、ダイズ、ペチュニア、トマト、およびタバコの植物細胞培養物もまた宿主として利用できる。
【0146】
しかしながら、脊椎動物細胞の場合が最も興味深く、そして培養物(組織培養物)中の脊椎動物細胞の増殖が定型的な操作法となっている。有用な哺乳類宿主細胞系統の例は、SV40で形質転換したサル腎CV1系統(COS−7,ATCC CRL1651);ヒト胚性腎系統(懸濁培養物中での生育のためにサブクローニングされた293または293細胞、Grahamら、J.Gen Virol.,36:59(1977));ベビーハムスター腎細胞(BHK,ATCC CCL10);チャイニーズハムスター卵巣細胞/−DHFR(CHO,Urlaubら、Proc.Natl.Acad.Sci.USA,77:4216(1980));マウスセルトーリ細胞(TM4,Mather,Biol.Reprod.,23:243−251(1980));サル腎細胞(CV1、ATCC CCL70);アフリカミドリザル腎細胞(VERO−76,ATCC CRL−1587);ヒト子宮頸癌腫細胞(HELA,ATCC CCL2);イヌ腎細胞(MDCK,ATCC CCL34);バッファローラット肝細胞(BRL3A,ATCC CRL1442);ヒト肺細胞(W138,ATCC CCL75);ヒト肝細胞(HepG2,HB8065);マウス乳腺腫瘍(MMT060562,ATCC CCL51);TRI細胞(Matherら、Annals N.Y.Acad.Sci.,383:44−68(1982));MRC5細胞;FS4細胞;およびヒトヘパトーマ系統(HepG2)である。
【0147】
宿主細胞は抗体産生のための上記した発現またはクローニングベクターで形質転換され、そして、プロモーターを誘導し、形質転換体を選択し、または所望の配列をコードする遺伝子を増幅するために適切であるように修飾された従来の栄養培地中で培養される。
【0148】
本発明の抗体の産生のために使用される宿主細胞は種々の培地中で培養してよい。市販されている培地、例えばHamのF10(Sigma)、最小必須培地((MEM)、(Sigma)、RPMI−1640(Sigma)、およびダルベッコの変性イーグル培地((DMEM)、Sigma)が宿主細胞を培養するために適している。さらにまた、Hamら、Meth.Enz.,58:44(1979),Barnesら、Anal.Biochem.,102:255(1980)、米国特許4,767,704;4,657,866;4,927,762;4,560,655;または5,122,469;WO90/03430;WO87/00195;または米国特許30,985に記載されている培地のいずれかを宿主細胞のための培養培地として使用してよい。これらの培地の何れにも、必要に応じてホルモンおよび/または他の成長因子(例えばインスリン、トランスフェリン、または表皮成長因子)、塩類(例えば塩化ナトリウム、カルシウム、マグネシウム、およびリン酸塩)、緩衝液(例えばHEPES)、ヌクレオチド(例えばアデノシンおよびチミジン)、抗生物質(例えばGENTAMYCINTM)、微量元素(マイクロモル範囲の再終濃度において通常は存在する無機化合物として定義される)、およびグルコースまたは同等のエネルギー源を補充してよい。何れかの他の必要な補充物もまた、当業者に知られる適切な濃度において包含させてよい。培養条件、例えば温度、pH等は発現のために選択された宿主細胞と共に以前に使用されていたものであり、そして当業者の知る通りである。
【0149】
組み換え手法を使用する場合、抗体はペリプラズム空間内に細胞内で産生されるか、または培地中に直接分泌され得る。抗体が細胞内で産生される場合、第1工程として、宿主細胞または溶解細胞の何れかである粒状破砕物を例えば遠心分離または限外濾過により除去する。抗体を培地中に分泌させる場合、そのような発現系に由来する上澄みは一般的には先ず市販のタンパク質濃縮フィルター、例えばAmiconまたはMillipore Pellicon限外濾過装置を用いて濃縮される。プロテアーゼ阻害剤、例えばPMSFを上記した工程の何れかに包含させることによりタンパク質分解を阻害してよく、そして抗生物質を包含させることにより有害な夾雑体の生育を防止してよい。
【0150】
細胞から調製した抗体組成物は例えばヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、およびアフィニティークロマトグラフィーを用いて精製することができ、アフィニティークロマトグラフィーが好ましい精製手法である。アフィニティーリガンドとしてのプロテインAの適当性は抗体中に存在する何れかの免疫グロブリンFcドメインの種およびアイソタイプに依存している。プロテインAはヒトγ1、γ2、またはγ4重鎖に基づいた抗体を精製するために使用できる(Lindmarkら、J.Immunol.Meth.,62:1−13(1983))。プロテインGは全てのマウスアイソタイプに対して、そしてヒトγ3に対して推奨されている(Gussら、EMBO J.,5:1567−1575(1986))。アフィニティーリガンドを連結するマトリックスは最も頻繁にはアガロースであるが、他のマトリックスも使用される。機械的に安定なマトリックス、例えば制御細孔ガラスまたはポリ(スチレンジビニル)ベンゼンはアガロースを用いて達成できるよりも高速な流量および短いプロセシング時間を可能にする。抗体がCH3ドメインを含む場合、Bakerbond ABXTM樹脂(J.T.Baker,Phillipsburg,N.J.)が精製のために有用である。タンパク質精製のための他の手法、例えばイオン交換カラム上の分画、エタノール沈殿、逆相HPLC、シリカ上のクロマトグラフィー、ヘパリンSEPHAROSETM上のクロマトグラフィー、アニオンまたはカチオン交換樹脂(例えばポリアスパラギン酸カラム)の上のクロマトグラフィー、等電点電気泳動、SDS−PAGE、および硫酸アンモニウム沈殿もまた回収すべき抗体に応じて使用される。
【0151】
他の組み換えポリペプチド、例えばCYTL1およびCYTL1変異体は同様の操作法により製造できる。
【0152】
2.アゴニスト抗CYTL1抗体または他のCYTL1アゴニストを得るためのスクリーニング
CYTL1のアゴニスト抗体および他のアゴニストは伝統的(競合的)結合アッセイまたは活性アッセイにおいて同定できる。
【0153】
CYTL1アゴニストを得るためのスクリーニングアッセイはuPARと結合または複合体形成するか、または別様にCYTL1とuPARとの相互作用を妨害し、そしてuPARへのuPAの結合を競合的に阻害できる化合物を同定するために設計してよい。本明細書に記載するスクリーニングアッセイは化学ライブラリの高スループットスクリーニングに適したアッセイを包含し、低分子薬剤候補を同定するために特に適するものとなっている。一般的に、結合アッセイおよび活性アッセイが与えられる。
【0154】
アッセイは種々のフォーマット、例えば限定しないが、タンパク質−タンパク質結合アッセイ、生化学的スクリーニングアッセイ、イムノアッセイ、および細胞系アッセイにおいて実施することができ、これらは当該分野で良く特徴付けられている。
【0155】
全てのアッセイは、それらが候補CYTL1アゴニストをuPA、uPARおよびCYTL1に、これら2成分が相互作用を起こすために十分な条件下および時間に渡り接触させることを必要とする点において共通である。結合アッセイにおいては、相互作用は結合であり、そして形成された複合体を単離するか、または反応混合物中で検出することができる。特定の実施形態においては、uPAまたはuPARポリペプチドまたは候補アゴニストの何れかを固相上、例えばマイクロプレート上に、共有結合または非共有結合的な連結により固定化する。アッセイはアンカリングされた成分を含有するコーティングされた表面のような固定化成分に、検出可能な標識で標識されていてよい非固定化成分を添加することにより実施する。反応が終了した時点で、非反応成分を例えば洗浄により除去し、そして固体表面上にアンカリングされた複合体を検出する。元来固定化されていない成分が検出可能な標識を担持している場合は、表面上に固定化された標識の検出は複合体化が起こったことを示している。元来固定化されていない成分が標識を担持していない場合は、複合体化は例えば固定化された複合体に特異的に結合する標識された抗体を用いて検出できる。
【0156】
好ましくは、CYTL1拮抗剤を、それらがuPAの生物学的活性を阻害する能力、例えば腫瘍の形成または転移を媒介するか、または血管形成、例えば限定しないが腫瘍の血管形成を誘導または支援するuPAの能力に基づいて、同定するか、または追加的に試験する。
【0157】
当然ながら、本明細書において特に考察しているスクリーニングアッセイは例示に過ぎない。スクリーニングされる拮抗剤候補(例えばポリペプチド、ペプチド、非ペプチド有機低分子、核酸等)の型に応じて選択できる種々の他のアッセイは当業者に良く知られており、そして、本発明の目的のために等しく適当である。
【0158】
3.医薬組成物
CYTL1およびCYTL1アゴニスト、例えばアゴニストCYTL1抗体は、uPAの産生または機能に関連する種々の障害、例えば種々のタイプの腫瘍または癌、例えば腫瘍または癌の転移、および望ましくない血管形成、例えば限定しないが、腫瘍および癌の血管形成を特徴とする疾患および障害の処置のために、医薬組成物の形態において投与できる。
【0159】
抗体フラグメントを使用する場合は、標的タンパク質の結合ドメインに特異的に結合する最も小さい阻害性のフラグメントが好ましい。例えば、抗体の可変領域配列に基づいて、標的タンパク質配列に結合する能力を保持しているペプチド分子を設計することができる。そのようなペプチドは化学合成および/または組み換えDNA技術による産生が可能である。例えばMarascoらProc.Natl.Acad.Sci.USA,90:7889−7893(1993)を参照。
【0160】
活性成分はまた、例えばコアセルベーション手法により、または界面重合により調製されたマイクロカプセル、例えばそれぞれ、ヒドロキシメチルセルロースまたはゼラチン−マイクロカプセルおよびポリ−(メチルメタクリレート)(poly−(methylmethacylate))マイクロカプセル中に、コロイド状薬物送達システム(例えばリポソーム、アルブミン微小球、マイクロエマルジョン、ナノ粒子、およびナノカプセル)中、またはマクロエマルジョン中に封入してよい。そのような手法は上記したRemington’s Pharmaceutical Sciencesに開示されている。
【0161】
インビボの投与のために使用されることになる製剤は滅菌されていなければならない。これは滅菌濾過膜を通す濾過により容易に達成される。
【0162】
除放性調製物を調製してよい。除放性調製物の適当な例は抗体を含有する固体疎水性重合体の半透過性マトリックスを包含し、このマトリックスは形状付与された物品、例えばフィルムまたはマイクロカプセルの形態にある。除放性マトリックスの例はポリエステル、ヒドロゲル(例えばポリ(2−ヒドロキシエチル−メタクリレート)、またはポリ(ビニルアルコール))、ポリラクチド(米国特許3,773,919)、L−グルタミン酸とγエチル−L−グルタメートの共重合体、非分解性エチレン−酢酸ビニル、分解性乳酸−グリコール酸共重合体、例えばLUPRON DEPOTTM(乳酸−グリコール酸共重合体および酢酸ロイプロリドよりなる注射可能な微小球)、およびポリ−D−(−)−3−ヒドロキシ酪酸を包含する。エチレン−酢酸ビニルおよび乳酸−グリコール酸のような重合体は100日間よりも長い期間に渡り分子を放出できるのに対し、特定のヒドロゲルはより短い期間タンパク質を放出する。カプセル化された抗生物質が長時間身体内に残存する場合、それらは37℃において水分に曝露される結果として変性または凝集を起こし、その結果生物学的活性の損失および免疫原性変化の可能性が生じる場合がある。関与する機序に応じて安定化のための合理的な方策を講じることができる。例えば凝集の機序がチオ−ジスルフィド交換を介した分子間S−S結合の形成であることが発見されれば、安定化は、スルフィドリル残基を修飾すること、酸性の溶液から凍結乾燥すること、水分含有量を調節すること、適切な添加剤を使用すること、および特定の重合体マトリックス組成物を開発することにより、達成してよい。
【0163】
本発明における製剤はまた、処置すべき特定の適応症のために必要に応じて1つより多い活性化合物、好ましくは相互に悪影響を及ぼさない補完的活性を有するものを含有してよい。そのような分子は意図する目的のために有効な量において組み合わせて適宜存在させるか、或いは別個に製剤化し、そして同時に、または何れかの順序において連続して投与してよい。
【0164】
例えば、本発明のCYTL1およびCYTL1アゴニストは1つ以上の追加的治療薬、例えば抗血管形成剤、抗新生物性組成物、化学療法剤および/または細胞毒性剤と組み合わせて投与してよい。
【0165】
以下の実施例は説明目的のみのために提示しており、そして如何様にも本発明の範囲を限定する意図はない。
【0166】
本明細書において引用する全ての特許および参考文献は参照により全体が本明細書に組み込まれる。
【実施例】
【0167】
材料および方法
材料−組み換えヒトuPAR、uPAおよびuPA−ATFはR&D Systems(Minneapolis,MN)より入手した。Pro−uPAはCortex Biochem(SanLeandro,CA)より入手した。ブタ粘膜由来の低分子量ヘパリンはSigma Chemical Company(St.Louis,MO)から購入した。
【0168】
インサイチュハイブリダイゼーション−薄膜切片のインサイチュハイブリダイゼーションのために、PCRプライマーを設計し、NM_001081106[5’−CCCACCTGCTACTCTCGGATG−3’(配列番号3)および5’−GGCAGGTCTAACAGTGGCACTAA−3’(配列番号4)]のnt.111−560から伸長するマウスCYTL1の449bpのフラグメント、またはNM_018659[5’−TCCCCCGACCTGCTACTC−3’(配列番号5)および5’−CCTGTGAGGGTCATGGCTCTGGC−3’(配列番号6)]のnt.94−525から伸長するヒトCYTL1の431bpのフラグメントを増幅した。プライマーは27ヌクレオチドのT7またはT3RNAポリメラーゼ開始部位をコードする伸長部を包含することにより、増幅産物からのそれぞれセンスまたはアンチセンスプローブのインビトロの転写を可能とした。
【0169】
ホルマリン固定パラフィン包埋5μm切片を脱パラフィン処理し、37℃で30分間4μg/mlのプロテイナーゼK中で脱タンパク質処理し、そして以前に報告されている通りインサイチュハイブリダイゼーション用にさらに処理した(Jubbら(2006)Methods in Molecular Biology 326,255−264)。33P−UTP標識のセンスおよびアンチセンスのプローブを一夜55℃において切片にハイブリダイズさせた。未ハイブリダイズのプローブはRNase処理およびストリンジェントな洗浄により除去した。スライドガラスを段階濃度エタノールで脱水し、NTB核トラックエマルジョン(Eastman Kodak)中に浸積し、4℃で4週間露光し、次に現像し、そしてヘマトキシリンおよびエオシンで逆染色した。
【0170】
ATDC5細胞成熟アッセイ−ATDC5細胞はRIKEN Cellbank,Japanより入手し、そして、5%ウシ胎児血清および2mM L−グルタミンを添加したダルベッコの変性イーグル培地(DMEM)およびHamのF12培地の50:50混合物中に維持した。細胞を12穴プレートの各ウェル当たり105個の細胞となるように播種し、コンフルエントとなるまで生育させ、次に10μg/ml組み換えウシインスリンおよび50μg/mlアスコルビン酸で処理することにより軟骨細胞分化を誘導した。成熟化中、種々の時点において、RNAを3連のウェルから採取した。第11日に、一部の細胞を0.1ng/mlのIL−1βで処理し、そしてさらに24時間の後RNAを採取した。
【0171】
コラーゲン誘導関節炎−7〜8週齢の雄性DBA/1JマウスをThe Jackson Laboratory(Bar Harbor,ME)から入手し、そしてAmerican Association of Laboratory Animal Careガイドラインに従って維持した。全ての実験操作法はGenentechのInstitutional Animal Care and Use Committeeにより認可されている。コラーゲン誘導関節炎は第0日および第21日に完全フロイントアジュバント中のウシII型コラーゲンの注射により標準的プロトコル(Barckら(2004)Arthritis and rheumatism 50(10),3377−3386)に従って誘導した。全RNAは臨床疾患のある手掌から、そして健常DBA/1J対照マウスの手掌から抽出した。
【0172】
GeneLogic BioExpressデータ−市販のデータベースBioExpress(GeneLogic,Gaithersburg,MD)を用いてコラーゲン誘導関節炎の間の遺伝子発現のクエリを行った。試料は製造元(Affymetrix,Santa Clara,CA,USA)のプロトコルに従ってAffymetrix U133A GeneChip上で分析した。
【0173】
CYTL1の発現および精製−ミツバチメリチンシグナルペプチドと共にFastBacバキュロウィルス系(Invitrogen,Carlsbad,CA)を用いて昆虫細胞中にN末端ポリヒスチジンタグを有するヒトCYTL1を発現させた。タンパク質はNi++−セファロース6FFカラム(Amersham Biosciences,Piscataway,NJ)上で精製した。タグは組み換えエンテロキナーゼ(Novagen,Madison,WI)で切断し、そしてタグおよび酵素をNi++−セファロースおよびEKaptureアガロース(Novagen)を用いて枯渇させた。CYTL1はさらに塩勾配溶離によりヘパリンカラム(HiTrap His,Amersham)から精製し、次にCentricon遠心濃縮器(Millipore,Billerica,MA)を用いて3kDaのMWカットオフで濃縮した。濾過のフロースルーは表面プラズモン共鳴および細胞系アッセイにおける緩衝液対照として使用するために温存しておいた。
【0174】
発現クローニング−pCMV−SPORT系ベクター中に約14,000の完全長ヒトcDNAを含有する発現ライブラリをOrigene(Rockville,MD)から入手した。ヒト胎盤アルカリホスファターゼ(AP)融合タンパク質(proteinsl);TACI−APおよびヒトおよびネズミCYTL1−APをpRK5系ベクターを用いて一過性トランスフェクションにより293細胞中で発現させた。細胞馴化培地を酵素活性に関してアッセイし、そして必要に応じて希釈することにより活性を等しくした。24穴プレート中のCOS細胞をFuGene6(Roche)を用いて発現コンストラクトのプール物でトランスフェクトした。48時間後、接着細胞を洗浄し、次に0.1%BSAを添加したCYTL1−APまたはTACI−APを含有する細胞馴化培地を用いて室温で30分間インキュベートした。洗浄後、結合した融合タンパク質を10%中性緩衝ホルマリンと共にインキュベートすることにより表面に交差結合させた。90分間65℃でプレートをインキュベートすることにより内因性のアルカリホスファターゼを不活性化し、次にウエスタンブルー安定化基質(Promega,Madison,WI)を用いてアルカリホスファターゼの活性を検出した。
【0175】
uPAの不活性化−室温で2時間10mMジイソプロピルフルオロホスフェート(DFP)で処理することによりuPAのプロテアーゼ活性を非可逆的に阻害した。過剰なDFPは十分な透析により除去した。完全な不活性化はuPA基質S−2444(diaPharma,West Chester,OH)を用いることにより確認した。
【0176】
細胞表面結合アッセイ−図の説明に示す通りCOSまたはCHO細胞を完全長のヒトまたはマウスuPARをコードする発現ベクターでトランスフェクトした。24時間後、トランスフェクション体を24穴プレートに移した。さらに24時間の後、細胞を洗浄し、次に「発現クローニング」の下に詳述したとおりCYTL1−APを含有する細胞馴化培地を用いてプローブした。結合アルカリホスファターゼ活性の定量のために、BluePhos(KPL,Gaithersburg,MD)を検出試薬として使用した。競合アッセイのためには記載濃度のプロ−uPA、uPA−ATF、DFP−uPAまたは精製されたCYTL1を、uPARトランスフェクト細胞とのインキュベーションの直前にCYTL1−AP細胞馴化培地に添加した。
【0177】
RNA抽出および定量的リアルタイムPCR(qRT−PCR)−全RNAはRNeasy miniprepキット(Qiagen,Germantown,MD)を用いて抽出し、光学密度により定量し、そしてOmniscript RT(Qiagen)を用いて逆転写した。形成したcDNAを7500リアルタイムPCRシステム(Applied Biosystems,Foster City,CA)においてQuantiTectプローブを用いて2連でアッセイした。増幅されたシグナルを記載の通りβ−アクチンまたはGAPDHのものに対して標準化した。プライマーおよびプローブは表1に列挙する通りである。
【0178】
表面プラズモン共鳴−表面プラズモン共鳴実験は10μl/分の流量においてHBS−P緩衝液(50mM HEPES pH7.4、150mM NaCl、0.005%界面活性剤)を用いてBIAcore 3000(BIAcore Inc.,Uppsala,Sweden)により実施した。組み換えヒトuPARを標準的なアミンカップリング化学を用いてCM5センサーチップ上に固定化した。慨すれば、新しく混合したN−ヒドロキシスクシンイミドおよび1−エチル−3−(3−ジメチルアミノプロピル)−カルボジイミド塩酸塩の8分間注入により表面を活性化し、uPARを10mM酢酸ナトリウムpH4.5中20μg/mlで注入し、そして未反応のエステルを1Mエタノールアミンでクエンチした。空のフローセルおよびカップリングしたBSAを有するフローセルを対照として使用した。平衡結合分析のために種々の濃度の精製CYTL1および等価な希釈度の緩衝液を全フローセルに渡って注入した。緩衝液単独への応答をCYTL1応答から差し引き、そして平衡時のピーク応答を記録した。希釈および計測は3回実施した。BIAEvalソフトウエア(BIAcore)を用いてラングミュアの方程式を使用して平衡結合データをフィットさせた。CYTL1とプロ−uPAとの間の競合を示すために、飽和濃度のプロ−uPAの注入の前、および直後に、uPARカップリングフローセルにCYTL1を注入した。
【0179】
二次構造予測−CYTL1種相同体はBLASTおよびBLAT検索によりESTおよびゲノム配列から同定し、そしてClustalXを用いてアラインした。JPredウエブサイトを介したJnetニューラルネットワーク系アルゴリズム(33)への入力として多数種アライメントを使用することにより、二次構造予測を生じさせた。ソース配列:ヒトNM018659,AY359101;チンパンジー(パン・トログロダイト)ゲノム予測:GenScanのchr4_2.3;マカクサル(リーサス・マカク、マカラ・マラッタ)ゲノム予測:Genscanのchr5.4.000.a;ウシ(ボス・タウルス)BE683329、BE683328、BE683327;マウスNM_001081106;ラットCK476529、CK363591、CK359536;ポッサム(モノデルフィス・ドメスチカ)ゲノム予測:GenScanのc_619.34;ニワトリ(ガルス・ガルス)BX931297、アフリカツメガエル(ラビス)EB474543、EB474068、EC277031;P.オリブ(バスタード・ハリブー、パラリクチス・オリバセウス)CX285777、CX285687;ハリブー(ヒポグロスス・ヒポグロスス)EB036635;フグ(ラブライプス)BLATにより同定されたゲノム配列−chrUn:189、159、975〜189、160、957;メダカ(オリジアス・ラチプス)AU241696、AU241459、AU177768;サケ(サルモ・サラー)EG847194、EG833037、EG802555;マス(オンコラインクス・ミキス)BX889777、BX866010;キュウリウオ−オスメルス・モルダックスEL55175;ミノウ(ピメファレス・プロメラス)DT309382、DT347709、DT106962;ゼブラフィッシュ(ゼブラフィッシュ、ダニオ・レリオ)EE214359、EE213198、EB957495;ゼブラフィッシュ2(ダニオレリオ−第2のCYTL1相同体)AL913033、EH466946、EH441042、AL913034。
【0180】
結果
CYTL1のインビボの発現−ホールマウントインサイチュハイブリダイゼーションによれば軟骨原基に相当する段階において肋骨、脊椎および長骨を包含するマウス胚中の発生中の骨格エレメントの種々のものにおけるCYTL1の発現が明らかになっている(図1)。薄膜切片のインサイチュハイブリダイゼーションによれば、主に軟骨におけるCYTL1の発現が明らかにされており、関節軟骨の表面層において特に高値の発現がある。CYTL1発現はまた、気管の軟骨輪においても観察されている。高レベルの発現は腱と骨との間の接触部位の周囲における腱鞘において観察されている。CYTL1発現は発生中の長骨の肥大性の軟骨細胞中、そしてさらに、動脈(静脈では観察されない)内皮細胞中にも観察され、第17日胚において大動脈中に先ず検出され、そして成体マウスにおいては筋肉動脈全体に渡って拡張している。
【0181】
軟骨細胞成熟化の間のCYTL1の発現の過程をクローンマウス前軟骨細胞系統ATDC5を用いて検討した(図2)。ATDC5細胞はインスリンに応答して分化し、遺伝子発現における十分特性化された変化を有している。CYTL1は未分化細胞において検出可能ではなかったが、分化の開始後は、CYTL1発現は第4日までには検出され、早期軟骨細胞マーカーコラーゲンIIの発現を伴っていた。CYTL1発現は第12日まで増大し続け、成熟軟骨細胞のマーカーであるアグレカンの増大した発現を伴っていた。
【0182】
発現のモジュレーション−多くのサイトカインが炎症において重要な調節の役割を果たしている(Heinrichら(2003)Biochem.J.374(Pt1),1−20)ため、本発明者らは実験的関節炎疾患のマウスモデルの経過の間のCYTL1発現を調べた。コラーゲン誘導関節炎(CIA)モデルから得られたマイクロアレイデータは、マウス関節中のCYTL1の発現は疾患においてダウンレギュレートされ、臨床評点および炎症サイトカイン、特にIL−1βの産生のピークと一致していることを示している(図3a)。別の実験においてマウスの手掌から抽出されたRNAの定量的RT−PCR分析によればCIAにおけるCYTL1発現のダウンレギュレーションが確認された(図3b)。IL−1βはCIAの進行において重要な役割を果たし(Williams,R.O.(2004)Methods in Molecular Medicine 98,207−216)、軟骨破壊に必須のメディエーターであり(Zwerinaら(2007)Proc.Natl.Acad.Sci.USA,104(28),11742−11747)、そして軟骨細胞に対して直接作用することにより数種の軟骨細胞成熟化関連の遺伝子のダウンレギュレーションを誘導する(LeFebvreら(1990)Biochimica et biophysica acta 1052(3),366−378)。CIAにおけるCYTL1のダウンレギュレーションがIL−1βの直接の作用により説明できるかどうかを試験するために、本発明者らは分化したATDC5軟骨細胞をIL−1βに曝露し、そしてqRT−PCRにより遺伝子発現をアッセイした。本発明者らは0.1ng/mlのIL−1βに応答したCYTL1発現の10分の1への低減を観察した(図3c)。逆に、uPAR発現はIL−1β処置によりアップレギュレートされた。
【0183】
受容体の同定−CYTL1はより早期には数種の両親媒性のα−ヘリックスの二次構造の予測に基づいて4ヘリックスサイトカインとして説明されていた(Liuら(2000)Genomics 65(3),283−292)。より詳細な折畳みの分析のために、本発明者らはまず、GenBANKの網羅的PsiBLASTクエリにより全ての既知のCYTL1オーソログの進化的に多様な収集物を作成し(Altschulら(1997)Nucl.Acids Res.25(17),3389−3402)、配列をClustalXとアラインし(Thompsonら(1997)Nucl.Acids Res.25(24),4876−4882)、そして次にPsiPREDとJNetアルゴリズムの両方を使用することにより(Jones,D.T.(1999)J Mol Biol 292(2),195−202;Cuff,J.A.,and Barton,G.J.(2000)Proteins 40(3),502−511)、正確な二次構造の予測を発生させた(補足図1)。短鎖サイトカインのヘリックスA〜Cに合致する長さと間隔を有する3つのα−ヘリックス(Rozwarskiら(1994)Structure 2(3),159−173)が高い信頼性で予測され、それと共に、GM−CSF、IL−3およびIL−13のような短鎖サイトカインにおいて観察されるようなα−ヘリックスおよび2つの短いβ−鎖の追加的な単一ターンが伴っている。CYTL1と4ヘリックスサイトカインファミリーとの間の関連性はそのゲノム機構の分析により強化される。4ヘリックスサイトカインは構造的特徴と比較してエキソン境界のステレオタイプの位置付けを示しており、全接合部が第0相にあり、これはCYTL1により共有されている特性である。しかしながら、その予測された二次構造においては、CYTL1は、第4番目、即ちDヘリックスが非存在であるか、極度にトランケーションされているそのC末端領域におけるヘリックスサイトカインパターンから劇的に逸脱している。このヘリックスはヘリックスサイトカインのそれらの高親和性受容体への結合のために必須であるため(Clackson,T.,and Wells,J.A.(1995)Science(New York,N.Y 267(5196),383−386;Boulanger,M.J.,およびGarcia,K.C.(2004)Advances in Protein Chemistry 68,107−146)、これは、CYTL1が恐らくは無関連の受容体ファミリーにとって欠如している受容体結合の相違した様式を採用している可能性を示唆していた。
【0184】
本発明者らは、発現クローニングの手順を用いてCYTL1の受容体に関する偏りのない検索を実施した。CYTL1−アルカリホスファターゼ融合タンパク質(CYTL1−AP)を用いて、完全長のヒトクローンのライブラリに由来するプールされた発現コンストラクトでトランスフェクトされたCOS細胞をプローブした。本発明者らはCYTL1−APに結合する能力を細胞に付与する単一のプールを同定し、次にこのプールを構成している個々のクローンをスクリーニングすることにより原因となる単一のクローンを同定した(図4a)。このクローンの配列決定によりそれがuPARをコードしていることが判明した。uPARの発現は、酵素活性により計測した場合、CYTL1−APと等しい濃度で使用した対照アルカリホスファターゼ融合タンパク質TACI−AP(膜貫通活性化因子カルシウムモジュレーターおよびシクロフィリンリガンド相互作用物質)に対して結合をもたらさなかった。同様の結果がCHO細胞に対して観察された。uPARに構造的に関連しているC4.4Aの発現はCYTL1−APに結合する能力をもたらさなかった(データ示さず)。ネズミuPARでトランスフェクトしたCOS細胞はマウスCYTL1−AP融合タンパク質に結合することができ(図4b)、CYTL1とuPARとの間の相互作用の進化論的保存を示していた。種間交差性の結合は殆ど、或いは全く観察されなかった。
【0185】
過剰量の精製されたCYTL1は濃度依存的な態様においてuPARトランスフェクト細胞へのCYTL1−APの結合を阻害することができた(図4c)。相同競合を推定すれば、3つの独立した実験で得られた細胞表面におけるuPARへのCYTL1の親和性は約1.5μM±0.5μM(SEM)である。
【0186】
uPARは多くの細胞表面および細胞外のタンパク質、例えばuPA、αvおよびβ1インテグリン、ビトロネクチン、uPAR−関連タンパク質(uPARAP/エンド180)およびIGF−IIR(Ploug,M.(2003)Current Pharmaceutical Design 9(19),1499−1528)と相互作用することがわかっている。CYTL1がuPARと直接、そしてアクセサリータンパク質を必要とすることなく相互作用するかどうかを調べるために、本発明者らはBIAcore3000を用いて表面プラズモン共鳴(SPR)分析を実施した。精製されたCYTL1は固定化されたuPARに特異的に結合した(図5)。同様の結合は哺乳類細胞および細菌において発現されたCYTL1でも観察された(データ示さず)。相互作用の速度論的動態は極めて急速であり、そして正確に定量することはできなかった。平衡結合の計測値は固定化uPAR上、および、対照フローセル上に種々の濃度のCYTL1を注入することにより得た(図5c)。単一部位結合に関するラングミュアの方程式はデータに良好にフィットし、1.1μM±0.06μMのCYTL1とuPARとの相互作用に関する親和性定数の計測値が得られた。固定化されたuPARの密度を5倍範囲に渡って変動させても親和性の相違は観察されなかった。
【0187】
グリコサミノグリカンは数種のサイトカインのそれらの受容体への結合に大きく影響することが知られている。本発明者らによるCYTL1タンパク質の精製はCYTL1とヘパリンとの間の相互作用を利用しており、そのため本発明者らはこの相互作用をさらに特性化することを試みた。CYTL1およびヘパリンを単独および組み合わせて、固定化されたuPAR上に注入した(図5d)。ヘパリンおよびCYTL1単独では各々、一過性の結合を示し(88RUおよび112RU);CYTL1とヘパリンを共に注入した場合、超相加的作用(353RU)が観察され、ヘパリンがCYTL1のuPARへの結合を増強していることが示された。
【0188】
競合的結合−uPARはuPAに対する細胞表面受容体である。uPARへの結合に関してCYTL1がuPAと競合するかどうか明らかにするため、uPARでトランスフェクトした細胞を種々の濃度のタンパク質分解的に不活性なプロ酵素(プロ−uPA)、受容体結合アミノ末端フラグメント(uPA−ATF)またはジイソプロピルフルオロホスフェートで処理することにより不活性化された完全長uPA(DFP−uPA)の存在下でCYTL1APと共にインキュベートした。uPAの3形態全てがuPAR発現細胞の表面への結合に関してCYTL1−APと競合し(図6a〜c)、結合の50%阻害は競合物質10nM〜40nMで達成された。この競合が追加のタンパク質の非存在下、および嵩高のアルカリホスファターゼタグの非存在下で起こるかどうかを試験するために、SPR分析を再度使用した(図6d)。CYTL1を固定化uPAR上に注入し、282応答単位(RU)の結合を生じさせるようにした。次にプロ−uPAを高濃度で注入することにより50%超の受容体占有状態を達成した。CYTL1の2回目の注入は大きく減衰した結合(95R)をもたらし、プロ−uPAによる受容体占有がCYTL1への結合のために使用できる受容体の量を低減したことを示していた。低pHによるuPARからのプロ−uPAの解離の後、CYTL1に結合する能力は回復した。
【0189】
考察
CYTL1は軟骨細胞および動脈内皮細胞により主に組織限定的様式において産生される分泌タンパク質である。その予測二次構造およびゲノム機構において、CYTL1は4α−ヘリックス造血性サイトカインに部分的類似性を保有している。僅かな例外を除き、このタンパク質ファミリーの全メンバーは、フィブロネクチン3型(Fn3)様のサイトカイン結合モジュールの保存された対により区別できる細胞表面受容体の進化論的に保存されたファミリーに結合する(Bazan,J.F.(1990)Proc.Natl.Acad.Sci.USA,87(18),6934−6938)。これらの受容体はそれらの細胞内伸長部を介してJAKファミリーのタンパク質チロシンキナーゼおよびSTAT転写因子と会合し、これを介して、細胞外リガンド結合により駆動される受容体オリゴマー化の後にシグナルがトリガーされる。この原理の3つの例外であるM−CSF、SCFおよびFlt3Lは、第2のクラスのPDGFR様受容体について「欠損」しており、これは自身のリガンドを免疫グロブリン(Ig)様ドメインに結合させ、そして細胞質チロシンキナーゼドメインを介してシグナルを発生するものである。CYTL1に関する受容体のスクリーニングにおいては従来サイトカイン受容体は同定されていない。同定された受容体であるuPARは上記ファミリーの何れにも構造的に無関連であり、そしてむしろ、GPIアンカーにより原形質膜に結合されているヘビ神経毒中に構造的に初めて同定されたドメインである3つのLy6−uPAR(LU)様モジュール(Ploug,M.,and Ellis,V.(1994)FEBS Lett 349(2),163−168;Barinkaら(2006)J Mol Biol 363(2),482−495;Llinasら(2005)Embo J 24(9),1655−1663)で大部分が構成されている。4ヘリックスサイトカインとLUモジュラータンパク質との間では、これらのドメインの折畳み関連物はTGF−βに対するシグナリング受容体のサイトカイン結合セグメント、アクチビンおよび骨形態発生タンパク質(BMP)を含む(Gyetkoら(1994)J.Clin.Invest.93(4),1380−1387)にも関わらず、相互作用は報告されていない。CYTL1が実際に4ヘリックスサイトカインであれば、これはLU折畳み型に関する相互作用の全く新規なクラスとなる。
【0190】
サイトカインは典型的には一連の受容体に逐次的に結合して細胞内ホスホリル化のカスケードをトリガーする高親和性のシグナリング複合体を形成することによりそれらの機能を呈する。しかしなお、uPARがCYTL1に対する真のシグナリング受容体であるかどうか、または、それがCYTL1結合タンパク質としてより正確に説明されるかどうかは不明である。細胞内および膜貫通ドメインを欠いているため、uPARの単独の作用はシグナリング受容体としては機能できない。しかしながら、多くのシグナリング経路がuPARのライゲーションの後に活性化されることがわかっており、タンパク質キナーゼSrc、JAK−1、Hck、FAKおよびERK1/2の活性化が報告されている(Webbら、J.Cell Biol.,152(4),741−752;Tangら(1998)J Biol Chem 273(29),18268−18272;Resnatiら、Embo J.,15(7),1572−1582;Dumlerら(1998)J.Biol.Chem.,273(1),315−321)。シグナルトランスダクションの機序は、インテグリンに関する役割が強く示唆されているものの、なお不明である。uPARとインテグリンα3β1との間の直接でuPA依存性の相互作用(Weiら(2001)Mol.Biol.Cell 12(10),2975−2986)が間接的相互作用を有するものとして示されており、その場合、ビトロネクチンはuPA依存的な様式でuPARに結合し、そして次にインテグリンをライゲーションする(Madsenら(2007)J.Cell Biol.177(5),927−939)。uPARへのCYTL1の結合は、CYTL1が他の膜貫通鎖にuPARを架橋できれば、或いは、それがインテグリンまたは他のシグナリング分子とのuPARの相互作用を局所的に調節すれば、同様の様式で細胞内シグナルトランスダクションをもたらす可能性がある。
【0191】
本発明者らは、CYTL1がプロ−uPA、uPA−ATFおよびDFP−不活性化uPAと競合しながらuPARに結合することを明らかにしている。従ってCYTL1はuPARとのuPAの相互作用の拮抗剤として機能できる。uPA−uPAR相互作用はプロ−uPAを切断して成熟uPAを形成するヘプシンおよびマトリプターゼのような細胞表面活性化因子にプロ−uPAを接触させる作用を有する。uPARへの結合はまた、成熟uPAの活性を増強する。uPARの結合に関して競合することにより、CYTL1は細胞関連uPA活性を低減すると考えられる。uPA−uPAR相互作用の競合的阻害剤の生物学的効果は多くの系において検討されている。uPAのタンパク質分解的に不活性化された突然変異体の型によりトランスフェクトされた腫瘍細胞は低減した転移能力を示し(Crowlyら(1993)Proc.Natl.Acad.Sci.USA,90(11),5021−5025)、uPA−ATFは微小血管内皮細胞による管形成(Croonら(1999)Am J Pathol 154(6),1731−1742)および網膜損傷後の血管形成(Le Gatら(2003)Gene Ther 10(25),2098−2103)を阻害する。uPAと競合しながらuPARに結合することにより、軟骨誘導CYTL1は同様の効果を媒介する場合があり、uPAR発現細胞が侵襲する能力を低減し、そして軟骨の一体性を維持することを支援する。
【0192】
健常な関節においてはuPARの発現は殆どないが;関節炎の間は、浸潤白血球がuPAおよびuPARの両方を発現し、そして軟骨細胞および滑膜細胞の両方が炎症性サイトカインに応答してuPARを発現し(Bussoら(1997)Ann Rheum Dis 56(9),550−557;Guiducciら(2005)Clin Exp Rheumatol 23(3),364−372;Schwabら(2001)Histochem Cell Biol 115(4),317−323)、そして軟骨の破壊的な血管新生がuPAR発現内皮細胞が侵襲するに従い発生する。uPAR拮抗剤の潜在的な効果はコラーゲン誘導関節炎を有するマウスへのuPA−ATFの全身送達により明らかにされており、これは疾患の発生率および重症度、および軟骨血管新生の程度を低減している(Apparaillyら(2002)Gene Therapy 9(3),192−200)。炎症の非存在下においては、CYTL1は関節軟骨において強力に発現されるが、その発現は炎症シグナルに応答して劇的に低減される。uPA拮抗剤活性のこの消失は細胞侵襲および細胞外マトリックスのリモデリングを促進する場合がある。現在実施中のCYTL1遺伝子欠損トランスジェニックマウスの表現型分析がインビボの役割の明確化に役立つはずである。
【0193】
骨ホメオスタシスにおけるuPARの役割は、uPAR欠損マウスの研究を介して最近明らかにされた(Furlanら(2007)J Bone Miner Res 22(9),1387−1396)。uPAR発現の非存在下において、骨鉱物質密度は上昇し、そして骨容量は低減している。増強された骨芽細胞活性および骨表面への破骨細胞接着の不全がこの表現型を説明していると考えられる。これは軟骨におけるCYTL1の発現が隣接する骨におけるマトリックスリモデリングに影響する機序となっている。
【0194】
BIAcore分析により定量した場合のuPARへのCYTL1の結合は中間的な親和性、即ち約1μMを有する。これはサイトカインのそれらの細胞表面受容体に対する親和性の典型的な範囲より有意に低値である。なお同定されていないCYTL1に対する共受容体が存在する可能性があり、それはシグナルトランスダクションの機序を与えることに加えて、細胞表面におけるuPARに対するCYTL1の親和性を増大させ、これによりそれをより強力なuPA−uPAR相互作用拮抗剤とする可能性がある。共受容体の非存在下においても、CYTL1はuPAと効果的に競合するために十分高い局所濃度に到達することも想像される。大規模遺伝子発現試験(Kumarら(2001)Osteoarthritis and Cartilage/OARS,Osteoarthritis Research Society 9(7),641−653)から得られたデータの慎重な分析によれば、CYTL1はmRNAレベルで豊富に表われ、その傾向は他の如何なるサイトカインよりも高いことがわかった。このことはCYTL1が銀染色2Dゲル中で容易に検出される関節軟骨のプロテオーム分析(Hermanssonら(2004)J Biol Chem 279(42),43514−43521)においてタンパク質レベルで当てはまると考えられる。CYTL1とヘパリンとの間の相互作用はCYTL1の局所濃度がブーストされると考えられる機序となっている。ヘパリンが生理学的結合相手である可能性は低いが、軟骨の豊富な高度硫酸化グリコサミノグリカンはCYTL1に結合してそれを発現部位の近傍に局在化させる可能性がある。さらにまた、ヘパリンにより達成されたuPARへのCYTL1の結合の強化は、細胞外マトリックスまたは細胞表面の何れかにおけるグリコサミノグリカンがuPARへのCYTL1の親和性を増強している可能性があることを示唆している。
【0195】
本明細書の作成の間、CYTL1の別の研究が発表された(Kimら(2007)J Biol Chem In press.PMID:17644814)。著者らはCYTL1が間葉細胞の軟骨細胞への分化を促進することを発見している。本発明者らは軟骨細胞に分化しているATDC5細胞へのCYTL1APの結合を観察しており(補足図2a)、そして本発明者らはqRT−PCRによるこれらの細胞によるuPARの発現を示している(補足図2b)。
【0196】
CYTL1の分子作用が4ヘリックスサイトカインファミリーのメンバーとして非典型的であるとすれば、CYTL1が真にこのファミリーのメンバーであるかどうかという疑問は未解決のままであり、そして構造的特性化なくしては解答が得られないことになる。ファミリーの多くのメンバーが多面発現性であり、異なる細胞コンパートメントにおいて極めて異なる機能的役割を果たしていることに留意すべきである。例えばレプチンは全身的には脂質代謝の調節物質として、そして局所的には造血(Wautersら(2000)Eur.J.Endocrin.143(3),293−311)および調節性T細胞発生(De Rosaら(2007)Immunity 26(2),241−255)を調節するという両方の作用を有している。同様に、CD34+造血先祖細胞により発現されるCYTL1はより多くの従来のサイトカイン様機能を保持しているが、軟骨および血管壁においては極めて異なる機能を採用している可能性もある。
【技術分野】
【0001】
(発明の分野)
本発明はウロキナーゼ型プラスミノーゲン活性化因子(uPA)のその受容体uPARへの結合を阻害するための方法、および/またはuPAの生物学的活性を阻害するための方法に関する。本発明はさらに、腫瘍の形成または転移、血管形成、例えば腫瘍血管形成を阻害するための方法、およびCYTL1アゴニストを同定するためのスクリーニングアッセイに関する。
【背景技術】
【0002】
(発明の背景)
ウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)は如何なる既知の造血系サイトカイン受容体にも構造的に類似していない。これは広範な種類の遊走性の細胞型により発現されるグリコシルホスファチジルイノシトール(GPI)アンカリング細胞表面タンパク質である(非特許文献1;非特許文献2;非特許文献3)。uPARは2つの異なる作用を有しており、即ち、第1にはそれが不活性な前ウロキナーゼ型プラスミノーゲン活性化因子(プロ−uPA)を運搬して細胞表面プロテアーゼに接近させ、それにより分解されてウロキナーゼ型プラスミノーゲン活性化因子(uPA)となり、これは細胞表面に係留されて残存し(非特許文献4)、そこでそれはセリンプロテアーゼカスケードを開始し、細胞周囲のタンパク質分解をもたらすことになり(非特許文献5;非特許文献6);そして第2にそれは細胞外マトリックス成分ビトロネクチンに結合し(非特許文献7)、これが次に細胞表面インテグリンに結合する(非特許文献8)。細胞外マトリックスへの接着とその分解の両方に影響することにより、uPAのuPARへの結合は、uPAR欠損マウスにおける好中球リクルートメントの不全により顕在化される通り、細胞の遊走において重要な役割を果たしている(非特許文献9;非特許文献10)。
【0003】
uPA−uPAR相互作用の競合的阻害剤の生物学的効果は多くの系において検討されている。uPAのタンパク質分解的に不活性な突然変異体でトランスフェクトされた腫瘍細胞は腫瘍転移の能力の低減を示しており(非特許文献11)、uPA−ATFは微小血管内皮細胞による管形成(非特許文献12)および網膜傷害後の血管形成(非特許文献13)を阻害する。
【0004】
CYTL1(サイトカイン様1)は新規の分泌タンパク質を同定して分析するための大規模な研究の一部としてクローニングされており(非特許文献14)、そして米国特許出願公開20050037465において「PRO4425」として開示されている。これは造血幹先祖細胞の希少なCD34+サブセットにおいて高度に発現される転写物の産物であるC17と同一であることがわかっている(非特許文献15)。CYTL1特異的cDNAの原料組織の分析、プロテオーム研究(非特許文献16)、および大規模な遺伝子発現分析(非特許文献17;非特許文献18)によれば軟骨および骨における豊富なCYTL1発現が示されている。それと対応して、ヒトにおいてCYTL1をコードする遺伝子(NM_018659)は骨と軟骨の発達に関与を示唆されている遺伝子がリッチ化されている領域(4p16−15)である染色体4:5,067,217−5,072,098にマッピングされている(非特許文献18;非特許文献19;非特許文献20;非特許文献21)。両親媒性の分析によれば4つのα−ヘリックスの存在が示唆されている(非特許文献15)。CYTL1のサイトカイン様性質は造血系の4つのα−ヘリックス束のサイトカインを連想させるパターンにおけるCYTL1鎖中の両親媒性のα−ヘリックスの予測に基づいている(非特許文献22)。本ファミリーのメンバーは有意な配列の類似性の非存在下に十分に保存されたコア折畳み部を有しており(非特許文献23)、そしてこのタンパク質の構造は、特殊化された膜貫通受容体のある集団とのそれらの相互作用を指向する傾向がある(非特許文献24)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Pepperら(1993)J Cell Biol 122(3),673−684
【非特許文献2】Danoら(1999)Apmis 107(1),120−127
【非特許文献3】Gyetkoら、(1994)J Clin Invest 93(4),1380−1387
【非特許文献4】Cubellisら(1986)J Biol Chem 261(34),15819−15822
【非特許文献5】Danoら、(2005)Thrombosis and Haemostasis 93(4),676−681
【非特許文献6】Ploug,M.(2003)Current Pharmaceutical Design 9(19),1499−1528
【非特許文献7】Weiら(1994)J Biol Chem 269(51),32380−32388
【非特許文献8】Madsenら(2007)J Cell Biol 177(5),927−939
【非特許文献9】Rijneveldら(2002)J Immunol 168(7),3507−3511
【非特許文献10】Gyetkoら(2000)J Immunol 165(3),1513−1519
【非特許文献11】Crowlyら(1993)Proc Natl Acad Sci USA 90(11),5021−5025
【非特許文献12】Kroonら(1999)Am J Pathol 154(6),1731−1742
【非特許文献13】L Gatら(2003)Gene Ther 10(25),2098−2103
【非特許文献14】Clarkら(2003)Genome research 13(10),2265−2270
【非特許文献15】Liuら(2000)Genomics 65(3),283−292(2)
【非特許文献16】Hermanssonら、(2004)J Biol Chem 279(42),43514−43521
【非特許文献17】Kumarら(2001)Osteoarthritis and cartilage/OARS,Osteoarthritis Research Society 9(7),641−653
【非特許文献18】Yagerら(2004)Genomics 84(3),524−535
【非特許文献19】Mangionら(1999)American Journal of Human Genetics 65(1),151−157
【非特許文献20】Polymeropoulosら(1996)Genomics 35(1),1−5
【非特許文献21】Shiangら(1994)Cell 78(2),335−342
【非特許文献22】Bazan,J.F.(1990)Immunology today 11(10),350−354
【非特許文献23】Hillら、(2002)J Mol Biol 322(1),205−233
【非特許文献24】Sprang and Bazan,(1993)Current Opinion in Structural Biology 3(6),815−827
【発明の概要】
【課題を解決するための手段】
【0006】
上記において考察したとおり、uPAとその受容体uPARとの間の相互作用は、セリンプロテアーゼカスケードの開始を細胞膜に局在化させること、そして細胞表面受容体と細胞外マトリックスとの間の会合をモジュレートすることの両方により、種々の細胞型の遊走において重要な役割を果たしている。本発明は少なくとも部分的には、uPAR結合に関してuPAと競合するuPARに対する追加的なリガンドとしての分泌サイトカイン様タンパク質CYTL1/C17の同定に基づいている。
【0007】
1つの態様において、本発明はサイトカイン様1(CYTL1)ポリペプチドまたはそのアゴニストにウロキナーゼ型プラスミノーゲン活性化因子(uPA)およびウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)を含む混合物を接触させることを含むuPAとuPARとの相互作用を阻害する方法に関する。
【0008】
別の態様において、本発明は有効量のCYTL1またはそのアゴニストにインビボでウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)およびウロキナーゼ型プラスミノーゲン活性化因子(uPA)を発現する細胞を接触させることを含むuPAの生物学的活性を阻害する方法に関する。
【0009】
さらに別の態様において、本発明は、哺乳類被験体における腫瘍形成または腫瘍転移を阻害するための方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む方法に関する。
【0010】
さらなる態様において、本発明は、哺乳類被験体における血管形成を阻害するための方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む方法に関する。
【0011】
またさらに別の態様において、本発明は(a)ウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)およびCYT1またはそのアゴニストを含有する混合物を候補拮抗剤とともにインキュベートすること、および(b)uPARへのuPAまたはそのアゴニストの結合を競合的に阻害する候補拮抗剤の能力を計測すること、を含むウロキナーゼ型プラスミノーゲン活性化因子(uPA)の拮抗剤のスクリーニングの方法に関する。
【0012】
全ての態様において、アゴニストは例えば、ポリペプチド、ペプチド、非ペプチド性低分子、またはアゴニストCYTL1抗体またはそのフラグメントであってよい。
【0013】
全ての態様において、ポリペプチドは例えばCYTL1変異体(variant)、例えば配列番号2のCYTL1のようなネイティブ配列CYTL1分子の配列と少なくとも約60%、または65%、または70%、または75%、または80%、または85%、または90%、または95%、または98%、または99%のアミノ酸配列同一性を有するCYTL1変異体であってよい。
【0014】
全ての態様において、アゴニストCYTL1抗体または抗体フラグメントは好ましくはモノクローナルであり、そしてキメラ、ヒト化、またはヒト型であることができる。
【0015】
全ての態様において、抗体フラグメントは例えばFab、Fab’、F(ab’)2、およびFvフラグメント;ダイアボディ;直鎖状抗体;単鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体を包含する。
【0016】
全ての態様において、哺乳類被験体は好ましくはヒト患者である。
【0017】
全ての態様において、腫瘍または癌は例えば、癌腫、リンパ腫、芽腫、肉腫、および白血病、例えば、限定しないが、乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択されてよい。
【図面の簡単な説明】
【0018】
【図1】CYTL1のインビボ発現。CYTL1 mRNA発現は(a〜d)ホールマウント標本および(e〜p)薄片に対して実施したインサイチュのハイブリダイゼーションにより分析した。全ての組織は特段の記載が無い限りマウスおよび成人より入手した。(e〜j)においては、上パネルは暗視野像であり、下パネルは相当する明視野像である。a)第12日胚の脊椎骨、b)第13日胚の手掌、c)第12日胚の気管および肺、d)第12日胚の肋骨、e)腱鞘(腱(T)、中足骨(M))、f)気管、g)肺動脈、h)第18日胚中足骨、i)表面軟骨層におけるCYTL1発現を示す(h)における関節表面のより高い倍率、およびj)ヒト気管。白色のバーは100□mを示す。
【図2】軟骨細胞分化の間のCYTL1の発現。ATDC5前軟骨細胞をコンフルエントとなるまで生育させ、次にインスリンおよびアスコルビン酸で処置して分化を促進した。記載した時点において、RNAを採取し、そしてTaqManにより遺伝子発現をアッセイした。記載した結果はGAPDHに対して標準化(normalized)されており、そして観察された最大値に尺度を合わせてある。丸印−コラーゲンII、三角印−アグレカン、四角印−CYTL1。結果は3つの独立した実験の代表である。
【図3】CIAにおけるCYTL1のダウンレギュレーション。(A)IL−1βおよびCYTL1の発現レベルはCIAプロトコル開始してから種々の日数の後にマウスの関節から抽出したRNAのマイクロアレイ分析により計測した。(B)RNAは健常マウスの手掌およびCIAを有するマウスの炎症手掌から抽出した。CYTL1発現はqRT−PCRでアッセイし、そしてβ−アクチンに対して標準化した。平均およびSEM(N=5対照/3CIA)。(C)軟骨細胞の分化は11日間ATDC5細胞において誘導した。次に細胞を0.1ng/mlのIL−1β(黒色バー)または培地単独(白色バー)で24時間処置した。RNAを採取しqRT−PCRでアッセイし、そしてβ−アクチンで標準化した。4つの独立した実験に由来する標準化したデータの平均およびSEM。「相対的発現」とは対照と比較した場合の発現である。
【図4】uPARはCYTL1に対する受容体である。(A)擬似トランスフェクトされたCOS細胞およびuPAR発現コンストラクトでトランスフェクトされたCOS細胞をCYTL1−APまたは対照であるTACI−APとともにインキュベートし、そして結合したAP活性を染料付着により検出した。(B)ヒトおよびマウスCYTL1−APを未トランスフェクトのCOS細胞(白色バー)、またはネズミ(斜線バー)またはヒト(黒色バー)のuPARでトランスフェクトしたCOS細胞とともにインキュベートした。結合したPA活性を比色酵素アッセイにより計測した。(C)uPARトランスフェクトした細胞をCYTL1−APおよび記載濃度の精製CYTL1とともにインキュベートした。CYTL1−APの結合は比色酵素アッセイにより計測し、そしてバックグラウンド(未トランスフェクト細胞)を差し引いた後の精製されたCYTL1非存在下に観察された結合のパーセンテージとして表示した。
【図5】CYTL1−uPAR相互作用の表面プラズモン共鳴分析。記載した濃度における精製されたCYTL1をA)固定化uPAR、B)対照のフローセル上に注入した。C)平衡結合分析−ピークuPAR依存性応答を注入したCYTL1の濃度に対してプロットする。曲線は読み取り値の3つの独立したセットに対するラングミュア方程式の最良フィットを示す。D)以下に示すもの、即ちi)CYTL1(1μm)、ii)CYTL1(1μm)+ヘパリン(160μg/ml)、iii)緩衝液単独、およびiv)ヘパリン(160μg/ml)を固定化したuPAR上に注入した。各条件の3つの独立した調製物の重複応答曲線(対照フローセル差し引き後)を、記載した平衡時の平均の応答とともに示す。
【図6】CYTL1およびuPAは競合してuPARに結合する。CYTL1−APを記載濃度のA)uPA−ATF、B)プロ−uPAまたはC)DFP不活性化uPAの存在下にuPARトランスフェクト細胞と共にインキュベートした。細胞表面結合CYTL1−APを酵素アッセイにより計測し、そして競合物質非存在下の結合パーセントとして表示した。示された曲線によれば、単一の結合部位に対する可逆的競合が推定される。R2値は0.998、0.987および0.880である。D)組み換えヒトuPAR(4776RU)をBIAcoreセンサーチップの1つのフローセル中に固定化した。CYTL1(1μm)の3つの同一注入を80μg/mlプロ−uPAの注入の前後、およびプロ−uPAの解離(洗浄)後に実施した。対照フローセル(BSA固定化、5157RU)の差し引き後の平衡応答を各CYTL1注入に関して記載する。
【図7A】図7Aおよび図7B:ネイティブ配列のヒトPRO4425(CYTL1)cDNAのヌクレオチド配列(配列番号1)およびネイティブヒトPRO4425(CYTL1)ポリペプチドの推定アミノ酸配列(配列番号2)。
【図7B】図7Aおよび図7B:ネイティブ配列のヒトPRO4425(CYTL1)cDNAのヌクレオチド配列(配列番号1)およびネイティブヒトPRO4425(CYTL1)ポリペプチドの推定アミノ酸配列(配列番号2)。
【図8】CYTL1−APを用いてプローブしたアビジンコーティングプレート上に固定化した酵素的にビオチン化されたuPARを示し、結合はAP基質を用いて検出した。CYTL1−APは他のタンパク質の非存在下でuPARに直接結合する。
【図9】CYTL1−APを用いてプローブしたアビジンコーティングプレート上に固定化した酵素的にビオチン化されたuPARを示し、結合はAP基質を用いて検出した。uPARへのCYTL1−APの結合は抗iPARポリクローナルにより阻害される。
【図10】CYTL1−AP融合タンパク質を用いたBIAcore試験。
【図11】CYTL1はPC−3細胞によるマトリゲル侵襲を阻害する。
【図12】CYTL1はuPA依存性細胞増殖を阻害する。
【図13】CYTL1の存在下で成育させた軟骨細胞において低減したコラーゲンII発現が観察される。
【図14】CYTL1KOマウスは肉視的には正常な軟骨および骨を発達させている。
【図15】CYTL1KOマウスは正常な骨鉱物質密度を生じている。
【図16】CYTL1KOマウスは重症度の低い関節炎を有するように観察される。
【図17】組み換えヒトCYTL1に関する精製スキーム。
【図18】CYTL1の安定性を最適化するためのスクリーニング緩衝液。
【図19】CYTL1の安定性を最適化するためのスクリーニング緩衝液。
【図20】CYTL1の安定性を最適化するためのスクリーニング緩衝液。
【図21】CYTL1の安定性を最適化するためのスクリーニング緩衝液。
【図22】マウスCYTL1を精製するための精製スキーム。
【図23】CYTL1結晶回折の結果。
【図24】補足図1。CYTL1の二次構造の推定およびゲノム構造。CYTL1種の相同体のアライメントを示しており、推定二次構造は下部に示す通りであり、H:ヘリックス、E:延長コンホメーション(β−鎖)を示す。「Jnet Rel」とは各残基における推定に関する信頼性を示す(9=最良)。3つのヘリックスが容易に推定され、それに伴い、短いα−ヘリックス(僅か4残基のみ信頼性をもって推定)およびβ−鎖がABループ中に、そして短いβ−鎖がC−Dループ中に在り、これは多くの短鎖サイトカインに特徴的である。注目すべきはC末端領域におけるプロリン残基であり、これはα−ヘリックスの形成とは相反する可能性がある。ヒトCYTL1のエキソン境界に相当する位置は矢印で示す通りであり、矢印上方の数字はエキソン境界相およびイントロンの長さを示す。4ヘリックスサイトカインファミリーにおいて、エキソン境界は特徴的にはヘリックスAの後、ヘリックスBの前、そしてヘリックスCの後に観察され、全てがゼロ相であり、これはCYTL1に関して観察される通りである。
【図25】補足図2。CYTL1のATDC5細胞への結合。A)材料および方法においてトランスフェクトされたCOS細胞に関して記載した通り種々の時点においてATDC5細胞をプローブするためにマウスCYTL1−APを用いた。成熟軟骨細胞の形態を有する細胞への融合タンパク質の結合は第4日より観察され、より後期の時点においてはさらに広範化した。B)ATDC5成熟化の間の種々の時点においてqRT−PCRにより遺伝子発現をアッセイした。記載した結果はGAPDHに対して標準化されており、そして観察された最大値に尺度を合わせてある。丸印−コラーゲンII、三角印−uPAR、四角印−CYTL1。
【図26】表1。qRT−PCRに関するプライマーおよびプローブ。全プローブはFAMレポーター染料およびTAMRAクエンチャーで標識した。
【発明を実施するための形態】
【0019】
I.定義
特段の記載が無い限り、本明細書で使用する専門技術用語は本発明が属する技術における通常の技術者が一般的に理解するものと同様の意味を有する。例えばSingletonら、Dictionary of Microbiology and Molecular Biology第2版、J.Wiley&Sons(New York,NY 1994);Sambrookら、Molecular Cloning,A Laboratory Manual,Cold Springs Harbor Press(Cold Springs Harbor,NY 1989)を参照。本発明の目的のためには以下の用語を下記記載の通り定義する。
【0020】
「ウロキナーゼ型プラスミノーゲン活性化因子」即ち「u−PA」という用語は本明細書においては、431アミノ酸のヒト前プロu−PA(NP_002649;Moroi and Aoki,J.Biol.Chem.251(19),5956−5965(1976))および相当する313アミノ酸の成熟ヒトポリペプチドを21アミノ酸シグナル配列を伴うか伴うことなく(Roldanら、EMBO J.1990;9:467-474)包含するネイティブ配列のu−PAポリペプチド、および、オルタナティブスプライシングされた、そして、対立遺伝子の変異体およびアイソフォーム、並びにその可溶性形態のような全ての天然に存在する変異体を包含する非ヒト哺乳類のそのネイティブ配列相同体を指す。
【0021】
「CYTL1」、「サイトカイン様1」および「PRO4425」という用語は本明細書においては互換的に使用され、そして種々の原料から、例えばヒト組織型から、または他の原料から単離してよく、または組み換えまたは合成方法により製造してよい。「CYTL1ポリペプチド」または「PRO4425ポリペプチド」に言及する本明細書における全ての開示内容はポリペプチドの各々を個別に、または総括して指すものとする。例えば調製、精製、誘導体化、対応または対抗する抗体の形成、投与、含有組成物、関連疾患の処置等の記載は、個別に本発明の各ポリペプチドに関する。「CYTL1ポリペプチド」または「PRO4425ポリペプチド」という用語はまた、本明細書に開示したCYTL1/PRO4425ポリペプチドの変異体を包含する。
【0022】
「ネイティブ配列CYTL1/PRO4425ポリペプチド」は天然から誘導された相当するCYTL1ポリペプチドと同じアミノ酸配列を有するポリペプチドを含む。そのようなネイティブ配列CYTL1ポリペプチドは天然より単離することができ、或いは組み換えまたは合成手段により製造できる。「ネイティブ配列CYTL1ポリペプチド」という用語は特記すれば特定のCYTL1ポリペプチドの天然に存在するトランケーションされた、または分泌された形態(例えば細胞外ドメイン配列)、ポリペプチドの天然に存在する変異体型(例えばオルタナティブスプライシング型)および天然に存在する対立遺伝子変異体を包含する。本発明の種々の実施形態において、本明細書に開示したネイティブ配列CYTL1ポリペプチドは添付図面に示す完全長アミノ酸配列を含む成熟または完全長のネイティブ配列ポリペプチドである。開始および終止コドンは図中太字および下線で示す。しかしながら、添付の図7に開示するCYTL1ポリペプチドは1により示されるメチオニンで開始するように記載されているが、図7中のアミノ酸の1位より上流または下流の何れかに位置する他のメチオニン残基をCYTL1ポリペプチドに関する開始アミノ酸残基として使用してもよいことが考えられ、かつ可能である。
【0023】
本明細書に開示したCYTL1ポリペプチドの「シグナルペプチド」の概ねの位置は図7に示す通りである。しかしながら、留意すべきは、シグナルペプチドのC末端境界は、シグナルペプチドのC末端境界がその型のアミノ酸配列エレメントを同定するために当該分野で定型的に使用されている基準に従って同定してよい、本発明において当初同定されたシグナルペプチドC末端境界の何れかの側において、殆どは約5アミノ酸を超えないが、変動する場合がある(例えばNielsenら、Prot.Eng.10:1−6(1997)およびvon Heinjeら、Nucl.Acids.Res.14:4683−4690(1986))。さらにまた、同様に認識されていることとして、一部の場合においては、分泌されたポリペプチドからのシグナル配列の切断は完全に均一ではなく、1つより多い分泌物質種をもたらす。シグナルペプチドが本発明において同定されたシグナルペプチドのC末端境界の何れかの側において約5アミノ酸を超えない範囲で切断されるこの成熟ポリペプチド、およびそれらをコードするポリヌクレオチドは本発明の意図するものである。
【0024】
「CYTL1変異体」は、本明細書に開示した完全長ネイティブ配列CYTL1ポリペプチド配列と少なくとも約80%アミノ酸配列同一性を有する上述および後述の通り定義される活性なCYTL1ポリペプチド、本明細書に開示したシグナルペプチドを欠いたCYTL1ポリペプチド配列、本明細書に開示したシグナルペプチドを伴うか伴わないCYTL1ポリペプチドの細胞外ドメイン、または本明細書に開示した完全長CYTL1ポリペプチドの何れかの他のフラグメントを意味する。そのようなCYTL1ポリペプチド変異体は例えば、完全長ネイティブアミノ酸配列のNまたはC末端においてアミノ酸残基1つ以上が付加または欠失しているCYTL1ポリペプチドを包含する。通常は、CYTL1ポリペプチド変異体は、少なくとも約80%アミノ酸配列同一性、或いは少なくとも約81%アミノ酸配列同一性、或いは少なくとも約82%アミノ酸配列同一性、或いは少なくとも約83%アミノ酸配列同一性、或いは少なくとも約84%アミノ酸配列同一性、或いは少なくとも約85%アミノ酸配列同一性、或いは少なくとも約86%アミノ酸配列同一性、或いは少なくとも約87%アミノ酸配列同一性、或いは少なくとも約88%アミノ酸配列同一性、或いは少なくとも約89%アミノ酸配列同一性、或いは少なくとも約90%アミノ酸配列同一性、或いは少なくとも約91%アミノ酸配列同一性、或いは少なくとも約92%アミノ酸配列同一性、或いは少なくとも約93%アミノ酸配列同一性、或いは少なくとも約94%アミノ酸配列同一性、或いは少なくとも約95%アミノ酸配列同一性、或いは少なくとも約96%アミノ酸配列同一性、或いは少なくとも約97%アミノ酸配列同一性、或いは少なくとも約98%アミノ酸配列同一性および或いは少なくとも約99%アミノ酸配列同一性を、本明細書に開示した完全長ネイティブ配列CYTL1ポリペプチド配列、本明細書に開示したシグナルペプチドを欠いたCYTL1ポリペプチド配列、本明細書に開示したシグナルペプチドを伴うか伴わないCYTL1ポリペプチドの細胞外ドメイン、または本明細書に開示した完全長CYTL1ポリペプチド配列の何れかの他の特に定義されたフラグメントに対して有することになる。通常は、CYTL1変異体ポリペプチドは少なくとも約10アミノ酸長、或いは少なくとも約20アミノ酸長、或いは少なくとも約30アミノ酸長、或いは少なくとも約40アミノ酸長、或いは少なくとも約50アミノ酸長、或いは少なくとも約60アミノ酸長、或いは少なくとも約70アミノ酸長、或いは少なくとも約80アミノ酸長、或いは少なくとも約90アミノ酸長、或いは少なくとも約100アミノ酸長、或いは少なくとも約150アミノ酸長、或いは少なくとも約200アミノ酸長、或いは少なくとも約300アミノ酸長、またはそれより長いものである。
【0025】
本発明において同定されたCYTL1ポリペプチド配列に関する「パーセント(%)アミノ酸配列同一性」は、配列をアラインし、そして、最大パーセント配列同一性を達成するために必要に応じてギャップを導入した後、そして、如何なる保存的置換も配列同一性の部分として考慮しない場合に、特定のCYTL1ポリペプチド配列におけるアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセンテージとして定義される。パーセントアミノ酸配列同一性を決定する目的のためのアライメントは、当業者の能力の範囲内の種々の方法において、例えば公に入手できるコンピュータソフトウエア、例えばBLAST、BLAST−2、ALIGNまたはMegalign(DNASTAR)ソフトウエアを用いて達成できる。当業者であれば比較すべき配列の完全長に渡って最大アライメントを達成するために必要な何れかのアルゴリズムを包含するアライメントを計測するための適切なパラメータを決定できる。しかしながら本発明の目的のためには、%アミノ酸配列同一性の値は配列比較コンピュータプログラムALIGN−2を用いて得られ、その場合、ALIGN−2プログラムに関する完全なソースコードは後に示す表Iに記載する通りである。ALIGN−2配列比較コンピュータプログラムはGenentech,Inc.の著作であり、そして後に示す表Iに記載するソースコードは米国著作権局(Washington D.C.,20559)においてユーザマニュアルと共にファイルされており、そこで米国著作権登録番号TXU510087の下に登録されている。ALIGN−2プログラムはGenentech,Inc.,South San Francisco,Calif.から公的に入手するか、または後に示す表Iに記載するソースコードからコンパイルしてよい。ALIGN−2プログラムはUNIX(登録商標)のオペレーティングシステム、好ましくはデジタルUNIX(登録商標)V4.0D上の使用のためにコンパイルしなければならない。全ての配列比較パラメータはALIGN−2プログラムにより設定され、変動しない。
【0026】
アミノ酸配列比較のためにALIGN−2を使用する状況において、所定のアミノ酸配列Aの、所定のアミノ酸配列Bへの、Bとの、またはBに対抗した%アミノ酸配列同一性(これは言い換えれば、所定のアミノ酸配列Bへの、Bとの、またはBに対抗した特定の%アミノ酸配列同一性を有するか、または含んでいる所定のアミノ酸配列Aと表現できる)は以下の通り計算される。
【0027】
100×(分数X/Y)
式中、Xは配列アライメントプログラムALIGN−2によりそのプログラムのAおよびBのアライメントにおいて同一マッチとして採点されたアミノ酸残基の数であり、そして式中、YはBにおけるアミノ酸残基の総数である。当然ながら、アミノ酸配列Aの長さがアミノ酸配列Bの長さと等しくない場合、AのBへの%アミノ酸配列同一性はBのAへの%アミノ酸配列同一性と等しくならない。この方法を用いた%アミノ酸配列同一性の計算の例として、表2および3は「CYTL1」と表記されたアミノ酸配列への「比較タンパク質」と表記されたアミノ酸配列の%アミノ酸配列同一性をどのようにして計算するかを示しており、ここで、「CYTL1」は目的の仮説的CYTL1ポリペプチドのアミノ酸配列を示し、「比較タンパク質」は目的の「CYTL1」ポリペプチドを比較すべき相手であるポリペプチドのアミノ酸配列を示し、そして「X」、「Y」および「Z」は各々異なる仮説的アミノ酸残基を示す。
【0028】
特段の記載が無い限り、本明細書において使用する全ての%アミノ酸配列同一性の値はALIGN−2コンピュータプログラムを用いてこの直前に記載した段落において説明した通りに得られる。しかしながら、%アミノ酸配列同一性の値はまた、WU−BLAST−2コンピュータプログラムを用いて後述する通りに得てもよい(Altschulら、Methods in Enzymology 266:460−480(1996))。WU−BLAST−2検索パラメータの大部分はデフォルト値に設定される。デフォルト値に設定されないもの、即ち調節可能なパラメータは、以下の値、即ち:オーバーラップスパン=1、オーバーラップ割合=0.125、ワード閾値(T)=11、および採点マトリックス=BLOSUM62で設定される。WU−BLAST−2を使用する場合、%アミノ酸配列同一性の値は(a)WU−BLAST−2により決定されたものとしての、ネイティブCYTL1ポリペプチドから誘導された配列を有する目的のCYTL1ポリペプチドのアミノ酸配列と目的の比較アミノ酸配列(即ちCYTL1変異体ポリペプチドであってよい目的のCYTL1ポリペプチドが比較される相手となる配列)との間のマッチする同一アミノ酸残基の数を(b)目的のCYTL1ポリペプチドのアミノ酸残基の総数で割ることにより求められる。例えば、「アミノ酸配列Bへの少なくとも80%のアミノ酸配列同一性を有するアミノ酸配列Aを含むポリペプチド」という文言において、アミノ酸配列Aは目的の比較アミノ酸配列であり、そしてアミノ酸配列Bは目的のCYTL1ポリペプチドのアミノ酸配列である。
【0029】
パーセントアミノ酸配列同一性はまた、配列比較プログラムNCBI−BLAST2(Altschulら、Nucleic Acids Res.25:3389−3402(1997))を用いて決定してもよい。NCBI−BLAST2配列比較プログラムはhttp://www.ncbi.nlm.nih.govからダウンロードしてよく、或いは別様にNational Institute of Health,Bethesda,Mdから得てよい。NCBI−BLAST2はいくつかの検索パラメータを使用しており、この場合、これらの検索パラメータの全てはデフォルト値、例えばアンマスク=有り、鎖=全て、予測発生度=10、最小の低複雑度長=15/5、多重通過e値=0.01、多重通過の定数=25、最終ギャップ付きアライメントに関するドロップオフ=25および採点マトリックス=BLOSUM62に設定されている。
【0030】
アミノ酸配列比較のためにNCBI−BLAST2を使用する状況において、所定のアミノ酸配列Aの、所定のアミノ酸配列Bへの、Bとの、またはBに対抗した%アミノ酸配列同一性(これは言い換えれば、所定のアミノ酸配列Bへの、Bとの、またはBに対抗した特定の%アミノ酸配列同一性を有するか、または含んでいる所定のアミノ酸配列Aと表現できる)は以下の通り計算される。
【0031】
100×(分数X/Y)
式中、Xは配列アライメントプログラムNCBI−BLAST2によりそのプログラムのAおよびBのアライメントにおいて同一マッチとして採点されたアミノ酸残基の数であり、そして式中、YはBにおけるアミノ酸残基の総数である。当然ながら、アミノ酸配列Aの長さがアミノ酸配列Bの長さと等しくない場合、AのBへの%アミノ酸配列同一性はBのAへの%アミノ酸配列同一性と等しくならない。
【0032】
「単離された」とは、本明細書に開示した種々のポリペプチドを説明するために用いられる場合、自身の天然の環境の成分から同定および分離および/または回収されたポリペプチドを意味する。自身の天然の環境の夾雑成分はポリペプチドに関する診断または治療用途を典型的には妨害する物質であり、そして酵素、ホルモン、および他のタンパク質性または非タンパク質性の溶質を包含してよい。好ましい実施形態においては、ポリペプチドは(1)スピニングカップシークエネーターの使用によりN末端または内部のアミノ酸配列の少なくとも15残基を得るために十分な程度にまで、または(2)クーマシーブルーまたは、好ましくは銀染色を用いた非還元または還元条件下のSDS−PAGEによる均一性にまで、精製されることになる。単離されたポリペプチドは、CYTL1ポリペプチドの少なくとも1つの天然の環境の成分が存在しないことから、組み換え細胞内部のインサイチュのポリペプチドを包含する。しかしながら通常は単離されたポリペプチドは少なくとも1つの精製工程により調製されることになる。
【0033】
「単離された」CYTL1ポリペプチドをコードする核酸または他のポリペプチドをコードする核酸はポリペプチドをコードする核酸の天然の原料において自身が通常会合している少なくとも1つの夾雑核酸分子から同定され、そして分離される核酸分子である。単離されたポリペプチドをコードする核酸分子は自身が天然に存在する形態またはセッティングとは異なる。従って単離されたポリペプチドをコードする核酸分子は、自身が天然の細胞中に存在することから特定のポリペプチドをコードする核酸分子からは区別される。しかしながら、単離されたポリペプチドをコードする核酸分子は例えば天然の細胞の場合とは異なる染色体位置に核酸分子があるポリペプチドを通常発現する細胞中に含有されたポリペプチドをコードする核酸分子を包含する。
【0034】
「対照配列」という用語は特定の宿主生物中の作動可能に連結したコード配列の発現のために必要なDNA配列を指す。原核生物に適する対照配列は例えばプロモーター、場合によりオペレーター配列、およびリボソーム結合部位を包含する。真核生物の細胞はプロモーター、ポリアデニル化シグナル、およびエンハンサーを利用することが知られている。
【0035】
核酸はそれが別の核酸配列と機能的関係を有するように位置づけられれば「作動可能に連結」されている。例えば、プレ配列または分泌リーダーに関するDNAは、それがポリペプチドの分泌に関与するプレタンパク質として発現されればポリペプチドに関するDNAに作動可能に連結しており、プロモーターまたはエンハンサーはそれが配列の転写に影響すればコード配列に作動可能に連結しており、或いはリボソーム結合部位はそれが翻訳を促進するように位置づけられていればコード配列に作動可能に連結している。一般的に「作動可能に連結」とは連結されるべきDNA配列が近接しており、そして分泌リーダーの場合は、近接し、そして読み相にあることを意味している。しかしながら、エンハンサーは近接している必要はない。連結は好都合な制限部位におけるライゲーションにより達成される。そのような部位が存在しない場合は、合成のオリゴヌクレオチドアダプターまたはリンカーを従来の慣行に従って使用する。
【0036】
ハイブリダイゼーション反応の「ストリンジェンシー」は当業者により容易に決定され得、そして一般的にはプローブ長、洗浄温度、および塩濃度に依存した実験的計算である。一般的に、より長いプローブは適切なアニーリングのためにより高温を必要とするのに対し、より短いプローブはより低い温度を必要とする。ハイブリダイゼーションは一般的に、相補鎖がそれらの融点より低値の環境に存在する場合に再アニーリングする変性DNAの能力に依存している。プローブとハイブリダイズ可能な配列との間の所望の相同性の程度が高値であるほど、使用できる相対的な温度は高値となる。その結果、より高値の相対的温度は反応条件をよりストリンジェントとする傾向があるのに対し、より低値の温度のそのような傾向は低い。ハイブリダイゼーション反応のストリンジェンシーのさらに詳細な説明および解説に関しては、Ausubelら、Current Protocols in Molecular Biology,Wiley Interscience Publishers,(1995)を参照。
【0037】
「ストリンジェントな条件」または「高ストリンジェンシー条件」とは、本明細書において定義される場合は、以下のもの、即ち:(1)洗浄のために低イオン強度および高温を使用、例えば、0.015M塩化ナトリウム/0.0015Mクエン酸ナトリウム/0.1%ドデシル硫酸ナトリウムを50℃で使用;(2)ハイブリダイゼーション中に変性剤、例えばホルムアミド、例えば50%(v/v)ホルムアミド+0.1%ウシ血清アルブミン/0.1%Ficoll/0.1%ポリビニルピロリドン/50mMリン酸ナトリウム緩衝液pH6.5+750mM塩化ナトリウム、75mMクエン酸ナトリウムを42℃で使用;または(3)50%ホルムアミド、5xSSC(0.75MNaCl、0.075Mクエン酸ナトリウム)、50mMリン酸ナトリウム(pH6.8)、0.1%ピロリン酸ナトリウム、5xDenhardt溶液、超音波処理サケ精子DNA(50μg/ml)、0.1%SDS、および10%デキストランスルフェートを42℃で使用し、そして洗浄は42℃において0.2xSSC(塩化ナトリウム/クエン酸ナトリウム)および50%ホルムアミド中55℃、ついで高ストリンジェンシー洗浄を0.1xSSC+EDTAを用いて55℃で実施、により同定してよい。
【0038】
「中ストリンジェント条件」はSambrookら、Molecular Cloning:A Laboratory Manual,New York:Cold Spring Harbor Press,1989に記載の通り同定してよく、そして、上記したものよりも低ストリンジェントな洗浄溶液およびハイブリダイゼーションの条件(例えば温度、イオン強度および%SDS)の使用を包含する。中ストリンジェント条件の例は20%ホルムアミド、5xSSC(150mMNaCl、15mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5xDenhardt溶液、10%デキストランスルフェート、および20mg/ml変性粉砕サケ精子DNAを含む溶液中37℃で一夜インキュベーション、ついで約37〜50℃において1xSSC中フィルター洗浄である。プローブ長等のような要素に適合するために必要に応じて温度、イオン強度などを調節する方法は当業者の知る通りである。
【0039】
「タグ付けされたエピトープ」という用語は本明細書において使用する場合、「タグポリペプチド」に融合したCYTL1ポリペプチドを含むキメラポリペプチドを指す。タグポリペプチドはそれに対抗して抗体が形成できるエピトープをもたらすために十分な残基を有するが、なお、自身が融合する相手であるポリペプチドの活性を妨害しない程度に十分短い。タグポリペプチドはさらに抗体が他のエピトープと実質的に交差反応しないように十分独特である。適当なタグポリペプチドは一般的に少なくとも6つのアミノ酸残基、そして通常は約8〜50アミノ酸残基(好ましくは約10〜20アミノ酸残基)を有する。
【0040】
本発明の目的のための「活性な」または「活性」とは、ネイティブまたは天然に存在するCYTL1の生物学的および/または免疫学的な活性を保持しているCYTL1ポリペプチドの型を指し、その場合、「生物学的」活性はネイティブまたは天然に存在するCYTL1により保有される抗原性のエピトープに対抗する抗体の産生を誘導する能力以外のネイティブまたは天然に存在するCYTL1により誘発される生物学的機能(阻害性または促進性の何れか)を指し、そして「免疫学的」活性はネイティブまたは天然に存在するCYTL1により保有される抗原性のエピトープに対抗する抗体の産生を誘導する能力を指す。本発明の目的のためには、好ましい生物学的活性はウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)に結合し、uPARへのuPAの結合を競合的に阻害し、および/またはuPAの生物学的活性を阻害する能力を包含する。
【0041】
「uPAの生物学的活性」という用語は最も広範な意味において使用され、そして限定しないが、uPARに結合する能力、細胞遊走のモジュレーション、細胞膜へのセリンプロテアーゼカスケードの開始の局在化、細胞表面受容体と細胞外マトリックスとの間の会合のモジュレーション、腫瘍転移の媒介への関与、および血管形成活性を包含する。
【0042】
「アゴニスト」という用語は本明細書においては元も広範な意味において使用される。CYTL1アゴニストは伏在する機序に関わらずネイティブ配列のCYTL1により媒介される生物学的活性を模倣する何れかの分子である。本発明の目的のためには生物学的活性は好ましくは上記定義したuPA生物学的活性を阻害する能力である。CYTL1アゴニストの例は限定しないが、アゴニスト抗CYTL1抗体、ペプチドおよび非ペプチド性有機低分子を包含する。
【0043】
「抗体」という用語は本明細書においては、最も広範な意味において使用され、そして特に、所望の生物学的活性を呈する限りにおいて、インタクトな(intact)抗体、モノクローナル抗体、ポリクローナル抗体、少なくとも2つのインタクトな抗体から形成された多重特異性抗体(例えば二重特異性抗体)、および抗体フラグメントを網羅している。
【0044】
「モノクローナル抗体」という用語は本明細書においては、実質的に均質な抗体の集団から得られた抗体を指し、即ち、集団を構成する個々の抗体は、少量で存在する場合がある可能な天然に存在する突然変異を除き、同一である。モノクローナル抗体は高度に特異的であり、単一の抗原部位に対するものである。さらにまた、異なる決定基(エピトープ)に対する異なる抗体を包含するポリクローナル抗体調製物とは対照的に、各モノクローナル抗体は抗原上の単一の決定基に対するものである。それらの特異性に加えて、モノクローナル抗体は、それらが他の抗体による夾雑を被らずに合成される点において好都合である。「モノクローナル」という修飾語は抗体の実質的に均質な集団から得られるものとしての抗体の特徴を示しており、そして何れかの特定の方法による抗体の産生を要求するものとみなしてはならない。例えば本発明に従って使用するべきモノクローナル抗体はKohlerら、Nature,256:495(1975)により最初に報告されたハイブリドーマ法により作成してよく、或いは組み換えDNA法により作成してよい(例えば米国特許4,816,567参照)。「モノクローナル抗体」はまた例えばClacksonら、Nature,352:624−628(1991)およびMarksら、J.Mol.Biol.,222:581−597(1991)に記載の手法を用いてファージ抗体ライブラリから単離してよい。
【0045】
抗体は特に、所望の生物学的活性を呈する限り、重鎖および/または軽鎖の一部分が特定の種から誘導されるか、または特定の抗体クラスまたはサブクラスに属する抗体の相当する配列と同一または相同であるが、鎖の残余は別の種から誘導されるか、または別の抗体クラスまたはサブクラスに属する抗体の相当する配列と同一または相同である、「キメラ」抗体、並びにそのような抗体のフラグメントを包含する(米国特許4,816,567;およびMorrisonら、Proc.Natl.Acad.Sci.USA,81:6851−6855(1984))。本明細書において目的とされるキメラ抗体は非ヒト霊長類(例えば旧世界猿、類人猿等)およびヒトの定常領域配列から誘導した可変ドメイン抗原結合配列を含む霊長類化抗体を包含する。
【0046】
「抗体フラグメント」はインタクトな抗体の一部分、好ましくはその抗原結合または可変領域を含むものを含む。抗体フラグメントの例はFab、Fab’、F(ab’)2、およびFvフラグメント;ダイアボディ;直鎖状抗体;単鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体を包含する。
【0047】
「インタクトな」抗体は抗原結合可変領域並びに軽鎖定常ドメイン(CL)および重鎖定常ドメインCH1、CH2およびCH3を含むものである。定常ドメインはネイティブ配列定常ドメイン(例えばヒトネイティブ配列定常ドメイン)またはそのアミノ酸配列変異体であってよい。好ましくは、インタクトな抗体は1つ以上のエフェクター機能を有する。
【0048】
非ヒト(例えばげっ歯類)抗体の「ヒト化」型は非ヒト免疫グロブリンから誘導した最小配列を含有するキメラ抗体である。大部分において、ヒト化抗体は、レシピエントの超可変領域に由来する残基が所望の特異性、親和性、および能力を有するマウス、ラット、ウサギまたは非ヒト霊長類のような非ヒト種(ドナー抗体)の超可変領域に由来する残基により置き換えられているヒト免疫グロブリン(レシピエント抗体)である。一部の場合においては、ヒト免疫グロブリンのフレームワーク領域(FR)残基は相当する非ヒト残基により置き換えられている。さらにまた、ヒト化抗体はレシピエント抗体中、またはドナー抗体中には存在しない残基を含んでよい。そのような修飾は抗体の性能をさらに精鋭化するために行われる。一般的に、ヒト化抗体は、少なくとも1つ、そして典型的には2つの可変ドメイン(Fab、Fab’、F(ab’)2、Fabc、Fv)の実質的に全てを含むことになり、その場合、超可変ループの全てまたは実質的に全てが非ヒト免疫グロブリンのものに相当し、そしてFRの全てまたは実質的に全てがヒト免疫グロブリン配列のものである。ヒト化抗体は場合によりやはり免疫グロブリン定常領域の少なくとも一部分(Fc)、典型的にはヒト免疫グロブリンのものを含むことになる。さらに詳細な説明はJonesら、Nature 321:522−525(1986);Riechmannら、Nature 332:323−329(1988);およびPresta,Curr.Op.Struct.Biol.2:593−596(1992)を参照。
【0049】
「超可変領域」という用語は本明細書において使用する場合、配列において超可変であり、および/または構造的に定義されたループを形成する抗体可変ドメインの領域を指す。超可変領域は「相補性決定領域」即ち「CDR」に由来するアミノ酸残基(即ち軽鎖可変ドメインの残基24〜34、50〜56、および89〜97、および重鎖可変ドメインの残基31〜35、50〜65、および95〜102;Kabatら、Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,MD.(1991))および/または「超可変ループ」に由来する残基(即ち軽鎖可変ドメインの残基26〜32、50〜52、および91〜96および重鎖可変ドメインの残基26〜32、53〜55、および96〜101;Chothia and Lesk J.Mol.Biol.196:901−917(1987))を含む。両方の場合において、可変ドメイン残基は後述においてさらに詳細に考察する通り、上記したKabatらに従ってナンバリングされる。「フレームワーク」即ち「FR」残基は本明細書において定義する通り超可変領域における残基以外の可変ドメイン残基である。
【0050】
「親抗体」または「野生型」抗体は本明細書に開示した抗体変異体と比較した場合に1つ以上のアミノ酸配列改変を欠いているアミノ酸配列を含む抗体である。即ち、親抗体は一般的に本明細書に開示した抗体変異体の相当する超可変領域のアミノ酸配列とはアミノ酸配列が異なる少なくとも1つの超可変領域を有する。親ポリペプチドはネイティブ配列(即ち天然に存在する)抗体(天然に存在する対立遺伝子変異体を包含する)、または天然に存在する配列の既存のアミノ酸配列修飾(例えば挿入、欠失および/または他の改変)を有する抗体を含んでよい。本開示全体に渡って、「野生型」、「WT」、「wt」および「親」または「親の」抗体は、互換的に使用される。
【0051】
本明細書において使用する場合、「抗体変異体」または「変異体抗体」とは、親抗体のアミノ酸配列とは異なるアミノ酸配列を有する抗体を指す。好ましくは抗体変異体は天然には存在しないアミノ酸配列を有する重鎖可変ドメインまたは軽鎖可変ドメインを含む。そのような変異体は必然的に親抗体と100%未満の配列の同一性または類似性を有する。好ましい実施形態においては、抗体変異体は親抗体の重鎖または軽鎖の何れかの可変ドメインのアミノ酸配列と約75%〜100%未満、より好ましくは約80%〜100%未満、より好ましくは約85%〜100%未満、より好ましくは約90%〜100%未満、そして最も好ましくは約95%〜100%未満のアミノ酸配列の同一性または類似性のアミノ酸配列を有することになる。抗体変異体は一般的に、1つ以上の自身の超可変領域の中、またはそれに隣接して1つ以上のアミノ酸改変を含むものである。
【0052】
「アミノ酸改変」とは所定のアミノ酸配列のアミノ酸配列における変化を指す。例示される改変は挿入、置換および欠失を包含する。「アミノ酸置換」は所定のアミノ酸配列における別の異なるアミノ酸残基による既存のアミノ酸残基の置き換えを指す。
【0053】
「置き換え」アミノ酸残基はアミノ酸配列において別のアミノ酸残基を置き換えるか置換するアミノ酸残基を指す。置き換え残基は天然に存在する、または天然に存在しないアミノ酸残基であってよい。
【0054】
「アミノ酸挿入」は所定のアミノ酸配列内部への1つ以上のアミノ酸残基の導入を指す。アミノ酸挿入は「ペプチド挿入」を包含し得、この場合は、ペプチド結合で連結された2つ以上のアミノ酸残基を含むペプチドが所定のアミノ酸配列内部に導入される。アミノ酸挿入にペプチドの挿入が関与する場合、挿入されるペプチドは、天然には存在しないアミノ酸配列を有するようにランダム突然変異誘発により形成してよい。「超可変領域に隣接する」アミノ酸改変とは、超可変領域のN末端および/またはC末端における1つ以上のアミノ酸残基の導入または置換であって、挿入または置き換えのアミノ酸残基の少なくとも1つが当該超可変領域のN末端またはC末端のアミノ酸残基とペプチド結合を形成するようなものを指す。
【0055】
「天然に存在するアミノ酸残基」とはアラニン(Ala);アルギニン(Arg);アスパラギン(Asn);アスパラギン酸(Asp);システイン(Cys);グルタミン(Gln);グルタミン酸(Glu);グリシン(Gly);ヒスチジン(His);イソロイシン(Ile):ロイシン(Leu);リジン(Lys);メチオニン(Met);フェニルアラニン(Phe);プロリン(Pro);セリン(Ser);スレオニン(Thr);トリプトファン(Trp);チロシン(Tyr);およびバリン(Val)よりなる群から一般的に選択される遺伝子コードによりコードされるものである。
【0056】
「天然に存在しないアミノ酸残基」とは本明細書においては、ポリペプチド鎖中で隣接するアミノ酸残基に共有結合することができる上記列挙した天然に存在するアミノ酸残基以外のアミノ酸残基である。天然に存在しないアミノ酸残基の例はノルロイシン、オルニチン、ノルバリン、ホモセリンおよび他のアミノ酸残基類縁体、例えばEllmanら、Meth.Enzym.202:301−336(1991)に記載されているものを包含する。そのような天然に存在しないアミノ酸残基を形成するためには、Norenら、Science 244:182(1989)および上記したEllmanらの操作法を使用できる。慨すれば、これらの操作法では天然に存在しないアミノ酸残基を用いてサプレッサーtRNAを化学的に活性化し、その後、RNAのインビトロの転写および翻訳を行う。
【0057】
本開示全体に渡り、Kabat,E.A.ら、Sequences of Proteins of Immunological Interest(National Institutes of Health,Bethesda,Md.(1987)および(1991)のナンバリングシステムを参照することとする。これらの解説書において、Kabatは各サブクラスに関する抗体の多くのアミノ酸配列を列挙し、そしてそのサブクラスにおける各残基位置に関して最も一般的に生じているアミノ酸を列挙している。Kabatは列挙された配列における各アミノ酸に残基番号を割り付けるための方法を使用しており、そして残基番号を割り付けるためのこの方法は本分野において標準的なものとなっている。Kabatナンバリングスキームは本明細書において後述する通りである。本発明の目的のために、Kabatの解説書に包含されない候補抗体アミノ酸配列に残基番号を割り付けるためには、以下の工程に従う。一般的に、候補配列はKabatの何れかの免疫グロブリン配列または何れかのコンセンサス配列とアラインされる。アライメントは手作業によるか、または一般的に受け入れられているコンピュータプログラムを用いてコンピュータにより実施してよく、そのようなプログラムの例はAlign2プログラムである。アライメントは大部分のFab配列にとって共通である一部のアミノ酸残基を用いることにより容易にしてよい。例えば軽鎖および重鎖は各々典型的には、同じ残基番号を有する2つのシステインを有し;VLドメインにおいては2つのシステインは典型的には残基番号23および88にあり、そしてVHドメインにおいては2つのシステイン残基は典型的には22および92と番号付けされている。フレームワーク残基は常時ではないが一般的には概ね同じ数の残基を有するが、CDRの大きさは変動することになる。例えば自身がアラインされるKabatの配列におけるCDRよりも長い候補配列に由来するCDRの場合、典型的には接尾辞を残基番号に付加することにより追加の残基の挿入を示す(例えば図1Bの残基100abc参照)。例えば残基34および36に関してKabat配列とアラインするが、それらの間に残基35とアラインすべき残基を有しない候補配列の場合、番号35は単に残基に割り付けられない。
【0058】
本明細書において使用する場合、「高親和性」を有する抗体はナノモル(nM)範囲かそれより良好なKD、即ち解離定数を有する抗体である。「ナノモル範囲かそれより良好」なKDはXnMと表記してよく、ここでXは約10未満の数である。
【0059】
「フィラメント状ファージ」という用語は自身の表面上に非相同ポリペプチドをディスプレイできるウィルス粒子を指し、そして限定しないがf1、fd、Pf1、およびM13を包含する。フィラメント状ファージはテトラサイクリンのような選択可能なマーカーを含有してよい(例えば「fd−tet」)。種々のフィラメント状ファージディスプレイ系が当業者に良く知られている(例えばZacherら、Gene9:127−140(1980),Smithら、Science 228:1315−1317(1985);およびParmley and Smith,Gene 73:305−318(1988)参照)。
【0060】
「パニング」という用語は標的に対する高い親和性および特異性を有する抗体のようなファージ担持化合物の同定および単離におけるスクリーニングプロセスの多数回実施を指す。
【0061】
「処置する」、「処置」および「療法」という用語は本明細書において使用する場合、治癒的療法、予防的療法、および予防療法を指す。連続処置または投与は1日以上の処置における中断を伴わない少なくとも毎日を基準とした処置を指す。間歇処置または投与、または間歇的様式における処置または投与は連続的ではなく、むしろ周期的な性質にある処置を指す。
【0062】
「哺乳類」という用語は本明細書において使用する場合、哺乳類に分類される何れかの哺乳類、例えばヒト、高等な非ヒト霊長類、げっ歯類、家畜および牧場動物、例えばウシ、ウマ、イヌおよびネコを指す。本発明の好ましい実施形態においては哺乳類はヒトである。
【0063】
1つ以上の別の治療薬「と組み合わせた」投与は、同時(併用)および何れかの順序の連続投与を包含する。
【0064】
「有効量」とは有益な、または所望の治療(予防を包含)結果を起こすために十分な量である。有効量は1回以上の投与において投与できる。
【0065】
本明細書において使用する場合、「細胞」、「細胞系統」および「細胞培養物」という表現は互換的に使用され、そして全てのそのような表記は子孫を包含する。即ち、「形質転換体」および「形質転換された細胞」という単語は初期の対象細胞および形質転換の回数に関わらずそれより誘導された培養物を包含する。さらにまた、全ての子孫は意図的または偶発的な突然変異に起因してDNA含有量において厳密には同一とならない場合がある。「子孫」という用語は元来形質転換された細胞または細胞系統の後の各世代の全ての新生仔を指す。初回形質転換された細胞においてスクリーニングの対象となるものと同じ機能または生物学的活性を有する突然変異体子孫が包含される。異なる表記が意図される場合は、それは文脈から明らかになるはずである。
【0066】
「癌」および「癌性の」という用語は典型的には調節されない細胞の成育を特徴とする哺乳類における生理学的状態を指す、または説明するものである。癌の例は癌腫、リンパ腫、芽腫、肉腫、および白血病を包含するがこれらに限定されない。そのような癌のより特定的な例は、限定しないが、乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌を包含する。
【0067】
「制御配列」という用語は特定の宿主生物において作動可能に連結したコード配列の発現のために必要なDNA配列を指す。原核生物に適する制御配列は例えばプロモーター、場合によりオペレーター配列、およびリボソーム結合部位を包含する。真核生物の細胞はプロモーター、ポリアデニル化シグナルおよびエンハンサーを利用することが知られている。
【0068】
核酸はそれが別の核酸配列と機能的関連性に有るようにおかれた場合に「作動可能に連結」している。例えば、プレ配列または分泌リーダーに関するDNAは、それがポリペプチドの分泌に関与するプレタンパク質として発現される場合には、そのポリペプチドに関するDNAに作動可能に連結しており;プロモーターまたはエンハンサーはそれが配列の転写に影響する場合にコード配列に作動可能に連結しており;或いは、リボソーム結合部位はそれが翻訳を促進するように位置づけられていればコード配列に作動可能に連結している。一般的に「作動可能に連結」とは連結されるべきDNA配列が近接しており、そして分泌リーダーの場合は、近接し、そして読み相にあることを意味している。しかしながら、エンハンサーは近接している必要はない。連結は好都合な制限部位におけるライゲーションにより達成される。そのような部位が存在しない場合は、合成のオリゴヌクレオチドアダプターまたはリンカーを従来の慣行に従って使用する。
【0069】
「低分子」とは約1000ダルトン未満、好ましくは約500ダルトン未満の分子量を有するものとして本明細書においては定義される。
【0070】
「抗血管形成剤」とは血管の発生をブロック、またはある程度まで妨害する化合物を指す。抗血管形成因子は例えば血管形成を促進することに関与する成長因子または成長因子受容体に結合する低分子または抗体であってよい。本発明において好ましい抗血管形成因子はベバシズマブ(AVASTIN(登録商標))のような血管内皮成長因子(VEGF)に結合する抗体である。
【0071】
「抗新生物性組成物」という用語は少なくとも1つの活性な治療薬、例えば「抗癌剤」を含む癌を処置する場合に有用な組成物を指す。治療薬(抗癌剤)の例は、限定しないが例えば、化学療法剤、成長阻害剤、細胞毒性剤、放射線療法において使用される薬剤、抗血管形成剤、アポトーシス剤、抗チューブリン剤、および癌を処置するための他の薬剤、例えば抗HER−2抗体、抗CD20抗体、表皮成長因子受容体(EGFR)拮抗剤(例えばチロシンキナーゼ阻害剤)、HER1/EGFR阻害剤(例えばエルロチニブ(TarcevaTM)、血小板誘導成長因子阻害剤(例えばGleevecTM(イマチニブメシレート))、COX−2阻害剤(例えばセレコキシブ)、インターフェロン、サイトカイン、以下の標的ErbB2、ErbB3、ErbB4、PDGFR−β、BlyS、APRIL、BCMAまたはVEGF受容体の1つ以上に結合する拮抗剤(例えば中和抗体)、TRAIL/Apo2、および他の生物活性および有機化学の薬剤等を包含する。これらの組み合わせもまた本発明に包含される。
【0072】
本明細書において使用する「細胞毒性剤」という用語は、細胞の機能を阻害または防止する、および/または、細胞の破壊を誘発する物質を指す。該用語は放射性同位体(例えばAt211、I131、I125、Y90、Re186、Re188、Sm153、Bi212、P32およびLuの放射性同位体)、化学療法剤、および毒素、例えば低分子毒素または細菌、カビ、植物または動物起源の酵素的に活性な毒素、およびそのフラグメントおよび/または変異体を包含することを意図している。
【0073】
「化学療法剤」は癌の処置において有用な化合物である。(登録商標)化学療法剤の例はアルキル化剤、例えばチオテパおよびCYTOXANシクロホスファミド;アルキルスルホネート、例えばブスルファン、イムプロスルファンおよびピポスルファン;アジリジン、例えばベンゾドパ、カルボクオン、メツレドパ、およびウレドパ;エチレンイミンおよびメチラメラミン、例えばアルトレタミン、トリエチレンメラミン、トリエチレンホスホルアミド、トリエチレンチオホスホルアミドおよびトリメチローロメラミン;TLK 286(TELCYTATM);アセトゲニン(特にブラタシンおよびブラタシノン);デルタ−9−テトラヒドロカンナビノール(ドロナビノール、MARINOL(登録商標));β−ラパコン;ラパコール;コルヒチン;ベツリン酸;カンプトテシン(例えば合成の類縁体トポテカン(HYCAMTIN(登録商標))、CPT−11(イリノテカン、CAMPTOSAR(登録商標))、アセチルカンプトテシン、スコポレクチン、および9−アミノカンプトテシン);ブリオスタチン;カリスタチン;CC−1065(例えばそのアドゼレシン、カルゼレシンおよびビゼレシン合成類縁体);ポドフィロトキシン;ポドフィリン酸;テニポシド;クリプトフィシン(特にクリプトフィシン1およびクリプトフィシン8);ドラスタチン;デュオカルマイシン(例えば合成類縁体、KW−2189およびCB1−TM1);エレウセロビン;パンクラチスタチン;サルコジクチン;スポンジスタチン;窒素マスタード、例えばクロラムブシル、クロルナファジン、コロホスファミド、エストラムスチン、イフォスファミド、メクロレタミン、塩酸メクロレタミンオキシド、メルファラン、ノベムビシン、フェネステリン、プレドニムスチン、トロフォスファミド、ウラシルマスタード;ニトロソ尿素、例えばカルムスチン、クロロゾトシン、フォテムスチン、ロムスチン、ニムスチン、およびラニムヌスチン;ビスホスホネート、例えばクロドロネート;抗生物質、例えばエネジン抗生物質(例えばカリケアマイシン、特にカリケアマイシンγ1IおよびカリケアマイシンオメガI1(例えばAgnew、Chem Intl.編Engl.、33:183−186(1994)参照)およびアントラサイクリン、例えばアナマイシン、AD32、アルカルビシン、ダウノルビシン、デキスラゾキサン、DX−52−1、エピルビシン、GPX−100、イダルビシン、KRN5500、メノガリル、ジネミシン、例えばジネミシンA、エスペラミシン、ネオカルジノスタチン発色団および関連の発色タンパク質エネジン抗生物質(antiobiotic)発色団、アクラシノマイシン、アクチノマイシン、オースラマイシン、アザセリン、ブレオマイシン、カクチノマイシン、カラビシン、カルミノマイシン、カルジノフィリン、クロモマイシニス、ダクチノマイシン、デトルビシン、6−ジアゾ−5−オキソ−L−ノルロイシン、ADRIAMYCIN(登録商標)ドキソルビシン(例えばモルホリノ−ドキソルビシン、シアノモルホリノ−ドキソルビシン、2−ピロリノ−ドキソルビシン、リポソームドキソルビシン、およびデオキシドキソルビシン)、エソルビシン、マルセロマイシン、マイトマイシン、例えばマイトマイシンC、ミコフェノール酸、ノガラマイシン、オリボマイシン、ペプロマイシン、ポトフィロマイシン、ピューロマイシン、ケラマイシン、ロドルビシン、ストレプトニグリン、ストレプトゾシン、ツベルシジン、ウベニメックス、ジノスタチン、およびゾルビシン;葉酸類縁体、例えばデノプテリン、プテロプテリン、およびトリメトレキセート;プリン類縁体、例えばフルダラビン、6−メルカプトプリン、チアミプリン、およびチオグアニン;ピリミジン類縁体、例えばアンシタビン、アザシチジン、6−アザウリジン、カルモフル、シタラビン、ジデオキシウリジン、ドキシフルリジン、エノシタビン、およびフロクスウリジン;アンドロゲン、例えばカルステロン、ドロモスタノロンプロピオネート、エピチオスタノール、メピチオスタン、およびテストラクトン;抗アドレナール、例えばアミノグルテチミド、ミトタン、およびトリロスタン;葉酸補給剤、例えばフォリン酸(ロイコボリン);アセグラトン;抗フォレート抗新生物性剤、例えばALIMTA7、LY231514ペメトレクスド、ジヒドロフォレートレダクターゼ阻害剤、例えばメトトレキセート、代謝拮抗剤、例えば5−フルオロウラシル(5−FU)およびそのプロドラッグ、例えばUFT、S−1およびカペシタビン、およびチミジレートシンターゼ阻害剤およびグリシンアミドリボヌクレオチドホルミルトランスフェラーゼ阻害剤、例えばラルチトレキセド(TOMUDEXTM、TDX);ジヒドロピリミジンデヒドロゲナーゼの阻害剤、例えばエニルウラシル;アルドホスファミドグリコシド;アミノレブリン酸;アムサクリン;ベストラブシル;ビサントレン;エダトラキセート;デフォファミン;デメコルチン;ジアジコン;エルホルニチン;酢酸エリプチニウム;エポチロン;エトグルシド;硝酸ガリウム;ヒドロキシ尿素;レンチナン;ロニダイニン;マイタンシノイド、例えばマイタンシンおよびアンサミトシン;ミトグアゾン;ミトキサントロン;モピダンモール;ニトラエリン;ペントスタチン;フェナメット;ピラルビシン;ロソキサントロン;2−エチルヒドラジド;プロカルバジン;PSK7多糖類複合体(JHS Natural Products,Eugene,OR);ラゾキサン;リゾキシン;シゾフィラン;スピロゲルマニウム;テヌアゾン酸;トリアジコン;2,2’,2’’−トリクロロトリエチルアミン;トリコテセン(特にT−2毒素、ベラキュリンA、ロリジンAおよびアングイジン);ウレタン;ビンデシン(ELDISINE(登録商標)、FILDESIN(登録商標));ダカルバジン;マンノムスチン;ミトブロニトール;ミトラクトール;ピポブロマン;ガシトシン;アラビノシド(「Ara−C」);シクロホスファミド;チオテパ;タキソイドおよびタキサン、例えばTAXOL(登録商標)パクリタキセル(Bristol−Myers Squibb Oncology,Princeton,N.J.)、ABRAXANETMCremophor非含有、パクリタキセルのアルブミン操作ナノ粒子製剤(American Pharmaceutical Partners,Schaumberg,Illinois)、およびTAXOTERE(登録商標)ドセタキセル(Rho^ne−Poulenc Rorer,Antony,France);クロラムブシル;ゲムシタビン(GEMZAR(登録商標));6−チオグアニン;メルカプトプリン;白金;白金類縁体または白金系類縁体、例えばシスプラチン、オキサリプラチンおよびカルボプラチン;ビンブラスチン(VELBAN(登録商標));エトポシド(VP−16);イフォスファミド;ミトキサントロン;ビンクリスチン(ONCOVIN(登録商標));ビンカアルカロイド;ビノレルビン(NAVELBINE(登録商標));ノバントロン;エダトレキセート;ダウノマイシン;アミノプテリン;キセロダ;イバンドロネート;トポイソメラーゼ阻害剤RFS2000;ジフルオロメチルオルニチン(DMFO);レチノイド、例えばレチノイン酸;上記何れかの製薬上許容しうる塩、酸または誘導体;並びに上記2つ以上の組み合わせ、例えばCHOP、即ちシクロホスファミド、ドキソルビシン、ビンクリスチン、およびプレドニゾロンの複合療法の略語、およびFOLFOX、即ち5−FUおよびロイコボリンと組み合わせたオキサリプラチン(ELOXATINTM)を用いた処置計画の略語を包含する。
【0074】
この定義に同様に包含されるものは、腫瘍に対するホルモンの作用を調節または阻害するように作用する抗ホルモン剤、例えば抗エストロゲンおよび選択的エストロゲン受容体モジュレーター(SERM)、例えばタモキシフェン(例えばNOLVADEX(登録商標)タモキシフェン)、ラロキシフェン、ドロロキシフェン、4−ヒドロキシタモキシフェン、トリオキシフェン、ケオキシフェン、LY117018、オナプリストン、およびFARESTON(登録商標)トレミフェン;副腎におけるエストロゲン産生を調節する酵素アロマターゼを阻害するアロマターゼ阻害剤、例えば4(5)−イミダゾール、アミノグルテチミド、MEGASE(登録商標)酢酸メゲストロール、AROMASIN(登録商標)エキセメスタン、ホルメスタニー、ファドロゾール、RIVISOR(登録商標)ボロゾール、FEMARA(登録商標)レトロゾール、およびARIMIDEX(登録商標)アナストロゾール;および抗アンドロゲン、例えばフルタミド、ニルタミド、ビカルタミド、ロイプロリド、およびゴセレリン;並びにトロキサシタビン(1,3−ジオキソランヌクレオシドシトシン類縁体);アンチセンスオリゴヌクレオチド、特に異常な細胞増殖への関与を示唆されているシグナリング経路における遺伝子の発現を阻害するもの、例えばPKC−α、Raf、H−Ras、および表皮成長因子受容体(EGF−R);ワクチン、例えば遺伝子療法ワクチン、例えばALLOVECTIN(登録商標)ワクチン、LEUVECTIN(登録商標)ワクチン、およびVAXID(登録商標)ワクチン;PROLEUKIN(登録商標)rIL−2;LURTOTECAN(登録商標)トポイソメラーゼ1阻害剤;ABARELIX(登録商標)rmRH;および上記何れかの製薬上許容しうる塩、酸または誘導体を包含する。
【0075】
「代謝拮抗物質化学療法剤」は代謝産物に構造的には同様であるが生産的な様式では身体により使用され得ない薬剤である。多くの代謝拮抗物質化学療法剤は核酸、RNAおよびDNAの産生を妨害する。代謝拮抗物質化学療法剤の例は、ゲムシタビン(GEMZAR(登録商標))、5−フルオロウラシル(5−FU)、カペシタビン(XELODATM)、6−メルカプトプリン、メトトレキセート、6−チオグアニン、ペメトレクスド、ラルチトレクスド、アラビノシルシトシンARA−Cシタラビン(CYTOSAR−U(登録商標))、ダカルバジン(DTIC−DOME(登録商標))、アゾシトシン、デオキシシトシン、ピリミジン、フルダラビン(FLUDARA(登録商標))、クラドラビン、2−デオキシ−D−グルコース等を包含する。好ましい代謝拮抗物質化学療法剤はゲムシタビンである。
【0076】
「ゲムシタビン」即ち「2’−デオキシ−2’,2’−ジフルオロシチジン一塩酸塩(b異性体)」は抗腫瘍活性を示すヌクレオシド類縁体である。ゲムシタビンHClに関する実験式はC9H11F2N3O4 A HClである。ゲムシタビンHClは商標GEMZAR(登録商標)の下にEli Lillyから販売されている。
【0077】
「白金系化学療法剤」は分子の一体的部分として白金を含有する有機化合物を含む。白金系化学療法剤の例はカルボプラチン、シスプラチン、およびオキサリプラチンを包含する。
【0078】
「白金系化学療法」とは、場合により1つ以上の他の化学療法剤と組み合わせた1つ以上の白金系化学療法剤を用いた療法を意図する。
【0079】
II.発明を実施するための形態
本発明の実施は特段の記載が無い限り当業者の技量の範囲内の分子生物学(組み換え手法を含む)、微生物学、細胞生物学、生物化学および免疫学の従来の手法を使用することになる。そのような手法は文献、例えば「Molecular Cloning:A Laboratory Manual」,第2版(Sambrookら、1989);「Oligonucleotide Synthesis」(M.J.Gait編、1984);「Animal Cell Culture」(R.I.Freshney編、1987);「Methods in Enzymology」(Academic Press,Inc.);「Handbook of Experimental Immunology」,第4版(D.M.Weir&C.C.Blackwell編、Blackwell Science Inc.,1987);「GeneTransfer Vectors for Mammalian Cells」(J.M.Miller&M.P.Calos編、1987);「Current Protocols in Molecular Biology」(F.M.Ausubelら編、1987);「PCR:The Polymerase Chain Reaction」(Mullisら編、1994);および「Current Protocols in Immunology」(J.E.Coliganら編、1991)において詳細に説明されている。
【0080】
1.アゴニスト抗CYTL1抗体の調製
抗CYTL1抗体は組み換えDNA技術の手法を包含する当該分野で知られた方法により製造できる。
【0081】
i)抗原の調製
場合により他の分子にコンジュゲートされた可溶性抗原またはそのフラグメントは抗体を形成するための免疫原として使用できる。膜貫通分子、例えば受容体に関しては、これらのフラグメント(例えば受容体の細胞外ドメイン)を免疫原として使用できる。或いは、膜貫通分子を発現する細胞を免疫原として使用できる。そのような細胞は天然原料(例えば癌細胞系統)から誘導することができ、或いは膜貫通分子を発現するように組み換え手法により形質転換されている細胞であってよい。抗体を調製するために有用な他の抗原およびその形態は当業者の知る通りである。
【0082】
(ii)ポリクローナル抗体
ポリクローナル抗体は好ましくは該当する抗原およびアジュバントの多数回の皮下(sc)または腹腔内(ip)注射により動物中で育成される。2官能性または誘導体化剤、例えばマレイミドベンゾイルスルホスクシンイミドエステル(システイン残基を介したコンジュゲーション)、N−ヒドロキシスクシンイミド(リジン残基を介する)、グルタルアルデヒド、無水コハク酸、SOCl2またはR1N=C=NR、ただし式中RおよびR1は異なるアルキル基であるもの、を用いて、免疫すべき種において免疫原性であるタンパク質、例えばキーホールリンペットヘモシアニン、血清アルブミン、ウシサイログロブリン、または大豆トリプシン阻害剤に該当する抗原をコンジュゲートすることが有用である場合がある。
【0083】
動物は100μgまたは5μgのタンパク質またはコンジュゲート(それぞれウサギまたはマウスに関する)を3容量のフロイント完全アジュバントと混合し、溶液を多数部位において皮内注射することにより抗原、免疫原性コンジュゲート、または誘導体に対抗して免疫される。1ヵ月後、多数部位における皮下注射によりフロイントの完全アジュバント中のペプチドまたはコンジュゲートの元の量の1/5〜1/10を用いて動物をブーストする。7〜14日後、動物から出血させ、そして血清の抗体力価をアッセイする。動物は力価が平衡に達するまでブーストする。好ましくは、同じ抗原であるが、異なるタンパク質に、および/または異なる交差結合試薬を介してコンジュゲートされているコンジュゲートで動物をブーストする。コンジュゲートはまた、タンパク質融合物として組み換え細胞培養物中に作成できる。さらにまた、ミョウバンのような凝集剤を適宜使用して免疫応答を増強する。
【0084】
(iii)モノクローナル抗体
モノクローナル抗体はKohlerら、Nature,256:495(1975)により最初に記載されたハイブリドーマ法を用いて作成してよく、或いは、組み換えDNA法(米国特許4,816,567)により作成してよい。ハイブリドーマ法においては、マウスまたは他の適切な宿主動物、例えばハムスターまたはマカクサルを上記した通り免疫することにより、免疫のために使用されるタンパク質に特異的に結合することになる抗体を産生するか産生生産できるリンパ球を生じさせる。或いは、リンパ球はインビトロで免疫してよい。次にリンパ球を適切な融合剤、例えばポリエチレングリコールを用いて骨髄腫細胞と融合させることによりハイブリドーマ細胞を形成する(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。
【0085】
このようにして調製されたハイブリドーマ細胞は、未融合の親骨髄腫細胞の成育または生存を阻害する1つ以上の物質を好ましくは含有する適当な培養培地中に播種して生育させる。例えば、親骨髄腫細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠いている場合、ハイブリドーマのための培養培地は典型的にはヒポキサンチン、アミノプテリン、およびチミジンを包含することになり(HAT培地)、これらの物質はHGPRT欠損細胞の成育を防止する。
【0086】
好ましい骨髄腫細胞は、効率的に融合し、選択された抗体産生細胞による抗体の安定な高レベル産生を支援し、そしてHAT培地のような培地に対して感受性であるものである。それらのうち、好ましい骨髄腫細胞系統はネズミ骨髄腫系統、例えばSalk Institute Cell Distribution Center,San Diego,Calif.USAから入手可能なMOPC−21およびMPC−11マウス腫瘍、および、American Type Culture Collection,Rockville,Md.USAから入手可能なSP−2またはX63−Ag8−653細胞から誘導したものである。ヒト骨髄腫およびマウス−ヒトヘテロ骨髄腫細胞系統もまた、ヒトモノクローナル抗体の産生に関して記載されている(Kozbor,J.Immunol.,133:3001(1984);Brodeurら、Monoclonal Antibody Production Techniques and Applications,pp.51−63(Marcel Dekker,Inc.,New York,1987))。
【0087】
ハイブリドーマ細胞を成育させている培養培地は抗原に対するモノクローナル抗体の産生に関してアッセイされる。好ましくは、ハイブリドーマ細胞により産生されたモノクローナル抗体の結合特異性は、免疫沈降により、またはインビトロの結合アッセイ、例えばラジオイムノアッセイ(RIA)または酵素結合免疫吸着アッセイ(ELISA)により、測定される。
【0088】
所望の特異性、親和性、および/または活性の抗体を産生するハイブリドーマ細胞が同定された後、クローンは限界希釈操作法によりサブクローニングし、そして標準的な方法により生育させてよい(Goding,Monoclonal Antibodies:Principles and Practice,pp.59−103(Academic Press,1986))。この目的のための適当な培養培地は例えばD−MEMまたはRPMI−1640培地を包含する。さらにまたハイブリドーマ細胞は動物中の腹水腫瘍としてインビボで成育させてよい。
【0089】
サブクローンにより分泌されたモノクローナル抗体は例えばプロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、またはアフィニティークロマトグラフィーのような従来の免疫グロブリン精製操作法により培養培地、腹水、または血清から適当に分離される。
【0090】
モノクローナル抗体をコードするDNAは従来の操作法を用いて容易に単離して配列決定される(例えばモノクローナル抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合できるオリゴヌクレオチドプローブを使用することによる)。ハイブリドーマ細胞はそのようなDNAの好ましい原料として供される。単離された後、DNAは発現ベクター内に入れてよく、そしてこれを次に宿主細胞、例えばE.coli細胞、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、または別様には免疫グロブリンタンパク質を産生しない骨髄腫細胞内にトランスフェクトすることにより、組み換え宿主細胞内のモノクローナル抗体の合成を達成できる。抗体の組み換え産生は後述においてより詳細に説明する。
【0091】
別の実施形態においては、抗体または抗体フラグメントはMcCaffertyら、Nature,348:552−554(1990)に記載の手法を用いて作成された抗体ファージライブラリから単離できる。
【0092】
Clacksonら、Nature,352:624−628(1991)およびMarksら、J.Mol.Biol.,222:581−597(1991)はファージライブラリを用いたそれぞれネズミおよびヒトの抗体の単離を記載している。その後の出版物は鎖シャフリングによる高親和性(nM範囲)のヒト抗体の産生(Marksら、Bio/Technology,10:779−783(1992))並びに極めて大型のファージライブラリを構築するための方策としてのコンビナトリアルな感染およびインビボの組み換え(Waterhouseら、Nuc.Acids.Res.,21:2265−2266(1993))を記載している。即ち、これらの手法はモノクローナル抗体の単離のための伝統的なモノクローナル抗体ハイブリドーマ手法の実行可能な代替法である。
【0093】
DNAはまた、相同ネズミ配列の代わりにヒト重鎖および軽鎖定常ドメインに関するコード配列を代替とすること(米国特許4,816,567;Morrisonら、Proc.Natl Acad.Sci.USA,81:6851(1984))によるか、または、非免疫グロブリンポリペプチドに関するコード配列の全てまたは部分を免疫グロブリンコード配列に共有結合することにより、修飾してよい。
【0094】
典型的には、そのような非免疫グロブリンポリペプチドは抗体の定常ドメインの代替とされるか、またはそれらは、抗体の1つの抗原複合化部位の可変ドメインの代替とされ、これにより抗原に対する特異性を有する1つの抗原複合化部位および異なる抗原に対する特異性を有する別の抗原複合化部位を含むキメラ2価抗体を作製する。
【0095】
(iv)ヒト化およびヒト抗体
ヒト化抗体は非ヒトである原料から自身内に導入された1つ以上のアミノ酸残基を有する。これらの非ヒトアミノ酸残基はしばしば「インポート」残基と称され、これは典型的には「インポート」可変ドメインから取り出される。ヒト化は本質的にはWinterらの方法(Jonesら、Nature,321:522−525(1986);Riechmannら、Nature,332:323−327(1988);Verhoeyenら、Science,239:1534−1536(1988))に従って、ヒト抗体の相当する配列に対してげっ歯類のCDRまたはCDR配列を置換することにより実施することができる。従って、そのような「ヒト化」抗体は実質的にはインタクトなヒト可変ドメインに満たない部分が非ヒト種由来の相当する配列により置換されているキメラ抗体(米国特許4,816,567)である。実際には、ヒト化抗体は典型的には一部のCDR残基および可能性として一部のFR残基がげっ歯類抗体中の類似の部位に由来する残基により置換されているヒト抗体である。
【0096】
ヒト化抗体を作成する場合に使用するべき軽鎖および重鎖の両方のヒト可変ドメインの選択は抗原性を低減するためには極めて重要である。所謂「ベストフィット」法に従って、げっ歯類抗体の可変ドメインの配列を既知ヒト可変ドメイン配列の完全なライブラリに対してスクリーニングする。次にげっ歯類のものと最も近似しているヒト配列をヒト化抗体のためのヒトフレームワーク(FR)として許容する(Simsら、J.Immunol.,151:2296(1993);Chothiaら、J.Mol.Biol.,196:901(1987))。別の方法は軽鎖または重鎖の特定のサブグループの全ヒト抗体のコンセンサス配列から誘導された特定のフレームワークを使用する。同じフレームワークをいくつかの異なるヒト化抗体に対して使用してよい(Carterら、Proc.Natl.Acad Sci.USA,89:4285(1992);Prestaら、J.Immnol.,151:2623(1993))。
【0097】
さらにまた、抗原に対する高親和性および他の望ましい生物学的特性を保持して抗体をヒト化することが重要である。この目標を達成するためには、好ましい方法に従って、親およびヒト化配列の3次元モデルを用いて親配列および種々の概念的なヒト化産物の分析のプロセスによりヒト化抗体を調製する。3次元免疫グロブリンモデルは公的に入手可能であり、そして当業者に良く知られている。選択された候補免疫グロブリン配列の推定の3次元コンホメーション構造を図示して表示できるコンピュータプログラムが使用可能である。これらの表示物の検討により候補免疫グロブリン配列の機能における残基の考えられる役割の分析、即ち、候補免疫グロブリンがその抗原に結合する能力に影響する残基の分析が可能となる。このような方法により、FR残基を選択し、そして、所望の抗体特性、例えば標的抗原に対する増大した親和性を達成できるようにレシピエントおよびインポート配列と組み合わせることができる。一般的に、CDR残基は、抗原結合に対して影響を与えることにおいては、直接、そして最も大きく関与している。
【0098】
或いは、内因性の免疫グロブリンの産生の非存在下においてヒト抗体の完全なレパートリーの産生が免疫により可能となるトランスジェニック動物(例えばマウス)を作成することが、現在可能である。例えば、キメラおよび生殖細胞系統の突然変異体マウスにおける抗体重鎖連結領域(JH)遺伝子のホモ接合欠失は内因性抗体産生の完全な阻害をもたらすことが記載されている。そのような生殖細胞系統突然変異体マウスにおけるヒト生殖細胞系統免疫グロブリン遺伝子アレイの転移は、抗原攻撃時のヒト抗体の産生をもたらすことになる。例えばJakobovitsら、Proc.Natl.Acad.Sci.USA,90:2551(1993);Jakobovitsら、Nature,362:255−258(1993);Bruggermannら、Year in Immuno.,7:33(1993);およびDuchosalら、Nature,355:258(1992)を参照。ヒト抗体はまたファージディスプレイライブラリから誘導できる(Hoogenboomら、J.Mol.Biol.,227:381(1991);Marksら、J.MoL Biol.,222:581−597(1991);Vaughanら、Nature Biotech.,14:309(1996))。抗体ファージディスプレイライブラリからのヒト抗体の形成を以下にさらに説明する。
【0099】
(v)抗体フラグメント
種々の手法が抗体フラグメントの産生のために開発されている。伝統的にはこれらのフラグメントはインタクトな抗体のタンパク質分解性消化を介して誘導されている(例えばMorimotoら、Journal of Biochemical and Biophysical Methods,24:107−117(1992)およびBrennanら、Science,229:81(1985)参照)。しかしながらこれらのフラグメントは現在では組み換え宿主細胞により直接産生できる。例えば抗体フラグメントは上記考察した抗体ファージライブラリから単離できる。或いはFab’−SHフラグメントをE.coliから直接回収し、そして化学的にカップリングさせることによりF(ab’)2フラグメントを形成できる(Carterら、Bio/Technology,10:163−167(1992))。後述する実施例において説明する通り別の実施形態においては、F(ab’)2はF(ab’)2分子の組み立てを促進するためにロイシンジッパーGCN4を用いて形成される。別の手順によれば、F(ab’)2フラグメントは組み換え宿主細胞培養物から直接単離できる。抗体フラグメントの産生のための他の手法は当業者に良く知られている。他の実施形態においては選択される抗体は単鎖Fvフラグメント(scFv)である。WO93/16185参照。
【0100】
(vi)多重特異性抗体
多重特異性抗体はエピトープが通常は異なる抗原に由来する少なくとも2つの異なるエピトープに対する結合特異性を有する。そのような分子は通常は2つの異なるエピトープに結合するのみとなる(即ち二重特異性抗体、BsAb)が、三重特異性抗体のような追加的特異性を有する抗体も本明細書において使用する場合にこの表現に包含される。
【0101】
二重特異性抗体を作成するための方法は当該分野において知られている。完全長二重特異性抗体の伝統的な産生は2つの鎖が異なる特異性を有する2つの免疫グロブリン重鎖−軽鎖の対の同時発現に基づいている(Millsteinら、Nature,305:537−539(1983))。免疫グロブリン重鎖および軽鎖のランダムな組み合わせのために、これらのハイブリドーマ(クアドローマ)は10種の異なる抗体分子の潜在的混合物を産生し、そのうち僅か1つのみが正しい二重特異性の構造を有する。通常はアフィニティークロマトグラフィー工程により行われる正しい分子の精製はむしろ負担が大きく、そして生成物の収率は低値である。同様の操作法がWO93/08829、およびTrauneckerら、EMBO J.,10:3655−3659(1991)に開示されている。異なる手順によれば、所望の結合特異性を有する抗体可変ドメイン(抗体−抗原複合化部位)を免疫グロブリン定常ドメイン配列に融合させる。融合は好ましくはヒンジ、CH2、およびCH3領域の少なくとも一部分を含む免疫グロブリン重鎖定常ドメインと行う。軽鎖結合のために必要な部位を含有する第1の重鎖定常領域(CH1)が融合物の少なくとも1つにおいて存在することが好ましい。免疫グロブリン重鎖融合物、および所望により免疫グロブリン軽鎖をコードするDNAを別個の発現ベクター内に挿入し、そして適当な宿主生物内に同時トランスフェクトする。これにより、構築において使用される3つのポリペプチド鎖の等しくない比率が最適な収率をもたらす場合の実施形態において3つのポリペプチドフラグメントの相互の割合を調節する場合に、大きい柔軟性がもたらされる。しかしながら、等しい比率における少なくとも2つのポリペプチド鎖の発現が高い収率をもたらす場合、または比率が特に重要ではない場合に、1つの発現ベクター中に2つまたは3つ全てのポリペプチド鎖に関するコード配列を挿入することが可能である。
【0102】
この手順の好ましい実施形態において、二重特異性抗体は、1つのアームにおいて第1の結合特異性を有するハイブリッド免疫グロブリン重鎖、および別のアームにおいてハイブリッド免疫グロブリン重鎖−軽鎖対(第2の結合特異性を与える)を有している。この非対称の構造は、二重特異性分子の半分のみにおける免疫グロブリン軽鎖の存在が容易な分離方法を可能とすることから、望ましくない免疫グロブリン鎖の組み合わせからの所望の二重特異性の化合物の分離を容易にする。この手順はWO94/04690に開示されている。二重特異性抗体の形成はさらに詳細には例えばSureshら、Methods in Enzymology,121:210(1986)を参照。
【0103】
WO96/27011に記載されている別の手順に従えば、抗体分子の対の間の界面は、組み換え細胞培養物から回収されるヘテロ二量体のパーセントを最大限とするように操作できる。好ましい界面は抗体定常ドメインのCH3ドメインの少なくとも一部分を含む。この方法においては、第1の抗体分子の界面に由来する1つ以上の小型アミノ酸側鎖をより大型の側鎖(例えばチロシンまたはトリプトファン)と置き換える。大型側鎖と同一または同様のサイズの代償分の「空洞」は、大型アミノ酸側鎖をより小さいもの(例えばアラニンまたはスレオニン)で置き換えることにより第2の抗体分子の界面上に形成される。これによりホモ二量体のような他の望ましくない最終生成物を超えてヘテロ二量体の収率を増大させるための機序が与えられる。
【0104】
二重特異性抗体は交差結合または「ヘテロコンジュゲート」抗体を包含する。例えばヘテロコンジュゲートにおける抗体の1つをアビジンに、もう1つをビオチンにカップリングできる。そのような抗体は例えば、望ましくない細胞に免疫系細胞をターゲティングするため(米国特許4,676,980)およびHIV感染症の処置のため(WO91/00360、WO92/200373)に提案されている。ヘテロコンジュゲート抗体は何れかの好都合な交差結合法を用いて作製してよい。適当な交差結合剤は当該分野で良く知られており、そして多くの交差結合手法と共に米国特許4,676,980に開示されている。
【0105】
抗体フラグメントから二重特異性抗体を形成するための手法もまた、文献に記載されている。例えば二重特異性抗体は化学結合を用いて調製できる。Brennanら、Science,229:81(1985)はインタクトな抗体をタンパク質分解的に切断することによりF(ab’)2フラグメントを形成する操作法を記載している。これらのフラグメントをジチオール錯化剤(dithiol complexing agent)である亜ヒ酸ナトリウムの存在下に還元することにより近接したジチオールを安定化させ、そして分子内のジスルフィド形成を防止する。次に形成されたFab’フラグメントをチオニトロベンゾエート(TNB)誘導体に変換する。次にFab’−TNB誘導体の一方をメルカプトエチルアミンを用いた還元によりFab’−チオールに再変換し、そして等モル量の他のFab’−TNB誘導体と混合することにより二重特異性抗体を形成する。生成した二重特異性抗体は酵素の選択的固定化のための薬剤として使用できる。
【0106】
Fab’−SHフラグメントはまたE.coliから直接回収することができ、そして化学的にカップリングすることにより二重特異性抗体を形成できる。Shalabyら、J.Exp.Med.,175:217−225(1992)は完全にヒト化された二重特異性抗体F(ab’)2分子の産生を記載している。各Fab’フラグメントをE.coliから別個に分泌させ、そしてインビトロの指向的化学カップリングに付すことにより二重特異性抗体を形成している。
【0107】
組み換え細胞培養物から直接二重特異性抗体フラグメントを作成して単離するための種々の手法もまた記載されている。例えば、二重特異性抗体はロイシンジッパーを用いて産生されている。Kostelnyら、J.Immunol.,148(5):1547−1553(1992)。FosおよびJunタンパク質に由来するロイシンジッパーペプチド(pepbdes)を遺伝子融合により2つの異なる抗体のFab’部分に連結している。抗体ホモ二量体をヒンジ領域で還元することにより単量体を形成し、そして次に再酸化して抗体ヘテロ二量体を形成している。この方法はまた、抗体ホモ二量体の産生のためにも利用できる。Hollingerら、Proc.Nati.Acad.Sci.USA,90:6444−6448(1993)の記載している「ダイアボディ」技術は二重特異性抗体フラグメントを作成するための代替的な機序を与えている。フラグメントは同じ鎖上の2ドメインの間の対形成を可能にするには短すぎるリンカーにより軽鎖可変ドメイン(VL)に結合した重鎖可変ドメイン(VH)を含む。従って、1つのフラグメントのVHおよびVLドメインは別のフラグメントの相補なVLおよびVHドメインと強制的に対形成させられ、これにより2つの抗原結合部位が形成される。単鎖Fv(sFv)二量体の使用により二重特異性抗体フラグメントを形成するための別の方策もまた報告されている。Gruberら、J.Immunol.,152:5368(1994)を参照。
【0108】
2より多い結合価を有する抗体もまた意図される。例えば三重特異性抗体を調製できる。Tuftら、J.Immunol.,147:60(1991)。
【0109】
(vii)エフェクター機能の操作
抗体の有効性を増強するように、エフェクター機能に関して本発明の抗体を修飾することが望ましい場合がある。例えば、システイン残基をFc領域に導入することによりこの領域における鎖間ジスルフィド結合の形成を可能にしてよい。このようにして形成されたホモ二量体抗体は向上した内在化能力および/または増大した補体媒介細胞殺傷および抗体依存性細胞毒性(ADCC)を有する場合がある。Caronら、J.Exp Med.,176:1191−1195(1992)およびShopes,B.J.Immunol.,148:2918−2922(1992)を参照。増強された抗腫瘍活性を有するホモ二量体抗体はまた、Wolffら、Cancer Research,53:2560−2565(1993)において記載される通りヘテロ2官能性交差結合剤を用いて調製してよい。或いは、抗体は二重のFc領域を有するように操作することができ、そしてこれにより、増強された補体溶解およびADCC能力を有する場合がある。Stevensonら、Anti−Cancer Drug Design 3:219−230(1989)を参照。
【0110】
(viii)抗体サルベージ受容体結合エピトープ融合物
本発明の特定の実施形態においては、例えば腫瘍浸透性を増大させるためにインタクトな抗体よりもむしろ抗体フラグメントを使用することが望ましい場合がある。そのような場合、抗体フラグメントはその血清中半減期を増大させるために修飾することが望ましい場合がある。これは例えば抗体フラグメント内へのサルベージ受容体結合エピトープの取り込みにより達成してよい(例えば抗体フラグメント内の適切な領域の突然変異により、或いは、エピトープをペプチドタグ内に取り込ませ、次にこれを、例えばDNAまたはペプチド合成により、抗体フラグメントに何れかの末端または中央部において融合する)。
【0111】
サルベージ受容体結合エピトープは好ましくは、Fcドメインの1つまたは2つのループに由来するアミノ酸残基の何れか1つ以上が抗体フラグメントの類似の位置に転移する領域を構築する。さらに好ましくは、Fcドメインの1つまたは2つのループに由来する残基3つ以上が転移する。なおさらに好ましくは、エピトープはFc領域のCH2ドメイン(例えばIgGのもの)から取り出し、そして、抗体のCH1、CH3、またはVH領域、または1つより多くのそのような領域に転移させる。或いは、エピトープをFc領域のCH2ドメインから取り出し、そして抗体フラグメントのCL領域またはVL領域、または両方に転移させる。
【0112】
(ix)抗体の他の共有結合的修飾
抗体の共有結合的修飾は本発明の範囲内に包含される。それらは適宜、化学合成によるか、または酵素的または化学的な抗体切断により行ってよい。抗体の共有結合的修飾の他の型は、選択された側鎖またはNまたはC末端残基と反応できる有機の誘導体化剤に抗体のターゲティングされたアミノ酸残基を反応させることにより分子内に導入される。共有結合的修飾の例は参照により本明細書に組み込まれる米国特許5,534,615に記載されている。抗体の共有結合的修飾の好ましい型は米国特許4,640,835;4,496,689;4,301,144;4,670,417;4,791,192または4,179,337に記載されている様式において、種々の非タンパク質性重合体、例えばポリエチレングリコール、ポリプロピレングリコール、またはポリオキシアルキレンの1つに抗体を連結させることを含む。
【0113】
(x)合成抗体ファージライブラリからの抗体の形成
好ましい実施形態においては、本発明はユニークなファージディスプレイの手順を用いて新規な抗体を形成および選択するための方法を提供する。該手順においては、単一のフレームワーク鋳型に基づく合成抗体ファージライブラリの形成、可変ドメイン内部の十分な多様性の設計、多様化された可変ドメインを有するポリペプチドのディスプレイ、抗原をターゲティングするための高親和性を有する候補抗体の選択、および選択された抗体の単離を行う。
【0114】
ファージディスプレイの方法の詳細は例えば参照により全体が本明細書に組み込まれる2003年12月11日に公開されたWO03/102157に記載されている。
【0115】
1つの態様において、本発明において使用される抗体ライブラリは抗体可変ドメインの少なくとも1つのCDRにおいて溶媒接触性および/または高多様性の位置を突然変異させることにより形成できる。CDRの一部または全てを本明細書に記載した方法を用いて突然変異させることができる。一部の実施形態においては、多様な抗体ライブラリは、単一のライブラリを形成するためにCDRH1、CDRH2およびCDRH3における位置を突然変異させることによるか、または単一のライブラリを形成するためにCDRL3およびCDRH3における位置を突然変異させることによるか、または単一のライブラリを形成するためにCDRL3およびCDRH1、CDRH2およびCDRH3における位置を突然変異させることにより形成することが好ましい場合がある。
【0116】
抗体可変ドメインのライブラリは例えばCDRH1、CDRH2およびCDRH3の溶媒接触性および/または高多様性の位置において突然変異を有するように形成できる。別のライブラリはCDRL1、CDRL2およびCDRL3において突然変異を有するように形成できる。これらのライブラリはまた、所望の親和性を有するバインダーを形成するために相互に組み合わせて使用できる。例えば標的抗原への結合のための重鎖ライブラリの選択を1回以上行った後、軽鎖ライブラリを重鎖バインダーの集団内に入れ換え、選択回数をさらに重ねることによりバインダーの親和性を増大させることができる。
【0117】
好ましくは、重鎖配列の可変領域のCDRH3領域において変異体アミノ酸により元のアミノ酸を置換することによりライブラリを形成する。得られたライブラリは複数の抗体配列を含有することができ、そこにおいては配列の多様性は重鎖配列のCDRH3領域に主に存在する。
【0118】
1つの態様において、ライブラリはヒト化抗体4D5配列、またはヒト化抗体4D5配列のフレームワークアミノ酸の配列との関連で形成される。好ましくはライブラリはDVKコドンセットによりコードされるアミノ酸による重鎖の少なくとも残基95〜100aの置換により形成され、その場合、DVKコドンセットはこれらの位置の各々に対して変異体アミノ酸のセットをコードするために使用される。これらの置換を形成するために有用なオリゴヌクレオチドのセットの例は配列(DVK)7を含む。一部の実施形態においては、ライブラリはDVKおよびNNKコドンセットの両方でコードされたアミノ酸による残基95〜100aの置換により形成する。これらの置換を形成するために有用なオリゴヌクレオチドのセットの例は配列(DVK)6(NNK)を含む。別の実施形態においては、ライブラリはDVKおよびNNKコドンセットの両方でコードされたアミノ酸による少なくとも残基95〜100aの置換により形成する。これらの置換を形成するために有用なオリゴヌクレオチドのセットの例は配列(DVK)5(NNK)を含む。これらの置換を形成するために有用なオリゴヌクレオチドセットの別の例は配列(NNK)6を含む。適当なオリゴヌクレオチド配列の他の例は本明細書に記載する基準に従って当業者が決定できる。
【0119】
別の実施形態においては、異なるCDRH3設計を利用することにより高親和性バインダーを単離し、そして種々のエピトープに対するバインダーを単離する。このライブラリにおいて形成されるCDRH3の長さの範囲は11〜13アミノ酸であるが、これとは異なる長さも形成することができる。H3の多様性はNNK、DVKおよびNVKコドンセット、並びにNおよび/またはC末端におけるより限定された多様性を使用することにより拡張できる。
【0120】
多様性はまた、CDRH1およびCDRH2において形成することもできる。CDR−H1およびH2の多様性の設計は以前の設計よりも天然の多様性により緊密にマッチしている多様性に着目した変更を伴って記載した天然の抗体レパートリーを模倣するためのターゲティングの方策に従う。
【0121】
CDRH3における多様性に関しては、異なる長さのH3を有するように多数のライブラリを別個に構築し、そして次に、抗原をターゲティングするためのバインダーを求める選別のために組み合わせることができる。多数のライブラリをプールし、そして明細書中前述および後述する固体支持体選択および溶液分類方法を用いて分類することができる。マルチソーティングの方策を使用してよい。例えば1つの変法では固体に結合した標的に対する分類を行い、その後、融合ポリペプチド上に存在する場合があるタグ(例えば抗gDタグ)に対する分類を行い、そしてその後、固体に結合した標的に対する別の分類を行う。或いは、ライブラリを固体表面に結合した標的に対して先ず分類することができ、次に溶出したバインダーを標的抗原の漸減濃度における溶液相結合を用いて分類する。異なる分類方法の組み合わせを利用することは高度に発現される配列のみの選択の最小限化を可能にし、そして、多くの異なる高親和性クローンの選択を可能にする。
【0122】
標的抗原に対する高親和性バインダーをライブラリから単離できる。H1/H2領域における限定された多様性は縮重性を約104〜105倍低下させ、そしてより高値のH3多様性を可能にすれば、より高親和性のバインダーが得られる。CDRH3における異なる型の多様性を有するライブラリを利用することにより(例えばDVKまたはNVTを利用することにより)、標的抗原の異なるエピトープに結合する場合があるバインダーの単離が可能となる。
【0123】
上記した通りプールされたライブラリから単離されたバインダーのうち、親和性は軽鎖において限定された多様性をもたらすことによりさらに向上する場合があることが発見された。本実施形態において軽鎖の多様性は以下の通り、即ちCDRL1において:アミノ酸位置28はRDTによりコードされ;アミノ酸位置29はRKTによりコードされ;アミノ酸位置30はRVWによりコードされ;アミノ酸位置31はANWによりコードされ;アミノ酸位置32はTHTによりコードされ;場合により、アミノ酸位置33はCTGによりコードされ;CDRL2において:アミノ酸位置50はKBGによりコードされ;アミノ酸位置53はAVCによりコードされ;そして場合により、アミノ酸位置55はGMAによりコードされ;CDRL3において:アミノ酸位置91はTMTまたはSRTまたは両方によりコードされ;アミノ酸位置92はDMCによりコードされ;アミノ酸位置93はRVTによりコードされ;アミノ酸位置94はNHTによりコードされ;そしてアミノ酸位置96はTWTまたはYKGまたは両方によりコードされるように、形成される。
【0124】
別の実施形態においては、CDRH1、CDRH2およびCDRH3領域における多様性を有するライブラリを形成する。本実施形態においてCDRH3における多様性は種々の長さのH3領域を用いて、そして主にコドンセットXYZおよびNNKまたはNNSを用いて形成する。ライブラリは個々のオリゴヌクレオチドを用いて形成してプールすることができ、或いはオリゴヌクレオチドをプールしてライブラリのサブセットを形成することができる。本実施形態のライブラリは固体に結合した標的に対抗して分類できる。多数の分類から単離されたクローンはELISAアッセイを用いて特異性および親和性に関してスクリーニングできる。特異性に関しては、クローンは所望の標的抗原並びに他の非標的抗原に対抗してスクリーニングできる。次に標的抗原に対するこのようなバインダーを溶液結合競合ELISAアッセイまたはスポット競合アッセイにおいて親和性に関してスクリーニングできる。高親和性のバインダーは上記した通り調製したXYZコドンセットを利用してライブラリから単離できる。これらのバインダーは細胞培養物中において高収率で抗体または抗原結合フラグメントとして容易に製造できる。
【0125】
一部の実施形態においては、CDRH3領域における長さにおいてより大きい多様性を有するライブラリを形成することが望ましい場合がある。例えば約7〜19アミノ酸の範囲のCDRH3領域を有するライブラリを形成することが望ましい場合がある。
【0126】
これらの実施形態のライブラリから単離された高親和性バインダーは高収率で細菌および真核生物細胞の培養物中において容易に製造される。ベクターはgDタグ、ウィルス被膜タンパク質成分配列のような配列を容易に除去するように、および/または、高収率で完全長抗体または抗原結合フラグメントの産生を可能にする定常領域配列において付加するために設計できる。
【0127】
CDRH3において突然変異を有するライブラリは他のCDR、例えばCDRL1、CDRL2、CDRL3、CDRH1および/またはCDRH2の変異体型を含有するライブラリと組み合わせることができる。即ち、例えば、1つの実施形態において、CDRH3ライブラリを、所定のコドンセットを用いて28、29、30、31、および/または32位において変異体アミノ酸を有するヒト化4D5抗体配列との関連で形成されたCDRL3ライブラリと組み合わせる。別の実施形態においては、CDRH3への突然変異を有するライブラリは、変異体CDRH1および/またはCDRH2重鎖可変ドメインを含むライブラリと組み合わせることができる。1つの実施形態において、CDRH1ライブラリは28、30、31、32および33位において変異体アミノ酸を有するヒト化抗体4D5配列を有するように形成される。CDRH2ライブラリは所定のコドンセットを用いて50、52、53、54、56および58位において変異体アミノ酸を有するヒト化抗体4D5配列を有するように形成される。
【0128】
(xi)抗体突然変異体
抗体突然変異を形成するためには、1つ以上のアミノ酸改変(例えば置換)を親抗体の1つ以上の超可変領域に導入する。或いは、または追加的に、フレームワーク領域残基の1つ以上の改変(例えば置換)を親抗体に導入してよく、その場合、これらは第2の哺乳類種に由来する抗原に対する抗体突然変異体の結合親和性の向上をもたらす。修飾すべきフレームワーク領域の残基の例は、直接抗原に非共有結合的に結合し(Amitら、(1986)Science 233:747−753);CDRのコンホメーションに相互作用/影響し(Chothiaら、(1987)J.Mol.Biol.196:901−917);および/またはVL−VH界面に関与する(EP239 400B1)ものを包含する。特定の実施形態においては、1つ以上のそのようなフレームワーク領域残基の修飾は第2の哺乳類種に由来する抗原に対する抗体の結合親和性の増強をもたらす。例えば、約1〜約5フレームワーク残基を本発明の本実施形態において改変してよい。場合により、これは、超可変領域の残基の何れも改変されていない場合であっても、前臨床治験における使用のために適する抗体突然変異体をもたらすためには十分である場合がある。しかしながら通常は、抗体突然変異体は追加的な超可変領域の改変を含むことになる。
【0129】
改変される超可変領域残基は、特に親抗体の元の結合親和性がランダムに産生された抗体突然変異体が容易にスクリーニングされ得るようなものである場合には、ランダムに変更してよい。
【0130】
そのような抗体突然変異体を形成するための1つの有用な操作法は「アラニンスキャニング突然変異誘発」と称されている(Cunningham and Wells(1989)Science 244:1081−1085)。ここでは1つ以上の超可変領域残基をアラニンまたはポリアラニン残基により置き換えることにより第2の哺乳類種に由来する抗原とのアミノ酸の相互作用に影響を及ぼす。次に置換に対して機能的感受性を示している超可変領域残基を、置換部位において、またはそれに対して追加の、または他の突然変異を導入することにより純化する。即ち、アミノ酸配列変異を導入するための部位は予め決定されているが、突然変異の性質自体は予め決定されている必要はない。この方法で産生したala突然変異体を本明細書に記載する通りそれらの生物学的活性に関してスクリーニングする。
【0131】
通常は「好ましい置換」の見出しの下に以下に示すもののような保存的置換により開始する。そのような置換が生物学的活性(例えば結合親和性)の変化をもたらす場合、次に、以下の表において「例示される置換」と命名されている、またはアミノ酸クラスを参照して後述においてさらに説明する通り、より実質的な変化を導入し、そして生成物をスクリーニングする。
【0132】
【表2】
【0133】
抗体の生物学的特性におけるより実質的な修飾は以下、即ち:(a)例えばシートまたはヘリックス型のコンホメーションとしての置換の領域におけるポリペプチド骨格の構造、(b)標的部位における分子の電荷または疎水性、または(c)側鎖の嵩、を維持することに対する自身の作用において有意に異なっている置換を選択することにより達成される。天然に存在する残基は共通の側鎖の特性、即ち:
(1)疎水性:ノルロイシン、met、ala、val、leu、ile;
(2)中性親水性:cys、ser、thr、asn、gln;
(3)酸性:asp、glu;
(4)塩基性:his、lys、arg;
(5)鎖の配向に影響を与える残基:gly、pro;および、
(6)芳香族:trp、tyr、phe
に基づいて群に分割される。
【0134】
非保存的置換はこれらのクラスの1つのメンバーを別のクラスに交換することを包含することになる。
【0135】
別の実施形態においては、修飾のために選択される部位はファージディスプレイを用いてアフィニティー成熟化される(上記参照)。
【0136】
アミノ酸配列突然変異体をコードする核酸分子は当該分野で知られる種々の方法により調製される。これらの方法は限定しないがオリゴヌクレオチド媒介(または部位指向性)突然変異誘発、PCR突然変異誘発、およびより早期に調製された突然変異体または親抗体の非突然変異体型のカセット突然変異誘発を包含する。突然変異体を作成するための好ましい方法は部位指向性突然変異誘発である(例えばKunkel(1985)Proc.Natl.Acad.Sci.USA 82:488参照)。
【0137】
特定の実施形態においては、抗体突然変異体は単一の超可変領域残基が置換されているのみとなる。他の実施形態においては、親抗体の1つ以上の超可変領域残基が置換されることになり、例えば約2〜約10超可変領域置換となる。
【0138】
通常は、向上した生物学的特性を有する抗体突然変異体は親抗体の重鎖または軽鎖可変ドメインの何れかのアミノ酸配列と少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、そして最も好ましくは少なくとも95%のアミノ酸配列同一性または類似性を有するアミノ酸配列を有することになる。この配列に関する同一性または類似性は、最大パーセント配列同一性を達成するために、配列をアラインし、そして必要に応じてギャップを導入した後に、親抗体残基と同一(即ち同じ残基)または類似(即ち共通の側鎖特性に基づいた同じ群に由来するアミノ酸残基、上記参照)である候補配列中のアミノ酸残基のパーセントとして本明細書においては定義される。可変ドメインの外部の抗体配列内へのN末端、C末端、または内部の伸長、欠失、または挿入の何れも、配列の同一性または類似性に影響するものとみなされない。
【0139】
抗体突然変異体の産生の後、親抗体と比較した場合のその分子の生物学的活性を測定する。上記した通り、ここでは抗体の結合親和性および/または他の生物学的活性の測定を行ってよい。本発明の好ましい実施形態においては、抗体突然変異体のパネルを調製し、そして抗原またはそのフラグメントに対する結合親和性に関してスクリーニングする。この初回スクリーニングから選択された1つ以上の抗体突然変異体を、場合により1つ以上の別の生物学的活性アッセイに付すことにより、増強された結合親和性を有する抗体突然変異体が実際に、例えば前臨床試験のために有用であることを確認する。
【0140】
このようにして選択された抗体突然変異体は、多くの場合抗体の意図する用途に応じてさらに修飾に付してよい。そのような修飾では、アミノ酸配列をさらに改変し、非相同ポリペプチドに融合し、および/または後述の通り操作する共有結合的修飾を行ってよい。アミノ酸配列改変に関しては、例示される修飾は上記の通り操作される。例えば抗体突然変異体の適切なコンホメーションの維持に関与する何れかのシステイン残基もまた、一般的にはセリンで置換することにより、分子の酸化安定性を向上させ、そして異常な交差結合を防止してよい。逆に、システインの結合を抗体に付加することによりその安定性を向上させてよい(特に抗体がFvフラグメントのような抗体フラグメントである場合)。別の型のアミノ酸突然変異体は改変されたグリコシル化パターンを有する。これは、抗体中に観察される1つ以上の炭水化物部分を欠失させることにより、および/または、抗体中に存在しない1つ以上のグリコシル化部位を付加することにより、達成してよい。抗体のグリコシル化は典型的にはN連結またはO連結の何れかである。N連結はアスパラギン残基の側鎖への炭水化物部分の連結を指す。Xがプロリン以外の何れかのアミノ酸であるトリペプチド配列であるアスパラギン−X−セリンおよびアスパラギン−X−スレオニンは、アスパラギン側鎖への炭水化物部分の酵素的連結のための認識配列である。即ち、ポリペプチド中のこれらのトリペプチド配列の何れかの存在は、潜在的なグリコシル部位を生じさせる。O連結グリコシル化は糖類であるN−アセチルガラクトサミン、ガラクトース、またはキシロースのうちの1つの、最も一般的にはセリンまたはスレオニンであるが5−ヒドロキシプロリンまたは5−ヒドロキシリジンでもよいヒドロキシアミノ酸への連結を指す。抗体へのグリコシル化部位の付加は、アミノ酸配列をそれが上記トリペプチド配列のうち1つ以上を含有するように改変することにより、好都合に達成される(N連結グリコシル化部位の場合)。改変はまた、元の抗体の配列に対する1つ以上のセリンまたはスレオニン残基の付加または置換により行ってもよい(O連結グリコシル化部位の場合)。
【0141】
同じ型の操作法および改変を用いることによりネイティブ配列CYTL1ポリペプチドのアゴニストとして作用するCYTL1変異体を作成できる。
【0142】
(xii)抗体および他のポリペプチドの組み換え産生
抗体の組み換え産生のためには、それをコードする核酸を単離し、そして後のクローニング(DNAの増幅)または発現のために複製可能なベクター内に挿入する。モノクローナル抗体をコードするDNAは従来の操作法を用いて(例えば抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合できるオリゴヌクレオチドプローブを使用することにより)容易に単離および配列決定される。多くのベクターが入手可能である。ベクター成分は一般的に、限定しないが、以下のもの、即ちシグナル配列、複製起点、1つ以上のマーカー遺伝子、エンハンサーエレメント、プロモーターおよび転写終止配列(例えば参照により本明細書に組み込まれる米国特許5,534,615記載)のうちの1つ以上を包含する。
【0143】
本発明におけるベクター中でのDNAのクローニングまたは発現のための適当な宿主細胞は、上記した原核生物、酵母、または他の高等な真核生物の細胞である。この目的のための適当な原核生物は、真正細菌、例えばグラム陰性またはグラム陽性の生物、例えば腸内細菌科、例えばエシェリシア、例えばE.coli、エンテロバクター、エルウィニア、クレブシエラ、プロテウス、サルモネラ、例えばサルモネラ・チフィムリウム、セラチア、例えばセラチア・マルセスカンス、およびシゲラ、並びにバチルス、例えばB.サブチルスおよびB.リチェニホルミス(例えば1989年4月12日公開のDD266,710に開示されているB.リチェニホルミス41P)、シュードモナス、例えばP.アエルギノーサ、およびストレプトマイセスを包含する。1つの好ましいE.coliクローニング宿主は、E.coli 294(ATCC31,446)であるが、他の菌株、例えばE.coli B、E.coli X1776(ATCC31,537)、およびE.coli W3110(ATCC27,325)も適している。これらの例は限定的なものではなく、例示に過ぎない。
【0144】
原核生物に加えて、真核生物の微生物、例えば糸状カビまたは酵母も抗体をコードするベクターのための適当なクローニングまたは発現の宿主である。サッカロマイセス・セレビシアエ、即ち一般的にはパン酵母が、下等な真核生物宿主微生物の中で最も一般的に使用されている。しかしながら多くの他の属、種、および系統が本発明において一般的に入手可能であり、そして有用であり、例えばスキゾサッカロマイセス・ポンベ;クルイベロマイセス宿主、例えばK.ラクチス、K.フラジリス(ATCC12,424)、K.ブルガリクス(ATCC16,045)、K.ウイケラミ(ATCC24,178)、K.ワルチ(ATCC56,500)、K.ドロソフィラルム(ATCC36,906)、K.サーモトレランス、およびK.マルキシアヌス;ヤロウイニア(EP402,226);ピチア・パストリス(EP183,070);カンジダ;トリコデルマ・リーシア(EP244,234);ニューロスポラ・クラッサ;シュワニオマイセス、例えばシュワニオマイセス・オクシデンタリス;および糸状カビ、例えばニューロスポラ、ペニシリウム、トリポクラジウム、およびアスペルギルス宿主、例えばA.ニヅランスおよびA.ナイジャーが挙げられる。
【0145】
グリコシル化抗体の発現のための適当な宿主細胞は多細胞生物から誘導される。無脊椎動物細胞の例は植物および昆虫の細胞を包含する。多くのバキュロウィルス菌株および変異体およびスポドプテラ・フルギペルダ(イモムシ)、アエデス・アエギプチ(カ)、アエデス・アルボピクツス(カ)、ドロソフィリア・メラノガスター(ミバエ)、およびボンビックス・モリのような宿主に由来する相当する許容性昆虫宿主細胞が同定されている。トランスフェクションのための種々のウィルス菌株、例えばオートグラファ・カリフォルニカNPVのL−1変異体およびボンビックス・モリNPVのBm−5菌株が公的に入手可能であり、そしてそのようなウィルスは、特にスポドプテラ・フルギペルダ細胞のトランスフェクションのために、本発明に従ってここでウィルスとして使用してよい。綿、トウモロコシ、バレイショ、ダイズ、ペチュニア、トマト、およびタバコの植物細胞培養物もまた宿主として利用できる。
【0146】
しかしながら、脊椎動物細胞の場合が最も興味深く、そして培養物(組織培養物)中の脊椎動物細胞の増殖が定型的な操作法となっている。有用な哺乳類宿主細胞系統の例は、SV40で形質転換したサル腎CV1系統(COS−7,ATCC CRL1651);ヒト胚性腎系統(懸濁培養物中での生育のためにサブクローニングされた293または293細胞、Grahamら、J.Gen Virol.,36:59(1977));ベビーハムスター腎細胞(BHK,ATCC CCL10);チャイニーズハムスター卵巣細胞/−DHFR(CHO,Urlaubら、Proc.Natl.Acad.Sci.USA,77:4216(1980));マウスセルトーリ細胞(TM4,Mather,Biol.Reprod.,23:243−251(1980));サル腎細胞(CV1、ATCC CCL70);アフリカミドリザル腎細胞(VERO−76,ATCC CRL−1587);ヒト子宮頸癌腫細胞(HELA,ATCC CCL2);イヌ腎細胞(MDCK,ATCC CCL34);バッファローラット肝細胞(BRL3A,ATCC CRL1442);ヒト肺細胞(W138,ATCC CCL75);ヒト肝細胞(HepG2,HB8065);マウス乳腺腫瘍(MMT060562,ATCC CCL51);TRI細胞(Matherら、Annals N.Y.Acad.Sci.,383:44−68(1982));MRC5細胞;FS4細胞;およびヒトヘパトーマ系統(HepG2)である。
【0147】
宿主細胞は抗体産生のための上記した発現またはクローニングベクターで形質転換され、そして、プロモーターを誘導し、形質転換体を選択し、または所望の配列をコードする遺伝子を増幅するために適切であるように修飾された従来の栄養培地中で培養される。
【0148】
本発明の抗体の産生のために使用される宿主細胞は種々の培地中で培養してよい。市販されている培地、例えばHamのF10(Sigma)、最小必須培地((MEM)、(Sigma)、RPMI−1640(Sigma)、およびダルベッコの変性イーグル培地((DMEM)、Sigma)が宿主細胞を培養するために適している。さらにまた、Hamら、Meth.Enz.,58:44(1979),Barnesら、Anal.Biochem.,102:255(1980)、米国特許4,767,704;4,657,866;4,927,762;4,560,655;または5,122,469;WO90/03430;WO87/00195;または米国特許30,985に記載されている培地のいずれかを宿主細胞のための培養培地として使用してよい。これらの培地の何れにも、必要に応じてホルモンおよび/または他の成長因子(例えばインスリン、トランスフェリン、または表皮成長因子)、塩類(例えば塩化ナトリウム、カルシウム、マグネシウム、およびリン酸塩)、緩衝液(例えばHEPES)、ヌクレオチド(例えばアデノシンおよびチミジン)、抗生物質(例えばGENTAMYCINTM)、微量元素(マイクロモル範囲の再終濃度において通常は存在する無機化合物として定義される)、およびグルコースまたは同等のエネルギー源を補充してよい。何れかの他の必要な補充物もまた、当業者に知られる適切な濃度において包含させてよい。培養条件、例えば温度、pH等は発現のために選択された宿主細胞と共に以前に使用されていたものであり、そして当業者の知る通りである。
【0149】
組み換え手法を使用する場合、抗体はペリプラズム空間内に細胞内で産生されるか、または培地中に直接分泌され得る。抗体が細胞内で産生される場合、第1工程として、宿主細胞または溶解細胞の何れかである粒状破砕物を例えば遠心分離または限外濾過により除去する。抗体を培地中に分泌させる場合、そのような発現系に由来する上澄みは一般的には先ず市販のタンパク質濃縮フィルター、例えばAmiconまたはMillipore Pellicon限外濾過装置を用いて濃縮される。プロテアーゼ阻害剤、例えばPMSFを上記した工程の何れかに包含させることによりタンパク質分解を阻害してよく、そして抗生物質を包含させることにより有害な夾雑体の生育を防止してよい。
【0150】
細胞から調製した抗体組成物は例えばヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、およびアフィニティークロマトグラフィーを用いて精製することができ、アフィニティークロマトグラフィーが好ましい精製手法である。アフィニティーリガンドとしてのプロテインAの適当性は抗体中に存在する何れかの免疫グロブリンFcドメインの種およびアイソタイプに依存している。プロテインAはヒトγ1、γ2、またはγ4重鎖に基づいた抗体を精製するために使用できる(Lindmarkら、J.Immunol.Meth.,62:1−13(1983))。プロテインGは全てのマウスアイソタイプに対して、そしてヒトγ3に対して推奨されている(Gussら、EMBO J.,5:1567−1575(1986))。アフィニティーリガンドを連結するマトリックスは最も頻繁にはアガロースであるが、他のマトリックスも使用される。機械的に安定なマトリックス、例えば制御細孔ガラスまたはポリ(スチレンジビニル)ベンゼンはアガロースを用いて達成できるよりも高速な流量および短いプロセシング時間を可能にする。抗体がCH3ドメインを含む場合、Bakerbond ABXTM樹脂(J.T.Baker,Phillipsburg,N.J.)が精製のために有用である。タンパク質精製のための他の手法、例えばイオン交換カラム上の分画、エタノール沈殿、逆相HPLC、シリカ上のクロマトグラフィー、ヘパリンSEPHAROSETM上のクロマトグラフィー、アニオンまたはカチオン交換樹脂(例えばポリアスパラギン酸カラム)の上のクロマトグラフィー、等電点電気泳動、SDS−PAGE、および硫酸アンモニウム沈殿もまた回収すべき抗体に応じて使用される。
【0151】
他の組み換えポリペプチド、例えばCYTL1およびCYTL1変異体は同様の操作法により製造できる。
【0152】
2.アゴニスト抗CYTL1抗体または他のCYTL1アゴニストを得るためのスクリーニング
CYTL1のアゴニスト抗体および他のアゴニストは伝統的(競合的)結合アッセイまたは活性アッセイにおいて同定できる。
【0153】
CYTL1アゴニストを得るためのスクリーニングアッセイはuPARと結合または複合体形成するか、または別様にCYTL1とuPARとの相互作用を妨害し、そしてuPARへのuPAの結合を競合的に阻害できる化合物を同定するために設計してよい。本明細書に記載するスクリーニングアッセイは化学ライブラリの高スループットスクリーニングに適したアッセイを包含し、低分子薬剤候補を同定するために特に適するものとなっている。一般的に、結合アッセイおよび活性アッセイが与えられる。
【0154】
アッセイは種々のフォーマット、例えば限定しないが、タンパク質−タンパク質結合アッセイ、生化学的スクリーニングアッセイ、イムノアッセイ、および細胞系アッセイにおいて実施することができ、これらは当該分野で良く特徴付けられている。
【0155】
全てのアッセイは、それらが候補CYTL1アゴニストをuPA、uPARおよびCYTL1に、これら2成分が相互作用を起こすために十分な条件下および時間に渡り接触させることを必要とする点において共通である。結合アッセイにおいては、相互作用は結合であり、そして形成された複合体を単離するか、または反応混合物中で検出することができる。特定の実施形態においては、uPAまたはuPARポリペプチドまたは候補アゴニストの何れかを固相上、例えばマイクロプレート上に、共有結合または非共有結合的な連結により固定化する。アッセイはアンカリングされた成分を含有するコーティングされた表面のような固定化成分に、検出可能な標識で標識されていてよい非固定化成分を添加することにより実施する。反応が終了した時点で、非反応成分を例えば洗浄により除去し、そして固体表面上にアンカリングされた複合体を検出する。元来固定化されていない成分が検出可能な標識を担持している場合は、表面上に固定化された標識の検出は複合体化が起こったことを示している。元来固定化されていない成分が標識を担持していない場合は、複合体化は例えば固定化された複合体に特異的に結合する標識された抗体を用いて検出できる。
【0156】
好ましくは、CYTL1拮抗剤を、それらがuPAの生物学的活性を阻害する能力、例えば腫瘍の形成または転移を媒介するか、または血管形成、例えば限定しないが腫瘍の血管形成を誘導または支援するuPAの能力に基づいて、同定するか、または追加的に試験する。
【0157】
当然ながら、本明細書において特に考察しているスクリーニングアッセイは例示に過ぎない。スクリーニングされる拮抗剤候補(例えばポリペプチド、ペプチド、非ペプチド有機低分子、核酸等)の型に応じて選択できる種々の他のアッセイは当業者に良く知られており、そして、本発明の目的のために等しく適当である。
【0158】
3.医薬組成物
CYTL1およびCYTL1アゴニスト、例えばアゴニストCYTL1抗体は、uPAの産生または機能に関連する種々の障害、例えば種々のタイプの腫瘍または癌、例えば腫瘍または癌の転移、および望ましくない血管形成、例えば限定しないが、腫瘍および癌の血管形成を特徴とする疾患および障害の処置のために、医薬組成物の形態において投与できる。
【0159】
抗体フラグメントを使用する場合は、標的タンパク質の結合ドメインに特異的に結合する最も小さい阻害性のフラグメントが好ましい。例えば、抗体の可変領域配列に基づいて、標的タンパク質配列に結合する能力を保持しているペプチド分子を設計することができる。そのようなペプチドは化学合成および/または組み換えDNA技術による産生が可能である。例えばMarascoらProc.Natl.Acad.Sci.USA,90:7889−7893(1993)を参照。
【0160】
活性成分はまた、例えばコアセルベーション手法により、または界面重合により調製されたマイクロカプセル、例えばそれぞれ、ヒドロキシメチルセルロースまたはゼラチン−マイクロカプセルおよびポリ−(メチルメタクリレート)(poly−(methylmethacylate))マイクロカプセル中に、コロイド状薬物送達システム(例えばリポソーム、アルブミン微小球、マイクロエマルジョン、ナノ粒子、およびナノカプセル)中、またはマクロエマルジョン中に封入してよい。そのような手法は上記したRemington’s Pharmaceutical Sciencesに開示されている。
【0161】
インビボの投与のために使用されることになる製剤は滅菌されていなければならない。これは滅菌濾過膜を通す濾過により容易に達成される。
【0162】
除放性調製物を調製してよい。除放性調製物の適当な例は抗体を含有する固体疎水性重合体の半透過性マトリックスを包含し、このマトリックスは形状付与された物品、例えばフィルムまたはマイクロカプセルの形態にある。除放性マトリックスの例はポリエステル、ヒドロゲル(例えばポリ(2−ヒドロキシエチル−メタクリレート)、またはポリ(ビニルアルコール))、ポリラクチド(米国特許3,773,919)、L−グルタミン酸とγエチル−L−グルタメートの共重合体、非分解性エチレン−酢酸ビニル、分解性乳酸−グリコール酸共重合体、例えばLUPRON DEPOTTM(乳酸−グリコール酸共重合体および酢酸ロイプロリドよりなる注射可能な微小球)、およびポリ−D−(−)−3−ヒドロキシ酪酸を包含する。エチレン−酢酸ビニルおよび乳酸−グリコール酸のような重合体は100日間よりも長い期間に渡り分子を放出できるのに対し、特定のヒドロゲルはより短い期間タンパク質を放出する。カプセル化された抗生物質が長時間身体内に残存する場合、それらは37℃において水分に曝露される結果として変性または凝集を起こし、その結果生物学的活性の損失および免疫原性変化の可能性が生じる場合がある。関与する機序に応じて安定化のための合理的な方策を講じることができる。例えば凝集の機序がチオ−ジスルフィド交換を介した分子間S−S結合の形成であることが発見されれば、安定化は、スルフィドリル残基を修飾すること、酸性の溶液から凍結乾燥すること、水分含有量を調節すること、適切な添加剤を使用すること、および特定の重合体マトリックス組成物を開発することにより、達成してよい。
【0163】
本発明における製剤はまた、処置すべき特定の適応症のために必要に応じて1つより多い活性化合物、好ましくは相互に悪影響を及ぼさない補完的活性を有するものを含有してよい。そのような分子は意図する目的のために有効な量において組み合わせて適宜存在させるか、或いは別個に製剤化し、そして同時に、または何れかの順序において連続して投与してよい。
【0164】
例えば、本発明のCYTL1およびCYTL1アゴニストは1つ以上の追加的治療薬、例えば抗血管形成剤、抗新生物性組成物、化学療法剤および/または細胞毒性剤と組み合わせて投与してよい。
【0165】
以下の実施例は説明目的のみのために提示しており、そして如何様にも本発明の範囲を限定する意図はない。
【0166】
本明細書において引用する全ての特許および参考文献は参照により全体が本明細書に組み込まれる。
【実施例】
【0167】
材料および方法
材料−組み換えヒトuPAR、uPAおよびuPA−ATFはR&D Systems(Minneapolis,MN)より入手した。Pro−uPAはCortex Biochem(SanLeandro,CA)より入手した。ブタ粘膜由来の低分子量ヘパリンはSigma Chemical Company(St.Louis,MO)から購入した。
【0168】
インサイチュハイブリダイゼーション−薄膜切片のインサイチュハイブリダイゼーションのために、PCRプライマーを設計し、NM_001081106[5’−CCCACCTGCTACTCTCGGATG−3’(配列番号3)および5’−GGCAGGTCTAACAGTGGCACTAA−3’(配列番号4)]のnt.111−560から伸長するマウスCYTL1の449bpのフラグメント、またはNM_018659[5’−TCCCCCGACCTGCTACTC−3’(配列番号5)および5’−CCTGTGAGGGTCATGGCTCTGGC−3’(配列番号6)]のnt.94−525から伸長するヒトCYTL1の431bpのフラグメントを増幅した。プライマーは27ヌクレオチドのT7またはT3RNAポリメラーゼ開始部位をコードする伸長部を包含することにより、増幅産物からのそれぞれセンスまたはアンチセンスプローブのインビトロの転写を可能とした。
【0169】
ホルマリン固定パラフィン包埋5μm切片を脱パラフィン処理し、37℃で30分間4μg/mlのプロテイナーゼK中で脱タンパク質処理し、そして以前に報告されている通りインサイチュハイブリダイゼーション用にさらに処理した(Jubbら(2006)Methods in Molecular Biology 326,255−264)。33P−UTP標識のセンスおよびアンチセンスのプローブを一夜55℃において切片にハイブリダイズさせた。未ハイブリダイズのプローブはRNase処理およびストリンジェントな洗浄により除去した。スライドガラスを段階濃度エタノールで脱水し、NTB核トラックエマルジョン(Eastman Kodak)中に浸積し、4℃で4週間露光し、次に現像し、そしてヘマトキシリンおよびエオシンで逆染色した。
【0170】
ATDC5細胞成熟アッセイ−ATDC5細胞はRIKEN Cellbank,Japanより入手し、そして、5%ウシ胎児血清および2mM L−グルタミンを添加したダルベッコの変性イーグル培地(DMEM)およびHamのF12培地の50:50混合物中に維持した。細胞を12穴プレートの各ウェル当たり105個の細胞となるように播種し、コンフルエントとなるまで生育させ、次に10μg/ml組み換えウシインスリンおよび50μg/mlアスコルビン酸で処理することにより軟骨細胞分化を誘導した。成熟化中、種々の時点において、RNAを3連のウェルから採取した。第11日に、一部の細胞を0.1ng/mlのIL−1βで処理し、そしてさらに24時間の後RNAを採取した。
【0171】
コラーゲン誘導関節炎−7〜8週齢の雄性DBA/1JマウスをThe Jackson Laboratory(Bar Harbor,ME)から入手し、そしてAmerican Association of Laboratory Animal Careガイドラインに従って維持した。全ての実験操作法はGenentechのInstitutional Animal Care and Use Committeeにより認可されている。コラーゲン誘導関節炎は第0日および第21日に完全フロイントアジュバント中のウシII型コラーゲンの注射により標準的プロトコル(Barckら(2004)Arthritis and rheumatism 50(10),3377−3386)に従って誘導した。全RNAは臨床疾患のある手掌から、そして健常DBA/1J対照マウスの手掌から抽出した。
【0172】
GeneLogic BioExpressデータ−市販のデータベースBioExpress(GeneLogic,Gaithersburg,MD)を用いてコラーゲン誘導関節炎の間の遺伝子発現のクエリを行った。試料は製造元(Affymetrix,Santa Clara,CA,USA)のプロトコルに従ってAffymetrix U133A GeneChip上で分析した。
【0173】
CYTL1の発現および精製−ミツバチメリチンシグナルペプチドと共にFastBacバキュロウィルス系(Invitrogen,Carlsbad,CA)を用いて昆虫細胞中にN末端ポリヒスチジンタグを有するヒトCYTL1を発現させた。タンパク質はNi++−セファロース6FFカラム(Amersham Biosciences,Piscataway,NJ)上で精製した。タグは組み換えエンテロキナーゼ(Novagen,Madison,WI)で切断し、そしてタグおよび酵素をNi++−セファロースおよびEKaptureアガロース(Novagen)を用いて枯渇させた。CYTL1はさらに塩勾配溶離によりヘパリンカラム(HiTrap His,Amersham)から精製し、次にCentricon遠心濃縮器(Millipore,Billerica,MA)を用いて3kDaのMWカットオフで濃縮した。濾過のフロースルーは表面プラズモン共鳴および細胞系アッセイにおける緩衝液対照として使用するために温存しておいた。
【0174】
発現クローニング−pCMV−SPORT系ベクター中に約14,000の完全長ヒトcDNAを含有する発現ライブラリをOrigene(Rockville,MD)から入手した。ヒト胎盤アルカリホスファターゼ(AP)融合タンパク質(proteinsl);TACI−APおよびヒトおよびネズミCYTL1−APをpRK5系ベクターを用いて一過性トランスフェクションにより293細胞中で発現させた。細胞馴化培地を酵素活性に関してアッセイし、そして必要に応じて希釈することにより活性を等しくした。24穴プレート中のCOS細胞をFuGene6(Roche)を用いて発現コンストラクトのプール物でトランスフェクトした。48時間後、接着細胞を洗浄し、次に0.1%BSAを添加したCYTL1−APまたはTACI−APを含有する細胞馴化培地を用いて室温で30分間インキュベートした。洗浄後、結合した融合タンパク質を10%中性緩衝ホルマリンと共にインキュベートすることにより表面に交差結合させた。90分間65℃でプレートをインキュベートすることにより内因性のアルカリホスファターゼを不活性化し、次にウエスタンブルー安定化基質(Promega,Madison,WI)を用いてアルカリホスファターゼの活性を検出した。
【0175】
uPAの不活性化−室温で2時間10mMジイソプロピルフルオロホスフェート(DFP)で処理することによりuPAのプロテアーゼ活性を非可逆的に阻害した。過剰なDFPは十分な透析により除去した。完全な不活性化はuPA基質S−2444(diaPharma,West Chester,OH)を用いることにより確認した。
【0176】
細胞表面結合アッセイ−図の説明に示す通りCOSまたはCHO細胞を完全長のヒトまたはマウスuPARをコードする発現ベクターでトランスフェクトした。24時間後、トランスフェクション体を24穴プレートに移した。さらに24時間の後、細胞を洗浄し、次に「発現クローニング」の下に詳述したとおりCYTL1−APを含有する細胞馴化培地を用いてプローブした。結合アルカリホスファターゼ活性の定量のために、BluePhos(KPL,Gaithersburg,MD)を検出試薬として使用した。競合アッセイのためには記載濃度のプロ−uPA、uPA−ATF、DFP−uPAまたは精製されたCYTL1を、uPARトランスフェクト細胞とのインキュベーションの直前にCYTL1−AP細胞馴化培地に添加した。
【0177】
RNA抽出および定量的リアルタイムPCR(qRT−PCR)−全RNAはRNeasy miniprepキット(Qiagen,Germantown,MD)を用いて抽出し、光学密度により定量し、そしてOmniscript RT(Qiagen)を用いて逆転写した。形成したcDNAを7500リアルタイムPCRシステム(Applied Biosystems,Foster City,CA)においてQuantiTectプローブを用いて2連でアッセイした。増幅されたシグナルを記載の通りβ−アクチンまたはGAPDHのものに対して標準化した。プライマーおよびプローブは表1に列挙する通りである。
【0178】
表面プラズモン共鳴−表面プラズモン共鳴実験は10μl/分の流量においてHBS−P緩衝液(50mM HEPES pH7.4、150mM NaCl、0.005%界面活性剤)を用いてBIAcore 3000(BIAcore Inc.,Uppsala,Sweden)により実施した。組み換えヒトuPARを標準的なアミンカップリング化学を用いてCM5センサーチップ上に固定化した。慨すれば、新しく混合したN−ヒドロキシスクシンイミドおよび1−エチル−3−(3−ジメチルアミノプロピル)−カルボジイミド塩酸塩の8分間注入により表面を活性化し、uPARを10mM酢酸ナトリウムpH4.5中20μg/mlで注入し、そして未反応のエステルを1Mエタノールアミンでクエンチした。空のフローセルおよびカップリングしたBSAを有するフローセルを対照として使用した。平衡結合分析のために種々の濃度の精製CYTL1および等価な希釈度の緩衝液を全フローセルに渡って注入した。緩衝液単独への応答をCYTL1応答から差し引き、そして平衡時のピーク応答を記録した。希釈および計測は3回実施した。BIAEvalソフトウエア(BIAcore)を用いてラングミュアの方程式を使用して平衡結合データをフィットさせた。CYTL1とプロ−uPAとの間の競合を示すために、飽和濃度のプロ−uPAの注入の前、および直後に、uPARカップリングフローセルにCYTL1を注入した。
【0179】
二次構造予測−CYTL1種相同体はBLASTおよびBLAT検索によりESTおよびゲノム配列から同定し、そしてClustalXを用いてアラインした。JPredウエブサイトを介したJnetニューラルネットワーク系アルゴリズム(33)への入力として多数種アライメントを使用することにより、二次構造予測を生じさせた。ソース配列:ヒトNM018659,AY359101;チンパンジー(パン・トログロダイト)ゲノム予測:GenScanのchr4_2.3;マカクサル(リーサス・マカク、マカラ・マラッタ)ゲノム予測:Genscanのchr5.4.000.a;ウシ(ボス・タウルス)BE683329、BE683328、BE683327;マウスNM_001081106;ラットCK476529、CK363591、CK359536;ポッサム(モノデルフィス・ドメスチカ)ゲノム予測:GenScanのc_619.34;ニワトリ(ガルス・ガルス)BX931297、アフリカツメガエル(ラビス)EB474543、EB474068、EC277031;P.オリブ(バスタード・ハリブー、パラリクチス・オリバセウス)CX285777、CX285687;ハリブー(ヒポグロスス・ヒポグロスス)EB036635;フグ(ラブライプス)BLATにより同定されたゲノム配列−chrUn:189、159、975〜189、160、957;メダカ(オリジアス・ラチプス)AU241696、AU241459、AU177768;サケ(サルモ・サラー)EG847194、EG833037、EG802555;マス(オンコラインクス・ミキス)BX889777、BX866010;キュウリウオ−オスメルス・モルダックスEL55175;ミノウ(ピメファレス・プロメラス)DT309382、DT347709、DT106962;ゼブラフィッシュ(ゼブラフィッシュ、ダニオ・レリオ)EE214359、EE213198、EB957495;ゼブラフィッシュ2(ダニオレリオ−第2のCYTL1相同体)AL913033、EH466946、EH441042、AL913034。
【0180】
結果
CYTL1のインビボの発現−ホールマウントインサイチュハイブリダイゼーションによれば軟骨原基に相当する段階において肋骨、脊椎および長骨を包含するマウス胚中の発生中の骨格エレメントの種々のものにおけるCYTL1の発現が明らかになっている(図1)。薄膜切片のインサイチュハイブリダイゼーションによれば、主に軟骨におけるCYTL1の発現が明らかにされており、関節軟骨の表面層において特に高値の発現がある。CYTL1発現はまた、気管の軟骨輪においても観察されている。高レベルの発現は腱と骨との間の接触部位の周囲における腱鞘において観察されている。CYTL1発現は発生中の長骨の肥大性の軟骨細胞中、そしてさらに、動脈(静脈では観察されない)内皮細胞中にも観察され、第17日胚において大動脈中に先ず検出され、そして成体マウスにおいては筋肉動脈全体に渡って拡張している。
【0181】
軟骨細胞成熟化の間のCYTL1の発現の過程をクローンマウス前軟骨細胞系統ATDC5を用いて検討した(図2)。ATDC5細胞はインスリンに応答して分化し、遺伝子発現における十分特性化された変化を有している。CYTL1は未分化細胞において検出可能ではなかったが、分化の開始後は、CYTL1発現は第4日までには検出され、早期軟骨細胞マーカーコラーゲンIIの発現を伴っていた。CYTL1発現は第12日まで増大し続け、成熟軟骨細胞のマーカーであるアグレカンの増大した発現を伴っていた。
【0182】
発現のモジュレーション−多くのサイトカインが炎症において重要な調節の役割を果たしている(Heinrichら(2003)Biochem.J.374(Pt1),1−20)ため、本発明者らは実験的関節炎疾患のマウスモデルの経過の間のCYTL1発現を調べた。コラーゲン誘導関節炎(CIA)モデルから得られたマイクロアレイデータは、マウス関節中のCYTL1の発現は疾患においてダウンレギュレートされ、臨床評点および炎症サイトカイン、特にIL−1βの産生のピークと一致していることを示している(図3a)。別の実験においてマウスの手掌から抽出されたRNAの定量的RT−PCR分析によればCIAにおけるCYTL1発現のダウンレギュレーションが確認された(図3b)。IL−1βはCIAの進行において重要な役割を果たし(Williams,R.O.(2004)Methods in Molecular Medicine 98,207−216)、軟骨破壊に必須のメディエーターであり(Zwerinaら(2007)Proc.Natl.Acad.Sci.USA,104(28),11742−11747)、そして軟骨細胞に対して直接作用することにより数種の軟骨細胞成熟化関連の遺伝子のダウンレギュレーションを誘導する(LeFebvreら(1990)Biochimica et biophysica acta 1052(3),366−378)。CIAにおけるCYTL1のダウンレギュレーションがIL−1βの直接の作用により説明できるかどうかを試験するために、本発明者らは分化したATDC5軟骨細胞をIL−1βに曝露し、そしてqRT−PCRにより遺伝子発現をアッセイした。本発明者らは0.1ng/mlのIL−1βに応答したCYTL1発現の10分の1への低減を観察した(図3c)。逆に、uPAR発現はIL−1β処置によりアップレギュレートされた。
【0183】
受容体の同定−CYTL1はより早期には数種の両親媒性のα−ヘリックスの二次構造の予測に基づいて4ヘリックスサイトカインとして説明されていた(Liuら(2000)Genomics 65(3),283−292)。より詳細な折畳みの分析のために、本発明者らはまず、GenBANKの網羅的PsiBLASTクエリにより全ての既知のCYTL1オーソログの進化的に多様な収集物を作成し(Altschulら(1997)Nucl.Acids Res.25(17),3389−3402)、配列をClustalXとアラインし(Thompsonら(1997)Nucl.Acids Res.25(24),4876−4882)、そして次にPsiPREDとJNetアルゴリズムの両方を使用することにより(Jones,D.T.(1999)J Mol Biol 292(2),195−202;Cuff,J.A.,and Barton,G.J.(2000)Proteins 40(3),502−511)、正確な二次構造の予測を発生させた(補足図1)。短鎖サイトカインのヘリックスA〜Cに合致する長さと間隔を有する3つのα−ヘリックス(Rozwarskiら(1994)Structure 2(3),159−173)が高い信頼性で予測され、それと共に、GM−CSF、IL−3およびIL−13のような短鎖サイトカインにおいて観察されるようなα−ヘリックスおよび2つの短いβ−鎖の追加的な単一ターンが伴っている。CYTL1と4ヘリックスサイトカインファミリーとの間の関連性はそのゲノム機構の分析により強化される。4ヘリックスサイトカインは構造的特徴と比較してエキソン境界のステレオタイプの位置付けを示しており、全接合部が第0相にあり、これはCYTL1により共有されている特性である。しかしながら、その予測された二次構造においては、CYTL1は、第4番目、即ちDヘリックスが非存在であるか、極度にトランケーションされているそのC末端領域におけるヘリックスサイトカインパターンから劇的に逸脱している。このヘリックスはヘリックスサイトカインのそれらの高親和性受容体への結合のために必須であるため(Clackson,T.,and Wells,J.A.(1995)Science(New York,N.Y 267(5196),383−386;Boulanger,M.J.,およびGarcia,K.C.(2004)Advances in Protein Chemistry 68,107−146)、これは、CYTL1が恐らくは無関連の受容体ファミリーにとって欠如している受容体結合の相違した様式を採用している可能性を示唆していた。
【0184】
本発明者らは、発現クローニングの手順を用いてCYTL1の受容体に関する偏りのない検索を実施した。CYTL1−アルカリホスファターゼ融合タンパク質(CYTL1−AP)を用いて、完全長のヒトクローンのライブラリに由来するプールされた発現コンストラクトでトランスフェクトされたCOS細胞をプローブした。本発明者らはCYTL1−APに結合する能力を細胞に付与する単一のプールを同定し、次にこのプールを構成している個々のクローンをスクリーニングすることにより原因となる単一のクローンを同定した(図4a)。このクローンの配列決定によりそれがuPARをコードしていることが判明した。uPARの発現は、酵素活性により計測した場合、CYTL1−APと等しい濃度で使用した対照アルカリホスファターゼ融合タンパク質TACI−AP(膜貫通活性化因子カルシウムモジュレーターおよびシクロフィリンリガンド相互作用物質)に対して結合をもたらさなかった。同様の結果がCHO細胞に対して観察された。uPARに構造的に関連しているC4.4Aの発現はCYTL1−APに結合する能力をもたらさなかった(データ示さず)。ネズミuPARでトランスフェクトしたCOS細胞はマウスCYTL1−AP融合タンパク質に結合することができ(図4b)、CYTL1とuPARとの間の相互作用の進化論的保存を示していた。種間交差性の結合は殆ど、或いは全く観察されなかった。
【0185】
過剰量の精製されたCYTL1は濃度依存的な態様においてuPARトランスフェクト細胞へのCYTL1−APの結合を阻害することができた(図4c)。相同競合を推定すれば、3つの独立した実験で得られた細胞表面におけるuPARへのCYTL1の親和性は約1.5μM±0.5μM(SEM)である。
【0186】
uPARは多くの細胞表面および細胞外のタンパク質、例えばuPA、αvおよびβ1インテグリン、ビトロネクチン、uPAR−関連タンパク質(uPARAP/エンド180)およびIGF−IIR(Ploug,M.(2003)Current Pharmaceutical Design 9(19),1499−1528)と相互作用することがわかっている。CYTL1がuPARと直接、そしてアクセサリータンパク質を必要とすることなく相互作用するかどうかを調べるために、本発明者らはBIAcore3000を用いて表面プラズモン共鳴(SPR)分析を実施した。精製されたCYTL1は固定化されたuPARに特異的に結合した(図5)。同様の結合は哺乳類細胞および細菌において発現されたCYTL1でも観察された(データ示さず)。相互作用の速度論的動態は極めて急速であり、そして正確に定量することはできなかった。平衡結合の計測値は固定化uPAR上、および、対照フローセル上に種々の濃度のCYTL1を注入することにより得た(図5c)。単一部位結合に関するラングミュアの方程式はデータに良好にフィットし、1.1μM±0.06μMのCYTL1とuPARとの相互作用に関する親和性定数の計測値が得られた。固定化されたuPARの密度を5倍範囲に渡って変動させても親和性の相違は観察されなかった。
【0187】
グリコサミノグリカンは数種のサイトカインのそれらの受容体への結合に大きく影響することが知られている。本発明者らによるCYTL1タンパク質の精製はCYTL1とヘパリンとの間の相互作用を利用しており、そのため本発明者らはこの相互作用をさらに特性化することを試みた。CYTL1およびヘパリンを単独および組み合わせて、固定化されたuPAR上に注入した(図5d)。ヘパリンおよびCYTL1単独では各々、一過性の結合を示し(88RUおよび112RU);CYTL1とヘパリンを共に注入した場合、超相加的作用(353RU)が観察され、ヘパリンがCYTL1のuPARへの結合を増強していることが示された。
【0188】
競合的結合−uPARはuPAに対する細胞表面受容体である。uPARへの結合に関してCYTL1がuPAと競合するかどうか明らかにするため、uPARでトランスフェクトした細胞を種々の濃度のタンパク質分解的に不活性なプロ酵素(プロ−uPA)、受容体結合アミノ末端フラグメント(uPA−ATF)またはジイソプロピルフルオロホスフェートで処理することにより不活性化された完全長uPA(DFP−uPA)の存在下でCYTL1APと共にインキュベートした。uPAの3形態全てがuPAR発現細胞の表面への結合に関してCYTL1−APと競合し(図6a〜c)、結合の50%阻害は競合物質10nM〜40nMで達成された。この競合が追加のタンパク質の非存在下、および嵩高のアルカリホスファターゼタグの非存在下で起こるかどうかを試験するために、SPR分析を再度使用した(図6d)。CYTL1を固定化uPAR上に注入し、282応答単位(RU)の結合を生じさせるようにした。次にプロ−uPAを高濃度で注入することにより50%超の受容体占有状態を達成した。CYTL1の2回目の注入は大きく減衰した結合(95R)をもたらし、プロ−uPAによる受容体占有がCYTL1への結合のために使用できる受容体の量を低減したことを示していた。低pHによるuPARからのプロ−uPAの解離の後、CYTL1に結合する能力は回復した。
【0189】
考察
CYTL1は軟骨細胞および動脈内皮細胞により主に組織限定的様式において産生される分泌タンパク質である。その予測二次構造およびゲノム機構において、CYTL1は4α−ヘリックス造血性サイトカインに部分的類似性を保有している。僅かな例外を除き、このタンパク質ファミリーの全メンバーは、フィブロネクチン3型(Fn3)様のサイトカイン結合モジュールの保存された対により区別できる細胞表面受容体の進化論的に保存されたファミリーに結合する(Bazan,J.F.(1990)Proc.Natl.Acad.Sci.USA,87(18),6934−6938)。これらの受容体はそれらの細胞内伸長部を介してJAKファミリーのタンパク質チロシンキナーゼおよびSTAT転写因子と会合し、これを介して、細胞外リガンド結合により駆動される受容体オリゴマー化の後にシグナルがトリガーされる。この原理の3つの例外であるM−CSF、SCFおよびFlt3Lは、第2のクラスのPDGFR様受容体について「欠損」しており、これは自身のリガンドを免疫グロブリン(Ig)様ドメインに結合させ、そして細胞質チロシンキナーゼドメインを介してシグナルを発生するものである。CYTL1に関する受容体のスクリーニングにおいては従来サイトカイン受容体は同定されていない。同定された受容体であるuPARは上記ファミリーの何れにも構造的に無関連であり、そしてむしろ、GPIアンカーにより原形質膜に結合されているヘビ神経毒中に構造的に初めて同定されたドメインである3つのLy6−uPAR(LU)様モジュール(Ploug,M.,and Ellis,V.(1994)FEBS Lett 349(2),163−168;Barinkaら(2006)J Mol Biol 363(2),482−495;Llinasら(2005)Embo J 24(9),1655−1663)で大部分が構成されている。4ヘリックスサイトカインとLUモジュラータンパク質との間では、これらのドメインの折畳み関連物はTGF−βに対するシグナリング受容体のサイトカイン結合セグメント、アクチビンおよび骨形態発生タンパク質(BMP)を含む(Gyetkoら(1994)J.Clin.Invest.93(4),1380−1387)にも関わらず、相互作用は報告されていない。CYTL1が実際に4ヘリックスサイトカインであれば、これはLU折畳み型に関する相互作用の全く新規なクラスとなる。
【0190】
サイトカインは典型的には一連の受容体に逐次的に結合して細胞内ホスホリル化のカスケードをトリガーする高親和性のシグナリング複合体を形成することによりそれらの機能を呈する。しかしなお、uPARがCYTL1に対する真のシグナリング受容体であるかどうか、または、それがCYTL1結合タンパク質としてより正確に説明されるかどうかは不明である。細胞内および膜貫通ドメインを欠いているため、uPARの単独の作用はシグナリング受容体としては機能できない。しかしながら、多くのシグナリング経路がuPARのライゲーションの後に活性化されることがわかっており、タンパク質キナーゼSrc、JAK−1、Hck、FAKおよびERK1/2の活性化が報告されている(Webbら、J.Cell Biol.,152(4),741−752;Tangら(1998)J Biol Chem 273(29),18268−18272;Resnatiら、Embo J.,15(7),1572−1582;Dumlerら(1998)J.Biol.Chem.,273(1),315−321)。シグナルトランスダクションの機序は、インテグリンに関する役割が強く示唆されているものの、なお不明である。uPARとインテグリンα3β1との間の直接でuPA依存性の相互作用(Weiら(2001)Mol.Biol.Cell 12(10),2975−2986)が間接的相互作用を有するものとして示されており、その場合、ビトロネクチンはuPA依存的な様式でuPARに結合し、そして次にインテグリンをライゲーションする(Madsenら(2007)J.Cell Biol.177(5),927−939)。uPARへのCYTL1の結合は、CYTL1が他の膜貫通鎖にuPARを架橋できれば、或いは、それがインテグリンまたは他のシグナリング分子とのuPARの相互作用を局所的に調節すれば、同様の様式で細胞内シグナルトランスダクションをもたらす可能性がある。
【0191】
本発明者らは、CYTL1がプロ−uPA、uPA−ATFおよびDFP−不活性化uPAと競合しながらuPARに結合することを明らかにしている。従ってCYTL1はuPARとのuPAの相互作用の拮抗剤として機能できる。uPA−uPAR相互作用はプロ−uPAを切断して成熟uPAを形成するヘプシンおよびマトリプターゼのような細胞表面活性化因子にプロ−uPAを接触させる作用を有する。uPARへの結合はまた、成熟uPAの活性を増強する。uPARの結合に関して競合することにより、CYTL1は細胞関連uPA活性を低減すると考えられる。uPA−uPAR相互作用の競合的阻害剤の生物学的効果は多くの系において検討されている。uPAのタンパク質分解的に不活性化された突然変異体の型によりトランスフェクトされた腫瘍細胞は低減した転移能力を示し(Crowlyら(1993)Proc.Natl.Acad.Sci.USA,90(11),5021−5025)、uPA−ATFは微小血管内皮細胞による管形成(Croonら(1999)Am J Pathol 154(6),1731−1742)および網膜損傷後の血管形成(Le Gatら(2003)Gene Ther 10(25),2098−2103)を阻害する。uPAと競合しながらuPARに結合することにより、軟骨誘導CYTL1は同様の効果を媒介する場合があり、uPAR発現細胞が侵襲する能力を低減し、そして軟骨の一体性を維持することを支援する。
【0192】
健常な関節においてはuPARの発現は殆どないが;関節炎の間は、浸潤白血球がuPAおよびuPARの両方を発現し、そして軟骨細胞および滑膜細胞の両方が炎症性サイトカインに応答してuPARを発現し(Bussoら(1997)Ann Rheum Dis 56(9),550−557;Guiducciら(2005)Clin Exp Rheumatol 23(3),364−372;Schwabら(2001)Histochem Cell Biol 115(4),317−323)、そして軟骨の破壊的な血管新生がuPAR発現内皮細胞が侵襲するに従い発生する。uPAR拮抗剤の潜在的な効果はコラーゲン誘導関節炎を有するマウスへのuPA−ATFの全身送達により明らかにされており、これは疾患の発生率および重症度、および軟骨血管新生の程度を低減している(Apparaillyら(2002)Gene Therapy 9(3),192−200)。炎症の非存在下においては、CYTL1は関節軟骨において強力に発現されるが、その発現は炎症シグナルに応答して劇的に低減される。uPA拮抗剤活性のこの消失は細胞侵襲および細胞外マトリックスのリモデリングを促進する場合がある。現在実施中のCYTL1遺伝子欠損トランスジェニックマウスの表現型分析がインビボの役割の明確化に役立つはずである。
【0193】
骨ホメオスタシスにおけるuPARの役割は、uPAR欠損マウスの研究を介して最近明らかにされた(Furlanら(2007)J Bone Miner Res 22(9),1387−1396)。uPAR発現の非存在下において、骨鉱物質密度は上昇し、そして骨容量は低減している。増強された骨芽細胞活性および骨表面への破骨細胞接着の不全がこの表現型を説明していると考えられる。これは軟骨におけるCYTL1の発現が隣接する骨におけるマトリックスリモデリングに影響する機序となっている。
【0194】
BIAcore分析により定量した場合のuPARへのCYTL1の結合は中間的な親和性、即ち約1μMを有する。これはサイトカインのそれらの細胞表面受容体に対する親和性の典型的な範囲より有意に低値である。なお同定されていないCYTL1に対する共受容体が存在する可能性があり、それはシグナルトランスダクションの機序を与えることに加えて、細胞表面におけるuPARに対するCYTL1の親和性を増大させ、これによりそれをより強力なuPA−uPAR相互作用拮抗剤とする可能性がある。共受容体の非存在下においても、CYTL1はuPAと効果的に競合するために十分高い局所濃度に到達することも想像される。大規模遺伝子発現試験(Kumarら(2001)Osteoarthritis and Cartilage/OARS,Osteoarthritis Research Society 9(7),641−653)から得られたデータの慎重な分析によれば、CYTL1はmRNAレベルで豊富に表われ、その傾向は他の如何なるサイトカインよりも高いことがわかった。このことはCYTL1が銀染色2Dゲル中で容易に検出される関節軟骨のプロテオーム分析(Hermanssonら(2004)J Biol Chem 279(42),43514−43521)においてタンパク質レベルで当てはまると考えられる。CYTL1とヘパリンとの間の相互作用はCYTL1の局所濃度がブーストされると考えられる機序となっている。ヘパリンが生理学的結合相手である可能性は低いが、軟骨の豊富な高度硫酸化グリコサミノグリカンはCYTL1に結合してそれを発現部位の近傍に局在化させる可能性がある。さらにまた、ヘパリンにより達成されたuPARへのCYTL1の結合の強化は、細胞外マトリックスまたは細胞表面の何れかにおけるグリコサミノグリカンがuPARへのCYTL1の親和性を増強している可能性があることを示唆している。
【0195】
本明細書の作成の間、CYTL1の別の研究が発表された(Kimら(2007)J Biol Chem In press.PMID:17644814)。著者らはCYTL1が間葉細胞の軟骨細胞への分化を促進することを発見している。本発明者らは軟骨細胞に分化しているATDC5細胞へのCYTL1APの結合を観察しており(補足図2a)、そして本発明者らはqRT−PCRによるこれらの細胞によるuPARの発現を示している(補足図2b)。
【0196】
CYTL1の分子作用が4ヘリックスサイトカインファミリーのメンバーとして非典型的であるとすれば、CYTL1が真にこのファミリーのメンバーであるかどうかという疑問は未解決のままであり、そして構造的特性化なくしては解答が得られないことになる。ファミリーの多くのメンバーが多面発現性であり、異なる細胞コンパートメントにおいて極めて異なる機能的役割を果たしていることに留意すべきである。例えばレプチンは全身的には脂質代謝の調節物質として、そして局所的には造血(Wautersら(2000)Eur.J.Endocrin.143(3),293−311)および調節性T細胞発生(De Rosaら(2007)Immunity 26(2),241−255)を調節するという両方の作用を有している。同様に、CD34+造血先祖細胞により発現されるCYTL1はより多くの従来のサイトカイン様機能を保持しているが、軟骨および血管壁においては極めて異なる機能を採用している可能性もある。
【特許請求の範囲】
【請求項1】
ウロキナーゼ型プラスミノーゲン活性化因子(uPA)とウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)との相互作用を阻害する方法であって、サイトカイン様1(CYTL1)ポリペプチドまたはそのアゴニストにuPAおよびuPARを含む混合物を接触させることを含む、方法。
【請求項2】
前記接触がインビトロで行われる、請求項1記載の方法。
【請求項3】
前記接触がインビボで行われる、請求項1記載の方法。
【請求項4】
前記混合物がuPAおよびuPARを発現する細胞を含む、請求項1記載の方法。
【請求項5】
前記細胞が癌細胞である、請求項4記載の方法。
【請求項6】
前記癌が癌腫、リンパ腫、芽腫、肉腫、および白血病よりなる群から選択される、請求項5記載の方法。
【請求項7】
前記癌が乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択される、請求項5記載の方法。
【請求項8】
前記アゴニストがCYTL1フラグメント、CYTL1アミノ酸変異体、アゴニスト抗CYTL1抗体およびそのフラグメント、uPARに結合できる、ペプチドおよび非ペプチド性低分子よりなる群から選択される、請求項1記載の方法。
【請求項9】
ウロキナーゼ型プラスミノーゲン活性化因子(uPA)の生物学的活性を阻害する方法であって、有効量のCYTL1またはそのアゴニストにインビボでウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)およびuPAを発現する細胞を接触させることを含む、方法。
【請求項10】
前記細胞が癌細胞である、請求項9記載の方法。
【請求項11】
前記癌が癌腫、リンパ腫、芽腫、肉腫、および白血病よりなる群から選択される、請求項10記載の方法。
【請求項12】
前記癌が乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択される、請求項10記載の方法。
【請求項13】
前記細胞が内皮細胞である、請求項9記載の方法。
【請求項14】
前記アゴニストがCYTL1フラグメント、CYTL1アミノ酸変異体、アゴニスト抗CYTL1抗体およびそのフラグメント、uPARに結合できる、ペプチドおよび非ペプチド性低分子よりなる群から選択される、請求項9記載の方法。
【請求項15】
哺乳類被験体における腫瘍形成または腫瘍転移を阻害するための方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む、方法。
【請求項16】
前記哺乳類被験体がヒト患者である、請求項15記載の方法。
【請求項17】
前記腫瘍が癌である、請求項16記載の方法。
【請求項18】
前記癌が癌腫、リンパ腫、芽腫、肉腫、および白血病よりなる群から選択される、請求項18記載の方法。
【請求項19】
前記癌が乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択される、請求項18記載の方法。
【請求項20】
前記アゴニストがCYTL1フラグメント、CYTL1アミノ酸変異体、アゴニスト抗CYTL1抗体およびそのフラグメント、uPARに結合できる、ペプチドおよび非ペプチド性低分子よりなる群から選択される、請求項16記載の方法。
【請求項21】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約70%の同一性を有する、請求項20記載の方法。
【請求項22】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約80%の同一性を有する、請求項20記載の方法。
【請求項23】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約85%の同一性を有する、請求項20記載の方法。
【請求項24】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約90%の同一性を有する、請求項20記載の方法。
【請求項25】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約95%の同一性を有する、請求項20記載の方法。
【請求項26】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約99%の同一性を有する、請求項20記載の方法。
【請求項27】
前記CYTL1変異体が配列番号2のCYTL1ポリペプチドの天然に存在する変異体である、請求項20記載の方法。
【請求項28】
前記アゴニストが抗CYTL1抗体またはそのフラグメントである、請求項20記載の方法。
【請求項29】
前記抗体がモノクローナル抗体またはそのフラグメントである、請求項28記載の方法。
【請求項30】
前記モノクローナル抗体がキメラ抗体、ヒト化抗体またはヒト抗体である、請求項29記載の方法。
【請求項31】
前記抗体フラグメントがFab、Fab’、F(ab’)2、およびFvフラグメント;ダイアボディ;直鎖状抗体;単鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体よりなる群から選択される、請求項29記載の方法。
【請求項32】
哺乳類被験体における血管形成を阻害するための方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む、方法。
【請求項33】
前記哺乳類被験体がヒト患者である、請求項32記載の方法。
【請求項34】
前記血管形成が腫瘍血管形成である、請求項33記載の方法。
【請求項35】
前記腫瘍が癌腫、リンパ腫、芽腫、肉腫、および白血病よりなる群から選択される、請求項34記載の方法。
【請求項36】
前記腫瘍が乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択される、請求項34記載の方法。
【請求項37】
前記アゴニストがCYTL1フラグメント、CYTL1アミノ酸変異体、アゴニスト抗CYTL1抗体およびそのフラグメント、uPARに結合できる、ペプチドおよび非ペプチド性低分子よりなる群から選択される、請求項33記載の方法。
【請求項38】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約70%の同一性を有する、請求項37記載の方法。
【請求項39】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約80%の同一性を有する、請求項37記載の方法。
【請求項40】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約85%の同一性を有する、請求項37記載の方法。
【請求項41】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約90%の同一性を有する、請求項37記載の方法。
【請求項42】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約95%の同一性を有する、請求項37記載の方法。
【請求項43】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約99%の同一性を有する、請求項37記載の方法。
【請求項44】
前記CYTL1変異体が配列番号2のCYTL1ポリペプチドの天然に存在する変異体である、請求項37記載の方法。
【請求項45】
前記アゴニストが抗CYTL1抗体またはそのフラグメントである、請求項37記載の方法。
【請求項46】
前記抗体がモノクローナル抗体またはそのフラグメントである、請求項45記載の方法。
【請求項47】
前記モノクローナル抗体がキメラ抗体、ヒト化抗体またはヒト抗体である、請求項46記載の方法。
【請求項48】
前記抗体フラグメントがFab、Fab’、F(ab’)2、およびFvフラグメント;ダイアボディ;直鎖状抗体;単鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体よりなる群から選択される、請求項46記載の方法。
【請求項49】
ウロキナーゼ型プラスミノーゲン活性化因子(uPA)の拮抗剤のスクリーニングの方法であって、(a)ウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)およびCYTL1またはそのアゴニストを含有する混合物を候補拮抗剤とともにインキュベートすること、および(b)該uPARへの該uPAまたはそのアゴニストの結合を競合的に阻害する該候補拮抗剤の能力を計測すること、を含む、方法。
【請求項50】
哺乳類被験体における網膜血管新生を低減する方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む、方法。
【請求項51】
哺乳類被験体における関節炎の発生率または重症度を低減する方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む、方法。
【請求項1】
ウロキナーゼ型プラスミノーゲン活性化因子(uPA)とウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)との相互作用を阻害する方法であって、サイトカイン様1(CYTL1)ポリペプチドまたはそのアゴニストにuPAおよびuPARを含む混合物を接触させることを含む、方法。
【請求項2】
前記接触がインビトロで行われる、請求項1記載の方法。
【請求項3】
前記接触がインビボで行われる、請求項1記載の方法。
【請求項4】
前記混合物がuPAおよびuPARを発現する細胞を含む、請求項1記載の方法。
【請求項5】
前記細胞が癌細胞である、請求項4記載の方法。
【請求項6】
前記癌が癌腫、リンパ腫、芽腫、肉腫、および白血病よりなる群から選択される、請求項5記載の方法。
【請求項7】
前記癌が乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択される、請求項5記載の方法。
【請求項8】
前記アゴニストがCYTL1フラグメント、CYTL1アミノ酸変異体、アゴニスト抗CYTL1抗体およびそのフラグメント、uPARに結合できる、ペプチドおよび非ペプチド性低分子よりなる群から選択される、請求項1記載の方法。
【請求項9】
ウロキナーゼ型プラスミノーゲン活性化因子(uPA)の生物学的活性を阻害する方法であって、有効量のCYTL1またはそのアゴニストにインビボでウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)およびuPAを発現する細胞を接触させることを含む、方法。
【請求項10】
前記細胞が癌細胞である、請求項9記載の方法。
【請求項11】
前記癌が癌腫、リンパ腫、芽腫、肉腫、および白血病よりなる群から選択される、請求項10記載の方法。
【請求項12】
前記癌が乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択される、請求項10記載の方法。
【請求項13】
前記細胞が内皮細胞である、請求項9記載の方法。
【請求項14】
前記アゴニストがCYTL1フラグメント、CYTL1アミノ酸変異体、アゴニスト抗CYTL1抗体およびそのフラグメント、uPARに結合できる、ペプチドおよび非ペプチド性低分子よりなる群から選択される、請求項9記載の方法。
【請求項15】
哺乳類被験体における腫瘍形成または腫瘍転移を阻害するための方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む、方法。
【請求項16】
前記哺乳類被験体がヒト患者である、請求項15記載の方法。
【請求項17】
前記腫瘍が癌である、請求項16記載の方法。
【請求項18】
前記癌が癌腫、リンパ腫、芽腫、肉腫、および白血病よりなる群から選択される、請求項18記載の方法。
【請求項19】
前記癌が乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択される、請求項18記載の方法。
【請求項20】
前記アゴニストがCYTL1フラグメント、CYTL1アミノ酸変異体、アゴニスト抗CYTL1抗体およびそのフラグメント、uPARに結合できる、ペプチドおよび非ペプチド性低分子よりなる群から選択される、請求項16記載の方法。
【請求項21】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約70%の同一性を有する、請求項20記載の方法。
【請求項22】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約80%の同一性を有する、請求項20記載の方法。
【請求項23】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約85%の同一性を有する、請求項20記載の方法。
【請求項24】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約90%の同一性を有する、請求項20記載の方法。
【請求項25】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約95%の同一性を有する、請求項20記載の方法。
【請求項26】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約99%の同一性を有する、請求項20記載の方法。
【請求項27】
前記CYTL1変異体が配列番号2のCYTL1ポリペプチドの天然に存在する変異体である、請求項20記載の方法。
【請求項28】
前記アゴニストが抗CYTL1抗体またはそのフラグメントである、請求項20記載の方法。
【請求項29】
前記抗体がモノクローナル抗体またはそのフラグメントである、請求項28記載の方法。
【請求項30】
前記モノクローナル抗体がキメラ抗体、ヒト化抗体またはヒト抗体である、請求項29記載の方法。
【請求項31】
前記抗体フラグメントがFab、Fab’、F(ab’)2、およびFvフラグメント;ダイアボディ;直鎖状抗体;単鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体よりなる群から選択される、請求項29記載の方法。
【請求項32】
哺乳類被験体における血管形成を阻害するための方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む、方法。
【請求項33】
前記哺乳類被験体がヒト患者である、請求項32記載の方法。
【請求項34】
前記血管形成が腫瘍血管形成である、請求項33記載の方法。
【請求項35】
前記腫瘍が癌腫、リンパ腫、芽腫、肉腫、および白血病よりなる群から選択される、請求項34記載の方法。
【請求項36】
前記腫瘍が乳癌、前立腺癌、結腸癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、ヘパトーマ、結腸直腸癌、子宮内膜癌腫、唾液腺癌腫、腎臓癌、外陰部癌、甲状腺癌、肝臓癌腫、および種々のタイプの頭頚部癌よりなる群から選択される、請求項34記載の方法。
【請求項37】
前記アゴニストがCYTL1フラグメント、CYTL1アミノ酸変異体、アゴニスト抗CYTL1抗体およびそのフラグメント、uPARに結合できる、ペプチドおよび非ペプチド性低分子よりなる群から選択される、請求項33記載の方法。
【請求項38】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約70%の同一性を有する、請求項37記載の方法。
【請求項39】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約80%の同一性を有する、請求項37記載の方法。
【請求項40】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約85%の同一性を有する、請求項37記載の方法。
【請求項41】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約90%の同一性を有する、請求項37記載の方法。
【請求項42】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約95%の同一性を有する、請求項37記載の方法。
【請求項43】
前記CYTL1変異体が配列番号2のCYTL1のアミノ酸配列に少なくとも約99%の同一性を有する、請求項37記載の方法。
【請求項44】
前記CYTL1変異体が配列番号2のCYTL1ポリペプチドの天然に存在する変異体である、請求項37記載の方法。
【請求項45】
前記アゴニストが抗CYTL1抗体またはそのフラグメントである、請求項37記載の方法。
【請求項46】
前記抗体がモノクローナル抗体またはそのフラグメントである、請求項45記載の方法。
【請求項47】
前記モノクローナル抗体がキメラ抗体、ヒト化抗体またはヒト抗体である、請求項46記載の方法。
【請求項48】
前記抗体フラグメントがFab、Fab’、F(ab’)2、およびFvフラグメント;ダイアボディ;直鎖状抗体;単鎖抗体分子;および抗体フラグメントから形成された多重特異性抗体よりなる群から選択される、請求項46記載の方法。
【請求項49】
ウロキナーゼ型プラスミノーゲン活性化因子(uPA)の拮抗剤のスクリーニングの方法であって、(a)ウロキナーゼ型プラスミノーゲン活性化因子受容体(uPAR)およびCYTL1またはそのアゴニストを含有する混合物を候補拮抗剤とともにインキュベートすること、および(b)該uPARへの該uPAまたはそのアゴニストの結合を競合的に阻害する該候補拮抗剤の能力を計測すること、を含む、方法。
【請求項50】
哺乳類被験体における網膜血管新生を低減する方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む、方法。
【請求項51】
哺乳類被験体における関節炎の発生率または重症度を低減する方法であって、有効量のCYTL1またはそのアゴニストを該被験体に投与することを含む、方法。
【図1】

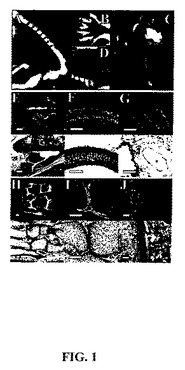
【図2】
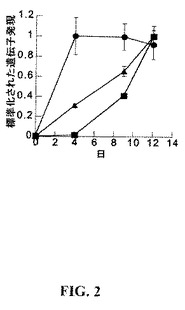
【図3】
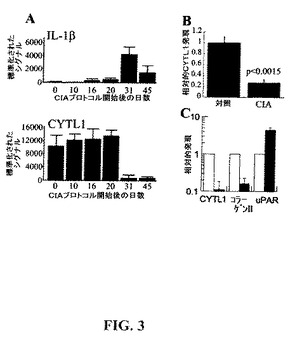
【図4】
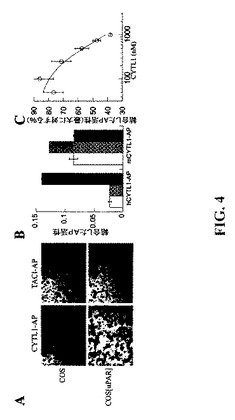
【図5】
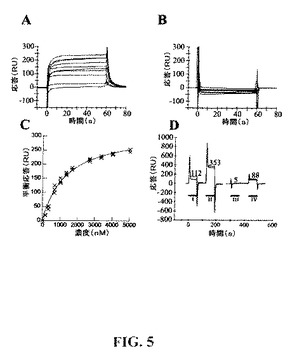
【図6】
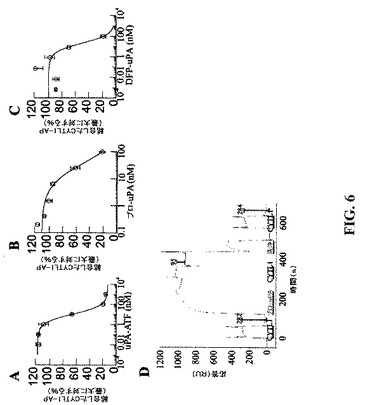
【図7A】
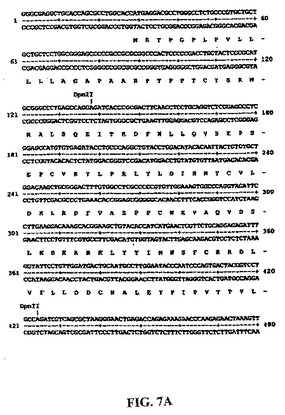
【図7B】

【図8】
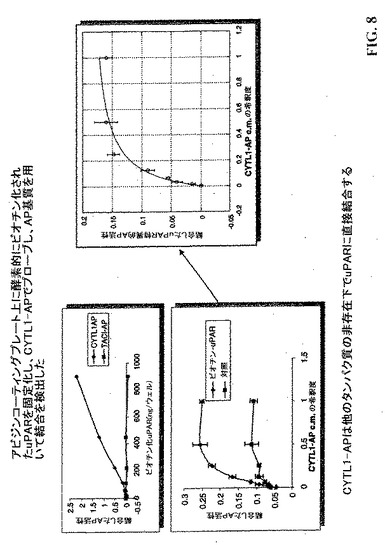
【図9】
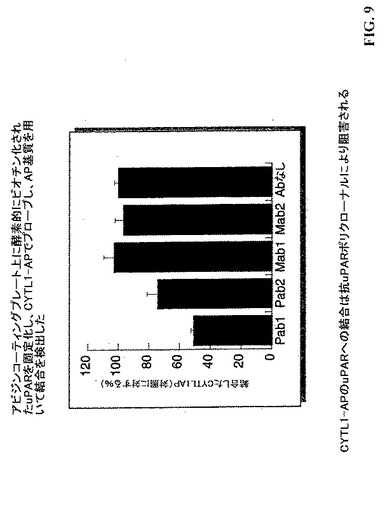
【図10】
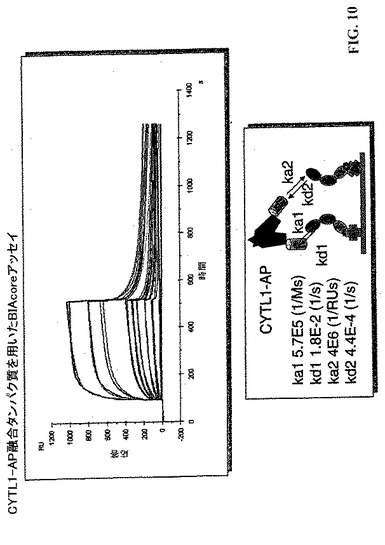
【図11】
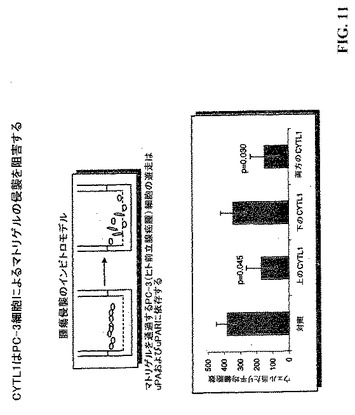
【図12】
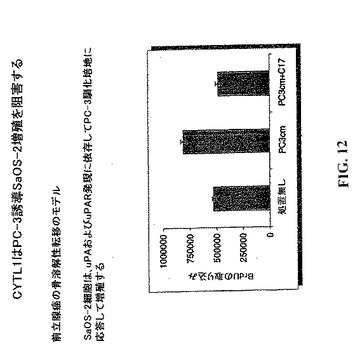
【図13】
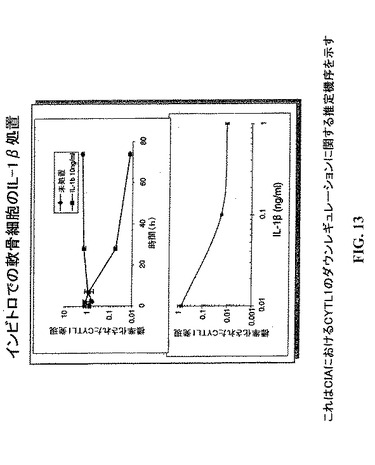
【図16】
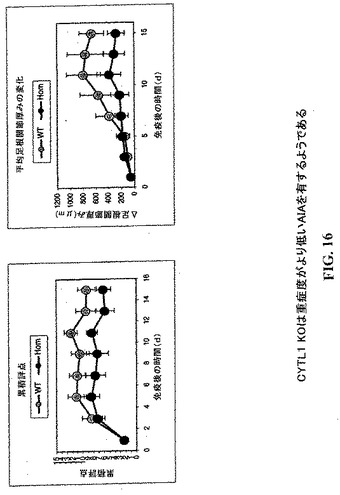
【図17】
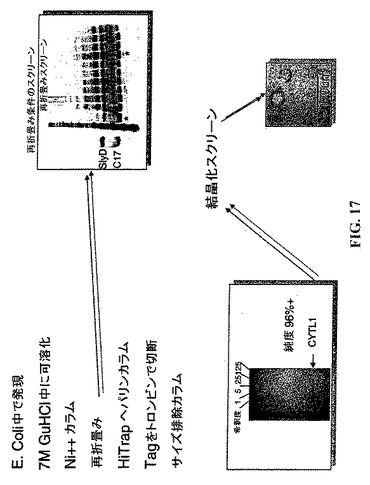
【図19】
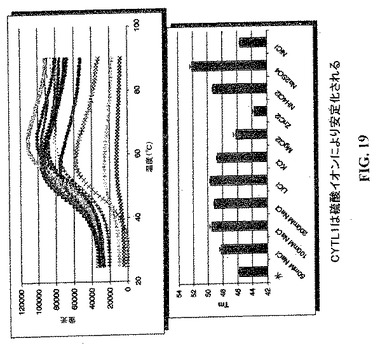
【図21】
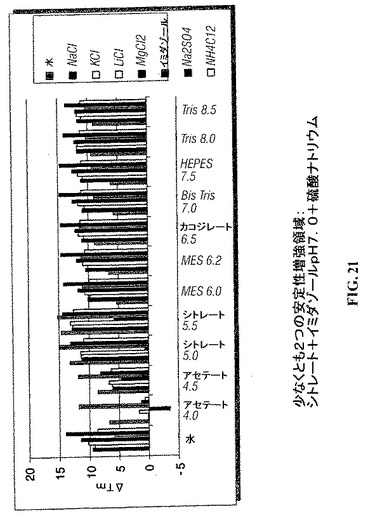
【図23】
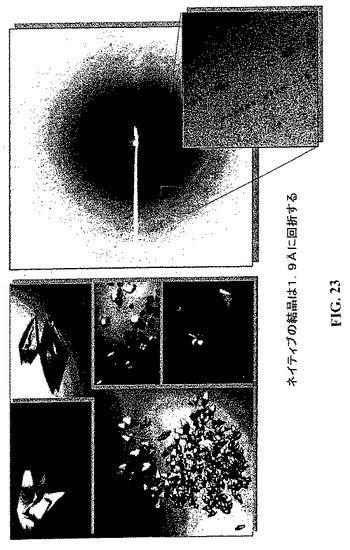
【図25】
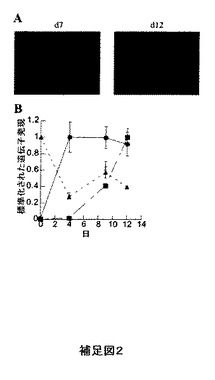
【図26】

【図14】
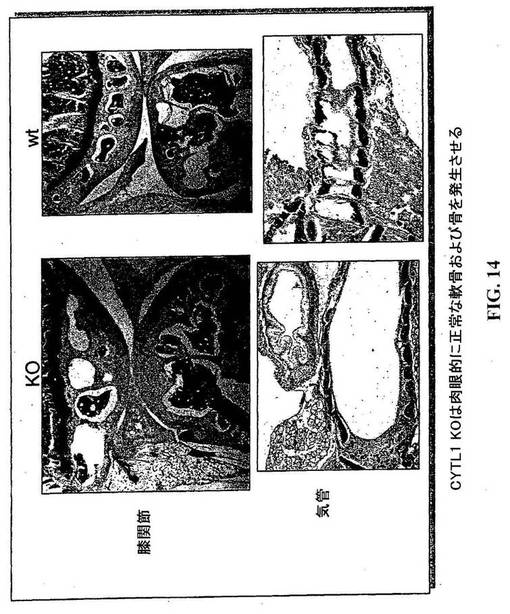
【図15】
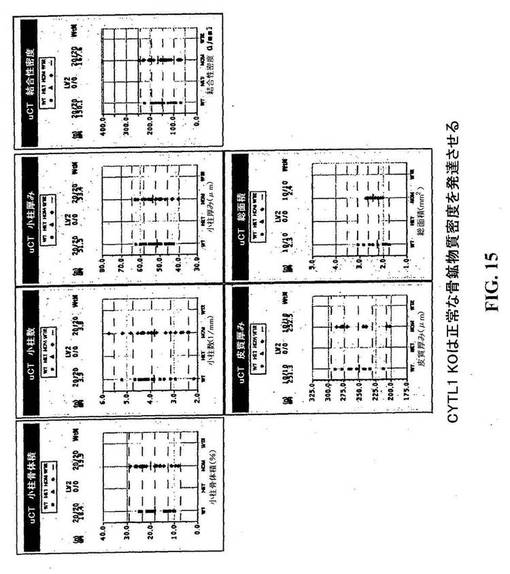
【図18】
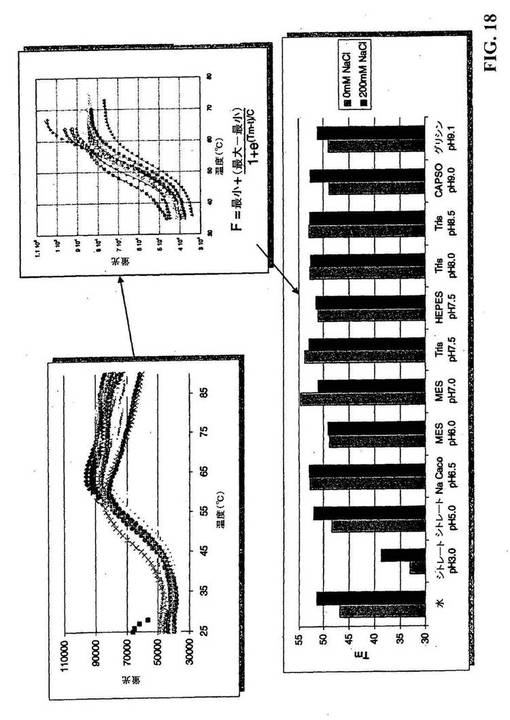
【図20】
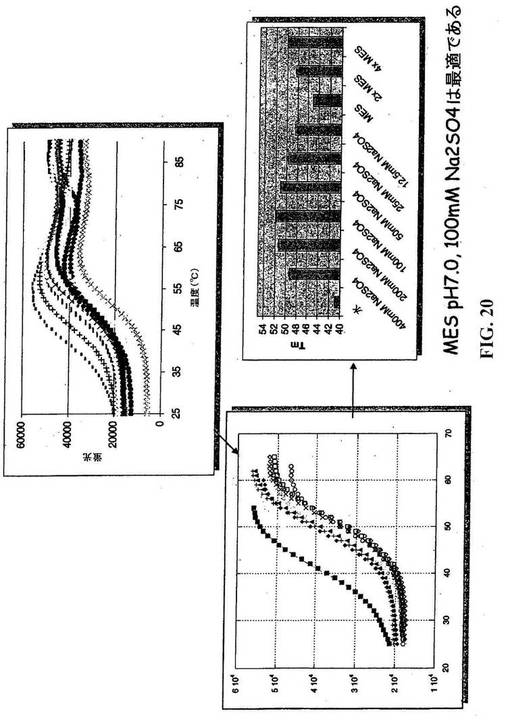
【図22】
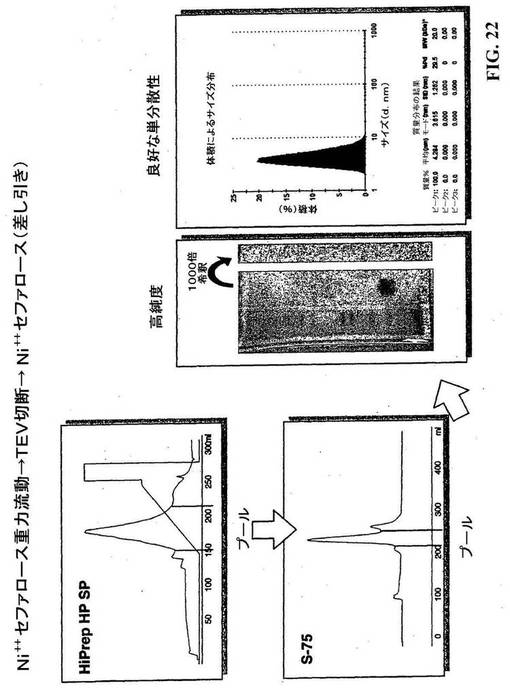
【図24】
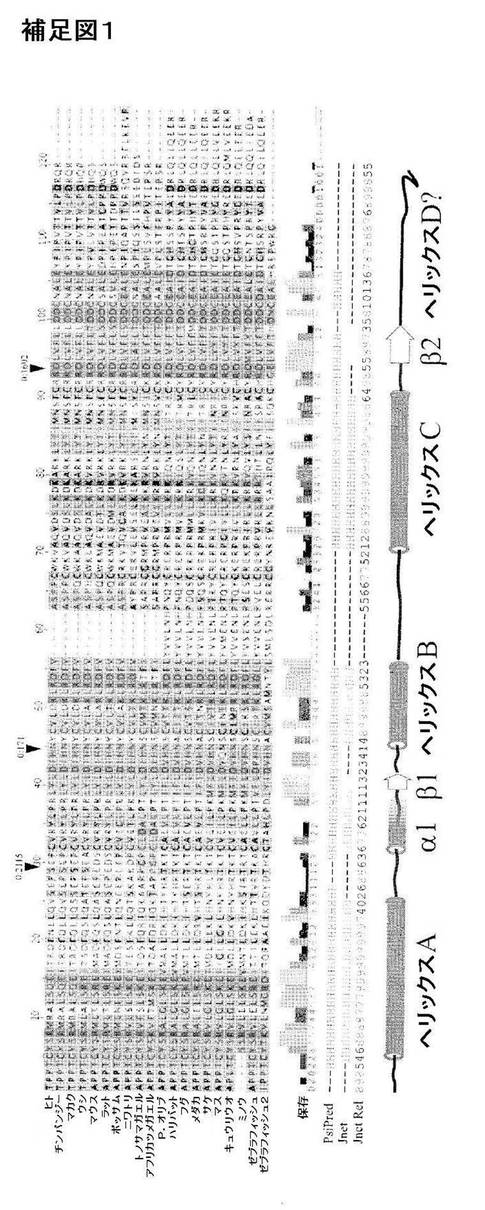
【図2】
【図3】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図16】
【図17】
【図19】
【図21】
【図23】
【図25】
【図26】
【図14】
【図15】
【図18】
【図20】
【図22】
【図24】
【公表番号】特表2011−503531(P2011−503531A)
【公表日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願番号】特願2010−531254(P2010−531254)
【出願日】平成20年10月23日(2008.10.23)
【国際出願番号】PCT/US2008/081018
【国際公開番号】WO2009/055613
【国際公開日】平成21年4月30日(2009.4.30)
【出願人】(509012625)ジェネンテック, インコーポレイテッド (357)
【Fターム(参考)】
【公表日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願日】平成20年10月23日(2008.10.23)
【国際出願番号】PCT/US2008/081018
【国際公開番号】WO2009/055613
【国際公開日】平成21年4月30日(2009.4.30)
【出願人】(509012625)ジェネンテック, インコーポレイテッド (357)
【Fターム(参考)】
[ Back to top ]