
エオキシン形成の修飾物質の同定方法
生物システムにおける14,15-LTC4(エオキシンC4;EoxC4)、14,15-LTD4(エオキシンD4;EoxD4)、又は14,15-LTE4(エオキシンE4;EoxE4)の形成を調節する化合物を同定する方法。抗炎症効果を有する化合物を同定する方法で、該方法は生物システムにおける14,15-LTC4(エオキシンC4;EoxC4)、14,15-LTD4(エオキシンD4;EoxD4)、又は14,15-LTE4(エオキシンE4;EoxE4)の形成及び/又は活性に対する効果について化合物を試験することを含む。(i)本発明の方法によって抗炎症又は骨量減少低減化合物を同定し;(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせることを含む抗炎症組成物又は骨量減少低減組成物を製造する方法。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は薬学的に有用な化合物を同定するための方法に関する。本発明は更に炎症疾患又は炎症全般の治療に有用な化合物を同定するための方法に関する。本発明はまた抗炎症組成物を製造する方法及び薬学的に有用な化合物を使用する方法にも関する。
【背景技術】
【0002】
その性質が炎症性である多くの疾患/障害がある。炎症症状の現在の治療法に伴う主な問題の一つは効能の欠如及び/又は副作用(実際のもの又はそのように思われているもの)の発生である。
喘息は、先進工業国の成人人口の6%〜8%で発症している慢性の炎症性疾患である。子供における発生率はさらに高く、殆どの国では10%に近い。喘息は、15才以下の子供の入院原因の最も一般的なものである。
喘息の治療法は症状の重症度に基づく。軽度の症例では治療しないか、又はβ-アゴニストの吸入で治療されるのみである。より重症の喘息を患っている患者は、定期的に抗炎症化合物を用いて治療するのが典型的である。
【0003】
現在の維持療法(主としてコルチコステロイド類の吸入)には、少なくともある程度のリスクがあると思われるため、喘息の治療が不十分な場合がかなりある。これらには、不要な罹患及び死亡に至る、子供の成長遅延及び骨ミネラル密度の損失の危険性が含まれる。ステロイド類の代替法として、ロイコトリエンレセプターアンタゴニスト(LTRas)が開発されている。これらの薬剤は経口的に与えることができるが、吸入されたステロイド類に比べてあまり有効ではなく、通常、気道炎症を満足に抑制できない。
この要因が組み合わさって、喘息の全患者の少なくとも50%が満足のいく治療を受けていない。
【0004】
治療が不十分であるという同様のパターンがアレルギー疾患に関連して存在し、多くの一般的症状を治療するのに薬剤を利用できるが、明白な副作用を考慮して使用されていない。鼻炎、結膜炎及び皮膚炎はアレルギー要素を有しうるが、潜在するアレルギーがない場合でも生じる場合がある。実際、このクラスの非アレルギー性症状は、多くの症例で治療が困難である。
【0005】
慢性閉塞性肺疾患(COPD)は、世界の人口の6%〜8%が罹患している一般的な疾患である。疾患は致死的となりうるおそれがあり、この病気の罹患率及び死亡率はかなりのものである。現在、COPDの経過を変えることができる既知の薬物学的治療法は存在しない。
【0006】
挙げることのできる他の炎症性疾患には次のものがある:
(a)特発性肺線維症(これはCOPDよりも一般的ではないが、非常に悪い予後を伴う重篤な疾患である。治癒的治療法は存在しない);
(b)炎症性大腸炎(高罹患率の疾患群。今日、このような疾患では対症療法が利用できるに過ぎない);及び
(c)関節リウマチ及び骨関節炎(関節のありふれた無能化炎症性疾患。現在では、このような症状への対応に利用できる治癒的治療法は存在せず、また適度に効果的な対症療法のみである)。
【0007】
炎症はまた痛みの一般的な原因である。炎症痛は多くの理由、例えば感染、手術又は他の外傷により生じるおそれがある。更に、数種の悪性腫瘍は、患者の総体的な症状に加わる炎症要素を有することが知られている。
よって、新規な及び/又は代替の抗炎症治療法が、上述した患者群の全てにとって有益となる。特に、実際の又は知覚される副作用を生じないで喘息のような炎症性疾患を治療できる有効な抗炎症剤に対して真のかつ実質的に満たされていないニーズがある。
【0008】
哺乳類のリポキシゲナーゼは、アラキドン酸の酸素化を触媒する構造的に関連した酵素のファミリーである。3種類のヒトリポキシゲナーゼが知られており、これらは炭素位置5、12及び151におけるアラキドン酸への分子酸素の挿入を触媒する。よって、酵素は、それぞれ5-、12-及び15-リポキシゲナーゼと称されている。
リポキシゲナーゼの作用によって生成されるアラキドン酸の代謝産物は、炎症誘発効果を含む明白な病態生理学的活性を有していることが知られている。
【0009】
ロイコトリエン生合成における重要な酵素は、2段階反応でアラキドン酸をロイコトリエン(LT)A4に転換する5-リポキシゲナーゼである。この代謝物は酵素的にロイコトリエンB4(LTB4)まで加水分解されるか、又はロイコトリエンC4(LTC4)シンターゼによってグルタチオンとコンジュゲートさせられてLTC4を形成し、これがついでγグルタミル-トランスペプチダーゼ及びジペプチダーゼの連続的作用によって切断されロイコトリエンD4(LTD4)とロイコトリエンE4(LTE4)をそれぞれ生じうる。LTC4とその代謝物は炎症誘発剤及び強力な気管支収縮剤2である。これらの代謝産物の作用並びにそれらを形成する生物学的プロセスを阻害する薬剤の開発に多大な努力がなされている。この目的のために開発された薬剤には、5-リポキシゲナーゼインヒビターと、これまでに述べたように、ロイコトリエンレセプターアンタゴニスト(LTRas)が含まれる。FLAP(Five Lipoxygenase Activating Protein)のインヒビターもまた研究されている。
【0010】
アラキドン酸を代謝する他のクラスの酵素はシクロオキシゲナーゼ類である。このプロセスにより生成されるアラキドン酸代謝産物には、プロスタグランジン類、トロンボキサン類、及びプロスタサイクリンが含まれ、それらの全てが生理学的及び病態生理学的活性を有している。特に、プロスタグランジンPGE2は、熱及び痛みをまた誘発する強力な炎症誘発媒介物である。従って、「NSAIDs」(非ステロイド性抗炎症剤)及び「コキシブ(coxibs)」(選択的シクロオキシゲナーゼ-2インヒビター)を含む、PGE2の生成を阻害する多くの薬剤が開発されている。これらのクラスの化合物は、主として一種又は数種のシクロオキシゲナーゼの阻害を介して作用する。またレセプターアンタゴニストが研究されている。
【0011】
よって、一般に、アラキドン酸代謝産物の生成を阻止可能な薬剤は、炎症の治療に有用であると思われる。
【0012】
例えば免疫抑制性患者、例えば化学療法、移植拒絶を防止するための治療を受けている患者又はAIDS患者において炎症誘発剤はまた有用でありうる。
5-リポキシゲナーゼに加えて、12-及び15-リポキシゲナーゼがまたヒトの体で発現される。
12-リポキシゲナーゼ(12-LO)はヒト血小板中で高度に発現されるがこの酵素の生物学的な役割については殆ど知られていない1。
【0013】
15-リポキシゲナーゼ(15-LO)1型(ヒト15-LO:EC1.13.11.33)は非ヘム鉄を含む75-kDaのタンパク質である。15-LOはアラキドン酸(20:4)のC-15又はリノール酸(18:2)のC-13に分子酸素を導入して15S-ヒドロペルオキシ-5,8,11-cis-13-trans-エイコサテトラエン酸(15-HPETE)又は13S-ヒドロペルオキシ-9-cis-13-trans-オクタデカジエン酸(13-HPODE)をそれぞれ形成する。少量の12S-ヒドロペルオキシ-5,8,14-cis-10-trans-エイコサテトラエン酸(12-HPETE)がまた15-LO酵素によってアラキドン酸から形成される。簡単に述べると、触媒反応は不飽和脂肪酸のcis-cis-1,4-ペンタジエン構造(20:4又は18:2)のC-3における水素除去を含んでおり、炭素ラジカルとFe3+からFe2+への鉄の還元を生じる(以下のスキーム1参照)。
【0014】
ラジカルはC-5に移動し、そこで分子酸素と反応してペルオキシ基を形成する。ペルオキシ基による鉄からFe3+への酸化は、プロトン化されて最終の脂質ヒドロペルオキシドを生じるペルオキシ陰イオンを生じる。
【0015】
ウサギ酵素15-LOはクローニングされ結晶化され、基質結合部位が、その基部がPhe353、Met419、Ile418及びIle593の側鎖によって定まる深い疎水性ポケット22であることが見出されている。脂肪酸基質は(酵素の表面で)Arg403によって繋留され位置せしめられている。変異原性研究により、疎水性ポケットのサイズの変動が酵素の位置特異性に影響を及ぼすことが立証されている。よって、 15-LOのポケットの拡大が12-リポキシゲネーション産物(12-HPETE)の量を増大させた。15-LO中の鉄はヒスチジンとC末端イソロイシンのカルボキシ基に配位されている。
【0016】
15(S)-HETEへの還元を受けることに加えて、15(S)-HPETEはまた脱水を受けて14,15-オキシド-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTA4)を生じる。このアリルエポキシドはジヒドロキシ酸8(R,S),15(S)-LTB4又は14(R,S),15(S)-LTB4に加水分解されうる11-12。また、基質として15-HPETEを用いて、15-LOは二重の二原子酸素付加産物5(S),15(S)-DHETE,8(S,R),15(S)-DHETE及び14(R,S),15(S)-DHETEの形成をまた触媒する。
【0017】
15(S)-リポキシゲナーゼ(15-LO)は遊離脂肪酸を代謝するばかりでなく、膜に存在しているポリエン脂肪酸を攻撃することもできる。該酵素はミトコンドリア膜の崩壊を通して網状赤血球の成熟において重要な役割を担っていることが予想された3。しかしながら、該酵素はケラチノサイトや眼水晶体の成熟のような、細胞の分化においてより一般的な役割を担いうる4。15-LOは主として網状赤血球、好酸球、皮膚、単球/マクロファージ及び気道上皮において発現される5。インターロイキン-4(IL-4)及びインターロイキン-13(IL-13)は培養された単球及び上皮細胞における15-LOの発現を誘導するが6、ある種の細胞への損傷が15-LOの活性を増加させることが報告されている(Green FA. Transformation of 5-HETE by activated keratinocyte 15-lipoxygenase and the activation mechanism. Lipids 25, 618-623 (1990))。
【0018】
5-リポキシゲナーゼ及び12-リポキシゲナーゼとは異なり、15-LOはウサギ及びヒト細胞においてのみ発現されると思われる1。ヒト15-LOのマウスオルソログは白血球型の12-LOであると思われる1。しかしながら、二つの酵素活性の調節はかなり異なっているようである7。
15-LOは20年以上も前に発見され、均一になるまで精製される最初の15-LOはウサギ網状赤血球から取り出された8。該酵素のヒト型は網状赤血球及び上皮細胞からクローニングされており、免疫学的研究から、該酵素は網状赤血球、好酸球及び気道上皮において同一である9。15-LOの第二の型(2型)はヒト毛根からのcDNAライブラリーからクローニングされ、続く該酵素の発現とアミノ酸解析により、この15-LOの2型酵素は先に開示された15-LOの1型とおよそ40%同一であることが証明された10。
【0019】
15-LOは、主として抗炎症効果を有していると思われるリポキシンの形成に関与していると思われるが13、15-LO誘導産物の5-オキソ-15-ヒドロキシ-6,8,1,13-エイコサテトラエン酸はヒト好酸球に対する強力な走化性剤であると報告されている14。ヒトにおける15-LO経路の生理学的及び病態生理学的役割は十分には特徴付けされていない。
【0020】
ロイコトリエン類のような生物システム中で生じることが知られているアラキドン酸代謝物に基づいて、多くの異なったアナログが合成されている。例えば14,15-LTC4のようなLTC4の合成アナログは気管支収縮を誘導しなかった。Drazen, J.M., Lewis, R. A., Austin, K.F., Toda, M., Brion, F., Marfat, A.及びCorey, E.J.: Contractile activities of structural analogs of leukotrienes C and D: necessity of a hydrophobic region. Proc. Natl. Acad. Sci. 78, 3195-3198 (1981)。
【発明の概要】
【0021】
我々は炎症誘発効果を有するアラキドン酸代謝物の生物システム中における産生を同定した。該代謝物は15-リポキシゲナーゼの作用を介して、可溶型グルタチオンSトランスフェラーゼ及び/又はロイコトリエンC4シンターゼの作用を介して形成されうる。該代謝物はエオキシンC4、エオキシンD4及びエオキシンE4と命名される。我々は、抗炎症化合物及びエオキシンC4、D4及びE4形成を阻害する化合物を同定するためのスクリーニング法を提供する。
【0022】
我々は、ホジキン細胞株L123615が15-リポキシゲナーゼ(15-LO)の1型活性を有しており、15-LO経路を介してアラキドン酸をエオキシン類に転換することを示す。L1236中におけるエオキシンC4、D4及びE4の存在及び形成を明らかにした後、我々は、これらエオキシン類が他のヒト細胞及び組織、特に好中球において形成されるが、また鼻ポリープ組織及び樹状細胞(実施例2)によっても形成されることをまた示す。我々はまた広範囲の細胞型が15-LOを発現することを示す。興味深いことに、炎症疾患又は炎症プロセスを含む疾患に罹患している患者の細胞、例えばホジキンリンパ腫の患者のリンパ節中のリード・シュテルンベルク細胞、サルコイドーシス患者の肺バイオプシーの類上皮細胞、関節リウマチの滑膜細胞及びアルツハイマー病の患者のニューロン(実施例3)は、コントロール細胞と比較して15-LOの増大した発現を示した。
【0023】
これらの新しい代謝物の生合成の立証と、関連する酵素の同定に加えて、エオキシン類の生物学的効果をまた記載する。例えば、実施例6aには、エオキシン類C4、D4及びE4は内皮細胞の透過を引き起こす点でヒスタミンより約100倍強力であることが示され(炎症のインビトロモデル)、5-LO経路を通して形成された姉妹代謝物LTC4に匹敵する作用強度を示す。例えば、エオキシン類C4、D4及びE4はコンフルエントなウシ又はヒト内皮細胞を有するモデル系において10-7及び10-8Mにて有意な血漿漏出を誘導する。炎症の他のモデルでは、 EoxC4及びEoxD4はGM-CSFで刺激された好中球による酸素フリーラジカル生成を主に誘導したが、EoxE4はIL-5で刺激された好酸球による酸素フリーラジカル生成を主に誘導した。我々はまたエオキシン類C4、D4及びE4の形成を阻止する化合物を実証した。
【0024】
我々は、それぞれエオキシンC4(EoxC4)、エオキシンD4(EoxD4)及びエオキシンE4(EoxE4)と命名されたエオキシン:14,15-LTC4、14,15-LTD4、及び14,15-LTE4と命名されたアラキドン酸からの3種の15-リポキシゲナーゼ誘導代謝物を同定する。
アラキドン酸からの15-LO誘導システイニル含有産物エオキシンC4、D4及びE4の形成は真核又は原核細胞において以前には決して記載されていない。
【0025】
RP-HPLCを使用して代謝物を単離し、代謝物の構造を陰イオンエレクトロスプレータンデム質量分析法によって決定した。エオキシン類は282nmで最大UV吸光度を示すコンジュゲートトリエン構造を有している。我々は、これらの代謝物が、アラキドン酸が15-LOで触媒された転換により14,15-オキシド-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTA4;エオキシンA4)になり、これがグルタチオンとコンジュゲートさせられて14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTC4;エオキシンC4)になり、これが続いて14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTD4;エオキシンD4)及び14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTE4;エオキシンE4)まで代謝されることによって形成されると考える。
【0026】
14,15-LTA4(エオキシンA4)からの14,15-LTD4(エオキシンD4)の形成に関与する酵素は、実施例4において検討されるように、LTC4-シンターゼ及び/又は可溶型グルタチオンS-トランスフェラーゼ(GST)、特にGSTM1、及びGSTP1であると考えられる。
示唆される経路を以下に示す:
【発明を実施するための形態】
【0027】
本発明の第一の態様は、生物システムにおけるエオキシンC4,エオキシンD4,又はエオキシンE4の形成を調節する化合物を同定する方法であって、15-LO活性又はGSTM1-1、GSTP1-1及び/又はロイコトリエンC4シンターゼ活性に対する効果について化合物を試験することを含む方法を提供する。
本発明の第二の態様は、炎症調節効果又は骨量減少調節(典型的には骨量減少低減)効果を有する化合物を同定する方法であって、生物システムにおけるエオキシンC4,エオキシンD4又はエオキシンE4の形成及び/又は活性に対する効果について化合物を試験することを含む方法を提供する。
【0028】
「形成を調節する」には、エオキシンC4、D4又はE4の形成及び/又は活性を阻害又は促進することが含まれる。典型的には、該方法は、EoxC4、D4又はE4の形成及び/又は活性を阻害するための化合物を同定するために使用しうる。かかる化合物はまた骨量減少低減化合物であると考えられる。該化合物は典型的にはEoxC4、EoxD4及びEoxE4の形成及び/又は活性を阻害しうる。
該方法は、あるいは、EoxC4、EoxD4又はEoxE4の形成及び/又は活性を促進するための化合物を同定するために使用することができる。かかる化合物は炎症誘発性であると考えられる。かかる化合物は典型的にはEoxC4、EoxD4及びEoxE4の形成を促進しうる。
【0029】
本発明の第一の態様の方法は、生物システムにおけるEoxC4、EoxD4又はEoxE4の形成又は活性に対する効果について化合物を試験する工程を更に含みうる。
生物システムは典型的には15-LO、グルタチオンSトランスフェラーゼ(典型的には可溶型グルタチオンSトランスフェラーゼ、例えばGSTM1-1又はGSTP1-1)及び/又はロイコトリエンC4シンターゼ活性を含む。
【0030】
化合物は、以下で更に検討するように、細胞画分又は単離酵素又は酵素群を含む系において試験することができる。あるいは、本発明の第一又は第二の態様の方法は、ホールセルに対する試験化合物の効果を試験することによって実施することができる。例えば、該方法は、エキソビボヒト細胞又は細胞株を使用して実施することができる。例えば、ヒト好酸球又は肥満細胞(例えば実施例8に記載したような、例えば臍帯血由来の肥満細胞)又はL1236細胞株を使用することができるが、例えば15-LO及び/又はGSTのような必要な酵素を発現する(内因性又は組換えの何れでも)ヒト鼻ポリープ組織及び/又は樹状細胞又は任意の他の細胞又は細胞株もまた使用することができる。しかしながら、この種の代謝経路に関与する酵素が同じ位置にある必要はない。様々なアラキドン酸代謝物の形成におけるよく知られた現象は経細胞生合成、つまり最初の酵素工程が一つの細胞型において触媒され、中間体が他の細胞型に搬出されて、そこで第二の酵素工程が実施されるものである。例えばエオキシンA4の生合成は一細胞型において生じる一方、エオキシンA4からエオキシンC4への転換(及び場合によってはエオキシンC4からエオキシンD4及びエオキシンE4への転換も)は第二の細胞型において生じうる。よって、試験は、エオキシン類の生合成のための補完酵素能力を持つ2つの細胞型の同時インキュベーションで実施することもまたできる。
【0031】
当業者には明らかなように、アッセイ手順の前又はその一部として細胞を刺激し;又は例えば実施例に記載しているように、アッセイにアラキドン酸を提供することが必要である。実施例8において検討するように、LTC4、プロスタグランジンD2又はIL-5によって好酸球を刺激してEoxC4を産生させることができる。15-LO-1の発現を誘導するためには例えばIL-4を使用して肥満細胞を刺激し;ついで実施例8で検討するようにアッセイにおいてアラキドン酸を提供することが望ましい場合がある。他の細胞はエオキシン類を放出することができるために刺激を必要としない場合がある。例えば、実施例8において検討されるように、ヒト鼻ポリープ細胞はエオキシン類を自発的に放出することができると考えられる。
【0032】
当業者には明らかなように、エオキシンC4の形成に対する効果が試験されているならば、エオキシンC4の代謝物の生合成に関与する現在の酵素(例えばg-グルタミル-トランスペプチダーゼ又はジペプチダーゼ)を有していることは必要ではない。同様に、15-LO活性に対する効果が試験されている場合、15-LO(エオキシンA4)の代謝物の生合成に関与する現在の酵素、例えばLTC4シンターゼ又は可溶型グルタチオンSトランスフェラーゼ、例えばGSTM1-1又はGSTP1-1を有していることは必要ではない。当業者には明らかなように、一又は限られた数の酵素が存在している系と、より多くの酵素(例えばアラキドン酸からのエオキシン合成に関与する酵素の全て、例えば15-LO;LTC4シンターゼ又は可溶型GST、例えばGSTM1-1又はGSTP1-1)が存在する系の双方において化合物を試験することが望ましい場合がある。当業者にはよく知られているように、単一又は限られた酵素系での試験は構造-機能関係を樹立するのに有用でありうる一方;複雑な系での試験は化合物がインビボで有用である可能性があるかどうかを樹立するのに有用でありうる。
【0033】
上述のように、ある化合物がエオキシンC4、エオキシンD4又はエオキシンE4の形成を阻害することができるかどうかを試験する適切な方法は、実施例7に例証する。しかしながら、別法として、他の細胞株は、それらがエオキシンを生産する酵素的機構を有しているならば使用することができ、細胞型に応じた手順の変更は当業者には明らかである。
エオキシン類は、例えばHPLC、ガスクロマトグラフィー、質量分析法、UV分光法及び異なった免疫学的方法、例えばEIA及びRIAによって検出し定量することができる。
【0034】
現在、上記のように、15-LO酵素の2つの型がクローニングされている。2型15-LOは1型15-LOとおよそ40%同一である。1型15-LOとは異なり、リノール酸は2型15-LOのための乏しい基質である。現在の知識では、阻害のための好ましい酵素はよって15-LOの1型であるが、15-LOの2型を、エオキシン類C4、D4及びE4の形成に至る代謝経路における酵素として除外することはできない。
15-LOの触媒活性は例えば記載されているようにして測定することができる9。該酵素は、当該試験化合物の存在下又は不在下で15-(s)-HETEへのアラキドン酸(基質)の転換又は13-ヒドロキシオクタデカジエン酸(13-HODE)へのリノール酸(基質)の転換を評価することによってアッセイすることができる。化合物が存在している場合における15-(S)-HETE又は13-HODEの形成の減少は酵素活性における減少を例証する。そのような化合物は、エオキシン類C4、D4及びE4の形成において最初の酵素段階を阻止する。かかる化合物はインビボで抗炎症作用を有していると考えられる。
【0035】
本発明の第一の態様の一側面では、ヒト15-LOの触媒活性を試験するためのアッセイは実施例5に記載されているように考えられる。ここに示したアッセイは、ジフェニル-1-ピレニルホスホン(DPPP)をホスフィンオキシドに酸化する脂質ヒドロペルオキシドの能力を利用する。ジフェニル-1-ピレニルホスホンは酸化されるまでは非蛍光化合物である。DPPPの酸化は蛍光化合物を生じ、これは温度に感受性がなく、少なくとも60分間安定である。
【0036】
本発明の第一の態様の他の側面では、問題の試験化合物の存在下又は不在下で、15-(S)-HETEへのアラキドン酸又は13-ヒドロキシオクタデカジエン酸(13-HODE)へのリノール酸(基質)の転換を決定するために細胞可溶化物においてアッセイを実施する。
上記の新規の15-リポキシゲナーゼで触媒されたアラキドン酸代謝物の形成における重要な酵素の他のものは可溶型GSTである。実施例4に示すように、GSTM1-1、GSTP1-1及び/又はLTC4シンターゼはエオキシン類C4、D4及びE4の形成に関与しうる。よって、本発明の第一態様の実施態様では、GSTM1-1、GSTP1-1及び/又はLTC4シンターゼの触媒活性に対する試験化合物の効果が評価される。GSTM1-1又はGSTP1-1の触媒活性のアッセイは本質的に実施例4に記載されているようにして実施することができる。
【0037】
本発明の第二の態様の実施態様では、実施例6aに記載されたもののように、炎症のインビトロモデルにおいてエオキシンC4、D4又はE4で誘導した内皮細胞の透過を調節するその能力について試験化合物をアッセイする。かかる試験はまた本発明の第一の態様の方法との関連で実施することもできる。
化合物は、例えば炎症のウサギモデルのように15-LOが存在している動物又は炎症のエキソビボモデルにおいて又は15-LOを発現するトランスジェニック動物(例えばマウス又はラット)において試験することができる。化合物は炎症のヒトエキソビボモデルにおいて、例えばヒト末梢血又はヒト臍帯血について、又はヒト末梢血もしくはヒト臍帯血から単離した細胞について、例えば白血球、例えば好中球、好酸球、樹状細胞又は肥満細胞について試験することができる。かかる試験の例は実施例の例えば実施例6bに記載される。該化合物はまた例えばUS2003/0175680に記載されているように、幹細胞又は他の骨形成細胞前駆体を骨細胞に分化させる能力について試験することができる。
【0038】
試験される化合物は、例えば上で検討したように、化合物の構造が15-LOの基質結合部位と比較され、又はそれに基づいて設計もしくは修飾される構造ベースのスクリーニング法に基づいて選択することができる。出発化合物は最初は15-LO酵素活性に対する効果について(例えば実施例5に記載されているアッセイを使用して)スクリーニングすることによって選択することができ;ついで、構造と比較し;例えばエオキシンC4、D4又はE4形成に対する効果について更なるモデル化及び/又は合成及び評価によってついで試験することができる更なる化合物を設計するための基礎として使用される。
【0039】
(例えば実施例5に記載されているような)DPPPを使用する15-LO活性に対する効果の評価は、分光光度アッセイを使用することができ、直ぐに実施でき、例えば高スループット試験に非常に適したマイクロタイタープレート方式に使用することができることを意味する。アラキドン酸又は15-HPETE又は他のアラキドン酸代謝物を基質として使用する評価は、15-LO活性をインビボで調節する化合物の能力を確認するのに有用でありうる。
化合物にはまた当業者によく知られているように他の試験、例えば毒性学又は代謝試験を施すことができる。
【0040】
本発明は、EoxC4、EoxD4及びEoxE4の一又は複数の形成又は活性を調節、例えば亢進又は阻害するのに有用な薬剤を同定しようとする際に使用されるスクリーニングアッセイを提供することが理解される。該方法において同定される化合物はそれ自体薬剤として有用であり得、あるいはそれらはより効能のある化合物の設計と合成のためのリード化合物を表しうる。
該化合物は上記の化合物を同定する方法の各々に対して薬物様化合物又は薬物様化合物の開発のためのリード化合物でありうる。上記方法は、当業者によく知られているように、薬学的化合物又は薬物の開発におけるスクリーニングアッセイとして有用でありうることが理解される。
【0041】
「薬物様化合物」という用語は、当業者によく知られており、医学における使用に適する特性、例えば、医薬品における有効成分としての特性を有する化合物の意味を含んでいてよい。従って、例えば、薬物様化合物は、有機化学の技術によるか、それより好ましくはないが、分子生物学又は生化学の技術により合成される分子であってよく、5000ダルトン未満であってよい小分子であることが好ましい。薬物様化合物は、特定のタンパク質又はタンパク質群との選択的な相互作用の特徴を更に示し、生物利用及び/又は細胞膜を透過可能でありうるが、これらの特徴が必須ではないことは理解されるであろう。
「リード化合物」という用語は、同様に当業者によく知られており、それ自体薬物としての使用に適さない(例えば、その所期の標的に対して弱い効力を示すにすぎず、その作用が非選択的で、不安定で、溶解性に乏しく、合成が困難又は不十分な生物学的利用能を有する)が、その化合物がより望ましい特性を有する他の化合物の設計の出発点を与えるものであるという意味を含みうる。
【0042】
高スループット操作が可能なスクリーニングアッセイが特に好ましいことが理解される。上で検討したように、DPPPを使用する酵素アッセイ法を使用することができる。
適切な場合には、炎症又はインビボでのエオキシンC4、D4又はE4の形成又は活性を調節しうる化合物を同定することは望ましいであろうことは理解される。よって、本発明の方法で使用される試薬と条件は、例えば15-LOと基質の間の相互作用が、ヒト15ーLOと天然に生じる基質(例えばアラキドン酸)との間のものに類似するように選択することができることは理解されるであろう。当業者には明らかなように、異なったアッセイ系を用いて化合物を評価することができ、その幾つかにおいてエオキシン形成に関与する特定の酵素に対する効果に対する特異性又はアッセイの簡便性を(例えば単一酵素系を使用して)最適化されうる一方、他のものでは、インビボ関連性を、例えばホールセル中の化合物の効果を評価することによって最適にすることができる。
【0043】
本発明のスクリーニングアッセイは炎症疾患又は炎症プロセスを含む疾患の治療において有用な化合物の探索に有用である。インビトロ及び/又はインビボスクリーンイングをベースとして選択した化合物を、炎症疾患又は炎症プロセスを含む疾患における抗炎症効果について更に試験することができる。同様に、本発明のスクリーニングアッセイは骨量減少の治療又は防止/骨形成亢進に有用である化合物の探索に有用である。インビトロ及び/又はインビボスクリーニングをベースにして選択された化合物は、骨量減少の患者又はその危険のある患者又は炎症プロセスを含む疾患における骨量減少/骨形成亢進効果について更に試験することができる。
【0044】
「炎症」という用語は、上で述べたもののような身体外傷、感染、慢性疾患によって誘発されうる局所的又は全身性保護反応、及び/又は外部刺激に対する化学的及び/又は生理学的反応(例えばアレルギー反応の一部として)によって特徴付けられる任意の症状を含むと当業者には理解される。有害な薬剤と傷ついた組織の双方を破壊し、希釈し又は隔離する作用をするあらゆるそのような応答は、例えば発熱、腫れ、痛み、発赤、血管拡張及び/又は血流増加、白血球の罹患領域への侵入、機能喪失及び/又は炎症症状に伴うことが知られている任意の他の徴候に顕れうる。
よって、「炎症」という用語は、任意の炎症性疾患、障害又は症状自体、それを伴う炎症要素を持つ任意の症状、及び/又はとりわけ急性、慢性、潰瘍、特異、アレルギー性及び壊死性炎症、及び当業者に知られている炎症の他の形態を含む、徴候として炎症によって特徴付けられる任意の症状を含むものとまた理解される。よって、その用語はまた本発明の目的に対して、炎症痛及び/又は発熱を含む。
【0045】
エオキシンC4、D4又はE4の形成又は活性を減少させる化合物は炎症疾患又は炎症プロセスを含む疾患を治療するのに有用である。従って、本発明のスクリーニング法は、喘息、慢性閉塞性肺疾患(COPD)、肺線維症、アレルギー性疾患、鼻炎、炎症性腸疾患、潰瘍、炎症性痛覚、発熱、アテローム性動脈硬化症、冠動脈疾患、脈管炎、膵炎、関節炎、変形性関節炎、関節リウマチ、結膜炎、虹彩炎、強膜炎、ブドウ膜炎、創傷、皮膚炎、湿疹、乾癬、発作、糖尿病、自己免疫疾患、アルツハイマー病、多発性硬化症、サルコイドーシス、アテローム性動脈硬化症、高血圧(例えばAnning PB等 (March 2005) Elevated endothelial nitric oxide bioactivity and resistance to angiotensin-dependent hypertension in 12/15-lipoxygenase knockout mice. Am J Pathol. 166(3):653-62を参照)、ホジキンリンパ腫及び他の悪性腫瘍、又は炎症成分を有する任意の他の疾患の治療に有用であり得る化合物を同定するのに有用である。
【0046】
本発明のスクリーニング法は、またUS2003/0175680において検討されているように、例えば骨粗鬆症、骨関節炎、パジェット病又は歯周病に伴う骨量減少の治療又は予防に有用である化合物を同定するのに有用であると考えられる。よって、本発明のスクリーニング法は、患者における骨ミネラル密度の増加、並びに骨折の発生の低減及び/又は治癒において有用であり得る化合物を同定するのにも有用であり得る。
該化合物は上述の症状の治療的及び/又は予防的処置の双方において有用であり得る。
【0047】
エオキシンC4、D4又はE4の形成又は活性を減少させる化合物は、例えば免疫抑制性患者、例えば化学療法又は移植の拒絶を減少させる治療を受けている患者において炎症応答の促進が望ましい場合がある患者を治療するのに有用であると考えられる。例えばEoxC4、EoxD4又はEoxE4は、炎症応答の促進が望ましい患者を治療するのに有用であると考えられる。
【0048】
従って、本発明の更なる態様は、治療的有効量のEoxC4,EoxD4又はEoxE4を患者に投与することを含む、炎症反応の促進を必要とする患者の治療方法を提供する。本発明の更なる態様は、炎症反応の促進を必要とする患者の治療のための医薬の製造におけるEoxC4,EoxD4又はEoxE4の使用を提供する。
【0049】
本発明のスクリーニング法を実施する結果として、例えば実施例7に示されるように、典型的にはペプチド又は低分子でありうる化合物が同定される。該化合物は、薬学的使用、例えば患者、例えば動物(典型的には15-LOを発現する哺乳動物、例えばマウス、ラット、ウサギ、イヌ又はサル、好ましくはウサギ又は他の動物(例えばトランスジェニック動物))又はヒトにおけるインビボ治験での使用のために製剤化することができる。
【0050】
本発明の第三の態様は、抗炎症組成物又は骨量減少低減組成物を製造する方法であって、
(i)本発明の第二の態様の方法によって抗炎症又は骨量減少低減化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法を提供する。
本発明の第四の態様は、エオキシン形成調節組成物を製造する方法であって、
(i)本発明の第一の態様の方法によってエオキシン形成調節化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法を提供する。
【0051】
該組成物は典型的には有効量の化合物を患者に供給するものである。「有効量」という用語は、治療された患者に治療効果を付与する化合物の量を意味する。効果は客観的(すなわち、ある試験又はマーカーで測定可能)でありうるか、又は主観的(すなわち、患者が効果の顕れを示すか又は効果を感じる)でありうる。
該組成物は、経口的、静脈内、皮下的、口腔的、経直腸的、皮膚的、経鼻的、経気管的、経気管支的、舌下的、任意の他の非経口経路によって又は吸入を介して、 薬学的に許容可能な投薬形態で投与されることが意図され、又はそのようにして投与されるのに適したものでありうる。該組成物は、例えば経口投与のための錠剤、カプセル剤又はエリキシル剤、直腸投与のための坐薬、非経口又は筋肉内投与のための滅菌液又は懸濁液等々でありうる。該組成物は吸入用のスプレー又は注射用の液体であり得る。かかる組成物は標準的な及び/又は認められている薬学的実務に従って調製することができる。
【0052】
該組成物はまた(例えばNSAIDs、コキシブ類、副腎皮質ステロイド類、鎮痛剤、5-リポキシゲナーゼ阻害剤、FLAP阻害剤(5-リポキシゲナーゼ活性化タンパク質)、及びロイコトリエンレセプターアンタゴニスト(LTRas)、及び/又は炎症の治療に有用な他の治療剤から選択される)ここで定義された炎症の治療に有用な他の治療剤を含みうる。
あるいは、該組成物はまた骨量減少の低減又は骨形成の増加に有用な他の治療剤、例えばビスホスホネート、エストロゲン、選択的エストロゲンレセプター調節因子(SERM)、カルシトニン(calcinonin)又はタンパク質同化薬を含みうる。かかる治療剤の例は、例えばUS2003/0175680の、例えば段落[0065]で検討されている。
かかる組合せ製品は他の治療剤との関連で同定された化合物の投与をもたらし、よって、その製剤の少なくとも一つが同定されたものを含み、少なくとも一つが他の治療剤を含む別個の製剤として提供することができ、又は併用調製物(つまり同定された化合物と他の治療剤を含む単一製剤として提供)として提供(つまり製剤化)されうる。
【0053】
化合物は様々な用量で投与できる。経口、肺及び局所用量は、1日・kg体重当たり約0.01(mg/kg/日)から約100mg/kg/日、好ましくは約0.01から約10mg/kg/日、より好ましくは約0.1から約5.0mg/kg/日の範囲であり得る。例えば、経口投与では、組成物は典型的には約0.01mgから約500mg、好ましくは約1mgから約100mgの活性成分を含む。経静脈的には、最も好ましい用量は一定速度の注入の間、約0.001から約10mg/kg/時間の範囲である。有利には、化合物は単一の一日用量で投与することができ、又は一日の全用量を、毎日2回、3回又は4回の分割量で投与してもよい。
【0054】
とにかく、医師又は当業者であれば、個々の患者に最も適し、投与経路、治療されるべき症状のタイプと重症度、並びに処置される特定の患者の種、年齢、体重、性別、腎機能、肝機能及び応答によって変わり得る実際の用量を決定することができるであろう。上記の用量は平均的な場合の例である;より高い又は低い用量範囲が有利となる個々の場合ももちろん有り得、それも本発明の範囲内である。例えば実施例7はエオキシン類の形成を阻害可能な化合物が同定される方法を示している。
次に、本発明を次の非限定的な図面及び実施例を参照しながら更に説明する。
【図面の簡単な説明】
【0055】
【図1】アラキドン酸(40μM)と共に10分間インキュベーションした後のL1236細胞によって形成された236nmでのUV吸光度を持つ産物のRP-HPLCクロマトグラム。挿入:それぞれ15(S)-HETE及び12(S)-HETEで共溶出した物質のUVスペクトル。
【図2a】アラキドン酸(40μM)と共に10分間インキュベーションした後のL1236細胞によって形成された280nmでのUV吸光度を持つ産物のRP-HPLCクロマトグラム。1-5の数字は合成標準の保持時間に対応する:1)8(R),15(S)-LTB4;2)8(S),15(S)-DHETE;3)8(S),15(S)-LTB4,4)8(R),15(S)-DHETE及び5)14(R),15(S)-DHETE。
【図2b】ピークI-IV下で溶出する代謝物のUVスペクトル。
【図3】ピークIIの物質の[M-H]-イオンの400eV(Elab)CIDスペクトル。上方パネル:LTC4;中央パネル:ピークII;下方パネル:合成14,15-LTC4。
【図4】ピークIVの物質の[M-H]-イオンの400eV(Elab)CIDスペクトル。上方パネル:LTD4;中央パネル:ピークIV;下方パネル:合成14,15-LTD4。
【図5】14,15-LTA4(2μM)と共に30分インキュベートした後にL1236細胞によって生成された産物のRP-HPLCクロマトグラム。1、3及び5の数は、それぞれ8(R),15(S)-LTB4;8(s),15(S)-LTB4、及び14(R),15(S)-LTB4に対する合成標準の保持時間に対応する。ピークI-IVは図2のピークI-IVと同じ保持時間及びUVスペクトルを有していた。挿入:ピークV下で溶出する物質のUVスペクトル。
【図6】ピークVの物質の[M-H]-イオンの400eV(Elab)CIDスペクトル。上方パネル:LTE4;中央パネル:ピークV;下方パネル:合成14,15-LTE4。
【図7】15-LO経路及び14,15-ロイコトリエン類(エオキシン類)の生合成のための示唆された経路の概観。
【図8a】アラキドン酸と共になされたL1236細胞のインキュベーション。a)40μMのアラキドン酸と共に行ったインキュベーション後の14,15-LTC4及び14,15-LTD4の形成の時間経過。
【図8b】アラキドン酸と共になされたL1236細胞のインキュベーション。b)示されている様々な濃度のアラキドン酸と共に行った5分のインキュベーション後の14,15-LTC4及び14,15-LTD4の形成の用量応答曲線。
【図9a】アラキドン酸(20μM)と共に5分間なされたインキュベーション後にヒト好酸球細胞によって生成された産物のRP-HPLCクロマトグラム。a)形成された産物のHPLCクロマトグラム。エオキシンC4、LTC4、8,15-DHETEs(図2参照)及び14(R),15(S)-DHETEに対する合成標準との共溶出が示されている。
【図9b】アラキドン酸(20μM)と共に5分間なされたインキュベーション後にヒト好酸球細胞によって生成された産物のRP-HPLCクロマトグラム。b)合成エオシンC4と共溶出するピーク下の物質のUVスペクトル。
【図10】a)エオシンC4と共溶出する物質、b)合成エオシンC4及びc)合成LTC4のポジティブイオンMS/MSスペクトル。
【図11】好酸球を5、10、20及び40μMのアラキドン酸と共に、a)2分、b)5分及びc)10分インキュベートした。2回の別個の実験の平均。
【図12a】切り取ったヒト鼻ポリープを40μMのアラキドン酸と共に5分間インキュベートした後にエオキシンC4(14,15-LTC4)及び一連の8,15-DHETE及び14,15-DHETE代謝物を形成。
【図12b】合成エオキシンC4(14,15-LTC4)と共溶出するピークの物質のUVスペクトル。
【図13a】ヒト樹状細胞を40μMのアラキドン酸と共に5分間インキュベートした後にエオキシンC4(14,15-LTC4)及び一連の8,15-DHETE及び14,15-DHETE代謝物を形成。
【図13b】合成エオキシンC4(14,15-LTC4)と共溶出するピークの物質のUVスペクトル。
【図14】免疫染色。ホジキンリンパ腫の患者からのリンパ節における15-LOの発現。ヒト15-LOの特異的ペプチド配列に対して産生されたポリクローナル15-リピキシゲナーゼ抗血清と続いて二次ペルオキシダーゼコンジュゲート抗体を用いた。リード・シュテルンベルク細胞における強い染色が観察された(図の中央部分の多核細胞)。
【図15】免疫染色。サルコイドーシスの患者からの肺バイオプシーにおける15-LOの発現。ヒト15-LOの特異的ペプチド配列に対して産生されたポリクローナル15-リピキシゲナーゼ抗血清と続いて二次ペルオキシダーゼコンジュゲート抗体を用いた。類上皮細胞(肉芽腫)における強い染色が観察された。
【図16】免疫染色。関節リウマチ患者のA)滑膜細胞及びB)軟骨細胞、C)アルツハイマー患者の脳皮質からの錐体細胞、及びD)星状細胞腫検体からの星状膠細胞の検体における15-LOの発現。
【図17a】生成されたGSTM1-1によるA)エオキシンA4(14,15-LTA4)からエオキシンC4(14,15-LTC4)への代謝。
【図17b】生成されたGSTM1-1によるB)LTA4からLTC4への代謝。
【図18】様々な可溶型グルタチオンS-トランスフェラーゼによるエオキシンA4(14,15-LTA4)からエオキシンC4(14,15-LTC4)への代謝。
【図19】好中球の細胞成分分画をEoxA4(40μM)と共に5分間インキュベートした後に形成された生成物のHPLCクロマトグラム。A)上清及びB)膜分画。
【図20】EoxA4(10μM)(上方パネル)LTA4(10mM)(中央パネル)又は両方(下方パネル)と共に37℃で5分間インキュベートした後のヒト好酸球に形成された生成物のHPLCクロマトグラム。
【図21】エオキシンC4及びLTC4の形成に対する5-リポキシゲナーゼ阻害剤(BW70C)及びLTC4シンターゼ阻害剤(MK886)の効果。37℃で5分間のA23187(1μM)及びアラキドン酸(10μM)での刺激の前に阻害剤の不在下又は存在下で2分間プレインキュベートした。
【図22】コンフルエントなウシ内皮細胞(最大効果70%)を持つモデル系における減少した抵抗として測定した血漿漏出に対するエオキシンC4(14,15-LTC4)、エオキシンD4(14,15-LTD4)、エオキシンE4(14,15-LTE4)及びLTC4の効果。
【図23a】GM-CSFで刺激された好中球によるエオキシン誘導酸化的代謝。測定されたパラメータを示す動的化学発光曲線。a.酸素フリーラジカルを生産するように刺激された(塗り潰し記号)又はGM-CSF処理された後に刺激された(白抜き記号)精製血液好中球。「BLX-26」はBLX-926を示すために使用する。
【図23b】GM-CSFで刺激された好中球によるエオキシン誘導酸化的代謝。測定されたパラメータを示す動的化学発光曲線。b:EoxD4(BLX-926)はGM-CSF刺激好中球によって酸素フリーラジカルを誘導した。結果は全ラジカル生成を示し、コントロール(刺激細胞)の%として、また平均+/−標準偏差、n=13として表している。
【図23c】GM-CSFで刺激された好中球によるエオキシン誘導酸化的代謝。測定されたパラメータを示す動的化学発光曲線。c:散乱したプロットは、EoxD4(BLX-926)がGM-CSF刺激好中球により酸素フリーラジカルを誘導したことを示している。結果は1分当たりのカウント(CPM×106),n=13として測定された全ラジカル生成を示す。線は基準線(k=1)を示し、その線の上の観測は全て刺激に対するレスポンダーであることを示す。
【図24a】EoxE4(BLX-927)。測定されたパラメータを示す動的化学発光曲線。a.酸素フリーラジカルを生産するように刺激された(塗り潰し記号)又はIL-5処理された後に刺激された(白抜き記号)精製血液好酸球。「BLX-27」はBLX-927を示すために使用する。
【図24b】EoxE4(BLX-927)。測定されたパラメータを示す動的化学発光曲線。b:刺激及びEoxD4(BLX-927)はIL-5刺激好酸球によって酸素フリーラジカルを誘導した。結果は全ラジカル生成を示し、コントロール(刺激細胞)の%として、また平均+/−標準偏差、n=6として表している。
【図24c】EoxE4(BLX-927)。測定されたパラメータを示す動的化学発光曲線。c:散乱したプロットは、EoxD4(BLX-927)がIL-5刺激好酸球により酸素フリーラジカルを誘導したことを示している。結果は1分当たりのカウント(CPM×106),n=6として測定された全ラジカル生成を示す。線は基準線(k=1)を示し、その線の上の観測は全て刺激に対するレスポンダーであることを示す。
【図25a】実施例7及び図27及び28で試験した化合物の構造。
【図25b】実施例7及び図27及び28で試験した化合物の構造。
【図26】エオキシンC4(14,15-LTC4)及びエオキシンD4(14,15-LTD4)の形成に対するロイコトリエン阻害剤ETYAの効果。
【図27a】エオキシンC4の形成に対する試験化合物の効果。
【図27b】エオキシンC4の形成に対する試験化合物の効果。
【図28a】エオキシンC4及び15-HETEの形成に対する試験化合物の効果の比較。a)化合物A、B、C、D及びE。
【図28b】エオキシンC4及び15-HETEの形成に対する試験化合物の効果の比較。b)化合物D、F及びG。
【図29】単離されたヒト好酸球における新規な15-リポキシゲナーゼ産物の形成。単離された好酸球(10×106細胞)をリン酸緩衝生理食塩水(1ml)中に再懸濁させ、2分間プレインキュベートした後、37℃で5分間アラキドン酸(10μM)で刺激した。3容量のメタノールを加えて反応を終結させた。蒸発後、試料を移動相に再懸濁させ、移動相としてAcN:MeOH:H20:HAc(29:19:72:0.8体積)pH5.6を使用するRP-HPLC-UV系中での分析を行った。
【図30a】新規な15-リポキシゲナーゼ産物の液体クロマトグラフィー-質量分析。パネルA:合成14,15-LTC4及びLTC4標準と比較した好酸球により形成された代謝物IのLC-MS/MSスペクトル。上方パネルは、37℃で5分間のアラキドン酸(10μM)付与後に好酸球に形成された代謝物の626.3[M+H]+の生成物イオンスキャンのスペクトルを示す。中央パネルは合成14,15-LTC4標準のスペクトルであり、下方パネルは合成LTC4標準のスペクトルである。
【図30b】新規な15-リポキシゲナーゼ産物の液体クロマトグラフィー-質量分析。パネルB:14,15-LTD4及びLTD4合成標準と比較した好酸球により形成された代謝物IIのLC-MS/MSスペクトル。上方パネルは、37℃で60分間好酸球と共にインキュベートした14,15-LTC4(10μM)から形成された代謝物の497.2[M+H]+の生成物イオンスキャンのスペクトルを示す。中央パネルは合成14,15-LTC4標準のスペクトルであり、下方パネルは合成LTC4標準のスペクトルである。
【図30c】新規な15-リポキシゲナーゼ産物の液体クロマトグラフィー-質量分析。パネルC:14,15-LTE4及びLTE4合成標準と比較した好酸球により形成された代謝物IIIのLC-MS/MSスペクトル。上方パネルは、37℃で60分間の好酸球と共にインキュベートした14,15-LTC4(10μM)から形成された代謝物の440.2[M+H]+の生成物イオンスキャンのスペクトルを示す。中央パネルは合成14,15-LTC4標準のスペクトルであり、下方パネルは合成LTC4標準のスペクトルである。
【図31】衝突は14,15-ロイコトリエン類をシステイニルロイコトリエンから区別する解離断片を誘導した。陽イオンモードのMS/MS生成物イオンスペクトルにおけるシステイニルロイコトリエン類と14,15-ロイコトリエン類の顕著な差はm/z189及び205である。これらの断片イオンの推定起点を示す。
【図32a】14,15-ロイコトリエン類によって誘導された血管透過性の変化。パネルA:14,15-LTC4、14,15-LTD4及び14,15-LTE4、LTC4(10-8M)及びヒスタミン(10-5M)に応答した内皮細胞障壁機能における急速な変化の解析。分析は内皮細胞単層の各側に電極を配した経内皮電気抵抗(TEER)の連続的記録によって達成した。このモデルの最大効果は-70%の抵抗変化である。5回のうち2回の典型的な実験が実証される。各値は3通りの測定の平均値である。
【図32b】14,15-ロイコトリエン類によって誘導された血管透過性の変化。パネルB:LTD4(10-8M)と比較した、14,15-LTD4(それぞれ10-8M及び10-7M)によって引き起こされる内皮細胞障壁機能の変化の時間経過。
【図33】14,15-ロイコトリエン類(エオキシン類)の形成に対して提案する代謝経路。
【実施例】
【0056】
実施例1:14,15-LTC4、14,15-LTD4、及び14,15-LTE4(つまりエオキシンC4、エオキシンD4及びエオキシンE4)の生合成の証明。
材料と方法
合成14,15-オキシド-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTA4)、14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,10(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTC4)、14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTD4)、14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTE4)、8(R),15(S)-ジヒドロキシ-5,9,11,13(Z,E,E,E)-エイコサテトラエン酸(8(R),15(S)-LTB4)、8(S),15(S)-ジヒドロキシ-5,9,11,13(Z,E,Z,E)-エイコサテトラエン酸(8(S),15(S)-DHETE)、8(S),15(S)-ジヒドロキシ-5,9,11,13(Z,E,E,E)-エイコサテトラエン酸(8(S),15(S)-LTB4)、8(R),15(S)-ジヒドロキシ-5,9,11,13(Z,E,Z,E)-エイコサテトラエン酸(8(R),15(S)-DHETE),14(R),15(S)-ジヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14(R),15(S)-DHETE又は14(R),15(S)-LTB4)、15(S)-ヒドロキシ-5,8,11,13(Z,Z,Z,E)-エイコサテトラエン酸(15-HETE)、12(S)-ヒドロキシ-5,8,10,14エイコサテトラエン酸(Z,Z,E,Z)(12(S)-HETE)及び13(S)-ヒドロキシオクタデカ-9,11(Z,E)-ジエン酸(13(S)-HODE)はBiomole社(Plymouth Meeting, USA)から入手した。細胞培養培地RPMI 1640、仔ウシ血清、ペニシリン、ストレプロマイシン及びグルタミンはGibco,BRLから購入した。HPLC溶媒はRathburnケミカルズ社 (Walkerburn, United Kingdom)から得た。
【0057】
L1236細胞株
細胞株L1236は混合細胞性サブタイプのホジキンリンパ腫(HL)に罹患した患者の末梢血から元々は生産した15。細胞は37℃で5%のCO2の雰囲気下で懸濁させて増殖させた。培養培地は、10%の仔ウシ血清及び2mMのグルタミンを補填した100U/mlのペニシリンと100mg/mlのストレプトマイシンを含むRPMI1640であった。
【0058】
実験条件及びHPLC分析
細胞懸濁液を200×gで5分間、遠心分離し、ペレットをPBS中に懸濁させた。PBSで2回洗浄した後、細胞を10-20×106細胞/mlの濃度でPBS中に溶解させた。試料を37℃で2分間前もって温めた後、アラキドン酸、14,15-LTA4又は他の示された代謝物を添加し、続いて示された期間、インキュベートした。インキュベーションは3容量の氷冷メタノールの添加により終結させた。沈殿したタンパク質と細胞断片を遠心分離によって除去し、過剰の水とエタノールを減圧下で蒸発させた。残留物をメタノールに溶解させ、m試験管に移し、窒素流下で乾固させた。続いて、残留物を以下に記載するクロマトグラフィー溶剤で再構成させた。試料をオクタデシル逆相カラム(Chromabond, C18, 100mm, Macherey-Nagal, Duren, Germany)に充填し、0.4ml/分の流量で溶出させた。移動相は、システイニル含有アラキドン酸代謝物及びジヒドロキシ酸又はモノヒドロキシ酸それぞれの分析に対して、アセトニトリル/メタノール/水/酢酸(体積で28:18:72:0.8、pHは30%NaOHで5.6に調整)又はメタノール/水/酢酸(体積で72.5:27.5:0.01)であった。溶出した代謝物を、プログラム可能なWaters991ダイオードアレイ分光光度計を使用して検出し定量した。
【0059】
陰イオンエレクトロスプレータンデム質量分析
スペクトルを、記載されたように16、衝突ガスとしてXeを用いて400eVの実験室フレーム衝突エネルギーでハイブリッド磁気セクター/飛行時間型タンデム機器に記録した。
【0060】
結果
アラキドン酸と共にL1236細胞を10分間インキュベートすると、主要産物が得られ、これを合成15(S)-HETEと共に同時にクロマトグラフィーにかけた(図1)。また、合成12(S)-HETEと共溶出した236nmの最大UV吸光度のマイナーなピークを観察した。これらのピークの物質のUVスペクトルはそれぞれ15-HETE及び12-HETEに対して報告されたスペクトルと一致している。15-HETE対12HETEの比は約9:1であり、これはこれらの産物の15-LOの1型に触媒された形成と一致している5。これらの結果は、細胞が15-LOの1型の活性を有しているが5-リポキシゲナーゼ活性を有していないことを証明している。リノール酸は優れた基質であり、L1236細胞はリノール酸を13-HODEに効率的に代謝したが(データは示さず)、これはこれらの細胞における15-LOの1型の発現に一致している。超音波処理細胞を抗15-LOウサギポリクローナル抗体を用いたウェスタンブロットが続くSDS/PAGEによって分析すると、網状赤血球15-LOと共遊走した免疫反応性バンドが現れた(データは示さず)。
【0061】
236nmのUVを吸収するピークに加えて、約270nmの最大UV吸光度と5-7分の保持時間を持つ多数のピークがクロマトグラムに観察された。アセトニトリルベースの移動相を使用してこれら産物を分離した。図2aはアラキドン酸と共に10分のインキュベーションを行った後に形成された生成物の典型的なRP-HPLCクロマトグラムを示している。コンジュゲートトリエンスペクトルと272nmに最大UV吸光度を含む5つのピーク(1-5)が観察された。これらのピークはそれぞれ8(R),15(S)-LTB4;8(S),15(S)-DHETE;8(S),15(S)-LTB4;8(R),15(S)-DHETE及び14(R),15(S)-DHETEの合成標準に対応する溶出時間を有していた。これら生成物のパターンはアラキドン酸と共にインキュベートしたヒト気道上皮について報告されたものと殆ど同一であった17。しかしながら、クロマトグラム中の2つの更なる主要なピークはまたコンジュゲートトリエンスペクトルを有していたが、282nm(II及びIV)に最大UV吸光度を有していた(図2b)。溶出物質を集め、陰イオンエレクトロスプレータンデム質量分析法によって分析した。図3はピークII(中央パネル)からの物質の[M-H]イオンの衝突解離(CID)スペクトルを示す。質量スペクトルは、LTC4に見出されなかったイオンm/z306の存在を除いて、624に分子イオンを持ち、LTC4(上方パネル)と殆ど同一であった。これらの細胞における観察された15-LO活性に鑑みると、質量スペクトルは、代謝物が14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12-エイコサテトラエン酸(14,15-LTC4)であることを示している。生成物が確かに14,15-LTC4であったことを確認するために、合成14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸を調製した。この合成代謝物の質量スペクトル(図3、下方パネル)はピークIIの物質のスペクトルと同一であった。他の主要な代謝物ピークIVの[M-H]-イオンの対応するCIDスペクトルを図4に示す。この質量スペクトルは、LTD4に見出されなかったイオン250及び251の存在を除いて、同じ分子イオンm/z495を持つLTD4のスペクトルと非常に類似しており、代謝物が14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12-エイコサテトラエン酸(14,15-LTD4)であることを示している。合成14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸の[M-H]-イオンの対応するCIDスペクトルは代謝物IVのスペクトルとまた同一であった(図4、下方パネル)。更に、合成14,15-LTC4及び14,15-LTD4はRP-HPLCでそれぞれ代謝物II及びIVと正確に共溶出した。合成14,15-LTC4と共に細胞をインキュベートすると14,15-LTD4が形成された(データは示さず)。また、コンジュゲートトリエンスペクトルと278nmに最大UV吸光度を持つ二つの他のマイナーなピークが観察された(図2a及びb、ピークI及びIII)。ピークIIIの物質を陰イオンエレクトロスプレータンデム質量分析法によって分析した。この代謝物のCIDスペクトルは14,15-LTD4のCIDスペクトルと同一であった(データは示さず)。この代謝物の正確な構造は確認されなかった。しかしながら、この代謝物のCIDスペクトル及びUVプロファイルは、スペクトルが14,15-LTD4のスペクトルと比較して4nm低色素的にシフトしているので、オールトランストリエン構造と一致している。従って、ピークIIIの物質はおそらくは8-trans-14,15-LTD4である。ピークIの物質は質量スペクトルを得るには不十分であったが、8-trans-14,15-LTD4の形成に沿って、該代謝物は8-trans-14,15-LTC4である可能性が非常に高い。更に、細胞を合成14,15-LTC4又は14,15-LTD4と共にインキュベートするとそれぞれ代謝物I及びIIIが形成され、これは、親14,15-LTC4及び14,15-LTD4から想定されたオールトランストリエンが形成されたことを裏付けている(データは示さず)。
【0062】
14,15-LTC4及び14,15-LTD4の形成機構及び二重結合構造を解明するために、細胞を2μMの合成14,15-オキシド-5,8,10,12(Z,Z,EE)-エイコサテトラエン酸(14,15-LTA4)と共に30分インキュベートした。図5は細胞によって産生された生成物のHPLCクロマトグラムを示している。272nmに最大UV吸光度とコンジュゲートトリエンスペクトルを有する3つのピークが観察された(ピーク1、3及び5)。これらの生成物はまた細胞をアラキドン酸と共にインキュベートしたときにも形成され(図2a)、生成物をそれぞれ合成8(R),15(S)-LTB4;8(S),15(S)-LTB4及び14(R),15(S)-LTB4と同時にクロマトグラフィーにかけた。コンジュゲートトリエンスペクトルと282nmに最大UV吸光度を持つ3つのピークが観察された(ピークII、IV及びV)。代謝物II及びIVは、HPLCでそれぞれ合成14,15-LTC4及び14,15-LTD4と同一の保持時間を有しており、これらの代謝物が14,15-LTC4及び14,15-LTD4であったことを裏付けている。ピークVの物質はコンジュゲートトリエンを含み、282nmに最大吸光度を有していた。この代謝物は、細胞をアラキドン酸と共にインキュベートしたときは(図2a)、二重二原子酸素添加産物8,15-DHETEとのHPLCでの共溶出のために、検出されなかった。この後者の生成物は14,15-LTA4からは形成されないので、代謝物VはHPLCクロマトグラムで可視でき、二重二原子酸素添加産物の後ろに隠れてはいない。細胞を合成14,15-LTD4と共にインキュベートしたとくに代謝物Vがまた形成された(データは示さず)。合成14,15-LTD4と共に細胞をインキュベートした後に形成されたピークVの物質を陰イオンエレクトロスプレータンデム質量分析法によって分析した。図6は、ピークVの物質の質量スペクトルが合成14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12-エイコサテトラエン酸(14,15-LTE4)のスペクトルと同一であったことを示している。代謝物VはまたHPLCで合成14,15-LTE4と同じ保持時間を有していた。また、より少量の8-trans-14,15-LTC4(ピークI)及び8-trans-14,15-LTD4(ピークIII)が14,15-LTA4と共にインキュベートした後に細胞によって生産された。
【0063】
15-LO経路及び14,15-LTC4、14,15-LTD4及び14,15-LTE4の生合成のための示唆された経路の概観を図7に示す。
アラキドン酸(40μM)と共になされたインキュベーション後の14,15-LTC4及び14,15-LTD4の形成の時間経過を図8aに示す。実質的な量の14,15-LTC4、37pmol/106細胞が、アラキドン酸と共に30秒インキュベートした後に既に10×106細胞によって生産された。14,15-LTC4の最大レベルが5分のインキュベーション後に観察され、その後14,15-LTC4のレベルは時間と共に減少した。測定可能な量の14,15-LTD4がアラキドン酸と共にインキュベートした2分後に観察され、レベルは時間と共に増加し、30分後に最大レベルに達した。8,15-DHETEはこのHPLC系において14,15-LTE4と同時にクロマトグラフィーにかけたので、この実験において14,15-LTE4のレベルを測定することができなかった。アラキドン酸からの14,15-ロイコトリエン類の形成についての用量応答曲線は、有意な量の14,15-LTC4及び14,15-LTD4が5μMのアラキドン酸濃度で既に形成されたことを示している(5分のインキュベーション時間;図8b)。これらの代謝物のレベルはアラキドン酸濃度と共に増加し、40μMのアラキドン酸においてプラトーに達した。
14,15-LTC4;14,15-LTD4及び14,15-LTE4はそれぞれエオキシンC4、エオキシンD4及びエオキシンE4と命名した。
【0064】
実施例2:他の細胞及び組織におけるエオキシンC4(14,15-LTC4)の生合成の証明
材料と方法
ヒト好酸球の単離
静脈血(100ml)を、健常な志願者又は好酸球増加症候群(HES)を含む好酸球増加症、慢性リンパ性白血病(CLL)及びある種の炎症疾患の患者からから採血した。400×Gで15分間、最初の遠心分離を実施した。上相を捨て下相を集め、室温で30分間、Dextran(商標)沈降を施した。沈降後、白血球細胞を含む上相を400×Gで10分間、一回遠心分離した。ペレットを溶解バッファーに懸濁させ、室温で30分インキュベートして混入した赤血球を除去した。インキュベーション後、Lymphoprep(商標)を用いて密度勾配遠心分離を実施した。好中球と好酸球を含む多形核(PMN)画分を集め、磁気細胞分離(MACS)を施した。好酸球は、磁気ミクロビーズに結合させたCD16抗体を使用するネガティブ選択によって単離した。純度は、形態学的検査とメイ・グリューネバルト-ギムザ溶液での染色によって確認した。純度は常に95%を越えていることが見出された。
【0065】
細胞のインキュベーション
単離された好酸球(5−10×106/試料)をリン酸緩衝生理食塩水(1ml/試料)に再懸濁させ、37℃でイオノフォアA23187(1μM)、アラキドン酸(5、10、20又は40μM)又は双方で2、5又は10分、刺激する前に37℃で2分間、プレインキュベートした。反応を3容量のメタノールの添加により終結させた。
【0066】
15-リポキシゲナーゼ生成物の分析
細胞片及びタンパク質を除去するために、遠心分離を実施した。上清を減圧下で蒸発させ、残留物をメタノールに溶解させ、試験管に移した。ついで、試料を窒素流下で蒸発乾固させ、最後に200μlの移動相(アセトニトリル:メタノール:H2O:酢酸、29:19:72:0.8、pHは5.6にNH3で調整)に再懸濁させた。抽出された試料に、記載されたようにして逆相HPLC系での分析を施した。簡単に述べると、試料をオクタデシル逆相カラム(Chromabond C18,Macherey-Nagel,Duren,Germany)に充填し、溶出流量は1.2ml/分であった。溶出した化合物を、Waters1996HPLCシステムに連結したWaters996光ダイオードアレイ分光光度計を利用して検出し定量した。
【0067】
質量分析
質量分析を実施例1に記載されるようにして実施した。
結果
ヒト好酸球中でのエオキシンC4の生合成
HPLCによって分析した場合、コンジュゲートトリエンスペクトルを持つ一つのピーク(最大吸光度282nm)がアラキドン酸と共に行ったインキュベーション後に観察された(図9a及びb)。生成物を合成14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(EoxC4)と共にクロマトグラフィーにかけた。得られた生成物の同一性を確保するために、タンデム質量分析を実施した。代謝物のMS/MSスペクトルはEoxC4のものと本質的に同一であった(図10)。ホジキン細胞株L1236に対して示された生成物プロファイルとは異なり、好酸球は直ぐにはEoxC4をEoxD4及びEoxE4に転換しなかった。
【0068】
15-リポキシゲナーゼの発現に加えて、好酸球はまた5-リポキシゲナーゼを発現し、これがロイコトリエン類を生成する。用量応答実験を、EoxC4生成の実験条件を最適化するために実施した。またEoxC4及びLTC4の形成の時間経過を調べた(図11)。しかしながら、エオキシンD4もエオキシンE4も5分のインキュベーション後に検出されなかった。
総括すると、該結果は、好酸球が外因性アラキドン酸をEoxC4に直ぐに転換させることを証明している。
【0069】
ヒト鼻ポリープ組織におけるエオキシンC4(14,15-LTC4)の生合成
気道上皮源は鼻ポリープの外科検体である。切り取ったヒト鼻ポリープをアラキドン酸(40μM)と共に5分間インキュベートすると、14,15-LTC4及び一連の8,15-DHETE及び14,15-DHETE代謝物が形成された(図12a)。合成14,15-LTC4と共に共溶出した物質のUVスペクトルは特徴的なトリエンスペクトルを示した(図12b)。
【0070】
ヒト樹状細胞におけるエオキシンC4(14,15-LTC4)の生合成
成長因子とインターロイキン-4の存在下でのバフィーコート細胞(白血球)の分離と培養によって樹状細胞を得た。単離細胞は樹状細胞の特徴的な機能と形態を示した。これらの細胞をアラキドン酸(40μM)と共に5分間インキュベートすると、主として8,15-DHETE及び14,15-DHETEの形成に至ったが、有意な量の14,15-LTC4も形成された(図13a)。14,15-LTC4の同定はまたUV分光法に基づいた(図13b)。
【0071】
実施例3:15-LOを発現する細胞の同定
気道呼吸器上皮、好酸球、肥満細胞、及びマクロファージにおける先に記載された15-LO発現に加えて、本発明者等は、(ヒト15-LOに対して産生された抗体を使用して)数種の正常なまた病理組織における15-LOの発現を証明した。15-LOはホジキンリンパ腫(HL)の患者からのリンパ節において発現することが見出された。HLの13/15例において、15-LOの発現がリード・シュテルンベルク細胞において検出された(図14)。非ホジキンリンパ腫の何れの例でも染色はなかった。サルコイドーシスの患者からのバイオプシーは、類上皮細胞(肉芽腫)における15-LOの強い染色を裏付けた。結果は、全ての類上皮細胞が15-LOを発現することを示唆している。類上皮細胞はクローン病、TBC及び数タイプの血管炎の患者の肉芽腫において見出される。15-LOを発現することが見出された他の細胞型は、関節リウマチ患者からの滑膜細胞(図16a)及び軟骨細胞(図16b)、アルツハイマー患者の脳皮質からの錐体細胞(図16c)、及び星状細胞腫検体からの星状膠細胞(図16d)(正常組織ではない)を含んでいた。
総括すると、該結果は、15-LOと、この経路を通して形成された代謝物がこれらの疾患の病態生理に関与していることを示している。
【0072】
実施例4:L1236細胞においてエオキシンA4(14,15-LTA4)をエオキシンC4(14,15-LTC4)に転換する酵素の同定
方法
細胞培養
細胞培養は実施例1に記載されているようにして実施した。
ヒト好酸球の単離
ヒト好酸球の単離は実施例2に記載されているようにして実施した。
RT-PCR分析
好酸球からのmRNAの単離と続くRT-PCR分析を標準的な方法によって実施した。PCR反応に用いたプライマーは、ミクロソームグルタチオントランスフェラーゼI(MGSTI)、ミクロソームグルタチオントランスフェラーゼII(MGSTII)、ミクロソームグルタチオントランスフェラーゼIII(MGSTIII)及びβ-アクチン(陽性コントロール)
相補的であった。
【0073】
細胞のインキュベーション
単離された好酸球(5-10×106/試料)をリン酸緩衝生理食塩水(1ml/試料)に再懸濁させ、37℃で5分間、EoxA4(10μM)、LTA4(10μM)又は双方で刺激する前に37℃で2分間、プレインキュベートした。あるいは、阻害剤の研究では、好酸球を、イオノフォアA23187(1μM)及びアラキドン酸(40μM)で37℃で5分間刺激する前にBW70C(1μM)又はMK886(10μM)の存在下で37℃で2分間、プレインキュベートした。反応はメタノール3容量の添加によって終結させた。代謝物は、Waters1996HPLCシステムに連結したWaters996光ダイオードアレイ分光光度計を利用して検出し定量した。
【0074】
酵素研究
EoxA4からEoxC4への転換におけるグルタチオントランスフェラーゼの可能な関連を探究するために、組換え酵素(1μg)をEoxA4(100μM)及びグルタチオン(5mM)と共に37℃で5分間インキュベートした。反応を1容量の停止液(AcN:MeOH:HAc,体積50:50:1)の添加により終結させ、EoxC4を、Waters1996HPLCシステムに連結したWaters996光ダイオードアレイ分光光度計を利用して検出し定量した。
【0075】
結果
ホジキン細胞株L1236においてEoxA4をEoxC4に転換する酵素の単離、精製及び特徴付け
L1236細胞を標準的な手順によって細胞成分分画し、酵素活性を含む細胞質画分をグルタチオンセファロースカラムに充填した。結合物質をグルタチオンバッファー勾配によって溶出させた。溶出された画分を酵素活性について試験し、SDS-PAGEゲルで実施した。28kDのおよそのMwを持つ一つの単一バンドを得た。アミノ酸解析(PAC)、cDNA配列解析及びBLAST検索により、得られたバンドの同定は可溶型グルタチオントランスフェラーゼM1b-1b(GSTM1b-1b)であった。組換えGSTM1b-1bはEoxA4をEoxC4に転換したが、同一条件下ではLTA4をLTC4に代謝しなかった(図17A及びB)。
【0076】
ヒト可溶型グルタチオントランスフェラーゼファミリーのスクリーニング
GSTM1b-1bはヒト可溶型グルタチオントランスフェラーゼファミリーのメンバーであるので19、EoxA4をEoxC4へ転換する可溶型グルタチオントランスフェラーゼの能力を調べた。GSTM1b-1bに加えて、GSTP1-1の二つのアイソフォーム(Val105及びIle105)がEoxA4をEoxC4に転換する大きな能力をまた有していたことが証明された(図18)。
【0077】
好酸球におけるEoxC4シンターゼ活性の局在化
可溶型グルタチオントランスフェラーゼが好酸球におけるEoxC4形成に関与するかどうかを調べるために、単離されたヒト好酸球の細胞成分分画を調製し、EoxA4と共にインキュベートした。L1236とは異なり、EoxC4シンターゼ活性は好中球の膜画分に殆ど完全に局在化した(図19)。
【0078】
好酸球における「酵素X」の特徴付け
好酸球中のEoxC4シンターゼ活性は膜画分にあったので、本発明者等はこの点における膜結合性グルタチオントランスフェラーゼの関与を調べた。
MAPEG(エイコサノイド及びグルタチオン代謝における膜結合型タンパク質(Membrane Associated Proteins in Eicosanoid and Glutathione metabolism))スーパーファミリー21のメンバーはLTC4シンターゼ、FLAP(5-リポキシゲナーゼ活性化タンパク質)、PGEシンターゼ、MGSTI(ミクロソームグルタチオントランスフェラーゼI)、MGSTII及びMGSTIII(3)である。PCR解析は、好酸球はMGSTII及びMGSTIIIをコードするmRNAを有する一方、MGSTIをコードするmRNAは観察されなかったことを明らかにした。先の研究から、好酸球がFLAPとLTC4シンターゼを有していることは知られている。
【0079】
EoxA4及びグルタチオンと共にMAPEGファミリーメンバーをインキュベートすることにより、LTC4シンターゼだけがEoxA4をEoxC4に転換する能力を有していることが示された(データは示さず)。
LTC4シンターゼは伝統的にLTA4からグルタチオン含有ロイコトリエン(LT)C4を生合成する際の重要な酵素であることが知られている。LTA4は5-リポキシゲナーゼ経路を介して合成される。好酸球は5-リポキシゲナーゼ及び15-リポキシゲナーゼ活性を共に有しており、LTC4シンターゼがEoxA4をEoxC4へ転換することができる唯一のMAPEGファミリーメンバーであるという結果は、LTC4シンターゼが二重の活性:LTC4及びEoxC4合成を備えていることを示唆している。LTC4シンターゼがEoxC4形成においてもまた重要な酵素であるという仮説を試験するために、単離した好酸球をEoxA4(10μM)、LTA4(10μM)、又は双方と共に37℃で5分間インキュベートした(図20)。
【0080】
好酸球をEoxA4と共にインキュベーションするとEoxC4が生合成される一方、好酸球をLTA4と共にインキュベーションするとLTC4を生じる。細胞を双方の基質と共にインキュベートすると、LTC4とEoxC4がおよそ9:1の比で生産された。これはこれらの基質に対するLTC4シンターゼの過去に報告された親和性と一致している20。
LTC4シンターゼがまた好酸球においてEoxC4シンターゼとしてまた作用するという知見を更に裏付けるために、発明者等は阻害研究を実施した。
【0081】
BW70Cは5-リポキシゲナーゼ阻害剤で、MK886はFLAP阻害剤であり、これらは高い(10μM)濃度で使用される場合、LTC4シンターゼをまたブロックする(図21)。BW70Cの存在下でのプレインキュベーションがLTC4ピークを完全に消滅させ、MK886の存在下でのプレインキュベーションがまたEoxC4合成を阻止した。これらの結果は、LTC4シンターゼが好酸球中でのEoxC4形成における重要な酵素であることを更に裏付けている。
【0082】
実施例5:ヒト15-リポキシゲナーゼ(15-LO)によって形成される脂質ヒドロペルオキシドの検出のためのアッセイ
ここに記載されたアッセイは、ジフェニル-1-ピレニルホスホン(DPPP)をホスフィンオキシドに酸化する脂質ヒドロペルオキシドの能力を利用する(以下のスキームを参照のこと)。ジフェニル-1-ピレニルホスホンは酸化されるまでは非蛍光化合物である。DPPPの酸化は蛍光化合物を生じ、これは温度に感受性がなく、少なくとも60分間安定である。
【0083】
【0084】
材料
ジフェニル-1-ピレニルホスホン(DPPP)(D-7894)はMolecular Probesから購入している。コール酸はSigmaから、リノール酸はBiomolから、PBSはGibco Life Technologiesから購入している。蛍光はMolecular Devices製の二重走査型マイクロプレート分光蛍光光度計Spectramax Geminiを用いて測定する。Spectramax Gemini分光蛍光光度計はキセノンランプと二つのホログラフィック回折格子モノクロメータを使用する。384ウェルまでのマイクロプレートを分光蛍光光度計と共に使用することができる。蛍光の検出は、ウェル中の試料の深さ全体に亘ってウェルの上面から達成される。
【0085】
全ての試薬は、フルオロフォア(DPPP)及び基質、リノール酸を除いてそのまま使用できる濃度で提供される。DPPPは粉末として提供され、-20℃で保存されなければならず、光から十分に保護されなければならない。リノール酸はエタノール中の100mM原液として供給される。酵素調製物は凍結アリコートとして提供される。
【0086】
酵素はバキュロウイルスを使用して発現され、スポドプテラ・フルギペルダ細胞(Sf9)から部分的に精製され、トリス-HCl(pH8.0)、75mMのNaCl及び5%グリセロールで送達される。精製された酵素は-70℃で少なくとも5ヶ月まで安定である。3回までの凍結解凍サイクルは酵素活性に影響を与えないと思われる。しかしながら、酵素の凍結解凍サイクルは、必ずしも必要ではないなら、避けなければならない。酵素を凍結し実験の直前でPBSに希釈する。希釈した酵素は同じ日中に使用されなければならない。
【0087】
DPPP(粉末)を、0.25mMの最終濃度までエタノール及び2-ブタノール(1:1の比)に溶解させた。この溶液をボルテックスして混合し、続いて+4℃で超音波水浴で10秒間超音波処理してDPPPを完全に溶解させた。この溶液は実験が実施される日には新鮮でなければならない。該溶液は遮光されなければならない。
【0088】
脂質ヒドロペルオキシド検出のためのジフェニル-1-ピレニルホスフィン法
アッセイは室温(20-22℃)で96ウェルの全て黒のプレートで実施される。各ウェルは、500μMのコール酸と15-LO酵素(プレミックス)を補填したダルベッコのリン酸緩衝生理食塩水(PBS)からなる45μlを含む。阻害剤(1μl)又はビヒクル(DMSO)を、基質である5μlの1mMリノール酸の添加の5-15分前に添加する。ついで、プレートを室温で20分間インキュベートする。酵素反応を50μlのメタノールの添加によって終結させる。プレートは、脂質ヒドロペルオキシドの有意な損失なしに60分までの間、室温で保存することができる。50μlの25μM DPPP溶液を加え、プレートを室温でインキュベートする。30-60分後、蛍光は、355nmの励起波長と380nmの発光波長を使用して読み取ることができる。
水溶液中でのDPPPの低い自動酸化のため、ブランクの一つは酵素反応のものと同じ比でバッファー、メタノール及びDPPPを含まなければならない。
【0089】
実施例6:エオキシンC4、エオキシンD4及びエオキシンE4(14,15-LTC4、14-15-LTD4、14-15-LTE4)の生物学的効果の証明
実施例6a:内皮細胞障壁機能の評価
材料と方法
内皮細胞アッセイ
ウシ大動脈内皮細胞(BAEC)及びヒト臍帯静脈内皮細胞(HUVEC)を単離しウシ血清又はヒト血清と共に培養した。第一から第五継代のBAEC又はHUVECを、短時間(2分)のトリプシン-EDTA処理(0.25%のトリプシン/0.01%のEDTA)によって剥離させ、3.0μmの孔径のポリカーボネートフィルター(Tissue Culture inserts, 10mm; NUNC, Roskilde, Denmark)上又は目視検査のために0.2μmの孔径のAnopore無機膜(濡れると透明)に再び蒔いた。細胞分化を促進させ、ECの付着を向上させるために、フィルターを50μlのBiomatrix I(167μg/ml)で前処理し、空気乾燥させた。ECをフィルター当たり2×105細胞の密度で播種し、空気中に5%CO2の加湿雰囲気下、37℃で培養培地中でインキュベートした。透明な0.2μmの孔径のフィルターで単層に亘る電気抵抗を測定し、毎日顕微鏡で観察して評価して、ECをコンフルエントな単層になるまで増殖させた。これらの実験条件下で播種後7日で最大の抵抗が得られ、ついでその日にEC単層を実験に使用した。
【0090】
EC障壁機能の測定
様々なアラキドン酸代謝物に応答した内皮細胞(EC)障壁機能の迅速な変化を、EC単層の各側の電極での経内皮電気抵抗(TEER)の連続的記録によって分析した18。TEERの測定では、ECを有するフィルターインサートを、繰り返し測定の間に互いのかつフィルターインサートに対しての電極の正確な位置決めを達成するように変更された抵抗測定チャンバー(Endohm-12, World Precision Instruments, Sarasota, FL, USA)に移した。チャンバー(下方コンパートメント)及びフィルターインサート(上方コンパートメント)にそれぞれ2ml及び400μlの培養培地を満たした。全ての測定は細胞培養インキュベーター中に配した装置で37℃にて行った。電気抵抗の直接の読みが抵抗チャンバーの電極に接続したオーム計(Evom, World Precision Instruments, Sarasota, FL, US)によって与えられた。個々のEC単層の電気抵抗は、ECの播種前に測定したBiomatrixでコートされた対応する裸のフィルターの抵抗を減算することによって得られた。よって、TEER値はEC単層の抵抗を表し、隣接する内皮細胞間の密着結合の発生に依存する。
【0091】
結果
14,15-システイニル-含有ロイコトリエン類(エオキシン類)の生物学的効果
血管透過性の増加は炎症のマーカーであり、システイニル含有ロイコトリエン類がヒスタミンとブラジキニンと同じ形で特異的血漿漏出を引き起こすことであることはよく知られている。5-リポキシゲナーゼ誘導システイニル含有ロイコトリエン類とは異なり、EoxC4とその代謝物はモルモット肺実質性条片又は回腸への収縮効果を有していないように思われる。EoxC4とその代謝物EoxD4及びEoxE4の血漿漏出を引き起こす能力を探索するために、内皮細胞障壁機能における迅速な変化を登録するためのモデル系を構築した。発明者等は、3種の14,15-システイニル含有ロイコトリエン類が全て血管透過性を増大させたことを見出した。化合物の10-7及び10-8Mの濃度はTEERの有意な変化を誘導した。比較すると、EoxC4、D4及びE4は最も強力な5-リポキシゲナーゼ誘導ロイコトリエン類より約5倍から10倍弱かったがこの点ではヒスタミンより100倍以上強かった(図22)。
【0092】
実施例6b:白血球の酸化的代謝に対するエオキシン類の効果
好中球及び好酸球による酸化的代謝は様々な薬剤、例えば酢酸ミリスチン酸ホルボール(PMA)、血小板活性化因子(PAF)、血清処理ザイモサン(STZ)、免疫グロブリン又はロイコトリエン類(LTB4)によって、またサイトカイン、例えばIL-5及びGM-CSFによって誘導できる。薬物刺激(プライミング)は、細胞を活性化しないで、刺激に対する細胞の増加した応答性をもたらすプロセスを意味する。細胞の薬物刺激状態を生じる機構はなお不確かであるが、造血性サイトカインIL-3、IL-5及びGM-CSFは好酸球と好中球をインビトロで薬物刺激することが知られている。しかしながら、IL-5だけはヒト血液好酸球の選択的活性化因子であることが示されており、IL-3及びGM-CSFは好酸球と好中球の双方に作用しうる。
【0093】
材料と方法
好中球及び好酸球顆粒球を、Percoll勾配及び磁気細胞分離システム(MACS)を使用して末梢全血から精製した。
顆粒球にIL-5(10.5ng/mL)又はGM-CSF(10.5ng/mL)の何れかで37℃で30分薬物刺激を与え、続いてIL-5(100ng/mL)、GM-CSF(100ng/mL)、LTB4(10-7M)又はエオキシン(10-8-10-6Mで刺激した。酸化的代謝をルシゲニン増強化学発光(CL)アッセイによって測定した。
使用した試験化合物はEoxC4(BLX-925)、EoxD4(BLX-926)、EoxE4(BLX-927)であった。
【0094】
結果
非薬物刺激好中球によるエオキシン誘導酸化的代謝
精製した好中球を、好中球に対する異なったエオキシン類の刺激効果との比較のために、酸化的代謝の二つの既知の活性化因子であるサイトカインGM-CSF及びLTB4を用いて刺激した。双方の活性化因子が非薬物刺激好中球による酸素フリーラジカル生成を誘導したが、何れのエオキシン類も非薬物刺激血液好中球による酸素フリーラジカル生成を誘導しなかった。
【0095】
一つの代表的な実験を示す動態曲線(図23a)は、非薬物刺激細胞と比較した場合のサイトカイン薬物刺激細胞の刺激後の向上したラジカル生成を示している。この向上したラジカル生成は、異なったドナーの全結果を平均値として示している図23bに示されるように用量依存パターンを示した。我々は異なった血液ドナー間でEoxD4(BLX-926)に対する強い個々の用量応答性を経験したので、全ての個々のデータをプロットしScatterプロットで示した(図23c)。このようにすることによって、EoxD4(BLX-926)が、異なったドナーに対して異なった最適な用量で、用量依存的な形で薬物刺激細胞によるラジカル生成を誘導したことが明らかになった。13の内、7-10の検体がEoxD4(BLX-926)に対するレスポンダーであった。
類似のパターンがまたEoxC4(BLX-925)に対して見られた。この化合物は用量依存的な形でGM-CSF薬物刺激好中球によるラジカル生成を誘導した(13の内、7-8の検体がEoxC4に対するレスポンダーであった;データは示さず)。
【0096】
非薬物刺激好酸球によるエオキシン誘導酸化的代謝
3種全てのエオキシン類が非薬物刺激好酸球による酸素フリーラジカルの少量の生成を誘導した。しかしながら、酸素フリーラジカルの生成量は、好酸球による酸化的代謝の他の既知の活性化因子、この場合は好酸球特異的サイトカインIL-5と比較した場合にかなり少ない(図24a)。
IL-5薬物刺激好酸球によるエオキシン誘導酸化的代謝
EoxE4(BLX-27と命名)は、図24に示されるように、IL-5薬物刺激好酸球によるラジカル生成の広い用量関連誘導を示し、該エオキシンが試験したものの中で好酸球による酸化的代謝の最良の誘導因子であるかも知れないことを示している。6の検体の内、3がEoxE4に対してレスポンダーであった。
結論
EoxC4(BLX-925)及びEoxD4(BLX-926)はGM-CSF薬物刺激好中球による酸素フリーラジカル生成を主に誘導したが、EoxE4(BLX-927)はIL-5薬物刺激好酸球による酸素フリーラジカル生成を主に誘導した。
【0097】
実施例7:エオキシンC4の形成に対する15-LO阻害の効果
ホジキン細胞株L1236をアラキドン酸(40μM)で5分間刺激した。L1236細胞はアラキドン酸をEoxC4、EoxD4及び一連の8,15-DHETE及び14,15-DHETE代謝物を効果的に転換した。
材料と方法
細胞インキュベーション
ホジキン細胞株L1236(10×106細胞/試料)を1mlのPBSバッファーに懸濁させ、試験物質(1、10又は50μM)の添加前に37℃で2分予め温めた。試験物質と共に5分間プレインキュベートした後、アラキドン酸(40μM)を加えた。インキュベーションを2分後に1容量のメタノールを添加して停止させた。各試験物質濃度は二組をインキュベートした。
【0098】
固相抽出
細胞及び細胞片を試料から遠心分離(2600rpm、6分)によって除去した。上清を水で希釈し、ついで洗浄し平衡にした抽出カートリッジOasis HLB 1cc 10mg(Waters, Sweden)に移した。代謝物をメタノールで溶出させ、HPLC系に注入する前に水で希釈した。
HPLC分析
RP-HPLCをNova Pak C18カラム(2.1×150mm, Waters AB, Sweden)で実施した。最初の移動相は、0.4ml/分の流量での100%A(0.05%(8.7mM)酢酸、pHをアンモニウムで5.6に調整)からなる。線形勾配を5分後に20分の36%B(60:40のアセトニトリル:メタノール)まで開始した。ついで、移動相は、更に20分は64:36のA:Bの均一濃度に維持された。カラム流出液はダイオードアレイ検出(PDA 996, Waters AB, Sweden)を使用してモニターし、UVスペクトルを200-340nmの間で獲得した。
【0099】
試験化合物
図25はEoxC4の形成を阻害した様々な化合物の構造を示している。幾つかのものはWO2004/080999に記載されている。1-フェニルチオカルバモイル-1H-ピラゾール-3-カルボン酸(2-クロロフェニル)-アミド(化合物A)は実施例47に記載されている。ピラゾール-1,3-ジカルボン酸1-(ベンゾ[1,3]ジオキソール-5-イルアミド)3-(2-クロロ-4-フルオロフェニルアミド(化合物B)は実施例64に記載されている。6-{[3-(2-クロロ-4-フルオロフェニルカルバモイル)ピラゾール-1-カルボニル]-アミノ}-ヘキサン酸エチルエステル(化合物C)は実施例65に記載されている。ピラゾール-1,3-ジカルボン酸3-(2-クロロ-4-フルオロフェニルアミド)1-ペンチル-アミド(化合物D)は実施例66に記載されている。ピラゾール-1,3-ジカルボン酸3-(2-クロロ-4-フルオロフェニルアミド)1-ブチルアミド(化合物E)はWO2004/080999(イソシアン酸ブチルがイソシアン酸ペンチルの代わりに用いられている特に実施例66を参照)に記載の方法と同様にして調製される。
【0100】
N'-(2-(エトキシイミノ)-1-フェニルエチリデン)ベンゾヒドラジド(化合物F)は次のようにして調製することができる:
(a)安息香酸(2-ヒドロキシイミノ-1-フェニルエチリデン)ヒドラジド
安息香酸ヒドラジド(1から1.2当量)をアルコール性溶媒(例えばEtOH又はMeOH)に溶解させた。2滴の酸(例えば濃硫酸又は氷酢酸)を加えた後、2-イソニトロソアセトフェノン(1当量)を滴下して加えた。反応混合物を(例えば上昇温度(例えば50-85℃)で)撹拌した後、濾過によって形成された固形物を集め、場合によっては(例えばEtOH/水からの)粗物質を再結晶して副題化合物を得た。
【0101】
(b)N'-(2-(エトキシイミノ)-1-フェニルエチリデン)ベンゾヒドラジド
ジメチルスルホキシド中の安息香酸(2-ヒドロキシイミノ-1-フェニルエチリデン)ヒドラジド(1当量;上の工程(a)を参照)及びKOH(1から1.2当量)の溶液に臭化エチル(少なくとも1当量)を加えた。反応は所定の期間(例えば6日)上昇温度(例えば45℃)で撹拌した。水を加え、混合物をジクロロメタンで抽出した。組み合わせた有機相をNaOH(2M)と水で洗浄した。有機相を乾燥させ(Na2SO4)、真空で蒸発させ、場合によっては再結晶化させて(例えばEtOH/水から)表題化合物を得た。
【0102】
3-クロロ-N'-(2-(ヒドロキシイミノ)-1-(3-クロロフェニル)エチリデン)ベンゾヒドラジド(化合物G)は次のようにして調製することができる:
(a)2-(3-クロロフェニル)-2-オキソアセトアルデヒドオキシム
亜硝酸イソアミル(1.83g、15.6mmol)を氷冷エタノール性ナトリウムエトキシド(0.9M、18.3mL、16.5mmol)に溶解させた。冷却された液に3-クロロアセトフェノン(14.9mmol)を滴下して加え、反応を室温に達するようにし、撹拌を16時間続けた。反応の過程で形成された固形物を濾過し、ジエチルエーテルで洗浄し、水に溶解させ、水溶液を氷酢酸で酸性化した。得られた結晶性固形物を濾過し、EtOH/水から再結晶化して副題化合物を得た。
【0103】
(b)3-クロロ-N'-(2-(ヒドロキシイミノ)-1-(3-クロロフェニル)エチリデン)ベンゾヒドラジド
3-クロロ安息香酸ヒドラジド(1から1.2当量)をEtOHに溶解し、それに2滴の濃硫酸と2-(3-クロロフェニル)-2-オキソアセトアルデヒドオキシム(1当量;上の工程(a)を参照)を加えた。反応を一晩50℃に加熱し、濾過し、EtOHで洗浄し、EtOH/水から再結晶化して表題化合物を得た。
【0104】
結果
ヒト細胞株L1236におけるEoxC4形成の阻害
10-4−4Mの濃度のエイコサテトラエン酸(ETYA)は、EoxC4、EoxD4(図26)及び他の15-LO誘導生成物を完全に阻止した。これらの生成物の形成はまた10-5-5MのETYAの存在下で減少した。これらの結果は、リポキシゲナーゼ阻害剤ETYAがEoxC4及びその代謝物の形成を阻止できることを示している。
EoxC4及び15-HETEの形成に対する化合物の効果
L1236細胞を2分間、示された物質(1μM)と共にプレインキュベートされ、続いて5分間、アラキドン酸(12.5μM)と共にインキュベートした。インキュベーションと抽出は上に記載されているようにして実施された。全ての試験化合物(図25に記載のもの)はEoxC4の形成を阻害した(図27)。また、阻害剤は同様の度合いでEoxC4及び15-HETEの合成を阻止した(図28)。
【0105】
実施例8:ヒト好酸球及び肥満細胞において15-リポキシゲナーゼ-1経路を介して生産された炎症誘発性アラキドン酸代謝物の同定
略語
CBMC 臍帯血肥満細胞
EC 内皮細胞
EIA エンザイムイムノアッセイ
HETE ヒドロキシルエイコサテトラエン酸
HPLC 高速エキタイクロマトグラフィー
IL インターロイキン
LO リポキシゲナーゼ
LT ロイコトリエン
MS 質量分析法
PG プロスタグランジン
【0106】
ヒト好酸球は豊富な量の15-リポキシゲナーゼ-1(15-LO-1)を含んでいる。しかしながら、ヒトにおける15-LO-1の生物学的役割は不確かである。アラキドン酸と共に5分間好酸球をインキュベーションすると、282nmで最大吸光度を有しロイコトリエン(LT)C4よりもRP-HPLCでの保持時間が短い主要生成物の形成に至る。陽イオンエレクトロスプレータンデム質量分析法での分析によって未知の好酸球代謝物を14,15-LTC4と同定した。この化合物は好酸球中において14,15-LTD4及び14,15-LTE4に代謝されうる。また臍帯血由来肥満細胞及び外科的に除去された鼻ポリープは14,15-LTC4を生産した。好酸球をアラキドン酸と共にインキュベーションするとLTC4よりも14,15-ロイコトリエンC4の生産の方が優勢になる一方、カルシウムイオノフォアでの刺激では専らLTC4が形成された。14,15-システイニル含有ロイコトリエン類は、炎症の特徴であるインビトロでの内皮細胞(EC)単層の血管透過性の増大を誘導することができる。このモデル系において、14,15-ロイコトリエン類はLTC4及びLTD4と殆ど同じく強力であったがヒスタミンより100倍強かった。好酸球は、LTC4、プロスタグランジンD2又はインターロイキン-5での刺激後に14,15-LTC4を生産し、これは、14,15-LTC4がアラキドン酸の内因性プールから合成され得ることを証明している。総括すると、本報告は、特にヒト好酸球とまたヒト肥満細胞及び鼻ポリープにおける炎症誘発性14,15-ロイコトリエン類の形成を記載する。好酸球はこれらの新たに特徴付けた代謝物の豊富な供給源であるので、5-リポキシゲナーゼ誘導ロイコトリエン類との混同を避けるために、我々はそれぞれ14,15-ロイコトリエンC4、D4及びE4の代わりにエオキシン類C4、D4及びE4という名称を示唆する。
【0107】
序
哺乳動物のリポキシゲナーゼは、喘息及び他の炎症疾患の病因に関係がある構造的に関連した脂質過酸化酵素のファミリーに属する。5-リポキシゲナーゼ(5-LO)はロイコトリエン合成における重要な酵素であり、この酵素はアラキドン酸のロイコトリエン(LT)A4への転換を触媒する。この化合物は酵素的にLTB4まで加水分解されるか又はグルタチオンとコンジュゲートされてLTC4を形成し、これが次にLTD4及びLTE4まで代謝されうる。ロイコトリエンC4とその代謝物は炎症誘発性薬剤及び強力な気管支収縮剤である(1)。5-LOとは異なり、ヒト15-リポキシゲナーゼ(15-LO-1)の生物学的機能についてはあまり知られていない。ヒト気道内皮細胞、好酸球及び肥満細胞及び樹状細胞のサブセットは高い量の15-LO-1を構成的に発現する(2-6)。ヒト15-LOの第二の形態もまた存在し、これは15-LO-2と命名される(7)。この酵素と15-LO-1の間のアミノ酸類似性は40%だけである(3)。インターロイキン(IL)-4及び/又はIL-13は培養された上皮細胞、単球及び肥満細胞における15-LO-1の発現を誘導し(4,8,9)、よってこれらのサイトカインはインビボでの15-LO-1発現の重要な生理学的/病態生理学的調節因子として作用しうる。
15-リポキシゲナーゼ-1は気管支喘息の病因に関与していることが示唆されている。15-LO-1の量は、コントロール患者と比較して喘息又は慢性気管支炎の患者の気管支粘膜下層において有意に増加している。15-リポキシゲナーゼ-1はアラキドン酸を15-ヒドロキシ-エイコサテトラエン酸(15-HETE)に転換し、これはインビボ並びにインビトロでの様々な細胞及び組織における15-LO活性のマーカーとして使用されている。ヒト気管上皮細胞、切り出したヒト気管支及びヒト肺ホモジネートは主としてアラキドン酸を15-HETEに転換する。喘息患者からの肺検体は非喘息患者の検体よりも多くの15-HETEを生産し(12-14)、15-HETEはアトピー性及び非アトピー性喘息患者からの刺激されたヒト肺組織における主なアラキドン酸代謝物であった(15)。増加した量の15-HETEは、抗原曝露アトピー性喘息患者の気管支肺胞洗浄液(BALF)(16)、喘息患者の痰試料(17)及び健常なコントロール患者からの鼻分泌物(18)に見出された。持続性気道好酸球増加症の深刻な喘息患者は健常な被験者又は好酸球増加症のない喘息患者と比較してBALF中の高いレベルの15-HETEを有している(14)。15-HETEのレベルは基底下(sub-basement)厚と相関することが見出され、15-LOが気道リモデリングに関連しているかも知れないことを示している(14)。
【0108】
15-LO経路を介して生産される最も豊富なアラキドン酸代謝物は15(S)-HETE)であり、これは15(S)-ヒドロペルオキシ-エイコサテトラエン酸(15(S)-HPETE)から形成される。15-HETEへの還元を受けることに加えて、15-HPETEはまた脱水を受けて14,15-エポキシ-エイコサテトラエン酸(14,15-LTA4)を形成する(19,20)。他の15-リポキシゲナーゼ生成物の5-オキソ-15-ヒドロキシ-6,8,11,13-エイコサテトラエン酸はヒト好酸球に対する強力な走化性剤であると報告されている(21)。しかしながら、15-LO-1はリポキシン類及びレゾルビン類(resolvins)の形成に関与しうるので、15-LO生成物は炎症疾患において保護的役割を有しうるという証拠がまたある(22)。総括すると、ヒトにおける15-LO経路の生理学的及び病態生理学的役割は確かではない。
【0109】
この報告では、我々は、14,15-ロイコトリエンC4、D4及びE4の形成及びヒトにおいて15-LO-1に対して炎症誘発性役割を示す生物学的効果を記載する。好酸球はこれらの新たに特徴付けられた代謝物のかかる豊富な供給源であるので、5-リポキシゲナーゼ誘導ロイコトリエン類との混同を避けるために、我々はそれぞれ14,15-ロイコトリエンC4、D4及びE4の代わりにエオキシン類C4、D4及びE4という名称を示唆する。
【0110】
結果
単離されたヒト好酸球における新規な15-リポキシゲナーゼ生成物の形成
単離されたヒト好酸球をアラキドン酸と共に5分間インキュベートしておよそ9:1の比で15(S)-HETE及び12(S)-HETEが形成され(データは示さず)、これは15-LO-1の活性と一致している(2)。約270nmに最大UV吸光度を持ち5-7分の間の保持時間を有する多くのピークが同じクロマトグラムで観察された。これらのより極性の代謝物をよりよく分離するために、アセトニトリルベースの移動HPLC相を使用した。コンジュゲートトリエンスペクトルと272nmでの最大吸光度を含む5つのピークが同定された(図29)。これらの5つのピークの物質の保持時間は、二重酸素添加生成物8(S),15(S)-DHETE及び8(R),15(S)-DHETE、14,15-オキシド-ロイコトリエンA4から誘導された2つの8(S),15(S)-LTB4及び8(R),15(S)-LTB4(19,20)及び14(R),15(S)-DHETEの保持時間に対応した。形成された生成物のパターンは、アラキドン酸と共にインキュベートされたヒト気道上皮細胞に対して報告されているものと同様であった(5)。コンジュゲートしたトリエンであるが282nmに最大の特徴的なUVスペクトルを持つ少量のロイコトリエン(LT)C4がまた観察された。これらのよく知られたアラキドン酸誘導代謝物に加えて、コンジュゲートされたトリエンスペクトルを有し、282nmに最大UV吸光度を有するより極性の主要な代謝物(代謝物1)がクロマトグラムに観察された(インサート,図29)。
【0111】
LC-MS/MSによる代謝物1の同定
代謝物1を更に分析するために、外因性アラキドン酸と共にインキュベートしたヒト好酸球の試料に、陽イオンエレクトロスプレータンデム質量分析を使用するLC-MS/MSによる分析を施した。得られたイオンスキャンスペクトルはLTC4のスペクトル中にも存在する数個の断片を含んでいた(図30A)。好酸球における既知の高い15-LO活性と併せてこの知見は、代謝物1が14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTC4)でありうることを示している。この仮説を確認するために、合成14,15-LTC4を調製し(Biomol Inc, USA)、代謝物1から得られた衝突誘導解離スペクトルを、合成14,15-LTC4に対して得られたスペクトルと比較した(図30A)。試料を、プロトン化された14,15-LTC4と等価な626.2m/zでの生成物イオンスキャンで分析した。クロマトグラムは標準14,15-LTC4と同じ保持時間のピークを含んでおり、質量スペクトルを比較すると、合成14,15-LTC4と同一であった。
【0112】
ヒト好酸球における14,15-LTC4の代謝
好酸球が14,15-LTC4を14,15-LTD4及び14,15-LTE4に代謝し得るかどうかを分析するために、好酸球を、37℃で60分間、合成14,15-LTC4又は14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTD4)と共にインキュベートした。形成された代謝物を、14,15-LTD4に対しては497.2m/z、14,15-LTE4に対しては440.2m/zの生成物イオンスキャンで分析した。14,15-LTC4から形成された生成物(代謝物2)の保持時間と質量スペクトルは合成14,15-LTD4と同一であり(図30B)、14,15-LTD4とのインキュベーション後に形成された生成物(代謝物3)は合成14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTE4)と同一の保持時間と質量スペクトルを有していた(図30C)。37℃での60分のインキュベーションの間、14,15-LTC4及び14,15-LTD4の非酵素的代謝はなかった(データは示さず)。
基本的には、14,15-ロイコトリエン類と対応する5-リポキシゲナーゼ誘導システイニル-ロイコトリエン類の生成物イオン質量スペクトルは殆ど同一である。しかしながら、14,15-ロイコトリエン類は205m/zで断片を形成する一方、5-リポキシゲナーゼ誘導システイニル-ロイコトリエン類における対応する断片は189m/zにおいてである(図31)。これらの衝突誘導解離生成物は、おそらくは、システイニル基が位置しており、14,15-ロイコトリエン類では炭素14、ロイコトリエン類に対しては炭素6である炭素の隣での切断によって形成される。
【0113】
好酸球における14,15-LTC4とLTC4の形成間の比較
ヒト好酸球は豊富な量の15-リポキシゲナーゼ及び5-リポキシゲナーゼ双方を含んでいる。15-リポキシゲナーゼ及び5-リポキシゲナーゼ経路が最も活性である条件を調べるために、好酸球をアラキドン酸又はカルシウムイオノフォアA23187の何れか、又は双方と共にインキュベートした。細胞を37℃で5分間インキュベートし、RP-HPLCを用いて14,15-LTC4及びLTC4の量を分析した。インキュベーションの5分後では好酸球には14,15-LTC4又はLTC4の代謝は殆どない(データは示さず)。従って、これらの代謝物の測定は、それぞれ15-リポキシゲナーゼ誘導システイニル含有14,15-ロイコトリエン類及び5-リポキシゲナーゼ誘導システイニル含有ロイコトリエン類の全形成を反映していると考えられる。外因性アラキドン酸との好酸球のインキュベーションにより14,15-LTC4が優先的に形成されたが、LTC4もまた幾つかのインキュベーションで形成された(表1)。カルシウムイオノフォアA23187の細胞の活性化はLTC4の形成を専ら誘導したが、イオノフォアA23187及びアラキドン酸双方での好酸球の刺激により14,15-LTC4とLTC4の双方が形成された。同様の量のこれらの代謝物が幾つかのインキュベーションで生産されたが、LTC4のレベルは、ある試料では更に高かった。総括すると、アラキドン酸と共に細胞をインキュベートすると、14,15-LTC4の形成が優勢になり、細胞のカルシウム活性化は5-LOをトリガーし、5-LO誘導ロイコトリエン類の形成を強く優勢にする。
【0114】
表1:好酸球中における14,15-LTC4及びLTC4の生合成。好酸球を健常なドナー又は患者から単離した。細胞をリン酸緩衝生理食塩水(1ml)に懸濁させ、37℃で5分間、アラキドン酸(10μM)、イオノフォアA23187(1μM)又は双方と共にインキュベートし、ついで上清を逆相HPLC分析にかけた。14,15-LTC4及びLTC4の相対量をpmol/107好酸球として計算した。
【0115】
14,15-ロイコトリエン類の生物学的効果
血漿漏出に至る血管透過性の増加は炎症の特徴であり、システイニル含有ロイコトリエン類がヒスタミン及びブラジキニンと同様な形でこの効果に寄与しうることはよく知られている(23)。14,15-LTC4とその代謝物14,15-LTD4及び14,15-LTE4が血管透過性における改変を誘導する能力を探究するために、ヒト内皮細胞(EC)単層を通しての電気抵抗変化に基づく確立されたインビトロ透過性アッセイを使用した。このモデルにおける最大刺激で誘導されたEC抵抗変化はおよそ-70%の低下に達する。用量応答曲線は、ヒスタミンの最大効果がこのモデル系において10-5Mで得られたことを示している(データは示さず)。全ての14,15-ロイコトリエン類がEC単層透過性において最大の増加を誘導することができた(図32A)。LTC4及びLTD4より些か強力ではないが、14,15-ロイコトリエン類はこの点でヒスタミンより約100倍強力であった(図32A)。14,15-LTD4は10-7MでEC透過性の最大増加を誘導したが、EC透過性の有意な増加がまた10-8Mで観察された(図32B)。14,15-ロイコトリエン類及びLTD4に応答したEC透過性の変化の動態は、ヒスタミン及びLTC4のような直接的に作用するアゴニストに対して観察されたパターンに類似している(図32B)(24)。他方ではLTB4はEC障壁機能に影響を及ぼすために顆粒球の存在を必要とし、LTB4の効果はこのモデル系では観察されなかった。
【0116】
好酸球における14,15-LTC4生合成のレセプター媒介活性化
14,15-LTC4形成の可能なレセプター媒介活性化を、アレルゲン誘導肥満細胞メディエーターLTC4及びプロスタグランジン(PG)D2並びに強力な好酸球活性化因子であるIL-5を使用することによって調べた。単離された好酸球(10×106/試料)を37℃で5分間、LTC4(1μM)、PGD2(1μM)又はIL-5(10ng/ml)と共にインキュベートした。ロイコトリエンC4、PGD2及びIL-5は全て5分のインキュベーション後に好酸球からの14,15-LTC4の放出を誘導した(表2)。これらの結果はレセプター媒介メカニズムを介した好酸球におけるアラキドン酸の内因性プールからの14,15-LTC4の形成を実証している。
【0117】
表2:好酸球における14,15-LTC4生合成のレセプター媒介活性化。健常な志願者又は好酸球増加症と診断された患者から単離されたヒト好酸球を1mlのPBSに懸濁させ、LTC4(1μM)、PGD2(1μM)又はIL-5(10ng/ml)の存在下で37℃で5分間インキュベートした。コントロール、LTC4、及びPGD2、n=4。IL-5、n=2。14,15-LTC4の量はEIAによって分析した。
【0118】
臍帯血誘導肥満細胞(CBMCMNC)における14,15-LTC4及び14,15-LTD4の生合成
肥満細胞は喘息の病態生理学において重要な役割を担っている。最近、我々は、インターロイキン(IL-4)がヒト臍帯血誘導肥満細胞においてインビトロで15-LO-1の発現を誘導し、肥満細胞がインビボで15-LO-1を発現することを報告した(4)。従って、肥満細胞が14,15-ロイコトリエン類を生産できるかを調べることは興味深かった。IL-4で薬物刺激された肥満細胞をアラキドン酸(10μM)と共に5分間インキュベートした。上清を集め、14,15-LTC4に対してEIAを使用して分析した。これらの条件下で、ヒト肥満細胞は0.8+/−0.6pmol/106細胞(n=3)を放出し、アラキドン酸刺激の後に、免疫反応性の14,15-LTC4を合成し放出する能力を示している。
【0119】
14,15-LTC4の同一性は更にLC-MS/MS単一反応モニタリングSRMで証明された。よって、アラキドン酸と共にインキュベートした肥満細胞が、14,15-LTC4のSRM遷移に対するものと同じ保持時間と典型的な断片パターンを有する代謝物を生じた(データは示さず)。また、肥満細胞試料を特異的14,15-LTD4 SRM法で分析して、4,15-LTC4が細胞によって更に代謝されるかどうかを調べた。確かに、合成14,15-LTD4と同一の断片化パターンと保持時間を有する代謝物が証明された。総括すると、肥満細胞は14,15-LTC4と14,15-LTD4の双方を生産する能力を有している。
【0120】
ヒト鼻ポリープにとる14,15-LTC4の自然の放出
鼻ポリープ症は、好酸球のような炎症細胞による組織浸潤からなる多因子性疾患である(25)。14,15-LTC4がヒト鼻ポリープによって合成されうるかを調べるために、無傷の外科的に除去した鼻ポリープを洗浄し、プロテアーゼ阻害剤(Complete MINI(登録商標))及びインドメタシンを含むPBS中で37℃で5又は30分、インキュベートした。ついで、インキュベーション培地を、14,15-LTC4に対してEIAを使用して分析した。その結果は、鼻ポリープが自然に14,15-LTC4(それぞれ5及び30分のインキュベーション後に、10.1±0.007及び7.18±1.18pmolの14,15-LTC4/gポリープ組織(n=2))を放出することを裏付けている。経時的な14,15-LTC4の減少レベルは14,15-LTD4及び14,15-LTE4への更なる代謝のためであり、これは14,15-LTC4に対して産生された抗体によっては検出されない(14,15-LTD4及び14,15-LTE4に対して1%未満の交差反応性、材料と方法を参照)。
【0121】
エオキシン類-14,15-ロイコトリエン類に対して示唆された新規な名前
ロイコトリエンという名前は、5-リポキシゲナーゼ生成物に伝統的に関連している。アラキドン酸において14,15-エポキシドを介して形成される15-リポキシゲナーゼ代謝物に対する機能的名前は、それは5-リポキシゲナーゼ経路を介して形成されないが、14,15-ロイコトリエンであった。好酸球はこの報告において記載された新規な15-リポキシゲナーゼ生成物に対するリッチな供給源であるので、5-LO誘導生成物との混同を避けるために、我々は、それぞれ14,15-LTC4、14,15-LTD4及び14,15-LTE4の代わりに、エオキシン(Eox)C4、EoxD4及びEoxE4という名を示唆する(図33)。
【0122】
検討
ヒト気道上皮細胞及び好酸球は高い量の15-LO-1を発現する(3,5,6)。また、ヒト肥満細胞、マクロファージ及び樹状細胞は15-LO-1を発現する(26+Gullilsson)。この研究分野での多くの研究にかかわらず、赤血球新生の間の酵素の機能を除いて、この酵素の生物学的役割は不確かである(2,3)。本報告では、我々は、ヒト好酸球、特に臍帯血誘導肥満細胞及び鼻ポリープがアラキドン酸を15-LO-1経路を介して、コンジュゲートされたトリエンスペクトルと282nmでの最大UV吸光度を有する代謝物に転換させる能力を有していることを記載する(図29)。好酸球及び肥満細胞において形成されたこれらの代謝物の構造は陽イオンタンデム質量分析法及びRP-HPLC/UV-分光法によって決定し、合成14,15-LTC4、14,15-LTD4及び14,15-LTE4と同一であることが見出された(図30及び31)。好酸球は主として14,15-LTC4を生産し、それぞれ合成14,15-LTC4及び14,15-LTD4と共にこれらの細胞のインキュベーションによって証明されるように、この代謝物は更に14,15-LTD4及び14,15-LTE4に代謝されうる。合成14,15-LTC4は様々な生物システムにおいて研究され(例えば、Drazen等(1981) Proc Natl Acad Sci 78, 3195-3198を参照)、14,15-LTA4は上皮細胞及び好酸球において形成されることは先に見出されているけれども(例えば文献32を参照)、システイニル含有14,15-ロイコトリエン類の生合成がヒト細胞及び組織中でアラキドン酸から生じることは決して証明されていない。
【0123】
外因的なアラキドン酸と共に好酸球をインキュベーションすると、14,15-LTC4の生成がLTC4より優性になり(表1)、代わりにカルシウムイオノフォアで細胞内カルシウムレベルが増加すると、5-LOを誘導し、ただLTC4だけを生産する。よって、5-LO及び15-LOロイコトリエンの生産の調節は異なり、排出される主要な代謝物にどの代謝物がなるかは刺激に依存する。この点で、14,15-ロイコトリエン経路はプロスタグランジン経路に似ており、遊離のアラキドン酸が利用できる場合に直ぐに生じ、カルシウム流に独立している。
【0124】
5-リポキシゲナーゼ誘導システイニル含有ロイコトリエン類とは異なり、14,15-LTC4とその代謝物はモルモット肺実質性条片又は回腸への収縮効果を有していないように思われる(27)。しかしながら、我々は、ここで、コンフルエントなヒト内皮細胞を持つモデル系において、3種全ての14,15-ロイコトリエン類が炎症のマーカーである血管漏出を増加させたことを立証する。14,15-ロイコトリエン類は10-7及び10-8Mで増大したEC透過性を誘導し、システイニル含有ロイコトリエン類と殆ど同じほど強力で、この点ではヒスタミンより100倍以上強かった(図32)。この研究では、我々は、内皮細胞障壁機能のメディエーター誘導混乱の解析のためにインビトロモデル系を使用した。TEERの連続的測定により、EC透過性の急速な変化の検出が可能になり、培養された内皮細胞単層の障壁能力の感度のよい指標であることが過去に示されている(24,28)。ヒスタミンやシステイニルロイコトリエン類のような炎症メディエーターの透過性増大活性は以前から知られている。これらのメディエーターは、細胞内Ca2+移動及び細胞骨格再配置の惹起を通してEC障壁機能にその効果を作用させ(29)、傍細胞 間隙形成及び亢進された巨大分子透過性を生じる。同様に、14,15-ロイコトリエン類は同様な方法で血管透過性の増加を刺激する。
【0125】
LTC4、PGD2及びIL-5でのヒト好酸球のレセプター媒介活性化がまた14,15-ロイコトリエン類の生産と放出を惹起した(表2)。よって、14,15-ロイコトリエン類は好酸球中のアラキドン酸の内因性プールから製造することができる。LTC4及びPGD2は、共にアラキドン酸誘導化合物であり、抗原投与時に肥満細胞によって合成され放出されることが知られており、活性化された肥満細胞は、14,15-ロイコトリエン類自体の生産に加えて、好酸球におけるこれらの生成物の生合成を刺激することができる。LTC4及びPGD2に加えて、IL-5は14,15-LTC4の内因性生合成を活性化することができる。IL-5は、その生産、活性化及び組織補充を含む好酸球の幾つかの側面を調節することに決定的に関与したサイトカインである。
【0126】
鼻ポリープ形成の原因は知られていないが、アラキドン酸代謝物は、特にアスピリン過敏症副鼻腔炎/喘息徴候を持つ患者において、病因において重要であると思われる。鼻ポリープ症の主要な特徴は活性化された好酸球の蓄積であると考えられ、これは組織内の炎症性細胞の28%までを占める(30)。従って、インビトロでインキュベートされたヒト鼻ポリープが14,15-ロイコトリエン類を生産し放出するかどうかを調べることは興味深かった。確かに、14,15-LTC4は鼻ポリープによってインキュベーション培地中に自然に放出された。活性化された好酸球は鼻ポリープ中の14,15-LTC4の主要なソースであると思われる。しかしながら、気道内皮細胞は、これらの細胞は多量の15-LOを含んでいることが知られているので、所定の役割を担っていることは除外できない(文献2-6を参照)。
好酸球はこれらの新規代謝物のリッチな供給源であるので、我々は、5-LO経路を介して生産された化合物との混同を避けるために、14,15-ロイコトリエンの代わりにエオキシンなる名前を示唆する。よって、14,15-ロイコトリエンA4、C4、D4及びE4はそれぞれエオキシン(Eox)A4、EoxC4、EoxD4及びEoxE4で置き換えられる(図33)。エオキシン類はヒト細胞においてアラキドン酸から生産されることは決して報告されていない。しかしながら、ヒト好塩基球は外因性14,15-ロイコトリエンA4を14,15-ロイコトリエンC4に転換することが先に見出されている(33)。
【0127】
総括すると、この報告は、好酸球及び肥満細胞によって生産される新規な炎症誘発性のアラキドン酸代謝物の形成、並びにこれらの代謝物の生合成に対する様々な生理学的トリガーの同定を記載する。よって、炎症性気道中の活性化された肥満細胞とTh2細胞がLTC4、PGD2及びIL-5を排出し、これが近くの好酸球を誘導してエオキシン類を生産させ放出し、血漿漏出を生じるということができる。更に、アレルギー性炎症に加えて、遊離のアラキドン酸が高量で存在する非アレルギー性炎症においてもまた、エオキシン生産が起こり、炎症プロセスに寄与することが推測できる。
多くの報告が、コントロール被験者と比較して喘息患者の気道組織における15-LO-1の増加した量と活性を実証している。健常な喫煙者は、特に慢性気管支炎の喫煙者の場合、気管支バイオプシー中での増加した15-LO-1の発現を示している。この点で、我々は、15-LO-1経路が好酸球及び肥満細胞において炎症誘発性の14,15-ロイコトリエン類(エオキシン類)を生産することができることを示しており、15-LO-1の阻害がヒトにおける喘息及び鼻炎の治療のための魅力的な標的であり得ることを示している。
【0128】
材料と方法
バキュテナー(登録商標)採血管はベクトン・ディキンソン(Rutherford, NJ)から得た。モノクローナルマウス抗ヒトCD16抗体にコンジュゲートされた常磁性ミクロビーズ、磁気スタンド及びLS+磁気分離カラムはMiltenyi Biotec(Bergisch Gladbach, Germany)から得た。組換えヒト(rh)IL-4はIn Vitro Sweden AB(Sweden)から購入した。14,15-LTC4 EIAキットはCayman Chemical(Ann Arbor, MI)から購入した。イオノフォアA23187はCalbiochem-Boehring(La Jolla, CA)から購入した。インドメタシンはSigma-Aldrichから購入した。全ての溶媒はHPLCグレードであった。アラキドン酸はNu-Chek prepから購入した。合成14,15-エポキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTA4)、14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTC4)、14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTD4)、14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTE4)、8(R),15(S)-ジヒドロキシ-5,9,11,13(Z,E,E,E)-エイコサテトラエン酸(8(R),15(S)-LTB4)、8(S),15(S)-ジヒドロキシ-5,9,11,13(Z,E,Z,E)-エイコサテトラエン酸(8(S),15(S)-DHETE)、8(S),15(S)-ジヒドロキシ-5,9,11,13(Z,E,E,E)-エイコサテトラエン酸(8(S),15(S)-LTB4)、8(R),15(S)-ジヒドロキシ-5,9,11,13(Z,E,Z,E)-エイコサテトラエン酸(8(R),15(S)-DHETE)、14(R),15(S)-ジヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14(R),15(S)-DHETE又は14(R),15(S)-LTB4)、15(S)-ヒドロキシ-5,8,11,13(Z,Z,Z,E)-エイコサテトラエン酸(15-HETE)、LTC4、LTD4及びLTE4は、Biomol Inc (Plymouth Meeting, USA)から購入した。
【0129】
ヒト好酸球の単離
静脈血(100ml)を、健常な志願者又は好酸球増加症候群(HES)を含む好酸球増加症、慢性リンパ性白血病(CLL)及びある種の炎症疾患の患者からから採血した。400×gで15分間の最初の遠心分離を実施した。上相を捨て下相を集め、20℃で30分間、Dextran(商標)沈降を施した。沈降後、白血球細胞を含む上相を400×gで10分間、一回遠心分離した。ペレットを溶解バッファー(100mMのNH4Cl、10mMのTRIS-HCl、pH7.4)に懸濁させ、室温で30分インキュベートして混入した赤血球を除去した。インキュベーション後、Lymphoprep(商標)を用いて密度勾配遠心分離を実施した。好中球と好酸球を含む多形核(PMN)画分を集め、磁気細胞分離(MACS)を施した。好酸球は、磁気ミクロビーズ(登録商標)に結合させたCD16抗体を使用するネガティブ選択によって単離した。純度は、形態学的検査とメイ・グリューネバルト-ギムザ溶液での染色によって確認した。純度は常に95%を越えていることが見出された。
臍帯血誘導肥満細胞(CBMCMNC)の調製及びポリープの単離
臍帯血誘導肥満細胞(CBMCMNC)を記載されたようにして樹立した(4)。ポリープは外科的に除去し、4℃で最大1時間、PBS中に維持した。ポリープを回りの組織から分離し、Ca2+及びMg2+なしのPBSで洗浄した。洗浄した濡れたポリープの重さは0.7-1gであった。
【0130】
細胞インキュベーション
好酸球
単離された好酸球(1−20×106/試料)をリン酸緩衝生理食塩水(1ml/試料)に再懸濁させ、37℃で示された時間の間、イオノフォアA23187(1μM)/アラキドン酸(1又は10μM)/A23187プラスアラキドン酸/PGD2(1μM)/LTC4(1μM)/IL-5(10ng/ml)/14,15-LTC4(10μM)又は14,15-LTD4(10μM)又は双方で2、5又は10分、刺激する前に37℃で2分間(インドメタシンの存在下又は不在下で、1μM)、プレインキュベートした。インキュベーションは1容量のメタノールの添加によって終結させた(RP-HPLCによる分析のため)。エンザイムイムノアッセイ(EIA)による続く分析のために、反応を、1200×g、5分の遠心分離前に0℃で急速冷却して終結させた。上清を凍結管に移し、分析するまで-70℃で保存した。
【0131】
肥満細胞
インキュベーション前に、CBMC細胞にIL-4(10ng/ml)を用いて4日間薬物刺激を施した。機械的な除去及びPBSでの洗浄後に、細胞をPBSに再懸濁させ、インドメタシン(1μM)と共に又はそれなしで37℃で2分間プレインキュベートした。CBMC細胞を続いて37℃でアラキドン酸と共に示された期間の間、インキュベートした。インキュベーションは1容量のメタノールの添加によって(LC-MS分析)又は0℃に急速冷却することによって終結させ、続いてEIAによって分析した。
ヒト鼻ポリープ
洗浄したヒトの無傷のポリープを、1mlの全容量中にPBS、プロテアーゼ阻害剤(Complete MINI(登録商標))及びインドメタシン(1μM)を含む清浄な試験管中に移した。試料を37℃でそれぞれ5分及び30分、インキュベートした。反応を0℃に急速冷却することによって終結させ、続いて1200×g、5分、遠心分離した。上清を凍結管に移し、EIAによる分析まで-70℃で保存した。
【0132】
15-リポキシゲナーゼ生成物の分析
逆相-高速液体クロマトグラフィー-UV(RP-HPLC-UV)
細胞片及びタンパク質を除去するために、1200×gで5分間、遠心分離を実施した。上清を減圧下で蒸発させ、残留物をメタノールに溶解させ、試験管に移した。ついで、試料を窒素流下で蒸発乾固させ、最後に200μlの移動相(ロイコトリエン類及び14,15-ロイコトリエン類の分析に対してはアセトニトリル:メタノール:H2O:酢酸、29:19:72:0.8、pHは5.6にNH3で調整、及びモノヒドロキシ酸の分析に対してはメタノール:水:トリフルオロ酢酸、70:30:0.007)に再懸濁させた。抽出された試料に、記載されたようにして逆相HPLC系での分析を施した。簡単に述べると、試料をオクタデシル逆相カラム(Chromabond 18C,Macherey-Nagel,Duren,Germany)に充填し、溶出流量は1.2ml/分であった。溶出した化合物を、Waters1996HPLCシステムに連結したWaters996光ダイオードアレイ分光光度計を利用して検出し定量した。
【0133】
液体クロマトグラフィー-質量分析解析(LC-MS/MS)
インキュベーションの終結後、試料を遠心分離し(1400×g、6分)、上清を水で希釈して最大25%のメタノールを含ませ、洗浄及び平衡抽出カートリッジOasis HLB 1cc 10mg(Waters AB, Sweden)に移した。カラムを水で洗浄し、200μlのメタノールで溶出させて代謝物を回収した。
液体クロマトグラフィー-質量分析はTSQ・Quantum・Ultraトリプル四極子質量分析計に連結したSurveyor・MS(Thermo Electron Corporation, Sweden)で実施した。注入前に1容量の水を試料に加えた。逆相LCは、400μl/分に一定に保たれた流量でZorbax・Eclipse・Plus3.5μM・C18カラム2.1×50mm(ChromTech, Sweden)を用いて実施した。移動相Aは水中2%のアセトニトリルと0.1%の酢酸からなり、移動相Bは水中80%のアセトニトリルと0.1%の酢酸から構成されている。使用した全ての水はMilliQグレードであった。90%のAで1.5分、均一濃度で開始して、100%のBに達する6.5分の線形勾配を続けた。該系を100%のBで4分洗浄し、ついで90%のAで4分間平衡にした。14,15-ロイコトリエン類を約0.5分、ロイコトリエン類からクロマトグラフィー的に分離した。質量分析計はポジティブモードでエレクトロスプレー大気圧イオン化源を使用して操作した。スプレー電圧は4500Vに設定し、キャピラリー温度は375℃であり、シース及び補助ガスはそれぞれ40及び5(任意単位)で最適であった。スキマーオフセットは10Vで、チューブレンズは121Vであった。
【0134】
好酸球代謝物に対するLC-MS/MS
LC-MS/MSを使用して、プロトン化14,15-LTC4、14,15-LTD4及び14,15-LTE4の生成物イオンスキャンを実施した。626.2m/zの14,15-LTC4を22eVの衝突エネルギーで断片化し、497.2m/zの14,15-LTD4及び440.2m/zの14,15-LTE4は共に18eVで断片化した。
肥満細胞代謝物に対するLC-MS/MS
14,15-LTC4及び14,15-LTD4は単一反応モニタリングSRMによって検出した。SRMスキャンで使用された遷移は、14,15-LTC4に対して626m/z→279m/z、14,15-LTD4に対して497m/z→205m/zであり、双方とも20eVの衝突エネルギーであった。14,15-LTC4に対して選択される遷移はまたLTC4に対して存在していたが、これらの化合物はクロマトグラフィー的にはおよそ0.5分で分離した。497m/z→205m/zの遷移は14,15-LTD4に対して特異的であり、よってLTD4には適用できなかった。
【0135】
14,15-LTC4に対するエンザイムイムノアッセイ(EIA)の特徴付け
14,15-LTC4に対するエンザイムイムノアッセイ(EIA)は少量の14,15-LTC4を定量するためにCayman Chemical (Ann Arbor, MI)によって開発された。この方法は、14,15-LTC4の定量においてRP-HPLCよりおよそ千倍感度が良かった。EIA交差反応性は製造者によって決定された。14,15-LTD4及び14,15-LTE4に対する交差反応性はそれぞれ0.9%及び0.04%であった。ロイコトリエンC4、ロイコトリエンD4、ロイコトリエンE4、アラキドン酸、12-HETE、15-HETE、5(S),6(S)-ジHETE、5(S),15(S)-ジHETE、8(S),15(S)-ジHETE及び14,15-ジHETEの交差反応性は0.01%未満であった。
上清をEIAバッファーで希釈し、分析を製造者のプロトコルに従って実施した。EIA定量からの光学密度(OD)を用いて次の関係に従って%B/B0を計算した:
%B/B0=(試料OD-NSB OD)/(ゼロ標準OD-NSB OD)×100
7.8-1000pg/ml(0.012-1.6pmol/ml)14,15-LTC4の範囲の標準曲線を作成し、試料中の14,15-LTC4の濃度を決定するために使用した。
【0136】
内皮細胞障壁機能の評価
ヒト臍帯静脈内皮細胞 (HUVEC)を単離し、過去に記載されているようにして培養した。細胞を3μmの孔径のポリカーボネートフィルター(Tissue Culture inserts, 10 mm; NUNC, Roskilde, Denmark)に2×105細胞の密度で播種し、コンフルエントな単層まで増殖させた。内皮細胞(EC)単層の障壁機能に変化を誘導するアラキドン酸代謝物の能力は、経内皮電気抵抗(TEER)の連続的記録によって分析した。TEERの測定では、ECを有するフィルターインサートを、繰り返し測定の間に互いのかつフィルターインサートに対しての電極の正確な位置決めを達成するように変更された抵抗測定チャンバー(Endohm-12, World Precision Instruments, Sarasota, FL, USA)に移した。チャンバー(下方コンパートメント)及びフィルターインサート(上方コンパートメント)にそれぞれ2ml及び400μlの培養培地を満たした。全ての測定は細胞培養インキュベーター中に配した装置で37℃にて行った。個々のEC単層の電気抵抗は、ECの播種前に測定したBiomatrixでコートされた対応する裸のフィルターの抵抗を減算することによって得られた。よって、TEER値はEC単層だけの抵抗を表す。
【0137】
1.Claesson, H. E.及びDahlen, S. E. (1999) J Intern Med 245, 205-27。
2.Kuhn, H., Walther, M.及びKuban, R. J. (2002) Prostaglandins Other Lipid Mediat 68-69, 263-90。
3.Kuhn, H.及びO'Donnell, V. B. (2006) Prog Lipid Res 45, 334-56。
4.Gulliksson, M., Brunnstrom, A., Johannesson, M., Backman, L., Nilsson, G., Harvima, I. T., Dahlen, B., Kumlin, M.及びClaesson, H-E. (2007) Biochem Biopys Acta in press.
5.Hunter, J. A., Finkbeiner, W. E., Nadel, J. A., Goetzl, E. J.及びHoltzman, M. J. (1985) Proc Natl Acad Sci U S A 82, 4633-7。
6.Turk, J., Maas, R. L., Brash, A. R., Roberts, L. J., 2nd及びOates, J. A. (1982) J Biol Chem 257, 7068-76。
7.Brash, A. R., Boeglin, W. E.及びChang, M. S. (1997) Proc Natl Acad Sci U S A 94, 6148-52。
8.Conrad, D. J., Kuhn, H., Mulkins, M., Highland, E.及びSigal, E. (1992) Proc Natl Acad Sci U S A 89, 217-21。
9.Brinckmann, R., Topp, M. S., Zalan, I., Heydeck, D., Ludwig, P., Kuhn, H., Berdel, W. E.及びHabenicht, J. R. (1996) Biochem J 318 ( Pt 1), 305-12。
10.Shannon, V. R., Chanez, P., Bousquet, J.及びHoltzman, M. J. (1993) Am Rev Respir Dis 147, 1024-8。
11.Bradding, P., Redington, A. E., Djukanovic, R., Conrad, D. J.及びHolgate, S. T. (1995) Am J Respir Crit Care Med 151, 1201-4。
12.Hamberg, M., Hedqvist, P.及びRadegran, K. (1980) Acta Physiol Scand 110, 219-21。
13.Kumlin, M., Ohlson, E., Bjorck, T., Hamberg, M., Granstrom, E., Dahlen, B., Zetterstrom, O.及びDahlen, S. E. (1991) Adv Prostaglandin Thromboxane Leukot Res 21A, 441-4。
14.Chu, H. W., Balzar, S., Westcott, J. Y., Trudeau, J. B., Sun, Y., Conrad, D. J.及びWenzel, S. E. (2002) Clin Exp Allergy 32, 1558-65。
15.Dahlen, S. E., Hansson, G., Hedqvist, P., Bjorck, T., Granstrom, E.及びDahlen, B. (1983) Proc Natl Acad Sci U S A 80, 1712-6。
16.Murray, J. J., Tonnel, A. B., Brash, A. R., Roberts, L. J., 2nd, Gosset, P., Workman, R., Capron, A.及びOates, J. A. (1985) Trans Assoc Am Physicians 98, 275-80。
17.Profita, M., Sala, A., Riccobono, L., Paterno, A., Mirabella, A., Bonanno, A., Guerrera, D., Pace, E., Bonsignore, G., Bousquet, J.及びVignola, A. M. (2000) J Allergy Clin Immunol 105, 711-6。
18.Ramis, I., Rosello-Catafau, J., Bulbena, O., Picado, C.及びGelpi, E. (1989) J Chromatogr 496, 416-22。
19.Jubiz, W., Radmark, O., Lindgren, J. A., Malmsten, C.及びSamuelsson, B. (1981) Biochem Biophys Res Commun 99, 976-86。
20.Maas, R. L.及びBrash, A. R. (1983) Proc Natl Acad Sci U S A 80, 2884-8。
21.Schwenk, U., Morita, E., Engel, R.及びSchroder, J. M. (1992) J Biol Chem 267, 12482-8。
22.Serhan, C. N.及びSavill, J. (2005) Nat Immunol 6, 1191-7。
23.Raud, J., Lindbom, L.及びHedqvist, P. (1992) Acta Physiol Scand 146, 545-6。
24.Gautam, N., Hedqvist, P. & Lindbom, L. (1998) Br J Pharmacol 125, 1109-14。
25.Bachert, C., Gevaert, P., Holtappels, G.及びvan Cauwenberge, P. (2002) Curr Allergy Asthma Rep 2, 481-7。
26.Spanbroek, R., Hildner, M., Kohler, A., Muller, A., Zintl, F., Kuhn, H., Radmark, O., Samuelsson, B.及びHabenicht, A. J. (2001) Proc Natl Acad Sci U S A 98, 5152-7。
27.Lewis, R. A., Drazen, J. M., Austen, K. F., Toda, M., Brion, F., Marfat, A.及びCorey, E. J. (1981) Proc Natl Acad Sci U S A 78, 4579-83。
28.Gautam, N., Herwald, H., Hedqvist, P.及びLindbom, L. (2000) J Exp Med 191, 1829-39。
29.Huang, A. J., Manning, J. E., Bandak, T. M., Ratau, M. C., Hanser, K. R.及びSilverstein, S. C. (1993) J Cell Biol 120, 1371-80。
30.Ferguson, B. J. (2000) Laryngoscope 110, 799-813。
31.Holtzman, M. J., Hansbrough, J. R., Rosen, G. D.及びTurk, J. (1988) Biochim Biophys Acta 963, 401-13。
32.Turk, J., Rand, T. H., Maas, R. L., Lawson, J. A., Brash, A. R., Roberts, L. J., 2nd, Colley, D. G.及びOates, J. A. (1983) Biochim Biophys Acta 750, 78-90。
33.Sok, D. E., Han, C. O., Shieh, W. R., Zhou, B. N.及びSih, C. J. (1982) Biochem Biophys Res Commun 104, 1363-70。
【0138】
文献(実施例8のもの以外)
1.Funk CD. The molecular biology of mammalian lipoxygenases and the quest for eicosanoid functions using lipoxygenase-deficient mice [Review]. Biochimica et Biophysica Acta - Lipids & Lipid Metabolism 1996; 1304(1):65-84。
2.Claesson HE, Dahlen SE. Asthma and leukotrienes: antileukotrienes as novel anti-asthmatic drugs [Review]. Journal of Internal Medicine 1999; 245(3):205-227。
3.Rapoport SM及びSchewe T. The maturational breakdown of mitochondria in reticulocytes. Biochim. Biophys. Acta 1986; 864:471-495。
4.Vanleyen K, Duvoisin RM, Engelhardt H, Wiedmann M. A function for lipoxygenase in programmed organelle degradation. Nature 1998; 395(6700):392-395。
5.Ford-Hutchinson AW. Arachidonate 15-lipoxygenase; characteristics and potential biological significance. Eicosanoids 1991; 4:65-74。
6.Conrad DJ, Kuhn H, Mulkins M等 Specific inflammatory cytokines regulate the expression of human monocyte 15-Lipoxygenase. Proc Natl Acad Sci USA 1992; 89(1):217-221。
7.Holtzman MJ, Hansbrough, R. J., Rosen, G. D.及びTurk, J. Uptake, release and novel species-dependent oxygenation of arachidonic acid in human and animal airway epithelial cells. Biochim. Biophys. Acta 1988; 963:401-413。
8.Rapoport SM, Schewe,T, Wiesner, R., Halangk, W., Ludwig, P., Janicke-Hohne, M., Tannert, C., Hiebsch, C.及びKlat, D. The lipoxygenase of reticulocytes. Purification, characterisation and biological dynamics of the lipoxygenase: its identity with the respiratory inhibitors of the reticulocyte. Eur. J. Biochem 1979; 96:545-561。
9.Sigal E, Grunberger, D., Highland, E., Gross, C., Dixon, R. A., Craik, C. S. Expression of cloned human reticulocyte 15-lipoxygenase and immunological evidence that 15-lipoxygenase of different cell types are related. J. Biol. Chem. 1990; 265:5113-5120。
10.Brash AR, Boeglin WE, Chang MS. Discovery of a second 15s-lipoxygenase in humans. Proc Natl Acad Sci USA 1997; 94(12):6148-6152。
11.Maas RL, Brash, A. R.及びOates, J. A. A second pathway of leukotriene biosynthesis in porcine leukocytes. Proc. Natl. Acad. Sci. 1981; 78:5523-5527。
12.Lundberg U, Malmsten C, Samuelsson B. Transformation of 15-hydroperoxy-5,9,11,13-eicosatetraenoic acid into novel leukotrienes. FEBS Lett 1981; 126:127-132。
13.Serhan CN, Haeggstrom, J. Z.及びLeslie, C. C. Lipid mediator networks in cell signaling: update and impact och cytokines. The Faseb journal 1996; 10:1147-1158。
14.Schwenk U, Morita E, Engel R, Schroder JM. Identification of 5-oxo-15-hydroxy-6,8,11,13-eicosatetraenoic acid as a novel and potent human eosinophil chemotactic eicosanoid. J Biol Chem 1992; 267(18):12482-12488.
15.Wolf W, Kapp U, Bohlen H, Kornacker M, Schoch C, Stahl B, Mucke S, von Kalle C, Fonatsch C, Schaefer H-E, Hansmann M-L and Diehl V: Peripheral blood mononuclear cells of a patient with advanced Hodgkin's lymphoma give rise to permanently growing Hodgkin-Reed Sternberg cells. Blood, 1996; Vol 87 (8) 3418-3428。
16.Griffiths W, Yang Y, Sjovall J及びLindgren JA: Electrospray tandem mass spectrometry of cysteinyl leukotrienes. Rapid Commun. Mass Spectrom. 1996; 10:1054-10709。
17.Hunter JA, Finkbeiner WE, Nadel EJ等 Predominant generation of 15-lipoxygenase metabolites of arachidonic acid by epithelial cells from human trachea. Proc. Natl. Acad. Sci. USA 1985; 82:4633-4637。
18.Gautam N, Hedqvist P及びLindbom L (1998). Kinetics of leukocyte-induced changes in endothelial barrier function. Br J Pharmacol 125: 1109-1114。
19.Mannervik B及びDanielsson UH (1988). グルタチオントランスフェラーゼs: structure and catalytic activity. CRC Crit Rev Biochem 3, 283-337。
20.Izumi Takashi等(1988). Solubilization and partial purification of leukotriene C4 synthase from guinea-pig lung: a ミクロソーム enzyme with high specificity towards 5,6-epoxide leukotriene A4. Biochem Biophys Acta 959, 305-315。
21.Jakobsson P-J, Morgenstern R, Mancini J, Ford-Hutchinson A及びPersson B (2000). Membrane-associated proteins in eicosanoid and glutathione metabolism. Am J Respir Crit Care Med 161, 20-24。
22.Gillmor, S.A., Villasenor, A., Fletterick, R., Sigal, E.及びBrowner, M.F., 1997, The structure of mammalian 15-lipoxygenase reveals similarity to the lipases and the determinants of substrate specificty Nature Structural Biology, vol 4, issue 12, 1003-1009。
【技術分野】
【0001】
本発明は薬学的に有用な化合物を同定するための方法に関する。本発明は更に炎症疾患又は炎症全般の治療に有用な化合物を同定するための方法に関する。本発明はまた抗炎症組成物を製造する方法及び薬学的に有用な化合物を使用する方法にも関する。
【背景技術】
【0002】
その性質が炎症性である多くの疾患/障害がある。炎症症状の現在の治療法に伴う主な問題の一つは効能の欠如及び/又は副作用(実際のもの又はそのように思われているもの)の発生である。
喘息は、先進工業国の成人人口の6%〜8%で発症している慢性の炎症性疾患である。子供における発生率はさらに高く、殆どの国では10%に近い。喘息は、15才以下の子供の入院原因の最も一般的なものである。
喘息の治療法は症状の重症度に基づく。軽度の症例では治療しないか、又はβ-アゴニストの吸入で治療されるのみである。より重症の喘息を患っている患者は、定期的に抗炎症化合物を用いて治療するのが典型的である。
【0003】
現在の維持療法(主としてコルチコステロイド類の吸入)には、少なくともある程度のリスクがあると思われるため、喘息の治療が不十分な場合がかなりある。これらには、不要な罹患及び死亡に至る、子供の成長遅延及び骨ミネラル密度の損失の危険性が含まれる。ステロイド類の代替法として、ロイコトリエンレセプターアンタゴニスト(LTRas)が開発されている。これらの薬剤は経口的に与えることができるが、吸入されたステロイド類に比べてあまり有効ではなく、通常、気道炎症を満足に抑制できない。
この要因が組み合わさって、喘息の全患者の少なくとも50%が満足のいく治療を受けていない。
【0004】
治療が不十分であるという同様のパターンがアレルギー疾患に関連して存在し、多くの一般的症状を治療するのに薬剤を利用できるが、明白な副作用を考慮して使用されていない。鼻炎、結膜炎及び皮膚炎はアレルギー要素を有しうるが、潜在するアレルギーがない場合でも生じる場合がある。実際、このクラスの非アレルギー性症状は、多くの症例で治療が困難である。
【0005】
慢性閉塞性肺疾患(COPD)は、世界の人口の6%〜8%が罹患している一般的な疾患である。疾患は致死的となりうるおそれがあり、この病気の罹患率及び死亡率はかなりのものである。現在、COPDの経過を変えることができる既知の薬物学的治療法は存在しない。
【0006】
挙げることのできる他の炎症性疾患には次のものがある:
(a)特発性肺線維症(これはCOPDよりも一般的ではないが、非常に悪い予後を伴う重篤な疾患である。治癒的治療法は存在しない);
(b)炎症性大腸炎(高罹患率の疾患群。今日、このような疾患では対症療法が利用できるに過ぎない);及び
(c)関節リウマチ及び骨関節炎(関節のありふれた無能化炎症性疾患。現在では、このような症状への対応に利用できる治癒的治療法は存在せず、また適度に効果的な対症療法のみである)。
【0007】
炎症はまた痛みの一般的な原因である。炎症痛は多くの理由、例えば感染、手術又は他の外傷により生じるおそれがある。更に、数種の悪性腫瘍は、患者の総体的な症状に加わる炎症要素を有することが知られている。
よって、新規な及び/又は代替の抗炎症治療法が、上述した患者群の全てにとって有益となる。特に、実際の又は知覚される副作用を生じないで喘息のような炎症性疾患を治療できる有効な抗炎症剤に対して真のかつ実質的に満たされていないニーズがある。
【0008】
哺乳類のリポキシゲナーゼは、アラキドン酸の酸素化を触媒する構造的に関連した酵素のファミリーである。3種類のヒトリポキシゲナーゼが知られており、これらは炭素位置5、12及び151におけるアラキドン酸への分子酸素の挿入を触媒する。よって、酵素は、それぞれ5-、12-及び15-リポキシゲナーゼと称されている。
リポキシゲナーゼの作用によって生成されるアラキドン酸の代謝産物は、炎症誘発効果を含む明白な病態生理学的活性を有していることが知られている。
【0009】
ロイコトリエン生合成における重要な酵素は、2段階反応でアラキドン酸をロイコトリエン(LT)A4に転換する5-リポキシゲナーゼである。この代謝物は酵素的にロイコトリエンB4(LTB4)まで加水分解されるか、又はロイコトリエンC4(LTC4)シンターゼによってグルタチオンとコンジュゲートさせられてLTC4を形成し、これがついでγグルタミル-トランスペプチダーゼ及びジペプチダーゼの連続的作用によって切断されロイコトリエンD4(LTD4)とロイコトリエンE4(LTE4)をそれぞれ生じうる。LTC4とその代謝物は炎症誘発剤及び強力な気管支収縮剤2である。これらの代謝産物の作用並びにそれらを形成する生物学的プロセスを阻害する薬剤の開発に多大な努力がなされている。この目的のために開発された薬剤には、5-リポキシゲナーゼインヒビターと、これまでに述べたように、ロイコトリエンレセプターアンタゴニスト(LTRas)が含まれる。FLAP(Five Lipoxygenase Activating Protein)のインヒビターもまた研究されている。
【0010】
アラキドン酸を代謝する他のクラスの酵素はシクロオキシゲナーゼ類である。このプロセスにより生成されるアラキドン酸代謝産物には、プロスタグランジン類、トロンボキサン類、及びプロスタサイクリンが含まれ、それらの全てが生理学的及び病態生理学的活性を有している。特に、プロスタグランジンPGE2は、熱及び痛みをまた誘発する強力な炎症誘発媒介物である。従って、「NSAIDs」(非ステロイド性抗炎症剤)及び「コキシブ(coxibs)」(選択的シクロオキシゲナーゼ-2インヒビター)を含む、PGE2の生成を阻害する多くの薬剤が開発されている。これらのクラスの化合物は、主として一種又は数種のシクロオキシゲナーゼの阻害を介して作用する。またレセプターアンタゴニストが研究されている。
【0011】
よって、一般に、アラキドン酸代謝産物の生成を阻止可能な薬剤は、炎症の治療に有用であると思われる。
【0012】
例えば免疫抑制性患者、例えば化学療法、移植拒絶を防止するための治療を受けている患者又はAIDS患者において炎症誘発剤はまた有用でありうる。
5-リポキシゲナーゼに加えて、12-及び15-リポキシゲナーゼがまたヒトの体で発現される。
12-リポキシゲナーゼ(12-LO)はヒト血小板中で高度に発現されるがこの酵素の生物学的な役割については殆ど知られていない1。
【0013】
15-リポキシゲナーゼ(15-LO)1型(ヒト15-LO:EC1.13.11.33)は非ヘム鉄を含む75-kDaのタンパク質である。15-LOはアラキドン酸(20:4)のC-15又はリノール酸(18:2)のC-13に分子酸素を導入して15S-ヒドロペルオキシ-5,8,11-cis-13-trans-エイコサテトラエン酸(15-HPETE)又は13S-ヒドロペルオキシ-9-cis-13-trans-オクタデカジエン酸(13-HPODE)をそれぞれ形成する。少量の12S-ヒドロペルオキシ-5,8,14-cis-10-trans-エイコサテトラエン酸(12-HPETE)がまた15-LO酵素によってアラキドン酸から形成される。簡単に述べると、触媒反応は不飽和脂肪酸のcis-cis-1,4-ペンタジエン構造(20:4又は18:2)のC-3における水素除去を含んでおり、炭素ラジカルとFe3+からFe2+への鉄の還元を生じる(以下のスキーム1参照)。
【0014】
ラジカルはC-5に移動し、そこで分子酸素と反応してペルオキシ基を形成する。ペルオキシ基による鉄からFe3+への酸化は、プロトン化されて最終の脂質ヒドロペルオキシドを生じるペルオキシ陰イオンを生じる。
【0015】
ウサギ酵素15-LOはクローニングされ結晶化され、基質結合部位が、その基部がPhe353、Met419、Ile418及びIle593の側鎖によって定まる深い疎水性ポケット22であることが見出されている。脂肪酸基質は(酵素の表面で)Arg403によって繋留され位置せしめられている。変異原性研究により、疎水性ポケットのサイズの変動が酵素の位置特異性に影響を及ぼすことが立証されている。よって、 15-LOのポケットの拡大が12-リポキシゲネーション産物(12-HPETE)の量を増大させた。15-LO中の鉄はヒスチジンとC末端イソロイシンのカルボキシ基に配位されている。
【0016】
15(S)-HETEへの還元を受けることに加えて、15(S)-HPETEはまた脱水を受けて14,15-オキシド-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTA4)を生じる。このアリルエポキシドはジヒドロキシ酸8(R,S),15(S)-LTB4又は14(R,S),15(S)-LTB4に加水分解されうる11-12。また、基質として15-HPETEを用いて、15-LOは二重の二原子酸素付加産物5(S),15(S)-DHETE,8(S,R),15(S)-DHETE及び14(R,S),15(S)-DHETEの形成をまた触媒する。
【0017】
15(S)-リポキシゲナーゼ(15-LO)は遊離脂肪酸を代謝するばかりでなく、膜に存在しているポリエン脂肪酸を攻撃することもできる。該酵素はミトコンドリア膜の崩壊を通して網状赤血球の成熟において重要な役割を担っていることが予想された3。しかしながら、該酵素はケラチノサイトや眼水晶体の成熟のような、細胞の分化においてより一般的な役割を担いうる4。15-LOは主として網状赤血球、好酸球、皮膚、単球/マクロファージ及び気道上皮において発現される5。インターロイキン-4(IL-4)及びインターロイキン-13(IL-13)は培養された単球及び上皮細胞における15-LOの発現を誘導するが6、ある種の細胞への損傷が15-LOの活性を増加させることが報告されている(Green FA. Transformation of 5-HETE by activated keratinocyte 15-lipoxygenase and the activation mechanism. Lipids 25, 618-623 (1990))。
【0018】
5-リポキシゲナーゼ及び12-リポキシゲナーゼとは異なり、15-LOはウサギ及びヒト細胞においてのみ発現されると思われる1。ヒト15-LOのマウスオルソログは白血球型の12-LOであると思われる1。しかしながら、二つの酵素活性の調節はかなり異なっているようである7。
15-LOは20年以上も前に発見され、均一になるまで精製される最初の15-LOはウサギ網状赤血球から取り出された8。該酵素のヒト型は網状赤血球及び上皮細胞からクローニングされており、免疫学的研究から、該酵素は網状赤血球、好酸球及び気道上皮において同一である9。15-LOの第二の型(2型)はヒト毛根からのcDNAライブラリーからクローニングされ、続く該酵素の発現とアミノ酸解析により、この15-LOの2型酵素は先に開示された15-LOの1型とおよそ40%同一であることが証明された10。
【0019】
15-LOは、主として抗炎症効果を有していると思われるリポキシンの形成に関与していると思われるが13、15-LO誘導産物の5-オキソ-15-ヒドロキシ-6,8,1,13-エイコサテトラエン酸はヒト好酸球に対する強力な走化性剤であると報告されている14。ヒトにおける15-LO経路の生理学的及び病態生理学的役割は十分には特徴付けされていない。
【0020】
ロイコトリエン類のような生物システム中で生じることが知られているアラキドン酸代謝物に基づいて、多くの異なったアナログが合成されている。例えば14,15-LTC4のようなLTC4の合成アナログは気管支収縮を誘導しなかった。Drazen, J.M., Lewis, R. A., Austin, K.F., Toda, M., Brion, F., Marfat, A.及びCorey, E.J.: Contractile activities of structural analogs of leukotrienes C and D: necessity of a hydrophobic region. Proc. Natl. Acad. Sci. 78, 3195-3198 (1981)。
【発明の概要】
【0021】
我々は炎症誘発効果を有するアラキドン酸代謝物の生物システム中における産生を同定した。該代謝物は15-リポキシゲナーゼの作用を介して、可溶型グルタチオンSトランスフェラーゼ及び/又はロイコトリエンC4シンターゼの作用を介して形成されうる。該代謝物はエオキシンC4、エオキシンD4及びエオキシンE4と命名される。我々は、抗炎症化合物及びエオキシンC4、D4及びE4形成を阻害する化合物を同定するためのスクリーニング法を提供する。
【0022】
我々は、ホジキン細胞株L123615が15-リポキシゲナーゼ(15-LO)の1型活性を有しており、15-LO経路を介してアラキドン酸をエオキシン類に転換することを示す。L1236中におけるエオキシンC4、D4及びE4の存在及び形成を明らかにした後、我々は、これらエオキシン類が他のヒト細胞及び組織、特に好中球において形成されるが、また鼻ポリープ組織及び樹状細胞(実施例2)によっても形成されることをまた示す。我々はまた広範囲の細胞型が15-LOを発現することを示す。興味深いことに、炎症疾患又は炎症プロセスを含む疾患に罹患している患者の細胞、例えばホジキンリンパ腫の患者のリンパ節中のリード・シュテルンベルク細胞、サルコイドーシス患者の肺バイオプシーの類上皮細胞、関節リウマチの滑膜細胞及びアルツハイマー病の患者のニューロン(実施例3)は、コントロール細胞と比較して15-LOの増大した発現を示した。
【0023】
これらの新しい代謝物の生合成の立証と、関連する酵素の同定に加えて、エオキシン類の生物学的効果をまた記載する。例えば、実施例6aには、エオキシン類C4、D4及びE4は内皮細胞の透過を引き起こす点でヒスタミンより約100倍強力であることが示され(炎症のインビトロモデル)、5-LO経路を通して形成された姉妹代謝物LTC4に匹敵する作用強度を示す。例えば、エオキシン類C4、D4及びE4はコンフルエントなウシ又はヒト内皮細胞を有するモデル系において10-7及び10-8Mにて有意な血漿漏出を誘導する。炎症の他のモデルでは、 EoxC4及びEoxD4はGM-CSFで刺激された好中球による酸素フリーラジカル生成を主に誘導したが、EoxE4はIL-5で刺激された好酸球による酸素フリーラジカル生成を主に誘導した。我々はまたエオキシン類C4、D4及びE4の形成を阻止する化合物を実証した。
【0024】
我々は、それぞれエオキシンC4(EoxC4)、エオキシンD4(EoxD4)及びエオキシンE4(EoxE4)と命名されたエオキシン:14,15-LTC4、14,15-LTD4、及び14,15-LTE4と命名されたアラキドン酸からの3種の15-リポキシゲナーゼ誘導代謝物を同定する。
アラキドン酸からの15-LO誘導システイニル含有産物エオキシンC4、D4及びE4の形成は真核又は原核細胞において以前には決して記載されていない。
【0025】
RP-HPLCを使用して代謝物を単離し、代謝物の構造を陰イオンエレクトロスプレータンデム質量分析法によって決定した。エオキシン類は282nmで最大UV吸光度を示すコンジュゲートトリエン構造を有している。我々は、これらの代謝物が、アラキドン酸が15-LOで触媒された転換により14,15-オキシド-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTA4;エオキシンA4)になり、これがグルタチオンとコンジュゲートさせられて14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTC4;エオキシンC4)になり、これが続いて14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTD4;エオキシンD4)及び14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTE4;エオキシンE4)まで代謝されることによって形成されると考える。
【0026】
14,15-LTA4(エオキシンA4)からの14,15-LTD4(エオキシンD4)の形成に関与する酵素は、実施例4において検討されるように、LTC4-シンターゼ及び/又は可溶型グルタチオンS-トランスフェラーゼ(GST)、特にGSTM1、及びGSTP1であると考えられる。
示唆される経路を以下に示す:
【発明を実施するための形態】
【0027】
本発明の第一の態様は、生物システムにおけるエオキシンC4,エオキシンD4,又はエオキシンE4の形成を調節する化合物を同定する方法であって、15-LO活性又はGSTM1-1、GSTP1-1及び/又はロイコトリエンC4シンターゼ活性に対する効果について化合物を試験することを含む方法を提供する。
本発明の第二の態様は、炎症調節効果又は骨量減少調節(典型的には骨量減少低減)効果を有する化合物を同定する方法であって、生物システムにおけるエオキシンC4,エオキシンD4又はエオキシンE4の形成及び/又は活性に対する効果について化合物を試験することを含む方法を提供する。
【0028】
「形成を調節する」には、エオキシンC4、D4又はE4の形成及び/又は活性を阻害又は促進することが含まれる。典型的には、該方法は、EoxC4、D4又はE4の形成及び/又は活性を阻害するための化合物を同定するために使用しうる。かかる化合物はまた骨量減少低減化合物であると考えられる。該化合物は典型的にはEoxC4、EoxD4及びEoxE4の形成及び/又は活性を阻害しうる。
該方法は、あるいは、EoxC4、EoxD4又はEoxE4の形成及び/又は活性を促進するための化合物を同定するために使用することができる。かかる化合物は炎症誘発性であると考えられる。かかる化合物は典型的にはEoxC4、EoxD4及びEoxE4の形成を促進しうる。
【0029】
本発明の第一の態様の方法は、生物システムにおけるEoxC4、EoxD4又はEoxE4の形成又は活性に対する効果について化合物を試験する工程を更に含みうる。
生物システムは典型的には15-LO、グルタチオンSトランスフェラーゼ(典型的には可溶型グルタチオンSトランスフェラーゼ、例えばGSTM1-1又はGSTP1-1)及び/又はロイコトリエンC4シンターゼ活性を含む。
【0030】
化合物は、以下で更に検討するように、細胞画分又は単離酵素又は酵素群を含む系において試験することができる。あるいは、本発明の第一又は第二の態様の方法は、ホールセルに対する試験化合物の効果を試験することによって実施することができる。例えば、該方法は、エキソビボヒト細胞又は細胞株を使用して実施することができる。例えば、ヒト好酸球又は肥満細胞(例えば実施例8に記載したような、例えば臍帯血由来の肥満細胞)又はL1236細胞株を使用することができるが、例えば15-LO及び/又はGSTのような必要な酵素を発現する(内因性又は組換えの何れでも)ヒト鼻ポリープ組織及び/又は樹状細胞又は任意の他の細胞又は細胞株もまた使用することができる。しかしながら、この種の代謝経路に関与する酵素が同じ位置にある必要はない。様々なアラキドン酸代謝物の形成におけるよく知られた現象は経細胞生合成、つまり最初の酵素工程が一つの細胞型において触媒され、中間体が他の細胞型に搬出されて、そこで第二の酵素工程が実施されるものである。例えばエオキシンA4の生合成は一細胞型において生じる一方、エオキシンA4からエオキシンC4への転換(及び場合によってはエオキシンC4からエオキシンD4及びエオキシンE4への転換も)は第二の細胞型において生じうる。よって、試験は、エオキシン類の生合成のための補完酵素能力を持つ2つの細胞型の同時インキュベーションで実施することもまたできる。
【0031】
当業者には明らかなように、アッセイ手順の前又はその一部として細胞を刺激し;又は例えば実施例に記載しているように、アッセイにアラキドン酸を提供することが必要である。実施例8において検討するように、LTC4、プロスタグランジンD2又はIL-5によって好酸球を刺激してEoxC4を産生させることができる。15-LO-1の発現を誘導するためには例えばIL-4を使用して肥満細胞を刺激し;ついで実施例8で検討するようにアッセイにおいてアラキドン酸を提供することが望ましい場合がある。他の細胞はエオキシン類を放出することができるために刺激を必要としない場合がある。例えば、実施例8において検討されるように、ヒト鼻ポリープ細胞はエオキシン類を自発的に放出することができると考えられる。
【0032】
当業者には明らかなように、エオキシンC4の形成に対する効果が試験されているならば、エオキシンC4の代謝物の生合成に関与する現在の酵素(例えばg-グルタミル-トランスペプチダーゼ又はジペプチダーゼ)を有していることは必要ではない。同様に、15-LO活性に対する効果が試験されている場合、15-LO(エオキシンA4)の代謝物の生合成に関与する現在の酵素、例えばLTC4シンターゼ又は可溶型グルタチオンSトランスフェラーゼ、例えばGSTM1-1又はGSTP1-1を有していることは必要ではない。当業者には明らかなように、一又は限られた数の酵素が存在している系と、より多くの酵素(例えばアラキドン酸からのエオキシン合成に関与する酵素の全て、例えば15-LO;LTC4シンターゼ又は可溶型GST、例えばGSTM1-1又はGSTP1-1)が存在する系の双方において化合物を試験することが望ましい場合がある。当業者にはよく知られているように、単一又は限られた酵素系での試験は構造-機能関係を樹立するのに有用でありうる一方;複雑な系での試験は化合物がインビボで有用である可能性があるかどうかを樹立するのに有用でありうる。
【0033】
上述のように、ある化合物がエオキシンC4、エオキシンD4又はエオキシンE4の形成を阻害することができるかどうかを試験する適切な方法は、実施例7に例証する。しかしながら、別法として、他の細胞株は、それらがエオキシンを生産する酵素的機構を有しているならば使用することができ、細胞型に応じた手順の変更は当業者には明らかである。
エオキシン類は、例えばHPLC、ガスクロマトグラフィー、質量分析法、UV分光法及び異なった免疫学的方法、例えばEIA及びRIAによって検出し定量することができる。
【0034】
現在、上記のように、15-LO酵素の2つの型がクローニングされている。2型15-LOは1型15-LOとおよそ40%同一である。1型15-LOとは異なり、リノール酸は2型15-LOのための乏しい基質である。現在の知識では、阻害のための好ましい酵素はよって15-LOの1型であるが、15-LOの2型を、エオキシン類C4、D4及びE4の形成に至る代謝経路における酵素として除外することはできない。
15-LOの触媒活性は例えば記載されているようにして測定することができる9。該酵素は、当該試験化合物の存在下又は不在下で15-(s)-HETEへのアラキドン酸(基質)の転換又は13-ヒドロキシオクタデカジエン酸(13-HODE)へのリノール酸(基質)の転換を評価することによってアッセイすることができる。化合物が存在している場合における15-(S)-HETE又は13-HODEの形成の減少は酵素活性における減少を例証する。そのような化合物は、エオキシン類C4、D4及びE4の形成において最初の酵素段階を阻止する。かかる化合物はインビボで抗炎症作用を有していると考えられる。
【0035】
本発明の第一の態様の一側面では、ヒト15-LOの触媒活性を試験するためのアッセイは実施例5に記載されているように考えられる。ここに示したアッセイは、ジフェニル-1-ピレニルホスホン(DPPP)をホスフィンオキシドに酸化する脂質ヒドロペルオキシドの能力を利用する。ジフェニル-1-ピレニルホスホンは酸化されるまでは非蛍光化合物である。DPPPの酸化は蛍光化合物を生じ、これは温度に感受性がなく、少なくとも60分間安定である。
【0036】
本発明の第一の態様の他の側面では、問題の試験化合物の存在下又は不在下で、15-(S)-HETEへのアラキドン酸又は13-ヒドロキシオクタデカジエン酸(13-HODE)へのリノール酸(基質)の転換を決定するために細胞可溶化物においてアッセイを実施する。
上記の新規の15-リポキシゲナーゼで触媒されたアラキドン酸代謝物の形成における重要な酵素の他のものは可溶型GSTである。実施例4に示すように、GSTM1-1、GSTP1-1及び/又はLTC4シンターゼはエオキシン類C4、D4及びE4の形成に関与しうる。よって、本発明の第一態様の実施態様では、GSTM1-1、GSTP1-1及び/又はLTC4シンターゼの触媒活性に対する試験化合物の効果が評価される。GSTM1-1又はGSTP1-1の触媒活性のアッセイは本質的に実施例4に記載されているようにして実施することができる。
【0037】
本発明の第二の態様の実施態様では、実施例6aに記載されたもののように、炎症のインビトロモデルにおいてエオキシンC4、D4又はE4で誘導した内皮細胞の透過を調節するその能力について試験化合物をアッセイする。かかる試験はまた本発明の第一の態様の方法との関連で実施することもできる。
化合物は、例えば炎症のウサギモデルのように15-LOが存在している動物又は炎症のエキソビボモデルにおいて又は15-LOを発現するトランスジェニック動物(例えばマウス又はラット)において試験することができる。化合物は炎症のヒトエキソビボモデルにおいて、例えばヒト末梢血又はヒト臍帯血について、又はヒト末梢血もしくはヒト臍帯血から単離した細胞について、例えば白血球、例えば好中球、好酸球、樹状細胞又は肥満細胞について試験することができる。かかる試験の例は実施例の例えば実施例6bに記載される。該化合物はまた例えばUS2003/0175680に記載されているように、幹細胞又は他の骨形成細胞前駆体を骨細胞に分化させる能力について試験することができる。
【0038】
試験される化合物は、例えば上で検討したように、化合物の構造が15-LOの基質結合部位と比較され、又はそれに基づいて設計もしくは修飾される構造ベースのスクリーニング法に基づいて選択することができる。出発化合物は最初は15-LO酵素活性に対する効果について(例えば実施例5に記載されているアッセイを使用して)スクリーニングすることによって選択することができ;ついで、構造と比較し;例えばエオキシンC4、D4又はE4形成に対する効果について更なるモデル化及び/又は合成及び評価によってついで試験することができる更なる化合物を設計するための基礎として使用される。
【0039】
(例えば実施例5に記載されているような)DPPPを使用する15-LO活性に対する効果の評価は、分光光度アッセイを使用することができ、直ぐに実施でき、例えば高スループット試験に非常に適したマイクロタイタープレート方式に使用することができることを意味する。アラキドン酸又は15-HPETE又は他のアラキドン酸代謝物を基質として使用する評価は、15-LO活性をインビボで調節する化合物の能力を確認するのに有用でありうる。
化合物にはまた当業者によく知られているように他の試験、例えば毒性学又は代謝試験を施すことができる。
【0040】
本発明は、EoxC4、EoxD4及びEoxE4の一又は複数の形成又は活性を調節、例えば亢進又は阻害するのに有用な薬剤を同定しようとする際に使用されるスクリーニングアッセイを提供することが理解される。該方法において同定される化合物はそれ自体薬剤として有用であり得、あるいはそれらはより効能のある化合物の設計と合成のためのリード化合物を表しうる。
該化合物は上記の化合物を同定する方法の各々に対して薬物様化合物又は薬物様化合物の開発のためのリード化合物でありうる。上記方法は、当業者によく知られているように、薬学的化合物又は薬物の開発におけるスクリーニングアッセイとして有用でありうることが理解される。
【0041】
「薬物様化合物」という用語は、当業者によく知られており、医学における使用に適する特性、例えば、医薬品における有効成分としての特性を有する化合物の意味を含んでいてよい。従って、例えば、薬物様化合物は、有機化学の技術によるか、それより好ましくはないが、分子生物学又は生化学の技術により合成される分子であってよく、5000ダルトン未満であってよい小分子であることが好ましい。薬物様化合物は、特定のタンパク質又はタンパク質群との選択的な相互作用の特徴を更に示し、生物利用及び/又は細胞膜を透過可能でありうるが、これらの特徴が必須ではないことは理解されるであろう。
「リード化合物」という用語は、同様に当業者によく知られており、それ自体薬物としての使用に適さない(例えば、その所期の標的に対して弱い効力を示すにすぎず、その作用が非選択的で、不安定で、溶解性に乏しく、合成が困難又は不十分な生物学的利用能を有する)が、その化合物がより望ましい特性を有する他の化合物の設計の出発点を与えるものであるという意味を含みうる。
【0042】
高スループット操作が可能なスクリーニングアッセイが特に好ましいことが理解される。上で検討したように、DPPPを使用する酵素アッセイ法を使用することができる。
適切な場合には、炎症又はインビボでのエオキシンC4、D4又はE4の形成又は活性を調節しうる化合物を同定することは望ましいであろうことは理解される。よって、本発明の方法で使用される試薬と条件は、例えば15-LOと基質の間の相互作用が、ヒト15ーLOと天然に生じる基質(例えばアラキドン酸)との間のものに類似するように選択することができることは理解されるであろう。当業者には明らかなように、異なったアッセイ系を用いて化合物を評価することができ、その幾つかにおいてエオキシン形成に関与する特定の酵素に対する効果に対する特異性又はアッセイの簡便性を(例えば単一酵素系を使用して)最適化されうる一方、他のものでは、インビボ関連性を、例えばホールセル中の化合物の効果を評価することによって最適にすることができる。
【0043】
本発明のスクリーニングアッセイは炎症疾患又は炎症プロセスを含む疾患の治療において有用な化合物の探索に有用である。インビトロ及び/又はインビボスクリーンイングをベースとして選択した化合物を、炎症疾患又は炎症プロセスを含む疾患における抗炎症効果について更に試験することができる。同様に、本発明のスクリーニングアッセイは骨量減少の治療又は防止/骨形成亢進に有用である化合物の探索に有用である。インビトロ及び/又はインビボスクリーニングをベースにして選択された化合物は、骨量減少の患者又はその危険のある患者又は炎症プロセスを含む疾患における骨量減少/骨形成亢進効果について更に試験することができる。
【0044】
「炎症」という用語は、上で述べたもののような身体外傷、感染、慢性疾患によって誘発されうる局所的又は全身性保護反応、及び/又は外部刺激に対する化学的及び/又は生理学的反応(例えばアレルギー反応の一部として)によって特徴付けられる任意の症状を含むと当業者には理解される。有害な薬剤と傷ついた組織の双方を破壊し、希釈し又は隔離する作用をするあらゆるそのような応答は、例えば発熱、腫れ、痛み、発赤、血管拡張及び/又は血流増加、白血球の罹患領域への侵入、機能喪失及び/又は炎症症状に伴うことが知られている任意の他の徴候に顕れうる。
よって、「炎症」という用語は、任意の炎症性疾患、障害又は症状自体、それを伴う炎症要素を持つ任意の症状、及び/又はとりわけ急性、慢性、潰瘍、特異、アレルギー性及び壊死性炎症、及び当業者に知られている炎症の他の形態を含む、徴候として炎症によって特徴付けられる任意の症状を含むものとまた理解される。よって、その用語はまた本発明の目的に対して、炎症痛及び/又は発熱を含む。
【0045】
エオキシンC4、D4又はE4の形成又は活性を減少させる化合物は炎症疾患又は炎症プロセスを含む疾患を治療するのに有用である。従って、本発明のスクリーニング法は、喘息、慢性閉塞性肺疾患(COPD)、肺線維症、アレルギー性疾患、鼻炎、炎症性腸疾患、潰瘍、炎症性痛覚、発熱、アテローム性動脈硬化症、冠動脈疾患、脈管炎、膵炎、関節炎、変形性関節炎、関節リウマチ、結膜炎、虹彩炎、強膜炎、ブドウ膜炎、創傷、皮膚炎、湿疹、乾癬、発作、糖尿病、自己免疫疾患、アルツハイマー病、多発性硬化症、サルコイドーシス、アテローム性動脈硬化症、高血圧(例えばAnning PB等 (March 2005) Elevated endothelial nitric oxide bioactivity and resistance to angiotensin-dependent hypertension in 12/15-lipoxygenase knockout mice. Am J Pathol. 166(3):653-62を参照)、ホジキンリンパ腫及び他の悪性腫瘍、又は炎症成分を有する任意の他の疾患の治療に有用であり得る化合物を同定するのに有用である。
【0046】
本発明のスクリーニング法は、またUS2003/0175680において検討されているように、例えば骨粗鬆症、骨関節炎、パジェット病又は歯周病に伴う骨量減少の治療又は予防に有用である化合物を同定するのに有用であると考えられる。よって、本発明のスクリーニング法は、患者における骨ミネラル密度の増加、並びに骨折の発生の低減及び/又は治癒において有用であり得る化合物を同定するのにも有用であり得る。
該化合物は上述の症状の治療的及び/又は予防的処置の双方において有用であり得る。
【0047】
エオキシンC4、D4又はE4の形成又は活性を減少させる化合物は、例えば免疫抑制性患者、例えば化学療法又は移植の拒絶を減少させる治療を受けている患者において炎症応答の促進が望ましい場合がある患者を治療するのに有用であると考えられる。例えばEoxC4、EoxD4又はEoxE4は、炎症応答の促進が望ましい患者を治療するのに有用であると考えられる。
【0048】
従って、本発明の更なる態様は、治療的有効量のEoxC4,EoxD4又はEoxE4を患者に投与することを含む、炎症反応の促進を必要とする患者の治療方法を提供する。本発明の更なる態様は、炎症反応の促進を必要とする患者の治療のための医薬の製造におけるEoxC4,EoxD4又はEoxE4の使用を提供する。
【0049】
本発明のスクリーニング法を実施する結果として、例えば実施例7に示されるように、典型的にはペプチド又は低分子でありうる化合物が同定される。該化合物は、薬学的使用、例えば患者、例えば動物(典型的には15-LOを発現する哺乳動物、例えばマウス、ラット、ウサギ、イヌ又はサル、好ましくはウサギ又は他の動物(例えばトランスジェニック動物))又はヒトにおけるインビボ治験での使用のために製剤化することができる。
【0050】
本発明の第三の態様は、抗炎症組成物又は骨量減少低減組成物を製造する方法であって、
(i)本発明の第二の態様の方法によって抗炎症又は骨量減少低減化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法を提供する。
本発明の第四の態様は、エオキシン形成調節組成物を製造する方法であって、
(i)本発明の第一の態様の方法によってエオキシン形成調節化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法を提供する。
【0051】
該組成物は典型的には有効量の化合物を患者に供給するものである。「有効量」という用語は、治療された患者に治療効果を付与する化合物の量を意味する。効果は客観的(すなわち、ある試験又はマーカーで測定可能)でありうるか、又は主観的(すなわち、患者が効果の顕れを示すか又は効果を感じる)でありうる。
該組成物は、経口的、静脈内、皮下的、口腔的、経直腸的、皮膚的、経鼻的、経気管的、経気管支的、舌下的、任意の他の非経口経路によって又は吸入を介して、 薬学的に許容可能な投薬形態で投与されることが意図され、又はそのようにして投与されるのに適したものでありうる。該組成物は、例えば経口投与のための錠剤、カプセル剤又はエリキシル剤、直腸投与のための坐薬、非経口又は筋肉内投与のための滅菌液又は懸濁液等々でありうる。該組成物は吸入用のスプレー又は注射用の液体であり得る。かかる組成物は標準的な及び/又は認められている薬学的実務に従って調製することができる。
【0052】
該組成物はまた(例えばNSAIDs、コキシブ類、副腎皮質ステロイド類、鎮痛剤、5-リポキシゲナーゼ阻害剤、FLAP阻害剤(5-リポキシゲナーゼ活性化タンパク質)、及びロイコトリエンレセプターアンタゴニスト(LTRas)、及び/又は炎症の治療に有用な他の治療剤から選択される)ここで定義された炎症の治療に有用な他の治療剤を含みうる。
あるいは、該組成物はまた骨量減少の低減又は骨形成の増加に有用な他の治療剤、例えばビスホスホネート、エストロゲン、選択的エストロゲンレセプター調節因子(SERM)、カルシトニン(calcinonin)又はタンパク質同化薬を含みうる。かかる治療剤の例は、例えばUS2003/0175680の、例えば段落[0065]で検討されている。
かかる組合せ製品は他の治療剤との関連で同定された化合物の投与をもたらし、よって、その製剤の少なくとも一つが同定されたものを含み、少なくとも一つが他の治療剤を含む別個の製剤として提供することができ、又は併用調製物(つまり同定された化合物と他の治療剤を含む単一製剤として提供)として提供(つまり製剤化)されうる。
【0053】
化合物は様々な用量で投与できる。経口、肺及び局所用量は、1日・kg体重当たり約0.01(mg/kg/日)から約100mg/kg/日、好ましくは約0.01から約10mg/kg/日、より好ましくは約0.1から約5.0mg/kg/日の範囲であり得る。例えば、経口投与では、組成物は典型的には約0.01mgから約500mg、好ましくは約1mgから約100mgの活性成分を含む。経静脈的には、最も好ましい用量は一定速度の注入の間、約0.001から約10mg/kg/時間の範囲である。有利には、化合物は単一の一日用量で投与することができ、又は一日の全用量を、毎日2回、3回又は4回の分割量で投与してもよい。
【0054】
とにかく、医師又は当業者であれば、個々の患者に最も適し、投与経路、治療されるべき症状のタイプと重症度、並びに処置される特定の患者の種、年齢、体重、性別、腎機能、肝機能及び応答によって変わり得る実際の用量を決定することができるであろう。上記の用量は平均的な場合の例である;より高い又は低い用量範囲が有利となる個々の場合ももちろん有り得、それも本発明の範囲内である。例えば実施例7はエオキシン類の形成を阻害可能な化合物が同定される方法を示している。
次に、本発明を次の非限定的な図面及び実施例を参照しながら更に説明する。
【図面の簡単な説明】
【0055】
【図1】アラキドン酸(40μM)と共に10分間インキュベーションした後のL1236細胞によって形成された236nmでのUV吸光度を持つ産物のRP-HPLCクロマトグラム。挿入:それぞれ15(S)-HETE及び12(S)-HETEで共溶出した物質のUVスペクトル。
【図2a】アラキドン酸(40μM)と共に10分間インキュベーションした後のL1236細胞によって形成された280nmでのUV吸光度を持つ産物のRP-HPLCクロマトグラム。1-5の数字は合成標準の保持時間に対応する:1)8(R),15(S)-LTB4;2)8(S),15(S)-DHETE;3)8(S),15(S)-LTB4,4)8(R),15(S)-DHETE及び5)14(R),15(S)-DHETE。
【図2b】ピークI-IV下で溶出する代謝物のUVスペクトル。
【図3】ピークIIの物質の[M-H]-イオンの400eV(Elab)CIDスペクトル。上方パネル:LTC4;中央パネル:ピークII;下方パネル:合成14,15-LTC4。
【図4】ピークIVの物質の[M-H]-イオンの400eV(Elab)CIDスペクトル。上方パネル:LTD4;中央パネル:ピークIV;下方パネル:合成14,15-LTD4。
【図5】14,15-LTA4(2μM)と共に30分インキュベートした後にL1236細胞によって生成された産物のRP-HPLCクロマトグラム。1、3及び5の数は、それぞれ8(R),15(S)-LTB4;8(s),15(S)-LTB4、及び14(R),15(S)-LTB4に対する合成標準の保持時間に対応する。ピークI-IVは図2のピークI-IVと同じ保持時間及びUVスペクトルを有していた。挿入:ピークV下で溶出する物質のUVスペクトル。
【図6】ピークVの物質の[M-H]-イオンの400eV(Elab)CIDスペクトル。上方パネル:LTE4;中央パネル:ピークV;下方パネル:合成14,15-LTE4。
【図7】15-LO経路及び14,15-ロイコトリエン類(エオキシン類)の生合成のための示唆された経路の概観。
【図8a】アラキドン酸と共になされたL1236細胞のインキュベーション。a)40μMのアラキドン酸と共に行ったインキュベーション後の14,15-LTC4及び14,15-LTD4の形成の時間経過。
【図8b】アラキドン酸と共になされたL1236細胞のインキュベーション。b)示されている様々な濃度のアラキドン酸と共に行った5分のインキュベーション後の14,15-LTC4及び14,15-LTD4の形成の用量応答曲線。
【図9a】アラキドン酸(20μM)と共に5分間なされたインキュベーション後にヒト好酸球細胞によって生成された産物のRP-HPLCクロマトグラム。a)形成された産物のHPLCクロマトグラム。エオキシンC4、LTC4、8,15-DHETEs(図2参照)及び14(R),15(S)-DHETEに対する合成標準との共溶出が示されている。
【図9b】アラキドン酸(20μM)と共に5分間なされたインキュベーション後にヒト好酸球細胞によって生成された産物のRP-HPLCクロマトグラム。b)合成エオシンC4と共溶出するピーク下の物質のUVスペクトル。
【図10】a)エオシンC4と共溶出する物質、b)合成エオシンC4及びc)合成LTC4のポジティブイオンMS/MSスペクトル。
【図11】好酸球を5、10、20及び40μMのアラキドン酸と共に、a)2分、b)5分及びc)10分インキュベートした。2回の別個の実験の平均。
【図12a】切り取ったヒト鼻ポリープを40μMのアラキドン酸と共に5分間インキュベートした後にエオキシンC4(14,15-LTC4)及び一連の8,15-DHETE及び14,15-DHETE代謝物を形成。
【図12b】合成エオキシンC4(14,15-LTC4)と共溶出するピークの物質のUVスペクトル。
【図13a】ヒト樹状細胞を40μMのアラキドン酸と共に5分間インキュベートした後にエオキシンC4(14,15-LTC4)及び一連の8,15-DHETE及び14,15-DHETE代謝物を形成。
【図13b】合成エオキシンC4(14,15-LTC4)と共溶出するピークの物質のUVスペクトル。
【図14】免疫染色。ホジキンリンパ腫の患者からのリンパ節における15-LOの発現。ヒト15-LOの特異的ペプチド配列に対して産生されたポリクローナル15-リピキシゲナーゼ抗血清と続いて二次ペルオキシダーゼコンジュゲート抗体を用いた。リード・シュテルンベルク細胞における強い染色が観察された(図の中央部分の多核細胞)。
【図15】免疫染色。サルコイドーシスの患者からの肺バイオプシーにおける15-LOの発現。ヒト15-LOの特異的ペプチド配列に対して産生されたポリクローナル15-リピキシゲナーゼ抗血清と続いて二次ペルオキシダーゼコンジュゲート抗体を用いた。類上皮細胞(肉芽腫)における強い染色が観察された。
【図16】免疫染色。関節リウマチ患者のA)滑膜細胞及びB)軟骨細胞、C)アルツハイマー患者の脳皮質からの錐体細胞、及びD)星状細胞腫検体からの星状膠細胞の検体における15-LOの発現。
【図17a】生成されたGSTM1-1によるA)エオキシンA4(14,15-LTA4)からエオキシンC4(14,15-LTC4)への代謝。
【図17b】生成されたGSTM1-1によるB)LTA4からLTC4への代謝。
【図18】様々な可溶型グルタチオンS-トランスフェラーゼによるエオキシンA4(14,15-LTA4)からエオキシンC4(14,15-LTC4)への代謝。
【図19】好中球の細胞成分分画をEoxA4(40μM)と共に5分間インキュベートした後に形成された生成物のHPLCクロマトグラム。A)上清及びB)膜分画。
【図20】EoxA4(10μM)(上方パネル)LTA4(10mM)(中央パネル)又は両方(下方パネル)と共に37℃で5分間インキュベートした後のヒト好酸球に形成された生成物のHPLCクロマトグラム。
【図21】エオキシンC4及びLTC4の形成に対する5-リポキシゲナーゼ阻害剤(BW70C)及びLTC4シンターゼ阻害剤(MK886)の効果。37℃で5分間のA23187(1μM)及びアラキドン酸(10μM)での刺激の前に阻害剤の不在下又は存在下で2分間プレインキュベートした。
【図22】コンフルエントなウシ内皮細胞(最大効果70%)を持つモデル系における減少した抵抗として測定した血漿漏出に対するエオキシンC4(14,15-LTC4)、エオキシンD4(14,15-LTD4)、エオキシンE4(14,15-LTE4)及びLTC4の効果。
【図23a】GM-CSFで刺激された好中球によるエオキシン誘導酸化的代謝。測定されたパラメータを示す動的化学発光曲線。a.酸素フリーラジカルを生産するように刺激された(塗り潰し記号)又はGM-CSF処理された後に刺激された(白抜き記号)精製血液好中球。「BLX-26」はBLX-926を示すために使用する。
【図23b】GM-CSFで刺激された好中球によるエオキシン誘導酸化的代謝。測定されたパラメータを示す動的化学発光曲線。b:EoxD4(BLX-926)はGM-CSF刺激好中球によって酸素フリーラジカルを誘導した。結果は全ラジカル生成を示し、コントロール(刺激細胞)の%として、また平均+/−標準偏差、n=13として表している。
【図23c】GM-CSFで刺激された好中球によるエオキシン誘導酸化的代謝。測定されたパラメータを示す動的化学発光曲線。c:散乱したプロットは、EoxD4(BLX-926)がGM-CSF刺激好中球により酸素フリーラジカルを誘導したことを示している。結果は1分当たりのカウント(CPM×106),n=13として測定された全ラジカル生成を示す。線は基準線(k=1)を示し、その線の上の観測は全て刺激に対するレスポンダーであることを示す。
【図24a】EoxE4(BLX-927)。測定されたパラメータを示す動的化学発光曲線。a.酸素フリーラジカルを生産するように刺激された(塗り潰し記号)又はIL-5処理された後に刺激された(白抜き記号)精製血液好酸球。「BLX-27」はBLX-927を示すために使用する。
【図24b】EoxE4(BLX-927)。測定されたパラメータを示す動的化学発光曲線。b:刺激及びEoxD4(BLX-927)はIL-5刺激好酸球によって酸素フリーラジカルを誘導した。結果は全ラジカル生成を示し、コントロール(刺激細胞)の%として、また平均+/−標準偏差、n=6として表している。
【図24c】EoxE4(BLX-927)。測定されたパラメータを示す動的化学発光曲線。c:散乱したプロットは、EoxD4(BLX-927)がIL-5刺激好酸球により酸素フリーラジカルを誘導したことを示している。結果は1分当たりのカウント(CPM×106),n=6として測定された全ラジカル生成を示す。線は基準線(k=1)を示し、その線の上の観測は全て刺激に対するレスポンダーであることを示す。
【図25a】実施例7及び図27及び28で試験した化合物の構造。
【図25b】実施例7及び図27及び28で試験した化合物の構造。
【図26】エオキシンC4(14,15-LTC4)及びエオキシンD4(14,15-LTD4)の形成に対するロイコトリエン阻害剤ETYAの効果。
【図27a】エオキシンC4の形成に対する試験化合物の効果。
【図27b】エオキシンC4の形成に対する試験化合物の効果。
【図28a】エオキシンC4及び15-HETEの形成に対する試験化合物の効果の比較。a)化合物A、B、C、D及びE。
【図28b】エオキシンC4及び15-HETEの形成に対する試験化合物の効果の比較。b)化合物D、F及びG。
【図29】単離されたヒト好酸球における新規な15-リポキシゲナーゼ産物の形成。単離された好酸球(10×106細胞)をリン酸緩衝生理食塩水(1ml)中に再懸濁させ、2分間プレインキュベートした後、37℃で5分間アラキドン酸(10μM)で刺激した。3容量のメタノールを加えて反応を終結させた。蒸発後、試料を移動相に再懸濁させ、移動相としてAcN:MeOH:H20:HAc(29:19:72:0.8体積)pH5.6を使用するRP-HPLC-UV系中での分析を行った。
【図30a】新規な15-リポキシゲナーゼ産物の液体クロマトグラフィー-質量分析。パネルA:合成14,15-LTC4及びLTC4標準と比較した好酸球により形成された代謝物IのLC-MS/MSスペクトル。上方パネルは、37℃で5分間のアラキドン酸(10μM)付与後に好酸球に形成された代謝物の626.3[M+H]+の生成物イオンスキャンのスペクトルを示す。中央パネルは合成14,15-LTC4標準のスペクトルであり、下方パネルは合成LTC4標準のスペクトルである。
【図30b】新規な15-リポキシゲナーゼ産物の液体クロマトグラフィー-質量分析。パネルB:14,15-LTD4及びLTD4合成標準と比較した好酸球により形成された代謝物IIのLC-MS/MSスペクトル。上方パネルは、37℃で60分間好酸球と共にインキュベートした14,15-LTC4(10μM)から形成された代謝物の497.2[M+H]+の生成物イオンスキャンのスペクトルを示す。中央パネルは合成14,15-LTC4標準のスペクトルであり、下方パネルは合成LTC4標準のスペクトルである。
【図30c】新規な15-リポキシゲナーゼ産物の液体クロマトグラフィー-質量分析。パネルC:14,15-LTE4及びLTE4合成標準と比較した好酸球により形成された代謝物IIIのLC-MS/MSスペクトル。上方パネルは、37℃で60分間の好酸球と共にインキュベートした14,15-LTC4(10μM)から形成された代謝物の440.2[M+H]+の生成物イオンスキャンのスペクトルを示す。中央パネルは合成14,15-LTC4標準のスペクトルであり、下方パネルは合成LTC4標準のスペクトルである。
【図31】衝突は14,15-ロイコトリエン類をシステイニルロイコトリエンから区別する解離断片を誘導した。陽イオンモードのMS/MS生成物イオンスペクトルにおけるシステイニルロイコトリエン類と14,15-ロイコトリエン類の顕著な差はm/z189及び205である。これらの断片イオンの推定起点を示す。
【図32a】14,15-ロイコトリエン類によって誘導された血管透過性の変化。パネルA:14,15-LTC4、14,15-LTD4及び14,15-LTE4、LTC4(10-8M)及びヒスタミン(10-5M)に応答した内皮細胞障壁機能における急速な変化の解析。分析は内皮細胞単層の各側に電極を配した経内皮電気抵抗(TEER)の連続的記録によって達成した。このモデルの最大効果は-70%の抵抗変化である。5回のうち2回の典型的な実験が実証される。各値は3通りの測定の平均値である。
【図32b】14,15-ロイコトリエン類によって誘導された血管透過性の変化。パネルB:LTD4(10-8M)と比較した、14,15-LTD4(それぞれ10-8M及び10-7M)によって引き起こされる内皮細胞障壁機能の変化の時間経過。
【図33】14,15-ロイコトリエン類(エオキシン類)の形成に対して提案する代謝経路。
【実施例】
【0056】
実施例1:14,15-LTC4、14,15-LTD4、及び14,15-LTE4(つまりエオキシンC4、エオキシンD4及びエオキシンE4)の生合成の証明。
材料と方法
合成14,15-オキシド-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTA4)、14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,10(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTC4)、14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTD4)、14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTE4)、8(R),15(S)-ジヒドロキシ-5,9,11,13(Z,E,E,E)-エイコサテトラエン酸(8(R),15(S)-LTB4)、8(S),15(S)-ジヒドロキシ-5,9,11,13(Z,E,Z,E)-エイコサテトラエン酸(8(S),15(S)-DHETE)、8(S),15(S)-ジヒドロキシ-5,9,11,13(Z,E,E,E)-エイコサテトラエン酸(8(S),15(S)-LTB4)、8(R),15(S)-ジヒドロキシ-5,9,11,13(Z,E,Z,E)-エイコサテトラエン酸(8(R),15(S)-DHETE),14(R),15(S)-ジヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14(R),15(S)-DHETE又は14(R),15(S)-LTB4)、15(S)-ヒドロキシ-5,8,11,13(Z,Z,Z,E)-エイコサテトラエン酸(15-HETE)、12(S)-ヒドロキシ-5,8,10,14エイコサテトラエン酸(Z,Z,E,Z)(12(S)-HETE)及び13(S)-ヒドロキシオクタデカ-9,11(Z,E)-ジエン酸(13(S)-HODE)はBiomole社(Plymouth Meeting, USA)から入手した。細胞培養培地RPMI 1640、仔ウシ血清、ペニシリン、ストレプロマイシン及びグルタミンはGibco,BRLから購入した。HPLC溶媒はRathburnケミカルズ社 (Walkerburn, United Kingdom)から得た。
【0057】
L1236細胞株
細胞株L1236は混合細胞性サブタイプのホジキンリンパ腫(HL)に罹患した患者の末梢血から元々は生産した15。細胞は37℃で5%のCO2の雰囲気下で懸濁させて増殖させた。培養培地は、10%の仔ウシ血清及び2mMのグルタミンを補填した100U/mlのペニシリンと100mg/mlのストレプトマイシンを含むRPMI1640であった。
【0058】
実験条件及びHPLC分析
細胞懸濁液を200×gで5分間、遠心分離し、ペレットをPBS中に懸濁させた。PBSで2回洗浄した後、細胞を10-20×106細胞/mlの濃度でPBS中に溶解させた。試料を37℃で2分間前もって温めた後、アラキドン酸、14,15-LTA4又は他の示された代謝物を添加し、続いて示された期間、インキュベートした。インキュベーションは3容量の氷冷メタノールの添加により終結させた。沈殿したタンパク質と細胞断片を遠心分離によって除去し、過剰の水とエタノールを減圧下で蒸発させた。残留物をメタノールに溶解させ、m試験管に移し、窒素流下で乾固させた。続いて、残留物を以下に記載するクロマトグラフィー溶剤で再構成させた。試料をオクタデシル逆相カラム(Chromabond, C18, 100mm, Macherey-Nagal, Duren, Germany)に充填し、0.4ml/分の流量で溶出させた。移動相は、システイニル含有アラキドン酸代謝物及びジヒドロキシ酸又はモノヒドロキシ酸それぞれの分析に対して、アセトニトリル/メタノール/水/酢酸(体積で28:18:72:0.8、pHは30%NaOHで5.6に調整)又はメタノール/水/酢酸(体積で72.5:27.5:0.01)であった。溶出した代謝物を、プログラム可能なWaters991ダイオードアレイ分光光度計を使用して検出し定量した。
【0059】
陰イオンエレクトロスプレータンデム質量分析
スペクトルを、記載されたように16、衝突ガスとしてXeを用いて400eVの実験室フレーム衝突エネルギーでハイブリッド磁気セクター/飛行時間型タンデム機器に記録した。
【0060】
結果
アラキドン酸と共にL1236細胞を10分間インキュベートすると、主要産物が得られ、これを合成15(S)-HETEと共に同時にクロマトグラフィーにかけた(図1)。また、合成12(S)-HETEと共溶出した236nmの最大UV吸光度のマイナーなピークを観察した。これらのピークの物質のUVスペクトルはそれぞれ15-HETE及び12-HETEに対して報告されたスペクトルと一致している。15-HETE対12HETEの比は約9:1であり、これはこれらの産物の15-LOの1型に触媒された形成と一致している5。これらの結果は、細胞が15-LOの1型の活性を有しているが5-リポキシゲナーゼ活性を有していないことを証明している。リノール酸は優れた基質であり、L1236細胞はリノール酸を13-HODEに効率的に代謝したが(データは示さず)、これはこれらの細胞における15-LOの1型の発現に一致している。超音波処理細胞を抗15-LOウサギポリクローナル抗体を用いたウェスタンブロットが続くSDS/PAGEによって分析すると、網状赤血球15-LOと共遊走した免疫反応性バンドが現れた(データは示さず)。
【0061】
236nmのUVを吸収するピークに加えて、約270nmの最大UV吸光度と5-7分の保持時間を持つ多数のピークがクロマトグラムに観察された。アセトニトリルベースの移動相を使用してこれら産物を分離した。図2aはアラキドン酸と共に10分のインキュベーションを行った後に形成された生成物の典型的なRP-HPLCクロマトグラムを示している。コンジュゲートトリエンスペクトルと272nmに最大UV吸光度を含む5つのピーク(1-5)が観察された。これらのピークはそれぞれ8(R),15(S)-LTB4;8(S),15(S)-DHETE;8(S),15(S)-LTB4;8(R),15(S)-DHETE及び14(R),15(S)-DHETEの合成標準に対応する溶出時間を有していた。これら生成物のパターンはアラキドン酸と共にインキュベートしたヒト気道上皮について報告されたものと殆ど同一であった17。しかしながら、クロマトグラム中の2つの更なる主要なピークはまたコンジュゲートトリエンスペクトルを有していたが、282nm(II及びIV)に最大UV吸光度を有していた(図2b)。溶出物質を集め、陰イオンエレクトロスプレータンデム質量分析法によって分析した。図3はピークII(中央パネル)からの物質の[M-H]イオンの衝突解離(CID)スペクトルを示す。質量スペクトルは、LTC4に見出されなかったイオンm/z306の存在を除いて、624に分子イオンを持ち、LTC4(上方パネル)と殆ど同一であった。これらの細胞における観察された15-LO活性に鑑みると、質量スペクトルは、代謝物が14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12-エイコサテトラエン酸(14,15-LTC4)であることを示している。生成物が確かに14,15-LTC4であったことを確認するために、合成14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸を調製した。この合成代謝物の質量スペクトル(図3、下方パネル)はピークIIの物質のスペクトルと同一であった。他の主要な代謝物ピークIVの[M-H]-イオンの対応するCIDスペクトルを図4に示す。この質量スペクトルは、LTD4に見出されなかったイオン250及び251の存在を除いて、同じ分子イオンm/z495を持つLTD4のスペクトルと非常に類似しており、代謝物が14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12-エイコサテトラエン酸(14,15-LTD4)であることを示している。合成14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸の[M-H]-イオンの対応するCIDスペクトルは代謝物IVのスペクトルとまた同一であった(図4、下方パネル)。更に、合成14,15-LTC4及び14,15-LTD4はRP-HPLCでそれぞれ代謝物II及びIVと正確に共溶出した。合成14,15-LTC4と共に細胞をインキュベートすると14,15-LTD4が形成された(データは示さず)。また、コンジュゲートトリエンスペクトルと278nmに最大UV吸光度を持つ二つの他のマイナーなピークが観察された(図2a及びb、ピークI及びIII)。ピークIIIの物質を陰イオンエレクトロスプレータンデム質量分析法によって分析した。この代謝物のCIDスペクトルは14,15-LTD4のCIDスペクトルと同一であった(データは示さず)。この代謝物の正確な構造は確認されなかった。しかしながら、この代謝物のCIDスペクトル及びUVプロファイルは、スペクトルが14,15-LTD4のスペクトルと比較して4nm低色素的にシフトしているので、オールトランストリエン構造と一致している。従って、ピークIIIの物質はおそらくは8-trans-14,15-LTD4である。ピークIの物質は質量スペクトルを得るには不十分であったが、8-trans-14,15-LTD4の形成に沿って、該代謝物は8-trans-14,15-LTC4である可能性が非常に高い。更に、細胞を合成14,15-LTC4又は14,15-LTD4と共にインキュベートするとそれぞれ代謝物I及びIIIが形成され、これは、親14,15-LTC4及び14,15-LTD4から想定されたオールトランストリエンが形成されたことを裏付けている(データは示さず)。
【0062】
14,15-LTC4及び14,15-LTD4の形成機構及び二重結合構造を解明するために、細胞を2μMの合成14,15-オキシド-5,8,10,12(Z,Z,EE)-エイコサテトラエン酸(14,15-LTA4)と共に30分インキュベートした。図5は細胞によって産生された生成物のHPLCクロマトグラムを示している。272nmに最大UV吸光度とコンジュゲートトリエンスペクトルを有する3つのピークが観察された(ピーク1、3及び5)。これらの生成物はまた細胞をアラキドン酸と共にインキュベートしたときにも形成され(図2a)、生成物をそれぞれ合成8(R),15(S)-LTB4;8(S),15(S)-LTB4及び14(R),15(S)-LTB4と同時にクロマトグラフィーにかけた。コンジュゲートトリエンスペクトルと282nmに最大UV吸光度を持つ3つのピークが観察された(ピークII、IV及びV)。代謝物II及びIVは、HPLCでそれぞれ合成14,15-LTC4及び14,15-LTD4と同一の保持時間を有しており、これらの代謝物が14,15-LTC4及び14,15-LTD4であったことを裏付けている。ピークVの物質はコンジュゲートトリエンを含み、282nmに最大吸光度を有していた。この代謝物は、細胞をアラキドン酸と共にインキュベートしたときは(図2a)、二重二原子酸素添加産物8,15-DHETEとのHPLCでの共溶出のために、検出されなかった。この後者の生成物は14,15-LTA4からは形成されないので、代謝物VはHPLCクロマトグラムで可視でき、二重二原子酸素添加産物の後ろに隠れてはいない。細胞を合成14,15-LTD4と共にインキュベートしたとくに代謝物Vがまた形成された(データは示さず)。合成14,15-LTD4と共に細胞をインキュベートした後に形成されたピークVの物質を陰イオンエレクトロスプレータンデム質量分析法によって分析した。図6は、ピークVの物質の質量スペクトルが合成14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12-エイコサテトラエン酸(14,15-LTE4)のスペクトルと同一であったことを示している。代謝物VはまたHPLCで合成14,15-LTE4と同じ保持時間を有していた。また、より少量の8-trans-14,15-LTC4(ピークI)及び8-trans-14,15-LTD4(ピークIII)が14,15-LTA4と共にインキュベートした後に細胞によって生産された。
【0063】
15-LO経路及び14,15-LTC4、14,15-LTD4及び14,15-LTE4の生合成のための示唆された経路の概観を図7に示す。
アラキドン酸(40μM)と共になされたインキュベーション後の14,15-LTC4及び14,15-LTD4の形成の時間経過を図8aに示す。実質的な量の14,15-LTC4、37pmol/106細胞が、アラキドン酸と共に30秒インキュベートした後に既に10×106細胞によって生産された。14,15-LTC4の最大レベルが5分のインキュベーション後に観察され、その後14,15-LTC4のレベルは時間と共に減少した。測定可能な量の14,15-LTD4がアラキドン酸と共にインキュベートした2分後に観察され、レベルは時間と共に増加し、30分後に最大レベルに達した。8,15-DHETEはこのHPLC系において14,15-LTE4と同時にクロマトグラフィーにかけたので、この実験において14,15-LTE4のレベルを測定することができなかった。アラキドン酸からの14,15-ロイコトリエン類の形成についての用量応答曲線は、有意な量の14,15-LTC4及び14,15-LTD4が5μMのアラキドン酸濃度で既に形成されたことを示している(5分のインキュベーション時間;図8b)。これらの代謝物のレベルはアラキドン酸濃度と共に増加し、40μMのアラキドン酸においてプラトーに達した。
14,15-LTC4;14,15-LTD4及び14,15-LTE4はそれぞれエオキシンC4、エオキシンD4及びエオキシンE4と命名した。
【0064】
実施例2:他の細胞及び組織におけるエオキシンC4(14,15-LTC4)の生合成の証明
材料と方法
ヒト好酸球の単離
静脈血(100ml)を、健常な志願者又は好酸球増加症候群(HES)を含む好酸球増加症、慢性リンパ性白血病(CLL)及びある種の炎症疾患の患者からから採血した。400×Gで15分間、最初の遠心分離を実施した。上相を捨て下相を集め、室温で30分間、Dextran(商標)沈降を施した。沈降後、白血球細胞を含む上相を400×Gで10分間、一回遠心分離した。ペレットを溶解バッファーに懸濁させ、室温で30分インキュベートして混入した赤血球を除去した。インキュベーション後、Lymphoprep(商標)を用いて密度勾配遠心分離を実施した。好中球と好酸球を含む多形核(PMN)画分を集め、磁気細胞分離(MACS)を施した。好酸球は、磁気ミクロビーズに結合させたCD16抗体を使用するネガティブ選択によって単離した。純度は、形態学的検査とメイ・グリューネバルト-ギムザ溶液での染色によって確認した。純度は常に95%を越えていることが見出された。
【0065】
細胞のインキュベーション
単離された好酸球(5−10×106/試料)をリン酸緩衝生理食塩水(1ml/試料)に再懸濁させ、37℃でイオノフォアA23187(1μM)、アラキドン酸(5、10、20又は40μM)又は双方で2、5又は10分、刺激する前に37℃で2分間、プレインキュベートした。反応を3容量のメタノールの添加により終結させた。
【0066】
15-リポキシゲナーゼ生成物の分析
細胞片及びタンパク質を除去するために、遠心分離を実施した。上清を減圧下で蒸発させ、残留物をメタノールに溶解させ、試験管に移した。ついで、試料を窒素流下で蒸発乾固させ、最後に200μlの移動相(アセトニトリル:メタノール:H2O:酢酸、29:19:72:0.8、pHは5.6にNH3で調整)に再懸濁させた。抽出された試料に、記載されたようにして逆相HPLC系での分析を施した。簡単に述べると、試料をオクタデシル逆相カラム(Chromabond C18,Macherey-Nagel,Duren,Germany)に充填し、溶出流量は1.2ml/分であった。溶出した化合物を、Waters1996HPLCシステムに連結したWaters996光ダイオードアレイ分光光度計を利用して検出し定量した。
【0067】
質量分析
質量分析を実施例1に記載されるようにして実施した。
結果
ヒト好酸球中でのエオキシンC4の生合成
HPLCによって分析した場合、コンジュゲートトリエンスペクトルを持つ一つのピーク(最大吸光度282nm)がアラキドン酸と共に行ったインキュベーション後に観察された(図9a及びb)。生成物を合成14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(EoxC4)と共にクロマトグラフィーにかけた。得られた生成物の同一性を確保するために、タンデム質量分析を実施した。代謝物のMS/MSスペクトルはEoxC4のものと本質的に同一であった(図10)。ホジキン細胞株L1236に対して示された生成物プロファイルとは異なり、好酸球は直ぐにはEoxC4をEoxD4及びEoxE4に転換しなかった。
【0068】
15-リポキシゲナーゼの発現に加えて、好酸球はまた5-リポキシゲナーゼを発現し、これがロイコトリエン類を生成する。用量応答実験を、EoxC4生成の実験条件を最適化するために実施した。またEoxC4及びLTC4の形成の時間経過を調べた(図11)。しかしながら、エオキシンD4もエオキシンE4も5分のインキュベーション後に検出されなかった。
総括すると、該結果は、好酸球が外因性アラキドン酸をEoxC4に直ぐに転換させることを証明している。
【0069】
ヒト鼻ポリープ組織におけるエオキシンC4(14,15-LTC4)の生合成
気道上皮源は鼻ポリープの外科検体である。切り取ったヒト鼻ポリープをアラキドン酸(40μM)と共に5分間インキュベートすると、14,15-LTC4及び一連の8,15-DHETE及び14,15-DHETE代謝物が形成された(図12a)。合成14,15-LTC4と共に共溶出した物質のUVスペクトルは特徴的なトリエンスペクトルを示した(図12b)。
【0070】
ヒト樹状細胞におけるエオキシンC4(14,15-LTC4)の生合成
成長因子とインターロイキン-4の存在下でのバフィーコート細胞(白血球)の分離と培養によって樹状細胞を得た。単離細胞は樹状細胞の特徴的な機能と形態を示した。これらの細胞をアラキドン酸(40μM)と共に5分間インキュベートすると、主として8,15-DHETE及び14,15-DHETEの形成に至ったが、有意な量の14,15-LTC4も形成された(図13a)。14,15-LTC4の同定はまたUV分光法に基づいた(図13b)。
【0071】
実施例3:15-LOを発現する細胞の同定
気道呼吸器上皮、好酸球、肥満細胞、及びマクロファージにおける先に記載された15-LO発現に加えて、本発明者等は、(ヒト15-LOに対して産生された抗体を使用して)数種の正常なまた病理組織における15-LOの発現を証明した。15-LOはホジキンリンパ腫(HL)の患者からのリンパ節において発現することが見出された。HLの13/15例において、15-LOの発現がリード・シュテルンベルク細胞において検出された(図14)。非ホジキンリンパ腫の何れの例でも染色はなかった。サルコイドーシスの患者からのバイオプシーは、類上皮細胞(肉芽腫)における15-LOの強い染色を裏付けた。結果は、全ての類上皮細胞が15-LOを発現することを示唆している。類上皮細胞はクローン病、TBC及び数タイプの血管炎の患者の肉芽腫において見出される。15-LOを発現することが見出された他の細胞型は、関節リウマチ患者からの滑膜細胞(図16a)及び軟骨細胞(図16b)、アルツハイマー患者の脳皮質からの錐体細胞(図16c)、及び星状細胞腫検体からの星状膠細胞(図16d)(正常組織ではない)を含んでいた。
総括すると、該結果は、15-LOと、この経路を通して形成された代謝物がこれらの疾患の病態生理に関与していることを示している。
【0072】
実施例4:L1236細胞においてエオキシンA4(14,15-LTA4)をエオキシンC4(14,15-LTC4)に転換する酵素の同定
方法
細胞培養
細胞培養は実施例1に記載されているようにして実施した。
ヒト好酸球の単離
ヒト好酸球の単離は実施例2に記載されているようにして実施した。
RT-PCR分析
好酸球からのmRNAの単離と続くRT-PCR分析を標準的な方法によって実施した。PCR反応に用いたプライマーは、ミクロソームグルタチオントランスフェラーゼI(MGSTI)、ミクロソームグルタチオントランスフェラーゼII(MGSTII)、ミクロソームグルタチオントランスフェラーゼIII(MGSTIII)及びβ-アクチン(陽性コントロール)
相補的であった。
【0073】
細胞のインキュベーション
単離された好酸球(5-10×106/試料)をリン酸緩衝生理食塩水(1ml/試料)に再懸濁させ、37℃で5分間、EoxA4(10μM)、LTA4(10μM)又は双方で刺激する前に37℃で2分間、プレインキュベートした。あるいは、阻害剤の研究では、好酸球を、イオノフォアA23187(1μM)及びアラキドン酸(40μM)で37℃で5分間刺激する前にBW70C(1μM)又はMK886(10μM)の存在下で37℃で2分間、プレインキュベートした。反応はメタノール3容量の添加によって終結させた。代謝物は、Waters1996HPLCシステムに連結したWaters996光ダイオードアレイ分光光度計を利用して検出し定量した。
【0074】
酵素研究
EoxA4からEoxC4への転換におけるグルタチオントランスフェラーゼの可能な関連を探究するために、組換え酵素(1μg)をEoxA4(100μM)及びグルタチオン(5mM)と共に37℃で5分間インキュベートした。反応を1容量の停止液(AcN:MeOH:HAc,体積50:50:1)の添加により終結させ、EoxC4を、Waters1996HPLCシステムに連結したWaters996光ダイオードアレイ分光光度計を利用して検出し定量した。
【0075】
結果
ホジキン細胞株L1236においてEoxA4をEoxC4に転換する酵素の単離、精製及び特徴付け
L1236細胞を標準的な手順によって細胞成分分画し、酵素活性を含む細胞質画分をグルタチオンセファロースカラムに充填した。結合物質をグルタチオンバッファー勾配によって溶出させた。溶出された画分を酵素活性について試験し、SDS-PAGEゲルで実施した。28kDのおよそのMwを持つ一つの単一バンドを得た。アミノ酸解析(PAC)、cDNA配列解析及びBLAST検索により、得られたバンドの同定は可溶型グルタチオントランスフェラーゼM1b-1b(GSTM1b-1b)であった。組換えGSTM1b-1bはEoxA4をEoxC4に転換したが、同一条件下ではLTA4をLTC4に代謝しなかった(図17A及びB)。
【0076】
ヒト可溶型グルタチオントランスフェラーゼファミリーのスクリーニング
GSTM1b-1bはヒト可溶型グルタチオントランスフェラーゼファミリーのメンバーであるので19、EoxA4をEoxC4へ転換する可溶型グルタチオントランスフェラーゼの能力を調べた。GSTM1b-1bに加えて、GSTP1-1の二つのアイソフォーム(Val105及びIle105)がEoxA4をEoxC4に転換する大きな能力をまた有していたことが証明された(図18)。
【0077】
好酸球におけるEoxC4シンターゼ活性の局在化
可溶型グルタチオントランスフェラーゼが好酸球におけるEoxC4形成に関与するかどうかを調べるために、単離されたヒト好酸球の細胞成分分画を調製し、EoxA4と共にインキュベートした。L1236とは異なり、EoxC4シンターゼ活性は好中球の膜画分に殆ど完全に局在化した(図19)。
【0078】
好酸球における「酵素X」の特徴付け
好酸球中のEoxC4シンターゼ活性は膜画分にあったので、本発明者等はこの点における膜結合性グルタチオントランスフェラーゼの関与を調べた。
MAPEG(エイコサノイド及びグルタチオン代謝における膜結合型タンパク質(Membrane Associated Proteins in Eicosanoid and Glutathione metabolism))スーパーファミリー21のメンバーはLTC4シンターゼ、FLAP(5-リポキシゲナーゼ活性化タンパク質)、PGEシンターゼ、MGSTI(ミクロソームグルタチオントランスフェラーゼI)、MGSTII及びMGSTIII(3)である。PCR解析は、好酸球はMGSTII及びMGSTIIIをコードするmRNAを有する一方、MGSTIをコードするmRNAは観察されなかったことを明らかにした。先の研究から、好酸球がFLAPとLTC4シンターゼを有していることは知られている。
【0079】
EoxA4及びグルタチオンと共にMAPEGファミリーメンバーをインキュベートすることにより、LTC4シンターゼだけがEoxA4をEoxC4に転換する能力を有していることが示された(データは示さず)。
LTC4シンターゼは伝統的にLTA4からグルタチオン含有ロイコトリエン(LT)C4を生合成する際の重要な酵素であることが知られている。LTA4は5-リポキシゲナーゼ経路を介して合成される。好酸球は5-リポキシゲナーゼ及び15-リポキシゲナーゼ活性を共に有しており、LTC4シンターゼがEoxA4をEoxC4へ転換することができる唯一のMAPEGファミリーメンバーであるという結果は、LTC4シンターゼが二重の活性:LTC4及びEoxC4合成を備えていることを示唆している。LTC4シンターゼがEoxC4形成においてもまた重要な酵素であるという仮説を試験するために、単離した好酸球をEoxA4(10μM)、LTA4(10μM)、又は双方と共に37℃で5分間インキュベートした(図20)。
【0080】
好酸球をEoxA4と共にインキュベーションするとEoxC4が生合成される一方、好酸球をLTA4と共にインキュベーションするとLTC4を生じる。細胞を双方の基質と共にインキュベートすると、LTC4とEoxC4がおよそ9:1の比で生産された。これはこれらの基質に対するLTC4シンターゼの過去に報告された親和性と一致している20。
LTC4シンターゼがまた好酸球においてEoxC4シンターゼとしてまた作用するという知見を更に裏付けるために、発明者等は阻害研究を実施した。
【0081】
BW70Cは5-リポキシゲナーゼ阻害剤で、MK886はFLAP阻害剤であり、これらは高い(10μM)濃度で使用される場合、LTC4シンターゼをまたブロックする(図21)。BW70Cの存在下でのプレインキュベーションがLTC4ピークを完全に消滅させ、MK886の存在下でのプレインキュベーションがまたEoxC4合成を阻止した。これらの結果は、LTC4シンターゼが好酸球中でのEoxC4形成における重要な酵素であることを更に裏付けている。
【0082】
実施例5:ヒト15-リポキシゲナーゼ(15-LO)によって形成される脂質ヒドロペルオキシドの検出のためのアッセイ
ここに記載されたアッセイは、ジフェニル-1-ピレニルホスホン(DPPP)をホスフィンオキシドに酸化する脂質ヒドロペルオキシドの能力を利用する(以下のスキームを参照のこと)。ジフェニル-1-ピレニルホスホンは酸化されるまでは非蛍光化合物である。DPPPの酸化は蛍光化合物を生じ、これは温度に感受性がなく、少なくとも60分間安定である。
【0083】
【0084】
材料
ジフェニル-1-ピレニルホスホン(DPPP)(D-7894)はMolecular Probesから購入している。コール酸はSigmaから、リノール酸はBiomolから、PBSはGibco Life Technologiesから購入している。蛍光はMolecular Devices製の二重走査型マイクロプレート分光蛍光光度計Spectramax Geminiを用いて測定する。Spectramax Gemini分光蛍光光度計はキセノンランプと二つのホログラフィック回折格子モノクロメータを使用する。384ウェルまでのマイクロプレートを分光蛍光光度計と共に使用することができる。蛍光の検出は、ウェル中の試料の深さ全体に亘ってウェルの上面から達成される。
【0085】
全ての試薬は、フルオロフォア(DPPP)及び基質、リノール酸を除いてそのまま使用できる濃度で提供される。DPPPは粉末として提供され、-20℃で保存されなければならず、光から十分に保護されなければならない。リノール酸はエタノール中の100mM原液として供給される。酵素調製物は凍結アリコートとして提供される。
【0086】
酵素はバキュロウイルスを使用して発現され、スポドプテラ・フルギペルダ細胞(Sf9)から部分的に精製され、トリス-HCl(pH8.0)、75mMのNaCl及び5%グリセロールで送達される。精製された酵素は-70℃で少なくとも5ヶ月まで安定である。3回までの凍結解凍サイクルは酵素活性に影響を与えないと思われる。しかしながら、酵素の凍結解凍サイクルは、必ずしも必要ではないなら、避けなければならない。酵素を凍結し実験の直前でPBSに希釈する。希釈した酵素は同じ日中に使用されなければならない。
【0087】
DPPP(粉末)を、0.25mMの最終濃度までエタノール及び2-ブタノール(1:1の比)に溶解させた。この溶液をボルテックスして混合し、続いて+4℃で超音波水浴で10秒間超音波処理してDPPPを完全に溶解させた。この溶液は実験が実施される日には新鮮でなければならない。該溶液は遮光されなければならない。
【0088】
脂質ヒドロペルオキシド検出のためのジフェニル-1-ピレニルホスフィン法
アッセイは室温(20-22℃)で96ウェルの全て黒のプレートで実施される。各ウェルは、500μMのコール酸と15-LO酵素(プレミックス)を補填したダルベッコのリン酸緩衝生理食塩水(PBS)からなる45μlを含む。阻害剤(1μl)又はビヒクル(DMSO)を、基質である5μlの1mMリノール酸の添加の5-15分前に添加する。ついで、プレートを室温で20分間インキュベートする。酵素反応を50μlのメタノールの添加によって終結させる。プレートは、脂質ヒドロペルオキシドの有意な損失なしに60分までの間、室温で保存することができる。50μlの25μM DPPP溶液を加え、プレートを室温でインキュベートする。30-60分後、蛍光は、355nmの励起波長と380nmの発光波長を使用して読み取ることができる。
水溶液中でのDPPPの低い自動酸化のため、ブランクの一つは酵素反応のものと同じ比でバッファー、メタノール及びDPPPを含まなければならない。
【0089】
実施例6:エオキシンC4、エオキシンD4及びエオキシンE4(14,15-LTC4、14-15-LTD4、14-15-LTE4)の生物学的効果の証明
実施例6a:内皮細胞障壁機能の評価
材料と方法
内皮細胞アッセイ
ウシ大動脈内皮細胞(BAEC)及びヒト臍帯静脈内皮細胞(HUVEC)を単離しウシ血清又はヒト血清と共に培養した。第一から第五継代のBAEC又はHUVECを、短時間(2分)のトリプシン-EDTA処理(0.25%のトリプシン/0.01%のEDTA)によって剥離させ、3.0μmの孔径のポリカーボネートフィルター(Tissue Culture inserts, 10mm; NUNC, Roskilde, Denmark)上又は目視検査のために0.2μmの孔径のAnopore無機膜(濡れると透明)に再び蒔いた。細胞分化を促進させ、ECの付着を向上させるために、フィルターを50μlのBiomatrix I(167μg/ml)で前処理し、空気乾燥させた。ECをフィルター当たり2×105細胞の密度で播種し、空気中に5%CO2の加湿雰囲気下、37℃で培養培地中でインキュベートした。透明な0.2μmの孔径のフィルターで単層に亘る電気抵抗を測定し、毎日顕微鏡で観察して評価して、ECをコンフルエントな単層になるまで増殖させた。これらの実験条件下で播種後7日で最大の抵抗が得られ、ついでその日にEC単層を実験に使用した。
【0090】
EC障壁機能の測定
様々なアラキドン酸代謝物に応答した内皮細胞(EC)障壁機能の迅速な変化を、EC単層の各側の電極での経内皮電気抵抗(TEER)の連続的記録によって分析した18。TEERの測定では、ECを有するフィルターインサートを、繰り返し測定の間に互いのかつフィルターインサートに対しての電極の正確な位置決めを達成するように変更された抵抗測定チャンバー(Endohm-12, World Precision Instruments, Sarasota, FL, USA)に移した。チャンバー(下方コンパートメント)及びフィルターインサート(上方コンパートメント)にそれぞれ2ml及び400μlの培養培地を満たした。全ての測定は細胞培養インキュベーター中に配した装置で37℃にて行った。電気抵抗の直接の読みが抵抗チャンバーの電極に接続したオーム計(Evom, World Precision Instruments, Sarasota, FL, US)によって与えられた。個々のEC単層の電気抵抗は、ECの播種前に測定したBiomatrixでコートされた対応する裸のフィルターの抵抗を減算することによって得られた。よって、TEER値はEC単層の抵抗を表し、隣接する内皮細胞間の密着結合の発生に依存する。
【0091】
結果
14,15-システイニル-含有ロイコトリエン類(エオキシン類)の生物学的効果
血管透過性の増加は炎症のマーカーであり、システイニル含有ロイコトリエン類がヒスタミンとブラジキニンと同じ形で特異的血漿漏出を引き起こすことであることはよく知られている。5-リポキシゲナーゼ誘導システイニル含有ロイコトリエン類とは異なり、EoxC4とその代謝物はモルモット肺実質性条片又は回腸への収縮効果を有していないように思われる。EoxC4とその代謝物EoxD4及びEoxE4の血漿漏出を引き起こす能力を探索するために、内皮細胞障壁機能における迅速な変化を登録するためのモデル系を構築した。発明者等は、3種の14,15-システイニル含有ロイコトリエン類が全て血管透過性を増大させたことを見出した。化合物の10-7及び10-8Mの濃度はTEERの有意な変化を誘導した。比較すると、EoxC4、D4及びE4は最も強力な5-リポキシゲナーゼ誘導ロイコトリエン類より約5倍から10倍弱かったがこの点ではヒスタミンより100倍以上強かった(図22)。
【0092】
実施例6b:白血球の酸化的代謝に対するエオキシン類の効果
好中球及び好酸球による酸化的代謝は様々な薬剤、例えば酢酸ミリスチン酸ホルボール(PMA)、血小板活性化因子(PAF)、血清処理ザイモサン(STZ)、免疫グロブリン又はロイコトリエン類(LTB4)によって、またサイトカイン、例えばIL-5及びGM-CSFによって誘導できる。薬物刺激(プライミング)は、細胞を活性化しないで、刺激に対する細胞の増加した応答性をもたらすプロセスを意味する。細胞の薬物刺激状態を生じる機構はなお不確かであるが、造血性サイトカインIL-3、IL-5及びGM-CSFは好酸球と好中球をインビトロで薬物刺激することが知られている。しかしながら、IL-5だけはヒト血液好酸球の選択的活性化因子であることが示されており、IL-3及びGM-CSFは好酸球と好中球の双方に作用しうる。
【0093】
材料と方法
好中球及び好酸球顆粒球を、Percoll勾配及び磁気細胞分離システム(MACS)を使用して末梢全血から精製した。
顆粒球にIL-5(10.5ng/mL)又はGM-CSF(10.5ng/mL)の何れかで37℃で30分薬物刺激を与え、続いてIL-5(100ng/mL)、GM-CSF(100ng/mL)、LTB4(10-7M)又はエオキシン(10-8-10-6Mで刺激した。酸化的代謝をルシゲニン増強化学発光(CL)アッセイによって測定した。
使用した試験化合物はEoxC4(BLX-925)、EoxD4(BLX-926)、EoxE4(BLX-927)であった。
【0094】
結果
非薬物刺激好中球によるエオキシン誘導酸化的代謝
精製した好中球を、好中球に対する異なったエオキシン類の刺激効果との比較のために、酸化的代謝の二つの既知の活性化因子であるサイトカインGM-CSF及びLTB4を用いて刺激した。双方の活性化因子が非薬物刺激好中球による酸素フリーラジカル生成を誘導したが、何れのエオキシン類も非薬物刺激血液好中球による酸素フリーラジカル生成を誘導しなかった。
【0095】
一つの代表的な実験を示す動態曲線(図23a)は、非薬物刺激細胞と比較した場合のサイトカイン薬物刺激細胞の刺激後の向上したラジカル生成を示している。この向上したラジカル生成は、異なったドナーの全結果を平均値として示している図23bに示されるように用量依存パターンを示した。我々は異なった血液ドナー間でEoxD4(BLX-926)に対する強い個々の用量応答性を経験したので、全ての個々のデータをプロットしScatterプロットで示した(図23c)。このようにすることによって、EoxD4(BLX-926)が、異なったドナーに対して異なった最適な用量で、用量依存的な形で薬物刺激細胞によるラジカル生成を誘導したことが明らかになった。13の内、7-10の検体がEoxD4(BLX-926)に対するレスポンダーであった。
類似のパターンがまたEoxC4(BLX-925)に対して見られた。この化合物は用量依存的な形でGM-CSF薬物刺激好中球によるラジカル生成を誘導した(13の内、7-8の検体がEoxC4に対するレスポンダーであった;データは示さず)。
【0096】
非薬物刺激好酸球によるエオキシン誘導酸化的代謝
3種全てのエオキシン類が非薬物刺激好酸球による酸素フリーラジカルの少量の生成を誘導した。しかしながら、酸素フリーラジカルの生成量は、好酸球による酸化的代謝の他の既知の活性化因子、この場合は好酸球特異的サイトカインIL-5と比較した場合にかなり少ない(図24a)。
IL-5薬物刺激好酸球によるエオキシン誘導酸化的代謝
EoxE4(BLX-27と命名)は、図24に示されるように、IL-5薬物刺激好酸球によるラジカル生成の広い用量関連誘導を示し、該エオキシンが試験したものの中で好酸球による酸化的代謝の最良の誘導因子であるかも知れないことを示している。6の検体の内、3がEoxE4に対してレスポンダーであった。
結論
EoxC4(BLX-925)及びEoxD4(BLX-926)はGM-CSF薬物刺激好中球による酸素フリーラジカル生成を主に誘導したが、EoxE4(BLX-927)はIL-5薬物刺激好酸球による酸素フリーラジカル生成を主に誘導した。
【0097】
実施例7:エオキシンC4の形成に対する15-LO阻害の効果
ホジキン細胞株L1236をアラキドン酸(40μM)で5分間刺激した。L1236細胞はアラキドン酸をEoxC4、EoxD4及び一連の8,15-DHETE及び14,15-DHETE代謝物を効果的に転換した。
材料と方法
細胞インキュベーション
ホジキン細胞株L1236(10×106細胞/試料)を1mlのPBSバッファーに懸濁させ、試験物質(1、10又は50μM)の添加前に37℃で2分予め温めた。試験物質と共に5分間プレインキュベートした後、アラキドン酸(40μM)を加えた。インキュベーションを2分後に1容量のメタノールを添加して停止させた。各試験物質濃度は二組をインキュベートした。
【0098】
固相抽出
細胞及び細胞片を試料から遠心分離(2600rpm、6分)によって除去した。上清を水で希釈し、ついで洗浄し平衡にした抽出カートリッジOasis HLB 1cc 10mg(Waters, Sweden)に移した。代謝物をメタノールで溶出させ、HPLC系に注入する前に水で希釈した。
HPLC分析
RP-HPLCをNova Pak C18カラム(2.1×150mm, Waters AB, Sweden)で実施した。最初の移動相は、0.4ml/分の流量での100%A(0.05%(8.7mM)酢酸、pHをアンモニウムで5.6に調整)からなる。線形勾配を5分後に20分の36%B(60:40のアセトニトリル:メタノール)まで開始した。ついで、移動相は、更に20分は64:36のA:Bの均一濃度に維持された。カラム流出液はダイオードアレイ検出(PDA 996, Waters AB, Sweden)を使用してモニターし、UVスペクトルを200-340nmの間で獲得した。
【0099】
試験化合物
図25はEoxC4の形成を阻害した様々な化合物の構造を示している。幾つかのものはWO2004/080999に記載されている。1-フェニルチオカルバモイル-1H-ピラゾール-3-カルボン酸(2-クロロフェニル)-アミド(化合物A)は実施例47に記載されている。ピラゾール-1,3-ジカルボン酸1-(ベンゾ[1,3]ジオキソール-5-イルアミド)3-(2-クロロ-4-フルオロフェニルアミド(化合物B)は実施例64に記載されている。6-{[3-(2-クロロ-4-フルオロフェニルカルバモイル)ピラゾール-1-カルボニル]-アミノ}-ヘキサン酸エチルエステル(化合物C)は実施例65に記載されている。ピラゾール-1,3-ジカルボン酸3-(2-クロロ-4-フルオロフェニルアミド)1-ペンチル-アミド(化合物D)は実施例66に記載されている。ピラゾール-1,3-ジカルボン酸3-(2-クロロ-4-フルオロフェニルアミド)1-ブチルアミド(化合物E)はWO2004/080999(イソシアン酸ブチルがイソシアン酸ペンチルの代わりに用いられている特に実施例66を参照)に記載の方法と同様にして調製される。
【0100】
N'-(2-(エトキシイミノ)-1-フェニルエチリデン)ベンゾヒドラジド(化合物F)は次のようにして調製することができる:
(a)安息香酸(2-ヒドロキシイミノ-1-フェニルエチリデン)ヒドラジド
安息香酸ヒドラジド(1から1.2当量)をアルコール性溶媒(例えばEtOH又はMeOH)に溶解させた。2滴の酸(例えば濃硫酸又は氷酢酸)を加えた後、2-イソニトロソアセトフェノン(1当量)を滴下して加えた。反応混合物を(例えば上昇温度(例えば50-85℃)で)撹拌した後、濾過によって形成された固形物を集め、場合によっては(例えばEtOH/水からの)粗物質を再結晶して副題化合物を得た。
【0101】
(b)N'-(2-(エトキシイミノ)-1-フェニルエチリデン)ベンゾヒドラジド
ジメチルスルホキシド中の安息香酸(2-ヒドロキシイミノ-1-フェニルエチリデン)ヒドラジド(1当量;上の工程(a)を参照)及びKOH(1から1.2当量)の溶液に臭化エチル(少なくとも1当量)を加えた。反応は所定の期間(例えば6日)上昇温度(例えば45℃)で撹拌した。水を加え、混合物をジクロロメタンで抽出した。組み合わせた有機相をNaOH(2M)と水で洗浄した。有機相を乾燥させ(Na2SO4)、真空で蒸発させ、場合によっては再結晶化させて(例えばEtOH/水から)表題化合物を得た。
【0102】
3-クロロ-N'-(2-(ヒドロキシイミノ)-1-(3-クロロフェニル)エチリデン)ベンゾヒドラジド(化合物G)は次のようにして調製することができる:
(a)2-(3-クロロフェニル)-2-オキソアセトアルデヒドオキシム
亜硝酸イソアミル(1.83g、15.6mmol)を氷冷エタノール性ナトリウムエトキシド(0.9M、18.3mL、16.5mmol)に溶解させた。冷却された液に3-クロロアセトフェノン(14.9mmol)を滴下して加え、反応を室温に達するようにし、撹拌を16時間続けた。反応の過程で形成された固形物を濾過し、ジエチルエーテルで洗浄し、水に溶解させ、水溶液を氷酢酸で酸性化した。得られた結晶性固形物を濾過し、EtOH/水から再結晶化して副題化合物を得た。
【0103】
(b)3-クロロ-N'-(2-(ヒドロキシイミノ)-1-(3-クロロフェニル)エチリデン)ベンゾヒドラジド
3-クロロ安息香酸ヒドラジド(1から1.2当量)をEtOHに溶解し、それに2滴の濃硫酸と2-(3-クロロフェニル)-2-オキソアセトアルデヒドオキシム(1当量;上の工程(a)を参照)を加えた。反応を一晩50℃に加熱し、濾過し、EtOHで洗浄し、EtOH/水から再結晶化して表題化合物を得た。
【0104】
結果
ヒト細胞株L1236におけるEoxC4形成の阻害
10-4−4Mの濃度のエイコサテトラエン酸(ETYA)は、EoxC4、EoxD4(図26)及び他の15-LO誘導生成物を完全に阻止した。これらの生成物の形成はまた10-5-5MのETYAの存在下で減少した。これらの結果は、リポキシゲナーゼ阻害剤ETYAがEoxC4及びその代謝物の形成を阻止できることを示している。
EoxC4及び15-HETEの形成に対する化合物の効果
L1236細胞を2分間、示された物質(1μM)と共にプレインキュベートされ、続いて5分間、アラキドン酸(12.5μM)と共にインキュベートした。インキュベーションと抽出は上に記載されているようにして実施された。全ての試験化合物(図25に記載のもの)はEoxC4の形成を阻害した(図27)。また、阻害剤は同様の度合いでEoxC4及び15-HETEの合成を阻止した(図28)。
【0105】
実施例8:ヒト好酸球及び肥満細胞において15-リポキシゲナーゼ-1経路を介して生産された炎症誘発性アラキドン酸代謝物の同定
略語
CBMC 臍帯血肥満細胞
EC 内皮細胞
EIA エンザイムイムノアッセイ
HETE ヒドロキシルエイコサテトラエン酸
HPLC 高速エキタイクロマトグラフィー
IL インターロイキン
LO リポキシゲナーゼ
LT ロイコトリエン
MS 質量分析法
PG プロスタグランジン
【0106】
ヒト好酸球は豊富な量の15-リポキシゲナーゼ-1(15-LO-1)を含んでいる。しかしながら、ヒトにおける15-LO-1の生物学的役割は不確かである。アラキドン酸と共に5分間好酸球をインキュベーションすると、282nmで最大吸光度を有しロイコトリエン(LT)C4よりもRP-HPLCでの保持時間が短い主要生成物の形成に至る。陽イオンエレクトロスプレータンデム質量分析法での分析によって未知の好酸球代謝物を14,15-LTC4と同定した。この化合物は好酸球中において14,15-LTD4及び14,15-LTE4に代謝されうる。また臍帯血由来肥満細胞及び外科的に除去された鼻ポリープは14,15-LTC4を生産した。好酸球をアラキドン酸と共にインキュベーションするとLTC4よりも14,15-ロイコトリエンC4の生産の方が優勢になる一方、カルシウムイオノフォアでの刺激では専らLTC4が形成された。14,15-システイニル含有ロイコトリエン類は、炎症の特徴であるインビトロでの内皮細胞(EC)単層の血管透過性の増大を誘導することができる。このモデル系において、14,15-ロイコトリエン類はLTC4及びLTD4と殆ど同じく強力であったがヒスタミンより100倍強かった。好酸球は、LTC4、プロスタグランジンD2又はインターロイキン-5での刺激後に14,15-LTC4を生産し、これは、14,15-LTC4がアラキドン酸の内因性プールから合成され得ることを証明している。総括すると、本報告は、特にヒト好酸球とまたヒト肥満細胞及び鼻ポリープにおける炎症誘発性14,15-ロイコトリエン類の形成を記載する。好酸球はこれらの新たに特徴付けた代謝物の豊富な供給源であるので、5-リポキシゲナーゼ誘導ロイコトリエン類との混同を避けるために、我々はそれぞれ14,15-ロイコトリエンC4、D4及びE4の代わりにエオキシン類C4、D4及びE4という名称を示唆する。
【0107】
序
哺乳動物のリポキシゲナーゼは、喘息及び他の炎症疾患の病因に関係がある構造的に関連した脂質過酸化酵素のファミリーに属する。5-リポキシゲナーゼ(5-LO)はロイコトリエン合成における重要な酵素であり、この酵素はアラキドン酸のロイコトリエン(LT)A4への転換を触媒する。この化合物は酵素的にLTB4まで加水分解されるか又はグルタチオンとコンジュゲートされてLTC4を形成し、これが次にLTD4及びLTE4まで代謝されうる。ロイコトリエンC4とその代謝物は炎症誘発性薬剤及び強力な気管支収縮剤である(1)。5-LOとは異なり、ヒト15-リポキシゲナーゼ(15-LO-1)の生物学的機能についてはあまり知られていない。ヒト気道内皮細胞、好酸球及び肥満細胞及び樹状細胞のサブセットは高い量の15-LO-1を構成的に発現する(2-6)。ヒト15-LOの第二の形態もまた存在し、これは15-LO-2と命名される(7)。この酵素と15-LO-1の間のアミノ酸類似性は40%だけである(3)。インターロイキン(IL)-4及び/又はIL-13は培養された上皮細胞、単球及び肥満細胞における15-LO-1の発現を誘導し(4,8,9)、よってこれらのサイトカインはインビボでの15-LO-1発現の重要な生理学的/病態生理学的調節因子として作用しうる。
15-リポキシゲナーゼ-1は気管支喘息の病因に関与していることが示唆されている。15-LO-1の量は、コントロール患者と比較して喘息又は慢性気管支炎の患者の気管支粘膜下層において有意に増加している。15-リポキシゲナーゼ-1はアラキドン酸を15-ヒドロキシ-エイコサテトラエン酸(15-HETE)に転換し、これはインビボ並びにインビトロでの様々な細胞及び組織における15-LO活性のマーカーとして使用されている。ヒト気管上皮細胞、切り出したヒト気管支及びヒト肺ホモジネートは主としてアラキドン酸を15-HETEに転換する。喘息患者からの肺検体は非喘息患者の検体よりも多くの15-HETEを生産し(12-14)、15-HETEはアトピー性及び非アトピー性喘息患者からの刺激されたヒト肺組織における主なアラキドン酸代謝物であった(15)。増加した量の15-HETEは、抗原曝露アトピー性喘息患者の気管支肺胞洗浄液(BALF)(16)、喘息患者の痰試料(17)及び健常なコントロール患者からの鼻分泌物(18)に見出された。持続性気道好酸球増加症の深刻な喘息患者は健常な被験者又は好酸球増加症のない喘息患者と比較してBALF中の高いレベルの15-HETEを有している(14)。15-HETEのレベルは基底下(sub-basement)厚と相関することが見出され、15-LOが気道リモデリングに関連しているかも知れないことを示している(14)。
【0108】
15-LO経路を介して生産される最も豊富なアラキドン酸代謝物は15(S)-HETE)であり、これは15(S)-ヒドロペルオキシ-エイコサテトラエン酸(15(S)-HPETE)から形成される。15-HETEへの還元を受けることに加えて、15-HPETEはまた脱水を受けて14,15-エポキシ-エイコサテトラエン酸(14,15-LTA4)を形成する(19,20)。他の15-リポキシゲナーゼ生成物の5-オキソ-15-ヒドロキシ-6,8,11,13-エイコサテトラエン酸はヒト好酸球に対する強力な走化性剤であると報告されている(21)。しかしながら、15-LO-1はリポキシン類及びレゾルビン類(resolvins)の形成に関与しうるので、15-LO生成物は炎症疾患において保護的役割を有しうるという証拠がまたある(22)。総括すると、ヒトにおける15-LO経路の生理学的及び病態生理学的役割は確かではない。
【0109】
この報告では、我々は、14,15-ロイコトリエンC4、D4及びE4の形成及びヒトにおいて15-LO-1に対して炎症誘発性役割を示す生物学的効果を記載する。好酸球はこれらの新たに特徴付けられた代謝物のかかる豊富な供給源であるので、5-リポキシゲナーゼ誘導ロイコトリエン類との混同を避けるために、我々はそれぞれ14,15-ロイコトリエンC4、D4及びE4の代わりにエオキシン類C4、D4及びE4という名称を示唆する。
【0110】
結果
単離されたヒト好酸球における新規な15-リポキシゲナーゼ生成物の形成
単離されたヒト好酸球をアラキドン酸と共に5分間インキュベートしておよそ9:1の比で15(S)-HETE及び12(S)-HETEが形成され(データは示さず)、これは15-LO-1の活性と一致している(2)。約270nmに最大UV吸光度を持ち5-7分の間の保持時間を有する多くのピークが同じクロマトグラムで観察された。これらのより極性の代謝物をよりよく分離するために、アセトニトリルベースの移動HPLC相を使用した。コンジュゲートトリエンスペクトルと272nmでの最大吸光度を含む5つのピークが同定された(図29)。これらの5つのピークの物質の保持時間は、二重酸素添加生成物8(S),15(S)-DHETE及び8(R),15(S)-DHETE、14,15-オキシド-ロイコトリエンA4から誘導された2つの8(S),15(S)-LTB4及び8(R),15(S)-LTB4(19,20)及び14(R),15(S)-DHETEの保持時間に対応した。形成された生成物のパターンは、アラキドン酸と共にインキュベートされたヒト気道上皮細胞に対して報告されているものと同様であった(5)。コンジュゲートしたトリエンであるが282nmに最大の特徴的なUVスペクトルを持つ少量のロイコトリエン(LT)C4がまた観察された。これらのよく知られたアラキドン酸誘導代謝物に加えて、コンジュゲートされたトリエンスペクトルを有し、282nmに最大UV吸光度を有するより極性の主要な代謝物(代謝物1)がクロマトグラムに観察された(インサート,図29)。
【0111】
LC-MS/MSによる代謝物1の同定
代謝物1を更に分析するために、外因性アラキドン酸と共にインキュベートしたヒト好酸球の試料に、陽イオンエレクトロスプレータンデム質量分析を使用するLC-MS/MSによる分析を施した。得られたイオンスキャンスペクトルはLTC4のスペクトル中にも存在する数個の断片を含んでいた(図30A)。好酸球における既知の高い15-LO活性と併せてこの知見は、代謝物1が14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTC4)でありうることを示している。この仮説を確認するために、合成14,15-LTC4を調製し(Biomol Inc, USA)、代謝物1から得られた衝突誘導解離スペクトルを、合成14,15-LTC4に対して得られたスペクトルと比較した(図30A)。試料を、プロトン化された14,15-LTC4と等価な626.2m/zでの生成物イオンスキャンで分析した。クロマトグラムは標準14,15-LTC4と同じ保持時間のピークを含んでおり、質量スペクトルを比較すると、合成14,15-LTC4と同一であった。
【0112】
ヒト好酸球における14,15-LTC4の代謝
好酸球が14,15-LTC4を14,15-LTD4及び14,15-LTE4に代謝し得るかどうかを分析するために、好酸球を、37℃で60分間、合成14,15-LTC4又は14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTD4)と共にインキュベートした。形成された代謝物を、14,15-LTD4に対しては497.2m/z、14,15-LTE4に対しては440.2m/zの生成物イオンスキャンで分析した。14,15-LTC4から形成された生成物(代謝物2)の保持時間と質量スペクトルは合成14,15-LTD4と同一であり(図30B)、14,15-LTD4とのインキュベーション後に形成された生成物(代謝物3)は合成14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTE4)と同一の保持時間と質量スペクトルを有していた(図30C)。37℃での60分のインキュベーションの間、14,15-LTC4及び14,15-LTD4の非酵素的代謝はなかった(データは示さず)。
基本的には、14,15-ロイコトリエン類と対応する5-リポキシゲナーゼ誘導システイニル-ロイコトリエン類の生成物イオン質量スペクトルは殆ど同一である。しかしながら、14,15-ロイコトリエン類は205m/zで断片を形成する一方、5-リポキシゲナーゼ誘導システイニル-ロイコトリエン類における対応する断片は189m/zにおいてである(図31)。これらの衝突誘導解離生成物は、おそらくは、システイニル基が位置しており、14,15-ロイコトリエン類では炭素14、ロイコトリエン類に対しては炭素6である炭素の隣での切断によって形成される。
【0113】
好酸球における14,15-LTC4とLTC4の形成間の比較
ヒト好酸球は豊富な量の15-リポキシゲナーゼ及び5-リポキシゲナーゼ双方を含んでいる。15-リポキシゲナーゼ及び5-リポキシゲナーゼ経路が最も活性である条件を調べるために、好酸球をアラキドン酸又はカルシウムイオノフォアA23187の何れか、又は双方と共にインキュベートした。細胞を37℃で5分間インキュベートし、RP-HPLCを用いて14,15-LTC4及びLTC4の量を分析した。インキュベーションの5分後では好酸球には14,15-LTC4又はLTC4の代謝は殆どない(データは示さず)。従って、これらの代謝物の測定は、それぞれ15-リポキシゲナーゼ誘導システイニル含有14,15-ロイコトリエン類及び5-リポキシゲナーゼ誘導システイニル含有ロイコトリエン類の全形成を反映していると考えられる。外因性アラキドン酸との好酸球のインキュベーションにより14,15-LTC4が優先的に形成されたが、LTC4もまた幾つかのインキュベーションで形成された(表1)。カルシウムイオノフォアA23187の細胞の活性化はLTC4の形成を専ら誘導したが、イオノフォアA23187及びアラキドン酸双方での好酸球の刺激により14,15-LTC4とLTC4の双方が形成された。同様の量のこれらの代謝物が幾つかのインキュベーションで生産されたが、LTC4のレベルは、ある試料では更に高かった。総括すると、アラキドン酸と共に細胞をインキュベートすると、14,15-LTC4の形成が優勢になり、細胞のカルシウム活性化は5-LOをトリガーし、5-LO誘導ロイコトリエン類の形成を強く優勢にする。
【0114】
表1:好酸球中における14,15-LTC4及びLTC4の生合成。好酸球を健常なドナー又は患者から単離した。細胞をリン酸緩衝生理食塩水(1ml)に懸濁させ、37℃で5分間、アラキドン酸(10μM)、イオノフォアA23187(1μM)又は双方と共にインキュベートし、ついで上清を逆相HPLC分析にかけた。14,15-LTC4及びLTC4の相対量をpmol/107好酸球として計算した。
【0115】
14,15-ロイコトリエン類の生物学的効果
血漿漏出に至る血管透過性の増加は炎症の特徴であり、システイニル含有ロイコトリエン類がヒスタミン及びブラジキニンと同様な形でこの効果に寄与しうることはよく知られている(23)。14,15-LTC4とその代謝物14,15-LTD4及び14,15-LTE4が血管透過性における改変を誘導する能力を探究するために、ヒト内皮細胞(EC)単層を通しての電気抵抗変化に基づく確立されたインビトロ透過性アッセイを使用した。このモデルにおける最大刺激で誘導されたEC抵抗変化はおよそ-70%の低下に達する。用量応答曲線は、ヒスタミンの最大効果がこのモデル系において10-5Mで得られたことを示している(データは示さず)。全ての14,15-ロイコトリエン類がEC単層透過性において最大の増加を誘導することができた(図32A)。LTC4及びLTD4より些か強力ではないが、14,15-ロイコトリエン類はこの点でヒスタミンより約100倍強力であった(図32A)。14,15-LTD4は10-7MでEC透過性の最大増加を誘導したが、EC透過性の有意な増加がまた10-8Mで観察された(図32B)。14,15-ロイコトリエン類及びLTD4に応答したEC透過性の変化の動態は、ヒスタミン及びLTC4のような直接的に作用するアゴニストに対して観察されたパターンに類似している(図32B)(24)。他方ではLTB4はEC障壁機能に影響を及ぼすために顆粒球の存在を必要とし、LTB4の効果はこのモデル系では観察されなかった。
【0116】
好酸球における14,15-LTC4生合成のレセプター媒介活性化
14,15-LTC4形成の可能なレセプター媒介活性化を、アレルゲン誘導肥満細胞メディエーターLTC4及びプロスタグランジン(PG)D2並びに強力な好酸球活性化因子であるIL-5を使用することによって調べた。単離された好酸球(10×106/試料)を37℃で5分間、LTC4(1μM)、PGD2(1μM)又はIL-5(10ng/ml)と共にインキュベートした。ロイコトリエンC4、PGD2及びIL-5は全て5分のインキュベーション後に好酸球からの14,15-LTC4の放出を誘導した(表2)。これらの結果はレセプター媒介メカニズムを介した好酸球におけるアラキドン酸の内因性プールからの14,15-LTC4の形成を実証している。
【0117】
表2:好酸球における14,15-LTC4生合成のレセプター媒介活性化。健常な志願者又は好酸球増加症と診断された患者から単離されたヒト好酸球を1mlのPBSに懸濁させ、LTC4(1μM)、PGD2(1μM)又はIL-5(10ng/ml)の存在下で37℃で5分間インキュベートした。コントロール、LTC4、及びPGD2、n=4。IL-5、n=2。14,15-LTC4の量はEIAによって分析した。
【0118】
臍帯血誘導肥満細胞(CBMCMNC)における14,15-LTC4及び14,15-LTD4の生合成
肥満細胞は喘息の病態生理学において重要な役割を担っている。最近、我々は、インターロイキン(IL-4)がヒト臍帯血誘導肥満細胞においてインビトロで15-LO-1の発現を誘導し、肥満細胞がインビボで15-LO-1を発現することを報告した(4)。従って、肥満細胞が14,15-ロイコトリエン類を生産できるかを調べることは興味深かった。IL-4で薬物刺激された肥満細胞をアラキドン酸(10μM)と共に5分間インキュベートした。上清を集め、14,15-LTC4に対してEIAを使用して分析した。これらの条件下で、ヒト肥満細胞は0.8+/−0.6pmol/106細胞(n=3)を放出し、アラキドン酸刺激の後に、免疫反応性の14,15-LTC4を合成し放出する能力を示している。
【0119】
14,15-LTC4の同一性は更にLC-MS/MS単一反応モニタリングSRMで証明された。よって、アラキドン酸と共にインキュベートした肥満細胞が、14,15-LTC4のSRM遷移に対するものと同じ保持時間と典型的な断片パターンを有する代謝物を生じた(データは示さず)。また、肥満細胞試料を特異的14,15-LTD4 SRM法で分析して、4,15-LTC4が細胞によって更に代謝されるかどうかを調べた。確かに、合成14,15-LTD4と同一の断片化パターンと保持時間を有する代謝物が証明された。総括すると、肥満細胞は14,15-LTC4と14,15-LTD4の双方を生産する能力を有している。
【0120】
ヒト鼻ポリープにとる14,15-LTC4の自然の放出
鼻ポリープ症は、好酸球のような炎症細胞による組織浸潤からなる多因子性疾患である(25)。14,15-LTC4がヒト鼻ポリープによって合成されうるかを調べるために、無傷の外科的に除去した鼻ポリープを洗浄し、プロテアーゼ阻害剤(Complete MINI(登録商標))及びインドメタシンを含むPBS中で37℃で5又は30分、インキュベートした。ついで、インキュベーション培地を、14,15-LTC4に対してEIAを使用して分析した。その結果は、鼻ポリープが自然に14,15-LTC4(それぞれ5及び30分のインキュベーション後に、10.1±0.007及び7.18±1.18pmolの14,15-LTC4/gポリープ組織(n=2))を放出することを裏付けている。経時的な14,15-LTC4の減少レベルは14,15-LTD4及び14,15-LTE4への更なる代謝のためであり、これは14,15-LTC4に対して産生された抗体によっては検出されない(14,15-LTD4及び14,15-LTE4に対して1%未満の交差反応性、材料と方法を参照)。
【0121】
エオキシン類-14,15-ロイコトリエン類に対して示唆された新規な名前
ロイコトリエンという名前は、5-リポキシゲナーゼ生成物に伝統的に関連している。アラキドン酸において14,15-エポキシドを介して形成される15-リポキシゲナーゼ代謝物に対する機能的名前は、それは5-リポキシゲナーゼ経路を介して形成されないが、14,15-ロイコトリエンであった。好酸球はこの報告において記載された新規な15-リポキシゲナーゼ生成物に対するリッチな供給源であるので、5-LO誘導生成物との混同を避けるために、我々は、それぞれ14,15-LTC4、14,15-LTD4及び14,15-LTE4の代わりに、エオキシン(Eox)C4、EoxD4及びEoxE4という名を示唆する(図33)。
【0122】
検討
ヒト気道上皮細胞及び好酸球は高い量の15-LO-1を発現する(3,5,6)。また、ヒト肥満細胞、マクロファージ及び樹状細胞は15-LO-1を発現する(26+Gullilsson)。この研究分野での多くの研究にかかわらず、赤血球新生の間の酵素の機能を除いて、この酵素の生物学的役割は不確かである(2,3)。本報告では、我々は、ヒト好酸球、特に臍帯血誘導肥満細胞及び鼻ポリープがアラキドン酸を15-LO-1経路を介して、コンジュゲートされたトリエンスペクトルと282nmでの最大UV吸光度を有する代謝物に転換させる能力を有していることを記載する(図29)。好酸球及び肥満細胞において形成されたこれらの代謝物の構造は陽イオンタンデム質量分析法及びRP-HPLC/UV-分光法によって決定し、合成14,15-LTC4、14,15-LTD4及び14,15-LTE4と同一であることが見出された(図30及び31)。好酸球は主として14,15-LTC4を生産し、それぞれ合成14,15-LTC4及び14,15-LTD4と共にこれらの細胞のインキュベーションによって証明されるように、この代謝物は更に14,15-LTD4及び14,15-LTE4に代謝されうる。合成14,15-LTC4は様々な生物システムにおいて研究され(例えば、Drazen等(1981) Proc Natl Acad Sci 78, 3195-3198を参照)、14,15-LTA4は上皮細胞及び好酸球において形成されることは先に見出されているけれども(例えば文献32を参照)、システイニル含有14,15-ロイコトリエン類の生合成がヒト細胞及び組織中でアラキドン酸から生じることは決して証明されていない。
【0123】
外因的なアラキドン酸と共に好酸球をインキュベーションすると、14,15-LTC4の生成がLTC4より優性になり(表1)、代わりにカルシウムイオノフォアで細胞内カルシウムレベルが増加すると、5-LOを誘導し、ただLTC4だけを生産する。よって、5-LO及び15-LOロイコトリエンの生産の調節は異なり、排出される主要な代謝物にどの代謝物がなるかは刺激に依存する。この点で、14,15-ロイコトリエン経路はプロスタグランジン経路に似ており、遊離のアラキドン酸が利用できる場合に直ぐに生じ、カルシウム流に独立している。
【0124】
5-リポキシゲナーゼ誘導システイニル含有ロイコトリエン類とは異なり、14,15-LTC4とその代謝物はモルモット肺実質性条片又は回腸への収縮効果を有していないように思われる(27)。しかしながら、我々は、ここで、コンフルエントなヒト内皮細胞を持つモデル系において、3種全ての14,15-ロイコトリエン類が炎症のマーカーである血管漏出を増加させたことを立証する。14,15-ロイコトリエン類は10-7及び10-8Mで増大したEC透過性を誘導し、システイニル含有ロイコトリエン類と殆ど同じほど強力で、この点ではヒスタミンより100倍以上強かった(図32)。この研究では、我々は、内皮細胞障壁機能のメディエーター誘導混乱の解析のためにインビトロモデル系を使用した。TEERの連続的測定により、EC透過性の急速な変化の検出が可能になり、培養された内皮細胞単層の障壁能力の感度のよい指標であることが過去に示されている(24,28)。ヒスタミンやシステイニルロイコトリエン類のような炎症メディエーターの透過性増大活性は以前から知られている。これらのメディエーターは、細胞内Ca2+移動及び細胞骨格再配置の惹起を通してEC障壁機能にその効果を作用させ(29)、傍細胞 間隙形成及び亢進された巨大分子透過性を生じる。同様に、14,15-ロイコトリエン類は同様な方法で血管透過性の増加を刺激する。
【0125】
LTC4、PGD2及びIL-5でのヒト好酸球のレセプター媒介活性化がまた14,15-ロイコトリエン類の生産と放出を惹起した(表2)。よって、14,15-ロイコトリエン類は好酸球中のアラキドン酸の内因性プールから製造することができる。LTC4及びPGD2は、共にアラキドン酸誘導化合物であり、抗原投与時に肥満細胞によって合成され放出されることが知られており、活性化された肥満細胞は、14,15-ロイコトリエン類自体の生産に加えて、好酸球におけるこれらの生成物の生合成を刺激することができる。LTC4及びPGD2に加えて、IL-5は14,15-LTC4の内因性生合成を活性化することができる。IL-5は、その生産、活性化及び組織補充を含む好酸球の幾つかの側面を調節することに決定的に関与したサイトカインである。
【0126】
鼻ポリープ形成の原因は知られていないが、アラキドン酸代謝物は、特にアスピリン過敏症副鼻腔炎/喘息徴候を持つ患者において、病因において重要であると思われる。鼻ポリープ症の主要な特徴は活性化された好酸球の蓄積であると考えられ、これは組織内の炎症性細胞の28%までを占める(30)。従って、インビトロでインキュベートされたヒト鼻ポリープが14,15-ロイコトリエン類を生産し放出するかどうかを調べることは興味深かった。確かに、14,15-LTC4は鼻ポリープによってインキュベーション培地中に自然に放出された。活性化された好酸球は鼻ポリープ中の14,15-LTC4の主要なソースであると思われる。しかしながら、気道内皮細胞は、これらの細胞は多量の15-LOを含んでいることが知られているので、所定の役割を担っていることは除外できない(文献2-6を参照)。
好酸球はこれらの新規代謝物のリッチな供給源であるので、我々は、5-LO経路を介して生産された化合物との混同を避けるために、14,15-ロイコトリエンの代わりにエオキシンなる名前を示唆する。よって、14,15-ロイコトリエンA4、C4、D4及びE4はそれぞれエオキシン(Eox)A4、EoxC4、EoxD4及びEoxE4で置き換えられる(図33)。エオキシン類はヒト細胞においてアラキドン酸から生産されることは決して報告されていない。しかしながら、ヒト好塩基球は外因性14,15-ロイコトリエンA4を14,15-ロイコトリエンC4に転換することが先に見出されている(33)。
【0127】
総括すると、この報告は、好酸球及び肥満細胞によって生産される新規な炎症誘発性のアラキドン酸代謝物の形成、並びにこれらの代謝物の生合成に対する様々な生理学的トリガーの同定を記載する。よって、炎症性気道中の活性化された肥満細胞とTh2細胞がLTC4、PGD2及びIL-5を排出し、これが近くの好酸球を誘導してエオキシン類を生産させ放出し、血漿漏出を生じるということができる。更に、アレルギー性炎症に加えて、遊離のアラキドン酸が高量で存在する非アレルギー性炎症においてもまた、エオキシン生産が起こり、炎症プロセスに寄与することが推測できる。
多くの報告が、コントロール被験者と比較して喘息患者の気道組織における15-LO-1の増加した量と活性を実証している。健常な喫煙者は、特に慢性気管支炎の喫煙者の場合、気管支バイオプシー中での増加した15-LO-1の発現を示している。この点で、我々は、15-LO-1経路が好酸球及び肥満細胞において炎症誘発性の14,15-ロイコトリエン類(エオキシン類)を生産することができることを示しており、15-LO-1の阻害がヒトにおける喘息及び鼻炎の治療のための魅力的な標的であり得ることを示している。
【0128】
材料と方法
バキュテナー(登録商標)採血管はベクトン・ディキンソン(Rutherford, NJ)から得た。モノクローナルマウス抗ヒトCD16抗体にコンジュゲートされた常磁性ミクロビーズ、磁気スタンド及びLS+磁気分離カラムはMiltenyi Biotec(Bergisch Gladbach, Germany)から得た。組換えヒト(rh)IL-4はIn Vitro Sweden AB(Sweden)から購入した。14,15-LTC4 EIAキットはCayman Chemical(Ann Arbor, MI)から購入した。イオノフォアA23187はCalbiochem-Boehring(La Jolla, CA)から購入した。インドメタシンはSigma-Aldrichから購入した。全ての溶媒はHPLCグレードであった。アラキドン酸はNu-Chek prepから購入した。合成14,15-エポキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTA4)、14(R)-グルタチオニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTC4)、14(R)-システイニル-グリシル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTD4)、14(R)-システイニル-15(S)-ヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14,15-LTE4)、8(R),15(S)-ジヒドロキシ-5,9,11,13(Z,E,E,E)-エイコサテトラエン酸(8(R),15(S)-LTB4)、8(S),15(S)-ジヒドロキシ-5,9,11,13(Z,E,Z,E)-エイコサテトラエン酸(8(S),15(S)-DHETE)、8(S),15(S)-ジヒドロキシ-5,9,11,13(Z,E,E,E)-エイコサテトラエン酸(8(S),15(S)-LTB4)、8(R),15(S)-ジヒドロキシ-5,9,11,13(Z,E,Z,E)-エイコサテトラエン酸(8(R),15(S)-DHETE)、14(R),15(S)-ジヒドロキシ-5,8,10,12(Z,Z,E,E)-エイコサテトラエン酸(14(R),15(S)-DHETE又は14(R),15(S)-LTB4)、15(S)-ヒドロキシ-5,8,11,13(Z,Z,Z,E)-エイコサテトラエン酸(15-HETE)、LTC4、LTD4及びLTE4は、Biomol Inc (Plymouth Meeting, USA)から購入した。
【0129】
ヒト好酸球の単離
静脈血(100ml)を、健常な志願者又は好酸球増加症候群(HES)を含む好酸球増加症、慢性リンパ性白血病(CLL)及びある種の炎症疾患の患者からから採血した。400×gで15分間の最初の遠心分離を実施した。上相を捨て下相を集め、20℃で30分間、Dextran(商標)沈降を施した。沈降後、白血球細胞を含む上相を400×gで10分間、一回遠心分離した。ペレットを溶解バッファー(100mMのNH4Cl、10mMのTRIS-HCl、pH7.4)に懸濁させ、室温で30分インキュベートして混入した赤血球を除去した。インキュベーション後、Lymphoprep(商標)を用いて密度勾配遠心分離を実施した。好中球と好酸球を含む多形核(PMN)画分を集め、磁気細胞分離(MACS)を施した。好酸球は、磁気ミクロビーズ(登録商標)に結合させたCD16抗体を使用するネガティブ選択によって単離した。純度は、形態学的検査とメイ・グリューネバルト-ギムザ溶液での染色によって確認した。純度は常に95%を越えていることが見出された。
臍帯血誘導肥満細胞(CBMCMNC)の調製及びポリープの単離
臍帯血誘導肥満細胞(CBMCMNC)を記載されたようにして樹立した(4)。ポリープは外科的に除去し、4℃で最大1時間、PBS中に維持した。ポリープを回りの組織から分離し、Ca2+及びMg2+なしのPBSで洗浄した。洗浄した濡れたポリープの重さは0.7-1gであった。
【0130】
細胞インキュベーション
好酸球
単離された好酸球(1−20×106/試料)をリン酸緩衝生理食塩水(1ml/試料)に再懸濁させ、37℃で示された時間の間、イオノフォアA23187(1μM)/アラキドン酸(1又は10μM)/A23187プラスアラキドン酸/PGD2(1μM)/LTC4(1μM)/IL-5(10ng/ml)/14,15-LTC4(10μM)又は14,15-LTD4(10μM)又は双方で2、5又は10分、刺激する前に37℃で2分間(インドメタシンの存在下又は不在下で、1μM)、プレインキュベートした。インキュベーションは1容量のメタノールの添加によって終結させた(RP-HPLCによる分析のため)。エンザイムイムノアッセイ(EIA)による続く分析のために、反応を、1200×g、5分の遠心分離前に0℃で急速冷却して終結させた。上清を凍結管に移し、分析するまで-70℃で保存した。
【0131】
肥満細胞
インキュベーション前に、CBMC細胞にIL-4(10ng/ml)を用いて4日間薬物刺激を施した。機械的な除去及びPBSでの洗浄後に、細胞をPBSに再懸濁させ、インドメタシン(1μM)と共に又はそれなしで37℃で2分間プレインキュベートした。CBMC細胞を続いて37℃でアラキドン酸と共に示された期間の間、インキュベートした。インキュベーションは1容量のメタノールの添加によって(LC-MS分析)又は0℃に急速冷却することによって終結させ、続いてEIAによって分析した。
ヒト鼻ポリープ
洗浄したヒトの無傷のポリープを、1mlの全容量中にPBS、プロテアーゼ阻害剤(Complete MINI(登録商標))及びインドメタシン(1μM)を含む清浄な試験管中に移した。試料を37℃でそれぞれ5分及び30分、インキュベートした。反応を0℃に急速冷却することによって終結させ、続いて1200×g、5分、遠心分離した。上清を凍結管に移し、EIAによる分析まで-70℃で保存した。
【0132】
15-リポキシゲナーゼ生成物の分析
逆相-高速液体クロマトグラフィー-UV(RP-HPLC-UV)
細胞片及びタンパク質を除去するために、1200×gで5分間、遠心分離を実施した。上清を減圧下で蒸発させ、残留物をメタノールに溶解させ、試験管に移した。ついで、試料を窒素流下で蒸発乾固させ、最後に200μlの移動相(ロイコトリエン類及び14,15-ロイコトリエン類の分析に対してはアセトニトリル:メタノール:H2O:酢酸、29:19:72:0.8、pHは5.6にNH3で調整、及びモノヒドロキシ酸の分析に対してはメタノール:水:トリフルオロ酢酸、70:30:0.007)に再懸濁させた。抽出された試料に、記載されたようにして逆相HPLC系での分析を施した。簡単に述べると、試料をオクタデシル逆相カラム(Chromabond 18C,Macherey-Nagel,Duren,Germany)に充填し、溶出流量は1.2ml/分であった。溶出した化合物を、Waters1996HPLCシステムに連結したWaters996光ダイオードアレイ分光光度計を利用して検出し定量した。
【0133】
液体クロマトグラフィー-質量分析解析(LC-MS/MS)
インキュベーションの終結後、試料を遠心分離し(1400×g、6分)、上清を水で希釈して最大25%のメタノールを含ませ、洗浄及び平衡抽出カートリッジOasis HLB 1cc 10mg(Waters AB, Sweden)に移した。カラムを水で洗浄し、200μlのメタノールで溶出させて代謝物を回収した。
液体クロマトグラフィー-質量分析はTSQ・Quantum・Ultraトリプル四極子質量分析計に連結したSurveyor・MS(Thermo Electron Corporation, Sweden)で実施した。注入前に1容量の水を試料に加えた。逆相LCは、400μl/分に一定に保たれた流量でZorbax・Eclipse・Plus3.5μM・C18カラム2.1×50mm(ChromTech, Sweden)を用いて実施した。移動相Aは水中2%のアセトニトリルと0.1%の酢酸からなり、移動相Bは水中80%のアセトニトリルと0.1%の酢酸から構成されている。使用した全ての水はMilliQグレードであった。90%のAで1.5分、均一濃度で開始して、100%のBに達する6.5分の線形勾配を続けた。該系を100%のBで4分洗浄し、ついで90%のAで4分間平衡にした。14,15-ロイコトリエン類を約0.5分、ロイコトリエン類からクロマトグラフィー的に分離した。質量分析計はポジティブモードでエレクトロスプレー大気圧イオン化源を使用して操作した。スプレー電圧は4500Vに設定し、キャピラリー温度は375℃であり、シース及び補助ガスはそれぞれ40及び5(任意単位)で最適であった。スキマーオフセットは10Vで、チューブレンズは121Vであった。
【0134】
好酸球代謝物に対するLC-MS/MS
LC-MS/MSを使用して、プロトン化14,15-LTC4、14,15-LTD4及び14,15-LTE4の生成物イオンスキャンを実施した。626.2m/zの14,15-LTC4を22eVの衝突エネルギーで断片化し、497.2m/zの14,15-LTD4及び440.2m/zの14,15-LTE4は共に18eVで断片化した。
肥満細胞代謝物に対するLC-MS/MS
14,15-LTC4及び14,15-LTD4は単一反応モニタリングSRMによって検出した。SRMスキャンで使用された遷移は、14,15-LTC4に対して626m/z→279m/z、14,15-LTD4に対して497m/z→205m/zであり、双方とも20eVの衝突エネルギーであった。14,15-LTC4に対して選択される遷移はまたLTC4に対して存在していたが、これらの化合物はクロマトグラフィー的にはおよそ0.5分で分離した。497m/z→205m/zの遷移は14,15-LTD4に対して特異的であり、よってLTD4には適用できなかった。
【0135】
14,15-LTC4に対するエンザイムイムノアッセイ(EIA)の特徴付け
14,15-LTC4に対するエンザイムイムノアッセイ(EIA)は少量の14,15-LTC4を定量するためにCayman Chemical (Ann Arbor, MI)によって開発された。この方法は、14,15-LTC4の定量においてRP-HPLCよりおよそ千倍感度が良かった。EIA交差反応性は製造者によって決定された。14,15-LTD4及び14,15-LTE4に対する交差反応性はそれぞれ0.9%及び0.04%であった。ロイコトリエンC4、ロイコトリエンD4、ロイコトリエンE4、アラキドン酸、12-HETE、15-HETE、5(S),6(S)-ジHETE、5(S),15(S)-ジHETE、8(S),15(S)-ジHETE及び14,15-ジHETEの交差反応性は0.01%未満であった。
上清をEIAバッファーで希釈し、分析を製造者のプロトコルに従って実施した。EIA定量からの光学密度(OD)を用いて次の関係に従って%B/B0を計算した:
%B/B0=(試料OD-NSB OD)/(ゼロ標準OD-NSB OD)×100
7.8-1000pg/ml(0.012-1.6pmol/ml)14,15-LTC4の範囲の標準曲線を作成し、試料中の14,15-LTC4の濃度を決定するために使用した。
【0136】
内皮細胞障壁機能の評価
ヒト臍帯静脈内皮細胞 (HUVEC)を単離し、過去に記載されているようにして培養した。細胞を3μmの孔径のポリカーボネートフィルター(Tissue Culture inserts, 10 mm; NUNC, Roskilde, Denmark)に2×105細胞の密度で播種し、コンフルエントな単層まで増殖させた。内皮細胞(EC)単層の障壁機能に変化を誘導するアラキドン酸代謝物の能力は、経内皮電気抵抗(TEER)の連続的記録によって分析した。TEERの測定では、ECを有するフィルターインサートを、繰り返し測定の間に互いのかつフィルターインサートに対しての電極の正確な位置決めを達成するように変更された抵抗測定チャンバー(Endohm-12, World Precision Instruments, Sarasota, FL, USA)に移した。チャンバー(下方コンパートメント)及びフィルターインサート(上方コンパートメント)にそれぞれ2ml及び400μlの培養培地を満たした。全ての測定は細胞培養インキュベーター中に配した装置で37℃にて行った。個々のEC単層の電気抵抗は、ECの播種前に測定したBiomatrixでコートされた対応する裸のフィルターの抵抗を減算することによって得られた。よって、TEER値はEC単層だけの抵抗を表す。
【0137】
1.Claesson, H. E.及びDahlen, S. E. (1999) J Intern Med 245, 205-27。
2.Kuhn, H., Walther, M.及びKuban, R. J. (2002) Prostaglandins Other Lipid Mediat 68-69, 263-90。
3.Kuhn, H.及びO'Donnell, V. B. (2006) Prog Lipid Res 45, 334-56。
4.Gulliksson, M., Brunnstrom, A., Johannesson, M., Backman, L., Nilsson, G., Harvima, I. T., Dahlen, B., Kumlin, M.及びClaesson, H-E. (2007) Biochem Biopys Acta in press.
5.Hunter, J. A., Finkbeiner, W. E., Nadel, J. A., Goetzl, E. J.及びHoltzman, M. J. (1985) Proc Natl Acad Sci U S A 82, 4633-7。
6.Turk, J., Maas, R. L., Brash, A. R., Roberts, L. J., 2nd及びOates, J. A. (1982) J Biol Chem 257, 7068-76。
7.Brash, A. R., Boeglin, W. E.及びChang, M. S. (1997) Proc Natl Acad Sci U S A 94, 6148-52。
8.Conrad, D. J., Kuhn, H., Mulkins, M., Highland, E.及びSigal, E. (1992) Proc Natl Acad Sci U S A 89, 217-21。
9.Brinckmann, R., Topp, M. S., Zalan, I., Heydeck, D., Ludwig, P., Kuhn, H., Berdel, W. E.及びHabenicht, J. R. (1996) Biochem J 318 ( Pt 1), 305-12。
10.Shannon, V. R., Chanez, P., Bousquet, J.及びHoltzman, M. J. (1993) Am Rev Respir Dis 147, 1024-8。
11.Bradding, P., Redington, A. E., Djukanovic, R., Conrad, D. J.及びHolgate, S. T. (1995) Am J Respir Crit Care Med 151, 1201-4。
12.Hamberg, M., Hedqvist, P.及びRadegran, K. (1980) Acta Physiol Scand 110, 219-21。
13.Kumlin, M., Ohlson, E., Bjorck, T., Hamberg, M., Granstrom, E., Dahlen, B., Zetterstrom, O.及びDahlen, S. E. (1991) Adv Prostaglandin Thromboxane Leukot Res 21A, 441-4。
14.Chu, H. W., Balzar, S., Westcott, J. Y., Trudeau, J. B., Sun, Y., Conrad, D. J.及びWenzel, S. E. (2002) Clin Exp Allergy 32, 1558-65。
15.Dahlen, S. E., Hansson, G., Hedqvist, P., Bjorck, T., Granstrom, E.及びDahlen, B. (1983) Proc Natl Acad Sci U S A 80, 1712-6。
16.Murray, J. J., Tonnel, A. B., Brash, A. R., Roberts, L. J., 2nd, Gosset, P., Workman, R., Capron, A.及びOates, J. A. (1985) Trans Assoc Am Physicians 98, 275-80。
17.Profita, M., Sala, A., Riccobono, L., Paterno, A., Mirabella, A., Bonanno, A., Guerrera, D., Pace, E., Bonsignore, G., Bousquet, J.及びVignola, A. M. (2000) J Allergy Clin Immunol 105, 711-6。
18.Ramis, I., Rosello-Catafau, J., Bulbena, O., Picado, C.及びGelpi, E. (1989) J Chromatogr 496, 416-22。
19.Jubiz, W., Radmark, O., Lindgren, J. A., Malmsten, C.及びSamuelsson, B. (1981) Biochem Biophys Res Commun 99, 976-86。
20.Maas, R. L.及びBrash, A. R. (1983) Proc Natl Acad Sci U S A 80, 2884-8。
21.Schwenk, U., Morita, E., Engel, R.及びSchroder, J. M. (1992) J Biol Chem 267, 12482-8。
22.Serhan, C. N.及びSavill, J. (2005) Nat Immunol 6, 1191-7。
23.Raud, J., Lindbom, L.及びHedqvist, P. (1992) Acta Physiol Scand 146, 545-6。
24.Gautam, N., Hedqvist, P. & Lindbom, L. (1998) Br J Pharmacol 125, 1109-14。
25.Bachert, C., Gevaert, P., Holtappels, G.及びvan Cauwenberge, P. (2002) Curr Allergy Asthma Rep 2, 481-7。
26.Spanbroek, R., Hildner, M., Kohler, A., Muller, A., Zintl, F., Kuhn, H., Radmark, O., Samuelsson, B.及びHabenicht, A. J. (2001) Proc Natl Acad Sci U S A 98, 5152-7。
27.Lewis, R. A., Drazen, J. M., Austen, K. F., Toda, M., Brion, F., Marfat, A.及びCorey, E. J. (1981) Proc Natl Acad Sci U S A 78, 4579-83。
28.Gautam, N., Herwald, H., Hedqvist, P.及びLindbom, L. (2000) J Exp Med 191, 1829-39。
29.Huang, A. J., Manning, J. E., Bandak, T. M., Ratau, M. C., Hanser, K. R.及びSilverstein, S. C. (1993) J Cell Biol 120, 1371-80。
30.Ferguson, B. J. (2000) Laryngoscope 110, 799-813。
31.Holtzman, M. J., Hansbrough, J. R., Rosen, G. D.及びTurk, J. (1988) Biochim Biophys Acta 963, 401-13。
32.Turk, J., Rand, T. H., Maas, R. L., Lawson, J. A., Brash, A. R., Roberts, L. J., 2nd, Colley, D. G.及びOates, J. A. (1983) Biochim Biophys Acta 750, 78-90。
33.Sok, D. E., Han, C. O., Shieh, W. R., Zhou, B. N.及びSih, C. J. (1982) Biochem Biophys Res Commun 104, 1363-70。
【0138】
文献(実施例8のもの以外)
1.Funk CD. The molecular biology of mammalian lipoxygenases and the quest for eicosanoid functions using lipoxygenase-deficient mice [Review]. Biochimica et Biophysica Acta - Lipids & Lipid Metabolism 1996; 1304(1):65-84。
2.Claesson HE, Dahlen SE. Asthma and leukotrienes: antileukotrienes as novel anti-asthmatic drugs [Review]. Journal of Internal Medicine 1999; 245(3):205-227。
3.Rapoport SM及びSchewe T. The maturational breakdown of mitochondria in reticulocytes. Biochim. Biophys. Acta 1986; 864:471-495。
4.Vanleyen K, Duvoisin RM, Engelhardt H, Wiedmann M. A function for lipoxygenase in programmed organelle degradation. Nature 1998; 395(6700):392-395。
5.Ford-Hutchinson AW. Arachidonate 15-lipoxygenase; characteristics and potential biological significance. Eicosanoids 1991; 4:65-74。
6.Conrad DJ, Kuhn H, Mulkins M等 Specific inflammatory cytokines regulate the expression of human monocyte 15-Lipoxygenase. Proc Natl Acad Sci USA 1992; 89(1):217-221。
7.Holtzman MJ, Hansbrough, R. J., Rosen, G. D.及びTurk, J. Uptake, release and novel species-dependent oxygenation of arachidonic acid in human and animal airway epithelial cells. Biochim. Biophys. Acta 1988; 963:401-413。
8.Rapoport SM, Schewe,T, Wiesner, R., Halangk, W., Ludwig, P., Janicke-Hohne, M., Tannert, C., Hiebsch, C.及びKlat, D. The lipoxygenase of reticulocytes. Purification, characterisation and biological dynamics of the lipoxygenase: its identity with the respiratory inhibitors of the reticulocyte. Eur. J. Biochem 1979; 96:545-561。
9.Sigal E, Grunberger, D., Highland, E., Gross, C., Dixon, R. A., Craik, C. S. Expression of cloned human reticulocyte 15-lipoxygenase and immunological evidence that 15-lipoxygenase of different cell types are related. J. Biol. Chem. 1990; 265:5113-5120。
10.Brash AR, Boeglin WE, Chang MS. Discovery of a second 15s-lipoxygenase in humans. Proc Natl Acad Sci USA 1997; 94(12):6148-6152。
11.Maas RL, Brash, A. R.及びOates, J. A. A second pathway of leukotriene biosynthesis in porcine leukocytes. Proc. Natl. Acad. Sci. 1981; 78:5523-5527。
12.Lundberg U, Malmsten C, Samuelsson B. Transformation of 15-hydroperoxy-5,9,11,13-eicosatetraenoic acid into novel leukotrienes. FEBS Lett 1981; 126:127-132。
13.Serhan CN, Haeggstrom, J. Z.及びLeslie, C. C. Lipid mediator networks in cell signaling: update and impact och cytokines. The Faseb journal 1996; 10:1147-1158。
14.Schwenk U, Morita E, Engel R, Schroder JM. Identification of 5-oxo-15-hydroxy-6,8,11,13-eicosatetraenoic acid as a novel and potent human eosinophil chemotactic eicosanoid. J Biol Chem 1992; 267(18):12482-12488.
15.Wolf W, Kapp U, Bohlen H, Kornacker M, Schoch C, Stahl B, Mucke S, von Kalle C, Fonatsch C, Schaefer H-E, Hansmann M-L and Diehl V: Peripheral blood mononuclear cells of a patient with advanced Hodgkin's lymphoma give rise to permanently growing Hodgkin-Reed Sternberg cells. Blood, 1996; Vol 87 (8) 3418-3428。
16.Griffiths W, Yang Y, Sjovall J及びLindgren JA: Electrospray tandem mass spectrometry of cysteinyl leukotrienes. Rapid Commun. Mass Spectrom. 1996; 10:1054-10709。
17.Hunter JA, Finkbeiner WE, Nadel EJ等 Predominant generation of 15-lipoxygenase metabolites of arachidonic acid by epithelial cells from human trachea. Proc. Natl. Acad. Sci. USA 1985; 82:4633-4637。
18.Gautam N, Hedqvist P及びLindbom L (1998). Kinetics of leukocyte-induced changes in endothelial barrier function. Br J Pharmacol 125: 1109-1114。
19.Mannervik B及びDanielsson UH (1988). グルタチオントランスフェラーゼs: structure and catalytic activity. CRC Crit Rev Biochem 3, 283-337。
20.Izumi Takashi等(1988). Solubilization and partial purification of leukotriene C4 synthase from guinea-pig lung: a ミクロソーム enzyme with high specificity towards 5,6-epoxide leukotriene A4. Biochem Biophys Acta 959, 305-315。
21.Jakobsson P-J, Morgenstern R, Mancini J, Ford-Hutchinson A及びPersson B (2000). Membrane-associated proteins in eicosanoid and glutathione metabolism. Am J Respir Crit Care Med 161, 20-24。
22.Gillmor, S.A., Villasenor, A., Fletterick, R., Sigal, E.及びBrowner, M.F., 1997, The structure of mammalian 15-lipoxygenase reveals similarity to the lipases and the determinants of substrate specificty Nature Structural Biology, vol 4, issue 12, 1003-1009。
【特許請求の範囲】
【請求項1】
生物システムにおける14,15-LTC4(エオキシンC4;EoxC4)、14,15-LTD4(エオキシンD4;EoxD4)、又は14,15-LTE4(エオキシンE4;EoxE4)の形成を調節する化合物を同定する方法であって、15リポキシゲナーゼ(15-LO)活性又はグルタチオンSトランスフェラーゼM1-1(GSTM1-1),グルタチオンSトランスフェラーゼP1-1(GSTP1-1)及び/又はロイコトリエンC4シンターゼ活性に対する効果について化合物を試験することを含む方法。
【請求項2】
炎症調節効果又は骨量減少調節効果を有する化合物を同定する方法であって、生物システムにおけるEoxC4,EoxD4又はEoxE4の形成及び/又は活性に対する効果について化合物を試験することを含む方法。
【請求項3】
該方法は、抗炎症効果又は骨量減少低減効果を有する化合物を同定するためのものであり、生物システムにおけるEoxC4,EoxD4又はEoxE4の形成及び/又は活性に対する阻害効果について化合物を試験することを含む請求項2に記載の方法。
【請求項4】
生物システムにおけるEoxC4,EoxD4又はEoxE4の形成又は活性に対する効果について化合物を試験する工程を更に含む請求項1に記載の方法。
【請求項5】
生物システムが、15-LO,グルタチオンSトランスフェラーゼ(例えばGSTM1-1又はGSTP1-1)及び/又はロイコトリエンC4シンターゼ活性を含む請求項2から4の何れか一項に記載の方法。
【請求項6】
化合物を、細胞画分又は単離酵素を含む系において試験する請求項1から5の何れか一項に記載の方法。
【請求項7】
化合物を、細胞を含む系において試験する請求項1から5の何れか一項に記載の方法。
【請求項8】
化合物を、細胞培養を含む系において試験する請求項7に記載の方法。
【請求項9】
細胞がL1236細胞又は一次好酸球白血球又は肥満細胞である請求項7又は8に記載の方法。
【請求項10】
細胞をアラキドン酸で刺激し又はアラキドン酸と共にインキュベートする請求項7に記載の方法。
【請求項11】
化合物を、組織又は器官を含む系において試験する請求項7に記載の方法。
【請求項12】
化合物を、動物を含む系において試験する請求項7に記載の方法。
【請求項13】
化合物を、炎症のインビトロモデルにおいて内皮細胞のエオキシン誘導透過に対する阻害効果;又は白血球によるエオキシン誘導酸素フリーラジカル生成について試験する請求項7に記載の方法。
【請求項14】
化合物を、炎症疾患又は炎症プロセスを含む疾患における抗炎症効果について更に試験する請求項1から13の何れか一項に記載の方法。
【請求項15】
炎症疾患が、喘息、慢性閉塞性肺疾患(COPD)、肺線維症、アレルギー性疾患、鼻炎、炎症性腸疾患、潰瘍、炎症性痛覚、発熱、アテローム性動脈硬化症、冠動脈疾患、脈管炎、膵炎、関節炎、変形性関節炎、関節リウマチ、結膜炎、虹彩炎、強膜炎、ブドウ膜炎、創傷、皮膚炎、湿疹、乾癬、発作、糖尿病、自己免疫疾患、アルツハイマー病、多発性硬化症、サルコイドーシス、アテローム性動脈硬化症、高血圧、ホジキンリンパ腫又は他の悪性腫瘍である、請求項14に記載の方法。
【請求項16】
化合物を、骨粗鬆症、骨関節炎、多発性骨髄腫、パジェット病又は歯周病における骨量減少低減効果について更に試験する請求項1から13の何れか一項に記載の方法。
【請求項17】
抗炎症組成物を製造する方法であって、
(i)請求項2又は請求項2に従属する請求項3から15の何れか一項に記載の方法によって抗炎症化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法。
【請求項18】
エオキシン形成調節組成物を製造する方法であって、
(i)請求項1又は請求項1に従属する請求項3から16の何れか一項に記載の方法によってエオキシン形成調節化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法。
【請求項19】
組成物を、炎症の治療に有用な更なる治療剤と組み合わせる工程を更に含む請求項17又は18に記載の方法。
【請求項20】
骨量減少低減組成物を製造する方法であって、
(i)請求項2又は請求項2に従属する請求項3から16の何れか一項に記載の方法によって骨量減少低減化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法。
【請求項21】
組成物を、骨量減少の治療に有用な更なる治療剤と組み合わせる工程を更に含む請求項20に記載の方法。
【請求項22】
炎症調節効果又は骨量減少調節効果を有する化合物を同定するため、又は生物システムにおけるEoxC4,EoxD4,又はEoxE4の形成を調節する化合物を同定するための方法における、L1236細胞又は細胞培養物、あるいは15リポキシゲナーゼと、グルタチオンSトランスフェラーゼM1-1(GSTM1-1),グルタチオンSトランスフェラーゼP1-1(GSTP1-1)及びロイコトリエンC4シンターゼの少なくとも一つを発現する他の細胞の使用。
【請求項23】
治療的有効量のEoxC4,EoxD4又はEoxE4を患者に投与することを含む、炎症反応の促進を必要とする患者の治療方法。
【請求項24】
炎症反応の促進を必要とする患者の治療のための医薬の製造におけるEoxC4,EoxD4又はEoxE4の使用。
【請求項25】
医薬に使用するためのEoxC4,EoxD4又はEoxE4。
【請求項1】
生物システムにおける14,15-LTC4(エオキシンC4;EoxC4)、14,15-LTD4(エオキシンD4;EoxD4)、又は14,15-LTE4(エオキシンE4;EoxE4)の形成を調節する化合物を同定する方法であって、15リポキシゲナーゼ(15-LO)活性又はグルタチオンSトランスフェラーゼM1-1(GSTM1-1),グルタチオンSトランスフェラーゼP1-1(GSTP1-1)及び/又はロイコトリエンC4シンターゼ活性に対する効果について化合物を試験することを含む方法。
【請求項2】
炎症調節効果又は骨量減少調節効果を有する化合物を同定する方法であって、生物システムにおけるEoxC4,EoxD4又はEoxE4の形成及び/又は活性に対する効果について化合物を試験することを含む方法。
【請求項3】
該方法は、抗炎症効果又は骨量減少低減効果を有する化合物を同定するためのものであり、生物システムにおけるEoxC4,EoxD4又はEoxE4の形成及び/又は活性に対する阻害効果について化合物を試験することを含む請求項2に記載の方法。
【請求項4】
生物システムにおけるEoxC4,EoxD4又はEoxE4の形成又は活性に対する効果について化合物を試験する工程を更に含む請求項1に記載の方法。
【請求項5】
生物システムが、15-LO,グルタチオンSトランスフェラーゼ(例えばGSTM1-1又はGSTP1-1)及び/又はロイコトリエンC4シンターゼ活性を含む請求項2から4の何れか一項に記載の方法。
【請求項6】
化合物を、細胞画分又は単離酵素を含む系において試験する請求項1から5の何れか一項に記載の方法。
【請求項7】
化合物を、細胞を含む系において試験する請求項1から5の何れか一項に記載の方法。
【請求項8】
化合物を、細胞培養を含む系において試験する請求項7に記載の方法。
【請求項9】
細胞がL1236細胞又は一次好酸球白血球又は肥満細胞である請求項7又は8に記載の方法。
【請求項10】
細胞をアラキドン酸で刺激し又はアラキドン酸と共にインキュベートする請求項7に記載の方法。
【請求項11】
化合物を、組織又は器官を含む系において試験する請求項7に記載の方法。
【請求項12】
化合物を、動物を含む系において試験する請求項7に記載の方法。
【請求項13】
化合物を、炎症のインビトロモデルにおいて内皮細胞のエオキシン誘導透過に対する阻害効果;又は白血球によるエオキシン誘導酸素フリーラジカル生成について試験する請求項7に記載の方法。
【請求項14】
化合物を、炎症疾患又は炎症プロセスを含む疾患における抗炎症効果について更に試験する請求項1から13の何れか一項に記載の方法。
【請求項15】
炎症疾患が、喘息、慢性閉塞性肺疾患(COPD)、肺線維症、アレルギー性疾患、鼻炎、炎症性腸疾患、潰瘍、炎症性痛覚、発熱、アテローム性動脈硬化症、冠動脈疾患、脈管炎、膵炎、関節炎、変形性関節炎、関節リウマチ、結膜炎、虹彩炎、強膜炎、ブドウ膜炎、創傷、皮膚炎、湿疹、乾癬、発作、糖尿病、自己免疫疾患、アルツハイマー病、多発性硬化症、サルコイドーシス、アテローム性動脈硬化症、高血圧、ホジキンリンパ腫又は他の悪性腫瘍である、請求項14に記載の方法。
【請求項16】
化合物を、骨粗鬆症、骨関節炎、多発性骨髄腫、パジェット病又は歯周病における骨量減少低減効果について更に試験する請求項1から13の何れか一項に記載の方法。
【請求項17】
抗炎症組成物を製造する方法であって、
(i)請求項2又は請求項2に従属する請求項3から15の何れか一項に記載の方法によって抗炎症化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法。
【請求項18】
エオキシン形成調節組成物を製造する方法であって、
(i)請求項1又は請求項1に従属する請求項3から16の何れか一項に記載の方法によってエオキシン形成調節化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法。
【請求項19】
組成物を、炎症の治療に有用な更なる治療剤と組み合わせる工程を更に含む請求項17又は18に記載の方法。
【請求項20】
骨量減少低減組成物を製造する方法であって、
(i)請求項2又は請求項2に従属する請求項3から16の何れか一項に記載の方法によって骨量減少低減化合物を同定し;
(ii)該化合物を薬学的に許容可能な賦形剤又は担体と組み合わせる
ことを含む方法。
【請求項21】
組成物を、骨量減少の治療に有用な更なる治療剤と組み合わせる工程を更に含む請求項20に記載の方法。
【請求項22】
炎症調節効果又は骨量減少調節効果を有する化合物を同定するため、又は生物システムにおけるEoxC4,EoxD4,又はEoxE4の形成を調節する化合物を同定するための方法における、L1236細胞又は細胞培養物、あるいは15リポキシゲナーゼと、グルタチオンSトランスフェラーゼM1-1(GSTM1-1),グルタチオンSトランスフェラーゼP1-1(GSTP1-1)及びロイコトリエンC4シンターゼの少なくとも一つを発現する他の細胞の使用。
【請求項23】
治療的有効量のEoxC4,EoxD4又はEoxE4を患者に投与することを含む、炎症反応の促進を必要とする患者の治療方法。
【請求項24】
炎症反応の促進を必要とする患者の治療のための医薬の製造におけるEoxC4,EoxD4又はEoxE4の使用。
【請求項25】
医薬に使用するためのEoxC4,EoxD4又はEoxE4。
【図1】

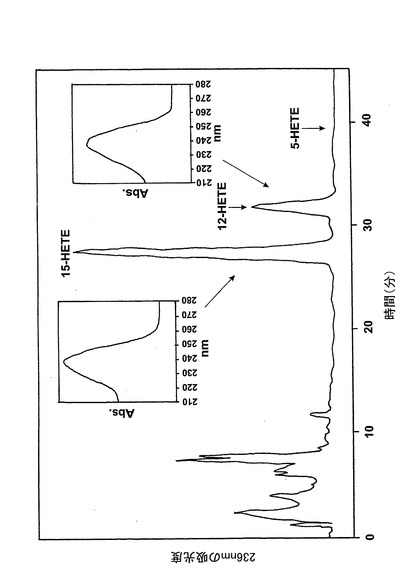
【図2a】
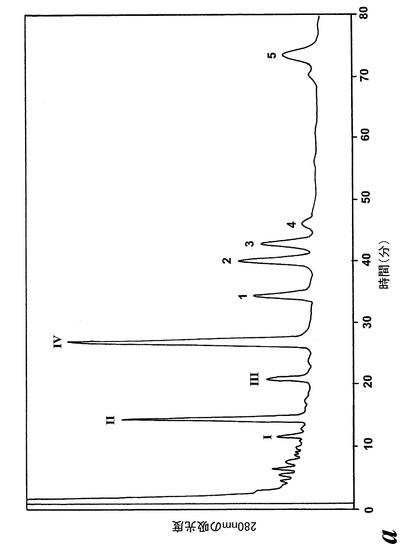
【図2b】
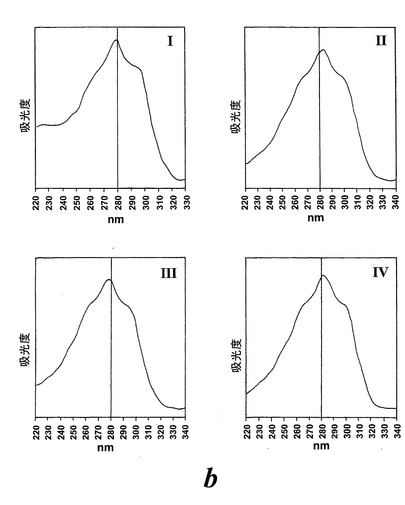
【図3】
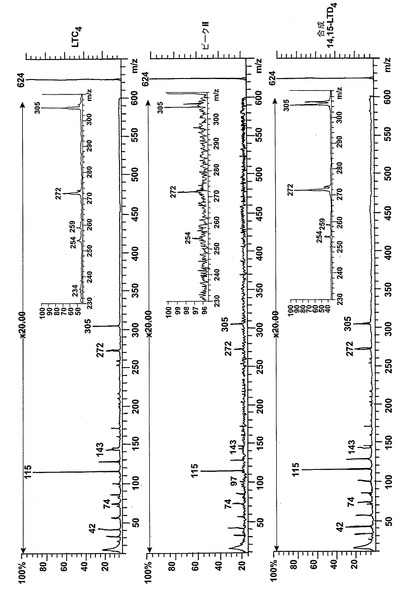
【図4】
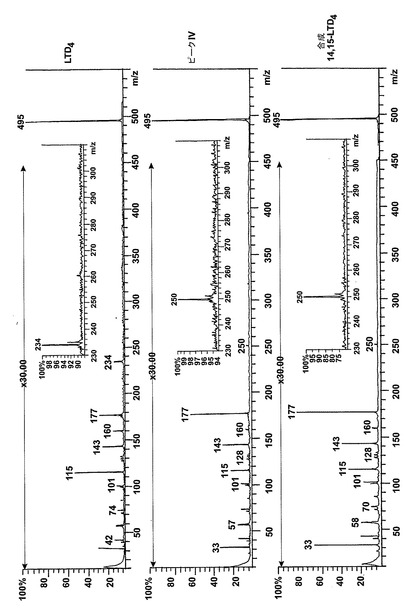
【図5】
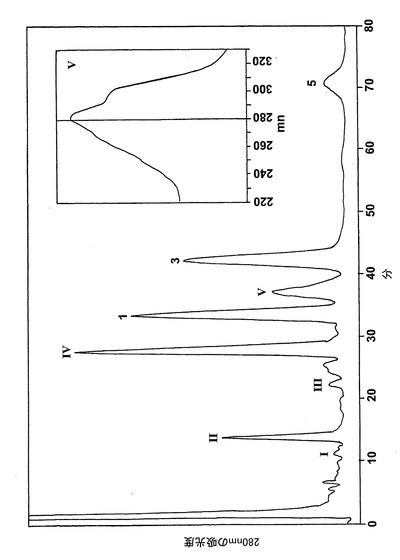
【図6】
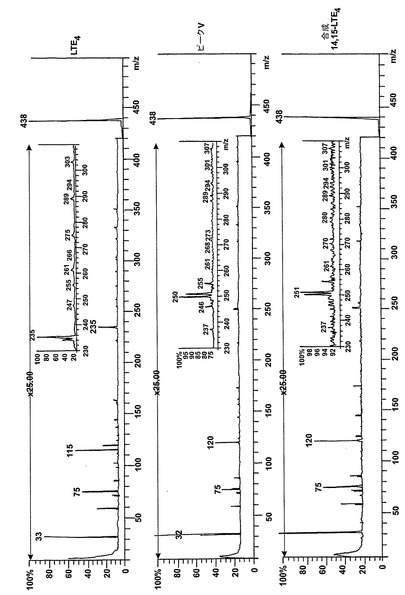
【図7】
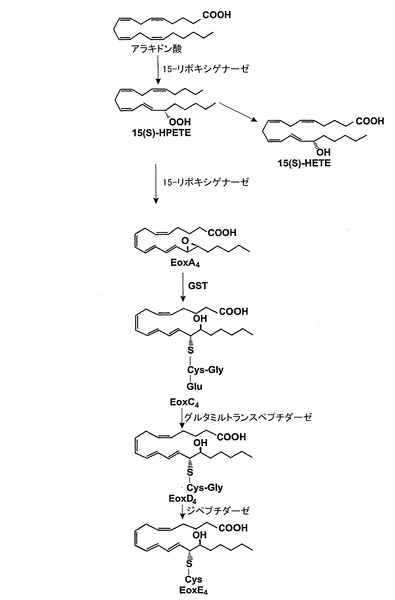
【図8a】
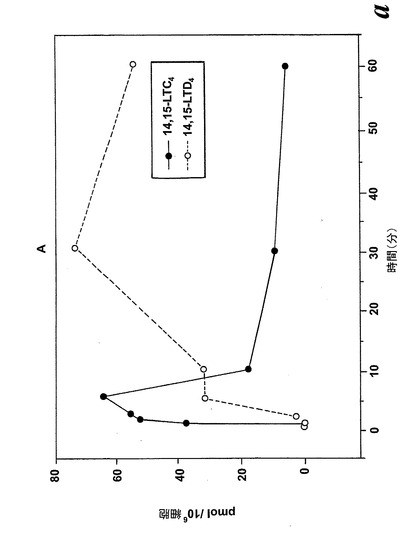
【図8b】
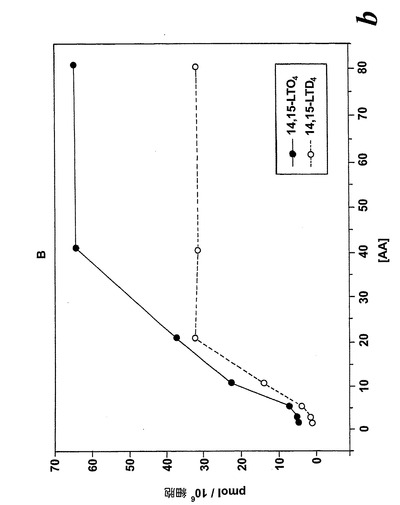
【図9a】
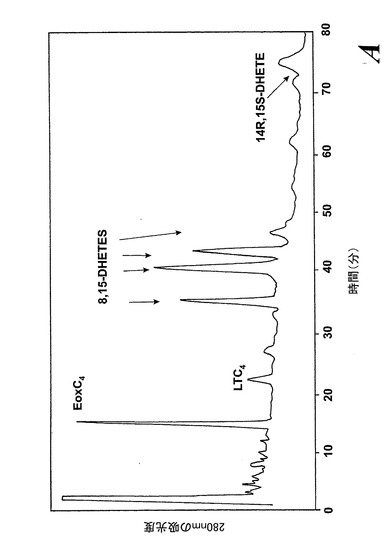
【図9b】
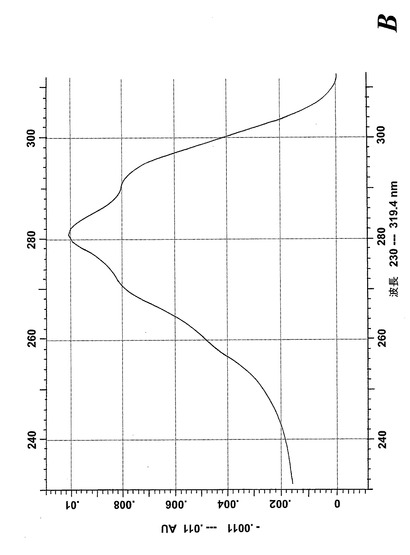
【図10】
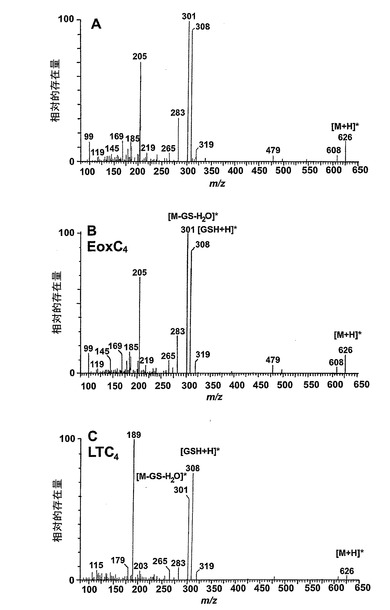
【図11】
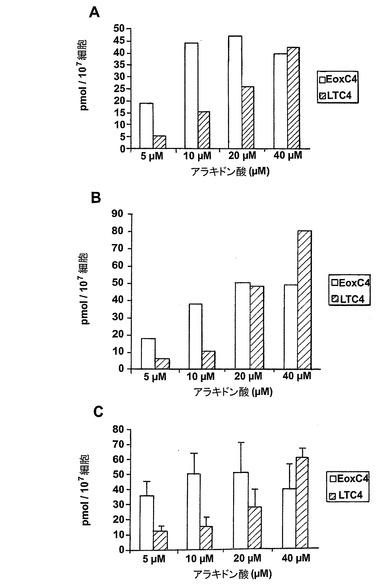
【図12a】
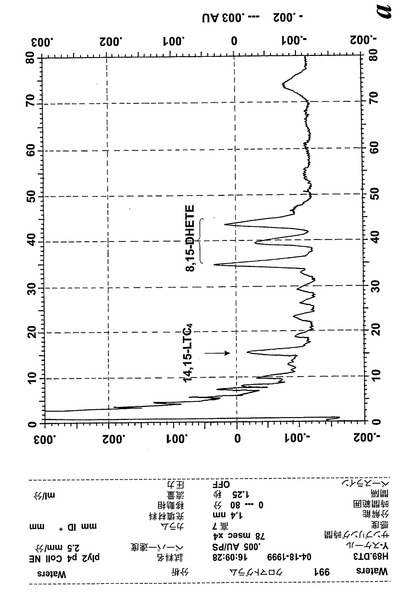
【図12b】
【図13a】
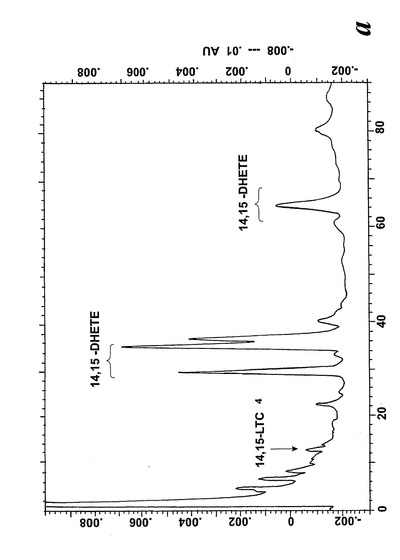
【図13b】
【図14】
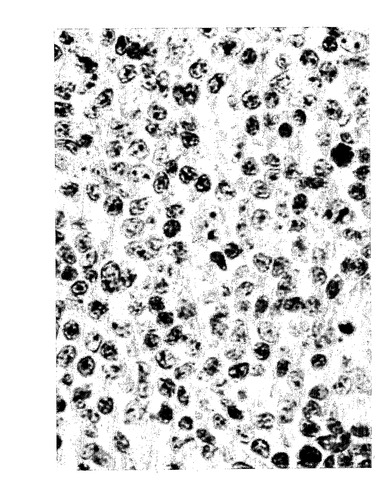
【図15】
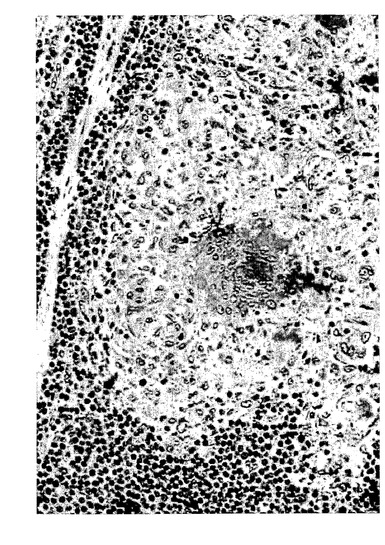
【図16】
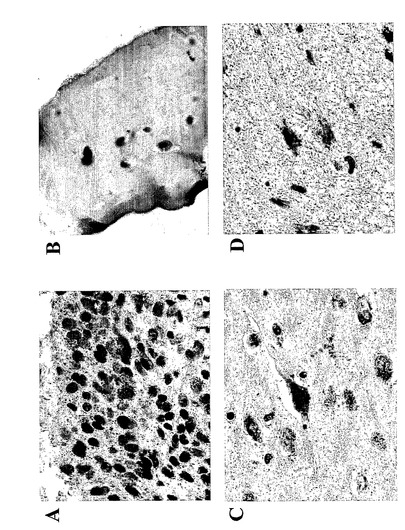
【図17a】
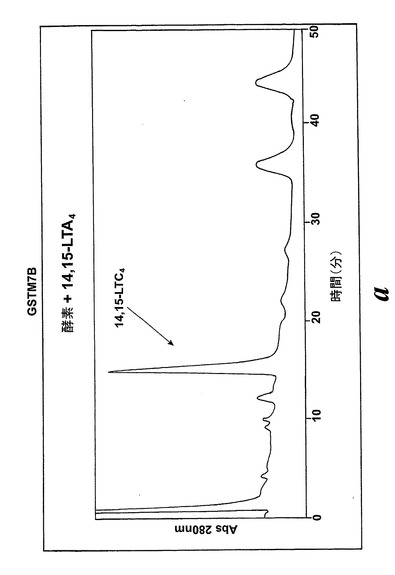
【図17b】
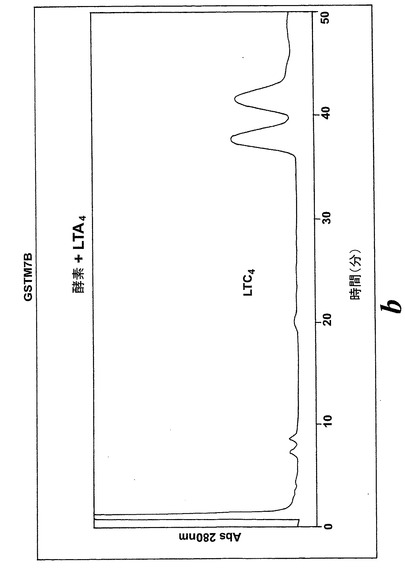
【図18】

【図19】
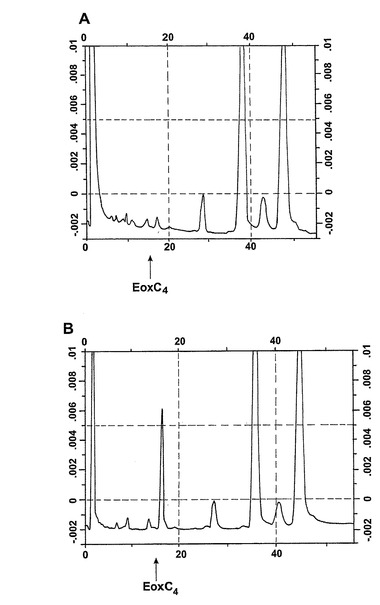
【図20】
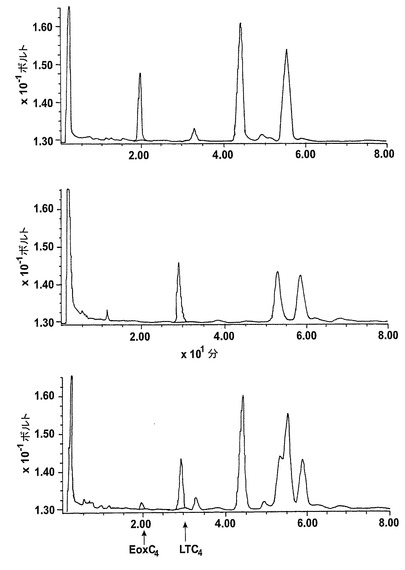
【図21】
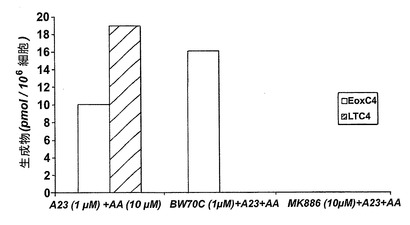
【図22】
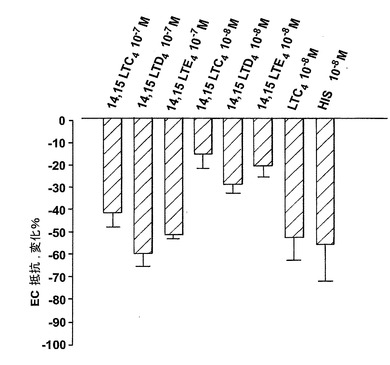
【図23a】
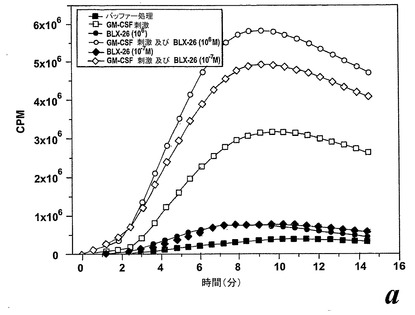
【図23b】
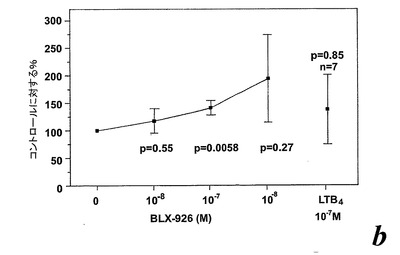
【図23c】
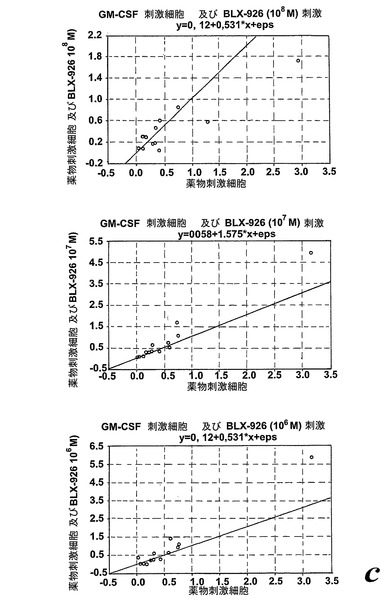
【図24a】
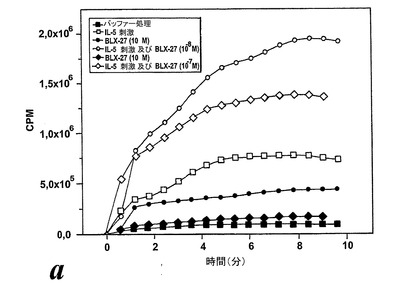
【図24b】
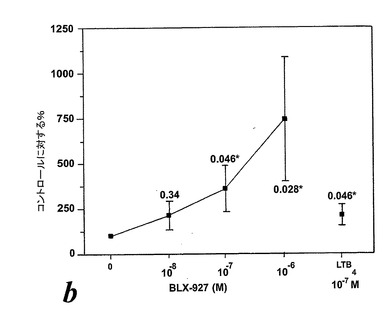
【図24c】
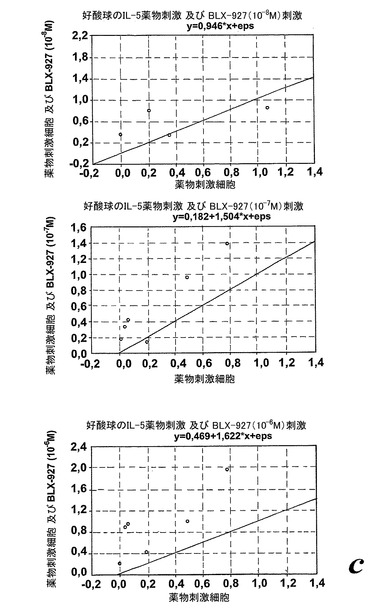
【図25a】
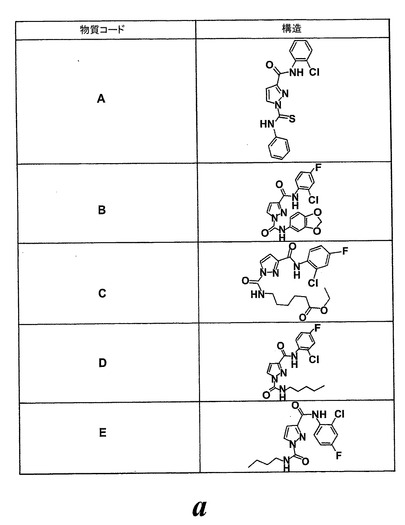
【図25b】
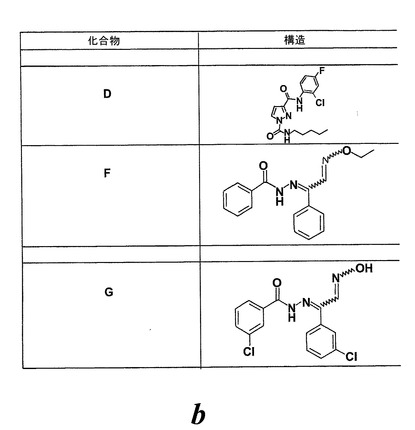
【図26】
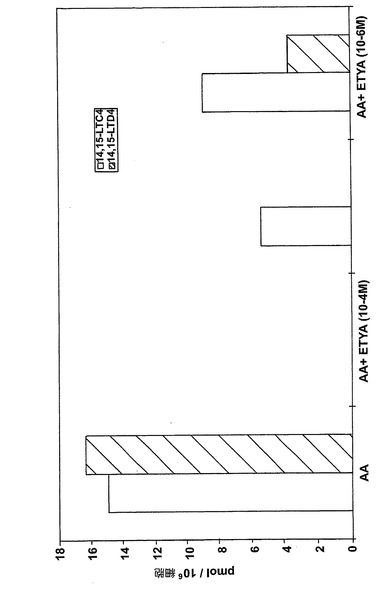
【図27a】
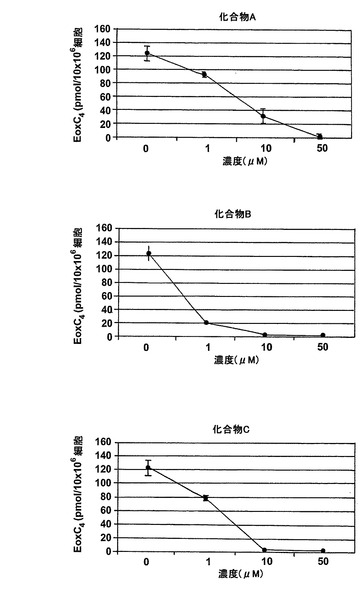
【図27b】
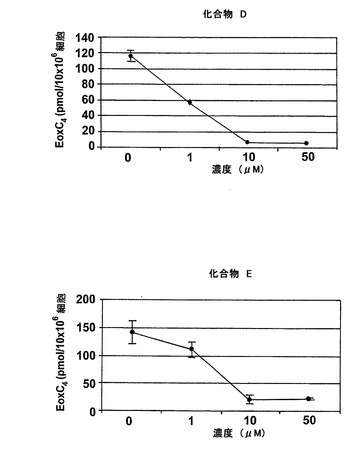
【図28a】
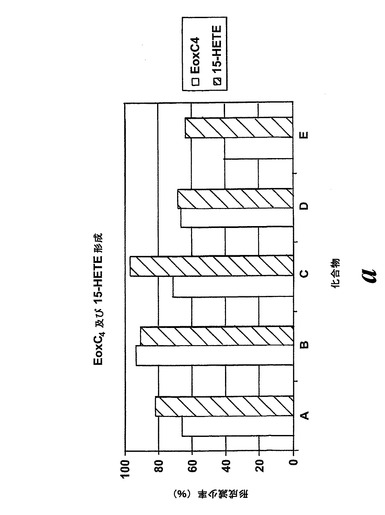
【図28b】
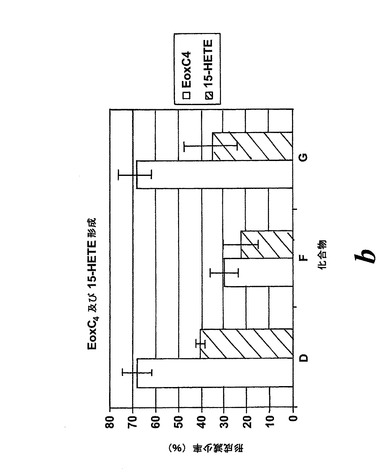
【図29】
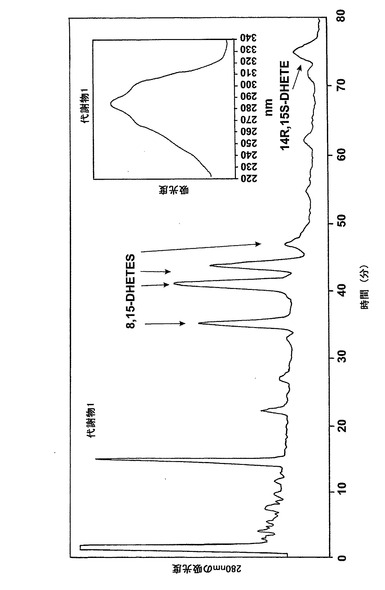
【図30a】
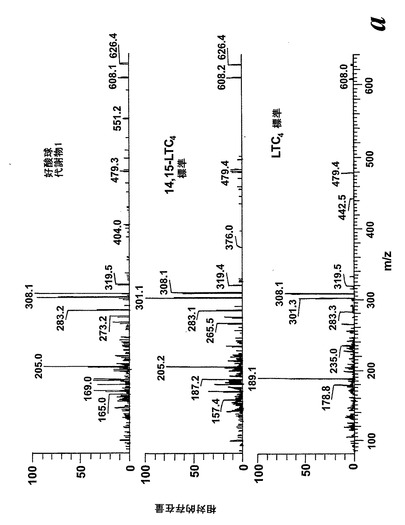
【図30b】
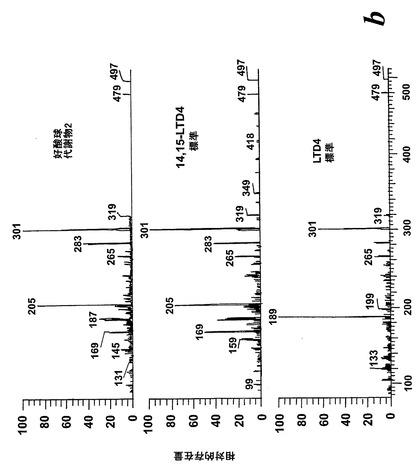
【図30c】
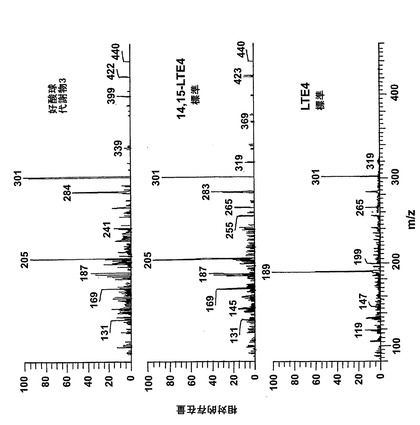
【図31】
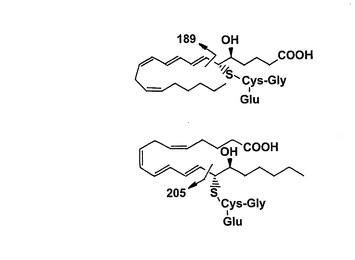
【図32a】
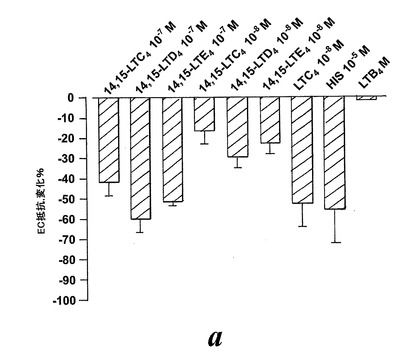
【図32b】
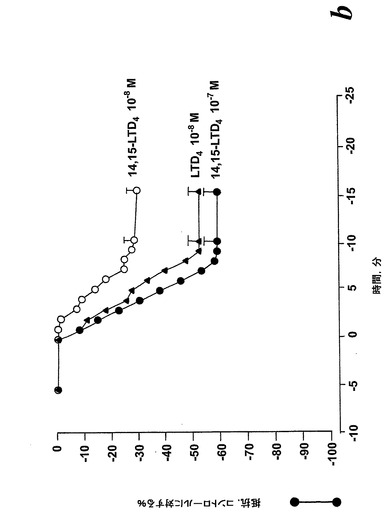
【図32c】
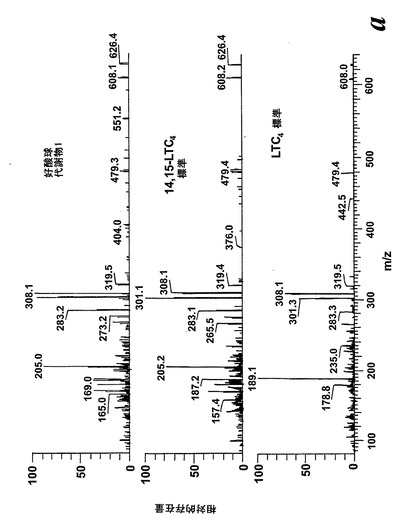
【図33】
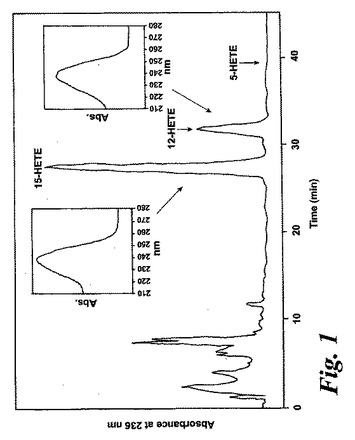
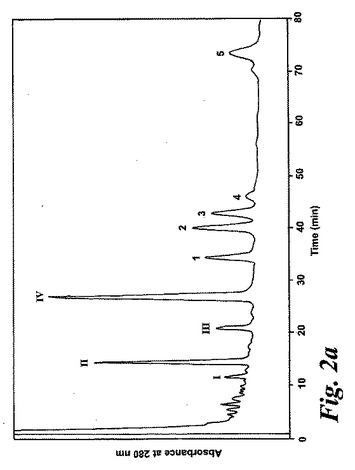
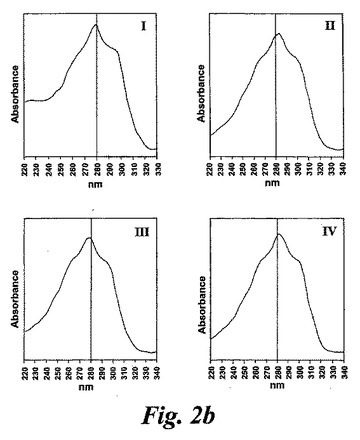
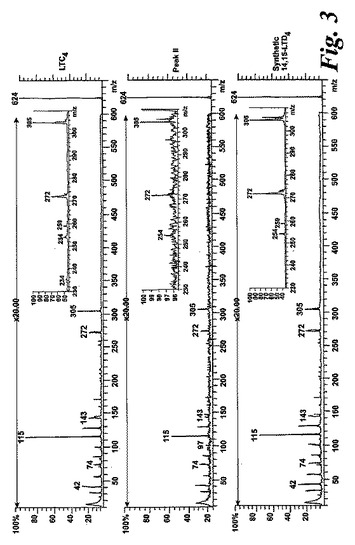
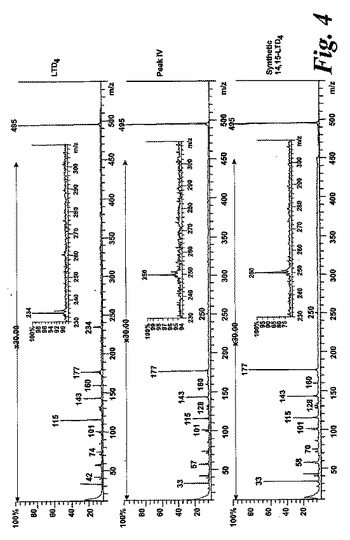
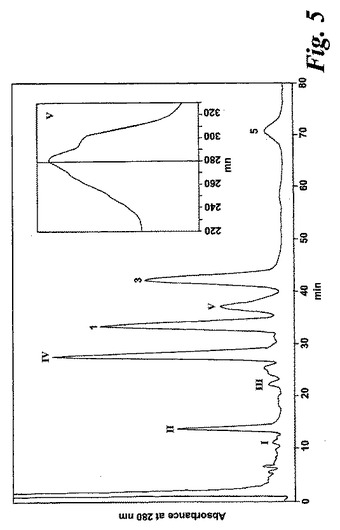
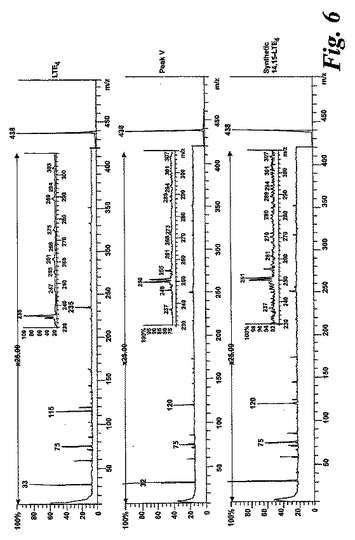
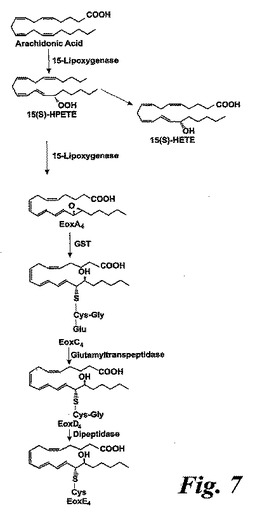
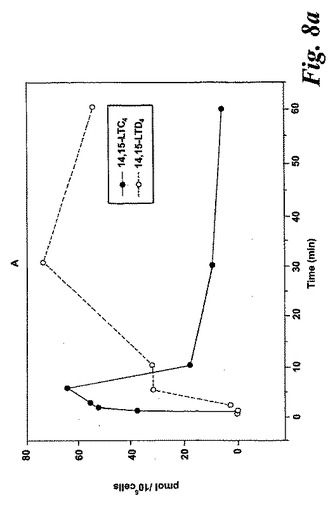
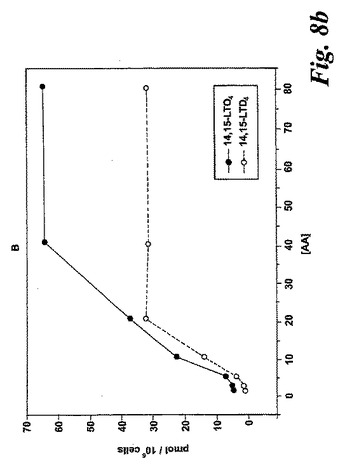
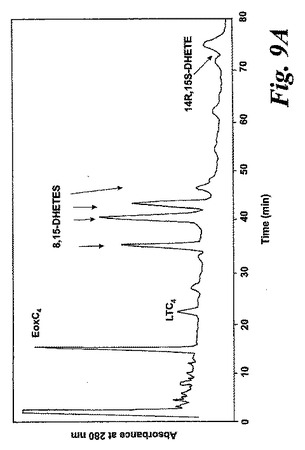
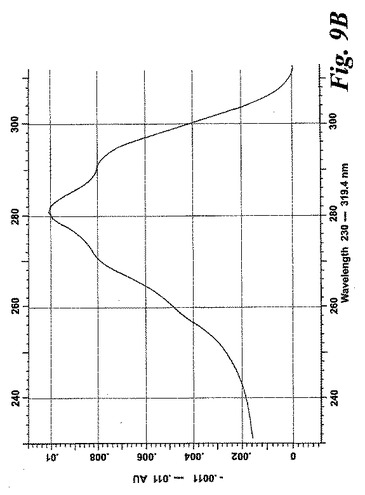
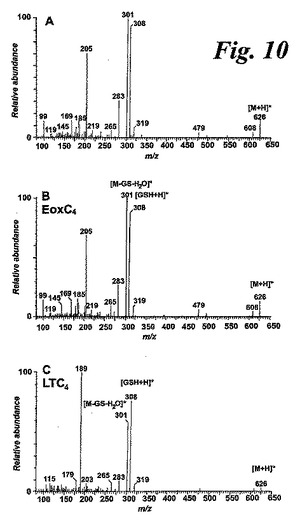
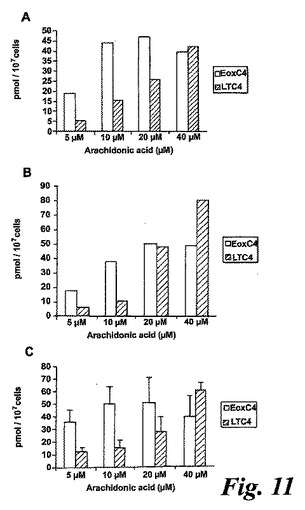
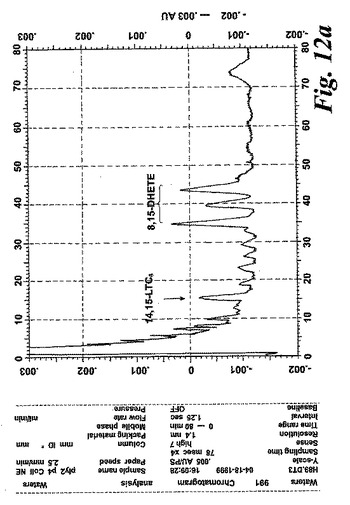
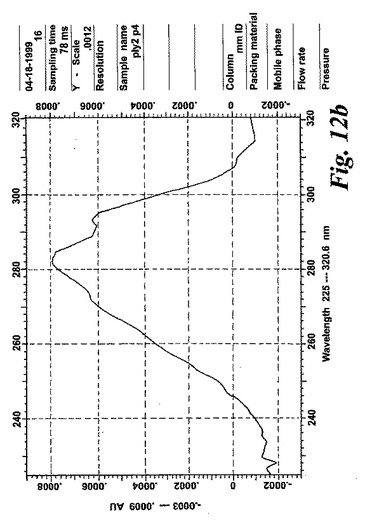
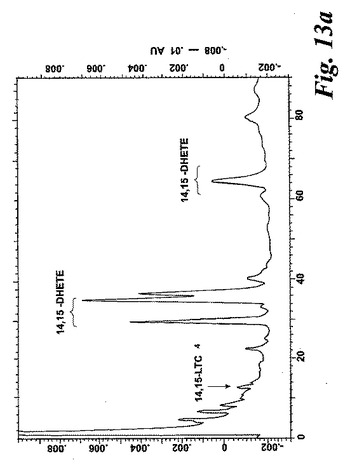
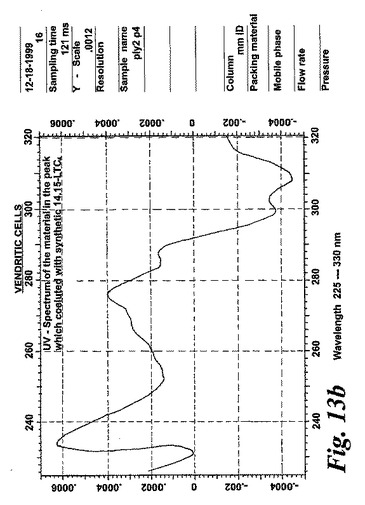
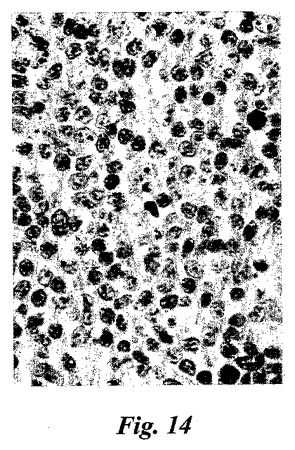
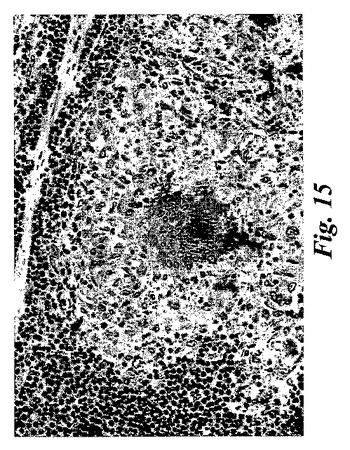
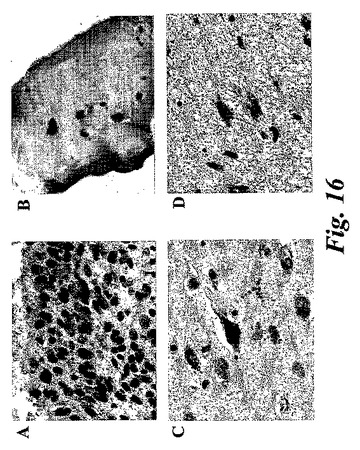
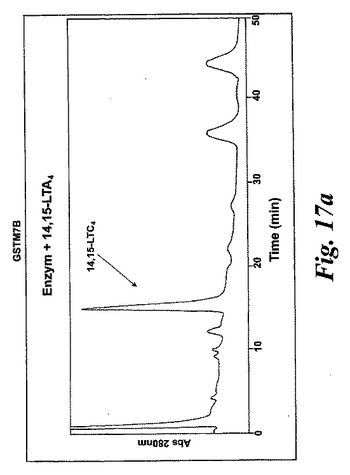
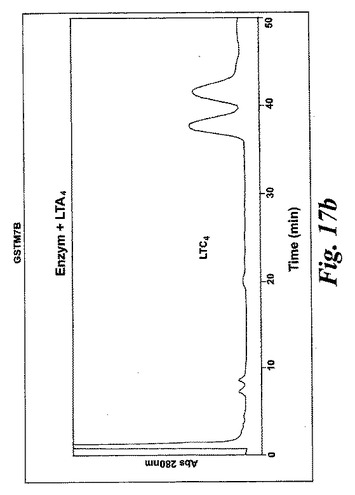
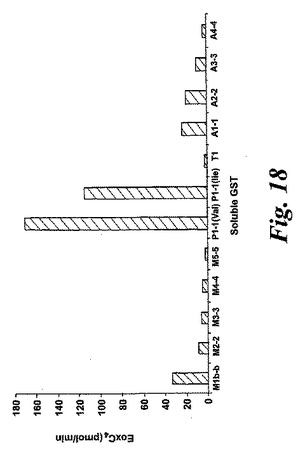
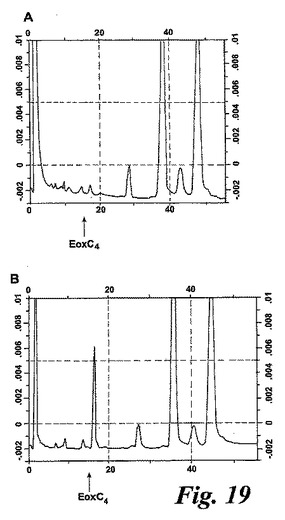
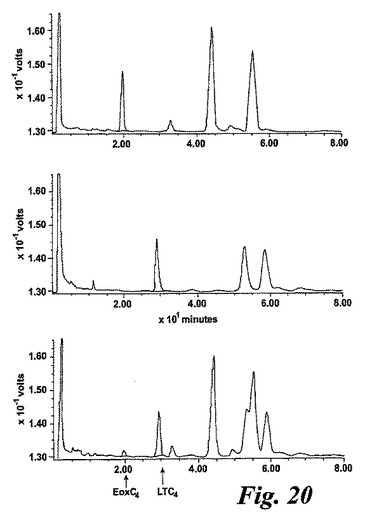
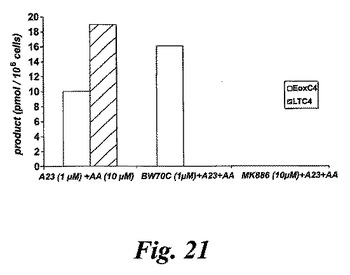
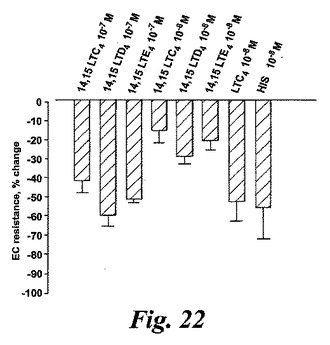
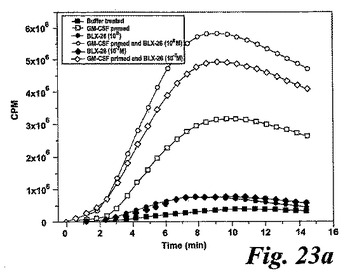
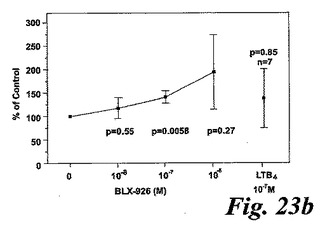
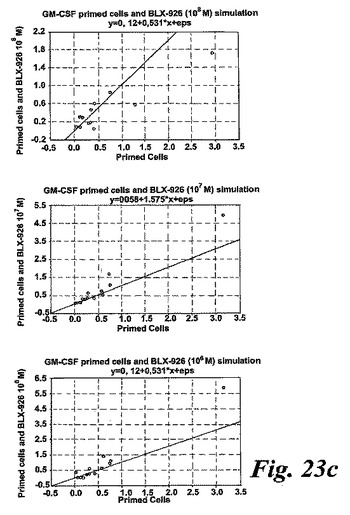
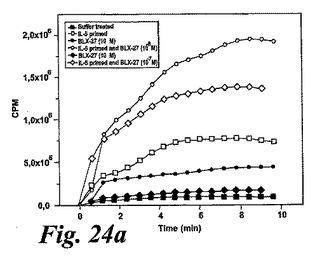
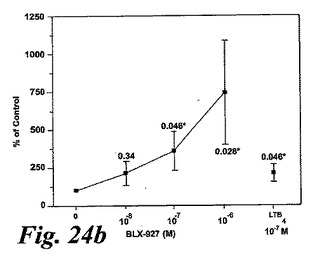
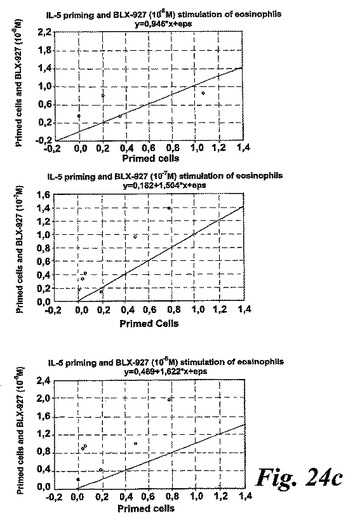
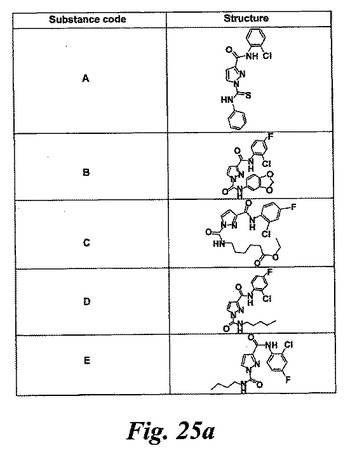
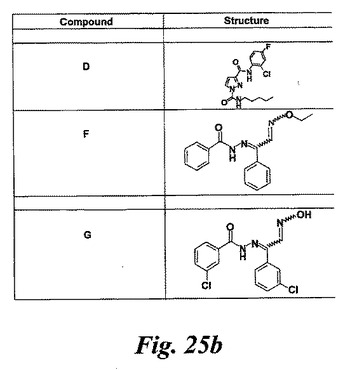
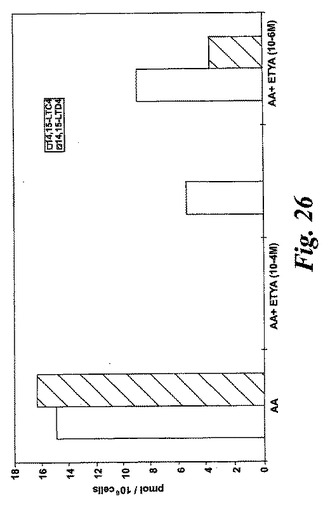
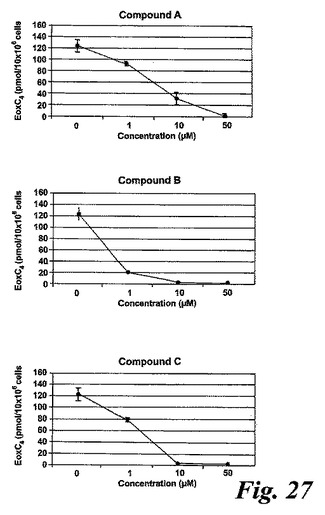
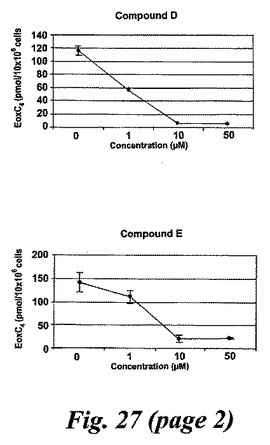
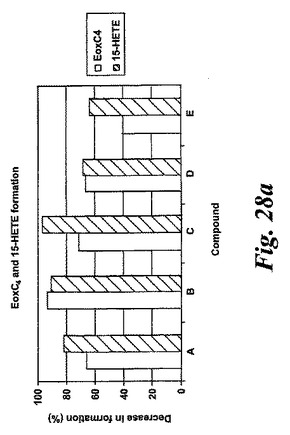
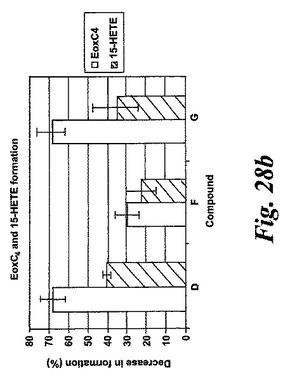
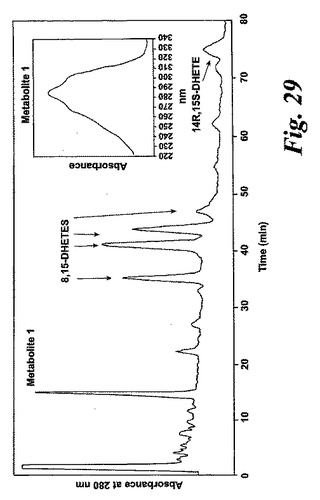
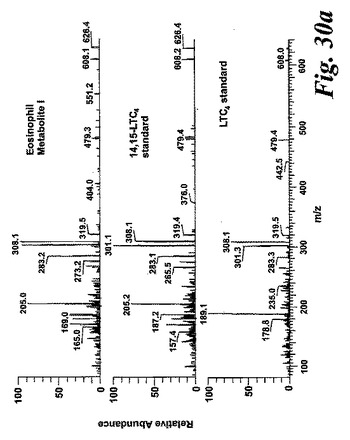
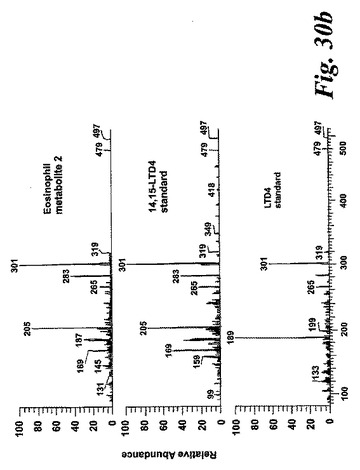
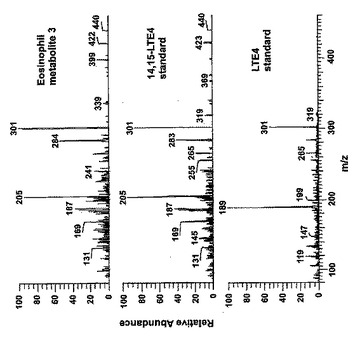
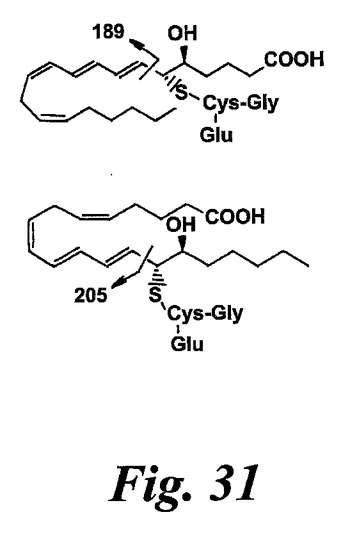
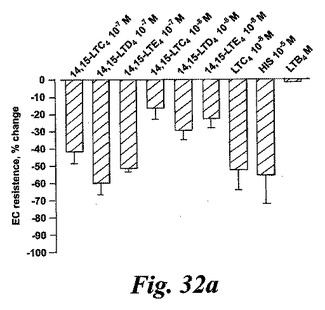
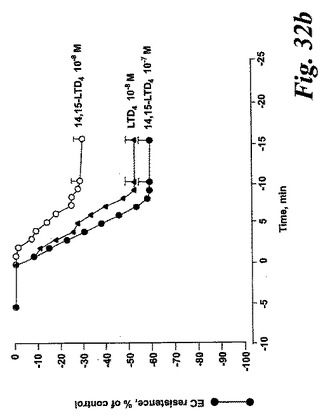
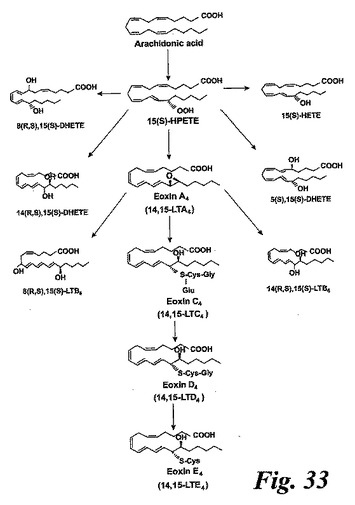
【図2a】
【図2b】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8a】
【図8b】
【図9a】
【図9b】
【図10】
【図11】
【図12a】
【図12b】
【図13a】
【図13b】
【図14】
【図15】
【図16】
【図17a】
【図17b】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23a】
【図23b】
【図23c】
【図24a】
【図24b】
【図24c】
【図25a】
【図25b】
【図26】
【図27a】
【図27b】
【図28a】
【図28b】
【図29】
【図30a】
【図30b】
【図30c】
【図31】
【図32a】
【図32b】
【図32c】
【図33】
【公表番号】特表2009−540856(P2009−540856A)
【公表日】平成21年11月26日(2009.11.26)
【国際特許分類】
【出願番号】特願2009−517391(P2009−517391)
【出願日】平成19年6月27日(2007.6.27)
【国際出願番号】PCT/GB2007/002394
【国際公開番号】WO2008/001079
【国際公開日】平成20年1月3日(2008.1.3)
【出願人】(505347558)バイオリポックス エービー (26)
【Fターム(参考)】
【公表日】平成21年11月26日(2009.11.26)
【国際特許分類】
【出願日】平成19年6月27日(2007.6.27)
【国際出願番号】PCT/GB2007/002394
【国際公開番号】WO2008/001079
【国際公開日】平成20年1月3日(2008.1.3)
【出願人】(505347558)バイオリポックス エービー (26)
【Fターム(参考)】
[ Back to top ]