
エストロン3硫酸トランスポーター活性の阻害剤
【課題】 細胞内に取り込まれる必要がなく、ドラッグデリバリー性に優れ、副作用がきわめて少ないエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤を提供すること。
【解決手段】 ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸からえらばれる1種以上の成分をエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤の有効成分とする。
【解決手段】 ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸からえらばれる1種以上の成分をエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤の有効成分とする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、エストロン3硫酸(エストロン−3−サルフェート;estrone-3-sulfate)トランスポーターのトランスポーター活性の阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤に関する。
【背景技術】
【0002】
細胞内外の物質輸送を行う膜タンパク質等はトランスポーターと呼ばれ、脂質二重膜中に埋め込まれているトランスポーターに特定の分子が結合すると、コンフォメーションが変化して物質が取込みあるいは排出輸送されることが知られている。近年、かかるトランスポーター、例えば有機アニオン性物質を輸送するOAT1等の有機アニオントランスポーター,有機カチオン性物質を輸送するOCT1等の有機カチオントランスポーター,ペプチド性物質を輸送するPEPT1等のペプチドトランスポーターなどの遺伝子が相次いで単離・同定されている。これらトランスポーター遺伝子は、全身の正常組織・臓器に偏在しているものもあるが、腎臓,肝臓,脳などといった特定の組織・臓器に局在するものも知られている。
【0003】
一方、エストロゲンは雌性動物に発情現象を起こす、いわゆる女性ホルモンであり、天然に存在するエストロン(E1),エストラジオール(E2),エストリオール(E3),エステトロール(E4)と、それらと同様の生物活性を有する合成エストロゲンに分類される。スチルベステロールなどの合成型の一部を除いて、エストロゲンはステロイド構造を有する。エストロン0.1μgのもつ生物学的作用を1IU(国際単位)とし、これがエストロゲンの単位として用いられている。分泌源は主として卵巣の卵胞及び黄体であるが、妊娠時の胎児胎盤系、副腎、精巣などからも分泌される。卵巣からのエストロゲンの分泌は、下垂体前葉より分泌される性腺刺激ホルモンにより支配される(下行性調節)が、逆にエストロゲンによる間脳下垂体系へのフィードバック作用も認められ(上行性調節)、両者の相互関係により性周期が成立するといわれている。
【0004】
さらに、エストロゲンはその受容体(レセプター)を介して作用し、エストロゲンの作用は標的組織である間脳−下垂体前葉−性器及び乳腺のみならず全身に及び、主な生理作用は、子宮内膜の増殖,子宮筋の発育,第二次性徴の発現,月経周期の成立の媒介,妊娠時の母体変化の惹起,乳腺管の増殖分泌促進などである。エストロゲンは主として肝臓において代謝をうけ、エストロン3硫酸等の抱合型エストロゲンとなり尿中に排泄される。臨床的には、無月経や月経異常の治療,月経の人為的移動,更年期障害,前立腺癌や乳癌に対するホルモン療法,骨粗鬆症などに対して用いられている。
【0005】
乳癌は、発癌時点からエストロゲン受容体システムの異常作動があり、初期はエストロゲン依存性の増殖を続けるが、次第にこの制御を離れた増殖となる。このことから、乳癌の発生と進展にエストロゲンとその受容体が深く関与していると考えられているが、その機序は未だ明らかではない。乳癌は乳腺の上皮組織の末梢乳管や腺房上皮から発生する悪性腫瘍で、欧米では女性の癌死亡の第一位である。また、日本においても、近年の食生活の欧米化、生活様式の変化などに伴い乳がんによる死亡率は上昇傾向にある。好発年齢は40歳代で、リスク因子として、乳癌の既往・家族歴,未婚,高齢初産,早期初潮と晩期閉経,肥満,放射線被爆,高脂肪食,良性乳腺疾患の既往などを挙げることができる。リンパ節転移は腋窩,鎖骨下,胸骨傍に頻度が高く、転移個数は予後と相関する。血行性転移は骨,肺,肝に多い。主症状は乳房腫瘤であり、表面不整,硬,境界不明瞭で可動性が少ない。
【0006】
乳癌の2/3は、エストロゲン依存性である(非特許文献1)、すなわち、細胞増殖がエストロゲンによって制御されている。エストロゲンの生物活性型がエストラジオールであり、2つの主要経路を経て合成される(非特許文献2、非特許文献3)。1つは、アロマターゼがアンドロゲンをエストロゲンに変換するアロマターゼ経路であり、もう1つは、ステロイドスルファターゼがエストロン3硫酸をエストロンに変換するスルファターゼ経路である。患者乳癌組織では、スルファターゼの活性はアロマターゼの活性と比べて50〜200倍であるため、スルファターゼ経路は、エストロン3硫酸由来生物活性エストロゲンの主要な供給源である(非特許文献3、非特許文献4)。また、エストロン3硫酸の循環血漿濃度は、非抱合エストロゲンの場合と比べて約10〜20倍であり、エストロン3硫酸の半減期はエストラジオールの半減期よりも長い(非特許文献5〜7)。したがって、エストロン3硫酸は、エストロン3硫酸自体があまり生物活性を示さなくても、活性エストロゲンの貯蔵庫として、乳癌の進行に重要な役割を果たすと考えられている。
【0007】
エストロン3硫酸による乳癌細胞増殖の刺激には、下記の一連のプロセスが関わっている:エストロン3硫酸の細胞への取り込み、エストロゲンスルファターゼによる脱硫酸化、I型の17β−ヒドロキシステロイドデヒドロゲナーゼによるエストロンのエストラジオールへの変換、核エストロゲン受容体への結合、及び遺伝子転写の制御(非特許文献8〜11)。エストロン3硫酸に対する乳癌細胞の応答に関与している酵素や受容体についてよく調べてみたが、エストロン3硫酸などのリガンドが細胞膜を通過する輸送メカニズムはわかっていない。エストロン3硫酸のlogP値は1.4であると報告されており、その値はエストロンの値(4.4)やエストラジオールの値(3.9)より著しく低いため、エストロン及びエストラジオールが拡散性であるにも関わらず、拡散によって細胞膜を通過することは容易ではないであろう。(非特許文献12)。エストロン3硫酸とともにインキュベーションした後のMCF−7細胞の細胞質画分中で、エストロン3硫酸はインタクトな形で検出されており(非特許文献13)、エストロゲン作用を示していた(非特許文献4、非特許文献14、非特許文献15)。したがって、ホルモン依存性増殖を示す乳癌細胞へのエストロン3硫酸の供給には、特異的トランスポーターが関与していると思われる。
【0008】
有機アニオントランスポーター(OAT、SLC22A)(非特許文献16)ファミリー及び有機アニオン輸送ペプチド(OATP、SLC21A、SLCO)(非特許文献17、非特許文献18)ファミリーのメンバーのなかには、エストロン3硫酸を輸送するものがあることが既に知られている。それらは主として肝臓、腎臓、脳、腸、その他の部位で発現するが、乳癌細胞中での発現は報告されていない。エストロン3硫酸に対するそれらのKm値は0.05μM〜59μMの範囲内である。女性では、エストロン3硫酸の血漿中レベルは1〜10nMの範囲で変動するため(非特許文献19)、これらの又はその他のトランスポーターは、ホルモン依存性増殖の第一歩としての、エストロン3硫酸の癌細胞への取り込みにおいて、ある役割を果たしていると思われる。
【0009】
最近、Pizzagalli et al.が、ヒトの乳腺におけるステロイドサルフェートトランスポーターであるOATP−Bの発現を示唆している(非特許文献20)。しかし、エストロゲン依存性MCF−7細胞及びT−47D細胞中でOATP−B mRNAが検出されなかったのに対し、OATP−D及びOATP−E mRNAはこれらの細胞中で発見されている(非特許文献21)。OATP−D及びOATP−Eは、その遺伝子をHEK293細胞中でトランジェントに発現させた場合にエストロン3硫酸に対する輸送活性を示すため(非特許文献22)、癌細胞におけるエストロン3硫酸の輸送に関与していると思われる。しかし、エストロゲン依存性細胞においてトランスポーターが媒介するエストロン3硫酸の取り込みは、これまで調べられていなかったが、肝臓、胎盤及び脳等のほかの組織における取り込みは報告されている。乳癌細胞の増殖メカニズムを充分に理解するためには、かかる細胞へのエストロゲンの供給メカニズムを解明しなくてはならない。
【0010】
【非特許文献1】N Engl J Med 302: 78-90, 1980
【非特許文献2】J Clin Endocrinol Metab 57: 1125-1128, 1983
【非特許文献3】Breast Cancer Res Treat 7: 35-44, 1986
【非特許文献4】Ann N Y Acad Sci 464: 126-137, 1986
【非特許文献5】J Clin Endocrinol Metab 81: 1460-1464, 1996
【非特許文献6】J Biol Chem 236: 1043-1050, 1961
【非特許文献7】J Clin Invest 51: 1020-1033, 1972
【非特許文献8】Endocrinology 138: 863-870, 1997
【非特許文献9】Mol Endocrinol 11: 77-86, 1997
【非特許文献10】Endocrinology 123: 1281-1287, 1988
【非特許文献11】Breast Cancer 9: 296-302, 2002
【非特許文献12】J Clin Endocrinol Metab 59: 1128-1132, 1984
【非特許文献13】Endocrinology 106: 1079-1086, 1980
【非特許文献14】J Clin Endocrinol Metab 81: 1460-1464, 1996
【非特許文献15】J Steroid Biochem Mol Biol 73: 225-235, 2000
【非特許文献16】Pflugers Arch 440: 337-350, 2000
【非特許文献17】Biochim Biophys Acta 1609: 1-18, 2003
【非特許文献18】Pflugers Arch 447: 653-665, 2003
【非特許文献19】Hormone Res 27: 61-68, 1987
【非特許文献20】J Clin Endocrinol Metab 88: 3902-3912, 2003
【非特許文献21】J Clin Endocrinol Metab 88: 3902-3912, 2003
【非特許文献22】Pharm Res 18: 1262-1269, 2001
【発明の開示】
【発明が解決しようとする課題】
【0011】
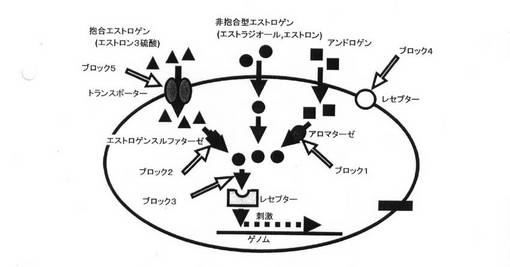
乳癌細胞におけるエストロゲンとその受容体との結合によるシグナル伝達をアップストリームにブロックする乳癌細胞の増殖阻害については、図1に示されるように、以下の方法(ブロック1〜4)が知られている。
【0012】
ブロック1(アンドロゲンからのエストロゲンの合成阻害)
ファドロゾール:閉経後、エストロゲン合成の律速酵素として働くアロマターゼを選択的に阻害して、アンドロゲンからエストロゲンへの変換を妨げることにより、生体内のエストロゲン濃度を低下させ、乳ガンの増殖を抑制する。
【0013】
ブロック2(エストロゲンレセプターでの競合阻害)
タモキシフェン、トレミフェン:乳ガン組織などのエストロゲン受容体に対し、エストロゲンと競合的に阻害し、抗エストロゲン作用を示すことによって抗乳ガン作用を発揮する。
【0014】
ブロック3(エストロン硫酸からのエストロゲン合成阻害)
抱合型エストロゲンは細胞内でエストロゲンサルファターゼによって活性型となり、ガン細胞増殖を促進する。この脱抱合酵素を阻害する。
【0015】
ブロック4(膜タンパクに対する抗体を利用した細胞障害作用)
ハーセプチン:乳ガン細胞にはHER2タンパクが高発現するものがある。このHER2と抗体の結合を利用し、抗体依存性細胞障害作用による抗腫瘍効果を発揮する。
【0016】
乳癌細胞の増殖阻害における、上記ブロック1〜4の方法では、アロマターゼ阻害剤やエストロゲンサルファターゼ阻害剤、あるいは、タモキシフェンやトレミフェン等のエストロゲン競合阻害剤が乳癌細胞内に取り込まれないと抗腫瘍効果が発揮されないことからドラッグデリバリーの面で、また、細胞内に取り込まれると副作用の面で、それぞれ問題なしとはいえない。本発明の課題は、細胞内に取り込まれる必要がなく、ドラッグデリバリー性に優れ、副作用がきわめて少ないエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤を提供することにある。
【課題を解決するための手段】
【0017】
本発明者らは、上記課題を解決するために鋭意研究し、乳癌細胞の増殖阻害の方法として、以下のブロック5(図1参照)について検討した。
ブロック5(トランスポーター活性の阻害)
エストロゲン感受性の乳ガン細胞に対し、エストロゲンの供給源であるエストロン3硫酸の細胞内取込みを阻害することで抗腫瘍効果が現れる。
【0018】
そして、ヒト乳癌培養細胞MCF−7細胞株をエストロン3硫酸と、被検物質としてのブロモスルフォフタレイン(Bromosulfophtalein;BSP)の存在下に培養した時、細胞表面に発現するエストロン3硫酸トランスポーターを介してのエストロン3硫酸の細胞内への取込みが阻害され、MCF−7細胞株の増殖が抑制されることを見い出し、本発明を完成するに至った。
【0019】
すなわち本発明は、(1)ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とするエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や、(2)ブロモスルフォフタレインを有効成分とするエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や、(3)ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とする乳癌細胞増殖抑制剤や、(4)ブロモスルフォフタレインを有効成分とする乳癌細胞増殖抑制剤や、(5)ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とする乳癌治療剤や、(6)ブロモスルフォフタレインを有効成分とする乳癌治療剤に関する。
【発明の効果】
【0020】
本発明によると、アロマターゼ阻害剤やエストロゲンサルファターゼ阻害剤、タモキシフェンやトレミフェン等のエストロゲン競合阻害剤と異なり、細胞内に取り込まれる必要がなく、ドラッグデリバリー性に優れ、副作用がきわめて少ないエストロン3硫酸トランスポーターの阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤を提供することができる。
【発明を実施するための最良の形態】
【0021】
本発明のエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤としては、ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸を挙げることができるが、中でもブロモスルフォフタレインを好適に例示することができる。また、これらは1種単独又は2種以上を併用することができる。
【0022】
本発明のエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤(以下「本件阻害剤」という場合がある)は、エストロン3硫酸トランスポーターを同定する上で重要である。エストロン3硫酸トランスポーターの候補トランスポーターとしては、SLCトランスポーター,OAT1,OAT2,OAT3,OAT4,OATP−A,OATP−B,OATP−C,OATP−D,OATP−E,OATP−F,OATP−8,NTCP,MRPs,BCRPなどを好適に挙げることができる。同定する方法としては、候補エストロン3硫酸トランスポーターを細胞表面に発現する細胞を培養し、培養した細胞内への候補エストロン3硫酸の取り込みを本件阻害剤の存在下において測定し、本件阻害剤による細胞内へのエストロン3硫酸のトランスポーター活性の阻害の程度を測定・評価する方法や、候補エストロン3硫酸トランスポーターを細胞表面に発現する細胞から単離した細胞膜画分に、エストロン3硫酸と本件阻害剤とを接触させ、本件阻害剤によるエストロン3硫酸の細胞膜画分への特異的結合の阻害の程度を測定・評価する方法や、候補エストロン3硫酸トランスポーターを細胞表面に発現する細胞から細胞膜画分を単離し、該細胞膜画分から小胞を作製して単離細胞膜小胞とし、該単離細胞膜小胞にエストロン3硫酸と本件阻害剤とを接触させ、本件阻害剤による小胞内へのエストロン3硫酸の取込み量もしくは取込み量の阻害の程度を測定・評価する方法を挙げることができる。上記エストロン3硫酸と本件阻害剤との共存下の細胞の培養は、使用細胞の増殖用培地を用い、通常の細胞培養条件下、(本件阻害剤非存在下の場合に)少なくとも細胞増殖が有意に認められるまで行うことが望ましく、上記エストロン3硫酸と本件阻害剤との細胞膜画分との接触や、上記エストロン3硫酸と本件阻害剤との単離細胞膜小胞との接触は、使用細胞の増殖用培地や適当な緩衝液中でインキュベーションすることにより行うことができる。
【0023】
上記本件阻害剤による細胞内へのエストロン3硫酸のトランスポーター活性の阻害の程度を測定・評価する方法としては、細胞内のエストロン3硫酸濃度を測定・評価する方法や、細胞増殖の程度を測定・評価する方法を挙げることができる。後述するMCF−7細胞株、KPL−1,MKL−F又はそれら細胞株由来の細胞株等のヒト乳癌培養細胞株を用いる場合は、細胞内の本件阻害剤濃度を測定・評価する方法と、細胞増殖の程度を測定・評価する方法とを利用することができるが、候補エストロン3硫酸トランスポーターを発現する形質転換細胞を用いる場合は、細胞内の本件阻害剤濃度を測定・評価する方法を利用することができる。また、候補エストロン3硫酸トランスポーター遺伝子をアフリカツメガエル卵母細胞等に発現させ、2本の微小電極を用いて、トランスポーターを介した基質輸送に伴って生じる膜電位の変化を検出(電流を測定)する電気生理学的方法により測定・評価することもできる。
【0024】
エストロン3硫酸トランスポーターを細胞表面に発現する細胞から細胞膜画分を単離する方法としては、F. Pietri-Rouxelら(Eur. J. Biochem., 247, 1174-1179, 1997)の方法など、従来公知の方法を用いることができ、また、細胞膜画分から小胞を作製して単離細胞膜小胞とする方法としては、J.E. Leverら(J. Biol. Chem., 252, 1990-1997 (1977))の方法など、従来公知の方法を用いることができる。そしてまた、本件阻害剤によるエストロン3硫酸の細胞膜画分への特異的結合の阻害の程度を測定する方法としては、放射性標識エストロン3硫酸を用いた定常状態結合阻害から平衡解離定数KDを測定する方法を例示することができ、本件阻害剤による単離細胞膜小胞内へのエストロン3硫酸の取込み量もしくは取込み量の阻害の程度を測定する方法としては、I. Tamaiら(Biochim. Biophys. Acta, 1512, 273-284 (2001))記載の方法など、従来公知の手法を細胞内エストロン3硫酸の測定方法として用いることができる。
【0025】
上記エストロン3硫酸トランスポーターを細胞表面に発現する細胞としては、候補エストロン3硫酸トランスポーターを発現する形質転換細胞を使用することができる。かかる形質転換細胞の作製に使用される前記SLCトランスポーター,OAT1,OAT2,OAT3,OAT4,OATP−A,OATP−B,OATP−C,OATP−D,OATP−E,OATP−F,OATP−8,NTCP,MRPs,BCRPをコードする遺伝子の由来としては特に制限されるものではなく、例えばヒト,イヌ,ウシ,ウマ,ヤギ,ヒツジ,サル,ブタ,ウサギ,ラット,マウス等を挙げることができるが、ヒト由来が好ましい。
【0026】
上記のようなトランスポーターををコードする遺伝子やcDNAを細胞に発現させる方法としては特に限定されないが、Davisら(BASIC METHODS IN MOLECULAR BIOLOGY, 1986)及びSambrookら(MOLECULAR CLONING: A LABORATORY MANUAL, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989)などの多くの標準的な実験室マニュアルに記載される方法、例えば、リン酸カルシウムトランスフェクション,DEAE−デキストラン媒介トランスフェクション,トランスベクション(transvection)マイクロインジェクション,カチオン性脂質媒介トランスフェクション,エレクトロポレーション,形質導入,スクレープローディング(scrape loading),弾丸導入(ballistic introduction),感染等の遺伝子導入法により、トランスポーター遺伝子を宿主細胞へ導入することにより発現させることができるが、トランスポーター遺伝子を含む発現ベクターを上記宿主細胞に導入することが好ましく、中でもトランスポーター遺伝子を含む発現ベクターを宿主細胞に感染させることにより、トランスポーターを細胞に発現させることがより好ましい。
【0027】
上記発現ベクターとしては、例えば、非分裂細胞を含む全ての細胞(血球系以外)での一過性発現に用いられるアデノウイルスベクター(Science, 252, 431-434, 1991)や、分裂細胞での長期発現に用いられるレトロウイルスベクター(Microbiology and Immunology, 158, 1-23, 1992)や、非病原性、非分裂細胞にも導入可能で、長期発現に用いられるアデノ随伴ウイルスベクター(Curr. Top. Microbiol. Immunol., 158, 97-129, 1992)、SV40のようなパポバウイルスベクター、ワクシニアウイルスベクター等のウイルスベクターの他、リポソーム等を具体的に挙げることができるが、これらに限定されるものではない。これらの中でも、高い効率で細胞における遺伝子発現が可能なアデノウイルスベクターが特に好ましい。また、これらの発現ベクターへのトランスポーター遺伝子の導入は常法によって行うことができ、例えばこれら発現ベクター中の適当なプロモーターの下流にトランスポーター遺伝子等を挿入することにより発現ベクターを構築することができる。また、発現ベクターはトランスポーター遺伝子及びマーカー遺伝子に加え、トランスポーターの細胞当たりの発現量を規格化するためのIRES(mRNA内部のリボソーム結合サイト)の他、エンハンサー,ターミネーター等発現を調節する制御配列を含んでいてもよい。
【0028】
トランスポーターを発現する宿主細胞としては、ドロソフィラS2,スポドプテラSf9等の昆虫細胞,Vero細胞,HeLa細胞,CHO細胞,WI−38細胞,BHK細胞,COS−7細胞,MDCK細胞,C127細胞,HKG細胞,ヒト腎細胞株,CV−1細胞,LLC−MK2細胞,MDBK細胞,MRC−5細胞,Caco−2細胞,HT29細胞,ヒトリンパ芽球細胞,アフリカツメガエル等の卵母細胞や、これらのdhfr欠損株,HGPRT欠損株,ウアバイン耐性株等を挙げることができる。具体的には、CHO−K1(チャイニーズハムスター卵巣細胞:ATCC CCL61),BHK(ハムスター腎細胞:ATCC CCL10),COS−7(CV−1 Origin,SV−40細胞:ATCC CRL1651),Vero細胞(アフリカミドリザル腎細胞:ATCC CCL81),ヒトリンパ芽球細胞(IM−9,ATCC CCL159)を例示することができる。
【0029】
トランスポーターは細胞膜表面に発現する膜タンパク質であるため、エストロン3硫酸トランスポーターに対する中和抗体は細胞外から選択的に標的を阻害する可能性があり、このことから本件阻害剤によりエストロン3硫酸トランスポーターを同定することができると、エストロン3硫酸トランスポーターに対する中和抗体を作製することができる。特異的に認識する抗体としては、モノクローナル抗体,ポリクローナル抗体,一本鎖抗体,ヒト化抗体,キメラ抗体,2つのエピトープを同時に認識することができる二機能性抗体等を例示することができる。これら抗体は、慣用のプロトコールを用いて、動物(好ましくはヒト以外)にエストロン3硫酸トランスポーターを膜表面に発現している細胞やその細胞膜画分を投与することにより産生され、例えばモノクローナル抗体の調製には、連続細胞系の培養物により産生される抗体をもたらす、ハイブリドーマ法(Nature 256, 495-497, 1975),トリオーマ法,ヒトB細胞ハイブリドーマ法(Immunology Today 4, 72, 1983)及びEBV−ハイブリドーマ法(MONOCLONAL ANTIBODIES AND CANCER THERAPY, pp.77-96, Alan R.Liss, Inc.,1985)など任意の方法を用いることができる。
【0030】
本発明の乳癌細胞増殖抑制剤は、エストロン3硫酸トランスポーターの同定用として、また乳癌細胞の増殖を抑制しうることから乳癌治療剤として用いることができる。同定する方法としては、候補エストロン3硫酸トランスポーターを細胞表面に発現する細胞を、本発明の乳癌細胞増殖抑制剤の存在下において培養し、細胞増殖の程度を測定・評価する方法を挙げることができる。
【0031】
本発明の乳癌治療剤は、乳癌の予防薬としても用いることができ、医薬用の治療剤として用いる場合は、薬学的に許容される通常の担体、結合剤、安定化剤、賦形剤、希釈剤、pH緩衝剤、崩壊剤、可溶化剤、溶解補助剤、等張剤などの各種調剤用配合成分を添加することができる。これら治療剤は、経口的又は非経口的に投与することができ、例えば粉末、顆粒、錠剤、カプセル剤、シロップ剤、懸濁液等の剤型で経口的に投与することができ、あるいは、例えば溶液、乳剤、懸濁液等の剤型にしたものを注射の型で非経口投与することができる。非経口投与する場合、局所に直接投与することができる。経口的に投与する製剤の場合、薬理学的に許容される担体としては、慣用の各種有機あるいは無機担体物質が用いられ、例えば錠剤には乳糖、デンプン等の賦形剤、タルク、ステアリン酸マグネシウム等の滑沢剤、ヒドロキシプロピルセルロース、ポリビニルピロリドン等の結合剤、カルボキシメチルセルロース等の崩壊剤等を配合することができ、懸濁液製剤には生理的食塩水アルコール等の溶剤、ポリエチレングリコール、プロピレングリコール等の溶解補助剤、ステアリルトリエタノールアミン、ラウリル硫酸ナトリウム、レシチン等の懸濁化剤、グリセリン、D−マンニトール等の等張化剤、リン酸塩、酢酸塩、クエン酸塩等の緩衝剤などを配合することができる。また必要に応じて、防腐剤、抗酸化剤、着色剤、甘味剤などの製剤添加物を配合することもできる。非経口的に投与する製剤の場合、蒸留水、生理的食塩水等の水溶性溶剤、サリチル酸ナトリウム等の溶解補助剤、塩化ナトリウム、グリセリン、D−マンニトール等の等張化剤、ヒト血清アルブミン等の安定化剤、メチルパラベン等の保存剤、ベンジルアルコール等の局麻剤を配合することができる。治療薬の投与量は、患者の体重や年齢、投与形態、症状等により適宜選定することができるが、例えば成人に投与する場合、有効成分を通常1回量として約0.001〜500mg、好ましくは1〜50mgを1日1回〜3回投与するのが望ましい。
【0032】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。
【実施例1】
【0033】
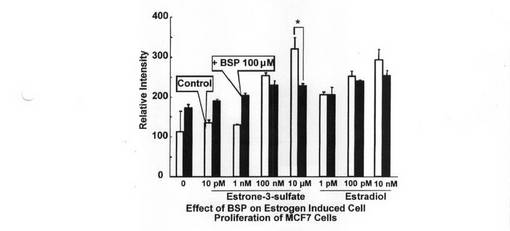
MCF−7細胞(ATCC−HTB22)のエストロン3硫酸による細胞増殖促進効果の抑制が、エストロン3硫酸の細胞内蓄積を抑えることで達成されるか否かを検討した。MCF−7細胞を8×104個/cm2の濃度で96ウェル プレートに播種し1日培養した後、10pM〜10μMの各種濃度のエストロン3硫酸(Sigma Chemicals社製)と100μMのブロモスルフォフタレイン(BSP:Sigma Chemicals社製)、及び、1pM〜10nMの各種濃度のエストラジオール(estradiol:Sigma Chemicals社製)と100μMのBSPを含む培地をそれぞれ添加し、さらに3日間培養した。その後、細胞量を測定した。エストロン3硫酸及びエストラジオールによる細胞増殖促進効果の結果を図2に示す。
【0034】
図2から明らかなように、ポジティブコントロールであるエストロン3硫酸単独存在下及びエストラジオール単独存在下においては、細胞増殖促進効果が観察された(open column)。一方、エストロン3硫酸又はエストラジオールに、100μMのBSPを同時添加することによって共存下に培養すると、エストロン3硫酸低濃度条件において細胞増殖促進効果が見られるが、これは、BSP自身あるいはその代謝物によるエストロゲン活性であると考えられる。この効果はエストロゲン濃度が増加するに従い、相対的に低くなった。興味深いことに、エストロン3硫酸10μM添加時、BSPによる細胞増殖効果は阻害されたのに対し、エストラジオール添加による効果には変化しなかった。したがって、エストロン3硫酸の細胞内取込みはBSPにより阻害され、脂溶性の高いエストラジオールは阻害されないため、細胞増殖効果にも変化がなかったと考えられた。以上のことから、細胞増殖におけるエストロン3硫酸の取り込みトランスポーターの重要性が示唆され、そのトランスポーターを阻害することによる細胞増殖抑制の可能性が示された。
【実施例2】
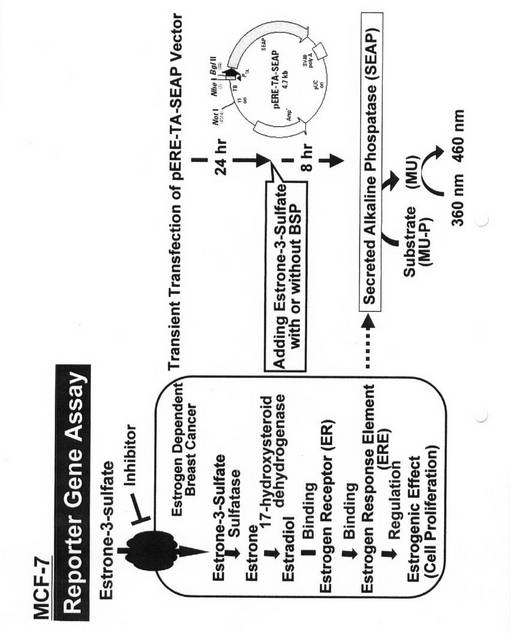
【0035】
MCF−7細胞(ATCC−HTB22)におけるエストロン3硫酸によるERE(エストロゲンレスポンスエレメント)活性の誘導がBSPにより抑制しうるか否かを検討した。MCF−7細胞を、図3に示すpERE−TA−SEAPベクター(BD Biosciencesから購入)で一過的にトランスフェクションした。このpERE−TA−SEAPベクターは、エストロゲンが結合するDNA配列であるERE配列を上流に持つSEAP(Secreted Alkaline phospatase)蛋白質発現ベクターで、エストロゲンが存在すると、SEAP蛋白質合成がEREを介して促進され、SEAP活性が上昇する。また、SEAPはリン酸結合を切断する酵素で、この酵素の基質をMU−P(methyl umbelliferone phosphate)とし、生じたMU(methyl umbelliferone)を定量することにより、SEAP活性を測定することができる。
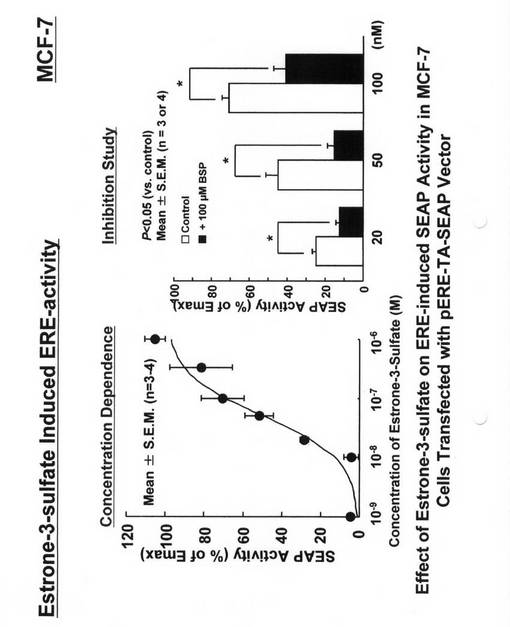
【0036】
pERE−TA−SEAPベクター(BD Biosciencesから購入)キットのマニュアルに沿ってトランスフェクションしたMCF−7細胞を8×104個/cm2の濃度で96ウェル プレートに播種し1日培養した後、10−9〜10−6Mの各種濃度のエストロン3硫酸(Sigma Chemicals社製)を添加し、さらに8時間培養したのちに、SEAP活性を測定した。結果を図4(左)に示す。その結果、pERE−TA−SEAPベクターで一過的にトランスフェクションしたMCF−7細胞は、添加したエストロン3硫酸の濃度依存的にSEAP活性を示した。次に、pERE−TA−SEAPベクター(BD Biosciencesから購入)キットのマニュアルに沿ってトランスフェクションしたMCF−7細胞を8×104個/cm2の濃度で96ウェル プレートに播種し1日培養した後、20nM,50nM,100nMの各種濃度のエストロン3硫酸と、100μMのブロモスルフォフタレイン(BSP:Sigma Chemicals社製)を含む培地又は含まない培地をそれぞれ添加し、さらに8時間培養したのちに、SEAP活性を測定した。結果を図4(右)に示す。その結果、pERE−TA−SEAPベクターで一過的にトランスフェクションしたMCF−7細胞は、100μMのBSPの添加により、エストロン3硫酸添加により誘導されるSEAP活性を抑制した。
【実施例3】
【0037】
MCF−7細胞(ATCC−HTB22)におけるエストロン3硫酸によるエストロゲン依存性の細胞増殖がBSPによりエストロン3硫酸トランスポーター特異的に抑制しうるか否かを検討した。エストロンはエストロン3硫酸と異なり、エストロン3硫酸トランスポーターを介さずに細胞内に取り込まれることから、BSPはエストロン依存性の細胞増殖を抑制することができないことが予想された(図5参照)。MCF−7細胞を8×103個/0.32cm2の濃度で96ウェル プレートに播種し1日培養した後、10−12〜10−7Mの各種濃度のエストロン(Sigma Chemicals社製)と、0μM,30μM,100μMのブロモスルフォフタレイン(BSP:Sigma Chemicals社製)を含む培地をそれぞれ添加し、さらに2日間培養したのちに、細胞増殖を測定した。結果を図6(左)に示す。その結果、30μM,100μMのBSPを添加しても、エストロン依存性の細胞増殖を抑制することができなかった。次に、エストロゲン依存性の細胞増殖がBSPによりエストロン3硫酸トランスポーター特異的に抑制しうることを確かめた。MCF−7細胞を8×103個/0.32cm2の濃度で96ウェル プレートに播種し1日培養した後、10−9〜10−5Mの各種濃度のエストロン3硫酸と、0μM,30μM,100μMのBSPを含む培地をそれぞれ添加し、さらに2日間培養したのちに、細胞増殖を測定した。結果を図6(右)に示す。その結果、30μM,100μMのBSPの添加により、エストロン3硫酸依存性の細胞増殖を抑制することができることが確認された。
【実施例4】
【0038】
1.材料と方法
(材料)
[3H]エストロン3硫酸、アンモニア塩(1702.0GBq/mmol)及び[3H]デヒドロエピアンドロステロンサルフェート(DHEAS、2926.7GBq/mmol)を、PerkinElmer Life Science Products, Inc.社より購入した。金沢大学がん研究所の佐々木琢磨教授より、T−47D細胞の供与を受けた。ウシ胎仔血清(FCS)は、Invitrogen Corp.社より入手した。その他の試薬はすべてSigma Chemicals社及びWako Pure Chemical Industries社より購入した。
【0039】
(T−47D細胞増殖アッセイ)
フェノールレッド及び10%FCSを含むRPMI1640培地(Sigma Chemicals社製)にT−47D細胞を入れ、加湿したインキュベーター内で、37℃、5%CO2にて通常どおり増殖させた。増殖アッセイのため、FCSを0.5%のデキストラン炭(DCC)とともに4℃で一晩インキュベーションし、その後培地を取り出してDCCを濾過し(0.2μm)、ステロイドホルモンを除去した。この処理を3回繰り返した。その後、2.5%のDCC処理FCSを含むフェノールレッド非含有RPMI1640培地を入れた96ウェルプレートに、8,000個/ウェルの密度でT−47D細胞を播種した。播種の1日後、水中又はジメチルスルフォキシド(DMSO、0.1%)中で、原液からの段階的濃度にてエストラジオール又はエストロン3硫酸を添加した。ネガティブコントロールは、溶媒のみであった。播種後、所定の日数で、該細胞をトリプシン処理し、細胞数を数えた。
【0040】
(エストロン3硫酸の取り込み活性の阻害)
文献(Tamai et al., 2001)記載のとおり、輸送実験を行った。15cmのディッシュでT−47D細胞を培養した後、該細胞を採取し、125mMのNaCl、4.8mMのKCl、5.6mMのD−グルコース、1.2mMのCaCl2、1.2mMのKH2PO4、1.2mMのMgSO4及び25mMのHepesを含み、pH7.4に調整された輸送培地に懸濁した。輸送培地中の細胞懸濁液及び放射標識を施した被験化合物を含む溶液を、20分間37℃で個別にインキュベーションして、それらを混合することにより輸送を開始させた。適時、混合液を150μLずつ取り除き、シリコーンオイル(SH550、Toray Dow Corning Co.社製)混合液の層、及び密度1.03の流動パラフィン(Wako Pure Chemical Industries社製)を通しての遠心濾過によって、輸送培地から細胞を分離した。各細胞ペレットを3NのKOH中で可溶化し、HClで中和した。その後、液体シンチレーション流体としてクリアゾル−1(Nacalai tesque社製)を用いた液体シンチレーションカウンターで、関連する放射能を測定した。
【0041】
(逆転写PCR法)
T−47D細胞におけるOATP及びOATトランスポーターの発現をRT−PCR法で調べた。オリゴ(dT)プライマー(Invitrogen Corp.社製)を用いて、一本鎖cDNAを構築した。下記の特異的プライマーを使用した。
OATP−A:
(F)5’-AAACAAGCTGCCCACATAGG-3’(配列番号1)
(R)3’-CAGCAAGACAAGCTGACAGA-5’(配列番号2)
OATP−B:
(F)5’-CCTGCCGCTCTTCTTTATCGG-3’(配列番号3)
(R)3’-ACCAGATGGCTGCACGTTG-5’(配列番号4)
OATP−C:
(F)5’-CACTTGGAGGCACCTCACA-3’(配列番号5)
(R)3’-ACAAGCCCAAGTAGACCCTT-5’(配列番号6)
OATP―D:
(F)5’-CAGGCCATGCTCTCCGAAA-3’(配列番号7)
(R)3’-AGCCACCACTGCAATCTCC-5’(配列番号8)
OATP−E:
(F)5’-CCCTGGGAATCCAGTGGATTG-3’(配列番号9)
(R)3’-AGCAGGCTATGGCAAAGAAGAG-5’(配列番号10)
OATP−F:
(F)5’-GGAAATTCCTCAGGCATAGTGG-3’(配列番号11)
(R)3’-CTGGGATTCCTGCAAGAACTC-5’(配列番号12)
OATP8:
(F)5’-GGGAATCATAACCATTCCTACGG-3’(配列番号13)
(R)3’-GAGGATTTGCATCCTGCTAGAC-5’(配列番号14)
OAT1:
(F)5’-CTGATGGCTTCTCACAACAC-3’(配列番号15)
(R)3’-CCGACTCAATGAAGAACCAG-5’(配列番号16)
OAT2:
(F)5’-GCTGGTTTTACCATCATCGT-3’(配列番号17)
(R)3’-GACTCAGGCCGTAATAGGAG-5’(配列番号18)
OAT3:
(F)5’-AAGTGACCTGTTCCGGATAC-3’(配列番号19)
(R)3’-CCATACCTGTTTGCCTGATG-5’(配列番号20)
OAT4:
(F)5’-GGCGTTATCTCCATTGCTTC-3’(配列番号21)
(R)3’-GAGATTGGAACCCAGTCTCT-5’(配列番号22)
デオキシヌクレオチド及びタグポリメラーゼ(Takara Bio Inc.社製)の存在下で、94℃で30秒、58℃(OATP)又は56℃(OAT)で30秒、72℃で30秒、最終的な伸長が72℃で10分間というサイクルを35回繰返して、反応を行った。1%アガロースゲル(w/v)電気泳動でPCR産物を分析し、かかるゲルを臭化エチジウムで染色して、バンドを視覚化した。
【0042】
(分析方法)
標準物質としてのウシ血清アルブミン(Bradford, 1976)とBioRadタンパク質アッセイキット(Hercules社製)を使用し、Bradfordの方法にしたがって、細胞タンパク質含有量を測定した。飽和性輸送の動態学的パラメータを見積もるために、KaleidaGraphTM(Synergy社製)を用いた非線形最少二乗回帰分析によって下記の方程式に取り込み速度(v)を適用した。
v=Vmax*s/(Km+s)
ここで、v及びsはそれぞれ取り込み速度と基質濃度であり、KmとVmaxはそれぞれ半飽和濃度(ミカエリス定数)と最大輸送速度を示す。データはすべて平均値±S.E.M.で表した。有意さの基準としてP<0.05を用いたStudent’s t testにより統計学的分析を行った。細胞取り込み量を取り込み培地中の被験化合物の濃度で割ることにより、細胞対培地比を得た。
【0043】
2.結果
(T−47D細胞の増殖に対するエストラジオール及びエストロン3硫酸の作用)
T−47D細胞の増殖に対するエストロン3硫酸の作用を解明するため、かかる細胞をエストラジオール及びエストロン3硫酸で処理した。エストロゲン依存性MCF−7細胞を用いた従来の研究(Billich et al., 2000)と一致して、T−47D細胞の増殖は、溶媒だけで処理した細胞と比較して、1pMのエストラジオールと100nMのエストロン3硫酸の両方によって刺激されていた(図3A)。播種から6日後に評価したところ、エストロゲン依存性の刺激は、濃度依存的に増大していた(図3B)。非線形回帰分析により、エストラジオール及びエストロン3硫酸の50%有効濃度(EC50)値を、それぞれ約22.5fM及び17.1nMと評価した。
【0044】
(T−47D細胞によるエストロン3硫酸の取り込み)
図4は、非標識1mMエストロン3硫酸の存在下又は不在下でのT−47D細胞による[3H]エストロン3硫酸(7.1nM)取り込みの経時変化を示す。[3H]エストロン3硫酸の取り込みは、20分にわたって直線的に増大した。非標識1mMエストロン3硫酸の存在下では、[3H]エストロン3硫酸の取り込み量の有意な増大は、35分間観察されなかった(図4)。これは、単なる拡散によってエストロン3硫酸がT−47D細胞に取り込まれることは、ほとんどないことを示唆している。したがって、以下の実験では、T−47D細胞によるエストロン3硫酸の取り込みを10分にわたって動態学的に分析した。さらに、細胞膜への吸着及び単なる拡散による取り込みを考慮に入れるため、非標識エストロン3硫酸の存在下での取り込み(非飽和性取り込み)を差し引いた後の外見上の取り込みを分析した。
【0045】
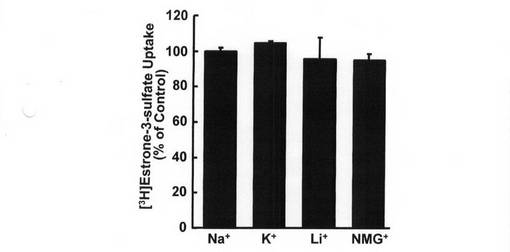
エストロン3硫酸の輸送機序を特徴づけるため、Na+を各種カチオンに置換して調べてみた(図5)。Na+を、K+、Li+、又はN−メチルグルカミン+と置換した場合、[3H]エストロン3硫酸の取り込みは、Na+存在下の場合と同等であった。これは、Na+非依存性輸送機序を示唆するものである。
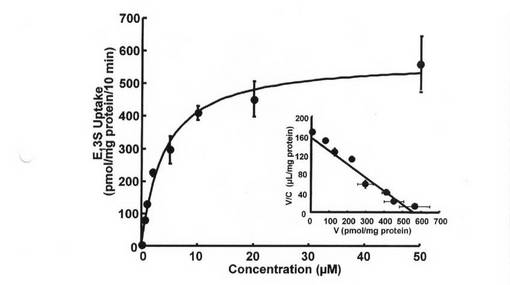
T−47D細胞によるエストロン3硫酸取り込みの動態学的パラメータを得るため、エストロン3硫酸取り込みの濃度依存性を調べた。図6に示すように、1mMのエストロン3硫酸の存在下での値を差し引いた後のエストロン3硫酸の取り込みは、飽和性であった。Eadie-Hofsteeプロット分析では単一の直線が得られており、これは、単一の飽和性機序が動態学的に関与していることを示唆している。非線形回帰分析では、Km及びVmaxの値(平均値±S.E.M.)はそれぞれ3.9±0.78μM及び576.3±32.9pmol/mgタンパク質/10minであった。
【0046】
(エストロン3硫酸の取り込み活性の阻害)
輸送システムの基質特異性を特徴づけるため、T−47D細胞による[3H]エストロン3硫酸の取り込みに対する、ホルモン、その抱合代謝産物、及び各種有機化合物の阻害作用を調べた。結果を(表1)に示す。表1中、対照(%)は、対照におけるエストロン3硫酸の取り込み量を100%としたときの割合で示され、各値は平均±S.E.M.(n=4)で表されれ、*P<0.05は対照との比較(Student’ test)。
【0047】
【表1】

【0048】
その結果、非標識エストロン3硫酸それ自体が、最も強い阻害作用を示した(5μM及び50μMのコントロールに対し、それぞれ34.7%と6.6%)。DHEASの阻害作用は有意であった(50μMのコントロールに対し、44.8%)が、エストロン3硫酸の阻害作用よりも弱かった。グルクロニド抱合体であるエストラジオール−17β−グルクロニドは、50μMでは全く作用を示さなかった。非抱合ステロイドホルモンは、そのすべてが有意に[3H]エストロン3硫酸の取り込みを阻害した。しかし、それらの親和性は、硫酸抱合体、エストロン3硫酸及びDHEASの親和性よりも低かった。調べてみたその他の有機化合物のうち、ブロモスルフォフタレイン及びタウロコール酸は強い阻害活性を示し、100μMでの阻害作用は、それぞれコントロールの18.9%と69.3%であった。その他の有機化合物の中で、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンは、ブロモスルフォフタレインに比較して少し弱いながらも阻害作用を示した。一方、サリチル酸は、全く作用を示さなかった。
【0049】
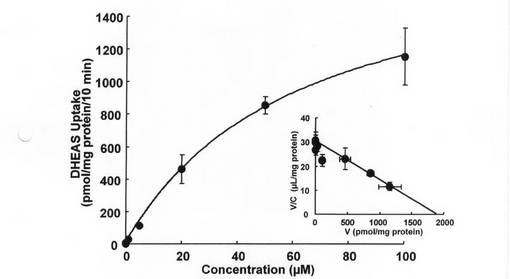
閉経後の女性においては、アンドロゲン及びその硫酸抱合体は、活性エストロゲンの供給源として、乳癌の進行に重要な役割を果たしている。DHEASは、多くの閉経後患者で高濃度(約1μM)で存在することから、主要な前駆物質である(J Clin Endocrinol Metab 54: 22-26., 1982)。そのため、DHEASが特異的輸送機序を介して輸送されるかどうかを調べてみた。DHEASの取り込みは、30分にわたって直線的に増大し、5mMの非標識DHEASの存在下で有意に減少した(データは図示せず)。5mMの非標識DHEASの存在下での取り込み(非特異的取り込み)を差し引いた後のDHEASの取り込みは、飽和性であった(図7)。DHEASのEadie-Hofstee分析では、エストロン3硫酸の場合と同様に単一の直線が得られており(図7、挿入図)、Km及びVmaxの値を計算したところ、それぞれ62.5±21.1μM及び1893±315.7pmol/mgタンパク質/10minであった。
【0050】
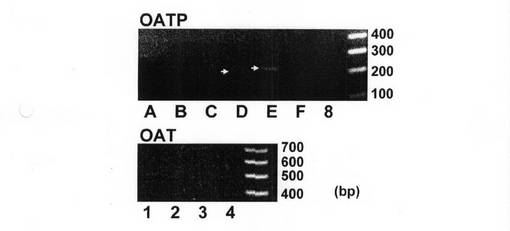
T−47D細胞におけるエストロン3硫酸及びDHEASの取り込みを仲介するトランスポーターを同定するため、エストロン3硫酸を輸送する可能性があるOATP及びOATトランスポーターの発現を、RT−PCR分析で測定した。特異的プライマーを用いることにより、テストしたOATP及びOATのうちOATP−D及びOATP−Eの発現がT−47D細胞中で検出されたが(図8)、RT反応を含まないPCRではバンドは全く検出されなかった(データは図示せず)。したがって、OATP−D及びOATP−Eは、エストロゲン依存性乳癌細胞中でエストロン3硫酸及びDHEASの取り込みを仲介する候補物質であるのかもしれない。
【0051】
3.考察
エストロン3硫酸は主要な循環エストロゲンであり、エストロゲン依存性乳癌の進行に関与している(J Steroid Biochem 34: 155-163, 1989)。腫瘍細胞内で、ステロイドスルファターゼにより脱硫酸化され、続いて生物活性エストロゲンに変換される。乳癌細胞の細胞質中でステロイドスルファターゼが検出されているため(Breast Cancer 6: 331-337, 1999)、エストロン3硫酸は、脱硫酸化の前に細胞膜を通過して細胞内に輸送されたに違いない。エストロン3硫酸の親水性を鑑みると、エストロン3硫酸の内在化は、特異的な輸送機序により仲介されるようである。これに対し、疎水性非抱合ステロイドホルモンは単に細胞内に拡散するにすぎないようである(J Clin Endocrinol Metab 59: 1128-1132, 1984)。そのため、本研究では、エストロゲン依存性増殖を示すモデル細胞と見なされているT−47D乳癌細胞への、エストロン3硫酸及びDHEASの取り込みを仲介する特異的な輸送システムの有無を調べてみた。
【0052】
最初に、細胞増殖のエストロゲン依存性を確認した。エストロン3硫酸、又はエストラジオールでの処理により、T−47D細胞の増殖が増大し、エストロン3硫酸のEC50の値が17.1nMとなった。この値は、エストロン3硫酸の生理学的血漿濃度に近い。エストロン3硫酸自体が、エストロゲン受容体での低い直接的生物反応を誘導するため(Endocrinology 138: 863-870, 1997)、かかる結果は、細胞膜を通過してエストロン3硫酸が内在化し、続いて細胞中で活性非抱合エストロゲンに変換されることを示唆している。したがって、本研究は、エストロゲン活性を解明する第一歩として、T−47D細胞によるエストロゲンの硫酸抱合体の取り込みに焦点を当てている。
【0053】
T−47D細胞によるエストロン3硫酸取り込みの経時的変化は、高濃度のエストロン3硫酸(1mM)の存在下では取り込みが有意に減少することから、取り込みが特異的トランスポーターによって仲介されることを示唆している(図4)。エストロン3硫酸の特異的な取り込みは、Na+を、Li+、K+、又はN−メチルグルカミン+と置換しても影響を受けない(図5)。OATP及びOATファミリーのメンバーは、Na+非依存性エストロン3硫酸取り込みトランスポーターの候補物質である。T−47D細胞によるエストロン3硫酸取り込みのKm値を計算したところ、3.9±0.78μMであった(図6)。これは、OATP及びOATの既知のKm値範囲である0.05μM〜59μMと一致する(Pharm Res 18: 1262-1269, 2001; J Biol Chem 274: 13675-13680, 1999; J Biol Chem 275: 4507-4512, 2000)。
【0054】
輸送機序をさらに特徴づけ、関与するトランスポーターを同定するために、T−47D細胞によるエストロン3硫酸の取り込みに対する阻害について研究した。エストロン3硫酸及びDHEASなどのステロイドホルモンの硫酸抱合体の阻害作用が、非抱合ステロイドホルモンの阻害作用よりも強力であったのに対し、グルクロニド抱合体には阻害作用が全くなかった。したがって、硫酸部分がトランスポーターに対する高親和性を与えているのかもしれず、その一方、グルクロニド部分はほとんど認識されていないのかもしれない。硫酸抱合体は、非抱合ステロイド又はグルクロニド抱合体と比較してイオン化定数(pKa)値が低いため、これらの結果は、トランスポーターによる基質の認識にはアニオン性部分が不可欠であることを示唆している。ステロイドホルモンの硫酸抱合体の血漿濃度は、非抱合ホルモン又はそれらのグルクロニド抱合体と比較して高いため、かかるトランスポーターは、乳房腫瘍へのエストロゲンの供給に対し、効果的な作用を及ぼしているはずである。さらに、T−47D細胞ではDHEASの取り込みも飽和性であり、Km値を計算したところ62.5±21.1μMであり、これはエストロン3硫酸の値(3.9μM、図6及び7)よりもかなり高いものである。エストロン3硫酸とDHEASの血漿濃度は、それぞれ約1〜10nMと1μMであるため(Hormone Res 27: 61-68, 1987; J Clin Endocrinol Metab 54: 22-26, 1982)、乳癌細胞は、飽和することなく効果的にそれらを取り込むのかもしれない。
【0055】
OATP、OAT3及びOAT4のメンバーには、ステロイドホルモンの硫酸抱合体を輸送できるものがある(Biochim Biophys Acta 1609: 1-18, 2003, Pflugers Arch 447: 653-665, 2003; Breast Cancer 6: 331-337, 1999; J Biol Chem 274: 13675-13680, 1999)。そのなかでも、OATP−B及びOAT4は、それぞれKm値が9.04μMと1.01μMの硫酸抱合体を認識することができるが、その一方で、ステロイドホルモンのグルクロニド抱合体に対する親和性は皆無である(Pharm Res 18: 1262-1269, 2001; J Biol Chem 275: 4507-4512, 2000)。T−47D細胞中で、[3H]エストロン3硫酸の取り込みがグルクロニド抱合体によって阻害されなかったため、OATP−B又はOAT4の機能的特質は、ステロイド抱合体の基質認識に関しては、T−47D細胞における取り込み機序と一致している。その一方で、OATP−A、OATP−C、OATP8、及びOAT3は、硫酸抱合体とグルクロニド抱合体の両方を輸送するため、これらのトランスポーターが、T−47D細胞中でのエストロン3硫酸の輸送に関与している可能性は低いと思われる(Biochim Biophys Acta 1609: 1-18, 2003, Pflugers Arch 447: 653-665, 2003; J Biol Chem 274: 13675-13680, 1999)。鍵となるトランスポーターの性質に関する洞察を得るために、各種ヒトOATP及びOATの発現をRT−PCR分析で調べてみた(図8)。発現が検出されたのはOATP−DとOATP−Eだけであり、これは、以前の報告(J Clin Endocrinol Metab 88: 3902-3912, 2003)と一致するものであった。OATP−DとOATP−Eは、HEK293細胞中で一過性に発現した際にエストロン3硫酸に対する輸送活性を示したため(Biochem Biophys Res Commun 273: 251-260, 2000)、T−47D細胞中で細胞膜を通過してのエストロン3硫酸の取り込みに関与しているのかもしれない。しかし、それらの機能的特質がよくわかっていないため、T−47D細胞によるエストロン3硫酸の取り込みに関与するトランスポーターを本研究で同定することはできなかった。BCRP(ABCG2)及びOSTalpha−OSTbetaが、基質としてエストロン3硫酸を受容できることも、報告されている(Breast Cancer 9: 296-302, 2002; Proc Natl Acad Sci USA 95: 15665-15670, 1998; Biochem Biophys Res Commun 298: 41-452002; Mol Pharmacol 64: 610-618, 2003; J Biol Chem 278: 27473-27482, 2003)。したがって、OATPとOAT以外のトランスポーターも、T−47D細胞におけるエストロン3硫酸の取り込みに関与している可能性がある。
【0056】
上記実施例により、エストロン3硫酸が、特異的輸送機序を介してエストロゲン依存性T−47D細胞により取り込まれることが、はっきりと実証された。これは、主要な循環エストロゲン、エストロン3硫酸及びDHEASが、特異的なトランスポーターを介してエストロゲン依存性乳癌細胞に供給されることを初めて実証するものである。T−47D細胞中のトランスポーターを同定するためにはさらなる研究が必要だが、そのようなトランスポーター分子は、エストロゲン依存性乳癌治療のための新しい分子標的となるであろう。
【図面の簡単な説明】
【0057】
【図1】乳癌の分子ターゲットを示す模式図である。
【図2】BSPによるエストロン3硫酸の乳癌細胞への取込み阻害を示す図である。
【図3】乳癌細胞へ取り込まれたエストロン3硫酸によるERE(エストロゲンレスポンスエレメント)活性の誘導を示す模式図、及びレポータージーンアッセイに用いられるpERE−TA−SEAPベクターの模式図である。
【図4】pERE−TA−SEAPベクターで一過的にトランスフェクションしたMCF−7細胞におけるERE誘導SEAP(Secreted Alkaline phospatase)活性に及ぼすエストロン3硫酸の効果を示す図である。
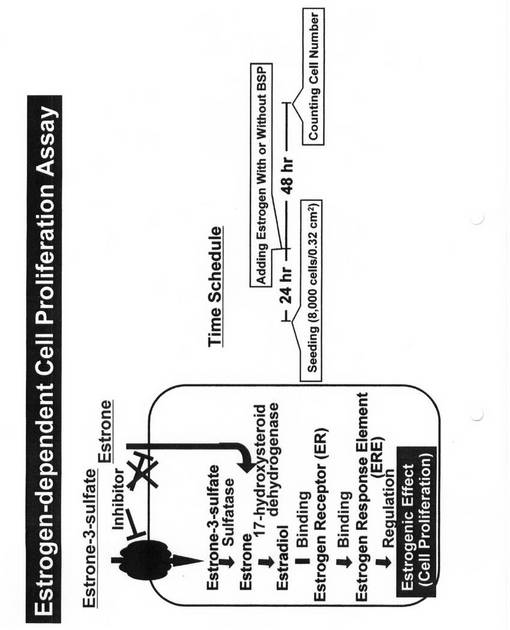
【図5】エストロゲン依存性細胞増殖アッセイの模式図、及びそのタイムスケジュールを示す図である。
【図6】MCF−7細胞にエストロゲン依存性細胞増殖に及ぼすブロモスルフォフタレインの効果を示す図である。
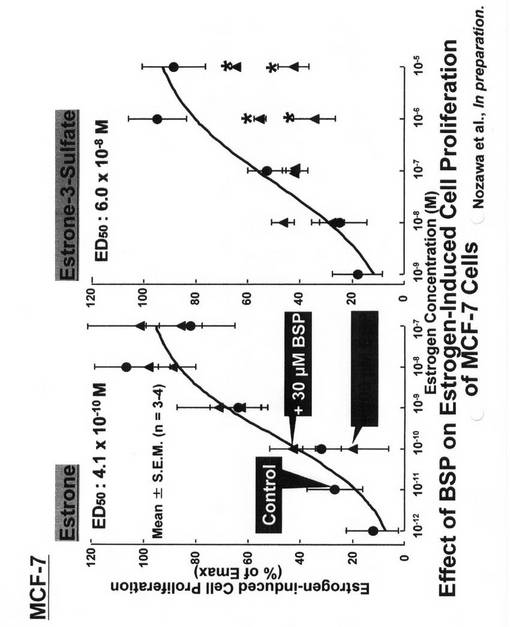
【図7】エストロゲンによるT−47D乳癌細胞増殖刺激の経時変化(a)及び濃度依存性(b)を示す図である。(a)T−47D細胞を密度8,000個/ウェルで播種し、培養した。播種の24時間後(第1日)、エストラジオール(1pM、▲)、エストロン3硫酸(100nM、●)、DMSO(0.1%、△)又は水(○)を添加した。所定日数経過後、T−47D細胞をトリプシン処理し、計数した。(b)濃度範囲がそれぞれ10−16〜10−11M、又は10−11〜10−6Mの各種濃度のエストラジオール(▲)又はエストロン3硫酸(●)の存在下で、T−47D細胞を増殖させた。播種6日後に細胞数を数えた。各値は平均値±S.E.M.を示す(n=3)。
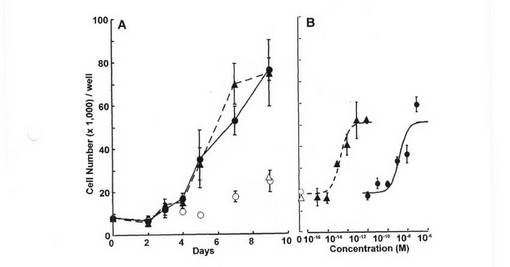
【図8】T−47D細胞による[3H]エストロン3硫酸取り込みの経時変化を示す図である。培養したT−47D細胞を、[3H]エストロン3硫酸(7.1nM)を含み、さらに1mMの非標識エストロン3硫酸を含む培地(○)又は含まない培地(●)で、37℃で35分間インキュベーションした。各値は平均値±S.E.M.を示す(n=4)。エラーバーが表示されていない場合、それらが記号よりも小さいことを示す。
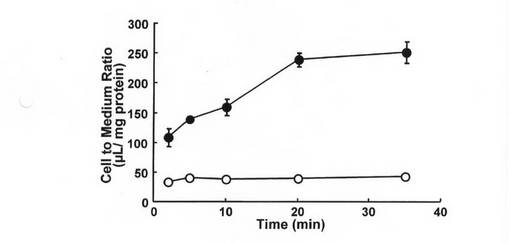
【図9】T−47D細胞による[3H]エストロン3硫酸の取り込みに対する細胞外カチオンの作用を示す図である。 細胞外Na+の存在下又は不在下で、[3H]エストロン3硫酸(7.1nM)の取り込み速度を10分間にわたって測定した。細胞外Na+を、K+、Li+、又はN−メチルグルカミン+(NMG+)と置換した。各値は平均値±S.E.M.を示す(n=4)。
【図10】T−47D細胞によるエストロン3硫酸取り込みの濃度依存性を示す図である。5.1nMから50μMまで、各種濃度のエストロン3硫酸の取り込みを、37℃で10分間測定した。過剰な非標識エストロン3硫酸(1mM)の存在下での取り込みを差し引いた後、飽和性取り込みを得て、非線形最少二乗分析による速度定数の評価に使用した。挿入図は、エストロン3硫酸の飽和性取り込みのEadie-Hofsteeプロットを示す。各値は平均値±S.E.M.を示す(n=4)。エラーバーが表示されていない場合、それらが記号よりも小さいことを示す。
【図11】T−47D細胞によるDHEAS取り込みの濃度依存性を示す図である。10.3nMから100μMまで、各種濃度のDHEASの取り込みを、37℃で10分間測定した。過剰な非標識DHEAS(5mM)の存在下での取り込みを差し引いた後、飽和性取り込みを得て、非線形最少二乗分析による速度定数の評価に使用した。挿入図は、DHEASの飽和性取り込みのEadie-Hofsteeプロットを示す。各値は平均値±S.E.M.を示す(n=4)。エラーバーが表示されていない場合、それらが記号よりも小さいことを示す。
【図12】T−47D細胞におけるOATP及びOATトランスポーターの発現を示す図である。T−47D細胞から得たmRNAを用いて、RT−PCR分析を行った。材料と手法に記載のとおりに反応を行った。矢印は、OATP−D及びOATP−Eの特異的なバンドを示す。
【技術分野】
【0001】
本発明は、エストロン3硫酸(エストロン−3−サルフェート;estrone-3-sulfate)トランスポーターのトランスポーター活性の阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤に関する。
【背景技術】
【0002】
細胞内外の物質輸送を行う膜タンパク質等はトランスポーターと呼ばれ、脂質二重膜中に埋め込まれているトランスポーターに特定の分子が結合すると、コンフォメーションが変化して物質が取込みあるいは排出輸送されることが知られている。近年、かかるトランスポーター、例えば有機アニオン性物質を輸送するOAT1等の有機アニオントランスポーター,有機カチオン性物質を輸送するOCT1等の有機カチオントランスポーター,ペプチド性物質を輸送するPEPT1等のペプチドトランスポーターなどの遺伝子が相次いで単離・同定されている。これらトランスポーター遺伝子は、全身の正常組織・臓器に偏在しているものもあるが、腎臓,肝臓,脳などといった特定の組織・臓器に局在するものも知られている。
【0003】
一方、エストロゲンは雌性動物に発情現象を起こす、いわゆる女性ホルモンであり、天然に存在するエストロン(E1),エストラジオール(E2),エストリオール(E3),エステトロール(E4)と、それらと同様の生物活性を有する合成エストロゲンに分類される。スチルベステロールなどの合成型の一部を除いて、エストロゲンはステロイド構造を有する。エストロン0.1μgのもつ生物学的作用を1IU(国際単位)とし、これがエストロゲンの単位として用いられている。分泌源は主として卵巣の卵胞及び黄体であるが、妊娠時の胎児胎盤系、副腎、精巣などからも分泌される。卵巣からのエストロゲンの分泌は、下垂体前葉より分泌される性腺刺激ホルモンにより支配される(下行性調節)が、逆にエストロゲンによる間脳下垂体系へのフィードバック作用も認められ(上行性調節)、両者の相互関係により性周期が成立するといわれている。
【0004】
さらに、エストロゲンはその受容体(レセプター)を介して作用し、エストロゲンの作用は標的組織である間脳−下垂体前葉−性器及び乳腺のみならず全身に及び、主な生理作用は、子宮内膜の増殖,子宮筋の発育,第二次性徴の発現,月経周期の成立の媒介,妊娠時の母体変化の惹起,乳腺管の増殖分泌促進などである。エストロゲンは主として肝臓において代謝をうけ、エストロン3硫酸等の抱合型エストロゲンとなり尿中に排泄される。臨床的には、無月経や月経異常の治療,月経の人為的移動,更年期障害,前立腺癌や乳癌に対するホルモン療法,骨粗鬆症などに対して用いられている。
【0005】
乳癌は、発癌時点からエストロゲン受容体システムの異常作動があり、初期はエストロゲン依存性の増殖を続けるが、次第にこの制御を離れた増殖となる。このことから、乳癌の発生と進展にエストロゲンとその受容体が深く関与していると考えられているが、その機序は未だ明らかではない。乳癌は乳腺の上皮組織の末梢乳管や腺房上皮から発生する悪性腫瘍で、欧米では女性の癌死亡の第一位である。また、日本においても、近年の食生活の欧米化、生活様式の変化などに伴い乳がんによる死亡率は上昇傾向にある。好発年齢は40歳代で、リスク因子として、乳癌の既往・家族歴,未婚,高齢初産,早期初潮と晩期閉経,肥満,放射線被爆,高脂肪食,良性乳腺疾患の既往などを挙げることができる。リンパ節転移は腋窩,鎖骨下,胸骨傍に頻度が高く、転移個数は予後と相関する。血行性転移は骨,肺,肝に多い。主症状は乳房腫瘤であり、表面不整,硬,境界不明瞭で可動性が少ない。
【0006】
乳癌の2/3は、エストロゲン依存性である(非特許文献1)、すなわち、細胞増殖がエストロゲンによって制御されている。エストロゲンの生物活性型がエストラジオールであり、2つの主要経路を経て合成される(非特許文献2、非特許文献3)。1つは、アロマターゼがアンドロゲンをエストロゲンに変換するアロマターゼ経路であり、もう1つは、ステロイドスルファターゼがエストロン3硫酸をエストロンに変換するスルファターゼ経路である。患者乳癌組織では、スルファターゼの活性はアロマターゼの活性と比べて50〜200倍であるため、スルファターゼ経路は、エストロン3硫酸由来生物活性エストロゲンの主要な供給源である(非特許文献3、非特許文献4)。また、エストロン3硫酸の循環血漿濃度は、非抱合エストロゲンの場合と比べて約10〜20倍であり、エストロン3硫酸の半減期はエストラジオールの半減期よりも長い(非特許文献5〜7)。したがって、エストロン3硫酸は、エストロン3硫酸自体があまり生物活性を示さなくても、活性エストロゲンの貯蔵庫として、乳癌の進行に重要な役割を果たすと考えられている。
【0007】
エストロン3硫酸による乳癌細胞増殖の刺激には、下記の一連のプロセスが関わっている:エストロン3硫酸の細胞への取り込み、エストロゲンスルファターゼによる脱硫酸化、I型の17β−ヒドロキシステロイドデヒドロゲナーゼによるエストロンのエストラジオールへの変換、核エストロゲン受容体への結合、及び遺伝子転写の制御(非特許文献8〜11)。エストロン3硫酸に対する乳癌細胞の応答に関与している酵素や受容体についてよく調べてみたが、エストロン3硫酸などのリガンドが細胞膜を通過する輸送メカニズムはわかっていない。エストロン3硫酸のlogP値は1.4であると報告されており、その値はエストロンの値(4.4)やエストラジオールの値(3.9)より著しく低いため、エストロン及びエストラジオールが拡散性であるにも関わらず、拡散によって細胞膜を通過することは容易ではないであろう。(非特許文献12)。エストロン3硫酸とともにインキュベーションした後のMCF−7細胞の細胞質画分中で、エストロン3硫酸はインタクトな形で検出されており(非特許文献13)、エストロゲン作用を示していた(非特許文献4、非特許文献14、非特許文献15)。したがって、ホルモン依存性増殖を示す乳癌細胞へのエストロン3硫酸の供給には、特異的トランスポーターが関与していると思われる。
【0008】
有機アニオントランスポーター(OAT、SLC22A)(非特許文献16)ファミリー及び有機アニオン輸送ペプチド(OATP、SLC21A、SLCO)(非特許文献17、非特許文献18)ファミリーのメンバーのなかには、エストロン3硫酸を輸送するものがあることが既に知られている。それらは主として肝臓、腎臓、脳、腸、その他の部位で発現するが、乳癌細胞中での発現は報告されていない。エストロン3硫酸に対するそれらのKm値は0.05μM〜59μMの範囲内である。女性では、エストロン3硫酸の血漿中レベルは1〜10nMの範囲で変動するため(非特許文献19)、これらの又はその他のトランスポーターは、ホルモン依存性増殖の第一歩としての、エストロン3硫酸の癌細胞への取り込みにおいて、ある役割を果たしていると思われる。
【0009】
最近、Pizzagalli et al.が、ヒトの乳腺におけるステロイドサルフェートトランスポーターであるOATP−Bの発現を示唆している(非特許文献20)。しかし、エストロゲン依存性MCF−7細胞及びT−47D細胞中でOATP−B mRNAが検出されなかったのに対し、OATP−D及びOATP−E mRNAはこれらの細胞中で発見されている(非特許文献21)。OATP−D及びOATP−Eは、その遺伝子をHEK293細胞中でトランジェントに発現させた場合にエストロン3硫酸に対する輸送活性を示すため(非特許文献22)、癌細胞におけるエストロン3硫酸の輸送に関与していると思われる。しかし、エストロゲン依存性細胞においてトランスポーターが媒介するエストロン3硫酸の取り込みは、これまで調べられていなかったが、肝臓、胎盤及び脳等のほかの組織における取り込みは報告されている。乳癌細胞の増殖メカニズムを充分に理解するためには、かかる細胞へのエストロゲンの供給メカニズムを解明しなくてはならない。
【0010】
【非特許文献1】N Engl J Med 302: 78-90, 1980
【非特許文献2】J Clin Endocrinol Metab 57: 1125-1128, 1983
【非特許文献3】Breast Cancer Res Treat 7: 35-44, 1986
【非特許文献4】Ann N Y Acad Sci 464: 126-137, 1986
【非特許文献5】J Clin Endocrinol Metab 81: 1460-1464, 1996
【非特許文献6】J Biol Chem 236: 1043-1050, 1961
【非特許文献7】J Clin Invest 51: 1020-1033, 1972
【非特許文献8】Endocrinology 138: 863-870, 1997
【非特許文献9】Mol Endocrinol 11: 77-86, 1997
【非特許文献10】Endocrinology 123: 1281-1287, 1988
【非特許文献11】Breast Cancer 9: 296-302, 2002
【非特許文献12】J Clin Endocrinol Metab 59: 1128-1132, 1984
【非特許文献13】Endocrinology 106: 1079-1086, 1980
【非特許文献14】J Clin Endocrinol Metab 81: 1460-1464, 1996
【非特許文献15】J Steroid Biochem Mol Biol 73: 225-235, 2000
【非特許文献16】Pflugers Arch 440: 337-350, 2000
【非特許文献17】Biochim Biophys Acta 1609: 1-18, 2003
【非特許文献18】Pflugers Arch 447: 653-665, 2003
【非特許文献19】Hormone Res 27: 61-68, 1987
【非特許文献20】J Clin Endocrinol Metab 88: 3902-3912, 2003
【非特許文献21】J Clin Endocrinol Metab 88: 3902-3912, 2003
【非特許文献22】Pharm Res 18: 1262-1269, 2001
【発明の開示】
【発明が解決しようとする課題】
【0011】
乳癌細胞におけるエストロゲンとその受容体との結合によるシグナル伝達をアップストリームにブロックする乳癌細胞の増殖阻害については、図1に示されるように、以下の方法(ブロック1〜4)が知られている。
【0012】
ブロック1(アンドロゲンからのエストロゲンの合成阻害)
ファドロゾール:閉経後、エストロゲン合成の律速酵素として働くアロマターゼを選択的に阻害して、アンドロゲンからエストロゲンへの変換を妨げることにより、生体内のエストロゲン濃度を低下させ、乳ガンの増殖を抑制する。
【0013】
ブロック2(エストロゲンレセプターでの競合阻害)
タモキシフェン、トレミフェン:乳ガン組織などのエストロゲン受容体に対し、エストロゲンと競合的に阻害し、抗エストロゲン作用を示すことによって抗乳ガン作用を発揮する。
【0014】
ブロック3(エストロン硫酸からのエストロゲン合成阻害)
抱合型エストロゲンは細胞内でエストロゲンサルファターゼによって活性型となり、ガン細胞増殖を促進する。この脱抱合酵素を阻害する。
【0015】
ブロック4(膜タンパクに対する抗体を利用した細胞障害作用)
ハーセプチン:乳ガン細胞にはHER2タンパクが高発現するものがある。このHER2と抗体の結合を利用し、抗体依存性細胞障害作用による抗腫瘍効果を発揮する。
【0016】
乳癌細胞の増殖阻害における、上記ブロック1〜4の方法では、アロマターゼ阻害剤やエストロゲンサルファターゼ阻害剤、あるいは、タモキシフェンやトレミフェン等のエストロゲン競合阻害剤が乳癌細胞内に取り込まれないと抗腫瘍効果が発揮されないことからドラッグデリバリーの面で、また、細胞内に取り込まれると副作用の面で、それぞれ問題なしとはいえない。本発明の課題は、細胞内に取り込まれる必要がなく、ドラッグデリバリー性に優れ、副作用がきわめて少ないエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤を提供することにある。
【課題を解決するための手段】
【0017】
本発明者らは、上記課題を解決するために鋭意研究し、乳癌細胞の増殖阻害の方法として、以下のブロック5(図1参照)について検討した。
ブロック5(トランスポーター活性の阻害)
エストロゲン感受性の乳ガン細胞に対し、エストロゲンの供給源であるエストロン3硫酸の細胞内取込みを阻害することで抗腫瘍効果が現れる。
【0018】
そして、ヒト乳癌培養細胞MCF−7細胞株をエストロン3硫酸と、被検物質としてのブロモスルフォフタレイン(Bromosulfophtalein;BSP)の存在下に培養した時、細胞表面に発現するエストロン3硫酸トランスポーターを介してのエストロン3硫酸の細胞内への取込みが阻害され、MCF−7細胞株の増殖が抑制されることを見い出し、本発明を完成するに至った。
【0019】
すなわち本発明は、(1)ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とするエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や、(2)ブロモスルフォフタレインを有効成分とするエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や、(3)ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とする乳癌細胞増殖抑制剤や、(4)ブロモスルフォフタレインを有効成分とする乳癌細胞増殖抑制剤や、(5)ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とする乳癌治療剤や、(6)ブロモスルフォフタレインを有効成分とする乳癌治療剤に関する。
【発明の効果】
【0020】
本発明によると、アロマターゼ阻害剤やエストロゲンサルファターゼ阻害剤、タモキシフェンやトレミフェン等のエストロゲン競合阻害剤と異なり、細胞内に取り込まれる必要がなく、ドラッグデリバリー性に優れ、副作用がきわめて少ないエストロン3硫酸トランスポーターの阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤を提供することができる。
【発明を実施するための最良の形態】
【0021】
本発明のエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤や乳癌細胞増殖抑制剤や乳癌治療剤としては、ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸を挙げることができるが、中でもブロモスルフォフタレインを好適に例示することができる。また、これらは1種単独又は2種以上を併用することができる。
【0022】
本発明のエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤(以下「本件阻害剤」という場合がある)は、エストロン3硫酸トランスポーターを同定する上で重要である。エストロン3硫酸トランスポーターの候補トランスポーターとしては、SLCトランスポーター,OAT1,OAT2,OAT3,OAT4,OATP−A,OATP−B,OATP−C,OATP−D,OATP−E,OATP−F,OATP−8,NTCP,MRPs,BCRPなどを好適に挙げることができる。同定する方法としては、候補エストロン3硫酸トランスポーターを細胞表面に発現する細胞を培養し、培養した細胞内への候補エストロン3硫酸の取り込みを本件阻害剤の存在下において測定し、本件阻害剤による細胞内へのエストロン3硫酸のトランスポーター活性の阻害の程度を測定・評価する方法や、候補エストロン3硫酸トランスポーターを細胞表面に発現する細胞から単離した細胞膜画分に、エストロン3硫酸と本件阻害剤とを接触させ、本件阻害剤によるエストロン3硫酸の細胞膜画分への特異的結合の阻害の程度を測定・評価する方法や、候補エストロン3硫酸トランスポーターを細胞表面に発現する細胞から細胞膜画分を単離し、該細胞膜画分から小胞を作製して単離細胞膜小胞とし、該単離細胞膜小胞にエストロン3硫酸と本件阻害剤とを接触させ、本件阻害剤による小胞内へのエストロン3硫酸の取込み量もしくは取込み量の阻害の程度を測定・評価する方法を挙げることができる。上記エストロン3硫酸と本件阻害剤との共存下の細胞の培養は、使用細胞の増殖用培地を用い、通常の細胞培養条件下、(本件阻害剤非存在下の場合に)少なくとも細胞増殖が有意に認められるまで行うことが望ましく、上記エストロン3硫酸と本件阻害剤との細胞膜画分との接触や、上記エストロン3硫酸と本件阻害剤との単離細胞膜小胞との接触は、使用細胞の増殖用培地や適当な緩衝液中でインキュベーションすることにより行うことができる。
【0023】
上記本件阻害剤による細胞内へのエストロン3硫酸のトランスポーター活性の阻害の程度を測定・評価する方法としては、細胞内のエストロン3硫酸濃度を測定・評価する方法や、細胞増殖の程度を測定・評価する方法を挙げることができる。後述するMCF−7細胞株、KPL−1,MKL−F又はそれら細胞株由来の細胞株等のヒト乳癌培養細胞株を用いる場合は、細胞内の本件阻害剤濃度を測定・評価する方法と、細胞増殖の程度を測定・評価する方法とを利用することができるが、候補エストロン3硫酸トランスポーターを発現する形質転換細胞を用いる場合は、細胞内の本件阻害剤濃度を測定・評価する方法を利用することができる。また、候補エストロン3硫酸トランスポーター遺伝子をアフリカツメガエル卵母細胞等に発現させ、2本の微小電極を用いて、トランスポーターを介した基質輸送に伴って生じる膜電位の変化を検出(電流を測定)する電気生理学的方法により測定・評価することもできる。
【0024】
エストロン3硫酸トランスポーターを細胞表面に発現する細胞から細胞膜画分を単離する方法としては、F. Pietri-Rouxelら(Eur. J. Biochem., 247, 1174-1179, 1997)の方法など、従来公知の方法を用いることができ、また、細胞膜画分から小胞を作製して単離細胞膜小胞とする方法としては、J.E. Leverら(J. Biol. Chem., 252, 1990-1997 (1977))の方法など、従来公知の方法を用いることができる。そしてまた、本件阻害剤によるエストロン3硫酸の細胞膜画分への特異的結合の阻害の程度を測定する方法としては、放射性標識エストロン3硫酸を用いた定常状態結合阻害から平衡解離定数KDを測定する方法を例示することができ、本件阻害剤による単離細胞膜小胞内へのエストロン3硫酸の取込み量もしくは取込み量の阻害の程度を測定する方法としては、I. Tamaiら(Biochim. Biophys. Acta, 1512, 273-284 (2001))記載の方法など、従来公知の手法を細胞内エストロン3硫酸の測定方法として用いることができる。
【0025】
上記エストロン3硫酸トランスポーターを細胞表面に発現する細胞としては、候補エストロン3硫酸トランスポーターを発現する形質転換細胞を使用することができる。かかる形質転換細胞の作製に使用される前記SLCトランスポーター,OAT1,OAT2,OAT3,OAT4,OATP−A,OATP−B,OATP−C,OATP−D,OATP−E,OATP−F,OATP−8,NTCP,MRPs,BCRPをコードする遺伝子の由来としては特に制限されるものではなく、例えばヒト,イヌ,ウシ,ウマ,ヤギ,ヒツジ,サル,ブタ,ウサギ,ラット,マウス等を挙げることができるが、ヒト由来が好ましい。
【0026】
上記のようなトランスポーターををコードする遺伝子やcDNAを細胞に発現させる方法としては特に限定されないが、Davisら(BASIC METHODS IN MOLECULAR BIOLOGY, 1986)及びSambrookら(MOLECULAR CLONING: A LABORATORY MANUAL, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989)などの多くの標準的な実験室マニュアルに記載される方法、例えば、リン酸カルシウムトランスフェクション,DEAE−デキストラン媒介トランスフェクション,トランスベクション(transvection)マイクロインジェクション,カチオン性脂質媒介トランスフェクション,エレクトロポレーション,形質導入,スクレープローディング(scrape loading),弾丸導入(ballistic introduction),感染等の遺伝子導入法により、トランスポーター遺伝子を宿主細胞へ導入することにより発現させることができるが、トランスポーター遺伝子を含む発現ベクターを上記宿主細胞に導入することが好ましく、中でもトランスポーター遺伝子を含む発現ベクターを宿主細胞に感染させることにより、トランスポーターを細胞に発現させることがより好ましい。
【0027】
上記発現ベクターとしては、例えば、非分裂細胞を含む全ての細胞(血球系以外)での一過性発現に用いられるアデノウイルスベクター(Science, 252, 431-434, 1991)や、分裂細胞での長期発現に用いられるレトロウイルスベクター(Microbiology and Immunology, 158, 1-23, 1992)や、非病原性、非分裂細胞にも導入可能で、長期発現に用いられるアデノ随伴ウイルスベクター(Curr. Top. Microbiol. Immunol., 158, 97-129, 1992)、SV40のようなパポバウイルスベクター、ワクシニアウイルスベクター等のウイルスベクターの他、リポソーム等を具体的に挙げることができるが、これらに限定されるものではない。これらの中でも、高い効率で細胞における遺伝子発現が可能なアデノウイルスベクターが特に好ましい。また、これらの発現ベクターへのトランスポーター遺伝子の導入は常法によって行うことができ、例えばこれら発現ベクター中の適当なプロモーターの下流にトランスポーター遺伝子等を挿入することにより発現ベクターを構築することができる。また、発現ベクターはトランスポーター遺伝子及びマーカー遺伝子に加え、トランスポーターの細胞当たりの発現量を規格化するためのIRES(mRNA内部のリボソーム結合サイト)の他、エンハンサー,ターミネーター等発現を調節する制御配列を含んでいてもよい。
【0028】
トランスポーターを発現する宿主細胞としては、ドロソフィラS2,スポドプテラSf9等の昆虫細胞,Vero細胞,HeLa細胞,CHO細胞,WI−38細胞,BHK細胞,COS−7細胞,MDCK細胞,C127細胞,HKG細胞,ヒト腎細胞株,CV−1細胞,LLC−MK2細胞,MDBK細胞,MRC−5細胞,Caco−2細胞,HT29細胞,ヒトリンパ芽球細胞,アフリカツメガエル等の卵母細胞や、これらのdhfr欠損株,HGPRT欠損株,ウアバイン耐性株等を挙げることができる。具体的には、CHO−K1(チャイニーズハムスター卵巣細胞:ATCC CCL61),BHK(ハムスター腎細胞:ATCC CCL10),COS−7(CV−1 Origin,SV−40細胞:ATCC CRL1651),Vero細胞(アフリカミドリザル腎細胞:ATCC CCL81),ヒトリンパ芽球細胞(IM−9,ATCC CCL159)を例示することができる。
【0029】
トランスポーターは細胞膜表面に発現する膜タンパク質であるため、エストロン3硫酸トランスポーターに対する中和抗体は細胞外から選択的に標的を阻害する可能性があり、このことから本件阻害剤によりエストロン3硫酸トランスポーターを同定することができると、エストロン3硫酸トランスポーターに対する中和抗体を作製することができる。特異的に認識する抗体としては、モノクローナル抗体,ポリクローナル抗体,一本鎖抗体,ヒト化抗体,キメラ抗体,2つのエピトープを同時に認識することができる二機能性抗体等を例示することができる。これら抗体は、慣用のプロトコールを用いて、動物(好ましくはヒト以外)にエストロン3硫酸トランスポーターを膜表面に発現している細胞やその細胞膜画分を投与することにより産生され、例えばモノクローナル抗体の調製には、連続細胞系の培養物により産生される抗体をもたらす、ハイブリドーマ法(Nature 256, 495-497, 1975),トリオーマ法,ヒトB細胞ハイブリドーマ法(Immunology Today 4, 72, 1983)及びEBV−ハイブリドーマ法(MONOCLONAL ANTIBODIES AND CANCER THERAPY, pp.77-96, Alan R.Liss, Inc.,1985)など任意の方法を用いることができる。
【0030】
本発明の乳癌細胞増殖抑制剤は、エストロン3硫酸トランスポーターの同定用として、また乳癌細胞の増殖を抑制しうることから乳癌治療剤として用いることができる。同定する方法としては、候補エストロン3硫酸トランスポーターを細胞表面に発現する細胞を、本発明の乳癌細胞増殖抑制剤の存在下において培養し、細胞増殖の程度を測定・評価する方法を挙げることができる。
【0031】
本発明の乳癌治療剤は、乳癌の予防薬としても用いることができ、医薬用の治療剤として用いる場合は、薬学的に許容される通常の担体、結合剤、安定化剤、賦形剤、希釈剤、pH緩衝剤、崩壊剤、可溶化剤、溶解補助剤、等張剤などの各種調剤用配合成分を添加することができる。これら治療剤は、経口的又は非経口的に投与することができ、例えば粉末、顆粒、錠剤、カプセル剤、シロップ剤、懸濁液等の剤型で経口的に投与することができ、あるいは、例えば溶液、乳剤、懸濁液等の剤型にしたものを注射の型で非経口投与することができる。非経口投与する場合、局所に直接投与することができる。経口的に投与する製剤の場合、薬理学的に許容される担体としては、慣用の各種有機あるいは無機担体物質が用いられ、例えば錠剤には乳糖、デンプン等の賦形剤、タルク、ステアリン酸マグネシウム等の滑沢剤、ヒドロキシプロピルセルロース、ポリビニルピロリドン等の結合剤、カルボキシメチルセルロース等の崩壊剤等を配合することができ、懸濁液製剤には生理的食塩水アルコール等の溶剤、ポリエチレングリコール、プロピレングリコール等の溶解補助剤、ステアリルトリエタノールアミン、ラウリル硫酸ナトリウム、レシチン等の懸濁化剤、グリセリン、D−マンニトール等の等張化剤、リン酸塩、酢酸塩、クエン酸塩等の緩衝剤などを配合することができる。また必要に応じて、防腐剤、抗酸化剤、着色剤、甘味剤などの製剤添加物を配合することもできる。非経口的に投与する製剤の場合、蒸留水、生理的食塩水等の水溶性溶剤、サリチル酸ナトリウム等の溶解補助剤、塩化ナトリウム、グリセリン、D−マンニトール等の等張化剤、ヒト血清アルブミン等の安定化剤、メチルパラベン等の保存剤、ベンジルアルコール等の局麻剤を配合することができる。治療薬の投与量は、患者の体重や年齢、投与形態、症状等により適宜選定することができるが、例えば成人に投与する場合、有効成分を通常1回量として約0.001〜500mg、好ましくは1〜50mgを1日1回〜3回投与するのが望ましい。
【0032】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。
【実施例1】
【0033】
MCF−7細胞(ATCC−HTB22)のエストロン3硫酸による細胞増殖促進効果の抑制が、エストロン3硫酸の細胞内蓄積を抑えることで達成されるか否かを検討した。MCF−7細胞を8×104個/cm2の濃度で96ウェル プレートに播種し1日培養した後、10pM〜10μMの各種濃度のエストロン3硫酸(Sigma Chemicals社製)と100μMのブロモスルフォフタレイン(BSP:Sigma Chemicals社製)、及び、1pM〜10nMの各種濃度のエストラジオール(estradiol:Sigma Chemicals社製)と100μMのBSPを含む培地をそれぞれ添加し、さらに3日間培養した。その後、細胞量を測定した。エストロン3硫酸及びエストラジオールによる細胞増殖促進効果の結果を図2に示す。
【0034】
図2から明らかなように、ポジティブコントロールであるエストロン3硫酸単独存在下及びエストラジオール単独存在下においては、細胞増殖促進効果が観察された(open column)。一方、エストロン3硫酸又はエストラジオールに、100μMのBSPを同時添加することによって共存下に培養すると、エストロン3硫酸低濃度条件において細胞増殖促進効果が見られるが、これは、BSP自身あるいはその代謝物によるエストロゲン活性であると考えられる。この効果はエストロゲン濃度が増加するに従い、相対的に低くなった。興味深いことに、エストロン3硫酸10μM添加時、BSPによる細胞増殖効果は阻害されたのに対し、エストラジオール添加による効果には変化しなかった。したがって、エストロン3硫酸の細胞内取込みはBSPにより阻害され、脂溶性の高いエストラジオールは阻害されないため、細胞増殖効果にも変化がなかったと考えられた。以上のことから、細胞増殖におけるエストロン3硫酸の取り込みトランスポーターの重要性が示唆され、そのトランスポーターを阻害することによる細胞増殖抑制の可能性が示された。
【実施例2】
【0035】
MCF−7細胞(ATCC−HTB22)におけるエストロン3硫酸によるERE(エストロゲンレスポンスエレメント)活性の誘導がBSPにより抑制しうるか否かを検討した。MCF−7細胞を、図3に示すpERE−TA−SEAPベクター(BD Biosciencesから購入)で一過的にトランスフェクションした。このpERE−TA−SEAPベクターは、エストロゲンが結合するDNA配列であるERE配列を上流に持つSEAP(Secreted Alkaline phospatase)蛋白質発現ベクターで、エストロゲンが存在すると、SEAP蛋白質合成がEREを介して促進され、SEAP活性が上昇する。また、SEAPはリン酸結合を切断する酵素で、この酵素の基質をMU−P(methyl umbelliferone phosphate)とし、生じたMU(methyl umbelliferone)を定量することにより、SEAP活性を測定することができる。
【0036】
pERE−TA−SEAPベクター(BD Biosciencesから購入)キットのマニュアルに沿ってトランスフェクションしたMCF−7細胞を8×104個/cm2の濃度で96ウェル プレートに播種し1日培養した後、10−9〜10−6Mの各種濃度のエストロン3硫酸(Sigma Chemicals社製)を添加し、さらに8時間培養したのちに、SEAP活性を測定した。結果を図4(左)に示す。その結果、pERE−TA−SEAPベクターで一過的にトランスフェクションしたMCF−7細胞は、添加したエストロン3硫酸の濃度依存的にSEAP活性を示した。次に、pERE−TA−SEAPベクター(BD Biosciencesから購入)キットのマニュアルに沿ってトランスフェクションしたMCF−7細胞を8×104個/cm2の濃度で96ウェル プレートに播種し1日培養した後、20nM,50nM,100nMの各種濃度のエストロン3硫酸と、100μMのブロモスルフォフタレイン(BSP:Sigma Chemicals社製)を含む培地又は含まない培地をそれぞれ添加し、さらに8時間培養したのちに、SEAP活性を測定した。結果を図4(右)に示す。その結果、pERE−TA−SEAPベクターで一過的にトランスフェクションしたMCF−7細胞は、100μMのBSPの添加により、エストロン3硫酸添加により誘導されるSEAP活性を抑制した。
【実施例3】
【0037】
MCF−7細胞(ATCC−HTB22)におけるエストロン3硫酸によるエストロゲン依存性の細胞増殖がBSPによりエストロン3硫酸トランスポーター特異的に抑制しうるか否かを検討した。エストロンはエストロン3硫酸と異なり、エストロン3硫酸トランスポーターを介さずに細胞内に取り込まれることから、BSPはエストロン依存性の細胞増殖を抑制することができないことが予想された(図5参照)。MCF−7細胞を8×103個/0.32cm2の濃度で96ウェル プレートに播種し1日培養した後、10−12〜10−7Mの各種濃度のエストロン(Sigma Chemicals社製)と、0μM,30μM,100μMのブロモスルフォフタレイン(BSP:Sigma Chemicals社製)を含む培地をそれぞれ添加し、さらに2日間培養したのちに、細胞増殖を測定した。結果を図6(左)に示す。その結果、30μM,100μMのBSPを添加しても、エストロン依存性の細胞増殖を抑制することができなかった。次に、エストロゲン依存性の細胞増殖がBSPによりエストロン3硫酸トランスポーター特異的に抑制しうることを確かめた。MCF−7細胞を8×103個/0.32cm2の濃度で96ウェル プレートに播種し1日培養した後、10−9〜10−5Mの各種濃度のエストロン3硫酸と、0μM,30μM,100μMのBSPを含む培地をそれぞれ添加し、さらに2日間培養したのちに、細胞増殖を測定した。結果を図6(右)に示す。その結果、30μM,100μMのBSPの添加により、エストロン3硫酸依存性の細胞増殖を抑制することができることが確認された。
【実施例4】
【0038】
1.材料と方法
(材料)
[3H]エストロン3硫酸、アンモニア塩(1702.0GBq/mmol)及び[3H]デヒドロエピアンドロステロンサルフェート(DHEAS、2926.7GBq/mmol)を、PerkinElmer Life Science Products, Inc.社より購入した。金沢大学がん研究所の佐々木琢磨教授より、T−47D細胞の供与を受けた。ウシ胎仔血清(FCS)は、Invitrogen Corp.社より入手した。その他の試薬はすべてSigma Chemicals社及びWako Pure Chemical Industries社より購入した。
【0039】
(T−47D細胞増殖アッセイ)
フェノールレッド及び10%FCSを含むRPMI1640培地(Sigma Chemicals社製)にT−47D細胞を入れ、加湿したインキュベーター内で、37℃、5%CO2にて通常どおり増殖させた。増殖アッセイのため、FCSを0.5%のデキストラン炭(DCC)とともに4℃で一晩インキュベーションし、その後培地を取り出してDCCを濾過し(0.2μm)、ステロイドホルモンを除去した。この処理を3回繰り返した。その後、2.5%のDCC処理FCSを含むフェノールレッド非含有RPMI1640培地を入れた96ウェルプレートに、8,000個/ウェルの密度でT−47D細胞を播種した。播種の1日後、水中又はジメチルスルフォキシド(DMSO、0.1%)中で、原液からの段階的濃度にてエストラジオール又はエストロン3硫酸を添加した。ネガティブコントロールは、溶媒のみであった。播種後、所定の日数で、該細胞をトリプシン処理し、細胞数を数えた。
【0040】
(エストロン3硫酸の取り込み活性の阻害)
文献(Tamai et al., 2001)記載のとおり、輸送実験を行った。15cmのディッシュでT−47D細胞を培養した後、該細胞を採取し、125mMのNaCl、4.8mMのKCl、5.6mMのD−グルコース、1.2mMのCaCl2、1.2mMのKH2PO4、1.2mMのMgSO4及び25mMのHepesを含み、pH7.4に調整された輸送培地に懸濁した。輸送培地中の細胞懸濁液及び放射標識を施した被験化合物を含む溶液を、20分間37℃で個別にインキュベーションして、それらを混合することにより輸送を開始させた。適時、混合液を150μLずつ取り除き、シリコーンオイル(SH550、Toray Dow Corning Co.社製)混合液の層、及び密度1.03の流動パラフィン(Wako Pure Chemical Industries社製)を通しての遠心濾過によって、輸送培地から細胞を分離した。各細胞ペレットを3NのKOH中で可溶化し、HClで中和した。その後、液体シンチレーション流体としてクリアゾル−1(Nacalai tesque社製)を用いた液体シンチレーションカウンターで、関連する放射能を測定した。
【0041】
(逆転写PCR法)
T−47D細胞におけるOATP及びOATトランスポーターの発現をRT−PCR法で調べた。オリゴ(dT)プライマー(Invitrogen Corp.社製)を用いて、一本鎖cDNAを構築した。下記の特異的プライマーを使用した。
OATP−A:
(F)5’-AAACAAGCTGCCCACATAGG-3’(配列番号1)
(R)3’-CAGCAAGACAAGCTGACAGA-5’(配列番号2)
OATP−B:
(F)5’-CCTGCCGCTCTTCTTTATCGG-3’(配列番号3)
(R)3’-ACCAGATGGCTGCACGTTG-5’(配列番号4)
OATP−C:
(F)5’-CACTTGGAGGCACCTCACA-3’(配列番号5)
(R)3’-ACAAGCCCAAGTAGACCCTT-5’(配列番号6)
OATP―D:
(F)5’-CAGGCCATGCTCTCCGAAA-3’(配列番号7)
(R)3’-AGCCACCACTGCAATCTCC-5’(配列番号8)
OATP−E:
(F)5’-CCCTGGGAATCCAGTGGATTG-3’(配列番号9)
(R)3’-AGCAGGCTATGGCAAAGAAGAG-5’(配列番号10)
OATP−F:
(F)5’-GGAAATTCCTCAGGCATAGTGG-3’(配列番号11)
(R)3’-CTGGGATTCCTGCAAGAACTC-5’(配列番号12)
OATP8:
(F)5’-GGGAATCATAACCATTCCTACGG-3’(配列番号13)
(R)3’-GAGGATTTGCATCCTGCTAGAC-5’(配列番号14)
OAT1:
(F)5’-CTGATGGCTTCTCACAACAC-3’(配列番号15)
(R)3’-CCGACTCAATGAAGAACCAG-5’(配列番号16)
OAT2:
(F)5’-GCTGGTTTTACCATCATCGT-3’(配列番号17)
(R)3’-GACTCAGGCCGTAATAGGAG-5’(配列番号18)
OAT3:
(F)5’-AAGTGACCTGTTCCGGATAC-3’(配列番号19)
(R)3’-CCATACCTGTTTGCCTGATG-5’(配列番号20)
OAT4:
(F)5’-GGCGTTATCTCCATTGCTTC-3’(配列番号21)
(R)3’-GAGATTGGAACCCAGTCTCT-5’(配列番号22)
デオキシヌクレオチド及びタグポリメラーゼ(Takara Bio Inc.社製)の存在下で、94℃で30秒、58℃(OATP)又は56℃(OAT)で30秒、72℃で30秒、最終的な伸長が72℃で10分間というサイクルを35回繰返して、反応を行った。1%アガロースゲル(w/v)電気泳動でPCR産物を分析し、かかるゲルを臭化エチジウムで染色して、バンドを視覚化した。
【0042】
(分析方法)
標準物質としてのウシ血清アルブミン(Bradford, 1976)とBioRadタンパク質アッセイキット(Hercules社製)を使用し、Bradfordの方法にしたがって、細胞タンパク質含有量を測定した。飽和性輸送の動態学的パラメータを見積もるために、KaleidaGraphTM(Synergy社製)を用いた非線形最少二乗回帰分析によって下記の方程式に取り込み速度(v)を適用した。
v=Vmax*s/(Km+s)
ここで、v及びsはそれぞれ取り込み速度と基質濃度であり、KmとVmaxはそれぞれ半飽和濃度(ミカエリス定数)と最大輸送速度を示す。データはすべて平均値±S.E.M.で表した。有意さの基準としてP<0.05を用いたStudent’s t testにより統計学的分析を行った。細胞取り込み量を取り込み培地中の被験化合物の濃度で割ることにより、細胞対培地比を得た。
【0043】
2.結果
(T−47D細胞の増殖に対するエストラジオール及びエストロン3硫酸の作用)
T−47D細胞の増殖に対するエストロン3硫酸の作用を解明するため、かかる細胞をエストラジオール及びエストロン3硫酸で処理した。エストロゲン依存性MCF−7細胞を用いた従来の研究(Billich et al., 2000)と一致して、T−47D細胞の増殖は、溶媒だけで処理した細胞と比較して、1pMのエストラジオールと100nMのエストロン3硫酸の両方によって刺激されていた(図3A)。播種から6日後に評価したところ、エストロゲン依存性の刺激は、濃度依存的に増大していた(図3B)。非線形回帰分析により、エストラジオール及びエストロン3硫酸の50%有効濃度(EC50)値を、それぞれ約22.5fM及び17.1nMと評価した。
【0044】
(T−47D細胞によるエストロン3硫酸の取り込み)
図4は、非標識1mMエストロン3硫酸の存在下又は不在下でのT−47D細胞による[3H]エストロン3硫酸(7.1nM)取り込みの経時変化を示す。[3H]エストロン3硫酸の取り込みは、20分にわたって直線的に増大した。非標識1mMエストロン3硫酸の存在下では、[3H]エストロン3硫酸の取り込み量の有意な増大は、35分間観察されなかった(図4)。これは、単なる拡散によってエストロン3硫酸がT−47D細胞に取り込まれることは、ほとんどないことを示唆している。したがって、以下の実験では、T−47D細胞によるエストロン3硫酸の取り込みを10分にわたって動態学的に分析した。さらに、細胞膜への吸着及び単なる拡散による取り込みを考慮に入れるため、非標識エストロン3硫酸の存在下での取り込み(非飽和性取り込み)を差し引いた後の外見上の取り込みを分析した。
【0045】
エストロン3硫酸の輸送機序を特徴づけるため、Na+を各種カチオンに置換して調べてみた(図5)。Na+を、K+、Li+、又はN−メチルグルカミン+と置換した場合、[3H]エストロン3硫酸の取り込みは、Na+存在下の場合と同等であった。これは、Na+非依存性輸送機序を示唆するものである。
T−47D細胞によるエストロン3硫酸取り込みの動態学的パラメータを得るため、エストロン3硫酸取り込みの濃度依存性を調べた。図6に示すように、1mMのエストロン3硫酸の存在下での値を差し引いた後のエストロン3硫酸の取り込みは、飽和性であった。Eadie-Hofsteeプロット分析では単一の直線が得られており、これは、単一の飽和性機序が動態学的に関与していることを示唆している。非線形回帰分析では、Km及びVmaxの値(平均値±S.E.M.)はそれぞれ3.9±0.78μM及び576.3±32.9pmol/mgタンパク質/10minであった。
【0046】
(エストロン3硫酸の取り込み活性の阻害)
輸送システムの基質特異性を特徴づけるため、T−47D細胞による[3H]エストロン3硫酸の取り込みに対する、ホルモン、その抱合代謝産物、及び各種有機化合物の阻害作用を調べた。結果を(表1)に示す。表1中、対照(%)は、対照におけるエストロン3硫酸の取り込み量を100%としたときの割合で示され、各値は平均±S.E.M.(n=4)で表されれ、*P<0.05は対照との比較(Student’ test)。
【0047】
【表1】
【0048】
その結果、非標識エストロン3硫酸それ自体が、最も強い阻害作用を示した(5μM及び50μMのコントロールに対し、それぞれ34.7%と6.6%)。DHEASの阻害作用は有意であった(50μMのコントロールに対し、44.8%)が、エストロン3硫酸の阻害作用よりも弱かった。グルクロニド抱合体であるエストラジオール−17β−グルクロニドは、50μMでは全く作用を示さなかった。非抱合ステロイドホルモンは、そのすべてが有意に[3H]エストロン3硫酸の取り込みを阻害した。しかし、それらの親和性は、硫酸抱合体、エストロン3硫酸及びDHEASの親和性よりも低かった。調べてみたその他の有機化合物のうち、ブロモスルフォフタレイン及びタウロコール酸は強い阻害活性を示し、100μMでの阻害作用は、それぞれコントロールの18.9%と69.3%であった。その他の有機化合物の中で、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンは、ブロモスルフォフタレインに比較して少し弱いながらも阻害作用を示した。一方、サリチル酸は、全く作用を示さなかった。
【0049】
閉経後の女性においては、アンドロゲン及びその硫酸抱合体は、活性エストロゲンの供給源として、乳癌の進行に重要な役割を果たしている。DHEASは、多くの閉経後患者で高濃度(約1μM)で存在することから、主要な前駆物質である(J Clin Endocrinol Metab 54: 22-26., 1982)。そのため、DHEASが特異的輸送機序を介して輸送されるかどうかを調べてみた。DHEASの取り込みは、30分にわたって直線的に増大し、5mMの非標識DHEASの存在下で有意に減少した(データは図示せず)。5mMの非標識DHEASの存在下での取り込み(非特異的取り込み)を差し引いた後のDHEASの取り込みは、飽和性であった(図7)。DHEASのEadie-Hofstee分析では、エストロン3硫酸の場合と同様に単一の直線が得られており(図7、挿入図)、Km及びVmaxの値を計算したところ、それぞれ62.5±21.1μM及び1893±315.7pmol/mgタンパク質/10minであった。
【0050】
T−47D細胞におけるエストロン3硫酸及びDHEASの取り込みを仲介するトランスポーターを同定するため、エストロン3硫酸を輸送する可能性があるOATP及びOATトランスポーターの発現を、RT−PCR分析で測定した。特異的プライマーを用いることにより、テストしたOATP及びOATのうちOATP−D及びOATP−Eの発現がT−47D細胞中で検出されたが(図8)、RT反応を含まないPCRではバンドは全く検出されなかった(データは図示せず)。したがって、OATP−D及びOATP−Eは、エストロゲン依存性乳癌細胞中でエストロン3硫酸及びDHEASの取り込みを仲介する候補物質であるのかもしれない。
【0051】
3.考察
エストロン3硫酸は主要な循環エストロゲンであり、エストロゲン依存性乳癌の進行に関与している(J Steroid Biochem 34: 155-163, 1989)。腫瘍細胞内で、ステロイドスルファターゼにより脱硫酸化され、続いて生物活性エストロゲンに変換される。乳癌細胞の細胞質中でステロイドスルファターゼが検出されているため(Breast Cancer 6: 331-337, 1999)、エストロン3硫酸は、脱硫酸化の前に細胞膜を通過して細胞内に輸送されたに違いない。エストロン3硫酸の親水性を鑑みると、エストロン3硫酸の内在化は、特異的な輸送機序により仲介されるようである。これに対し、疎水性非抱合ステロイドホルモンは単に細胞内に拡散するにすぎないようである(J Clin Endocrinol Metab 59: 1128-1132, 1984)。そのため、本研究では、エストロゲン依存性増殖を示すモデル細胞と見なされているT−47D乳癌細胞への、エストロン3硫酸及びDHEASの取り込みを仲介する特異的な輸送システムの有無を調べてみた。
【0052】
最初に、細胞増殖のエストロゲン依存性を確認した。エストロン3硫酸、又はエストラジオールでの処理により、T−47D細胞の増殖が増大し、エストロン3硫酸のEC50の値が17.1nMとなった。この値は、エストロン3硫酸の生理学的血漿濃度に近い。エストロン3硫酸自体が、エストロゲン受容体での低い直接的生物反応を誘導するため(Endocrinology 138: 863-870, 1997)、かかる結果は、細胞膜を通過してエストロン3硫酸が内在化し、続いて細胞中で活性非抱合エストロゲンに変換されることを示唆している。したがって、本研究は、エストロゲン活性を解明する第一歩として、T−47D細胞によるエストロゲンの硫酸抱合体の取り込みに焦点を当てている。
【0053】
T−47D細胞によるエストロン3硫酸取り込みの経時的変化は、高濃度のエストロン3硫酸(1mM)の存在下では取り込みが有意に減少することから、取り込みが特異的トランスポーターによって仲介されることを示唆している(図4)。エストロン3硫酸の特異的な取り込みは、Na+を、Li+、K+、又はN−メチルグルカミン+と置換しても影響を受けない(図5)。OATP及びOATファミリーのメンバーは、Na+非依存性エストロン3硫酸取り込みトランスポーターの候補物質である。T−47D細胞によるエストロン3硫酸取り込みのKm値を計算したところ、3.9±0.78μMであった(図6)。これは、OATP及びOATの既知のKm値範囲である0.05μM〜59μMと一致する(Pharm Res 18: 1262-1269, 2001; J Biol Chem 274: 13675-13680, 1999; J Biol Chem 275: 4507-4512, 2000)。
【0054】
輸送機序をさらに特徴づけ、関与するトランスポーターを同定するために、T−47D細胞によるエストロン3硫酸の取り込みに対する阻害について研究した。エストロン3硫酸及びDHEASなどのステロイドホルモンの硫酸抱合体の阻害作用が、非抱合ステロイドホルモンの阻害作用よりも強力であったのに対し、グルクロニド抱合体には阻害作用が全くなかった。したがって、硫酸部分がトランスポーターに対する高親和性を与えているのかもしれず、その一方、グルクロニド部分はほとんど認識されていないのかもしれない。硫酸抱合体は、非抱合ステロイド又はグルクロニド抱合体と比較してイオン化定数(pKa)値が低いため、これらの結果は、トランスポーターによる基質の認識にはアニオン性部分が不可欠であることを示唆している。ステロイドホルモンの硫酸抱合体の血漿濃度は、非抱合ホルモン又はそれらのグルクロニド抱合体と比較して高いため、かかるトランスポーターは、乳房腫瘍へのエストロゲンの供給に対し、効果的な作用を及ぼしているはずである。さらに、T−47D細胞ではDHEASの取り込みも飽和性であり、Km値を計算したところ62.5±21.1μMであり、これはエストロン3硫酸の値(3.9μM、図6及び7)よりもかなり高いものである。エストロン3硫酸とDHEASの血漿濃度は、それぞれ約1〜10nMと1μMであるため(Hormone Res 27: 61-68, 1987; J Clin Endocrinol Metab 54: 22-26, 1982)、乳癌細胞は、飽和することなく効果的にそれらを取り込むのかもしれない。
【0055】
OATP、OAT3及びOAT4のメンバーには、ステロイドホルモンの硫酸抱合体を輸送できるものがある(Biochim Biophys Acta 1609: 1-18, 2003, Pflugers Arch 447: 653-665, 2003; Breast Cancer 6: 331-337, 1999; J Biol Chem 274: 13675-13680, 1999)。そのなかでも、OATP−B及びOAT4は、それぞれKm値が9.04μMと1.01μMの硫酸抱合体を認識することができるが、その一方で、ステロイドホルモンのグルクロニド抱合体に対する親和性は皆無である(Pharm Res 18: 1262-1269, 2001; J Biol Chem 275: 4507-4512, 2000)。T−47D細胞中で、[3H]エストロン3硫酸の取り込みがグルクロニド抱合体によって阻害されなかったため、OATP−B又はOAT4の機能的特質は、ステロイド抱合体の基質認識に関しては、T−47D細胞における取り込み機序と一致している。その一方で、OATP−A、OATP−C、OATP8、及びOAT3は、硫酸抱合体とグルクロニド抱合体の両方を輸送するため、これらのトランスポーターが、T−47D細胞中でのエストロン3硫酸の輸送に関与している可能性は低いと思われる(Biochim Biophys Acta 1609: 1-18, 2003, Pflugers Arch 447: 653-665, 2003; J Biol Chem 274: 13675-13680, 1999)。鍵となるトランスポーターの性質に関する洞察を得るために、各種ヒトOATP及びOATの発現をRT−PCR分析で調べてみた(図8)。発現が検出されたのはOATP−DとOATP−Eだけであり、これは、以前の報告(J Clin Endocrinol Metab 88: 3902-3912, 2003)と一致するものであった。OATP−DとOATP−Eは、HEK293細胞中で一過性に発現した際にエストロン3硫酸に対する輸送活性を示したため(Biochem Biophys Res Commun 273: 251-260, 2000)、T−47D細胞中で細胞膜を通過してのエストロン3硫酸の取り込みに関与しているのかもしれない。しかし、それらの機能的特質がよくわかっていないため、T−47D細胞によるエストロン3硫酸の取り込みに関与するトランスポーターを本研究で同定することはできなかった。BCRP(ABCG2)及びOSTalpha−OSTbetaが、基質としてエストロン3硫酸を受容できることも、報告されている(Breast Cancer 9: 296-302, 2002; Proc Natl Acad Sci USA 95: 15665-15670, 1998; Biochem Biophys Res Commun 298: 41-452002; Mol Pharmacol 64: 610-618, 2003; J Biol Chem 278: 27473-27482, 2003)。したがって、OATPとOAT以外のトランスポーターも、T−47D細胞におけるエストロン3硫酸の取り込みに関与している可能性がある。
【0056】
上記実施例により、エストロン3硫酸が、特異的輸送機序を介してエストロゲン依存性T−47D細胞により取り込まれることが、はっきりと実証された。これは、主要な循環エストロゲン、エストロン3硫酸及びDHEASが、特異的なトランスポーターを介してエストロゲン依存性乳癌細胞に供給されることを初めて実証するものである。T−47D細胞中のトランスポーターを同定するためにはさらなる研究が必要だが、そのようなトランスポーター分子は、エストロゲン依存性乳癌治療のための新しい分子標的となるであろう。
【図面の簡単な説明】
【0057】
【図1】乳癌の分子ターゲットを示す模式図である。
【図2】BSPによるエストロン3硫酸の乳癌細胞への取込み阻害を示す図である。
【図3】乳癌細胞へ取り込まれたエストロン3硫酸によるERE(エストロゲンレスポンスエレメント)活性の誘導を示す模式図、及びレポータージーンアッセイに用いられるpERE−TA−SEAPベクターの模式図である。
【図4】pERE−TA−SEAPベクターで一過的にトランスフェクションしたMCF−7細胞におけるERE誘導SEAP(Secreted Alkaline phospatase)活性に及ぼすエストロン3硫酸の効果を示す図である。
【図5】エストロゲン依存性細胞増殖アッセイの模式図、及びそのタイムスケジュールを示す図である。
【図6】MCF−7細胞にエストロゲン依存性細胞増殖に及ぼすブロモスルフォフタレインの効果を示す図である。
【図7】エストロゲンによるT−47D乳癌細胞増殖刺激の経時変化(a)及び濃度依存性(b)を示す図である。(a)T−47D細胞を密度8,000個/ウェルで播種し、培養した。播種の24時間後(第1日)、エストラジオール(1pM、▲)、エストロン3硫酸(100nM、●)、DMSO(0.1%、△)又は水(○)を添加した。所定日数経過後、T−47D細胞をトリプシン処理し、計数した。(b)濃度範囲がそれぞれ10−16〜10−11M、又は10−11〜10−6Mの各種濃度のエストラジオール(▲)又はエストロン3硫酸(●)の存在下で、T−47D細胞を増殖させた。播種6日後に細胞数を数えた。各値は平均値±S.E.M.を示す(n=3)。
【図8】T−47D細胞による[3H]エストロン3硫酸取り込みの経時変化を示す図である。培養したT−47D細胞を、[3H]エストロン3硫酸(7.1nM)を含み、さらに1mMの非標識エストロン3硫酸を含む培地(○)又は含まない培地(●)で、37℃で35分間インキュベーションした。各値は平均値±S.E.M.を示す(n=4)。エラーバーが表示されていない場合、それらが記号よりも小さいことを示す。
【図9】T−47D細胞による[3H]エストロン3硫酸の取り込みに対する細胞外カチオンの作用を示す図である。 細胞外Na+の存在下又は不在下で、[3H]エストロン3硫酸(7.1nM)の取り込み速度を10分間にわたって測定した。細胞外Na+を、K+、Li+、又はN−メチルグルカミン+(NMG+)と置換した。各値は平均値±S.E.M.を示す(n=4)。
【図10】T−47D細胞によるエストロン3硫酸取り込みの濃度依存性を示す図である。5.1nMから50μMまで、各種濃度のエストロン3硫酸の取り込みを、37℃で10分間測定した。過剰な非標識エストロン3硫酸(1mM)の存在下での取り込みを差し引いた後、飽和性取り込みを得て、非線形最少二乗分析による速度定数の評価に使用した。挿入図は、エストロン3硫酸の飽和性取り込みのEadie-Hofsteeプロットを示す。各値は平均値±S.E.M.を示す(n=4)。エラーバーが表示されていない場合、それらが記号よりも小さいことを示す。
【図11】T−47D細胞によるDHEAS取り込みの濃度依存性を示す図である。10.3nMから100μMまで、各種濃度のDHEASの取り込みを、37℃で10分間測定した。過剰な非標識DHEAS(5mM)の存在下での取り込みを差し引いた後、飽和性取り込みを得て、非線形最少二乗分析による速度定数の評価に使用した。挿入図は、DHEASの飽和性取り込みのEadie-Hofsteeプロットを示す。各値は平均値±S.E.M.を示す(n=4)。エラーバーが表示されていない場合、それらが記号よりも小さいことを示す。
【図12】T−47D細胞におけるOATP及びOATトランスポーターの発現を示す図である。T−47D細胞から得たmRNAを用いて、RT−PCR分析を行った。材料と手法に記載のとおりに反応を行った。矢印は、OATP−D及びOATP−Eの特異的なバンドを示す。
【特許請求の範囲】
【請求項1】
ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とするエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤。
【請求項2】
ブロモスルフォフタレインを有効成分とするエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤。
【請求項3】
ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とする乳癌細胞増殖抑制剤。
【請求項4】
ブロモスルフォフタレインを有効成分とする乳癌細胞増殖抑制剤。
【請求項5】
ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とする乳癌治療剤。
【請求項6】
ブロモスルフォフタレインを有効成分とする乳癌治療剤。
【請求項1】
ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とするエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤。
【請求項2】
ブロモスルフォフタレインを有効成分とするエストロン3硫酸トランスポーターのトランスポーター活性の阻害剤。
【請求項3】
ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とする乳癌細胞増殖抑制剤。
【請求項4】
ブロモスルフォフタレインを有効成分とする乳癌細胞増殖抑制剤。
【請求項5】
ブロモスルフォフタレイン,デヒドロエピアンドロステロンサルフェート,デヒドロエピアンドロステロン,タウロコール酸、pアミノ馬尿酸、テトラエチルアンモニウム、プロベネシド及びベンジルペニシリンからえらばれる1種以上の成分を有効成分とする乳癌治療剤。
【請求項6】
ブロモスルフォフタレインを有効成分とする乳癌治療剤。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公開番号】特開2006−176427(P2006−176427A)
【公開日】平成18年7月6日(2006.7.6)
【国際特許分類】
【出願番号】特願2004−370290(P2004−370290)
【出願日】平成16年12月21日(2004.12.21)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成16年8月24日 The American Society for Pharmacology and Experimental Therapeutics発行の「The Journal of Pharmacology and Experimental Therapeutics,Vol.311,No.3,」に発表
【出願人】(502133066)株式会社ジェノメンブレン (7)
【Fターム(参考)】
【公開日】平成18年7月6日(2006.7.6)
【国際特許分類】
【出願日】平成16年12月21日(2004.12.21)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成16年8月24日 The American Society for Pharmacology and Experimental Therapeutics発行の「The Journal of Pharmacology and Experimental Therapeutics,Vol.311,No.3,」に発表
【出願人】(502133066)株式会社ジェノメンブレン (7)
【Fターム(参考)】
[ Back to top ]