
エタノール生産におけるトランスヒドロゲナーゼ遺伝子の増大した発現及びその使用
本発明は、トランスヒドロゲナーゼ遺伝子の増大した発現を有する単離又は組換えエタノール生産性細菌及び調製の方法を提供する。本発明はまた、この細菌及び対応するキットを使用してエタノールを生産する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願)
本出願は、それらの開示全体が参照により本明細書に組み込まれる、2009年5月15日出願の米国特許仮出願第61/178,672号、2009年8月19日出願の米国特許仮出願第61/235,340号、及び2010年1月4日出願の米国特許仮出願第61/292,094号の優先権を主張する。
【0002】
(政府支援研究)
本発明は、米国エネルギー省(U.S.Department of Energy)によって与えられた、契約番号第US DOE FG02−96ER20222号、第DE−FG36−08GO88142号、及び第DE−FC36−GO17058号の下での米国政府の支援を得て行われた。政府は本発明に一定の権利を有する。
【背景技術】
【0003】
リグノセルロースバイオマスからの糖類を基質として使用して多種多様な発酵産物を作ることができる(9、13、16、37)。しかし、発酵の前に、炭水化物ポリマーであるセルロース及びヘミセルロースを、化学的工程と酵素的工程の組合せを用いて可溶性糖類に変換しなければならない(38、41)。化学的工程は、微生物触媒の代謝に負の影響を及ぼす、アルコール類、酸及びアルデヒド類などの少量の生成物の混合物を生じる副反応を伴う。アルコール類(カテコール、シリンゴール等)は、細胞膜の透過性を上げることによって作用し、毒性は分子の疎水性とよく相関することが示されている(46)。有機酸(酢酸、ギ酸等)は、中性形態で膜を横断し、細胞質内でイオン化して、プロトン駆動力を崩壊させることによって増殖を阻害すると考えられる(31、45)。アルデヒド類の阻害機構はより複雑である。アルデヒド類は、直接の物理的作用及び代謝作用に加えて、多くの細胞成分と反応して生成物を形成し得る(26、34)。全体として、化学的前処理からのこれらの少量の生成物は、細胞増殖を遅延させ、バイオマス由来の糖類の発酵を減速させ得る(10、30)。
【0004】
フルフラール(五炭糖類の脱水生成物)は特に重要である。フルフラールはリグノセルロース分解の天然生成物である。フルフラールは、酸性条件下でのセルロースバイオマスの解重合の間に五炭糖類の脱水によっても形成される(21)。この化合物はヘミセルロースシロップの毒性に対する重要な寄与因子であり、他の化合物の毒性を増大する(44)。ヘミセルロースの希酸加水分解物中のフルフラール含量は毒性と関連づけられてきた(22)。石灰の添加(pH10)によるフルフラールの除去は加水分解物を容易に発酵可能にしたが、フルフラールを再添加すると毒性が回復した(21)。フルフラールはまた、ヘミセルロースの酸加水分解物中に存在することが公知の他の化合物の毒性を増強することも示されている(44〜46)。フルフラールは、DNAの構造と配列を変化させ(3、17)、解糖酵素を阻害して(6)、糖代謝を遅延させる(11)ことが報告されている。
【0005】
これらの阻害剤の存在下で機能する発酵生物の能力が広く探求されてきた。アルギン酸塩へのサッカロミセス・セレビシエ(Saccharomyces cerevisiae)の封入は保護的であり、ヘミセルロースの酸加水分解物における発酵を改善することが示された(36)。S.セレビシエの株は、加水分解物阻害剤に対して改善された耐性を有することがこれまでに記述されている(1、19、28)。大腸菌(Escherichia coli)(7)、S.セレビシエ(2)及び他の微生物(4)は、フルフラールをより毒性の低い生成物、フルフリルアルコールに還元することを触媒する酵素を含むことが示された(46)。大腸菌では、フルフラールレダクターゼ活性はNADPH依存性であると思われる(7)。NADPH依存性のフルフラールレダクターゼは大腸菌から精製されたが、その他のものも存在し得る。5−ヒドロキシメチルフルフラール(六炭糖類の脱水生成物)を還元することができるNADPH依存性酵素がS.セレビシエにおいて特徴づけられ、ADH6遺伝子と同定された(33)。
【発明の概要】
【発明が解決しようとする課題】
【0006】
従って、エタノール生産性微生物によるフルフラールに対する耐性を高めることができれば、これらの微生物によるエタノール生産の増大をもたらすだろう。
【課題を解決するための手段】
【0007】
本発明は、大規模な燃料生産のための生物を提供する。特に、本発明は、増大したフルフラールの存在下で増殖し、エタノールを生産することができる細菌を提供する。
【0008】
本発明は、参照細菌と比較して少なくとも1つのトランスヒドロゲナーゼ遺伝子の増大した発現を有する、単離されたか又は組換えたエタノール生産性細菌を提供する。一実施形態では、トランスヒドロゲナーゼ遺伝子はpntA及びpntBである。
【0009】
本発明はまた、参照細菌と比較してpntA及びpntBの増大した発現を有する、単離された細菌又は組換え細菌を提供する。
【0010】
一実施形態では、細菌は、参照細菌と比較して増大したフルフラール耐性を有する。
【0011】
別の実施形態では、細菌は野生型細菌である。
【0012】
別の実施形態では、細菌はエタノール生産性である。
【0013】
別の実施形態では、細菌は参照細菌と比較して増大したエタノール生産を示す。
【0014】
別の実施形態では、細菌は、参照細菌と比較してフルフラールの存在下で増大したエタノール生産を示す。
【0015】
別の実施形態では、細菌は参照細菌と比較して増大した増殖を有する。
【0016】
別の実施形態では、細菌は、参照細菌と比較してフルフラールの存在下で増大した増殖を有する。
【0017】
別の実施形態では、細菌は、約0.025%フルフラール〜約0.15%フルフラールの濃度のフルフラールの存在下で増大した増殖を有する。
【0018】
別の実施形態では、細菌は、参照細菌と比較して増大した増殖及び増大したエタノール生産を有する。
【0019】
別の実施形態では、細菌は、参照細菌と比較して加水分解物の存在下で増大した増殖を有する。
【0020】
別の実施形態では、細菌は加水分解物の存在下で増大した増殖を有し、加水分解物は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス又はセルロースバイオマスを含む生成物に由来する。
【0021】
別の実施形態では、pntA及びpntB遺伝子の発現は、pntA及びpntB遺伝子の発現を調節するプロモータを修飾又は付加することによって増大又は変化する。
【0022】
別の実施形態では、pntA及びpntB遺伝子の発現は、遺伝子を異なる調節タンパク質の制御下又は付加的な調節タンパク質の制御下に置くことによって増大又は変化する。
【0023】
別の実施形態では、細菌は、嫌気又は微好気条件下で主発酵生成物としてエタノールを生産することができる。
【0024】
別の実施形態では、細菌は、グラム陰性細菌及びグラム陽性細菌から成る群より選択される。
【0025】
別の実施形態では、細菌は、グラム陰性細菌及びグラム陽性細菌から成る群より選択され、グラム陰性細菌は、エシェリキア属(Escherichia)、ザイモモナス属(Zymomonas)、アシネトバクター属(Acinetobacter)、グルコノバクター属(Gluconobacter)、ジオバクター属(Geobacter)、シュワネラ属(Shewanella)、サルモネラ属(Salmonella)、エンテロバクター属(Enterobacter)及びクレブシエラ属(Klebsiella)から成る群より選択される。
【0026】
別の実施形態では、細菌は、グラム陰性細菌及びグラム陽性細菌から成る群より選択され、グラム陽性細菌は、バチルス属(Bacillus)、クロストリジウム属(Clostridium)、コリネバクテリウム属(Corynebacterium)、乳酸杆菌属(Lactobacillus)、ラクトコッカス属(Lactococcus)、オエノコッカス属(Oenococcus)、連鎖球菌属(Streptococcus)及びユーバクテリウム属(Eubacterium)から成る群より選択される。
【0027】
別の実施形態では、細菌は大腸菌(Escherichia coli)である。
【0028】
別の実施形態では、細菌はクレブシエラ・オキシトカ(Klebsiella oxytoca)である。
【0029】
本発明は、参照細菌と比較してPntA及びPntBタンパク質の活性が増大している、単離された細菌又は組換え細菌を提供する。
【0030】
本発明は、参照細菌と比較してPntA及びPntBタンパク質の活性が増大しており、且つ参照細菌と比較して増大したフルフラール耐性を有する、単離された細菌又は組換え細菌を提供する。
【0031】
本発明はまた、参照細菌と比較してpntA及びpntB遺伝子の発現が増大しており、且つ参照細菌と比較して増大したフルフラール耐性を有する、単離された細菌又は組換え細菌を提供する。
【0032】
本発明はまた、参照細菌と比較してPntA及びPntB遺伝子の発現又はPntA及びPntBポリペプチドの活性が増大しており、参照細菌と比較してフルフラール耐性が増大した、エタノールを生産することができ、並びに
(a)細菌の候補株をフルフラールの存在下で増殖させること;及び
(b)フルフラールの存在下でエタノールを生産する細菌を選択すること
を含む工程によって調製される、単離された細菌又は組換え細菌を提供する。
【0033】
本発明はまた、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖を本発明の単離された細菌又は組換え細菌のいずれかと接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含む、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産する方法を提供する。
【0034】
さらに、本発明は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖を本発明の単離された細菌又は組換え細菌と接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含む、フルフラールの存在下でバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産する方法を提供する。
【0035】
加えて、本発明は、本発明の方法によって生産されるエタノールを提供する。
【0036】
本発明はまた、本発明の単離された細菌又は組換え細菌を含むキットを提供する。
【図面の簡単な説明】
【0037】
【図1】図1A〜Bは、フルフラールの存在下で種々のレベルのsthA遺伝子発現を有するプラスミドを保持する様々なLY180株の増殖。24時間後(図1A)及び48時間後(図1B)の細胞密度(光学密度)とフルフラール濃度の関係を表すグラフを示す。記号:(■)、挿入物を有さないpTrc99a対照;(▲)、pTrc99a−sthAの順方向非誘導sthA発現;(▲)、0.01mM IPTGで誘導したpTrc99a−sthAの順方向sthA発現;及び(◆)、0.1mM IPTGで誘導したpTrc99a−sthAの順方向sthA発現。
【図2】図2A〜Bは、フルフラールの存在下で種々のレベルのpntA及びpntB遺伝子発現を有するプラスミドを保持する様々なLY180株の増殖。24時間後(図2A)及び48時間後(図2B)の細胞密度(光学密度)とフルフラール濃度の関係を表すグラフを示す。記号:(■)、挿入物を有さないpTrc99a対照;(▲)、pTrc99a−pntABの順方向非誘導pntAB発現;(▲)、0.01mM IPTGで誘導したpTrc99a−pntABの順方向pntAB発現;及び(◆)、0.1mM IPTGで誘導したpTrc99a−pntABの順方向pntAB発現。
【図3】図3A〜Bは、フルフラール耐性へのトランスヒドロゲナーゼ(SthA及びPntAB)の増大した発現の影響を示す。培養物を、50g/lのキシロース及び1.0g/lのフルフラールを含むAM1最少培地中で48時間増殖させた。空のベクタを対照として使用した。接種の前に誘導物質を添加した。細胞密度(550nmでの光学密度に関して)を、24時間後の約0.05%フルフラール濃度(図3A)及び48時間後の約0.10%フルフラール濃度(図3B)での各々の株について示す。対照株を白い棒グラフで示している。種々のレベルのpntA発現を有するプラスミドを保持する株を斜線の棒グラフで示す。種々のレベルのpntAB遺伝子発現を保持する株を黒い棒グラフで示す。
【図4】図4A〜Bは、pntAの核酸配列(図4A)及びアミノ酸配列(図4B)を示す。
【図5】図5A〜Bは、pntBの核酸配列(図5A)及びアミノ酸配列(図5B)を示す。
【図6】図6A〜Bは、0.5g/lフルフラール抗原での抗原投与後のLY180における転写及び調節の変化を示す。有意に変動した調節遺伝子をNCAによって同定し、ヌル分布に比して0.05をP値のカットオフとした。増大した活性を伴う調節因子を実線の枠で示し、減少した活性を伴う調節因子を破線の枠で示す。混合活性を示した調節因子は明灰色で示している(DcuR)。図6Aは部分的な調節因子−遺伝子応答地図を示す。2倍より大きく変動した代表的な遺伝子を示しており、中実(点々)の枠は増大した発現(減少した発現)を有する遺伝子を示す。実線は連結した調節因子による活性化を示し、破線は抑制を示す。DcuRについては変動の方向が不明確であり、この調節因子は明灰色で示している。図6Bは、フルフラールによる増殖阻害の機構を説明するモデルを示す。フルフラールの添加は、NADPHについての基本的生合成反応と競合する2つのNADPH依存性オキシドレダクターゼ(YqhD及びDkgA)を誘導する。硫酸塩の同化はシステイン当たり4つのNADPHを必要とする。二次的な結果は、ストリンジェント因子SF並びに生合成調節因子ArgR、PurR及びRutRの変動によって証明されるように、含硫アミノ酸の枯渇と、停止した翻訳及び多くの非含硫ビルディングブロック中間体の蓄積から、より一般的なストレス応答への事象のカスケードとを含む。
【図7】図7は、ヒスチジン経路におけるhisG以外の遺伝子のフルフラール添加による発現増大を示す。変化の割合を括弧内に示す。略語:ACR、アミノイミダゾールカルボキサミドリボヌクレオチド。
【図8】図8は、硫黄のシステイン及びメチオニンへの同化に関与する遺伝子のフルフラールによる発現増大を示す。比較のためにアスパラギン酸からのトレオニン及びイソロイシンの合成のための経路を含める。1.5倍又はそれ以上上方調節された遺伝子をプラスの記号で示す。1.5倍又はそれ以上下方調節された遺伝子をマイナスの記号で示す。その他はすべてプラス又はマイナス記号のない黒字で示している。
【図9】図9は、1g/lフルフラールの存在下での増殖への培地添加物の影響を示す。48時間の培養後に培養物を比較する(AM1培地、50g/lキシロース、37℃)。図9Aは個々のアミノ酸(各々0.1mM)の添加を示す。図9Bはアミノ酸(各々0.5mM)の添加を示す。図9Cはシステインの添加を示す。図9Dは代替硫黄源の添加を示す。
【図10】図10は、表1であり、細菌株、プラスミド及びプライマを示す。
【図11】図11は、表2であり、機能群によって分類した、0.5g/lフルフラールでの処理に対するLY180の応答が2倍より大きく変動した遺伝子を示す。
【図12】図12は、表3であり、フルフラール(0.5g/l)の添加に応答して5倍又はそれ以上の発現比の変化が生じた遺伝子を示す。これらの遺伝子の一部は、対照実験で水の添加に対する応答も2倍より大きく変化した(星印で示している)。
【図13】図13は、表4であり、NCAによって決定したとき、ヌル分布に比してLY180のフルフラール応答が有意に(P<0.05)変動した調節因子を示す。
【図14】図14は、嫌気的増殖及び発酵への5−HMFの効果である。キシロース(100g/lキシロース)を添加したAM1無機塩培地で細胞を増殖させた。A.1.0g/ lの5−HMFで増殖中の細胞質量;B.1.0g/lの5−HMFで発酵中のエタノール生産;C.発酵中の5−HMF(1.0g/l)の還元;D.2.5g/lの5−HMFで増殖中の細胞質量;E.2.5g/lの5−HMFで発酵中のエタノール生産;F.発酵中の5−HMF(2.5g/l 5−HMF)の還元。比較のために5−HMFなしでの並行発酵をA及びBに含めている(破線)。すべてのデータは標準偏差と共に平均値としてプロットしている(n=3)。すべてについての記号:□、LY180;及び●、EMFR9。
【図15】図15は、5−HMFのインビトロでの還元及び5−HMF耐性へのYqhD及びDkgAの効果。A.インビトロでの5−HMF還元に対する比活性。溶解細胞抽出物(2mMのNADPH、20mMの5−HMF)において活性を測定した。B.EMFR9(耐性変異株)の細胞収率へのプラスミドからのyqhD及びdkgA発現の影響。実験は、50g/lのキシロース及び1.0g/lの5−HMFを含むAM1培地での試験管培養物において実施した(48時間培養)。プラスミドの維持のためにカナマイシンを含めることは5−HMF耐性を減少することに留意されたい。誘導株(Ind.)は0.1mMのIPTGを添加して増殖させた。C.LY180(親株)の細胞収率へのyqhD及びdkgA欠失の影響。実験は、50g/lのキシロース及び2.5g/lの5−HMFを含むAM1培地での試験管培養物において実施した(48時間培養)。すべてのデータは標準偏差と共に平均値としてプロットしている(n=4)。
【図16】図16は、5−HMF耐性へのプラスミドからのpntAB発現の影響。実験は、指示されているように50g/lのキシロース及び5−HMFを含むAM1培地でBioscreen C増殖曲線分析器を使用して実施した。すべてのデータは標準偏差とと共に平均値としてプロットしている(n=10)。明瞭さのために連結点は省略した。A.添加物なし;B.0.9g/lの5−HMFを添加;及びC.1.8g/lの5−HMFを添加。すべてについての記号:△、LY180(pTrc99a−対照);○、非誘導LY180(pTrc99a−pntAB);●、0.01mMのIPTGで誘導したLY180(pTrc99a−pntAB)。
【図17】図17は、LY180の5−HMF耐性へのL−システインの効果。実験は、50g/lのキシロース及び5−HMFを含むAM1培地での試験管培養物において実施した(24時間培養)。培養物に、指示されているようにろ過滅菌したL−システインを添加した。すべてのデータは標準偏差と共に平均値としてプロットしている(n=4)。A.1.0g/lの5−HMF;B.2.0g/lの5−HMF。
【発明を実施するための形態】
【0038】
(I.定義)
本明細書で使用される、「単離された」は、他の細菌による汚染がないことを意味する。単離された細菌は、その単離細菌の特性及び機能を妨げない少量の他の細菌の存在下で存在し得る。単離された細菌は、一般に少なくとも30%、40%、50%、60%、70%、80%、85%、90%、95%、98%又は99%純粋である。好ましくは、本発明による単離された細菌は少なくとも98%又は少なくとも99%純粋である。
【0039】
本明細書で使用される、「細菌」は、「非組換え細菌」、「組換え細菌」及び「突然変異細菌」を含む。
【0040】
本明細書で使用される、「非組換え細菌」は、異種ポリヌクレオチド配列を含まず、本発明の組成物及び方法を用いたさらなる修飾に適する、例えば遺伝子操作、例えば異種ポリヌクレオチド配列を組み込むことができる、例えばトランスフェクトすることができる、細菌細胞を含む。この用語は、最初にトランスフェクトされた細胞の子孫を包含することが意図されている。特定の実施形態では、細胞はグラム陰性細菌細胞及びグラム陽性細菌細胞である。
【0041】
細菌に関して本明細書で使用される「組換え体」は、遺伝子操作に適する若しくは遺伝子操作に供される、又は異種ポリヌクレオチド配列が組み込まれている、又は天然ポリヌクレオチド配列が変異若しくは欠失するように処理された細菌細胞を意味する。
【0042】
細菌に関して本明細書で使用される「突然変異体」は、本明細書中以下で定義される参照細菌と同一ではない細菌細胞を意味する。
【0043】
「突然変異」細菌は「組換え」細菌を含む。
【0044】
本明細書で使用される、「エタノール生産性」は、炭水化物から主発酵生成物としてエタノールを生産する細菌の能力を意味する。この用語は、天然に生じるエタノール生産性生物及び天然に生じた又は誘導された突然変異を有するエタノール生産性生物を包含することが意図されている。
【0045】
「非エタノール生産性」という用語は、細菌が炭水化物から主発酵生成物としてエタノールを生産することができないことを意味する。この用語は、総非ガス発酵生成物の40%未満を含有している微量発酵生成物としてエタノールを生産する微生物を包含することが意図されている。
【0046】
本明細書で使用される「エタノール生産」は、炭水化物からの主発酵生成物としてのエタノールの生産を意味する。
【0047】
本明細書で使用される「エタノールを生産することができる」は、本明細書で定義されるように「エタノール生産」できることを意味する。
【0048】
「発酵する」及び「発酵」という用語は、複合糖類の分解又は解重合並びにその糖残基のエタノール、酢酸エステル及びコハク酸エステルへの生物変換を意味する。これらの用語は、エタノールが炭水化物から生産される、特に発酵の主生成物として生産される酵素的過程(例えば細胞又は無細胞の、例えば溶解生成物又は精製ポリペプチド混合物の)を包含することが意図されている。
【0049】
「主発酵生成物」及び「主要発酵生成物」という用語は、本明細書では交換可能に使用され、総非ガス生成物の約50%より多くを含有している非ガス性の発酵生成物を包含することが意図されている。主発酵生成物は最も大量の非ガス生成物である。本発明のある特定の実施形態では、主発酵生成物はエタノールである。
【0050】
本明細書で使用される「微量発酵生成物」という用語は、総非ガス生成物の40%未満を含有する非ガス性の発酵生成物を包含することが意図されている。本発明のある実施形態では、微量発酵生成物はエタノールである。
【0051】
「同時糖化発酵」又は「SSF」という用語は、複合糖の同時分解又は解重合及び発酵によるその糖残基のエタノールへの生物変換反応のための1又はそれ以上の組換え宿主(又は、精製又は非精製抽出物を含む、その抽出物)の使用を包含することが意図されている。SSFは、バイオマスを、細菌によって最終的にエタノールに変換可能な多糖類に分解するために使用できる周知の工程である。天然で起こる場合のバイオマスの分解を反映して、SSFは、リグノセルロースなどの糖源を、最終的にエタノールに変換できる単糖類に分解するためにエタノール生産性細菌(又はそれに由来する酵素)の活性と真菌(又は真菌から抽出されたセルラーゼなどの酵素)の活性を組み合わせる。SSF反応は、典型的には、高価な真菌酵素の使用を最適化するために酸性pHで実施される。
【0052】
「糖」という用語は、糖分子(複数も)を含む何れの炭水化物源もを包含することが意図されている。そのような糖は、本発明の生成物及び方法による、解重合(所望により)並びにその後の発酵によるアセトアルデヒドへの及びその後のエタノールへの生物変換のための糖類の可能性のある供給源である。糖の供給源は、大部分の植物における燃料貯蔵の主要形態であるデンプン、植物の堅い細胞壁並びに繊維性及び木質組織の主たる細胞外構造成分であるヘミセルロース及びセルロースを含む。この用語は、単糖(simple sugars)とも呼ばれる単糖類(monosaccharides)、オリゴ糖類及び多糖類を包含することが意図されている。ある特定の実施形態では、糖は、例えばグルコース、キシロース、アラビノース、マンノース、ガラクトース、スクロース及びラクトースを含む。他の実施形態では、糖はグルコースである。
【0053】
本明細書で使用される場合、「PntAB」は、ピリジンヌクレオチドトランスヒドロゲナーゼを意味する。pntA及びpntBとしても知られるpntABは、PntABトランスヒドロゲナーゼに対応する遺伝子を指し、一方PntABという用語はpntAB遺伝子の産物を示す。
【0054】
pntA及びpntB遺伝子に対応するアミノ酸配列及び核酸配列をそれぞれ図4A〜4B及び図5A〜5Bに示す。
【0055】
本明細書で使用される場合、「SthA」は、細胞質トランスヒドロゲナーゼを意味する。sthAはSthAトランスヒドロゲナーゼに対応する遺伝子を指し、一方SthAという用語はsthA遺伝子の産物を示す。
【0056】
本明細書で使用される場合、「突然変異核酸分子」又は「突然変異遺伝子」は、そのポリペプチド又は変異株によってコードされ得るポリペプチドがそのポリペプチド又は野生型核酸分子若しくは遺伝子によってコードされるポリペプチドとは異なる活性又は特性を示すように、少なくとも1つの変化(例えば置換、挿入、欠失)を含むヌクレオチド配列を有する核酸分子又は遺伝子を包含することが意図されている。
【0057】
核酸分子又は遺伝子に関して本明細書で用いられる「突然変異」は、核酸若しくは遺伝子の変化、挿入若しくは欠失、又は核酸若しくは遺伝子の発現レベルの増大若しくは減少を意味し、発現の増大又は減少は、それぞれ、核酸分子又は遺伝子によってコードされ得るポリペプチドの発現の増大又は減少を生じさせる。突然変異はまた、そのポリペプチド又はその変異株によってコードされ得るポリペプチドがそのポリペプチド又は野生型核酸分子若しくは遺伝子によってコードされるポリペプチドとは異なる活性又は特性を示すように、少なくとも1つの変化(例えば置換、挿入、欠失)を含むヌクレオチド配列を有する核酸分子又は遺伝子を意味する。
【0058】
本明細書で使用される、「突然変異タンパク質」又は「突然変異タンパク質又はアミノ酸配列」は、ポリペプチド又は突然変異アミノ酸配列によってコードされ得るポリペプチドがそのポリペプチド又は野生型アミノ酸配列によってコードされるポリペプチドとは異なる活性又は特性を示すように、少なくとも1つの変化(例えば置換、挿入、欠失)を含むアミノ酸配列を包含することが意図されている。
【0059】
タンパク質又はアミノ酸配列に関して本明細書で使用される、「突然変異」は、アミノ酸配列のアミノ酸の変化、挿入若しくは欠失、又はアミノ酸配列の発現レベルの増大若しくは減少を意味し、発現の増大又は減少は、アミノ酸配列によってコードされ得るポリペプチドの発現の増大又は減少を生じさせる。突然変異はまた、そのポリペプチド又は変異株によってコードされ得るポリペプチドがそのポリペプチド又は野生型アミノ酸配列によってコードされるポリペプチドとは異なる活性又は特性を示すように、少なくとも1つの変化(例えば置換、挿入、欠失)を含むアミノ酸配列を有するタンパク質又はアミノ酸配列を意味する。
【0060】
本明細書で使用される「断片」又は「サブ配列」は、親若しくは参照核酸配列若しくはアミノ酸配列の一部、又は親若しくは参照配列、ポリペプチド若しくは遺伝子の生物学的機能又は特性をコードするか又は保持する、ポリペプチド若しくは遺伝子の一部を包含することが意図されている。
【0061】
「突然変異」細菌は、本明細書中上記で定義した「突然変異」を含む細菌を包含する。
【0062】
本明細書で使用される「参照物」又は「参照細菌」は、少なくとも、野生型細菌及び親細菌を包含する。
【0063】
本明細書で使用される「野生型」は、生物又は菌株の典型的な形態、例えば突然突然変異の不在下で天然に生じる細菌、遺伝子又は特徴を意味する。「野生型」は、天然集団における最も一般的な表現型を示す。野生型は、遺伝子型及び表現型についての参照物の標準である。
【0064】
本明細書で使用される「親」又は「親細菌」は、目的の細菌を生じる細菌を示す。
【0065】
本明細書で使用される「遺伝子」は、酵素又は他のポリペプチド分子の合成を誘導できる、例えばコード配列、例えばポリペプチド、そのサブ配列をコードする隣接するオープンリーディングフレーム(ORF)を含み得る、又は生物においてそれ自体が機能性であり得る核酸である。生物中の遺伝子は、本明細書で定義されるオペロンにクラスター化することができ、オペロンは遺伝子間DNAによって他の遺伝子及び/又はオペロンから切り離されている。オペロン内に含まれる個々の遺伝子は、個々の遺伝子の間の遺伝子間DNAなしでオーバーラップし得る。加えて、「遺伝子」という用語は、選択された目的のための特定遺伝子を包含することが意図されている。遺伝子は、宿主細胞にとって内因性であり得るか又は、例えばエピソームに保持されたプラスミド若しくはゲノムに安定に組み込まれたプラスミド(若しくはその断片)として、宿主細胞に組換え導入され得る。異種遺伝子は、細胞に導入される、その細胞にとって天然ではない遺伝子である。
【0066】
「核酸」という用語は、ポリペプチド、そのサブ配列をコードするオープンリーディングフレームを含み、非コード調節配列及びイントロンをさらに含み得る核酸分子、例えばポリヌクレオチドを包含することが意図されている。加えて、この用語は、機能的遺伝子座に位置づけられる1つ又はそれ以上の遺伝子を包含することが意図されている。加えて、この用語は、選択された目的のための特定遺伝子を包含することが意図されている。一実施形態では、遺伝子という用語は、pntA及びpntBを含むがこれらに限定されない、トランスヒドロゲナーゼをコードする任意の遺伝子を包含する。一実施形態では、遺伝子又はポリヌクレオチドセグメントは、炭水化物のエタノールへの生物変換反応における少なくとも1つの工程に関与する。生物中の遺伝子は、本明細書で定義されるオペロンにクラスター化することができ、オペロンは遺伝子間DNAによって他の遺伝子及び/又はオペロンから切り離されている。
【0067】
本明細書で使用される「増加している」又は「増加する」又は「増加した」は、例えば、参照細菌と比較してpntA及びpntB遺伝子の発現増加を有する細菌において、pntA及びpntB遺伝子の発現レベルと比較して、少なくとも5%、例えば5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、99、100%又はそれ以上増加していることを示す。
【0068】
本明細書で使用される「増加している」又は「増加する」又は「増加した」はまた、例えば、参照細菌と比較してpntA及びpntB遺伝子の発現増加を有する細菌において、pntA及びpntB遺伝子の発現レベルと比較して、少なくとも1倍、例えば1、2、3、4、5、6、7、8、9、10、15、20、30、40、50、60、70、80、90、100、200、500、1000倍又はそれ以上の増加を意味する。
【0069】
本明細書で使用される場合、「減少している」又は「減少する」又は「減少した」は、例えば、参照細菌と比較して、細菌におけるpntA及びpntB遺伝子の発現レベルの減少と比較して、少なくとも5%、例えば5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、99又は100%減少していることを示す。
【0070】
本明細書で使用される場合、「減少している」又は「減少する」又は「減少した」はまた、例えば、参照細菌と比較して、細菌におけるpntA及びpntB遺伝子の発現レベルと比較して、少なくとも1倍、例えば1、2、3、4、5、6、7、8、9、10、15、20、30、40、50、60、70、80、90、100、200、500、1000倍又はそれ以上の減少を意味する。
【0071】
「減少した」又は「縮小した」も、活性、発現等の検出可能なレベルが存在しない、例えばpntA及びpntB遺伝子の発現の検出可能なレベルが存在しない又はPntA及びPntBタンパク質の検出可能な活性が存在しないように排除されていることを意味する。
【0072】
本明細書で使用される「活性」は、遺伝子の活性、例えば遺伝子の転写のレベルを示す。「活性」はまた、mRNAの活性、例えばmRNAの翻訳のレベルを示す。「活性」はまた、タンパク質、例えばPntA及びPntBの活性を示す。
【0073】
「活性の増大」は、活性の速度及び/又はレベルの増大を含む。
【0074】
本明細書で使用される「pntA及びpntBの発現」におけるような「発現」は、pntA及びpntB遺伝子のタンパク質産物の発現を示す。本明細書で使用される場合、「pntA及びpntBの発現」におけるような「発現」はまた、pntA及び/又はpntB遺伝子に対応するmRNA転写産物の検出可能なレベルの発現を示す。
【0075】
発現レベルに関連して「変化している」は、目的とする遺伝子、mRNA又はタンパク質、例えばpntA及び/又はpntB遺伝子の発現が減少していることを意味する。
【0076】
本明細書で使用される「発現されない」は、目的とする遺伝子又はmRNA、例えばpntA及び/又はpntB遺伝子の産物の検出可能なレベルが存在しないことを意味する。
【0077】
本明細書で使用される「排除する」は、検出不能なレベルへの減少を意味する。
【0078】
本明細書で使用される「フルフラールの耐性」は、フルフラールの存在下で、例えば0.1g/l又はそれ以上(例えば0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2.0、2.5、3.0g/l又はそれ以上)の濃度のフルフラールの存在下で増殖するか又はエタノールを生産する、エタノール生産性細菌の能力を意味する。フルフラールの耐性はまた、野生型細菌又は親細菌による増殖又はエタノール生産のレベルと比較して増大しているレベルで、フルフラールの存在下で増殖するか又はエタノールを生産するエタノール生産性細胞の能力を意味する。
【0079】
本明細書で使用されるフルフラールの存在に適用されるときの「の存在下で」は、少なくとも0.1g/l又はそれ以上(例えば0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2.0、2.5、3.0g/l又はそれ以上)のフルフラールの存在下に細菌を保持することを意味する。
【0080】
本明細書で使用されるフルフラールの不在に適用されるときの「の不在下で」は、検出可能でないレベルを含む、0.1g/l又はそれ以下のフルフラールを含有する培地中に細菌を保持することを意味する。
【0081】
本明細書で使用される「増殖」は、経時的な細胞の数又は質量の、本明細書で定義される増大を意味する。
【0082】
本明細書で使用される「ヘミセルロース加水分解物」は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス又はセルロースバイオマスに由来する加水分解物を含むが、これに限定されない。
【0083】
本明細書で使用される「に由来する」は、それを起源とすることを意味する。
【0084】
「グラム陰性細菌細胞」という用語は、当該技術分野で広く認められているこの用語の定義を包含することが意図されている。例示的なグラム陰性細菌は、アシネトバクター属、グルコノバクター属、ザイモモナス属、エシェリキア属、ジオバクター属、シュワネラ属、サルモネラ属、エンテロバクター属及びクレブシエラ属を含む。
【0085】
「グラム陽性細菌」という用語は、当該技術分野で広く認められているこの用語の定義を包含することが意図されている。例示的なグラム陽性細菌は、バチルス属、クロストリジウム属、コリネバクテリウム属、乳酸杆菌属、ラクトコッカス属、オエノコッカス属、連鎖球菌属及びユーバクテリウム属を含む。
【0086】
「アミノ酸」という用語は、タンパク質において通常生じる20のα−アミノ酸を包含することが意図されている。塩基性荷電アミノ酸は、アルギニン、アスパラギン、グルタミン、ヒスチジン及びリシンを含む。中性荷電アミノ酸は、アラニン、システイン、グリシン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、プロリン、セリン、トレオニン、トリプトファン、チロシン及びバリンを含む。酸性アミノ酸は、アスパラギン酸及びグルタミン酸を含む。
【0087】
本明細書で使用される「選択すること」は、同定された細菌がフルフラールの存在下でエタノールを生産することを確認する工程を示す。
【0088】
本明細書で使用される「同定すること」は、細菌を評価し、その細菌がフルフラールの存在下でエタノールを生産することを確認する工程を示す。
【0089】
本明細書で使用される「フルフラールの濃度を増大すること」は、0〜5g/lの増加、例えば1μg/lの増加、1mg/lの増加又は1g/lの増加を意味する。
【0090】
本開示において、「含む(comprises)」、「含んでいる(comprising)」、「含有する(containing)」及び「有する(having)」等は、米国特許法においてそれらに帰せられている制約のない意味を有し、「包含する(includes)」、「包含している(including)」等を意味する。
【0091】
本明細書で使用される「トランスヒドロゲナーゼ」という用語は、ニコチンアミドアデニンジヌクレオチド補因子NAD(H)とNADP(H)の間の還元性等価物の相互変換を触媒する酵素を示す。
【0092】
本明細書で使用される「トランスヒドロゲナーゼ遺伝子」という用語は、その産物がトランスヒドロゲナーゼである遺伝子を示す。
【0093】
本明細書で使用される「5−ヒドロキシメチルフルフラール」又は「5−HMF」という用語は、構造:
【0094】
【化1】

【0095】
を有する糖類の脱水に由来する有機化合物を意味することが意図されている。
【0096】
II.細菌
本発明は、エタノールの形成のために糖類を分解するのに適した細菌に関する。この細菌は、特にフルフラール及び/又は5−HMFを含む培地において、改善されたエタノール生産能力を有する。改善されたエタノール生産の能力は、糖消化及び発酵の間のフルフラール及び/又は5−HMFに対する細胞の耐性を増大するトランスヒドロゲナーゼ遺伝子の選択された増大した発現に関連する。
【0097】
従って、本発明は、参照細菌と比較して少なくとも1つのトランスヒドロゲナーゼ遺伝子の増大した発現を有するエタノール生産性細菌を提供する。本発明の1つの態様では、細菌は単離された細菌である。本発明の別の態様では、細菌は組換え細菌である。本発明のさらに別の態様では、トランスヒドロゲナーゼ遺伝子はpntA及びpntB遺伝子を含む。
【0098】
本発明はまた、pntA及びpntB遺伝子の増大した発現を有する単離された細菌又は組換え細菌を提供する。本発明の1つの態様では、本発明の単離された細菌は野生型細菌を含む。
【0099】
本発明の細菌はそれらの増大した増殖を特徴とし得る。本発明の細菌は、さらに、増大した濃度のフルフラール及び/又は5−HMF中で増殖するそれらの能力を特徴とする。従って、本発明の他の態様では、単離された細菌又は組換え細菌は、参照細菌と比較して増大した増殖、又は参照細菌と比較して増大したフルフラール及び/若しくは5−HMF耐性、又は参照細菌と比較してフルフラール及び/若しくは5−HMFの存在下での増大した増殖、又はフルフラール及び/若しくは5−HMFの存在下での、例えば約0.025%フルフラール〜約0.15%フルフラールのフルフラール濃度、若しくは約0.025%5−HMF〜約0.15%5−HMFの5−HMF濃度での増大した増殖を有する。
【0100】
本発明の細菌はまた、糖源から主発酵生成物としてエタノールを生産するそれらの能力を特徴とし得る。フルフラール及び5−HMFは、通常、セルロース消化及びエタノールへの発酵の間エタノール生産性細菌の増殖を阻害するが、本発明の細菌は、高濃度のフルフラール及び/又は5−HMF中でエタノールを生産することができる。従って、本発明の他の態様は、エタノール生産性である、又は参照細菌と比較して増大したエタノール生産を示す、又は参照細菌と比較してフルフラール及び/若しくは5−HMFの存在下で増大したエタノール生産を示す、又は嫌気若しくは微好気条件下で主発酵生成物としてエタノールを生産することができる、又は参照細菌と比較してフルフラール及び/若しくは5−HMFの存在下で増大した増殖及び増大したエタノール生産を示す細菌を含む。
【0101】
本発明のさらなる態様は、参照細菌と比較して加水分解物の存在下で増大した増殖を有する細菌を含む。本発明は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス又はセルロースバイオマスに由来する加水分解物を含むがこれに限定されない様々な加水分解物を提供する。
【0102】
参照細菌と。例えば、プロモータを、異なるプロモータによるプロモータの置換、あるいは、例えば核酸を挿入する、置換する、重複させる若しくは除去することによる又はプロモータにおいて調節エレメント若しくはモチーフを挿入する、置換する、重複させる若しくは除去することによるプロモータの修飾を含むがこれらに限定されない、当該技術分野で一般に認められている方法によって変化させる。従って、本発明の1つの態様では、本発明のpntA及びpntB遺伝子などのトランスヒドロゲナーゼ遺伝子の発現を、参照細菌と比較して遺伝子発現を調節するプロモータを修飾する又は付加することを含むがこれらに限定されない、当該技術分野で公知の方法によって増大させる。
【0103】
本発明は、pntA及びpntB遺伝子を参照細菌と比較して異なる調節タンパク質の制御下に置くか又は付加的な調節タンパク質の制御下に置くことを含むがこれに限定されない、当該技術分野で公知の方法によって、pntA及びpntB遺伝子の調節を変化させる方法を提供する。一実施形態では、調節タンパク質は抑制因子である。選択的な実施形態では、調節タンパク質は誘導因子である。
【0104】
本発明の細菌は、非組換え又は組換えでよい。本発明の細菌は、グラム陰性細菌及びグラム陽性細菌から成る群より選択され、グラム陰性細菌は、アシネトバクター属、グルコノバクター属、ザイモモナス属、エシェリキア属、ジオバクター属、シュワネラ属、サルモネラ属、エンテロバクター属及びクレブシエラ属から成る群より選択され、そしてグラム陽性細菌は、バチルス属、クロストリジウム属、コリネバクテリウム属、乳酸杆菌属、ラクトコッカス属、オエノコッカス属、連鎖球菌属及びユーバクテリウム属から成る群より選択される。一態様では、本発明の細菌は大腸菌であり、別の態様では、本発明の細菌はクレブシエラ・オキシトカである。
【0105】
本発明の細菌のpntA及びpntB遺伝子などの増大したトランスヒドロゲナーゼ遺伝子に関連するタンパク質活性を増大又は変化できることは当業者に理解される。例えば、遺伝子産物中のアミノ酸を、天然に生じる遺伝子産物と比較して遺伝子産物の機能に実質的な影響を及ぼさずにある程度まで置換するか、付加するか又は欠失させることができる。従って、本発明はまた、PntA及びPntBタンパク質の活性が参照細菌と比較して増大している又は変化している、単離された細菌又は組換え細菌を提供する。一実施形態では、そのような細菌は、参照細菌と比較して増大したフルフラール及び/又は5−HMF耐性を有する。
【0106】
本発明は、上記で論じたように、例えばpntA及びpntB遺伝子などのトランスヒドロゲナーゼ遺伝子の増大した又は変化した発現を有して、増大した濃度のフルフラール中で増殖し続けることができる細菌を提供する。従って、本発明はさらに、参照細菌と比較してpntA及びpntB遺伝子の発現が増大又は変化しており、且つ参照細菌と比較して増大したフルフラール及び/又は5−HMF耐性を有する、単離された細菌又は組換え細菌を提供する。発現は、pntA又はpntB遺伝子の修飾(例えば、遺伝子をコードする配列において核酸又はアミノ酸を挿入するか、置換するか又は除去することによる)を含むがこれに限定されない、当該技術分野で公知の方法によって増大させるか又は変化させる。
【0107】
本発明はまた、参照細菌と比較してpntA及びpntB遺伝子の発現又はPntA及びPntBポリペプチドの活性が増大しており、参照細菌と比較してフルフラール及び/又は5−HMF耐性が高く、エタノールを生産することができ、並びに(a)細菌の候補株をフルフラール及び/又は5−HMFの存在下で増殖させること;及び(b)フルフラール及び/又は5−HMFの存在下でエタノールを生産する細菌を選択することを含む工程によって調製される、単離された細菌又は組換え細菌を提供する。
【0108】
本発明はさらに、フルフラール及び/又は5−HMFの存在下でのエタノール生産のために糖類を発酵させるのに適した微生物を提供する。従って、本発明は、微生物にとって内因性であるか又は、例えばエピソームに保持されたプラスミド若しくはゲノムに安定に組み込まれたプラスミド(若しくはそのフラグメント)として微生物に組換え導入されるトランスヒドロゲナーゼ遺伝子の増大した発現を有する微生物を提供する。
【0109】
III.製造方法
本発明はさらに、フルフラール及び/又は5−HMFの存在下での、例えば増大したフルフラール及び/又は5−HMFの存在下でのエタノール生産のために糖類を発酵させるのに適した細菌を生産する方法に関する。従って、本発明は、参照細菌と比較してpntA及びpntB遺伝子の発現又はPntABポリペプチドの活性が増大しており、参照細菌と比較してフルフラール及び/又は5−HMF耐性が増大しており、細菌はエタノールを生産することができ、細菌はフルフラール及び/又は5−HMFの存在下で細菌の候補株を増殖させることと、フルフラール及び/又は5−HMFの存在下でエタノールを生産する細菌を選択することとを含む工程によって調製される、単離された細菌又は組換え細菌を提供する。
【0110】
組換えエタノール生産性微生物を製造する方法は、分子生物学の技術分野において公知である。適切な材料及び方法並びに組換え微生物は、例えば、参照により本明細書に組み込まれる米国特許第7,026,152号明細書、同第6,849,434号明細書、同第6,333,181号明細書、同第5,821,093号明細書、同第5,482,846号明細書、同第5,424,202号明細書、同第5,028,539号明細書、同第5,000,000号明細書、同第5,487,989号明細書、同第5,554,520号明細書及び同第5,162,516号明細書並びに国際公開第WO2003/025117号に記載されており、本発明を実施する際に使用できる。本明細書で述べられている本発明の細菌は、当業者によって日常的に実施され、本願で示す実施例において明らかにされる方法によって製造できる。
【0111】
IV.エタノールを生産するための方法
本発明の細菌は、エタノールの生産のために糖類を分解するのに適している。従って、本発明は、上述した本発明の単離された細菌又は組換え細菌と供給源を接触させ、それによって供給源からエタノールを生産することを含む、供給源からエタノールを生産する方法を提供する。本発明の特定の実施形態では、供給源は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス若しくはオリゴ糖源、又はそれらの何れかの組合せから成る群より選択できる。
【0112】
本発明の細菌は、増大した濃度のフルフラール及び/又は5−HMFの存在下で糖類からエタノールを生産するために使用しできる。従って、本発明は、フルフラール及び/又は5−HMFの存在下で、供給源を上述した本発明の単離された細菌又は組換え細菌と接触させ、それによって供給源からエタノールを生産することを含む、供給源からエタノールを生産する方法を提供する。本発明の特定の実施形態では、供給源は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス若しくはオリゴ糖源、又はそれらの何れかの組合せから成る群より選択できる。
【0113】
本発明の微生物は、嫌気条件下でのエタノール生産を特徴とする。野生型大腸菌は、嫌気的増殖の間にエタノールと酢酸塩を1:1の割合で生産する。増殖の定常期の間に、野生型大腸菌は主要生成物として乳酸塩を生産し、全発酵生成物中のエタノールの割合は約20%である。これらのすべての発酵における生成物は様々な酸を含み、それ故混合酸発酵という用語を導く。
【0114】
一般的には、産生宿主細胞株の最良増殖速度及び培養物によって生成される酵素のための触媒条件を促進するための最適pH及び温度を提供する発酵条件が選択される(Doran et al., (1993) Biotechnol. Progress. 9:533-538)。様々な例示的発酵条件が米国特許第5,487,989号明細書及び同第5,554,520号明細書に開示されている。非限定的な例として、約25〜約40℃の範囲の温度及び約4.5〜8.0の範囲のpHを含む条件を選択できる。例えば、この参照により明確に本明細書に組み込まれる、米国特許第5,424,202号明細書及び同第5,916,787号明細書を参照されたい。
【0115】
本発明は、トランスヒドロゲナーゼの増大した発現を有する単離された細菌若しくは組換え細菌、又は参照細菌と比較してpntA及びpntB遺伝子の増大した発現を有する単離された細菌若しくは組換え細菌を提供する。この細菌は、エタノールを生産するため、特に、例えばバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖などの供給源からエタノールを生産するために使用できる。従って、本発明は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源を本発明の細菌と接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含む、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産するための方法を提供する。そのような生産は、フルフラール及び/又は5−HMFの存在下又は不在下で起こり得る。
【0116】
本発明の方法に従って、本明細書で述べられている細菌は、本発明の細菌でセルロースバイオマス又はオリゴ糖源を単糖に分解又は解重合し、それによってオリゴ糖源などのバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマスからエタノールを生産する。その後、細菌は、トランスヒドロゲナーゼ遺伝子の増大した発現によって又は、特にそれらが担持するpntA及びpntB遺伝子の増大した発現によって、より単純な糖類を発酵によってエタノールへと異化する。同時に起こる、複合糖のより小さな糖残基への解重合とそれに続く発酵のこのプロセスは、同時糖化発酵(SSF)と称される。
【0117】
現在、リグノセルロースなどの複合糖の変換は非常に複雑な多工程方法である。例えば、リグノセルロースを、最初に酸加水分解を用いて分解又は解重合しなければならない。この後に、固体から液体を分離し、その後これらの生成物を洗浄して、無毒化し、さらに解重合できるセルロースを生じさせて、最後に、適切なエタノール生産性宿主細胞によって発酵させる工程が続く。これに対し、トウモロコシの発酵ははるかに簡単であり、基本的に一工程方法で、エタノール生産性宿主による即時生物変換反応のためにアミラーゼを使用してトウモロコシデンプンを分解することができる。
【0118】
本発明の細菌及び方法が、リグノセルロースを発酵させるためのより効率的な工程の使用を提供することは当業者に認識される。例えば、本発明の方法は、酸加水分解を完全に回避する方法を包含することが意図されている。さらに、本発明の微生物は、好都合にも増大した濃度の毒素フルフラールの存在下で糖類を発酵させることができる。
【0119】
本発明の1つの利点は、これまで十分に活用されていなかったサッカリド源を使用できることである。その結果として、多くの複合サッカリド基質を、解重合並びに本発明の組換え細菌及び方法を使用したその後の発酵のための出発源として使用することができる。理想的には、再生可能資源をSSF工程において使用できる。オフィスから出る混合古紙は好ましい基質であり(Brooks et al.,(1995)Biotechnol.Progress.11:619-625;Ingram et al.,(1995)米国特許第5,424,202号明細書)、酸で前処理したバガス(Doran et al.,(1994)Biotech.Bioeng.44:240-247)又は高度に精製された結晶セルロース(Doran et al.,(1993)Biotechnol.Progress.9:533-538)よりもはるかに容易に消化される。エンドグルカナーゼ及びエキソグルカナーゼの両方のグルカナーゼがセルロース結合ドメインを含み、これらの酵素は、遠心分離を用いて消化されなかったセルロース残基を採取することにより、その後の発酵のために容易に再生利用できる(Brooks et al.,(1995)Biotechnol.Progress.11:619-625)。より高いリグニン含量を有する基質に関しては再生利用工程の数が限られるかもしれないが、そのようなアプローチは精製セルロースに関して良好に機能する。本発明の範囲内である他の基質源は、あらゆる種類の加工又は非加工植物材料、例えば、刈り取った芝、トウモロコシの皮、トウモロコシの穂軸、葉柄、葉、繊維、パルプ、麻、おがくず、新聞紙等を含む。
【0120】
本発明はまた、上述した本発明の単離された細菌又は組換え細菌を含むキットを提供する。このキットは、随意に例えば本明細書で述べる方法及び工程に従ってエタノールを生産するための指示書のような、使用のための指示書を提供する。そのような指示書は、随意に増大した濃度のフルフラール中でエタノールを生産することを記述してもよい。一実施形態では、キットは糖源を含む。
【実施例】
【0121】
本発明を以下の実施例によってさらに説明するが、それらは限定と解釈されるべきではない。実施例全体を通じて、特に明記されない限り以下の材料及び方法を使用する。
【0122】
材料及び方法
LY168株は、ヘミセルロース加水分解物における糖類の発酵に関してこれまでに記述されている。LY180(NRRL B−50239)を生じさせる基質範囲を改善するためにいくつかの変更を加えた(ラクトース利用の回復、エンドグルカナーゼの組込み及びセロビオース利用の組込み)。これらの菌株についての関連する特徴を以下の表1に示す。表1に示す組込みのために使用した線状フラグメントはGenBankに寄託されている。
【0123】
【表1】
【0124】
LY180株をLBグルコースアンピシリンプレートで一晩増殖させ、各々を使用してAM1キシロースの試験管に3〜4ODまで接種した。この培養物を直ちに使用して、4mLのAM1 5%キシロース、アンピシリン及び表示の濃度のフルフラールを含む13の100mmの蓋付き試験管に、0.05の初期ODまで接種した。培養物を37℃の水浴で増殖させ、24及び48時間後にOD 500nmでの読み取りを行った。
【実施例1】
【0125】
sthAは、主としてNADPH酸化の方向における機能を促進する動力学的特性を備えた細胞質トランスヒドロゲナーゼである(52)。大腸菌の細胞質トランスヒドロゲナーゼSthA遺伝子をpTrc99aにクローニングし、配列決定によって確認した。LY180株を、図1A〜1Bに示すようにこれらのプラスミドで改変した。クローニングした遺伝子の機能性をインビトロアッセイによって確認した。AM1、アンピシリン及び5%キシロース中で改変したLY180株の増殖を24時間後及び48時間後に比較した。光学密度を漸増フルフラール濃度に対してプロットした。空のベクタを対照として使用した。接種の前にインデューサを添加した。
【0126】
0.1mMのIPTGでの誘導すると、活性が約1.0nmol/分/mgタンパク質から18nmol/分/mgタンパク質に増大することが認められた。プラスミドからのsthA遺伝子の発現は、IPTGでの誘導の有無に関わらずフルフラール耐性を変化させなかった(図1)。図1A〜1Bの結果は、24時間後(図1A)及び48時間後(図2B)に、それぞれ、すべてのLY180株がsthA遺伝子の発現レベルに関わりなく漸増フルフラール濃度で実質的に同様の増殖の減少を示したことを明らかにする。sthA遺伝子の過剰発現はフルフラール耐性に影響を及ぼさないと見られた。
【実施例2】
【0127】
PntABは、発酵増殖の間機能しないと考えられているプロトン転位トランスヒドロゲナーゼであるが、潜在的にNADPHのプールを増大させることができる(52)。実施例2及び3では、大腸菌の細胞質トランスヒドロゲナーゼPntABをpTrc99aにクローニングし、配列決定によって確認した。天然のリボソーム結合部位及びrho依存性ターミネータを有するPntABを、HindIII消化可能末端を有するプライマを使用して大腸菌株LY180ゲノムDNAからポリメラーゼ連鎖反応(PCR)で増幅した。PCR産物とpTrc99aをHindIII消化し、Qiagen Qiaprep Spin Miniprep Kitを用いて精製して、T4 Quick DNAリガーゼを使用して連結した。生じたベクタをTOP10F’に形質転換した後、プラスミドを抽出し、LY180を形質転換するために使用した。方向をPCR分析によって確認した。LY180株を、図2〜3に示すようにこれらのプラスミドで改変した。空のベクタを対照として使用した。接種の前にインデューサを添加した。pntA及びpntB遺伝子の発現増大を有するプラスミドを担持するLY180株の増殖を比較した(図2A〜2B)。24及び48時間後の光学密度を漸増フルフラール濃度に対してプロットした。24時間後、0.05%のフルフラール濃度で、pTrc99a−pntAB正プラスミドを担持するLY180株は、試験したその他の菌株のいずれについても2倍以上の光学細胞密度測定値を示した(図2A)。48時間後、pTrc99a−pntAB正プラスミドを担持するLY180株は、0.10%のフルフラール濃度で持続的な細胞増殖を示したが、その他のすべての菌株は実質的に増殖を停止していた(図2B)。これらの結果は、選択されたpntA及びpntB過剰発現がフルフラール耐性を増大させることを指示する。
【実施例3】
【0128】
様々なレベルのトランスヒドロゲナーゼsthA及びpntAB遺伝子発現増大を有するプラスミドを担持するLY180株の増殖の結果を、それぞれ、24時間後と48時間後に0.05%及び0.10%フルフラール濃度で比較した(図3A〜3B)。24時間後、0.05%のフルフラール濃度で、pTrc99a−pntAB正プラスミドを担持するLY180株は、試験したその他の菌株のいずれについても約2倍の光学細胞密度を示した(図3A)。48時間後、0.10%のフルフラール濃度で、pTrc99a−pntAB正プラスミドを担持するLY180株は、試験したその他の菌株のいずれについても約7倍の光学細胞密度を示し、試験したその他の菌株は、そのようなフルフラール濃度で、あったとしてもごくわずかな増殖を示した。この場合も、これらの結果は、選択されたpntA及びpntB過剰発現がフルフラール耐性を増大させることを明らかにしている。
【0129】
非誘導プラスミドからのpntABの漏洩発現は、1g/l フルフラールの存在下で部分的に増殖を回復した(図3)。この酵素のより高いレベルを発現させるためにIPTGを添加すると、フルフラールに対する耐性を排除し、フルフラールの不在下での細胞の増殖も阻害した。これらの結果に基づき、フルフラールは、生合成のために必要なNADPHの供給を枯渇させることによって増殖を阻害する。硫酸塩同化のためにシステイン当たり4当量という大量のNADPHを必要とし、発酵の間のキシロースからのNADPH生産のための経路が限られていることから、含硫アミノ酸の生産はNADPHに関するフルフラールレダクターゼからの競合を最も受けやすかった。
【実施例4】
【0130】
菌株、培地及び増殖条件
エタノール生産性菌株を、固体培地については20g/lキシロースを添加し、発酵実験については50g/lキシロース又はそれ以上を添加したAM1無機塩培地に保持した(22)。大腸菌株LY180(48、60)はKO11の誘導体であり、この試験のための出発点として使用した。大腸菌W(ATCC9637)は、当初大腸菌Bの誘導体であると報告された、KO11株の親であることに留意されたい(29)。
【0131】
発酵は、2N KOHの自動添加を用いて先に述べられているように(100g/lキシロース、37℃、150rpm、pH6.5)実施した(23)。
【0132】
先に記述されているように、総容量4mLのAM1及び50g/lのろ過滅菌したキシロースを含むスタンディングチューブ中で増殖を測定することによってフルフラール耐性を試験した(48)。チューブを37℃で培養し、24時間後と48時間後に測定した。報告されている数値は4回の測定の平均である。
【0133】
菌株の構築
大腸菌トランスヒドロゲナーゼ遺伝子を、隣接するHindIII部位を提供するプライマによりBioRad iCycler(Hercules,CA)を用いてLY180株ゲノムDNAから増幅した(リボソーム結合部位、コード領域、及び200bpのターミネータ領域)(25)。HindIIIで消化した後、生成物をHindIII消化したpTrc99a(ベクタ)に連結し、大腸菌TOP10F’(Carlsbad,CA)に形質転換した。QiaPrep Spin Mini Prep Kit(Valencia,CA)を用いてプラスミドを精製した。制限酵素での消化及びポリメラーゼ連鎖反応によって遺伝子の配向性を確認した(図10の表1)。
【0134】
マイクロアレイ分析
培養物を小さな発酵槽で670mg dcw/lの密度に増殖させた。初期試料を取り出し、対照として使用した。50g/l保存水溶液から直ちにフルフラールを添加して(最終濃度0.5g/l)、2回目の試料採取の前に15分間培養を続けた。試料をエタノール−ドライアイス浴中で急速に冷却し、4℃で遠心分離によって採取し、Qiagen RNA Laterに再懸濁して、−80℃で保存した。Qiagen RNeasy Mini Kitを用いてRNAを抽出し、DNase Iで処理して、フェノール/クロロホルム抽出及びエタノール沈殿によって精製した。大腸菌K12のために設計されたプローブを使用したマイクロアレイ比較のために、RNAをNimbleGen(Madison,WI)に送付した。各々の試料は、4つの発酵槽からのプールされた材料から成っていた。完全な実験を2回実施して、データを平均した(8つの発酵槽)。ArrayStar(DNA Star,Madison,WI)及びSimPhenyソフトウエア(Genomatica Inc.,San Diego,CA)を使用してデータを解析した。発現比を2つのプールされたデータセットの平均として提示する。対照として水だけを添加した実験では、54の遺伝子(>トランスクリプトームの2%)が水の添加後に2倍より大きく変化することが認められた。これらのうち8つだけがフルフラール添加によって影響を受け、フルフラール送達手順の転写への影響がフルフラールの作用に比べて無視し得るものであることを指示した。
【0135】
ネットワーク成分解析
ネットワーク成分解析(Network Component Analysis;NCA)は、発現比及び公知の調節関係から転写因子活性比を計算するものであり、これを先に記述されているように実施した(45、46、56)。関連性ファイルをRegulon DB及びEcoCyc(4、14)に従って更新した。「緊縮調節因子」のレギュロンを、MOPSグルコース中のBW25113の中間対数増殖の間のセリンヒドロキサメート処理によるセリン飢餓に対する5分間応答の分析により、先に記述されているように定義した(12)。有意に変化した調節活性を有する調節因子を、ヌル分布との比較によって及び0.05のP値カットオフを使用して同定した。
【0136】
フルフラールのトランスクリプトーム効果
活発に増殖している細胞において0.5g/lフルフラールの添加前と添加の15分後にメッセージレベルを比較した。水対照も比較のために含めた。小さな群の遺伝子の発現がフルフラールに応答して5倍より大きく変化していることが認められた(図12の表3)。よりストリンジェントでない測定基準(2倍又はそれ以上)を使用すると、約400の遺伝子(トランスクリプトームの10%)についての発現レベルが、水対照又はフルフラール添加前の培養物と比較して、フルフラール添加に応答して変化した。これらの変化した遺伝子の分布は機能群の間で大きく異なっており、フルフラールの作用機構に関する有用な洞察を提供した(図11の表2)。大部分の機能群では、遺伝子成員の10%未満の発現レベルが2倍又はそれ以上変化した。この低頻度の変化を有する群は、補因子、炭素化合物、調節、高分子合成(細胞構造、DNA、脂質、転写及び翻訳)、及びその他(ファージ、推定上/IS、調節及び未分類/不明)を含んだ。4つの群(細胞プロセス、中枢代謝、エネルギー及び輸送)では成員遺伝子の10%〜20%についての発現レベルが変化した。中枢代謝、エネルギー及び輸送に関連する、影響を受けた遺伝子の大部分は、フルフラール添加時に発現が増大した。これらの変化は、利用可能であり得る付加的な化合物を捕捉し、代謝する、及びエネルギー産生のための炭素流を増大させる機会を提供する。細胞プロセスに関係する変化した遺伝子の多くは運動性及び化学走性に関与するが、LY180株は非運動性である(データは示していない)。
【0137】
以前の報告(48)と一致して、フルフラールのNADPH特異的還元を触媒することができる2つのオキシドレダクターゼ、YqhD及びDkgAは、先に報告されているように、フルフラールの添加によって親において5倍以上上方調節された(mRNA)。これら2つの遺伝子はフルフラール耐性変異株(EMFR9)では沈黙化し、フルフラール耐性の増大を生じさせた。
【0138】
2つの機能群における成員遺伝子の20%以上についての発現レベルが、フルフラール、アミノ酸及びヌクレオチドの添加によって変化した。これらの群では、変化した遺伝子の2/3以上が、フルフラールの添加によって2倍又はそれ以上低減した。プリン、ピリミジン及びあらゆるファミリーのアミノ酸の生合成に影響を及ぼす個々の遺伝子についての発現レベルは、フルフラールの添加によって2倍又はそれ以上低減した。ヌクレオチド代謝における1個の遺伝子及びアミノ酸代謝に関与する8個の遺伝子だけが、少なくとも2倍のフルフラール依存性発現増大を示した。合わせて考慮すると、これらの変化は、フルフラールの添加時に観察される生合成及び増殖の全般的な減少と一致する。
【0139】
調節活性へのフルフラールの効果
フルフラールに対する細胞応答の全体的見解を提供するためにネットワーク成分解析(NCA)を用いた。この解析は、トランスクリプトームデータから、摂動活性を有する調節因子を同定するために公知の調節ネットワーク構造を利用する(46、56)。この解析に含まれた60の調節因子のうちで、22がフルフラール負荷後に有意に変化した発現を有すると同定され、有意性はランダムネットワークに対して評価した(図6A)。システイン及びメチオニン生合成の調節因子(CysB及びMetJ)並びにアミノ酸のリプレッサ(ArgR)及びヌクレオチド生合成のリプレッサ(PurR)は、フルフラール添加によって有意に影響された。緊縮応答(アミノ酸及び炭素飢餓の間、資源を増殖から転用すること)の集合的なインジケータである緊縮調節因子も、停滞した生合成及び多くの中間体の過剰と一致する活性化を示す。合わせて考慮すると、これらの結果は、多くのアミノ酸及び生合成中間体のプールがフルフラール添加によって変化していたことを示す。システイン及びメチオニン生合成に関与する遺伝子の発現は増大したが、大部分の他の生合成経路の発現減少したという事実は、フルフラール負荷から生じる初期事象としてのシステイン及びメチオニンプールの枯渇と一致する。
【0140】
一般的なストレス応答についてのシグナルとして働くσ因子、RpoSもフルフラールの添加によって影響され、細胞がストレス誘導物質の存在を認識することを示した。大腸菌ゲノムの10%までが何らかの方法でRpoSによって調節されるので(51、59)、特異的阻害応答を決定することは困難である。発現増大を有するRpoS調節性遺伝子の例は、poxB(ピルビン酸塩の酢酸塩とCO2への変換)(64)及びotsA(浸透圧ストレス応答の間のトレハロース産生)(55)を含む。
【0141】
中枢代謝における数多くの遺伝子(aceB、aceE、sdhC、dctA、cyoA、fumA)の発現増大及び嫌気条件下で活性であることが一般に知られている遺伝子、fumBの下方調節によって証明されたように、ArcAはNCAによって有意に下方調節された。フルフラール耐性へのグリセロール添加の影響を検討した。しかし、グリセロール(1.0〜20g/l)の添加はフルフラール耐性に影響を及ぼさなかった(データは含めていない)。fis及びcrpを含むいくつかの他の調節因子は、NCAによって有意に変化することが認められた。
【0142】
ヒスチジンもフルフラールの添加によって制限され得る。His調節因子(ヒスチジニル−tRNA)の制御下の遺伝子(hisA、hisB、hisC、hisD、hisF、hisH及びhisI)は、フルフラールの添加後、2倍未満であるが、一般に増大した(図7)。ヒスチジン生合成の2つの最終工程は、発酵に関連する高いNADH/NAD+比によって減速できる反応である、NAD+のNADHへの還元を含む。
【0143】
硫黄のアミノ酸への同化に関与する遺伝子の発現へのフルフラールの効果
硫黄のシステイン及びメチオニンへの同化に関与する多くの遺伝子(cysC、cysH、cysI、cysM、cysN、cysQ、metA、metB、metC、metL、sbp、tauA、tauB、tauC及びtauD)は2倍又はそれ以上増大した。これらはいくつかの機能群内に散らばっている(図11の表2):アミノ酸、中枢代謝、調節及び輸送。硫黄の同化に関与する多くの付加的な遺伝子も2倍未満上方調節されて、幅広いフルフラール応答を明らかにするために含めている(図8)。硫黄はAM1培地中で硫酸塩として供給され、4分子のNADPHを必要とするエネルギー消費型反応である、組込みのために硫化水素のレベルに還元されねばならない。これらの遺伝子の発現のフルフラール誘導性増大は、アミノ酸、プリン及びピリミジンの生合成に関与する多くの他の遺伝子について認められる減少とは著しい対照を成している(図11の表2)。タウリン輸送遺伝子(tauABCD;硫黄の選択的供給源)、硫酸塩結合性輸送タンパク質(sbp)、及び多くのシステイン生合成遺伝子の転写活性化因子(cbl)の発現は、添加されたフルフラールに応答して5倍を超えて増大した(図12の表3)。合わせて考慮すると、これらの結果は、フルフラールの添加が、この経路における高いNADPH必要量に関連した含硫アミノ酸(システイン及びメチオニン)の細胞内欠乏を生じさせることを明らかにしている。
【0144】
フルフラール耐性へのアミノ酸添加物の影響
20アミノ酸すべてを、AM1無機塩培地におけるLY180の増殖を改善するそれらの能力に関して個別に試験した(図9A)。個々のアミノ酸の細胞含量におおよそ基づいて0.1mMの濃度を選択した(50)。5つのアミノ酸だけが、この低濃度で供給した場合にフルフラール耐性を改善した:システイン>メチオニン>セリン、アルギニン>ヒスチジン。2つの含硫アミノ酸は、フルフラール耐性のために明らかに最も有益であった。5倍高い濃度(0.5mM)で供給した場合は、すべてのアミノ酸がある程度まで有益であった(図9B)。しかし、システインが最も有効であることは変わらず、次いでセリン、メチオニン及びアルギニンであった。0.05mMのシステイン濃度では、LY180は、総細胞硫黄量にほぼ等しい、1g/lフルフラールの存在下で0.8g/lの密度に増殖することが可能であった(図4C)。0.01mMシステインではフルフラール耐性の測定可能な改善は認められなかった。
【0145】
システインの防御濃度(0.05mM)は1g/lフルフラール(10mM)の濃度よりも200倍低かった。無機塩培地中のフルフラールはその特徴的なスペクトルによって容易に定量され(Martinez et al.,2000)、ごくわずかな化学的反応性と一致して、37℃で48時間の培養の間不変のままであった。
【0146】
ヒスチジン生合成に関与する大部分の遺伝子は、2倍未満であるが、フルフラール添加に応答して増大した(図7)。発酵の間のヒスチジンの新規生合成は、嫌気的増殖においては高いNADH/NAD+比によって及び生合成の2つの最終工程ではNAD+のさらなる還元のための必要量によって抑制される。同様に、セリン生合成における最初の関与工程もNAD+の還元を含み、発酵の間妨げられる。セリンを増大させることは、H2Sからの還元硫黄をシステインに組み込む効率を高める。アルギニン生合成に関与する遺伝子(argA、argB、argC、argD、car A、carB及びargG)は、一般にフルフラールの添加後2倍以上減少した。アルギニンデカルボキシラーゼをコードするspeAの発現レベルはフルフラールの添加によって増大した。アルギニンの分解は、生合成のための有用な中間体及び補因子を提供する。
【0147】
フルフラール耐性への選択的硫黄源の影響
フルフラールの添加は増殖を阻害し、硫黄の同化に関与する遺伝子の転写を増大させた。選択的硫黄化合物、タウリンの取込み及び組込みに関与する遺伝子(tauABC及びtauD)は、最大の発現増大を有する10遺伝子に含まれた(図12の表3)。tau遺伝子は、典型的には硫黄飢餓の間だけ発現される(57)。システインはフルフラール阻害を取り除くのに有効であったので、これらの遺伝子の発現増大はフルフラールによる含硫アミノ酸のプールの減少から生じる。フルフラールは、硫酸塩からの還元硫黄(H2S)のアベイラビリティを制限することによって又は還元硫黄のシステインへの組込みを阻害することによって、含硫アミノ酸の生合成を阻害する。
【0148】
1g/lフルフラールを含むAM1培地での増殖の間に、還元の異なるレベルで代謝に入る選択的な硫黄源(L−システイン、D−システイン、タウリン及びチオ硫酸ナトリウム)の作用(図9D)を比較した。システインへの同化の前に硫酸塩を完全に還元するためには4分子のNADPHと2つのレダクターゼ酵素(CysH及びCysIJ)が必要であることに留意しなければならない。L−システイン、D−システイン及びチオ硫酸塩は両方のレダクターゼ酵素を回避し、すべてがフルフラール阻害を取り除くのに有効であった。D−システインは直接組み込まれることができず、最初にH2Sに異化される(49)。チオ硫酸塩はまた、CysMによる組込みのための還元硫黄の供給源としても働く(47)。タウリンは細胞質内で亜硫酸塩に異化され、システインへの同化の前に亜硫酸レダクターゼ(CysIJ及び3分子のNADPH)によって還元されねばならない(54)。システイン及びチオ硫酸塩と異なり、タウリンは1g/lフルフラールによる増殖の阻害を防ぐのに有効ではなかった。
【0149】
選択的硫黄源に関するこれらの結果は、フルフラールが、還元硫黄のシステインへの組込みを阻害することによってではなく、硫酸塩からの還元H2Sの生産を制限することによって増殖を阻害するように働くことを示す。硫黄源としてのタウリンに関しては、フルフラールは、フルフラールレダクターゼYqhD(8μM)よりも高いNADPHに対するKm(80μM)(53)を有する、亜硫酸レダクターゼ(CysIJ)のレベルで作用しなければならない(48)。硫黄源としての硫酸塩に関しては、硫黄同化のより早い工程でのフルフラールのさらなる作用を除外することはできない。
【実施例5】
【0150】
エタノール生産性細菌における5−ヒドロキシメチルフルフラール耐性の増大
ヘミセルロース加水分解物中に存在するフラン阻害剤を耐容する生物触媒の能力は、再生可能な化学物質の生産のために重要である。この実施例は、エタノール生産性大腸菌(E.coli)LY180のフルフラール耐性変異株であるEMFR9が、5−ヒドロキシメチルフルフラール(5−HMF)に対する耐性も獲得していることを明らかにする。フラン耐性は、NADPHに対して低いKmを有する2つのフランレダクターゼ、yqhD及びdkgAのより低い発現から生じる。フラン耐性はまた、NADPH/NADHトランスヒドロゲナーゼ(pntAB)をコードするプラスミドを付加することによっても増大した。
【0151】
材料及び方法
菌株、培地及び増殖条件
この試験で使用した菌株及びプラスミドは以前に記述されている(70及び23)。これらは、LY180(大腸菌のエタノール生産性誘導体)、EMFR9(LY180のフルフラール耐性誘導体)、LY180ΔyqhD、LY180ΔdkgA、LY180ΔyqhDΔdkgA、yqhDを含有するpLOI4301を含む。dkgAを含有するプラスミドpLOI4303(48)、及びpntABを含有するpLOI4316(70)も使用した。培養物を、20g/lキシロース(固体培地)、50g/lキシロース(Bioscreen C増殖アナライザ及び試験管培養物)、又は100g/l(pH制御発酵)を含むAM1最小培地(23)において37℃で増殖させた。
【0152】
HMFに対する耐性を、指示されているようにAM1 4ml及び5−HMFを含む13×100mmの閉管を用いて試験した。適切な場合は、プラスミドの保持のための抗生物質を含ませた。管に、0.05OD550nmの初期密度で接種した。48時間の培養(60rpm)後にSpectronic 20D+分光光度計(Thermo,Waltham,MA)を用いて増殖を測定した。フラン耐性へのpntABの作用を調べるために、各ウエルにつき400μlのAM1(及び5−HMF又はフルフラール)を含むマルチウォールプレートに上記のように接種した。Bioscreen C増殖アナライザ(Oy Growth Curves,Helsinki,Finland)を用いてOD(420〜580nmの帯域幅)を72時間測定した。
【0153】
発酵実験のために、LY180及びEMFR9の種培養物を、AM1培地350mlを含む小さな発酵槽で一晩増殖させた(37℃、200rpm)。2NのKOHの自動添加によって培養液をpH6.5に保持した。中間対数増殖期に達したとき、実験発酵槽に0.1 OD550nmの初期細胞密度(33mg乾燥細胞重量/l)で接種した。細胞質量(OD550nm)及びフランレベルを先に述べられているように(20)12時間の間隔で観測した。
【0154】
インビボでのフランの還元を、pH制御発酵槽を用いて測定した。培養物が約1OD550nmに達したとき、10%w/v保存溶液を使用してフランを添加した。細胞質量及び5−HMFを0、15、30及び60分後に測定した。
【0155】
インビトロでのフランの還元
AM1及び0.1mMのIPTGを含む培養試験管(13×100mm)を0.05OD550nmに接種し、37℃で培養した。これらを1〜2OD550nmの密度で採取した。細胞ペレットを100mMのリン酸カリウム緩衝液(pH7.0)で1回洗浄して、10OD550nmの密度で緩衝液に再懸濁した。試料(1ml)を、Lysing Matrix Bを含む2ml試験管に添加し、FastPrep−24(MP Biomedicals,Solon,OH)を用いて破壊した(20秒間)。NADPHのフラン依存性酸化を、DU800分光光度計(Beckman Coulter,Fullerton,CA)を用いて340nmで測定した。反応物(総容量200μl;37℃)は、粗抽出物50μL、0.2mMのNADPH及び20mMの5−HMFを含んでいた。BCAアッセイ(Thermo Scientific,Rockford,IL)を用いてタンパク質を測定した。
【0156】
統計分析
データを平均±SD(n≧3)として提示する。Graphpad Prismソフトウエア(La Jolla,CA)を使用して統計比較(両側スチューデントt検定)を行った。
【0157】
結果
EMFR9株は5−HMFに対する増大した耐性を示す
フルフラール耐性変異株、EMFR9中に存在する突然変異は、5−HMFに対する耐性も増大させた(図14)。1.0g/Lの5−HMFで、EMFR9による増殖及びエタノール生産は、5−HMFの不在下でのLY180(親)のものに等しかった(図14A、14B)。5−HMFは、細胞収率又はエタノール収率に有害な影響を及ぼさずに発酵の最初の24時間にEMFR9によって速やかに代謝された。LY180は1.0g/lの5−HMFによって完全に阻害されたが、5−HMFレベルは培養の間緩やかに減少した(図14A、14B及び14C)。接種しない場合は減少が認められず(データは示していない)、これが代謝作用の結果であることを確認した。
【0158】
EMFR9に関しては、2.5gの5−HMF/lを含めることによってエタノール生産及び増殖は減速したが、それでも進行して96時間後に完了した(図14D、14E及び14F)。このより高いレベルの5−HMFに関する細胞収率及びエタノール収率は、5−HMFなしでのLY180と同等であった。5−HMFのレベルはEMFR9によって急速且つ完全に減少した。LY180に関しては、5−HMFの代謝は遅く且つ不完全であった(図14F)。
【0159】
5−HMF耐性へのYqhD及びDkgAの作用
EMFR9におけるフルフラール耐性は、2つのNADPH依存性オキシドレダクターゼ、YqhD及びDkgAのサイレンシングから生じることが以前に明らかにされた(23)。これらの活性をコードする遺伝子をpCR2.1 TOPOにクローニングし、EMFR9に形質転換して、0.1mMのIPTGで誘導した。細胞を採取し、破壊して、5−HMFレダクターゼ活性に関して試験した(図15A)。プラスミドからの個別のyqhD及びdkgAの発現は、NADPHの5−HMF依存性酸化の速度の5倍増大を生じさせ、YqhD及びDkgAが5−HMFを基質として利用することを確認した。
【0160】
プラスミドからのyqhD及びdkgAの個別発現は、5−HMFに対するEMFR9の耐性を減少した(図15B)。プラスミドの維持のためにカナマイシン(12.5mg/l)を添加すると、すべての株において5−HMF耐性が減少し、この実験では5−HMFのより低い濃度(1.0g/l)の使用を必要とした。プラスミドpCR2.1は、IPTGの不在下でクローニングした遺伝子の発現に関して露出性である(71)。yqhDの非誘導発現でさえも、5−HMFに対するEMFR9の感受性を回復するのに十分であった。5−HMFによる増殖阻害はyqhD誘導によってさらに増大した。dkgAの発現はそれほど有効ではなく、EMFR9において5−HMF感受性を回復するには誘導が必要であった。これら2つのオキシドレダクターゼの間の有効性の差は、DkgAについての23μMと比較して、NADPHに対するYqhDのより低い見かけKm(8μM)と一致する(23)。
【0161】
LY180からのyqhDの欠失は、耐性を2.5g/l 5−HMFに増大した(図15C)。マーカが染色体内に残存する欠失はそれよりも有効でなかったが、yqhDの不活性化はすべての場合に5−HMF耐性のために有益であることを確認した。
【0162】
NADPHのアベイラビリティを高めることは5−HMF耐性を増大させた
NADPHのアベイラビリティを高めるためにプロトン転位トランスヒドロゲナーゼpntAB(68)をLY180において過剰発現させた(図16)。阻害剤の不在下では(図16A)、ベクタを含むLY180(対照)及びLY180(pTrc99a pntAB)の両方が同じ速度で増殖した。IPTG(0.01mM)でのLY180(pTrc99a−pntA)の誘導は、5−HMFの不在下では有害であった。しかし、非誘導LY180(pTrc099a pntAB)は、5−HMF(0.9g/l及び1.8g/l)の存在下でベクタ対照よりも速やかに増殖した(図16B及び16C)。pntABの同様の効果がフルフラールに関して以前に認められている(23)。従って両方のフランによる増殖の阻害はフランの還元から生じると思われ、生合成のために必要なNADPHのプールを枯渇させる。加えて、pntABの過剰発現は、フルフラールの不在下でさえも、72時間後に全体的な増殖の増大を導き、生合成がこれらの条件下でNADPHによって制限され得ることを指示した。
【0163】
硫黄の同化及びシステインの生合成はNADPHの特に高い必要量を有する。システインを添加することは大腸菌LY180においてフルフラール耐性を増大させることが以前に示されているが(70)、5−HMF耐性に関してはそれより有益でないことが認められた(図17)。LY180の増殖は1g/lの5−HMFによって部分的に阻害され、100μMのシステインを添加することによって完全に回復した(図17A)。2.5g/lの5−HMFの存在下での増殖は、100μM又は1000μMシステインによって回復しなかった(図17B)。フルフラールと異なり、システインの添加は5−HMFについてのMICを増大しなかった。
【0164】
参考文献
1. Almeida, J.R.M., T. Modig, A. Petersson, B. Hahn-Hagerdal, G. Liden, and M.F. Gorwa-Grauslund. 2007. Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates by Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 82:340-349.
2. Almeida, J.R.M, A. Roder, T. Modig, B. Laddan, G. Liden, and M.F. Gorwa-Grauslund. 2008. NADH- vs. NADPH-coupled reduction of 5-hydroxymethyl furfural (HMF) and its implications on product distribution in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 78:939-945.
3. Barciszewski, J., G.E. Siboska, B.O. Pedersen, B.F.C. Clark, and S.I.S. Ratten. 1997. A mechanism for the in vivo formation of N-6-furfuryladenine, kinetin, as a secondary oxidative damage product of DNA. FEBS letters. 414:457-460.
4. Boopathy, R., H. Bokang, and L. Daniels. 1993. Biotransformation of furfural and 5-hydroxymethyl furfural by enteric bacteria. J. Indust. Microbiol. 11:147-150.
5. Datsenko, K.A., and B.L. Wanner. 2000. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. PNAS. 97:6640-6645.
6. Gorsich, S.W., B.S. Dien, N.N. Nichols, P.J. Slinger, Z.L. Liu, and C.D. Skory. 2006. Tolerance to furfural-induced stress is associated with pentose phosphate pathway genes ZWF1, GND1, RPE1, and TKL1, in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 71:339-349.
7. Gutierrez, T., L.O. Ingram, and J.F. Preston. 2006. Purification and characterization of a furfural reductase (FFR) from Escherichia coli strain LY01-An enzyme important in the detoxification of furfural during ethanol production. J. Bacteriol. 121:154-164.
8. Habrych, M., S. Rodriguez, and J. Stewart. 2002. Purification and identification of an Escherichia coli Beta-Keto Ester Reductase as 2,5-diketo-D-gluconate reductase YqhE. Biotechnol. Prog. 18:257-261.
9. Hahn-Hagerdal, B., M. Galbe, M.F. Gorwa-Grauslund, G. Liden, and G. Zacchi. 2006. Bio-ethanol - the fuel of tomorrow from the residues of today. Trends Biotechnol. 24:549-556.
10. Horvath, I.S., M.J. Taherzadeh, C. Niklasson, and G. Liden. 2001. Effects of furfural on anaerobic continuous cultivation of Saccharomyces cerevisiae. Biotechnol. Bioeng. 75:540-549.
11. Hristozova, Ts., A. Angelov, B. Tzvetkova, D. Paskaleva, V. Gotcheva, S. Gargova, and K. Pavlova. 2006. Effect of furfural on carbon metabolism key enzymes of lactose-assimilating yeasts. Enzyme Microbiol. Technol. 39:1108-1112.
12. Jantama, K., X. Zhang, J.C. Moore, K.T. Shanmugam, S.A. Svoronos, and L.O. Ingram. 2008. Eliminating side products and increasing succinate yields in engineered strains of Escherichia coli C. Biotech. Bioeng. 30:881-893.
13. Jarboe, L.R., T.B. Grabar, L.P. Yomano, K.T. Shanmugan, and L.O. Ingram. 2007. Development of ethanologenic bacteria. Adv. Biochem. Engin/Biotechnol. 108:237-261.
14. Jeudy, S., V. Monchois, C. Maza, JM Claverie, C. Abergel. 2006. Crystal structure of Escherichia coli DkgA, a broad-specificity aldo-keto reductase. Proteins. 62:302-307.
15. Karp, P.D., I.M. Keseler, A. Shearer, M. Latendresse, M. Krummenacker, S.M. Paley, I. Paulsen, J. Collado-Vides, S. Gama-Castro, M. Peralta-Gil, A. Santos-Zavaleta, M.I. Penaloza-Spinola, C. Bonavides-Martinez, and J. Ingraham. 2007. Multidimensional annotation of the Escherichia coli K12 genome. Nucleic Acids Res. 35:7577-7590.
16. Katahira, S., A. Mizuike, H. Fukuda, and A. Kondo. 2006. Ethanol fermentation from lignocellulosic hydrolysate by a recombinant xylose and cellooligosaccharide-assimilating yeast strain. Appl. Microbiol. Biotechnol. 72:1136-1143.
17. Khan, Q.A., F.A. Shamsi, and S.M. Hadi (1995) Mutagenicity of furfural in plasmid DNA. Cancer Lett. 89:95-99.
18. Ko, J., I. Kim, S. Yoo, B. Min, K. Kim, and C. Park. 2005. Conversion of methylglyoxal to acetol by Escherichia coli aldo-keto reductases. J. Bacteriol. 187:5782-5789.
19. Martin, C., M. Marcet, O. Almazan, and L.J. Jonsson. 2007. Adaptation of a recombinant xylose-utilizing Saccharomyces cerevisiae strain to a sugarcane bagasse hydrolysate with high content of fermentation inhibitors. Bioresour. Technol. 98:1767-1773.
20. Martinez, A., M.E. Rodriguez, S.W. York, J.F. Preston, and L.O. Ingram. 2000. Use of UV absorbance to monitor furans in dilute acid hydrolysates of biomass. Biotechnol Prog. 16:637-641.
21. Martinez, A., M.E. Rodriguez, M.L. Wells, S.W. York, J.F. Preston, and L.O. Ingram. 2001. Detoxification of dilute acid hydrolysates of lignocellulose with lime. Biotechnol. Prog. 17:287-293.
22. Martinez, A., M.E. Rodriguez, S.W. York, J.F. Preston and L. O. Ingram. 2000. Effects of Ca(OH)2 treatments ("overliming") on the composition and toxicity of bagasse hemicellulose hydrolysates. Biotechnol. Bioengin. 69(5): 526-536.
23. Martinez, A., T.B. Grabar, K.T. Shanmugam, L.P. Yomano, S.W. York, and L.O. Ingram. 2007. Low salt medium for lactate and ethanol production by recombinant Escherichia coli B. Biotechnol. Lett. 29:397-404.
24. McCue, L.A., W. Thompson, C.S. Carmack, M.P. Ryan, J.S. Liu, V. Derbyshire, and C.E. Lawrence. 2001. Phylogenetic footprinting of transcription factor binding sites in proteobacterial genomes. Nucleic Acids Res. 29:774-782.
25. Miller, J.H.. 1992. A short course in bacterial genetics. CSHL Press. Plainview, New York.
26. Modig, T., G. Liden, and M.J. Taherzadeh. 2002. Inhibition effects of furfural on alcohol dehydrogenase, aldehyde dehydrogenase, and pyruvate dehydrogenase. Biochem. J. 363:769-776.
27. Nakamura, C.E., and Whited GM. 2003. Metabolic engineering for the microbial production of 1,3-propandiol. Current Opinion in Biotechnology. 14:454-459.
28. Nilsson, A., M.F. Gorwa-Grauslund, B. Hahn-Hagerdal, and G. Liden. 2005. Cofactor dependence in furan reduction by Saccharomyces cerevisiae in fermentation of acid-hydrolyzed lignocellulose. Appl. Environ. Microbiol. 71:7866-7871
29. Ohta, K., D.S. Beall, K.T. Shanmugam, and L.O. Ingram. 1991. Genetic improvement of Escherichia coli for ethanol production: chromosomal integration of Zymomonas mobilis genes encoding pyruvate decarboxylase and alcohol dehydrogenase II. Appl. Environ. Microbiol. 57:893-900.
30. Palmqvist, E., and B. Hahn-Hagerdal. 2000. Fermentation of lignocellulosic hydrolysates. I: inhibition and detoxification. Bioresour. Technol. 74:17-24.
31. Palmqvist, E., and B. Hahn-Hagerdal. 2000. Fermentation of lignocellulosic hydrolysates. II: inhibitors and mechanisms of inhibition. Bioresour. Technol. 74:25-33.
32. Perez, J.M., F.A. Arenas, G.A. Pradenas, J.M. Sandoval, and C.C. Vasquez. 2008. Escherichia coli YqhD exhibits aldehyde reductase activity and protects from the harmful effect of lipid peroxidation-derived aldehydes. J. Biol. Chem. 283:7346-7353.
33. Petersson, A., J.R.M. Almeida, T. Modig, K. Karhumaa, B. Hahn-Hagerdal, M.F. Gorwa-Grauslund, and G. Liden. 2006. A 5-hydroxymethyl furfural reducing enzyme encoded by Saccharomyces cerevisiae ADH6 gene conveys HMF tolerance. YEAST. 23:455-464.
34. Singh, N.P., and A. Khan. 1995. Acetaldehyde: Genotoxicity and cytotoxicity in human lymphocytes. Mutat. Res 337:9-17.
35. Sulzenbacher, G., K. Alvarez, R. Heuvel, C. Versluis, S. Spinelli, V. Campanacci, C. Valencia, C. Cambillau, H. Eklund, and M. Tegoni. 2004. Crystal structure of E. coli alcohol dehydrogenase YqhD: Evidence of a covalently modified NADP coenzyme. J. Mol. Biol. 342:489-502.
36. Talebnia, F., and M.J. Taherzadeh. 2006. In situ detoxification and continuous cultivation of dilute-acid hydrolysate to ethanol by encapsulated S-cerevisiae. J Biotechnol. 125:377-384.
37. Tokiwa, Y., and B.P.Calabia. 2008. Biological production of functional chemicals from renewable resources. Can. J. Chem. 86:548-555.
38. Um, B.H., M.N. Karim, and L.L. Henk 2003. Effect of sulfuric and phosphoric acid pretreatments on enzymatic hydrolysis of corn stover. Appl Biochem Biotechnol. 105-108:115-125.
39. White, D. 2000. The Physiology and Biochemistry of Prokaryotes. 2nd edition. Oxford University Press. New York, NY.
40. Almeida, J.R.M., M. Betilsson, M.F. Gorwa-Grauslund, S. Gorsich, and G. Liden. 2009. Metabolic effects of furaldehydes and impacts on biotechnological processes. Appl. Microbiol. Biotechnol. 82:625-638.
41. Amann, E., B. Ochs, and K.J. Abel. 1988. Tightly regulated tac promoter vectors for the expression of unfused and fused proteins in Escherichia coli. Gene 69:301-315.
42. Gama-Castro, S., J.J. Veronica, M. Peralta-Gil, A. Santos-Zavaleta, M.I. Penaloza-Spindola, B. Contreras-Moreira, J. Segura-Salazar, L. Muniz-Rascado, I. Martinez-Flores, H. Salgado, C. Bonavides-Martinez, C. Abreu-Goodger, C. Rodriguez-Penagos, J. Miranda-Rios, E. Morett, E. Merino, A.M. Huerta and J. Collado-Vides. 2008 RegulonDB (version 6.0): gene regulation model of Escherichia coli K-12 beyond transcription, active (experimental) annotated promoters and Textpresso navigation. Nucl. Acids Res., 36:D120-D124.
43. Gralla, J.D. 2005. Escherichia coli ribosomal RNA transcription: regulatory roles for ppGpp, NTPs, architectural proteins and a polymerase-binding protein. Molecular Microbiology. 55:973-977.
44. Gyaneshwar, P., O. Paliy, J. McAuliffe, D.L. Papham, M.I. Jordan, and S. Kustu. 2005. Sulfur and nitrogen limitation in Escherichia coli K-12: Specific homeostatic responses. J. Bacteriol. 187:1074-1090.
45. Hyduke, D.H., L.R. Jarboe, L.M. Tran, K.J.Y. Chou, and J.C. Liao. 2007. Integrated network analysis identifies nitric oxide response networks and dihydroxyacid dehydratase as a crucial target in Escherichia coli. PNAS. 104:8484-8489.
46. Liao, J.C., R. Boscolo, Y.L. Yang, L.M. Tran, C. Sabatti, and V.P. Roychowdhury. 2003. Network Component Analysis: Reconstruction of regulatory signals in biological systems. PNAS. 100:15522-15527.
47. Maier, T.H. 2003. Semisynthetic production of L-alpha-amino acids by metabolic engineering of the cysteine-biosynthetic pathway. Nature Biotechnol. 4:422-427.
48. Miller, E.M., L.R. Jarboe, L.P. Yomano, S.W. York, K.T. Shanmugam and L.O. Ingram. 2009. Silencing of NADPH-dependent oxidoreductases (yqhD and dkgA) in furfural-resistant ethanologenic Escherichia coli. Appl. Environ. Microbiol. IN PRESS.
49. Nagasawa T., T. Ishi, H. Kumagai, and H. Yamada. 1985. D-cysteine desulfhydrase of Escherichia coli - purification and characterization. Eur. J. Biochem. 153:541-551.
50. Neidhardt, F.C. Escherichia coli and Salmonella Typhimurium. 1987. vol. 1. American Society for Microbiology. Washington, DC.
51. Patten, C.L., M.G. Kirchhof, M.R. Schertzberg, R.A. Morton, and H.E. Schellhorn. 2004. Microarray analysis of RpoS-mediated gene expression in Escherichia coli K-12. Mol. Genet. Genomics. 272(5):580-591.
52. Sauer, U., F. Canonaco, S. Heri, A Perrenoud, E. Fisher. 2004. The soluble and membrane-bound transhydrogenases UdhA and PntAB have divergent functions in NADPH metabolism of Escherichia coli. J. Biol. Chem. 279:6613-6619.
53. Schomburg I., A. Chang, O. Hofmann, C. Ebeling, F. Ehrentreich, D. Schomburg. 2002. BRENDA: a resource for enzyme data and metabolic information. Trends Biochem. Sci. 27(1):54-6.
54. Siegel, L.M., P.S. Davis. 1974. Reduced Nicotinamide Adenine Dinucleotide Phosphate-Sulfite Reductase of Enterobacteria. J. Biol. Chem. 249:1587-1598.
55. Strom, A.R., and I. Kaasen. 1993. Trehalose metabolism in Escherichia coli:stress protection and stress regulation of gene expression. Mol. Microbiol. 8:205-210.
56. Tran, L. M., M. P. Brynildsen, K. C. Kao, J. K. Suen, and J. C. Liao. 2005. gNCA: a framework for determining transcription factor activity based on transcriptome: identifiability and numerical implementation. Metab Eng 7:128-41.
57. van der Ploeg, J.R., M.A. Weiss, E. Saller, H. Nahimoto, N. Saito, M.A. Kertesz, T. Leisinger. 1996. Identification of sulfate starvation-regulated genes in Escherichia coli: a gene cluster involved in the utilization of taurine as a sulfur source. J. Bacteriol. 178:5438-5446.
58. Vemuri, G.N., E. Altman, D.P. Sangurdekar, A.B. Khodursky, M.A. Eiteman. 2006. Overflow metabolism in Escherichia coli during steady-state growth: Transcriptional regulation and effect of the redox ratio. Appl. Environ. Microbiol. 72:3653-3661.
59. Weber, H., T. Polen, J. Heuveling, V.F. Wendish, and R. Hengge. 2005. Genome-wide analysis of the general stress response network in Escherichia coli: sigma S-dependent genes, promoters, and sigma factor selectivity. J. Bacteriol. 187:1591-1603.
60. Yomano, L.P., S.W. York, K.T. Shanmugam, and L.O. Ingram. 2009. Deletion of methylglyoxal synthase gene (mgsA) increased sugar co-metabolism in ethanol-producing Escherichia coli. Biotechnol. Lett. IN PRESS.
61. Zaldivar, J., and L.O. Ingram. 1999. Effect of organic acids on the growth and fermentation of ethanologenic Escherichia coli LY01. Biotechnol. Bioeng. 66: 203-210
62. Zaldivar, J., A. Martinez, and L.O. Ingram. 1999. Effect of selected aldehydes on the growth and fermentation of ethanologenic Escherichia coli. Biotechnol. Bioeng. 65: 24-33.
63. Zaldivar, J., A. Martinez, and L.O. Ingram. 2000. Effect of alcohol compounds found in hemicellulose hydrolysate on the growth and fermentation of ethanologenic Escherichia coli. Biotechnol. Bioeng. 68:524-530.
64. Zhang, T.F., and L.P. Hager. 1987. A single-step large scale purification of pyruvate oxidase. Arch. Biochem. Biophys. 257:485-48.
65. Almeida JRM, Roder A, Modig T, Laadan B, Liden G, Gorwa-Grauslund MF (2008) NADH- vs. NADPH-coupled reduction of 5-hydroxymethyl furfural (HMF) and its implications on product distribution in Saccharomyces cerevisiae. Appl Microbiol Biotechnol 78:939-945.
66. Cheng KK, Cai BY, Zhang JA, Ling HZ, Zhou YH, Ge JP, Xu JM (2008) Sugarcane bagasse hemicellulose hydrolysate for ethanol production by acid recovery process. Biochem Eng J 38:105-109.
67. Heer D, Heine D, Sauer U (2009) Resistance of Saccharomyces cerevisiae to high furfural concentration is based on NADPH-dependent reduction by at least two oxidoreductases. Appl Environ Microbiol doi:10.1128/AEM.01649-9.
68. Keseler IM, Bonavides-Martinez C, Collado-Vides J, Gama-Castro S, Gunsalus RP, Johnson DA, Krummenacker M, Nolan LM, Paley S, Paulsen IT, Peralta-Gil M, Santos-Zavaleta A, Shearer AG, Karp PD (2009) EcoCyc: a comprehensive view of Escherichia coli biology. Nucleic Acids Res 37:D464-70.
69. Liu ZL, Moon J (2009) A novel NADPH-dependent aldehyde reductase gene from Saccharomyces cerevisiae NRRL Y-12632 involved in the detoxification of aldehyde inhibitors derived from lignocellulosic biomass conversion. Gene 446:1-10.
70. Miller EN, Jarboe LR, Turner PC, Pharkya P, Yomano LP, York SW, Nunn D, Shanmugam KT, Ingram LO (2009a) Furfural inhibits growth by limiting sulfur assimilation in ethanologenic Escherichia coli strain LY180. Appl Environ Microbiol 75:6132-6141.
71. Purvis JE, Yomano LP, Ingram LO (2005) Enhanced trehalose production improves growth of Escherichia coli under osmotic stress. Appl Environ Microbiol 71:3761-3769.
72. Taherzadeh MJ, Gustafsson L, Niklasson C, Liden G (2000) Physiological effects of 5-hydroxymethylfurfural on Saccharomyces cerevisiae. Appl Microbiol Biotechnol 53:701-708.
73. Um BH, Karim MN, Henk LL (2003) Effect of sulfuric and phosphoric acid pretreatments on enzymatic hydrolysis of corn stover. Appl Biochem Biotechnol 105:115-125.
74. Wyman CE, Dale BE, Elander RT, Holtzapple M, Ladisch MR, Lee YY (2005) Comparative sugar recovery data from laboratory scale application of leading pretreatment technologies to corn stover. Bioresour Technol, 96:2026-2032.
【0165】
参照による組み込み
本明細書で特定された、全ての出版物、特許出願及び特許は、その全体が参照により本明細書に明確に組み込まれている。
【0166】
均等物
当業者は、通常の実験を実施するだけで、本明細書に記載される本発明の特定の実施形態の多くの均等物を認識又は確認することができる。そのような均等物は、本明細書に包含されることが意図されている。
【技術分野】
【0001】
(関連出願)
本出願は、それらの開示全体が参照により本明細書に組み込まれる、2009年5月15日出願の米国特許仮出願第61/178,672号、2009年8月19日出願の米国特許仮出願第61/235,340号、及び2010年1月4日出願の米国特許仮出願第61/292,094号の優先権を主張する。
【0002】
(政府支援研究)
本発明は、米国エネルギー省(U.S.Department of Energy)によって与えられた、契約番号第US DOE FG02−96ER20222号、第DE−FG36−08GO88142号、及び第DE−FC36−GO17058号の下での米国政府の支援を得て行われた。政府は本発明に一定の権利を有する。
【背景技術】
【0003】
リグノセルロースバイオマスからの糖類を基質として使用して多種多様な発酵産物を作ることができる(9、13、16、37)。しかし、発酵の前に、炭水化物ポリマーであるセルロース及びヘミセルロースを、化学的工程と酵素的工程の組合せを用いて可溶性糖類に変換しなければならない(38、41)。化学的工程は、微生物触媒の代謝に負の影響を及ぼす、アルコール類、酸及びアルデヒド類などの少量の生成物の混合物を生じる副反応を伴う。アルコール類(カテコール、シリンゴール等)は、細胞膜の透過性を上げることによって作用し、毒性は分子の疎水性とよく相関することが示されている(46)。有機酸(酢酸、ギ酸等)は、中性形態で膜を横断し、細胞質内でイオン化して、プロトン駆動力を崩壊させることによって増殖を阻害すると考えられる(31、45)。アルデヒド類の阻害機構はより複雑である。アルデヒド類は、直接の物理的作用及び代謝作用に加えて、多くの細胞成分と反応して生成物を形成し得る(26、34)。全体として、化学的前処理からのこれらの少量の生成物は、細胞増殖を遅延させ、バイオマス由来の糖類の発酵を減速させ得る(10、30)。
【0004】
フルフラール(五炭糖類の脱水生成物)は特に重要である。フルフラールはリグノセルロース分解の天然生成物である。フルフラールは、酸性条件下でのセルロースバイオマスの解重合の間に五炭糖類の脱水によっても形成される(21)。この化合物はヘミセルロースシロップの毒性に対する重要な寄与因子であり、他の化合物の毒性を増大する(44)。ヘミセルロースの希酸加水分解物中のフルフラール含量は毒性と関連づけられてきた(22)。石灰の添加(pH10)によるフルフラールの除去は加水分解物を容易に発酵可能にしたが、フルフラールを再添加すると毒性が回復した(21)。フルフラールはまた、ヘミセルロースの酸加水分解物中に存在することが公知の他の化合物の毒性を増強することも示されている(44〜46)。フルフラールは、DNAの構造と配列を変化させ(3、17)、解糖酵素を阻害して(6)、糖代謝を遅延させる(11)ことが報告されている。
【0005】
これらの阻害剤の存在下で機能する発酵生物の能力が広く探求されてきた。アルギン酸塩へのサッカロミセス・セレビシエ(Saccharomyces cerevisiae)の封入は保護的であり、ヘミセルロースの酸加水分解物における発酵を改善することが示された(36)。S.セレビシエの株は、加水分解物阻害剤に対して改善された耐性を有することがこれまでに記述されている(1、19、28)。大腸菌(Escherichia coli)(7)、S.セレビシエ(2)及び他の微生物(4)は、フルフラールをより毒性の低い生成物、フルフリルアルコールに還元することを触媒する酵素を含むことが示された(46)。大腸菌では、フルフラールレダクターゼ活性はNADPH依存性であると思われる(7)。NADPH依存性のフルフラールレダクターゼは大腸菌から精製されたが、その他のものも存在し得る。5−ヒドロキシメチルフルフラール(六炭糖類の脱水生成物)を還元することができるNADPH依存性酵素がS.セレビシエにおいて特徴づけられ、ADH6遺伝子と同定された(33)。
【発明の概要】
【発明が解決しようとする課題】
【0006】
従って、エタノール生産性微生物によるフルフラールに対する耐性を高めることができれば、これらの微生物によるエタノール生産の増大をもたらすだろう。
【課題を解決するための手段】
【0007】
本発明は、大規模な燃料生産のための生物を提供する。特に、本発明は、増大したフルフラールの存在下で増殖し、エタノールを生産することができる細菌を提供する。
【0008】
本発明は、参照細菌と比較して少なくとも1つのトランスヒドロゲナーゼ遺伝子の増大した発現を有する、単離されたか又は組換えたエタノール生産性細菌を提供する。一実施形態では、トランスヒドロゲナーゼ遺伝子はpntA及びpntBである。
【0009】
本発明はまた、参照細菌と比較してpntA及びpntBの増大した発現を有する、単離された細菌又は組換え細菌を提供する。
【0010】
一実施形態では、細菌は、参照細菌と比較して増大したフルフラール耐性を有する。
【0011】
別の実施形態では、細菌は野生型細菌である。
【0012】
別の実施形態では、細菌はエタノール生産性である。
【0013】
別の実施形態では、細菌は参照細菌と比較して増大したエタノール生産を示す。
【0014】
別の実施形態では、細菌は、参照細菌と比較してフルフラールの存在下で増大したエタノール生産を示す。
【0015】
別の実施形態では、細菌は参照細菌と比較して増大した増殖を有する。
【0016】
別の実施形態では、細菌は、参照細菌と比較してフルフラールの存在下で増大した増殖を有する。
【0017】
別の実施形態では、細菌は、約0.025%フルフラール〜約0.15%フルフラールの濃度のフルフラールの存在下で増大した増殖を有する。
【0018】
別の実施形態では、細菌は、参照細菌と比較して増大した増殖及び増大したエタノール生産を有する。
【0019】
別の実施形態では、細菌は、参照細菌と比較して加水分解物の存在下で増大した増殖を有する。
【0020】
別の実施形態では、細菌は加水分解物の存在下で増大した増殖を有し、加水分解物は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス又はセルロースバイオマスを含む生成物に由来する。
【0021】
別の実施形態では、pntA及びpntB遺伝子の発現は、pntA及びpntB遺伝子の発現を調節するプロモータを修飾又は付加することによって増大又は変化する。
【0022】
別の実施形態では、pntA及びpntB遺伝子の発現は、遺伝子を異なる調節タンパク質の制御下又は付加的な調節タンパク質の制御下に置くことによって増大又は変化する。
【0023】
別の実施形態では、細菌は、嫌気又は微好気条件下で主発酵生成物としてエタノールを生産することができる。
【0024】
別の実施形態では、細菌は、グラム陰性細菌及びグラム陽性細菌から成る群より選択される。
【0025】
別の実施形態では、細菌は、グラム陰性細菌及びグラム陽性細菌から成る群より選択され、グラム陰性細菌は、エシェリキア属(Escherichia)、ザイモモナス属(Zymomonas)、アシネトバクター属(Acinetobacter)、グルコノバクター属(Gluconobacter)、ジオバクター属(Geobacter)、シュワネラ属(Shewanella)、サルモネラ属(Salmonella)、エンテロバクター属(Enterobacter)及びクレブシエラ属(Klebsiella)から成る群より選択される。
【0026】
別の実施形態では、細菌は、グラム陰性細菌及びグラム陽性細菌から成る群より選択され、グラム陽性細菌は、バチルス属(Bacillus)、クロストリジウム属(Clostridium)、コリネバクテリウム属(Corynebacterium)、乳酸杆菌属(Lactobacillus)、ラクトコッカス属(Lactococcus)、オエノコッカス属(Oenococcus)、連鎖球菌属(Streptococcus)及びユーバクテリウム属(Eubacterium)から成る群より選択される。
【0027】
別の実施形態では、細菌は大腸菌(Escherichia coli)である。
【0028】
別の実施形態では、細菌はクレブシエラ・オキシトカ(Klebsiella oxytoca)である。
【0029】
本発明は、参照細菌と比較してPntA及びPntBタンパク質の活性が増大している、単離された細菌又は組換え細菌を提供する。
【0030】
本発明は、参照細菌と比較してPntA及びPntBタンパク質の活性が増大しており、且つ参照細菌と比較して増大したフルフラール耐性を有する、単離された細菌又は組換え細菌を提供する。
【0031】
本発明はまた、参照細菌と比較してpntA及びpntB遺伝子の発現が増大しており、且つ参照細菌と比較して増大したフルフラール耐性を有する、単離された細菌又は組換え細菌を提供する。
【0032】
本発明はまた、参照細菌と比較してPntA及びPntB遺伝子の発現又はPntA及びPntBポリペプチドの活性が増大しており、参照細菌と比較してフルフラール耐性が増大した、エタノールを生産することができ、並びに
(a)細菌の候補株をフルフラールの存在下で増殖させること;及び
(b)フルフラールの存在下でエタノールを生産する細菌を選択すること
を含む工程によって調製される、単離された細菌又は組換え細菌を提供する。
【0033】
本発明はまた、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖を本発明の単離された細菌又は組換え細菌のいずれかと接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含む、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産する方法を提供する。
【0034】
さらに、本発明は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖を本発明の単離された細菌又は組換え細菌と接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含む、フルフラールの存在下でバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産する方法を提供する。
【0035】
加えて、本発明は、本発明の方法によって生産されるエタノールを提供する。
【0036】
本発明はまた、本発明の単離された細菌又は組換え細菌を含むキットを提供する。
【図面の簡単な説明】
【0037】
【図1】図1A〜Bは、フルフラールの存在下で種々のレベルのsthA遺伝子発現を有するプラスミドを保持する様々なLY180株の増殖。24時間後(図1A)及び48時間後(図1B)の細胞密度(光学密度)とフルフラール濃度の関係を表すグラフを示す。記号:(■)、挿入物を有さないpTrc99a対照;(▲)、pTrc99a−sthAの順方向非誘導sthA発現;(▲)、0.01mM IPTGで誘導したpTrc99a−sthAの順方向sthA発現;及び(◆)、0.1mM IPTGで誘導したpTrc99a−sthAの順方向sthA発現。
【図2】図2A〜Bは、フルフラールの存在下で種々のレベルのpntA及びpntB遺伝子発現を有するプラスミドを保持する様々なLY180株の増殖。24時間後(図2A)及び48時間後(図2B)の細胞密度(光学密度)とフルフラール濃度の関係を表すグラフを示す。記号:(■)、挿入物を有さないpTrc99a対照;(▲)、pTrc99a−pntABの順方向非誘導pntAB発現;(▲)、0.01mM IPTGで誘導したpTrc99a−pntABの順方向pntAB発現;及び(◆)、0.1mM IPTGで誘導したpTrc99a−pntABの順方向pntAB発現。
【図3】図3A〜Bは、フルフラール耐性へのトランスヒドロゲナーゼ(SthA及びPntAB)の増大した発現の影響を示す。培養物を、50g/lのキシロース及び1.0g/lのフルフラールを含むAM1最少培地中で48時間増殖させた。空のベクタを対照として使用した。接種の前に誘導物質を添加した。細胞密度(550nmでの光学密度に関して)を、24時間後の約0.05%フルフラール濃度(図3A)及び48時間後の約0.10%フルフラール濃度(図3B)での各々の株について示す。対照株を白い棒グラフで示している。種々のレベルのpntA発現を有するプラスミドを保持する株を斜線の棒グラフで示す。種々のレベルのpntAB遺伝子発現を保持する株を黒い棒グラフで示す。
【図4】図4A〜Bは、pntAの核酸配列(図4A)及びアミノ酸配列(図4B)を示す。
【図5】図5A〜Bは、pntBの核酸配列(図5A)及びアミノ酸配列(図5B)を示す。
【図6】図6A〜Bは、0.5g/lフルフラール抗原での抗原投与後のLY180における転写及び調節の変化を示す。有意に変動した調節遺伝子をNCAによって同定し、ヌル分布に比して0.05をP値のカットオフとした。増大した活性を伴う調節因子を実線の枠で示し、減少した活性を伴う調節因子を破線の枠で示す。混合活性を示した調節因子は明灰色で示している(DcuR)。図6Aは部分的な調節因子−遺伝子応答地図を示す。2倍より大きく変動した代表的な遺伝子を示しており、中実(点々)の枠は増大した発現(減少した発現)を有する遺伝子を示す。実線は連結した調節因子による活性化を示し、破線は抑制を示す。DcuRについては変動の方向が不明確であり、この調節因子は明灰色で示している。図6Bは、フルフラールによる増殖阻害の機構を説明するモデルを示す。フルフラールの添加は、NADPHについての基本的生合成反応と競合する2つのNADPH依存性オキシドレダクターゼ(YqhD及びDkgA)を誘導する。硫酸塩の同化はシステイン当たり4つのNADPHを必要とする。二次的な結果は、ストリンジェント因子SF並びに生合成調節因子ArgR、PurR及びRutRの変動によって証明されるように、含硫アミノ酸の枯渇と、停止した翻訳及び多くの非含硫ビルディングブロック中間体の蓄積から、より一般的なストレス応答への事象のカスケードとを含む。
【図7】図7は、ヒスチジン経路におけるhisG以外の遺伝子のフルフラール添加による発現増大を示す。変化の割合を括弧内に示す。略語:ACR、アミノイミダゾールカルボキサミドリボヌクレオチド。
【図8】図8は、硫黄のシステイン及びメチオニンへの同化に関与する遺伝子のフルフラールによる発現増大を示す。比較のためにアスパラギン酸からのトレオニン及びイソロイシンの合成のための経路を含める。1.5倍又はそれ以上上方調節された遺伝子をプラスの記号で示す。1.5倍又はそれ以上下方調節された遺伝子をマイナスの記号で示す。その他はすべてプラス又はマイナス記号のない黒字で示している。
【図9】図9は、1g/lフルフラールの存在下での増殖への培地添加物の影響を示す。48時間の培養後に培養物を比較する(AM1培地、50g/lキシロース、37℃)。図9Aは個々のアミノ酸(各々0.1mM)の添加を示す。図9Bはアミノ酸(各々0.5mM)の添加を示す。図9Cはシステインの添加を示す。図9Dは代替硫黄源の添加を示す。
【図10】図10は、表1であり、細菌株、プラスミド及びプライマを示す。
【図11】図11は、表2であり、機能群によって分類した、0.5g/lフルフラールでの処理に対するLY180の応答が2倍より大きく変動した遺伝子を示す。
【図12】図12は、表3であり、フルフラール(0.5g/l)の添加に応答して5倍又はそれ以上の発現比の変化が生じた遺伝子を示す。これらの遺伝子の一部は、対照実験で水の添加に対する応答も2倍より大きく変化した(星印で示している)。
【図13】図13は、表4であり、NCAによって決定したとき、ヌル分布に比してLY180のフルフラール応答が有意に(P<0.05)変動した調節因子を示す。
【図14】図14は、嫌気的増殖及び発酵への5−HMFの効果である。キシロース(100g/lキシロース)を添加したAM1無機塩培地で細胞を増殖させた。A.1.0g/ lの5−HMFで増殖中の細胞質量;B.1.0g/lの5−HMFで発酵中のエタノール生産;C.発酵中の5−HMF(1.0g/l)の還元;D.2.5g/lの5−HMFで増殖中の細胞質量;E.2.5g/lの5−HMFで発酵中のエタノール生産;F.発酵中の5−HMF(2.5g/l 5−HMF)の還元。比較のために5−HMFなしでの並行発酵をA及びBに含めている(破線)。すべてのデータは標準偏差と共に平均値としてプロットしている(n=3)。すべてについての記号:□、LY180;及び●、EMFR9。
【図15】図15は、5−HMFのインビトロでの還元及び5−HMF耐性へのYqhD及びDkgAの効果。A.インビトロでの5−HMF還元に対する比活性。溶解細胞抽出物(2mMのNADPH、20mMの5−HMF)において活性を測定した。B.EMFR9(耐性変異株)の細胞収率へのプラスミドからのyqhD及びdkgA発現の影響。実験は、50g/lのキシロース及び1.0g/lの5−HMFを含むAM1培地での試験管培養物において実施した(48時間培養)。プラスミドの維持のためにカナマイシンを含めることは5−HMF耐性を減少することに留意されたい。誘導株(Ind.)は0.1mMのIPTGを添加して増殖させた。C.LY180(親株)の細胞収率へのyqhD及びdkgA欠失の影響。実験は、50g/lのキシロース及び2.5g/lの5−HMFを含むAM1培地での試験管培養物において実施した(48時間培養)。すべてのデータは標準偏差と共に平均値としてプロットしている(n=4)。
【図16】図16は、5−HMF耐性へのプラスミドからのpntAB発現の影響。実験は、指示されているように50g/lのキシロース及び5−HMFを含むAM1培地でBioscreen C増殖曲線分析器を使用して実施した。すべてのデータは標準偏差とと共に平均値としてプロットしている(n=10)。明瞭さのために連結点は省略した。A.添加物なし;B.0.9g/lの5−HMFを添加;及びC.1.8g/lの5−HMFを添加。すべてについての記号:△、LY180(pTrc99a−対照);○、非誘導LY180(pTrc99a−pntAB);●、0.01mMのIPTGで誘導したLY180(pTrc99a−pntAB)。
【図17】図17は、LY180の5−HMF耐性へのL−システインの効果。実験は、50g/lのキシロース及び5−HMFを含むAM1培地での試験管培養物において実施した(24時間培養)。培養物に、指示されているようにろ過滅菌したL−システインを添加した。すべてのデータは標準偏差と共に平均値としてプロットしている(n=4)。A.1.0g/lの5−HMF;B.2.0g/lの5−HMF。
【発明を実施するための形態】
【0038】
(I.定義)
本明細書で使用される、「単離された」は、他の細菌による汚染がないことを意味する。単離された細菌は、その単離細菌の特性及び機能を妨げない少量の他の細菌の存在下で存在し得る。単離された細菌は、一般に少なくとも30%、40%、50%、60%、70%、80%、85%、90%、95%、98%又は99%純粋である。好ましくは、本発明による単離された細菌は少なくとも98%又は少なくとも99%純粋である。
【0039】
本明細書で使用される、「細菌」は、「非組換え細菌」、「組換え細菌」及び「突然変異細菌」を含む。
【0040】
本明細書で使用される、「非組換え細菌」は、異種ポリヌクレオチド配列を含まず、本発明の組成物及び方法を用いたさらなる修飾に適する、例えば遺伝子操作、例えば異種ポリヌクレオチド配列を組み込むことができる、例えばトランスフェクトすることができる、細菌細胞を含む。この用語は、最初にトランスフェクトされた細胞の子孫を包含することが意図されている。特定の実施形態では、細胞はグラム陰性細菌細胞及びグラム陽性細菌細胞である。
【0041】
細菌に関して本明細書で使用される「組換え体」は、遺伝子操作に適する若しくは遺伝子操作に供される、又は異種ポリヌクレオチド配列が組み込まれている、又は天然ポリヌクレオチド配列が変異若しくは欠失するように処理された細菌細胞を意味する。
【0042】
細菌に関して本明細書で使用される「突然変異体」は、本明細書中以下で定義される参照細菌と同一ではない細菌細胞を意味する。
【0043】
「突然変異」細菌は「組換え」細菌を含む。
【0044】
本明細書で使用される、「エタノール生産性」は、炭水化物から主発酵生成物としてエタノールを生産する細菌の能力を意味する。この用語は、天然に生じるエタノール生産性生物及び天然に生じた又は誘導された突然変異を有するエタノール生産性生物を包含することが意図されている。
【0045】
「非エタノール生産性」という用語は、細菌が炭水化物から主発酵生成物としてエタノールを生産することができないことを意味する。この用語は、総非ガス発酵生成物の40%未満を含有している微量発酵生成物としてエタノールを生産する微生物を包含することが意図されている。
【0046】
本明細書で使用される「エタノール生産」は、炭水化物からの主発酵生成物としてのエタノールの生産を意味する。
【0047】
本明細書で使用される「エタノールを生産することができる」は、本明細書で定義されるように「エタノール生産」できることを意味する。
【0048】
「発酵する」及び「発酵」という用語は、複合糖類の分解又は解重合並びにその糖残基のエタノール、酢酸エステル及びコハク酸エステルへの生物変換を意味する。これらの用語は、エタノールが炭水化物から生産される、特に発酵の主生成物として生産される酵素的過程(例えば細胞又は無細胞の、例えば溶解生成物又は精製ポリペプチド混合物の)を包含することが意図されている。
【0049】
「主発酵生成物」及び「主要発酵生成物」という用語は、本明細書では交換可能に使用され、総非ガス生成物の約50%より多くを含有している非ガス性の発酵生成物を包含することが意図されている。主発酵生成物は最も大量の非ガス生成物である。本発明のある特定の実施形態では、主発酵生成物はエタノールである。
【0050】
本明細書で使用される「微量発酵生成物」という用語は、総非ガス生成物の40%未満を含有する非ガス性の発酵生成物を包含することが意図されている。本発明のある実施形態では、微量発酵生成物はエタノールである。
【0051】
「同時糖化発酵」又は「SSF」という用語は、複合糖の同時分解又は解重合及び発酵によるその糖残基のエタノールへの生物変換反応のための1又はそれ以上の組換え宿主(又は、精製又は非精製抽出物を含む、その抽出物)の使用を包含することが意図されている。SSFは、バイオマスを、細菌によって最終的にエタノールに変換可能な多糖類に分解するために使用できる周知の工程である。天然で起こる場合のバイオマスの分解を反映して、SSFは、リグノセルロースなどの糖源を、最終的にエタノールに変換できる単糖類に分解するためにエタノール生産性細菌(又はそれに由来する酵素)の活性と真菌(又は真菌から抽出されたセルラーゼなどの酵素)の活性を組み合わせる。SSF反応は、典型的には、高価な真菌酵素の使用を最適化するために酸性pHで実施される。
【0052】
「糖」という用語は、糖分子(複数も)を含む何れの炭水化物源もを包含することが意図されている。そのような糖は、本発明の生成物及び方法による、解重合(所望により)並びにその後の発酵によるアセトアルデヒドへの及びその後のエタノールへの生物変換のための糖類の可能性のある供給源である。糖の供給源は、大部分の植物における燃料貯蔵の主要形態であるデンプン、植物の堅い細胞壁並びに繊維性及び木質組織の主たる細胞外構造成分であるヘミセルロース及びセルロースを含む。この用語は、単糖(simple sugars)とも呼ばれる単糖類(monosaccharides)、オリゴ糖類及び多糖類を包含することが意図されている。ある特定の実施形態では、糖は、例えばグルコース、キシロース、アラビノース、マンノース、ガラクトース、スクロース及びラクトースを含む。他の実施形態では、糖はグルコースである。
【0053】
本明細書で使用される場合、「PntAB」は、ピリジンヌクレオチドトランスヒドロゲナーゼを意味する。pntA及びpntBとしても知られるpntABは、PntABトランスヒドロゲナーゼに対応する遺伝子を指し、一方PntABという用語はpntAB遺伝子の産物を示す。
【0054】
pntA及びpntB遺伝子に対応するアミノ酸配列及び核酸配列をそれぞれ図4A〜4B及び図5A〜5Bに示す。
【0055】
本明細書で使用される場合、「SthA」は、細胞質トランスヒドロゲナーゼを意味する。sthAはSthAトランスヒドロゲナーゼに対応する遺伝子を指し、一方SthAという用語はsthA遺伝子の産物を示す。
【0056】
本明細書で使用される場合、「突然変異核酸分子」又は「突然変異遺伝子」は、そのポリペプチド又は変異株によってコードされ得るポリペプチドがそのポリペプチド又は野生型核酸分子若しくは遺伝子によってコードされるポリペプチドとは異なる活性又は特性を示すように、少なくとも1つの変化(例えば置換、挿入、欠失)を含むヌクレオチド配列を有する核酸分子又は遺伝子を包含することが意図されている。
【0057】
核酸分子又は遺伝子に関して本明細書で用いられる「突然変異」は、核酸若しくは遺伝子の変化、挿入若しくは欠失、又は核酸若しくは遺伝子の発現レベルの増大若しくは減少を意味し、発現の増大又は減少は、それぞれ、核酸分子又は遺伝子によってコードされ得るポリペプチドの発現の増大又は減少を生じさせる。突然変異はまた、そのポリペプチド又はその変異株によってコードされ得るポリペプチドがそのポリペプチド又は野生型核酸分子若しくは遺伝子によってコードされるポリペプチドとは異なる活性又は特性を示すように、少なくとも1つの変化(例えば置換、挿入、欠失)を含むヌクレオチド配列を有する核酸分子又は遺伝子を意味する。
【0058】
本明細書で使用される、「突然変異タンパク質」又は「突然変異タンパク質又はアミノ酸配列」は、ポリペプチド又は突然変異アミノ酸配列によってコードされ得るポリペプチドがそのポリペプチド又は野生型アミノ酸配列によってコードされるポリペプチドとは異なる活性又は特性を示すように、少なくとも1つの変化(例えば置換、挿入、欠失)を含むアミノ酸配列を包含することが意図されている。
【0059】
タンパク質又はアミノ酸配列に関して本明細書で使用される、「突然変異」は、アミノ酸配列のアミノ酸の変化、挿入若しくは欠失、又はアミノ酸配列の発現レベルの増大若しくは減少を意味し、発現の増大又は減少は、アミノ酸配列によってコードされ得るポリペプチドの発現の増大又は減少を生じさせる。突然変異はまた、そのポリペプチド又は変異株によってコードされ得るポリペプチドがそのポリペプチド又は野生型アミノ酸配列によってコードされるポリペプチドとは異なる活性又は特性を示すように、少なくとも1つの変化(例えば置換、挿入、欠失)を含むアミノ酸配列を有するタンパク質又はアミノ酸配列を意味する。
【0060】
本明細書で使用される「断片」又は「サブ配列」は、親若しくは参照核酸配列若しくはアミノ酸配列の一部、又は親若しくは参照配列、ポリペプチド若しくは遺伝子の生物学的機能又は特性をコードするか又は保持する、ポリペプチド若しくは遺伝子の一部を包含することが意図されている。
【0061】
「突然変異」細菌は、本明細書中上記で定義した「突然変異」を含む細菌を包含する。
【0062】
本明細書で使用される「参照物」又は「参照細菌」は、少なくとも、野生型細菌及び親細菌を包含する。
【0063】
本明細書で使用される「野生型」は、生物又は菌株の典型的な形態、例えば突然突然変異の不在下で天然に生じる細菌、遺伝子又は特徴を意味する。「野生型」は、天然集団における最も一般的な表現型を示す。野生型は、遺伝子型及び表現型についての参照物の標準である。
【0064】
本明細書で使用される「親」又は「親細菌」は、目的の細菌を生じる細菌を示す。
【0065】
本明細書で使用される「遺伝子」は、酵素又は他のポリペプチド分子の合成を誘導できる、例えばコード配列、例えばポリペプチド、そのサブ配列をコードする隣接するオープンリーディングフレーム(ORF)を含み得る、又は生物においてそれ自体が機能性であり得る核酸である。生物中の遺伝子は、本明細書で定義されるオペロンにクラスター化することができ、オペロンは遺伝子間DNAによって他の遺伝子及び/又はオペロンから切り離されている。オペロン内に含まれる個々の遺伝子は、個々の遺伝子の間の遺伝子間DNAなしでオーバーラップし得る。加えて、「遺伝子」という用語は、選択された目的のための特定遺伝子を包含することが意図されている。遺伝子は、宿主細胞にとって内因性であり得るか又は、例えばエピソームに保持されたプラスミド若しくはゲノムに安定に組み込まれたプラスミド(若しくはその断片)として、宿主細胞に組換え導入され得る。異種遺伝子は、細胞に導入される、その細胞にとって天然ではない遺伝子である。
【0066】
「核酸」という用語は、ポリペプチド、そのサブ配列をコードするオープンリーディングフレームを含み、非コード調節配列及びイントロンをさらに含み得る核酸分子、例えばポリヌクレオチドを包含することが意図されている。加えて、この用語は、機能的遺伝子座に位置づけられる1つ又はそれ以上の遺伝子を包含することが意図されている。加えて、この用語は、選択された目的のための特定遺伝子を包含することが意図されている。一実施形態では、遺伝子という用語は、pntA及びpntBを含むがこれらに限定されない、トランスヒドロゲナーゼをコードする任意の遺伝子を包含する。一実施形態では、遺伝子又はポリヌクレオチドセグメントは、炭水化物のエタノールへの生物変換反応における少なくとも1つの工程に関与する。生物中の遺伝子は、本明細書で定義されるオペロンにクラスター化することができ、オペロンは遺伝子間DNAによって他の遺伝子及び/又はオペロンから切り離されている。
【0067】
本明細書で使用される「増加している」又は「増加する」又は「増加した」は、例えば、参照細菌と比較してpntA及びpntB遺伝子の発現増加を有する細菌において、pntA及びpntB遺伝子の発現レベルと比較して、少なくとも5%、例えば5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、99、100%又はそれ以上増加していることを示す。
【0068】
本明細書で使用される「増加している」又は「増加する」又は「増加した」はまた、例えば、参照細菌と比較してpntA及びpntB遺伝子の発現増加を有する細菌において、pntA及びpntB遺伝子の発現レベルと比較して、少なくとも1倍、例えば1、2、3、4、5、6、7、8、9、10、15、20、30、40、50、60、70、80、90、100、200、500、1000倍又はそれ以上の増加を意味する。
【0069】
本明細書で使用される場合、「減少している」又は「減少する」又は「減少した」は、例えば、参照細菌と比較して、細菌におけるpntA及びpntB遺伝子の発現レベルの減少と比較して、少なくとも5%、例えば5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、99又は100%減少していることを示す。
【0070】
本明細書で使用される場合、「減少している」又は「減少する」又は「減少した」はまた、例えば、参照細菌と比較して、細菌におけるpntA及びpntB遺伝子の発現レベルと比較して、少なくとも1倍、例えば1、2、3、4、5、6、7、8、9、10、15、20、30、40、50、60、70、80、90、100、200、500、1000倍又はそれ以上の減少を意味する。
【0071】
「減少した」又は「縮小した」も、活性、発現等の検出可能なレベルが存在しない、例えばpntA及びpntB遺伝子の発現の検出可能なレベルが存在しない又はPntA及びPntBタンパク質の検出可能な活性が存在しないように排除されていることを意味する。
【0072】
本明細書で使用される「活性」は、遺伝子の活性、例えば遺伝子の転写のレベルを示す。「活性」はまた、mRNAの活性、例えばmRNAの翻訳のレベルを示す。「活性」はまた、タンパク質、例えばPntA及びPntBの活性を示す。
【0073】
「活性の増大」は、活性の速度及び/又はレベルの増大を含む。
【0074】
本明細書で使用される「pntA及びpntBの発現」におけるような「発現」は、pntA及びpntB遺伝子のタンパク質産物の発現を示す。本明細書で使用される場合、「pntA及びpntBの発現」におけるような「発現」はまた、pntA及び/又はpntB遺伝子に対応するmRNA転写産物の検出可能なレベルの発現を示す。
【0075】
発現レベルに関連して「変化している」は、目的とする遺伝子、mRNA又はタンパク質、例えばpntA及び/又はpntB遺伝子の発現が減少していることを意味する。
【0076】
本明細書で使用される「発現されない」は、目的とする遺伝子又はmRNA、例えばpntA及び/又はpntB遺伝子の産物の検出可能なレベルが存在しないことを意味する。
【0077】
本明細書で使用される「排除する」は、検出不能なレベルへの減少を意味する。
【0078】
本明細書で使用される「フルフラールの耐性」は、フルフラールの存在下で、例えば0.1g/l又はそれ以上(例えば0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2.0、2.5、3.0g/l又はそれ以上)の濃度のフルフラールの存在下で増殖するか又はエタノールを生産する、エタノール生産性細菌の能力を意味する。フルフラールの耐性はまた、野生型細菌又は親細菌による増殖又はエタノール生産のレベルと比較して増大しているレベルで、フルフラールの存在下で増殖するか又はエタノールを生産するエタノール生産性細胞の能力を意味する。
【0079】
本明細書で使用されるフルフラールの存在に適用されるときの「の存在下で」は、少なくとも0.1g/l又はそれ以上(例えば0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2.0、2.5、3.0g/l又はそれ以上)のフルフラールの存在下に細菌を保持することを意味する。
【0080】
本明細書で使用されるフルフラールの不在に適用されるときの「の不在下で」は、検出可能でないレベルを含む、0.1g/l又はそれ以下のフルフラールを含有する培地中に細菌を保持することを意味する。
【0081】
本明細書で使用される「増殖」は、経時的な細胞の数又は質量の、本明細書で定義される増大を意味する。
【0082】
本明細書で使用される「ヘミセルロース加水分解物」は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス又はセルロースバイオマスに由来する加水分解物を含むが、これに限定されない。
【0083】
本明細書で使用される「に由来する」は、それを起源とすることを意味する。
【0084】
「グラム陰性細菌細胞」という用語は、当該技術分野で広く認められているこの用語の定義を包含することが意図されている。例示的なグラム陰性細菌は、アシネトバクター属、グルコノバクター属、ザイモモナス属、エシェリキア属、ジオバクター属、シュワネラ属、サルモネラ属、エンテロバクター属及びクレブシエラ属を含む。
【0085】
「グラム陽性細菌」という用語は、当該技術分野で広く認められているこの用語の定義を包含することが意図されている。例示的なグラム陽性細菌は、バチルス属、クロストリジウム属、コリネバクテリウム属、乳酸杆菌属、ラクトコッカス属、オエノコッカス属、連鎖球菌属及びユーバクテリウム属を含む。
【0086】
「アミノ酸」という用語は、タンパク質において通常生じる20のα−アミノ酸を包含することが意図されている。塩基性荷電アミノ酸は、アルギニン、アスパラギン、グルタミン、ヒスチジン及びリシンを含む。中性荷電アミノ酸は、アラニン、システイン、グリシン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、プロリン、セリン、トレオニン、トリプトファン、チロシン及びバリンを含む。酸性アミノ酸は、アスパラギン酸及びグルタミン酸を含む。
【0087】
本明細書で使用される「選択すること」は、同定された細菌がフルフラールの存在下でエタノールを生産することを確認する工程を示す。
【0088】
本明細書で使用される「同定すること」は、細菌を評価し、その細菌がフルフラールの存在下でエタノールを生産することを確認する工程を示す。
【0089】
本明細書で使用される「フルフラールの濃度を増大すること」は、0〜5g/lの増加、例えば1μg/lの増加、1mg/lの増加又は1g/lの増加を意味する。
【0090】
本開示において、「含む(comprises)」、「含んでいる(comprising)」、「含有する(containing)」及び「有する(having)」等は、米国特許法においてそれらに帰せられている制約のない意味を有し、「包含する(includes)」、「包含している(including)」等を意味する。
【0091】
本明細書で使用される「トランスヒドロゲナーゼ」という用語は、ニコチンアミドアデニンジヌクレオチド補因子NAD(H)とNADP(H)の間の還元性等価物の相互変換を触媒する酵素を示す。
【0092】
本明細書で使用される「トランスヒドロゲナーゼ遺伝子」という用語は、その産物がトランスヒドロゲナーゼである遺伝子を示す。
【0093】
本明細書で使用される「5−ヒドロキシメチルフルフラール」又は「5−HMF」という用語は、構造:
【0094】
【化1】
【0095】
を有する糖類の脱水に由来する有機化合物を意味することが意図されている。
【0096】
II.細菌
本発明は、エタノールの形成のために糖類を分解するのに適した細菌に関する。この細菌は、特にフルフラール及び/又は5−HMFを含む培地において、改善されたエタノール生産能力を有する。改善されたエタノール生産の能力は、糖消化及び発酵の間のフルフラール及び/又は5−HMFに対する細胞の耐性を増大するトランスヒドロゲナーゼ遺伝子の選択された増大した発現に関連する。
【0097】
従って、本発明は、参照細菌と比較して少なくとも1つのトランスヒドロゲナーゼ遺伝子の増大した発現を有するエタノール生産性細菌を提供する。本発明の1つの態様では、細菌は単離された細菌である。本発明の別の態様では、細菌は組換え細菌である。本発明のさらに別の態様では、トランスヒドロゲナーゼ遺伝子はpntA及びpntB遺伝子を含む。
【0098】
本発明はまた、pntA及びpntB遺伝子の増大した発現を有する単離された細菌又は組換え細菌を提供する。本発明の1つの態様では、本発明の単離された細菌は野生型細菌を含む。
【0099】
本発明の細菌はそれらの増大した増殖を特徴とし得る。本発明の細菌は、さらに、増大した濃度のフルフラール及び/又は5−HMF中で増殖するそれらの能力を特徴とする。従って、本発明の他の態様では、単離された細菌又は組換え細菌は、参照細菌と比較して増大した増殖、又は参照細菌と比較して増大したフルフラール及び/若しくは5−HMF耐性、又は参照細菌と比較してフルフラール及び/若しくは5−HMFの存在下での増大した増殖、又はフルフラール及び/若しくは5−HMFの存在下での、例えば約0.025%フルフラール〜約0.15%フルフラールのフルフラール濃度、若しくは約0.025%5−HMF〜約0.15%5−HMFの5−HMF濃度での増大した増殖を有する。
【0100】
本発明の細菌はまた、糖源から主発酵生成物としてエタノールを生産するそれらの能力を特徴とし得る。フルフラール及び5−HMFは、通常、セルロース消化及びエタノールへの発酵の間エタノール生産性細菌の増殖を阻害するが、本発明の細菌は、高濃度のフルフラール及び/又は5−HMF中でエタノールを生産することができる。従って、本発明の他の態様は、エタノール生産性である、又は参照細菌と比較して増大したエタノール生産を示す、又は参照細菌と比較してフルフラール及び/若しくは5−HMFの存在下で増大したエタノール生産を示す、又は嫌気若しくは微好気条件下で主発酵生成物としてエタノールを生産することができる、又は参照細菌と比較してフルフラール及び/若しくは5−HMFの存在下で増大した増殖及び増大したエタノール生産を示す細菌を含む。
【0101】
本発明のさらなる態様は、参照細菌と比較して加水分解物の存在下で増大した増殖を有する細菌を含む。本発明は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス又はセルロースバイオマスに由来する加水分解物を含むがこれに限定されない様々な加水分解物を提供する。
【0102】
参照細菌と。例えば、プロモータを、異なるプロモータによるプロモータの置換、あるいは、例えば核酸を挿入する、置換する、重複させる若しくは除去することによる又はプロモータにおいて調節エレメント若しくはモチーフを挿入する、置換する、重複させる若しくは除去することによるプロモータの修飾を含むがこれらに限定されない、当該技術分野で一般に認められている方法によって変化させる。従って、本発明の1つの態様では、本発明のpntA及びpntB遺伝子などのトランスヒドロゲナーゼ遺伝子の発現を、参照細菌と比較して遺伝子発現を調節するプロモータを修飾する又は付加することを含むがこれらに限定されない、当該技術分野で公知の方法によって増大させる。
【0103】
本発明は、pntA及びpntB遺伝子を参照細菌と比較して異なる調節タンパク質の制御下に置くか又は付加的な調節タンパク質の制御下に置くことを含むがこれに限定されない、当該技術分野で公知の方法によって、pntA及びpntB遺伝子の調節を変化させる方法を提供する。一実施形態では、調節タンパク質は抑制因子である。選択的な実施形態では、調節タンパク質は誘導因子である。
【0104】
本発明の細菌は、非組換え又は組換えでよい。本発明の細菌は、グラム陰性細菌及びグラム陽性細菌から成る群より選択され、グラム陰性細菌は、アシネトバクター属、グルコノバクター属、ザイモモナス属、エシェリキア属、ジオバクター属、シュワネラ属、サルモネラ属、エンテロバクター属及びクレブシエラ属から成る群より選択され、そしてグラム陽性細菌は、バチルス属、クロストリジウム属、コリネバクテリウム属、乳酸杆菌属、ラクトコッカス属、オエノコッカス属、連鎖球菌属及びユーバクテリウム属から成る群より選択される。一態様では、本発明の細菌は大腸菌であり、別の態様では、本発明の細菌はクレブシエラ・オキシトカである。
【0105】
本発明の細菌のpntA及びpntB遺伝子などの増大したトランスヒドロゲナーゼ遺伝子に関連するタンパク質活性を増大又は変化できることは当業者に理解される。例えば、遺伝子産物中のアミノ酸を、天然に生じる遺伝子産物と比較して遺伝子産物の機能に実質的な影響を及ぼさずにある程度まで置換するか、付加するか又は欠失させることができる。従って、本発明はまた、PntA及びPntBタンパク質の活性が参照細菌と比較して増大している又は変化している、単離された細菌又は組換え細菌を提供する。一実施形態では、そのような細菌は、参照細菌と比較して増大したフルフラール及び/又は5−HMF耐性を有する。
【0106】
本発明は、上記で論じたように、例えばpntA及びpntB遺伝子などのトランスヒドロゲナーゼ遺伝子の増大した又は変化した発現を有して、増大した濃度のフルフラール中で増殖し続けることができる細菌を提供する。従って、本発明はさらに、参照細菌と比較してpntA及びpntB遺伝子の発現が増大又は変化しており、且つ参照細菌と比較して増大したフルフラール及び/又は5−HMF耐性を有する、単離された細菌又は組換え細菌を提供する。発現は、pntA又はpntB遺伝子の修飾(例えば、遺伝子をコードする配列において核酸又はアミノ酸を挿入するか、置換するか又は除去することによる)を含むがこれに限定されない、当該技術分野で公知の方法によって増大させるか又は変化させる。
【0107】
本発明はまた、参照細菌と比較してpntA及びpntB遺伝子の発現又はPntA及びPntBポリペプチドの活性が増大しており、参照細菌と比較してフルフラール及び/又は5−HMF耐性が高く、エタノールを生産することができ、並びに(a)細菌の候補株をフルフラール及び/又は5−HMFの存在下で増殖させること;及び(b)フルフラール及び/又は5−HMFの存在下でエタノールを生産する細菌を選択することを含む工程によって調製される、単離された細菌又は組換え細菌を提供する。
【0108】
本発明はさらに、フルフラール及び/又は5−HMFの存在下でのエタノール生産のために糖類を発酵させるのに適した微生物を提供する。従って、本発明は、微生物にとって内因性であるか又は、例えばエピソームに保持されたプラスミド若しくはゲノムに安定に組み込まれたプラスミド(若しくはそのフラグメント)として微生物に組換え導入されるトランスヒドロゲナーゼ遺伝子の増大した発現を有する微生物を提供する。
【0109】
III.製造方法
本発明はさらに、フルフラール及び/又は5−HMFの存在下での、例えば増大したフルフラール及び/又は5−HMFの存在下でのエタノール生産のために糖類を発酵させるのに適した細菌を生産する方法に関する。従って、本発明は、参照細菌と比較してpntA及びpntB遺伝子の発現又はPntABポリペプチドの活性が増大しており、参照細菌と比較してフルフラール及び/又は5−HMF耐性が増大しており、細菌はエタノールを生産することができ、細菌はフルフラール及び/又は5−HMFの存在下で細菌の候補株を増殖させることと、フルフラール及び/又は5−HMFの存在下でエタノールを生産する細菌を選択することとを含む工程によって調製される、単離された細菌又は組換え細菌を提供する。
【0110】
組換えエタノール生産性微生物を製造する方法は、分子生物学の技術分野において公知である。適切な材料及び方法並びに組換え微生物は、例えば、参照により本明細書に組み込まれる米国特許第7,026,152号明細書、同第6,849,434号明細書、同第6,333,181号明細書、同第5,821,093号明細書、同第5,482,846号明細書、同第5,424,202号明細書、同第5,028,539号明細書、同第5,000,000号明細書、同第5,487,989号明細書、同第5,554,520号明細書及び同第5,162,516号明細書並びに国際公開第WO2003/025117号に記載されており、本発明を実施する際に使用できる。本明細書で述べられている本発明の細菌は、当業者によって日常的に実施され、本願で示す実施例において明らかにされる方法によって製造できる。
【0111】
IV.エタノールを生産するための方法
本発明の細菌は、エタノールの生産のために糖類を分解するのに適している。従って、本発明は、上述した本発明の単離された細菌又は組換え細菌と供給源を接触させ、それによって供給源からエタノールを生産することを含む、供給源からエタノールを生産する方法を提供する。本発明の特定の実施形態では、供給源は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス若しくはオリゴ糖源、又はそれらの何れかの組合せから成る群より選択できる。
【0112】
本発明の細菌は、増大した濃度のフルフラール及び/又は5−HMFの存在下で糖類からエタノールを生産するために使用しできる。従って、本発明は、フルフラール及び/又は5−HMFの存在下で、供給源を上述した本発明の単離された細菌又は組換え細菌と接触させ、それによって供給源からエタノールを生産することを含む、供給源からエタノールを生産する方法を提供する。本発明の特定の実施形態では、供給源は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス若しくはオリゴ糖源、又はそれらの何れかの組合せから成る群より選択できる。
【0113】
本発明の微生物は、嫌気条件下でのエタノール生産を特徴とする。野生型大腸菌は、嫌気的増殖の間にエタノールと酢酸塩を1:1の割合で生産する。増殖の定常期の間に、野生型大腸菌は主要生成物として乳酸塩を生産し、全発酵生成物中のエタノールの割合は約20%である。これらのすべての発酵における生成物は様々な酸を含み、それ故混合酸発酵という用語を導く。
【0114】
一般的には、産生宿主細胞株の最良増殖速度及び培養物によって生成される酵素のための触媒条件を促進するための最適pH及び温度を提供する発酵条件が選択される(Doran et al., (1993) Biotechnol. Progress. 9:533-538)。様々な例示的発酵条件が米国特許第5,487,989号明細書及び同第5,554,520号明細書に開示されている。非限定的な例として、約25〜約40℃の範囲の温度及び約4.5〜8.0の範囲のpHを含む条件を選択できる。例えば、この参照により明確に本明細書に組み込まれる、米国特許第5,424,202号明細書及び同第5,916,787号明細書を参照されたい。
【0115】
本発明は、トランスヒドロゲナーゼの増大した発現を有する単離された細菌若しくは組換え細菌、又は参照細菌と比較してpntA及びpntB遺伝子の増大した発現を有する単離された細菌若しくは組換え細菌を提供する。この細菌は、エタノールを生産するため、特に、例えばバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖などの供給源からエタノールを生産するために使用できる。従って、本発明は、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源を本発明の細菌と接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含む、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産するための方法を提供する。そのような生産は、フルフラール及び/又は5−HMFの存在下又は不在下で起こり得る。
【0116】
本発明の方法に従って、本明細書で述べられている細菌は、本発明の細菌でセルロースバイオマス又はオリゴ糖源を単糖に分解又は解重合し、それによってオリゴ糖源などのバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマスからエタノールを生産する。その後、細菌は、トランスヒドロゲナーゼ遺伝子の増大した発現によって又は、特にそれらが担持するpntA及びpntB遺伝子の増大した発現によって、より単純な糖類を発酵によってエタノールへと異化する。同時に起こる、複合糖のより小さな糖残基への解重合とそれに続く発酵のこのプロセスは、同時糖化発酵(SSF)と称される。
【0117】
現在、リグノセルロースなどの複合糖の変換は非常に複雑な多工程方法である。例えば、リグノセルロースを、最初に酸加水分解を用いて分解又は解重合しなければならない。この後に、固体から液体を分離し、その後これらの生成物を洗浄して、無毒化し、さらに解重合できるセルロースを生じさせて、最後に、適切なエタノール生産性宿主細胞によって発酵させる工程が続く。これに対し、トウモロコシの発酵ははるかに簡単であり、基本的に一工程方法で、エタノール生産性宿主による即時生物変換反応のためにアミラーゼを使用してトウモロコシデンプンを分解することができる。
【0118】
本発明の細菌及び方法が、リグノセルロースを発酵させるためのより効率的な工程の使用を提供することは当業者に認識される。例えば、本発明の方法は、酸加水分解を完全に回避する方法を包含することが意図されている。さらに、本発明の微生物は、好都合にも増大した濃度の毒素フルフラールの存在下で糖類を発酵させることができる。
【0119】
本発明の1つの利点は、これまで十分に活用されていなかったサッカリド源を使用できることである。その結果として、多くの複合サッカリド基質を、解重合並びに本発明の組換え細菌及び方法を使用したその後の発酵のための出発源として使用することができる。理想的には、再生可能資源をSSF工程において使用できる。オフィスから出る混合古紙は好ましい基質であり(Brooks et al.,(1995)Biotechnol.Progress.11:619-625;Ingram et al.,(1995)米国特許第5,424,202号明細書)、酸で前処理したバガス(Doran et al.,(1994)Biotech.Bioeng.44:240-247)又は高度に精製された結晶セルロース(Doran et al.,(1993)Biotechnol.Progress.9:533-538)よりもはるかに容易に消化される。エンドグルカナーゼ及びエキソグルカナーゼの両方のグルカナーゼがセルロース結合ドメインを含み、これらの酵素は、遠心分離を用いて消化されなかったセルロース残基を採取することにより、その後の発酵のために容易に再生利用できる(Brooks et al.,(1995)Biotechnol.Progress.11:619-625)。より高いリグニン含量を有する基質に関しては再生利用工程の数が限られるかもしれないが、そのようなアプローチは精製セルロースに関して良好に機能する。本発明の範囲内である他の基質源は、あらゆる種類の加工又は非加工植物材料、例えば、刈り取った芝、トウモロコシの皮、トウモロコシの穂軸、葉柄、葉、繊維、パルプ、麻、おがくず、新聞紙等を含む。
【0120】
本発明はまた、上述した本発明の単離された細菌又は組換え細菌を含むキットを提供する。このキットは、随意に例えば本明細書で述べる方法及び工程に従ってエタノールを生産するための指示書のような、使用のための指示書を提供する。そのような指示書は、随意に増大した濃度のフルフラール中でエタノールを生産することを記述してもよい。一実施形態では、キットは糖源を含む。
【実施例】
【0121】
本発明を以下の実施例によってさらに説明するが、それらは限定と解釈されるべきではない。実施例全体を通じて、特に明記されない限り以下の材料及び方法を使用する。
【0122】
材料及び方法
LY168株は、ヘミセルロース加水分解物における糖類の発酵に関してこれまでに記述されている。LY180(NRRL B−50239)を生じさせる基質範囲を改善するためにいくつかの変更を加えた(ラクトース利用の回復、エンドグルカナーゼの組込み及びセロビオース利用の組込み)。これらの菌株についての関連する特徴を以下の表1に示す。表1に示す組込みのために使用した線状フラグメントはGenBankに寄託されている。
【0123】
【表1】
【0124】
LY180株をLBグルコースアンピシリンプレートで一晩増殖させ、各々を使用してAM1キシロースの試験管に3〜4ODまで接種した。この培養物を直ちに使用して、4mLのAM1 5%キシロース、アンピシリン及び表示の濃度のフルフラールを含む13の100mmの蓋付き試験管に、0.05の初期ODまで接種した。培養物を37℃の水浴で増殖させ、24及び48時間後にOD 500nmでの読み取りを行った。
【実施例1】
【0125】
sthAは、主としてNADPH酸化の方向における機能を促進する動力学的特性を備えた細胞質トランスヒドロゲナーゼである(52)。大腸菌の細胞質トランスヒドロゲナーゼSthA遺伝子をpTrc99aにクローニングし、配列決定によって確認した。LY180株を、図1A〜1Bに示すようにこれらのプラスミドで改変した。クローニングした遺伝子の機能性をインビトロアッセイによって確認した。AM1、アンピシリン及び5%キシロース中で改変したLY180株の増殖を24時間後及び48時間後に比較した。光学密度を漸増フルフラール濃度に対してプロットした。空のベクタを対照として使用した。接種の前にインデューサを添加した。
【0126】
0.1mMのIPTGでの誘導すると、活性が約1.0nmol/分/mgタンパク質から18nmol/分/mgタンパク質に増大することが認められた。プラスミドからのsthA遺伝子の発現は、IPTGでの誘導の有無に関わらずフルフラール耐性を変化させなかった(図1)。図1A〜1Bの結果は、24時間後(図1A)及び48時間後(図2B)に、それぞれ、すべてのLY180株がsthA遺伝子の発現レベルに関わりなく漸増フルフラール濃度で実質的に同様の増殖の減少を示したことを明らかにする。sthA遺伝子の過剰発現はフルフラール耐性に影響を及ぼさないと見られた。
【実施例2】
【0127】
PntABは、発酵増殖の間機能しないと考えられているプロトン転位トランスヒドロゲナーゼであるが、潜在的にNADPHのプールを増大させることができる(52)。実施例2及び3では、大腸菌の細胞質トランスヒドロゲナーゼPntABをpTrc99aにクローニングし、配列決定によって確認した。天然のリボソーム結合部位及びrho依存性ターミネータを有するPntABを、HindIII消化可能末端を有するプライマを使用して大腸菌株LY180ゲノムDNAからポリメラーゼ連鎖反応(PCR)で増幅した。PCR産物とpTrc99aをHindIII消化し、Qiagen Qiaprep Spin Miniprep Kitを用いて精製して、T4 Quick DNAリガーゼを使用して連結した。生じたベクタをTOP10F’に形質転換した後、プラスミドを抽出し、LY180を形質転換するために使用した。方向をPCR分析によって確認した。LY180株を、図2〜3に示すようにこれらのプラスミドで改変した。空のベクタを対照として使用した。接種の前にインデューサを添加した。pntA及びpntB遺伝子の発現増大を有するプラスミドを担持するLY180株の増殖を比較した(図2A〜2B)。24及び48時間後の光学密度を漸増フルフラール濃度に対してプロットした。24時間後、0.05%のフルフラール濃度で、pTrc99a−pntAB正プラスミドを担持するLY180株は、試験したその他の菌株のいずれについても2倍以上の光学細胞密度測定値を示した(図2A)。48時間後、pTrc99a−pntAB正プラスミドを担持するLY180株は、0.10%のフルフラール濃度で持続的な細胞増殖を示したが、その他のすべての菌株は実質的に増殖を停止していた(図2B)。これらの結果は、選択されたpntA及びpntB過剰発現がフルフラール耐性を増大させることを指示する。
【実施例3】
【0128】
様々なレベルのトランスヒドロゲナーゼsthA及びpntAB遺伝子発現増大を有するプラスミドを担持するLY180株の増殖の結果を、それぞれ、24時間後と48時間後に0.05%及び0.10%フルフラール濃度で比較した(図3A〜3B)。24時間後、0.05%のフルフラール濃度で、pTrc99a−pntAB正プラスミドを担持するLY180株は、試験したその他の菌株のいずれについても約2倍の光学細胞密度を示した(図3A)。48時間後、0.10%のフルフラール濃度で、pTrc99a−pntAB正プラスミドを担持するLY180株は、試験したその他の菌株のいずれについても約7倍の光学細胞密度を示し、試験したその他の菌株は、そのようなフルフラール濃度で、あったとしてもごくわずかな増殖を示した。この場合も、これらの結果は、選択されたpntA及びpntB過剰発現がフルフラール耐性を増大させることを明らかにしている。
【0129】
非誘導プラスミドからのpntABの漏洩発現は、1g/l フルフラールの存在下で部分的に増殖を回復した(図3)。この酵素のより高いレベルを発現させるためにIPTGを添加すると、フルフラールに対する耐性を排除し、フルフラールの不在下での細胞の増殖も阻害した。これらの結果に基づき、フルフラールは、生合成のために必要なNADPHの供給を枯渇させることによって増殖を阻害する。硫酸塩同化のためにシステイン当たり4当量という大量のNADPHを必要とし、発酵の間のキシロースからのNADPH生産のための経路が限られていることから、含硫アミノ酸の生産はNADPHに関するフルフラールレダクターゼからの競合を最も受けやすかった。
【実施例4】
【0130】
菌株、培地及び増殖条件
エタノール生産性菌株を、固体培地については20g/lキシロースを添加し、発酵実験については50g/lキシロース又はそれ以上を添加したAM1無機塩培地に保持した(22)。大腸菌株LY180(48、60)はKO11の誘導体であり、この試験のための出発点として使用した。大腸菌W(ATCC9637)は、当初大腸菌Bの誘導体であると報告された、KO11株の親であることに留意されたい(29)。
【0131】
発酵は、2N KOHの自動添加を用いて先に述べられているように(100g/lキシロース、37℃、150rpm、pH6.5)実施した(23)。
【0132】
先に記述されているように、総容量4mLのAM1及び50g/lのろ過滅菌したキシロースを含むスタンディングチューブ中で増殖を測定することによってフルフラール耐性を試験した(48)。チューブを37℃で培養し、24時間後と48時間後に測定した。報告されている数値は4回の測定の平均である。
【0133】
菌株の構築
大腸菌トランスヒドロゲナーゼ遺伝子を、隣接するHindIII部位を提供するプライマによりBioRad iCycler(Hercules,CA)を用いてLY180株ゲノムDNAから増幅した(リボソーム結合部位、コード領域、及び200bpのターミネータ領域)(25)。HindIIIで消化した後、生成物をHindIII消化したpTrc99a(ベクタ)に連結し、大腸菌TOP10F’(Carlsbad,CA)に形質転換した。QiaPrep Spin Mini Prep Kit(Valencia,CA)を用いてプラスミドを精製した。制限酵素での消化及びポリメラーゼ連鎖反応によって遺伝子の配向性を確認した(図10の表1)。
【0134】
マイクロアレイ分析
培養物を小さな発酵槽で670mg dcw/lの密度に増殖させた。初期試料を取り出し、対照として使用した。50g/l保存水溶液から直ちにフルフラールを添加して(最終濃度0.5g/l)、2回目の試料採取の前に15分間培養を続けた。試料をエタノール−ドライアイス浴中で急速に冷却し、4℃で遠心分離によって採取し、Qiagen RNA Laterに再懸濁して、−80℃で保存した。Qiagen RNeasy Mini Kitを用いてRNAを抽出し、DNase Iで処理して、フェノール/クロロホルム抽出及びエタノール沈殿によって精製した。大腸菌K12のために設計されたプローブを使用したマイクロアレイ比較のために、RNAをNimbleGen(Madison,WI)に送付した。各々の試料は、4つの発酵槽からのプールされた材料から成っていた。完全な実験を2回実施して、データを平均した(8つの発酵槽)。ArrayStar(DNA Star,Madison,WI)及びSimPhenyソフトウエア(Genomatica Inc.,San Diego,CA)を使用してデータを解析した。発現比を2つのプールされたデータセットの平均として提示する。対照として水だけを添加した実験では、54の遺伝子(>トランスクリプトームの2%)が水の添加後に2倍より大きく変化することが認められた。これらのうち8つだけがフルフラール添加によって影響を受け、フルフラール送達手順の転写への影響がフルフラールの作用に比べて無視し得るものであることを指示した。
【0135】
ネットワーク成分解析
ネットワーク成分解析(Network Component Analysis;NCA)は、発現比及び公知の調節関係から転写因子活性比を計算するものであり、これを先に記述されているように実施した(45、46、56)。関連性ファイルをRegulon DB及びEcoCyc(4、14)に従って更新した。「緊縮調節因子」のレギュロンを、MOPSグルコース中のBW25113の中間対数増殖の間のセリンヒドロキサメート処理によるセリン飢餓に対する5分間応答の分析により、先に記述されているように定義した(12)。有意に変化した調節活性を有する調節因子を、ヌル分布との比較によって及び0.05のP値カットオフを使用して同定した。
【0136】
フルフラールのトランスクリプトーム効果
活発に増殖している細胞において0.5g/lフルフラールの添加前と添加の15分後にメッセージレベルを比較した。水対照も比較のために含めた。小さな群の遺伝子の発現がフルフラールに応答して5倍より大きく変化していることが認められた(図12の表3)。よりストリンジェントでない測定基準(2倍又はそれ以上)を使用すると、約400の遺伝子(トランスクリプトームの10%)についての発現レベルが、水対照又はフルフラール添加前の培養物と比較して、フルフラール添加に応答して変化した。これらの変化した遺伝子の分布は機能群の間で大きく異なっており、フルフラールの作用機構に関する有用な洞察を提供した(図11の表2)。大部分の機能群では、遺伝子成員の10%未満の発現レベルが2倍又はそれ以上変化した。この低頻度の変化を有する群は、補因子、炭素化合物、調節、高分子合成(細胞構造、DNA、脂質、転写及び翻訳)、及びその他(ファージ、推定上/IS、調節及び未分類/不明)を含んだ。4つの群(細胞プロセス、中枢代謝、エネルギー及び輸送)では成員遺伝子の10%〜20%についての発現レベルが変化した。中枢代謝、エネルギー及び輸送に関連する、影響を受けた遺伝子の大部分は、フルフラール添加時に発現が増大した。これらの変化は、利用可能であり得る付加的な化合物を捕捉し、代謝する、及びエネルギー産生のための炭素流を増大させる機会を提供する。細胞プロセスに関係する変化した遺伝子の多くは運動性及び化学走性に関与するが、LY180株は非運動性である(データは示していない)。
【0137】
以前の報告(48)と一致して、フルフラールのNADPH特異的還元を触媒することができる2つのオキシドレダクターゼ、YqhD及びDkgAは、先に報告されているように、フルフラールの添加によって親において5倍以上上方調節された(mRNA)。これら2つの遺伝子はフルフラール耐性変異株(EMFR9)では沈黙化し、フルフラール耐性の増大を生じさせた。
【0138】
2つの機能群における成員遺伝子の20%以上についての発現レベルが、フルフラール、アミノ酸及びヌクレオチドの添加によって変化した。これらの群では、変化した遺伝子の2/3以上が、フルフラールの添加によって2倍又はそれ以上低減した。プリン、ピリミジン及びあらゆるファミリーのアミノ酸の生合成に影響を及ぼす個々の遺伝子についての発現レベルは、フルフラールの添加によって2倍又はそれ以上低減した。ヌクレオチド代謝における1個の遺伝子及びアミノ酸代謝に関与する8個の遺伝子だけが、少なくとも2倍のフルフラール依存性発現増大を示した。合わせて考慮すると、これらの変化は、フルフラールの添加時に観察される生合成及び増殖の全般的な減少と一致する。
【0139】
調節活性へのフルフラールの効果
フルフラールに対する細胞応答の全体的見解を提供するためにネットワーク成分解析(NCA)を用いた。この解析は、トランスクリプトームデータから、摂動活性を有する調節因子を同定するために公知の調節ネットワーク構造を利用する(46、56)。この解析に含まれた60の調節因子のうちで、22がフルフラール負荷後に有意に変化した発現を有すると同定され、有意性はランダムネットワークに対して評価した(図6A)。システイン及びメチオニン生合成の調節因子(CysB及びMetJ)並びにアミノ酸のリプレッサ(ArgR)及びヌクレオチド生合成のリプレッサ(PurR)は、フルフラール添加によって有意に影響された。緊縮応答(アミノ酸及び炭素飢餓の間、資源を増殖から転用すること)の集合的なインジケータである緊縮調節因子も、停滞した生合成及び多くの中間体の過剰と一致する活性化を示す。合わせて考慮すると、これらの結果は、多くのアミノ酸及び生合成中間体のプールがフルフラール添加によって変化していたことを示す。システイン及びメチオニン生合成に関与する遺伝子の発現は増大したが、大部分の他の生合成経路の発現減少したという事実は、フルフラール負荷から生じる初期事象としてのシステイン及びメチオニンプールの枯渇と一致する。
【0140】
一般的なストレス応答についてのシグナルとして働くσ因子、RpoSもフルフラールの添加によって影響され、細胞がストレス誘導物質の存在を認識することを示した。大腸菌ゲノムの10%までが何らかの方法でRpoSによって調節されるので(51、59)、特異的阻害応答を決定することは困難である。発現増大を有するRpoS調節性遺伝子の例は、poxB(ピルビン酸塩の酢酸塩とCO2への変換)(64)及びotsA(浸透圧ストレス応答の間のトレハロース産生)(55)を含む。
【0141】
中枢代謝における数多くの遺伝子(aceB、aceE、sdhC、dctA、cyoA、fumA)の発現増大及び嫌気条件下で活性であることが一般に知られている遺伝子、fumBの下方調節によって証明されたように、ArcAはNCAによって有意に下方調節された。フルフラール耐性へのグリセロール添加の影響を検討した。しかし、グリセロール(1.0〜20g/l)の添加はフルフラール耐性に影響を及ぼさなかった(データは含めていない)。fis及びcrpを含むいくつかの他の調節因子は、NCAによって有意に変化することが認められた。
【0142】
ヒスチジンもフルフラールの添加によって制限され得る。His調節因子(ヒスチジニル−tRNA)の制御下の遺伝子(hisA、hisB、hisC、hisD、hisF、hisH及びhisI)は、フルフラールの添加後、2倍未満であるが、一般に増大した(図7)。ヒスチジン生合成の2つの最終工程は、発酵に関連する高いNADH/NAD+比によって減速できる反応である、NAD+のNADHへの還元を含む。
【0143】
硫黄のアミノ酸への同化に関与する遺伝子の発現へのフルフラールの効果
硫黄のシステイン及びメチオニンへの同化に関与する多くの遺伝子(cysC、cysH、cysI、cysM、cysN、cysQ、metA、metB、metC、metL、sbp、tauA、tauB、tauC及びtauD)は2倍又はそれ以上増大した。これらはいくつかの機能群内に散らばっている(図11の表2):アミノ酸、中枢代謝、調節及び輸送。硫黄の同化に関与する多くの付加的な遺伝子も2倍未満上方調節されて、幅広いフルフラール応答を明らかにするために含めている(図8)。硫黄はAM1培地中で硫酸塩として供給され、4分子のNADPHを必要とするエネルギー消費型反応である、組込みのために硫化水素のレベルに還元されねばならない。これらの遺伝子の発現のフルフラール誘導性増大は、アミノ酸、プリン及びピリミジンの生合成に関与する多くの他の遺伝子について認められる減少とは著しい対照を成している(図11の表2)。タウリン輸送遺伝子(tauABCD;硫黄の選択的供給源)、硫酸塩結合性輸送タンパク質(sbp)、及び多くのシステイン生合成遺伝子の転写活性化因子(cbl)の発現は、添加されたフルフラールに応答して5倍を超えて増大した(図12の表3)。合わせて考慮すると、これらの結果は、フルフラールの添加が、この経路における高いNADPH必要量に関連した含硫アミノ酸(システイン及びメチオニン)の細胞内欠乏を生じさせることを明らかにしている。
【0144】
フルフラール耐性へのアミノ酸添加物の影響
20アミノ酸すべてを、AM1無機塩培地におけるLY180の増殖を改善するそれらの能力に関して個別に試験した(図9A)。個々のアミノ酸の細胞含量におおよそ基づいて0.1mMの濃度を選択した(50)。5つのアミノ酸だけが、この低濃度で供給した場合にフルフラール耐性を改善した:システイン>メチオニン>セリン、アルギニン>ヒスチジン。2つの含硫アミノ酸は、フルフラール耐性のために明らかに最も有益であった。5倍高い濃度(0.5mM)で供給した場合は、すべてのアミノ酸がある程度まで有益であった(図9B)。しかし、システインが最も有効であることは変わらず、次いでセリン、メチオニン及びアルギニンであった。0.05mMのシステイン濃度では、LY180は、総細胞硫黄量にほぼ等しい、1g/lフルフラールの存在下で0.8g/lの密度に増殖することが可能であった(図4C)。0.01mMシステインではフルフラール耐性の測定可能な改善は認められなかった。
【0145】
システインの防御濃度(0.05mM)は1g/lフルフラール(10mM)の濃度よりも200倍低かった。無機塩培地中のフルフラールはその特徴的なスペクトルによって容易に定量され(Martinez et al.,2000)、ごくわずかな化学的反応性と一致して、37℃で48時間の培養の間不変のままであった。
【0146】
ヒスチジン生合成に関与する大部分の遺伝子は、2倍未満であるが、フルフラール添加に応答して増大した(図7)。発酵の間のヒスチジンの新規生合成は、嫌気的増殖においては高いNADH/NAD+比によって及び生合成の2つの最終工程ではNAD+のさらなる還元のための必要量によって抑制される。同様に、セリン生合成における最初の関与工程もNAD+の還元を含み、発酵の間妨げられる。セリンを増大させることは、H2Sからの還元硫黄をシステインに組み込む効率を高める。アルギニン生合成に関与する遺伝子(argA、argB、argC、argD、car A、carB及びargG)は、一般にフルフラールの添加後2倍以上減少した。アルギニンデカルボキシラーゼをコードするspeAの発現レベルはフルフラールの添加によって増大した。アルギニンの分解は、生合成のための有用な中間体及び補因子を提供する。
【0147】
フルフラール耐性への選択的硫黄源の影響
フルフラールの添加は増殖を阻害し、硫黄の同化に関与する遺伝子の転写を増大させた。選択的硫黄化合物、タウリンの取込み及び組込みに関与する遺伝子(tauABC及びtauD)は、最大の発現増大を有する10遺伝子に含まれた(図12の表3)。tau遺伝子は、典型的には硫黄飢餓の間だけ発現される(57)。システインはフルフラール阻害を取り除くのに有効であったので、これらの遺伝子の発現増大はフルフラールによる含硫アミノ酸のプールの減少から生じる。フルフラールは、硫酸塩からの還元硫黄(H2S)のアベイラビリティを制限することによって又は還元硫黄のシステインへの組込みを阻害することによって、含硫アミノ酸の生合成を阻害する。
【0148】
1g/lフルフラールを含むAM1培地での増殖の間に、還元の異なるレベルで代謝に入る選択的な硫黄源(L−システイン、D−システイン、タウリン及びチオ硫酸ナトリウム)の作用(図9D)を比較した。システインへの同化の前に硫酸塩を完全に還元するためには4分子のNADPHと2つのレダクターゼ酵素(CysH及びCysIJ)が必要であることに留意しなければならない。L−システイン、D−システイン及びチオ硫酸塩は両方のレダクターゼ酵素を回避し、すべてがフルフラール阻害を取り除くのに有効であった。D−システインは直接組み込まれることができず、最初にH2Sに異化される(49)。チオ硫酸塩はまた、CysMによる組込みのための還元硫黄の供給源としても働く(47)。タウリンは細胞質内で亜硫酸塩に異化され、システインへの同化の前に亜硫酸レダクターゼ(CysIJ及び3分子のNADPH)によって還元されねばならない(54)。システイン及びチオ硫酸塩と異なり、タウリンは1g/lフルフラールによる増殖の阻害を防ぐのに有効ではなかった。
【0149】
選択的硫黄源に関するこれらの結果は、フルフラールが、還元硫黄のシステインへの組込みを阻害することによってではなく、硫酸塩からの還元H2Sの生産を制限することによって増殖を阻害するように働くことを示す。硫黄源としてのタウリンに関しては、フルフラールは、フルフラールレダクターゼYqhD(8μM)よりも高いNADPHに対するKm(80μM)(53)を有する、亜硫酸レダクターゼ(CysIJ)のレベルで作用しなければならない(48)。硫黄源としての硫酸塩に関しては、硫黄同化のより早い工程でのフルフラールのさらなる作用を除外することはできない。
【実施例5】
【0150】
エタノール生産性細菌における5−ヒドロキシメチルフルフラール耐性の増大
ヘミセルロース加水分解物中に存在するフラン阻害剤を耐容する生物触媒の能力は、再生可能な化学物質の生産のために重要である。この実施例は、エタノール生産性大腸菌(E.coli)LY180のフルフラール耐性変異株であるEMFR9が、5−ヒドロキシメチルフルフラール(5−HMF)に対する耐性も獲得していることを明らかにする。フラン耐性は、NADPHに対して低いKmを有する2つのフランレダクターゼ、yqhD及びdkgAのより低い発現から生じる。フラン耐性はまた、NADPH/NADHトランスヒドロゲナーゼ(pntAB)をコードするプラスミドを付加することによっても増大した。
【0151】
材料及び方法
菌株、培地及び増殖条件
この試験で使用した菌株及びプラスミドは以前に記述されている(70及び23)。これらは、LY180(大腸菌のエタノール生産性誘導体)、EMFR9(LY180のフルフラール耐性誘導体)、LY180ΔyqhD、LY180ΔdkgA、LY180ΔyqhDΔdkgA、yqhDを含有するpLOI4301を含む。dkgAを含有するプラスミドpLOI4303(48)、及びpntABを含有するpLOI4316(70)も使用した。培養物を、20g/lキシロース(固体培地)、50g/lキシロース(Bioscreen C増殖アナライザ及び試験管培養物)、又は100g/l(pH制御発酵)を含むAM1最小培地(23)において37℃で増殖させた。
【0152】
HMFに対する耐性を、指示されているようにAM1 4ml及び5−HMFを含む13×100mmの閉管を用いて試験した。適切な場合は、プラスミドの保持のための抗生物質を含ませた。管に、0.05OD550nmの初期密度で接種した。48時間の培養(60rpm)後にSpectronic 20D+分光光度計(Thermo,Waltham,MA)を用いて増殖を測定した。フラン耐性へのpntABの作用を調べるために、各ウエルにつき400μlのAM1(及び5−HMF又はフルフラール)を含むマルチウォールプレートに上記のように接種した。Bioscreen C増殖アナライザ(Oy Growth Curves,Helsinki,Finland)を用いてOD(420〜580nmの帯域幅)を72時間測定した。
【0153】
発酵実験のために、LY180及びEMFR9の種培養物を、AM1培地350mlを含む小さな発酵槽で一晩増殖させた(37℃、200rpm)。2NのKOHの自動添加によって培養液をpH6.5に保持した。中間対数増殖期に達したとき、実験発酵槽に0.1 OD550nmの初期細胞密度(33mg乾燥細胞重量/l)で接種した。細胞質量(OD550nm)及びフランレベルを先に述べられているように(20)12時間の間隔で観測した。
【0154】
インビボでのフランの還元を、pH制御発酵槽を用いて測定した。培養物が約1OD550nmに達したとき、10%w/v保存溶液を使用してフランを添加した。細胞質量及び5−HMFを0、15、30及び60分後に測定した。
【0155】
インビトロでのフランの還元
AM1及び0.1mMのIPTGを含む培養試験管(13×100mm)を0.05OD550nmに接種し、37℃で培養した。これらを1〜2OD550nmの密度で採取した。細胞ペレットを100mMのリン酸カリウム緩衝液(pH7.0)で1回洗浄して、10OD550nmの密度で緩衝液に再懸濁した。試料(1ml)を、Lysing Matrix Bを含む2ml試験管に添加し、FastPrep−24(MP Biomedicals,Solon,OH)を用いて破壊した(20秒間)。NADPHのフラン依存性酸化を、DU800分光光度計(Beckman Coulter,Fullerton,CA)を用いて340nmで測定した。反応物(総容量200μl;37℃)は、粗抽出物50μL、0.2mMのNADPH及び20mMの5−HMFを含んでいた。BCAアッセイ(Thermo Scientific,Rockford,IL)を用いてタンパク質を測定した。
【0156】
統計分析
データを平均±SD(n≧3)として提示する。Graphpad Prismソフトウエア(La Jolla,CA)を使用して統計比較(両側スチューデントt検定)を行った。
【0157】
結果
EMFR9株は5−HMFに対する増大した耐性を示す
フルフラール耐性変異株、EMFR9中に存在する突然変異は、5−HMFに対する耐性も増大させた(図14)。1.0g/Lの5−HMFで、EMFR9による増殖及びエタノール生産は、5−HMFの不在下でのLY180(親)のものに等しかった(図14A、14B)。5−HMFは、細胞収率又はエタノール収率に有害な影響を及ぼさずに発酵の最初の24時間にEMFR9によって速やかに代謝された。LY180は1.0g/lの5−HMFによって完全に阻害されたが、5−HMFレベルは培養の間緩やかに減少した(図14A、14B及び14C)。接種しない場合は減少が認められず(データは示していない)、これが代謝作用の結果であることを確認した。
【0158】
EMFR9に関しては、2.5gの5−HMF/lを含めることによってエタノール生産及び増殖は減速したが、それでも進行して96時間後に完了した(図14D、14E及び14F)。このより高いレベルの5−HMFに関する細胞収率及びエタノール収率は、5−HMFなしでのLY180と同等であった。5−HMFのレベルはEMFR9によって急速且つ完全に減少した。LY180に関しては、5−HMFの代謝は遅く且つ不完全であった(図14F)。
【0159】
5−HMF耐性へのYqhD及びDkgAの作用
EMFR9におけるフルフラール耐性は、2つのNADPH依存性オキシドレダクターゼ、YqhD及びDkgAのサイレンシングから生じることが以前に明らかにされた(23)。これらの活性をコードする遺伝子をpCR2.1 TOPOにクローニングし、EMFR9に形質転換して、0.1mMのIPTGで誘導した。細胞を採取し、破壊して、5−HMFレダクターゼ活性に関して試験した(図15A)。プラスミドからの個別のyqhD及びdkgAの発現は、NADPHの5−HMF依存性酸化の速度の5倍増大を生じさせ、YqhD及びDkgAが5−HMFを基質として利用することを確認した。
【0160】
プラスミドからのyqhD及びdkgAの個別発現は、5−HMFに対するEMFR9の耐性を減少した(図15B)。プラスミドの維持のためにカナマイシン(12.5mg/l)を添加すると、すべての株において5−HMF耐性が減少し、この実験では5−HMFのより低い濃度(1.0g/l)の使用を必要とした。プラスミドpCR2.1は、IPTGの不在下でクローニングした遺伝子の発現に関して露出性である(71)。yqhDの非誘導発現でさえも、5−HMFに対するEMFR9の感受性を回復するのに十分であった。5−HMFによる増殖阻害はyqhD誘導によってさらに増大した。dkgAの発現はそれほど有効ではなく、EMFR9において5−HMF感受性を回復するには誘導が必要であった。これら2つのオキシドレダクターゼの間の有効性の差は、DkgAについての23μMと比較して、NADPHに対するYqhDのより低い見かけKm(8μM)と一致する(23)。
【0161】
LY180からのyqhDの欠失は、耐性を2.5g/l 5−HMFに増大した(図15C)。マーカが染色体内に残存する欠失はそれよりも有効でなかったが、yqhDの不活性化はすべての場合に5−HMF耐性のために有益であることを確認した。
【0162】
NADPHのアベイラビリティを高めることは5−HMF耐性を増大させた
NADPHのアベイラビリティを高めるためにプロトン転位トランスヒドロゲナーゼpntAB(68)をLY180において過剰発現させた(図16)。阻害剤の不在下では(図16A)、ベクタを含むLY180(対照)及びLY180(pTrc99a pntAB)の両方が同じ速度で増殖した。IPTG(0.01mM)でのLY180(pTrc99a−pntA)の誘導は、5−HMFの不在下では有害であった。しかし、非誘導LY180(pTrc099a pntAB)は、5−HMF(0.9g/l及び1.8g/l)の存在下でベクタ対照よりも速やかに増殖した(図16B及び16C)。pntABの同様の効果がフルフラールに関して以前に認められている(23)。従って両方のフランによる増殖の阻害はフランの還元から生じると思われ、生合成のために必要なNADPHのプールを枯渇させる。加えて、pntABの過剰発現は、フルフラールの不在下でさえも、72時間後に全体的な増殖の増大を導き、生合成がこれらの条件下でNADPHによって制限され得ることを指示した。
【0163】
硫黄の同化及びシステインの生合成はNADPHの特に高い必要量を有する。システインを添加することは大腸菌LY180においてフルフラール耐性を増大させることが以前に示されているが(70)、5−HMF耐性に関してはそれより有益でないことが認められた(図17)。LY180の増殖は1g/lの5−HMFによって部分的に阻害され、100μMのシステインを添加することによって完全に回復した(図17A)。2.5g/lの5−HMFの存在下での増殖は、100μM又は1000μMシステインによって回復しなかった(図17B)。フルフラールと異なり、システインの添加は5−HMFについてのMICを増大しなかった。
【0164】
参考文献
1. Almeida, J.R.M., T. Modig, A. Petersson, B. Hahn-Hagerdal, G. Liden, and M.F. Gorwa-Grauslund. 2007. Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates by Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 82:340-349.
2. Almeida, J.R.M, A. Roder, T. Modig, B. Laddan, G. Liden, and M.F. Gorwa-Grauslund. 2008. NADH- vs. NADPH-coupled reduction of 5-hydroxymethyl furfural (HMF) and its implications on product distribution in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 78:939-945.
3. Barciszewski, J., G.E. Siboska, B.O. Pedersen, B.F.C. Clark, and S.I.S. Ratten. 1997. A mechanism for the in vivo formation of N-6-furfuryladenine, kinetin, as a secondary oxidative damage product of DNA. FEBS letters. 414:457-460.
4. Boopathy, R., H. Bokang, and L. Daniels. 1993. Biotransformation of furfural and 5-hydroxymethyl furfural by enteric bacteria. J. Indust. Microbiol. 11:147-150.
5. Datsenko, K.A., and B.L. Wanner. 2000. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. PNAS. 97:6640-6645.
6. Gorsich, S.W., B.S. Dien, N.N. Nichols, P.J. Slinger, Z.L. Liu, and C.D. Skory. 2006. Tolerance to furfural-induced stress is associated with pentose phosphate pathway genes ZWF1, GND1, RPE1, and TKL1, in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 71:339-349.
7. Gutierrez, T., L.O. Ingram, and J.F. Preston. 2006. Purification and characterization of a furfural reductase (FFR) from Escherichia coli strain LY01-An enzyme important in the detoxification of furfural during ethanol production. J. Bacteriol. 121:154-164.
8. Habrych, M., S. Rodriguez, and J. Stewart. 2002. Purification and identification of an Escherichia coli Beta-Keto Ester Reductase as 2,5-diketo-D-gluconate reductase YqhE. Biotechnol. Prog. 18:257-261.
9. Hahn-Hagerdal, B., M. Galbe, M.F. Gorwa-Grauslund, G. Liden, and G. Zacchi. 2006. Bio-ethanol - the fuel of tomorrow from the residues of today. Trends Biotechnol. 24:549-556.
10. Horvath, I.S., M.J. Taherzadeh, C. Niklasson, and G. Liden. 2001. Effects of furfural on anaerobic continuous cultivation of Saccharomyces cerevisiae. Biotechnol. Bioeng. 75:540-549.
11. Hristozova, Ts., A. Angelov, B. Tzvetkova, D. Paskaleva, V. Gotcheva, S. Gargova, and K. Pavlova. 2006. Effect of furfural on carbon metabolism key enzymes of lactose-assimilating yeasts. Enzyme Microbiol. Technol. 39:1108-1112.
12. Jantama, K., X. Zhang, J.C. Moore, K.T. Shanmugam, S.A. Svoronos, and L.O. Ingram. 2008. Eliminating side products and increasing succinate yields in engineered strains of Escherichia coli C. Biotech. Bioeng. 30:881-893.
13. Jarboe, L.R., T.B. Grabar, L.P. Yomano, K.T. Shanmugan, and L.O. Ingram. 2007. Development of ethanologenic bacteria. Adv. Biochem. Engin/Biotechnol. 108:237-261.
14. Jeudy, S., V. Monchois, C. Maza, JM Claverie, C. Abergel. 2006. Crystal structure of Escherichia coli DkgA, a broad-specificity aldo-keto reductase. Proteins. 62:302-307.
15. Karp, P.D., I.M. Keseler, A. Shearer, M. Latendresse, M. Krummenacker, S.M. Paley, I. Paulsen, J. Collado-Vides, S. Gama-Castro, M. Peralta-Gil, A. Santos-Zavaleta, M.I. Penaloza-Spinola, C. Bonavides-Martinez, and J. Ingraham. 2007. Multidimensional annotation of the Escherichia coli K12 genome. Nucleic Acids Res. 35:7577-7590.
16. Katahira, S., A. Mizuike, H. Fukuda, and A. Kondo. 2006. Ethanol fermentation from lignocellulosic hydrolysate by a recombinant xylose and cellooligosaccharide-assimilating yeast strain. Appl. Microbiol. Biotechnol. 72:1136-1143.
17. Khan, Q.A., F.A. Shamsi, and S.M. Hadi (1995) Mutagenicity of furfural in plasmid DNA. Cancer Lett. 89:95-99.
18. Ko, J., I. Kim, S. Yoo, B. Min, K. Kim, and C. Park. 2005. Conversion of methylglyoxal to acetol by Escherichia coli aldo-keto reductases. J. Bacteriol. 187:5782-5789.
19. Martin, C., M. Marcet, O. Almazan, and L.J. Jonsson. 2007. Adaptation of a recombinant xylose-utilizing Saccharomyces cerevisiae strain to a sugarcane bagasse hydrolysate with high content of fermentation inhibitors. Bioresour. Technol. 98:1767-1773.
20. Martinez, A., M.E. Rodriguez, S.W. York, J.F. Preston, and L.O. Ingram. 2000. Use of UV absorbance to monitor furans in dilute acid hydrolysates of biomass. Biotechnol Prog. 16:637-641.
21. Martinez, A., M.E. Rodriguez, M.L. Wells, S.W. York, J.F. Preston, and L.O. Ingram. 2001. Detoxification of dilute acid hydrolysates of lignocellulose with lime. Biotechnol. Prog. 17:287-293.
22. Martinez, A., M.E. Rodriguez, S.W. York, J.F. Preston and L. O. Ingram. 2000. Effects of Ca(OH)2 treatments ("overliming") on the composition and toxicity of bagasse hemicellulose hydrolysates. Biotechnol. Bioengin. 69(5): 526-536.
23. Martinez, A., T.B. Grabar, K.T. Shanmugam, L.P. Yomano, S.W. York, and L.O. Ingram. 2007. Low salt medium for lactate and ethanol production by recombinant Escherichia coli B. Biotechnol. Lett. 29:397-404.
24. McCue, L.A., W. Thompson, C.S. Carmack, M.P. Ryan, J.S. Liu, V. Derbyshire, and C.E. Lawrence. 2001. Phylogenetic footprinting of transcription factor binding sites in proteobacterial genomes. Nucleic Acids Res. 29:774-782.
25. Miller, J.H.. 1992. A short course in bacterial genetics. CSHL Press. Plainview, New York.
26. Modig, T., G. Liden, and M.J. Taherzadeh. 2002. Inhibition effects of furfural on alcohol dehydrogenase, aldehyde dehydrogenase, and pyruvate dehydrogenase. Biochem. J. 363:769-776.
27. Nakamura, C.E., and Whited GM. 2003. Metabolic engineering for the microbial production of 1,3-propandiol. Current Opinion in Biotechnology. 14:454-459.
28. Nilsson, A., M.F. Gorwa-Grauslund, B. Hahn-Hagerdal, and G. Liden. 2005. Cofactor dependence in furan reduction by Saccharomyces cerevisiae in fermentation of acid-hydrolyzed lignocellulose. Appl. Environ. Microbiol. 71:7866-7871
29. Ohta, K., D.S. Beall, K.T. Shanmugam, and L.O. Ingram. 1991. Genetic improvement of Escherichia coli for ethanol production: chromosomal integration of Zymomonas mobilis genes encoding pyruvate decarboxylase and alcohol dehydrogenase II. Appl. Environ. Microbiol. 57:893-900.
30. Palmqvist, E., and B. Hahn-Hagerdal. 2000. Fermentation of lignocellulosic hydrolysates. I: inhibition and detoxification. Bioresour. Technol. 74:17-24.
31. Palmqvist, E., and B. Hahn-Hagerdal. 2000. Fermentation of lignocellulosic hydrolysates. II: inhibitors and mechanisms of inhibition. Bioresour. Technol. 74:25-33.
32. Perez, J.M., F.A. Arenas, G.A. Pradenas, J.M. Sandoval, and C.C. Vasquez. 2008. Escherichia coli YqhD exhibits aldehyde reductase activity and protects from the harmful effect of lipid peroxidation-derived aldehydes. J. Biol. Chem. 283:7346-7353.
33. Petersson, A., J.R.M. Almeida, T. Modig, K. Karhumaa, B. Hahn-Hagerdal, M.F. Gorwa-Grauslund, and G. Liden. 2006. A 5-hydroxymethyl furfural reducing enzyme encoded by Saccharomyces cerevisiae ADH6 gene conveys HMF tolerance. YEAST. 23:455-464.
34. Singh, N.P., and A. Khan. 1995. Acetaldehyde: Genotoxicity and cytotoxicity in human lymphocytes. Mutat. Res 337:9-17.
35. Sulzenbacher, G., K. Alvarez, R. Heuvel, C. Versluis, S. Spinelli, V. Campanacci, C. Valencia, C. Cambillau, H. Eklund, and M. Tegoni. 2004. Crystal structure of E. coli alcohol dehydrogenase YqhD: Evidence of a covalently modified NADP coenzyme. J. Mol. Biol. 342:489-502.
36. Talebnia, F., and M.J. Taherzadeh. 2006. In situ detoxification and continuous cultivation of dilute-acid hydrolysate to ethanol by encapsulated S-cerevisiae. J Biotechnol. 125:377-384.
37. Tokiwa, Y., and B.P.Calabia. 2008. Biological production of functional chemicals from renewable resources. Can. J. Chem. 86:548-555.
38. Um, B.H., M.N. Karim, and L.L. Henk 2003. Effect of sulfuric and phosphoric acid pretreatments on enzymatic hydrolysis of corn stover. Appl Biochem Biotechnol. 105-108:115-125.
39. White, D. 2000. The Physiology and Biochemistry of Prokaryotes. 2nd edition. Oxford University Press. New York, NY.
40. Almeida, J.R.M., M. Betilsson, M.F. Gorwa-Grauslund, S. Gorsich, and G. Liden. 2009. Metabolic effects of furaldehydes and impacts on biotechnological processes. Appl. Microbiol. Biotechnol. 82:625-638.
41. Amann, E., B. Ochs, and K.J. Abel. 1988. Tightly regulated tac promoter vectors for the expression of unfused and fused proteins in Escherichia coli. Gene 69:301-315.
42. Gama-Castro, S., J.J. Veronica, M. Peralta-Gil, A. Santos-Zavaleta, M.I. Penaloza-Spindola, B. Contreras-Moreira, J. Segura-Salazar, L. Muniz-Rascado, I. Martinez-Flores, H. Salgado, C. Bonavides-Martinez, C. Abreu-Goodger, C. Rodriguez-Penagos, J. Miranda-Rios, E. Morett, E. Merino, A.M. Huerta and J. Collado-Vides. 2008 RegulonDB (version 6.0): gene regulation model of Escherichia coli K-12 beyond transcription, active (experimental) annotated promoters and Textpresso navigation. Nucl. Acids Res., 36:D120-D124.
43. Gralla, J.D. 2005. Escherichia coli ribosomal RNA transcription: regulatory roles for ppGpp, NTPs, architectural proteins and a polymerase-binding protein. Molecular Microbiology. 55:973-977.
44. Gyaneshwar, P., O. Paliy, J. McAuliffe, D.L. Papham, M.I. Jordan, and S. Kustu. 2005. Sulfur and nitrogen limitation in Escherichia coli K-12: Specific homeostatic responses. J. Bacteriol. 187:1074-1090.
45. Hyduke, D.H., L.R. Jarboe, L.M. Tran, K.J.Y. Chou, and J.C. Liao. 2007. Integrated network analysis identifies nitric oxide response networks and dihydroxyacid dehydratase as a crucial target in Escherichia coli. PNAS. 104:8484-8489.
46. Liao, J.C., R. Boscolo, Y.L. Yang, L.M. Tran, C. Sabatti, and V.P. Roychowdhury. 2003. Network Component Analysis: Reconstruction of regulatory signals in biological systems. PNAS. 100:15522-15527.
47. Maier, T.H. 2003. Semisynthetic production of L-alpha-amino acids by metabolic engineering of the cysteine-biosynthetic pathway. Nature Biotechnol. 4:422-427.
48. Miller, E.M., L.R. Jarboe, L.P. Yomano, S.W. York, K.T. Shanmugam and L.O. Ingram. 2009. Silencing of NADPH-dependent oxidoreductases (yqhD and dkgA) in furfural-resistant ethanologenic Escherichia coli. Appl. Environ. Microbiol. IN PRESS.
49. Nagasawa T., T. Ishi, H. Kumagai, and H. Yamada. 1985. D-cysteine desulfhydrase of Escherichia coli - purification and characterization. Eur. J. Biochem. 153:541-551.
50. Neidhardt, F.C. Escherichia coli and Salmonella Typhimurium. 1987. vol. 1. American Society for Microbiology. Washington, DC.
51. Patten, C.L., M.G. Kirchhof, M.R. Schertzberg, R.A. Morton, and H.E. Schellhorn. 2004. Microarray analysis of RpoS-mediated gene expression in Escherichia coli K-12. Mol. Genet. Genomics. 272(5):580-591.
52. Sauer, U., F. Canonaco, S. Heri, A Perrenoud, E. Fisher. 2004. The soluble and membrane-bound transhydrogenases UdhA and PntAB have divergent functions in NADPH metabolism of Escherichia coli. J. Biol. Chem. 279:6613-6619.
53. Schomburg I., A. Chang, O. Hofmann, C. Ebeling, F. Ehrentreich, D. Schomburg. 2002. BRENDA: a resource for enzyme data and metabolic information. Trends Biochem. Sci. 27(1):54-6.
54. Siegel, L.M., P.S. Davis. 1974. Reduced Nicotinamide Adenine Dinucleotide Phosphate-Sulfite Reductase of Enterobacteria. J. Biol. Chem. 249:1587-1598.
55. Strom, A.R., and I. Kaasen. 1993. Trehalose metabolism in Escherichia coli:stress protection and stress regulation of gene expression. Mol. Microbiol. 8:205-210.
56. Tran, L. M., M. P. Brynildsen, K. C. Kao, J. K. Suen, and J. C. Liao. 2005. gNCA: a framework for determining transcription factor activity based on transcriptome: identifiability and numerical implementation. Metab Eng 7:128-41.
57. van der Ploeg, J.R., M.A. Weiss, E. Saller, H. Nahimoto, N. Saito, M.A. Kertesz, T. Leisinger. 1996. Identification of sulfate starvation-regulated genes in Escherichia coli: a gene cluster involved in the utilization of taurine as a sulfur source. J. Bacteriol. 178:5438-5446.
58. Vemuri, G.N., E. Altman, D.P. Sangurdekar, A.B. Khodursky, M.A. Eiteman. 2006. Overflow metabolism in Escherichia coli during steady-state growth: Transcriptional regulation and effect of the redox ratio. Appl. Environ. Microbiol. 72:3653-3661.
59. Weber, H., T. Polen, J. Heuveling, V.F. Wendish, and R. Hengge. 2005. Genome-wide analysis of the general stress response network in Escherichia coli: sigma S-dependent genes, promoters, and sigma factor selectivity. J. Bacteriol. 187:1591-1603.
60. Yomano, L.P., S.W. York, K.T. Shanmugam, and L.O. Ingram. 2009. Deletion of methylglyoxal synthase gene (mgsA) increased sugar co-metabolism in ethanol-producing Escherichia coli. Biotechnol. Lett. IN PRESS.
61. Zaldivar, J., and L.O. Ingram. 1999. Effect of organic acids on the growth and fermentation of ethanologenic Escherichia coli LY01. Biotechnol. Bioeng. 66: 203-210
62. Zaldivar, J., A. Martinez, and L.O. Ingram. 1999. Effect of selected aldehydes on the growth and fermentation of ethanologenic Escherichia coli. Biotechnol. Bioeng. 65: 24-33.
63. Zaldivar, J., A. Martinez, and L.O. Ingram. 2000. Effect of alcohol compounds found in hemicellulose hydrolysate on the growth and fermentation of ethanologenic Escherichia coli. Biotechnol. Bioeng. 68:524-530.
64. Zhang, T.F., and L.P. Hager. 1987. A single-step large scale purification of pyruvate oxidase. Arch. Biochem. Biophys. 257:485-48.
65. Almeida JRM, Roder A, Modig T, Laadan B, Liden G, Gorwa-Grauslund MF (2008) NADH- vs. NADPH-coupled reduction of 5-hydroxymethyl furfural (HMF) and its implications on product distribution in Saccharomyces cerevisiae. Appl Microbiol Biotechnol 78:939-945.
66. Cheng KK, Cai BY, Zhang JA, Ling HZ, Zhou YH, Ge JP, Xu JM (2008) Sugarcane bagasse hemicellulose hydrolysate for ethanol production by acid recovery process. Biochem Eng J 38:105-109.
67. Heer D, Heine D, Sauer U (2009) Resistance of Saccharomyces cerevisiae to high furfural concentration is based on NADPH-dependent reduction by at least two oxidoreductases. Appl Environ Microbiol doi:10.1128/AEM.01649-9.
68. Keseler IM, Bonavides-Martinez C, Collado-Vides J, Gama-Castro S, Gunsalus RP, Johnson DA, Krummenacker M, Nolan LM, Paley S, Paulsen IT, Peralta-Gil M, Santos-Zavaleta A, Shearer AG, Karp PD (2009) EcoCyc: a comprehensive view of Escherichia coli biology. Nucleic Acids Res 37:D464-70.
69. Liu ZL, Moon J (2009) A novel NADPH-dependent aldehyde reductase gene from Saccharomyces cerevisiae NRRL Y-12632 involved in the detoxification of aldehyde inhibitors derived from lignocellulosic biomass conversion. Gene 446:1-10.
70. Miller EN, Jarboe LR, Turner PC, Pharkya P, Yomano LP, York SW, Nunn D, Shanmugam KT, Ingram LO (2009a) Furfural inhibits growth by limiting sulfur assimilation in ethanologenic Escherichia coli strain LY180. Appl Environ Microbiol 75:6132-6141.
71. Purvis JE, Yomano LP, Ingram LO (2005) Enhanced trehalose production improves growth of Escherichia coli under osmotic stress. Appl Environ Microbiol 71:3761-3769.
72. Taherzadeh MJ, Gustafsson L, Niklasson C, Liden G (2000) Physiological effects of 5-hydroxymethylfurfural on Saccharomyces cerevisiae. Appl Microbiol Biotechnol 53:701-708.
73. Um BH, Karim MN, Henk LL (2003) Effect of sulfuric and phosphoric acid pretreatments on enzymatic hydrolysis of corn stover. Appl Biochem Biotechnol 105:115-125.
74. Wyman CE, Dale BE, Elander RT, Holtzapple M, Ladisch MR, Lee YY (2005) Comparative sugar recovery data from laboratory scale application of leading pretreatment technologies to corn stover. Bioresour Technol, 96:2026-2032.
【0165】
参照による組み込み
本明細書で特定された、全ての出版物、特許出願及び特許は、その全体が参照により本明細書に明確に組み込まれている。
【0166】
均等物
当業者は、通常の実験を実施するだけで、本明細書に記載される本発明の特定の実施形態の多くの均等物を認識又は確認することができる。そのような均等物は、本明細書に包含されることが意図されている。
【特許請求の範囲】
【請求項1】
参照細菌と比較して少なくとも1つのトランスヒドロゲナーゼ遺伝子の発現が増大している、単離又は組換えエタノール生産性細菌。
【請求項2】
トランスヒドロゲナーゼ遺伝子がpntA及びpntBである、請求項1に記載の細菌。
【請求項3】
当該細菌が、参照細菌と比較してpntA及びpntB遺伝子の増大した発現を有する、単離又は組換え細菌。
【請求項4】
当該細菌が、参照細菌と比較してフルフラール耐性が増大している、請求項3に記載の細菌。
【請求項5】
参照細菌が野生型細菌である、請求項3に記載の細菌。
【請求項6】
当該細菌がエタノール生産性である、請求項3に記載の細菌。
【請求項7】
当該細菌が参照細菌と比較してエタノール生産性が増大している、請求項3に記載の細菌。
【請求項8】
当該細菌が、参照細菌と比較してフルフラール又は5−HMFの存在下で増大したエタノール生産性を示す、請求項3に記載の細菌。
【請求項9】
当該細菌が参照細菌と比較して増殖性が増大している、請求項3に記載の細菌。
【請求項10】
当該細菌が、参照細菌と比較してフルフラール又は5−HMFの存在下に増殖性が増大している、請求項3に記載の細菌。
【請求項11】
当該細菌が、約0.025%フルフラール〜約0.15%フルフラールの濃度で5−HMFの存在下に増殖性が増大している、請求項3に記載の細菌。
【請求項12】
当該細菌が、約0.025%フルフラール〜約0.15%フルフラールの濃度でフルフラールの存在下に増殖性が増大している、請求項3に記載の細菌。
【請求項13】
当該細菌が、参照細菌と比較して増殖性及びエタノール生産性が増大している、請求項3に記載の細菌。
【請求項14】
当該細菌が、参照細菌と比較して加水分解物の存在下で増殖性が増大している、請求項3に記載の細菌。
【請求項15】
加水分解物が、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス又はセルロースバイオマスを含む生成物に由来する、請求項14に記載の細菌。
【請求項16】
当該発現が、pntA及びpntB遺伝子の発現を調節するプロモータ又は調節タンパク質を修飾する又は付加することによって増大する、請求項3に記載の細菌。
【請求項17】
当該細菌が、嫌気又は微好気条件下で主発酵生成物としてエタノールを生産することができる、請求項3に記載の細菌。
【請求項18】
当該細菌が、グラム陰性細菌及びグラム陽性細菌から成る群より選択される、請求項3に記載の細菌。
【請求項19】
グラム陰性細菌が、エシェリキア属(Escherichia)、アシネトバクター属(Acinetobacter)、ザイモモナス属(Zymomonas)、グルコノバクター属(Gluconobacter)、ジオバクター属(Geobacter)、シュワネラ属(Shewanella)、サルモネラ属(Salmonella)、エンテロバクター属(Enterobacter)及びクレブシエラ属(Klebsiella)から成る群より選択される、請求項18に記載の細菌。
【請求項20】
グラム陽性細菌が、バチルス属(Bacillus)、クロストリジウム属(Clostridium)、コリネバクテリウム属(Corynebacterium)、乳酸杆菌属(Lactobacillus)、ラクトコッカス属(Lactococcus)、オエノコッカス属(Oenococcus)、連鎖球菌属(Streptococcus)及びユーバクテリウム属(Eubacterium)から成る群より選択される、請求項18に記載の細菌。
【請求項21】
細菌が大腸菌(Escherichia coli)である、請求項3に記載の細菌。
【請求項22】
細菌がクレブシエラ・オキシトカ(Klebsiella oxytoca)である、請求項3に記載の細菌。
【請求項23】
参照細菌と比較してPntA及びPntBタンパク質の活性が増大している、単離又は組換え細菌。
【請求項24】
当該細菌が、参照細菌と比較してフルフラール耐性を増大している、請求項23に記載の細菌。
【請求項25】
当該細菌が、参照細菌と比較して5−HMF耐性を増大している、請求項23に記載の細菌。
【請求項26】
参照細菌と比較してpntA及びpntB遺伝子の発現が増大しており、且つ細菌が参照細菌と比較してフルフラール耐性を増大している、単離又は組換え細菌。
【請求項27】
参照細菌と比較してpntA及びpntB遺伝子の発現又はPntA及びPntBポリペプチドの活性が増大しており、参照細菌と比較してフルフラール又は5−HMFの耐性が増大しており、当該細菌がエタノールを生産することができ、並びに当該細菌が:
(a)細菌の候補株をフルフラール又は5−HMFの存在下で増殖させること;及び
(b)フルフラール又は5−HMFの存在下でエタノールを生産する細菌を選択すること:
を含む工程によって調製される、単離又は組換え細菌。
【請求項28】
バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖を、請求項1〜26のいずれか一項に記載の単離又は組換え細菌と接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含有してなる、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産する方法。
【請求項29】
バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖を、請求項1〜26のいずれか一項に記載の単離された又は組換え細菌と接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含有してなる、フルフラールの存在下でバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産する方法。
【請求項30】
請求項28に記載の方法によって生産されるエタノール。
【請求項31】
請求項29に記載の方法によって生産されるエタノール。
【請求項32】
請求項1〜27のいずれか一項に記載の単離又は組換え細菌を含むキット。
【請求項1】
参照細菌と比較して少なくとも1つのトランスヒドロゲナーゼ遺伝子の発現が増大している、単離又は組換えエタノール生産性細菌。
【請求項2】
トランスヒドロゲナーゼ遺伝子がpntA及びpntBである、請求項1に記載の細菌。
【請求項3】
当該細菌が、参照細菌と比較してpntA及びpntB遺伝子の増大した発現を有する、単離又は組換え細菌。
【請求項4】
当該細菌が、参照細菌と比較してフルフラール耐性が増大している、請求項3に記載の細菌。
【請求項5】
参照細菌が野生型細菌である、請求項3に記載の細菌。
【請求項6】
当該細菌がエタノール生産性である、請求項3に記載の細菌。
【請求項7】
当該細菌が参照細菌と比較してエタノール生産性が増大している、請求項3に記載の細菌。
【請求項8】
当該細菌が、参照細菌と比較してフルフラール又は5−HMFの存在下で増大したエタノール生産性を示す、請求項3に記載の細菌。
【請求項9】
当該細菌が参照細菌と比較して増殖性が増大している、請求項3に記載の細菌。
【請求項10】
当該細菌が、参照細菌と比較してフルフラール又は5−HMFの存在下に増殖性が増大している、請求項3に記載の細菌。
【請求項11】
当該細菌が、約0.025%フルフラール〜約0.15%フルフラールの濃度で5−HMFの存在下に増殖性が増大している、請求項3に記載の細菌。
【請求項12】
当該細菌が、約0.025%フルフラール〜約0.15%フルフラールの濃度でフルフラールの存在下に増殖性が増大している、請求項3に記載の細菌。
【請求項13】
当該細菌が、参照細菌と比較して増殖性及びエタノール生産性が増大している、請求項3に記載の細菌。
【請求項14】
当該細菌が、参照細菌と比較して加水分解物の存在下で増殖性が増大している、請求項3に記載の細菌。
【請求項15】
加水分解物が、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス又はセルロースバイオマスを含む生成物に由来する、請求項14に記載の細菌。
【請求項16】
当該発現が、pntA及びpntB遺伝子の発現を調節するプロモータ又は調節タンパク質を修飾する又は付加することによって増大する、請求項3に記載の細菌。
【請求項17】
当該細菌が、嫌気又は微好気条件下で主発酵生成物としてエタノールを生産することができる、請求項3に記載の細菌。
【請求項18】
当該細菌が、グラム陰性細菌及びグラム陽性細菌から成る群より選択される、請求項3に記載の細菌。
【請求項19】
グラム陰性細菌が、エシェリキア属(Escherichia)、アシネトバクター属(Acinetobacter)、ザイモモナス属(Zymomonas)、グルコノバクター属(Gluconobacter)、ジオバクター属(Geobacter)、シュワネラ属(Shewanella)、サルモネラ属(Salmonella)、エンテロバクター属(Enterobacter)及びクレブシエラ属(Klebsiella)から成る群より選択される、請求項18に記載の細菌。
【請求項20】
グラム陽性細菌が、バチルス属(Bacillus)、クロストリジウム属(Clostridium)、コリネバクテリウム属(Corynebacterium)、乳酸杆菌属(Lactobacillus)、ラクトコッカス属(Lactococcus)、オエノコッカス属(Oenococcus)、連鎖球菌属(Streptococcus)及びユーバクテリウム属(Eubacterium)から成る群より選択される、請求項18に記載の細菌。
【請求項21】
細菌が大腸菌(Escherichia coli)である、請求項3に記載の細菌。
【請求項22】
細菌がクレブシエラ・オキシトカ(Klebsiella oxytoca)である、請求項3に記載の細菌。
【請求項23】
参照細菌と比較してPntA及びPntBタンパク質の活性が増大している、単離又は組換え細菌。
【請求項24】
当該細菌が、参照細菌と比較してフルフラール耐性を増大している、請求項23に記載の細菌。
【請求項25】
当該細菌が、参照細菌と比較して5−HMF耐性を増大している、請求項23に記載の細菌。
【請求項26】
参照細菌と比較してpntA及びpntB遺伝子の発現が増大しており、且つ細菌が参照細菌と比較してフルフラール耐性を増大している、単離又は組換え細菌。
【請求項27】
参照細菌と比較してpntA及びpntB遺伝子の発現又はPntA及びPntBポリペプチドの活性が増大しており、参照細菌と比較してフルフラール又は5−HMFの耐性が増大しており、当該細菌がエタノールを生産することができ、並びに当該細菌が:
(a)細菌の候補株をフルフラール又は5−HMFの存在下で増殖させること;及び
(b)フルフラール又は5−HMFの存在下でエタノールを生産する細菌を選択すること:
を含む工程によって調製される、単離又は組換え細菌。
【請求項28】
バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖を、請求項1〜26のいずれか一項に記載の単離又は組換え細菌と接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含有してなる、バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産する方法。
【請求項29】
バイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖を、請求項1〜26のいずれか一項に記載の単離された又は組換え細菌と接触させ、それによってバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産することを含有してなる、フルフラールの存在下でバイオマス、ヘミセルロースバイオマス、リグノセルロースバイオマス、セルロースバイオマス又はオリゴ糖源からエタノールを生産する方法。
【請求項30】
請求項28に記載の方法によって生産されるエタノール。
【請求項31】
請求項29に記載の方法によって生産されるエタノール。
【請求項32】
請求項1〜27のいずれか一項に記載の単離又は組換え細菌を含むキット。
【図1】

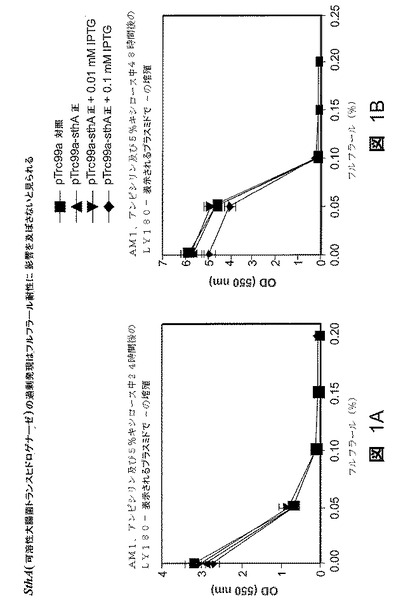
【図2】
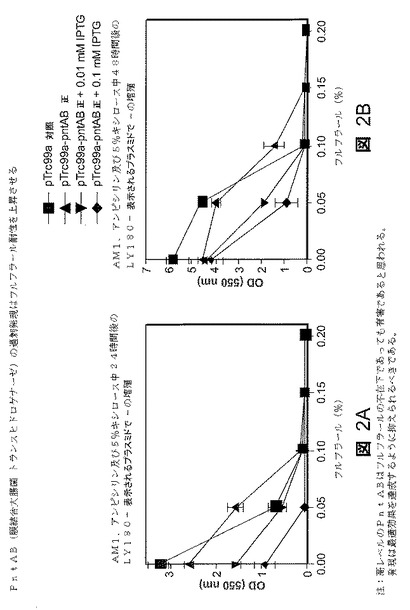
【図3】
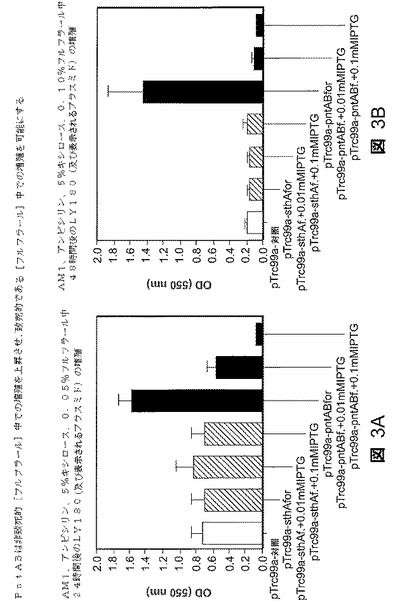
【図4】

【図5】

【図6A】
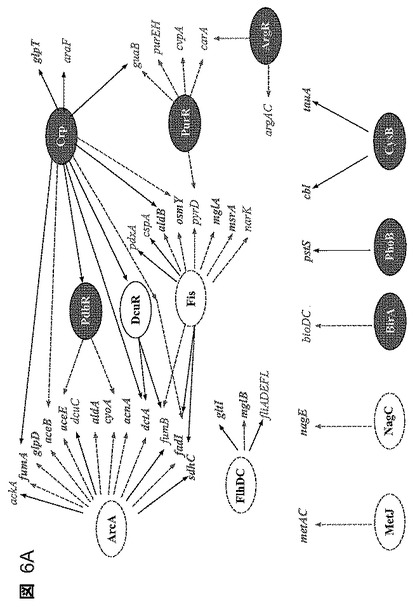
【図6B】
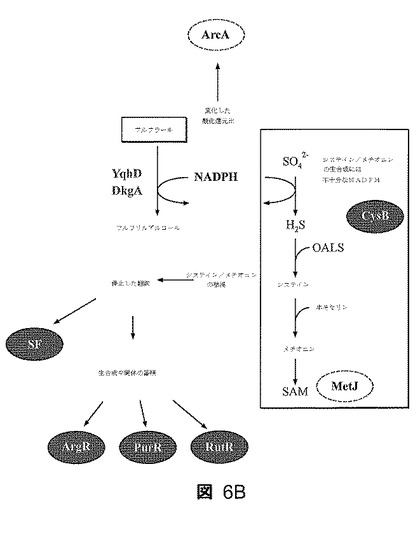
【図7】

【図8】
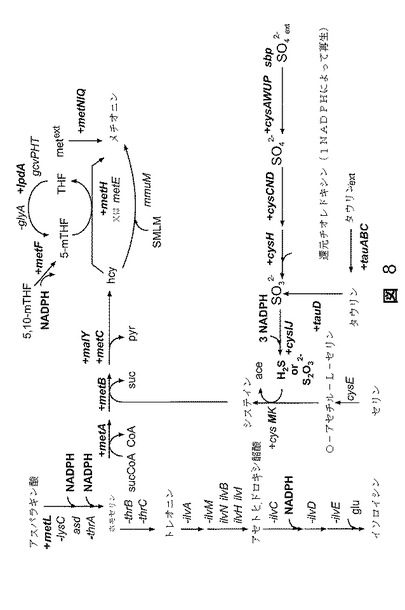
【図9−1】
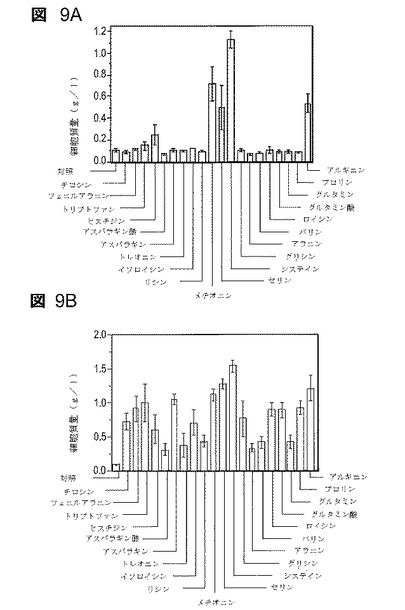
【図9−2】
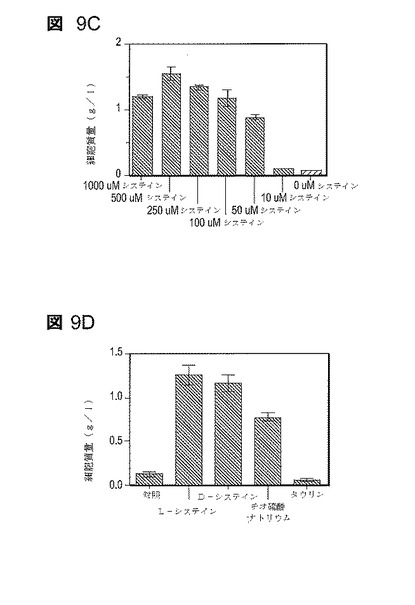
【図10】
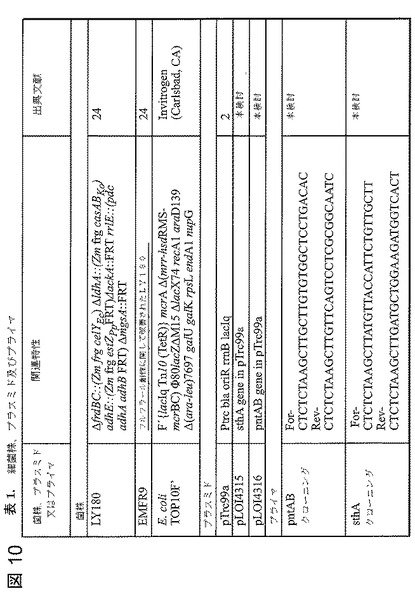
【図11−1】

【図11−2】

【図12】
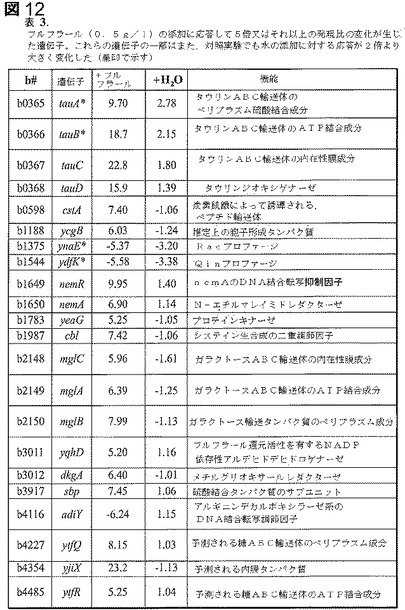
【図13】
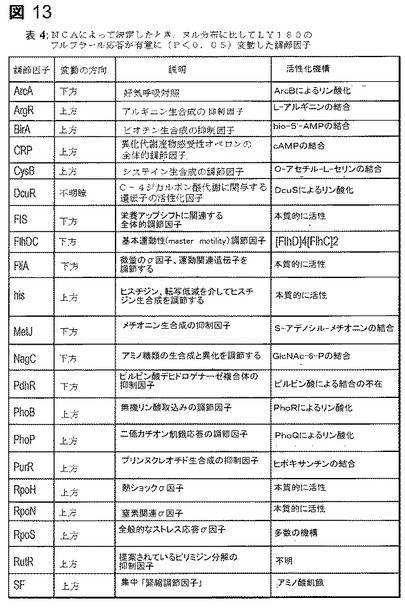
【図14−1】
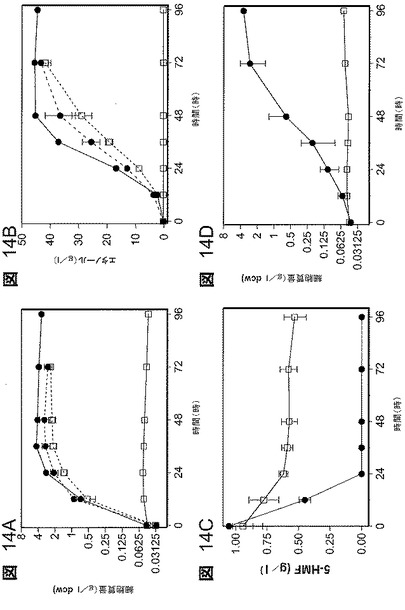
【図14−2】
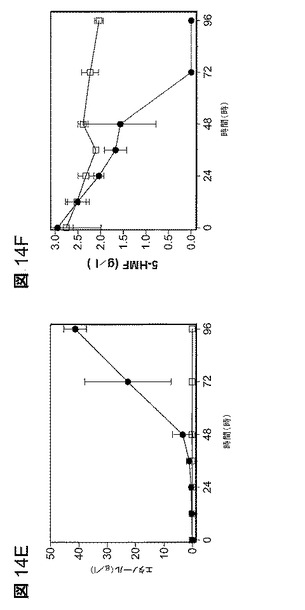
【図15】
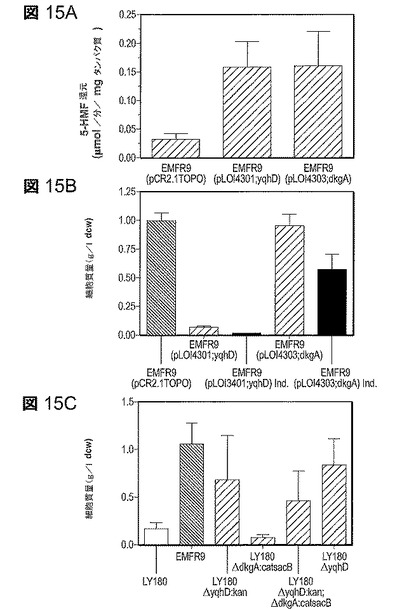
【図16】
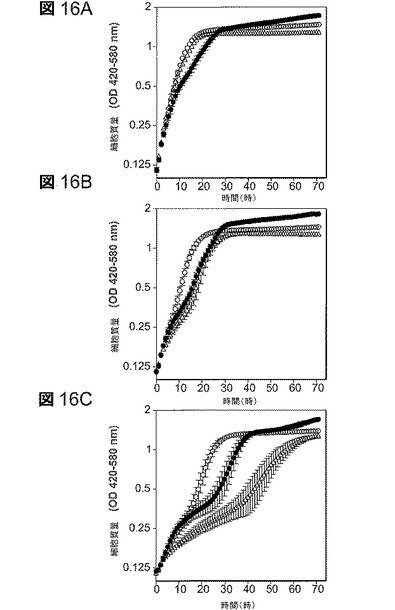
【図17】
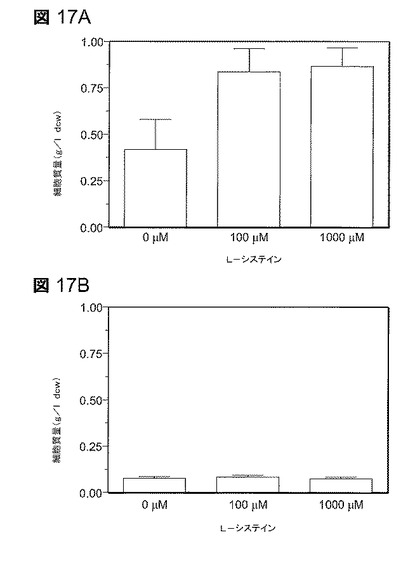
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図7】
【図8】
【図9−1】
【図9−2】
【図10】
【図11−1】
【図11−2】
【図12】
【図13】
【図14−1】
【図14−2】
【図15】
【図16】
【図17】
【公表番号】特表2012−526552(P2012−526552A)
【公表日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願番号】特願2012−510788(P2012−510788)
【出願日】平成22年3月15日(2010.3.15)
【国際出願番号】PCT/US2010/000769
【国際公開番号】WO2010/132079
【国際公開日】平成22年11月18日(2010.11.18)
【出願人】(506040249)ユニバーシティー オブ フロリダ リサーチ ファンデーション, インク. (3)
【Fターム(参考)】
【公表日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願日】平成22年3月15日(2010.3.15)
【国際出願番号】PCT/US2010/000769
【国際公開番号】WO2010/132079
【国際公開日】平成22年11月18日(2010.11.18)
【出願人】(506040249)ユニバーシティー オブ フロリダ リサーチ ファンデーション, インク. (3)
【Fターム(参考)】
[ Back to top ]