
エピジェネティック制御の小分子モジュレーター及びそれらの治療適用
開示される事項は、転写因子の機能を調節するための方法および組成物に関する、特にエピジェネティックレギュレーター(ヒストン修飾酵素)を特異的な DNA プロモーターにリクルートする転写因子である。標的とされる転写因子には、筋細胞増強因子(MEF2), フォークヘッド/ウイングドヘリックス 転写因子 FOXP3 および 転写因子 GATA3が含まれるが、これらに限定されない。また、開示される事項は、MEF2及びその関連因子〔ヒストン デアセチラーゼ (HDACs), p300/CBP および Cabin 1を含むが、これらに限定されない〕の小分子モジュレーター及びその治療適用である。
【発明の詳細な説明】
【発明の説明】
【0001】
[関連出願の相互参照]
この出願は、2008年11月5日に出願された米国特許仮出願第61/111,689号, および2009年9月29日に出願された米国特許仮出願第61/246,934号の利益を主張する出願である。上記の仮出願は、本明細書中に参照によって援用される。
【0002】
[連邦政府により援助された研究および開発に関する記載]
本発明は、NIH R21AI49905, R01HL076334, および RC1DA028790の助成による部分的な援助のもとでなされた。従って、米国政府は、特定の権利を有している。
【0003】
[発明の分野]
本発明は、一般に分子医学の分野に関する。特に、本発明は、転写因子の機能を調節するための方法および組成物に関する、特にエピジェネティックレギュレーター(ヒストン修飾酵素)を特異的な DNA プロモーターにリクルートする転写因子に関する。標的とされる転写因子には、筋細胞増強因子(MEF2), フォークヘッド/ウイングドヘリックス 転写因子 FOXP3 および 転写因子 GATA3が含まれるが、これらに限定されない。特に、本発明は、MEF2及びその関連因子〔ヒストン デアセチラーゼ (HDACs), p300/CBP および Cabin 1を含むが、これらに限定されない〕の小分子のモジュレーター及びその治療への適用に関する。
【0004】
[発明の背景]
本発明は、特定の疾患と関連する転写因子の機能を調節する小分子の使用に関する。転写因子は、特定のDNA配列に結合し、直接的に又は関連するタンパク質〔例えば、コアクチベータおよびコリプレッサー〕をとおして、又はヒストン 修飾酵素〔例えば、ヒストン アセチルトランスフェラーゼ (HATs) および ヒストン デアセチラーゼ (HDACs)〕をリクルートすることにより遺伝子発現を制御するタンパク質である。転写因子は、適切なレベルの遺伝子発現を保証することにより多くの生物学的プロセスに重要な役割を果たしている。また、転写を制御する能力が異常に修飾される場合には特定の病的状態と関連づけすることができる。特定の転写因子の機能を選択的に調節できる小分子を同定し、開発することによって、潜在的に新しい治療適用を導入することができる。本発明は、二つの基本的なアイデアに基づく。一つは、特定の転写因子(例えば、MEF2, FOXP3 および GATA3)に結合し、転写のコアクチベータ および コリプレッサーとの相互作用を調節する小分子を開発することである。もう一つは、HAT (例えば、p300およびCBP) およびHDACs および他のヒストン修飾酵素(例えば、ヒストン メチルトランスフェラーゼ, およびデメチラーゼ, DNAメチルトランスフェラーゼ)のリクルートおよびクロマチンの特定の領域に対するクロマチンリモデリング機構をブロックする小分子を開発することである。
【0005】
特に、本発明は、筋肉, 免疫および神経系の発生および適応応答に重要な役割を担う筋細胞エンハンサー因子-2 (MEF2: myocyte enhancer factor-2)(Flavell et al., 2006; Kim et al., 2008; Mao et al., 1999; McKinsey et al., 2002; Pan et al., 2004; Potthoff and Olson, 2007; Youn and Liu, 2000; Youn et al., 1999)に関する。MEF2は、心筋細胞における肥大応答(hypertrophic responses)の重要な制御因子と指摘された。病理学的な刺激により誘導された心臓肥大は、多くの形態の心血管疾患において心不全を誘発しえる。
【0006】
一般にMEF2は、四つのメンバー(MEF2A, MEF2B, MEF2C および MEF2D)の転写因子のファミリーを規定する。それらの機能の重要性は、マウスおよびショウジョウバエの遺伝学を使用することにより詳細に実証された(Potthoff and Olson, 2007)。最初に同定された骨格筋におけるMEF2は、筋原性のベーシック ヘリックス・ループ・ヘリックス 転写因子(例えば、MyoD)と共に筋形成を促進し、維持する(Molkentin and Olson, 1996)。MEF2A(MEF2ファミリーのメンバーの一つ)は、最近このタンパク質が冠動脈疾患 (CAD) および 心筋梗塞 (MI)と関連することから「心臓発作遺伝子(heart attack gene)」であるとされた(Wang et al., 2003)。これらの所見は、ヒトの心臓疾患におけるMEF2の重要な役割の基礎をなす(Kim et al., 2008; Wei et al., 2008; Zhang et al., 2002)。
【0007】
MEF2は、現在では多くの他の細胞タイプの基本転写因子として知られる。一例を挙げると, MEF2は、リンパ系の発生においてカルシウムシグナル伝達を媒介する重要な転写因子の一つである(Pan et al., 2004; Youn and Liu, 2000; Youn et al., 1999)。MEF2は、サイトカイン発現 および 免疫応答を制御する。また、MEF2は、ニューロンの生存 および シナプスのリモデリングの基礎的な転写プログラムを制御する(Chen and Cepko, 2009; Flavell et al., 2006; Flavell and Greenberg, 2008; Flavell et al., 2008; Mao et al., 1999; Morrow et al., 2008; Shalizi et al,, 2006; Shalizi and Bonni, 2005; Yang et al., 2009)。これらの観察は、MEF2 機能を調節する小分子が心肥大および心不全, 自己免疫疾患および移植拒絶, 神経変性疾患および学習および記憶の病理学的な障害に治療効果を有することを示唆する(Fischer et al., 2007; Stefanko et al., 2009)。
【0008】
細胞の内側のMEF2の作用には、三つの別個の工程が含まれる: (i) 転写抑制; (ii) カルシウム依存性の脱抑制;および (iii) 転写活性化。
【0009】
MEF2による転写抑制は、内因性の又は関連するヒストンデアセチラーゼ (HDAC) 活性を有する種々の転写コリプレッサーとの結合に依存する。T細胞中でMEF2はCabin 1と結合し、次にクラス IのHDACs(例えば、 HDAC 1, HADC2 および HDAC3)と結合する(Youn and Liu, 2000)。筋肉細胞において、MEF2は、直接的にクラス II HDACs(例えば、HDAC4, HDAC5, および HDAC9)と結合し、筋肉の発生 および 適応応答に関与する特異的な遺伝子の発現を阻害する(Chan et al., 2003; Gregoire et al., 2006; Gregoire and Yang, 2005; McKinsey et al., 2001, 2002; Miska et al., 1999; Sparrow et al., 1999)。HDAC5またはHDAC9がノックアウトされたマウスは肥大性刺激への感受性の増加を示し、心臓肥大におけるクラス II HDACsの重要な役割が示唆される(Potthoff and Olson, 2007)。これらの及び他のデータは、MEF2およびクラス II ヒストン デアセチラーゼ (HDACs)(特に、HDAC5およびHDAC9)が心筋細胞における肥大性シグナルの重要なメディエータであることを示す。多くのデータは、MEF2/クラス II HDAC 経路が心臓肥大の潜在的な治療上の標的であることを示唆する。
【0010】
特定のカルシウムシグナルへの応答において、Cabinl および HDACsは、MEF2から除去される(Potthoff and Olson, 2007)。次に、MEF2は、コアクチベータ(例えば、p300 および CBP)をリクルートし、種々の転写のアクチベータ および コアクチベータと結合することにより別個のプログラムを開始させる(McKinsey et al., 2001; Sartorelli et al., 1997; Slepak et al., 2001; Wei et al., 2008)。p300の小さな増加が、MEF2依存性の心肥大を誘発させることに必要十分であることが示された。MEF2は、よく特徴が調べれたMADS-ボックスおよびMEF2-特異的ドメインからなる高度に保存されたN末端領域 (残基2-93)を有する(Shore and Sharrocks, 1995)。MADS-ボックス/MEF2ドメインは機能が著しく豊富であり、DNA結合, 二量体化, およびCabin 1, クラス IIa HDACs および p300/CBPを含む無数のMEF2の転写パートナーとのタンパク質間相互作用を媒介する(McKinsey et al., 2001, 2002)。MEF2におけるMADS-ボックス/MEF2 ドメインが、クラス II HDACs および Cabin1に保存された小さいモチーフと結合するため必要十分であることが示された。また、p300 および CBPのCH3ドメインは、MADS-ボックスMEF2ドメインと結合することが示された。
【0011】
当該技術において利用可能な様々な細胞プロセスにおけるMEF2の関与について広範な知識が存在するにもかかわらず、これまで適切な分子ツールを欠いていることからその知識を利用することは不可能であった。特に、如何にして小分子によりMEF2の活性を調節するかについては長年にわたる挑戦であり、それはMEF2が明らかな酵素活性のない相対的に小さい転写因子であるからであり;MEF2の主な機能は特定のDNAと結合し、転写コレギュレータ(例えば、Cabin 1, クラス II HDACs および p300/CBP)を特異的なプロモーターにリクルートすることである。この様式の作用は、一般に薬にできないと考えられており、少なくとも小分子による標的化は非常に困難である。このような分子の発見または創造によって、この分野のさらなる進捗が促進され、MEF2-関連疾患(炎症, 自己免疫疾患, 神経変性疾患, 癌, および心血管疾患)のための新しい機構に基づく、また構造に基づく治療適用が導かれえる。
【0012】
また、本出願に記載される発明は、他の転写因子(例えば、フォークヘッド/ウイングドヘリックス転写因子 FOXP3)の活性を調節することに拡張できる(Bennett et al., 2001; Fontenot et al., 2003; Hori et al., 2003; Wu et al., 2006; Zheng and Rudensky, 2007)。FOXP3は、制御性T細胞(Tregs)の発生および機能に重要な鍵となる転写因子である。Tregsは免疫系の過剰な活性化を抑制するため必要とされるT細胞の特殊な集団であり、変異によるFOXP3の機能喪失や他の機構によって胎性の自己免疫疾患(例えば、IPEX)が誘発され、他方でFOXP3の発現又はその活性の増強によって抑制機能を付与することができる。FOXP3の機能の上昇は自己免疫疾患 および 移植拒絶の治療に有益でありえるが、他方でFOXP3 活性のダウンレギュレーションを戦略的に使用して免疫に基づく抗腫瘍療法を開発することができる(Zuo et al., 2007a; Zuo et al., 2007b)。以上のとおり、FOXP3と結合し、そのコリプレッサー および コアクチベータとの相互作用を調節する小分子は、自己免疫疾患, 移植拒絶および癌治療における治療適用を有するだろう。
【0013】
MEF2と同様にFOXP3の機能は、HAT (例えば、TIP60) および HDACs (クラス I および クラス II HDACsを含む)を含む転写コレギュレータにより厳格に制御される(Li et al., 2007)。FOXP3と結合し、ヒストン修飾酵素 およびクロマチンリモデリング機構などのエピジェネティックレギュレーターを含むコレギュレータとの相互作用をブロックする本発明に記載された方法により小分子を開発できるだろう。
【0014】
同じように、転写コレギュレータを標的とすることは、同様な挑戦である。
【0015】
転写因子の転写コレギュレータ(例えば、MEF2, クラス II HDACs)は、最良の試験群である。
【0016】
ヒストン デアセチラーゼ (HDACs) (ECナンバー 3.5.1)は、ヒストンにおけるc-N-アセチル リジン アミノ酸からアセチル基を除去する酵素のクラスである。その作用は、ヒストン アセチルトランスフェラーゼ (HATs)の作用と逆である。HDACs タンパク質は、現在では多数の非-ヒストンタンパク質も含まれる彼らの標的というよりむしろ彼らの活性を正確に記載するため、リジン デアセチラーゼ (KDAC)とも称される。
【0017】
彼らの名称から示唆されるように、HDACsの主な機能の一つは、アセチル基をヒストンタンパク質から除去することである。ヒストンタンパク質は、クロマチンの主要なタンパク質成分である。彼らは、周囲にDNAが巻きつくスプールとして働き、遺伝子制御 および DNAパッケージングに重要な役割を担っている。ヒストンタンパク質は、リジン および アルギニンアミノ酸に存在するアミン基のため通常陽性に荷電した尾部を有する。これらの陽性の荷電は、ヒストン尾部がDNAバックボーンにおける陰性に荷電したリン酸基との相互作用や結合を援助する。
【0018】
DNA と ヒストンの間の結合は、転写因子がDNAにアクセスする能力を制御する重要な制御機構として作用する。DNAおよびヒストンの間の強い結合は、転写因子によるアクセスを制限し、遺伝子の転写を抑制する(Morrison et al., 2007)。ヒストンまたはDNAの修飾によって、それらの結合の強さを変化させ、転写活性を調節することができる(Morrison et al., 2007)。メチル, リン酸, またはアセチル部分を共有結合で付加することによって、ヌクレオソームの状態が変化し、結果的に転写に影響することが示されている。アセチル化は、ヒストンにおける保存されたN 末端リジン残基のε-アミノ基へのアセチル基の付加から生じる。アセチル基のヒストンへの付加は陽性に荷電したヒストンタンパク質 および 陰性に荷電したDNAのリン酸バックボーンの間の引力を減少させ、よりリラックスし、アクセス可能なクロマチン構造を生じる。HATsは、ヒストンアセチル化を促進するので、転写アクチベータであると信じられている。逆に, HDACsは、アセチル基をヒストンから除去するため貢献し、それによって転写が抑制される。従って、局所性のクロマチン構造 および 遺伝子発現を主に支配するのはHATs および HDACs活性の間の相互作用である。HDACsは、クロマチンの脱アセチルをとおして全体的な遺伝子転写を変化させる。HDACsは直接的にDNA配列に結合せず、遺伝子の認識を標的とする付加的な因子を必要とすることに注意すべきである(Morrison et al., 2007)。
【0019】
機能およびDNA配列の類似性に基づくHDAC タンパク質の4 つの認められたサブタイプ (クラス I - IV)が存在する。最初の二つのサブタイプは、活性がトリコスタチン A (TSA)により阻害される「古典的」HDACsと考えられる。クラス I HDACsはHDACs 1, 2, 3 および 8を含み、これらはユビキタスに発現される(Zhang and Olsen, 2000)。クラス II HDACsは、4, 5, 7 および 9を含むIIaおよびHDAC6 および HDAC10を含むIIbの二つのサブグループを有する。クラス IIaは、カルボキシル-末端の触媒ドメイン、また転写因子の筋細胞増強因子 2 (MEF2) ファミリーのメンバーとの相互作用を媒介するアミノ-末端の伸展部で共通の構造上の構成を共有する。また、HDACsは細胞内局在が異なり、一般にクラス Iは核に認められ、クラス IIbは大抵が細胞質に局在し、クラス IIaは核および細胞質の間をシャトルする。クラス IのHDACsとは異なり、クラス IIaのHDACsは組織限定的であり、特に高いレベルの発現を心臓, 骨格筋, および脳で示す(Zhang et al., 2002)。クラス IIIはTSAにより影響されないNAD+依存性タンパク質のファミリーであり、クラス IVはそれ自身の非定型のカテゴリーであると考えられる。HDAC11 は、クラスV にグループ化される。
【0020】
HDACsは細胞プロセスにおいて重要な役割を有するので、HDAC インヒビター (HDACi)の医学的な適用は研究が活発な領域である。しかしながら、医学におけるHDACiの多くの使用は、基本的な機構の知識とは無関係に発見された。例えば、精神医学および神経学において、バルプロ酸を精神安定剤(mood stabilizer)および抗癲癇剤として用いてきた長い歴史がある。バルプロ酸の抗痙攣薬特性は、抗痙攣薬として調査された幾つかの他の化合物のビヒクルとして使用された際に偶然発見された。後にバルプロ酸はHDACiと同定された。近年、HDACiは、神経変性疾患のための緩和剤(mitigator)または治療剤として活発に研究されている。癌治療のためのHDACiを開発する多くの努力がなされてきた。例えば、ボリノスタット (SARA)は、最近皮膚の T-細胞 リンパ腫 (CTCL)の治療のため承認された。CTCLの臨床評価中の別の剤は、環状デプシペプチドの天然物 FK228 (Romidepsin)であり、これはクラス IのHDACsの強力なインヒビターである。加えて、臨床試験は、HIVの感染者の潜在的なプールにおいてバルプロ酸の効果を研究している。HDACiの薬適用に大きな関心があるにもかかわらず、これらの化合物が働く正確な機構はまだよく理解されていない。このような事情から、これらの仕事は、大部分は憶測や試行錯誤により導かれた。
【0021】
HDACiを使用する際に特に問題となる事項の一つは、HDAC 活性を阻害する既知の小分子の多くがHDACsの触媒活性を標的とすることで機能するよう設計されたことである。活性部位は多数の異なるHDACアイソフォームにより共有された保存された特徴であるので、アイソフォーム-選択的な HDACiを同定することは本来困難である。従って、多くのHDACiは、特異性が低く、HDACの特定の種を特異的に標的とする能力がない。例えば、トリコスタチン A (TSA)は、現在知られている最も強力な可逆的なHDACiであり、低ナノモル範囲のIC50を有する;また、TSAはヒドロキサム酸基及びフェニル基との五-炭素原子リンカーを有し、HDACの活性部位に適合する至適なコンホメーションを有する(de Ruijter et al., 2003; Somoza et al., 2004)。全てのHDACsは、TSAによる阻害に凡そ等しく感受性であると考えられる(de Ruijter et al., 2003)。
【0022】
以上のように、HDACsの機能を阻害し、関連する転写因子の活性を調節する小分子を発見することに関する主要な障害は、HDAC酵素の活性部位に結合する能力をとおして同定され、評価されるHDACiの発見 および 最適化に最新技術が焦点を合わせていることである。典型的には、これらの HDACiは、一般構造式R-L-Zを有する。式中のRはタンパク質表面認識基であり、短い脂肪性リンカーLを介して活性部位の亜鉛原子と結合するZn2+ -キレート化基Zに連結される。既知のHDACiにおいて特徴づけられた最も共通するキレート基(Z)は、ヒドロキサム酸 (TSA, ボリノスタット, LAQ824, ベリノスタット), チオール誘導体 (FK228, ラルガゾール) または求電子性ケトン (トラポキシンA)である。Zn2+のような金属カチオンに緊密に結合する係る基の潜在的な欠点は、それらが特定のタンパク質に対して十分な選択性を欠いていることから、様々な副作用を生じる可能性があることである。
【0023】
別のクラスの既知のHDACiは、オルト-アミノアニリド (2-アミノアニリド)部分を備え(feature)るベンズアミドであり、MS-725, MGCD0103, ピメロイルアニリド オルト-アミノアニリド (PAOA)および化合物 106 (N'-(2 アミノフェニル)-N7-p-トリルヘプタンジアミド)を含み、これはフリートライヒ失調症およびハンチントン舞踏病を含む神経変性疾患の潜在的な治療剤として調査された(Chou et al., 2008; Herman et al., 2006; Paris et al., 2008; Rai et al., 2008; Thomas et al., 2008; Wong et al., 2003)。このクラスのHDACiの作用の詳細な分子機構は知られていなく、o-アミノアニリド基のHDAC活性部位の亜鉛原子への結合にこれらの分子が関与することが前提とされたが、このような結合モチーフに関して直接的な証拠を欠いている。そのうえ、係る分子によってHDAC 機能の阻害が関係する生物活性が呈されることが認められたが、他の非選択的なHDACiは類似する活性を示さなかった。例えば、化合物106はHDAC阻害は弱いが、フラタキシンの誘導に関して非常に活性であることが示され、他方で密接に関連するHDACi(例えば、SAHA)はこのタイプの活性を有していなかった。結果的には、このクラスの化合物の作用に関する分子機構が提示されていないことから、それらの治療上の潜在的な能力を最適化することが阻まれてきた。
【0024】
様々な HDAC アイソフォームの良好に制御された生物学的役割に起因する幾つかの付加的な要素によって、HDACiのデザインのための更なる挑戦が従来のアプローチに課される。実験によって、アセチル化したヒストンの量がHDACiの存在下で増加することが示された。しかしなから、DNAに結合した転写因子によるHATs および HDACsのリクルートによって、複数のタンパク質の転写調節性の複合体の形成が生じ、従属的な遺伝子の一群(arrays)に細胞タイプ特異性およびシグナル依存的な制御が付与される。無差別にHDACsを阻害するHDACiを用いることは、モンキーランチを複雑で繊細なバランスのとれた機械に投げ入れることに似ており、このことからHDACiが関与する試行の多くで観察された多数の望ましくない副作用が説明される。
【0025】
また、HDACiを医学的に使用する研究において直面する問題は、エピジェネティック制御を研究する研究者により共有されている。エピジェネティック制御は、永久にDNA 配列を変化させることのない遺伝性の遺伝子発現パターンの確立である。この制御は、細胞機能を制御する重要な機構として出現した。
【0026】
エピジェネティック制御の変化は、多くの疾患(特に、癌)の特徴である。これらの疾患の治療における薬物として開発される小分子(表現型スクリーニングを介して見つけられた)は、細胞プロセスのエピジェネティックコントロールを調節することにより作用する。エピジェネティック制御自体の基礎的な機構は、非常に興味がもたれる領域である。同時に、小分子のエピジェネティックレギュレーターの探索は、創薬のため非常に有望な領域となっている。
【0027】
エピジェネティック制御はDNA (例えば、DNA シトシン メチルトランスフェラーゼ)またはタンパク質(例えば、HATS, HDACs, ヒストンメチラーゼ, および ヒストンデメチラーゼ)に作用する酵素によるクロマチン構造の化学的な修飾をとおして大部分が達成されるので、この関連において、DNA転写の制御におけるHDACsの役割をエピジェネティック制御のコンポーネントとして認めることができる。HDACiと類似して、エピジェネティックレギュレーターの現行の化学的なモジュレーターの大多数は、これらの酵素を別の細胞の役割を有する複数の酵素によりしばしば共有される触媒部位に結合することにより阻害する。
【0028】
転写因子(例えば、MEF2)の役割に関する最近の進歩及びその幾つかの主要な疾患において関係性があるにもかかわらず、MEF2の機能を調節する能力のある小分子を同定することは可能ではなかった。このような分子によって、この分野のさらなる進捗が促進され、MEF2-関連疾患(炎症, 自己免疫疾患, 神経変性疾患, 癌, および心血管疾患)のための新しい機構に基づく、また構造に基づく治療適用が導かれえる。
【0029】
以上のとおり、この領域の研究における一般的な問題は、特異的なエピジェネティック制御因子の酵素またはタンパク質を標的としえる小分子を欠いていることである。また、更なる問題は、全体の酵素またはタンパク質のクラスをとおして広域の活性を有することに起因する欠点がなく、必要とされる選択性が与えられる様式で、係る小分子をデザインし, 評価し 最適化する方法を欠いていることである。係る分子は、エピジェネティック制御の基礎的な機構を研究する分子ツールとして、同様に、治療上の介入を目標とする治療剤として重要な適用を有する。
【0030】
[発明の概要]
広範な生化学的な研究及び構造研究(Guo et al., 2007; Han et al., 2005; Han et al., 2003)により、予想外にも発明者は小分子を用いて活性を調節することを可能とするMEF2のユニークな構造上の特性を発見した。この発見は、従来の知識に反することから本発明の核心部分であり、この発見によって新しい可能性のドアが開かれる。この予想外の発見に基づき、発明者は、転写因子とそれらのコファクターとの間の境界のインヒビターを介した相互作用をブロックすることにより細胞プロセスを調節する一般的な戦略を発明した。それゆえ、本発明は、ファクターコファクター複合体の境界の表面の部位に結合し、複合体の形成を破壊する能力のある小分子モジュレーターを同定する, スクリーニングする, アッセイする, および合成するための方法も提供する。
【0031】
以上のとおり、本発明は、これまで「薬になりえない(undrugable)」転写因子(例えば、MEF2)を標的とする長年にわたる課題を解決した。
【0032】
HDACiが直面する特定の問題に関して、本発明はHDAC機能の小分子モジュレーターの開発のための代替的なアプローチを提供する。特定のHDACの活性部位を標的とするよりむしろ、本発明は、転写を制御するため必要とされる関連する転写因子とHDACとの結合部位を標的とする。異なる HDAC サブタイプは異なる転写因子に対し異なる結合表面を有することから、これらのタンパク質間相互作用の境界(interface)を標的とすることにより保存された活性部位に関連して生じる特異性の問題が解消される。
【0033】
特に、本発明は、転写因子 MEF2 および クラス IIa HDACsの間の相互作用を標的とする。MEF2の活性は、標的の遺伝子発現を抑制する特異的なプロモーターにおいてMEF2と結合するクラス IIa HDACsにより制御される(Potthoff and Olson, 2007)。また、様々な癌の治療のため開発される幾つかのHDACsの小分子インヒビター(HDACi)によって、MEF2のディレギュレーション(deregulation)およびHDACs活性が関与する疾患(心肥大, 神経変性障害, および免疫機能不全を含む)の治療の可能性が示された(Morrison et al., 2007; Paris et al., 2008)。これらの観察は、クラス IIa HDAC:MEF2 相互作用をブロックする小分子がメンバー特異的なHDACiとして類似する臨床上の利益を提供しえることを示唆する(Guo et al., 2007; Han et al., 2005; Han et al., 2003)。
【0034】
クラス IIa HDACsは、筋肉, ニューロン および T細胞においてMEF2と密接に作用する。プロモーターを標的とするためクラス IIa HDACsはDNAに結合しないが、その相互作用はDNA-結合型のMEF2に依存する。この相互作用は、クラス IIa HDACsに保存されるが、他のHDACsでは保存されていない短い両親媒性のヘリックスにより媒介され、これがMEF2 のMADS-ボックス/MEF2 ドメインにおける疎水性の溝と結合する(Guo et al., 2007; Han et al., 2005; Han et al., 2003)。このようなリガンド/レセプター様の結合機構が存在することは、小分子を使用してクラス IIa HDACsのMEF2-特異的なプロモーターへのリクルートをブロックすることが可能であろうことを示唆する(Guo et al., 2007; Han et al., 2005; Han et al., 2003)。
【0035】
本発明は、MEF2と結合する小分子が転写コレギュレータ(例えば、Cabin1, クラス IIa HADCS および p300/CBP)のリクルートにおいてその活性を調節できるとの体系的な構造 および 生化学的な研究に基づく(Guo et al., 2007; Han et al., 2005; Han et al., 2003)。これらのコレギュレータの多くは、クロマチンを修飾する内因性の機能(例えば、HDACs, p300 および CBP)またはクロマチン修飾の酵素および機構をリクルートする能力を有する(mSin3AはCabin1, HP1, CtBPと結合し、14-3-3はクラス IIa HDACsと結合する)。従って、MEF2と結合し、他の転写コレギュレータとの相互作用を調節する小分子は、MEF2が重要な調節的な役割を担う組織におけるエピジェネティックモジュレーターとして貢献できる。それゆえ、これらの小分子を使用して、MEF2-依存的な遺伝子発現の活性の制御が異常な疾患を治療することができる。調節不全(dysregulation)は、MEF2及びその関連する因子の遺伝的な変異, MEF2機能の減少または過度な活性化を導く減少した又は過剰なシグナル, MEF2と結合する、また相互作用するコファクターの異常な過小または過剰な発現から生じえる。MEF2-結合小分子の潜在的な臨床適用には、筋肉系, 免疫系および神経系の疾患が含まれるが、これらに限定されない。筋肉系においては、心肥大, 筋線維タイプのリモデリング(muscle fiber type remodelin), およびMEF2機能の不均衡から生じる他の筋肉関連疾患である。免疫系においては、種々の自己免疫疾患又は過剰な又は非常に僅かなMEF2-依存的な遺伝子発現から生じる免疫不全である。また、MEF2-結合小分子を使用して、移植拒絶を予防するため制御性T細胞の機能や全体の免疫応答を操作することができる。MEF2-依存的な遺伝子発現は密接にシナプスのリモデリングやニューロンの生存と関連するので、MEF2 結合小分子を使用して制御がはずれたMEF2機能から生じる種々の神経変性疾患(例えば、アルツハイマー病やハンチントン舞踏病など), 自閉症, 精神医学的な障害, および学習および記憶の障害を治療することができる。
【0036】
本発明の小分子モジュレーターは転写因子及びそのコファクターのタンパク質間相互作用の境界(protein-protein interaction interface)に局在する結合部位を標的とすることにより機能するので、それらも境界のインヒビターとして本出願において参照される。
【0037】
本発明の基礎的な原則を説明したので、我々は次に本発明の様々な側面 および 態様を以下に要約する:
第一の側面において、本発明は、境界のインヒビターの候補をスクリーニングする, 同定する, または最適化するためのアッセイを提供する。この本発明の側面と一致するアッセイは、一般に境界のインヒビターの候補を、各々のMEF2 単量体のベータ鎖S1, S2 およびS3 およびヘリックスH2およびCabin 1およびHDAC4およびHDAC 9からの短いヘリックスモチーフを含むタンパク質間の境界により規定される分子表面を含んでいる評価するエレメント(evaluating element)と接触させる工程を含むだろう。幾つかの好適な態様において、評価するエレメントは、候補インヒビターについての情報を提供するレポートエレメントと作動可能に連結される。好適な態様において、前記アッセイは、MEF2と結合し、その転写コレギュレータへの結合を調節する小分子の迅速でハイスループットなスクリーニングや最適化を許容する細胞に基づくルシフェラーゼアッセイである。このアッセイは、発明者の研究室によるMEF2 複合体の構造および生化学の10年以上の研究に基づき開発された。本発明は、最初に結晶研究により同定され、更に本発明の構造に基づく変異の研究により実証されたタンパク質間の境界がMEF2-結合小分子に高度に特異的で敏感なスクリーニングのための分子的な基礎として貢献できるだろうことを主張する。他の態様において、前記アッセイの評価するエレメントは、本発明により同定されたタンパク質間の境界に基づくプルダウン, 免疫共沈降 または蛍光クエンチングおよび異方性(anisotropy)または任意のインビトロ結合アッセイおよび 細胞に基づくルシフェラーゼ レポーター アッセイ, トランスジェニック レポーターアッセイを含むが、これらに限定されない理学的なアッセイで実行されえる。
【0038】
第二側面において、本発明は、筋肉, 免疫系および神経系を含むが、これらに限定されない種々の哺乳類の組織におけるMEF2-依存的な転写のエピジェネティックモジュレーターとして有用な化合物も提供する。幾つかの好適な態様において、この本発明の側面と一致する化合物は、以前に公開された化合物(Chou et al., 2008; Herman et al., 2006; Paris et al., 2008; Rai et al., 2008; Thomas et al., 2008; Wong et al., 2003), ピメロイルアニリド オルソアミノアニリドPAOA, および商業的に利用可能なホモログ スベロイルアニリド オルソアミノアニリド(suberoylanilide orthoaminoanilide), またはBML-210, 同様に、それらの構造的に関連する誘導体を含みえるが、これらに限定されない。
【0039】
第三の側面において、本発明は、表 1における結晶構造座標で表されるMEF2へのBML-210結合の結晶構造により樹立された構造上のフレームワークから由来するMEF2におけるBML-210 結合部位により規定された分子フレームワーク(molecular framework)も提供する。我々は、BML-210をDNAと結合したMEF2A (1-78) 二量体と共結晶化した。結晶を2.4Aで回折し、MEF2A (1-78): DNA複合体を探索モデルとして用いた分子置換で構造を解析した(Santelli and Richmond, 2000)。BML-210は、拡張したコンホメーションに適合し、MEF2の疎水性ポケットに結合する(図 1d)。分子の一端でフェニルアミド基は、Leu66, Leu67, Thr70, Leu66' (プライム記号は他の単量体からの残基を示す) および Thr70'を含む幾つかの疎水性残基で周囲をとりまかれる。また、この端でのアミド基は、所定の位置にあり、それぞれThr70 および Thr70'との水素結合相互作用に関与する。BML-210の他の端で環連結の電子密度(ring-link electron density)は、Asn73, Gln56', Asp61' および Asp63'で周囲をとりまかれるより親水性の環境にある。この領域は、オルト-アミノアニリド基に対応する。ここでオルトアミノアニリド部分とそのアミド基とでMEF2の残基と広範なファンデルワールス接触や潜在的な水素結合相互作用を作る(図 1d)。オクタンジアミドのメチレン基は、二つのMEF2 分子のヘリックスH2の間に適切に合い、主として疎水性の性質のMEF2残基の主鎖 および 側鎖と多数の接触を作る(図 1d)。MEF2に結合したBML-210の結晶構造に基づき、我々はBML 210 および PAOAと接触する又は近くに近接して接触する表面残基を使用して、実験および計算に基づくアプローチによりMEF2に対する小分子のデザイン および スクリーニングを導くことできることを主張する。これらの残基は、暴露されるMEF2の鎖 S1, S2 および S3 および ヘリックスH2における全ての残基を含む。この構造上のフレームを本発明に記載のハイスループットで特異的な高感度アッセイと組み合せることで、BML210 およびPAOA誘導体および新規の分子骨格(molecular scaffolds)に基づく分子を含むMEF2-結合小分子の迅速なデザインおよび最適化が許容される。本発明において開示されたMEF2に結合したBML-210の結晶構造に加えて本出願に記載される小分子 MEF2-結合部位を規定することで、BML-210に及び他のベンズアミド含有HDACインヒビターに存在するオルト-アミノアニリド部分のための有望な結合部位が初めて示された。特に、この結合部位は、このクラスのHDACインヒビターに関して以前に仮定されたHDAC 酵素の活性部位と異なる。
【0040】
第四の側面において、本発明は、上記の第三の側面における分子フレームワークにより規定された境界の結合部位への結合に有用なMEF2-結合小分子を提供する。一態様において、提供される分子は、MEF2と結合するBML-210の結晶構造から由来する同定された結合部位から由来する一般構造式を有するMEF2-結合小分子である。提供される小分子は、記載された構造上のフォールドと結合するため設計され、潜在的に高い親和性および選択性でMEF2と結合するだろう。この本発明の側面の態様において提供される化合物は、一般式 Ra-L Rbの化合物を含み、この化合物はMEF2 結合部位に結合し、式中:
Ra は、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(S02)-を含む基から選択されるMEF2結合部位の疎水性領域と結合する認識基であり、式中:
Rc および Rd は、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される、
L は、20 炭素原子までの鎖からなるリンカーであり(但し、三炭素原子までが酸素, 窒素 またはイオウ原子で置換でき、更にアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミド, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)、
Rbは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(S02)-を含む基から選択されるMEF2結合部位の親水性領域と結合する認識基であり、式中:
Rc および Rd は、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される。
【0041】
好適な態様において、提供される化合物は、一般構造式Ar1-L1-L2-L3-Ar2を有し、式中:
Ar1 および Ar2 は、独立にベンゼン, ナフタレン, ピリジン, ピリミジン, ピラジン, キノリン, イソキノリン, ピロール, フラン, チオフェン, イミダゾール, ピラゾール, オキサゾール, チアゾール, イソキサゾール, インドール, ベンズイミダゾール, ベンゾチアゾール, ベンゾキサゾールからなる群から選択される芳香環である(但し、前記芳香環は、水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アミノ, アルキルアミノ, ジアルキルアミノ, アリールアミノ, ヘテロアリール アミノ, ヒドロキシ, またはハロからなる群から選択される七置換基までを含み得る)。また、前記置換基は、共に連結して、12 原子までの環を形成できる、
L1 および L3 は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2 は、10 炭素原子までの鎖からなる群から選択される連結基であり(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【0042】
更なる好適な態様において、提供される化合物は、次の一般式を有する:
【化1】

【0043】
式中:
R1 - R10は、独立に次のものからなる群から選択される
L1 および L3 は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2 は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【0044】
更なる好適な態様において、提供される化合物は、次のリストの化合物から選択される(図 2)。
【0045】
これらの化合物は、インビトロやインビボでMEF2との結合において広範囲の親和性を示し、上記のとおり筋肉, 免疫および神経系においてMEF2に基づく療法を開発するためのリード化合物として使用できるだろう。実際、前記化合物は、インビトロおよびマウスモデルで制御性T細胞機能の促進に効果を示し、そのため自己免疫疾患や移植拒絶を予防するための可能性のあるリード薬である。
【0046】
また、提供されたMEF2-結合分子を調製するための方法が本発明において提供される。
【0047】
第五の側面において、本発明は、MEF2-依存的な転写のディレギュレーションから生じる疾患を治療するため有用な化合物 および 組成物も提供する。本発明のこの側面と一致する組成物は、一または二以上のMEF2及びそのコファクターの結合をブロックする能力がある化合物を含む。提供される化合物 および 組成物は、転写因子及びそれらのリクルートしたヒストン-修飾酵素の相互作用と関連するエピジェネティック制御の調節が関与する治療用に使用できる。特に, 提供されるMEF2-結合小分子を使用して、MEF2-依存的な転写の調節解除から生じる様々な疾患を治療でき、これにはMEF2機能の不均衡から生じる心肥大, 筋線維タイプのリモデリング, および他の筋肉関連疾患; 過剰な又は非常に僅かなMEF2-依存的な遺伝子発現から生じる自己免疫疾患又は免疫不全, および移植拒絶; 制御からはずれたMEF2機能から生じる様々な神経変性疾患(例えば、アルツハイマー型の疾患およびハンチントン疾患など), 自閉症, 精神医学的な障害, および学習および記憶の障害が含まれるが、これらに限定されない。この本発明の態様の更なる適用は、FOXP3-依存的な転写の調節解除(deregulation)から生じる様々な疾患を治療するため使用できるFOXP3 転写因子の機能を調節する小分子が関与する。これらの疾患には、自己免疫疾患, 移植拒絶 および 癌が含まれるが、これらに限定されない。
【0048】
本発明の第六の態様は、薬:MEF2複合体の調製、また構造決定のための結晶化が関与する。このプロトコールは、MEF2と結合する化合物の高品質の結晶を得るためMEF2Aの特定のタンパク質断片(2-78)および緩衝液条件の範囲を規定する。この複合体の調製および結晶化の方法は、現在および将来のMEF2と結合する小分子の構造上の特徴に、また前記構造を使用してリード化合物の至適化を導くため必須である。
【0049】
本発明の前述の及び他の特性、それらを得て、用いる様式は、以下の記載を参照し、添付される図面を考慮することでより明らかとなり、理解されるだろう。前記図面は本発明の典型的な態様のみを記載しており、従って、その範囲は限定されない。
【0050】
本発明の他の側面および利点は、以下の記載および特許請求の範囲から明らかになるであろう。
【図面の簡単な説明】
【0051】
【図1a】図1はMEF2および同定されたリガンド結合部位の結晶構造の三つの異なる表現(a-c)を示す; (d) 2.4Aの解像度でMEF2A(1-78)と結合したBML-210を示しており、BML-210との結合の相互作用に関与するアミノ酸が含まれる。
【図1b】図1はMEF2および同定されたリガンド結合部位の結晶構造の三つの異なる表現(a-c)を示す; (d) 2.4Aの解像度でMEF2A(1-78)と結合したBML-210を示しており、BML-210との結合の相互作用に関与するアミノ酸が含まれる。
【図1c】図1はMEF2および同定されたリガンド結合部位の結晶構造の三つの異なる表現(a-c)を示す; (d) 2.4Aの解像度でMEF2A(1-78)と結合したBML-210を示しており、BML-210との結合の相互作用に関与するアミノ酸が含まれる。
【図1d】図1はMEF2および同定されたリガンド結合部位の結晶構造の三つの異なる表現(a-c)を示す; (d) 2.4Aの解像度でMEF2A(1-78)と結合したBML-210を示しており、BML-210との結合の相互作用に関与するアミノ酸が含まれる。
【図2】図2は、BML-210, PAOA および関連する例示のMEF2-結合分子の構造を示す。
【図3a】図3は、HDACiに関するMEF2-HDAC ルシフェラーゼレポーターアッセイの模式図を示す。
【図3b】図3は、HDACiに関するMEF2-HDAC ルシフェラーゼレポーターアッセイの模式図を示す。
【図4】図4は、BML-210での処理の24 hrs後のMEF2 HDAC ルシフェラーゼ レポーター アッセイにおけるルシフェラーゼ活性に関して標準化した応答を示す。ルシフェラーゼ活性は、用量依存的な様式で減少した。
【図5a】(a) BiacoreアッセイにおいてBML-210とインキュベーション後にHDACと結合するMEF2の競合阻害を示す; (b) BML-210およびHDAC9がMEF2上でオーバーラップする領域に結合することを示している構造分析。
【図5b】(a) BiacoreアッセイにおいてBML-210とインキュベーション後にHDACと結合するMEF2の競合阻害を示す; (b) BML-210およびHDAC9がMEF2上でオーバーラップする領域に結合することを示している構造分析。
【0052】
[詳細な記載]
本発明の様々な側面を要約したので、我々は次に更に本発明を説明するため様々な例示の態様を詳細に記載する。
【0053】
HDACsの特異的な境界のインヒビター及びその使用
上述のとおり、エピジェネティックレギュレーターの研究を難しくしたことは、多くが別個の機能を有する多数の酵素アイソフォームが存在するとの事実である。例えば、四つのHDAC酵素スーパーファミリーのうち、クラス I (HDAC 1, 2, 3, および 8) および クラス II (HDAC 4, 5, 6, 7, 9, および 10)は癌に関与する二つの主要なクラスである。各々の個々のメンバーのユニークな機能は分子生物学アプローチ(例えば、ノックアウトまたはノックダウン)により研究できるにもかかわらず、それらの薬治療における役割は評価することができない。というのも、多くの現在のHDAC インヒビターは、多くのHDACs (クラス I および II)に共通の触媒ドメインを標的とするからである。事実、おそらく異なる HDAC アイソフォームの活性に影響することにより、所与のHDACインヒビターが異なる細胞条件において逆に影響しているようにみえる結果を誘導することがしばしば観察される。以上のように、現行のHDACiがアイソフォーム特異性を欠くことは、特定のHDACsの薬の影響および個々の機能に関する合理的な機構的な研究の主要な障壁を代表する事項である。また、多数のHDACsまたは小数のサブセット又はそれどころか特定のメンバーを阻害することが薬理学的に有益であるかどうかは明らかではない。特定の臨床適用(例えば、癌治療)は前者を必要とし、他方で他のもの(例えば、神経変性および炎症)は後者から利益をえるだろう。
【0054】
現在利用可能なクラス I および クラス II HDACsのインヒビターは、彼等の化学的な構造の重要な特性に基づいて六つのカテゴリーにグループ化できる:
(1) ヒドロキサム酸;
(2) チオール含有分子;
(3) 求電子性のケトン(electrophilic ketones);
(4) 短鎖脂肪酸(short-chain fatty acids);
(5) ベンズアミド(オルトアミノアニリド);および
(6) 環状のデプシペプチド。
【0055】
これらの分子(ヒドロキサム酸, チオール, および求電子性ケトンを含む)の幾つかがHDAC 活性部位の亜鉛イオンに結合することで働くことは確認されているが、他のカテゴリーの阻害機構ははっきりしていない。異なる HDAC インヒビターは、異なるクラスのHDACsにおいて異なる活性を示した。例えば、ヒドロキサム酸のカテゴリーのメンバー(例えば、TSA および SAHA)は、全クラス I および クラス IIのHDACsにわたる強力なインヒビターであるが、幾つかのメンバーのベンズアミドファミリー〔例えば、ピメロイルアニリド オルト アミノアニリド (PAOA), MS-275, CI-994, MGCD-0103〕, および環状ペプチドのファミリー(例えば、FK228)は特定のHDACメンバー又はサブクラスに対し適度(modest)の選択性を示した。しかしながら、一般的にHDACインヒビターの分子機構の理解を欠いていることが理由で、この選択性が如何に達成されるかは明らかではない。結果的に、幾つかのHDACインヒビターにおける選択性の経験的な観察によって、サブタイプ特異的なHDACインヒビターの合理的なデザインを導くことはできなかった。幾つかのクラスI (HDAC8)およびクラスII (HDAC4およびHDAC7)メンバーの触媒ドメイン及びそれらの異なるインヒビターとの複合体の結晶構造が解析され、これらの構造およびクラスIおよびクラスII HDACsの構造に基づく配列アラインメントによって、活性部位が高度に保存され、周囲の領域に軽微な差のみが示されることが示唆される。従って、彼らも選択性の潜在的な機構についての見識を提供することに失敗している。この背景から、高度に保存された触媒ドメインに対するサブタイプ特異的なインヒビターの合理的なデザインは、非常に挑戦的であると広く考えられた。
【0056】
HDACsがしばしば特異的なコレギュレータおよび他のクロマチン修飾酵素との大きなマルチ-タンパク質複合体に存在するとの観察に基づいて、本発明の発明者は機構に基づくアプローチにより新規のHDACに基づく小分子エピジェネティックレギュレーターを同定し、最適化するためのよい道筋が提供されるだろうと考えた。つまり、ドラッグデザインの従来のアプローチに支配的な酵素の活性部位を標的とするのではなく、その代わりに、HDACs及びそれらの関連性のある機能的なパートナーの間のタンパク質間相互作用を標的とする小分子が開発されるだろう。
【0057】
従って、一側面において、本発明は、HDACが関連する転写因子と結合することをブロックすることによりHDAC 機能を調節する方法を提供する。一般的に、本発明のこの側面による方法は、前記転写因子を前記転写因子 および HDACの間の境界の表面内の部位に選択的に結合する能力のある境界のインヒビターと接触させて、このHDACの触媒活性を前記転写因子のDNA-結合部位の近傍に局在するタンパク質部位で機能することを妨げる工程を有する。
【0058】
前記HDACは、古典的なHDAC又はそのHDACのサブセットの任意のHDACアイソフォームであってもよい。例示のHDACには、HDAC 4, 5, 6, 7, 9 および 10,又はその任意のサブセットが含まれえる。好適な態様において、HDACは、クラス IIa HDACsである。また、更なる好適な態様において、HDACは、HDAC4またはHDAC9の何れである。
【0059】
転写因子は、HDACと結合することが知られる任意の転写因子であってもよい。例示の転写因子には、MEF2, FOXP3, GATA3, および Cabinsが含まれえるが、それらに限定されない。好適な態様において、転写因子はMEF2である。
【0060】
境界のインヒビター(interfacial inhibitor)は、小分子であってもよい。
【0061】
上記に記載される方法は、インビボまたはインビトロで、HDACs および 対応する転写因子の機能および機構に関する情報を得るための研究設定に使用しえる。また、これらを疾患を治療するための臨床設定に使用しえる。
【0062】
本発明の方法で治療しえる疾患は、一般にMEF2機能の不均衡から生じる心肥大, 筋線維タイプのリモデリング,および他の筋肉関連疾患; 過剰な又は非常に僅かなMEF2-依存性の遺伝子発現から生じる自己免疫疾患又は免疫不全; 制御からはずれたMEF2機能から生じる様々な神経変性疾患(例えば、Friedrich's 運動失調, アルツハイマー病およびハンチントン病など), 自閉症, 精神医学的な障害, および学習および記憶の障害が関与する疾患である。例示の疾患には、神経変性疾患, 心臓疾患, 自己免疫疾患, 炎症, および 癌が含まれえるが、これらに限定されない。
【0063】
臨床に使用する場合、本発明のこの側面に基づく方法は、一般的にHDACの転写因子への結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。
【0064】
異なる臨床に使用する場合、本発明のこの側面に基づく方法は、一般的にCabin1のMEF2への結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。Cabin1は、カルシニュリン依存的な転写プログラムの転写コリプレッサーであり、T細胞およびニューロン細胞で高度に発現される。
【0065】
さらに異なる臨床設定に使用する場合、本発明のこの側面に基づく方法は、一般的にp300のMEF2への結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。
【0066】
さらに異なる臨床設定に使用する場合、本発明のこの側面に基づく方法は、一般的にCBPのMEF2への結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。
【0067】
一般的な設定において、本発明のこの側面に基づく方法は、一般的にMEF2と結合する任意の転写コレギュレータの結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。
【0068】
上記で表されるように、HDACは、任意の古典的なHDAC又はそのHDACのサブセットであってもよい。遮断薬(blocking agent)は、前記遮断薬がHDAC および 転写因子の間の境界の表面に局在する部位に選択的に結合する能力がある限り、小分子, ラセン状ペプチドミメティックス,又はその組み合わせであってもよい。好適な態様において、転写因子はMEF2である。
【0069】
境界のインヒビターを開発するための方法およびツール
本発明の別の側面において、HDACおよび転写因子の間の境界部位に結合して、その相互作用をブロックする能力のある境界のインヒビターを同定するためのアッセイも提供する。
【0070】
本発明のこの側面におけるアッセイは、一般にMEF2二量体における構造エレメントの第一の群および構造エレメントの第二の群の間の境界により規定されるMEF2 二量体における境界の結合部位を含む評価エレメント(evaluation element)に試験化合物を導入する工程を有する。構造エレメントの第一の群は、各々のMEF2 単量体のベータ鎖S1, S2 およびS3,およびヘリックスH2を含む〔二次的な構造エレメントおよび対応する残基の範囲は、Han等(Han et al,, Nature 2003)に記載されたとおりである〕。第二の群は、Cabin 1, HDAC4, HDAC9, HDAC5, HDAC7, p300 および CBPの短いヘリックスモチーフを含む。また、評価エレメントは、試験化合物との結合または非結合に関する情報を報告するためのレポーターエレメントと作動可能に連結(operatively coupled)されてもよい。
【0071】
好適な態様において、本出願においてベイトと融合させた結合ドメイン, プレイと融合させた活性化ドメイン, およびレポーター遺伝子;並びにレポーターシグナルレベルを決定することを含むtwo-hybridシステムに基づくアッセイが開示される。前記結合ドメインはGAL4 DNA と融合させたMEF2D(GAL4-MEF2)を含む; 前記活性化ドメインはVP-16と融合させたHDAC4のMEF2結合モチーフ(HDAC 4-VP16)を含む。レポーター遺伝子はGAL4-駆動性のレポータープラスミド(GAL4Luc)であり、全てが宿主細胞に保持される。
【0072】
本発明のこの好適な態様は、転写因子(例えば、MEF2)と結合し、その転写コレギュレータへの結合を調節する小分子の迅速でハイスループットなスクリーニングや最適化を許容する細胞に基づくルシフェラーゼアッセイである。前記アッセイは、発明者の研究室によるMEF2 複合体の構造および生化学についての10年以上にわたる研究に基づき開発された。この例示の態様は、最初に結晶研究により同定され、更に本発明の構造に基づく変異の研究により実証されたタンパク質間の境界がMEF2-結合小分子に高度に特異的で敏感なスクリーニングのための分子的な基礎として貢献できるだろうことに基づく。
【0073】
各々のMEF2 単量体のベータ鎖S1, S2 およびS3 およびヘリックスH2およびCabin 1およびHDAC4およびHDAC 9からの短いヘリックスモチーフを含むこのタンパク質間の境界に基づく任意のアッセイは、本発明の範囲内であると考えられる。他の例示的なアッセイの実施態様は、任意の理学的なアッセイ技術を含みえる(例えば、本発明により同定されたタンパク質間の境界に基づく、プルダウン, 共免疫沈降; ルシフェラーゼ レポーター アッセイ,およびトランスジェニック レポーター アッセイを含みえるが、これらに限定されない蛍光に基づく結合アッセイおよび機能的なアッセイを含みえるが、これらに限定されない)。
【0074】
特に、提供される化合物が実際に細胞の内側でクラス IIa HDACの結合を崩壊させることできるかどうかを確認するため、クロマチン免疫沈降(ChIP)アッセイを含む幾つかのタイプのアッセイを使用できる。HDAC4 プラスミド構築物は、一過性にHela細胞にトランスフェクトされる。MEF2媒介性プロモーターにおけるHDAC4の存在(occupation)は、試験される化合物および緩衝剤のコントロールの存在下で、適切な特異的な抗体およびPCRプライマーを用いたChIPにより検出される。ChIP分析の代替的なアプローチとして蛍光イメージングに基づく方法も使用できる。GFP-融合MEF2C および HDAC4をHeLaまたはC2C12 細胞にトランスフェクトして、それらの相互作用が研究された。単独で発現させた場合、GFP-HDAC4は拡散により細胞質に局在し、他方でGFP-MEF2も拡散パターンで核に局在する。共発現させた場合、HDAC4 および MEF2は核の内側に点状ボディー(punctate bodies)を形成する。これらの点状の核ボディー(punctate nuclear bodies)の性質は未知であるが、それらの形成は明らかにMEF2:HDAC4の相互作用に依存的である。というのも、機能的なMEF2-結合モチーフを欠いているHDAC4 変異体はMEF2の核ボディーの標的化に失敗するからである。最終的に、mRNAのプロファイリング(マイクロアレイ)および結合位置(ChIP-on-chip)によるMEF2標的遺伝子のゲノムワイドな分析によって、更に静止または活性化(例えば、カルシウム シグナルの興奮)状態におけるMEF2-依存的な抑制または活性化に大きい反応を示す周知のMEF2標的遺伝子を選択することによりこの方法を促進することができる。この方法を用いて、これらの遺伝子がクラス II HDACsまたは他のMEF2コリプレッサー(例えば、Cabin 1)により潜在的に制御されるかどうかを、ChIPを用いてクラス IIa HDACのプロモーターの存在を検出すること、薬処理に際する又はHDAC 4, 5, 7または9のsiRNA ノックダウン(siRNAによる発現変化が最初に評価される)後の発現の変化をモニターすることにより確認することができる。全体的に、この方法を使用して、様々な濃度の提供される化合物の存在下で、マイクロアレイを用いて遺伝子発現を分析すること、ゲノムワイドでクラス IIa HDACsの結合を検出することにより、化合物をゲノムワイドで評価することができる。
【0075】
アッセイの実施を促進するため、本発明はMEF2-結合小分子を探索するためのハイスループットで高度に感受性で特異的なスクリーニングのプラットホーム(screen platform)も提供する。このプラットホームは、GAL4-駆動性のレポータープラスミド (GAL4Luc), GAL4 DNA-結合ドメインと融合させたMEF2D(GAL4-MEF2), VP-16と融合させたHDAC4のMEF2-結合モチーフ(HDAC4-VP 16), VP-16と融合させたGAL4 DNA-結合ドメイン(ポジティブコントロール)並びにネガティブコントロールおよびポジティブコントロールとして様々な化合物を含んでいる安定的に形質転換された細胞株を含む。上記の安定な細胞株, プラスミド, およびコントロール化合物からなるキットは、新しいMEF2-結合分子を探索する使用者によるスクリーニングのため、また存在するリーディング化合物を最適化するため作出できる。また、MEF2D をMEF2A, MEF2BおよびMEF2Cにスイッチすることにより、この方法を使用してMEF2 ファミリーのアイソフォームに選択的に結合する化合物を探索することができる。係る化合物を使用して疾患における特定のMEF2ファミリーメンバーの機能や関与を研究することができる、またMEF2-関連疾患の診断剤の開発や特異的な治療剤の同定のため使用することができる。
【0076】
別の側面において、本発明は、制御性の及び機能的な複合体のターゲティングによるサブタイプ特異的なHDAC インヒビター/モジュレーターを同定するための方法も提供する。本発明のこの側面に基づく方法は、通常は以下の工程を含む: (1) 機能上重要な境界を含む構造または下部構造(substructure)を解析すること(solving); (2) 現存の又は新しい潜在的なHDACインヒビターから選択される検査分子を計算でドッキングさせることによりドッキング分析を解析した構造に適用すること; (3) 所望のHDAC複合体の間のタンパク質間相互作用を崩壊させることができる化合物をスクリーニングするためのアッセイを開発すること; (4) 工程(3)において同定された化合物の特徴を調べること; (5) 前記化合物を計算で最適化すること;および (6) 最適化された化合物を合成し、前記化合物を工程 (3)のアッセイを用いて確認すること(validating)。
【0077】
工程 1に関して、構造は、探索モデルとして現存の構造を用いる分子置換により解析できる。必要に応じて、実験の段階は、MADまたはMIRで得られてもよい。工程 2に関して、ドッキングは、標準のパッケージ(例えば、AutoDock)を用いて行なうことができる。工程 3は上記記載のとおりであり、大抵は哺乳類のtwo-hybridアッセイに基づく。工程の残りは化合物の性質に依存して当該技術分野において既知の方法を用いてなされる。
【0078】
別の側面において、本発明は、HDAC および 転写因子の間の結合をブロックするための遮断薬として有用な化合物も提供する。HDACは、任意の古典的なHDAC又はそのサブセットであってもよい。転写因子は、HDACと結合することが知られる任意の転写因子であってもよく、MEF2, FOXP3 および GATA3が含まれるが、これらに限定されない。
【0079】
好適な態様において、転写因子はMEF2である。
【0080】
本発明のこの側面による化合物は、小さい有機分子およびラセン状のペプチドミメティックスを含む。
【0081】
MEF2-結合小分子は、開示されたMEF2 結合部位をガイドとして利用することにより本出願において提供される方法を用いて同定できる。提供される小分子は、MEF2と結合するBML-210の結晶構造から明らかとされた同定された結合部位から由来する一般構造式を有する。提供される小分子は、記載された構造上のフォールドと結合するため設計され、潜在的に高い親和性および選択性でMEF2と結合するだろう。このアプローチを用いて強力で選択的な化合物を同定するため、当該技術分野において既知の方法を利用でき、この方法にはリード化合物を同定し、続いてリード化合物の最適化するためのコンピュータを利用した構造に基づくデザインとインシリコスクリーニングとの組み合わせ, コンビナトリアルライブラリデザインとハイスループットスクリーニングとの組み合わせ, およびフラグメントベースのドラッグディスカバリー(fragment-based drug discovery)が含まれるが、これらに限定されない。
【0082】
好適な態様において、本発明のもとで提供される化合物は、MEF2結合部位に結合する化合物を含み、次の一般式を有する:
Ra-L-Rb, 式中で:
Ra は、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(S02)-を含む基から選択されるMEF2結合部位の疎水性領域と結合する認識基であり、式中:
Rc および Rd は、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される。
【0083】
L は、20 炭素原子までの鎖からなるリンカーであり(但し、三炭素原子までが酸素, 窒素 またはイオウ原子で置換でき、更にアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミド, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)、
Rbは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(S02)-を含む基から選択されるMEF2結合部位の親水性領域と結合する認識基であり、式中:
Rc および Rd は、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される。
【0084】
好適な態様において、提供される化合物は、一般構造式Ar1-L1-L2-L3-Ar2を有し、式中:
Ar1 および Ar2 は、独立にベンゼン, ナフタレン, ピリジン, ピリミジン, ピラジン, キノリン, イソキノリン, ピロール, フラン, チオフェン, イミダゾール, ピラゾール, オキサゾール, チアゾール, イソキサゾール, インドール; ベンズイミダゾール, ベンゾチアゾール, ベンゾキサゾールからなる群から選択される芳香環である(但し、前記芳香環は、水素, アルキル, アルケニル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アミノ, アルキルアミノ, ジアルキルアミノ, アリールアミノ, ヘテロアリール アミノ, ヒドロキシ, またはハロからなる群から選択される七置換基までを含み得る)。また、前記置換基は、共に連結して、12 原子までの環を形成できる、
L1 および L3 は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2 は、10 炭素原子までの鎖からなる群から選択される連結基であり(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【0085】
更なる好適な態様において、提供される化合物は、次の一般式を有する:
【化2】
【0086】
式中で:
R1 - R10は、独立に次のものからなる群から選択される
L1 および L3 は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2 は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【0087】
更なる好適な態様において、提供される化合物は、次のリストの化合物から選択される(図 2)。
【化3】
【0088】
本発明の様々な側面および派生の完全な理解を更に促進するため、次の実例となる例が提供される。
【例】
【0089】
[例1] サブタイプ特異的なHDACインヒビター/モジュレーターを同定するための戦略
本発明は、サブタイプ-特異的なHDACインヒビター/モジュレーターとして作用しえるリード化合物を同定するための新規の戦略を提供する。前記戦略は、幾つかの反復する工程からなる:
工程 1: 所望のHDAC複合体に関して、HDACの関連する調節タンパク質に結合するHDACの構造または下部構造が解析される又はHDACの機能上重要な境界を含む構造または下部構造が得られる。
【0090】
工程 2: 前記構造は、タンパク質間の境界に結合しえる現存のまたは新しい潜在的なHDACインヒビターのドッキング分析のため使用される。仮想的なスクリーニングは、機能的なデータ(例えば、HDACインヒビターが標的のHDAC複合体が関与する細胞プロセス(cellular processes)において効果を示す場合、またHDAC インヒビターが活性部位の阻害以外の機構をとおして作用するように思われる場合)により導かれる。
【0091】
工程 3: また、構造や関連する生化学的な情報を使用して、意図するタンパク質間の境界を破壊することができる化合物をスクリーニングするため使用できるアッセイの開発が導かれる。
【0092】
工程 4: 一度このようなリード薬が見つけられたら、タンパク質標的との複合体は、工程 1において樹立された構造研究システムを用いて特徴が調べられる。
【0093】
工程 5: 構造の情報と組み合わせて関連する化学の技術を使用して、高度な親和性および特異性を有する標的タンパク質と結合できるアナログ(analogs)のデザインが導かれる。使用される方法は、工程 2の方法と類似する。
【0094】
工程 6: 設計されたアナログが合成され、工程 3において樹立されたアッセイにより分析される。最終的に、最適化された化合物は、親化合物の効果を模倣し、高い強度および低い非特異性/副作用を有しえるかどうかを検査するためのインビボ試験のため使用される。
【0095】
上記の機構に基づくアプローチを利用して、発明者は、以前の既知のHDAC インヒビター(PAOA)がMEF2インヒビター(MEF2i)として作用することによりクラス IIa HDACsの機能を特異的に破壊することができるリード化合物として同定されることを実証した。そのうえ、PAOAの代表的な構造上のアナログも、本出願において記載された方法で設計され、合成され、評価された。そして、幾つかの強力なMEF2iが同定され(図 2)、このアプローチの更なる側面が実証された。
【0096】
[例2] 治療に適用されるサブタイプ-特異的なHDACインヒビターの使用
クラス IIaのHDACsは、ニューロンの生存/シナプスの形成, T細胞の選択/活性化, および筋肉のリモデリングに重大な役割を担う。これらの活性の調節不全は、神経変性(neurodegeneration), 炎症および心肥大を含む幾つかの疾患において指摘される。癌治療のため開発された幾つかのHDACiは、これらの障害に対し有益な効果を示した。これらのHDAC インヒビターの非特異的な性質によりこれらの疾患における臨床適用が妨げられる。しかしながら、これらの観察によって、観察された治療効果がクラス IIaのHDACsと関連するかどうか、またクラス IIaのHDAC機能の選択的な破壊がこれらの疾患を治療するための実行可能な戦略であるかどうかの魅力的な質問が提起される。これらの問いに対応するため、クラス IIaのHDACsの機能を特異的に破壊できる小分子が必要とされる。
【0097】
既知のMEF2およびクラス IIaのHDACsの細胞機能や筋肉, 免疫およびニューロン系における幾つかのHDACインヒビターの効果に基づき、我々は、MEF2機能の不均衡から生じる心肥大, 筋線維タイプのリモデリング,および他の筋肉関連疾患; 過剰な又は非常に僅かなMEF2-依存的な遺伝子発現から生じる自己免疫疾患又は免疫不全,および移植拒絶; 制御からはずれたMEF2機能から生じる様々な神経変性疾患(例えば、Friedrich's 運動失調, アルツハイマー病およびハンチントン病など), 自閉症, 精神医学的な障害, および学習および記憶の障害を治療するため使用できるだろう本発明の方法および化合物を提唱する。
【0098】
上記の様々なヒト疾患を治療するため開発されたMEF2-結合分子は、経口的に, 筋肉内に, 腹腔内に, 皮下に, 静脈内に注射で投与できるだろう。他の送達法も可能であり、正確なプロトコールは治療されるコンディションに依存するだろう。用量は特定の臨床適用に応じて変動するであろうが、化合物がインビトロで効果を示す標準的なアッセイはインビトロでは約0.1 - 10 uMであり、動物モデルの研究では1-10 mg/KG体重である。
【0099】
図 2に示される本発明のもとで提供される好適な化合物は、インビトロやインビボでMEF2との結合において広範囲の親和性を示し、上記のとおり筋肉, 免疫および神経系においてMEF2に基づく療法を開発するためのリード化合物として貢献できるだろう。実際、前記化合物は、インビトロおよびマウスモデルで調節性のT細胞機能の促進に効果を示し、そのため自己免疫疾患や移植拒絶を予防するための可能性のあるリード薬である。例えば、MEF2-結合分子NKL30の0.15μMによって、インビボでの活性の抑制を増強することにより調節性のT細胞機能が明らかに非常に増強された。恒常的な増殖アッセイのマウスのモデルにおいて、静脈内注射により投与された 1 mg/KG体重の前記化合物は、インビボでTreg 機能を非常に増強させた。これらのデータは、本発明により提供されたMEF2-結合分子が自己免疫疾患を治療するため、また移植拒絶を予防するため使用できるだろうことを強く示唆する。
【0100】
[例3] 機能的な調節のためのクラス IIaのHDACsのターゲティング
他のHDACsと比較して、クラス IIaファミリーは、機能 および 制御の幾つかの側面においてユニークである。第一に、クラス IIa HDACsは、選択的に筋肉, 脳 および T細胞中で発現し、これらの組織におけるそれらの機能と一致する。第二に、クラス IIaのHDACsの活性は、クラス IIaのHDACsが発現される組織における優勢な二次メッセンジャーであるカルシウム シグナルにより厳格に制御される。第三に、クラス IIのHDACsは、ユニークな特性をこのサブクラスのHDACsに与える触媒ドメインのN端の大きな制御ドメインを含む。N末端の調節領域は、種々のタンパク質と相互作用するドメインおよびモチーフを含み、これらにはクラスII HDACsのカルシウム応答性を制御するもの(例えば、CaM, CaMKおよび14-3-3),およびクラスIIのHDACsを特異的なプロモーターを標的とさせるもの(例えば、MEF2およびBCL-6),および他のエピジェネティックレギュレーターおよびエフェクター(例えば、クラスIのHDACs, CtBPおよびHP-1、これはクラスIIaのHDACsと協調的に機能する)が含まれる。幾つかのこれらの複合体の構造は、特異的な破壊の潜在的な標的として調査された。
【0101】
クラス IIaのHDACsの制御および機能に関与する多くの複合体のなかで、生化学および構造の点で最もよく調べられたものはMEF2 複合体である。MEF2は、クラス IIaのHDACsと同じ発現パターンを有する配列特異的な転写因子(MEF2A-D)のファミリーであり、神経変性, 炎症および心臓疾患にも関与する。MEF2ファミリーの転写因子は高度に保存されたMADS-ボックス/MEF2S ドメインと称されるN末端領域を共有し、これがDNA結合, 二量体化並びに種々の転写因子およびコレギュレータとのタンパク質間相互作用を媒介する。クラス IIaのHDACsは、脱アセチルに関して特異的なクロマチン領域を標的とするためDNAには結合せず、MEF2との相互作用に依存している。この相互作用をブロックすることが可能性がある方式として選択されてクラス IIaのHDACsの機能が破壊される。
【0102】
クラス IIaのHDACsおよびMEF2の間の相互作用が広範な機能的および生化学的な分析の対象とされ、クラス IIaのHDACsにおいて保存された短配列モチーフ(MEF2-結合モチーフ)およびMEF2のMADS-ボックス/MEF2Sドメインが彼らの結合に必要十分であることが明らかとされた。体系的な構造研究や生物物理学的な研究をMEF2 および クラス IIaのHDACsの間の相互作用や類似するMEF2-結合モチーフを含む関連する転写抑制(Cabin1)について行った。MEF2-結合モチーフが短い両親媒性のヘリックス構造に適合し、MEF2のMADS- ボックス/MEF2ドメイン上の疎水性の溝と結合することが結晶構造により明らかとされた。このようなリガンド/レセプター様の結合機構が存在することは、小分子を使用してクラス IIa HDACsのMEF2-特異的なプロモーターへのリクルートをブロックすることが可能であろうことを示唆する(refs Han Nature and 2005)。
【0103】
[例4] サブ-クラス特異的なHDAC阻害アッセイの開発
一連のMEF2-依存的なルシフェラーゼ レポーター アッセイと一過性にトランスフェクトされたMEF2D, HDAC4 および コアクチベータ p300を使用して、最初に様々な化合物をスクリーニングした。しかし、これらのアッセイは、シグナルが弱く、高頻度で偽陽性を呈した(おそらく、MEF2の複合体の転写活性化機構や内因性の因子からの干渉が原因であろう)。これらの観察をとおして、細胞の内側のHDAC4 および MEF2の間の分子の相互作用を繰り返すことができる高度に感受性で特異的なアッセイが必須であることが発見された。この問題を解決するため、発明者は、HDAC4 および MEF2Dの間の相互作用を内因性の因子からの干渉を最小限で検出する能力のある哺乳類のtwo-hybrid系を発明した(図 3)。
【0104】
このアッセイ系において、MEF2DはGAL4 DNA結合ドメインと融合され(GAL4-MEF2D)、HDAC4のMEF2-結合モチーフ(aa 155-220)がVP-16と融合された(HDAC4-VP 16)。双方の構築物 および GAL4-駆動性のレポータープラスチド(GAL4Luc)で一過性にトランスフェクトされたHela 細胞がGAL4-VP 16のポジティブコントロールにより産生されるものと匹敵する強いシグナルを生じ、他方でHDAC4との結合に欠陥のあることが前に示されたMEF2D 変異体 Leu67Asp (L67D)がレポーターの活性化に失敗することが予備的な分析により示された(データ示さず)。全てのルシフェラーゼ レポーター アッセイにおけるタンパク質の発現レベルをウエスタンブロットで確認した。
【0105】
上記の構造上の見識やMEF2D L67D 変異体をきっかけに、我々は前にインビトロでHDAC4:MEF2の相互作用を破壊することが示されたHDAC4の変異を導入した。これらの変異も細胞に基づくアッセイにおいてルシフェラーゼ シグナルを減少させた(図 3b)。最も興味深い事項は、インビトロで〜60%までMEF2 の結合を弱める(野生型および変異体のHDAC4によるMEF2結合のKdは、それぞれ0.47μM および 0.81μMである)HDCA4のVal180Lysの変異は、細胞に基づくアッセイにおいてルシフェラーゼのシグナルを部分的に減少させた(図 3b)。これらの観察は、哺乳類のtwo-hybridアッセイからのシグナルがHDAC4 および MEF2の間の分子相互作用と非常によく相関することを示す。これらの結果によって、HDAC4:MEF2相互作用の構造モデルに関して更なる支持が提供されるのみならず、細胞の内側のHADC4:MEF2相互作用を検出するための敏感で特異的な方法も確認される。
【0106】
[例5] MEF2/HDAC 相互作用の選択的なインヒビターの同定
スクリーニングの複雑さを減らすため、現存のHDACインヒビターにおける多数の機能的なデータを利用した。これらのインヒビターの多くが触媒ドメインを標的とするが、細胞 に基づくヒストンアセチル化アッセイにより発見された幾つかの化合物はクラス IIaのHDACsのMEF2への結合を含むHDAC機能の他の側面に影響しえる。この事項を踏まえ、我々は、HDAC9:MEF2 複合体の結晶構造から由来するファルマコフォアモデルを用いた小分子データベースに対する仮想的なスクリーニング(3D ドッキング)を行なった。この探索により新規の標的を生じなかったにもかかわらず、MEF2の疎水性ポケットが特定の長さのリンカーにより連結された二つの芳香族環を有する化合物を好むことが示された。この結果は、MEF2二量体がHDAC9のフェニルアラニンを結合できる二対称性関連部位(two symmetry-related sites)を含むことを示している結晶解析と一致する(refs Han et Nature and JMB)。以上のことから、我々は、このような構造上の特性を有する既知のHDACインヒビターを探索し、哺乳類のtwo-hybridアッセイを用いてHDAC4:MEF2 相互作用における彼らの効果を試験した。
【0107】
哺乳類のtwo-hybridアッセイを用いて選択されたHDAC インヒビターのプールのスクリーニングによって、前に研究された化合物PAOA(図 2)が用量依存的な様式でレポーター シグナルを阻害することが明らかとされた(図 4)。PAOAはHDAC4-VP 16の発現に影響しなかったが、GAL4-VP16により駆動されるレポーター シグナルを5.6 倍まで10μMで減少させ(データ示さず)、この化合物の我々の実験条件下でのルシフェラーゼ活性の発現の非特異的な阻害を示している。しかしながら、同じ濃度のPAOAは、GAL4-MEF2DおよびHDAC4- VP16により駆動されるレポーターシグナルを約 26 倍まで減少させ、PAOAが通常の阻害効果をこえてHDAC4:MEF2 相互作用を破壊する特異的な効果を有することを示唆している。対照的に、亜鉛活性部位を標的とする強力なHDACインヒビターのトリコスタチン A (TSA)は、GAL4-VP 16 および GAL4-MEF2D/HDAC4-VP 16により駆動されるレポーター シグナルに類似する阻害効果を示した(データ示さず)。これらの結果は、PAOA(しかし、TSAではない)がHDAC4 および MEF2Dの間の相互作用を破壊できることを示唆する。
【0108】
HDAC4:MEF2 相互作用におけるPAOAのIC50は、哺乳類のtwo-hybridアッセイに基づくと約5μMであり、ヒストンアセチル化阻害アッセイを用いて決定したものと類似する。MEF2と結合するHDAC4に関するKdは、前に0.47μMであると決定された。我々がHDAC4の平衡濃度が我々のアッセイ条件下で〜0.5μMであると仮定する場合、PAOAのMEF2への結合に関するKdは5μMであると見積もられた。しかしながら、フリーのHDAC4濃度が細胞に基づくアッセイにおいて低い場合、見積ったKdはより大きいだろう。
【0109】
PAOAがHDAC4と競合してMEF2 と結合するかどうかについてもインビトロでBiacore T-100での表面プラズモン共鳴 (SPR)を用いて評価した。HDAC4 (aa 155-220)をCM5 センサーチップに固定化し、精製 MEF2A (1-95)を分析物として使用した。様々な濃度でのHDAC4へのMEF2Aの結合によって、一連の明確なセンサーグラムが得られた(データ示さず)。PAOAの濃度を増加させてMEF2Aとインキュベーションしたところ、固定化された HDAC4への結合の用量依存的な減少が示された(図 5)。競合的な結合反応は複雑であり、BML-210のMEF2への直接の結合が機器の検出限界をこえたことがBiacoreのデータの分析から示された。これらの技術的な制限から、定量的な結合定数を得ることは困難である。それにもかかわらず、予備的なデータはBML-210が実際にインビトロでHDAC4と競合的にMEF2と結合することを示唆している。
【0110】
当初PAOAはヒストンのアセチル化を選択的に誘導するがチューブリンでは誘導しない化合物のグループの一部として発見され、このような特徴はおそらくチューブリン特異的なHDACであるHDAC6以外のHDACsの阻害によるものであろう。PAOAは、インビトロでHDAC4と競合的にMEF2と結合する。右の挿入図は、Biacoreによるアッセイを記載する。HDAC4: 赤色のヘリックス; MEF2: 緑色のクロス, デアセチラーゼ。この選択性の分子的な基礎は知られていないが、HDAC6はクラス lIb サブファミリーに属し、クラス IIaに保存されるMEF2-結合モチーフを有していなく、機能するためMEF2を必要とするようにみえることに注目すべきである。最近、PAOA及びその誘導体は、フリートライヒ失調症においてフラタキシンの発現を増強することが示された。この機構にはヒストンアセチル化の誘導が関与するようにみえるが、強力であるが特異性が低いHDACインヒビター(例えば、TSA および SAHA)はフラタキシン発現に影響を示さず、しかし細胞中のトータルのヒストンアセチル化はPAOAよりも高いレベルで誘導できる。これらの観察は、PAOA及びその誘導体がフラタキシンのサイレンシングに関与する特異的なHDACまたはHDAC複合体を阻害するユニークな機能を有することを示唆する。さらにまた、PAOAの作用の分子的な基礎は、更に本発明に開示されたMEF2に結合したBML 210の結晶構造により明らかにできる。本発明において記載された小分子のMEF2-結合部位を規定することに加えて、この構造によりBML-210に、同様に、PAOA及び他のベンズアミド含有HDACインヒビターに存在するオルト-アミノアニリド部分に関する有望な結合部位が初めて示された。特に、この結合部位は、このクラスのHDACインヒビターに関して以前に仮定されたHDAC 酵素の活性部位と異なる。
【0111】
[例5] DNA上のMEF2に結合したBML-210の複合体の調製およびBML-210:MEF2:DNA 複合体の結晶および複合体構造の原子の詳細
MEF2に結合したBML-210の詳細な相互作用を特徴づけるため、またより強力なMEF2-結合分子のデザインを導く構造上の情報を使用するため、我々はDNA上のMEF2に結合したBML-210の結晶構造を決定した。MEF2A 1-78をコードする遺伝子をMEF2AFLからPCR増幅で作出し、pET30b 発現ベクターにクローニングした。タンパク質をE. coli株BL21(DE3)pLysSで25C, 一晩で発現し、連続的なクロマトグラフィー工程をSp-セファロース, およびゲル濾過で4Cで250mM NaC1, 10mM Hepes(pH7.6), 1mM EDTA, 1 mM DTTで精製を行い0.6mg/lの最終収量を得た。オリゴヌクレオチド (Santelli and Richmond, 2000)をIDT DNA technologiesから購入し、MonoQ FPLC カラムを用いて精製し、透析し、凍結乾燥し、サーマルサイクラーを用いてアニーリングした。タンパク質サンプルの1/10容量の10mM BML 210をタンパク質サンプルに0.5mg/mlで添加して凡そ17mg/ml(0.9mM)に濃縮し、DNA二重鎖(DNA duplex)をトレイにセットする前に1:1の比で添加した(10% DMSO終濃度)。プレート様の結晶をハンギングドロップ水蒸気拡散法の条件を18Cで 24% PEG4000, 140mM NaCl, 5mM MgCl2, 10mM CaCl2, 0.004%NaN3, 3.3% グリセロール, 50mM TrisHCl (pH 5.8 - pH8.18)を用いて得た。薬物濃度の結晶をpH 8.18で得た。結晶は、2.4Åの解像度で回折され、空間群 P1 (a=41.567Å, b=61.622Å c=61.478Å α=114.12° β=89.99° γ=89.95°)に属する。構造は、探索モデルとして1TQE.pdbを用いた分子置換により解析された(refs Richmond 2000)。最終的なモデルは、Rfree 26%およびRw 23%を有する。座標は、(11.1_001_nr_nh_bml.pdb)に添付された。
【0112】
[例6] 構造に基づくデザインおよび化学的な方法を用いたリード化合物の最適化
機能的なスクリーニングにより発見された多くのHDACインヒビターは、マイクロモルまたは更にミリモルの範囲のIC50の適度(modest)な強度を有する。リード化合物の最適化は、典型的には化学構造および構造活性相関(SAR)の研究の体系的な修飾により行った。しかしながら、標的および化合物と標的の間の詳細な結合相互作用を知ることがなければ、このような経験的なアプローチはしばしば労働集約的であり、有効性は限定的である。このような状況は一連のアナログがフリートライヒ失調症を治療するため使用しえる強力な化合物を探索するため合成されたPAOAの場合に事実である。幾つかのPAOAのアナログは親化合物よりも高い活性を示したにもかかわらず、効果は非常に控えめ(very modest)であり、また改善の機構は明らかではなかった。
【0113】
特に、これらのPAOA誘導体がフラタキシン発現を活性化する能力は、HDAC阻害活性と相関しない。例えば、幾つかの誘導体は、ヒストン脱アセチル阻害アッセイにおいて非常に弱いが、フラタキシン誘導において非常に活性であった。PAOAがMEF2と結合し、クラス IIa HDACsのリクルートをブロックするとの我々の予備的な所見によって、これらの興味ある結果の潜在的な分子機構が提供される。クラス IIaのHDACは、デアセチラーゼ活性と非依存性に転写を抑制できる。例えば、全C末端触媒ドメインを欠いているHDAC9 の天然のスプライシングバリアント(MITRとしても知られている)は、MEF2-依存的な遺伝子発現の強力な転写リプレッサーであり、この機構はおそらく他のエピジェネティックエフェクター(例えば、HP1 および CtBP)をリクルートすることによるものである。その意味で、クラス IIaのHDACsの触媒ドメインを標的とする小分子インヒビターは、これらのタンパク質を完全にエピジェネティックにサイレンシングする可能性を排除できない。この事項によって、フラタキシン発現の再活性化におけるTSA および SAHAの無効性が説明されえる。他方で、PAOAは、HDAC活性および他の転写リプレッサーのリクルートを共にブロックできる。MEF2 は転写アクチベータ(例えば、CBP/p300)をリクルートすることによる遺伝子活性化に関与するが、PAOAのMEF2依存的な遺伝子発現における主作用はクラスIIaのHDACsや他の転写リプレッサーのサイレンシング効果を緩和することであることを現在のデータは示唆している。
【0114】
上記の分析およびBML 210:MEF2:DNAの開示された構造に依存することにより、またPAOAのMEF2Aへのドッキングを解釈することにより、この薬分子が好ましく実験的に観察された電子密度に適合できることが示され、MEF2結合に関する新しいPAOA様化合物のデザインのためのICM-ドッキング プロトコール (Molsoft L.L.C)が比較的に有効であることを実証している。このドッキング アプローチを使用して、潜在的に大きな親和性および選択性を有する新しいPAOAアナログをデザインし、分析することができる。構造の幾つかの特性を以下に高い親和性でMEF2と結合しえる新しい分子のデザインに使用される原理を説明するため簡単に説明する。
【0115】
BML-210は、拡大したコンホメーションに適合し、MEF2の疎水性ポケットに結合する(図 1)。また、このポケットはクラス IIaのHDACs および Cabin 1において保存されたMEF2-結合モチーフの結合部位である。電子密度の一端が単純な芳香環と類似し、Leu66, Leu67, Thr70, Leu66', Leu67' および Thr70'(プライム記号は他の単量体からの残基を示す)を含む幾つかの疎水性残基によりとりまかれているので、我々はこの密度をフェニル基に割り当てた。また、この端でのカルボニル基は、所定の位置にあり、Thr70'との水素結合相互作用に関与する(図 1)。
【0116】
新しく最適化されたMEF2-結合小分子の構造に基づく設計は、幾つかのMEF2の既知の構造を用いて達成することができる。我々は、今回三つのMEF2複合体の結晶構造を解析した。その二つは、それぞれCabin1 および HDAC9のMEF2-結合モチーフから由来するペプチドを含み(Guo et al., 2007; Han et al., 2005; Han et al., 2003)、他方で三つめは本出願中で記載されるBML-210複合体であり、如何に小分子がMEF2に結合できるかを示している最初の構造である。全ての三つの複合体において、天然のペプチドであろうと合成の分子であろうと小分子リガンドは、MEF2 二量体の表面の深い溝に結合する(図 1)。Cabin l および HDAC9が、類似する別個のタンパク質間相互作用によりMEF2に結合することを我々の前の研究は示す(図 1)。興味深いことに、BML-210は、MEF2への結合において天然リガンドの幾つかの点を模倣するようにみえる。例えば、PAOAのフェニル環のLeu66, Leu67, Thr70, Leu66', Leu67' および Thr70'により形成された中心の疎水性ポケットへの結合は、HDAC9におけるLeu147の結合を暗示(reminiscent)する(図 1)。三つの構造の詳細な分析により、我々は幾つかの別々の疎水性ポケット, 水素結合のドナーおよびアクセプター, および幾つかの荷電した残基を含む小分子結合を探索しえるMEF2の溝の様々な構造上の特性を同定した。我々は、この構造上の情報の利用を計画して、選択的にMEF2と結合できる新しい系列の小分子をデザインし、至適化する。
【0117】
[例7] MEF2/HDAC相互作用の小分子インヒビターの構造に基づくデザイン
結晶構造をガイドとして用いて、新しいMEF2-活性小分子を同定するための基礎を提供できる多数のPAOAアナログを設計した。一群のアナログを設計して、PAOA構造の二つの一般的なエレメントを探索した。一つ目は、リンカーの長さと強剛性(rigidity)である。二つ目は、官能基及びその二つの芳香族の結合単位におけるポジションである。リンカーの電子密度は複数のコンホメーションに適合することを示し、この領域中でのPAOA および MEF2の間の非至適な結合を示唆している。他方HDAC9において、Lys144 および Val143の脂肪族側鎖は、MEF2の溝を充填して広範なファンデルワールスの接触および水素結合を適切に確立する。これらの天然の相互作用を模倣するため/改善するためPAOAリンカーに導入した官能基の結合親和性を増強しえる。設計した化合物を上記のドッキング分析に供して、精力的に好ましくないものが除去される。残っている分子を標準技術を用いて合成し、前に記載されたインビトロおよびインビボ分析に供した。
【0118】
最初の系列の潜在的なインヒビター(図 2)を合成し、異なる化合物が哺乳類のtwo-hybrid アッセイのレポーターシグナルを阻害する能力において有意に異なる活性を示すことが既に認められた。最も興味深いことに、これらの一つの化合物 4がPAOAと類似する活性を示すが、GAL-VP 16により駆動されるコントロールシグナルに影響せず、この新しい誘導体がPAOAよりも特異的であることを示唆している。PAOAにおけるオルト-アミノアニリド部分は、前にクラス I および クラス II HDAGsの活性部位に結合できる亜鉛キレート基であることが想定されたが、この作用様式の直接の証拠はまだ得られていない。化合物 4において、この亜鉛キレート基は、アミノ基をメタ位に移動することにより除去された。さらに、この誘導体は、PAOAと同等に活性であり、非特異的な影響が低く、我々のアッセイ条件下で、細胞の内側で観察されたPAOAの効果は触媒活性を阻害するよりむしろ主にMEF2:HDAC4の相互作用を破壊する能力によりことが示唆される。
【0119】
[例8] 化合物 10の合成
提供される化合物は、当該技術分野において既知の方法を適合させることにより調製できる。例えば、化合物 10の合成は:
【化4】
【0120】
次の工程により調製された:
工程 1: ピメリン酸(1 equiv) および 3-ブロモアニリン(1 equiv)をフラスコに添加し、130 ℃で一晩撹拌した。反応混合物をEtOAcに希釈し、10%水酸化カリウムで抽出した。水層をconc.HClでpH〜2に酸性化し、酢酸エチルで抽出した。有機層を減圧下で減少させ、アセトニトリル/水で再結晶化した。
【0121】
工程 2. ジクロロメタン中にフェニレンジアミンを含む溶液に(Boc)2O (1 equiv)をrtで添加し、混合物を一晩撹拌した。反応物を減圧下で濃縮し、酢酸エチルで希釈し、水および塩水で三回抽出した。有機層を減圧下で減少させ、クロロホルム/ヘキサンで再結晶化した。
【0122】
工程 3. 工程 1の産物(100 mg)をDMSO (3 mL)に溶解し、この溶液にHunigの塩基(1 equiv), HBTU (1 equiv), および工程 2のモノ保護フェニレンジアミン産物を添加した。生じた溶液を一晩rtで撹拌した。溶液を酢酸エチルで希釈し、塩水で三回抽出した。有機層を減圧下で減少させ、カラムクロマトグラフィー (ヘキサン/酢酸エチル勾配)で精製した。単離された産物をジクロロメタンに溶解し、℃に冷却し、トリフルオロ酢酸 (1 mL)で処置した。溶液をrtに温め、一晩撹拌した。反応を重炭酸ナトリウムで中和し、減圧下で濃縮した。生じた固形物を酢酸エチルに溶解し、飽和塩化ナトリウム溶液で抽出した。有機層を硫酸マグネシウムで乾燥し、濾過し、減圧下で濃縮し、カラム クロマトグラフィー (ヘキサン/酢酸エチル および ジクロロメタン/メタノール 勾配)で精製した。減圧下で溶媒を蒸発させ、純粋な産物 10が提供され、構造および純度がNMR分光学で検証された。
【0123】
本発明が特定の例示の態様および例に記載されるが、本出願に開示された態様が説明の目的でのみ記載され、様々な修飾および変更が請求項に記載される本発明の精神および範囲から逸脱することなく、当業者によりなされることが理解されるだろう。
【表1−1】
【表1−2】
【表1−3】
【表1−4】
【表1−5】
【表1−6】
【表1−7】
【表1−8】
【表1−9】
【表1−10】
【表1−11】
【表1−12】
【表1−13】
【表1−14】
【表1−15】
【表1−16】
【表1−17】
【表1−18】
【表1−19】
【表1−20】
【表1−21】
【表1−22】
【表1−23】
【表1−24】
【表1−25】
【表1−26】
【表1−27】
【表1−28】
【表1−29】
【表1−30】
【表1−31】
【表1−32】
【表1−33】
【表1−34】
【表1−35】
【表1−36】
【表1−37】
【表1−38】
【表1−39】
【表1−40】
【表1−41】
【表1−42】
【表1−43】
【表1−44】
【表1−45】
【表1−46】
【表1−47】
【表1−48】
【表1−49】
【表1−50】
【表1−51】
【表1−52】
【表1−53】
【表1−54】
【表1−55】
【表1−56】
【表1−57】
【表1−58】
【表1−59】
【表1−60】
【表1−61】
【表1−62】
【表1−63】
【表1−64】
【0124】
[参照文献]
本出願中で参照される全ての引用は、その全体が参照によって援用される。
【0125】
Bennett, C.L., Christie, J., Ramsdell, F., Brunkow, M.E., Ferguson, P.J., Whitesell, L., Kelly, T.E., Saulsbury, F.T., Chance, P.F., and Ochs, H.D. (2001). The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nature genetics 2 7, 20-21.
Chan, J.K., Sun, L., Yang, X.J., Zhu, G., and Wu, Z. (2003). Functional characterization of an amino-terminal region of HDAC4 that possesses MEF2 binding and transcriptional repressive activity. The Journal of biological chemistry 278, 23515-23521.
Chen, B., and Cepko, C.L. (2009). HDAC4 regulates neuronal survival in normal and diseased retinas. Science (New York, NY 323, 256-259.
Chou, C.J., Herman, D., and Gottesfeld, J.M. (2008), Pimelic diphenylamide 106 is a slow, tight-binding inhibitor of class l histone deacetylases. The Journal of biological chemistry 283, 35402-35409.
de Ruijter, A.J., van Gennip, A.H., Caron, H.N., Kemp, S., and van Kuilenburg, A.B. (2003). Histone deacetylases (HDACs): characterization of the classical HDAC family. The Biochemical journal 370, 737-749.
Fischer, A., Sananbenesi, F., Wang, X., Dobbin, M., and Tsai, L.H. (2007). Recovery of learning and memory is associated with chromatin remodelling. Nature 447, 178-182.
Flavell, S.W., Cowan, C.W., Kim, T.K., Greer, P.L., Lin, Y., Paradis, S., Griffith, E.C., Hu, L.S., Chen, C., and Greenberg, M.E. (2006). Activity-dependent regulation of MEF2 transcription factors suppresses excitatory synapse number. Science (New York, NY 311, 1008-1012.
Flavell, S.W., and Greenberg, M.E. (2008). Signaling mechanisms linking neuronal activity to gene expression and plasticity of the nervous system. Annual review of neuroscience 31, 563-590,
Flavell, S.W., Kim, T.K,, Gray, J.M., Harmin, D.A,, Hemberg, M., Hong, E.J., Markenscoff Papadimitriou, E., Bear, D.M., and Greenberg, M.E. (2008). Genome-wide analysis of MEF22 transcriptional program reveals synaptic target genes and neuronal activity-dependent polyadenylation site selection. Neuron 60, 1022-1038.
Fontenot, J.D., Gavin, M.A., and Rudensky, A.Y. (2003). Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nature immunology 4, 3 3 0-3 3 6.
Gregoire, S., Tremblay, A.M., Xiao, L., Yang, Q., Ma, K., Nie, J., Mao, Z., Wu, Z., Giguere, V., and Yang, X.J. (2006). Control of MEF2 transcriptional activity by coordinated phosphorylation and sumoylation. The Journal of biological chemistry 281, 4423-4433.
Gregoire, S., and Yang, X.J. (2005). Association with class IIa histone deacetylases upregulates the sumoylation of MEF2 transcription factors. Molecular and cellular biology 25, 2273-2287.
Guo, L., Han, A., Bates, D.L., Cao, J., and Chen, L. (2007). Crystal structure of a conserved N-terminal domain of histone deacetylase 4 reveals functional insights into glutamine-rich domains. Proceedings of the National Academy of Sciences of the United States of America 104, 4297-4302.
Han, A., He, J., Wu, Y., Liu, J.O., and Chen, L. (2005). Mechanism of recruitment of class TI histone deacetylases by myocyte enhancer factor-2. Journal of molecular biology 345, 91-102.
Han, A., Pan, F., Stroud, J.C., Youn, H.D., Liu, J.O., and Chen, L. (2003). Sequence specific recruitment of transcriptional co-repressor Cabin l by myocyte enhancer factor-2. Nature 422, 730-734.
Herman, D., Jenssen, K., Burnett, R., Soragni, E., Perlman, S.L., and Gottesfeld, J.M. (2006). Histone deacetylase inhibitors reverse gene silencing in Friedreich's ataxia. Nature chemical biology 2, 551-558.
Hori, S., Nomura, T., and Sakaguchi, S. (2003). Control of regulatory T cell development by the transcription factor Foxp3. Science (New York, NY 299, 1057-1061.
Kim, Y., Phan, D., van Rooij, E., Wang, D.Z., MoAnally, J., Qi, X., Richardson, J.A., Hill, J.A., Bassel-Duby, R., and Olson, EX (2008). The MEF2D transcription factor mediates stress-dependent cardiac remodeling in mice. J Clin Invest 118, 124-132.
Li, B., Samanta, A., Song, X., Iacono, K.T., Brennan, P., Chatila, T.A., Roncador, G., Banham, A.H., Riley, J.L., Wang, Q., et al. (2007). FOXP3 is a homo oligomer and a component of a supramolecular regulatory complex disabled in the human XLAAD/IPEX autoimmune disease. International immunology 19, 825-835.
Mao, Z., Bonni, A., Xia, F,, Nadal-Vicens, M., and Greenberg, M.E. (1999). Neuronal activity-dependent cell survival mediated by transcription factor MEF2. Science (New York, NY 286,785-790.
McKinsey, T.A., Zhang, C.L., and Olson, E.N. (2001). Control of muscle development by dueling HATs and HDACs. Current opinion in genetics & development 11, 497-504.
McKinsey, T.A., Zhang, C.L., and Olson, E.N. (2002), MEF2: a calcium-dependent regulator of cell division, differentiation and death, Trends Biochem Sci 27, 40-47.
Miska, E.A., Karlsson, C., Langley, E., Nielsen, S.J., Pines, J., and Kouzarides, T.(1999). HDAC4 deacetylase associates with and represses the MEF2 transcription factor. The EMBO journal 18, 5099-5107.
Molkentin, J.D., and Olson, E.N. (1996). Combinatorial control of muscle development by basic helix-loop-helix and MARS-box transcription factors. Proceedings of the National Academy of Sciences of the United States of America 93, 9366- 9373.
Morrison, B.E., Majdzadeh, N., and D'Mello, S.R. (2007). Histone deacetylases: focus on the nervous system. Cell Mol Life Sci 64, 2258-2269.
Morrow, E.M., Yoo, S.Y., Flavell, S.W., Kim, T.K., Lin, Y., Hill, R.S., Mukaddes, N.M., Balkhy, S., Gascon, G., Hashmi, A., et al. (2008). Identifying autism loci and genes by tracing recent shared ancestry. Science (New York, NY 321, 218-223.
Pan, F., Ye, Z., Cheng, L., and Liu, J.O. (2004). Myocyte enhancer factor 2 mediates calcium-dependent transcription of the interleukin-2 gene in T lymphocytes: a calcium signaling module that is distinct from but collaborates with the nuclear factor of activated T cells (NFAT). The Journal of biological hemistry 279, 14477-14480.
Paris, M., Porcelloni, M., Binaschi, M., and Fattori, D. (2008). Histone deacetylase inhibitors: from bench to clinic. Journal of medicinal chemistry 51, 1505- 1529.
Potthoff, M.J., and Olson, E.N. (2007). MEF2: a central regulator of diverse developmental programs. Development (Cambridge, England) 134, 4131- 4140.
Rai, M., Soragni, E., Jenssen, K., Burnett, R., Herman, D., Coppola, G., Geschwind, D.H., Gottesfeld, J.M., and Pandolfo, M. (2008). HDAC inhibitors correct frataxin deficiency in a Friedreich ataxia mouse model. PloS one 3, eI958.
Santelli, E., and Richmond, T.J. (2000). Crystal structure of MEF2A core bound to DNA at 1.5 A resolution. Journal of molecular biology 297, 437-449.
Sartorelli, V., Huang, J., Hamamori, Y., and Kedes, L. (1997). Molecular mechanisms of myogenic coactivation by p300: direct interaction with the activation domain of MyoD and with the MADS box of MEF2C. Molecular and cellular biology 17, 1010-1026.
Shalizi, A., Gaudilliere, B., Yuan, Z., Stegmuller, J., Shirogane, T., Ge, Q., Tan, Y., Schulman, B., Harper, J.W., and Bonni, A. (2006). A calcium-regulated MEF2 sumoylation switch controls postsynaptic differentiation. Science (New York, NY 311, 1012-1017.
Shalizi, A.K., and Bonni, A. (2005). Brawn for Brains: The Role of MEF2 Proteins in the Developing Nervous System. Current topics in developmental biology 69, 239-266.
Shore, P., and Sharrocks, A.D. (1995). The MADS-box family of transcription factors. Eur J Biochem 229, 1-13.
Slepak, T.I., Webster, K.A., Zang, J., Prentice, H., O'Dowd, A., Hicks, M.N., and Bishopric, N.H. (2001). Control of cardiac-specific transcription by p300 through myocyte enhancer factor-2D. The Journal of biological chemistry 276, 7575-7585.
Somoza, J.R., Skene, R.J., Katz, B.A., Mol, C., Ho, J.D., Jennings, A.J., Luong, C., Arvai, A., Buggy, J.J., Chi, E., et al. (2004). Structural snapshots of human HDAC8 provide insights into the class I histone deacetylases. Structure 12, 1325-1334.
Sparrow, D.B., Miska, E.A., Langley, E., Reynaud-Deonauth, S., Kotecha, S., Towers, N., Spohr, G., Kouzarides, T., and Mohun, T.J. (1999). MEF-2 function is modified by a novel co-repressor, MITR. The EMBO journal 18, 5085-5098.
Stefanko, D.P., Barrett, R.M., Ly, A.R., Reolon, G.K., and Wood, M.A. (2009). Modulation of long-term memory for object recognition via HDAC inhibition. Proceedings of the National Academy of Sciences of the United States of America 106,9447-9452.
Thomas, E.A., Coppola, G., Desplats, P.A., Tang, B., Soragni, E., Burnett, R., Gao, F., Fitzgerald, K.M., Borok, J.F., Herman, D., et al. (2008). The HDAC inhibitor 4b ameliorates the disease phenotype and transcriptional abnormalities in Huntington's disease transgenic mice. Proceedings of the National Academy of Sciences of the United States of America 105, 15564-15569.
Wang, L., Fan, C., Topol, S.E., Topol, E.J., and Wang, Q. (2003). Mutation of MEF2A in an inherited disorder with features of coronary artery disease. Science (New York, NY 302, 1578-1581.
Wei, J.Q., Shehadeh, L.A., Mitrani, J.M., Pessanha, M., Slepak, TI, Webster, K.A,, and Bishopric, N.H. (2008). Quantitative control of adaptive cardiac hypertrophy by acetyltransferase p300. Circulation 118, 934-946.
Wong, J.C., Hong, R., and Schreiber, S.L. (2003). Structural biasing elements for in cell histone deacetylase paralog selectivity. Journal of the American Chemical Society 125, 5586-5587.
Wu, Y., Borde, M., Heissmeyer, V., Feuerer, M., Lapan, A.D., Stroud, J.C., Bates, D.L., Guo, L., Han, A., Ziegler, S.F., et al. (2006). FOXP3 controls regulatory T cell function through cooperation with NFAT. Cell 126, 375-387.
Yang, Q., She, H., Gearing, M., Colla, B., Lee, M., Shacka, J.J., and Mao, Z. (2009). Regulation of neuronal survival factor MEF2D by chaperone-mediated autophagy. Science (New York, NY 323, 124-127.
Youn, H.D., and Liu, J.O. (2000). Cabin1 represses MEF2-dependent Nur77 expression and T cell apoptosis by controlling association of histone deacetylases and acetylases with MEF2. Immunity 13, 85-94.
Youn, H.D., Sun, L., Prywes, R., and Liu, J.O. (1999). Apoptosis of T cells mediated by Ca2+-induced release of the transcription factor MEF2. Science (New York, NY 286, 790-793.
Zhang, C.L., McKinsey, T.A., Chang, S., Antos, C.L., Hill, J.A., and Olson, E.N. (2002). Class II histone deacetylases act as signal-responsive repressors of cardiac hypertrophy. Cell 110, 479-488.
Zheng, Y., and Rudensky, A.Y. (2007). Foxp3 in control of the regulatory T cell lineage. Nature immunology 8,457-462.
Zuo, T., Liu, R., Zhang, H., Chang, X., Liu, Y., Wang, L., Zheng, P., and Liu, Y.(2007a). FOXP3 is a novel transcriptional repressor for the breast cancer oncogene SKP2. J Clin Invest.
Zuo, T., Wang, L., Morrison, C., Chang, X., Zhang, H., Li, W., Liu, Y., Wang, Y., Liu, X., Chan, M.W., et al. (2007b). FOXP3 is an X-linked breast cancer suppressor gene and an important repressor of the HER-21ErbB2 oncogene. Cell 129,1275-1286.
【発明の説明】
【0001】
[関連出願の相互参照]
この出願は、2008年11月5日に出願された米国特許仮出願第61/111,689号, および2009年9月29日に出願された米国特許仮出願第61/246,934号の利益を主張する出願である。上記の仮出願は、本明細書中に参照によって援用される。
【0002】
[連邦政府により援助された研究および開発に関する記載]
本発明は、NIH R21AI49905, R01HL076334, および RC1DA028790の助成による部分的な援助のもとでなされた。従って、米国政府は、特定の権利を有している。
【0003】
[発明の分野]
本発明は、一般に分子医学の分野に関する。特に、本発明は、転写因子の機能を調節するための方法および組成物に関する、特にエピジェネティックレギュレーター(ヒストン修飾酵素)を特異的な DNA プロモーターにリクルートする転写因子に関する。標的とされる転写因子には、筋細胞増強因子(MEF2), フォークヘッド/ウイングドヘリックス 転写因子 FOXP3 および 転写因子 GATA3が含まれるが、これらに限定されない。特に、本発明は、MEF2及びその関連因子〔ヒストン デアセチラーゼ (HDACs), p300/CBP および Cabin 1を含むが、これらに限定されない〕の小分子のモジュレーター及びその治療への適用に関する。
【0004】
[発明の背景]
本発明は、特定の疾患と関連する転写因子の機能を調節する小分子の使用に関する。転写因子は、特定のDNA配列に結合し、直接的に又は関連するタンパク質〔例えば、コアクチベータおよびコリプレッサー〕をとおして、又はヒストン 修飾酵素〔例えば、ヒストン アセチルトランスフェラーゼ (HATs) および ヒストン デアセチラーゼ (HDACs)〕をリクルートすることにより遺伝子発現を制御するタンパク質である。転写因子は、適切なレベルの遺伝子発現を保証することにより多くの生物学的プロセスに重要な役割を果たしている。また、転写を制御する能力が異常に修飾される場合には特定の病的状態と関連づけすることができる。特定の転写因子の機能を選択的に調節できる小分子を同定し、開発することによって、潜在的に新しい治療適用を導入することができる。本発明は、二つの基本的なアイデアに基づく。一つは、特定の転写因子(例えば、MEF2, FOXP3 および GATA3)に結合し、転写のコアクチベータ および コリプレッサーとの相互作用を調節する小分子を開発することである。もう一つは、HAT (例えば、p300およびCBP) およびHDACs および他のヒストン修飾酵素(例えば、ヒストン メチルトランスフェラーゼ, およびデメチラーゼ, DNAメチルトランスフェラーゼ)のリクルートおよびクロマチンの特定の領域に対するクロマチンリモデリング機構をブロックする小分子を開発することである。
【0005】
特に、本発明は、筋肉, 免疫および神経系の発生および適応応答に重要な役割を担う筋細胞エンハンサー因子-2 (MEF2: myocyte enhancer factor-2)(Flavell et al., 2006; Kim et al., 2008; Mao et al., 1999; McKinsey et al., 2002; Pan et al., 2004; Potthoff and Olson, 2007; Youn and Liu, 2000; Youn et al., 1999)に関する。MEF2は、心筋細胞における肥大応答(hypertrophic responses)の重要な制御因子と指摘された。病理学的な刺激により誘導された心臓肥大は、多くの形態の心血管疾患において心不全を誘発しえる。
【0006】
一般にMEF2は、四つのメンバー(MEF2A, MEF2B, MEF2C および MEF2D)の転写因子のファミリーを規定する。それらの機能の重要性は、マウスおよびショウジョウバエの遺伝学を使用することにより詳細に実証された(Potthoff and Olson, 2007)。最初に同定された骨格筋におけるMEF2は、筋原性のベーシック ヘリックス・ループ・ヘリックス 転写因子(例えば、MyoD)と共に筋形成を促進し、維持する(Molkentin and Olson, 1996)。MEF2A(MEF2ファミリーのメンバーの一つ)は、最近このタンパク質が冠動脈疾患 (CAD) および 心筋梗塞 (MI)と関連することから「心臓発作遺伝子(heart attack gene)」であるとされた(Wang et al., 2003)。これらの所見は、ヒトの心臓疾患におけるMEF2の重要な役割の基礎をなす(Kim et al., 2008; Wei et al., 2008; Zhang et al., 2002)。
【0007】
MEF2は、現在では多くの他の細胞タイプの基本転写因子として知られる。一例を挙げると, MEF2は、リンパ系の発生においてカルシウムシグナル伝達を媒介する重要な転写因子の一つである(Pan et al., 2004; Youn and Liu, 2000; Youn et al., 1999)。MEF2は、サイトカイン発現 および 免疫応答を制御する。また、MEF2は、ニューロンの生存 および シナプスのリモデリングの基礎的な転写プログラムを制御する(Chen and Cepko, 2009; Flavell et al., 2006; Flavell and Greenberg, 2008; Flavell et al., 2008; Mao et al., 1999; Morrow et al., 2008; Shalizi et al,, 2006; Shalizi and Bonni, 2005; Yang et al., 2009)。これらの観察は、MEF2 機能を調節する小分子が心肥大および心不全, 自己免疫疾患および移植拒絶, 神経変性疾患および学習および記憶の病理学的な障害に治療効果を有することを示唆する(Fischer et al., 2007; Stefanko et al., 2009)。
【0008】
細胞の内側のMEF2の作用には、三つの別個の工程が含まれる: (i) 転写抑制; (ii) カルシウム依存性の脱抑制;および (iii) 転写活性化。
【0009】
MEF2による転写抑制は、内因性の又は関連するヒストンデアセチラーゼ (HDAC) 活性を有する種々の転写コリプレッサーとの結合に依存する。T細胞中でMEF2はCabin 1と結合し、次にクラス IのHDACs(例えば、 HDAC 1, HADC2 および HDAC3)と結合する(Youn and Liu, 2000)。筋肉細胞において、MEF2は、直接的にクラス II HDACs(例えば、HDAC4, HDAC5, および HDAC9)と結合し、筋肉の発生 および 適応応答に関与する特異的な遺伝子の発現を阻害する(Chan et al., 2003; Gregoire et al., 2006; Gregoire and Yang, 2005; McKinsey et al., 2001, 2002; Miska et al., 1999; Sparrow et al., 1999)。HDAC5またはHDAC9がノックアウトされたマウスは肥大性刺激への感受性の増加を示し、心臓肥大におけるクラス II HDACsの重要な役割が示唆される(Potthoff and Olson, 2007)。これらの及び他のデータは、MEF2およびクラス II ヒストン デアセチラーゼ (HDACs)(特に、HDAC5およびHDAC9)が心筋細胞における肥大性シグナルの重要なメディエータであることを示す。多くのデータは、MEF2/クラス II HDAC 経路が心臓肥大の潜在的な治療上の標的であることを示唆する。
【0010】
特定のカルシウムシグナルへの応答において、Cabinl および HDACsは、MEF2から除去される(Potthoff and Olson, 2007)。次に、MEF2は、コアクチベータ(例えば、p300 および CBP)をリクルートし、種々の転写のアクチベータ および コアクチベータと結合することにより別個のプログラムを開始させる(McKinsey et al., 2001; Sartorelli et al., 1997; Slepak et al., 2001; Wei et al., 2008)。p300の小さな増加が、MEF2依存性の心肥大を誘発させることに必要十分であることが示された。MEF2は、よく特徴が調べれたMADS-ボックスおよびMEF2-特異的ドメインからなる高度に保存されたN末端領域 (残基2-93)を有する(Shore and Sharrocks, 1995)。MADS-ボックス/MEF2ドメインは機能が著しく豊富であり、DNA結合, 二量体化, およびCabin 1, クラス IIa HDACs および p300/CBPを含む無数のMEF2の転写パートナーとのタンパク質間相互作用を媒介する(McKinsey et al., 2001, 2002)。MEF2におけるMADS-ボックス/MEF2 ドメインが、クラス II HDACs および Cabin1に保存された小さいモチーフと結合するため必要十分であることが示された。また、p300 および CBPのCH3ドメインは、MADS-ボックスMEF2ドメインと結合することが示された。
【0011】
当該技術において利用可能な様々な細胞プロセスにおけるMEF2の関与について広範な知識が存在するにもかかわらず、これまで適切な分子ツールを欠いていることからその知識を利用することは不可能であった。特に、如何にして小分子によりMEF2の活性を調節するかについては長年にわたる挑戦であり、それはMEF2が明らかな酵素活性のない相対的に小さい転写因子であるからであり;MEF2の主な機能は特定のDNAと結合し、転写コレギュレータ(例えば、Cabin 1, クラス II HDACs および p300/CBP)を特異的なプロモーターにリクルートすることである。この様式の作用は、一般に薬にできないと考えられており、少なくとも小分子による標的化は非常に困難である。このような分子の発見または創造によって、この分野のさらなる進捗が促進され、MEF2-関連疾患(炎症, 自己免疫疾患, 神経変性疾患, 癌, および心血管疾患)のための新しい機構に基づく、また構造に基づく治療適用が導かれえる。
【0012】
また、本出願に記載される発明は、他の転写因子(例えば、フォークヘッド/ウイングドヘリックス転写因子 FOXP3)の活性を調節することに拡張できる(Bennett et al., 2001; Fontenot et al., 2003; Hori et al., 2003; Wu et al., 2006; Zheng and Rudensky, 2007)。FOXP3は、制御性T細胞(Tregs)の発生および機能に重要な鍵となる転写因子である。Tregsは免疫系の過剰な活性化を抑制するため必要とされるT細胞の特殊な集団であり、変異によるFOXP3の機能喪失や他の機構によって胎性の自己免疫疾患(例えば、IPEX)が誘発され、他方でFOXP3の発現又はその活性の増強によって抑制機能を付与することができる。FOXP3の機能の上昇は自己免疫疾患 および 移植拒絶の治療に有益でありえるが、他方でFOXP3 活性のダウンレギュレーションを戦略的に使用して免疫に基づく抗腫瘍療法を開発することができる(Zuo et al., 2007a; Zuo et al., 2007b)。以上のとおり、FOXP3と結合し、そのコリプレッサー および コアクチベータとの相互作用を調節する小分子は、自己免疫疾患, 移植拒絶および癌治療における治療適用を有するだろう。
【0013】
MEF2と同様にFOXP3の機能は、HAT (例えば、TIP60) および HDACs (クラス I および クラス II HDACsを含む)を含む転写コレギュレータにより厳格に制御される(Li et al., 2007)。FOXP3と結合し、ヒストン修飾酵素 およびクロマチンリモデリング機構などのエピジェネティックレギュレーターを含むコレギュレータとの相互作用をブロックする本発明に記載された方法により小分子を開発できるだろう。
【0014】
同じように、転写コレギュレータを標的とすることは、同様な挑戦である。
【0015】
転写因子の転写コレギュレータ(例えば、MEF2, クラス II HDACs)は、最良の試験群である。
【0016】
ヒストン デアセチラーゼ (HDACs) (ECナンバー 3.5.1)は、ヒストンにおけるc-N-アセチル リジン アミノ酸からアセチル基を除去する酵素のクラスである。その作用は、ヒストン アセチルトランスフェラーゼ (HATs)の作用と逆である。HDACs タンパク質は、現在では多数の非-ヒストンタンパク質も含まれる彼らの標的というよりむしろ彼らの活性を正確に記載するため、リジン デアセチラーゼ (KDAC)とも称される。
【0017】
彼らの名称から示唆されるように、HDACsの主な機能の一つは、アセチル基をヒストンタンパク質から除去することである。ヒストンタンパク質は、クロマチンの主要なタンパク質成分である。彼らは、周囲にDNAが巻きつくスプールとして働き、遺伝子制御 および DNAパッケージングに重要な役割を担っている。ヒストンタンパク質は、リジン および アルギニンアミノ酸に存在するアミン基のため通常陽性に荷電した尾部を有する。これらの陽性の荷電は、ヒストン尾部がDNAバックボーンにおける陰性に荷電したリン酸基との相互作用や結合を援助する。
【0018】
DNA と ヒストンの間の結合は、転写因子がDNAにアクセスする能力を制御する重要な制御機構として作用する。DNAおよびヒストンの間の強い結合は、転写因子によるアクセスを制限し、遺伝子の転写を抑制する(Morrison et al., 2007)。ヒストンまたはDNAの修飾によって、それらの結合の強さを変化させ、転写活性を調節することができる(Morrison et al., 2007)。メチル, リン酸, またはアセチル部分を共有結合で付加することによって、ヌクレオソームの状態が変化し、結果的に転写に影響することが示されている。アセチル化は、ヒストンにおける保存されたN 末端リジン残基のε-アミノ基へのアセチル基の付加から生じる。アセチル基のヒストンへの付加は陽性に荷電したヒストンタンパク質 および 陰性に荷電したDNAのリン酸バックボーンの間の引力を減少させ、よりリラックスし、アクセス可能なクロマチン構造を生じる。HATsは、ヒストンアセチル化を促進するので、転写アクチベータであると信じられている。逆に, HDACsは、アセチル基をヒストンから除去するため貢献し、それによって転写が抑制される。従って、局所性のクロマチン構造 および 遺伝子発現を主に支配するのはHATs および HDACs活性の間の相互作用である。HDACsは、クロマチンの脱アセチルをとおして全体的な遺伝子転写を変化させる。HDACsは直接的にDNA配列に結合せず、遺伝子の認識を標的とする付加的な因子を必要とすることに注意すべきである(Morrison et al., 2007)。
【0019】
機能およびDNA配列の類似性に基づくHDAC タンパク質の4 つの認められたサブタイプ (クラス I - IV)が存在する。最初の二つのサブタイプは、活性がトリコスタチン A (TSA)により阻害される「古典的」HDACsと考えられる。クラス I HDACsはHDACs 1, 2, 3 および 8を含み、これらはユビキタスに発現される(Zhang and Olsen, 2000)。クラス II HDACsは、4, 5, 7 および 9を含むIIaおよびHDAC6 および HDAC10を含むIIbの二つのサブグループを有する。クラス IIaは、カルボキシル-末端の触媒ドメイン、また転写因子の筋細胞増強因子 2 (MEF2) ファミリーのメンバーとの相互作用を媒介するアミノ-末端の伸展部で共通の構造上の構成を共有する。また、HDACsは細胞内局在が異なり、一般にクラス Iは核に認められ、クラス IIbは大抵が細胞質に局在し、クラス IIaは核および細胞質の間をシャトルする。クラス IのHDACsとは異なり、クラス IIaのHDACsは組織限定的であり、特に高いレベルの発現を心臓, 骨格筋, および脳で示す(Zhang et al., 2002)。クラス IIIはTSAにより影響されないNAD+依存性タンパク質のファミリーであり、クラス IVはそれ自身の非定型のカテゴリーであると考えられる。HDAC11 は、クラスV にグループ化される。
【0020】
HDACsは細胞プロセスにおいて重要な役割を有するので、HDAC インヒビター (HDACi)の医学的な適用は研究が活発な領域である。しかしながら、医学におけるHDACiの多くの使用は、基本的な機構の知識とは無関係に発見された。例えば、精神医学および神経学において、バルプロ酸を精神安定剤(mood stabilizer)および抗癲癇剤として用いてきた長い歴史がある。バルプロ酸の抗痙攣薬特性は、抗痙攣薬として調査された幾つかの他の化合物のビヒクルとして使用された際に偶然発見された。後にバルプロ酸はHDACiと同定された。近年、HDACiは、神経変性疾患のための緩和剤(mitigator)または治療剤として活発に研究されている。癌治療のためのHDACiを開発する多くの努力がなされてきた。例えば、ボリノスタット (SARA)は、最近皮膚の T-細胞 リンパ腫 (CTCL)の治療のため承認された。CTCLの臨床評価中の別の剤は、環状デプシペプチドの天然物 FK228 (Romidepsin)であり、これはクラス IのHDACsの強力なインヒビターである。加えて、臨床試験は、HIVの感染者の潜在的なプールにおいてバルプロ酸の効果を研究している。HDACiの薬適用に大きな関心があるにもかかわらず、これらの化合物が働く正確な機構はまだよく理解されていない。このような事情から、これらの仕事は、大部分は憶測や試行錯誤により導かれた。
【0021】
HDACiを使用する際に特に問題となる事項の一つは、HDAC 活性を阻害する既知の小分子の多くがHDACsの触媒活性を標的とすることで機能するよう設計されたことである。活性部位は多数の異なるHDACアイソフォームにより共有された保存された特徴であるので、アイソフォーム-選択的な HDACiを同定することは本来困難である。従って、多くのHDACiは、特異性が低く、HDACの特定の種を特異的に標的とする能力がない。例えば、トリコスタチン A (TSA)は、現在知られている最も強力な可逆的なHDACiであり、低ナノモル範囲のIC50を有する;また、TSAはヒドロキサム酸基及びフェニル基との五-炭素原子リンカーを有し、HDACの活性部位に適合する至適なコンホメーションを有する(de Ruijter et al., 2003; Somoza et al., 2004)。全てのHDACsは、TSAによる阻害に凡そ等しく感受性であると考えられる(de Ruijter et al., 2003)。
【0022】
以上のように、HDACsの機能を阻害し、関連する転写因子の活性を調節する小分子を発見することに関する主要な障害は、HDAC酵素の活性部位に結合する能力をとおして同定され、評価されるHDACiの発見 および 最適化に最新技術が焦点を合わせていることである。典型的には、これらの HDACiは、一般構造式R-L-Zを有する。式中のRはタンパク質表面認識基であり、短い脂肪性リンカーLを介して活性部位の亜鉛原子と結合するZn2+ -キレート化基Zに連結される。既知のHDACiにおいて特徴づけられた最も共通するキレート基(Z)は、ヒドロキサム酸 (TSA, ボリノスタット, LAQ824, ベリノスタット), チオール誘導体 (FK228, ラルガゾール) または求電子性ケトン (トラポキシンA)である。Zn2+のような金属カチオンに緊密に結合する係る基の潜在的な欠点は、それらが特定のタンパク質に対して十分な選択性を欠いていることから、様々な副作用を生じる可能性があることである。
【0023】
別のクラスの既知のHDACiは、オルト-アミノアニリド (2-アミノアニリド)部分を備え(feature)るベンズアミドであり、MS-725, MGCD0103, ピメロイルアニリド オルト-アミノアニリド (PAOA)および化合物 106 (N'-(2 アミノフェニル)-N7-p-トリルヘプタンジアミド)を含み、これはフリートライヒ失調症およびハンチントン舞踏病を含む神経変性疾患の潜在的な治療剤として調査された(Chou et al., 2008; Herman et al., 2006; Paris et al., 2008; Rai et al., 2008; Thomas et al., 2008; Wong et al., 2003)。このクラスのHDACiの作用の詳細な分子機構は知られていなく、o-アミノアニリド基のHDAC活性部位の亜鉛原子への結合にこれらの分子が関与することが前提とされたが、このような結合モチーフに関して直接的な証拠を欠いている。そのうえ、係る分子によってHDAC 機能の阻害が関係する生物活性が呈されることが認められたが、他の非選択的なHDACiは類似する活性を示さなかった。例えば、化合物106はHDAC阻害は弱いが、フラタキシンの誘導に関して非常に活性であることが示され、他方で密接に関連するHDACi(例えば、SAHA)はこのタイプの活性を有していなかった。結果的には、このクラスの化合物の作用に関する分子機構が提示されていないことから、それらの治療上の潜在的な能力を最適化することが阻まれてきた。
【0024】
様々な HDAC アイソフォームの良好に制御された生物学的役割に起因する幾つかの付加的な要素によって、HDACiのデザインのための更なる挑戦が従来のアプローチに課される。実験によって、アセチル化したヒストンの量がHDACiの存在下で増加することが示された。しかしなから、DNAに結合した転写因子によるHATs および HDACsのリクルートによって、複数のタンパク質の転写調節性の複合体の形成が生じ、従属的な遺伝子の一群(arrays)に細胞タイプ特異性およびシグナル依存的な制御が付与される。無差別にHDACsを阻害するHDACiを用いることは、モンキーランチを複雑で繊細なバランスのとれた機械に投げ入れることに似ており、このことからHDACiが関与する試行の多くで観察された多数の望ましくない副作用が説明される。
【0025】
また、HDACiを医学的に使用する研究において直面する問題は、エピジェネティック制御を研究する研究者により共有されている。エピジェネティック制御は、永久にDNA 配列を変化させることのない遺伝性の遺伝子発現パターンの確立である。この制御は、細胞機能を制御する重要な機構として出現した。
【0026】
エピジェネティック制御の変化は、多くの疾患(特に、癌)の特徴である。これらの疾患の治療における薬物として開発される小分子(表現型スクリーニングを介して見つけられた)は、細胞プロセスのエピジェネティックコントロールを調節することにより作用する。エピジェネティック制御自体の基礎的な機構は、非常に興味がもたれる領域である。同時に、小分子のエピジェネティックレギュレーターの探索は、創薬のため非常に有望な領域となっている。
【0027】
エピジェネティック制御はDNA (例えば、DNA シトシン メチルトランスフェラーゼ)またはタンパク質(例えば、HATS, HDACs, ヒストンメチラーゼ, および ヒストンデメチラーゼ)に作用する酵素によるクロマチン構造の化学的な修飾をとおして大部分が達成されるので、この関連において、DNA転写の制御におけるHDACsの役割をエピジェネティック制御のコンポーネントとして認めることができる。HDACiと類似して、エピジェネティックレギュレーターの現行の化学的なモジュレーターの大多数は、これらの酵素を別の細胞の役割を有する複数の酵素によりしばしば共有される触媒部位に結合することにより阻害する。
【0028】
転写因子(例えば、MEF2)の役割に関する最近の進歩及びその幾つかの主要な疾患において関係性があるにもかかわらず、MEF2の機能を調節する能力のある小分子を同定することは可能ではなかった。このような分子によって、この分野のさらなる進捗が促進され、MEF2-関連疾患(炎症, 自己免疫疾患, 神経変性疾患, 癌, および心血管疾患)のための新しい機構に基づく、また構造に基づく治療適用が導かれえる。
【0029】
以上のとおり、この領域の研究における一般的な問題は、特異的なエピジェネティック制御因子の酵素またはタンパク質を標的としえる小分子を欠いていることである。また、更なる問題は、全体の酵素またはタンパク質のクラスをとおして広域の活性を有することに起因する欠点がなく、必要とされる選択性が与えられる様式で、係る小分子をデザインし, 評価し 最適化する方法を欠いていることである。係る分子は、エピジェネティック制御の基礎的な機構を研究する分子ツールとして、同様に、治療上の介入を目標とする治療剤として重要な適用を有する。
【0030】
[発明の概要]
広範な生化学的な研究及び構造研究(Guo et al., 2007; Han et al., 2005; Han et al., 2003)により、予想外にも発明者は小分子を用いて活性を調節することを可能とするMEF2のユニークな構造上の特性を発見した。この発見は、従来の知識に反することから本発明の核心部分であり、この発見によって新しい可能性のドアが開かれる。この予想外の発見に基づき、発明者は、転写因子とそれらのコファクターとの間の境界のインヒビターを介した相互作用をブロックすることにより細胞プロセスを調節する一般的な戦略を発明した。それゆえ、本発明は、ファクターコファクター複合体の境界の表面の部位に結合し、複合体の形成を破壊する能力のある小分子モジュレーターを同定する, スクリーニングする, アッセイする, および合成するための方法も提供する。
【0031】
以上のとおり、本発明は、これまで「薬になりえない(undrugable)」転写因子(例えば、MEF2)を標的とする長年にわたる課題を解決した。
【0032】
HDACiが直面する特定の問題に関して、本発明はHDAC機能の小分子モジュレーターの開発のための代替的なアプローチを提供する。特定のHDACの活性部位を標的とするよりむしろ、本発明は、転写を制御するため必要とされる関連する転写因子とHDACとの結合部位を標的とする。異なる HDAC サブタイプは異なる転写因子に対し異なる結合表面を有することから、これらのタンパク質間相互作用の境界(interface)を標的とすることにより保存された活性部位に関連して生じる特異性の問題が解消される。
【0033】
特に、本発明は、転写因子 MEF2 および クラス IIa HDACsの間の相互作用を標的とする。MEF2の活性は、標的の遺伝子発現を抑制する特異的なプロモーターにおいてMEF2と結合するクラス IIa HDACsにより制御される(Potthoff and Olson, 2007)。また、様々な癌の治療のため開発される幾つかのHDACsの小分子インヒビター(HDACi)によって、MEF2のディレギュレーション(deregulation)およびHDACs活性が関与する疾患(心肥大, 神経変性障害, および免疫機能不全を含む)の治療の可能性が示された(Morrison et al., 2007; Paris et al., 2008)。これらの観察は、クラス IIa HDAC:MEF2 相互作用をブロックする小分子がメンバー特異的なHDACiとして類似する臨床上の利益を提供しえることを示唆する(Guo et al., 2007; Han et al., 2005; Han et al., 2003)。
【0034】
クラス IIa HDACsは、筋肉, ニューロン および T細胞においてMEF2と密接に作用する。プロモーターを標的とするためクラス IIa HDACsはDNAに結合しないが、その相互作用はDNA-結合型のMEF2に依存する。この相互作用は、クラス IIa HDACsに保存されるが、他のHDACsでは保存されていない短い両親媒性のヘリックスにより媒介され、これがMEF2 のMADS-ボックス/MEF2 ドメインにおける疎水性の溝と結合する(Guo et al., 2007; Han et al., 2005; Han et al., 2003)。このようなリガンド/レセプター様の結合機構が存在することは、小分子を使用してクラス IIa HDACsのMEF2-特異的なプロモーターへのリクルートをブロックすることが可能であろうことを示唆する(Guo et al., 2007; Han et al., 2005; Han et al., 2003)。
【0035】
本発明は、MEF2と結合する小分子が転写コレギュレータ(例えば、Cabin1, クラス IIa HADCS および p300/CBP)のリクルートにおいてその活性を調節できるとの体系的な構造 および 生化学的な研究に基づく(Guo et al., 2007; Han et al., 2005; Han et al., 2003)。これらのコレギュレータの多くは、クロマチンを修飾する内因性の機能(例えば、HDACs, p300 および CBP)またはクロマチン修飾の酵素および機構をリクルートする能力を有する(mSin3AはCabin1, HP1, CtBPと結合し、14-3-3はクラス IIa HDACsと結合する)。従って、MEF2と結合し、他の転写コレギュレータとの相互作用を調節する小分子は、MEF2が重要な調節的な役割を担う組織におけるエピジェネティックモジュレーターとして貢献できる。それゆえ、これらの小分子を使用して、MEF2-依存的な遺伝子発現の活性の制御が異常な疾患を治療することができる。調節不全(dysregulation)は、MEF2及びその関連する因子の遺伝的な変異, MEF2機能の減少または過度な活性化を導く減少した又は過剰なシグナル, MEF2と結合する、また相互作用するコファクターの異常な過小または過剰な発現から生じえる。MEF2-結合小分子の潜在的な臨床適用には、筋肉系, 免疫系および神経系の疾患が含まれるが、これらに限定されない。筋肉系においては、心肥大, 筋線維タイプのリモデリング(muscle fiber type remodelin), およびMEF2機能の不均衡から生じる他の筋肉関連疾患である。免疫系においては、種々の自己免疫疾患又は過剰な又は非常に僅かなMEF2-依存的な遺伝子発現から生じる免疫不全である。また、MEF2-結合小分子を使用して、移植拒絶を予防するため制御性T細胞の機能や全体の免疫応答を操作することができる。MEF2-依存的な遺伝子発現は密接にシナプスのリモデリングやニューロンの生存と関連するので、MEF2 結合小分子を使用して制御がはずれたMEF2機能から生じる種々の神経変性疾患(例えば、アルツハイマー病やハンチントン舞踏病など), 自閉症, 精神医学的な障害, および学習および記憶の障害を治療することができる。
【0036】
本発明の小分子モジュレーターは転写因子及びそのコファクターのタンパク質間相互作用の境界(protein-protein interaction interface)に局在する結合部位を標的とすることにより機能するので、それらも境界のインヒビターとして本出願において参照される。
【0037】
本発明の基礎的な原則を説明したので、我々は次に本発明の様々な側面 および 態様を以下に要約する:
第一の側面において、本発明は、境界のインヒビターの候補をスクリーニングする, 同定する, または最適化するためのアッセイを提供する。この本発明の側面と一致するアッセイは、一般に境界のインヒビターの候補を、各々のMEF2 単量体のベータ鎖S1, S2 およびS3 およびヘリックスH2およびCabin 1およびHDAC4およびHDAC 9からの短いヘリックスモチーフを含むタンパク質間の境界により規定される分子表面を含んでいる評価するエレメント(evaluating element)と接触させる工程を含むだろう。幾つかの好適な態様において、評価するエレメントは、候補インヒビターについての情報を提供するレポートエレメントと作動可能に連結される。好適な態様において、前記アッセイは、MEF2と結合し、その転写コレギュレータへの結合を調節する小分子の迅速でハイスループットなスクリーニングや最適化を許容する細胞に基づくルシフェラーゼアッセイである。このアッセイは、発明者の研究室によるMEF2 複合体の構造および生化学の10年以上の研究に基づき開発された。本発明は、最初に結晶研究により同定され、更に本発明の構造に基づく変異の研究により実証されたタンパク質間の境界がMEF2-結合小分子に高度に特異的で敏感なスクリーニングのための分子的な基礎として貢献できるだろうことを主張する。他の態様において、前記アッセイの評価するエレメントは、本発明により同定されたタンパク質間の境界に基づくプルダウン, 免疫共沈降 または蛍光クエンチングおよび異方性(anisotropy)または任意のインビトロ結合アッセイおよび 細胞に基づくルシフェラーゼ レポーター アッセイ, トランスジェニック レポーターアッセイを含むが、これらに限定されない理学的なアッセイで実行されえる。
【0038】
第二側面において、本発明は、筋肉, 免疫系および神経系を含むが、これらに限定されない種々の哺乳類の組織におけるMEF2-依存的な転写のエピジェネティックモジュレーターとして有用な化合物も提供する。幾つかの好適な態様において、この本発明の側面と一致する化合物は、以前に公開された化合物(Chou et al., 2008; Herman et al., 2006; Paris et al., 2008; Rai et al., 2008; Thomas et al., 2008; Wong et al., 2003), ピメロイルアニリド オルソアミノアニリドPAOA, および商業的に利用可能なホモログ スベロイルアニリド オルソアミノアニリド(suberoylanilide orthoaminoanilide), またはBML-210, 同様に、それらの構造的に関連する誘導体を含みえるが、これらに限定されない。
【0039】
第三の側面において、本発明は、表 1における結晶構造座標で表されるMEF2へのBML-210結合の結晶構造により樹立された構造上のフレームワークから由来するMEF2におけるBML-210 結合部位により規定された分子フレームワーク(molecular framework)も提供する。我々は、BML-210をDNAと結合したMEF2A (1-78) 二量体と共結晶化した。結晶を2.4Aで回折し、MEF2A (1-78): DNA複合体を探索モデルとして用いた分子置換で構造を解析した(Santelli and Richmond, 2000)。BML-210は、拡張したコンホメーションに適合し、MEF2の疎水性ポケットに結合する(図 1d)。分子の一端でフェニルアミド基は、Leu66, Leu67, Thr70, Leu66' (プライム記号は他の単量体からの残基を示す) および Thr70'を含む幾つかの疎水性残基で周囲をとりまかれる。また、この端でのアミド基は、所定の位置にあり、それぞれThr70 および Thr70'との水素結合相互作用に関与する。BML-210の他の端で環連結の電子密度(ring-link electron density)は、Asn73, Gln56', Asp61' および Asp63'で周囲をとりまかれるより親水性の環境にある。この領域は、オルト-アミノアニリド基に対応する。ここでオルトアミノアニリド部分とそのアミド基とでMEF2の残基と広範なファンデルワールス接触や潜在的な水素結合相互作用を作る(図 1d)。オクタンジアミドのメチレン基は、二つのMEF2 分子のヘリックスH2の間に適切に合い、主として疎水性の性質のMEF2残基の主鎖 および 側鎖と多数の接触を作る(図 1d)。MEF2に結合したBML-210の結晶構造に基づき、我々はBML 210 および PAOAと接触する又は近くに近接して接触する表面残基を使用して、実験および計算に基づくアプローチによりMEF2に対する小分子のデザイン および スクリーニングを導くことできることを主張する。これらの残基は、暴露されるMEF2の鎖 S1, S2 および S3 および ヘリックスH2における全ての残基を含む。この構造上のフレームを本発明に記載のハイスループットで特異的な高感度アッセイと組み合せることで、BML210 およびPAOA誘導体および新規の分子骨格(molecular scaffolds)に基づく分子を含むMEF2-結合小分子の迅速なデザインおよび最適化が許容される。本発明において開示されたMEF2に結合したBML-210の結晶構造に加えて本出願に記載される小分子 MEF2-結合部位を規定することで、BML-210に及び他のベンズアミド含有HDACインヒビターに存在するオルト-アミノアニリド部分のための有望な結合部位が初めて示された。特に、この結合部位は、このクラスのHDACインヒビターに関して以前に仮定されたHDAC 酵素の活性部位と異なる。
【0040】
第四の側面において、本発明は、上記の第三の側面における分子フレームワークにより規定された境界の結合部位への結合に有用なMEF2-結合小分子を提供する。一態様において、提供される分子は、MEF2と結合するBML-210の結晶構造から由来する同定された結合部位から由来する一般構造式を有するMEF2-結合小分子である。提供される小分子は、記載された構造上のフォールドと結合するため設計され、潜在的に高い親和性および選択性でMEF2と結合するだろう。この本発明の側面の態様において提供される化合物は、一般式 Ra-L Rbの化合物を含み、この化合物はMEF2 結合部位に結合し、式中:
Ra は、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(S02)-を含む基から選択されるMEF2結合部位の疎水性領域と結合する認識基であり、式中:
Rc および Rd は、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される、
L は、20 炭素原子までの鎖からなるリンカーであり(但し、三炭素原子までが酸素, 窒素 またはイオウ原子で置換でき、更にアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミド, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)、
Rbは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(S02)-を含む基から選択されるMEF2結合部位の親水性領域と結合する認識基であり、式中:
Rc および Rd は、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される。
【0041】
好適な態様において、提供される化合物は、一般構造式Ar1-L1-L2-L3-Ar2を有し、式中:
Ar1 および Ar2 は、独立にベンゼン, ナフタレン, ピリジン, ピリミジン, ピラジン, キノリン, イソキノリン, ピロール, フラン, チオフェン, イミダゾール, ピラゾール, オキサゾール, チアゾール, イソキサゾール, インドール, ベンズイミダゾール, ベンゾチアゾール, ベンゾキサゾールからなる群から選択される芳香環である(但し、前記芳香環は、水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アミノ, アルキルアミノ, ジアルキルアミノ, アリールアミノ, ヘテロアリール アミノ, ヒドロキシ, またはハロからなる群から選択される七置換基までを含み得る)。また、前記置換基は、共に連結して、12 原子までの環を形成できる、
L1 および L3 は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2 は、10 炭素原子までの鎖からなる群から選択される連結基であり(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【0042】
更なる好適な態様において、提供される化合物は、次の一般式を有する:
【化1】
【0043】
式中:
R1 - R10は、独立に次のものからなる群から選択される
L1 および L3 は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2 は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【0044】
更なる好適な態様において、提供される化合物は、次のリストの化合物から選択される(図 2)。
【0045】
これらの化合物は、インビトロやインビボでMEF2との結合において広範囲の親和性を示し、上記のとおり筋肉, 免疫および神経系においてMEF2に基づく療法を開発するためのリード化合物として使用できるだろう。実際、前記化合物は、インビトロおよびマウスモデルで制御性T細胞機能の促進に効果を示し、そのため自己免疫疾患や移植拒絶を予防するための可能性のあるリード薬である。
【0046】
また、提供されたMEF2-結合分子を調製するための方法が本発明において提供される。
【0047】
第五の側面において、本発明は、MEF2-依存的な転写のディレギュレーションから生じる疾患を治療するため有用な化合物 および 組成物も提供する。本発明のこの側面と一致する組成物は、一または二以上のMEF2及びそのコファクターの結合をブロックする能力がある化合物を含む。提供される化合物 および 組成物は、転写因子及びそれらのリクルートしたヒストン-修飾酵素の相互作用と関連するエピジェネティック制御の調節が関与する治療用に使用できる。特に, 提供されるMEF2-結合小分子を使用して、MEF2-依存的な転写の調節解除から生じる様々な疾患を治療でき、これにはMEF2機能の不均衡から生じる心肥大, 筋線維タイプのリモデリング, および他の筋肉関連疾患; 過剰な又は非常に僅かなMEF2-依存的な遺伝子発現から生じる自己免疫疾患又は免疫不全, および移植拒絶; 制御からはずれたMEF2機能から生じる様々な神経変性疾患(例えば、アルツハイマー型の疾患およびハンチントン疾患など), 自閉症, 精神医学的な障害, および学習および記憶の障害が含まれるが、これらに限定されない。この本発明の態様の更なる適用は、FOXP3-依存的な転写の調節解除(deregulation)から生じる様々な疾患を治療するため使用できるFOXP3 転写因子の機能を調節する小分子が関与する。これらの疾患には、自己免疫疾患, 移植拒絶 および 癌が含まれるが、これらに限定されない。
【0048】
本発明の第六の態様は、薬:MEF2複合体の調製、また構造決定のための結晶化が関与する。このプロトコールは、MEF2と結合する化合物の高品質の結晶を得るためMEF2Aの特定のタンパク質断片(2-78)および緩衝液条件の範囲を規定する。この複合体の調製および結晶化の方法は、現在および将来のMEF2と結合する小分子の構造上の特徴に、また前記構造を使用してリード化合物の至適化を導くため必須である。
【0049】
本発明の前述の及び他の特性、それらを得て、用いる様式は、以下の記載を参照し、添付される図面を考慮することでより明らかとなり、理解されるだろう。前記図面は本発明の典型的な態様のみを記載しており、従って、その範囲は限定されない。
【0050】
本発明の他の側面および利点は、以下の記載および特許請求の範囲から明らかになるであろう。
【図面の簡単な説明】
【0051】
【図1a】図1はMEF2および同定されたリガンド結合部位の結晶構造の三つの異なる表現(a-c)を示す; (d) 2.4Aの解像度でMEF2A(1-78)と結合したBML-210を示しており、BML-210との結合の相互作用に関与するアミノ酸が含まれる。
【図1b】図1はMEF2および同定されたリガンド結合部位の結晶構造の三つの異なる表現(a-c)を示す; (d) 2.4Aの解像度でMEF2A(1-78)と結合したBML-210を示しており、BML-210との結合の相互作用に関与するアミノ酸が含まれる。
【図1c】図1はMEF2および同定されたリガンド結合部位の結晶構造の三つの異なる表現(a-c)を示す; (d) 2.4Aの解像度でMEF2A(1-78)と結合したBML-210を示しており、BML-210との結合の相互作用に関与するアミノ酸が含まれる。
【図1d】図1はMEF2および同定されたリガンド結合部位の結晶構造の三つの異なる表現(a-c)を示す; (d) 2.4Aの解像度でMEF2A(1-78)と結合したBML-210を示しており、BML-210との結合の相互作用に関与するアミノ酸が含まれる。
【図2】図2は、BML-210, PAOA および関連する例示のMEF2-結合分子の構造を示す。
【図3a】図3は、HDACiに関するMEF2-HDAC ルシフェラーゼレポーターアッセイの模式図を示す。
【図3b】図3は、HDACiに関するMEF2-HDAC ルシフェラーゼレポーターアッセイの模式図を示す。
【図4】図4は、BML-210での処理の24 hrs後のMEF2 HDAC ルシフェラーゼ レポーター アッセイにおけるルシフェラーゼ活性に関して標準化した応答を示す。ルシフェラーゼ活性は、用量依存的な様式で減少した。
【図5a】(a) BiacoreアッセイにおいてBML-210とインキュベーション後にHDACと結合するMEF2の競合阻害を示す; (b) BML-210およびHDAC9がMEF2上でオーバーラップする領域に結合することを示している構造分析。
【図5b】(a) BiacoreアッセイにおいてBML-210とインキュベーション後にHDACと結合するMEF2の競合阻害を示す; (b) BML-210およびHDAC9がMEF2上でオーバーラップする領域に結合することを示している構造分析。
【0052】
[詳細な記載]
本発明の様々な側面を要約したので、我々は次に更に本発明を説明するため様々な例示の態様を詳細に記載する。
【0053】
HDACsの特異的な境界のインヒビター及びその使用
上述のとおり、エピジェネティックレギュレーターの研究を難しくしたことは、多くが別個の機能を有する多数の酵素アイソフォームが存在するとの事実である。例えば、四つのHDAC酵素スーパーファミリーのうち、クラス I (HDAC 1, 2, 3, および 8) および クラス II (HDAC 4, 5, 6, 7, 9, および 10)は癌に関与する二つの主要なクラスである。各々の個々のメンバーのユニークな機能は分子生物学アプローチ(例えば、ノックアウトまたはノックダウン)により研究できるにもかかわらず、それらの薬治療における役割は評価することができない。というのも、多くの現在のHDAC インヒビターは、多くのHDACs (クラス I および II)に共通の触媒ドメインを標的とするからである。事実、おそらく異なる HDAC アイソフォームの活性に影響することにより、所与のHDACインヒビターが異なる細胞条件において逆に影響しているようにみえる結果を誘導することがしばしば観察される。以上のように、現行のHDACiがアイソフォーム特異性を欠くことは、特定のHDACsの薬の影響および個々の機能に関する合理的な機構的な研究の主要な障壁を代表する事項である。また、多数のHDACsまたは小数のサブセット又はそれどころか特定のメンバーを阻害することが薬理学的に有益であるかどうかは明らかではない。特定の臨床適用(例えば、癌治療)は前者を必要とし、他方で他のもの(例えば、神経変性および炎症)は後者から利益をえるだろう。
【0054】
現在利用可能なクラス I および クラス II HDACsのインヒビターは、彼等の化学的な構造の重要な特性に基づいて六つのカテゴリーにグループ化できる:
(1) ヒドロキサム酸;
(2) チオール含有分子;
(3) 求電子性のケトン(electrophilic ketones);
(4) 短鎖脂肪酸(short-chain fatty acids);
(5) ベンズアミド(オルトアミノアニリド);および
(6) 環状のデプシペプチド。
【0055】
これらの分子(ヒドロキサム酸, チオール, および求電子性ケトンを含む)の幾つかがHDAC 活性部位の亜鉛イオンに結合することで働くことは確認されているが、他のカテゴリーの阻害機構ははっきりしていない。異なる HDAC インヒビターは、異なるクラスのHDACsにおいて異なる活性を示した。例えば、ヒドロキサム酸のカテゴリーのメンバー(例えば、TSA および SAHA)は、全クラス I および クラス IIのHDACsにわたる強力なインヒビターであるが、幾つかのメンバーのベンズアミドファミリー〔例えば、ピメロイルアニリド オルト アミノアニリド (PAOA), MS-275, CI-994, MGCD-0103〕, および環状ペプチドのファミリー(例えば、FK228)は特定のHDACメンバー又はサブクラスに対し適度(modest)の選択性を示した。しかしながら、一般的にHDACインヒビターの分子機構の理解を欠いていることが理由で、この選択性が如何に達成されるかは明らかではない。結果的に、幾つかのHDACインヒビターにおける選択性の経験的な観察によって、サブタイプ特異的なHDACインヒビターの合理的なデザインを導くことはできなかった。幾つかのクラスI (HDAC8)およびクラスII (HDAC4およびHDAC7)メンバーの触媒ドメイン及びそれらの異なるインヒビターとの複合体の結晶構造が解析され、これらの構造およびクラスIおよびクラスII HDACsの構造に基づく配列アラインメントによって、活性部位が高度に保存され、周囲の領域に軽微な差のみが示されることが示唆される。従って、彼らも選択性の潜在的な機構についての見識を提供することに失敗している。この背景から、高度に保存された触媒ドメインに対するサブタイプ特異的なインヒビターの合理的なデザインは、非常に挑戦的であると広く考えられた。
【0056】
HDACsがしばしば特異的なコレギュレータおよび他のクロマチン修飾酵素との大きなマルチ-タンパク質複合体に存在するとの観察に基づいて、本発明の発明者は機構に基づくアプローチにより新規のHDACに基づく小分子エピジェネティックレギュレーターを同定し、最適化するためのよい道筋が提供されるだろうと考えた。つまり、ドラッグデザインの従来のアプローチに支配的な酵素の活性部位を標的とするのではなく、その代わりに、HDACs及びそれらの関連性のある機能的なパートナーの間のタンパク質間相互作用を標的とする小分子が開発されるだろう。
【0057】
従って、一側面において、本発明は、HDACが関連する転写因子と結合することをブロックすることによりHDAC 機能を調節する方法を提供する。一般的に、本発明のこの側面による方法は、前記転写因子を前記転写因子 および HDACの間の境界の表面内の部位に選択的に結合する能力のある境界のインヒビターと接触させて、このHDACの触媒活性を前記転写因子のDNA-結合部位の近傍に局在するタンパク質部位で機能することを妨げる工程を有する。
【0058】
前記HDACは、古典的なHDAC又はそのHDACのサブセットの任意のHDACアイソフォームであってもよい。例示のHDACには、HDAC 4, 5, 6, 7, 9 および 10,又はその任意のサブセットが含まれえる。好適な態様において、HDACは、クラス IIa HDACsである。また、更なる好適な態様において、HDACは、HDAC4またはHDAC9の何れである。
【0059】
転写因子は、HDACと結合することが知られる任意の転写因子であってもよい。例示の転写因子には、MEF2, FOXP3, GATA3, および Cabinsが含まれえるが、それらに限定されない。好適な態様において、転写因子はMEF2である。
【0060】
境界のインヒビター(interfacial inhibitor)は、小分子であってもよい。
【0061】
上記に記載される方法は、インビボまたはインビトロで、HDACs および 対応する転写因子の機能および機構に関する情報を得るための研究設定に使用しえる。また、これらを疾患を治療するための臨床設定に使用しえる。
【0062】
本発明の方法で治療しえる疾患は、一般にMEF2機能の不均衡から生じる心肥大, 筋線維タイプのリモデリング,および他の筋肉関連疾患; 過剰な又は非常に僅かなMEF2-依存性の遺伝子発現から生じる自己免疫疾患又は免疫不全; 制御からはずれたMEF2機能から生じる様々な神経変性疾患(例えば、Friedrich's 運動失調, アルツハイマー病およびハンチントン病など), 自閉症, 精神医学的な障害, および学習および記憶の障害が関与する疾患である。例示の疾患には、神経変性疾患, 心臓疾患, 自己免疫疾患, 炎症, および 癌が含まれえるが、これらに限定されない。
【0063】
臨床に使用する場合、本発明のこの側面に基づく方法は、一般的にHDACの転写因子への結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。
【0064】
異なる臨床に使用する場合、本発明のこの側面に基づく方法は、一般的にCabin1のMEF2への結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。Cabin1は、カルシニュリン依存的な転写プログラムの転写コリプレッサーであり、T細胞およびニューロン細胞で高度に発現される。
【0065】
さらに異なる臨床設定に使用する場合、本発明のこの側面に基づく方法は、一般的にp300のMEF2への結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。
【0066】
さらに異なる臨床設定に使用する場合、本発明のこの側面に基づく方法は、一般的にCBPのMEF2への結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。
【0067】
一般的な設定において、本発明のこの側面に基づく方法は、一般的にMEF2と結合する任意の転写コレギュレータの結合をブロックする能力のある遮断薬の薬学的な有効量を患者に投与する工程を含む。
【0068】
上記で表されるように、HDACは、任意の古典的なHDAC又はそのHDACのサブセットであってもよい。遮断薬(blocking agent)は、前記遮断薬がHDAC および 転写因子の間の境界の表面に局在する部位に選択的に結合する能力がある限り、小分子, ラセン状ペプチドミメティックス,又はその組み合わせであってもよい。好適な態様において、転写因子はMEF2である。
【0069】
境界のインヒビターを開発するための方法およびツール
本発明の別の側面において、HDACおよび転写因子の間の境界部位に結合して、その相互作用をブロックする能力のある境界のインヒビターを同定するためのアッセイも提供する。
【0070】
本発明のこの側面におけるアッセイは、一般にMEF2二量体における構造エレメントの第一の群および構造エレメントの第二の群の間の境界により規定されるMEF2 二量体における境界の結合部位を含む評価エレメント(evaluation element)に試験化合物を導入する工程を有する。構造エレメントの第一の群は、各々のMEF2 単量体のベータ鎖S1, S2 およびS3,およびヘリックスH2を含む〔二次的な構造エレメントおよび対応する残基の範囲は、Han等(Han et al,, Nature 2003)に記載されたとおりである〕。第二の群は、Cabin 1, HDAC4, HDAC9, HDAC5, HDAC7, p300 および CBPの短いヘリックスモチーフを含む。また、評価エレメントは、試験化合物との結合または非結合に関する情報を報告するためのレポーターエレメントと作動可能に連結(operatively coupled)されてもよい。
【0071】
好適な態様において、本出願においてベイトと融合させた結合ドメイン, プレイと融合させた活性化ドメイン, およびレポーター遺伝子;並びにレポーターシグナルレベルを決定することを含むtwo-hybridシステムに基づくアッセイが開示される。前記結合ドメインはGAL4 DNA と融合させたMEF2D(GAL4-MEF2)を含む; 前記活性化ドメインはVP-16と融合させたHDAC4のMEF2結合モチーフ(HDAC 4-VP16)を含む。レポーター遺伝子はGAL4-駆動性のレポータープラスミド(GAL4Luc)であり、全てが宿主細胞に保持される。
【0072】
本発明のこの好適な態様は、転写因子(例えば、MEF2)と結合し、その転写コレギュレータへの結合を調節する小分子の迅速でハイスループットなスクリーニングや最適化を許容する細胞に基づくルシフェラーゼアッセイである。前記アッセイは、発明者の研究室によるMEF2 複合体の構造および生化学についての10年以上にわたる研究に基づき開発された。この例示の態様は、最初に結晶研究により同定され、更に本発明の構造に基づく変異の研究により実証されたタンパク質間の境界がMEF2-結合小分子に高度に特異的で敏感なスクリーニングのための分子的な基礎として貢献できるだろうことに基づく。
【0073】
各々のMEF2 単量体のベータ鎖S1, S2 およびS3 およびヘリックスH2およびCabin 1およびHDAC4およびHDAC 9からの短いヘリックスモチーフを含むこのタンパク質間の境界に基づく任意のアッセイは、本発明の範囲内であると考えられる。他の例示的なアッセイの実施態様は、任意の理学的なアッセイ技術を含みえる(例えば、本発明により同定されたタンパク質間の境界に基づく、プルダウン, 共免疫沈降; ルシフェラーゼ レポーター アッセイ,およびトランスジェニック レポーター アッセイを含みえるが、これらに限定されない蛍光に基づく結合アッセイおよび機能的なアッセイを含みえるが、これらに限定されない)。
【0074】
特に、提供される化合物が実際に細胞の内側でクラス IIa HDACの結合を崩壊させることできるかどうかを確認するため、クロマチン免疫沈降(ChIP)アッセイを含む幾つかのタイプのアッセイを使用できる。HDAC4 プラスミド構築物は、一過性にHela細胞にトランスフェクトされる。MEF2媒介性プロモーターにおけるHDAC4の存在(occupation)は、試験される化合物および緩衝剤のコントロールの存在下で、適切な特異的な抗体およびPCRプライマーを用いたChIPにより検出される。ChIP分析の代替的なアプローチとして蛍光イメージングに基づく方法も使用できる。GFP-融合MEF2C および HDAC4をHeLaまたはC2C12 細胞にトランスフェクトして、それらの相互作用が研究された。単独で発現させた場合、GFP-HDAC4は拡散により細胞質に局在し、他方でGFP-MEF2も拡散パターンで核に局在する。共発現させた場合、HDAC4 および MEF2は核の内側に点状ボディー(punctate bodies)を形成する。これらの点状の核ボディー(punctate nuclear bodies)の性質は未知であるが、それらの形成は明らかにMEF2:HDAC4の相互作用に依存的である。というのも、機能的なMEF2-結合モチーフを欠いているHDAC4 変異体はMEF2の核ボディーの標的化に失敗するからである。最終的に、mRNAのプロファイリング(マイクロアレイ)および結合位置(ChIP-on-chip)によるMEF2標的遺伝子のゲノムワイドな分析によって、更に静止または活性化(例えば、カルシウム シグナルの興奮)状態におけるMEF2-依存的な抑制または活性化に大きい反応を示す周知のMEF2標的遺伝子を選択することによりこの方法を促進することができる。この方法を用いて、これらの遺伝子がクラス II HDACsまたは他のMEF2コリプレッサー(例えば、Cabin 1)により潜在的に制御されるかどうかを、ChIPを用いてクラス IIa HDACのプロモーターの存在を検出すること、薬処理に際する又はHDAC 4, 5, 7または9のsiRNA ノックダウン(siRNAによる発現変化が最初に評価される)後の発現の変化をモニターすることにより確認することができる。全体的に、この方法を使用して、様々な濃度の提供される化合物の存在下で、マイクロアレイを用いて遺伝子発現を分析すること、ゲノムワイドでクラス IIa HDACsの結合を検出することにより、化合物をゲノムワイドで評価することができる。
【0075】
アッセイの実施を促進するため、本発明はMEF2-結合小分子を探索するためのハイスループットで高度に感受性で特異的なスクリーニングのプラットホーム(screen platform)も提供する。このプラットホームは、GAL4-駆動性のレポータープラスミド (GAL4Luc), GAL4 DNA-結合ドメインと融合させたMEF2D(GAL4-MEF2), VP-16と融合させたHDAC4のMEF2-結合モチーフ(HDAC4-VP 16), VP-16と融合させたGAL4 DNA-結合ドメイン(ポジティブコントロール)並びにネガティブコントロールおよびポジティブコントロールとして様々な化合物を含んでいる安定的に形質転換された細胞株を含む。上記の安定な細胞株, プラスミド, およびコントロール化合物からなるキットは、新しいMEF2-結合分子を探索する使用者によるスクリーニングのため、また存在するリーディング化合物を最適化するため作出できる。また、MEF2D をMEF2A, MEF2BおよびMEF2Cにスイッチすることにより、この方法を使用してMEF2 ファミリーのアイソフォームに選択的に結合する化合物を探索することができる。係る化合物を使用して疾患における特定のMEF2ファミリーメンバーの機能や関与を研究することができる、またMEF2-関連疾患の診断剤の開発や特異的な治療剤の同定のため使用することができる。
【0076】
別の側面において、本発明は、制御性の及び機能的な複合体のターゲティングによるサブタイプ特異的なHDAC インヒビター/モジュレーターを同定するための方法も提供する。本発明のこの側面に基づく方法は、通常は以下の工程を含む: (1) 機能上重要な境界を含む構造または下部構造(substructure)を解析すること(solving); (2) 現存の又は新しい潜在的なHDACインヒビターから選択される検査分子を計算でドッキングさせることによりドッキング分析を解析した構造に適用すること; (3) 所望のHDAC複合体の間のタンパク質間相互作用を崩壊させることができる化合物をスクリーニングするためのアッセイを開発すること; (4) 工程(3)において同定された化合物の特徴を調べること; (5) 前記化合物を計算で最適化すること;および (6) 最適化された化合物を合成し、前記化合物を工程 (3)のアッセイを用いて確認すること(validating)。
【0077】
工程 1に関して、構造は、探索モデルとして現存の構造を用いる分子置換により解析できる。必要に応じて、実験の段階は、MADまたはMIRで得られてもよい。工程 2に関して、ドッキングは、標準のパッケージ(例えば、AutoDock)を用いて行なうことができる。工程 3は上記記載のとおりであり、大抵は哺乳類のtwo-hybridアッセイに基づく。工程の残りは化合物の性質に依存して当該技術分野において既知の方法を用いてなされる。
【0078】
別の側面において、本発明は、HDAC および 転写因子の間の結合をブロックするための遮断薬として有用な化合物も提供する。HDACは、任意の古典的なHDAC又はそのサブセットであってもよい。転写因子は、HDACと結合することが知られる任意の転写因子であってもよく、MEF2, FOXP3 および GATA3が含まれるが、これらに限定されない。
【0079】
好適な態様において、転写因子はMEF2である。
【0080】
本発明のこの側面による化合物は、小さい有機分子およびラセン状のペプチドミメティックスを含む。
【0081】
MEF2-結合小分子は、開示されたMEF2 結合部位をガイドとして利用することにより本出願において提供される方法を用いて同定できる。提供される小分子は、MEF2と結合するBML-210の結晶構造から明らかとされた同定された結合部位から由来する一般構造式を有する。提供される小分子は、記載された構造上のフォールドと結合するため設計され、潜在的に高い親和性および選択性でMEF2と結合するだろう。このアプローチを用いて強力で選択的な化合物を同定するため、当該技術分野において既知の方法を利用でき、この方法にはリード化合物を同定し、続いてリード化合物の最適化するためのコンピュータを利用した構造に基づくデザインとインシリコスクリーニングとの組み合わせ, コンビナトリアルライブラリデザインとハイスループットスクリーニングとの組み合わせ, およびフラグメントベースのドラッグディスカバリー(fragment-based drug discovery)が含まれるが、これらに限定されない。
【0082】
好適な態様において、本発明のもとで提供される化合物は、MEF2結合部位に結合する化合物を含み、次の一般式を有する:
Ra-L-Rb, 式中で:
Ra は、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(S02)-を含む基から選択されるMEF2結合部位の疎水性領域と結合する認識基であり、式中:
Rc および Rd は、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される。
【0083】
L は、20 炭素原子までの鎖からなるリンカーであり(但し、三炭素原子までが酸素, 窒素 またはイオウ原子で置換でき、更にアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミド, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)、
Rbは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(S02)-を含む基から選択されるMEF2結合部位の親水性領域と結合する認識基であり、式中:
Rc および Rd は、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される。
【0084】
好適な態様において、提供される化合物は、一般構造式Ar1-L1-L2-L3-Ar2を有し、式中:
Ar1 および Ar2 は、独立にベンゼン, ナフタレン, ピリジン, ピリミジン, ピラジン, キノリン, イソキノリン, ピロール, フラン, チオフェン, イミダゾール, ピラゾール, オキサゾール, チアゾール, イソキサゾール, インドール; ベンズイミダゾール, ベンゾチアゾール, ベンゾキサゾールからなる群から選択される芳香環である(但し、前記芳香環は、水素, アルキル, アルケニル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アミノ, アルキルアミノ, ジアルキルアミノ, アリールアミノ, ヘテロアリール アミノ, ヒドロキシ, またはハロからなる群から選択される七置換基までを含み得る)。また、前記置換基は、共に連結して、12 原子までの環を形成できる、
L1 および L3 は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2 は、10 炭素原子までの鎖からなる群から選択される連結基であり(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【0085】
更なる好適な態様において、提供される化合物は、次の一般式を有する:
【化2】
【0086】
式中で:
R1 - R10は、独立に次のものからなる群から選択される
L1 および L3 は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2 は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【0087】
更なる好適な態様において、提供される化合物は、次のリストの化合物から選択される(図 2)。
【化3】
【0088】
本発明の様々な側面および派生の完全な理解を更に促進するため、次の実例となる例が提供される。
【例】
【0089】
[例1] サブタイプ特異的なHDACインヒビター/モジュレーターを同定するための戦略
本発明は、サブタイプ-特異的なHDACインヒビター/モジュレーターとして作用しえるリード化合物を同定するための新規の戦略を提供する。前記戦略は、幾つかの反復する工程からなる:
工程 1: 所望のHDAC複合体に関して、HDACの関連する調節タンパク質に結合するHDACの構造または下部構造が解析される又はHDACの機能上重要な境界を含む構造または下部構造が得られる。
【0090】
工程 2: 前記構造は、タンパク質間の境界に結合しえる現存のまたは新しい潜在的なHDACインヒビターのドッキング分析のため使用される。仮想的なスクリーニングは、機能的なデータ(例えば、HDACインヒビターが標的のHDAC複合体が関与する細胞プロセス(cellular processes)において効果を示す場合、またHDAC インヒビターが活性部位の阻害以外の機構をとおして作用するように思われる場合)により導かれる。
【0091】
工程 3: また、構造や関連する生化学的な情報を使用して、意図するタンパク質間の境界を破壊することができる化合物をスクリーニングするため使用できるアッセイの開発が導かれる。
【0092】
工程 4: 一度このようなリード薬が見つけられたら、タンパク質標的との複合体は、工程 1において樹立された構造研究システムを用いて特徴が調べられる。
【0093】
工程 5: 構造の情報と組み合わせて関連する化学の技術を使用して、高度な親和性および特異性を有する標的タンパク質と結合できるアナログ(analogs)のデザインが導かれる。使用される方法は、工程 2の方法と類似する。
【0094】
工程 6: 設計されたアナログが合成され、工程 3において樹立されたアッセイにより分析される。最終的に、最適化された化合物は、親化合物の効果を模倣し、高い強度および低い非特異性/副作用を有しえるかどうかを検査するためのインビボ試験のため使用される。
【0095】
上記の機構に基づくアプローチを利用して、発明者は、以前の既知のHDAC インヒビター(PAOA)がMEF2インヒビター(MEF2i)として作用することによりクラス IIa HDACsの機能を特異的に破壊することができるリード化合物として同定されることを実証した。そのうえ、PAOAの代表的な構造上のアナログも、本出願において記載された方法で設計され、合成され、評価された。そして、幾つかの強力なMEF2iが同定され(図 2)、このアプローチの更なる側面が実証された。
【0096】
[例2] 治療に適用されるサブタイプ-特異的なHDACインヒビターの使用
クラス IIaのHDACsは、ニューロンの生存/シナプスの形成, T細胞の選択/活性化, および筋肉のリモデリングに重大な役割を担う。これらの活性の調節不全は、神経変性(neurodegeneration), 炎症および心肥大を含む幾つかの疾患において指摘される。癌治療のため開発された幾つかのHDACiは、これらの障害に対し有益な効果を示した。これらのHDAC インヒビターの非特異的な性質によりこれらの疾患における臨床適用が妨げられる。しかしながら、これらの観察によって、観察された治療効果がクラス IIaのHDACsと関連するかどうか、またクラス IIaのHDAC機能の選択的な破壊がこれらの疾患を治療するための実行可能な戦略であるかどうかの魅力的な質問が提起される。これらの問いに対応するため、クラス IIaのHDACsの機能を特異的に破壊できる小分子が必要とされる。
【0097】
既知のMEF2およびクラス IIaのHDACsの細胞機能や筋肉, 免疫およびニューロン系における幾つかのHDACインヒビターの効果に基づき、我々は、MEF2機能の不均衡から生じる心肥大, 筋線維タイプのリモデリング,および他の筋肉関連疾患; 過剰な又は非常に僅かなMEF2-依存的な遺伝子発現から生じる自己免疫疾患又は免疫不全,および移植拒絶; 制御からはずれたMEF2機能から生じる様々な神経変性疾患(例えば、Friedrich's 運動失調, アルツハイマー病およびハンチントン病など), 自閉症, 精神医学的な障害, および学習および記憶の障害を治療するため使用できるだろう本発明の方法および化合物を提唱する。
【0098】
上記の様々なヒト疾患を治療するため開発されたMEF2-結合分子は、経口的に, 筋肉内に, 腹腔内に, 皮下に, 静脈内に注射で投与できるだろう。他の送達法も可能であり、正確なプロトコールは治療されるコンディションに依存するだろう。用量は特定の臨床適用に応じて変動するであろうが、化合物がインビトロで効果を示す標準的なアッセイはインビトロでは約0.1 - 10 uMであり、動物モデルの研究では1-10 mg/KG体重である。
【0099】
図 2に示される本発明のもとで提供される好適な化合物は、インビトロやインビボでMEF2との結合において広範囲の親和性を示し、上記のとおり筋肉, 免疫および神経系においてMEF2に基づく療法を開発するためのリード化合物として貢献できるだろう。実際、前記化合物は、インビトロおよびマウスモデルで調節性のT細胞機能の促進に効果を示し、そのため自己免疫疾患や移植拒絶を予防するための可能性のあるリード薬である。例えば、MEF2-結合分子NKL30の0.15μMによって、インビボでの活性の抑制を増強することにより調節性のT細胞機能が明らかに非常に増強された。恒常的な増殖アッセイのマウスのモデルにおいて、静脈内注射により投与された 1 mg/KG体重の前記化合物は、インビボでTreg 機能を非常に増強させた。これらのデータは、本発明により提供されたMEF2-結合分子が自己免疫疾患を治療するため、また移植拒絶を予防するため使用できるだろうことを強く示唆する。
【0100】
[例3] 機能的な調節のためのクラス IIaのHDACsのターゲティング
他のHDACsと比較して、クラス IIaファミリーは、機能 および 制御の幾つかの側面においてユニークである。第一に、クラス IIa HDACsは、選択的に筋肉, 脳 および T細胞中で発現し、これらの組織におけるそれらの機能と一致する。第二に、クラス IIaのHDACsの活性は、クラス IIaのHDACsが発現される組織における優勢な二次メッセンジャーであるカルシウム シグナルにより厳格に制御される。第三に、クラス IIのHDACsは、ユニークな特性をこのサブクラスのHDACsに与える触媒ドメインのN端の大きな制御ドメインを含む。N末端の調節領域は、種々のタンパク質と相互作用するドメインおよびモチーフを含み、これらにはクラスII HDACsのカルシウム応答性を制御するもの(例えば、CaM, CaMKおよび14-3-3),およびクラスIIのHDACsを特異的なプロモーターを標的とさせるもの(例えば、MEF2およびBCL-6),および他のエピジェネティックレギュレーターおよびエフェクター(例えば、クラスIのHDACs, CtBPおよびHP-1、これはクラスIIaのHDACsと協調的に機能する)が含まれる。幾つかのこれらの複合体の構造は、特異的な破壊の潜在的な標的として調査された。
【0101】
クラス IIaのHDACsの制御および機能に関与する多くの複合体のなかで、生化学および構造の点で最もよく調べられたものはMEF2 複合体である。MEF2は、クラス IIaのHDACsと同じ発現パターンを有する配列特異的な転写因子(MEF2A-D)のファミリーであり、神経変性, 炎症および心臓疾患にも関与する。MEF2ファミリーの転写因子は高度に保存されたMADS-ボックス/MEF2S ドメインと称されるN末端領域を共有し、これがDNA結合, 二量体化並びに種々の転写因子およびコレギュレータとのタンパク質間相互作用を媒介する。クラス IIaのHDACsは、脱アセチルに関して特異的なクロマチン領域を標的とするためDNAには結合せず、MEF2との相互作用に依存している。この相互作用をブロックすることが可能性がある方式として選択されてクラス IIaのHDACsの機能が破壊される。
【0102】
クラス IIaのHDACsおよびMEF2の間の相互作用が広範な機能的および生化学的な分析の対象とされ、クラス IIaのHDACsにおいて保存された短配列モチーフ(MEF2-結合モチーフ)およびMEF2のMADS-ボックス/MEF2Sドメインが彼らの結合に必要十分であることが明らかとされた。体系的な構造研究や生物物理学的な研究をMEF2 および クラス IIaのHDACsの間の相互作用や類似するMEF2-結合モチーフを含む関連する転写抑制(Cabin1)について行った。MEF2-結合モチーフが短い両親媒性のヘリックス構造に適合し、MEF2のMADS- ボックス/MEF2ドメイン上の疎水性の溝と結合することが結晶構造により明らかとされた。このようなリガンド/レセプター様の結合機構が存在することは、小分子を使用してクラス IIa HDACsのMEF2-特異的なプロモーターへのリクルートをブロックすることが可能であろうことを示唆する(refs Han Nature and 2005)。
【0103】
[例4] サブ-クラス特異的なHDAC阻害アッセイの開発
一連のMEF2-依存的なルシフェラーゼ レポーター アッセイと一過性にトランスフェクトされたMEF2D, HDAC4 および コアクチベータ p300を使用して、最初に様々な化合物をスクリーニングした。しかし、これらのアッセイは、シグナルが弱く、高頻度で偽陽性を呈した(おそらく、MEF2の複合体の転写活性化機構や内因性の因子からの干渉が原因であろう)。これらの観察をとおして、細胞の内側のHDAC4 および MEF2の間の分子の相互作用を繰り返すことができる高度に感受性で特異的なアッセイが必須であることが発見された。この問題を解決するため、発明者は、HDAC4 および MEF2Dの間の相互作用を内因性の因子からの干渉を最小限で検出する能力のある哺乳類のtwo-hybrid系を発明した(図 3)。
【0104】
このアッセイ系において、MEF2DはGAL4 DNA結合ドメインと融合され(GAL4-MEF2D)、HDAC4のMEF2-結合モチーフ(aa 155-220)がVP-16と融合された(HDAC4-VP 16)。双方の構築物 および GAL4-駆動性のレポータープラスチド(GAL4Luc)で一過性にトランスフェクトされたHela 細胞がGAL4-VP 16のポジティブコントロールにより産生されるものと匹敵する強いシグナルを生じ、他方でHDAC4との結合に欠陥のあることが前に示されたMEF2D 変異体 Leu67Asp (L67D)がレポーターの活性化に失敗することが予備的な分析により示された(データ示さず)。全てのルシフェラーゼ レポーター アッセイにおけるタンパク質の発現レベルをウエスタンブロットで確認した。
【0105】
上記の構造上の見識やMEF2D L67D 変異体をきっかけに、我々は前にインビトロでHDAC4:MEF2の相互作用を破壊することが示されたHDAC4の変異を導入した。これらの変異も細胞に基づくアッセイにおいてルシフェラーゼ シグナルを減少させた(図 3b)。最も興味深い事項は、インビトロで〜60%までMEF2 の結合を弱める(野生型および変異体のHDAC4によるMEF2結合のKdは、それぞれ0.47μM および 0.81μMである)HDCA4のVal180Lysの変異は、細胞に基づくアッセイにおいてルシフェラーゼのシグナルを部分的に減少させた(図 3b)。これらの観察は、哺乳類のtwo-hybridアッセイからのシグナルがHDAC4 および MEF2の間の分子相互作用と非常によく相関することを示す。これらの結果によって、HDAC4:MEF2相互作用の構造モデルに関して更なる支持が提供されるのみならず、細胞の内側のHADC4:MEF2相互作用を検出するための敏感で特異的な方法も確認される。
【0106】
[例5] MEF2/HDAC 相互作用の選択的なインヒビターの同定
スクリーニングの複雑さを減らすため、現存のHDACインヒビターにおける多数の機能的なデータを利用した。これらのインヒビターの多くが触媒ドメインを標的とするが、細胞 に基づくヒストンアセチル化アッセイにより発見された幾つかの化合物はクラス IIaのHDACsのMEF2への結合を含むHDAC機能の他の側面に影響しえる。この事項を踏まえ、我々は、HDAC9:MEF2 複合体の結晶構造から由来するファルマコフォアモデルを用いた小分子データベースに対する仮想的なスクリーニング(3D ドッキング)を行なった。この探索により新規の標的を生じなかったにもかかわらず、MEF2の疎水性ポケットが特定の長さのリンカーにより連結された二つの芳香族環を有する化合物を好むことが示された。この結果は、MEF2二量体がHDAC9のフェニルアラニンを結合できる二対称性関連部位(two symmetry-related sites)を含むことを示している結晶解析と一致する(refs Han et Nature and JMB)。以上のことから、我々は、このような構造上の特性を有する既知のHDACインヒビターを探索し、哺乳類のtwo-hybridアッセイを用いてHDAC4:MEF2 相互作用における彼らの効果を試験した。
【0107】
哺乳類のtwo-hybridアッセイを用いて選択されたHDAC インヒビターのプールのスクリーニングによって、前に研究された化合物PAOA(図 2)が用量依存的な様式でレポーター シグナルを阻害することが明らかとされた(図 4)。PAOAはHDAC4-VP 16の発現に影響しなかったが、GAL4-VP16により駆動されるレポーター シグナルを5.6 倍まで10μMで減少させ(データ示さず)、この化合物の我々の実験条件下でのルシフェラーゼ活性の発現の非特異的な阻害を示している。しかしながら、同じ濃度のPAOAは、GAL4-MEF2DおよびHDAC4- VP16により駆動されるレポーターシグナルを約 26 倍まで減少させ、PAOAが通常の阻害効果をこえてHDAC4:MEF2 相互作用を破壊する特異的な効果を有することを示唆している。対照的に、亜鉛活性部位を標的とする強力なHDACインヒビターのトリコスタチン A (TSA)は、GAL4-VP 16 および GAL4-MEF2D/HDAC4-VP 16により駆動されるレポーター シグナルに類似する阻害効果を示した(データ示さず)。これらの結果は、PAOA(しかし、TSAではない)がHDAC4 および MEF2Dの間の相互作用を破壊できることを示唆する。
【0108】
HDAC4:MEF2 相互作用におけるPAOAのIC50は、哺乳類のtwo-hybridアッセイに基づくと約5μMであり、ヒストンアセチル化阻害アッセイを用いて決定したものと類似する。MEF2と結合するHDAC4に関するKdは、前に0.47μMであると決定された。我々がHDAC4の平衡濃度が我々のアッセイ条件下で〜0.5μMであると仮定する場合、PAOAのMEF2への結合に関するKdは5μMであると見積もられた。しかしながら、フリーのHDAC4濃度が細胞に基づくアッセイにおいて低い場合、見積ったKdはより大きいだろう。
【0109】
PAOAがHDAC4と競合してMEF2 と結合するかどうかについてもインビトロでBiacore T-100での表面プラズモン共鳴 (SPR)を用いて評価した。HDAC4 (aa 155-220)をCM5 センサーチップに固定化し、精製 MEF2A (1-95)を分析物として使用した。様々な濃度でのHDAC4へのMEF2Aの結合によって、一連の明確なセンサーグラムが得られた(データ示さず)。PAOAの濃度を増加させてMEF2Aとインキュベーションしたところ、固定化された HDAC4への結合の用量依存的な減少が示された(図 5)。競合的な結合反応は複雑であり、BML-210のMEF2への直接の結合が機器の検出限界をこえたことがBiacoreのデータの分析から示された。これらの技術的な制限から、定量的な結合定数を得ることは困難である。それにもかかわらず、予備的なデータはBML-210が実際にインビトロでHDAC4と競合的にMEF2と結合することを示唆している。
【0110】
当初PAOAはヒストンのアセチル化を選択的に誘導するがチューブリンでは誘導しない化合物のグループの一部として発見され、このような特徴はおそらくチューブリン特異的なHDACであるHDAC6以外のHDACsの阻害によるものであろう。PAOAは、インビトロでHDAC4と競合的にMEF2と結合する。右の挿入図は、Biacoreによるアッセイを記載する。HDAC4: 赤色のヘリックス; MEF2: 緑色のクロス, デアセチラーゼ。この選択性の分子的な基礎は知られていないが、HDAC6はクラス lIb サブファミリーに属し、クラス IIaに保存されるMEF2-結合モチーフを有していなく、機能するためMEF2を必要とするようにみえることに注目すべきである。最近、PAOA及びその誘導体は、フリートライヒ失調症においてフラタキシンの発現を増強することが示された。この機構にはヒストンアセチル化の誘導が関与するようにみえるが、強力であるが特異性が低いHDACインヒビター(例えば、TSA および SAHA)はフラタキシン発現に影響を示さず、しかし細胞中のトータルのヒストンアセチル化はPAOAよりも高いレベルで誘導できる。これらの観察は、PAOA及びその誘導体がフラタキシンのサイレンシングに関与する特異的なHDACまたはHDAC複合体を阻害するユニークな機能を有することを示唆する。さらにまた、PAOAの作用の分子的な基礎は、更に本発明に開示されたMEF2に結合したBML 210の結晶構造により明らかにできる。本発明において記載された小分子のMEF2-結合部位を規定することに加えて、この構造によりBML-210に、同様に、PAOA及び他のベンズアミド含有HDACインヒビターに存在するオルト-アミノアニリド部分に関する有望な結合部位が初めて示された。特に、この結合部位は、このクラスのHDACインヒビターに関して以前に仮定されたHDAC 酵素の活性部位と異なる。
【0111】
[例5] DNA上のMEF2に結合したBML-210の複合体の調製およびBML-210:MEF2:DNA 複合体の結晶および複合体構造の原子の詳細
MEF2に結合したBML-210の詳細な相互作用を特徴づけるため、またより強力なMEF2-結合分子のデザインを導く構造上の情報を使用するため、我々はDNA上のMEF2に結合したBML-210の結晶構造を決定した。MEF2A 1-78をコードする遺伝子をMEF2AFLからPCR増幅で作出し、pET30b 発現ベクターにクローニングした。タンパク質をE. coli株BL21(DE3)pLysSで25C, 一晩で発現し、連続的なクロマトグラフィー工程をSp-セファロース, およびゲル濾過で4Cで250mM NaC1, 10mM Hepes(pH7.6), 1mM EDTA, 1 mM DTTで精製を行い0.6mg/lの最終収量を得た。オリゴヌクレオチド (Santelli and Richmond, 2000)をIDT DNA technologiesから購入し、MonoQ FPLC カラムを用いて精製し、透析し、凍結乾燥し、サーマルサイクラーを用いてアニーリングした。タンパク質サンプルの1/10容量の10mM BML 210をタンパク質サンプルに0.5mg/mlで添加して凡そ17mg/ml(0.9mM)に濃縮し、DNA二重鎖(DNA duplex)をトレイにセットする前に1:1の比で添加した(10% DMSO終濃度)。プレート様の結晶をハンギングドロップ水蒸気拡散法の条件を18Cで 24% PEG4000, 140mM NaCl, 5mM MgCl2, 10mM CaCl2, 0.004%NaN3, 3.3% グリセロール, 50mM TrisHCl (pH 5.8 - pH8.18)を用いて得た。薬物濃度の結晶をpH 8.18で得た。結晶は、2.4Åの解像度で回折され、空間群 P1 (a=41.567Å, b=61.622Å c=61.478Å α=114.12° β=89.99° γ=89.95°)に属する。構造は、探索モデルとして1TQE.pdbを用いた分子置換により解析された(refs Richmond 2000)。最終的なモデルは、Rfree 26%およびRw 23%を有する。座標は、(11.1_001_nr_nh_bml.pdb)に添付された。
【0112】
[例6] 構造に基づくデザインおよび化学的な方法を用いたリード化合物の最適化
機能的なスクリーニングにより発見された多くのHDACインヒビターは、マイクロモルまたは更にミリモルの範囲のIC50の適度(modest)な強度を有する。リード化合物の最適化は、典型的には化学構造および構造活性相関(SAR)の研究の体系的な修飾により行った。しかしながら、標的および化合物と標的の間の詳細な結合相互作用を知ることがなければ、このような経験的なアプローチはしばしば労働集約的であり、有効性は限定的である。このような状況は一連のアナログがフリートライヒ失調症を治療するため使用しえる強力な化合物を探索するため合成されたPAOAの場合に事実である。幾つかのPAOAのアナログは親化合物よりも高い活性を示したにもかかわらず、効果は非常に控えめ(very modest)であり、また改善の機構は明らかではなかった。
【0113】
特に、これらのPAOA誘導体がフラタキシン発現を活性化する能力は、HDAC阻害活性と相関しない。例えば、幾つかの誘導体は、ヒストン脱アセチル阻害アッセイにおいて非常に弱いが、フラタキシン誘導において非常に活性であった。PAOAがMEF2と結合し、クラス IIa HDACsのリクルートをブロックするとの我々の予備的な所見によって、これらの興味ある結果の潜在的な分子機構が提供される。クラス IIaのHDACは、デアセチラーゼ活性と非依存性に転写を抑制できる。例えば、全C末端触媒ドメインを欠いているHDAC9 の天然のスプライシングバリアント(MITRとしても知られている)は、MEF2-依存的な遺伝子発現の強力な転写リプレッサーであり、この機構はおそらく他のエピジェネティックエフェクター(例えば、HP1 および CtBP)をリクルートすることによるものである。その意味で、クラス IIaのHDACsの触媒ドメインを標的とする小分子インヒビターは、これらのタンパク質を完全にエピジェネティックにサイレンシングする可能性を排除できない。この事項によって、フラタキシン発現の再活性化におけるTSA および SAHAの無効性が説明されえる。他方で、PAOAは、HDAC活性および他の転写リプレッサーのリクルートを共にブロックできる。MEF2 は転写アクチベータ(例えば、CBP/p300)をリクルートすることによる遺伝子活性化に関与するが、PAOAのMEF2依存的な遺伝子発現における主作用はクラスIIaのHDACsや他の転写リプレッサーのサイレンシング効果を緩和することであることを現在のデータは示唆している。
【0114】
上記の分析およびBML 210:MEF2:DNAの開示された構造に依存することにより、またPAOAのMEF2Aへのドッキングを解釈することにより、この薬分子が好ましく実験的に観察された電子密度に適合できることが示され、MEF2結合に関する新しいPAOA様化合物のデザインのためのICM-ドッキング プロトコール (Molsoft L.L.C)が比較的に有効であることを実証している。このドッキング アプローチを使用して、潜在的に大きな親和性および選択性を有する新しいPAOAアナログをデザインし、分析することができる。構造の幾つかの特性を以下に高い親和性でMEF2と結合しえる新しい分子のデザインに使用される原理を説明するため簡単に説明する。
【0115】
BML-210は、拡大したコンホメーションに適合し、MEF2の疎水性ポケットに結合する(図 1)。また、このポケットはクラス IIaのHDACs および Cabin 1において保存されたMEF2-結合モチーフの結合部位である。電子密度の一端が単純な芳香環と類似し、Leu66, Leu67, Thr70, Leu66', Leu67' および Thr70'(プライム記号は他の単量体からの残基を示す)を含む幾つかの疎水性残基によりとりまかれているので、我々はこの密度をフェニル基に割り当てた。また、この端でのカルボニル基は、所定の位置にあり、Thr70'との水素結合相互作用に関与する(図 1)。
【0116】
新しく最適化されたMEF2-結合小分子の構造に基づく設計は、幾つかのMEF2の既知の構造を用いて達成することができる。我々は、今回三つのMEF2複合体の結晶構造を解析した。その二つは、それぞれCabin1 および HDAC9のMEF2-結合モチーフから由来するペプチドを含み(Guo et al., 2007; Han et al., 2005; Han et al., 2003)、他方で三つめは本出願中で記載されるBML-210複合体であり、如何に小分子がMEF2に結合できるかを示している最初の構造である。全ての三つの複合体において、天然のペプチドであろうと合成の分子であろうと小分子リガンドは、MEF2 二量体の表面の深い溝に結合する(図 1)。Cabin l および HDAC9が、類似する別個のタンパク質間相互作用によりMEF2に結合することを我々の前の研究は示す(図 1)。興味深いことに、BML-210は、MEF2への結合において天然リガンドの幾つかの点を模倣するようにみえる。例えば、PAOAのフェニル環のLeu66, Leu67, Thr70, Leu66', Leu67' および Thr70'により形成された中心の疎水性ポケットへの結合は、HDAC9におけるLeu147の結合を暗示(reminiscent)する(図 1)。三つの構造の詳細な分析により、我々は幾つかの別々の疎水性ポケット, 水素結合のドナーおよびアクセプター, および幾つかの荷電した残基を含む小分子結合を探索しえるMEF2の溝の様々な構造上の特性を同定した。我々は、この構造上の情報の利用を計画して、選択的にMEF2と結合できる新しい系列の小分子をデザインし、至適化する。
【0117】
[例7] MEF2/HDAC相互作用の小分子インヒビターの構造に基づくデザイン
結晶構造をガイドとして用いて、新しいMEF2-活性小分子を同定するための基礎を提供できる多数のPAOAアナログを設計した。一群のアナログを設計して、PAOA構造の二つの一般的なエレメントを探索した。一つ目は、リンカーの長さと強剛性(rigidity)である。二つ目は、官能基及びその二つの芳香族の結合単位におけるポジションである。リンカーの電子密度は複数のコンホメーションに適合することを示し、この領域中でのPAOA および MEF2の間の非至適な結合を示唆している。他方HDAC9において、Lys144 および Val143の脂肪族側鎖は、MEF2の溝を充填して広範なファンデルワールスの接触および水素結合を適切に確立する。これらの天然の相互作用を模倣するため/改善するためPAOAリンカーに導入した官能基の結合親和性を増強しえる。設計した化合物を上記のドッキング分析に供して、精力的に好ましくないものが除去される。残っている分子を標準技術を用いて合成し、前に記載されたインビトロおよびインビボ分析に供した。
【0118】
最初の系列の潜在的なインヒビター(図 2)を合成し、異なる化合物が哺乳類のtwo-hybrid アッセイのレポーターシグナルを阻害する能力において有意に異なる活性を示すことが既に認められた。最も興味深いことに、これらの一つの化合物 4がPAOAと類似する活性を示すが、GAL-VP 16により駆動されるコントロールシグナルに影響せず、この新しい誘導体がPAOAよりも特異的であることを示唆している。PAOAにおけるオルト-アミノアニリド部分は、前にクラス I および クラス II HDAGsの活性部位に結合できる亜鉛キレート基であることが想定されたが、この作用様式の直接の証拠はまだ得られていない。化合物 4において、この亜鉛キレート基は、アミノ基をメタ位に移動することにより除去された。さらに、この誘導体は、PAOAと同等に活性であり、非特異的な影響が低く、我々のアッセイ条件下で、細胞の内側で観察されたPAOAの効果は触媒活性を阻害するよりむしろ主にMEF2:HDAC4の相互作用を破壊する能力によりことが示唆される。
【0119】
[例8] 化合物 10の合成
提供される化合物は、当該技術分野において既知の方法を適合させることにより調製できる。例えば、化合物 10の合成は:
【化4】
【0120】
次の工程により調製された:
工程 1: ピメリン酸(1 equiv) および 3-ブロモアニリン(1 equiv)をフラスコに添加し、130 ℃で一晩撹拌した。反応混合物をEtOAcに希釈し、10%水酸化カリウムで抽出した。水層をconc.HClでpH〜2に酸性化し、酢酸エチルで抽出した。有機層を減圧下で減少させ、アセトニトリル/水で再結晶化した。
【0121】
工程 2. ジクロロメタン中にフェニレンジアミンを含む溶液に(Boc)2O (1 equiv)をrtで添加し、混合物を一晩撹拌した。反応物を減圧下で濃縮し、酢酸エチルで希釈し、水および塩水で三回抽出した。有機層を減圧下で減少させ、クロロホルム/ヘキサンで再結晶化した。
【0122】
工程 3. 工程 1の産物(100 mg)をDMSO (3 mL)に溶解し、この溶液にHunigの塩基(1 equiv), HBTU (1 equiv), および工程 2のモノ保護フェニレンジアミン産物を添加した。生じた溶液を一晩rtで撹拌した。溶液を酢酸エチルで希釈し、塩水で三回抽出した。有機層を減圧下で減少させ、カラムクロマトグラフィー (ヘキサン/酢酸エチル勾配)で精製した。単離された産物をジクロロメタンに溶解し、℃に冷却し、トリフルオロ酢酸 (1 mL)で処置した。溶液をrtに温め、一晩撹拌した。反応を重炭酸ナトリウムで中和し、減圧下で濃縮した。生じた固形物を酢酸エチルに溶解し、飽和塩化ナトリウム溶液で抽出した。有機層を硫酸マグネシウムで乾燥し、濾過し、減圧下で濃縮し、カラム クロマトグラフィー (ヘキサン/酢酸エチル および ジクロロメタン/メタノール 勾配)で精製した。減圧下で溶媒を蒸発させ、純粋な産物 10が提供され、構造および純度がNMR分光学で検証された。
【0123】
本発明が特定の例示の態様および例に記載されるが、本出願に開示された態様が説明の目的でのみ記載され、様々な修飾および変更が請求項に記載される本発明の精神および範囲から逸脱することなく、当業者によりなされることが理解されるだろう。
【表1−1】
【表1−2】
【表1−3】
【表1−4】
【表1−5】
【表1−6】
【表1−7】
【表1−8】
【表1−9】
【表1−10】
【表1−11】
【表1−12】
【表1−13】
【表1−14】
【表1−15】
【表1−16】
【表1−17】
【表1−18】
【表1−19】
【表1−20】
【表1−21】
【表1−22】
【表1−23】
【表1−24】
【表1−25】
【表1−26】
【表1−27】
【表1−28】
【表1−29】
【表1−30】
【表1−31】
【表1−32】
【表1−33】
【表1−34】
【表1−35】
【表1−36】
【表1−37】
【表1−38】
【表1−39】
【表1−40】
【表1−41】
【表1−42】
【表1−43】
【表1−44】
【表1−45】
【表1−46】
【表1−47】
【表1−48】
【表1−49】
【表1−50】
【表1−51】
【表1−52】
【表1−53】
【表1−54】
【表1−55】
【表1−56】
【表1−57】
【表1−58】
【表1−59】
【表1−60】
【表1−61】
【表1−62】
【表1−63】
【表1−64】
【0124】
[参照文献]
本出願中で参照される全ての引用は、その全体が参照によって援用される。
【0125】
Bennett, C.L., Christie, J., Ramsdell, F., Brunkow, M.E., Ferguson, P.J., Whitesell, L., Kelly, T.E., Saulsbury, F.T., Chance, P.F., and Ochs, H.D. (2001). The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nature genetics 2 7, 20-21.
Chan, J.K., Sun, L., Yang, X.J., Zhu, G., and Wu, Z. (2003). Functional characterization of an amino-terminal region of HDAC4 that possesses MEF2 binding and transcriptional repressive activity. The Journal of biological chemistry 278, 23515-23521.
Chen, B., and Cepko, C.L. (2009). HDAC4 regulates neuronal survival in normal and diseased retinas. Science (New York, NY 323, 256-259.
Chou, C.J., Herman, D., and Gottesfeld, J.M. (2008), Pimelic diphenylamide 106 is a slow, tight-binding inhibitor of class l histone deacetylases. The Journal of biological chemistry 283, 35402-35409.
de Ruijter, A.J., van Gennip, A.H., Caron, H.N., Kemp, S., and van Kuilenburg, A.B. (2003). Histone deacetylases (HDACs): characterization of the classical HDAC family. The Biochemical journal 370, 737-749.
Fischer, A., Sananbenesi, F., Wang, X., Dobbin, M., and Tsai, L.H. (2007). Recovery of learning and memory is associated with chromatin remodelling. Nature 447, 178-182.
Flavell, S.W., Cowan, C.W., Kim, T.K., Greer, P.L., Lin, Y., Paradis, S., Griffith, E.C., Hu, L.S., Chen, C., and Greenberg, M.E. (2006). Activity-dependent regulation of MEF2 transcription factors suppresses excitatory synapse number. Science (New York, NY 311, 1008-1012.
Flavell, S.W., and Greenberg, M.E. (2008). Signaling mechanisms linking neuronal activity to gene expression and plasticity of the nervous system. Annual review of neuroscience 31, 563-590,
Flavell, S.W., Kim, T.K,, Gray, J.M., Harmin, D.A,, Hemberg, M., Hong, E.J., Markenscoff Papadimitriou, E., Bear, D.M., and Greenberg, M.E. (2008). Genome-wide analysis of MEF22 transcriptional program reveals synaptic target genes and neuronal activity-dependent polyadenylation site selection. Neuron 60, 1022-1038.
Fontenot, J.D., Gavin, M.A., and Rudensky, A.Y. (2003). Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nature immunology 4, 3 3 0-3 3 6.
Gregoire, S., Tremblay, A.M., Xiao, L., Yang, Q., Ma, K., Nie, J., Mao, Z., Wu, Z., Giguere, V., and Yang, X.J. (2006). Control of MEF2 transcriptional activity by coordinated phosphorylation and sumoylation. The Journal of biological chemistry 281, 4423-4433.
Gregoire, S., and Yang, X.J. (2005). Association with class IIa histone deacetylases upregulates the sumoylation of MEF2 transcription factors. Molecular and cellular biology 25, 2273-2287.
Guo, L., Han, A., Bates, D.L., Cao, J., and Chen, L. (2007). Crystal structure of a conserved N-terminal domain of histone deacetylase 4 reveals functional insights into glutamine-rich domains. Proceedings of the National Academy of Sciences of the United States of America 104, 4297-4302.
Han, A., He, J., Wu, Y., Liu, J.O., and Chen, L. (2005). Mechanism of recruitment of class TI histone deacetylases by myocyte enhancer factor-2. Journal of molecular biology 345, 91-102.
Han, A., Pan, F., Stroud, J.C., Youn, H.D., Liu, J.O., and Chen, L. (2003). Sequence specific recruitment of transcriptional co-repressor Cabin l by myocyte enhancer factor-2. Nature 422, 730-734.
Herman, D., Jenssen, K., Burnett, R., Soragni, E., Perlman, S.L., and Gottesfeld, J.M. (2006). Histone deacetylase inhibitors reverse gene silencing in Friedreich's ataxia. Nature chemical biology 2, 551-558.
Hori, S., Nomura, T., and Sakaguchi, S. (2003). Control of regulatory T cell development by the transcription factor Foxp3. Science (New York, NY 299, 1057-1061.
Kim, Y., Phan, D., van Rooij, E., Wang, D.Z., MoAnally, J., Qi, X., Richardson, J.A., Hill, J.A., Bassel-Duby, R., and Olson, EX (2008). The MEF2D transcription factor mediates stress-dependent cardiac remodeling in mice. J Clin Invest 118, 124-132.
Li, B., Samanta, A., Song, X., Iacono, K.T., Brennan, P., Chatila, T.A., Roncador, G., Banham, A.H., Riley, J.L., Wang, Q., et al. (2007). FOXP3 is a homo oligomer and a component of a supramolecular regulatory complex disabled in the human XLAAD/IPEX autoimmune disease. International immunology 19, 825-835.
Mao, Z., Bonni, A., Xia, F,, Nadal-Vicens, M., and Greenberg, M.E. (1999). Neuronal activity-dependent cell survival mediated by transcription factor MEF2. Science (New York, NY 286,785-790.
McKinsey, T.A., Zhang, C.L., and Olson, E.N. (2001). Control of muscle development by dueling HATs and HDACs. Current opinion in genetics & development 11, 497-504.
McKinsey, T.A., Zhang, C.L., and Olson, E.N. (2002), MEF2: a calcium-dependent regulator of cell division, differentiation and death, Trends Biochem Sci 27, 40-47.
Miska, E.A., Karlsson, C., Langley, E., Nielsen, S.J., Pines, J., and Kouzarides, T.(1999). HDAC4 deacetylase associates with and represses the MEF2 transcription factor. The EMBO journal 18, 5099-5107.
Molkentin, J.D., and Olson, E.N. (1996). Combinatorial control of muscle development by basic helix-loop-helix and MARS-box transcription factors. Proceedings of the National Academy of Sciences of the United States of America 93, 9366- 9373.
Morrison, B.E., Majdzadeh, N., and D'Mello, S.R. (2007). Histone deacetylases: focus on the nervous system. Cell Mol Life Sci 64, 2258-2269.
Morrow, E.M., Yoo, S.Y., Flavell, S.W., Kim, T.K., Lin, Y., Hill, R.S., Mukaddes, N.M., Balkhy, S., Gascon, G., Hashmi, A., et al. (2008). Identifying autism loci and genes by tracing recent shared ancestry. Science (New York, NY 321, 218-223.
Pan, F., Ye, Z., Cheng, L., and Liu, J.O. (2004). Myocyte enhancer factor 2 mediates calcium-dependent transcription of the interleukin-2 gene in T lymphocytes: a calcium signaling module that is distinct from but collaborates with the nuclear factor of activated T cells (NFAT). The Journal of biological hemistry 279, 14477-14480.
Paris, M., Porcelloni, M., Binaschi, M., and Fattori, D. (2008). Histone deacetylase inhibitors: from bench to clinic. Journal of medicinal chemistry 51, 1505- 1529.
Potthoff, M.J., and Olson, E.N. (2007). MEF2: a central regulator of diverse developmental programs. Development (Cambridge, England) 134, 4131- 4140.
Rai, M., Soragni, E., Jenssen, K., Burnett, R., Herman, D., Coppola, G., Geschwind, D.H., Gottesfeld, J.M., and Pandolfo, M. (2008). HDAC inhibitors correct frataxin deficiency in a Friedreich ataxia mouse model. PloS one 3, eI958.
Santelli, E., and Richmond, T.J. (2000). Crystal structure of MEF2A core bound to DNA at 1.5 A resolution. Journal of molecular biology 297, 437-449.
Sartorelli, V., Huang, J., Hamamori, Y., and Kedes, L. (1997). Molecular mechanisms of myogenic coactivation by p300: direct interaction with the activation domain of MyoD and with the MADS box of MEF2C. Molecular and cellular biology 17, 1010-1026.
Shalizi, A., Gaudilliere, B., Yuan, Z., Stegmuller, J., Shirogane, T., Ge, Q., Tan, Y., Schulman, B., Harper, J.W., and Bonni, A. (2006). A calcium-regulated MEF2 sumoylation switch controls postsynaptic differentiation. Science (New York, NY 311, 1012-1017.
Shalizi, A.K., and Bonni, A. (2005). Brawn for Brains: The Role of MEF2 Proteins in the Developing Nervous System. Current topics in developmental biology 69, 239-266.
Shore, P., and Sharrocks, A.D. (1995). The MADS-box family of transcription factors. Eur J Biochem 229, 1-13.
Slepak, T.I., Webster, K.A., Zang, J., Prentice, H., O'Dowd, A., Hicks, M.N., and Bishopric, N.H. (2001). Control of cardiac-specific transcription by p300 through myocyte enhancer factor-2D. The Journal of biological chemistry 276, 7575-7585.
Somoza, J.R., Skene, R.J., Katz, B.A., Mol, C., Ho, J.D., Jennings, A.J., Luong, C., Arvai, A., Buggy, J.J., Chi, E., et al. (2004). Structural snapshots of human HDAC8 provide insights into the class I histone deacetylases. Structure 12, 1325-1334.
Sparrow, D.B., Miska, E.A., Langley, E., Reynaud-Deonauth, S., Kotecha, S., Towers, N., Spohr, G., Kouzarides, T., and Mohun, T.J. (1999). MEF-2 function is modified by a novel co-repressor, MITR. The EMBO journal 18, 5085-5098.
Stefanko, D.P., Barrett, R.M., Ly, A.R., Reolon, G.K., and Wood, M.A. (2009). Modulation of long-term memory for object recognition via HDAC inhibition. Proceedings of the National Academy of Sciences of the United States of America 106,9447-9452.
Thomas, E.A., Coppola, G., Desplats, P.A., Tang, B., Soragni, E., Burnett, R., Gao, F., Fitzgerald, K.M., Borok, J.F., Herman, D., et al. (2008). The HDAC inhibitor 4b ameliorates the disease phenotype and transcriptional abnormalities in Huntington's disease transgenic mice. Proceedings of the National Academy of Sciences of the United States of America 105, 15564-15569.
Wang, L., Fan, C., Topol, S.E., Topol, E.J., and Wang, Q. (2003). Mutation of MEF2A in an inherited disorder with features of coronary artery disease. Science (New York, NY 302, 1578-1581.
Wei, J.Q., Shehadeh, L.A., Mitrani, J.M., Pessanha, M., Slepak, TI, Webster, K.A,, and Bishopric, N.H. (2008). Quantitative control of adaptive cardiac hypertrophy by acetyltransferase p300. Circulation 118, 934-946.
Wong, J.C., Hong, R., and Schreiber, S.L. (2003). Structural biasing elements for in cell histone deacetylase paralog selectivity. Journal of the American Chemical Society 125, 5586-5587.
Wu, Y., Borde, M., Heissmeyer, V., Feuerer, M., Lapan, A.D., Stroud, J.C., Bates, D.L., Guo, L., Han, A., Ziegler, S.F., et al. (2006). FOXP3 controls regulatory T cell function through cooperation with NFAT. Cell 126, 375-387.
Yang, Q., She, H., Gearing, M., Colla, B., Lee, M., Shacka, J.J., and Mao, Z. (2009). Regulation of neuronal survival factor MEF2D by chaperone-mediated autophagy. Science (New York, NY 323, 124-127.
Youn, H.D., and Liu, J.O. (2000). Cabin1 represses MEF2-dependent Nur77 expression and T cell apoptosis by controlling association of histone deacetylases and acetylases with MEF2. Immunity 13, 85-94.
Youn, H.D., Sun, L., Prywes, R., and Liu, J.O. (1999). Apoptosis of T cells mediated by Ca2+-induced release of the transcription factor MEF2. Science (New York, NY 286, 790-793.
Zhang, C.L., McKinsey, T.A., Chang, S., Antos, C.L., Hill, J.A., and Olson, E.N. (2002). Class II histone deacetylases act as signal-responsive repressors of cardiac hypertrophy. Cell 110, 479-488.
Zheng, Y., and Rudensky, A.Y. (2007). Foxp3 in control of the regulatory T cell lineage. Nature immunology 8,457-462.
Zuo, T., Liu, R., Zhang, H., Chang, X., Liu, Y., Wang, L., Zheng, P., and Liu, Y.(2007a). FOXP3 is a novel transcriptional repressor for the breast cancer oncogene SKP2. J Clin Invest.
Zuo, T., Wang, L., Morrison, C., Chang, X., Zhang, H., Li, W., Liu, Y., Wang, Y., Liu, X., Chan, M.W., et al. (2007b). FOXP3 is an X-linked breast cancer suppressor gene and an important repressor of the HER-21ErbB2 oncogene. Cell 129,1275-1286.
【特許請求の範囲】
【請求項1】
細胞プロセスを調節する方法であって、調節される細胞プロセスと関係する標的の転写因子またはコファクターを選択することと;前記転写因子またはコファクターを前記転写因子またはコファクターおよび結合パートナーの間の境界に局在する結合部位と選択的に結合する能力のある境界のインヒビターと接触させることと,それにより前記転写因子またはコファクターと前記結合パートナーの間の相互作用を破壊することとを含む方法。
【請求項2】
請求項1に記載の方法であって、前記転写コファクターがHDAC酵素である方法。
【請求項3】
請求項2に記載の方法であって、前記HDAC 酵素がクラス I および クラスIIのHDACsからなる群から選択される方法。
【請求項4】
請求項2に記載の方法であって、前記HDAC 酵素がクラスIIaのHDACsからなる群から選択される方法。
【請求項5】
請求項2に記載の方法であって、前記境界の結合部位が前記HDAC酵素およびMEF2の間の境界により規定される方法。
【請求項6】
請求項2に記載の方法であって、前記境界の結合部位が前記HDAC酵素および前記HDACを標的のクロマチン領域にリクルートするコファクターの間の境界により規定される方法。
【請求項7】
請求項1に記載の方法であって、前記転写因子がMEF2, FOXP3からなる群から選択される方法。
【請求項8】
請求項1に記載の方法であって、前記細胞プロセスが転写の調節不全と関係する方法。
【請求項9】
請求項1に記載の方法であって、前記境界のインヒビターがMEF2の境界の結合部位と結合する能力のある一般式Ra-L-Rbを有している化合物であり、式中で、
Raは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(SO2)-を含む基から選択されるMEF2結合部位の疎水性領域と結合する認識基であり、式中の:
RcおよびRdは、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択され、
L は、20 炭素原子までの鎖からなるリンカーであり(但し、三炭素原子までが酸素, 窒素 またはイオウ原子で置換でき、更にアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミド, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)、
Rbは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(SO2)-を含む基から選択されるMEF2結合部位の親水性領域と結合する認識基であり、式中の:
RcおよびRdは、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される方法。
【請求項10】
請求項1に記載の方法であって、前記境界のインヒビターがMEF2の境界の結合部位と結合する能力のある一般式Ar1-L1-L2-L3-Ar2を有している化合物であり、式中で、
Ar1およびAr2は、独立にベンゼン, ナフタレン, ピリジン, ピリミジン, ピラジン, キノリン, イソキノリン, ピロール, フラン, チオフェン, イミダゾール, ピラゾール, オキサゾール, チアゾール, イソキサゾール, インドール, ベンズイミダゾール, ベンゾチアゾール, ベンゾキサゾールからなる群から選択される芳香環であり(但し、前記芳香環は、水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アミノ, アルキルアミノ, ジアルキルアミノ, アリールアミノ, ヘテロアリール アミノ, ヒドロキシ, またはハロからなる群から選択される七置換基までを含み得る)、 前記置換基は共に連結して12 原子までの環を形成でき、
L1およびL3は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)方法。
【請求項11】
請求項1に記載の方法であって、前記境界のインヒビターが以下の一般式を有している化合物であり:
【化1】
R1 - R10は、独立に次のものからなる群から選択され、
L1およびL3は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)方法。
【請求項12】
請求項1に記載の方法であって、前記境界のインヒビターが以下の化合物からなる群から選択される式を有しているMEF2の境界の結合部位と結合する能力のある化合物である方法。
【化2】
【請求項13】
転写因子依存的な調節不全と関係する疾患を罹患している患者を治療する方法であって、薬理学的な有効量の境界のインヒビターを前記患者に投与することを含み、前記境界のインヒビターは標的の転写因子又はそのコファクターに選択的に結合する能力があるものであり、前記転写因子またはコファクターは前記疾患のプロセスと関係することが知られる方法。
【請求項14】
請求項10に記載の方法であって、前記疾患は、炎症, 自己免疫疾患, 神経変性疾患, 癌, および心血管疾患から選択される方法。
【請求項15】
請求項10に記載の方法であって、前記転写因子がHDACsを標的DNA領域にリクルートする方法。
【請求項16】
請求項12に記載の方法であって、前記転写因子がMEF2, FOXP3又はその組み合わせから選択される方法。
【請求項17】
請求項12に記載の方法であって、前記転写コファクターがHDACsのメンバーである方法。
【請求項18】
請求項12に記載の方法であって、前記転写コファクターがクラス I および クラスIIのHDACsからなる群から選択される方法。
【請求項19】
請求項12に記載の方法であって、前記HDACの調節解除と関係する疾患は、癌, 神経変性疾患, 心疾患(heart disease),又はその組み合わせからなる群から選択される方法。
【請求項20】
選択された転写因子またはコファクターとその結合パートナーとの間の相互作用を調節するための境界のインヒビターを同定し、最適化する方法であって、以下を含む方法:
a. 前記転写因子またはコファクター,又はその機能上重要な境界を含んでいる部分の結晶構造を表している相対座標のセットを得ること(ここで、前記座標は、前記転写因子またはコファクター,又はその機能上重要な境界を含んでいる部分がその結合パートナーまたはリード化合物と複合体の状態にある場合の前記転写因子またはコファクターにより推測されるコンホメーションを表す);および
b. 前記境界を表している該座標の部分に基づきインヒビターを設計すること。
【請求項21】
請求項17に記載の方法であって、前記転写因子がMEF2またはFOXP3である方法。
【請求項22】
請求項17に記載の方法であって、前記転写コファクターがクラス I および クラスIIのHDACからなる群から選択される方法。
【請求項23】
請求項17に記載の方法であって、前記座標は、表 1に記載の各々のMEF2 単量体のベータ鎖S1, S2 およびS3 およびヘリックスH2を含む方法。
【請求項24】
請求項17に記載の方法であって、前記座標は、表 1に記載の座標である方法。
【請求項25】
境界のインヒビターとしての使用に適切なリード化合物をスクリーニングする, 同定する, または最適化するためのアッセイ方法であって、以下を含むアッセイ方法:
転写因子またはコファクターの境界の結合部位を表している構造を有している分子フレームワークを含む評価するエレメントを提供すること;
試験化合物を前記評価するエレメントと接触させること;および
前記試験化合物を評価するエレメントからの既定の応答によりリード化合物として同定すること。
【請求項26】
請求項25に記載のアッセイ方法であって、前記評価するエレメントは、選択された転写因子またはコファクターと融合させた結合ドメイン, 前記転写因子またはコファクターの結合パートナーと融合させた活性化ドメイン, およびレポーター遺伝子を有しているtwo-hybridアッセイシステムであり、前記レポーターのシグナルが既定のレベル未満に低下する場合、前記試験化合物はリード化合物として同定されるアッセイ方法。
【請求項27】
請求項26に記載のアッセイ方法であって、前記転写因子はMEF2Dのメンバーであり、前記結合パートナーはHDACsのメンバーであるアッセイ方法。
【請求項28】
請求項26に記載のアッセイ方法であって、前記転写因子はMEF2Dであり、前記結合パートナーはHDAC4又はその相互作用する部分であり、該相互作用する部分はMEF2Dと結合するHDAC4のセグメントを含むアッセイ方法。
【請求項29】
請求項25に記載のアッセイシステムであって、前記転写因子は、MEF2, FOXP3, およびHDACsを標的DNA領域にリクルートする他の類似の転写因子から選択されるアッセイシステム。
【請求項30】
請求項26に記載のアッセイを行なうためのキットであって、前記融合させた結合ドメインおよび作用ドメイン(acting domain)をコードしているベクターを含むキット。
【請求項31】
選択されたHDACおよび転写因子の間の結合を阻害するための有用な化合物であって、前記化合物は、以下からなる群から選択される任意の構造から選択される構造式を有している化合物。
【化3】
【請求項32】
請求項31に記載の化合物であって、前記化合物は、NEF2の境界のインヒビターとして有用である化合物。
【請求項33】
請求項1に記載の方法であって、前記境界のインヒビターがMEF2の境界の結合部位と結合する能力のある一般式Ra-L-Rbを有している化合物であり、式中で、
Raは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(SO2)-を含む基から選択されるMEF2結合部位の疎水性領域と結合する認識基であり、式中の:
RcおよびRdは、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択され、
L は、20 炭素原子までの鎖からなるリンカーであり(但し、三炭素原子までが酸素, 窒素 またはイオウ原子で置換でき、更にアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミド, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)、
Rbは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(SO2)-を含む基から選択されるMEF2結合部位の親水性領域と結合する認識基であり、式中の:
RcおよびRdは、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される方法。
【請求項34】
MEF2の境界の結合部位と結合する能力のある一般式Ar1-L1-L2-L3-Ar2を有しているMEF2の境界のインヒビターとして有用な化合物であって、式中で:
Ar1およびAr2は、独立にベンゼン, ナフタレン, ピリジン, ピリミジン, ピラジン, キノリン, イソキノリン, ピロール, フラン, チオフェン, イミダゾール, ピラゾール, オキサゾール, チアゾール, イソキサゾール, インドール; ベンズイミダゾール, ベンゾチアゾール, ベンゾキサゾールからなる群から選択される芳香環であり(但し、前記芳香環は、水素, アルキル, アルケニル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アミノ, アルキルアミノ, ジアルキルアミノ, アリールアミノ, ヘテロアリール アミノ, ヒドロキシ, またはハロからなる群から選択される七置換基までを含み得る)、 前記置換基は共に連結して12 原子までの環を形成でき、
L1およびL3は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) またはS(=0)2-からなる群から選択される連結基であり、
L2は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)化合物。
【請求項35】
MEF2の境界のインヒビターとして有用な化合物であって、以下の一般式を有している化合物:
【化4】
式中で:
R1 - R10は、独立に次のものからなる群から選択され、
L1およびL3は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【請求項36】
HDACs および NEF2の間の相互作用を調節するため有用な組成物であって、請求項 31, 33, 34, 35に記載の化合物,又はその組み合わせを含む組成物。
【請求項1】
細胞プロセスを調節する方法であって、調節される細胞プロセスと関係する標的の転写因子またはコファクターを選択することと;前記転写因子またはコファクターを前記転写因子またはコファクターおよび結合パートナーの間の境界に局在する結合部位と選択的に結合する能力のある境界のインヒビターと接触させることと,それにより前記転写因子またはコファクターと前記結合パートナーの間の相互作用を破壊することとを含む方法。
【請求項2】
請求項1に記載の方法であって、前記転写コファクターがHDAC酵素である方法。
【請求項3】
請求項2に記載の方法であって、前記HDAC 酵素がクラス I および クラスIIのHDACsからなる群から選択される方法。
【請求項4】
請求項2に記載の方法であって、前記HDAC 酵素がクラスIIaのHDACsからなる群から選択される方法。
【請求項5】
請求項2に記載の方法であって、前記境界の結合部位が前記HDAC酵素およびMEF2の間の境界により規定される方法。
【請求項6】
請求項2に記載の方法であって、前記境界の結合部位が前記HDAC酵素および前記HDACを標的のクロマチン領域にリクルートするコファクターの間の境界により規定される方法。
【請求項7】
請求項1に記載の方法であって、前記転写因子がMEF2, FOXP3からなる群から選択される方法。
【請求項8】
請求項1に記載の方法であって、前記細胞プロセスが転写の調節不全と関係する方法。
【請求項9】
請求項1に記載の方法であって、前記境界のインヒビターがMEF2の境界の結合部位と結合する能力のある一般式Ra-L-Rbを有している化合物であり、式中で、
Raは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(SO2)-を含む基から選択されるMEF2結合部位の疎水性領域と結合する認識基であり、式中の:
RcおよびRdは、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択され、
L は、20 炭素原子までの鎖からなるリンカーであり(但し、三炭素原子までが酸素, 窒素 またはイオウ原子で置換でき、更にアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミド, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)、
Rbは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(SO2)-を含む基から選択されるMEF2結合部位の親水性領域と結合する認識基であり、式中の:
RcおよびRdは、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される方法。
【請求項10】
請求項1に記載の方法であって、前記境界のインヒビターがMEF2の境界の結合部位と結合する能力のある一般式Ar1-L1-L2-L3-Ar2を有している化合物であり、式中で、
Ar1およびAr2は、独立にベンゼン, ナフタレン, ピリジン, ピリミジン, ピラジン, キノリン, イソキノリン, ピロール, フラン, チオフェン, イミダゾール, ピラゾール, オキサゾール, チアゾール, イソキサゾール, インドール, ベンズイミダゾール, ベンゾチアゾール, ベンゾキサゾールからなる群から選択される芳香環であり(但し、前記芳香環は、水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アミノ, アルキルアミノ, ジアルキルアミノ, アリールアミノ, ヘテロアリール アミノ, ヒドロキシ, またはハロからなる群から選択される七置換基までを含み得る)、 前記置換基は共に連結して12 原子までの環を形成でき、
L1およびL3は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)方法。
【請求項11】
請求項1に記載の方法であって、前記境界のインヒビターが以下の一般式を有している化合物であり:
【化1】
R1 - R10は、独立に次のものからなる群から選択され、
L1およびL3は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)方法。
【請求項12】
請求項1に記載の方法であって、前記境界のインヒビターが以下の化合物からなる群から選択される式を有しているMEF2の境界の結合部位と結合する能力のある化合物である方法。
【化2】
【請求項13】
転写因子依存的な調節不全と関係する疾患を罹患している患者を治療する方法であって、薬理学的な有効量の境界のインヒビターを前記患者に投与することを含み、前記境界のインヒビターは標的の転写因子又はそのコファクターに選択的に結合する能力があるものであり、前記転写因子またはコファクターは前記疾患のプロセスと関係することが知られる方法。
【請求項14】
請求項10に記載の方法であって、前記疾患は、炎症, 自己免疫疾患, 神経変性疾患, 癌, および心血管疾患から選択される方法。
【請求項15】
請求項10に記載の方法であって、前記転写因子がHDACsを標的DNA領域にリクルートする方法。
【請求項16】
請求項12に記載の方法であって、前記転写因子がMEF2, FOXP3又はその組み合わせから選択される方法。
【請求項17】
請求項12に記載の方法であって、前記転写コファクターがHDACsのメンバーである方法。
【請求項18】
請求項12に記載の方法であって、前記転写コファクターがクラス I および クラスIIのHDACsからなる群から選択される方法。
【請求項19】
請求項12に記載の方法であって、前記HDACの調節解除と関係する疾患は、癌, 神経変性疾患, 心疾患(heart disease),又はその組み合わせからなる群から選択される方法。
【請求項20】
選択された転写因子またはコファクターとその結合パートナーとの間の相互作用を調節するための境界のインヒビターを同定し、最適化する方法であって、以下を含む方法:
a. 前記転写因子またはコファクター,又はその機能上重要な境界を含んでいる部分の結晶構造を表している相対座標のセットを得ること(ここで、前記座標は、前記転写因子またはコファクター,又はその機能上重要な境界を含んでいる部分がその結合パートナーまたはリード化合物と複合体の状態にある場合の前記転写因子またはコファクターにより推測されるコンホメーションを表す);および
b. 前記境界を表している該座標の部分に基づきインヒビターを設計すること。
【請求項21】
請求項17に記載の方法であって、前記転写因子がMEF2またはFOXP3である方法。
【請求項22】
請求項17に記載の方法であって、前記転写コファクターがクラス I および クラスIIのHDACからなる群から選択される方法。
【請求項23】
請求項17に記載の方法であって、前記座標は、表 1に記載の各々のMEF2 単量体のベータ鎖S1, S2 およびS3 およびヘリックスH2を含む方法。
【請求項24】
請求項17に記載の方法であって、前記座標は、表 1に記載の座標である方法。
【請求項25】
境界のインヒビターとしての使用に適切なリード化合物をスクリーニングする, 同定する, または最適化するためのアッセイ方法であって、以下を含むアッセイ方法:
転写因子またはコファクターの境界の結合部位を表している構造を有している分子フレームワークを含む評価するエレメントを提供すること;
試験化合物を前記評価するエレメントと接触させること;および
前記試験化合物を評価するエレメントからの既定の応答によりリード化合物として同定すること。
【請求項26】
請求項25に記載のアッセイ方法であって、前記評価するエレメントは、選択された転写因子またはコファクターと融合させた結合ドメイン, 前記転写因子またはコファクターの結合パートナーと融合させた活性化ドメイン, およびレポーター遺伝子を有しているtwo-hybridアッセイシステムであり、前記レポーターのシグナルが既定のレベル未満に低下する場合、前記試験化合物はリード化合物として同定されるアッセイ方法。
【請求項27】
請求項26に記載のアッセイ方法であって、前記転写因子はMEF2Dのメンバーであり、前記結合パートナーはHDACsのメンバーであるアッセイ方法。
【請求項28】
請求項26に記載のアッセイ方法であって、前記転写因子はMEF2Dであり、前記結合パートナーはHDAC4又はその相互作用する部分であり、該相互作用する部分はMEF2Dと結合するHDAC4のセグメントを含むアッセイ方法。
【請求項29】
請求項25に記載のアッセイシステムであって、前記転写因子は、MEF2, FOXP3, およびHDACsを標的DNA領域にリクルートする他の類似の転写因子から選択されるアッセイシステム。
【請求項30】
請求項26に記載のアッセイを行なうためのキットであって、前記融合させた結合ドメインおよび作用ドメイン(acting domain)をコードしているベクターを含むキット。
【請求項31】
選択されたHDACおよび転写因子の間の結合を阻害するための有用な化合物であって、前記化合物は、以下からなる群から選択される任意の構造から選択される構造式を有している化合物。
【化3】
【請求項32】
請求項31に記載の化合物であって、前記化合物は、NEF2の境界のインヒビターとして有用である化合物。
【請求項33】
請求項1に記載の方法であって、前記境界のインヒビターがMEF2の境界の結合部位と結合する能力のある一般式Ra-L-Rbを有している化合物であり、式中で、
Raは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(SO2)-を含む基から選択されるMEF2結合部位の疎水性領域と結合する認識基であり、式中の:
RcおよびRdは、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択され、
L は、20 炭素原子までの鎖からなるリンカーであり(但し、三炭素原子までが酸素, 窒素 またはイオウ原子で置換でき、更にアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミド, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)、
Rbは、低級アルキル, アルケニル, アルキニル, アリール, ヘテロアリール, アルコキシ, アリールオキシ, アルキルチオ, アリールチオ, またはRcRdNC(=O)-, RcRdN(SO2)-を含む基から選択されるMEF2結合部位の親水性領域と結合する認識基であり、式中の:
RcおよびRdは、独立に水素, アルキル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アルキルアミノ, ジアルキルアミノ, アリールアミノ またはヘテロアリール アミノからなる群から選択される方法。
【請求項34】
MEF2の境界の結合部位と結合する能力のある一般式Ar1-L1-L2-L3-Ar2を有しているMEF2の境界のインヒビターとして有用な化合物であって、式中で:
Ar1およびAr2は、独立にベンゼン, ナフタレン, ピリジン, ピリミジン, ピラジン, キノリン, イソキノリン, ピロール, フラン, チオフェン, イミダゾール, ピラゾール, オキサゾール, チアゾール, イソキサゾール, インドール; ベンズイミダゾール, ベンゾチアゾール, ベンゾキサゾールからなる群から選択される芳香環であり(但し、前記芳香環は、水素, アルキル, アルケニル, アルケニル, アルキニル, アルコキシ, アリール, ヘテロアリール, アミノ, アルキルアミノ, ジアルキルアミノ, アリールアミノ, ヘテロアリール アミノ, ヒドロキシ, またはハロからなる群から選択される七置換基までを含み得る)、 前記置換基は共に連結して12 原子までの環を形成でき、
L1およびL3は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) またはS(=0)2-からなる群から選択される連結基であり、
L2は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)化合物。
【請求項35】
MEF2の境界のインヒビターとして有用な化合物であって、以下の一般式を有している化合物:
【化4】
式中で:
R1 - R10は、独立に次のものからなる群から選択され、
L1およびL3は、独立にアミノ, アルキルアミノ, アリールアミノ, オキサ, ケト, NHC (=0), NR (C=O), S(=0) または-S(=0)2-からなる群から選択される連結基であり、
L2は、10 炭素原子までの鎖からなる群から選択される連結基である(但し、三原子までが酸素, 窒素 またはイオウ原子で置換でき、更にこれらの原子はアルキル, アルケニル, アルキニル, アリール, ヘテロアリール, ベンゾ, ヒドロキシ, アルコキシ, アリールオキシ, オキサ, ケト, アミノ, スルホンアミド, またはフルオロからなる群から選択される置換基を含むことができる)。
【請求項36】
HDACs および NEF2の間の相互作用を調節するため有用な組成物であって、請求項 31, 33, 34, 35に記載の化合物,又はその組み合わせを含む組成物。
【図1a】

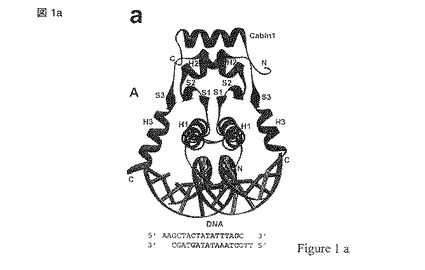
【図1b】
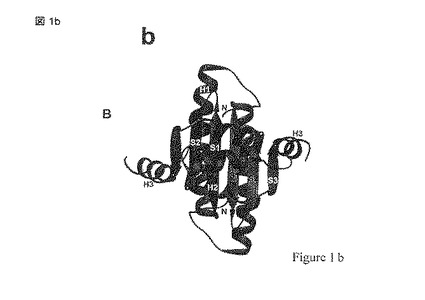
【図1c】
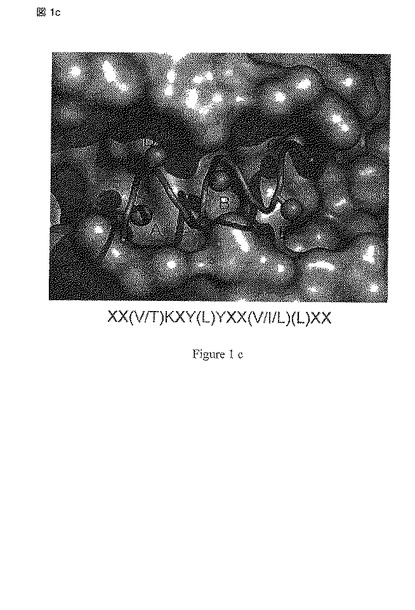
【図1d】

【図2】
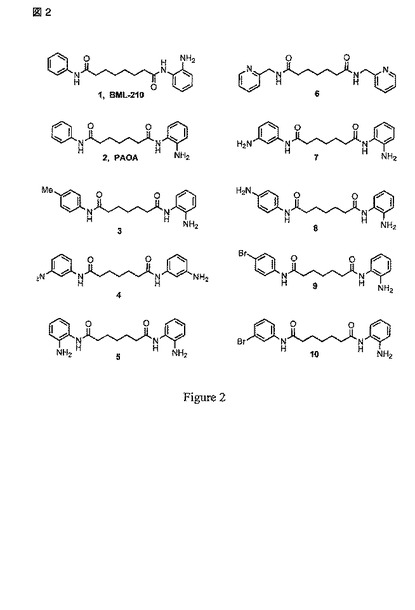
【図3a】
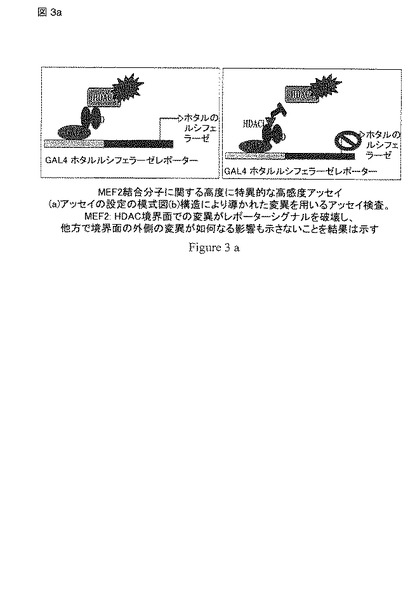
【図3b】
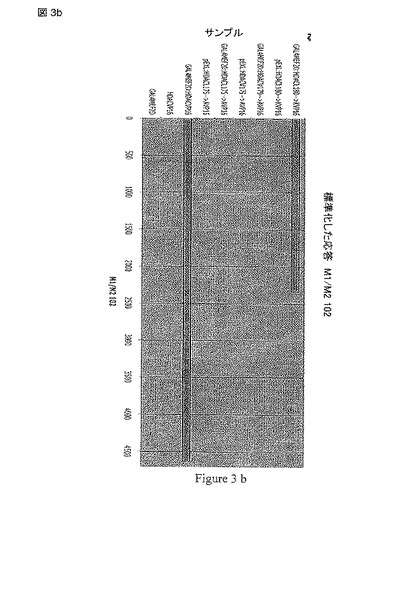
【図4】
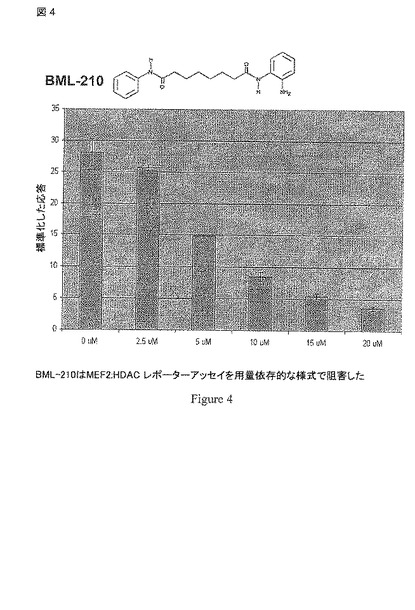
【図5a】
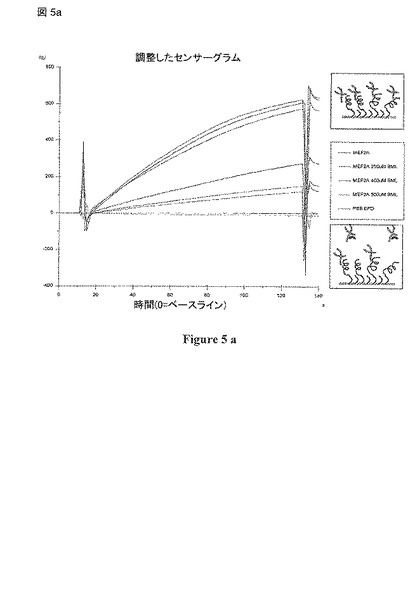
【図5b】

【図1b】
【図1c】
【図1d】
【図2】
【図3a】
【図3b】
【図4】
【図5a】
【図5b】
【公表番号】特表2012−508245(P2012−508245A)
【公表日】平成24年4月5日(2012.4.5)
【国際特許分類】
【出願番号】特願2011−535674(P2011−535674)
【出願日】平成21年11月5日(2009.11.5)
【国際出願番号】PCT/US2009/063456
【国際公開番号】WO2010/054126
【国際公開日】平成22年5月14日(2010.5.14)
【出願人】(301040556)ユニヴァーシティー オブ サザン カリフォルニア (15)
【Fターム(参考)】
【公表日】平成24年4月5日(2012.4.5)
【国際特許分類】
【出願日】平成21年11月5日(2009.11.5)
【国際出願番号】PCT/US2009/063456
【国際公開番号】WO2010/054126
【国際公開日】平成22年5月14日(2010.5.14)
【出願人】(301040556)ユニヴァーシティー オブ サザン カリフォルニア (15)
【Fターム(参考)】
[ Back to top ]