
エルゴチオナーゼ、およびエルゴチオネインの定量方法
【課題】エルゴチオネインの特異的な定量に有用なエルゴチオナーゼ、それをコードする遺伝子、該遺伝子を含む形質転換体、該エルゴチオナーゼの製造方法、該エルゴチオナーゼを利用したエルゴチオネインの定量方法を提供することを課題とする。
【解決手段】エルゴチオナーゼ生産能を有するバークホルデリア・スピーシーズHME13株を見出し、当該株より、エルゴチオナーゼを精製した結果、L-エルゴチオネインに特異的に作用することが明らかになった。また本酵素をコードする遺伝子をクローニングし、大腸菌により高発現させ、本酵素を容易にかつ大量調製することが可能となった。さらに、本酵素を用いて特異的かつ簡便にL-エルゴチオネインを定量可能なことが明らかになった。
【解決手段】エルゴチオナーゼ生産能を有するバークホルデリア・スピーシーズHME13株を見出し、当該株より、エルゴチオナーゼを精製した結果、L-エルゴチオネインに特異的に作用することが明らかになった。また本酵素をコードする遺伝子をクローニングし、大腸菌により高発現させ、本酵素を容易にかつ大量調製することが可能となった。さらに、本酵素を用いて特異的かつ簡便にL-エルゴチオネインを定量可能なことが明らかになった。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、エルゴチオネインの特異的な定量に有用な酵素であるエルゴチオナーゼ、それをコードする遺伝子、該遺伝子を含む形質転換体、該酵素の製造方法、該酵素を利用したエルゴチオネインの定量方法に関する。
【背景技術】
【0002】
エルゴチオネインは、1909年にライ麦角菌(Claviceps purpurea)より単離、構造決定されたアミノ酸の1種であり、その後、幅広い動物、植物に存在することが明らかとなったが、動物、植物などはエルゴチオネイン合成能を有しておらず、一部のカビ、キノコ類のみが生産し、動物、植物は、吸収、食餌によりこれを取り込んでいると考えられている。
【0003】
エルゴチオネインは、光化学傷害に対する保護作用、活性酸素種(Reactive Oxygen Species、ROS)や活性窒素種(Reactive Nitrogen Species, RNS)の消去やこれらによる酸化ストレスの低減など優れた抗酸化活性を有し、一方で、高い水溶性、熱・酸素に対する安定性を有するため、抗酸化物質として、アンチエイジング素材、美肌成分、生体成分、食料品などの酸化防止剤として、その利用が期待されている。
【0004】
また、ヒトにおいて、有機カチオントランスポーター(organic cation transporter)によりエルゴチオネインが細胞内に特異的に取り込まれること、その変異がクローン病のリスクと高い相関があること、リューマチ患者の赤血球にエルゴチオネイン含量が高いことなどから、自己免疫疾患との関係が指摘されている。
【0005】
このような背景より、食品、化粧品、臓器、血液など生体材料中のエルゴチオネインを特異的に、かつ、簡便に測定する方法が求められていたが(特許文献1)、これまでに報告されている比色法、高圧液体クロマトグラフィー(HPLC)を用いた方法では、特異性、簡便性において問題があった。具体的には、2,2’-dithiopyridineを用いた比色法では夾雑するチオール化合物の影響を除くために、サンプルを予めアルカリ条件下、銅触媒により酸化する必要があり、本前処理を行った場合でも非特異的に発色を完全には抑制できない(非特許文献1)。また、HPLCを用いた定量では、本化合物の逆相カラムへの保持が非常に弱いこと、非特異的な吸収波長(254 nm)で検出する必要があることから、Thiopropyl-Sepharoseにより前処理する(非特許文献2)、カラムを2本タンデムに連結する、検出に質量分析器を利用する、など複雑な操作や高価な機械が必要であった(非特許文献3)。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2009-192383
【非特許文献】
【0007】
【非特許文献1】Biochem. J., 139, 221-235 (1974)
【非特許文献2】Chem. Pharm. Bull., 26, 3772-3778 (1978)
【非特許文献3】Int. J. Med. Mushrooms, 8, 215-222 (2006)
【発明の概要】
【発明が解決しようとする課題】
【0008】
上記課題に鑑み、本発明者らは、より簡便かつ特異的にエルゴチオネインを定量するために、酵素を用いた特異的なエルゴチオネイン定量方法の開発を検討した。
【0009】
エルゴチオネインに特異的に作用する酵素として、エルゴチオネインを炭素源、エネルギー源として利用可能な大腸菌W株より、L-エルゴチオネインに作用し、チオウロカン酸(3-(2,3-ジヒドロ-2-チオキソ-1H-イミダゾール-4-イル)プロペン酸)およびトリメチルアミンを生成する(式1)に示すエルゴチオナーゼが知られている(非特許文献4:J. Biol. Chem., 237, 874-881 (1962))。
【0010】
式1
【化1】

【0011】
本発明者らは、エルゴチオナーゼを用いて、エルゴチオネインの簡便かつ特異的な定量が可能か検討するために、以下の参考例1に示すように、この大腸菌W株よりエルゴチオナーゼの調製を試みたが、本菌株にエルゴチオナーゼ活性を検出できなかった。
【0012】
そこで本発明者らは、エルゴチオネインの簡便かつ特異的な定量に利用可能なエルゴチオナーゼを見出すために、エルゴチオネインを窒素源として利用可能な微生物のスクリーニングを土壌(高知県のいち動物公園)および研究室保有の微生物保存株より行った。その結果、土壌よりエルゴチオナーゼ生産能を有する微生物を見出した。最も高い活性を示したバークホルデリア・スピーシーズ(Burkholderia sp.)HME13株より、エルゴチオナーゼを精製し、その酵素科学的性質を解析した結果、本酵素がL-エルゴチオネインに特異的に作用することが明らかになった。さらに、本酵素をコードする遺伝子をクローニングし、大腸菌により高発現させ、本酵素を容易にかつ大量調製することが可能となった。さらに、本酵素を用いてL-エルゴチオネインの定量を検討した結果、特異的かつ簡便にL-エルゴチオネインを定量可能なことが明らかになり、本発明を完成した。
【0013】
なお、非特許文献5(J. Bacteriol., 74, 381-385 (1957))では、研究室保存のバチルス・サチルス(Bacillus subtilis)、バチルス・ポリミキサ(Bacillus polymyxa)、大腸菌(Escherichia coli)、イシェリヒア・フロインディー(Escherichia freundii)、アエロバクター・アエロゲネス(Aerobacter aerogenes)、ストレプトコッカス・フェーカリス(Streptococcus faecalis)、ストレプトコッカス・リキファッシェンス(Streptococcus liquifaciens)、ミクロコッカス・パイロゲネス・バラエティ・アウレウス(Micrococcus pyogenes var. aureus)、セラチア・マルセッセンス(Serratia marcescens)、プロテウス・バルガリス(Proteus vulgaris)、シュードモナス・エルギノーザ(Pseudomonas aeruginosa)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、ナイセリア・カタルラリス(Neisseria catarrhalis)、アルカリゲネス・フェーカリス(Alcaligenes faecalis)は、エルゴチオネインを資化できず、土壌よりエルゴチオネイン資化性を持つ菌として単離したアルカリゲネス・フェーカリス(Alcaligenes faecalis)のみがエルゴチオネインを代謝可能であったことを報告しており、エルゴチオネイン資化性を有する微生物が自然界において希にしか存在しないことを示している。
【0014】
即ち本発明は、エルゴチオネインの特異的な定量に有用なエルゴチオナーゼ、それをコードする遺伝子、該遺伝子を含む形質転換体、該エルゴチオナーゼの製造方法、該エルゴチオナーゼを利用したエルゴチオネインの定量方法に関する。
【0015】
本発明は、より具体的には以下の〔1〕〜〔14〕を提供するものである。
〔1〕 次の(1)〜(2)に示す理化学的性状を有するエルゴチオナーゼ;
(1)作用:L-エルゴチオネインに作用し、チオウロカン酸(3-(2,3-ジヒドロ-2-チオキソ-1H-イミダゾール-4-イル)プロペン酸)およびトリメチルアミンを生成する、および
(2)基質特異性:L-エルゴチオネインに作用し、D-, L-ヒスチジン、D-, L-チロシン、D-, L-フェニルアラニンに作用しない。
〔2〕 さらに付加的に、次の(3)〜(8)に示す理化学的性状を有する〔1〕に記載のエルゴチオナーゼ;
(3)至適pH:7.0〜9.5、
(4)至適温度:50〜65℃、
(5)分子量:ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動による分子量が45,000〜55,000、ゲルろ過による分子量が200,000〜260,000。
(6)熱安定性: 60℃、30分で90%以上の活性を保持、
(7)pH安定性: pH 6.5〜11.0、30分で80%以上の活性を保持、および
(8)阻害剤: p-ヒドロキシ水銀安息香酸(p-hydroxymercuribenzoate)、ヨード酢酸(iodoacetate)、および/またはL-システイン(L-cysteine)存在下において80%以上の活性を保持。
〔3〕 バークホルデリア属、もしくはシュードモナス属に属する微生物に由来する、〔1〕または〔2〕に記載のエルゴチオナーゼ。
〔4〕 前記バークホルデリア属、もしくはシュードモナス属に属する微生物が、下記の群から選択されるいずれかの微生物であることを特徴とする〔3〕に記載のエルゴチオナーゼ;
バークホルデリア・エスピー NITE P-861(Burkholderia sp. NITE P-861)、および
シュードモナス・エスピー (Pseudomonas sp.)。
〔5〕 下記(a)〜(e)のいずれかに記載のエルゴチオナーゼ活性を有するタンパク質;
(a)配列番号:3に記載のアミノ酸配列を含むタンパク質、
(b)配列番号:4に記載された塩基配列を含むポリヌクレオチドによってコードされるタンパク質、
(c)配列番号:3に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含むタンパク質、
(d)配列番号:4に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質、または
(e)配列番号:3に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列を含むタンパク質。
〔6〕 〔5〕に記載のタンパク質をコードするポリヌクレオチド。
〔7〕 〔1〕〜〔4〕のいずれかに記載のエルゴチオナーゼをコードするポリヌクレオチドまたは〔6〕に記載のポリヌクレオチドを保持したベクター。
〔8〕 〔7〕に記載のベクターにより形質転換された形質転換体。
〔9〕 前記宿主が大腸菌である〔8〕に記載の形質転換体。
〔10〕 バークホルデリア属、もしくはシュードモナス属に属し、エルゴチオナーゼ活性を有する微生物、もしくは〔8〕または〔9〕に記載の形質転換体を培養する工程と、培養物よりエルゴチオナーゼを回収する工程を含むエルゴチオナーゼの製造方法。
〔11〕 エルゴチオネインに作用してチオウロカン酸を生成する能力を有する酵素活性物質を、被検対象に接触させる工程を含む、エルゴチオネインの定量方法。
〔12〕 前記酵素活性物質が、〔1〕〜〔4〕のいずれかに記載のエルゴチオナーゼ、〔5〕に記載のエルゴチオナーゼ活性を有するタンパク質、当該エルゴチオナーゼまたはエルゴチオナーゼ活性を有するタンパク質を生産する微生物、およびその処理物からなる群から選択される、〔11〕に記載のエルゴチオネインの定量方法。
〔13〕 エルゴチオナーゼおよび/または酵素活性物質を含むことを特徴とするエルゴチオネイン定量試薬、およびキット。
〔14〕 チオウロカン酸およびトリメチルアミンにエルゴチオナーゼおよび/または酵素活性物質を作用させることを特徴とする、エルゴチオネインの製造方法。
【発明の効果】
【0016】
本発明により、エルゴチオネインに特異的に作用するエルゴチオナーゼにより、特異的かつ簡便にエルゴチオネインを定量する方法が提供された。また、エルゴチオナーゼが提供された。さらに、エルゴチオナーゼをコードするDNAを単離し、当該酵素を高発現する組換え菌が提供された。本発明の方法は、エルゴチオナーゼの高い特異性を利用して、簡便にエルゴチオネインを定量できるため、簡易な方法が望まれる食品検査、臨床検査等において有用である。本発明によって、エルゴチオネインを特異的かつ簡便に定量することが可能となった。
【図面の簡単な説明】
【0017】
【図1】分子系統樹を示す図である。
【図2】エルゴチオナーゼの至適温度を検討した結果を示す図である。
【図3】エルゴチオナーゼの熱安定性について検討した結果を示す図である。
【図4】エルゴチオナーゼの至適pHについて検討した結果を示す図である。
【図5】エルゴチオナーゼの安定性に与えるpHの影響について検討した結果を示す図である。
【図6】エルゴチオナーゼによるエルゴチオネインの定量結果を示す図である。
【発明を実施するための形態】
【0018】
本発明は、次の(1)〜(2)に示す理化学的性状を有するエルゴチオナーゼを提供する;
(1)作用:L-エルゴチオネインに作用し、チオウロカン酸およびトリメチルアミンを生成する、および
(2)基質特異性:L-エルゴチオネインに作用し、D-, L-ヒスチジン、D-, L-チロシン、D-, L-フェニルアラニンに作用しない。
【0019】
本発明者らが見出したエルゴチオナーゼは、好ましくは前記性状(1)〜(2)に加えて、さらに次の(3)〜(8)に示す理化学的性状を有する;
(3)至適pH:7.0〜9.5、
(4)至適温度:50〜65℃、
(5)分子量:ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動(以下、SDS-PAGEと略す)による分子量が45,000〜55,000、ゲルろ過による分子量が200,000〜260,000、
(6)熱安定性: 60℃、30分で90%以上の活性を保持、
(7)pH安定性: pH 6.5〜11.0、30分で80%以上の活性を保持、および
(8)阻害剤: p-ヒドロキシ水銀安息香酸(p-hydroxymercuribenzoate)、ヨード酢酸(iodoacetate、グリセルアルデヒド3リン酸デヒドロゲナーゼ阻害剤)、L-システイン(L-cysteine)存在下において80%以上の活性を保持。
【0020】
なお上記(5)に記載の分子量において、より好ましくは、SDS-PAGEによる分子量が47,000〜51,000、ゲルろ過による分子量が230,000である。さらに好ましくは、SDS-PAGEによる分子量が49,000である。またアミノ酸配列から予測される分子量は、約55,000である。なお本発明者らが見出したエルゴチオナーゼは、活性状態では4量体(テトラマー)であると考えられる。
【0021】
本発明のエルゴチオナーゼは、動物、植物等特に限定されないが、例えば、微生物から得ることができる。本発明における好ましい微生物として、例えば、バークホルデリア属もしくはシュードモナス属に属する微生物を挙げることができる。これらバークホルデリア属もしくはシュードモナス属に属する微生物は、エルゴチオネインに作用し、チオウロカン酸を生成する。
本発明におけるエルゴチオネインは好ましくは、L-エルゴチオネインである。
【0022】
バークホルデリア属に属する微生物として、具体的には次の微生物を示すことができる。この微生物は、本発明において特異的にエルゴチオネインに作用し、チオウロカン酸を生成する好ましい微生物である。
バークホルデリア・エスピー HME 13(Burkholderia sp. HME 13)
【0023】
このバークホルデリア・エスピー HME 13は、以下のとおり寄託されている。以下に寄託を特定する内容を記載する。
(1)寄託機関の名称・あて名:
名称:独立行政法人製品評価技術基盤機構
あて名:日本国千葉県木更津市かずさ鎌足2−5−8(郵便番号292−0818)
(2)受託日(寄託日):2010年1月20日
(3)受託番号:
バークホルデリア・エスピー HME 13 (Burkholderia sp. HME 13) ;受託番号 NITE P-861
【0024】
また本発明において、エルゴチオネインに作用し、チオウロカン酸およびトリメチルアミンを生成する能力を有する微生物は、これらの寄託菌株のみならず、種々の微生物資源(microbiological resource)からも見出すことができる。微生物資源には、自然環境から単離される微生物群、微生物寄託機関に保存された微生物菌株などが含まれる。例えば本発明において、エルゴチオネインに作用し、チオウロカン酸およびトリメチルアミンを生成する能力を有する微生物として、好ましくは次のようなバークホルデリア属やシュードモナス属に属する微生物が挙げられる。
シュードモナス・エスピー(Pseudomonas sp.)
【0025】
即ち本発明は、バークホルデリア属もしくはシュードモナス属に属する微生物が下記の群から選択されるいずれかの微生物であることを特徴とするエルゴチオナーゼに関する;
バークホルデリア・エスピー NITE P-861(Burkholderia sp. NITE P-861)、および
シュードモナス・エスピー (Pseudomonas sp.)。
【0026】
本発明において、エルゴチオナーゼ活性(本明細書では単に「活性」、あるいは「本酵素活性」と記載する場合がある)とは、L-エルゴチオネインを基質として、チオウロカン酸およびトリメチルアミンを生成する酵素活性を言う。
【0027】
本発明では、本酵素活性を有する微生物であれば、任意の微生物を用いることができる。本発明に用いることができる微生物は、エルゴチオネインに作用しチオウロカン酸およびトリメチルアミンを生成する能力や作用を有することを確認することにより得ることができる。
【0028】
目的の能力や作用を有する微生物を選択するため、例えば、予め適当な培地中で被験微生物を培養し、生育した微生物、もしくは菌体処理物をエルゴチオネインと接触、反応させる。その結果、チオウロカン酸またはトリメチルアミンの生成が確認できれば、当該微生物を選択し本発明に用いることができる。
【0029】
このとき、被験微生物の培養時に、培地中にエルゴチオネインを加えておく、もしくはエルゴチオネインを単一窒素源として加えておけば、エルゴチオナーゼの誘導あるいは製造が期待できる。
【0030】
チオウロカン酸が生成されたか否かは、例えば以下の方法により測定することができる。
0.25 mM エルゴチオネイン、10 mM ピロリン酸ナトリウム緩衝液(pH 9.0)およびび酵素活性物質を含む 1 mLの反応液を用いて30℃で反応し、チオウロカン酸生成に由来する311 nmの吸光度の増加を測定する。1 Uは、上記条件下で1分間に1μmolのチオウロカン酸生成を触媒する酵素量とし、チオウロカン酸の分子吸光係数(ε=18,000)より計算する。
【0031】
例えば、微生物寄託機関に保存されたバークホルデリア属やシュードモナス属微生物を対象として、上記選択方法によって、目的の作用を有する微生物を選択することができる。
寄託された微生物は、例えば次のような寄託機関から分譲を受けることができる;
生物遺伝資源センター(NBRC)、
理化学研究所(JCM)、
東京農業大学菌株保存室(IAM)、
American Type Culture Collection(ATCC)、および
Deutsche Sammlung von Mikroorganismen(DSM)。
【0032】
あるいはリボソーマルDNA(rDNA)の塩基配列情報に基づいて、本発明に利用しうる微生物を選択することもできる。一般にrDNAの塩基配列情報は、生物間の遺伝学的な近さを表す指標として有用であることが広く知られている。特に16S rRNAをコードするDNA(以下16S rDNAと記載する)は、微生物の遺伝的な同一性あるいは類似性を決定するためのツールとして活用されている。16S rRNAは、全生物に存在しており、生物間で構造の違いは見られるが、アライメント可能な程度に配列が保存されている。16S rRNAの生物間水平伝播の頻度は低いので、分類に使用する方法が汎用されている。
【0033】
具体的には、小サブユニットリボソームRNA(SSU rRNA)は、タンパク質合成の場であるリボソームを構成する核酸分子の1つである。その塩基配列の長さは、多少の違いはあるものの、細菌からヒトまで起源は同じものとみなされている。したがって、rRNAの塩基配列は進化的保存性が高く、生物の系統解析において最も頻繁に利用されている(Microbiol. Rev., 58, 1-9 (1994))。原核生物におけるSSU rRNA は約1500塩基の16S rRNAである。原核生物の分類と同定に16S rRNA の塩基配列を利用する系統分類が一般に用いられている(ASM News, 65, 752-757 (1999)、生物工学実験書 p113 日本生物工学会編, 培風館)。
【0034】
本発明者らが見出した上記の菌株は、16S rDNA中に配列番号:5に示す塩基配列を含んでいた。塩基配列情報データベースを対象として、これらの塩基配列情報を検索すれば、既に16S rDNAが決定された微生物の中から、その16S rDNA中に同一性の高い塩基配列を含む菌株を見つけ出すことができる。例えばDNA Data Bank of Japan(DDBJ)などの公共データベースサービス機関では、16S rRNA(Prokaryotes)の塩基配列情報を対象とする検索サービスが提供されている。こうして見出された微生物のうち、特にバークホルデリア属やシュードモナス属に属する微生物を選択することによって、本発明に利用しうる微生物とすることができる。
【0035】
これら微生物は、醗酵学の分野で公知の方法に従って培養することができる。培地としては炭素源、窒素源、無機物およびその他の栄養素を適量含有する培地ならば、合成培地または天然培地のいずれでも使用することができる。培地は、液体培地または固体培地を使用することができる。
【0036】
具体的には、炭素源として、次に示すような一般的な炭素源より、使用する微生物の資化性を考慮して、適宜一種または二種以上選択して使用する。
糖類:グルコース、フルクトース、マルトース、スクロース、ラクトースなど
デンプン、
デンプン加水分解物、
廃糖蜜、
アルコール:グリセロール、メタノール、エタノールなど
糖アルコール、
有機酸:酢酸、グルコン酸、ピルビン酸、α-ケトグルタル酸、クエン酸など
脂肪類:パーム油、大豆油、オリーブオイルなど
炭化水素類:ノルマルパラフィンなど
【0037】
窒素源としては、次に示すような一般的な窒素源の中から、使用する微生物の資化性を考慮して、適宜一種または二種以上選択して使用する。
有機窒素化合物:例えば、肉エキス、ペプトン、酵母エキス、麦芽エキス、大豆加水分解物、ミルクカゼイン、カザミノ酸、廃糖蜜、アスパラギン、グルタミン、あるいはグルタミン酸などのアミノ酸類やコーンスティープリカー、NZアミンなど
無機窒素化合物:アンモニア、硝酸アンモニウム、硫酸アンモニウムなど、尿素、硝酸塩など
また、エルゴチオナーゼの誘導を目的として、エルゴチオネインを単一の窒素源として使用することができる。
【0038】
さらに、無機塩として微量のマグネシウム、マンガン、カリウム、カルシウム、ナトリウム、銅、亜鉛などの塩や各種ビタミンなどの微量栄養素を添加することもできる。また、必要に応じて植物油、界面活性剤、シリコンなどの消泡剤を培養液中に添加することもできる。
【0039】
培養は前記培地成分を含有する液体培地中で、通常の培養方法を用いて行うことができる。例えば次のような培養方法を利用することができる。
振とう培養(shaking culture)
通気攪拌培養(aeration-agitation culture)
連続培養(continuous culture)
流加培養(feeding culture, fed batch culture)
【0040】
培養条件は、微生物の種類、培養の種類、培養方法により適宜選択することができる。利用する菌株が増殖し、エルゴチオナーゼを生成しうる条件であれば特に制限はない。一般的な培養条件として次のような条件を示すことができる。
培養開始時のpHを4から10、好ましくは6から8に調節
15から70℃、好ましくは20から40℃の温度条件下で培養
培養時間はエルゴチオナーゼ活性を有する菌体を得ることができれば特に制限されない。通常は1日から14日、好ましくは1日から7日培養する。
【0041】
本発明において、微生物菌体の処理物とは、菌体を目的とする酵素活性が維持される条件で処理したものを意味する。酵素活性の維持とは、生菌体で得られる酵素活性の一般的には20%以上、通常30%以上、好ましくは50%以上、より好ましくは70%以上の活性を維持することを言う。生菌体とその処理物の酵素活性は、生菌体の酵素活性と、同量の生菌体から得られた処理物の酵素活性を比較することによって定量的に比較することができる。本発明における処理物は、生菌体よりも高い酵素活性を有する場合もある。
【0042】
このような処理物を得る方法としては、凍結乾燥、有機溶媒による脱水乾燥、菌体の自己消化、酵素活性画分の抽出、超音波処理などを利用することができる。このような処理方法によって、例えば、凍結乾燥菌体、アセトン乾燥菌体、菌体自己消化物、菌体抽出物、菌体磨砕物、菌体の超音波処理物等を得ることができる。
【0043】
これらの処理は、単独で、あるいは複数の処理を重複して適用することもできる。例えば適当な培養液で微生物を培養後、菌体を破砕して、遠心分離等により無細胞抽出液を調製することができる。菌体は、機械的な破砕、超音波、高圧処理、酵素消化、自己消化などにより破砕することができる。無細胞抽出液は本発明における菌体の処理物に含まれる。
【0044】
さらに本発明においては、当該菌体から当該反応を触媒する酵素を部分精製したものや完全に精製したものなどを得ることができる。
すなわち本発明は、本発明によって提供されたエルゴチオナーゼ活性を有する微生物の菌体から精製することができるエルゴチオナーゼに関する。
【0045】
本発明のエルゴチオナーゼは、上記本発明の微生物、あるいはその無細胞抽出液などから精製することができる。具体的には、次のような精製工程を適宜組み合わせることによって、無細胞抽出液から実質的に純粋なエルゴチオナーゼを得ることができる;
硫安、アセトンなどを用いた溶解度による分割、
イオン交換クロマトグラフィー、
疎水クロマトグラフィー、
2',5'-ADP セファロースやブルーもしくはレッドセファロースなどを用いたアフィニティクロマトグラフィー、あるいは
ゲルろ過。
【0046】
例えば、微生物を集菌後、次のような精製工程を経て、本発明のエルゴチオナーゼを実質的に純粋な酵素として単離することができる;
超音波による菌体の破砕と無細胞抽出液の調製、
硫安分画によるタンパク画分の分離、
DEAE-トヨパールを用いたイオン交換クロマトグラフィー、および
ブチル-トヨパールを用いた疎水クロマトグラフィーによるエルゴチオナーゼ活性分画の単離。
【0047】
本発明において、「実質的に純粋」とは、当該酵素が、当該酵素以外の生物学的な高分子化合物や化学物質を実質的に含まないことを言う。当該酵素以外の生物学的な高分子化合物とは、例えば菌体や培養液に由来するタンパク、糖類、脂質、核酸類が含まれる。本発明における実質的に純粋な酵素は、少なくとも75%以上、通常80%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましくは95%以上、あるいは99%以上の純度を有する。酵素の純度を決定する方法は公知である。例えば、各種のクロマトグラフィーや、ポリアクリルアミドゲル電気泳動などによって蛋白質の純度を決定する方法が周知である。
【0048】
あるいは本発明によって、上記理化学的性状(1)〜(2)を有する単離されたエルゴチオナーゼが提供される。本発明において、単離されたエルゴチオナーゼとは、それが存在する天然の環境から分離されて存在していることを意味する。したがって、菌体から分離されたエルゴチオナーゼは、本発明における単離されたエルゴチオナーゼに含まれる。
【0049】
本発明における実質的に純粋なエルゴチオナーゼ、あるいは単離されたエルゴチオナーゼは、精製、あるいは単離された後に、酵素組成物とすることもできる。例えば、精製、あるいは単離されたエルゴチオナーゼを適当な担体と配合することによって、本発明のエルゴチオナーゼを含む酵素組成物を調製することができる。
【0050】
担体としては、アルブミンなどの不活性蛋白質、ショ糖などの糖類あるいは糖アルコールなどを利用することができる。酵素組成物は、液状であることもできるし、乾燥状態であることもできる。酵素と担体を含む水溶液を凍結乾燥することによって得られる酵素組成物は、本発明における好ましい組成物の一つである。
【0051】
本発明のエルゴチオナーゼは、以下の方法によりその活性を定量することができる。すなわち、次の組成の反応液中(1 mL)で、30℃で反応し、チオウロカン酸生成に由来する311 nm の吸光度の増加を測定する。
0.25 mM エルゴチオネイン
10 mM ピロリン酸ナトリウム緩衝液 (pH 9.0) および
酵素
1 Uは、上記条件下で1分間に1μmol のチオウロカン酸生成を触媒する酵素量とし、チオウロカン酸の分子吸光係数(ε=18,000)より計算する。
【0052】
また本発明は、下記(a)から(e)のいずれかに記載のエルゴチオナーゼ活性を有するタンパク質を提供する;
(a)配列番号:3に記載のアミノ酸配列を含むタンパク質、
(b)配列番号:4に記載された塩基配列を含むポリヌクレオチドによってコードされるタンパク質、
(c)配列番号:3に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含むタンパク質、
(d)配列番号:4に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質、
(e)配列番号:3に記載のアミノ酸配列と80%以上の相同性を有するアミノ酸配列を含むタンパク質。
【0053】
本発明において、ポリヌクレオチドは、DNAやRNA等の天然のポリヌクレオチドに加え、人工的なヌクレオチド誘導体を含む人工的な分子であることもできる。また本発明のポリヌクレオチドは、DNA-RNAのキメラ分子であることもできる。
【0054】
本発明のエルゴチオナーゼ活性を有するタンパク質にはシグナル配列が含まれていない。
本発明のエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドは、例えば配列番号:4に示す塩基配列を含む。配列番号:4に示す塩基配列は、配列番号:3に示すアミノ酸配列を含むタンパク質をコードしており、このアミノ酸配列を含むタンパク質は、本発明によるエルゴチオナーゼの好ましい態様を構成する。
【0055】
本発明のエルゴチオナーゼ活性を有するタンパク質には、本発明のエルゴチオナーゼをコードするポリヌクレオチドまたは本発明のエルゴチオナーゼ活性を有するタンパク質のホモログも含まれる。
【0056】
本発明のエルゴチオナーゼをコードするポリヌクレオチドのホモログとは、配列番号:3に記載のアミノ酸配列に1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列を含み、かつ、配列番号:3に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質を意味する。本発明において、配列番号:3に記載のアミノ酸配列からなるタンパク質と機能的に同等とは、当該タンパク質が前記理化学的性質(1)〜(2)を有することを意味する。改変されるアミノ酸の数は、改変後のタンパク質が、前記理化学的性質を有している限り、特に制限はないが、一般的には、50アミノ酸以内、好ましくは30アミノ酸以内、より好ましくは10アミノ酸以内(例えば、5アミノ酸以内、3アミノ酸以内)である。あるいは、アミノ酸配列全体のうち、例えば20%以下、具体的には10%以下(例えば、10、9、8、7、6、5、4、3、2、または1%以下)のアミノ酸残基の改変は許容される。すなわち、配列番号:3に記載のアミノ酸配列と、好ましくは80%以上、より好ましくは90%以上(例えば、90、91、92、93、94、95、96、97、98、99%以上)の相同性を有するアミノ酸配列を含むタンパク質も、本発明のエルゴチオナーゼ活性を有するタンパク質に含まれる。
【0057】
一般にタンパク質の機能の維持のためには、置換するアミノ酸は、置換前のアミノ酸と類似の性質を有するアミノ酸であることが好ましい。このようなアミノ酸残基の置換は、保存的置換と呼ばれている。例えば、Ala、Val、Leu、Ile、Pro、Met、Phe、Trpは、共に非極性アミノ酸に分類されるため、互いに似た性質を有する。また、非荷電性アミノ酸としては、Gly、Ser、Thr、Cys、Tyr、Asn、Glnが挙げられる。また、酸性アミノ酸としては、AspおよびGluが挙げられる。また、塩基性アミノ酸としては、Lys、Arg、Hisが挙げられる。これらの各グループ内のアミノ酸置換は許容される。
【0058】
そして当業者であれば、配列番号:4に記載の塩基配列を含むポリヌクレオチドに部位特異的変異導入法(Nucleic Acid Res. 10,pp.6487 (1982) , Methods in Enzymol.100,pp.448 (1983), Molecular Cloning 2ndEdt., Cold Spring Harbor Laboratory Press (1989) , PCR A Practical Approach IRL Press pp.200 (1991) などを用いて、適宜置換、欠失、挿入、および/または付加変異を導入することによりポリヌクレオチドのホモログを得ることが可能である。そのホモログをコードするポリヌクレオチドを宿主に導入して発現させることにより、配列番号:3に記載のアミノ酸配列からなるタンパク質のホモログを得ることが可能である。
【0059】
また、本発明におけるポリヌクレオチドのホモログは、配列番号:4に示される塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズできるポリヌクレオチドであって、かつ、前記理化学的性質(1)〜(2)を有するタンパク質をコードするポリヌクレオチドも含む。ストリンジェントな条件でハイブリダイズできるポリヌクレオチドとは、配列番号:4に記載の塩基配列中の任意の少なくとも20個、好ましくは少なくとも30個、例えば40、60または100個の連続した配列を一つまたは複数選択したDNAをプローブDNAとし、例えばECL direct nucleic acid labeling and detection system (GEヘルスケア・ジャパン社製)を用いて、マニュアルに記載の条件(wash:42℃、0.5x SSCを含むprimary wash buffer)において、ハイブリダイズするポリヌクレオチドを指す。
【0060】
より具体的な「ストリンジェントな条件」とは、例えば、通常、42℃、2×SSC、0.1%SDSの条件であり、好ましくは50℃、2×SSC 、0.1%SDSの条件であり、さらに「高度にストリンジェントな条件」とは例えば、65℃、0.1×SSCおよび0.1%SDSの条件であるが、これらの条件に特に制限されない。ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度や塩濃度など複数の要素が考えられ、当業者であればこれら要素を適宜選択することで最適なストリンジェンシーを実現することが可能である。
【0061】
さらに、本発明におけるポリヌクレオチドのホモログは、配列番号:3に示されるアミノ酸配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは90%以上(90、91、92、93、94、95、96、97、98、または99%以上)のホモロジーを有するタンパク質をコードするポリヌクレオチドを含む。タンパク質のホモロジー検索は、例えばSWISS-PROT、PIR、DADなどのタンパク質のアミノ酸配列に関するデータベースやDDBJ、EMBL、あるいはGene-BankなどのDNA配列に関するデータベース、DNA配列を基にした予想アミノ酸配列に関するデータベースなどを対象に、BLAST、FASTAなどのプログラムを利用して、例えば、インターネットを通じて行うことができる。
【0062】
また、本発明におけるタンパク質には、配列番号:3に記載のアミノ酸配列にシグナル配列を含めた配列番号:1に記載のアミノ酸配列を含むタンパク質も含まれる。
また、本発明におけるポリヌクレオチドには、配列番号:4に記載の塩基配列にシグナル配列を含めた配列番号:2に記載の塩基配列を含むポリヌクレオチドも含まれる。
【0063】
また本発明は、本発明のエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドに関する。本発明のエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドは、例えば、以下のような方法によって単離することができる。
【0064】
配列番号:4に記載の塩基配列を基にPCR用のプライマーを設計し、酵素生産株の染色体DNAもしくは、cDNAライブラリーを鋳型としてPCRを行うことにより本発明のポリヌクレオチドを得ることができる。
【0065】
さらに、得られたDNA断片をプローブとして、酵素生産株の染色体DNAの制限酵素消化物をファージ、プラスミドなどに導入し、大腸菌を形質転換して得られたライブラリーやcDNAライブラリーを利用して、コロニーハイブリダイゼーション、プラークハイブリダイゼーションなどにより、本発明のポリヌクレオチドを得ることができる。
【0066】
また、PCRにより得られたDNA断片の塩基配列を解析し、得られた配列から、既知のDNAの外側に伸長させるためのPCRプライマーを設計し、酵素生産株の染色体DNAを適当な制限酵素で消化後、自己環化反応によりDNAを鋳型として逆PCRを行うことにより(Genetics 120, 621-623 (1988))、また、RACE法(Rapid Amplification of cDNA End、「PCR実験マニュアル」p25-33, HBJ出版局)などにより本発明のポリヌクレオチドを得ることも可能である。
なお本発明のポリヌクレオチドには、以上のような方法によってクローニングされたゲノムDNA、あるいはcDNAの他、合成によって得られたDNAが含まれる。
【0067】
また本発明は、上記本発明のポリヌクレオチドが挿入された発現ベクターを提供する。例えば、上述のようにして単離されたエルゴチオナーゼをコードするポリヌクレオチド、またはエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドを公知の発現ベクターに挿入することにより、エルゴチオナーゼ発現ベクターが提供される。
【0068】
また本発明は、本発明の発現ベクターにより形質転換された形質転換体に関する。この発現ベクターで形質転換した形質転換体を培養することにより、本発明のエルゴチオナーゼを形質転換体(組換え体)から得ることができる。
【0069】
本発明においてエルゴチオナーゼを発現させるために、形質転換の対象となる微生物は、エルゴチオナーゼを有するポリペプチドをコードするポリヌクレオチド、またはエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドを含む発現(組換え)ベクターにより形質転換され、エルゴチオナーゼ活性を発現することができる生物であれば特に制限はない。利用可能な微生物としては、例えば以下のような微生物を示すことができる。
エシェリヒア(Escherichia)属
バチルス(Bacillus)属
シュードモナス(Pseudomonas)属
セラチア(Serratia)属
ブレビバクテリウム(Brevibacterium)属
コリネバクテリイウム(Corynebacterium)属
ストレプトコッカス(Streptococcus)属
ラクトバチルス(Lactobacillus)属など宿主ベクター系の開発されている細菌
ロドコッカス(Rhodococcus)属
ストレプトマイセス(Streptomyces)属など宿主ベクター系の開発されている放線菌
サッカロマイセス(Saccharomyces)属
クライベロマイセス(Kluyveromyces)属
シゾサッカロマイセス(Schizosaccharomyces)属
チゴサッカロマイセス(Zygosaccharomyces)属
ヤロウイア(Yarrowia)属
トリコスポロン(Trichosporon)属
ロドスポリジウム(Rhodosporidium)属
ピキア(Pichia)属
キャンディダ(Candida)属などの宿主ベクター系の開発されている酵母
ノイロスポラ(Neurospora)属
アスペルギルス(Aspergillus)属
セファロスポリウム(Cephalosporium)属
トリコデルマ(Trichoderma)属などの宿主ベクター系の開発されているカビ
【0070】
形質転換体の作製のための手順および宿主に適合した組換えベクターの構築は、分子生物学、生物工学、遺伝子工学の分野において慣用されている技術に準じて行うことができる(例えば、Sambrookら、モレキュラー・クローニング、Cold Spring Harbor Laboratories)。微生物中などにおいて、本発明のエルゴチオナーゼ遺伝子を発現させるためには、まず微生物中において安定に存在するプラスミドベクターやファージベクター中にこのDNAを導入し、その遺伝情報を転写・翻訳させる必要がある。
【0071】
そのためには、転写・翻訳を制御するユニットにあたるプロモーターを本発明のDNA鎖の5'-側上流に、より好ましくはターミネーターを3'-側下流に、それぞれ組み込めばよい。このプロモーター、ターミネーターとしては、宿主として利用する微生物中において機能することが知られているプロモーター、ターミネーターを用いる必要がある。これら各種微生物において利用可能なベクター、プロモーター、ターミネータ−などに関して「微生物学基礎講座8遺伝子工学・共立出版」、特に酵母に関しては、Adv. Biochem. Eng. 43, 75-102 (1990)、Yeast 8, 423-488 (1992)、などに詳細に記述されている。
【0072】
例えばエシェリヒア属、特に大腸菌エシェリヒア・コリ(Escherichia coli)においては、プラスミドベクターとして、pBR、pUC系プラスミドを利用でき、lac(β−ガラクトシダーゼ)、trp(トリプトファンオペロン)、tac、 trc (lac、trpの融合)、λファージ PL、PRなどに由来するプロモーターなどが利用できる。また、ターミネーターとしては、trpA由来、ファージ由来、rrnBリボソーマルRNA由来のターミネーターなどを用いることができる。これらの中で、市販のpSE420(Invitrogen製)のマルチクローニングサイトを一部改変したベクターpSE420D(特開2000-189170に記載)が好適に利用できる。
【0073】
バチルス属においては、ベクターとしてpUB110系プラスミド、pC194系プラスミドなどが利用可能であり、染色体にインテグレートすることもできる。また、プロモーター、ターミネーターとしてapr(アルカリプロテアーゼ)、npr(中性プロテアーゼ)、amy(α−アミラーゼ)などが利用できる。
【0074】
シュードモナス属においては、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・セパシア(Pseudomonas cepacia)などで宿主ベクター系が開発されている。トルエン化合物の分解に関与するプラスミドTOLプラスミドを基本にした広宿主域ベクター(RSF1010などに由来する自律的複製に必要な遺伝子を含む)pKT240などが利用可能であり、プロモーター、ターミネーターとして、リパーゼ(特開平5-284973)遺伝子などが利用できる。
【0075】
ブレビバクテリウム属、特に、ブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)においては、pAJ43(Gene 39, 281 (1985))などのプラスミドベクターが利用可能である。プロモーター、ターミネーターとしては、大腸菌で使用されているプロモーター、ターミネーターがそのまま利用可能である。
【0076】
コリネバクテリウム属、特にコリネバクテリウム・グルタミカム(Corynebacterium glutamicum)においては、pCS11(特開昭57-183799)、pCB101(Mol. Gen. Genet. 196, 175 (1984)などのプラスミドベクターが利用可能である。
【0077】
ストレプトコッカス(Streptococcus)属においては、pHV1301(FEMS Microbiol. Lett. 26, 239 (1985)、pGK1(Appl. Environ. Microbiol. 50, 94 (1985))などがプラスミドベクターとして利用可能である。
【0078】
ラクトバチルス(Lactobacillus)属においては、ストレプトコッカス属用に開発されたpAMβ1(J. Bacteriol. 137, 614 (1979))などが利用可能であり、プロモーターとして大腸菌で利用されているものが利用可能である。
【0079】
ロドコッカス(Rhodococcus)属においては、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)から単離されたプラスミドベクターが使用可能である (J. Gen. Microbiol. 138,1003 (1992) )。
【0080】
ストレプトマイセス(Streptomyces)属においては、HopwoodらのGenetic Manipulation of Streptomyces: A Laboratory Manual Cold Spring Harbor Laboratories (1985)に記載の方法に従って、プラスミドを構築することができる。特に、ストレプトマイセス・リビダンス(Streptomyces lividans)においては、pIJ486 (Mol. Gen. Genet. 203, 468-478, 1986)、pKC1064(Gene 103,97-99 (1991) )、pUWL-KS (Gene 165,149-150 (1995) )が使用できる。また、ストレプトマイセス・バージニア(Streptomyces virginiae)においても、同様のプラスミドを使用することができる(Actinomycetol. 11, 46-53 (1997) )。
【0081】
サッカロマイセス(Saccharomyces)属、特にサッカロマイセス・セレビジアエ(Saccharomyces cerevisiae)においては、YRp系、YEp系、YCp系、YIp系プラスミドが利用可能であり、染色体内に多コピー存在するリボソームDNAとの相同組換えを利用したインテグレーションベクター(EP 537456など)は、多コピーで遺伝子を導入でき、かつ安定に遺伝子を保持できるため極めて有用である。また、ADH(アルコール脱水素酵素)、GAPDH(グリセルアルデヒド−3−リン酸脱水素酵素)、PHO(酸性フォスファターゼ)、GAL(β−ガラクトシダーゼ)、PGK(ホスホグリセレートキナーゼ)、ENO(エノラーゼ)などのプロモーター、ターミネーターが利用可能である。
【0082】
クライベロマイセス属、特にクライベロマイセス・ラクティス(Kluyveromyces lactis)においては、サッカロマイセス・セレビジアエ由来2μm系プラスミド、pKD1系プラスミド(J. Bacteriol. 145, 382-390 (1981))、キラー活性に関与するpGKl1由来プラスミド、クライベロマイセス属における自律増殖遺伝子KARS系プラスミド、リボソームDNAなどとの相同組換えにより染色体中にインテグレート可能なベクタープラスミド(EP 537456など)などが利用可能である。また、ADH、PGKなどに由来するプロモーター、ターミネーターが利用可能である。
【0083】
シゾサッカロマイセス(Schizosaccharomyces)属においては、シゾサッカロマイセス・ポンベ由来のARS (自律複製に関与する遺伝子)およびびサッカロマイセス・セレビジアエ由来の栄養要求性を相補する選択マーカーを含むプラスミドベクターが利用可能である(Mol. Cell. Biol. 6, 80 (1986))。また、シゾサッカロマイセス・ポンベ由来のADHプロモーターなどが利用できる(EMBO J. 6, 729 (1987))。特に、pAUR224は、タカラバイオから市販されており容易に利用できる。
【0084】
チゴサッカロマイセス属(Zygosaccharomyces)においては、チゴサッカロマイセス・ロウキシ(Zygosaccharomyces rouxii)由来のpSB3(Nucleic Acids Res. 13, 4267 (1985))などに由来するプラスミドベクターが利用可能であり、サッカロマイセス・セレビジアエ由来 PHO5 プロモーターや、チゴサッカロマイセス・ロウキシ由来 GAP-Zr(グリセルアルデヒド−3−リン酸脱水素酵素)のプロモーター(Agri. Biol. Chem. 54, 2521 (1990))などが利用可能である。
【0085】
ピキア(Pichia)属においては、ピキア・アンガスタ(旧名:ハンゼヌラ・ポリモルファ(Hansenula polymorpha))において宿主ベクター系が開発されている。ベクターとしては、ピキア・アンガスタ由来自律複製に関与する遺伝子(HARS1、HARS2)も利用可能であるが、比較的不安定であるため、染色体への多コピーインテグレーションが有効である(Yeast 7, 431-443 (1991))。また、メタノールなどで誘導されるAOX(アルコールオキシダーゼ)、FDH(ギ酸脱水素酵素)のプロモーターなどが利用可能である。また、ピキア・パストリス(Pichia pastoris)などにピキア由来自律複製に関与する遺伝子 (PARS1、 PARS2)などを利用した宿主ベクター系が開発されており(Mol. Cell. Biol. 5, 3376 (1985))、高濃度培養とメタノールで誘導可能なAOXなど強いプロモーターが利用できる(Nucleic Acids Res. 15, 3859 (1987))。
【0086】
キャンディダ(Candida)属においては、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・アルビカンス(Candida albicans)、キャンディダ・トロピカリス(Candida tropicalis)、キャンディダ・ウチルス(Candida utilis)などにおいて宿主ベクター系が開発されている。キャンディダ・マルトーサにおいてはキャンディダ・マルトーサ由来ARSがクローニングされ(Agri. Biol. Chem. 51, 51, 1587 (1987))、これを利用したベクターが開発されている。また、キャンディダ・ウチルスにおいては、染色体インテグレートタイプのベクターは強力なプロモーターが開発されている(特開平 08-173170)。
【0087】
アスペルギルス(Aspergillus)属においては、アスペルギルス・ニガー (Aspergillus niger) 、アスペルギルス・オリジー (Aspergillus oryzae) などがカビの中で最もよく研究されており、プラスミドや染色体へのインテグレーションが利用可能であり、菌体外プロテアーゼやアミラーゼ由来のプロモーターが利用可能である(Trends in Biotechnology 7, 283-287 (1989))。
【0088】
トリコデルマ(Trichoderma)属においては、トリコデルマ・リーゼイ(Trichoderma reesei)を利用したホストベクター系が開発され、菌体外セルラーゼ遺伝子由来プロモーターなどが利用できる(Biotechnology 7, 596-603 (1989))。
【0089】
また、微生物以外でも、植物、動物において様々な宿主・ベクター系が開発されており、特に蚕を用いた昆虫(Nature 315, 592-594 (1985))や菜種、トウモロコシ、ジャガイモなどの植物中に大量に異種タンパク質を発現させる系が開発されており、好適に利用できる。
【0090】
本発明において使用するエルゴチオナーゼ生産能を有する微生物は、エルゴチオナーゼ生産能を有する全ての菌株、突然変異株、変種、遺伝子操作技術の利用により作成された本発明の酵素生産能を獲得した形質転換株を含む。
【0091】
また本発明は、エルゴチオナーゼの製造方法を提供する。本製造方法には、バークホルデリア属もしくはシュードモナス属に属しエルゴチオナーゼ活性を有する微生物、もしくは上記本発明の形質転換体を培養する工程と、培養物よりエルゴチオナーゼを回収する工程が含まれる。具体的な培養工程および回収工程は、上述のとおりである。
【0092】
また本発明は、エルゴチオネインに作用してチオウロカン酸を生成する能力を有する酵素活性物質(enzymatic active materials)を、被検対象に接触させる工程を含む、エルゴチオネインの定量方法に関する。
本発明のエルゴチオネインの定量方法は、エルゴチオナーゼ活性を有する酵素活性物質を、この酵素活性物質によってエルゴチオネインからチオウロカン酸を生成することが可能な条件下で、被検対象に接触させる工程を含む。
【0093】
本発明において被検対象としては、下記(1)〜(2)を挙げることができる。
(1)被験者(クローン病患者を含む)および臨床検体等から単離されたあらゆる生体由来物質(組織、臓器、体液、血液、細胞等)、および
(2)エルゴチオネイン、エルゴチオネイン含有物質、またはその原料や処理物(例えば、一般食品、健康食品、美容食品等の食品、化粧品、生体材料、カビ類、キノコ類、マイコバクテリア等)。
【0094】
本発明において、酵素活性物質とは、酵素活性を有する物質を指し、例えばエルゴチオナーゼ活性を有する物質を挙げることができる。具体的には、エルゴチオネインに作用してチオウロカン酸およびトリメチルアミンを生成する能力を有する物質であり、より好ましくは微生物に由来する物質であって、当該能力が維持された物質である。
【0095】
本発明の酵素活性物質は、例えば、本発明のエルゴチオナーゼ(エルゴチオナーゼ活性を有するタンパク質を含む)、当該エルゴチオナーゼを生産する微生物、およびその処理物からなる群から選択することができる。より具体的には、例えば次の要素が、エルゴチオナーゼ活性を維持する限り、本発明の酵素活性物質に含まれる。
i) エルゴチオネインに作用し、チオウロカン酸およびトリメチルアミンを生成する能力を有する微生物
ii) i)の微生物の菌体、菌体培養物、菌体もしくは菌体培養物の処理物(菌体もしくは菌体培養物の乾燥物や濃縮物、菌体培養液から菌体を除去した上清、菌体自体の乾燥物や濃縮物等)
iii) i)またはii)から単離された物質
なお本発明における酵素活性物質は、精製された標品である必要はない。
【0096】
本発明において、微生物は、培養物であることもできるし、菌体であることもできる。培養物は、微生物の生存と増殖を支持する培地とともに当該微生物が存在する状態を言う。例えば微生物を含む液体培地(培養液)は、培養物である。一方、本発明においては、微生物そのものを指す用語として「菌体」(microbial cell)を用いる。菌体は、培養物から回収された微生物細胞を指す。例えば遠心分離によって液体培地から微生物の菌体を回収することができる。培養物から回収された菌体を洗浄することによって、菌体表面の培地成分を除くこともできる。
【0097】
さらに別の態様においては、本発明における酵素活性物質は、上記i)の微生物から単離することができるエルゴチオナーゼを含む。また当該エルゴチオナーゼを担体に固定化したものも、エルゴチオナーゼ活性を維持する限り、酵素活性物質に含まれる。
【0098】
本発明において酵素活性物質は、そのまま、あるいは固定化して基質化合物と接触させることができる。微生物菌体あるいはその処理物を固定化するための種々の方法が公知である。例えば、以下のような方法が微生物やその処理物の固定化方法として公知である。
ポリアクリルアミド法、
含硫多糖ゲル法(κ-カラギーナンゲル法など)、
アルギン酸ゲル法、
寒天ゲル法、
イオン交換樹脂法
あるいは、限外ろ過膜等を用いたメンブレンバイオリアクター中で、微生物、その処理物、あるいは酵素を基質化合物と接触させて反応させることもできる。
【0099】
本発明におけるエルゴチオネイン定量方法は、微生物を酵素が誘導される適切な前記培養条件において培養を行い、得られた培養液、あるいは培養液から採取した菌体や該菌体処理物、並びに酵素を、上記被検対象に接触させ、インキュベートすることにより行うことができる。例えば、エルゴチオナーゼ活性により生成されたチオウロカン酸を上述の方法で定量することで、被検対象に含有されていたエルゴチオネインの定量を行うことができる。具体的には、チオウロカン酸の生成量(311 nmの吸光度の増加)を測定し、被検物質中のエルゴチオネイン濃度を計算することによって行われる。実際に測定する際には、例えば検量線を作成し、検量線の直線性を確認の上で、最終生成されたチオウロカン酸生成に由来する311 nm の吸光度の値から、被検対象に含有されていたエルゴチオネインの定量を行うという手法をとることが可能である。
【0100】
反応条件としては、例えば以下の条件を示すことができる。
pH 5.0から11.0、好ましくはpH 7.0から9.5、
温度25から80℃、好ましくは25から60℃
【0101】
すなわち本発明は、下記(1)〜(2)の工程を含むエルゴチオネインの定量方法を提供する;
(1) エルゴチオネインに作用してチオウロカン酸およびトリメチルアミンを生成する能力を有する酵素活性物質(enzymatic active materials)を、被検対象に接触させる工程、および
(2) 生成されたチオウロカン酸またはトリメチルアミンを定量する工程。
【0102】
また本発明は、エルゴチオナーゼまたは酵素活性物質を含むことを特徴とするエルゴチオネイン定量試薬およびキットを提供する。本定量試薬およびキットにより、エルゴチオネインの定量を行うことができる。
【0103】
本発明の定量試薬やキットにおいては、有効成分であるエルゴチオナーゼまたは酵素活性物質以外に、例えば、滅菌水、生理食塩水、植物油、界面活性剤、脂質、溶解補助剤、緩衝剤、タンパク質安定剤(BSAやゼラチンなど)、保存剤、ブロッキング溶液、反応溶液、反応停止液、試料を処理するための試薬等が必要に応じて混合されていてもよい。さらにキットには、その他、使用方法を記載した指示書等をパッケージしておくこともできる。
【0104】
また本発明は、エルゴチオネインの製造方法を提供する。本製造方法には、チオウロカン酸およびトリメチルアミンにエルゴチオナーゼまたは酵素活性物質を作用させる工程が含まれる。
【0105】
なお本明細書において引用されたすべての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0106】
以下、実施例により本発明を詳細に説明するが、本発明はこれに限定されない。
【0107】
〔参考例1〕
非特許文献4(J. Biol. Chem., 237, 874-881 (1962))に記載の培地を用い、大腸菌W株を0.02% 硫酸マグネシウム・7水和物、0.2% クエン酸、1% K2HPO4、0.5% グルコース及び5 mM エルゴチオネインを含有する培地(以下、エルゴチオネイン培地, pH 7.2)で37℃、5日間培養したが菌の生育は見られなかった。
【0108】
LB培地(1% Bacto-Trypton(Difco製)、0.5% Bacto-Yeast extract(Difco製)、1% 塩化ナトリウム、pH 7.2)で培養後、菌体を生理食塩水で洗浄し、エルゴチオネイン培地で懸濁し、さらに37℃、5日間誘導培養した後に菌を回収し、超音波破砕後の無細胞抽出液を用いて酵素活性の測定を行ったが、エルゴチオナーゼ活性は検出されなかった。
【0109】
[エルゴチオナーゼ活性の測定]
0.25 mM エルゴチオネイン、10 mM ピロリン酸ナトリウム緩衝液(pH 9.0)および酵素を含む1 mLの反応液を用いて30℃で反応し、チオウロカン酸生成に由来する311 nmの吸光度の増加を測定した。1 U は、上記条件下で1分間に1μmolのチオウロカン酸生成を触媒する酵素量とし、チオウロカン酸の分子吸光係数(ε=18,000)より計算した。
【0110】
〔実施例1〕スクリーニング
エルゴチオネイン培地(5 mM エルゴチオネイン、0.02% 硫酸マグネシウム・7水和物、0.2% クエン酸、1% 無水リン酸水素二カリウム、pH7.2)を1 mLずつ分注し、オートクレーブにより加熱滅菌した。ここに高知県立のいち動物公園(高知県香南市野市町)から採取した土壌試料を小スパチュラで1サジ加え、30℃、4日間、振盪培養した。この培養液5μLを、0.1 mg/L シクロヘキシミドを含むエルゴチオネイン培地に植菌し、30℃、2日間、振盪培養した。この培養液を100倍希釈し、20μLを0.1 mg/Lシクロヘキシミドを含むエルゴチオネイン平板培地に塗布し、30℃で静置培養後、コロニーを単離した。その結果、エルゴチオネイン資化性細菌が単離された。
【0111】
〔実施例2〕HME13の同定
単離された資化性細菌(菌株)のうちHME13の微生物学的特徴は、表1にまとめたとおりである。
【0112】
【表1】
【0113】
続いて16S-rDNAの解析によりHME13を同定した。16S-rDNAの塩基配列は、以下の各プロトコールに基づいて解析した。DNA抽出:InstaGene Matrix (BIO-RAD)、PCR:PrimeSTAR HS DNA Polymerase(タカラバイオ)、サイクルシークエンス:BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems)、シークエンス:ABI PRISM 3130 xl Genetic Analyzer System (Applied Biosystems)、配列決定:ChromasPro 1.4 (Technelysium Pty Ltd.)。
得られた塩基配列を配列番号:5に示した。配列番号:5の塩基配列を元に、アポロンDB-BA4.0(テクノスルガ・ラボ)を用いて、Blast検索を行った結果、次の菌株の16S-rDNA配列にそれぞれ99.4%の相同性を示した。
Burkholderia arboris R-24201
B. stabilis LMG14294
【0114】
得られた結果を元に分子系統樹を作成した結果を図1に示した。クラスター解析の結果、HME13は、Burkholderia属に含まれると考えられるが既知種とは異なることが示唆されたので、HME13菌をバークホルデリア・エスピー (Burkholderia sp.) と同定した。
【0115】
〔実施例3〕 酵素反応
実施例1に記載のエルゴチオネイン培地で一晩振盪培養し、遠心分離にて得られた菌体を10 mM Sodium pyrophosphate (pH9.0), 1mM PMSFに懸濁し、超音波破砕した。破砕液を遠心分離し、未破砕菌体および菌体の残渣を除いた粗酵素液を得た。0.25 mM エルゴチオネインを含む10 mM Sodium pyrophosphate (pH 9.0)に本粗酵素液20μLを加え、30℃で保持しながら311 nmの吸光度を測定してチオウロカン酸の増加を追跡することで、酵素活性を測定した。また、タンパク質濃度をProtein assay CBB solution(ナカライテスク)にて定量した。エルゴチオナーゼ1Uは、1分間に1μmolのチオウロカン酸を生成する酵素量とした。結果を表2に示した。
【0116】
【表2】
【0117】
〔実施例4〕 HME13からのエルゴチオナーゼの精製
エルゴチオネイン培地1 mLにBurkholderia sp. HME13を植菌し、30℃、一晩振盪培養し、得られた前培養液100μLを500 mLのエルゴチオネイン培地に植菌し、30℃、45時間振盪培養した。培養液を遠心分離し、得られた菌体を1 mM PMSF を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) に懸濁し、超音波により菌体を破砕した。菌体破砕液を35-65% 硫安分画を行い、得られた酵素を10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で2回透析した後、20 mM Tris-HCl (pH 7.2)で平衡化したDEAE-トヨパールカラム (250 mL-Resin)にアプライし、塩化ナトリウムにより段階的に溶出した。活性画分を回収し、濃縮後、35% 飽和硫安を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で透析し、35% 飽和硫安を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で平衡化したButyl-トヨパールカラム (200 mL-Resin)にアプライし、硫安濃度を段階的に下げて酵素を溶出した。精製の要約を表3(HME13からのエルゴチオナーゼの精製)に示した。
【0118】
【表3】
【0119】
続いてHME13から精製されたエルゴチオナーゼの理化学的特徴を明らかにした。
【0120】
(1) 至適温度:
HME13から精製されたエルゴチオナーゼの至適温度を検討した。0.25 mMエルゴチオネイン、10 mM リン酸ナトリウム緩衝液(pH7.2)、23 ng 精製酵素を含む全量1.0 mlの溶液を反応液とし、25℃〜80℃まで5℃間隔でインキュベートし、反応液中に生成するチオウロカン酸の吸収(311 nm)の増加を測定した。結果を図2に示す。この結果から、HME13から精製されたエルゴチオナーゼの至適温度は65℃であることが確認された。また50-65℃で最大活性の80%以上が維持された。
【0121】
(2) 熱安定性:
次に、HME13から精製されたエルゴチオナーゼの熱安定性について検討した。0.1 mg /ml 精製酵素溶液を4,20,30,40,50,60,65,70,80℃で30分間加温した。その後、0.25 mM エルゴチオネイン,10 mMリン酸ナトリウム緩衝液(pH7.2),各温度で処理された精製酵素47 ngを含む全量1.0 mlの溶液を反応液とし、30℃で反応液中に生成するチオウロカン酸の吸収(311 nm)の増加を測定した。結果を図3に示す。この結果から、HME13から精製されたエルゴチオナーゼは60℃まで約90%の活性を維持し、70℃以上の温度では急速に活性を失うことが確認された。
【0122】
(3) 至適pH:
さらに、HME13から精製されたエルゴチオナーゼの至適pHについて検討した。緩衝液を変えた他は(1)と同じ組成の反応液にエルゴチオナーゼ(47 ng/mL)を加えて反応を開始し、30℃で活性を測定した。実験に使用した緩衝液と、そのpHは次のとおりである。
100 mM酢酸ナトリウム緩衝液:pH5.0−6.5
100 mMリン酸ナトリウム緩衝液:pH6.5−7.5
100 mM Tris塩酸緩衝液:pH7.5−8.5
100 mMグリシン水酸化ナトリウム緩衝液:pH8.5−11.0
【0123】
生成するチオウロカン酸の吸収(311 nm)を測定した。結果を図4に示す。この結果から、HME13から精製されたエルゴチオナーゼの至適pHは8.0であることが確認された。またpH 7.0-9.5 で最大活性の80%以上が維持された。
【0124】
(4) pH安定性:
次に、HME13から精製されたエルゴチオナーゼの安定性に与えるpHの影響について検討した。各緩衝液に溶解したエルゴチオナーゼ(0.1mg/mL)を30℃で30分間プレインキュベーションした。次いで各pHで処理されたエルゴチオナーゼ47 ngを(1)と同じ組成の反応液に加えて反応を開始し、30℃で生成するチオウロカン酸の吸収(311 nm)を測定した。結果を図5に示す。この結果から、HME13から精製されたエルゴチオナーゼはpH 6.5-11.0で30℃, 30分間処理後に活性の80%以上を維持することが確認された。
【0125】
(5) 基質特異性:
HME13から精製されたエルゴチオナーゼの基質特異性について検討した。基質を変えた他は(1)と同じ組成の反応液にエルゴチオナーゼ(47 ng/mL)を加えて反応を開始し、30℃で活性を測定した。実験に使用した基質は表5のとおりである。エルゴチオネインを基質とした場合には生成するチオウロカン酸の吸収(311 nm)を測定した。ヒスチジンを基質とした場合には生成するウロカニン酸の吸収(277 nm)を測定した。チロシンを基質とした場合には生成するp-ヒドロキシケイ皮酸の吸収(315 nm)を測定した。フェニルアラニンを基質とした場合には生成するトランス-ケイ皮酸の吸収(290 nm)を測定した。この結果、エルゴチオナーゼはエルゴチオネイン以外のいずれのアミノ酸にも作用しなかった(表4)。
【0126】
【表4】
【0127】
(6) 阻害剤の影響:
HME13から精製されたエルゴチオナーゼの阻害剤の影響について検討した。各阻害剤を加えた他は(1)と同じ組成の反応液にエルゴチオナーゼ(47 ng/mL)を加えて反応を開始し、30℃で生成するチオウロカン酸の吸収(311 nm)を測定した。但し、p-ヒドロキシ水銀安息香酸については、エルゴチオネインを除く反応液を30℃で20分間加温後、エルゴチオネインを加えて反応を開始した。実験に使用した阻害剤は表5のとおりである。この結果、エルゴチオナーゼは検討した各阻害剤によりほとんど影響を受けず、特にp-ヒドロキシ水銀安息香酸(p-hydroxymercuribenzoate)、ヨード酢酸(iodoacetate)、L-システイン(L-cysteine)存在下で80%以上の活性を保持していた点において、非特許文献4に記載の大腸菌W株由来エルゴチオナーゼと異なっていた(表5)。
【0128】
【表5】
【0129】
〔実施例5〕 エルゴチオナーゼの部分アミノ酸配列
実施例4で得られた酵素を用いて、プロテインシーケンサー(HT492 アプライドバイオシステムズ社)によりN末端アミノ酸配列を解析した。アミノ酸配列を配列番号:6に示した。
【0130】
〔実施例6〕 エルゴチオナーゼ遺伝子のコア領域のクローニング
N末端アミノ酸配列を元にBLAST検索した結果、完全に一致するアミノ酸配列が得られた。得られた配列は次のとおりである。
Burkholderia sp.383 由来 Bcep18194_B3018 タンパク質のアミノ酸配列の一部
Burkholderia pseudomarei 1106a 由来 BURPS1106A_2045 タンパク質のアミノ酸配列の一部
Burkholderia pseudomarei 1710b 由来 BURPS1710b_2187 タンパク質のアミノ酸配列の一部
Burkholderia pseudomarei K96243 由来 BPSL1676 タンパク質のアミノ酸配列の一部
【0131】
これら菌株のゲノムデータベース情報を基にそれぞれセンスプライマー、アンチセンスプライマーを合成した。それぞれの塩基配列を配列番号:7、8に示した。
配列番号:7:CGACGTGCACACGCTCGCCGCGCT
配列番号:8:CCCGACGCGAGCTGCACGAGATGCTT
【0132】
プライマー各10 pmol、dNTP 10 nmol、Burkholderia sp. HME13由来染色体DNA 140 ng、Ex-Taq用緩衝液(タカラバイオ製)、Ex-Taq 1.25 U(タカラバイオ製)を含む50μLの反応液を用い、94℃、1分の後、98℃、10秒 、68℃、2分を30サイクル、最後に72℃、10分、GeneAmp PCR System 9700(アプライド・バイオシステムズ製)を用いて行った。得られたDNA断片の塩基配列を解析し、決定されたエルゴチオナーゼ遺伝子の塩基配列を配列番号:2に、予想されるアミノ酸配列を配列番号:1に示した。予想されるアミノ酸配列をSignalP v3.0 (http://www.cbs.dtu.dk/services/SignalP/) で解析したところ、MQRFLMRSAIALALFAAVPAFA(配列番号:9)がシグナル配列であり、成熟型酵素では切断されると予測され、精製酵素のN末端アミノ酸配列解析結果と一致した。シグナル配列が切断された場合のエルゴチオナーゼのアミノ酸配列を配列番号:3に、塩基配列を配列番号:4に示す。
【0133】
〔実施例7〕 エルゴチオナーゼ遺伝子のクローニング
エルゴチオナーゼの構造遺伝子配列を基にORFのみをクローニングするためのプライマー(配列番号:10、11)を合成した。プライマーを各10 pmol、dNTP 10 nmol、Burkholderia sp. HME13由来染色体DNA 144 ng、Ex-Taq用緩衝液(タカラバイオ製)、Ex-Taq 1.25 U(タカラバイオ製)を含む50μLの反応液を用い、94℃、1分の後、98℃、10秒、68℃、1分を25サイクル、最後に72℃、10分、GeneAmp PCR System 9700(アプライド・バイオシステムズ製)を用いて行った。
配列番号:10:GGAATTCCATATGCAACGCTTCCTCATGCGGAGC
配列番号:11:GGGAAGCTTTCAGTCGTGATACGCGCGCAGCAG
【0134】
得られたDNA断片を制限酵素NdeI、HindIIIで2重消化し、アガロースゲル電気泳動を行い、目的とするバンドの部分を切り出し精製した。得られたDNA断片を、NdeI、HindIIIで2重消化したpET21a(+) (Novagen製)とLigation-convenience kit (ニッポンジーン製)を用いて、ライゲーションし、大腸菌DH5α株を形質転換した。
【0135】
得られた形質転換株を、アンピシリンを含むLB培地で生育させ、生育した形質転換体からPlasmidを抽出し、エルゴチオナーゼ遺伝子が挿入されていることとが確認できたプラスミドをpET21a(+)/ERTaseとした。pET21a(+)/ERTaseで大腸菌 Rosetta 2 (DE3) 株 (Novagen製)を形質転換し、エルゴチオナーゼ発現株を得た。
【0136】
〔実施例8〕 組換えエルゴチオナーゼの活性評価
エルゴチオナーゼを発現するプラスミドpET21a(+)/ERTaseで形質転換された大腸菌Rosetta 2 (DE3) 株をアンピシリンとクロラムフェニコールを含む液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに3時間培養を行った。
【0137】
菌体を遠心分離により集菌した後、1.0 mM PMSF、0.01% 2-メルカプトエタノールを含む10 mM ピロリン酸ナトリウム緩衝液(pH 7.2)に懸濁し、密閉式超音波破砕装置UCD-200TM(コスモバイオ製)を用いて菌体を破砕した。菌体破砕液を遠心分離し、その上清を菌体抽出液として回収した。また該遺伝子を含まない大腸菌Rosetta 2 (DE3)株をLB培地で終夜培養し、0.1 mM IPTG添加後さらに3時間培養した菌体を同様に破砕して、菌体抽出液を回収した。それぞれの菌体抽出液を用いてエルゴチオナーゼ活性を測定した。結果を表6に示した。活性測定の方法は実施例3と同様とした。
【0138】
【表6】
【0139】
〔実施例9〕 組換えエルゴチオナーゼのN末端アミノ酸配列の決定
エルゴチオナーゼを発現するプラスミドで形質転換された大腸菌Rosetta2 (DE3) 株を、1 mM PMSFを含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) に懸濁し、超音波により菌体を破砕した。菌体破砕液を30-50% 硫安分画を行い、得られた酵素を10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で2回透析した後、20mM Tris-HCl (pH 7.2)で平衡化したDEAE-トヨパールカラムにアプライし、塩化ナトリウムにより段階的に溶出した。活性画分を回収し、濃縮後、30% 飽和硫安を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で透析し、30% 飽和硫安を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で平衡化したButyl-トヨパールカラム (200mL-Resin)にアプライし、硫安濃度を段階的に下げて酵素を溶出した。精製の要約を表7(組換えエルゴチオナーゼの精製)に示した。
【0140】
【表7】
【0141】
得られた酵素を用いて、プロテインシーケンサー(HT492 アプライドバイオシステムズ社)によりN末端アミノ酸配列を解析した。アミノ酸配列を配列番号:12に示した。
配列番号:12:DVTLDGRSVTPESIARI
【0142】
組換えエルゴチオナーゼのN末端アミノ酸配列は、親株のエルゴチオナーゼと同様にN末端のシグナル配列が切断されていた。
【0143】
〔実施例10〕 シグナル配列を除いた組換えエルゴチオナーゼの活性評価
エルゴチオナーゼのシグナル配列を除いたORFのみをクローニングするためのプライマー(配列番号:13、14)を合成した。プライマーを各15 pmol、dNTP 20 nmol、pET21a(+)/ERTase plasmid、KOD FX用緩衝液(TOYOBO製)、KOD FX 1U(TOYOBO製)を含む50μLの反応液を用い、94℃、2分の後、98℃、10秒、74℃、1分30秒を5サイクル、98℃、10秒、72℃、1分30秒を20サイクル、最後に68℃、7分、GeneAmp PCR System 9700(アプライド・バイオシステムズ製)を用いて行った。
配列番号:13:GGAATTCCATATGGACGTCACGCTCGACGGCCGGT
配列番号:14:GGGAAGCTTTCAGTCGTGATACGCGCGCAGCAG
【0144】
得られたDNA断片を制限酵素NdeI、HindIIIで2重消化し、アガロースゲル電気泳動を行い、目的とするバンドの部分を切り出し精製した。得られたDNA断片を、NdeI、HindIIIで2重消化したpET21a(+) (Novagen製)とLigation-convenience kit (ニッポンジーン製)を用いて、ライゲーションし、大腸菌DH5α株を形質転換した。
【0145】
得られた形質転換株を、アンピシリンを含むLB培地で生育させ、生育した形質転換体からPlasmidを抽出し、エルゴチオナーゼ遺伝子が挿入されていることとが確認できたプラスミドをpET21a(+)/ERTase-sigとした。pET21a(+)/ERTase-sigで大腸菌 Rosetta 2 (DE3) 株 (Novagen製)を形質転換し、シグナル配列を除いたエルゴチオナーゼ発現株を得た。
【0146】
シグナル配列を含むエルゴチオナーゼを発現するプラスミドpET21a(+)/ERTaseで形質転換された大腸菌Rosetta 2 (DE3) 株とシグナル配列を含まないエルゴチオナーゼを発現するプラスミドpET21a(+)/ERTase-sigで形質転換された大腸菌Rosetta 2 (DE3) 株をアンピシリンとクロラムフェニコールを含む液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに3時間培養を行った。
【0147】
菌体を遠心分離により集菌した後、1.0 mM PMSF、0.01% 2-メルカプトエタノールを含む10 mM ピロリン酸ナトリウム緩衝液(pH 7.2)に懸濁し、密閉式超音波破砕装置UCD-200TM(コスモバイオ製)を用いて菌体を破砕した。菌体破砕液を遠心分離し、その上清を菌体抽出液として回収した。また該遺伝子を含まない大腸菌Rosetta 2 (DE3)株をLB培地で終夜培養し、0.1 mM IPTG添加後さらに3時間培養した菌体を同様に破砕して、菌体抽出液を回収した。それぞれの菌体抽出液を用いてエルゴチオナーゼ活性を測定した。結果を表8に示した。活性測定の方法は実施例3と同様とした。
【0148】
【表8】
【0149】
〔実施例11〕 エルゴチオナーゼによるエルゴチオネインの定量
0〜500μg/mLエルゴチオネイン、10mM リン酸ナトリウム緩衝液(pH7.2)、1.9 mg/mL精製酵素を含む全量1.0 mLの溶液を反応液とし、30℃で1時間インキュベートし、2N HClを50μL加えて反応を停止し、反応液中に生成するチオウロカン酸の吸収(311 nm)の増加を測定した。結果を図6に示す。この結果から、HME13から精製されたエルゴチオナーゼを用いて0-500μg/mLの範囲でエルゴチオネインを定量可能であることが確認された。
【産業上の利用可能性】
【0150】
本発明は、エルゴチオナーゼの利用による、エルゴチオネインの特異的かつ簡便な定量方法を提供する。本発明によるエルゴチオネインの定量は、例えば、クローン病の診断法として有用である。
【技術分野】
【0001】
本発明は、エルゴチオネインの特異的な定量に有用な酵素であるエルゴチオナーゼ、それをコードする遺伝子、該遺伝子を含む形質転換体、該酵素の製造方法、該酵素を利用したエルゴチオネインの定量方法に関する。
【背景技術】
【0002】
エルゴチオネインは、1909年にライ麦角菌(Claviceps purpurea)より単離、構造決定されたアミノ酸の1種であり、その後、幅広い動物、植物に存在することが明らかとなったが、動物、植物などはエルゴチオネイン合成能を有しておらず、一部のカビ、キノコ類のみが生産し、動物、植物は、吸収、食餌によりこれを取り込んでいると考えられている。
【0003】
エルゴチオネインは、光化学傷害に対する保護作用、活性酸素種(Reactive Oxygen Species、ROS)や活性窒素種(Reactive Nitrogen Species, RNS)の消去やこれらによる酸化ストレスの低減など優れた抗酸化活性を有し、一方で、高い水溶性、熱・酸素に対する安定性を有するため、抗酸化物質として、アンチエイジング素材、美肌成分、生体成分、食料品などの酸化防止剤として、その利用が期待されている。
【0004】
また、ヒトにおいて、有機カチオントランスポーター(organic cation transporter)によりエルゴチオネインが細胞内に特異的に取り込まれること、その変異がクローン病のリスクと高い相関があること、リューマチ患者の赤血球にエルゴチオネイン含量が高いことなどから、自己免疫疾患との関係が指摘されている。
【0005】
このような背景より、食品、化粧品、臓器、血液など生体材料中のエルゴチオネインを特異的に、かつ、簡便に測定する方法が求められていたが(特許文献1)、これまでに報告されている比色法、高圧液体クロマトグラフィー(HPLC)を用いた方法では、特異性、簡便性において問題があった。具体的には、2,2’-dithiopyridineを用いた比色法では夾雑するチオール化合物の影響を除くために、サンプルを予めアルカリ条件下、銅触媒により酸化する必要があり、本前処理を行った場合でも非特異的に発色を完全には抑制できない(非特許文献1)。また、HPLCを用いた定量では、本化合物の逆相カラムへの保持が非常に弱いこと、非特異的な吸収波長(254 nm)で検出する必要があることから、Thiopropyl-Sepharoseにより前処理する(非特許文献2)、カラムを2本タンデムに連結する、検出に質量分析器を利用する、など複雑な操作や高価な機械が必要であった(非特許文献3)。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2009-192383
【非特許文献】
【0007】
【非特許文献1】Biochem. J., 139, 221-235 (1974)
【非特許文献2】Chem. Pharm. Bull., 26, 3772-3778 (1978)
【非特許文献3】Int. J. Med. Mushrooms, 8, 215-222 (2006)
【発明の概要】
【発明が解決しようとする課題】
【0008】
上記課題に鑑み、本発明者らは、より簡便かつ特異的にエルゴチオネインを定量するために、酵素を用いた特異的なエルゴチオネイン定量方法の開発を検討した。
【0009】
エルゴチオネインに特異的に作用する酵素として、エルゴチオネインを炭素源、エネルギー源として利用可能な大腸菌W株より、L-エルゴチオネインに作用し、チオウロカン酸(3-(2,3-ジヒドロ-2-チオキソ-1H-イミダゾール-4-イル)プロペン酸)およびトリメチルアミンを生成する(式1)に示すエルゴチオナーゼが知られている(非特許文献4:J. Biol. Chem., 237, 874-881 (1962))。
【0010】
式1
【化1】
【0011】
本発明者らは、エルゴチオナーゼを用いて、エルゴチオネインの簡便かつ特異的な定量が可能か検討するために、以下の参考例1に示すように、この大腸菌W株よりエルゴチオナーゼの調製を試みたが、本菌株にエルゴチオナーゼ活性を検出できなかった。
【0012】
そこで本発明者らは、エルゴチオネインの簡便かつ特異的な定量に利用可能なエルゴチオナーゼを見出すために、エルゴチオネインを窒素源として利用可能な微生物のスクリーニングを土壌(高知県のいち動物公園)および研究室保有の微生物保存株より行った。その結果、土壌よりエルゴチオナーゼ生産能を有する微生物を見出した。最も高い活性を示したバークホルデリア・スピーシーズ(Burkholderia sp.)HME13株より、エルゴチオナーゼを精製し、その酵素科学的性質を解析した結果、本酵素がL-エルゴチオネインに特異的に作用することが明らかになった。さらに、本酵素をコードする遺伝子をクローニングし、大腸菌により高発現させ、本酵素を容易にかつ大量調製することが可能となった。さらに、本酵素を用いてL-エルゴチオネインの定量を検討した結果、特異的かつ簡便にL-エルゴチオネインを定量可能なことが明らかになり、本発明を完成した。
【0013】
なお、非特許文献5(J. Bacteriol., 74, 381-385 (1957))では、研究室保存のバチルス・サチルス(Bacillus subtilis)、バチルス・ポリミキサ(Bacillus polymyxa)、大腸菌(Escherichia coli)、イシェリヒア・フロインディー(Escherichia freundii)、アエロバクター・アエロゲネス(Aerobacter aerogenes)、ストレプトコッカス・フェーカリス(Streptococcus faecalis)、ストレプトコッカス・リキファッシェンス(Streptococcus liquifaciens)、ミクロコッカス・パイロゲネス・バラエティ・アウレウス(Micrococcus pyogenes var. aureus)、セラチア・マルセッセンス(Serratia marcescens)、プロテウス・バルガリス(Proteus vulgaris)、シュードモナス・エルギノーザ(Pseudomonas aeruginosa)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、ナイセリア・カタルラリス(Neisseria catarrhalis)、アルカリゲネス・フェーカリス(Alcaligenes faecalis)は、エルゴチオネインを資化できず、土壌よりエルゴチオネイン資化性を持つ菌として単離したアルカリゲネス・フェーカリス(Alcaligenes faecalis)のみがエルゴチオネインを代謝可能であったことを報告しており、エルゴチオネイン資化性を有する微生物が自然界において希にしか存在しないことを示している。
【0014】
即ち本発明は、エルゴチオネインの特異的な定量に有用なエルゴチオナーゼ、それをコードする遺伝子、該遺伝子を含む形質転換体、該エルゴチオナーゼの製造方法、該エルゴチオナーゼを利用したエルゴチオネインの定量方法に関する。
【0015】
本発明は、より具体的には以下の〔1〕〜〔14〕を提供するものである。
〔1〕 次の(1)〜(2)に示す理化学的性状を有するエルゴチオナーゼ;
(1)作用:L-エルゴチオネインに作用し、チオウロカン酸(3-(2,3-ジヒドロ-2-チオキソ-1H-イミダゾール-4-イル)プロペン酸)およびトリメチルアミンを生成する、および
(2)基質特異性:L-エルゴチオネインに作用し、D-, L-ヒスチジン、D-, L-チロシン、D-, L-フェニルアラニンに作用しない。
〔2〕 さらに付加的に、次の(3)〜(8)に示す理化学的性状を有する〔1〕に記載のエルゴチオナーゼ;
(3)至適pH:7.0〜9.5、
(4)至適温度:50〜65℃、
(5)分子量:ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動による分子量が45,000〜55,000、ゲルろ過による分子量が200,000〜260,000。
(6)熱安定性: 60℃、30分で90%以上の活性を保持、
(7)pH安定性: pH 6.5〜11.0、30分で80%以上の活性を保持、および
(8)阻害剤: p-ヒドロキシ水銀安息香酸(p-hydroxymercuribenzoate)、ヨード酢酸(iodoacetate)、および/またはL-システイン(L-cysteine)存在下において80%以上の活性を保持。
〔3〕 バークホルデリア属、もしくはシュードモナス属に属する微生物に由来する、〔1〕または〔2〕に記載のエルゴチオナーゼ。
〔4〕 前記バークホルデリア属、もしくはシュードモナス属に属する微生物が、下記の群から選択されるいずれかの微生物であることを特徴とする〔3〕に記載のエルゴチオナーゼ;
バークホルデリア・エスピー NITE P-861(Burkholderia sp. NITE P-861)、および
シュードモナス・エスピー (Pseudomonas sp.)。
〔5〕 下記(a)〜(e)のいずれかに記載のエルゴチオナーゼ活性を有するタンパク質;
(a)配列番号:3に記載のアミノ酸配列を含むタンパク質、
(b)配列番号:4に記載された塩基配列を含むポリヌクレオチドによってコードされるタンパク質、
(c)配列番号:3に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含むタンパク質、
(d)配列番号:4に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質、または
(e)配列番号:3に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列を含むタンパク質。
〔6〕 〔5〕に記載のタンパク質をコードするポリヌクレオチド。
〔7〕 〔1〕〜〔4〕のいずれかに記載のエルゴチオナーゼをコードするポリヌクレオチドまたは〔6〕に記載のポリヌクレオチドを保持したベクター。
〔8〕 〔7〕に記載のベクターにより形質転換された形質転換体。
〔9〕 前記宿主が大腸菌である〔8〕に記載の形質転換体。
〔10〕 バークホルデリア属、もしくはシュードモナス属に属し、エルゴチオナーゼ活性を有する微生物、もしくは〔8〕または〔9〕に記載の形質転換体を培養する工程と、培養物よりエルゴチオナーゼを回収する工程を含むエルゴチオナーゼの製造方法。
〔11〕 エルゴチオネインに作用してチオウロカン酸を生成する能力を有する酵素活性物質を、被検対象に接触させる工程を含む、エルゴチオネインの定量方法。
〔12〕 前記酵素活性物質が、〔1〕〜〔4〕のいずれかに記載のエルゴチオナーゼ、〔5〕に記載のエルゴチオナーゼ活性を有するタンパク質、当該エルゴチオナーゼまたはエルゴチオナーゼ活性を有するタンパク質を生産する微生物、およびその処理物からなる群から選択される、〔11〕に記載のエルゴチオネインの定量方法。
〔13〕 エルゴチオナーゼおよび/または酵素活性物質を含むことを特徴とするエルゴチオネイン定量試薬、およびキット。
〔14〕 チオウロカン酸およびトリメチルアミンにエルゴチオナーゼおよび/または酵素活性物質を作用させることを特徴とする、エルゴチオネインの製造方法。
【発明の効果】
【0016】
本発明により、エルゴチオネインに特異的に作用するエルゴチオナーゼにより、特異的かつ簡便にエルゴチオネインを定量する方法が提供された。また、エルゴチオナーゼが提供された。さらに、エルゴチオナーゼをコードするDNAを単離し、当該酵素を高発現する組換え菌が提供された。本発明の方法は、エルゴチオナーゼの高い特異性を利用して、簡便にエルゴチオネインを定量できるため、簡易な方法が望まれる食品検査、臨床検査等において有用である。本発明によって、エルゴチオネインを特異的かつ簡便に定量することが可能となった。
【図面の簡単な説明】
【0017】
【図1】分子系統樹を示す図である。
【図2】エルゴチオナーゼの至適温度を検討した結果を示す図である。
【図3】エルゴチオナーゼの熱安定性について検討した結果を示す図である。
【図4】エルゴチオナーゼの至適pHについて検討した結果を示す図である。
【図5】エルゴチオナーゼの安定性に与えるpHの影響について検討した結果を示す図である。
【図6】エルゴチオナーゼによるエルゴチオネインの定量結果を示す図である。
【発明を実施するための形態】
【0018】
本発明は、次の(1)〜(2)に示す理化学的性状を有するエルゴチオナーゼを提供する;
(1)作用:L-エルゴチオネインに作用し、チオウロカン酸およびトリメチルアミンを生成する、および
(2)基質特異性:L-エルゴチオネインに作用し、D-, L-ヒスチジン、D-, L-チロシン、D-, L-フェニルアラニンに作用しない。
【0019】
本発明者らが見出したエルゴチオナーゼは、好ましくは前記性状(1)〜(2)に加えて、さらに次の(3)〜(8)に示す理化学的性状を有する;
(3)至適pH:7.0〜9.5、
(4)至適温度:50〜65℃、
(5)分子量:ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動(以下、SDS-PAGEと略す)による分子量が45,000〜55,000、ゲルろ過による分子量が200,000〜260,000、
(6)熱安定性: 60℃、30分で90%以上の活性を保持、
(7)pH安定性: pH 6.5〜11.0、30分で80%以上の活性を保持、および
(8)阻害剤: p-ヒドロキシ水銀安息香酸(p-hydroxymercuribenzoate)、ヨード酢酸(iodoacetate、グリセルアルデヒド3リン酸デヒドロゲナーゼ阻害剤)、L-システイン(L-cysteine)存在下において80%以上の活性を保持。
【0020】
なお上記(5)に記載の分子量において、より好ましくは、SDS-PAGEによる分子量が47,000〜51,000、ゲルろ過による分子量が230,000である。さらに好ましくは、SDS-PAGEによる分子量が49,000である。またアミノ酸配列から予測される分子量は、約55,000である。なお本発明者らが見出したエルゴチオナーゼは、活性状態では4量体(テトラマー)であると考えられる。
【0021】
本発明のエルゴチオナーゼは、動物、植物等特に限定されないが、例えば、微生物から得ることができる。本発明における好ましい微生物として、例えば、バークホルデリア属もしくはシュードモナス属に属する微生物を挙げることができる。これらバークホルデリア属もしくはシュードモナス属に属する微生物は、エルゴチオネインに作用し、チオウロカン酸を生成する。
本発明におけるエルゴチオネインは好ましくは、L-エルゴチオネインである。
【0022】
バークホルデリア属に属する微生物として、具体的には次の微生物を示すことができる。この微生物は、本発明において特異的にエルゴチオネインに作用し、チオウロカン酸を生成する好ましい微生物である。
バークホルデリア・エスピー HME 13(Burkholderia sp. HME 13)
【0023】
このバークホルデリア・エスピー HME 13は、以下のとおり寄託されている。以下に寄託を特定する内容を記載する。
(1)寄託機関の名称・あて名:
名称:独立行政法人製品評価技術基盤機構
あて名:日本国千葉県木更津市かずさ鎌足2−5−8(郵便番号292−0818)
(2)受託日(寄託日):2010年1月20日
(3)受託番号:
バークホルデリア・エスピー HME 13 (Burkholderia sp. HME 13) ;受託番号 NITE P-861
【0024】
また本発明において、エルゴチオネインに作用し、チオウロカン酸およびトリメチルアミンを生成する能力を有する微生物は、これらの寄託菌株のみならず、種々の微生物資源(microbiological resource)からも見出すことができる。微生物資源には、自然環境から単離される微生物群、微生物寄託機関に保存された微生物菌株などが含まれる。例えば本発明において、エルゴチオネインに作用し、チオウロカン酸およびトリメチルアミンを生成する能力を有する微生物として、好ましくは次のようなバークホルデリア属やシュードモナス属に属する微生物が挙げられる。
シュードモナス・エスピー(Pseudomonas sp.)
【0025】
即ち本発明は、バークホルデリア属もしくはシュードモナス属に属する微生物が下記の群から選択されるいずれかの微生物であることを特徴とするエルゴチオナーゼに関する;
バークホルデリア・エスピー NITE P-861(Burkholderia sp. NITE P-861)、および
シュードモナス・エスピー (Pseudomonas sp.)。
【0026】
本発明において、エルゴチオナーゼ活性(本明細書では単に「活性」、あるいは「本酵素活性」と記載する場合がある)とは、L-エルゴチオネインを基質として、チオウロカン酸およびトリメチルアミンを生成する酵素活性を言う。
【0027】
本発明では、本酵素活性を有する微生物であれば、任意の微生物を用いることができる。本発明に用いることができる微生物は、エルゴチオネインに作用しチオウロカン酸およびトリメチルアミンを生成する能力や作用を有することを確認することにより得ることができる。
【0028】
目的の能力や作用を有する微生物を選択するため、例えば、予め適当な培地中で被験微生物を培養し、生育した微生物、もしくは菌体処理物をエルゴチオネインと接触、反応させる。その結果、チオウロカン酸またはトリメチルアミンの生成が確認できれば、当該微生物を選択し本発明に用いることができる。
【0029】
このとき、被験微生物の培養時に、培地中にエルゴチオネインを加えておく、もしくはエルゴチオネインを単一窒素源として加えておけば、エルゴチオナーゼの誘導あるいは製造が期待できる。
【0030】
チオウロカン酸が生成されたか否かは、例えば以下の方法により測定することができる。
0.25 mM エルゴチオネイン、10 mM ピロリン酸ナトリウム緩衝液(pH 9.0)およびび酵素活性物質を含む 1 mLの反応液を用いて30℃で反応し、チオウロカン酸生成に由来する311 nmの吸光度の増加を測定する。1 Uは、上記条件下で1分間に1μmolのチオウロカン酸生成を触媒する酵素量とし、チオウロカン酸の分子吸光係数(ε=18,000)より計算する。
【0031】
例えば、微生物寄託機関に保存されたバークホルデリア属やシュードモナス属微生物を対象として、上記選択方法によって、目的の作用を有する微生物を選択することができる。
寄託された微生物は、例えば次のような寄託機関から分譲を受けることができる;
生物遺伝資源センター(NBRC)、
理化学研究所(JCM)、
東京農業大学菌株保存室(IAM)、
American Type Culture Collection(ATCC)、および
Deutsche Sammlung von Mikroorganismen(DSM)。
【0032】
あるいはリボソーマルDNA(rDNA)の塩基配列情報に基づいて、本発明に利用しうる微生物を選択することもできる。一般にrDNAの塩基配列情報は、生物間の遺伝学的な近さを表す指標として有用であることが広く知られている。特に16S rRNAをコードするDNA(以下16S rDNAと記載する)は、微生物の遺伝的な同一性あるいは類似性を決定するためのツールとして活用されている。16S rRNAは、全生物に存在しており、生物間で構造の違いは見られるが、アライメント可能な程度に配列が保存されている。16S rRNAの生物間水平伝播の頻度は低いので、分類に使用する方法が汎用されている。
【0033】
具体的には、小サブユニットリボソームRNA(SSU rRNA)は、タンパク質合成の場であるリボソームを構成する核酸分子の1つである。その塩基配列の長さは、多少の違いはあるものの、細菌からヒトまで起源は同じものとみなされている。したがって、rRNAの塩基配列は進化的保存性が高く、生物の系統解析において最も頻繁に利用されている(Microbiol. Rev., 58, 1-9 (1994))。原核生物におけるSSU rRNA は約1500塩基の16S rRNAである。原核生物の分類と同定に16S rRNA の塩基配列を利用する系統分類が一般に用いられている(ASM News, 65, 752-757 (1999)、生物工学実験書 p113 日本生物工学会編, 培風館)。
【0034】
本発明者らが見出した上記の菌株は、16S rDNA中に配列番号:5に示す塩基配列を含んでいた。塩基配列情報データベースを対象として、これらの塩基配列情報を検索すれば、既に16S rDNAが決定された微生物の中から、その16S rDNA中に同一性の高い塩基配列を含む菌株を見つけ出すことができる。例えばDNA Data Bank of Japan(DDBJ)などの公共データベースサービス機関では、16S rRNA(Prokaryotes)の塩基配列情報を対象とする検索サービスが提供されている。こうして見出された微生物のうち、特にバークホルデリア属やシュードモナス属に属する微生物を選択することによって、本発明に利用しうる微生物とすることができる。
【0035】
これら微生物は、醗酵学の分野で公知の方法に従って培養することができる。培地としては炭素源、窒素源、無機物およびその他の栄養素を適量含有する培地ならば、合成培地または天然培地のいずれでも使用することができる。培地は、液体培地または固体培地を使用することができる。
【0036】
具体的には、炭素源として、次に示すような一般的な炭素源より、使用する微生物の資化性を考慮して、適宜一種または二種以上選択して使用する。
糖類:グルコース、フルクトース、マルトース、スクロース、ラクトースなど
デンプン、
デンプン加水分解物、
廃糖蜜、
アルコール:グリセロール、メタノール、エタノールなど
糖アルコール、
有機酸:酢酸、グルコン酸、ピルビン酸、α-ケトグルタル酸、クエン酸など
脂肪類:パーム油、大豆油、オリーブオイルなど
炭化水素類:ノルマルパラフィンなど
【0037】
窒素源としては、次に示すような一般的な窒素源の中から、使用する微生物の資化性を考慮して、適宜一種または二種以上選択して使用する。
有機窒素化合物:例えば、肉エキス、ペプトン、酵母エキス、麦芽エキス、大豆加水分解物、ミルクカゼイン、カザミノ酸、廃糖蜜、アスパラギン、グルタミン、あるいはグルタミン酸などのアミノ酸類やコーンスティープリカー、NZアミンなど
無機窒素化合物:アンモニア、硝酸アンモニウム、硫酸アンモニウムなど、尿素、硝酸塩など
また、エルゴチオナーゼの誘導を目的として、エルゴチオネインを単一の窒素源として使用することができる。
【0038】
さらに、無機塩として微量のマグネシウム、マンガン、カリウム、カルシウム、ナトリウム、銅、亜鉛などの塩や各種ビタミンなどの微量栄養素を添加することもできる。また、必要に応じて植物油、界面活性剤、シリコンなどの消泡剤を培養液中に添加することもできる。
【0039】
培養は前記培地成分を含有する液体培地中で、通常の培養方法を用いて行うことができる。例えば次のような培養方法を利用することができる。
振とう培養(shaking culture)
通気攪拌培養(aeration-agitation culture)
連続培養(continuous culture)
流加培養(feeding culture, fed batch culture)
【0040】
培養条件は、微生物の種類、培養の種類、培養方法により適宜選択することができる。利用する菌株が増殖し、エルゴチオナーゼを生成しうる条件であれば特に制限はない。一般的な培養条件として次のような条件を示すことができる。
培養開始時のpHを4から10、好ましくは6から8に調節
15から70℃、好ましくは20から40℃の温度条件下で培養
培養時間はエルゴチオナーゼ活性を有する菌体を得ることができれば特に制限されない。通常は1日から14日、好ましくは1日から7日培養する。
【0041】
本発明において、微生物菌体の処理物とは、菌体を目的とする酵素活性が維持される条件で処理したものを意味する。酵素活性の維持とは、生菌体で得られる酵素活性の一般的には20%以上、通常30%以上、好ましくは50%以上、より好ましくは70%以上の活性を維持することを言う。生菌体とその処理物の酵素活性は、生菌体の酵素活性と、同量の生菌体から得られた処理物の酵素活性を比較することによって定量的に比較することができる。本発明における処理物は、生菌体よりも高い酵素活性を有する場合もある。
【0042】
このような処理物を得る方法としては、凍結乾燥、有機溶媒による脱水乾燥、菌体の自己消化、酵素活性画分の抽出、超音波処理などを利用することができる。このような処理方法によって、例えば、凍結乾燥菌体、アセトン乾燥菌体、菌体自己消化物、菌体抽出物、菌体磨砕物、菌体の超音波処理物等を得ることができる。
【0043】
これらの処理は、単独で、あるいは複数の処理を重複して適用することもできる。例えば適当な培養液で微生物を培養後、菌体を破砕して、遠心分離等により無細胞抽出液を調製することができる。菌体は、機械的な破砕、超音波、高圧処理、酵素消化、自己消化などにより破砕することができる。無細胞抽出液は本発明における菌体の処理物に含まれる。
【0044】
さらに本発明においては、当該菌体から当該反応を触媒する酵素を部分精製したものや完全に精製したものなどを得ることができる。
すなわち本発明は、本発明によって提供されたエルゴチオナーゼ活性を有する微生物の菌体から精製することができるエルゴチオナーゼに関する。
【0045】
本発明のエルゴチオナーゼは、上記本発明の微生物、あるいはその無細胞抽出液などから精製することができる。具体的には、次のような精製工程を適宜組み合わせることによって、無細胞抽出液から実質的に純粋なエルゴチオナーゼを得ることができる;
硫安、アセトンなどを用いた溶解度による分割、
イオン交換クロマトグラフィー、
疎水クロマトグラフィー、
2',5'-ADP セファロースやブルーもしくはレッドセファロースなどを用いたアフィニティクロマトグラフィー、あるいは
ゲルろ過。
【0046】
例えば、微生物を集菌後、次のような精製工程を経て、本発明のエルゴチオナーゼを実質的に純粋な酵素として単離することができる;
超音波による菌体の破砕と無細胞抽出液の調製、
硫安分画によるタンパク画分の分離、
DEAE-トヨパールを用いたイオン交換クロマトグラフィー、および
ブチル-トヨパールを用いた疎水クロマトグラフィーによるエルゴチオナーゼ活性分画の単離。
【0047】
本発明において、「実質的に純粋」とは、当該酵素が、当該酵素以外の生物学的な高分子化合物や化学物質を実質的に含まないことを言う。当該酵素以外の生物学的な高分子化合物とは、例えば菌体や培養液に由来するタンパク、糖類、脂質、核酸類が含まれる。本発明における実質的に純粋な酵素は、少なくとも75%以上、通常80%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましくは95%以上、あるいは99%以上の純度を有する。酵素の純度を決定する方法は公知である。例えば、各種のクロマトグラフィーや、ポリアクリルアミドゲル電気泳動などによって蛋白質の純度を決定する方法が周知である。
【0048】
あるいは本発明によって、上記理化学的性状(1)〜(2)を有する単離されたエルゴチオナーゼが提供される。本発明において、単離されたエルゴチオナーゼとは、それが存在する天然の環境から分離されて存在していることを意味する。したがって、菌体から分離されたエルゴチオナーゼは、本発明における単離されたエルゴチオナーゼに含まれる。
【0049】
本発明における実質的に純粋なエルゴチオナーゼ、あるいは単離されたエルゴチオナーゼは、精製、あるいは単離された後に、酵素組成物とすることもできる。例えば、精製、あるいは単離されたエルゴチオナーゼを適当な担体と配合することによって、本発明のエルゴチオナーゼを含む酵素組成物を調製することができる。
【0050】
担体としては、アルブミンなどの不活性蛋白質、ショ糖などの糖類あるいは糖アルコールなどを利用することができる。酵素組成物は、液状であることもできるし、乾燥状態であることもできる。酵素と担体を含む水溶液を凍結乾燥することによって得られる酵素組成物は、本発明における好ましい組成物の一つである。
【0051】
本発明のエルゴチオナーゼは、以下の方法によりその活性を定量することができる。すなわち、次の組成の反応液中(1 mL)で、30℃で反応し、チオウロカン酸生成に由来する311 nm の吸光度の増加を測定する。
0.25 mM エルゴチオネイン
10 mM ピロリン酸ナトリウム緩衝液 (pH 9.0) および
酵素
1 Uは、上記条件下で1分間に1μmol のチオウロカン酸生成を触媒する酵素量とし、チオウロカン酸の分子吸光係数(ε=18,000)より計算する。
【0052】
また本発明は、下記(a)から(e)のいずれかに記載のエルゴチオナーゼ活性を有するタンパク質を提供する;
(a)配列番号:3に記載のアミノ酸配列を含むタンパク質、
(b)配列番号:4に記載された塩基配列を含むポリヌクレオチドによってコードされるタンパク質、
(c)配列番号:3に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含むタンパク質、
(d)配列番号:4に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質、
(e)配列番号:3に記載のアミノ酸配列と80%以上の相同性を有するアミノ酸配列を含むタンパク質。
【0053】
本発明において、ポリヌクレオチドは、DNAやRNA等の天然のポリヌクレオチドに加え、人工的なヌクレオチド誘導体を含む人工的な分子であることもできる。また本発明のポリヌクレオチドは、DNA-RNAのキメラ分子であることもできる。
【0054】
本発明のエルゴチオナーゼ活性を有するタンパク質にはシグナル配列が含まれていない。
本発明のエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドは、例えば配列番号:4に示す塩基配列を含む。配列番号:4に示す塩基配列は、配列番号:3に示すアミノ酸配列を含むタンパク質をコードしており、このアミノ酸配列を含むタンパク質は、本発明によるエルゴチオナーゼの好ましい態様を構成する。
【0055】
本発明のエルゴチオナーゼ活性を有するタンパク質には、本発明のエルゴチオナーゼをコードするポリヌクレオチドまたは本発明のエルゴチオナーゼ活性を有するタンパク質のホモログも含まれる。
【0056】
本発明のエルゴチオナーゼをコードするポリヌクレオチドのホモログとは、配列番号:3に記載のアミノ酸配列に1もしくは複数のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列を含み、かつ、配列番号:3に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質を意味する。本発明において、配列番号:3に記載のアミノ酸配列からなるタンパク質と機能的に同等とは、当該タンパク質が前記理化学的性質(1)〜(2)を有することを意味する。改変されるアミノ酸の数は、改変後のタンパク質が、前記理化学的性質を有している限り、特に制限はないが、一般的には、50アミノ酸以内、好ましくは30アミノ酸以内、より好ましくは10アミノ酸以内(例えば、5アミノ酸以内、3アミノ酸以内)である。あるいは、アミノ酸配列全体のうち、例えば20%以下、具体的には10%以下(例えば、10、9、8、7、6、5、4、3、2、または1%以下)のアミノ酸残基の改変は許容される。すなわち、配列番号:3に記載のアミノ酸配列と、好ましくは80%以上、より好ましくは90%以上(例えば、90、91、92、93、94、95、96、97、98、99%以上)の相同性を有するアミノ酸配列を含むタンパク質も、本発明のエルゴチオナーゼ活性を有するタンパク質に含まれる。
【0057】
一般にタンパク質の機能の維持のためには、置換するアミノ酸は、置換前のアミノ酸と類似の性質を有するアミノ酸であることが好ましい。このようなアミノ酸残基の置換は、保存的置換と呼ばれている。例えば、Ala、Val、Leu、Ile、Pro、Met、Phe、Trpは、共に非極性アミノ酸に分類されるため、互いに似た性質を有する。また、非荷電性アミノ酸としては、Gly、Ser、Thr、Cys、Tyr、Asn、Glnが挙げられる。また、酸性アミノ酸としては、AspおよびGluが挙げられる。また、塩基性アミノ酸としては、Lys、Arg、Hisが挙げられる。これらの各グループ内のアミノ酸置換は許容される。
【0058】
そして当業者であれば、配列番号:4に記載の塩基配列を含むポリヌクレオチドに部位特異的変異導入法(Nucleic Acid Res. 10,pp.6487 (1982) , Methods in Enzymol.100,pp.448 (1983), Molecular Cloning 2ndEdt., Cold Spring Harbor Laboratory Press (1989) , PCR A Practical Approach IRL Press pp.200 (1991) などを用いて、適宜置換、欠失、挿入、および/または付加変異を導入することによりポリヌクレオチドのホモログを得ることが可能である。そのホモログをコードするポリヌクレオチドを宿主に導入して発現させることにより、配列番号:3に記載のアミノ酸配列からなるタンパク質のホモログを得ることが可能である。
【0059】
また、本発明におけるポリヌクレオチドのホモログは、配列番号:4に示される塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズできるポリヌクレオチドであって、かつ、前記理化学的性質(1)〜(2)を有するタンパク質をコードするポリヌクレオチドも含む。ストリンジェントな条件でハイブリダイズできるポリヌクレオチドとは、配列番号:4に記載の塩基配列中の任意の少なくとも20個、好ましくは少なくとも30個、例えば40、60または100個の連続した配列を一つまたは複数選択したDNAをプローブDNAとし、例えばECL direct nucleic acid labeling and detection system (GEヘルスケア・ジャパン社製)を用いて、マニュアルに記載の条件(wash:42℃、0.5x SSCを含むprimary wash buffer)において、ハイブリダイズするポリヌクレオチドを指す。
【0060】
より具体的な「ストリンジェントな条件」とは、例えば、通常、42℃、2×SSC、0.1%SDSの条件であり、好ましくは50℃、2×SSC 、0.1%SDSの条件であり、さらに「高度にストリンジェントな条件」とは例えば、65℃、0.1×SSCおよび0.1%SDSの条件であるが、これらの条件に特に制限されない。ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度や塩濃度など複数の要素が考えられ、当業者であればこれら要素を適宜選択することで最適なストリンジェンシーを実現することが可能である。
【0061】
さらに、本発明におけるポリヌクレオチドのホモログは、配列番号:3に示されるアミノ酸配列と少なくとも70%、好ましくは少なくとも80%、より好ましくは90%以上(90、91、92、93、94、95、96、97、98、または99%以上)のホモロジーを有するタンパク質をコードするポリヌクレオチドを含む。タンパク質のホモロジー検索は、例えばSWISS-PROT、PIR、DADなどのタンパク質のアミノ酸配列に関するデータベースやDDBJ、EMBL、あるいはGene-BankなどのDNA配列に関するデータベース、DNA配列を基にした予想アミノ酸配列に関するデータベースなどを対象に、BLAST、FASTAなどのプログラムを利用して、例えば、インターネットを通じて行うことができる。
【0062】
また、本発明におけるタンパク質には、配列番号:3に記載のアミノ酸配列にシグナル配列を含めた配列番号:1に記載のアミノ酸配列を含むタンパク質も含まれる。
また、本発明におけるポリヌクレオチドには、配列番号:4に記載の塩基配列にシグナル配列を含めた配列番号:2に記載の塩基配列を含むポリヌクレオチドも含まれる。
【0063】
また本発明は、本発明のエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドに関する。本発明のエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドは、例えば、以下のような方法によって単離することができる。
【0064】
配列番号:4に記載の塩基配列を基にPCR用のプライマーを設計し、酵素生産株の染色体DNAもしくは、cDNAライブラリーを鋳型としてPCRを行うことにより本発明のポリヌクレオチドを得ることができる。
【0065】
さらに、得られたDNA断片をプローブとして、酵素生産株の染色体DNAの制限酵素消化物をファージ、プラスミドなどに導入し、大腸菌を形質転換して得られたライブラリーやcDNAライブラリーを利用して、コロニーハイブリダイゼーション、プラークハイブリダイゼーションなどにより、本発明のポリヌクレオチドを得ることができる。
【0066】
また、PCRにより得られたDNA断片の塩基配列を解析し、得られた配列から、既知のDNAの外側に伸長させるためのPCRプライマーを設計し、酵素生産株の染色体DNAを適当な制限酵素で消化後、自己環化反応によりDNAを鋳型として逆PCRを行うことにより(Genetics 120, 621-623 (1988))、また、RACE法(Rapid Amplification of cDNA End、「PCR実験マニュアル」p25-33, HBJ出版局)などにより本発明のポリヌクレオチドを得ることも可能である。
なお本発明のポリヌクレオチドには、以上のような方法によってクローニングされたゲノムDNA、あるいはcDNAの他、合成によって得られたDNAが含まれる。
【0067】
また本発明は、上記本発明のポリヌクレオチドが挿入された発現ベクターを提供する。例えば、上述のようにして単離されたエルゴチオナーゼをコードするポリヌクレオチド、またはエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドを公知の発現ベクターに挿入することにより、エルゴチオナーゼ発現ベクターが提供される。
【0068】
また本発明は、本発明の発現ベクターにより形質転換された形質転換体に関する。この発現ベクターで形質転換した形質転換体を培養することにより、本発明のエルゴチオナーゼを形質転換体(組換え体)から得ることができる。
【0069】
本発明においてエルゴチオナーゼを発現させるために、形質転換の対象となる微生物は、エルゴチオナーゼを有するポリペプチドをコードするポリヌクレオチド、またはエルゴチオナーゼ活性を有するタンパク質をコードするポリヌクレオチドを含む発現(組換え)ベクターにより形質転換され、エルゴチオナーゼ活性を発現することができる生物であれば特に制限はない。利用可能な微生物としては、例えば以下のような微生物を示すことができる。
エシェリヒア(Escherichia)属
バチルス(Bacillus)属
シュードモナス(Pseudomonas)属
セラチア(Serratia)属
ブレビバクテリウム(Brevibacterium)属
コリネバクテリイウム(Corynebacterium)属
ストレプトコッカス(Streptococcus)属
ラクトバチルス(Lactobacillus)属など宿主ベクター系の開発されている細菌
ロドコッカス(Rhodococcus)属
ストレプトマイセス(Streptomyces)属など宿主ベクター系の開発されている放線菌
サッカロマイセス(Saccharomyces)属
クライベロマイセス(Kluyveromyces)属
シゾサッカロマイセス(Schizosaccharomyces)属
チゴサッカロマイセス(Zygosaccharomyces)属
ヤロウイア(Yarrowia)属
トリコスポロン(Trichosporon)属
ロドスポリジウム(Rhodosporidium)属
ピキア(Pichia)属
キャンディダ(Candida)属などの宿主ベクター系の開発されている酵母
ノイロスポラ(Neurospora)属
アスペルギルス(Aspergillus)属
セファロスポリウム(Cephalosporium)属
トリコデルマ(Trichoderma)属などの宿主ベクター系の開発されているカビ
【0070】
形質転換体の作製のための手順および宿主に適合した組換えベクターの構築は、分子生物学、生物工学、遺伝子工学の分野において慣用されている技術に準じて行うことができる(例えば、Sambrookら、モレキュラー・クローニング、Cold Spring Harbor Laboratories)。微生物中などにおいて、本発明のエルゴチオナーゼ遺伝子を発現させるためには、まず微生物中において安定に存在するプラスミドベクターやファージベクター中にこのDNAを導入し、その遺伝情報を転写・翻訳させる必要がある。
【0071】
そのためには、転写・翻訳を制御するユニットにあたるプロモーターを本発明のDNA鎖の5'-側上流に、より好ましくはターミネーターを3'-側下流に、それぞれ組み込めばよい。このプロモーター、ターミネーターとしては、宿主として利用する微生物中において機能することが知られているプロモーター、ターミネーターを用いる必要がある。これら各種微生物において利用可能なベクター、プロモーター、ターミネータ−などに関して「微生物学基礎講座8遺伝子工学・共立出版」、特に酵母に関しては、Adv. Biochem. Eng. 43, 75-102 (1990)、Yeast 8, 423-488 (1992)、などに詳細に記述されている。
【0072】
例えばエシェリヒア属、特に大腸菌エシェリヒア・コリ(Escherichia coli)においては、プラスミドベクターとして、pBR、pUC系プラスミドを利用でき、lac(β−ガラクトシダーゼ)、trp(トリプトファンオペロン)、tac、 trc (lac、trpの融合)、λファージ PL、PRなどに由来するプロモーターなどが利用できる。また、ターミネーターとしては、trpA由来、ファージ由来、rrnBリボソーマルRNA由来のターミネーターなどを用いることができる。これらの中で、市販のpSE420(Invitrogen製)のマルチクローニングサイトを一部改変したベクターpSE420D(特開2000-189170に記載)が好適に利用できる。
【0073】
バチルス属においては、ベクターとしてpUB110系プラスミド、pC194系プラスミドなどが利用可能であり、染色体にインテグレートすることもできる。また、プロモーター、ターミネーターとしてapr(アルカリプロテアーゼ)、npr(中性プロテアーゼ)、amy(α−アミラーゼ)などが利用できる。
【0074】
シュードモナス属においては、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・セパシア(Pseudomonas cepacia)などで宿主ベクター系が開発されている。トルエン化合物の分解に関与するプラスミドTOLプラスミドを基本にした広宿主域ベクター(RSF1010などに由来する自律的複製に必要な遺伝子を含む)pKT240などが利用可能であり、プロモーター、ターミネーターとして、リパーゼ(特開平5-284973)遺伝子などが利用できる。
【0075】
ブレビバクテリウム属、特に、ブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)においては、pAJ43(Gene 39, 281 (1985))などのプラスミドベクターが利用可能である。プロモーター、ターミネーターとしては、大腸菌で使用されているプロモーター、ターミネーターがそのまま利用可能である。
【0076】
コリネバクテリウム属、特にコリネバクテリウム・グルタミカム(Corynebacterium glutamicum)においては、pCS11(特開昭57-183799)、pCB101(Mol. Gen. Genet. 196, 175 (1984)などのプラスミドベクターが利用可能である。
【0077】
ストレプトコッカス(Streptococcus)属においては、pHV1301(FEMS Microbiol. Lett. 26, 239 (1985)、pGK1(Appl. Environ. Microbiol. 50, 94 (1985))などがプラスミドベクターとして利用可能である。
【0078】
ラクトバチルス(Lactobacillus)属においては、ストレプトコッカス属用に開発されたpAMβ1(J. Bacteriol. 137, 614 (1979))などが利用可能であり、プロモーターとして大腸菌で利用されているものが利用可能である。
【0079】
ロドコッカス(Rhodococcus)属においては、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)から単離されたプラスミドベクターが使用可能である (J. Gen. Microbiol. 138,1003 (1992) )。
【0080】
ストレプトマイセス(Streptomyces)属においては、HopwoodらのGenetic Manipulation of Streptomyces: A Laboratory Manual Cold Spring Harbor Laboratories (1985)に記載の方法に従って、プラスミドを構築することができる。特に、ストレプトマイセス・リビダンス(Streptomyces lividans)においては、pIJ486 (Mol. Gen. Genet. 203, 468-478, 1986)、pKC1064(Gene 103,97-99 (1991) )、pUWL-KS (Gene 165,149-150 (1995) )が使用できる。また、ストレプトマイセス・バージニア(Streptomyces virginiae)においても、同様のプラスミドを使用することができる(Actinomycetol. 11, 46-53 (1997) )。
【0081】
サッカロマイセス(Saccharomyces)属、特にサッカロマイセス・セレビジアエ(Saccharomyces cerevisiae)においては、YRp系、YEp系、YCp系、YIp系プラスミドが利用可能であり、染色体内に多コピー存在するリボソームDNAとの相同組換えを利用したインテグレーションベクター(EP 537456など)は、多コピーで遺伝子を導入でき、かつ安定に遺伝子を保持できるため極めて有用である。また、ADH(アルコール脱水素酵素)、GAPDH(グリセルアルデヒド−3−リン酸脱水素酵素)、PHO(酸性フォスファターゼ)、GAL(β−ガラクトシダーゼ)、PGK(ホスホグリセレートキナーゼ)、ENO(エノラーゼ)などのプロモーター、ターミネーターが利用可能である。
【0082】
クライベロマイセス属、特にクライベロマイセス・ラクティス(Kluyveromyces lactis)においては、サッカロマイセス・セレビジアエ由来2μm系プラスミド、pKD1系プラスミド(J. Bacteriol. 145, 382-390 (1981))、キラー活性に関与するpGKl1由来プラスミド、クライベロマイセス属における自律増殖遺伝子KARS系プラスミド、リボソームDNAなどとの相同組換えにより染色体中にインテグレート可能なベクタープラスミド(EP 537456など)などが利用可能である。また、ADH、PGKなどに由来するプロモーター、ターミネーターが利用可能である。
【0083】
シゾサッカロマイセス(Schizosaccharomyces)属においては、シゾサッカロマイセス・ポンベ由来のARS (自律複製に関与する遺伝子)およびびサッカロマイセス・セレビジアエ由来の栄養要求性を相補する選択マーカーを含むプラスミドベクターが利用可能である(Mol. Cell. Biol. 6, 80 (1986))。また、シゾサッカロマイセス・ポンベ由来のADHプロモーターなどが利用できる(EMBO J. 6, 729 (1987))。特に、pAUR224は、タカラバイオから市販されており容易に利用できる。
【0084】
チゴサッカロマイセス属(Zygosaccharomyces)においては、チゴサッカロマイセス・ロウキシ(Zygosaccharomyces rouxii)由来のpSB3(Nucleic Acids Res. 13, 4267 (1985))などに由来するプラスミドベクターが利用可能であり、サッカロマイセス・セレビジアエ由来 PHO5 プロモーターや、チゴサッカロマイセス・ロウキシ由来 GAP-Zr(グリセルアルデヒド−3−リン酸脱水素酵素)のプロモーター(Agri. Biol. Chem. 54, 2521 (1990))などが利用可能である。
【0085】
ピキア(Pichia)属においては、ピキア・アンガスタ(旧名:ハンゼヌラ・ポリモルファ(Hansenula polymorpha))において宿主ベクター系が開発されている。ベクターとしては、ピキア・アンガスタ由来自律複製に関与する遺伝子(HARS1、HARS2)も利用可能であるが、比較的不安定であるため、染色体への多コピーインテグレーションが有効である(Yeast 7, 431-443 (1991))。また、メタノールなどで誘導されるAOX(アルコールオキシダーゼ)、FDH(ギ酸脱水素酵素)のプロモーターなどが利用可能である。また、ピキア・パストリス(Pichia pastoris)などにピキア由来自律複製に関与する遺伝子 (PARS1、 PARS2)などを利用した宿主ベクター系が開発されており(Mol. Cell. Biol. 5, 3376 (1985))、高濃度培養とメタノールで誘導可能なAOXなど強いプロモーターが利用できる(Nucleic Acids Res. 15, 3859 (1987))。
【0086】
キャンディダ(Candida)属においては、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・アルビカンス(Candida albicans)、キャンディダ・トロピカリス(Candida tropicalis)、キャンディダ・ウチルス(Candida utilis)などにおいて宿主ベクター系が開発されている。キャンディダ・マルトーサにおいてはキャンディダ・マルトーサ由来ARSがクローニングされ(Agri. Biol. Chem. 51, 51, 1587 (1987))、これを利用したベクターが開発されている。また、キャンディダ・ウチルスにおいては、染色体インテグレートタイプのベクターは強力なプロモーターが開発されている(特開平 08-173170)。
【0087】
アスペルギルス(Aspergillus)属においては、アスペルギルス・ニガー (Aspergillus niger) 、アスペルギルス・オリジー (Aspergillus oryzae) などがカビの中で最もよく研究されており、プラスミドや染色体へのインテグレーションが利用可能であり、菌体外プロテアーゼやアミラーゼ由来のプロモーターが利用可能である(Trends in Biotechnology 7, 283-287 (1989))。
【0088】
トリコデルマ(Trichoderma)属においては、トリコデルマ・リーゼイ(Trichoderma reesei)を利用したホストベクター系が開発され、菌体外セルラーゼ遺伝子由来プロモーターなどが利用できる(Biotechnology 7, 596-603 (1989))。
【0089】
また、微生物以外でも、植物、動物において様々な宿主・ベクター系が開発されており、特に蚕を用いた昆虫(Nature 315, 592-594 (1985))や菜種、トウモロコシ、ジャガイモなどの植物中に大量に異種タンパク質を発現させる系が開発されており、好適に利用できる。
【0090】
本発明において使用するエルゴチオナーゼ生産能を有する微生物は、エルゴチオナーゼ生産能を有する全ての菌株、突然変異株、変種、遺伝子操作技術の利用により作成された本発明の酵素生産能を獲得した形質転換株を含む。
【0091】
また本発明は、エルゴチオナーゼの製造方法を提供する。本製造方法には、バークホルデリア属もしくはシュードモナス属に属しエルゴチオナーゼ活性を有する微生物、もしくは上記本発明の形質転換体を培養する工程と、培養物よりエルゴチオナーゼを回収する工程が含まれる。具体的な培養工程および回収工程は、上述のとおりである。
【0092】
また本発明は、エルゴチオネインに作用してチオウロカン酸を生成する能力を有する酵素活性物質(enzymatic active materials)を、被検対象に接触させる工程を含む、エルゴチオネインの定量方法に関する。
本発明のエルゴチオネインの定量方法は、エルゴチオナーゼ活性を有する酵素活性物質を、この酵素活性物質によってエルゴチオネインからチオウロカン酸を生成することが可能な条件下で、被検対象に接触させる工程を含む。
【0093】
本発明において被検対象としては、下記(1)〜(2)を挙げることができる。
(1)被験者(クローン病患者を含む)および臨床検体等から単離されたあらゆる生体由来物質(組織、臓器、体液、血液、細胞等)、および
(2)エルゴチオネイン、エルゴチオネイン含有物質、またはその原料や処理物(例えば、一般食品、健康食品、美容食品等の食品、化粧品、生体材料、カビ類、キノコ類、マイコバクテリア等)。
【0094】
本発明において、酵素活性物質とは、酵素活性を有する物質を指し、例えばエルゴチオナーゼ活性を有する物質を挙げることができる。具体的には、エルゴチオネインに作用してチオウロカン酸およびトリメチルアミンを生成する能力を有する物質であり、より好ましくは微生物に由来する物質であって、当該能力が維持された物質である。
【0095】
本発明の酵素活性物質は、例えば、本発明のエルゴチオナーゼ(エルゴチオナーゼ活性を有するタンパク質を含む)、当該エルゴチオナーゼを生産する微生物、およびその処理物からなる群から選択することができる。より具体的には、例えば次の要素が、エルゴチオナーゼ活性を維持する限り、本発明の酵素活性物質に含まれる。
i) エルゴチオネインに作用し、チオウロカン酸およびトリメチルアミンを生成する能力を有する微生物
ii) i)の微生物の菌体、菌体培養物、菌体もしくは菌体培養物の処理物(菌体もしくは菌体培養物の乾燥物や濃縮物、菌体培養液から菌体を除去した上清、菌体自体の乾燥物や濃縮物等)
iii) i)またはii)から単離された物質
なお本発明における酵素活性物質は、精製された標品である必要はない。
【0096】
本発明において、微生物は、培養物であることもできるし、菌体であることもできる。培養物は、微生物の生存と増殖を支持する培地とともに当該微生物が存在する状態を言う。例えば微生物を含む液体培地(培養液)は、培養物である。一方、本発明においては、微生物そのものを指す用語として「菌体」(microbial cell)を用いる。菌体は、培養物から回収された微生物細胞を指す。例えば遠心分離によって液体培地から微生物の菌体を回収することができる。培養物から回収された菌体を洗浄することによって、菌体表面の培地成分を除くこともできる。
【0097】
さらに別の態様においては、本発明における酵素活性物質は、上記i)の微生物から単離することができるエルゴチオナーゼを含む。また当該エルゴチオナーゼを担体に固定化したものも、エルゴチオナーゼ活性を維持する限り、酵素活性物質に含まれる。
【0098】
本発明において酵素活性物質は、そのまま、あるいは固定化して基質化合物と接触させることができる。微生物菌体あるいはその処理物を固定化するための種々の方法が公知である。例えば、以下のような方法が微生物やその処理物の固定化方法として公知である。
ポリアクリルアミド法、
含硫多糖ゲル法(κ-カラギーナンゲル法など)、
アルギン酸ゲル法、
寒天ゲル法、
イオン交換樹脂法
あるいは、限外ろ過膜等を用いたメンブレンバイオリアクター中で、微生物、その処理物、あるいは酵素を基質化合物と接触させて反応させることもできる。
【0099】
本発明におけるエルゴチオネイン定量方法は、微生物を酵素が誘導される適切な前記培養条件において培養を行い、得られた培養液、あるいは培養液から採取した菌体や該菌体処理物、並びに酵素を、上記被検対象に接触させ、インキュベートすることにより行うことができる。例えば、エルゴチオナーゼ活性により生成されたチオウロカン酸を上述の方法で定量することで、被検対象に含有されていたエルゴチオネインの定量を行うことができる。具体的には、チオウロカン酸の生成量(311 nmの吸光度の増加)を測定し、被検物質中のエルゴチオネイン濃度を計算することによって行われる。実際に測定する際には、例えば検量線を作成し、検量線の直線性を確認の上で、最終生成されたチオウロカン酸生成に由来する311 nm の吸光度の値から、被検対象に含有されていたエルゴチオネインの定量を行うという手法をとることが可能である。
【0100】
反応条件としては、例えば以下の条件を示すことができる。
pH 5.0から11.0、好ましくはpH 7.0から9.5、
温度25から80℃、好ましくは25から60℃
【0101】
すなわち本発明は、下記(1)〜(2)の工程を含むエルゴチオネインの定量方法を提供する;
(1) エルゴチオネインに作用してチオウロカン酸およびトリメチルアミンを生成する能力を有する酵素活性物質(enzymatic active materials)を、被検対象に接触させる工程、および
(2) 生成されたチオウロカン酸またはトリメチルアミンを定量する工程。
【0102】
また本発明は、エルゴチオナーゼまたは酵素活性物質を含むことを特徴とするエルゴチオネイン定量試薬およびキットを提供する。本定量試薬およびキットにより、エルゴチオネインの定量を行うことができる。
【0103】
本発明の定量試薬やキットにおいては、有効成分であるエルゴチオナーゼまたは酵素活性物質以外に、例えば、滅菌水、生理食塩水、植物油、界面活性剤、脂質、溶解補助剤、緩衝剤、タンパク質安定剤(BSAやゼラチンなど)、保存剤、ブロッキング溶液、反応溶液、反応停止液、試料を処理するための試薬等が必要に応じて混合されていてもよい。さらにキットには、その他、使用方法を記載した指示書等をパッケージしておくこともできる。
【0104】
また本発明は、エルゴチオネインの製造方法を提供する。本製造方法には、チオウロカン酸およびトリメチルアミンにエルゴチオナーゼまたは酵素活性物質を作用させる工程が含まれる。
【0105】
なお本明細書において引用されたすべての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0106】
以下、実施例により本発明を詳細に説明するが、本発明はこれに限定されない。
【0107】
〔参考例1〕
非特許文献4(J. Biol. Chem., 237, 874-881 (1962))に記載の培地を用い、大腸菌W株を0.02% 硫酸マグネシウム・7水和物、0.2% クエン酸、1% K2HPO4、0.5% グルコース及び5 mM エルゴチオネインを含有する培地(以下、エルゴチオネイン培地, pH 7.2)で37℃、5日間培養したが菌の生育は見られなかった。
【0108】
LB培地(1% Bacto-Trypton(Difco製)、0.5% Bacto-Yeast extract(Difco製)、1% 塩化ナトリウム、pH 7.2)で培養後、菌体を生理食塩水で洗浄し、エルゴチオネイン培地で懸濁し、さらに37℃、5日間誘導培養した後に菌を回収し、超音波破砕後の無細胞抽出液を用いて酵素活性の測定を行ったが、エルゴチオナーゼ活性は検出されなかった。
【0109】
[エルゴチオナーゼ活性の測定]
0.25 mM エルゴチオネイン、10 mM ピロリン酸ナトリウム緩衝液(pH 9.0)および酵素を含む1 mLの反応液を用いて30℃で反応し、チオウロカン酸生成に由来する311 nmの吸光度の増加を測定した。1 U は、上記条件下で1分間に1μmolのチオウロカン酸生成を触媒する酵素量とし、チオウロカン酸の分子吸光係数(ε=18,000)より計算した。
【0110】
〔実施例1〕スクリーニング
エルゴチオネイン培地(5 mM エルゴチオネイン、0.02% 硫酸マグネシウム・7水和物、0.2% クエン酸、1% 無水リン酸水素二カリウム、pH7.2)を1 mLずつ分注し、オートクレーブにより加熱滅菌した。ここに高知県立のいち動物公園(高知県香南市野市町)から採取した土壌試料を小スパチュラで1サジ加え、30℃、4日間、振盪培養した。この培養液5μLを、0.1 mg/L シクロヘキシミドを含むエルゴチオネイン培地に植菌し、30℃、2日間、振盪培養した。この培養液を100倍希釈し、20μLを0.1 mg/Lシクロヘキシミドを含むエルゴチオネイン平板培地に塗布し、30℃で静置培養後、コロニーを単離した。その結果、エルゴチオネイン資化性細菌が単離された。
【0111】
〔実施例2〕HME13の同定
単離された資化性細菌(菌株)のうちHME13の微生物学的特徴は、表1にまとめたとおりである。
【0112】
【表1】
【0113】
続いて16S-rDNAの解析によりHME13を同定した。16S-rDNAの塩基配列は、以下の各プロトコールに基づいて解析した。DNA抽出:InstaGene Matrix (BIO-RAD)、PCR:PrimeSTAR HS DNA Polymerase(タカラバイオ)、サイクルシークエンス:BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems)、シークエンス:ABI PRISM 3130 xl Genetic Analyzer System (Applied Biosystems)、配列決定:ChromasPro 1.4 (Technelysium Pty Ltd.)。
得られた塩基配列を配列番号:5に示した。配列番号:5の塩基配列を元に、アポロンDB-BA4.0(テクノスルガ・ラボ)を用いて、Blast検索を行った結果、次の菌株の16S-rDNA配列にそれぞれ99.4%の相同性を示した。
Burkholderia arboris R-24201
B. stabilis LMG14294
【0114】
得られた結果を元に分子系統樹を作成した結果を図1に示した。クラスター解析の結果、HME13は、Burkholderia属に含まれると考えられるが既知種とは異なることが示唆されたので、HME13菌をバークホルデリア・エスピー (Burkholderia sp.) と同定した。
【0115】
〔実施例3〕 酵素反応
実施例1に記載のエルゴチオネイン培地で一晩振盪培養し、遠心分離にて得られた菌体を10 mM Sodium pyrophosphate (pH9.0), 1mM PMSFに懸濁し、超音波破砕した。破砕液を遠心分離し、未破砕菌体および菌体の残渣を除いた粗酵素液を得た。0.25 mM エルゴチオネインを含む10 mM Sodium pyrophosphate (pH 9.0)に本粗酵素液20μLを加え、30℃で保持しながら311 nmの吸光度を測定してチオウロカン酸の増加を追跡することで、酵素活性を測定した。また、タンパク質濃度をProtein assay CBB solution(ナカライテスク)にて定量した。エルゴチオナーゼ1Uは、1分間に1μmolのチオウロカン酸を生成する酵素量とした。結果を表2に示した。
【0116】
【表2】
【0117】
〔実施例4〕 HME13からのエルゴチオナーゼの精製
エルゴチオネイン培地1 mLにBurkholderia sp. HME13を植菌し、30℃、一晩振盪培養し、得られた前培養液100μLを500 mLのエルゴチオネイン培地に植菌し、30℃、45時間振盪培養した。培養液を遠心分離し、得られた菌体を1 mM PMSF を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) に懸濁し、超音波により菌体を破砕した。菌体破砕液を35-65% 硫安分画を行い、得られた酵素を10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で2回透析した後、20 mM Tris-HCl (pH 7.2)で平衡化したDEAE-トヨパールカラム (250 mL-Resin)にアプライし、塩化ナトリウムにより段階的に溶出した。活性画分を回収し、濃縮後、35% 飽和硫安を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で透析し、35% 飽和硫安を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で平衡化したButyl-トヨパールカラム (200 mL-Resin)にアプライし、硫安濃度を段階的に下げて酵素を溶出した。精製の要約を表3(HME13からのエルゴチオナーゼの精製)に示した。
【0118】
【表3】
【0119】
続いてHME13から精製されたエルゴチオナーゼの理化学的特徴を明らかにした。
【0120】
(1) 至適温度:
HME13から精製されたエルゴチオナーゼの至適温度を検討した。0.25 mMエルゴチオネイン、10 mM リン酸ナトリウム緩衝液(pH7.2)、23 ng 精製酵素を含む全量1.0 mlの溶液を反応液とし、25℃〜80℃まで5℃間隔でインキュベートし、反応液中に生成するチオウロカン酸の吸収(311 nm)の増加を測定した。結果を図2に示す。この結果から、HME13から精製されたエルゴチオナーゼの至適温度は65℃であることが確認された。また50-65℃で最大活性の80%以上が維持された。
【0121】
(2) 熱安定性:
次に、HME13から精製されたエルゴチオナーゼの熱安定性について検討した。0.1 mg /ml 精製酵素溶液を4,20,30,40,50,60,65,70,80℃で30分間加温した。その後、0.25 mM エルゴチオネイン,10 mMリン酸ナトリウム緩衝液(pH7.2),各温度で処理された精製酵素47 ngを含む全量1.0 mlの溶液を反応液とし、30℃で反応液中に生成するチオウロカン酸の吸収(311 nm)の増加を測定した。結果を図3に示す。この結果から、HME13から精製されたエルゴチオナーゼは60℃まで約90%の活性を維持し、70℃以上の温度では急速に活性を失うことが確認された。
【0122】
(3) 至適pH:
さらに、HME13から精製されたエルゴチオナーゼの至適pHについて検討した。緩衝液を変えた他は(1)と同じ組成の反応液にエルゴチオナーゼ(47 ng/mL)を加えて反応を開始し、30℃で活性を測定した。実験に使用した緩衝液と、そのpHは次のとおりである。
100 mM酢酸ナトリウム緩衝液:pH5.0−6.5
100 mMリン酸ナトリウム緩衝液:pH6.5−7.5
100 mM Tris塩酸緩衝液:pH7.5−8.5
100 mMグリシン水酸化ナトリウム緩衝液:pH8.5−11.0
【0123】
生成するチオウロカン酸の吸収(311 nm)を測定した。結果を図4に示す。この結果から、HME13から精製されたエルゴチオナーゼの至適pHは8.0であることが確認された。またpH 7.0-9.5 で最大活性の80%以上が維持された。
【0124】
(4) pH安定性:
次に、HME13から精製されたエルゴチオナーゼの安定性に与えるpHの影響について検討した。各緩衝液に溶解したエルゴチオナーゼ(0.1mg/mL)を30℃で30分間プレインキュベーションした。次いで各pHで処理されたエルゴチオナーゼ47 ngを(1)と同じ組成の反応液に加えて反応を開始し、30℃で生成するチオウロカン酸の吸収(311 nm)を測定した。結果を図5に示す。この結果から、HME13から精製されたエルゴチオナーゼはpH 6.5-11.0で30℃, 30分間処理後に活性の80%以上を維持することが確認された。
【0125】
(5) 基質特異性:
HME13から精製されたエルゴチオナーゼの基質特異性について検討した。基質を変えた他は(1)と同じ組成の反応液にエルゴチオナーゼ(47 ng/mL)を加えて反応を開始し、30℃で活性を測定した。実験に使用した基質は表5のとおりである。エルゴチオネインを基質とした場合には生成するチオウロカン酸の吸収(311 nm)を測定した。ヒスチジンを基質とした場合には生成するウロカニン酸の吸収(277 nm)を測定した。チロシンを基質とした場合には生成するp-ヒドロキシケイ皮酸の吸収(315 nm)を測定した。フェニルアラニンを基質とした場合には生成するトランス-ケイ皮酸の吸収(290 nm)を測定した。この結果、エルゴチオナーゼはエルゴチオネイン以外のいずれのアミノ酸にも作用しなかった(表4)。
【0126】
【表4】
【0127】
(6) 阻害剤の影響:
HME13から精製されたエルゴチオナーゼの阻害剤の影響について検討した。各阻害剤を加えた他は(1)と同じ組成の反応液にエルゴチオナーゼ(47 ng/mL)を加えて反応を開始し、30℃で生成するチオウロカン酸の吸収(311 nm)を測定した。但し、p-ヒドロキシ水銀安息香酸については、エルゴチオネインを除く反応液を30℃で20分間加温後、エルゴチオネインを加えて反応を開始した。実験に使用した阻害剤は表5のとおりである。この結果、エルゴチオナーゼは検討した各阻害剤によりほとんど影響を受けず、特にp-ヒドロキシ水銀安息香酸(p-hydroxymercuribenzoate)、ヨード酢酸(iodoacetate)、L-システイン(L-cysteine)存在下で80%以上の活性を保持していた点において、非特許文献4に記載の大腸菌W株由来エルゴチオナーゼと異なっていた(表5)。
【0128】
【表5】
【0129】
〔実施例5〕 エルゴチオナーゼの部分アミノ酸配列
実施例4で得られた酵素を用いて、プロテインシーケンサー(HT492 アプライドバイオシステムズ社)によりN末端アミノ酸配列を解析した。アミノ酸配列を配列番号:6に示した。
【0130】
〔実施例6〕 エルゴチオナーゼ遺伝子のコア領域のクローニング
N末端アミノ酸配列を元にBLAST検索した結果、完全に一致するアミノ酸配列が得られた。得られた配列は次のとおりである。
Burkholderia sp.383 由来 Bcep18194_B3018 タンパク質のアミノ酸配列の一部
Burkholderia pseudomarei 1106a 由来 BURPS1106A_2045 タンパク質のアミノ酸配列の一部
Burkholderia pseudomarei 1710b 由来 BURPS1710b_2187 タンパク質のアミノ酸配列の一部
Burkholderia pseudomarei K96243 由来 BPSL1676 タンパク質のアミノ酸配列の一部
【0131】
これら菌株のゲノムデータベース情報を基にそれぞれセンスプライマー、アンチセンスプライマーを合成した。それぞれの塩基配列を配列番号:7、8に示した。
配列番号:7:CGACGTGCACACGCTCGCCGCGCT
配列番号:8:CCCGACGCGAGCTGCACGAGATGCTT
【0132】
プライマー各10 pmol、dNTP 10 nmol、Burkholderia sp. HME13由来染色体DNA 140 ng、Ex-Taq用緩衝液(タカラバイオ製)、Ex-Taq 1.25 U(タカラバイオ製)を含む50μLの反応液を用い、94℃、1分の後、98℃、10秒 、68℃、2分を30サイクル、最後に72℃、10分、GeneAmp PCR System 9700(アプライド・バイオシステムズ製)を用いて行った。得られたDNA断片の塩基配列を解析し、決定されたエルゴチオナーゼ遺伝子の塩基配列を配列番号:2に、予想されるアミノ酸配列を配列番号:1に示した。予想されるアミノ酸配列をSignalP v3.0 (http://www.cbs.dtu.dk/services/SignalP/) で解析したところ、MQRFLMRSAIALALFAAVPAFA(配列番号:9)がシグナル配列であり、成熟型酵素では切断されると予測され、精製酵素のN末端アミノ酸配列解析結果と一致した。シグナル配列が切断された場合のエルゴチオナーゼのアミノ酸配列を配列番号:3に、塩基配列を配列番号:4に示す。
【0133】
〔実施例7〕 エルゴチオナーゼ遺伝子のクローニング
エルゴチオナーゼの構造遺伝子配列を基にORFのみをクローニングするためのプライマー(配列番号:10、11)を合成した。プライマーを各10 pmol、dNTP 10 nmol、Burkholderia sp. HME13由来染色体DNA 144 ng、Ex-Taq用緩衝液(タカラバイオ製)、Ex-Taq 1.25 U(タカラバイオ製)を含む50μLの反応液を用い、94℃、1分の後、98℃、10秒、68℃、1分を25サイクル、最後に72℃、10分、GeneAmp PCR System 9700(アプライド・バイオシステムズ製)を用いて行った。
配列番号:10:GGAATTCCATATGCAACGCTTCCTCATGCGGAGC
配列番号:11:GGGAAGCTTTCAGTCGTGATACGCGCGCAGCAG
【0134】
得られたDNA断片を制限酵素NdeI、HindIIIで2重消化し、アガロースゲル電気泳動を行い、目的とするバンドの部分を切り出し精製した。得られたDNA断片を、NdeI、HindIIIで2重消化したpET21a(+) (Novagen製)とLigation-convenience kit (ニッポンジーン製)を用いて、ライゲーションし、大腸菌DH5α株を形質転換した。
【0135】
得られた形質転換株を、アンピシリンを含むLB培地で生育させ、生育した形質転換体からPlasmidを抽出し、エルゴチオナーゼ遺伝子が挿入されていることとが確認できたプラスミドをpET21a(+)/ERTaseとした。pET21a(+)/ERTaseで大腸菌 Rosetta 2 (DE3) 株 (Novagen製)を形質転換し、エルゴチオナーゼ発現株を得た。
【0136】
〔実施例8〕 組換えエルゴチオナーゼの活性評価
エルゴチオナーゼを発現するプラスミドpET21a(+)/ERTaseで形質転換された大腸菌Rosetta 2 (DE3) 株をアンピシリンとクロラムフェニコールを含む液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに3時間培養を行った。
【0137】
菌体を遠心分離により集菌した後、1.0 mM PMSF、0.01% 2-メルカプトエタノールを含む10 mM ピロリン酸ナトリウム緩衝液(pH 7.2)に懸濁し、密閉式超音波破砕装置UCD-200TM(コスモバイオ製)を用いて菌体を破砕した。菌体破砕液を遠心分離し、その上清を菌体抽出液として回収した。また該遺伝子を含まない大腸菌Rosetta 2 (DE3)株をLB培地で終夜培養し、0.1 mM IPTG添加後さらに3時間培養した菌体を同様に破砕して、菌体抽出液を回収した。それぞれの菌体抽出液を用いてエルゴチオナーゼ活性を測定した。結果を表6に示した。活性測定の方法は実施例3と同様とした。
【0138】
【表6】
【0139】
〔実施例9〕 組換えエルゴチオナーゼのN末端アミノ酸配列の決定
エルゴチオナーゼを発現するプラスミドで形質転換された大腸菌Rosetta2 (DE3) 株を、1 mM PMSFを含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) に懸濁し、超音波により菌体を破砕した。菌体破砕液を30-50% 硫安分画を行い、得られた酵素を10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で2回透析した後、20mM Tris-HCl (pH 7.2)で平衡化したDEAE-トヨパールカラムにアプライし、塩化ナトリウムにより段階的に溶出した。活性画分を回収し、濃縮後、30% 飽和硫安を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で透析し、30% 飽和硫安を含む10 mM ピロリン酸ナトリウム緩衝液 (pH 7.2) で平衡化したButyl-トヨパールカラム (200mL-Resin)にアプライし、硫安濃度を段階的に下げて酵素を溶出した。精製の要約を表7(組換えエルゴチオナーゼの精製)に示した。
【0140】
【表7】
【0141】
得られた酵素を用いて、プロテインシーケンサー(HT492 アプライドバイオシステムズ社)によりN末端アミノ酸配列を解析した。アミノ酸配列を配列番号:12に示した。
配列番号:12:DVTLDGRSVTPESIARI
【0142】
組換えエルゴチオナーゼのN末端アミノ酸配列は、親株のエルゴチオナーゼと同様にN末端のシグナル配列が切断されていた。
【0143】
〔実施例10〕 シグナル配列を除いた組換えエルゴチオナーゼの活性評価
エルゴチオナーゼのシグナル配列を除いたORFのみをクローニングするためのプライマー(配列番号:13、14)を合成した。プライマーを各15 pmol、dNTP 20 nmol、pET21a(+)/ERTase plasmid、KOD FX用緩衝液(TOYOBO製)、KOD FX 1U(TOYOBO製)を含む50μLの反応液を用い、94℃、2分の後、98℃、10秒、74℃、1分30秒を5サイクル、98℃、10秒、72℃、1分30秒を20サイクル、最後に68℃、7分、GeneAmp PCR System 9700(アプライド・バイオシステムズ製)を用いて行った。
配列番号:13:GGAATTCCATATGGACGTCACGCTCGACGGCCGGT
配列番号:14:GGGAAGCTTTCAGTCGTGATACGCGCGCAGCAG
【0144】
得られたDNA断片を制限酵素NdeI、HindIIIで2重消化し、アガロースゲル電気泳動を行い、目的とするバンドの部分を切り出し精製した。得られたDNA断片を、NdeI、HindIIIで2重消化したpET21a(+) (Novagen製)とLigation-convenience kit (ニッポンジーン製)を用いて、ライゲーションし、大腸菌DH5α株を形質転換した。
【0145】
得られた形質転換株を、アンピシリンを含むLB培地で生育させ、生育した形質転換体からPlasmidを抽出し、エルゴチオナーゼ遺伝子が挿入されていることとが確認できたプラスミドをpET21a(+)/ERTase-sigとした。pET21a(+)/ERTase-sigで大腸菌 Rosetta 2 (DE3) 株 (Novagen製)を形質転換し、シグナル配列を除いたエルゴチオナーゼ発現株を得た。
【0146】
シグナル配列を含むエルゴチオナーゼを発現するプラスミドpET21a(+)/ERTaseで形質転換された大腸菌Rosetta 2 (DE3) 株とシグナル配列を含まないエルゴチオナーゼを発現するプラスミドpET21a(+)/ERTase-sigで形質転換された大腸菌Rosetta 2 (DE3) 株をアンピシリンとクロラムフェニコールを含む液体LB培地で終夜30℃培養し、0.1 mM IPTGを加え、さらに3時間培養を行った。
【0147】
菌体を遠心分離により集菌した後、1.0 mM PMSF、0.01% 2-メルカプトエタノールを含む10 mM ピロリン酸ナトリウム緩衝液(pH 7.2)に懸濁し、密閉式超音波破砕装置UCD-200TM(コスモバイオ製)を用いて菌体を破砕した。菌体破砕液を遠心分離し、その上清を菌体抽出液として回収した。また該遺伝子を含まない大腸菌Rosetta 2 (DE3)株をLB培地で終夜培養し、0.1 mM IPTG添加後さらに3時間培養した菌体を同様に破砕して、菌体抽出液を回収した。それぞれの菌体抽出液を用いてエルゴチオナーゼ活性を測定した。結果を表8に示した。活性測定の方法は実施例3と同様とした。
【0148】
【表8】
【0149】
〔実施例11〕 エルゴチオナーゼによるエルゴチオネインの定量
0〜500μg/mLエルゴチオネイン、10mM リン酸ナトリウム緩衝液(pH7.2)、1.9 mg/mL精製酵素を含む全量1.0 mLの溶液を反応液とし、30℃で1時間インキュベートし、2N HClを50μL加えて反応を停止し、反応液中に生成するチオウロカン酸の吸収(311 nm)の増加を測定した。結果を図6に示す。この結果から、HME13から精製されたエルゴチオナーゼを用いて0-500μg/mLの範囲でエルゴチオネインを定量可能であることが確認された。
【産業上の利用可能性】
【0150】
本発明は、エルゴチオナーゼの利用による、エルゴチオネインの特異的かつ簡便な定量方法を提供する。本発明によるエルゴチオネインの定量は、例えば、クローン病の診断法として有用である。
【特許請求の範囲】
【請求項1】
次の(1)〜(2)に示す理化学的性状を有するエルゴチオナーゼ;
(1)作用:L-エルゴチオネインに作用し、チオウロカン酸(3-(2,3-ジヒドロ-2-チオキソ-1H-イミダゾール-4-イル)プロペン酸)およびトリメチルアミンを生成する、および
(2)基質特異性:L-エルゴチオネインに作用し、D-, L-ヒスチジン、D-, L-チロシン、D-, L-フェニルアラニンに作用しない。
【請求項2】
さらに付加的に、次の(3)〜(8)に示す理化学的性状を有する請求項1に記載のエルゴチオナーゼ;
(3)至適pH:7.0〜9.5、
(4)至適温度:50〜65℃、
(5)分子量:ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動による分子量が45,000〜55,000、ゲルろ過による分子量が200,000〜260,000。
(6)熱安定性: 60℃、30分で90%以上の活性を保持、
(7)pH安定性: pH 6.5〜11.0、30分で80%以上の活性を保持、および
(8)阻害剤: p-ヒドロキシ水銀安息香酸(p-hydroxymercuribenzoate)、ヨード酢酸(iodoacetate)、および/またはL-システイン(L-cysteine)存在下において80%以上の活性を保持。
【請求項3】
バークホルデリア属、もしくはシュードモナス属に属する微生物に由来する、請求項1または2に記載のエルゴチオナーゼ。
【請求項4】
前記バークホルデリア属、もしくはシュードモナス属に属する微生物が、下記の群から選択されるいずれかの微生物であることを特徴とする請求項3に記載のエルゴチオナーゼ;
バークホルデリア・エスピー NITE P-861(Burkholderia sp. NITE P-861)、および
シュードモナス・エスピー (Pseudomonas sp.)。
【請求項5】
下記(a)〜(e)のいずれかに記載のエルゴチオナーゼ活性を有するタンパク質;
(a)配列番号:3に記載のアミノ酸配列を含むタンパク質、
(b)配列番号:4に記載された塩基配列を含むポリヌクレオチドによってコードされるタンパク質、
(c)配列番号:3に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含むタンパク質、
(d)配列番号:4に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質、または
(e)配列番号:3に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列を含むタンパク質。
【請求項6】
請求項5に記載のタンパク質をコードするポリヌクレオチド。
【請求項7】
請求項1〜4のいずれかに記載のエルゴチオナーゼをコードするポリヌクレオチドまたは請求項6に記載のポリヌクレオチドを保持したベクター。
【請求項8】
請求項7に記載のベクターにより形質転換された形質転換体。
【請求項9】
前記宿主が大腸菌である請求項8に記載の形質転換体。
【請求項10】
バークホルデリア属、もしくはシュードモナス属に属し、エルゴチオナーゼ活性を有する微生物、もしくは請求項8または9に記載の形質転換体を培養する工程と、培養物よりエルゴチオナーゼを回収する工程を含むエルゴチオナーゼの製造方法。
【請求項11】
エルゴチオネインに作用してチオウロカン酸を生成する能力を有する酵素活性物質を、被検対象に接触させる工程を含む、エルゴチオネインの定量方法。
【請求項12】
前記酵素活性物質が、請求項1〜4のいずれかに記載のエルゴチオナーゼ、請求項5に記載のエルゴチオナーゼ活性を有するタンパク質、当該エルゴチオナーゼまたはエルゴチオナーゼ活性を有するタンパク質を生産する微生物、およびその処理物からなる群から選択される、請求項11に記載のエルゴチオネインの定量方法。
【請求項13】
エルゴチオナーゼおよび/または酵素活性物質を含むことを特徴とするエルゴチオネイン定量試薬、およびキット。
【請求項14】
チオウロカン酸およびトリメチルアミンにエルゴチオナーゼおよび/または酵素活性物質を作用させることを特徴とする、エルゴチオネインの製造方法。
【請求項1】
次の(1)〜(2)に示す理化学的性状を有するエルゴチオナーゼ;
(1)作用:L-エルゴチオネインに作用し、チオウロカン酸(3-(2,3-ジヒドロ-2-チオキソ-1H-イミダゾール-4-イル)プロペン酸)およびトリメチルアミンを生成する、および
(2)基質特異性:L-エルゴチオネインに作用し、D-, L-ヒスチジン、D-, L-チロシン、D-, L-フェニルアラニンに作用しない。
【請求項2】
さらに付加的に、次の(3)〜(8)に示す理化学的性状を有する請求項1に記載のエルゴチオナーゼ;
(3)至適pH:7.0〜9.5、
(4)至適温度:50〜65℃、
(5)分子量:ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動による分子量が45,000〜55,000、ゲルろ過による分子量が200,000〜260,000。
(6)熱安定性: 60℃、30分で90%以上の活性を保持、
(7)pH安定性: pH 6.5〜11.0、30分で80%以上の活性を保持、および
(8)阻害剤: p-ヒドロキシ水銀安息香酸(p-hydroxymercuribenzoate)、ヨード酢酸(iodoacetate)、および/またはL-システイン(L-cysteine)存在下において80%以上の活性を保持。
【請求項3】
バークホルデリア属、もしくはシュードモナス属に属する微生物に由来する、請求項1または2に記載のエルゴチオナーゼ。
【請求項4】
前記バークホルデリア属、もしくはシュードモナス属に属する微生物が、下記の群から選択されるいずれかの微生物であることを特徴とする請求項3に記載のエルゴチオナーゼ;
バークホルデリア・エスピー NITE P-861(Burkholderia sp. NITE P-861)、および
シュードモナス・エスピー (Pseudomonas sp.)。
【請求項5】
下記(a)〜(e)のいずれかに記載のエルゴチオナーゼ活性を有するタンパク質;
(a)配列番号:3に記載のアミノ酸配列を含むタンパク質、
(b)配列番号:4に記載された塩基配列を含むポリヌクレオチドによってコードされるタンパク質、
(c)配列番号:3に記載のアミノ酸配列において、1もしくは複数のアミノ酸が置換、欠失、挿入、および/または付加したアミノ酸配列を含むタンパク質、
(d)配列番号:4に記載された塩基配列からなるDNAと高度にストリンジェントな条件下でハイブリダイズするポリヌクレオチドによってコードされるタンパク質、または
(e)配列番号:3に記載のアミノ酸配列と80%以上の同一性を有するアミノ酸配列を含むタンパク質。
【請求項6】
請求項5に記載のタンパク質をコードするポリヌクレオチド。
【請求項7】
請求項1〜4のいずれかに記載のエルゴチオナーゼをコードするポリヌクレオチドまたは請求項6に記載のポリヌクレオチドを保持したベクター。
【請求項8】
請求項7に記載のベクターにより形質転換された形質転換体。
【請求項9】
前記宿主が大腸菌である請求項8に記載の形質転換体。
【請求項10】
バークホルデリア属、もしくはシュードモナス属に属し、エルゴチオナーゼ活性を有する微生物、もしくは請求項8または9に記載の形質転換体を培養する工程と、培養物よりエルゴチオナーゼを回収する工程を含むエルゴチオナーゼの製造方法。
【請求項11】
エルゴチオネインに作用してチオウロカン酸を生成する能力を有する酵素活性物質を、被検対象に接触させる工程を含む、エルゴチオネインの定量方法。
【請求項12】
前記酵素活性物質が、請求項1〜4のいずれかに記載のエルゴチオナーゼ、請求項5に記載のエルゴチオナーゼ活性を有するタンパク質、当該エルゴチオナーゼまたはエルゴチオナーゼ活性を有するタンパク質を生産する微生物、およびその処理物からなる群から選択される、請求項11に記載のエルゴチオネインの定量方法。
【請求項13】
エルゴチオナーゼおよび/または酵素活性物質を含むことを特徴とするエルゴチオネイン定量試薬、およびキット。
【請求項14】
チオウロカン酸およびトリメチルアミンにエルゴチオナーゼおよび/または酵素活性物質を作用させることを特徴とする、エルゴチオネインの製造方法。
【図1】

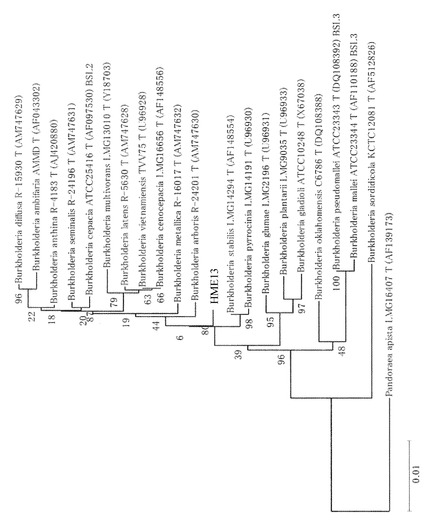
【図2】
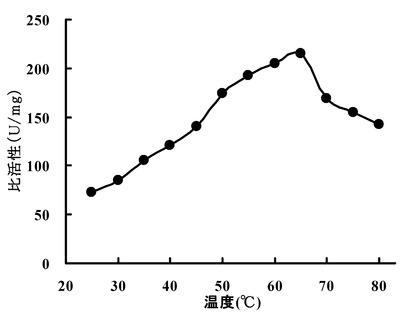
【図3】
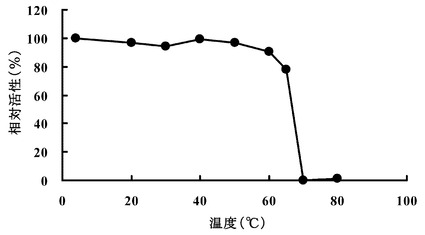
【図4】
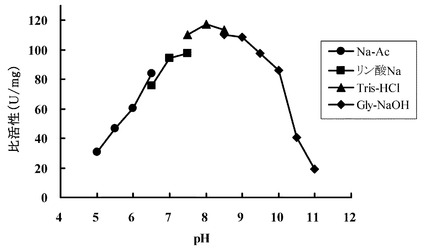
【図5】
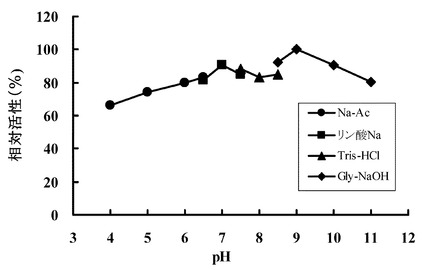
【図6】
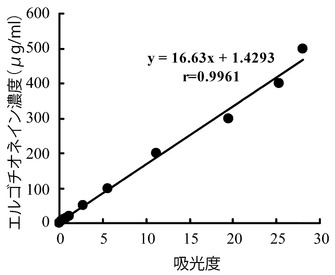
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−65591(P2012−65591A)
【公開日】平成24年4月5日(2012.4.5)
【国際特許分類】
【出願番号】特願2010−213226(P2010−213226)
【出願日】平成22年9月24日(2010.9.24)
【出願人】(000002901)株式会社ダイセル (1,236)
【出願人】(504174180)国立大学法人高知大学 (174)
【Fターム(参考)】
【公開日】平成24年4月5日(2012.4.5)
【国際特許分類】
【出願日】平成22年9月24日(2010.9.24)
【出願人】(000002901)株式会社ダイセル (1,236)
【出願人】(504174180)国立大学法人高知大学 (174)
【Fターム(参考)】
[ Back to top ]