
エンドサイトーシス依存性DNA取り込み抑制剤、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染阻害剤、ウイルス感染予防剤、及びウイルス感染促進剤、並びに、遺伝子マーカー、モノクローナル抗体、ハイブリドーマ、及びそれらの使用方法。
【課題】エンドサイトーシス依存性DNA取り込み抑制剤、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染阻害剤、ウイルス感染予防剤及びウイルス感染促進剤、並びに、遺伝子マーカー、モノクローナル抗体、ハイブリドーマ及びそれらの使用方法を提供すること。
【解決手段】DNaseXタンパク質の機能を増強することによって、エンドサイトーシス依存性のDNAの取り込みを抑制することができる。ここで、DNaseXタンパク質の機能を増強する因子としては、DNaseX遺伝子発現ベクター等である。一方、DNaseXタンパク質の機能を抑制することによって、エンドサイトーシス依存性のDNAの取り込みを促進することができる。ここで、DNaseXタンパク質の機能を抑制する因子としては、例えば、DNaseX遺伝子を標的とするsiRNA等である。また、DNaseX遺伝子のhmRNA、mRNA、タンパク質等の発現レベルによって、ウイルス感染に対する易感染性予測やウイルス感染後の病状推定を行うこともできる。
【解決手段】DNaseXタンパク質の機能を増強することによって、エンドサイトーシス依存性のDNAの取り込みを抑制することができる。ここで、DNaseXタンパク質の機能を増強する因子としては、DNaseX遺伝子発現ベクター等である。一方、DNaseXタンパク質の機能を抑制することによって、エンドサイトーシス依存性のDNAの取り込みを促進することができる。ここで、DNaseXタンパク質の機能を抑制する因子としては、例えば、DNaseX遺伝子を標的とするsiRNA等である。また、DNaseX遺伝子のhmRNA、mRNA、タンパク質等の発現レベルによって、ウイルス感染に対する易感染性予測やウイルス感染後の病状推定を行うこともできる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、エンドサイトーシス依存性DNA取り込み抑制剤、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染阻害剤、ウイルス感染予防剤、及びウイルス感染促進剤、並びに、遺伝子マーカー、モノクローナル抗体、ハイブリドーマ、及びそれらの使用方法に関する。
【背景技術】
【0002】
DNaseXは、DNaseI様DNaseとして同定された初めての哺乳動物DNaseである。DNaseXは、そのC末端に疎水性ドメイン(HD)を有することが分かっており(非特許文献1参照)、この疎水性ドメインは各種の哺乳動物間で保存されている(非特許文献2参照)。
【0003】
近年、DNaseタンパク質の機能を用いて、癌及びその前駆病変(例えば、良性腫瘍、悪性腫瘍、癌腫、肉腫等)を検出する方法や、癌及びその前駆病変等の疾患を診断する方法が開発された(非特許文献3参照)。しかしながら、DNaseXタンパク質の詳細な機能は未だ分かっていなかった。
【非特許文献1】Shiokawa, D., and Tanuma, S., Biochemistry, 40, 143-152 (2001)
【非特許文献2】Shiokawa, D., Shika, Y., Saito, K., Yamazaki, K. and Tanuma, S., Biochem. J., 392, 511-517 (2005)
【非特許文献3】特表2006―510363号公報
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明はエンドサイトーシス依存性DNA取り込み抑制剤、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染阻害剤、ウイルス感染予防剤、ウイルス感染促進剤、及び新規な疾患モデル動物、並びに、遺伝子マーカー、モノクローナル抗体、ハイブリドーマ、及びそれらの使用方法を提供することを目的とする。
【課題を解決するための手段】
【0005】
本発明者らは、RD細胞(rhabdomyosarcoma)をin vitroで分化させた筋管細胞は、未分化のRD細胞より外来lacZ遺伝子をエンドサイトーシスにより効率よく取り込むが、その遺伝子産物であるβガラクトシダーゼの発現は10%以下であり、強く抑制されることを見出した。DNaseXタンパク質が生体の筋組織及びRD細胞由来の筋管細胞において高く発現していることや、さらに、未分化の筋管細胞でDNaseXタンパク質を強制的に発現させると、エンドサイトーシスによる遺伝子の取り込みが強く抑制され、一方、RD細胞由来の筋管細胞でsiRNAによってDNaseX遺伝子の発現を抑制すると、エンドサイトーシスによる遺伝子の取り込みが促進することを見出した。こうして、発明者らは、外来遺伝子のエンドサイトーシスに対するDNaseXタンパク質の機能を明らかにした。一方、本発明者らは、ヒトDNaseXタンパク質の一部の配列であるオリゴペプチドを用い、ヒトDNaseXタンパク質を特異的に認識する抗体を作製し、本発明を完成するに至った。
【0006】
すなわち、本発明にかかるDNA取り込み抑制剤は、DNaseXタンパク質の機能を増強するエンドサイトーシス依存性DNA取り込み抑制剤であることを特徴とする。なお、前記DNA取り込み抑制剤は、DNaseXタンパク質の発現を増強するDNA取り込み抑制剤であってもよい。
【0007】
また、本発明にかかるウイルス感染阻害剤は、DNaseXタンパク質の機能を増強するウイルス感染阻害剤であることを特徴とする。なお、前記ウイルス感染阻害剤は、DNaseXタンパク質の発現を増強するウイルス感染阻害剤であってもよい。また、本発明にかかる細胞に対するウイルス感染阻害方法は、当該細胞内でDNaseXタンパク質の機能を増強することを特徴とする。その一手段として、DNaseXタンパク質の発現を増強してもよい。
【0008】
さらに、本発明にかかるウイルス感染予防剤又はウイルス感染に対するワクチンは、前記ウイルス感染阻害剤を含有することを特徴とする。また、本発明にかかるヒト又はヒト以外の脊椎動物に対するウイルス感染予防方法は、当該動物内でDNaseXタンパク質の機能を増強することを特徴とする。その一手段として、DNaseXタンパク質の発現を増強してもよい。
【0009】
また、本発明にかかる発現ベクターは、DNaseXタンパク質を強制発現させる発現ベクターであることを特徴とする。
【0010】
さらに、本発明にかかる細胞は、DNaseXタンパク質の機能が増強された細胞であることを特徴とする。ここで、前記細胞は、DNaseXタンパク質を強制発現させる発現ベクターを有していてもよい。
【0011】
また、本発明にかかるヒト又はヒト以外の脊椎動物は、体の一部又は全部で、DNaseXタンパク質が異所的に又は過剰に機能が増強していることを特徴とする。ここで、前記脊椎動物は、DNaseXタンパク質を強制発現させる発現ベクターを細胞に導入することにより作製されてもよい。また、前記脊椎動物は、DNaseXタンパク質を強制発現させる発現ベクターによって作製されたトランスジェニック脊椎動物であってもよい。
【0012】
さらに、本発明にかかるスクリーニング方法は、DNaseXタンパク質の機能を増強する化合物をスクリーニングすることを特徴とする。
【0013】
本発明のモノクローナル抗体は、RSLLHTAAAFDFPTSFQL(配列番号3)の一部又は全部をエピトープとする。本発明のハイブリドーマは、このモノクローナル抗体を産生する。本発明のハイブリドーマは、寄託受領番号FERM AP-21282であることが好ましい。本発明のモノクローナル抗体は、この寄託受領番号FERM AP-21282のハイブリドーマから産生されるものでもよい。
【0014】
また、本発明の検出方法は、上記いずれかのモノクローナル抗体を用いて、DNaseXタンパク質を検出する検出方法である。
【0015】
また、本発明にかかるDNA取り込み促進剤は、DNaseXタンパク質の機能を抑制するエンドサイトーシス依存性DNA取り込み促進剤であることを特徴とする。なお、前記DNA取り込み促進剤は、上記いずれかのモノクローナル抗体を有効成分として含有してもよく、DNaseXタンパク質の発現を抑制してもよい。
【0016】
さらに、本発明にかかるウイルス感染促進剤は、DNaseXタンパク質の機能を抑制するウイルス感染促進剤であることを特徴とする。なお、前記ウイルス感染促進剤は、上記いずれかのモノクローナル抗体を有効成分として含有してもよく、DNaseXタンパク質の発現を抑制してもよい。また、前記ウイルス感染促進剤は、DNaseX遺伝子に対するsiRNAを有効成分として含有することを特徴とする。ここで、前記siRNAは、配列番号32を有していることが好ましい。また、本発明にかかる細胞又はヒト又はヒト以外の脊椎動物に対するウイルス感染促進方法は、当該細胞又は当該ヒト又はヒト以外の脊椎動物内でDNaseXタンパク質の機能を抑制することを特徴とする。その一手段として、DNaseXタンパク質の発現を抑制してもよい。
【0017】
また、本発明にかかる発現ベクターは、DNaseX遺伝子に対するsiRNAを発現する発現ベクターであることを特徴とする。
【0018】
さらに、本発明にかかる細胞は、内在性DNaseXタンパク質の機能が抑制されている細胞であることを特徴とする。ここで、前記細胞は、DNaseX遺伝子に対するsiRNAを発現する発現ベクターを有することが好ましい。
【0019】
また、本発明にかかるヒト又はヒト以外の脊椎動物は、体の一部又は全部で、内在性DNaseXタンパク質の機能が抑制されていることを特徴とする。ここで、前記脊椎動物は、DNaseX遺伝子のノックダウン又はノックアウトによって作製されていてもよい。
【0020】
さらに、発明にかかるスクリーニング方法は、DNaseXタンパク質の機能を抑制する化合物をスクリーニングすることを特徴とする。上記いずれかのモノクローナル抗体を用いて、スクリーニングしてもよい。
【0021】
なお、本明細書で「エンドサイトーシス依存性DNA取り込み」というのは、エンドサイトーシスに依存して、細胞内にほぼ分解されずにDNAを取り込むことをいい、特に、DNAが細胞内に取り込まれる際に、DNaseXタンパク質によって分解されないことを意味する。また、ウイルスの宿主への感染とは、ウイルスが宿主に接触してDNAを宿主内に導入する段階のことをいい、ゲノム内にDNAが挿入されたり、細胞内でウイルスDNAが増幅したり、ウイルスが増殖したり、それによって様々な現象が引き起こされたりするのは、感染後に生じることである。
【0022】
また、本発明の遺伝子マーカーは、DNaseX遺伝子関連物質である。特に、ウイルスに対する易感染性予測マーカー、又はウイルス感染後の病状推定マーカーであることを特徴としてもよい。前記ウイルスが、インフルエンザウイルス、肝炎ウイルス、HIVウイルス、ATLウイルス、エボラウイルス、黄熱病ウイルス、狂犬病ウイルス、サイトメガロウイルス、SARSウイルス、JCウイルス(進行性多巣性白質脳症)、水痘・帯状疱疹ウイルス、コクサッキーA16型・エンテロウイルス71ウイルス(手足口病)、デング熱ウイルス、パルボウイルス(伝染性紅斑)、EBウイルス(伝染性単核球症)、天然痘ウイルス、風疹ウイルス、ポリオウイルス、麻疹ウイルス、アデノウイルス(プール熱)、マールブルグウイルス(フィロウイルス科)、クリミア・コンゴ出血熱ウイルス、ラッサウイルス、フニンウイルス等の新大陸アレナウイルス、ハンタウイルス、リフトバレー熱ウイルス、ムンプスウイルス(流行性耳下腺炎)、ウエストナイルウイルス、単純ヘルペスウイルス、コクサッキーウイルス、ロタウイルス、ノロウイルス、チクングニアウイルス(アルファウイルス属、トガウイルス科)からなる群から選ばれてもよい。前記遺伝子関連物質がDNA、hnRNA、mRNA、及びタンパク質からなる群から選ばれることが好ましい。
【0023】
さらに、本発明のキットは、ウイルスに対する易感染性予測又はウイルス感染後の病状推定のためのキットであって、DNaseXタンパク質に対する抗体又はDNaseXcDNAを増幅するためのPCR用プライマーを含有する。前記抗体がモノクローナル抗体であってもよい。このモノクローナル抗体が、配列番号3の一部又は全部をエピトープとしてもよい。さらに、DNaseXタンパク質の検出には、抗原抗体反応を利用する全ての方法が可能であるが、特に免疫クロマトグラフ測定法を利用していることが好ましい。
【発明の効果】
【0024】
本発明によればエンドサイトーシス依存性DNA取り込み抑制剤、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染阻害剤、ウイルス感染予防剤、ウイルス感染促進剤、及び新規な疾患モデル動物、並びに遺伝子マーカー、モノクローナル抗体、ハイブリドーマ、及びそれらの使用方法を提供することができる。
【発明を実施するための最良の形態】
【0025】
以下、上記知見に基づき完成した本発明の実施の形態を、実施例を挙げながら詳細に説明する。実施の形態及び実施例に特に説明がない場合には、J. Sambrook, E. F. Fritsch & T. Maniatis (Ed.), Molecular cloning, a laboratory manual (3rd edition), Cold Spring Harbor Press, Cold Spring Harbor, New York (2001); F. M. Ausubel, R. Brent, R. E. Kingston, D. D. Moore, J.G. Seidman, J. A. Smith, K. Struhl (Ed.), Current Protocols in Molecular Biology, John Wiley & Sons Ltd.等の標準的なプロトコール集に記載の方法、あるいはそれを修飾したり、改変した方法を用いる。また、市販の試薬キットや測定装置を用いている場合には、特に説明が無い場合、それらに添付のプロトコールを用いる。
【0026】
なお、本発明の目的、特徴、利点、及びそのアイデアは、本明細書の記載により、当業者には明らかであり、本明細書の記載から、当業者であれば、容易に本発明を再現できる。以下に記載された発明の実施の形態及び具体的に実施例等は、本発明の好ましい実施態様を示すものであり、例示又は説明のために示されているのであって、本発明をそれらに限定するものではない。本明細書で開示されている本発明の意図並びに範囲内で、本明細書の記載に基づき、様々な改変並びに修飾ができることは、当業者にとって明らかである。
【0027】
==本発明の薬剤と使用方法==
本発明者らが明らかにしたように、DNaseXタンパク質は、GPI結合性膜タンパク質であり、外来DNAのエンドサイトーシス依存性取り込みを抑制する機能を有する。
【0028】
(1)エンドサイトーシス依存性DNA取り込み抑制剤
従って、DNaseXタンパク質の機能を増強する物質は、エンドサイトーシス依存性DNA取り込み抑制剤として有用である。エンドサイトーシスによってDNAが細胞内に取り込まれる場合としては、例えば、ウイルスが細胞に感染した場合等が挙げられる。そこで、DNaseXタンパク質の機能を増強する物質を有効成分として含有する薬剤は、ウイルス感染を阻害できるウイルス感染阻害剤や、ウイルス感染予防剤や、ウイルス感染に対するワクチンとして有用である。なお、ウイルスの種類は、特に限定されない。
【0029】
ここで、「DNaseXタンパク質の機能を増強する」とは、細胞全体でDNaseXタンパク質の機能が増強されれば良く、例えばDNaseXタンパク質分子の活性が増強されることにより機能が増強されるだけでなく、細胞内でDNaseXタンパク質分子の量が増加することにより機能が増強されてもよい。
【0030】
DNaseXタンパク質の機能を増強するための方法は特に限定されないが、例えば、DNaseXタンパク質を直接細胞に導入する方法やDNaseXタンパク質の機能を増強する物質を細胞に投与する方法が考えられる。
【0031】
DNaseXタンパク質を細胞に導入するための方法としては特に限定されず、Protein Transduction Domains (PTD) 融合タンパク質としてTATやVP22との融合タンパク質を用いてもよく、BioPorterTM、ChariotTM等のタンパク質導入試薬を用いてもよい。
【0032】
DNaseXタンパク質の機能を増強する物質としては特に限定されず、DNaseXタンパク質の活性を増強する物質、内在DNaseX遺伝子の発現レベルを亢進させる物質、外来DNaseX遺伝子を強制発現させる発現ベクター等、様々な物質が含まれるが外来DNaseX遺伝子を強制発現させる発現ベクターであることが好ましい。
【0033】
内在DNaseX遺伝子の発現レベルを亢進させる物質としては、例えば、相同組換えによってゲノム中のDNaseX遺伝子のプロモーターをより強力なプロモーターで置換することができる相同組換え用ベクターや、DNaseX遺伝子のプロモーターを活性化するタンパク質をコードする遺伝子の発現ベクター等を用いることができる。
【0034】
ここで、内在DNaseX遺伝子とは、ゲノム中の本来のDNaseX遺伝子座に存在するDNaseX遺伝子のことをいい、外来DNaseX遺伝子とは、細胞外に由来し、ゲノム中の本来のDNaseX遺伝子座以外の場所に存在するDNaseX遺伝子のことをいう。
【0035】
外来DNaseX遺伝子を強制発現させる発現ベクターは、導入する宿主細胞で機能するプロモーターを備えていれば、どんなベクターでもよく、プラスミドベクターでも、ウイルスベクターでもよい。また、遺伝子導入に際しては、遺伝子を導入後、発現ベクターを染色体の外におく一過的発現(transient expression)であっても、発現ベクターが染色体に組み込まれた永久発現(permanent expression)であってもよい。遺伝子導入は、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法、マイクロインジェクション法、ウイルス感染法、その他様々なDDS(drug delivery system)等、その方法は特に限定されない。
【0036】
(2)エンドサイトーシス依存性DNA取り込み促進剤
一方、DNaseXタンパク質の機能を抑制する物質を有効成分として含有する薬剤は、ウイルス感染を促進することができる。従って、DNaseXタンパク質の機能を抑制する物質は、エンドサイトーシス依存性DNA取り込み促進剤として有用であり、DNaseXタンパク質の機能を抑制する物質を有効成分として含有する薬剤は、ウイルス感染を促進できるウイルス感染促進剤として有用である。ここで、「DNaseXタンパク質の機能を抑制する」とは、細胞全体でDNaseXタンパク質の機能が抑制されれば良く、例えばDNaseXタンパク質分子の活性が低下することにより機能が抑制されるだけでなく、細胞内でDNaseXタンパク質分子の量が減少することにより機能が抑制されてもよい。
【0037】
DNaseXタンパク質の機能を抑制するための方法は特に限定されないが、例えば、細胞内にあるDNaseXタンパク質を分解する方法やDNaseXタンパク質の機能を抑制する物質を細胞に投与する方法等が考えられる。
【0038】
細胞内にあるDNaseXタンパク質を分解する方法は特に限定されないが、例えば、相同組換えによって、ゲノム内のDNaseX遺伝子を、ユビキチンの標的配列を有したDNaseXタンパク質をコードする融合DNaseX遺伝子に置換し、外来遺伝子によってユビキチンを供給する方法等が考えられる。
【0039】
なお、DNaseXタンパク質の機能を抑制する物質は、例えば、DNaseXタンパク質に特異的に結合し、その機能を特異的に抑制する抗体やアプタマーや、化合物等であってもよい。この抗体は、ポリクローナル抗体でもモノクローナル抗体でもよいが、モノクローナル抗体であることが好ましく、下記のようにして作製された、RSLLHTAAAFDFPTSFQLの一部又は全部をエピトープとするモノクローナル抗体であってもよい。また、DNaseXタンパク質の機能を抑制するために、内在DNaseX遺伝子の発現を抑制してもよく、例えば、DNaseX遺伝子のプロモーターを抑制する物質やDNaseX遺伝子の発現を標的にする、siRNA、miRNA、アンチセンスRNA等を用いてもよく、相同組換え等を用いてゲノム中のDNaseX遺伝子を破壊してもよい。
【0040】
ここで、用いられる細胞は、培養細胞であっても、脊椎動物個体中の細胞であってもよい。
【0041】
==疾患モデル動物及びその有用性==
(1)疾患モデル動物の作製
本発明の疾患モデル動物は、脊椎動物個体に対し、体の全部又は一部でDNaseXタンパク質の機能を異所的に又は過剰に増強させる処理を施すこと、又は体の全部又は一部でDNaseXタンパク質の機能を抑制する処理を施すことにより作製することができる。
【0042】
脊椎動物個体において、DNaseXタンパク質の機能を増強するための方法は特に限定されないが、例えば、DNaseXタンパク質を個体に直接に導入する方法やDNaseXタンパク質の機能を増強する物質を個体に投与する方法が考えられる。投与方法は、投与する物質によって適宜使い分ければよく、経口投与でも非経口投与でもよい。
【0043】
DNaseXタンパク質を個体に導入するための方法としては特に限定されず、Protein Transduction Domains (PTD) 融合タンパク質としてTATやVP22との融合タンパク質を用いてもよく、BioPorterTM、ChariotTM等のタンパク質導入試薬を用いてもよく、DNaseXタンパク質を注入してもよい。
【0044】
DNaseXタンパク質の機能を増強する物質としては特に限定されず、DNaseXタンパク質の活性を増強する物質、内在DNaseX遺伝子の発現レベルを亢進させる物質、外来DNaseX遺伝子を強制発現させる発現ベクター等、様々な物質が含まれる。特にDNA等は、リポフェクション法、エレクトロポレーション法、マイクロインジェクション法、ウイルス感染法等、常法に従って投与すればよい。
【0045】
内在DNaseX遺伝子の発現レベルを亢進させる物質としては、例えば、相同組換えによってゲノム中のDNaseX遺伝子のプロモーターをより強力なプロモーターで置換することができる相同組換え用ベクターや、DNaseX遺伝子のプロモーターを活性化するタンパク質をコードする遺伝子の発現ベクター等を用いることができる。
【0046】
外来DNaseX遺伝子を強制発現させる発現ベクターは、導入する個体で機能するプロモーターを備えていれば、どんなベクターでもよく、プラスミドベクターでも、ウイルスベクターでもよい。また、遺伝子導入の系としては、発現ベクターを染色体の外におく一過的発現(transient expression)の系や、発現ベクターが染色体に組み込まれた永久発現(permanent expression)の系を用いてもよい。
【0047】
なお、予め培養細胞や培養組織においてDNaseXタンパク質の機能を増強させ、それらの細胞や組織を脊椎動物個体に移植することにより、体の全部又は一部でDNaseXタンパク質の機能を増強させてもよいが、体の全部でDNaseXタンパク質の機能を増強させるためには、遺伝子操作によることが好ましく、外来DNaseX遺伝子を強制発現させる発現ベクターを用いて、トランスジェニック脊椎動物を作製することがより好ましい。
【0048】
また、脊椎動物個体においてDNaseXタンパク質の機能を抑制するための方法は特に限定されないが、例えば、DNaseXタンパク質の機能を抑制する物質を個体に投与する方法等が考えられる。投与方法は、投与する物質によって適宜使い分ければよいのであって、経口投与でも非経口投与でもよい。
【0049】
なお、DNaseXタンパク質の機能を抑制する物質は、例えば、DNaseXタンパク質に特異的に結合し、その機能を特異的に抑制する抗体やアプタマーや、化合物等であってもよい。その抗体は、ポリクローナル抗体でもモノクローナル抗体でもよいが、モノクローナル抗体であることが好ましく、下記のようにして作製された、RSLLHTAAAFDFPTSFQLの一部又は全部をエピトープとするモノクローナル抗体であってもよい。また、DNaseXタンパク質の機能を抑制するために、内在DNaseX遺伝子の発現を抑制してもよく、例えば、DNaseX遺伝子のプロモーターを抑制する物質やDNaseX遺伝子の発現を標的にする、siRNA、miRNA、アンチセンスRNA等を用いてもよく、相同組換え等を用いてゲノム中のDNaseX遺伝子を破壊してもよい。特にDNAやRNA等では、リポフェクション法、エレクトロポレーション法、マイクロインジェクション法、ウイルス感染法等、常法に従って行えばよい。
【0050】
なお、予め培養細胞や培養組織においてDNaseXタンパク質の機能を抑制し、それらの細胞や組織を脊椎動物個体に移植することにより、体の全部又は一部でDNaseXタンパク質の機能を抑制してもよいが、体の全部でDNaseXタンパク質の機能を抑制するためには、遺伝子操作によることが好ましく、ノックアウト脊椎動物やノックダウン脊椎動物を作製することがより好ましい。例えば、相同組換え等を用いた内在DNaseX遺伝子における変異(欠失、置換、付加、挿入等を含む)の導入による変異体(条件突然変異体も含む)の作製、ドミナント・ネガティブ変異をもつ変異タンパク質を有するトランスジェニック動物の作製、アンチセンスRNAやRNAi、又はそれらをコードするDNAを用いたノックダウン動物の作製をすればよい。
【0051】
なお、疾患モデル動物としては、例えば、マウス、ラット等のヒト以外の脊椎動物であることが好ましい。
【0052】
(2)疾患モデル動物の有用性
DNaseXタンパク質の機能を増強させた動物は、ウイルス等に感染しにくくなるため、感染症抵抗性を示すモデルとして有用である。一方、DNaseXタンパク質のDNaseX活性を抑制した動物は、ウイルス等に感染しやすくなるため、易感染性を示すモデルとして有用である。ここで、感染症とは、ウイルス等の病原体が感染して増殖し、病的症状を発症する疾患のことをいう。
【0053】
==スクリーニング方法==
DNaseXタンパク質の機能を増強する化合物をスクリーニングすることにより、エンドサイトーシス依存性DNA取り込み抑制剤、ウイルス感染阻害剤、ウイルス感染予防剤及びワクチン等に用いられる化合物を得ることができる。
【0054】
具体的な一例として、DNaseX遺伝子を発現する培養細胞、培養組織、培養器官等に、スクリーニングの対象となる物質を投与する。そして、投与前後で、その培養物におけるDNaseX遺伝子の転写物やタンパク質の発現量を測定する。投与前後のDNaseX遺伝子の発現量を比較し、投与後にDNaseX遺伝子の発現量が増強しているかどうかを調べることにより、感染症に対して抵抗性を有する物質を同定することができる。
【0055】
一方、DNaseXタンパク質の機能を抑制する化合物をスクリーニングすることにより、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染促進剤等に用いられる化合物を得ることができる。 例えば、DNaseX遺伝子を発現する培養細胞、培養組織、培養器官等に、スクリーニングの対象となる物質を投与する。そして、投与前後で、その培養物におけるDNaseX遺伝子の転写物やタンパク質の発現量を測定する。投与前後のDNaseXタンパク質の発現量を比較し、投与後にDNaseXタンパク質の発現量が低下しているかどうかを調べることにより、感染症に対して予防できる物質や感染症を治療することができる物質を同定することができる。
【0056】
DNaseX遺伝子の発現量は、転写物の量を測定するRT-PCTを用いたり、タンパク質の発現量を測定するELISA法、RIA法、ウエスタンブロッティング法、免疫組織化学染色等を利用したりして、常法に従って測定することができる。なお、タンパク質の発現量を検出するための抗体は、ポリクローナル抗体でもモノクローナル抗体でもよいが、モノクローナル抗体であることが好ましく、下記のようにして作製された、RSLLHTAAAFDFPTSFQLの一部又は全部をエピトープとするモノクローナル抗体であってもよい。
【0057】
DNaseXタンパク質の機能を測定する方法は、DNaseXタンパク質の発現を測定する以外にも、DNAの単位時間当たりの分解量等を測定してもよく、特に限定されない。
【0058】
なお、培養細胞等の代わりに、上記「疾患モデル動物」を用いて、DNaseXタンパク質の機能を増強する化合物又は抑制する化合物をスクリーニングしてもよい。スクリーニング方法は、上述と同様の方法を用いればよい。
【0059】
==DNaseXタンパク質及びDNA==
なお、本発明にかかるDNaseXタンパク質は、以下の(a)又は(b)のタンパク質である。
(a)ヒトDNaseXタンパク質(配列番号1のアミノ酸配列を有するタンパク質)及びそのホモログ。
(b)(a)に記載のタンパク質のアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、又は付加したアミノ酸配列からなり、DNaseX活性を有するタンパク質。
【0060】
これらのタンパク質を合成するには、そのタンパク質をコードするDNAを用いて、発現ベクター等を構築し、細胞あるいは個体に導入してもよい。発現ベクターは、導入する細胞及び個体で機能するプロモーターを備えていれば、どんなベクターでもよく、プラスミドベクターでも、ウイルスベクターでもよい。また、in vitro 転写/翻訳系で合成してもよく、あるいは、そのタンパク質を化学合成してもよい。
【0061】
上記の(a)又は(b)のタンパク質をコードするDNAとしては、そのタンパク質のアミノ酸配列よりコドン表を用いて作成された配列を有するDNAでもよいが、以下の(a)又は(b)のDNAが特に好ましい。
(a)ヒトDNaseX遺伝子のcDNA(配列番号2の塩基配列を有するDNA、Accession No. X90392)及びDNaseX遺伝子ホモログのcDNA。
(b)(a)に記載のcDNAの塩基配列において、1もしくは数個の塩基が欠失、置換、又は付加した塩基配列を有し、DNaseX活性を有するタンパク質をコードするDNA。
【0062】
なお、これらのDNAを得るには、そのDNA配列を有するクローンが含まれるゲノムライブラリーやcDNAライブラリーをスクリーニングしてもよく、そのDNA配列を有する生物種のゲノムやRNAを用いてPCRやRACE法等で増幅してもよく、これらを組み合わせてもよい。また、そのDNAを化学合成してもよい。
【0063】
==抗ヒトDNaseXモノクローナル抗体及びその有用性===
本発明の抗ヒトDNaseXモノクローナル抗体は、ハイブリドーマにより産生されるモノクローナル抗体である。本発明のモノクローナル抗体は、公知の技術を用いて作製することができる。
【0064】
(1)抗ヒトDNaseXモノクローナル抗体の作製
本発明のモノクローナル抗体の作製には、DNaseXタンパク質のうち抗原性を有するタンパク質を免疫原として用いることが好ましい。抗原性を有するタンパク質としては、例えば、ヒトDNaseXの237-254位に対応するペプチド(RSLLHTAAAFDFPTSFQL(配列番号3)が挙げられる。このペプチドは、化学合成によって作製することができる。なお、免疫原には、DNaseXタンパク質を発現している細胞そのものを用いてもよい。
【0065】
免疫に用いる動物は、細胞融合に使用する親細胞との適合性を考慮して選択するのが好ましい。このような動物としては、例えば、マウス、ラット、モルモット、ウサギ等が挙げられる。
【0066】
上記動物に免疫原を投与する方法は、当業者に公知の方法を用いて行えばよいが、免疫原とフロイント完全アジュバント等のアジュバントを併せて投与することが好ましい。
【0067】
上記動物に免疫原を投与し、その動物の血清中の抗体価が上昇したのを確認した後に、免疫動物から脾臓を取り出し、脾臓から脾細胞を調製し、以下の細胞融合を行う。
【0068】
細胞融合は、当業者に公知の方法を用いて行うことができる。ここで、細胞融合に使用する親細胞としては、X63-Ag8-653、NS-1、NS-0/1、SP2O等のミエローマ細胞が好ましい。また、脾細胞とミエローマ細胞との細胞数比は任意に設定することができるが、例えば、ミエローマ細胞に対して脾細胞を10倍とするのが好ましい。細胞融合に用いる培養液としては、例えば、前記ミエローマ細胞株の増殖に好適なRPMI1640培養液、MEM培養液等が挙げられる。なお、PEGは、分子量4000の溶液であることが好ましい。
【0069】
細胞融合後は、HAT培地を用いてハイブリドーマをスクリーニングし、限界希釈法によって抗体を産生するハイブリドーマをクローニングすればよい。また、スクリーニングは、ヒトDNaseXタンパク質を目的とした、ELISA法、ウエスタンブロッティング法、免疫組織化学染色を行えばよい。
【0070】
このようにしてスクリーニングされ、クローン化されたハイブリドーマから産生されるモノクローナル抗体は、ヒトDNaseXタンパク質に対して特異的に反応する。モノクローナル抗体を産生するハイブリドーマは、公知の方法を用いて培養することができる。そして、その培養上清から、モノクローナル抗体を得ることができる。又は、ハイブリドーマをマウス等の動物の腹腔内に投与し、その動物の腹水から、モノクローナル抗体を得てもよい。
【0071】
本発明のモノクローナル抗体としては、例えば、独立行政法人産業技術総合研究所特許生物寄託センターに2007年3月30日に受託受領番号FERM AP-21282で寄託されたhDNase X 1B1 Hybridomaが挙げられる。
【0072】
(2)抗ヒトDNaseXモノクローナル抗体の有用性
(i)易感染性予測マーカー
一般に、ウイルスは、細胞表面に吸着し、その細胞自身が持っているエンドサイトーシス機構によって、エンドソーム小胞として細胞内に取り込まれることが知られている。これより、細胞のエンドサイトーシス機構を抑制すれば、ウイルス感染は阻害され、一方、細胞のエンドサイトーシス機構を促進すれば、ウイルス感染は助長される。
【0073】
以下の実施例に示すように、RD細胞由来の筋管細胞にDNaseXタンパク質を強制的に発現させると、エンドサイトーシスによる遺伝子の取り込みは抑制され、一方、siRNAによってDNaseX遺伝子の発現を抑制すると、エンドサイトーシスによる遺伝子の取り込みは促進する。従って、DNaseX遺伝子の発現レベルによって、ウイルスの細胞への感染のしやすさを予測することができ、DNaseXの遺伝子関連物質はウイルスに対する易感染性予測マーカーとして利用できる。なお、DNaseXの遺伝子関連物質は、DNAであっても、RNA(hnRNA、mRNA)であっても、タンパク質であってもよい。
【0074】
具体的には、例えば、多数の健常人のDNaseXの遺伝子関連物質のレベルを測定し、正常なレベルの範囲を設定し、あるヒト又はヒト以外の脊椎動物におけるDNaseXの遺伝子関連物質のレベルが、その範囲より多かったり少なかったりすれば、その脊椎動物は、通常よりウイルスに感染し難かったり、感染しやすかったりする。これにより、例えば、投与する抗ウイルス剤の強さや量を決めることも可能になる。
【0075】
(ii)ウイルス感染後の病状推定マーカー
一般に、ウイルスが細胞に感染すると、そのウイルスは細胞内で大量に増殖し、細胞本来の生理機能が破錠したり細胞膜の破壊が起きたりして、宿主細胞は死に至る場合がある。また、ウイルスによっては、短期間で大量のウイルスを作って直ちに宿主細胞を殺すのではなく、むしろ宿主細胞へのダメージが少なくなるよう、少量のウイルスを長期間にわたって持続的に産生(持続感染)する場合もある。
【0076】
上記(i)の通り、RD細胞由来の筋管細胞にDNaseXタンパク質を強制的に発現させると、エンドサイトーシスによる遺伝子の取り込みは抑制される。一方、siRNAによってDNaseX遺伝子の発現を抑制すると、エンドサイトーシスによる遺伝子の取り込みは促進する。このように、細胞におけるDNaseX量は、ウイルスに感染したヒト又はヒト以外の脊椎動物の中で、ウイルスが増殖する時の細胞抵抗性を決めると考えられる。従って、ウイルス感染脊椎動物のDNaseX遺伝子の発現レベルより、その動物がどのように病状推移するか、例えば、軽症で済みそうか重症に移行しそうか、等を推定することができ、DNaseXの遺伝子関連物質は病状推移推定マーカーとして利用できる。なお、DNaseXの遺伝子関連物質は、DNAであっても、RNA(hnRNA、mRNA)であっても、タンパク質であってもよい。
【0077】
(iii)対象ウイルス
上記(i)及び(ii)のウイルスとしては、例えば、インフルエンザウイルス、肝炎ウイルス、HIVウイルス、ATLウイルス、エボラウイルス、黄熱病ウイルス、狂犬病ウイルス、サイトメガロウイルス、SARSウイルス、JCウイルス、水痘・帯状疱疹ウイルス、コクサッキーA16型・エンテロウイルス71ウイルス、デング熱ウイルス、パルボウイルス、EBウイルス、天然痘ウイルス、風疹ウイルス、ポリオウイルス、麻疹ウイルス、アデノウイルス、マールブルグウイルス、フィロウイルス、クリミア・コンゴ出血熱ウイルス、ラッサウイルス、フニンウイルス、新大陸アレナウイルス、ハンタウイルス、リフトバレー熱ウイルス、ムンプスウイルス、ウエストナイルウイルス、単純ヘルペスウイルス、コクサッキーウイルス、ロタウイルス、ノロウイルス、チクングニアウイルス、トガウイルス等が挙げられるが、これらに限定されない。
【0078】
==キット==
上記のように、ヒト又はヒト以外の脊椎動物のDNaseX遺伝子の発現レベルより、易感染性予測又はウイルス感染後の病状推定をすることができる。従って、DNaseX遺伝子の発現レベルを測定できるキットは、ウイルスに対する易感染性予測又はウイルス感染後の病状推定に有用である。このキットは、DNaseX遺伝子の発現レベルを測定するために、DNaseXタンパク質に対する抗体、あるいはDNaseXのcDNAを増幅するためのPCR用プライマーを含有することが好ましい。なお、この抗体はポリクローナル抗体であってもモノクローナル抗体であってもよく、モノクローナル抗体が、RSLLHTAAAFDFPTSFQLの一部又は全部をエピトープとしてもよい。なお、簡便さの点から、このキットは、免疫クロマトグラフ測定法等の装置状になっていることが好ましいが抗原抗体反応を利用した方法であれば特に限定されない。
【実施例】
【0079】
以下、本発明の実施例について詳細に述べる。
<実施例1:新規モノクローナル抗体の作製と特異性の確認>
本実施例で作製したモノクローナル抗体は、実施例2において使用した。
【0080】
(1)新規抗ヒトDNaseXモノクローナル抗体の作製
本法では、ヒトDNaseXの237-254位に対応するペプチド(RSLLHTAAAFDFPTSFQL(配列番号3))のN末端にCysを付加した合成ペプチドとチオグロブリンとをN末端のCysによって結合したペプチドを免疫原として用いた。まず、6週齢雌BDF1マウスに、上記抗原50μgを完全フロイントアジュバントと共に皮下注射して初回免疫を行った。初回免疫から2週後、3週後、4週後に、上記抗原50μgを不完全フロイントアジュバントと共に皮下注射して追加免疫を行った。最後に、上記抗原50μgを静脈注射して最終免疫を行った。最終免疫から4日後、上記マウスから脾臓を取り出して脾細胞を調製し、ポリエチレングリコールを用いて、調製した脾細胞とX63-Ag8-653マウスミエローマ細胞(Kinebuchi, M., Ide, T., Lupin, D., Tamatani, T., Miyasaka, M., Matsuura, A., Nagai, Y., Kikuchi, K. and Uede, T., J. Immunol., 146, 3721-3728 (1991))とを細胞融合した。その後、HAT培地を用いてハイブリドーマをスクリーニングし、限界希釈法によってクローニングを行い、最終的にクローン化したハイブリドーマ株1B1を得た(寄託受領番号:FERM AP-21282)。なお、1B1ハイブリドーマから産生される抗ヒトDNaseXモノクローナル抗体のサブクラスはIgG1/κであった。
【0081】
作製した抗ヒトDNaseXモノクローナル抗体の反応性は、ウエスタンブロッティング法によって確認した。まず、C末端にMyc-Hisタグを結合させたヒトDNase(DNaseI、DNaseX、DNaseγ、DNAS1L2)をコードする各cDNAを発現する発現ベクター(phDNaseX)を構築した(Shiokawa et al. 2005, Biochem J., vol.392, p.511-517.)。各発現ベクターをCHO-K1細胞に導入し、各タンパク質を発現した細胞より調製した細胞抽出物を用いてウエスタンブロッティングを行った(図1A)。また、HeLa S3細胞、ヒト胎児性横紋筋肉腫細胞(以下、RD細胞)、COS-7細胞、及びCHO-K1細胞の各細胞より調製した細胞抽出物を用いてウエスタンブロッティングを行った(図1B)。さらに、各動物種のDNaseXcDNAをpcDNA3.1 myc-His B(Invitrogen社)のEco RVサイトへサブクローニングして、DNaseX発現ベクターを構築した。なお、cDNAの作製に用いられたプライマー配列は以下の通りである。
サルDNaseX(センスプライマー):5’-CCACCATGCACTATCCAACTGCACTC-3'(配列番号5)
(アンチセンスプライマー):5’- GGTGGCAGGGCACAGTTGAGG-3’(配列番号6)
ブタDNaseX(センスプライマー):5’-CCACCATGGATTCCTCTGGAGGGTT CC -3’(配列番号7)
(アンチセンスプライマー):5’-GGCCGCCAGGCCCAGCTGTGG-3’(配列番号8)
ウシDNaseX(センスプライマー):5’-CCACCATGCATTCCTCTGGAGGGTTCC-3’(配列番号9)
(アンチセンスプライマー):5’-GGCCACCAGGCCCAGCTGGGG-3'(配列番号10)
マウスDNaseX(センスプライマー):5’-CCACCATGCCCTTTGGACAACCTGGA-3'(配列番号11)
(アンチセンスプライマー):5’-GTCCAGCTGAGATGGCAGGAG-3’(配列番号12)
ラットDNaseX(センスプライマー):5’-CCACCATGCCCTCTGGACAGCCTGTA-3'(配列番号13)
(アンチセンスプライマー):5’-GCCCAGCTGAGATAGTGACAG-3’(配列番号14)
ハムスターDNaseX(センスプライマー):5’-CCACCATGCCATATATGGCCATGCAT-3’(配列番号15)
(アンチセンスプライマー):5’-GCTCAGTTGAGATGGCAAAAG-3’(配列番号16)
【0082】
上記の方法によって構築した各DNaseX発現ベクター1μgに対し、TNT QuickCoupled Transcription/Translation System(Promega社)のTNT Quick Master Mixに加え、50μlの反応液で1時間、30℃でインキュベートした。得られた各DNaseXタンパク質を含む一定量の反応液を用いてウエスタンブロッティング法を行った(図1C)。なおコントロールには、挿入DNAのない発現ベクターに対し、上記in vitro転写/翻訳系によって同様に得られた等量の反応液を用いた。また、ウエスタンブロッティング法に用いた抗GAPDHモノクローナル抗体(MAB374)はChemicon社から、抗Myc tag抗体(R950-25)はInvitrogen社から購入した。
【0083】
その結果、作製した抗ヒトDNaseXモノクローナル抗体は、DNaseXに対して特異的に反応し(図1A)、DNaseXを発現することが知られている細胞(HeLa S3細胞、RD細胞)に対しても特異的に反応し(図1B)、ヒトDNaseXに対して特異的に反応した(図1C)。なお、ヒトDNaseXタンパク質は35kDaのバンドとして検出された。
【0084】
(2)新規抗ヒトDNaseIモノクローナル抗体の作製
上記(1)と同様の方法を用いて、抗ヒトDNaseIモノクローナル抗体を作製し、最終的にクローン化したハイブリドーマ株10A2を得た。なお、免疫原には、ヒトDNaseIの245-262位に対応するペプチド(GAVVPDSALPFNFQAAYG(配列番号4))のN末端にCysを付加した合成ペプチドとチオグロブリンとを結合させたペプチドを用いた。得られた抗ヒトDNaseIモノクローナル抗体のサブクラスはIgG1/κであった。
【0085】
作製した抗ヒトDNaseIモノクローナル抗体の特異性を、ウエスタンブロッティング法によって確認した。ここで、ウエスタンブロッティング法に用いた細胞抽出液は、(1)と同様である。なお、各動物種のDNaseIタンパク質発現ベクターを構築するにあたり、cDNAの作製に用いたプライマー配列は以下の通りである。
ブタDNaseI(センスプライマー):5’-CCACCATGAGGGCGGCCAGGCTGATG-3’(配列番号17)
(アンチセンスプライマー):5’-GGCTCTCTTCAGTGTCACCTC-3’(配列番号18)
マウスDNaseI(センスプライマー):5’-CCACCATGCGGTACACAGGGCTAATG-3’(配列番号19)
(アンチセンスプライマー):5’-GATTTTTCTGAGTGTCACCTC-3’(配列番号20)
【0086】
上記の方法によって構築した各DNaseI発現ベクターに対し、TNT QuickCoupled Transcription/Translation System(Promega社)を用いて各DNaseIタンパク質を作製し、ウエスタンブロッティング法を行った(図2B)。コントロールには、挿入DNAのない発現ベクターを用いた。また、ウエスタンブロッティング法で用いた抗Myc tag抗体(R950-25)はInvitrogen社から購入した。
その結果、作製した抗ヒトDNaseIモノクローナル抗体は、DNaseIに対して特異的に反応し(図2A)、ヒトDNaseIに対して特異的に反応した(図2B)。
【0087】
<実施例2:エンドサイトーシス依存性DNA取り込み>
本実施例では、DNaseXタンパク質の機能増強又は機能抑制が、それぞれエンドサイトーシス依存性DNA取り込みの抑制又は増強をもたらすことを示す。
【0088】
2−1 材料
(1)細胞の調製
HeLa S3細胞、COS-7細胞、及びCHO-KI細胞は、10%胎仔ウシ血清(FCS)、100単位/mlペニシリン及び0.1mg/mlストレプトマイシン含有ダルベッコ改変イーグル培地(DMEM)で培養した。また、RD細胞は、増殖培地(GM)(20%FCS、100単位/mlペニシリン及び0.1mg/mlストレプトマイシン含有DMEM)で培養した。なお、RD細胞は、ヒューマンサイエンス研究資源バンクから入手した。
【0089】
(2)抗体
抗ヒトDNaseXモノクローナル抗体及び抗ヒトDNaseIモノクローナル抗体は、実施例1で作製したものを使用した。
また、マウス抗SERCA-1モノクローナル抗体(Ve121G9)はLab vision社から、抗GAPDHモノクローナル抗体(MAB374)はChemicon社から、抗Myc tag抗体(R950-25)はInvitrogen社から購入した。さらに、ヤギ抗GM130(P-20)ポリクローナル抗体はSanta Cruz Biotechnology社から、ウサギ抗マウスFas(341289)ポリクローナル抗体はCalbiochem社から、抗EEA1(PA1-063)抗体はAffinity BioReagents社から、抗LAMP-1(H228)抗体はSanta Cruz Biotechnology社から、抗カルレティキュリン(C4606)抗体及び抗pan-cadherin(C3678)抗体はSigma社から購入した。
【0090】
(3)発現ベクターの作製
i) hDNaseXΔC発現ベクター(phDNaseXΔC)
PCRによって以下のプライマーを用いてhDNaseXΔC cDNAを合成し、このcDNAをpcDNA3.1 myc-His B(Invitrogen社)(挿入配列がコードするペプチドのC末端にMycタグとHisタグを付加し得る)のEco RVサイトへサブクローニングして、hDNaseXΔC発現ベクターを構築した。
hDNaseXΔC(センスプタイマー):5’-CCACCATGCACTACCCAACTGCACTC--3'(センスプライマー)(配列番号21)
(アンチセンスプライマー): 5'-CTGGCTCAGCTTCAGCTCCAC-3'(配列番号22)
【0091】
ii) mFasΔC発現ベクター(pmFasΔC)
PCRによって以下のプライマーを用いてmFasΔC cDNAを合成し、このcDNAをpcDNA3.1 myc-His B(Invitrogen社)のEco RVサイトへサブクローニングして、mFasΔC発現ベクターを構築した。
mFasΔC(センスプライマー): 5'-CCACCATGCTGTGGATCTGGGCTGTC-3'(配列番号23)
(アンチセンスプライマー): 5’-TTCCTGGATTGTCATGTCTTCAGC-3’(配列番号24)
【0092】
iii) hDNase I-HD発現ベクター(phDNase I-HD)
PCRによって以下のプライマーを用いて、hDNase Iと疎水性ドメイン(HD)を含むDNase Xの外部末端領域(アミノ酸配列256-284位)をコードするcDNAを作製し、このcDNAをpcDNA3.1 myc-His B(Invitrogen社)のEco RVサイトへサブクローニングして、hDNase I-HD発現ベクターを構築した。
hDNase I(センスプライマー):5’-CCACCATGAGGGGCATGAAGCTGCTG-3’(配列番号25)
(アンチセンスプライマー):5'-CTTCAGCATCACCTCCACTGG-3'(配列番号26)
HD(センスプライマー):5’-CTGAGCCAGGCGCACAGCGTC-3’(配列番号27)
(アンチセンスプライマー):5'-GGCAGCAGGGCACAGCTGAGG-3'(配列番号28)
【0093】
iv) hDNase X H148A発現ベクター(phDNase X H148A)
phDNase X H148Aは、LA PCR in vitro mutagenesisキット(タカラバイオ株式会社)を用いた部位特異的突然変異誘発法によって148番目のHisをAlaに置換し、phDNase Xと同様に作製した。
【0094】
v) hDNaseI-myc-His発現ベクター(phDNaseI-myc-His)、hDNaseX-myc-His発現ベクター(phDNaseX-myc-His)、hDNase γ-myc-His発現ベクター(phDNase γ-myc-His)、hDNAS1L2-myc-His発現ベクター(phDNAS1L2-myc-His)、及びcDNA 3.1/myc-His/lacZ(placZ)発現ベクター(pcDNA 3.1/myc-His/lacZ(placZ))
hDNaseI(phDNase I-myc-His)、hNaseX(phDNase X-myc-His)、DNaseγ(phDNase γ-myc-His)、及びhDNAS1L2(phDNAS1L2-myc-His)は、Shiokawaらの方法(Shiokawa, D., and Tanuma, S. Biochemistry, 2001, vol.40, p.143-152)に従って作製した。また、pcDNA 3.1/myc-His/lacZ(placZ)(βガラクトシダーゼ発現ベクター)は、Invitrogen社から購入した。
【0095】
2-2 アッセイ法
(1)筋管細胞への分化誘導方法
継代培養したRD細胞を、タイプIコラーゲンでコートされた培養皿(Iwaki社製)に播種し、GM培地で培養した。次に、GM培地を分化培地(DM)(2%ウマ血清、100単位/mlペニシリン及び0.1mg/mlストレプトマイシン含有DMEM)に変更して、培地交換をしながらRD細胞を培養し、筋管細胞へ分化誘導した。
【0096】
(2)ウエスタンブロッティング法
ウエスタンブロッティング法に用いた各抗体の希釈濃度は、抗ヒトDNaseXモノクローナル抗体は500ng/ml、抗DNaseIモノクローナル抗体は100ng/ml、抗Myc tag抗体は1000倍希釈、抗GAPDHモノクローナル抗体は250倍希釈、マウス抗SERCA-1モノクローナル抗体は100倍希釈、ウサギ抗マウスFasポリクローナル抗体は2000倍希釈、抗pan-cadherin抗体は400倍希釈であった。
【0097】
(3)免疫組織化学染色
ホルマリンで固定したサンプルをパラフィンで包埋し、10μmの切片を作製した。なお、パラフィン包埋された正常ヒト組織は、BioChain社から購入した。
免疫組織化学染色は、Cell and Tissue Staining kit(HRP-DAB)(R&D Systems社)を用いて行った。まず、組織切片を乗せたスライドをAntigen Retrieval Dewax Solution(BioChain社)で処理し、この組織切片に、3μg/mlの抗DNaseXモノクローナル抗体をのせて4℃、オーバーナイトでインキュベートした。次に、これらの組織切片にContrast BLUE Solution(KPL)をのせて対比染色を行った。なお、これらの画像は、CK40 light microscope system(Olympus)を用いて記録した。
【0098】
(4)間接免疫蛍光染色
免疫蛍光染色による解析は、従来の方法(Shiokawa et al., Biochem. J., 2005, vol. 392, p.511-517)を改良して行った。具体的には、以下の通りである。
まず、培養皿の底に敷いたカバーグラス上で細胞を培養した。培養後、これらの細胞を3.5%ホルムアルデヒドで固定し、Cytonin solution(R&D Systems社)を添加して、室温で30分間インキュベートした。次に、一次抗体の希釈液をカバーグラス上の細胞に滴下し、4℃、オーバーナイトでインキュベートした。ここで、各一次抗体は、抗ヒトDNaseXモノクローナル抗体は3μg/ml、抗DNase Iモノクローナル抗体は1μg/ml、抗カルレティキュリン抗体は2000倍希釈、マウス抗SERCA-1モノクローナル抗体は50倍希釈、ヤギ抗GM130(P-20)ポリクローナル抗体は50倍希釈、ウサギ抗マウスFas(341289)ポリクローナル抗体は100倍希釈、抗pan-cadherin抗体は400倍希釈、抗EEA1(PA1-063)抗体は1000倍希釈、抗LAMP-1(H228)抗体は100倍希釈の濃度に調製して使用した。
【0099】
次に、細胞をDPBS(ダルベッコ改変リン酸緩衝生理食塩水;GIBCO)で洗浄し、二次抗体の希釈液をカバーガラス上の細胞に滴下し、室温で30分間インキュベートした。なお、二次抗体(Alexa 488ニワトリ抗マウスIgG抗体(Molecular Probes社)、Alexa 594ロバ抗ヤギIgG抗体(Molecular Probes社)、Alexa 594ヤギ抗ウサギIgG抗体(Molecular Probes社))は2000倍希釈の濃度に調製して使用した。
【0100】
また、Zenon Tricolor Mouse IgG1 Labeling Kit #2(Molecular Probes社)を用いて、抗ヒトDNaseXモノクローナル抗体とマウス抗SERCA-1モノクローナル抗体との二重染色を行った。なお、抗ヒトDNaseXモノクローナル抗体はAlexa488で、マウス抗SERCA-1モノクローナル抗体はAlexa594で標識したものを使用した。これらの抗体の希釈液をカバーガラス上の細胞に滴下し、室温で90分間インキュベートした。
【0101】
最後に、細胞をDPBSで洗浄し、100nM Hoechst 33342で対比染色し、蛍光顕微鏡(BX60、オリンパス株式会社)や共焦点レーザー顕微鏡(TCS SP2、ライカマイクロシステムズ株式会社)で観察した。
【0102】
(5)PI-PLC(ホスファチジルイノシトール特異性ホスホリパーゼ)による処理
免疫組織化学染色には、カバーグラス上の細胞を、DPBSで2回リンスし、5 unit/ml Bacillus thuringiensisホスファチジルイノシトール特異性ホスホリパーゼ(PI-PLC)(Sigma社)含有Leibovidz培養培地で37℃、30分間インキュベートした後、(4)の通りに固定し、抗体で処理した。
【0103】
また、PI-PLC処理後に、細胞と上清を分画するためには、付着している細胞をDPBSでリンスし、1.5mlチューブに回収し、このチューブに5 unit/ml PI-PLCを100μl入れて37℃で20分間インキュベートした後、10分間遠心分離(2000×g)をし、上清を別なチューブ(Sup fraction)に回収した。また、チューブ内の細胞ペレットをDPBSで遠心分離にて2回洗浄し、Leibovidz培地(cell fraction)500μlに再懸濁した。
【0104】
(6)TUNELアッセイ
カバーグラス上の細胞を、placZ-Transome IV複合体存在下で30分間インキュベートした。次に、これらの細胞をDPBSで2回洗浄し、3.7%ホルムアルデヒド含有DPBSをのせて室温で10分間固定した。次に、lacZプラスミドに生じる3’-OH末端を、Flow TACS in situ TUNEL Detection kit(R&D Systems社)を用いたTUNEL法によって検出した。なお、アッセイ全体は、FITC結合ストレプトアビジンをAlexa488結合ストレプトアビジン(Molecular Probes社)に変更した以外は、キット製造業者のプロトコールに従って行った。最後に、これらの細胞をDPBSで3回洗浄し、10μM Hoeschst 33342含有DPBSをのせて、エンドサイトーシスを起こしたDNAを室温で10分間対比染色させ、蛍光顕微鏡で観察した。
【0105】
(7)蛍光顕微鏡によるDNA画像のキャプチャー
細胞核及びエンドサイトーシスを起こしたDNAを、Hoechst 33342で対比染色した。核DNAの観察には2種類のNDフィルター(U-ND6及びU-ND25)(オリンパス株式会社)を用いて、一方、エンドサイトーシスを起こしたDNAの観察には、NDフィルターを用いないで、それぞれのDNA画像を観察した。
【0106】
2-3 DNaseXタンパク質によるエンドサイトーシス依存性DNA取り込み抑制
(1)DNaseXタンパク質の発現
DNaseXタンパク質の発現がRD細胞の筋肉への分化段階に関連していること示すために、免疫組織化学染色を用いて、ヒト組織におけるDNaseXタンパク質の発現を調べたところ、図3Aに示すように、心筋細胞及び骨格筋線維では、DNaseXタンパク質の強いシグナルが得られたが、脳ではシグナルが検出できなかった。このように、DNaseXタンパク質は、筋細胞に強く発現している。
次に、低濃度のマイトジェン条件下でRD細胞を培養すると、筋管が形成される(図3B)ので、この系を用いて、DNaseXタンパク質の発現の筋肉分化における変化をウエスタンブロッティング法で調べたところ、図3Cに示すように、DNaseXタンパク質の発現は分化誘導3日目に増加し、分化誘導5日目にはさらに増加した。このDNaseXタンパク質の発現の変化は、筋肉分化マーカー(SERCA-1:筋小胞体Ca2+-ATPase1)の発現と同様の挙動を示した(図3C)。
このように、DNaseXタンパク質の発現は、未分化筋細胞では弱く、分化するにつれ強くなる。
【0107】
(2)DNaseXタンパク質の細胞内分布の検討
次に、6日間低濃度のマイトジェン条件下で培養して分化させた筋管細胞に対し、抗DNaseX抗体及び抗カルレティキュリン抗体、抗SERCA-1抗体、又は抗GM130抗体で二重染色し、共焦点レーザー顕微鏡を用いてDNaseXタンパク質が膜に分布していることを示した。
【0108】
図3D(浸透化処理有り)に示すように、DNaseXタンパク質は、カルレティキュリンやSERCA-1と共に小胞体(ER)と筋小胞体に局在化していた。また、ゴルジ体のマーカー(GM130)との2重染色でもシグナルが重なり、DNaseXタンパク質がゴルジ体に存在していることが認められた。
さらに、浸透化処理無しで、免疫蛍光染色を行ったところ、図3E(蛍光顕微鏡像)及びF(共焦点レーザー顕微鏡像)に示すように、同様に分化した筋管細胞膜に局在することが観察され、DNaseXタンパク質は細胞外の膜表面に存在した。
このように、DNaseXタンパク質は、分化したRD由来の筋管において分泌経路に局在していることから、タンパク質合成後、細胞膜に送達されることが結論される。
【0109】
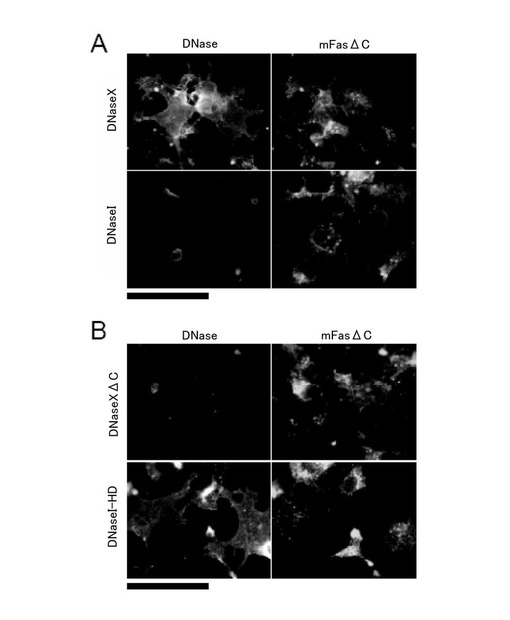
(3)細胞表面におけるDNaseXタンパク質の局在性
DNaseXタンパク質の細胞膜における局在を確認するために、Fugene6 (Roche)を用いて、DNaseX発現ベクターとDNaseI発現ベクターをCOS-7細胞に導入し、免疫蛍光染色を用いて各DNaseの細胞内での分布を観察した。なお、マウスFas抗原の突然変異型(mFasΔC)は、膜貫通タンパクとして細胞表面に発現していることが知られているので、細胞膜における観察を容易にするために、mFasΔCを同時にCOS-7細胞に導入した。
図4Aに示すように、DNaseXタンパク質は、細胞膜に局在して発現し、mFasΔCと同様な分布を示していた(図4A上段)。一方、コントロールとして用いたDNaseIタンパク質は、細胞膜に発現していなかった(図4A下段)。
次に、疎水性のC端ドメインを欠失するDNaseXΔCの発現ベクターとDNaseXのC端ドメインを有するDNaseI-HDの発現ベクターを用いて、同様の実験を行ったところ、図4Bに示すように、DNaseXΔCタンパク質は膜に局在せず、DNaseI-HDタンパク質は膜に局在した。このことから、疎水性のC端ドメインは、DNaseXタンパク質が膜に局在するために必須の領域であることが結論される。
【0110】
(4)DNaseXタンパク質の膜結合性
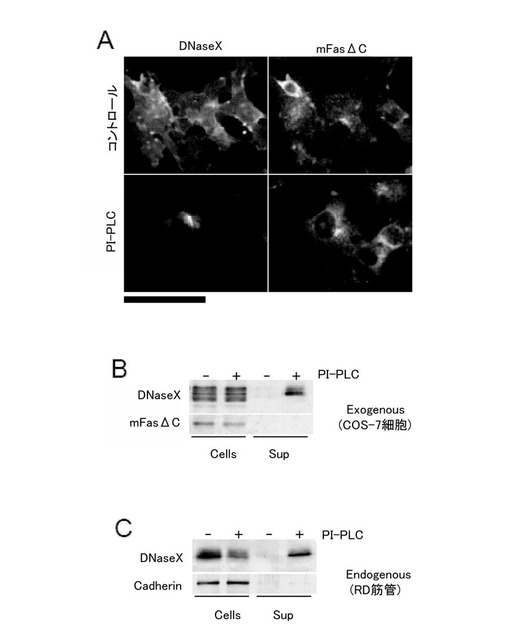
ここでは、GPIアンカーを解離する酵素として知られているPI-PLC(ホスファチジルイノシトール特異性ホスホリパーゼ)を用いて、以下の実験を行ない、DNaseXタンパク質がGPIアンカーで細胞膜に結合していることを示す。
【0111】
まず、Fugene6(Roche)を用いて、DNaseX発現ベクターとmFasΔC発現ベクターを同時にCOS-7細胞に導入し、PI-PLC処理をした後、免疫蛍光染色を用いてDNaseXの細胞膜への分布を観察した(図5A下段)。コントロールとして、PI-PLCの処理をしない細胞におけるDNaseXの細胞膜への分布を観察した(図5A上段)。図5Aに示すように、PI-PLCの処理を行うと、膜貫通タンパク(mFasΔC)の発現量はコントロールと比べて変化しなかったが、DNaseXタンパク質の発現量はコントロールと比べて減少した。
【0112】
さらに、DNaseX発現ベクターとmFasΔC発現ベクターを導入したCOS-7細胞に対し、PI-PLCの処置を行った後に、細胞と上清を分画して、DNaseXタンパク質の存在を、ウエスタンブロッティング法によって調べたところ、図5Bに示すように、PI-PLC処理によって、膜貫通タンパク(mFasΔC)は培養上清中に放出されなかったが、DNaseXタンパク質は培養上清中に放出された。
【0113】
また、RD細胞由来の筋管細胞を用いて、PI-PLC処理後、同様にウエスタンブロッティングを行ったところ、内在性のDNaseXにおいても、上記と同様の結果が得られた(図5C)。なお、ここでは、コントロールとして、細胞間接着分子である膜タンパク質カドヘリンを用いた。
【0114】
このように、DNaseXタンパク質は、細胞の外側で、GPIアンカーによって細胞膜に結合して存在していることが結論される。
【0115】
(5)RD細胞の分化段階とエンドサイトーシス依存性遺伝子導入効率の比較
ここでは、RD細胞が分化するに従い、DNaseXタンパク質によるエンドサイトーシス依存性DNA取り込み阻害が向上することを示す。
【0116】
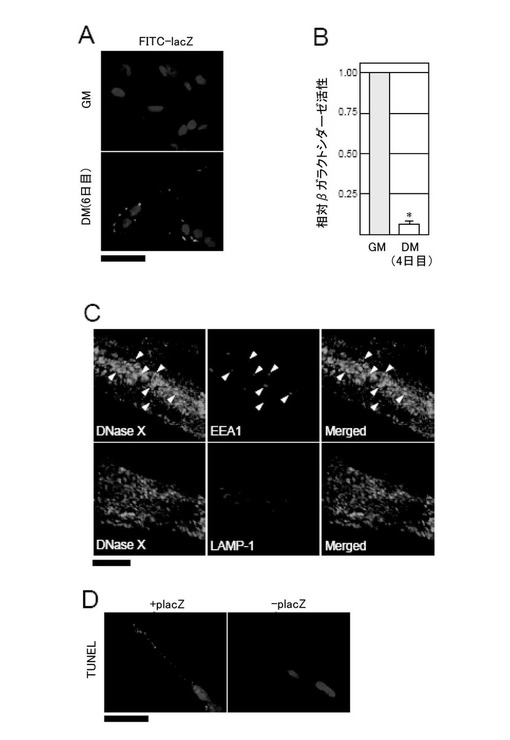
まず、エンドサイトーシスによる遺伝子導入効率を評価するために、人工的に作製したαヘリックスのカチオン型ペプチドによる遺伝子導入を利用した(Niidome, T., Ohmori, N., Ichinose, A., Wada, A., Mihara, H., Hirayama, T., Aoyagi, H., J. Biol. Chem., 272, 15307-15312 (1997), Ohmori, N., Niidome, T., Kiyota, T., Lee, S., Sugihara, G., Wada, A. Hirayama, T. and Aoyagi, H., Biochem. Biophys. Res. Commun., 245, 259-265 (1998))。未分化RD細胞及びRD細胞を6日間分化させた筋管細胞を直径60mmの培養皿で培養し、あらかじめカチオン性ペプチドLLKLLLKLWKKLLKLLK(配列番号29)(遺伝子導入試薬Transome IV(和光純薬工業株式会社))と複合体をつくらせたFITC標識のLacZ cDNAベクター(FITC-lacZ)を導入した。なお、 FITC-lacZは、FITC標識のプライマー(5’-FITC-GAC GGATCGGGAGATCTCCCG-3’(センスプライマー)(配列番号30)、5'-FITC-CGAAAGGAGCGGGCGCTAGGG-3’(アンチセンスプライマー)(配列番号31)を用い、placZを鋳型にしてPCR法を用いて作製した。24時間後、これらの細胞を、DPBSで2回洗浄し、3.7%ホルムアルデヒドで固定(室温、10分間)した。その後、これらの細胞をDPBSで3回洗浄し、100nM Hoeschst 33342で核を染色した後、対比染色して、蛍光顕微鏡にてこれらの細胞を観察した。図6A(上図:未分化RD細胞、下図:筋管細胞)に示したように、FITCのシグナルは、未分化の細胞より筋管細胞で多く観察された。このことは、筋管細胞は、未分化の細胞と比べて、蛍光標識をした遺伝子(FITC-lacZ)を多く取り込むことを示す。
【0117】
次に、同様に、Transome IVと複合体をつくらせたFITC-lacZ又はインサートを有しない空のベクターを各細胞に導入し、BetaGro Assay System(Promega株式会社)を用いてβガラクトシダーゼ活性を測定した。なお、pFITC-lacZで得られた測定値から、空のベクターで得られた測定値(バックグラウンドの値と考えられる)を引くことにより、実際の値を算出した。図6Bに示したように、筋管細胞は、未分化の細胞と比べて、蛍光標識をした遺伝子(FITC-lacZ)を多く取り込んだにもかかわらず、未分化の細胞と比べて、10%以下しかβガラクトシダーゼを発現していなかった。
【0118】
次に、分化した筋管細胞において、エンドサイトーシス小胞(初期、後期)とDNaseXタンパク質との関係を調べるため、抗DNaseX抗体及び抗EEA1抗体あるいは抗LAMP-1抗体と二重染色したところ、図6Cに示す通り、DNaseXタンパク質は、初期エンドソームマーカーEEA1と同じ場所に検出されたが、成熟リソソームマーカーLAMP-1と同じ場所には検出されなかった。
【0119】
さらに、細胞内で分解されたDNA断片をin situで検出するために、筋管細胞にLacZ cDNA発現ベクター(placZ)をトランスフェクトし、TUNEL法を用いて、断片化されたDNA3'末端ヒドロキシル基を検出した(Gavrieli et al., J. Cell Biol., 1992, vol.119, p.493-501)ところ、図6Dに示すように、placZを導入した筋管細胞では、placZを導入しない筋管細胞に対し、TUNELに対してより強い陽性反応を示した。このことは、筋管細胞において、取り込まれたplacZが、細胞内で分解されていることを示す。
【0120】
このように、筋管細胞は、未分化なRD細胞に比べ、DNaseXタンパク質が多量に存在すること、エンドサイトーシス活性は高いものの、取り込まれた遺伝子は分解されやすいことを示している。
【0121】
(6)DNaseXタンパク質の過剰発現
本実施例では、細胞内でDNaseXタンパク質を過剰発現させると、細胞のエンドサイトーシス依存性DNA取り込み能が低下することを示す。
【0122】
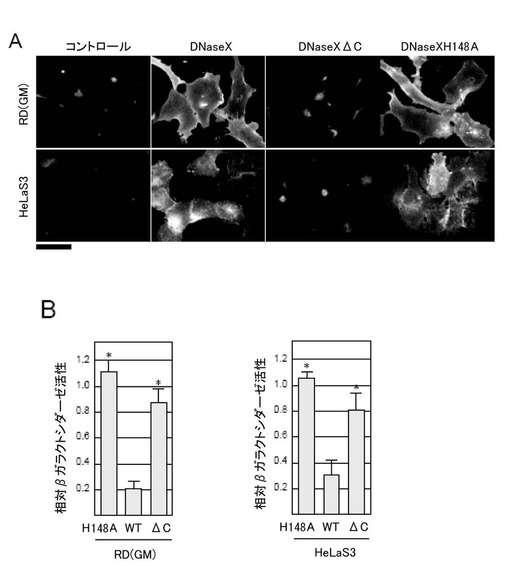
まず、Fugene6 (Roche)を用いて、本来DNaseXの発現がほとんど認められない未分化RD細胞及びHeLaS3細胞に、野生型のDNaseXを発現するphDNaseX、C端の疎水性領域を欠失したDNaseXを発現するphDNaseXΔC、活性部位に点突然変異を有するDNaseXを発現するphDNaseXH148Aの各発現ベクターを導入した。コントロールの細胞には、空のベクターを導入した。
【0123】
抗DNaseX抗体を用いた免疫蛍光染色によってDNaseXタンパク質の発現を観察したところ、HeLa S3細胞及び未分化のRD細胞の細胞表面において、phDNaseX、及びphDNaseXH148Aを導入した場合、細胞膜上にDNaseXタンパク質の発現が認められた(図7A)。一方、C末端疎水性ペプチドを欠損している欠失変異体phDNaseXΔCを導入した場合では、細胞膜でのDNaseXの発現が認められなかった(図7A)。
【0124】
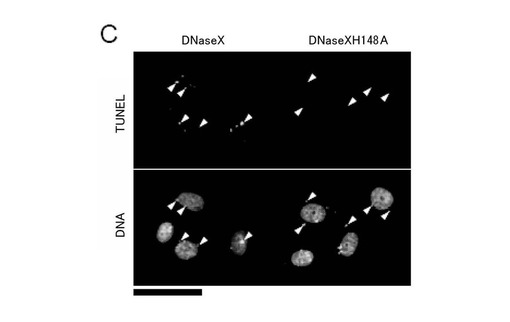
また、(5)と同様にしてエンドサイトーシスによる遺伝子導入の影響を調べたところ、図7Bに示すように、野生型のDNaseXタンパク質を過剰発現させた場合、両方の細胞において、βガラクトシダーゼ活性を阻害するが、C末端を欠失させるか、活性部位に点突然変異を導入することによってβガラクトシダーゼの活性阻害は減少した(図7B)。
【0125】
さらに、TUNEL法を用いて、HeLa S3細胞に導入されたDNAの切断を検出したところphDNaseXを導入した細胞は、陽性シグナルが検出されたが、phDNaseXH148Aを導入した細胞は、陽性シグナルが検出できなかった(図7C)。
【0126】
これらの実験結果から、細胞内でDNaseXタンパク質を過剰発現させると、DNaseXタンパク質依存的に細胞のエンドサイトーシス依存性DNA取り込みを抑制できることが結論される。
【0127】
(7)DNaseXタンパク質の発現抑制
本実施例では、細胞内でDNaseXタンパク質を発現抑制すると、細胞のエンドサイトーシス依存性DNA取り込み能が向上することを示す。
(1)RNA
ヒトDNaseX遺伝子(配列番号2)に対するsiRNA及びncRNA (ネガティブコントロールとして使用するRNA duplex)は、Invitrogen社から購入した。なお、本実施例にて用いるsiRNA(small interfering RNA)は、標的配列5'-CCTGCTTCGAGAACTCAATCGATTT-3(配列番号32)に対応する。
(2)siRNAを用いたDNaseX遺伝子の発現抑制
まず、継代培養しているRD細胞を60mm培養皿に播種し、DMEM培地で培養した。次に、培地を分化培地(DM)に変更し、Lipofectamine 2000 transfection regent(Invitrogen社)を用いて、120pmolのsiRNAをRD細胞にトランスフェクトした。その後も培地交換を行ながら、分化培地で4日間RD細胞の培養を行った。各細胞に対し、抗DNaseX抗体、抗SERCA-1抗体、抗GADPH抗体を用いてウエスタンブロッティング法を行った。ここで、SERCA-1は筋管細胞への分化マーカーであり、GADPHはポジティブコントロールである。なお、コントロールとして、何もトランスフェクトしないで、他の処理は同様に行った細胞を用いた。
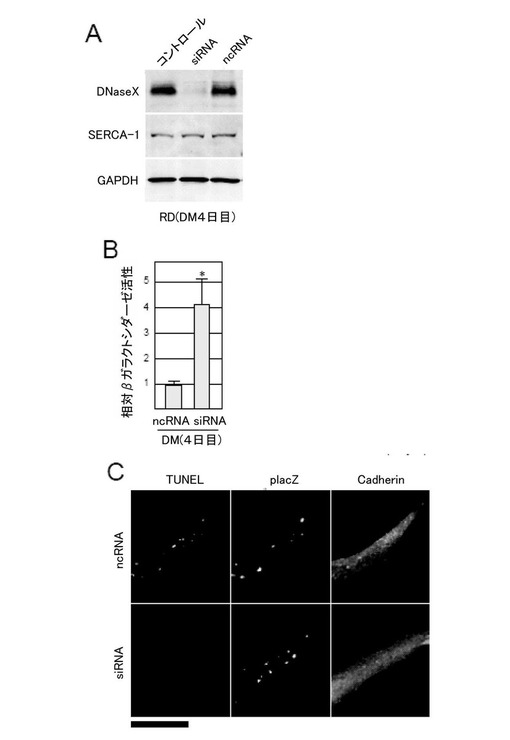
【0128】
図8Aに示すように、DNaseX遺伝子の発現は、ヒトDNaseX遺伝子に対して特異的に反応するsiRNAを導入した細胞において有意に減少した。一方、ncRNAを導入した細胞は、DNaseX遺伝子の発現に変化はなかった。この結果は、用いたsiRNAが、DNaseX遺伝子の発現を抑制できることを示す。また、いずれの場合もSERCA-1の発現が検出されることから、DNaseXは、RD細胞の筋肉分化に不可欠な要素ではないことが示される。
【0129】
さらに、siRNAを導入したRD細胞由来の筋管細胞において、(5)と同様にしてエンドサイトーシスによる遺伝子導入の影響を調べたところ、siRNAを用いてDNaseX遺伝子をノックダウンすると、分化した筋管細胞において、4倍以上も導入遺伝子の発現が促進された(図8B)。さらに、TUNEL法を用いて、分化した筋管細胞に導入されたプラスミドの切断を検出したところ、siRNAが導入された筋管細胞では、陽性反応を示すシグナルは、ほとんど認められなかった(図8C)。
【0130】
このように、細胞内でDNaseXタンパク質を発現抑制すると、細胞のエンドサイトーシス依存性DNA取り込みは促進されることが結論される。
【図面の簡単な説明】
【0131】
【図1】本発明の一実施例において、新規抗ヒトDNaseXモノクローナル抗体の高い特異性をウエスタンブロッティング法で示した図である。
【図2】本発明の一実施例において、新規抗ヒトDNaseIモノクローナル抗体の高い特異性をウエスタンブロッティング法で示した図である。
【図3−1】本発明の一実施例において、ヒト筋肉組織及びRD細胞におけるDNaseXの発現を示す図である。(A)免疫組織化学染色(スケール:100μm)、(B)位相差顕微鏡像(スケール:100μm)、(C)ウエスタンブロッティング法(コントロール:GAPDH)による。
【図3−2】本発明の一実施例において、ヒト筋肉組織及びRD細胞におけるDNaseXの発現を示す図である。(D)共焦点レーザー顕微鏡像(スケール:50μm)、(E)蛍光顕微鏡像(スケール:50μm)、(F)共焦点レーザー顕微鏡像(スケール:50μm)による。
【図4】本発明の一実施例において、DNaseXのC端の疎水性領域が膜に結合するために必要であることを示す図である。なお、mFasΔCはマウスFasタンパク質のC端が欠失した変異体である。(A)蛍光顕微鏡像(スケール:50μm)、(B)蛍光顕微鏡像(スケール:50μm)による。
【図5】本発明の一実施例において、PI-PLC処理による細胞表面からのDNaseXの解離を示す図である。(A)蛍光顕微鏡像(スケール:50μm)、(B)(C)ともウエスタンブロッティング法による。(+)5単位/ml PI-PLC存在下、(-)PI-PLC非存在下を表す。(Cells)細胞中(Sup)培養上清中のDNaseXの存在を調べた。
【図6】本発明の一実施例において、増殖性RD細胞及び分化した筋管細胞におけるエンドサイトーシス依存性遺伝子取り込みを示す図である。(A)蛍光顕微鏡像(スケール:50μm)(C)共焦点レーザー顕微鏡像(スケール:10μm)、(D)蛍光顕微鏡像(スケール:50μm)による。GMは増殖性RD細胞を、DMは分化筋管細胞を示す。(B)lacZの活性を示す(N=5、*=p<0.001(t検定))。
【図7−1】本発明の一実施例において、細胞内でDNaseXタンパク質を過剰発現させると、細胞のエンドサイトーシス依存性DNA取り込み能が低下することを示す図である。(A)蛍光顕微鏡像(スケール:50μm)による。(B)lacZの活性を示す(N=6、*=p<0.001(t検定))。
【図7−2】本発明の一実施例において、細胞内でDNaseXタンパク質を過剰発現させると、細胞のエンドサイトーシス依存性DNA取り込み能が低下することを示す図である。(C)蛍光顕微鏡像(スケール:50μm)による。
【図8】本発明の一実施例において、細胞内でDNaseXタンパク質を発現抑制すると、細胞のエンドサイトーシス依存性DNA取り込み能が向上することを示す図である。(A)ウエスタンブロッティング法(C)蛍光顕微鏡像(スケール:50μm)による。(B)lacZの活性を示す(N=6、*=p<0.001(t検定))。
【技術分野】
【0001】
本発明は、エンドサイトーシス依存性DNA取り込み抑制剤、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染阻害剤、ウイルス感染予防剤、及びウイルス感染促進剤、並びに、遺伝子マーカー、モノクローナル抗体、ハイブリドーマ、及びそれらの使用方法に関する。
【背景技術】
【0002】
DNaseXは、DNaseI様DNaseとして同定された初めての哺乳動物DNaseである。DNaseXは、そのC末端に疎水性ドメイン(HD)を有することが分かっており(非特許文献1参照)、この疎水性ドメインは各種の哺乳動物間で保存されている(非特許文献2参照)。
【0003】
近年、DNaseタンパク質の機能を用いて、癌及びその前駆病変(例えば、良性腫瘍、悪性腫瘍、癌腫、肉腫等)を検出する方法や、癌及びその前駆病変等の疾患を診断する方法が開発された(非特許文献3参照)。しかしながら、DNaseXタンパク質の詳細な機能は未だ分かっていなかった。
【非特許文献1】Shiokawa, D., and Tanuma, S., Biochemistry, 40, 143-152 (2001)
【非特許文献2】Shiokawa, D., Shika, Y., Saito, K., Yamazaki, K. and Tanuma, S., Biochem. J., 392, 511-517 (2005)
【非特許文献3】特表2006―510363号公報
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明はエンドサイトーシス依存性DNA取り込み抑制剤、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染阻害剤、ウイルス感染予防剤、ウイルス感染促進剤、及び新規な疾患モデル動物、並びに、遺伝子マーカー、モノクローナル抗体、ハイブリドーマ、及びそれらの使用方法を提供することを目的とする。
【課題を解決するための手段】
【0005】
本発明者らは、RD細胞(rhabdomyosarcoma)をin vitroで分化させた筋管細胞は、未分化のRD細胞より外来lacZ遺伝子をエンドサイトーシスにより効率よく取り込むが、その遺伝子産物であるβガラクトシダーゼの発現は10%以下であり、強く抑制されることを見出した。DNaseXタンパク質が生体の筋組織及びRD細胞由来の筋管細胞において高く発現していることや、さらに、未分化の筋管細胞でDNaseXタンパク質を強制的に発現させると、エンドサイトーシスによる遺伝子の取り込みが強く抑制され、一方、RD細胞由来の筋管細胞でsiRNAによってDNaseX遺伝子の発現を抑制すると、エンドサイトーシスによる遺伝子の取り込みが促進することを見出した。こうして、発明者らは、外来遺伝子のエンドサイトーシスに対するDNaseXタンパク質の機能を明らかにした。一方、本発明者らは、ヒトDNaseXタンパク質の一部の配列であるオリゴペプチドを用い、ヒトDNaseXタンパク質を特異的に認識する抗体を作製し、本発明を完成するに至った。
【0006】
すなわち、本発明にかかるDNA取り込み抑制剤は、DNaseXタンパク質の機能を増強するエンドサイトーシス依存性DNA取り込み抑制剤であることを特徴とする。なお、前記DNA取り込み抑制剤は、DNaseXタンパク質の発現を増強するDNA取り込み抑制剤であってもよい。
【0007】
また、本発明にかかるウイルス感染阻害剤は、DNaseXタンパク質の機能を増強するウイルス感染阻害剤であることを特徴とする。なお、前記ウイルス感染阻害剤は、DNaseXタンパク質の発現を増強するウイルス感染阻害剤であってもよい。また、本発明にかかる細胞に対するウイルス感染阻害方法は、当該細胞内でDNaseXタンパク質の機能を増強することを特徴とする。その一手段として、DNaseXタンパク質の発現を増強してもよい。
【0008】
さらに、本発明にかかるウイルス感染予防剤又はウイルス感染に対するワクチンは、前記ウイルス感染阻害剤を含有することを特徴とする。また、本発明にかかるヒト又はヒト以外の脊椎動物に対するウイルス感染予防方法は、当該動物内でDNaseXタンパク質の機能を増強することを特徴とする。その一手段として、DNaseXタンパク質の発現を増強してもよい。
【0009】
また、本発明にかかる発現ベクターは、DNaseXタンパク質を強制発現させる発現ベクターであることを特徴とする。
【0010】
さらに、本発明にかかる細胞は、DNaseXタンパク質の機能が増強された細胞であることを特徴とする。ここで、前記細胞は、DNaseXタンパク質を強制発現させる発現ベクターを有していてもよい。
【0011】
また、本発明にかかるヒト又はヒト以外の脊椎動物は、体の一部又は全部で、DNaseXタンパク質が異所的に又は過剰に機能が増強していることを特徴とする。ここで、前記脊椎動物は、DNaseXタンパク質を強制発現させる発現ベクターを細胞に導入することにより作製されてもよい。また、前記脊椎動物は、DNaseXタンパク質を強制発現させる発現ベクターによって作製されたトランスジェニック脊椎動物であってもよい。
【0012】
さらに、本発明にかかるスクリーニング方法は、DNaseXタンパク質の機能を増強する化合物をスクリーニングすることを特徴とする。
【0013】
本発明のモノクローナル抗体は、RSLLHTAAAFDFPTSFQL(配列番号3)の一部又は全部をエピトープとする。本発明のハイブリドーマは、このモノクローナル抗体を産生する。本発明のハイブリドーマは、寄託受領番号FERM AP-21282であることが好ましい。本発明のモノクローナル抗体は、この寄託受領番号FERM AP-21282のハイブリドーマから産生されるものでもよい。
【0014】
また、本発明の検出方法は、上記いずれかのモノクローナル抗体を用いて、DNaseXタンパク質を検出する検出方法である。
【0015】
また、本発明にかかるDNA取り込み促進剤は、DNaseXタンパク質の機能を抑制するエンドサイトーシス依存性DNA取り込み促進剤であることを特徴とする。なお、前記DNA取り込み促進剤は、上記いずれかのモノクローナル抗体を有効成分として含有してもよく、DNaseXタンパク質の発現を抑制してもよい。
【0016】
さらに、本発明にかかるウイルス感染促進剤は、DNaseXタンパク質の機能を抑制するウイルス感染促進剤であることを特徴とする。なお、前記ウイルス感染促進剤は、上記いずれかのモノクローナル抗体を有効成分として含有してもよく、DNaseXタンパク質の発現を抑制してもよい。また、前記ウイルス感染促進剤は、DNaseX遺伝子に対するsiRNAを有効成分として含有することを特徴とする。ここで、前記siRNAは、配列番号32を有していることが好ましい。また、本発明にかかる細胞又はヒト又はヒト以外の脊椎動物に対するウイルス感染促進方法は、当該細胞又は当該ヒト又はヒト以外の脊椎動物内でDNaseXタンパク質の機能を抑制することを特徴とする。その一手段として、DNaseXタンパク質の発現を抑制してもよい。
【0017】
また、本発明にかかる発現ベクターは、DNaseX遺伝子に対するsiRNAを発現する発現ベクターであることを特徴とする。
【0018】
さらに、本発明にかかる細胞は、内在性DNaseXタンパク質の機能が抑制されている細胞であることを特徴とする。ここで、前記細胞は、DNaseX遺伝子に対するsiRNAを発現する発現ベクターを有することが好ましい。
【0019】
また、本発明にかかるヒト又はヒト以外の脊椎動物は、体の一部又は全部で、内在性DNaseXタンパク質の機能が抑制されていることを特徴とする。ここで、前記脊椎動物は、DNaseX遺伝子のノックダウン又はノックアウトによって作製されていてもよい。
【0020】
さらに、発明にかかるスクリーニング方法は、DNaseXタンパク質の機能を抑制する化合物をスクリーニングすることを特徴とする。上記いずれかのモノクローナル抗体を用いて、スクリーニングしてもよい。
【0021】
なお、本明細書で「エンドサイトーシス依存性DNA取り込み」というのは、エンドサイトーシスに依存して、細胞内にほぼ分解されずにDNAを取り込むことをいい、特に、DNAが細胞内に取り込まれる際に、DNaseXタンパク質によって分解されないことを意味する。また、ウイルスの宿主への感染とは、ウイルスが宿主に接触してDNAを宿主内に導入する段階のことをいい、ゲノム内にDNAが挿入されたり、細胞内でウイルスDNAが増幅したり、ウイルスが増殖したり、それによって様々な現象が引き起こされたりするのは、感染後に生じることである。
【0022】
また、本発明の遺伝子マーカーは、DNaseX遺伝子関連物質である。特に、ウイルスに対する易感染性予測マーカー、又はウイルス感染後の病状推定マーカーであることを特徴としてもよい。前記ウイルスが、インフルエンザウイルス、肝炎ウイルス、HIVウイルス、ATLウイルス、エボラウイルス、黄熱病ウイルス、狂犬病ウイルス、サイトメガロウイルス、SARSウイルス、JCウイルス(進行性多巣性白質脳症)、水痘・帯状疱疹ウイルス、コクサッキーA16型・エンテロウイルス71ウイルス(手足口病)、デング熱ウイルス、パルボウイルス(伝染性紅斑)、EBウイルス(伝染性単核球症)、天然痘ウイルス、風疹ウイルス、ポリオウイルス、麻疹ウイルス、アデノウイルス(プール熱)、マールブルグウイルス(フィロウイルス科)、クリミア・コンゴ出血熱ウイルス、ラッサウイルス、フニンウイルス等の新大陸アレナウイルス、ハンタウイルス、リフトバレー熱ウイルス、ムンプスウイルス(流行性耳下腺炎)、ウエストナイルウイルス、単純ヘルペスウイルス、コクサッキーウイルス、ロタウイルス、ノロウイルス、チクングニアウイルス(アルファウイルス属、トガウイルス科)からなる群から選ばれてもよい。前記遺伝子関連物質がDNA、hnRNA、mRNA、及びタンパク質からなる群から選ばれることが好ましい。
【0023】
さらに、本発明のキットは、ウイルスに対する易感染性予測又はウイルス感染後の病状推定のためのキットであって、DNaseXタンパク質に対する抗体又はDNaseXcDNAを増幅するためのPCR用プライマーを含有する。前記抗体がモノクローナル抗体であってもよい。このモノクローナル抗体が、配列番号3の一部又は全部をエピトープとしてもよい。さらに、DNaseXタンパク質の検出には、抗原抗体反応を利用する全ての方法が可能であるが、特に免疫クロマトグラフ測定法を利用していることが好ましい。
【発明の効果】
【0024】
本発明によればエンドサイトーシス依存性DNA取り込み抑制剤、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染阻害剤、ウイルス感染予防剤、ウイルス感染促進剤、及び新規な疾患モデル動物、並びに遺伝子マーカー、モノクローナル抗体、ハイブリドーマ、及びそれらの使用方法を提供することができる。
【発明を実施するための最良の形態】
【0025】
以下、上記知見に基づき完成した本発明の実施の形態を、実施例を挙げながら詳細に説明する。実施の形態及び実施例に特に説明がない場合には、J. Sambrook, E. F. Fritsch & T. Maniatis (Ed.), Molecular cloning, a laboratory manual (3rd edition), Cold Spring Harbor Press, Cold Spring Harbor, New York (2001); F. M. Ausubel, R. Brent, R. E. Kingston, D. D. Moore, J.G. Seidman, J. A. Smith, K. Struhl (Ed.), Current Protocols in Molecular Biology, John Wiley & Sons Ltd.等の標準的なプロトコール集に記載の方法、あるいはそれを修飾したり、改変した方法を用いる。また、市販の試薬キットや測定装置を用いている場合には、特に説明が無い場合、それらに添付のプロトコールを用いる。
【0026】
なお、本発明の目的、特徴、利点、及びそのアイデアは、本明細書の記載により、当業者には明らかであり、本明細書の記載から、当業者であれば、容易に本発明を再現できる。以下に記載された発明の実施の形態及び具体的に実施例等は、本発明の好ましい実施態様を示すものであり、例示又は説明のために示されているのであって、本発明をそれらに限定するものではない。本明細書で開示されている本発明の意図並びに範囲内で、本明細書の記載に基づき、様々な改変並びに修飾ができることは、当業者にとって明らかである。
【0027】
==本発明の薬剤と使用方法==
本発明者らが明らかにしたように、DNaseXタンパク質は、GPI結合性膜タンパク質であり、外来DNAのエンドサイトーシス依存性取り込みを抑制する機能を有する。
【0028】
(1)エンドサイトーシス依存性DNA取り込み抑制剤
従って、DNaseXタンパク質の機能を増強する物質は、エンドサイトーシス依存性DNA取り込み抑制剤として有用である。エンドサイトーシスによってDNAが細胞内に取り込まれる場合としては、例えば、ウイルスが細胞に感染した場合等が挙げられる。そこで、DNaseXタンパク質の機能を増強する物質を有効成分として含有する薬剤は、ウイルス感染を阻害できるウイルス感染阻害剤や、ウイルス感染予防剤や、ウイルス感染に対するワクチンとして有用である。なお、ウイルスの種類は、特に限定されない。
【0029】
ここで、「DNaseXタンパク質の機能を増強する」とは、細胞全体でDNaseXタンパク質の機能が増強されれば良く、例えばDNaseXタンパク質分子の活性が増強されることにより機能が増強されるだけでなく、細胞内でDNaseXタンパク質分子の量が増加することにより機能が増強されてもよい。
【0030】
DNaseXタンパク質の機能を増強するための方法は特に限定されないが、例えば、DNaseXタンパク質を直接細胞に導入する方法やDNaseXタンパク質の機能を増強する物質を細胞に投与する方法が考えられる。
【0031】
DNaseXタンパク質を細胞に導入するための方法としては特に限定されず、Protein Transduction Domains (PTD) 融合タンパク質としてTATやVP22との融合タンパク質を用いてもよく、BioPorterTM、ChariotTM等のタンパク質導入試薬を用いてもよい。
【0032】
DNaseXタンパク質の機能を増強する物質としては特に限定されず、DNaseXタンパク質の活性を増強する物質、内在DNaseX遺伝子の発現レベルを亢進させる物質、外来DNaseX遺伝子を強制発現させる発現ベクター等、様々な物質が含まれるが外来DNaseX遺伝子を強制発現させる発現ベクターであることが好ましい。
【0033】
内在DNaseX遺伝子の発現レベルを亢進させる物質としては、例えば、相同組換えによってゲノム中のDNaseX遺伝子のプロモーターをより強力なプロモーターで置換することができる相同組換え用ベクターや、DNaseX遺伝子のプロモーターを活性化するタンパク質をコードする遺伝子の発現ベクター等を用いることができる。
【0034】
ここで、内在DNaseX遺伝子とは、ゲノム中の本来のDNaseX遺伝子座に存在するDNaseX遺伝子のことをいい、外来DNaseX遺伝子とは、細胞外に由来し、ゲノム中の本来のDNaseX遺伝子座以外の場所に存在するDNaseX遺伝子のことをいう。
【0035】
外来DNaseX遺伝子を強制発現させる発現ベクターは、導入する宿主細胞で機能するプロモーターを備えていれば、どんなベクターでもよく、プラスミドベクターでも、ウイルスベクターでもよい。また、遺伝子導入に際しては、遺伝子を導入後、発現ベクターを染色体の外におく一過的発現(transient expression)であっても、発現ベクターが染色体に組み込まれた永久発現(permanent expression)であってもよい。遺伝子導入は、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法、マイクロインジェクション法、ウイルス感染法、その他様々なDDS(drug delivery system)等、その方法は特に限定されない。
【0036】
(2)エンドサイトーシス依存性DNA取り込み促進剤
一方、DNaseXタンパク質の機能を抑制する物質を有効成分として含有する薬剤は、ウイルス感染を促進することができる。従って、DNaseXタンパク質の機能を抑制する物質は、エンドサイトーシス依存性DNA取り込み促進剤として有用であり、DNaseXタンパク質の機能を抑制する物質を有効成分として含有する薬剤は、ウイルス感染を促進できるウイルス感染促進剤として有用である。ここで、「DNaseXタンパク質の機能を抑制する」とは、細胞全体でDNaseXタンパク質の機能が抑制されれば良く、例えばDNaseXタンパク質分子の活性が低下することにより機能が抑制されるだけでなく、細胞内でDNaseXタンパク質分子の量が減少することにより機能が抑制されてもよい。
【0037】
DNaseXタンパク質の機能を抑制するための方法は特に限定されないが、例えば、細胞内にあるDNaseXタンパク質を分解する方法やDNaseXタンパク質の機能を抑制する物質を細胞に投与する方法等が考えられる。
【0038】
細胞内にあるDNaseXタンパク質を分解する方法は特に限定されないが、例えば、相同組換えによって、ゲノム内のDNaseX遺伝子を、ユビキチンの標的配列を有したDNaseXタンパク質をコードする融合DNaseX遺伝子に置換し、外来遺伝子によってユビキチンを供給する方法等が考えられる。
【0039】
なお、DNaseXタンパク質の機能を抑制する物質は、例えば、DNaseXタンパク質に特異的に結合し、その機能を特異的に抑制する抗体やアプタマーや、化合物等であってもよい。この抗体は、ポリクローナル抗体でもモノクローナル抗体でもよいが、モノクローナル抗体であることが好ましく、下記のようにして作製された、RSLLHTAAAFDFPTSFQLの一部又は全部をエピトープとするモノクローナル抗体であってもよい。また、DNaseXタンパク質の機能を抑制するために、内在DNaseX遺伝子の発現を抑制してもよく、例えば、DNaseX遺伝子のプロモーターを抑制する物質やDNaseX遺伝子の発現を標的にする、siRNA、miRNA、アンチセンスRNA等を用いてもよく、相同組換え等を用いてゲノム中のDNaseX遺伝子を破壊してもよい。
【0040】
ここで、用いられる細胞は、培養細胞であっても、脊椎動物個体中の細胞であってもよい。
【0041】
==疾患モデル動物及びその有用性==
(1)疾患モデル動物の作製
本発明の疾患モデル動物は、脊椎動物個体に対し、体の全部又は一部でDNaseXタンパク質の機能を異所的に又は過剰に増強させる処理を施すこと、又は体の全部又は一部でDNaseXタンパク質の機能を抑制する処理を施すことにより作製することができる。
【0042】
脊椎動物個体において、DNaseXタンパク質の機能を増強するための方法は特に限定されないが、例えば、DNaseXタンパク質を個体に直接に導入する方法やDNaseXタンパク質の機能を増強する物質を個体に投与する方法が考えられる。投与方法は、投与する物質によって適宜使い分ければよく、経口投与でも非経口投与でもよい。
【0043】
DNaseXタンパク質を個体に導入するための方法としては特に限定されず、Protein Transduction Domains (PTD) 融合タンパク質としてTATやVP22との融合タンパク質を用いてもよく、BioPorterTM、ChariotTM等のタンパク質導入試薬を用いてもよく、DNaseXタンパク質を注入してもよい。
【0044】
DNaseXタンパク質の機能を増強する物質としては特に限定されず、DNaseXタンパク質の活性を増強する物質、内在DNaseX遺伝子の発現レベルを亢進させる物質、外来DNaseX遺伝子を強制発現させる発現ベクター等、様々な物質が含まれる。特にDNA等は、リポフェクション法、エレクトロポレーション法、マイクロインジェクション法、ウイルス感染法等、常法に従って投与すればよい。
【0045】
内在DNaseX遺伝子の発現レベルを亢進させる物質としては、例えば、相同組換えによってゲノム中のDNaseX遺伝子のプロモーターをより強力なプロモーターで置換することができる相同組換え用ベクターや、DNaseX遺伝子のプロモーターを活性化するタンパク質をコードする遺伝子の発現ベクター等を用いることができる。
【0046】
外来DNaseX遺伝子を強制発現させる発現ベクターは、導入する個体で機能するプロモーターを備えていれば、どんなベクターでもよく、プラスミドベクターでも、ウイルスベクターでもよい。また、遺伝子導入の系としては、発現ベクターを染色体の外におく一過的発現(transient expression)の系や、発現ベクターが染色体に組み込まれた永久発現(permanent expression)の系を用いてもよい。
【0047】
なお、予め培養細胞や培養組織においてDNaseXタンパク質の機能を増強させ、それらの細胞や組織を脊椎動物個体に移植することにより、体の全部又は一部でDNaseXタンパク質の機能を増強させてもよいが、体の全部でDNaseXタンパク質の機能を増強させるためには、遺伝子操作によることが好ましく、外来DNaseX遺伝子を強制発現させる発現ベクターを用いて、トランスジェニック脊椎動物を作製することがより好ましい。
【0048】
また、脊椎動物個体においてDNaseXタンパク質の機能を抑制するための方法は特に限定されないが、例えば、DNaseXタンパク質の機能を抑制する物質を個体に投与する方法等が考えられる。投与方法は、投与する物質によって適宜使い分ければよいのであって、経口投与でも非経口投与でもよい。
【0049】
なお、DNaseXタンパク質の機能を抑制する物質は、例えば、DNaseXタンパク質に特異的に結合し、その機能を特異的に抑制する抗体やアプタマーや、化合物等であってもよい。その抗体は、ポリクローナル抗体でもモノクローナル抗体でもよいが、モノクローナル抗体であることが好ましく、下記のようにして作製された、RSLLHTAAAFDFPTSFQLの一部又は全部をエピトープとするモノクローナル抗体であってもよい。また、DNaseXタンパク質の機能を抑制するために、内在DNaseX遺伝子の発現を抑制してもよく、例えば、DNaseX遺伝子のプロモーターを抑制する物質やDNaseX遺伝子の発現を標的にする、siRNA、miRNA、アンチセンスRNA等を用いてもよく、相同組換え等を用いてゲノム中のDNaseX遺伝子を破壊してもよい。特にDNAやRNA等では、リポフェクション法、エレクトロポレーション法、マイクロインジェクション法、ウイルス感染法等、常法に従って行えばよい。
【0050】
なお、予め培養細胞や培養組織においてDNaseXタンパク質の機能を抑制し、それらの細胞や組織を脊椎動物個体に移植することにより、体の全部又は一部でDNaseXタンパク質の機能を抑制してもよいが、体の全部でDNaseXタンパク質の機能を抑制するためには、遺伝子操作によることが好ましく、ノックアウト脊椎動物やノックダウン脊椎動物を作製することがより好ましい。例えば、相同組換え等を用いた内在DNaseX遺伝子における変異(欠失、置換、付加、挿入等を含む)の導入による変異体(条件突然変異体も含む)の作製、ドミナント・ネガティブ変異をもつ変異タンパク質を有するトランスジェニック動物の作製、アンチセンスRNAやRNAi、又はそれらをコードするDNAを用いたノックダウン動物の作製をすればよい。
【0051】
なお、疾患モデル動物としては、例えば、マウス、ラット等のヒト以外の脊椎動物であることが好ましい。
【0052】
(2)疾患モデル動物の有用性
DNaseXタンパク質の機能を増強させた動物は、ウイルス等に感染しにくくなるため、感染症抵抗性を示すモデルとして有用である。一方、DNaseXタンパク質のDNaseX活性を抑制した動物は、ウイルス等に感染しやすくなるため、易感染性を示すモデルとして有用である。ここで、感染症とは、ウイルス等の病原体が感染して増殖し、病的症状を発症する疾患のことをいう。
【0053】
==スクリーニング方法==
DNaseXタンパク質の機能を増強する化合物をスクリーニングすることにより、エンドサイトーシス依存性DNA取り込み抑制剤、ウイルス感染阻害剤、ウイルス感染予防剤及びワクチン等に用いられる化合物を得ることができる。
【0054】
具体的な一例として、DNaseX遺伝子を発現する培養細胞、培養組織、培養器官等に、スクリーニングの対象となる物質を投与する。そして、投与前後で、その培養物におけるDNaseX遺伝子の転写物やタンパク質の発現量を測定する。投与前後のDNaseX遺伝子の発現量を比較し、投与後にDNaseX遺伝子の発現量が増強しているかどうかを調べることにより、感染症に対して抵抗性を有する物質を同定することができる。
【0055】
一方、DNaseXタンパク質の機能を抑制する化合物をスクリーニングすることにより、エンドサイトーシス依存性DNA取り込み促進剤、ウイルス感染促進剤等に用いられる化合物を得ることができる。 例えば、DNaseX遺伝子を発現する培養細胞、培養組織、培養器官等に、スクリーニングの対象となる物質を投与する。そして、投与前後で、その培養物におけるDNaseX遺伝子の転写物やタンパク質の発現量を測定する。投与前後のDNaseXタンパク質の発現量を比較し、投与後にDNaseXタンパク質の発現量が低下しているかどうかを調べることにより、感染症に対して予防できる物質や感染症を治療することができる物質を同定することができる。
【0056】
DNaseX遺伝子の発現量は、転写物の量を測定するRT-PCTを用いたり、タンパク質の発現量を測定するELISA法、RIA法、ウエスタンブロッティング法、免疫組織化学染色等を利用したりして、常法に従って測定することができる。なお、タンパク質の発現量を検出するための抗体は、ポリクローナル抗体でもモノクローナル抗体でもよいが、モノクローナル抗体であることが好ましく、下記のようにして作製された、RSLLHTAAAFDFPTSFQLの一部又は全部をエピトープとするモノクローナル抗体であってもよい。
【0057】
DNaseXタンパク質の機能を測定する方法は、DNaseXタンパク質の発現を測定する以外にも、DNAの単位時間当たりの分解量等を測定してもよく、特に限定されない。
【0058】
なお、培養細胞等の代わりに、上記「疾患モデル動物」を用いて、DNaseXタンパク質の機能を増強する化合物又は抑制する化合物をスクリーニングしてもよい。スクリーニング方法は、上述と同様の方法を用いればよい。
【0059】
==DNaseXタンパク質及びDNA==
なお、本発明にかかるDNaseXタンパク質は、以下の(a)又は(b)のタンパク質である。
(a)ヒトDNaseXタンパク質(配列番号1のアミノ酸配列を有するタンパク質)及びそのホモログ。
(b)(a)に記載のタンパク質のアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、又は付加したアミノ酸配列からなり、DNaseX活性を有するタンパク質。
【0060】
これらのタンパク質を合成するには、そのタンパク質をコードするDNAを用いて、発現ベクター等を構築し、細胞あるいは個体に導入してもよい。発現ベクターは、導入する細胞及び個体で機能するプロモーターを備えていれば、どんなベクターでもよく、プラスミドベクターでも、ウイルスベクターでもよい。また、in vitro 転写/翻訳系で合成してもよく、あるいは、そのタンパク質を化学合成してもよい。
【0061】
上記の(a)又は(b)のタンパク質をコードするDNAとしては、そのタンパク質のアミノ酸配列よりコドン表を用いて作成された配列を有するDNAでもよいが、以下の(a)又は(b)のDNAが特に好ましい。
(a)ヒトDNaseX遺伝子のcDNA(配列番号2の塩基配列を有するDNA、Accession No. X90392)及びDNaseX遺伝子ホモログのcDNA。
(b)(a)に記載のcDNAの塩基配列において、1もしくは数個の塩基が欠失、置換、又は付加した塩基配列を有し、DNaseX活性を有するタンパク質をコードするDNA。
【0062】
なお、これらのDNAを得るには、そのDNA配列を有するクローンが含まれるゲノムライブラリーやcDNAライブラリーをスクリーニングしてもよく、そのDNA配列を有する生物種のゲノムやRNAを用いてPCRやRACE法等で増幅してもよく、これらを組み合わせてもよい。また、そのDNAを化学合成してもよい。
【0063】
==抗ヒトDNaseXモノクローナル抗体及びその有用性===
本発明の抗ヒトDNaseXモノクローナル抗体は、ハイブリドーマにより産生されるモノクローナル抗体である。本発明のモノクローナル抗体は、公知の技術を用いて作製することができる。
【0064】
(1)抗ヒトDNaseXモノクローナル抗体の作製
本発明のモノクローナル抗体の作製には、DNaseXタンパク質のうち抗原性を有するタンパク質を免疫原として用いることが好ましい。抗原性を有するタンパク質としては、例えば、ヒトDNaseXの237-254位に対応するペプチド(RSLLHTAAAFDFPTSFQL(配列番号3)が挙げられる。このペプチドは、化学合成によって作製することができる。なお、免疫原には、DNaseXタンパク質を発現している細胞そのものを用いてもよい。
【0065】
免疫に用いる動物は、細胞融合に使用する親細胞との適合性を考慮して選択するのが好ましい。このような動物としては、例えば、マウス、ラット、モルモット、ウサギ等が挙げられる。
【0066】
上記動物に免疫原を投与する方法は、当業者に公知の方法を用いて行えばよいが、免疫原とフロイント完全アジュバント等のアジュバントを併せて投与することが好ましい。
【0067】
上記動物に免疫原を投与し、その動物の血清中の抗体価が上昇したのを確認した後に、免疫動物から脾臓を取り出し、脾臓から脾細胞を調製し、以下の細胞融合を行う。
【0068】
細胞融合は、当業者に公知の方法を用いて行うことができる。ここで、細胞融合に使用する親細胞としては、X63-Ag8-653、NS-1、NS-0/1、SP2O等のミエローマ細胞が好ましい。また、脾細胞とミエローマ細胞との細胞数比は任意に設定することができるが、例えば、ミエローマ細胞に対して脾細胞を10倍とするのが好ましい。細胞融合に用いる培養液としては、例えば、前記ミエローマ細胞株の増殖に好適なRPMI1640培養液、MEM培養液等が挙げられる。なお、PEGは、分子量4000の溶液であることが好ましい。
【0069】
細胞融合後は、HAT培地を用いてハイブリドーマをスクリーニングし、限界希釈法によって抗体を産生するハイブリドーマをクローニングすればよい。また、スクリーニングは、ヒトDNaseXタンパク質を目的とした、ELISA法、ウエスタンブロッティング法、免疫組織化学染色を行えばよい。
【0070】
このようにしてスクリーニングされ、クローン化されたハイブリドーマから産生されるモノクローナル抗体は、ヒトDNaseXタンパク質に対して特異的に反応する。モノクローナル抗体を産生するハイブリドーマは、公知の方法を用いて培養することができる。そして、その培養上清から、モノクローナル抗体を得ることができる。又は、ハイブリドーマをマウス等の動物の腹腔内に投与し、その動物の腹水から、モノクローナル抗体を得てもよい。
【0071】
本発明のモノクローナル抗体としては、例えば、独立行政法人産業技術総合研究所特許生物寄託センターに2007年3月30日に受託受領番号FERM AP-21282で寄託されたhDNase X 1B1 Hybridomaが挙げられる。
【0072】
(2)抗ヒトDNaseXモノクローナル抗体の有用性
(i)易感染性予測マーカー
一般に、ウイルスは、細胞表面に吸着し、その細胞自身が持っているエンドサイトーシス機構によって、エンドソーム小胞として細胞内に取り込まれることが知られている。これより、細胞のエンドサイトーシス機構を抑制すれば、ウイルス感染は阻害され、一方、細胞のエンドサイトーシス機構を促進すれば、ウイルス感染は助長される。
【0073】
以下の実施例に示すように、RD細胞由来の筋管細胞にDNaseXタンパク質を強制的に発現させると、エンドサイトーシスによる遺伝子の取り込みは抑制され、一方、siRNAによってDNaseX遺伝子の発現を抑制すると、エンドサイトーシスによる遺伝子の取り込みは促進する。従って、DNaseX遺伝子の発現レベルによって、ウイルスの細胞への感染のしやすさを予測することができ、DNaseXの遺伝子関連物質はウイルスに対する易感染性予測マーカーとして利用できる。なお、DNaseXの遺伝子関連物質は、DNAであっても、RNA(hnRNA、mRNA)であっても、タンパク質であってもよい。
【0074】
具体的には、例えば、多数の健常人のDNaseXの遺伝子関連物質のレベルを測定し、正常なレベルの範囲を設定し、あるヒト又はヒト以外の脊椎動物におけるDNaseXの遺伝子関連物質のレベルが、その範囲より多かったり少なかったりすれば、その脊椎動物は、通常よりウイルスに感染し難かったり、感染しやすかったりする。これにより、例えば、投与する抗ウイルス剤の強さや量を決めることも可能になる。
【0075】
(ii)ウイルス感染後の病状推定マーカー
一般に、ウイルスが細胞に感染すると、そのウイルスは細胞内で大量に増殖し、細胞本来の生理機能が破錠したり細胞膜の破壊が起きたりして、宿主細胞は死に至る場合がある。また、ウイルスによっては、短期間で大量のウイルスを作って直ちに宿主細胞を殺すのではなく、むしろ宿主細胞へのダメージが少なくなるよう、少量のウイルスを長期間にわたって持続的に産生(持続感染)する場合もある。
【0076】
上記(i)の通り、RD細胞由来の筋管細胞にDNaseXタンパク質を強制的に発現させると、エンドサイトーシスによる遺伝子の取り込みは抑制される。一方、siRNAによってDNaseX遺伝子の発現を抑制すると、エンドサイトーシスによる遺伝子の取り込みは促進する。このように、細胞におけるDNaseX量は、ウイルスに感染したヒト又はヒト以外の脊椎動物の中で、ウイルスが増殖する時の細胞抵抗性を決めると考えられる。従って、ウイルス感染脊椎動物のDNaseX遺伝子の発現レベルより、その動物がどのように病状推移するか、例えば、軽症で済みそうか重症に移行しそうか、等を推定することができ、DNaseXの遺伝子関連物質は病状推移推定マーカーとして利用できる。なお、DNaseXの遺伝子関連物質は、DNAであっても、RNA(hnRNA、mRNA)であっても、タンパク質であってもよい。
【0077】
(iii)対象ウイルス
上記(i)及び(ii)のウイルスとしては、例えば、インフルエンザウイルス、肝炎ウイルス、HIVウイルス、ATLウイルス、エボラウイルス、黄熱病ウイルス、狂犬病ウイルス、サイトメガロウイルス、SARSウイルス、JCウイルス、水痘・帯状疱疹ウイルス、コクサッキーA16型・エンテロウイルス71ウイルス、デング熱ウイルス、パルボウイルス、EBウイルス、天然痘ウイルス、風疹ウイルス、ポリオウイルス、麻疹ウイルス、アデノウイルス、マールブルグウイルス、フィロウイルス、クリミア・コンゴ出血熱ウイルス、ラッサウイルス、フニンウイルス、新大陸アレナウイルス、ハンタウイルス、リフトバレー熱ウイルス、ムンプスウイルス、ウエストナイルウイルス、単純ヘルペスウイルス、コクサッキーウイルス、ロタウイルス、ノロウイルス、チクングニアウイルス、トガウイルス等が挙げられるが、これらに限定されない。
【0078】
==キット==
上記のように、ヒト又はヒト以外の脊椎動物のDNaseX遺伝子の発現レベルより、易感染性予測又はウイルス感染後の病状推定をすることができる。従って、DNaseX遺伝子の発現レベルを測定できるキットは、ウイルスに対する易感染性予測又はウイルス感染後の病状推定に有用である。このキットは、DNaseX遺伝子の発現レベルを測定するために、DNaseXタンパク質に対する抗体、あるいはDNaseXのcDNAを増幅するためのPCR用プライマーを含有することが好ましい。なお、この抗体はポリクローナル抗体であってもモノクローナル抗体であってもよく、モノクローナル抗体が、RSLLHTAAAFDFPTSFQLの一部又は全部をエピトープとしてもよい。なお、簡便さの点から、このキットは、免疫クロマトグラフ測定法等の装置状になっていることが好ましいが抗原抗体反応を利用した方法であれば特に限定されない。
【実施例】
【0079】
以下、本発明の実施例について詳細に述べる。
<実施例1:新規モノクローナル抗体の作製と特異性の確認>
本実施例で作製したモノクローナル抗体は、実施例2において使用した。
【0080】
(1)新規抗ヒトDNaseXモノクローナル抗体の作製
本法では、ヒトDNaseXの237-254位に対応するペプチド(RSLLHTAAAFDFPTSFQL(配列番号3))のN末端にCysを付加した合成ペプチドとチオグロブリンとをN末端のCysによって結合したペプチドを免疫原として用いた。まず、6週齢雌BDF1マウスに、上記抗原50μgを完全フロイントアジュバントと共に皮下注射して初回免疫を行った。初回免疫から2週後、3週後、4週後に、上記抗原50μgを不完全フロイントアジュバントと共に皮下注射して追加免疫を行った。最後に、上記抗原50μgを静脈注射して最終免疫を行った。最終免疫から4日後、上記マウスから脾臓を取り出して脾細胞を調製し、ポリエチレングリコールを用いて、調製した脾細胞とX63-Ag8-653マウスミエローマ細胞(Kinebuchi, M., Ide, T., Lupin, D., Tamatani, T., Miyasaka, M., Matsuura, A., Nagai, Y., Kikuchi, K. and Uede, T., J. Immunol., 146, 3721-3728 (1991))とを細胞融合した。その後、HAT培地を用いてハイブリドーマをスクリーニングし、限界希釈法によってクローニングを行い、最終的にクローン化したハイブリドーマ株1B1を得た(寄託受領番号:FERM AP-21282)。なお、1B1ハイブリドーマから産生される抗ヒトDNaseXモノクローナル抗体のサブクラスはIgG1/κであった。
【0081】
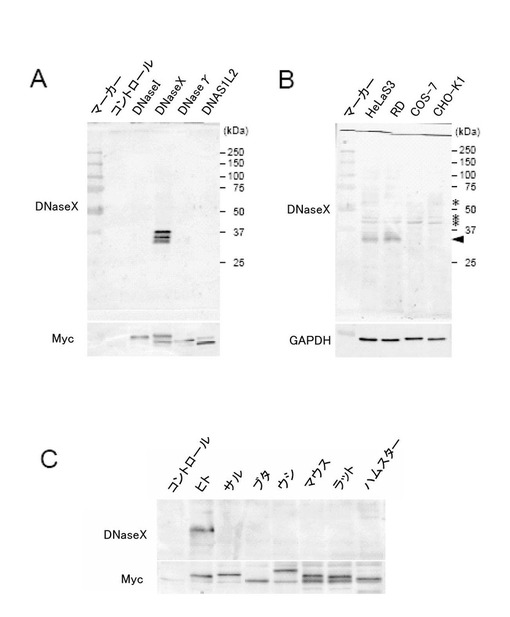
作製した抗ヒトDNaseXモノクローナル抗体の反応性は、ウエスタンブロッティング法によって確認した。まず、C末端にMyc-Hisタグを結合させたヒトDNase(DNaseI、DNaseX、DNaseγ、DNAS1L2)をコードする各cDNAを発現する発現ベクター(phDNaseX)を構築した(Shiokawa et al. 2005, Biochem J., vol.392, p.511-517.)。各発現ベクターをCHO-K1細胞に導入し、各タンパク質を発現した細胞より調製した細胞抽出物を用いてウエスタンブロッティングを行った(図1A)。また、HeLa S3細胞、ヒト胎児性横紋筋肉腫細胞(以下、RD細胞)、COS-7細胞、及びCHO-K1細胞の各細胞より調製した細胞抽出物を用いてウエスタンブロッティングを行った(図1B)。さらに、各動物種のDNaseXcDNAをpcDNA3.1 myc-His B(Invitrogen社)のEco RVサイトへサブクローニングして、DNaseX発現ベクターを構築した。なお、cDNAの作製に用いられたプライマー配列は以下の通りである。
サルDNaseX(センスプライマー):5’-CCACCATGCACTATCCAACTGCACTC-3'(配列番号5)
(アンチセンスプライマー):5’- GGTGGCAGGGCACAGTTGAGG-3’(配列番号6)
ブタDNaseX(センスプライマー):5’-CCACCATGGATTCCTCTGGAGGGTT CC -3’(配列番号7)
(アンチセンスプライマー):5’-GGCCGCCAGGCCCAGCTGTGG-3’(配列番号8)
ウシDNaseX(センスプライマー):5’-CCACCATGCATTCCTCTGGAGGGTTCC-3’(配列番号9)
(アンチセンスプライマー):5’-GGCCACCAGGCCCAGCTGGGG-3'(配列番号10)
マウスDNaseX(センスプライマー):5’-CCACCATGCCCTTTGGACAACCTGGA-3'(配列番号11)
(アンチセンスプライマー):5’-GTCCAGCTGAGATGGCAGGAG-3’(配列番号12)
ラットDNaseX(センスプライマー):5’-CCACCATGCCCTCTGGACAGCCTGTA-3'(配列番号13)
(アンチセンスプライマー):5’-GCCCAGCTGAGATAGTGACAG-3’(配列番号14)
ハムスターDNaseX(センスプライマー):5’-CCACCATGCCATATATGGCCATGCAT-3’(配列番号15)
(アンチセンスプライマー):5’-GCTCAGTTGAGATGGCAAAAG-3’(配列番号16)
【0082】
上記の方法によって構築した各DNaseX発現ベクター1μgに対し、TNT QuickCoupled Transcription/Translation System(Promega社)のTNT Quick Master Mixに加え、50μlの反応液で1時間、30℃でインキュベートした。得られた各DNaseXタンパク質を含む一定量の反応液を用いてウエスタンブロッティング法を行った(図1C)。なおコントロールには、挿入DNAのない発現ベクターに対し、上記in vitro転写/翻訳系によって同様に得られた等量の反応液を用いた。また、ウエスタンブロッティング法に用いた抗GAPDHモノクローナル抗体(MAB374)はChemicon社から、抗Myc tag抗体(R950-25)はInvitrogen社から購入した。
【0083】
その結果、作製した抗ヒトDNaseXモノクローナル抗体は、DNaseXに対して特異的に反応し(図1A)、DNaseXを発現することが知られている細胞(HeLa S3細胞、RD細胞)に対しても特異的に反応し(図1B)、ヒトDNaseXに対して特異的に反応した(図1C)。なお、ヒトDNaseXタンパク質は35kDaのバンドとして検出された。
【0084】
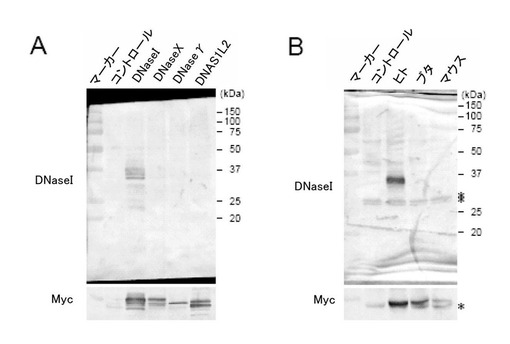
(2)新規抗ヒトDNaseIモノクローナル抗体の作製
上記(1)と同様の方法を用いて、抗ヒトDNaseIモノクローナル抗体を作製し、最終的にクローン化したハイブリドーマ株10A2を得た。なお、免疫原には、ヒトDNaseIの245-262位に対応するペプチド(GAVVPDSALPFNFQAAYG(配列番号4))のN末端にCysを付加した合成ペプチドとチオグロブリンとを結合させたペプチドを用いた。得られた抗ヒトDNaseIモノクローナル抗体のサブクラスはIgG1/κであった。
【0085】
作製した抗ヒトDNaseIモノクローナル抗体の特異性を、ウエスタンブロッティング法によって確認した。ここで、ウエスタンブロッティング法に用いた細胞抽出液は、(1)と同様である。なお、各動物種のDNaseIタンパク質発現ベクターを構築するにあたり、cDNAの作製に用いたプライマー配列は以下の通りである。
ブタDNaseI(センスプライマー):5’-CCACCATGAGGGCGGCCAGGCTGATG-3’(配列番号17)
(アンチセンスプライマー):5’-GGCTCTCTTCAGTGTCACCTC-3’(配列番号18)
マウスDNaseI(センスプライマー):5’-CCACCATGCGGTACACAGGGCTAATG-3’(配列番号19)
(アンチセンスプライマー):5’-GATTTTTCTGAGTGTCACCTC-3’(配列番号20)
【0086】
上記の方法によって構築した各DNaseI発現ベクターに対し、TNT QuickCoupled Transcription/Translation System(Promega社)を用いて各DNaseIタンパク質を作製し、ウエスタンブロッティング法を行った(図2B)。コントロールには、挿入DNAのない発現ベクターを用いた。また、ウエスタンブロッティング法で用いた抗Myc tag抗体(R950-25)はInvitrogen社から購入した。
その結果、作製した抗ヒトDNaseIモノクローナル抗体は、DNaseIに対して特異的に反応し(図2A)、ヒトDNaseIに対して特異的に反応した(図2B)。
【0087】
<実施例2:エンドサイトーシス依存性DNA取り込み>
本実施例では、DNaseXタンパク質の機能増強又は機能抑制が、それぞれエンドサイトーシス依存性DNA取り込みの抑制又は増強をもたらすことを示す。
【0088】
2−1 材料
(1)細胞の調製
HeLa S3細胞、COS-7細胞、及びCHO-KI細胞は、10%胎仔ウシ血清(FCS)、100単位/mlペニシリン及び0.1mg/mlストレプトマイシン含有ダルベッコ改変イーグル培地(DMEM)で培養した。また、RD細胞は、増殖培地(GM)(20%FCS、100単位/mlペニシリン及び0.1mg/mlストレプトマイシン含有DMEM)で培養した。なお、RD細胞は、ヒューマンサイエンス研究資源バンクから入手した。
【0089】
(2)抗体
抗ヒトDNaseXモノクローナル抗体及び抗ヒトDNaseIモノクローナル抗体は、実施例1で作製したものを使用した。
また、マウス抗SERCA-1モノクローナル抗体(Ve121G9)はLab vision社から、抗GAPDHモノクローナル抗体(MAB374)はChemicon社から、抗Myc tag抗体(R950-25)はInvitrogen社から購入した。さらに、ヤギ抗GM130(P-20)ポリクローナル抗体はSanta Cruz Biotechnology社から、ウサギ抗マウスFas(341289)ポリクローナル抗体はCalbiochem社から、抗EEA1(PA1-063)抗体はAffinity BioReagents社から、抗LAMP-1(H228)抗体はSanta Cruz Biotechnology社から、抗カルレティキュリン(C4606)抗体及び抗pan-cadherin(C3678)抗体はSigma社から購入した。
【0090】
(3)発現ベクターの作製
i) hDNaseXΔC発現ベクター(phDNaseXΔC)
PCRによって以下のプライマーを用いてhDNaseXΔC cDNAを合成し、このcDNAをpcDNA3.1 myc-His B(Invitrogen社)(挿入配列がコードするペプチドのC末端にMycタグとHisタグを付加し得る)のEco RVサイトへサブクローニングして、hDNaseXΔC発現ベクターを構築した。
hDNaseXΔC(センスプタイマー):5’-CCACCATGCACTACCCAACTGCACTC--3'(センスプライマー)(配列番号21)
(アンチセンスプライマー): 5'-CTGGCTCAGCTTCAGCTCCAC-3'(配列番号22)
【0091】
ii) mFasΔC発現ベクター(pmFasΔC)
PCRによって以下のプライマーを用いてmFasΔC cDNAを合成し、このcDNAをpcDNA3.1 myc-His B(Invitrogen社)のEco RVサイトへサブクローニングして、mFasΔC発現ベクターを構築した。
mFasΔC(センスプライマー): 5'-CCACCATGCTGTGGATCTGGGCTGTC-3'(配列番号23)
(アンチセンスプライマー): 5’-TTCCTGGATTGTCATGTCTTCAGC-3’(配列番号24)
【0092】
iii) hDNase I-HD発現ベクター(phDNase I-HD)
PCRによって以下のプライマーを用いて、hDNase Iと疎水性ドメイン(HD)を含むDNase Xの外部末端領域(アミノ酸配列256-284位)をコードするcDNAを作製し、このcDNAをpcDNA3.1 myc-His B(Invitrogen社)のEco RVサイトへサブクローニングして、hDNase I-HD発現ベクターを構築した。
hDNase I(センスプライマー):5’-CCACCATGAGGGGCATGAAGCTGCTG-3’(配列番号25)
(アンチセンスプライマー):5'-CTTCAGCATCACCTCCACTGG-3'(配列番号26)
HD(センスプライマー):5’-CTGAGCCAGGCGCACAGCGTC-3’(配列番号27)
(アンチセンスプライマー):5'-GGCAGCAGGGCACAGCTGAGG-3'(配列番号28)
【0093】
iv) hDNase X H148A発現ベクター(phDNase X H148A)
phDNase X H148Aは、LA PCR in vitro mutagenesisキット(タカラバイオ株式会社)を用いた部位特異的突然変異誘発法によって148番目のHisをAlaに置換し、phDNase Xと同様に作製した。
【0094】
v) hDNaseI-myc-His発現ベクター(phDNaseI-myc-His)、hDNaseX-myc-His発現ベクター(phDNaseX-myc-His)、hDNase γ-myc-His発現ベクター(phDNase γ-myc-His)、hDNAS1L2-myc-His発現ベクター(phDNAS1L2-myc-His)、及びcDNA 3.1/myc-His/lacZ(placZ)発現ベクター(pcDNA 3.1/myc-His/lacZ(placZ))
hDNaseI(phDNase I-myc-His)、hNaseX(phDNase X-myc-His)、DNaseγ(phDNase γ-myc-His)、及びhDNAS1L2(phDNAS1L2-myc-His)は、Shiokawaらの方法(Shiokawa, D., and Tanuma, S. Biochemistry, 2001, vol.40, p.143-152)に従って作製した。また、pcDNA 3.1/myc-His/lacZ(placZ)(βガラクトシダーゼ発現ベクター)は、Invitrogen社から購入した。
【0095】
2-2 アッセイ法
(1)筋管細胞への分化誘導方法
継代培養したRD細胞を、タイプIコラーゲンでコートされた培養皿(Iwaki社製)に播種し、GM培地で培養した。次に、GM培地を分化培地(DM)(2%ウマ血清、100単位/mlペニシリン及び0.1mg/mlストレプトマイシン含有DMEM)に変更して、培地交換をしながらRD細胞を培養し、筋管細胞へ分化誘導した。
【0096】
(2)ウエスタンブロッティング法
ウエスタンブロッティング法に用いた各抗体の希釈濃度は、抗ヒトDNaseXモノクローナル抗体は500ng/ml、抗DNaseIモノクローナル抗体は100ng/ml、抗Myc tag抗体は1000倍希釈、抗GAPDHモノクローナル抗体は250倍希釈、マウス抗SERCA-1モノクローナル抗体は100倍希釈、ウサギ抗マウスFasポリクローナル抗体は2000倍希釈、抗pan-cadherin抗体は400倍希釈であった。
【0097】
(3)免疫組織化学染色
ホルマリンで固定したサンプルをパラフィンで包埋し、10μmの切片を作製した。なお、パラフィン包埋された正常ヒト組織は、BioChain社から購入した。
免疫組織化学染色は、Cell and Tissue Staining kit(HRP-DAB)(R&D Systems社)を用いて行った。まず、組織切片を乗せたスライドをAntigen Retrieval Dewax Solution(BioChain社)で処理し、この組織切片に、3μg/mlの抗DNaseXモノクローナル抗体をのせて4℃、オーバーナイトでインキュベートした。次に、これらの組織切片にContrast BLUE Solution(KPL)をのせて対比染色を行った。なお、これらの画像は、CK40 light microscope system(Olympus)を用いて記録した。
【0098】
(4)間接免疫蛍光染色
免疫蛍光染色による解析は、従来の方法(Shiokawa et al., Biochem. J., 2005, vol. 392, p.511-517)を改良して行った。具体的には、以下の通りである。
まず、培養皿の底に敷いたカバーグラス上で細胞を培養した。培養後、これらの細胞を3.5%ホルムアルデヒドで固定し、Cytonin solution(R&D Systems社)を添加して、室温で30分間インキュベートした。次に、一次抗体の希釈液をカバーグラス上の細胞に滴下し、4℃、オーバーナイトでインキュベートした。ここで、各一次抗体は、抗ヒトDNaseXモノクローナル抗体は3μg/ml、抗DNase Iモノクローナル抗体は1μg/ml、抗カルレティキュリン抗体は2000倍希釈、マウス抗SERCA-1モノクローナル抗体は50倍希釈、ヤギ抗GM130(P-20)ポリクローナル抗体は50倍希釈、ウサギ抗マウスFas(341289)ポリクローナル抗体は100倍希釈、抗pan-cadherin抗体は400倍希釈、抗EEA1(PA1-063)抗体は1000倍希釈、抗LAMP-1(H228)抗体は100倍希釈の濃度に調製して使用した。
【0099】
次に、細胞をDPBS(ダルベッコ改変リン酸緩衝生理食塩水;GIBCO)で洗浄し、二次抗体の希釈液をカバーガラス上の細胞に滴下し、室温で30分間インキュベートした。なお、二次抗体(Alexa 488ニワトリ抗マウスIgG抗体(Molecular Probes社)、Alexa 594ロバ抗ヤギIgG抗体(Molecular Probes社)、Alexa 594ヤギ抗ウサギIgG抗体(Molecular Probes社))は2000倍希釈の濃度に調製して使用した。
【0100】
また、Zenon Tricolor Mouse IgG1 Labeling Kit #2(Molecular Probes社)を用いて、抗ヒトDNaseXモノクローナル抗体とマウス抗SERCA-1モノクローナル抗体との二重染色を行った。なお、抗ヒトDNaseXモノクローナル抗体はAlexa488で、マウス抗SERCA-1モノクローナル抗体はAlexa594で標識したものを使用した。これらの抗体の希釈液をカバーガラス上の細胞に滴下し、室温で90分間インキュベートした。
【0101】
最後に、細胞をDPBSで洗浄し、100nM Hoechst 33342で対比染色し、蛍光顕微鏡(BX60、オリンパス株式会社)や共焦点レーザー顕微鏡(TCS SP2、ライカマイクロシステムズ株式会社)で観察した。
【0102】
(5)PI-PLC(ホスファチジルイノシトール特異性ホスホリパーゼ)による処理
免疫組織化学染色には、カバーグラス上の細胞を、DPBSで2回リンスし、5 unit/ml Bacillus thuringiensisホスファチジルイノシトール特異性ホスホリパーゼ(PI-PLC)(Sigma社)含有Leibovidz培養培地で37℃、30分間インキュベートした後、(4)の通りに固定し、抗体で処理した。
【0103】
また、PI-PLC処理後に、細胞と上清を分画するためには、付着している細胞をDPBSでリンスし、1.5mlチューブに回収し、このチューブに5 unit/ml PI-PLCを100μl入れて37℃で20分間インキュベートした後、10分間遠心分離(2000×g)をし、上清を別なチューブ(Sup fraction)に回収した。また、チューブ内の細胞ペレットをDPBSで遠心分離にて2回洗浄し、Leibovidz培地(cell fraction)500μlに再懸濁した。
【0104】
(6)TUNELアッセイ
カバーグラス上の細胞を、placZ-Transome IV複合体存在下で30分間インキュベートした。次に、これらの細胞をDPBSで2回洗浄し、3.7%ホルムアルデヒド含有DPBSをのせて室温で10分間固定した。次に、lacZプラスミドに生じる3’-OH末端を、Flow TACS in situ TUNEL Detection kit(R&D Systems社)を用いたTUNEL法によって検出した。なお、アッセイ全体は、FITC結合ストレプトアビジンをAlexa488結合ストレプトアビジン(Molecular Probes社)に変更した以外は、キット製造業者のプロトコールに従って行った。最後に、これらの細胞をDPBSで3回洗浄し、10μM Hoeschst 33342含有DPBSをのせて、エンドサイトーシスを起こしたDNAを室温で10分間対比染色させ、蛍光顕微鏡で観察した。
【0105】
(7)蛍光顕微鏡によるDNA画像のキャプチャー
細胞核及びエンドサイトーシスを起こしたDNAを、Hoechst 33342で対比染色した。核DNAの観察には2種類のNDフィルター(U-ND6及びU-ND25)(オリンパス株式会社)を用いて、一方、エンドサイトーシスを起こしたDNAの観察には、NDフィルターを用いないで、それぞれのDNA画像を観察した。
【0106】
2-3 DNaseXタンパク質によるエンドサイトーシス依存性DNA取り込み抑制
(1)DNaseXタンパク質の発現
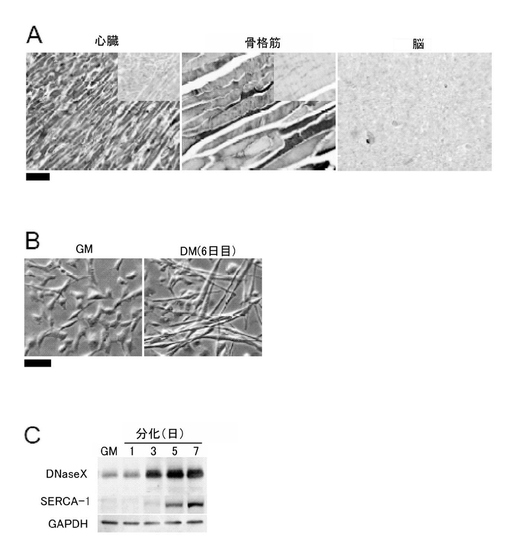
DNaseXタンパク質の発現がRD細胞の筋肉への分化段階に関連していること示すために、免疫組織化学染色を用いて、ヒト組織におけるDNaseXタンパク質の発現を調べたところ、図3Aに示すように、心筋細胞及び骨格筋線維では、DNaseXタンパク質の強いシグナルが得られたが、脳ではシグナルが検出できなかった。このように、DNaseXタンパク質は、筋細胞に強く発現している。
次に、低濃度のマイトジェン条件下でRD細胞を培養すると、筋管が形成される(図3B)ので、この系を用いて、DNaseXタンパク質の発現の筋肉分化における変化をウエスタンブロッティング法で調べたところ、図3Cに示すように、DNaseXタンパク質の発現は分化誘導3日目に増加し、分化誘導5日目にはさらに増加した。このDNaseXタンパク質の発現の変化は、筋肉分化マーカー(SERCA-1:筋小胞体Ca2+-ATPase1)の発現と同様の挙動を示した(図3C)。
このように、DNaseXタンパク質の発現は、未分化筋細胞では弱く、分化するにつれ強くなる。
【0107】
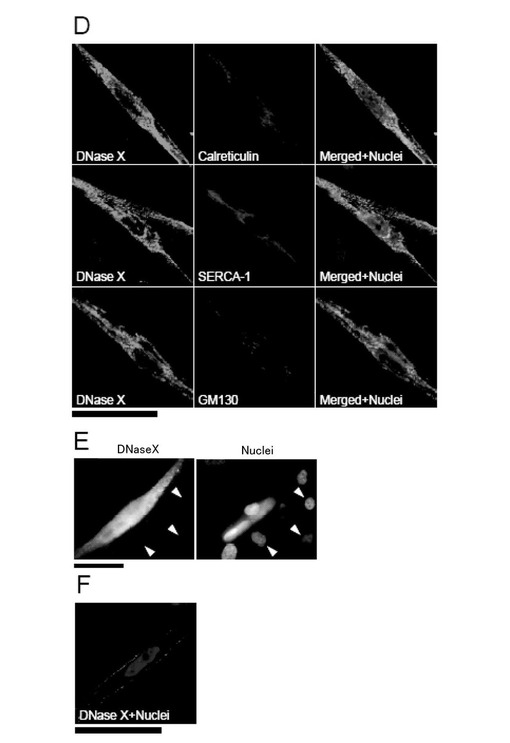
(2)DNaseXタンパク質の細胞内分布の検討
次に、6日間低濃度のマイトジェン条件下で培養して分化させた筋管細胞に対し、抗DNaseX抗体及び抗カルレティキュリン抗体、抗SERCA-1抗体、又は抗GM130抗体で二重染色し、共焦点レーザー顕微鏡を用いてDNaseXタンパク質が膜に分布していることを示した。
【0108】
図3D(浸透化処理有り)に示すように、DNaseXタンパク質は、カルレティキュリンやSERCA-1と共に小胞体(ER)と筋小胞体に局在化していた。また、ゴルジ体のマーカー(GM130)との2重染色でもシグナルが重なり、DNaseXタンパク質がゴルジ体に存在していることが認められた。
さらに、浸透化処理無しで、免疫蛍光染色を行ったところ、図3E(蛍光顕微鏡像)及びF(共焦点レーザー顕微鏡像)に示すように、同様に分化した筋管細胞膜に局在することが観察され、DNaseXタンパク質は細胞外の膜表面に存在した。
このように、DNaseXタンパク質は、分化したRD由来の筋管において分泌経路に局在していることから、タンパク質合成後、細胞膜に送達されることが結論される。
【0109】
(3)細胞表面におけるDNaseXタンパク質の局在性
DNaseXタンパク質の細胞膜における局在を確認するために、Fugene6 (Roche)を用いて、DNaseX発現ベクターとDNaseI発現ベクターをCOS-7細胞に導入し、免疫蛍光染色を用いて各DNaseの細胞内での分布を観察した。なお、マウスFas抗原の突然変異型(mFasΔC)は、膜貫通タンパクとして細胞表面に発現していることが知られているので、細胞膜における観察を容易にするために、mFasΔCを同時にCOS-7細胞に導入した。
図4Aに示すように、DNaseXタンパク質は、細胞膜に局在して発現し、mFasΔCと同様な分布を示していた(図4A上段)。一方、コントロールとして用いたDNaseIタンパク質は、細胞膜に発現していなかった(図4A下段)。
次に、疎水性のC端ドメインを欠失するDNaseXΔCの発現ベクターとDNaseXのC端ドメインを有するDNaseI-HDの発現ベクターを用いて、同様の実験を行ったところ、図4Bに示すように、DNaseXΔCタンパク質は膜に局在せず、DNaseI-HDタンパク質は膜に局在した。このことから、疎水性のC端ドメインは、DNaseXタンパク質が膜に局在するために必須の領域であることが結論される。
【0110】
(4)DNaseXタンパク質の膜結合性
ここでは、GPIアンカーを解離する酵素として知られているPI-PLC(ホスファチジルイノシトール特異性ホスホリパーゼ)を用いて、以下の実験を行ない、DNaseXタンパク質がGPIアンカーで細胞膜に結合していることを示す。
【0111】
まず、Fugene6(Roche)を用いて、DNaseX発現ベクターとmFasΔC発現ベクターを同時にCOS-7細胞に導入し、PI-PLC処理をした後、免疫蛍光染色を用いてDNaseXの細胞膜への分布を観察した(図5A下段)。コントロールとして、PI-PLCの処理をしない細胞におけるDNaseXの細胞膜への分布を観察した(図5A上段)。図5Aに示すように、PI-PLCの処理を行うと、膜貫通タンパク(mFasΔC)の発現量はコントロールと比べて変化しなかったが、DNaseXタンパク質の発現量はコントロールと比べて減少した。
【0112】
さらに、DNaseX発現ベクターとmFasΔC発現ベクターを導入したCOS-7細胞に対し、PI-PLCの処置を行った後に、細胞と上清を分画して、DNaseXタンパク質の存在を、ウエスタンブロッティング法によって調べたところ、図5Bに示すように、PI-PLC処理によって、膜貫通タンパク(mFasΔC)は培養上清中に放出されなかったが、DNaseXタンパク質は培養上清中に放出された。
【0113】
また、RD細胞由来の筋管細胞を用いて、PI-PLC処理後、同様にウエスタンブロッティングを行ったところ、内在性のDNaseXにおいても、上記と同様の結果が得られた(図5C)。なお、ここでは、コントロールとして、細胞間接着分子である膜タンパク質カドヘリンを用いた。
【0114】
このように、DNaseXタンパク質は、細胞の外側で、GPIアンカーによって細胞膜に結合して存在していることが結論される。
【0115】
(5)RD細胞の分化段階とエンドサイトーシス依存性遺伝子導入効率の比較
ここでは、RD細胞が分化するに従い、DNaseXタンパク質によるエンドサイトーシス依存性DNA取り込み阻害が向上することを示す。
【0116】
まず、エンドサイトーシスによる遺伝子導入効率を評価するために、人工的に作製したαヘリックスのカチオン型ペプチドによる遺伝子導入を利用した(Niidome, T., Ohmori, N., Ichinose, A., Wada, A., Mihara, H., Hirayama, T., Aoyagi, H., J. Biol. Chem., 272, 15307-15312 (1997), Ohmori, N., Niidome, T., Kiyota, T., Lee, S., Sugihara, G., Wada, A. Hirayama, T. and Aoyagi, H., Biochem. Biophys. Res. Commun., 245, 259-265 (1998))。未分化RD細胞及びRD細胞を6日間分化させた筋管細胞を直径60mmの培養皿で培養し、あらかじめカチオン性ペプチドLLKLLLKLWKKLLKLLK(配列番号29)(遺伝子導入試薬Transome IV(和光純薬工業株式会社))と複合体をつくらせたFITC標識のLacZ cDNAベクター(FITC-lacZ)を導入した。なお、 FITC-lacZは、FITC標識のプライマー(5’-FITC-GAC GGATCGGGAGATCTCCCG-3’(センスプライマー)(配列番号30)、5'-FITC-CGAAAGGAGCGGGCGCTAGGG-3’(アンチセンスプライマー)(配列番号31)を用い、placZを鋳型にしてPCR法を用いて作製した。24時間後、これらの細胞を、DPBSで2回洗浄し、3.7%ホルムアルデヒドで固定(室温、10分間)した。その後、これらの細胞をDPBSで3回洗浄し、100nM Hoeschst 33342で核を染色した後、対比染色して、蛍光顕微鏡にてこれらの細胞を観察した。図6A(上図:未分化RD細胞、下図:筋管細胞)に示したように、FITCのシグナルは、未分化の細胞より筋管細胞で多く観察された。このことは、筋管細胞は、未分化の細胞と比べて、蛍光標識をした遺伝子(FITC-lacZ)を多く取り込むことを示す。
【0117】
次に、同様に、Transome IVと複合体をつくらせたFITC-lacZ又はインサートを有しない空のベクターを各細胞に導入し、BetaGro Assay System(Promega株式会社)を用いてβガラクトシダーゼ活性を測定した。なお、pFITC-lacZで得られた測定値から、空のベクターで得られた測定値(バックグラウンドの値と考えられる)を引くことにより、実際の値を算出した。図6Bに示したように、筋管細胞は、未分化の細胞と比べて、蛍光標識をした遺伝子(FITC-lacZ)を多く取り込んだにもかかわらず、未分化の細胞と比べて、10%以下しかβガラクトシダーゼを発現していなかった。
【0118】
次に、分化した筋管細胞において、エンドサイトーシス小胞(初期、後期)とDNaseXタンパク質との関係を調べるため、抗DNaseX抗体及び抗EEA1抗体あるいは抗LAMP-1抗体と二重染色したところ、図6Cに示す通り、DNaseXタンパク質は、初期エンドソームマーカーEEA1と同じ場所に検出されたが、成熟リソソームマーカーLAMP-1と同じ場所には検出されなかった。
【0119】
さらに、細胞内で分解されたDNA断片をin situで検出するために、筋管細胞にLacZ cDNA発現ベクター(placZ)をトランスフェクトし、TUNEL法を用いて、断片化されたDNA3'末端ヒドロキシル基を検出した(Gavrieli et al., J. Cell Biol., 1992, vol.119, p.493-501)ところ、図6Dに示すように、placZを導入した筋管細胞では、placZを導入しない筋管細胞に対し、TUNELに対してより強い陽性反応を示した。このことは、筋管細胞において、取り込まれたplacZが、細胞内で分解されていることを示す。
【0120】
このように、筋管細胞は、未分化なRD細胞に比べ、DNaseXタンパク質が多量に存在すること、エンドサイトーシス活性は高いものの、取り込まれた遺伝子は分解されやすいことを示している。
【0121】
(6)DNaseXタンパク質の過剰発現
本実施例では、細胞内でDNaseXタンパク質を過剰発現させると、細胞のエンドサイトーシス依存性DNA取り込み能が低下することを示す。
【0122】
まず、Fugene6 (Roche)を用いて、本来DNaseXの発現がほとんど認められない未分化RD細胞及びHeLaS3細胞に、野生型のDNaseXを発現するphDNaseX、C端の疎水性領域を欠失したDNaseXを発現するphDNaseXΔC、活性部位に点突然変異を有するDNaseXを発現するphDNaseXH148Aの各発現ベクターを導入した。コントロールの細胞には、空のベクターを導入した。
【0123】
抗DNaseX抗体を用いた免疫蛍光染色によってDNaseXタンパク質の発現を観察したところ、HeLa S3細胞及び未分化のRD細胞の細胞表面において、phDNaseX、及びphDNaseXH148Aを導入した場合、細胞膜上にDNaseXタンパク質の発現が認められた(図7A)。一方、C末端疎水性ペプチドを欠損している欠失変異体phDNaseXΔCを導入した場合では、細胞膜でのDNaseXの発現が認められなかった(図7A)。
【0124】
また、(5)と同様にしてエンドサイトーシスによる遺伝子導入の影響を調べたところ、図7Bに示すように、野生型のDNaseXタンパク質を過剰発現させた場合、両方の細胞において、βガラクトシダーゼ活性を阻害するが、C末端を欠失させるか、活性部位に点突然変異を導入することによってβガラクトシダーゼの活性阻害は減少した(図7B)。
【0125】
さらに、TUNEL法を用いて、HeLa S3細胞に導入されたDNAの切断を検出したところphDNaseXを導入した細胞は、陽性シグナルが検出されたが、phDNaseXH148Aを導入した細胞は、陽性シグナルが検出できなかった(図7C)。
【0126】
これらの実験結果から、細胞内でDNaseXタンパク質を過剰発現させると、DNaseXタンパク質依存的に細胞のエンドサイトーシス依存性DNA取り込みを抑制できることが結論される。
【0127】
(7)DNaseXタンパク質の発現抑制
本実施例では、細胞内でDNaseXタンパク質を発現抑制すると、細胞のエンドサイトーシス依存性DNA取り込み能が向上することを示す。
(1)RNA
ヒトDNaseX遺伝子(配列番号2)に対するsiRNA及びncRNA (ネガティブコントロールとして使用するRNA duplex)は、Invitrogen社から購入した。なお、本実施例にて用いるsiRNA(small interfering RNA)は、標的配列5'-CCTGCTTCGAGAACTCAATCGATTT-3(配列番号32)に対応する。
(2)siRNAを用いたDNaseX遺伝子の発現抑制
まず、継代培養しているRD細胞を60mm培養皿に播種し、DMEM培地で培養した。次に、培地を分化培地(DM)に変更し、Lipofectamine 2000 transfection regent(Invitrogen社)を用いて、120pmolのsiRNAをRD細胞にトランスフェクトした。その後も培地交換を行ながら、分化培地で4日間RD細胞の培養を行った。各細胞に対し、抗DNaseX抗体、抗SERCA-1抗体、抗GADPH抗体を用いてウエスタンブロッティング法を行った。ここで、SERCA-1は筋管細胞への分化マーカーであり、GADPHはポジティブコントロールである。なお、コントロールとして、何もトランスフェクトしないで、他の処理は同様に行った細胞を用いた。
【0128】
図8Aに示すように、DNaseX遺伝子の発現は、ヒトDNaseX遺伝子に対して特異的に反応するsiRNAを導入した細胞において有意に減少した。一方、ncRNAを導入した細胞は、DNaseX遺伝子の発現に変化はなかった。この結果は、用いたsiRNAが、DNaseX遺伝子の発現を抑制できることを示す。また、いずれの場合もSERCA-1の発現が検出されることから、DNaseXは、RD細胞の筋肉分化に不可欠な要素ではないことが示される。
【0129】
さらに、siRNAを導入したRD細胞由来の筋管細胞において、(5)と同様にしてエンドサイトーシスによる遺伝子導入の影響を調べたところ、siRNAを用いてDNaseX遺伝子をノックダウンすると、分化した筋管細胞において、4倍以上も導入遺伝子の発現が促進された(図8B)。さらに、TUNEL法を用いて、分化した筋管細胞に導入されたプラスミドの切断を検出したところ、siRNAが導入された筋管細胞では、陽性反応を示すシグナルは、ほとんど認められなかった(図8C)。
【0130】
このように、細胞内でDNaseXタンパク質を発現抑制すると、細胞のエンドサイトーシス依存性DNA取り込みは促進されることが結論される。
【図面の簡単な説明】
【0131】
【図1】本発明の一実施例において、新規抗ヒトDNaseXモノクローナル抗体の高い特異性をウエスタンブロッティング法で示した図である。
【図2】本発明の一実施例において、新規抗ヒトDNaseIモノクローナル抗体の高い特異性をウエスタンブロッティング法で示した図である。
【図3−1】本発明の一実施例において、ヒト筋肉組織及びRD細胞におけるDNaseXの発現を示す図である。(A)免疫組織化学染色(スケール:100μm)、(B)位相差顕微鏡像(スケール:100μm)、(C)ウエスタンブロッティング法(コントロール:GAPDH)による。
【図3−2】本発明の一実施例において、ヒト筋肉組織及びRD細胞におけるDNaseXの発現を示す図である。(D)共焦点レーザー顕微鏡像(スケール:50μm)、(E)蛍光顕微鏡像(スケール:50μm)、(F)共焦点レーザー顕微鏡像(スケール:50μm)による。
【図4】本発明の一実施例において、DNaseXのC端の疎水性領域が膜に結合するために必要であることを示す図である。なお、mFasΔCはマウスFasタンパク質のC端が欠失した変異体である。(A)蛍光顕微鏡像(スケール:50μm)、(B)蛍光顕微鏡像(スケール:50μm)による。
【図5】本発明の一実施例において、PI-PLC処理による細胞表面からのDNaseXの解離を示す図である。(A)蛍光顕微鏡像(スケール:50μm)、(B)(C)ともウエスタンブロッティング法による。(+)5単位/ml PI-PLC存在下、(-)PI-PLC非存在下を表す。(Cells)細胞中(Sup)培養上清中のDNaseXの存在を調べた。
【図6】本発明の一実施例において、増殖性RD細胞及び分化した筋管細胞におけるエンドサイトーシス依存性遺伝子取り込みを示す図である。(A)蛍光顕微鏡像(スケール:50μm)(C)共焦点レーザー顕微鏡像(スケール:10μm)、(D)蛍光顕微鏡像(スケール:50μm)による。GMは増殖性RD細胞を、DMは分化筋管細胞を示す。(B)lacZの活性を示す(N=5、*=p<0.001(t検定))。
【図7−1】本発明の一実施例において、細胞内でDNaseXタンパク質を過剰発現させると、細胞のエンドサイトーシス依存性DNA取り込み能が低下することを示す図である。(A)蛍光顕微鏡像(スケール:50μm)による。(B)lacZの活性を示す(N=6、*=p<0.001(t検定))。
【図7−2】本発明の一実施例において、細胞内でDNaseXタンパク質を過剰発現させると、細胞のエンドサイトーシス依存性DNA取り込み能が低下することを示す図である。(C)蛍光顕微鏡像(スケール:50μm)による。
【図8】本発明の一実施例において、細胞内でDNaseXタンパク質を発現抑制すると、細胞のエンドサイトーシス依存性DNA取り込み能が向上することを示す図である。(A)ウエスタンブロッティング法(C)蛍光顕微鏡像(スケール:50μm)による。(B)lacZの活性を示す(N=6、*=p<0.001(t検定))。
【特許請求の範囲】
【請求項1】
DNaseXタンパク質の機能を増強するエンドサイトーシス依存性DNA取り込み抑制剤。
【請求項2】
DNaseXタンパク質の発現を増強することを特徴とする請求項1に記載のエンドサイトーシス依存性DNA取り込み抑制剤。
【請求項3】
DNaseXタンパク質の機能を増強するウイルス感染阻害剤。
【請求項4】
DNaseXタンパク質の発現を増強することを特徴とする請求項3に記載のウイルス感染阻害剤。
【請求項5】
請求項3に記載のウイルス感染阻害剤を含有するウイルス感染予防剤。
【請求項6】
請求項5に記載のウイルス感染阻害剤を含有するウイルス感染に対するワクチン。
【請求項7】
DNaseXタンパク質を強制発現させる発現ベクター。
【請求項8】
DNaseXタンパク質の機能が増強された細胞。
【請求項9】
DNaseXタンパク質を強制発現させる発現ベクターを有する請求項7に記載の細胞。
【請求項10】
体の一部又は全部で、DNaseXタンパク質が異所的に又は過剰に機能が増強していることを特徴とする脊椎動物。
【請求項11】
DNaseXタンパク質を強制発現させる発現ベクターを細胞に導入することにより作製された請求項10に記載の脊椎動物。
【請求項12】
DNaseXタンパク質を強制発現させる発現ベクターによって作製されたトランスジェニック脊椎動物である請求項11に記載の脊椎動物。
【請求項13】
DNaseXタンパク質の機能を増強する化合物をスクリーニングするスクリーニング方法。
【請求項14】
配列番号3の一部又は全部をエピトープとするモノクローナル抗体。
【請求項15】
請求項14に記載のモノクローナル抗体を産生するハイブリドーマ。
【請求項16】
寄託受領番号FERM AP-21282である請求項15に記載のハイブリドーマ。
【請求項17】
請求項16に記載のハイブリドーマから産出されるモノクローナル抗体。
【請求項18】
請求項14又は17のいずれかに記載のモノクローナル抗体を用いて、DNaseXタンパク質を検出する検出方法。
【請求項19】
DNaseXタンパク質の機能を抑制するエンドサイトーシス依存性DNA取り込み促進剤。
【請求項20】
請求項17に記載のモノクローナル抗体を有効成分として含有するエンドサイトーシス依存性DNA取り込み促進剤。
【請求項21】
DNaseXタンパク質の発現を抑制することを特徴とする請求項19に記載のエンドサイトーシス依存性DNA取り込み促進剤。
【請求項22】
DNaseXタンパク質の機能を抑制するウイルス感染促進剤。
【請求項23】
請求項17に記載のモノクローナル抗体を有効成分として含有するウイルス感染促進剤。
【請求項24】
DNaseXタンパク質の発現を抑制することを特徴とする請求項22に記載のウイルス感染促進剤。
【請求項25】
DNaseX遺伝子に対するsiRNAを有効成分として含有することを特徴とする請求項24に記載のウイルス感染促進剤。
【請求項26】
前記siRNAが、配列番号32を有することを特徴とする請求項25に記載のウイルス感染促進剤。
【請求項27】
DNaseX遺伝子に対するsiRNAを発現する発現ベクター。
【請求項28】
内在性DNaseXタンパク質の機能が抑制されている細胞。
【請求項29】
DNaseX遺伝子に対するsiRNAを発現する発現ベクターを有することを特徴とする請求項28に記載の細胞。
【請求項30】
体の一部又は全部で、内在性DNaseXタンパク質の機能が抑制されている脊椎動物。
【請求項31】
DNaseX遺伝子のノックダウン又はノックアウトによって作製された請求項30に記載の脊椎動物。
【請求項32】
DNaseXタンパク質の機能を抑制する化合物をスクリーニングするスクリーニング方法。
【請求項33】
請求項32に記載の検出方法を用いて、DNaseXタンパク質の機能を抑制する化合物をスクリーニングするスクリーニング方法。
【請求項34】
DNaseX遺伝子関連物質である、遺伝子マーカー。
【請求項35】
ウイルスに対する易感染性予測マーカー、又はウイルス感染後の病状推定マーカーであることを特徴とする請求項34に記載の遺伝子マーカー。
【請求項36】
前記ウイルスが、インフルエンザウイルス、肝炎ウイルス、HIVウイルス、ATLウイルス、エボラウイルス、黄熱病ウイルス、狂犬病ウイルス、サイトメガロウイルス、SARSウイルス、JCウイルス(進行性多巣性白質脳症)、水痘・帯状疱疹ウイルス、コクサッキーA16型・エンテロウイルス71ウイルス(手足口病)、デング熱ウイルス、パルボウイルス(伝染性紅斑)、EBウイルス(伝染性単核球症)、天然痘ウイルス、風疹ウイルス、ポリオウイルス、麻疹ウイルス、アデノウイルス(プール熱)、マールブルグウイルス(フィロウイルス科)、クリミア・コンゴ出血熱ウイルス、ラッサウイルス、フニンウイルス等の新大陸アレナウイルス、ハンタウイルス、リフトバレー熱ウイルス、ムンプスウイルス(流行性耳下腺炎)、ウエストナイルウイルス、単純ヘルペスウイルス、コクサッキーウイルス、ロタウイルス、ノロウイルス、チクングニアウイルス(アルファウイルス属、トガウイルス科)からなる群から選ばれることを特徴とする請求項35に記載の遺伝子マーカー。
【請求項37】
前記遺伝子関連物質がDNA、hnRNA、mRNA、及びタンパク質からなる群から選ばれることを特徴とする請求項34〜36のいずれかに記載の遺伝子マーカー。
【請求項38】
ウイルスに対する易感染性予測又はウイルス感染後の病状推定のためのキットであって、DNaseXタンパク質に対する抗体又はDNaseXcDNAを増幅するためのPCR用プライマーを含有することを特徴とするキット。
【請求項39】
前記抗体がモノクローナル抗体であることを特徴とする請求項38に記載のキット。
【請求項40】
前記モノクローナル抗体が、配列番号3の一部又は全部をエピトープとすることを特徴とする請求項39に記載のキット。
【請求項41】
DNaseXタンパク質を抗原抗体反応を利用した方法で検出することを特徴とする請求項40に記載のキット。
【請求項42】
前記抗原抗体反応を利用した方法が免疫クロマトグラフ測定法であることを特徴とする請求項41に記載のキット。
【請求項1】
DNaseXタンパク質の機能を増強するエンドサイトーシス依存性DNA取り込み抑制剤。
【請求項2】
DNaseXタンパク質の発現を増強することを特徴とする請求項1に記載のエンドサイトーシス依存性DNA取り込み抑制剤。
【請求項3】
DNaseXタンパク質の機能を増強するウイルス感染阻害剤。
【請求項4】
DNaseXタンパク質の発現を増強することを特徴とする請求項3に記載のウイルス感染阻害剤。
【請求項5】
請求項3に記載のウイルス感染阻害剤を含有するウイルス感染予防剤。
【請求項6】
請求項5に記載のウイルス感染阻害剤を含有するウイルス感染に対するワクチン。
【請求項7】
DNaseXタンパク質を強制発現させる発現ベクター。
【請求項8】
DNaseXタンパク質の機能が増強された細胞。
【請求項9】
DNaseXタンパク質を強制発現させる発現ベクターを有する請求項7に記載の細胞。
【請求項10】
体の一部又は全部で、DNaseXタンパク質が異所的に又は過剰に機能が増強していることを特徴とする脊椎動物。
【請求項11】
DNaseXタンパク質を強制発現させる発現ベクターを細胞に導入することにより作製された請求項10に記載の脊椎動物。
【請求項12】
DNaseXタンパク質を強制発現させる発現ベクターによって作製されたトランスジェニック脊椎動物である請求項11に記載の脊椎動物。
【請求項13】
DNaseXタンパク質の機能を増強する化合物をスクリーニングするスクリーニング方法。
【請求項14】
配列番号3の一部又は全部をエピトープとするモノクローナル抗体。
【請求項15】
請求項14に記載のモノクローナル抗体を産生するハイブリドーマ。
【請求項16】
寄託受領番号FERM AP-21282である請求項15に記載のハイブリドーマ。
【請求項17】
請求項16に記載のハイブリドーマから産出されるモノクローナル抗体。
【請求項18】
請求項14又は17のいずれかに記載のモノクローナル抗体を用いて、DNaseXタンパク質を検出する検出方法。
【請求項19】
DNaseXタンパク質の機能を抑制するエンドサイトーシス依存性DNA取り込み促進剤。
【請求項20】
請求項17に記載のモノクローナル抗体を有効成分として含有するエンドサイトーシス依存性DNA取り込み促進剤。
【請求項21】
DNaseXタンパク質の発現を抑制することを特徴とする請求項19に記載のエンドサイトーシス依存性DNA取り込み促進剤。
【請求項22】
DNaseXタンパク質の機能を抑制するウイルス感染促進剤。
【請求項23】
請求項17に記載のモノクローナル抗体を有効成分として含有するウイルス感染促進剤。
【請求項24】
DNaseXタンパク質の発現を抑制することを特徴とする請求項22に記載のウイルス感染促進剤。
【請求項25】
DNaseX遺伝子に対するsiRNAを有効成分として含有することを特徴とする請求項24に記載のウイルス感染促進剤。
【請求項26】
前記siRNAが、配列番号32を有することを特徴とする請求項25に記載のウイルス感染促進剤。
【請求項27】
DNaseX遺伝子に対するsiRNAを発現する発現ベクター。
【請求項28】
内在性DNaseXタンパク質の機能が抑制されている細胞。
【請求項29】
DNaseX遺伝子に対するsiRNAを発現する発現ベクターを有することを特徴とする請求項28に記載の細胞。
【請求項30】
体の一部又は全部で、内在性DNaseXタンパク質の機能が抑制されている脊椎動物。
【請求項31】
DNaseX遺伝子のノックダウン又はノックアウトによって作製された請求項30に記載の脊椎動物。
【請求項32】
DNaseXタンパク質の機能を抑制する化合物をスクリーニングするスクリーニング方法。
【請求項33】
請求項32に記載の検出方法を用いて、DNaseXタンパク質の機能を抑制する化合物をスクリーニングするスクリーニング方法。
【請求項34】
DNaseX遺伝子関連物質である、遺伝子マーカー。
【請求項35】
ウイルスに対する易感染性予測マーカー、又はウイルス感染後の病状推定マーカーであることを特徴とする請求項34に記載の遺伝子マーカー。
【請求項36】
前記ウイルスが、インフルエンザウイルス、肝炎ウイルス、HIVウイルス、ATLウイルス、エボラウイルス、黄熱病ウイルス、狂犬病ウイルス、サイトメガロウイルス、SARSウイルス、JCウイルス(進行性多巣性白質脳症)、水痘・帯状疱疹ウイルス、コクサッキーA16型・エンテロウイルス71ウイルス(手足口病)、デング熱ウイルス、パルボウイルス(伝染性紅斑)、EBウイルス(伝染性単核球症)、天然痘ウイルス、風疹ウイルス、ポリオウイルス、麻疹ウイルス、アデノウイルス(プール熱)、マールブルグウイルス(フィロウイルス科)、クリミア・コンゴ出血熱ウイルス、ラッサウイルス、フニンウイルス等の新大陸アレナウイルス、ハンタウイルス、リフトバレー熱ウイルス、ムンプスウイルス(流行性耳下腺炎)、ウエストナイルウイルス、単純ヘルペスウイルス、コクサッキーウイルス、ロタウイルス、ノロウイルス、チクングニアウイルス(アルファウイルス属、トガウイルス科)からなる群から選ばれることを特徴とする請求項35に記載の遺伝子マーカー。
【請求項37】
前記遺伝子関連物質がDNA、hnRNA、mRNA、及びタンパク質からなる群から選ばれることを特徴とする請求項34〜36のいずれかに記載の遺伝子マーカー。
【請求項38】
ウイルスに対する易感染性予測又はウイルス感染後の病状推定のためのキットであって、DNaseXタンパク質に対する抗体又はDNaseXcDNAを増幅するためのPCR用プライマーを含有することを特徴とするキット。
【請求項39】
前記抗体がモノクローナル抗体であることを特徴とする請求項38に記載のキット。
【請求項40】
前記モノクローナル抗体が、配列番号3の一部又は全部をエピトープとすることを特徴とする請求項39に記載のキット。
【請求項41】
DNaseXタンパク質を抗原抗体反応を利用した方法で検出することを特徴とする請求項40に記載のキット。
【請求項42】
前記抗原抗体反応を利用した方法が免疫クロマトグラフ測定法であることを特徴とする請求項41に記載のキット。
【図1】


【図2】
【図3−1】
【図3−2】
【図4】
【図5】
【図6】
【図7−1】
【図7−2】
【図8】
【図2】
【図3−1】
【図3−2】
【図4】
【図5】
【図6】
【図7−1】
【図7−2】
【図8】
【公開番号】特開2008−253188(P2008−253188A)
【公開日】平成20年10月23日(2008.10.23)
【国際特許分類】
【出願番号】特願2007−98925(P2007−98925)
【出願日】平成19年4月4日(2007.4.4)
【出願人】(803000115)学校法人東京理科大学 (545)
【出願人】(399032282)株式会社 免疫生物研究所 (14)
【Fターム(参考)】
【公開日】平成20年10月23日(2008.10.23)
【国際特許分類】
【出願日】平成19年4月4日(2007.4.4)
【出願人】(803000115)学校法人東京理科大学 (545)
【出願人】(399032282)株式会社 免疫生物研究所 (14)
【Fターム(参考)】
[ Back to top ]