
エンドリシン活性を有する新規ポリペプチド及びその使用
本発明は、クロストリジウム・ディフィシレの細胞に特異的に結合し及び/又はそれを溶解させることができる、配列番号1のアミノ酸配列を含む単離したポリペプチド、又はそのフラグメント、バリアント、誘導体又は融合体、及びそのフラグメント、バリアント、誘導体又は融合体がクロストリジウム・ディフィシレのバクテリオファージの天然リシンではないという条件で、それを製造する手段を提供する。本発明は、クロストリジウム・ディフィシレの細胞など細菌細胞を殺細胞し、それの感染に関連する疾患及び状態を診断、治療、そして予防する方法を更に提供する。本発明は、また、当該方法に使用するための診断キットを提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、クロストリジウム・ディフィシレ(Clostridium difficile)のバクテリオファージからのエンドリシン由来新規ポリペプチド及びそれをコードする核酸分子、並びにその組成物に関する。本発明は、クロストリジウム・ディフィシレなどの微生物細胞に関連する病態及び疾患の診断及び治療における当該ポリペプチド及び核酸分子の使用をも提供する。特に、本発明は、クロストリジウム・ディフィシレのバクテリオファージΦCD27由来エンドリシン活性を有する新規ポリペプチド及びその使用を提供する。
【背景技術】
【0002】
クロストリジウム・ディフィシレに関連する高まりつつある問題、特に、抗生物質使用にしばしば関連する院内感染におけるその役割は十分裏付けられている(非特許文献1)。C.ディフィシレは、加熱、乾燥及び消毒剤に耐性を有する胞子形成能を有する嫌気性グラム陽性菌である。非塩素系の洗浄剤への露出は、実際に胞子形成を増加させるという幾つかの証拠がある。これらの特性は、この生物が病院環境中に生き残る能力に寄与し、それにより患者に感染する能力を有する病原体の貯蔵庫を維持する。C.ディフィシレ関連疾患(CDAD)は、その増大する速度及び深刻さの双方を伴い、UKにおいても世界的にも高まりつつある問題である。イングランド及びウェールズでは、C.ディフィシレに関連する死亡が1999年の975人から2004年の2,247人まで増加した。CDADの届出は、1999年の1000人から2000年の15,000人、そして2003年の35,500人まで増加した(非特許文献2)。上記のヒトの健康への脅威に加えて、C.ディフィシレが、動物、特に子ウシ及びヒツジなどの家畜における罹患及び死亡の重大な原因でもあることを銘記すべきである。従って、ヒトにおけるこの問題に対処する方法に関する本明細書での開示は、同様に家畜対象にも当てはまると読み取るべきである。
【0003】
特に重大な進展は、当初カナダとUSAにおける、しかし現在はUKと幾つかの他のヨーロッパ諸国で著しい、C.ディフィシレの高毒性株の出現である。C.ディフィシレリボタイプ027と特定されたこの新菌株は、174例及び19名の死亡にかかわる大流行において2003年にUKで検出された。2006年までに75の病院からC.ディフィシレリボタイプ027の450の別のUK分離株が認められた(非特許文献1)。
【0004】
C.ディフィシレは土壌及び動物の腸管に広く分布している。それは、3%のヒト成人並びに80%の健常新生児及び小児の便から培養することができる(非特許文献1)。病原性能力は、C.ディフィシレの強力な毒素を産生する能力に関連する;2つの主要な特徴的毒素は、308kDaの外毒素の毒素A(TcdA)及び270kDaの細胞毒素の毒素B(TcdB)であり、これらはアミノ酸レベルで63%の相同性を共有する(非特許文献3)。これらの毒素をコード化する遺伝子は、病原性島PaLoc(非特許文献4)に関係し、そして菌株はこれらの2つの毒素を産生するそれらの能力の点で異なる。他の毒性因子も関与していると考えられ、そして別の2成分系毒素CDTが明らかにされている(非特許文献5、6)。
【0005】
毒性C.ディフィシレ株の病原性能力は、胃腸管(GIT)ミクロフローラが損なわれ又はアンバランスになっている場合に成り立ち、そしてこれは抗生物質治療の一般的な帰結である。このように、病院環境はヒトの病気を蔓延させ引き起こすC.ディフィシレの理想的な環境である(非特許文献1)。
【0006】
CDADは、C.ディフィシレの病原性株が十分強い地位をGITミクロフローラ内で
獲得すると起こり、宿主上皮を損傷する1つ又は複数の毒素を産生する。GITミクロフローラは病原性微生物に対する重要なバリアであり、宿主と有利な様式で相互作用しながらホメオスタシス平衡に維持される、凡そ500から1000の異なる種の複合群落を形成する。古典的な抗生物質治療は、程度の差はるが無差別的であり、それはGITの微生物群落の微妙なバランスを損なう可能性がある。従来の抗生物質治療又は他の要因の結果として、正常なミクロフローラの破壊はCDADの発症の主な要因である。
【0007】
それ故、複合GITミクロフローラの保護能力を損傷しない、C.ディフィシレの制御に対する新しい処置法及び取り組み方法の必要性が高まっている。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Kuijper, E. J., Coignard, B. & Tull, P. (2006) Clin Microbiol Infect 12 Suppl 6, 2-18.
【非特許文献2】Anonymous (2006) Health Statistics Quarterly 30, 56-60.
【非特許文献3】Rupnik, M., Dupuy, B., Fairweather, N. F., Gerding, D. N., Johnson, S., Just, I., Lyerly, D. M., Popoff, M. R., Rood, J. I., Sonenshein, A. L., Thelestam, M., Wren, B. W., Wilkins, T. D. & von Eichel-Streiber, C. (2005) J Med Microbiol 54, 113-7.
【非特許文献4】Braun, V., Hundsberger, T., Leukel, P., Sauerborn, M. & von Eichel-Streiber, C. (1996) Gene 181, 29-38.
【非特許文献5】Goncalves, C., Decre, D., Barbut, F., Burghoffer, B. & Petit, J. C. (2004) J Clin Microbiol 42, 1933-9.
【非特許文献6】Popoff, M. R., Rubin, E. J., Gill, D. M. & Boquet, P. (1988) Infect Immun 56, 2299-306.
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明の第1の態様は、クロストリジウム・ディフィシレの細胞に特異的に結合し及び/又はそれを溶解させることができる、配列番号1のアミノ酸配列を含む単離ポリペプチド、又はそのフラグメント、バリアント、誘導体若しくは融合体を提供することである。
【課題を解決するための手段】
【0010】
下記のアミノ酸配列は、クロストリジウム・ディフィシレのバクテリオファージΦCD27の野生型(即ち、天然産)エンドリシンの配列である。
【化1】
【0011】
NCBI受入番号YP_002290910及びACH91325をも参照されたい。
【0012】
1つの実施態様では、ポリペプチドは、クロストリジウム・ディフィシレ(ΦCD27以外の)のバクテリオファージの天然リシンではない。従って、本発明の第1の態様は、配列番号1のアミノ酸配列を含み又はそれから成る単離したポリペプチド、及びその非天然型フラグメント、バリアント、誘導体若しくは融合体を提供する。
【発明を実施するための形態】
【0013】
本明細書で使用される用語「アミノ酸」は、遺伝的にコード化された20の標準アミノ酸及びそれらの対応する「D」型の立体異性体(天然の「L」型と比較して)、ω−アミノ酸及び他の天然アミノ酸、特殊アミノ酸(例えば、α,α−ジ置換アミノ酸、N−アルキルアミノ酸など)並びに化学誘導アミノ酸(下記を参照)を含む。
【0014】
従って、アミノ酸を、「アラニン」又は「Ala」又は「A」のように特別に羅列する場合、この用語は特に明示しない限りL−アラニン及びD−アラニンの両方を指す。所望の機能特性がそのポリペプチドにより保持される限り、他の特殊アミノ酸も本発明のポリペプチドの好適な成分であり得る。例示したペプチドに対して、コード化されたアミノ酸残基は、それぞれ、必要に応じて標準アミノ酸の慣用名に対応する単一文字記号表示によって表わされる。
【0015】
好ましくは、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は、L−アミノ酸を含み又はそれから成る。
【0016】
「単離した」により、本発明のポリペプチド、特にバクテリオファージΦCD27の野生型エンドリシンが天然に見出されるもの以外の形態で提供されることを意味する。好ましくは、ポリペプチドはインタクトバクテリオファージを含まないで提供される。
【0017】
1つの実施態様では、単離した形態で与えられるバクテリオファージΦCD27[配列番号1]の天然エンドリシンである。
【0018】
先行技術において公知のクロストリジウム・ディフィシレバクテリオファージの他の天然リシンは、本発明の第1の態様に包含されない。特に、クロストリジウム・ディフィシレバクテリオファージの以下のリシンは、本発明の第1の態様の範囲から明確に除外される:
(a)バクテリオファージΦCD119のリシン(lysin);
(b)バクテリオファージΦC2のリシン;及び
(c)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン。
【0019】
例えば、以下の公知のタンパク質(それらのNCBI受入番号を参照することにより定義される):
PhiC2推定エンドリシン: YP_001110754;
CD630ファージエンドリシン: YP_001087453;
phiCD119推定リシン: YP_529586;
QCD−32g58仮想タンパク質: ZP_01803398;
QCD−32g58仮想タンパク質: ZP_01803228;
は、本発明の第1の態様の範囲から明確に除外される。
【0020】
1つの実施態様では、本発明の第1の態様のポリペプチドは、配列番号1のアミノ酸配列を含む。例えば、ポリペプチドは、配列番号1のアミノ酸配列から成ることができる。
【0021】
しかしながら、本発明の第1の態様は、クロストリジウム・ディフィシレの細胞に特異的に結合し及び/又はそれを溶解させることができる配列番号1のアミノ酸のフラグメント、バリアント、誘導体及び融合体にも及ぶ。
【0022】
「クロストリジウム・ディフィシレの細胞に特異的に結合することができる」により、ポリペプチドがクロストリジウム・ディフィシレの細胞に優先的に結合することができることを意味する。しかし、当然のことながら、当該ポリペプチドは1つ又はそれ以上の更なる細胞型にも優先的に結合することができる。好ましくは、クロストリジウム種の細胞に排他的に結合する。当該細胞結合活性は、当技術分野で周知の方法を用いて測定することができる。
【0023】
「クロストリジウム・ディフィシレの細胞を溶解させることができる」により、ポリペプチド、又はそのフラグメント、バリアント、誘導体若しくは融合体が、バクテリオファージΦCD27の野生型エンドリシンが細菌性細胞を溶解させる能力(少なくとも一部)を保持することを意味する。当然のことながら、当該溶解活性は、全細胞型に対する非特異的細胞毒性活性よりむしろ細胞特異的(例えば、クロストリジウム・ディフィシレの細胞に対して)であるべきである。当該細胞溶解活性は、以下の実施例に詳述される方法(Loessner et al. [37]をも参照されたい、その開示内容は参照することにより本明細書に組み入れられている)など当技術分野で周知の方法を用いて測定することができる。好ましくは、ポリペプチドがクロストリジウム・ディフィシレの細胞を溶解させる能力は新鮮細胞を用いて測定される。
【0024】
好ましい実施態様では、ポリペプチドがクロストリジウム・ディフィシレの細胞を溶解させる能力は、11204菌株の細胞を用いて測定される。
【0025】
当業者には当然のことながら、ポリペプチド、又はそのフラグメント、バリアント、誘導体若しくは融合体は、細菌細胞を溶解させるバクテリオファージΦCD27の野生型エンドリシンの全能力を保持する必要はない。むしろ、該ポリペプチド、フラグメント、バリアント、誘導体又はその融合体は、細菌細胞を溶解するバクテリオファージΦCD27の野生型エンドリシンのただ単に少なくとも10%の能力を保持する必要があるだけである。しかしながら、好ましくは、ポリペプチド、フラグメント、バリアント、誘導体又はその融合体は、少なくとも20%の、例えば、少なくとも30%、40%、50%、60%、70%、80%、90%、100%、150%、200%又はそれ以上の、細菌細胞を溶解させるバクテリオファージΦCD27の野生型エンドリシンの能力を示す。
【0026】
それ故に、本発明の第1の態様の実施態様では、ポリペプチドは、クロストリジウム・ディフィシレの細胞を溶解させることができる、配列番号1のアミノ酸配列のフラグメントを含み又はそれから成る。
【0027】
多くのバクテリオファージ・エンドリシンが2つの異なるドメイン(例えば、Sheehan et al., 1996, FEMS Microbiology Letters 140:23-28を参照されたい、その開示内容は参照することにより本明細書に組み入れられている)から成ることは十分立証されている。1つは細胞壁分解に関与する触媒ドメインであり、これらは幾つかの異なる形態で存在することが知られている。他のドメインは、細胞表面モチーフを認識する細胞壁結合ドメインであり、その標的細胞へのエンドリシンの付着を可能にする。
【0028】
酵素ドメインは、同じタイプの溶解活性を共有する溶菌酵素の他の類似領域に対するそのアミノ酸相同性によって同定することができる。バクテリオファージΦCD27のエンドリシンの場合、酵素ドメインはN−アセチルムラモイル−L−アラニンアミダーゼと同定されており、それはエンドリシンのアミノ末端領域を占有する(これは公知酵素ドメインによる配列番号1の並列解析によって、例えば、NCBICDD検索ツールを用いて確認することができる;Marchler-Bauer & Bryant, 2004, Nuc. Acids Res. 32[W]:327-331を参照されたい。その開示内容は、参照することにより本明細書に組み入れられている)。細胞壁結合ドメインは、エンドリシンのカルボキシ末端領域を占有すると考えられる。
【0029】
1つの実施態様では、酵素ドメインは配列番号1の1から175のアミノ酸内に含まれる。従って、酵素ドメインを含むフラグメントは、アミノ酸1、5、10、15、20、25、30、35、40、45、50、60、70、80、90又は100のいずれかから開始し、そしてアミノ酸175、170、165、160、155、150、145、140、135、130、125、120、115、110又は105のいずれかで終了する配列番号1の配列から成ると考えられる。例えば、酵素ドメインを含むフラグメントは、配列番号1の10から140までのアミノ酸、又は配列番号1の25から155までのアミノ酸、又は上記の開始及び終点の他の可能な順列のいずれかからなると考えられる。
【0030】
1つの実施態様では、細胞壁結合ドメインは配列番号1の175から270のアミノ酸内に含まれる。従って、細胞壁結合ドメインを含むフラグメントは、アミノ酸175、180、185、190、195、200、205、210、215、220のいずれかから開始し、そしてアミノ酸270、265、260、255、250、245、240、235、230又は225のいずれかで終了する配列番号1の配列から成ると考えられる。例えば、細胞壁結合ドメインを含むフラグメントは、配列番号1の195から265までのアミノ酸、又は配列番号1の180から240までのアミノ酸、又は上記の開始及び終点の他の可能な順列のいずれかからなると考えられる。
【0031】
本発明の第1の態様のポリペプチドは、好ましくは酵素ドメイン及び細胞壁結合ドメインの両方に対応する、配列番号1のアミノ酸配列の1つ又はそれ以上のフラグメントを含み又はそれから成る。
【0032】
しかしながら、当業者には当然のことながら、配列番号1の細胞壁結合ドメインは、代わりにクロストリジウム・ディフィシレの細胞を溶解させることができる別の起源からの酵素(溶解)ドメインに融合するか又はそうでなければ結合してもよい。キメラ的なリシンの製造は、Sheehan et al., 1996, FEMS Microbiology Letters 140:23-28に記載されており、その開示内容は参照することにより本明細書に組み入れられている。従って、別の実施態様では、本発明の第1の態様のポリペプチドは、細胞壁結合ドメインに対応する配列番号1のアミノ酸配列の1つ又はそれ以上のフラグメントを含み又はそれから成ることができる。
【0033】
フラグメントは、配列番号1の少なくとも50の隣接アミノ酸、例えば、配列番号1の、少なくとも60、70、80、90、100、110、120、130、140、150、160、170、175、180、190、200、210、220、230、240、250、260又は265の隣接アミノ酸を含み又はそれから成ることができる。
【0034】
別の実施態様では、本発明の第1の態様のポリペプチドは、クロストリジウム・ディフィシレの細胞を溶解させることができる、配列番号1のアミノ酸配列のバリアント、又はそのフラグメントを含み又はそれから成ることができる。
【0035】
ポリペプチドの「バリアント」により、配列番号1のアミノ酸配列に関連して保存性であれ又は非保存性であれ、挿入、欠失及び/又は置換を含む。特に、バリアントポリペプチドは、非天然バリアントであってもよい。
【0036】
例えば、ポリペプチドは、配列番号1のアミノ酸配列に対して少なくとも60%の同一性、より好ましくは該配列に対して少なくとも70%又は80%又は85%又は90%の同一性、そして最も好ましくは該アミノ酸配列に対して少なくとも95%、96%、97%、98%又は99%の同一性を有するアミノ酸配列を含んでもよい。
【0037】
当然のことながら、上記の配列同一性は配列番号1のアミノ酸配列の全長にわたり、又はその一部にわたってもよい。しかしながら、好ましくは、配列同一性は、配列番号1のアミノ酸配列の少なくとも50のアミノ酸にわたり、例えば、その中の少なくとも60、70、80、90、100、110、120、130、140、150、160、170、180、190、200、210、220、230、240、250、260又はそれ以上のアミノ酸にわたる。
【0038】
同一性パーセントは、当技術分野で周知の方法、例えば、ExPASyファシリティウェブサイトでのLALIGNプログラム(Huang and Miller, Adv. Appl. Math. (1991) 12:337-357、その開示内容は参照することにより本明細書に組み入れられている) :
www.ch.embnet.org/software/LALIGN_form.html
を用いて、パラメーターとしてグローバルアラインメントオプション、スコアリングマトリックススBLOSUM62、開口ギャップペナルティ−14、拡張ギャップペナルティ−4を用いて測定することができる。
【0039】
或いはまた、2ポリペプチド間の配列同一性パーセントは、適切なコンピュータプログラム、例えば、AlignX, Vector NTI Advance 10 (Invitrogen Corporationから) 又はGAPプログラム(University of Wisconsin Genetic Computing Groupから)を用いて測定してもよい。
【0040】
当然のことながら、同一性パーセントは、その配列が最適にアラインメントされているポリペプチドに関して計算される。
【0041】
配列番号1のアミノ酸配列のフラグメント及びバリアントは、当技術分野で周知のタンパク質工学及び部位特異的変異誘発の方法を用いて作製されてもよい(例えば、Molecular Cloning: a Laboratory Manual, 3rd edition(「分子クローニング:実験室マニュアル第3版」), Sambrook & Russell, 2001, Cold Spring Harbor Laboratory Pressを参照されたい。その開示内容は参照することにより本明細書に組み入れられている)。
【0042】
当業者には当然のことながら、本発明のポリペプチド、又はそのフラグメント、バリアント若しくは融合体は、修飾又は誘導体化された1つ又はそれ以上のアミノ酸を含んでもよい。従って、ポリペプチドは、配列番号1のアミノ酸配列の誘導体、又はそのフラグメント若しくはバリアントを含み又はそれから成ることができる。
【0043】
1つ又はそれ以上のアミノ酸の化学誘導体は、官能性側鎖基との反応により得ることができる。当該誘導体化分子は、例えば、遊離アミノ基が誘導体化されて、アミン塩酸塩、p−トルエンスルホニル基、カルボキシベンゾキシ基、t−ブチルオキシカルボニル基、クロロアセチル基又はホルミル基を形成するそれらの分子を含む。遊離カルボキシル基は、誘導体化して塩、メチル及びエチルエステル又は他のタイプのエステル及びヒドラジドを形成してもよい。遊離のヒドロキシル基は、誘導体化してO−アシル又はO−アルキル誘導体を形成してもよい。化学誘導体としては、20の標準アミノ酸の天然アミノ酸誘導体を含むそれらのペプチドも含まれる。例えば:4−ヒドロキシプロリンは、プロリンと置換してもよい;5−ヒドロキシリジンは、リジンと置換してもよい;3−メチルヒスチジンは、ヒスチジンと置換してもよい;ホモセリンはセリンと、そしてオルニチンはリジンと置換してもよい。誘導体は、また、必要活性が維持される限り、1つ又はそれ以上の付加又は欠失を含むペプチドをも含む。含まれる他の修飾は、アミド化、アミノ末端アシル化(例えば、アセチル化又はチオグリコール酸アミド化)、末端カルボキシアミド化(例えば、アンモニア又はメチルアミンによる)、などの末端修飾である。
【0044】
当業者には更に当然のことながら、ペプチド模倣的化合物も有用であり得る。従って、「ポリペプチド」により、エンドリシン活性を呈するペプチド模倣的化合物を含む。用語「ペプチド模倣的」は、治療薬として特定のポリペプチドの立体配座及び所望の特徴を模倣した化合物を指す。
【0045】
例えば、本明細書に記載のポリペプチドは、アミノ酸残基がペプチド(−CO−NH−)結合によって結合される分子だけでなく、ペプチド結合が逆向きの分子も含む。当該逆反転ペプチド模倣体は、当技術分野で公知の方法、例えば、Meziere et al. (1997) J. Immunol. 159, 3230-3237に記載の方法などを用いて作製してもよく、その開示内容は参照することにより本明細書に組み入れられている。CO−NHの代わりにNH−CO結合を含む当該逆反転ペプチド模擬体は、タンパク質分解に対して更にいっそう抵抗性である。或いはまた、本発明のポリペプチドは、1つ又はそれ以上のアミノ酸残基が、通常のアミド結合の代わりに−γ(CH2NH)−結合によって結合したペプチド模倣的化合物であってもよい。
【0046】
当然のことながら、ポリペプチドは、細胞外タンパク質分解消化に対する感受性を低減するのを助けるために、そのN−又はC−末端において、例えば、アミド化により都合よくブロックしてもよい。
【0047】
上記のように、D−アミノ酸及びN−メチルアミノ酸などの様々な非コード又は修飾アミノ酸は、本発明のポリペプチドを修飾するのに使用してもよい。加えて、推定生物活性立体配座は、環化などの共有結合修飾により、又はラクタムの組み込み若しくは他のタイプの架橋によって安定化してもよい。ジスルフィド、スルフィド及びアルキレン架橋を含む、環状ホモデチックペプチド及び環状ヘテロデチックペプチドの合成方法は、米国特許第5643872号に開示されている。環化方法の他の例は、米国特許第6008058号にて検討され開示されており、それらの文書における関連開示内容は、参照することにより本明細書に組み入れられている。環状の安定化ペプチド模倣的化合物の合成への更なる取り組みは、閉環メタセシス(RCM)である。
【0048】
要約すれば、周知のような末端修飾は、プロテイナーゼ消化による感受性を低減させ、その結果、溶液中、特にプロテアーゼが存在すると考えられる生体液中のペプチドの半減期を延長するのに有用である。ポリペプチド環化は、また、有用な修飾であり、環化によって形成される安定構造に因りそして環状ペプチドで認められる生物活性の点で好ましい。
【0049】
従って、1つの実施態様では、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は環状である。しかしながら、好ましい実施態様では、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は線状である。
【0050】
本発明の第1の態様の更なる実施態様では、ポリペプチドは配列番号1のアミノ酸配列の融合体、又はそのフラグメント、バリアント若しくは誘導体を含み又はそれから成る。
【0051】
ポリペプチドの「融合体」は、その他のポリペプチドに融合したポリペプチドを含む。例えば、ポリペプチドは、配列番号1のアミノ酸配列の内部に及び/又はN−及び/又はC−末端に挿入された1つ又はそれ以上の更なるアミノ酸、又はそのフラグメント、バリアント若しくは誘導体を含んでもよい。
【0052】
従って上記のように、1つの実施態様では、本発明の第1の態様のポリペプチドは、異なる起源の酵素ドメインがそれに融合した、細胞壁結合ドメイン(又はその細胞壁結合活性を保持するようなドメイン配列のバリアント)から成る配列番号1のフラグメントを含む。
【0053】
他の好適な酵素ドメインの例は:
L−アラノイル−D−グルタミン酸エンドペプチダーゼ;D−グルタミル−m−DAPエンドペプチダーゼ;ペプチド間架橋特異的エンドペプチダーゼ;N−アセチル−β−D−グルコサミニダーゼ(=ムラモイルヒドロラーゼ);N−アセチル−β−D−ムラミダーゼ(=リゾチーム);溶菌性トランスグリコシラーゼを含む。
【0054】
また、他の起源からのN−アセチルムラモイル−L−アラニンアミダーゼも利用可能である(Loessner, 2005, Current Opinion in Microbiology 8: 480-487を参照されたい。その開示内容は参照することにより本明細書に組み入れられている)。
【0055】
例えば、該ポリペプチドは、該ポリペプチドの精製を容易にするために、グルタチオン−S−トランスフェラーゼ(GST)又はタンパク質Aのようなポリペプチドに融合してもよい。当該融合体の例は当業者に周知である。同様に、該ポリペプチドは、His6などのオリゴヒスチジンタグに、又は周知のMycタグエピトープなどの抗体によって認識されるエピトープに融合してもよい。該ポリペプチドのいずれのフラグメント、バリアント又は誘導体に対する融合体も、本発明の範囲内に含まれる。当然のことながら、所望の特性、即ち、エンドリシン活性を保持する融合体(又はそのバリアント若しくは誘導体)が好ましい。融合体が本明細書に記載の方法に使用するのに好適なものであれば、それも特に好ましい。
【0056】
例えば、融合体は、本発明の該ポリペプチドに所望の特性を賦与する更なる部分を含んでもよい;例えば、その部分はポリペプチドを検出又は単離し、ポリペプチドの細胞内取り込みを促進し、又は細胞からのタンパク質の分泌を導くのに有用と考えられる。その部分は、当業者に周知の、例えば、ビオチン部分構造、放射性部分構造、蛍光部分構造、例えば、小分子蛍光プローブ又は緑色蛍光タンパク質(GFP)蛍光プローブであってもよい。その部分構造は、当業者に周知の免疫原性タグ、例えば、Mycタグであってもよく、又は当業者に周知の、ポリペプチドの細胞内取り込みを促進することができる親油性分子若しくはポリペプチドドメインであってもよい。
【0057】
当業者には当然のことながら、本発明のポリペプチドは、また、上記ポリペプチドの薬学的に許容される酸又は塩基付加塩をも含む。本発明において有用な、上記の塩基化合物の薬学的に許容される酸付加塩を製造するために使用される酸は、非毒性酸付加塩、即ち、塩酸塩、臭化水素酸塩、ヨウ化水素酸塩、硝酸塩、硫酸塩、重硫酸塩、リン酸塩、過リン酸塩、酢酸塩、乳酸塩、クエン酸塩、酸性クエン酸塩、酒石酸塩、酸性酒石酸塩、コハク酸塩、マレイン酸塩、フマル酸塩、グルコン酸塩、糖酸塩、安息香酸塩、メタンスルホン酸塩、エタンスルホン酸塩、ベンゼンスルホン酸塩、p−トルエンスルホン酸塩、及びとりわけ、パモエート[即ち、1,1′−メチレン−ビス−(2−ヒドロキシ−3−ナフトエート)]塩などの、薬学的に許容されるアニオンを含有する塩を形成する酸である。
【0058】
薬学的に許容される塩基付加塩も、また、ポリペプチドの薬学的に許容される塩の形態を生成するために使用してもよい。本来酸性の本発明化合物の、薬学的に許容される塩基塩を製造する試薬として使用してもよい化学塩基は、当該化合物と非毒性塩基塩を形成する塩基である。当該非毒性塩基塩は、アルカリ金属カチオン(例えば、カリウム及びナトリウム)及びアルカリ土類金属カチオン(例えば、カルシウム及びマグネシウム)、N−メチルグルカミン(メグルミン)などの、アンモニウム又は水溶性アミン付加塩、並びに、とりわけ、薬学的に許容される有機アミンの低級アルカノールアンモニウム及び他の塩基塩などの当該薬学的に許容されるカチオンから由来する塩を含むが、これらに限定されるものではない。
【0059】
ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は、また、貯蔵のために凍結乾燥し、そして使用前に好適な担体中に再構成することもできる。いずれの好適な凍結乾燥方法(例えば、噴霧乾燥、ケーキ乾燥)及び/又は再構成方法も採用することができる。当業者には当然のことながら、凍結乾燥及び再構成は様々な程度の活性ロスをもたらし、そした使用レベルはそれを補うために上方へ調整しなければならない。好ましくは、凍結乾燥(フリーズドライ)ポリペプチドは、再水和した場合、その活性(凍結乾燥する前)の、最高で約20%、又は最高で約25%、又は最高で約30%、又は最高で約35%、又は最高で約40%、又は最高で約45%、又は最高で約50%を失う。
【0060】
本発明のポリペプチドの本質的な特徴は、クロストリジウム・ディフィシレの細胞の溶解能力である。好ましくは、ポリペプチドはクロストリジウム・ディフィシレの複数の株の細胞を溶解することができる。例えば、ポリペプチドは、配列番号1のΦCD27リシンによって溶解するクロストリジウム・ディフィシレの1つ又はそれ以上の菌株を溶解することができる(下の表1を参照)。
【0061】
当然のことながら、本発明のポリペプチドは、また、バシラス種(例えば、バシラス・セレウス(Bacillus cereus)、枯草菌及び/又は炭疽菌)、他のクロストリジウム種(例えば、クロストリジウム・ビフェルメンタンス(Clostridium bifermentas))及び/又はリステリア種(例えば、リステリア・イバノビイ(Listeria ivanovii))など他の細菌種の細胞を溶解することができる。
【0062】
1つの実施態様では、本発明のポリペプチドは、健常な腸生理を維持するのに有用な細菌を実質的に溶菌することができない。例えば、ポリペプチドが、クロストリジウム・レプツム(Clostridium leptum)、 クロストリジウム・ネキシル(Clostridium nexile)、クロストリジウム・コッコイデス(Clostridium coccoides)、クロストリジウム・イノクーム(Clostridium innocuum)、クロストリジウム・ラモスム(Clostridium ramosum)及び/又はアネロコッカス・ヒドロジェナリス(Anaerococcus hydrogenalis)の細胞を溶解しない場合、それは有益である。
【0063】
最も好ましくは、本発明のポリペプチドは、カナダ、US及び現在全ヨーロッパで出現しているクロストリジウム・ディフィシレの高病原性株である、クロストリジウム・ディフィシレ株リボタイプ027の細胞を溶解することができる。例えば、ポリペプチドは、クロストリジウム・ディフィシレリボタイプ027の細胞に対して、配列番号1のポリペプチドの少なくとも10%、例えば、少なくとも20%、30%、40%、50%、60%、70%、80%、90%、100%又はそれ以上の溶解活性を示すことができる。ポリペプチドは、クロストリジウム・ディフィシレリボタイプ027の細胞に対して、配列番号1のポリペプチドより大きい溶解活性、例えば、少なくとも110%、120%、130%、140%、150%、160%、170%、180%、190%、200%、250%、300%、500%又はそれ以上の溶解活性さえも示すことができる。
【0064】
有利なことには、ポリペプチドは、病原性細菌の細胞を選択的に、即ち、非病原性細菌よりも大幅に溶解させることができる。
【0065】
本発明の第1の態様において使用するポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体の製造方法は、当技術分野で周知である。好都合なことには、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は、組み換えポリペプチドであり又はそれを含む。
【0066】
このように、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体をコード化する核酸分子(又はポリヌクレオチド)は、好適な宿主及びそれから得られるポリペプチドに発現することができる。当該組み換えポリペプチドの好適な生産方法は、当技術分野で周知である(例えば、Sambrook & Russell, 2000, Molecular Cloning, A Laboratory Manual, Third Edition(「分子クローニング、実験室マニュアル、第3版」), Cold Spring Harbor, New Yorkを参照されたい。その文書の関連する開示内容は、参照することにより本明細書に組み入れられている)。
【0067】
簡潔にいうと、発現ベクターは、適切な宿主内で核酸分子によりコード化されるポリペプチドを発現することができる核酸分子を含んで、構成することができる。
【0068】
核酸分子、特にDNAを、例えば、相補的凝集性末端経由で、操作可能的にベクターに結合する様々な方法が開発されている。例えば、相補的ホモポリマー領域は、ベクターDNAに挿入するようにDNAセグメントに付加することができる。次いで、ベクター及びDNAセグメントは、相補的ホモポリマーの尾部間で水素結合により接続し、組み換えDNA分子を形成する。
【0069】
1つ又はそれ以上の制限部位を含む合成リンカーは、DNAセグメントをベクターへ接続する代替法を提供する。例えば、エンドヌクレアーゼ制限消化により生成するDNAセグメントは、3′−5′−エキソヌクレアーゼ活性を有する突出3′−1本鎖末端を除去し、そしてそれらのポリメラーゼ活性を有する陥凹3′−末端を埋める酵素である、バクテリオファージT4DNAポリメラーゼ又は大腸菌DNAポリメラーゼIで処理される。
【0070】
従って、これらの活性の組み合わせは平滑末端DNAセグメントを産生する。次いで、平滑末端セグメントは、バクテリオファージT4DNAリガーゼなどの、平滑末端DNA分子の連結反応を触媒することができる酵素の存在下に、大過剰モルのリンカー分子とインキュベートされる。このように、反応の生成物は、それらの末端に高分子リンカー配列を持っているDNAセグメントである。次いで、これらのDNAセグメントは好適な制限酵素により切断され、そしてDNAセグメントの末端と適合する末端を生成する酵素で切断されている発現ベクターに連結される。
【0071】
DNA(又はレトロウイルスベクター、RNA)は、次いで、好適な宿主に発現してポリペプチドを産生する。このように、ポリペプチドをコード化するDNAは公知の方法に従って使用され、本明細書に含まれる教示を勘案して適切に修正されて発現ベクターを構築し、そのベクターは、次いで、本発明の化合物又はその結合部分を発現及び産生するように、適切な宿主細胞を形質転換するために使用することができる。当該技術は当技術分野で周知である。
【0072】
ポリペプチドをコード化するDNA(又はレトロウイルスベクターの場合には、RNA)は、適切な宿主への導入のために多種多様な他のDNA配列に結合してもよい。相手のDNAは、宿主の性質、宿主へのDNAの導入様式、及びエピソームの保持又は統合が求められるかどうかによって決まる。
【0073】
一般的に、DNAは、適正な方向に及び発現のための正しいリーディングフレームにおいて、プラスミドなどの発現ベクターに挿入される。必要に応じて、DNAは、所望の宿主によって認識される適切な転写及び翻訳調節制御ヌクレオチド配列に結合し得るが、当該制御は一般的に発現ベクター中で実現可能である。次いで、ベクターは標準的方法を介して宿主に導入される。一般的に、全ての宿主がベクターにより形質転換されるわけではない。従って、形質転換宿主細胞を選択する必要性がある。1つの選択方法は、抗生物質耐性などの、形質転換細胞中の選択可能な形質をコードするあらゆる必要な制御要素によって、発現ベクターにDNA配列を組み込むことを含む。或いはまた、当該選択可能な形質の遺伝子は、所望の宿主細胞を同時形質転換するのに使用される別のベクター上にあり得る。
【0074】
発現ベクターにより形質転換されている宿主細胞は、次いで、十分な時間そしてポリペプチドの発現を可能にする本明細書に開示の教示に照らして当業者に公知の適切な条件下で培養し、次いでこれを回収することができる。
【0075】
多くの発現系が知られており、細菌(大腸菌、枯草菌)、酵母(例えば、サッカロミセス・セレビジエ)、糸状菌(例えば、アスペルギルス)、植物細胞、動物細胞及び昆虫細胞が挙げられる。
【0076】
代表的なベクターは、典型的には、ベクターが他の非原核生物細胞型における発現に使用される場合でも、原核生物中の増殖のためのColE1複製起点(ori)などの原核生物レプリコンを含む。ベクターは、また、それによって形質転換した大腸菌などの細菌性宿主細胞における遺伝子の発現(転写及び転座)を導くことができる、原核生物プロモーターなどの適切なプロモーターをも含むことができる。
【0077】
代表的な原核生物ベクタープラスミドは、Biorad Laboratories (Richmond, CA, USA) から入手できるpUC18、pUC19、pBR322及びpBR329、及びPharmacia(Piscataway, NJ, USA) から入手できるpTrc99及びpKK223−3である。
【0078】
代表的な哺乳動物細胞ベクタープラスミドは、Pharmacia, Piscataway(NJ, USA) から入手できるpSVLである。このベクターは、クローン化遺伝子の発現を推進するSV40後期プロモーターを用いるが、発現の最高レベルはCOS−1細胞などのT抗原産生細胞において、見出されている。
【0079】
誘導型哺乳動物ベクターの例は、Pharmaciaから同様に入手できるpMSGである。このベクターは、クローン化遺伝子の発現を推進するために、マウス哺乳動物腫瘍ウイルスの長い末端反復のグルココルチコイド誘導プロモーターを用いる。
【0080】
他のベクター及び発現系は、様々な宿主細胞を用いて使用するために当技術分野で周知である。
【0081】
宿主細胞は原核細胞であっても又は真核細胞であってもよい。細菌細胞は、好ましい原核宿主細胞であり、そして一般的には、例えば、Bethesda Research Laboratories Inc.(Bethesda, MD, USA)から入手できる大腸菌DH5株、及びAmerican Type Culture Collection (ATCC) (Rockville, MD, USA) (No. ATCC 31343)から入手できるRR1などの大腸菌の1菌株である。好ましい真核宿主細胞は、酵母、昆虫及び哺乳動物細胞、好ましくはマウス、ラット、サル又はヒト線維芽細胞及び腎細胞系などの脊椎動物細胞である。酵母宿主細胞は、Stratagene Cloning Systems(La Jolla, CA 92037, USA)から一般に入手できるYPH499、YPH500及びYPH501を含む。好ましい哺乳動物宿主細胞は、ヒト胚性腎細胞であるCRL1658及び293細胞のようなATCCから入手できるチャイニーズハムスター卵巣(CHO)細胞を含む。好ましい昆虫細胞はバキュロウイルス発現ベクターで形質転換し得るSf9細胞である。
【0082】
宿主細胞を培養しそして組み換えタンパク質を分離する方法は、当技術分野でで周知である。当然のことながら、宿主細胞によっては、産生される本発明のポリペプチドは異なってもよい。例えば、酵母又は細菌細胞などの特定の細胞は、違った方法で翻訳後修飾されると考えられる本発明の化合物の形態の産生をもたらす可能性のある、異なる翻訳後修飾系を有しないか又は有する。
【0083】
本発明のポリペプチドは、また、ウサギ網状赤血球溶血液又はコムギ胚芽溶解物(Promegaから入手できる)などの市販のインビトロ翻訳系を用いてインビトロで製造してもよい。好ましくは、翻訳系はウサギ網状赤血球溶血液である。都合の良いことには、翻訳系は、TNT転写−翻訳系(Promega)などの転写系に連結されてもよい。この系は、翻訳と同じ反応でコード化DNAポリヌクレオチドからの好適なmRNA転写物を産生する利点を有する。
【0084】
CS Bio Company Inc.(Menlo Park, USA)から入手できるもののような、自動ポリペプチド合成機も、また、使用することができる。
【0085】
従って、本発明の第2の態様は、本発明の第1の態様のポリペプチドをコード化する単離した核酸分子を提供することである。
【0086】
核酸分子はDNA(例えば、cDNA)又はRNAであってもよい。
【0087】
好ましい実施態様では、核酸分子は図3に示されるヌクレオチド配列[配列番号2]を含み又はそれから成る。
【0088】
本発明の第3の態様は、本発明の第2の態様の核酸分子を含むベクターを提供することである。1つの実施態様では、ベクターは発現ベクターである。好ましくは、ベクターはpET15b及びpACYC184から成る群から選択される。
【0089】
当業者には当然のことながら、発現ベクターの選択は宿主細胞の選択により決定してもよい。従って、乳酸連鎖球菌における本発明のポリペプチドの発現では、2成分制御系をコード化するnisR 及びnisK遺伝子をも発現する乳酸連鎖球菌のバックグランド株を用いて、本発明のポリペプチドがnisAオペロンのプロモーターの制御下に発現する、ナイシン(nisin)発現系を使用することができる。この系の下での発現は正に調節され、そして外因性ナイシンの供給により誘導される(Ruyter at el., 1996, Applied and Environmental Microbiology 62:3662-3667を参照されたい。その開示内容は、参照することにより本明細書に組み入れられている)。
【0090】
代わりの実施態様では、ナイシンの全生合成遺伝子クラスターは、インデューサーがその細胞によって合成される場合と同じ宿主細胞内にもたらされる。
【0091】
更なる代わりの実施態様では、本発明のポリペプチドは、プラスミド系か又は染色体に組み込まれた系を用いて、ラクトース異化オペロンの制御下に乳酸連鎖球菌に発現してもよい(例えば、Payne et al., 1996, FEMS Microbiology Letters 136: 19-24及びvan Rooijen et al., 1992, Journal of Bacteriology 174: 2273-2280を参照されたい。その開示内容は、参照することにより本明細書に組み入れられている)。
【0092】
本発明の第4の態様は、本発明の第2の態様の核酸分子又は本発明の第3の態様のベクターを含む宿主細胞を提供することである。1つの実施態様では、宿主細胞は微生物細胞、例えば、細菌細胞である。好ましくは、宿主細胞は非病原性である。
【0093】
例えば、宿主細胞は、大腸菌、ラクトコッカス種、バクテロイデス種、ラクトバシラス種、エンテロコッカス種及びバシラス種から成る群から選択してもよい。
【0094】
好ましい実施態様では、宿主細胞は乳酸連鎖球菌の細胞である。
【0095】
或いはまた、宿主細胞は酵母細胞、例えば、サッカロミセス種であってもよい。
【0096】
本発明の第5の態様は、ポリペプチドを発現させる条件下に、本発明の第2の態様の核酸分子又は本発明の第3の態様のベクターを含む宿主細胞集団を培養し、それからポリペプチドを単離することを含む、本発明のポリペプチドを製造する方法を提供することである。
【0097】
本発明の第6の態様は、以下のもの:
(a)本発明の第1の態様のポリペプチド;
(b)本発明の第2の態様の核酸分子;
(c)本発明の第3態様のベクター;
(d)本発明の第4態様の宿主;及び/又は
(e)本発明の第1の態様のポリペプチドを発現することができるバクテリオファージ;
及び薬学的に許容される担体、賦形剤又は医薬品添加剤を含む、医薬組成物を提供することである。
【0098】
本明細書で使用される「医薬組成物」は、本発明の方法に使用するための治療上有効な製剤を意味する。
【0099】
本明細書で使用される「治療的有効量」、又は「有効量」、又は「治療的に有効な」は、所定の病態及び投与レジメンに対し治療的効果を与える量を指す。これは、所要の添加剤及び賦形剤、即ち、担体又は投与賦形剤と併せて、所望の治療的効果をもたらすように算出される活性物質の所定量である。更に、宿主の活動性、機能及び反応性の臨床的に重大な欠如を減少させ、そして最も好ましくはそれを予防するのに十分な量を意味することを意図している。或いはまた、治療的有効量は、宿主の臨床的に重大な病態の改善をもたらすのに十分な量である。当業者には当然のことながら、化合物の量はその比活性度によって変動してもよい。好適な用量は、所要の賦形剤と併せて所望の治療効果をもたらすように算出される活性組成物の所定量を含むことができる。本発明の組成物を製造するための方法及び使用では、活性成分の治療的有効量が供給される。治療的有効量は、当技術分野で周知のように、年齢、体重、性別、病態、合併症、他の疾患など、患者の特性を基準にして通常の技量の医療又は獣医従事者により決定することができる。
【0100】
本発明の1つの実施態様では、医薬組成物は、本発明の第1の態様のポリペプチドを含む。
【0101】
ポリペプチドは、使用されるポリペプチドの有効性/毒性に応じて種々の濃度で製剤化し得る。好ましくは、製剤は、0.1μMと1mMの間、より好ましくは1μMと100μMの間、5μMと50μMの間、10μMと50μMの間、20μMと40μMの間、及び最も好ましくは約30μMの濃度でポリペプチドを含む。インビトロ適用の場合は、製剤は類似濃度のポリペプチドを含むことができる(しかし、当然のことながら、より高濃度も使用してもよい)。
【0102】
従って、この医薬製剤は、当該細胞により感染し又は感染を起こし易い患者において、クロストリジウム・ディフィシレの細胞増殖を少なくとも部分的に阻害するのに十分な量のポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体を含むことができる。好ましくは、医薬製剤は、患者においてクロストリジウム・ディフィシレの細胞を死滅させるのに十分な量のポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体を含む。
【0103】
当業者には当然のことながら、本発明のポリペプチドは、一般的に、所定の投与経路及び標準的薬剤治療(例えば、Remington: The Science and Practice of Pharmacy, 19th edition(「薬学の科学及び実践、第19版」), 1995, Ed. Alfonso Gennaro, Mack Publishing Company, Pennsylvania, USAを参照されたい。その開示内容は、参照することにより本明細書に組み入れられている)に関して選択される好適な医薬品添加剤、賦形剤又は担体との混合物で投与される。
【0104】
例えば、ポリペプチドは、即時性、遅延性又は放出制御の適用のために、着香剤又は着色剤を含んでもよい錠剤、カプセル剤、腔坐剤、エリキシル剤、水剤又は懸濁剤の形態で、経口、口腔内又は舌下で投与することができる。ポリペプチドは、直接注射(例えば胃腸管へ)を経て投与してもよい。
【0105】
しかしながら、好ましくは、ポリペプチド及びその医薬組成物は、経口投与用である。
【0106】
好適な錠剤処方は、微結晶セルロース、乳糖、クエン酸ナトリウム、炭酸カルシウム、第二リン酸カルシウム及びグリシンなどの医薬品添加剤、澱粉(好ましくは、トウモロコシ、ジャガイモ又はタピオカ澱粉)、澱粉グリコール酸ナトリウム、クロスカルメロースナトリウム、及び特定の複合珪酸塩などの崩壊剤、及びポリビニルピロリドン、ヒドロキシル−プロピルメチルセルロース(HPMC)、ヒドロキシ−プロピルセルロース(HPC)、蔗糖、ゼラチン及びアラビアゴムなどの造粒結合剤を含んでもよい。加えて、ステアリン酸マグネシウム、ステアリン酸、グリセリルベヘナート及びタルクなどの滑沢剤が含まれ得る。
【0107】
類似タイプの固形組成物も、ゼラチンカプセル剤の充填剤として用いてもよい。この関連の好ましい医薬品添加剤は、ラクトース、澱粉、セルロース、乳糖又は高分子量ポリエチレングリコールをも含むことができる。水性懸濁剤及び/又はエリキシル剤では、ポリペプチドは種々の甘味剤又は着香剤、着色料又は色素と、乳化剤及び/又は懸濁剤と及び水、エタノール、プロピレングリコール及びグリセリンなどの希釈剤、及びその組み合わせと併用してもよい。
【0108】
ポリペプチドは、非経口で、例えば、静脈内、関節内、動脈内、腹腔内、髄腔内、心室内、胸骨内、頭蓋内、筋肉内若しくは皮下に投与してもよく、又は注入法で投与してもよい。それらは、他の物質、例えば、血液と等張溶液にするのに十分な塩又はグルコースを含有してもよい滅菌水溶液の形態での使用に最も適している。水溶液は、必要に応じて、好適に緩衝化(好ましくは3から9のpHに)すべきである。無菌状態での好適な非経口製剤の調剤は、当業者に周知の標準的調剤技術により容易に達成し得る。
【0109】
非経口投与に好適な製剤は、抗酸化剤、緩衝剤、静菌薬、及び製剤を対象とする受容者の血液と等張にする溶質を含有してもよい水溶性又は非水溶性無菌注射液;及び懸濁剤及び濃稠化剤を含有してもよい水溶性又は非水溶性滅菌懸濁液を含む。製剤は、1回用量又は多回用量容器、例えば、密封アンプル及びバイアルで提供してもよく、そして使用直前に滅菌液体担体、例えば、注射用蒸留水の添加だけを必要とする凍結乾燥(凍乾)状態で保存してもよい。即時注射液及び懸濁液は、前記の種類の滅菌粉末、顆粒及び錠剤から調剤してもよい。
【0110】
ヒトの患者への経口及び非経口投与では、ポリペプチドの日用量レベルは、通常成人当り1から100mg(即ち、約0.015から15mg/kg)であち、単回用量又は分割用量で投与される。例えば、3mg/kgのような、1〜10mg/kgの用量が使用されてよい。
【0111】
本発明の別の実施態様では、医薬組成物は、ポリペプチドそのものを含まないで、代わりに該ポリペプチドを発現することができる核酸分子を含む。好適な核酸分子、発現ベクター、及び宿主は詳細に上述されている。
【0112】
例えば、組み換えプロバイオティックを使用してもよい(LAB株、例えば、乳酸連鎖球菌又はラクトバシラス種)。
【0113】
本発明の更なる実施態様では、医薬組成物は、本発明の第1の態様のポリペプチドを発現することができるバクテリオファージを含む。例えば、本発明の第1の態様のポリペプチドを送達するために、野生型バクテリオファージΦCD27を使用してもよい。当該バクテリオファージに基づく治療を実施する方法は、当技術分野において周知である(例えば、Watanabe et al., 2007, Antimicrobial Agents & Chemotherapy 51:446-452を参照されたい)。
【0114】
このように、本明細書に記載の細菌感染症の処置では、本発明のポリペプチドは、同種の(cognate)タンパク質として、核酸構築物、ベクター又は同種のタンパク質を発現する宿主細胞として、同種のタンパク質(バクテリオファージを含む)を発現する生物の一部として、又はその細菌標的とリシンの接触を達成するために、それがC. ディフィシレのような病原性細菌であれ、又は本明細書に更に記載される別の病原体若しくは潜在的病原体であれ、当技術分野で公知のその他簡便な方法によって投与してもよい。
【0115】
理想的には、タンパク質は保護された形態で胃腸管に送達される。これは当技術分野で公知の多種多様の方法によって達成され得る。例えば、適切な用量のリシンは、胃の酸性状態は切り抜けるが、腸に入るにつれてタンパク質を放出する形態でマイクロカプセル化される。送達は、乳酸連鎖球菌、ラクトバシラス種、ビフィドバクテリウム種又はバクテロイデスを含むがこれらに限定されない、胃腸管通過を切り抜ける非病原性微生物によって行われる。当業者は、本明細書に開示されるリシンなどの活性化合物の胃腸管送達のためのそのような手段に使用できる選択肢について熟知している。これらの手段は、細胞内産生、secA分泌又は別の分泌経路による分泌、及び制御溶解による送達を含む。好ましくは、タンパク質は一度に全て放出されるのではなく、投与されたボーラスが胃腸管を横断するにつれて漸増的に放出される。或いはまた、リシンは、胃腸管における適切な位置で又は適切なシグナルの受信時にリシンを発現する、良性細菌の一部として導入される。本明細書に開示の好ましい実施態様では、非病原性ラクトコッカスが、胃腸管の特定の位置に到達時にΦCD27リシンを発現するように処理される。発現シグナルは、pH感受性プロモーター、又はこの目的のために当技術分野で公知の別の手段によって規定することができる。
【0116】
送達の他の手段としては、以下:
(a)国際公開公報第2006/111553号(ポリ尿素及び他の多層カプセル材);
(b)国際公開公報第2006/111570号及び欧州特許第1715739号(シクロデキストリンカプセル化);
(c)国際公開公報第2006/100308号及び欧州特許第1742728号(酵母及び他の微生物細胞カプセル化技術);
(d)米国特許第5153182号、欧州特許第1499183号及び国際公開公報第03/092378号;米国特許第6831070号(腸細胞発現による治療遺伝子産物送達);
(e)米国特許第7202236号(放出調節用医薬製剤);
(f)米国特許第5762904号(本発明のリシンを送達するために改変し得る、重合リポソームを用いるワクチンの経口デリバリー);
(g)米国特許第7195906号(本発明のリシンを発現させるために改変し得るビフィドバクテリウム);及び
(h)本明細書に引用された文献;
が含まれ、これらの全ては、本発明の新規な送達方法及び組成物を達成するために、当業者が本発明の開示内容を活用するのを可能にする目的で、参照することにより本明細書に組み入れられている。
【0117】
このように、本発明の医薬組成物の好ましい実施態様では、ポリペプチド、それをコード化する核酸分子などはマイクロカプセル化される(例えば、シクロデキストリン若しくは脂質二重層などの化学エンベロープ内で、又は処理したラクトコッカス細胞などの生存又は非生存微生物細胞内で)。このような方法で、ポリペプチド、核酸分子などは、胃腸管の作用部位への途中で胃の酸性状態に対して保護することができる。
【0118】
本発明の第7の態様は、医薬に使用するための、本発明の第1の態様のポリペプチド又は本発明の第6の態様の医薬組成物を提供することである。
【0119】
本発明の第8の態様は、患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制する薬剤の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。
【0120】
当然のことながら、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を示すポリペプチドは、必ずしもクロストリジウム・ディフィシレのバクテリオファージから由来する必要はない。例えば、ポリペプチドは以下の群:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン;
から選択することができる。
【0121】
或いはまた、ポリペプチドは、クロストリジウム・ビフェルメンタンス又はクロストリジウム・ソルデリ(Clostridium sordelli)などの異なるクロストリジウム種のバクテリオファージから由来(例えば、コード化)してもよい。
【0122】
しかしながら、好ましい実施態様では、ポリペプチドは、クロストリジウム・ディフィシレのバクテリオファージから由来する。
【0123】
従って、本発明の第8の態様の使用は、本発明の第1の態様のポリペプチドに限定されるものではなく、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するいずれのポリペプチドの使用をも包含する (Goh et al., 2007, Microbiology 153:676-685に記載のΦC2のリシンを含む。その開示内容は、参照することにより本明細書に組み入れられている)。
【0124】
本発明の関連する態様は、患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するための、クロストリジウム・ディフィシルのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。
【0125】
本発明の更なる態様は、患者中の微生物細胞に関連する疾患又は病態の治療又は予防のための薬剤の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。本発明の関連する態様は、患者中の微生物細胞に関連する疾患又は病態を治療又は予防するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。
【0126】
「患者中の微生物細胞に関連する疾患又は病態」には、クロストリジウム・ディフィシレによる患者の感染から生じ又は拮抗する疾患及び病態が包含される。当該疾患及び病態には、クロストリジウム・ディフィシレ関連疾患(CDAD)が含まれる。
【0127】
本発明の上記の使用の1つの実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、本発明の第1の態様のポリペプチドであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触により溶解を起こし易い他の細菌細胞から成る群から選択されるポリペプチドである(下記の表1と2を参照)。
【0128】
好ましくは、微生物細胞は、クロストリジウム・ディフィシレを含み又はそれから成る。このように、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、クロストリジウム・ディフィシレ細胞による感染に関連する疾患及び病態(クロストリジウム・ディフィシレ関連疾患、CDADなど)を治療又は予防するのに使用することができる。
【0129】
最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレリボタイプ027細胞を含み又はそれから成る。
【0130】
従って、本発明は更に以下の方法:
(a)患者中の微生物細胞を殺滅し及び/又はその増殖を阻害/抑制するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを患者に投与することを含む方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法;
(b)患者において微生物細胞に関連する疾患又は病態を治療又は予防するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若バクテリオファージを患者に投与することを含む方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法;
を提供する。
【0131】
本発明の上記の方法の1つの実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは本発明の第1の態様のポリペプチドであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触により溶解を起こし易い他の細菌細胞から成る群から選択されるポリペプチドである(下記の表1と2を参照)。好ましくは、微生物細胞は、クロストリジウム・ディフィシレ細胞を含み又はそれから成る。このように、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、クロストリジウム・ディフィシレ細胞による感染に関連する疾患及び病態(クロストリジウム・ディフィシレ関連疾患、CDADなど)を治療又は予防するのに使用することができる。最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレリボタイプ027の細胞を含み又はそれから成る。
【0132】
当業者には更に当然のことながら、本発明の使用及び方法は医学及び獣医学分野の両方で有用性を有する。従って、本薬剤はヒト及び非ヒト動物(ウマ、ウシ、イヌ及びネコなど)の両方の治療に使用することができる。しかしながら、好ましくは、患者はヒトである。
【0133】
「治療」には、患者の治療的及び予防的治療法が含まれる。用語「予防的」は、患者又は対象におけるクロストリジウム・ディフィシレによる感染の可能性を防止するか低減する、本明細書に記載のポリペプチド又は製剤の使用を包含するように用いられる。
【0134】
上記のように、用語「有効量」は、治療する疾患又は病態の好ましい変化をもたらすのに使用することができる、本発明のポリペプチドの濃度又は量を記載するために本明細書で用いられるが、その変化が寛解、好ましい生理的結果、治療する病状又は病態の好転又は減弱、惹起している病態又は病状の可能性の予防又は低減であるかは、治療する疾患又は病態によって決まる。
【0135】
当然のことながら、本明細書に記載の薬剤は、1つ又はそれ以上の更なる治療薬との併用で患者に投与してもよい。
【0136】
例えば、本明細書に記載の薬剤は:
(a)1つ又はそれ以上の従来の抗生物質治療(β−ラクタム、アミノグリコシド及び/又はキノロン);
(b)1つ又はそれ以上の更なるリシン、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージ;
(c)1つ又はそれ以上の抗生物質、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくは細菌;及び/又は
(d)腸内のクロストリジウム・ディフィシレ細胞の細菌溶菌時に放出される毒素を中和する療法;
と併用して投与してもよい。好適な中和療法は、抗生物質(Babcock et al., 2006, Infect. Immun. 74:6339-6347を参照されたい)及びトレバマー(tolevamer)などの毒素吸収剤(Barker et al., 2006, Aliment. Pharmacol. Ther. 24:1525-1534を参照されたい)を含んでもよい。
【0137】
本発明の更なる態様は、インビトロ及び/又はエクスビボで微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。例えば、エンドリシン活性を有する該ポリペプチドは、そのような細菌細胞で汚染され易い病院、キッチンなどの表面を清浄化するのに使用することができる。
【0138】
好ましくは、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、本発明の第1の態様のポリペプチドであって、その微生物細胞が、クロストリジウム・ディフィシレ及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択されるポリペプチドである(下記の表1及び2を参照)。例えば、微生物細胞は、クロストリジウム・ディフィシレ細胞を含み又はそれから成ってよい。最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレ・リボタイプ027の細胞を含み又はそれから成る。
【0139】
本発明の更なる態様は、試料中の微生物細胞の存在を検出するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性及び/又は細胞結合特異性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを含むキットであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択されるキットを提供することである。
【0140】
好ましい実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、本発明の第1の態様のポリペプチドであって、微生物細胞がクロストリジウム・ディフィシレ及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞からなる群から選択されるポリペプチドである(下記の表1及び2を参照)。例えば、微生物細胞は、クロストリジウム・ディフィシレ細胞を含み及びそれから成ってよい。最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレリボタイプ027の細胞を含み又はそれから成る。
【0141】
本発明のキットの更なる実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、マルチウェルプレートなどの好適な表面に固定化される。
【0142】
キットは、組織試料、細胞培養試料及び拭い液由来の細胞試料(例えば、微生物細胞による汚染を試験する表面から採取される)などの、いずれの好適な細胞試料と併せて使用してもよい。
【0143】
場合により、キットは、陰性対照試料(例えば、クロストリジウム・ディフィシレ細胞を試験するための細胞型を含まない)及び/又は陽性対照試料(試験するための細胞を含む)を更に含む。
【0144】
本発明の関連する態様は、以下の使用及び方法:
(a)クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、微生物細胞に関連する疾患又は病態の診断薬の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用;
(b)クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、微生物細胞に関連する疾患又は病態を診断するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用;
(c)インビトロ及び/又はエクスビボで試料中の微生物細胞の存在を検出するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用;及び
(d)患者中の微生物細胞に関連する疾患又は病態を診断するための、試験すべき患者からの細胞試料をクロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージに接触させ、そして試料中の細胞がそれにより溶解されるかどうかを測定することを含む方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法;
を提供することである。
【0145】
本発明の上記の使用及び方法の1つの実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、本発明の第1の態様のポリペプチドであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択されるポリペプチドである(下記の表1及び2を参照)。好ましくは、微生物細胞は、クロストリジウム・ディフィシレ細胞を含み又はそれから成る。このように、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、クロストリジウム・ディフィシレ細胞による感染に関連する疾患及び病態(クロストリジウム・ディフィシレ関連疾患、CDADなど)を診断するのに使用することができる。最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレ・リボタイプ027の細胞を含み又はそれから成る。
【0146】
当該診断のための使用及び方法では、細胞の溶解は当技術分野で周知の方法を用いて検出し得る。例えば、ATPのレベルは、細胞溶解の指標として測定し得る。
【0147】
本発明の上記の使用及び方法の他の実施態様では、ポリペプチドは、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合ドメインを含み又はそれから成る。検出を可能にするために、このようなポリペプチドは、磁気ビーズに融合され又は好適なレポーター(例えば、緑色蛍光タンパク質)を含む融合タンパク質として使用してもよい。
【0148】
当該診断方法は、リステリアエンドリシンなど他の系由来のエンドリシンにて十分確立されている(例えば、Loessner et al., 2002, Mol Microbiol 44, 335-49; Kretzer et al., 2007, Applied Environ. Microbiol. 73:1992-2000を参照されたい。その開示内容は、参照することにより本明細書に組み入れられている; 好適な分析法も、また、例えばProfos, Germany [www.profos.de/content/view/164/69/lang,en/でそれらのウェブサイトを参照されたい]から市販されている) 。
【0149】
本発明の典型的な実施態様は、以下の図を参照しながら、以下の非限定的実施例にて記載される。
【図面の簡単な説明】
【0150】
【図1】ΦCD27の電子顕微鏡写真。試料は飽和酢酸ウラニル中でネガティブ染色された。
【図2】予測されたORFを示すΦCD27ゲノム地図。矢印は転写の方向を示す。提案された機能モジュールは、BLAST結果並びにΦCD119、ΦC2、及びC.ディフィシレ株630プロファージの公表配列との類似性に基づいて表示される。
【図3】ΦCD27リシンのヌクレオチド配列、配列番号2。
【図4a−1】公表C.ディフィシレのバクテリオファージ(ΦC2(32);ΦCD119(31))、又はプロファージ(配列ゲノムからのCD630プロファージ1及び2(36))配列による、ΦCD27 ヌクレオチド配列のアラインメント。AlignX、Vector NTI Advance 10, Invitrogenにより行なわれたアラインメント。ΦCD27アミノ酸配列は配列番号2である。
【図4a−2】図4a−1の続きである。
【図4a−3】図4a−2の続きである。
【図4b】公表C.ディフィシレのバクテリオファージ(ΦC2(32);ΦCD119(31))、又はプロファージ(配列ゲノムからのCD630プロファージ1及び2(36))配列による、ΦCD27推定アミノ酸配列のアラインメント。AlignX、Vector NTI Advance 10, Invitrogenにより行なわれたアラインメント。ΦCD27アミノ酸配列は配列番号2である。
【図5】pET15bベクター(Novagen)のクローニング部位。
【図6a】(a)ΦCD27リシンを発現する大腸菌からの粗タンパク質溶解産物のゲル解析。レーン1は、SeeBlueマーカー(Invitrogen、サイズ191、97、64、51、39、28及び19kDa)、レーン2〜5は、BL21(DE3)pET15bΦCD27L総蛋白質抽出物。レーン2〜4は、IPTG−2で3時間誘導した抽出物及び20mMのTriS−HCl、pH8、50mMのNaClで抽出した3、プロテアーゼ阻害剤(Roche Complete mini EDTA-free)を含む3及び変性緩衝液(8Mの尿素、0.1MのNaH2PO4、0.01MのTriS−HCl、pH8.0)で抽出した4。レーン5は20mMのTriS−HCl、pH8、50mMのNaClで抽出した非誘導対照。レーン6及び7は、20mMのTriS−HCl、pH8、50mMのNaClで抽出したBL21(DE3)pET15bΦCD630L1総蛋白質抽出物、レーン6は、IPTGで3時間誘導しただけの(a)6×His抗体による(b)ゲルのウェスタン解析。
【図6b】(a)ΦCD27リシンを発現する大腸菌からの粗タンパク質溶解産物のゲル解析。レーン1は、SeeBlueマーカー(Invitrogen、サイズ191、97、64、51、39、28及び19kDa)、レーン2〜5は、BL21(DE3)pET15bΦCD27L総蛋白質抽出物。レーン2〜4は、IPTG−2で3時間誘導した抽出物及び20mMのTriS−HCl、pH8、50mMのNaClで抽出した3、プロテアーゼ阻害剤(Roche Complete mini EDTA-free)を含む3及び変性緩衝液(8Mの尿素、0.1MのNaH2PO4、0.01MのTriS−HCl、pH8.0)で抽出した4。レーン5は20mMのTriS−HCl、pH8、50mMのNaClで抽出した非誘導対照。レーン6及び7は、20mMのTriS−HCl、pH8、50mMのNaClで抽出したBL21(DE3)pET15bΦCD630L1総蛋白質抽出物、レーン6は、IPTGで3時間誘導しただけの(a)6×His抗体による(b)ゲルのウェスタン解析。
【図7】NiNTAカラム精製Hisタグ化ΦCD27リシンのゲル解析。レーン1は、SeeBlueマーカー(Invitrogen、サイズ191、97、64、51、39、28及び19kDa)、レーン2〜5は、IPTGで誘導後のBL21(DE3)pET15bΦCD27L総蛋白質抽出物、レーン1は、粗溶解産物、レーン2は、カラム流入、レーン3は、一次洗浄溶離液、レーン4は、洗浄二次溶離液、レーン5は、一次溶出液(E1、1ml)、レーン6は、二次溶出液(E2)。
【図8】エンド対数に増殖し、液体窒素中で急速冷凍し、次いでPBS中に再懸濁したC. ディフィシレ11204の細胞によるBioscreen溶解試験。ΦCD27リシン及びCD630リシンは大腸菌に発現し、そしてNiNTAカラムでHisタグを用いて精製された(図6を参照)。270μlの細胞をE1抽出物の30μlの希釈液に加えた。値は二重複分析の平均値+/−標準偏差である。CD630L1抽出物による細胞溶解は、緩衝液だけの対照に見られるものと同等であった。
【図9】エンド対数に増殖し、4℃で遠心分離により採取し、次いでPBS中に再懸濁したC. ディフィシレ11204の細胞によるBioscreen溶解試験は、1〜1.5の間のODを与えた。ΦCD27リシンは大腸菌に発現し、そしてNiNTAカラムでHisタグを用いて精製された(図6を参照)。270μlの細胞を溶離緩衝液で希釈した溶出液1(E1)の30μl試料に加えて、1分析当り10.5μgから0.35ngの範囲の濃度を与えた。新鮮細胞の使用は、緩衝液だけの対照において有意に少ない溶解を与えた。70ng未満のNiNTA精製タンパク質で緩衝液だけの対照と差が見られなかった。
【図10a】活性のスペクトルを試験するためにC.ディフィシル細胞に添加されたΦCD27リシンのBioscreen溶解試験。細胞を大腸菌から産生した3.5μgNiNTA精製タンパク質(E1)とインキュベートした。試験した30株のうち、宿主株12727及びΦCD27非感受性株11208及び高毒性リボタイプ027R23613を含み、全てが感受性であった。インキュベーションは緩衝液(B)か又はリシン(L)を用いて二重複でなされた。
【図10b】活性のスペクトルを試験するためにC.ディフィシル細胞に添加されたΦCD27リシンのBioscreen溶解試験。細胞を大腸菌から産生した3.5μgNiNTA精製タンパク質(E1)とインキュベートした。試験した30株のうち、宿主株12727及びΦCD27非感受性株11208及び高毒性リボタイプ027R23613を含み、全てが感受性であった。インキュベーションは緩衝液(B)か又はリシン(L)を用いて二重複でなされた。
【図10c】活性のスペクトルを試験するためにC.ディフィシル細胞に添加されたΦCD27リシンのBioscreen溶解試験。細胞を大腸菌から産生した3.5μgNiNTA精製タンパク質(E1)とインキュベートした。試験した30株のうち、宿主株12727及びΦCD27非感受性株11208及び高毒性リボタイプ027R23613を含み、全てが感受性であった。インキュベーションは緩衝液(B)か又はリシン(L)を用いて二重複でなされた。
【図11a−1】クロストリジウム種及び一般的な腸細菌に対するΦCD27リシンの活性。細胞を後期安定相で採取し、PBS中で再懸濁し、大腸菌から産生した7μgNiNTA精製タンパク質(E1)とインキュベートした。結果は、二重複分析の平均値+/−標準偏差である。ΦCD27リシンは大部分の種において細胞溶解を誘導しなかった(a及び表2を参照)。例外として(b及び表2を参照)、クロストリジウム・ビフェルメンタンスの迅速溶解、バシラス・セレウスの溶解、そして長い誘導期を有するB.サチリス、及びリステリア・イバノビイ(b)に対する微弱作用が含まれる。
【図11a−2】図11a−1の続きである。
【図11a−3】図11a−2の続きである。
【図11a−4】図11a−3の続きである。
【図11b−1】クロストリジウム種及び一般的な腸細菌に対するΦCD27リシンの活性。細胞を後期安定相で採取し、PBS中で再懸濁し、大腸菌から産生した7μgNiNTA精製タンパク質(E1)とインキュベートした。結果は、二重複分析の平均値+/−標準偏差である。ΦCD27リシンは大部分の種において細胞溶解を誘導しなかった(a及び表2を参照)。例外として(b及び表2を参照)、クロストリジウム・ビフェルメンタンスの迅速溶解、バシラス・セレウスの溶解、そして長い誘導期を有するB.サチリス、及びリステリア・イバノビイ(b)に対する微弱作用が含まれる。
【図11b−2】図11b−1の続きである。
【図11b−3】図11b−2の続きである。
【図11b−4】図11b−3の続きである。
【図12】ΦCD27リシン活性のpHプロファイル。C.ディフィシレ11204細胞を一定の範囲のpHに調整したPBSで再懸濁し、そして大腸菌から産生したNi−NTA精製リシンE1の活性を前と同様にbioscreenで測定した。
【図13A】(a)ΦCD27リシンを発現する乳酸連鎖球菌由来の粗タンパク質溶解物のゲル分析。レーン1及び10は、SeeBlue(Invitrogen、サイズ191、97、64、51,39、28及び19kDa)、レーン2〜5は、5時間(2、4)誘導又は非誘導(3、5)のphiCD27LpUK200HIS(2、3)又は空ベクターpUK200HIS対照(4、5)を含有する乳酸連鎖球菌UKLC10。レーン6〜9は、全て4時間誘導した(レーン当り10μg)phiCD27LpET15b(6、8、9)又は空ベクター対照(7)を含有する大腸菌BL21(DE3)。全タンパク質を、レーン8(20mMのリン酸ナトリウム、pH8)及び9(50mMのTrisHCl、pH7.5)を除いて20mMのTris−HCl、pH8、50mMのNaClで抽出した。(a)6×His抗体による(b)ゲルのウェスタン分析。
【図13B】(a)ΦCD27リシンを発現する乳酸連鎖球菌由来の粗タンパク質溶解物のゲル分析。レーン1及び10は、SeeBlue(Invitrogen、サイズ191、97、64、51,39、28及び19kDa)、レーン2〜5は、5時間(2、4)誘導又は非誘導(3、5)のphiCD27LpUK200HIS(2、3)又は空ベクターpUK200HIS対照(4、5)を含有する乳酸連鎖球菌UKLC10。レーン6〜9は、全て4時間誘導した(レーン当り10μg)phiCD27LpET15b(6、8、9)又は空ベクター対照(7)を含有する大腸菌BL21(DE3)。全タンパク質を、レーン8(20mMのリン酸ナトリウム、pH8)及び9(50mMのTrisHCl、pH7.5)を除いて20mMのTris−HCl、pH8、50mMのNaClで抽出した。(a)6×His抗体による(b)ゲルのウェスタン分析。
【図14】エンプティーベクター対照からの抽出物と比較した、C.ディフィシレ株11204の新鮮細胞とインキュベートしたphiCD27リシン発現大腸菌及び乳酸連鎖球菌由来の粗タンパク質抽出物のBioscreen分析。50μgタンパク質を各分析で使用し、結果は二重複分析の平均値+/−標準偏差である。
【図15】Zeba緩衝液交換カラム(Pierce)を20mMのリン酸ナトリウム、pH6.0に通した分割量と比較した、元の抽出物の活性を示す大腸菌産生Ni−NTA精製リシンE1のBioscreen分析。リシン及び緩衝液対照をC.ディフィシレ株11204の急速冷凍細胞とインキュベートした。結果は二重複分析の平均値+/−標準偏差である。
【図16】LM4−CD27L(レーン2)及びLM4−CD27LE(レーン3)の粗細胞抽出物のSDS−PAGE、及びHisタグ化タンパク質を強調表示する対応のウェスタンブロット。タンパク質を20mMのリン酸ナトリウム、pH6.0で抽出し、そして10μg分割量をMOPS緩衝液(Invitrogen)中の10%Bis-TrisNuPageゲルで電気泳動した。レーン1は、SeeBlueマーカー。
【図17】中期対数まで増殖し、次いで液体窒素中で急速冷凍したC.ディフィシレ株11204細胞の溶解を示すBioscreen分析。細胞を10μgNiNTA精製E1(溶出液1)又は対照としての溶出緩衝液とインキュベートした。
【実施例】
【0151】
背景
抗菌薬としての細菌ウイルスの利用で、近年多少のルネサンスを経験している。一部において、これは、薬剤耐性病原体の継続的出現に続く従来型抗生物質の代替を見出す必要性を反映している。最近のレビューはこの可能性を強調しているが、バクテリオファージの使用に内在する限界をも強調している(7、8)。
【0152】
一般的に、バクテリオファージは顕著な系統差を示すことから、それらは限定範囲の個々の菌株に対して活性であることを意味する。バクテリオファージとその細菌宿主間の相互作用の動力学は、バクテリオファージ攻撃に抵抗性の宿主変異体の即時選択を含む。他の関連する問題は、宿主生菌によるバクテリオファージ製剤の汚染可能性、及びバクテリオファージが遺伝子流動及び毒性因子の拡大に寄与する可能性を含む(9)。バクテリオファージによる毒素遺伝子の輸送は特に十分立証されており、その例はコレラ毒素(10)、ボツリヌス毒素(9)、志賀毒素(11)及びジフテリア毒素(9)を含む。これらの限定にもかかわらず、バクテリオファージは、マウスモデルにおける大腸菌(12)、黄色ブドウ球菌(13)及びバンコマイシン耐性エンテロコッカス・フェシウム(14)を制御するのに実験的に使用されている。バクテリオファージ療法はニワトリにおけるカンピロバクター(15)及び大腸菌(16)の制御のために研究中である。クロストリジウムに関して、ハムスターモデルにおけるC.ディフィシレを標的とする研究が報告されている(17)。更に、FDAは、最近全食品におけるリステリア制御のためのバクテリオファージ(LISTEX(登録商標), EBI Food Safety) へのGRAS承認を更に拡大した(18)。
【0153】
インタクト・バクテリオファージの使用に加えて、抗菌薬としてバクテリオファージ・エンドリシスを使用する可能性がある。バクテリオファージ・ライフサイクルの最終段階は、新たに複製されたインタクト・バクテリオファージ粒子のプールを放出するための細菌宿主細胞の溶解を含む。一般的に、これは、膜破壊性ホリンの厳密な時限産生が、そのペプチドグリカン標的に細胞壁分解性エンドリシンが接近することを可能にする、2段階過程によって達成される。エンドリシン酵素は分泌されず、ホリンの作用により及び細胞壁を分解するそれ自体の能力により細胞から放出される。一度放出された場合、エンドリシンは細胞の外側からペプチドグリカンを攻撃することができ、この現象は初期のバクテリオファージ研究の時から観察されている:それを「外部からの溶解」と称する。最も特徴的なバクテリオファージ・エンドリシンの構造は、触媒ドメイン及び特徴的な細胞壁結合ドメイン(CBD)を有するモジュール構造である。触媒ドメインは変化することができ、そしてほとんどの場合それはアミダーゼか又はムラミダーゼである。CBDは細胞壁表面上の糖モチーフを認識するレクチン様能力を有し、そして関係する特異性の変動は、エンドリシンに特異的な分類群へのそれらの特徴的なターゲッティングをもたらす(19、20)。
【0154】
Gassonらは、新しい抗菌薬として及びモデル系としてのリステリア及びクロストリジウムを用いた新しい検出技術の基礎の両方で、バクテリオファージ・エンドリシンの開発の先駆者となった(21)。その後、標的抗菌薬としてのエンドリシンの可能性が、炭疽菌(23)、肺炎連鎖球菌(24)及びエンテロコッカス・フェカーリスを標的とする公表例により広く認識されている(22)。リステリアに関して、重要な更なる研究が Martin Loessner at ETH, Switzerlandにより行なわれている(19、20)。加えて、クロストリジウム・パーフリンジェンスに対して活性なエンドリシンが特徴付けられている(26)。
【0155】
新しいバクテリオファージ・リシンの特徴付け及びその使用方法
テンペレート・バクテリオファージΦCD27は、クロストリジウム・ディフィシレ菌株保存株NCTC12727から単離された。他の25のC.ディフィシレ株に対してΦCD27を試験し、そしてタイプ株11204を含む他の4株に対して有効なことが示された。バクテリオファージ・ゲノムDNAを抽出して配列決定し、そしてBLAST検索によりエンドリシン配列が同定された。その配列は、公表されたC.ディフィシレのバクテリオファージ・エンドリシン(ΦCD119、ΦC2、配列決定されたC.ディフィシレCD630におけるプロファージ1及び2)と明瞭なアミノ酸及びヌクレオチド相同性を示した。リシンはpET15bへサブクローニングされ、そして6×Hisタグにより大腸菌に発現された。リシンはニッケルカラムで一部精製され、そしてファージ感受性及び非感受性株の両方を溶解することが、37℃でインキュベーション時の光学密度の減少により証明された。試験した30株のうち、毒性リボタイプ027株を含む全てが溶解することを示した。一連の菌属からの多数の他の細菌は、リシンに対して感受性を示さなかった。しかしながら若干の活性がC.ビフェルメンタンス、C.ソルデリ、セレウス菌、枯草菌に対して認められ、そしてリステリア・イバノビイに対して非常にわずかな活性が認められた。部分精製リシンの比活性は、C.ディフィシレ株によって変動した。従って、本明細書に開示されたリシンは、C.ディフィシレ病原性の治療及び検出に対する新しい強力な武器となる。
【0156】
本明細書で同定されそして特徴付けられたリシンは、C.ディフィシレ感染症、及びヒト及び動物の他の細菌感染症を治療するために使用できる新しい組成物である。本発明によれば、ΦCD27リシンは、当技術分野で公知の方法によって生産してもよい。それは、この目的のために増殖したウイルスから使用のために単離してもよい。しかしながら、好ましくは、それは本明細書に開示した組み換え法により、そして当業者に公知の代替法により生産される。この分子の関連サブポーションについて、細菌に特異的に結合し、それらの細菌を溶解するそれらの能力が特徴付けられた。これらの分子サブポーションは、天然分子として別々に又は同時に生産され、そして使用されてもよい。
【0157】
ΦCD27リシンの発見、クローニング及び活性
溶解物の生産及び活性分析は、記載のように行われた(27)。C.ディフィシレ株NCTC12727(the Health Protection Agency, Colindale, London から入手可能−糞便から単離され、1992年にS. Tabaqchali, St. Bart's Hospital, London により寄託)は、BHI+C(ビタミンK(10μl、0.5容量%/l)、ヘミン(5mg/l)、レザズリン(1mg/l)及びL−システイン(0.5g/l)を補充したBHI(Oxoid) )中、37℃で嫌気的に24時間増殖させた。バクテリオファージ生産をマイトマイシンC(Sigma)により3μg/mlの最終濃度で24時間誘導した。培養液を4℃において4000×gで20分間遠心分離し、上清を0.45μmフィルターユニット(Millipore) を通して濾過し、そして4℃で保存した。150μlの一夜C.ディフィシレBHI+C培養液を加えたBHI軟寒天(0.75%)を重層化したBHIプレート(1.5%寒天)上に、上清の25μl部をスポットし、そして37℃で一夜嫌気的にインキュベートした。培養液(表1を参照)を2重複で試験し、12727上清からの明瞭なプラーク形成を、4株−C.ディフィシレ11204(タイプ株)、11205、11207及び11209で確認した。菌株11204からのプラークは、滅菌パスツールピペットで250μlBHI+C中へ採取し、4℃で一夜インキュベートした。バクテリオファージ−ΦCD27の存在は電子顕微鏡で確認したが、それがカウドウイルス目に属することを示した(28)(図1)。総数で25のC.ディフィシレ株がマイトマイシンCで誘導され、その上清は25株全てに対して交差試験した。ΦCD27は、この方法で発見された唯一のプラーク形成単位であった。C.ディフィシレからのバクテリオファージ発見が稀なことはこれまでの刊行物にも示されており、それには94分離株から2バクテリオファージ産生株(29)又は56分離株から3産生株(30)が見出される。
【0158】
力価を増加させるために、100μlのプラーク溶出液を100μlのC.ディフィシレ24時間培養液と5mlBHI軟寒天中で混合し、BHI寒天上に塗布した。37℃での一夜の嫌気性培養は近似融合溶菌を与え、そして5mlBHI+Cで2時間の溶離は、2×106pfu/mlの力価を与えた。力価は、11204の液体培養での連続インキュベーションにより増加し、初期から中期対数期まで25mlBHI+C培養における細胞を増殖させ、バクテリオファージ:細胞の比を少なくとも4:1にする光学密度(OD)を与えた。この方法は、細菌懸濁液の完全透明化をもたらし、そして2代継代は2.5×1011pfu/mlの力価を与えた。DNA抽出のために、OD0.3の細胞をc.7.の感染の多重度まで濾過溶解物でインキュベートした。3時間のインキュベーションは完全溶解をもたらし、上清を上記のように採取して濾過し、そして2つの50ml部をQiagen λ midikit (Qiagen)中で使用した結果、c.160μgのバクテリオファージのゲノムDNA収量を得た。
【0159】
バクテリオファージΦCD27ゲノムのシークエンシング及びアセンブリーを、Phred-Phrapプログラムを用いてBiochemistry DNA Sequencing Facility (University of Cambridge, UK)により実施した。循環ゲノムは50,930bpであり、75の提案されたオープン・リーディング・フレーム(orf)を含んだ(図2)。これらの多くは、C.ディフィシレのバクテリオファージΦCD119(31)及びΦC2(32)からのものを含み、同定したバクテリオファージORFに顕著な相同性を示した。ORFは、BITS(Harpenden)を経て行なわれたArtemis (33)及びBlastP 検索(34,35)により解析した。提案されたΦCD27リシン配列は816bpであり、N−アセチルムラモイル−L−アラニンアミダーゼに相同性を示す271アミノ酸予測タンパク質をコードした。ヌクレトチド及びアミノ酸配列(図3)の両方は、C.ディフィシレのバクテリオファージ及びプロファージ(図4)の公表配列に合っていたが、ΦC2に対してその最大相同性(95.9%ヌクレオチド及びアミノ酸同一性)を有した。
【0160】
ΦCD27リシン配列はプライマーを用いてゲノムDNAから増幅され、初めのMet残基(プライマーCD27L_NDE、5’−TTA CAT ATG AAA ATA TGT ATA ACA GTA GG[配列番号3]、Sigma Genosys)周囲のNdeI(CATATG)部位を、そしてコード配列(プライマーCD27L_XHO、5’−CAA CCA CCT CGA GTT GAT AAC[配列番号4]の下流にXhoI部位(CTCGAG)を作製し、発現ベクターpET15b(Novagen)におけるサブクローニングを促進させた。増幅は、1×Phusion緩衝液、200μMのdNTP、0.5μMの各プライマー、200ngのゲノムDNA鋳型を含む50μlの反応液中、高性能PhusionDNAポリメラーゼ(0.02U/μl、Finnzymes)により実施した。増幅条件は、98℃で30秒間の初期変性、その後に30サイクルの変性(90℃で10秒)、アニーリング(58℃で30秒)及び伸長(90℃で10秒)、次いで72℃で5分間の最終伸長が続いた。平滑末端PCR産物はSureClean (Bioline)を用いて精製し、そして72℃で20分間インキュベートして、1×AmpliTaq緩衝液、0.2mMのdATP及び1UのAmpliTaqDNAポリメラーゼ(Applied Biosystems)を含む50μlの反応液中に3’A−オーバーハングがもたらされた。生成物をSureCleanで精製し、次いでTAクローニングキット(Invitrogen)を用いてpCR2.1に結合された。ライゲーション生成物は、TOP10化学的コンピーテント大腸菌(Invitrogen)中へ形質転換して、陽性菌を100μg/mlアンピシリンを補充したL寒天で選択し、そしてブルー・ホワイト選択のために40mg/mlX−ガル溶液の40μlで重層化した。プラスミドDNAをプラスミドミニキット(Qiagen) を用いて抽出し、挿入部分をベクタープライマー及びBigDye v3.1シークエンシングキット(Applied Biosystems)を用いて配列決定した。リシン原配列と100%配列相同性を示すが、付加したNdeI及びXhoI部位を有するクローンは挿入部分を放出するのが限定的であった。これはゲル精製され(Qiaex II, Qiagen)、そしてFast-Link DNAリガーゼ (Epicentre)を用いて、リシン配列がIPTG誘導性lacオペレーターによる高発現T7プロモーターの制御下に6−ヒスチジンタグの下流に発現するように、pET15bへと結合された。ライゲーション産物は、先ず配列確認のために TOP10細胞へ、次いでタンパク質発現のために化学的コンピーテントBL−21(DE3)細胞 (Invitrogen)へ形質転換された。配列決定したC.ディフィシレ(36)のプロファージ1からのリシン配列を、 ベクターpUC57へとGenscript Corp. (Piscataway, USA)によって合成し、そしてプライマーCD630L1_NDE(5’−TGC TCA TAT GAA AAT AGG TAT AAA TTG)[配列番号5]、及びXhoI部位を含む特定のベクターDNAでリシンを増幅したM13前方(5’−GTA AAA CGA CGG CCA GT)[配列番号6]を用いて、同様にHis−タグ化発現のためにサブクローニングした。
【0161】
His−タグ化リシンは、メーカーにより示唆されたように、OD6000.4まで100μg/mlアンピシリンにより10mlLブロス中に増殖したBL21(DE3)細胞に発現し、次いで0.5mMのIPTG(Melford Biosciences)で3〜4時間誘導した。細胞を4000×gで遠心分離することにより採取し、次いで1mlの懸濁液(20mMTris−HCl、pH8、50mMのNaCl)中4℃で20分間再懸濁し、そして2mlスクリューキャップチューブに移した。粗タンパク質溶解物は、FastPrep FP120 細胞破壊器(Savant)中の0.1mm酸洗浄ガラスビーズ(Sigma)を用いて、バースト間に5〜10分間氷上でインキュベートしながら4×30秒のバースト(速度10)での細胞破壊によって得られた。破壊細片を13,000×gで4℃にて20分間遠心分離によりペレット化し、上清を4℃で保存した。粗溶解物は、IPTG誘導無しで増殖したリシンを含む細胞から、及び誘導の有り無しで増殖したpET15b空ベクターを含む細胞からも製造された。タンパク質含量をBradford試薬(Bio Rad)を用いて測定し、10μg部をMOPS緩衝液(Invitrogen)中10%NuPage Novex Bis Trisゲルで電気泳動した。His−タグ化リシンの存在は、抗Hisタグモノクローナル抗体(Novagen)を用いてウェスタンブロットにより確認した。タンパク質はNuPage緩衝液 (Invitrogen)を用いてPVDF膜に移され、そして検出は二次抗体として抗マウスIgGを用いてQiagen (Qiaexpress検出及び分析便覧)に記載のように行なわれ、そして比色検出はSigma FastBCIP/NBTアルカリ性ホスファターゼ基質でなされた。これにより、IPTG誘導溶解物におけるc.33kDaのHisタグ化バンドの高発現及び非誘導溶解物の低発現を証明した(図6)。
【0162】
粗溶解物によるC.ディフィシレの11204及び11207株細胞のリシスは、Loessner ら(37)によって記載された方法を用いて評価した。11204株細胞は、エンド対数期まで増殖し、1.8ml分割量をスクリューキャップチューブ(13,000×g、2分)中への遠心分離によって採取し、そしてペレットは液体窒素中で急速冷凍して−20℃で保存した。ペレットは900μlの20mMのTris−HCl、pH8中氷上で再懸濁し、そして100μlの粗タンパク質溶解物を含むキュベットに加えて、その後OD600の低下を読み込み前に混合して1時間モニターした。この系で、C.ディフィシレ細胞は緩衝系中で一定量の溶解を示したが、ΦCD27リシン粗抽出物による溶解はより急速で多大であった。しかしながら、誘導pET15bエンプティーベクター粗溶解物によるその後の試験は同等の溶解を示し、大腸菌リゾチームの活性を示唆した。この問題を回避するために、ΦCD27及びCD630L1リシンをQiagenNiNTAキットでアフィニティー精製した。BL−21(DE3)細胞は、100μg/mlのアンピシリンを含む250mlのLブロス中でOD6000.6まで増殖し、次いで1mMの最終濃度でIPTGにより5時間誘導した。細胞を4000×gで4℃にて20分間遠心分離することにより採取して、−20℃で保存した。タンパク質を自然条件で精製し、そして精製はNuPageゲル分析によって確認した(図7)。この方法は大部分がリシンの部分精製タンパク質を産生し、その収量は第1のΦCD27溶出液(E1)では2.3mg総タンパク質であり、第2(E2)では0.5mgであった。E1溶出液の希釈液のインキュベーションは、同様に調製した細胞からの溶出液に比べて11204株細胞の急速溶解を示したが、pET15b空ベクターを発現した。しかしながら、CD630L1E1溶出液は11204株を溶解せずに、そしてΦCD27リシンとの相乗効果は見られなかった。
【0163】
溶解分析は、Bioscreen C (Labsystems)及び溶出緩衝液(EB, Qiagen)でのNiNTA部分精製リシン抽出物を用いたマルチウェルプレート中で継続した。初期分析は、分光光度計分析におけるように、30μlのEB及び270μlの細胞の全容積中の約(c.)7μgのタンパク質を用いた。分析は氷上にセットアップし、次いで37℃に予熱されたBioscreen Cに移し、そしてプログラムは以下のように実施した:600nmの光学密度でサンプリング前に10秒振盪して2分毎にサンプリングした。各分析は、緩衝液だけの2ウェル及びリシンの2ウェルで実施し、全部で4ウェルに同じ細菌細胞懸濁液から植菌した。この系では、感受性株のリシンウェル中の溶解は迅速であり−その差は5分以内で顕著であった。しかしながら、リシン誘導溶解よりもはるかに低速にもかかわらず、緩衝液だけの対照での溶解も顕著であった(図8)。
【0164】
C.ディフィシレ及び他の細菌細胞の両方は、エンド対数まで増殖して、凍結せずに氷上で採取し、次いで可及的速やかに分析した場合、緩衝液だけの溶解は減少し又は全体的に欠如しており(図9)、他の種全ての溶解は、クロストリジウム・ビフェルメンタンス、クロストリジウム・ソルデリ、バシラス・セレウスを注目すべき例外として欠如しており、そして枯草菌及びリステリア・イバノビイイで若干欠如していた(図11、表2)。胃腸管ミクロフローラのATリッチクロストリジウム様成分の代表である更なる菌株について、ΦCD27リシンに対する感受性を試験した。表3に示されるように、試験したいずれもリシンに感受性ではなかった。
【0165】
新鮮細胞の使用は、12分までの顕著な遅延を伴う、C.ディフィシレへの溶解の迅速開始の程度は小さかった(図9)。全てのC.ディフィシレ株を第2NiNTAカラムから単離した3.5μgのリシンを用いて再試験した(第1精製と同等のリシスを示すことを試験した;図10)。新鮮又は凍結細胞を用いたいずれの場合にも、感受性プロファイルは、リシンに対して明らかな感受性示す30株全てで同様であった(表1)。
【0166】
ΦCD27リシンのpHプロファイルはを、感受性株11204を用いて試験した−活性は、pH4.5、5.8、6.5、7.0、7.3(PBSの通常のpH)、7.6及び8.3で試験して、かなりのpH範囲内でほとんど変動を示さなかった。希釈系列は、300μl分析において10.5μgタンパク質での活性が最大であったが、良好な溶解は3.5μg及び0.7μgでも見られた。しかしながら、0.35μgは緩衝液対照をわずかに下回る反応を示し、より低い量では45分分析内で溶解を示さなかった。
【0167】
胃腸管へのΦCD27リシンの送達は、物理的カプセル化又は乳酸菌の1メンバーなどの組み換え片利共生微生物の使用により達成することができた。乳酸連鎖球菌はこの点で可能性を確立しており、それ故この種におけるΦCD27リシンのサブクローニング及び発現が実証された。ΦCD27リシン配列をベクターpUK200Hisへサブクローニングした。これは、NcoIによるpUK200の制限、末端充填、次いでナイシン誘導性プロモーターから下流の6−ヒスチジンタグ(AGT CAT CAC CAT CAC CAT CAC GC)[配列番号7]をコード化するオリゴマーの挿入により構築される、nisA翻訳融合プラスミドpUK200の誘導体である(38)。再循環した場合、これはサブクローニングのためのNcoI部位を再作製した(Hornら、未発表)。ベクターpUK200Hisは、NcoIで制限され、T4DNAポリメラーゼ(Promega)で末端充填されて、nisAプロモーターの制御下翻訳融合のための最初のATGコドンを作製した。phicd27l配列は、pCR2.1(上記)においてサブクローニングされたCD27L−NDE…CD27L−XHO PCR産物から増幅された。プライマーCD27LCOD2_F(5’−AAA ATA TGT ATA ACA GTA GGA CAC)[配列番号8]及びM13前方(5’−GTA AAA CGA CGG CCA GT)[配列番号9]は、第2コドンAAAからの完全配列、及びある種のベクター配列を増幅し、リシンコード配列のすぐ後にEcoRI部位を与えた。増幅は上記のようであったが、アニーリング温度は56℃であった。PCR産物及びNcoI-カットの、末端充填pUK200Hisベクターは、EcoRIにより制限され、そして相互に連結されてnisAプロモーターの制御下にHisタグ化翻訳融合体が作製された。連結反応産物は、配列検証のためにエレクトロコンピテント大腸菌株MC1022に形質転換され、陽性形質転換体をクロラムフェニコール(15μg/ml)で選択した。次いで精製プラスミド調製品をエレクトロコンピテント乳酸連鎖球菌株FI10676に形質転換し、そして5μg/mlクロラムフェニコールを補充したGM17寒天で選択した。
【0168】
pUK200His−phiCD27L又はpUK200His空ベクター制御を発現する乳酸連鎖球菌株は、30℃静置で5μg/mlのクロラムフェニコールにより10mlのGM17ブロス中で増殖した。100μlの一夜培養液を使用して予熱ブロスを植菌した結果、培養液は中間期対数(OD6000.5)まで増殖した。発現は30℃で5時間1ng/mlのナイシンで誘導し、粗タンパク質溶解物は、20mMのTris−HCl、pH8.5、50mMのNaCl中に大腸菌で記載のように産生した。ΦCD27リシンの乳酸菌発現の証明が、タンパク質ゲル解析で示された(図13)。乳酸連鎖球菌に発現したΦCD27リシンに対するクロストリジウム・ディフィシレ株11204の感受性が、図14に示されるように粗タンパク質抽出物を用いて証明された。
【0169】
【表1】
【0170】
【表2】
【0171】
【表3】
【0172】
【表4】
【0173】
【表5】
【0174】
上記に加えて、健常腸生理を維持するために、望ましくは有害でない片利共生生物株を代表する多くの追加菌株は、本発明のリシンとの接触により害されないことが知られている。ΦCD27に対して試験した以下のクロストリジウム種の全て、DSMZからの全てにおいて、全てが溶解をもたらさなかった。これらの菌株は、Eckbergら(2005) Science 308 1635- 及び補充論文、及び Kikuchi ら (2002) Microbiol. Immunol. 46, 353 及び引用文献を参考にして、ヒトの腸に通常見られる主要クロストリジウム菌株群の代表であることを基準に、特異的に選択された。
【0175】
【表6】
【0176】
全てのC.ディフィシレ株を、同じ方法で大腸菌に発現したC.ディフィシレ株630のプロファージ1リシンに対して試験し、その結果CD630L1リシンは溶解をもたらさなかった。
【0177】
細胞生存性
細胞生存性に対するphiCD27の効果を測定するために、反復分析が前還元(pre-reduced)緩衝液及び培地を用いて嫌気性条件下に設定された。細胞はエンド対数で増殖され、嫌気性条件下で遠心分離により採取し、次いでPBS緩衝液中pH7.3で再懸濁した。PBS中でc.1×108細胞からc.1×103細胞まで10倍希釈を行なった;各分析において細胞数の評価が可能なように、それらの10μl分割量を0時点でBHI寒天上にスポットした。分析は2回ずつとし、100μgの部分精製エンドリシン(E1)か又は等量(50μl)の緩衝液(EB)及び300μlの最終量までの細胞を含有した。連続的な穏やかな振盪による2時間のインキュベーション後、PBS中の10倍連続希釈で30μlの試料を採取した;これらの希釈液の10μl分割量をBHI寒天上にスポットし、各2回ずつのペアの1つから残りの270μl分析量を細胞計数可能なように塗布した。
【0178】
0時点でc.1×108細胞を含む分析では、2時間のインキュベーション後1対数の低下を示したが、一方で1×107細胞又は1×106細胞が加えられた分析では、緩衝液対照に比べて2対数の低下を示した。低めの初期細胞数による分析では、リシンはより有効で、1×105細胞を植菌した分析からわずか4生存コロニーが回収されただけで、1×104細胞以下の分析では生存細胞は残っていなかった。
【0179】
次いで上記の生存性分析を、Ni−NTA溶出緩衝液(EB)を20mMのpH6リン酸緩衝液(NP)で置き換えるために、2mlのZeba Desaltスピンカラム(Pierce)を用いて、緩衝液交換にかけていたE1の400μlの分割量を用いて繰り返した。NP緩衝液中のリシンは、クロストリジウム・ディフィシレ11204の凍結細胞に対して元のNiNTAE1と同等の活性を示した(図15)。50μgのE1−NP又はNP緩衝液対照及びc.1×106細胞を用いて、上記の生存性分析を繰り返した;リシンによる2時間インキュベーションは緩衝液対照に比べて3対数の低下をもたらした。
【0180】
上記データはその後公表科学論文原稿(Mayer et al., 2008, J. Bacteriol. 190:6734-6740)の基礎となったが、その開示内容は、参照により本明細書に組み込まれている。
【0181】
ドメイン交換
スプライス重複PCRによるΦCD27Lエンドリシン上への新しい酵素ドメインの設計
リステリア・モノサイトゲネスに対して活性なバクテリオファージΦLM4からのエンドリシンLM−4は、宿主細胞の有効な溶解をもたらすことが示された(英国特許公開公報第2255561(B)号)。エンドリシンは864bpの長さで、pfam2557、VanY、蛋白質の始部のD−アラニル−D−アラニルカルボキシペプチダーゼ及びCOG5632、全配列にわたりN−アセチルムラモイル−L−アラニンアミダーゼ(NCBI Blast)に相同性を示す287アミノ酸のタンパク質を与えた。提案酵素活性ドメイン(EAD)をコード化する配列の前半は、スプライス重複伸長PCRにより、CD27L細胞壁結合ドメイン(CBD、Asn180から最終のArg270)か又は全270アミノ酸酵素の上流に挿入された。LM4酵素ドメインは、LM4のATGのNdeI部位を作製するために、プライマーLM4Nde5’−GGA TGA TTA CAT ATG GCA TTA ACA G[配列番号10]、及び2つのスプライス重複プラマーの1つ:LM4 EAD−CD27L EAD−CBDを与えるために、CD27L酵素の最初の15ヌクレオチドに適合する尾部を有するLM4配列の、ヌクレオチド439〜453、Thr147からAsn151を表わす、LM4−スプライス−CD27LE 5’−TAT ACA TAT TTT CAT GTT TTG TGT CGC AGT[配列番号11];又は、LM4 EAD−CD27L CBDを与えるために、Asn180からArg270のCD27Lの提案C末端結合ドメインに適合する尾部を有するLM4配列のヌクレオチド439〜453、Thr147からAsn151を表わす、LM4−スプライス−CD27L 5’−TTT AAC TCC CTC ATT GTT TTG TGT CGC AGT[配列番号12]、を用いてプラスミドpFI567 (Payne et al., 1996 FEMS Microbiology Letters 136: 19-24)からPCRによって増幅した。同様に、CD27L全配列又はCBDは、ベクター、T7T 5’−GCT AGT TAT TGC TCA GCG G[配列番号13]からのプライマー、及びLM4 EAD配列末端−そこでプライマーの最後の20ntがMet1からのCD27L配列の始めをコード化する、全配列に対する、CD27LEスプライスLM4 5’−ACT GCG ACA CAA AAC ATG AAA ATA TGT ATA ACA GT[配列番号14];及びプライマーの最後の16ntがAsn180からのCD27L配列の提案CBDをコード化する、CBDだけに対するCD27LスプライスLM4 5’− CT GCG ACA CAA AAC AAT GAG GGA GTT AAA C[配列番号15]に適合する尾部を有したスプライシングプライマー、を用いてΦCD27L−pET15bから増幅した。PCRを、元の鋳型に100%適合を与えるスプライシングプライマー部分に適合するように5サイクルのアニーリング温度を用い、次いで20サイクルは全スプライシングプライマーに適合するアニーリング温度にて、メーカー推奨の条件でPhusion (Finnzymes)により実施した。生成物は、SureClean (Bioline) を用いて精製し、50μlの容積に再懸濁した。これらの鋳型を100倍希釈し、そして1μl分割量を、2配列のスプライシングを可能にするアニーリング温度(54℃)で、元の外側プライマー−LM4Nde及びT7T−を用いてPCR反応に使用した。最終生成物をSureCleanで精製し、NdeI及びXhoIで制限し、そしてHis−タグ化LM4−CD27LE及びLM4−CD27Lを生産するために、pET15bにサブクローニングした。次いでこれらのプラスミドは大腸菌に形質転換され、それらの配列が確認された。
【0182】
複合酵素の粗抽出物及びNiNTA精製抽出物の両方を生成して、SDS−PAGE及びウェスタンブロット法で解析し、そして前述のように分析した(図16参照)。His−タグ化LM4−CD27LE及びLM4−CD27Lの両方は、粗抽出物中に高レベルで存在した。C.ディフィシレ11204の凍結細胞をPBS緩衝液中、pH5.8でインキュベートした場合、10μgのNiNTA精製抽出物は、緩衝液対照に比べて迅速溶解をもたらし(図17参照)、LM4−CD27LEは天然のCD27Lと同様の溶解速度を示した。細胞希釈剤としてpH7.3でPBS緩衝液を用いて等価活性が見られた。
【0183】
生存性分析において、LM4−CD27LE及びLM4−CD27Lの両方のNiNTA精製溶出液は生存数の低下をもたらした(図17参照)。50μgNiNTAE1を用いた、c.1×104細胞を含む分析は、緩衝液対照に比べて2時間インキュベーション後に少なくとも1対数の減少を示した。この低下は天然酵素で見られたほどに大きくはなく、他の酵素ドメインの添加は、C.ディフィシレを死滅させる能力を有する活性な新しい酵素を生産することができるという原理を証明した。
【0184】
野生型LM4及びドメインを交換したリシンのヌクレオチド及びアミノ酸配列
【化2】
【0185】
【化3】
【0186】
【化4】
【0187】
参考文献
1.Kuijper, E. J., Coignard, B. & Tull, P. (2006) Clin Microbiol Infect 12 Suppl 6, 2-18.
2.Anonymous (2006) Health Statistics Quarterly 30, 56-60.
3. Rupnik, M., Dupuy, B., Fairweather, N. F., Gerding, D. N., Johnson, S., Just, I., Lyerly, D. M., Popoff, M. R., Rood, J. I., Sonenshein, A. L., Thelestam, M., Wren, B. W., Wilkins, T. D. & von Eichel-Streiber, C. (2005) J Med Microbiol 54, 113-7.
4. Braun, V., Hundsberger, T., Leukel, P., Sauerborn, M. & von Eichel-Streiber, C. (1996) Gene 181, 29-38.
5. Goncalves, C., Decre, D., Barbut, F., Burghoffer, B. & Petit, J. C. (2004) J Clin Microbiol 42, 1933-9.
6. Popoff, M. R., Rubin, E. J., Gill, D. M. & Boquet, P. (1988) Infect Immun
56, 2299-306.
7. Skurnik, M. & Strauch, E. (2006) Int J Med Microbiol 296, 5-14.
8. Projan, S. (2004) Nat Biotechnol 22, 167-8.
9. Brussow, H., Canchaya, C. & Hardt, W. D. (2004) Microbiol Mol Biol Rev 68, 560-602.
10. Davis, B. M. & Waldor, M. K. (2003) Curr Opin Microbiol 6, 35-42.
11. Strauch, E., Schaudinn, C. & Beutin, L. (2004) Infect Immun 72, 7030-9.
12. Chibani-Chennoufi, S., Sidoti, J., Bruttin, A., Kutter, E., Sarker, S. & Brussow, H. (2004) Antimicrob Agents Chemother 48, 2558-69.
13. Matsuzaki, S., Yasuda, M., Nishikawa, H., Kuroda, M., Ujihara, T., Shuin,
T., Shen, Y., Jin, Z., Fujimoto, S., Nasimuzzaman, M. D., Wakiguchi, H., Sugihara, S., Sugiura, T., Koda, S., Muraoka, A. & Imai, S. (2003) J Infect Dis 187, 613-24.
14. Biswas, B., Adhya, S., Washart, P., Paul, B., Trostel, A. N., Powell, B.,
Carlton, R. & Merril, C. R. (2002) Infect Immun 70, 204-10.
15. Loc Carrillo, C., Atterbury, R. J., el-Shibiny, A., Connerton, P. L., Dillon, E., Scott, A. & Connerton, I. F. (2005) Appl Environ Microbiol 71, 6554-63.16. Huff, W. E., Huff, G. R., Rath, N. C., Balog, J. M. & Donoghue, A. M. (2004) Poult Sci 83, 1944-7.
17. Ramesh, V., Fralick, J. A. & Rolfe, R. D. (1999) Anaerobe 5, 69-78.
18. Wray, T. (2007) National Provisioner 5th July
19. Loessner, M. J., Kramer, K., Ebel, F. & Scherer, S. (2002) Mol Microbiol 44, 335-49.
20. Loessner, M. J. (2005) Curr Opin Microbiol 8, 480-7.
21. Gasson (1995-2003) Patents GB 2255561 B (1995); AU 650737B (1994); US 5763251 (1998); US 6083684 (2000); CA 2066387 (2003); EP 0510907B (2003).
22. Fischetti, V. A. (2005) Trends Microbiol 13, 491-6.
23. Schuch, R., Nelson, D. & Fischetti, V. A. (2002) Nature 418, 884-9.
24. Loeffler, J. M., Djurkovic, S. & Fischetti, V. A. (2003) Infect Immun 71,
6199-204.
25. Yoong, P., Schuch, R., Nelson, D. & Fischetti, V. A. (2004) J Bacteriol 186, 4808-12.
26. Zimmer, M., Vukov, N., Scherer, S. & Loessner, M. J. (2002) Appl Environ Microbiol 68, 5311-7.
27. Sell, T. L., Schaberg, D. R. & Fekety, F. R. (1983) J Clin Microbiol 17, 1148-52.
28. Nelson, D. (2004) J Bacteriol 186, 7029-31.
29. Mahony, D. E., Bell, P. D. & Easterbrook, K. B. (1985) J Clin Microbiol 21, 251-4.
30. Goh, S., Riley, T. V. & Chang, B. J. (2005) Appl Environ Microbiol 71, 1079-83.
31. Govind, R., Fralick, J. A. & Rolfe, R. D. (2006) J Bacteriol 188, 2568-77.
32. Goh, S., Ong, P. F., Song, K. P., Riley, T. V. & Chang, B. J. (2007) Microbiology 153, 676-85.
33. Rutherford, K., J. Parkhill, J. Crook, T. Horsnell, P. Rice, M-A. Rajandream and B. Barrell . (2000) Bioinformatics 16, 944-945.
34. Altschul, S. F., Thomas L. Madden, Alejandro A. Schaffer, & Jinghui Zhang, Z. Z., Webb Miller, and David J. Lipman (1997) Nucleic Acids Res. 25, 3389-3402
35. Schaffer, A. A., L. Aravind, Thomas L. Madden, Sergei , Shavirin, J. L. S., Yuri I. Wolf, Eugene V. Koonin, and & Altschul, S. F. (2001) Nucleic Acids Res. 29, 2994-3005.
36. Sebaihia, M., Wren, B. W., Mullany, P., Fairweather, N. F., Minton, N., Stabler, R., Thomson, N. R., Roberts, A. P., Cerdeno-Tarraga, A. M., Wang, H., Holden, M. T., Wright, A., Churcher, C., Quail, M. A., Baker, S., Bason, N., Brooks, K., Chillingworth, T., Cronin, A., Davis, P., Dowd, L., Fraser, A., Feltwell,
T., Hance, Z., Holroyd, S., Jagels, K., Moule, S., Mungall, K., Price, C., Rabbinowitsch, E., Sharp, S., Simmonds, M., Stevens, K., Unwin, L., Whithead, S., Dupuy, B., Dougan, G., Barrell, B. & Parkhill, J. (2006) Nat Genet 38, 779-86.
37. Loessner, M. J., Wendlinger, G. & Scherer, S. (1995) Mol Microbiol 16, 1231-41.
38. Wegmann, U., Klein, J. R., Drumm, I., Kuipers, O. P. & Henrich, B. (1999) Appl Environ Microbiol 65, 4729-33.
【技術分野】
【0001】
本発明は、クロストリジウム・ディフィシレ(Clostridium difficile)のバクテリオファージからのエンドリシン由来新規ポリペプチド及びそれをコードする核酸分子、並びにその組成物に関する。本発明は、クロストリジウム・ディフィシレなどの微生物細胞に関連する病態及び疾患の診断及び治療における当該ポリペプチド及び核酸分子の使用をも提供する。特に、本発明は、クロストリジウム・ディフィシレのバクテリオファージΦCD27由来エンドリシン活性を有する新規ポリペプチド及びその使用を提供する。
【背景技術】
【0002】
クロストリジウム・ディフィシレに関連する高まりつつある問題、特に、抗生物質使用にしばしば関連する院内感染におけるその役割は十分裏付けられている(非特許文献1)。C.ディフィシレは、加熱、乾燥及び消毒剤に耐性を有する胞子形成能を有する嫌気性グラム陽性菌である。非塩素系の洗浄剤への露出は、実際に胞子形成を増加させるという幾つかの証拠がある。これらの特性は、この生物が病院環境中に生き残る能力に寄与し、それにより患者に感染する能力を有する病原体の貯蔵庫を維持する。C.ディフィシレ関連疾患(CDAD)は、その増大する速度及び深刻さの双方を伴い、UKにおいても世界的にも高まりつつある問題である。イングランド及びウェールズでは、C.ディフィシレに関連する死亡が1999年の975人から2004年の2,247人まで増加した。CDADの届出は、1999年の1000人から2000年の15,000人、そして2003年の35,500人まで増加した(非特許文献2)。上記のヒトの健康への脅威に加えて、C.ディフィシレが、動物、特に子ウシ及びヒツジなどの家畜における罹患及び死亡の重大な原因でもあることを銘記すべきである。従って、ヒトにおけるこの問題に対処する方法に関する本明細書での開示は、同様に家畜対象にも当てはまると読み取るべきである。
【0003】
特に重大な進展は、当初カナダとUSAにおける、しかし現在はUKと幾つかの他のヨーロッパ諸国で著しい、C.ディフィシレの高毒性株の出現である。C.ディフィシレリボタイプ027と特定されたこの新菌株は、174例及び19名の死亡にかかわる大流行において2003年にUKで検出された。2006年までに75の病院からC.ディフィシレリボタイプ027の450の別のUK分離株が認められた(非特許文献1)。
【0004】
C.ディフィシレは土壌及び動物の腸管に広く分布している。それは、3%のヒト成人並びに80%の健常新生児及び小児の便から培養することができる(非特許文献1)。病原性能力は、C.ディフィシレの強力な毒素を産生する能力に関連する;2つの主要な特徴的毒素は、308kDaの外毒素の毒素A(TcdA)及び270kDaの細胞毒素の毒素B(TcdB)であり、これらはアミノ酸レベルで63%の相同性を共有する(非特許文献3)。これらの毒素をコード化する遺伝子は、病原性島PaLoc(非特許文献4)に関係し、そして菌株はこれらの2つの毒素を産生するそれらの能力の点で異なる。他の毒性因子も関与していると考えられ、そして別の2成分系毒素CDTが明らかにされている(非特許文献5、6)。
【0005】
毒性C.ディフィシレ株の病原性能力は、胃腸管(GIT)ミクロフローラが損なわれ又はアンバランスになっている場合に成り立ち、そしてこれは抗生物質治療の一般的な帰結である。このように、病院環境はヒトの病気を蔓延させ引き起こすC.ディフィシレの理想的な環境である(非特許文献1)。
【0006】
CDADは、C.ディフィシレの病原性株が十分強い地位をGITミクロフローラ内で
獲得すると起こり、宿主上皮を損傷する1つ又は複数の毒素を産生する。GITミクロフローラは病原性微生物に対する重要なバリアであり、宿主と有利な様式で相互作用しながらホメオスタシス平衡に維持される、凡そ500から1000の異なる種の複合群落を形成する。古典的な抗生物質治療は、程度の差はるが無差別的であり、それはGITの微生物群落の微妙なバランスを損なう可能性がある。従来の抗生物質治療又は他の要因の結果として、正常なミクロフローラの破壊はCDADの発症の主な要因である。
【0007】
それ故、複合GITミクロフローラの保護能力を損傷しない、C.ディフィシレの制御に対する新しい処置法及び取り組み方法の必要性が高まっている。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Kuijper, E. J., Coignard, B. & Tull, P. (2006) Clin Microbiol Infect 12 Suppl 6, 2-18.
【非特許文献2】Anonymous (2006) Health Statistics Quarterly 30, 56-60.
【非特許文献3】Rupnik, M., Dupuy, B., Fairweather, N. F., Gerding, D. N., Johnson, S., Just, I., Lyerly, D. M., Popoff, M. R., Rood, J. I., Sonenshein, A. L., Thelestam, M., Wren, B. W., Wilkins, T. D. & von Eichel-Streiber, C. (2005) J Med Microbiol 54, 113-7.
【非特許文献4】Braun, V., Hundsberger, T., Leukel, P., Sauerborn, M. & von Eichel-Streiber, C. (1996) Gene 181, 29-38.
【非特許文献5】Goncalves, C., Decre, D., Barbut, F., Burghoffer, B. & Petit, J. C. (2004) J Clin Microbiol 42, 1933-9.
【非特許文献6】Popoff, M. R., Rubin, E. J., Gill, D. M. & Boquet, P. (1988) Infect Immun 56, 2299-306.
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明の第1の態様は、クロストリジウム・ディフィシレの細胞に特異的に結合し及び/又はそれを溶解させることができる、配列番号1のアミノ酸配列を含む単離ポリペプチド、又はそのフラグメント、バリアント、誘導体若しくは融合体を提供することである。
【課題を解決するための手段】
【0010】
下記のアミノ酸配列は、クロストリジウム・ディフィシレのバクテリオファージΦCD27の野生型(即ち、天然産)エンドリシンの配列である。
【化1】
【0011】
NCBI受入番号YP_002290910及びACH91325をも参照されたい。
【0012】
1つの実施態様では、ポリペプチドは、クロストリジウム・ディフィシレ(ΦCD27以外の)のバクテリオファージの天然リシンではない。従って、本発明の第1の態様は、配列番号1のアミノ酸配列を含み又はそれから成る単離したポリペプチド、及びその非天然型フラグメント、バリアント、誘導体若しくは融合体を提供する。
【発明を実施するための形態】
【0013】
本明細書で使用される用語「アミノ酸」は、遺伝的にコード化された20の標準アミノ酸及びそれらの対応する「D」型の立体異性体(天然の「L」型と比較して)、ω−アミノ酸及び他の天然アミノ酸、特殊アミノ酸(例えば、α,α−ジ置換アミノ酸、N−アルキルアミノ酸など)並びに化学誘導アミノ酸(下記を参照)を含む。
【0014】
従って、アミノ酸を、「アラニン」又は「Ala」又は「A」のように特別に羅列する場合、この用語は特に明示しない限りL−アラニン及びD−アラニンの両方を指す。所望の機能特性がそのポリペプチドにより保持される限り、他の特殊アミノ酸も本発明のポリペプチドの好適な成分であり得る。例示したペプチドに対して、コード化されたアミノ酸残基は、それぞれ、必要に応じて標準アミノ酸の慣用名に対応する単一文字記号表示によって表わされる。
【0015】
好ましくは、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は、L−アミノ酸を含み又はそれから成る。
【0016】
「単離した」により、本発明のポリペプチド、特にバクテリオファージΦCD27の野生型エンドリシンが天然に見出されるもの以外の形態で提供されることを意味する。好ましくは、ポリペプチドはインタクトバクテリオファージを含まないで提供される。
【0017】
1つの実施態様では、単離した形態で与えられるバクテリオファージΦCD27[配列番号1]の天然エンドリシンである。
【0018】
先行技術において公知のクロストリジウム・ディフィシレバクテリオファージの他の天然リシンは、本発明の第1の態様に包含されない。特に、クロストリジウム・ディフィシレバクテリオファージの以下のリシンは、本発明の第1の態様の範囲から明確に除外される:
(a)バクテリオファージΦCD119のリシン(lysin);
(b)バクテリオファージΦC2のリシン;及び
(c)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン。
【0019】
例えば、以下の公知のタンパク質(それらのNCBI受入番号を参照することにより定義される):
PhiC2推定エンドリシン: YP_001110754;
CD630ファージエンドリシン: YP_001087453;
phiCD119推定リシン: YP_529586;
QCD−32g58仮想タンパク質: ZP_01803398;
QCD−32g58仮想タンパク質: ZP_01803228;
は、本発明の第1の態様の範囲から明確に除外される。
【0020】
1つの実施態様では、本発明の第1の態様のポリペプチドは、配列番号1のアミノ酸配列を含む。例えば、ポリペプチドは、配列番号1のアミノ酸配列から成ることができる。
【0021】
しかしながら、本発明の第1の態様は、クロストリジウム・ディフィシレの細胞に特異的に結合し及び/又はそれを溶解させることができる配列番号1のアミノ酸のフラグメント、バリアント、誘導体及び融合体にも及ぶ。
【0022】
「クロストリジウム・ディフィシレの細胞に特異的に結合することができる」により、ポリペプチドがクロストリジウム・ディフィシレの細胞に優先的に結合することができることを意味する。しかし、当然のことながら、当該ポリペプチドは1つ又はそれ以上の更なる細胞型にも優先的に結合することができる。好ましくは、クロストリジウム種の細胞に排他的に結合する。当該細胞結合活性は、当技術分野で周知の方法を用いて測定することができる。
【0023】
「クロストリジウム・ディフィシレの細胞を溶解させることができる」により、ポリペプチド、又はそのフラグメント、バリアント、誘導体若しくは融合体が、バクテリオファージΦCD27の野生型エンドリシンが細菌性細胞を溶解させる能力(少なくとも一部)を保持することを意味する。当然のことながら、当該溶解活性は、全細胞型に対する非特異的細胞毒性活性よりむしろ細胞特異的(例えば、クロストリジウム・ディフィシレの細胞に対して)であるべきである。当該細胞溶解活性は、以下の実施例に詳述される方法(Loessner et al. [37]をも参照されたい、その開示内容は参照することにより本明細書に組み入れられている)など当技術分野で周知の方法を用いて測定することができる。好ましくは、ポリペプチドがクロストリジウム・ディフィシレの細胞を溶解させる能力は新鮮細胞を用いて測定される。
【0024】
好ましい実施態様では、ポリペプチドがクロストリジウム・ディフィシレの細胞を溶解させる能力は、11204菌株の細胞を用いて測定される。
【0025】
当業者には当然のことながら、ポリペプチド、又はそのフラグメント、バリアント、誘導体若しくは融合体は、細菌細胞を溶解させるバクテリオファージΦCD27の野生型エンドリシンの全能力を保持する必要はない。むしろ、該ポリペプチド、フラグメント、バリアント、誘導体又はその融合体は、細菌細胞を溶解するバクテリオファージΦCD27の野生型エンドリシンのただ単に少なくとも10%の能力を保持する必要があるだけである。しかしながら、好ましくは、ポリペプチド、フラグメント、バリアント、誘導体又はその融合体は、少なくとも20%の、例えば、少なくとも30%、40%、50%、60%、70%、80%、90%、100%、150%、200%又はそれ以上の、細菌細胞を溶解させるバクテリオファージΦCD27の野生型エンドリシンの能力を示す。
【0026】
それ故に、本発明の第1の態様の実施態様では、ポリペプチドは、クロストリジウム・ディフィシレの細胞を溶解させることができる、配列番号1のアミノ酸配列のフラグメントを含み又はそれから成る。
【0027】
多くのバクテリオファージ・エンドリシンが2つの異なるドメイン(例えば、Sheehan et al., 1996, FEMS Microbiology Letters 140:23-28を参照されたい、その開示内容は参照することにより本明細書に組み入れられている)から成ることは十分立証されている。1つは細胞壁分解に関与する触媒ドメインであり、これらは幾つかの異なる形態で存在することが知られている。他のドメインは、細胞表面モチーフを認識する細胞壁結合ドメインであり、その標的細胞へのエンドリシンの付着を可能にする。
【0028】
酵素ドメインは、同じタイプの溶解活性を共有する溶菌酵素の他の類似領域に対するそのアミノ酸相同性によって同定することができる。バクテリオファージΦCD27のエンドリシンの場合、酵素ドメインはN−アセチルムラモイル−L−アラニンアミダーゼと同定されており、それはエンドリシンのアミノ末端領域を占有する(これは公知酵素ドメインによる配列番号1の並列解析によって、例えば、NCBICDD検索ツールを用いて確認することができる;Marchler-Bauer & Bryant, 2004, Nuc. Acids Res. 32[W]:327-331を参照されたい。その開示内容は、参照することにより本明細書に組み入れられている)。細胞壁結合ドメインは、エンドリシンのカルボキシ末端領域を占有すると考えられる。
【0029】
1つの実施態様では、酵素ドメインは配列番号1の1から175のアミノ酸内に含まれる。従って、酵素ドメインを含むフラグメントは、アミノ酸1、5、10、15、20、25、30、35、40、45、50、60、70、80、90又は100のいずれかから開始し、そしてアミノ酸175、170、165、160、155、150、145、140、135、130、125、120、115、110又は105のいずれかで終了する配列番号1の配列から成ると考えられる。例えば、酵素ドメインを含むフラグメントは、配列番号1の10から140までのアミノ酸、又は配列番号1の25から155までのアミノ酸、又は上記の開始及び終点の他の可能な順列のいずれかからなると考えられる。
【0030】
1つの実施態様では、細胞壁結合ドメインは配列番号1の175から270のアミノ酸内に含まれる。従って、細胞壁結合ドメインを含むフラグメントは、アミノ酸175、180、185、190、195、200、205、210、215、220のいずれかから開始し、そしてアミノ酸270、265、260、255、250、245、240、235、230又は225のいずれかで終了する配列番号1の配列から成ると考えられる。例えば、細胞壁結合ドメインを含むフラグメントは、配列番号1の195から265までのアミノ酸、又は配列番号1の180から240までのアミノ酸、又は上記の開始及び終点の他の可能な順列のいずれかからなると考えられる。
【0031】
本発明の第1の態様のポリペプチドは、好ましくは酵素ドメイン及び細胞壁結合ドメインの両方に対応する、配列番号1のアミノ酸配列の1つ又はそれ以上のフラグメントを含み又はそれから成る。
【0032】
しかしながら、当業者には当然のことながら、配列番号1の細胞壁結合ドメインは、代わりにクロストリジウム・ディフィシレの細胞を溶解させることができる別の起源からの酵素(溶解)ドメインに融合するか又はそうでなければ結合してもよい。キメラ的なリシンの製造は、Sheehan et al., 1996, FEMS Microbiology Letters 140:23-28に記載されており、その開示内容は参照することにより本明細書に組み入れられている。従って、別の実施態様では、本発明の第1の態様のポリペプチドは、細胞壁結合ドメインに対応する配列番号1のアミノ酸配列の1つ又はそれ以上のフラグメントを含み又はそれから成ることができる。
【0033】
フラグメントは、配列番号1の少なくとも50の隣接アミノ酸、例えば、配列番号1の、少なくとも60、70、80、90、100、110、120、130、140、150、160、170、175、180、190、200、210、220、230、240、250、260又は265の隣接アミノ酸を含み又はそれから成ることができる。
【0034】
別の実施態様では、本発明の第1の態様のポリペプチドは、クロストリジウム・ディフィシレの細胞を溶解させることができる、配列番号1のアミノ酸配列のバリアント、又はそのフラグメントを含み又はそれから成ることができる。
【0035】
ポリペプチドの「バリアント」により、配列番号1のアミノ酸配列に関連して保存性であれ又は非保存性であれ、挿入、欠失及び/又は置換を含む。特に、バリアントポリペプチドは、非天然バリアントであってもよい。
【0036】
例えば、ポリペプチドは、配列番号1のアミノ酸配列に対して少なくとも60%の同一性、より好ましくは該配列に対して少なくとも70%又は80%又は85%又は90%の同一性、そして最も好ましくは該アミノ酸配列に対して少なくとも95%、96%、97%、98%又は99%の同一性を有するアミノ酸配列を含んでもよい。
【0037】
当然のことながら、上記の配列同一性は配列番号1のアミノ酸配列の全長にわたり、又はその一部にわたってもよい。しかしながら、好ましくは、配列同一性は、配列番号1のアミノ酸配列の少なくとも50のアミノ酸にわたり、例えば、その中の少なくとも60、70、80、90、100、110、120、130、140、150、160、170、180、190、200、210、220、230、240、250、260又はそれ以上のアミノ酸にわたる。
【0038】
同一性パーセントは、当技術分野で周知の方法、例えば、ExPASyファシリティウェブサイトでのLALIGNプログラム(Huang and Miller, Adv. Appl. Math. (1991) 12:337-357、その開示内容は参照することにより本明細書に組み入れられている) :
www.ch.embnet.org/software/LALIGN_form.html
を用いて、パラメーターとしてグローバルアラインメントオプション、スコアリングマトリックススBLOSUM62、開口ギャップペナルティ−14、拡張ギャップペナルティ−4を用いて測定することができる。
【0039】
或いはまた、2ポリペプチド間の配列同一性パーセントは、適切なコンピュータプログラム、例えば、AlignX, Vector NTI Advance 10 (Invitrogen Corporationから) 又はGAPプログラム(University of Wisconsin Genetic Computing Groupから)を用いて測定してもよい。
【0040】
当然のことながら、同一性パーセントは、その配列が最適にアラインメントされているポリペプチドに関して計算される。
【0041】
配列番号1のアミノ酸配列のフラグメント及びバリアントは、当技術分野で周知のタンパク質工学及び部位特異的変異誘発の方法を用いて作製されてもよい(例えば、Molecular Cloning: a Laboratory Manual, 3rd edition(「分子クローニング:実験室マニュアル第3版」), Sambrook & Russell, 2001, Cold Spring Harbor Laboratory Pressを参照されたい。その開示内容は参照することにより本明細書に組み入れられている)。
【0042】
当業者には当然のことながら、本発明のポリペプチド、又はそのフラグメント、バリアント若しくは融合体は、修飾又は誘導体化された1つ又はそれ以上のアミノ酸を含んでもよい。従って、ポリペプチドは、配列番号1のアミノ酸配列の誘導体、又はそのフラグメント若しくはバリアントを含み又はそれから成ることができる。
【0043】
1つ又はそれ以上のアミノ酸の化学誘導体は、官能性側鎖基との反応により得ることができる。当該誘導体化分子は、例えば、遊離アミノ基が誘導体化されて、アミン塩酸塩、p−トルエンスルホニル基、カルボキシベンゾキシ基、t−ブチルオキシカルボニル基、クロロアセチル基又はホルミル基を形成するそれらの分子を含む。遊離カルボキシル基は、誘導体化して塩、メチル及びエチルエステル又は他のタイプのエステル及びヒドラジドを形成してもよい。遊離のヒドロキシル基は、誘導体化してO−アシル又はO−アルキル誘導体を形成してもよい。化学誘導体としては、20の標準アミノ酸の天然アミノ酸誘導体を含むそれらのペプチドも含まれる。例えば:4−ヒドロキシプロリンは、プロリンと置換してもよい;5−ヒドロキシリジンは、リジンと置換してもよい;3−メチルヒスチジンは、ヒスチジンと置換してもよい;ホモセリンはセリンと、そしてオルニチンはリジンと置換してもよい。誘導体は、また、必要活性が維持される限り、1つ又はそれ以上の付加又は欠失を含むペプチドをも含む。含まれる他の修飾は、アミド化、アミノ末端アシル化(例えば、アセチル化又はチオグリコール酸アミド化)、末端カルボキシアミド化(例えば、アンモニア又はメチルアミンによる)、などの末端修飾である。
【0044】
当業者には更に当然のことながら、ペプチド模倣的化合物も有用であり得る。従って、「ポリペプチド」により、エンドリシン活性を呈するペプチド模倣的化合物を含む。用語「ペプチド模倣的」は、治療薬として特定のポリペプチドの立体配座及び所望の特徴を模倣した化合物を指す。
【0045】
例えば、本明細書に記載のポリペプチドは、アミノ酸残基がペプチド(−CO−NH−)結合によって結合される分子だけでなく、ペプチド結合が逆向きの分子も含む。当該逆反転ペプチド模倣体は、当技術分野で公知の方法、例えば、Meziere et al. (1997) J. Immunol. 159, 3230-3237に記載の方法などを用いて作製してもよく、その開示内容は参照することにより本明細書に組み入れられている。CO−NHの代わりにNH−CO結合を含む当該逆反転ペプチド模擬体は、タンパク質分解に対して更にいっそう抵抗性である。或いはまた、本発明のポリペプチドは、1つ又はそれ以上のアミノ酸残基が、通常のアミド結合の代わりに−γ(CH2NH)−結合によって結合したペプチド模倣的化合物であってもよい。
【0046】
当然のことながら、ポリペプチドは、細胞外タンパク質分解消化に対する感受性を低減するのを助けるために、そのN−又はC−末端において、例えば、アミド化により都合よくブロックしてもよい。
【0047】
上記のように、D−アミノ酸及びN−メチルアミノ酸などの様々な非コード又は修飾アミノ酸は、本発明のポリペプチドを修飾するのに使用してもよい。加えて、推定生物活性立体配座は、環化などの共有結合修飾により、又はラクタムの組み込み若しくは他のタイプの架橋によって安定化してもよい。ジスルフィド、スルフィド及びアルキレン架橋を含む、環状ホモデチックペプチド及び環状ヘテロデチックペプチドの合成方法は、米国特許第5643872号に開示されている。環化方法の他の例は、米国特許第6008058号にて検討され開示されており、それらの文書における関連開示内容は、参照することにより本明細書に組み入れられている。環状の安定化ペプチド模倣的化合物の合成への更なる取り組みは、閉環メタセシス(RCM)である。
【0048】
要約すれば、周知のような末端修飾は、プロテイナーゼ消化による感受性を低減させ、その結果、溶液中、特にプロテアーゼが存在すると考えられる生体液中のペプチドの半減期を延長するのに有用である。ポリペプチド環化は、また、有用な修飾であり、環化によって形成される安定構造に因りそして環状ペプチドで認められる生物活性の点で好ましい。
【0049】
従って、1つの実施態様では、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は環状である。しかしながら、好ましい実施態様では、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は線状である。
【0050】
本発明の第1の態様の更なる実施態様では、ポリペプチドは配列番号1のアミノ酸配列の融合体、又はそのフラグメント、バリアント若しくは誘導体を含み又はそれから成る。
【0051】
ポリペプチドの「融合体」は、その他のポリペプチドに融合したポリペプチドを含む。例えば、ポリペプチドは、配列番号1のアミノ酸配列の内部に及び/又はN−及び/又はC−末端に挿入された1つ又はそれ以上の更なるアミノ酸、又はそのフラグメント、バリアント若しくは誘導体を含んでもよい。
【0052】
従って上記のように、1つの実施態様では、本発明の第1の態様のポリペプチドは、異なる起源の酵素ドメインがそれに融合した、細胞壁結合ドメイン(又はその細胞壁結合活性を保持するようなドメイン配列のバリアント)から成る配列番号1のフラグメントを含む。
【0053】
他の好適な酵素ドメインの例は:
L−アラノイル−D−グルタミン酸エンドペプチダーゼ;D−グルタミル−m−DAPエンドペプチダーゼ;ペプチド間架橋特異的エンドペプチダーゼ;N−アセチル−β−D−グルコサミニダーゼ(=ムラモイルヒドロラーゼ);N−アセチル−β−D−ムラミダーゼ(=リゾチーム);溶菌性トランスグリコシラーゼを含む。
【0054】
また、他の起源からのN−アセチルムラモイル−L−アラニンアミダーゼも利用可能である(Loessner, 2005, Current Opinion in Microbiology 8: 480-487を参照されたい。その開示内容は参照することにより本明細書に組み入れられている)。
【0055】
例えば、該ポリペプチドは、該ポリペプチドの精製を容易にするために、グルタチオン−S−トランスフェラーゼ(GST)又はタンパク質Aのようなポリペプチドに融合してもよい。当該融合体の例は当業者に周知である。同様に、該ポリペプチドは、His6などのオリゴヒスチジンタグに、又は周知のMycタグエピトープなどの抗体によって認識されるエピトープに融合してもよい。該ポリペプチドのいずれのフラグメント、バリアント又は誘導体に対する融合体も、本発明の範囲内に含まれる。当然のことながら、所望の特性、即ち、エンドリシン活性を保持する融合体(又はそのバリアント若しくは誘導体)が好ましい。融合体が本明細書に記載の方法に使用するのに好適なものであれば、それも特に好ましい。
【0056】
例えば、融合体は、本発明の該ポリペプチドに所望の特性を賦与する更なる部分を含んでもよい;例えば、その部分はポリペプチドを検出又は単離し、ポリペプチドの細胞内取り込みを促進し、又は細胞からのタンパク質の分泌を導くのに有用と考えられる。その部分は、当業者に周知の、例えば、ビオチン部分構造、放射性部分構造、蛍光部分構造、例えば、小分子蛍光プローブ又は緑色蛍光タンパク質(GFP)蛍光プローブであってもよい。その部分構造は、当業者に周知の免疫原性タグ、例えば、Mycタグであってもよく、又は当業者に周知の、ポリペプチドの細胞内取り込みを促進することができる親油性分子若しくはポリペプチドドメインであってもよい。
【0057】
当業者には当然のことながら、本発明のポリペプチドは、また、上記ポリペプチドの薬学的に許容される酸又は塩基付加塩をも含む。本発明において有用な、上記の塩基化合物の薬学的に許容される酸付加塩を製造するために使用される酸は、非毒性酸付加塩、即ち、塩酸塩、臭化水素酸塩、ヨウ化水素酸塩、硝酸塩、硫酸塩、重硫酸塩、リン酸塩、過リン酸塩、酢酸塩、乳酸塩、クエン酸塩、酸性クエン酸塩、酒石酸塩、酸性酒石酸塩、コハク酸塩、マレイン酸塩、フマル酸塩、グルコン酸塩、糖酸塩、安息香酸塩、メタンスルホン酸塩、エタンスルホン酸塩、ベンゼンスルホン酸塩、p−トルエンスルホン酸塩、及びとりわけ、パモエート[即ち、1,1′−メチレン−ビス−(2−ヒドロキシ−3−ナフトエート)]塩などの、薬学的に許容されるアニオンを含有する塩を形成する酸である。
【0058】
薬学的に許容される塩基付加塩も、また、ポリペプチドの薬学的に許容される塩の形態を生成するために使用してもよい。本来酸性の本発明化合物の、薬学的に許容される塩基塩を製造する試薬として使用してもよい化学塩基は、当該化合物と非毒性塩基塩を形成する塩基である。当該非毒性塩基塩は、アルカリ金属カチオン(例えば、カリウム及びナトリウム)及びアルカリ土類金属カチオン(例えば、カルシウム及びマグネシウム)、N−メチルグルカミン(メグルミン)などの、アンモニウム又は水溶性アミン付加塩、並びに、とりわけ、薬学的に許容される有機アミンの低級アルカノールアンモニウム及び他の塩基塩などの当該薬学的に許容されるカチオンから由来する塩を含むが、これらに限定されるものではない。
【0059】
ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は、また、貯蔵のために凍結乾燥し、そして使用前に好適な担体中に再構成することもできる。いずれの好適な凍結乾燥方法(例えば、噴霧乾燥、ケーキ乾燥)及び/又は再構成方法も採用することができる。当業者には当然のことながら、凍結乾燥及び再構成は様々な程度の活性ロスをもたらし、そした使用レベルはそれを補うために上方へ調整しなければならない。好ましくは、凍結乾燥(フリーズドライ)ポリペプチドは、再水和した場合、その活性(凍結乾燥する前)の、最高で約20%、又は最高で約25%、又は最高で約30%、又は最高で約35%、又は最高で約40%、又は最高で約45%、又は最高で約50%を失う。
【0060】
本発明のポリペプチドの本質的な特徴は、クロストリジウム・ディフィシレの細胞の溶解能力である。好ましくは、ポリペプチドはクロストリジウム・ディフィシレの複数の株の細胞を溶解することができる。例えば、ポリペプチドは、配列番号1のΦCD27リシンによって溶解するクロストリジウム・ディフィシレの1つ又はそれ以上の菌株を溶解することができる(下の表1を参照)。
【0061】
当然のことながら、本発明のポリペプチドは、また、バシラス種(例えば、バシラス・セレウス(Bacillus cereus)、枯草菌及び/又は炭疽菌)、他のクロストリジウム種(例えば、クロストリジウム・ビフェルメンタンス(Clostridium bifermentas))及び/又はリステリア種(例えば、リステリア・イバノビイ(Listeria ivanovii))など他の細菌種の細胞を溶解することができる。
【0062】
1つの実施態様では、本発明のポリペプチドは、健常な腸生理を維持するのに有用な細菌を実質的に溶菌することができない。例えば、ポリペプチドが、クロストリジウム・レプツム(Clostridium leptum)、 クロストリジウム・ネキシル(Clostridium nexile)、クロストリジウム・コッコイデス(Clostridium coccoides)、クロストリジウム・イノクーム(Clostridium innocuum)、クロストリジウム・ラモスム(Clostridium ramosum)及び/又はアネロコッカス・ヒドロジェナリス(Anaerococcus hydrogenalis)の細胞を溶解しない場合、それは有益である。
【0063】
最も好ましくは、本発明のポリペプチドは、カナダ、US及び現在全ヨーロッパで出現しているクロストリジウム・ディフィシレの高病原性株である、クロストリジウム・ディフィシレ株リボタイプ027の細胞を溶解することができる。例えば、ポリペプチドは、クロストリジウム・ディフィシレリボタイプ027の細胞に対して、配列番号1のポリペプチドの少なくとも10%、例えば、少なくとも20%、30%、40%、50%、60%、70%、80%、90%、100%又はそれ以上の溶解活性を示すことができる。ポリペプチドは、クロストリジウム・ディフィシレリボタイプ027の細胞に対して、配列番号1のポリペプチドより大きい溶解活性、例えば、少なくとも110%、120%、130%、140%、150%、160%、170%、180%、190%、200%、250%、300%、500%又はそれ以上の溶解活性さえも示すことができる。
【0064】
有利なことには、ポリペプチドは、病原性細菌の細胞を選択的に、即ち、非病原性細菌よりも大幅に溶解させることができる。
【0065】
本発明の第1の態様において使用するポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体の製造方法は、当技術分野で周知である。好都合なことには、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体は、組み換えポリペプチドであり又はそれを含む。
【0066】
このように、ポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体をコード化する核酸分子(又はポリヌクレオチド)は、好適な宿主及びそれから得られるポリペプチドに発現することができる。当該組み換えポリペプチドの好適な生産方法は、当技術分野で周知である(例えば、Sambrook & Russell, 2000, Molecular Cloning, A Laboratory Manual, Third Edition(「分子クローニング、実験室マニュアル、第3版」), Cold Spring Harbor, New Yorkを参照されたい。その文書の関連する開示内容は、参照することにより本明細書に組み入れられている)。
【0067】
簡潔にいうと、発現ベクターは、適切な宿主内で核酸分子によりコード化されるポリペプチドを発現することができる核酸分子を含んで、構成することができる。
【0068】
核酸分子、特にDNAを、例えば、相補的凝集性末端経由で、操作可能的にベクターに結合する様々な方法が開発されている。例えば、相補的ホモポリマー領域は、ベクターDNAに挿入するようにDNAセグメントに付加することができる。次いで、ベクター及びDNAセグメントは、相補的ホモポリマーの尾部間で水素結合により接続し、組み換えDNA分子を形成する。
【0069】
1つ又はそれ以上の制限部位を含む合成リンカーは、DNAセグメントをベクターへ接続する代替法を提供する。例えば、エンドヌクレアーゼ制限消化により生成するDNAセグメントは、3′−5′−エキソヌクレアーゼ活性を有する突出3′−1本鎖末端を除去し、そしてそれらのポリメラーゼ活性を有する陥凹3′−末端を埋める酵素である、バクテリオファージT4DNAポリメラーゼ又は大腸菌DNAポリメラーゼIで処理される。
【0070】
従って、これらの活性の組み合わせは平滑末端DNAセグメントを産生する。次いで、平滑末端セグメントは、バクテリオファージT4DNAリガーゼなどの、平滑末端DNA分子の連結反応を触媒することができる酵素の存在下に、大過剰モルのリンカー分子とインキュベートされる。このように、反応の生成物は、それらの末端に高分子リンカー配列を持っているDNAセグメントである。次いで、これらのDNAセグメントは好適な制限酵素により切断され、そしてDNAセグメントの末端と適合する末端を生成する酵素で切断されている発現ベクターに連結される。
【0071】
DNA(又はレトロウイルスベクター、RNA)は、次いで、好適な宿主に発現してポリペプチドを産生する。このように、ポリペプチドをコード化するDNAは公知の方法に従って使用され、本明細書に含まれる教示を勘案して適切に修正されて発現ベクターを構築し、そのベクターは、次いで、本発明の化合物又はその結合部分を発現及び産生するように、適切な宿主細胞を形質転換するために使用することができる。当該技術は当技術分野で周知である。
【0072】
ポリペプチドをコード化するDNA(又はレトロウイルスベクターの場合には、RNA)は、適切な宿主への導入のために多種多様な他のDNA配列に結合してもよい。相手のDNAは、宿主の性質、宿主へのDNAの導入様式、及びエピソームの保持又は統合が求められるかどうかによって決まる。
【0073】
一般的に、DNAは、適正な方向に及び発現のための正しいリーディングフレームにおいて、プラスミドなどの発現ベクターに挿入される。必要に応じて、DNAは、所望の宿主によって認識される適切な転写及び翻訳調節制御ヌクレオチド配列に結合し得るが、当該制御は一般的に発現ベクター中で実現可能である。次いで、ベクターは標準的方法を介して宿主に導入される。一般的に、全ての宿主がベクターにより形質転換されるわけではない。従って、形質転換宿主細胞を選択する必要性がある。1つの選択方法は、抗生物質耐性などの、形質転換細胞中の選択可能な形質をコードするあらゆる必要な制御要素によって、発現ベクターにDNA配列を組み込むことを含む。或いはまた、当該選択可能な形質の遺伝子は、所望の宿主細胞を同時形質転換するのに使用される別のベクター上にあり得る。
【0074】
発現ベクターにより形質転換されている宿主細胞は、次いで、十分な時間そしてポリペプチドの発現を可能にする本明細書に開示の教示に照らして当業者に公知の適切な条件下で培養し、次いでこれを回収することができる。
【0075】
多くの発現系が知られており、細菌(大腸菌、枯草菌)、酵母(例えば、サッカロミセス・セレビジエ)、糸状菌(例えば、アスペルギルス)、植物細胞、動物細胞及び昆虫細胞が挙げられる。
【0076】
代表的なベクターは、典型的には、ベクターが他の非原核生物細胞型における発現に使用される場合でも、原核生物中の増殖のためのColE1複製起点(ori)などの原核生物レプリコンを含む。ベクターは、また、それによって形質転換した大腸菌などの細菌性宿主細胞における遺伝子の発現(転写及び転座)を導くことができる、原核生物プロモーターなどの適切なプロモーターをも含むことができる。
【0077】
代表的な原核生物ベクタープラスミドは、Biorad Laboratories (Richmond, CA, USA) から入手できるpUC18、pUC19、pBR322及びpBR329、及びPharmacia(Piscataway, NJ, USA) から入手できるpTrc99及びpKK223−3である。
【0078】
代表的な哺乳動物細胞ベクタープラスミドは、Pharmacia, Piscataway(NJ, USA) から入手できるpSVLである。このベクターは、クローン化遺伝子の発現を推進するSV40後期プロモーターを用いるが、発現の最高レベルはCOS−1細胞などのT抗原産生細胞において、見出されている。
【0079】
誘導型哺乳動物ベクターの例は、Pharmaciaから同様に入手できるpMSGである。このベクターは、クローン化遺伝子の発現を推進するために、マウス哺乳動物腫瘍ウイルスの長い末端反復のグルココルチコイド誘導プロモーターを用いる。
【0080】
他のベクター及び発現系は、様々な宿主細胞を用いて使用するために当技術分野で周知である。
【0081】
宿主細胞は原核細胞であっても又は真核細胞であってもよい。細菌細胞は、好ましい原核宿主細胞であり、そして一般的には、例えば、Bethesda Research Laboratories Inc.(Bethesda, MD, USA)から入手できる大腸菌DH5株、及びAmerican Type Culture Collection (ATCC) (Rockville, MD, USA) (No. ATCC 31343)から入手できるRR1などの大腸菌の1菌株である。好ましい真核宿主細胞は、酵母、昆虫及び哺乳動物細胞、好ましくはマウス、ラット、サル又はヒト線維芽細胞及び腎細胞系などの脊椎動物細胞である。酵母宿主細胞は、Stratagene Cloning Systems(La Jolla, CA 92037, USA)から一般に入手できるYPH499、YPH500及びYPH501を含む。好ましい哺乳動物宿主細胞は、ヒト胚性腎細胞であるCRL1658及び293細胞のようなATCCから入手できるチャイニーズハムスター卵巣(CHO)細胞を含む。好ましい昆虫細胞はバキュロウイルス発現ベクターで形質転換し得るSf9細胞である。
【0082】
宿主細胞を培養しそして組み換えタンパク質を分離する方法は、当技術分野でで周知である。当然のことながら、宿主細胞によっては、産生される本発明のポリペプチドは異なってもよい。例えば、酵母又は細菌細胞などの特定の細胞は、違った方法で翻訳後修飾されると考えられる本発明の化合物の形態の産生をもたらす可能性のある、異なる翻訳後修飾系を有しないか又は有する。
【0083】
本発明のポリペプチドは、また、ウサギ網状赤血球溶血液又はコムギ胚芽溶解物(Promegaから入手できる)などの市販のインビトロ翻訳系を用いてインビトロで製造してもよい。好ましくは、翻訳系はウサギ網状赤血球溶血液である。都合の良いことには、翻訳系は、TNT転写−翻訳系(Promega)などの転写系に連結されてもよい。この系は、翻訳と同じ反応でコード化DNAポリヌクレオチドからの好適なmRNA転写物を産生する利点を有する。
【0084】
CS Bio Company Inc.(Menlo Park, USA)から入手できるもののような、自動ポリペプチド合成機も、また、使用することができる。
【0085】
従って、本発明の第2の態様は、本発明の第1の態様のポリペプチドをコード化する単離した核酸分子を提供することである。
【0086】
核酸分子はDNA(例えば、cDNA)又はRNAであってもよい。
【0087】
好ましい実施態様では、核酸分子は図3に示されるヌクレオチド配列[配列番号2]を含み又はそれから成る。
【0088】
本発明の第3の態様は、本発明の第2の態様の核酸分子を含むベクターを提供することである。1つの実施態様では、ベクターは発現ベクターである。好ましくは、ベクターはpET15b及びpACYC184から成る群から選択される。
【0089】
当業者には当然のことながら、発現ベクターの選択は宿主細胞の選択により決定してもよい。従って、乳酸連鎖球菌における本発明のポリペプチドの発現では、2成分制御系をコード化するnisR 及びnisK遺伝子をも発現する乳酸連鎖球菌のバックグランド株を用いて、本発明のポリペプチドがnisAオペロンのプロモーターの制御下に発現する、ナイシン(nisin)発現系を使用することができる。この系の下での発現は正に調節され、そして外因性ナイシンの供給により誘導される(Ruyter at el., 1996, Applied and Environmental Microbiology 62:3662-3667を参照されたい。その開示内容は、参照することにより本明細書に組み入れられている)。
【0090】
代わりの実施態様では、ナイシンの全生合成遺伝子クラスターは、インデューサーがその細胞によって合成される場合と同じ宿主細胞内にもたらされる。
【0091】
更なる代わりの実施態様では、本発明のポリペプチドは、プラスミド系か又は染色体に組み込まれた系を用いて、ラクトース異化オペロンの制御下に乳酸連鎖球菌に発現してもよい(例えば、Payne et al., 1996, FEMS Microbiology Letters 136: 19-24及びvan Rooijen et al., 1992, Journal of Bacteriology 174: 2273-2280を参照されたい。その開示内容は、参照することにより本明細書に組み入れられている)。
【0092】
本発明の第4の態様は、本発明の第2の態様の核酸分子又は本発明の第3の態様のベクターを含む宿主細胞を提供することである。1つの実施態様では、宿主細胞は微生物細胞、例えば、細菌細胞である。好ましくは、宿主細胞は非病原性である。
【0093】
例えば、宿主細胞は、大腸菌、ラクトコッカス種、バクテロイデス種、ラクトバシラス種、エンテロコッカス種及びバシラス種から成る群から選択してもよい。
【0094】
好ましい実施態様では、宿主細胞は乳酸連鎖球菌の細胞である。
【0095】
或いはまた、宿主細胞は酵母細胞、例えば、サッカロミセス種であってもよい。
【0096】
本発明の第5の態様は、ポリペプチドを発現させる条件下に、本発明の第2の態様の核酸分子又は本発明の第3の態様のベクターを含む宿主細胞集団を培養し、それからポリペプチドを単離することを含む、本発明のポリペプチドを製造する方法を提供することである。
【0097】
本発明の第6の態様は、以下のもの:
(a)本発明の第1の態様のポリペプチド;
(b)本発明の第2の態様の核酸分子;
(c)本発明の第3態様のベクター;
(d)本発明の第4態様の宿主;及び/又は
(e)本発明の第1の態様のポリペプチドを発現することができるバクテリオファージ;
及び薬学的に許容される担体、賦形剤又は医薬品添加剤を含む、医薬組成物を提供することである。
【0098】
本明細書で使用される「医薬組成物」は、本発明の方法に使用するための治療上有効な製剤を意味する。
【0099】
本明細書で使用される「治療的有効量」、又は「有効量」、又は「治療的に有効な」は、所定の病態及び投与レジメンに対し治療的効果を与える量を指す。これは、所要の添加剤及び賦形剤、即ち、担体又は投与賦形剤と併せて、所望の治療的効果をもたらすように算出される活性物質の所定量である。更に、宿主の活動性、機能及び反応性の臨床的に重大な欠如を減少させ、そして最も好ましくはそれを予防するのに十分な量を意味することを意図している。或いはまた、治療的有効量は、宿主の臨床的に重大な病態の改善をもたらすのに十分な量である。当業者には当然のことながら、化合物の量はその比活性度によって変動してもよい。好適な用量は、所要の賦形剤と併せて所望の治療効果をもたらすように算出される活性組成物の所定量を含むことができる。本発明の組成物を製造するための方法及び使用では、活性成分の治療的有効量が供給される。治療的有効量は、当技術分野で周知のように、年齢、体重、性別、病態、合併症、他の疾患など、患者の特性を基準にして通常の技量の医療又は獣医従事者により決定することができる。
【0100】
本発明の1つの実施態様では、医薬組成物は、本発明の第1の態様のポリペプチドを含む。
【0101】
ポリペプチドは、使用されるポリペプチドの有効性/毒性に応じて種々の濃度で製剤化し得る。好ましくは、製剤は、0.1μMと1mMの間、より好ましくは1μMと100μMの間、5μMと50μMの間、10μMと50μMの間、20μMと40μMの間、及び最も好ましくは約30μMの濃度でポリペプチドを含む。インビトロ適用の場合は、製剤は類似濃度のポリペプチドを含むことができる(しかし、当然のことながら、より高濃度も使用してもよい)。
【0102】
従って、この医薬製剤は、当該細胞により感染し又は感染を起こし易い患者において、クロストリジウム・ディフィシレの細胞増殖を少なくとも部分的に阻害するのに十分な量のポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体を含むことができる。好ましくは、医薬製剤は、患者においてクロストリジウム・ディフィシレの細胞を死滅させるのに十分な量のポリペプチド、又はそのフラグメント、バリアント、融合体若しくは誘導体を含む。
【0103】
当業者には当然のことながら、本発明のポリペプチドは、一般的に、所定の投与経路及び標準的薬剤治療(例えば、Remington: The Science and Practice of Pharmacy, 19th edition(「薬学の科学及び実践、第19版」), 1995, Ed. Alfonso Gennaro, Mack Publishing Company, Pennsylvania, USAを参照されたい。その開示内容は、参照することにより本明細書に組み入れられている)に関して選択される好適な医薬品添加剤、賦形剤又は担体との混合物で投与される。
【0104】
例えば、ポリペプチドは、即時性、遅延性又は放出制御の適用のために、着香剤又は着色剤を含んでもよい錠剤、カプセル剤、腔坐剤、エリキシル剤、水剤又は懸濁剤の形態で、経口、口腔内又は舌下で投与することができる。ポリペプチドは、直接注射(例えば胃腸管へ)を経て投与してもよい。
【0105】
しかしながら、好ましくは、ポリペプチド及びその医薬組成物は、経口投与用である。
【0106】
好適な錠剤処方は、微結晶セルロース、乳糖、クエン酸ナトリウム、炭酸カルシウム、第二リン酸カルシウム及びグリシンなどの医薬品添加剤、澱粉(好ましくは、トウモロコシ、ジャガイモ又はタピオカ澱粉)、澱粉グリコール酸ナトリウム、クロスカルメロースナトリウム、及び特定の複合珪酸塩などの崩壊剤、及びポリビニルピロリドン、ヒドロキシル−プロピルメチルセルロース(HPMC)、ヒドロキシ−プロピルセルロース(HPC)、蔗糖、ゼラチン及びアラビアゴムなどの造粒結合剤を含んでもよい。加えて、ステアリン酸マグネシウム、ステアリン酸、グリセリルベヘナート及びタルクなどの滑沢剤が含まれ得る。
【0107】
類似タイプの固形組成物も、ゼラチンカプセル剤の充填剤として用いてもよい。この関連の好ましい医薬品添加剤は、ラクトース、澱粉、セルロース、乳糖又は高分子量ポリエチレングリコールをも含むことができる。水性懸濁剤及び/又はエリキシル剤では、ポリペプチドは種々の甘味剤又は着香剤、着色料又は色素と、乳化剤及び/又は懸濁剤と及び水、エタノール、プロピレングリコール及びグリセリンなどの希釈剤、及びその組み合わせと併用してもよい。
【0108】
ポリペプチドは、非経口で、例えば、静脈内、関節内、動脈内、腹腔内、髄腔内、心室内、胸骨内、頭蓋内、筋肉内若しくは皮下に投与してもよく、又は注入法で投与してもよい。それらは、他の物質、例えば、血液と等張溶液にするのに十分な塩又はグルコースを含有してもよい滅菌水溶液の形態での使用に最も適している。水溶液は、必要に応じて、好適に緩衝化(好ましくは3から9のpHに)すべきである。無菌状態での好適な非経口製剤の調剤は、当業者に周知の標準的調剤技術により容易に達成し得る。
【0109】
非経口投与に好適な製剤は、抗酸化剤、緩衝剤、静菌薬、及び製剤を対象とする受容者の血液と等張にする溶質を含有してもよい水溶性又は非水溶性無菌注射液;及び懸濁剤及び濃稠化剤を含有してもよい水溶性又は非水溶性滅菌懸濁液を含む。製剤は、1回用量又は多回用量容器、例えば、密封アンプル及びバイアルで提供してもよく、そして使用直前に滅菌液体担体、例えば、注射用蒸留水の添加だけを必要とする凍結乾燥(凍乾)状態で保存してもよい。即時注射液及び懸濁液は、前記の種類の滅菌粉末、顆粒及び錠剤から調剤してもよい。
【0110】
ヒトの患者への経口及び非経口投与では、ポリペプチドの日用量レベルは、通常成人当り1から100mg(即ち、約0.015から15mg/kg)であち、単回用量又は分割用量で投与される。例えば、3mg/kgのような、1〜10mg/kgの用量が使用されてよい。
【0111】
本発明の別の実施態様では、医薬組成物は、ポリペプチドそのものを含まないで、代わりに該ポリペプチドを発現することができる核酸分子を含む。好適な核酸分子、発現ベクター、及び宿主は詳細に上述されている。
【0112】
例えば、組み換えプロバイオティックを使用してもよい(LAB株、例えば、乳酸連鎖球菌又はラクトバシラス種)。
【0113】
本発明の更なる実施態様では、医薬組成物は、本発明の第1の態様のポリペプチドを発現することができるバクテリオファージを含む。例えば、本発明の第1の態様のポリペプチドを送達するために、野生型バクテリオファージΦCD27を使用してもよい。当該バクテリオファージに基づく治療を実施する方法は、当技術分野において周知である(例えば、Watanabe et al., 2007, Antimicrobial Agents & Chemotherapy 51:446-452を参照されたい)。
【0114】
このように、本明細書に記載の細菌感染症の処置では、本発明のポリペプチドは、同種の(cognate)タンパク質として、核酸構築物、ベクター又は同種のタンパク質を発現する宿主細胞として、同種のタンパク質(バクテリオファージを含む)を発現する生物の一部として、又はその細菌標的とリシンの接触を達成するために、それがC. ディフィシレのような病原性細菌であれ、又は本明細書に更に記載される別の病原体若しくは潜在的病原体であれ、当技術分野で公知のその他簡便な方法によって投与してもよい。
【0115】
理想的には、タンパク質は保護された形態で胃腸管に送達される。これは当技術分野で公知の多種多様の方法によって達成され得る。例えば、適切な用量のリシンは、胃の酸性状態は切り抜けるが、腸に入るにつれてタンパク質を放出する形態でマイクロカプセル化される。送達は、乳酸連鎖球菌、ラクトバシラス種、ビフィドバクテリウム種又はバクテロイデスを含むがこれらに限定されない、胃腸管通過を切り抜ける非病原性微生物によって行われる。当業者は、本明細書に開示されるリシンなどの活性化合物の胃腸管送達のためのそのような手段に使用できる選択肢について熟知している。これらの手段は、細胞内産生、secA分泌又は別の分泌経路による分泌、及び制御溶解による送達を含む。好ましくは、タンパク質は一度に全て放出されるのではなく、投与されたボーラスが胃腸管を横断するにつれて漸増的に放出される。或いはまた、リシンは、胃腸管における適切な位置で又は適切なシグナルの受信時にリシンを発現する、良性細菌の一部として導入される。本明細書に開示の好ましい実施態様では、非病原性ラクトコッカスが、胃腸管の特定の位置に到達時にΦCD27リシンを発現するように処理される。発現シグナルは、pH感受性プロモーター、又はこの目的のために当技術分野で公知の別の手段によって規定することができる。
【0116】
送達の他の手段としては、以下:
(a)国際公開公報第2006/111553号(ポリ尿素及び他の多層カプセル材);
(b)国際公開公報第2006/111570号及び欧州特許第1715739号(シクロデキストリンカプセル化);
(c)国際公開公報第2006/100308号及び欧州特許第1742728号(酵母及び他の微生物細胞カプセル化技術);
(d)米国特許第5153182号、欧州特許第1499183号及び国際公開公報第03/092378号;米国特許第6831070号(腸細胞発現による治療遺伝子産物送達);
(e)米国特許第7202236号(放出調節用医薬製剤);
(f)米国特許第5762904号(本発明のリシンを送達するために改変し得る、重合リポソームを用いるワクチンの経口デリバリー);
(g)米国特許第7195906号(本発明のリシンを発現させるために改変し得るビフィドバクテリウム);及び
(h)本明細書に引用された文献;
が含まれ、これらの全ては、本発明の新規な送達方法及び組成物を達成するために、当業者が本発明の開示内容を活用するのを可能にする目的で、参照することにより本明細書に組み入れられている。
【0117】
このように、本発明の医薬組成物の好ましい実施態様では、ポリペプチド、それをコード化する核酸分子などはマイクロカプセル化される(例えば、シクロデキストリン若しくは脂質二重層などの化学エンベロープ内で、又は処理したラクトコッカス細胞などの生存又は非生存微生物細胞内で)。このような方法で、ポリペプチド、核酸分子などは、胃腸管の作用部位への途中で胃の酸性状態に対して保護することができる。
【0118】
本発明の第7の態様は、医薬に使用するための、本発明の第1の態様のポリペプチド又は本発明の第6の態様の医薬組成物を提供することである。
【0119】
本発明の第8の態様は、患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制する薬剤の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。
【0120】
当然のことながら、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を示すポリペプチドは、必ずしもクロストリジウム・ディフィシレのバクテリオファージから由来する必要はない。例えば、ポリペプチドは以下の群:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン;
から選択することができる。
【0121】
或いはまた、ポリペプチドは、クロストリジウム・ビフェルメンタンス又はクロストリジウム・ソルデリ(Clostridium sordelli)などの異なるクロストリジウム種のバクテリオファージから由来(例えば、コード化)してもよい。
【0122】
しかしながら、好ましい実施態様では、ポリペプチドは、クロストリジウム・ディフィシレのバクテリオファージから由来する。
【0123】
従って、本発明の第8の態様の使用は、本発明の第1の態様のポリペプチドに限定されるものではなく、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するいずれのポリペプチドの使用をも包含する (Goh et al., 2007, Microbiology 153:676-685に記載のΦC2のリシンを含む。その開示内容は、参照することにより本明細書に組み入れられている)。
【0124】
本発明の関連する態様は、患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するための、クロストリジウム・ディフィシルのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。
【0125】
本発明の更なる態様は、患者中の微生物細胞に関連する疾患又は病態の治療又は予防のための薬剤の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。本発明の関連する態様は、患者中の微生物細胞に関連する疾患又は病態を治療又は予防するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。
【0126】
「患者中の微生物細胞に関連する疾患又は病態」には、クロストリジウム・ディフィシレによる患者の感染から生じ又は拮抗する疾患及び病態が包含される。当該疾患及び病態には、クロストリジウム・ディフィシレ関連疾患(CDAD)が含まれる。
【0127】
本発明の上記の使用の1つの実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、本発明の第1の態様のポリペプチドであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触により溶解を起こし易い他の細菌細胞から成る群から選択されるポリペプチドである(下記の表1と2を参照)。
【0128】
好ましくは、微生物細胞は、クロストリジウム・ディフィシレを含み又はそれから成る。このように、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、クロストリジウム・ディフィシレ細胞による感染に関連する疾患及び病態(クロストリジウム・ディフィシレ関連疾患、CDADなど)を治療又は予防するのに使用することができる。
【0129】
最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレリボタイプ027細胞を含み又はそれから成る。
【0130】
従って、本発明は更に以下の方法:
(a)患者中の微生物細胞を殺滅し及び/又はその増殖を阻害/抑制するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを患者に投与することを含む方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法;
(b)患者において微生物細胞に関連する疾患又は病態を治療又は予防するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若バクテリオファージを患者に投与することを含む方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法;
を提供する。
【0131】
本発明の上記の方法の1つの実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは本発明の第1の態様のポリペプチドであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触により溶解を起こし易い他の細菌細胞から成る群から選択されるポリペプチドである(下記の表1と2を参照)。好ましくは、微生物細胞は、クロストリジウム・ディフィシレ細胞を含み又はそれから成る。このように、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、クロストリジウム・ディフィシレ細胞による感染に関連する疾患及び病態(クロストリジウム・ディフィシレ関連疾患、CDADなど)を治療又は予防するのに使用することができる。最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレリボタイプ027の細胞を含み又はそれから成る。
【0132】
当業者には更に当然のことながら、本発明の使用及び方法は医学及び獣医学分野の両方で有用性を有する。従って、本薬剤はヒト及び非ヒト動物(ウマ、ウシ、イヌ及びネコなど)の両方の治療に使用することができる。しかしながら、好ましくは、患者はヒトである。
【0133】
「治療」には、患者の治療的及び予防的治療法が含まれる。用語「予防的」は、患者又は対象におけるクロストリジウム・ディフィシレによる感染の可能性を防止するか低減する、本明細書に記載のポリペプチド又は製剤の使用を包含するように用いられる。
【0134】
上記のように、用語「有効量」は、治療する疾患又は病態の好ましい変化をもたらすのに使用することができる、本発明のポリペプチドの濃度又は量を記載するために本明細書で用いられるが、その変化が寛解、好ましい生理的結果、治療する病状又は病態の好転又は減弱、惹起している病態又は病状の可能性の予防又は低減であるかは、治療する疾患又は病態によって決まる。
【0135】
当然のことながら、本明細書に記載の薬剤は、1つ又はそれ以上の更なる治療薬との併用で患者に投与してもよい。
【0136】
例えば、本明細書に記載の薬剤は:
(a)1つ又はそれ以上の従来の抗生物質治療(β−ラクタム、アミノグリコシド及び/又はキノロン);
(b)1つ又はそれ以上の更なるリシン、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージ;
(c)1つ又はそれ以上の抗生物質、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくは細菌;及び/又は
(d)腸内のクロストリジウム・ディフィシレ細胞の細菌溶菌時に放出される毒素を中和する療法;
と併用して投与してもよい。好適な中和療法は、抗生物質(Babcock et al., 2006, Infect. Immun. 74:6339-6347を参照されたい)及びトレバマー(tolevamer)などの毒素吸収剤(Barker et al., 2006, Aliment. Pharmacol. Ther. 24:1525-1534を参照されたい)を含んでもよい。
【0137】
本発明の更なる態様は、インビトロ及び/又はエクスビボで微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用を提供することである。例えば、エンドリシン活性を有する該ポリペプチドは、そのような細菌細胞で汚染され易い病院、キッチンなどの表面を清浄化するのに使用することができる。
【0138】
好ましくは、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、本発明の第1の態様のポリペプチドであって、その微生物細胞が、クロストリジウム・ディフィシレ及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択されるポリペプチドである(下記の表1及び2を参照)。例えば、微生物細胞は、クロストリジウム・ディフィシレ細胞を含み又はそれから成ってよい。最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレ・リボタイプ027の細胞を含み又はそれから成る。
【0139】
本発明の更なる態様は、試料中の微生物細胞の存在を検出するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性及び/又は細胞結合特異性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを含むキットであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択されるキットを提供することである。
【0140】
好ましい実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、本発明の第1の態様のポリペプチドであって、微生物細胞がクロストリジウム・ディフィシレ及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞からなる群から選択されるポリペプチドである(下記の表1及び2を参照)。例えば、微生物細胞は、クロストリジウム・ディフィシレ細胞を含み及びそれから成ってよい。最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレリボタイプ027の細胞を含み又はそれから成る。
【0141】
本発明のキットの更なる実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、マルチウェルプレートなどの好適な表面に固定化される。
【0142】
キットは、組織試料、細胞培養試料及び拭い液由来の細胞試料(例えば、微生物細胞による汚染を試験する表面から採取される)などの、いずれの好適な細胞試料と併せて使用してもよい。
【0143】
場合により、キットは、陰性対照試料(例えば、クロストリジウム・ディフィシレ細胞を試験するための細胞型を含まない)及び/又は陽性対照試料(試験するための細胞を含む)を更に含む。
【0144】
本発明の関連する態様は、以下の使用及び方法:
(a)クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、微生物細胞に関連する疾患又は病態の診断薬の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用;
(b)クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、微生物細胞に関連する疾患又は病態を診断するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用;
(c)インビトロ及び/又はエクスビボで試料中の微生物細胞の存在を検出するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用;及び
(d)患者中の微生物細胞に関連する疾患又は病態を診断するための、試験すべき患者からの細胞試料をクロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージに接触させ、そして試料中の細胞がそれにより溶解されるかどうかを測定することを含む方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法;
を提供することである。
【0145】
本発明の上記の使用及び方法の1つの実施態様では、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、本発明の第1の態様のポリペプチドであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択されるポリペプチドである(下記の表1及び2を参照)。好ましくは、微生物細胞は、クロストリジウム・ディフィシレ細胞を含み又はそれから成る。このように、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドは、クロストリジウム・ディフィシレ細胞による感染に関連する疾患及び病態(クロストリジウム・ディフィシレ関連疾患、CDADなど)を診断するのに使用することができる。最も好ましくは、微生物細胞は、クロストリジウム・ディフィシレ・リボタイプ027の細胞を含み又はそれから成る。
【0146】
当該診断のための使用及び方法では、細胞の溶解は当技術分野で周知の方法を用いて検出し得る。例えば、ATPのレベルは、細胞溶解の指標として測定し得る。
【0147】
本発明の上記の使用及び方法の他の実施態様では、ポリペプチドは、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合ドメインを含み又はそれから成る。検出を可能にするために、このようなポリペプチドは、磁気ビーズに融合され又は好適なレポーター(例えば、緑色蛍光タンパク質)を含む融合タンパク質として使用してもよい。
【0148】
当該診断方法は、リステリアエンドリシンなど他の系由来のエンドリシンにて十分確立されている(例えば、Loessner et al., 2002, Mol Microbiol 44, 335-49; Kretzer et al., 2007, Applied Environ. Microbiol. 73:1992-2000を参照されたい。その開示内容は、参照することにより本明細書に組み入れられている; 好適な分析法も、また、例えばProfos, Germany [www.profos.de/content/view/164/69/lang,en/でそれらのウェブサイトを参照されたい]から市販されている) 。
【0149】
本発明の典型的な実施態様は、以下の図を参照しながら、以下の非限定的実施例にて記載される。
【図面の簡単な説明】
【0150】
【図1】ΦCD27の電子顕微鏡写真。試料は飽和酢酸ウラニル中でネガティブ染色された。
【図2】予測されたORFを示すΦCD27ゲノム地図。矢印は転写の方向を示す。提案された機能モジュールは、BLAST結果並びにΦCD119、ΦC2、及びC.ディフィシレ株630プロファージの公表配列との類似性に基づいて表示される。
【図3】ΦCD27リシンのヌクレオチド配列、配列番号2。
【図4a−1】公表C.ディフィシレのバクテリオファージ(ΦC2(32);ΦCD119(31))、又はプロファージ(配列ゲノムからのCD630プロファージ1及び2(36))配列による、ΦCD27 ヌクレオチド配列のアラインメント。AlignX、Vector NTI Advance 10, Invitrogenにより行なわれたアラインメント。ΦCD27アミノ酸配列は配列番号2である。
【図4a−2】図4a−1の続きである。
【図4a−3】図4a−2の続きである。
【図4b】公表C.ディフィシレのバクテリオファージ(ΦC2(32);ΦCD119(31))、又はプロファージ(配列ゲノムからのCD630プロファージ1及び2(36))配列による、ΦCD27推定アミノ酸配列のアラインメント。AlignX、Vector NTI Advance 10, Invitrogenにより行なわれたアラインメント。ΦCD27アミノ酸配列は配列番号2である。
【図5】pET15bベクター(Novagen)のクローニング部位。
【図6a】(a)ΦCD27リシンを発現する大腸菌からの粗タンパク質溶解産物のゲル解析。レーン1は、SeeBlueマーカー(Invitrogen、サイズ191、97、64、51、39、28及び19kDa)、レーン2〜5は、BL21(DE3)pET15bΦCD27L総蛋白質抽出物。レーン2〜4は、IPTG−2で3時間誘導した抽出物及び20mMのTriS−HCl、pH8、50mMのNaClで抽出した3、プロテアーゼ阻害剤(Roche Complete mini EDTA-free)を含む3及び変性緩衝液(8Mの尿素、0.1MのNaH2PO4、0.01MのTriS−HCl、pH8.0)で抽出した4。レーン5は20mMのTriS−HCl、pH8、50mMのNaClで抽出した非誘導対照。レーン6及び7は、20mMのTriS−HCl、pH8、50mMのNaClで抽出したBL21(DE3)pET15bΦCD630L1総蛋白質抽出物、レーン6は、IPTGで3時間誘導しただけの(a)6×His抗体による(b)ゲルのウェスタン解析。
【図6b】(a)ΦCD27リシンを発現する大腸菌からの粗タンパク質溶解産物のゲル解析。レーン1は、SeeBlueマーカー(Invitrogen、サイズ191、97、64、51、39、28及び19kDa)、レーン2〜5は、BL21(DE3)pET15bΦCD27L総蛋白質抽出物。レーン2〜4は、IPTG−2で3時間誘導した抽出物及び20mMのTriS−HCl、pH8、50mMのNaClで抽出した3、プロテアーゼ阻害剤(Roche Complete mini EDTA-free)を含む3及び変性緩衝液(8Mの尿素、0.1MのNaH2PO4、0.01MのTriS−HCl、pH8.0)で抽出した4。レーン5は20mMのTriS−HCl、pH8、50mMのNaClで抽出した非誘導対照。レーン6及び7は、20mMのTriS−HCl、pH8、50mMのNaClで抽出したBL21(DE3)pET15bΦCD630L1総蛋白質抽出物、レーン6は、IPTGで3時間誘導しただけの(a)6×His抗体による(b)ゲルのウェスタン解析。
【図7】NiNTAカラム精製Hisタグ化ΦCD27リシンのゲル解析。レーン1は、SeeBlueマーカー(Invitrogen、サイズ191、97、64、51、39、28及び19kDa)、レーン2〜5は、IPTGで誘導後のBL21(DE3)pET15bΦCD27L総蛋白質抽出物、レーン1は、粗溶解産物、レーン2は、カラム流入、レーン3は、一次洗浄溶離液、レーン4は、洗浄二次溶離液、レーン5は、一次溶出液(E1、1ml)、レーン6は、二次溶出液(E2)。
【図8】エンド対数に増殖し、液体窒素中で急速冷凍し、次いでPBS中に再懸濁したC. ディフィシレ11204の細胞によるBioscreen溶解試験。ΦCD27リシン及びCD630リシンは大腸菌に発現し、そしてNiNTAカラムでHisタグを用いて精製された(図6を参照)。270μlの細胞をE1抽出物の30μlの希釈液に加えた。値は二重複分析の平均値+/−標準偏差である。CD630L1抽出物による細胞溶解は、緩衝液だけの対照に見られるものと同等であった。
【図9】エンド対数に増殖し、4℃で遠心分離により採取し、次いでPBS中に再懸濁したC. ディフィシレ11204の細胞によるBioscreen溶解試験は、1〜1.5の間のODを与えた。ΦCD27リシンは大腸菌に発現し、そしてNiNTAカラムでHisタグを用いて精製された(図6を参照)。270μlの細胞を溶離緩衝液で希釈した溶出液1(E1)の30μl試料に加えて、1分析当り10.5μgから0.35ngの範囲の濃度を与えた。新鮮細胞の使用は、緩衝液だけの対照において有意に少ない溶解を与えた。70ng未満のNiNTA精製タンパク質で緩衝液だけの対照と差が見られなかった。
【図10a】活性のスペクトルを試験するためにC.ディフィシル細胞に添加されたΦCD27リシンのBioscreen溶解試験。細胞を大腸菌から産生した3.5μgNiNTA精製タンパク質(E1)とインキュベートした。試験した30株のうち、宿主株12727及びΦCD27非感受性株11208及び高毒性リボタイプ027R23613を含み、全てが感受性であった。インキュベーションは緩衝液(B)か又はリシン(L)を用いて二重複でなされた。
【図10b】活性のスペクトルを試験するためにC.ディフィシル細胞に添加されたΦCD27リシンのBioscreen溶解試験。細胞を大腸菌から産生した3.5μgNiNTA精製タンパク質(E1)とインキュベートした。試験した30株のうち、宿主株12727及びΦCD27非感受性株11208及び高毒性リボタイプ027R23613を含み、全てが感受性であった。インキュベーションは緩衝液(B)か又はリシン(L)を用いて二重複でなされた。
【図10c】活性のスペクトルを試験するためにC.ディフィシル細胞に添加されたΦCD27リシンのBioscreen溶解試験。細胞を大腸菌から産生した3.5μgNiNTA精製タンパク質(E1)とインキュベートした。試験した30株のうち、宿主株12727及びΦCD27非感受性株11208及び高毒性リボタイプ027R23613を含み、全てが感受性であった。インキュベーションは緩衝液(B)か又はリシン(L)を用いて二重複でなされた。
【図11a−1】クロストリジウム種及び一般的な腸細菌に対するΦCD27リシンの活性。細胞を後期安定相で採取し、PBS中で再懸濁し、大腸菌から産生した7μgNiNTA精製タンパク質(E1)とインキュベートした。結果は、二重複分析の平均値+/−標準偏差である。ΦCD27リシンは大部分の種において細胞溶解を誘導しなかった(a及び表2を参照)。例外として(b及び表2を参照)、クロストリジウム・ビフェルメンタンスの迅速溶解、バシラス・セレウスの溶解、そして長い誘導期を有するB.サチリス、及びリステリア・イバノビイ(b)に対する微弱作用が含まれる。
【図11a−2】図11a−1の続きである。
【図11a−3】図11a−2の続きである。
【図11a−4】図11a−3の続きである。
【図11b−1】クロストリジウム種及び一般的な腸細菌に対するΦCD27リシンの活性。細胞を後期安定相で採取し、PBS中で再懸濁し、大腸菌から産生した7μgNiNTA精製タンパク質(E1)とインキュベートした。結果は、二重複分析の平均値+/−標準偏差である。ΦCD27リシンは大部分の種において細胞溶解を誘導しなかった(a及び表2を参照)。例外として(b及び表2を参照)、クロストリジウム・ビフェルメンタンスの迅速溶解、バシラス・セレウスの溶解、そして長い誘導期を有するB.サチリス、及びリステリア・イバノビイ(b)に対する微弱作用が含まれる。
【図11b−2】図11b−1の続きである。
【図11b−3】図11b−2の続きである。
【図11b−4】図11b−3の続きである。
【図12】ΦCD27リシン活性のpHプロファイル。C.ディフィシレ11204細胞を一定の範囲のpHに調整したPBSで再懸濁し、そして大腸菌から産生したNi−NTA精製リシンE1の活性を前と同様にbioscreenで測定した。
【図13A】(a)ΦCD27リシンを発現する乳酸連鎖球菌由来の粗タンパク質溶解物のゲル分析。レーン1及び10は、SeeBlue(Invitrogen、サイズ191、97、64、51,39、28及び19kDa)、レーン2〜5は、5時間(2、4)誘導又は非誘導(3、5)のphiCD27LpUK200HIS(2、3)又は空ベクターpUK200HIS対照(4、5)を含有する乳酸連鎖球菌UKLC10。レーン6〜9は、全て4時間誘導した(レーン当り10μg)phiCD27LpET15b(6、8、9)又は空ベクター対照(7)を含有する大腸菌BL21(DE3)。全タンパク質を、レーン8(20mMのリン酸ナトリウム、pH8)及び9(50mMのTrisHCl、pH7.5)を除いて20mMのTris−HCl、pH8、50mMのNaClで抽出した。(a)6×His抗体による(b)ゲルのウェスタン分析。
【図13B】(a)ΦCD27リシンを発現する乳酸連鎖球菌由来の粗タンパク質溶解物のゲル分析。レーン1及び10は、SeeBlue(Invitrogen、サイズ191、97、64、51,39、28及び19kDa)、レーン2〜5は、5時間(2、4)誘導又は非誘導(3、5)のphiCD27LpUK200HIS(2、3)又は空ベクターpUK200HIS対照(4、5)を含有する乳酸連鎖球菌UKLC10。レーン6〜9は、全て4時間誘導した(レーン当り10μg)phiCD27LpET15b(6、8、9)又は空ベクター対照(7)を含有する大腸菌BL21(DE3)。全タンパク質を、レーン8(20mMのリン酸ナトリウム、pH8)及び9(50mMのTrisHCl、pH7.5)を除いて20mMのTris−HCl、pH8、50mMのNaClで抽出した。(a)6×His抗体による(b)ゲルのウェスタン分析。
【図14】エンプティーベクター対照からの抽出物と比較した、C.ディフィシレ株11204の新鮮細胞とインキュベートしたphiCD27リシン発現大腸菌及び乳酸連鎖球菌由来の粗タンパク質抽出物のBioscreen分析。50μgタンパク質を各分析で使用し、結果は二重複分析の平均値+/−標準偏差である。
【図15】Zeba緩衝液交換カラム(Pierce)を20mMのリン酸ナトリウム、pH6.0に通した分割量と比較した、元の抽出物の活性を示す大腸菌産生Ni−NTA精製リシンE1のBioscreen分析。リシン及び緩衝液対照をC.ディフィシレ株11204の急速冷凍細胞とインキュベートした。結果は二重複分析の平均値+/−標準偏差である。
【図16】LM4−CD27L(レーン2)及びLM4−CD27LE(レーン3)の粗細胞抽出物のSDS−PAGE、及びHisタグ化タンパク質を強調表示する対応のウェスタンブロット。タンパク質を20mMのリン酸ナトリウム、pH6.0で抽出し、そして10μg分割量をMOPS緩衝液(Invitrogen)中の10%Bis-TrisNuPageゲルで電気泳動した。レーン1は、SeeBlueマーカー。
【図17】中期対数まで増殖し、次いで液体窒素中で急速冷凍したC.ディフィシレ株11204細胞の溶解を示すBioscreen分析。細胞を10μgNiNTA精製E1(溶出液1)又は対照としての溶出緩衝液とインキュベートした。
【実施例】
【0151】
背景
抗菌薬としての細菌ウイルスの利用で、近年多少のルネサンスを経験している。一部において、これは、薬剤耐性病原体の継続的出現に続く従来型抗生物質の代替を見出す必要性を反映している。最近のレビューはこの可能性を強調しているが、バクテリオファージの使用に内在する限界をも強調している(7、8)。
【0152】
一般的に、バクテリオファージは顕著な系統差を示すことから、それらは限定範囲の個々の菌株に対して活性であることを意味する。バクテリオファージとその細菌宿主間の相互作用の動力学は、バクテリオファージ攻撃に抵抗性の宿主変異体の即時選択を含む。他の関連する問題は、宿主生菌によるバクテリオファージ製剤の汚染可能性、及びバクテリオファージが遺伝子流動及び毒性因子の拡大に寄与する可能性を含む(9)。バクテリオファージによる毒素遺伝子の輸送は特に十分立証されており、その例はコレラ毒素(10)、ボツリヌス毒素(9)、志賀毒素(11)及びジフテリア毒素(9)を含む。これらの限定にもかかわらず、バクテリオファージは、マウスモデルにおける大腸菌(12)、黄色ブドウ球菌(13)及びバンコマイシン耐性エンテロコッカス・フェシウム(14)を制御するのに実験的に使用されている。バクテリオファージ療法はニワトリにおけるカンピロバクター(15)及び大腸菌(16)の制御のために研究中である。クロストリジウムに関して、ハムスターモデルにおけるC.ディフィシレを標的とする研究が報告されている(17)。更に、FDAは、最近全食品におけるリステリア制御のためのバクテリオファージ(LISTEX(登録商標), EBI Food Safety) へのGRAS承認を更に拡大した(18)。
【0153】
インタクト・バクテリオファージの使用に加えて、抗菌薬としてバクテリオファージ・エンドリシスを使用する可能性がある。バクテリオファージ・ライフサイクルの最終段階は、新たに複製されたインタクト・バクテリオファージ粒子のプールを放出するための細菌宿主細胞の溶解を含む。一般的に、これは、膜破壊性ホリンの厳密な時限産生が、そのペプチドグリカン標的に細胞壁分解性エンドリシンが接近することを可能にする、2段階過程によって達成される。エンドリシン酵素は分泌されず、ホリンの作用により及び細胞壁を分解するそれ自体の能力により細胞から放出される。一度放出された場合、エンドリシンは細胞の外側からペプチドグリカンを攻撃することができ、この現象は初期のバクテリオファージ研究の時から観察されている:それを「外部からの溶解」と称する。最も特徴的なバクテリオファージ・エンドリシンの構造は、触媒ドメイン及び特徴的な細胞壁結合ドメイン(CBD)を有するモジュール構造である。触媒ドメインは変化することができ、そしてほとんどの場合それはアミダーゼか又はムラミダーゼである。CBDは細胞壁表面上の糖モチーフを認識するレクチン様能力を有し、そして関係する特異性の変動は、エンドリシンに特異的な分類群へのそれらの特徴的なターゲッティングをもたらす(19、20)。
【0154】
Gassonらは、新しい抗菌薬として及びモデル系としてのリステリア及びクロストリジウムを用いた新しい検出技術の基礎の両方で、バクテリオファージ・エンドリシンの開発の先駆者となった(21)。その後、標的抗菌薬としてのエンドリシンの可能性が、炭疽菌(23)、肺炎連鎖球菌(24)及びエンテロコッカス・フェカーリスを標的とする公表例により広く認識されている(22)。リステリアに関して、重要な更なる研究が Martin Loessner at ETH, Switzerlandにより行なわれている(19、20)。加えて、クロストリジウム・パーフリンジェンスに対して活性なエンドリシンが特徴付けられている(26)。
【0155】
新しいバクテリオファージ・リシンの特徴付け及びその使用方法
テンペレート・バクテリオファージΦCD27は、クロストリジウム・ディフィシレ菌株保存株NCTC12727から単離された。他の25のC.ディフィシレ株に対してΦCD27を試験し、そしてタイプ株11204を含む他の4株に対して有効なことが示された。バクテリオファージ・ゲノムDNAを抽出して配列決定し、そしてBLAST検索によりエンドリシン配列が同定された。その配列は、公表されたC.ディフィシレのバクテリオファージ・エンドリシン(ΦCD119、ΦC2、配列決定されたC.ディフィシレCD630におけるプロファージ1及び2)と明瞭なアミノ酸及びヌクレオチド相同性を示した。リシンはpET15bへサブクローニングされ、そして6×Hisタグにより大腸菌に発現された。リシンはニッケルカラムで一部精製され、そしてファージ感受性及び非感受性株の両方を溶解することが、37℃でインキュベーション時の光学密度の減少により証明された。試験した30株のうち、毒性リボタイプ027株を含む全てが溶解することを示した。一連の菌属からの多数の他の細菌は、リシンに対して感受性を示さなかった。しかしながら若干の活性がC.ビフェルメンタンス、C.ソルデリ、セレウス菌、枯草菌に対して認められ、そしてリステリア・イバノビイに対して非常にわずかな活性が認められた。部分精製リシンの比活性は、C.ディフィシレ株によって変動した。従って、本明細書に開示されたリシンは、C.ディフィシレ病原性の治療及び検出に対する新しい強力な武器となる。
【0156】
本明細書で同定されそして特徴付けられたリシンは、C.ディフィシレ感染症、及びヒト及び動物の他の細菌感染症を治療するために使用できる新しい組成物である。本発明によれば、ΦCD27リシンは、当技術分野で公知の方法によって生産してもよい。それは、この目的のために増殖したウイルスから使用のために単離してもよい。しかしながら、好ましくは、それは本明細書に開示した組み換え法により、そして当業者に公知の代替法により生産される。この分子の関連サブポーションについて、細菌に特異的に結合し、それらの細菌を溶解するそれらの能力が特徴付けられた。これらの分子サブポーションは、天然分子として別々に又は同時に生産され、そして使用されてもよい。
【0157】
ΦCD27リシンの発見、クローニング及び活性
溶解物の生産及び活性分析は、記載のように行われた(27)。C.ディフィシレ株NCTC12727(the Health Protection Agency, Colindale, London から入手可能−糞便から単離され、1992年にS. Tabaqchali, St. Bart's Hospital, London により寄託)は、BHI+C(ビタミンK(10μl、0.5容量%/l)、ヘミン(5mg/l)、レザズリン(1mg/l)及びL−システイン(0.5g/l)を補充したBHI(Oxoid) )中、37℃で嫌気的に24時間増殖させた。バクテリオファージ生産をマイトマイシンC(Sigma)により3μg/mlの最終濃度で24時間誘導した。培養液を4℃において4000×gで20分間遠心分離し、上清を0.45μmフィルターユニット(Millipore) を通して濾過し、そして4℃で保存した。150μlの一夜C.ディフィシレBHI+C培養液を加えたBHI軟寒天(0.75%)を重層化したBHIプレート(1.5%寒天)上に、上清の25μl部をスポットし、そして37℃で一夜嫌気的にインキュベートした。培養液(表1を参照)を2重複で試験し、12727上清からの明瞭なプラーク形成を、4株−C.ディフィシレ11204(タイプ株)、11205、11207及び11209で確認した。菌株11204からのプラークは、滅菌パスツールピペットで250μlBHI+C中へ採取し、4℃で一夜インキュベートした。バクテリオファージ−ΦCD27の存在は電子顕微鏡で確認したが、それがカウドウイルス目に属することを示した(28)(図1)。総数で25のC.ディフィシレ株がマイトマイシンCで誘導され、その上清は25株全てに対して交差試験した。ΦCD27は、この方法で発見された唯一のプラーク形成単位であった。C.ディフィシレからのバクテリオファージ発見が稀なことはこれまでの刊行物にも示されており、それには94分離株から2バクテリオファージ産生株(29)又は56分離株から3産生株(30)が見出される。
【0158】
力価を増加させるために、100μlのプラーク溶出液を100μlのC.ディフィシレ24時間培養液と5mlBHI軟寒天中で混合し、BHI寒天上に塗布した。37℃での一夜の嫌気性培養は近似融合溶菌を与え、そして5mlBHI+Cで2時間の溶離は、2×106pfu/mlの力価を与えた。力価は、11204の液体培養での連続インキュベーションにより増加し、初期から中期対数期まで25mlBHI+C培養における細胞を増殖させ、バクテリオファージ:細胞の比を少なくとも4:1にする光学密度(OD)を与えた。この方法は、細菌懸濁液の完全透明化をもたらし、そして2代継代は2.5×1011pfu/mlの力価を与えた。DNA抽出のために、OD0.3の細胞をc.7.の感染の多重度まで濾過溶解物でインキュベートした。3時間のインキュベーションは完全溶解をもたらし、上清を上記のように採取して濾過し、そして2つの50ml部をQiagen λ midikit (Qiagen)中で使用した結果、c.160μgのバクテリオファージのゲノムDNA収量を得た。
【0159】
バクテリオファージΦCD27ゲノムのシークエンシング及びアセンブリーを、Phred-Phrapプログラムを用いてBiochemistry DNA Sequencing Facility (University of Cambridge, UK)により実施した。循環ゲノムは50,930bpであり、75の提案されたオープン・リーディング・フレーム(orf)を含んだ(図2)。これらの多くは、C.ディフィシレのバクテリオファージΦCD119(31)及びΦC2(32)からのものを含み、同定したバクテリオファージORFに顕著な相同性を示した。ORFは、BITS(Harpenden)を経て行なわれたArtemis (33)及びBlastP 検索(34,35)により解析した。提案されたΦCD27リシン配列は816bpであり、N−アセチルムラモイル−L−アラニンアミダーゼに相同性を示す271アミノ酸予測タンパク質をコードした。ヌクレトチド及びアミノ酸配列(図3)の両方は、C.ディフィシレのバクテリオファージ及びプロファージ(図4)の公表配列に合っていたが、ΦC2に対してその最大相同性(95.9%ヌクレオチド及びアミノ酸同一性)を有した。
【0160】
ΦCD27リシン配列はプライマーを用いてゲノムDNAから増幅され、初めのMet残基(プライマーCD27L_NDE、5’−TTA CAT ATG AAA ATA TGT ATA ACA GTA GG[配列番号3]、Sigma Genosys)周囲のNdeI(CATATG)部位を、そしてコード配列(プライマーCD27L_XHO、5’−CAA CCA CCT CGA GTT GAT AAC[配列番号4]の下流にXhoI部位(CTCGAG)を作製し、発現ベクターpET15b(Novagen)におけるサブクローニングを促進させた。増幅は、1×Phusion緩衝液、200μMのdNTP、0.5μMの各プライマー、200ngのゲノムDNA鋳型を含む50μlの反応液中、高性能PhusionDNAポリメラーゼ(0.02U/μl、Finnzymes)により実施した。増幅条件は、98℃で30秒間の初期変性、その後に30サイクルの変性(90℃で10秒)、アニーリング(58℃で30秒)及び伸長(90℃で10秒)、次いで72℃で5分間の最終伸長が続いた。平滑末端PCR産物はSureClean (Bioline)を用いて精製し、そして72℃で20分間インキュベートして、1×AmpliTaq緩衝液、0.2mMのdATP及び1UのAmpliTaqDNAポリメラーゼ(Applied Biosystems)を含む50μlの反応液中に3’A−オーバーハングがもたらされた。生成物をSureCleanで精製し、次いでTAクローニングキット(Invitrogen)を用いてpCR2.1に結合された。ライゲーション生成物は、TOP10化学的コンピーテント大腸菌(Invitrogen)中へ形質転換して、陽性菌を100μg/mlアンピシリンを補充したL寒天で選択し、そしてブルー・ホワイト選択のために40mg/mlX−ガル溶液の40μlで重層化した。プラスミドDNAをプラスミドミニキット(Qiagen) を用いて抽出し、挿入部分をベクタープライマー及びBigDye v3.1シークエンシングキット(Applied Biosystems)を用いて配列決定した。リシン原配列と100%配列相同性を示すが、付加したNdeI及びXhoI部位を有するクローンは挿入部分を放出するのが限定的であった。これはゲル精製され(Qiaex II, Qiagen)、そしてFast-Link DNAリガーゼ (Epicentre)を用いて、リシン配列がIPTG誘導性lacオペレーターによる高発現T7プロモーターの制御下に6−ヒスチジンタグの下流に発現するように、pET15bへと結合された。ライゲーション産物は、先ず配列確認のために TOP10細胞へ、次いでタンパク質発現のために化学的コンピーテントBL−21(DE3)細胞 (Invitrogen)へ形質転換された。配列決定したC.ディフィシレ(36)のプロファージ1からのリシン配列を、 ベクターpUC57へとGenscript Corp. (Piscataway, USA)によって合成し、そしてプライマーCD630L1_NDE(5’−TGC TCA TAT GAA AAT AGG TAT AAA TTG)[配列番号5]、及びXhoI部位を含む特定のベクターDNAでリシンを増幅したM13前方(5’−GTA AAA CGA CGG CCA GT)[配列番号6]を用いて、同様にHis−タグ化発現のためにサブクローニングした。
【0161】
His−タグ化リシンは、メーカーにより示唆されたように、OD6000.4まで100μg/mlアンピシリンにより10mlLブロス中に増殖したBL21(DE3)細胞に発現し、次いで0.5mMのIPTG(Melford Biosciences)で3〜4時間誘導した。細胞を4000×gで遠心分離することにより採取し、次いで1mlの懸濁液(20mMTris−HCl、pH8、50mMのNaCl)中4℃で20分間再懸濁し、そして2mlスクリューキャップチューブに移した。粗タンパク質溶解物は、FastPrep FP120 細胞破壊器(Savant)中の0.1mm酸洗浄ガラスビーズ(Sigma)を用いて、バースト間に5〜10分間氷上でインキュベートしながら4×30秒のバースト(速度10)での細胞破壊によって得られた。破壊細片を13,000×gで4℃にて20分間遠心分離によりペレット化し、上清を4℃で保存した。粗溶解物は、IPTG誘導無しで増殖したリシンを含む細胞から、及び誘導の有り無しで増殖したpET15b空ベクターを含む細胞からも製造された。タンパク質含量をBradford試薬(Bio Rad)を用いて測定し、10μg部をMOPS緩衝液(Invitrogen)中10%NuPage Novex Bis Trisゲルで電気泳動した。His−タグ化リシンの存在は、抗Hisタグモノクローナル抗体(Novagen)を用いてウェスタンブロットにより確認した。タンパク質はNuPage緩衝液 (Invitrogen)を用いてPVDF膜に移され、そして検出は二次抗体として抗マウスIgGを用いてQiagen (Qiaexpress検出及び分析便覧)に記載のように行なわれ、そして比色検出はSigma FastBCIP/NBTアルカリ性ホスファターゼ基質でなされた。これにより、IPTG誘導溶解物におけるc.33kDaのHisタグ化バンドの高発現及び非誘導溶解物の低発現を証明した(図6)。
【0162】
粗溶解物によるC.ディフィシレの11204及び11207株細胞のリシスは、Loessner ら(37)によって記載された方法を用いて評価した。11204株細胞は、エンド対数期まで増殖し、1.8ml分割量をスクリューキャップチューブ(13,000×g、2分)中への遠心分離によって採取し、そしてペレットは液体窒素中で急速冷凍して−20℃で保存した。ペレットは900μlの20mMのTris−HCl、pH8中氷上で再懸濁し、そして100μlの粗タンパク質溶解物を含むキュベットに加えて、その後OD600の低下を読み込み前に混合して1時間モニターした。この系で、C.ディフィシレ細胞は緩衝系中で一定量の溶解を示したが、ΦCD27リシン粗抽出物による溶解はより急速で多大であった。しかしながら、誘導pET15bエンプティーベクター粗溶解物によるその後の試験は同等の溶解を示し、大腸菌リゾチームの活性を示唆した。この問題を回避するために、ΦCD27及びCD630L1リシンをQiagenNiNTAキットでアフィニティー精製した。BL−21(DE3)細胞は、100μg/mlのアンピシリンを含む250mlのLブロス中でOD6000.6まで増殖し、次いで1mMの最終濃度でIPTGにより5時間誘導した。細胞を4000×gで4℃にて20分間遠心分離することにより採取して、−20℃で保存した。タンパク質を自然条件で精製し、そして精製はNuPageゲル分析によって確認した(図7)。この方法は大部分がリシンの部分精製タンパク質を産生し、その収量は第1のΦCD27溶出液(E1)では2.3mg総タンパク質であり、第2(E2)では0.5mgであった。E1溶出液の希釈液のインキュベーションは、同様に調製した細胞からの溶出液に比べて11204株細胞の急速溶解を示したが、pET15b空ベクターを発現した。しかしながら、CD630L1E1溶出液は11204株を溶解せずに、そしてΦCD27リシンとの相乗効果は見られなかった。
【0163】
溶解分析は、Bioscreen C (Labsystems)及び溶出緩衝液(EB, Qiagen)でのNiNTA部分精製リシン抽出物を用いたマルチウェルプレート中で継続した。初期分析は、分光光度計分析におけるように、30μlのEB及び270μlの細胞の全容積中の約(c.)7μgのタンパク質を用いた。分析は氷上にセットアップし、次いで37℃に予熱されたBioscreen Cに移し、そしてプログラムは以下のように実施した:600nmの光学密度でサンプリング前に10秒振盪して2分毎にサンプリングした。各分析は、緩衝液だけの2ウェル及びリシンの2ウェルで実施し、全部で4ウェルに同じ細菌細胞懸濁液から植菌した。この系では、感受性株のリシンウェル中の溶解は迅速であり−その差は5分以内で顕著であった。しかしながら、リシン誘導溶解よりもはるかに低速にもかかわらず、緩衝液だけの対照での溶解も顕著であった(図8)。
【0164】
C.ディフィシレ及び他の細菌細胞の両方は、エンド対数まで増殖して、凍結せずに氷上で採取し、次いで可及的速やかに分析した場合、緩衝液だけの溶解は減少し又は全体的に欠如しており(図9)、他の種全ての溶解は、クロストリジウム・ビフェルメンタンス、クロストリジウム・ソルデリ、バシラス・セレウスを注目すべき例外として欠如しており、そして枯草菌及びリステリア・イバノビイイで若干欠如していた(図11、表2)。胃腸管ミクロフローラのATリッチクロストリジウム様成分の代表である更なる菌株について、ΦCD27リシンに対する感受性を試験した。表3に示されるように、試験したいずれもリシンに感受性ではなかった。
【0165】
新鮮細胞の使用は、12分までの顕著な遅延を伴う、C.ディフィシレへの溶解の迅速開始の程度は小さかった(図9)。全てのC.ディフィシレ株を第2NiNTAカラムから単離した3.5μgのリシンを用いて再試験した(第1精製と同等のリシスを示すことを試験した;図10)。新鮮又は凍結細胞を用いたいずれの場合にも、感受性プロファイルは、リシンに対して明らかな感受性示す30株全てで同様であった(表1)。
【0166】
ΦCD27リシンのpHプロファイルはを、感受性株11204を用いて試験した−活性は、pH4.5、5.8、6.5、7.0、7.3(PBSの通常のpH)、7.6及び8.3で試験して、かなりのpH範囲内でほとんど変動を示さなかった。希釈系列は、300μl分析において10.5μgタンパク質での活性が最大であったが、良好な溶解は3.5μg及び0.7μgでも見られた。しかしながら、0.35μgは緩衝液対照をわずかに下回る反応を示し、より低い量では45分分析内で溶解を示さなかった。
【0167】
胃腸管へのΦCD27リシンの送達は、物理的カプセル化又は乳酸菌の1メンバーなどの組み換え片利共生微生物の使用により達成することができた。乳酸連鎖球菌はこの点で可能性を確立しており、それ故この種におけるΦCD27リシンのサブクローニング及び発現が実証された。ΦCD27リシン配列をベクターpUK200Hisへサブクローニングした。これは、NcoIによるpUK200の制限、末端充填、次いでナイシン誘導性プロモーターから下流の6−ヒスチジンタグ(AGT CAT CAC CAT CAC CAT CAC GC)[配列番号7]をコード化するオリゴマーの挿入により構築される、nisA翻訳融合プラスミドpUK200の誘導体である(38)。再循環した場合、これはサブクローニングのためのNcoI部位を再作製した(Hornら、未発表)。ベクターpUK200Hisは、NcoIで制限され、T4DNAポリメラーゼ(Promega)で末端充填されて、nisAプロモーターの制御下翻訳融合のための最初のATGコドンを作製した。phicd27l配列は、pCR2.1(上記)においてサブクローニングされたCD27L−NDE…CD27L−XHO PCR産物から増幅された。プライマーCD27LCOD2_F(5’−AAA ATA TGT ATA ACA GTA GGA CAC)[配列番号8]及びM13前方(5’−GTA AAA CGA CGG CCA GT)[配列番号9]は、第2コドンAAAからの完全配列、及びある種のベクター配列を増幅し、リシンコード配列のすぐ後にEcoRI部位を与えた。増幅は上記のようであったが、アニーリング温度は56℃であった。PCR産物及びNcoI-カットの、末端充填pUK200Hisベクターは、EcoRIにより制限され、そして相互に連結されてnisAプロモーターの制御下にHisタグ化翻訳融合体が作製された。連結反応産物は、配列検証のためにエレクトロコンピテント大腸菌株MC1022に形質転換され、陽性形質転換体をクロラムフェニコール(15μg/ml)で選択した。次いで精製プラスミド調製品をエレクトロコンピテント乳酸連鎖球菌株FI10676に形質転換し、そして5μg/mlクロラムフェニコールを補充したGM17寒天で選択した。
【0168】
pUK200His−phiCD27L又はpUK200His空ベクター制御を発現する乳酸連鎖球菌株は、30℃静置で5μg/mlのクロラムフェニコールにより10mlのGM17ブロス中で増殖した。100μlの一夜培養液を使用して予熱ブロスを植菌した結果、培養液は中間期対数(OD6000.5)まで増殖した。発現は30℃で5時間1ng/mlのナイシンで誘導し、粗タンパク質溶解物は、20mMのTris−HCl、pH8.5、50mMのNaCl中に大腸菌で記載のように産生した。ΦCD27リシンの乳酸菌発現の証明が、タンパク質ゲル解析で示された(図13)。乳酸連鎖球菌に発現したΦCD27リシンに対するクロストリジウム・ディフィシレ株11204の感受性が、図14に示されるように粗タンパク質抽出物を用いて証明された。
【0169】
【表1】
【0170】
【表2】
【0171】
【表3】
【0172】
【表4】
【0173】
【表5】
【0174】
上記に加えて、健常腸生理を維持するために、望ましくは有害でない片利共生生物株を代表する多くの追加菌株は、本発明のリシンとの接触により害されないことが知られている。ΦCD27に対して試験した以下のクロストリジウム種の全て、DSMZからの全てにおいて、全てが溶解をもたらさなかった。これらの菌株は、Eckbergら(2005) Science 308 1635- 及び補充論文、及び Kikuchi ら (2002) Microbiol. Immunol. 46, 353 及び引用文献を参考にして、ヒトの腸に通常見られる主要クロストリジウム菌株群の代表であることを基準に、特異的に選択された。
【0175】
【表6】
【0176】
全てのC.ディフィシレ株を、同じ方法で大腸菌に発現したC.ディフィシレ株630のプロファージ1リシンに対して試験し、その結果CD630L1リシンは溶解をもたらさなかった。
【0177】
細胞生存性
細胞生存性に対するphiCD27の効果を測定するために、反復分析が前還元(pre-reduced)緩衝液及び培地を用いて嫌気性条件下に設定された。細胞はエンド対数で増殖され、嫌気性条件下で遠心分離により採取し、次いでPBS緩衝液中pH7.3で再懸濁した。PBS中でc.1×108細胞からc.1×103細胞まで10倍希釈を行なった;各分析において細胞数の評価が可能なように、それらの10μl分割量を0時点でBHI寒天上にスポットした。分析は2回ずつとし、100μgの部分精製エンドリシン(E1)か又は等量(50μl)の緩衝液(EB)及び300μlの最終量までの細胞を含有した。連続的な穏やかな振盪による2時間のインキュベーション後、PBS中の10倍連続希釈で30μlの試料を採取した;これらの希釈液の10μl分割量をBHI寒天上にスポットし、各2回ずつのペアの1つから残りの270μl分析量を細胞計数可能なように塗布した。
【0178】
0時点でc.1×108細胞を含む分析では、2時間のインキュベーション後1対数の低下を示したが、一方で1×107細胞又は1×106細胞が加えられた分析では、緩衝液対照に比べて2対数の低下を示した。低めの初期細胞数による分析では、リシンはより有効で、1×105細胞を植菌した分析からわずか4生存コロニーが回収されただけで、1×104細胞以下の分析では生存細胞は残っていなかった。
【0179】
次いで上記の生存性分析を、Ni−NTA溶出緩衝液(EB)を20mMのpH6リン酸緩衝液(NP)で置き換えるために、2mlのZeba Desaltスピンカラム(Pierce)を用いて、緩衝液交換にかけていたE1の400μlの分割量を用いて繰り返した。NP緩衝液中のリシンは、クロストリジウム・ディフィシレ11204の凍結細胞に対して元のNiNTAE1と同等の活性を示した(図15)。50μgのE1−NP又はNP緩衝液対照及びc.1×106細胞を用いて、上記の生存性分析を繰り返した;リシンによる2時間インキュベーションは緩衝液対照に比べて3対数の低下をもたらした。
【0180】
上記データはその後公表科学論文原稿(Mayer et al., 2008, J. Bacteriol. 190:6734-6740)の基礎となったが、その開示内容は、参照により本明細書に組み込まれている。
【0181】
ドメイン交換
スプライス重複PCRによるΦCD27Lエンドリシン上への新しい酵素ドメインの設計
リステリア・モノサイトゲネスに対して活性なバクテリオファージΦLM4からのエンドリシンLM−4は、宿主細胞の有効な溶解をもたらすことが示された(英国特許公開公報第2255561(B)号)。エンドリシンは864bpの長さで、pfam2557、VanY、蛋白質の始部のD−アラニル−D−アラニルカルボキシペプチダーゼ及びCOG5632、全配列にわたりN−アセチルムラモイル−L−アラニンアミダーゼ(NCBI Blast)に相同性を示す287アミノ酸のタンパク質を与えた。提案酵素活性ドメイン(EAD)をコード化する配列の前半は、スプライス重複伸長PCRにより、CD27L細胞壁結合ドメイン(CBD、Asn180から最終のArg270)か又は全270アミノ酸酵素の上流に挿入された。LM4酵素ドメインは、LM4のATGのNdeI部位を作製するために、プライマーLM4Nde5’−GGA TGA TTA CAT ATG GCA TTA ACA G[配列番号10]、及び2つのスプライス重複プラマーの1つ:LM4 EAD−CD27L EAD−CBDを与えるために、CD27L酵素の最初の15ヌクレオチドに適合する尾部を有するLM4配列の、ヌクレオチド439〜453、Thr147からAsn151を表わす、LM4−スプライス−CD27LE 5’−TAT ACA TAT TTT CAT GTT TTG TGT CGC AGT[配列番号11];又は、LM4 EAD−CD27L CBDを与えるために、Asn180からArg270のCD27Lの提案C末端結合ドメインに適合する尾部を有するLM4配列のヌクレオチド439〜453、Thr147からAsn151を表わす、LM4−スプライス−CD27L 5’−TTT AAC TCC CTC ATT GTT TTG TGT CGC AGT[配列番号12]、を用いてプラスミドpFI567 (Payne et al., 1996 FEMS Microbiology Letters 136: 19-24)からPCRによって増幅した。同様に、CD27L全配列又はCBDは、ベクター、T7T 5’−GCT AGT TAT TGC TCA GCG G[配列番号13]からのプライマー、及びLM4 EAD配列末端−そこでプライマーの最後の20ntがMet1からのCD27L配列の始めをコード化する、全配列に対する、CD27LEスプライスLM4 5’−ACT GCG ACA CAA AAC ATG AAA ATA TGT ATA ACA GT[配列番号14];及びプライマーの最後の16ntがAsn180からのCD27L配列の提案CBDをコード化する、CBDだけに対するCD27LスプライスLM4 5’− CT GCG ACA CAA AAC AAT GAG GGA GTT AAA C[配列番号15]に適合する尾部を有したスプライシングプライマー、を用いてΦCD27L−pET15bから増幅した。PCRを、元の鋳型に100%適合を与えるスプライシングプライマー部分に適合するように5サイクルのアニーリング温度を用い、次いで20サイクルは全スプライシングプライマーに適合するアニーリング温度にて、メーカー推奨の条件でPhusion (Finnzymes)により実施した。生成物は、SureClean (Bioline) を用いて精製し、50μlの容積に再懸濁した。これらの鋳型を100倍希釈し、そして1μl分割量を、2配列のスプライシングを可能にするアニーリング温度(54℃)で、元の外側プライマー−LM4Nde及びT7T−を用いてPCR反応に使用した。最終生成物をSureCleanで精製し、NdeI及びXhoIで制限し、そしてHis−タグ化LM4−CD27LE及びLM4−CD27Lを生産するために、pET15bにサブクローニングした。次いでこれらのプラスミドは大腸菌に形質転換され、それらの配列が確認された。
【0182】
複合酵素の粗抽出物及びNiNTA精製抽出物の両方を生成して、SDS−PAGE及びウェスタンブロット法で解析し、そして前述のように分析した(図16参照)。His−タグ化LM4−CD27LE及びLM4−CD27Lの両方は、粗抽出物中に高レベルで存在した。C.ディフィシレ11204の凍結細胞をPBS緩衝液中、pH5.8でインキュベートした場合、10μgのNiNTA精製抽出物は、緩衝液対照に比べて迅速溶解をもたらし(図17参照)、LM4−CD27LEは天然のCD27Lと同様の溶解速度を示した。細胞希釈剤としてpH7.3でPBS緩衝液を用いて等価活性が見られた。
【0183】
生存性分析において、LM4−CD27LE及びLM4−CD27Lの両方のNiNTA精製溶出液は生存数の低下をもたらした(図17参照)。50μgNiNTAE1を用いた、c.1×104細胞を含む分析は、緩衝液対照に比べて2時間インキュベーション後に少なくとも1対数の減少を示した。この低下は天然酵素で見られたほどに大きくはなく、他の酵素ドメインの添加は、C.ディフィシレを死滅させる能力を有する活性な新しい酵素を生産することができるという原理を証明した。
【0184】
野生型LM4及びドメインを交換したリシンのヌクレオチド及びアミノ酸配列
【化2】
【0185】
【化3】
【0186】
【化4】
【0187】
参考文献
1.Kuijper, E. J., Coignard, B. & Tull, P. (2006) Clin Microbiol Infect 12 Suppl 6, 2-18.
2.Anonymous (2006) Health Statistics Quarterly 30, 56-60.
3. Rupnik, M., Dupuy, B., Fairweather, N. F., Gerding, D. N., Johnson, S., Just, I., Lyerly, D. M., Popoff, M. R., Rood, J. I., Sonenshein, A. L., Thelestam, M., Wren, B. W., Wilkins, T. D. & von Eichel-Streiber, C. (2005) J Med Microbiol 54, 113-7.
4. Braun, V., Hundsberger, T., Leukel, P., Sauerborn, M. & von Eichel-Streiber, C. (1996) Gene 181, 29-38.
5. Goncalves, C., Decre, D., Barbut, F., Burghoffer, B. & Petit, J. C. (2004) J Clin Microbiol 42, 1933-9.
6. Popoff, M. R., Rubin, E. J., Gill, D. M. & Boquet, P. (1988) Infect Immun
56, 2299-306.
7. Skurnik, M. & Strauch, E. (2006) Int J Med Microbiol 296, 5-14.
8. Projan, S. (2004) Nat Biotechnol 22, 167-8.
9. Brussow, H., Canchaya, C. & Hardt, W. D. (2004) Microbiol Mol Biol Rev 68, 560-602.
10. Davis, B. M. & Waldor, M. K. (2003) Curr Opin Microbiol 6, 35-42.
11. Strauch, E., Schaudinn, C. & Beutin, L. (2004) Infect Immun 72, 7030-9.
12. Chibani-Chennoufi, S., Sidoti, J., Bruttin, A., Kutter, E., Sarker, S. & Brussow, H. (2004) Antimicrob Agents Chemother 48, 2558-69.
13. Matsuzaki, S., Yasuda, M., Nishikawa, H., Kuroda, M., Ujihara, T., Shuin,
T., Shen, Y., Jin, Z., Fujimoto, S., Nasimuzzaman, M. D., Wakiguchi, H., Sugihara, S., Sugiura, T., Koda, S., Muraoka, A. & Imai, S. (2003) J Infect Dis 187, 613-24.
14. Biswas, B., Adhya, S., Washart, P., Paul, B., Trostel, A. N., Powell, B.,
Carlton, R. & Merril, C. R. (2002) Infect Immun 70, 204-10.
15. Loc Carrillo, C., Atterbury, R. J., el-Shibiny, A., Connerton, P. L., Dillon, E., Scott, A. & Connerton, I. F. (2005) Appl Environ Microbiol 71, 6554-63.16. Huff, W. E., Huff, G. R., Rath, N. C., Balog, J. M. & Donoghue, A. M. (2004) Poult Sci 83, 1944-7.
17. Ramesh, V., Fralick, J. A. & Rolfe, R. D. (1999) Anaerobe 5, 69-78.
18. Wray, T. (2007) National Provisioner 5th July
19. Loessner, M. J., Kramer, K., Ebel, F. & Scherer, S. (2002) Mol Microbiol 44, 335-49.
20. Loessner, M. J. (2005) Curr Opin Microbiol 8, 480-7.
21. Gasson (1995-2003) Patents GB 2255561 B (1995); AU 650737B (1994); US 5763251 (1998); US 6083684 (2000); CA 2066387 (2003); EP 0510907B (2003).
22. Fischetti, V. A. (2005) Trends Microbiol 13, 491-6.
23. Schuch, R., Nelson, D. & Fischetti, V. A. (2002) Nature 418, 884-9.
24. Loeffler, J. M., Djurkovic, S. & Fischetti, V. A. (2003) Infect Immun 71,
6199-204.
25. Yoong, P., Schuch, R., Nelson, D. & Fischetti, V. A. (2004) J Bacteriol 186, 4808-12.
26. Zimmer, M., Vukov, N., Scherer, S. & Loessner, M. J. (2002) Appl Environ Microbiol 68, 5311-7.
27. Sell, T. L., Schaberg, D. R. & Fekety, F. R. (1983) J Clin Microbiol 17, 1148-52.
28. Nelson, D. (2004) J Bacteriol 186, 7029-31.
29. Mahony, D. E., Bell, P. D. & Easterbrook, K. B. (1985) J Clin Microbiol 21, 251-4.
30. Goh, S., Riley, T. V. & Chang, B. J. (2005) Appl Environ Microbiol 71, 1079-83.
31. Govind, R., Fralick, J. A. & Rolfe, R. D. (2006) J Bacteriol 188, 2568-77.
32. Goh, S., Ong, P. F., Song, K. P., Riley, T. V. & Chang, B. J. (2007) Microbiology 153, 676-85.
33. Rutherford, K., J. Parkhill, J. Crook, T. Horsnell, P. Rice, M-A. Rajandream and B. Barrell . (2000) Bioinformatics 16, 944-945.
34. Altschul, S. F., Thomas L. Madden, Alejandro A. Schaffer, & Jinghui Zhang, Z. Z., Webb Miller, and David J. Lipman (1997) Nucleic Acids Res. 25, 3389-3402
35. Schaffer, A. A., L. Aravind, Thomas L. Madden, Sergei , Shavirin, J. L. S., Yuri I. Wolf, Eugene V. Koonin, and & Altschul, S. F. (2001) Nucleic Acids Res. 29, 2994-3005.
36. Sebaihia, M., Wren, B. W., Mullany, P., Fairweather, N. F., Minton, N., Stabler, R., Thomson, N. R., Roberts, A. P., Cerdeno-Tarraga, A. M., Wang, H., Holden, M. T., Wright, A., Churcher, C., Quail, M. A., Baker, S., Bason, N., Brooks, K., Chillingworth, T., Cronin, A., Davis, P., Dowd, L., Fraser, A., Feltwell,
T., Hance, Z., Holroyd, S., Jagels, K., Moule, S., Mungall, K., Price, C., Rabbinowitsch, E., Sharp, S., Simmonds, M., Stevens, K., Unwin, L., Whithead, S., Dupuy, B., Dougan, G., Barrell, B. & Parkhill, J. (2006) Nat Genet 38, 779-86.
37. Loessner, M. J., Wendlinger, G. & Scherer, S. (1995) Mol Microbiol 16, 1231-41.
38. Wegmann, U., Klein, J. R., Drumm, I., Kuipers, O. P. & Henrich, B. (1999) Appl Environ Microbiol 65, 4729-33.
【特許請求の範囲】
【請求項1】
クロストリジウム・ディフィシレの細胞に特異的に結合し及び/又はそれを溶解させることができる、配列番号1のアミノ酸配列を含む単離したポリペプチド、又はそのフラグメント、バリアント、誘導体若しくは融合体。
【請求項2】
ポリペプチドが、クロストリジウム・ディフィシレ株630(CD630)のバクテリオファージΦCD119、バクテリオファージΦC2又はプロファージ1若しくは2のリシンではない、請求項1に記載のポリペプチド。
【請求項3】
請求項1又は2に記載のポリペプチドであって、そのフラグメント、バリアント、誘導体又は融合体が、配列番号1のアミノ酸配列と少なくとも60%同一性を示すポリペプチド。
【請求項4】
請求項1〜3のいずれか1項に記載のポリペプチドであって、そのフラグメント、バリアント、誘導体又は融合体が、クロストリジウム・ディフィシレのバクテリオファージの天然リシンではないポリペプチド。
【請求項5】
クロストリジウム・ディフィシレの細胞に特異的に結合することができる、請求項1〜4のいずれか1項に記載の単離したポリペプチド。
【請求項6】
クロストリジウム・ディフィシレの細胞を溶解させることができる、請求項1〜5のいずれか1項に記載の単離したポリペプチド。
【請求項7】
クロストリジウム・ディフィシレの細胞に特異的に結合し及び/又はそれを溶解させることができる、請求項1〜6のいずれか1項に記載の単離したポリペプチド。
【請求項8】
配列番号1のアミノ酸配列を含む、請求項1〜7のいずれか1項に記載の単離したポリペプチド。
【請求項9】
配列番号1のアミノ酸配列から成る、請求項8に記載の単離したポリペプチド。
【請求項10】
配列番号1のアミノ酸配列のフラグメントを含み又はそれから成る、請求項1〜9のいずれか1項に記載の単離したポリペプチド。
【請求項11】
請求項10に記載の単離したポリペプチドであって、フラグメントが、配列番号1の少なくとも50の隣接アミノ酸、例えば、配列番号1の少なくとも60、70、80、90、100、110、120、130、140、150、160、170、180、190、200、210、220、230、240、250、260又は265の隣接アミノ酸を含むポリペプチド。
【請求項12】
フラグメントが配列番号1の酵素(細胞溶解)ドメインを含み又はそれから成る、請求項10又は11に記載の単離したポリペプチド。
【請求項13】
フラグメントが配列番号1の細胞壁結合ドメインを含み又はそれから成る、請求項10〜12のいずれか1項に記載の単離したポリペプチド。
【請求項14】
配列番号1のアミノ酸配列のバリアント、又はそのフラグメントを含み又はそれから成る、請求項1〜13のいずれか1項に記載の単離したポリペプチド。
【請求項15】
請求項14に記載の単離したポリペプチドであって、バリアントが、配列番号1のアミノ酸配列、又はそのフラグメントに対して少なくとも60%同一性、より好ましくは該配列に対して少なくとも70%又は80%又は85%又は90%同一性、そして最も好ましくは該アミノ酸配列に対して少なくとも95%、96%、97%、98%又は99%同一性を有するアミノ酸配列を含み又はそれから成るポリペプチド。
【請求項16】
配列番号1のアミノ酸配列の誘導体、又はそのフラグメント若しくはバリアントを含み又はそれから成る、請求項1〜15のいずれか1項に記載の単離したポリペプチド。
【請求項17】
配列番号1のアミノ酸配列の融合体、又はそのフラグメント、バリアント若しくは誘導体を含み又はそれから成る、請求項1〜16のいずれか1項に記載の単離したポリペプチド。
【請求項18】
配列番号1のアミノ酸配列、又はそのフラグメント、バリアント若しくは誘導体のN−及び/又はC−末端に挿入された1つ又はそれ以上の更なるアミノ酸を含み又はそれから成る、請求項17に記載の単離したポリペプチド。
【請求項19】
配列番号1の細胞壁結合ドメイン及び配列番号1中のものと異なる酵素(細胞溶解)ドメインを含み又はそれから成る、請求項17又は18に記載の単離したポリペプチド。
【請求項20】
ポリペプチドが、クロストリジウム・ディフィシレの複数株の細胞を溶解させることができる、請求項1〜19のいずれか1項に記載の単離したポリペプチド。
【請求項21】
ポリペプチドが、バシラス種(例えば、バシラス・セレウス、枯草菌及び炭疽菌)、他のクロストリジウム種(例えば、クロストリジウム・ソルデリ及びクロストリジウム・ビフェルメンタンス)及びリステリア種(例えば、リステリア・イバノビイ)の細胞から成る群から選択される1つ又はそれ以上の細胞型を溶解させることができる、請求項1〜20のいずれか1項に記載の単離したポリペプチド。
【請求項22】
ポリペプチドが、クロストリジウム・レプツム、 クロストリジウム・ネキシル、クロストリジウム・コッコイデス、クロストリジウム・イノクーム、クロストリジウム・ラモスム及び/又はアネロコッカス・ヒドロジェナリスの細胞を溶解しない、請求項1〜21のいずれか1項に記載の単離したポリペプチド。
【請求項23】
ポリペプチドがクロストリジウム・ディフィシレリボタイプ027の細胞を溶解することができる、請求項1〜22のいずれか1項に記載の単離したポリペプチド。
【請求項24】
ポリペプチドが、クロストリジウム・ディフィシレリボタイプ027の細胞に対して配列番号1のポリペプチドの少なくとも10%、例えば、少なくとも20%、30%、40%、50%、60%、70%、80%、90%、100%又はそれ以上の溶解活性を示す、請求項23に記載の単離したポリペプチド。
【請求項25】
ポリペプチドが、クロストリジウム・ディフィシレリボタイプ027の細胞に対して配列番号1のポリペプチドの少なくとも100%、例えば、少なくとも、120%、130%、140%、150%、160%、170%、180%、190%、200%、250%、300%、500%又はそれ以上の溶解活性を示す、請求項24に記載の単離したポリペプチド。
【請求項26】
ポリペプチドが病原菌の細胞を選択的に溶解することができる、請求項1〜25のいずれか1項に記載の単離したポリペプチド。
【請求項27】
ポリペプチドが組み換えポリペプチドである、請求項1〜26のいずれか1項に記載の単離したポリペプチド。
【請求項28】
請求項1〜27のいずれか1項に記載のポリペプチドをコード化する単離した核酸分子。
【請求項29】
核酸分子が配列番号2の核酸配列を含み又はそれから成る、請求項28に記載の核酸分子。
【請求項30】
請求項28又は29に記載の核酸分子を含むベクター。
【請求項31】
ベクターが発現ベクターである、請求項30に記載のベクター。
【請求項32】
ベクターがpET15b及びpACYC184から成る群から選択される、請求項30又は31に記載のベクター。
【請求項33】
請求項28又は29に記載の核酸分子又は請求項30又は31に記載のベクターを含む宿主細胞。
【請求項34】
宿主細胞が請求項1〜27のいずれか1項に記載のポリペプチドを発現することができる、請求項33に記載の宿主細胞。
【請求項35】
宿主細胞が微生物細胞である、請求項33又は34に記載の宿主細胞。
【請求項36】
宿主細胞が細菌細胞である、請求項33〜35のいずれか1項に記載の宿主細胞。
【請求項37】
宿主細胞が非病原性である、請求項35又は36に記載の宿主細胞。
【請求項38】
宿主細胞が、大腸菌、ラクトコッカス種、バクテロイデス種、ラクトバシラス種、エンテロコッカス種及びバシラス種の細胞から成る群から選択される、請求項33〜37のいずれか1項に記載の宿主細胞。
【請求項39】
宿主細胞が乳酸連鎖球菌細胞である、請求項38に記載の宿主細胞。
【請求項40】
ポリペプチドを発現する条件下で請求項28又は29に記載の核酸分子、又は請求項30〜32のいずれか1項に記載のベクターを含む宿主細胞集団を培養し、そこからポリペプチドを単離することを含む、請求項1〜27のいずれか1項に記載のポリペプチドを生産する方法。
【請求項41】
以下のもの:
(a)請求項1〜27のいずれか1項に記載のポリペプチド;
(b)請求項28又は29に記載の核酸分子;
(c)請求項30〜32のいずれか1項に記載のベクター;
(d)請求項33〜39のいずれか1項に記載の宿主;及び/又は
(e)本発明の第1の態様に記載のポリペプチドを発現することができるバクテリオファージ;
及び薬学的に許容される担体、賦形剤又は医薬品添加剤を含む医薬組成物。
【請求項42】
請求項1〜27のいずれか1項に記載のポリペプチドを含む、請求項41に記載の医薬組成物。
【請求項43】
経口投与のための、請求項41又は42に記載の医薬組成物。
【請求項44】
ポリペプチドがマイクロカプセル化された、請求項41〜43のいずれか1項に記載の医薬組成物。
【請求項45】
ポリペプチドを胃腸管に送達することができる、請求項41〜44のいずれか1項に記載の医薬組成物。
【請求項46】
請求項28又は29に記載の核酸分子及び/又は請求項30〜32のいずれか1項に記載のベクターを含む、請求項41〜45のいずれか1項に記載の医薬組成物。
【請求項47】
請求項33〜39のいずれか1項に記載の宿主細胞を含む、請求項41〜46のいずれか1項に記載の医薬組成物。
【請求項48】
請求項1〜27のいずれか1項に記載のポリペプチドを発現するように、そして胃腸管内の所定の場所に到達時に該ポリペプチドを放出するように、遺伝子組み換えした病原性細菌宿主細胞を含む、請求項47に記載の医薬組成物。
【請求項49】
請求項1〜27のいずれか1項に記載のポリペプチドを発現することができるバクテリオファージを含む、請求項41〜48のいずれか1項に記載の医薬組成物。
【請求項50】
組成物が胃腸管内でポリペプチドの持続性又は徐放性放出を可能にする、請求項41〜49のいずれか1項に記載の医薬組成物。
【請求項51】
医薬に使用するための、請求項1〜27のいずれか1項に記載のポリペプチド又は請求項41〜50のいずれか1項に記載の医薬組成物。
【請求項52】
患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するための薬剤の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞が、クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項53】
患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制に使用するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、ポリペプチド又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージ。
【請求項54】
患者中の微生物細胞に関連する疾患又は病態の治療又は予防のための薬剤製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項55】
患者中の微生物細胞に関連する疾患又は病態の治療又は予防に使用するのための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、ポリペプチド又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージ。
【請求項56】
請求項52〜55のいずれか1項に記載の使用であって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン;
から成る群から選択される使用。
【請求項57】
請求項52〜56のいずれか1項に記載の使用であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞が、クロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項58】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項52〜57のいずれか1項に記載の使用。
【請求項59】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項58に記載の使用。
【請求項60】
クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを患者に投与することを含む、患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制する方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項61】
クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを患者に投与することを含む、患者中の微生物細胞に関連する疾患又は病態を治療又は予防する方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項62】
請求項60又は61に記載の方法であって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン;
から成る群から選択される方法。
【請求項63】
請求項60〜62のいずれか1項に記載の方法であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項64】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項60〜63のいずれか1項に記載の方法。
【請求項65】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項64に記載の方法。
【請求項66】
インビトロ及び/又はエクスビボで微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項67】
請求項66に記載の使用であって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630のプロファージ1及び2のリシン;
から成る群から選択される使用。
【請求項68】
請求項66又は67に記載の使用であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項69】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項66〜68のいずれか1項に記載の使用。
【請求項70】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項69に記載の使用。
【請求項71】
クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを含む、試料中の微生物細胞の存在を検出するためのキットであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択されるキット。
【請求項72】
請求項71に記載のキットであって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630のプロファージ1及び2のリシン;
から成る群から選択されるキット。
【請求項73】
請求項71又は72に記載のキットであって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択されるキット。
【請求項74】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項71〜73のいずれか1項に記載のキット。
【請求項75】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項74に記載のキット。
【請求項76】
ポリペプチドが表面に固定化されている、請求項71〜75のいずれか1項に記載のキット。
【請求項77】
試料が細胞試料である、請求項71〜76のいずれか1項に記載のキット。
【請求項78】
試料が微生物細胞による汚染の試験をするために表面から採取された拭い液由来である、請求項71〜77のいずれか1項に記載のキット。
【請求項79】
陰性対照試料を更に含む、請求項71〜78のいずれか1項に記載のキット。
【請求項80】
陽性対照試料を更に含む、請求項71〜79のいずれか1項に記載のキット。
【請求項81】
クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、微生物細胞に関連する疾患又は病態の診断薬の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用。
【請求項82】
クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される微生物細胞に関連する疾患又は病態診断に使用するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージ。
【請求項83】
インビトロ及び/又はエクスビボで試料中の微生物細胞の存在を検出するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項84】
請求項81〜83のいずれか1項に記載の使用であって、細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン;
から成る群から選択される使用。
【請求項85】
請求項81〜84のいずれか1項に記載の使用であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチドが請求項1〜24のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項86】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項81〜85のいずれか1項に記載の使用。
【請求項87】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項86に記載の使用。
【請求項88】
クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを用いて試験するために患者の細胞試料を接触させ、そして試料中の細胞がそれにより溶解されるかどうかを測定することを含む、患者中の微生物細胞に関連する疾患又は病態を診断する方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項89】
請求項88に記載の方法であって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630のプロファージ1及び2のリシン;
から成る群から選択される方法。
【請求項90】
請求項88又は89に記載の方法であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項91】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項88〜90のいずれか1項に記載の方法。
【請求項92】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項91に記載の方法。
【請求項93】
実質的に実施例に関連して本明細書に記載した通りの、単離したポリペプチド。
【請求項94】
実質的に実施例に関連した本明細書に記載した通りの単離した核酸分子。
【請求項95】
実質的に実施例に関連して本明細書に記載した通りのベクター。
【請求項96】
実質的に実施例に関連して本明細書に記載した通りの宿主細胞。
【請求項97】
実質的に実施例に関連して本明細書に記載した通りの、請求項1〜27のいずれか1項に記載したポリペプチドを製造する方法。
【請求項98】
実質的に実施例に関連して本明細書に記載した通りの、医薬組成物。
【請求項99】
実質的に実施例に関連して本明細書に記載した通りの、微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するためのポリペプチドの使用。
【請求項100】
実質的に実施例に関連して本明細書に記載した通りの、微生物細胞に関連する疾患又は病態を治療又は予防するための薬剤製造におけるポリペプチドの使用。
【請求項101】
実質的に実施例に関連して本明細書に記載した通りの、試料中の微生物細胞の存在を検出するためのキット。
【請求項102】
実質的に実施例に関連して本明細書に記載した通りの、微生物細胞に関連する疾患又は病態の診断薬の製造におけるポリペプチドの使用。
【請求項103】
実質的に実施例に関連して本明細書に記載した通りの、試料中の微生物細胞の存在を検出するためのポリペプチドの使用。
【請求項1】
クロストリジウム・ディフィシレの細胞に特異的に結合し及び/又はそれを溶解させることができる、配列番号1のアミノ酸配列を含む単離したポリペプチド、又はそのフラグメント、バリアント、誘導体若しくは融合体。
【請求項2】
ポリペプチドが、クロストリジウム・ディフィシレ株630(CD630)のバクテリオファージΦCD119、バクテリオファージΦC2又はプロファージ1若しくは2のリシンではない、請求項1に記載のポリペプチド。
【請求項3】
請求項1又は2に記載のポリペプチドであって、そのフラグメント、バリアント、誘導体又は融合体が、配列番号1のアミノ酸配列と少なくとも60%同一性を示すポリペプチド。
【請求項4】
請求項1〜3のいずれか1項に記載のポリペプチドであって、そのフラグメント、バリアント、誘導体又は融合体が、クロストリジウム・ディフィシレのバクテリオファージの天然リシンではないポリペプチド。
【請求項5】
クロストリジウム・ディフィシレの細胞に特異的に結合することができる、請求項1〜4のいずれか1項に記載の単離したポリペプチド。
【請求項6】
クロストリジウム・ディフィシレの細胞を溶解させることができる、請求項1〜5のいずれか1項に記載の単離したポリペプチド。
【請求項7】
クロストリジウム・ディフィシレの細胞に特異的に結合し及び/又はそれを溶解させることができる、請求項1〜6のいずれか1項に記載の単離したポリペプチド。
【請求項8】
配列番号1のアミノ酸配列を含む、請求項1〜7のいずれか1項に記載の単離したポリペプチド。
【請求項9】
配列番号1のアミノ酸配列から成る、請求項8に記載の単離したポリペプチド。
【請求項10】
配列番号1のアミノ酸配列のフラグメントを含み又はそれから成る、請求項1〜9のいずれか1項に記載の単離したポリペプチド。
【請求項11】
請求項10に記載の単離したポリペプチドであって、フラグメントが、配列番号1の少なくとも50の隣接アミノ酸、例えば、配列番号1の少なくとも60、70、80、90、100、110、120、130、140、150、160、170、180、190、200、210、220、230、240、250、260又は265の隣接アミノ酸を含むポリペプチド。
【請求項12】
フラグメントが配列番号1の酵素(細胞溶解)ドメインを含み又はそれから成る、請求項10又は11に記載の単離したポリペプチド。
【請求項13】
フラグメントが配列番号1の細胞壁結合ドメインを含み又はそれから成る、請求項10〜12のいずれか1項に記載の単離したポリペプチド。
【請求項14】
配列番号1のアミノ酸配列のバリアント、又はそのフラグメントを含み又はそれから成る、請求項1〜13のいずれか1項に記載の単離したポリペプチド。
【請求項15】
請求項14に記載の単離したポリペプチドであって、バリアントが、配列番号1のアミノ酸配列、又はそのフラグメントに対して少なくとも60%同一性、より好ましくは該配列に対して少なくとも70%又は80%又は85%又は90%同一性、そして最も好ましくは該アミノ酸配列に対して少なくとも95%、96%、97%、98%又は99%同一性を有するアミノ酸配列を含み又はそれから成るポリペプチド。
【請求項16】
配列番号1のアミノ酸配列の誘導体、又はそのフラグメント若しくはバリアントを含み又はそれから成る、請求項1〜15のいずれか1項に記載の単離したポリペプチド。
【請求項17】
配列番号1のアミノ酸配列の融合体、又はそのフラグメント、バリアント若しくは誘導体を含み又はそれから成る、請求項1〜16のいずれか1項に記載の単離したポリペプチド。
【請求項18】
配列番号1のアミノ酸配列、又はそのフラグメント、バリアント若しくは誘導体のN−及び/又はC−末端に挿入された1つ又はそれ以上の更なるアミノ酸を含み又はそれから成る、請求項17に記載の単離したポリペプチド。
【請求項19】
配列番号1の細胞壁結合ドメイン及び配列番号1中のものと異なる酵素(細胞溶解)ドメインを含み又はそれから成る、請求項17又は18に記載の単離したポリペプチド。
【請求項20】
ポリペプチドが、クロストリジウム・ディフィシレの複数株の細胞を溶解させることができる、請求項1〜19のいずれか1項に記載の単離したポリペプチド。
【請求項21】
ポリペプチドが、バシラス種(例えば、バシラス・セレウス、枯草菌及び炭疽菌)、他のクロストリジウム種(例えば、クロストリジウム・ソルデリ及びクロストリジウム・ビフェルメンタンス)及びリステリア種(例えば、リステリア・イバノビイ)の細胞から成る群から選択される1つ又はそれ以上の細胞型を溶解させることができる、請求項1〜20のいずれか1項に記載の単離したポリペプチド。
【請求項22】
ポリペプチドが、クロストリジウム・レプツム、 クロストリジウム・ネキシル、クロストリジウム・コッコイデス、クロストリジウム・イノクーム、クロストリジウム・ラモスム及び/又はアネロコッカス・ヒドロジェナリスの細胞を溶解しない、請求項1〜21のいずれか1項に記載の単離したポリペプチド。
【請求項23】
ポリペプチドがクロストリジウム・ディフィシレリボタイプ027の細胞を溶解することができる、請求項1〜22のいずれか1項に記載の単離したポリペプチド。
【請求項24】
ポリペプチドが、クロストリジウム・ディフィシレリボタイプ027の細胞に対して配列番号1のポリペプチドの少なくとも10%、例えば、少なくとも20%、30%、40%、50%、60%、70%、80%、90%、100%又はそれ以上の溶解活性を示す、請求項23に記載の単離したポリペプチド。
【請求項25】
ポリペプチドが、クロストリジウム・ディフィシレリボタイプ027の細胞に対して配列番号1のポリペプチドの少なくとも100%、例えば、少なくとも、120%、130%、140%、150%、160%、170%、180%、190%、200%、250%、300%、500%又はそれ以上の溶解活性を示す、請求項24に記載の単離したポリペプチド。
【請求項26】
ポリペプチドが病原菌の細胞を選択的に溶解することができる、請求項1〜25のいずれか1項に記載の単離したポリペプチド。
【請求項27】
ポリペプチドが組み換えポリペプチドである、請求項1〜26のいずれか1項に記載の単離したポリペプチド。
【請求項28】
請求項1〜27のいずれか1項に記載のポリペプチドをコード化する単離した核酸分子。
【請求項29】
核酸分子が配列番号2の核酸配列を含み又はそれから成る、請求項28に記載の核酸分子。
【請求項30】
請求項28又は29に記載の核酸分子を含むベクター。
【請求項31】
ベクターが発現ベクターである、請求項30に記載のベクター。
【請求項32】
ベクターがpET15b及びpACYC184から成る群から選択される、請求項30又は31に記載のベクター。
【請求項33】
請求項28又は29に記載の核酸分子又は請求項30又は31に記載のベクターを含む宿主細胞。
【請求項34】
宿主細胞が請求項1〜27のいずれか1項に記載のポリペプチドを発現することができる、請求項33に記載の宿主細胞。
【請求項35】
宿主細胞が微生物細胞である、請求項33又は34に記載の宿主細胞。
【請求項36】
宿主細胞が細菌細胞である、請求項33〜35のいずれか1項に記載の宿主細胞。
【請求項37】
宿主細胞が非病原性である、請求項35又は36に記載の宿主細胞。
【請求項38】
宿主細胞が、大腸菌、ラクトコッカス種、バクテロイデス種、ラクトバシラス種、エンテロコッカス種及びバシラス種の細胞から成る群から選択される、請求項33〜37のいずれか1項に記載の宿主細胞。
【請求項39】
宿主細胞が乳酸連鎖球菌細胞である、請求項38に記載の宿主細胞。
【請求項40】
ポリペプチドを発現する条件下で請求項28又は29に記載の核酸分子、又は請求項30〜32のいずれか1項に記載のベクターを含む宿主細胞集団を培養し、そこからポリペプチドを単離することを含む、請求項1〜27のいずれか1項に記載のポリペプチドを生産する方法。
【請求項41】
以下のもの:
(a)請求項1〜27のいずれか1項に記載のポリペプチド;
(b)請求項28又は29に記載の核酸分子;
(c)請求項30〜32のいずれか1項に記載のベクター;
(d)請求項33〜39のいずれか1項に記載の宿主;及び/又は
(e)本発明の第1の態様に記載のポリペプチドを発現することができるバクテリオファージ;
及び薬学的に許容される担体、賦形剤又は医薬品添加剤を含む医薬組成物。
【請求項42】
請求項1〜27のいずれか1項に記載のポリペプチドを含む、請求項41に記載の医薬組成物。
【請求項43】
経口投与のための、請求項41又は42に記載の医薬組成物。
【請求項44】
ポリペプチドがマイクロカプセル化された、請求項41〜43のいずれか1項に記載の医薬組成物。
【請求項45】
ポリペプチドを胃腸管に送達することができる、請求項41〜44のいずれか1項に記載の医薬組成物。
【請求項46】
請求項28又は29に記載の核酸分子及び/又は請求項30〜32のいずれか1項に記載のベクターを含む、請求項41〜45のいずれか1項に記載の医薬組成物。
【請求項47】
請求項33〜39のいずれか1項に記載の宿主細胞を含む、請求項41〜46のいずれか1項に記載の医薬組成物。
【請求項48】
請求項1〜27のいずれか1項に記載のポリペプチドを発現するように、そして胃腸管内の所定の場所に到達時に該ポリペプチドを放出するように、遺伝子組み換えした病原性細菌宿主細胞を含む、請求項47に記載の医薬組成物。
【請求項49】
請求項1〜27のいずれか1項に記載のポリペプチドを発現することができるバクテリオファージを含む、請求項41〜48のいずれか1項に記載の医薬組成物。
【請求項50】
組成物が胃腸管内でポリペプチドの持続性又は徐放性放出を可能にする、請求項41〜49のいずれか1項に記載の医薬組成物。
【請求項51】
医薬に使用するための、請求項1〜27のいずれか1項に記載のポリペプチド又は請求項41〜50のいずれか1項に記載の医薬組成物。
【請求項52】
患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するための薬剤の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞が、クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項53】
患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制に使用するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、ポリペプチド又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージ。
【請求項54】
患者中の微生物細胞に関連する疾患又は病態の治療又は予防のための薬剤製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項55】
患者中の微生物細胞に関連する疾患又は病態の治療又は予防に使用するのための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、ポリペプチド又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージ。
【請求項56】
請求項52〜55のいずれか1項に記載の使用であって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン;
から成る群から選択される使用。
【請求項57】
請求項52〜56のいずれか1項に記載の使用であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞が、クロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項58】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項52〜57のいずれか1項に記載の使用。
【請求項59】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項58に記載の使用。
【請求項60】
クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを患者に投与することを含む、患者中の微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制する方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項61】
クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを患者に投与することを含む、患者中の微生物細胞に関連する疾患又は病態を治療又は予防する方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項62】
請求項60又は61に記載の方法であって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン;
から成る群から選択される方法。
【請求項63】
請求項60〜62のいずれか1項に記載の方法であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項64】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項60〜63のいずれか1項に記載の方法。
【請求項65】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項64に記載の方法。
【請求項66】
インビトロ及び/又はエクスビボで微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項67】
請求項66に記載の使用であって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630のプロファージ1及び2のリシン;
から成る群から選択される使用。
【請求項68】
請求項66又は67に記載の使用であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項69】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項66〜68のいずれか1項に記載の使用。
【請求項70】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項69に記載の使用。
【請求項71】
クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを含む、試料中の微生物細胞の存在を検出するためのキットであって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択されるキット。
【請求項72】
請求項71に記載のキットであって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630のプロファージ1及び2のリシン;
から成る群から選択されるキット。
【請求項73】
請求項71又は72に記載のキットであって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択されるキット。
【請求項74】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項71〜73のいずれか1項に記載のキット。
【請求項75】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項74に記載のキット。
【請求項76】
ポリペプチドが表面に固定化されている、請求項71〜75のいずれか1項に記載のキット。
【請求項77】
試料が細胞試料である、請求項71〜76のいずれか1項に記載のキット。
【請求項78】
試料が微生物細胞による汚染の試験をするために表面から採取された拭い液由来である、請求項71〜77のいずれか1項に記載のキット。
【請求項79】
陰性対照試料を更に含む、請求項71〜78のいずれか1項に記載のキット。
【請求項80】
陽性対照試料を更に含む、請求項71〜79のいずれか1項に記載のキット。
【請求項81】
クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される、微生物細胞に関連する疾患又は病態の診断薬の製造における、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用。
【請求項82】
クロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される微生物細胞に関連する疾患又は病態診断に使用するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージ。
【請求項83】
インビトロ及び/又はエクスビボで試料中の微生物細胞の存在を検出するための、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージの使用であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項84】
請求項81〜83のいずれか1項に記載の使用であって、細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630(CD630)のプロファージ1及び2のリシン;
から成る群から選択される使用。
【請求項85】
請求項81〜84のいずれか1項に記載の使用であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞壁結合活性及び/又は細胞溶解活性を有するポリペプチドが請求項1〜24のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される使用。
【請求項86】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項81〜85のいずれか1項に記載の使用。
【請求項87】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項86に記載の使用。
【請求項88】
クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチド、又はそれを発現することができる核酸分子、ベクター、宿主細胞若しくはバクテリオファージを用いて試験するために患者の細胞試料を接触させ、そして試料中の細胞がそれにより溶解されるかどうかを測定することを含む、患者中の微生物細胞に関連する疾患又は病態を診断する方法であって、微生物細胞がクロストリジウム・ディフィシレ細胞及び該エンドリシンによる溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項89】
請求項88に記載の方法であって、細胞溶解活性を有するポリペプチドが以下のもの:
(a)バクテリオファージΦCD27のリシン;
(b)バクテリオファージΦCD119のリシン;
(c)バクテリオファージΦC2のリシン;及び
(d)クロストリジウム・ディフィシレ株630のプロファージ1及び2のリシン;
から成る群から選択される方法。
【請求項90】
請求項88又は89に記載の方法であって、クロストリジウム・ディフィシレのバクテリオファージ由来エンドリシンの細胞溶解活性を有するポリペプチドが請求項1〜27のいずれか1項に記載のポリペプチドであり、そして微生物細胞がクロストリジウム・ディフィシレ細胞及び配列番号1のポリペプチドとの接触時に溶解を起こし易い他の細菌細胞から成る群から選択される方法。
【請求項91】
微生物細胞がクロストリジウム・ディフィシレ細胞である、請求項88〜90のいずれか1項に記載の方法。
【請求項92】
微生物細胞がクロストリジウム・ディフィシレ・リボタイプ027細胞である、請求項91に記載の方法。
【請求項93】
実質的に実施例に関連して本明細書に記載した通りの、単離したポリペプチド。
【請求項94】
実質的に実施例に関連した本明細書に記載した通りの単離した核酸分子。
【請求項95】
実質的に実施例に関連して本明細書に記載した通りのベクター。
【請求項96】
実質的に実施例に関連して本明細書に記載した通りの宿主細胞。
【請求項97】
実質的に実施例に関連して本明細書に記載した通りの、請求項1〜27のいずれか1項に記載したポリペプチドを製造する方法。
【請求項98】
実質的に実施例に関連して本明細書に記載した通りの、医薬組成物。
【請求項99】
実質的に実施例に関連して本明細書に記載した通りの、微生物細胞を殺細胞し及び/又はその増殖を阻害/抑制するためのポリペプチドの使用。
【請求項100】
実質的に実施例に関連して本明細書に記載した通りの、微生物細胞に関連する疾患又は病態を治療又は予防するための薬剤製造におけるポリペプチドの使用。
【請求項101】
実質的に実施例に関連して本明細書に記載した通りの、試料中の微生物細胞の存在を検出するためのキット。
【請求項102】
実質的に実施例に関連して本明細書に記載した通りの、微生物細胞に関連する疾患又は病態の診断薬の製造におけるポリペプチドの使用。
【請求項103】
実質的に実施例に関連して本明細書に記載した通りの、試料中の微生物細胞の存在を検出するためのポリペプチドの使用。
【図1】

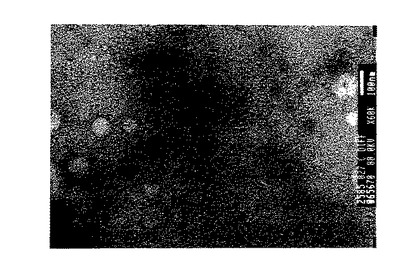
【図2】
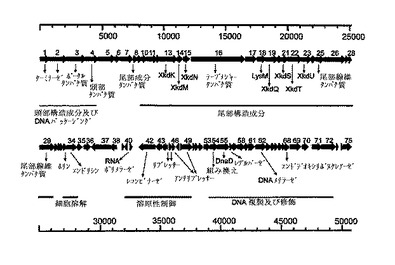
【図3】

【図4a−1】
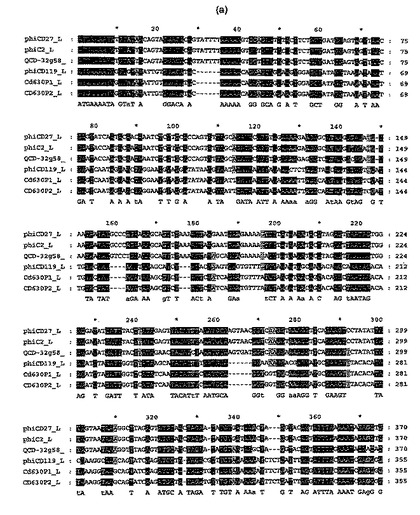
【図4a−2】
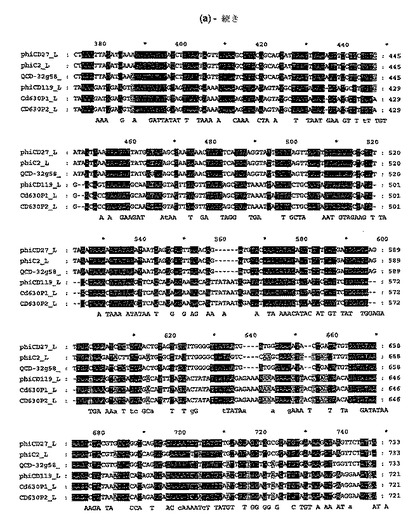
【図4a−3】

【図4b】

【図5】

【図6a】
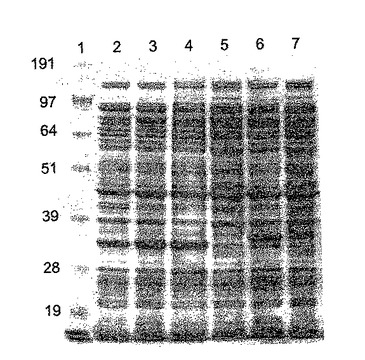
【図6b】
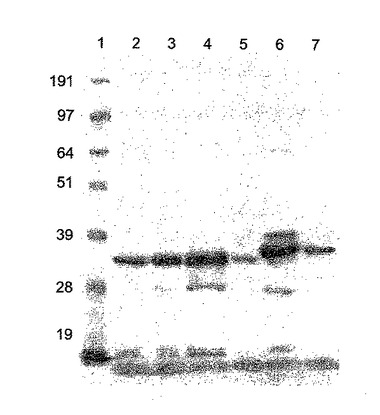
【図7】
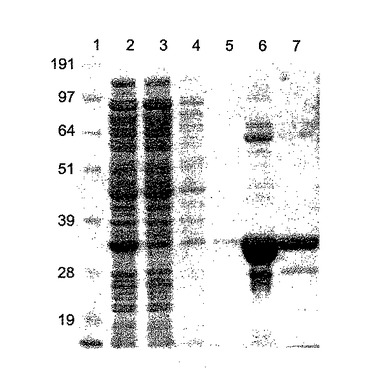
【図8】
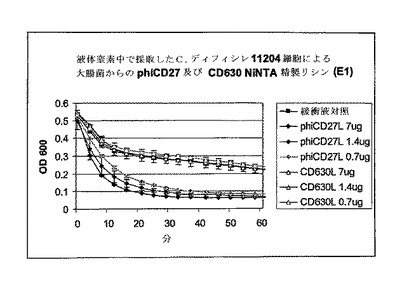
【図9】
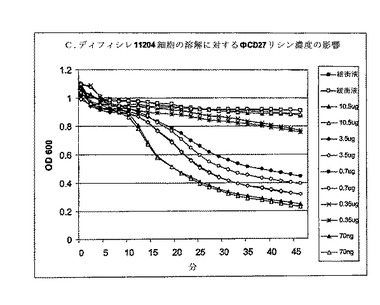
【図10a】
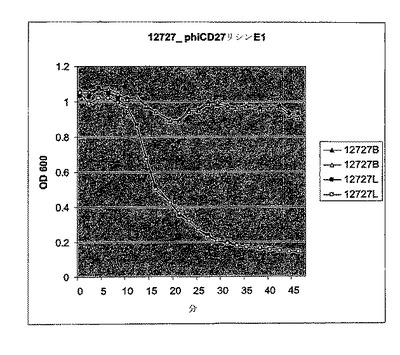
【図10b】
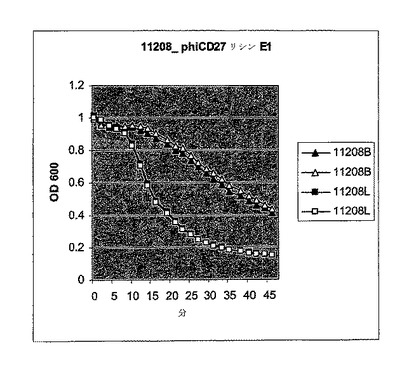
【図10c】
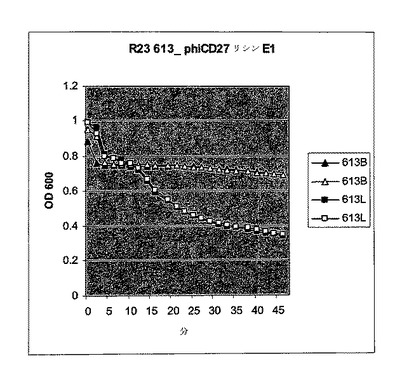
【図11a−1】
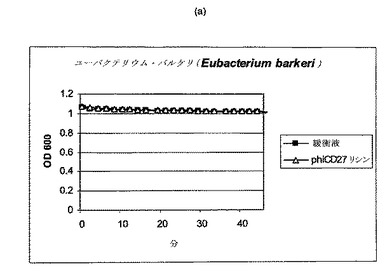
【図11a−2】
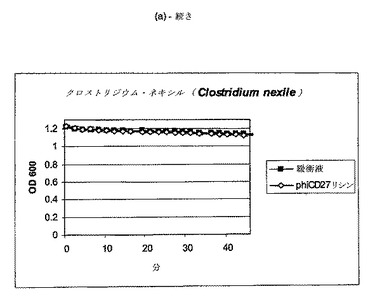
【図11a−3】
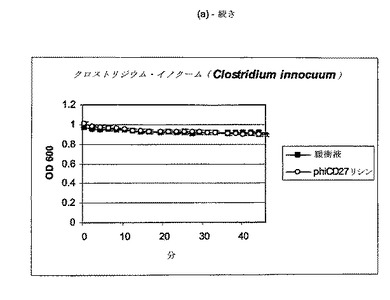
【図11a−4】
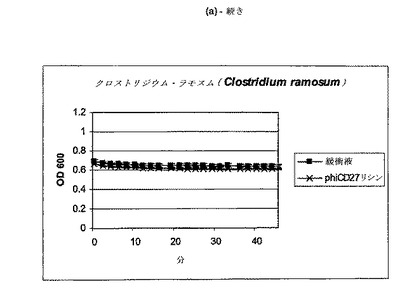
【図11b−1】
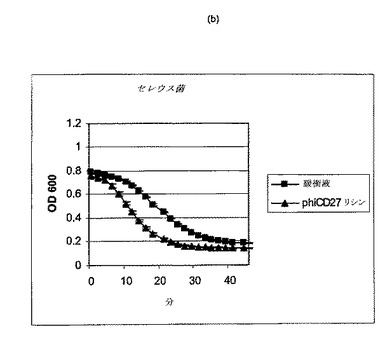
【図11b−2】
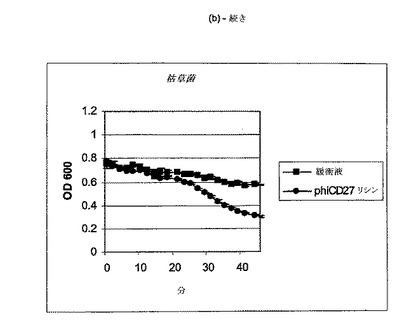
【図11b−3】
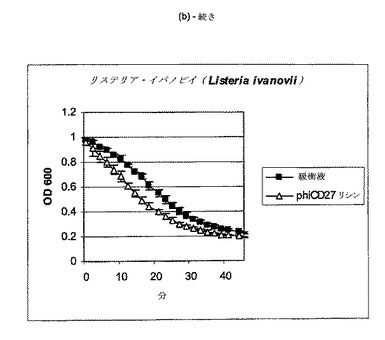
【図11b−4】
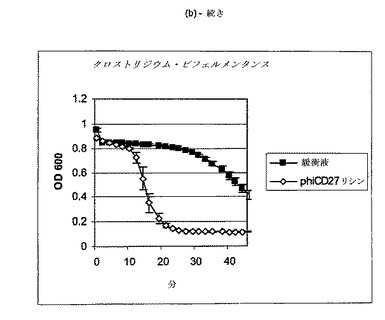
【図12】
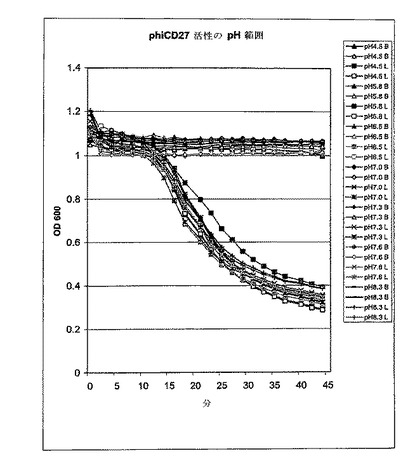
【図13A】
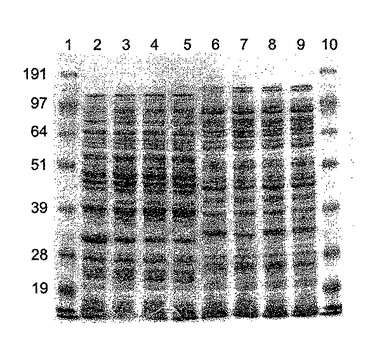
【図13B】
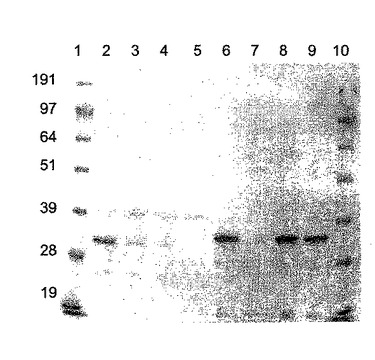
【図14】
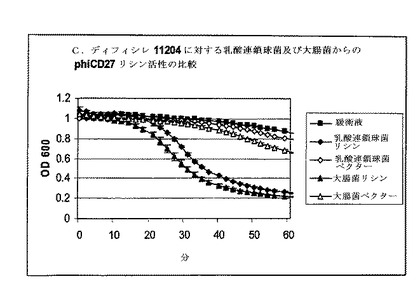
【図15】
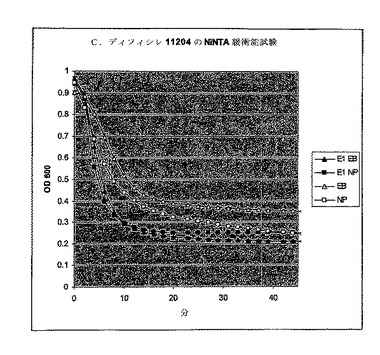
【図16】
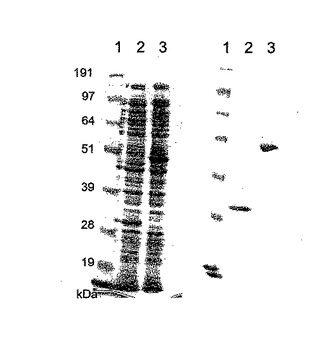
【図17】
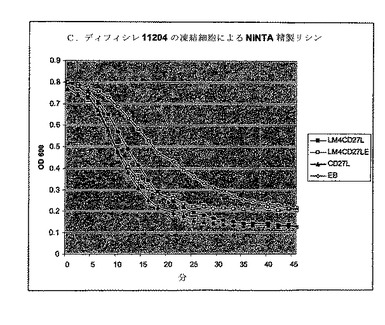
【図2】
【図3】
【図4a−1】
【図4a−2】
【図4a−3】
【図4b】
【図5】
【図6a】
【図6b】
【図7】
【図8】
【図9】
【図10a】
【図10b】
【図10c】
【図11a−1】
【図11a−2】
【図11a−3】
【図11a−4】
【図11b−1】
【図11b−2】
【図11b−3】
【図11b−4】
【図12】
【図13A】
【図13B】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2011−504366(P2011−504366A)
【公表日】平成23年2月10日(2011.2.10)
【国際特許分類】
【出願番号】特願2010−534544(P2010−534544)
【出願日】平成20年11月24日(2008.11.24)
【国際出願番号】PCT/GB2008/003923
【国際公開番号】WO2009/068858
【国際公開日】平成21年6月4日(2009.6.4)
【出願人】(500060766)プラント バイオサイエンス リミティド (1)
【Fターム(参考)】
【公表日】平成23年2月10日(2011.2.10)
【国際特許分類】
【出願日】平成20年11月24日(2008.11.24)
【国際出願番号】PCT/GB2008/003923
【国際公開番号】WO2009/068858
【国際公開日】平成21年6月4日(2009.6.4)
【出願人】(500060766)プラント バイオサイエンス リミティド (1)
【Fターム(参考)】
[ Back to top ]