
エンバク中のアベナンスラミド濃度を増加させる方法
偽製麦を通してエンバク中のアベナンスラミド濃度を増加させる方法を開示する。最初に、エンバクを二次休眠の誘発又は増強に供し、次いで、高温で5日間まで製麦する。次いで、製麦したが発芽していないエンバクを乾燥させ、このままで使用する、或いは食品、飼料、栄養補助食品若しくはパーソナルケア製品及び成分を製造するためにさらに加工又は製粉する。非休眠エンバクを休眠状態及び偽製麦に適した状態にするための方法も提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アベナンスラミド、及びより具体的にはエンバク中のアベナンスラミド濃度を増加させる方法に関する。
【背景技術】
【0002】
エンバクは、アベナンスラミド、アベナコシラート(avenacosylate)、トリテルペンサポニン(アベナコシド)及びステロイドサポニン(アベナシン)を含むいくつかの群の生理活性ファイトケミカルを合成し貯蔵する能力において、穀物の中で独特である。
【0003】
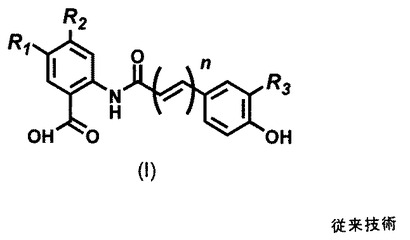
アベナンスラミドは、図1に示す一般式の、約30種の異なるN−アロイルアントラニル酸アルカロイドの群である。
【0004】
アベナンスラミドは、抗酸化物質であり、ヒト及び他の動物中で生物学的に利用可能であり、インビボで心血管の健康作用を有することが示されてきた。
【0005】
精製アベナンスラミド混合物を使用して、アベナンスラミドがハムスター及びヒト中で生物学的に利用可能であり、ビタミンCと相乗的に作用してLDL−コレステロール耐酸化を増強することが示されてきた。別の研究では、健康な成人におけるアベナンスラミド摂取の薬物動態が決定された。約60及び120mgの2種の経口投与量レベルで、精製アベナンスラミドを使用した無作為プラセボ対照三元交差試験を使用したところ、アベナンスラミドは生物学的に利用可能であり、血漿グルタチオンレベルによって測定されるように、用量依存的にヒトの酸化防御システムを増強した。観察された生理活性を得るために、エンムギふすま50g一給餌(a 50 g serving)などの食餌源送達系(dietary source delivery system)から、閾値レベルのアベナンスラミド、すなわち、約30〜60mgを得るには、約600〜1,200百万分率(ppm)の総アベナンスラミドを含むエンバク製品を要するだろう。これは、現存するエンバク品種又は現存するエンバク製品について現在記録されているものよりも実質的に高い濃度である。
【0006】
炎症が主な原因であることが知られている湿疹、ツタウルシ毒、虫さされ、日焼け及び帯状疱疹などの皮膚の状態のために、コロイド懸濁液中に通常調製されるオートミールが何百年もの間局所的に使用されてきたことは周知である。約10ppmのアベナンスラミドは、その局所的な抗刺激、抗掻痒及び抗炎症活性のために、現在、世界中のいくつかのパーソナル及びペットケア市場の製品中の中核有効成分を形成している。
【0007】
アベナンスラミドはまた、インビトロで、抗がん予防性を示した。約40ppmの合成アベナンスラミドC(図1;n=1、R1=OH、R2=H、R3=OH)が、ラット及びヒトの両胚大動脈血管壁培養液中で平滑筋細胞の増殖及び一酸化窒素産生を阻害することがインビトロで近年示された。このインビトロ系中4〜20ppmの濃度で、VAO-6ふすまから精製したアベナンスラミド混合物は、酸化LDL−コレステロールに対するこれらの細胞の「粘着性」を有意に減少させ、コレステロールに基づく動脈プラークの蓄積を促進する炎症誘発性シグナル分子の形成を阻害した。さらに、精製エンバクアベナンスラミド、特にアベナンスラミドCは、約20〜50ppmの濃度で処理した場合に結腸がん細胞系に対して抗炎症及び抗増殖活性を有するが、正常な細胞系には影響しないことが最近分かった。
【0008】
アベナンスラミドはまた、インビトロで強力な抗酸化物質でもある。アベナンスラミドは、西洋社会の罹患率及び死亡率の主因であるアテローム性動脈硬化症(心血管疾患)の特定の発症段階の阻害剤であることが示されてきた。ヒト大動脈上皮細胞培養液を使用したところ、エンムギふすまから抽出した個々のアベナンスラミド及び精製エンバクアベナンスラミド混合物は、抗アテローム生成及び抗炎症生理活性を示した。さらに最近では、ヒト大動脈単層細胞培養液におけるアベナンスラミドの抗炎症作用の機構は、アテローム発生の開始、進行及び併発における主要因である、lkBキナーゼ及びlkkタンパク質のリン酸化を阻害することによる核因子kB活性化の阻害に起因していた。
【0009】
アベナンスラミドは、これらの利点を引き出すにはあまりに低い濃度で現在のエンバク品種中に存在する。ヒト被検体を含む臨床的証拠から、単一用量の有効最小濃度は、1000〜3000ppmの抗酸化物質の範囲でなければならないだろう。
【0010】
しかしながら、北米及びスカンジナビアに及ぶエンバク品種は、一般的に、約4ppm〜約150ppmの総アベナンスラミドを含有し、これらの濃度は、遺伝子型、環境、作物年度及び立地に応じて広く変化し得る。遺伝子型/環境相互作用にかかわらず、穀粒の外層のアベナンスラミド含量は、デンプン質胚乳の含量よりも常に高く、アベナンスラミドが主としてふすま分画中に局在していることを示している。従来技術の栽培努力は、濃度を、乾燥穀粒中、出発濃度の約130%まで増加させることができることを示している。
【0011】
全粒穀粒中のアベナンスラミド濃度はまた、生理学的及び/又は機械的工程を通して増加させることもできる。製麦は、穀物粒を浸漬及び発芽させて、種々の最終目的のために穀粒の組成物を変化させる工程であり、数千年間実行されてきた。例えば製麦工程は、複合炭水化物、脂質及びタンパク質の分解をもたらし、これらの糖、脂肪酸及びアミノ酸の貯蔵源を、胚性植物のさらなる発育のために穀粒自体、及び穀粒を消費する生物の両方がより栄養的に利用できるようにする。
【0012】
例えば、アベナンスラミド総量が48時間の発芽期間中に150%に増加することが示された。20℃で10時間の水道水への浸麦中、総アベナンスラミドは、全粒エンバク穀粒中で、約90ppmから約110ppmまで、27%増加することも示された。総アベナンスラミド濃度の増加は、時間及び温度依存性であった。浸麦の温度を上昇させると、濃度は、10時間の浸麦中に約75ppmまで50%も増加した。20℃より上、例えば40℃に温度を上昇させる、又は浸麦を例えば48時間に延長しても、アベナンスラミドのさらなる増加はもたらされなかった。濃度はわずかしか増加せず、実質的な量の蓄積はもたらされなかったが、単に乾燥穀粒の水吸収を開始することによって、アベナンスラミド増加がもたらされる。
【0013】
製麦を通してアベナンスラミドを増加させるための現在の技術は、上で概説した所望の生理反応を引き出すのに十分高い濃度のアベナンスラミドを有する製麦製品を産生しないことが現在の研究から明らかである。さらに、特に2日を超える製麦時間による、これらの増加をもたらす製麦/発芽工程はまた、根、苗条及び部分的に枯渇した製麦穀粒を有する発芽したエンバク実生をもたらすので、発芽製品は、根、子葉鞘及び発芽した穀粒と関連する他の解剖学的修飾の存在のために、さらなる乾燥分画/製粉用途における使用を限定してしまうだろう。さらに、現在の技術は、主に被覆エンバク用に開発されてきたので、殻がまだ付着している発芽穀粒をもたらす。業界で広く使用されている衝撃又は圧縮空気殻除去などの機械的手段による製麦前の殻除去、並びに第2の「研磨」手順、すなわち脱皮エンバクの外側に位置するトリコームの除去は、無傷の穀粒の完全性を激しく損ない、製麦にとって望ましくない損傷した穀粒をもたらし得る。さらに、製麦製品の殻除去及び研磨は、製麦材料の柔軟性及び密度の変化によって複雑化される。
【0014】
根、苗条及び部分的に枯渇した製麦穀粒からなる発芽したエンバク新芽はまた、直接消費できる食品への取り込み又は成分のような焼成食品への取り込みに適した、押し麦フレーク、エンムギふすま又はエンバク粉などのいずれかの伝統的なエンバク製品に製粉するのが困難である。
【0015】
さらに、既に観察された強力な遺伝子型/環境相互作用があるとすれば、伝統的な育種技術を通して高アベナンスラミドエンバクを産生する可能性は、達成するのに何年もかかり得る長期的な解決策を意味する。
【0016】
そのため、心血管の健康を回復させるなどのアベナンスラミドの潜在的に有益な効果を認めることができるようにエンバク穀粒中のアベナンスラミド濃度の実質的な増加が必要とされている。
【発明の概要】
【課題を解決するための手段】
【0017】
本発明の一実施形態によると、休眠中のエンバクを偽製麦(false malting)に供し、アベナンスラミド濃度は増加しているが発芽はしていないエンバクをもたらすことによって、アベナンスラミド濃度が増加したエンバクを生産する方法が提供される。
【0018】
好ましくは、エンバクが非休眠性である場合、休眠が誘発される。
【0019】
好ましくは、エンバクが非休眠性である場合、休眠が増強される。
【0020】
好ましくは、エンバクは休眠を誘発又は増強するために嫌気的に浸麦される。
【0021】
好ましくは、エンバクは、休眠を誘発するために、偽製麦の前に乾熱処理される。
【0022】
好ましくは、エンバクは、30〜40℃で48〜72時間乾熱処理され、その後約70℃で144〜約168時間さらに乾熱処理される。
【0023】
好ましくは、エンバクは、エンバクを4〜40℃で12〜18時間、水に浸漬することによって嫌気的に浸麦される。
【0024】
好ましくは、エンバクは、カルシウムイオンを含む水に嫌気的に浸麦される。
【0025】
好ましくは、エンバクは、偽製麦のために、4〜40℃の温度で96〜120時間インキュベートされる。
【0026】
好ましくは、エンバクは、休眠、殻無しエンバクである。
【0027】
好ましくは、エンバク中の増加したアベナンスラミド濃度は、乾燥量基準で、750ppmより高い。
【0028】
好ましくは、エンバクが被覆されており、休眠エンバクである場合には、エンバクは殻除去される。
【0029】
好ましくは、エンバクは、乾燥量基準で約3〜10%の最終水分含量まで乾燥される。
【0030】
好ましくは、エンバクは、約35%の水分含量まで嫌気的に浸麦される。
【0031】
好ましくは、外側のふすま成分及び残留脱ふすま挽割は、摩耗製粉を使用して回収される。
【0032】
好ましくは、表皮剥離ふすま成分は、摩耗製粉前のエンバクの初期重量の3〜30%を含む。
【0033】
本発明のこれらの及び他の特徴は、添付の図面を参照して以下の説明からより明らかとなるだろう。
【図面の簡単な説明】
【0034】
【図1】アベナンスラミドの一般構造式を示す図である。
【図2】個々のアベナンスラミドの構造を説明する図である。
【図3】アベナンスラミド分画の典型的なHPLCプロファイルを説明する図である。
【図4】アベナンスラミド濃度を増加させる代表的な方法のステップを示す図である。
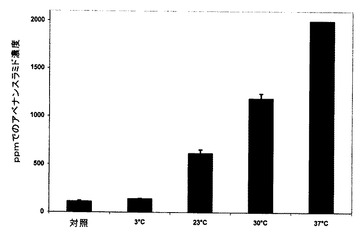
【図5】温度の、加熱処理され、嫌気的に浸麦された殻無しエンバクの総アベナンスラミド濃度への影響を示す図である。
【図6】37℃での偽製麦中の非休眠エンバクへのアベナンスラミドの蓄積の時間経過を示す図である。
【図7】Satake乾燥製粉によって決定される、未製麦穀粒中の総アベナンスラミドの分布を説明する図である。
【図8】Satake乾燥製粉によって決定される、偽製麦後の一品種(VAO-48)における穀粒中の総アベナンスラミドの分布を説明する図である。
【図9】Satake乾燥製粉によって決定される、偽製麦後の別の品種(VAO-22)における穀粒中の総アベナンスラミドの分布を説明する図である。
【発明を実施するための形態】
【0035】
本発明を実施するための本発明者等により考えられた最良の形態を含む本発明のいくつかの特定の実施形態についてここで詳細に言及する。これらの特定の実施形態の例は、添付の図面中に説明される。本発明は、これらの特定の実施形態と組み合わせて記載されるが、本発明を記載の実施形態に限定することを意図していないことが理解されよう。それどころか、添付の特許請求の範囲によって定義される本発明の精神及び範囲内に含まれ得る、代替、修正及び同等形態を包含することを意図している。以下の説明で、本発明の完全な理解をもたらすために、多数の特定の詳細が明らかにされる。本発明は、これらの特定の詳細のいくつか又は全てなしでも実施することができる。他の例では、本発明を不必要に不明瞭にしないために、周知の工程の操作は詳細には記載しなかった。
【0036】
本明細書及び添付の特許請求の範囲において、単数形「a」「an」及び「the」は、文脈が特に明確に指示しない限りは、複数の言及を含む。特に定義しない限りは、本明細書で使用する全ての技術的及び科学的用語は、本発明が属する技術分野の当業者に一般に理解されるのと同じ意味を有する。
【0037】
本発明は、1又は2以上の実施形態に関して記載されている。しかしながら、特許請求の範囲に定義される本発明の範囲から逸脱することなく、いくつかの変形及び修正をすることができることが、当業者に明らかであろう。
【0038】
一般的なエンバク(アベナ・サティバ(Avena sativa))は、穀物の種である。エンバクは、種々の用途を有している。ヒトが消費するためのエンバクは、ポリッジ、朝食用シリアル、クッキー及びスナックバーを作るために使用される。農業では、エンバクの最も一般的な用途の1つは、家畜の飼料である。エンバクは、ウマの食餌の大部分を構成し、ウシにも定期的に与えられている。エンバクはまた、イヌ及びニワトリの飼料のいくつかのブランドにも使用されている。エンバクは、世界中の種々の地域で2000年間栽培されてきた。エンバクは、食品として消費される前は、医療目的、エンバクがまだなお尊ばれている用途で使用されていた。エンバクは、スコットランド、グレートブリテン、ドイツ及びスカンジナビア諸国を含む多くの国々の人々にとっての主食であったので、ヨーロッパでのエンバクの栽培は広範であり、エンバクは重要な商業作物を構成した。
【0039】
殻、すなわち外穎及び内穎は、穀粒の発達中に重要な役割を果たす。殻は、発達中の穀粒に炭水化物を供給する。発達中の穀粒にとって、アミノ酸は主な窒素源であり、この大部分は外穎及び内穎によって与えられ得る。さらに、外穎及び内穎は外側の被覆であるので、小花及び穀粒を病原体及び昆虫による攻撃から保護することができる。
【0040】
大半のエンバクは、収穫される際、殻が付着している。例えばVAO-48など、無殻である品種、すなわち、殻が穀粒に緩く付着しており、コンバイン処理及び収穫中に圃場中に残される品種も存在する。
【0041】
休眠状態で母本から放出される成熟エンバク穀粒は、一次休眠として知られる、種皮強制休眠若しくは胚性休眠のいずれか、又は両方を示すことができる。
【0042】
「休眠」という用語は、通常は発芽に好ましい環境条件下でさえ、種子が一時的に発芽するのを妨げられている状態を説明することを意図している。これらの条件は、水、光、温度、ガス、機械的制限、種皮及びホルモン構造の複雑な組合せであり得る。種子休眠は、例えば、参照によりその内容が本明細書に組み込まれている、Genetic and Molecular Control of Seed Dormancy, B. Li and M.E. Foley, Trends in Plant Science, (1997) Vol.2 (10), pp.384-389に記載されてきた。
【0043】
休眠は、発芽を遅らせ、散布のための時間を許し、同時に全種子が発芽するのを防ぐ。発芽をずらすことは、いくつかの種子及び実生を、短期間の悪天候若しくは一過性の草食動物から損傷又は死をこうむることから保護する。
【0044】
「追熟」の期間後、光、温度、湿度などの環境的に好ましい条件下で、非休眠穀粒は、急速に推移し、発芽の開始がもたらされる。
【0045】
しかしながら、好ましくない環境条件に曝露された、部分的又は完全に追熟した非休眠穀粒は、「二次休眠」として知られる、誘発された休止状態を示し得る。
【0046】
「二次休眠」という用語は、いくつかの非休眠及び休眠後の種子が高温などの発芽にとって好ましくない条件に曝露された状態を説明することを意図している。「休眠」という用語は、一次休眠、例えば、それだけに限定されないが、種皮強制休眠又は真の胚性休眠;二次休眠、例えば、それだけに限定されないが、化学的に誘発される休眠、物理的に誘発される休眠又は他の任意の形の誘発される休眠、自然の二次休眠;及び他の任意の形の休眠を含む、休眠状態の任意の種子を含むことを意図している。
【0047】
多くの園芸植物は、その野生の祖先は休眠していたにもかかわらず、水があり、十分に暖かいとすぐに容易に発芽する種子を有する。これらの栽培植物は、休眠を欠く植物を栽培し維持した植物育種家及び庭師による選択圧の産生のために、休眠を欠く。
【0048】
例えば、以下に記載のエンバク種子の品種であるVAO-48など、逆に、長期の二次休眠を有するよう遺伝的に選択されてきた品種が存在する。
【0049】
製麦の技術分野の当業者によく知られた典型的な製麦工程中に実施されるように、エンバクが、浸漬又は浸麦によって処理され、発芽を可能にされた場合、アベナンスラミド濃度は、乾燥種子中の初期アベナンスラミド濃度に対して、時間依存的様式で、発芽実生中で増加する。アベナンスラミド濃度を増強するための短期間の製麦の有用性は、アベナンスラミドのわずかな増加によって限定され、延長された製麦期間、例えば4〜5日は、より高濃度のアベナンスラミドを含む発芽穀物製品をもたらすが、従来の製粉及び加工市場における有用性を減少させる。
【0050】
本発明の一実施形態によると、以下に記載の二次休眠の誘発及び偽製麦を通して大量のアベナンスラミドが蓄積される。二次休眠中のエンバクを製麦工程に供した場合、驚くべきことに、発芽しないが、アベナンスラミド濃度が25〜35倍まで劇的に増加し得る。
【0051】
言い換えれば、発芽が起こらない条件を作り出す予測可能な方法で二次休眠を利用することによって、発芽穀物と比較して、全粒穀粒の栄養価を変化させることができる。
【0052】
6日間までの休眠エンバクの浸漬及び発芽中、発芽の肉眼的視覚的指標、例えば、根の発生、分岐及び伸長、子葉鞘及び苗条の拡大及び伸長等が観察されていないという事実にもかかわらず、アベナンスラミドは産生され続ける。典型的な製麦手順を伴うが、休眠中の穀物が発芽しないこの処理を、以降、「偽製麦」と呼ぶ。
【0053】
「偽製麦」という用語は、当業者によって実施される製麦技術と類似又は同一の処理を説明することを意図している。しかしながら、種子は休眠、例えば二次休眠中であるので、偽製麦に供された種子は発芽しない。
【0054】
偽製麦は、穀物一般、特にエンバクの組成を変化させる新しい技術を表す。製麦中のエンバクの発芽の抑制は、多数の確立された食品、飼料及び産業利用の市場への取り込みに貢献するアベナンスラミド含量が増加した全粒エンバク製品をもたらす。
【0055】
アベナンスラミドの定量分析の方法
エンバク材料の質的及び量的なアベナンスラミド組成物の決定を以下に記載するように行った。エンバク試料、すなわち種子を、恒量まで37℃で、オーブン中で乾燥させ(約48時間)、分析まで−20℃で真空パックビニール袋中に貯蔵した。抽出前に市販のコーヒーミルを使用して種子を挽いた。抽出及び定量分析は、一般的に、2反復を使用して行った。
【0056】
抽出
還流酸性化80%エタノール(エタノール:水:氷酢酸、80:19.9:0.1(v/v/v))75mlに、挽いたエンバク試料10gを、抗酸化剤としての亜ジチオン酸ナトリウム5mgと共に、激しく攪拌しながら添加した。混合物を熱から離し、攪拌しながら室温で20分間冷却させた。次いで、全内容物を、フリットディスクを備えた目盛り付きガラスクロマトグラフィーカラムにデカントした。重力によって懸濁液を沈降させ、透明な上澄みを有する軽く圧縮された抽出物ベッド(ベッド容積=VbmL)を形成した。上澄みを重力流によって回収し、抽出ベッドを「浸出抽出」によって3×Vbの酸性化80%エタノールで溶出し、透明な黄緑色溶離液を得た。
【0057】
疎水性相互作用及び芳香族吸着クロマトグラフィーによるアベナンスラミドの精製
この抽出物から親油性成分を除去するため、Octyl Sepharose(商標) CL 4-Bクロマトグラフィービーズを添加し(抽出物g当たり0.5ml)、混合物を回転蒸発によって真空下40℃で濃縮乾固した。酸化を防ぐために、蒸留水を混合物に添加して、確実に乾燥中にアベナンスラミドが沈殿するようにした。乾燥混合物を酸性化50%エタノール(エタノール:水:氷酢酸、50:49.9:0.1(v/v/v))に再懸濁し、前もって重力圧縮し、酸性化50%エタノール(例えば、25mlの試料10gについて、最終ベッド容積Vb=30mL)にあらかじめ平衡化したOctyl Sepharose(商標)CL 4-Bを含有する目盛り付きガラスクロマトグラフィーカラムに定量的に移した。次いで、カラムを3×Vbの酸性化50%エタノールで溶出した。合わせた溶離液を回転蒸発によって真空下40℃で濃縮して本質的に脂質を含まない抽出物を得た。
【0058】
サポニン、フラボノイド配糖体、アルコール可溶性タンパク質及びペプチド、遊離糖、芳香族、有機物及びアミノ酸を除去するため、濃縮抽出物を小容積(試料10g当たり約3mlの酸性化40%エタノール(エタノール:水:氷酢酸、40:59.9:0.1(v/v/v)))に溶解し、Sephadex(商標)LH-20を使用してクロマトグラフィーによって精製した。溶液を、前もって重力圧縮し、酸性化40%エタノール(例えば、試料10gについて、最終ベッド容積Vb=25ml)にあらかじめ平衡化したSephadex(商標)LH-20を含有する目盛り付きガラスクロマトグラフィーカラムに定量的に移した。最初に、サポニン、遊離糖、アミノ酸等を、2×Vbの酸性化40%エタノールで溶出することによって除去した。吸収されたアベナンスラミドは、3×Vbの酸性化95%エタノール(エタノール:水:氷酢酸、95:4.9:0.1(v/v/v))で溶出することによって回収した。再度、酸化を防ぐための亜ジチオン酸ナトリウム5mgと共に蒸発前に水を添加した。溶離液を回転蒸発によって真空下40℃で濃縮乾固して精製アベナンスラミド分画を得た。
【0059】
アベナンスラミド分画のHPLC分析
第1の方法では、精製アベナンスラミド分画を50%エタノール(エタノール:水、50:50(v/v))5mlに溶解し、0.45μmフィルターを通して濾過し、HPLCにかけた。試料(10μl)は、Rheodyne(商標)インジェクターを使用して、C18ガードカラムを備えたCERA Column Cooler 250を使用して25℃に維持したC18逆相カラム(ODS Hypersil(商標)C18、5μm、4.6mm×250mm)に注入した。Thermo Separation Products(TSP)Spectra System P4000ポンプを使用してHPLC分析を行い、TSP SpectraSystem(商標)UV3000スペクトルスキャン検出器及びChromQuestソフトウェアを使用して330nmで監視した。流速は1分当たり0.8mlに維持した。HPLC用溶媒は、A:メタノール、B:水及びC:5%酢酸とした。溶媒の勾配(容積%)は、40A:55B:5Cからなり、40分で50A:45B:5Cに直線的に増加し、15分で80A:15B:5Cに直線的に増加し、次いで3分で100Aに達し、3分間維持した。溶媒の勾配は、3分で元の状態に戻し、4分間平衡化した。全ての主要アベナンスラミドのピークを、相対的保持時間及びUVスペクトル(240〜380nmを監視)の真正標準(authentic standards)との比較によって同定した。全ての微量アベナンスラミドは、HPLC−質量分析のみによって同定した。
【0060】
第2の方法では、第1の方法の溶媒系を以下のように変更した。HPLC用溶媒は、A:メタノール、B:水及びC:0.1Mリン酸とした。溶媒の勾配(容積%)は、45A:45B:10Cからなり、55分で60A:30B:10Cに直線的に増加し、次いで3分で100Aに達し、3分間維持した。溶媒の勾配は、3分で元の状態に戻し、4分間平衡化した。
【0061】
第3の方法では、試料(10μl)を30℃に維持したC18逆相カラム(Zorbax Stable(商標)結合C18、3.5μm、4.6mm×150mm)に注入した。HPLC用溶媒は、A:メタノール、B:水及びC:0.5Mギ酸とした。溶媒の勾配(容積%)は、45A:45B:10Cからなり、24分で55A:35B:10Cに直線的に増加し、9分で70A:20B:10Cに直線的に増加し、次いで3分で95A:5Cに達し、3分間維持した。溶媒の勾配は、3分で元の状態に戻し、3分間平衡化した。
【0062】
アベナンスラミド分画のHPLC質量分析
微量アベナンスラミドのピークを、Surveyor(商標)HPLC-UVダイオードアレイ検出器システムを備えたThermo Finnigan LCQ Advantage(商標)質量分析計を使用して、HPLC−MS−MSによって同定した(HPLC条件:Hypersil(商標)ODS、120Å、5μ、250×4.6mmカラム)。HPLC UVモニタリングは330nmで行った。1分当たり0.8mLの流速で、第1の方法で上記したのと同じ溶媒系を使用した。溶媒の勾配(容積%)は、40A:55B:5Cからなり、80分で60A:35B:5Cに直線的に増加し、5分で80A:15B:5Aに直線的に増加し、次いで3分で100Aに達し、3分間維持した。溶媒の勾配は、3分で元の状態に戻し、3分間平衡化した。MS−MS条件:エレクトロスプレーイオン化(ESI,electro-spray ionization、ネガティブモード)、電源電圧:4.5Kボルト、キャピラリー電圧:−10ボルト、キャピラリー温度:300℃、シースガス流量:80%満、補助ガス流量:20%最大(分割流無し)。
【0063】
アベナンスラミドの定量的推定
個々のアベナンスラミドは、外部真正アベナンスラミドA標準に対する330nmのピーク面積を決定することによって定量化し、アベナンスラミドA重量当量として表した。総アベナンスラミドは、アベナンスラミドA重量当量として計算した個々のアベナンスラミド量を合計することによって計算し、乾燥重量基準で百万分率(ppm)アベナンスラミドA当量として表した。
【0064】
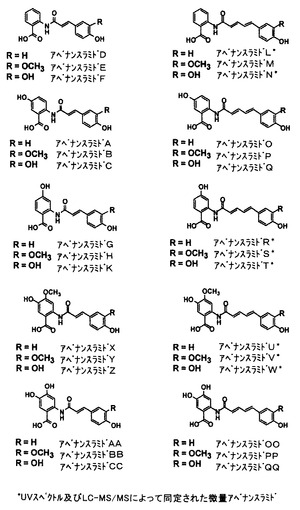
アベナンスラミドは、図1に示す一般式の、約30種の異なるN−アロイルアントラニル酸アルカロイドの群である。図2は、麦芽エンバク及び麦芽になっていないエンバクの両方で生じる個々のアベナンスラミドの構造を説明する。
【0065】
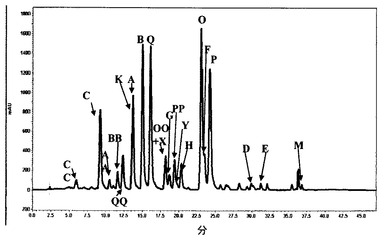
図3は、図2において麦芽にされ、命名を割り当てられた、上記第3の方法を使用したエンバクからのアベナンスラミド分画の典型的なHPLCプロファイルを説明する。
【0066】
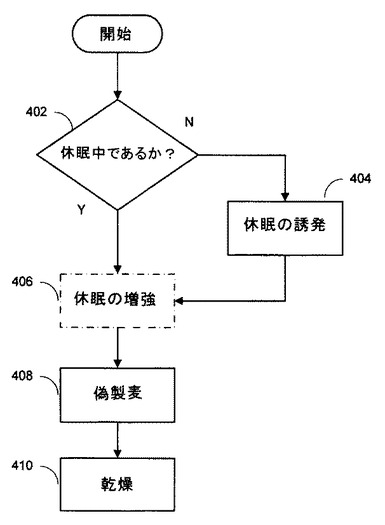
図4は、本発明の実施形態に従って、アベナンスラミド濃度を増加させる方法のステップを示す。
【0067】
エンバクが休眠中でない場合(402)、二次休眠が誘発され得る(404)。そうでなければ、エンバクは休眠、一般的には、自然の又は誘発された二次休眠中である。
【0068】
種子が休眠中だとしても、二次休眠中の種子は、経時的に休眠から覚醒し得るので、休眠を増強するための任意のステップ(406)が含まれ得る。
【0069】
二次休眠を誘発又は増強するための1つの非限定的方法は、エンバクを嫌気的に水和させる、又は焼戻しすることである。
【0070】
製麦は、一般的に、エンバクが特定の水分含量に達するまで、エンバクを水に浸麦することから始まる。浸麦は、一般的に、エンバクの空気への曝露が点在しており、エンバクが追加の酸素を得ることが可能になっている。
【0071】
エンバクの休眠は、嫌気的な浸麦を通して誘発(404)又は増強(406)され得る。嫌気的な浸麦では、エンバクは、例えば30℃より高い温度で12〜18時間、嫌気的に焼戻し又は水和される。
【0072】
次いで、エンバクは、偽製麦(408)、すなわち製麦と類似又は同一であるが発芽しない条件、例えば23〜37℃で96〜120時間供される。偽製麦後、エンバクは、貯蔵又はさらなる加工前に、例えば、35℃で24〜48時間乾燥される(410)。
【0073】
以下は、本発明の実施形態によるアベナンスラミド濃度の増加を示す非限定的実施例である。
【実施例1】
【0074】
休眠対非休眠エンバク
新しく収穫した休眠、殻無し、覆い無し(bald)播種エンバク育種系統(VAO-48)の種子試料及び非休眠、殻無し、覆い無し播種エンバク育種系統(VAO-2、品種AC GEHLとして現在登録されている)の種子試料を以下に記載のように製麦した。
【0075】
各々の育種系統約20gを、穏やかに攪拌しながら、室温で20分間、1%次亜塩素酸ナトリウム水溶液に液浸することによって手短に表面殺菌し、次いで、過剰な次亜塩素酸ナトリウム溶液を除去するために取り出し、徹底的にすすいだ。次いで、室温及び散乱光で4日間、湿った濾紙ディスク上の被覆したペトリ皿(150×15mm)中で種子を発芽させた。各々の育種系統の別の試料20gを対照として取っておいた。
【0076】
4日後、2つの試料の発芽率を決定した。次いで、発芽及び未発芽両方を含む種子を取り出し、室温で2日間、種子乾燥機中で乾燥させた。平均発芽率は、以下のとおりであった。
休眠、殻無し、覆い無し(VAO-48) 1%
非休眠、殻無し、覆い無し(VAO-22) 75%
【0077】
(偽)製麦前及び後両方の2つの試料の総アベナンスラミド含量を、上記の第1の方法を使用して決定し、表1に要約する。
【0078】
【表1】
【0079】
表1は、休眠エンバクが、驚くべきことに、偽製麦期間中にアベナンスラミドを蓄積したことを示している。
【0080】
休眠種子は、出発材料と同様の形態を有し、非休眠種子とは異なり、根、子葉鞘、並びに新生の葉及び苗条を有さなかった。
【実施例2】
【0081】
嫌気的浸麦及び休眠種子の貯蔵の休眠への影響
休眠エンバクは、貯蔵の条件(例えば、酸素濃度、温度、水分含量)に応じて、時間と共にその二次休眠から覚醒することが知られている。特に、低酸素濃度は、休眠を延長する、及び/又は発芽を阻害する。サブゼロ温度(例えば、−20℃)で貯蔵した種子は、長期間その遺伝的に伝達する形質の大半を保持することができることも、エンバク育種家及び種子胚芽プラズマ保存技術分野の当業者に知られている。しかしながら、製麦などの大規模工程に実用的であるためには、種子を休眠状態に維持するための高価な低温貯蔵に頼らず、休眠エンバクの一定供給が、一年中定期的に利用可能であるべきである。そのため、休眠エンバクの休眠を延長及び増強し、非休眠エンバクの二次休眠を誘発することが好ましい。
【0082】
室温で3週間、次いで−20℃で1週間貯蔵した、実施例1で使用した休眠エンバク系統(VAO-48)の種子を、約23℃で水道水に約48時間までの種々の時間浸した。小規模実験では、種子を、水で完全に満たし、Parafilmで密封した気密ペトリ皿に散布した(すなわち、嫌気的浸麦)。約2、4、6、8、12、24及び48時間後、種子を取り出し、室温で、種子乾燥機中で手短に風乾した。次いで、種子を表面殺菌し、散乱光中室温で4日間、上記のように製麦した。4日後、発芽率を上記のように決定した。
【0083】
さらなる実験では、より大量の種子を使用し、水道水で満たし、Parafilm(商標)で密封した気密Erlenmeyerフラスコ中で嫌気的浸麦を行った。フラスコ形状の頂上の「先細り」は、残気の排除を促進し、水位より下への種子の水浸を保証した。両実験の結果を表2に要約する。
【0084】
【表2】
【0085】
空気への曝露なしで、約8〜18時間一晩中種子を単に浸漬又は浸麦することによって、浸麦なしの約50%から嫌気的浸麦後10%未満まで発芽率がかなり下がったことが分かった。これは、8〜18時間の嫌気的浸麦によって発芽が実質的に減少し得ることを示している。浸麦温度を最適化するためのさらなる実験は、30℃〜32℃の温度で18時間が、3%未満の発芽(すなわち、97%を超える休眠)という一貫した発芽率をもたらすことを示した。
【0086】
種々の貯蔵期間後の休眠エンバク系統の休眠属性の保持を試験するために、室温で14週間まで種々の期間貯蔵した試料を使用して発芽実験を行った。さらに、控え試料を追加で一晩中嫌気的浸麦に供し、手短に風乾し、次いで同様の条件下で発芽を試験した。両時間経過実験では、種子を室温及び散乱光で、4日間ペトリ皿中で発芽させ、発芽率を計算した。両実験の結果を表3に示す。
【0087】
【表3】
【0088】
休眠エンバク系統の休眠は、貯蔵の最初の8〜10週間で増加し、次いで劇的に減少して、約90%の種子が発芽する。予想通り、16時間嫌気的に浸麦したエンバクは対応する非浸麦試料よりも、室温で8週間以上の貯蔵後により大きい休眠を示した。こうして室温での貯蔵で徐々に休眠から覚醒するエンバクの休眠を増強し、この工程により適したものにすることができる。
【実施例3】
【0089】
加熱前処理及び嫌気的浸麦の、非休眠、殻無しエンバクにおける休眠及びアベナンスラミド蓄積への影響
二次休眠を増強及び誘発する他の方法が存在する。エンバクの休眠を増強及び誘発する非限定的な代表的な方法は、30〜70℃の乾熱で2週間までの種々の期間エンバクを処理することである。この手順は通常、2段階で行う。第1段階は、約30℃の温度で数日間、種子の水分含量を約3%まで下げることを含み、その後、第2段階は約70℃で1週間までである。このような処理はまた、種子伝染性糸状菌及び細菌胞子を減少させるのに有効であることも示されてきた。この体制下で種子は損傷せず、いくつかの場合には、発芽率が事実上増加する。
【0090】
非休眠、殻無しエンバク系統(VAO-2)の種子(500g)を対流式乾燥器中37℃で72時間、次いで70℃で144時間加熱処理した。室温まで冷却後、加熱処理種子を小口試料に分割し、ある群(5反復)は32℃で18時間嫌気的に浸麦し、別の群は嫌気的に浸麦しなかった(5反復)。両群を上記のように表面殺菌し、次いで散乱光中室温で、4日間ペトリ皿(150×50mm)中で製麦した。製麦後、種子を、上記の第1の方法を使用して、発芽率(発芽%)及びアベナンスラミド含量について評価した(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)。結果を表4に要約する。
【0091】
【表4】
【0092】
上記のように、2段階加熱処理下でのエンバクの乾熱処理及び18時間の嫌気的浸麦の両処理は、別々に製麦中のエンバクの発芽を減少させた(すなわち、休眠を増加させた)。加熱処理の後に嫌気的浸麦と、共に使用した場合、発芽率は実質的に0%まで低下し、4日の製麦期間中、種子は休眠したままであった。さらに、加熱及び嫌気的浸麦の処理を組み合わせた4日の「偽製麦」したエンバクの総アベナンスラミド含量は、最初の77ppmから製麦後445ppmまで増加し、初期値に対して約480%(5.8倍)の増加を示した。
【0093】
休眠を誘発するための2段階乾熱処理工程が生存率の減少をもたらさなかったことに留意する。加熱処理エンバクを100ppmのジベレリン(例えば、GA3)存在下で5日間インキュベートした場合、発芽のほぼ完全な回復が観察された。
【実施例4】
【0094】
温度の、偽製麦中のアベナンスラミド蓄積への影響
伝統的な製麦では、発芽及び後の発酵のための種子のデンプン貯蔵物質の遊離糖への同時の加水分解を最大化するように最適温度が選択される。これらの目的のため、最適温度は通常、約25℃未満である。温度のアベナンスラミド蓄積への影響を研究するため、4種の異なる温度で、暗中で4日間偽製麦を行った。比較のため、散乱光下、室温でも偽製麦を行った。これらの研究のため、加熱前処理及び嫌気的浸麦によって休眠状態にした非休眠、殻無しエンバク品種を使用した。
【0095】
本工程のための4種の温度は、3℃、23℃(室温)、30℃及び37℃とし、暗中で偽製麦を行い、比較試料を23℃で、散乱光中で製麦した。3℃での製麦は実験室の冷蔵庫で行い、30℃及び37℃での製麦は制御温度対流式オーブンで行った。
【0096】
AC Baton殻無しエンバク(AC Baton、発芽率は95%を超える)の試料50gを使用した。種子は以下:上記実施例3のように37℃で72時間、その後70℃で144時間加熱処理し、室温(23℃)まで手短に冷却し、次いで32℃で18時間水道水に嫌気的に浸麦し、種子乾燥機中で約4時間風乾した。種子は、室温で1時間0.25%次亜塩素酸ナトリウム水溶液を使用するのみで上記のように表面殺菌し、次いで4つの小口試料に分割し、実施例3の制御温度対流式オーブンを使用して、4種の異なる温度で、暗中で96時間「偽製麦」した。
【0097】
製麦後、種子を発芽率(発芽%)について評価し、室温で一晩、種子乾燥機中で風乾し、アベナンスラミド含量を上記の第1の方法によって決定した(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)。3種の主要アベナンスラミド(アベナンスラミドA、B及びC)、他の全てのアベナンスラミド及び総アベナンスラミドの含量を計算し、未製麦材料中の対応する濃度と比較した。結果を表5に要約する。
【0098】
【表5】
*発芽%の値は、4日の発芽率を表す。括弧内のアベナンスラミドの値は、製麦前濃度を表す。
【0099】
比較の観点から、表5は、蓄積したアベナンスラミドA、B、C及び他の全てのアベナンスラミドの濃度が製麦温度の上昇と共に偽製麦中に増加したことを示している。使用した最も低い温度(3℃)は、未製麦材料中の114ppmから製麦後約142ppmまで(約25%又は1.25倍)の増加をもたらした一方で、最も高い温度(37℃)は、ほぼ2,000ppm(約1,650%又は17.5倍)の総アベナンスラミド濃度をもたらした。ここで使用した前処理体制下で、高温及び低温の両方が発芽をわずかに減少させた。
【0100】
温度の、加熱及び嫌気的浸麦手順によって休眠状態にした非休眠(95%を超える生存種子)、殻無しエンバクAC Batonを使用した96時間の偽製麦中に蓄積した総アベナンスラミドへの影響を図5に示す。単に最も熟練した製麦技術で使用される温度(通常10〜25℃)より上の温度で製麦することによって示される、対照(すなわち、未製麦)種子に対する劇的な増加を容易に見ることができる。
【実施例5】
【0101】
時間の、偽製麦中のアベナンスラミド蓄積への影響
偽製麦工程中のアベナンスラミド蓄積の時間経過を研究するために、非休眠、殻無しエンバク品種を使用して、種々の期間で工程を行い、蓄積したアベナンスラミド濃度を決定した。
【0102】
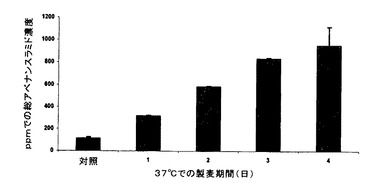
AC Baton殻無しエンバク(AC Baton、発芽率は95%を超える)の試料120gを使用した。種子を実施例3のように加熱処理し、室温(23℃)まで手短に冷却し、次いで32℃で18時間水道水に嫌気的に浸麦し、約4時間種子乾燥機中で風乾した。種子を実施例4のように表面殺菌し、次いで、湿ったペーパータオル上のアルミニウム箔で覆った浅いトレイ内で、暗中37℃で種々の期間「偽製麦」した。発芽率は、4日後に1%未満であった。各々の製麦期間後、種子の代表的な小口試料(約30g)を取り出し、室温で4時間乾燥させ(強制空気)、次いで37℃で約40時間乾燥させることによって恒量にし、上記の第1の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析するまで−20℃で貯蔵し、未製麦材料と比較した。結果を表6に要約し、図6に示す。
【0103】
【表6】
*括弧内のアベナンスラミドの値は、製麦前濃度を表す。
【0104】
表6及び図6のデータから、アベナンスラミドA、B、C及び総アベナンスラミドの濃度全てが偽製麦期間中に時間と共に増加し、増加の時間経過は3日までやや線形パターンに従い、4日で水平になることが分かる。総アベナンスラミドの4日全体の蓄積は、初期値に対して約740%(8.4倍)の増加を示した。
【実施例6】
【0105】
カルシウムイオン浸麦処理の、休眠エンバクVAO-48の偽製麦中のアベナンスラミド蓄積への影響
嫌気的浸麦段階中のCa+2イオンの存在が、その後の製麦段階中のアベナンスラミドの蓄積に何らかの影響を有するかどうかを調査するために、浸麦水に一連の増加するCa+2濃度を使用し、製麦後にアベナンスラミド含量を決定した。
【0106】
殻無し、無毛の休眠エンバクVAO-48の試料25gを、35℃で水又は種々の濃度のUSP−グレードCaCl2・(H2O)2溶液のいずれかに嫌気的に浸麦した。18時間の浸麦及び種子乾燥機中で約4時間風乾後、種子を上記のように表面殺菌し、次いで制御温度オーブン中、30℃、暗中で4日間、ペトリ皿(150×50mm)を使用して「偽製麦」した。全ての場合の発芽率は、4日後に1%未満であった。製麦後、種子を取り出し、室温で4時間乾燥させ(強制空気)、次いで37℃で約40時間乾燥させることによって恒量にし、上記の第3の方法によって(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、アベナンスラミド含量について分析するまで−20℃で貯蔵し、未製麦材料と比較した。結果を表7に要約する。
【0107】
表7に示すように、個々のアベナンスラミド濃度は未製麦種子中で非常に低く、製麦前の総アベナンスラミド含量はわずか約32ppmであった。カルシウムイオンを添加していない水への浸麦及び30℃で4日の製麦後、全アベナンスラミド濃度は、ずっと高く、439ppmの総アベナンスラミド濃度は、約13.7倍の増加を示した。浸麦水中のCa+2濃度の増加と共に、麦芽穀粒中のアベナンスラミド濃度は、増加傾向を示し、Ca+2を添加しなかった場合の約439ppmから1%Ca+2を添加した場合の約757ppm(約1.7倍)まで浸麦中に増加し、未製麦種子に対して1%Ca+2を添加した場合全体として約23.6倍増加した。これらの結果は、浸麦期間中のCa+2の添加が、その後の製麦中のアベナンスラミド蓄積を増強することを示している。
【0108】
【表7】
【実施例7】
【0109】
高濃度の塩化カルシウム浸麦処理の、非休眠エンバクVAO-22の偽製麦中のアベナンスラミド蓄積への影響
非休眠、殻無し、無毛のエンバク系統VAO-22及び浸麦水中でより高濃度のCa+2を実施例7で使用した。
【0110】
VAO-22殻無し、無毛、非休眠エンバク育種家の種子の試料1300gを使用した(4日後の発芽率84%)。次いで、種子を実施例5のように加熱処理した。未製麦種子を表す対照として試料25gを使用し、分析するまで−20℃で貯蔵した。加熱処理した材料の残りを1.0%又は2.0%CaCl2・(H2O)2のいずれかの中、35℃で18時間嫌気的に浸麦し、種子乾燥機中で約4時間風乾した。次いで、種子を上記のように表面殺菌し、制御温度オーブン中、暗中30℃で4日間、ペトリ皿(150×50mm)を使用して「偽製麦」した。製麦後、種子を取り出し、室温で4時間乾燥させ(強制空気)、次いで37℃で約40時間乾燥させることによって恒量にし、上記の第3の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析するまで−20℃で貯蔵し、未製麦材料と比較した。結果を表8に要約する。
【0111】
表8から分かるように、この品種のエンバクの個々の及び総アベナンスラミド濃度は、1及び2%の両濃度のCaCl2に浸麦した後、4日の偽製麦中に増加した。例えば、1%CaCl2に18時間嫌気的に浸麦し、30℃で4日間偽製麦した場合、アベナンスラミドBは、3.4ppmからほぼ80ppmまで(2,240%;初期値に対して23.4倍)増加した一方で、2%CaCl2では、濃度は103ppm(2,930%;初期値に対して30.3倍)に達した。アベナンスラミドCについては、対応する増加は、1%CaCl2で2.3ppmから65.2ppmまで(2,735%;初期値に対して28.4倍)、2%CaCl2で100.1ppmまで(4,252%;初期値に対して43.5倍)であった。他のアベナンスラミドも同様に増加し、総アベナンスラミド濃度は、1%及び2%CaCl2でそれぞれ2,307%及び3,190%(初期値に対して24.1倍及び31.9倍)の濃度を示した。これら2つの実施例のデータは、嫌気的浸麦ステップ中に添加した少なくとも2%までの濃度範囲のCaCl2の、その後の製麦エンバク中のアベナンスラミドの蓄積及び/又は保持を増強する役割を支持する。
【0112】
【表8】
【実施例8】
【0113】
偽製麦前及び後の休眠エンバクVAO48エンバクのSatake製粉の、ふすま及び脱ふすま挽割乾燥粉砕分画へのアベナンスラミド蓄積への影響
前述のように、偽製麦は、大部分は未製麦穀粒と本質的に同じであり、それゆえ、従来の乾燥摩耗製粉、例えばSatake製粉を通してふすま及び脱ふすま製品への加工に容易に使用可能なエンバク穀粒を生じる。
【0114】
殻無し、無毛、休眠エンバクVAO-48の試料(約380g)を、Satakeを使用して製粉し、以下の連続的な分画を得た。
0〜7(重量)%ふすま分画
7〜15(重量)%ふすま分画
15〜23(重量)%ふすま分画
23(重量)%を超える脱ふすま挽割
【0115】
全製粉分画は、直ちに氷上で冷却し、分析するまで−20℃で貯蔵した。
【0116】
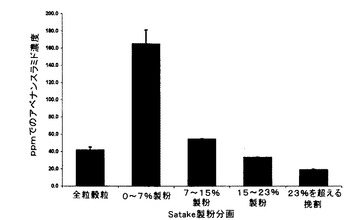
未製粉全粒種子及びSatake製粉分画の控え試料を、上記の第1の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析し、結果を表9及び図7に要約する。
【0117】
【表9】
【0118】
表9に示すように、未製麦VAO-48エンバクのアベナンスラミド含量は分画間でかなり変化し、最高濃度(全粒穀粒中の濃度のほぼ4倍)が最外側の(すなわち、0〜7%)ふすま層で示され、後続のふすま分画で内側に減少した。穀粒の23%を除去した後の残りの脱ふすま挽割は、全粒穀粒のアベナンスラミド含量の半分未満の最低アベナンスラミド濃度を有した。外層の除去の増加と共に含量が減少するこの傾向は、総アベナンスラミド含量においてのみならず、大部分において、個々のアベナンスラミドの各々の濃度でも見られた。
【0119】
実施例5における上記のように加熱処理し、使用するまで−20℃で貯蔵した同じ材料のさらなる試料(377g)を35℃で18時間、水道水に嫌気的に浸麦した。種子を実施例4のように表面殺菌し、次いで、制御温度オーブン中、30℃で4日間、「偽製麦」した。全ての場合において、発芽率は4日後に1%未満であった。製麦後、種子を取り出し、制御温度オーブン中37℃で恒量まで乾燥させ、製粉するまで−20℃で貯蔵した。以下のSatake製粉分画は上記のように調製した。
0〜10.7(重量)%ふすま分画
10.7〜20.6(重量)%ふすま分画
20.6(重量)%を超える脱ふすま挽割
【0120】
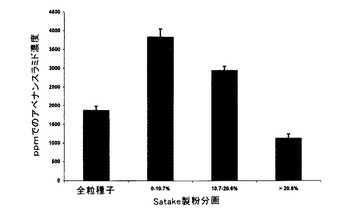
未製粉全粒製麦エンバク及び製粉分画の各々の控え試料を上記の第2の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析した。結果を表10及び図8に要約する。
【0121】
【表10】
【0122】
明白な減少の程度は、未製麦穀粒ほど劇的でなく、中間のふすま材料及び脱ふすま挽割の相対的含量は、未製麦材料中の同様の分画よりも比例的にわずかに高かったが、脱ふすまの程度が増加するにつれてアベナンスラミド濃度が減少する傾向は、回収した個々の分画で同様であった。0〜10.7%脱ふすま分画中に蓄積した総アベナンスラミドは、3,832ppmも高くに達し、未製麦穀粒からの同様の分画に対して2,218%(22.2倍)の増加及び全粒未製麦材料に対して91.5倍の増加を示した。
【0123】
0〜10.7%及び10.7〜20.6%のふすま分画を合わせて0〜20.6%の全体的な分画(すなわち、20.6%製粉収率)を形成した場合、材料は、約3,388ppmの総アベナンスラミド濃度を有する。
【0124】
これらの結果は、穀粒の外層が最高濃度の全アベナンスラミドを含有することを示している。
【実施例9】
【0125】
偽製麦前及び後の殻無し、無毛、非休眠エンバクVAO22エンバクのSatake製粉の、ふすま及び脱ふすま挽割乾燥製粉分画へのアベナンスラミド蓄積への影響
偽製麦ふすまの外層中のアベナンスラミド濃度は、全粒穀粒中の濃度に対して高いが、このふすまの収率(製麦穀粒の7〜10%から)は低い。穀粒の特徴的な細長い形状及び深く区切られた中央のしわの形態のために、ふすま及び胚乳の明確な分離が限定され、胚乳組織の混入が最小限のふすまの収率は、少ない脱ふすま及び低いふすま分画製粉収率に制限される。しかしながら、殻無し、無毛、非休眠系統VAO-22育種系統は、短い穀粒及び比較的浅いしわを有し、全体形状及び形態がコムギ穀粒に似ており、Satake摩耗製粉にはるかにより適したものになっている。
【0126】
VAO-22殻無し、無毛、非休眠エンバクの育種家の種子の実施例7で使用した材料のさらなる試料を使用した(4日後の発芽率84%)。次いで、種子を実施例5のように加熱処理した。試料25gを、未製麦種子を表す対照として使用し、分析するまで−20℃で貯蔵した。加熱処理した材料の残りは、水道水中に35℃で18時間嫌気的に浸麦した。次いで、種子を実施例5のように表面殺菌し、制御温度オーブン中、暗中30℃で4日間「偽製麦」した。製麦後、種子を取り出し、制御温度オーブン中37℃で恒量まで乾燥させ、製粉するまで−20℃で貯蔵した。以下のSatake製粉分画を上記のように調製した。
0〜10.1(重量)%ふすま分画
10.1〜20.1(重量)%ふすま分画
20.1(重量)%を超える脱ふすま挽割
【0127】
未製粉全粒製麦エンバク及び製粉分画の各々の控え試料を、上記の第3の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析した方法2を使用してHPLCによってアベナンスラミド含量について分析し、未製麦材料と比較した。結果を表11及び図9に要約する。
【0128】
【表11】
【0129】
表11に示す全粒製麦穀粒の総アベナンスラミド含量は、約42ppmから約1,100ppmまで増加し、未製麦穀粒に対して約2,560%(26.6倍)の増加を示した。0〜10.1%ふすま分画中のアベナンスラミド濃度は、ほぼ2,200ppmまで増加し、全粒未製麦穀粒中の濃度に対して、この材料中のアベナンスラミドにおいて5,170%の増加(52.7倍)を示した。同様の増加は、10.1〜20.1%ふすま分画中でも見られ、合わせて0〜20.1%の全体分画(20.1%製粉収率で)を形成すると、約2,183ppmの総アベナンスラミド濃度を有するだろう。
【0130】
これは、VAO-48エンバクを使用した表10に示す同様の結果よりも低いが、それでも約20%の製粉収率とかなりのアベナンスラミド増加を示す。しかしながら、このエンバクは、VAO-48材料で観察される同様の中間の製粉分画中のアベナンスラミド含量における急速な減少傾向を含まないことが明白である。
【0131】
未製麦及び偽製麦の殻無し、無毛エンバクのSatake製粉によって産生されるものなどの乾燥摩耗製粉分画を使用して、任意の所望の濃度及び同程度の脱ふすまから約3,000ppmもの高い総アベナンスラミド濃度のふすま混合物を製造し、穀粒内の局在に関連するタンパク質、β−グルカン、フィチン酸塩等の他の成分の組成における変化を最小限にすることができることも実施例8及び9から明白である。
【実施例10】
【0132】
30℃での4日の偽製麦後の被覆、非休眠エンバク品種へのアベナンスラミドの蓄積
殻無し、無毛エンバク品種を使用する経済的利点にもかかわらず、最終製品中の殻の包含が正当化され得る技術の潜在的用途、例えば、それだけに限定されないが、殻の除去が経済的であると判明していない、飼料用途、非食品用産業用分画用途が存在する。
【0133】
本発明で開発された技術を使用した被覆エンバクの製麦は、殻が製麦後に除去されているか、又は製品中にまだ存在しているかのいずれかである製品をもたらす。従来の殻除去及び研磨装置によって被覆エンバクの殻を除去しようとする試み及び毛(トリコーム)の除去は、穀粒へのある程度の機械的損傷及びその後の製麦工程中の複雑化をもたらす。
【0134】
それにもかかわらず、上記と同じ手順の下で商業用エンバクを慎重に殻除去し製麦することによって被覆エンバクを使用する試みがなされた。あるいは、全粒被覆エンバクも以下に記載のように試された。
【0135】
被覆、非休眠エンバクJordan実験用種子の試料約500gを使用した。種子を、殻がついたまま実施例5のように加熱処理し、実施例5のように表面殺菌した。小口試料80gを、未製麦種子を表す対照として使用し、分析するまで−20℃で貯蔵した。加熱処理した材料の残りは、水道水中に35℃で18時間嫌気的に浸麦した。次いで、種子を、0.25%次亜塩素酸ナトリウム水溶液を使用して室温で1時間、実施例5のように表面殺菌し、制御温度オーブン中、暗中30℃で4日間「偽製麦」した。製麦後、種子を取り出し、制御温度オーブン中37℃で恒量まで乾燥させ、−20℃で貯蔵した。全粒製麦エンバクの控え試料を、上記の第3の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析し、未製麦材料と比較した。
【0136】
被覆、非休眠エンバクAC Goslinの試料1,000gを使用した。種子を、殻がついたまま実施例5のように加熱処理し、穀粒に損傷を与えるのを避けるために小さなローラー殻除去機を使用して機械的に殻を除去し、実施例5のように表面殺菌した。小口試料25gを、未製麦種子を表す対照として使用し、分析するまで−20℃で貯蔵した。種子の残りは、水道水中に35℃で18時間嫌気的に浸麦し、種子乾燥機中室温で4時間簡手短に風乾し、0.25%次亜塩素酸ナトリウム水溶液を使用して室温で1時間、実施例5のように表面殺菌した。次いで、種子を、室温及び散乱光で4日間、湿った濾紙ディスク上の大きなペトリ皿(150×15mm)中で「偽製麦」した。製麦後、種子を取り出し、種子乾燥機中で4日間乾燥させ、分析するまで−20℃で貯蔵した。全粒製麦エンバクの控え試料を、上記の第1の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析し、未製麦材料と比較した。
【0137】
結果を表12に要約する。
【0138】
【表12】
【0139】
表12で分かるように、殻を有する未製麦無傷試料は、約57ppmの総アベナンスラミド含量を有していた。30℃で4日間製麦した場合、種子は、総計183.7ppmのアベナンスラミドを蓄積し、未製麦試料に対して約220%の増加を示した。
【技術分野】
【0001】
本発明は、アベナンスラミド、及びより具体的にはエンバク中のアベナンスラミド濃度を増加させる方法に関する。
【背景技術】
【0002】
エンバクは、アベナンスラミド、アベナコシラート(avenacosylate)、トリテルペンサポニン(アベナコシド)及びステロイドサポニン(アベナシン)を含むいくつかの群の生理活性ファイトケミカルを合成し貯蔵する能力において、穀物の中で独特である。
【0003】
アベナンスラミドは、図1に示す一般式の、約30種の異なるN−アロイルアントラニル酸アルカロイドの群である。
【0004】
アベナンスラミドは、抗酸化物質であり、ヒト及び他の動物中で生物学的に利用可能であり、インビボで心血管の健康作用を有することが示されてきた。
【0005】
精製アベナンスラミド混合物を使用して、アベナンスラミドがハムスター及びヒト中で生物学的に利用可能であり、ビタミンCと相乗的に作用してLDL−コレステロール耐酸化を増強することが示されてきた。別の研究では、健康な成人におけるアベナンスラミド摂取の薬物動態が決定された。約60及び120mgの2種の経口投与量レベルで、精製アベナンスラミドを使用した無作為プラセボ対照三元交差試験を使用したところ、アベナンスラミドは生物学的に利用可能であり、血漿グルタチオンレベルによって測定されるように、用量依存的にヒトの酸化防御システムを増強した。観察された生理活性を得るために、エンムギふすま50g一給餌(a 50 g serving)などの食餌源送達系(dietary source delivery system)から、閾値レベルのアベナンスラミド、すなわち、約30〜60mgを得るには、約600〜1,200百万分率(ppm)の総アベナンスラミドを含むエンバク製品を要するだろう。これは、現存するエンバク品種又は現存するエンバク製品について現在記録されているものよりも実質的に高い濃度である。
【0006】
炎症が主な原因であることが知られている湿疹、ツタウルシ毒、虫さされ、日焼け及び帯状疱疹などの皮膚の状態のために、コロイド懸濁液中に通常調製されるオートミールが何百年もの間局所的に使用されてきたことは周知である。約10ppmのアベナンスラミドは、その局所的な抗刺激、抗掻痒及び抗炎症活性のために、現在、世界中のいくつかのパーソナル及びペットケア市場の製品中の中核有効成分を形成している。
【0007】
アベナンスラミドはまた、インビトロで、抗がん予防性を示した。約40ppmの合成アベナンスラミドC(図1;n=1、R1=OH、R2=H、R3=OH)が、ラット及びヒトの両胚大動脈血管壁培養液中で平滑筋細胞の増殖及び一酸化窒素産生を阻害することがインビトロで近年示された。このインビトロ系中4〜20ppmの濃度で、VAO-6ふすまから精製したアベナンスラミド混合物は、酸化LDL−コレステロールに対するこれらの細胞の「粘着性」を有意に減少させ、コレステロールに基づく動脈プラークの蓄積を促進する炎症誘発性シグナル分子の形成を阻害した。さらに、精製エンバクアベナンスラミド、特にアベナンスラミドCは、約20〜50ppmの濃度で処理した場合に結腸がん細胞系に対して抗炎症及び抗増殖活性を有するが、正常な細胞系には影響しないことが最近分かった。
【0008】
アベナンスラミドはまた、インビトロで強力な抗酸化物質でもある。アベナンスラミドは、西洋社会の罹患率及び死亡率の主因であるアテローム性動脈硬化症(心血管疾患)の特定の発症段階の阻害剤であることが示されてきた。ヒト大動脈上皮細胞培養液を使用したところ、エンムギふすまから抽出した個々のアベナンスラミド及び精製エンバクアベナンスラミド混合物は、抗アテローム生成及び抗炎症生理活性を示した。さらに最近では、ヒト大動脈単層細胞培養液におけるアベナンスラミドの抗炎症作用の機構は、アテローム発生の開始、進行及び併発における主要因である、lkBキナーゼ及びlkkタンパク質のリン酸化を阻害することによる核因子kB活性化の阻害に起因していた。
【0009】
アベナンスラミドは、これらの利点を引き出すにはあまりに低い濃度で現在のエンバク品種中に存在する。ヒト被検体を含む臨床的証拠から、単一用量の有効最小濃度は、1000〜3000ppmの抗酸化物質の範囲でなければならないだろう。
【0010】
しかしながら、北米及びスカンジナビアに及ぶエンバク品種は、一般的に、約4ppm〜約150ppmの総アベナンスラミドを含有し、これらの濃度は、遺伝子型、環境、作物年度及び立地に応じて広く変化し得る。遺伝子型/環境相互作用にかかわらず、穀粒の外層のアベナンスラミド含量は、デンプン質胚乳の含量よりも常に高く、アベナンスラミドが主としてふすま分画中に局在していることを示している。従来技術の栽培努力は、濃度を、乾燥穀粒中、出発濃度の約130%まで増加させることができることを示している。
【0011】
全粒穀粒中のアベナンスラミド濃度はまた、生理学的及び/又は機械的工程を通して増加させることもできる。製麦は、穀物粒を浸漬及び発芽させて、種々の最終目的のために穀粒の組成物を変化させる工程であり、数千年間実行されてきた。例えば製麦工程は、複合炭水化物、脂質及びタンパク質の分解をもたらし、これらの糖、脂肪酸及びアミノ酸の貯蔵源を、胚性植物のさらなる発育のために穀粒自体、及び穀粒を消費する生物の両方がより栄養的に利用できるようにする。
【0012】
例えば、アベナンスラミド総量が48時間の発芽期間中に150%に増加することが示された。20℃で10時間の水道水への浸麦中、総アベナンスラミドは、全粒エンバク穀粒中で、約90ppmから約110ppmまで、27%増加することも示された。総アベナンスラミド濃度の増加は、時間及び温度依存性であった。浸麦の温度を上昇させると、濃度は、10時間の浸麦中に約75ppmまで50%も増加した。20℃より上、例えば40℃に温度を上昇させる、又は浸麦を例えば48時間に延長しても、アベナンスラミドのさらなる増加はもたらされなかった。濃度はわずかしか増加せず、実質的な量の蓄積はもたらされなかったが、単に乾燥穀粒の水吸収を開始することによって、アベナンスラミド増加がもたらされる。
【0013】
製麦を通してアベナンスラミドを増加させるための現在の技術は、上で概説した所望の生理反応を引き出すのに十分高い濃度のアベナンスラミドを有する製麦製品を産生しないことが現在の研究から明らかである。さらに、特に2日を超える製麦時間による、これらの増加をもたらす製麦/発芽工程はまた、根、苗条及び部分的に枯渇した製麦穀粒を有する発芽したエンバク実生をもたらすので、発芽製品は、根、子葉鞘及び発芽した穀粒と関連する他の解剖学的修飾の存在のために、さらなる乾燥分画/製粉用途における使用を限定してしまうだろう。さらに、現在の技術は、主に被覆エンバク用に開発されてきたので、殻がまだ付着している発芽穀粒をもたらす。業界で広く使用されている衝撃又は圧縮空気殻除去などの機械的手段による製麦前の殻除去、並びに第2の「研磨」手順、すなわち脱皮エンバクの外側に位置するトリコームの除去は、無傷の穀粒の完全性を激しく損ない、製麦にとって望ましくない損傷した穀粒をもたらし得る。さらに、製麦製品の殻除去及び研磨は、製麦材料の柔軟性及び密度の変化によって複雑化される。
【0014】
根、苗条及び部分的に枯渇した製麦穀粒からなる発芽したエンバク新芽はまた、直接消費できる食品への取り込み又は成分のような焼成食品への取り込みに適した、押し麦フレーク、エンムギふすま又はエンバク粉などのいずれかの伝統的なエンバク製品に製粉するのが困難である。
【0015】
さらに、既に観察された強力な遺伝子型/環境相互作用があるとすれば、伝統的な育種技術を通して高アベナンスラミドエンバクを産生する可能性は、達成するのに何年もかかり得る長期的な解決策を意味する。
【0016】
そのため、心血管の健康を回復させるなどのアベナンスラミドの潜在的に有益な効果を認めることができるようにエンバク穀粒中のアベナンスラミド濃度の実質的な増加が必要とされている。
【発明の概要】
【課題を解決するための手段】
【0017】
本発明の一実施形態によると、休眠中のエンバクを偽製麦(false malting)に供し、アベナンスラミド濃度は増加しているが発芽はしていないエンバクをもたらすことによって、アベナンスラミド濃度が増加したエンバクを生産する方法が提供される。
【0018】
好ましくは、エンバクが非休眠性である場合、休眠が誘発される。
【0019】
好ましくは、エンバクが非休眠性である場合、休眠が増強される。
【0020】
好ましくは、エンバクは休眠を誘発又は増強するために嫌気的に浸麦される。
【0021】
好ましくは、エンバクは、休眠を誘発するために、偽製麦の前に乾熱処理される。
【0022】
好ましくは、エンバクは、30〜40℃で48〜72時間乾熱処理され、その後約70℃で144〜約168時間さらに乾熱処理される。
【0023】
好ましくは、エンバクは、エンバクを4〜40℃で12〜18時間、水に浸漬することによって嫌気的に浸麦される。
【0024】
好ましくは、エンバクは、カルシウムイオンを含む水に嫌気的に浸麦される。
【0025】
好ましくは、エンバクは、偽製麦のために、4〜40℃の温度で96〜120時間インキュベートされる。
【0026】
好ましくは、エンバクは、休眠、殻無しエンバクである。
【0027】
好ましくは、エンバク中の増加したアベナンスラミド濃度は、乾燥量基準で、750ppmより高い。
【0028】
好ましくは、エンバクが被覆されており、休眠エンバクである場合には、エンバクは殻除去される。
【0029】
好ましくは、エンバクは、乾燥量基準で約3〜10%の最終水分含量まで乾燥される。
【0030】
好ましくは、エンバクは、約35%の水分含量まで嫌気的に浸麦される。
【0031】
好ましくは、外側のふすま成分及び残留脱ふすま挽割は、摩耗製粉を使用して回収される。
【0032】
好ましくは、表皮剥離ふすま成分は、摩耗製粉前のエンバクの初期重量の3〜30%を含む。
【0033】
本発明のこれらの及び他の特徴は、添付の図面を参照して以下の説明からより明らかとなるだろう。
【図面の簡単な説明】
【0034】
【図1】アベナンスラミドの一般構造式を示す図である。
【図2】個々のアベナンスラミドの構造を説明する図である。
【図3】アベナンスラミド分画の典型的なHPLCプロファイルを説明する図である。
【図4】アベナンスラミド濃度を増加させる代表的な方法のステップを示す図である。
【図5】温度の、加熱処理され、嫌気的に浸麦された殻無しエンバクの総アベナンスラミド濃度への影響を示す図である。
【図6】37℃での偽製麦中の非休眠エンバクへのアベナンスラミドの蓄積の時間経過を示す図である。
【図7】Satake乾燥製粉によって決定される、未製麦穀粒中の総アベナンスラミドの分布を説明する図である。
【図8】Satake乾燥製粉によって決定される、偽製麦後の一品種(VAO-48)における穀粒中の総アベナンスラミドの分布を説明する図である。
【図9】Satake乾燥製粉によって決定される、偽製麦後の別の品種(VAO-22)における穀粒中の総アベナンスラミドの分布を説明する図である。
【発明を実施するための形態】
【0035】
本発明を実施するための本発明者等により考えられた最良の形態を含む本発明のいくつかの特定の実施形態についてここで詳細に言及する。これらの特定の実施形態の例は、添付の図面中に説明される。本発明は、これらの特定の実施形態と組み合わせて記載されるが、本発明を記載の実施形態に限定することを意図していないことが理解されよう。それどころか、添付の特許請求の範囲によって定義される本発明の精神及び範囲内に含まれ得る、代替、修正及び同等形態を包含することを意図している。以下の説明で、本発明の完全な理解をもたらすために、多数の特定の詳細が明らかにされる。本発明は、これらの特定の詳細のいくつか又は全てなしでも実施することができる。他の例では、本発明を不必要に不明瞭にしないために、周知の工程の操作は詳細には記載しなかった。
【0036】
本明細書及び添付の特許請求の範囲において、単数形「a」「an」及び「the」は、文脈が特に明確に指示しない限りは、複数の言及を含む。特に定義しない限りは、本明細書で使用する全ての技術的及び科学的用語は、本発明が属する技術分野の当業者に一般に理解されるのと同じ意味を有する。
【0037】
本発明は、1又は2以上の実施形態に関して記載されている。しかしながら、特許請求の範囲に定義される本発明の範囲から逸脱することなく、いくつかの変形及び修正をすることができることが、当業者に明らかであろう。
【0038】
一般的なエンバク(アベナ・サティバ(Avena sativa))は、穀物の種である。エンバクは、種々の用途を有している。ヒトが消費するためのエンバクは、ポリッジ、朝食用シリアル、クッキー及びスナックバーを作るために使用される。農業では、エンバクの最も一般的な用途の1つは、家畜の飼料である。エンバクは、ウマの食餌の大部分を構成し、ウシにも定期的に与えられている。エンバクはまた、イヌ及びニワトリの飼料のいくつかのブランドにも使用されている。エンバクは、世界中の種々の地域で2000年間栽培されてきた。エンバクは、食品として消費される前は、医療目的、エンバクがまだなお尊ばれている用途で使用されていた。エンバクは、スコットランド、グレートブリテン、ドイツ及びスカンジナビア諸国を含む多くの国々の人々にとっての主食であったので、ヨーロッパでのエンバクの栽培は広範であり、エンバクは重要な商業作物を構成した。
【0039】
殻、すなわち外穎及び内穎は、穀粒の発達中に重要な役割を果たす。殻は、発達中の穀粒に炭水化物を供給する。発達中の穀粒にとって、アミノ酸は主な窒素源であり、この大部分は外穎及び内穎によって与えられ得る。さらに、外穎及び内穎は外側の被覆であるので、小花及び穀粒を病原体及び昆虫による攻撃から保護することができる。
【0040】
大半のエンバクは、収穫される際、殻が付着している。例えばVAO-48など、無殻である品種、すなわち、殻が穀粒に緩く付着しており、コンバイン処理及び収穫中に圃場中に残される品種も存在する。
【0041】
休眠状態で母本から放出される成熟エンバク穀粒は、一次休眠として知られる、種皮強制休眠若しくは胚性休眠のいずれか、又は両方を示すことができる。
【0042】
「休眠」という用語は、通常は発芽に好ましい環境条件下でさえ、種子が一時的に発芽するのを妨げられている状態を説明することを意図している。これらの条件は、水、光、温度、ガス、機械的制限、種皮及びホルモン構造の複雑な組合せであり得る。種子休眠は、例えば、参照によりその内容が本明細書に組み込まれている、Genetic and Molecular Control of Seed Dormancy, B. Li and M.E. Foley, Trends in Plant Science, (1997) Vol.2 (10), pp.384-389に記載されてきた。
【0043】
休眠は、発芽を遅らせ、散布のための時間を許し、同時に全種子が発芽するのを防ぐ。発芽をずらすことは、いくつかの種子及び実生を、短期間の悪天候若しくは一過性の草食動物から損傷又は死をこうむることから保護する。
【0044】
「追熟」の期間後、光、温度、湿度などの環境的に好ましい条件下で、非休眠穀粒は、急速に推移し、発芽の開始がもたらされる。
【0045】
しかしながら、好ましくない環境条件に曝露された、部分的又は完全に追熟した非休眠穀粒は、「二次休眠」として知られる、誘発された休止状態を示し得る。
【0046】
「二次休眠」という用語は、いくつかの非休眠及び休眠後の種子が高温などの発芽にとって好ましくない条件に曝露された状態を説明することを意図している。「休眠」という用語は、一次休眠、例えば、それだけに限定されないが、種皮強制休眠又は真の胚性休眠;二次休眠、例えば、それだけに限定されないが、化学的に誘発される休眠、物理的に誘発される休眠又は他の任意の形の誘発される休眠、自然の二次休眠;及び他の任意の形の休眠を含む、休眠状態の任意の種子を含むことを意図している。
【0047】
多くの園芸植物は、その野生の祖先は休眠していたにもかかわらず、水があり、十分に暖かいとすぐに容易に発芽する種子を有する。これらの栽培植物は、休眠を欠く植物を栽培し維持した植物育種家及び庭師による選択圧の産生のために、休眠を欠く。
【0048】
例えば、以下に記載のエンバク種子の品種であるVAO-48など、逆に、長期の二次休眠を有するよう遺伝的に選択されてきた品種が存在する。
【0049】
製麦の技術分野の当業者によく知られた典型的な製麦工程中に実施されるように、エンバクが、浸漬又は浸麦によって処理され、発芽を可能にされた場合、アベナンスラミド濃度は、乾燥種子中の初期アベナンスラミド濃度に対して、時間依存的様式で、発芽実生中で増加する。アベナンスラミド濃度を増強するための短期間の製麦の有用性は、アベナンスラミドのわずかな増加によって限定され、延長された製麦期間、例えば4〜5日は、より高濃度のアベナンスラミドを含む発芽穀物製品をもたらすが、従来の製粉及び加工市場における有用性を減少させる。
【0050】
本発明の一実施形態によると、以下に記載の二次休眠の誘発及び偽製麦を通して大量のアベナンスラミドが蓄積される。二次休眠中のエンバクを製麦工程に供した場合、驚くべきことに、発芽しないが、アベナンスラミド濃度が25〜35倍まで劇的に増加し得る。
【0051】
言い換えれば、発芽が起こらない条件を作り出す予測可能な方法で二次休眠を利用することによって、発芽穀物と比較して、全粒穀粒の栄養価を変化させることができる。
【0052】
6日間までの休眠エンバクの浸漬及び発芽中、発芽の肉眼的視覚的指標、例えば、根の発生、分岐及び伸長、子葉鞘及び苗条の拡大及び伸長等が観察されていないという事実にもかかわらず、アベナンスラミドは産生され続ける。典型的な製麦手順を伴うが、休眠中の穀物が発芽しないこの処理を、以降、「偽製麦」と呼ぶ。
【0053】
「偽製麦」という用語は、当業者によって実施される製麦技術と類似又は同一の処理を説明することを意図している。しかしながら、種子は休眠、例えば二次休眠中であるので、偽製麦に供された種子は発芽しない。
【0054】
偽製麦は、穀物一般、特にエンバクの組成を変化させる新しい技術を表す。製麦中のエンバクの発芽の抑制は、多数の確立された食品、飼料及び産業利用の市場への取り込みに貢献するアベナンスラミド含量が増加した全粒エンバク製品をもたらす。
【0055】
アベナンスラミドの定量分析の方法
エンバク材料の質的及び量的なアベナンスラミド組成物の決定を以下に記載するように行った。エンバク試料、すなわち種子を、恒量まで37℃で、オーブン中で乾燥させ(約48時間)、分析まで−20℃で真空パックビニール袋中に貯蔵した。抽出前に市販のコーヒーミルを使用して種子を挽いた。抽出及び定量分析は、一般的に、2反復を使用して行った。
【0056】
抽出
還流酸性化80%エタノール(エタノール:水:氷酢酸、80:19.9:0.1(v/v/v))75mlに、挽いたエンバク試料10gを、抗酸化剤としての亜ジチオン酸ナトリウム5mgと共に、激しく攪拌しながら添加した。混合物を熱から離し、攪拌しながら室温で20分間冷却させた。次いで、全内容物を、フリットディスクを備えた目盛り付きガラスクロマトグラフィーカラムにデカントした。重力によって懸濁液を沈降させ、透明な上澄みを有する軽く圧縮された抽出物ベッド(ベッド容積=VbmL)を形成した。上澄みを重力流によって回収し、抽出ベッドを「浸出抽出」によって3×Vbの酸性化80%エタノールで溶出し、透明な黄緑色溶離液を得た。
【0057】
疎水性相互作用及び芳香族吸着クロマトグラフィーによるアベナンスラミドの精製
この抽出物から親油性成分を除去するため、Octyl Sepharose(商標) CL 4-Bクロマトグラフィービーズを添加し(抽出物g当たり0.5ml)、混合物を回転蒸発によって真空下40℃で濃縮乾固した。酸化を防ぐために、蒸留水を混合物に添加して、確実に乾燥中にアベナンスラミドが沈殿するようにした。乾燥混合物を酸性化50%エタノール(エタノール:水:氷酢酸、50:49.9:0.1(v/v/v))に再懸濁し、前もって重力圧縮し、酸性化50%エタノール(例えば、25mlの試料10gについて、最終ベッド容積Vb=30mL)にあらかじめ平衡化したOctyl Sepharose(商標)CL 4-Bを含有する目盛り付きガラスクロマトグラフィーカラムに定量的に移した。次いで、カラムを3×Vbの酸性化50%エタノールで溶出した。合わせた溶離液を回転蒸発によって真空下40℃で濃縮して本質的に脂質を含まない抽出物を得た。
【0058】
サポニン、フラボノイド配糖体、アルコール可溶性タンパク質及びペプチド、遊離糖、芳香族、有機物及びアミノ酸を除去するため、濃縮抽出物を小容積(試料10g当たり約3mlの酸性化40%エタノール(エタノール:水:氷酢酸、40:59.9:0.1(v/v/v)))に溶解し、Sephadex(商標)LH-20を使用してクロマトグラフィーによって精製した。溶液を、前もって重力圧縮し、酸性化40%エタノール(例えば、試料10gについて、最終ベッド容積Vb=25ml)にあらかじめ平衡化したSephadex(商標)LH-20を含有する目盛り付きガラスクロマトグラフィーカラムに定量的に移した。最初に、サポニン、遊離糖、アミノ酸等を、2×Vbの酸性化40%エタノールで溶出することによって除去した。吸収されたアベナンスラミドは、3×Vbの酸性化95%エタノール(エタノール:水:氷酢酸、95:4.9:0.1(v/v/v))で溶出することによって回収した。再度、酸化を防ぐための亜ジチオン酸ナトリウム5mgと共に蒸発前に水を添加した。溶離液を回転蒸発によって真空下40℃で濃縮乾固して精製アベナンスラミド分画を得た。
【0059】
アベナンスラミド分画のHPLC分析
第1の方法では、精製アベナンスラミド分画を50%エタノール(エタノール:水、50:50(v/v))5mlに溶解し、0.45μmフィルターを通して濾過し、HPLCにかけた。試料(10μl)は、Rheodyne(商標)インジェクターを使用して、C18ガードカラムを備えたCERA Column Cooler 250を使用して25℃に維持したC18逆相カラム(ODS Hypersil(商標)C18、5μm、4.6mm×250mm)に注入した。Thermo Separation Products(TSP)Spectra System P4000ポンプを使用してHPLC分析を行い、TSP SpectraSystem(商標)UV3000スペクトルスキャン検出器及びChromQuestソフトウェアを使用して330nmで監視した。流速は1分当たり0.8mlに維持した。HPLC用溶媒は、A:メタノール、B:水及びC:5%酢酸とした。溶媒の勾配(容積%)は、40A:55B:5Cからなり、40分で50A:45B:5Cに直線的に増加し、15分で80A:15B:5Cに直線的に増加し、次いで3分で100Aに達し、3分間維持した。溶媒の勾配は、3分で元の状態に戻し、4分間平衡化した。全ての主要アベナンスラミドのピークを、相対的保持時間及びUVスペクトル(240〜380nmを監視)の真正標準(authentic standards)との比較によって同定した。全ての微量アベナンスラミドは、HPLC−質量分析のみによって同定した。
【0060】
第2の方法では、第1の方法の溶媒系を以下のように変更した。HPLC用溶媒は、A:メタノール、B:水及びC:0.1Mリン酸とした。溶媒の勾配(容積%)は、45A:45B:10Cからなり、55分で60A:30B:10Cに直線的に増加し、次いで3分で100Aに達し、3分間維持した。溶媒の勾配は、3分で元の状態に戻し、4分間平衡化した。
【0061】
第3の方法では、試料(10μl)を30℃に維持したC18逆相カラム(Zorbax Stable(商標)結合C18、3.5μm、4.6mm×150mm)に注入した。HPLC用溶媒は、A:メタノール、B:水及びC:0.5Mギ酸とした。溶媒の勾配(容積%)は、45A:45B:10Cからなり、24分で55A:35B:10Cに直線的に増加し、9分で70A:20B:10Cに直線的に増加し、次いで3分で95A:5Cに達し、3分間維持した。溶媒の勾配は、3分で元の状態に戻し、3分間平衡化した。
【0062】
アベナンスラミド分画のHPLC質量分析
微量アベナンスラミドのピークを、Surveyor(商標)HPLC-UVダイオードアレイ検出器システムを備えたThermo Finnigan LCQ Advantage(商標)質量分析計を使用して、HPLC−MS−MSによって同定した(HPLC条件:Hypersil(商標)ODS、120Å、5μ、250×4.6mmカラム)。HPLC UVモニタリングは330nmで行った。1分当たり0.8mLの流速で、第1の方法で上記したのと同じ溶媒系を使用した。溶媒の勾配(容積%)は、40A:55B:5Cからなり、80分で60A:35B:5Cに直線的に増加し、5分で80A:15B:5Aに直線的に増加し、次いで3分で100Aに達し、3分間維持した。溶媒の勾配は、3分で元の状態に戻し、3分間平衡化した。MS−MS条件:エレクトロスプレーイオン化(ESI,electro-spray ionization、ネガティブモード)、電源電圧:4.5Kボルト、キャピラリー電圧:−10ボルト、キャピラリー温度:300℃、シースガス流量:80%満、補助ガス流量:20%最大(分割流無し)。
【0063】
アベナンスラミドの定量的推定
個々のアベナンスラミドは、外部真正アベナンスラミドA標準に対する330nmのピーク面積を決定することによって定量化し、アベナンスラミドA重量当量として表した。総アベナンスラミドは、アベナンスラミドA重量当量として計算した個々のアベナンスラミド量を合計することによって計算し、乾燥重量基準で百万分率(ppm)アベナンスラミドA当量として表した。
【0064】
アベナンスラミドは、図1に示す一般式の、約30種の異なるN−アロイルアントラニル酸アルカロイドの群である。図2は、麦芽エンバク及び麦芽になっていないエンバクの両方で生じる個々のアベナンスラミドの構造を説明する。
【0065】
図3は、図2において麦芽にされ、命名を割り当てられた、上記第3の方法を使用したエンバクからのアベナンスラミド分画の典型的なHPLCプロファイルを説明する。
【0066】
図4は、本発明の実施形態に従って、アベナンスラミド濃度を増加させる方法のステップを示す。
【0067】
エンバクが休眠中でない場合(402)、二次休眠が誘発され得る(404)。そうでなければ、エンバクは休眠、一般的には、自然の又は誘発された二次休眠中である。
【0068】
種子が休眠中だとしても、二次休眠中の種子は、経時的に休眠から覚醒し得るので、休眠を増強するための任意のステップ(406)が含まれ得る。
【0069】
二次休眠を誘発又は増強するための1つの非限定的方法は、エンバクを嫌気的に水和させる、又は焼戻しすることである。
【0070】
製麦は、一般的に、エンバクが特定の水分含量に達するまで、エンバクを水に浸麦することから始まる。浸麦は、一般的に、エンバクの空気への曝露が点在しており、エンバクが追加の酸素を得ることが可能になっている。
【0071】
エンバクの休眠は、嫌気的な浸麦を通して誘発(404)又は増強(406)され得る。嫌気的な浸麦では、エンバクは、例えば30℃より高い温度で12〜18時間、嫌気的に焼戻し又は水和される。
【0072】
次いで、エンバクは、偽製麦(408)、すなわち製麦と類似又は同一であるが発芽しない条件、例えば23〜37℃で96〜120時間供される。偽製麦後、エンバクは、貯蔵又はさらなる加工前に、例えば、35℃で24〜48時間乾燥される(410)。
【0073】
以下は、本発明の実施形態によるアベナンスラミド濃度の増加を示す非限定的実施例である。
【実施例1】
【0074】
休眠対非休眠エンバク
新しく収穫した休眠、殻無し、覆い無し(bald)播種エンバク育種系統(VAO-48)の種子試料及び非休眠、殻無し、覆い無し播種エンバク育種系統(VAO-2、品種AC GEHLとして現在登録されている)の種子試料を以下に記載のように製麦した。
【0075】
各々の育種系統約20gを、穏やかに攪拌しながら、室温で20分間、1%次亜塩素酸ナトリウム水溶液に液浸することによって手短に表面殺菌し、次いで、過剰な次亜塩素酸ナトリウム溶液を除去するために取り出し、徹底的にすすいだ。次いで、室温及び散乱光で4日間、湿った濾紙ディスク上の被覆したペトリ皿(150×15mm)中で種子を発芽させた。各々の育種系統の別の試料20gを対照として取っておいた。
【0076】
4日後、2つの試料の発芽率を決定した。次いで、発芽及び未発芽両方を含む種子を取り出し、室温で2日間、種子乾燥機中で乾燥させた。平均発芽率は、以下のとおりであった。
休眠、殻無し、覆い無し(VAO-48) 1%
非休眠、殻無し、覆い無し(VAO-22) 75%
【0077】
(偽)製麦前及び後両方の2つの試料の総アベナンスラミド含量を、上記の第1の方法を使用して決定し、表1に要約する。
【0078】
【表1】
【0079】
表1は、休眠エンバクが、驚くべきことに、偽製麦期間中にアベナンスラミドを蓄積したことを示している。
【0080】
休眠種子は、出発材料と同様の形態を有し、非休眠種子とは異なり、根、子葉鞘、並びに新生の葉及び苗条を有さなかった。
【実施例2】
【0081】
嫌気的浸麦及び休眠種子の貯蔵の休眠への影響
休眠エンバクは、貯蔵の条件(例えば、酸素濃度、温度、水分含量)に応じて、時間と共にその二次休眠から覚醒することが知られている。特に、低酸素濃度は、休眠を延長する、及び/又は発芽を阻害する。サブゼロ温度(例えば、−20℃)で貯蔵した種子は、長期間その遺伝的に伝達する形質の大半を保持することができることも、エンバク育種家及び種子胚芽プラズマ保存技術分野の当業者に知られている。しかしながら、製麦などの大規模工程に実用的であるためには、種子を休眠状態に維持するための高価な低温貯蔵に頼らず、休眠エンバクの一定供給が、一年中定期的に利用可能であるべきである。そのため、休眠エンバクの休眠を延長及び増強し、非休眠エンバクの二次休眠を誘発することが好ましい。
【0082】
室温で3週間、次いで−20℃で1週間貯蔵した、実施例1で使用した休眠エンバク系統(VAO-48)の種子を、約23℃で水道水に約48時間までの種々の時間浸した。小規模実験では、種子を、水で完全に満たし、Parafilmで密封した気密ペトリ皿に散布した(すなわち、嫌気的浸麦)。約2、4、6、8、12、24及び48時間後、種子を取り出し、室温で、種子乾燥機中で手短に風乾した。次いで、種子を表面殺菌し、散乱光中室温で4日間、上記のように製麦した。4日後、発芽率を上記のように決定した。
【0083】
さらなる実験では、より大量の種子を使用し、水道水で満たし、Parafilm(商標)で密封した気密Erlenmeyerフラスコ中で嫌気的浸麦を行った。フラスコ形状の頂上の「先細り」は、残気の排除を促進し、水位より下への種子の水浸を保証した。両実験の結果を表2に要約する。
【0084】
【表2】
【0085】
空気への曝露なしで、約8〜18時間一晩中種子を単に浸漬又は浸麦することによって、浸麦なしの約50%から嫌気的浸麦後10%未満まで発芽率がかなり下がったことが分かった。これは、8〜18時間の嫌気的浸麦によって発芽が実質的に減少し得ることを示している。浸麦温度を最適化するためのさらなる実験は、30℃〜32℃の温度で18時間が、3%未満の発芽(すなわち、97%を超える休眠)という一貫した発芽率をもたらすことを示した。
【0086】
種々の貯蔵期間後の休眠エンバク系統の休眠属性の保持を試験するために、室温で14週間まで種々の期間貯蔵した試料を使用して発芽実験を行った。さらに、控え試料を追加で一晩中嫌気的浸麦に供し、手短に風乾し、次いで同様の条件下で発芽を試験した。両時間経過実験では、種子を室温及び散乱光で、4日間ペトリ皿中で発芽させ、発芽率を計算した。両実験の結果を表3に示す。
【0087】
【表3】
【0088】
休眠エンバク系統の休眠は、貯蔵の最初の8〜10週間で増加し、次いで劇的に減少して、約90%の種子が発芽する。予想通り、16時間嫌気的に浸麦したエンバクは対応する非浸麦試料よりも、室温で8週間以上の貯蔵後により大きい休眠を示した。こうして室温での貯蔵で徐々に休眠から覚醒するエンバクの休眠を増強し、この工程により適したものにすることができる。
【実施例3】
【0089】
加熱前処理及び嫌気的浸麦の、非休眠、殻無しエンバクにおける休眠及びアベナンスラミド蓄積への影響
二次休眠を増強及び誘発する他の方法が存在する。エンバクの休眠を増強及び誘発する非限定的な代表的な方法は、30〜70℃の乾熱で2週間までの種々の期間エンバクを処理することである。この手順は通常、2段階で行う。第1段階は、約30℃の温度で数日間、種子の水分含量を約3%まで下げることを含み、その後、第2段階は約70℃で1週間までである。このような処理はまた、種子伝染性糸状菌及び細菌胞子を減少させるのに有効であることも示されてきた。この体制下で種子は損傷せず、いくつかの場合には、発芽率が事実上増加する。
【0090】
非休眠、殻無しエンバク系統(VAO-2)の種子(500g)を対流式乾燥器中37℃で72時間、次いで70℃で144時間加熱処理した。室温まで冷却後、加熱処理種子を小口試料に分割し、ある群(5反復)は32℃で18時間嫌気的に浸麦し、別の群は嫌気的に浸麦しなかった(5反復)。両群を上記のように表面殺菌し、次いで散乱光中室温で、4日間ペトリ皿(150×50mm)中で製麦した。製麦後、種子を、上記の第1の方法を使用して、発芽率(発芽%)及びアベナンスラミド含量について評価した(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)。結果を表4に要約する。
【0091】
【表4】
【0092】
上記のように、2段階加熱処理下でのエンバクの乾熱処理及び18時間の嫌気的浸麦の両処理は、別々に製麦中のエンバクの発芽を減少させた(すなわち、休眠を増加させた)。加熱処理の後に嫌気的浸麦と、共に使用した場合、発芽率は実質的に0%まで低下し、4日の製麦期間中、種子は休眠したままであった。さらに、加熱及び嫌気的浸麦の処理を組み合わせた4日の「偽製麦」したエンバクの総アベナンスラミド含量は、最初の77ppmから製麦後445ppmまで増加し、初期値に対して約480%(5.8倍)の増加を示した。
【0093】
休眠を誘発するための2段階乾熱処理工程が生存率の減少をもたらさなかったことに留意する。加熱処理エンバクを100ppmのジベレリン(例えば、GA3)存在下で5日間インキュベートした場合、発芽のほぼ完全な回復が観察された。
【実施例4】
【0094】
温度の、偽製麦中のアベナンスラミド蓄積への影響
伝統的な製麦では、発芽及び後の発酵のための種子のデンプン貯蔵物質の遊離糖への同時の加水分解を最大化するように最適温度が選択される。これらの目的のため、最適温度は通常、約25℃未満である。温度のアベナンスラミド蓄積への影響を研究するため、4種の異なる温度で、暗中で4日間偽製麦を行った。比較のため、散乱光下、室温でも偽製麦を行った。これらの研究のため、加熱前処理及び嫌気的浸麦によって休眠状態にした非休眠、殻無しエンバク品種を使用した。
【0095】
本工程のための4種の温度は、3℃、23℃(室温)、30℃及び37℃とし、暗中で偽製麦を行い、比較試料を23℃で、散乱光中で製麦した。3℃での製麦は実験室の冷蔵庫で行い、30℃及び37℃での製麦は制御温度対流式オーブンで行った。
【0096】
AC Baton殻無しエンバク(AC Baton、発芽率は95%を超える)の試料50gを使用した。種子は以下:上記実施例3のように37℃で72時間、その後70℃で144時間加熱処理し、室温(23℃)まで手短に冷却し、次いで32℃で18時間水道水に嫌気的に浸麦し、種子乾燥機中で約4時間風乾した。種子は、室温で1時間0.25%次亜塩素酸ナトリウム水溶液を使用するのみで上記のように表面殺菌し、次いで4つの小口試料に分割し、実施例3の制御温度対流式オーブンを使用して、4種の異なる温度で、暗中で96時間「偽製麦」した。
【0097】
製麦後、種子を発芽率(発芽%)について評価し、室温で一晩、種子乾燥機中で風乾し、アベナンスラミド含量を上記の第1の方法によって決定した(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)。3種の主要アベナンスラミド(アベナンスラミドA、B及びC)、他の全てのアベナンスラミド及び総アベナンスラミドの含量を計算し、未製麦材料中の対応する濃度と比較した。結果を表5に要約する。
【0098】
【表5】
*発芽%の値は、4日の発芽率を表す。括弧内のアベナンスラミドの値は、製麦前濃度を表す。
【0099】
比較の観点から、表5は、蓄積したアベナンスラミドA、B、C及び他の全てのアベナンスラミドの濃度が製麦温度の上昇と共に偽製麦中に増加したことを示している。使用した最も低い温度(3℃)は、未製麦材料中の114ppmから製麦後約142ppmまで(約25%又は1.25倍)の増加をもたらした一方で、最も高い温度(37℃)は、ほぼ2,000ppm(約1,650%又は17.5倍)の総アベナンスラミド濃度をもたらした。ここで使用した前処理体制下で、高温及び低温の両方が発芽をわずかに減少させた。
【0100】
温度の、加熱及び嫌気的浸麦手順によって休眠状態にした非休眠(95%を超える生存種子)、殻無しエンバクAC Batonを使用した96時間の偽製麦中に蓄積した総アベナンスラミドへの影響を図5に示す。単に最も熟練した製麦技術で使用される温度(通常10〜25℃)より上の温度で製麦することによって示される、対照(すなわち、未製麦)種子に対する劇的な増加を容易に見ることができる。
【実施例5】
【0101】
時間の、偽製麦中のアベナンスラミド蓄積への影響
偽製麦工程中のアベナンスラミド蓄積の時間経過を研究するために、非休眠、殻無しエンバク品種を使用して、種々の期間で工程を行い、蓄積したアベナンスラミド濃度を決定した。
【0102】
AC Baton殻無しエンバク(AC Baton、発芽率は95%を超える)の試料120gを使用した。種子を実施例3のように加熱処理し、室温(23℃)まで手短に冷却し、次いで32℃で18時間水道水に嫌気的に浸麦し、約4時間種子乾燥機中で風乾した。種子を実施例4のように表面殺菌し、次いで、湿ったペーパータオル上のアルミニウム箔で覆った浅いトレイ内で、暗中37℃で種々の期間「偽製麦」した。発芽率は、4日後に1%未満であった。各々の製麦期間後、種子の代表的な小口試料(約30g)を取り出し、室温で4時間乾燥させ(強制空気)、次いで37℃で約40時間乾燥させることによって恒量にし、上記の第1の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析するまで−20℃で貯蔵し、未製麦材料と比較した。結果を表6に要約し、図6に示す。
【0103】
【表6】
*括弧内のアベナンスラミドの値は、製麦前濃度を表す。
【0104】
表6及び図6のデータから、アベナンスラミドA、B、C及び総アベナンスラミドの濃度全てが偽製麦期間中に時間と共に増加し、増加の時間経過は3日までやや線形パターンに従い、4日で水平になることが分かる。総アベナンスラミドの4日全体の蓄積は、初期値に対して約740%(8.4倍)の増加を示した。
【実施例6】
【0105】
カルシウムイオン浸麦処理の、休眠エンバクVAO-48の偽製麦中のアベナンスラミド蓄積への影響
嫌気的浸麦段階中のCa+2イオンの存在が、その後の製麦段階中のアベナンスラミドの蓄積に何らかの影響を有するかどうかを調査するために、浸麦水に一連の増加するCa+2濃度を使用し、製麦後にアベナンスラミド含量を決定した。
【0106】
殻無し、無毛の休眠エンバクVAO-48の試料25gを、35℃で水又は種々の濃度のUSP−グレードCaCl2・(H2O)2溶液のいずれかに嫌気的に浸麦した。18時間の浸麦及び種子乾燥機中で約4時間風乾後、種子を上記のように表面殺菌し、次いで制御温度オーブン中、30℃、暗中で4日間、ペトリ皿(150×50mm)を使用して「偽製麦」した。全ての場合の発芽率は、4日後に1%未満であった。製麦後、種子を取り出し、室温で4時間乾燥させ(強制空気)、次いで37℃で約40時間乾燥させることによって恒量にし、上記の第3の方法によって(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、アベナンスラミド含量について分析するまで−20℃で貯蔵し、未製麦材料と比較した。結果を表7に要約する。
【0107】
表7に示すように、個々のアベナンスラミド濃度は未製麦種子中で非常に低く、製麦前の総アベナンスラミド含量はわずか約32ppmであった。カルシウムイオンを添加していない水への浸麦及び30℃で4日の製麦後、全アベナンスラミド濃度は、ずっと高く、439ppmの総アベナンスラミド濃度は、約13.7倍の増加を示した。浸麦水中のCa+2濃度の増加と共に、麦芽穀粒中のアベナンスラミド濃度は、増加傾向を示し、Ca+2を添加しなかった場合の約439ppmから1%Ca+2を添加した場合の約757ppm(約1.7倍)まで浸麦中に増加し、未製麦種子に対して1%Ca+2を添加した場合全体として約23.6倍増加した。これらの結果は、浸麦期間中のCa+2の添加が、その後の製麦中のアベナンスラミド蓄積を増強することを示している。
【0108】
【表7】
【実施例7】
【0109】
高濃度の塩化カルシウム浸麦処理の、非休眠エンバクVAO-22の偽製麦中のアベナンスラミド蓄積への影響
非休眠、殻無し、無毛のエンバク系統VAO-22及び浸麦水中でより高濃度のCa+2を実施例7で使用した。
【0110】
VAO-22殻無し、無毛、非休眠エンバク育種家の種子の試料1300gを使用した(4日後の発芽率84%)。次いで、種子を実施例5のように加熱処理した。未製麦種子を表す対照として試料25gを使用し、分析するまで−20℃で貯蔵した。加熱処理した材料の残りを1.0%又は2.0%CaCl2・(H2O)2のいずれかの中、35℃で18時間嫌気的に浸麦し、種子乾燥機中で約4時間風乾した。次いで、種子を上記のように表面殺菌し、制御温度オーブン中、暗中30℃で4日間、ペトリ皿(150×50mm)を使用して「偽製麦」した。製麦後、種子を取り出し、室温で4時間乾燥させ(強制空気)、次いで37℃で約40時間乾燥させることによって恒量にし、上記の第3の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析するまで−20℃で貯蔵し、未製麦材料と比較した。結果を表8に要約する。
【0111】
表8から分かるように、この品種のエンバクの個々の及び総アベナンスラミド濃度は、1及び2%の両濃度のCaCl2に浸麦した後、4日の偽製麦中に増加した。例えば、1%CaCl2に18時間嫌気的に浸麦し、30℃で4日間偽製麦した場合、アベナンスラミドBは、3.4ppmからほぼ80ppmまで(2,240%;初期値に対して23.4倍)増加した一方で、2%CaCl2では、濃度は103ppm(2,930%;初期値に対して30.3倍)に達した。アベナンスラミドCについては、対応する増加は、1%CaCl2で2.3ppmから65.2ppmまで(2,735%;初期値に対して28.4倍)、2%CaCl2で100.1ppmまで(4,252%;初期値に対して43.5倍)であった。他のアベナンスラミドも同様に増加し、総アベナンスラミド濃度は、1%及び2%CaCl2でそれぞれ2,307%及び3,190%(初期値に対して24.1倍及び31.9倍)の濃度を示した。これら2つの実施例のデータは、嫌気的浸麦ステップ中に添加した少なくとも2%までの濃度範囲のCaCl2の、その後の製麦エンバク中のアベナンスラミドの蓄積及び/又は保持を増強する役割を支持する。
【0112】
【表8】
【実施例8】
【0113】
偽製麦前及び後の休眠エンバクVAO48エンバクのSatake製粉の、ふすま及び脱ふすま挽割乾燥粉砕分画へのアベナンスラミド蓄積への影響
前述のように、偽製麦は、大部分は未製麦穀粒と本質的に同じであり、それゆえ、従来の乾燥摩耗製粉、例えばSatake製粉を通してふすま及び脱ふすま製品への加工に容易に使用可能なエンバク穀粒を生じる。
【0114】
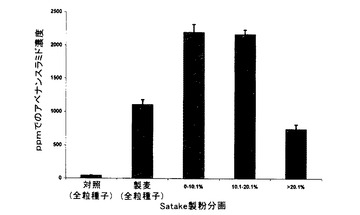
殻無し、無毛、休眠エンバクVAO-48の試料(約380g)を、Satakeを使用して製粉し、以下の連続的な分画を得た。
0〜7(重量)%ふすま分画
7〜15(重量)%ふすま分画
15〜23(重量)%ふすま分画
23(重量)%を超える脱ふすま挽割
【0115】
全製粉分画は、直ちに氷上で冷却し、分析するまで−20℃で貯蔵した。
【0116】
未製粉全粒種子及びSatake製粉分画の控え試料を、上記の第1の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析し、結果を表9及び図7に要約する。
【0117】
【表9】
【0118】
表9に示すように、未製麦VAO-48エンバクのアベナンスラミド含量は分画間でかなり変化し、最高濃度(全粒穀粒中の濃度のほぼ4倍)が最外側の(すなわち、0〜7%)ふすま層で示され、後続のふすま分画で内側に減少した。穀粒の23%を除去した後の残りの脱ふすま挽割は、全粒穀粒のアベナンスラミド含量の半分未満の最低アベナンスラミド濃度を有した。外層の除去の増加と共に含量が減少するこの傾向は、総アベナンスラミド含量においてのみならず、大部分において、個々のアベナンスラミドの各々の濃度でも見られた。
【0119】
実施例5における上記のように加熱処理し、使用するまで−20℃で貯蔵した同じ材料のさらなる試料(377g)を35℃で18時間、水道水に嫌気的に浸麦した。種子を実施例4のように表面殺菌し、次いで、制御温度オーブン中、30℃で4日間、「偽製麦」した。全ての場合において、発芽率は4日後に1%未満であった。製麦後、種子を取り出し、制御温度オーブン中37℃で恒量まで乾燥させ、製粉するまで−20℃で貯蔵した。以下のSatake製粉分画は上記のように調製した。
0〜10.7(重量)%ふすま分画
10.7〜20.6(重量)%ふすま分画
20.6(重量)%を超える脱ふすま挽割
【0120】
未製粉全粒製麦エンバク及び製粉分画の各々の控え試料を上記の第2の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析した。結果を表10及び図8に要約する。
【0121】
【表10】
【0122】
明白な減少の程度は、未製麦穀粒ほど劇的でなく、中間のふすま材料及び脱ふすま挽割の相対的含量は、未製麦材料中の同様の分画よりも比例的にわずかに高かったが、脱ふすまの程度が増加するにつれてアベナンスラミド濃度が減少する傾向は、回収した個々の分画で同様であった。0〜10.7%脱ふすま分画中に蓄積した総アベナンスラミドは、3,832ppmも高くに達し、未製麦穀粒からの同様の分画に対して2,218%(22.2倍)の増加及び全粒未製麦材料に対して91.5倍の増加を示した。
【0123】
0〜10.7%及び10.7〜20.6%のふすま分画を合わせて0〜20.6%の全体的な分画(すなわち、20.6%製粉収率)を形成した場合、材料は、約3,388ppmの総アベナンスラミド濃度を有する。
【0124】
これらの結果は、穀粒の外層が最高濃度の全アベナンスラミドを含有することを示している。
【実施例9】
【0125】
偽製麦前及び後の殻無し、無毛、非休眠エンバクVAO22エンバクのSatake製粉の、ふすま及び脱ふすま挽割乾燥製粉分画へのアベナンスラミド蓄積への影響
偽製麦ふすまの外層中のアベナンスラミド濃度は、全粒穀粒中の濃度に対して高いが、このふすまの収率(製麦穀粒の7〜10%から)は低い。穀粒の特徴的な細長い形状及び深く区切られた中央のしわの形態のために、ふすま及び胚乳の明確な分離が限定され、胚乳組織の混入が最小限のふすまの収率は、少ない脱ふすま及び低いふすま分画製粉収率に制限される。しかしながら、殻無し、無毛、非休眠系統VAO-22育種系統は、短い穀粒及び比較的浅いしわを有し、全体形状及び形態がコムギ穀粒に似ており、Satake摩耗製粉にはるかにより適したものになっている。
【0126】
VAO-22殻無し、無毛、非休眠エンバクの育種家の種子の実施例7で使用した材料のさらなる試料を使用した(4日後の発芽率84%)。次いで、種子を実施例5のように加熱処理した。試料25gを、未製麦種子を表す対照として使用し、分析するまで−20℃で貯蔵した。加熱処理した材料の残りは、水道水中に35℃で18時間嫌気的に浸麦した。次いで、種子を実施例5のように表面殺菌し、制御温度オーブン中、暗中30℃で4日間「偽製麦」した。製麦後、種子を取り出し、制御温度オーブン中37℃で恒量まで乾燥させ、製粉するまで−20℃で貯蔵した。以下のSatake製粉分画を上記のように調製した。
0〜10.1(重量)%ふすま分画
10.1〜20.1(重量)%ふすま分画
20.1(重量)%を超える脱ふすま挽割
【0127】
未製粉全粒製麦エンバク及び製粉分画の各々の控え試料を、上記の第3の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析した方法2を使用してHPLCによってアベナンスラミド含量について分析し、未製麦材料と比較した。結果を表11及び図9に要約する。
【0128】
【表11】
【0129】
表11に示す全粒製麦穀粒の総アベナンスラミド含量は、約42ppmから約1,100ppmまで増加し、未製麦穀粒に対して約2,560%(26.6倍)の増加を示した。0〜10.1%ふすま分画中のアベナンスラミド濃度は、ほぼ2,200ppmまで増加し、全粒未製麦穀粒中の濃度に対して、この材料中のアベナンスラミドにおいて5,170%の増加(52.7倍)を示した。同様の増加は、10.1〜20.1%ふすま分画中でも見られ、合わせて0〜20.1%の全体分画(20.1%製粉収率で)を形成すると、約2,183ppmの総アベナンスラミド濃度を有するだろう。
【0130】
これは、VAO-48エンバクを使用した表10に示す同様の結果よりも低いが、それでも約20%の製粉収率とかなりのアベナンスラミド増加を示す。しかしながら、このエンバクは、VAO-48材料で観察される同様の中間の製粉分画中のアベナンスラミド含量における急速な減少傾向を含まないことが明白である。
【0131】
未製麦及び偽製麦の殻無し、無毛エンバクのSatake製粉によって産生されるものなどの乾燥摩耗製粉分画を使用して、任意の所望の濃度及び同程度の脱ふすまから約3,000ppmもの高い総アベナンスラミド濃度のふすま混合物を製造し、穀粒内の局在に関連するタンパク質、β−グルカン、フィチン酸塩等の他の成分の組成における変化を最小限にすることができることも実施例8及び9から明白である。
【実施例10】
【0132】
30℃での4日の偽製麦後の被覆、非休眠エンバク品種へのアベナンスラミドの蓄積
殻無し、無毛エンバク品種を使用する経済的利点にもかかわらず、最終製品中の殻の包含が正当化され得る技術の潜在的用途、例えば、それだけに限定されないが、殻の除去が経済的であると判明していない、飼料用途、非食品用産業用分画用途が存在する。
【0133】
本発明で開発された技術を使用した被覆エンバクの製麦は、殻が製麦後に除去されているか、又は製品中にまだ存在しているかのいずれかである製品をもたらす。従来の殻除去及び研磨装置によって被覆エンバクの殻を除去しようとする試み及び毛(トリコーム)の除去は、穀粒へのある程度の機械的損傷及びその後の製麦工程中の複雑化をもたらす。
【0134】
それにもかかわらず、上記と同じ手順の下で商業用エンバクを慎重に殻除去し製麦することによって被覆エンバクを使用する試みがなされた。あるいは、全粒被覆エンバクも以下に記載のように試された。
【0135】
被覆、非休眠エンバクJordan実験用種子の試料約500gを使用した。種子を、殻がついたまま実施例5のように加熱処理し、実施例5のように表面殺菌した。小口試料80gを、未製麦種子を表す対照として使用し、分析するまで−20℃で貯蔵した。加熱処理した材料の残りは、水道水中に35℃で18時間嫌気的に浸麦した。次いで、種子を、0.25%次亜塩素酸ナトリウム水溶液を使用して室温で1時間、実施例5のように表面殺菌し、制御温度オーブン中、暗中30℃で4日間「偽製麦」した。製麦後、種子を取り出し、制御温度オーブン中37℃で恒量まで乾燥させ、−20℃で貯蔵した。全粒製麦エンバクの控え試料を、上記の第3の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析し、未製麦材料と比較した。
【0136】
被覆、非休眠エンバクAC Goslinの試料1,000gを使用した。種子を、殻がついたまま実施例5のように加熱処理し、穀粒に損傷を与えるのを避けるために小さなローラー殻除去機を使用して機械的に殻を除去し、実施例5のように表面殺菌した。小口試料25gを、未製麦種子を表す対照として使用し、分析するまで−20℃で貯蔵した。種子の残りは、水道水中に35℃で18時間嫌気的に浸麦し、種子乾燥機中室温で4時間簡手短に風乾し、0.25%次亜塩素酸ナトリウム水溶液を使用して室温で1時間、実施例5のように表面殺菌した。次いで、種子を、室温及び散乱光で4日間、湿った濾紙ディスク上の大きなペトリ皿(150×15mm)中で「偽製麦」した。製麦後、種子を取り出し、種子乾燥機中で4日間乾燥させ、分析するまで−20℃で貯蔵した。全粒製麦エンバクの控え試料を、上記の第1の方法を使用して(アベナンスラミドA当量μg/乾燥重量g=アベナンスラミドA ppm)、HPLCによってアベナンスラミド含量について分析し、未製麦材料と比較した。
【0137】
結果を表12に要約する。
【0138】
【表12】
【0139】
表12で分かるように、殻を有する未製麦無傷試料は、約57ppmの総アベナンスラミド含量を有していた。30℃で4日間製麦した場合、種子は、総計183.7ppmのアベナンスラミドを蓄積し、未製麦試料に対して約220%の増加を示した。
【特許請求の範囲】
【請求項1】
休眠中のエンバクを提供するステップと、
前記エンバクを、アベナンスラミド濃度が増加しているが発芽はしていないエンバクをもたらす偽製麦に供するステップと
を含む、アベナンスラミド濃度が増加したエンバクを生産する方法。
【請求項2】
エンバクが非休眠性であり、休眠中のエンバクを提供するステップが、前記休眠を誘発させるステップをさらに含む、請求項1に記載の方法。
【請求項3】
エンバクが休眠性であり、休眠中のエンバクを提供するステップが、前記休眠を増強させるステップをさらに含む、請求項1に記載の方法。
【請求項4】
休眠を誘発又は増強するために、エンバクを嫌気的に浸麦するステップをさらに含む、請求項1に記載の方法。
【請求項5】
偽製麦が、エンバクを4〜40℃の温度で96〜120時間インキュベートするステップを含む、請求項1に記載の方法。
【請求項6】
嫌気的浸麦が、エンバクを4〜40℃で12〜18時間、水に浸漬するステップを含む、請求項4に記載の方法。
【請求項7】
嫌気的浸麦が、カルシウムイオンを含む水中である、請求項4に記載の方法。
【請求項8】
休眠を誘発するために偽製麦前にエンバクを乾熱処理するステップをさらに含む、請求項1に記載の方法。
【請求項9】
エンバクが休眠、殻無しエンバクである、請求項1に記載の方法。
【請求項10】
エンバク中の増加したアベナンスラミド濃度が、乾燥量基準で、750ppmより高い、請求項1に記載の方法。
【請求項11】
エンバクが30〜40℃で48〜72時間乾熱処理され、その後約70℃で144〜約168時間さらに乾熱処理される、請求項8に記載の方法。
【請求項12】
エンバクが被覆されており、休眠エンバクであり、前記エンバクの殻を除去するステップをさらに含む、請求項1に記載の方法。
【請求項13】
請求項1に従って生産されるエンバク。
【請求項14】
請求項13に記載のエンバクから生産される栄養補助食品、食品、飼料又は工業製品。
【請求項15】
食品製品を製造するための、請求項1に従って生産されたエンバクの使用。
【請求項16】
エンバクが、乾燥量基準で約3〜10%の最終水分含量まで乾燥される、請求項8に記載の方法。
【請求項17】
エンバクが、約35%の水分含量まで嫌気的に浸麦される、請求項4に記載の方法。
【請求項18】
外側のふすま成分及び残留脱ふすま挽割を回収するために摩耗製粉を使用して、請求項1に従って生産されたエンバクから得られるエンバクふすま及び脱ふすまエンバク挽割。
【請求項19】
表皮剥離ふすま成分が、摩耗製粉前のエンバクの初期重量の3〜30%を含む、請求項18に記載のエンバクふすま及び脱ふすまエンバク挽割。
【請求項20】
脱ふすま挽割が、摩耗製粉前のエンバクの初期重量の97〜70%を含む、請求項18に記載のエンバクふすま及び脱ふすまエンバク挽割。
【請求項1】
休眠中のエンバクを提供するステップと、
前記エンバクを、アベナンスラミド濃度が増加しているが発芽はしていないエンバクをもたらす偽製麦に供するステップと
を含む、アベナンスラミド濃度が増加したエンバクを生産する方法。
【請求項2】
エンバクが非休眠性であり、休眠中のエンバクを提供するステップが、前記休眠を誘発させるステップをさらに含む、請求項1に記載の方法。
【請求項3】
エンバクが休眠性であり、休眠中のエンバクを提供するステップが、前記休眠を増強させるステップをさらに含む、請求項1に記載の方法。
【請求項4】
休眠を誘発又は増強するために、エンバクを嫌気的に浸麦するステップをさらに含む、請求項1に記載の方法。
【請求項5】
偽製麦が、エンバクを4〜40℃の温度で96〜120時間インキュベートするステップを含む、請求項1に記載の方法。
【請求項6】
嫌気的浸麦が、エンバクを4〜40℃で12〜18時間、水に浸漬するステップを含む、請求項4に記載の方法。
【請求項7】
嫌気的浸麦が、カルシウムイオンを含む水中である、請求項4に記載の方法。
【請求項8】
休眠を誘発するために偽製麦前にエンバクを乾熱処理するステップをさらに含む、請求項1に記載の方法。
【請求項9】
エンバクが休眠、殻無しエンバクである、請求項1に記載の方法。
【請求項10】
エンバク中の増加したアベナンスラミド濃度が、乾燥量基準で、750ppmより高い、請求項1に記載の方法。
【請求項11】
エンバクが30〜40℃で48〜72時間乾熱処理され、その後約70℃で144〜約168時間さらに乾熱処理される、請求項8に記載の方法。
【請求項12】
エンバクが被覆されており、休眠エンバクであり、前記エンバクの殻を除去するステップをさらに含む、請求項1に記載の方法。
【請求項13】
請求項1に従って生産されるエンバク。
【請求項14】
請求項13に記載のエンバクから生産される栄養補助食品、食品、飼料又は工業製品。
【請求項15】
食品製品を製造するための、請求項1に従って生産されたエンバクの使用。
【請求項16】
エンバクが、乾燥量基準で約3〜10%の最終水分含量まで乾燥される、請求項8に記載の方法。
【請求項17】
エンバクが、約35%の水分含量まで嫌気的に浸麦される、請求項4に記載の方法。
【請求項18】
外側のふすま成分及び残留脱ふすま挽割を回収するために摩耗製粉を使用して、請求項1に従って生産されたエンバクから得られるエンバクふすま及び脱ふすまエンバク挽割。
【請求項19】
表皮剥離ふすま成分が、摩耗製粉前のエンバクの初期重量の3〜30%を含む、請求項18に記載のエンバクふすま及び脱ふすまエンバク挽割。
【請求項20】
脱ふすま挽割が、摩耗製粉前のエンバクの初期重量の97〜70%を含む、請求項18に記載のエンバクふすま及び脱ふすまエンバク挽割。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2012−521746(P2012−521746A)
【公表日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願番号】特願2012−501095(P2012−501095)
【出願日】平成22年3月25日(2010.3.25)
【国際出願番号】PCT/CA2010/000458
【国際公開番号】WO2010/108277
【国際公開日】平成22年9月30日(2010.9.30)
【出願人】(511223187)
【氏名又は名称原語表記】HER MAJESTY THE QUEEN In Right of Canada as represented BY THE MINISTER OF AGRICULTURE AND AGRI−FOOD
【Fターム(参考)】
【公表日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願日】平成22年3月25日(2010.3.25)
【国際出願番号】PCT/CA2010/000458
【国際公開番号】WO2010/108277
【国際公開日】平成22年9月30日(2010.9.30)
【出願人】(511223187)
【氏名又は名称原語表記】HER MAJESTY THE QUEEN In Right of Canada as represented BY THE MINISTER OF AGRICULTURE AND AGRI−FOOD
【Fターム(参考)】
[ Back to top ]