
エンプリンとS100A9の結合の阻害を指標とする慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法

【課題】S100A9の新規受容体を標的とする慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法の提供。
【解決手段】本発明は、慢性炎症抑制剤又は癌転移抑制剤の候補物質がエンプリンとS100A9又はS100A8/A9との結合を有意に阻害する場合に、当該候補物質は慢性炎症又は癌転移を有意に抑制させると評価する、慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法、を提供する。
【解決手段】本発明は、慢性炎症抑制剤又は癌転移抑制剤の候補物質がエンプリンとS100A9又はS100A8/A9との結合を有意に阻害する場合に、当該候補物質は慢性炎症又は癌転移を有意に抑制させると評価する、慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法、を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、S100A9の新規受容体であるエンプリンを標的とする慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法を提供する。
【背景技術】
【0002】
過剰増殖や乾癬においてアップレギュレーションされるタンパク質としてS100A8およびS100A9が知られる。S100A8およびS100A9は、20を超えるメンバーから構成されるEF−ハンドカルシウム結合S100タンパク質ファミリーに属する(非特許文献1:Marenholz I et al., Biochem Biophys Res Commun (2004) 322:1111-1122)。どちらのタンパク質も好中球、活性化単球、およびマクロファージによって分泌され、それらの細胞の化学走性分子として機能し、炎症性細胞の漸増に関する正のフィードバックループに関与する(非特許文献2:Roth J et al., Trends Immunol (2003) 24:155-158)。S100A8およびS100A9陽性骨髄細胞は、炎症領域内に浸潤する最初の細胞である(非特許文献3:Odink K et al., Nature (1987) 330:80-82)。慢性関節リウマチ(非特許文献4:Liao H et al., Arthritis Rheum (2004) 50:3792-3803)、多発性硬化症(非特許文献5:Bogumil T et al., Neurosci Lett (1998) 247:195-197)、クローン病(非特許文献6:Lugering N, et al., Digestion (1995) 56:406-414)、および結合組織疾患(非特許文献7:Kuruto R, et al., J Biochem (Tokyo) (1990) 108:650-653)を含む多数のヒト炎症性疾患で高いS100A8およびS100A9血清レベルが観察されている。従って、S100A8およびS100A9は、炎症の誘導および伝播に重要な役割を担うと考えられている。
【0003】
上皮細胞中でS100A8と100A9が果たす生物学的機能について、本発明者は以前、外因性S100A8とS100A9が複合体(S100A8/A9)(別名:カルプロテクチン)を形成することで正常表皮角化細胞(NHEK)を刺激して乾癬性病変などにおいて発現亢進される炎症性サイトカインを産生させ、さらにS100A8/A9誘導性サイトカインがNHEK中でのS100A8およびS100A9の産生および分泌を刺激することを明らかにした(非特許文献8:J Cell Biochem. 2007 Nov 28, Epub ahead of print)。さらに、S100A8/A9自体がNHEKの増殖を増強することも見出した。これらの結果は、主要メディエーターとしてS100A8/A9が関与するNHEKの増殖と炎症の正のフィードバック機構の存在を明らかにした。即ち、S100A8/A9が炎症性サイトカインの産生を誘導して炎症性疾患を惹起し、その炎症が細胞増殖を誘導し、さらには細胞増殖が炎症を誘導するといったスパイラルを形成し、増殖・炎症が連鎖する持続性皮膚炎症性疾患、例えばアトピー性皮膚炎や乾癬などの原因となることが示唆された。
【0004】
S100A8/A9により引き起こされる慢性炎症の負のサイクル形成を阻止するためには、S100A8およびA9のレセプターの同定が必要と考えられる。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Biochem Biophys Res Commun (2004) 322:1111-1122
【非特許文献2】Trends Immunol (2003) 24:155-158
【非特許文献3】Nature (1987) 330:80-82
【非特許文献4】Arthritis Rheum (2004) 50:3792-3803
【非特許文献5】Neurosci Lett (1998) 247:195-197
【非特許文献6】Digestion (1995) 56:406-414
【非特許文献7】J Biochem (Tokyo) (1990) 108:650-653
【非特許文献8】J Cell Biochem. (2008) 104:453-464
【非特許文献9】Nature Cell Biol. (2006) 8(12): 1369-1375
【非特許文献10】Hum Genet (2002) 111:310-313
【非特許文献11】Morrison TB et al., Biotechniques (1998) 24:954-958, 960, 962
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、S100A9の新規受容体を標的とする慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法を提供する。
【課題を解決するための手段】
【0007】
S100タンパク質ファミリーの受容体として知られているRAGE (Receptor for advanced glycation endoproducts)は、S100A9とも結合することが本発明者らにより確認された。しかしながら、両者の結合による信号系の存在は、中和抗体、siRNA試験のいずれによっても確認できなかった。
【0008】
そこで、本発明者らが、培養ケラチノサイトからS100A8及び/又はA9と結合するタンパク質を単離してLC/MS/MS解析にかけ、S100A8/A9結合タンパク質の同定を試みた結果、S100A9のレセプター候補が多数発見された。その中から、免疫グロブリンスーパーファミリーのメンバーであり、且つ膜貫通型の糖タンパク質であるエンプリン(Emmprin)(The extracellular matrix metalloproteinase inducer)(別名バシジン又はCD147)に着目したところ、エンプリンの発現の抑制によりS100A9によるサイトカイン誘導やマトリックスメタロプロテアーゼ誘導が顕著に低下することが分かった。また、免疫染色の結果は、S100A9及びエンプリンがアトピー性皮膚炎や乾癬に罹患している患者の表皮、そして、浸潤するメラノーマ細胞において強発現していることを示した。
【0009】
従って、エンプリンがS100A9のレセプターであること、そして、これらの間の結合を阻害することで、慢性炎症の抑制、更には癌の転移を抑制することが見出され、本発明が完成するに至った。
【0010】
従って、本願は以下の発明を包含する:
(1)慢性炎症抑制剤又は癌転移抑制剤の候補物質がエンプリンとS100A9又はS100A8/A9との結合を有意に阻害する場合に、当該候補物質は慢性炎症又は癌転移を有意に抑制させると評価する、慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法。
(2)慢性炎症抑制剤又は癌転移抑制剤の候補物質の存在下、エンプリンとS100A9又はS100A8/A9とをインキュベートし、エンプリンとS100A9又はS100A8/A9との結合を阻害する物質を慢性炎症抑制剤又は癌転移抑制剤として選定することを含んで成る、(1)に記載の方法。
(3)エンプリンが固体支持体に固相化されている、(1)又は(2)に記載の方法。
(4)前記結合の阻害がELISA法により決定される、(1)〜(3)のいずれかに記載の方法。
(5)エンプリンとS100A9との結合を阻害する薬剤を含んで成る、慢性炎症抑制剤又は癌転移抑制剤。
(6)前記薬剤が、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物を一種又は二種以上含む、(6)に記載の慢性炎症抑制剤又は癌転移抑制剤。
(7)エンプリンとS100A9との結合を阻害する薬剤を被験者に投与することを含んで成る、慢性炎症又は癌転移の抑制方法。
(8)前記薬剤が、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物を一種又は二種以上含む、(9)に記載の方法。
【発明の効果】
【0011】
エンプリンは、マトリックスメタロプロテアーゼ(MMP)を誘導して癌の転移を誘導する。このように、エンプリンと悪性腫瘍との関係はよく知られている。また、S100A8/A9についても、転移の好発部位は、癌細胞が出すVEGF、TNF等の因子などと反応して、S100A8/A9を分泌し、これが癌細胞の転移を誘導することが知られている(非特許文献9:Nature Cell Biol. (2006) 8(12): 1369-1375)。本発明者らが培養ケラチノサイトをS100A8/A9で刺激したところ、癌の浸潤に関与するMMPは、当該刺激により発現が亢進されたものの、エンプリンをノックダウンした場合にはその発現が有意に抑制されることが確認された(結果は示さず)。従来、MMPの発現亢進はエンプリンの自己分泌ループによると考えられていたが、上記の結果は、MMPの発現はエンプリンのみでは亢進されず、S100A9刺激を経由することが必要であることを示している。
【0012】
癌細胞の転移に関与しているエンプリンがS100A9のレセプターとして機能していることは、本発明者らによって初めて見出された。実際、エンプリンの発現を抑制することでS100A9によるサイトカインやMMPの発現亢進は低下する(実施例)。そのため、エンプリンとS100A9の関係を考慮すれば、癌の転移とその悪性度について新たな解釈が可能になると考えられる。更に、エンプリンは、S100A9と同様にアトピー性皮膚炎や乾癬の患者の表皮上層やメラノーマ細胞の表皮において発現していたため、両者はアトピー性皮膚炎、乾癬、癌等の慢性炎症にも関与していることが予想される。従って、本発明によれば、エンプリンとS100A9との結合の阻害を指標とすることで、慢性炎症抑制剤又は癌転移抑制剤の探索が可能になる。
【図面の簡単な説明】
【0013】
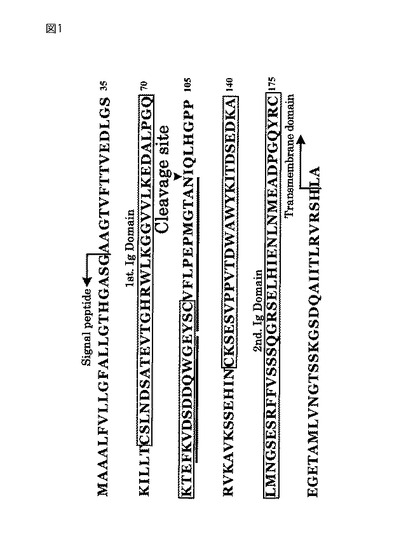
【図1】エンプリンの一次構造。
【図2】多次元キャピラリーLC/MS/MSを用いたタンパク質の網羅的解析法により同定されたS100A9の受容体候補タンパク質。
【図3】エンプリンsiRNAによるエンプリンの発現抑制効果。
【図4】エンプリンの発現抑制に伴うサイトカインの発現変化。
【図5】エンプリンの発現抑制に伴うMMPの発現変化。
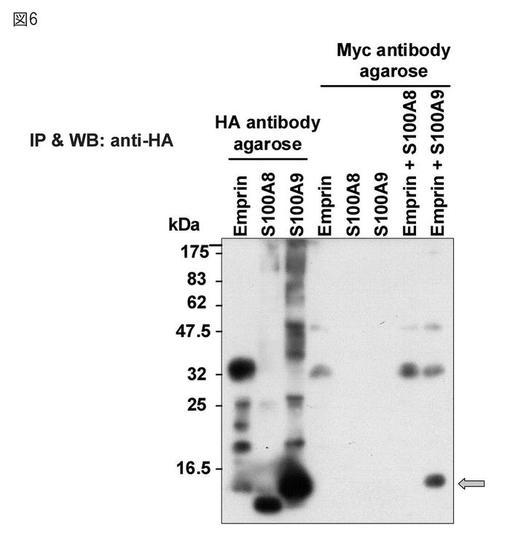
【図6】ウェスタンブロットによるエンプリン結合タンパク質の同定。矢印はS100A9のバンドを示す。
【図7】可溶性エンプリンによるMMP1の誘導効果。
【図8A】ヒト正常皮膚におけるエンプリン及びS100タンパク質の局在を示す免疫染色図。
【図8B】皮膚モデルにおけるエンプリン及びS100タンパク質の局在を示す免疫染色図。
【図8C】アトピー性皮膚疾患皮膚におけるエンプリン及びS100タンパク質の局在を示す免疫染色図。
【図8D】S100A8抗体、27E10、Dapiを用いて免疫染色したアトピー性皮膚炎及び乾癬の皮膚の比較。
【図8E】S100A9抗体、27E10、Dapiを用いて免疫染色したアトピー性皮膚炎及び乾癬の皮膚の比較。
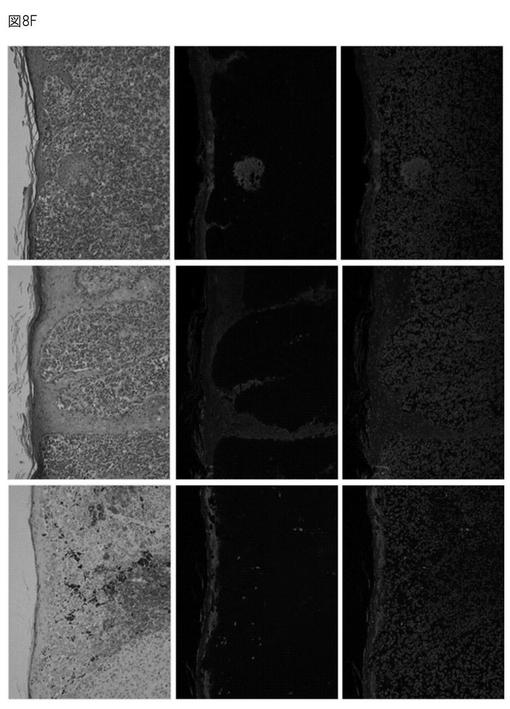
【図8F】メラノーマ組織におけるS100A9の局在を示す免疫染色図(上段はHE染色、中段はS100A9抗体による染色、下段はDAPI染色。左側、中央、右側の写真はそれぞれ異なるサンプルに由来する)。
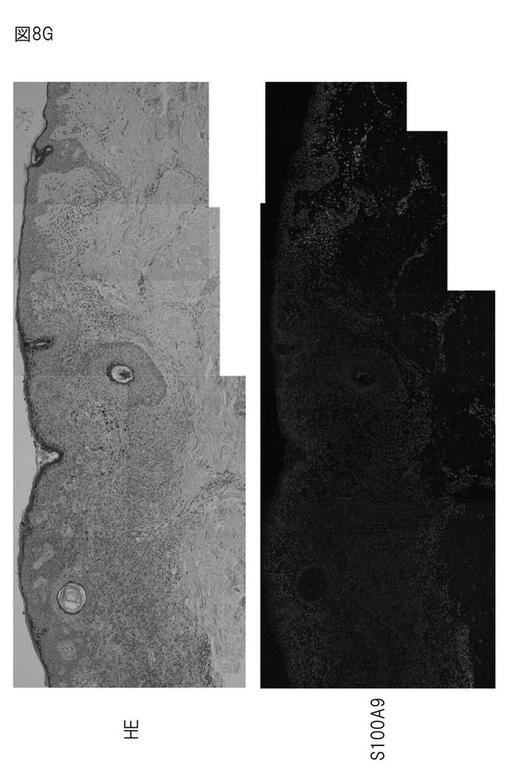
【図8G】悪性メラノーマにおけるS100A9の局在を示す免疫染色図。
【図8H】メラノーマ組織におけるエンプリンの局在を示す免疫染色図。
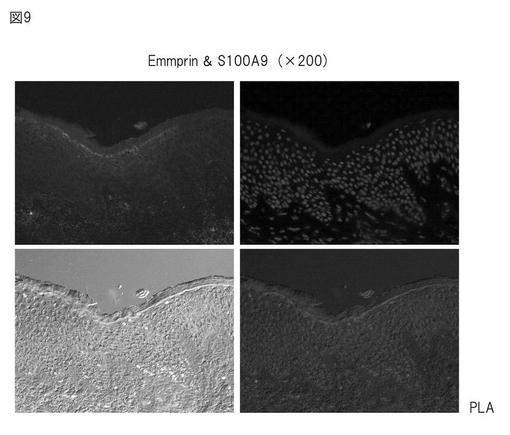
【図9】PLA(Proximity Ligation Assay) 法によるアトピー性皮膚におけるエンプリンとS100A9の相互作用の証明(200倍)。
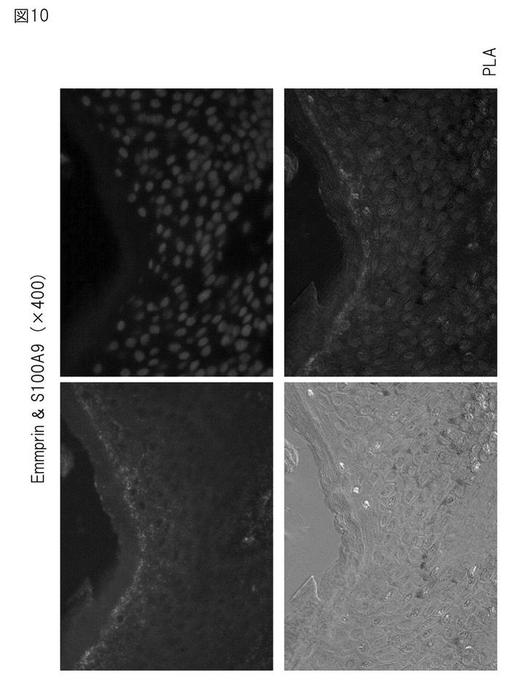
【図10】PLA法によるアトピー性皮膚におけるエンプリンとS100A9の相互作用の証明(400倍)。
【図11】S100A9とエンプリンとの結合を阻害する植物抽出物のスクリーニング結果。
【図12】ヨモギエキス、オドリコソウエキス、トウキエキスのS100A9−エンプリン結合阻害効果の比較。
【発明を実施するための形態】
【0014】
エンプリンは、2個のIgドメインを有する、1回膜貫通型の糖タンパク質であり、コラゲナーゼ(MMP−1)の発現亢進作用を有している。エンプリンヌルマウスには、精子形成、受精、感覚機能及び記憶機能、並びに混合リンパ球反応の欠損が見られる。エンプリンの一次構造を図1に、そしてエンプリンの全長配列を配列番号1に示す。MMP−1で切断されたIgドメイン1は、コラーゲンの発現を誘導する。
【0015】
本発明のスクリーニング方法は、特に限定されるものではないが、候補物質の存在下エンプリンとS100A9とをインキュベーションし、エンプリンとS100A9との結合を有意に阻害する候補薬剤をS100A9に起因する慢性炎症抑制剤又は癌転移抑制剤として選択することからなる。その評価基準として、例えばエンプリンとS100A9タンパク質との結合がコントロールを作用させた場合と比べ10%以上、又は20%以上、又は30%以上、又は50%以上、又は70%以上、又は100%阻害されていたなら慢性炎症又は癌転移を「有意に抑制する」、と判断してよい。
【0016】
S100A9は、上述のとおりS100A8と複合体を形成していることがあり、この複合体がエンプリンと結合することもある。従って、S100A8/A9とエンプリンとの結合を阻害する物質を慢性炎症抑制剤又は癌転移抑制剤としてスクリーニングしてもよい。
【0017】
エンプリンとS100A9との結合の阻害を検出する手段は特に限定されるわけではないが、ELISA法に基づきエンプリンとS100A9(又はS100A8/A9)との結合における検量線を作製し、この結合を阻害する分子、すなわち吸光度の低下する分子を慢性炎症抑制剤又は癌転移抑制剤の候補薬剤として検出することができる。良好な検出感度を確保する観点から、固体支持体に吸着される分子は、分子量が大きいエンプリンが好ましい。
【0018】
本明細書で使用する「慢性炎症」は、アトピー性皮膚炎、乾癬の他、癌なども包含する。また、本明細書で使用する「癌転移抑制」とは、癌細胞が原発巣である組織から、浸潤、血中およびリンパ管を通しての遊走、新たな組織への定着を経て、増殖を開始する過程のいずれかもしくは全てを抑制することを意味し、癌細胞の増殖抑制とは異なる。
【0019】
S100A8およびA9
S100A8およびA9のアミノ酸配列およびそれをコードするDNA配列は、例えばHum Genet (2002) 111:310-313(非特許文献10)に公開されている。本発明において使用できるS100A8およびA9は、通常ヒト由来の天然型、あるいは組み換えタンパク質であるが、活性を有すれば改変型、異種由来、もしくは非精製品を用いることができる。S100A8およびA9の組換タンパク質は、当業界で周知の方法に従い、例えば単離したまたはPCRにより合成したS100A8又はA9遺伝子 (cDNA) を例えばプラスミド、ウィルス等に挿入して発現ベクターを調製し、これを宿主細胞、例えば微生物、動物細胞又は植物細胞等の培養細胞に導入し、発現させることにより、大量調製することが可能である。
【0020】
S100A9は、水や培地、例えば表皮角化細胞の培養に適当な培地、例えば上記EpiLife(登録商標)培地に溶解し、本発明のスクリーニング系に添加する。添加量は、一概には規定できないが1ng/mlから1mg/ml程度、好ましくは10ng/mlから100μg/ml程度、より好ましくは100ng/mlから10μg/ml程度の濃度とする。S100A9またはS100A8/A9の添加は、好ましくは塩化カルシウムの存在下で行う。S100A9またはS100A8/A9の存在下でのインキュベーション時間、インキュベーション温度といった培養条件は特に制限されることはなく、好ましくは30〜37℃で1〜14時間、より好ましくは34〜37℃で2〜7時間、好ましくはCO25%の下でインキュベーションを行う。
【0021】
本発明の慢性炎症抑制剤は、S100A8/A9に起因する持続性皮膚炎症性疾患、例えばアトピー性皮膚炎、乾癬などの予防、治療といった改善等に有効な医薬品又は化粧品として利用できる。
【0022】
本発明のスクリーニング方法により得られた慢性炎症抑制剤又は癌転移抑制剤として、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物が挙げられる。特に、ヨモギエキスはS100A9とエンプリンとの結合を有意に阻害することが確認されているため(図12)、慢性炎症抑制剤又は癌転移抑制剤の好ましい活性成分であることが予想される。ここで、本発明で使用する各植物の植物体又はその抽出物は、各々の植物体の各種部位(花、花穂、果皮、果実、茎、葉、枝、枝葉、幹、樹皮、根茎、根皮、根、種子又は全草など)をそのまま又は乾燥したものを粉砕して乾燥粉末としたもの、あるいはそのまま又は乾燥・粉砕後、溶媒で抽出したものである。
【0023】
抽出物の場合、抽出に用いられる抽出溶媒は通常抽出に用いられる溶媒であれば何でもよく、特にメタノール、エタノールあるいは1,3−ブチレングリコール等のアルコール類、含水アルコール類、アセトン、酢酸エチルエステル等の有機溶媒を単独あるいは組み合わせて用いることができ、このうち特に、アルコール類、含水アルコール類が好ましく、特にメタノール、エタノール、1,3−ブチレングリコール、含水エタノールまたは含水1,3−ブチレングリコールが好ましい。また前記溶媒は、室温乃至溶媒の沸点以下の温度で用いることが好ましい。
【0024】
抽出方法は特に制限されるものはないが、通常、常温から、常圧下での溶媒の沸点の範囲であれば良く、抽出後は濾過又はイオン交換樹脂を用い、吸着・脱色・精製して溶液状、ペースト状、ゲル状、粉末状とすれば良い。更に多くの場合は、そのままの状態で利用できるが、必要ならば、その効果に影響のない範囲で更に脱臭、脱色等の精製処理を加えても良く、脱臭・脱色等の精製処理手段としては、活性炭カラム等を用いれば良く、抽出物質により一般的に適用される通常の手段を任意に選択して行えば良い。
【0025】
植物体の抽出部位として、ヨモギの場合、葉が、トウキの場合、根が、オドリコソウの場合、茎、葉、花が考えられるが、抽出部位はこれらに限定されない。
【0026】
上記溶媒で抽出して得られた抽出物をそのまま、あるいは例えば凍結乾燥などにより濃縮したエキスを使用でき、また必要であれば吸着法、例えばイオン交換樹脂を用いて不純物を除去したものや、ポーラスポリマー(例えばアンバーライトXAD−2)のカラムにて吸着させた後、所望の溶媒で溶出し、さらに濃縮したものも使用することができる。
【0027】
本発明の慢性炎症抑制剤又は癌転移抑制剤は、前記植物体又はその抽出物の一種または二種以上からなるものであることが好ましいが、本発明の効果を損なわない範囲において、他の種々の成分を含有することができる。また、本発明の慢性炎症抑制剤又は癌転移抑制剤は、その使用目的に合わせて用量、用法、剤型を適宜決定することが可能である。例えば、本発明の慢性炎症抑制剤の投与形態は、経口、非経口、外用等であってよい。剤型としては、例えば錠剤、粉剤、カプセル剤、顆粒剤、エキス剤、シロップ剤等の経口投与剤、又は注射剤、点滴剤、若しくは坐剤等の非経口投与剤軟膏、クリーム、乳液、ローション、パック、浴用剤等の外用剤を挙げることができる。
【0028】
本発明の慢性炎症抑制剤又は癌転移抑制剤の上記エキス成分の配合量は、用途に応じて適宜決定できるが、一般には阻害剤全量中、乾燥物として0.0001〜20.0質量%、好ましくは0.0001〜10.0質量%である。ヨモギエキス、オドリコソウエキス、トウキエキスは濃度依存的に慢性炎症又は癌転移を抑制することが考えられる。
【0029】
また、慢性炎症抑制剤中又は癌転移抑制剤中には、上記薬剤以外に、例えば、通常の食品や医薬品に使用される賦形剤、防湿剤、防腐剤、強化剤、増粘剤、乳化剤、酸化防止剤、甘味料、酸味料、調味料、着色料、香料等、化粧品等に通常用いられる美白剤、保湿剤、油性成分、紫外線吸収剤、界面活性剤、増粘剤、アルコール類、粉末成分、色剤、水性成分、水、各種皮膚栄養剤等を必要に応じて適宜配合することができる。
【0030】
さらに、本発明の慢性炎症抑制剤又は癌転移抑制剤を皮膚外用剤として使用する場合、皮膚外用剤に慣用の助剤、例えばエデト酸二ナトリウム、エデト酸三ナトリウム、クエン酸ナトリウム、ポリリン酸ナトリウム、メタリン酸ナトリウム、グルコン酸等の金属封鎖剤、カフェイン、タンニン、ベラパミル、トラネキサム酸およびその誘導体、甘草抽出物、グラブリジン、カリンの果実の熱水抽出物、各種生薬、酢酸トコフェロール、グリチルリチン酸およびその誘導体またはその塩等の薬剤、ビタミンC、アスコルビン酸リン酸マグネシウム、アスコルビン酸グルコシド、アルブチン、コウジ酸等の美白剤、グルコース、フルクトース、マンノース、ショ糖、トレハロース等の糖類、レチノイン酸、レチノール、酢酸レチノール、パルミチン酸レチノール等のビタミンA類なども適宜配合することができる。
【0031】
以下、具体例を挙げて、本発明を更に具体的に説明する。なお、本発明はこれにより限定されるものではない。
【実施例】
【0032】
1.新規S100A9受容体候補のスクリーニング
培養ケラチノサイトから回収したタンパク質混合物とGST融合S100A9又はS100A8/A9タンパク質とを混合した後、このサンプルについてキャピラリーLC/MS/MSによるタンパク質の網羅的解析法を行った。
【0033】
LC/MS/MS解析
内径100μm、長さ120mmの未充填のキャピラリーカラム(New Objetive社製)のテーパー状の出口側の端部に、充填剤を保持するために、シリカ製のフリットを作製した。得られたキャピラリーカラムに、平均粒径が5μmのオクタデシル化シリカ型充填剤Aqua C18(Phenomenex社製)を、高さが100mmとなるように充填し、分析用逆相キャピラリーカラムを得た。
【0034】
内径250μm、長さ150mmの未充填のキャピラリーカラム(Agilent社製)の出口側の端部に、充填剤を保持するために、シリカ製のフリットを作製した。得られたキャピラリーカラムの出口側に、平均粒径が5μmのカチオン交換樹脂型充填剤Partisphere SCX resins(Whatman社製)と、平均粒径が5μmのアニオン交換樹脂型充填剤PolyWAX LP(PolyLC社製)を質量比2:1で混合したもの、入口側に、平均粒径が5μmのオクタデシル化シリカ型充填剤Aqua C18(Phenomenex社製)を、それぞれ高さが25mmとなるように充填し、トラップ用逆相キャピラリーカラム及びSCX−WAX混合キャピラリーカラムからなる二相型キャピラリーカラムを得た。
【0035】
なお、分析用逆相キャピラリーカラム及び二相型キャピラリーカラムを作製する際には、高圧窒素ガス及び加圧型充填容器を用いて、スラリー充填法により充填剤を充填した。
【0036】
続いて、6ステップのMudPIT型分析(二次元HPLC/ESI MS/MS)により、上記タンパク質混合物を分析した。
【0037】
まず、ペプチドを約4μg含む上清を、加圧法により、二相型キャピラリーカラムにロードした後、試料溶液の10倍以上の体積の移動相A(水、アセトニトリル及びギ酸の体積比95:5:0.1の混合液;pH〜2.6)を用いて、洗浄、脱塩した。この二相型キャピラリーカラム10を、貫通孔型ユニオン(Upchurch Scientific社製)(不図示)を介して、分析用逆相キャピラリーカラム20と接続した。次に、内径が100μmのキャピラリーを配管として用いたNanospace SI−2型HPLC装置(資生堂社製)に接続した。このとき、トラップ用逆相キャピラリーカラム11は、SCX−WAX混合キャピラリーカラム12及び分析用逆相キャピラリーカラム20の上流側に配置した。
【0038】
移動相としては、移動相A、移動相B(水、アセトニトリル及びギ酸の体積比20:80:0.1の混合液)、移動相C(500mMの酢酸アンモニウムを含む移動相A;pH〜6.8)を用い、ペプチドの溶出法は、矩形状に加える移動相Cの体積%をステップ毎に漸増させた、計6ステップのグラジエント溶出法とした。
【0039】
ステップ1のグラジエントプロファイルは、5分間移動相Aを流し、次の5分間で移動相Bの比率を0体積%から15体積%まで増加させ、次の60分間で移動相Bの比率を45体積%まで増加させ、次の10分間で移動相Bの比率を75体積%まで増加させた後、この比率で5分間流すものである。
【0040】
ステップ2〜6のグラジエントプロファイルは、1分間移動相Aを流し、次の4分間移動相Cの比率をX[体積%]として流し、次の5分間で移動相Cの比率を0体積%から15体積%まで増加させ、次の60分間で移動相Cの比率を45体積%まで増加させ、次の10分間で移動相Cの比率を75体積%まで増加させた後、この比率で5分間流すものである。このとき、ポンプの送液の流速を250μL/分とし、抵抗型キャピラリーによるスプリットにより、カラムの流速を300〜400nL/分に調整した。
【0041】
また、ESI MS/MSを測定する際には、イオントラップ型質量分析計LCQ−Deca(Thermo Fisher Scientific社製)を用いた。このとき、分析用逆相キャピラリーカラムから溶出されたペプチドは、スプリットすることなく、質量分析計に直接導入した。
【0042】
なお、質量電荷比(m/z)が400〜1400のフルスキャンMSスペクトル測定1回及びデータ依存型MS/MSスペクトル測定3回を、各ステップを通じて繰り返した。このとき、標準化解裂エネルギーは35%とした。また、マイクロスキャンは、MSスペクトル測定及びMS/MS測定ともに3とした。さらに、動的排除設定は、リピートカウント1、リピート期間0.50分、排除リストサイズ25、排除期間10.00分とした。
【0043】
得られたMS/MSスペクトルは、Bioworksソフトウェア(Thermo Fisher Scientific社製)上で動くSEQUESTアルゴリズムにより、非冗長ヒトデータベース(ftp://ftp.ncbi.nih.gov/blast/db/FASTA/nr.gz、2007/2/8版)に対して、検索した。
【0044】
その結果、図2に列記ような受容体候補タンパク質が同定された。これらのタンパク質の中から、バシジン(エンプリン)をS100A9の新規受容体として以下の実験を行った。
【0045】
2.siRNAによるエンプリンの発現抑制
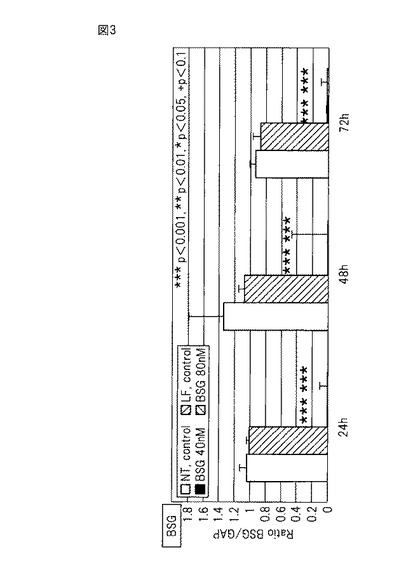
エンプリンの発現抑制のために、RNAiMaxを用いて、エンプリンsiRNA(Santa Cruz: sc-35298)を、終濃度40 nM又は80 nMとなるように増殖期の培養ケラチノサイトにトランスフェクションした(図3中の「BSG」)。コントロールとして、何も添加していないもの(図3中の「NT」(non-treated control))およびヒト遺伝子のいずれの部分とも相同性を有していないcontrol siRNA-A (Santa Cruz Biotechnology, Inc., sc-3707)(図3中の「LF」)を使用した。尚、トランスフェクションは、培養培地を増殖因子を含まない基礎培地に交換してから行った。トランスフェクションから24時間後にS100A9で増殖ケラチノサイトを刺激し、さらに24時間経過後にRNAを採取した。その結果、図3に示すとおり、エンプリンsiRNAをトランスフェクションした場合、24、48、72時間後には、上記コントロールを用いた場合のエンプリンの発現量と比較して1〜3%まで発現が抑制された。
【0046】
3.エンプリンの発現抑制によるサイトカイン及びMMPの発現変化
IL−8(CXCL−8)、TNFα、IL1−F9、CXCL−1は、S100A8/A9の添加により、ケラチノサイトでの発現が亢進されることが明らかとなっている(前掲J Cell Biochem. (2008) 104:453-464)(非特許文献8)。S100A9の添加によってもIL−8(CXCL−8)、TNFα、IL1−F9及びCXCL−1の発現が亢進されるか、また、エンプリンの発現抑制がこれらのサイトカインの発現にどのような影響を与えるかについてリアルタイム定量PCRにより検討した。同様の方法により、S100A9刺激がMMP−1及びMMP−10の発現に及ぼす影響についても検討した。
【0047】
リアルタイム定量PCR
EpiLife(商標)-KG2(Cascade Biologies社)中で培養した増殖期のNHEKを、2 mMの塩化カルシウム、S100A8またはS100A9(各10μg/ml)を含有又は非含有の同培地に置換し、3時間にわたり曝露させ、MagNA(商標) Pure mRNA抽出キットおよびMagNA Pure(商標)機器(Roche Diagnostics社、日本国、東京都)を用いてmRNAを抽出した。得られたmRNAは、SuperScript(商標) II(Invitrogen Corporation社、米国、カリフォルニア州、カールズバッド)を用いて逆転写させた。リアルタイム定量PCRは、製造業者の取扱説明書にしたがってLightCycler FastStart DNA master SYBR green Iキット(Roche Diagnostics社)を用いてLightCycler高速サーマルサイクラーシステム上で実施した。典型的な反応条件は、10分間の活性化ステップ、それに続く95℃で15秒の変性、60℃で10秒のアニーリング、72℃で10秒の伸長からなるサイクル40回であった。使用したプライマーは、下記の表1に示す。各プライマーの最終濃度は20μlの総反応容量中で0.2〜0.25μMとした。グリセルアルデヒド−3−リン酸脱水素酵素(GAPDH)遺伝子を対照遺伝子として使用した。増幅させたフラグメントの特異性は融解曲線分析によって確認した。各遺伝子の発現レベルは、LightCycler分析用ソフトウエアを用いて定量分析した(非特許文献11:Morrison TB et al., Biotechniques (1998)24:954-958, 960, 962)。目的のmRNA量は、A8/control siRNA-A (santa Cruz: sc-37007)(A8/LF)のmRNAの量に対する比率として表した。
【0048】
【表1】

【0049】
図4に示すとおり、S100A9を添加した試料(A9/LF)は、全てのサイトカインの発現を誘導した。一方、エンプリンsiRNAを添加した試料(A9/siRNA)は、全てのサイトカイン発現量が有意に低下した。これは、エンプリンsiRNAによってエンプリンの発現が抑制された場合、S100A9でサイトカインの発現を刺激してもサイトカインの発現が抑制されることを明確に示している。
【0050】
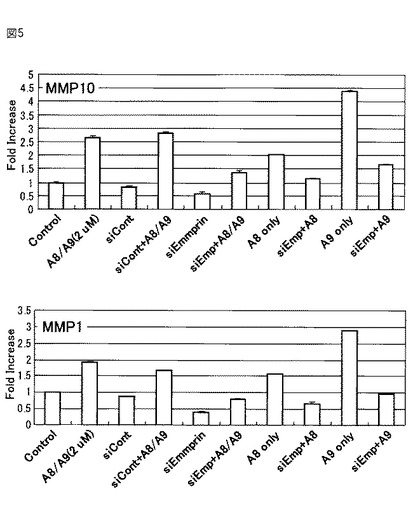
MMPについても、S100A9を添加した場合、発現が亢進されるのに対し、エンプリンがノックダウンされている場合、S100A9を添加しても発現が有意に抑制されることが明らかとなった(図5)。
【0051】
4.エンプリンとS100タンパク質との結合試験
1)エンプリン細胞外ドメインの作製
[方法]
細胞: ヒト胎児腎細胞株 (HEK293) は ATCC 社より購入したものを使用し、 培養ヒト正常線維芽細胞 OUMS-24は、難波正義博士により単離されたものを使用した。 HEK293 と OUMS-24 は、 Gibco 社の DMEM/F12 培地 (最終濃度が 10% となるように牛胎児血清を添加) を用いて培養した。
【0052】
2)エンプリン細胞外ドメイン発現コンストラクト:
CMV イントロンプロモーター (CMVi) を導入した PDNR 1r ベクター (プロモーターレスドナーベクター;Clontech 社) を構築し、CMVi の下流にヒトエンプリン細胞外ドメイン (C末にmyc-HA-Flag-6Hisタグが付加されている) をコードするcDNAを挿入した(pCMVi-exEmmp: エンプリン細胞外ドメイン発現ドナーベクター)。挿入cDNAの塩基配列は DNA シークエンサーにより正しいことを確認済みである。
【0053】
3)エンプリン細胞外ドメインの細胞外への分泌:
pCMVi-exEmmp を、FuGENE-HD (Roche社) トランスフェクション試薬を用いて HEK293 に導入し、48時間後に培養上清を回収した。培養上清に Sigma 社の抗 HA tag 抗体共有結合担体を添加し、4℃で3時間振盪混和した。その後、5000 rpm、1分間の遠心分離を行い、沈降してきた担体結合タンパク質を酸性バッファーにより溶出した。溶出サンプルを12% の SDS-PAGE を用いて電気泳動した後、PVDF 膜にエレクトロブロットして、CST 社の抗 HA tag 抗体を用いてウエスタンブロットを行い、エンプリン細胞外ドメインが分泌されていることを確認した。
【0054】
4)エンプリン細胞外ドメイン発現アデノウィルス (Ad-exEmmp):
pCMVi-exEmmp のアデノウィルスベクターへの変換は、アデノウィルス作製キット(Adeno-X-expression system: Clontech 社)を使用して行った。
【0055】
5)エンプリン細胞外ドメインの大量精製:
Ad-exEmmp (20 MOI)を培養ヒト正常線維芽細胞 OUMS-24 (10 cm dish x 20)に感染させた。感染させる時期は、OUMS-24 が高密度状態になった時とした。これは、高密度培養により接触阻止が惹起された細胞では細胞分裂が起こらず、細胞内に存在するアデノウィルス由来エピソーム含量の低下が抑制され、その結果、アデノウィルスによる標的遺伝子発現が極めて長期間(2-3週間)持続するからである。しかも、OUMS-24 は無血清培養が可能であることより、長期に渡って培養上清中に分泌された組み換えタンパク質を、血清を含まない状態で回収することができる。感染操作後、24時間培養して無血清培地 DMEM/F12 (フェノールレッド不含) に置換する。3日の間隔で液換えを行い、その度に培養上清を回収して4℃で保存する (タンパク質の安定性に応じて保存条件を変える)。この操作を30日間行った。約2Lの回収培養上清について、80%飽和硫安条件で得られた沈殿を50mlの純水に溶かし、その後、純水に対して透析することで硫安を除いた。 透析後、目的の組み換えタンパク質は、抗 HA tag 抗体共有結合担体充填カラム(sigma 社)を用いて回収した。
【0056】
6)エンプリン結合タンパク質の同定:
エンプリンがS100タンパク質の新規レセプターであることを確認するべく、免疫沈降及びウェスタンブロットによりエンプリン結合タンパク質の同定を行った。本実験において、エンプリンはC末にmyc-HA-Flag-6Hisタグが付加されているものを使用した。また、S100タンパク質としてS100A8及びS100A9タンパク質を使用した。これらのタンパク質がコードされているプラスミド(C末にHAタグが付加されている)をそれぞれHEK293細胞にトランスフェクションし、その培養上清からそれぞれのタンパク質を単離して使用した。
【0057】
エンプリンとS100A8及びS100A9タンパク質の結合解析のために、それぞれのタンパク質が含まれる培養上清を混合して反応させた後、HA抗体及びMyc抗体を用いて免疫沈降を行った。ウェスタンブロットの結果を図6に示す。エンプリンとS100A8とを混合した試料については、32kDa付近にエンプリンのバンドのみが確認された(「Emprin + S100A8」)。一方、エンプリンとS100A9タンパク質とを混合した試料では(「Emprin + S100A9」)、47.5kDa付近にそれらの結合を示すバンドが確認された。以上の結果から、エンプリンはS100A9タンパク質の新規レセプター候補であることが明らかとなった。
【0058】
5.可溶性エンプリンのMMP発現に及ぼす影響
従来、MMPの発現亢進は、エンプリンの細胞外ドメインがMMPにより分解され、放出された可溶性のエンプリンが細胞表面に存在する受容体としてのエンプリンに結合し、MMPの産生を促すというエンプリンの自己分泌(オートクリン)に起因すると考えられていた。従って、可溶性エンプリンが実際にMMPの発現を亢進させるか否かについて検討した。
【0059】
可溶性エンプリンとして上記方法により精製したエンプリン細胞外ドメインを用い、これをケラチノサイトに添加したところ、0.025、0.25、2.5μMのいずれの濃度でもMMP−1誘導効果はほとんど見られなかった。また、S100A9単独ではMMP−1の発現が顕著に亢進されたのに対し、可溶性エンプリンとS100A9とを一緒にケラチノサイトに添加した場合、S100A9によるMMP1発現亢進効果は有意に抑制された。結果を図7に示す。可溶性エンプリンとS100A9とが共存した場合にMMPの発現が抑制されたのは、MMP産生を誘導するS100A9が、可溶性エンプリンに捕捉され、両者が結合体したことによるものと考えられる。これらの結果から、従来提唱されていたエンプリンの自己分泌によるメカニズムより、S100A9刺激がエンプリンを通じてMMPの発現を亢進させるというメカニズムの方が合理的と思われる。結果は示さないが、可溶性エンプリンはMMP−10、TNFα、IL−8の発現も有意に抑制した。
【0060】
6.表皮におけるエンプリンの局在
1)免疫染色
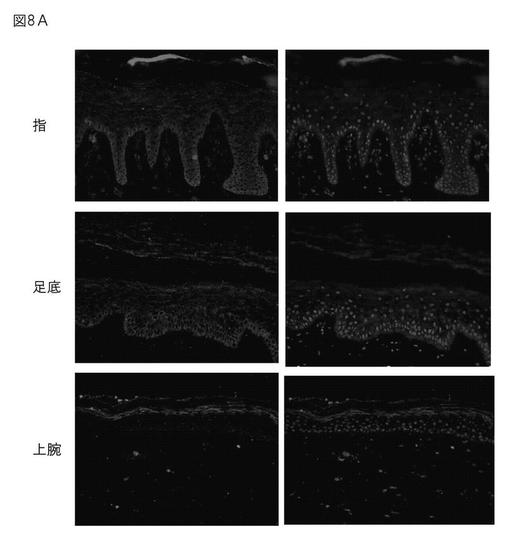
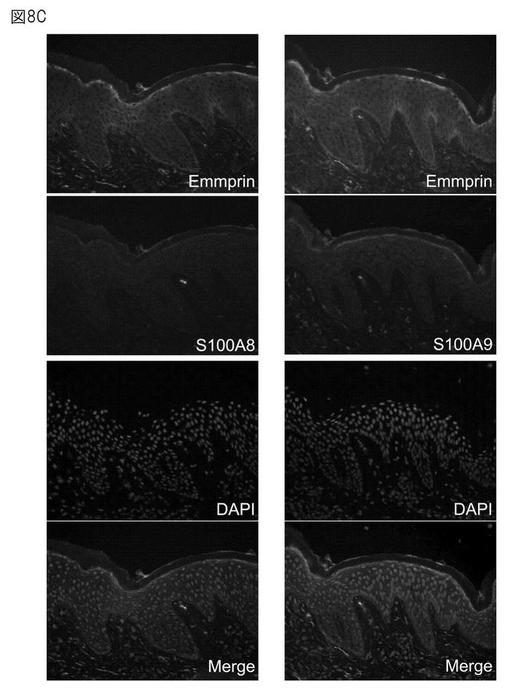
エンプリンがヒト表皮に存在するか、また、S100タンパク質と同一局在を示すかについて、免疫染色により確認した。結果を図8A〜Cに示す。エンプリンは、正常表皮、皮膚モデル、アトピー性皮膚炎(AD)の皮膚のいずれでも顆粒層で多く発現している。また、S100タンパク質もエンプリン付近で発現していた。
【0061】
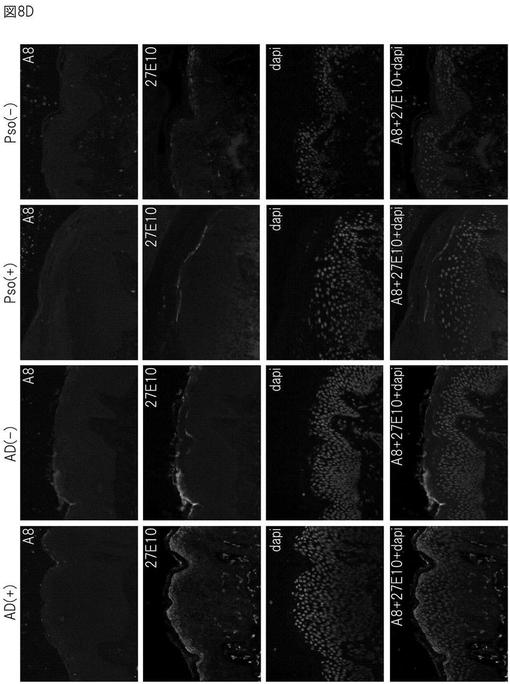
更に、アトピー性皮膚炎の皮膚では、エンプリンが高発現しており、S100A8及びS100A9タンパク質の発現も亢進されていることが明らかとなった。また、S100A8/A9複合体を特異的に結合する27E10抗体を用いた免疫染色の結果は、乾癬(Pso)の皮膚と比較した場合、アトピー性皮膚疾患の皮膚の顆粒層でS100A8/A9複合体が高発現していることを示している(図8D及び図8E)。
【0062】
免疫染色の結果によると、正常表皮では、S100A8、A9、エンプリンはほとんど発現していない。しかし、メラノーマ組織におけるS100A9の発現について免疫染色により確認したところ、いずれのサンプルでも表皮側にS100A9の強発現が認められた(図8F、左側、中央、右側の写真)。また、正常部位ではS100A9の発現はほとんど認められないのに対し、悪性メラノーマ(Clark's level III)では、メラノーマ細胞の浸潤に対応するように基底層直上の表皮にS100A9が発現していることが確認された(図8G)。一方、同じ腫瘍塊でも、母斑組織では表皮側にS100A9の発現は認められなかった。
【0063】
S100A9抗体とエンプリン(CD147)抗体を用いて免疫染色した部位について、エンプリン抗体に代えてメラノーマ特異的抗体(HMB45)を用いて染色したところ、メラノーマ特異的抗体により染色される部位がエンプリン抗体のものと重複していた(図8H)。この結果により、エンプリンは浸潤するメラノーマ細胞において発現していることが確認された。
【0064】
2)アトピー皮膚におけるエンプリンとS100A9の相互作用の証明
エンプリンとS100A9タンパク質とが、単に結合しているだけでなく、実際に相互作用していることをPLA (Proximity Ligation Assay) 法により確認した。PLA法によれば、DNAプローブで標識された2種類の抗体を用い、蛍光色素をラベルした相補的DNAをハイブリダイズさせることで、それらのタンパク質が相互作用しているか否かを明らかにすることができる。PLA法は、通常の免疫染色と比較してはるかに高感度である。
【0065】
Olink社のDuolink in situ PLAキットを用い、相互作用試験を行った。アトピー患者から得られた患部皮膚組織を、4% パラホルムアルデヒドで固定後、通常の方法でパラフィンに包埋した。 4μmで細切後、キシレン処理、エタノール処理を経てPBSにて洗浄し、ブロッキング後、一次抗体(以下の表1参照)と4℃で一晩反応させた。PBSで洗浄後、PLAプローブ(以下の表1参照)と、37℃で2時間反応させた。
【0066】
【表2】
【0067】
洗浄後、DNAプローブとハイブリダイゼーションを行い、TBS-Tで洗浄し、リガーゼを加えて37℃で15分インキュベートし、プローブを融合させた。ポリメラーゼを加え、37℃で90分インキュベートし、ライゲートしたDNAプローブの増幅を行った。Detection kit 613 (Olink社) を用いて蛍光色素をラベルし、顕微鏡観察を行った。結果を図9及び図10に示す。
【0068】
エンプリンとS100A9抗体とを用いてPLA法を行った場合、有棘層から顆粒層付近に強い陽性反応が認められた(図9)。これは、エンプリンとS100A9とが相互作用をしていることを示すものである。一方、S100A8についても、顆粒層付近に陽性反応が認められたが、これはS100A9とダイマーを形成した結果によるものと考えられる(結果は示さない)。
【0069】
7.エンプリン細胞外ドメインへのS100A8、S100A9タンパク質の結合を阻害する薬剤のスクリーニング
1)リコンビナントS100A8、S100A9の調製とビオチン化:
ヒトS100A8、S100A9をGST融合タンパク質として大腸菌で産生させ、グルタチオン共有結合担体によるアフィニティークロマトグラフィーで精製した。その後、GSTを切断・除去した。精製したS100A8、S100A9タンパク質のビオチン化については次の方法をとる。各精製タンパク質濃度に対して3倍モル量の Biotin-(AC5)2Sulfo-OSu (Dojindo社) を混合した。室温で 2 時間反応させた後に Nap-5 (GE Healthcare社) により未反応のビオチン化試薬を除いた。
【0070】
2)エンプリン細胞外ドメインへのS100A8、S100A9タンパク質の結合を阻害する薬剤のスクリーニング:
リコンビナントエンプリン細胞外ドメイン(図1中のsignal peptideの後から、transmembrane domainの前に相当)を96 well プレート (Pierce社) のウェル上に結合させる。ウェルを洗浄後、非特異的吸着を抑えるため、各ウェルは5%BSAその他のブロッキング剤で処理する。次に試験する薬剤(対象としては、溶媒のみ)をウェル内に添加して室温で 1 時間インキュベートする。ウェルを洗浄後、リコンビナントS100A9w(エンプリンの全長配列)を加え室温で1時間インキュベートする。さらにHRP標識した抗 S100A9抗体を同ウェル内に添加して反応させる。再度洗浄し、発色基質 (オルソフェニレンジアミン) を添加して ELISAリーダーで吸光度 (O.D.492 nm)を測定する。この操作により、まずはエンプリンとS100A9(又はS100A8/A9)との結合における検量線を作製する。次にこの結合を阻害する分子のスクリーニングを行う。上記のアッセイ系に薬剤を添加し、吸光度の低下するものを候補薬剤とする。
【0071】
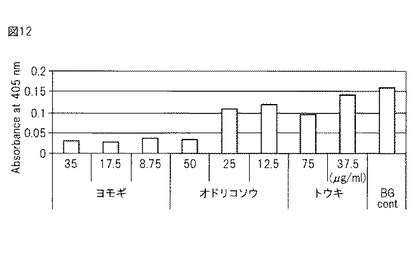
種々の植物抽出物を上記スクリーニング方法にかけたところ、ヨモギエキス、トウキエキス、オドリコソウエキスがコントロールよりも有意にエンプリンとS100A9との結合を阻害することが明らかとなった(図11)。中でも、ヨモギエキスは強い阻害効果を示した(図12)。
【技術分野】
【0001】
本発明は、S100A9の新規受容体であるエンプリンを標的とする慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法を提供する。
【背景技術】
【0002】
過剰増殖や乾癬においてアップレギュレーションされるタンパク質としてS100A8およびS100A9が知られる。S100A8およびS100A9は、20を超えるメンバーから構成されるEF−ハンドカルシウム結合S100タンパク質ファミリーに属する(非特許文献1:Marenholz I et al., Biochem Biophys Res Commun (2004) 322:1111-1122)。どちらのタンパク質も好中球、活性化単球、およびマクロファージによって分泌され、それらの細胞の化学走性分子として機能し、炎症性細胞の漸増に関する正のフィードバックループに関与する(非特許文献2:Roth J et al., Trends Immunol (2003) 24:155-158)。S100A8およびS100A9陽性骨髄細胞は、炎症領域内に浸潤する最初の細胞である(非特許文献3:Odink K et al., Nature (1987) 330:80-82)。慢性関節リウマチ(非特許文献4:Liao H et al., Arthritis Rheum (2004) 50:3792-3803)、多発性硬化症(非特許文献5:Bogumil T et al., Neurosci Lett (1998) 247:195-197)、クローン病(非特許文献6:Lugering N, et al., Digestion (1995) 56:406-414)、および結合組織疾患(非特許文献7:Kuruto R, et al., J Biochem (Tokyo) (1990) 108:650-653)を含む多数のヒト炎症性疾患で高いS100A8およびS100A9血清レベルが観察されている。従って、S100A8およびS100A9は、炎症の誘導および伝播に重要な役割を担うと考えられている。
【0003】
上皮細胞中でS100A8と100A9が果たす生物学的機能について、本発明者は以前、外因性S100A8とS100A9が複合体(S100A8/A9)(別名:カルプロテクチン)を形成することで正常表皮角化細胞(NHEK)を刺激して乾癬性病変などにおいて発現亢進される炎症性サイトカインを産生させ、さらにS100A8/A9誘導性サイトカインがNHEK中でのS100A8およびS100A9の産生および分泌を刺激することを明らかにした(非特許文献8:J Cell Biochem. 2007 Nov 28, Epub ahead of print)。さらに、S100A8/A9自体がNHEKの増殖を増強することも見出した。これらの結果は、主要メディエーターとしてS100A8/A9が関与するNHEKの増殖と炎症の正のフィードバック機構の存在を明らかにした。即ち、S100A8/A9が炎症性サイトカインの産生を誘導して炎症性疾患を惹起し、その炎症が細胞増殖を誘導し、さらには細胞増殖が炎症を誘導するといったスパイラルを形成し、増殖・炎症が連鎖する持続性皮膚炎症性疾患、例えばアトピー性皮膚炎や乾癬などの原因となることが示唆された。
【0004】
S100A8/A9により引き起こされる慢性炎症の負のサイクル形成を阻止するためには、S100A8およびA9のレセプターの同定が必要と考えられる。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Biochem Biophys Res Commun (2004) 322:1111-1122
【非特許文献2】Trends Immunol (2003) 24:155-158
【非特許文献3】Nature (1987) 330:80-82
【非特許文献4】Arthritis Rheum (2004) 50:3792-3803
【非特許文献5】Neurosci Lett (1998) 247:195-197
【非特許文献6】Digestion (1995) 56:406-414
【非特許文献7】J Biochem (Tokyo) (1990) 108:650-653
【非特許文献8】J Cell Biochem. (2008) 104:453-464
【非特許文献9】Nature Cell Biol. (2006) 8(12): 1369-1375
【非特許文献10】Hum Genet (2002) 111:310-313
【非特許文献11】Morrison TB et al., Biotechniques (1998) 24:954-958, 960, 962
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、S100A9の新規受容体を標的とする慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法を提供する。
【課題を解決するための手段】
【0007】
S100タンパク質ファミリーの受容体として知られているRAGE (Receptor for advanced glycation endoproducts)は、S100A9とも結合することが本発明者らにより確認された。しかしながら、両者の結合による信号系の存在は、中和抗体、siRNA試験のいずれによっても確認できなかった。
【0008】
そこで、本発明者らが、培養ケラチノサイトからS100A8及び/又はA9と結合するタンパク質を単離してLC/MS/MS解析にかけ、S100A8/A9結合タンパク質の同定を試みた結果、S100A9のレセプター候補が多数発見された。その中から、免疫グロブリンスーパーファミリーのメンバーであり、且つ膜貫通型の糖タンパク質であるエンプリン(Emmprin)(The extracellular matrix metalloproteinase inducer)(別名バシジン又はCD147)に着目したところ、エンプリンの発現の抑制によりS100A9によるサイトカイン誘導やマトリックスメタロプロテアーゼ誘導が顕著に低下することが分かった。また、免疫染色の結果は、S100A9及びエンプリンがアトピー性皮膚炎や乾癬に罹患している患者の表皮、そして、浸潤するメラノーマ細胞において強発現していることを示した。
【0009】
従って、エンプリンがS100A9のレセプターであること、そして、これらの間の結合を阻害することで、慢性炎症の抑制、更には癌の転移を抑制することが見出され、本発明が完成するに至った。
【0010】
従って、本願は以下の発明を包含する:
(1)慢性炎症抑制剤又は癌転移抑制剤の候補物質がエンプリンとS100A9又はS100A8/A9との結合を有意に阻害する場合に、当該候補物質は慢性炎症又は癌転移を有意に抑制させると評価する、慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法。
(2)慢性炎症抑制剤又は癌転移抑制剤の候補物質の存在下、エンプリンとS100A9又はS100A8/A9とをインキュベートし、エンプリンとS100A9又はS100A8/A9との結合を阻害する物質を慢性炎症抑制剤又は癌転移抑制剤として選定することを含んで成る、(1)に記載の方法。
(3)エンプリンが固体支持体に固相化されている、(1)又は(2)に記載の方法。
(4)前記結合の阻害がELISA法により決定される、(1)〜(3)のいずれかに記載の方法。
(5)エンプリンとS100A9との結合を阻害する薬剤を含んで成る、慢性炎症抑制剤又は癌転移抑制剤。
(6)前記薬剤が、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物を一種又は二種以上含む、(6)に記載の慢性炎症抑制剤又は癌転移抑制剤。
(7)エンプリンとS100A9との結合を阻害する薬剤を被験者に投与することを含んで成る、慢性炎症又は癌転移の抑制方法。
(8)前記薬剤が、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物を一種又は二種以上含む、(9)に記載の方法。
【発明の効果】
【0011】
エンプリンは、マトリックスメタロプロテアーゼ(MMP)を誘導して癌の転移を誘導する。このように、エンプリンと悪性腫瘍との関係はよく知られている。また、S100A8/A9についても、転移の好発部位は、癌細胞が出すVEGF、TNF等の因子などと反応して、S100A8/A9を分泌し、これが癌細胞の転移を誘導することが知られている(非特許文献9:Nature Cell Biol. (2006) 8(12): 1369-1375)。本発明者らが培養ケラチノサイトをS100A8/A9で刺激したところ、癌の浸潤に関与するMMPは、当該刺激により発現が亢進されたものの、エンプリンをノックダウンした場合にはその発現が有意に抑制されることが確認された(結果は示さず)。従来、MMPの発現亢進はエンプリンの自己分泌ループによると考えられていたが、上記の結果は、MMPの発現はエンプリンのみでは亢進されず、S100A9刺激を経由することが必要であることを示している。
【0012】
癌細胞の転移に関与しているエンプリンがS100A9のレセプターとして機能していることは、本発明者らによって初めて見出された。実際、エンプリンの発現を抑制することでS100A9によるサイトカインやMMPの発現亢進は低下する(実施例)。そのため、エンプリンとS100A9の関係を考慮すれば、癌の転移とその悪性度について新たな解釈が可能になると考えられる。更に、エンプリンは、S100A9と同様にアトピー性皮膚炎や乾癬の患者の表皮上層やメラノーマ細胞の表皮において発現していたため、両者はアトピー性皮膚炎、乾癬、癌等の慢性炎症にも関与していることが予想される。従って、本発明によれば、エンプリンとS100A9との結合の阻害を指標とすることで、慢性炎症抑制剤又は癌転移抑制剤の探索が可能になる。
【図面の簡単な説明】
【0013】
【図1】エンプリンの一次構造。
【図2】多次元キャピラリーLC/MS/MSを用いたタンパク質の網羅的解析法により同定されたS100A9の受容体候補タンパク質。
【図3】エンプリンsiRNAによるエンプリンの発現抑制効果。
【図4】エンプリンの発現抑制に伴うサイトカインの発現変化。
【図5】エンプリンの発現抑制に伴うMMPの発現変化。
【図6】ウェスタンブロットによるエンプリン結合タンパク質の同定。矢印はS100A9のバンドを示す。
【図7】可溶性エンプリンによるMMP1の誘導効果。
【図8A】ヒト正常皮膚におけるエンプリン及びS100タンパク質の局在を示す免疫染色図。
【図8B】皮膚モデルにおけるエンプリン及びS100タンパク質の局在を示す免疫染色図。
【図8C】アトピー性皮膚疾患皮膚におけるエンプリン及びS100タンパク質の局在を示す免疫染色図。
【図8D】S100A8抗体、27E10、Dapiを用いて免疫染色したアトピー性皮膚炎及び乾癬の皮膚の比較。
【図8E】S100A9抗体、27E10、Dapiを用いて免疫染色したアトピー性皮膚炎及び乾癬の皮膚の比較。
【図8F】メラノーマ組織におけるS100A9の局在を示す免疫染色図(上段はHE染色、中段はS100A9抗体による染色、下段はDAPI染色。左側、中央、右側の写真はそれぞれ異なるサンプルに由来する)。
【図8G】悪性メラノーマにおけるS100A9の局在を示す免疫染色図。
【図8H】メラノーマ組織におけるエンプリンの局在を示す免疫染色図。
【図9】PLA(Proximity Ligation Assay) 法によるアトピー性皮膚におけるエンプリンとS100A9の相互作用の証明(200倍)。
【図10】PLA法によるアトピー性皮膚におけるエンプリンとS100A9の相互作用の証明(400倍)。
【図11】S100A9とエンプリンとの結合を阻害する植物抽出物のスクリーニング結果。
【図12】ヨモギエキス、オドリコソウエキス、トウキエキスのS100A9−エンプリン結合阻害効果の比較。
【発明を実施するための形態】
【0014】
エンプリンは、2個のIgドメインを有する、1回膜貫通型の糖タンパク質であり、コラゲナーゼ(MMP−1)の発現亢進作用を有している。エンプリンヌルマウスには、精子形成、受精、感覚機能及び記憶機能、並びに混合リンパ球反応の欠損が見られる。エンプリンの一次構造を図1に、そしてエンプリンの全長配列を配列番号1に示す。MMP−1で切断されたIgドメイン1は、コラーゲンの発現を誘導する。
【0015】
本発明のスクリーニング方法は、特に限定されるものではないが、候補物質の存在下エンプリンとS100A9とをインキュベーションし、エンプリンとS100A9との結合を有意に阻害する候補薬剤をS100A9に起因する慢性炎症抑制剤又は癌転移抑制剤として選択することからなる。その評価基準として、例えばエンプリンとS100A9タンパク質との結合がコントロールを作用させた場合と比べ10%以上、又は20%以上、又は30%以上、又は50%以上、又は70%以上、又は100%阻害されていたなら慢性炎症又は癌転移を「有意に抑制する」、と判断してよい。
【0016】
S100A9は、上述のとおりS100A8と複合体を形成していることがあり、この複合体がエンプリンと結合することもある。従って、S100A8/A9とエンプリンとの結合を阻害する物質を慢性炎症抑制剤又は癌転移抑制剤としてスクリーニングしてもよい。
【0017】
エンプリンとS100A9との結合の阻害を検出する手段は特に限定されるわけではないが、ELISA法に基づきエンプリンとS100A9(又はS100A8/A9)との結合における検量線を作製し、この結合を阻害する分子、すなわち吸光度の低下する分子を慢性炎症抑制剤又は癌転移抑制剤の候補薬剤として検出することができる。良好な検出感度を確保する観点から、固体支持体に吸着される分子は、分子量が大きいエンプリンが好ましい。
【0018】
本明細書で使用する「慢性炎症」は、アトピー性皮膚炎、乾癬の他、癌なども包含する。また、本明細書で使用する「癌転移抑制」とは、癌細胞が原発巣である組織から、浸潤、血中およびリンパ管を通しての遊走、新たな組織への定着を経て、増殖を開始する過程のいずれかもしくは全てを抑制することを意味し、癌細胞の増殖抑制とは異なる。
【0019】
S100A8およびA9
S100A8およびA9のアミノ酸配列およびそれをコードするDNA配列は、例えばHum Genet (2002) 111:310-313(非特許文献10)に公開されている。本発明において使用できるS100A8およびA9は、通常ヒト由来の天然型、あるいは組み換えタンパク質であるが、活性を有すれば改変型、異種由来、もしくは非精製品を用いることができる。S100A8およびA9の組換タンパク質は、当業界で周知の方法に従い、例えば単離したまたはPCRにより合成したS100A8又はA9遺伝子 (cDNA) を例えばプラスミド、ウィルス等に挿入して発現ベクターを調製し、これを宿主細胞、例えば微生物、動物細胞又は植物細胞等の培養細胞に導入し、発現させることにより、大量調製することが可能である。
【0020】
S100A9は、水や培地、例えば表皮角化細胞の培養に適当な培地、例えば上記EpiLife(登録商標)培地に溶解し、本発明のスクリーニング系に添加する。添加量は、一概には規定できないが1ng/mlから1mg/ml程度、好ましくは10ng/mlから100μg/ml程度、より好ましくは100ng/mlから10μg/ml程度の濃度とする。S100A9またはS100A8/A9の添加は、好ましくは塩化カルシウムの存在下で行う。S100A9またはS100A8/A9の存在下でのインキュベーション時間、インキュベーション温度といった培養条件は特に制限されることはなく、好ましくは30〜37℃で1〜14時間、より好ましくは34〜37℃で2〜7時間、好ましくはCO25%の下でインキュベーションを行う。
【0021】
本発明の慢性炎症抑制剤は、S100A8/A9に起因する持続性皮膚炎症性疾患、例えばアトピー性皮膚炎、乾癬などの予防、治療といった改善等に有効な医薬品又は化粧品として利用できる。
【0022】
本発明のスクリーニング方法により得られた慢性炎症抑制剤又は癌転移抑制剤として、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物が挙げられる。特に、ヨモギエキスはS100A9とエンプリンとの結合を有意に阻害することが確認されているため(図12)、慢性炎症抑制剤又は癌転移抑制剤の好ましい活性成分であることが予想される。ここで、本発明で使用する各植物の植物体又はその抽出物は、各々の植物体の各種部位(花、花穂、果皮、果実、茎、葉、枝、枝葉、幹、樹皮、根茎、根皮、根、種子又は全草など)をそのまま又は乾燥したものを粉砕して乾燥粉末としたもの、あるいはそのまま又は乾燥・粉砕後、溶媒で抽出したものである。
【0023】
抽出物の場合、抽出に用いられる抽出溶媒は通常抽出に用いられる溶媒であれば何でもよく、特にメタノール、エタノールあるいは1,3−ブチレングリコール等のアルコール類、含水アルコール類、アセトン、酢酸エチルエステル等の有機溶媒を単独あるいは組み合わせて用いることができ、このうち特に、アルコール類、含水アルコール類が好ましく、特にメタノール、エタノール、1,3−ブチレングリコール、含水エタノールまたは含水1,3−ブチレングリコールが好ましい。また前記溶媒は、室温乃至溶媒の沸点以下の温度で用いることが好ましい。
【0024】
抽出方法は特に制限されるものはないが、通常、常温から、常圧下での溶媒の沸点の範囲であれば良く、抽出後は濾過又はイオン交換樹脂を用い、吸着・脱色・精製して溶液状、ペースト状、ゲル状、粉末状とすれば良い。更に多くの場合は、そのままの状態で利用できるが、必要ならば、その効果に影響のない範囲で更に脱臭、脱色等の精製処理を加えても良く、脱臭・脱色等の精製処理手段としては、活性炭カラム等を用いれば良く、抽出物質により一般的に適用される通常の手段を任意に選択して行えば良い。
【0025】
植物体の抽出部位として、ヨモギの場合、葉が、トウキの場合、根が、オドリコソウの場合、茎、葉、花が考えられるが、抽出部位はこれらに限定されない。
【0026】
上記溶媒で抽出して得られた抽出物をそのまま、あるいは例えば凍結乾燥などにより濃縮したエキスを使用でき、また必要であれば吸着法、例えばイオン交換樹脂を用いて不純物を除去したものや、ポーラスポリマー(例えばアンバーライトXAD−2)のカラムにて吸着させた後、所望の溶媒で溶出し、さらに濃縮したものも使用することができる。
【0027】
本発明の慢性炎症抑制剤又は癌転移抑制剤は、前記植物体又はその抽出物の一種または二種以上からなるものであることが好ましいが、本発明の効果を損なわない範囲において、他の種々の成分を含有することができる。また、本発明の慢性炎症抑制剤又は癌転移抑制剤は、その使用目的に合わせて用量、用法、剤型を適宜決定することが可能である。例えば、本発明の慢性炎症抑制剤の投与形態は、経口、非経口、外用等であってよい。剤型としては、例えば錠剤、粉剤、カプセル剤、顆粒剤、エキス剤、シロップ剤等の経口投与剤、又は注射剤、点滴剤、若しくは坐剤等の非経口投与剤軟膏、クリーム、乳液、ローション、パック、浴用剤等の外用剤を挙げることができる。
【0028】
本発明の慢性炎症抑制剤又は癌転移抑制剤の上記エキス成分の配合量は、用途に応じて適宜決定できるが、一般には阻害剤全量中、乾燥物として0.0001〜20.0質量%、好ましくは0.0001〜10.0質量%である。ヨモギエキス、オドリコソウエキス、トウキエキスは濃度依存的に慢性炎症又は癌転移を抑制することが考えられる。
【0029】
また、慢性炎症抑制剤中又は癌転移抑制剤中には、上記薬剤以外に、例えば、通常の食品や医薬品に使用される賦形剤、防湿剤、防腐剤、強化剤、増粘剤、乳化剤、酸化防止剤、甘味料、酸味料、調味料、着色料、香料等、化粧品等に通常用いられる美白剤、保湿剤、油性成分、紫外線吸収剤、界面活性剤、増粘剤、アルコール類、粉末成分、色剤、水性成分、水、各種皮膚栄養剤等を必要に応じて適宜配合することができる。
【0030】
さらに、本発明の慢性炎症抑制剤又は癌転移抑制剤を皮膚外用剤として使用する場合、皮膚外用剤に慣用の助剤、例えばエデト酸二ナトリウム、エデト酸三ナトリウム、クエン酸ナトリウム、ポリリン酸ナトリウム、メタリン酸ナトリウム、グルコン酸等の金属封鎖剤、カフェイン、タンニン、ベラパミル、トラネキサム酸およびその誘導体、甘草抽出物、グラブリジン、カリンの果実の熱水抽出物、各種生薬、酢酸トコフェロール、グリチルリチン酸およびその誘導体またはその塩等の薬剤、ビタミンC、アスコルビン酸リン酸マグネシウム、アスコルビン酸グルコシド、アルブチン、コウジ酸等の美白剤、グルコース、フルクトース、マンノース、ショ糖、トレハロース等の糖類、レチノイン酸、レチノール、酢酸レチノール、パルミチン酸レチノール等のビタミンA類なども適宜配合することができる。
【0031】
以下、具体例を挙げて、本発明を更に具体的に説明する。なお、本発明はこれにより限定されるものではない。
【実施例】
【0032】
1.新規S100A9受容体候補のスクリーニング
培養ケラチノサイトから回収したタンパク質混合物とGST融合S100A9又はS100A8/A9タンパク質とを混合した後、このサンプルについてキャピラリーLC/MS/MSによるタンパク質の網羅的解析法を行った。
【0033】
LC/MS/MS解析
内径100μm、長さ120mmの未充填のキャピラリーカラム(New Objetive社製)のテーパー状の出口側の端部に、充填剤を保持するために、シリカ製のフリットを作製した。得られたキャピラリーカラムに、平均粒径が5μmのオクタデシル化シリカ型充填剤Aqua C18(Phenomenex社製)を、高さが100mmとなるように充填し、分析用逆相キャピラリーカラムを得た。
【0034】
内径250μm、長さ150mmの未充填のキャピラリーカラム(Agilent社製)の出口側の端部に、充填剤を保持するために、シリカ製のフリットを作製した。得られたキャピラリーカラムの出口側に、平均粒径が5μmのカチオン交換樹脂型充填剤Partisphere SCX resins(Whatman社製)と、平均粒径が5μmのアニオン交換樹脂型充填剤PolyWAX LP(PolyLC社製)を質量比2:1で混合したもの、入口側に、平均粒径が5μmのオクタデシル化シリカ型充填剤Aqua C18(Phenomenex社製)を、それぞれ高さが25mmとなるように充填し、トラップ用逆相キャピラリーカラム及びSCX−WAX混合キャピラリーカラムからなる二相型キャピラリーカラムを得た。
【0035】
なお、分析用逆相キャピラリーカラム及び二相型キャピラリーカラムを作製する際には、高圧窒素ガス及び加圧型充填容器を用いて、スラリー充填法により充填剤を充填した。
【0036】
続いて、6ステップのMudPIT型分析(二次元HPLC/ESI MS/MS)により、上記タンパク質混合物を分析した。
【0037】
まず、ペプチドを約4μg含む上清を、加圧法により、二相型キャピラリーカラムにロードした後、試料溶液の10倍以上の体積の移動相A(水、アセトニトリル及びギ酸の体積比95:5:0.1の混合液;pH〜2.6)を用いて、洗浄、脱塩した。この二相型キャピラリーカラム10を、貫通孔型ユニオン(Upchurch Scientific社製)(不図示)を介して、分析用逆相キャピラリーカラム20と接続した。次に、内径が100μmのキャピラリーを配管として用いたNanospace SI−2型HPLC装置(資生堂社製)に接続した。このとき、トラップ用逆相キャピラリーカラム11は、SCX−WAX混合キャピラリーカラム12及び分析用逆相キャピラリーカラム20の上流側に配置した。
【0038】
移動相としては、移動相A、移動相B(水、アセトニトリル及びギ酸の体積比20:80:0.1の混合液)、移動相C(500mMの酢酸アンモニウムを含む移動相A;pH〜6.8)を用い、ペプチドの溶出法は、矩形状に加える移動相Cの体積%をステップ毎に漸増させた、計6ステップのグラジエント溶出法とした。
【0039】
ステップ1のグラジエントプロファイルは、5分間移動相Aを流し、次の5分間で移動相Bの比率を0体積%から15体積%まで増加させ、次の60分間で移動相Bの比率を45体積%まで増加させ、次の10分間で移動相Bの比率を75体積%まで増加させた後、この比率で5分間流すものである。
【0040】
ステップ2〜6のグラジエントプロファイルは、1分間移動相Aを流し、次の4分間移動相Cの比率をX[体積%]として流し、次の5分間で移動相Cの比率を0体積%から15体積%まで増加させ、次の60分間で移動相Cの比率を45体積%まで増加させ、次の10分間で移動相Cの比率を75体積%まで増加させた後、この比率で5分間流すものである。このとき、ポンプの送液の流速を250μL/分とし、抵抗型キャピラリーによるスプリットにより、カラムの流速を300〜400nL/分に調整した。
【0041】
また、ESI MS/MSを測定する際には、イオントラップ型質量分析計LCQ−Deca(Thermo Fisher Scientific社製)を用いた。このとき、分析用逆相キャピラリーカラムから溶出されたペプチドは、スプリットすることなく、質量分析計に直接導入した。
【0042】
なお、質量電荷比(m/z)が400〜1400のフルスキャンMSスペクトル測定1回及びデータ依存型MS/MSスペクトル測定3回を、各ステップを通じて繰り返した。このとき、標準化解裂エネルギーは35%とした。また、マイクロスキャンは、MSスペクトル測定及びMS/MS測定ともに3とした。さらに、動的排除設定は、リピートカウント1、リピート期間0.50分、排除リストサイズ25、排除期間10.00分とした。
【0043】
得られたMS/MSスペクトルは、Bioworksソフトウェア(Thermo Fisher Scientific社製)上で動くSEQUESTアルゴリズムにより、非冗長ヒトデータベース(ftp://ftp.ncbi.nih.gov/blast/db/FASTA/nr.gz、2007/2/8版)に対して、検索した。
【0044】
その結果、図2に列記ような受容体候補タンパク質が同定された。これらのタンパク質の中から、バシジン(エンプリン)をS100A9の新規受容体として以下の実験を行った。
【0045】
2.siRNAによるエンプリンの発現抑制
エンプリンの発現抑制のために、RNAiMaxを用いて、エンプリンsiRNA(Santa Cruz: sc-35298)を、終濃度40 nM又は80 nMとなるように増殖期の培養ケラチノサイトにトランスフェクションした(図3中の「BSG」)。コントロールとして、何も添加していないもの(図3中の「NT」(non-treated control))およびヒト遺伝子のいずれの部分とも相同性を有していないcontrol siRNA-A (Santa Cruz Biotechnology, Inc., sc-3707)(図3中の「LF」)を使用した。尚、トランスフェクションは、培養培地を増殖因子を含まない基礎培地に交換してから行った。トランスフェクションから24時間後にS100A9で増殖ケラチノサイトを刺激し、さらに24時間経過後にRNAを採取した。その結果、図3に示すとおり、エンプリンsiRNAをトランスフェクションした場合、24、48、72時間後には、上記コントロールを用いた場合のエンプリンの発現量と比較して1〜3%まで発現が抑制された。
【0046】
3.エンプリンの発現抑制によるサイトカイン及びMMPの発現変化
IL−8(CXCL−8)、TNFα、IL1−F9、CXCL−1は、S100A8/A9の添加により、ケラチノサイトでの発現が亢進されることが明らかとなっている(前掲J Cell Biochem. (2008) 104:453-464)(非特許文献8)。S100A9の添加によってもIL−8(CXCL−8)、TNFα、IL1−F9及びCXCL−1の発現が亢進されるか、また、エンプリンの発現抑制がこれらのサイトカインの発現にどのような影響を与えるかについてリアルタイム定量PCRにより検討した。同様の方法により、S100A9刺激がMMP−1及びMMP−10の発現に及ぼす影響についても検討した。
【0047】
リアルタイム定量PCR
EpiLife(商標)-KG2(Cascade Biologies社)中で培養した増殖期のNHEKを、2 mMの塩化カルシウム、S100A8またはS100A9(各10μg/ml)を含有又は非含有の同培地に置換し、3時間にわたり曝露させ、MagNA(商標) Pure mRNA抽出キットおよびMagNA Pure(商標)機器(Roche Diagnostics社、日本国、東京都)を用いてmRNAを抽出した。得られたmRNAは、SuperScript(商標) II(Invitrogen Corporation社、米国、カリフォルニア州、カールズバッド)を用いて逆転写させた。リアルタイム定量PCRは、製造業者の取扱説明書にしたがってLightCycler FastStart DNA master SYBR green Iキット(Roche Diagnostics社)を用いてLightCycler高速サーマルサイクラーシステム上で実施した。典型的な反応条件は、10分間の活性化ステップ、それに続く95℃で15秒の変性、60℃で10秒のアニーリング、72℃で10秒の伸長からなるサイクル40回であった。使用したプライマーは、下記の表1に示す。各プライマーの最終濃度は20μlの総反応容量中で0.2〜0.25μMとした。グリセルアルデヒド−3−リン酸脱水素酵素(GAPDH)遺伝子を対照遺伝子として使用した。増幅させたフラグメントの特異性は融解曲線分析によって確認した。各遺伝子の発現レベルは、LightCycler分析用ソフトウエアを用いて定量分析した(非特許文献11:Morrison TB et al., Biotechniques (1998)24:954-958, 960, 962)。目的のmRNA量は、A8/control siRNA-A (santa Cruz: sc-37007)(A8/LF)のmRNAの量に対する比率として表した。
【0048】
【表1】
【0049】
図4に示すとおり、S100A9を添加した試料(A9/LF)は、全てのサイトカインの発現を誘導した。一方、エンプリンsiRNAを添加した試料(A9/siRNA)は、全てのサイトカイン発現量が有意に低下した。これは、エンプリンsiRNAによってエンプリンの発現が抑制された場合、S100A9でサイトカインの発現を刺激してもサイトカインの発現が抑制されることを明確に示している。
【0050】
MMPについても、S100A9を添加した場合、発現が亢進されるのに対し、エンプリンがノックダウンされている場合、S100A9を添加しても発現が有意に抑制されることが明らかとなった(図5)。
【0051】
4.エンプリンとS100タンパク質との結合試験
1)エンプリン細胞外ドメインの作製
[方法]
細胞: ヒト胎児腎細胞株 (HEK293) は ATCC 社より購入したものを使用し、 培養ヒト正常線維芽細胞 OUMS-24は、難波正義博士により単離されたものを使用した。 HEK293 と OUMS-24 は、 Gibco 社の DMEM/F12 培地 (最終濃度が 10% となるように牛胎児血清を添加) を用いて培養した。
【0052】
2)エンプリン細胞外ドメイン発現コンストラクト:
CMV イントロンプロモーター (CMVi) を導入した PDNR 1r ベクター (プロモーターレスドナーベクター;Clontech 社) を構築し、CMVi の下流にヒトエンプリン細胞外ドメイン (C末にmyc-HA-Flag-6Hisタグが付加されている) をコードするcDNAを挿入した(pCMVi-exEmmp: エンプリン細胞外ドメイン発現ドナーベクター)。挿入cDNAの塩基配列は DNA シークエンサーにより正しいことを確認済みである。
【0053】
3)エンプリン細胞外ドメインの細胞外への分泌:
pCMVi-exEmmp を、FuGENE-HD (Roche社) トランスフェクション試薬を用いて HEK293 に導入し、48時間後に培養上清を回収した。培養上清に Sigma 社の抗 HA tag 抗体共有結合担体を添加し、4℃で3時間振盪混和した。その後、5000 rpm、1分間の遠心分離を行い、沈降してきた担体結合タンパク質を酸性バッファーにより溶出した。溶出サンプルを12% の SDS-PAGE を用いて電気泳動した後、PVDF 膜にエレクトロブロットして、CST 社の抗 HA tag 抗体を用いてウエスタンブロットを行い、エンプリン細胞外ドメインが分泌されていることを確認した。
【0054】
4)エンプリン細胞外ドメイン発現アデノウィルス (Ad-exEmmp):
pCMVi-exEmmp のアデノウィルスベクターへの変換は、アデノウィルス作製キット(Adeno-X-expression system: Clontech 社)を使用して行った。
【0055】
5)エンプリン細胞外ドメインの大量精製:
Ad-exEmmp (20 MOI)を培養ヒト正常線維芽細胞 OUMS-24 (10 cm dish x 20)に感染させた。感染させる時期は、OUMS-24 が高密度状態になった時とした。これは、高密度培養により接触阻止が惹起された細胞では細胞分裂が起こらず、細胞内に存在するアデノウィルス由来エピソーム含量の低下が抑制され、その結果、アデノウィルスによる標的遺伝子発現が極めて長期間(2-3週間)持続するからである。しかも、OUMS-24 は無血清培養が可能であることより、長期に渡って培養上清中に分泌された組み換えタンパク質を、血清を含まない状態で回収することができる。感染操作後、24時間培養して無血清培地 DMEM/F12 (フェノールレッド不含) に置換する。3日の間隔で液換えを行い、その度に培養上清を回収して4℃で保存する (タンパク質の安定性に応じて保存条件を変える)。この操作を30日間行った。約2Lの回収培養上清について、80%飽和硫安条件で得られた沈殿を50mlの純水に溶かし、その後、純水に対して透析することで硫安を除いた。 透析後、目的の組み換えタンパク質は、抗 HA tag 抗体共有結合担体充填カラム(sigma 社)を用いて回収した。
【0056】
6)エンプリン結合タンパク質の同定:
エンプリンがS100タンパク質の新規レセプターであることを確認するべく、免疫沈降及びウェスタンブロットによりエンプリン結合タンパク質の同定を行った。本実験において、エンプリンはC末にmyc-HA-Flag-6Hisタグが付加されているものを使用した。また、S100タンパク質としてS100A8及びS100A9タンパク質を使用した。これらのタンパク質がコードされているプラスミド(C末にHAタグが付加されている)をそれぞれHEK293細胞にトランスフェクションし、その培養上清からそれぞれのタンパク質を単離して使用した。
【0057】
エンプリンとS100A8及びS100A9タンパク質の結合解析のために、それぞれのタンパク質が含まれる培養上清を混合して反応させた後、HA抗体及びMyc抗体を用いて免疫沈降を行った。ウェスタンブロットの結果を図6に示す。エンプリンとS100A8とを混合した試料については、32kDa付近にエンプリンのバンドのみが確認された(「Emprin + S100A8」)。一方、エンプリンとS100A9タンパク質とを混合した試料では(「Emprin + S100A9」)、47.5kDa付近にそれらの結合を示すバンドが確認された。以上の結果から、エンプリンはS100A9タンパク質の新規レセプター候補であることが明らかとなった。
【0058】
5.可溶性エンプリンのMMP発現に及ぼす影響
従来、MMPの発現亢進は、エンプリンの細胞外ドメインがMMPにより分解され、放出された可溶性のエンプリンが細胞表面に存在する受容体としてのエンプリンに結合し、MMPの産生を促すというエンプリンの自己分泌(オートクリン)に起因すると考えられていた。従って、可溶性エンプリンが実際にMMPの発現を亢進させるか否かについて検討した。
【0059】
可溶性エンプリンとして上記方法により精製したエンプリン細胞外ドメインを用い、これをケラチノサイトに添加したところ、0.025、0.25、2.5μMのいずれの濃度でもMMP−1誘導効果はほとんど見られなかった。また、S100A9単独ではMMP−1の発現が顕著に亢進されたのに対し、可溶性エンプリンとS100A9とを一緒にケラチノサイトに添加した場合、S100A9によるMMP1発現亢進効果は有意に抑制された。結果を図7に示す。可溶性エンプリンとS100A9とが共存した場合にMMPの発現が抑制されたのは、MMP産生を誘導するS100A9が、可溶性エンプリンに捕捉され、両者が結合体したことによるものと考えられる。これらの結果から、従来提唱されていたエンプリンの自己分泌によるメカニズムより、S100A9刺激がエンプリンを通じてMMPの発現を亢進させるというメカニズムの方が合理的と思われる。結果は示さないが、可溶性エンプリンはMMP−10、TNFα、IL−8の発現も有意に抑制した。
【0060】
6.表皮におけるエンプリンの局在
1)免疫染色
エンプリンがヒト表皮に存在するか、また、S100タンパク質と同一局在を示すかについて、免疫染色により確認した。結果を図8A〜Cに示す。エンプリンは、正常表皮、皮膚モデル、アトピー性皮膚炎(AD)の皮膚のいずれでも顆粒層で多く発現している。また、S100タンパク質もエンプリン付近で発現していた。
【0061】
更に、アトピー性皮膚炎の皮膚では、エンプリンが高発現しており、S100A8及びS100A9タンパク質の発現も亢進されていることが明らかとなった。また、S100A8/A9複合体を特異的に結合する27E10抗体を用いた免疫染色の結果は、乾癬(Pso)の皮膚と比較した場合、アトピー性皮膚疾患の皮膚の顆粒層でS100A8/A9複合体が高発現していることを示している(図8D及び図8E)。
【0062】
免疫染色の結果によると、正常表皮では、S100A8、A9、エンプリンはほとんど発現していない。しかし、メラノーマ組織におけるS100A9の発現について免疫染色により確認したところ、いずれのサンプルでも表皮側にS100A9の強発現が認められた(図8F、左側、中央、右側の写真)。また、正常部位ではS100A9の発現はほとんど認められないのに対し、悪性メラノーマ(Clark's level III)では、メラノーマ細胞の浸潤に対応するように基底層直上の表皮にS100A9が発現していることが確認された(図8G)。一方、同じ腫瘍塊でも、母斑組織では表皮側にS100A9の発現は認められなかった。
【0063】
S100A9抗体とエンプリン(CD147)抗体を用いて免疫染色した部位について、エンプリン抗体に代えてメラノーマ特異的抗体(HMB45)を用いて染色したところ、メラノーマ特異的抗体により染色される部位がエンプリン抗体のものと重複していた(図8H)。この結果により、エンプリンは浸潤するメラノーマ細胞において発現していることが確認された。
【0064】
2)アトピー皮膚におけるエンプリンとS100A9の相互作用の証明
エンプリンとS100A9タンパク質とが、単に結合しているだけでなく、実際に相互作用していることをPLA (Proximity Ligation Assay) 法により確認した。PLA法によれば、DNAプローブで標識された2種類の抗体を用い、蛍光色素をラベルした相補的DNAをハイブリダイズさせることで、それらのタンパク質が相互作用しているか否かを明らかにすることができる。PLA法は、通常の免疫染色と比較してはるかに高感度である。
【0065】
Olink社のDuolink in situ PLAキットを用い、相互作用試験を行った。アトピー患者から得られた患部皮膚組織を、4% パラホルムアルデヒドで固定後、通常の方法でパラフィンに包埋した。 4μmで細切後、キシレン処理、エタノール処理を経てPBSにて洗浄し、ブロッキング後、一次抗体(以下の表1参照)と4℃で一晩反応させた。PBSで洗浄後、PLAプローブ(以下の表1参照)と、37℃で2時間反応させた。
【0066】
【表2】
【0067】
洗浄後、DNAプローブとハイブリダイゼーションを行い、TBS-Tで洗浄し、リガーゼを加えて37℃で15分インキュベートし、プローブを融合させた。ポリメラーゼを加え、37℃で90分インキュベートし、ライゲートしたDNAプローブの増幅を行った。Detection kit 613 (Olink社) を用いて蛍光色素をラベルし、顕微鏡観察を行った。結果を図9及び図10に示す。
【0068】
エンプリンとS100A9抗体とを用いてPLA法を行った場合、有棘層から顆粒層付近に強い陽性反応が認められた(図9)。これは、エンプリンとS100A9とが相互作用をしていることを示すものである。一方、S100A8についても、顆粒層付近に陽性反応が認められたが、これはS100A9とダイマーを形成した結果によるものと考えられる(結果は示さない)。
【0069】
7.エンプリン細胞外ドメインへのS100A8、S100A9タンパク質の結合を阻害する薬剤のスクリーニング
1)リコンビナントS100A8、S100A9の調製とビオチン化:
ヒトS100A8、S100A9をGST融合タンパク質として大腸菌で産生させ、グルタチオン共有結合担体によるアフィニティークロマトグラフィーで精製した。その後、GSTを切断・除去した。精製したS100A8、S100A9タンパク質のビオチン化については次の方法をとる。各精製タンパク質濃度に対して3倍モル量の Biotin-(AC5)2Sulfo-OSu (Dojindo社) を混合した。室温で 2 時間反応させた後に Nap-5 (GE Healthcare社) により未反応のビオチン化試薬を除いた。
【0070】
2)エンプリン細胞外ドメインへのS100A8、S100A9タンパク質の結合を阻害する薬剤のスクリーニング:
リコンビナントエンプリン細胞外ドメイン(図1中のsignal peptideの後から、transmembrane domainの前に相当)を96 well プレート (Pierce社) のウェル上に結合させる。ウェルを洗浄後、非特異的吸着を抑えるため、各ウェルは5%BSAその他のブロッキング剤で処理する。次に試験する薬剤(対象としては、溶媒のみ)をウェル内に添加して室温で 1 時間インキュベートする。ウェルを洗浄後、リコンビナントS100A9w(エンプリンの全長配列)を加え室温で1時間インキュベートする。さらにHRP標識した抗 S100A9抗体を同ウェル内に添加して反応させる。再度洗浄し、発色基質 (オルソフェニレンジアミン) を添加して ELISAリーダーで吸光度 (O.D.492 nm)を測定する。この操作により、まずはエンプリンとS100A9(又はS100A8/A9)との結合における検量線を作製する。次にこの結合を阻害する分子のスクリーニングを行う。上記のアッセイ系に薬剤を添加し、吸光度の低下するものを候補薬剤とする。
【0071】
種々の植物抽出物を上記スクリーニング方法にかけたところ、ヨモギエキス、トウキエキス、オドリコソウエキスがコントロールよりも有意にエンプリンとS100A9との結合を阻害することが明らかとなった(図11)。中でも、ヨモギエキスは強い阻害効果を示した(図12)。
【特許請求の範囲】
【請求項1】
慢性炎症抑制剤又は癌転移抑制剤の候補物質がエンプリンとS100A9又はS100A8/A9との結合を有意に阻害する場合に、当該候補物質は慢性炎症又は癌転移を有意に抑制させると評価する、慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法。
【請求項2】
慢性炎症抑制剤又は癌転移抑制剤をスクリーニングする方法であって、慢性炎症抑制剤又は癌転移抑制剤の候補物質の存在下、エンプリンとS100A9又はS100A8/A9とをインキュベートし、エンプリンとS100A9又はS100A8/A9との結合を阻害する物質を慢性炎症抑制剤又は癌転移抑制剤として選定することを含んで成る、請求項1に記載の方法。
【請求項3】
エンプリンが固体支持体に固相化されている、請求項1又は2に記載の方法。
【請求項4】
前記結合の阻害がELISA法により決定される、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
エンプリンとS100A9との結合を阻害する薬剤を含んで成る、慢性炎症抑制剤又は癌転移抑制剤。
【請求項6】
前記薬剤が、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物を一種又は二種以上含む、請求項5に記載の慢性炎症抑制剤又は癌転移抑制剤。
【請求項7】
エンプリンとS100A9との結合を阻害する薬剤を被験者に投与することを含んで成る、慢性炎症又は癌転移の抑制方法。
【請求項8】
前記薬剤が、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物を一種又は二種以上含む、請求項7に記載の方法。
【請求項1】
慢性炎症抑制剤又は癌転移抑制剤の候補物質がエンプリンとS100A9又はS100A8/A9との結合を有意に阻害する場合に、当該候補物質は慢性炎症又は癌転移を有意に抑制させると評価する、慢性炎症抑制剤又は癌転移抑制剤のスクリーニング方法。
【請求項2】
慢性炎症抑制剤又は癌転移抑制剤をスクリーニングする方法であって、慢性炎症抑制剤又は癌転移抑制剤の候補物質の存在下、エンプリンとS100A9又はS100A8/A9とをインキュベートし、エンプリンとS100A9又はS100A8/A9との結合を阻害する物質を慢性炎症抑制剤又は癌転移抑制剤として選定することを含んで成る、請求項1に記載の方法。
【請求項3】
エンプリンが固体支持体に固相化されている、請求項1又は2に記載の方法。
【請求項4】
前記結合の阻害がELISA法により決定される、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
エンプリンとS100A9との結合を阻害する薬剤を含んで成る、慢性炎症抑制剤又は癌転移抑制剤。
【請求項6】
前記薬剤が、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物を一種又は二種以上含む、請求項5に記載の慢性炎症抑制剤又は癌転移抑制剤。
【請求項7】
エンプリンとS100A9との結合を阻害する薬剤を被験者に投与することを含んで成る、慢性炎症又は癌転移の抑制方法。
【請求項8】
前記薬剤が、ヨモギ、トウキ及びオドリコソウから成る群から選択される植物体又はその抽出物を一種又は二種以上含む、請求項7に記載の方法。
【図1】

【図2】

【図3】
【図4】

【図5】
【図7】

【図11】

【図12】
【図6】
【図8A】
【図8B】

【図8C】
【図8D】
【図8E】

【図8F】
【図8G】
【図8H】

【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図7】
【図11】
【図12】
【図6】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図8G】
【図8H】
【図9】
【図10】
【公開番号】特開2011−47932(P2011−47932A)
【公開日】平成23年3月10日(2011.3.10)
【国際特許分類】
【出願番号】特願2010−174038(P2010−174038)
【出願日】平成22年8月2日(2010.8.2)
【出願人】(000001959)株式会社資生堂 (1,748)
【Fターム(参考)】
【公開日】平成23年3月10日(2011.3.10)
【国際特許分類】
【出願日】平成22年8月2日(2010.8.2)
【出願人】(000001959)株式会社資生堂 (1,748)
【Fターム(参考)】
[ Back to top ]