
オイル生成能を有する微生物を用いた脂肪酸アルキルエステルの製造方法
本発明は、オイル生成能を有する微生物を利用した脂肪酸アルキルエステル(fatty acid alkyl ester)の製造方法に関し、より詳細にはオイル生成能を有する微生物を培養して、オイルを大量蓄積した後、生成されたオイルを微生物内で自己分解して、遊離脂肪酸を生成した後、アルキルエステル化することを特徴とする脂肪酸アルキルエステルの製造方法に関する。本発明に係る方法は、代謝工学的アプローチを介して、微生物が産生する代表的なオイルであるトリアシルグリセロール(Triacyl glycerol)のような微生物内に蓄積されたオイルを高い転換効率で脂肪酸アルキルエステルに転換でき、最近バイオディーゼルの効用価値が確認された脂肪酸アルキルエステルの産業的生産方法で役立つ。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、オイル(oil)生成能を有する微生物を利用した脂肪酸アルキルエステル(fatty acid alkyl ester)の製造方法に関し、より詳細にはオイル生成能を有する微生物を培養して、オイルを大量蓄積した後、生成されたオイルを微生物内で自己分解して、遊離脂肪酸を生成した後、アルキルエステル化することを特徴とする脂肪酸アルキルエステルの製造方法に関する。
【背景技術】
【0002】
最近、油の高価格と環境問題から微生物を利用したバイオ燃料生産が大きな関心をよんでいる。最近、バイオディーゼルが軽油に代わり、または軽油と混合してディーゼルエンジンに使用できる代替燃料として浮上し、市場規模が急速に増加している。ヨーロッパ連合(EU)では2008年一年間660万トンのバイオディーゼルが生産され、55億ユーロの市場規模となっている(Biodiesel Market, Frost & Sullivan)。また、米国でも2006年一年間3億ガロンのバイオディーゼルが生産された(Biodiesel Market, Global Industry Analysts Inc, 2006. 5)。
【0003】
バイオディーゼルは、燃焼率が良く、有毒ガスの排出が低く、軽油より発熱量が10%程度低く、発火点はより高いため、運送と保存により安全な長所を有している。バイオディーゼルは、主に動植物の脂肪成分を軽油と類似の特性を有するように加工したり、植物性油脂(ぬか、廃食用油、大豆油、菜種油等)とアルコールを反応させて製造した。しかし、この場合、大量生産が難しい短所がある。従って、軽油の代替燃料に適するバイオディーゼルを微生物を利用して大量生産すると、原油輸入代替効果及び温室ガス排出減少による環境的効果等を齎すと期待された。
【0004】
一方、オイルは、微生物が、炭素源は豊富であるが他の成長因子(窒素、燐、酸素、硫黄等)が不足する時、細胞内で合成して蓄積するエネルギー貯蔵物質であり、成長環境が変わって制限された成長因子が再提供されると、蓄積しておいたオイルを分解してエネルギー源として使用するようになる。オイルは、生産する微生物の種類や、供給される化学物質の種類、培養条件等の変化によって100種類以上の単量体が構成要素となることが知られている。
【0005】
最近、サトウキビ等の植物性脂肪酸を利用して、アルコールを加えて脂肪酸アルキルエステル(fatty acid alkylester)を生産する技術が開発され、これは現在バイオディーゼル燃料として使われている。また、遊離脂肪酸をエステル化する方法は、ヨーロッパ公開特許127104A号、ヨーロッパ公開特許184740A号及び米国登録特許4,164,506号等に開示されているが、開示された特許では脂肪酸及び脂肪酸トリグリセリドの混合物とメタノールを共に加熱して、エステル化反応を行った。また、油脂から脂肪酸アルキルエステルの収率を高めるための方法として、エステル交換反応の結果として得られたグリセリンから遊離脂肪酸(free fatty acid)を分離した後、分離した遊離脂肪酸をエステル化する方法がヨーロッパ公開特許708813A号に開示されている。
【0006】
しかし、このような方法では多量の脂肪酸又は遊離脂肪酸が得られ難く、現在最も多く使われている植物性脂肪酸は、植物の成長期間が長く、代謝工学的アプローチが多少難しいため、脂肪酸蓄積の増大及びこれを利用した生産性の増加が困難である。
【0007】
そこで、本発明者等は代謝工学的アプローチを介して生産性が増加した、バイオディーゼルとして使用できる脂肪酸アルキルエステルを高効率で提供できる新しい方法を開発しようと鋭意努力した結果、代謝工学的方法でオイル生成能を有する微生物を利用して、オイル生成を最大化した後、これを微生物内で自己分解させて遊離脂肪酸を生産して、アルキルエステル化することによって、高効率で脂肪酸アルキルエステルを製造する可能性があることを確認して、本発明を完成するようになった。
【発明の概要】
【0008】
本発明の目的は、代謝工学的アプローチを介して生産性が増加した、バイオディーゼルとして使用できる脂肪酸アルキルエステルを高効率で大量生産できる新しい方法を提供することである。
【0009】
前記目的を達成するために、本発明は下記の工程を含む脂肪酸アルキルエステルの製造方法を提供する:
(a)オイル生成能を有する微生物を培養してオイルを生成する工程、
(b)前記生成されたオイルを含有する微生物においてオイルの自己分解を誘導して、遊離脂肪酸を生成する工程、及び
(c)前記生成された遊離脂肪酸にアルコールを添加し、反応させて脂肪酸アルキルエステルを製造する工程。
【0010】
本発明はまた、下記工程を含む、オイル生成能とリパーゼをコードする遺伝子を有する微生物を利用した脂肪酸メチルエステルの製造方法を提供する:
(a)オイル生成能とリパーゼをコードする遺伝子を有する微生物を培養して、オイルを生成させる工程、
(b)前記生成されたオイルを含有する微生物においてリパーゼによるオイルの自己分解を誘導して、遊離脂肪酸を生成させる工程、及び
(c)前記生成された遊離脂肪酸にメタノールを添加し、反応させて脂肪酸メチルエステルを製造する工程。
【0011】
本発明の他の特徴及び具現例は、以下の詳細な説明及び添付された特許請求範囲からより一層明白になる。
【図面の簡単な説明】
【0012】
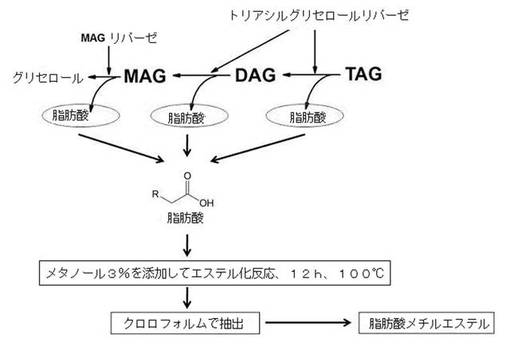
【図1】ロドコッカス・オパカスPD630菌株を利用して代表的オイル成分であるトリアシルグリセロール(Triacylglycerol、TAG)から脂肪酸メチルエステル(fatty acid methyl ester)を高効率で製造する工程を示した図である。
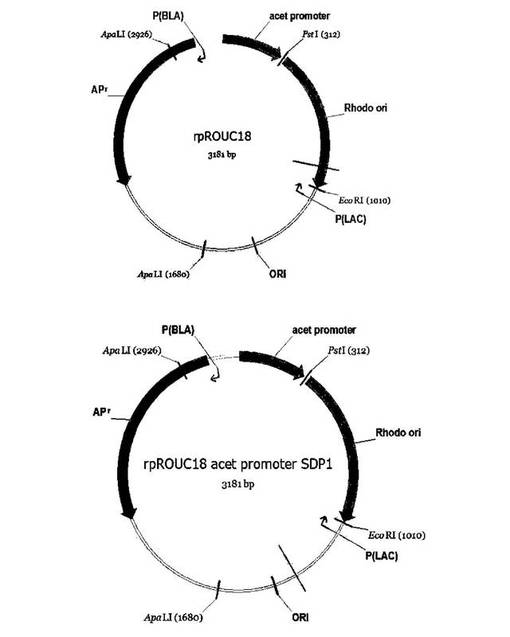
【図2】組換えベクターrpROUC18(a)及びrpROUC18_KM(b)の遺伝子地図を示した図である。
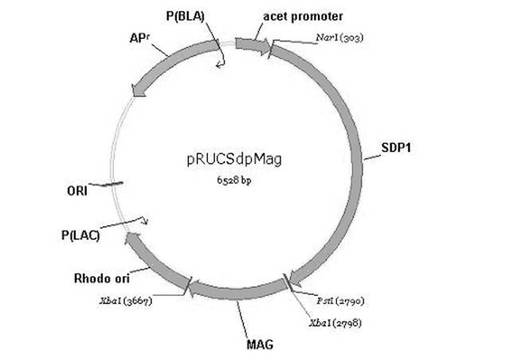
【図3】sdp1及びMSMEG_0220遺伝子を含む組換えベクターpRUCSdpMagの遺伝子地図を示した図である。
【図4】表1の情報から作られたプラスミドの種類とその情報を示した図である。
【図5】遊離脂肪酸に対するガスクロマトグラフィー(Gas chromatography)分析結果を示した図である。
【図6】脂肪酸メチルエステルに対するガスクロマトグラフィー分析結果を示した図である。
【図7】脂肪酸メチルエステルに対するガスクロマトグラフィー分析結果を示した図である。
【発明を実施するための形態】
【0013】
他の方式で定義されない限り、本明細書において使用されたあらゆる技術的・科学的用語は、本発明が属する技術分野に熟練した専門家によって通常理解されるものと同じ意味を有する。通常、本明細書において使用された命名法は、本技術分野において周知であり、しかも汎用されるものである。
本発明の詳細な説明などにおいて使用される主な用語の定義は、下記の通りである。
【0014】
本願において、「オイル(oil)」とは、微生物が炭素源は豊富であるが、他の成長因子(窒素、燐、酸素、硫黄等)が不足する時、細胞内で合成して蓄積するエネルギー貯蔵物質をいい、これは脂肪酸の前駆体として加水分解によって遊離脂肪酸とグリセロールに分離する。
【0015】
本願において、「脂肪酸」とは、鎖状の飽和又は不飽和モノカルボン酸である。これは、炭素鎖の長さ及び飽和の可否によって分類され、オイル、即ち、脂肪が加水分解して生じる脂肪酸を特に「遊離脂肪酸」という。
【0016】
一観点において、本発明はオイル生成能を有する微生物を利用した脂肪酸アルキルエステルの製造方法に関する。より具体的には、本発明による次の工程を含むことができる;
(a)オイル生成能を有する微生物を培養してオイルを生成する工程、
(b)前記生成されたオイルを含有する微生物においてオイルの自己分解を誘導して、遊離脂肪酸を生成する工程、及び
(c)前記生成された遊離脂肪酸にアルコールを添加し、反応させて脂肪酸アルキルエステルを製造する工程。
本発明は下記の工程を含む脂肪酸アルキルエステルの製造方法に関する:
(a)オイル生成能を有する微生物を培養してオイルを生成する工程、
(b)前記生成されたオイルを含有する微生物においてオイルの自己分解を誘導して、遊離脂肪酸を生成する工程、及び
(c)前記生成された遊離脂肪酸にアルコールを添加し、反応させて脂肪酸アルキルエステルを製造する工程。
【0017】
本発明において、前記オイルは、トリアシルグリセロール(TAG)、ジアシルグリセロール(diacylglycerol、DAG)、モノアシルグリセロール(monoacylglycerol、MAG)、リン脂質(phospholipid)、ステロール脂質(sterol lipid)、スフィンゴ脂質(sphingolipid)、サッカロ脂質(saccharolipid)、プレノール脂質(prenol lipid)及びポリケチド(polyketide)等であってもよいが、微生物で産生されるオイルであれば、これに限定されない。
【0018】
ここで、前記オイルの分解によって生成される遊離脂肪酸は、飽和または不飽和脂肪であってもよく、不飽和脂肪酸は脂肪酸の炭素鎖において一つ以上の二重結合を有する脂肪酸をいい、例示的にオレイン酸(oleic acid)、リノール酸(linoleic acid)、リノレン酸(linolenic acid)、パルミトレイン酸(palmitoleic acid)、リシノール酸(ricinoleic acid)、バクセン酸(vaccenic acid)、カドレン酸(gadoleic acid)、アラキドン酸(arachidonic acid)、EPA(5,8,11,14,17-eicosapentaenoic acid)、エルカ酸(erucic acid)、DHA(4,7,10,13,16,19-docosahexaenoic acid)等が挙げられる。また、飽和脂肪酸は、炭素鎖に二重結合を持たない脂肪酸をいい、例示的に酪酸(butyric acid)、カプロン酸(caproic acid)、カプリル酸(caprylic acid)、カプリン酸(capric acid)、ラウリン酸(lauric acid)、ミリスチン酸(myristic acid)、パルミチン酸(palmitic acid)、ステアリン酸(stearic acid)、アラキジン酸(arachidic acid)、ベヘン酸(behenic acid)、リグノセンリン酸(lignoceric acid)等が挙げられる。前記脂肪酸は置換基で修飾されたものであってもよく、例示的に芳香族環基、エポキシ基、シアノ基、及びハロゲン基からなる群から選択される置換基で修飾されたものが用いられるが、これに限定されない。
【0019】
本発明において、前記遊離脂肪酸の前駆体になるオイルは、オイル生成能を有する微生物によって生成されることを特徴とし、例えば、アエロモナス(Aeromonas)属微生物、アクロモバクター(Achromobacter)属微生物、アシドボラックス・デラフィルディ(Acidovorax delafieldii)、アシドボラックス・ファシリス(Acidovorax facilis)、アシネトバクター(Acinetobacter)属微生物、アクチノマイセス(Actinomyces)属微生物、アエロモナス(Aeromonas)属微生物、アルカリゲネス(Alcaligenes)属微生物、アルテロモナス(Alteromonas)属微生物、アルソルニア(Althornia)属微生物、アプラノキトリウム(Aplanochytrium)属微生物、アスペルギルス(Aspergillus)属微生物、アモエボバクター(Amoebobacter)属微生物、アファノカプサ属微生物(Aphanocapsa sp.)、アファノテーケ属微生物(Aphanothece sp.)、アクアスピリラム・オートトロフィカム(Aquaspirillum autotrophicum)、アゾリゾビウム・カウリノダンス(Azorhizobium caulinodans)、アゾスピリラム属微生物(Azospirillum sp.)、アゾスピリラム(Azospirillum)属微生物、アゾトバクター(Azotobacter)属微生物、バシラス(Bacillus)属微生物、ベギアトア(Beggiatoa)属微生物、ベイジェリンキア(Beijerinckia)属微生物、ベネケア(Beneckea)属微生物、ブラケスレア(Blakeslea)属微生物、ボルデテラ・パーツシス(Bordetella pertussis)、ブラディリゾビウム・ジャポニカム(Bradyrhizobium japonicum)、カリオファノン・ラタム(Caryophanon latum)、カウロバクター(Caulobacter)属微生物、クロログロエア(Chlorogloea)属微生物、クロマチウム(Chromatium)属微生物、クロモバクテリウム(Chromobacterium)属微生物、クロストリジウム(Clostridium)属微生物、コマモナス(Comamonas)属微生物、コリネバクテリウム(Corynebacterium)属微生物、クリプテコジニウム(Crypthecodinium)属微生物、シアノバクテリア(Cyanobacteria)属微生物、デルキシア(Derxia)属微生物、デスルホネマ(Desulfonema)属微生物、デスルホサルシナ・バリアビリス(Desulfosarcina variabilis)、デスルホビブリオ・サポボランス(Desulfovibrio sapovorans)、エクトチオロドスピラ(Ectothiorhodospira)属微生物、エリナ(Elina)属微生物、エントモフトラ(Entomophthora)属微生物、フェロバシラス・フェロキシダンス(Ferrobacillus ferroxidans)、フラボバクテリウム属微生物(Flavobacterium sp.)、ヘモフィルス・インフルエンザ(Haemophilus influenzae)、ハロバクテリウム(Halobacterium)属微生物、ヒドロクラスラタス・クラスラタス(Hydroclathratus clathratus)、ヒドロゲノモナス・ファシリス(Hydrogenomonas facilis)、ヒドロゲノファガ(Hydrogenophaga)属微生物、ハイフォミクロビウム(Hyphomicrobium)属微生物、イリオバクター・デラフィエルディ(Ilyobacter delafieldii)、ジャポノキトリウム(Japonochytrium)属微生物、ラブリス・モナクス(Labrys monachus)、ランプロシスティス・ロゼオペルシシーナ(Lamprocystis roseopersicina)、ランプロペディア・ハイアリーナ(Lampropedia hyalina)、レジオネラ属微生物(Legionella sp.)、レプトスリックス・ディスコホラス(Leptothrix discophorus)、メチロバクテリウム(Methylobacterium)属微生物、メチロシナス(Methylosinus)属微生物、ミクロコッカス(Micrococcus)属微生物、モルティエレラ(Mortierella)属微生物、マイコバクテリウム(Mycobacterium)属微生物、ニトロバクター(Nitrobacter)属微生物、ノカルディア(Nocardia)属微生物、パラコッカス・デントリフィカンス(Paracoccus dentrificans)、オシラトリア(Oscillatoria)、ペニシリウム・シクロピウム(Penicillium cyclopium)、フォトバクテリウム(Photobacterium)属微生物、フィサルム・ポリセファラム(Physarum ploycephalum)、フィコミセス(Phycomyces)属微生物、シュードモナス(Pseudomonas)属微生物、ピシウム(Pythium)属微生物、ラルストニア(Ralstonia)属微生物、リゾビウム(Rhizobium)属微生物、ロドバシラス(Rhodobacillus)属微生物、ロドバクター(Rhodobacter)属微生物、ロドコッカス(Rhodococcus)属微生物、ロドシクルス(Rhodocyclus)属微生物、ロドミクロビウム・バニエリ(Rhodomicrobium vannielii)、ロドシュードモナス(Rhodopseudomonas)属微生物、ロドスピリラム(Rhodospirillum)属微生物、シゾチトリウム(Schizochytrium)属微生物、スフィンゴモナス・パウシモビリス(Sphingomonas paucimobilis)、スピリラム(Spirillum)属微生物、スピルリナ(Spirulina)属微生物、スタフィロコッカス(Staphylococcus)属微生物、ステラ(Stella)属微生物、ストレプトマイセス(Streptomyces)属微生物、シントロフォモナス・ウォルフェイ(Syntrophomonas wolfei)、サーモフィリック・シアノバクテリア(Thermophilic cyanobacteria)、サーマス・サーモフィルス(Thermus thermophilus)、チオバシラスA2(Thiobacillus A2)、チオバシラス(Thiobacillus)属微生物、チオカプサ(Thiocapsa)属微生物、トラウストキトリウム(Thraustochytrium)属微生物、チオシスティス・バイオラセア(Thiocystis violacea)、ビブリオ・パラハエモリティカス(Vibrio parahaemolyticus)、キサントバクター・オートトロフィカス(Xanthobacter autotrophicus)、キサントモナス・マルトフィリア(Xanthomonas maltophilia)、ズーグロエア(Zoogloea)属微生物及びオイル生成能を有する酵素をコードする遺伝子に形質転換された微生物等が挙げられるが、オイルを産生できる微生物であれば、前記に制限されず本発明に係る方法が適用されることは明らかである。
【0020】
本発明において、前記工程(a)の培養は、菌体増殖のための1段階培養及びオイル生成のための2段階培養を含むが、この時、オイル生成のための培養はオイル生成量を向上するために窒素源が制限された培地で行うことを特徴とする。
【0021】
微生物によって生成されたオイルは微生物において自己分解するが、この時、前記工程(b)の自己分解は、リパーゼによって行われる。このようなリパーゼとしては、トリアシルグリセロールリパーゼ(triacyl glycerol lipase、EC: 3.1.1.34, 3.1.1.13)、モノアシルグリセロールリパーゼ(monoacyl glycerol lipase、EC: 3.1.1.23)、リソホスホポリパーゼ(lysophospholipase、EC: 3.1.1.5)等を利用してもよい。
【0022】
この時、望ましくは、前記オイル生成能を有する微生物にリパーゼをコードする遺伝子を導入または増幅することができ、さらに望ましくは基質に反応して活性化できるリパーゼ遺伝子を導入したことを特徴とする。本発明の一実施形態では、微生物のオイル自己分解のために、リパーゼ遺伝子、即ち、配列番号5及び8のリパーゼ遺伝子を導入した菌株を利用した。それと共に、本発明の他の実施形態では、配列番号17、18及び19のトリアシルグリセロールリパーゼ遺伝子または配列番号20のモノアシルグリセロールリパーゼ遺伝子を各々または組み合わせて導入した菌株を利用した。
【0023】
本発明において、前記工程(c)で添加されるアルコールは、1次、2次及び3次構造の全てのアルコールを利用してもよいが、望ましくは炭素数1乃至8のアルコールまたはこの中の2以上の混合物を利用でき、さらに望ましくは、メタノール(methanol)を利用することを特徴とする。
【0024】
また、前記工程(c)の反応は、80〜120℃で1〜24時間行うことを特徴とする。また、有機溶媒を追加で添加して行うことを特徴とし、望ましくはクロロホルムを添加して行われる。
【0025】
本発明の一実施形態では、オイル生成能を有する微生物として、ロドコッカス・オパカスPD630菌株を使用し、菌体増殖のための1段階培養とオイル生成のための2段階培養で分けて行った。前記オイル生成のための2段階培養は、窒素源を制限した培地を使ってオイルの生成を誘導した。この時、生成されたオイルの自己分解のために、前記菌株にアセトアミドによって活性化するリパーゼ遺伝子を導入し、微生物にアセトアミドを加えて活性化したリパーゼを利用してin vivoで約0.27g/Lの遊離脂肪酸を生成した。
【0026】
また、前記取得された遊離脂肪酸溶液は、凍結乾燥して水を取り除いた後、クロロホルムを添加し、H2SO4を含むメタノールを添加した後、100℃で12時間反応させて、水を添加した後、有機溶媒層を分離して、遊離脂肪酸メチルエステルを取得した。生成された遊離脂肪酸メチルエステル(Free fatty acid methyl ester)の濃度は、0.2g/Lであり、これは遊離脂肪酸から遊離脂肪酸メチルエステルへの転換が行われたことを意味する(図1)。そこで、本発明に係る方法を利用することによって、より容易でかつ親環境的方法で脂肪酸メチルエステルを高効率に産生できることが明らかになり、従来の軽油等を代替できるバイオディーゼルの生産に非常に有用な方法であることが確認された。
【0027】
他の観点において、本発明は、下記工程を含む、オイル生成能とリパーゼをコードする遺伝子を有する微生物を利用した脂肪酸メチルエステルの製造方法に関する:
(a)オイル生成能とリパーゼをコードする遺伝子を有する微生物を培養して、オイルを生成させる工程、
(b)前記生成されたオイルを含有する微生物においてリパーゼによるオイルの自己分解を誘導して、遊離脂肪酸を生成させる工程、及び
(c)前記生成された遊離脂肪酸にメタノールを添加し、反応させて脂肪酸メチルエステルを製造する工程。
【0028】
本発明の他の実施形態では、トリアシルグリセロールリパーゼ遺伝子またはモノアシルグリセロールリパーゼ遺伝子を各々または組み合わせて導入した菌株を利用した後、トリアシルグリセロールリパーゼ遺伝子またはモノアシルグリセロールリパーゼ遺伝子だけを導入した場合より、前記二つのリパーゼ遺伝子を共に導入させた場合、同量のブドウ糖からより多量の遊離脂肪酸を生産し、より優秀な効率で脂肪酸メチルエステルを取得する可能性があることを確認した。従って、本発明において、望ましくは、トリアシルグリセロールリパーゼ遺伝子及びモノアシルグリセロールリパーゼ遺伝子を共に導入して用いられる。
【0029】
一方、本発明の実施形態では特定培地と培養方法だけを例示したが、文献に報告されたように、ホエー(whey)、CSL(corn steep liquor)等の糖化液と異なる培地を使うこと、及び流加培養(fed-batch culture)、連続培養等の多様な方法を使うこと(Lee et al., Bioprocess Biosyst. Eng., 26: 63, 2003; Lee et al., Appl. Microbiol. Biotechnol., 58: 663, 2002; Lee et al., Biotechnol. Lett., 25: 111, 2003; Lee et al., Appl. Microbiol. Biotechnol., 54: 23, 2000; Lee et al., Biotechnol. Bioeng., 72: 41, 2001)も当業界において通常の知識を有する者には自明な事項である。
【実施例】
【0030】
以下、本発明を実施例を挙げて詳述する。これらの実施例は単に本発明をより具体的に説明するためのものであり、本発明の範囲がこれらの実施例に制限されないことは当業者において通常の知識を有する者にとって自明である。
【0031】
特に、下記実施例では、ロドコッカス・オパカスPD630を宿主微生物として利用した方法についてだけ示されているが、オイル生成能を有すると知られた微生物またはオイル生成能を有するように形質転換させた微生物であれば、本発明に係る方法を適用できることは本明細書に開示された内容から当業界において通常の知識を有する者には自明である。
【0032】
また、下記実施例では、遊離脂肪酸のエステル化過程において、アルコールとしてメタノールを使ったが、その他アルコールを使って、エステル化反応を行うことによって、多様な種類の脂肪酸アルキルエステルを製造できることも当業界において通常の知識を有する者においては自明である。
【0033】
《実施例1:オイルを自己分解させる遺伝子を導入したロドコッカス・オパカスPD630の組換え菌株の製造(1)》
1−1.プラスミドpRUCSdpの作製
pUC18プラスミド(Phamacia、Biotech, Uppsala, Sweden)から図2の遺伝子地図を有するrpROUC18(配列番号1)及びrpROUC18_KM(配列番号2)組換えベクターを製造した後、下記のように遺伝子切片を導入した。
【0034】
まず、シロイヌナズナ(Arabidopsis thaliana col.)のゲノムDNAを鋳型とし、合成された配列番号3と4のプライマーでPCRを行い、トリアシルグリセロールリパーゼをコードするsdp1遺伝子切片を作製した。
配列番号3:5’−TATAGGCGCCATGGATATAAGTAATGAGGC−3’
配列番号4:5’−TGTCCTGCAGCTAAGCATCTATAACACTAC−3’
【0035】
次に、前記製造されたsdp1切片をrpROUC18プラスミド(Phamacia、Biotech, Uppsala, Sweden)に制限酵素(NarI及びPstI)を処理した後、T4DNAリガーゼを処理して、制限酵素で切断されたsdp1切片(配列番号5)とpUC18プラスミドを接合させることによって、組換えプラスミドであるpRUCSdpを作製した。
【0036】
1−2.プラスミドpRUCSdpMagの作製
マイコバクテリウム・スメグマティス(Mycobacterium smegmatis)(KCTC9108)のゲノムDNAを鋳型とし、合成された配列番号6と7のプライマーでPCRを行い、モノアシルグリセロールリパーゼをコードするMSMEG_0220遺伝子切片を作製した。
配列番号6:5’−TATATCTAGAACAACGGGGAGGACAACCGAATGGTGAGCAGCACCCGCAGTGAACAC−3’
配列番号7:5’−TATATCTAGATCACAGATGACTCACGATCCATGAG−3’
【0037】
次に、前記製造されたMSMEG_0220切片をpRUCSdpプラスミドに制限酵素(XbaI)を処理した後、T4DNAリガーゼを処理して、制限酵素で切断されたMSMEG_0220切片(配列番号8)とpRUCSdpプラスミドを接合させることによって、図3の組換えプラスミドであるpRUCSdpMagを作製した。その次に、製造した前記組換えプラスミドであるpRUCSdpMagをロドコッカス・オパカスPD630DSM44193菌株(DSMZ(Deutsche Sammlung von Mikroorganismen und Zellkulturen), Germany)に導入して、アセトアミド(acetamide)により活性を有するように作製されたリパーゼ遺伝子が導入された組換え菌株を製造した。
【0038】
《実施例2:オイルを自己分解させる遺伝子を導入したロドコッカス・オパカスPD630の組換え菌株の製造(2)》
2−1.プラスミドの作製
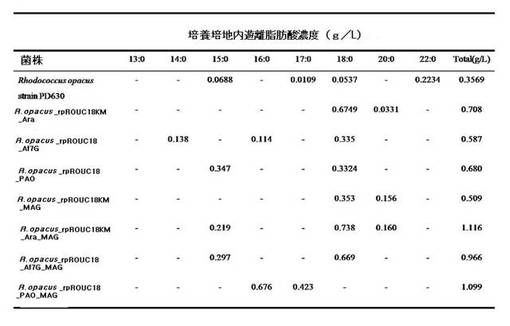
下記提示されたプライマーを使って、表1の明示された条件と遺伝子鋳型からトリアシルグリセロールリパーゼとモノアシルグリセロールリパーゼを実施例1−1のrpROUC18プラスミド及びrpROUC18_KMプラスミドに導入する方式によって、図4に示されたように多様な種類のプラスミドを作製した。この時、トリアシルグリセロールリパーゼとモノアシルグリセロールリパーゼを共に導入した。表1は、各々の制限酵素の種類と遺伝子の由来等が記載されており、この他にも多様な種の遺伝子を導入してもよい。
【0039】
実施例1に示したように、rpROUC18プラスミドには、アセトアミドに誘導されるプロモーターを使って、希望する時期に導入させた遺伝子を機能できるようにし、この時、アセトアミドで誘導させる場合には0.5%(w/w)の濃度を用いた。
ARAT_fプライマー(配列番号9)
5’−TATATTCCATGGGGAGGACAACATATAAGTAATGAGGCTAGT−3’
ARAT−rプライマー(配列番号10)
5’−CCGCCTGCAGCTAAGCATCTATAACACTAC−3’
ATAG7_fプライマー(配列番号11)
5’−TATTGACGTCGACAACGGGGAGGACAACCGAATGGAACGCGGATCCACTTG−3’
ATAG7−rプライマー(配列番号12)
5’−CTTGTACTAAGTCCCGGGTTAGTGGACGACCTCGAAGC−3’
Mlip2_fプライマー(配列番号13)
5’−TATTGGCGCCGACAACGGGGAGGACAACCGAATGGTGAGCAGCACCCGCAGTGAA−3’
Mlip2_rプライマー(配列番号14)
5’−CCACGATGGACACGTTGTACTAAGTCTGCAGTCACAGATGACTCACGATCC−3'
PAO_fプライマー(配列番号15)
5’−TATAGACGTCATGAAGAAGAAGTCTCTGCTCCCC−3’
PAO_rプライマー(配列番号16)
5’−TCGAaagcttCTACAGGCTGGCGTTCTTCA−3’
【0040】
【表1】
【0041】
この時、図4のrpROUC18KM_AraプラスミドとrpROUC18KM_Ara_MAGプラスミドに導入されたシロイヌナズナ(Arabidopsis thaliana)TAGリパーゼ遺伝子切片は、配列番号17の塩基配列を有し、rpROUC18KM_Af7GプラスミドとrpROUC18KM_Af7G_MAGプラスミドに導入されたアスペルギルス・フミガータスTAG7Gリパーゼ遺伝子切片は、配列番号18の塩基配列を有し、rpROUC18KM_PAOプラスミドとrpROUC18KM_PAO_MAGプラスミドに導入されたシュードモナス・アエルギノーザ(Pseudomonas aeruginosa)TAGリパーゼ遺伝子切片は、配列番号19の塩基配列を有し、rpROUC18KM_MAGプラスミド、RPROUC18KM_ARA_MAGプラスミド、rpROUC18KM_Af7G_MAGプラスミド及びrpROUC18KM_PAO_MAGプラスミドに導入されたM.スメグナティス(M. smegmatis)MAGリパーゼ遺伝子切片は配列番号20の塩基配列を有する。
【0042】
《実施例3:ロドコッカス・オパカスPD630の組換え菌株を利用した脂肪酸メチルエステルの産生》
3−1:実施例1のロドコッカス・オパカスPD630の組換え菌株を利用した遊離脂肪酸(fatty acid)の産生(1)
実施例1のアセトアミドにより活性を有するように作製されたリパーゼ遺伝子が導入されたロドコッカス・オパカスPD630の組換え菌株を培養するに当たり、培地内に窒素源(nitrogen)を制限して、オイル産生のために2段階培養を行った。
【0043】
先ず、1段階培養では、実施例1の組換え菌株を100mLのNB(nutrient broth)を含有した250mLフラスコで30℃、250rpmで24時間培養した。
【0044】
前記培養液を6000rpmで10分間遠心分離して菌体を回収し、2段階培養に使用されるMSM培地で細胞に残っているNB培地性分を洗浄した後、再度6000rpmで10分間遠心分離して菌体を回収して、100mLのMSM培地に懸濁させた。MSM培地(pH7.0)の組成は、下記の通りである。蒸溜水1L当りKH2PO4 0.8g、Na2HPO4 5.58g、(NH4)2SO4 0.1g、MgSO47H2O 0.12g、FeSO45H2O 0.5mg、MnSO45H2O 1.54mg、H3BO3 2.86mg、CuSO45H2O 0.039mg、CoCl26H2O 0.041mg、ZnCl2 0.021mg、Na2MoO42H2O 0.025mg、CaCl22H2O 11.6mgが含まれていている。
【0045】
100mLのMSM培地に懸濁させた菌株は、炭素源として20g/Lのグルコースを添加した後、30℃、250rpmで24時間培養した。その次に、顕微鏡でリアルタイムモニターして、菌株内のオイル蓄積をチェックした後、遊離脂肪酸を生成するようにリパーゼを活性化させるために培養液に0.5%(w/v)アセトアミドを入れて30℃で48時間培養した。
【0046】
培養終了後、培養液を6000rpmで10分間遠心分離して細胞を回収した。回収した細胞を蒸溜水で一度洗った後、100℃の乾燥器で24時間乾燥した。
乾燥された細胞は、キャピラリーカラムが装着されたAgilent6890Nシリーズガスクロマトグラフィー装置(Chiraldex G-TA of Astec, USA)を利用して、ガスクロマトグラフィー分析を行って、細胞内の合成された遊離脂肪酸含有量を測定した。2段階フラスコ培養結果、遊離脂肪酸は、0.27g/Lの濃度で生成されたことを確認した。
【0047】
3−2:実施例2のロドコッカス・オパカスPD630の組換え菌株を利用した遊離脂肪酸の生産(2)
実施例2のアセトアミドにより活性を有するように作製されたリパーゼ遺伝子が導入されたロドコッカス・オパカスPD630の組換え菌株を培養するに当たり、培地内に窒素源を制限して、オイル産生のための2段階培養を行った。
【0048】
先ず、1段階培養では、実施例2の組換え菌株を各々200mLのTSB(Tryptic soy broth)を含有した250mLのフラスコで30℃、200rpmで16時間培養した。
【0049】
前記培養液を3000rpmで30分間遠心分離して菌体を回収して、2段階培養に使用されるMSM培地で細胞に残っているTSB培地性分を洗浄した後、再度3000rpmで30分間遠心分離して菌体を回収して、200mLのMSM培地に懸濁した。MSM培地(pH7.0)の組成は下記の通りである。蒸溜水1L当りKH2PO4 0.8g、Na2HPO4 5.58g、(NH4)2SO4 0.1g、MgSO47H2O 0.12g、FeSO45H2O 1.0mg、MnSO45H2O 3.08mg、H3BO3 5.72mg、CuSO45H2O 0.078mg、CoCl26H2O 0.082mg、ZnCl2 0.042mg、Na2MoO42H2O 0.050mg、CaCl22H2O 23.2mgが含まれている。
【0050】
200mLのMSM培地に懸濁させた菌株は、炭素源として20g/Lのグルコースを添加した後、30℃、200rpmで48時間培養した。その次に、顕微鏡でリアルタイムモニターして、菌株内のオイル蓄積をチェックした後、遊離脂肪酸を生成するようにリパーゼを活性化させるために培養液に0.5%(w/v)アセトアミドを入れて30℃で24時間培養した。
【0051】
培養終了後、培養液を3000rpmで30分間遠心分離して細胞を回収した。上澄液は−45℃、10mmトール(Torr)で48時間凍結乾燥した。そして、回収した細胞は、蒸溜水で一度洗い落とした後、80℃の乾燥器で24時間乾燥した。各々の物質は0.1gずつ取って脂肪酸分析のために、Microbial Identification System(Microbial ID, Inc., Network, Del., USA)に提示された方法でカスクロマトグラフィーサンプルを準備した。
【0052】
準備されたサンプルは、毛管カラムが装着されたAgilent6890Nシリーズガスクロマトグラフィー装置(Chiraldex G-TA of Astec, USA)を利用して、ガスクロマトグラフィー分析を行って、細胞内の合成された遊離脂肪酸含有量を測定した。
【0053】
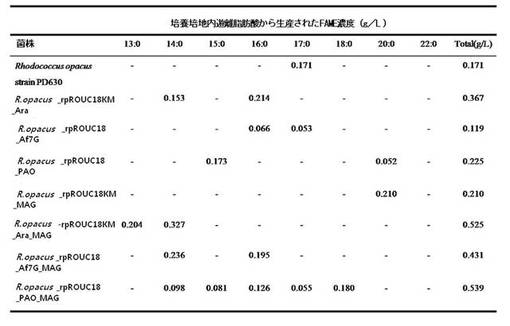
このように2段階フラスコ培養結果、細胞内での遊離脂肪酸含有量に比べて上澄液の凍結乾燥物内の遊離脂肪酸は顕著に多量であり、そこで遊離脂肪酸は細胞外に分泌されることが分かった。図5は、上澄液の凍結建造物内の遊離脂肪酸を測定した結果で、遊離脂肪酸が多様な長さの遊離脂肪酸の混合体として生成されたことを確認することができた。また、トリアシルグリセロールリパーゼだけを導入させた場合より、モノアシルグリセロールリパーゼを共に導入させた場合、同量のブドウ糖からさらに多量の遊離脂肪酸を産生することが分かった。
【0054】
3−3:遊離脂肪酸から脂肪酸メチルエステルへの転換
実施例3−1で得られた乾燥菌株にクロロホルム2mLを添加して、3%(v/v)H2SO4を含むメタノール1mLを添加した後、この混合物を100℃で12時間反応させた。
【0055】
反応終了後、混合物を常温まで冷まして、混合物に蒸溜水1mLを添加して5分間激しく混ぜて、有機溶媒(chloroform)層と水(水溶液)層を分離させて、10,000rpmで10分間遠心分離して有機溶媒層だけを採取して、毛管カラムが装着されたAgilent6890Nシリーズガスクロマトグラフィー装置(Chiraldex G-TA of Astec, USA)を利用してガスクロマトグラフィー分析を行って、生成された脂肪酸メチルエステル濃度を測定した。
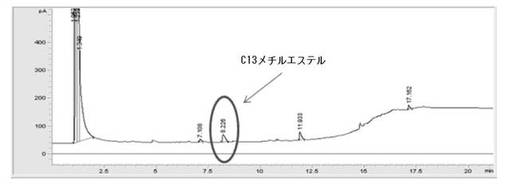
【0056】
その結果、図6に示したように、C13脂肪酸メチルエステルを確認でき、これは、0.2g/Lで取得された。これは、遊離脂肪酸から遊離脂肪酸メチルエステルロへの転換がなされたことを意味する。
また、実施例3−2で得られた上澄液に前記同様にメタノールを添加して反応させた後、生成された脂肪酸メチルエステルを測定した。
【0057】
その結果、図7に示したように、脂肪酸メチルエステルへ転換されたことを確認でき、特にトリアシルグリセロールリパーゼ(TAGリパーゼ)またはモノアシルグリセロールリパーゼ(MAGリパーゼ)だけを導入した場合に比べて、二つのリパーゼを共に導入させた場合、顕著に多量の脂肪酸メチルエステルが取得されることを確認することができた。このように本発明に係る微生物を利用した方法は、そのままバイオ燃料生産に使われるほどの生産効率を有し、本発明の脂肪酸メチルエステル生産効率は、既に公示された脂肪酸アルキルエステル生産に比べて、顕著に増加したことを確認することができた。
【産業上の利用可能性】
【0058】
以上説明したように、本発明に係る方法は、代謝工学的アプローチを介して、微生物が産生する代表的なオイルであるトリアシルグリセロール(Triacyl glycerol)のような微生物内に蓄積されたオイルを高い転換効率で脂肪酸アルキルエステルに転換でき、最近バイオディーゼルの利用価値が確認された脂肪酸アルキルエステルの産業的生産方法で役立つ。
【0059】
以上、本発明の内容の特定の部分を詳述したが、当業界における通常の知識を持った者にとって、このような具体的な記述は単なる好適な実施態様に過ぎず、これにより本発明の範囲が制限されることはないという点は明らかである。よって、本発明の実質的な範囲は特許請求の範囲とこれらの等価物により定義されると言える。
【技術分野】
【0001】
本発明は、オイル(oil)生成能を有する微生物を利用した脂肪酸アルキルエステル(fatty acid alkyl ester)の製造方法に関し、より詳細にはオイル生成能を有する微生物を培養して、オイルを大量蓄積した後、生成されたオイルを微生物内で自己分解して、遊離脂肪酸を生成した後、アルキルエステル化することを特徴とする脂肪酸アルキルエステルの製造方法に関する。
【背景技術】
【0002】
最近、油の高価格と環境問題から微生物を利用したバイオ燃料生産が大きな関心をよんでいる。最近、バイオディーゼルが軽油に代わり、または軽油と混合してディーゼルエンジンに使用できる代替燃料として浮上し、市場規模が急速に増加している。ヨーロッパ連合(EU)では2008年一年間660万トンのバイオディーゼルが生産され、55億ユーロの市場規模となっている(Biodiesel Market, Frost & Sullivan)。また、米国でも2006年一年間3億ガロンのバイオディーゼルが生産された(Biodiesel Market, Global Industry Analysts Inc, 2006. 5)。
【0003】
バイオディーゼルは、燃焼率が良く、有毒ガスの排出が低く、軽油より発熱量が10%程度低く、発火点はより高いため、運送と保存により安全な長所を有している。バイオディーゼルは、主に動植物の脂肪成分を軽油と類似の特性を有するように加工したり、植物性油脂(ぬか、廃食用油、大豆油、菜種油等)とアルコールを反応させて製造した。しかし、この場合、大量生産が難しい短所がある。従って、軽油の代替燃料に適するバイオディーゼルを微生物を利用して大量生産すると、原油輸入代替効果及び温室ガス排出減少による環境的効果等を齎すと期待された。
【0004】
一方、オイルは、微生物が、炭素源は豊富であるが他の成長因子(窒素、燐、酸素、硫黄等)が不足する時、細胞内で合成して蓄積するエネルギー貯蔵物質であり、成長環境が変わって制限された成長因子が再提供されると、蓄積しておいたオイルを分解してエネルギー源として使用するようになる。オイルは、生産する微生物の種類や、供給される化学物質の種類、培養条件等の変化によって100種類以上の単量体が構成要素となることが知られている。
【0005】
最近、サトウキビ等の植物性脂肪酸を利用して、アルコールを加えて脂肪酸アルキルエステル(fatty acid alkylester)を生産する技術が開発され、これは現在バイオディーゼル燃料として使われている。また、遊離脂肪酸をエステル化する方法は、ヨーロッパ公開特許127104A号、ヨーロッパ公開特許184740A号及び米国登録特許4,164,506号等に開示されているが、開示された特許では脂肪酸及び脂肪酸トリグリセリドの混合物とメタノールを共に加熱して、エステル化反応を行った。また、油脂から脂肪酸アルキルエステルの収率を高めるための方法として、エステル交換反応の結果として得られたグリセリンから遊離脂肪酸(free fatty acid)を分離した後、分離した遊離脂肪酸をエステル化する方法がヨーロッパ公開特許708813A号に開示されている。
【0006】
しかし、このような方法では多量の脂肪酸又は遊離脂肪酸が得られ難く、現在最も多く使われている植物性脂肪酸は、植物の成長期間が長く、代謝工学的アプローチが多少難しいため、脂肪酸蓄積の増大及びこれを利用した生産性の増加が困難である。
【0007】
そこで、本発明者等は代謝工学的アプローチを介して生産性が増加した、バイオディーゼルとして使用できる脂肪酸アルキルエステルを高効率で提供できる新しい方法を開発しようと鋭意努力した結果、代謝工学的方法でオイル生成能を有する微生物を利用して、オイル生成を最大化した後、これを微生物内で自己分解させて遊離脂肪酸を生産して、アルキルエステル化することによって、高効率で脂肪酸アルキルエステルを製造する可能性があることを確認して、本発明を完成するようになった。
【発明の概要】
【0008】
本発明の目的は、代謝工学的アプローチを介して生産性が増加した、バイオディーゼルとして使用できる脂肪酸アルキルエステルを高効率で大量生産できる新しい方法を提供することである。
【0009】
前記目的を達成するために、本発明は下記の工程を含む脂肪酸アルキルエステルの製造方法を提供する:
(a)オイル生成能を有する微生物を培養してオイルを生成する工程、
(b)前記生成されたオイルを含有する微生物においてオイルの自己分解を誘導して、遊離脂肪酸を生成する工程、及び
(c)前記生成された遊離脂肪酸にアルコールを添加し、反応させて脂肪酸アルキルエステルを製造する工程。
【0010】
本発明はまた、下記工程を含む、オイル生成能とリパーゼをコードする遺伝子を有する微生物を利用した脂肪酸メチルエステルの製造方法を提供する:
(a)オイル生成能とリパーゼをコードする遺伝子を有する微生物を培養して、オイルを生成させる工程、
(b)前記生成されたオイルを含有する微生物においてリパーゼによるオイルの自己分解を誘導して、遊離脂肪酸を生成させる工程、及び
(c)前記生成された遊離脂肪酸にメタノールを添加し、反応させて脂肪酸メチルエステルを製造する工程。
【0011】
本発明の他の特徴及び具現例は、以下の詳細な説明及び添付された特許請求範囲からより一層明白になる。
【図面の簡単な説明】
【0012】
【図1】ロドコッカス・オパカスPD630菌株を利用して代表的オイル成分であるトリアシルグリセロール(Triacylglycerol、TAG)から脂肪酸メチルエステル(fatty acid methyl ester)を高効率で製造する工程を示した図である。
【図2】組換えベクターrpROUC18(a)及びrpROUC18_KM(b)の遺伝子地図を示した図である。
【図3】sdp1及びMSMEG_0220遺伝子を含む組換えベクターpRUCSdpMagの遺伝子地図を示した図である。
【図4】表1の情報から作られたプラスミドの種類とその情報を示した図である。
【図5】遊離脂肪酸に対するガスクロマトグラフィー(Gas chromatography)分析結果を示した図である。
【図6】脂肪酸メチルエステルに対するガスクロマトグラフィー分析結果を示した図である。
【図7】脂肪酸メチルエステルに対するガスクロマトグラフィー分析結果を示した図である。
【発明を実施するための形態】
【0013】
他の方式で定義されない限り、本明細書において使用されたあらゆる技術的・科学的用語は、本発明が属する技術分野に熟練した専門家によって通常理解されるものと同じ意味を有する。通常、本明細書において使用された命名法は、本技術分野において周知であり、しかも汎用されるものである。
本発明の詳細な説明などにおいて使用される主な用語の定義は、下記の通りである。
【0014】
本願において、「オイル(oil)」とは、微生物が炭素源は豊富であるが、他の成長因子(窒素、燐、酸素、硫黄等)が不足する時、細胞内で合成して蓄積するエネルギー貯蔵物質をいい、これは脂肪酸の前駆体として加水分解によって遊離脂肪酸とグリセロールに分離する。
【0015】
本願において、「脂肪酸」とは、鎖状の飽和又は不飽和モノカルボン酸である。これは、炭素鎖の長さ及び飽和の可否によって分類され、オイル、即ち、脂肪が加水分解して生じる脂肪酸を特に「遊離脂肪酸」という。
【0016】
一観点において、本発明はオイル生成能を有する微生物を利用した脂肪酸アルキルエステルの製造方法に関する。より具体的には、本発明による次の工程を含むことができる;
(a)オイル生成能を有する微生物を培養してオイルを生成する工程、
(b)前記生成されたオイルを含有する微生物においてオイルの自己分解を誘導して、遊離脂肪酸を生成する工程、及び
(c)前記生成された遊離脂肪酸にアルコールを添加し、反応させて脂肪酸アルキルエステルを製造する工程。
本発明は下記の工程を含む脂肪酸アルキルエステルの製造方法に関する:
(a)オイル生成能を有する微生物を培養してオイルを生成する工程、
(b)前記生成されたオイルを含有する微生物においてオイルの自己分解を誘導して、遊離脂肪酸を生成する工程、及び
(c)前記生成された遊離脂肪酸にアルコールを添加し、反応させて脂肪酸アルキルエステルを製造する工程。
【0017】
本発明において、前記オイルは、トリアシルグリセロール(TAG)、ジアシルグリセロール(diacylglycerol、DAG)、モノアシルグリセロール(monoacylglycerol、MAG)、リン脂質(phospholipid)、ステロール脂質(sterol lipid)、スフィンゴ脂質(sphingolipid)、サッカロ脂質(saccharolipid)、プレノール脂質(prenol lipid)及びポリケチド(polyketide)等であってもよいが、微生物で産生されるオイルであれば、これに限定されない。
【0018】
ここで、前記オイルの分解によって生成される遊離脂肪酸は、飽和または不飽和脂肪であってもよく、不飽和脂肪酸は脂肪酸の炭素鎖において一つ以上の二重結合を有する脂肪酸をいい、例示的にオレイン酸(oleic acid)、リノール酸(linoleic acid)、リノレン酸(linolenic acid)、パルミトレイン酸(palmitoleic acid)、リシノール酸(ricinoleic acid)、バクセン酸(vaccenic acid)、カドレン酸(gadoleic acid)、アラキドン酸(arachidonic acid)、EPA(5,8,11,14,17-eicosapentaenoic acid)、エルカ酸(erucic acid)、DHA(4,7,10,13,16,19-docosahexaenoic acid)等が挙げられる。また、飽和脂肪酸は、炭素鎖に二重結合を持たない脂肪酸をいい、例示的に酪酸(butyric acid)、カプロン酸(caproic acid)、カプリル酸(caprylic acid)、カプリン酸(capric acid)、ラウリン酸(lauric acid)、ミリスチン酸(myristic acid)、パルミチン酸(palmitic acid)、ステアリン酸(stearic acid)、アラキジン酸(arachidic acid)、ベヘン酸(behenic acid)、リグノセンリン酸(lignoceric acid)等が挙げられる。前記脂肪酸は置換基で修飾されたものであってもよく、例示的に芳香族環基、エポキシ基、シアノ基、及びハロゲン基からなる群から選択される置換基で修飾されたものが用いられるが、これに限定されない。
【0019】
本発明において、前記遊離脂肪酸の前駆体になるオイルは、オイル生成能を有する微生物によって生成されることを特徴とし、例えば、アエロモナス(Aeromonas)属微生物、アクロモバクター(Achromobacter)属微生物、アシドボラックス・デラフィルディ(Acidovorax delafieldii)、アシドボラックス・ファシリス(Acidovorax facilis)、アシネトバクター(Acinetobacter)属微生物、アクチノマイセス(Actinomyces)属微生物、アエロモナス(Aeromonas)属微生物、アルカリゲネス(Alcaligenes)属微生物、アルテロモナス(Alteromonas)属微生物、アルソルニア(Althornia)属微生物、アプラノキトリウム(Aplanochytrium)属微生物、アスペルギルス(Aspergillus)属微生物、アモエボバクター(Amoebobacter)属微生物、アファノカプサ属微生物(Aphanocapsa sp.)、アファノテーケ属微生物(Aphanothece sp.)、アクアスピリラム・オートトロフィカム(Aquaspirillum autotrophicum)、アゾリゾビウム・カウリノダンス(Azorhizobium caulinodans)、アゾスピリラム属微生物(Azospirillum sp.)、アゾスピリラム(Azospirillum)属微生物、アゾトバクター(Azotobacter)属微生物、バシラス(Bacillus)属微生物、ベギアトア(Beggiatoa)属微生物、ベイジェリンキア(Beijerinckia)属微生物、ベネケア(Beneckea)属微生物、ブラケスレア(Blakeslea)属微生物、ボルデテラ・パーツシス(Bordetella pertussis)、ブラディリゾビウム・ジャポニカム(Bradyrhizobium japonicum)、カリオファノン・ラタム(Caryophanon latum)、カウロバクター(Caulobacter)属微生物、クロログロエア(Chlorogloea)属微生物、クロマチウム(Chromatium)属微生物、クロモバクテリウム(Chromobacterium)属微生物、クロストリジウム(Clostridium)属微生物、コマモナス(Comamonas)属微生物、コリネバクテリウム(Corynebacterium)属微生物、クリプテコジニウム(Crypthecodinium)属微生物、シアノバクテリア(Cyanobacteria)属微生物、デルキシア(Derxia)属微生物、デスルホネマ(Desulfonema)属微生物、デスルホサルシナ・バリアビリス(Desulfosarcina variabilis)、デスルホビブリオ・サポボランス(Desulfovibrio sapovorans)、エクトチオロドスピラ(Ectothiorhodospira)属微生物、エリナ(Elina)属微生物、エントモフトラ(Entomophthora)属微生物、フェロバシラス・フェロキシダンス(Ferrobacillus ferroxidans)、フラボバクテリウム属微生物(Flavobacterium sp.)、ヘモフィルス・インフルエンザ(Haemophilus influenzae)、ハロバクテリウム(Halobacterium)属微生物、ヒドロクラスラタス・クラスラタス(Hydroclathratus clathratus)、ヒドロゲノモナス・ファシリス(Hydrogenomonas facilis)、ヒドロゲノファガ(Hydrogenophaga)属微生物、ハイフォミクロビウム(Hyphomicrobium)属微生物、イリオバクター・デラフィエルディ(Ilyobacter delafieldii)、ジャポノキトリウム(Japonochytrium)属微生物、ラブリス・モナクス(Labrys monachus)、ランプロシスティス・ロゼオペルシシーナ(Lamprocystis roseopersicina)、ランプロペディア・ハイアリーナ(Lampropedia hyalina)、レジオネラ属微生物(Legionella sp.)、レプトスリックス・ディスコホラス(Leptothrix discophorus)、メチロバクテリウム(Methylobacterium)属微生物、メチロシナス(Methylosinus)属微生物、ミクロコッカス(Micrococcus)属微生物、モルティエレラ(Mortierella)属微生物、マイコバクテリウム(Mycobacterium)属微生物、ニトロバクター(Nitrobacter)属微生物、ノカルディア(Nocardia)属微生物、パラコッカス・デントリフィカンス(Paracoccus dentrificans)、オシラトリア(Oscillatoria)、ペニシリウム・シクロピウム(Penicillium cyclopium)、フォトバクテリウム(Photobacterium)属微生物、フィサルム・ポリセファラム(Physarum ploycephalum)、フィコミセス(Phycomyces)属微生物、シュードモナス(Pseudomonas)属微生物、ピシウム(Pythium)属微生物、ラルストニア(Ralstonia)属微生物、リゾビウム(Rhizobium)属微生物、ロドバシラス(Rhodobacillus)属微生物、ロドバクター(Rhodobacter)属微生物、ロドコッカス(Rhodococcus)属微生物、ロドシクルス(Rhodocyclus)属微生物、ロドミクロビウム・バニエリ(Rhodomicrobium vannielii)、ロドシュードモナス(Rhodopseudomonas)属微生物、ロドスピリラム(Rhodospirillum)属微生物、シゾチトリウム(Schizochytrium)属微生物、スフィンゴモナス・パウシモビリス(Sphingomonas paucimobilis)、スピリラム(Spirillum)属微生物、スピルリナ(Spirulina)属微生物、スタフィロコッカス(Staphylococcus)属微生物、ステラ(Stella)属微生物、ストレプトマイセス(Streptomyces)属微生物、シントロフォモナス・ウォルフェイ(Syntrophomonas wolfei)、サーモフィリック・シアノバクテリア(Thermophilic cyanobacteria)、サーマス・サーモフィルス(Thermus thermophilus)、チオバシラスA2(Thiobacillus A2)、チオバシラス(Thiobacillus)属微生物、チオカプサ(Thiocapsa)属微生物、トラウストキトリウム(Thraustochytrium)属微生物、チオシスティス・バイオラセア(Thiocystis violacea)、ビブリオ・パラハエモリティカス(Vibrio parahaemolyticus)、キサントバクター・オートトロフィカス(Xanthobacter autotrophicus)、キサントモナス・マルトフィリア(Xanthomonas maltophilia)、ズーグロエア(Zoogloea)属微生物及びオイル生成能を有する酵素をコードする遺伝子に形質転換された微生物等が挙げられるが、オイルを産生できる微生物であれば、前記に制限されず本発明に係る方法が適用されることは明らかである。
【0020】
本発明において、前記工程(a)の培養は、菌体増殖のための1段階培養及びオイル生成のための2段階培養を含むが、この時、オイル生成のための培養はオイル生成量を向上するために窒素源が制限された培地で行うことを特徴とする。
【0021】
微生物によって生成されたオイルは微生物において自己分解するが、この時、前記工程(b)の自己分解は、リパーゼによって行われる。このようなリパーゼとしては、トリアシルグリセロールリパーゼ(triacyl glycerol lipase、EC: 3.1.1.34, 3.1.1.13)、モノアシルグリセロールリパーゼ(monoacyl glycerol lipase、EC: 3.1.1.23)、リソホスホポリパーゼ(lysophospholipase、EC: 3.1.1.5)等を利用してもよい。
【0022】
この時、望ましくは、前記オイル生成能を有する微生物にリパーゼをコードする遺伝子を導入または増幅することができ、さらに望ましくは基質に反応して活性化できるリパーゼ遺伝子を導入したことを特徴とする。本発明の一実施形態では、微生物のオイル自己分解のために、リパーゼ遺伝子、即ち、配列番号5及び8のリパーゼ遺伝子を導入した菌株を利用した。それと共に、本発明の他の実施形態では、配列番号17、18及び19のトリアシルグリセロールリパーゼ遺伝子または配列番号20のモノアシルグリセロールリパーゼ遺伝子を各々または組み合わせて導入した菌株を利用した。
【0023】
本発明において、前記工程(c)で添加されるアルコールは、1次、2次及び3次構造の全てのアルコールを利用してもよいが、望ましくは炭素数1乃至8のアルコールまたはこの中の2以上の混合物を利用でき、さらに望ましくは、メタノール(methanol)を利用することを特徴とする。
【0024】
また、前記工程(c)の反応は、80〜120℃で1〜24時間行うことを特徴とする。また、有機溶媒を追加で添加して行うことを特徴とし、望ましくはクロロホルムを添加して行われる。
【0025】
本発明の一実施形態では、オイル生成能を有する微生物として、ロドコッカス・オパカスPD630菌株を使用し、菌体増殖のための1段階培養とオイル生成のための2段階培養で分けて行った。前記オイル生成のための2段階培養は、窒素源を制限した培地を使ってオイルの生成を誘導した。この時、生成されたオイルの自己分解のために、前記菌株にアセトアミドによって活性化するリパーゼ遺伝子を導入し、微生物にアセトアミドを加えて活性化したリパーゼを利用してin vivoで約0.27g/Lの遊離脂肪酸を生成した。
【0026】
また、前記取得された遊離脂肪酸溶液は、凍結乾燥して水を取り除いた後、クロロホルムを添加し、H2SO4を含むメタノールを添加した後、100℃で12時間反応させて、水を添加した後、有機溶媒層を分離して、遊離脂肪酸メチルエステルを取得した。生成された遊離脂肪酸メチルエステル(Free fatty acid methyl ester)の濃度は、0.2g/Lであり、これは遊離脂肪酸から遊離脂肪酸メチルエステルへの転換が行われたことを意味する(図1)。そこで、本発明に係る方法を利用することによって、より容易でかつ親環境的方法で脂肪酸メチルエステルを高効率に産生できることが明らかになり、従来の軽油等を代替できるバイオディーゼルの生産に非常に有用な方法であることが確認された。
【0027】
他の観点において、本発明は、下記工程を含む、オイル生成能とリパーゼをコードする遺伝子を有する微生物を利用した脂肪酸メチルエステルの製造方法に関する:
(a)オイル生成能とリパーゼをコードする遺伝子を有する微生物を培養して、オイルを生成させる工程、
(b)前記生成されたオイルを含有する微生物においてリパーゼによるオイルの自己分解を誘導して、遊離脂肪酸を生成させる工程、及び
(c)前記生成された遊離脂肪酸にメタノールを添加し、反応させて脂肪酸メチルエステルを製造する工程。
【0028】
本発明の他の実施形態では、トリアシルグリセロールリパーゼ遺伝子またはモノアシルグリセロールリパーゼ遺伝子を各々または組み合わせて導入した菌株を利用した後、トリアシルグリセロールリパーゼ遺伝子またはモノアシルグリセロールリパーゼ遺伝子だけを導入した場合より、前記二つのリパーゼ遺伝子を共に導入させた場合、同量のブドウ糖からより多量の遊離脂肪酸を生産し、より優秀な効率で脂肪酸メチルエステルを取得する可能性があることを確認した。従って、本発明において、望ましくは、トリアシルグリセロールリパーゼ遺伝子及びモノアシルグリセロールリパーゼ遺伝子を共に導入して用いられる。
【0029】
一方、本発明の実施形態では特定培地と培養方法だけを例示したが、文献に報告されたように、ホエー(whey)、CSL(corn steep liquor)等の糖化液と異なる培地を使うこと、及び流加培養(fed-batch culture)、連続培養等の多様な方法を使うこと(Lee et al., Bioprocess Biosyst. Eng., 26: 63, 2003; Lee et al., Appl. Microbiol. Biotechnol., 58: 663, 2002; Lee et al., Biotechnol. Lett., 25: 111, 2003; Lee et al., Appl. Microbiol. Biotechnol., 54: 23, 2000; Lee et al., Biotechnol. Bioeng., 72: 41, 2001)も当業界において通常の知識を有する者には自明な事項である。
【実施例】
【0030】
以下、本発明を実施例を挙げて詳述する。これらの実施例は単に本発明をより具体的に説明するためのものであり、本発明の範囲がこれらの実施例に制限されないことは当業者において通常の知識を有する者にとって自明である。
【0031】
特に、下記実施例では、ロドコッカス・オパカスPD630を宿主微生物として利用した方法についてだけ示されているが、オイル生成能を有すると知られた微生物またはオイル生成能を有するように形質転換させた微生物であれば、本発明に係る方法を適用できることは本明細書に開示された内容から当業界において通常の知識を有する者には自明である。
【0032】
また、下記実施例では、遊離脂肪酸のエステル化過程において、アルコールとしてメタノールを使ったが、その他アルコールを使って、エステル化反応を行うことによって、多様な種類の脂肪酸アルキルエステルを製造できることも当業界において通常の知識を有する者においては自明である。
【0033】
《実施例1:オイルを自己分解させる遺伝子を導入したロドコッカス・オパカスPD630の組換え菌株の製造(1)》
1−1.プラスミドpRUCSdpの作製
pUC18プラスミド(Phamacia、Biotech, Uppsala, Sweden)から図2の遺伝子地図を有するrpROUC18(配列番号1)及びrpROUC18_KM(配列番号2)組換えベクターを製造した後、下記のように遺伝子切片を導入した。
【0034】
まず、シロイヌナズナ(Arabidopsis thaliana col.)のゲノムDNAを鋳型とし、合成された配列番号3と4のプライマーでPCRを行い、トリアシルグリセロールリパーゼをコードするsdp1遺伝子切片を作製した。
配列番号3:5’−TATAGGCGCCATGGATATAAGTAATGAGGC−3’
配列番号4:5’−TGTCCTGCAGCTAAGCATCTATAACACTAC−3’
【0035】
次に、前記製造されたsdp1切片をrpROUC18プラスミド(Phamacia、Biotech, Uppsala, Sweden)に制限酵素(NarI及びPstI)を処理した後、T4DNAリガーゼを処理して、制限酵素で切断されたsdp1切片(配列番号5)とpUC18プラスミドを接合させることによって、組換えプラスミドであるpRUCSdpを作製した。
【0036】
1−2.プラスミドpRUCSdpMagの作製
マイコバクテリウム・スメグマティス(Mycobacterium smegmatis)(KCTC9108)のゲノムDNAを鋳型とし、合成された配列番号6と7のプライマーでPCRを行い、モノアシルグリセロールリパーゼをコードするMSMEG_0220遺伝子切片を作製した。
配列番号6:5’−TATATCTAGAACAACGGGGAGGACAACCGAATGGTGAGCAGCACCCGCAGTGAACAC−3’
配列番号7:5’−TATATCTAGATCACAGATGACTCACGATCCATGAG−3’
【0037】
次に、前記製造されたMSMEG_0220切片をpRUCSdpプラスミドに制限酵素(XbaI)を処理した後、T4DNAリガーゼを処理して、制限酵素で切断されたMSMEG_0220切片(配列番号8)とpRUCSdpプラスミドを接合させることによって、図3の組換えプラスミドであるpRUCSdpMagを作製した。その次に、製造した前記組換えプラスミドであるpRUCSdpMagをロドコッカス・オパカスPD630DSM44193菌株(DSMZ(Deutsche Sammlung von Mikroorganismen und Zellkulturen), Germany)に導入して、アセトアミド(acetamide)により活性を有するように作製されたリパーゼ遺伝子が導入された組換え菌株を製造した。
【0038】
《実施例2:オイルを自己分解させる遺伝子を導入したロドコッカス・オパカスPD630の組換え菌株の製造(2)》
2−1.プラスミドの作製
下記提示されたプライマーを使って、表1の明示された条件と遺伝子鋳型からトリアシルグリセロールリパーゼとモノアシルグリセロールリパーゼを実施例1−1のrpROUC18プラスミド及びrpROUC18_KMプラスミドに導入する方式によって、図4に示されたように多様な種類のプラスミドを作製した。この時、トリアシルグリセロールリパーゼとモノアシルグリセロールリパーゼを共に導入した。表1は、各々の制限酵素の種類と遺伝子の由来等が記載されており、この他にも多様な種の遺伝子を導入してもよい。
【0039】
実施例1に示したように、rpROUC18プラスミドには、アセトアミドに誘導されるプロモーターを使って、希望する時期に導入させた遺伝子を機能できるようにし、この時、アセトアミドで誘導させる場合には0.5%(w/w)の濃度を用いた。
ARAT_fプライマー(配列番号9)
5’−TATATTCCATGGGGAGGACAACATATAAGTAATGAGGCTAGT−3’
ARAT−rプライマー(配列番号10)
5’−CCGCCTGCAGCTAAGCATCTATAACACTAC−3’
ATAG7_fプライマー(配列番号11)
5’−TATTGACGTCGACAACGGGGAGGACAACCGAATGGAACGCGGATCCACTTG−3’
ATAG7−rプライマー(配列番号12)
5’−CTTGTACTAAGTCCCGGGTTAGTGGACGACCTCGAAGC−3’
Mlip2_fプライマー(配列番号13)
5’−TATTGGCGCCGACAACGGGGAGGACAACCGAATGGTGAGCAGCACCCGCAGTGAA−3’
Mlip2_rプライマー(配列番号14)
5’−CCACGATGGACACGTTGTACTAAGTCTGCAGTCACAGATGACTCACGATCC−3'
PAO_fプライマー(配列番号15)
5’−TATAGACGTCATGAAGAAGAAGTCTCTGCTCCCC−3’
PAO_rプライマー(配列番号16)
5’−TCGAaagcttCTACAGGCTGGCGTTCTTCA−3’
【0040】
【表1】
【0041】
この時、図4のrpROUC18KM_AraプラスミドとrpROUC18KM_Ara_MAGプラスミドに導入されたシロイヌナズナ(Arabidopsis thaliana)TAGリパーゼ遺伝子切片は、配列番号17の塩基配列を有し、rpROUC18KM_Af7GプラスミドとrpROUC18KM_Af7G_MAGプラスミドに導入されたアスペルギルス・フミガータスTAG7Gリパーゼ遺伝子切片は、配列番号18の塩基配列を有し、rpROUC18KM_PAOプラスミドとrpROUC18KM_PAO_MAGプラスミドに導入されたシュードモナス・アエルギノーザ(Pseudomonas aeruginosa)TAGリパーゼ遺伝子切片は、配列番号19の塩基配列を有し、rpROUC18KM_MAGプラスミド、RPROUC18KM_ARA_MAGプラスミド、rpROUC18KM_Af7G_MAGプラスミド及びrpROUC18KM_PAO_MAGプラスミドに導入されたM.スメグナティス(M. smegmatis)MAGリパーゼ遺伝子切片は配列番号20の塩基配列を有する。
【0042】
《実施例3:ロドコッカス・オパカスPD630の組換え菌株を利用した脂肪酸メチルエステルの産生》
3−1:実施例1のロドコッカス・オパカスPD630の組換え菌株を利用した遊離脂肪酸(fatty acid)の産生(1)
実施例1のアセトアミドにより活性を有するように作製されたリパーゼ遺伝子が導入されたロドコッカス・オパカスPD630の組換え菌株を培養するに当たり、培地内に窒素源(nitrogen)を制限して、オイル産生のために2段階培養を行った。
【0043】
先ず、1段階培養では、実施例1の組換え菌株を100mLのNB(nutrient broth)を含有した250mLフラスコで30℃、250rpmで24時間培養した。
【0044】
前記培養液を6000rpmで10分間遠心分離して菌体を回収し、2段階培養に使用されるMSM培地で細胞に残っているNB培地性分を洗浄した後、再度6000rpmで10分間遠心分離して菌体を回収して、100mLのMSM培地に懸濁させた。MSM培地(pH7.0)の組成は、下記の通りである。蒸溜水1L当りKH2PO4 0.8g、Na2HPO4 5.58g、(NH4)2SO4 0.1g、MgSO47H2O 0.12g、FeSO45H2O 0.5mg、MnSO45H2O 1.54mg、H3BO3 2.86mg、CuSO45H2O 0.039mg、CoCl26H2O 0.041mg、ZnCl2 0.021mg、Na2MoO42H2O 0.025mg、CaCl22H2O 11.6mgが含まれていている。
【0045】
100mLのMSM培地に懸濁させた菌株は、炭素源として20g/Lのグルコースを添加した後、30℃、250rpmで24時間培養した。その次に、顕微鏡でリアルタイムモニターして、菌株内のオイル蓄積をチェックした後、遊離脂肪酸を生成するようにリパーゼを活性化させるために培養液に0.5%(w/v)アセトアミドを入れて30℃で48時間培養した。
【0046】
培養終了後、培養液を6000rpmで10分間遠心分離して細胞を回収した。回収した細胞を蒸溜水で一度洗った後、100℃の乾燥器で24時間乾燥した。
乾燥された細胞は、キャピラリーカラムが装着されたAgilent6890Nシリーズガスクロマトグラフィー装置(Chiraldex G-TA of Astec, USA)を利用して、ガスクロマトグラフィー分析を行って、細胞内の合成された遊離脂肪酸含有量を測定した。2段階フラスコ培養結果、遊離脂肪酸は、0.27g/Lの濃度で生成されたことを確認した。
【0047】
3−2:実施例2のロドコッカス・オパカスPD630の組換え菌株を利用した遊離脂肪酸の生産(2)
実施例2のアセトアミドにより活性を有するように作製されたリパーゼ遺伝子が導入されたロドコッカス・オパカスPD630の組換え菌株を培養するに当たり、培地内に窒素源を制限して、オイル産生のための2段階培養を行った。
【0048】
先ず、1段階培養では、実施例2の組換え菌株を各々200mLのTSB(Tryptic soy broth)を含有した250mLのフラスコで30℃、200rpmで16時間培養した。
【0049】
前記培養液を3000rpmで30分間遠心分離して菌体を回収して、2段階培養に使用されるMSM培地で細胞に残っているTSB培地性分を洗浄した後、再度3000rpmで30分間遠心分離して菌体を回収して、200mLのMSM培地に懸濁した。MSM培地(pH7.0)の組成は下記の通りである。蒸溜水1L当りKH2PO4 0.8g、Na2HPO4 5.58g、(NH4)2SO4 0.1g、MgSO47H2O 0.12g、FeSO45H2O 1.0mg、MnSO45H2O 3.08mg、H3BO3 5.72mg、CuSO45H2O 0.078mg、CoCl26H2O 0.082mg、ZnCl2 0.042mg、Na2MoO42H2O 0.050mg、CaCl22H2O 23.2mgが含まれている。
【0050】
200mLのMSM培地に懸濁させた菌株は、炭素源として20g/Lのグルコースを添加した後、30℃、200rpmで48時間培養した。その次に、顕微鏡でリアルタイムモニターして、菌株内のオイル蓄積をチェックした後、遊離脂肪酸を生成するようにリパーゼを活性化させるために培養液に0.5%(w/v)アセトアミドを入れて30℃で24時間培養した。
【0051】
培養終了後、培養液を3000rpmで30分間遠心分離して細胞を回収した。上澄液は−45℃、10mmトール(Torr)で48時間凍結乾燥した。そして、回収した細胞は、蒸溜水で一度洗い落とした後、80℃の乾燥器で24時間乾燥した。各々の物質は0.1gずつ取って脂肪酸分析のために、Microbial Identification System(Microbial ID, Inc., Network, Del., USA)に提示された方法でカスクロマトグラフィーサンプルを準備した。
【0052】
準備されたサンプルは、毛管カラムが装着されたAgilent6890Nシリーズガスクロマトグラフィー装置(Chiraldex G-TA of Astec, USA)を利用して、ガスクロマトグラフィー分析を行って、細胞内の合成された遊離脂肪酸含有量を測定した。
【0053】
このように2段階フラスコ培養結果、細胞内での遊離脂肪酸含有量に比べて上澄液の凍結乾燥物内の遊離脂肪酸は顕著に多量であり、そこで遊離脂肪酸は細胞外に分泌されることが分かった。図5は、上澄液の凍結建造物内の遊離脂肪酸を測定した結果で、遊離脂肪酸が多様な長さの遊離脂肪酸の混合体として生成されたことを確認することができた。また、トリアシルグリセロールリパーゼだけを導入させた場合より、モノアシルグリセロールリパーゼを共に導入させた場合、同量のブドウ糖からさらに多量の遊離脂肪酸を産生することが分かった。
【0054】
3−3:遊離脂肪酸から脂肪酸メチルエステルへの転換
実施例3−1で得られた乾燥菌株にクロロホルム2mLを添加して、3%(v/v)H2SO4を含むメタノール1mLを添加した後、この混合物を100℃で12時間反応させた。
【0055】
反応終了後、混合物を常温まで冷まして、混合物に蒸溜水1mLを添加して5分間激しく混ぜて、有機溶媒(chloroform)層と水(水溶液)層を分離させて、10,000rpmで10分間遠心分離して有機溶媒層だけを採取して、毛管カラムが装着されたAgilent6890Nシリーズガスクロマトグラフィー装置(Chiraldex G-TA of Astec, USA)を利用してガスクロマトグラフィー分析を行って、生成された脂肪酸メチルエステル濃度を測定した。
【0056】
その結果、図6に示したように、C13脂肪酸メチルエステルを確認でき、これは、0.2g/Lで取得された。これは、遊離脂肪酸から遊離脂肪酸メチルエステルロへの転換がなされたことを意味する。
また、実施例3−2で得られた上澄液に前記同様にメタノールを添加して反応させた後、生成された脂肪酸メチルエステルを測定した。
【0057】
その結果、図7に示したように、脂肪酸メチルエステルへ転換されたことを確認でき、特にトリアシルグリセロールリパーゼ(TAGリパーゼ)またはモノアシルグリセロールリパーゼ(MAGリパーゼ)だけを導入した場合に比べて、二つのリパーゼを共に導入させた場合、顕著に多量の脂肪酸メチルエステルが取得されることを確認することができた。このように本発明に係る微生物を利用した方法は、そのままバイオ燃料生産に使われるほどの生産効率を有し、本発明の脂肪酸メチルエステル生産効率は、既に公示された脂肪酸アルキルエステル生産に比べて、顕著に増加したことを確認することができた。
【産業上の利用可能性】
【0058】
以上説明したように、本発明に係る方法は、代謝工学的アプローチを介して、微生物が産生する代表的なオイルであるトリアシルグリセロール(Triacyl glycerol)のような微生物内に蓄積されたオイルを高い転換効率で脂肪酸アルキルエステルに転換でき、最近バイオディーゼルの利用価値が確認された脂肪酸アルキルエステルの産業的生産方法で役立つ。
【0059】
以上、本発明の内容の特定の部分を詳述したが、当業界における通常の知識を持った者にとって、このような具体的な記述は単なる好適な実施態様に過ぎず、これにより本発明の範囲が制限されることはないという点は明らかである。よって、本発明の実質的な範囲は特許請求の範囲とこれらの等価物により定義されると言える。
【特許請求の範囲】
【請求項1】
次の工程を含む、オイル生成能を有する微生物を利用した脂肪酸アルキルエステルの製造方法:
(a)オイル生成能を有する微生物を培養してオイルを生成する工程、
(b)前記生成されたオイルを含有する微生物においてオイルの自己分解を誘導して、遊離脂肪酸を生成する工程、及び
(c)前記生成された遊離脂肪酸にアルコールを添加し、反応させて脂肪酸アルキルエステルを製造する工程。
【請求項2】
前記オイルはトリアシルグリセロール(TAG)、ジアシルグリセロール(DAG)、モノアシルグリセロール(MAG)、リン脂質、ステロール脂質、スフィンゴ脂質、サッカロ脂質、プレノール脂質及びポリケチドからなる群より選択される請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項3】
前記オイル生成能を有する微生物はアエロモナス属微生物、アクロモバクター属微生物、アシドボラックス・デラフィルディ、アシドボラックス・ファシリス、アシネトバクター属微生物、アクチノマイセス属微生物、アエロモナス属微生物、アルカリゲネス属微生物、アルテロモナス属微生物、アルソルニア属微生物、アプラノキトリウム属微生物、アスペルギルス属微生物、アモエボバクター属微生物、アファノカプサ属微生物、アファノテーケ属微生物、アクアスピリラム・オートトロフィカム、アゾリゾビウム・カウリノダンス、アゾスピリラム属微生物、アゾスピリラム属微生物、アゾトバクター属微生物、バシラス属微生物、ベギアトア属微生物、ベイジェリンキア属微生物、ベネケア属微生物、ブラケスレア属微生物、ボルデテラ・パーツシス、ブラディリゾビウム・ジャポニカム、カリオファノン・ラタム、カウロバクター属微生物、クロログロエア属微生物、クロマチウム属微生物、クロモバクテリウム属微生物、クロストリジウム属微生物、コマモナス属微生物、コリネバクテリウム属微生物、クリプテコジニウム属微生物、シアノバクテリア属微生物、デルキシア属微生物、デスルホネマ属微生物、デスルホサルシナ・バリアビリス、デスルホビブリオ・サポボランス、エクトチオロドスピラ属微生物、エリナ属微生物、エントモフトラ属微生物、フェロバシラス・フェロキシダンス、フラボバクテリウム属微生物、ヘモフィルス・インフルエンザ、ハロバクテリウム属微生物、ヒドロクラスラタス・クラスラタス、ヒドロゲノモナス・ファシリス、ヒドロゲノファガ属微生物、ハイフォミクロビウム属微生物、イリオバクター・デラフィエルディ、ジャポノキトリウム属微生物、ラブリス・モナクス、ランプロシスティス・ロゼオペルシシーナ、ランプロペディア・ハイアリーナ、レジオネラ属微生物、レプトスリックス・ディスコホラス、メチロバクテリウム属微生物、メチロシナス属微生物、ミクロコッカス属微生物、モルティエレラ属微生物、マイコバクテリウム属微生物、ニトロバクター属微生物、ノカルディア属微生物、パラコッカス・デントリフィカンス、オシラトリア、ペニシリウム・シクロピウム、フォトバクテリウム属微生物、フィサルム・ポリセファラム、フィコミセス属微生物、シュードモナス属微生物、ピシウム属微生物、ラルストニア属微生物、リゾビウム属微生物、ロドバシラス属微生物、ロドバクター属微生物、ロドコッカス属微生物、ロドシクルス属微生物、ロドミクロビウム・バニエリ、ロドシュードモナス属微生物、ロドスピリラム属微生物、シゾチトリウム属微生物、スフィンゴモナス・パウシモビリス、スピリラム属微生物、スピルリナ属微生物、スタフィロコッカス属微生物、ステラ属微生物、ストレプトマイセス属微生物、シントロフォモナス・ウォルフェイ、サーモフィリック・シアノバクテリア、サーマス・サーモフィルス、チオバシラスA2、チオバシラス属微生物、チオカプサ属微生物、トラウストキトリウム属微生物、チオシスティス・バイオラセア、ビブリオ・パラハエモリティカス、キサントバクター・オートトロフィカス、キサントモナス・マルトフィリア、ズーグロエア属微生物及びオイル生成能を有する酵素をコードする遺伝子により形質転換された微生物からなる群より選択される微生物である請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項4】
前記工程(a)の培養は、窒素源が制限された培地で行う請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項5】
前記遊離脂肪酸は飽和又は不飽和脂肪酸である請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項6】
前記遊離脂肪酸はオレイン酸、リノール酸、リノレン酸、パルミトレイン酸、リシノール酸、バクセン酸、カドレン酸、アラキドン酸、EPA(5,8,11,14,17-eicosapentaenoic acid)、エルカ酸、DHA(4,7,10,13,16,19-docosahexaenoic acid)酪酸、カプロン酸、カプリル酸、カプリン酸、ラウリン酸、ミリスチン酸、パルミチン酸、ステアリン酸、アラキジン酸、ベヘン酸及びリグノセンリン酸からなる群より選択される請求項5に記載の脂肪酸アルキルエステルの製造方法。
【請求項7】
前記遊離脂肪酸は芳香族環基、エポキシ基、シアノ基及びハロゲン基からなる群から選択される置換基で置換されたものである請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項8】
前記工程(b)の自己分解は、リパーゼによって行われる請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項9】
前記リパーゼはトリアシルグリセロールリパーゼ、モノアシルグリセロールリパーゼ及びリソホスホポリパーゼのいずれか一つ以上である請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項10】
前記オイル生成能を有する微生物はリパーゼをコードする遺伝子が導入または増幅された微生物である請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項11】
前記オイル生成能を有する微生物はロドコッカス・オパカスである請求項10に記載の脂肪酸アルキルエステルの製造方法。
【請求項12】
前記オイル生成能を有する微生物はトリアシルグリセロールリパーゼ、モノアシルグリセロールリパーゼ及びリソホスホポリパーゼからなる群より選択される二つ以上のリパーゼをコードする遺伝子が導入された微生物である請求項10に記載の脂肪酸アルキルエステルの製造方法。
【請求項13】
前記リパーゼをコードする遺伝子は配列番号5、8、17、18、19及び20からなる群より選択されるいずれか一つの塩基配列を有する遺伝子である請求項10に記載の脂肪酸アルキルエステルの製造方法。
【請求項14】
前記工程(c)の反応は、80〜120℃で1〜24時間行う請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項15】
次の工程を含む、オイル生成能とリパーゼをコードする遺伝子を有する微生物を利用した脂肪酸メチルエステルの製造方法:
(a)オイル生成能とリパーゼをコードする遺伝子を有する微生物を培養して、オイルを生成させる工程、
(b)前記生成されたオイルを含有する微生物においてリパーゼによるオイルの自己分解を誘導して、遊離脂肪酸を生成させる工程、及び
(c)前記生成された遊離脂肪酸にメタノールを添加し、反応させて脂肪酸メチルエステルを製造する工程。
【請求項16】
前記微生物はトリアシルグリセロールリパーゼをコードする遺伝子及びモノアシルグリセロールリパーゼをコードする遺伝子が導入された微生物である請求項15に記載脂肪酸メチルエステルの製造方法。
【請求項17】
前記トリアシルグリセロールリパーゼをコードする遺伝子は配列番号5、17、18、及び19からなる群より選択されるいずれか一つの塩基配列を有する遺伝子であり、前記モノアシルグリセロールリパーゼをコードする遺伝子は配列番号8または20の塩基配列を有する遺伝子である請求項16に記載の脂肪酸メチルエステルの製造方法。
【請求項1】
次の工程を含む、オイル生成能を有する微生物を利用した脂肪酸アルキルエステルの製造方法:
(a)オイル生成能を有する微生物を培養してオイルを生成する工程、
(b)前記生成されたオイルを含有する微生物においてオイルの自己分解を誘導して、遊離脂肪酸を生成する工程、及び
(c)前記生成された遊離脂肪酸にアルコールを添加し、反応させて脂肪酸アルキルエステルを製造する工程。
【請求項2】
前記オイルはトリアシルグリセロール(TAG)、ジアシルグリセロール(DAG)、モノアシルグリセロール(MAG)、リン脂質、ステロール脂質、スフィンゴ脂質、サッカロ脂質、プレノール脂質及びポリケチドからなる群より選択される請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項3】
前記オイル生成能を有する微生物はアエロモナス属微生物、アクロモバクター属微生物、アシドボラックス・デラフィルディ、アシドボラックス・ファシリス、アシネトバクター属微生物、アクチノマイセス属微生物、アエロモナス属微生物、アルカリゲネス属微生物、アルテロモナス属微生物、アルソルニア属微生物、アプラノキトリウム属微生物、アスペルギルス属微生物、アモエボバクター属微生物、アファノカプサ属微生物、アファノテーケ属微生物、アクアスピリラム・オートトロフィカム、アゾリゾビウム・カウリノダンス、アゾスピリラム属微生物、アゾスピリラム属微生物、アゾトバクター属微生物、バシラス属微生物、ベギアトア属微生物、ベイジェリンキア属微生物、ベネケア属微生物、ブラケスレア属微生物、ボルデテラ・パーツシス、ブラディリゾビウム・ジャポニカム、カリオファノン・ラタム、カウロバクター属微生物、クロログロエア属微生物、クロマチウム属微生物、クロモバクテリウム属微生物、クロストリジウム属微生物、コマモナス属微生物、コリネバクテリウム属微生物、クリプテコジニウム属微生物、シアノバクテリア属微生物、デルキシア属微生物、デスルホネマ属微生物、デスルホサルシナ・バリアビリス、デスルホビブリオ・サポボランス、エクトチオロドスピラ属微生物、エリナ属微生物、エントモフトラ属微生物、フェロバシラス・フェロキシダンス、フラボバクテリウム属微生物、ヘモフィルス・インフルエンザ、ハロバクテリウム属微生物、ヒドロクラスラタス・クラスラタス、ヒドロゲノモナス・ファシリス、ヒドロゲノファガ属微生物、ハイフォミクロビウム属微生物、イリオバクター・デラフィエルディ、ジャポノキトリウム属微生物、ラブリス・モナクス、ランプロシスティス・ロゼオペルシシーナ、ランプロペディア・ハイアリーナ、レジオネラ属微生物、レプトスリックス・ディスコホラス、メチロバクテリウム属微生物、メチロシナス属微生物、ミクロコッカス属微生物、モルティエレラ属微生物、マイコバクテリウム属微生物、ニトロバクター属微生物、ノカルディア属微生物、パラコッカス・デントリフィカンス、オシラトリア、ペニシリウム・シクロピウム、フォトバクテリウム属微生物、フィサルム・ポリセファラム、フィコミセス属微生物、シュードモナス属微生物、ピシウム属微生物、ラルストニア属微生物、リゾビウム属微生物、ロドバシラス属微生物、ロドバクター属微生物、ロドコッカス属微生物、ロドシクルス属微生物、ロドミクロビウム・バニエリ、ロドシュードモナス属微生物、ロドスピリラム属微生物、シゾチトリウム属微生物、スフィンゴモナス・パウシモビリス、スピリラム属微生物、スピルリナ属微生物、スタフィロコッカス属微生物、ステラ属微生物、ストレプトマイセス属微生物、シントロフォモナス・ウォルフェイ、サーモフィリック・シアノバクテリア、サーマス・サーモフィルス、チオバシラスA2、チオバシラス属微生物、チオカプサ属微生物、トラウストキトリウム属微生物、チオシスティス・バイオラセア、ビブリオ・パラハエモリティカス、キサントバクター・オートトロフィカス、キサントモナス・マルトフィリア、ズーグロエア属微生物及びオイル生成能を有する酵素をコードする遺伝子により形質転換された微生物からなる群より選択される微生物である請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項4】
前記工程(a)の培養は、窒素源が制限された培地で行う請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項5】
前記遊離脂肪酸は飽和又は不飽和脂肪酸である請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項6】
前記遊離脂肪酸はオレイン酸、リノール酸、リノレン酸、パルミトレイン酸、リシノール酸、バクセン酸、カドレン酸、アラキドン酸、EPA(5,8,11,14,17-eicosapentaenoic acid)、エルカ酸、DHA(4,7,10,13,16,19-docosahexaenoic acid)酪酸、カプロン酸、カプリル酸、カプリン酸、ラウリン酸、ミリスチン酸、パルミチン酸、ステアリン酸、アラキジン酸、ベヘン酸及びリグノセンリン酸からなる群より選択される請求項5に記載の脂肪酸アルキルエステルの製造方法。
【請求項7】
前記遊離脂肪酸は芳香族環基、エポキシ基、シアノ基及びハロゲン基からなる群から選択される置換基で置換されたものである請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項8】
前記工程(b)の自己分解は、リパーゼによって行われる請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項9】
前記リパーゼはトリアシルグリセロールリパーゼ、モノアシルグリセロールリパーゼ及びリソホスホポリパーゼのいずれか一つ以上である請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項10】
前記オイル生成能を有する微生物はリパーゼをコードする遺伝子が導入または増幅された微生物である請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項11】
前記オイル生成能を有する微生物はロドコッカス・オパカスである請求項10に記載の脂肪酸アルキルエステルの製造方法。
【請求項12】
前記オイル生成能を有する微生物はトリアシルグリセロールリパーゼ、モノアシルグリセロールリパーゼ及びリソホスホポリパーゼからなる群より選択される二つ以上のリパーゼをコードする遺伝子が導入された微生物である請求項10に記載の脂肪酸アルキルエステルの製造方法。
【請求項13】
前記リパーゼをコードする遺伝子は配列番号5、8、17、18、19及び20からなる群より選択されるいずれか一つの塩基配列を有する遺伝子である請求項10に記載の脂肪酸アルキルエステルの製造方法。
【請求項14】
前記工程(c)の反応は、80〜120℃で1〜24時間行う請求項1に記載の脂肪酸アルキルエステルの製造方法。
【請求項15】
次の工程を含む、オイル生成能とリパーゼをコードする遺伝子を有する微生物を利用した脂肪酸メチルエステルの製造方法:
(a)オイル生成能とリパーゼをコードする遺伝子を有する微生物を培養して、オイルを生成させる工程、
(b)前記生成されたオイルを含有する微生物においてリパーゼによるオイルの自己分解を誘導して、遊離脂肪酸を生成させる工程、及び
(c)前記生成された遊離脂肪酸にメタノールを添加し、反応させて脂肪酸メチルエステルを製造する工程。
【請求項16】
前記微生物はトリアシルグリセロールリパーゼをコードする遺伝子及びモノアシルグリセロールリパーゼをコードする遺伝子が導入された微生物である請求項15に記載脂肪酸メチルエステルの製造方法。
【請求項17】
前記トリアシルグリセロールリパーゼをコードする遺伝子は配列番号5、17、18、及び19からなる群より選択されるいずれか一つの塩基配列を有する遺伝子であり、前記モノアシルグリセロールリパーゼをコードする遺伝子は配列番号8または20の塩基配列を有する遺伝子である請求項16に記載の脂肪酸メチルエステルの製造方法。
【図1】

【図2】
【図3】
【図4】

【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2012−533292(P2012−533292A)
【公表日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願番号】特願2012−520549(P2012−520549)
【出願日】平成22年7月19日(2010.7.19)
【国際出願番号】PCT/KR2010/004701
【国際公開番号】WO2011/008058
【国際公開日】平成23年1月20日(2011.1.20)
【出願人】(502318478)コリア アドバンスド インスティチュート オブ サイエンス アンド テクノロジィ (27)
【Fターム(参考)】
【公表日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願日】平成22年7月19日(2010.7.19)
【国際出願番号】PCT/KR2010/004701
【国際公開番号】WO2011/008058
【国際公開日】平成23年1月20日(2011.1.20)
【出願人】(502318478)コリア アドバンスド インスティチュート オブ サイエンス アンド テクノロジィ (27)
【Fターム(参考)】
[ Back to top ]