
オキシドレダクターゼ活性を有するポリペプチドおよびその使用
本発明は、配列番号3に示されるアミノ酸配列もしくは配列番号4のヌクレオチド配列によってコードされたアミノ酸配列、または配列番号3の配列と45%以上の配列同一性を有するその変異体ポリペプチドを含む、オキシドレダクターゼ活性を有するポリペプチドに関する。本発明はまた、2,5−フラン−ジカルボン酸(FDCA)の生成または5−ヒドロキシメチル−2−フランカルボン酸(HMF酸)の生成のための方法に関する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、オキシドレダクターゼ活性を有するポリペプチドに関し、このポリペプチドをコードする遺伝子を含むポリヌクレオチド配列に関する。本発明は、さらに2,5−フラン−ジカルボン酸(FDCA)の生成に関する。さらに、これらのポリペプチドを生成するのに適した、本発明によるポリヌクレオチドによって形質転換された細胞も、本発明に含まれ、これらの細胞はまた、ヒドロキシメチルフルフラール(HMF)のFDCAへの生体内変換のために使用することもできる。
【0002】
[背景技術]
2,5−フラン−ジカルボン酸(FDCA)は、PETなどのポリエステルの生成においてテレフタレートの生体ベースの代替物になる可能性が大きい。そのため、また他の理由から、FDCAは、バイオマスからの高付加価値化学物質(Top Value−Added Chemicals)に関するDOEレポートにおいてトップ12優先化学物質の1つと選定された(Top Value−Added Chemicals from Biomass、Volume I−Results of screening for potential Candidates from Sugars and Synthesis gas、 Department of Energy(USA)、2004)。この化合物は、酸性条件下でヘキソース糖を加熱することにより生成され得る5−ヒドロキシメチルフルフラール(HMF)の酸化によって得ることができる。DOEレポートの27ページに、FDCAのいくつかの潜在的な有用性が開示されている。これらの有用性には、コハク酸、2,5−ビス(アミノメチル)−テトラヒドロフラン、2,5−ジヒドロキシメチル−テトラヒドロフラン、2,5−ジヒドロキシメチルフラン、および2,5−フランジカルバルデヒドを生成する基質としての役割が含まれる。C6糖の化学的酸化脱水によるFDCAの生成およびFDCAの用途はよく知られており、26ページの表13には技術的な障害も提示されているが、生体内変換(おそらく酵素的変換)については、その状況は不明であった。
【0003】
FDCAの酵素的調製の方法は、国際公開2009/023174号パンフレットに開示されている。この開示では、ヒドロキシメチルフルフラール類がクロロペルオキシドレダクターゼにより酸化され、ヒドロキシメチルフルフラールのC1位置にカルボン酸基を有する生成物、具体的にはホルミルフランカルボン酸すなわちFFCAが過酸化水素により酸化された。その結果を、例えば図1に示す。別の実施形態では、酸化基質の存在下でHMFをオキシドレダクターゼと接触させ、それによってHMFをジホルミルフランまたはホルミルフランカルボン酸の少なくとも1つに酸化する。
【0004】
国際公開2009/023174号パンフレットにある公知の方法の欠点は、この反応が過酸化水素を必要とすること、および形成される生成物がFDCAと2つの混入副産物、ヒドロキシメチルフランカルボン酸(HmFCA)およびホルミルフランカルボン酸(FFCA)との混合物であることである。その結果、HMFからFDCAへの収率は低下し、実質的に精製された形でFDCAを得るために回収ステップを追加する必要がある。
【0005】
データベースEBIにおいて、「バークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)PsJNの第1染色体の完全配列」の中で、グルコース−メタノール−コリンオキシドレダクターゼとの相同性から推定されたUniprot B2T4R9、577個のアミノ酸配列が同定された。
【0006】
データベースEBIにおいて、「バークホルデリア・フィマツム(Burkholderia phymatum)STM815のプラスミド1の完全配列」の中で、グルコース−メタノール−コリンオキシドレダクターゼとの相同性から推定された、Uniprot B2JSZ0、576個のアミノ酸配列が同定された。
【0007】
Deurzen,M.P.Jら、J.Carbohydrate Chemistry 16(3)、299〜309(1997)では、5−ヒドロキシメチルフルフラールのクロロペルオキシダーゼ触媒による酸化が開示されている。この反応では、フラン−2,5−ジカルボキサルデヒド(FDC)への選択性が60〜74%である。副産物は、5−ヒドロキシメチル−2−フランカルボン酸(HFCA)および5−ホルミルフラン−2−カルボン酸(FFCA)であった。
【0008】
[発明の概要]
本発明の目的は、分子状酸素をレドックス反応に使用することができるオキシドレダクターゼを提供することである。別の目的は、幅広い反応スペクトルを有するオキシドレダクターゼを提供することである。別の目的は、高い部位特異性を有するオキシドレダクターゼを提供することである。本発明のさらなる目的は、実質量の副生成物を回避することができる、FDCAを生成するための方法を提供することである。本発明のさらなる目的は、実質量の副生成物を回避することができる、5−ヒドロキシメチル−2−フランカルボン酸(HMF酸)を生成するための方法を提供することである。本発明によってこれらの目的の1つまたは複数が達成される。
【0009】
本発明は、オキシドレダクターゼ活性を有するポリペプチドをコードするポリヌクレオチドを提供する。本発明のポリヌクレオチドは、オキシドレダクターゼ活性、具体的にはHmfHオキシドレダクターゼ活性を有するポリペプチドを通常はコードする。
【0010】
したがって、本発明によれば、配列番号3に示されるアミノ酸配列もしくは配列番号4のヌクレオチド配列によってコードされたアミノ酸配列もしくは配列番号3の配列と45%以上の配列同一性もしくは45%超の配列同一性を有するその変異体ポリペプチドを含む、オキシドレダクターゼ活性を有するポリペプチドが提供される。
【0011】
これらのオキシドレダクターゼポリペプチドは、付加価値化合物FDCAを生成するための、化学的酸化ルートの代わりとなる酵素的代替ルートを提供し、温和な反応条件(30℃、pH7)を可能にして、かつ無用な生成物を少なくする。このオキシドレダクターゼは、真正のオキシダーゼである。すなわち、分子状酸素のみがこの酸化反応に必要であり、高価な補因子を再生する必要がない。この酵素は幅広い反応特異性および部位特異性を示し、アルコール基およびアルデヒド基の両方を、これらの基がフラン骨格のC2位置またはC5位置のいずれにあるかにかかわらず、最終的にカルボン酸へ酸化する。
【0012】
さらに、本発明によれば、本発明のポリヌクレオチド配列を組み込んでいる発現ベクターなどのベクター、および本発明によるポリペプチド、ポリヌクレオチド、またはベクターを含む細胞が提供される。
【0013】
本発明は、オキシドレダクターゼ活性を有するポリペプチドを製造するための方法をさらに提供し、この方法は、前記ポリペプチドの発現を可能にする条件下で本発明の細胞を培養すること、ならびに場合によっては、この発現ポリペプチドおよびこのような方法によって得られるポリペプチドを回収することを含む。
【0014】
本発明の別の実施形態は、強健な全細胞生体触媒を使用して、フルクトースリッチな(リッチにされた)流加からFDCAを生成する統合的方法に関する。
【図面の簡単な説明】
【0015】
【図1】HmfHオキシドレダクターゼによって触媒される酸化の反応スキームを示す図である。
【図2】HmfH発現ベクターpJT’hmfHのプラスミドマップを示す図である。Ptac’、tacプロモーター;rep、pRO1600複製タンパク質をコードする遺伝子;gmR、ゲンタマイシン耐性遺伝子;bla、β‐ラクタマーゼ;pRO1600−ori、P.プチダ(P.putida)のための複製開始点;pUC ori、大腸菌(E.coli)のための複製開始点。
【図3A】C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成を示す概略図である。
【図3B】C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成を示す概略図である。
【図3C】C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成を示す概略図である。
【図3D】C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成を示す概略図である。
【図4】P.プチダ(P.putida)S12 pJT’hmfHの粗細胞抽出物中でのHMFからのFDCA、HMF−アルコール(H−oh)、およびHMF−酸(H−酸)の形成を示す図である。
【図5】P.プチダ(P.putida)_pJT’hmfHの流加回分培養法におけるHMF酸およびFDCAの形成を示す図である。左側矢印は、HMF流加の開始を示す;右側矢印は、流加へのグリセロールの追加を示す。
【図6】P.プチダ(P.putida)_pJT’hmfHの流加回分培養法におけるHMF酸およびFDCAの形成を示す図である。矢印は、HMFとグリセロールを合わせた流加の開始を示す。
【図7A】7aは、実施例VIIIの流加回分発酵におけるHMF酸、FDCA、およびバイオマスの濃度を示す図である。7bは、実施例VIIIの流加回分発酵におけるグリセロールおよびHMFの流加速度を示す図である。
【図7B】7aは、実施例VIIIの流加回分発酵におけるHMF酸、FDCA、およびバイオマスの濃度を示す図である。7bは、実施例VIIIの流加回分発酵におけるグリセロールおよびHMFの流加速度を示す図である。
【0016】
[配列リストの簡単な説明]
配列番号1は、プライマーFN23のDNA配列:5’−CGGAATTCCACATGACAAGGGGAGACCG−3’を示す。下線の配列は、EcoRI制限部位を示す。
配列番号2は、プライマーFN24のDNA配列:5’−CGGAATTCGCTTCGGTCTTCAACTCGGATG−3’を示す。下線の配列は、EcoRI制限部位を示す。
配列番号3は、HmfHのアミノ酸配列を示す。
配列番号4は、hmfHのコード配列を示す。
【0017】
[発明の詳細な説明]
本明細書および添付の特許請求の範囲を通して、単語「含む(comprise)」および「含む(include)」、ならびに「含む(comprises)」、「含んでいる(comprising)」、および「含んでいる(including)」などの語尾変化語は、包含的であると解釈されるべきである。すなわち、これらの単語には、文脈の枠内で、具体的に列挙されない他のエレメントまたは整数も包含され得ることを示す意図がある。
【0018】
本明細書において使用する冠詞「a」および「an」は、この冠詞の文法上の目的語が1つまたは2以上である(すなわち1つまたは少なくとも1つである)ことを指す。例として、「1つのエレメント(an element)」は1つのエレメントまたは2つ以上のエレメントを意味することができる。
【0019】
本明細書における幅広い反応スペクトルとは、オキシドレダクターゼが多くの様々な基質、例えばHMF、HMF酸、HMFアルコール、フルフラール、フルフリルアルコールに対して触媒として作用することができることを意味する。部位特異性とは、オキシドレダクターゼが特定のC原子のみを酸化することを意味する。
【0020】
オキシドレダクターゼポリペプチド
オキシドレダクターゼ活性を有する本発明によるポリペプチドは、配列番号3に示されるアミノ酸配列もしくは配列番号4のヌクレオチド配列によってコードされたアミノ酸配列または配列番号3に示される配列と少なくとも45%の配列同一性を有するその変異体ポリペプチドを含む。
【0021】
一実施形態では、変異体核酸分子は、配列番号4に示されるヌクレオチド配列に少なくとも66%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上の相同性の実質的に相同なヌクレオチド配列を含む、タンパク質をコードするヌクレオチド配列を含む。別の実施形態では、変異体タンパク質は、配列番号3に示されるアミノ酸配列に少なくとも45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上の相同性の実質的に相同なアミノ酸配列を含む。
【0022】
本発明の一実施形態は、(a)配列番号4に示されるヌクレオチド配列;(b)配列番号4の逆相補鎖であるポリヌクレオチドと選択的にハイブリダイズするヌクレオチド配列;(c)配列番号4のヌクレオチド配列と少なくとも66%以上の配列同一性を有するヌクレオチド配列;(d)少なくとも約100ヌクレオチド長である、(a)、(b)、または(c)で定義されるヌクレオチド配列のフラグメント;(e)(a)、(b)、(c)、または(d)のいずれか1つに定義される配列に、遺伝子コードの結果として縮重した配列;(f)(a)、(b)、(c)、(d)、もしくは(e)に定義されるヌクレオチド配列の逆相補鎖であるヌクレオチド配列、またはポリペプチドをコードするヌクレオチド配列を含むポリヌクレオチド、および関連ポリペプチドに対するポリヌクレオチドである。
【0023】
一実施形態は、GC含量が56%以上、58%以上、または58〜65%であり得るこのポリヌクレオチドを含む核酸構築物である。さらなる実施形態は、このポリヌクレオチド配列を組み込んでいるベクター、または核酸構築物である。
【0024】
用語「相同性」、「配列同一性」などは、本明細書では互換的に使用される。本発明の目的のために、2つのアミノ酸配列間または2つの核酸配列間で共通する配列同一性の程度を決定するために、最適な比較を目指した配列アライメントが本明細書において定義される(例えば第2のアミノ酸配列または核酸配列との最適アライメントのために、第1のアミノ酸配列または核酸配列の配列の中にギャップを導入することができる)。このようなアライメントは、比較される配列の全長にわたって実行することができる。あるいは、このアライメントは、より短い比較鎖長、例えば約20、約50、約100以上の核酸/塩基またはアミノ酸にわたって実行することができる。
【0025】
次いで、対応するアミノ酸位置またはヌクレオチド位置にあるアミノ酸残基またはヌクレオチドを比較する。第1の配列中の位置が第2の配列中の対応する位置と同じアミノ酸残基またはヌクレオチドによって占められている場合に、これらの分子はその位置において同一である。配列間で共通する同一性の程度は、2つの配列間の同一率によって通常表現され、配列間で共通する同一位置の数の関数である(すなわち、同一率(%)=同一位置の数/位置(すなわち、重なり位置)の総数×100)。比較される2つの配列は同じまたは実質的に同じ長さであることが好ましい。
【0026】
2つの配列間の相同性を決定するために、数種類の異なるコンピュータプログラムが入手可能であるという事実は、当業者ならば承知しているであろう。例えば、2つの配列間の配列の比較および同一率の決定は、数学アルゴリズムを用いて達成することができる。好ましい実施形態では、2つのアミノ酸配列間の同一率は、Accelrys GCGソフトウェアパッケージ(http://www.accelrys.com/products/gcg/から入手可能)の中のGAPプログラムに組み込まれているNeedlemanおよびWunsch(J.Mol.Biol.(48):444〜453(1970))のアルゴリズムによって、Blossom62マトリックスまたはPAM250マトリックスのいずれか、ならびに16、14、12、10、8、または4のギャップ重みおよび1、2、3、4、5、または6の鎖長重みを使用して決定される。これらすべての異なるパラメータにより、若干異なる結果が得られること、しかし2つの配列の全体的な同一率は、異なるアルゴリズムを用いる場合にもあまり変わらないことは、当業者ならば認識するであろう。
【0027】
さらに別の実施形態では、2つのヌクレオチド配列間の同一率は、Accelrys GCGソフトウェアパッケージ(http://www.accelrys.com/products/gcg/から入手可能)の中のGAPプログラムによって、NWSgapdna.CMPマトリックスならびに40、50、60、70、または80のギャップ重みおよび1、2、3、4、5、または6の鎖長重みを使用して決定される。別の実施形態では、2つのアミノ酸配列間またはヌクレオチド配列間の同一率は、ALIGNプログラム(バージョン2.0)(GenestreamサーバーIGH Montpellier France http://vega.igh.cnrs.fr/bin/align−guess.cgiの配列データを用いてALIGN Queryから入手可能)の中に組み込まれているE.MeyersおよびW.Miller(CABIOS、4:11〜17(1989))のアルゴリズムによって、PAM120重み残基表、12のギャップ長ペナルティ、および4のギャップペナルティを使用して決定される。
【0028】
本発明の核酸配列およびタンパク質配列を「クエリー配列」として使用して、例えば他のファミリーメンバーまたは関連配列を同定するために公開データベースの検索を行うことができる。このような検索は、Altschulら、(1990)J.Mol.Biol.215:403〜10のBLASTnおよびBLASTxのプログラム(バージョン2.0)を使用して行なうことができる。BLASTヌクレオチド検索を、NBLASTプログラム(スコア=100、ワード長=12)で行って、本発明のオキシドレダクターゼ核酸分子に相同なヌクレオチド配列を得ることができる。BLASTタンパク質検索を、BLASTxプログラム(スコア=50、ワード長=3)で行って、本発明のオキシドレダクターゼタンパク質分子に相同なアミノ酸配列を得ることができる。比較のためのギャップアライメントを得るために、Altschulら、(1997)Nucleic Acids Res.25(17):3389〜3402に記載のGapped BLASTを利用することができる。BLASTおよびGapped BLASTプログラムを利用する場合、それぞれのプログラム(例えばBLASTxおよびBLASTn)のデフォルトパラメーターを使用することができる。National Center for Biotechnology Informationのホームページ(http://www.ncbi.nlm.nih.gov/)を参照されたい。
【0029】
本明細書において使用する用語「選択的にハイブリダイズする(selectively hybridizing, hybridizes selctively)」および類似の用語は、少なくとも66%、少なくとも70%、少なくとも75%、少なくとも80%、より好ましくは少なくとも85%、さらにより好ましくは少なくとも90%、好ましくは少なくとも95%、より好ましくは少なくとも98%、またはより好ましくは少なくとも99%の相同性を互いに有するヌクレオチド配列が、通常互いにハイブリダイズした状態を保つハイブリダイゼーションおよび洗浄の条件を記載する意図がある。すなわち、このようなハイブリダイズする配列は、少なくとも45%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65、少なくとも70%、少なくとも75%、少なくとも80%、より好ましくは少なくとも85%、さらにより好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも98%、またはより好ましくは少なくとも99%の配列同一性を共有し得る。
【0030】
このようなハイブリダイゼーション条件の好ましい非限定例では、約45℃にて6×塩化ナトリウム/クエン酸ナトリウム(SSC)中でハイブリダイゼーションを行った後、約50℃、好ましくは約55℃、好ましくは約60℃、さらにより好ましくは約65℃にて、1×SSC、0.1%SDS中で1回または複数回洗浄する。
【0031】
高ストリンジェント条件としては、例えば、約68℃にて、5×SSC/5×Denhardt溶液/1.0%SDS中でハイブリダイズし、室温にて、0.2×SSC/0.1%SDS中で洗浄することが挙げられる。あるいは、洗浄は42℃で行ってもよい。
【0032】
当業者であれば、ストリンジェントおよび高ストリンジェントハイブリダイゼーション条件として、いずれの条件を適用すべきかを知っていよう。そのような条件に関するさらなる手引きは、当技術分野では容易に入手可能であり、例えば、Sambrookら、1989、Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Press、N.Y.;およびAusubelら(編者)、1995、Current Protocols in Molecular Biology、(John Wiley & Sons、N.Y.)が挙げられる。
【0033】
もちろん、ポリA配列(mRNAの3’末端ポリ(A)トラクトなど)またはT(またはU)残基の相補的ストレッチに対してのみハイブリダイズするポリヌクレオチドは、ポリ(A)ストレッチまたはその相補体(例えば、実際には任意の二本鎖cDNAクローン)を含有するいかなる核酸分子にもハイブリダイズするため、本発明の核酸の一部に特異的にハイブリダイズさせるために使用される本発明のポリヌクレオチドには含まれない。
【0034】
典型的なアプローチでは、他の生物、例えば細菌から、具体的には微生物マユハキタケ科(Trichomaceae)から、例えばバークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)などのバークホルデリア属(Burkholderia)から構築された遺伝子ライブラリーをスクリーニングすることができる。
【0035】
例えば、相同なオキシドレダクターゼポリヌクレオチドについて、サザンブロット分析により、バークホルデリア(Burkholderia)株をスクリーニングすることができる。本発明の相同なDNA制限フラグメントが検出されたなら、当業者にはよく知られた標準的手法を用いて、適切な菌株から同じ大きさの染色体フラグメントを基に遺伝子ライブラリーを構築することができる。あるいは、微生物が真核生物である場合、オキシドレダクターゼHmfHのmRNA転写物はノーザンハイブリダイゼーションによって同定することができ、この転写物が同定されたなら、真核微生物から単離された全RNAを使用してcDNAライブラリーを作製することができる。
【0036】
相同遺伝子配列は、例えば、本明細書において教示したヌクレオチド配列を基に設計された2つの縮重オリゴヌクレオチドプライマープールを使用してPCRを行うことにより単離することができる。
【0037】
この反応のテンプレートは、本発明のポリヌクレオチドを発現すると知られているかまたは推定される菌株由来の全染色体DNAとすることができる。PCR生成物をサブクローニングし、増幅された配列が新規のオキシドレダクターゼの核酸配列またはその機能的等価物の配列であることを確認するために配列を決定することができる。
【0038】
あるいは、この反応のテンプレートは、本発明のポリヌクレオチドを発現すると知られているかまたは推定される菌株から調製されたmRNAの逆転写により得られたcDNAとすることができる。PCR生成物をサブクローニングし、増幅された配列が新規のオキシドレダクターゼの核酸配列またはその機能的等価物の配列であることを確認するために配列を決定することができる。
【0039】
PCRフラグメントは、その後、様々な公知の方法により、完全長cDNAクローンを単離するために使用することができる。例えば、増幅フラグメントを標識し、バクテリオファージまたはコスミドcDNAライブラリーをスクリーニングするために使用することができる。あるいは、ゲノムライブラリーをスクリーニングするために、この標識フラグメントを使用することができる。
【0040】
PCR技術はまた、他の生物から完全長cDNA配列を単離するために使用することができる。例えば、適切な細胞源または組織源から標準的手法によりRNAを単離することができる。第1鎖の合成をプライミングするための、増幅フラグメントのほとんどの5’末端に対して特異的なオリゴヌクレオチドプライマーを使用して、RNA上で逆転写反応を行うことができる。
【0041】
次いで、得られたRNA/DNAハイブリッドに、標準ターミナルトランスフェラーゼ反応を使用して「尾」を付け(例えばグアニンで)、そのハイブリッドをRNase Hにより消化した後、第2鎖の合成を(例えば、ポリ−Cプライマーにより)プライミングすることができる。こうして、増幅フラグメントの上流のcDNAを容易に単離することができる。有用なクローニング戦略のレビューには、上記のSambrookらおよびAusubelらを参照されたい。
【0042】
本発明の別の態様は、クローニングベクターおよび発現ベクターを含めた、オキシドレダクターゼタンパク質またはその機能的等価物をコードする本発明のポリヌクレオチドを含むベクター、ならびに例えば本発明のポリヌクレオチドの発現が起こる条件下で、適切な宿主細胞においてこのようなベクターを増殖、形質転換または形質移入する方法に関する。本明細書において使用する用語「ベクター」は、別の核酸分子に連結して、その核酸分子を輸送することができる核酸分子を指す。
【0043】
本発明のポリヌクレオチドは、組換え複製可能ベクター、例えばクローニングベクターまたは発現ベクターに組み込むこむことができる。このベクターを使用して、適合性宿主細胞中で核酸を複製してもよい。こうしてさらなる実施形態では、本発明は、本発明のポリヌクレオチドを複製可能ベクターに導入すること、ベクターを適合性宿主細胞に導入すること、およびベクターの複製をもたらす条件下で宿主細胞を増殖させることによって、本発明のポリペプチドを作製する方法を提供する。ベクターは、宿主細胞から回収してもよい。適切な宿主細胞は以下に記載がある。
【0044】
本発明の発現カセットまたはポリヌクレオチドが挿入されているベクターは、組換えDNA手法に便利に供することができる任意のベクターであってもよく、ベクターの選択は、ベクターが導入されることになる宿主細胞に依存することが多い。
【0045】
本発明によるベクターは、自律的に複製するベクター、すなわち、その複製が染色体の複製に依存しない、染色体外の実体として存在するベクター、例えばプラスミドであってもよい。あるいは、このベクターは、宿主細胞に導入された場合、宿主細胞ゲノムに組み込まれ、それを組み込んだ染色体と一緒に複製されるものであってもよい。
【0046】
ベクターのタイプの1つは「プラスミド」であり、追加のDNAセグメントを連結することができる環状二本鎖DNAループを指す。ベクターの別のタイプはウイルスベクターであり、このベクターではウイルスゲノムに追加のDNAセグメントを連結することができる。ある種のベクターは、導入された宿主細胞中で自己複製することができる(例えば、細菌性複製起点を有する細菌ベクターおよびエピソーム性哺乳動物ベクター)。他のベクター(例えば、非エピソーム性哺乳動物ベクター)は、宿主細胞に導入されると同時に宿主細胞のゲノムに組み込まれ、その結果、宿主ゲノムと一緒に複製される。さらに、ある種のベクターは、作動可能に連結された遺伝子の発現を指示することができる。本明細書では、このようなベクターを「発現ベクター」という。一般に、組換えDNA技術において有用な発現ベクターは、プラスミドの形態である場合が多い。プラスミドはベクターの最も一般的に使用される形態であるので、用語「プラスミド」および「ベクター」は、本明細書において互換的に使用することができる。しかしながら、本発明は、機能的に等価な、コスミド、ウイルスベクター(例えば、複製欠損レトロウイルス、アデノウイルス、およびアデノ随伴ウイルス)、およびファージベクターなどの他の形態の発現ベクターを包含することが意図される。
【0047】
本発明によるベクターは、例えばRNAの生成のために、または宿主細胞の形質移入もしくは形質転換のためにインビトロで使用してもよい。
【0048】
本発明のベクターは、2個以上、例えば3個、4個、または5個の本発明のポリヌクレオチドを、例えば過剰発現のために含んでもよい。
【0049】
本発明の組換え発現ベクターは、宿主細胞中で核酸を発現するのに適した形態で本発明の核酸を含む。このことは、組換え発現ベクターが、発現のために使用される宿主細胞に基づき選択され、発現される核酸配列に作動可能に連結された1つまたは複数の調節配列を含むことを意味する。
【0050】
発現ベクターなどのベクターにおいて、「作動可能に連結された」は、目的のヌクレオチド配列が、(例えば、インビトロ転写/翻訳系、またはベクターが宿主細胞に導入される場合は宿主細胞において)ヌクレオチド配列の発現を可能にするように、調節配列に連結されていることを意味することが意図される。すなわち、用語「作動可能に連結された」は、記載の成分がその意図される様式で機能することを可能にする位置関係で並置されることを意味する。コード配列に「作動可能に連結された」プロモーター、エンハンサー、または他の発現調節シグナルなどの調節配列を、コード配列の発現が制御配列に適合する条件下で達成されるように位置させるか、または意図した目的のために協調して機能するように、例えば、転写がプロモーターで始まり、ポリペプチドをコードするDNA配列を通って進行するようにその配列を配置する。
【0051】
用語「調節配列」または「制御配列」は、プロモーター、エンハンサー、および他の発現調節エレメント(例えばポリアデニル化シグナル)を含むことが意図される。このような調節配列は、例えば、Goeddel;Gene Expression Technology:Methods in Enzymology 185、Academic Press、San Diego、CA(1990)に記載されている。
【0052】
用語「調節配列」または「制御配列」には、多くのタイプの宿主細胞においてヌクレオチド配列の構成的な発現を指示する配列、および特定の宿主細胞においてのみヌクレオチド配列の発現を指示する配列(例えば、組織特異的調節配列)が含まれる。
【0053】
したがって、所与の宿主細胞に対するベクターまたは発現構築物は、第1の発明のポリペプチドをコードする配列のコード鎖に関して、5’末端から3’末端に向かって連続した順序で互いに作動的に連結された次のエレメントを含んでもよい。(1)所与の宿主細胞において、ポリペプチドをコードするヌクレオチド配列の転写を指示することができるプロモーター配列;(2)場合によっては、所与の宿主細胞から培地へのポリペプチドの分泌を指示することができるシグナル配列;(3)セロビオヒドロラーゼ活性を有するポリペプチドの成熟型および好ましくは活性型をコードする本発明のDNA配列;好ましくはさらに(4)ポリペプチドをコードするヌクレオチド配列の下流にある、転写を終結させることができる転写終結領域(ターミネーター)。
【0054】
本発明によるヌクレオチド配列の下流に、1つまたは複数の転写終結部位(例えば、ターミネーター)を含有する3’非翻訳領域が存在し得る。ターミネーターの起源はそれほど重要ではない。例えば、ターミネーターは、ポリペプチドをコードするDNA配列にとって本来のものにすることができる。しかしながら、酵母宿主細胞の中では酵母のターミネーターを使用すること、また、糸状菌宿主細胞の中では糸状菌のターミネーターを使用することが好ましい。ターミネーターが宿主細胞(この中でポリペプチドをコードするヌクレオチド配列が発現されることになる)にとって内因性のものであることがより好ましい。転写領域では、翻訳のためのリボソーム結合部位が存在してもよい。構築物によって発現される成熟転写物のコード部分には、始点には翻訳開始のAUG、また翻訳されるポリペプチドの終点には適切な場所に終止コドンが含まれる。
【0055】
さらに、本発明のポリヌクレオチドの発現は、異種の制御領域、例えばプロモーター、分泌リーダー、および/またはターミネーターの領域を選ぶことによって増強し得る。これらの制御領域は、発現の増強、所望の場合には、発現宿主からの目的タンパク質の分泌レベルの増強、および/または本発明のポリペプチド発現の誘導制御の実現を促進させ得る。
【0056】
発現ベクターの設計は、形質変換される宿主細胞の選択、所望タンパク質の発現レベル等の要因に依存し得ることは、当業者ならば認識されよう。発現ベクターなど、本発明のベクターを宿主細胞に導入し、それによって本明細書に記載の核酸がコードするタンパク質またはペプチド(例えば、オキシドレダクターゼタンパク質、オキシドレダクターゼタンパク質の突然変異形態、オキシドレダクターゼタンパク質のフラグメント、変異体、または機能的等価物、融合タンパク質など)を生成させることができる。
【0057】
組換え発現ベクターなど、本発明のベクターは、原核細胞または真核細胞の中でオキシドレダクターゼタンパク質を発現するように設計することができる。例えば、オキシドレダクターゼタンパク質は、大腸菌(E.coli)などの細菌細胞、昆虫細胞(バキュロウイルス発現ベクターを使用して)、糸状菌、酵母細胞、または哺乳類細胞の中で発現することができる。適切な宿主細胞は、Goeddel、Gene Expression Technology:Methods in Enzymology 185、Academic Press、San Diego、 CA(1990)の中にさらに取り上げられている。適切な宿主の代表例は本明細書の以下に記載がある。
【0058】
上記宿主細胞のための適切な培地および条件は、当技術分野で知られている。
【0059】
上述の用語「制御配列」または「調節配列」は、本明細書では、ポリペプチドの発現に必要および/または有利となり得る成分を少なくとも1つ含むと定義される。いずれの制御配列も、ポリペプチドをコードする本発明の核酸配列にとって本来のものであっても、または外来のものであってもよい。このような制御配列としては、プロモーター、リーダー、最適な翻訳開始配列(Kozak、1991、J.Biol.Chem.266:19867〜19870に記載)、分泌シグナル配列、プロペプチド配列、ポリアデニル化配列、転写ターミネーターを挙げ得るが、これらに限定されるものではない。最低限でも、制御配列はプロモーターならびに転写および翻訳の終止シグナルを通常含んでいる。
【0060】
安定に形質転換された微生物とは、1つまたは複数のDNAフラグメントが導入されており、増殖培養において、その導入分子を維持し、複製し、かつ隔離するような微生物である。安定な形質転換は、多重もしくは単一の染色体組込みまたはプラスミドベクターなどの染色体外エレメントによる可能性がある。プラスミドベクターは、特定のDNAフラグメントによってコードされたポリペプチドの発現を指示することができる。
【0061】
発現は、構成的であってもよく、または特定のポリペプチドをコードする機能的に関連するDNAフラグメントを高レベルで転写させることができる誘導性(または抑制性)プロモーターによって調節されてもよい。
【0062】
オキシドレダクターゼの単離
オキシドレダクターゼまたはオキシドレダクターゼを発現するDNA物質は、生物、好ましくはオキシドレダクターゼを発現する微生物から単離し得る。微生物はHMFを利用できることが好ましいが、これは必須ではない。微生物は、カプリアビダス属(Cupriavidus)、バークホルデリア属(Burkholderia)、ブラディリゾビウム属(Bradyhrizobium)、メチロバクテリウム属(Methylobacterium);カプリアビダス・バシレンシス(Cupriavidus basisliensis)、バークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)、ブラディリゾビウム・ジャポニクム(Bradyhrizobium japonicum)、メチロバクテリウム・ラデイオトレランス(Methylobacterium radiotolerans)、カプリアビダス・バシスレンシス(Cupriavidus basisliensis)HMF14、バークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)PsJN、ブラディリゾビウム・ジャポニクム(Bradyhrizobium japonicum)USDA110、メチロバクテリウム・ラデイオトレランス(Methylobacterium radiotolerans)JCM2831からなる群から選択されることが好ましい。
【0063】
本発明において有用な最も好ましいオキシドレダクターゼは、HMFを利用するものであり、カプリアビダス・バシレンシス(Cupriavidus basisliensis)HMF14から単離されたオキシドレダクターゼである。このオキシドレダクターゼは、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約(Budapest Treaty on International Recognition of the Deposits of Microorganisms for the Purpose of Patent Procedures)に従ってドイツ細胞バンク(DSMZ)に寄託されている:カプリアビダス・バシレンシスHMF14=DSM22875、寄託日:2009年8月19日、寄託者TNO、Schoemakerstraat 97、2628VK Delft、Netherlands。
【0064】
したがって、我々はHMF利用細菌であるカプリアビダス・バシレンシス株HMF14を分離し、HMF分解経路に関与する遺伝子を同定した。これらの遺伝子の1つ(本明細書における定義はhmfH)は、579個のアミノ酸からなる62kDaのFAD依存性オキシドレダクターゼをコードしており、このオキシドレダクターゼはフルフリルアルコール、フルフラール、HMF、および5−ヒドロキシメチルフロン酸を酸化することが見出された。これらの分子中のC2およびC5のアルコール基/アルデヒド基は、レダクターゼ活性の存在は有利かもしれないが、レダクターゼをコードする核酸構築物の追加を必要とすることなく、酸化された(図1を参照されたい)。
【0065】
このようにして、本発明は、ポリペプチド、例えばオキシドレダクターゼ(EC1.1+EC1.2の活性)活性を有する酵素をコードするポリヌクレオチドを提供する。本明細書では、酵素はポリペプチドのサブクラスである。
【0066】
酸化反応
本明細書では、酸化反応は、本発明のオキシドレダクターゼおよび1つまたは複数の補酵素(本明細書の下記に記載)の存在下でのフラン化合物とオキシダントとの1回または複数回の反応である。この酸化反応は、生成物を生じる1回の酸化反応ステップ(例えばHMF−酸のFDCAへの酸化)を含むことができる。あるいは、この酸化反応は、2回以上の酸化反応ステップを含むことができ、各ステップで中間体が生まれ、最後の中間体が最終生成物である(例えばHMFのFDCAへの酸化)。酸化反応の例を図1に示す。
【0067】
1つの酸化反応は2,5−フランジカルボン酸(FDCA)の生成であり、この反応では、オキシドレダクターゼ触媒および1つまたは複数の補酵素の存在下でのオキシダントとの反応によって、FDCAの1つまたは複数のフラン前駆体がFDCAに変換される。ここで、オキシドレダクターゼ触媒は本発明によるポリペプチドを含む。FDCAのフラン前駆体は、5−ヒドロキシメチルフルフラール(HMF)、2,5−ジヒドロキシメチルフラン(HMFアルコール)、および5−ヒドロキシメチル−2−フランカルボン酸(HMF酸)からなる群から選択することができ、好ましくは、フラン前駆体はHMFである。HMFは、従来の方法で酸の存在下で加熱することにより1つまたは複数のヘキソース糖から得てもよい。ヘキソース糖はバイオマスから得てもよい。この酸化反応はまた、5−ヒドロキシメチル−2−フランカルボン酸(HMF酸)を生成するための方法であってもよく、この反応では、オキシドレダクターゼ触媒および1つまたは複数の補酵素の存在下でのオキシダントとの反応によって、HMF酸の1つまたは複数のフラン前駆体がHMF酸に変換される。ここで、オキシドレダクターゼ触媒は本発明によるポリペプチドを含む。一実施形態では、HMF酸のフラン前駆体は、5−ヒドロキシメチルフルフラール(HMF)および2,5−ジヒドロキシメチルフラン(HMFアルコール)から選択される群から選択される。他の酸化方法も可能である。
【0068】
酸化反応は、比較的温和な温度、すなわち10〜80℃、より好ましくは20〜45℃、最も好ましくはおよそ25〜40℃にて行うことが好ましい。反応中のpHは、好ましくはpH3〜8、より好ましくはおよそpH7である。反応時間は、大気中酸素または純酸素を使用して、6〜18時間であり、酵素はより長時間活性であることが好ましい。
【0069】
反応器は任意の適切な(通気型)バイオリアクターとすることができる。反応器は、バッチで、連続的に、または好ましくは流加回分形式で操作することができる。
【0070】
FDCA、HMF−酸などのような酸化生成物は、冷却/再結晶および結晶化した酸化生成物、例えば結晶化FDCAの分離によって反応混合物から回収することができる。しかしながら、当技術分野で知られているように、酸沈殿および溶媒抽出などの他の回収方法が適切であるが、これらに限定されるものではない。
【0071】
場合によっては、反応は補酵素の存在下で行われる。補酵素は、ニコチンアミドアデニンジヌクレオチド(NAD+)および/またはフラビンアデニンジヌクレオチド(FAD)および/またはピロロキノリンキノロン(PQQ)であってもよい。デヒドロゲナーゼ活性、例えばその細胞抽出物において酸化反応が起こるかまたは存在する細胞の中に見出される内因性の脱水素活性との相乗効果が本発明の酸化反応で見出された。モノマーの1つが、記載のFDCAである、1つまたは複数のモノマーからポリマーを生成するための方法。
【0072】
[オキシダント]
本発明による反応中のオキシダントは、任意のオキシダント、好ましくは酸素であってもよい。酸素の最も経済的な供給源は空気である。このことは、空気が大気から容易に得られ、無料、無毒性、かつ反応後に除去する必要もない点で有利である。あるいは、分子状酸素放出系を利用してもよい。酸素発生系は、当技術分野で開示された様々な酸素発生系から原則として選択し得る。例えば、反応混合物の中にすでに存在するカタラーゼ酵素を利用して、過酸化水素から酸素を発生させ得る。
【0073】
[フラン化合物]
本明細書では、フラン化合物は、2,5−フラン−ジカルボン酸またはその前駆体に酸化され得るフラン基を有する任意の化合物であると理解される。好ましいフラン化合物としては、ヒドロキシメチルフルフラール(HMF)、ヒドロキシメチルフランカルボン酸(HMF酸)、2,5−ジヒドロキシメチルフラン(HMFアルコール)が挙げられる。フラン環または任意もしくはその置換可能な側鎖は、例えばOH、C1〜C10アルキル、アルキル、アリル、アリール、またはRO−エーテル部分、および環式基により、フラン環の任意の可能な位置で置換されていてもよい。
【0074】
[オキシドレダクターゼの発現]
酵素の発現のために利用される厳密な手法に関係なく、当技術分野で公知の方法を用いて、これらの酵素をコードする遺伝子を別の宿主細胞に導入して、その発現を移行させることができると考えられる。本明細書において定義する遺伝エレメントには、関連酵素を発現させるかまたはその発現を調節するタンパク質、特に酵素、アポタンパク質、またはアンチセンスRNAなどの生成物のための発現可能コード配列を有する核酸(一般にはDNAまたはRNA)が含まれる。発現されたタンパク質は、酵素として機能するか、酵素活性を抑制もしくは抑制解除するか、または酵素の発現を制御することができる。これらの発現可能配列をコードする組換えDNAは、染色体にある(例えば相同遺伝子組換えによって宿主細胞染色体へ組み込まれている)か、または染色体外にある(例えば、1つまたは複数のプラスミド、コスミド、および自己複製可能な他のベクターによって保持されている)ことができる。本発明による、宿主細胞を形質転換するために利用される組換えDNAには、構造遺伝子および転写因子に加えて、タンパク質、アポタンパク質、またはアンチセンスRNAのためのコード配列の発現または抑制除去を制御するように作用する、プロモーター、リプレッサー、およびエンハンサーを含む発現調節配列が含まれ得ることが理解される。例えば、このような制御配列は、野生型の宿主細胞に挿入されて、宿主細胞ゲノム中にすでにコードされていた、選択された酵素の過剰発現を促進することができる。あるいは、その代わりに染色体外にコードされた酵素の合成を制御するために使用することができる。
【0075】
組換えDNAは、宿主細胞への遺伝エレメントの移入を媒介する、プラスミド、コスミド、ファージ、酵母人工染色体、または他のベクターを含むが、これらに限定されない、任意の手段によって宿主細胞へ導入することができる。これらのベクターは、ベクターの複製およびベクターが保持する遺伝エレメントを制御するシス作用性の制御エレメントと一緒に、複製起点を含むことができる。選択可能マーカーは、遺伝エレメントが導入された宿主細胞の同定を助けるためにベクター上に存在させることができる。
【0076】
遺伝エレメントを宿主細胞に導入する手段(例えばクローニング)は、当業者にはよく知られている。染色体外の多重コピープラスミドベクターを利用して、本発明による遺伝エレメントを挿入することができる。宿主細胞への遺伝エレメントのプラスミド運搬導入には、制限酵素によりプラスミドベクターを最初に切断した後、プラスミドと本発明による標的酵素類をコードする遺伝エレメントとを連結することが含まれる。この連結組換えプラスミドの再環状化と同時に、感染(例えばλファージへのパッケージング)またはプラスミド移入のための他の手法(例えば電気穿孔、ミクロ注入など)が宿主細胞へプラスミドを移入するために利用される。宿主細胞への遺伝エレメントの挿入に適したプラスミドは、当業者にはよく知られている。
【0077】
他の遺伝子クローニング法には、染色体への遺伝物質の直接的な組込みが含まれるが、これに限定されるものではない。これは、宿主染色体の相同DNA配列によって挟まれた非複製プラスミド上に本明細書に記載の遺伝エレメントをクローニングすること、および前記組換えプラスミドを宿主に形質転換すると同時に、DNA組換えによって染色体へ遺伝エレメントが導入され得ることを含む様々な手段によって実現できる。組込みDNAフラグメントが抗生物質耐性などの選択可能マーカーを含有している場合に、このような組換え株を回収することができる。あるいは、遺伝エレメントは、非複製プラスミドを使用することなく、宿主細胞の染色体へ直接導入することができる。これは、宿主染色体の相同DNA配列も含有する、本発明による遺伝エレメントのDNAフラグメントを合成的に生成することによって達成できる。これらの合成DNAフラグメントが選択可能マーカーをさらに含有する場合にも、遺伝エレメントを宿主染色体に挿入することができる。
【0078】
[宿主細胞]
本発明の別の実施形態は、本発明によるポリペプチド、ポリヌクレオチド、核酸構築物、またはベクターを含む細胞である。宿主細胞は、ポリペプチド、ポリヌクレオチド、核酸構築物、またはベクターが適切に発現され得る細胞である。
【0079】
オキシドレダクターゼは、宿主細胞中で好都合に発現され得る。本発明による宿主細胞は任意の宿主細胞でよい。この細胞は、原核細胞、真核細胞、植物細胞、または動物細胞であってもよい。このような細胞では、1つまたは複数の遺伝子が、完全もしくは部分的に、欠落、ノックアウト、または破壊されていることがあり、場合によっては、1つまたは複数の遺伝子はプロテアーゼをコードしている。一実施形態によれば、本発明による宿主細胞は真核生物の宿主細胞である。真核細胞は、哺乳類、昆虫、植物、菌類、または藻類の細胞であることが好ましい。好ましい哺乳類細胞としては、例えばチャイニーズハムスター卵巣(CHO)細胞、COS細胞、293細胞、PerC6細胞、およびハイブリドーマが挙げられる。好ましい昆虫細胞としては、例えばSf9細胞およびSf21細胞ならびにそれらの派生細胞が挙げられる。より好ましくは、真核細胞は、菌類細胞、すなわちカンジダ属(Candida)、ハンゼヌラ属(Hansenula)、クリベロミセス属(Kluyveromyces)、ピチア属(Pichia)、サッカロミセス属(Saccharomyces)、シゾサッカロミセス属(Schizosaccharomyces)、またはヤロウイア属(Yarrowia)の菌株等の酵母細胞である。より好ましくは、クリベロミセス・ラクチス(Kluyveromyces lactis)、S.セルビシエ(S.cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ヤロウイア・リポリティカ(Yarrowia lipolytica)、およびピチア・パストリス(Pichia pastoris)由来の細胞または糸状菌細胞。最も好ましくは、真核細胞は糸状菌細胞である。
【0080】
「糸状菌」は、真菌亜門および卵菌亜門のすべての糸状形態を含む(Hawksworthら、Ainsworth and Bisby’s Dictionary of The Fungi、第8版、1995、CAB International、University Press、Cambridge、UKによる定義)。糸状菌は、キチン、セルロース、グルカン、キトサン、マンナン、および他の複合多糖で構成される菌糸壁を特徴とする。栄養成長は菌糸の伸長によるものであり、炭素異化反応は偏性好気性である。糸状菌株としては、アクレモニウム属(Acremonium)、アガリクス属(Agaricus)、アスペルギルス属(Aspergillus)、アウレオバシディウム属(Aureobasidium)、クリソスポリウム属(Chrysosporium)、コプリナス属(Coprinus)、クリプトコックス属(Cryptococcus)、フィリバシジウム属(Filibasidium)、フザリウム属(Fusarium)、フミコラ属(Humicola)、マグナポルテ属(Magnaporthe)、ムコール属(Mucor)、ミセリオフトーラ属(Myceliophthora)、ネオカリマスティクス属(Neocallimastix)、ニューロスポラ属(Neurospora)、ペシロミセス属(Paecilomyces)、ペニシリウム属(Penicillium)、ピロミセス属(Piromyces)、パネロカエテ属(Panerochaete)、プロイロータス属(Pleurotus)、シゾフィラム属(Schizophyllum)、タラロミセス属(Talaromyces)、サーモアスカス属(Thermoascus)、チエラビア属(Thielavia)、トリポクラジウム属(Tolypocladium)、およびトリコデルマ属(Trichoderma)の菌株が挙げられるが、これらに限定されるものではない。
【0081】
好ましい糸状菌細胞は、アスペルギルス属(Aspergillus)、クリソスポリウム属(Chrysosporium)、ペニシリウム属(Penicillium)、タラロミセス属(Talaromyces)、またはトリコデルマ属(Trichoderma)の菌種、最も好ましくは、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ソジェ(Aspergillus sojae)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、タラロミセス・エメルソニ(Talaromyces emersonii)、アスペルギルス・オリゼ(Aspergillus oryzae)、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)、トリコデルマ・リーゼイ(Trichoderma reesei)、またはペニシリウム・クリゾゲヌム(Penicillium chrysogenum)の菌種に属する。本発明による宿主細胞がアスペルギルス属(Aspergillus)宿主細胞である場合、宿主細胞にはCBS513.88、CBS124.903、またはその派生細胞が好ましい。
【0082】
別の実施形態によれば、本発明による宿主細胞は原核細胞である。好ましくは、原核生物の宿主細胞は細菌細胞である。用語「細菌細胞」は、グラム陰性菌およびグラム陽性菌の両方を含む。適切な細菌は、例えば大腸菌属(Escherichia)、アナベーナ属(Anabaena)、コーロバクター属(Caulobacter)、グルコノバクター属(Gluconobacter)、ロドバクター属(Rhodobacter)、シュードモナス属(Pseudomonas)、パラコッカス属(Paracoccus)、バチルス属(Bacillus)、ブレビバクテリウム属(Brevibacterium)、コリネバクテリウム属(Corynebacterium)、リゾビウム属(Rhizobium)(シノリゾビウム属(Sinorhizobium))、フラボバクテリウム属(Flavobacterium)、クレブシェラ属(Klebsiella)、エンテロバクター属(Enterobacter)、ラクトバチルス属(Lactobacillus)、ラクトコッカス属(Lactococcus)、メチロバクテリウム属(Methylobacterium)、スタフィロコッカス属(Staphylococcus)、またはストレプトミセス属(Streptomyces)から選択することができる。好ましくは、細菌細胞は、B.スブチリス(B.subtilis)、B.アミロリケファシエンス(B.amyloliquefaciens)、B.リケニフォルミス(B.licheniformis)、B.プンチス(B.puntis)、B.メガテリウム(B.megaterium)、B.ハロデュランス(B.halodurans)、B.プミルス(B.pumilus)、グルコノバクター・オキシダンス(Gluconobacter oxydans)、コーロバクター・クレセンタス(Caulobacter crescentus)CB15、メチロバクテリウム・エキストロクエンス(Methylobacterium extorquens)、ロドバクター・スフェロイデス(Rhodobacter sphaeroides)、シュードモナス・ゼアキサンチニファシエンス(Pseudomonas zeaxanthinifaciens)、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・プチダ(Pseudomonas putida)S12、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)、大腸菌(E.coli)、C.グルタミクム(C.glutamicum)、スタフィロコッカス・カルノーサス(Staphylococcus carnosus)、ストレプトミセス・リビダンス(Streptomyces lividans)、シノリゾビウム・メリロティ(Sinorhizobium melioti)、およびリゾビウム・ラジオバクター(Rhizobium radiobacter)からなる群から選択される。
【0083】
糸状菌のいくつかの菌株、例えばアスペルギルス・ニガー(Aspergillus niger)CBS513.88、アスペルギルス・オリゼ(Aspergillus oryzae)ATCC20423、IFO4177、ATCC1011、ATCC9576、ATCC14488〜14491、ATCC11601、ATCC12892、P.クリソゲヌム(chrysogenum)CBS455.95、ペニシリウム・シトリヌム(Penicillium citrinum)ATCC38065、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)P2、タラロミセス・エメルソニ(Talaromyces emersonii)CBS124.902、アクレモニウム・クリソゲヌム(Acremonium chrysogenum)ATCC36225またはATCC48272、トリコデルマ・リーゼイ(Trichoderma reesei)ATCC26921またはATCC56765またはATCC26921、アスペルギルス・ソジェ(Aspergillus sojae)ATCC11906、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)ATCC44006は、米国微生物系統保存機関(American Type Culture Collection(ATCC))、ドイツ微生物細胞培養コレクション(Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ))、オランダ微生物株保存センター(Centraalbureau Voor Schimmelcultures (CBS))、およびAgricultural Research Service Patent Culture Collection,Northern Regional Research Center(NRRL)などのいくつかの微生物株保存機関において公的に容易に入手できる。それらの派生株もまた使用し得る。
【0084】
別の実施形態によれば、本発明による宿主細胞は原核細胞である。好ましくは、原核宿主細胞は細菌細胞である。用語「細菌細胞」は、グラム陰性菌およびグラム陽性菌の両方を含む。適切な細菌は、例えば大腸菌属(Escherichia)、アナベーナ属(Anabaena)、コーロバクター属(Caulobacter)、グルコノバクター属(Gluconobacter)、ロドバクター属(Rhodobacter)、シュードモナス属(Pseudomonas)、パラコッカス属(Paracoccus)、バチルス属(Bacillus)、ブレビバクテリウム属(Brevibacterium)、コリネバクテリウム属(Corynebacterium)、リゾビウム属(Rhizobium)(シノリゾビウム属(Sinorhizobium))、フラボバクテリウム属(Flavobacterium)、クレブシェラ属(Klebsiella)、エンテロバクター属(Enterobacter)、ラクトバチルス属(Lactobacillus)、ラクトコッカス属(Lactococcus)、メチロバクテリウム属(Methylobacterium)、スタフィロコッカス属(Staphylococcus)、またはストレプトミセス属(Streptomyces)から選択することができる。好ましくは、細菌細胞は、B.スブチリス(B.subtilis)、B.アミロリケファシエンス(B.amyloliquefaciens)、B.リケニフォルミス(B.licheniformis)、B.プンチス(B.puntis)、B.メガテリウム(B.megaterium)、B.ハロデュランス(B.halodurans)、B.プミルス(B.pumilus)、G.オキシダンス(G.oxydans)、コーロバクター・クレセンタス(Caulobacter crescentus)CB15、メチロバクテリウム・エキストロクエンス(Methylobacterium extorquens)、ロドバクター・スフェロイデス(Rhodobacter sphaeroides)、シュードモナス・プチダ(Pseudomonas putida)、パラコッカス・ゼアキサンチニファシエンス(Paracoccus zeaxanthinifaciens)、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)、大腸菌(E.coli)、C.グルタミクム(C.glutamicum)、スタフィロコッカス・カルノーサス(Staphylococcus carnosus)、ストレプトミセス・リビダンス(Streptomyces lividans)、シノリゾビウム・メリロティ(Sinorhizobium melioti)、およびリゾビウム・ラジオバクター(Rhizobium radiobacter)からなる群から選択される。
【0085】
本発明による宿主細胞中で生成される化合物の特定用途のために、その用途に応じて宿主細胞の選択がなされ得る。例えば、本発明による宿主細胞中で生成される化合物が、食品用途で使用されることになる場合、宿主細胞はサッカロミセス・セレビシア(Saccharomyces cerevisiae)などの食用生物から選択してもよい。特定用途には、食品、(動物)飼料、医薬、作物保護などの農業、および/またはパーソナルケアの用途が含まれるが、これらに限定されものではない。
【0086】
さらに、本発明は、オキシドレダクターゼ活性を有するポリペプチドを製造するための方法であって、前記ポリペプチドの発現を可能にする条件下で、本発明による細胞を培養すること、および場合によっては、発現されたポリペプチドを回収することを含む方法に関し、その方法によって得られるポリペプチドに関する。
【0087】
[流加原料]
フルクタンが本来豊富な農業作物(例えばトビナンブールまたはチコリルート)は、従来の加水分解および熱化学(の組合せ)処理によってHMFに富んだ流加原料に変換し得る。フルクトースからHMFを生成する技術は確立しており、確固としたものである。さらにグルコースに富んだ流加原料も利用することができるが、HMFの熱化学的形成はフルクトースからの方がより効率的に起こる。したがって、グルコースイソメラーゼを使用してグルコースをフルクトースに変換するために、追加の酵素ステップを含むことができる。後者のプロセスは、加水分解されたデンプンから高フルクトースコーンシロップ(HFCS)を生成するために食品産業において確立されている。食品用途との競合を回避するならば、リグノセルロース水解物が、HMF/FDCAを生成するための好ましい流加原料になるであろう。
【0088】
[生体内変換]
HMFのFDCAへの生体内変換については、本明細書において前述したカプリアビダス・バシレンシス(Cupriavidus basilensis)HMFオキシドレダクターゼを発現する強健な全細胞生体触媒(遊離細胞または固定化細胞)を使用する。このプロセスでは、全細胞生体触媒は酵素触媒にまさるいくつかの利点を有している。すなわち、HMFオキシドレダクターゼが高反応性基質による化学的不活性化から保護され、かつHMFから(おそらく)対応する一塩基酸に導き、次いでこの一塩基酸がHmfHによって二価酸に変換される最初の2つの酸化ステップにおいて、宿主固有のデヒドロゲナーゼが、HMFオキシドレダクターゼを助けることができる。補因子再生を確実にするために、追加の処置を要求し得ることが好ましい。全細胞生体触媒は、HMF流加を最小限で処理できるものであるべきである。すなわち、低pH、高温、および流加原料の加水分解/熱化学による変換中に生成される有毒化合物(その中に基質がある)に対する耐性があることが好ましい。シュードモナス・プチダ(Pseudomonas putida)S12は、種々の化学的ストレス要因に対する耐性、比較的広いpH領域、およびHMFオキシドレダクターゼを助けてHMFのFDCAへのより効率的な生体内変換を生じさせる固有な(indiginous)デヒドロゲナーゼの存在の観点から、適切な宿主生物としての資格があると見なし得る。
【0089】
全細胞生体触媒の代わりに、全細胞生体触媒と同様に、細胞溶解物、精製酵素、または単独もしくは酵素混合物としての固定化(immobolized)酵素を使用してもよい。
【0090】
[生成物およびバイオマスの回収]
生体内変換の後、細胞は確立した方法によってブロスから分離し、再使用してもよい。FDCAは酸沈殿によって無細胞ブロスから回収し、適切な有機溶媒中に高温で再溶解してもよい。溶解後に、FDCAは、酸沈殿および溶媒抽出、または当技術分野で公知の他の精製法によって、所望の場合には二価酸の形態にて高純度で回収してもよい。
【0091】
[FDCAの用途]
FDCAはポリエステルの製造において、テレフタレート(terephtalate)の代わりとして使用し得る。FDCAはまた、極めて多様な高価値化合物のための基質として使用し得る。例えば、コハク酸、2,5−ビス(アミノメチル)−テトラヒドロフラン、2,5−ジヒドロキシメチル−テトラヒドロフラン、2,5−ジヒドロキシメチルフラン、および2,5−フランジカルバルデヒドを製造するための公知の基質である。FDCAは、コーティングの製造において、例えばアルキド樹脂および熱可塑性コーティングの中に使用してもよい。FDCAはまた、バイオ燃料におけるキシレン等価物として、また溶媒として使用してもよい。
【0092】
FDCAはエステル化してもよく、このエステルは可塑剤として使用し得る。FDCAはそのジオールに変換してもよく、このジオールはPET様ポリエステルおよびポリウレタンの中に使用し得る。さらに、FDCAはそのジアミンに変換してもよく、このジアミンは連鎖延長剤として使用し得る。またこのジアミンはジイソシアネートに変換してもよく、このジイソシアネートはポリウレタンの製造に使用することができる。本発明による方法によって、FDCAおよびFDCAから製造される生成物は、リグノセルロースバイオマスを含むバイオマスから生体内変換を介して作ることができる。
【0093】
[実施例]
[一般的方法]
菌株およびプラスミド カプリアビダス・バシレンシス(Cupriavidus basilensis)HMF14は、DSMZ:カプリアビダス・バシレンシス(Cupriavidus basilensis)HMF14=DSM22875、寄託日:2009年8月19日に寄託されており、唯一の炭素源としてフランを使用することができる土壌分離菌である。シュードモナス・プチダ(Pseudomonas putida)S12(ATCC700801)をHMFオキシドレダクターゼ発現のための宿主として使用した。大腸菌(Escherichia coli)DH5a(Invitrogen)を一般のクローニング目的に使用した。pUCP22由来の大腸菌(E.coli)−P.プチダ(P.putida)シャトルプラスミドpJT’mcs(未公表)を、構成的tacプロモーターの制御下でHMFオキシドレダクターゼを発現させるために使用した。大腸菌(E.coli)における複製については、pUC複製起点を使用する;P.プチダ(P.putida)における複製については、pRO1600複製起点を使用する。hmfH遺伝子の発現は構成的tacプロモーターにより促進される。大腸菌(E.coli)の抗生物質選択(アンピシリン抵抗性)のために、β―ラクタマーゼマーカー遺伝子(bla)を使用する。P.プチダ(P.putida)の抗生物質選択のために、ゲンタマイシンアセチルトランスフェラーゼマーカー遺伝子(gmR)を使用する。
【0094】
HmfH発現ベクターpJT’hmfHのプラスミドマップを図2に示す。Ptac’、tacプロモーター;rep、広範囲の宿主領域複製起点;gmR、ゲンタマイシン耐性遺伝子;bla、β‐ラクタマーゼ;pUC ori、大腸菌(E.coli)のための複製起点。
【0095】
培地および培養条件 ミネラル塩培地(MM)を特定培地として使用した。MMには次のものが含有されていた(脱イオン水1リットル当たり):指定の炭素源と共に添加された、K2HPO4 3.88g、NaH2HPO4 1.63g、(NH4)2SO4 2.0g、MgCl2・6H2O 0.1g、EDTA 10mg、ZnSO4・7H2O 2mg、CaCl2・2H2O 1mg、FeSO4・7H2O 5mg、Na2MoO4・2H2O 0.2mg、CuSO4・5H2O 0.2mg、CoCl2・6H2O 0.4mg、およびMnCl2・2H2O 1mg。ルリアブロス(L−ブロス:10g/lバクトトリプトン(Bacto trypton)(ディフコ(Difco)社)、5g/l酵母抽出物(ディフコ社)、5g/l NaCl)を、P.プチダ(P.putida)S12および派生菌株、C.バシレンシス(C.basilensis)HMF14および大腸菌(E.coli)DH5aおよび派生菌を調製するための完全培地として使用した。固体のL−ブロスは2%(w/v)寒天(ディフコ社)を使用して凝固させた。
【0096】
流加回分培養実験では、最初の回分段階は、次の組成を有する適合ミネラル塩培地1l中で行なわれた:K2HPO4 3.88g、NaH2HPO4 1.63g、(NH4)2SO4 2.0g、MgCl2・6H2O 0.2g、EDTA 20mg、ZnSO4・7H2O 4mg、CaCl2・2H2O 2mg、FeSO4・7H2O 10mg、Na2MoO4・2H2O 0.4mg、CuSO4・5H2O 0.4mg、CoCl2・6H2O 0.8mg、MnCl2・2H2O 2mg、10mg/Lゲンタマイシン、および100mMグリセロール。最初のグリセロールが枯渇した後、流加を開始し、培養物中に制限基質としてグリセロールを維持しながら、最大の増殖が得られるように制御した。流加溶液には、(1l当たり):グリセロール368.4gおよび10g/l MgCl2・6H2O、ならびに12.6g/l HMFが含有されていた。
【0097】
抗生物質:アンピシリン(amp)を大腸菌(E.coli)に対して100μg/ml添加した。ゲンタマイシン(gm)をP.プチダ(P.putida)S12に対して、ルリアブロス中に30μg/ml、ミネラル塩培地中に10μg/ml添加した。抗生物質はシグマアルドリッチ(Sigma−Aldrich)社から購入した。
【0098】
培養:P.プチダ(P.putida)およびC.バシレンシス(C.basilensis)は30℃にて培養した。大腸菌(E.coli)は37℃にて培養した。MM上での振とうフラスコ実験は、水平振とうインキュベーター中でボストンボトル(Boston bottle)(Alltech applied sciences BV;Breda,The Netherlands)を用いて行った。L−ブロス上の振とうフラスコ実験は、水平振とうインキュベーター中で、綿栓を詰めたエルレンマイヤーフラスコを用いて行った。流加回分実験は、BioFlo110コントローラーを使用して、1l発酵槽(ニュー・ブランズウィック・サイエンティフィック(New Brunswick Scientific)社)中で行った。最初の回分発酵は、40mMグリセロールおよび2mMグルコースを添加したMM100ml中での一晩前培養物から得られた洗浄細胞で開始した。最初の撹拌速度を200rpmに設定し、M+W Istruments D−5111マスフローコントローラを使用して空気を1lmin−1でヘッドスペースに供給した。InProモデル6900プローブ(Mettler Toledo BV;Tiel、The Netherlands)を使用して溶存酸素圧(DO)を連続的にモニターし、最大1000rpmまで撹拌速度を自動調整することによって30%空気飽和を維持した。最大撹拌速度に達したとき、0.2lmin−1の流量で空気を精製酸素と交換し、最大撹拌速度を800rpmに設定した。最初の回分段階の間は、25%NH4OHの自動添加によってpHを7.0に維持し、流加段階の間は、10mM NaOHの自動添加によってpHを一定に保った。温度は30℃に保った。
【0099】
アッセイおよび分析法:細菌培養の細胞乾燥重量(CDW)は、平底96穴マイクロプレート(Greiner)を使用し、Biowave Cell Density Meter(WPA社)またはμQuant MQX200汎用マイクロプレート分光光度計(Biotek)を用いて、600nm(OD600)での光学密度を測定することにより決定した。OD600 1.0は、P.プチダ(P.putida)については、0.56g CDW/L(Biowave)または1.4g CDW/L(μQuant)に相当する。
【0100】
HPLC分析:FDCA、HMF、HMF−アルコール、およびHMF−酸は、230nmに設定されたダイオードアレー検出器を使用して、RP−HPLC(Agilent 1100システム)により分析した。使用したカラムは、Zorbax Eclipse XDB−C8(孔径80Å、表面積180m2/g、Agilent)であり、25℃で操作した。溶離液として、1%アセトニトリルと共に20mM KH2P04(pH2またはpH6)中のアセトニトリルにグラディエントをかけて使用し、流量1.2ml/分で、アセトニトリルを3.5分で0〜5%まで、2.5分で5〜40%まで増加させた。
【0101】
細胞抽出物の調製:HMFオキシドレダクターゼを発現する、野生型C.バシレンシス(C.basilensis)HMF14またはP.プチダ(P.putida)S12の形質転換細胞の細胞抽出物は、12mMコハク酸(C.バシレンシス(C.basilensis)HMF14、OD600およそ1.5)または20mMグルコース(P.プチダ(P.putida)S12、OD600およそ4)のいずれかを添加したMMを使用した、指数増殖期後半の培養物50mlから調製した。培養物を遠心分離によって採取し、アッセイ緩衝液3ml中に再懸濁した。細胞は、Branson sonifier(パルスモードでのマイクロチップ、出力設定3、およびパーセントデューティー(duty)サイクル設定40%;3サイクルの超音波処理:45秒パルスおよび15秒休止)またはSonics Vibra−Cell(Sonics&Materials、USA)(5mm先細マイクロチップ;パルスモード設定1分(0,5秒パルス、2秒休止)のいずれかを使用して、超音波処理により破壊した。超音波処理の後、8228×gにて4℃で3分間遠心分離してデブリを除去した。上清は、PD10ゲル濾過カラム(GE healthcare)を使用して脱塩し、HMFオキシドレダクターゼアッセイのための細胞抽出物として使用した。タンパク質濃度はBradford試薬(シグマアルドリッチ社)を使用して測定した。
【0102】
HMFオキシドレダクターゼアッセイ:HMFオキシドレダクターゼアッセイは、C.バシレンシス(C.basilensis)HMF14 HMFオキシドレダクターゼを発現する、野生型C.バシレンシス(C.basilensis)HMF14またはP.プチダ(P.putida)S12の形質転換細胞の細胞抽出物で行った。ネガティブ対照として、hmfH遺伝子にトランスポゾン挿入がある、野生型P.プチダ(P.putida)S12またはC.バシレンシス(C.basilensis)HMF14変異体を使用した。細胞抽出物を、酸素負荷条件下で30℃にてフルフラール、フルフリルアルコール、HMF、またはHMF−酸と共にインキュベートした。反応混合物は、細胞抽出液1ml、酸素飽和MM976μl、2mMフラビンアデニンジヌクレオチド(FAD)溶液20μl、および基質(フルフラール、フルフリルアルコール、HMF、またはHMF酸)の0.5Mストック4μlを含有した。サンプルを設定間隔で抜き取り、最終濃度1MまでHClを添加して反応を直ちに停止させた。サンプル中の基質および生成物の濃度はHPLCによって測定した。酸素欠乏対照として、連続窒素ガス流によって反応混合物の様々な成分を酸素欠乏状態にし、同反応混合物を、ゴム栓付きヘッドスペースバイアル中で窒素ガス下インキュベートした。
【0103】
試薬:分析用標準のFDCAは、Immunosource B.V.(Halle−Zoersel、Belgium)から購入した。5−ヒドロキシメチル−フロン酸(HMF酸)は、Matrix Scientific(Columbia SC、United States)から購入した。この化合物は高度にエステル化されていることが見出された。使用の直前に、エステル化されたHMF酸の10mM溶液を2M H2S04中で2時間沸騰させて冷却し、50mMリン酸緩衝液を加えた後、NaOHによりpH 7.0に調整した。他のすべての試薬は、シグマアルドリッチChemie B.V.(Zwijndrecht、The Netherlands)から購入した。
【0104】
分子および遺伝子の手法:ゲノムDNAは、DNeasy tissue kit(QIAGEN)を用いて単離した。プラスミドDNAは、QIAprep spin miniprep kit(QIAGEN)を用いて単離した。アガローストラップDNAフラグメントは、QIAEXII gel extraction kit(QIAGEN)を用いて単離した。
【0105】
PCR反応は、製造業者の説明書に従ってAccuprime Pfxポリメラーゼ(Invitrogen)を用いて行った。C.バシレンシス(C.basilensis)HMF14のゲノムDNAからHMFオキシドレダクターゼを増幅するために使用したプライマーは、FN23:5’−CGGAATTCCACATGACAAGGGGAGACCG−3’(配列番号1)およびFN24:5’−CGGAATTCGCTTCGGTCTTCAACTCGGATG−3’(配列番号2)であった。下線の配列はEcoRI制限部位を示す。
【0106】
プラスミドDNAは、Gene Pulserエレクトロポレーションデバイス(BioRad)を用いて、エレクトロコンピテント(electrocompetent)細胞へ導入した。
【0107】
トランスポゾンに隣接する染色体DNAは、当技術分野で公知の標準的方法{Ausubel,F.M.ら、Current protocols in molecular biology(Green publishing association、New York;1987)}によって同定し、完全遺伝子座の配列をプライマーウォーキング(primer walking)によって得た。オリゴヌクレオチド合成およびDNA塩基配列決定は、MWG Biotech AG(Germany)によって行った。
【0108】
他の標準分子生物学手法は、SambrookおよびRussel{Sambrook,J.、Russel,D.W.Molecular cloning;a laboratory manual(Cold spring Harbor Laboratoy Press、New York:2001)に従って行った。
【0109】
[実施例I]
[C.バシレンシス(C.basilensis)HMF14の細胞抽出物におけるFDCA生成]
12mMコハク酸および3mM HMFが添加されたMM上で増殖した、対数増殖期後半の前培養物(OD600およそ1.5)から得られたC.バシレンシス(C.basilensis)の細胞抽出物を、HMFまたはHMF酸のいずれかと共にインキュベートすると、FDCAの形成が観察された。
【0110】
基質としてHMFを使用すると、HMF−酸およびHMF−アルコールの迅速な一時的蓄積がFDCA形成と同時に観察された。脱塩細胞抽出物中では、粗細胞抽出物中と比較して、HMF−酸濃度の減少速度は低下し、FDCA生成は遅くなった。FDCAが粗細胞抽出物中と同様に脱塩細胞抽出物中でも生成したので、HMFのFDCAへの変換には補因子は関与していないと考えられた。しかしながら、粗細胞抽出物由来の補因子または他の低分子量成分が、FDCA形成に関して相乗効果を有するように見えた。基質としてHMF−酸を加えると、FDCAの即時形成が粗細胞抽出物中および脱塩細胞抽出物中の両方で観察された。さらに、FDCAが嫌気条件下で生成しなかったので、酸素の存在がFDCA形成には必要であることが実証された。HMF酸のFDCAへの化学量論的変換が観察された。
【0111】
[実施例II]
[HMFオキシドレダクターゼをコードするhmfH遺伝子の単離および特徴づけ]
hmfH遺伝子は、FAD依存性グルコース−メタノール−コリン(GMC)オキシドレダクターゼファミリーに属する62 317 Daのタンパク質をコードすることが見出された。この酵素にはフラン−ジカルボン酸(FDCA)へHMF−酸を酸化する能力があることが示された。さらに、この酵素は、基質としてHMFも受け入れ、HMFをHMF−酸を介してFDCAに酸化した。非重複性NCBIデータベース中でHmfHと最も高い相同性を有するものが、バークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)PsJNのGMCオキシドレダクターゼであることがわかった(座位タグBphyt_2180;562個のアミノ酸ストレッチにわたって68%の同一性)。C.バシレンシス(C.basilensis)HMF14のHMF/フルフラールオペロンの配列データに基づいて、他の潜在的なHMF/フルフラールディグレーダー(degrader)が同定された。選択された菌株は、唯一の炭素源としてHMFまたはフルフラールのいずれかを含むミネラル塩培地上での増殖について試験された。HMF上で増殖できた菌株は、HmfHと45〜68%の同一性があるオキシドレダクターゼをコードするhmfHオルソログを有していた。1菌株(バークホルデリア・ゼノボランス(Burkholderia xenovorans LB400)は、そのオキシドレダクターゼがHmfHと44%の同一性があったが、HMFを利用することができなかった。
【0112】
図3に、C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成の概略図を示す。色は図1における酵素活性に対応する。矢印の下の太数字(x/y)は、yアミノ酸ストレッチ中の対応するC.バシレンシス(C.basilensis)HMF14タンパク質との同一性(x)の割合を示す。オルソロガス遺伝子は、National Center for Biotechnology Informationの非重複性タンパク質データベースに対するBLASTxホモロジー検索によって同定された。HmfAと少なくとも50%の同一性がある酵素をコードするhmfAオルソログがあると同時に、hmfA、B、C、D、およびEに対するオルソログが単一ゲノム中にある場合に、フルフラールクラスターに対するヒットは妥当であると定めた。同じ基準をhmfFおよびhmfGのオルソログを定義するために用いたが、他方hmfHオルソログに対する基準としてはHmfHとの40%の同一性を用いた。イタリック体の数字は、指定菌株のゲノム座位タグを示す。白矢印は、代謝機能を有しない遺伝子を示す。C:唯一の炭素源としてフルフラールまたはHMF(3mM)のいずれかを含むミネラル塩培地上での試験菌株の増殖表現型の概要。ND:測定していない。
【0113】
これらの図では、HMFオキシドレダクターゼHmfHをコードする翻訳領域のヌクレオチド配列(図4)および推定上のアミノ酸配列(図3)が示されている。
【0114】
[実施例III]
[P.プチダ(P.putida)S12中のhmfHのクローニングおよびコードされたオキシドレダクターゼの発現]
HMFオキシドレダクターゼをコードするhmfH遺伝子を、発現ベクターpJT’mcsにクローニングした。C.バシレンシス(C.basilensis)HMF14ゲノムDNA上でプライマーFN23およびFN24を用いて得られたPCRフラグメントを、EcoRI(Fermentas)により消化した。この消化フラグメントを、EcoRI消化およびFastAP(Fermentas)処理のpJT’mcsベクターに連結し、HmfH発現プラスミドpJT’hmfHを得た。hmfH挿入の方向が正しいことを、当技術分野で公知の制御消化およびヌクレオチド配列決定によって確認した。HMFオキシドレダクターゼの発現は、MM+20mMグルコースおよび10mg/lゲンタマイシン中で増殖したP.プチダ(P.putida)_pJT’hmfH(OD600およそ4)の細胞抽出物において、基質としてHMFまたはHMF酸を使用したときにFDCAが形成されたことによって実証された(表1)。
【0115】
【表1】

【0116】
これらの結果により、FDCAの形成が酸素存在下でのみ起こり、hmfHにコードされたオキシドレダクターゼによって触媒されることが確認された。HMFを基質として用いた場合には、脱塩細胞抽出物中および粗細胞抽出物中の両方においてHMF−酸の一時的な蓄積が観察された。脱塩細胞抽出物(低分子量補因子が除去された)中のHMF−酸の形成により、HmfHが、HMF−酸をFDCAへ酸化するだけでなく、HMFをそのモノカルボン酸形へ酸化できることも示された。
【0117】
HmfHに加えて、P.プチダ(P.putida)S12に固有で見かけ上非特異的なデヒドロゲナーゼもまた、その還元等量が脱塩によって除去されなければ、HMFからHMF酸を形成することができる(表2)。アルデヒドレダクターゼ/アルコールデヒドロゲナーゼ活性はNADH/NAD+依存であるが、アルデヒドデヒドロゲナーゼ活性は2つの人工電子伝達体、PMSおよびDCPIPを組み合せることによって維持することができる。ここで、PMSは最初の電子担体であり、DCPIPは最終電子受容体である。同様な活性は、C.バシレンシス(C.basilensis)HMF14の細胞抽出物中でも観察された。
【0118】
【表2】
【0119】
P.プチダ(P.putida)S12 pJT’hmfHの粗細胞抽出物中の、HMFからFDCA、HMF−アルコール(H−oh)およびHMF−酸(H−酸)の形成を図4に示す。図4は、P.プチダ(P.putida)S12 pJT’hmfH細胞抽出物中の、内因性デヒドロゲナーゼおよびHmfHの相乗効果を説明する。最初に、アルデヒドレダクターゼがおそらく触媒したHMF−アルコールの形成と同時にHMFの迅速な減少が観察された。同時に、HMF−酸(おそらくアルデヒドデヒドロゲナーゼを介して)およびFDCA(HmfHを介して)が蓄積し始めた。HMF−アルコールは、形成直後に、おそらくHMF、HMF−酸およびFDCAへの再酸化を通して消失した。HMF−酸のみが、25時間のタイムスパンの中で、FDCAに(ほとんど)完全に酸化される前に顕著な蓄積を示した。これらの結果により、HMF−酸のFDCAへの酸化がこの無細胞系における律速段階であったことが示唆される。
【0120】
[実施例IV]
[HmfHを発現するP.プチダ(P.putida)S12によるHMFのFDCAへの全細胞変換]
P.プチダ(P.putida)S12の内因性デヒドロゲナーゼが、HMFからHMF−酸を生成する追加の手段となって、HMFのFDCAへの酸化においてHmfHと相乗的に作用することができるので、HMFのFDCAへの全細胞生体内変換法には、酵素法にまさる利点がある場合がある。この可能性を調べるために、静止細胞、増殖細胞、および破壊細胞を、HMFからFDCAの生成について試験した。
【0121】
振とうフラスコ培養を、1lエルレンマイヤーフラスコを用いて、10mg/Lゲンタマイシンを添加したMM+40mMグリセロールおよび2mMグルコース150ml中で行った。細胞を対数増殖期の終わり(OD600およそ4)に収集して、洗浄し、19.4g/l K2HP04、8.15g/l NaH2HP04、40mMグリセロールおよび10mg/lゲンタマイシンを添加したMM中に濃縮した。濃縮細胞懸濁液(CDW1.65gに相当)のアリコート(10ml)を、250mlのエルレンマイヤーフラスコ中でHMFと共にインキュベートし、FDCAの分析のために一定間隔でサンプルを採取した。静止細胞については、グリセロールを再懸濁培地から除去し、破壊細胞は、超音波処理によって得た。
【0122】
これまでの観察のように、HMFがHMF−アルコールおよびHMF酸に迅速に変換された後、HMF酸の減少およびそれと同時のFDCAの形成が起こった。試験したHMFの2つの濃度について、FDCAの生成速度およびHMF酸の減少速度の両方を表IV−aに示す。
【0123】
【表3】
【0124】
表3では、FDCA生成がHmfHを発現するP.プチダ(P.putida)S12の増殖細胞でより効率的であったことが示されている。全体的なFDCAの生成速度、変換効率、およびHMF−酸の減少速度は、増殖細胞のインキュベーションにおいてより高いものであった。さらに、FDCAの生成速度はHMFの初期濃度に依存しており、この系は飽和していないことが示唆された。
【0125】
[実施例V]
[HmfHを発現するP.プチダ(P.putida)S12を用いた、流加回分によるHMFからのFDCAの生成]
FDCAの生成は、HmfHを発現するP.プチダ(P.putida)S12の増殖細胞で最も効率的であることが見出された。そこで、P.プチダ(P.putida)S12 pJT’hmfHの全細胞によるFDCAの生成を実証するために流加回分実験を行った。
【0126】
初期段階では、P.プチダ(P.putida)_pJT’hmfHを、グリセロールの流加を含むMM上で13gCDW/lの細胞密度になるまで培養した。この段階の終わりに、HMF流加を開始し、0.8mmol/l/時の速度でHMFを加えた。グリセロールを一緒に流加しないと、HMF−酸およびFDCAは0.41mmol/l/時の同じ速度で形成された(図5を参照されたい)。次いで、0.21M HMFおよび4Mグリセロールを含有する溶液に流加を取り替え、およそ5ml/時で流加した。このようにグリセロールを一緒に流加すると、HMF−酸の濃度は迅速に低下したが、FDCAの形成はほぼ同じ速度で続いた(図5)。総量44,6mmolのHMFが流加回分に加えられた時点で、この実験を停止した。結果はFDCAへのほとんど完全な変換(すなわち96.5%の効率)を示した。
【0127】
同様な実験で、0.7mmol/l/時の平均速度でグリセロールをHMFと共に流加した。この流加回転培養では、HMF酸中間体の蓄積は観察されず、HMFはすべてFDCAに直接変換された(図6)。
【0128】
[実施例VI]
[発酵ブロスからのFDCAの精製]
pH1.0でのFDCAの溶解度は、水中で約1.5g/lであることがわかった。この特性を有利に利用して、発酵ブロスからFDCA生成物を回収した。遠心分離(9500×gで5分間)により細胞を除去した後に、室温で連続して撹拌しながらpH1になるまで、96%H2S04約10mlを透明ブロス100mlに加え、FDCAを沈殿させた。この沈殿を8228×gで10分間遠心分離して回収し、風乾したペレットを60℃でメタノール中に再融解した。予熱された0.22μmフィルターでろ過して溶解しないデブリを除去した後すぐに、そのろ液を純度分析のためにHPLCに供した。上記手順によって発酵ブロスから回収したFDCAの純度は、約65%であった。FDCAのさらなる精製は当技術分野で公知の方法により行うことができる。
【0129】
[実施例VII]
[HmfHを発現するP.プチダ(P.putida)S12を用いた、流加回分によるHMFからのFDCAの生成]
第2の流加回分によるFDCA生成実験は、P.プチダ(P.putida)S12 pJT’hmfHの全細胞による、FDCAのタイターおよび生産性を向上させるために行った。
【0130】
最初の回分段階では、P.プチダ(P.putida)_pJT’hmfHを、最初のグリセロールが23.2時間の発酵後(OD600およそ8)に枯渇するまで、MM上で培養した。この時点で、バイオマス増加が約0.45gCDW/l/時になる速度でグリセロール流加を開始した。50.7時間の発酵後に、バイオマス増加が約0.045gCDW/l/時になる速度までグリセロール流加を低下させた。123.4時間の発酵後に、バイオマス増加が約0.25gCDW/l/時になる速度に発酵の終了までグリセロール流加を増加させた。この期間にわたり、HMF流加を行った。26.7時間の発酵後に、0.65mmol/時/gCDWの速度でHMF流加を開始し、HMF酸およびFDCAの両方の蓄積をもたらした。33.3時間の発酵後に、HMF流加速度を0.28mmol/時/gCDWに低下させて、72.8時間の発酵後に、0.09mmol/時/gCDWへとさらに低下させ、発酵の117.4時間後に停止した。HMF流加の減少は、発酵槽中のHMF酸濃度の漸減をもたらしたが、FDCA濃度の増加は継続した。HMF酸がもはや検出できなくなった時点で、発酵を停止した。この時点で、188mmolのHMFを発酵槽に加えると、30.8g/l濃度で約182mmolのFDCAの生成(すなわち97%の効率)がもたらされた。図7aに、P.プチダ(P.putida)_pJT’hmfHの流加回分培養中のバイオマス、HMF酸、およびFDCAの形成を示す。図7bに、同じ培養におけるグリセロールおよびHMFの流加速度を示す。
【0131】
[実施例VIII]
[発酵ブロスからのFDCAの精製]
pH0.5でのFDCAの溶解度は、水中で約0.4g/lであることがわかった。沈殿の後に、この濃度が溶液中に残るので、実施例VIIIで得られるタイターの増加は、沈殿による精製に起因する生成物の損失を顕著に減少させる。
【0132】
遠心分離(9500×gで15分間)による細胞の除去後に、透明ブロス500mlを3分間沸騰させてタンパク質を沈殿させ、9500×gで20分間遠心分離した。4℃で連続して撹拌しながらpH0.5になるまで、96%H2S04約50mlを透明上清に加え、FDCAを沈殿させた。この沈殿を8228×gで20分間遠心分離して回収し、水250mlで1回洗浄して、再び8228×gで20分間遠心分離した。得られたペレットを風乾した後、テトラヒドロフラン(THF)約1200ml中に30℃で溶解した。ろ過により溶解しないデブリを除去した後すぐに、発酵ブロス中で最初79%純度のFDCAが高純度(>99%)の乾燥FDCA粉末11.8gになるまで透明THF溶液を50℃で真空蒸発させた。必要があれば、当技術分野で公知の方法を使用して、FDCAの精製をさらに最適にすることができる。
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、オキシドレダクターゼ活性を有するポリペプチドに関し、このポリペプチドをコードする遺伝子を含むポリヌクレオチド配列に関する。本発明は、さらに2,5−フラン−ジカルボン酸(FDCA)の生成に関する。さらに、これらのポリペプチドを生成するのに適した、本発明によるポリヌクレオチドによって形質転換された細胞も、本発明に含まれ、これらの細胞はまた、ヒドロキシメチルフルフラール(HMF)のFDCAへの生体内変換のために使用することもできる。
【0002】
[背景技術]
2,5−フラン−ジカルボン酸(FDCA)は、PETなどのポリエステルの生成においてテレフタレートの生体ベースの代替物になる可能性が大きい。そのため、また他の理由から、FDCAは、バイオマスからの高付加価値化学物質(Top Value−Added Chemicals)に関するDOEレポートにおいてトップ12優先化学物質の1つと選定された(Top Value−Added Chemicals from Biomass、Volume I−Results of screening for potential Candidates from Sugars and Synthesis gas、 Department of Energy(USA)、2004)。この化合物は、酸性条件下でヘキソース糖を加熱することにより生成され得る5−ヒドロキシメチルフルフラール(HMF)の酸化によって得ることができる。DOEレポートの27ページに、FDCAのいくつかの潜在的な有用性が開示されている。これらの有用性には、コハク酸、2,5−ビス(アミノメチル)−テトラヒドロフラン、2,5−ジヒドロキシメチル−テトラヒドロフラン、2,5−ジヒドロキシメチルフラン、および2,5−フランジカルバルデヒドを生成する基質としての役割が含まれる。C6糖の化学的酸化脱水によるFDCAの生成およびFDCAの用途はよく知られており、26ページの表13には技術的な障害も提示されているが、生体内変換(おそらく酵素的変換)については、その状況は不明であった。
【0003】
FDCAの酵素的調製の方法は、国際公開2009/023174号パンフレットに開示されている。この開示では、ヒドロキシメチルフルフラール類がクロロペルオキシドレダクターゼにより酸化され、ヒドロキシメチルフルフラールのC1位置にカルボン酸基を有する生成物、具体的にはホルミルフランカルボン酸すなわちFFCAが過酸化水素により酸化された。その結果を、例えば図1に示す。別の実施形態では、酸化基質の存在下でHMFをオキシドレダクターゼと接触させ、それによってHMFをジホルミルフランまたはホルミルフランカルボン酸の少なくとも1つに酸化する。
【0004】
国際公開2009/023174号パンフレットにある公知の方法の欠点は、この反応が過酸化水素を必要とすること、および形成される生成物がFDCAと2つの混入副産物、ヒドロキシメチルフランカルボン酸(HmFCA)およびホルミルフランカルボン酸(FFCA)との混合物であることである。その結果、HMFからFDCAへの収率は低下し、実質的に精製された形でFDCAを得るために回収ステップを追加する必要がある。
【0005】
データベースEBIにおいて、「バークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)PsJNの第1染色体の完全配列」の中で、グルコース−メタノール−コリンオキシドレダクターゼとの相同性から推定されたUniprot B2T4R9、577個のアミノ酸配列が同定された。
【0006】
データベースEBIにおいて、「バークホルデリア・フィマツム(Burkholderia phymatum)STM815のプラスミド1の完全配列」の中で、グルコース−メタノール−コリンオキシドレダクターゼとの相同性から推定された、Uniprot B2JSZ0、576個のアミノ酸配列が同定された。
【0007】
Deurzen,M.P.Jら、J.Carbohydrate Chemistry 16(3)、299〜309(1997)では、5−ヒドロキシメチルフルフラールのクロロペルオキシダーゼ触媒による酸化が開示されている。この反応では、フラン−2,5−ジカルボキサルデヒド(FDC)への選択性が60〜74%である。副産物は、5−ヒドロキシメチル−2−フランカルボン酸(HFCA)および5−ホルミルフラン−2−カルボン酸(FFCA)であった。
【0008】
[発明の概要]
本発明の目的は、分子状酸素をレドックス反応に使用することができるオキシドレダクターゼを提供することである。別の目的は、幅広い反応スペクトルを有するオキシドレダクターゼを提供することである。別の目的は、高い部位特異性を有するオキシドレダクターゼを提供することである。本発明のさらなる目的は、実質量の副生成物を回避することができる、FDCAを生成するための方法を提供することである。本発明のさらなる目的は、実質量の副生成物を回避することができる、5−ヒドロキシメチル−2−フランカルボン酸(HMF酸)を生成するための方法を提供することである。本発明によってこれらの目的の1つまたは複数が達成される。
【0009】
本発明は、オキシドレダクターゼ活性を有するポリペプチドをコードするポリヌクレオチドを提供する。本発明のポリヌクレオチドは、オキシドレダクターゼ活性、具体的にはHmfHオキシドレダクターゼ活性を有するポリペプチドを通常はコードする。
【0010】
したがって、本発明によれば、配列番号3に示されるアミノ酸配列もしくは配列番号4のヌクレオチド配列によってコードされたアミノ酸配列もしくは配列番号3の配列と45%以上の配列同一性もしくは45%超の配列同一性を有するその変異体ポリペプチドを含む、オキシドレダクターゼ活性を有するポリペプチドが提供される。
【0011】
これらのオキシドレダクターゼポリペプチドは、付加価値化合物FDCAを生成するための、化学的酸化ルートの代わりとなる酵素的代替ルートを提供し、温和な反応条件(30℃、pH7)を可能にして、かつ無用な生成物を少なくする。このオキシドレダクターゼは、真正のオキシダーゼである。すなわち、分子状酸素のみがこの酸化反応に必要であり、高価な補因子を再生する必要がない。この酵素は幅広い反応特異性および部位特異性を示し、アルコール基およびアルデヒド基の両方を、これらの基がフラン骨格のC2位置またはC5位置のいずれにあるかにかかわらず、最終的にカルボン酸へ酸化する。
【0012】
さらに、本発明によれば、本発明のポリヌクレオチド配列を組み込んでいる発現ベクターなどのベクター、および本発明によるポリペプチド、ポリヌクレオチド、またはベクターを含む細胞が提供される。
【0013】
本発明は、オキシドレダクターゼ活性を有するポリペプチドを製造するための方法をさらに提供し、この方法は、前記ポリペプチドの発現を可能にする条件下で本発明の細胞を培養すること、ならびに場合によっては、この発現ポリペプチドおよびこのような方法によって得られるポリペプチドを回収することを含む。
【0014】
本発明の別の実施形態は、強健な全細胞生体触媒を使用して、フルクトースリッチな(リッチにされた)流加からFDCAを生成する統合的方法に関する。
【図面の簡単な説明】
【0015】
【図1】HmfHオキシドレダクターゼによって触媒される酸化の反応スキームを示す図である。
【図2】HmfH発現ベクターpJT’hmfHのプラスミドマップを示す図である。Ptac’、tacプロモーター;rep、pRO1600複製タンパク質をコードする遺伝子;gmR、ゲンタマイシン耐性遺伝子;bla、β‐ラクタマーゼ;pRO1600−ori、P.プチダ(P.putida)のための複製開始点;pUC ori、大腸菌(E.coli)のための複製開始点。
【図3A】C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成を示す概略図である。
【図3B】C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成を示す概略図である。
【図3C】C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成を示す概略図である。
【図3D】C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成を示す概略図である。
【図4】P.プチダ(P.putida)S12 pJT’hmfHの粗細胞抽出物中でのHMFからのFDCA、HMF−アルコール(H−oh)、およびHMF−酸(H−酸)の形成を示す図である。
【図5】P.プチダ(P.putida)_pJT’hmfHの流加回分培養法におけるHMF酸およびFDCAの形成を示す図である。左側矢印は、HMF流加の開始を示す;右側矢印は、流加へのグリセロールの追加を示す。
【図6】P.プチダ(P.putida)_pJT’hmfHの流加回分培養法におけるHMF酸およびFDCAの形成を示す図である。矢印は、HMFとグリセロールを合わせた流加の開始を示す。
【図7A】7aは、実施例VIIIの流加回分発酵におけるHMF酸、FDCA、およびバイオマスの濃度を示す図である。7bは、実施例VIIIの流加回分発酵におけるグリセロールおよびHMFの流加速度を示す図である。
【図7B】7aは、実施例VIIIの流加回分発酵におけるHMF酸、FDCA、およびバイオマスの濃度を示す図である。7bは、実施例VIIIの流加回分発酵におけるグリセロールおよびHMFの流加速度を示す図である。
【0016】
[配列リストの簡単な説明]
配列番号1は、プライマーFN23のDNA配列:5’−CGGAATTCCACATGACAAGGGGAGACCG−3’を示す。下線の配列は、EcoRI制限部位を示す。
配列番号2は、プライマーFN24のDNA配列:5’−CGGAATTCGCTTCGGTCTTCAACTCGGATG−3’を示す。下線の配列は、EcoRI制限部位を示す。
配列番号3は、HmfHのアミノ酸配列を示す。
配列番号4は、hmfHのコード配列を示す。
【0017】
[発明の詳細な説明]
本明細書および添付の特許請求の範囲を通して、単語「含む(comprise)」および「含む(include)」、ならびに「含む(comprises)」、「含んでいる(comprising)」、および「含んでいる(including)」などの語尾変化語は、包含的であると解釈されるべきである。すなわち、これらの単語には、文脈の枠内で、具体的に列挙されない他のエレメントまたは整数も包含され得ることを示す意図がある。
【0018】
本明細書において使用する冠詞「a」および「an」は、この冠詞の文法上の目的語が1つまたは2以上である(すなわち1つまたは少なくとも1つである)ことを指す。例として、「1つのエレメント(an element)」は1つのエレメントまたは2つ以上のエレメントを意味することができる。
【0019】
本明細書における幅広い反応スペクトルとは、オキシドレダクターゼが多くの様々な基質、例えばHMF、HMF酸、HMFアルコール、フルフラール、フルフリルアルコールに対して触媒として作用することができることを意味する。部位特異性とは、オキシドレダクターゼが特定のC原子のみを酸化することを意味する。
【0020】
オキシドレダクターゼポリペプチド
オキシドレダクターゼ活性を有する本発明によるポリペプチドは、配列番号3に示されるアミノ酸配列もしくは配列番号4のヌクレオチド配列によってコードされたアミノ酸配列または配列番号3に示される配列と少なくとも45%の配列同一性を有するその変異体ポリペプチドを含む。
【0021】
一実施形態では、変異体核酸分子は、配列番号4に示されるヌクレオチド配列に少なくとも66%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上の相同性の実質的に相同なヌクレオチド配列を含む、タンパク質をコードするヌクレオチド配列を含む。別の実施形態では、変異体タンパク質は、配列番号3に示されるアミノ酸配列に少なくとも45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上の相同性の実質的に相同なアミノ酸配列を含む。
【0022】
本発明の一実施形態は、(a)配列番号4に示されるヌクレオチド配列;(b)配列番号4の逆相補鎖であるポリヌクレオチドと選択的にハイブリダイズするヌクレオチド配列;(c)配列番号4のヌクレオチド配列と少なくとも66%以上の配列同一性を有するヌクレオチド配列;(d)少なくとも約100ヌクレオチド長である、(a)、(b)、または(c)で定義されるヌクレオチド配列のフラグメント;(e)(a)、(b)、(c)、または(d)のいずれか1つに定義される配列に、遺伝子コードの結果として縮重した配列;(f)(a)、(b)、(c)、(d)、もしくは(e)に定義されるヌクレオチド配列の逆相補鎖であるヌクレオチド配列、またはポリペプチドをコードするヌクレオチド配列を含むポリヌクレオチド、および関連ポリペプチドに対するポリヌクレオチドである。
【0023】
一実施形態は、GC含量が56%以上、58%以上、または58〜65%であり得るこのポリヌクレオチドを含む核酸構築物である。さらなる実施形態は、このポリヌクレオチド配列を組み込んでいるベクター、または核酸構築物である。
【0024】
用語「相同性」、「配列同一性」などは、本明細書では互換的に使用される。本発明の目的のために、2つのアミノ酸配列間または2つの核酸配列間で共通する配列同一性の程度を決定するために、最適な比較を目指した配列アライメントが本明細書において定義される(例えば第2のアミノ酸配列または核酸配列との最適アライメントのために、第1のアミノ酸配列または核酸配列の配列の中にギャップを導入することができる)。このようなアライメントは、比較される配列の全長にわたって実行することができる。あるいは、このアライメントは、より短い比較鎖長、例えば約20、約50、約100以上の核酸/塩基またはアミノ酸にわたって実行することができる。
【0025】
次いで、対応するアミノ酸位置またはヌクレオチド位置にあるアミノ酸残基またはヌクレオチドを比較する。第1の配列中の位置が第2の配列中の対応する位置と同じアミノ酸残基またはヌクレオチドによって占められている場合に、これらの分子はその位置において同一である。配列間で共通する同一性の程度は、2つの配列間の同一率によって通常表現され、配列間で共通する同一位置の数の関数である(すなわち、同一率(%)=同一位置の数/位置(すなわち、重なり位置)の総数×100)。比較される2つの配列は同じまたは実質的に同じ長さであることが好ましい。
【0026】
2つの配列間の相同性を決定するために、数種類の異なるコンピュータプログラムが入手可能であるという事実は、当業者ならば承知しているであろう。例えば、2つの配列間の配列の比較および同一率の決定は、数学アルゴリズムを用いて達成することができる。好ましい実施形態では、2つのアミノ酸配列間の同一率は、Accelrys GCGソフトウェアパッケージ(http://www.accelrys.com/products/gcg/から入手可能)の中のGAPプログラムに組み込まれているNeedlemanおよびWunsch(J.Mol.Biol.(48):444〜453(1970))のアルゴリズムによって、Blossom62マトリックスまたはPAM250マトリックスのいずれか、ならびに16、14、12、10、8、または4のギャップ重みおよび1、2、3、4、5、または6の鎖長重みを使用して決定される。これらすべての異なるパラメータにより、若干異なる結果が得られること、しかし2つの配列の全体的な同一率は、異なるアルゴリズムを用いる場合にもあまり変わらないことは、当業者ならば認識するであろう。
【0027】
さらに別の実施形態では、2つのヌクレオチド配列間の同一率は、Accelrys GCGソフトウェアパッケージ(http://www.accelrys.com/products/gcg/から入手可能)の中のGAPプログラムによって、NWSgapdna.CMPマトリックスならびに40、50、60、70、または80のギャップ重みおよび1、2、3、4、5、または6の鎖長重みを使用して決定される。別の実施形態では、2つのアミノ酸配列間またはヌクレオチド配列間の同一率は、ALIGNプログラム(バージョン2.0)(GenestreamサーバーIGH Montpellier France http://vega.igh.cnrs.fr/bin/align−guess.cgiの配列データを用いてALIGN Queryから入手可能)の中に組み込まれているE.MeyersおよびW.Miller(CABIOS、4:11〜17(1989))のアルゴリズムによって、PAM120重み残基表、12のギャップ長ペナルティ、および4のギャップペナルティを使用して決定される。
【0028】
本発明の核酸配列およびタンパク質配列を「クエリー配列」として使用して、例えば他のファミリーメンバーまたは関連配列を同定するために公開データベースの検索を行うことができる。このような検索は、Altschulら、(1990)J.Mol.Biol.215:403〜10のBLASTnおよびBLASTxのプログラム(バージョン2.0)を使用して行なうことができる。BLASTヌクレオチド検索を、NBLASTプログラム(スコア=100、ワード長=12)で行って、本発明のオキシドレダクターゼ核酸分子に相同なヌクレオチド配列を得ることができる。BLASTタンパク質検索を、BLASTxプログラム(スコア=50、ワード長=3)で行って、本発明のオキシドレダクターゼタンパク質分子に相同なアミノ酸配列を得ることができる。比較のためのギャップアライメントを得るために、Altschulら、(1997)Nucleic Acids Res.25(17):3389〜3402に記載のGapped BLASTを利用することができる。BLASTおよびGapped BLASTプログラムを利用する場合、それぞれのプログラム(例えばBLASTxおよびBLASTn)のデフォルトパラメーターを使用することができる。National Center for Biotechnology Informationのホームページ(http://www.ncbi.nlm.nih.gov/)を参照されたい。
【0029】
本明細書において使用する用語「選択的にハイブリダイズする(selectively hybridizing, hybridizes selctively)」および類似の用語は、少なくとも66%、少なくとも70%、少なくとも75%、少なくとも80%、より好ましくは少なくとも85%、さらにより好ましくは少なくとも90%、好ましくは少なくとも95%、より好ましくは少なくとも98%、またはより好ましくは少なくとも99%の相同性を互いに有するヌクレオチド配列が、通常互いにハイブリダイズした状態を保つハイブリダイゼーションおよび洗浄の条件を記載する意図がある。すなわち、このようなハイブリダイズする配列は、少なくとも45%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65、少なくとも70%、少なくとも75%、少なくとも80%、より好ましくは少なくとも85%、さらにより好ましくは少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも98%、またはより好ましくは少なくとも99%の配列同一性を共有し得る。
【0030】
このようなハイブリダイゼーション条件の好ましい非限定例では、約45℃にて6×塩化ナトリウム/クエン酸ナトリウム(SSC)中でハイブリダイゼーションを行った後、約50℃、好ましくは約55℃、好ましくは約60℃、さらにより好ましくは約65℃にて、1×SSC、0.1%SDS中で1回または複数回洗浄する。
【0031】
高ストリンジェント条件としては、例えば、約68℃にて、5×SSC/5×Denhardt溶液/1.0%SDS中でハイブリダイズし、室温にて、0.2×SSC/0.1%SDS中で洗浄することが挙げられる。あるいは、洗浄は42℃で行ってもよい。
【0032】
当業者であれば、ストリンジェントおよび高ストリンジェントハイブリダイゼーション条件として、いずれの条件を適用すべきかを知っていよう。そのような条件に関するさらなる手引きは、当技術分野では容易に入手可能であり、例えば、Sambrookら、1989、Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Press、N.Y.;およびAusubelら(編者)、1995、Current Protocols in Molecular Biology、(John Wiley & Sons、N.Y.)が挙げられる。
【0033】
もちろん、ポリA配列(mRNAの3’末端ポリ(A)トラクトなど)またはT(またはU)残基の相補的ストレッチに対してのみハイブリダイズするポリヌクレオチドは、ポリ(A)ストレッチまたはその相補体(例えば、実際には任意の二本鎖cDNAクローン)を含有するいかなる核酸分子にもハイブリダイズするため、本発明の核酸の一部に特異的にハイブリダイズさせるために使用される本発明のポリヌクレオチドには含まれない。
【0034】
典型的なアプローチでは、他の生物、例えば細菌から、具体的には微生物マユハキタケ科(Trichomaceae)から、例えばバークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)などのバークホルデリア属(Burkholderia)から構築された遺伝子ライブラリーをスクリーニングすることができる。
【0035】
例えば、相同なオキシドレダクターゼポリヌクレオチドについて、サザンブロット分析により、バークホルデリア(Burkholderia)株をスクリーニングすることができる。本発明の相同なDNA制限フラグメントが検出されたなら、当業者にはよく知られた標準的手法を用いて、適切な菌株から同じ大きさの染色体フラグメントを基に遺伝子ライブラリーを構築することができる。あるいは、微生物が真核生物である場合、オキシドレダクターゼHmfHのmRNA転写物はノーザンハイブリダイゼーションによって同定することができ、この転写物が同定されたなら、真核微生物から単離された全RNAを使用してcDNAライブラリーを作製することができる。
【0036】
相同遺伝子配列は、例えば、本明細書において教示したヌクレオチド配列を基に設計された2つの縮重オリゴヌクレオチドプライマープールを使用してPCRを行うことにより単離することができる。
【0037】
この反応のテンプレートは、本発明のポリヌクレオチドを発現すると知られているかまたは推定される菌株由来の全染色体DNAとすることができる。PCR生成物をサブクローニングし、増幅された配列が新規のオキシドレダクターゼの核酸配列またはその機能的等価物の配列であることを確認するために配列を決定することができる。
【0038】
あるいは、この反応のテンプレートは、本発明のポリヌクレオチドを発現すると知られているかまたは推定される菌株から調製されたmRNAの逆転写により得られたcDNAとすることができる。PCR生成物をサブクローニングし、増幅された配列が新規のオキシドレダクターゼの核酸配列またはその機能的等価物の配列であることを確認するために配列を決定することができる。
【0039】
PCRフラグメントは、その後、様々な公知の方法により、完全長cDNAクローンを単離するために使用することができる。例えば、増幅フラグメントを標識し、バクテリオファージまたはコスミドcDNAライブラリーをスクリーニングするために使用することができる。あるいは、ゲノムライブラリーをスクリーニングするために、この標識フラグメントを使用することができる。
【0040】
PCR技術はまた、他の生物から完全長cDNA配列を単離するために使用することができる。例えば、適切な細胞源または組織源から標準的手法によりRNAを単離することができる。第1鎖の合成をプライミングするための、増幅フラグメントのほとんどの5’末端に対して特異的なオリゴヌクレオチドプライマーを使用して、RNA上で逆転写反応を行うことができる。
【0041】
次いで、得られたRNA/DNAハイブリッドに、標準ターミナルトランスフェラーゼ反応を使用して「尾」を付け(例えばグアニンで)、そのハイブリッドをRNase Hにより消化した後、第2鎖の合成を(例えば、ポリ−Cプライマーにより)プライミングすることができる。こうして、増幅フラグメントの上流のcDNAを容易に単離することができる。有用なクローニング戦略のレビューには、上記のSambrookらおよびAusubelらを参照されたい。
【0042】
本発明の別の態様は、クローニングベクターおよび発現ベクターを含めた、オキシドレダクターゼタンパク質またはその機能的等価物をコードする本発明のポリヌクレオチドを含むベクター、ならびに例えば本発明のポリヌクレオチドの発現が起こる条件下で、適切な宿主細胞においてこのようなベクターを増殖、形質転換または形質移入する方法に関する。本明細書において使用する用語「ベクター」は、別の核酸分子に連結して、その核酸分子を輸送することができる核酸分子を指す。
【0043】
本発明のポリヌクレオチドは、組換え複製可能ベクター、例えばクローニングベクターまたは発現ベクターに組み込むこむことができる。このベクターを使用して、適合性宿主細胞中で核酸を複製してもよい。こうしてさらなる実施形態では、本発明は、本発明のポリヌクレオチドを複製可能ベクターに導入すること、ベクターを適合性宿主細胞に導入すること、およびベクターの複製をもたらす条件下で宿主細胞を増殖させることによって、本発明のポリペプチドを作製する方法を提供する。ベクターは、宿主細胞から回収してもよい。適切な宿主細胞は以下に記載がある。
【0044】
本発明の発現カセットまたはポリヌクレオチドが挿入されているベクターは、組換えDNA手法に便利に供することができる任意のベクターであってもよく、ベクターの選択は、ベクターが導入されることになる宿主細胞に依存することが多い。
【0045】
本発明によるベクターは、自律的に複製するベクター、すなわち、その複製が染色体の複製に依存しない、染色体外の実体として存在するベクター、例えばプラスミドであってもよい。あるいは、このベクターは、宿主細胞に導入された場合、宿主細胞ゲノムに組み込まれ、それを組み込んだ染色体と一緒に複製されるものであってもよい。
【0046】
ベクターのタイプの1つは「プラスミド」であり、追加のDNAセグメントを連結することができる環状二本鎖DNAループを指す。ベクターの別のタイプはウイルスベクターであり、このベクターではウイルスゲノムに追加のDNAセグメントを連結することができる。ある種のベクターは、導入された宿主細胞中で自己複製することができる(例えば、細菌性複製起点を有する細菌ベクターおよびエピソーム性哺乳動物ベクター)。他のベクター(例えば、非エピソーム性哺乳動物ベクター)は、宿主細胞に導入されると同時に宿主細胞のゲノムに組み込まれ、その結果、宿主ゲノムと一緒に複製される。さらに、ある種のベクターは、作動可能に連結された遺伝子の発現を指示することができる。本明細書では、このようなベクターを「発現ベクター」という。一般に、組換えDNA技術において有用な発現ベクターは、プラスミドの形態である場合が多い。プラスミドはベクターの最も一般的に使用される形態であるので、用語「プラスミド」および「ベクター」は、本明細書において互換的に使用することができる。しかしながら、本発明は、機能的に等価な、コスミド、ウイルスベクター(例えば、複製欠損レトロウイルス、アデノウイルス、およびアデノ随伴ウイルス)、およびファージベクターなどの他の形態の発現ベクターを包含することが意図される。
【0047】
本発明によるベクターは、例えばRNAの生成のために、または宿主細胞の形質移入もしくは形質転換のためにインビトロで使用してもよい。
【0048】
本発明のベクターは、2個以上、例えば3個、4個、または5個の本発明のポリヌクレオチドを、例えば過剰発現のために含んでもよい。
【0049】
本発明の組換え発現ベクターは、宿主細胞中で核酸を発現するのに適した形態で本発明の核酸を含む。このことは、組換え発現ベクターが、発現のために使用される宿主細胞に基づき選択され、発現される核酸配列に作動可能に連結された1つまたは複数の調節配列を含むことを意味する。
【0050】
発現ベクターなどのベクターにおいて、「作動可能に連結された」は、目的のヌクレオチド配列が、(例えば、インビトロ転写/翻訳系、またはベクターが宿主細胞に導入される場合は宿主細胞において)ヌクレオチド配列の発現を可能にするように、調節配列に連結されていることを意味することが意図される。すなわち、用語「作動可能に連結された」は、記載の成分がその意図される様式で機能することを可能にする位置関係で並置されることを意味する。コード配列に「作動可能に連結された」プロモーター、エンハンサー、または他の発現調節シグナルなどの調節配列を、コード配列の発現が制御配列に適合する条件下で達成されるように位置させるか、または意図した目的のために協調して機能するように、例えば、転写がプロモーターで始まり、ポリペプチドをコードするDNA配列を通って進行するようにその配列を配置する。
【0051】
用語「調節配列」または「制御配列」は、プロモーター、エンハンサー、および他の発現調節エレメント(例えばポリアデニル化シグナル)を含むことが意図される。このような調節配列は、例えば、Goeddel;Gene Expression Technology:Methods in Enzymology 185、Academic Press、San Diego、CA(1990)に記載されている。
【0052】
用語「調節配列」または「制御配列」には、多くのタイプの宿主細胞においてヌクレオチド配列の構成的な発現を指示する配列、および特定の宿主細胞においてのみヌクレオチド配列の発現を指示する配列(例えば、組織特異的調節配列)が含まれる。
【0053】
したがって、所与の宿主細胞に対するベクターまたは発現構築物は、第1の発明のポリペプチドをコードする配列のコード鎖に関して、5’末端から3’末端に向かって連続した順序で互いに作動的に連結された次のエレメントを含んでもよい。(1)所与の宿主細胞において、ポリペプチドをコードするヌクレオチド配列の転写を指示することができるプロモーター配列;(2)場合によっては、所与の宿主細胞から培地へのポリペプチドの分泌を指示することができるシグナル配列;(3)セロビオヒドロラーゼ活性を有するポリペプチドの成熟型および好ましくは活性型をコードする本発明のDNA配列;好ましくはさらに(4)ポリペプチドをコードするヌクレオチド配列の下流にある、転写を終結させることができる転写終結領域(ターミネーター)。
【0054】
本発明によるヌクレオチド配列の下流に、1つまたは複数の転写終結部位(例えば、ターミネーター)を含有する3’非翻訳領域が存在し得る。ターミネーターの起源はそれほど重要ではない。例えば、ターミネーターは、ポリペプチドをコードするDNA配列にとって本来のものにすることができる。しかしながら、酵母宿主細胞の中では酵母のターミネーターを使用すること、また、糸状菌宿主細胞の中では糸状菌のターミネーターを使用することが好ましい。ターミネーターが宿主細胞(この中でポリペプチドをコードするヌクレオチド配列が発現されることになる)にとって内因性のものであることがより好ましい。転写領域では、翻訳のためのリボソーム結合部位が存在してもよい。構築物によって発現される成熟転写物のコード部分には、始点には翻訳開始のAUG、また翻訳されるポリペプチドの終点には適切な場所に終止コドンが含まれる。
【0055】
さらに、本発明のポリヌクレオチドの発現は、異種の制御領域、例えばプロモーター、分泌リーダー、および/またはターミネーターの領域を選ぶことによって増強し得る。これらの制御領域は、発現の増強、所望の場合には、発現宿主からの目的タンパク質の分泌レベルの増強、および/または本発明のポリペプチド発現の誘導制御の実現を促進させ得る。
【0056】
発現ベクターの設計は、形質変換される宿主細胞の選択、所望タンパク質の発現レベル等の要因に依存し得ることは、当業者ならば認識されよう。発現ベクターなど、本発明のベクターを宿主細胞に導入し、それによって本明細書に記載の核酸がコードするタンパク質またはペプチド(例えば、オキシドレダクターゼタンパク質、オキシドレダクターゼタンパク質の突然変異形態、オキシドレダクターゼタンパク質のフラグメント、変異体、または機能的等価物、融合タンパク質など)を生成させることができる。
【0057】
組換え発現ベクターなど、本発明のベクターは、原核細胞または真核細胞の中でオキシドレダクターゼタンパク質を発現するように設計することができる。例えば、オキシドレダクターゼタンパク質は、大腸菌(E.coli)などの細菌細胞、昆虫細胞(バキュロウイルス発現ベクターを使用して)、糸状菌、酵母細胞、または哺乳類細胞の中で発現することができる。適切な宿主細胞は、Goeddel、Gene Expression Technology:Methods in Enzymology 185、Academic Press、San Diego、 CA(1990)の中にさらに取り上げられている。適切な宿主の代表例は本明細書の以下に記載がある。
【0058】
上記宿主細胞のための適切な培地および条件は、当技術分野で知られている。
【0059】
上述の用語「制御配列」または「調節配列」は、本明細書では、ポリペプチドの発現に必要および/または有利となり得る成分を少なくとも1つ含むと定義される。いずれの制御配列も、ポリペプチドをコードする本発明の核酸配列にとって本来のものであっても、または外来のものであってもよい。このような制御配列としては、プロモーター、リーダー、最適な翻訳開始配列(Kozak、1991、J.Biol.Chem.266:19867〜19870に記載)、分泌シグナル配列、プロペプチド配列、ポリアデニル化配列、転写ターミネーターを挙げ得るが、これらに限定されるものではない。最低限でも、制御配列はプロモーターならびに転写および翻訳の終止シグナルを通常含んでいる。
【0060】
安定に形質転換された微生物とは、1つまたは複数のDNAフラグメントが導入されており、増殖培養において、その導入分子を維持し、複製し、かつ隔離するような微生物である。安定な形質転換は、多重もしくは単一の染色体組込みまたはプラスミドベクターなどの染色体外エレメントによる可能性がある。プラスミドベクターは、特定のDNAフラグメントによってコードされたポリペプチドの発現を指示することができる。
【0061】
発現は、構成的であってもよく、または特定のポリペプチドをコードする機能的に関連するDNAフラグメントを高レベルで転写させることができる誘導性(または抑制性)プロモーターによって調節されてもよい。
【0062】
オキシドレダクターゼの単離
オキシドレダクターゼまたはオキシドレダクターゼを発現するDNA物質は、生物、好ましくはオキシドレダクターゼを発現する微生物から単離し得る。微生物はHMFを利用できることが好ましいが、これは必須ではない。微生物は、カプリアビダス属(Cupriavidus)、バークホルデリア属(Burkholderia)、ブラディリゾビウム属(Bradyhrizobium)、メチロバクテリウム属(Methylobacterium);カプリアビダス・バシレンシス(Cupriavidus basisliensis)、バークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)、ブラディリゾビウム・ジャポニクム(Bradyhrizobium japonicum)、メチロバクテリウム・ラデイオトレランス(Methylobacterium radiotolerans)、カプリアビダス・バシスレンシス(Cupriavidus basisliensis)HMF14、バークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)PsJN、ブラディリゾビウム・ジャポニクム(Bradyhrizobium japonicum)USDA110、メチロバクテリウム・ラデイオトレランス(Methylobacterium radiotolerans)JCM2831からなる群から選択されることが好ましい。
【0063】
本発明において有用な最も好ましいオキシドレダクターゼは、HMFを利用するものであり、カプリアビダス・バシレンシス(Cupriavidus basisliensis)HMF14から単離されたオキシドレダクターゼである。このオキシドレダクターゼは、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約(Budapest Treaty on International Recognition of the Deposits of Microorganisms for the Purpose of Patent Procedures)に従ってドイツ細胞バンク(DSMZ)に寄託されている:カプリアビダス・バシレンシスHMF14=DSM22875、寄託日:2009年8月19日、寄託者TNO、Schoemakerstraat 97、2628VK Delft、Netherlands。
【0064】
したがって、我々はHMF利用細菌であるカプリアビダス・バシレンシス株HMF14を分離し、HMF分解経路に関与する遺伝子を同定した。これらの遺伝子の1つ(本明細書における定義はhmfH)は、579個のアミノ酸からなる62kDaのFAD依存性オキシドレダクターゼをコードしており、このオキシドレダクターゼはフルフリルアルコール、フルフラール、HMF、および5−ヒドロキシメチルフロン酸を酸化することが見出された。これらの分子中のC2およびC5のアルコール基/アルデヒド基は、レダクターゼ活性の存在は有利かもしれないが、レダクターゼをコードする核酸構築物の追加を必要とすることなく、酸化された(図1を参照されたい)。
【0065】
このようにして、本発明は、ポリペプチド、例えばオキシドレダクターゼ(EC1.1+EC1.2の活性)活性を有する酵素をコードするポリヌクレオチドを提供する。本明細書では、酵素はポリペプチドのサブクラスである。
【0066】
酸化反応
本明細書では、酸化反応は、本発明のオキシドレダクターゼおよび1つまたは複数の補酵素(本明細書の下記に記載)の存在下でのフラン化合物とオキシダントとの1回または複数回の反応である。この酸化反応は、生成物を生じる1回の酸化反応ステップ(例えばHMF−酸のFDCAへの酸化)を含むことができる。あるいは、この酸化反応は、2回以上の酸化反応ステップを含むことができ、各ステップで中間体が生まれ、最後の中間体が最終生成物である(例えばHMFのFDCAへの酸化)。酸化反応の例を図1に示す。
【0067】
1つの酸化反応は2,5−フランジカルボン酸(FDCA)の生成であり、この反応では、オキシドレダクターゼ触媒および1つまたは複数の補酵素の存在下でのオキシダントとの反応によって、FDCAの1つまたは複数のフラン前駆体がFDCAに変換される。ここで、オキシドレダクターゼ触媒は本発明によるポリペプチドを含む。FDCAのフラン前駆体は、5−ヒドロキシメチルフルフラール(HMF)、2,5−ジヒドロキシメチルフラン(HMFアルコール)、および5−ヒドロキシメチル−2−フランカルボン酸(HMF酸)からなる群から選択することができ、好ましくは、フラン前駆体はHMFである。HMFは、従来の方法で酸の存在下で加熱することにより1つまたは複数のヘキソース糖から得てもよい。ヘキソース糖はバイオマスから得てもよい。この酸化反応はまた、5−ヒドロキシメチル−2−フランカルボン酸(HMF酸)を生成するための方法であってもよく、この反応では、オキシドレダクターゼ触媒および1つまたは複数の補酵素の存在下でのオキシダントとの反応によって、HMF酸の1つまたは複数のフラン前駆体がHMF酸に変換される。ここで、オキシドレダクターゼ触媒は本発明によるポリペプチドを含む。一実施形態では、HMF酸のフラン前駆体は、5−ヒドロキシメチルフルフラール(HMF)および2,5−ジヒドロキシメチルフラン(HMFアルコール)から選択される群から選択される。他の酸化方法も可能である。
【0068】
酸化反応は、比較的温和な温度、すなわち10〜80℃、より好ましくは20〜45℃、最も好ましくはおよそ25〜40℃にて行うことが好ましい。反応中のpHは、好ましくはpH3〜8、より好ましくはおよそpH7である。反応時間は、大気中酸素または純酸素を使用して、6〜18時間であり、酵素はより長時間活性であることが好ましい。
【0069】
反応器は任意の適切な(通気型)バイオリアクターとすることができる。反応器は、バッチで、連続的に、または好ましくは流加回分形式で操作することができる。
【0070】
FDCA、HMF−酸などのような酸化生成物は、冷却/再結晶および結晶化した酸化生成物、例えば結晶化FDCAの分離によって反応混合物から回収することができる。しかしながら、当技術分野で知られているように、酸沈殿および溶媒抽出などの他の回収方法が適切であるが、これらに限定されるものではない。
【0071】
場合によっては、反応は補酵素の存在下で行われる。補酵素は、ニコチンアミドアデニンジヌクレオチド(NAD+)および/またはフラビンアデニンジヌクレオチド(FAD)および/またはピロロキノリンキノロン(PQQ)であってもよい。デヒドロゲナーゼ活性、例えばその細胞抽出物において酸化反応が起こるかまたは存在する細胞の中に見出される内因性の脱水素活性との相乗効果が本発明の酸化反応で見出された。モノマーの1つが、記載のFDCAである、1つまたは複数のモノマーからポリマーを生成するための方法。
【0072】
[オキシダント]
本発明による反応中のオキシダントは、任意のオキシダント、好ましくは酸素であってもよい。酸素の最も経済的な供給源は空気である。このことは、空気が大気から容易に得られ、無料、無毒性、かつ反応後に除去する必要もない点で有利である。あるいは、分子状酸素放出系を利用してもよい。酸素発生系は、当技術分野で開示された様々な酸素発生系から原則として選択し得る。例えば、反応混合物の中にすでに存在するカタラーゼ酵素を利用して、過酸化水素から酸素を発生させ得る。
【0073】
[フラン化合物]
本明細書では、フラン化合物は、2,5−フラン−ジカルボン酸またはその前駆体に酸化され得るフラン基を有する任意の化合物であると理解される。好ましいフラン化合物としては、ヒドロキシメチルフルフラール(HMF)、ヒドロキシメチルフランカルボン酸(HMF酸)、2,5−ジヒドロキシメチルフラン(HMFアルコール)が挙げられる。フラン環または任意もしくはその置換可能な側鎖は、例えばOH、C1〜C10アルキル、アルキル、アリル、アリール、またはRO−エーテル部分、および環式基により、フラン環の任意の可能な位置で置換されていてもよい。
【0074】
[オキシドレダクターゼの発現]
酵素の発現のために利用される厳密な手法に関係なく、当技術分野で公知の方法を用いて、これらの酵素をコードする遺伝子を別の宿主細胞に導入して、その発現を移行させることができると考えられる。本明細書において定義する遺伝エレメントには、関連酵素を発現させるかまたはその発現を調節するタンパク質、特に酵素、アポタンパク質、またはアンチセンスRNAなどの生成物のための発現可能コード配列を有する核酸(一般にはDNAまたはRNA)が含まれる。発現されたタンパク質は、酵素として機能するか、酵素活性を抑制もしくは抑制解除するか、または酵素の発現を制御することができる。これらの発現可能配列をコードする組換えDNAは、染色体にある(例えば相同遺伝子組換えによって宿主細胞染色体へ組み込まれている)か、または染色体外にある(例えば、1つまたは複数のプラスミド、コスミド、および自己複製可能な他のベクターによって保持されている)ことができる。本発明による、宿主細胞を形質転換するために利用される組換えDNAには、構造遺伝子および転写因子に加えて、タンパク質、アポタンパク質、またはアンチセンスRNAのためのコード配列の発現または抑制除去を制御するように作用する、プロモーター、リプレッサー、およびエンハンサーを含む発現調節配列が含まれ得ることが理解される。例えば、このような制御配列は、野生型の宿主細胞に挿入されて、宿主細胞ゲノム中にすでにコードされていた、選択された酵素の過剰発現を促進することができる。あるいは、その代わりに染色体外にコードされた酵素の合成を制御するために使用することができる。
【0075】
組換えDNAは、宿主細胞への遺伝エレメントの移入を媒介する、プラスミド、コスミド、ファージ、酵母人工染色体、または他のベクターを含むが、これらに限定されない、任意の手段によって宿主細胞へ導入することができる。これらのベクターは、ベクターの複製およびベクターが保持する遺伝エレメントを制御するシス作用性の制御エレメントと一緒に、複製起点を含むことができる。選択可能マーカーは、遺伝エレメントが導入された宿主細胞の同定を助けるためにベクター上に存在させることができる。
【0076】
遺伝エレメントを宿主細胞に導入する手段(例えばクローニング)は、当業者にはよく知られている。染色体外の多重コピープラスミドベクターを利用して、本発明による遺伝エレメントを挿入することができる。宿主細胞への遺伝エレメントのプラスミド運搬導入には、制限酵素によりプラスミドベクターを最初に切断した後、プラスミドと本発明による標的酵素類をコードする遺伝エレメントとを連結することが含まれる。この連結組換えプラスミドの再環状化と同時に、感染(例えばλファージへのパッケージング)またはプラスミド移入のための他の手法(例えば電気穿孔、ミクロ注入など)が宿主細胞へプラスミドを移入するために利用される。宿主細胞への遺伝エレメントの挿入に適したプラスミドは、当業者にはよく知られている。
【0077】
他の遺伝子クローニング法には、染色体への遺伝物質の直接的な組込みが含まれるが、これに限定されるものではない。これは、宿主染色体の相同DNA配列によって挟まれた非複製プラスミド上に本明細書に記載の遺伝エレメントをクローニングすること、および前記組換えプラスミドを宿主に形質転換すると同時に、DNA組換えによって染色体へ遺伝エレメントが導入され得ることを含む様々な手段によって実現できる。組込みDNAフラグメントが抗生物質耐性などの選択可能マーカーを含有している場合に、このような組換え株を回収することができる。あるいは、遺伝エレメントは、非複製プラスミドを使用することなく、宿主細胞の染色体へ直接導入することができる。これは、宿主染色体の相同DNA配列も含有する、本発明による遺伝エレメントのDNAフラグメントを合成的に生成することによって達成できる。これらの合成DNAフラグメントが選択可能マーカーをさらに含有する場合にも、遺伝エレメントを宿主染色体に挿入することができる。
【0078】
[宿主細胞]
本発明の別の実施形態は、本発明によるポリペプチド、ポリヌクレオチド、核酸構築物、またはベクターを含む細胞である。宿主細胞は、ポリペプチド、ポリヌクレオチド、核酸構築物、またはベクターが適切に発現され得る細胞である。
【0079】
オキシドレダクターゼは、宿主細胞中で好都合に発現され得る。本発明による宿主細胞は任意の宿主細胞でよい。この細胞は、原核細胞、真核細胞、植物細胞、または動物細胞であってもよい。このような細胞では、1つまたは複数の遺伝子が、完全もしくは部分的に、欠落、ノックアウト、または破壊されていることがあり、場合によっては、1つまたは複数の遺伝子はプロテアーゼをコードしている。一実施形態によれば、本発明による宿主細胞は真核生物の宿主細胞である。真核細胞は、哺乳類、昆虫、植物、菌類、または藻類の細胞であることが好ましい。好ましい哺乳類細胞としては、例えばチャイニーズハムスター卵巣(CHO)細胞、COS細胞、293細胞、PerC6細胞、およびハイブリドーマが挙げられる。好ましい昆虫細胞としては、例えばSf9細胞およびSf21細胞ならびにそれらの派生細胞が挙げられる。より好ましくは、真核細胞は、菌類細胞、すなわちカンジダ属(Candida)、ハンゼヌラ属(Hansenula)、クリベロミセス属(Kluyveromyces)、ピチア属(Pichia)、サッカロミセス属(Saccharomyces)、シゾサッカロミセス属(Schizosaccharomyces)、またはヤロウイア属(Yarrowia)の菌株等の酵母細胞である。より好ましくは、クリベロミセス・ラクチス(Kluyveromyces lactis)、S.セルビシエ(S.cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ヤロウイア・リポリティカ(Yarrowia lipolytica)、およびピチア・パストリス(Pichia pastoris)由来の細胞または糸状菌細胞。最も好ましくは、真核細胞は糸状菌細胞である。
【0080】
「糸状菌」は、真菌亜門および卵菌亜門のすべての糸状形態を含む(Hawksworthら、Ainsworth and Bisby’s Dictionary of The Fungi、第8版、1995、CAB International、University Press、Cambridge、UKによる定義)。糸状菌は、キチン、セルロース、グルカン、キトサン、マンナン、および他の複合多糖で構成される菌糸壁を特徴とする。栄養成長は菌糸の伸長によるものであり、炭素異化反応は偏性好気性である。糸状菌株としては、アクレモニウム属(Acremonium)、アガリクス属(Agaricus)、アスペルギルス属(Aspergillus)、アウレオバシディウム属(Aureobasidium)、クリソスポリウム属(Chrysosporium)、コプリナス属(Coprinus)、クリプトコックス属(Cryptococcus)、フィリバシジウム属(Filibasidium)、フザリウム属(Fusarium)、フミコラ属(Humicola)、マグナポルテ属(Magnaporthe)、ムコール属(Mucor)、ミセリオフトーラ属(Myceliophthora)、ネオカリマスティクス属(Neocallimastix)、ニューロスポラ属(Neurospora)、ペシロミセス属(Paecilomyces)、ペニシリウム属(Penicillium)、ピロミセス属(Piromyces)、パネロカエテ属(Panerochaete)、プロイロータス属(Pleurotus)、シゾフィラム属(Schizophyllum)、タラロミセス属(Talaromyces)、サーモアスカス属(Thermoascus)、チエラビア属(Thielavia)、トリポクラジウム属(Tolypocladium)、およびトリコデルマ属(Trichoderma)の菌株が挙げられるが、これらに限定されるものではない。
【0081】
好ましい糸状菌細胞は、アスペルギルス属(Aspergillus)、クリソスポリウム属(Chrysosporium)、ペニシリウム属(Penicillium)、タラロミセス属(Talaromyces)、またはトリコデルマ属(Trichoderma)の菌種、最も好ましくは、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ソジェ(Aspergillus sojae)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、タラロミセス・エメルソニ(Talaromyces emersonii)、アスペルギルス・オリゼ(Aspergillus oryzae)、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)、トリコデルマ・リーゼイ(Trichoderma reesei)、またはペニシリウム・クリゾゲヌム(Penicillium chrysogenum)の菌種に属する。本発明による宿主細胞がアスペルギルス属(Aspergillus)宿主細胞である場合、宿主細胞にはCBS513.88、CBS124.903、またはその派生細胞が好ましい。
【0082】
別の実施形態によれば、本発明による宿主細胞は原核細胞である。好ましくは、原核生物の宿主細胞は細菌細胞である。用語「細菌細胞」は、グラム陰性菌およびグラム陽性菌の両方を含む。適切な細菌は、例えば大腸菌属(Escherichia)、アナベーナ属(Anabaena)、コーロバクター属(Caulobacter)、グルコノバクター属(Gluconobacter)、ロドバクター属(Rhodobacter)、シュードモナス属(Pseudomonas)、パラコッカス属(Paracoccus)、バチルス属(Bacillus)、ブレビバクテリウム属(Brevibacterium)、コリネバクテリウム属(Corynebacterium)、リゾビウム属(Rhizobium)(シノリゾビウム属(Sinorhizobium))、フラボバクテリウム属(Flavobacterium)、クレブシェラ属(Klebsiella)、エンテロバクター属(Enterobacter)、ラクトバチルス属(Lactobacillus)、ラクトコッカス属(Lactococcus)、メチロバクテリウム属(Methylobacterium)、スタフィロコッカス属(Staphylococcus)、またはストレプトミセス属(Streptomyces)から選択することができる。好ましくは、細菌細胞は、B.スブチリス(B.subtilis)、B.アミロリケファシエンス(B.amyloliquefaciens)、B.リケニフォルミス(B.licheniformis)、B.プンチス(B.puntis)、B.メガテリウム(B.megaterium)、B.ハロデュランス(B.halodurans)、B.プミルス(B.pumilus)、グルコノバクター・オキシダンス(Gluconobacter oxydans)、コーロバクター・クレセンタス(Caulobacter crescentus)CB15、メチロバクテリウム・エキストロクエンス(Methylobacterium extorquens)、ロドバクター・スフェロイデス(Rhodobacter sphaeroides)、シュードモナス・ゼアキサンチニファシエンス(Pseudomonas zeaxanthinifaciens)、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・プチダ(Pseudomonas putida)S12、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)、大腸菌(E.coli)、C.グルタミクム(C.glutamicum)、スタフィロコッカス・カルノーサス(Staphylococcus carnosus)、ストレプトミセス・リビダンス(Streptomyces lividans)、シノリゾビウム・メリロティ(Sinorhizobium melioti)、およびリゾビウム・ラジオバクター(Rhizobium radiobacter)からなる群から選択される。
【0083】
糸状菌のいくつかの菌株、例えばアスペルギルス・ニガー(Aspergillus niger)CBS513.88、アスペルギルス・オリゼ(Aspergillus oryzae)ATCC20423、IFO4177、ATCC1011、ATCC9576、ATCC14488〜14491、ATCC11601、ATCC12892、P.クリソゲヌム(chrysogenum)CBS455.95、ペニシリウム・シトリヌム(Penicillium citrinum)ATCC38065、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)P2、タラロミセス・エメルソニ(Talaromyces emersonii)CBS124.902、アクレモニウム・クリソゲヌム(Acremonium chrysogenum)ATCC36225またはATCC48272、トリコデルマ・リーゼイ(Trichoderma reesei)ATCC26921またはATCC56765またはATCC26921、アスペルギルス・ソジェ(Aspergillus sojae)ATCC11906、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)ATCC44006は、米国微生物系統保存機関(American Type Culture Collection(ATCC))、ドイツ微生物細胞培養コレクション(Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ))、オランダ微生物株保存センター(Centraalbureau Voor Schimmelcultures (CBS))、およびAgricultural Research Service Patent Culture Collection,Northern Regional Research Center(NRRL)などのいくつかの微生物株保存機関において公的に容易に入手できる。それらの派生株もまた使用し得る。
【0084】
別の実施形態によれば、本発明による宿主細胞は原核細胞である。好ましくは、原核宿主細胞は細菌細胞である。用語「細菌細胞」は、グラム陰性菌およびグラム陽性菌の両方を含む。適切な細菌は、例えば大腸菌属(Escherichia)、アナベーナ属(Anabaena)、コーロバクター属(Caulobacter)、グルコノバクター属(Gluconobacter)、ロドバクター属(Rhodobacter)、シュードモナス属(Pseudomonas)、パラコッカス属(Paracoccus)、バチルス属(Bacillus)、ブレビバクテリウム属(Brevibacterium)、コリネバクテリウム属(Corynebacterium)、リゾビウム属(Rhizobium)(シノリゾビウム属(Sinorhizobium))、フラボバクテリウム属(Flavobacterium)、クレブシェラ属(Klebsiella)、エンテロバクター属(Enterobacter)、ラクトバチルス属(Lactobacillus)、ラクトコッカス属(Lactococcus)、メチロバクテリウム属(Methylobacterium)、スタフィロコッカス属(Staphylococcus)、またはストレプトミセス属(Streptomyces)から選択することができる。好ましくは、細菌細胞は、B.スブチリス(B.subtilis)、B.アミロリケファシエンス(B.amyloliquefaciens)、B.リケニフォルミス(B.licheniformis)、B.プンチス(B.puntis)、B.メガテリウム(B.megaterium)、B.ハロデュランス(B.halodurans)、B.プミルス(B.pumilus)、G.オキシダンス(G.oxydans)、コーロバクター・クレセンタス(Caulobacter crescentus)CB15、メチロバクテリウム・エキストロクエンス(Methylobacterium extorquens)、ロドバクター・スフェロイデス(Rhodobacter sphaeroides)、シュードモナス・プチダ(Pseudomonas putida)、パラコッカス・ゼアキサンチニファシエンス(Paracoccus zeaxanthinifaciens)、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)、大腸菌(E.coli)、C.グルタミクム(C.glutamicum)、スタフィロコッカス・カルノーサス(Staphylococcus carnosus)、ストレプトミセス・リビダンス(Streptomyces lividans)、シノリゾビウム・メリロティ(Sinorhizobium melioti)、およびリゾビウム・ラジオバクター(Rhizobium radiobacter)からなる群から選択される。
【0085】
本発明による宿主細胞中で生成される化合物の特定用途のために、その用途に応じて宿主細胞の選択がなされ得る。例えば、本発明による宿主細胞中で生成される化合物が、食品用途で使用されることになる場合、宿主細胞はサッカロミセス・セレビシア(Saccharomyces cerevisiae)などの食用生物から選択してもよい。特定用途には、食品、(動物)飼料、医薬、作物保護などの農業、および/またはパーソナルケアの用途が含まれるが、これらに限定されものではない。
【0086】
さらに、本発明は、オキシドレダクターゼ活性を有するポリペプチドを製造するための方法であって、前記ポリペプチドの発現を可能にする条件下で、本発明による細胞を培養すること、および場合によっては、発現されたポリペプチドを回収することを含む方法に関し、その方法によって得られるポリペプチドに関する。
【0087】
[流加原料]
フルクタンが本来豊富な農業作物(例えばトビナンブールまたはチコリルート)は、従来の加水分解および熱化学(の組合せ)処理によってHMFに富んだ流加原料に変換し得る。フルクトースからHMFを生成する技術は確立しており、確固としたものである。さらにグルコースに富んだ流加原料も利用することができるが、HMFの熱化学的形成はフルクトースからの方がより効率的に起こる。したがって、グルコースイソメラーゼを使用してグルコースをフルクトースに変換するために、追加の酵素ステップを含むことができる。後者のプロセスは、加水分解されたデンプンから高フルクトースコーンシロップ(HFCS)を生成するために食品産業において確立されている。食品用途との競合を回避するならば、リグノセルロース水解物が、HMF/FDCAを生成するための好ましい流加原料になるであろう。
【0088】
[生体内変換]
HMFのFDCAへの生体内変換については、本明細書において前述したカプリアビダス・バシレンシス(Cupriavidus basilensis)HMFオキシドレダクターゼを発現する強健な全細胞生体触媒(遊離細胞または固定化細胞)を使用する。このプロセスでは、全細胞生体触媒は酵素触媒にまさるいくつかの利点を有している。すなわち、HMFオキシドレダクターゼが高反応性基質による化学的不活性化から保護され、かつHMFから(おそらく)対応する一塩基酸に導き、次いでこの一塩基酸がHmfHによって二価酸に変換される最初の2つの酸化ステップにおいて、宿主固有のデヒドロゲナーゼが、HMFオキシドレダクターゼを助けることができる。補因子再生を確実にするために、追加の処置を要求し得ることが好ましい。全細胞生体触媒は、HMF流加を最小限で処理できるものであるべきである。すなわち、低pH、高温、および流加原料の加水分解/熱化学による変換中に生成される有毒化合物(その中に基質がある)に対する耐性があることが好ましい。シュードモナス・プチダ(Pseudomonas putida)S12は、種々の化学的ストレス要因に対する耐性、比較的広いpH領域、およびHMFオキシドレダクターゼを助けてHMFのFDCAへのより効率的な生体内変換を生じさせる固有な(indiginous)デヒドロゲナーゼの存在の観点から、適切な宿主生物としての資格があると見なし得る。
【0089】
全細胞生体触媒の代わりに、全細胞生体触媒と同様に、細胞溶解物、精製酵素、または単独もしくは酵素混合物としての固定化(immobolized)酵素を使用してもよい。
【0090】
[生成物およびバイオマスの回収]
生体内変換の後、細胞は確立した方法によってブロスから分離し、再使用してもよい。FDCAは酸沈殿によって無細胞ブロスから回収し、適切な有機溶媒中に高温で再溶解してもよい。溶解後に、FDCAは、酸沈殿および溶媒抽出、または当技術分野で公知の他の精製法によって、所望の場合には二価酸の形態にて高純度で回収してもよい。
【0091】
[FDCAの用途]
FDCAはポリエステルの製造において、テレフタレート(terephtalate)の代わりとして使用し得る。FDCAはまた、極めて多様な高価値化合物のための基質として使用し得る。例えば、コハク酸、2,5−ビス(アミノメチル)−テトラヒドロフラン、2,5−ジヒドロキシメチル−テトラヒドロフラン、2,5−ジヒドロキシメチルフラン、および2,5−フランジカルバルデヒドを製造するための公知の基質である。FDCAは、コーティングの製造において、例えばアルキド樹脂および熱可塑性コーティングの中に使用してもよい。FDCAはまた、バイオ燃料におけるキシレン等価物として、また溶媒として使用してもよい。
【0092】
FDCAはエステル化してもよく、このエステルは可塑剤として使用し得る。FDCAはそのジオールに変換してもよく、このジオールはPET様ポリエステルおよびポリウレタンの中に使用し得る。さらに、FDCAはそのジアミンに変換してもよく、このジアミンは連鎖延長剤として使用し得る。またこのジアミンはジイソシアネートに変換してもよく、このジイソシアネートはポリウレタンの製造に使用することができる。本発明による方法によって、FDCAおよびFDCAから製造される生成物は、リグノセルロースバイオマスを含むバイオマスから生体内変換を介して作ることができる。
【0093】
[実施例]
[一般的方法]
菌株およびプラスミド カプリアビダス・バシレンシス(Cupriavidus basilensis)HMF14は、DSMZ:カプリアビダス・バシレンシス(Cupriavidus basilensis)HMF14=DSM22875、寄託日:2009年8月19日に寄託されており、唯一の炭素源としてフランを使用することができる土壌分離菌である。シュードモナス・プチダ(Pseudomonas putida)S12(ATCC700801)をHMFオキシドレダクターゼ発現のための宿主として使用した。大腸菌(Escherichia coli)DH5a(Invitrogen)を一般のクローニング目的に使用した。pUCP22由来の大腸菌(E.coli)−P.プチダ(P.putida)シャトルプラスミドpJT’mcs(未公表)を、構成的tacプロモーターの制御下でHMFオキシドレダクターゼを発現させるために使用した。大腸菌(E.coli)における複製については、pUC複製起点を使用する;P.プチダ(P.putida)における複製については、pRO1600複製起点を使用する。hmfH遺伝子の発現は構成的tacプロモーターにより促進される。大腸菌(E.coli)の抗生物質選択(アンピシリン抵抗性)のために、β―ラクタマーゼマーカー遺伝子(bla)を使用する。P.プチダ(P.putida)の抗生物質選択のために、ゲンタマイシンアセチルトランスフェラーゼマーカー遺伝子(gmR)を使用する。
【0094】
HmfH発現ベクターpJT’hmfHのプラスミドマップを図2に示す。Ptac’、tacプロモーター;rep、広範囲の宿主領域複製起点;gmR、ゲンタマイシン耐性遺伝子;bla、β‐ラクタマーゼ;pUC ori、大腸菌(E.coli)のための複製起点。
【0095】
培地および培養条件 ミネラル塩培地(MM)を特定培地として使用した。MMには次のものが含有されていた(脱イオン水1リットル当たり):指定の炭素源と共に添加された、K2HPO4 3.88g、NaH2HPO4 1.63g、(NH4)2SO4 2.0g、MgCl2・6H2O 0.1g、EDTA 10mg、ZnSO4・7H2O 2mg、CaCl2・2H2O 1mg、FeSO4・7H2O 5mg、Na2MoO4・2H2O 0.2mg、CuSO4・5H2O 0.2mg、CoCl2・6H2O 0.4mg、およびMnCl2・2H2O 1mg。ルリアブロス(L−ブロス:10g/lバクトトリプトン(Bacto trypton)(ディフコ(Difco)社)、5g/l酵母抽出物(ディフコ社)、5g/l NaCl)を、P.プチダ(P.putida)S12および派生菌株、C.バシレンシス(C.basilensis)HMF14および大腸菌(E.coli)DH5aおよび派生菌を調製するための完全培地として使用した。固体のL−ブロスは2%(w/v)寒天(ディフコ社)を使用して凝固させた。
【0096】
流加回分培養実験では、最初の回分段階は、次の組成を有する適合ミネラル塩培地1l中で行なわれた:K2HPO4 3.88g、NaH2HPO4 1.63g、(NH4)2SO4 2.0g、MgCl2・6H2O 0.2g、EDTA 20mg、ZnSO4・7H2O 4mg、CaCl2・2H2O 2mg、FeSO4・7H2O 10mg、Na2MoO4・2H2O 0.4mg、CuSO4・5H2O 0.4mg、CoCl2・6H2O 0.8mg、MnCl2・2H2O 2mg、10mg/Lゲンタマイシン、および100mMグリセロール。最初のグリセロールが枯渇した後、流加を開始し、培養物中に制限基質としてグリセロールを維持しながら、最大の増殖が得られるように制御した。流加溶液には、(1l当たり):グリセロール368.4gおよび10g/l MgCl2・6H2O、ならびに12.6g/l HMFが含有されていた。
【0097】
抗生物質:アンピシリン(amp)を大腸菌(E.coli)に対して100μg/ml添加した。ゲンタマイシン(gm)をP.プチダ(P.putida)S12に対して、ルリアブロス中に30μg/ml、ミネラル塩培地中に10μg/ml添加した。抗生物質はシグマアルドリッチ(Sigma−Aldrich)社から購入した。
【0098】
培養:P.プチダ(P.putida)およびC.バシレンシス(C.basilensis)は30℃にて培養した。大腸菌(E.coli)は37℃にて培養した。MM上での振とうフラスコ実験は、水平振とうインキュベーター中でボストンボトル(Boston bottle)(Alltech applied sciences BV;Breda,The Netherlands)を用いて行った。L−ブロス上の振とうフラスコ実験は、水平振とうインキュベーター中で、綿栓を詰めたエルレンマイヤーフラスコを用いて行った。流加回分実験は、BioFlo110コントローラーを使用して、1l発酵槽(ニュー・ブランズウィック・サイエンティフィック(New Brunswick Scientific)社)中で行った。最初の回分発酵は、40mMグリセロールおよび2mMグルコースを添加したMM100ml中での一晩前培養物から得られた洗浄細胞で開始した。最初の撹拌速度を200rpmに設定し、M+W Istruments D−5111マスフローコントローラを使用して空気を1lmin−1でヘッドスペースに供給した。InProモデル6900プローブ(Mettler Toledo BV;Tiel、The Netherlands)を使用して溶存酸素圧(DO)を連続的にモニターし、最大1000rpmまで撹拌速度を自動調整することによって30%空気飽和を維持した。最大撹拌速度に達したとき、0.2lmin−1の流量で空気を精製酸素と交換し、最大撹拌速度を800rpmに設定した。最初の回分段階の間は、25%NH4OHの自動添加によってpHを7.0に維持し、流加段階の間は、10mM NaOHの自動添加によってpHを一定に保った。温度は30℃に保った。
【0099】
アッセイおよび分析法:細菌培養の細胞乾燥重量(CDW)は、平底96穴マイクロプレート(Greiner)を使用し、Biowave Cell Density Meter(WPA社)またはμQuant MQX200汎用マイクロプレート分光光度計(Biotek)を用いて、600nm(OD600)での光学密度を測定することにより決定した。OD600 1.0は、P.プチダ(P.putida)については、0.56g CDW/L(Biowave)または1.4g CDW/L(μQuant)に相当する。
【0100】
HPLC分析:FDCA、HMF、HMF−アルコール、およびHMF−酸は、230nmに設定されたダイオードアレー検出器を使用して、RP−HPLC(Agilent 1100システム)により分析した。使用したカラムは、Zorbax Eclipse XDB−C8(孔径80Å、表面積180m2/g、Agilent)であり、25℃で操作した。溶離液として、1%アセトニトリルと共に20mM KH2P04(pH2またはpH6)中のアセトニトリルにグラディエントをかけて使用し、流量1.2ml/分で、アセトニトリルを3.5分で0〜5%まで、2.5分で5〜40%まで増加させた。
【0101】
細胞抽出物の調製:HMFオキシドレダクターゼを発現する、野生型C.バシレンシス(C.basilensis)HMF14またはP.プチダ(P.putida)S12の形質転換細胞の細胞抽出物は、12mMコハク酸(C.バシレンシス(C.basilensis)HMF14、OD600およそ1.5)または20mMグルコース(P.プチダ(P.putida)S12、OD600およそ4)のいずれかを添加したMMを使用した、指数増殖期後半の培養物50mlから調製した。培養物を遠心分離によって採取し、アッセイ緩衝液3ml中に再懸濁した。細胞は、Branson sonifier(パルスモードでのマイクロチップ、出力設定3、およびパーセントデューティー(duty)サイクル設定40%;3サイクルの超音波処理:45秒パルスおよび15秒休止)またはSonics Vibra−Cell(Sonics&Materials、USA)(5mm先細マイクロチップ;パルスモード設定1分(0,5秒パルス、2秒休止)のいずれかを使用して、超音波処理により破壊した。超音波処理の後、8228×gにて4℃で3分間遠心分離してデブリを除去した。上清は、PD10ゲル濾過カラム(GE healthcare)を使用して脱塩し、HMFオキシドレダクターゼアッセイのための細胞抽出物として使用した。タンパク質濃度はBradford試薬(シグマアルドリッチ社)を使用して測定した。
【0102】
HMFオキシドレダクターゼアッセイ:HMFオキシドレダクターゼアッセイは、C.バシレンシス(C.basilensis)HMF14 HMFオキシドレダクターゼを発現する、野生型C.バシレンシス(C.basilensis)HMF14またはP.プチダ(P.putida)S12の形質転換細胞の細胞抽出物で行った。ネガティブ対照として、hmfH遺伝子にトランスポゾン挿入がある、野生型P.プチダ(P.putida)S12またはC.バシレンシス(C.basilensis)HMF14変異体を使用した。細胞抽出物を、酸素負荷条件下で30℃にてフルフラール、フルフリルアルコール、HMF、またはHMF−酸と共にインキュベートした。反応混合物は、細胞抽出液1ml、酸素飽和MM976μl、2mMフラビンアデニンジヌクレオチド(FAD)溶液20μl、および基質(フルフラール、フルフリルアルコール、HMF、またはHMF酸)の0.5Mストック4μlを含有した。サンプルを設定間隔で抜き取り、最終濃度1MまでHClを添加して反応を直ちに停止させた。サンプル中の基質および生成物の濃度はHPLCによって測定した。酸素欠乏対照として、連続窒素ガス流によって反応混合物の様々な成分を酸素欠乏状態にし、同反応混合物を、ゴム栓付きヘッドスペースバイアル中で窒素ガス下インキュベートした。
【0103】
試薬:分析用標準のFDCAは、Immunosource B.V.(Halle−Zoersel、Belgium)から購入した。5−ヒドロキシメチル−フロン酸(HMF酸)は、Matrix Scientific(Columbia SC、United States)から購入した。この化合物は高度にエステル化されていることが見出された。使用の直前に、エステル化されたHMF酸の10mM溶液を2M H2S04中で2時間沸騰させて冷却し、50mMリン酸緩衝液を加えた後、NaOHによりpH 7.0に調整した。他のすべての試薬は、シグマアルドリッチChemie B.V.(Zwijndrecht、The Netherlands)から購入した。
【0104】
分子および遺伝子の手法:ゲノムDNAは、DNeasy tissue kit(QIAGEN)を用いて単離した。プラスミドDNAは、QIAprep spin miniprep kit(QIAGEN)を用いて単離した。アガローストラップDNAフラグメントは、QIAEXII gel extraction kit(QIAGEN)を用いて単離した。
【0105】
PCR反応は、製造業者の説明書に従ってAccuprime Pfxポリメラーゼ(Invitrogen)を用いて行った。C.バシレンシス(C.basilensis)HMF14のゲノムDNAからHMFオキシドレダクターゼを増幅するために使用したプライマーは、FN23:5’−CGGAATTCCACATGACAAGGGGAGACCG−3’(配列番号1)およびFN24:5’−CGGAATTCGCTTCGGTCTTCAACTCGGATG−3’(配列番号2)であった。下線の配列はEcoRI制限部位を示す。
【0106】
プラスミドDNAは、Gene Pulserエレクトロポレーションデバイス(BioRad)を用いて、エレクトロコンピテント(electrocompetent)細胞へ導入した。
【0107】
トランスポゾンに隣接する染色体DNAは、当技術分野で公知の標準的方法{Ausubel,F.M.ら、Current protocols in molecular biology(Green publishing association、New York;1987)}によって同定し、完全遺伝子座の配列をプライマーウォーキング(primer walking)によって得た。オリゴヌクレオチド合成およびDNA塩基配列決定は、MWG Biotech AG(Germany)によって行った。
【0108】
他の標準分子生物学手法は、SambrookおよびRussel{Sambrook,J.、Russel,D.W.Molecular cloning;a laboratory manual(Cold spring Harbor Laboratoy Press、New York:2001)に従って行った。
【0109】
[実施例I]
[C.バシレンシス(C.basilensis)HMF14の細胞抽出物におけるFDCA生成]
12mMコハク酸および3mM HMFが添加されたMM上で増殖した、対数増殖期後半の前培養物(OD600およそ1.5)から得られたC.バシレンシス(C.basilensis)の細胞抽出物を、HMFまたはHMF酸のいずれかと共にインキュベートすると、FDCAの形成が観察された。
【0110】
基質としてHMFを使用すると、HMF−酸およびHMF−アルコールの迅速な一時的蓄積がFDCA形成と同時に観察された。脱塩細胞抽出物中では、粗細胞抽出物中と比較して、HMF−酸濃度の減少速度は低下し、FDCA生成は遅くなった。FDCAが粗細胞抽出物中と同様に脱塩細胞抽出物中でも生成したので、HMFのFDCAへの変換には補因子は関与していないと考えられた。しかしながら、粗細胞抽出物由来の補因子または他の低分子量成分が、FDCA形成に関して相乗効果を有するように見えた。基質としてHMF−酸を加えると、FDCAの即時形成が粗細胞抽出物中および脱塩細胞抽出物中の両方で観察された。さらに、FDCAが嫌気条件下で生成しなかったので、酸素の存在がFDCA形成には必要であることが実証された。HMF酸のFDCAへの化学量論的変換が観察された。
【0111】
[実施例II]
[HMFオキシドレダクターゼをコードするhmfH遺伝子の単離および特徴づけ]
hmfH遺伝子は、FAD依存性グルコース−メタノール−コリン(GMC)オキシドレダクターゼファミリーに属する62 317 Daのタンパク質をコードすることが見出された。この酵素にはフラン−ジカルボン酸(FDCA)へHMF−酸を酸化する能力があることが示された。さらに、この酵素は、基質としてHMFも受け入れ、HMFをHMF−酸を介してFDCAに酸化した。非重複性NCBIデータベース中でHmfHと最も高い相同性を有するものが、バークホルデリア・フィトフィルマンス(Burkholderia phytofirmans)PsJNのGMCオキシドレダクターゼであることがわかった(座位タグBphyt_2180;562個のアミノ酸ストレッチにわたって68%の同一性)。C.バシレンシス(C.basilensis)HMF14のHMF/フルフラールオペロンの配列データに基づいて、他の潜在的なHMF/フルフラールディグレーダー(degrader)が同定された。選択された菌株は、唯一の炭素源としてHMFまたはフルフラールのいずれかを含むミネラル塩培地上での増殖について試験された。HMF上で増殖できた菌株は、HmfHと45〜68%の同一性があるオキシドレダクターゼをコードするhmfHオルソログを有していた。1菌株(バークホルデリア・ゼノボランス(Burkholderia xenovorans LB400)は、そのオキシドレダクターゼがHmfHと44%の同一性があったが、HMFを利用することができなかった。
【0112】
図3に、C.バシレンシス(C.basilensis)HMF14(A)ならびにフルフラールおよび/またはHMFを利用する可能性があると同定された他種(B)における、フルフラールおよびHMFの代謝関連遺伝子の遺伝子構成の概略図を示す。色は図1における酵素活性に対応する。矢印の下の太数字(x/y)は、yアミノ酸ストレッチ中の対応するC.バシレンシス(C.basilensis)HMF14タンパク質との同一性(x)の割合を示す。オルソロガス遺伝子は、National Center for Biotechnology Informationの非重複性タンパク質データベースに対するBLASTxホモロジー検索によって同定された。HmfAと少なくとも50%の同一性がある酵素をコードするhmfAオルソログがあると同時に、hmfA、B、C、D、およびEに対するオルソログが単一ゲノム中にある場合に、フルフラールクラスターに対するヒットは妥当であると定めた。同じ基準をhmfFおよびhmfGのオルソログを定義するために用いたが、他方hmfHオルソログに対する基準としてはHmfHとの40%の同一性を用いた。イタリック体の数字は、指定菌株のゲノム座位タグを示す。白矢印は、代謝機能を有しない遺伝子を示す。C:唯一の炭素源としてフルフラールまたはHMF(3mM)のいずれかを含むミネラル塩培地上での試験菌株の増殖表現型の概要。ND:測定していない。
【0113】
これらの図では、HMFオキシドレダクターゼHmfHをコードする翻訳領域のヌクレオチド配列(図4)および推定上のアミノ酸配列(図3)が示されている。
【0114】
[実施例III]
[P.プチダ(P.putida)S12中のhmfHのクローニングおよびコードされたオキシドレダクターゼの発現]
HMFオキシドレダクターゼをコードするhmfH遺伝子を、発現ベクターpJT’mcsにクローニングした。C.バシレンシス(C.basilensis)HMF14ゲノムDNA上でプライマーFN23およびFN24を用いて得られたPCRフラグメントを、EcoRI(Fermentas)により消化した。この消化フラグメントを、EcoRI消化およびFastAP(Fermentas)処理のpJT’mcsベクターに連結し、HmfH発現プラスミドpJT’hmfHを得た。hmfH挿入の方向が正しいことを、当技術分野で公知の制御消化およびヌクレオチド配列決定によって確認した。HMFオキシドレダクターゼの発現は、MM+20mMグルコースおよび10mg/lゲンタマイシン中で増殖したP.プチダ(P.putida)_pJT’hmfH(OD600およそ4)の細胞抽出物において、基質としてHMFまたはHMF酸を使用したときにFDCAが形成されたことによって実証された(表1)。
【0115】
【表1】
【0116】
これらの結果により、FDCAの形成が酸素存在下でのみ起こり、hmfHにコードされたオキシドレダクターゼによって触媒されることが確認された。HMFを基質として用いた場合には、脱塩細胞抽出物中および粗細胞抽出物中の両方においてHMF−酸の一時的な蓄積が観察された。脱塩細胞抽出物(低分子量補因子が除去された)中のHMF−酸の形成により、HmfHが、HMF−酸をFDCAへ酸化するだけでなく、HMFをそのモノカルボン酸形へ酸化できることも示された。
【0117】
HmfHに加えて、P.プチダ(P.putida)S12に固有で見かけ上非特異的なデヒドロゲナーゼもまた、その還元等量が脱塩によって除去されなければ、HMFからHMF酸を形成することができる(表2)。アルデヒドレダクターゼ/アルコールデヒドロゲナーゼ活性はNADH/NAD+依存であるが、アルデヒドデヒドロゲナーゼ活性は2つの人工電子伝達体、PMSおよびDCPIPを組み合せることによって維持することができる。ここで、PMSは最初の電子担体であり、DCPIPは最終電子受容体である。同様な活性は、C.バシレンシス(C.basilensis)HMF14の細胞抽出物中でも観察された。
【0118】
【表2】
【0119】
P.プチダ(P.putida)S12 pJT’hmfHの粗細胞抽出物中の、HMFからFDCA、HMF−アルコール(H−oh)およびHMF−酸(H−酸)の形成を図4に示す。図4は、P.プチダ(P.putida)S12 pJT’hmfH細胞抽出物中の、内因性デヒドロゲナーゼおよびHmfHの相乗効果を説明する。最初に、アルデヒドレダクターゼがおそらく触媒したHMF−アルコールの形成と同時にHMFの迅速な減少が観察された。同時に、HMF−酸(おそらくアルデヒドデヒドロゲナーゼを介して)およびFDCA(HmfHを介して)が蓄積し始めた。HMF−アルコールは、形成直後に、おそらくHMF、HMF−酸およびFDCAへの再酸化を通して消失した。HMF−酸のみが、25時間のタイムスパンの中で、FDCAに(ほとんど)完全に酸化される前に顕著な蓄積を示した。これらの結果により、HMF−酸のFDCAへの酸化がこの無細胞系における律速段階であったことが示唆される。
【0120】
[実施例IV]
[HmfHを発現するP.プチダ(P.putida)S12によるHMFのFDCAへの全細胞変換]
P.プチダ(P.putida)S12の内因性デヒドロゲナーゼが、HMFからHMF−酸を生成する追加の手段となって、HMFのFDCAへの酸化においてHmfHと相乗的に作用することができるので、HMFのFDCAへの全細胞生体内変換法には、酵素法にまさる利点がある場合がある。この可能性を調べるために、静止細胞、増殖細胞、および破壊細胞を、HMFからFDCAの生成について試験した。
【0121】
振とうフラスコ培養を、1lエルレンマイヤーフラスコを用いて、10mg/Lゲンタマイシンを添加したMM+40mMグリセロールおよび2mMグルコース150ml中で行った。細胞を対数増殖期の終わり(OD600およそ4)に収集して、洗浄し、19.4g/l K2HP04、8.15g/l NaH2HP04、40mMグリセロールおよび10mg/lゲンタマイシンを添加したMM中に濃縮した。濃縮細胞懸濁液(CDW1.65gに相当)のアリコート(10ml)を、250mlのエルレンマイヤーフラスコ中でHMFと共にインキュベートし、FDCAの分析のために一定間隔でサンプルを採取した。静止細胞については、グリセロールを再懸濁培地から除去し、破壊細胞は、超音波処理によって得た。
【0122】
これまでの観察のように、HMFがHMF−アルコールおよびHMF酸に迅速に変換された後、HMF酸の減少およびそれと同時のFDCAの形成が起こった。試験したHMFの2つの濃度について、FDCAの生成速度およびHMF酸の減少速度の両方を表IV−aに示す。
【0123】
【表3】
【0124】
表3では、FDCA生成がHmfHを発現するP.プチダ(P.putida)S12の増殖細胞でより効率的であったことが示されている。全体的なFDCAの生成速度、変換効率、およびHMF−酸の減少速度は、増殖細胞のインキュベーションにおいてより高いものであった。さらに、FDCAの生成速度はHMFの初期濃度に依存しており、この系は飽和していないことが示唆された。
【0125】
[実施例V]
[HmfHを発現するP.プチダ(P.putida)S12を用いた、流加回分によるHMFからのFDCAの生成]
FDCAの生成は、HmfHを発現するP.プチダ(P.putida)S12の増殖細胞で最も効率的であることが見出された。そこで、P.プチダ(P.putida)S12 pJT’hmfHの全細胞によるFDCAの生成を実証するために流加回分実験を行った。
【0126】
初期段階では、P.プチダ(P.putida)_pJT’hmfHを、グリセロールの流加を含むMM上で13gCDW/lの細胞密度になるまで培養した。この段階の終わりに、HMF流加を開始し、0.8mmol/l/時の速度でHMFを加えた。グリセロールを一緒に流加しないと、HMF−酸およびFDCAは0.41mmol/l/時の同じ速度で形成された(図5を参照されたい)。次いで、0.21M HMFおよび4Mグリセロールを含有する溶液に流加を取り替え、およそ5ml/時で流加した。このようにグリセロールを一緒に流加すると、HMF−酸の濃度は迅速に低下したが、FDCAの形成はほぼ同じ速度で続いた(図5)。総量44,6mmolのHMFが流加回分に加えられた時点で、この実験を停止した。結果はFDCAへのほとんど完全な変換(すなわち96.5%の効率)を示した。
【0127】
同様な実験で、0.7mmol/l/時の平均速度でグリセロールをHMFと共に流加した。この流加回転培養では、HMF酸中間体の蓄積は観察されず、HMFはすべてFDCAに直接変換された(図6)。
【0128】
[実施例VI]
[発酵ブロスからのFDCAの精製]
pH1.0でのFDCAの溶解度は、水中で約1.5g/lであることがわかった。この特性を有利に利用して、発酵ブロスからFDCA生成物を回収した。遠心分離(9500×gで5分間)により細胞を除去した後に、室温で連続して撹拌しながらpH1になるまで、96%H2S04約10mlを透明ブロス100mlに加え、FDCAを沈殿させた。この沈殿を8228×gで10分間遠心分離して回収し、風乾したペレットを60℃でメタノール中に再融解した。予熱された0.22μmフィルターでろ過して溶解しないデブリを除去した後すぐに、そのろ液を純度分析のためにHPLCに供した。上記手順によって発酵ブロスから回収したFDCAの純度は、約65%であった。FDCAのさらなる精製は当技術分野で公知の方法により行うことができる。
【0129】
[実施例VII]
[HmfHを発現するP.プチダ(P.putida)S12を用いた、流加回分によるHMFからのFDCAの生成]
第2の流加回分によるFDCA生成実験は、P.プチダ(P.putida)S12 pJT’hmfHの全細胞による、FDCAのタイターおよび生産性を向上させるために行った。
【0130】
最初の回分段階では、P.プチダ(P.putida)_pJT’hmfHを、最初のグリセロールが23.2時間の発酵後(OD600およそ8)に枯渇するまで、MM上で培養した。この時点で、バイオマス増加が約0.45gCDW/l/時になる速度でグリセロール流加を開始した。50.7時間の発酵後に、バイオマス増加が約0.045gCDW/l/時になる速度までグリセロール流加を低下させた。123.4時間の発酵後に、バイオマス増加が約0.25gCDW/l/時になる速度に発酵の終了までグリセロール流加を増加させた。この期間にわたり、HMF流加を行った。26.7時間の発酵後に、0.65mmol/時/gCDWの速度でHMF流加を開始し、HMF酸およびFDCAの両方の蓄積をもたらした。33.3時間の発酵後に、HMF流加速度を0.28mmol/時/gCDWに低下させて、72.8時間の発酵後に、0.09mmol/時/gCDWへとさらに低下させ、発酵の117.4時間後に停止した。HMF流加の減少は、発酵槽中のHMF酸濃度の漸減をもたらしたが、FDCA濃度の増加は継続した。HMF酸がもはや検出できなくなった時点で、発酵を停止した。この時点で、188mmolのHMFを発酵槽に加えると、30.8g/l濃度で約182mmolのFDCAの生成(すなわち97%の効率)がもたらされた。図7aに、P.プチダ(P.putida)_pJT’hmfHの流加回分培養中のバイオマス、HMF酸、およびFDCAの形成を示す。図7bに、同じ培養におけるグリセロールおよびHMFの流加速度を示す。
【0131】
[実施例VIII]
[発酵ブロスからのFDCAの精製]
pH0.5でのFDCAの溶解度は、水中で約0.4g/lであることがわかった。沈殿の後に、この濃度が溶液中に残るので、実施例VIIIで得られるタイターの増加は、沈殿による精製に起因する生成物の損失を顕著に減少させる。
【0132】
遠心分離(9500×gで15分間)による細胞の除去後に、透明ブロス500mlを3分間沸騰させてタンパク質を沈殿させ、9500×gで20分間遠心分離した。4℃で連続して撹拌しながらpH0.5になるまで、96%H2S04約50mlを透明上清に加え、FDCAを沈殿させた。この沈殿を8228×gで20分間遠心分離して回収し、水250mlで1回洗浄して、再び8228×gで20分間遠心分離した。得られたペレットを風乾した後、テトラヒドロフラン(THF)約1200ml中に30℃で溶解した。ろ過により溶解しないデブリを除去した後すぐに、発酵ブロス中で最初79%純度のFDCAが高純度(>99%)の乾燥FDCA粉末11.8gになるまで透明THF溶液を50℃で真空蒸発させた。必要があれば、当技術分野で公知の方法を使用して、FDCAの精製をさらに最適にすることができる。
【特許請求の範囲】
【請求項1】
オキシドレダクターゼ活性を有するポリペプチドであって、配列番号3に示されるアミノ酸配列もしくは配列番号4のヌクレオチド配列によってコードされたアミノ酸配列または配列番号3の配列と少なくとも45%以上の配列同一性を有するその変異体ポリペプチドを含むポリペプチド。
【請求項2】
(a)配列番号4に示される前記ヌクレオチド配列;
(b)配列番号4の逆相補鎖であるポリヌクレオチドと選択的にハイブリダイズするヌクレオチド配列;
(c)配列番号4の前記ヌクレオチド配列と少なくとも66%以上の配列同一性を有するヌクレオチド配列;
(d)少なくとも約100ヌクレオチド長である、(a)または(b)で定義されるヌクレオチド配列のフラグメント;
(e)(a)、(b)、(c)、または(d)のいずれか1つに定義される配列に、遺伝子コードの結果として縮重した配列;
(f)(a)、(b)、(c)、(d)、または(e)に定義されるヌクレオチド配列の逆相補鎖であるヌクレオチド配列
を含むポリペプチド。
【請求項3】
請求項1に記載のポリペプチドをコードするポリヌクレオチド。
【請求項4】
請求項2または3のいずれか一項に記載のポリヌクレオチドを含む核酸構築物。
【請求項5】
請求項2〜3のいずれか一項に記載のポリヌクレオチド配列または請求項4に記載の核酸構築物を挿入するベクター。
【請求項6】
請求項1に記載のポリペプチド、請求項2もしくは3に記載のポリヌクレオチド、請求項4に記載の核酸構築物、または請求項5に記載のベクターを含む細胞。
【請求項7】
前記細胞が、大腸菌属(Escherichia)、アナベーナ属(Anabaena)、コーロバクター属(Caulobacter)、グルコノバクター属(Gluconobacter)、ロドバクター属(Rhodobacter)、シュードモナス属(Pseudomonas)、パラコッカス属(Paracoccus)、バチルス属(Bacillus)、ブレビバクテリウム属(Brevibacterium)、コリネバクテリウム属(Corynebacterium)、リゾビウム属(Rhizobium)(シノリゾビウム属(Sinorhizobium))、フラボバクテリウム属(Flavobacterium)、クレブシェラ属(Klebsiella)、エンテロバクター属(Enterobacter)、ラクトバチルス属(Lactobacillus)、ラクトコッカス属(Lactococcus)、メチロバクテリウム属(Methylobacterium)、スタフィロコッカス属(Staphylococcus)、もしくはストレプトミセス属(Streptomyces);またはB.スブチリス種(B.subtilis)、B.アミロリケファシエンス種(B.amyloliquefaciens)、B.リケニフォルミス種(B.licheniformis)、B.プンチス種(B.puntis)、B.メガテリウム種(B.megaterium)、B.ハロデュランス種(B.halodurans)、B.プミルス種(B.pumilus)、G.オキシダンス種(G.oxydans)、コーロバクター・クレセンタス(Caulobacter crescentus)CB15種、メチロバクテリウム・エキストロクエンス種(Methylobacterium extorquens)、ロドバクター・スフェロイデス種(Rhodobacter sphaeroides)、シュードモナス・プチダ種(Pseudomonas putida)、パラコッカス・ゼアキサンチニファシエンス種(Paracoccus zeaxanthinifaciens)、パラコッカス・デニトリフィカンス種(Paracoccus denitrificans)、大腸菌種(E.coli)、C.グルタミクム種(C.glutamicum)、スタフィロコッカス・カルノーサス種(Staphylococcus carnosus)、ストレプトミセス・リビダンス種(Streptomyces lividans)、シノリゾビウム・メリオティ種(Sinorhizobium melioti)、およびリゾビウム・ラジオバクター種(Rhizobium radiobacter)の群から選択される細菌である、請求項6に記載の細胞。
【請求項8】
前記細胞が、カンジダ属(Candida)、ハンゼヌラ属(Hansenula)、クリベロミセス属(Kluyveromyces)、ピチア属(Pichia)、サッカロミセス属(Saccharomyces)、シゾサッカロミセス属(Schizosaccharomyces)、もしくはヤロウイア属(Yarrowia);またはクリベロミセス・ラクチス種(Kluyveromyces lactis)、S.セルビシエ種(S.cerevisiae)、ハンゼヌラ・ポリモルファ種(Hansenula polymorpha)、ヤロウィア・リポリティカ種(Yarrowia lipolytica)、およびピチア・パストリス種(Pichia pastoris)からなる群からの酵母細胞、あるいはアスペルギルス属(Aspergillus)、クリソスポリウム属(Chrysosporium)、ペニシリウム属(Penicillium)、タラロミセス属(Talaromyces)、もしくはトリコデルマ属(Trichoderma);または、アスペルギルス・ニガー種(Aspergillus niger)、アスペルギルス・アワモリ種(Aspergillus awamori)、アスペルギルス・フォエティダス種(Aspergillus foetidus)、アスペルギルス・ソジェ種(Aspergillus sojae)、アスペルギルス・フミガーツス種(Aspergillus fumigatus)、タラロミセス・エメルソニ種(Talaromyces emersonii)、アスペルギルス・オリゼ種(Aspergillus oryzae)、クリソスポリウム・ラクノウェンス種(Chrysosporium lucknowense)、トリコデルマ・リーゼイ種(Trichoderma reesei)、もしくはペニシリウム・クリゾゲヌム種(Penicillium chrysogenum)の群からの糸状菌細胞である、請求項6に記載の細胞。
【請求項9】
オキシドレダクターゼ活性を有するポリペプチドを製造するための方法であって、前記ポリペプチドの発現を可能にする条件下で、請求項6〜8のいずれか一項に記載の細胞を培養することと、場合によっては、発現されたポリペプチドを回収することとを含む方法。
【請求項10】
請求項9に記載の方法によって得られるポリペプチド。
【請求項11】
2,5−フランジカルボン酸(FDCA)を生成するための方法であって、オキシドレダクターゼ触媒および場合によっては1つまたは複数の補酵素の存在下でのオキシダントとの反応によって、FDCAの1つまたは複数のフラン前駆体がFDCAに変換され、前記オキシドレダクターゼ触媒が請求項1または請求項10に記載のポリペプチドを含む方法。
【請求項12】
FDCAの前記フラン前駆体が、5−ヒドロキシメチルフルフラール(HMF)、2,5−ジヒドロキシメチルフラン(HMFアルコール)、および5−ヒドロキシメチル−2−フランカルボン酸(HMF酸)からなる群から選択される群から選択され、好ましくは前記フラン前駆体がHMFである、請求項11に記載の方法。
【請求項13】
前記前駆体がHMFであり、酸の存在下で加熱することにより1つまたは複数のヘキソース糖からHMFが得られ、好ましくは前記1つまたは複数のヘキソース糖がバイオマスから得られる、請求項15に記載の方法。
【請求項14】
5−ハイドロキシメチル−2−フランカルボン酸(HMF酸)を生成するための方法であって、HMF酸の1つまたは複数のフラン前駆体が、オキシドレダクターゼ触媒および場合によっては1つまたは複数の補酵素補因子の存在下でのオキシダントとの反応によってHMF酸に変換され、前記オキシドレダクターゼ触媒が請求項1または請求項12に記載のポリペプチドを含む方法。
【請求項15】
HMF酸の前記フラン前駆体が、5−ヒドロキシメチルフルフラール(HMF)および2,5−ジヒドロキシメチルフラン(HMFアルコール)から選択される群から選択される、請求項14に記載の方法。
【請求項16】
前記補酵素が、ニコチンアミドアデニンジヌクレオチド(NAD+)および/またはピロロキノリンキノロン(PQQ)および/またはフラビンアデニンジヌクレオチド(FAD)である、請求項11または15のいずれか一項に記載の方法。
【請求項17】
前記オキシドレダクターゼ触媒が、請求項6〜8のいずれか一項に記載の細胞の無細胞抽出物である、請求項11〜16のいずれか一項に記載の方法。
【請求項18】
前記オキシドレダクターゼ触媒が、請求項6〜8のいずれか一項に記載の細胞である全細胞生体触媒である、請求項11〜17のいずれか一項に記載の方法。
【請求項19】
1つまたは複数のモノマーからポリマーを生成するための方法であって、前記モノマーの1つが請求項18に記載のFDCAである方法。
【請求項1】
オキシドレダクターゼ活性を有するポリペプチドであって、配列番号3に示されるアミノ酸配列もしくは配列番号4のヌクレオチド配列によってコードされたアミノ酸配列または配列番号3の配列と少なくとも45%以上の配列同一性を有するその変異体ポリペプチドを含むポリペプチド。
【請求項2】
(a)配列番号4に示される前記ヌクレオチド配列;
(b)配列番号4の逆相補鎖であるポリヌクレオチドと選択的にハイブリダイズするヌクレオチド配列;
(c)配列番号4の前記ヌクレオチド配列と少なくとも66%以上の配列同一性を有するヌクレオチド配列;
(d)少なくとも約100ヌクレオチド長である、(a)または(b)で定義されるヌクレオチド配列のフラグメント;
(e)(a)、(b)、(c)、または(d)のいずれか1つに定義される配列に、遺伝子コードの結果として縮重した配列;
(f)(a)、(b)、(c)、(d)、または(e)に定義されるヌクレオチド配列の逆相補鎖であるヌクレオチド配列
を含むポリペプチド。
【請求項3】
請求項1に記載のポリペプチドをコードするポリヌクレオチド。
【請求項4】
請求項2または3のいずれか一項に記載のポリヌクレオチドを含む核酸構築物。
【請求項5】
請求項2〜3のいずれか一項に記載のポリヌクレオチド配列または請求項4に記載の核酸構築物を挿入するベクター。
【請求項6】
請求項1に記載のポリペプチド、請求項2もしくは3に記載のポリヌクレオチド、請求項4に記載の核酸構築物、または請求項5に記載のベクターを含む細胞。
【請求項7】
前記細胞が、大腸菌属(Escherichia)、アナベーナ属(Anabaena)、コーロバクター属(Caulobacter)、グルコノバクター属(Gluconobacter)、ロドバクター属(Rhodobacter)、シュードモナス属(Pseudomonas)、パラコッカス属(Paracoccus)、バチルス属(Bacillus)、ブレビバクテリウム属(Brevibacterium)、コリネバクテリウム属(Corynebacterium)、リゾビウム属(Rhizobium)(シノリゾビウム属(Sinorhizobium))、フラボバクテリウム属(Flavobacterium)、クレブシェラ属(Klebsiella)、エンテロバクター属(Enterobacter)、ラクトバチルス属(Lactobacillus)、ラクトコッカス属(Lactococcus)、メチロバクテリウム属(Methylobacterium)、スタフィロコッカス属(Staphylococcus)、もしくはストレプトミセス属(Streptomyces);またはB.スブチリス種(B.subtilis)、B.アミロリケファシエンス種(B.amyloliquefaciens)、B.リケニフォルミス種(B.licheniformis)、B.プンチス種(B.puntis)、B.メガテリウム種(B.megaterium)、B.ハロデュランス種(B.halodurans)、B.プミルス種(B.pumilus)、G.オキシダンス種(G.oxydans)、コーロバクター・クレセンタス(Caulobacter crescentus)CB15種、メチロバクテリウム・エキストロクエンス種(Methylobacterium extorquens)、ロドバクター・スフェロイデス種(Rhodobacter sphaeroides)、シュードモナス・プチダ種(Pseudomonas putida)、パラコッカス・ゼアキサンチニファシエンス種(Paracoccus zeaxanthinifaciens)、パラコッカス・デニトリフィカンス種(Paracoccus denitrificans)、大腸菌種(E.coli)、C.グルタミクム種(C.glutamicum)、スタフィロコッカス・カルノーサス種(Staphylococcus carnosus)、ストレプトミセス・リビダンス種(Streptomyces lividans)、シノリゾビウム・メリオティ種(Sinorhizobium melioti)、およびリゾビウム・ラジオバクター種(Rhizobium radiobacter)の群から選択される細菌である、請求項6に記載の細胞。
【請求項8】
前記細胞が、カンジダ属(Candida)、ハンゼヌラ属(Hansenula)、クリベロミセス属(Kluyveromyces)、ピチア属(Pichia)、サッカロミセス属(Saccharomyces)、シゾサッカロミセス属(Schizosaccharomyces)、もしくはヤロウイア属(Yarrowia);またはクリベロミセス・ラクチス種(Kluyveromyces lactis)、S.セルビシエ種(S.cerevisiae)、ハンゼヌラ・ポリモルファ種(Hansenula polymorpha)、ヤロウィア・リポリティカ種(Yarrowia lipolytica)、およびピチア・パストリス種(Pichia pastoris)からなる群からの酵母細胞、あるいはアスペルギルス属(Aspergillus)、クリソスポリウム属(Chrysosporium)、ペニシリウム属(Penicillium)、タラロミセス属(Talaromyces)、もしくはトリコデルマ属(Trichoderma);または、アスペルギルス・ニガー種(Aspergillus niger)、アスペルギルス・アワモリ種(Aspergillus awamori)、アスペルギルス・フォエティダス種(Aspergillus foetidus)、アスペルギルス・ソジェ種(Aspergillus sojae)、アスペルギルス・フミガーツス種(Aspergillus fumigatus)、タラロミセス・エメルソニ種(Talaromyces emersonii)、アスペルギルス・オリゼ種(Aspergillus oryzae)、クリソスポリウム・ラクノウェンス種(Chrysosporium lucknowense)、トリコデルマ・リーゼイ種(Trichoderma reesei)、もしくはペニシリウム・クリゾゲヌム種(Penicillium chrysogenum)の群からの糸状菌細胞である、請求項6に記載の細胞。
【請求項9】
オキシドレダクターゼ活性を有するポリペプチドを製造するための方法であって、前記ポリペプチドの発現を可能にする条件下で、請求項6〜8のいずれか一項に記載の細胞を培養することと、場合によっては、発現されたポリペプチドを回収することとを含む方法。
【請求項10】
請求項9に記載の方法によって得られるポリペプチド。
【請求項11】
2,5−フランジカルボン酸(FDCA)を生成するための方法であって、オキシドレダクターゼ触媒および場合によっては1つまたは複数の補酵素の存在下でのオキシダントとの反応によって、FDCAの1つまたは複数のフラン前駆体がFDCAに変換され、前記オキシドレダクターゼ触媒が請求項1または請求項10に記載のポリペプチドを含む方法。
【請求項12】
FDCAの前記フラン前駆体が、5−ヒドロキシメチルフルフラール(HMF)、2,5−ジヒドロキシメチルフラン(HMFアルコール)、および5−ヒドロキシメチル−2−フランカルボン酸(HMF酸)からなる群から選択される群から選択され、好ましくは前記フラン前駆体がHMFである、請求項11に記載の方法。
【請求項13】
前記前駆体がHMFであり、酸の存在下で加熱することにより1つまたは複数のヘキソース糖からHMFが得られ、好ましくは前記1つまたは複数のヘキソース糖がバイオマスから得られる、請求項15に記載の方法。
【請求項14】
5−ハイドロキシメチル−2−フランカルボン酸(HMF酸)を生成するための方法であって、HMF酸の1つまたは複数のフラン前駆体が、オキシドレダクターゼ触媒および場合によっては1つまたは複数の補酵素補因子の存在下でのオキシダントとの反応によってHMF酸に変換され、前記オキシドレダクターゼ触媒が請求項1または請求項12に記載のポリペプチドを含む方法。
【請求項15】
HMF酸の前記フラン前駆体が、5−ヒドロキシメチルフルフラール(HMF)および2,5−ジヒドロキシメチルフラン(HMFアルコール)から選択される群から選択される、請求項14に記載の方法。
【請求項16】
前記補酵素が、ニコチンアミドアデニンジヌクレオチド(NAD+)および/またはピロロキノリンキノロン(PQQ)および/またはフラビンアデニンジヌクレオチド(FAD)である、請求項11または15のいずれか一項に記載の方法。
【請求項17】
前記オキシドレダクターゼ触媒が、請求項6〜8のいずれか一項に記載の細胞の無細胞抽出物である、請求項11〜16のいずれか一項に記載の方法。
【請求項18】
前記オキシドレダクターゼ触媒が、請求項6〜8のいずれか一項に記載の細胞である全細胞生体触媒である、請求項11〜17のいずれか一項に記載の方法。
【請求項19】
1つまたは複数のモノマーからポリマーを生成するための方法であって、前記モノマーの1つが請求項18に記載のFDCAである方法。
【図1】

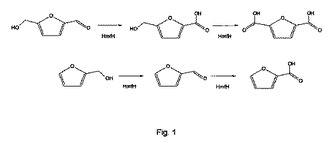
【図2】
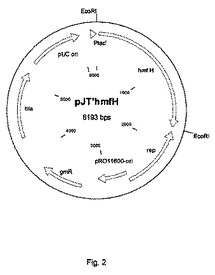
【図3A】
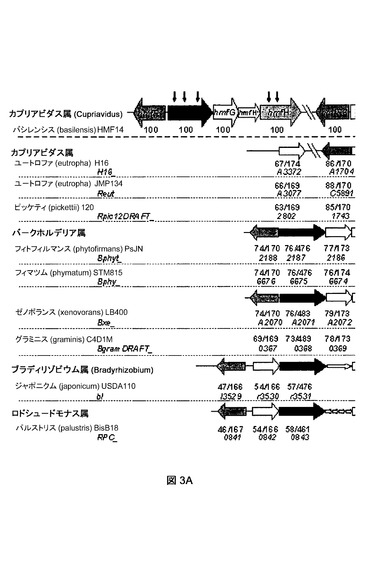
【図3B】

【図3C】
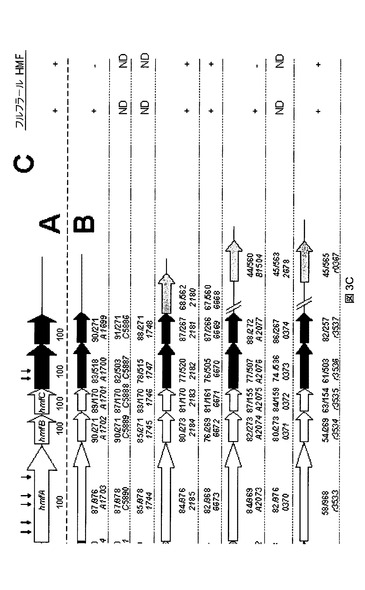
【図3D】
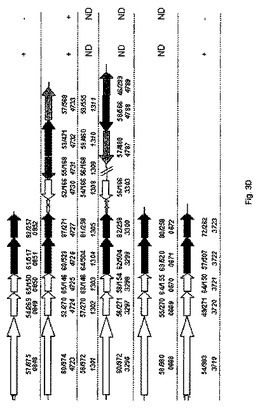
【図4】
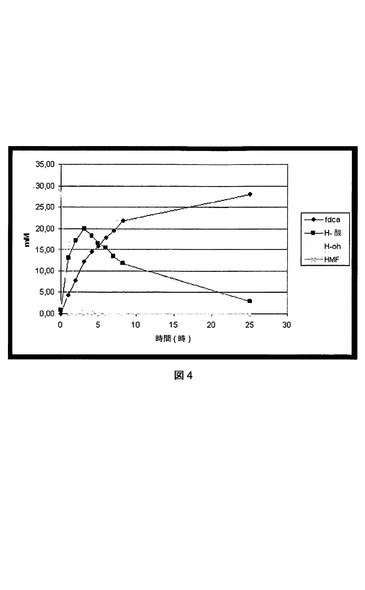
【図5】
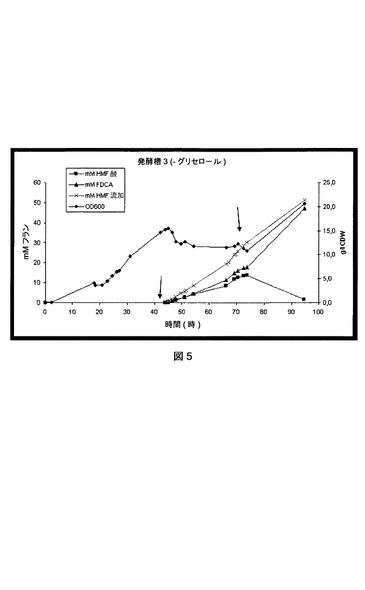
【図6】
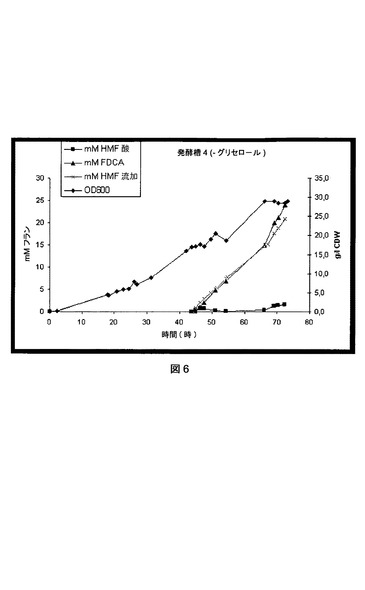
【図7A】
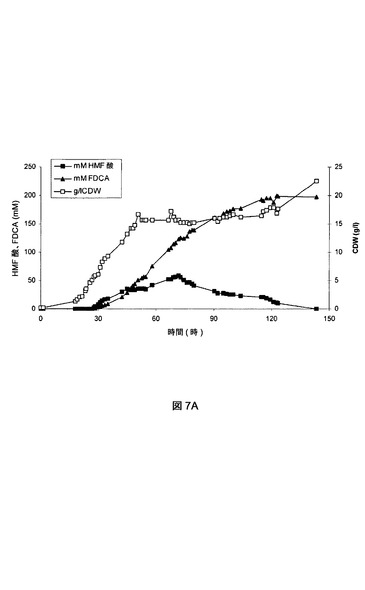
【図7B】
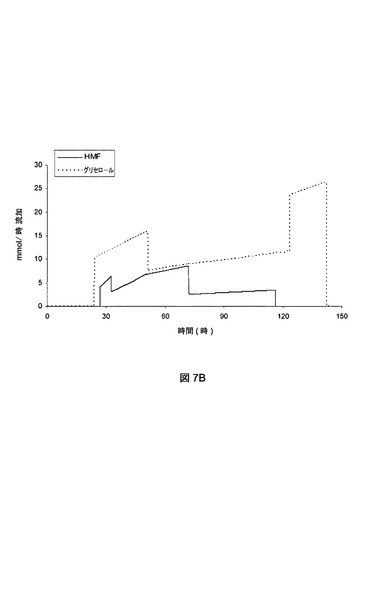
【図2】
【図3A】
【図3B】
【図3C】
【図3D】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【公表番号】特表2013−503614(P2013−503614A)
【公表日】平成25年2月4日(2013.2.4)
【国際特許分類】
【出願番号】特願2012−527325(P2012−527325)
【出願日】平成22年9月2日(2010.9.2)
【国際出願番号】PCT/EP2010/062896
【国際公開番号】WO2011/026913
【国際公開日】平成23年3月10日(2011.3.10)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成25年2月4日(2013.2.4)
【国際特許分類】
【出願日】平成22年9月2日(2010.9.2)
【国際出願番号】PCT/EP2010/062896
【国際公開番号】WO2011/026913
【国際公開日】平成23年3月10日(2011.3.10)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]