
オクロバクトラム・インターメディウムのリポ多糖類および哺乳動物の免疫刺激剤としてのそれらの使用
本発明は、オクロバクトラム・インターメディウム株 LMG3306由来のリポ多糖類(LPS)の単離、精製および特性化、ならびに哺乳動物の免疫刺激剤としてのそれらの使用、敗血症の処置および/または予防のための医薬化合物、ならびに免疫抑制動物における、およびリーシュマニア症に対するワクチン用アジュバントの製造方法に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、オクロバクトラム・インターメディウム(Ochrobactrum intermedium)株LMG3306由来のリポ多糖類(LPS)の単離、精製および特性化、ならびに哺乳動物の免疫刺激剤としてのそれらの使用、敗血症の処置および/または予防のための医薬化合物、ならびに免疫抑制動物における、およびリーシュマニア症に対するワクチン用アジュバントの製造方法に関する。
【背景技術】
【0002】
背景技術
オクロバクトラム属の細菌は、プロテオバクテリア門のアルファ2下位分類に含まれる。それらは、重症もしくは免疫不全患者または留置カテーテルを有する患者でのみ病原性となることが知られている主に土壌に生息する細菌である。オクロバクトラムが、髄膜炎、骨髄炎、菌血症、および敗血症を引き起こし得るような状況において、これらの細菌は、それら自体の慢性感染を確立することはできず、カテーテル除去後に正常な宿主から除去される(Cieslak, T. J., C. J. Drabick, and M. L. Robb. 1996. Pyogenic infections due to Ochrobactrum anthropi. Clin. Infect. Dis. 22:845−847.)
【0003】
グラム陰性菌の外膜(OM)の主成分である全てのリポ多糖類の組成は、2−ケト−3−デオキシオクツロソン酸(KDO)と称される特定の糖類を介して脂質Aと称される脂質成分に結合された多糖を含むという一般原則に従うため(Rietschel, E. T., Schade, U., Jensen, M., Wollenweber, H. W., Luderitz, O. and Greisman, S. G. 1982. Bacterial endotoxins:chemical structure, biological activity and role in septicaemia. Scand. J. Infect. Dis. Suppl. 3:8)、全てのLPS分子が同一の生物学的効果を有すると推定されている。
【0004】
しかしながら、この概念は、いくつかのLPS種のサイトカイン合成を生じる能力における大きな差異を示す研究によって修正され、この作用は、それらの脂質A部分の構造上の特徴と関係している(Netea, M. G., van Deuren, M., Kullberg, B. J., Cavaillon, J. M. and van der Meer, J. W. M. 2002. Does the shape of lipid A determine the interaction of LPS with Toll−like receptors? Trends Immunol. 23:135.)。
【0005】
グラム陰性通性細胞内細菌であるレジオネラニューモフィラは、そのLPSの低い内毒素性および化学構造の両方に関してブルセラ菌と類似であり、腸内細菌のLPSによって引き起こされる内毒素性より顕著に低い(Zahringer, U., Knirel, Y. A., Lindner, B., Helbig, J. H., Sonesson, A., Marre, R. and Rietschel, E. T. 1995. The lipopolysaccharide of Legionella pneumophila serogroup 1 (strain Philadelphia 1): chemical structure and biological significance. Prog. Clin. Biol. Res. 392:113. 26)。
【0006】
オクロバクトラム・アンスロピ属は、既知のブルセラ菌の最も近縁種である(Velasco et al. International Journal of Systematic Bacteriology 48 (1998) 759−768)。それらの近縁の系統発生的関係にも関わらず、ブルセラ・アボルタスとオクロバクトラムは、OM特性が顕著に異なり、これらの大きな差異は、LPSにおけるわずかな変化によって少なくとも一部分引き起こされる(Velasco et al. Infection and Immunity 68 (2000)3210−3218)。
【0007】
オクロバクトラム・インターメディウム LMG3301由来のLPSの完全なコア−脂質A骨格およびオクロバクトラム・アンスロピ LMG3331由来のLPSのO型鎖が、決定された(Velasco et al. Carbohydrates Research 306 (1996)123−126; Velasco et al. Carbohydrates Research 306 (1998)283−290; Velasco et al. Infection and Immunity 68 (2000)3210−3218)。
【0008】
オクロバクトラム・インターメディウム LMG3306株は、Velascoらにより発表されたが(International Journal of Systematic Bacteriology 48 (1998) 759−768)、それにもかかわらず、オクロバクトラム・インターメディウム LMG3306由来のLPSは、未だ報告されていない。
【0009】
一方、米国だけでも毎年およそ900,000症例の敗血症が発生し、その結果、およそ210,000人が死亡し、約17億ドルの費用がかかっている。敗血症は、罹患率が上昇しつつあり、死亡率27−48%の疾患である。敗血症性ショックは、敗血症の結果起こり、高い頻度で死に至る。敗血症罹患中に感染と共に生じる重篤な炎症は、いくつかの治療的介入の目標であった。それにもかかわらず、一般的な抗炎症剤を用いる20年以上の臨床試験での敗血症の炎症面は、この手法があまり成功しなかったことを示した。
【0010】
敗血症に至る病理学的機序は、複雑であって、理解が難しい。敗血症は、全身性炎症反応、その後の代償性抗炎症性反応を伴う。それらのバランスは宿主の生存に重要である。さらに、敗血症の動物モデルは、ヒト患者における敗血症のこの複雑な性質を容易には模倣し得ない。
【0011】
近年、敗血症性ショックのほとんどの影響がエンドトキシン(LPS)を循環させる大規模な放出によるものと考えられているため、トール様受容体(TLR)アンタゴニストによるそれらの作用を処理するいくつかの試みがなされている。従って、TLR4アンタゴニストとして作用するLPSの脂質A部分の構造類縁体であるエリトラン(Eritoran)は、フェーズIIの開発段階にあり、また重症敗血症患者の死亡率の低下を示した。さらに、TAK−242はTLR4シグナル伝達阻害剤であり、それは、炎症性サイトカインレベルを低下させ、また高いIL−6レベルを有する重症敗血症患者の下位集団における死亡率を低下させる。
【0012】
今日でも、利用可能なワクチンがない多くの感染性疾患が存在する。これらの場合のいくつかにおいては、失敗の原因は、正確かつ適当な免疫反応を誘導するための正確なアジュバントの欠如にある。細胞内細菌、ウイルスおよびほとんどの原生動物のようないくつかの感染源に関して、防御免疫反応はIFNγ産生に特徴付けられるTヘルパー1型(TH1)のものである。対照的に、ほとんどの蠕虫に対する防御反応は、IL−4産生を特徴とする2型(TH2)のものである(Fresno, M., M. Kopf, and L. Rivas. 1997. Cytokines and infectious disease. Immunol Today 18:56−58)。多くのアジュバントの毒性およびTh1誘導の悪化(exarcebated)により、ヒトワクチン剤形に利用可能なアジュバントはほとんどない。実際にアジュバントではないミョウバンが、ヒトワクチン剤形で最も使用されている。近年、トール様受容体(TLR)のアゴニストが、大きな関心を集めている(Hoffman, Nature Reviews Drug Discovery 4, 879, 2005)(Kwissa; Expert Vaccine Rev., 6, G73, 2007)。いくつかの抗原は、種々のTLRのいくつかのアゴニストと共に、現在分析されている。
【0013】
最後に、既に承認されているものを含む何れかのワクチン剤形の主な問題の1つは、それらが免疫抑制された対象においてほとんど作用しないことである。その他に、それらのうちのいくつかは、患者に副次的影響を有する(Kwissa, et al 2007)。
【0014】
オクロバクトラム・インターメディウム LMG 3301由来の野生型deep−rough LPS (O型鎖が存在せず、外核部分からのいくつかの単糖を欠くLPS) の完全なコア脂質A骨格は、Velascoらの、Carbohydrates Research 306 (1998)283−290に記載され;オクロバクトラム・アンスロピ LMG 3331 LPS由来の野生型smooth LPS由来のO型鎖は、Velascoらの、Carbohydrates Research 306 (1996)123−126に発表され;完全LPSの分子量は、Velascoらの、Infection and Immunity 68 (2000)3210−3218に報告されている。
【0015】
オクロバクトラム・インターメディウム LMG 3306株は、Velascoらによって発表されているが(International Journal of Systematic Bacteriology 48 (1998) 759−768)、それにも関わらず、オクロバクトラム・インターメディウム LMG 3306由来の野生型smooth LPSは未だ報告されていない。
【発明の概要】
【0016】
発明の概要
今日、オクロバクトラム・インターメディウム株 LMG 3306のリポ多糖類(IM)が、敗血症の処置および/または予防に有用であり、ならびに免疫抑制された動物およびリーシュマニア症患者におけるワクチン用アジュバントとして有用であることを実証した。
【0017】
故に、本発明の第一局面は、敗血症の処置および/または予防、ならびに免疫抑制された動物およびリーシュマニア症患者に対するワクチン用アジュバントのための医薬の製造における、オクロバクトラム・インターメディウム株 LMG 3306のリポ多糖類(IM)の使用に関する。
【0018】
本発明の一態様は、敗血症性ショックおよび内毒素ショックの処置および/または予防のための医薬の製造における、オクロバクトラム・インターメディウム株LMG 3306(IM)のリポ多糖類の使用を提供する。
【0019】
本発明の他の態様は、ヒトおよび動物の両方における感染症に対するワクチン用アジュバントとしてのオクロバクトラム・インターメディウム株 LMG 3306のリポ多糖類(IM)の使用を提供する。
【0020】
本発明の特定の態様は、免疫抑制されたヒトを含む動物における、感染症の処置および/または予防のための医薬の製造におけるIMの使用を提供する。
【0021】
本発明により、オクロバクトラム・インターメディウム株 LMG 3306のリポ多糖について記載されている使用は、ヒトおよび動物の両方に適用可能である。故に、本発明を通じて、動物における使用に関する記載は、ヒトもまた包含する。
【0022】
まとめると、哺乳類の免疫刺激剤は、敗血症および内毒素血症を処置および予防し、免疫抑制された動物におけるワクチン用アジュバント特性を有し、また足蹠リーシュマニア症における実験マウスモデルでの防御および治療効果を有する。
【0023】
IM(オクロバクトラム・インターメディウム株 LMG3306由来のリポ多糖類)は、
−マクロファージにおいてIL−12産生を誘導すること、
−本発明のLPSによって誘導されたTNFのレベルは、腸のLPSよりはるかに低く、病理学的効果を有さないこと、
−本発明のLPSはTヘルパー1(TH1)活性化およびγ−IFN産生を誘導すること、
を含む、免疫刺激剤として広範な活性を有することが証明された。
【0024】
本化合物は、哺乳動物における免疫応答を刺激する能力に関係している。故に、本発明は、オクロバクトラム・インターメディウム株 LMG3306 リポ多糖類および所望により1種またはそれ以上の薬学的に許容される賦形剤を含んでいてよい免疫刺激組成物を提供する。好ましくは、免疫応答の調節とは、ヒトの免疫能を増強することである。
【0025】
本発明は、敗血症および内毒素血症を予防および治療する化合物としての哺乳動物の免疫刺激剤であるIMの使用に関する。本発明は、特に、正常動物および実験的に免疫抑制された動物におけるワクチン効果を改善するためのアジュバントとしての使用に関する。この点で、そのアジュバント特性の例として、本発明は、特に、実験的なリーシュマニア感染症のためのワクチン用アジュバントとしての、およびリーシュマニア症、主に足蹠リーシュマニア症の実験マウスモデルを用いる皮膚リーシュマニア症の治療および防御効果としての、IMの使用に関する。
【0026】
本発明は、哺乳動物の免疫刺激剤として医薬分野で用いるための化合物としての、オクロバクトラム・インターメディウム株 LMG3306由来のLPSの製造方法に関する。
【0027】
本発明の化合物は、ウシ、ブタ、乳牛およびマウスで行う実験的野外試験における免疫刺激剤として特異的な活性を発現する:
−本発明の化合物と合わせて特定のワクチン接種後のIBRウイルスに対する中和抗体の増加の影響
−マクロファージ分化の誘導
−マクロファージにおけるTNFの誘導
−マクロファージにおけるIL−12の誘導
−脾臓リンパ球によるIFN−γ産生の誘導
−TLR4およびTLR2受容体による免疫刺激作用
−T細胞の刺激に対する用量依存的な影響。
【0028】
従って、本発明は、オクロバクトラム・インターメディウム株 LMG3306 リポ多糖類(LPS)の、哺乳動物における免疫応答を刺激し得る能力に関する。故に、本発明は、オクロバクトラム・インターメディウム株 LMG3306 リポ多糖類および所望により1種またはそれ以上の薬学的に許容される賦形剤を含んでいてよい免疫刺激組成物を提供する。好ましくは、免疫応答の調節とは、哺乳動物の免疫能を増強することである。
【0029】
本発明はまた、オクロバクトラム・インターメディウム株 LMG3306 LPSまたはそれを含む医薬組成物を哺乳動物に投与することを含む、哺乳動物における免疫応答を調節する方法に関する。故に、本発明はまた、哺乳動物に一般的な細胞性免疫を誘導するための医薬の製造における免疫刺激組成物の使用に関する。
【0030】
本発明はまた、オクロバクトラム・インターメディウム LMG3306のLPSを哺乳動物に投与することを含む、哺乳動物におけるマクロファージの分化誘導、TNF産生、IL−12産生、脾臓リンパ球によるIFN−γ産生、およびウイルス特異的CD8およびCD4 T−細胞の活性化のための医薬の製造における本発明の免疫刺激組成物の使用に関する。
【0031】
他の態様において、本発明の免疫刺激組成物は、ワクチン中に含まれる特定のウイルスおよび/または細菌に対してより免疫性にする(よりよい免疫化)ためのワクチン用アジュバントとして使用される。
【0032】
これらの致死未満量または他の低毒素LPSによる内毒素ショックの予防に関するほとんどの研究が、予め接種が必要であることを示している(Hatao, F., et al., The induction of super − resistance using synthetic lipopolysaccharide receptor agonist rescues fatal endotoxemia in rats without excessive immunosuppression. Shock2005; 23 (4): 365−70)。本発明者らは、驚くべきことに、オクロバクトラム・インターメディウム株 LMG3306のリポ多糖が、前もって脱感作期間を要せず、オクロバクトラム・インターメディウム株 LMG3306のリポ多糖類と毒性作用を保護したLPSを合わせた接種のみを要することを見出した。
【0033】
また、ほとんどの公表データで、インビトロまたは細胞株でのTNFの抑制とインビボでの脱感作作用との関係を見出されている(例えば、Lehner, MD, et al, lipopolysaccharide and Highly Purified Lipoteichoic Acid Via Different Toll −Like receptors Independent of Paracrine Mediators. 2,001. J. Immunol. 166: 5161−5167を参照)。しかしながら、本発明者らは、オクロバクトラム・インターメディウム株 LMG3306のリポ多糖が、インビボでLPSの作用を保護するが、マクロファージ株でも腹腔マクロファージまたは脾臓細胞においてもTNFまたはIL−12の分泌を低下させないことを見出した。インビボでの予備的結果はまた、オクロバクトラム・インターメディウム株 LMG 3306のリポ多糖(IM)が、何らかの機序で、LPSにより誘導されるTNFレベルを大幅に減少させ得ることを示唆する。何れの場合も、TNFレベルと保護に直接的な関係はない。これらの結果は、驚くべきことである。
【0034】
IL−10またはTGF−βの増大した産生の脱感作作用に起因すると考える他の仮説がある(Randow, F. et al., Mechanism of endotoxin desensitization: involvement of 35 interleukin 10 and transforming growth factor beta. J. Exp. Med 1995. 181, 1887−1892)。しかしながら、IL−10が、IMによりインビトロでほとんど誘導されないという事実により、そのような場合はなく、実際に、何かが腹腔マクロファージまたは脾臓細胞中でのIL−10の産生を阻害し(けれども、それは有意ではない)、LPSにより誘導されるIL−10のレベルは、C57BL/6マウス由来の脾臓細胞の場合を除いて、IMにより顕著に増大されないようである。
【0035】
他の興味のある局面は、病原体間の免疫応答を調節するIMの能力である。従って、リーシュマニア属の主な感染系を用いて、IMが、CpGと同様の方法ではなく、免疫源性リーシュマニア抗原抽出物の作用を増強することが明らかになった。CpGは、TLR9とのそれらの結合を介して、Th1応答を増強する(それは、リーシュマニア属および多くの他の細菌のような病原体から保護する)公知の免疫調節剤であり、免疫調節剤およびワクチンアジュバントとして特許が取得されている。しかしながら、IMは、インビボでTh1/Th2 プロフィールを変化させず、故に、CpG以上に悪化させる。このことは、IMが、体液性(Th2)および細胞性(Th1)の両方が防御に重要な役割を果たす感染における良好なCpGアジュバントであり得ることを示唆し得る。防御反応だけでなく、IMが防御を直接誘導し得て、すなわち、リーシュマニア属による感染に治療効果を有することを示すことを証明したことはおそらくより重要である。
【0036】
本明細書および特許請求の範囲の全体にわたって、用語“含む”および該用語の変形は、他の技術的特徴、付加物、成分または工程を除外することを意図しない。さらなる発明の目的、利点および特徴は、当該記載の検討により当業者に明らかになり得るか、または本発明の実施により習得され得る。以下の実施例および図面は、説明を目的として提供され、それらは、本発明を限定することを意図しない。さらに、本発明は、本明細書に記載の特定かつ好ましい態様の全ての可能性のある組み合わせを包含する。
【図面の簡単な説明】
【0037】
【図1】図1は、IMの使用による、大腸菌のLPSによって引き起こされる毒性作用の予防を示す。5匹のマウスの群を、LPS(1.75mg)またはIM(オクロバクトラム・インターメディウムのLPS)0.6μg/Kg(用量60)もしくは3μg/Kg(用量300)単独、またはそれらの組合せで処理し、生存を評価した。
【図2】図2は、LPS内毒素ショックの処置を示す。5匹の動物群をLPSで処理する(2mg/動物)。24時間後、それらにIM(オクロバクトラム・インターメディウムのLPS)0.6μg/Kg(用量60)または3μg/Kg(用量300)を注入し、生存を評価した。
【図3】図3は、大腸菌性腹膜炎の予防を示す。5匹のマウスの群に大腸菌(コロニー形成のための1E7ユニット、CFU)を注入し;24時間後に、動物にIM(オクロバクトラム・インターメディウムのLPS)0.6μg/Kg(用量60)または6μg/Kg(用量600)を注入し、生存(上パネル)および臨床症状(下パネル)を評価した。
【0038】
【図4】図4は、大腸菌性腹膜炎の治療を示す。5匹のマウスの群に大腸菌(コロニー形成のための1E8ユニット、CFU)を注入し;4時間後、動物にIM(オクロバクトラム・インターメディウムのLPS)0.6μg/Kg(用量600)を注入し、生存(上パネル)および臨床症状(下パネル)を評価した。
【図5】図5は、リーシュマニア症に対する防御免疫および作用経路の免疫図の一般的特徴を示す。図に示す通り、寄生虫とマクロファージの相互作用は免疫反応を生じ、それが主にTh1 リンパ球活性化に基づくとき、寄生虫に対する反応は有効かつ成功裏であるが、主にTh2の応答であるときは、感染が現れ得る。
【図6】図6は、リーシュマニア症に対するワクチンにおいて、可能性のあるアジュバント IM25(オクロバクトラム・インターメディウム LMG3306のリポ多糖類)を評価するための免疫化プロトコールを示す。5匹のマウスの群に、0日、15日および30日目にSLA抗原単独またはCpGまたはIMをワクチン接種した。60日目に、各動物の足蹠に1000プロマスチゴート(promastigote)を注入し、病変部の変化および免疫パラメーターを評価した。
【0039】
【図7】図7は、SLAおよびSLA−CpG(上パネル)またはアジュバントとしてSLA−IMをワクチン接種した動物における病変の変化を示す。感染による足蹠の腫れを定量する。
【図8】図8は、SLAおよびSLA−CpG SLA−IM 対 対照(PBS)をワクチン接種した動物の足蹠における傷の巨視的状態を示す。図中、6週間の感染期間中の、対照群の動物、1匹のSLAをワクチン接種した動物、1匹のSLA−CpGをワクチン接種した動物および6匹のSLA−IMをワクチン接種した動物の写真が含まれる。
【図9】図9は、脾臓およびリンパ節(DLN)中の寄生虫数の定量化を示し、SLAでのワクチン接種は、対照群に対して該臓器中で計数された寄生虫数の如何なる減少も生じないが、SLA−CpGは、その数をおよそ2log低下し、SLA−IM群でも同様の低下を示す結果が得られた。
【0040】
【図10】図10は、屠殺した動物の脾臓由来の細胞(単離され、インビトロでSLAで刺激された細胞)におけるTh1応答(IFN−γ)および/またはTh2(IL−4)の評価の結果を図示する。
【図11】図11は、特異的抗体アイソタイプIgG2a(Th1)およびIgG1(Th2)に関して、リーシュマニア属レベルの分析結果によって変わるTh1/Th2応答の評価を示す。
【図12】図12は、リーシュマニア症に実験的に感染させた動物における、IMの治療効果を評価するプロトコールを示す。
【図13】図13(A)は、IMをワクチン接種した動物における病変部の評価を示す。感染による足蹠の腫れを定量する。(B)は、対照群(PBS)と比較して、IMをワクチン接種した動物の足蹠病変部の巨視的状態を示す。6週間の感染中の、1匹の対照群の動物、1匹のCpGをワクチン接種した動物、および全ての6匹のSLA−IMをワクチン接種した動物の写真が含まれる。(C)は、7週後の脾臓およびリンパ節(DLN)中の寄生虫数の定量化を示す。
【0041】
【図14】図14は、IgG2a/IgG1比を含む、IgG1およびIgG2a抗体に対する、PBS対照サンプル、CPGサンプルおよびIMサンプルの比較表を示す。
【図15】図15は、免疫抑制された動物におけるアジュバントRTの役割を分析するために用いるプロトコールの概要を示す。5匹の動物群を、デキサメサゾンで免疫抑制し、SLA、SLA−CpGまたはSLA−IMをワクチン接種した。24時間後、動物を感染させ、その後8日目に再感染させた。60日目に屠殺後、病変部の変化および免疫パラメーターを評価した。
【図16】図16は、マウスの脾臓細胞における、SLAをワクチン接種した後のIMのアジュバント作用を示す。脾臓細胞を、抗原SLA(10g/ml)の存在下または不存在下で72時間培養し、細胞増殖を、72時間の間のDNAへのトリチウム化チミジンの挿入により評価する。結果は、各実験において6匹の動物を用いてトリプリケートで2回実験した平均を示す。
【0042】
【図17】図17は、ワクチン接種した動物由来の脾臓細胞によるIL−4産生への影響を示す。脾臓細胞を種々の動物から単離し、SLA(10g/ml)で刺激し、ELISAによりIL−4の分泌を分析した。
【図18】図18は、ワクチン接種し、デキサメサゾン(DEX)で処理したマウスの脾臓細胞によるIL−4およびIFN−γの効果を示す。
【図19】図19は、ワクチン接種した動物におけるSLAに対する細胞応答を示す。SLAのインビトロ添加後17日目での、IFN−γに対する特異的応答およびSLAに対する応答を示す。IFN−γ分泌(Th1)/IL−4(Th2)の比を示す。
【0043】
【図20】図20は、生マクロファージにおける、大腸菌LPSと比較した、免疫刺激化合物の添加後のJ774細胞の分化を示す。本発明の免疫刺激化合物は、J774マクロファージの分化の用量依存的阻害をもたらす。この作用は、J774または生マクロファージにおける重量/容量に基づく大腸菌由来のLPSで観察されたものよりもおよそ500倍低い(データ示さず)。
【図21】図21は、刺激していないマクロファージは、ELISAにより検出可能なTNFを合成しないことを示す。0.1−10μg/ml用量の本発明の免疫刺激組成物は、J774 マクロファージ(図21a)ならびに生細胞(データ示さず)において、用量依存的に4000pg/mlまでの有意なTNF産生反応レベルを誘導する。本発明の免疫刺激剤の効果は非常に有意であるが、大腸菌由来のLPSよりおよそ500倍低かった(図21b)。
【0044】
【図22】図22は、本発明の免疫刺激組成物が、マウスのBalb/cおよびC57B16株両方由来のマクロファージにおけるTNF産生を誘導するが、同じ濃度で大腸菌LPSより有効性が低いことを示す。
【図23】図23は、10mμg/mlの用量の本発明の免疫刺激剤は、C57Bl/6およびBalb/cの両方由来の腹膜マクロファージにおける、多量のIL−12を誘導することを示す。
【図24】図24は、コンカナバリンA(ConA)刺激が、両株のマウス由来の脾臓細胞による多量のIFN−γ分泌を誘導することを示し、IFNγの産生により特徴付けられるTh1応答を刺激することを示唆する。1−10μg/ml用量の本発明の免疫刺激組成物は、C57Bl/6およびBalb/cの両方由来の脾臓リンパ球の多量のIFNγを誘導する。
【0045】
【図25】図25は、本発明の免疫刺激組成物が、C57BI/6由来の腹腔マクロファージにおけるTNFを誘導するが、マクロファージにおけるTLR−2欠失は単に部分的に活性を阻害するのみで、TLR4の欠損は強く誘導を低下させることを示す。
【図26】図26は、本発明の免疫刺激組成物が、C57BI/6由来の腹腔マクロファージにおいてIL−12を誘導するが、マクロファージにおけるTLR−2の欠失は、単に部分的に活性を阻害するのみで、TLR4の欠損は強く誘導を低下させることを示す。
【図27】図27は、コンカナバリンA刺激がIFN−γ分泌を誘導し、それは、C57Bl/6 TLR2−/−由来の脾臓細胞で非常に低下し、C57Bl/6 TLR4−/− 細胞では、10mgの免疫刺激化合物を用いたとき、ほぼ完全に消失することを示す。
【図28】図28は、B6 C57マウス由来の脾臓細胞をリンパ球性脈絡髄膜炎ウイルスで感染させた後、異なる刺激;左から右へ:非ペプチド;LCMVNP396ペプチド(ペプチドNP396);培地のみ;PMA+イオノマイシン;異なる濃度の免疫刺激化合物、の添加によって誘導されたT細胞応答性CD4(図28a)またはCD8(図28b)を示す。
【0046】
特定の態様/例の詳細な説明
用語“免疫刺激組成物”は、本明細書で広義に定義され、該製品を接種された哺乳動物における免疫応答を刺激し得る投与可能な形態の任意の生物学的物質を意味する。
【0047】
本明細書で用いる用語“精製した”は、それがLPSと関係するとき、LPSが、それに限定されないが、上記の方法のような分画法または精製法により、種々の他の成分を除去され、組成物が実質的にその発現される生物学的活性を保持しないことを示す。用語“実質的に精製した”を用いるとき、この意味は、LPSが該組成物の主な非溶媒成分を形成する組成物を意味し得る。例えば、“実質的に精製したLPS”は、溶液または組成物の非溶媒成分の約50%以上、約60%以上、約70%以上、約80%以上、約90%以上、約95%以上が本発明のLPSであることを示す。
【0048】
本発明の好ましい態様に従い、免疫刺激組成物は、非溶媒成分は、0.2%以上の他の化合物(DNA、RNA、タンパク質、グルカン、脂質等)が混入してない、実質的に精製された形態のオクロバクトラム・インターメディウム株 LMG3306のLPSを含む。
【0049】
“免疫学的有効量”は、哺乳動物における検出可能な細胞性または体液性免疫応答を誘導するのに十分な免疫源の量を意味する。
【0050】
本明細書で用いる“防御免疫応答”は、特定の病原体により引き起こされる感染または疾患を予防または遅延する細胞性または体液性免疫応答を意味する。
【0051】
本発明の化合物は、賦形剤、pHおよび濃度に基づき常套的製造技術を用いて製造され得る。
【0052】
本発明の態様に従い、リポ多糖類は、オクロバクトラム・インターメディウム LMG3306から以下の工程で抽出される:
a)オクロバクトラム・インターメディウム LMG3306の不活化培養物を遠心し;
b)得られた沈殿物を再懸濁して生理懸濁液とし;
c)精製水およびポリエチレングリコールを用いて透析し;
d)4倍量のメタノールおよび1%の酢酸ナトリウム入り飽和メタノールで沈殿させ;
e)凍結乾燥させて、粗LPSを得る。
【0053】
好ましい態様に従い、一旦、粗LPSを凍結乾燥させて、それを以下の工程で精製する:
1. 粗LPSを、要すれば超音波を用いて、緩衝液(10mM Tris−HCl pH7,5)に溶解し;
2. プロテイナーゼKを添加し、室温でインキュベートし;
3. 精製したLPSを超遠心分離により集め;
4. サンプルを凍結乾燥させる。
【0054】
本発明の免疫刺激化合物の製造方法の好ましい態様のより詳細な記載を、以下のフローチャートで表わす:
オクロバクトラム・インターメディウム LMG3306 LPの抽出法のフローチャート:
【表1】
【0055】
製造法は、以下の工程を含む:
i)ワーキングシード細菌(LMG3306株)の解凍
接種前準備:
1. Triptycase Soy Broth(TSB)を1L準備する
2. 60mlのTSBを含むチューブを準備する
3. ワーキングシード凍結バイアルから種菌(pearl)を取り出し、60mlのTSBを含むチューブにそれを加える
4. 37±0.1℃で24時間、振とうしながらインキュベートする
5. 前接種源の制御:同定および純度制御:コロニーの形態および培養物の顕微鏡的形態の確認。
【0056】
ii)制御発酵槽中の増殖:
接種物製造
1. スクロース 2‰を添加した培養培地TSBを準備する
2. スクロース 2‰を添加した5LのTSB(pH6.75±0.15)を含む接種ボトルを準備する
3. 培養培地を含む接種ボトル中で前接種源を接種する
4. 37±0.1℃で24時間、振とうしながらインキュベートする
5. 接種源の制御:同定および純度制御:コロニーの形態および培養物の顕微鏡的形態の確認。
【0057】
培養
1. 発酵装置を浄化および滅菌する
2. スクロース 2‰を添加した400LのTSBを準備する
3. スクロース 2‰を添加し、pH6.75±0.15に合わせた400LのTSBを発酵装置に入れる
4. 培養培地を含む発酵装置に接種源を入れる
5. 37±0.1℃で24時間、最大50rpmで振とうしながらインキュベートする
6. 培養物の制御:同定および純度制御:コロニーの形態および培養物の顕微鏡的形態の確認。
【0058】
不活性化
1. 滅菌導管により不活性化タンクに培養物を移送する
2. ホルムアルデヒド溶液を、終濃度0.4%まで添加し、37℃で10rpmで振とうしながら3日間インキュベートする。
【0059】
遠心分離
1. 発酵槽を連続遠心分離機に接続し、発酵槽の内容物を遠心する
2. 10リットル容量のプラスチックボトルに不活化培養物を入れ、使用まで4℃で貯蔵する。
【0060】
iii)以下に記載の通り、LPS抽出法を行う:
1−オクロバクトラム・インターメディウム LMG3306の不活化培養物を遠心する。上清を除去する。
2−得られた沈殿物を再懸濁し生理懸濁液とする:200g/l 15mM NaCl。
3−65℃で、水溶液に90%でフェノールを添加する。
4−65℃で20分間振とうする。混合物を10℃まで冷却する。
5−9000xgで20分間遠心する。4℃で24時間維持する。
6−初めに純水で、次いでポリエチレングリコールで透析する。
7−4倍量のメタノールおよび1%の酢酸ナトリウム含有飽和メタノールで沈殿させる。
8−9000xgで20分間遠心する。上清を除去する。抽出物を純水で再懸濁し、振とうする。
9−工程7を繰り返す。
10−サンプルを凍結乾燥して、粗LPSを得る。
【0061】
iv)以下の方法によるLPSの精製:
1. 超音波を用いて、100mgの粗LPSを20(5mg/ml)の緩衝液(10mM Tris−HCl pH7,5)に溶解する。5−10mgのLPSを対照として使用する。
2. 0.5mlのプロテイナーゼK(1mgのLPS当たり、50μg/mlまたは5μgのプロテイナーゼ)を添加する。室温で12時間、撹拌および軽く振とうしながらインキュベートする。
3. 純粋なLPSを超遠心(100.000×g、6時間)により集める。沈殿物を純水に再懸濁する。
4. サンプルを凍結乾燥させる。
【0062】
v)バルク溶液を以下の通り製造してLPS濃度を計測:
材料および反応材
1. 1,25 N H2SO4:H2O DD中、6,66mlの市販の反応材で100mlとする。
2. 0,042N 過ヨウ素酸:100mlの0,125N H2SO4中、0,48gの過ヨウ素酸(パラ過ヨウ素酸、H15IO6)
3. 0,5N HCl中、2% 亜ヒ酸ナトリウム(200ml H2Oに8,3ml HCl濃縮物)
4. チオバルビツール酸(Sigma):0,3% pH2(H2O DD中に溶解、50℃)。15日以内に使用する。
5. ジメチルスルホキシド
6. 標準:
Kdo(分子量=255,1)(Sigma、−20℃で貯蔵)
デオキシリボース(分子量=136)(Sigma、−20℃で貯蔵)
7. 上記のバッチから定量したLPS
8. ビー玉(marble)が合致するガラス管(第二クロム混合物または等量物で浄化)。
9. 分光光度計。
【0063】
方法
1. Kdo標準(0.05μg)、デオキシリボース標準(0.025μg)、上記のバッチから定量したLPS(0.2μg)およびブランク反応材(200μl H2O)を含むチューブを準備し、
a)サンプルが抽出したLPSであるとき、凍結乾燥させ、0.2μgを計量し、1つのチューブに準備する。
b)サンプルが最終生成物のとき、200μlを含むチューブを準備する。
チューブにH2Oを加えて200μlとする。
2. サンプルを含むチューブを氷上に置き、20μlの1.25 N H2SO4を各チューブに加え、それらをビー玉で栓をして正確に20分間沸騰させる。氷上で冷却する。この工程は、標準サンプルには行わない。
3. 0.25mlの過ヨウ素酸をサンプルおよび標準サンプルに迅速に加え、振とうし、それらを室温で20分間インキュベートする(最初のチューブから計る)。過ヨウ素酸を加えるため、チューブをH2SO4が添加されたときと同じ順にしておく。測定されたKdoの量は、Kdoの可溶化によって、また加水分解条件中のその分解によっても変わるため、加水分解時間を最適化しなければならない(20分は、オクロバクトラム・インターメディウム LMG3306 LPSにとって十分である)。
4. 3と同じ順のチューブを、0.5mlの亜ヒ酸ナトリウムを添加し、ボルテックスして振とうし、最初のチューブへの添加から2分間待つ(試薬の添加により現れる黄色が、この時消失する)。
5. 各チューブに前と同じ順で2mlのチオバルビツール酸を迅速に添加する。それらをビー玉で栓をして20分間沸騰させる。
6. 各チューブに正確に1mlのDMSO(混濁の出現を避ける)を添加し、その後、それらを、水道水を入れたトレー中で冷却する。
7. 室温で30分間インキュベートする。ランプを安定化させるために、初めの30分間、分光光度計を点けておく。
8.各チューブを552nmおよび536nm(Kdoおよびデオキシリボースそれぞれの最大吸収)で読み出す。プラスチックトレーを用い得る。
【0064】
測定
Mk552=Abs552/μmolkを、Mk536、Md552 y Md536と同様に計算する
【化1】
k:KDO;d:デオキシリボース;Abs:吸光度。
【0065】
vi)以下に記載の最終プロセスまでのバッチの均質性:
定量化プロセスのこの均質性確認法は、製品製造中、EMEA/CVMP/598/99“プロセスバリデーションのガイダンスノート”に従って2回用いられる:
−オクロバクトラム・インターメディウム LMG3306不活性化培養物からのLPS抽出の終わりに、その濃度を知るために用いる。
−最終生成物中のLPSの量を調べるために用いる。
【0066】
該確認法は、抽出段階の終わりに適用されるプロセスである。分析法で用いられる量は、最終製品中で定量される量よりもはるかに多い。抽出されたLPSの濃度を知るためにこの方法を用いる。
【0067】
LPS定量化は、その成分の1つであるKdoの定量化で行う。Kdoは、LPS分子の一部であり、特定のLPSに常に一定量存在するが、異なる細菌LPSでは量が変わる。オクロバクトラム・インターメディウムにおいて、LMG3306は、各LPS分子について2分子のKdoである。
【0068】
定量化は、分光光度法によって行われる。
【0069】
Kdoの定量化は、サンプルが抽出LPSであるとき、LPSの抽出および精製工程の最後に残る残渣デオキシリボース(DNA成分)が、Kdoの最大吸収波長(552nm)での吸光度によるアッセイを妨げ得るため、明確ではない。
【0070】
標準Kdoを用いるKdoの定量化の確認は、Kdoが線形的に正確かつ正しく定量されることが証明され得る。LPSサンプルがデオキシリボース残渣の混入を含むとき、該方法の明確性の欠如は、補正式を用いて解決される。この式は、この干渉がサンプルに存在するとき、得られた理論的Kdo値をデオキシリボース干渉を除去する実数値に合わせる。
【0071】
デオキシリボース定量の直線性は、この成分が単独で直線性を示すだけでなく、LPSの汚染物質としても直線性を示すことを意味する。
【0072】
この方法が使用される毎に、異なる濃度の標準Kdoを含むサンプル、異なる濃度のデオキシリボースを含むサンプル、異なる濃度の上記で定量したLPSを含むサンプル、および異なる濃度の試験サンプルを含むサンプルを用い、またそれらは全て、デオキシリボース最大吸収波長(552nm)で測定され得る。
【0073】
以下の工程を行って結果を得る:
−標準Kdoを含む標準直線:直線性を試験する。
−標準デオキシリボースを含む標準直線:直線性を試験する。
−上記の通り定量したLPSを含む直線。方法をチェックする。直線性を試験する。
−標準Kdo、上記の通り定量したLPSおよび試験サンプルについて552nmでの吸光度、ならびに標準デオキシリボースについて536nmでの吸光度を分母として、各サンプルについて得られた吸光度値の商を計算する。
【0074】
式を適用する際、選択された値が標準直線内である限り、商値が標準Kdoおよび標準デオキシリボースの濃度から得られた値に最も近い試験サンプルの濃度値を用いる。値が直線内でない場合、最も近い値を変わりに用いる。
【0075】
式の適用は、試験サンプル中のKdoマイクロモルを決定する。
【0076】
本発明はさらに、本明細書で提供するLPS免疫刺激化合物を1種以上の薬学的に許容される担体または賦形剤と混合して含む医薬組成物を提供する。適当な担体は、投与経路と処置されるべき状態によって変わり得る。本発明の医薬組成物を、抗原に対する免疫応答を増大するか、または免疫系の細胞の活性を増強するためにアジュバントとして用い得るか、またはいくつかの例において、対象における特定の状態を予防または処置するために予防用または治療用組成物として用い得ることが理解され得る。
【0077】
故に、一態様において、本発明は、本発明のLPSおよび薬学的に許容される担体を含む医薬組成物を提供し、ここで、LPSは、免疫応答を調節するのに有効な量、所望の免疫応答を刺激し、疾患または状態を処置または阻害するという意味で所望の効果を達成するのに必要とされる化合物量で存在する。本医薬組成物はまた、抗原と共に投与されるとき、アジュバントとして作用する。
【0078】
本発明の免疫刺激組成物は、広範な投与量および広範な比率で投与されたとき、強力な免疫刺激効果を示す。好ましい態様によれば、免疫刺激組成物は、0.5ないし120μg/mlのLPSを含む。より好ましくは、組成物は、0.5ないし10μg/mlのLPSを含む。
【0079】
好ましい態様に従い、リポ多糖類は、ミセル相が4℃で1年以上安定である均質な懸濁液形態である。
【0080】
好ましくは、本発明の免疫刺激組成物のpHは、2−12である。
【0081】
ワクチン用量と共に投与された免疫刺激剤の量は、一般的に、それがワクチンと共に、典型的なワクチンにおける重大な副作用のない免疫保護応答を引き起こす量として選択される。かかる量は、どの特定の免疫源が使用され、かつそれらがどのように提示されるかによって大きく変わり得る。一般に、各用量は、約1−1000μgの抗原および0.1−200μgの免疫刺激剤を含み、最も典型的には、約2−100μgの抗原および0.5−150μgの免疫刺激剤を含み、好ましくは約5−50μgの抗原調製物および1−75μgの免疫刺激剤を含むと予期される。もちろん、投与された投与量は、年齢、体重、もしあれば併用処置剤の種類、ならびに投与される抗原および免疫刺激剤の性質によって変わり得る。
【0082】
本発明の免疫刺激組成物の投与経路は、一般的な免疫刺激剤を送達する何れかの好適な経路であり得る。しかしながら、免疫刺激組成物は、好ましくは筋肉内または皮下経路を介して非経腸的に投与される。より好ましくは、免疫刺激組成物は、皮下投与される。要すれば、他の投与経路も用いられ得て、故に本発明の組成物の例は、懸濁液、シロップまたはエリキシルのような、開口部用、例えば経口、経鼻、経肛門、経膣等の投与用液体調製物;および、滅菌懸濁液またはエマルジョンのような非経腸投与、皮下投与、皮内投与、筋肉内投与または静脈内投与(例えば、注射可能投与)用調製物を含む。そのような組成物において、本発明の免疫刺激組成物は、好適な担体、希釈剤、または滅菌水、生理的食塩水、グルコースなどのような賦形剤と混合され得る。
【0083】
適当な免疫保護剤およびかかる用量の非毒性用量は、当業者に容易に決定され得て、すなわち、本発明の製品中に含まれる本発明の菌株の適当な免疫保護かつ非毒性量は、有効量のLPSの範囲内であり得る。適用に推奨される最低の年齢はなく、実際に1日で行ういくつかの実験が示される。
【0084】
もちろん、必要ならば投与は適当な間隔を空けて繰り返され得る。
好ましくは、投与スケジュールは以下の通りである:
・皮下経路により0.1μg/Kg体重(b.w.)の第一投与。
・15日後の同じ投与量での第二投与。
さらなる投与:6ヶ月−1年の適用がより推奨される(0.1μg/Kg b.w.皮下経路)。
【実施例】
【0085】
実施例1.内毒素血症の処置および予防
これに関して、本発明者らは、Balb/cマウスの急性モデルにおいて大腸菌由来のLPSを用いる。
第一に、本発明者らは、6日間で100%死亡率をもたらす大腸菌LPSの用量を決定する(致死量100)。
大腸菌株O111:B4のLPSを、SIGMA社(St. Louis, MO, USA)から購入した。全ての実験において、LPSの希釈剤としてリン酸塩緩衝液を用いる。全ての場合において、用いるビークルはリン酸緩衝生理食塩水(PBS)であった。生存実験用のLPSの適当用量は、LPSの増大用量をメスBALB/cに腹腔内(IP)投与したときの用量応答曲線により決定した。
【0086】
内毒素血症の致死作用の予防:
内毒素血症を有するメスBALB/cマウスを、LPS(大腸菌 O111:B4)暴露の前に24時間、2つの異なる用量のIM、0.6μg/Kg(用量60)または3μg/Kg(用量300)でIP処理するか、または処理しなかった。LPSは、マウス1匹当たり1.75mgをIP投与した。
敗血症の場合に等しい急速輸液を供するために、全ての注入は、同量であった。
【0087】
IMは両用量で、大腸菌のLPSの内毒素ショックによる動物のLPS関連死を完全に予防した。さらに、IM処置した動物は、敗血症性ショックに関連した何らの徴候も示さなかった(図1)。用いたLPSの用量は、IMの用量よりもはるかに多かった。故に、IM(オクロバクトラム・インターメディウム 3306のLPS)は、LPSと競合され得るとは考えられない。
【0088】
内毒素致死性ショックの処置:
敗血症の主な問題は、通常、その過程が始まるときに、その予防ではなく、むしろ処置である。この試験のために、5匹のマウスを、LPS IM 0.6μg/Kg(用量60)または3μg/Kg(用量300)IPで処置の24時間後、動物1匹当たり2mgのLPS(大腸菌 O111:B4)でIP(腹腔内)感染させた。IMは、LPS誘導性死亡に保護効果を有する。故に、プラセボ処置後72時間で、全ての動物が死亡する。しかしながら、IM処置は、20%の動物の生存をもたらし、それらの死を顕著に遅延させたこのことは、IMが、内毒素ショックを有するマウスを部分的に処置し得ることを示す(図2).
【0089】
驚くことに、IM処置した動物は、プラセボ群においてもより低レベルの血液凝固を示した。この事実は、過剰な凝固の問題が敗血症の現象と関係するため非常に重要であり、実際、この条件に最近承認された化合物のみが凝固の阻害作用を有する。
【0090】
細菌性腹膜炎:
オクロバクトラム・インターメディウム LMG 3306のリポ多糖類を、敗血症の他のモデルである大腸菌誘導性腹膜炎誘導性敗血症で試験した。
【0091】
大腸菌性致死性腹膜炎の処置:
この実験に関して、メスC57BL/6マウスを、動物を、LPS IM、0.6μg/Kg(用量60)または3μg/Kg(用量300)、または単一のIP投与を用いるプラセボで処置した24または4時間後に、107または108の生存O26:B6 大腸菌コロニー形成単位(CFU)を接種した。
【0092】
生存および病理的症状(摂食障害、死亡率、脱毛、choquexia、身体活動など)をその後に評価した。
大腸菌注入の24時間後、107 CFUを最初接種した何匹かのマウスが死に(それらの20%)、かつ敗血症の病的症状を示す(0から5段階で平均3)。
IMでの処置は、用量応答的に動物の死を防ぎ、敗血症の病的症状を完全に回復させた(図3)。
【0093】
108 CFUの大腸菌の用量が用いられたとき、全ての動物は24時間で死亡する;故に、それらは大腸菌の摂取の4時間後にIMで処理された。それにも関わらず、その時、マウスは何れも死んでおらず、病理的症状の平均スコアは非常に高かった(5段階のうち4)。全ての大腸菌感染マウスが、14時間で死んだ。それにも関わらず、IM 3μg/Kg(用量300)は、ほとんどの動物の死を防ぎ(80%生存率)、臨床的重症度を徐々に低下させた(図4)。
【0094】
リーシュマニア症の実験的感染におけるアジュバント効果
足蹠感染におけるアジュバント効果:
この抗原は、Th1保護応答を活性化し得ない(図5)ため、SLA(可溶性リーシュマニア抗原)抗原抽出物が、主なリーシュマニア原虫での実験的感染に対する保護効果をもたらさないことは、よく知られている。しかし、SLAが、CpGとしてアジュバントと結合するとき(米国特許第7521063号)(TLR9による免疫応答を刺激し、Th1応答を増大させるためのワクチン用アジュバント)(Heeg, Int. J. Microbiol, 298, 33, 2008)、TLR9(トール様受容体)を介して保護的Th1(IFN−γ)の活性化をもたらし得る(米国特許第6890542号)。
【0095】
従って、このモデルは、実験的なリーシュマニア症に対するワクチン接種用のアジュバントとしてのCPGのよく確立された使用と比較してIMを試験するのに非常に有用である。
図6で、本発明者らは、この目的のためのプロトコールを示す。
【0096】
基本的に、SLA抗原は、単独で、CPGと共に、またはIMと共に、0、15および30日目に接種される。
【0097】
60日目で、1,000寄生虫(promastigote)を各マウスの足蹠に接種し、病変および免疫学的パラメーターを観察する。感染の進行は肉眼で観察され得て、寄生虫数と関係する。図7に示す通り、摂取後、対照群での感染が確認される。SLA抗原は、何らかの保護効果を生じないが、SLA+CPGとSLA+IM群の両方で顕著に病変を縮小する。
【0098】
さらに、感染後6週の巨視的結果を図8に示し、SLA単独で処理した動物の足蹠形と比較して、SLA−CpGまたはSLA−IMで処理した動物の病変の顕著な縮小を証明する。
【0099】
さらに、寄生虫数を、足蹠および脾臓で定量した。感染後7週で、動物を屠殺し、脾臓およびドレインリンパ節(DLN)に存在する寄生虫を決定した。SLA免疫は、IMおよびCpGとは対照的に、寄生虫数の減少をもたらさない。CpGは、2logに近い数を減少させ、IMは、少なくとも、全6匹の動物のうち4匹で同様の結果を示した(図9)。
【0100】
簡単には、SLAは単独では保護しないが、CpGまたはIMとの組合せは感染に対する保護効果を有する。
屠殺した動物の脾臓細胞を集め、SLAで“インビトロ”で刺激し、Th1(IFN−γ)および/またはTh2(IL−4)応答を評価した。
【0101】
感受性Balb/cモデルにおけるリーシュマニア属の主な感染症は、IFN−γレベルよりもIL−4レベルを比較的増強することがよく記載されている。SLAワクチン接種はこのパターンを修正しないが、CpGとのその組合せは、IFN−γの産生によって定量化されたTh1応答を増大させ、それ故に、IL−4の相対的発現を低下させた。従って、IFN−γ(応答Th1)/IL−4(応答Th2)比は、顕著に増加した(図10)。
【0102】
驚くことに、IMは、CpGと同様のより強い保護作用を有するが、免疫学的機序は異なる。その保護は、両方のパラメーターが同時に増大するため(図10)、IFN−γ(応答Th1)/IL−4(応答Th2)の比と厳密に関連しない。
【0103】
応答Th1/Th2を決定する第二の方法は、リーシュマニア症−特有のアイソタイプ抗体IgG2a(Th1)およびIgG1(Th2)のレベルを分析することである。
【0104】
機序は明らかではないが、IgG2aレベルは、リーシュマニア症からの保護と関連する。SLAで処理しないか、またはそれのみで処理した動物において、IgG2aレベルは検出されなかった。しかしながら、IgG2aレベルは、CpGまたはIMで処理した動物で検出された。故に、IgG2a/IgG1比は、SLA−CpおよびSLA−IM群の動物と対照的に、非処理動物またはSLAでのみ処理した動物でより低い(図11)。
【0105】
足蹠実験マウスモデルにおけるIMの予防および治療効果:
図12において、リーシュマニア症を有する進行性実験的完成におけるIMの治療効果を評価するためのプロトコールを示す。
【0106】
これに関し、IMを、2つの異なる用量でリーシュマニア原虫と同時に接種した:
1)IMF−1、2および3群は、0.6μg/Kg(18ng/マウス)を受容した。
2)IMF−4、5および6は、1.2μg/Kg(36ng/マウス)用量を受容した。
結果は、IMが、リーシュマニア属による主な感染症から、CpGで得られた効力と同様の効力で保護し得ることを示す(図13A)。
【0107】
IMで処理した動物は、顕著な病変を示さなかった(図13B)。感染の7週間後、動物を屠殺し、脾臓およびリンパ節(LN)中の寄生虫数を定量した。同様の結果が、CpGおよびIM群で得られ、リンパ節中で顕著に減少し(10.000倍)、脾臓において寄生虫は見られなかった(図13C)。
【0108】
最後に、上記の実験に従って、CpG群は、抗体IgG2a/IgG1レベルの比に顕著な変化を示す。さらに、IM用量は、得られた上記の結果と一致して、この比に顕著な変化を示さない(図14)。
【0109】
最終的な結論として、本発明者らは、IMがCpGと同様の特性を有するが、Th1/Th2またはIgG2a/IgG1のプロフィールおよび比が変化しないため、免疫機序における最終結果が異なることを見出した。本発明において、IMは、両方の(CpGで実証される通り、Th1のみではない)応答の活性化による、より良好なアジュバントであり得る。
【0110】
IM(オクロバクトラム・インターメディウムのLPS)は、免疫抑制された動物におけるワクチンアジュバント特性を有する:
IMは、抗原としてSLAを用いて、リーシュマニア原虫感染に対してワクチンアジュバント作用を有することが既報である。
【0111】
この抗原は、免疫源性が低く、より重要なことには、防御作用であるTh1応答(IgG2a応答およびIFN−γ産生により特徴付けられる)ではなく、主にTh2応答(IgG1抗体の優位およびIL−4産生により特徴付けられる)をもたらす。
【0112】
免疫抑制されたマウスに対するアジュバント作用を試験するために、Balb/cメスを、動物1匹当たりコルチコステロイド・デキサメサゾ(Dex)20μg IPで処理し、24時間後に、動物に、SLAを単独(10μg/動物)で、またはIM、0.6μg/Kg(用量60)または3μg/Kg(用量300)、もしくはTh1応答をもたらすことが既報の化合物であるCpGと共に接種した(用いたプロトコールについては図15を参照のこと)。マウスに、1週間後にSLA単独を再び接種し、17日目に動物を屠殺し、脾臓細胞を集め、血清中の抗SLA抗体力価を評価した。
【0113】
図16に示す通り、特定の抗SLA抗体は、Dex単独の対照ではなくSLAを注入された全ての動物で検出されたが、ほとんどの抗体は、IgG1アイソタイプであった。記載される通り、CpGアジュバントは、IgG2a/IgG1比を増加させ、より重要なことには、IMは、インビボでのTh2からTh1への切り替えを表すこの変更をCPGよりも良好にもたらした。このことは、IMが、免疫抑制された動物においてアジュバント活性を有し、より重要なことには、免疫抑制された動物においてさえも好ましいTh1応答を維持し続けることを示した。
【0114】
加えて、異なる動物由来の脾臓細胞を得て、インビトロでSLAで刺激し、増殖およびIFN−γ(リーシュマニア症に対する保護に必要なTh1サイトカイン)を測定した。図7に示す通り、SLAは、Dex処理動物において増殖応答をもたらした。CpG処理動物は、改善した増殖応答を有した。興味深いことに、IMで処理した動物もまた、用量依存的に、SLAよりも良好な増殖応答を有する(図17)。分裂促進物質に対する応答の相違は、全ての動物で観察されなかった(データ示さず)。
【0115】
本発明者らは、“インビトロ”での脾臓細胞のSLA処置後の脾臓細胞の上清に放出されたIFN−γ(Th1サイトカイン)およびIL−4(Th2サイトカイン)を調べた。図18に示す通り、ワクチン接種していないDex対照動物は、インビトロでのSLA暴露によりIL−4またはIFN−γの産生が検出不可能であった。SLA接種された動物において、“インビトロ”でのSLA暴露は、IL−4産生を生じるが、IFN−γは非常に少なく、同じ動物の血清におけるIgG1/IgG2aデータと一致する。
【0116】
CpG処理は、IL−4およびIFN−γの比をほとんど変えることなく、両方の増加をもたらした(図19)。興味深いことに、IMのLPSは、両サイトカインを増加させるが、IL−4よりもIFN−γが高い割合で増加した。これは、とりわけ、3μg/Kg(用量300)で処理した動物で明らかであり、IFN−γ/IL−4比が10倍増加した(図19)。
【0117】
実施例2.製造方法
免疫刺激性LPSを、オクロバクトラム・インターメディウム株 LMG3306から上記の製造法に従って得た。
1. 上記の工程(ii)(製造方法)の発酵バルク抗原を得るための方法
2. 上記の工程(iii)(製造方法)のLPS抽出方法
3. 上記の工程(iv)および(v)(製造方法)の、LPSの精製およびLPS量の測定。
【0118】
生成物を、4℃で24時間までにバッチの均質性を維持するために、予め滅菌し、連続撹拌システムを供するステンレススチール製タンク中で貯蔵した。生成物を、滅菌ガス管システムを用いて、該タンクから容器詰めする場所へ移した。
【0119】
容器詰めを、100Bバックグラウンドを用いてクラス100Aの滅菌環境で行った。ゴム栓およびカプセル化を、同じ場所で行った。
【0120】
最終生成物を含むバイアルを、それらにラベルが付され包装されるまで4℃で貯蔵した。ラベル添付および包装を、別の場所で行い、そして各時間にその場所へ入れるのは、最終生成物およびその調整材料の1つのバッチのみである。販売までの最終貯蔵を、5±3℃で行った。
【0121】
実施例3.組成:
【表2】
【0122】
実施例4.Kdo試験:
本実験の主な目的は、上記のKdoの比色法によって、LPS1ml当たりのナノモル数の測定値の再現性を確認することである。
試験すべきパラメーターは、
1−アッセイ内の正確性
2−アッセイ間の正確性
3−バッチ間の正確性
である。
【0123】
1−アッセイ内の正確性の結果
このアッセイにおいて、本発明の生成物の1つのバッチ(A003A)を用い、3つのサンプルを、サンプル1つ当たり10回同様の操作を繰り返して、抗体力価を測定した。次いで、各サンプルの3つの希釈物(1/2、1/4および1/8)を、希釈物1つ当たり10回同様の操作を繰り返して、該アッセイの再現性を確認した。
【0124】
【表3】
【表4】
【0125】
結果は、試験が再現性であることを示す; VCは10%未満である。
【表5】
【表6】
【表7】
【表8】
【0126】
本発明者らは、各希釈物についてVCを調べた。得られた結果は、試験が再現性であることを示す; VCは10%未満である。
【0127】
2−アッセイ間の正確性
主に実験者の操作による変動を調べるため、本発明の製品の同じバッチを、3日の異なる日に滴定した。各日、全ての必要な溶液を新鮮に調製し、生成物サンプルを実験の最初から最終日まで4℃で貯蔵した。
このアッセイにおいて、本発明の生成物の1つのバッチ(A003A)を、異なる3日に滴定し、3つの異なるサンプルをサンプル毎に10個の複製物を用いて行った。
【0128】
【表9】
【表10】
【0129】
得られた結果は、試験の再現性を示す; VCは10%未満である。
【0130】
3−バッチ間の正確性
バッチ製造の変動を調べる:
このアッセイにおいて、本発明の製造物の3つのバッチ(A001A、A002AおよびA003A)を、滴定した。各バッチの3つのサンプルをサンプル毎に10個の複製物を用いて行った。
【表11】
【表12】
【0131】
Kdo活性の顕著な相違は、アッセイ内、アッセイ間および経時的アッセイにおいて観察されなかった。
【0132】
実施例5.ウイルス不活性化ワクチン用アジュバントとしての免疫源性試験
野外試験
この野外試験を、血清−陰性のウシにおける、常套的に不活性化したワクチン接種による免疫化によりもたらされる、血清、ウシヘルペスウイルス1(BHV−1)または感染性ウシ鼻気管炎ウイルス(IBR)に対する抗体全体および中和抗体に対する免疫応答(inmunitary answer)への免疫刺激剤の効力を評価するために行った。
【0133】
材料および方法
1.ウシ群の選択
試験群の選択のために、乳牛の他の5つの群からの120頭の動物を予め試験した。これらの5つの群中に血清−陽性の動物がいたため、サンプルを6つの群に分けた;試験した動物は、血清−陰性であって、この群を選択した。
【0134】
血清検査を、IBRウイルスに対する全抗体数を検出するELISA試験で行った。
【0135】
2.プロトコールの実施:処理、ワクチン接種およびサンプル収集
2.1.処理およびワクチン接種
試験を、IBRウイルスに感染していない22頭の血清−陰性の若い雌牛で行った。新しい動物を試験中に群に加えなかった。
【0136】
若い雌牛を以下の群に分けた:
−群1.−13頭の若い雌牛を、試験0日目および7日目に5mlの免疫刺激化合物(6μg/mL)で処理し、7日目および28日目に、不活性化IBRウイルスを含む5mlの承認された市販のワクチンをワクチン接種した。両製品を、首の筋肉内深部に注射した。
−群2.−6頭の若い雌牛を、7日目および28日目に、同様のワクチンおよび同様の方法でワクチン接種して免疫化した。
−群3.−3頭の若い雌牛を対照群とした;それらは、ウイルスが感染しなかった個とを確認するために、処理または免疫化しなかった。
【0137】
2.2.サンプル収集
各動物の血液サンプルを、試験0日目、7日目、28日目および58日目に得た。
【0138】
試験開発スキーム
【表13】
【0139】
3.ウシヘルペスウイルス(BHV−1)中和血清抗体の検出
I.ウシヘルペスウイルス1中和抗体力価を、標準ウイルス中和試験により決定した;100 DI50CT/25μlのウシ腎臓由来のよく確立された細胞株である“Georgia bovine kidney”(GBK)のColorado株(Prof. Marcelino Alvarez, Dpto. Sanidad Animal, Facultad de Veterinaria, Universidad de Leon, Espanaにより提供される)を用いて行われ、各血清を4回試験した。
【0140】
血清を純粋な血清から滴定した。血清をウイルス(25μlの希釈した血清および25μlのウイルス懸濁液)と混合して希釈し、その抗体力価を、Spearman−Kaurber法(TRIMMED SPEARMAN−KARBER (TSK) PROGRAM VERSION 1.5, ECOLOGICAL MONITORING RESEARCH DIVISION ENVIRONMENTAL MONITORING SYSTEMS LABORATORY, U. S. ENVIRONMENTAL PROTECTION AGENCY CINCINNATI, OHIO 45268)を用いて見出した。50%保護する血清の最も高い希釈倍率の逆数として表される。
【0141】
II.2つの実験群において、平均対数抗体力価が見出され、それはその逆対数の逆数として表される。血清転換を、2つの血清収集毎間の抗体の4倍増加として定義する。
【0142】
III.異なる試験日に処理した、また処理しなかった動物群について得られた平均対数抗体力価間の統計的に関連する相違の存在を確認するために、統計分析(Kinskall−Wallis non−parameter test)を、Windows−environment EpiInfoソフトウェアを用いて行った。
【0143】
結果および考察
I.試験中に血清転換した動物はいなかった。
II.7日目および28日目での血清転換率は、処理群および非処理群のそれぞれで92.3%および83.3%であり、28日目および58日目では、処理群および非処理群のそれぞれで69.2%および16.7%であった。
【0144】
両方の場合において、血清転換率は、非処理動物群よりも処理動物群で高かった;しかし、それらの群間の相違は、28日目と58日目での血清転換率でのみ統計的に有意であった。
【0145】
III.28日目において、IBR中和抗体の平均対数抗体力価は、処理動物群で3.50であって、非処理動物群で得られた力価2.37よりも顕著に高かった。
【0146】
58日目において、IBR中和抗体の平均対数抗体力価は、処理動物群で12であって、非処理動物群で得られた力価4.8よりも顕著に高かった。
【0147】
処理動物群で得られたIBRウイルス血清抗体力価は、非処理動物群より高く、28日目および58日目でそれぞれ1.5および2.5であった。
【0148】
結論
1.得られた結果により示される通り、免疫刺激化合物の投与は、ワクチン接種した動物における中和抗体および保護レベルとしての体液性応答を顕著に増大させた;故に、それは、ワクチン接種によって誘導される免疫応答の実現に重要な役割を果たした。
2.その適用は、
−その低廉な価格のために、ウシに適用される免疫化パターンとして推奨され、
−基本的に、動物が、ウシを太らせる場所のようなストレス下の状況にあるとき、これらの結果が子牛に乳を飲ませて草を食べさせる状況下で行われた他の野外試験で見出された他の臨床的および生産的結果を裏付けるために、
推奨される。
【0149】
実施例6.マクロファージ分化の誘導
多くの細菌種由来のLPSを含む、トール様受容体(TLR)リガンドのような免疫調節分子の特徴の1つは、増殖活性の減少に関係するマクロファージ細胞系の形態変化を誘導するそれらの能力である。
【0150】
このことを研究するために、J774(クローンJ774A168)および生264.7マウスマクロファージ細胞株を、2mM L−グルタミン(Sigma)、抗生物質、ゲンタマイシンおよび5%FCSを添加したRPMI1640培地(GIBCO, Gran Island, NY)中で培養した。実験のために、細胞を、0.5%FCSおよび異なる用量の本発明の免疫刺激化合物または大腸菌由来のLPSを添加したRPMI中で培養した。培養物を、37℃で72時間インキュベートし、細胞増殖を、培養の最後の16時間での(3H)チミジン(New England Nuclear, Boston, MA)のDNAへの挿入により評価した。細胞を、1μCiの(3H)チミジンを用いてパルス処理し、自動細胞ハーベスターを用いてガラス繊維フィルター中に捕捉した。放射性物質の挿入を、液体シンチレーション分光計中で測定した。該アッセイを、トリプリケート培養で行った。
【0151】
本発明の免疫刺激組成物は、生マクロファージにおける大腸菌LPSと比較して、図20に示す通り、J774細胞の分化を誘導する。マクロファージの分化は、通常、細胞増殖の減少と関係する。本発明の免疫刺激組成物は、J774マクロファージの分化の用量依存的阻害を誘導する。この効果は、J774または生マクロファージにおいて、大腸菌由来のLPSの重量/volで観察された効果のおよそ500倍未満である。
【0152】
実施例7.マクロファージにおけるTNFの誘導:
TLR リガンドの活性を実験するための最も高感度のアッセイの1つは、マクロファージまたはマクロファージ細胞株により産生されるTNFサイトカインを研究することである。この試験のために、J774(クローンJ774A168)および生264.7(ATCC TIB−71から購入)マウスマクロファージ細胞株を、2mM L−グルタミン(Sigma)、抗生物質、ゲンタマイシンおよび5% FCSを添加したRPMI1640培地(GIBCO, Gran Island, NY)中で培養した。実験のために、細胞を、0.5%FCSおよび異なる用量の本発明の免疫刺激化合物または大腸菌由来のLPSを添加したRPMI中で培養した。培養物を、37℃、5% CO2下で24時間インキュベートし、上清を集めた。TNFを、2部位サンドイッチELISA(Endogen, Woburn, MA)により検出した。
【0153】
非刺激マクロファージは、ELISAによって検出可能なTNFを合成しない。用量0.1−10μg/mlの本発明の免疫刺激組成物は、用量依存的に、J774マクロファージ(図21a)ならびに生細胞において、4000pg/mlまでの顕著なTNF産生反応レベルをもたらす。本発明の免疫刺激組成物の効果は顕著であるが、大腸菌由来のLPSのおよそ500倍未満であった(図21b).
【0154】
さらに、腹膜マクロファージを、10%チオグリコレート溶液(1ml;Difco Laboratories)の単回の腹膜注入の4日後に、BALB/cまたはC57Bl/6、12齢マウスから腹腔洗浄法により単離した。Th1応答を増大する傾向にあるマウス株であるC57Bl/6からのマクロファージおよび対照的にTh2応答に主に作用すると考えられるBalb/cからのマクロファージを、本発明緒免疫刺激組成物で処理した。細胞(1.5×106/ウェル)を、示した量のLPS(026.B6 大腸菌血清型、Sigma)または本発明の免疫刺激組成物の存在下または不存在下で、新鮮なRPMI/0.5% FCSでコーティングした12ウェルの平底プレート中に1時間接着させた。培養を、37℃、5%CO2下で24時間インキュベートし、上清を集めた。TNFを、2部位サンドイッチELISA(Endogen, Woburn, MA)により検出した。
【0155】
本発明の免疫刺激組成物は、当量濃度の大腸菌LPSよりも低いが、マウスBalb/cおよびC57B16株の両方由来のマクロファージでTNF産生を誘導する(図22)。本発明の免疫刺激物質は、当量濃度で大腸菌LPSがTNFを誘導するものの500倍未満の効力であることが結論づけられ得る。故に、本発明の免疫刺激組成物は、免疫刺激用量のTNFを誘導し得るが、毒性となり得る過剰用量ではなく、大腸菌由来のLPSにより誘導される内毒素ショックに関与する。
【0156】
実施例8.マクロファージにおけるIL−12の誘導:
IL−12は、TCR リガンドを含む多くの刺激に応答してマクロファージにより主に分泌されるサイトカインである。IL−12は、IL−12がTh1表現型へのTヘルパー(Th)分化を制御するため、最も重要なサイトカインの1つである。IL−12は、p35/p40タンパク質鎖からなる70kDのヘテロ二量体である。
【0157】
これを試験するため、腹膜マクロファージを、10%チオグリコレート溶液(1ml;Difco Laboratories)の単回の腹膜注入の4日後に、BALB/cまたはC57Bl/6、12齢マウスから腹腔洗浄法により単離した。細胞(1.5×106/ウェル)を、示した量のLPS(026.B6 大腸菌血清型、Sigma)または本発明の免疫刺激組成物の存在下または不存在下で、新鮮なRPMI/0.5% FCSでコーティングした12ウェルの平底プレート中に1時間接着させた。培養を、37℃、5%CO2下で24時間インキュベートし、上清を集めた。Il−12をELISAにより検出した。用量10μg/mlの本発明の免疫刺激物質は、C57Bl/6およびBalb/cの両方由来の腹膜マクロファージにおいて、多量のIL−12を誘導する(図23)。
【0158】
実施例9.脾臓リンパ球によるIFN−γ産生の誘導:
脾臓細胞(SC)懸濁液を、マウスから製造した。SCを、蒸留水を含む低張の溶解液により赤血球を除き、5%FCS、2mM L−グルタミン、ペニシリン(100U/ml)およびストレプトマイシン(100ng/ml)(GIBCO Laboratories, Grand Island, NY)を含むRPMI−1640完全培地中に懸濁した。
【0159】
脾臓細胞(0.4x106細胞/ウェル)を、96ウェル平底培養プレート(Costar, Cambridge, MA)中、250μlの培養培地(RPMI1640、10% FCS、2mM L−グルタミン、5x10−5M 2−メルカプトエタノール、100U/mlペニシリン、0.1μg/mlストレプトマイシン)中、37℃で5%CO2雰囲気下で培養した。細胞を、示した量の免疫刺激化合物の存在下または不存在下、コンカナバリンA(ConA)(10ng/ml)を用いて活性化した。
【0160】
マウスIFN−γを、ConAの存在下または不存在下で、脾臓細胞の培養物から得られた24時間の上清で測定した。それらを、製造業者の指示に従い、特定のサンドイッチELISAマウスMiniKit(Endogen)によりアッセイした。図24に示す通り、ConA刺激は、両方のマウス株由来の脾臓細胞による多量のIFN−γ分泌を誘導し、IFNγの産生により特徴付けられるTh1応答を刺激することが示唆される。
【0161】
実施例10.TLR4およびTLR2受容体による免疫刺激作用:
本発明の免疫刺激物質により用いられると推定される受容体を明らかにするため、TLR2またはTLR4受容体の何れかを欠失するマウス腹膜マクロファージまたは脾臓細胞を、上記のアッセイに用いた。
【0162】
腹膜マクロファージを、10%チオグリコレート溶液(1ml;Difco Laboratories)の単回の腹膜注入の4日後に、C57Bl/6またはC57Bl/6 tlr4−/−またはC57Bl/6 tlr2−/−(Sarna JR, Dyck RH, Whishaw IQ. 2000. The Dalila effect: C57BL6 mice barber whiskers by plucking. Behavioral Brain Research, 108(1):39−45. PubMedID: 10680755)、12齢マウスから腹腔洗浄法により単離した。細胞(1.5×106/ウェル)を、示した量のLPS(026.B6 大腸菌血清型、Sigma)または本発明の免疫刺激組成物の存在下または不存在下で、新鮮なRPMI/0.5% FCSでコーティングした12ウェルの平底プレート中に1時間接着させた。培養を、37℃、5%CO2下で24時間インキュベートし、上清を集めた。TNFおよびIL−12を、2部位サンドイッチELISAにより検出した。マクロファージにおけるTLR4の欠失は、本発明の免疫刺激物質のTNF(図25)およびIL−12(図26)を誘導する能力を大幅に低減させたが、TLR−2の欠失は、活性を一部分阻害したのみであった。
【0163】
脾臓細胞(SC)懸濁液を、マウスから製造した。SCを、蒸留水を含む低張の溶解液により赤血球を除き、5%FCS、2mM L−グルタミン、ペニシリン(100U/ml)およびストレプトマイシン(100ng/ml)(GIBCO Laboratories, Grand Island, NY)を含むRPMI−1640完全培地中に再懸濁した。
【0164】
脾臓細胞(0.4x106細胞/ウェル)を、96ウェル平底培養プレート(Costar, Cambridge, MA)中、250μlの培養培地(RPMI1640、10% FCS、2mM L−グルタミン、5x10−5M 2−メルカプトエタノール、100U/mlペニシリン、0.1μg/mlストレプトマイシン)中、37℃で5%CO2雰囲気下で培養した。細胞を、示した量の免疫刺激化合物の存在下または不存在下、コンカナバリンA(ConA)(10ng/ml)を用いて活性化した。
【0165】
マウスIFN−γを、ConA+本発明の免疫刺激組成物の存在下または不存在下で、脾臓細胞の培養物から得られた24時間の上清で測定した。それらを、製造業者の指示に従い、特定のサンドイッチELISAマウスMiniKit(Endogen)によりアッセイした。図27に示す通り、ConA刺激は、多量のIFN−γ分泌を誘導し、それは、C57Bl/6 tlr2−/−由来の脾臓細胞で大幅に低下し、C57Bl/6 tlr4−/−細胞を用いたとき、ほぼ完全に消失した。
【0166】
故に、本発明の免疫刺激物質は、主にTLR4受容体を介して作用し、部分的にTLR2を介して作用する。
【0167】
実施例11. T細胞は、オクロバクトラム・インターメディウム LMG3306 LPSにより用量依存的に刺激される。
本発明者らは、ある特定の実験を行い、LPSの免疫刺激能を実験する。
B6 C57マウスを、2x105 pfuのリンパ球性脈絡髄膜炎ウイルス(LCMV)で感染させ、7日後に、ウイルスによって誘導されたT細胞応答を起こしたマウスを選択し、マウスを屠殺し、それらの脾臓細胞を単離し、その後、異なる刺激剤で数時間エクスビボ刺激した。
【0168】
ペプチド特異的刺激を実験するために、脾臓細胞を、培地のみまたはLCMVからの免疫優勢H2b制限CTLエピトープの1つであるLCMV NP396 ペプチドを含む培地と共に、ブレフェルジンA(IFNγ分泌を避けるため)の存在下で6時間インキュベートした(Denis Hudrisier, Joeulle Riond, and Jean Edouard Gairin, Molecular and Functional Dissection of the H−2Db−Restricted Subdominant Cytotoxic T−Cell Response to Lymphocytic Choriomeningitis Virus, J Virol. 2001 March; 75(5): 2468−2471)。
【0169】
この時点でのマウスにおける全ての活性化T細胞を検出するために(ペプチド特異性から独立して)、脾臓細胞を、最適濃度のホルボールミリステートアセテート(PMA)+イオノマイシンの存在下または不存在下でインキュベートした。
【0170】
最後に、オクロバクトラム・インターメディウム LPSの刺激能を実験するために、化合物を異なる濃度:0.1μg/ml、1μg/ml、10μg/mlおよび100μg/mlで添加した。
【0171】
図28に示す通り、0.1および1μg/mlでの刺激は低かった。最大レベルの刺激には、10μg/ml以上の濃度で達した。これらの濃度で、ほとんどの活性化CD8およびCD4−T細胞は、少なくともT細胞刺激の“最適な”プロトコールと比較して、IFNγ発現が陽性であった。
【0172】
腸内細菌のそれとは異なる構造的特徴を有する脂質A部分を含む、ブルセラ菌種およびオクロバクテリウム・アンスロピ由来のLPSは、高濃度(10μg/ml)でのみ、TLR−4による生化学的シグナル伝達を誘発する。PMA/イオノマイシンが、CD8およびCD4 T−細胞の両方に毒性であったが、LPSは用いた最大濃度でも毒性ではなかったことが分かった。これらの細胞の大部分は、PMA/イオノマイシンの添加によりアポトーシスが起こり、結果として、T細胞数が遙かに少なかった。
【0173】
。
オクロバクトラム・インターメディウムのLPSは、より古典的な使用によいアジュバントとなるだけでなく、DNAワクチンによって誘導される免疫応答を増加させるため、多くの特性を達成する。
【0174】
1.最適用量のLPSを用いた本発明者らの実験で得られた結果は、DNAプラスミドにより誘導される免疫応答の、マウスで誘導される抗体応答を少なくとも増加させる改善である。
2.LPSは、インビトロで添加したとき、用量依存的にウイルス特異的CD8およびCD4 T−細胞の両方を活性化し得る。
3.PMA/イオノマイシン(常用される)は、脾臓細胞にアポトーシスを誘導するが、100μg/mlで添加したときでさえ、全ての濃度で毒性ではない。
【0175】
それは、形質転換が容易な非病原性細菌によって生産される。
【技術分野】
【0001】
本発明は、オクロバクトラム・インターメディウム(Ochrobactrum intermedium)株LMG3306由来のリポ多糖類(LPS)の単離、精製および特性化、ならびに哺乳動物の免疫刺激剤としてのそれらの使用、敗血症の処置および/または予防のための医薬化合物、ならびに免疫抑制動物における、およびリーシュマニア症に対するワクチン用アジュバントの製造方法に関する。
【背景技術】
【0002】
背景技術
オクロバクトラム属の細菌は、プロテオバクテリア門のアルファ2下位分類に含まれる。それらは、重症もしくは免疫不全患者または留置カテーテルを有する患者でのみ病原性となることが知られている主に土壌に生息する細菌である。オクロバクトラムが、髄膜炎、骨髄炎、菌血症、および敗血症を引き起こし得るような状況において、これらの細菌は、それら自体の慢性感染を確立することはできず、カテーテル除去後に正常な宿主から除去される(Cieslak, T. J., C. J. Drabick, and M. L. Robb. 1996. Pyogenic infections due to Ochrobactrum anthropi. Clin. Infect. Dis. 22:845−847.)
【0003】
グラム陰性菌の外膜(OM)の主成分である全てのリポ多糖類の組成は、2−ケト−3−デオキシオクツロソン酸(KDO)と称される特定の糖類を介して脂質Aと称される脂質成分に結合された多糖を含むという一般原則に従うため(Rietschel, E. T., Schade, U., Jensen, M., Wollenweber, H. W., Luderitz, O. and Greisman, S. G. 1982. Bacterial endotoxins:chemical structure, biological activity and role in septicaemia. Scand. J. Infect. Dis. Suppl. 3:8)、全てのLPS分子が同一の生物学的効果を有すると推定されている。
【0004】
しかしながら、この概念は、いくつかのLPS種のサイトカイン合成を生じる能力における大きな差異を示す研究によって修正され、この作用は、それらの脂質A部分の構造上の特徴と関係している(Netea, M. G., van Deuren, M., Kullberg, B. J., Cavaillon, J. M. and van der Meer, J. W. M. 2002. Does the shape of lipid A determine the interaction of LPS with Toll−like receptors? Trends Immunol. 23:135.)。
【0005】
グラム陰性通性細胞内細菌であるレジオネラニューモフィラは、そのLPSの低い内毒素性および化学構造の両方に関してブルセラ菌と類似であり、腸内細菌のLPSによって引き起こされる内毒素性より顕著に低い(Zahringer, U., Knirel, Y. A., Lindner, B., Helbig, J. H., Sonesson, A., Marre, R. and Rietschel, E. T. 1995. The lipopolysaccharide of Legionella pneumophila serogroup 1 (strain Philadelphia 1): chemical structure and biological significance. Prog. Clin. Biol. Res. 392:113. 26)。
【0006】
オクロバクトラム・アンスロピ属は、既知のブルセラ菌の最も近縁種である(Velasco et al. International Journal of Systematic Bacteriology 48 (1998) 759−768)。それらの近縁の系統発生的関係にも関わらず、ブルセラ・アボルタスとオクロバクトラムは、OM特性が顕著に異なり、これらの大きな差異は、LPSにおけるわずかな変化によって少なくとも一部分引き起こされる(Velasco et al. Infection and Immunity 68 (2000)3210−3218)。
【0007】
オクロバクトラム・インターメディウム LMG3301由来のLPSの完全なコア−脂質A骨格およびオクロバクトラム・アンスロピ LMG3331由来のLPSのO型鎖が、決定された(Velasco et al. Carbohydrates Research 306 (1996)123−126; Velasco et al. Carbohydrates Research 306 (1998)283−290; Velasco et al. Infection and Immunity 68 (2000)3210−3218)。
【0008】
オクロバクトラム・インターメディウム LMG3306株は、Velascoらにより発表されたが(International Journal of Systematic Bacteriology 48 (1998) 759−768)、それにもかかわらず、オクロバクトラム・インターメディウム LMG3306由来のLPSは、未だ報告されていない。
【0009】
一方、米国だけでも毎年およそ900,000症例の敗血症が発生し、その結果、およそ210,000人が死亡し、約17億ドルの費用がかかっている。敗血症は、罹患率が上昇しつつあり、死亡率27−48%の疾患である。敗血症性ショックは、敗血症の結果起こり、高い頻度で死に至る。敗血症罹患中に感染と共に生じる重篤な炎症は、いくつかの治療的介入の目標であった。それにもかかわらず、一般的な抗炎症剤を用いる20年以上の臨床試験での敗血症の炎症面は、この手法があまり成功しなかったことを示した。
【0010】
敗血症に至る病理学的機序は、複雑であって、理解が難しい。敗血症は、全身性炎症反応、その後の代償性抗炎症性反応を伴う。それらのバランスは宿主の生存に重要である。さらに、敗血症の動物モデルは、ヒト患者における敗血症のこの複雑な性質を容易には模倣し得ない。
【0011】
近年、敗血症性ショックのほとんどの影響がエンドトキシン(LPS)を循環させる大規模な放出によるものと考えられているため、トール様受容体(TLR)アンタゴニストによるそれらの作用を処理するいくつかの試みがなされている。従って、TLR4アンタゴニストとして作用するLPSの脂質A部分の構造類縁体であるエリトラン(Eritoran)は、フェーズIIの開発段階にあり、また重症敗血症患者の死亡率の低下を示した。さらに、TAK−242はTLR4シグナル伝達阻害剤であり、それは、炎症性サイトカインレベルを低下させ、また高いIL−6レベルを有する重症敗血症患者の下位集団における死亡率を低下させる。
【0012】
今日でも、利用可能なワクチンがない多くの感染性疾患が存在する。これらの場合のいくつかにおいては、失敗の原因は、正確かつ適当な免疫反応を誘導するための正確なアジュバントの欠如にある。細胞内細菌、ウイルスおよびほとんどの原生動物のようないくつかの感染源に関して、防御免疫反応はIFNγ産生に特徴付けられるTヘルパー1型(TH1)のものである。対照的に、ほとんどの蠕虫に対する防御反応は、IL−4産生を特徴とする2型(TH2)のものである(Fresno, M., M. Kopf, and L. Rivas. 1997. Cytokines and infectious disease. Immunol Today 18:56−58)。多くのアジュバントの毒性およびTh1誘導の悪化(exarcebated)により、ヒトワクチン剤形に利用可能なアジュバントはほとんどない。実際にアジュバントではないミョウバンが、ヒトワクチン剤形で最も使用されている。近年、トール様受容体(TLR)のアゴニストが、大きな関心を集めている(Hoffman, Nature Reviews Drug Discovery 4, 879, 2005)(Kwissa; Expert Vaccine Rev., 6, G73, 2007)。いくつかの抗原は、種々のTLRのいくつかのアゴニストと共に、現在分析されている。
【0013】
最後に、既に承認されているものを含む何れかのワクチン剤形の主な問題の1つは、それらが免疫抑制された対象においてほとんど作用しないことである。その他に、それらのうちのいくつかは、患者に副次的影響を有する(Kwissa, et al 2007)。
【0014】
オクロバクトラム・インターメディウム LMG 3301由来の野生型deep−rough LPS (O型鎖が存在せず、外核部分からのいくつかの単糖を欠くLPS) の完全なコア脂質A骨格は、Velascoらの、Carbohydrates Research 306 (1998)283−290に記載され;オクロバクトラム・アンスロピ LMG 3331 LPS由来の野生型smooth LPS由来のO型鎖は、Velascoらの、Carbohydrates Research 306 (1996)123−126に発表され;完全LPSの分子量は、Velascoらの、Infection and Immunity 68 (2000)3210−3218に報告されている。
【0015】
オクロバクトラム・インターメディウム LMG 3306株は、Velascoらによって発表されているが(International Journal of Systematic Bacteriology 48 (1998) 759−768)、それにも関わらず、オクロバクトラム・インターメディウム LMG 3306由来の野生型smooth LPSは未だ報告されていない。
【発明の概要】
【0016】
発明の概要
今日、オクロバクトラム・インターメディウム株 LMG 3306のリポ多糖類(IM)が、敗血症の処置および/または予防に有用であり、ならびに免疫抑制された動物およびリーシュマニア症患者におけるワクチン用アジュバントとして有用であることを実証した。
【0017】
故に、本発明の第一局面は、敗血症の処置および/または予防、ならびに免疫抑制された動物およびリーシュマニア症患者に対するワクチン用アジュバントのための医薬の製造における、オクロバクトラム・インターメディウム株 LMG 3306のリポ多糖類(IM)の使用に関する。
【0018】
本発明の一態様は、敗血症性ショックおよび内毒素ショックの処置および/または予防のための医薬の製造における、オクロバクトラム・インターメディウム株LMG 3306(IM)のリポ多糖類の使用を提供する。
【0019】
本発明の他の態様は、ヒトおよび動物の両方における感染症に対するワクチン用アジュバントとしてのオクロバクトラム・インターメディウム株 LMG 3306のリポ多糖類(IM)の使用を提供する。
【0020】
本発明の特定の態様は、免疫抑制されたヒトを含む動物における、感染症の処置および/または予防のための医薬の製造におけるIMの使用を提供する。
【0021】
本発明により、オクロバクトラム・インターメディウム株 LMG 3306のリポ多糖について記載されている使用は、ヒトおよび動物の両方に適用可能である。故に、本発明を通じて、動物における使用に関する記載は、ヒトもまた包含する。
【0022】
まとめると、哺乳類の免疫刺激剤は、敗血症および内毒素血症を処置および予防し、免疫抑制された動物におけるワクチン用アジュバント特性を有し、また足蹠リーシュマニア症における実験マウスモデルでの防御および治療効果を有する。
【0023】
IM(オクロバクトラム・インターメディウム株 LMG3306由来のリポ多糖類)は、
−マクロファージにおいてIL−12産生を誘導すること、
−本発明のLPSによって誘導されたTNFのレベルは、腸のLPSよりはるかに低く、病理学的効果を有さないこと、
−本発明のLPSはTヘルパー1(TH1)活性化およびγ−IFN産生を誘導すること、
を含む、免疫刺激剤として広範な活性を有することが証明された。
【0024】
本化合物は、哺乳動物における免疫応答を刺激する能力に関係している。故に、本発明は、オクロバクトラム・インターメディウム株 LMG3306 リポ多糖類および所望により1種またはそれ以上の薬学的に許容される賦形剤を含んでいてよい免疫刺激組成物を提供する。好ましくは、免疫応答の調節とは、ヒトの免疫能を増強することである。
【0025】
本発明は、敗血症および内毒素血症を予防および治療する化合物としての哺乳動物の免疫刺激剤であるIMの使用に関する。本発明は、特に、正常動物および実験的に免疫抑制された動物におけるワクチン効果を改善するためのアジュバントとしての使用に関する。この点で、そのアジュバント特性の例として、本発明は、特に、実験的なリーシュマニア感染症のためのワクチン用アジュバントとしての、およびリーシュマニア症、主に足蹠リーシュマニア症の実験マウスモデルを用いる皮膚リーシュマニア症の治療および防御効果としての、IMの使用に関する。
【0026】
本発明は、哺乳動物の免疫刺激剤として医薬分野で用いるための化合物としての、オクロバクトラム・インターメディウム株 LMG3306由来のLPSの製造方法に関する。
【0027】
本発明の化合物は、ウシ、ブタ、乳牛およびマウスで行う実験的野外試験における免疫刺激剤として特異的な活性を発現する:
−本発明の化合物と合わせて特定のワクチン接種後のIBRウイルスに対する中和抗体の増加の影響
−マクロファージ分化の誘導
−マクロファージにおけるTNFの誘導
−マクロファージにおけるIL−12の誘導
−脾臓リンパ球によるIFN−γ産生の誘導
−TLR4およびTLR2受容体による免疫刺激作用
−T細胞の刺激に対する用量依存的な影響。
【0028】
従って、本発明は、オクロバクトラム・インターメディウム株 LMG3306 リポ多糖類(LPS)の、哺乳動物における免疫応答を刺激し得る能力に関する。故に、本発明は、オクロバクトラム・インターメディウム株 LMG3306 リポ多糖類および所望により1種またはそれ以上の薬学的に許容される賦形剤を含んでいてよい免疫刺激組成物を提供する。好ましくは、免疫応答の調節とは、哺乳動物の免疫能を増強することである。
【0029】
本発明はまた、オクロバクトラム・インターメディウム株 LMG3306 LPSまたはそれを含む医薬組成物を哺乳動物に投与することを含む、哺乳動物における免疫応答を調節する方法に関する。故に、本発明はまた、哺乳動物に一般的な細胞性免疫を誘導するための医薬の製造における免疫刺激組成物の使用に関する。
【0030】
本発明はまた、オクロバクトラム・インターメディウム LMG3306のLPSを哺乳動物に投与することを含む、哺乳動物におけるマクロファージの分化誘導、TNF産生、IL−12産生、脾臓リンパ球によるIFN−γ産生、およびウイルス特異的CD8およびCD4 T−細胞の活性化のための医薬の製造における本発明の免疫刺激組成物の使用に関する。
【0031】
他の態様において、本発明の免疫刺激組成物は、ワクチン中に含まれる特定のウイルスおよび/または細菌に対してより免疫性にする(よりよい免疫化)ためのワクチン用アジュバントとして使用される。
【0032】
これらの致死未満量または他の低毒素LPSによる内毒素ショックの予防に関するほとんどの研究が、予め接種が必要であることを示している(Hatao, F., et al., The induction of super − resistance using synthetic lipopolysaccharide receptor agonist rescues fatal endotoxemia in rats without excessive immunosuppression. Shock2005; 23 (4): 365−70)。本発明者らは、驚くべきことに、オクロバクトラム・インターメディウム株 LMG3306のリポ多糖が、前もって脱感作期間を要せず、オクロバクトラム・インターメディウム株 LMG3306のリポ多糖類と毒性作用を保護したLPSを合わせた接種のみを要することを見出した。
【0033】
また、ほとんどの公表データで、インビトロまたは細胞株でのTNFの抑制とインビボでの脱感作作用との関係を見出されている(例えば、Lehner, MD, et al, lipopolysaccharide and Highly Purified Lipoteichoic Acid Via Different Toll −Like receptors Independent of Paracrine Mediators. 2,001. J. Immunol. 166: 5161−5167を参照)。しかしながら、本発明者らは、オクロバクトラム・インターメディウム株 LMG3306のリポ多糖が、インビボでLPSの作用を保護するが、マクロファージ株でも腹腔マクロファージまたは脾臓細胞においてもTNFまたはIL−12の分泌を低下させないことを見出した。インビボでの予備的結果はまた、オクロバクトラム・インターメディウム株 LMG 3306のリポ多糖(IM)が、何らかの機序で、LPSにより誘導されるTNFレベルを大幅に減少させ得ることを示唆する。何れの場合も、TNFレベルと保護に直接的な関係はない。これらの結果は、驚くべきことである。
【0034】
IL−10またはTGF−βの増大した産生の脱感作作用に起因すると考える他の仮説がある(Randow, F. et al., Mechanism of endotoxin desensitization: involvement of 35 interleukin 10 and transforming growth factor beta. J. Exp. Med 1995. 181, 1887−1892)。しかしながら、IL−10が、IMによりインビトロでほとんど誘導されないという事実により、そのような場合はなく、実際に、何かが腹腔マクロファージまたは脾臓細胞中でのIL−10の産生を阻害し(けれども、それは有意ではない)、LPSにより誘導されるIL−10のレベルは、C57BL/6マウス由来の脾臓細胞の場合を除いて、IMにより顕著に増大されないようである。
【0035】
他の興味のある局面は、病原体間の免疫応答を調節するIMの能力である。従って、リーシュマニア属の主な感染系を用いて、IMが、CpGと同様の方法ではなく、免疫源性リーシュマニア抗原抽出物の作用を増強することが明らかになった。CpGは、TLR9とのそれらの結合を介して、Th1応答を増強する(それは、リーシュマニア属および多くの他の細菌のような病原体から保護する)公知の免疫調節剤であり、免疫調節剤およびワクチンアジュバントとして特許が取得されている。しかしながら、IMは、インビボでTh1/Th2 プロフィールを変化させず、故に、CpG以上に悪化させる。このことは、IMが、体液性(Th2)および細胞性(Th1)の両方が防御に重要な役割を果たす感染における良好なCpGアジュバントであり得ることを示唆し得る。防御反応だけでなく、IMが防御を直接誘導し得て、すなわち、リーシュマニア属による感染に治療効果を有することを示すことを証明したことはおそらくより重要である。
【0036】
本明細書および特許請求の範囲の全体にわたって、用語“含む”および該用語の変形は、他の技術的特徴、付加物、成分または工程を除外することを意図しない。さらなる発明の目的、利点および特徴は、当該記載の検討により当業者に明らかになり得るか、または本発明の実施により習得され得る。以下の実施例および図面は、説明を目的として提供され、それらは、本発明を限定することを意図しない。さらに、本発明は、本明細書に記載の特定かつ好ましい態様の全ての可能性のある組み合わせを包含する。
【図面の簡単な説明】
【0037】
【図1】図1は、IMの使用による、大腸菌のLPSによって引き起こされる毒性作用の予防を示す。5匹のマウスの群を、LPS(1.75mg)またはIM(オクロバクトラム・インターメディウムのLPS)0.6μg/Kg(用量60)もしくは3μg/Kg(用量300)単独、またはそれらの組合せで処理し、生存を評価した。
【図2】図2は、LPS内毒素ショックの処置を示す。5匹の動物群をLPSで処理する(2mg/動物)。24時間後、それらにIM(オクロバクトラム・インターメディウムのLPS)0.6μg/Kg(用量60)または3μg/Kg(用量300)を注入し、生存を評価した。
【図3】図3は、大腸菌性腹膜炎の予防を示す。5匹のマウスの群に大腸菌(コロニー形成のための1E7ユニット、CFU)を注入し;24時間後に、動物にIM(オクロバクトラム・インターメディウムのLPS)0.6μg/Kg(用量60)または6μg/Kg(用量600)を注入し、生存(上パネル)および臨床症状(下パネル)を評価した。
【0038】
【図4】図4は、大腸菌性腹膜炎の治療を示す。5匹のマウスの群に大腸菌(コロニー形成のための1E8ユニット、CFU)を注入し;4時間後、動物にIM(オクロバクトラム・インターメディウムのLPS)0.6μg/Kg(用量600)を注入し、生存(上パネル)および臨床症状(下パネル)を評価した。
【図5】図5は、リーシュマニア症に対する防御免疫および作用経路の免疫図の一般的特徴を示す。図に示す通り、寄生虫とマクロファージの相互作用は免疫反応を生じ、それが主にTh1 リンパ球活性化に基づくとき、寄生虫に対する反応は有効かつ成功裏であるが、主にTh2の応答であるときは、感染が現れ得る。
【図6】図6は、リーシュマニア症に対するワクチンにおいて、可能性のあるアジュバント IM25(オクロバクトラム・インターメディウム LMG3306のリポ多糖類)を評価するための免疫化プロトコールを示す。5匹のマウスの群に、0日、15日および30日目にSLA抗原単独またはCpGまたはIMをワクチン接種した。60日目に、各動物の足蹠に1000プロマスチゴート(promastigote)を注入し、病変部の変化および免疫パラメーターを評価した。
【0039】
【図7】図7は、SLAおよびSLA−CpG(上パネル)またはアジュバントとしてSLA−IMをワクチン接種した動物における病変の変化を示す。感染による足蹠の腫れを定量する。
【図8】図8は、SLAおよびSLA−CpG SLA−IM 対 対照(PBS)をワクチン接種した動物の足蹠における傷の巨視的状態を示す。図中、6週間の感染期間中の、対照群の動物、1匹のSLAをワクチン接種した動物、1匹のSLA−CpGをワクチン接種した動物および6匹のSLA−IMをワクチン接種した動物の写真が含まれる。
【図9】図9は、脾臓およびリンパ節(DLN)中の寄生虫数の定量化を示し、SLAでのワクチン接種は、対照群に対して該臓器中で計数された寄生虫数の如何なる減少も生じないが、SLA−CpGは、その数をおよそ2log低下し、SLA−IM群でも同様の低下を示す結果が得られた。
【0040】
【図10】図10は、屠殺した動物の脾臓由来の細胞(単離され、インビトロでSLAで刺激された細胞)におけるTh1応答(IFN−γ)および/またはTh2(IL−4)の評価の結果を図示する。
【図11】図11は、特異的抗体アイソタイプIgG2a(Th1)およびIgG1(Th2)に関して、リーシュマニア属レベルの分析結果によって変わるTh1/Th2応答の評価を示す。
【図12】図12は、リーシュマニア症に実験的に感染させた動物における、IMの治療効果を評価するプロトコールを示す。
【図13】図13(A)は、IMをワクチン接種した動物における病変部の評価を示す。感染による足蹠の腫れを定量する。(B)は、対照群(PBS)と比較して、IMをワクチン接種した動物の足蹠病変部の巨視的状態を示す。6週間の感染中の、1匹の対照群の動物、1匹のCpGをワクチン接種した動物、および全ての6匹のSLA−IMをワクチン接種した動物の写真が含まれる。(C)は、7週後の脾臓およびリンパ節(DLN)中の寄生虫数の定量化を示す。
【0041】
【図14】図14は、IgG2a/IgG1比を含む、IgG1およびIgG2a抗体に対する、PBS対照サンプル、CPGサンプルおよびIMサンプルの比較表を示す。
【図15】図15は、免疫抑制された動物におけるアジュバントRTの役割を分析するために用いるプロトコールの概要を示す。5匹の動物群を、デキサメサゾンで免疫抑制し、SLA、SLA−CpGまたはSLA−IMをワクチン接種した。24時間後、動物を感染させ、その後8日目に再感染させた。60日目に屠殺後、病変部の変化および免疫パラメーターを評価した。
【図16】図16は、マウスの脾臓細胞における、SLAをワクチン接種した後のIMのアジュバント作用を示す。脾臓細胞を、抗原SLA(10g/ml)の存在下または不存在下で72時間培養し、細胞増殖を、72時間の間のDNAへのトリチウム化チミジンの挿入により評価する。結果は、各実験において6匹の動物を用いてトリプリケートで2回実験した平均を示す。
【0042】
【図17】図17は、ワクチン接種した動物由来の脾臓細胞によるIL−4産生への影響を示す。脾臓細胞を種々の動物から単離し、SLA(10g/ml)で刺激し、ELISAによりIL−4の分泌を分析した。
【図18】図18は、ワクチン接種し、デキサメサゾン(DEX)で処理したマウスの脾臓細胞によるIL−4およびIFN−γの効果を示す。
【図19】図19は、ワクチン接種した動物におけるSLAに対する細胞応答を示す。SLAのインビトロ添加後17日目での、IFN−γに対する特異的応答およびSLAに対する応答を示す。IFN−γ分泌(Th1)/IL−4(Th2)の比を示す。
【0043】
【図20】図20は、生マクロファージにおける、大腸菌LPSと比較した、免疫刺激化合物の添加後のJ774細胞の分化を示す。本発明の免疫刺激化合物は、J774マクロファージの分化の用量依存的阻害をもたらす。この作用は、J774または生マクロファージにおける重量/容量に基づく大腸菌由来のLPSで観察されたものよりもおよそ500倍低い(データ示さず)。
【図21】図21は、刺激していないマクロファージは、ELISAにより検出可能なTNFを合成しないことを示す。0.1−10μg/ml用量の本発明の免疫刺激組成物は、J774 マクロファージ(図21a)ならびに生細胞(データ示さず)において、用量依存的に4000pg/mlまでの有意なTNF産生反応レベルを誘導する。本発明の免疫刺激剤の効果は非常に有意であるが、大腸菌由来のLPSよりおよそ500倍低かった(図21b)。
【0044】
【図22】図22は、本発明の免疫刺激組成物が、マウスのBalb/cおよびC57B16株両方由来のマクロファージにおけるTNF産生を誘導するが、同じ濃度で大腸菌LPSより有効性が低いことを示す。
【図23】図23は、10mμg/mlの用量の本発明の免疫刺激剤は、C57Bl/6およびBalb/cの両方由来の腹膜マクロファージにおける、多量のIL−12を誘導することを示す。
【図24】図24は、コンカナバリンA(ConA)刺激が、両株のマウス由来の脾臓細胞による多量のIFN−γ分泌を誘導することを示し、IFNγの産生により特徴付けられるTh1応答を刺激することを示唆する。1−10μg/ml用量の本発明の免疫刺激組成物は、C57Bl/6およびBalb/cの両方由来の脾臓リンパ球の多量のIFNγを誘導する。
【0045】
【図25】図25は、本発明の免疫刺激組成物が、C57BI/6由来の腹腔マクロファージにおけるTNFを誘導するが、マクロファージにおけるTLR−2欠失は単に部分的に活性を阻害するのみで、TLR4の欠損は強く誘導を低下させることを示す。
【図26】図26は、本発明の免疫刺激組成物が、C57BI/6由来の腹腔マクロファージにおいてIL−12を誘導するが、マクロファージにおけるTLR−2の欠失は、単に部分的に活性を阻害するのみで、TLR4の欠損は強く誘導を低下させることを示す。
【図27】図27は、コンカナバリンA刺激がIFN−γ分泌を誘導し、それは、C57Bl/6 TLR2−/−由来の脾臓細胞で非常に低下し、C57Bl/6 TLR4−/− 細胞では、10mgの免疫刺激化合物を用いたとき、ほぼ完全に消失することを示す。
【図28】図28は、B6 C57マウス由来の脾臓細胞をリンパ球性脈絡髄膜炎ウイルスで感染させた後、異なる刺激;左から右へ:非ペプチド;LCMVNP396ペプチド(ペプチドNP396);培地のみ;PMA+イオノマイシン;異なる濃度の免疫刺激化合物、の添加によって誘導されたT細胞応答性CD4(図28a)またはCD8(図28b)を示す。
【0046】
特定の態様/例の詳細な説明
用語“免疫刺激組成物”は、本明細書で広義に定義され、該製品を接種された哺乳動物における免疫応答を刺激し得る投与可能な形態の任意の生物学的物質を意味する。
【0047】
本明細書で用いる用語“精製した”は、それがLPSと関係するとき、LPSが、それに限定されないが、上記の方法のような分画法または精製法により、種々の他の成分を除去され、組成物が実質的にその発現される生物学的活性を保持しないことを示す。用語“実質的に精製した”を用いるとき、この意味は、LPSが該組成物の主な非溶媒成分を形成する組成物を意味し得る。例えば、“実質的に精製したLPS”は、溶液または組成物の非溶媒成分の約50%以上、約60%以上、約70%以上、約80%以上、約90%以上、約95%以上が本発明のLPSであることを示す。
【0048】
本発明の好ましい態様に従い、免疫刺激組成物は、非溶媒成分は、0.2%以上の他の化合物(DNA、RNA、タンパク質、グルカン、脂質等)が混入してない、実質的に精製された形態のオクロバクトラム・インターメディウム株 LMG3306のLPSを含む。
【0049】
“免疫学的有効量”は、哺乳動物における検出可能な細胞性または体液性免疫応答を誘導するのに十分な免疫源の量を意味する。
【0050】
本明細書で用いる“防御免疫応答”は、特定の病原体により引き起こされる感染または疾患を予防または遅延する細胞性または体液性免疫応答を意味する。
【0051】
本発明の化合物は、賦形剤、pHおよび濃度に基づき常套的製造技術を用いて製造され得る。
【0052】
本発明の態様に従い、リポ多糖類は、オクロバクトラム・インターメディウム LMG3306から以下の工程で抽出される:
a)オクロバクトラム・インターメディウム LMG3306の不活化培養物を遠心し;
b)得られた沈殿物を再懸濁して生理懸濁液とし;
c)精製水およびポリエチレングリコールを用いて透析し;
d)4倍量のメタノールおよび1%の酢酸ナトリウム入り飽和メタノールで沈殿させ;
e)凍結乾燥させて、粗LPSを得る。
【0053】
好ましい態様に従い、一旦、粗LPSを凍結乾燥させて、それを以下の工程で精製する:
1. 粗LPSを、要すれば超音波を用いて、緩衝液(10mM Tris−HCl pH7,5)に溶解し;
2. プロテイナーゼKを添加し、室温でインキュベートし;
3. 精製したLPSを超遠心分離により集め;
4. サンプルを凍結乾燥させる。
【0054】
本発明の免疫刺激化合物の製造方法の好ましい態様のより詳細な記載を、以下のフローチャートで表わす:
オクロバクトラム・インターメディウム LMG3306 LPの抽出法のフローチャート:
【表1】
【0055】
製造法は、以下の工程を含む:
i)ワーキングシード細菌(LMG3306株)の解凍
接種前準備:
1. Triptycase Soy Broth(TSB)を1L準備する
2. 60mlのTSBを含むチューブを準備する
3. ワーキングシード凍結バイアルから種菌(pearl)を取り出し、60mlのTSBを含むチューブにそれを加える
4. 37±0.1℃で24時間、振とうしながらインキュベートする
5. 前接種源の制御:同定および純度制御:コロニーの形態および培養物の顕微鏡的形態の確認。
【0056】
ii)制御発酵槽中の増殖:
接種物製造
1. スクロース 2‰を添加した培養培地TSBを準備する
2. スクロース 2‰を添加した5LのTSB(pH6.75±0.15)を含む接種ボトルを準備する
3. 培養培地を含む接種ボトル中で前接種源を接種する
4. 37±0.1℃で24時間、振とうしながらインキュベートする
5. 接種源の制御:同定および純度制御:コロニーの形態および培養物の顕微鏡的形態の確認。
【0057】
培養
1. 発酵装置を浄化および滅菌する
2. スクロース 2‰を添加した400LのTSBを準備する
3. スクロース 2‰を添加し、pH6.75±0.15に合わせた400LのTSBを発酵装置に入れる
4. 培養培地を含む発酵装置に接種源を入れる
5. 37±0.1℃で24時間、最大50rpmで振とうしながらインキュベートする
6. 培養物の制御:同定および純度制御:コロニーの形態および培養物の顕微鏡的形態の確認。
【0058】
不活性化
1. 滅菌導管により不活性化タンクに培養物を移送する
2. ホルムアルデヒド溶液を、終濃度0.4%まで添加し、37℃で10rpmで振とうしながら3日間インキュベートする。
【0059】
遠心分離
1. 発酵槽を連続遠心分離機に接続し、発酵槽の内容物を遠心する
2. 10リットル容量のプラスチックボトルに不活化培養物を入れ、使用まで4℃で貯蔵する。
【0060】
iii)以下に記載の通り、LPS抽出法を行う:
1−オクロバクトラム・インターメディウム LMG3306の不活化培養物を遠心する。上清を除去する。
2−得られた沈殿物を再懸濁し生理懸濁液とする:200g/l 15mM NaCl。
3−65℃で、水溶液に90%でフェノールを添加する。
4−65℃で20分間振とうする。混合物を10℃まで冷却する。
5−9000xgで20分間遠心する。4℃で24時間維持する。
6−初めに純水で、次いでポリエチレングリコールで透析する。
7−4倍量のメタノールおよび1%の酢酸ナトリウム含有飽和メタノールで沈殿させる。
8−9000xgで20分間遠心する。上清を除去する。抽出物を純水で再懸濁し、振とうする。
9−工程7を繰り返す。
10−サンプルを凍結乾燥して、粗LPSを得る。
【0061】
iv)以下の方法によるLPSの精製:
1. 超音波を用いて、100mgの粗LPSを20(5mg/ml)の緩衝液(10mM Tris−HCl pH7,5)に溶解する。5−10mgのLPSを対照として使用する。
2. 0.5mlのプロテイナーゼK(1mgのLPS当たり、50μg/mlまたは5μgのプロテイナーゼ)を添加する。室温で12時間、撹拌および軽く振とうしながらインキュベートする。
3. 純粋なLPSを超遠心(100.000×g、6時間)により集める。沈殿物を純水に再懸濁する。
4. サンプルを凍結乾燥させる。
【0062】
v)バルク溶液を以下の通り製造してLPS濃度を計測:
材料および反応材
1. 1,25 N H2SO4:H2O DD中、6,66mlの市販の反応材で100mlとする。
2. 0,042N 過ヨウ素酸:100mlの0,125N H2SO4中、0,48gの過ヨウ素酸(パラ過ヨウ素酸、H15IO6)
3. 0,5N HCl中、2% 亜ヒ酸ナトリウム(200ml H2Oに8,3ml HCl濃縮物)
4. チオバルビツール酸(Sigma):0,3% pH2(H2O DD中に溶解、50℃)。15日以内に使用する。
5. ジメチルスルホキシド
6. 標準:
Kdo(分子量=255,1)(Sigma、−20℃で貯蔵)
デオキシリボース(分子量=136)(Sigma、−20℃で貯蔵)
7. 上記のバッチから定量したLPS
8. ビー玉(marble)が合致するガラス管(第二クロム混合物または等量物で浄化)。
9. 分光光度計。
【0063】
方法
1. Kdo標準(0.05μg)、デオキシリボース標準(0.025μg)、上記のバッチから定量したLPS(0.2μg)およびブランク反応材(200μl H2O)を含むチューブを準備し、
a)サンプルが抽出したLPSであるとき、凍結乾燥させ、0.2μgを計量し、1つのチューブに準備する。
b)サンプルが最終生成物のとき、200μlを含むチューブを準備する。
チューブにH2Oを加えて200μlとする。
2. サンプルを含むチューブを氷上に置き、20μlの1.25 N H2SO4を各チューブに加え、それらをビー玉で栓をして正確に20分間沸騰させる。氷上で冷却する。この工程は、標準サンプルには行わない。
3. 0.25mlの過ヨウ素酸をサンプルおよび標準サンプルに迅速に加え、振とうし、それらを室温で20分間インキュベートする(最初のチューブから計る)。過ヨウ素酸を加えるため、チューブをH2SO4が添加されたときと同じ順にしておく。測定されたKdoの量は、Kdoの可溶化によって、また加水分解条件中のその分解によっても変わるため、加水分解時間を最適化しなければならない(20分は、オクロバクトラム・インターメディウム LMG3306 LPSにとって十分である)。
4. 3と同じ順のチューブを、0.5mlの亜ヒ酸ナトリウムを添加し、ボルテックスして振とうし、最初のチューブへの添加から2分間待つ(試薬の添加により現れる黄色が、この時消失する)。
5. 各チューブに前と同じ順で2mlのチオバルビツール酸を迅速に添加する。それらをビー玉で栓をして20分間沸騰させる。
6. 各チューブに正確に1mlのDMSO(混濁の出現を避ける)を添加し、その後、それらを、水道水を入れたトレー中で冷却する。
7. 室温で30分間インキュベートする。ランプを安定化させるために、初めの30分間、分光光度計を点けておく。
8.各チューブを552nmおよび536nm(Kdoおよびデオキシリボースそれぞれの最大吸収)で読み出す。プラスチックトレーを用い得る。
【0064】
測定
Mk552=Abs552/μmolkを、Mk536、Md552 y Md536と同様に計算する
【化1】
k:KDO;d:デオキシリボース;Abs:吸光度。
【0065】
vi)以下に記載の最終プロセスまでのバッチの均質性:
定量化プロセスのこの均質性確認法は、製品製造中、EMEA/CVMP/598/99“プロセスバリデーションのガイダンスノート”に従って2回用いられる:
−オクロバクトラム・インターメディウム LMG3306不活性化培養物からのLPS抽出の終わりに、その濃度を知るために用いる。
−最終生成物中のLPSの量を調べるために用いる。
【0066】
該確認法は、抽出段階の終わりに適用されるプロセスである。分析法で用いられる量は、最終製品中で定量される量よりもはるかに多い。抽出されたLPSの濃度を知るためにこの方法を用いる。
【0067】
LPS定量化は、その成分の1つであるKdoの定量化で行う。Kdoは、LPS分子の一部であり、特定のLPSに常に一定量存在するが、異なる細菌LPSでは量が変わる。オクロバクトラム・インターメディウムにおいて、LMG3306は、各LPS分子について2分子のKdoである。
【0068】
定量化は、分光光度法によって行われる。
【0069】
Kdoの定量化は、サンプルが抽出LPSであるとき、LPSの抽出および精製工程の最後に残る残渣デオキシリボース(DNA成分)が、Kdoの最大吸収波長(552nm)での吸光度によるアッセイを妨げ得るため、明確ではない。
【0070】
標準Kdoを用いるKdoの定量化の確認は、Kdoが線形的に正確かつ正しく定量されることが証明され得る。LPSサンプルがデオキシリボース残渣の混入を含むとき、該方法の明確性の欠如は、補正式を用いて解決される。この式は、この干渉がサンプルに存在するとき、得られた理論的Kdo値をデオキシリボース干渉を除去する実数値に合わせる。
【0071】
デオキシリボース定量の直線性は、この成分が単独で直線性を示すだけでなく、LPSの汚染物質としても直線性を示すことを意味する。
【0072】
この方法が使用される毎に、異なる濃度の標準Kdoを含むサンプル、異なる濃度のデオキシリボースを含むサンプル、異なる濃度の上記で定量したLPSを含むサンプル、および異なる濃度の試験サンプルを含むサンプルを用い、またそれらは全て、デオキシリボース最大吸収波長(552nm)で測定され得る。
【0073】
以下の工程を行って結果を得る:
−標準Kdoを含む標準直線:直線性を試験する。
−標準デオキシリボースを含む標準直線:直線性を試験する。
−上記の通り定量したLPSを含む直線。方法をチェックする。直線性を試験する。
−標準Kdo、上記の通り定量したLPSおよび試験サンプルについて552nmでの吸光度、ならびに標準デオキシリボースについて536nmでの吸光度を分母として、各サンプルについて得られた吸光度値の商を計算する。
【0074】
式を適用する際、選択された値が標準直線内である限り、商値が標準Kdoおよび標準デオキシリボースの濃度から得られた値に最も近い試験サンプルの濃度値を用いる。値が直線内でない場合、最も近い値を変わりに用いる。
【0075】
式の適用は、試験サンプル中のKdoマイクロモルを決定する。
【0076】
本発明はさらに、本明細書で提供するLPS免疫刺激化合物を1種以上の薬学的に許容される担体または賦形剤と混合して含む医薬組成物を提供する。適当な担体は、投与経路と処置されるべき状態によって変わり得る。本発明の医薬組成物を、抗原に対する免疫応答を増大するか、または免疫系の細胞の活性を増強するためにアジュバントとして用い得るか、またはいくつかの例において、対象における特定の状態を予防または処置するために予防用または治療用組成物として用い得ることが理解され得る。
【0077】
故に、一態様において、本発明は、本発明のLPSおよび薬学的に許容される担体を含む医薬組成物を提供し、ここで、LPSは、免疫応答を調節するのに有効な量、所望の免疫応答を刺激し、疾患または状態を処置または阻害するという意味で所望の効果を達成するのに必要とされる化合物量で存在する。本医薬組成物はまた、抗原と共に投与されるとき、アジュバントとして作用する。
【0078】
本発明の免疫刺激組成物は、広範な投与量および広範な比率で投与されたとき、強力な免疫刺激効果を示す。好ましい態様によれば、免疫刺激組成物は、0.5ないし120μg/mlのLPSを含む。より好ましくは、組成物は、0.5ないし10μg/mlのLPSを含む。
【0079】
好ましい態様に従い、リポ多糖類は、ミセル相が4℃で1年以上安定である均質な懸濁液形態である。
【0080】
好ましくは、本発明の免疫刺激組成物のpHは、2−12である。
【0081】
ワクチン用量と共に投与された免疫刺激剤の量は、一般的に、それがワクチンと共に、典型的なワクチンにおける重大な副作用のない免疫保護応答を引き起こす量として選択される。かかる量は、どの特定の免疫源が使用され、かつそれらがどのように提示されるかによって大きく変わり得る。一般に、各用量は、約1−1000μgの抗原および0.1−200μgの免疫刺激剤を含み、最も典型的には、約2−100μgの抗原および0.5−150μgの免疫刺激剤を含み、好ましくは約5−50μgの抗原調製物および1−75μgの免疫刺激剤を含むと予期される。もちろん、投与された投与量は、年齢、体重、もしあれば併用処置剤の種類、ならびに投与される抗原および免疫刺激剤の性質によって変わり得る。
【0082】
本発明の免疫刺激組成物の投与経路は、一般的な免疫刺激剤を送達する何れかの好適な経路であり得る。しかしながら、免疫刺激組成物は、好ましくは筋肉内または皮下経路を介して非経腸的に投与される。より好ましくは、免疫刺激組成物は、皮下投与される。要すれば、他の投与経路も用いられ得て、故に本発明の組成物の例は、懸濁液、シロップまたはエリキシルのような、開口部用、例えば経口、経鼻、経肛門、経膣等の投与用液体調製物;および、滅菌懸濁液またはエマルジョンのような非経腸投与、皮下投与、皮内投与、筋肉内投与または静脈内投与(例えば、注射可能投与)用調製物を含む。そのような組成物において、本発明の免疫刺激組成物は、好適な担体、希釈剤、または滅菌水、生理的食塩水、グルコースなどのような賦形剤と混合され得る。
【0083】
適当な免疫保護剤およびかかる用量の非毒性用量は、当業者に容易に決定され得て、すなわち、本発明の製品中に含まれる本発明の菌株の適当な免疫保護かつ非毒性量は、有効量のLPSの範囲内であり得る。適用に推奨される最低の年齢はなく、実際に1日で行ういくつかの実験が示される。
【0084】
もちろん、必要ならば投与は適当な間隔を空けて繰り返され得る。
好ましくは、投与スケジュールは以下の通りである:
・皮下経路により0.1μg/Kg体重(b.w.)の第一投与。
・15日後の同じ投与量での第二投与。
さらなる投与:6ヶ月−1年の適用がより推奨される(0.1μg/Kg b.w.皮下経路)。
【実施例】
【0085】
実施例1.内毒素血症の処置および予防
これに関して、本発明者らは、Balb/cマウスの急性モデルにおいて大腸菌由来のLPSを用いる。
第一に、本発明者らは、6日間で100%死亡率をもたらす大腸菌LPSの用量を決定する(致死量100)。
大腸菌株O111:B4のLPSを、SIGMA社(St. Louis, MO, USA)から購入した。全ての実験において、LPSの希釈剤としてリン酸塩緩衝液を用いる。全ての場合において、用いるビークルはリン酸緩衝生理食塩水(PBS)であった。生存実験用のLPSの適当用量は、LPSの増大用量をメスBALB/cに腹腔内(IP)投与したときの用量応答曲線により決定した。
【0086】
内毒素血症の致死作用の予防:
内毒素血症を有するメスBALB/cマウスを、LPS(大腸菌 O111:B4)暴露の前に24時間、2つの異なる用量のIM、0.6μg/Kg(用量60)または3μg/Kg(用量300)でIP処理するか、または処理しなかった。LPSは、マウス1匹当たり1.75mgをIP投与した。
敗血症の場合に等しい急速輸液を供するために、全ての注入は、同量であった。
【0087】
IMは両用量で、大腸菌のLPSの内毒素ショックによる動物のLPS関連死を完全に予防した。さらに、IM処置した動物は、敗血症性ショックに関連した何らの徴候も示さなかった(図1)。用いたLPSの用量は、IMの用量よりもはるかに多かった。故に、IM(オクロバクトラム・インターメディウム 3306のLPS)は、LPSと競合され得るとは考えられない。
【0088】
内毒素致死性ショックの処置:
敗血症の主な問題は、通常、その過程が始まるときに、その予防ではなく、むしろ処置である。この試験のために、5匹のマウスを、LPS IM 0.6μg/Kg(用量60)または3μg/Kg(用量300)IPで処置の24時間後、動物1匹当たり2mgのLPS(大腸菌 O111:B4)でIP(腹腔内)感染させた。IMは、LPS誘導性死亡に保護効果を有する。故に、プラセボ処置後72時間で、全ての動物が死亡する。しかしながら、IM処置は、20%の動物の生存をもたらし、それらの死を顕著に遅延させたこのことは、IMが、内毒素ショックを有するマウスを部分的に処置し得ることを示す(図2).
【0089】
驚くことに、IM処置した動物は、プラセボ群においてもより低レベルの血液凝固を示した。この事実は、過剰な凝固の問題が敗血症の現象と関係するため非常に重要であり、実際、この条件に最近承認された化合物のみが凝固の阻害作用を有する。
【0090】
細菌性腹膜炎:
オクロバクトラム・インターメディウム LMG 3306のリポ多糖類を、敗血症の他のモデルである大腸菌誘導性腹膜炎誘導性敗血症で試験した。
【0091】
大腸菌性致死性腹膜炎の処置:
この実験に関して、メスC57BL/6マウスを、動物を、LPS IM、0.6μg/Kg(用量60)または3μg/Kg(用量300)、または単一のIP投与を用いるプラセボで処置した24または4時間後に、107または108の生存O26:B6 大腸菌コロニー形成単位(CFU)を接種した。
【0092】
生存および病理的症状(摂食障害、死亡率、脱毛、choquexia、身体活動など)をその後に評価した。
大腸菌注入の24時間後、107 CFUを最初接種した何匹かのマウスが死に(それらの20%)、かつ敗血症の病的症状を示す(0から5段階で平均3)。
IMでの処置は、用量応答的に動物の死を防ぎ、敗血症の病的症状を完全に回復させた(図3)。
【0093】
108 CFUの大腸菌の用量が用いられたとき、全ての動物は24時間で死亡する;故に、それらは大腸菌の摂取の4時間後にIMで処理された。それにも関わらず、その時、マウスは何れも死んでおらず、病理的症状の平均スコアは非常に高かった(5段階のうち4)。全ての大腸菌感染マウスが、14時間で死んだ。それにも関わらず、IM 3μg/Kg(用量300)は、ほとんどの動物の死を防ぎ(80%生存率)、臨床的重症度を徐々に低下させた(図4)。
【0094】
リーシュマニア症の実験的感染におけるアジュバント効果
足蹠感染におけるアジュバント効果:
この抗原は、Th1保護応答を活性化し得ない(図5)ため、SLA(可溶性リーシュマニア抗原)抗原抽出物が、主なリーシュマニア原虫での実験的感染に対する保護効果をもたらさないことは、よく知られている。しかし、SLAが、CpGとしてアジュバントと結合するとき(米国特許第7521063号)(TLR9による免疫応答を刺激し、Th1応答を増大させるためのワクチン用アジュバント)(Heeg, Int. J. Microbiol, 298, 33, 2008)、TLR9(トール様受容体)を介して保護的Th1(IFN−γ)の活性化をもたらし得る(米国特許第6890542号)。
【0095】
従って、このモデルは、実験的なリーシュマニア症に対するワクチン接種用のアジュバントとしてのCPGのよく確立された使用と比較してIMを試験するのに非常に有用である。
図6で、本発明者らは、この目的のためのプロトコールを示す。
【0096】
基本的に、SLA抗原は、単独で、CPGと共に、またはIMと共に、0、15および30日目に接種される。
【0097】
60日目で、1,000寄生虫(promastigote)を各マウスの足蹠に接種し、病変および免疫学的パラメーターを観察する。感染の進行は肉眼で観察され得て、寄生虫数と関係する。図7に示す通り、摂取後、対照群での感染が確認される。SLA抗原は、何らかの保護効果を生じないが、SLA+CPGとSLA+IM群の両方で顕著に病変を縮小する。
【0098】
さらに、感染後6週の巨視的結果を図8に示し、SLA単独で処理した動物の足蹠形と比較して、SLA−CpGまたはSLA−IMで処理した動物の病変の顕著な縮小を証明する。
【0099】
さらに、寄生虫数を、足蹠および脾臓で定量した。感染後7週で、動物を屠殺し、脾臓およびドレインリンパ節(DLN)に存在する寄生虫を決定した。SLA免疫は、IMおよびCpGとは対照的に、寄生虫数の減少をもたらさない。CpGは、2logに近い数を減少させ、IMは、少なくとも、全6匹の動物のうち4匹で同様の結果を示した(図9)。
【0100】
簡単には、SLAは単独では保護しないが、CpGまたはIMとの組合せは感染に対する保護効果を有する。
屠殺した動物の脾臓細胞を集め、SLAで“インビトロ”で刺激し、Th1(IFN−γ)および/またはTh2(IL−4)応答を評価した。
【0101】
感受性Balb/cモデルにおけるリーシュマニア属の主な感染症は、IFN−γレベルよりもIL−4レベルを比較的増強することがよく記載されている。SLAワクチン接種はこのパターンを修正しないが、CpGとのその組合せは、IFN−γの産生によって定量化されたTh1応答を増大させ、それ故に、IL−4の相対的発現を低下させた。従って、IFN−γ(応答Th1)/IL−4(応答Th2)比は、顕著に増加した(図10)。
【0102】
驚くことに、IMは、CpGと同様のより強い保護作用を有するが、免疫学的機序は異なる。その保護は、両方のパラメーターが同時に増大するため(図10)、IFN−γ(応答Th1)/IL−4(応答Th2)の比と厳密に関連しない。
【0103】
応答Th1/Th2を決定する第二の方法は、リーシュマニア症−特有のアイソタイプ抗体IgG2a(Th1)およびIgG1(Th2)のレベルを分析することである。
【0104】
機序は明らかではないが、IgG2aレベルは、リーシュマニア症からの保護と関連する。SLAで処理しないか、またはそれのみで処理した動物において、IgG2aレベルは検出されなかった。しかしながら、IgG2aレベルは、CpGまたはIMで処理した動物で検出された。故に、IgG2a/IgG1比は、SLA−CpおよびSLA−IM群の動物と対照的に、非処理動物またはSLAでのみ処理した動物でより低い(図11)。
【0105】
足蹠実験マウスモデルにおけるIMの予防および治療効果:
図12において、リーシュマニア症を有する進行性実験的完成におけるIMの治療効果を評価するためのプロトコールを示す。
【0106】
これに関し、IMを、2つの異なる用量でリーシュマニア原虫と同時に接種した:
1)IMF−1、2および3群は、0.6μg/Kg(18ng/マウス)を受容した。
2)IMF−4、5および6は、1.2μg/Kg(36ng/マウス)用量を受容した。
結果は、IMが、リーシュマニア属による主な感染症から、CpGで得られた効力と同様の効力で保護し得ることを示す(図13A)。
【0107】
IMで処理した動物は、顕著な病変を示さなかった(図13B)。感染の7週間後、動物を屠殺し、脾臓およびリンパ節(LN)中の寄生虫数を定量した。同様の結果が、CpGおよびIM群で得られ、リンパ節中で顕著に減少し(10.000倍)、脾臓において寄生虫は見られなかった(図13C)。
【0108】
最後に、上記の実験に従って、CpG群は、抗体IgG2a/IgG1レベルの比に顕著な変化を示す。さらに、IM用量は、得られた上記の結果と一致して、この比に顕著な変化を示さない(図14)。
【0109】
最終的な結論として、本発明者らは、IMがCpGと同様の特性を有するが、Th1/Th2またはIgG2a/IgG1のプロフィールおよび比が変化しないため、免疫機序における最終結果が異なることを見出した。本発明において、IMは、両方の(CpGで実証される通り、Th1のみではない)応答の活性化による、より良好なアジュバントであり得る。
【0110】
IM(オクロバクトラム・インターメディウムのLPS)は、免疫抑制された動物におけるワクチンアジュバント特性を有する:
IMは、抗原としてSLAを用いて、リーシュマニア原虫感染に対してワクチンアジュバント作用を有することが既報である。
【0111】
この抗原は、免疫源性が低く、より重要なことには、防御作用であるTh1応答(IgG2a応答およびIFN−γ産生により特徴付けられる)ではなく、主にTh2応答(IgG1抗体の優位およびIL−4産生により特徴付けられる)をもたらす。
【0112】
免疫抑制されたマウスに対するアジュバント作用を試験するために、Balb/cメスを、動物1匹当たりコルチコステロイド・デキサメサゾ(Dex)20μg IPで処理し、24時間後に、動物に、SLAを単独(10μg/動物)で、またはIM、0.6μg/Kg(用量60)または3μg/Kg(用量300)、もしくはTh1応答をもたらすことが既報の化合物であるCpGと共に接種した(用いたプロトコールについては図15を参照のこと)。マウスに、1週間後にSLA単独を再び接種し、17日目に動物を屠殺し、脾臓細胞を集め、血清中の抗SLA抗体力価を評価した。
【0113】
図16に示す通り、特定の抗SLA抗体は、Dex単独の対照ではなくSLAを注入された全ての動物で検出されたが、ほとんどの抗体は、IgG1アイソタイプであった。記載される通り、CpGアジュバントは、IgG2a/IgG1比を増加させ、より重要なことには、IMは、インビボでのTh2からTh1への切り替えを表すこの変更をCPGよりも良好にもたらした。このことは、IMが、免疫抑制された動物においてアジュバント活性を有し、より重要なことには、免疫抑制された動物においてさえも好ましいTh1応答を維持し続けることを示した。
【0114】
加えて、異なる動物由来の脾臓細胞を得て、インビトロでSLAで刺激し、増殖およびIFN−γ(リーシュマニア症に対する保護に必要なTh1サイトカイン)を測定した。図7に示す通り、SLAは、Dex処理動物において増殖応答をもたらした。CpG処理動物は、改善した増殖応答を有した。興味深いことに、IMで処理した動物もまた、用量依存的に、SLAよりも良好な増殖応答を有する(図17)。分裂促進物質に対する応答の相違は、全ての動物で観察されなかった(データ示さず)。
【0115】
本発明者らは、“インビトロ”での脾臓細胞のSLA処置後の脾臓細胞の上清に放出されたIFN−γ(Th1サイトカイン)およびIL−4(Th2サイトカイン)を調べた。図18に示す通り、ワクチン接種していないDex対照動物は、インビトロでのSLA暴露によりIL−4またはIFN−γの産生が検出不可能であった。SLA接種された動物において、“インビトロ”でのSLA暴露は、IL−4産生を生じるが、IFN−γは非常に少なく、同じ動物の血清におけるIgG1/IgG2aデータと一致する。
【0116】
CpG処理は、IL−4およびIFN−γの比をほとんど変えることなく、両方の増加をもたらした(図19)。興味深いことに、IMのLPSは、両サイトカインを増加させるが、IL−4よりもIFN−γが高い割合で増加した。これは、とりわけ、3μg/Kg(用量300)で処理した動物で明らかであり、IFN−γ/IL−4比が10倍増加した(図19)。
【0117】
実施例2.製造方法
免疫刺激性LPSを、オクロバクトラム・インターメディウム株 LMG3306から上記の製造法に従って得た。
1. 上記の工程(ii)(製造方法)の発酵バルク抗原を得るための方法
2. 上記の工程(iii)(製造方法)のLPS抽出方法
3. 上記の工程(iv)および(v)(製造方法)の、LPSの精製およびLPS量の測定。
【0118】
生成物を、4℃で24時間までにバッチの均質性を維持するために、予め滅菌し、連続撹拌システムを供するステンレススチール製タンク中で貯蔵した。生成物を、滅菌ガス管システムを用いて、該タンクから容器詰めする場所へ移した。
【0119】
容器詰めを、100Bバックグラウンドを用いてクラス100Aの滅菌環境で行った。ゴム栓およびカプセル化を、同じ場所で行った。
【0120】
最終生成物を含むバイアルを、それらにラベルが付され包装されるまで4℃で貯蔵した。ラベル添付および包装を、別の場所で行い、そして各時間にその場所へ入れるのは、最終生成物およびその調整材料の1つのバッチのみである。販売までの最終貯蔵を、5±3℃で行った。
【0121】
実施例3.組成:
【表2】
【0122】
実施例4.Kdo試験:
本実験の主な目的は、上記のKdoの比色法によって、LPS1ml当たりのナノモル数の測定値の再現性を確認することである。
試験すべきパラメーターは、
1−アッセイ内の正確性
2−アッセイ間の正確性
3−バッチ間の正確性
である。
【0123】
1−アッセイ内の正確性の結果
このアッセイにおいて、本発明の生成物の1つのバッチ(A003A)を用い、3つのサンプルを、サンプル1つ当たり10回同様の操作を繰り返して、抗体力価を測定した。次いで、各サンプルの3つの希釈物(1/2、1/4および1/8)を、希釈物1つ当たり10回同様の操作を繰り返して、該アッセイの再現性を確認した。
【0124】
【表3】
【表4】
【0125】
結果は、試験が再現性であることを示す; VCは10%未満である。
【表5】
【表6】
【表7】
【表8】
【0126】
本発明者らは、各希釈物についてVCを調べた。得られた結果は、試験が再現性であることを示す; VCは10%未満である。
【0127】
2−アッセイ間の正確性
主に実験者の操作による変動を調べるため、本発明の製品の同じバッチを、3日の異なる日に滴定した。各日、全ての必要な溶液を新鮮に調製し、生成物サンプルを実験の最初から最終日まで4℃で貯蔵した。
このアッセイにおいて、本発明の生成物の1つのバッチ(A003A)を、異なる3日に滴定し、3つの異なるサンプルをサンプル毎に10個の複製物を用いて行った。
【0128】
【表9】
【表10】
【0129】
得られた結果は、試験の再現性を示す; VCは10%未満である。
【0130】
3−バッチ間の正確性
バッチ製造の変動を調べる:
このアッセイにおいて、本発明の製造物の3つのバッチ(A001A、A002AおよびA003A)を、滴定した。各バッチの3つのサンプルをサンプル毎に10個の複製物を用いて行った。
【表11】
【表12】
【0131】
Kdo活性の顕著な相違は、アッセイ内、アッセイ間および経時的アッセイにおいて観察されなかった。
【0132】
実施例5.ウイルス不活性化ワクチン用アジュバントとしての免疫源性試験
野外試験
この野外試験を、血清−陰性のウシにおける、常套的に不活性化したワクチン接種による免疫化によりもたらされる、血清、ウシヘルペスウイルス1(BHV−1)または感染性ウシ鼻気管炎ウイルス(IBR)に対する抗体全体および中和抗体に対する免疫応答(inmunitary answer)への免疫刺激剤の効力を評価するために行った。
【0133】
材料および方法
1.ウシ群の選択
試験群の選択のために、乳牛の他の5つの群からの120頭の動物を予め試験した。これらの5つの群中に血清−陽性の動物がいたため、サンプルを6つの群に分けた;試験した動物は、血清−陰性であって、この群を選択した。
【0134】
血清検査を、IBRウイルスに対する全抗体数を検出するELISA試験で行った。
【0135】
2.プロトコールの実施:処理、ワクチン接種およびサンプル収集
2.1.処理およびワクチン接種
試験を、IBRウイルスに感染していない22頭の血清−陰性の若い雌牛で行った。新しい動物を試験中に群に加えなかった。
【0136】
若い雌牛を以下の群に分けた:
−群1.−13頭の若い雌牛を、試験0日目および7日目に5mlの免疫刺激化合物(6μg/mL)で処理し、7日目および28日目に、不活性化IBRウイルスを含む5mlの承認された市販のワクチンをワクチン接種した。両製品を、首の筋肉内深部に注射した。
−群2.−6頭の若い雌牛を、7日目および28日目に、同様のワクチンおよび同様の方法でワクチン接種して免疫化した。
−群3.−3頭の若い雌牛を対照群とした;それらは、ウイルスが感染しなかった個とを確認するために、処理または免疫化しなかった。
【0137】
2.2.サンプル収集
各動物の血液サンプルを、試験0日目、7日目、28日目および58日目に得た。
【0138】
試験開発スキーム
【表13】
【0139】
3.ウシヘルペスウイルス(BHV−1)中和血清抗体の検出
I.ウシヘルペスウイルス1中和抗体力価を、標準ウイルス中和試験により決定した;100 DI50CT/25μlのウシ腎臓由来のよく確立された細胞株である“Georgia bovine kidney”(GBK)のColorado株(Prof. Marcelino Alvarez, Dpto. Sanidad Animal, Facultad de Veterinaria, Universidad de Leon, Espanaにより提供される)を用いて行われ、各血清を4回試験した。
【0140】
血清を純粋な血清から滴定した。血清をウイルス(25μlの希釈した血清および25μlのウイルス懸濁液)と混合して希釈し、その抗体力価を、Spearman−Kaurber法(TRIMMED SPEARMAN−KARBER (TSK) PROGRAM VERSION 1.5, ECOLOGICAL MONITORING RESEARCH DIVISION ENVIRONMENTAL MONITORING SYSTEMS LABORATORY, U. S. ENVIRONMENTAL PROTECTION AGENCY CINCINNATI, OHIO 45268)を用いて見出した。50%保護する血清の最も高い希釈倍率の逆数として表される。
【0141】
II.2つの実験群において、平均対数抗体力価が見出され、それはその逆対数の逆数として表される。血清転換を、2つの血清収集毎間の抗体の4倍増加として定義する。
【0142】
III.異なる試験日に処理した、また処理しなかった動物群について得られた平均対数抗体力価間の統計的に関連する相違の存在を確認するために、統計分析(Kinskall−Wallis non−parameter test)を、Windows−environment EpiInfoソフトウェアを用いて行った。
【0143】
結果および考察
I.試験中に血清転換した動物はいなかった。
II.7日目および28日目での血清転換率は、処理群および非処理群のそれぞれで92.3%および83.3%であり、28日目および58日目では、処理群および非処理群のそれぞれで69.2%および16.7%であった。
【0144】
両方の場合において、血清転換率は、非処理動物群よりも処理動物群で高かった;しかし、それらの群間の相違は、28日目と58日目での血清転換率でのみ統計的に有意であった。
【0145】
III.28日目において、IBR中和抗体の平均対数抗体力価は、処理動物群で3.50であって、非処理動物群で得られた力価2.37よりも顕著に高かった。
【0146】
58日目において、IBR中和抗体の平均対数抗体力価は、処理動物群で12であって、非処理動物群で得られた力価4.8よりも顕著に高かった。
【0147】
処理動物群で得られたIBRウイルス血清抗体力価は、非処理動物群より高く、28日目および58日目でそれぞれ1.5および2.5であった。
【0148】
結論
1.得られた結果により示される通り、免疫刺激化合物の投与は、ワクチン接種した動物における中和抗体および保護レベルとしての体液性応答を顕著に増大させた;故に、それは、ワクチン接種によって誘導される免疫応答の実現に重要な役割を果たした。
2.その適用は、
−その低廉な価格のために、ウシに適用される免疫化パターンとして推奨され、
−基本的に、動物が、ウシを太らせる場所のようなストレス下の状況にあるとき、これらの結果が子牛に乳を飲ませて草を食べさせる状況下で行われた他の野外試験で見出された他の臨床的および生産的結果を裏付けるために、
推奨される。
【0149】
実施例6.マクロファージ分化の誘導
多くの細菌種由来のLPSを含む、トール様受容体(TLR)リガンドのような免疫調節分子の特徴の1つは、増殖活性の減少に関係するマクロファージ細胞系の形態変化を誘導するそれらの能力である。
【0150】
このことを研究するために、J774(クローンJ774A168)および生264.7マウスマクロファージ細胞株を、2mM L−グルタミン(Sigma)、抗生物質、ゲンタマイシンおよび5%FCSを添加したRPMI1640培地(GIBCO, Gran Island, NY)中で培養した。実験のために、細胞を、0.5%FCSおよび異なる用量の本発明の免疫刺激化合物または大腸菌由来のLPSを添加したRPMI中で培養した。培養物を、37℃で72時間インキュベートし、細胞増殖を、培養の最後の16時間での(3H)チミジン(New England Nuclear, Boston, MA)のDNAへの挿入により評価した。細胞を、1μCiの(3H)チミジンを用いてパルス処理し、自動細胞ハーベスターを用いてガラス繊維フィルター中に捕捉した。放射性物質の挿入を、液体シンチレーション分光計中で測定した。該アッセイを、トリプリケート培養で行った。
【0151】
本発明の免疫刺激組成物は、生マクロファージにおける大腸菌LPSと比較して、図20に示す通り、J774細胞の分化を誘導する。マクロファージの分化は、通常、細胞増殖の減少と関係する。本発明の免疫刺激組成物は、J774マクロファージの分化の用量依存的阻害を誘導する。この効果は、J774または生マクロファージにおいて、大腸菌由来のLPSの重量/volで観察された効果のおよそ500倍未満である。
【0152】
実施例7.マクロファージにおけるTNFの誘導:
TLR リガンドの活性を実験するための最も高感度のアッセイの1つは、マクロファージまたはマクロファージ細胞株により産生されるTNFサイトカインを研究することである。この試験のために、J774(クローンJ774A168)および生264.7(ATCC TIB−71から購入)マウスマクロファージ細胞株を、2mM L−グルタミン(Sigma)、抗生物質、ゲンタマイシンおよび5% FCSを添加したRPMI1640培地(GIBCO, Gran Island, NY)中で培養した。実験のために、細胞を、0.5%FCSおよび異なる用量の本発明の免疫刺激化合物または大腸菌由来のLPSを添加したRPMI中で培養した。培養物を、37℃、5% CO2下で24時間インキュベートし、上清を集めた。TNFを、2部位サンドイッチELISA(Endogen, Woburn, MA)により検出した。
【0153】
非刺激マクロファージは、ELISAによって検出可能なTNFを合成しない。用量0.1−10μg/mlの本発明の免疫刺激組成物は、用量依存的に、J774マクロファージ(図21a)ならびに生細胞において、4000pg/mlまでの顕著なTNF産生反応レベルをもたらす。本発明の免疫刺激組成物の効果は顕著であるが、大腸菌由来のLPSのおよそ500倍未満であった(図21b).
【0154】
さらに、腹膜マクロファージを、10%チオグリコレート溶液(1ml;Difco Laboratories)の単回の腹膜注入の4日後に、BALB/cまたはC57Bl/6、12齢マウスから腹腔洗浄法により単離した。Th1応答を増大する傾向にあるマウス株であるC57Bl/6からのマクロファージおよび対照的にTh2応答に主に作用すると考えられるBalb/cからのマクロファージを、本発明緒免疫刺激組成物で処理した。細胞(1.5×106/ウェル)を、示した量のLPS(026.B6 大腸菌血清型、Sigma)または本発明の免疫刺激組成物の存在下または不存在下で、新鮮なRPMI/0.5% FCSでコーティングした12ウェルの平底プレート中に1時間接着させた。培養を、37℃、5%CO2下で24時間インキュベートし、上清を集めた。TNFを、2部位サンドイッチELISA(Endogen, Woburn, MA)により検出した。
【0155】
本発明の免疫刺激組成物は、当量濃度の大腸菌LPSよりも低いが、マウスBalb/cおよびC57B16株の両方由来のマクロファージでTNF産生を誘導する(図22)。本発明の免疫刺激物質は、当量濃度で大腸菌LPSがTNFを誘導するものの500倍未満の効力であることが結論づけられ得る。故に、本発明の免疫刺激組成物は、免疫刺激用量のTNFを誘導し得るが、毒性となり得る過剰用量ではなく、大腸菌由来のLPSにより誘導される内毒素ショックに関与する。
【0156】
実施例8.マクロファージにおけるIL−12の誘導:
IL−12は、TCR リガンドを含む多くの刺激に応答してマクロファージにより主に分泌されるサイトカインである。IL−12は、IL−12がTh1表現型へのTヘルパー(Th)分化を制御するため、最も重要なサイトカインの1つである。IL−12は、p35/p40タンパク質鎖からなる70kDのヘテロ二量体である。
【0157】
これを試験するため、腹膜マクロファージを、10%チオグリコレート溶液(1ml;Difco Laboratories)の単回の腹膜注入の4日後に、BALB/cまたはC57Bl/6、12齢マウスから腹腔洗浄法により単離した。細胞(1.5×106/ウェル)を、示した量のLPS(026.B6 大腸菌血清型、Sigma)または本発明の免疫刺激組成物の存在下または不存在下で、新鮮なRPMI/0.5% FCSでコーティングした12ウェルの平底プレート中に1時間接着させた。培養を、37℃、5%CO2下で24時間インキュベートし、上清を集めた。Il−12をELISAにより検出した。用量10μg/mlの本発明の免疫刺激物質は、C57Bl/6およびBalb/cの両方由来の腹膜マクロファージにおいて、多量のIL−12を誘導する(図23)。
【0158】
実施例9.脾臓リンパ球によるIFN−γ産生の誘導:
脾臓細胞(SC)懸濁液を、マウスから製造した。SCを、蒸留水を含む低張の溶解液により赤血球を除き、5%FCS、2mM L−グルタミン、ペニシリン(100U/ml)およびストレプトマイシン(100ng/ml)(GIBCO Laboratories, Grand Island, NY)を含むRPMI−1640完全培地中に懸濁した。
【0159】
脾臓細胞(0.4x106細胞/ウェル)を、96ウェル平底培養プレート(Costar, Cambridge, MA)中、250μlの培養培地(RPMI1640、10% FCS、2mM L−グルタミン、5x10−5M 2−メルカプトエタノール、100U/mlペニシリン、0.1μg/mlストレプトマイシン)中、37℃で5%CO2雰囲気下で培養した。細胞を、示した量の免疫刺激化合物の存在下または不存在下、コンカナバリンA(ConA)(10ng/ml)を用いて活性化した。
【0160】
マウスIFN−γを、ConAの存在下または不存在下で、脾臓細胞の培養物から得られた24時間の上清で測定した。それらを、製造業者の指示に従い、特定のサンドイッチELISAマウスMiniKit(Endogen)によりアッセイした。図24に示す通り、ConA刺激は、両方のマウス株由来の脾臓細胞による多量のIFN−γ分泌を誘導し、IFNγの産生により特徴付けられるTh1応答を刺激することが示唆される。
【0161】
実施例10.TLR4およびTLR2受容体による免疫刺激作用:
本発明の免疫刺激物質により用いられると推定される受容体を明らかにするため、TLR2またはTLR4受容体の何れかを欠失するマウス腹膜マクロファージまたは脾臓細胞を、上記のアッセイに用いた。
【0162】
腹膜マクロファージを、10%チオグリコレート溶液(1ml;Difco Laboratories)の単回の腹膜注入の4日後に、C57Bl/6またはC57Bl/6 tlr4−/−またはC57Bl/6 tlr2−/−(Sarna JR, Dyck RH, Whishaw IQ. 2000. The Dalila effect: C57BL6 mice barber whiskers by plucking. Behavioral Brain Research, 108(1):39−45. PubMedID: 10680755)、12齢マウスから腹腔洗浄法により単離した。細胞(1.5×106/ウェル)を、示した量のLPS(026.B6 大腸菌血清型、Sigma)または本発明の免疫刺激組成物の存在下または不存在下で、新鮮なRPMI/0.5% FCSでコーティングした12ウェルの平底プレート中に1時間接着させた。培養を、37℃、5%CO2下で24時間インキュベートし、上清を集めた。TNFおよびIL−12を、2部位サンドイッチELISAにより検出した。マクロファージにおけるTLR4の欠失は、本発明の免疫刺激物質のTNF(図25)およびIL−12(図26)を誘導する能力を大幅に低減させたが、TLR−2の欠失は、活性を一部分阻害したのみであった。
【0163】
脾臓細胞(SC)懸濁液を、マウスから製造した。SCを、蒸留水を含む低張の溶解液により赤血球を除き、5%FCS、2mM L−グルタミン、ペニシリン(100U/ml)およびストレプトマイシン(100ng/ml)(GIBCO Laboratories, Grand Island, NY)を含むRPMI−1640完全培地中に再懸濁した。
【0164】
脾臓細胞(0.4x106細胞/ウェル)を、96ウェル平底培養プレート(Costar, Cambridge, MA)中、250μlの培養培地(RPMI1640、10% FCS、2mM L−グルタミン、5x10−5M 2−メルカプトエタノール、100U/mlペニシリン、0.1μg/mlストレプトマイシン)中、37℃で5%CO2雰囲気下で培養した。細胞を、示した量の免疫刺激化合物の存在下または不存在下、コンカナバリンA(ConA)(10ng/ml)を用いて活性化した。
【0165】
マウスIFN−γを、ConA+本発明の免疫刺激組成物の存在下または不存在下で、脾臓細胞の培養物から得られた24時間の上清で測定した。それらを、製造業者の指示に従い、特定のサンドイッチELISAマウスMiniKit(Endogen)によりアッセイした。図27に示す通り、ConA刺激は、多量のIFN−γ分泌を誘導し、それは、C57Bl/6 tlr2−/−由来の脾臓細胞で大幅に低下し、C57Bl/6 tlr4−/−細胞を用いたとき、ほぼ完全に消失した。
【0166】
故に、本発明の免疫刺激物質は、主にTLR4受容体を介して作用し、部分的にTLR2を介して作用する。
【0167】
実施例11. T細胞は、オクロバクトラム・インターメディウム LMG3306 LPSにより用量依存的に刺激される。
本発明者らは、ある特定の実験を行い、LPSの免疫刺激能を実験する。
B6 C57マウスを、2x105 pfuのリンパ球性脈絡髄膜炎ウイルス(LCMV)で感染させ、7日後に、ウイルスによって誘導されたT細胞応答を起こしたマウスを選択し、マウスを屠殺し、それらの脾臓細胞を単離し、その後、異なる刺激剤で数時間エクスビボ刺激した。
【0168】
ペプチド特異的刺激を実験するために、脾臓細胞を、培地のみまたはLCMVからの免疫優勢H2b制限CTLエピトープの1つであるLCMV NP396 ペプチドを含む培地と共に、ブレフェルジンA(IFNγ分泌を避けるため)の存在下で6時間インキュベートした(Denis Hudrisier, Joeulle Riond, and Jean Edouard Gairin, Molecular and Functional Dissection of the H−2Db−Restricted Subdominant Cytotoxic T−Cell Response to Lymphocytic Choriomeningitis Virus, J Virol. 2001 March; 75(5): 2468−2471)。
【0169】
この時点でのマウスにおける全ての活性化T細胞を検出するために(ペプチド特異性から独立して)、脾臓細胞を、最適濃度のホルボールミリステートアセテート(PMA)+イオノマイシンの存在下または不存在下でインキュベートした。
【0170】
最後に、オクロバクトラム・インターメディウム LPSの刺激能を実験するために、化合物を異なる濃度:0.1μg/ml、1μg/ml、10μg/mlおよび100μg/mlで添加した。
【0171】
図28に示す通り、0.1および1μg/mlでの刺激は低かった。最大レベルの刺激には、10μg/ml以上の濃度で達した。これらの濃度で、ほとんどの活性化CD8およびCD4−T細胞は、少なくともT細胞刺激の“最適な”プロトコールと比較して、IFNγ発現が陽性であった。
【0172】
腸内細菌のそれとは異なる構造的特徴を有する脂質A部分を含む、ブルセラ菌種およびオクロバクテリウム・アンスロピ由来のLPSは、高濃度(10μg/ml)でのみ、TLR−4による生化学的シグナル伝達を誘発する。PMA/イオノマイシンが、CD8およびCD4 T−細胞の両方に毒性であったが、LPSは用いた最大濃度でも毒性ではなかったことが分かった。これらの細胞の大部分は、PMA/イオノマイシンの添加によりアポトーシスが起こり、結果として、T細胞数が遙かに少なかった。
【0173】
。
オクロバクトラム・インターメディウムのLPSは、より古典的な使用によいアジュバントとなるだけでなく、DNAワクチンによって誘導される免疫応答を増加させるため、多くの特性を達成する。
【0174】
1.最適用量のLPSを用いた本発明者らの実験で得られた結果は、DNAプラスミドにより誘導される免疫応答の、マウスで誘導される抗体応答を少なくとも増加させる改善である。
2.LPSは、インビトロで添加したとき、用量依存的にウイルス特異的CD8およびCD4 T−細胞の両方を活性化し得る。
3.PMA/イオノマイシン(常用される)は、脾臓細胞にアポトーシスを誘導するが、100μg/mlで添加したときでさえ、全ての濃度で毒性ではない。
【0175】
それは、形質転換が容易な非病原性細菌によって生産される。
【特許請求の範囲】
【請求項1】
敗血症を処置および/または予防するための、哺乳動物において免疫応答を刺激し得るオクロバクトラム・インターメディウム LMG3306由来のリポ多糖類。
【請求項2】
オクロバクトラム・インターメディウム株 LMG3306を、トリプチケースソイ培地を用いて培養する工程を含む、請求項1記載のリポ多糖類を製造する方法。
【請求項3】
リポ多糖類の抽出工程:
a)オクロバクトラム・インターメディウム株LMG3306の不活化培養物を遠心し;
b)得られた沈殿物を再懸濁して生理懸濁液とし;
c)精製水およびポリエチレングリコールを用いて透析し;
d)4倍量のメタノールおよび1%の酢酸ナトリウム含有飽和メタノールを用いて沈殿させ;
e)凍結乾燥させて、粗LPSを得る工程
をさらに含む、請求項2記載の方法。
【請求項4】
粗LPSの精製工程:
a)所望により超音波を用いて、粗LPSを緩衝液(10mM Tris−HCl pH7,5)に溶解し;
b)プロテイナーゼKを添加して、室温でインキュベートし;
c)精製したLPSを超遠心分離により集め;
d)サンプルを凍結乾燥する工程
をさらに含む、請求項2記載の方法。
【請求項5】
請求項1記載のオクロバクトラム・インターメディウム LMG3306由来のリポ多糖類および所望により1個またはそれ以上の薬学的に許容される賦形剤を含む、免疫刺激組成物。
【請求項6】
オクロバクトラム・インターメディウム LMG3306の化学的に純粋な(非溶媒成分が、0.2%以上の他の化合物の混入を含まない)LPSを含むことを特徴とする、請求項5記載の組成物。
【請求項7】
オクロバクトラム・インターメディウム株 LMG3306由来のLPSを0.5−120μg/ml含むことを特徴とする、請求項5または6記載の組成物。
【請求項8】
リポ多糖類が、ミセル相が4℃で1年以上安定である均質な懸濁液形態で存在する、請求項5ないし7のいずれか一項記載の組成物。
【請求項9】
組成物のpHが2−12である、請求項5ないし8のいずれか一項記載の組成物。
【請求項10】
哺乳動物における免疫応答の調節のための医薬の製造における、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項11】
調節が、哺乳動物における免疫能力を増強することを含む、請求項10記載の使用。
【請求項12】
免疫応答の調節が、マクロファージの分化誘導、TNF産生、IL−12産生、脾臓リンパ球によるIFN−γ産生、および/またはウイルス特異的CD8およびCD4 T細胞の活性化からなる、請求項11記載の使用。
【請求項13】
調節が、対象における炎症性サイトカインの産生を阻害することを含む、請求項11記載の使用。
【請求項14】
炎症性サイトカインがIL−12である、請求項13記載の使用。
【請求項15】
ワクチンに含まれる特定のウイルスおよび/または細菌に対して免疫を強化するためのワクチン用アジュバントとしての、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項16】
敗血症の処置および/または予防のための医薬の製造における、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項17】
内毒素血症の処置および/または予防のための医薬の製造における、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項18】
感染症の処置および/または予防のための医薬の製造における、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項19】
免疫抑制動物における感染症の処置および/または予防のための医薬の製造における、請求項18記載の組成物の使用。
【請求項20】
リーシュマニア原虫感染に対するワクチンにおけるアジュバントとしての、請求項18または19記載の組成物の使用。
【請求項21】
リーシュマニア原虫の皮膚感染に対するワクチンにおけるアジュバントとしての、請求項20記載の組成物の使用。
【請求項1】
敗血症を処置および/または予防するための、哺乳動物において免疫応答を刺激し得るオクロバクトラム・インターメディウム LMG3306由来のリポ多糖類。
【請求項2】
オクロバクトラム・インターメディウム株 LMG3306を、トリプチケースソイ培地を用いて培養する工程を含む、請求項1記載のリポ多糖類を製造する方法。
【請求項3】
リポ多糖類の抽出工程:
a)オクロバクトラム・インターメディウム株LMG3306の不活化培養物を遠心し;
b)得られた沈殿物を再懸濁して生理懸濁液とし;
c)精製水およびポリエチレングリコールを用いて透析し;
d)4倍量のメタノールおよび1%の酢酸ナトリウム含有飽和メタノールを用いて沈殿させ;
e)凍結乾燥させて、粗LPSを得る工程
をさらに含む、請求項2記載の方法。
【請求項4】
粗LPSの精製工程:
a)所望により超音波を用いて、粗LPSを緩衝液(10mM Tris−HCl pH7,5)に溶解し;
b)プロテイナーゼKを添加して、室温でインキュベートし;
c)精製したLPSを超遠心分離により集め;
d)サンプルを凍結乾燥する工程
をさらに含む、請求項2記載の方法。
【請求項5】
請求項1記載のオクロバクトラム・インターメディウム LMG3306由来のリポ多糖類および所望により1個またはそれ以上の薬学的に許容される賦形剤を含む、免疫刺激組成物。
【請求項6】
オクロバクトラム・インターメディウム LMG3306の化学的に純粋な(非溶媒成分が、0.2%以上の他の化合物の混入を含まない)LPSを含むことを特徴とする、請求項5記載の組成物。
【請求項7】
オクロバクトラム・インターメディウム株 LMG3306由来のLPSを0.5−120μg/ml含むことを特徴とする、請求項5または6記載の組成物。
【請求項8】
リポ多糖類が、ミセル相が4℃で1年以上安定である均質な懸濁液形態で存在する、請求項5ないし7のいずれか一項記載の組成物。
【請求項9】
組成物のpHが2−12である、請求項5ないし8のいずれか一項記載の組成物。
【請求項10】
哺乳動物における免疫応答の調節のための医薬の製造における、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項11】
調節が、哺乳動物における免疫能力を増強することを含む、請求項10記載の使用。
【請求項12】
免疫応答の調節が、マクロファージの分化誘導、TNF産生、IL−12産生、脾臓リンパ球によるIFN−γ産生、および/またはウイルス特異的CD8およびCD4 T細胞の活性化からなる、請求項11記載の使用。
【請求項13】
調節が、対象における炎症性サイトカインの産生を阻害することを含む、請求項11記載の使用。
【請求項14】
炎症性サイトカインがIL−12である、請求項13記載の使用。
【請求項15】
ワクチンに含まれる特定のウイルスおよび/または細菌に対して免疫を強化するためのワクチン用アジュバントとしての、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項16】
敗血症の処置および/または予防のための医薬の製造における、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項17】
内毒素血症の処置および/または予防のための医薬の製造における、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項18】
感染症の処置および/または予防のための医薬の製造における、請求項5ないし9のいずれか一項記載の組成物の使用。
【請求項19】
免疫抑制動物における感染症の処置および/または予防のための医薬の製造における、請求項18記載の組成物の使用。
【請求項20】
リーシュマニア原虫感染に対するワクチンにおけるアジュバントとしての、請求項18または19記載の組成物の使用。
【請求項21】
リーシュマニア原虫の皮膚感染に対するワクチンにおけるアジュバントとしての、請求項20記載の組成物の使用。
【図1】

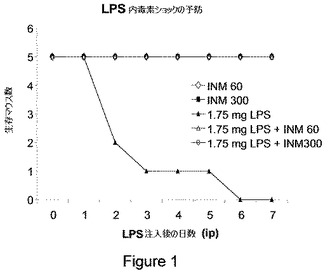
【図2】

【図3】
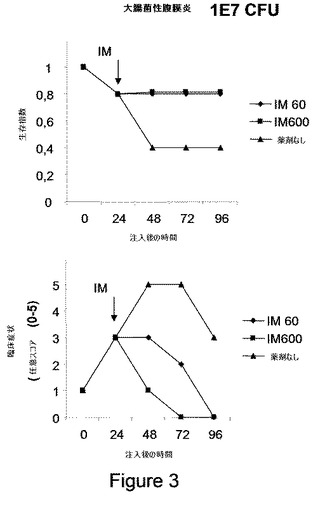
【図4】
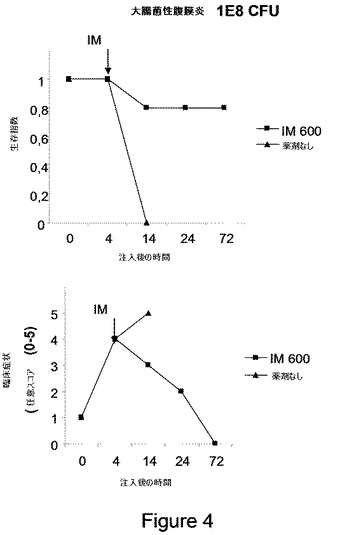
【図5】

【図6】
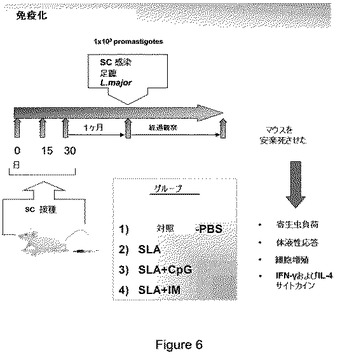
【図7】
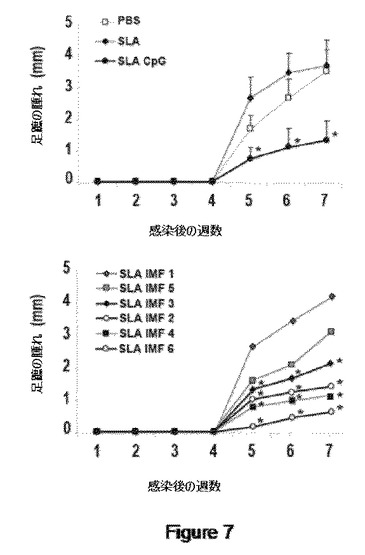
【図8】
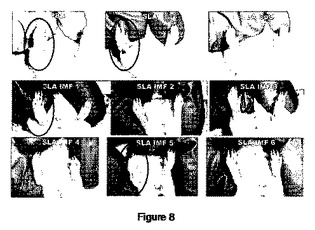
【図9】

【図10】
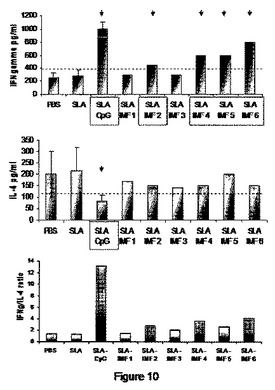
【図11】
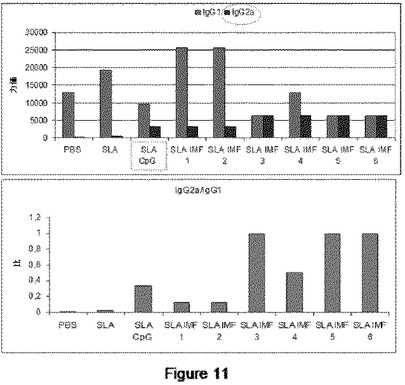
【図12】
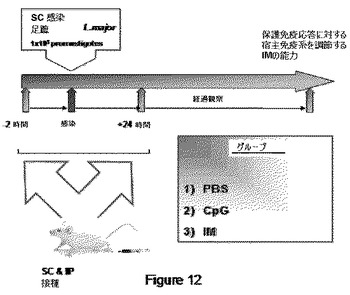
【図13】
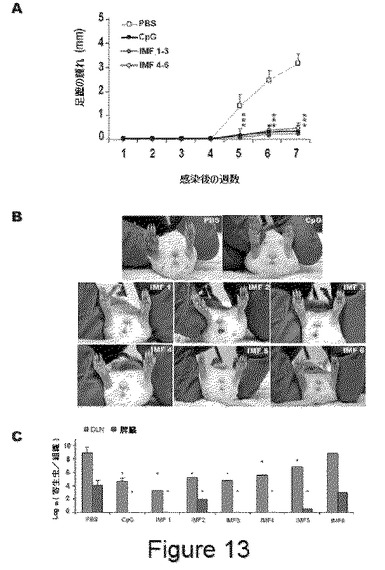
【図14】

【図15】
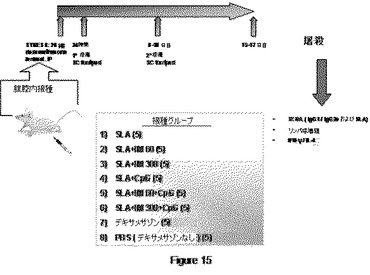
【図16】
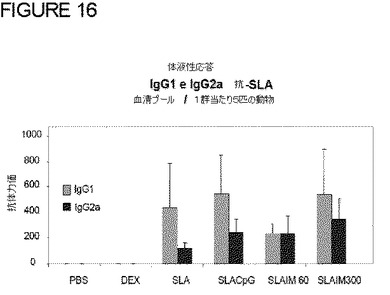
【図17】
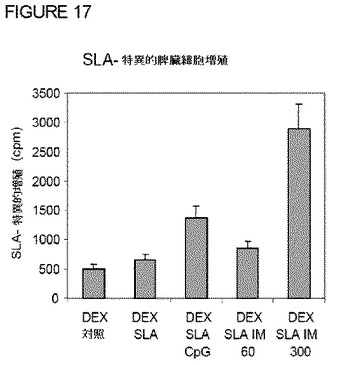
【図18】
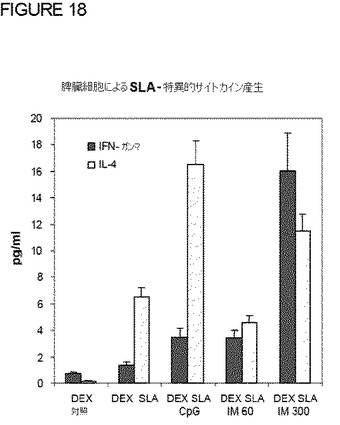
【図19】

【図20】
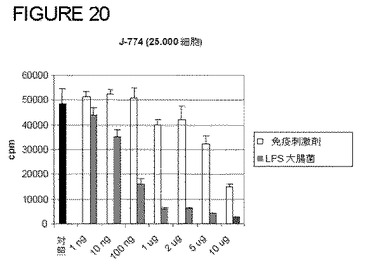
【図21】
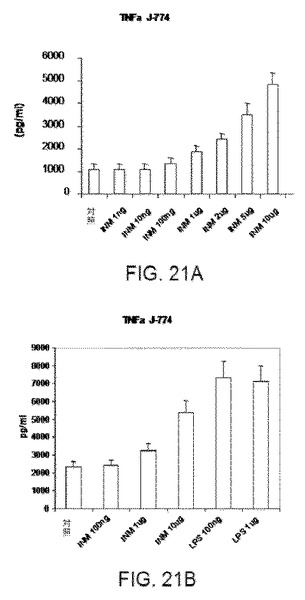
【図22】

【図23】

【図24】
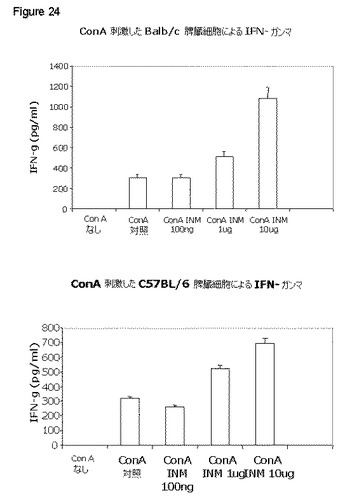
【図25】
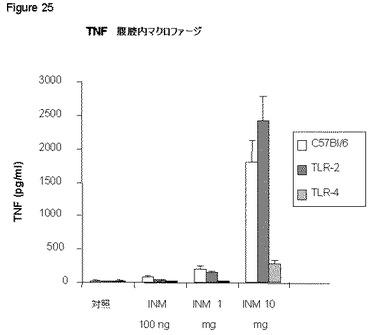
【図26】
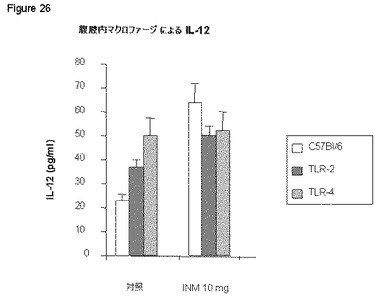
【図27】

【図28】
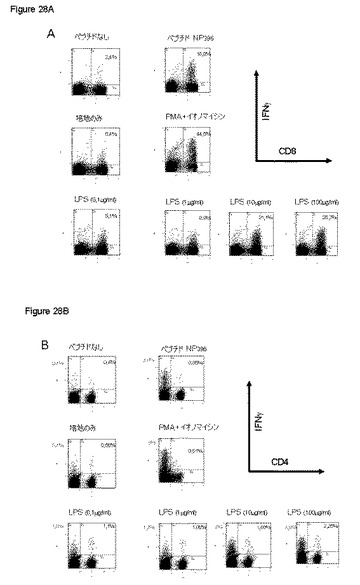
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【公表番号】特表2012−528805(P2012−528805A)
【公表日】平成24年11月15日(2012.11.15)
【国際特許分類】
【出願番号】特願2012−513476(P2012−513476)
【出願日】平成21年12月31日(2009.12.31)
【国際出願番号】PCT/EP2009/009342
【国際公開番号】WO2010/139352
【国際公開日】平成22年12月9日(2010.12.9)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.WINDOWS
【出願人】(511294279)ラボラトリオス・オベヘロ・ソシエダッド・アノニマ (1)
【氏名又は名称原語表記】LABORATORIOS OVEJERO, S.A.
【Fターム(参考)】
【公表日】平成24年11月15日(2012.11.15)
【国際特許分類】
【出願日】平成21年12月31日(2009.12.31)
【国際出願番号】PCT/EP2009/009342
【国際公開番号】WO2010/139352
【国際公開日】平成22年12月9日(2010.12.9)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.WINDOWS
【出願人】(511294279)ラボラトリオス・オベヘロ・ソシエダッド・アノニマ (1)
【氏名又は名称原語表記】LABORATORIOS OVEJERO, S.A.
【Fターム(参考)】
[ Back to top ]