
オステオポンチンsiRNA
【課題】オステオポンチンの亢進に起因する疾患の処置のための医薬等として有用なオステオポンチンのsiRNA及びそれを用いるスクリーニング方法を提供する。
【解決手段】オステオポンチンのためのsiRNA、それを含む組成物及び医薬、及びオステオポンチンと相互作用する物質及び相互作用を阻害又は促進する物質をスクリーニング方法。
【解決手段】オステオポンチンのためのsiRNA、それを含む組成物及び医薬、及びオステオポンチンと相互作用する物質及び相互作用を阻害又は促進する物質をスクリーニング方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、オステオポンチンの発現抑制ためのsiRNAに関するものである。
【背景技術】
【0002】
組織の構築には、細胞間の接着と同時に細胞―細胞外マトリックスとの接着が必要である。細胞―細胞外マトリックス間接着に関与する接着分子は、細胞骨格と細胞外を連結する役割を果たしており、細胞の分化、シグナル伝達に関与している。接着分子と細胞外マトリックスの接着機構を解析することにより、種々の疾患におけるメカニズムの解明、さらには治療につながることが期待されている。
【0003】
細胞外マトリックスの一種であるオステオポンチン(OPN)は、分子量約41kDaの分泌型酸性リン酸化糖タンパク質であり、分子中央部にはαv等のインテグリンとの接着に重要なGRGDS配列が存在し、その直後にはトロンビン開裂部位(R168S169)が存在する。また、GRGDS配列直後に存在するSVVYGLR配列はα9β1、α4β1、α4β7という炎症に関与するインテグリンと結合する。
【0004】
OPNは細胞接着、細胞遊走、一酸化窒素(NO)産生の制御、免疫系への関与など多彩な機能が報告されてきており、癌転移、リウマチ関節炎や多発性硬化症(EAE)などの慢性炎症疾患、自己免疫疾患など多くの難治疾患病態と関与することが示されている。
【0005】
特に、炎症性疾患に関しては、受容体からも推察されるように、直接的な関与が示唆されている。OPNの受容体であるα9インテグリンは好中球上に発現しており、α4インテグリンはリンパ球上に発現していることから、OPN機能を抑制することにより好中球を初めとする白血球の遊走を抑制することが考えられ、抗炎症作用が期待されている。
【0006】
事実、OPN欠損マウスは、腫瘍、リウマチ関節炎、多発性硬化症(EAE)、動脈硬化等の疾患に対し抵抗性を示すことが報告されている(非特許文献1〜4)。また、OPNに対する中和抗体を用いて、リウマチ関節炎が緩解した報告もある(特許文献3、非特許文献5)。
【0007】
そこで、OPN機能を阻害することにより、治療効果を期待することができると考え、新規分子特異的ノックダウン法であるRNAi(RNA干渉:RNA interference)法の治療への応用を検討した。
【0008】
RNAi法とは、短い干渉dsRNA(siRNA(small interfering RNA))を用いて、それが導入された細胞において、それと同じ配列を持つ特定遺伝子の発現を遺伝子レベルですみやかに発現抑制する技術である(非特許文献6及び7)。OPNのmRNA配列から、ターゲットとなる配列を複数選択し、そのsiRNAを合成し、OPNをノックダウンさせることにより疾患治癒に結びつけようと考えた。
【0009】
特許文献1には、オステオポンチンの部分をコードするRNAのIL−1βに関与する結合組織疾患の治療における使用が記載されている。しかし、siRNAとしての使用については、何ら開示されていない。
【0010】
特許文献2には、FGFR(線維芽細胞増殖因子レセプター:fibroblast growth factor receptors)のsiRNA及びFGFRがOPN遺伝子の発現を亢進していることが記載されている。しかし、OPNのsiRNAについては、何らの開示も示唆もない。
【0011】
特許文献3には、組換え抗オステオポンチン抗体及びそれを含む医薬が記載されている。しかし、OPNのsiRNAについては、何らの開示も示唆もない。
【0012】
また、二本鎖RNAをベクターを用いてインビトロで産生させ、これをRNaseIII核酸分解酵素ファミリーの一つである、dicerを用いて切断した、siRNAの集団からなる遺伝子操作用のキットが市販されている(SuperSilencing(登録商標)Human SPP1 siRNAキット(Secreted phosphoprotein 1(osteopontin, bone sialoprotein I, early T-lymphocyte activation 1))非特許文献8)。しかし、このキットは、遺伝子発現の研究のために、siRNAを用いて特定遺伝子の発現を阻害するためのものであり、siRNAの医薬用途については何ら開示されていない。また、上記製造方法によって得られたsiRNAは、分子特異的ではない短いRNAフラグメントをも含んでいることから、遺伝子ノックダウンの特異性が低いという欠点もあった。
【0013】
したがって、オステオポンチンの亢進に起因する疾患の処置のための医薬等として有用なOPNのsiRNAの開発が望まれていた。
【先行技術文献】
【特許文献】
【0014】
【特許文献1】特開平8−191693
【特許文献2】米国特許出願公開第2003/0143676A1号明細書
【特許文献3】国際公開第2003/02751号パンフレット
【非特許文献】
【0015】
【非特許文献1】Nemoto H, Rittling SR, Yoshitake H, Furuya K, Amagasa T, Tsuji K, Nifuji A, Denhardt DT, Noda M. Osteopontin deficiency reduces experimental tumor cell metastasis to bone and soft tissues. J Bone Miner Res. 16:652-9, 2001.
【非特許文献2】Yumoto K, Ishijima M, Rittling SR, Tsuji K, Tsuchiya Y, Kon S, Nifuji A, Uede T, Denhardt DT, Noda M. Osteopontin deficiency protects joints against destruction in anti-type II collagen antibody-induced arthritis in mice. Proc Natl Acad Sci U S A. 99:4556-4561, 2002.
【非特許文献3】Chabas D, Baranzini SE, Mitchell D, Bernard CC, Rittling SR, Denhardt DT, Sobel RA, Lock C, Karpuj M, Pedotti R, Heller R, Oksenberg JR, Steinman L. The influence of the proinflammatory cytokine, osteopontin, on autoimmune demyelinating disease.Science. 294:1731-5, 2001.
【非特許文献4】Matsui Y, Rittling SR, Okamoto H, Inobe M, Jia N, Shimizu T, Akino M, Sugawara T, Morimoto J, Kimura C, Kon S, Denhardt D, Kitabatake A, Uede T. Osteopontin Deficiency Attenuates Atherosclerosis in Female Apolipoprotein E-Deficient Mice. Arterioscler Thromb Vasc Biol. 23:1029-34, 2003.
【非特許文献5】Yamamoto N, Sakai F, Kon S, Morimoto J, Kimura C, Yamazaki H, Okazaki I, Seki N, Fujii T, Uede T. Essential role of the cryptic epitope SLAYGLR within osteopontin in a murine model of rheumatoid arthritis. J Clin Invest. 112:181-8, 2003.
【非特許文献6】Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391:806-11, 1998.
【非特許文献7】Elbashir SM, Harborth J, Lendeckel W, Yalcin A, Weber K, Tuschl T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature. 411:494-8, 2001.
【非特許文献8】SuperArray社、SuperSilencing(登録商標)製品説明、[平成16年3月22日検索]。インターネット<URL:http://www.superarray.com/sirnaqa.php>
【発明の概要】
【発明が解決しようとする課題】
【0016】
本発明の目的は、オステオポンチンのためのsiRNA、それを含む組成物及び医薬、及びオステオポンチンと相互作用する物質及び相互作用を阻害又は促進する物質をスクリーニングする方法を提供することである。
【課題を解決するための手段】
【0017】
本発明者らは、鋭意研究を行ったところ、オステオポンチンの発現を特異的に抑制するsiRNAを見出し、本発明を完成させたものである。したがって、本発明は、下記を提供する。
1.配列番号1、配列番号7又は配列番号21に示す配列からなるDNAに対応するmRNAの、20塩基〜30塩基の連続した任意の部分RNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体であって、オステオポンチンの発現を抑制するRNA。
2.配列番号1に示す配列からなるDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号298〜1140番から選ばれる連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体。
3.配列番号3、4、5若しくは6に示す配列からなるRNA、配列番号3、4、5若しくは6に示す配列と相補的な配列からなるRNA、若しくは配列番号3、4、5若しくは6に示す配列からなるRNAと配列番号3、4、5若しくは6に示す配列と相補的な配列からなるRNAとからなる二本鎖RNA、又はそれらの誘導体。
4.配列番号28、29若しくは30に示す配列からなるRNA、配列番号28、29若しくは30に示すと相補的な配列からなるRNA、若しくは配列番号28、29若しくは30に示す配列からなるRNAと配列番号28、29若しくは30に示すと相補的な配列からなるRNAとからなる二本鎖RNA、又はそれらの誘導体。
5.配列番号7に示す配列からなるDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号241〜1065番から選ばれる連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体。
6.配列番号9、10、11若しくは12に示す配列からなるRNA、配列番号9、10、11若しくは12に示す配列と相補的な配列からなるRNA、若しくは配列番号9、10、11若しくは12に示す配列からなるRNAと配列番号9、10、11若しくは12に示す配列と相補的な配列とからなるRNAからなる二本鎖RNA、又はそれらの誘導体。
7.配列番号21に示す配列からなるDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号1〜954番から選ばれる連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体。
8.配列番号23、24、25、26若しくは27に示す配列からなるRNA、配列番号23、24、25、26若しくは27に示す配列と相補的な配列からなるRNA、若しくは配列番号23、24、25、26若しくは27に示す配列からなるRNAと配列番号23、24、25、26若しくは27に示す配列と相補的な配列からなるRNAとからなる二本鎖RNA、又はそれらの誘導体。
9.上記1〜8の何れかに記載のRNAを含む、オステオポンチンの発現を抑制するためのオステオポンチン発現抑制剤。
10.上記1〜8の何れかに記載のRNAを含む、オステオポンチンの発現を抑制するための組成物。
11.上記1〜8の何れかに記載のRNAを有効成分として含む医薬。
12.オステオポンチンの発現を抑制するためのものである上記11記載の医薬。
13.上記1〜8の何れかに記載のRNAを有効成分として含む、オステオポンチンの亢進に起因する疾患の処置のための医薬。
14.上記オステオポンチンの亢進に起因する疾患が、腫瘍、肝炎、動脈硬化、多発性硬化症、関節炎、リウマチ及び/又は肺繊維症である、上記13に記載の医薬。
15.上記1〜8の何れかに記載のRNAを含む、siRNA発現ベクター。
16.被験物質と上記1〜8の何れかに記載のRNAとを接触させることを含む、オステオポンチンと相互作用する物質、又はオステオポンチン及びオステオポンチンと相互作用する物質の相互作用を促進又は阻害する物質をスクリーニングするための方法。
【図面の簡単な説明】
【0018】
【図1】マウスOPN siRNAのターゲット配列とその位置。
【図2】ヒトOPN siRNAのターゲット配列とその位置。
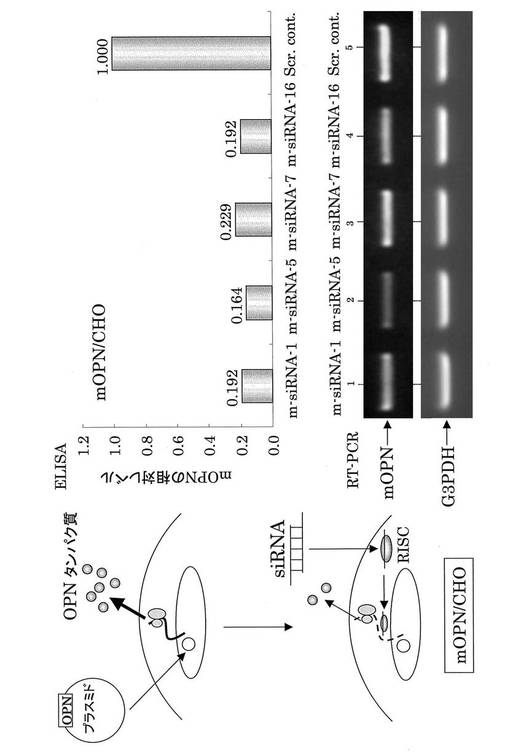
【図3】mOPN siRNAのノックダウン効果(マウスOPNを遺伝子導入したmOPN/CHO細胞を用いた。Scr contは、哺乳動物の細胞にはないことが確認されているsiRNAであり、siRNAのネガティブコントロールとして使用した。RISCはRNA-induced silencing complexを示す。)。mOPN siRNA は、mOPNタンパク質発現を抑制した。
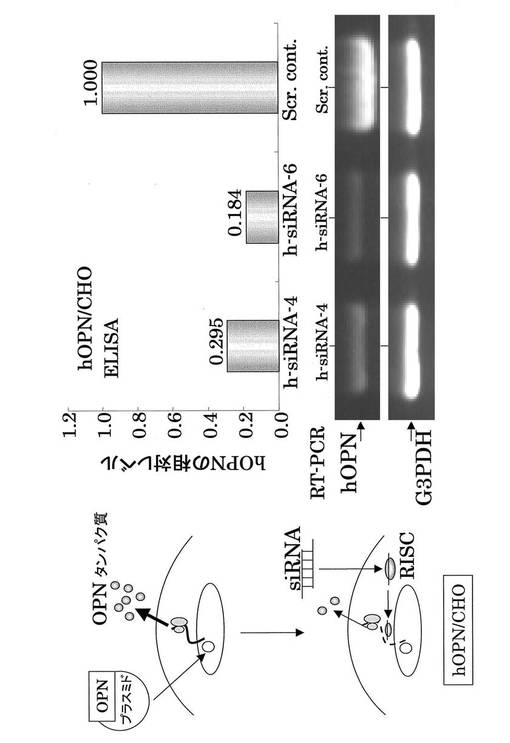
【図4】hOPN siRNAのノックダウン効果(ヒトOPNを遺伝子導入したhOPN/CHO細胞を用いている。Scr cont、RISCは図3と同義である)。hOPN siRNA はhOPNタンパク質発現を抑制した。
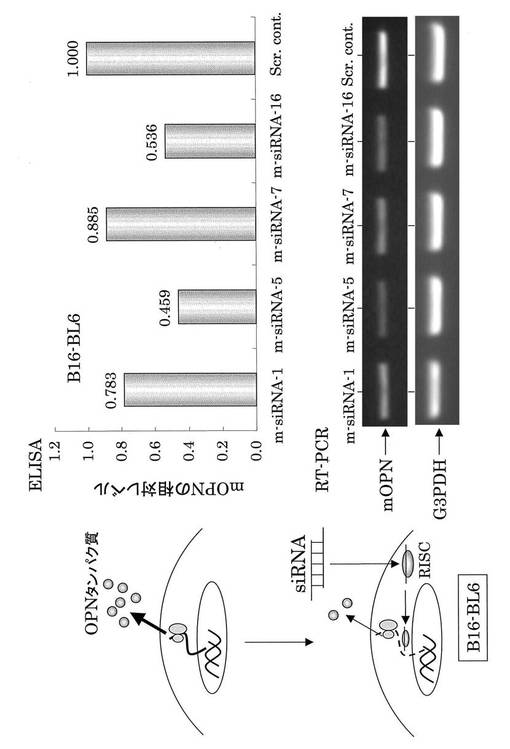
【図5】内在性OPNに対するsiRNAのノックダウン効果(細胞はマウス悪性メラノーマ細胞であるB16−BL6細胞株を用いている。Scr cont、RISCは図3と同義である)。mOPN siRNA は内在性mOPNの発現を抑制した。
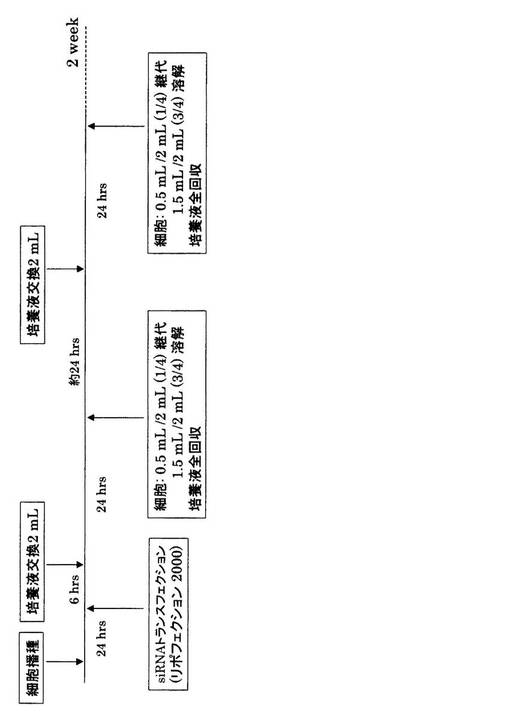
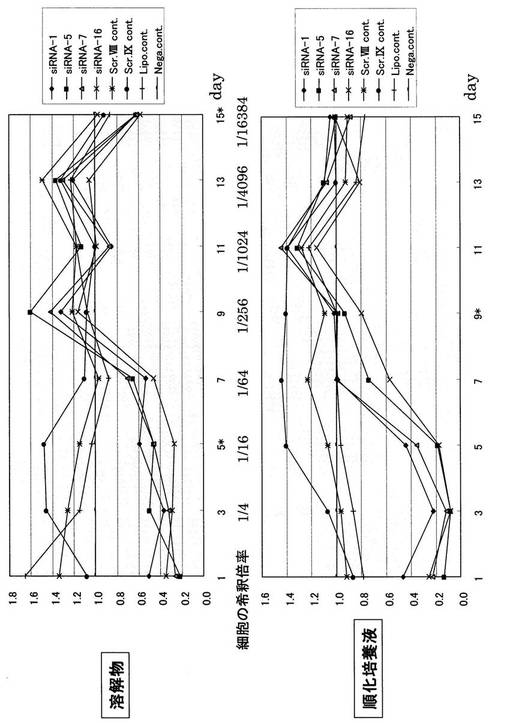
【図6】mOPN siRNAのRNAi効果持続時間解析実験のスケジュール。
【図7】mOPN siRNAのRNAi効果持続時間(Lipo. cont.は、siRNAを加えないでLipofectamine 2000を用いて遺伝子導入を行った結果である。Nega cont.は、遺伝子導入処理等を一切行わないで通常に培養していた状態である)。
【図8−1】肝炎誘発とmOPN siRNA による肝炎治療実験プロトコール。
【図8−2】肝炎誘発とmOPN siRNA による肝炎治療実験プロトコール。
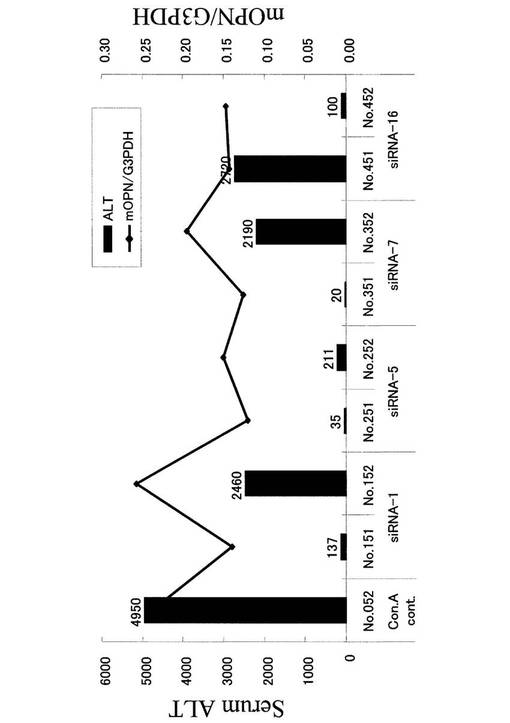
【図9】mOPN siRNAによる肝臓中OPNのノックダウン効果とALT値。
【図10】OPN siRNA投与による肝臓HE染色像
【図11】肝炎悪性度とOPN mRNA量の相関
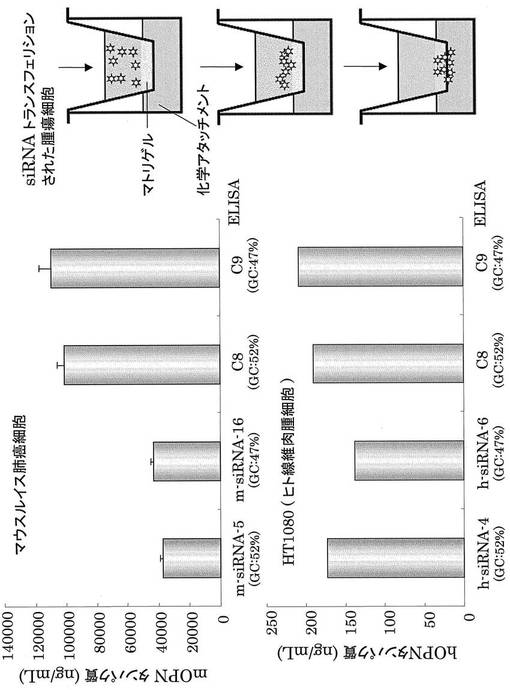
【図12】癌細胞に対するOPN siRNA のノックダウン効果。
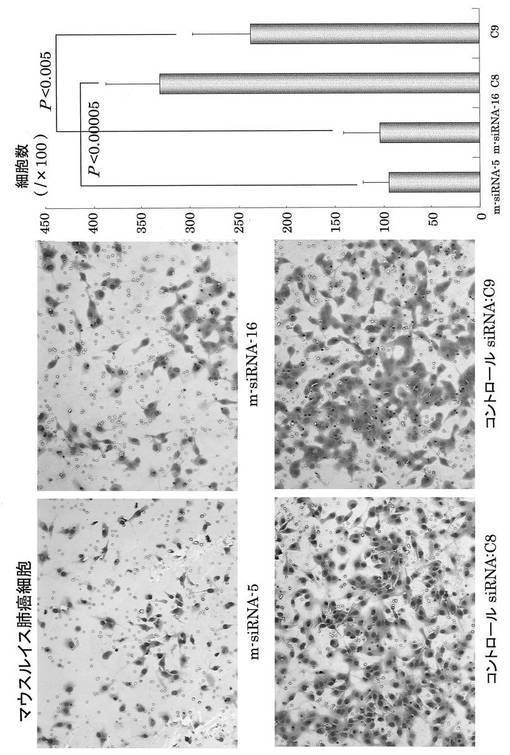
【図13】mOPN siRNA による癌細胞浸潤抑制効果。mOPN siRNAは肺癌細胞の浸潤を抑制した。
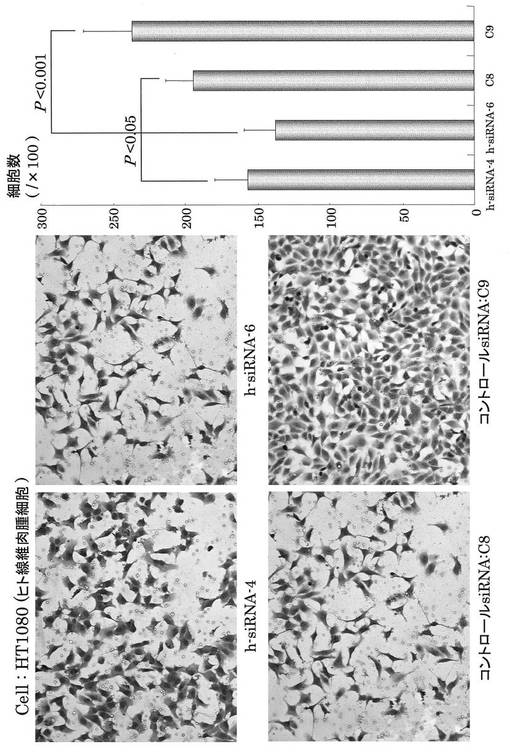
【図14】hOPN siRNAによる癌細胞浸潤抑制効果。hOPN siRNAはヒト線維肉腫の浸潤を抑制した。
【図15】OPN siRNA投与による肝炎治療効果(血清ALT値)。
【図16】蛍光siRNA(Cy5)を用いたFACS。
【図17】蛍光siRNA(Alexa Fluor 647)を用いたFACS。
【図18】siRNA−Cy5の細胞内局在。
【図19】siRNA−Alexa Fluor 647の細胞内局在。
【図20】siRNAの肝臓内リンパ球導入プロトコール。
【図21】肝臓内リンパ球のFACSによる分離・検出。
【図22】siRNAの肝臓内リンパ球への導入。
【図23】ラットOPN siRNAのターゲット配列とその位置。
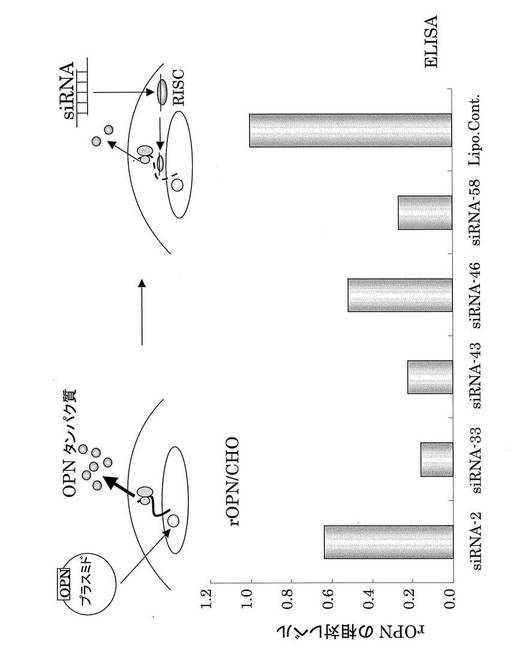
【図24】ラットOPN siRNAのノックダウン効果。rOPN siRNAはrOPNタンパク質発現を抑制した(RISCは図3と、Lipo. Cont.は図7のLipo. cont.と同義である)。
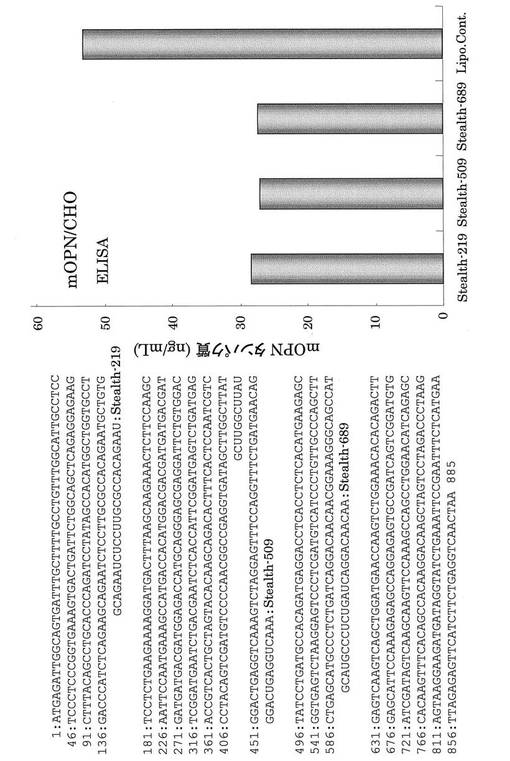
【図25】mOPN Stealth(商標)RNAiのターゲット配列とそのノックダウン効果。mOPN Stealth(商標)RNAiはmOPN タンパク質発現を抑制した。
【発明を実施するための形態】
【0019】
以下、本発明の実施の形態について詳細に説明する。
先ず、本明細書で使用する略称について説明する。
Aまたはa:アデニン
Tまたはt:チミン
Uまたはu:ウラシル
Gまたはg:グアニン
Cまたはc:シトシン
RNA:リボ核酸(通常、塩基としてA,U,G,Cを有するリボヌクレオチド単位から構成される。)
DNA:デオキシリボ核酸(通常、塩基としてA,T,G,Cを有するデオキシリボヌクレオチド単位からなる。)
DAPI:4‘,6−ジアミノ−2−フェニルインドール
ELISA:enzyme-linked immunosorbent assay
FACS:蛍光活性化セルソーター
FITC:フルオレセインイソチオシアネート
PE:フィコエリトリン(紅色の色素タンパク質)
Scr. RNA:スクランブルRNA。mRNA特異的なsiRNAとGC含量を揃えた非特異的なRNA。
Scr. cont.:対象となるsiRNAのコントロールとして用いるScr. RNA
「オステオポンチン」とは、分泌性リン酸タンパク質1(secreted phospho protein 1)とも呼ばれ、配列番号2(マウス)、配列番号8(ヒト)又は配列番号22(ラット)に示すアミノ酸配列からなるタンパク質であり、骨のマトリックスに含まれる接着タンパク質の一種である。オステオポンチンは、インテグリンと結合するRGD配列を有し、破骨細胞と骨芽細胞の両方を骨組織周辺に接着させる作用を有する。また、オステオポンチンは、強く負に荷電しており、ヒドロキシアパタイトを沈着させ、骨のカルシウムを保持する作用を有する。以下、オステオポンチンをOPNと略すこともある。
【0020】
マウス、ヒト及びラットのオステオポンチン遺伝子の塩基配列及びアミノ酸配列を、それぞれ配列番号1及び2、配列番号7及び8、並びに配列番号21及び22に示した。
【0021】
本発明はオステオポンチン遺伝子に対応するmRNAの部分配列であって、20塩基〜30塩基の連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体であって、オステオポンチンの発現を抑制するsiRNAに関する。
より具体的には本発明は、配列番号1に示す配列からなるマウスDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号298〜1140番から選ばれる連続した任意のRNA配列からなるセンスRNA、配列番号7に示す配列からなるヒトDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号241〜1065番から選ばれる連続した任意のRNA配列からなるセンスRNA、配列番号21に示す配列からなるラットDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号1〜954番から選ばれる連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体に関する。
なお、OPNの発現を制御するsiRNAは、通常OPN遺伝子の開始コドンから75塩基以上下流の最初の塩基配列AAを見つけ、当該AAを含む21〜23塩基、好ましくは21塩基であって、GC含量が50%前後の、OPNに対して特異的な配列を選択することによって得られる。特異的であるか否かは、例えばNCBI(http://www.ncbi.nlm.nih.gov/)のBLAST-searchにかけることによって確認することができる。
また、本発明の他の具体的な態様としては、上記各DNAの翻訳領域に対応するmRNAの部分配列であって、GCで始まる25塩基の連続した任意のRNA配列からなるセンスRNAと該センスRNAに相補的なアンチセンスRNAとからなる二本鎖RNAの誘導体であって、いわゆるステルス(商標)RNAiの構造を有するsiRNAが挙げられる(Stable plasma membrane levels of hCTR1 mediate cellular copper uptake. J Biol Chem. 2005 Mar 11;280(10):9635-9. Epub 2005 Jan 5. PMID: 15634665およびhttp://www.invitrogen.co.jp/products/molecular_biology/RNAi.shtml参照)。ステルス(商標)RNAiは、上記の21〜23塩基のsiRNAと同様にターゲット遺伝子を抑制し、かつ非特異的なインターフェロンストレス応答を抑制し、血清、細胞質中においても安定性が高いという特長を有する。
【0022】
本発明で相補的な配列からなるRNAとは、所定の塩基配列を有するRNAに対して、A:UおよびG:Cといった塩基対関係に基づいて、mRNAの配列に対応する配列を有する鎖を意味するものである。DNAの相補的二本鎖のうち、タンパク質をコードしている鎖、すなわち、mRNAと同一の配列を有する鎖がセンス鎖であり、センス鎖と塩基配列が相補的な関係にある配列を有する鎖がアンチセンス鎖である。
【0023】
本発明で誘導体とは、例えば、本発明のRNAの安定性を向上させるために、3’側にdTを数個、好ましくは2〜4個、より好ましくは2又は3個、最も好ましくは2個付加させた、3’オーバーハング型誘導体である。なお、dTの代わりにUを付加させても良い。
【0024】
また、本発明で誘導体とは、例えば、本発明のRNAの安定性を向上させるために、5’側にリン酸を付加させた誘導体である。
【0025】
また、本発明で誘導体とは、いわゆるタンデムタイプのsiRNA発現ベクターに組み込むための、2つのプロモーター、例えばU6プロモーター、センスRNA及びアンチセンスRNAを結合させた誘導体である。具体的には、U6プロモーター、センスRNA、5個のT、U6プロモーター、アンチセンスRNA、5個のTを結合させた誘導体が挙げられる。
【0026】
また、本発明で誘導体とは、いわゆるステムループタイプのsiRNA発現ベクターに組み込み、ショートヘアピンRNAを介して、siRNAを産生するための、プロモーター、例えばU6プロモーター、センスRNA、ループ配列及びアンチセンスRNAを結合させた誘導体である。具体的には、U6プロモーター、センスRNA、ループ配列、例えば、gtgtgctgtcc、アンチセンスRNAを結合させた誘導体が挙げられる。
また、本発明で誘導体とは、ステルス(商標)RNAi構造を有する誘導体である。
【0027】
siRNA(small interfering RNA)とは、20〜30bp、好ましくは21〜23bp、25bpまたは27bpからなる二本鎖RNAであり、RNA干渉(RNA interference: RNAi)を誘導することができる二本鎖RNAをいう。また、RNA干渉とは、二本鎖RNAにより、そのRNAと相同的な配列を有する遺伝子の発現が抑制される現象をいう。
【0028】
RNAiは、特定配列を有する遺伝子を破壊することができることから、遺伝子機能の解析や特定遺伝子の発現抑制に有用である。また、細胞への導入量が少量の割に比較的長時間効果が持続するという特徴を有する。
【0029】
本発明のsiRNAは、常法によって合成することができ、市販のDNA/RNAシンセサイザー、例えば、Applied Biosystems394型で行うことができる。
【0030】
本発明のRNAは、そのままか又は凍結乾燥させて、オステオポンチンの機序解明のための試薬として用いることができる。
【0031】
本発明のRNA又はその誘導体は、そのままもしくは公知の薬学的に許容される担体(賦形剤、増量剤、結合剤、滑沢剤などが含まれる)や慣用の添加剤などと混合して医薬組成物として調製することができる。当該医薬組成物は、調製する形態(錠剤、丸剤、カプセル剤、散剤、顆粒剤、シロップ剤などの経口投与剤;注射剤、点滴剤、外用剤、坐剤などの非経口投与剤)等に応じて経口投与または非経口投与することができる。また投与量は、有効成分の種類、投与経路、投与対象または患者の年齢、体重、症状などによって異なり一概に規定できないが、通常、1日投与用量として、数mg〜2g程度、好ましくは数十mg程度を、1日1〜数回にわけて投与することができる。
【0032】
siRNAベクターは、常法にしたがって調製することができる。タンデムタイプの場合は、PCRによってセンス、アンチセンス配列を含むプライマーによりプロモーター部分を増幅し、増幅断片を制限酵素で切断後、ベクターのプロモーター、例えばU6プロモーターの下流に挿入する。ステムループの場合、センス−ループ−アンチセンス配列を含むオリゴヌクレオチドを合成アニールさせ、ベクターのプロモーター、例えばU6プロモーターの下流に挿入する。
以下、実施例によって本発明を具体的に説明する。なお、実施例においてmOPNはマウス・オステオポンチンを、hOPNはヒト・オステオポンチンを、そしてrOPNはラット・オステオポンチンそれぞれ示す。
【実施例1】
【0033】
(1)オリゴヌクレオチドの調製
4種のリボヌクレオシド3’−ホスホロアミダイト(GLENリサーチ)を用いて、DNA自動合成機(Applied Biosystem Model394A)で配列番号3、4、5、6、9、10、11、12、23、24、25、26及び27に示す各配列の各オリゴリボヌクレオチド(RNA断片)を合成した。各RNA断片を1μmolスケールで合成した。合成終了後、合成したオリゴヌクレオチドが結合したCPG(Controlled Pore Glass)を濃アンモニア水:エタノール(3:1v/v)混液で室温2時間処理してオリゴリボヌクレオチドをCPG樹脂から切り出し、更に55℃で16時間加温した。
マウスOPNについて、RNAi designer(http://www.invitrogen.co.jp/primers/rnaidesign/)を用いて決定した配列である、配列番号28、29若しくは30に示す配列について、invitorgenの示す方法にしたがって、ステルス(商標)RNAiを調製した(Stable plasma membrane levels of hCTR1 mediate cellular copper uptake. J Biol Chem. 2005 Mar 11;280(10):9635-9. Epub 2005 Jan 5. PMID: 15634665およびhttp://www.invitrogen.co.jp/products/molecular_biology/RNAi.shtml参照)。
【0034】
溶媒を留去し、残渣に1mlの1M TBAF(テトラブチルアンモニウムフルオリド)/THF(テトラヒドロフラン)溶液を加え、37℃で16時間攪拌した。これに5mlの0.1Mトリエチルアンモニウムアセテート水溶液(pH7.0)を加えた後、C18(ウォーターズ社製)オープンカラムクロマトグラフィーを行なった(カラムサイズ1.5×12cm:5−40%アセトニトリル、50mMトリエチルアンモニウムビカルボネート水溶液の両溶媒を用いた濃度勾配により溶出した)。約30%濃度のアセトニトリルで溶出される末端リボヌクレオチド5’位の保護基であるジメトキシトリチルの発色を有するフラクションを集め、これに5mlの0.01N塩酸を加え、15分間攪拌した。0.1Nアンモニア水で中和し、水層を酢酸エチルで洗浄し、溶媒留去後、滅菌水1mlに溶解した。この画分中のオリゴリボヌクレオチドを逆相HPLCで分取後、さらにイオン交換HPLCで分取し、精製した。得られたオリゴヌクレオチドを後述の実験に供した。
【0035】
逆相およびイオン交換HPLCの条件は以下の通りであった:
逆相HPLC
カラム:μ−ボンダスフィアー(C−18)カラム、Ф3.9x150mm(ウォーターズ社製)
溶媒:A溶液 5% アセトニトリル/0.1M TEAA(トリエチルアンモニウムアセテート、pH7.0);
B溶液 25% アセトニトリル/0.1M TEAA(pH7.0)。
イオン交換HPLC
カラム:TSKgel DEAE 2SWカラム、4.6×250mm、東ソー(株)製
溶媒:A溶液 20% アセトニトリル;
B溶液 20% アセトニトリルを含む2M ギ酸アンモニウム。
【0036】
(2)OPNsiRNAの生物学的作用
〔方法〕
<培養細胞へのOPN siRNA導入方法>
上記の通り合成された各RNA断片をOPN siRNAとして、Lipofectamine 2000(Invitrogen, Carlsbad, CA)を用いるリポフェクション法により、24穴プレート中で増殖させた対数増殖期の培養細胞に導入した。
【0037】
F−12培地50μLにリポフェクタミン2000を2μLを加え、5分間放置した。別のチューブで、F−12培地50μLに各siRNA 0.8μgを加えた。両チューブの内容物を混ぜ、20分放置し、siRNA−リポフェクタミン2000複合体を調製した。細胞をF−12培地で洗浄後、F−12培地500μLを加え、さらにsiRNA−リポフェクタミン複合体を加え、これを24時間培養した。次いで、上清を回収し、細胞ペレットを、TNEバッファー(10mM Tris−HCl pH7.8、1.0% NP−40、0.15M NaCl、1mM EDTA)を用いて可溶化させた。ELISA(固相酵素免疫検定法)にて上清及び細胞ペレット可溶化物におけるOPN発現量を定量した。
【0038】
<ELISA及びRT−PCR>
ヒトOPN及びマウスOPNのsiRNAのRNAi効果をELISA法、RT−PCR法(逆転写−ポリメラーゼ連鎖反応法)及びリアルタイムPCR法を用いて検討した。
ヒトOPN、マウスOPNのELISAを用いた解析には、ヒトオステオポンチン測定キット及びマウスオステオポンチン測定キット(共に免疫生物研究所製)を使用した。
RT−PCR及びリアルタイムPCRには以下のプライマーを用いた。リアルタイムPCRは、LightCycler-FastStart DNA Master SYBR Green I(Roche Diagnostics、Mannheim、Germany)により作製したサンプルを用いて行った。定量は、グリセルアルデヒド三リン酸脱水素酵素(G3PDH)を標準とした標準比(Normalized Ratio)で行った。
また、ラットOPNのsiRNA及びマウスOPN stealth(商標)RNAのノックダウン効果を、上記と同様にELISAを用いて解析した。
【0039】
使用したプライマー
mOPN-forward: 5'-TCAAAGTCTAGGAGTTTCCAG-3'
mOPN-reverse: 5'-TTAGTTGACCTCAGAAGATGA-3'
hOPN-forward: 5'-CCATGAGAATTGCAGTGATTT-3'
hOPN-reverse: 5'-TTAATTGACCTCAGAAGATGC-3'
G3PDH-forward: 5'-ACCACAGTCCATGCCATCAC-3'
G3PDH-reverse: 5'-TCCACCACCCTGTTGCTGTA-3'
β-actin-forward: 5'-TGGAATCCTGTGGCATCCATGAAAC-3'
β-actin-reverse: 5'-TAAAACGCAGCTCAGTAACAGTCCG-3'
【0040】
<肝炎マウス作製>
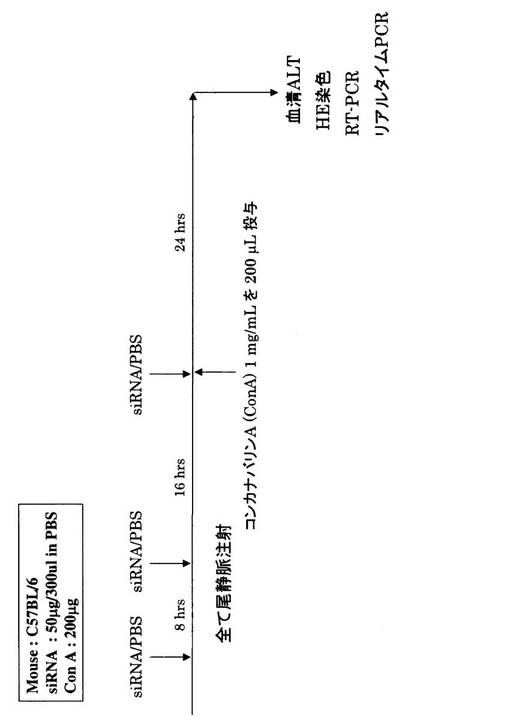
C57BL/6マウスにコンカナバリンA(ConA、Vector laboratories, Burlingame, CA)を尾静脈投与し(200μg/マウス)、肝炎モデルを作出した。ConA投与24時間後に、肝臓を摘出し、ALT(アラニンアミノトランスフェラーゼ;GPT)測定、HE(ヘマトキシリン−エオシン)染色、RNA抽出を行った。
【0041】
<肝炎治癒作用>
上記肝炎モデルにsiRNA50μg(300μl(図8−1)又は500μl(ハイドロダイナミクス法:図8−2))をマウスに尾静脈投与した。
siRNAを、ConA投与24時間前、16時間前及び同時の3回投与した。コントロールとして、PBS(リン酸緩衝生理食塩水)を上記と同条件で投与した(図8−1、図8−2)。
【0042】
<癌転移抑制作用/マトリゲルアッセイ>
癌浸潤能のインビトロアッセイは、マトリゲルインベージョンチャンバー(BD Biosciences, Bedford, MA)を用いて行った。マウスルイス肺癌細胞又はHT1080(ヒト線維肉腫細胞)(2.5×104細胞/ウェル)をチャンバー上層に接種し、下層には、5% FCS(ウシ胎仔血清)を含む培地を加え、22時間培養した。綿棒にてフィルター上層をこすり、上層部に残るマトリゲルや非浸潤細胞を取り去った。フィルター下層の浸潤した細胞をメタノールで固定し、ギムザで染色し、蒸留水で洗浄した。
【0043】
〔実験結果〕
<培養細胞におけるノックダウン効果>
mOPN siRNAのmOPNに対するRNAi効果を、mOPN遺伝子をCHO細胞に高発現させたトランスフェクタントであるmOPN/CHO細胞に種々のsiRNAを導入することにより解析した。RNAi効果の確認は、mOPNに対するELISAとRT−PCRによって行った。ELISAは、siRNA導入後の細胞可溶化物をサンプルとして用いた。その結果、m−siRNA−1、m−siRNA−5、m−siRNA−7及びm−siRNA−16の全てのsiRNAについて、mOPNのノックダウン効果が認められた(図3)。
【0044】
同様にhOPN siRNAのhOPNに対するRNAi効果を、hOPN/CHO細胞を用いて解析した。その結果、h−siRNA−4及びh−siRNA−6において、hOPNのノックダウン効果が認められた(図4)。
【0045】
本発明のsiRNAが、内在性OPN発現をも抑制するかどうかを、マウス悪性メラノーマ細胞株である、B16−BL6細胞を用いて解析した。細胞可溶化物をサンプルとして使用した結果、m−siRNA−5及びm−siRNA−16によって内在性OPN発現の抑制効果が認められた(図5)。
【0046】
次に、mOPN siRNAのRNAi効果持続期間について検討した。図6に示すようにmOPN/CHO細胞に遺伝子導入後、24時間後に上清と細胞3/4を回収した。残りの細胞1/4をさらに培養し、24時間後に培地を交換し、さらに24時間後に、培養上清及び細胞3/4を回収した。残りの細胞1/4をさらに培養し、同様な操作を繰り返し、上清及び細胞の回収を2週間行った。培養上清と細胞可溶化物におけるOPNの発現量をELISA法によって測定した。その結果、約1週間後においても、全てのsiRNAにおいて、ノックダウン効果が認められた(図7)。
【0047】
<OPNsiRANの治療効果>
次に、OPN siRNAを用いたRNAi医薬への可能性について検討するため、肝炎についてOPN機能抑制による治療効果を試験した。治癒効果検討方法の概略を図8−1及び図8−2に示した。すなわち、コンカナバリンA(ConA)200μgをC57BL/6マウスに尾静脈内投与することによって肝炎誘発モデルを作製した。siRNAをConA投与24時間前、16時間前、同時の3回、それぞれ50μgを尾静脈内に投与した。肝炎の評価は、ConA投与24時間後の血清ALT(アラニンアミノトランスフェラーゼ)値と肝臓組織のHE染色にて行った。OPNのRNAi効果は、RT−PCRとリアルタイムPCRによって評価した(図8−1)。
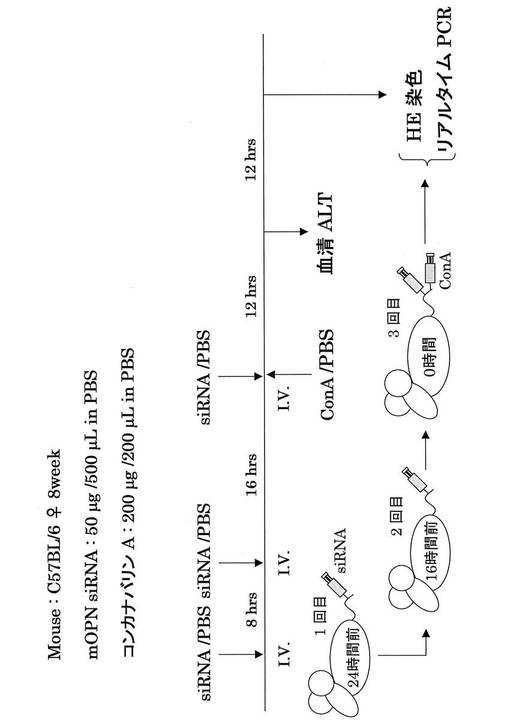
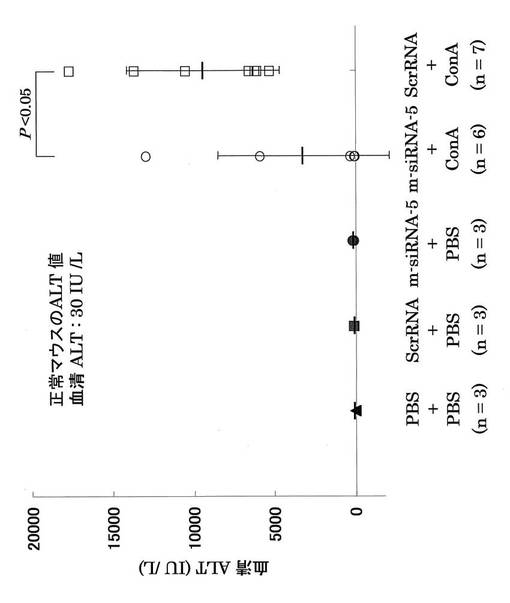
さらに、図8−2に示すプロトコールで、肝炎治療実験を行った。5群で実験を行った。すなわち、1群:siRNAの代わりにPBSとConAの代わりにPBS、2群:Scr siRNAとConAの代わりにPBS、3群:OPN siRNA−5とConAの代わりにPBS、4群:OPN siRNA−5とConA、5群:Scr siRNAとConA。ConA肝炎マウスに対してOPN siRNAを投与した4群では有意にALTの減少が見られた(図15)。
【0048】
肝臓におけるOPNノックダウン効果をRT−PCRによって検討した。その結果、siRNA処理群では、コントロール群と比較して低いOPN発現が見られた。一方、血清ALT値は、OPN抑制が認められた個体において、低値傾向が見られた(図9)。
【0049】
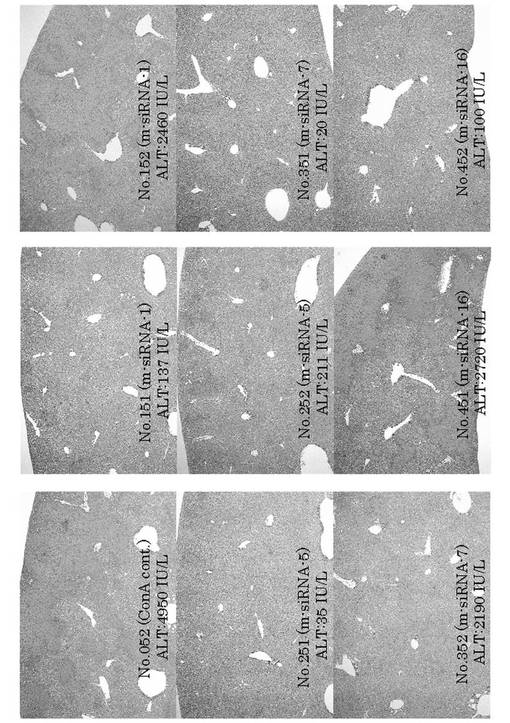
肝臓のHE染色を行った。OPN siRNAを投与していない個体は、肝臓に広範な壊死が観察された。一方、OPN siRNAを投与した個体は、肝臓内壊死が抑制されていた(図10)。
【0050】
OPNの発現量を定量化するために、リアルタイムPCRを行った。その際、ハウスキーピング遺伝子であるグリセルアルデヒド三リン酸脱水素酵素(G3PDH)の発現量を用いて標準化を行った。OPN発現量とALT値との相関を調べた結果、OPN量とALT値に強い相関が見られた(図11)。
【0051】
次に、癌転移のインビトロアッセイであるマトリゲル法を用いて、OPN siRNAによる癌転移抑制能を調べた。mOPN siRNA、hOPN siRNAの転移抑制能を調べるためにマウスルイス肺癌細胞 と HT1080(Human Fibrosarcoma cells)細胞をそれぞれ用いた。
【0052】
まず、mOPN、hOPNのノックダウン効果をそれぞれのOPN ELISA kitを用いて検討した。その結果、mOPN siRNAでは、m−siRNA−5及びm−siRNA−16の両方においてOPNのノックダウン効果が見られ、hOPN siRNAでは、h−siRNA−6においてノックダウン効果が観察された(図12)。C8、C9は、GC含量が一致するコントロールsiRNAを対照として使用しており、m−siRNA−5とh−siRNA−4のコントロールはC8、m−siRNA−16とh−siRNA−6のコントロールとしてC9を利用した。
【0053】
転移能の評価は、マトリゲル膜の裏側に浸潤した細胞を観測することにより検討した。siRNAを導入した細胞の浸潤能は、コントロールと比較して少ないことが分かった(図13、14)。特に、mOPN siRNAでは、m−siRNA−5が、hOPN siRNAではh−siRNA−6が強い浸潤抑制能を有していることが分かった(図13、14)。グラフは、浸潤している細胞を計測することにより定量化して、OPN siRNAによる浸潤抑制効果を確認した結果を示す。
【0054】
その結果、siRNAを導入した細胞の浸潤能は、コントロールと比較して明らかに抑制された(図12)。特に、mOPN siRNAでは、m−siRNA−5が、また、hOPN siRNAではh−siRNA−6がマトリゲルに対する細胞の浸潤を抑制し、これらが、強い浸潤抑制能を有していることが分かった(図13、図14)。
以上により、OPNノックダウン効果と浸潤抑制とが相関している結果を得ることができた。
【0055】
<siRNAのターゲット免疫細胞の同定>
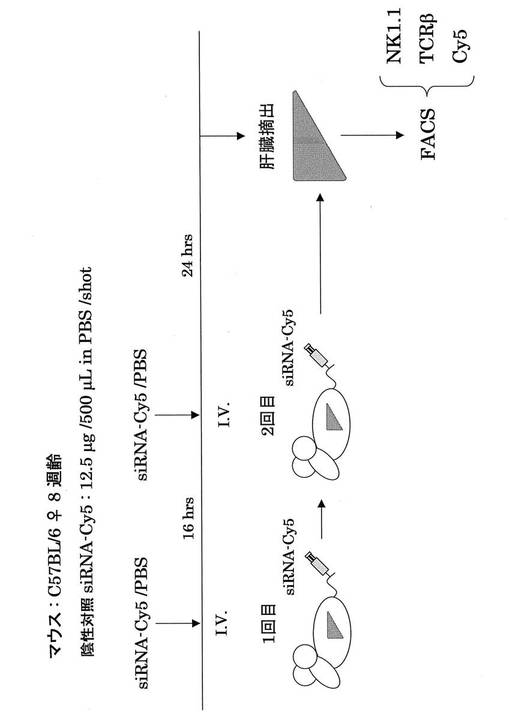
肝炎発症には、NKT細胞から分泌されるOPNが重要ということは既に報告されている(Diao H, Kon S, Iwabuchi K, Kimura C, Morimoto J, Ito D, Segawa T, Maeda M, Hamuro J, Nakayama T, Taniguchi M, Yagita H, Van Kaer L, Onoe K, Denhardt D, Rittling S, Uede T. Osteopontin as a mediator of NKT cell function in T cell-mediated liver diseases. Immunity. 2004 Oct;21(4):539-50.)。以上のことから、OPN siRNAの静脈内投与による肝炎治療効果は、NKT細胞から分泌されるOPNをOPN siRNAが抑制していると推察された。そこで、蛍光siRNAをマウス尾静脈に投与し、肝臓内の白血球のsiRNAの取り込みをFACSを用いて解析したところ、NKT細胞画分にsiRNAが取り込まれている結果を得ることができた。
【0056】
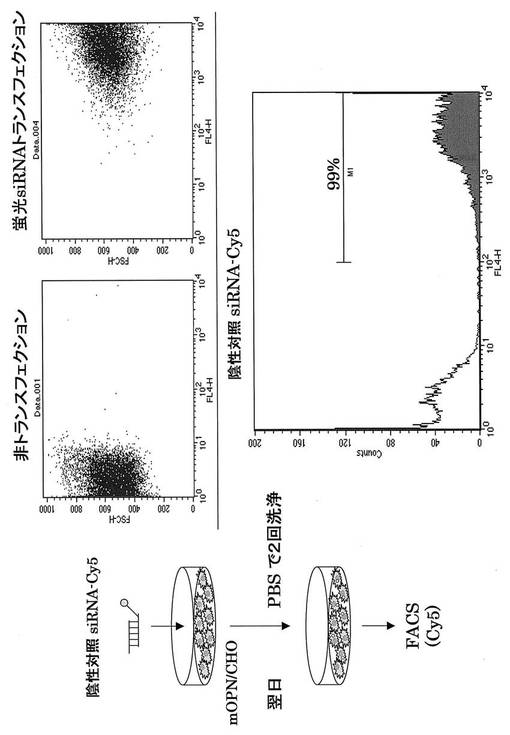
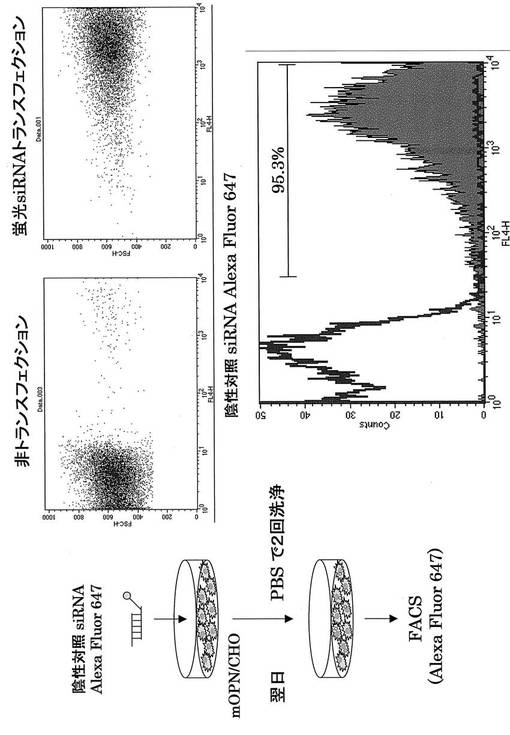
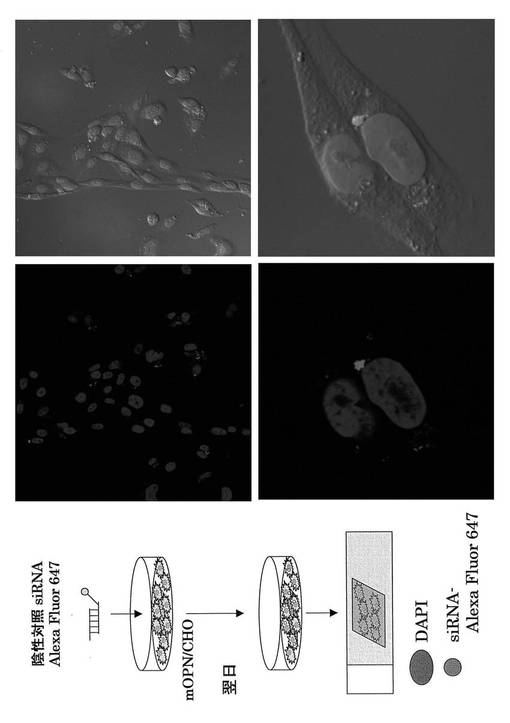
Alexa Fluor647またはCy5で蛍光標識してあるコントロールsiRNA (Qiagen)を使用して、OPN siRNAを投与した際の肝炎治療に関わる免疫細胞の同定を試みた。まず、それぞれの蛍光siRNAの遺伝子導入効率を検討するため、mOPN/CHO細胞に遺伝子導入し、FACS解析した。その結果、それぞれの蛍光siRNAは90%以上の導入効率を得ることができた(図16、17)。
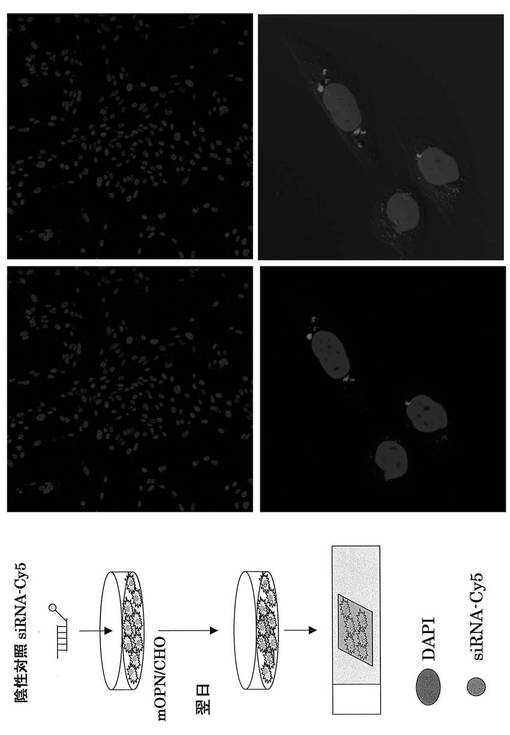
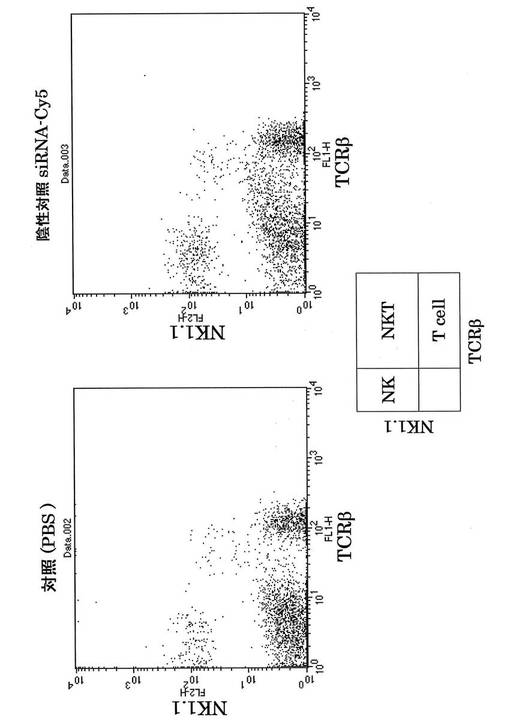
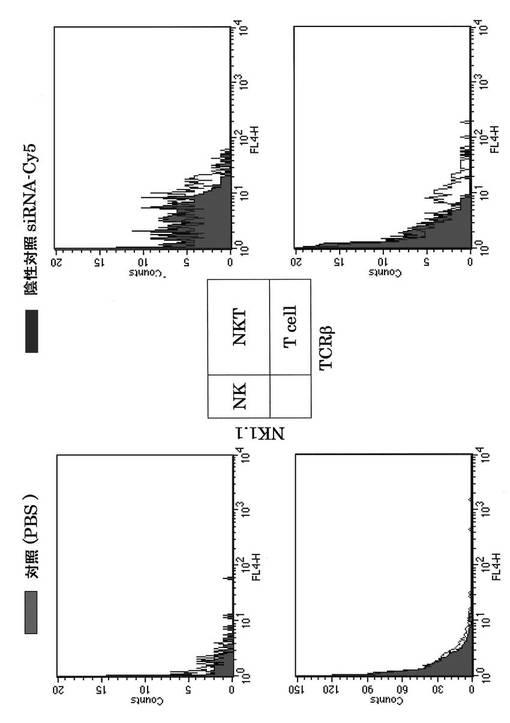
次にそれぞれの蛍光siRNAの細胞内局在を蛍光顕微鏡を用いて検討した。DAPIは核染色に用いた。その結果、核周囲の細胞質にsiRNAが局在することが分かり(図16、17)、これは報告と一致する(http://www1.qiagen.com/JP/geneXpression/NewsLetter/PDF/1027994_GExClubNL_5J.pdf)。Cy5標識siRNAを図20に示す方法にてマウスに導入し、肝臓内における免疫細胞のsiRNAのターゲット細胞同定を行った。PE標識抗NK1.1抗体とFITC標識抗TCRβ抗体にて展開し、NK細胞、T細胞、NKT細胞画分中の蛍光siRNAをFL4にて検出した。
その結果、siRNAは、特にNKT細胞とT細胞に多く取り込まれていることが分かった。すなわち、OPN siRNAの肝炎治療効果は、NKT細胞とT細胞が発現するOPNをノックダウンすることにより得られていることが示唆された(図21、22)。
<ラットOPN siRNAによるノックダウン効果とステルスsiRNA(invitrogen)によるマウスOPN siRNAのノックダウン効果>
上記と同様の方法によって、ラットOPN siRNAによるノックダウン効果とStealth(商標)siRNA(invitrogen)によるマウスOPN siRNAのノックダウン効果も確認された(図23〜25)。
【技術分野】
【0001】
本発明は、オステオポンチンの発現抑制ためのsiRNAに関するものである。
【背景技術】
【0002】
組織の構築には、細胞間の接着と同時に細胞―細胞外マトリックスとの接着が必要である。細胞―細胞外マトリックス間接着に関与する接着分子は、細胞骨格と細胞外を連結する役割を果たしており、細胞の分化、シグナル伝達に関与している。接着分子と細胞外マトリックスの接着機構を解析することにより、種々の疾患におけるメカニズムの解明、さらには治療につながることが期待されている。
【0003】
細胞外マトリックスの一種であるオステオポンチン(OPN)は、分子量約41kDaの分泌型酸性リン酸化糖タンパク質であり、分子中央部にはαv等のインテグリンとの接着に重要なGRGDS配列が存在し、その直後にはトロンビン開裂部位(R168S169)が存在する。また、GRGDS配列直後に存在するSVVYGLR配列はα9β1、α4β1、α4β7という炎症に関与するインテグリンと結合する。
【0004】
OPNは細胞接着、細胞遊走、一酸化窒素(NO)産生の制御、免疫系への関与など多彩な機能が報告されてきており、癌転移、リウマチ関節炎や多発性硬化症(EAE)などの慢性炎症疾患、自己免疫疾患など多くの難治疾患病態と関与することが示されている。
【0005】
特に、炎症性疾患に関しては、受容体からも推察されるように、直接的な関与が示唆されている。OPNの受容体であるα9インテグリンは好中球上に発現しており、α4インテグリンはリンパ球上に発現していることから、OPN機能を抑制することにより好中球を初めとする白血球の遊走を抑制することが考えられ、抗炎症作用が期待されている。
【0006】
事実、OPN欠損マウスは、腫瘍、リウマチ関節炎、多発性硬化症(EAE)、動脈硬化等の疾患に対し抵抗性を示すことが報告されている(非特許文献1〜4)。また、OPNに対する中和抗体を用いて、リウマチ関節炎が緩解した報告もある(特許文献3、非特許文献5)。
【0007】
そこで、OPN機能を阻害することにより、治療効果を期待することができると考え、新規分子特異的ノックダウン法であるRNAi(RNA干渉:RNA interference)法の治療への応用を検討した。
【0008】
RNAi法とは、短い干渉dsRNA(siRNA(small interfering RNA))を用いて、それが導入された細胞において、それと同じ配列を持つ特定遺伝子の発現を遺伝子レベルですみやかに発現抑制する技術である(非特許文献6及び7)。OPNのmRNA配列から、ターゲットとなる配列を複数選択し、そのsiRNAを合成し、OPNをノックダウンさせることにより疾患治癒に結びつけようと考えた。
【0009】
特許文献1には、オステオポンチンの部分をコードするRNAのIL−1βに関与する結合組織疾患の治療における使用が記載されている。しかし、siRNAとしての使用については、何ら開示されていない。
【0010】
特許文献2には、FGFR(線維芽細胞増殖因子レセプター:fibroblast growth factor receptors)のsiRNA及びFGFRがOPN遺伝子の発現を亢進していることが記載されている。しかし、OPNのsiRNAについては、何らの開示も示唆もない。
【0011】
特許文献3には、組換え抗オステオポンチン抗体及びそれを含む医薬が記載されている。しかし、OPNのsiRNAについては、何らの開示も示唆もない。
【0012】
また、二本鎖RNAをベクターを用いてインビトロで産生させ、これをRNaseIII核酸分解酵素ファミリーの一つである、dicerを用いて切断した、siRNAの集団からなる遺伝子操作用のキットが市販されている(SuperSilencing(登録商標)Human SPP1 siRNAキット(Secreted phosphoprotein 1(osteopontin, bone sialoprotein I, early T-lymphocyte activation 1))非特許文献8)。しかし、このキットは、遺伝子発現の研究のために、siRNAを用いて特定遺伝子の発現を阻害するためのものであり、siRNAの医薬用途については何ら開示されていない。また、上記製造方法によって得られたsiRNAは、分子特異的ではない短いRNAフラグメントをも含んでいることから、遺伝子ノックダウンの特異性が低いという欠点もあった。
【0013】
したがって、オステオポンチンの亢進に起因する疾患の処置のための医薬等として有用なOPNのsiRNAの開発が望まれていた。
【先行技術文献】
【特許文献】
【0014】
【特許文献1】特開平8−191693
【特許文献2】米国特許出願公開第2003/0143676A1号明細書
【特許文献3】国際公開第2003/02751号パンフレット
【非特許文献】
【0015】
【非特許文献1】Nemoto H, Rittling SR, Yoshitake H, Furuya K, Amagasa T, Tsuji K, Nifuji A, Denhardt DT, Noda M. Osteopontin deficiency reduces experimental tumor cell metastasis to bone and soft tissues. J Bone Miner Res. 16:652-9, 2001.
【非特許文献2】Yumoto K, Ishijima M, Rittling SR, Tsuji K, Tsuchiya Y, Kon S, Nifuji A, Uede T, Denhardt DT, Noda M. Osteopontin deficiency protects joints against destruction in anti-type II collagen antibody-induced arthritis in mice. Proc Natl Acad Sci U S A. 99:4556-4561, 2002.
【非特許文献3】Chabas D, Baranzini SE, Mitchell D, Bernard CC, Rittling SR, Denhardt DT, Sobel RA, Lock C, Karpuj M, Pedotti R, Heller R, Oksenberg JR, Steinman L. The influence of the proinflammatory cytokine, osteopontin, on autoimmune demyelinating disease.Science. 294:1731-5, 2001.
【非特許文献4】Matsui Y, Rittling SR, Okamoto H, Inobe M, Jia N, Shimizu T, Akino M, Sugawara T, Morimoto J, Kimura C, Kon S, Denhardt D, Kitabatake A, Uede T. Osteopontin Deficiency Attenuates Atherosclerosis in Female Apolipoprotein E-Deficient Mice. Arterioscler Thromb Vasc Biol. 23:1029-34, 2003.
【非特許文献5】Yamamoto N, Sakai F, Kon S, Morimoto J, Kimura C, Yamazaki H, Okazaki I, Seki N, Fujii T, Uede T. Essential role of the cryptic epitope SLAYGLR within osteopontin in a murine model of rheumatoid arthritis. J Clin Invest. 112:181-8, 2003.
【非特許文献6】Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391:806-11, 1998.
【非特許文献7】Elbashir SM, Harborth J, Lendeckel W, Yalcin A, Weber K, Tuschl T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature. 411:494-8, 2001.
【非特許文献8】SuperArray社、SuperSilencing(登録商標)製品説明、[平成16年3月22日検索]。インターネット<URL:http://www.superarray.com/sirnaqa.php>
【発明の概要】
【発明が解決しようとする課題】
【0016】
本発明の目的は、オステオポンチンのためのsiRNA、それを含む組成物及び医薬、及びオステオポンチンと相互作用する物質及び相互作用を阻害又は促進する物質をスクリーニングする方法を提供することである。
【課題を解決するための手段】
【0017】
本発明者らは、鋭意研究を行ったところ、オステオポンチンの発現を特異的に抑制するsiRNAを見出し、本発明を完成させたものである。したがって、本発明は、下記を提供する。
1.配列番号1、配列番号7又は配列番号21に示す配列からなるDNAに対応するmRNAの、20塩基〜30塩基の連続した任意の部分RNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体であって、オステオポンチンの発現を抑制するRNA。
2.配列番号1に示す配列からなるDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号298〜1140番から選ばれる連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体。
3.配列番号3、4、5若しくは6に示す配列からなるRNA、配列番号3、4、5若しくは6に示す配列と相補的な配列からなるRNA、若しくは配列番号3、4、5若しくは6に示す配列からなるRNAと配列番号3、4、5若しくは6に示す配列と相補的な配列からなるRNAとからなる二本鎖RNA、又はそれらの誘導体。
4.配列番号28、29若しくは30に示す配列からなるRNA、配列番号28、29若しくは30に示すと相補的な配列からなるRNA、若しくは配列番号28、29若しくは30に示す配列からなるRNAと配列番号28、29若しくは30に示すと相補的な配列からなるRNAとからなる二本鎖RNA、又はそれらの誘導体。
5.配列番号7に示す配列からなるDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号241〜1065番から選ばれる連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体。
6.配列番号9、10、11若しくは12に示す配列からなるRNA、配列番号9、10、11若しくは12に示す配列と相補的な配列からなるRNA、若しくは配列番号9、10、11若しくは12に示す配列からなるRNAと配列番号9、10、11若しくは12に示す配列と相補的な配列とからなるRNAからなる二本鎖RNA、又はそれらの誘導体。
7.配列番号21に示す配列からなるDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号1〜954番から選ばれる連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体。
8.配列番号23、24、25、26若しくは27に示す配列からなるRNA、配列番号23、24、25、26若しくは27に示す配列と相補的な配列からなるRNA、若しくは配列番号23、24、25、26若しくは27に示す配列からなるRNAと配列番号23、24、25、26若しくは27に示す配列と相補的な配列からなるRNAとからなる二本鎖RNA、又はそれらの誘導体。
9.上記1〜8の何れかに記載のRNAを含む、オステオポンチンの発現を抑制するためのオステオポンチン発現抑制剤。
10.上記1〜8の何れかに記載のRNAを含む、オステオポンチンの発現を抑制するための組成物。
11.上記1〜8の何れかに記載のRNAを有効成分として含む医薬。
12.オステオポンチンの発現を抑制するためのものである上記11記載の医薬。
13.上記1〜8の何れかに記載のRNAを有効成分として含む、オステオポンチンの亢進に起因する疾患の処置のための医薬。
14.上記オステオポンチンの亢進に起因する疾患が、腫瘍、肝炎、動脈硬化、多発性硬化症、関節炎、リウマチ及び/又は肺繊維症である、上記13に記載の医薬。
15.上記1〜8の何れかに記載のRNAを含む、siRNA発現ベクター。
16.被験物質と上記1〜8の何れかに記載のRNAとを接触させることを含む、オステオポンチンと相互作用する物質、又はオステオポンチン及びオステオポンチンと相互作用する物質の相互作用を促進又は阻害する物質をスクリーニングするための方法。
【図面の簡単な説明】
【0018】
【図1】マウスOPN siRNAのターゲット配列とその位置。
【図2】ヒトOPN siRNAのターゲット配列とその位置。
【図3】mOPN siRNAのノックダウン効果(マウスOPNを遺伝子導入したmOPN/CHO細胞を用いた。Scr contは、哺乳動物の細胞にはないことが確認されているsiRNAであり、siRNAのネガティブコントロールとして使用した。RISCはRNA-induced silencing complexを示す。)。mOPN siRNA は、mOPNタンパク質発現を抑制した。
【図4】hOPN siRNAのノックダウン効果(ヒトOPNを遺伝子導入したhOPN/CHO細胞を用いている。Scr cont、RISCは図3と同義である)。hOPN siRNA はhOPNタンパク質発現を抑制した。
【図5】内在性OPNに対するsiRNAのノックダウン効果(細胞はマウス悪性メラノーマ細胞であるB16−BL6細胞株を用いている。Scr cont、RISCは図3と同義である)。mOPN siRNA は内在性mOPNの発現を抑制した。
【図6】mOPN siRNAのRNAi効果持続時間解析実験のスケジュール。
【図7】mOPN siRNAのRNAi効果持続時間(Lipo. cont.は、siRNAを加えないでLipofectamine 2000を用いて遺伝子導入を行った結果である。Nega cont.は、遺伝子導入処理等を一切行わないで通常に培養していた状態である)。
【図8−1】肝炎誘発とmOPN siRNA による肝炎治療実験プロトコール。
【図8−2】肝炎誘発とmOPN siRNA による肝炎治療実験プロトコール。
【図9】mOPN siRNAによる肝臓中OPNのノックダウン効果とALT値。
【図10】OPN siRNA投与による肝臓HE染色像
【図11】肝炎悪性度とOPN mRNA量の相関
【図12】癌細胞に対するOPN siRNA のノックダウン効果。
【図13】mOPN siRNA による癌細胞浸潤抑制効果。mOPN siRNAは肺癌細胞の浸潤を抑制した。
【図14】hOPN siRNAによる癌細胞浸潤抑制効果。hOPN siRNAはヒト線維肉腫の浸潤を抑制した。
【図15】OPN siRNA投与による肝炎治療効果(血清ALT値)。
【図16】蛍光siRNA(Cy5)を用いたFACS。
【図17】蛍光siRNA(Alexa Fluor 647)を用いたFACS。
【図18】siRNA−Cy5の細胞内局在。
【図19】siRNA−Alexa Fluor 647の細胞内局在。
【図20】siRNAの肝臓内リンパ球導入プロトコール。
【図21】肝臓内リンパ球のFACSによる分離・検出。
【図22】siRNAの肝臓内リンパ球への導入。
【図23】ラットOPN siRNAのターゲット配列とその位置。
【図24】ラットOPN siRNAのノックダウン効果。rOPN siRNAはrOPNタンパク質発現を抑制した(RISCは図3と、Lipo. Cont.は図7のLipo. cont.と同義である)。
【図25】mOPN Stealth(商標)RNAiのターゲット配列とそのノックダウン効果。mOPN Stealth(商標)RNAiはmOPN タンパク質発現を抑制した。
【発明を実施するための形態】
【0019】
以下、本発明の実施の形態について詳細に説明する。
先ず、本明細書で使用する略称について説明する。
Aまたはa:アデニン
Tまたはt:チミン
Uまたはu:ウラシル
Gまたはg:グアニン
Cまたはc:シトシン
RNA:リボ核酸(通常、塩基としてA,U,G,Cを有するリボヌクレオチド単位から構成される。)
DNA:デオキシリボ核酸(通常、塩基としてA,T,G,Cを有するデオキシリボヌクレオチド単位からなる。)
DAPI:4‘,6−ジアミノ−2−フェニルインドール
ELISA:enzyme-linked immunosorbent assay
FACS:蛍光活性化セルソーター
FITC:フルオレセインイソチオシアネート
PE:フィコエリトリン(紅色の色素タンパク質)
Scr. RNA:スクランブルRNA。mRNA特異的なsiRNAとGC含量を揃えた非特異的なRNA。
Scr. cont.:対象となるsiRNAのコントロールとして用いるScr. RNA
「オステオポンチン」とは、分泌性リン酸タンパク質1(secreted phospho protein 1)とも呼ばれ、配列番号2(マウス)、配列番号8(ヒト)又は配列番号22(ラット)に示すアミノ酸配列からなるタンパク質であり、骨のマトリックスに含まれる接着タンパク質の一種である。オステオポンチンは、インテグリンと結合するRGD配列を有し、破骨細胞と骨芽細胞の両方を骨組織周辺に接着させる作用を有する。また、オステオポンチンは、強く負に荷電しており、ヒドロキシアパタイトを沈着させ、骨のカルシウムを保持する作用を有する。以下、オステオポンチンをOPNと略すこともある。
【0020】
マウス、ヒト及びラットのオステオポンチン遺伝子の塩基配列及びアミノ酸配列を、それぞれ配列番号1及び2、配列番号7及び8、並びに配列番号21及び22に示した。
【0021】
本発明はオステオポンチン遺伝子に対応するmRNAの部分配列であって、20塩基〜30塩基の連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体であって、オステオポンチンの発現を抑制するsiRNAに関する。
より具体的には本発明は、配列番号1に示す配列からなるマウスDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号298〜1140番から選ばれる連続した任意のRNA配列からなるセンスRNA、配列番号7に示す配列からなるヒトDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号241〜1065番から選ばれる連続した任意のRNA配列からなるセンスRNA、配列番号21に示す配列からなるラットDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号1〜954番から選ばれる連続した任意のRNA配列からなるセンスRNA、該センスRNAに相補的な配列からなるアンチセンスRNA、若しくは該センス配列と該アンチセンス配列とからなる二本鎖RNA、又はそれらの誘導体に関する。
なお、OPNの発現を制御するsiRNAは、通常OPN遺伝子の開始コドンから75塩基以上下流の最初の塩基配列AAを見つけ、当該AAを含む21〜23塩基、好ましくは21塩基であって、GC含量が50%前後の、OPNに対して特異的な配列を選択することによって得られる。特異的であるか否かは、例えばNCBI(http://www.ncbi.nlm.nih.gov/)のBLAST-searchにかけることによって確認することができる。
また、本発明の他の具体的な態様としては、上記各DNAの翻訳領域に対応するmRNAの部分配列であって、GCで始まる25塩基の連続した任意のRNA配列からなるセンスRNAと該センスRNAに相補的なアンチセンスRNAとからなる二本鎖RNAの誘導体であって、いわゆるステルス(商標)RNAiの構造を有するsiRNAが挙げられる(Stable plasma membrane levels of hCTR1 mediate cellular copper uptake. J Biol Chem. 2005 Mar 11;280(10):9635-9. Epub 2005 Jan 5. PMID: 15634665およびhttp://www.invitrogen.co.jp/products/molecular_biology/RNAi.shtml参照)。ステルス(商標)RNAiは、上記の21〜23塩基のsiRNAと同様にターゲット遺伝子を抑制し、かつ非特異的なインターフェロンストレス応答を抑制し、血清、細胞質中においても安定性が高いという特長を有する。
【0022】
本発明で相補的な配列からなるRNAとは、所定の塩基配列を有するRNAに対して、A:UおよびG:Cといった塩基対関係に基づいて、mRNAの配列に対応する配列を有する鎖を意味するものである。DNAの相補的二本鎖のうち、タンパク質をコードしている鎖、すなわち、mRNAと同一の配列を有する鎖がセンス鎖であり、センス鎖と塩基配列が相補的な関係にある配列を有する鎖がアンチセンス鎖である。
【0023】
本発明で誘導体とは、例えば、本発明のRNAの安定性を向上させるために、3’側にdTを数個、好ましくは2〜4個、より好ましくは2又は3個、最も好ましくは2個付加させた、3’オーバーハング型誘導体である。なお、dTの代わりにUを付加させても良い。
【0024】
また、本発明で誘導体とは、例えば、本発明のRNAの安定性を向上させるために、5’側にリン酸を付加させた誘導体である。
【0025】
また、本発明で誘導体とは、いわゆるタンデムタイプのsiRNA発現ベクターに組み込むための、2つのプロモーター、例えばU6プロモーター、センスRNA及びアンチセンスRNAを結合させた誘導体である。具体的には、U6プロモーター、センスRNA、5個のT、U6プロモーター、アンチセンスRNA、5個のTを結合させた誘導体が挙げられる。
【0026】
また、本発明で誘導体とは、いわゆるステムループタイプのsiRNA発現ベクターに組み込み、ショートヘアピンRNAを介して、siRNAを産生するための、プロモーター、例えばU6プロモーター、センスRNA、ループ配列及びアンチセンスRNAを結合させた誘導体である。具体的には、U6プロモーター、センスRNA、ループ配列、例えば、gtgtgctgtcc、アンチセンスRNAを結合させた誘導体が挙げられる。
また、本発明で誘導体とは、ステルス(商標)RNAi構造を有する誘導体である。
【0027】
siRNA(small interfering RNA)とは、20〜30bp、好ましくは21〜23bp、25bpまたは27bpからなる二本鎖RNAであり、RNA干渉(RNA interference: RNAi)を誘導することができる二本鎖RNAをいう。また、RNA干渉とは、二本鎖RNAにより、そのRNAと相同的な配列を有する遺伝子の発現が抑制される現象をいう。
【0028】
RNAiは、特定配列を有する遺伝子を破壊することができることから、遺伝子機能の解析や特定遺伝子の発現抑制に有用である。また、細胞への導入量が少量の割に比較的長時間効果が持続するという特徴を有する。
【0029】
本発明のsiRNAは、常法によって合成することができ、市販のDNA/RNAシンセサイザー、例えば、Applied Biosystems394型で行うことができる。
【0030】
本発明のRNAは、そのままか又は凍結乾燥させて、オステオポンチンの機序解明のための試薬として用いることができる。
【0031】
本発明のRNA又はその誘導体は、そのままもしくは公知の薬学的に許容される担体(賦形剤、増量剤、結合剤、滑沢剤などが含まれる)や慣用の添加剤などと混合して医薬組成物として調製することができる。当該医薬組成物は、調製する形態(錠剤、丸剤、カプセル剤、散剤、顆粒剤、シロップ剤などの経口投与剤;注射剤、点滴剤、外用剤、坐剤などの非経口投与剤)等に応じて経口投与または非経口投与することができる。また投与量は、有効成分の種類、投与経路、投与対象または患者の年齢、体重、症状などによって異なり一概に規定できないが、通常、1日投与用量として、数mg〜2g程度、好ましくは数十mg程度を、1日1〜数回にわけて投与することができる。
【0032】
siRNAベクターは、常法にしたがって調製することができる。タンデムタイプの場合は、PCRによってセンス、アンチセンス配列を含むプライマーによりプロモーター部分を増幅し、増幅断片を制限酵素で切断後、ベクターのプロモーター、例えばU6プロモーターの下流に挿入する。ステムループの場合、センス−ループ−アンチセンス配列を含むオリゴヌクレオチドを合成アニールさせ、ベクターのプロモーター、例えばU6プロモーターの下流に挿入する。
以下、実施例によって本発明を具体的に説明する。なお、実施例においてmOPNはマウス・オステオポンチンを、hOPNはヒト・オステオポンチンを、そしてrOPNはラット・オステオポンチンそれぞれ示す。
【実施例1】
【0033】
(1)オリゴヌクレオチドの調製
4種のリボヌクレオシド3’−ホスホロアミダイト(GLENリサーチ)を用いて、DNA自動合成機(Applied Biosystem Model394A)で配列番号3、4、5、6、9、10、11、12、23、24、25、26及び27に示す各配列の各オリゴリボヌクレオチド(RNA断片)を合成した。各RNA断片を1μmolスケールで合成した。合成終了後、合成したオリゴヌクレオチドが結合したCPG(Controlled Pore Glass)を濃アンモニア水:エタノール(3:1v/v)混液で室温2時間処理してオリゴリボヌクレオチドをCPG樹脂から切り出し、更に55℃で16時間加温した。
マウスOPNについて、RNAi designer(http://www.invitrogen.co.jp/primers/rnaidesign/)を用いて決定した配列である、配列番号28、29若しくは30に示す配列について、invitorgenの示す方法にしたがって、ステルス(商標)RNAiを調製した(Stable plasma membrane levels of hCTR1 mediate cellular copper uptake. J Biol Chem. 2005 Mar 11;280(10):9635-9. Epub 2005 Jan 5. PMID: 15634665およびhttp://www.invitrogen.co.jp/products/molecular_biology/RNAi.shtml参照)。
【0034】
溶媒を留去し、残渣に1mlの1M TBAF(テトラブチルアンモニウムフルオリド)/THF(テトラヒドロフラン)溶液を加え、37℃で16時間攪拌した。これに5mlの0.1Mトリエチルアンモニウムアセテート水溶液(pH7.0)を加えた後、C18(ウォーターズ社製)オープンカラムクロマトグラフィーを行なった(カラムサイズ1.5×12cm:5−40%アセトニトリル、50mMトリエチルアンモニウムビカルボネート水溶液の両溶媒を用いた濃度勾配により溶出した)。約30%濃度のアセトニトリルで溶出される末端リボヌクレオチド5’位の保護基であるジメトキシトリチルの発色を有するフラクションを集め、これに5mlの0.01N塩酸を加え、15分間攪拌した。0.1Nアンモニア水で中和し、水層を酢酸エチルで洗浄し、溶媒留去後、滅菌水1mlに溶解した。この画分中のオリゴリボヌクレオチドを逆相HPLCで分取後、さらにイオン交換HPLCで分取し、精製した。得られたオリゴヌクレオチドを後述の実験に供した。
【0035】
逆相およびイオン交換HPLCの条件は以下の通りであった:
逆相HPLC
カラム:μ−ボンダスフィアー(C−18)カラム、Ф3.9x150mm(ウォーターズ社製)
溶媒:A溶液 5% アセトニトリル/0.1M TEAA(トリエチルアンモニウムアセテート、pH7.0);
B溶液 25% アセトニトリル/0.1M TEAA(pH7.0)。
イオン交換HPLC
カラム:TSKgel DEAE 2SWカラム、4.6×250mm、東ソー(株)製
溶媒:A溶液 20% アセトニトリル;
B溶液 20% アセトニトリルを含む2M ギ酸アンモニウム。
【0036】
(2)OPNsiRNAの生物学的作用
〔方法〕
<培養細胞へのOPN siRNA導入方法>
上記の通り合成された各RNA断片をOPN siRNAとして、Lipofectamine 2000(Invitrogen, Carlsbad, CA)を用いるリポフェクション法により、24穴プレート中で増殖させた対数増殖期の培養細胞に導入した。
【0037】
F−12培地50μLにリポフェクタミン2000を2μLを加え、5分間放置した。別のチューブで、F−12培地50μLに各siRNA 0.8μgを加えた。両チューブの内容物を混ぜ、20分放置し、siRNA−リポフェクタミン2000複合体を調製した。細胞をF−12培地で洗浄後、F−12培地500μLを加え、さらにsiRNA−リポフェクタミン複合体を加え、これを24時間培養した。次いで、上清を回収し、細胞ペレットを、TNEバッファー(10mM Tris−HCl pH7.8、1.0% NP−40、0.15M NaCl、1mM EDTA)を用いて可溶化させた。ELISA(固相酵素免疫検定法)にて上清及び細胞ペレット可溶化物におけるOPN発現量を定量した。
【0038】
<ELISA及びRT−PCR>
ヒトOPN及びマウスOPNのsiRNAのRNAi効果をELISA法、RT−PCR法(逆転写−ポリメラーゼ連鎖反応法)及びリアルタイムPCR法を用いて検討した。
ヒトOPN、マウスOPNのELISAを用いた解析には、ヒトオステオポンチン測定キット及びマウスオステオポンチン測定キット(共に免疫生物研究所製)を使用した。
RT−PCR及びリアルタイムPCRには以下のプライマーを用いた。リアルタイムPCRは、LightCycler-FastStart DNA Master SYBR Green I(Roche Diagnostics、Mannheim、Germany)により作製したサンプルを用いて行った。定量は、グリセルアルデヒド三リン酸脱水素酵素(G3PDH)を標準とした標準比(Normalized Ratio)で行った。
また、ラットOPNのsiRNA及びマウスOPN stealth(商標)RNAのノックダウン効果を、上記と同様にELISAを用いて解析した。
【0039】
使用したプライマー
mOPN-forward: 5'-TCAAAGTCTAGGAGTTTCCAG-3'
mOPN-reverse: 5'-TTAGTTGACCTCAGAAGATGA-3'
hOPN-forward: 5'-CCATGAGAATTGCAGTGATTT-3'
hOPN-reverse: 5'-TTAATTGACCTCAGAAGATGC-3'
G3PDH-forward: 5'-ACCACAGTCCATGCCATCAC-3'
G3PDH-reverse: 5'-TCCACCACCCTGTTGCTGTA-3'
β-actin-forward: 5'-TGGAATCCTGTGGCATCCATGAAAC-3'
β-actin-reverse: 5'-TAAAACGCAGCTCAGTAACAGTCCG-3'
【0040】
<肝炎マウス作製>
C57BL/6マウスにコンカナバリンA(ConA、Vector laboratories, Burlingame, CA)を尾静脈投与し(200μg/マウス)、肝炎モデルを作出した。ConA投与24時間後に、肝臓を摘出し、ALT(アラニンアミノトランスフェラーゼ;GPT)測定、HE(ヘマトキシリン−エオシン)染色、RNA抽出を行った。
【0041】
<肝炎治癒作用>
上記肝炎モデルにsiRNA50μg(300μl(図8−1)又は500μl(ハイドロダイナミクス法:図8−2))をマウスに尾静脈投与した。
siRNAを、ConA投与24時間前、16時間前及び同時の3回投与した。コントロールとして、PBS(リン酸緩衝生理食塩水)を上記と同条件で投与した(図8−1、図8−2)。
【0042】
<癌転移抑制作用/マトリゲルアッセイ>
癌浸潤能のインビトロアッセイは、マトリゲルインベージョンチャンバー(BD Biosciences, Bedford, MA)を用いて行った。マウスルイス肺癌細胞又はHT1080(ヒト線維肉腫細胞)(2.5×104細胞/ウェル)をチャンバー上層に接種し、下層には、5% FCS(ウシ胎仔血清)を含む培地を加え、22時間培養した。綿棒にてフィルター上層をこすり、上層部に残るマトリゲルや非浸潤細胞を取り去った。フィルター下層の浸潤した細胞をメタノールで固定し、ギムザで染色し、蒸留水で洗浄した。
【0043】
〔実験結果〕
<培養細胞におけるノックダウン効果>
mOPN siRNAのmOPNに対するRNAi効果を、mOPN遺伝子をCHO細胞に高発現させたトランスフェクタントであるmOPN/CHO細胞に種々のsiRNAを導入することにより解析した。RNAi効果の確認は、mOPNに対するELISAとRT−PCRによって行った。ELISAは、siRNA導入後の細胞可溶化物をサンプルとして用いた。その結果、m−siRNA−1、m−siRNA−5、m−siRNA−7及びm−siRNA−16の全てのsiRNAについて、mOPNのノックダウン効果が認められた(図3)。
【0044】
同様にhOPN siRNAのhOPNに対するRNAi効果を、hOPN/CHO細胞を用いて解析した。その結果、h−siRNA−4及びh−siRNA−6において、hOPNのノックダウン効果が認められた(図4)。
【0045】
本発明のsiRNAが、内在性OPN発現をも抑制するかどうかを、マウス悪性メラノーマ細胞株である、B16−BL6細胞を用いて解析した。細胞可溶化物をサンプルとして使用した結果、m−siRNA−5及びm−siRNA−16によって内在性OPN発現の抑制効果が認められた(図5)。
【0046】
次に、mOPN siRNAのRNAi効果持続期間について検討した。図6に示すようにmOPN/CHO細胞に遺伝子導入後、24時間後に上清と細胞3/4を回収した。残りの細胞1/4をさらに培養し、24時間後に培地を交換し、さらに24時間後に、培養上清及び細胞3/4を回収した。残りの細胞1/4をさらに培養し、同様な操作を繰り返し、上清及び細胞の回収を2週間行った。培養上清と細胞可溶化物におけるOPNの発現量をELISA法によって測定した。その結果、約1週間後においても、全てのsiRNAにおいて、ノックダウン効果が認められた(図7)。
【0047】
<OPNsiRANの治療効果>
次に、OPN siRNAを用いたRNAi医薬への可能性について検討するため、肝炎についてOPN機能抑制による治療効果を試験した。治癒効果検討方法の概略を図8−1及び図8−2に示した。すなわち、コンカナバリンA(ConA)200μgをC57BL/6マウスに尾静脈内投与することによって肝炎誘発モデルを作製した。siRNAをConA投与24時間前、16時間前、同時の3回、それぞれ50μgを尾静脈内に投与した。肝炎の評価は、ConA投与24時間後の血清ALT(アラニンアミノトランスフェラーゼ)値と肝臓組織のHE染色にて行った。OPNのRNAi効果は、RT−PCRとリアルタイムPCRによって評価した(図8−1)。
さらに、図8−2に示すプロトコールで、肝炎治療実験を行った。5群で実験を行った。すなわち、1群:siRNAの代わりにPBSとConAの代わりにPBS、2群:Scr siRNAとConAの代わりにPBS、3群:OPN siRNA−5とConAの代わりにPBS、4群:OPN siRNA−5とConA、5群:Scr siRNAとConA。ConA肝炎マウスに対してOPN siRNAを投与した4群では有意にALTの減少が見られた(図15)。
【0048】
肝臓におけるOPNノックダウン効果をRT−PCRによって検討した。その結果、siRNA処理群では、コントロール群と比較して低いOPN発現が見られた。一方、血清ALT値は、OPN抑制が認められた個体において、低値傾向が見られた(図9)。
【0049】
肝臓のHE染色を行った。OPN siRNAを投与していない個体は、肝臓に広範な壊死が観察された。一方、OPN siRNAを投与した個体は、肝臓内壊死が抑制されていた(図10)。
【0050】
OPNの発現量を定量化するために、リアルタイムPCRを行った。その際、ハウスキーピング遺伝子であるグリセルアルデヒド三リン酸脱水素酵素(G3PDH)の発現量を用いて標準化を行った。OPN発現量とALT値との相関を調べた結果、OPN量とALT値に強い相関が見られた(図11)。
【0051】
次に、癌転移のインビトロアッセイであるマトリゲル法を用いて、OPN siRNAによる癌転移抑制能を調べた。mOPN siRNA、hOPN siRNAの転移抑制能を調べるためにマウスルイス肺癌細胞 と HT1080(Human Fibrosarcoma cells)細胞をそれぞれ用いた。
【0052】
まず、mOPN、hOPNのノックダウン効果をそれぞれのOPN ELISA kitを用いて検討した。その結果、mOPN siRNAでは、m−siRNA−5及びm−siRNA−16の両方においてOPNのノックダウン効果が見られ、hOPN siRNAでは、h−siRNA−6においてノックダウン効果が観察された(図12)。C8、C9は、GC含量が一致するコントロールsiRNAを対照として使用しており、m−siRNA−5とh−siRNA−4のコントロールはC8、m−siRNA−16とh−siRNA−6のコントロールとしてC9を利用した。
【0053】
転移能の評価は、マトリゲル膜の裏側に浸潤した細胞を観測することにより検討した。siRNAを導入した細胞の浸潤能は、コントロールと比較して少ないことが分かった(図13、14)。特に、mOPN siRNAでは、m−siRNA−5が、hOPN siRNAではh−siRNA−6が強い浸潤抑制能を有していることが分かった(図13、14)。グラフは、浸潤している細胞を計測することにより定量化して、OPN siRNAによる浸潤抑制効果を確認した結果を示す。
【0054】
その結果、siRNAを導入した細胞の浸潤能は、コントロールと比較して明らかに抑制された(図12)。特に、mOPN siRNAでは、m−siRNA−5が、また、hOPN siRNAではh−siRNA−6がマトリゲルに対する細胞の浸潤を抑制し、これらが、強い浸潤抑制能を有していることが分かった(図13、図14)。
以上により、OPNノックダウン効果と浸潤抑制とが相関している結果を得ることができた。
【0055】
<siRNAのターゲット免疫細胞の同定>
肝炎発症には、NKT細胞から分泌されるOPNが重要ということは既に報告されている(Diao H, Kon S, Iwabuchi K, Kimura C, Morimoto J, Ito D, Segawa T, Maeda M, Hamuro J, Nakayama T, Taniguchi M, Yagita H, Van Kaer L, Onoe K, Denhardt D, Rittling S, Uede T. Osteopontin as a mediator of NKT cell function in T cell-mediated liver diseases. Immunity. 2004 Oct;21(4):539-50.)。以上のことから、OPN siRNAの静脈内投与による肝炎治療効果は、NKT細胞から分泌されるOPNをOPN siRNAが抑制していると推察された。そこで、蛍光siRNAをマウス尾静脈に投与し、肝臓内の白血球のsiRNAの取り込みをFACSを用いて解析したところ、NKT細胞画分にsiRNAが取り込まれている結果を得ることができた。
【0056】
Alexa Fluor647またはCy5で蛍光標識してあるコントロールsiRNA (Qiagen)を使用して、OPN siRNAを投与した際の肝炎治療に関わる免疫細胞の同定を試みた。まず、それぞれの蛍光siRNAの遺伝子導入効率を検討するため、mOPN/CHO細胞に遺伝子導入し、FACS解析した。その結果、それぞれの蛍光siRNAは90%以上の導入効率を得ることができた(図16、17)。
次にそれぞれの蛍光siRNAの細胞内局在を蛍光顕微鏡を用いて検討した。DAPIは核染色に用いた。その結果、核周囲の細胞質にsiRNAが局在することが分かり(図16、17)、これは報告と一致する(http://www1.qiagen.com/JP/geneXpression/NewsLetter/PDF/1027994_GExClubNL_5J.pdf)。Cy5標識siRNAを図20に示す方法にてマウスに導入し、肝臓内における免疫細胞のsiRNAのターゲット細胞同定を行った。PE標識抗NK1.1抗体とFITC標識抗TCRβ抗体にて展開し、NK細胞、T細胞、NKT細胞画分中の蛍光siRNAをFL4にて検出した。
その結果、siRNAは、特にNKT細胞とT細胞に多く取り込まれていることが分かった。すなわち、OPN siRNAの肝炎治療効果は、NKT細胞とT細胞が発現するOPNをノックダウンすることにより得られていることが示唆された(図21、22)。
<ラットOPN siRNAによるノックダウン効果とステルスsiRNA(invitrogen)によるマウスOPN siRNAのノックダウン効果>
上記と同様の方法によって、ラットOPN siRNAによるノックダウン効果とStealth(商標)siRNA(invitrogen)によるマウスOPN siRNAのノックダウン効果も確認された(図23〜25)。
【特許請求の範囲】
【請求項1】
配列番号7に示す配列からなるDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号241〜1065番から選ばれる連続した任意のRNA配列と該センスRNAに相補的な配列とからなる二本鎖RNA。
【請求項2】
配列番号9、10、11又は12に示す配列からなるRNAと配列番号9、10、11又は12に示す配列と相補的な配列とからなるRNAからなる二本鎖RNA。
【請求項3】
請求項1又は2に記載の二本鎖RNAを含む、オステオポンチンの発現を抑制するための組成物。
【請求項4】
請求項1又は2に記載の二本鎖RNAを有効成分として含む、オステオポンチンの亢進に起因する疾患の処置のための医薬。
【請求項5】
上記オステオポンチンの亢進に起因する疾患が、腫瘍、肝炎、動脈硬化、多発性硬化症、関節炎、リウマチ及び/又は肺繊維症である、請求項4に記載の医薬。
【請求項1】
配列番号7に示す配列からなるDNAに対応するmRNAのAAで始まる21塩基のRNA配列であって、該DNAの配列番号241〜1065番から選ばれる連続した任意のRNA配列と該センスRNAに相補的な配列とからなる二本鎖RNA。
【請求項2】
配列番号9、10、11又は12に示す配列からなるRNAと配列番号9、10、11又は12に示す配列と相補的な配列とからなるRNAからなる二本鎖RNA。
【請求項3】
請求項1又は2に記載の二本鎖RNAを含む、オステオポンチンの発現を抑制するための組成物。
【請求項4】
請求項1又は2に記載の二本鎖RNAを有効成分として含む、オステオポンチンの亢進に起因する疾患の処置のための医薬。
【請求項5】
上記オステオポンチンの亢進に起因する疾患が、腫瘍、肝炎、動脈硬化、多発性硬化症、関節炎、リウマチ及び/又は肺繊維症である、請求項4に記載の医薬。
【図23】



【図1】

【図2】

【図3】
【図4】
【図5】
【図6】
【図7】
【図8−1】
【図8−2】
【図9】

【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図24】
【図25】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8−1】
【図8−2】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図24】
【図25】
【公開番号】特開2010−263917(P2010−263917A)
【公開日】平成22年11月25日(2010.11.25)
【国際特許分類】
【出願番号】特願2010−181658(P2010−181658)
【出願日】平成22年8月16日(2010.8.16)
【分割の表示】特願2005−114424(P2005−114424)の分割
【原出願日】平成17年4月12日(2005.4.12)
【出願人】(000195524)生化学工業株式会社 (143)
【出願人】(501416243)株式会社ジーンテクノサイエンス (9)
【Fターム(参考)】
【公開日】平成22年11月25日(2010.11.25)
【国際特許分類】
【出願日】平成22年8月16日(2010.8.16)
【分割の表示】特願2005−114424(P2005−114424)の分割
【原出願日】平成17年4月12日(2005.4.12)
【出願人】(000195524)生化学工業株式会社 (143)
【出願人】(501416243)株式会社ジーンテクノサイエンス (9)
【Fターム(参考)】
[ Back to top ]