
カイコの卵および眼の着色に関与する遺伝子およびその利用
【課題】 カイコのre遺伝子を同定すること、同定したre遺伝子を利用して、カイコの卵および眼の着色を改変すること、および、同定したre遺伝子を遺伝子マーカーとして利用して、遺伝子組換えカイコを選抜すること
【解決手段】 ポジショナルクローニングの手法などを利用して、re遺伝子を同定し、カイコの卵および眼における赤色の着色が、この遺伝子の機能が喪失によって標準型の色素が合成されないことにより生じていることを見出した。当該遺伝子を利用することにより、カイコの卵および眼の色の改変を行うことが可能であり、さらには、このような形質の変化を指標として、効率的に遺伝子組換えカイコを選抜することが可能であることを見出した。
【解決手段】 ポジショナルクローニングの手法などを利用して、re遺伝子を同定し、カイコの卵および眼における赤色の着色が、この遺伝子の機能が喪失によって標準型の色素が合成されないことにより生じていることを見出した。当該遺伝子を利用することにより、カイコの卵および眼の色の改変を行うことが可能であり、さらには、このような形質の変化を指標として、効率的に遺伝子組換えカイコを選抜することが可能であることを見出した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、カイコの卵および眼の着色に関与するred egg(re)遺伝子およびその利用に関する。
【背景技術】
【0002】
オモクローム系色素は、非常に多くの昆虫において、卵、眼、体表の色に関わっていることが報告されている(非特許文献1)。オモクローム系色素はトリプトファンから派生し、3-ヒドロキシキヌレニンを前駆体として酸化縮合により生合成される色素である(図1)(非特許文献2、3)。オモクロームに関与する遺伝子は、主にショウジョウバエの複眼変異系統の研究から見出されているが、3-ヒドロキシキヌレニンの合成と色素顆粒への取り込みに関する遺伝子については知見が多いものの、色素顆粒にとりこまれた後の経路に関する遺伝子は、ほとんど明らかにされていない。
【0003】
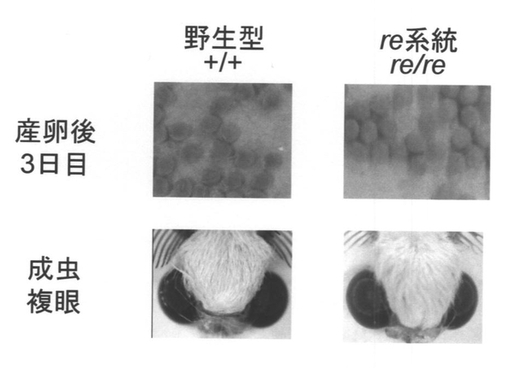
野生型(標準型)のカイコでは、オモクローム系色素によって産卵後72時間で卵は藤鼠色に着色するが、自然突然変異で見つかったカイコ赤卵突然変異系統red egg(re)は、劣性ホモの個体の卵が薄いオレンジに着色する(図2)。さらに時間が経つと、野生型の卵の色は濃くなり、reの劣性ホモ個体の卵は赤色になる。また、野生型の成虫の蛾の眼は黒色に着色するが、reの劣性ホモ個体の成虫の複眼は濃赤褐色に着色する(図2)。reでは、オモクローム系色素の前駆体である3-ヒドロキシキヌレニンは合成されており、色素顆粒に取り込みも行われているが、最終的に生じる色素を調べると、野生型の卵と複眼に存在する色素成分のいくつかが存在しないことが報告されていた(非特許文献4)。このため、reの原因遺伝子は、3-ヒドロキシキヌレニンを色素顆粒に取り込んだ後、最終的な色素の合成に関係していることが予想されていたが、その実体は不明であった。
【0004】
カイコでは、これまで3つのオモクローム関連変異体の原因遺伝子が同定されている。すなわち、キヌレニンを3-ヒドロキシキヌレニンに変換する遺伝子に欠損があるw-1、3-ヒドロキシキヌレニンを色素顆粒に取り込むためのトランスポーター遺伝子に欠損があるw-2とw-3であり、それぞれショウジョウバエのcinnabar、scarlet、whiteに対応している(特許文献1、非特許文献5〜7)。これら3つの変異体では、オモクローム系色素が合成されず、卵と複眼が白くなる。
【0005】
カイコの卵色眼色変異系統は、肉眼でも容易に判別できるため、カイコの遺伝子組換え個体を判別するマーカーとして着目されている。現在、w-1が遺伝子組換えマーカーとして利用されているが、w-1の卵色は母親の遺伝形質に依存するため、卵の時期では組換え個体の判別は付かない。さらに、w-1、w-2、w-3はすべて白色卵で未受精卵と同じ色であるため区別がつきにくい。
【0006】
一方、reは母性遺伝せず、さらに卵が赤くなるため、未受精卵とも、野生型の卵とも区別がつきやすい。reの原因遺伝子を同定することができれば、簡便に遺伝子組換え個体を判別する分子マーカーを作出することが可能となる。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許第4431739号公報
【非特許文献】
【0008】
【非特許文献1】Linzen, B., (1974) Advamces in Insect Physiology 10:117-246
【非特許文献2】Butenandt, A., W. Schafer, (1962) Ommochromes. New York: Academic Press. p13-33.
【非特許文献3】Reed, RD., LM. Nagy, (2005) Evol Dev 7:301-311
【非特許文献4】Kawase, S., H. Aruga, (1954) The Japanese journal of genetics 29:114-121
【非特許文献5】Quan, GX. et al., (2002) Mol Genet Genomics 267:1-9.
【非特許文献6】Komoto, N. et al., (2009) Insect Biochem Mol Biol 39:152-156.
【非特許文献7】Tatematsu, KI. et al., (2011) Positional cloning of silkworm white egg 2 (w-2) locus shows functional conservation and diversification of ABC transporters for pigmentation in insects. Genes Cells.(MID: 21294818)
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、このような状況に鑑みてなされたものであり、その目的は、カイコのre遺伝子を同定することにある。また、本発明の目的は、同定したre遺伝子を利用して、カイコの卵および眼の着色を改変することにある。さらなる本発明の目的は、同定したre遺伝子を遺伝子マーカーとして利用して、遺伝子組換えカイコを選抜することにある。
【課題を解決するための手段】
【0010】
本発明者らは、上記課題を解決すべく、まず、ポジショナルクローニングの手法によりre遺伝子が存在する領域の絞り込みを行った結果、202kbにまで絞り込むことに成功した。次いで、本発明者らは、絞り込まれた領域において5つの予測遺伝子を見出し、それらの卵における発現解析を行い、データベース上で正常にアノテーションされていないBGIBMGA003497-1が卵の着色に先立って発現していることを見出した。さらに、本発明者らは、BGIBMGA003497-1の構造の解析を行ったところ、re変異系統ではORF内部に重大な欠損が存在することが判明した。BGIBMGA003497-1は、12回膜貫通型の「Major Facilitator Superfamily」に属するトランスポーターをコードしていると考えられ、この欠損は、膜貫通領域に存在していた。以上から、本発明者らは、BGIBMGA003497-1がre遺伝子の本体であり、カイコの卵および眼における赤色の着色が、この遺伝子の機能が喪失によって標準型の色素が合成されないことにより生じていることを見出した。
【0011】
さらに、本発明者らは、当該遺伝子を利用することにより、カイコの卵および眼の色の改変を行うことが可能であり、さらには、このような形質の変化を指標として、効率的に遺伝子組換えカイコを選抜することが可能であることを見出した。
【0012】
本発明は、より詳しくは、以下の発明を提供するものである。
【0013】
[1] カイコの卵および眼を変色させる活性を有するタンパク質をコードする、下記(a)〜(e)のいずれかに記載のDNA。
(a)配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または3に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:1または3に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
(e)配列番号:1または3に記載の塩基配列からなるDNAと相同性が90%以上の塩基配列からなるDNA
【0014】
[2] カイコの卵および眼を変色させる活性を有するタンパク質をコードする、下記(a)〜(c)のいずれかに記載のDNA。
(a)[1]に記載のDNAの転写産物と相補的な二重鎖RNAをコードするDNA
(b)[1]に記載のDNAの転写産物と相補的なアンチセンスRNAをコードするDNA
(c)[1]に記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA
【0015】
[3] [1]または[2]に記載のDNAを含むベクター。
【0016】
[4] [1]または[2]に記載のDNAが導入された細胞。
【0017】
[5] [1]または[2]に記載のDNAが導入されたカイコ。
【0018】
[6] [5]に記載のカイコの卵、子孫またはクローン。
【0019】
[7] 卵および眼が変色されたカイコの生産方法であって、[1]または[2]に記載のDNAをカイコに導入することを特徴とする方法。
【0020】
[8] 遺伝子組換えカイコの生産方法であって、任意のDNAと[1]または[2]に記載のDNAをカイコに導入し、卵および眼が変色した個体を選抜することを特徴とする方法。
【発明の効果】
【0021】
本発明によって、カイコの卵および眼の着色に関与するre遺伝子が同定され、該遺伝子の染色体上の位置および構造が解明された。本発明により見出されたカイコのre遺伝子は、色素の合成に関与する蛋白質をコードし、その機能の有無により、卵および眼の色が変色する。本発明のre遺伝子を用いれば、カイコの卵や眼の色を変色させることが可能であり、当該遺伝子は、遺伝子組換え体の選抜のためのマーカーとして好適に利用することができる。
【0022】
現在最も早く組換え体の判別ができるEGFP等のマーカーでは、蛍光顕微鏡を用いて産卵後7日目の胚の眼の蛍光を観察することにより遺伝子組換え体を判別している。一方、本発明において同定されたre遺伝子を用いれば、産卵後3日目に遺伝子組換え体を選抜することが可能であり、従来よりも遺伝子組換え体の識別時期が4日間早い。しかも、この識別は肉眼で行うことが可能であり、高価な蛍光顕微鏡などを必要としない。
【0023】
また、肉眼で識別が可能な遺伝子マーカーとして、KMO遺伝子マーカーも用いられてはいるが、母性遺伝のために遺伝子組換え体の卵は全て着色され、卵では組換え個体の識別が不可能である。このように、本発明によれば、従来技術に比して、カイコの遺伝子組換え体の選抜を効率的に行うことができる。
【図面の簡単な説明】
【0024】
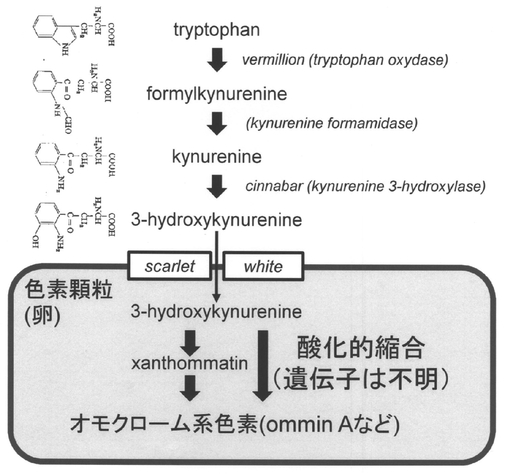
【図1】カイコの卵を着色するオモクロームの合成経路を示す図である。母親の体内で、トリプトファンからホルミルキヌレニン、ホルミルキヌレニンからキヌレニン、キヌレニンから3-ヒドロキシキヌレニンに変換される。3-ヒドロキシキヌレニンは卵に送られる。卵が発生する際、色素顆粒が作られ、scarlet、whiteから構成されるヘテロダイマーのトランスポーターを通過することにより、3-ヒドロキシキヌレニンが色素顆粒に取り込まれる。3-ヒドロキシキヌレニンのその後の色素合成反応は色素顆粒の中で起こる。カイコの卵の色素はommin Aなど複数の色素の混合物といわれている。3-ヒドロキシキヌレニンの変化までは、母親の体内で起きるため、cinnabarより上流の遺伝子への変異は母性遺伝する。遺伝子名はショウジョウバエの遺伝子名を記した。
【図2】野生型カイコとre系統カイコの卵と成虫複眼の写真である。
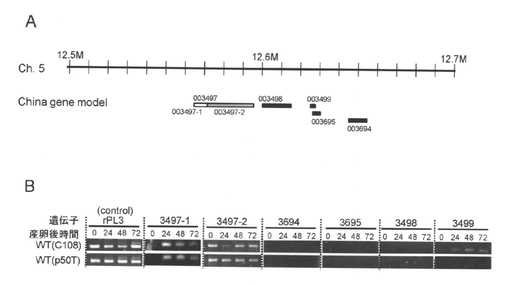
【図3】Aは、絞り込み領域内の予測遺伝子(China gene model)を示す図である。絞り込み領域内には、BGIBMGA003497、003498、003499、003494、003695が存在した。BGIBMGA003497は2つの遺伝子を融合して予測していることが推測された。003497-1は白の四角、003497-2はグレーの四角で示した。Bは、予測遺伝子の発現を示す写真である。C108、p50Tは野生型の遺伝子、rPL3はコントロール遺伝子である。
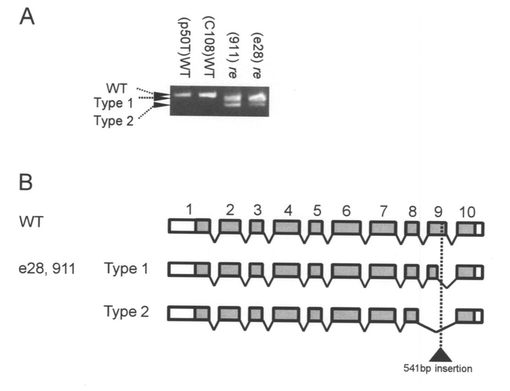
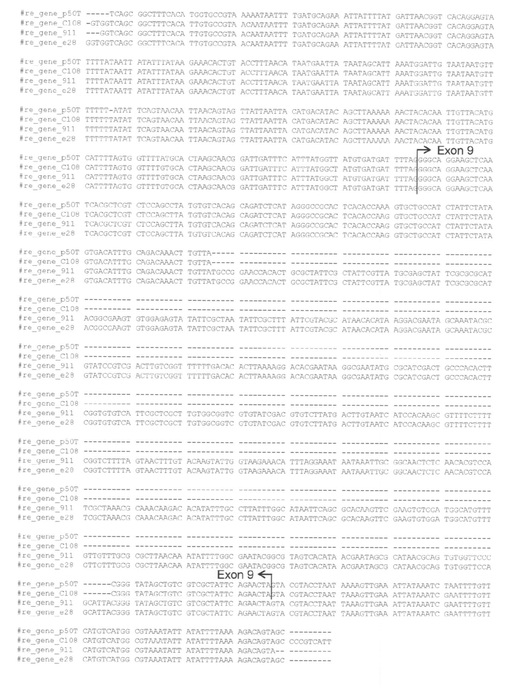
【図4】Aは、re遺伝子のRT-PCRの結果を示す写真である。産卵後24時間後の卵のcDNAを用いてre遺伝子を増幅すると、2種類のre系統(911、e28)では、野生型系統(p50T、C108)よりも短い2種類のPCR産物が増幅された。Bは、野生型系統(p50T、C108)とre系統(911、e28)の遺伝子構造の比較を示す図である。エキソンは四角で、イントロンは斜め線で表示した。黒い三角形は911とe28ではエキソン9に同一の541bpの配列の挿入(黒い三角形)が存在した。ORF領域はグレーで塗りつぶしている。
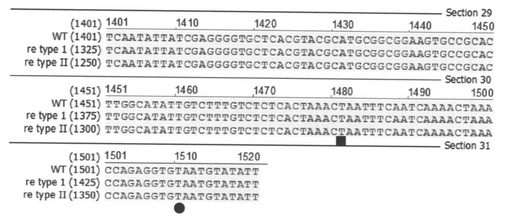
【図5A】野生型系統(WT)とre系統(re type I、re type II)のre遺伝子のcDNAのORF領域の整列図である。図中、*は、ORF開始コドン、黒丸は、WTの終止コドン、黒四角は、re type Iとre type IIの終止コドンを示す。
【図5B】図5Aの続きの図である。
【図5C】図5Bの続きの図である。
【図6】野生型系統(WT)とre系統(re type I、re type II)のORFのアミノ酸配列の整列図である。
【図7】野生型系統(p50T、C108)とre系統(911、e28)のBGIBMGA003497-1(re)遺伝子のexon 9近傍のゲノム配列の整列図である。re系統にはexon 9の中に541bpの挿入配列が存在する。
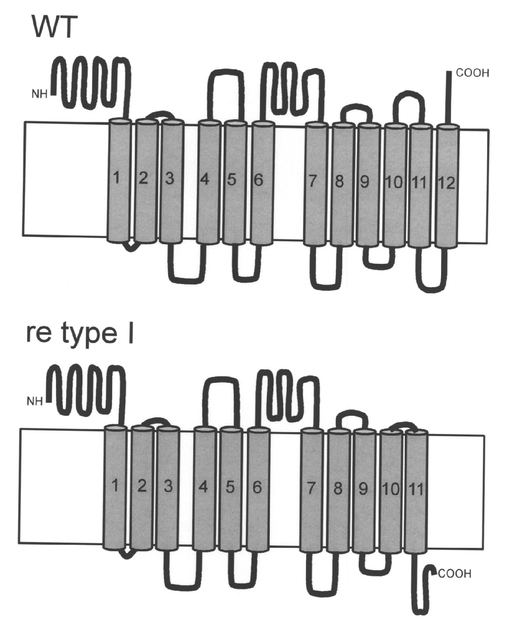
【図8】カイコの野生型系統とre系統のBGIBMGA003497-1(re)遺伝子の膜貫通ドメインの予測構造を示す図である。野生型の遺伝子では12回膜貫通構造と予測されるが、原因遺伝子と赤卵系統では、10個目の膜貫通ドメイン以降のアミノ酸配列が異なり、構造が変化している。
【図9】カイコのBGIBMGA003497-1(re)遺伝子と昆虫種のホモログについての、近隣接合法により作製した系統樹の図である。図9の昆虫の部分を拡大したものである。黒丸はカイコのBGIBMGA003497-1(re)遺伝子を示す。赤卵原因遺伝子のホモログは、ショウジョウバエ以外の昆虫で幅広い昆虫種に存在した。
【発明を実施するための形態】
【0025】
<カイコの卵および眼を変色させる活性を有するタンパク質をコードするDNA>
本発明は、カイコの卵および眼を変色させる活性を有する活性を有するタンパク質をコードするDNAを提供する。
【0026】
本発明者らにより同定された、カイコの卵および眼を変色させる活性を有するタンパク質をコードする野生型系統(P50T、C108)のre cDNAの塩基配列をそれぞれ配列番号:1、3に、これらDNAがコードするタンパク質のアミノ酸配列をそれぞれ配列番号:2、4に示す。
【0027】
ここで「カイコの卵および眼を変色させる活性」とは、典型的には、カイコの卵を藤鼠色に、成虫の眼を黒色に変色させる活性を意味する。
【0028】
本発明のDNAの1つの態様は、配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:1または3に記載の塩基配列のコード領域を含むDNA)である。
【0029】
現在の技術水準においては、当業者であれば、特定の野生型系統(例えば、P50T、C108)におけるre遺伝子の塩基配列情報が得られた場合、その塩基配列を改変し、そのコードするアミノ酸配列は異なるが、同様の活性を有するタンパク質をコードするDNAを取得することが可能である。また、自然界においても、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは起こり得ることである。従って、本発明は、これら野生型系統におけるタンパク質のアミノ酸配列(配列番号:2または4)において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなり、カイコの卵および眼を変色させる活性を有するタンパク質をコードするDNAをも含むものである。ここで「複数」とは、改変後のタンパク質がカイコの卵および眼を変色させる活性を維持する範囲における、アミノ酸の改変数であり、通常、50アミノ酸以内、好ましくは30アミノ酸以内、さらに好ましくは10アミノ酸以内(例えば、5アミノ酸以内、3アミノ酸以内、2アミノ酸)である。
【0030】
さらに、現在の技術水準においては、当業者であれば、特定の野生型系統(例えば、P50T、C108)からre遺伝子が得られた場合、そのDNAの塩基配列情報を利用して、カイコまたは他の生物から、同様の活性を有するタンパク質をコードするDNAを取得することが可能である。従って、本発明は、これら野生型系統(例えば、P50T、C108)におけるre遺伝子(配列番号:1または3)とストリンジェントな条件でハイブリダイズするDNAであって、カイコの卵および眼を変色させる活性を有するタンパク質をコードするDNAをも含むものである。
【0031】
こうして得られた変異DNAや相同DNAが、カイコの卵および眼を変色させる活性を有するタンパク質をコードするか否かは、例えば、被検DNAを、re変異系統(卵および眼が赤色となる系統)に導入し、卵が藤鼠色に変色するか否か、あるいは、成虫の眼が黒色に変色するか否かにより、判定することができる。
【0032】
本発明のDNAは、その導入により、カイコの卵および眼を変色させることが可能であるという意味において、カイコの卵および眼を変色させるための薬剤である。
【0033】
なお、上記した変異DNAを作製するための、DNAへの人為的な変異の導入は、例えば、部位特異的変異誘発(site-directed mutagenesis)法(Kramer, W. & Fritz, HJ., Methods Enzymol, 154:350-367, 1987)により行うことができる。
【0034】
また、上記した相同遺伝子を単離するための方法としては、例えば、ハイブリダイゼーション技術(Southern, E. M., Journal of Molecular Biology, 98:503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R. K., et al. Science, 230:1350-1354, 1985、Saiki, R. K. et al. Science, 239:487-491, 1988)が挙げられる。相同遺伝子をコードするDNAを単離するためには、通常ストリンジェントな条件下でハイブリダイゼーション反応を行なう。ストリンジェントなハイブリダイゼーション条件としては、6M尿素、0.4%SDS、0.5xSSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を例示できる。よりストリンジェンシーの高い条件、例えば、6M尿素、0.4%SDS、0.1xSSCの条件を用いれば、より相同性の高いDNAの単離を期待することができる。単離されたDNAは、核酸レベルあるいはアミノ酸配列レベルにおいて、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の相同性は、BLASTN(核酸レベル)やBLASTX(アミノ酸レベル)のプログラム(Altschul et al. J. Mol. Biol., 215:403-410, 1990)を利用して決定することができる。該プログラムは、KarlinおよびAltschulによるアルゴリズムBLAST(Proc. Natl. Acad. Sci. USA, 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA, 90:5873-5877, 1993)に基づいている。BLASTNによって塩基配列を解析する場合には、パラメーターは例えばscore=100、wordlength=12とする。また、BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えばscore=50、wordlength=3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res. 25:3389-3402, 1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0035】
本発明のreタンパク質をコードするDNAとしては、その形態に特に制限はなく、cDNAの他、ゲノムDNA、および化学合成DNAが含まれる。ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、カイコからゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、フォスミド、BAC、PACなどが利用できる)を作成し、これを展開して、re遺伝子(例えば、配列番号:1または3に記載のDNA)の塩基配列を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより調製することが可能である。また、re遺伝子に特異的なプライマーを作成し、これを利用したPCRを行うことによって調製することも可能である。また、cDNAは、例えば、鱗翅目昆虫から抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。
【0036】
<カイコのre遺伝子の発現を抑制するために用いるDNA>
また、本発明は、上記した本発明の野生型系統のre遺伝子の発現を抑制するために用いるDNAを提供する。本発明においては、re遺伝子の機能が喪失することにより、カイコの卵および眼が変色することが見出された。従って、re遺伝子の発現を抑制する活性を有するDNAの導入により、カイコの卵および眼を変色させることが可能である。この意味において、re遺伝子の発現を抑制するために用いるDNAは、カイコの卵および眼を変色させるための薬剤である。ここで「re遺伝子の発現の抑制」には、遺伝子の転写の抑制およびタンパク質への翻訳の抑制の双方が含まれる。また、「発現の抑制」には、発現の完全な停止のみならず発現の減少も含まれる。また、「カイコの卵および眼を変色させる活性」とは、典型的には、カイコの卵および成虫の眼を赤色に変色させる活性を意味する。被検DNAが、カイコの卵および成虫の眼を赤色に変色させる活性を有するか否かは、例えば、当該DNAを、野生型異系統(卵が藤鼠色および眼が黒色となる系統)に導入し、卵および成虫の眼が赤色に変色するか否かにより、判定することができる。
【0037】
re遺伝子の発現を抑制するために用いるDNAの一つの態様は、re遺伝子の転写産物と相補的なdsRNA(二重鎖RNA)をコードするDNAである。標的遺伝子配列と同一もしくは類似した配列を有するdsRNAを細胞内に導入することにより、導入した外来遺伝子および標的内因性遺伝子の発現がいずれも抑制される、RNAi(RNA干渉、RNA interference)と呼ばれる現象を引き起こすことができる。細胞に約40〜数百塩基対のdsRNAが導入されると、ヘリカーゼドメインを持つダイサー(Dicer)と呼ばれるRNaseIII様のヌクレアーゼが、ATP存在下で、dsRNAを3'末端から約21〜23塩基対ずつ切り出し、siRNA(short interference RNA)が生じる。このsiRNAに、特異的なタンパク質が結合して、ヌクレアーゼ複合体(RISC:RNA-induced silencing complex)が形成される。この複合体はsiRNAと同じ配列を認識して結合し、RNaseIII様の酵素活性によってsiRNAの中央部で標的遺伝子の転写産物(mRNA)を切断する。また、この経路とは別にsiRNAのアンチセンス鎖がmRNAに結合してRNA依存性RNAポリメラーゼ(RsRP)のプライマーとして作用し、dsRNAが合成される。このdsRNAが再びダイサーの基質となって、新たなsiRNAを生じて作用を増幅する経路も考えられている。
【0038】
本発明のdsRNAをコードするDNAは、標的遺伝子の転写産物(mRNA)のいずれかの領域に対するアンチセンスRNAをコードしたアンチセンスDNAと、該mRNAのいずれかの領域のセンスRNAをコードしたセンスDNAを含み、該アンチセンスDNAおよび該センスDNAより、それぞれアンチセンスRNAおよびセンスRNAを発現させることができる。また、これらのアンチセンスRNAおよびセンスRNAよりdsRNAを作成することができる。
【0039】
本発明のdsRNAの発現システムをベクター等に保持させる場合の構成としては、同一のベクターからアンチセンスRNAおよびセンスRNAを発現させる場合と、異なるベクターからそれぞれアンチセンスRNAとセンスRNAを発現させる場合がある。同一のベクターからアンチセンスRNAおよびセンスRNAを発現させる構成としては、例えば、アンチセンスDNAおよびセンスDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモーターを連結させたアンチセンスRNA発現カセットとセンスRNA発現カセットをそれぞれ構築し、これらカセットを同方向にあるいは逆方向にベクターに挿入する構成である。
【0040】
また、異なる鎖上に対向するように、アンチセンスDNAとセンスDNAとを逆向きに配置した発現システムを構成することもできる。この構成では、アンチセンスRNAコード鎖とセンスRNAコード鎖とが対となった一つの二本鎖DNAが備えられ、その両側にそれぞれの鎖からアンチセンスRNAとセンスRNAとを発現し得るようにプロモーターを対向して備える。この場合には、センスRNAとアンチセンスRNAの下流に余分な配列が付加されることを避けるために、それぞれの鎖(アンチセンスRNAコード鎖、センスRNAコード鎖)の3'末端にターミネーターをそれぞれ備えることが好ましい。このターミネーターは、A(アデニン)塩基を4つ以上連続させた配列などを用いることができる。また、このパリンドロームスタイルの発現システムでは、二つのプロモーターの種類は異なっていることが好ましい。
【0041】
また、異なるベクターからアンチセンスRNAおよびセンスRNAを発現させる構成としては、例えば、アンチセンスDNAおよびセンスDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモーターを連結させたアンチセンスRNA発現カセットとセンスRNA発現カセットとをそれぞれ構築し、これらカセットを異なるベクターに保持させる構成である。
【0042】
本発明に用いるdsRNAとしては、siRNAが好ましい。「siRNA」は、細胞内で毒性を示さない範囲の短鎖からなる二重鎖RNAを意味する。標的re遺伝子の発現を抑制することができ、かつ、毒性を示さなければ、その鎖長に特に制限はない。dsRNAの鎖長は、例えば、15〜500塩基対である。
【0043】
本発明のdsRNAをコードするDNAとしては、標的配列のインバーテッドリピートの間に適当な配列(イントロン配列が望ましい)を挿入し、ヘアピン構造を持つダブルストランドRNA(self-complementary 'hairpin' RNA(hpRNA))を作るようなコンストラクト(Smith, N.A., et al. Nature, 407:319, 2000、Wesley, S. V. et al. Plant J. 27:581, 2001、Piccin, A. et al. Nucleic Acids Res. 29:E55, 2001)を用いることが好ましい。
【0044】
本発明のdsRNAをコードするDNAは、標的re遺伝子の塩基配列と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の同一性は上述した手法(BLASTプログラム)により決定できる。
【0045】
dsRNAにおけるRNA同士が対合した二重鎖RNAの部分は、完全に対合しているものに限らず、ミスマッチ(対応する塩基が相補的でない)、バルジ(一方の鎖に対応する塩基がない)などにより不対合部分が含まれていてもよい。本発明においては、dsRNAにおけるRNA同士が対合する二重鎖RNA領域中に、バルジおよびミスマッチの両方が含まれていてもよい。
【0046】
re遺伝子の発現を抑制するために用いるDNAの他の態様は、re遺伝子の転写産物と相補的なアンチセンスRNAをコードするDNA(アンチセンスDNA)である。アンチセンスDNAが標的遺伝子の発現を抑制する作用としては、三重鎖形成による転写開始阻害、RNAポリメラーゼによって局部的に開状ループ構造がつくられた部位とのハイブリッド形成による転写抑制、合成の進みつつあるRNAとのハイブリッド形成による転写阻害、イントロンとエキソンとの接合点でのハイブリッド形成によるスプライシング抑制、スプライソソーム形成部位とのハイブリッド形成によるスプライシング抑制、mRNAとのハイブリッド形成による核から細胞質への移行抑制、キャッピング部位やポリ(A)付加部位とのハイブリッド形成によるスプライシング抑制、翻訳開始因子結合部位とのハイブリッド形成による翻訳開始抑制、開始コドン近傍のリボソーム結合部位とのハイブリッド形成による翻訳抑制、mRNAの翻訳領域やポリソーム結合部位とのハイブリッド形成によるペプチド鎖の伸長阻止、および核酸とタンパク質との相互作用部位とのハイブリッド形成による遺伝子発現抑制などが挙げられる。これらは、転写、スプライシング、または翻訳の過程を阻害して、標的遺伝子の発現を抑制する(平島および井上「新生化学実験講座2 核酸IV 遺伝子の複製と発現」,日本生化学会編,東京化学同人, pp.319-347, 1993)。本発明で用いられるアンチセンスDNAは、上記のいずれの作用で標的re遺伝子の発現を抑制してもよい。一つの態様として、標的遺伝子のmRNAの5'端近傍の非翻訳領域に相補的なアンチセンス配列を設計すれば、遺伝子の翻訳阻害に効果的であると考えられる。また、コード領域もしくは3'側の非翻訳領域に相補的な配列も使用し得る。このように、遺伝子の翻訳領域だけでなく非翻訳領域の配列のアンチセンス配列を含むDNAも、本発明で利用されるアンチセンスDNAに含まれる。使用されるアンチセンスDNAは、適当なプロモーターの下流に連結され、好ましくは3'側に転写終結シグナルを含む配列が連結される。
【0047】
アンチセンスDNAは、本発明のre遺伝子(例えば、配列番号:1または3に記載の塩基配列からなるDNA)の配列情報を基にホスホロチオネート法(Stein, Nucleic Acids Res., 16:3209-3221, 1988)などにより調製することが可能である。調製されたDNAは、後述する公知の方法で、カイコへ導入できる。アンチセンスDNAの配列は、カイコが持つ内因性のre遺伝子の転写産物と相補的な配列であることが好ましいが、遺伝子の発現を有効に阻害できる限り、完全に相補的でなくてもよい。転写されたRNAは、標的とする遺伝子の転写産物に対して好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の相補性を有する。効果的に標的遺伝子の発現を阻害するには、アンチセンスDNAの長さは、少なくとも15塩基以上であり、好ましくは100塩基以上であり、さらに好ましくは500塩基以上である。通常、用いられるアンチセンスDNAの長さは5kbよりも短く、好ましくは2.5kbよりも短い。
【0048】
re遺伝子の発現を抑制するために用いるDNAの他の態様は、re遺伝子の転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNAである。リボザイムには、グループIイントロン型や、RNasePに含まれるM1RNAのように400ヌクレオチド以上の大きさのものもあるが、ハンマーヘッド型やヘアピン型と呼ばれる40ヌクレオチド程度の活性ドメインを有するものもある(小泉誠および大塚栄子、蛋白質核酸酵素, 35:2191, 1990)。
【0049】
例えば、ハンマーヘッド型リボザイムの自己切断ドメインは、G13U14C15のC15の3'側を切断するが、活性にはU14が9位のAと塩基対を形成することが重要とされ、15位の塩基はCの他にAまたはUでも切断されることが示されている(Koizumi et. al., FEBS Lett. 228:225, 1988)。リボザイムの基質結合部を標的部位近傍のRNA配列と相補的になるように設計すれば、標的RNA中のUC、UUまたはUAという配列を認識する制限酵素的なRNA切断リボザイムを作出することが可能である(Koizumi et. al., FEBS Lett. 239:285, 1988、小泉誠および大塚栄子,蛋白質核酸酵素,35:2191, 1990、Koizumi et. al., Nucleic. Acids. Res. 17:7059, 1989)。
【0050】
また、ヘアピン型リボザイムも、本発明の目的のために有用である。ヘアピン型リボザイムは、例えばタバコリングスポットウイルスのサテライトRNAのマイナス鎖に見出される(Buzayan, Nature 323:349, 1986)。このリボザイムも、標的特異的なRNA切断を起こすように設計できることが示されている(Kikuchi and Sasaki, Nucleic Acids Res. 19:6751, 1992、菊池洋,化学と生物 30:112, 1992)。標的を切断できるよう設計されたリボザイムは、カイコ細胞中で転写されるようにアクチンプロモーターなどのプロモーターおよび転写終結配列に連結される。このような構成単位をタンデムに並べ、標的遺伝子内の複数の部位を切断できるようにして、より効果を高めることもできる(Yuyama et al., Biochem. Biophys. Res. Commun. 186:1271, 1992)。このようなリボザイムを用いて標的となるre遺伝子の転写産物を特異的に切断し、該遺伝子の発現を抑制することができる。
【0051】
<ベクター、形質転換細胞、形質転換カイコ>
本発明は、また、上記本発明のDNA(re遺伝子、およびre遺伝子の発現を抑制するためのDNA)を含むベクター、上記本発明のDNAが導入された細胞、上記本発明のDNAが導入されたカイコを提供する。
【0052】
上記本発明のDNAが導入される細胞としては、特に制限はなく、目的に応じて種々の細胞を用いることができる。例えば、組換えタンパク質生産の目的であれば、カイコ細胞の他、例えば、大腸菌、酵母、動物細胞、植物細胞などを用いることができる。遺伝子組換えカイコを製造する場合には、カイコ細胞が用いられる。
【0053】
本発明のベクターとしては、例えば、自律複製可能なベクターまたは染色体中に相同組換え可能なベクターを使用することができる。カイコ卵に対して本発明のDNAの導入を行う場合、例えば、カイコの発生初期卵へ、トランスポゾンをベクターとして注射する方法(Tamura,T. et al., (2000) Nature Biotechnology 18:81-84)に従って行うことができる。例えば、トランスポゾンの逆位末端反復配列(Handler AM. et al.,(1998) Proc. Natl. Acad. Sci. U.S.A. 95(13):7520-5)の間に上記DNAを挿入したベクターとともに、トランスポゾン転移酵素をコードするDNAを有するベクター(ヘルパーベクター)をカイコ卵に導入する。ヘルパーベクターとしては、pHA3PIG(Tamura,T. et al., (2000) Nature Biotechnology 18, 81-84)が挙げられるが、これに限定されるものではない。本発明におけるトランスポゾンとしては、piggyBacが好ましいが、これに限定されるものではなく、マリーナ(mariner)、ミノス(minos)等を用いることもできる(Shimizu,K. et al., (2000) Insect Mol. Biol., 9, 277-281;Wang W. et al.,(2000) Insect Mol Biol 9(2):145-55)。また、本発明では、バキュロウイルスベクターを使用することにより形質転換カイコを作出することも可能である(Yamao, M. et al., (1999) Genes Dev 13:511-516)。
【0054】
ベクターに用いるプロモーターとしては、例えば、アクチンプロモーター、AcNPVのhr5-IE1プロモーター、3xP3プロモーターなどを用いることができる。
【0055】
カイコ細胞へのベクターの導入は、例えば、リン酸カルシウム沈殿法、電気パルス穿孔法(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 9.1-9.9)、リポフェクタミン法(GIBCO-BRL社製)、マイクロインジェクション法などの公知の方法で行うことが可能である。
【0056】
一旦、染色体内に上記本発明のDNAが導入されたカイコが得られれば、該カイコから、卵、子孫、あるいはクローンを得て、それらを基にカイコを繁殖させることも可能である。本発明には、上記本発明のDNAが導入されたカイコおよびその卵、子孫およびクローンが含まれる。
【0057】
<卵および眼が変色されたカイコの生産方法>
本発明は、また、卵および眼が変色されたカイコの生産方法であって、上記本発明のDNA(re遺伝子、およびre遺伝子の発現を抑制するためのDNA)をカイコに導入することを特徴とする方法を提供する。
【0058】
本発明の方法の一つの態様は、野生型のre遺伝子をre変異系統のカイコに導入して、卵が藤鼠色に、成虫の眼が黒色に変色されたカイコを生産する方法である。
【0059】
本発明の方法の他の一つの態様は、re遺伝子の発現を抑制するためのDNAを野生型カイコに導入して、卵および眼が赤色に変色されたカイコを生産する方法である。
【0060】
<遺伝子組換えカイコの生産方法>
本発明は、また、遺伝子組換えカイコの生産方法であって、任意のDNAと上記本発明のDNA(re遺伝子、およびre遺伝子の発現を抑制するためのDNA)をカイコに導入し、卵および眼が変色した個体を選抜することを特徴とする方法を提供する。
【0061】
カイコに導入する「任意のDNA」としては、特に制限はない。例えば、医薬品や検査薬の原料となる有用タンパク質をコードするDNA、高機能シルクの生産に有用な蛋白質をコードするDNA、ヒトの病態のモデルカイコを作製するために有用なDNAが挙げられる。任意のDNAと上記本発明のDNAは、一つのベクターに配置してカイコに導入することが好ましいが、それぞれを異なるベクターに配置してカイコに導入することも可能である。任意のDNAと上記本発明のDNAは、ベクター上では、カイコにおいて発現を保証するプロモーター(例えば、アクチンプロモーター)に発現可能に連結されている。任意のDNAとre遺伝子が導入されたre変異系統カイコは、その卵が藤鼠色に変色し、成虫においては眼が黒色に変色する。また、任意のDNAとre遺伝子の発現を抑制するためのDNAが導入された野生型カイコは、その卵が赤色に変色し、成虫においては眼が赤色に変色する。この変色を指標に、遺伝子組換えカイコを選抜することができる。特に、卵においては、産卵後3日目には変色するため、従来よりも早期に、かつ、肉眼で、遺伝子組換え体を判別することが可能である。
【実施例】
【0062】
以下、実施例および比較例に基づいて本発明をより具体的に説明するが、本発明は以下の実施例に限定されるものではない。
【0063】
[実験材料と方法]
(1)カイコの飼育
カイコは人工飼料(日本農産工業)で25度〜27度で飼育した。C108系統は、研究室で累代飼育しているもの、911(re)系統は農業生物資源研究所内で累代飼育しているもの、e28系統はナショナルバイオリソースプロジェクトで累代飼育している系統(九州大学)を取り寄せて実験に用いた。
【0064】
(2)ポジショナルクローニング
reの表現型の責任領域のマッピングのため、C108(正常着色卵)と911(re)を交配して生まれたF1のオスのカイコと、911(re)のメスの交配を行った。この交配により生まれたBF1卵(正常着色282、赤卵114、計396)から孵化した1零幼虫からゲノムDNAをDNAzol(Invitrogen)を用いて抽出した。抽出したDNAに用いて、SNPマーカー(表1)を用い、文献(非特許文献7)と類似の方法で連鎖解析を行い、reの表現型の責任領域を絞り込んだ。SNPマーカーの一部は(Yamamoto, K. et al., (2008) Genome Biol 9:R21)で使用されているものである。
【0065】
【表1】

【0066】
(3)遺伝子の発現解析
P50T(正常着色卵、ゲノム解読系統)、C108(正常着色卵)、911(re変異系統)、e28(re変異系統)について、産下直後、24時間後、48時間後、72時間後の卵からtotal RNAをIsogen(NipponGene)を用いて抽出し、First-Strand cDNA Synthesis Kit(GE Healthcare)を用いてcDNAを合成した。RT-PCRは、絞り込み範囲内に存在した予測遺伝子BGIBMGA003497、003498、003499、003494、003695についてプライマー(表2)を設計し、total RNA 0.5ng分のcDNAを鋳型に用いて行った。PCRで増幅した断片の配列決定は、BigDye Ver.3.1(Applied Biosystems)とABI3130xl(Applied Biosystems)を用いて行い、配列解析はVector NTI ver.11(Invitrogen)を用いて行った。
【0067】
【表2】
【0068】
(4)膜貫通ドメイン予測
膜貫通ドメインはSOSUI(http://bp.nuap.nagoya-u.ac.jp/sosui/sosui_submit.html)プログラムで予測した。
【0069】
(5)分子系統解析
BGIBMGA003497-1と相同性が予測された100個の遺伝子について、MEGA5ソフトウェアを用い、アミノ酸配列のアラインメント及び近隣接合法で分子系統解析を行った。
【0070】
[実施例1] re遺伝子の同定
(1)ポジショナルクローニングによるre遺伝子の存在領域の絞り込み
reの責任領域の絞り込みのため、まず、C108(正常着色卵、+re/+re)のメスと911(re/re)のオスを掛け合わせ、F1を得た。F1のオス(re/+re)を911(re/re)のメスに掛け、戻し交配し、BF1個体(卵)を得た。BF1(卵)には正常着色卵と赤卵が存在した。BF1正常着色卵から生まれた幼虫282個体、BF1赤卵から生まれた幼虫114個体からそれぞれゲノムDNAを抽出し、5番染色体上に作成した40個のSNPマーカー(表1)を用いてジェノタイピングを行った。その結果、reの責任領域はSNPマーカーre-071119-R2_F1とre-080104R-R12_F1の間の202kbに絞り込まれた(表1、図3A)。
【0071】
(2)re遺伝子の候補の発現解析
ポジショナルクローニングより絞り込んだ領域には、5つの予測遺伝子BGIBMGA003497、003498、003499、003494、003695が存在した(図3A)。これらの配列をNCBIのデーターベースに対し検索を行ったところ、BGIBMGA003497の5’側はMajor Facilitator superfamilyに属する遺伝子に、3’側はChromodomain-helicase-DNA-binding proteinという全く異なる遺伝子にホモロジーが検出されたため、2つの遺伝子が融合して1つの遺伝子として誤って予測されていることが考えられた。そこで、BGIBMGA003497の5’側と3側を仮にBGIBMGA003497-1とBGIBMGA003497-2とした。reの責任領域範囲内の予測遺伝子の発現を調べるため、BGIBMGA003498、003499、003494、003695にはそれぞれ1セットずつ、BGIBMGA003497については、BGIBMGA003497-1とBGIBMGA003497-2に1セットずつ、プライマーを設計した。カイコの卵は、産卵後40時間頃に薄茶色に着色し始め、この時期には野生型系統とre系統を見分けにくいが、72時間頃になると、野生型は藤鼠色になるのに対し、re系統は薄オレンジ色または赤色を呈し、それ以降はそれぞれの色が濃くなる。このため、reの原因遺伝子は、主に産卵後72時間より前に働いていることが考えられた。そこで、標準型の卵色と複眼色を示すP50TとC108の2つの系統について産下直後、1日後、2日後、3日後の卵における遺伝子の発現をRT-PCRにより調べた(図3B)。その結果、BGIBMGA003499はC108で発現が見られたが、p50Tではその発現が非常に弱くなっていた。003498、003494、003695では発現を示すバンドが検出できなかった。BGIBMGA003497-2は発現が検出されたが、特に、p50T系統では一様に発現していた。興味深いことに、BGIBMGA003497-1は24~48時間後に発現がピークになり、72時間後には発現量が下がっており、卵の着色に先立って発現していると考えられた。BGIBMGA003497-1とBGIBMGA003497-2では発現パターンが異なることから、この二つは独立の遺伝子であることが考えられた。
【0072】
(3)BGIBMGA003497-1の構造の解析
BGIBMGA003497-1のORF配列を調べるため、カイコのESTライブラリーとゲノム配列にBlastN検索をかけたところ、アラタ体と神経由来のESTライブラリーから対応するESTが得られた。これらのEST配列を基にORF全長を野生型の卵およびre系統の卵(911系統とe28系統)のcDNAから単離し、遺伝子構造の比較を行った。P50T系統、C108系統共に、BGIBMGA003497-1は、495アミノ酸のORFをコードする転写産物が発現していた(図4、図5、図6)。911系統、e28系統では、BGIBMGA003497-1は正常な転写産物が検出されず、exon 9の一部を失ったType 1および、exon 9全体を失った転写産物Type 2の2種類の異常な転写産物が検出された(図4〜6、配列番号:5〜8)。Type 1では76塩基が、Type 2では100塩基が欠損しており、野生型とType 1、Type 2がコードする蛋白質のアミノ酸配列を比較すると、Type 1、Type 2では、欠損部位よりもC末側は野生型のアミノ酸配列よりも短く、また、フレームシフトの結果全く異なる配列になっていた(図6)。BGIBMGA003497-1は小さい溶質を通すトランスポーターのファミリーであるMajor Facilitator superfamilyに属する遺伝子にホモロジーを持っていた。Major Facilitator superfamilyは一般に12回膜貫通型もしくは14回膜貫通型の構造をとるため、BGIBMGA003497-1のコードする蛋白質の膜貫通ドメインをSOSUIプログラムで予測した。その結果、野生型の遺伝子では12回膜貫通型になると予測され、Type 1、Type 2の欠損は10番目と11番目の膜貫通部位の間及び、10番目の膜貫通部位に生じていることが判明した。re系統の転写産物がコードする蛋白質の膜貫通ドメインを予測すると、野生型とは異なり、11回膜貫通型構造をとることが予測された(図8)。以上から、re系統では、異常な構造のBGIBMGA003497-1蛋白質が発現していると考えられる。上述のように、reの責任領域範囲内の遺伝子では、卵の着色に先立って発現が上昇しているものは、BGIBMGA003497-1のみであることから、BGIBMGA003497-1がreの原因遺伝子であることが強く示唆された。
【0073】
また、C108、p50Tの野生型系統2種類と、911、e28のre系統2種類において、BGIBMGA003497-1のexon9近傍のゲノム配列を決定した。その結果、911、e28は共に、exon 9内部に541bpの配列が挿入しており、この挿入がスプライシング異常を引き起こしていることが考えられた(図7)。911、e28はでは同じ位置に同じ配列の挿入が見られたため、911はe28と由来が同じであることが強く示唆された。
(4)他の生物種におけるBGIBMGA003497-1の探索
他の生物種におけるBGIBMGA003497-1(re遺伝子)のホモログをNCBIのデーターベースでBlastP検索を行ったところ、脊椎動物から植物まで幅広い種でホモログが存在した。昆虫種で見ると、双翅目昆虫のアカイエカ、ハマダラカ、ネッタイシマカ、膜翅目昆虫のミツバチ、キョウソヤドリコバチ、オオアリ、ジャンプアリ、鞘翅目昆虫のコクヌストモドキ、シラミ目のコロモジラミ、半翅目昆虫のアブラムシでホモログが存在した(図9)。このことから、re遺伝子は、幅広い昆虫種でオモクローム合成系に寄与していることが考えられる。しかしながら、Blast検索で抽出されたホモログ(e-value>7e-28以上,ホモロジー25%以上)のもので、その機能が報告されているものは1つも存在しなかった。NCBIのデーターベースでBlastP検索でも、flybaseのBlast検索でも、ショウジョウバエ12種ではホモログが検出されなかった。このことから、ショウジョウバエ12種では、re遺伝子は失われている可能性が強く示唆される。
【産業上の利用可能性】
【0074】
本発明によって見出されたre遺伝子は、カイコの卵および眼の着色に関与するため、遺伝子組換え体の選抜のためのマーカーとして好適に利用することができる。本発明によれば、効率的に遺伝子組換え体を生産することができるため、本発明は、カイコにおける有用タンパク質生産や高機能シルク生産、カイコにおけるヒト病態モデルの作製などの幅広い利用が可能である。
【配列表フリーテキスト】
【0075】
配列番号9〜37
<223> 人工的に合成されたプライマーの配列
【技術分野】
【0001】
本発明は、カイコの卵および眼の着色に関与するred egg(re)遺伝子およびその利用に関する。
【背景技術】
【0002】
オモクローム系色素は、非常に多くの昆虫において、卵、眼、体表の色に関わっていることが報告されている(非特許文献1)。オモクローム系色素はトリプトファンから派生し、3-ヒドロキシキヌレニンを前駆体として酸化縮合により生合成される色素である(図1)(非特許文献2、3)。オモクロームに関与する遺伝子は、主にショウジョウバエの複眼変異系統の研究から見出されているが、3-ヒドロキシキヌレニンの合成と色素顆粒への取り込みに関する遺伝子については知見が多いものの、色素顆粒にとりこまれた後の経路に関する遺伝子は、ほとんど明らかにされていない。
【0003】
野生型(標準型)のカイコでは、オモクローム系色素によって産卵後72時間で卵は藤鼠色に着色するが、自然突然変異で見つかったカイコ赤卵突然変異系統red egg(re)は、劣性ホモの個体の卵が薄いオレンジに着色する(図2)。さらに時間が経つと、野生型の卵の色は濃くなり、reの劣性ホモ個体の卵は赤色になる。また、野生型の成虫の蛾の眼は黒色に着色するが、reの劣性ホモ個体の成虫の複眼は濃赤褐色に着色する(図2)。reでは、オモクローム系色素の前駆体である3-ヒドロキシキヌレニンは合成されており、色素顆粒に取り込みも行われているが、最終的に生じる色素を調べると、野生型の卵と複眼に存在する色素成分のいくつかが存在しないことが報告されていた(非特許文献4)。このため、reの原因遺伝子は、3-ヒドロキシキヌレニンを色素顆粒に取り込んだ後、最終的な色素の合成に関係していることが予想されていたが、その実体は不明であった。
【0004】
カイコでは、これまで3つのオモクローム関連変異体の原因遺伝子が同定されている。すなわち、キヌレニンを3-ヒドロキシキヌレニンに変換する遺伝子に欠損があるw-1、3-ヒドロキシキヌレニンを色素顆粒に取り込むためのトランスポーター遺伝子に欠損があるw-2とw-3であり、それぞれショウジョウバエのcinnabar、scarlet、whiteに対応している(特許文献1、非特許文献5〜7)。これら3つの変異体では、オモクローム系色素が合成されず、卵と複眼が白くなる。
【0005】
カイコの卵色眼色変異系統は、肉眼でも容易に判別できるため、カイコの遺伝子組換え個体を判別するマーカーとして着目されている。現在、w-1が遺伝子組換えマーカーとして利用されているが、w-1の卵色は母親の遺伝形質に依存するため、卵の時期では組換え個体の判別は付かない。さらに、w-1、w-2、w-3はすべて白色卵で未受精卵と同じ色であるため区別がつきにくい。
【0006】
一方、reは母性遺伝せず、さらに卵が赤くなるため、未受精卵とも、野生型の卵とも区別がつきやすい。reの原因遺伝子を同定することができれば、簡便に遺伝子組換え個体を判別する分子マーカーを作出することが可能となる。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許第4431739号公報
【非特許文献】
【0008】
【非特許文献1】Linzen, B., (1974) Advamces in Insect Physiology 10:117-246
【非特許文献2】Butenandt, A., W. Schafer, (1962) Ommochromes. New York: Academic Press. p13-33.
【非特許文献3】Reed, RD., LM. Nagy, (2005) Evol Dev 7:301-311
【非特許文献4】Kawase, S., H. Aruga, (1954) The Japanese journal of genetics 29:114-121
【非特許文献5】Quan, GX. et al., (2002) Mol Genet Genomics 267:1-9.
【非特許文献6】Komoto, N. et al., (2009) Insect Biochem Mol Biol 39:152-156.
【非特許文献7】Tatematsu, KI. et al., (2011) Positional cloning of silkworm white egg 2 (w-2) locus shows functional conservation and diversification of ABC transporters for pigmentation in insects. Genes Cells.(MID: 21294818)
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、このような状況に鑑みてなされたものであり、その目的は、カイコのre遺伝子を同定することにある。また、本発明の目的は、同定したre遺伝子を利用して、カイコの卵および眼の着色を改変することにある。さらなる本発明の目的は、同定したre遺伝子を遺伝子マーカーとして利用して、遺伝子組換えカイコを選抜することにある。
【課題を解決するための手段】
【0010】
本発明者らは、上記課題を解決すべく、まず、ポジショナルクローニングの手法によりre遺伝子が存在する領域の絞り込みを行った結果、202kbにまで絞り込むことに成功した。次いで、本発明者らは、絞り込まれた領域において5つの予測遺伝子を見出し、それらの卵における発現解析を行い、データベース上で正常にアノテーションされていないBGIBMGA003497-1が卵の着色に先立って発現していることを見出した。さらに、本発明者らは、BGIBMGA003497-1の構造の解析を行ったところ、re変異系統ではORF内部に重大な欠損が存在することが判明した。BGIBMGA003497-1は、12回膜貫通型の「Major Facilitator Superfamily」に属するトランスポーターをコードしていると考えられ、この欠損は、膜貫通領域に存在していた。以上から、本発明者らは、BGIBMGA003497-1がre遺伝子の本体であり、カイコの卵および眼における赤色の着色が、この遺伝子の機能が喪失によって標準型の色素が合成されないことにより生じていることを見出した。
【0011】
さらに、本発明者らは、当該遺伝子を利用することにより、カイコの卵および眼の色の改変を行うことが可能であり、さらには、このような形質の変化を指標として、効率的に遺伝子組換えカイコを選抜することが可能であることを見出した。
【0012】
本発明は、より詳しくは、以下の発明を提供するものである。
【0013】
[1] カイコの卵および眼を変色させる活性を有するタンパク質をコードする、下記(a)〜(e)のいずれかに記載のDNA。
(a)配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または3に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:1または3に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
(e)配列番号:1または3に記載の塩基配列からなるDNAと相同性が90%以上の塩基配列からなるDNA
【0014】
[2] カイコの卵および眼を変色させる活性を有するタンパク質をコードする、下記(a)〜(c)のいずれかに記載のDNA。
(a)[1]に記載のDNAの転写産物と相補的な二重鎖RNAをコードするDNA
(b)[1]に記載のDNAの転写産物と相補的なアンチセンスRNAをコードするDNA
(c)[1]に記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA
【0015】
[3] [1]または[2]に記載のDNAを含むベクター。
【0016】
[4] [1]または[2]に記載のDNAが導入された細胞。
【0017】
[5] [1]または[2]に記載のDNAが導入されたカイコ。
【0018】
[6] [5]に記載のカイコの卵、子孫またはクローン。
【0019】
[7] 卵および眼が変色されたカイコの生産方法であって、[1]または[2]に記載のDNAをカイコに導入することを特徴とする方法。
【0020】
[8] 遺伝子組換えカイコの生産方法であって、任意のDNAと[1]または[2]に記載のDNAをカイコに導入し、卵および眼が変色した個体を選抜することを特徴とする方法。
【発明の効果】
【0021】
本発明によって、カイコの卵および眼の着色に関与するre遺伝子が同定され、該遺伝子の染色体上の位置および構造が解明された。本発明により見出されたカイコのre遺伝子は、色素の合成に関与する蛋白質をコードし、その機能の有無により、卵および眼の色が変色する。本発明のre遺伝子を用いれば、カイコの卵や眼の色を変色させることが可能であり、当該遺伝子は、遺伝子組換え体の選抜のためのマーカーとして好適に利用することができる。
【0022】
現在最も早く組換え体の判別ができるEGFP等のマーカーでは、蛍光顕微鏡を用いて産卵後7日目の胚の眼の蛍光を観察することにより遺伝子組換え体を判別している。一方、本発明において同定されたre遺伝子を用いれば、産卵後3日目に遺伝子組換え体を選抜することが可能であり、従来よりも遺伝子組換え体の識別時期が4日間早い。しかも、この識別は肉眼で行うことが可能であり、高価な蛍光顕微鏡などを必要としない。
【0023】
また、肉眼で識別が可能な遺伝子マーカーとして、KMO遺伝子マーカーも用いられてはいるが、母性遺伝のために遺伝子組換え体の卵は全て着色され、卵では組換え個体の識別が不可能である。このように、本発明によれば、従来技術に比して、カイコの遺伝子組換え体の選抜を効率的に行うことができる。
【図面の簡単な説明】
【0024】
【図1】カイコの卵を着色するオモクロームの合成経路を示す図である。母親の体内で、トリプトファンからホルミルキヌレニン、ホルミルキヌレニンからキヌレニン、キヌレニンから3-ヒドロキシキヌレニンに変換される。3-ヒドロキシキヌレニンは卵に送られる。卵が発生する際、色素顆粒が作られ、scarlet、whiteから構成されるヘテロダイマーのトランスポーターを通過することにより、3-ヒドロキシキヌレニンが色素顆粒に取り込まれる。3-ヒドロキシキヌレニンのその後の色素合成反応は色素顆粒の中で起こる。カイコの卵の色素はommin Aなど複数の色素の混合物といわれている。3-ヒドロキシキヌレニンの変化までは、母親の体内で起きるため、cinnabarより上流の遺伝子への変異は母性遺伝する。遺伝子名はショウジョウバエの遺伝子名を記した。
【図2】野生型カイコとre系統カイコの卵と成虫複眼の写真である。
【図3】Aは、絞り込み領域内の予測遺伝子(China gene model)を示す図である。絞り込み領域内には、BGIBMGA003497、003498、003499、003494、003695が存在した。BGIBMGA003497は2つの遺伝子を融合して予測していることが推測された。003497-1は白の四角、003497-2はグレーの四角で示した。Bは、予測遺伝子の発現を示す写真である。C108、p50Tは野生型の遺伝子、rPL3はコントロール遺伝子である。
【図4】Aは、re遺伝子のRT-PCRの結果を示す写真である。産卵後24時間後の卵のcDNAを用いてre遺伝子を増幅すると、2種類のre系統(911、e28)では、野生型系統(p50T、C108)よりも短い2種類のPCR産物が増幅された。Bは、野生型系統(p50T、C108)とre系統(911、e28)の遺伝子構造の比較を示す図である。エキソンは四角で、イントロンは斜め線で表示した。黒い三角形は911とe28ではエキソン9に同一の541bpの配列の挿入(黒い三角形)が存在した。ORF領域はグレーで塗りつぶしている。
【図5A】野生型系統(WT)とre系統(re type I、re type II)のre遺伝子のcDNAのORF領域の整列図である。図中、*は、ORF開始コドン、黒丸は、WTの終止コドン、黒四角は、re type Iとre type IIの終止コドンを示す。
【図5B】図5Aの続きの図である。
【図5C】図5Bの続きの図である。
【図6】野生型系統(WT)とre系統(re type I、re type II)のORFのアミノ酸配列の整列図である。
【図7】野生型系統(p50T、C108)とre系統(911、e28)のBGIBMGA003497-1(re)遺伝子のexon 9近傍のゲノム配列の整列図である。re系統にはexon 9の中に541bpの挿入配列が存在する。
【図8】カイコの野生型系統とre系統のBGIBMGA003497-1(re)遺伝子の膜貫通ドメインの予測構造を示す図である。野生型の遺伝子では12回膜貫通構造と予測されるが、原因遺伝子と赤卵系統では、10個目の膜貫通ドメイン以降のアミノ酸配列が異なり、構造が変化している。
【図9】カイコのBGIBMGA003497-1(re)遺伝子と昆虫種のホモログについての、近隣接合法により作製した系統樹の図である。図9の昆虫の部分を拡大したものである。黒丸はカイコのBGIBMGA003497-1(re)遺伝子を示す。赤卵原因遺伝子のホモログは、ショウジョウバエ以外の昆虫で幅広い昆虫種に存在した。
【発明を実施するための形態】
【0025】
<カイコの卵および眼を変色させる活性を有するタンパク質をコードするDNA>
本発明は、カイコの卵および眼を変色させる活性を有する活性を有するタンパク質をコードするDNAを提供する。
【0026】
本発明者らにより同定された、カイコの卵および眼を変色させる活性を有するタンパク質をコードする野生型系統(P50T、C108)のre cDNAの塩基配列をそれぞれ配列番号:1、3に、これらDNAがコードするタンパク質のアミノ酸配列をそれぞれ配列番号:2、4に示す。
【0027】
ここで「カイコの卵および眼を変色させる活性」とは、典型的には、カイコの卵を藤鼠色に、成虫の眼を黒色に変色させる活性を意味する。
【0028】
本発明のDNAの1つの態様は、配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA(典型的には、配列番号:1または3に記載の塩基配列のコード領域を含むDNA)である。
【0029】
現在の技術水準においては、当業者であれば、特定の野生型系統(例えば、P50T、C108)におけるre遺伝子の塩基配列情報が得られた場合、その塩基配列を改変し、そのコードするアミノ酸配列は異なるが、同様の活性を有するタンパク質をコードするDNAを取得することが可能である。また、自然界においても、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは起こり得ることである。従って、本発明は、これら野生型系統におけるタンパク質のアミノ酸配列(配列番号:2または4)において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなり、カイコの卵および眼を変色させる活性を有するタンパク質をコードするDNAをも含むものである。ここで「複数」とは、改変後のタンパク質がカイコの卵および眼を変色させる活性を維持する範囲における、アミノ酸の改変数であり、通常、50アミノ酸以内、好ましくは30アミノ酸以内、さらに好ましくは10アミノ酸以内(例えば、5アミノ酸以内、3アミノ酸以内、2アミノ酸)である。
【0030】
さらに、現在の技術水準においては、当業者であれば、特定の野生型系統(例えば、P50T、C108)からre遺伝子が得られた場合、そのDNAの塩基配列情報を利用して、カイコまたは他の生物から、同様の活性を有するタンパク質をコードするDNAを取得することが可能である。従って、本発明は、これら野生型系統(例えば、P50T、C108)におけるre遺伝子(配列番号:1または3)とストリンジェントな条件でハイブリダイズするDNAであって、カイコの卵および眼を変色させる活性を有するタンパク質をコードするDNAをも含むものである。
【0031】
こうして得られた変異DNAや相同DNAが、カイコの卵および眼を変色させる活性を有するタンパク質をコードするか否かは、例えば、被検DNAを、re変異系統(卵および眼が赤色となる系統)に導入し、卵が藤鼠色に変色するか否か、あるいは、成虫の眼が黒色に変色するか否かにより、判定することができる。
【0032】
本発明のDNAは、その導入により、カイコの卵および眼を変色させることが可能であるという意味において、カイコの卵および眼を変色させるための薬剤である。
【0033】
なお、上記した変異DNAを作製するための、DNAへの人為的な変異の導入は、例えば、部位特異的変異誘発(site-directed mutagenesis)法(Kramer, W. & Fritz, HJ., Methods Enzymol, 154:350-367, 1987)により行うことができる。
【0034】
また、上記した相同遺伝子を単離するための方法としては、例えば、ハイブリダイゼーション技術(Southern, E. M., Journal of Molecular Biology, 98:503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R. K., et al. Science, 230:1350-1354, 1985、Saiki, R. K. et al. Science, 239:487-491, 1988)が挙げられる。相同遺伝子をコードするDNAを単離するためには、通常ストリンジェントな条件下でハイブリダイゼーション反応を行なう。ストリンジェントなハイブリダイゼーション条件としては、6M尿素、0.4%SDS、0.5xSSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を例示できる。よりストリンジェンシーの高い条件、例えば、6M尿素、0.4%SDS、0.1xSSCの条件を用いれば、より相同性の高いDNAの単離を期待することができる。単離されたDNAは、核酸レベルあるいはアミノ酸配列レベルにおいて、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の相同性は、BLASTN(核酸レベル)やBLASTX(アミノ酸レベル)のプログラム(Altschul et al. J. Mol. Biol., 215:403-410, 1990)を利用して決定することができる。該プログラムは、KarlinおよびAltschulによるアルゴリズムBLAST(Proc. Natl. Acad. Sci. USA, 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA, 90:5873-5877, 1993)に基づいている。BLASTNによって塩基配列を解析する場合には、パラメーターは例えばscore=100、wordlength=12とする。また、BLASTXによってアミノ酸配列を解析する場合には、パラメーターは例えばscore=50、wordlength=3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res. 25:3389-3402, 1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0035】
本発明のreタンパク質をコードするDNAとしては、その形態に特に制限はなく、cDNAの他、ゲノムDNA、および化学合成DNAが含まれる。ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、カイコからゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、フォスミド、BAC、PACなどが利用できる)を作成し、これを展開して、re遺伝子(例えば、配列番号:1または3に記載のDNA)の塩基配列を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより調製することが可能である。また、re遺伝子に特異的なプライマーを作成し、これを利用したPCRを行うことによって調製することも可能である。また、cDNAは、例えば、鱗翅目昆虫から抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。
【0036】
<カイコのre遺伝子の発現を抑制するために用いるDNA>
また、本発明は、上記した本発明の野生型系統のre遺伝子の発現を抑制するために用いるDNAを提供する。本発明においては、re遺伝子の機能が喪失することにより、カイコの卵および眼が変色することが見出された。従って、re遺伝子の発現を抑制する活性を有するDNAの導入により、カイコの卵および眼を変色させることが可能である。この意味において、re遺伝子の発現を抑制するために用いるDNAは、カイコの卵および眼を変色させるための薬剤である。ここで「re遺伝子の発現の抑制」には、遺伝子の転写の抑制およびタンパク質への翻訳の抑制の双方が含まれる。また、「発現の抑制」には、発現の完全な停止のみならず発現の減少も含まれる。また、「カイコの卵および眼を変色させる活性」とは、典型的には、カイコの卵および成虫の眼を赤色に変色させる活性を意味する。被検DNAが、カイコの卵および成虫の眼を赤色に変色させる活性を有するか否かは、例えば、当該DNAを、野生型異系統(卵が藤鼠色および眼が黒色となる系統)に導入し、卵および成虫の眼が赤色に変色するか否かにより、判定することができる。
【0037】
re遺伝子の発現を抑制するために用いるDNAの一つの態様は、re遺伝子の転写産物と相補的なdsRNA(二重鎖RNA)をコードするDNAである。標的遺伝子配列と同一もしくは類似した配列を有するdsRNAを細胞内に導入することにより、導入した外来遺伝子および標的内因性遺伝子の発現がいずれも抑制される、RNAi(RNA干渉、RNA interference)と呼ばれる現象を引き起こすことができる。細胞に約40〜数百塩基対のdsRNAが導入されると、ヘリカーゼドメインを持つダイサー(Dicer)と呼ばれるRNaseIII様のヌクレアーゼが、ATP存在下で、dsRNAを3'末端から約21〜23塩基対ずつ切り出し、siRNA(short interference RNA)が生じる。このsiRNAに、特異的なタンパク質が結合して、ヌクレアーゼ複合体(RISC:RNA-induced silencing complex)が形成される。この複合体はsiRNAと同じ配列を認識して結合し、RNaseIII様の酵素活性によってsiRNAの中央部で標的遺伝子の転写産物(mRNA)を切断する。また、この経路とは別にsiRNAのアンチセンス鎖がmRNAに結合してRNA依存性RNAポリメラーゼ(RsRP)のプライマーとして作用し、dsRNAが合成される。このdsRNAが再びダイサーの基質となって、新たなsiRNAを生じて作用を増幅する経路も考えられている。
【0038】
本発明のdsRNAをコードするDNAは、標的遺伝子の転写産物(mRNA)のいずれかの領域に対するアンチセンスRNAをコードしたアンチセンスDNAと、該mRNAのいずれかの領域のセンスRNAをコードしたセンスDNAを含み、該アンチセンスDNAおよび該センスDNAより、それぞれアンチセンスRNAおよびセンスRNAを発現させることができる。また、これらのアンチセンスRNAおよびセンスRNAよりdsRNAを作成することができる。
【0039】
本発明のdsRNAの発現システムをベクター等に保持させる場合の構成としては、同一のベクターからアンチセンスRNAおよびセンスRNAを発現させる場合と、異なるベクターからそれぞれアンチセンスRNAとセンスRNAを発現させる場合がある。同一のベクターからアンチセンスRNAおよびセンスRNAを発現させる構成としては、例えば、アンチセンスDNAおよびセンスDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモーターを連結させたアンチセンスRNA発現カセットとセンスRNA発現カセットをそれぞれ構築し、これらカセットを同方向にあるいは逆方向にベクターに挿入する構成である。
【0040】
また、異なる鎖上に対向するように、アンチセンスDNAとセンスDNAとを逆向きに配置した発現システムを構成することもできる。この構成では、アンチセンスRNAコード鎖とセンスRNAコード鎖とが対となった一つの二本鎖DNAが備えられ、その両側にそれぞれの鎖からアンチセンスRNAとセンスRNAとを発現し得るようにプロモーターを対向して備える。この場合には、センスRNAとアンチセンスRNAの下流に余分な配列が付加されることを避けるために、それぞれの鎖(アンチセンスRNAコード鎖、センスRNAコード鎖)の3'末端にターミネーターをそれぞれ備えることが好ましい。このターミネーターは、A(アデニン)塩基を4つ以上連続させた配列などを用いることができる。また、このパリンドロームスタイルの発現システムでは、二つのプロモーターの種類は異なっていることが好ましい。
【0041】
また、異なるベクターからアンチセンスRNAおよびセンスRNAを発現させる構成としては、例えば、アンチセンスDNAおよびセンスDNAの上流にそれぞれpolIII系のような短いRNAを発現し得るプロモーターを連結させたアンチセンスRNA発現カセットとセンスRNA発現カセットとをそれぞれ構築し、これらカセットを異なるベクターに保持させる構成である。
【0042】
本発明に用いるdsRNAとしては、siRNAが好ましい。「siRNA」は、細胞内で毒性を示さない範囲の短鎖からなる二重鎖RNAを意味する。標的re遺伝子の発現を抑制することができ、かつ、毒性を示さなければ、その鎖長に特に制限はない。dsRNAの鎖長は、例えば、15〜500塩基対である。
【0043】
本発明のdsRNAをコードするDNAとしては、標的配列のインバーテッドリピートの間に適当な配列(イントロン配列が望ましい)を挿入し、ヘアピン構造を持つダブルストランドRNA(self-complementary 'hairpin' RNA(hpRNA))を作るようなコンストラクト(Smith, N.A., et al. Nature, 407:319, 2000、Wesley, S. V. et al. Plant J. 27:581, 2001、Piccin, A. et al. Nucleic Acids Res. 29:E55, 2001)を用いることが好ましい。
【0044】
本発明のdsRNAをコードするDNAは、標的re遺伝子の塩基配列と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の配列の同一性を有する。配列の同一性は上述した手法(BLASTプログラム)により決定できる。
【0045】
dsRNAにおけるRNA同士が対合した二重鎖RNAの部分は、完全に対合しているものに限らず、ミスマッチ(対応する塩基が相補的でない)、バルジ(一方の鎖に対応する塩基がない)などにより不対合部分が含まれていてもよい。本発明においては、dsRNAにおけるRNA同士が対合する二重鎖RNA領域中に、バルジおよびミスマッチの両方が含まれていてもよい。
【0046】
re遺伝子の発現を抑制するために用いるDNAの他の態様は、re遺伝子の転写産物と相補的なアンチセンスRNAをコードするDNA(アンチセンスDNA)である。アンチセンスDNAが標的遺伝子の発現を抑制する作用としては、三重鎖形成による転写開始阻害、RNAポリメラーゼによって局部的に開状ループ構造がつくられた部位とのハイブリッド形成による転写抑制、合成の進みつつあるRNAとのハイブリッド形成による転写阻害、イントロンとエキソンとの接合点でのハイブリッド形成によるスプライシング抑制、スプライソソーム形成部位とのハイブリッド形成によるスプライシング抑制、mRNAとのハイブリッド形成による核から細胞質への移行抑制、キャッピング部位やポリ(A)付加部位とのハイブリッド形成によるスプライシング抑制、翻訳開始因子結合部位とのハイブリッド形成による翻訳開始抑制、開始コドン近傍のリボソーム結合部位とのハイブリッド形成による翻訳抑制、mRNAの翻訳領域やポリソーム結合部位とのハイブリッド形成によるペプチド鎖の伸長阻止、および核酸とタンパク質との相互作用部位とのハイブリッド形成による遺伝子発現抑制などが挙げられる。これらは、転写、スプライシング、または翻訳の過程を阻害して、標的遺伝子の発現を抑制する(平島および井上「新生化学実験講座2 核酸IV 遺伝子の複製と発現」,日本生化学会編,東京化学同人, pp.319-347, 1993)。本発明で用いられるアンチセンスDNAは、上記のいずれの作用で標的re遺伝子の発現を抑制してもよい。一つの態様として、標的遺伝子のmRNAの5'端近傍の非翻訳領域に相補的なアンチセンス配列を設計すれば、遺伝子の翻訳阻害に効果的であると考えられる。また、コード領域もしくは3'側の非翻訳領域に相補的な配列も使用し得る。このように、遺伝子の翻訳領域だけでなく非翻訳領域の配列のアンチセンス配列を含むDNAも、本発明で利用されるアンチセンスDNAに含まれる。使用されるアンチセンスDNAは、適当なプロモーターの下流に連結され、好ましくは3'側に転写終結シグナルを含む配列が連結される。
【0047】
アンチセンスDNAは、本発明のre遺伝子(例えば、配列番号:1または3に記載の塩基配列からなるDNA)の配列情報を基にホスホロチオネート法(Stein, Nucleic Acids Res., 16:3209-3221, 1988)などにより調製することが可能である。調製されたDNAは、後述する公知の方法で、カイコへ導入できる。アンチセンスDNAの配列は、カイコが持つ内因性のre遺伝子の転写産物と相補的な配列であることが好ましいが、遺伝子の発現を有効に阻害できる限り、完全に相補的でなくてもよい。転写されたRNAは、標的とする遺伝子の転写産物に対して好ましくは90%以上(例えば、95%、96%、97%、98%、99%以上)の相補性を有する。効果的に標的遺伝子の発現を阻害するには、アンチセンスDNAの長さは、少なくとも15塩基以上であり、好ましくは100塩基以上であり、さらに好ましくは500塩基以上である。通常、用いられるアンチセンスDNAの長さは5kbよりも短く、好ましくは2.5kbよりも短い。
【0048】
re遺伝子の発現を抑制するために用いるDNAの他の態様は、re遺伝子の転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNAである。リボザイムには、グループIイントロン型や、RNasePに含まれるM1RNAのように400ヌクレオチド以上の大きさのものもあるが、ハンマーヘッド型やヘアピン型と呼ばれる40ヌクレオチド程度の活性ドメインを有するものもある(小泉誠および大塚栄子、蛋白質核酸酵素, 35:2191, 1990)。
【0049】
例えば、ハンマーヘッド型リボザイムの自己切断ドメインは、G13U14C15のC15の3'側を切断するが、活性にはU14が9位のAと塩基対を形成することが重要とされ、15位の塩基はCの他にAまたはUでも切断されることが示されている(Koizumi et. al., FEBS Lett. 228:225, 1988)。リボザイムの基質結合部を標的部位近傍のRNA配列と相補的になるように設計すれば、標的RNA中のUC、UUまたはUAという配列を認識する制限酵素的なRNA切断リボザイムを作出することが可能である(Koizumi et. al., FEBS Lett. 239:285, 1988、小泉誠および大塚栄子,蛋白質核酸酵素,35:2191, 1990、Koizumi et. al., Nucleic. Acids. Res. 17:7059, 1989)。
【0050】
また、ヘアピン型リボザイムも、本発明の目的のために有用である。ヘアピン型リボザイムは、例えばタバコリングスポットウイルスのサテライトRNAのマイナス鎖に見出される(Buzayan, Nature 323:349, 1986)。このリボザイムも、標的特異的なRNA切断を起こすように設計できることが示されている(Kikuchi and Sasaki, Nucleic Acids Res. 19:6751, 1992、菊池洋,化学と生物 30:112, 1992)。標的を切断できるよう設計されたリボザイムは、カイコ細胞中で転写されるようにアクチンプロモーターなどのプロモーターおよび転写終結配列に連結される。このような構成単位をタンデムに並べ、標的遺伝子内の複数の部位を切断できるようにして、より効果を高めることもできる(Yuyama et al., Biochem. Biophys. Res. Commun. 186:1271, 1992)。このようなリボザイムを用いて標的となるre遺伝子の転写産物を特異的に切断し、該遺伝子の発現を抑制することができる。
【0051】
<ベクター、形質転換細胞、形質転換カイコ>
本発明は、また、上記本発明のDNA(re遺伝子、およびre遺伝子の発現を抑制するためのDNA)を含むベクター、上記本発明のDNAが導入された細胞、上記本発明のDNAが導入されたカイコを提供する。
【0052】
上記本発明のDNAが導入される細胞としては、特に制限はなく、目的に応じて種々の細胞を用いることができる。例えば、組換えタンパク質生産の目的であれば、カイコ細胞の他、例えば、大腸菌、酵母、動物細胞、植物細胞などを用いることができる。遺伝子組換えカイコを製造する場合には、カイコ細胞が用いられる。
【0053】
本発明のベクターとしては、例えば、自律複製可能なベクターまたは染色体中に相同組換え可能なベクターを使用することができる。カイコ卵に対して本発明のDNAの導入を行う場合、例えば、カイコの発生初期卵へ、トランスポゾンをベクターとして注射する方法(Tamura,T. et al., (2000) Nature Biotechnology 18:81-84)に従って行うことができる。例えば、トランスポゾンの逆位末端反復配列(Handler AM. et al.,(1998) Proc. Natl. Acad. Sci. U.S.A. 95(13):7520-5)の間に上記DNAを挿入したベクターとともに、トランスポゾン転移酵素をコードするDNAを有するベクター(ヘルパーベクター)をカイコ卵に導入する。ヘルパーベクターとしては、pHA3PIG(Tamura,T. et al., (2000) Nature Biotechnology 18, 81-84)が挙げられるが、これに限定されるものではない。本発明におけるトランスポゾンとしては、piggyBacが好ましいが、これに限定されるものではなく、マリーナ(mariner)、ミノス(minos)等を用いることもできる(Shimizu,K. et al., (2000) Insect Mol. Biol., 9, 277-281;Wang W. et al.,(2000) Insect Mol Biol 9(2):145-55)。また、本発明では、バキュロウイルスベクターを使用することにより形質転換カイコを作出することも可能である(Yamao, M. et al., (1999) Genes Dev 13:511-516)。
【0054】
ベクターに用いるプロモーターとしては、例えば、アクチンプロモーター、AcNPVのhr5-IE1プロモーター、3xP3プロモーターなどを用いることができる。
【0055】
カイコ細胞へのベクターの導入は、例えば、リン酸カルシウム沈殿法、電気パルス穿孔法(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 9.1-9.9)、リポフェクタミン法(GIBCO-BRL社製)、マイクロインジェクション法などの公知の方法で行うことが可能である。
【0056】
一旦、染色体内に上記本発明のDNAが導入されたカイコが得られれば、該カイコから、卵、子孫、あるいはクローンを得て、それらを基にカイコを繁殖させることも可能である。本発明には、上記本発明のDNAが導入されたカイコおよびその卵、子孫およびクローンが含まれる。
【0057】
<卵および眼が変色されたカイコの生産方法>
本発明は、また、卵および眼が変色されたカイコの生産方法であって、上記本発明のDNA(re遺伝子、およびre遺伝子の発現を抑制するためのDNA)をカイコに導入することを特徴とする方法を提供する。
【0058】
本発明の方法の一つの態様は、野生型のre遺伝子をre変異系統のカイコに導入して、卵が藤鼠色に、成虫の眼が黒色に変色されたカイコを生産する方法である。
【0059】
本発明の方法の他の一つの態様は、re遺伝子の発現を抑制するためのDNAを野生型カイコに導入して、卵および眼が赤色に変色されたカイコを生産する方法である。
【0060】
<遺伝子組換えカイコの生産方法>
本発明は、また、遺伝子組換えカイコの生産方法であって、任意のDNAと上記本発明のDNA(re遺伝子、およびre遺伝子の発現を抑制するためのDNA)をカイコに導入し、卵および眼が変色した個体を選抜することを特徴とする方法を提供する。
【0061】
カイコに導入する「任意のDNA」としては、特に制限はない。例えば、医薬品や検査薬の原料となる有用タンパク質をコードするDNA、高機能シルクの生産に有用な蛋白質をコードするDNA、ヒトの病態のモデルカイコを作製するために有用なDNAが挙げられる。任意のDNAと上記本発明のDNAは、一つのベクターに配置してカイコに導入することが好ましいが、それぞれを異なるベクターに配置してカイコに導入することも可能である。任意のDNAと上記本発明のDNAは、ベクター上では、カイコにおいて発現を保証するプロモーター(例えば、アクチンプロモーター)に発現可能に連結されている。任意のDNAとre遺伝子が導入されたre変異系統カイコは、その卵が藤鼠色に変色し、成虫においては眼が黒色に変色する。また、任意のDNAとre遺伝子の発現を抑制するためのDNAが導入された野生型カイコは、その卵が赤色に変色し、成虫においては眼が赤色に変色する。この変色を指標に、遺伝子組換えカイコを選抜することができる。特に、卵においては、産卵後3日目には変色するため、従来よりも早期に、かつ、肉眼で、遺伝子組換え体を判別することが可能である。
【実施例】
【0062】
以下、実施例および比較例に基づいて本発明をより具体的に説明するが、本発明は以下の実施例に限定されるものではない。
【0063】
[実験材料と方法]
(1)カイコの飼育
カイコは人工飼料(日本農産工業)で25度〜27度で飼育した。C108系統は、研究室で累代飼育しているもの、911(re)系統は農業生物資源研究所内で累代飼育しているもの、e28系統はナショナルバイオリソースプロジェクトで累代飼育している系統(九州大学)を取り寄せて実験に用いた。
【0064】
(2)ポジショナルクローニング
reの表現型の責任領域のマッピングのため、C108(正常着色卵)と911(re)を交配して生まれたF1のオスのカイコと、911(re)のメスの交配を行った。この交配により生まれたBF1卵(正常着色282、赤卵114、計396)から孵化した1零幼虫からゲノムDNAをDNAzol(Invitrogen)を用いて抽出した。抽出したDNAに用いて、SNPマーカー(表1)を用い、文献(非特許文献7)と類似の方法で連鎖解析を行い、reの表現型の責任領域を絞り込んだ。SNPマーカーの一部は(Yamamoto, K. et al., (2008) Genome Biol 9:R21)で使用されているものである。
【0065】
【表1】
【0066】
(3)遺伝子の発現解析
P50T(正常着色卵、ゲノム解読系統)、C108(正常着色卵)、911(re変異系統)、e28(re変異系統)について、産下直後、24時間後、48時間後、72時間後の卵からtotal RNAをIsogen(NipponGene)を用いて抽出し、First-Strand cDNA Synthesis Kit(GE Healthcare)を用いてcDNAを合成した。RT-PCRは、絞り込み範囲内に存在した予測遺伝子BGIBMGA003497、003498、003499、003494、003695についてプライマー(表2)を設計し、total RNA 0.5ng分のcDNAを鋳型に用いて行った。PCRで増幅した断片の配列決定は、BigDye Ver.3.1(Applied Biosystems)とABI3130xl(Applied Biosystems)を用いて行い、配列解析はVector NTI ver.11(Invitrogen)を用いて行った。
【0067】
【表2】
【0068】
(4)膜貫通ドメイン予測
膜貫通ドメインはSOSUI(http://bp.nuap.nagoya-u.ac.jp/sosui/sosui_submit.html)プログラムで予測した。
【0069】
(5)分子系統解析
BGIBMGA003497-1と相同性が予測された100個の遺伝子について、MEGA5ソフトウェアを用い、アミノ酸配列のアラインメント及び近隣接合法で分子系統解析を行った。
【0070】
[実施例1] re遺伝子の同定
(1)ポジショナルクローニングによるre遺伝子の存在領域の絞り込み
reの責任領域の絞り込みのため、まず、C108(正常着色卵、+re/+re)のメスと911(re/re)のオスを掛け合わせ、F1を得た。F1のオス(re/+re)を911(re/re)のメスに掛け、戻し交配し、BF1個体(卵)を得た。BF1(卵)には正常着色卵と赤卵が存在した。BF1正常着色卵から生まれた幼虫282個体、BF1赤卵から生まれた幼虫114個体からそれぞれゲノムDNAを抽出し、5番染色体上に作成した40個のSNPマーカー(表1)を用いてジェノタイピングを行った。その結果、reの責任領域はSNPマーカーre-071119-R2_F1とre-080104R-R12_F1の間の202kbに絞り込まれた(表1、図3A)。
【0071】
(2)re遺伝子の候補の発現解析
ポジショナルクローニングより絞り込んだ領域には、5つの予測遺伝子BGIBMGA003497、003498、003499、003494、003695が存在した(図3A)。これらの配列をNCBIのデーターベースに対し検索を行ったところ、BGIBMGA003497の5’側はMajor Facilitator superfamilyに属する遺伝子に、3’側はChromodomain-helicase-DNA-binding proteinという全く異なる遺伝子にホモロジーが検出されたため、2つの遺伝子が融合して1つの遺伝子として誤って予測されていることが考えられた。そこで、BGIBMGA003497の5’側と3側を仮にBGIBMGA003497-1とBGIBMGA003497-2とした。reの責任領域範囲内の予測遺伝子の発現を調べるため、BGIBMGA003498、003499、003494、003695にはそれぞれ1セットずつ、BGIBMGA003497については、BGIBMGA003497-1とBGIBMGA003497-2に1セットずつ、プライマーを設計した。カイコの卵は、産卵後40時間頃に薄茶色に着色し始め、この時期には野生型系統とre系統を見分けにくいが、72時間頃になると、野生型は藤鼠色になるのに対し、re系統は薄オレンジ色または赤色を呈し、それ以降はそれぞれの色が濃くなる。このため、reの原因遺伝子は、主に産卵後72時間より前に働いていることが考えられた。そこで、標準型の卵色と複眼色を示すP50TとC108の2つの系統について産下直後、1日後、2日後、3日後の卵における遺伝子の発現をRT-PCRにより調べた(図3B)。その結果、BGIBMGA003499はC108で発現が見られたが、p50Tではその発現が非常に弱くなっていた。003498、003494、003695では発現を示すバンドが検出できなかった。BGIBMGA003497-2は発現が検出されたが、特に、p50T系統では一様に発現していた。興味深いことに、BGIBMGA003497-1は24~48時間後に発現がピークになり、72時間後には発現量が下がっており、卵の着色に先立って発現していると考えられた。BGIBMGA003497-1とBGIBMGA003497-2では発現パターンが異なることから、この二つは独立の遺伝子であることが考えられた。
【0072】
(3)BGIBMGA003497-1の構造の解析
BGIBMGA003497-1のORF配列を調べるため、カイコのESTライブラリーとゲノム配列にBlastN検索をかけたところ、アラタ体と神経由来のESTライブラリーから対応するESTが得られた。これらのEST配列を基にORF全長を野生型の卵およびre系統の卵(911系統とe28系統)のcDNAから単離し、遺伝子構造の比較を行った。P50T系統、C108系統共に、BGIBMGA003497-1は、495アミノ酸のORFをコードする転写産物が発現していた(図4、図5、図6)。911系統、e28系統では、BGIBMGA003497-1は正常な転写産物が検出されず、exon 9の一部を失ったType 1および、exon 9全体を失った転写産物Type 2の2種類の異常な転写産物が検出された(図4〜6、配列番号:5〜8)。Type 1では76塩基が、Type 2では100塩基が欠損しており、野生型とType 1、Type 2がコードする蛋白質のアミノ酸配列を比較すると、Type 1、Type 2では、欠損部位よりもC末側は野生型のアミノ酸配列よりも短く、また、フレームシフトの結果全く異なる配列になっていた(図6)。BGIBMGA003497-1は小さい溶質を通すトランスポーターのファミリーであるMajor Facilitator superfamilyに属する遺伝子にホモロジーを持っていた。Major Facilitator superfamilyは一般に12回膜貫通型もしくは14回膜貫通型の構造をとるため、BGIBMGA003497-1のコードする蛋白質の膜貫通ドメインをSOSUIプログラムで予測した。その結果、野生型の遺伝子では12回膜貫通型になると予測され、Type 1、Type 2の欠損は10番目と11番目の膜貫通部位の間及び、10番目の膜貫通部位に生じていることが判明した。re系統の転写産物がコードする蛋白質の膜貫通ドメインを予測すると、野生型とは異なり、11回膜貫通型構造をとることが予測された(図8)。以上から、re系統では、異常な構造のBGIBMGA003497-1蛋白質が発現していると考えられる。上述のように、reの責任領域範囲内の遺伝子では、卵の着色に先立って発現が上昇しているものは、BGIBMGA003497-1のみであることから、BGIBMGA003497-1がreの原因遺伝子であることが強く示唆された。
【0073】
また、C108、p50Tの野生型系統2種類と、911、e28のre系統2種類において、BGIBMGA003497-1のexon9近傍のゲノム配列を決定した。その結果、911、e28は共に、exon 9内部に541bpの配列が挿入しており、この挿入がスプライシング異常を引き起こしていることが考えられた(図7)。911、e28はでは同じ位置に同じ配列の挿入が見られたため、911はe28と由来が同じであることが強く示唆された。
(4)他の生物種におけるBGIBMGA003497-1の探索
他の生物種におけるBGIBMGA003497-1(re遺伝子)のホモログをNCBIのデーターベースでBlastP検索を行ったところ、脊椎動物から植物まで幅広い種でホモログが存在した。昆虫種で見ると、双翅目昆虫のアカイエカ、ハマダラカ、ネッタイシマカ、膜翅目昆虫のミツバチ、キョウソヤドリコバチ、オオアリ、ジャンプアリ、鞘翅目昆虫のコクヌストモドキ、シラミ目のコロモジラミ、半翅目昆虫のアブラムシでホモログが存在した(図9)。このことから、re遺伝子は、幅広い昆虫種でオモクローム合成系に寄与していることが考えられる。しかしながら、Blast検索で抽出されたホモログ(e-value>7e-28以上,ホモロジー25%以上)のもので、その機能が報告されているものは1つも存在しなかった。NCBIのデーターベースでBlastP検索でも、flybaseのBlast検索でも、ショウジョウバエ12種ではホモログが検出されなかった。このことから、ショウジョウバエ12種では、re遺伝子は失われている可能性が強く示唆される。
【産業上の利用可能性】
【0074】
本発明によって見出されたre遺伝子は、カイコの卵および眼の着色に関与するため、遺伝子組換え体の選抜のためのマーカーとして好適に利用することができる。本発明によれば、効率的に遺伝子組換え体を生産することができるため、本発明は、カイコにおける有用タンパク質生産や高機能シルク生産、カイコにおけるヒト病態モデルの作製などの幅広い利用が可能である。
【配列表フリーテキスト】
【0075】
配列番号9〜37
<223> 人工的に合成されたプライマーの配列
【特許請求の範囲】
【請求項1】
カイコの卵および眼を変色させる活性を有するタンパク質をコードする、下記(a)〜(e)のいずれかに記載のDNA。
(a)配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または3に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:1または3に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
(e)配列番号:1または3に記載の塩基配列からなるDNAと相同性が90%以上の塩基配列からなるDNA
【請求項2】
カイコの卵および眼を変色させる活性を有するタンパク質をコードする、下記(a)〜(c)のいずれかに記載のDNA。
(a)請求項1に記載のDNAの転写産物と相補的な二重鎖RNAをコードするDNA
(b)請求項1に記載のDNAの転写産物と相補的なアンチセンスRNAをコードするDNA
(c)請求項1に記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA
【請求項3】
請求項1または2に記載のDNAを含むベクター。
【請求項4】
請求項1または2に記載のDNAが導入された細胞。
【請求項5】
請求項1または2に記載のDNAが導入されたカイコ。
【請求項6】
請求項5に記載のカイコの卵、子孫またはクローン。
【請求項7】
卵および眼が変色されたカイコの生産方法であって、請求項1または2に記載のDNAをカイコに導入することを特徴とする方法。
【請求項8】
遺伝子組換えカイコの生産方法であって、任意のDNAと請求項1または2に記載のDNAをカイコに導入し、卵および眼が変色した個体を選抜することを特徴とする方法。
【請求項1】
カイコの卵および眼を変色させる活性を有するタンパク質をコードする、下記(a)〜(e)のいずれかに記載のDNA。
(a)配列番号:2または4に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1または3に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2または4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号:1または3に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNA
(e)配列番号:1または3に記載の塩基配列からなるDNAと相同性が90%以上の塩基配列からなるDNA
【請求項2】
カイコの卵および眼を変色させる活性を有するタンパク質をコードする、下記(a)〜(c)のいずれかに記載のDNA。
(a)請求項1に記載のDNAの転写産物と相補的な二重鎖RNAをコードするDNA
(b)請求項1に記載のDNAの転写産物と相補的なアンチセンスRNAをコードするDNA
(c)請求項1に記載のDNAの転写産物を特異的に開裂するリボザイム活性を有するRNAをコードするDNA
【請求項3】
請求項1または2に記載のDNAを含むベクター。
【請求項4】
請求項1または2に記載のDNAが導入された細胞。
【請求項5】
請求項1または2に記載のDNAが導入されたカイコ。
【請求項6】
請求項5に記載のカイコの卵、子孫またはクローン。
【請求項7】
卵および眼が変色されたカイコの生産方法であって、請求項1または2に記載のDNAをカイコに導入することを特徴とする方法。
【請求項8】
遺伝子組換えカイコの生産方法であって、任意のDNAと請求項1または2に記載のDNAをカイコに導入し、卵および眼が変色した個体を選抜することを特徴とする方法。
【図1】

【図2】
【図3】
【図4】
【図5A】

【図5B】

【図5C】
【図6】

【図7】
【図8】
【図9】

【図2】
【図3】
【図4】
【図5A】
【図5B】
【図5C】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2012−205525(P2012−205525A)
【公開日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願番号】特願2011−72777(P2011−72777)
【出願日】平成23年3月29日(2011.3.29)
【国等の委託研究の成果に係る記載事項】(出願人による申告)農林水産省平成22年度アグリ・ゲノム研究の総合的な推進委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
【公開日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願日】平成23年3月29日(2011.3.29)
【国等の委託研究の成果に係る記載事項】(出願人による申告)農林水産省平成22年度アグリ・ゲノム研究の総合的な推進委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
[ Back to top ]