
カゼイン由来の抗菌ペプチドおよびそれを生成するLactobacillus株
本発明は、健康増進剤、特に抗菌剤の分野に全般的に関し、抗菌ペプチドおよび該抗菌ペプチドを与える細菌株を提供する。ある態様において、本発明は、その試料が2005年11月18日にスコットランドアバディーンのナショナル・コレクション・オブ・インダストリアル・アンド・マリン・バクテリアにアクセッション番号NCIMB41354の下で寄託されているLactobacillus acidophilus DPC6026株、または乳もしくは乳製品から抗菌活性を有するペプチドを製造できるその派生株もしくは変異株の生物学的に純粋な培養物を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、健康増進剤、特に抗菌剤の分野に全般的に関し、抗菌ペプチドおよび該抗菌ペプチドを与える細菌株を提供する。
【背景技術】
【0002】
発明の背景
病原菌による感染により、ヒトおよび動物の両方において重大な健康問題が起こり得る。特に、E.coli、E.sakazakii、Streptococcus mutansおよびListeria innocuaのような病原株による感染は、ひどく衰弱させ得る。病原菌に対する抗菌剤を供給する必要性があることが認識されている。
【0003】
病原菌の抗生物質に対する耐性の増加は、主要な問題点であり、新規薬物の探索が続いている(35)。多剤耐性のE.coli、Klebsiella spp.、およびE.sakazakiiの株は、驚くべき速度で増加しており、免疫無防備状態の宿主にしばしば関連して、該宿主にひどい影響を与え得る(6、14)。Enterobacter種は、それらがより古い抗菌剤に対する先天的な耐性を有し、かつそれらの院内感染との関連が増加しているために、近年、重要性が増加している病原体として認識されている(41)。30年より長きにわたって、E.sakazakiiは、新生児の髄膜炎の特有の症候群の原因として認識されている(20)。40〜80%の死亡率が報告されており、乳ベースの幼児用調合乳が感染の病原巣として働くという興味深い証拠が増加している((20)、(33)、(34))。E.sakazakii感染を制御するための現在の工業的な努力は、製造における衛生業務の改善および生物体についての最終製品の試験に焦点を当てている(34)。新規な抗菌ペプチドの候補を同定するためのある可能な経路は、乳タンパク質の探索にある。
【0004】
乳タンパク質は、生物活性ペプチドの豊富な供給源であり、アンギオテンシン−I変換酵素(ACE)阻害、オピオイド、免疫調節、抗血栓および抗菌活性を含む一連の健康増進特性を有すると報告されている(9)。生物活性ペプチドは、カゼインおよびホエーのような乳タンパク質にコード化された形で存在し、プロペプチドまたは成熟C末端ペプチドとして貯蔵され(13、19)、それらの放出にはタンパク質分解が必要である(11)。これらの機能的ペプチドのうち最も特徴付けられているものは、カゼインに由来し、心血管系、主にACE阻害における効果を有することが示されており、その結果、抗高血圧の特徴を有する(43)。MaruyamaおよびSuzuki(28)は、カゼインのトリプシン加水分解物が、ACEのin vitro活性を阻害したことを報告した。Pihlanto−Leppalaら(37)は、乳酸菌(LAB)で発酵させ、その後消化酵素で加水分解したホエーおよびカゼインタンパク質からのペプチドのACE阻害活性を研究した。同定されたペプチドは、αs1−カゼインf(142−147)、f(194−199)およびβ−カゼインf(108−113)、ならびにホエーからの2種のACE阻害ペプチドであった(37)。さらに、一連の抗菌ペプチドは、ウシのαs2カゼインから単離されている(23)。これらは、カソシジン(Casocidin)−1,f(164−179):KTKLTEEEKN RLNFLKKISQ RYQKFALPQY LKTVYQHQK、f(164−179):KKTKLTEEEK NRLNFL、およびf(187−207):QKFALPQYLK TVYQHQKAMK Qを含む。
【0005】
これらの3種のペプチドは、牛乳を酢酸および硫酸カルシウムで処理して混合物を形成し、次いでこれを加熱して沈殿物および上清を形成することから導かれる。上清は回収され、カチオン交換樹脂で処理される。ペプチドを、クロマトグラフィー手法を用いて溶出させ、活性ペプチドであるカソシジン−1を、消化プロテアーゼを用いて遊離させる。製造のコストは、これらの3つのペプチドの広い利用の1つの可能な制限である。特に、カソシジン−1は、Escherichia coliおよびStaphylococcus carnosusに対してin vitroで殺菌性であることが示されている(51)。
【0006】
しかし、カソシジン−Iを直接製造する細菌株は知られておらず、このことは、カソシジン−Iの製造が、製造コストを付加する酵素および化学物質の使用を必要とすることを意味する。さらなる情報は、「Antibiotic peptides from bovine milk」の表題のUS6579849に見出し得る。上記の特許にはこれらのペプチドの活性に対する制限は述べられていないが、Zuchtら、(FEBS Letters 372号(1995年)185〜188頁)は、これらのペプチドの阻害の範囲を記載し、グラム陰性細菌のみがこれらのペプチドにより影響を受けるようであり、このことは、グラム陽性菌の感染および汚染が、US6579849のペプチドでは治療し得ないことを意味する。
【0007】
LABを含む微生物起源のプロテイナーゼは、それらの特異性および活性により、カゼインからのペプチドの作製および抗菌ペプチドの遊離の可能性において主要な役割を演じる(29)。したがって、発酵の間、これらのプロテイナーゼを製造する微生物は、乳中で成長するために、カゼインおよびホエーのような乳タンパク質を分解できなければならない。LABによるカゼイン分解および分解産物のその後の利用は、複雑なタンパク質分解系を必要とする(9)。Lactococcus lactis(22、38)およびLactobacillus helveticus(27、50)のようなLABの高いタンパク質分解性、ならびに乳中での成長のために乳タンパク質を分解するそれらの必要性のために、生物活性ペプチドの作製のためのそれらのスターター培養物としての使用が報告されている(12、25、32、45)。非特許文献1(30)は、Lactobacillus helveticus PR4からのプロテイナーゼを用いて、6種の哺乳類からの乳のカゼインナトリウムから、抗菌活性を示す生物活性ペプチドを製造した。ヒトβ−カゼインf(184−210)に相当する抗菌ペプチドは、ヒトカゼインナトリウムの、L.helveticus PR4の部分精製プロテイナーゼを用いる加水分解により製造され、いくつかのグラム陽性細菌、およびE.coliに対しても抗菌活性を示した(30)。カゼイン分解の間に製造されるペプチドの特徴付けは、L.helveticus(11、49)について、およびL.casei(7)についてより少ない程度で記載されている。また、L.delbrueckii subsp.lactis ACA−DC 178の細胞壁結合プロテイナーゼが特徴付けされ、β−カゼインに対するその特異性が文書化されている(45)。
【0008】
カゼイン由来抗菌ペプチドはほとんど報告されておらず、記載されているものは、成熟C末端ペプチドに対する酵素の作用により得られ、このことは活性ペプチド配列の遊離を導く(43)。カゼイン起源の最初の抗菌ペプチドは、Hillら(17)により同定され、彼らは、カゼインをキモシンでpH6.4にてタンパク質分解した後に、カセシジンとして知られる抗菌糖ペプチドを単離した。1次アミノ酸構造R1PKHPIKHQGLPQEVLNENLLRF23を有するαs1−カゼインペプチド残基1−23である正に荷電した抗菌ペプチド、イスラシジン(17)は、グラム陽性およびグラム陰性細菌の両方に対して広い範囲の活性を有し(23)、ヒツジおよびウシの乳房炎感染を予防する(12)ことが示された。
【0009】
病原菌が時間の経過とともに、現存する抗菌化合物に対する耐性を発達させるという永続的な問題もある。よって、耐性菌により示される攻撃に継続的に対応するために、新規な抗菌ペプチドの開発に対する持続的な必要性が存在する。新規な抗菌化合物は、濃度の増加、投与回数の増加のようなより煩わしい投与法を必要とし得るか、あるいは狭い範囲の標的種に対してしか有効になり得ない場合がある。これらの状況であっても、新規な抗菌物質は、耐性の増加の問題のために、現存する化合物よりもまだ有利である。さらに、細菌の株を提供してこのような抗菌ペプチドを製造できることは、このことが、まず、該ペプチドの経済的な製造方法を提案でき、次に、該ペプチドをin situの状況で製造する手段を提供し得るので、非常に利益がある。
【非特許文献1】Minervini,F.ら、Applied and Environmental Microbiology:5297−5305(2003)
【発明の開示】
【発明が解決しようとする課題】
【0010】
(発明の目的)
本発明の目的の1つは、少なくとも1種の抗菌ペプチドを製造する細菌の新規な株を提供することである。少なくとも1種の抗菌ペプチドを提供することは、本発明のさらなる目的である。さらなる目的は、多剤耐性生物に対して用いるための抗菌ペプチドを提供することである。抗菌剤を含有する安全性が向上された食品を提供することも、本発明の目的である。ACE阻害の方法を提供することは、本発明のさらなる目的である。抗菌特性が向上された調合乳を提供することも、本発明の目的である。
【課題を解決するための手段】
【0011】
(発明の要旨)
ある実施形態において、本発明は、配列番号1(IKHQHPQE)、配列番号2(SDIPNPIGSENSEK)および配列番号3(VLNENLLR)からなる群より選択される1つまたは複数のペプチドを含む抗菌化合物を提供する。別の実施形態において、本発明は、その試料が2005年11月18日にアクセッション番号NCIMB 41354の下でスコットランドアバディーンのナショナル・コレクション・オブ・インダストリアル・アンド・マリン・バクテリア(National Collection of Industrial and Marine Bacteria)に寄託されているLactobacillus acidophilus DPC6026株、または乳もしくは乳製品から抗菌活性を有するペプチドを製造できるその派生株もしくは変異株の生物学的に純粋な培養物を提供する。
【0012】
本発明は、DPC6026により製造されるタンパク質も提供する。特に、本発明は、乳から抗菌ペプチドを製造できる1つまたは複数のペプチドを製造できる、DPC6026により製造される新規なタンパク質分解酵素タンパク質を提供する。これらのペプチドは、乳または乳製品からの配列番号1、配列番号2および配列番号3からなる群のうちの1つまたは複数に実質的に等しい配列を有するものであってよい。
【0013】
本発明は、乳または乳製品に、Lactobacillus acidophilus DPC6026株、または乳もしくは乳製品から抗菌活性を有するペプチドを製造できるその派生株もしくは変異株の生物学的に純粋な培養物、DPC6026培養物から得られる無細胞培養上清またはその画分、およびDPC6026により製造されるタンパク質であって、乳または乳製品から1つまたは抗菌ペプチドを製造可能であるタンパク質からなる群のうちの1つまたは複数を加えることを含む、乳または乳製品から抗菌ペプチドを製造する方法も提供する。
【0014】
本発明は、乳または乳製品から配列番号1、配列番号2または配列番号3のうちの少なくとも1つを製造可能であるタンパク質分解特性、触媒特性および/または酵素特性を有するDCP6026からの1つまたは複数種のタンパク質を製造する方法も提供する。
【0015】
請求項1に記載の抗菌化合物、請求項2に記載の培養物、請求項3に記載の上清および請求項4に記載のタンパク質からなる群より選択される1つまたは複数を含む抗菌組成物。
【0016】
本発明は、DPC6026株から得られる無細胞培養上清もしくはその画分、あるいは上記の1もしくは複数のペプチド、化合物、上清または株を適切な担体または希釈剤とともに含む抗菌組成物も提供する。
【0017】
本発明は、上記の化合物、ペプチド、株、タンパク質、上清または組成物の、微生物感染または微生物汚染の管理における使用も提供する。本発明の態様は、本発明の1つまたは複数の化合物、株、上清もしくは組成物を動物またはヒトに投与することを含む、微生物感染もしくは微生物汚染を予防または治療または制限する方法を提供する。微生物感染または微生物汚染は、Escherichia coli、E.sakazakii、Streptococcus mutans、Listeria innocua、Klebsiella sppおよびStaphylococcus carnosusからなる群の1もしくは複数種を原因とする汚染または感染であってよい。特に、感染または汚染は乳腺炎であってよい。
【0018】
別の実施形態において、感染または汚染は髄膜炎であってよく、新生児の乳腺炎であってよい。ある実施形態において、化合物、ペプチド、株、上清または組成物は、乳ベースの調合乳に供給してよい。つまり、本発明のペプチドは、幼児用調合乳製品の製造における組込まれた防御系として用い得る。1つまたは複数のペプチドは、特に、Enterobacter Sakazakii汚染に対して有効であり得る。
【0019】
構造、活性の範囲および効力の点での生物活性ペプチドの多様な性質のために(39)、これらのペプチドは、可能性のある新しい薬物設計の主な標的である。乳タンパク質由来の配列番号1(IKHQGLPQE)および配列番号3(VLNENLLR)のような抗菌ペプチドを、E.sakazakii 5920(ATCC12868)のような病原株に対する防御の組込まれた機構として用いる可能性は、幼児用調合乳製造において用いられるもののような粉乳の安全性を増進させる有用なアプローチを提供し得る。
【0020】
本発明は、候補の動物またはヒトを、本発明の1つまたは複数の組成物、化合物、上清、ペプチドまたは株で処置することを含む、アンギオテンシン−I変換酵素(ACE)の阻害、オピオイド調節、免疫調節および血栓症抑制の方法も提供する。
【0021】
本発明のペプチド、株、化合物、組成物および上清は、ウシおよびヒツジの乳房炎の治療に用いることもできる。配列番号1および配列番号3は、抗菌ペプチドであるイスラシジンの切断産物に類似する。配列番号2も用いてよい。イスラシジンは、マウスあたり10μg程度の低い用量で投与したときに、S.aureus、Streptococcus pyogenesおよびL.monocytogenesに対して強い防御効果を有すると記載され、ヒツジおよびウシの乳房炎の治療において以前に用いられたこともある。
【0022】
αs1−カゼインの最初の9残基はチーズの作製において蓄積し、いくつかのチーズに存在する苦味の原因であるので(Foxら、1995年、Chemistry of Structure−Function Relationships in Cheese(Malin, E. L.およびTunick, M. H.編)、Plenum Press、New York、59〜98頁)、本発明のペプチド、株、化合物、組成物および上清は、苦味チーズの製造における苦味に寄与するために用い得る(Pedersonら、1999年、J. Bacteriology 181号、4592〜4597頁)。
【0023】
本発明のペプチド、株、化合物、組成物および上清は、上記のような配列番号1、配列番号3および配列番号2の広い範囲の活性のために、食品または腐敗しやすい商品の、病原性グラム陰性およびグラム陽性細菌に対する防腐剤として用い得る。
【0024】
L.acidophilus DPC6026は、ブタの小腸から単離され、コークのDairy Products Research Centre(DPRC),Teagasc Moorepark,Fermoy,Co.に貯蔵されている新規な株である。イスラシジンまたはその断片の製造を示す細菌株は、以前に記載されていなかった。イスラシジンは、キモシンで処理したαs1−カゼインに由来した(HillらJ of Dairy research 1974年;41号:147頁)。このことは、L.acidophilus DPC6026は、その天然の状態で、酵素を必要とせずに3つのペプチドIKHQGLPQE、VLNENLLRおよびSDIPIGSENSEKを製造し、工業的なレベルで発酵プロセスの余分な経費を回避するので、経済的に有利である。イスラシジンのペプチド配列は、そのより長い鎖の長さのために、化学合成するには配列番号1、配列番号2および配列番号3よりも高価である。配列番号1、配列番号2、配列番号3はそれぞれ、より短い鎖長を有し、よって、化学合成がより安価である。
【0025】
イスラシジンαs1−カゼインf(1−23)は、溶液中で比較的安定な立体配置を維持するようである(Malinら、Journal of Protein Chemistry、20号、391〜404頁)。立体配置が安定なペプチドは、アレルゲンに特徴的であることが知られており、αs1−カゼインのこのセグメントは、アレルゲン性であることが知られている(Spuerginら、1996年 Allergy 51号、306〜312頁)。残基20の周囲のαs1−カゼインの領域は、ヒトIgEにより認識される重要なアレルゲン性エピトープを含むことが示唆されている(Spuerginら、1996年)。さらに、配列番号3は、ヒトIgEにより認識される重要なアレルゲン性エピトープを含むと考えられているαs1−カゼインの残基20を欠失しているので、アレルゲン性エピトープを提示し得ない(Spuerginら、1996年)。
【0026】
in vivoでの透過
イスラシジンは、in vivoで、Staphylococcus carnosusに対する防御効果を示すことが示されている。配列番号1および配列番号3は、病原性乳腺炎原因株に対して、in vivoでより大きい防御効果を示す。この理由は、より短い生物活性ペプチドが、よりよいin vivo効果を示すことであると仮定されている(Vermeirssenら、British Journal of Nutrition (2004年)、92号、357〜366頁)。
【0027】
αs1−カゼインf(1−9)(RPKHPIKHQ)は、自然発生高血圧ラット(SHR)において抗高血圧効果を有することが示されている(Saitoら、2000年、J. Dairy Science、83号、1434〜1440頁)。配列番号1は、最後の4アミノ酸(IKHQ)と100%の相同性を共有する。これは、ペプチドのC末端にプロリン、グルタミンおよびグルタミン酸も有する。ACE活性は、C末端トリペプチド領域でのプロリンの存在により、非常に遅くなる(Vermeirssenら、2004年、British Journal of Nutrition、92号、357〜366頁)。この事象は、配列番号1がACE阻害活性を有し得ることを示唆し、イスラシジンを2つのより短いペプチドに切断することの可能な利点を示唆する。
【0028】
ある状況下では、イスラシジン耐性の機構は、配列番号1によっても配列番号3によっても提示されない分子の「抗原性」領域により媒介されると仮定されてもいる。
【0029】
ラクトフェリシンBのような他のカチオン性抗菌ペプチドは、プロテアーゼを産生するので、Escherichia coliおよびStaphylococcus aureusのような耐性株を有することが知られており、このことは、E.coliおよびStaphylococcus aureusによるマゲイニンのような他のカチオン性抗菌ペプチドに対する主要な耐性機構として示唆されている(Ulvatneら、Journal of Antimicrobial Chemotherapy(2003年)50号、461〜467頁)。イスラシジンを本発明のペプチドの2つ、配列番号1および配列番号3に切断することは、イスラシジンに比較してより短いこれらの配列を切断できるプロテアーゼがより少ないので、耐性株の発生の可能性を低減させるようである(コンピュータプログラムhttp://ca.expasy.org/cgi−bin/peptidecutter/peptidecutter.plを用いて証明されるように)。
【発明を実施するための最良の形態】
【0030】
材料および方法
基質および化学薬品
ペプチドインドリシジンおよびセクロピンP1、ならびに酵素プロテイナーゼKはSigma(Sigma Aldrich Chemie、ドイツシュタインハイム)から購入した。ピマリシンは、Merck(Merck、ドイツダルムシュタット)から購入した。ペプチドイスラシジンは、Peptide Protein Research Ltd.,(英国フェアラム)により合成された。Maximum Recovery Diluent(MRD)は、Oxoid(Oxoid Ltd.,イングランドベイジングストーク)から購入した。ウシカゼインナトリウムは、Dairygold(アイルランドコークミッチェルズタウン)からであった。パルスフィールド保証アガロースは、Bio−Rad Laboratories(米国CA94547ハーキュリーズ)からであった。酵素Apa Iおよび低分子量PFGマーカー(NO350S)は、New England Biolabs(英国ハートフォードシャーSG4 OTY)から購入した。低融点アガロースは、Bio−Rad Laboratories(カリフォルニアリッチモンド)から購入した。
【0031】
微生物および培養条件
L.acidophilus DPC6026、L.johnsonii DPC6092およびL.salivarius DPC6027は、ブタの小腸から単離し(データは未公表)、アイルランドコークのTeagasc Dairy Products Research Centreの培養コレクションに貯蔵した。これらの株を、MRS培地(Oxoid Ltd、英国ベイジングストーク)中で嫌気的に24時間、37℃にて増殖させた。標準培養物は、10mlのMRS培地に、10μlの冷凍ストック(−80℃)を接種することにより調製し、次いで、37℃にて16〜24時間インキュベートした。NCIMB(ナショナル・コレクション・オブ・インダストリアル・アンド・マリン・バクテリア、スコットランドアバディーンAB24 3RY)から購入したL.innocua DPC3306、E.sakazakii 5920(ATCC 12868)、E.sakazakii 8272(NCTC8155)、ならびにE.coli DPC6053(Teagasc Dairy Products Research Centre、アイルランドコーク)を、試験株として用いた。
スキムミルク寒天(SMA)プレート上でのタンパク質分解活性の初期スクリーニング
ブタの腸起源の205種の細菌単離物ならびにヒト成人および幼児の糞便起源の55種の単離物(データは未公表)を、以前に記載されたようにして(36)SMAプレートを用いてタンパク質分解活性についてスクリーニングした。点在させた培養物の周囲の透明領域が、タンパク質分解を示した。直径4.0mm以上の領域を発生させる単離物を、さらなる研究のために選択した。
【0032】
高性能液体クロマトグラフィー(HPLC)
SMAプレートを用いてタンパク質分解活性を示す単離物を、カゼインナトリウム溶液(2.5%w/v)に接種し(1%w/v)、嫌気的に37℃にて一晩インキュベートした。次いで、Delta−Pak C18カラム(サイズ;600mm×7.5cm、Varian Chromatography Systems、米国カリフォルニアウォルナットクリーク)を用いる高性能液体クロマトグラフィーを、得られた発酵生成物に対して行った。移動相は、トリフルオロ酢酸(0.1%v/v)含有のアセトニトリルとHPLC等級の水(30%v/v)との2成分混合物であった。流速は、1ml/分であった。ブタ腸単離物および幼児糞便単離物によるカゼインの分解を、HP1100ダイオードアレイ検出器を用いて214nmでのUV吸収の測定により監視した。
【0033】
パルスフィールドゲル電気泳動
MRS培地からの15種の単離物のそれぞれからの高分子量DNAの調製は、プラグスライスの制限消化のために20Uの制限酵素ApaIを用いた以外は、Simpsonら(42)により以前に記載されたとおりであった。
株の種同定のための単離物の16S rDNA配列決定
ブタの腸、ならびに幼児ヒトおよび成人の糞便の15種の単離物は、著しいタンパク質分解活性を示すことが示され、個別の株の種同定のために16sリボソームDNA(rDNA)配列決定に付した(Lark Technologies Inc.,英国エセックス)。2つの16S rDNAプライマー、5’末端についてCO1(5’−AGTTTGATCCTGGCTCAG−3’)および3’末端についてCO2(5’−TACCTTGTTACGACT−3’)を用いて、約1.5kbの16S rDNA生成物を、以前に記載された(42)ポリメラーゼ連鎖反応(PCR)条件下で作製した。このPCR生成物を、CO1プライマー(Lark)を用いて部分的に配列決定した。BLASTプログラムを用いることにより得られた16S rDNA配列の比較により、株を特定の種に割り当てることが可能であった。通常、16S rDNA配列の類似性の値がBLASTプログラムを用いて97%を超えると、それらの株は同じ種に属すると考えられる(44)。
【0034】
GRAS株を用いる大規模発酵
タンパク質分解活性を示す15種の単離物のうち、3種の遺伝的に別個の株であるLactobacillus acidophilus DPC6026、L.salivarius DPC6027およびL.johnsonii DPC6092を、それらのタンパク質分解能力に基づいて、およびそれらが全て一般的に安全であるとみなされているので(GRAS)、さらなる発酵に個別に用いた。3つの別々の発酵を、3回ずつ行った。カゼインナトリウム基質(2.5%w/v)に、各株(1%w/v)を接種し、100rpmで混合しながら、0.1M NaOHの添加により維持する一定のpH7で、37℃にて24時間インキュベートした。発酵生成物を、次いで、80℃に加熱して培養物を不活性化し、その後、サイズ排除S1Y10 10kDaらせんカートリッジフィルタ(Millipore Ltd.,英国ハートフォードシャー)を通してろ過し、10kDa未満のペプチドを分離した。これらのペプチドを含む画分を凍結乾燥し、さらなる使用まで−20℃にて貯蔵した。
【0035】
発酵生成物の逆相HPLC分析
サイズが10kDa未満のペプチドを、狭い口径のカラム(Nucleosil C18、5mm×250mm:Varian Chromatography Systems、米国カリフォルニアウォルナットクリーク)、および214nmで作動するUV検出器を含むRP−HPLC逆相高性能液体クロマトグラフィーシステムを用いてカゼインナトリウム加水分解物から分離した。凍結乾燥粉末の一定量を、HPLC等級の蒸留水で希釈し、0.45μmのフィルタ(Millipore)を通してろ過し、30mg/mlの発酵生成物をカラムに添加した。移動相は、トリフルオロ酢酸(0.1%v/v)含有のアセトニトリルおよびHPLC等級の水(100%v/v)の2成分混合物であった。移動相中のアセトニトリル含量を、1ml/分の流速で、0から100%まで72分間で直線的に増加させた。ペプチドを、214nmの波長で作動する検出器を用いて検出した。回収した画分から、セントリバップコンソール(centrivap console)(Labconco Corporation、米国カンザスシティ)を用いる蒸発により溶媒を除去した。画分を、その後の抗菌活性のアッセイの前に、1mlの蒸留水に再溶解した。
【0036】
タンパク質濃度の決定
画分のタンパク質濃度は、Bioradタンパク質アッセイ法(24)を用いて決定した。595nmの吸光度を決定し、タンパク質濃度をmg/mlとして報告した。
【0037】
抗菌活性
ペプチドおよび合成した画分の抗菌活性の予備的特徴付けは、96ウェルアッセイでの成長阻害の測定(1、10)、および寒天拡散法の使用(16、39)を含んだ。96ウェルプレートアッセイは、各ペプチド画分単独の対照(セクロピンP1およびインドリシジン)とともに、ペプチド画分を含まない対照(増殖対照)を含んだ。細菌を含まない対照(無菌性対照)も含めた。プレートを37℃にて6時間インキュベートし、培養成長を1時間ごとに監視した。最小阻止濃度(MIC)を、マイクロタイタープレートリーダーでのOD600を記録することにより測定される、認識可能な成長がない最低濃度とした。ウェル拡散アッセイ(16、40)を用いて、精製および化学合成したペプチドの抗菌活性を検出した。これらのアッセイは、それぞれE.coli DPC6053、L.innocua DPC3306、E.sakazakii5920(ATCC12868)またはE.sakazakii8272(NCTC8155)の指示株の一晩培養物1mlを播種した脳心臓浸出物(BHI)、ルリアベルタニ(LB)または栄養寒天(NA)のいずれかで行った。直径4.6mmのウェルを、これらの寒天プレート中に切り取って入れ、各ウェル中に30μl(0.162mg/ml)のペプチド画分を入れた。プレートを4℃にて4時間貯蔵して、ペプチドの放射状の拡散を許容し、30℃にて嫌気的にまたは37℃にて好気的に24時間インキュベートし、阻害領域について調べた。ペプチドに対する株の感度を、ウェルの周囲の阻害領域の直径に従って評点した。実験は、3回行い、平均の領域サイズを計算した。E.coli DPC6053、E.sakazakii5920(ATCC12868)およびE.sakazakii8272(NCTC8155)についてイスラシジン、セクロピンP1、およびL.innocua DPC3306に対してインドリシジンを対照として用いた。
【0038】
抗菌ペプチドの精製、配列決定および合成
抗菌活性を示すタンパク質画分を、上記のRP−HPLC条件により再分画し、指示株E.coli DPC6053の成長を、市販のペプチドセクロピンP1と同様のタンパク質濃度で阻害した画分を回収して、337nmで作動するレーザおよび20kVの加速電圧で飛行時間型マトリクス支援レーザ脱離イオン化法(MALDI−TOF)質量分析計(PE Biosystems Voyager−DE STR Biospectrometry Workstation、Aberdeen Proteome Facility)を用いる質量分析(MS)により、ペプチド組成を分析した。各画分中のペプチドのアミノ酸配列を、誘導体化およびエドマン分解の後に決定した。これらのステップは、494Aプロテインシーケンサー(Applied Biosystems)で行った。抗菌活性を示す各画分中の同定された全てのペプチド(MS分析およびアミノ酸配列決定により)を、その後、Peptide Protein Research Ltd.,(英国フェアラム)により化学合成した。合成ペプチドの純度は、製造者により証明されるようにHPLC分析により決定して、95%を超えていた。
化学合成ペプチドおよび粗画分のプロテイナーゼKでの処理
化学合成ペプチドおよびE.coli DPC6053に対して抗菌活性を示す画分を、プロテイナーゼK(Sigma)に対する感受性について、プロテイナーゼK(2050U/ml)とペプチドまたは画分(0.554mg/ml)との1:1の容量比での4℃で6時間のインキュベーションにより、寒天ウェル拡散アッセイ(40)を用いて試験した。プレートを、37℃にて24時間インキュベートした。用いた陽性対照は、ペプチドイスラシジン(Peptide Protein Research Ltd.,)およびセクロピンP1(Sigma TM)であった。
【実施例】
【0039】
本研究の目的の1つは、哺乳動物の腸起源のLABのタンパク質分解能力を用いて放出されたウシカゼインから新規な生物活性ペプチドを発見することであった。ブタ小腸から得られた205種の単離物ならびにヒトの成人および幼児の糞便起源の55種の単離物を、本研究において用いた。
【0040】
タンパク質分解活性を有する単離物のスクリーニング
LAB寒天プレートでの260種の単離物全てを、基質としてSMAおよびカゼインナトリウムを用いてタンパク質分解活性についてスクリーニングした。ブタの腸起源の5種の単離物と、ヒトの幼児および成人の糞便起源の10種の単離物を、それらのタンパク質分解能力およびカゼインナトリウムから10kDa以下のペプチドを生じる能力に基づいて、選択した。15種全ての単離物は、タンパク質分解活性を示す直径サイズ4mm以上の領域サイズを生じ、15種全ての単離物の小規模カゼイン発酵生成物のHPLC分析により、最も複雑なペプチドプロファイルが示された。PFGEおよび16S rDNA配列決定の後に、選択された15種の単離物のうち、9種が乳酸桿菌の別個の種であると同定された(表1)。全てヒトの成人の糞便起源の5種の単離物は、Enterococcus faecalisと99〜100%の相同性を示し(BLASTプログラムを用いて)、よって、腸球菌は通常、安全とはみなされないので(GRAS)(21)、大規模発酵でのさらなる使用には適さないと考えられた。単離物3L7および33L1(ヒト幼児糞便単離物)は、それぞれE.hiraeおよびStaphylococcus epidermisと100%の相同性を示し(BLASTプログラム)、これらも、非GRAS状態のために、さらなる研究から排除した。9種の乳酸桿菌単離物のうちの3種は、PFGE(図1)および16S−rDNA配列決定により、L.acidophilus DPC6026、L.johnsonii DPC6092およびL.salivarius DPC6027と同定された。
【0041】
L.acidophilus DPC6026、L.salivarius DPC6027およびL.johnsonii DPC6092のカゼインナトリウム(2.5% w/v)での24時間後のインキュベーション、ならびにその後の10kDaの膜を通してのろ過の後に得られたペプチドプロファイルを、図2に示す。これらの3種の乳酸桿菌でのカゼインナトリウムの発酵は、カゼインナトリウム(図2A)のより低分子量のペプチド(図2B〜2D)への実質的な分解をもたらし、L.acidophilus DPC6026は、カゼインナトリウムのより小分子量のペプチドへの最大程度の分解を示した(表1)。L.acidophilus DPC6026は、得られたペプチドの60.74%が≦0.5kDa未満であり、17.49%が1〜0.5kDaの間のペプチドであり、11.57%が1〜2kDaの間のペプチドである発酵生成物を生じた。この株は、L.johnsonii DPC6092、L.salivarius DPC6027およびL.animalis DPC6028よりも0.5kDa〜2.0kDaの間のペプチドをより多く製造した(表1)。L.reuteri DPC6100、L.gasserri DPC6093、L.rhamnosus DPC6095およびL.brevi DPC6102により製造されたHPLCペプチドプロファイルは、選択した株と同程度のカゼインナトリウムの分解を示さず(表1)(HPLCプロファイルは示さず)、よって、さらなる研究のために選択しなかった。L.delbrueckii sp.bulgaricus DPC6104は、生物活性ペプチドの作製のためのその使用が以前に記載されているので、選択しなかった(11)。
【0042】
抗菌ペプチドの検出
Lactobacillus acidophilus DPC6026、L.salivarius DPC6027およびL.johnsonii DPC6092により製造されたカゼインナトリウム発酵生成物は、抗菌画分であった。しかし、L.acidophilus DPC6026から作製された画分のみを抗菌活性についてアッセイした。なぜなら、この株は、示したクロマトグラフィープロファイル(図2)から示されるように、3つの株のうちで最高のタンパク質分解活性により特徴付けられたからである。E.coli DPC6053、Listeria innocua DPC3306、E.sakazakii5920(ATCC12868)およびE.sakazakii8272(NCTC8155)の株に対する96ウェルプレートアッセイまたはウェル拡散アッセイのいずれかを用いて、粗発酵生成物の抗菌活性は検出されなかった。L.acidophilus DPC6026により製造されたカゼインナトリウム発酵生成物を、サイズ排除S1Y10 10kDaらせんカートリッジフィルタを通してろ過して、10kDa以下のペプチドを含む透過物を得た。L.acidophilus DPC6026により製造されたろ過カゼインナトリウム発酵生成物の72個の画分を、RP−HPLCにより回収して、E.coli DPC6053、E.sakazakii5920(ATCC12868)、E.sakazakii8272(NCTC8155)およびL.innocua DPC3306に対する抗菌活性について、ウェル拡散アッセイによりアッセイした。E.coli DPC6053およびL.innocua DPC3306に対するこれらの画分の抗菌活性は、96ウェルプレートアッセイ法を用いてOD600を測定することによってもアッセイした。3つの画分、A1−45、A1−49およびA1−54は、最も効力が高い抗菌活性を有し、それぞれ0.554mg/ml、0.5mg/mlおよび1.24mg/mlのペプチド濃度を有していた。これらの全ての画分は、試験株E.coli DPC6053に対して、それぞれこれらの濃度で阻害活性を示した。
【0043】
ペプチドの精製、配列決定および特徴付け
画分A1−45、A1−49およびA1−54中のペプチド混合物を、質量分析およびエドマン分解に付した。アミノ酸組成は、配列決定(Aberdeen Proteome Facility)によっても決定した。画分A1−45は、ペプチド配列IKHQGLPQE(配列番号1)を含むことが見出された(表2)。このペプチド配列は、αs1−カゼインf(21−29)に相当する。画分A1−49は、αs1−カゼインf(183−207)に相当するペプチド配列SDIPNPIGSENSEK(配列番号2)を含んでいた。A1−54は、αs1−カゼインf(30−37)に相当するペプチド配列VLNENLLR(配列番号3)を含むことが見出された。各ペプチドの推定および計算された質量を報告する(表2)。寒天ウェル拡散アッセイにより決定されたように、画分45に存在する配列番号1は、指示生物であるE.coli DPC6053を、0.05mMの濃度で阻害した(図3B)。このペプチドは、E.coli O157:H7派生株(E.coli DPC6054およびE.coli DPC6055)、E.sakazakii5920(ATCC12868)のような臨床対象の潜在的な病原細菌に対しても、同じ濃度(0.05mM)で阻害を示した(図3B)。画分54に存在する配列番号3は、指示生物であるE.coli DPC6053を0.22mMの濃度で(図3C)、また、E.sakazakii DPC6091のような潜在的病原株を同じ濃度で阻害したが(図3C)、配列番号2を含む画分A1−49は、Listeria innocua DPC3306に対してわずかな阻害活性しか示さず、E.coli DPC6053に対しては全く活性を示さなかった(表3)。
【0044】
ペプチドの化学合成
強い抗菌活性を示し、かつ広い範囲の活性を有していた画分から同定されたペプチドを、化学合成して、MALDI−TOF分析により同定された配列が、画分中の抗菌配列であるかを確認した。これらは、αs1−CN f(21−29)(配列番号1)、αs1−CN f(30−37)(配列番号3)およびαs1−CN f(183−207)(配列番号2)であった。化学合成ペプチド配列番号1および配列番号3は、未精製のペプチドを含む画分と同じ微生物を阻害した。合成ペプチド配列番号2(画分A1−49に存在)は、Listeria innocua DPC3306に対していくらかの阻害活性を示したが、E.coli DPC6053に対しては活性を示さなかった(表3)。
【0045】
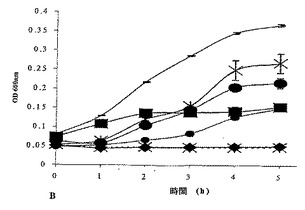
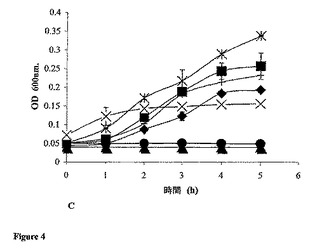
これらのペプチドの最小阻止濃度(MIC)を、イスラシジンを陽性対照として用いて決定した。イスラシジンは、E.coli DPC6053を、0.05mM〜1.9mMの範囲の濃度で阻害した(図4A)。イスラシジンについてのMICは、記載される実験条件下では0.059mMであると見出されたが、配列番号1についてのMICは、E.coli DPC6053の成長を阻害すると示された(図4B)0.078mMであり、配列番号3は、この微生物を0.22mM〜1.2mMの範囲の濃度で阻害した(図4C)。配列番号3について得られたMICは、0.22mMであった。つまり、本発明は、驚くべきことに、全てが抗菌特性を有する3種の新規ペプチドを製造した。イスラシジンは、最良の標準物である陽性対照とみなした。配列番号1は、イスラシジンに好ましく匹敵するが、配列番号2および配列番号3は、同じ濃度では、効果が1桁小さい。しかし、より大きい濃度が必要であるが、配列番号2および配列番号3はともに、抗菌特性を明らかに示していた。
【0046】
配列番号2および配列番号3に必要な濃度は、抗菌化合物として通常の濃度範囲内である。これらは、例えば、カソシジン−1、および望ましい組成物の濃度が0.1mg〜1mgであると示すUS6579849「Antibiotic peptides from bovine milk」に記載されるペプチドによく匹敵する。Florisら、Current Pharmaceutical Design、2003年、9号、1257〜1275頁は、イスラシジンが「比較的高濃度(0.1〜1mg/mL)ではあるが、乳酸桿菌および他のグラム陽性細菌のin vitro成長を阻害することが見出された」と述べている。
【0047】
アッセイ条件下では、全ての化学合成ペプチドおよび粗画分は、プロテイナーゼKで処理したときにそれらの抗菌活性を喪失した。コンピュータプログラムExpasy Peptide cutter(http://ca.expasy.org/cgi−bin/peptidecutter/peptidecutter.pl)を用いて、ペプチド配列IKHQGLPQE(配列番号1)、VLNENLLR(配列番号3)、SDIPIGSENSEK(配列番号2)が全て、トリプシンおよびキモトリプシンにより加水分解されることが見出された。
【0048】
考察
この研究において、カゼインナトリウムを、タンパク質分解株L.acidophilus DPC6026、L.salivarius DPC6027およびL.johnsonii DPC6092を用いてタンパク質分解に付した。L.acidophilus DPC6026株を、発酵で用いるために選択し、生じたペプチドを、E.coli JM109 DPC6053およびEnterobacter sakazakii DPC6090(ATCC12868)のような潜在的病原株に対する抗菌活性についてアッセイした。
【0049】
本研究で同定されたペプチド配列IKHQGLPQE(配列番号1)、VLNENLLR(配列番号3)およびSDIPIGSENSEK(配列番号2)は、抗菌活性について以前に報告されていない(8、47)。
【0050】
(Dziubaら、1999年(8))は、種々の供給源からの一連の生物活性ペプチドを含む生物活性データベースを参照している。データベースに含まれるペプチドは、抗菌ペプチド、アンギオテンシン−I変換酵素阻害ペプチド(ACE−I阻害ペプチド)、オピオイドペプチドおよびブラディキニン増強ペプチドである。行った実験で病原菌に対する最良の陽性対照として用いたペプチド、イスラシジン(ペプチドデータID 3035、名称イスラシジン、配列RPKHPIKHQGLPQEVLNENLLRP)は、このデータベースに詳述されている。
【0051】
(Wangら、2004年(47))は、抗菌ペプチドデータベースのみについて詳述するが、イスラシジンについてはわずかしか述べていない。
【0052】
研究の結論
牛乳タンパク質からの既知の抗菌ペプチドは、R1PKHPIKHQGLPQEVLNENLLRF23と決定された一次アミノ酸配列を有するイスラシジン、αs1−CN f1−23である(26)。このペプチドは、広い範囲の活性を有し(23)、配列番号1および配列番号3との高い程度の相同性のために、本研究において陽性対照として用いた。配列番号1は、イスラシジンと9残基が共通し、配列番号3はイスラシジンのC末端と8残基が共通している。ペプチド配列番号1は、0.05mMのMICを示したイスラシジン、ならびにより効力が高い商業的に入手可能なペプチドであるセクロピンP1およびインドリシジン(0.05mMのMIC)に匹敵する0.078mMのMICを示した。配列番号3は、E.coli DPC6053に対して、イスラシジンのMIC(0.05mM)に匹敵する0.22mMのMICを示した。配列番号1のペプチド(IKHQGLPQE)は、+2の正電荷を含み、疎水性(イソロイシン)末端および親水性(グルタメート)ドメインを有し、かつE.sakazakiiおよびE.coliのようなグラム陰性細菌に対して、グラム陽性細菌よりもよい活性を示す。
【0053】
抗菌ペプチドは、通常、5〜60の間のアミノ酸を有し、10kDa未満の分子質量を有し、通常、両親媒性であり、通常、広い範囲の活性を有する(Florisら、Current Pharmaceutical Design 2003年)。配列番号3(VLNENLLR)は、これらの特徴が共通している。
【0054】
ペプチドのカチオン性および疎水性の二重の特徴が、ペプチドと細菌膜との最初の相互作用に重要であることが知られている。正電荷は、細菌の外膜および細胞質膜との相互作用を促進する(Wuら、Journal of Biol. Chem.、1999年)。アルギニン(R)の強い正電荷が、Hancockら、1999年 Peptide Antibiotics, Antimicrobial agents and chemotherapy:1317〜1323頁に記載されるように、配列番号3(VLNENLLR)がグラム陰性細菌のリポ多糖領域と相互作用し、負に荷電した細胞質膜を透過することを可能にする。
【0055】
配列番号3はまた、疎水性領域(VL)および親水性領域(R)も有する。これは、また、L.innocuaのようなグラム陽性株に対するよりも病原性グラム陰性細菌に対して、より強力な活性を示す。また、化学合成ペプチドも、L.innocua DPC3306に対するよりもE.coli DPC6053のようなグラム陰性株に対してより強力な活性を示した。
【0056】
LABのプロテイナーゼは、β−CNおよびαs1−CNのペプチド結合の40%より多くを加水分解して、オリゴペプチドを生じることが示されている(22)。次いで、LABの複合ペプチダーゼは、これらのオリゴペプチドに対して作用する(31)。Minerviniらは、L.helveticus PR4からの部分精製プロテイナーゼを用いるACE阻害ペプチドの作製を報告した(30)。このグループは、グラム陽性およびグラム陰性細菌の両方に対して活性を示したヒトβ−CN f(184−210)からの抗菌ペプチドも同定した。β−CN由来のいくつかのカソキニンは、Lactobacillus helveticus CP790の細胞壁会合セリン型プロテイナーゼにより、遊離されている(49)。L.delbrueckii subsp.lactis ACA−DC178からの細胞壁結合プロテイナーゼは、β−CNから4つのペプチドを遊離させたが(45)、これらのペプチドの生物活性は報告されていなかった。
【0057】
L.acidophilus DPC6026は、カゼインに対するそのタンパク質分解能力、よって抗菌活性のような生物活性を有する多数のペプチドを作製するその能力を示した最初のアッセイに基づいて選択された。セクロピンP1は、ブタ小腸から単離されたペプチドであり、その活性の範囲が主にグラム陰性細菌に対するものであるので、対照として用いた(5)。
【0058】
配列番号1、配列番号2および配列番号3の化学合成ペプチド、ならびにこれらのペプチドを含む画分は、プロテイナーゼKで処理したときにその抗菌活性を失うことが見出された。トリプシンおよびキモトリプシンは、抗菌性粗画分で同定された化学合成ペプチドの全てを不活性化させた。Expasy Peptide cutterプログラム(http://ca.expasy.org/cgi−bin/peptidecutter/peptidecutter.pl)を用いて、ウシαs1−CNは、3つのエンドペプチダーゼおよびプロテイナーゼの組合せにより加水分解されて、抗菌ペプチド配列番号1および配列番号3を放出し得ることが見出された。プロリンエンドペプチダーゼは、ウシαs1−CNを、Pro20のC末端側で切断し、エンドペプチダーゼは、Glu29のC末端側で制限する。この酵素の組合せが、ウシαs1−CNから抗菌ペプチドIKHQGLPQE(配列番号1)を放出させるようである。プロリン特異的ペプチダーゼの存在が、カゼインからのプロリンに富むオリゴペプチドを分解するそれらの能力のために、LABの最適な成長に必要であり、プロリン特異的ペプチダーゼは、L.delbrueckii subsp bulgaricusから以前に単離されている(2)。ウシαs1−CNをArg37位で制限するArg−Cプロテイナーゼと、ウシαs1−CNをGlu29位で制限するエンドペプチダーゼとの組合せが、抗菌ペプチドVLNENLLR(配列番号3)を放出させるようである。
【0059】
理論に結び付けられることを望まないが、本明細書に記載される条件(0.1M NaOHで維持されたpH7、100rpm、37〜42℃)下で発酵に用いられたL.acidophilus DPC6026が、αs1−CNを、ペプチド配列番号1(IKHQGLPQE)および配列番号3(VLNENLLR)および配列番号2(SDIPIGSENSEK)に切断可能なArg−Cプロテイナーゼのような酵素(プロテイナーゼおよびペプチダーゼ)の新規な酵素の集団を生み出すと考えられている。この仮定は、コンピュータプログラムhttp://ca.expasy.org/cgi−bin/peptidecutter/peptidecutter.pl、およびαs1−CNを切断して配列番号1、配列番号2および配列番号3を製造できるという条件での酵素リストからの酵素を用いるαs1−Cnの切断分析に基づいている。
【0060】
ほとんどの抗菌ペプチドは、6〜50の間のアミノ酸残基を有する(48)。現在までに知られている最小の抗菌ペプチドは、ウシのラクトフェリシンから単離された配列R1RWQWRを有するヘキサペプチドと命名されている(46)。ほとんどが+2以上の正電荷を有し、疎水性および親水性の両方のドメインを有する両親媒性の構造に折り畳まれる(47)。これらの特徴により、グラム陰性細菌のリポ多糖領域と相互作用し、負に荷電した細胞質膜を透過することが可能になる(15)。
【0061】
配列データのみから、ペプチドの活性またはそれが形成し得る二次構造のいずれも予測することは困難であり得る(4)。ジスルフィド架橋を欠くペプチド(ペプチドIKHQGLPQE(配列番号1)およびVLNENLLR(配列番号3)のように)は、水中で無作為な構造を有し、膜に結合するかまたは自己凝集するときのみ、これらは構造を形成する(10)。しかし、これらのペプチドのいくつかの特徴は、作用機序が、カチオン性および中性の抗菌ペプチドについて以前に記載されている機構である(15、48)細胞質膜を介する自己促進型取り込みとそれに続く細胞質膜バリアとの干渉とを含み得ることを示唆する。
【0062】
特徴は、以下のことを含む(1)細菌の細胞質膜との最初の相互作用に重要なペプチドのカチオン性に寄与するアルギニンのような正に荷電したペプチド。(2)疎水性−これは、ペプチド配列番号1においてはアミノ酸イソロイシン、およびペプチド配列番号3においてはアミノ酸バリン(V)およびロイシン(L)によりもたらされる。本発明は、これらのペプチドの疎水性を増加させて、脂質アシル鎖と疎水性コアとの間の疎水性相互作用の増大によるこれらのペプチドの膜への結合を増加させることも提供する。
【0063】
結論
本研究は、L.acidophilus DPC6026株がカゼインからの抗菌ペプチドの作製に適し、該抗菌ペプチドのMIC濃度が商業的に合成されたペプチド(セクロピンP1)および既知の抗菌ペプチドであるイスラシジンに匹敵するので(22)、このLactobacillus株を用いて製造されたカゼインナトリウム発酵生成物を、抗菌性の利点を有する機能性食品の成分としてみなし得ることを示す。
【0064】
本研究は、ウシαs1−カゼインを、本研究で単離された広い範囲の抗菌ペプチドの前駆体としてみなすことができ、最終的には、これらのペプチドが、E.sakazakiiのような病原性細菌に対する組込まれた防御系として幼児調合乳製品に用いられる可能性を有し得ることも示す。
【0065】
Enterobacter sakazakii抑制効果を有するカゼインナトリウム粉末の製造方法の例
(1)カゼインナトリウム基質(5%w/v)に、(1%w/v)のL.acidophilus DPC6026を接種して、100rpmで混合しながら、0.1M NaOHの添加により維持する一定のpH7で、37℃にて24時間インキュベートしてよい。この発酵生成物を、次いで、80℃に加熱して培養物を不活性化し、これを次いで凍結乾燥して微細粉末を製造できる。
(2)カゼインナトリウム基質(5%w/v)に、(1%w/v)のL.acidophilus DPC6026を接種して、100rpmで混合しながら、0.1M NaOHの添加により維持する一定のpH7で、37℃にて24時間インキュベートしてよい。
(3)この発酵生成物を、次いで、80℃に加熱して培養物を不活性化し、その後、サイズ排除カラム10kDaを通してろ過して、10kDa未満のペプチドを含む粉末を作製してよい。この下流の処理により、活性ペプチドの活性が増大する。10kDa膜ろ過物を凍結乾燥し、−20℃で貯蔵して、真空包装する。
(4)上記のようにしてステップ(2)に従ってよいが、3kDa膜フィルタを用いて、ペプチド配列番号1および配列番号3の活性をさらに増大させてよい。
【0066】
上記の粉末は、10〜15%の濃度で乾燥乳ベースの幼児用調合乳に加えて、Enterobacter sakazakiiでの汚染およびこの病原菌を原因とするその後の大腸炎および髄膜炎を予防できる。
【0067】
上記のようにして製造されるEnterobacter sakazakii抑制粉末は、特に、カゼインベースの乳児用調合乳製品に加えた場合に、Enterobacter sakazakiiおよびその他の病原菌に対する内部の組込まれた防御機構を提供する。
【0068】
ある実施形態において、本発明のペプチドは、免疫無防備で早産の新生児の感染の顕著な原因であるEnterobacter sakazakii汚染に対する保護のために、乳ベースの幼児用調合乳の調製方法のための補助物質として別々または一緒に用い得る。
【0069】
本発明のペプチドは、αs1−CN(カゼイン)の最初の9残基が切断に耐性であり、チーズの熟成において苦味に寄与すると記載されているので(Pedersonら、1999年)、チーズ製造における苦味に寄与するために用いることができる。
【0070】
L.acidophilus DPC6026による、αs1−CN由来抗菌ペプチドの製造は、以前に報告されていない。作製されたペプチド配列は、種々の文献およびデータベース検索で見出されず、この点において新規であると考えられる。
【0071】
本発明に関連して本明細書で用いる場合、「含み/含む」の語および「有する/含有する」の語は、記載する特徴、整数、ステップまたは成分の存在が、1つまたは複数の他の特徴、整数、ステップ、成分もしくはそれらの群の存在または追加を排除しないことを明記するために用いられる。
【0072】
【数1】

【0073】
【数2】
【0074】
【数3】
【0075】
【数4】
表1:1未満〜20kDaの間のペプチドのサイズを生じる乳酸桿菌株を用いて導かれるカゼインナトリウムタンパク質由来加水分解画分
【0076】
【表1】
表2:ヒトの幼児および成人の糞便単離物、ならびにブタ小腸単離物の16S rDNA配列決定
【0077】
【表2】
DPC;Dairy Products Research Centre, Teagasc Moorepark,Fermoy,Co.アイルランド、コーク
a.Enterococcus種との相同性の百分率。
b.Lactobacillus種との相同性の百分率。
c.Staphylococcus種との相同性百分率。
【0078】
表3:L.acidophilus DPC6026により製造されたカゼインナトリウム加水分解物からの粗画分に含まれるペプチドの配列および対応するカゼイン(CN)断片
【0079】
【表3】
aアミノ酸について1文字記号を用いる。
b各ペプチドの推定質量について、平均質量を報告する。
【0080】
表4:カゼインナトリウム中でのL.acidophilus DPC6026の発酵に続いて合成された純粋ペプチドの阻害範囲
【0081】
【表4】
a.Dairy Products Research Centre、アイルランドコーク。
+++、直径2.0cmを超える阻害の領域が生じた。
++、直径1.5cmを超える阻害の領域が生じた。
+、直径1.0cmを超える阻害の領域が生じた。
−、領域は検出されず。
【図面の簡単な説明】
【0082】
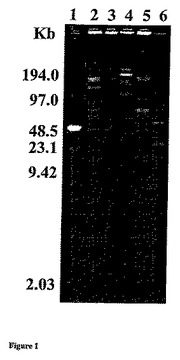
【図1】制限酵素ApaIについてのPFGEの巨視的な制限パターン。 パルス時間は、示すゲルについて1〜15秒であった。レーン1は、低分子量PFGマーカー(1% LMPアガロースに埋め込まれたラムダDNA−Hind III断片およびラムダコンカテマーの混合物)、レーン2〜6は、Lactobacillus acidophilus DPC6026、L.johnsonii DPC6092、L.salivarius DPC6027、L.animalis DPC6028およびL.delbrueckii sp.bulgaricus DPC6104を含んでいた。
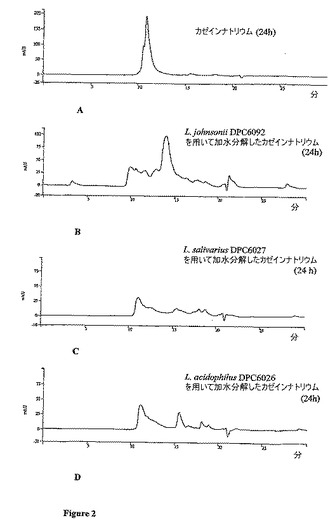
【図2】GRAS株のタンパク質分解活性。 ウシカゼインナトリウム(A)、Lactobacillus johnsonii DPC6092(B)、L.salivarius DPC6027(C)およびL.acidophilus DPC6026(D)での発酵後のそのカゼインナトリウム加水分解物の高性能液体クロマトグラフィー(HPLC)のペプチドプロファイル。HPLCは、214nmにて24時間監視した。試料30μlを、1ml/分の流速のDelta−Pak C−18(600mm×7.5cm)に添加した。タンパク質分解は、室温で行った。L.acidophilus DPC6026は、カゼインナトリウムのほぼ完全な分解を示した(D)。
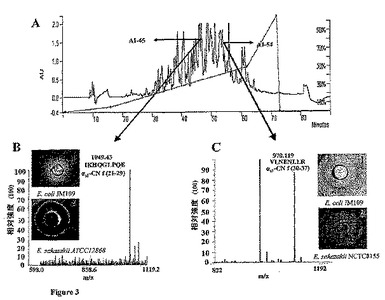
【図3】A.L.acidophilus DPC6026と24時間インキュベートしたpH7でのカゼインナトリウムの逆相高性能液体クロマトグラフィー(RP−HPLC)のクロマトグラム。矢印は、ペプチド画分A1−45およびA1−54の位置を示す。RP−HPLCは、室温で、材料および方法の部分に示す条件に従って行った。B.画分A1−45(基質としてのカゼインナトリウムおよびL.acidophilus DPC6026株を用いて単離した)の、ペプチドが検出されたm/z領域で記録された飛行時間型マトリクス支援レーザ脱離イオン化法(MALDI−TOF)スペクトル。ペプチド画分A1−45は、質量1049.43のペプチドを含有することが見出され、これは、配列決定の後に、αs1−CN f(21−29)に相当するIKHQGLPQEであるとわかった。1.L.acidophilus DPC6026でのカゼインナトリウム加水分解物の画分A1−45中で同定される化学合成ペプチドIKHQGLPQEによるE.coli DPC6053の阻害。寒天ウェル拡散アッセイ法を用いて、直径2.5cmの領域を記録した。2.L.acidophilus DPC6026でのカゼインナトリウム加水分解物の画分A1−45中で同定される化学合成ペプチドIKHQGLPQEによるEnterobacter sakazakii5920(ATCC128682)の阻害。寒天ウェル拡散アッセイ法を用いて、直径2.0cmの領域を記録した。C.画分A1−54(基質としてのカゼインナトリウムおよびL.acidophilus DPC6026株を用いて単離した)の、ペプチドが検出されたm/z領域で記録された飛行時間型マトリクス支援レーザ脱離イオン化法(MALDI−TOF)スペクトル。ペプチド画分A1−54は、質量970.119のペプチドを含有することが見出され、これは、配列決定の後に、αs1−CN f(30−37)に相当するVLNENLLRであるとわかった。1.L.acidophilus DPC6026でのカゼインナトリウム加水分解物の画分A1−54中で同定される化学合成ペプチドVLNENLLRによるE.coli DPC6053の阻害。寒天ウェル拡散アッセイ法を用いて、直径1.5cmの領域を記録した。2.L.acidophilus DPC6026でのカゼインナトリウム加水分解物の画分A1−54中で同定される化学合成ペプチドVLNENLLRによるEnterobacter sakazakii8272(NCTC8155)の阻害。寒天ウェル拡散アッセイ法を用いて、直径2.2cmの領域を記録した。
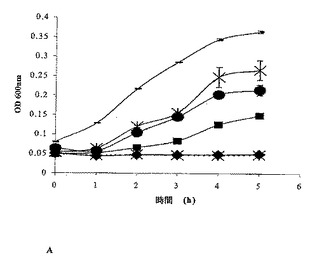
【図4A】A.96ウェルプレートアッセイを用いた、E.coli DPC6053(LB培地で1:10に希釈した一晩培養物)のOD600に対する種々の濃度のイスラシジンの影響。セクロピンP1を対照ペプチドとして用いる。記号:■、セクロピンP1(0.52mM)とインキュベートしたE.coli DPC6053。−、何も添加しないE.coli DPC6053対照。◆、イスラシジン(1.9mM)とインキュベートしたE.coli DPC6053。×、無菌性対照(何も添加しないLB培地)。●、0.23mMの濃度のイスラシジンとインキュベートしたE.coli DPC6053。X、イスラシジン(0.059mM)とインキュベートしたE.coli DPC6053の生存度。
【図4B】96ウェルプレートアッセイを用いた、E.coli DPC6053(LB培地で1:10に希釈した一晩培養物)のOD600に対する種々の濃度のペプチドIKHQGLPQEの影響。記号:−、何も添加しないE.coli DPC6053。X、IKHQGLPQE(0.078mM)とインキュベートしたE.coli DPC6053。■、セクロピンP1(0.52mM)とインキュベートしたE.coli DPC6053。●、IKHQGLPQE(0.15mM)とインキュベートしたE.coli DPC6053。・、IKHQGLPQE(0.31mM)とインキュベートしたE.coli DPC6053。◆、IKHQGLPQE(0.625mM)とインキュベートしたE.coli DPC6053。x、無菌性対照(何も添加しないLB培地)。
【図4C】96ウェルプレートアッセイを用いた、E.coli DPC6053(LB培地で1:10に希釈した一晩培養物)のOD600に対する種々の濃度のペプチドVLNENLLRの影響。記号:X、何も添加しないE.coli DPC6053。■、VLNENLLR(0.22mM)とインキュベートしたE.coli DPC6053。−、VLNENLLR(0.45mM)とインキュベートしたE.coli DPC6053。◆、VLNENLLR(1.2mM)とインキュベートしたE.coli DPC6053。X、セクロピンP1(0.52mM)とインキュベートしたE.coli DPC6053の生存度。●、VLNENLLR(2.5mM)とインキュベートしたE.coli DPC6053。▲、無菌性対照−何も添加しないLB培地。
【技術分野】
【0001】
発明の分野
本発明は、健康増進剤、特に抗菌剤の分野に全般的に関し、抗菌ペプチドおよび該抗菌ペプチドを与える細菌株を提供する。
【背景技術】
【0002】
発明の背景
病原菌による感染により、ヒトおよび動物の両方において重大な健康問題が起こり得る。特に、E.coli、E.sakazakii、Streptococcus mutansおよびListeria innocuaのような病原株による感染は、ひどく衰弱させ得る。病原菌に対する抗菌剤を供給する必要性があることが認識されている。
【0003】
病原菌の抗生物質に対する耐性の増加は、主要な問題点であり、新規薬物の探索が続いている(35)。多剤耐性のE.coli、Klebsiella spp.、およびE.sakazakiiの株は、驚くべき速度で増加しており、免疫無防備状態の宿主にしばしば関連して、該宿主にひどい影響を与え得る(6、14)。Enterobacter種は、それらがより古い抗菌剤に対する先天的な耐性を有し、かつそれらの院内感染との関連が増加しているために、近年、重要性が増加している病原体として認識されている(41)。30年より長きにわたって、E.sakazakiiは、新生児の髄膜炎の特有の症候群の原因として認識されている(20)。40〜80%の死亡率が報告されており、乳ベースの幼児用調合乳が感染の病原巣として働くという興味深い証拠が増加している((20)、(33)、(34))。E.sakazakii感染を制御するための現在の工業的な努力は、製造における衛生業務の改善および生物体についての最終製品の試験に焦点を当てている(34)。新規な抗菌ペプチドの候補を同定するためのある可能な経路は、乳タンパク質の探索にある。
【0004】
乳タンパク質は、生物活性ペプチドの豊富な供給源であり、アンギオテンシン−I変換酵素(ACE)阻害、オピオイド、免疫調節、抗血栓および抗菌活性を含む一連の健康増進特性を有すると報告されている(9)。生物活性ペプチドは、カゼインおよびホエーのような乳タンパク質にコード化された形で存在し、プロペプチドまたは成熟C末端ペプチドとして貯蔵され(13、19)、それらの放出にはタンパク質分解が必要である(11)。これらの機能的ペプチドのうち最も特徴付けられているものは、カゼインに由来し、心血管系、主にACE阻害における効果を有することが示されており、その結果、抗高血圧の特徴を有する(43)。MaruyamaおよびSuzuki(28)は、カゼインのトリプシン加水分解物が、ACEのin vitro活性を阻害したことを報告した。Pihlanto−Leppalaら(37)は、乳酸菌(LAB)で発酵させ、その後消化酵素で加水分解したホエーおよびカゼインタンパク質からのペプチドのACE阻害活性を研究した。同定されたペプチドは、αs1−カゼインf(142−147)、f(194−199)およびβ−カゼインf(108−113)、ならびにホエーからの2種のACE阻害ペプチドであった(37)。さらに、一連の抗菌ペプチドは、ウシのαs2カゼインから単離されている(23)。これらは、カソシジン(Casocidin)−1,f(164−179):KTKLTEEEKN RLNFLKKISQ RYQKFALPQY LKTVYQHQK、f(164−179):KKTKLTEEEK NRLNFL、およびf(187−207):QKFALPQYLK TVYQHQKAMK Qを含む。
【0005】
これらの3種のペプチドは、牛乳を酢酸および硫酸カルシウムで処理して混合物を形成し、次いでこれを加熱して沈殿物および上清を形成することから導かれる。上清は回収され、カチオン交換樹脂で処理される。ペプチドを、クロマトグラフィー手法を用いて溶出させ、活性ペプチドであるカソシジン−1を、消化プロテアーゼを用いて遊離させる。製造のコストは、これらの3つのペプチドの広い利用の1つの可能な制限である。特に、カソシジン−1は、Escherichia coliおよびStaphylococcus carnosusに対してin vitroで殺菌性であることが示されている(51)。
【0006】
しかし、カソシジン−Iを直接製造する細菌株は知られておらず、このことは、カソシジン−Iの製造が、製造コストを付加する酵素および化学物質の使用を必要とすることを意味する。さらなる情報は、「Antibiotic peptides from bovine milk」の表題のUS6579849に見出し得る。上記の特許にはこれらのペプチドの活性に対する制限は述べられていないが、Zuchtら、(FEBS Letters 372号(1995年)185〜188頁)は、これらのペプチドの阻害の範囲を記載し、グラム陰性細菌のみがこれらのペプチドにより影響を受けるようであり、このことは、グラム陽性菌の感染および汚染が、US6579849のペプチドでは治療し得ないことを意味する。
【0007】
LABを含む微生物起源のプロテイナーゼは、それらの特異性および活性により、カゼインからのペプチドの作製および抗菌ペプチドの遊離の可能性において主要な役割を演じる(29)。したがって、発酵の間、これらのプロテイナーゼを製造する微生物は、乳中で成長するために、カゼインおよびホエーのような乳タンパク質を分解できなければならない。LABによるカゼイン分解および分解産物のその後の利用は、複雑なタンパク質分解系を必要とする(9)。Lactococcus lactis(22、38)およびLactobacillus helveticus(27、50)のようなLABの高いタンパク質分解性、ならびに乳中での成長のために乳タンパク質を分解するそれらの必要性のために、生物活性ペプチドの作製のためのそれらのスターター培養物としての使用が報告されている(12、25、32、45)。非特許文献1(30)は、Lactobacillus helveticus PR4からのプロテイナーゼを用いて、6種の哺乳類からの乳のカゼインナトリウムから、抗菌活性を示す生物活性ペプチドを製造した。ヒトβ−カゼインf(184−210)に相当する抗菌ペプチドは、ヒトカゼインナトリウムの、L.helveticus PR4の部分精製プロテイナーゼを用いる加水分解により製造され、いくつかのグラム陽性細菌、およびE.coliに対しても抗菌活性を示した(30)。カゼイン分解の間に製造されるペプチドの特徴付けは、L.helveticus(11、49)について、およびL.casei(7)についてより少ない程度で記載されている。また、L.delbrueckii subsp.lactis ACA−DC 178の細胞壁結合プロテイナーゼが特徴付けされ、β−カゼインに対するその特異性が文書化されている(45)。
【0008】
カゼイン由来抗菌ペプチドはほとんど報告されておらず、記載されているものは、成熟C末端ペプチドに対する酵素の作用により得られ、このことは活性ペプチド配列の遊離を導く(43)。カゼイン起源の最初の抗菌ペプチドは、Hillら(17)により同定され、彼らは、カゼインをキモシンでpH6.4にてタンパク質分解した後に、カセシジンとして知られる抗菌糖ペプチドを単離した。1次アミノ酸構造R1PKHPIKHQGLPQEVLNENLLRF23を有するαs1−カゼインペプチド残基1−23である正に荷電した抗菌ペプチド、イスラシジン(17)は、グラム陽性およびグラム陰性細菌の両方に対して広い範囲の活性を有し(23)、ヒツジおよびウシの乳房炎感染を予防する(12)ことが示された。
【0009】
病原菌が時間の経過とともに、現存する抗菌化合物に対する耐性を発達させるという永続的な問題もある。よって、耐性菌により示される攻撃に継続的に対応するために、新規な抗菌ペプチドの開発に対する持続的な必要性が存在する。新規な抗菌化合物は、濃度の増加、投与回数の増加のようなより煩わしい投与法を必要とし得るか、あるいは狭い範囲の標的種に対してしか有効になり得ない場合がある。これらの状況であっても、新規な抗菌物質は、耐性の増加の問題のために、現存する化合物よりもまだ有利である。さらに、細菌の株を提供してこのような抗菌ペプチドを製造できることは、このことが、まず、該ペプチドの経済的な製造方法を提案でき、次に、該ペプチドをin situの状況で製造する手段を提供し得るので、非常に利益がある。
【非特許文献1】Minervini,F.ら、Applied and Environmental Microbiology:5297−5305(2003)
【発明の開示】
【発明が解決しようとする課題】
【0010】
(発明の目的)
本発明の目的の1つは、少なくとも1種の抗菌ペプチドを製造する細菌の新規な株を提供することである。少なくとも1種の抗菌ペプチドを提供することは、本発明のさらなる目的である。さらなる目的は、多剤耐性生物に対して用いるための抗菌ペプチドを提供することである。抗菌剤を含有する安全性が向上された食品を提供することも、本発明の目的である。ACE阻害の方法を提供することは、本発明のさらなる目的である。抗菌特性が向上された調合乳を提供することも、本発明の目的である。
【課題を解決するための手段】
【0011】
(発明の要旨)
ある実施形態において、本発明は、配列番号1(IKHQHPQE)、配列番号2(SDIPNPIGSENSEK)および配列番号3(VLNENLLR)からなる群より選択される1つまたは複数のペプチドを含む抗菌化合物を提供する。別の実施形態において、本発明は、その試料が2005年11月18日にアクセッション番号NCIMB 41354の下でスコットランドアバディーンのナショナル・コレクション・オブ・インダストリアル・アンド・マリン・バクテリア(National Collection of Industrial and Marine Bacteria)に寄託されているLactobacillus acidophilus DPC6026株、または乳もしくは乳製品から抗菌活性を有するペプチドを製造できるその派生株もしくは変異株の生物学的に純粋な培養物を提供する。
【0012】
本発明は、DPC6026により製造されるタンパク質も提供する。特に、本発明は、乳から抗菌ペプチドを製造できる1つまたは複数のペプチドを製造できる、DPC6026により製造される新規なタンパク質分解酵素タンパク質を提供する。これらのペプチドは、乳または乳製品からの配列番号1、配列番号2および配列番号3からなる群のうちの1つまたは複数に実質的に等しい配列を有するものであってよい。
【0013】
本発明は、乳または乳製品に、Lactobacillus acidophilus DPC6026株、または乳もしくは乳製品から抗菌活性を有するペプチドを製造できるその派生株もしくは変異株の生物学的に純粋な培養物、DPC6026培養物から得られる無細胞培養上清またはその画分、およびDPC6026により製造されるタンパク質であって、乳または乳製品から1つまたは抗菌ペプチドを製造可能であるタンパク質からなる群のうちの1つまたは複数を加えることを含む、乳または乳製品から抗菌ペプチドを製造する方法も提供する。
【0014】
本発明は、乳または乳製品から配列番号1、配列番号2または配列番号3のうちの少なくとも1つを製造可能であるタンパク質分解特性、触媒特性および/または酵素特性を有するDCP6026からの1つまたは複数種のタンパク質を製造する方法も提供する。
【0015】
請求項1に記載の抗菌化合物、請求項2に記載の培養物、請求項3に記載の上清および請求項4に記載のタンパク質からなる群より選択される1つまたは複数を含む抗菌組成物。
【0016】
本発明は、DPC6026株から得られる無細胞培養上清もしくはその画分、あるいは上記の1もしくは複数のペプチド、化合物、上清または株を適切な担体または希釈剤とともに含む抗菌組成物も提供する。
【0017】
本発明は、上記の化合物、ペプチド、株、タンパク質、上清または組成物の、微生物感染または微生物汚染の管理における使用も提供する。本発明の態様は、本発明の1つまたは複数の化合物、株、上清もしくは組成物を動物またはヒトに投与することを含む、微生物感染もしくは微生物汚染を予防または治療または制限する方法を提供する。微生物感染または微生物汚染は、Escherichia coli、E.sakazakii、Streptococcus mutans、Listeria innocua、Klebsiella sppおよびStaphylococcus carnosusからなる群の1もしくは複数種を原因とする汚染または感染であってよい。特に、感染または汚染は乳腺炎であってよい。
【0018】
別の実施形態において、感染または汚染は髄膜炎であってよく、新生児の乳腺炎であってよい。ある実施形態において、化合物、ペプチド、株、上清または組成物は、乳ベースの調合乳に供給してよい。つまり、本発明のペプチドは、幼児用調合乳製品の製造における組込まれた防御系として用い得る。1つまたは複数のペプチドは、特に、Enterobacter Sakazakii汚染に対して有効であり得る。
【0019】
構造、活性の範囲および効力の点での生物活性ペプチドの多様な性質のために(39)、これらのペプチドは、可能性のある新しい薬物設計の主な標的である。乳タンパク質由来の配列番号1(IKHQGLPQE)および配列番号3(VLNENLLR)のような抗菌ペプチドを、E.sakazakii 5920(ATCC12868)のような病原株に対する防御の組込まれた機構として用いる可能性は、幼児用調合乳製造において用いられるもののような粉乳の安全性を増進させる有用なアプローチを提供し得る。
【0020】
本発明は、候補の動物またはヒトを、本発明の1つまたは複数の組成物、化合物、上清、ペプチドまたは株で処置することを含む、アンギオテンシン−I変換酵素(ACE)の阻害、オピオイド調節、免疫調節および血栓症抑制の方法も提供する。
【0021】
本発明のペプチド、株、化合物、組成物および上清は、ウシおよびヒツジの乳房炎の治療に用いることもできる。配列番号1および配列番号3は、抗菌ペプチドであるイスラシジンの切断産物に類似する。配列番号2も用いてよい。イスラシジンは、マウスあたり10μg程度の低い用量で投与したときに、S.aureus、Streptococcus pyogenesおよびL.monocytogenesに対して強い防御効果を有すると記載され、ヒツジおよびウシの乳房炎の治療において以前に用いられたこともある。
【0022】
αs1−カゼインの最初の9残基はチーズの作製において蓄積し、いくつかのチーズに存在する苦味の原因であるので(Foxら、1995年、Chemistry of Structure−Function Relationships in Cheese(Malin, E. L.およびTunick, M. H.編)、Plenum Press、New York、59〜98頁)、本発明のペプチド、株、化合物、組成物および上清は、苦味チーズの製造における苦味に寄与するために用い得る(Pedersonら、1999年、J. Bacteriology 181号、4592〜4597頁)。
【0023】
本発明のペプチド、株、化合物、組成物および上清は、上記のような配列番号1、配列番号3および配列番号2の広い範囲の活性のために、食品または腐敗しやすい商品の、病原性グラム陰性およびグラム陽性細菌に対する防腐剤として用い得る。
【0024】
L.acidophilus DPC6026は、ブタの小腸から単離され、コークのDairy Products Research Centre(DPRC),Teagasc Moorepark,Fermoy,Co.に貯蔵されている新規な株である。イスラシジンまたはその断片の製造を示す細菌株は、以前に記載されていなかった。イスラシジンは、キモシンで処理したαs1−カゼインに由来した(HillらJ of Dairy research 1974年;41号:147頁)。このことは、L.acidophilus DPC6026は、その天然の状態で、酵素を必要とせずに3つのペプチドIKHQGLPQE、VLNENLLRおよびSDIPIGSENSEKを製造し、工業的なレベルで発酵プロセスの余分な経費を回避するので、経済的に有利である。イスラシジンのペプチド配列は、そのより長い鎖の長さのために、化学合成するには配列番号1、配列番号2および配列番号3よりも高価である。配列番号1、配列番号2、配列番号3はそれぞれ、より短い鎖長を有し、よって、化学合成がより安価である。
【0025】
イスラシジンαs1−カゼインf(1−23)は、溶液中で比較的安定な立体配置を維持するようである(Malinら、Journal of Protein Chemistry、20号、391〜404頁)。立体配置が安定なペプチドは、アレルゲンに特徴的であることが知られており、αs1−カゼインのこのセグメントは、アレルゲン性であることが知られている(Spuerginら、1996年 Allergy 51号、306〜312頁)。残基20の周囲のαs1−カゼインの領域は、ヒトIgEにより認識される重要なアレルゲン性エピトープを含むことが示唆されている(Spuerginら、1996年)。さらに、配列番号3は、ヒトIgEにより認識される重要なアレルゲン性エピトープを含むと考えられているαs1−カゼインの残基20を欠失しているので、アレルゲン性エピトープを提示し得ない(Spuerginら、1996年)。
【0026】
in vivoでの透過
イスラシジンは、in vivoで、Staphylococcus carnosusに対する防御効果を示すことが示されている。配列番号1および配列番号3は、病原性乳腺炎原因株に対して、in vivoでより大きい防御効果を示す。この理由は、より短い生物活性ペプチドが、よりよいin vivo効果を示すことであると仮定されている(Vermeirssenら、British Journal of Nutrition (2004年)、92号、357〜366頁)。
【0027】
αs1−カゼインf(1−9)(RPKHPIKHQ)は、自然発生高血圧ラット(SHR)において抗高血圧効果を有することが示されている(Saitoら、2000年、J. Dairy Science、83号、1434〜1440頁)。配列番号1は、最後の4アミノ酸(IKHQ)と100%の相同性を共有する。これは、ペプチドのC末端にプロリン、グルタミンおよびグルタミン酸も有する。ACE活性は、C末端トリペプチド領域でのプロリンの存在により、非常に遅くなる(Vermeirssenら、2004年、British Journal of Nutrition、92号、357〜366頁)。この事象は、配列番号1がACE阻害活性を有し得ることを示唆し、イスラシジンを2つのより短いペプチドに切断することの可能な利点を示唆する。
【0028】
ある状況下では、イスラシジン耐性の機構は、配列番号1によっても配列番号3によっても提示されない分子の「抗原性」領域により媒介されると仮定されてもいる。
【0029】
ラクトフェリシンBのような他のカチオン性抗菌ペプチドは、プロテアーゼを産生するので、Escherichia coliおよびStaphylococcus aureusのような耐性株を有することが知られており、このことは、E.coliおよびStaphylococcus aureusによるマゲイニンのような他のカチオン性抗菌ペプチドに対する主要な耐性機構として示唆されている(Ulvatneら、Journal of Antimicrobial Chemotherapy(2003年)50号、461〜467頁)。イスラシジンを本発明のペプチドの2つ、配列番号1および配列番号3に切断することは、イスラシジンに比較してより短いこれらの配列を切断できるプロテアーゼがより少ないので、耐性株の発生の可能性を低減させるようである(コンピュータプログラムhttp://ca.expasy.org/cgi−bin/peptidecutter/peptidecutter.plを用いて証明されるように)。
【発明を実施するための最良の形態】
【0030】
材料および方法
基質および化学薬品
ペプチドインドリシジンおよびセクロピンP1、ならびに酵素プロテイナーゼKはSigma(Sigma Aldrich Chemie、ドイツシュタインハイム)から購入した。ピマリシンは、Merck(Merck、ドイツダルムシュタット)から購入した。ペプチドイスラシジンは、Peptide Protein Research Ltd.,(英国フェアラム)により合成された。Maximum Recovery Diluent(MRD)は、Oxoid(Oxoid Ltd.,イングランドベイジングストーク)から購入した。ウシカゼインナトリウムは、Dairygold(アイルランドコークミッチェルズタウン)からであった。パルスフィールド保証アガロースは、Bio−Rad Laboratories(米国CA94547ハーキュリーズ)からであった。酵素Apa Iおよび低分子量PFGマーカー(NO350S)は、New England Biolabs(英国ハートフォードシャーSG4 OTY)から購入した。低融点アガロースは、Bio−Rad Laboratories(カリフォルニアリッチモンド)から購入した。
【0031】
微生物および培養条件
L.acidophilus DPC6026、L.johnsonii DPC6092およびL.salivarius DPC6027は、ブタの小腸から単離し(データは未公表)、アイルランドコークのTeagasc Dairy Products Research Centreの培養コレクションに貯蔵した。これらの株を、MRS培地(Oxoid Ltd、英国ベイジングストーク)中で嫌気的に24時間、37℃にて増殖させた。標準培養物は、10mlのMRS培地に、10μlの冷凍ストック(−80℃)を接種することにより調製し、次いで、37℃にて16〜24時間インキュベートした。NCIMB(ナショナル・コレクション・オブ・インダストリアル・アンド・マリン・バクテリア、スコットランドアバディーンAB24 3RY)から購入したL.innocua DPC3306、E.sakazakii 5920(ATCC 12868)、E.sakazakii 8272(NCTC8155)、ならびにE.coli DPC6053(Teagasc Dairy Products Research Centre、アイルランドコーク)を、試験株として用いた。
スキムミルク寒天(SMA)プレート上でのタンパク質分解活性の初期スクリーニング
ブタの腸起源の205種の細菌単離物ならびにヒト成人および幼児の糞便起源の55種の単離物(データは未公表)を、以前に記載されたようにして(36)SMAプレートを用いてタンパク質分解活性についてスクリーニングした。点在させた培養物の周囲の透明領域が、タンパク質分解を示した。直径4.0mm以上の領域を発生させる単離物を、さらなる研究のために選択した。
【0032】
高性能液体クロマトグラフィー(HPLC)
SMAプレートを用いてタンパク質分解活性を示す単離物を、カゼインナトリウム溶液(2.5%w/v)に接種し(1%w/v)、嫌気的に37℃にて一晩インキュベートした。次いで、Delta−Pak C18カラム(サイズ;600mm×7.5cm、Varian Chromatography Systems、米国カリフォルニアウォルナットクリーク)を用いる高性能液体クロマトグラフィーを、得られた発酵生成物に対して行った。移動相は、トリフルオロ酢酸(0.1%v/v)含有のアセトニトリルとHPLC等級の水(30%v/v)との2成分混合物であった。流速は、1ml/分であった。ブタ腸単離物および幼児糞便単離物によるカゼインの分解を、HP1100ダイオードアレイ検出器を用いて214nmでのUV吸収の測定により監視した。
【0033】
パルスフィールドゲル電気泳動
MRS培地からの15種の単離物のそれぞれからの高分子量DNAの調製は、プラグスライスの制限消化のために20Uの制限酵素ApaIを用いた以外は、Simpsonら(42)により以前に記載されたとおりであった。
株の種同定のための単離物の16S rDNA配列決定
ブタの腸、ならびに幼児ヒトおよび成人の糞便の15種の単離物は、著しいタンパク質分解活性を示すことが示され、個別の株の種同定のために16sリボソームDNA(rDNA)配列決定に付した(Lark Technologies Inc.,英国エセックス)。2つの16S rDNAプライマー、5’末端についてCO1(5’−AGTTTGATCCTGGCTCAG−3’)および3’末端についてCO2(5’−TACCTTGTTACGACT−3’)を用いて、約1.5kbの16S rDNA生成物を、以前に記載された(42)ポリメラーゼ連鎖反応(PCR)条件下で作製した。このPCR生成物を、CO1プライマー(Lark)を用いて部分的に配列決定した。BLASTプログラムを用いることにより得られた16S rDNA配列の比較により、株を特定の種に割り当てることが可能であった。通常、16S rDNA配列の類似性の値がBLASTプログラムを用いて97%を超えると、それらの株は同じ種に属すると考えられる(44)。
【0034】
GRAS株を用いる大規模発酵
タンパク質分解活性を示す15種の単離物のうち、3種の遺伝的に別個の株であるLactobacillus acidophilus DPC6026、L.salivarius DPC6027およびL.johnsonii DPC6092を、それらのタンパク質分解能力に基づいて、およびそれらが全て一般的に安全であるとみなされているので(GRAS)、さらなる発酵に個別に用いた。3つの別々の発酵を、3回ずつ行った。カゼインナトリウム基質(2.5%w/v)に、各株(1%w/v)を接種し、100rpmで混合しながら、0.1M NaOHの添加により維持する一定のpH7で、37℃にて24時間インキュベートした。発酵生成物を、次いで、80℃に加熱して培養物を不活性化し、その後、サイズ排除S1Y10 10kDaらせんカートリッジフィルタ(Millipore Ltd.,英国ハートフォードシャー)を通してろ過し、10kDa未満のペプチドを分離した。これらのペプチドを含む画分を凍結乾燥し、さらなる使用まで−20℃にて貯蔵した。
【0035】
発酵生成物の逆相HPLC分析
サイズが10kDa未満のペプチドを、狭い口径のカラム(Nucleosil C18、5mm×250mm:Varian Chromatography Systems、米国カリフォルニアウォルナットクリーク)、および214nmで作動するUV検出器を含むRP−HPLC逆相高性能液体クロマトグラフィーシステムを用いてカゼインナトリウム加水分解物から分離した。凍結乾燥粉末の一定量を、HPLC等級の蒸留水で希釈し、0.45μmのフィルタ(Millipore)を通してろ過し、30mg/mlの発酵生成物をカラムに添加した。移動相は、トリフルオロ酢酸(0.1%v/v)含有のアセトニトリルおよびHPLC等級の水(100%v/v)の2成分混合物であった。移動相中のアセトニトリル含量を、1ml/分の流速で、0から100%まで72分間で直線的に増加させた。ペプチドを、214nmの波長で作動する検出器を用いて検出した。回収した画分から、セントリバップコンソール(centrivap console)(Labconco Corporation、米国カンザスシティ)を用いる蒸発により溶媒を除去した。画分を、その後の抗菌活性のアッセイの前に、1mlの蒸留水に再溶解した。
【0036】
タンパク質濃度の決定
画分のタンパク質濃度は、Bioradタンパク質アッセイ法(24)を用いて決定した。595nmの吸光度を決定し、タンパク質濃度をmg/mlとして報告した。
【0037】
抗菌活性
ペプチドおよび合成した画分の抗菌活性の予備的特徴付けは、96ウェルアッセイでの成長阻害の測定(1、10)、および寒天拡散法の使用(16、39)を含んだ。96ウェルプレートアッセイは、各ペプチド画分単独の対照(セクロピンP1およびインドリシジン)とともに、ペプチド画分を含まない対照(増殖対照)を含んだ。細菌を含まない対照(無菌性対照)も含めた。プレートを37℃にて6時間インキュベートし、培養成長を1時間ごとに監視した。最小阻止濃度(MIC)を、マイクロタイタープレートリーダーでのOD600を記録することにより測定される、認識可能な成長がない最低濃度とした。ウェル拡散アッセイ(16、40)を用いて、精製および化学合成したペプチドの抗菌活性を検出した。これらのアッセイは、それぞれE.coli DPC6053、L.innocua DPC3306、E.sakazakii5920(ATCC12868)またはE.sakazakii8272(NCTC8155)の指示株の一晩培養物1mlを播種した脳心臓浸出物(BHI)、ルリアベルタニ(LB)または栄養寒天(NA)のいずれかで行った。直径4.6mmのウェルを、これらの寒天プレート中に切り取って入れ、各ウェル中に30μl(0.162mg/ml)のペプチド画分を入れた。プレートを4℃にて4時間貯蔵して、ペプチドの放射状の拡散を許容し、30℃にて嫌気的にまたは37℃にて好気的に24時間インキュベートし、阻害領域について調べた。ペプチドに対する株の感度を、ウェルの周囲の阻害領域の直径に従って評点した。実験は、3回行い、平均の領域サイズを計算した。E.coli DPC6053、E.sakazakii5920(ATCC12868)およびE.sakazakii8272(NCTC8155)についてイスラシジン、セクロピンP1、およびL.innocua DPC3306に対してインドリシジンを対照として用いた。
【0038】
抗菌ペプチドの精製、配列決定および合成
抗菌活性を示すタンパク質画分を、上記のRP−HPLC条件により再分画し、指示株E.coli DPC6053の成長を、市販のペプチドセクロピンP1と同様のタンパク質濃度で阻害した画分を回収して、337nmで作動するレーザおよび20kVの加速電圧で飛行時間型マトリクス支援レーザ脱離イオン化法(MALDI−TOF)質量分析計(PE Biosystems Voyager−DE STR Biospectrometry Workstation、Aberdeen Proteome Facility)を用いる質量分析(MS)により、ペプチド組成を分析した。各画分中のペプチドのアミノ酸配列を、誘導体化およびエドマン分解の後に決定した。これらのステップは、494Aプロテインシーケンサー(Applied Biosystems)で行った。抗菌活性を示す各画分中の同定された全てのペプチド(MS分析およびアミノ酸配列決定により)を、その後、Peptide Protein Research Ltd.,(英国フェアラム)により化学合成した。合成ペプチドの純度は、製造者により証明されるようにHPLC分析により決定して、95%を超えていた。
化学合成ペプチドおよび粗画分のプロテイナーゼKでの処理
化学合成ペプチドおよびE.coli DPC6053に対して抗菌活性を示す画分を、プロテイナーゼK(Sigma)に対する感受性について、プロテイナーゼK(2050U/ml)とペプチドまたは画分(0.554mg/ml)との1:1の容量比での4℃で6時間のインキュベーションにより、寒天ウェル拡散アッセイ(40)を用いて試験した。プレートを、37℃にて24時間インキュベートした。用いた陽性対照は、ペプチドイスラシジン(Peptide Protein Research Ltd.,)およびセクロピンP1(Sigma TM)であった。
【実施例】
【0039】
本研究の目的の1つは、哺乳動物の腸起源のLABのタンパク質分解能力を用いて放出されたウシカゼインから新規な生物活性ペプチドを発見することであった。ブタ小腸から得られた205種の単離物ならびにヒトの成人および幼児の糞便起源の55種の単離物を、本研究において用いた。
【0040】
タンパク質分解活性を有する単離物のスクリーニング
LAB寒天プレートでの260種の単離物全てを、基質としてSMAおよびカゼインナトリウムを用いてタンパク質分解活性についてスクリーニングした。ブタの腸起源の5種の単離物と、ヒトの幼児および成人の糞便起源の10種の単離物を、それらのタンパク質分解能力およびカゼインナトリウムから10kDa以下のペプチドを生じる能力に基づいて、選択した。15種全ての単離物は、タンパク質分解活性を示す直径サイズ4mm以上の領域サイズを生じ、15種全ての単離物の小規模カゼイン発酵生成物のHPLC分析により、最も複雑なペプチドプロファイルが示された。PFGEおよび16S rDNA配列決定の後に、選択された15種の単離物のうち、9種が乳酸桿菌の別個の種であると同定された(表1)。全てヒトの成人の糞便起源の5種の単離物は、Enterococcus faecalisと99〜100%の相同性を示し(BLASTプログラムを用いて)、よって、腸球菌は通常、安全とはみなされないので(GRAS)(21)、大規模発酵でのさらなる使用には適さないと考えられた。単離物3L7および33L1(ヒト幼児糞便単離物)は、それぞれE.hiraeおよびStaphylococcus epidermisと100%の相同性を示し(BLASTプログラム)、これらも、非GRAS状態のために、さらなる研究から排除した。9種の乳酸桿菌単離物のうちの3種は、PFGE(図1)および16S−rDNA配列決定により、L.acidophilus DPC6026、L.johnsonii DPC6092およびL.salivarius DPC6027と同定された。
【0041】
L.acidophilus DPC6026、L.salivarius DPC6027およびL.johnsonii DPC6092のカゼインナトリウム(2.5% w/v)での24時間後のインキュベーション、ならびにその後の10kDaの膜を通してのろ過の後に得られたペプチドプロファイルを、図2に示す。これらの3種の乳酸桿菌でのカゼインナトリウムの発酵は、カゼインナトリウム(図2A)のより低分子量のペプチド(図2B〜2D)への実質的な分解をもたらし、L.acidophilus DPC6026は、カゼインナトリウムのより小分子量のペプチドへの最大程度の分解を示した(表1)。L.acidophilus DPC6026は、得られたペプチドの60.74%が≦0.5kDa未満であり、17.49%が1〜0.5kDaの間のペプチドであり、11.57%が1〜2kDaの間のペプチドである発酵生成物を生じた。この株は、L.johnsonii DPC6092、L.salivarius DPC6027およびL.animalis DPC6028よりも0.5kDa〜2.0kDaの間のペプチドをより多く製造した(表1)。L.reuteri DPC6100、L.gasserri DPC6093、L.rhamnosus DPC6095およびL.brevi DPC6102により製造されたHPLCペプチドプロファイルは、選択した株と同程度のカゼインナトリウムの分解を示さず(表1)(HPLCプロファイルは示さず)、よって、さらなる研究のために選択しなかった。L.delbrueckii sp.bulgaricus DPC6104は、生物活性ペプチドの作製のためのその使用が以前に記載されているので、選択しなかった(11)。
【0042】
抗菌ペプチドの検出
Lactobacillus acidophilus DPC6026、L.salivarius DPC6027およびL.johnsonii DPC6092により製造されたカゼインナトリウム発酵生成物は、抗菌画分であった。しかし、L.acidophilus DPC6026から作製された画分のみを抗菌活性についてアッセイした。なぜなら、この株は、示したクロマトグラフィープロファイル(図2)から示されるように、3つの株のうちで最高のタンパク質分解活性により特徴付けられたからである。E.coli DPC6053、Listeria innocua DPC3306、E.sakazakii5920(ATCC12868)およびE.sakazakii8272(NCTC8155)の株に対する96ウェルプレートアッセイまたはウェル拡散アッセイのいずれかを用いて、粗発酵生成物の抗菌活性は検出されなかった。L.acidophilus DPC6026により製造されたカゼインナトリウム発酵生成物を、サイズ排除S1Y10 10kDaらせんカートリッジフィルタを通してろ過して、10kDa以下のペプチドを含む透過物を得た。L.acidophilus DPC6026により製造されたろ過カゼインナトリウム発酵生成物の72個の画分を、RP−HPLCにより回収して、E.coli DPC6053、E.sakazakii5920(ATCC12868)、E.sakazakii8272(NCTC8155)およびL.innocua DPC3306に対する抗菌活性について、ウェル拡散アッセイによりアッセイした。E.coli DPC6053およびL.innocua DPC3306に対するこれらの画分の抗菌活性は、96ウェルプレートアッセイ法を用いてOD600を測定することによってもアッセイした。3つの画分、A1−45、A1−49およびA1−54は、最も効力が高い抗菌活性を有し、それぞれ0.554mg/ml、0.5mg/mlおよび1.24mg/mlのペプチド濃度を有していた。これらの全ての画分は、試験株E.coli DPC6053に対して、それぞれこれらの濃度で阻害活性を示した。
【0043】
ペプチドの精製、配列決定および特徴付け
画分A1−45、A1−49およびA1−54中のペプチド混合物を、質量分析およびエドマン分解に付した。アミノ酸組成は、配列決定(Aberdeen Proteome Facility)によっても決定した。画分A1−45は、ペプチド配列IKHQGLPQE(配列番号1)を含むことが見出された(表2)。このペプチド配列は、αs1−カゼインf(21−29)に相当する。画分A1−49は、αs1−カゼインf(183−207)に相当するペプチド配列SDIPNPIGSENSEK(配列番号2)を含んでいた。A1−54は、αs1−カゼインf(30−37)に相当するペプチド配列VLNENLLR(配列番号3)を含むことが見出された。各ペプチドの推定および計算された質量を報告する(表2)。寒天ウェル拡散アッセイにより決定されたように、画分45に存在する配列番号1は、指示生物であるE.coli DPC6053を、0.05mMの濃度で阻害した(図3B)。このペプチドは、E.coli O157:H7派生株(E.coli DPC6054およびE.coli DPC6055)、E.sakazakii5920(ATCC12868)のような臨床対象の潜在的な病原細菌に対しても、同じ濃度(0.05mM)で阻害を示した(図3B)。画分54に存在する配列番号3は、指示生物であるE.coli DPC6053を0.22mMの濃度で(図3C)、また、E.sakazakii DPC6091のような潜在的病原株を同じ濃度で阻害したが(図3C)、配列番号2を含む画分A1−49は、Listeria innocua DPC3306に対してわずかな阻害活性しか示さず、E.coli DPC6053に対しては全く活性を示さなかった(表3)。
【0044】
ペプチドの化学合成
強い抗菌活性を示し、かつ広い範囲の活性を有していた画分から同定されたペプチドを、化学合成して、MALDI−TOF分析により同定された配列が、画分中の抗菌配列であるかを確認した。これらは、αs1−CN f(21−29)(配列番号1)、αs1−CN f(30−37)(配列番号3)およびαs1−CN f(183−207)(配列番号2)であった。化学合成ペプチド配列番号1および配列番号3は、未精製のペプチドを含む画分と同じ微生物を阻害した。合成ペプチド配列番号2(画分A1−49に存在)は、Listeria innocua DPC3306に対していくらかの阻害活性を示したが、E.coli DPC6053に対しては活性を示さなかった(表3)。
【0045】
これらのペプチドの最小阻止濃度(MIC)を、イスラシジンを陽性対照として用いて決定した。イスラシジンは、E.coli DPC6053を、0.05mM〜1.9mMの範囲の濃度で阻害した(図4A)。イスラシジンについてのMICは、記載される実験条件下では0.059mMであると見出されたが、配列番号1についてのMICは、E.coli DPC6053の成長を阻害すると示された(図4B)0.078mMであり、配列番号3は、この微生物を0.22mM〜1.2mMの範囲の濃度で阻害した(図4C)。配列番号3について得られたMICは、0.22mMであった。つまり、本発明は、驚くべきことに、全てが抗菌特性を有する3種の新規ペプチドを製造した。イスラシジンは、最良の標準物である陽性対照とみなした。配列番号1は、イスラシジンに好ましく匹敵するが、配列番号2および配列番号3は、同じ濃度では、効果が1桁小さい。しかし、より大きい濃度が必要であるが、配列番号2および配列番号3はともに、抗菌特性を明らかに示していた。
【0046】
配列番号2および配列番号3に必要な濃度は、抗菌化合物として通常の濃度範囲内である。これらは、例えば、カソシジン−1、および望ましい組成物の濃度が0.1mg〜1mgであると示すUS6579849「Antibiotic peptides from bovine milk」に記載されるペプチドによく匹敵する。Florisら、Current Pharmaceutical Design、2003年、9号、1257〜1275頁は、イスラシジンが「比較的高濃度(0.1〜1mg/mL)ではあるが、乳酸桿菌および他のグラム陽性細菌のin vitro成長を阻害することが見出された」と述べている。
【0047】
アッセイ条件下では、全ての化学合成ペプチドおよび粗画分は、プロテイナーゼKで処理したときにそれらの抗菌活性を喪失した。コンピュータプログラムExpasy Peptide cutter(http://ca.expasy.org/cgi−bin/peptidecutter/peptidecutter.pl)を用いて、ペプチド配列IKHQGLPQE(配列番号1)、VLNENLLR(配列番号3)、SDIPIGSENSEK(配列番号2)が全て、トリプシンおよびキモトリプシンにより加水分解されることが見出された。
【0048】
考察
この研究において、カゼインナトリウムを、タンパク質分解株L.acidophilus DPC6026、L.salivarius DPC6027およびL.johnsonii DPC6092を用いてタンパク質分解に付した。L.acidophilus DPC6026株を、発酵で用いるために選択し、生じたペプチドを、E.coli JM109 DPC6053およびEnterobacter sakazakii DPC6090(ATCC12868)のような潜在的病原株に対する抗菌活性についてアッセイした。
【0049】
本研究で同定されたペプチド配列IKHQGLPQE(配列番号1)、VLNENLLR(配列番号3)およびSDIPIGSENSEK(配列番号2)は、抗菌活性について以前に報告されていない(8、47)。
【0050】
(Dziubaら、1999年(8))は、種々の供給源からの一連の生物活性ペプチドを含む生物活性データベースを参照している。データベースに含まれるペプチドは、抗菌ペプチド、アンギオテンシン−I変換酵素阻害ペプチド(ACE−I阻害ペプチド)、オピオイドペプチドおよびブラディキニン増強ペプチドである。行った実験で病原菌に対する最良の陽性対照として用いたペプチド、イスラシジン(ペプチドデータID 3035、名称イスラシジン、配列RPKHPIKHQGLPQEVLNENLLRP)は、このデータベースに詳述されている。
【0051】
(Wangら、2004年(47))は、抗菌ペプチドデータベースのみについて詳述するが、イスラシジンについてはわずかしか述べていない。
【0052】
研究の結論
牛乳タンパク質からの既知の抗菌ペプチドは、R1PKHPIKHQGLPQEVLNENLLRF23と決定された一次アミノ酸配列を有するイスラシジン、αs1−CN f1−23である(26)。このペプチドは、広い範囲の活性を有し(23)、配列番号1および配列番号3との高い程度の相同性のために、本研究において陽性対照として用いた。配列番号1は、イスラシジンと9残基が共通し、配列番号3はイスラシジンのC末端と8残基が共通している。ペプチド配列番号1は、0.05mMのMICを示したイスラシジン、ならびにより効力が高い商業的に入手可能なペプチドであるセクロピンP1およびインドリシジン(0.05mMのMIC)に匹敵する0.078mMのMICを示した。配列番号3は、E.coli DPC6053に対して、イスラシジンのMIC(0.05mM)に匹敵する0.22mMのMICを示した。配列番号1のペプチド(IKHQGLPQE)は、+2の正電荷を含み、疎水性(イソロイシン)末端および親水性(グルタメート)ドメインを有し、かつE.sakazakiiおよびE.coliのようなグラム陰性細菌に対して、グラム陽性細菌よりもよい活性を示す。
【0053】
抗菌ペプチドは、通常、5〜60の間のアミノ酸を有し、10kDa未満の分子質量を有し、通常、両親媒性であり、通常、広い範囲の活性を有する(Florisら、Current Pharmaceutical Design 2003年)。配列番号3(VLNENLLR)は、これらの特徴が共通している。
【0054】
ペプチドのカチオン性および疎水性の二重の特徴が、ペプチドと細菌膜との最初の相互作用に重要であることが知られている。正電荷は、細菌の外膜および細胞質膜との相互作用を促進する(Wuら、Journal of Biol. Chem.、1999年)。アルギニン(R)の強い正電荷が、Hancockら、1999年 Peptide Antibiotics, Antimicrobial agents and chemotherapy:1317〜1323頁に記載されるように、配列番号3(VLNENLLR)がグラム陰性細菌のリポ多糖領域と相互作用し、負に荷電した細胞質膜を透過することを可能にする。
【0055】
配列番号3はまた、疎水性領域(VL)および親水性領域(R)も有する。これは、また、L.innocuaのようなグラム陽性株に対するよりも病原性グラム陰性細菌に対して、より強力な活性を示す。また、化学合成ペプチドも、L.innocua DPC3306に対するよりもE.coli DPC6053のようなグラム陰性株に対してより強力な活性を示した。
【0056】
LABのプロテイナーゼは、β−CNおよびαs1−CNのペプチド結合の40%より多くを加水分解して、オリゴペプチドを生じることが示されている(22)。次いで、LABの複合ペプチダーゼは、これらのオリゴペプチドに対して作用する(31)。Minerviniらは、L.helveticus PR4からの部分精製プロテイナーゼを用いるACE阻害ペプチドの作製を報告した(30)。このグループは、グラム陽性およびグラム陰性細菌の両方に対して活性を示したヒトβ−CN f(184−210)からの抗菌ペプチドも同定した。β−CN由来のいくつかのカソキニンは、Lactobacillus helveticus CP790の細胞壁会合セリン型プロテイナーゼにより、遊離されている(49)。L.delbrueckii subsp.lactis ACA−DC178からの細胞壁結合プロテイナーゼは、β−CNから4つのペプチドを遊離させたが(45)、これらのペプチドの生物活性は報告されていなかった。
【0057】
L.acidophilus DPC6026は、カゼインに対するそのタンパク質分解能力、よって抗菌活性のような生物活性を有する多数のペプチドを作製するその能力を示した最初のアッセイに基づいて選択された。セクロピンP1は、ブタ小腸から単離されたペプチドであり、その活性の範囲が主にグラム陰性細菌に対するものであるので、対照として用いた(5)。
【0058】
配列番号1、配列番号2および配列番号3の化学合成ペプチド、ならびにこれらのペプチドを含む画分は、プロテイナーゼKで処理したときにその抗菌活性を失うことが見出された。トリプシンおよびキモトリプシンは、抗菌性粗画分で同定された化学合成ペプチドの全てを不活性化させた。Expasy Peptide cutterプログラム(http://ca.expasy.org/cgi−bin/peptidecutter/peptidecutter.pl)を用いて、ウシαs1−CNは、3つのエンドペプチダーゼおよびプロテイナーゼの組合せにより加水分解されて、抗菌ペプチド配列番号1および配列番号3を放出し得ることが見出された。プロリンエンドペプチダーゼは、ウシαs1−CNを、Pro20のC末端側で切断し、エンドペプチダーゼは、Glu29のC末端側で制限する。この酵素の組合せが、ウシαs1−CNから抗菌ペプチドIKHQGLPQE(配列番号1)を放出させるようである。プロリン特異的ペプチダーゼの存在が、カゼインからのプロリンに富むオリゴペプチドを分解するそれらの能力のために、LABの最適な成長に必要であり、プロリン特異的ペプチダーゼは、L.delbrueckii subsp bulgaricusから以前に単離されている(2)。ウシαs1−CNをArg37位で制限するArg−Cプロテイナーゼと、ウシαs1−CNをGlu29位で制限するエンドペプチダーゼとの組合せが、抗菌ペプチドVLNENLLR(配列番号3)を放出させるようである。
【0059】
理論に結び付けられることを望まないが、本明細書に記載される条件(0.1M NaOHで維持されたpH7、100rpm、37〜42℃)下で発酵に用いられたL.acidophilus DPC6026が、αs1−CNを、ペプチド配列番号1(IKHQGLPQE)および配列番号3(VLNENLLR)および配列番号2(SDIPIGSENSEK)に切断可能なArg−Cプロテイナーゼのような酵素(プロテイナーゼおよびペプチダーゼ)の新規な酵素の集団を生み出すと考えられている。この仮定は、コンピュータプログラムhttp://ca.expasy.org/cgi−bin/peptidecutter/peptidecutter.pl、およびαs1−CNを切断して配列番号1、配列番号2および配列番号3を製造できるという条件での酵素リストからの酵素を用いるαs1−Cnの切断分析に基づいている。
【0060】
ほとんどの抗菌ペプチドは、6〜50の間のアミノ酸残基を有する(48)。現在までに知られている最小の抗菌ペプチドは、ウシのラクトフェリシンから単離された配列R1RWQWRを有するヘキサペプチドと命名されている(46)。ほとんどが+2以上の正電荷を有し、疎水性および親水性の両方のドメインを有する両親媒性の構造に折り畳まれる(47)。これらの特徴により、グラム陰性細菌のリポ多糖領域と相互作用し、負に荷電した細胞質膜を透過することが可能になる(15)。
【0061】
配列データのみから、ペプチドの活性またはそれが形成し得る二次構造のいずれも予測することは困難であり得る(4)。ジスルフィド架橋を欠くペプチド(ペプチドIKHQGLPQE(配列番号1)およびVLNENLLR(配列番号3)のように)は、水中で無作為な構造を有し、膜に結合するかまたは自己凝集するときのみ、これらは構造を形成する(10)。しかし、これらのペプチドのいくつかの特徴は、作用機序が、カチオン性および中性の抗菌ペプチドについて以前に記載されている機構である(15、48)細胞質膜を介する自己促進型取り込みとそれに続く細胞質膜バリアとの干渉とを含み得ることを示唆する。
【0062】
特徴は、以下のことを含む(1)細菌の細胞質膜との最初の相互作用に重要なペプチドのカチオン性に寄与するアルギニンのような正に荷電したペプチド。(2)疎水性−これは、ペプチド配列番号1においてはアミノ酸イソロイシン、およびペプチド配列番号3においてはアミノ酸バリン(V)およびロイシン(L)によりもたらされる。本発明は、これらのペプチドの疎水性を増加させて、脂質アシル鎖と疎水性コアとの間の疎水性相互作用の増大によるこれらのペプチドの膜への結合を増加させることも提供する。
【0063】
結論
本研究は、L.acidophilus DPC6026株がカゼインからの抗菌ペプチドの作製に適し、該抗菌ペプチドのMIC濃度が商業的に合成されたペプチド(セクロピンP1)および既知の抗菌ペプチドであるイスラシジンに匹敵するので(22)、このLactobacillus株を用いて製造されたカゼインナトリウム発酵生成物を、抗菌性の利点を有する機能性食品の成分としてみなし得ることを示す。
【0064】
本研究は、ウシαs1−カゼインを、本研究で単離された広い範囲の抗菌ペプチドの前駆体としてみなすことができ、最終的には、これらのペプチドが、E.sakazakiiのような病原性細菌に対する組込まれた防御系として幼児調合乳製品に用いられる可能性を有し得ることも示す。
【0065】
Enterobacter sakazakii抑制効果を有するカゼインナトリウム粉末の製造方法の例
(1)カゼインナトリウム基質(5%w/v)に、(1%w/v)のL.acidophilus DPC6026を接種して、100rpmで混合しながら、0.1M NaOHの添加により維持する一定のpH7で、37℃にて24時間インキュベートしてよい。この発酵生成物を、次いで、80℃に加熱して培養物を不活性化し、これを次いで凍結乾燥して微細粉末を製造できる。
(2)カゼインナトリウム基質(5%w/v)に、(1%w/v)のL.acidophilus DPC6026を接種して、100rpmで混合しながら、0.1M NaOHの添加により維持する一定のpH7で、37℃にて24時間インキュベートしてよい。
(3)この発酵生成物を、次いで、80℃に加熱して培養物を不活性化し、その後、サイズ排除カラム10kDaを通してろ過して、10kDa未満のペプチドを含む粉末を作製してよい。この下流の処理により、活性ペプチドの活性が増大する。10kDa膜ろ過物を凍結乾燥し、−20℃で貯蔵して、真空包装する。
(4)上記のようにしてステップ(2)に従ってよいが、3kDa膜フィルタを用いて、ペプチド配列番号1および配列番号3の活性をさらに増大させてよい。
【0066】
上記の粉末は、10〜15%の濃度で乾燥乳ベースの幼児用調合乳に加えて、Enterobacter sakazakiiでの汚染およびこの病原菌を原因とするその後の大腸炎および髄膜炎を予防できる。
【0067】
上記のようにして製造されるEnterobacter sakazakii抑制粉末は、特に、カゼインベースの乳児用調合乳製品に加えた場合に、Enterobacter sakazakiiおよびその他の病原菌に対する内部の組込まれた防御機構を提供する。
【0068】
ある実施形態において、本発明のペプチドは、免疫無防備で早産の新生児の感染の顕著な原因であるEnterobacter sakazakii汚染に対する保護のために、乳ベースの幼児用調合乳の調製方法のための補助物質として別々または一緒に用い得る。
【0069】
本発明のペプチドは、αs1−CN(カゼイン)の最初の9残基が切断に耐性であり、チーズの熟成において苦味に寄与すると記載されているので(Pedersonら、1999年)、チーズ製造における苦味に寄与するために用いることができる。
【0070】
L.acidophilus DPC6026による、αs1−CN由来抗菌ペプチドの製造は、以前に報告されていない。作製されたペプチド配列は、種々の文献およびデータベース検索で見出されず、この点において新規であると考えられる。
【0071】
本発明に関連して本明細書で用いる場合、「含み/含む」の語および「有する/含有する」の語は、記載する特徴、整数、ステップまたは成分の存在が、1つまたは複数の他の特徴、整数、ステップ、成分もしくはそれらの群の存在または追加を排除しないことを明記するために用いられる。
【0072】
【数1】
【0073】
【数2】
【0074】
【数3】
【0075】
【数4】
表1:1未満〜20kDaの間のペプチドのサイズを生じる乳酸桿菌株を用いて導かれるカゼインナトリウムタンパク質由来加水分解画分
【0076】
【表1】
表2:ヒトの幼児および成人の糞便単離物、ならびにブタ小腸単離物の16S rDNA配列決定
【0077】
【表2】
DPC;Dairy Products Research Centre, Teagasc Moorepark,Fermoy,Co.アイルランド、コーク
a.Enterococcus種との相同性の百分率。
b.Lactobacillus種との相同性の百分率。
c.Staphylococcus種との相同性百分率。
【0078】
表3:L.acidophilus DPC6026により製造されたカゼインナトリウム加水分解物からの粗画分に含まれるペプチドの配列および対応するカゼイン(CN)断片
【0079】
【表3】
aアミノ酸について1文字記号を用いる。
b各ペプチドの推定質量について、平均質量を報告する。
【0080】
表4:カゼインナトリウム中でのL.acidophilus DPC6026の発酵に続いて合成された純粋ペプチドの阻害範囲
【0081】
【表4】
a.Dairy Products Research Centre、アイルランドコーク。
+++、直径2.0cmを超える阻害の領域が生じた。
++、直径1.5cmを超える阻害の領域が生じた。
+、直径1.0cmを超える阻害の領域が生じた。
−、領域は検出されず。
【図面の簡単な説明】
【0082】
【図1】制限酵素ApaIについてのPFGEの巨視的な制限パターン。 パルス時間は、示すゲルについて1〜15秒であった。レーン1は、低分子量PFGマーカー(1% LMPアガロースに埋め込まれたラムダDNA−Hind III断片およびラムダコンカテマーの混合物)、レーン2〜6は、Lactobacillus acidophilus DPC6026、L.johnsonii DPC6092、L.salivarius DPC6027、L.animalis DPC6028およびL.delbrueckii sp.bulgaricus DPC6104を含んでいた。
【図2】GRAS株のタンパク質分解活性。 ウシカゼインナトリウム(A)、Lactobacillus johnsonii DPC6092(B)、L.salivarius DPC6027(C)およびL.acidophilus DPC6026(D)での発酵後のそのカゼインナトリウム加水分解物の高性能液体クロマトグラフィー(HPLC)のペプチドプロファイル。HPLCは、214nmにて24時間監視した。試料30μlを、1ml/分の流速のDelta−Pak C−18(600mm×7.5cm)に添加した。タンパク質分解は、室温で行った。L.acidophilus DPC6026は、カゼインナトリウムのほぼ完全な分解を示した(D)。
【図3】A.L.acidophilus DPC6026と24時間インキュベートしたpH7でのカゼインナトリウムの逆相高性能液体クロマトグラフィー(RP−HPLC)のクロマトグラム。矢印は、ペプチド画分A1−45およびA1−54の位置を示す。RP−HPLCは、室温で、材料および方法の部分に示す条件に従って行った。B.画分A1−45(基質としてのカゼインナトリウムおよびL.acidophilus DPC6026株を用いて単離した)の、ペプチドが検出されたm/z領域で記録された飛行時間型マトリクス支援レーザ脱離イオン化法(MALDI−TOF)スペクトル。ペプチド画分A1−45は、質量1049.43のペプチドを含有することが見出され、これは、配列決定の後に、αs1−CN f(21−29)に相当するIKHQGLPQEであるとわかった。1.L.acidophilus DPC6026でのカゼインナトリウム加水分解物の画分A1−45中で同定される化学合成ペプチドIKHQGLPQEによるE.coli DPC6053の阻害。寒天ウェル拡散アッセイ法を用いて、直径2.5cmの領域を記録した。2.L.acidophilus DPC6026でのカゼインナトリウム加水分解物の画分A1−45中で同定される化学合成ペプチドIKHQGLPQEによるEnterobacter sakazakii5920(ATCC128682)の阻害。寒天ウェル拡散アッセイ法を用いて、直径2.0cmの領域を記録した。C.画分A1−54(基質としてのカゼインナトリウムおよびL.acidophilus DPC6026株を用いて単離した)の、ペプチドが検出されたm/z領域で記録された飛行時間型マトリクス支援レーザ脱離イオン化法(MALDI−TOF)スペクトル。ペプチド画分A1−54は、質量970.119のペプチドを含有することが見出され、これは、配列決定の後に、αs1−CN f(30−37)に相当するVLNENLLRであるとわかった。1.L.acidophilus DPC6026でのカゼインナトリウム加水分解物の画分A1−54中で同定される化学合成ペプチドVLNENLLRによるE.coli DPC6053の阻害。寒天ウェル拡散アッセイ法を用いて、直径1.5cmの領域を記録した。2.L.acidophilus DPC6026でのカゼインナトリウム加水分解物の画分A1−54中で同定される化学合成ペプチドVLNENLLRによるEnterobacter sakazakii8272(NCTC8155)の阻害。寒天ウェル拡散アッセイ法を用いて、直径2.2cmの領域を記録した。
【図4A】A.96ウェルプレートアッセイを用いた、E.coli DPC6053(LB培地で1:10に希釈した一晩培養物)のOD600に対する種々の濃度のイスラシジンの影響。セクロピンP1を対照ペプチドとして用いる。記号:■、セクロピンP1(0.52mM)とインキュベートしたE.coli DPC6053。−、何も添加しないE.coli DPC6053対照。◆、イスラシジン(1.9mM)とインキュベートしたE.coli DPC6053。×、無菌性対照(何も添加しないLB培地)。●、0.23mMの濃度のイスラシジンとインキュベートしたE.coli DPC6053。X、イスラシジン(0.059mM)とインキュベートしたE.coli DPC6053の生存度。
【図4B】96ウェルプレートアッセイを用いた、E.coli DPC6053(LB培地で1:10に希釈した一晩培養物)のOD600に対する種々の濃度のペプチドIKHQGLPQEの影響。記号:−、何も添加しないE.coli DPC6053。X、IKHQGLPQE(0.078mM)とインキュベートしたE.coli DPC6053。■、セクロピンP1(0.52mM)とインキュベートしたE.coli DPC6053。●、IKHQGLPQE(0.15mM)とインキュベートしたE.coli DPC6053。・、IKHQGLPQE(0.31mM)とインキュベートしたE.coli DPC6053。◆、IKHQGLPQE(0.625mM)とインキュベートしたE.coli DPC6053。x、無菌性対照(何も添加しないLB培地)。
【図4C】96ウェルプレートアッセイを用いた、E.coli DPC6053(LB培地で1:10に希釈した一晩培養物)のOD600に対する種々の濃度のペプチドVLNENLLRの影響。記号:X、何も添加しないE.coli DPC6053。■、VLNENLLR(0.22mM)とインキュベートしたE.coli DPC6053。−、VLNENLLR(0.45mM)とインキュベートしたE.coli DPC6053。◆、VLNENLLR(1.2mM)とインキュベートしたE.coli DPC6053。X、セクロピンP1(0.52mM)とインキュベートしたE.coli DPC6053の生存度。●、VLNENLLR(2.5mM)とインキュベートしたE.coli DPC6053。▲、無菌性対照−何も添加しないLB培地。
【特許請求の範囲】
【請求項1】
配列番号1、配列番号2および配列番号3からなる群より選択される1つまたは複数のペプチドを含む抗菌化合物。
【請求項2】
試料が2005年11月18日にアクセッション番号NCIMB41354の下でスコットランド、アバディーンのナショナル・コレクション・オブ・インダストリアル・アンド・マリン・バクテリア(National Collection of Industrial and Marine Bacteria)に寄託されているLactobacillus acidophilus DPC6026株、または乳もしくは乳製品から抗菌活性を有するペプチドを製造できるその派生株もしくは変異株の生物学的に純粋な培養物。
【請求項3】
請求項2に記載の培養物から得られる無細胞培養上清またはその画分。
【請求項4】
DPC6026により製造される化合物であって、乳または乳製品から1つまたは複数の抗菌ペプチドを製造できる化合物。
【請求項5】
前記抗菌ペプチドが、配列番号1、配列番号2および配列番号3からなる群より選択される1つまたは複数である請求項4に記載の化合物。
【請求項6】
請求項1に記載の抗菌化合物、請求項2に記載の培養物、請求項3に記載の上清および請求項4または請求項5に記載の化合物からなる群より選択される1つまたは複数を含む抗菌組成物。
【請求項7】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物からなる群のうちの1つまたは複数の、微生物感染または微生物汚染の管理における使用。
【請求項8】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物からなる群のうちの1つまたは複数を、動物またはヒトに投与することを含む、微生物感染または微生物汚染を防止または治療または制限する方法。
【請求項9】
前記微生物感染または微生物汚染が、Escherichia coli、E.sakazakii、Streptococcus mutans、Listeria innocua、Klebsiella sppおよびStaphylococcus carnosusからなる群のうちの1つまたは複数を原因とする汚染または感染である請求項7に記載の使用または請求項8に記載の方法。
【請求項10】
前記感染または汚染が、乳腺炎である請求項7もしくは請求項9に記載の使用または請求項6に記載の方法。
【請求項11】
前記感染または汚染が、髄膜炎である請求項7もしくは請求項9に記載の使用または請求項8に記載の方法。
【請求項12】
前記化合物、株、上清または組成物が、乳をベースとする調合乳に供給される請求項11に記載の使用または方法。
【請求項13】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物の、アンギオテンシン−I変換酵素(ACE)の阻害、オピオイド調節、免疫調節および抗血栓症治療から選択される群のうちの1つまたは複数における使用。
【請求項14】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物を、動物またはヒトに投与することを含む、アンギオテンシン−I変換酵素(ACE)の阻害、オピオイド調節、免疫調節または血栓症抑制の方法。
【請求項15】
乳または乳製品に、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物および請求項6に記載の組成物からなる群のうちの1つまたは複数を加えることを含む、乳または乳製品から抗菌ペプチドを製造する方法。
【請求項16】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物および請求項6に記載の組成物の1つまたは複数を含む食品。
【請求項17】
前記食品が、幼児用調合乳、粉乳、ヨーグルト、チーズ、生菌飲料、乳児用食品調合乳、乳ベースの飲料および食品補助物質からなる群より選択される1つまたは複数である請求項16に記載の食品。
【請求項18】
治療有効量の、請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物のうちの1つまたは複数を含む医薬組成物。
【請求項19】
添付の図面および表を参照して本明細書に実質的に記載される化合物、株、上清、組成物、タンパク質、方法、使用、食品または医薬組成物。
【請求項1】
配列番号1、配列番号2および配列番号3からなる群より選択される1つまたは複数のペプチドを含む抗菌化合物。
【請求項2】
試料が2005年11月18日にアクセッション番号NCIMB41354の下でスコットランド、アバディーンのナショナル・コレクション・オブ・インダストリアル・アンド・マリン・バクテリア(National Collection of Industrial and Marine Bacteria)に寄託されているLactobacillus acidophilus DPC6026株、または乳もしくは乳製品から抗菌活性を有するペプチドを製造できるその派生株もしくは変異株の生物学的に純粋な培養物。
【請求項3】
請求項2に記載の培養物から得られる無細胞培養上清またはその画分。
【請求項4】
DPC6026により製造される化合物であって、乳または乳製品から1つまたは複数の抗菌ペプチドを製造できる化合物。
【請求項5】
前記抗菌ペプチドが、配列番号1、配列番号2および配列番号3からなる群より選択される1つまたは複数である請求項4に記載の化合物。
【請求項6】
請求項1に記載の抗菌化合物、請求項2に記載の培養物、請求項3に記載の上清および請求項4または請求項5に記載の化合物からなる群より選択される1つまたは複数を含む抗菌組成物。
【請求項7】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物からなる群のうちの1つまたは複数の、微生物感染または微生物汚染の管理における使用。
【請求項8】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物からなる群のうちの1つまたは複数を、動物またはヒトに投与することを含む、微生物感染または微生物汚染を防止または治療または制限する方法。
【請求項9】
前記微生物感染または微生物汚染が、Escherichia coli、E.sakazakii、Streptococcus mutans、Listeria innocua、Klebsiella sppおよびStaphylococcus carnosusからなる群のうちの1つまたは複数を原因とする汚染または感染である請求項7に記載の使用または請求項8に記載の方法。
【請求項10】
前記感染または汚染が、乳腺炎である請求項7もしくは請求項9に記載の使用または請求項6に記載の方法。
【請求項11】
前記感染または汚染が、髄膜炎である請求項7もしくは請求項9に記載の使用または請求項8に記載の方法。
【請求項12】
前記化合物、株、上清または組成物が、乳をベースとする調合乳に供給される請求項11に記載の使用または方法。
【請求項13】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物の、アンギオテンシン−I変換酵素(ACE)の阻害、オピオイド調節、免疫調節および抗血栓症治療から選択される群のうちの1つまたは複数における使用。
【請求項14】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物を、動物またはヒトに投与することを含む、アンギオテンシン−I変換酵素(ACE)の阻害、オピオイド調節、免疫調節または血栓症抑制の方法。
【請求項15】
乳または乳製品に、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物および請求項6に記載の組成物からなる群のうちの1つまたは複数を加えることを含む、乳または乳製品から抗菌ペプチドを製造する方法。
【請求項16】
請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物および請求項6に記載の組成物の1つまたは複数を含む食品。
【請求項17】
前記食品が、幼児用調合乳、粉乳、ヨーグルト、チーズ、生菌飲料、乳児用食品調合乳、乳ベースの飲料および食品補助物質からなる群より選択される1つまたは複数である請求項16に記載の食品。
【請求項18】
治療有効量の、請求項1に記載の化合物、請求項2に記載の株、請求項3に記載の上清、請求項4または請求項5に記載の化合物、および請求項6に記載の組成物のうちの1つまたは複数を含む医薬組成物。
【請求項19】
添付の図面および表を参照して本明細書に実質的に記載される化合物、株、上清、組成物、タンパク質、方法、使用、食品または医薬組成物。
【図1】

【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【公表番号】特表2009−516738(P2009−516738A)
【公表日】平成21年4月23日(2009.4.23)
【国際特許分類】
【出願番号】特願2008−541905(P2008−541905)
【出願日】平成18年11月20日(2006.11.20)
【国際出願番号】PCT/IE2006/000130
【国際公開番号】WO2007/057872
【国際公開日】平成19年5月24日(2007.5.24)
【出願人】(508152375)
【Fターム(参考)】
【公表日】平成21年4月23日(2009.4.23)
【国際特許分類】
【出願日】平成18年11月20日(2006.11.20)
【国際出願番号】PCT/IE2006/000130
【国際公開番号】WO2007/057872
【国際公開日】平成19年5月24日(2007.5.24)
【出願人】(508152375)
【Fターム(参考)】
[ Back to top ]