
カテプシンE特異的阻害剤
【課題】カテプシンEを特異的に阻害し、かつカテプシンEと類縁性の高いリソソーム酵素であるカテプシンDは阻害しない物質の提供。
【解決手段】特定の塩基配列を有するいずれかのDNA(可変領域)の3'末端に20〜40マーの全部または一部が二本鎖のDNA(テール領域)を結合したDNA複合体を提供する。また、可変領域と結合する末端と反対側のテール領域の末端がテールまたはループを形成しているDNA複合体を提供する。これらのDNA複合体は、カテプシンE阻害活性を有する。
【解決手段】特定の塩基配列を有するいずれかのDNA(可変領域)の3'末端に20〜40マーの全部または一部が二本鎖のDNA(テール領域)を結合したDNA複合体を提供する。また、可変領域と結合する末端と反対側のテール領域の末端がテールまたはループを形成しているDNA複合体を提供する。これらのDNA複合体は、カテプシンE阻害活性を有する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、カテプシンEに対して特異的な阻害作用を示すDNAおよびこのDNAを用いたカテプシンE阻害剤に関する。
【背景技術】
【0002】
カテプシンEは免疫担当細胞、とくに抗原提示細胞であるマクロファージ、樹状細胞などに限局的に存在する細胞内アスパラギン酸プロテアーゼである。山本らは、カテプシンE遺伝子欠損マウス(ノックアウトマウス)を作製し、本酵素の機能解析を進めてきた(Yamamoto, K. Cathepsin E and cathepsin D: biosynthesis, processing and subcellular location. Adv. Exp. Med. Biol., 362, 223-9 (1995)、非特許文献1)。カテプシンE欠損マウスは無菌環境下で飼育しても、全く異常を示さないが、通常環境下に移すとアトピー性皮膚炎様症状を示した。病理組織学的にもヒトにおけるアトピー性皮膚炎症状と酷似した症状を示した。カテプシンE欠損マウスから調製したマクロファージを用いた研究では、正常マクロファージに比べて、殺菌能、細胞生存率、遊走能などの細胞機能に著明な低下があることが示されている。このことは、カテプシンE欠損が宿主の免疫応答自体に低下や不調をもたらし、それが何らかの外的刺激によってアトピー性皮膚炎などの個体の病態を発症させていることを示唆している。事実、カテプシンE欠損マウスでは、慢性関節リウマチのモデルである実験的関節炎が誘導されると、野生型マウスに比べてその病態に著しい増悪が見られることが示されている。カテプシンE欠損が炎症性サイトカインの分泌パターンを変化させ、関節炎を増悪させることが考えられる。一方、正常マウス(DBA/1J)にコラーゲン関節炎を起こさせると、血清中のカテプシンE活性は対照マウスの約2.5倍(p<0.001)に上昇していることが示されている。(筑波 隆幸、山本 健二:アトピー性皮膚炎とカテプシンE. 日薬理誌(Folia Pharmacol. Jpn.) 122, 15-20 (2003)、非特許文献2)このことは、カテプシンE欠損という極端なケースを除けば、慢性関節リウマチなどの自己免疫疾患等においては、一般的にカテプシンEの活性増加が生じていると考えられる。このことは、慢性関節リウマチ患者の滑液中のカテプシンE活性が病態に比例して増大するという山本らの報告とよく一致する。
【0003】
一方、各種癌細胞を用いた実験から、カテプシンEが癌細胞表面のTRAIL (TNF-related apoptosis-inducing ligand)を特異的に切断遊離させ、TRAILとその受容体を介するシグナリングによって癌細胞をアポトーシスに誘導することが明らかにされている(山本ら:未発表)。TRAILはCD4+ T細胞、NK細胞、単球、樹状細胞、好中球などにも発現していることから、これらの細胞を介して癌細胞をアポトーシスに誘導する一方で、自らの細胞をもアポトーシスに誘導して宿主免疫系を破綻させる可能性も示唆されている。このことは、カテプシンEの過剰発現がTRAILを介するアポトーシス誘導機構を惹起させ、Th1型の免疫反応に破綻を誘導し、自己免疫疾患等の種々の疾患を生む可能性を強く示唆している。
【0004】
以上のような知見から、カテプシンE活性の増大は、自己免疫疾患等の様々な病態に繋がると考えられ、その際の過剰なカテプシンE活性を抑える薬剤はこうした病態の改善に大いに役立つものと期待される。カテプシンEに対する阻害剤については、これまで放線菌由来のペプスタチンA(Umezawa, H., Aoyagi, T., Morishima, H., Matsuzaki, M,.and Hamada, M.Pepstatin, a new pepsin inhibitor produced by Actinomycetes. J. Antibiot., 23, 259-62 (1970)、非特許文献3)と回虫から分離されたAscaris pepsin inhibitor(Kageyama, T. Molecular cloning, expression and characterization of an Ascaris inhibitor for pepsin and cathepsin E. Eur. J. Biochem., 253, 804-9 (1998)、非特許文献4)が知られている。
【非特許文献1】Yamamoto, K. Cathepsin E and cathepsin D: biosynthesis, processing and subcellular location. Adv. Exp. Med. Biol., 362, 223-9 (1995)
【非特許文献2】筑波 隆幸、山本 健二:アトピー性皮膚炎とカテプシンE. 日薬理誌(Folia Pharmacol. Jpn.) 122, 15-20 (2003)
【非特許文献3】Umezawa, H., Aoyagi, T., Morishima, H., Matsuzaki, M,.and Hamada, M.Pepstatin, a new pepsin inhibitor produced by Actinomycetes. J. Antibiot., 23, 259-62 (1970)
【非特許文献4】Kageyama, T. Molecular cloning, expression and characterization of an Ascaris inhibitor for pepsin and cathepsin E. Eur. J. Biochem., 253, 804-9 (1998)
【発明の開示】
【発明が解決しようとする課題】
【0005】
放線菌由来のペプスタチンAはアプパラギン酸プロテアーゼ群に共通の阻害剤で特異性がない。回虫ペプシン阻害剤(Ascaris pepsin inhibitor)はペプシンに対すると同様にカテプシンEを強く阻害し、他のアプパラギン酸プロテアーゼは阻害しないが、高分子タンパク質で細胞毒性が強いという欠点がある。そのため、カテプシンEに特異的な阻害剤として実用的なものは知られていないのが現状である。
【0006】
そこで本発明の目的は、カテプシンEを特異的に阻害し、かつカテプシンEと類縁性の高いリソソーム酵素であるカテプシンDは阻害しない、物質を提供することにある。
【課題を解決するための手段】
【0007】
本発明者らは、かねてより、「高速分子進化法」に基づき、従来からのDNAアプタマー調製法より格段に高速化された方法で上記阻害作用を有する物質を探査した。その結果、カテプシンDに対しては殆ど阻害活性を持たず、カテプシンEを特異的に阻害するDNAを見いだし、本発明を完成させた。
【0008】
本発明は、下記の何れかのDNAに関する。
(1)配列番号1〜20のいずれかの塩基配列を有するDNA、
(2)配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA、および
(3)配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA。
上記において、30マーの二本鎖DNAの一方の一本鎖は、配列番号21の塩基配列を有することができる。
【0009】
さらに本発明は、上記本発明のDNA(以下、可変領域という)の3'末端に20〜40マーの全部または一部が二本鎖のDNA(以下、テール領域という)を結合したDNA複合体に関する。
上記DNA複合体においては、テール領域が30マーの二本鎖DNAであることができ、さらに、二本鎖DNAであるテール領域の可変領域の3'末端に結合している一本鎖は、配列番号21の塩基配列を有することができる。
【0010】
さらに本発明は、上記本発明のDNA複合体を有効成分とするカテプシンE阻害剤に関する。
【発明の効果】
【0011】
本発明のDNA複合体は、カテプシンEに対して高い特異阻害活性を示し、かつカテプシンEと類縁性の高いリソソーム酵素であるカテプシンDは阻害せず、副作用等の点からも治療薬としての有用性が高いと期待できる。
【0012】
DNA複合体は分子の合成が容易である上に、生体内でRNAに較べ耐性がある。分子の改変も比較的容易であり、種々の修飾基の導入が可能である。さらに、得られた分子を手がかりにして、さらに高機能なDNA分子を進化させることも可能である。生体内にタンパク質ほど分解酵素が存在しないと考えられるので、寿命的にも有利な可能性がある。
【発明を実施するための最良の形態】
【0013】
本発明のDNAは、下記の何れかのDNAである。
(1)配列番号1〜20のいずれかの塩基配列を有するDNA、
(2)配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA、および
(3)配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA。
【0014】
配列番号1〜20で示される塩基配列を有するDNAは、いずれも以下の分子淘汰操作により得た。即ち、まずブロックシャフリング法でDNAライブラリーを作成し、それを用い、ターゲット分子であるカテプシンEに対する親和性で優れた分子を淘汰し、さらに、阻害活性で淘汰するという作業を行って、目的分子の淘汰を行い、上記DNAを得た。具体的には、以下の手順で行った。
【0015】
1)DNAライブラリーの作成
7種のアミノ酸に対応するコードをもつDNAライブラリーを作成した(表1参照)。このとき、ライブラリーの合成にはブロック合成法(YLBS法)を用いた。
【0016】
【表1】

【0017】
2)カテプシンE結合DNAアプタマーの淘汰
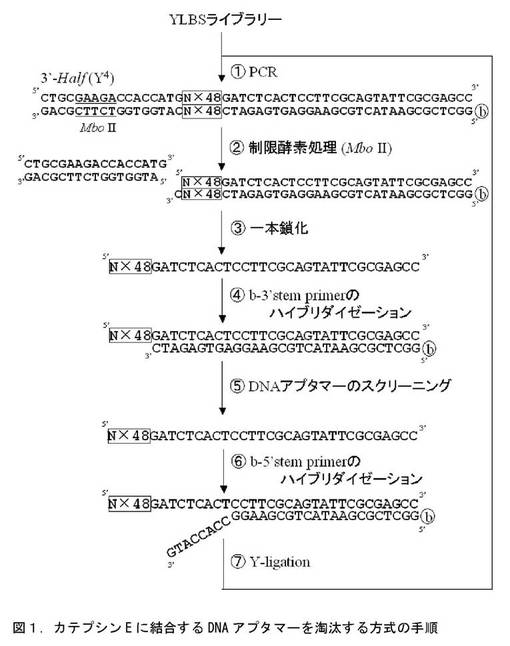
図1に示す操作手順に従って、カテプシンEに強く結合するDNAアプタマー群を淘汰した。
【0018】
3)SFリンク法により、カテプシンE阻害活性を持つ分子の淘汰
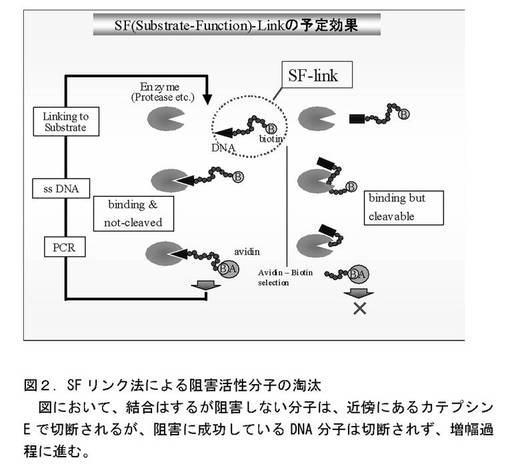
図2に示すSFリンク法により、結合はするが阻害活性のない分子は除き、カテプシンE阻害活性の高まったDNA分子群を得た。
【0019】
4)得られたDNA分子群について、適宜、結合強度、阻害強度、分子の分光学的特性などを解析した。
【0020】
上記分子淘汰操作(SFリンク法)を、条件を変えて複数回(6回)行い、カテプシンE阻害活性を持つ分子として、配列番号1〜20で示される塩基配列を有するDNAを得た。詳細は実施例に記載する。
【0021】
本明細書で言う「1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列」における「1から数個」の範囲は特には限定されないが、例えば、1から5個、好ましくは1から4個、より好ましくは1から3個、より好ましくは1から2個、さらに好ましくは1個を意味する。
【0022】
上記した「ストリンジェントな条件下でハイブリダイズする」とは、DNAをプローブとして使用し、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法、あるいはサザンブロットハイブリダイゼーション法等を用いることにより得られるDNAの塩基配列を意味し、例えば、コロニーあるいはプラーク由来のDNA又は該DNAの断片を固定化したフィルターを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2×SSC溶液(1×SSC溶液は、150mM塩化ナトリウム、15mMクエン酸ナトリウム)を用い、65℃条件下でフィルターを洗浄することにより同定できるDNA等を挙げることができる。ハイブリダイゼーションは、モレキュラークローニング第2版等に記載されている方法に準じて行うことができる。
【0023】
ストリンジェントな条件下でハイブリダイズするDNAとしては、プローブとして使用するDNAの塩基配列と一定以上の相同性を有するDNAが挙げられ、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは93%以上、特に好ましくは95%以上、最も好ましくは98%以上の相同性を有するDNAが挙げられる。
【0024】
本発明のDNAの取得方法は特に限定されない。本発明のDNAは、本明細書中の配列表の配列番号1から20に記載した塩基配列の情報に基づいて化学合成法(月刊「BIOINDUSTRY」1995 年8 月号参照)等により調製することができる。DNAの調製の操作は当業者に既知であり、例えば、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989(以下、モレキュラークローニング第2版と略す)、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)等に記載の方法に準じて行うことができる。
【0025】
本発明は、前述のように、(2)配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA、および(3)配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNAを包含する。配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有するDNAおよび配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有するDNAは、3'末端に30マーの二本鎖DNAを結合した複合体とした場合に、カテプシンE阻害活性を有するものであれば、本発明の範囲に含まれる。30マーの二本鎖DNAの一方の一本鎖は、配列番号21の塩基配列を有するものであることができる。
【0026】
3'末端に30マーの二本鎖DNAを結合したDNA複合体のカテプシンE阻害活性は、以下のように検定することができる。
(1)セレクションバッファー(50mM酢酸ナトリウム(pH4.5), 5mM MgCl2, 0.1M NaCl)86μLとカテプシンE溶液(3.36mM)3μLを混ぜたものを室温で5分間インキュベーション。
(2)この溶液にライブラリのPCR産物を一本鎖化したもの(非ビオチン鎖側、10pmol/μL)を1μL加え、室温で15分間インキュベーション。(ブランクとして水1μLを加えたものも同時に作製)
(3)この溶液にカテプシンE基質((株)ペプチド研 Code:3200-V)10μLを加え、40℃で10分間反応。
(4)10分後、すぐに氷中に移し、5%トリクロロ酢酸 1mLを加え反応を止める。
(5)この溶液を分光蛍光光度計で測定する(328nm Excitation, 396nm Emission)。カテプシンEの活性が高いもの程396nmのピークが高くなるので、ブランクとの値を比較することによって阻害活性が求まる。
【0027】
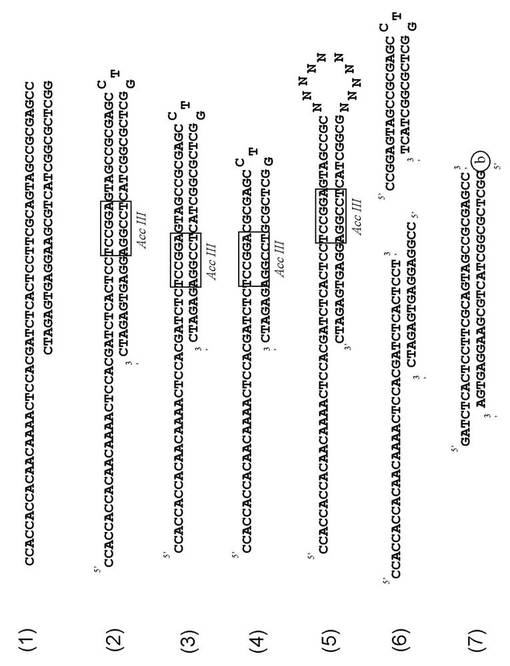
本発明は、上記配列番号1から20に記載したいずれかの塩基配列を有するDNA(可変領域)の3'末端に20〜40マーの全部または一部が二本鎖のDNA(テール領域)を結合したDNA複合体を包含する。テール領域の塩基数は、DNA複合体のカテプシンE阻害活性に影響するので、可変領域の種類に応じて、最適の塩基数を選択することが好ましい。テール領域の塩基配列も、適宜選択することができる。テール領域は、好ましくは25〜35マー、より好ましくは30マーの二本鎖DNAである。さらに、二本鎖DNAであるテール領域の、可変領域の3'末端に結合している一本鎖は、配列番号21の塩基配列を有することが好ましい。可変領域の3'末端に結合しているテール領域の鎖が配列番号21の塩基配列を有するDNA複合体の例(SF-R6#3 with tail)を図3の(1)に示す。
【0028】
図3の(1)に示すDNA複合体は、テール領域が2本のDNAがハイブリダイゼーションしているものである。それに対して、テール領域の2本のDNAは、テール領域の末端(可変領域と結合する末端と反対側の末端)で、テール(図3の(2)〜(4))、ループ(図3の(5))、または一部が一本鎖 (図3の(6)の左側のDNA複合体)を形成していてもよい。テールおよびループを形成する塩基数は特に制限はないが、例えば、3〜15の範囲であることができる。図3の(2)〜(4)に示す末端がループであるDNA複合体におけるループは、塩基数が3であり、CTGの塩基配列を有する。また、図3の(5)に示す末端がループであるDNA複合体におけるループは、塩基数が10である。このループの塩基配列はNで示され、NはA、T、G、Cの内どれか1つがランダムに出現していることを示す。図3の(6) に示すDNA複合体の部分的一本鎖(図中の左側のDNA複合体の右末端)の塩基数も特に制限はないが、例えば、1〜10の範囲であることができる。
【0029】
本発明のDNA複合体は、例えば、図3の(1)に示すDNA複合体のように、配列番号1から20に記載したいずれかの塩基配列を有するDNA(可変領域)の3'末端に、例えば、30マーの二本鎖のDNA(テール領域)を結合したDNA複合体であるが、それ以外に、図3の(2)〜(6)に示すDNA複合体のように、テール領域の末端にループを有するDNA複合体、及びテール領域の末端が部分的一本鎖であるDNA複合体であることもできる。
【0030】
本発明は、上記本発明のDNA複合体を有効成分とするカテプシンE阻害剤を包含する。本発明のDNA複合体を有効成分とするテプシンE阻害剤は、実験試薬として使用できる他、治療薬としても使用できる。
【0031】
例えば、カテプシンEの生理作用研究などにおいて、カテプシンE阻害剤としてペプスタチンAが使われている (Chain, B. M., Free, P., Medd, P., Swetman, C., Tabor, A. B. and Terrazzini, N.The expression and function of cathepsin E in dendritic cells. J. Immunol., 174, 1791-800 (2005). Cappiello, M. G., Wu, Z., Scott, B. B., McGeehan, G. M. and Harrison, R. K. Purification and characterization of recombinant human cathepsin E expressed in human kidney cell line 293. Protein Expr. Purif., 37, 53-60 (2004). )。しかし、ペプスタチンAはカテプシンDやペプシンも阻害するためにカテプシンE単独の効果を調べることが困難であった。それに対して本発明のDNA複合体は、カテプシンDを阻害しないので、カテプシンE単独の効果を調べることを可能にする。その結果、カテプシンEに起因する種々の病因を除く可能性がある。既に、インスリン依存性糖尿病や慢性関節リウマチの発症進展に、カテプシンE活性の亢進が見られる可能性が示唆されており (筑波 隆幸、山本 健二:アトピー性皮膚炎とカテプシンE. 日薬理誌(Folia Pharmacol. Jpn.) 122, 15-20 (2003).) 、カテプシンE特異的な阻害活性を持つ本発明のDNA複合体は、そのような疾患の治療薬となることが期待される。
【実施例】
【0032】
以下、本発明を実施例によりさらに詳細に説明する。
【0033】
例1
1)PCR 増幅
未淘汰のDNAライブラリをテンプレートとして以下に示す方法でPCR増幅した。
Taq ポリメラーゼ (Greiner, Tokyo, Japan) 1U
10 ×PCR バッファ 5μL
[670mM Tris-HCl, 166mM (NH4)2SO4, 4.5% TritonX100, 2mg/mLゼラチン]
MgCl2 (25mM) 5μL
dNTP (2.5mM) 4μL
テンプレート DNAライブラリ (1fmole以上)
プライマー b-3' stemプライマー(100pmol/μL) 0.5μL
5' partプライマー(100pmol/μL) 0.5μL
H2O 全体で50μL
【0034】
プログラム
90℃ 2分
90℃ 30秒
45℃ 1分
72℃ 30秒
90℃ 30秒、45℃ 1分及び72℃ 30秒を30サイクル行い、得られた増幅物は電気泳動法で確認し、エタノール沈殿で精製する。
【0035】
2)制限酵素処理 (Mbo II)
反応混合物:
Mbo II (Takara) 8U
10 × L Buffer 5μL
[100mM Tris-HCl (pH7.5), 100mM MgCl2, 10mM ジチオスレイトール]
DNA 50pmol
H2O 全体で 50μL
37℃で 1時間インキュベートする。
【0036】
3)一本鎖DNA の調製
a. DNAのストレプトアビジン磁気ビーズへの結合
制限酵素消化物 50μL
ストレプトアビジン磁気ビーズ 50μL
[Dynabeads M-280 Streptavidin (Dynal, Oslo, Norway)]
懸濁物を30分間振とうする。
磁石でビーズをチューブ底に固定し、上清を捨てる。
【0037】
b. 0.01% BSA水溶液による洗浄
100μL の 0.01% BSAを添加し、良く混合する。
磁石でビーズをチューブ底に固定し、上清を捨てる。
この操作を2回繰り返す。
【0038】
c. 二本鎖DNAの変性および非ビオチン標識一本鎖DNAの回収
室温で2分間50 μLの25% (w/v) アンモニア水で処理し、磁石でビーズをチューブ底に固定した後、上清を回収する。(非ビオチン標識一本鎖DNAは上清中に存在する)
この操作を2回繰り返す。
回収液を乾燥する。
【0039】
4)プライマーb-3'stemのハイブリダイゼーション
回収した ssDNAを 95μL のセレクションバッファ(以下参照)に溶解する。
5μL のb-3'stem プライマー溶液 (10pmol/μL)を添加する。
94℃(5 分) および 60℃(15分)の加熱処理でDNAをハイブリダイズする。
【0040】
5)DNA アプタマーの試験管内スクリーニング
材料:
1.セレクションバッファ:
50 mM 酢酸ナトリウム (pH 4.5)
100 mM NaCl
5 mM MgCl2
2.洗浄バッファ 1:
50 mM酢酸ナトリウム(pH 4.5)
0.5 M NaCl
3.洗浄バッファ2:
50 mM酢酸ナトリウム(pH 4.5)
1 M NaCl
4.溶出バッファ1:
50 mM酢酸ナトリウム(pH 4.5)
1 M NaCl
10 mM MgCl2
5.溶出バッファ2:
50 mM酢酸ナトリウム(pH 4.5)
2 M NaCl
10 mM MgCl2
【0041】
操作:
ステップ 1. セレクションバッファ中のDNA を固定化酵素と20-30 分間反応させる。
DNA: 100 μL (セレクションバッファ)
酵素: 5 μL (固定化操作および活性検定により調製したもの)
ここで、DNA : 酵素モル比 = 4 : 1 にする。
【0042】
ステップ 2. セレクションバッファによる洗浄
上記反応液に 200 μL のセレクションバッファを添加し、チューブを数回穏やかにタッピングする。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し捨てる。
以下の表2に記したようにこの操作を繰り返す。
【0043】
ステップ 3. 洗浄バッファ 1による洗浄
ステップ2の洗浄および遠心第5ラウンドの後、200 μL の洗浄バッファ 1を添加し、チューブを数回穏やかにタッピングする。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し捨てる。
以下の表2に記したようにこの操作を繰り返す。
【0044】
ステップ 4. 洗浄バッファ2による洗浄
このステップには二種類の操作が含まれる:
A) 200 μL の洗浄バッファ 2 を添加し、チューブを数回穏やかにタッピングする。 時々タッピングしながら室温で5分間反応させる。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し捨てる。
以下の表2に記したようにこの操作を繰り返す。
【0045】
B) 200 μL の洗浄バッファ 2 を添加し、チューブを数回穏やかにタッピングする。15000 rpm で1 分間遠心した後、上清を静かにピペットで吸い出し捨てる。
以下の表2に記したようにこの操作を繰り返す。
【0046】
ステップ 5. 溶出バッファ 1による溶出
最初に 37oCの恒温槽を準備する。100 μL の溶出バッファ 1 を添加して、チューブを数回穏やかにタッピングする。チューブを恒温槽に10分間浸す。2-5 分間間隔で穏やかにタッピングし、再び恒温槽に戻す。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し、きれいなチューブに回収する。
この操作を2回繰り返す。
【0047】
ステップ 6. 溶出バッファ2による溶出
最初に95℃の恒温槽を準備する。100 μL の溶出バッファ 2 を添加して、チューブを数回穏やかにタッピングする。チューブを恒温槽に5分間浸す。2分間間隔で穏やかにタッピングし、再び恒温槽に戻す。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し、きれいなチューブに回収する。
この操作を2回繰り返す。
【0048】
回収した上清からエタノール沈殿でDNAを精製する。
ただし、塩濃度が高い場合は、70%エタノールで2、3回ペレットを洗浄する。
【0049】
6)一本鎖DNA の調製 ( (3)と同様)
a. DNAのストレプトアビジン磁気ビーズへの結合
回収DNAに対し、以下のものを添加する。
水 10μL
ストレプトアビジン磁気ビーズ 10μL
[Dynabeads M-280 Streptavidin (Dynal, Oslo, Norway)]
懸濁物を30分間振とうする。
磁石でビーズをチューブ底に固定し、上清を捨てる。
【0050】
b. 0.01% BSA水溶液による洗浄
100μL の 0.01% BSAを添加し、良く混合する。
磁石でビーズをチューブ底に固定し、上清を捨てる。
この操作を2回繰り返す。
【0051】
c. 二本鎖DNAの変性および非ビオチン標識一本鎖DNAの回収
室温で2分間50 μLの25% (w/v) アンモニア水で処理し、磁石でビーズをチューブ底に固定した後、上清を回収する。(非ビオチン標識一本鎖DNAは上清中に存在する)
この操作を2回繰り返す。
回収液を乾燥する。
【0052】
7)ハイブリダイゼーション
回収した 一本鎖DNAを 2μL の水に溶解する。 1μL のプライマーb-5' stem溶液 (1pmol/μL)および5μL の 2×TRL バッファ[100mM Tris-HCl (pH 8.0), 20mM MgCl2, 20mg/L BSA, 2mM ヘキサアンミンコバルトクロライド (III) 、 50% ポリエチレングリコール 6000]を添加する。
94℃(5 分) および 60℃(15分)の加熱処理でDNAをハイブリダイゼーションする。
【0053】
8)Y-ライゲーション反応
1μL の1mM ATP および 2μL (50U)の T4 RNAリガーゼ (Takara)を添加し、25℃で一晩 (12時間)反応する。
ステップ1から8までの操作を反復する。
【0054】
【表2】
数字 (5, 10 ……) は、洗浄回数を示す。
各洗浄は 200μL のバッファを使用した。
SB-- セレクションバッファ
WB1-- 洗浄1
WB2-a --洗浄2 (室温5分間のインキュベーション)
WB2-b --洗浄2 (インキュベーション無し)
【0055】
ペプチド・DNA複合体の調製
以下のプロトコルに従って、ペプチド・DNA複合体を調製した。
(1)上記で作製したDNAライブラリを5'ビオチン標識プライマー(b-3h-Stem : GGCTCGCGAATACTGCGAAGGAGTGAGATC)(配列番号21)と5'チオール標識プライマー(SH-5sPartT7 : CTGCGAAGACCACCATG) (配列番号22)を用いてPCR法にて増幅する(各50pmole, 50μLのPCR2本)。
(2)100μLのストレプトアビジン磁気ビーズ懸濁液(2×B&W)を添加し、1時間振とうする。
(3)ビーズを25μLのアンモニア水(2分間)で2回抽出してチオール基標識鎖を回収後、乾燥する。
(4)二価試薬EMCS(N-[ε-マレイミドカプロイルオキシ]スクシンイミドエステル、1mg/mL-アセトニトリル)溶液を10μL添加し、37℃で1時間攪拌する。
(5)アセトニトリルをエバポレートした後、20μLの0.1Mリン酸緩衝液(pH7.0)に溶解し、ゲル濾過脱塩カラム(NAP5、Pharmacia)で脱塩する。
(6)回収した1mLの修飾DNA水溶液を乾燥後、1μLの1M重炭酸ナトリウム溶液、2μLの水及び6μLのペプチド水溶液(2mg/mL)(N末端ビオチン標識Pro Pro Thr Ile Phe Phe Arg Leu Lys)を添加し、37℃で一晩放置する。
(7)反応溶液をNAP5カラムで脱塩して、回収液を乾燥する。
(8)100μLの0.1Mリン酸緩衝液(pH7.0)に溶解し、100μLのストレプトアビジン磁気ビーズ懸濁液(2×B&W)を添加し、1時間振とうする。
(9)ビーズを25μLのアンモニア水(2分間)で2回洗浄後、25μLのアンモニア水(65℃、15分間)で2回抽出して、ペプチド結合DNAを回収する。
【0056】
淘汰試験方法
以下の方法に従って、淘汰試験を行った。
(1)ペプチド基質・DNA複合体とゲルビーズ固定化カテプシンEを(2:1)のモル比で混合した後、反応用バッファー中、40℃で10分間インキュベーションする。(反応用バッファー組成: 50mM 酢酸ナトリウム(pH4.5)、5mM MgCl2、0.1M NaCl)
(2)10分後、5%トリクロロ酢酸を投入して反応を止める。
(3)反応後の溶液に等量のストレプトアビジン磁気ビーズ懸濁液を投入し、攪拌しながら室温で1時間インキュベーションする。
(4)磁気ビーズを磁石で固定し、上清を廃棄する。残った磁気ビーズを0.01%BSA 200μLで2回洗浄する。
(5)磁気ビーズに25%アンモニア水25μLを投入し、60℃で15分インキュベーションする。その後、上清を回収する。この操作を2回行う。
(6)凍結乾燥機でアンモニア水を蒸発させた後、10μLの水で溶解したものをテンプレートとしてPCRを行う(94℃ 30秒、50℃ 1分及び72℃ 30秒を30サイクル行う)。
PCR条件
94℃ 2分
94℃ 30秒
50℃ 1分
72℃ 30秒
72℃ 5分
(7)PCR産物を元に次サイクル用のSFリンクを作製し、淘汰サイクルを回していく。
【0057】
カテプシンE阻害活性検定法
(1)セレクションバッファー(50mM酢酸ナトリウム(pH4.5), 5mM MgCl2, 0.1M NaCl)86μLとカテプシンE溶液(3.36mM)3μLを混ぜたものを室温で5分間インキュベーション。
(2)この溶液にライブラリのPCR産物を一本鎖化したもの(非ビオチン鎖側、10pmol/μL)を1μL加え、室温で15分間インキュベーション。(ブランクとして水1μLを加えたものも同時に作製)
(3)この溶液にカテプシンE基質((株)ペプチド研 Code:3200-V)10μLを加え、40℃で10分間反応。
(4)10分後、すぐに氷中に移し、5%トリクロロ酢酸 1mLを加え反応を止める。
(5)この溶液を分光蛍光光度計で測定する(328nm Excitation, 396nm Emission)。カテプシンEの活性が高いもの程396nmのピークが高くなるので、ブランクとの値を比較することによってライブラリの阻害活性が求まる。
【0058】
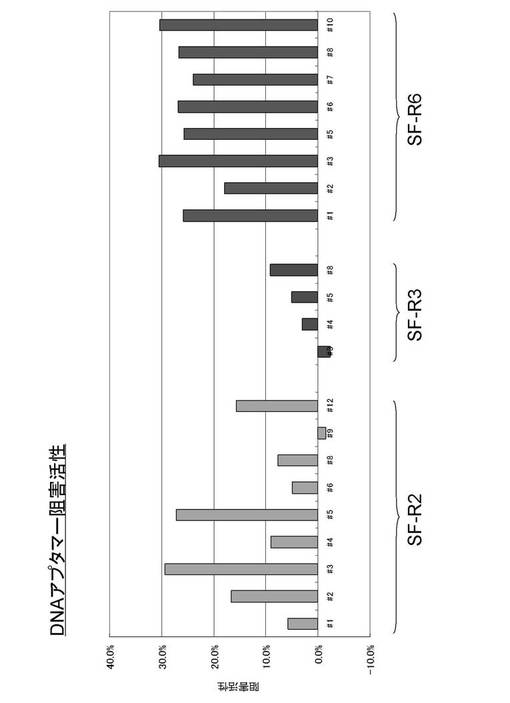
上記の方法に従い、カテプシンEに対する阻害活性で淘汰したDNAライブラリについて、カテプシンE阻害活性を検定し、カテプシンEに対する阻害活性を有するDNA複合体として、可変領域が配列番号1〜8で示される塩基配列を有するDNA(SF−R2の#1、2、3、4、5、6、8、9、12)、配列番号9〜11で示される塩基配列を有するDNA(SF−R3の#3、4、5、8)、配列番号12〜19で示される塩基配列を有するDNA(SF−R6の#1、2、3、5、6、7、8、10)を得た。これらのDNA複合体が有するカテプシンEに対する阻害活性の結果を図4に示す。
【0059】
図4に示すデータは全てテール領域付きの状態(DNA複合体)で測定したものである。具体的には、図3の(1)に描かれているような構造になっている。但し、図中の1本鎖領域がそれぞれのクローンによって異なる配列になっている。
【0060】
例2
SF-R6#3 DNA 作製法
1、PCR、制限酵素処理、一本鎖化によるSF6#3DNA作製
(1)SF-R6#3DNAのPCR増幅
Taq ポリメラーゼ(Greiner, Japan) 1U
10 × Buffer 5μL
[670mM Tris-HCl, 166mM (NH4)2SO4, 4.5% TritonX100, 2mg/mL ゼラチン]
MgCl2(25mM) 3μL
dNTP(2.5mM) 4μL
テンプレート SF-R6#3をクローン化したプラスミド溶液 1μL
プライマー b-3' stem プライマー (10pmol/μL) 1μL
5' part プライマー (10pmol/μL) 1μL
水で50μLに調整
b-3' stem プライマー配列:5' GGCTCGCGGCTACTGCGAAGGAGTGAGATC 3'(配列番号21)
(5'末端ビオチン化)
5' part プライマー配列:5' CTGCGAAGACCACCATG 3'(配列番号22)
【0061】
PCR条件
90℃, 2分
90℃, 30秒
60℃, 1分
72℃, 30秒
72℃, 5分
(上記90℃, 30秒から72℃, 30秒を30サイクル)
PCR後、エタノール沈殿により精製
【0062】
(2)制限酵素処理 (Mbo II)
エタノール沈殿後のペレット(約10 pmol)に対し、
Mbo II (TaKaRa, Japan) 10U
10 × L Buffer 1μL
[100mM Tris-HCl (pH7.5), 100mM MgCl2, 10mM ジチオスレイトール]
水で10μLに調整
を加え、37℃、1時間反応。
【0063】
(3)DNAの1本鎖化 1)
制限酵素反応後の溶液に対し、
ストレプトアビジンビーズ 50μL
[Dynabeads M-280 Streptavidin (Dynal, Norway)]
水 40μL
を加え、室温で振とうしながら30分間反応。
反応後、磁石を用いてビーズを固定し上清を取り除く。残ったビーズに0.01%BSA 100μLを加え、良く攪拌して洗浄する(2回繰り返す)。
洗浄後、25%アンモニア水50μLを加え、室温で2分振とう後、上清を回収(2回繰り返す)。
回収した上清は凍結乾燥機を用いて乾燥させておく。
【0064】
(4)3'stem プライマーのハイブリダイゼーション 2)
乾燥後のペレットに対し
3' stem プライマー (10pmol/μL) 1μL
(3' stem プライマー配列:5' GGCTCGCGGCTACTGCGAAGGAGTGAGATC 3')(配列番号21)
500mM リン酸バッファー (pH 7.4) 1μL
水で10μLに調整
を加え、
94℃, 5分後1℃/秒で冷却
60℃, 15分
の条件でアニーリング。
【0065】
【化1】
【0066】
例3
例2の(2)のMbo IIによる制限酵素処理を行わなかったこと以外は例2と同様の操作を行い、可変領域がSF−R6の#3L(配列番号20)であるDNA複合体を調製した。得られた可変領域がSF−R6の#3L(配列番号20)であるDNA複合体に関する情報を以下の表2にまとめて示す。尚、Kdは、SPR法による結合解離速度定数から求めた。カテプシンD活性阻害試験は、上記カテプシンE阻害活性検定法において、カテプシンE溶液に代えてカテプシンD溶液を用いて行った。
【0067】
【表3】
【0068】
可変領域がSF−R6の#3L(配列番号20)であるDNA複合体は、天然の強力な阻害剤ペプスタチンの活性と較べて、強度的には及ばないものの、特異性においては、逆に優れたものとなっている。
【0069】
例4
可変領域がSF−R6の#3であるDNA複合体(図3の(1)の複合体)に代わって、テール領域が図3の(2)〜(7)に示すDNA複合体をDNA合成により調製し、カテプシンEに対する阻害活性を求めた。各DNA複合体は、有機合成法により合成した。図3の(2)に示すDNA複合体は、85マーであり、配列番号24に示す。図3の(3)に示すDNA複合体は、74マーであり、配列番号25に示す。図3の(4)に示すDNA複合体は、64マーであり、配列番号26に示す。図3の(5)に示すDNA複合体は、83マーであり、配列番号27に示す。テール領域が図3の(6)の左側に示すDNA複合体は、図3の(2)に示すDNA複合体を制限酵素AccIIIで処理して得た。図3の(7)に示すDNA複合体は、各DNA鎖を合成し、ハイブリダイゼーションすることで得た。
【0070】
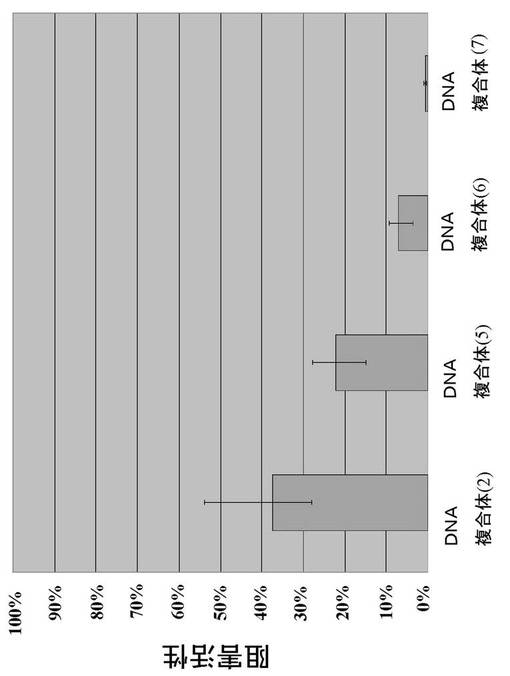
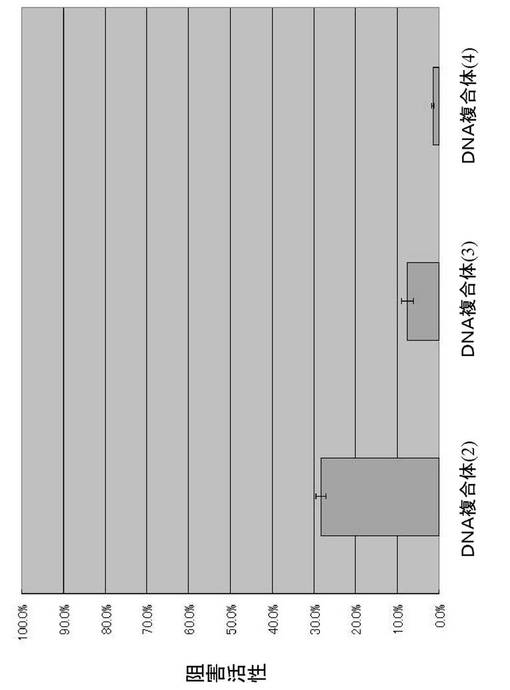
得られた各DNA複合体について、カテプシンEに対する阻害活性を求め、結果を図5および6に示す。テール領域によってカテプシンEに対する阻害活性が変化することが分かる。
【産業上の利用可能性】
【0071】
本発明は、カテプシンE特異的阻害DNA複合体の研究およびこれを用いた医薬の分野に有用である。
【図面の簡単な説明】
【0072】
【図1】カテプシンE結合DNAアプタマーの淘汰手順を示す。
【図2】SFリンク法によるカテプシンE阻害活性を持つ分子の淘汰の模式図。
【図3】本発明のDNA複合体の例。
【図4】例1で得られた本発明のDNA複合体のカテプシンEに対する阻害活性の結果。
【図5】例3で得られた本発明のDNA複合体(2)、(5)、(6)、(7)のカテプシンEに対する阻害活性の結果。
【図6】例3で得られた本発明のDNA複合体(2)、(3)、(4)のカテプシンEに対する阻害活性の結果。
【技術分野】
【0001】
本発明は、カテプシンEに対して特異的な阻害作用を示すDNAおよびこのDNAを用いたカテプシンE阻害剤に関する。
【背景技術】
【0002】
カテプシンEは免疫担当細胞、とくに抗原提示細胞であるマクロファージ、樹状細胞などに限局的に存在する細胞内アスパラギン酸プロテアーゼである。山本らは、カテプシンE遺伝子欠損マウス(ノックアウトマウス)を作製し、本酵素の機能解析を進めてきた(Yamamoto, K. Cathepsin E and cathepsin D: biosynthesis, processing and subcellular location. Adv. Exp. Med. Biol., 362, 223-9 (1995)、非特許文献1)。カテプシンE欠損マウスは無菌環境下で飼育しても、全く異常を示さないが、通常環境下に移すとアトピー性皮膚炎様症状を示した。病理組織学的にもヒトにおけるアトピー性皮膚炎症状と酷似した症状を示した。カテプシンE欠損マウスから調製したマクロファージを用いた研究では、正常マクロファージに比べて、殺菌能、細胞生存率、遊走能などの細胞機能に著明な低下があることが示されている。このことは、カテプシンE欠損が宿主の免疫応答自体に低下や不調をもたらし、それが何らかの外的刺激によってアトピー性皮膚炎などの個体の病態を発症させていることを示唆している。事実、カテプシンE欠損マウスでは、慢性関節リウマチのモデルである実験的関節炎が誘導されると、野生型マウスに比べてその病態に著しい増悪が見られることが示されている。カテプシンE欠損が炎症性サイトカインの分泌パターンを変化させ、関節炎を増悪させることが考えられる。一方、正常マウス(DBA/1J)にコラーゲン関節炎を起こさせると、血清中のカテプシンE活性は対照マウスの約2.5倍(p<0.001)に上昇していることが示されている。(筑波 隆幸、山本 健二:アトピー性皮膚炎とカテプシンE. 日薬理誌(Folia Pharmacol. Jpn.) 122, 15-20 (2003)、非特許文献2)このことは、カテプシンE欠損という極端なケースを除けば、慢性関節リウマチなどの自己免疫疾患等においては、一般的にカテプシンEの活性増加が生じていると考えられる。このことは、慢性関節リウマチ患者の滑液中のカテプシンE活性が病態に比例して増大するという山本らの報告とよく一致する。
【0003】
一方、各種癌細胞を用いた実験から、カテプシンEが癌細胞表面のTRAIL (TNF-related apoptosis-inducing ligand)を特異的に切断遊離させ、TRAILとその受容体を介するシグナリングによって癌細胞をアポトーシスに誘導することが明らかにされている(山本ら:未発表)。TRAILはCD4+ T細胞、NK細胞、単球、樹状細胞、好中球などにも発現していることから、これらの細胞を介して癌細胞をアポトーシスに誘導する一方で、自らの細胞をもアポトーシスに誘導して宿主免疫系を破綻させる可能性も示唆されている。このことは、カテプシンEの過剰発現がTRAILを介するアポトーシス誘導機構を惹起させ、Th1型の免疫反応に破綻を誘導し、自己免疫疾患等の種々の疾患を生む可能性を強く示唆している。
【0004】
以上のような知見から、カテプシンE活性の増大は、自己免疫疾患等の様々な病態に繋がると考えられ、その際の過剰なカテプシンE活性を抑える薬剤はこうした病態の改善に大いに役立つものと期待される。カテプシンEに対する阻害剤については、これまで放線菌由来のペプスタチンA(Umezawa, H., Aoyagi, T., Morishima, H., Matsuzaki, M,.and Hamada, M.Pepstatin, a new pepsin inhibitor produced by Actinomycetes. J. Antibiot., 23, 259-62 (1970)、非特許文献3)と回虫から分離されたAscaris pepsin inhibitor(Kageyama, T. Molecular cloning, expression and characterization of an Ascaris inhibitor for pepsin and cathepsin E. Eur. J. Biochem., 253, 804-9 (1998)、非特許文献4)が知られている。
【非特許文献1】Yamamoto, K. Cathepsin E and cathepsin D: biosynthesis, processing and subcellular location. Adv. Exp. Med. Biol., 362, 223-9 (1995)
【非特許文献2】筑波 隆幸、山本 健二:アトピー性皮膚炎とカテプシンE. 日薬理誌(Folia Pharmacol. Jpn.) 122, 15-20 (2003)
【非特許文献3】Umezawa, H., Aoyagi, T., Morishima, H., Matsuzaki, M,.and Hamada, M.Pepstatin, a new pepsin inhibitor produced by Actinomycetes. J. Antibiot., 23, 259-62 (1970)
【非特許文献4】Kageyama, T. Molecular cloning, expression and characterization of an Ascaris inhibitor for pepsin and cathepsin E. Eur. J. Biochem., 253, 804-9 (1998)
【発明の開示】
【発明が解決しようとする課題】
【0005】
放線菌由来のペプスタチンAはアプパラギン酸プロテアーゼ群に共通の阻害剤で特異性がない。回虫ペプシン阻害剤(Ascaris pepsin inhibitor)はペプシンに対すると同様にカテプシンEを強く阻害し、他のアプパラギン酸プロテアーゼは阻害しないが、高分子タンパク質で細胞毒性が強いという欠点がある。そのため、カテプシンEに特異的な阻害剤として実用的なものは知られていないのが現状である。
【0006】
そこで本発明の目的は、カテプシンEを特異的に阻害し、かつカテプシンEと類縁性の高いリソソーム酵素であるカテプシンDは阻害しない、物質を提供することにある。
【課題を解決するための手段】
【0007】
本発明者らは、かねてより、「高速分子進化法」に基づき、従来からのDNAアプタマー調製法より格段に高速化された方法で上記阻害作用を有する物質を探査した。その結果、カテプシンDに対しては殆ど阻害活性を持たず、カテプシンEを特異的に阻害するDNAを見いだし、本発明を完成させた。
【0008】
本発明は、下記の何れかのDNAに関する。
(1)配列番号1〜20のいずれかの塩基配列を有するDNA、
(2)配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA、および
(3)配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA。
上記において、30マーの二本鎖DNAの一方の一本鎖は、配列番号21の塩基配列を有することができる。
【0009】
さらに本発明は、上記本発明のDNA(以下、可変領域という)の3'末端に20〜40マーの全部または一部が二本鎖のDNA(以下、テール領域という)を結合したDNA複合体に関する。
上記DNA複合体においては、テール領域が30マーの二本鎖DNAであることができ、さらに、二本鎖DNAであるテール領域の可変領域の3'末端に結合している一本鎖は、配列番号21の塩基配列を有することができる。
【0010】
さらに本発明は、上記本発明のDNA複合体を有効成分とするカテプシンE阻害剤に関する。
【発明の効果】
【0011】
本発明のDNA複合体は、カテプシンEに対して高い特異阻害活性を示し、かつカテプシンEと類縁性の高いリソソーム酵素であるカテプシンDは阻害せず、副作用等の点からも治療薬としての有用性が高いと期待できる。
【0012】
DNA複合体は分子の合成が容易である上に、生体内でRNAに較べ耐性がある。分子の改変も比較的容易であり、種々の修飾基の導入が可能である。さらに、得られた分子を手がかりにして、さらに高機能なDNA分子を進化させることも可能である。生体内にタンパク質ほど分解酵素が存在しないと考えられるので、寿命的にも有利な可能性がある。
【発明を実施するための最良の形態】
【0013】
本発明のDNAは、下記の何れかのDNAである。
(1)配列番号1〜20のいずれかの塩基配列を有するDNA、
(2)配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA、および
(3)配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA。
【0014】
配列番号1〜20で示される塩基配列を有するDNAは、いずれも以下の分子淘汰操作により得た。即ち、まずブロックシャフリング法でDNAライブラリーを作成し、それを用い、ターゲット分子であるカテプシンEに対する親和性で優れた分子を淘汰し、さらに、阻害活性で淘汰するという作業を行って、目的分子の淘汰を行い、上記DNAを得た。具体的には、以下の手順で行った。
【0015】
1)DNAライブラリーの作成
7種のアミノ酸に対応するコードをもつDNAライブラリーを作成した(表1参照)。このとき、ライブラリーの合成にはブロック合成法(YLBS法)を用いた。
【0016】
【表1】
【0017】
2)カテプシンE結合DNAアプタマーの淘汰
図1に示す操作手順に従って、カテプシンEに強く結合するDNAアプタマー群を淘汰した。
【0018】
3)SFリンク法により、カテプシンE阻害活性を持つ分子の淘汰
図2に示すSFリンク法により、結合はするが阻害活性のない分子は除き、カテプシンE阻害活性の高まったDNA分子群を得た。
【0019】
4)得られたDNA分子群について、適宜、結合強度、阻害強度、分子の分光学的特性などを解析した。
【0020】
上記分子淘汰操作(SFリンク法)を、条件を変えて複数回(6回)行い、カテプシンE阻害活性を持つ分子として、配列番号1〜20で示される塩基配列を有するDNAを得た。詳細は実施例に記載する。
【0021】
本明細書で言う「1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列」における「1から数個」の範囲は特には限定されないが、例えば、1から5個、好ましくは1から4個、より好ましくは1から3個、より好ましくは1から2個、さらに好ましくは1個を意味する。
【0022】
上記した「ストリンジェントな条件下でハイブリダイズする」とは、DNAをプローブとして使用し、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法、あるいはサザンブロットハイブリダイゼーション法等を用いることにより得られるDNAの塩基配列を意味し、例えば、コロニーあるいはプラーク由来のDNA又は該DNAの断片を固定化したフィルターを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2×SSC溶液(1×SSC溶液は、150mM塩化ナトリウム、15mMクエン酸ナトリウム)を用い、65℃条件下でフィルターを洗浄することにより同定できるDNA等を挙げることができる。ハイブリダイゼーションは、モレキュラークローニング第2版等に記載されている方法に準じて行うことができる。
【0023】
ストリンジェントな条件下でハイブリダイズするDNAとしては、プローブとして使用するDNAの塩基配列と一定以上の相同性を有するDNAが挙げられ、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは93%以上、特に好ましくは95%以上、最も好ましくは98%以上の相同性を有するDNAが挙げられる。
【0024】
本発明のDNAの取得方法は特に限定されない。本発明のDNAは、本明細書中の配列表の配列番号1から20に記載した塩基配列の情報に基づいて化学合成法(月刊「BIOINDUSTRY」1995 年8 月号参照)等により調製することができる。DNAの調製の操作は当業者に既知であり、例えば、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989(以下、モレキュラークローニング第2版と略す)、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)等に記載の方法に準じて行うことができる。
【0025】
本発明は、前述のように、(2)配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA、および(3)配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNAを包含する。配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有するDNAおよび配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有するDNAは、3'末端に30マーの二本鎖DNAを結合した複合体とした場合に、カテプシンE阻害活性を有するものであれば、本発明の範囲に含まれる。30マーの二本鎖DNAの一方の一本鎖は、配列番号21の塩基配列を有するものであることができる。
【0026】
3'末端に30マーの二本鎖DNAを結合したDNA複合体のカテプシンE阻害活性は、以下のように検定することができる。
(1)セレクションバッファー(50mM酢酸ナトリウム(pH4.5), 5mM MgCl2, 0.1M NaCl)86μLとカテプシンE溶液(3.36mM)3μLを混ぜたものを室温で5分間インキュベーション。
(2)この溶液にライブラリのPCR産物を一本鎖化したもの(非ビオチン鎖側、10pmol/μL)を1μL加え、室温で15分間インキュベーション。(ブランクとして水1μLを加えたものも同時に作製)
(3)この溶液にカテプシンE基質((株)ペプチド研 Code:3200-V)10μLを加え、40℃で10分間反応。
(4)10分後、すぐに氷中に移し、5%トリクロロ酢酸 1mLを加え反応を止める。
(5)この溶液を分光蛍光光度計で測定する(328nm Excitation, 396nm Emission)。カテプシンEの活性が高いもの程396nmのピークが高くなるので、ブランクとの値を比較することによって阻害活性が求まる。
【0027】
本発明は、上記配列番号1から20に記載したいずれかの塩基配列を有するDNA(可変領域)の3'末端に20〜40マーの全部または一部が二本鎖のDNA(テール領域)を結合したDNA複合体を包含する。テール領域の塩基数は、DNA複合体のカテプシンE阻害活性に影響するので、可変領域の種類に応じて、最適の塩基数を選択することが好ましい。テール領域の塩基配列も、適宜選択することができる。テール領域は、好ましくは25〜35マー、より好ましくは30マーの二本鎖DNAである。さらに、二本鎖DNAであるテール領域の、可変領域の3'末端に結合している一本鎖は、配列番号21の塩基配列を有することが好ましい。可変領域の3'末端に結合しているテール領域の鎖が配列番号21の塩基配列を有するDNA複合体の例(SF-R6#3 with tail)を図3の(1)に示す。
【0028】
図3の(1)に示すDNA複合体は、テール領域が2本のDNAがハイブリダイゼーションしているものである。それに対して、テール領域の2本のDNAは、テール領域の末端(可変領域と結合する末端と反対側の末端)で、テール(図3の(2)〜(4))、ループ(図3の(5))、または一部が一本鎖 (図3の(6)の左側のDNA複合体)を形成していてもよい。テールおよびループを形成する塩基数は特に制限はないが、例えば、3〜15の範囲であることができる。図3の(2)〜(4)に示す末端がループであるDNA複合体におけるループは、塩基数が3であり、CTGの塩基配列を有する。また、図3の(5)に示す末端がループであるDNA複合体におけるループは、塩基数が10である。このループの塩基配列はNで示され、NはA、T、G、Cの内どれか1つがランダムに出現していることを示す。図3の(6) に示すDNA複合体の部分的一本鎖(図中の左側のDNA複合体の右末端)の塩基数も特に制限はないが、例えば、1〜10の範囲であることができる。
【0029】
本発明のDNA複合体は、例えば、図3の(1)に示すDNA複合体のように、配列番号1から20に記載したいずれかの塩基配列を有するDNA(可変領域)の3'末端に、例えば、30マーの二本鎖のDNA(テール領域)を結合したDNA複合体であるが、それ以外に、図3の(2)〜(6)に示すDNA複合体のように、テール領域の末端にループを有するDNA複合体、及びテール領域の末端が部分的一本鎖であるDNA複合体であることもできる。
【0030】
本発明は、上記本発明のDNA複合体を有効成分とするカテプシンE阻害剤を包含する。本発明のDNA複合体を有効成分とするテプシンE阻害剤は、実験試薬として使用できる他、治療薬としても使用できる。
【0031】
例えば、カテプシンEの生理作用研究などにおいて、カテプシンE阻害剤としてペプスタチンAが使われている (Chain, B. M., Free, P., Medd, P., Swetman, C., Tabor, A. B. and Terrazzini, N.The expression and function of cathepsin E in dendritic cells. J. Immunol., 174, 1791-800 (2005). Cappiello, M. G., Wu, Z., Scott, B. B., McGeehan, G. M. and Harrison, R. K. Purification and characterization of recombinant human cathepsin E expressed in human kidney cell line 293. Protein Expr. Purif., 37, 53-60 (2004). )。しかし、ペプスタチンAはカテプシンDやペプシンも阻害するためにカテプシンE単独の効果を調べることが困難であった。それに対して本発明のDNA複合体は、カテプシンDを阻害しないので、カテプシンE単独の効果を調べることを可能にする。その結果、カテプシンEに起因する種々の病因を除く可能性がある。既に、インスリン依存性糖尿病や慢性関節リウマチの発症進展に、カテプシンE活性の亢進が見られる可能性が示唆されており (筑波 隆幸、山本 健二:アトピー性皮膚炎とカテプシンE. 日薬理誌(Folia Pharmacol. Jpn.) 122, 15-20 (2003).) 、カテプシンE特異的な阻害活性を持つ本発明のDNA複合体は、そのような疾患の治療薬となることが期待される。
【実施例】
【0032】
以下、本発明を実施例によりさらに詳細に説明する。
【0033】
例1
1)PCR 増幅
未淘汰のDNAライブラリをテンプレートとして以下に示す方法でPCR増幅した。
Taq ポリメラーゼ (Greiner, Tokyo, Japan) 1U
10 ×PCR バッファ 5μL
[670mM Tris-HCl, 166mM (NH4)2SO4, 4.5% TritonX100, 2mg/mLゼラチン]
MgCl2 (25mM) 5μL
dNTP (2.5mM) 4μL
テンプレート DNAライブラリ (1fmole以上)
プライマー b-3' stemプライマー(100pmol/μL) 0.5μL
5' partプライマー(100pmol/μL) 0.5μL
H2O 全体で50μL
【0034】
プログラム
90℃ 2分
90℃ 30秒
45℃ 1分
72℃ 30秒
90℃ 30秒、45℃ 1分及び72℃ 30秒を30サイクル行い、得られた増幅物は電気泳動法で確認し、エタノール沈殿で精製する。
【0035】
2)制限酵素処理 (Mbo II)
反応混合物:
Mbo II (Takara) 8U
10 × L Buffer 5μL
[100mM Tris-HCl (pH7.5), 100mM MgCl2, 10mM ジチオスレイトール]
DNA 50pmol
H2O 全体で 50μL
37℃で 1時間インキュベートする。
【0036】
3)一本鎖DNA の調製
a. DNAのストレプトアビジン磁気ビーズへの結合
制限酵素消化物 50μL
ストレプトアビジン磁気ビーズ 50μL
[Dynabeads M-280 Streptavidin (Dynal, Oslo, Norway)]
懸濁物を30分間振とうする。
磁石でビーズをチューブ底に固定し、上清を捨てる。
【0037】
b. 0.01% BSA水溶液による洗浄
100μL の 0.01% BSAを添加し、良く混合する。
磁石でビーズをチューブ底に固定し、上清を捨てる。
この操作を2回繰り返す。
【0038】
c. 二本鎖DNAの変性および非ビオチン標識一本鎖DNAの回収
室温で2分間50 μLの25% (w/v) アンモニア水で処理し、磁石でビーズをチューブ底に固定した後、上清を回収する。(非ビオチン標識一本鎖DNAは上清中に存在する)
この操作を2回繰り返す。
回収液を乾燥する。
【0039】
4)プライマーb-3'stemのハイブリダイゼーション
回収した ssDNAを 95μL のセレクションバッファ(以下参照)に溶解する。
5μL のb-3'stem プライマー溶液 (10pmol/μL)を添加する。
94℃(5 分) および 60℃(15分)の加熱処理でDNAをハイブリダイズする。
【0040】
5)DNA アプタマーの試験管内スクリーニング
材料:
1.セレクションバッファ:
50 mM 酢酸ナトリウム (pH 4.5)
100 mM NaCl
5 mM MgCl2
2.洗浄バッファ 1:
50 mM酢酸ナトリウム(pH 4.5)
0.5 M NaCl
3.洗浄バッファ2:
50 mM酢酸ナトリウム(pH 4.5)
1 M NaCl
4.溶出バッファ1:
50 mM酢酸ナトリウム(pH 4.5)
1 M NaCl
10 mM MgCl2
5.溶出バッファ2:
50 mM酢酸ナトリウム(pH 4.5)
2 M NaCl
10 mM MgCl2
【0041】
操作:
ステップ 1. セレクションバッファ中のDNA を固定化酵素と20-30 分間反応させる。
DNA: 100 μL (セレクションバッファ)
酵素: 5 μL (固定化操作および活性検定により調製したもの)
ここで、DNA : 酵素モル比 = 4 : 1 にする。
【0042】
ステップ 2. セレクションバッファによる洗浄
上記反応液に 200 μL のセレクションバッファを添加し、チューブを数回穏やかにタッピングする。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し捨てる。
以下の表2に記したようにこの操作を繰り返す。
【0043】
ステップ 3. 洗浄バッファ 1による洗浄
ステップ2の洗浄および遠心第5ラウンドの後、200 μL の洗浄バッファ 1を添加し、チューブを数回穏やかにタッピングする。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し捨てる。
以下の表2に記したようにこの操作を繰り返す。
【0044】
ステップ 4. 洗浄バッファ2による洗浄
このステップには二種類の操作が含まれる:
A) 200 μL の洗浄バッファ 2 を添加し、チューブを数回穏やかにタッピングする。 時々タッピングしながら室温で5分間反応させる。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し捨てる。
以下の表2に記したようにこの操作を繰り返す。
【0045】
B) 200 μL の洗浄バッファ 2 を添加し、チューブを数回穏やかにタッピングする。15000 rpm で1 分間遠心した後、上清を静かにピペットで吸い出し捨てる。
以下の表2に記したようにこの操作を繰り返す。
【0046】
ステップ 5. 溶出バッファ 1による溶出
最初に 37oCの恒温槽を準備する。100 μL の溶出バッファ 1 を添加して、チューブを数回穏やかにタッピングする。チューブを恒温槽に10分間浸す。2-5 分間間隔で穏やかにタッピングし、再び恒温槽に戻す。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し、きれいなチューブに回収する。
この操作を2回繰り返す。
【0047】
ステップ 6. 溶出バッファ2による溶出
最初に95℃の恒温槽を準備する。100 μL の溶出バッファ 2 を添加して、チューブを数回穏やかにタッピングする。チューブを恒温槽に5分間浸す。2分間間隔で穏やかにタッピングし、再び恒温槽に戻す。卓上遠心機で30秒間遠心した後、上清を静かにピペットで吸い出し、きれいなチューブに回収する。
この操作を2回繰り返す。
【0048】
回収した上清からエタノール沈殿でDNAを精製する。
ただし、塩濃度が高い場合は、70%エタノールで2、3回ペレットを洗浄する。
【0049】
6)一本鎖DNA の調製 ( (3)と同様)
a. DNAのストレプトアビジン磁気ビーズへの結合
回収DNAに対し、以下のものを添加する。
水 10μL
ストレプトアビジン磁気ビーズ 10μL
[Dynabeads M-280 Streptavidin (Dynal, Oslo, Norway)]
懸濁物を30分間振とうする。
磁石でビーズをチューブ底に固定し、上清を捨てる。
【0050】
b. 0.01% BSA水溶液による洗浄
100μL の 0.01% BSAを添加し、良く混合する。
磁石でビーズをチューブ底に固定し、上清を捨てる。
この操作を2回繰り返す。
【0051】
c. 二本鎖DNAの変性および非ビオチン標識一本鎖DNAの回収
室温で2分間50 μLの25% (w/v) アンモニア水で処理し、磁石でビーズをチューブ底に固定した後、上清を回収する。(非ビオチン標識一本鎖DNAは上清中に存在する)
この操作を2回繰り返す。
回収液を乾燥する。
【0052】
7)ハイブリダイゼーション
回収した 一本鎖DNAを 2μL の水に溶解する。 1μL のプライマーb-5' stem溶液 (1pmol/μL)および5μL の 2×TRL バッファ[100mM Tris-HCl (pH 8.0), 20mM MgCl2, 20mg/L BSA, 2mM ヘキサアンミンコバルトクロライド (III) 、 50% ポリエチレングリコール 6000]を添加する。
94℃(5 分) および 60℃(15分)の加熱処理でDNAをハイブリダイゼーションする。
【0053】
8)Y-ライゲーション反応
1μL の1mM ATP および 2μL (50U)の T4 RNAリガーゼ (Takara)を添加し、25℃で一晩 (12時間)反応する。
ステップ1から8までの操作を反復する。
【0054】
【表2】
数字 (5, 10 ……) は、洗浄回数を示す。
各洗浄は 200μL のバッファを使用した。
SB-- セレクションバッファ
WB1-- 洗浄1
WB2-a --洗浄2 (室温5分間のインキュベーション)
WB2-b --洗浄2 (インキュベーション無し)
【0055】
ペプチド・DNA複合体の調製
以下のプロトコルに従って、ペプチド・DNA複合体を調製した。
(1)上記で作製したDNAライブラリを5'ビオチン標識プライマー(b-3h-Stem : GGCTCGCGAATACTGCGAAGGAGTGAGATC)(配列番号21)と5'チオール標識プライマー(SH-5sPartT7 : CTGCGAAGACCACCATG) (配列番号22)を用いてPCR法にて増幅する(各50pmole, 50μLのPCR2本)。
(2)100μLのストレプトアビジン磁気ビーズ懸濁液(2×B&W)を添加し、1時間振とうする。
(3)ビーズを25μLのアンモニア水(2分間)で2回抽出してチオール基標識鎖を回収後、乾燥する。
(4)二価試薬EMCS(N-[ε-マレイミドカプロイルオキシ]スクシンイミドエステル、1mg/mL-アセトニトリル)溶液を10μL添加し、37℃で1時間攪拌する。
(5)アセトニトリルをエバポレートした後、20μLの0.1Mリン酸緩衝液(pH7.0)に溶解し、ゲル濾過脱塩カラム(NAP5、Pharmacia)で脱塩する。
(6)回収した1mLの修飾DNA水溶液を乾燥後、1μLの1M重炭酸ナトリウム溶液、2μLの水及び6μLのペプチド水溶液(2mg/mL)(N末端ビオチン標識Pro Pro Thr Ile Phe Phe Arg Leu Lys)を添加し、37℃で一晩放置する。
(7)反応溶液をNAP5カラムで脱塩して、回収液を乾燥する。
(8)100μLの0.1Mリン酸緩衝液(pH7.0)に溶解し、100μLのストレプトアビジン磁気ビーズ懸濁液(2×B&W)を添加し、1時間振とうする。
(9)ビーズを25μLのアンモニア水(2分間)で2回洗浄後、25μLのアンモニア水(65℃、15分間)で2回抽出して、ペプチド結合DNAを回収する。
【0056】
淘汰試験方法
以下の方法に従って、淘汰試験を行った。
(1)ペプチド基質・DNA複合体とゲルビーズ固定化カテプシンEを(2:1)のモル比で混合した後、反応用バッファー中、40℃で10分間インキュベーションする。(反応用バッファー組成: 50mM 酢酸ナトリウム(pH4.5)、5mM MgCl2、0.1M NaCl)
(2)10分後、5%トリクロロ酢酸を投入して反応を止める。
(3)反応後の溶液に等量のストレプトアビジン磁気ビーズ懸濁液を投入し、攪拌しながら室温で1時間インキュベーションする。
(4)磁気ビーズを磁石で固定し、上清を廃棄する。残った磁気ビーズを0.01%BSA 200μLで2回洗浄する。
(5)磁気ビーズに25%アンモニア水25μLを投入し、60℃で15分インキュベーションする。その後、上清を回収する。この操作を2回行う。
(6)凍結乾燥機でアンモニア水を蒸発させた後、10μLの水で溶解したものをテンプレートとしてPCRを行う(94℃ 30秒、50℃ 1分及び72℃ 30秒を30サイクル行う)。
PCR条件
94℃ 2分
94℃ 30秒
50℃ 1分
72℃ 30秒
72℃ 5分
(7)PCR産物を元に次サイクル用のSFリンクを作製し、淘汰サイクルを回していく。
【0057】
カテプシンE阻害活性検定法
(1)セレクションバッファー(50mM酢酸ナトリウム(pH4.5), 5mM MgCl2, 0.1M NaCl)86μLとカテプシンE溶液(3.36mM)3μLを混ぜたものを室温で5分間インキュベーション。
(2)この溶液にライブラリのPCR産物を一本鎖化したもの(非ビオチン鎖側、10pmol/μL)を1μL加え、室温で15分間インキュベーション。(ブランクとして水1μLを加えたものも同時に作製)
(3)この溶液にカテプシンE基質((株)ペプチド研 Code:3200-V)10μLを加え、40℃で10分間反応。
(4)10分後、すぐに氷中に移し、5%トリクロロ酢酸 1mLを加え反応を止める。
(5)この溶液を分光蛍光光度計で測定する(328nm Excitation, 396nm Emission)。カテプシンEの活性が高いもの程396nmのピークが高くなるので、ブランクとの値を比較することによってライブラリの阻害活性が求まる。
【0058】
上記の方法に従い、カテプシンEに対する阻害活性で淘汰したDNAライブラリについて、カテプシンE阻害活性を検定し、カテプシンEに対する阻害活性を有するDNA複合体として、可変領域が配列番号1〜8で示される塩基配列を有するDNA(SF−R2の#1、2、3、4、5、6、8、9、12)、配列番号9〜11で示される塩基配列を有するDNA(SF−R3の#3、4、5、8)、配列番号12〜19で示される塩基配列を有するDNA(SF−R6の#1、2、3、5、6、7、8、10)を得た。これらのDNA複合体が有するカテプシンEに対する阻害活性の結果を図4に示す。
【0059】
図4に示すデータは全てテール領域付きの状態(DNA複合体)で測定したものである。具体的には、図3の(1)に描かれているような構造になっている。但し、図中の1本鎖領域がそれぞれのクローンによって異なる配列になっている。
【0060】
例2
SF-R6#3 DNA 作製法
1、PCR、制限酵素処理、一本鎖化によるSF6#3DNA作製
(1)SF-R6#3DNAのPCR増幅
Taq ポリメラーゼ(Greiner, Japan) 1U
10 × Buffer 5μL
[670mM Tris-HCl, 166mM (NH4)2SO4, 4.5% TritonX100, 2mg/mL ゼラチン]
MgCl2(25mM) 3μL
dNTP(2.5mM) 4μL
テンプレート SF-R6#3をクローン化したプラスミド溶液 1μL
プライマー b-3' stem プライマー (10pmol/μL) 1μL
5' part プライマー (10pmol/μL) 1μL
水で50μLに調整
b-3' stem プライマー配列:5' GGCTCGCGGCTACTGCGAAGGAGTGAGATC 3'(配列番号21)
(5'末端ビオチン化)
5' part プライマー配列:5' CTGCGAAGACCACCATG 3'(配列番号22)
【0061】
PCR条件
90℃, 2分
90℃, 30秒
60℃, 1分
72℃, 30秒
72℃, 5分
(上記90℃, 30秒から72℃, 30秒を30サイクル)
PCR後、エタノール沈殿により精製
【0062】
(2)制限酵素処理 (Mbo II)
エタノール沈殿後のペレット(約10 pmol)に対し、
Mbo II (TaKaRa, Japan) 10U
10 × L Buffer 1μL
[100mM Tris-HCl (pH7.5), 100mM MgCl2, 10mM ジチオスレイトール]
水で10μLに調整
を加え、37℃、1時間反応。
【0063】
(3)DNAの1本鎖化 1)
制限酵素反応後の溶液に対し、
ストレプトアビジンビーズ 50μL
[Dynabeads M-280 Streptavidin (Dynal, Norway)]
水 40μL
を加え、室温で振とうしながら30分間反応。
反応後、磁石を用いてビーズを固定し上清を取り除く。残ったビーズに0.01%BSA 100μLを加え、良く攪拌して洗浄する(2回繰り返す)。
洗浄後、25%アンモニア水50μLを加え、室温で2分振とう後、上清を回収(2回繰り返す)。
回収した上清は凍結乾燥機を用いて乾燥させておく。
【0064】
(4)3'stem プライマーのハイブリダイゼーション 2)
乾燥後のペレットに対し
3' stem プライマー (10pmol/μL) 1μL
(3' stem プライマー配列:5' GGCTCGCGGCTACTGCGAAGGAGTGAGATC 3')(配列番号21)
500mM リン酸バッファー (pH 7.4) 1μL
水で10μLに調整
を加え、
94℃, 5分後1℃/秒で冷却
60℃, 15分
の条件でアニーリング。
【0065】
【化1】
【0066】
例3
例2の(2)のMbo IIによる制限酵素処理を行わなかったこと以外は例2と同様の操作を行い、可変領域がSF−R6の#3L(配列番号20)であるDNA複合体を調製した。得られた可変領域がSF−R6の#3L(配列番号20)であるDNA複合体に関する情報を以下の表2にまとめて示す。尚、Kdは、SPR法による結合解離速度定数から求めた。カテプシンD活性阻害試験は、上記カテプシンE阻害活性検定法において、カテプシンE溶液に代えてカテプシンD溶液を用いて行った。
【0067】
【表3】
【0068】
可変領域がSF−R6の#3L(配列番号20)であるDNA複合体は、天然の強力な阻害剤ペプスタチンの活性と較べて、強度的には及ばないものの、特異性においては、逆に優れたものとなっている。
【0069】
例4
可変領域がSF−R6の#3であるDNA複合体(図3の(1)の複合体)に代わって、テール領域が図3の(2)〜(7)に示すDNA複合体をDNA合成により調製し、カテプシンEに対する阻害活性を求めた。各DNA複合体は、有機合成法により合成した。図3の(2)に示すDNA複合体は、85マーであり、配列番号24に示す。図3の(3)に示すDNA複合体は、74マーであり、配列番号25に示す。図3の(4)に示すDNA複合体は、64マーであり、配列番号26に示す。図3の(5)に示すDNA複合体は、83マーであり、配列番号27に示す。テール領域が図3の(6)の左側に示すDNA複合体は、図3の(2)に示すDNA複合体を制限酵素AccIIIで処理して得た。図3の(7)に示すDNA複合体は、各DNA鎖を合成し、ハイブリダイゼーションすることで得た。
【0070】
得られた各DNA複合体について、カテプシンEに対する阻害活性を求め、結果を図5および6に示す。テール領域によってカテプシンEに対する阻害活性が変化することが分かる。
【産業上の利用可能性】
【0071】
本発明は、カテプシンE特異的阻害DNA複合体の研究およびこれを用いた医薬の分野に有用である。
【図面の簡単な説明】
【0072】
【図1】カテプシンE結合DNAアプタマーの淘汰手順を示す。
【図2】SFリンク法によるカテプシンE阻害活性を持つ分子の淘汰の模式図。
【図3】本発明のDNA複合体の例。
【図4】例1で得られた本発明のDNA複合体のカテプシンEに対する阻害活性の結果。
【図5】例3で得られた本発明のDNA複合体(2)、(5)、(6)、(7)のカテプシンEに対する阻害活性の結果。
【図6】例3で得られた本発明のDNA複合体(2)、(3)、(4)のカテプシンEに対する阻害活性の結果。
【特許請求の範囲】
【請求項1】
下記の何れかのDNA。
(1)配列番号1〜20のいずれかの塩基配列を有するDNA、
(2)配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA、および
(3)配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA。
【請求項2】
30マーの二本鎖DNAの一方の一本鎖が、配列番号21の塩基配列を有する請求項1に記載のDNA。
【請求項3】
請求項1に記載のDNA(以下、可変領域という)の3'末端に20〜40マーの全部または一部が二本鎖のDNA(以下、テール領域という)を結合したDNA複合体。
【請求項4】
テール領域が30マーの二本鎖DNAである請求項3に記載のDNA複合体。
【請求項5】
二本鎖DNAであるテール領域の可変領域の3'末端に結合している一本鎖は、配列番号21の塩基配列を有する請求項3に記載のDNA複合体。
【請求項6】
可変領域と結合する末端と反対側のテール領域の末端がテールまたはループを形成している請求項3〜5のいずれか1項に記載のDNA複合体。
【請求項7】
請求項3〜6のいずれか1項に記載のDNA複合体を有効成分とするカテプシンE阻害剤。
【請求項1】
下記の何れかのDNA。
(1)配列番号1〜20のいずれかの塩基配列を有するDNA、
(2)配列番号1〜20のいずれかの塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA、および
(3)配列番号1〜20のいずれかの塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイスする塩基配列を有し、かつ3'末端に30マーの二本鎖DNAを結合した複合体はカテプシンE阻害活性を有するDNA。
【請求項2】
30マーの二本鎖DNAの一方の一本鎖が、配列番号21の塩基配列を有する請求項1に記載のDNA。
【請求項3】
請求項1に記載のDNA(以下、可変領域という)の3'末端に20〜40マーの全部または一部が二本鎖のDNA(以下、テール領域という)を結合したDNA複合体。
【請求項4】
テール領域が30マーの二本鎖DNAである請求項3に記載のDNA複合体。
【請求項5】
二本鎖DNAであるテール領域の可変領域の3'末端に結合している一本鎖は、配列番号21の塩基配列を有する請求項3に記載のDNA複合体。
【請求項6】
可変領域と結合する末端と反対側のテール領域の末端がテールまたはループを形成している請求項3〜5のいずれか1項に記載のDNA複合体。
【請求項7】
請求項3〜6のいずれか1項に記載のDNA複合体を有効成分とするカテプシンE阻害剤。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2007−49911(P2007−49911A)
【公開日】平成19年3月1日(2007.3.1)
【国際特許分類】
【出願番号】特願2005−235819(P2005−235819)
【出願日】平成17年8月16日(2005.8.16)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【出願人】(504190548)国立大学法人埼玉大学 (292)
【出願人】(504145342)国立大学法人九州大学 (960)
【Fターム(参考)】
【公開日】平成19年3月1日(2007.3.1)
【国際特許分類】
【出願日】平成17年8月16日(2005.8.16)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【出願人】(504190548)国立大学法人埼玉大学 (292)
【出願人】(504145342)国立大学法人九州大学 (960)
【Fターム(参考)】
[ Back to top ]