
カロテノイド化合物をコードする遺伝子
ユニークなカロテン産生生合成遺伝子クラスターが、パントエア・アグロメランス(Pantoea agglomerans)DC404株から単離され、クラスターの遺伝子構成はcrtE−idi−crtY−crtI−crtB−crtZである。このクラスター内に含有される遺伝子は、ゲラニルゲラニルピロリン酸(GGPP)合成酵素(CrtE)、イソペンテニルピロリン酸イソメラーゼ(Idi)、リコペンシクラーゼ(CrtY)、フィトエンデサチュラーゼ(CrtI)、フィトエンシンターゼ(CrtB)、およびβ−カロテン水酸化酵素(CrtZ)をコードする。遺伝子クラスター、遺伝子およびそれらの生成物は、ファルネシルピロリン酸のカロテノイドへの転換のために有用である。これらのDNAセグメントを含有するベクター、ベクターを含有する宿主細胞、および形質転換宿主生物体内で組換えDNA技術によってこれらの酵素を製造する方法が開示される。

【発明の詳細な説明】
【関連出願との関係】
【0001】
本願明細書は、2003年6月12日に出願された米国仮願第60/477,874号、および、2003年12月3日に出願された米国仮出願第60/527,083号の優先権の利益を主張する。
【技術分野】
【0002】
本発明は、分子生物学および微生物学の分野に関する。より具体的には、本発明は微生物のカロテノイド化合物産生に有用な酵素(例えばリコピン、β−カロテン、およびゼアキサンチン)をコードする、・アグロメランス(Pantoea agglomerans)から単離される核酸断片に関連する。
【背景技術】
【0003】
カロテノイドは、最も広く分布して構造的に多様性のあるクラスの天然色素の1つを代表し、淡黄色からオレンジから深紅色の色素色を生じる。カロテン産生性組織の顕著な例としては、ニンジン、トマト、赤ピーマン、およびラッパズイセンおよびマリーゴールドの花弁が挙げられる。カロテノイドは、全ての光合成生物体、ならびにいくつかの細菌および菌・カビ類によって合成される。これらの色素は、光合成、栄養、および光酸化損傷に対する保護おいて重要な機能を有する。例えば動物はカロテノイドを合成する能力を有さず、食物源を通じてこれらの栄養的に重要な化合物を得なくてはならない。
【0004】
自然界における600を越える異なるカロテノイドの存在にもかかわらず、工業的には、食物色素、動物飼料、医薬品、および化粧品のためにわずかなカロテノイドが使用されるのみである。これは生産の困難さが主因である。現在、工業目的で使用されるほとんどのカロテノイドは化学合成によって生産されるが、これらの化合物は化学的に製造することが非常に困難である(非特許文献1)。天然カロテノイドは、植物材料の抽出または微生物合成のいずれかによって得ることができる。しかし商業的なカロテノイド生産にはわずかな植物のみが広く使用され、これらの植物におけるカロテノイド合成の生産性は比較的低い。その結果、これらの植物から生成されるカロテノイドは非常に高価である。生合成の生産能力を増大させる一方法は、組換えDNA技術を応用することである(非特許文献2でレビューされている)。したがって非カロテン産生性細菌および酵母中でカロテノイドを生成することにより、最も適切で効率的な産生生物体の質、量、および選択のコントロールを可能にすることが望ましい。後者は、消費者にとって商業的生産経済(ひいては入手可能性)のために、特に重要である。
【0005】
構造的に最も一般的なカロテノイドは、炭素40個(C40)のテルペノイドである。しかし炭素原子30個のみのカロテノイド(C30、ジアポカロテノイド)が、いくつかの種で検出される。これらの各タイプのカロテノイドの生合成は、イソプレン生合成経路、およびその五炭素ユニバーサルイソプレン構成単位であるイソペンテニルピロリン酸(IPP)に由来する。この生合成経路は、2つの部分に分割できる。1)ファルネシルピロリン酸(FPP)の形成をもたらす上流イソプレン経路、および2)FPPを長鎖C30およびC40カロテン産生性化合物に転換する様々なcrt遺伝子を含んでなる下流カロテノイド生合成経路。この経路の双方の部分を図1に示す。
【0006】
典型的にフィトエンの形成が、C40カロテノイドの生合成にユニークな第1のステップを代表する(図1および2)。フィトエンそれ自体は無色のカロテノイドであり、イソペンテニルピロリン酸イソメラーゼ(遺伝子idiによってコードされる)によるIPPからジメチルアリルピロリン酸(DMAPP)への異性化を通じて生じる。この反応に、
一続きの3プレニル転移酵素反応が続き、そこではゲラニルピロリン酸(GPP)、ファルネシルピロリン酸(FPP)、およびゲラニルゲラニルピロリン酸(GGPP)が形成される。GGPP合成酵素をコードする遺伝子crtEが、この後者の反応を司る。最後に2個のGGPP分子が縮合して、フィトエン(PPPP)を形成する。この反応はフィトエンシンターゼ(遺伝子crtBによってコードされる)によって触媒される。
【0007】
リコピンはフィトエンから生成される「着色」カロテノイドである。リコピンは特徴的な赤色を熟したトマトに与え、食物色素として多くの用途を有する。それはまた、いくつかの細菌、菌・カビ類、および緑色植物におけるその他のカロテノイドの生合成の中間体でもある。リコピンは、遺伝子crtI(フィトエンデサチュラーゼをコードする)によって触媒される、8個の水素原子の除去による4つの逐次脱水素化反応を通じて、フィトエンから生合成的に調製される。この反応における中間物(Imtermediaries)は、フィトフルエン、ζ−カロテン、およびニューロスポレンである。
【0008】
リコピンシクラーゼ(CrtY)は、リコピンをβ−カロテンに転換する。β−カロテンは、カラースペクトルが黄色からオレンジの範囲である典型的なカロテンである。これは、マーガリンおよびバターの着色料として、ビタミンA生成源として、そして最近は特定種類の癌に対して防止効果の可能性がある化合物としての用途がある。
【0009】
β−カロテンは、β−カロテン水酸化酵素活性(crtZ遺伝子によってコードされる)によってもたらされる水酸化反応を通じて、ゼアキサンチンに転換する。例えばこれはトウモロコシの種子中に存在する黄色色素である。ゼアキサンチンは雌鶏または錦鯉の飼料内に含有され、それらの着色の重要な色素源である。
【0010】
フィトエン、リコピン、β−カロテン、およびゼアキサンチンの生成をつかさどるカロテノイド生合成遺伝子および酵素に加えて、(i)水素付加、(ii)脱水素化、(iii)環化、(iv)酸化、(v)エステル化/グリコシル化、またはこれらのプロセスのあらゆる組み合わせによって、C40化合物の分子内転換を可能にして、多数のその他の官能性付与カロテノイド化合物を生成する、その他の様々なcrt遺伝子が知られている。
【0011】
腸内細菌科内の細菌の多くは自然に着色しており、これらの生物体がカロテノイドを生成する能力が示される。さらにC40カロテノイド生合成は、かつて全てエルウィニア(Erwinia)属に所属するとされていた、細菌の大まかなグループに分類されていた生物体の小さなグループであるパントエア(Pantoea)属において、特に良く研究されている[エルウィニア(Erwinia)、ペクトバクテリウム(Pectobacterium)、ブレンネリア(Brenneria)gen.nov.、およびパントエア(Pantoea)を含んでなる4つの系統学的群に、かつての大きなエルウィニア(Erwinia)属が再分類された詳細については、非特許文献3を参照されたい]。例えば非特許文献4、非特許文献5などのいくつかのレビューが、カロテノイド色素生合成の遺伝学を考察している。crtEXYIBZをコードする遺伝子配列は、パントエア・アグロメランス(Pantoea agglomerans)(かつてE.ヘルビコラ(E.herbicola)EHO−10(ATCC番号39368)として知られていた)、P.アナナティス(P.ananatis)(かつてE.ウレドボラ(E.uredovora)20D3(ATCC番号19321)として知られていた)、P.ステワルティ(P.stewartii)(かつてE.ステワルティ(E.stewartii)(ATCC番号8200)として知られていた)、およびP.アグロメランスpv.ミレチア(P.agglomerans pv.milletiae)について入手できる(特許文献1、特許文献2、特許文献3、特許文献4、特許文献5、特許文献6、ジェンバンク(登録商標)登録番号M87280、D90087、AY166713、AB07
6662もまたそれぞれ参照されたい)。しかし既存の文献は、これらの詳しく特性決定されたパントエア(Pantoea)種にあまり関連しない生物体におけるcrtEXYIBZをコードする遺伝子配列、およびこれらの配列の遺伝子構成に関する限定的情報のみを提供する。
【0012】
したがって解決すべき問題は、パントエア・アグロメランス(Pantoea agglomerans)(ATCC番号39368)、P.アナナティス(P.ananatis)(ATCC番号19321)、P.ステワルティ(P.stewartii)(ATCC番号8200)、およびP.アグロメランスpv.ミレチア(P.agglomerans pv.milletiae)とより遠縁の生物体から、カロテノイド生合成酵素の全てまたは一部をコードする核酸配列を同定して、カロテノイド生合成経路のより良い理解のための研究を容易にし、これらの経路の操作のための遺伝的ツールを提供して、適切な遺伝子を適切な宿主中に導入し発現することで、カロテノイドを大量に合成する手段を提供することである。これは合成方法に勝るカロテノイド生成をもたらすであろう。
【0013】
本出願人は、パントエア・アグロメランス(Pantoea agglomerans)DC404株と称される黄色着色素細菌から、CrtE、Idi、CrtY、CrtI、CrtB、およびCrtZ酵素をコードするカロテノイド生合成経路中の6つのユニークな読み取り枠(ORF)を単離することで、既述の問題を解決した。またこれらの配列の遺伝子構成は新しい。
【0014】
【特許文献1】米国特許第5,656,472号明細書
【特許文献2】米国特許第5,545,816号明細書
【特許文献3】米国特許第5,530,189号明細書
【特許文献4】米国特許第5,530,188号明細書
【特許文献5】米国特許第5,429,939号明細書
【特許文献6】国際公開第02/079395A2号パンフレット
【非特許文献1】ネリス(Nelis)およびレーンハー(Leenheer)、Appl.Bacteriol.70:181〜191(1991)
【非特許文献2】ミサワ(Misawa)およびシマダ(Shimada)、J.Biotech.59:169〜181(1998)
【非特許文献3】ハウベン(Hauben)ら、Syst.Appl.Microbiol.21(3):384〜397(1998年8月)
【非特許文献4】G.アームストロング(Armstrong)、J.Bact.176:4795〜4802(1994)
【非特許文献5】G.アームストロング(Armstrong)、Annu.Rev.Microbiol.51:629−659(1997)
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明は、リコペン、β−カロテン、およびゼアキサンチンをはじめとする、様々なカロテノイドの合成に関与することが実証されたDC404株から単離される、6つの遺伝子を提供する。遺伝子は同一オペロン上にクラスター化され、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子を含む。crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子のDNA配列は、ORF1〜6および配列番号1、3、5、7、9、および11にそれぞれ対応する。
【0016】
したがって本発明は、カロテノイド生合成経路酵素をコードする単離された核酸分子を提供し、分子の遺伝子構成はcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子は
記載順にクラスター化され、crtZの転写はcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起きる。
【0017】
好ましい実施態様では、crtE−idi−crtY−crtI−crtB−crtZ遺伝子クラスターのカロテノイド生合成経路酵素の少なくとも1つは、配列番号2、4、6、8、10、および12よりなる群から選択されるアミノ酸配列によってコードされる。そしてさらに好ましい実施態様では、単離された酸分子は配列番号18に記載の配列を有するポリヌクレオチドを含んでなる。
【0018】
同様に本発明は、crtE−idi−crtY−crtI−crtB−crtZを含んでなる配列番号18で示される単離された核酸分子、または配列番号18と少なくとも95%の同一性を有する遺伝子または単離された核酸分子を提供し、単離された核酸分子は、全てのポリペプチドcrtE、idi、crtY、crtI、crtB、およびcrtZをコードする。
【0019】
さらに本発明は、
(a)配列番号2、4、6、8、10、および12よりなる群から選択されるアミノ酸配列をコードする単離された核酸分子、
(b)0.1×SSC、0.1%SDSで65℃、および2×SSC、0.1%SDSで洗浄後、0.1×SSC、0.1%SDSというハイブリダイゼーション条件下で(a)とハイブリダイズする単離された核酸分子、および
(c)(a)または(b)と相補的である単離された核酸分子
よりなる群から選択される、カロテノイド生合成経路酵素をコードする単離された核酸分子を提供する。
【0020】
本発明はさらに、細菌、酵母、糸状菌類、藻類、および植物における遺伝子の遺伝的発現に適した制御領域を含んでなる、本遺伝子および遺伝的キメラによってコードされるポリペプチド、ならびに該物質を含んでなる形質転換宿主を提供する。
【0021】
本発明は、
(a)本核酸分子ゲノムのライブラリーを探索し、
(b)本核酸分子とハイブリダイズするDNAクローンを同定し、
(c)ステップ(b)で同定されたクローンを含んでなるゲノム断片を配列決定することを含んでなる、カロテノイド生合成経路酵素をコードする核酸分子を得る方法を提供し、配列決定されたゲノム断片は、カロテノイド生合成酵素をコードする。
【0022】
同様に本発明は、
(a)本核酸配列の一部に対応する少なくとも1つのオリゴヌクレオチドプライマーを合成し、
(b)ステップ(a)のオリゴヌクレオチドプライマーを使用して、クローン化ベクター中に存在するインサートを増幅しすること
を含んでなる、カロテノイド生合成経路酵素をコードする核酸分子を得る方法を提供し、増幅されたインサートは、カロテノイド生合成経路酵素をコードするアミノ酸配列の一部をコードする。
【0023】
好ましい実施態様では、本発明は、
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、カロテノイド生合成経路酵素をコードする核酸分子
を含んでなる形質転換宿主細胞を提供し、
ここで、該分子の遺伝子構成はcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子は記載順にクラスター化し、crtZの転写はcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起き、
(b)適切な生育条件下でステップ(a)の宿主細胞を有効量の発酵性炭素基質に接触させることでカロテノイド化合物が生成することを含んでなるカロテノイド化合物の製造方法を提供する。
【0024】
同様に本発明は、カロテノイド化合物の生成に適した制御領域を含んでなる、本カロテノイド酵素をコードする一組の核酸分子および遺伝子キメラをさらに提供する。
【0025】
具体的な好ましい実施態様では、本発明は例えばメチロモナス(Methylomonas)16a(ATCCの称号PTA2402)のような高生育メタノトローフ細菌株などのC1代謝宿主におけるカロテノイド化合物の製造方法を提供し、C1代謝宿主は、
(a)メタンおよびメタノールよりなる群から選択されるC1炭素基質で生育し、
(b)機能性的エムデン−マイヤーホフ炭素経路を含んでなり、前記経路はピロリン酸依存性ホスホフルクトキナーゼ酵素をコードする遺伝子を含んでなる。
【0026】
さらに本発明は、生物体においてカロテノイド生合成が改変されるように、生物体中で配列番号1、3、5、7、9、および11よりなる群から選択される少なくとも1つのカロテノイド遺伝子を過剰発現するステップを含んでなる、生物体中でカロテノイド生合成を制御する方法を提供する。
【0027】
代案の実施態様では、本発明は、
(i)a)配列番号1、3、5、7、9、および11よりなる群から選択されるカロテノイド生合成経路酵素をコードする単離された核酸分子、
b)ステップ(a)の前記単離された核酸分子とハイブリダイズする第1のヌクレオチド断片集団、および
c)ステップ(a)の前記単離された核酸分子とハイブリッドしない第2のヌクレオチド断片集団
を含んでなるヌクレオチド配列の混合物を制限エンドヌクレアーゼで消化して、
制限酵素消化断片の混合物を生成させ、
(ii)前記制限酵素消化断片の混合物を変性し、
(iii)ステップ(ii)の変性した前記制限酵素消化断片の混合物をポリメラーゼと共にインキュベートし、
(iv)ステップ(ii)および(iii)を繰り返して、改変された生物学的活性を有するタンパク質をコードする突然変異カロテノイド遺伝子を生成させること
を含んでなる方法によって生成された、改変された生物学的活性を有するカロテノイド生合成経路酵素をコードする突然変異遺伝子を提供する。
【0028】
別の実施態様では、本発明は、配列番号16に記載の16s rDNA配列を含んでなるパントエア・アグロメランス(Pantoea agglomerans)DC404株を提供する。
【0029】
配列説明および生物学的寄託
本願明細書の一部を形成する以下の詳細な説明および添付の配列説明によって、本発明をより完全に理解できるであろう。
【0030】
以下の配列は、37C.F.R.1.821〜1.825(「ヌクレオチド配列および/またはアミノ酸配列開示を含む特許出願の要件−配列規則(Requirements
for Patent Applications Containing Nucleotide Sequences and/or Amino Acid Sequence Disclosures−the Sequence Rules)」)を満たし、世界知的所有権機関(WIPO)標準ST.25(1998)およびEPOおよびPCTの配列表要件(規則5.2および49.5(aの2)、および実施細則第208号および附属書C)に一致する。ヌクレオチドおよびアミノ酸配列データのために使用される記号および型式は、37C.F.R.§1.822で述べられる規則に従う。
【0031】
配列番号1〜12は、表1で同定される完全長の遺伝子またはタンパク質である。
【0032】
【表1】
【0033】
配列番号13〜15、および17は、プライマーHK12、JCR14、JCR15、およびTET−1FP−1をそれぞれコードするヌクレオチド配列である。
【0034】
配列番号16は、DC404株の16S rRNA遺伝子配列を提供する。
【0035】
配列番号18は、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子をコードするDC404株からのDNAの8,814bp断片のヌクレオチド配列である。
【0036】
配列番号19は、プライマーpWEB404Fのヌクレオチド配列である。
【0037】
配列番号20は、プライマーpWEB404Rのヌクレオチド配列である。
【0038】
出願人らは、「特許手続上の微生物の寄託の国際的承認に関するブダペスト条約」の条項に従って、以下の生物学的寄託を行った。
【0039】
【表2】
【0040】
ここでの用法では、「ATCC」は、米国VA20110−2209マナッサスのユニバーシティ・ブールヴァード10801のATCCに所在する米国微生物系統保存機関国際寄託局(American Type Culture Collection International Depository Authority)を指す。「国際寄託名」は、ATCCに寄託された培養物の登録番号である。
【0041】
列挙した寄託株は、表示された国際受託機関に少なくとも30年間保存され、それを開示する特許の付与時に一般に公開される。寄託株の利用可能性は、政府の行動によって付与された特許権を失墜させて主題発明を実施する認可とはみなされない。
【発明を実施するための最良の形態】
【0042】
本発明の遺伝子およびそれらの発現生成物は、様々なカロテノイド化合物を生成する能力を有する組換え生物体の作成のために有用である。CrtE、Idi、CrtY、CrtI、CrtB、およびCrtZをコードする核酸断片がパントエア・アグロメランス(Pantoea agglomerans)DC404株から単離され、当業者に良く知られているBLASTおよびFASTAアルゴリズムを使用して、ヌクレオチドおよびタンパク質配列を含む公共データベースとの比較によって同定されている。本発明の遺伝子および遺伝子産物は、カロテノイド化合物の改良または操作のために、多様なやり方で使用しても良い。これらの遺伝子を含んでなる遺伝子クラスターのユニークな遺伝子構成の結果として、さらなる利点があるかもしれない。
【0043】
カロテノイド化合物は化学的に作ることが非常に困難なので、これらの化合物の微生物による生成には一般的な実用的用途がある。(ネリス(Nelis)およびリーンハー(Leenheer)、前出)。ほとんどのカロテノイドは濃色を有し、天然色素または着色剤と見なすことができる。さらに多くのカロテノイドは強力な抗酸化特性を有するので、食餌中にこれらの化合物を含めることは健康的であると考えられる。良く知られている例はβ−カロテン、カンタキサンチン、およびアスタキサンチンである。さらにカロテノイドは、水産養殖で必要とされる要素である。カロテノイド着色はサケおよびエビの生物体の価値に対して極めて重要であるので、これらの水産養殖は本発明の特に有用な用途である(シャヒディ(Shahidi)F.、およびブラウン(Brown)J.A.、Critical reviews in Food Science、38(1):1〜67(1998))。最後にカロテノイドは、ステロイド、香料および香水、および可能な電気光学用途がある化合物の合成において中間体としての用途を有する。
【0044】
下の開示は、パントエア・アグロメランス(Pantoea agglomerans)DC404株からのカロテノイド合成遺伝子の単離、遺伝子工学によるこれらの遺伝子の修飾、および大腸菌(E.coli)、細菌、酵母、菌・カビ類、および高等植物中でのクローン化および発現に適した適合性プラスミド中へのそれらの挿入の詳細な説明を提供する。
【0045】
定義
本開示では、いくつかの用語および略語が使用される。以下の定義が提供される。
【0046】
「読み取り枠」はORFと略記される。
【0047】
「ポリメラーゼ連鎖反応」はPCRと略記される。
【0048】
「高速液体クロマトグラフィー」はHPLCと略記される。
【0049】
用語「イソプレノイド化合物」とは、形式的にイソプレン(2−メチルブタ−1,3−ジエン、CH2=C(CH3)CH=CH2)から誘導される化合物を指し、その骨格は分子中の反復として認識できる。これらの化合物は、イソペンテニルピロリン酸(IPP)から開始して、イソプレノイド経路を通じて生合成的に生成されてイソプレン単位の頭尾縮合によって形成され、例えば長さが炭素5、10、15、20、30、または40個であっても良い分子をもたらす。
【0050】
「カロテノイド生合成経路」という用語は、図1で示すように、本発明の上流イソプレノイド経路、および/または下流カロテノイド生合成経路のメンバーを含んでなる遺伝子を指す。
【0051】
「上流イソプレノイド経路」および「上流経路」という用語は区別なく使用され、ピルビン酸およびグリセルアルデヒド−3−リン酸からファルネシルピロリン酸(FPP)への転換に関与する酵素を指す。これらの酵素としては、「dxs」遺伝子(1−デオキシキシルロース−5−リン酸シンターゼをコードする)、「dxr」遺伝子(1−デオキシキシルロース−5−リン酸レダクトイソメラーゼをコードする)、「ispD」遺伝子(2C−メチル−D−エリスリトールシチジル転移酵素をコードする、ygbPとしても知られている)、「ispE」遺伝子(4−ジホスホシチジル−2−C−メチルエリスリトールキナーゼをコードする、ychBとしても知られている)、「ispF」遺伝子(2C−メチル−D−エリスリトール2,4−シクロ二リン酸シンターゼをコードする、ygbBとしても知られている)、「pyrG」遺伝子(CTPシンターゼをコードする)、ジメチルアリル二リン酸の形成に関与する「lytB」遺伝子、2−C−メチル−D−エリスリトール4−リン酸の合成に関与する「gcpE」遺伝子、「idi」遺伝子(IPPからジメチルアリルピロリン酸への分子内転換を司る)、およびイソプレノイド経路中の「ispA」遺伝子(ゲラニル転移酵素またはファルネシル二リン酸シンターゼをコードする)が挙げられるが、これに限定されるものではない。
【0052】
「Idi」という用語は、idi遺伝子によってコードされるイソペンテニル二リン酸イソメラーゼ(E.C.5.3.3.2)を指す。代表的なidi遺伝子は、配列番号3として提供される。
【0053】
用語「下流カロテノイド生合成経路」および「下流経路」は区別なく使用され、FPPを一連のカロテノイドに転換する酵素を指す。これらとしては(合成がC30カロテノイド生合成にユニークな第1のステップを代表する)ジアポフィトエン、または(合成がC40カロテノイド生合成にユニークな第1のステップを代表する)フィトエンのいずれかの即時の合成に関与する、遺伝子および遺伝子産物が挙げられる。様々なC30〜C40カロテノイド生成をもたらす全ての引き続く反応は、下流カロテノイド生合成経路に含まれる。これらの遺伝子および遺伝子産物には、crtM、crtN、crtN2、crtE、crtX、crtY、crtI、crtB、crtZ、crtW、crtO、crtA、crtC、crtD、crtF、およびcrtUをはじめとするが、これに限定されるものではない「crt」遺伝子の全てが含まれる。最後に「下流カロテノイド生合成酵素」という用語は、CrtM、CrtN、CrtN2、CrtE、CrtX、CrtY、
CrtI、CrtB、CrtZ、CrtW、CrtO、CrtA、CrtC、CrtD、CrtF、およびCrtUをはじめとするが、これに限定されるものではない、本下流経路中のあらゆる酵素を指す包括的用語である。
【0054】
本願明細書では、「カロテノイド化合物」という用語は、イソプレンから形式的に誘導される接合ポリエン炭素骨格を有する炭化水素のクラスとして定義される。このクラスの分子は、トリテルペン(C30ジアポカロテノイド)およびテトラテルペン(C40カロテノイド)およびそれらの酸素化誘導体から構成される。これらの分子は典型的に強力な光吸収特性を有し、長さがC200を超える範囲であっても良い。例えば長さがC35、C50、C60、C70、およびC80であるその他の「カロテノイド化合物」も知られている。
【0055】
「C30ジアポカロテノイド」は、2個の中央メチル基が1,6−位の関係にあり、残りの非末端メチル基が1,5−位の関係にあるように、イソプレノイド単位の配置が分子の中央で逆転するような様式で結合する6個のイソプレノイド単位からなる。全てのC30カロテノイドは、(i)水素付加、(ii)脱水素化、(iii)環化、(iv)酸化、(v)エステル化/グリコシル化、またはこれらのプロセスのいずれかの組み合わせによって、接合二重結合の長い中央鎖を有する非環式C30H42構造から形式的に誘導されても良い。
【0056】
「テトラテルペン」または「C40カロテノイド」は、2個の中央メチル基が1,6−位の関係にあり、残りの非末端メチル基が1,5−位の関係にあるように、イソプレノイド単位の配置が分子の中央で逆転するような様式で結合した8個のイソプレノイド単位からなる。全てのC40カロテノイドは、(i)水素付加、(ii)脱水素化、(iii)環化、(iv)酸化、(v)エステル化/グリコシル化、またはこれらのプロセスのいずれかの組み合わせによって、接合二重結合の長い中央鎖を有する非環式C40H56構造(下の式I)から形式的に誘導されても良い。このクラスはまた、炭素骨格(式I)の再配置、またはこの構造の一部の(形式的)除去から生じる特定の化合物も含む。
【0057】
【化1】
【0058】
便宜上、カロテノイドの式は、簡単な表記法で書かれることが多い(下の式IA)。
【0059】
【化2】
【0060】
式中、破線はイソプレノイド単位中の形式的区分を示す。
【0061】
「官能性付与された」または「官能性付与」という用語は、あらゆるカロテノイド主鎖
部分の(i)水素付加、(ii)脱水素化、(iii)環化、(iv)酸化、または(v)エステル化/グリコシル化を指す。この主鎖は、接合二重結合の長い中央鎖として定義される。官能性付与はまた、上のプロセスのあらゆる組み合わせによって生じても良い。
【0062】
「CrtE」という用語は、crtE遺伝子によってコードされ、トランス−トランス−ファルネシル二リン酸とイソペンテニル二リン酸をピロリン酸とゲラニルゲラニル二リン酸に転換する、ゲラニルゲラニルピロリン酸合成酵素(synthase)を指す。代表的なcrtE遺伝子は、配列番号1として提供される。
【0063】
「CrtY」という用語は、crtY遺伝子によってコードされ、リコペンをβ−カロテンに転換するリコペンシクラーゼ酵素を指す。代表的なcrtY遺伝子は、配列番号5として提供される。
【0064】
「CrtI」という用語は、crtI遺伝子によってコードされるフィトエンデサチュラーゼ酵素を指す。CrtIは4個の二重結合の導入により、フィトフルエン、β−カロテン、およびニューロスポレンの中間物を通じて、フィトエンをリコペンに転換する。代表的なcrtI遺伝子は、配列番号7として提供される。
【0065】
「CrtB」という用語は、プレフィトエン二リン酸からフィトエンへの反応を触媒するcrtB遺伝子によってコードされるフィトエンシンターゼ酵素を指す。代表的なcrtB遺伝子は、配列番号9として提供される。
【0066】
「CrtZ」という用語は、β−カロテンからゼアキサンチンへの水酸化反応を触媒するcrtZ遺伝子によってコードされる、β−カロテン水酸化酵素を指す。代表的なcrtZ遺伝子は、配列番号11として提供される。
【0067】
「CrtX」という用語は、crtX遺伝子によってコードされ、ゼアキサンチンをゼアキサンチン−β−ジグルコシドに転換する、ゼアキサンチングルコシル転移酵素を指す。
【0068】
「遺伝子構成」という用語は、ORFが単一核酸断片上に組織化される順序と方向を指す。
【0069】
本願明細書中では、3つの異なるDNAクラスターの遺伝子構成について述べられ、それぞれは下のように定義される。
1.「crtE−idi−crtY−crtI−crtB−crtZ」または「crtE−idi−crtYIBZ」という用語は、以下の遺伝子構成を有する分子を指す。crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子は述べられる順にクラスター化され、crtZの転写はcrtE、idi、crtY、crtI、およびcrtBと反対の方向で起きる。
2.「crtE−crtX−crtY−crtI−crtB−crtZ」または「crtEXYIBZ」という用語は、以下の遺伝子構成を有する分子を指す。crtE、crtX、crtY、crtI、crtB、およびcrtZ遺伝子は述べられる順にクラスター化され、crtZの転写はcrtE、crtX、crtY、crtI、およびcrtBと反対の方向で起きる。
3.「crtE−idi−crtX−crtY−crtI−crtB−crtZ」または「crtE−idi−crtXYIBZ」という用語は、以下の遺伝子構成を有する分子を指す。crtE、idi、crtX、crtY、crtI、crtB、およびcrtZ遺伝子は述べられる順にクラスター化され、crtZの転写はcrtE、idi、crtX、crtY、crtI、およびcrtBと反対の方向で起きる。
【0070】
「エムデン−マイヤーホフ経路」という用語は、グルコースおよびフルクトースなどのヘキソースをグリセルアルデヒド3−リン酸、ジヒドロキシアセトンリン酸、ホスホエノールピルビン酸、およびピルビン酸などの重要な細胞性三炭素中間体に転換するための一連の生化学的反応を指す。これらの反応は、ATPの形態の生化学的に有用なエネルギーの正味を生じて典型的に進行する。エムデン−マイヤーホフ経路にユニークな重要な酵素は、ホスホフルクトキナーゼおよびフルクトース1,6−ビスリン酸アルドラーゼである。
【0071】
「エントナー−ドゥドロフ経路」という用語は、生化学的に有用なエネルギーの正味の生成なしに、グルコースまたはフルクトースなどのヘキソースを重要な三炭素細胞性中間体ピルビン酸およびグリセルアルデヒド3−リン酸に転換するための一連の生化学的反応を指す。エントナー−ドゥドロフ経路にユニークな重要な酵素は、6−ホスホグルコン酸デヒドラターゼおよびケトデオキシホスホ−グルコン酸アルドラーゼである。
【0072】
「C1炭素基質」または「一炭素基質」という用語は、炭素−炭素結合が欠如するあらゆる炭素含有分子を指す。例はメタン、メタノール、ホルムアルデヒド、ギ酸、ホルマート、メチル化アミン(例えばモノ−、ジ−、およびトリ−メチルアミン)、メチル化チオール、および二酸化炭素である。
【0073】
「C1代謝個体」という用語は、そのエネルギーおよびバイオマスの唯一の供給源として一炭素基質を使用する能力を有する微生物を指す。C1代謝個体は、典型的にメチロトローフおよび/またはメタノトローフである。
【0074】
「メチロトローフ」という用語は、炭素−炭素結合を含有しない有機化合物を酸化できる生物体を意味する。メチロトローフがCH4を酸化できる場合、メチロトローフはまたメタノトローフでもある。
【0075】
「メタノトローフ」または「メタノトローフ細菌」という用語は、その主要な炭素およびエネルギー供給源としてメタンを利用できる原核生物を意味する。メタンから二酸化炭素への完全な酸化は、好気性分解経路によって起きる。本発明で有用なメタノトローフの典型的な例としては、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、およびメチロサイナス(Methylosinus)属が挙げられる(がこれに限定されるものではない)。
【0076】
「高生育メタノトローフ細菌株」という用語は、唯一の炭素およびエネルギー源としてメタンまたはメタノールで生育できる、機能的エムデンマイヤーホフ炭素流経路を有する細菌を指し、高速の生育および代謝されるC1基質1グラム当たりの高い細胞質量生成が得られる。ここで述べられる特定の「高生育メタノトローフ細菌株」は、「メチロモナス(Methylomonas)16a」、「16a」または「メチロモナス(Methylomonas)sp.16a」と称され、これらの用語は区別なく使用されてメチロモナス(Methylomonas)sp.16a(ATCC PTA−2402)株を指す(米国特許第6,689,601号明細書)。
【0077】
メチロモナス(Methylomonas)中の「crt遺伝子クラスター」という用語は、メチロモナス(Methylomonas)sp.16aの天然C30カロテノイド生合成経路において活性であるcrtN1、ald、およびcrtN2を含んでなる読み取り枠を指す。
【0078】
「CrtN1」という用語は、crtN1遺伝子によってコードされ、メチロモナス(Methylomonas)sp.16aの天然カロテノイド生合成経路において活性である酵素を指す。この遺伝子は、メチロモナス(Methylomonas)においてcrt遺伝子クラスター上に位置する第1の遺伝子である。
【0079】
「ALD」という用語は、ald遺伝子によってコードされ、メチロモナス(Methylomonas)sp.16aの天然カロテノイド生合成経路において活性である酵素を指す。この遺伝子は、メチロモナス(Methylomonas)においてcrt遺伝子クラスター上に位置する第2の遺伝子である。
【0080】
「CrtN3」という用語は、crtN3遺伝子によってコードされ、メチロモナス(Methylomonas)sp.16aの天然カロテノイド生合成に影響する酵素を指す。この遺伝子はcrt遺伝子クラスター内に位置せず、代わりにこの遺伝子はメチロモナス(Methylomonas)ゲノムの異なる遺伝子座内に存在する(国際公開第02/18617号パンフレット)。
【0081】
「色素なし」または「白色変異体」または「非着色株」という用語は、天然ピンク色素(例えばC30カロテノイド)を生成しないメチロモナス(Methylomonas)sp.16a細菌を指す。したがって細菌細胞の色はピンクとは対照的に、白色に見える。メチロモナス(Methylomonas)sp.16a白色変異体は、天然C30カロテノイド遺伝子の全てまたは一部を消去して遺伝子操作された。例えばメチロモナス(Methylomonas)sp.16a中のald/crtN1遺伝子または天然crt遺伝子クラスターを駆動するプロモーターのどちらかの中断は、C40カロテノイド生成によりふさわしい非着色(「白色」)変異体を作り出す(国際公開第02/18617号パンフレット)。
【0082】
「メチロモナス(Methylomonas)sp.16aMWM1000」または「MWM1100」という用語は、メチロモナス(Methylomonas)sp.16aに天然であるaldおよびcrtN1遺伝子の一部を除去して作り出された、非着色メタノトローフ細菌株を指す(国際公開第02/18617号パンフレット)。欠失は、MWM1000におけるC30カロテノイドの生成を中断させた。ald/crtN1欠失は「Δald/crtN1」と表される。
【0083】
ここでの用法では、「単離された核酸断片」は、場合により合成、非天然または修飾ヌクレオチド塩基を含有する一本鎖または二本鎖であるRNAまたはDNAのポリマーである。DNAポリマーの形態の単離された核酸断片は、1つまたはそれ以上のcDNA、ゲノムDNAまたは合成DNAの断片を含んでなっても良い。
【0084】
核酸分子は、適切な温度および溶液イオン強度条件下で、核酸分子の一本鎖形態がその他の核酸分子とアニールできる場合、cDNA、ゲノムDNA、またはRNAなどの別の核酸分子と「ハイブリダイズ可能」である。ハイブリダイズおよび洗浄条件については良く知られており、サムブルック(Sambrook)、J.、フリッチュ(Fritsch)、E.F.およびマニアティス(Maniatis)、T.「分子クローン化:実験室マニュアル(Molecular Cloning:A Laboratory Manual)」、第二版、コールド・スプリング・ハーバー・ラボラトリー(Cold Spring Harbor Laboratory)、ニューヨーク州コールド・スプリング・ハーバー(Cold Spring Harbor、NY)(1989)の特に第11章およびその表11.1で例証される。(以下「マニアティス(Maniatis)」)。温度およびイオン強度条件が、ハイブリダイズの「ストリンジェンシー」を定める。ストリンジェンシー条件は、(遠縁の生物からの相同的配列などの)中程度に類似の断
片をスクリーニングするため、そして(機能性酵素を複製する近縁の生物からの遺伝子などの)高度に類似した断片をスクリーニングするために調節できる。ハイブリダイズ後の洗浄が、ストリンジェンシー条件を決定する。1つの好ましい条件の組は、室温において6×SSC、0.5%SDSで15分間に始まり、次に45℃において2×SSC、0.5%SDSで30分間を反復し、次に50℃において0.2×SSC、0.5%SDSを30分間を2回反復する、一連の洗浄を使用する。より好ましいストリンジェンシー条件の組はより高い温度を使用し、そこでは洗浄は、最後の0.2×SSC、0.5%SDS中での2回の30分間の洗浄の温度を60℃に増大させること以外は上述したのと同一である。別の好ましい高度にストリンジェンシー条件の組は、65℃において0.1×SSC、0.1%SDS中での2回の最終洗浄を使用する。さらに別のストリンジェントな条件の組は、例えば0.1×SSC、0.1%SDSで65℃のハイブリダイズ、および2×SSC、0.1%SDSで洗浄後、0.1×SSC、0.1%SDSを含む。
【0085】
ハイブリダイズのストリンジェンシー次第で塩基間のミスマッチは可能であるが、ハイブリダイズは、2つの核酸が相補的配列を含有することを要求する。核酸がハイブリダイズする適切なストリンジェンシーは、技術分野で良く知られた変数である核酸の長さおよび相補性の程度に左右される。2つのヌクレオチド配列間の類似性または相同性の程度が高い程、これらの配列を有する核酸ハイブリッドのTm値は大きくなる。核酸ハイブリダイズの相対安定性(より高いTmに対応する)は、次の順で低下する。RNA:RNA、DNA:RNA、DNA:DNA。長さが100ヌクレオチドを超えるハイブリッドでは、Tmを計算する式が導かれている(サムブルック(Sambrook)ら、前述9.50〜9.51参照)。より短かい核酸、すなわちオリゴヌクレオチドによるハイブリダイズのためにはミスマッチの配置がより重要になり、オリゴヌクレオチドの長さがその特異性を決定する(サムブルック(Sambrook))ら、前述11.7〜11.8参照)。一実施態様では、ハイブリダイズ可能な核酸の長さは少なくともヌクレオチド約10個である。ハイブリダイズ可能な核酸の好ましい最小の長さは少なくともヌクレオチド約15個、より好ましくは少なくともヌクレオチド約20個、そして最も好ましくは長さが少なくともヌクレオチド約30個である。さらに当業者は、温度および洗浄液の塩濃度が、プローブの長さなどの要因次第で必要に応じて調節できることを認識する。
【0086】
アミノ酸またはヌクレオチド配列の「かなりの部分」とは、当業者による配列の手動評価によって、あるいはBLAST(「基礎的局在性整列化検索ツール(Basic Local Alignment Search Tool)」アルトシュル(Altschul)S.F.ら、J.Mol.Biol.215:403〜410(1993))などのアルゴリズムを使用したコンピュータ支援配列比較および同定によって、遺伝子またはポリペプチドの推定上の同定を得るのに十分なポリペプチドまたは遺伝子のヌクレオチド配列のアミノ酸配列を含んでなる部分である。推定的にポリペプチドまたは核酸配列が既知のタンパク質または遺伝子に相同的であると同定するためには、概して10個以上の隣接するアミノ酸または30個以上のヌクレオチド配列が必要である。さらにヌクレオチド配列に関して、20〜30個の隣接するヌクレオチドを含んでなる遺伝子特異的オリゴヌクレオチドプローブを配列依存遺伝子同定法(例えばサザンハイブリダイズ)および単離(例えば細菌コロニーまたはバクテリオファージ・プラークの原位置(in situ)ハイブリダイズ)において使用しても良い。さらにプライマーを含んでなる特定の核酸断片を得るために、塩基12〜15個の短いオリゴヌクレオチドをPCRで増幅プライマーとして使用しても良い。したがってヌクレオチド配列の「かなりの部分」は、配列を含んでなる核酸断片を特異的に同定および/または単離できるようにする十分な配列を含んでなる。本願明細書は、1つまたはそれ以上の特定の微生物タンパク質をコードする部分的または完全アミノ酸およびヌクレオチド配列を教示する。当業者はここで報告される配列の恩恵を被り、当業者に知られている目的のために、今や開示された配列の全てまたはかなりの部分を使用できる。したがって本発明は、添付の配列表で報告される完全な配列、
ならびに上で定義される配列のかなりの部分を含んでなる。
【0087】
「相補的」と言う用語は、互いにハイブリダイズできるヌクレオチド塩基間の関係について述べるために使用される。例えばDNAについて、アデノシンはチミンに相補的であり、シトシンはグアニンに相補的である。したがって本発明は添付の配列表で報告されるような完全な配列、ならびに実質的に類似した核酸配列に相補的である単離された核酸断片も含む。
【0088】
技術分野で既知の「パーセント同一性」と言う用語は、配列を比較して判定される2つ以上のポリペプチド配列または2つ以上のポリヌクレオチド配列の関係である。技術分野において「同一性」は、場合によってはこのような配列ストリング間の整合によって判定されるような、ポリペプチドまたはポリヌクレオチド配列間の配列関連性の程度も意味する。「同一性」および「類似性」は、1)「計算分子生物学(Computational Molecular Biology)(レスク(Lesk)A.M.編)、オックスフォードユニバーシティ(Oxford University)、ニューヨーク(NY)(1988)、2)「バイオコンピューティング:情報科学およびゲノムプロジェクト(Biocomputing:Informatics and Genome Projects)」(スミス(Smith)D.W.編)、アカデミック(Academic)、ニューヨーク(1993)、3)「配列データのコンピュータ分析(Computer Analysis of Sequence Data)」、第一部、(グリフィン(Griffin)A.M.、およびグリフィン(Griffin)H.G.編)、ヒュマーナ(Humana)、ニュージャージー(NJ)(1994)、4)「分子生物学における配列分析(Sequence Analysis in Molecular Biology)」(フォン・ハインェ(von Heinje)G.編)、アカデミック(1987)、5)「配列分析入門(Sequence Analysis Primer)」(グリブスコフ(Gribskov)M.およびデュヴルー(Devereux)J.編)、ストックトン(Stockton)、ニューヨーク(1991)で述べられたものをはじめとするがこれに限定されるものではない既知の方法によって容易に計算できる。同一性を判定する好ましい方法は、試験される配列間に最良の整合を与えるように設計される。同一性および類似性を判定する方法は、一般に入手できるコンピュータプログラムで体系化される。配列整合および同一性百分率の計算は、ウィスコンシンマディソンのDNASTAR(DNASTAR Inc.(Madison、WI))からのLASERGENEバイオインフォマティクス計算スイートのMegalignプログラムを使用して実施しても良い。Clustal整合法(ヒギンズ(Higgins)およびシャープ(Sharp)、CABIOS.5:151〜153(1989))を使用して、デフォルトのパラメータ(GAP PENALTY=10、GAP LENGTH PENALTY=10)で、配列の複数の整合を実施する。Clustal法を使用した対整合のデフォルトのパラメータは、KTUPLE 1、GAP PENALTY=3、WINDOW=5、およびDIAGONALS SAVED=5である。
【0089】
適切な核酸断片(本発明の単離ポリヌクレオチド)は、ここで報告するアミノ酸配列と少なくとも約70%同一の、好ましくは少なくとも約75%同一の、そしてより好ましくは少なくとも約80%同一のポリペプチドをコードする。好ましい核酸断片は、ここで報告するアミノ酸配列と約85%同一のアミノ酸配列をコードする。より好ましい核酸断片は、ここで報告するアミノ酸配列と少なくとも約90%同一のアミノ酸配列をコードする。最も好ましい核酸断片は、ここで報告するアミノ酸配列と少なくとも約95%同一のアミノ酸配列をコードする。適切な核酸断片は上の相同性を有するだけでなく、典型的に少なくとも50個のアミノ酸、好ましくは少なくとも100個のアミノ酸、より好ましくは少なくとも150個のアミノ酸、さらにより好ましくは少なくとも200個のアミノ酸、そして最も好ましくは少なくとも250個のアミノ酸を有するポリペプチドをコードする
。
【0090】
「コドン縮重」とは、コードされるポリペプチドのアミノ酸配列に影響することなく、ヌクレオチド配列の変動を可能にする遺伝コードにおける性質を指す。したがって本発明は、配列番号2、4、6、8、10、および12に記載される、本微生物のポリペプチドをコードするアミノ酸配列の全てまたはかなりの部分をコードするあらゆる核酸断片に関する。当業者は、任意のアミノ酸を特定化するためのヌクレオチドコドンの利用において、特定の宿主細胞によって示される「コドンバイアス」を十分承知している。したがって宿主細胞中における改善された発現のために遺伝子を合成する場合、コドン使用頻度が、宿主細胞の好ましいコドン使用頻度に近くなるようように、遺伝子をデザインすることが望ましい。
【0091】
「合成遺伝子」は、当業者に知られた手順を使用して化学的に合成されるオリゴヌクレオチド構成単位から構築できる。これらの構成単位をライゲートしアニールして遺伝子セグメントを形成し、次にそれを酵素的にアセンブルして遺伝子全体を構成しても良い。DNA配列に関連して「化学的に合成された」とは、構成要素ヌクレオチドが生体内で(in vitro)アセンブルされたことを意味する。DNAの手動化学合成は確立した手順を使用して達成されても良く、あるいはいくつかの市販の機器の1つを使用して自動化学合成を実施できる。したがって遺伝子をヌクレオチド配列の最適化に基づいて、最適な遺伝子発現のために調整し、宿主細胞のコドンバイアスを反映させることができる。当業者は、コドン利用が宿主によって好まれるコドンに偏っている場合の遺伝子発現成功の見込みの真価を認める。好ましいコドンの判定は、配列情報が利用できる宿主細胞から誘導された遺伝子の調査に基づくことができる。
【0092】
「遺伝子」とは、コード配列に先行する制御配列(5’非コード配列)およびその後ろの制御配列(3’非コード配列)を含めた特異性タンパクを発現する核酸断片を指す。「天然遺伝子」とは、それ自体の制御配列を有して自然界に見いだされる遺伝子を指す。「キメラ遺伝子」とは、自然界には一緒に見られない制御およびコード配列を含んでなる天然遺伝子でないあらゆる遺伝子を指す。したがってキメラ遺伝子は、異なる供給源から誘導される制御配列およびコード配列、あるいは同一供給源から誘導されるが、自然界に見られるのとは異なるやり方で配列された制御配列およびコード配列を含んでなっても良い。「内在性遺伝子」とは、生物のゲノム中の自然な部位にある天然遺伝子を指す。「外来性」遺伝子とは、常態では宿主生物に見られないが、遺伝子移入によって宿主生物中に導入される遺伝子を指す。外来遺伝子は、非天然生物に挿入された天然遺伝子、またはキメラ遺伝子を含んでなることができる。「導入遺伝子」とは、形質転換手順によってゲノム中に導入された遺伝子である。
【0093】
「コード配列」とは、特定のアミノ酸配列をコードするDNA配列を指す。「適切な調節配列」とは、コード配列の上流(5’非コード配列)、配列内、または下流(3’非コード配列)に位置して、転写、RNAプロセシングまたは安定性、または関連コード配列の翻訳に影響を及ぼすヌクレオチド配列を指す。調節配列は、プロモーター、翻訳リーダー配列、イントロン、ポリアデニル化認識配列、RNAプロセッシング部位、エフェクター結合部位、およびステム−ループ構造を含んでも良い。
【0094】
「プロモーター」とは、コード配列または機能性RNAの発現を調節できるDNA配列を指す。概してコード配列は、プロモーター配列に対して3’に位置する。プロモーターは、そっくりそのまま天然遺伝子から誘導されても良く、あるいは自然界に見られる異なるプロモーターから誘導される異なる要素からなっても良く、あるいは合成DNAセグメントを含んでなってさえ良い。異なるプロモーターは、異なる組織または細胞タイプ中で、あるいは異なる発達段階において、あるいは異なる環境または生理学的条件に呼応して
、遺伝子の発現を導いても良いことが当業者には理解される。ほとんどの細胞タイプ中でほとんどの場合に遺伝子の発現を引き起こすプロモーターは、一般に「構造プロモーター」と称される。ほとんどの場合、制御配列のはっきりした境界は完全に画定されていないので、異なる長さのDNA断片が同一のプロモーター活性を有しても良いこともさらに認識される。
【0095】
「3’非コード配列」は、コード配列の下流に位置するDNA配列を指し、mRNAプロセッシングまたは遺伝子発現に影響できる調節シグナルをコードするポリアデニル化認識配列(常態では真核細胞に限られる)およびその他の配列を含む。ポリアデニル化シグナルは、通常、mRNA前駆物質の3’末端へのポリアデニル酸トラクトの付加に影響することで特徴付られる。
【0096】
「RNA転写物」とは、DNA配列のRNAポリメラーゼ触媒転写から得られる生成物を指す。RNA転写物がDNA配列の完全な相補的コピーである場合、それは一次転写物と称され、あるいはそれは一次転写物の転写後プロセッシングから誘導されるRNA配列であるかもしれず、成熟RNAと称される。「メッセンジャーRNA」または「mRNA」とはイントロンがなく、細胞によってタンパク質に翻訳されることができるRNAを指す。「cDNA」とは、mRNAに対して相補的であり、それから誘導される二重鎖DNAを指す。「センスRNA」とは、mRNAを含み、細胞によってタンパク質に翻訳されることができるRNA転写物を指す。「アンチセンスRNA」とは、標的一次転写物またはmRNAの全部または一部に相補的であり、標的遺伝子の発現をブロックするRNA転写物を指す(米国特許第5,107,065号明細書、国際公開第99/28508号パンフレット)。アンチセンスRNAの相補性は、特定の遺伝子転写物のあらゆる部分、すなわち5’非コード配列、3’非コード配列、またはコード配列にあっても良い。「機能RNA」とは、翻訳されないがそれでもなお細胞過程に影響するアンチセンスRNA、リボザイムRNA、またはその他のRNAを指す。
【0097】
「作動的に結合した」と言う用語は、1つの機能が他方の機能によって影響される、単一核酸断片上の核酸配列のつながりを指す。例えばプロモーターはコード配列の発現に影響できる場合、そのコード配列と作動的に結合する(すなわちコード配列がプロモーターの転写調節の下にある)。コード配列は、センスまたはアンチセンスオリエンテーションで制御配列に作動的に結合できる。
【0098】
「発現」と言う用語は、ここでの用法では、本発明の核酸断片から誘導されるセンス(mRNA)またはアンチセンスRNAの転写および安定した蓄積を指す。発現はまた、mRNAのポリペプチドへの翻訳を指しても良い。
【0099】
「成熟」タンパク質とは、翻訳後処理されたポリペプチド、すなわちそれから一次翻訳生成物中に存在するあらゆるプレ−またはプロペプチドが除去されたものを指す。「前駆」タンパク質とは、mRNAの翻訳の一次生成物、すなわちプレ−またはプロペプチドがなおも存在するものを指す。プレ−またはプロペプチドは、細胞内局在化シグナルであっても良い(がそれに限定されない)。
【0100】
「シグナルペプチド」という用語は、分泌された成熟タンパク質に先行するアミノ末端ポリペプチドを指す。シグナルペプチドは成熟タンパク質から切断されるので、その中には存在しない。シグナルペプチドは、分泌されたタンパク質を細胞膜を越えて方向付け、転位置する機能を有する。シグナルペプチドはまた、シグナルタンパク質とも称される。
【0101】
「接合」とは、1つの細菌細胞(すなわち「供与体」)から別の(すなわち「受容体」)細菌細胞への一方向性のDNA転位(例えば細菌プラスミドからの)が起きる、特定の
タイプの形質転換を意味する。プロセスは直接的な細胞と細胞の接触を伴う。別の細菌細胞(すなわち「ヘルパー」)が存在して接合を容易にすることもある。
【0102】
「形質転換」とは、遺伝的に安定した遺伝形質をもたらす、宿主生物体中のゲノムへの核酸断片の転移を指す。形質転換核酸断片を含有する宿主生物体は、「遺伝子導入」、「組換え」または「形質転換」生物体と称される。
【0103】
「プラスミド」、「ベクター」、および「カセット」と言う用語は、細胞の中心的代謝の一部ではない遺伝子を運ぶことが多く、通常環状二本鎖DNA断片の形態である染色体外因子を指す。このような因子は、あらゆる供給源から誘導される一本鎖または二本鎖DNAまたはRNAの配列、ゲノム一体化配列、直鎖または環状のファージまたはヌクレオチド配列を自律的に複製するかもしれず、そこではいくつかのヌクレオチド配列が独自の構成に連結または組み換えされ、それは選択された遺伝子産物のために、適切な3’非翻訳配列と共にプロモーター断片およびDNA配列を細胞中に導入することができる。「形質転換カセット」とは、外来性遺伝子を含有し、外来遺伝子に加えて特定の宿主細胞の形質転換を容易にする因子を有する特定のベクターを指す。「発現カセット」とは、外来性遺伝子を含有し、外来性遺伝子に加えて外来性宿主におけるその遺伝子の促進された発現を可能にする因子を有する特定のベクターを指す。
【0104】
「改変された生物学的活性」と言う用語は、アッセイ方法によって測定できる、ヌクレオチド配列によってコードされるタンパク質に関連した活性を指し、活性は、天然配列に関連した活性を超え、またはそれ未満のいずれかである。「増強された生物学的活性」とは、天然配列に関連したものを超える改変された活性を指す。「低下した生物学的活性」とは、天然配列に関連したもの未満の改変された活性を指す。
【0105】
「配列分析ソフトウェア」と言う用語は、ヌクレオチドまたはアミノ酸配列の分析のために有用なあらゆるコンピュータアルゴリズムまたはソフトウェアプログラムを指す。「配列分析ソフトウェア」は、市販のものでも、あるいは独立して開発されても良い。典型的な配列分析ソフトウェアとしては、1.)ウィスコンシン州マディソンのジェネティック・コンピュータ・グループからのGCGパッケージプログラム(Wisconsin Package Version 9.0、Genetics Computer Group(GCG),Madison,WI)、2.)BLASTP、BLASTN、BLASTX(アルシュール(Altschul)ら著、J.Mol.Biol.215:403〜410(1990))、および3.)ウィスコンシン州マディソンのDNASTAR(DNASTAR,Inc.Madison、WI)からのDNASTAR、および4.)スミス−ウォーターマン・アルゴリズムを組み入れたFASTAプログラム(W.R.ピアースン(Pearson)著、Comput.Methods Genome Res.[Proc.Int.Symp.](1994)、1992年会議、111〜20、編集者:スハイ,サンドル(Suhai,Sandor)、Plenum、ニューヨーク州ニューヨーク(New York、NY))、および5.)メリーランド州ベセズダのインフォマックス(InforMax,Inc.(Bethesda、MD))からのベクターNTIプログラムが挙げられるが、これに限定されるものではない。本願明細書の文脈内では、配列分析ソフトウェアを分析のために使用する場合、分析結果は特に断りのない限り、言及されるプログラムの「デフォルト値」に基づくものと理解される。ここでの用法では、「デフォルト値」とは、最初に初期化されたときにソフトウェアに元々ロードされた、あらゆる値またはパラメータの組(製造業者によって設定される)を意味する。
【0106】
ここで使用される標準組換えDNAおよび分子クローン化技術は技術分野で良く知られており、マニアティス(Maniatis)(上述);シルハビー(Silhavy)T
.J.、ベンナン(Bennan)M.L.およびエンクイスト(Enquist)L.W.「遺伝子融合実験(Experiments with Gene Fusions)」コールド・スプリング・ハーバー・ラボラトリー:ニューヨーク州コールド・スプリング・ハーバー(1984);およびオースベル(Ausubel)、F.M.ら「分子生物学現代プロトコル(Current Protocols in Molecular Biology)」、グリーン・パブリッシング・アソシエーツ(Greene Publishing Assoc.)およびワイリー−インターサイエンス(Wiley−Interscience)による出版(1987)で述べられている。
【0107】
カロテノイド生成に関与する遺伝子
カロテノイド化合物の生合成に関与する酵素経路は、ピルビン酸およびグリセルアルデヒド−3−リン酸からファルネシルピロリン酸への転換を提供する上流イソプレノイド経路、およびジアポフィトエンまたはフィトエンいずれかの合成と、全ての引き続いて生成されるカロテノイドを提供する下流カロテノイド生合成経路の2つの部分に都合良く分けることができる(図1)。上流経路は多くの微生物に遍在性であり、これらの場合、所望のカロテノイド生合成のための下流経路を含んでなる遺伝子を導入することだけが必要かもしれない。2つの経路間の区分は、ファルネシルピロリン酸塩(FPP)の合成に関わる。FPPが天然に存在する場合、下流カロテノイド生合成経路の要素のみが必要である。しかしカロテノイド生成において下流経路カロテノイド遺伝子が効果的であるためには、宿主細胞が細胞内に適切なレベルのFPPを有することが必要なことが理解される。FPP合成が宿主細胞によって提供されない場合、FPPの生成に必要な遺伝子を導入することが必要である。これらの各経路について下で詳細に述べる。
【0108】
上流イソプレノイド経路
イソプレノイド生合成は、一般的なC5イソプレンサブユニットであるイソペンテニルピロリン酸塩(IPP)を生じる2つの経路のどちらかを通じて起きる。最初に良く知られている酢酸/メバロン酸経路を通じてIPPを合成しても良い。しかし最近の研究は、メバロン酸依存経路が全ての生物体で作動しているわけではないことを実証している。IPP生合成のための代案のメバロン酸非依存経路が、細菌および緑藻類および高等植物で特性決定されている(ホルバック(Horbach)ら、FEMS Microbiol.Lett.111:135〜140(1993);ローマー(Rohmer)ら、Biochem.295:517〜524(1993);シュベンダー(Schwender)ら、Biochem.316:73〜80(1996);アイゼンライヒ(Eisenreich)ら、Proc.Natl.Acad.Sci.USA 93:6431〜6436(1996))。
【0109】
メバロン酸非依存イソプレノイド経路中の多くのステップが知られている(図1)。例えばIPPの生成をもたらす代案の経路の最初のステップは、コール(Cole)ら(Nature 393:537〜544(1998))によって結核菌(Mycobacterium tuberculosis)において研究されている。経路の第1のステップは、D−1−デオキシキシルロース−5−リン酸として知られている五炭素化合物を生じる、2つの三炭素分子(ピルビン酸およびD−グリセルアルデヒド3−リン酸)の縮合を伴う。この反応はdxs遺伝子によってコードされるDxs酵素によって起きる。次にD−1−デオキシキシルロース−5−リン酸の異性化および還元から、2−C−メチル−D−エリスリトール−4−リン酸が生じる。異性化および還元プロセスに関与する酵素の1つは、遺伝子dxrによってコードされるD−1−デオキシキシルロース−5−リン酸レダクトイソメラーゼ(Dxr)である。2−C−メチル−D−エリスリトール−4−リン酸を引き続いて、注釈なしの遺伝子ygbP(コール(Cole)ら、前出)でコードされる酵素によって、CTP−依存反応において4−ジホスホシチジル−2C−メチル−D−エリスリトールに転換する。しかし最近ygbP遺伝子は、isp遺伝子クラスター
の一部としてispDと改名された(SwissProtein登録番号Q46893)。
【0110】
次にychB遺伝子によってコードされる酵素によって、4−ジホスホシチジル−2C−メチル−D−エリスリトールの第2位のヒドロキシ基をATP−依存反応においてリン酸化できる。この生成物は4−ジホスホシチジル−2C−メチル−D−エリスリトールをリン酸化して、4−ジホスホシチジル−2C−メチル−D−エリスリトール2−リン酸が得られる。ychB遺伝子もまたisp遺伝子クラスターの一部としてispEと改名された(SwissProtein登録番号P24209)。最後にygbB遺伝子の生成物が、CTP−依存様式で、4−ジホスホシチジル−2C−メチル−D−エリスリトール2−リン酸を2C−メチル−D−エリスリトール2,4−シクロ二リン酸に転換する。この遺伝子もまた最近改名されて、isp遺伝子クラスターに属する。具体的には、ygbB遺伝子の新しい名称はispF(SwissProtein登録番号P36663)である。pyrG遺伝子の生成物は、CTPシンターゼとしてこれらの反応において重要である。
【0111】
lytBおよびgcpE(およびおそらくはその他の)遺伝子によってコードされる酵素はイソペンテニルピロリン酸(IPP)およびジメチルアリルピロリン酸(DMAPP)の形成をもたらす反応に関与すると考えられる。IPPは、idi遺伝子によってコードされるイソペンテニル二リン酸イソメラーゼ(または「IPPイソメラーゼ」)を通じてDMAPPに異性化しても良いが、この酵素は生存には必須ではなく、2−C−メチル−D−エリスリトール4−リン酸(MEP)経路を使用するいくつかの細菌では不在でも良い。最近の証拠はMEP経路がIPPの前に分岐して、lytB遺伝子産物を通じてIPPとDMAPPを別個に生成することを示唆する。lytBノックアウト変異は、IPPおよびDMAPPの双方を補った培養液中以外では、大腸菌(E.coli)に対して致死性である。
【0112】
FPPの合成は、IPPのジメチルアリルピロリン酸(DMAPP)への異性化を通じて起きる。この反応に、ispAによって触媒される2つのプレニル転移酵素反応手順が続き、ゲラニルピロリン酸(GPP、炭素数10の分子)およびファルネシルピロリン酸(FPP、炭素数15の分子)の生成をそれぞれもたらす。
【0113】
下流カロテノイド生合成経路
上流イソプレノイド経路と下流カロテノイド経路の間の区分は、幾分主観的である。FPP合成はカロテン産生性および非カロテン産生性細菌の双方で一般的であるので、出願人は、C30ジアポカロテノイドまたはC40カロテノイドのどちらかの形成をもたらす分岐する2経路の化合物へのファルネシルピロリン酸(FPP)の転換を開始するための下流カロテノイド生合成経路中の第1のステップを考察する。
【0114】
C40経路中では、生合成経路の第1のステップは、IPPの添加によって、ファルネシルピロリン酸(FPP)をゲラニルゲラニルピロリン酸(GGPP)として知られている炭素数20の分子に転換するプレニル転移酵素反応で始まる。GGPP合成酵素をコードする遺伝子crtE(EC2.5.1.29)は、このプレニル転移酵素反応を司る。次に2個のGGPP分子の縮合反応が起きて、下流カロテノイド生合成経路の第1の炭素数40の分子であるフィトエン((7,8,11,12,7’,8’,11’,12’−ω−オクタヒドロ−ω,ω−カロテン、またはPPPP)を形成する。この酵素的反応はCrtB(フィトエンシンターゼ、EC2.5.1.−)によって触媒される。
【0115】
引き続く水素付加、脱水素、環化、酸化、またはこれらのプロセスのあらゆる組み合わせによって、化合物フィトエンからC40カロテノイドのスペクトルを生成した。例えば
「赤色」着色スペクトルを与えるリコピンは、遺伝子crtI(フィトエンデサチュラーゼをコードする)によって触媒される、8個の水素原子の除去による4つの逐次脱水素化反応を通じて、フィトエンから生成される(図2参照)。リコピンシクラーゼ(CrtY)は、リコピンをβ−カロテン(β,β−カロテン)に転換する。β−カロテンは、β−カロテン水酸化酵素(crtZ遺伝子によってコードされる)活性に起因する水酸化反応を通じて、ゼアキサンチン((3R,3’R)−β,β−カロテン−3,3’−ジオール)に転換される。
【0116】
フィトエン、リコペン、β−カロテン、およびゼアキサンチンを作り出す組み合わせで使用できるcrtE、crtY、crtI、crtB、およびcrtZに加えて、直鎖C40化合物の分子内転換を可能にし、多数のその他の官能性付与カロテノイド化合物を生成する様々なその他のcrt遺伝子が知られている。当業者は、公的に入手できる文献(例えばジェンバンク(登録商標))、特許文献、およびカロテノイドを生成する能力を有する微生物の実験的分析に従って、様々なその他のcrt遺伝子同定できるであろう。
例えば、
・ゼアキサンチンは、crtX遺伝子(EC2.4.1.−、例えばジェンバンク(登録商標)登録番号D90087、M87280、M90698)によってコードされるゼアキサンチングルコシル転移酵素によってゼアンキサンチン−β−グルコシドに転換できる。
・β−カロテンは、crtW(例えばジェンバンク(登録商標)登録番号AF218415、D45881、D58420、D58422、X86782、Y15112)、crtO(例えばジェンバンク(登録商標)登録番号X86782、Y15112)またはbktによってコードされるβ−カロテンケトラーゼによって、カンタキサンチンに転換できる。エキネノンはこの反応における中間体である。
・カンタキサンチンは、crtZ遺伝子によってコードされるβ−カロテン水酸化酵素によって、アスタキサンチンに転換できる。アドニルビンはこの反応における中間体である。
・ゼアキサンチンは、crtW、crtO、またはbktによってコードされるβ−カロテンケトラーゼによって、アスタキサンチンに転換できる。アドニキサンチンはこの反応における中間体である。
・スフェロイデンは、crtA(例えばジェンバンク(登録商標)登録番号AJ010302、Z11165、X52291)によってコードされるスフェロイデンモノオキシゲナーゼによって、スフェロイデノンに転換できる。
crtC(例えばジェンバンク(登録商標)登録番号AB034704、AF195122、AJ010302、AF287480、U73944、X52291、Z11165、Z21955)、crtD(例えばジェンバンク(登録商標)登録番号AJ010302、X63204、U73944、X52291、Z11165)、およびcrtF(例えばジェンバンク(登録商標)登録番号AB034704、AF288602、AJ010302、X52291、Z11165)遺伝子によってそれぞれコードされる、ヒドロキシニューロスポレンシンターゼ、メトキシニューロスポレンデサチュラーゼ、およびヒドロキシニューロスポレン−O−メチル転移酵素の逐次作用によって、ニューロスポレンはスフェロイデンに転換でき、リコペンはスピリロキサンチンに転換できる。
・β−カロテンは、crtU(例えばジェンバンク(登録商標)登録番号AF047490、AF121947、AF139916、AF195507、AF272737、AF372617、AJ133724、AJ224683、D26095、U38550、X89897、Y15115)によってコードされる、β−カロテンデサチュラーゼによってイソレニエラテンに転換できる。
【0117】
これらの例は制限を意図せず、このC40下流カロテノイド生合成経路内には、その他の多くのカロテノイド遺伝子および生成物が存在する。したがって宿主生物体中で十分な
FPP源が利用できれば、本発明の方法を使用して、場合により植物、動物、および/または細菌源から単離されたあらゆるその他の知られているcrt遺伝子に加えて、ここに示すcrtE、crtY、crtI、crtB、およびcrtZ遺伝子の様々な組み合わせを使用することで、無数の異なるカロテノイドおよびカロテノイド誘導体が作成できる。
【0118】
本発明の有用な生成物は、アンテラキサンチン、アドニルビン、アドニキサンチン、アスタキサンチン、カンタキサンチン、カプソルブリン、β−クリプトキサンチン、α−カロテン、β−カロテン、ε−カロテン、エキネノン、3−ヒドロキシエキネノン、3’−ヒドロキシエキネノン、α−カロテン、4−ケト−γ−カロテン、ζ−カロテン、α−クリプトキサンチン、デオキシフレキシキサンチン、ジアトキサンチン、7,8−ジデヒドロアスタキサンチン、フコキサンチン、フコキサンチノール、イソレニエラテン、ラクツカキサンチン、ルテイン、リコピン、ミキソバクトン、ネオキサンチン、ニューロスポレン、ヒドロキシニューロスポレン、ペリジニン、フィトエン、ロドピン、ロドピングルコシド、4−ケト−ルビキサンチン、シホナキサンチン、スフェロイデン、スフェロイデノン、スピリロキサンチン、4−ケト−トルレン、3−ヒドロキシ−4−ケト−トルレン、ウリオリド、ウリオリドアセタート、ビオラキサンチン、ゼアキサンチン−β−ジグルコシド、およびゼアキサンチンをはじめとするが、これに限定されるものではない、ここで定義されるあらゆるカロテノイド化合物を含むことが構想された。さらに本発明は、ヒドロキシ−、メトキシ−、オキソ−、エポキシ−、カルボキシ−、またはアルデヒド酸官能基、グリコシドエステル、またはスルフェートを作り出す、これらの分子の誘導化を包含する。
【0119】
上流イソプレノイド経路と下流カロテノイド生合成経路との相互作用
全体的なイソプレノイド生合成を促進させることによって、カロテノイド生成を促進する多様な研究が試みられている。特にidiの上方制御は、カロテノイド生成に劇的に影響することが実証されている。例えばカジワラ(Kajiwara)ら(Biochem.J.324:421〜426(1997))は、「IPPイソメラーゼは、原核生物大腸菌(E.coli)のイソプレノイド生合成において影響力が大きいステップを形成し、代謝遺伝子操作による工業的に有用なイソプレノイドの効率的生成の可能性がある」ことを最初に実証した。具体的には、外来性に発現されるIPPイソメラーゼが、エルウィニア(Erwinia)カロテノイド生合成遺伝子クラスターを含んでなる大腸菌(E.coli)において、対照と比べて3.6〜4.5倍高いレベルのリコピン生成を可能にし、同様に1.5〜2.7倍高いレベルのβ−カロテンおよび1.7〜2.1倍高いレベルのフィトエンが生成された。
【0120】
ワング(Wang)らの引き続く研究(Biotech.Bioengineering 62(2):235〜241(1999))は、大腸菌(E.coli)idi遺伝子、アーキオグロバス・フルギダス(Archaeoglobus fulgidus)gps遺伝子、およびアグロバクテリウム・オーランチアカム(Agrobacterium aurantiacum)crtBIYZW遺伝子クラスターで形質転換した大腸菌(E.coli)において、50倍高いアスタキサンチン生成をもたらした。GGPP合成における最後のステップが、カロテノイド生成における第1の速度制御ステップであり、第2の速度制御ステップはIPP異性化であると結論された。そして、最後にアルブレヒト(Albrecht)ら(Biotech.Letters 21:791〜795(1999))は、大腸菌(E.coli)における内在性dxsおよびdxr遺伝子および外来性idi遺伝子(ファフィア・ロドジマ(Phaffia rhodozyma)からの)の過剰発現が、カロテン産生を3.5倍まで刺激できることを見いだした。
【0121】
したがって大腸菌(E.coli)およびその他の細菌における、工業的に価値あるカ
ロテノイド生成の最大化に向けた代謝操作法は、上流イソプレノイド経路中の流束および律速ステップ、ならびに下流カロテノイド生合成経路における発現レベルを注意深く考慮すべきである。上流イソプレノイド経路中の律速遺伝子(例えばidi)の過剰発現は、カロテン産生を劇的に増大できる。
【0122】
P.アグロメランス(P.agglomerans)DC404株のカロテノイド生合成遺伝子および酵素の配列同定
C40カロテノイド生合成経路に関与する遺伝子産物をコードする多様なヌクレオチド配列が、DC404株から単離されている。例えばORFの1および3〜6は、下流カロテノイド生合成経路中のcrtE、Y、I、B、およびZ遺伝子をコードし(図1および2参照)、それらの酵素生成物は、着色カロテノイド、リコペン、β−カロテン、およびゼアキサンチンの生成をもたらす。ORF2は、上流イソプレノイド経路においてidi遺伝子をコードする。これらの6個のORFは、crtE−idi−crtY−crtI−crtB−crtZの遺伝子構成を有する1つの核酸断片(配列番号18)上に構成される。crtE−idi−crtY−crtI−crtB遺伝子は、オペロン内で作動的に結合されているように見え、他方crtZ遺伝子は、反対方向に転写される。
【0123】
DC404株から単離された遺伝子の組全体(crtE−idi−crtY−crtI−crtB−crtZ)は、ここで1つの配列(配列番号18)内に開示される。カロテノイド化合物の生成ために、この遺伝子クラスターをベクターに入れて、微生物宿主中で発現させる。当業者はコードされた全てのポリペプチドが発現して酵素的に活性ならば、その有用性に影響することなく、このような遺伝子クラスターに少数の核酸置換、付加、および欠失(特定の宿主細胞発現のための好ましいコドンの置換などの)があっても良いことを認識するであろう。したがってcrtE−idi−crtY−crtI−crtB−crtZを含んでなる配列番号18で示される単離された核酸分子、配列番号18と少なくとも95%の同一性を有する遺伝子または単離された核酸分子を提供することは本発明の範囲内であり、単離された核酸分子は、crtE、idi、crtY、crtI、crtB、およびcrtZの全てのポリペプチドをコードする。
【0124】
スミス−ウォーターマン位置合わせアルゴリズム(W.R.ピアソン(Pearson)著、Comput.Methods Genome Res.[Proc.Int.Symp.](1994)、1992年会議、111〜20、編集者:スハイ,サンドル(Suhai,Sandor)、Plenum、New York、NY)を使用して、crtEヌクレオチド塩基および推定されるアミノ酸配列(ORF1)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸301個の長さにわたり、ここで報告するCrtEのアミノ酸配列と約65%同一であることが明らかにされる。より好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtEコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtEの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0125】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、idiヌクレオチド塩基および推定されるアミノ酸配列(ORF2)と、公共データベースとの比較から、最も類似した既知の配列が、アミノ酸349個の長さにわたり、ここで報告するIdiのアミノ酸配列と約67%同一であることが明らかにされる。より好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいidiコード核酸配列は、活性タンパク質をコードするもので、ここで報告するidiの核酸配列と少なくとも約70%〜80
%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0126】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、crtYヌクレオチド塩基および推定されるアミノ酸配列(ORF3)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸389個の長さにわたり、ここで報告するCrtYのアミノ酸配列と約61%同一であることが明らかにされる。より好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtYコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtYの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0127】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、crtIヌクレオチド塩基および推定されるアミノ酸配列(ORF4)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸492の長さにわたり、ここで報告するCrtIのアミノ酸配列と約82%同一であることが明らかにされる。好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtIコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtIの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0128】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、crtBヌクレオチド塩基および推定されるアミノ酸配列(ORF5)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸308個の長さにわたり、ここで報告するCrtBのアミノ酸配列と約67%同一であることが明らかにされる。より好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtBコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtBの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0129】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、crtZヌクレオチド塩基および推定されるアミノ酸配列(ORF6)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸185個の長さにわたり、ここで報告するCrtZのアミノ酸配列と約73%同一であることが明らかにされる。好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtZコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtZの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0130】
相同体の単離
本発明のC40カロテノイド生合成経路の各核酸断片を使用して、同一またはその他の微生物の(または植物)種から相同タンパク質をコードする遺伝子を単離しても良い。配
列依存プロトコルを使用した相同遺伝子の単離は、技術分野で良く知られている。配列依存プロトコルの例としては、以下が挙げられるが、これに限定されるものではない。1.)核酸ハイブリダイズ法、2.)様々な核酸増幅技術の使用によって例証されるようなDNAおよびRNA増幅法[例えばポリメラーゼ連鎖反応(PCR)、ミュリス(Mullis)ら、米国特許第4,683,202号明細書;リガーゼ連鎖反応(LCR)、タボール(Tabor)S.ら、Proc.Natl.Acad.Sci.USA82:1074(1985);または鎖置換増幅(SDA)、ウォーカー(Walker)ら、Proc.Natl.Acad.Sci.USA、89:392(1992)]、および3.)相補によるライブラリー構築およびスクリーニング法。
【0131】
例えば、ここで述べるようなC40カロテノイド生合成経路のものと類似したタンパク質またはポリペプチドをコードする遺伝子は、当業者に良く知られている方法を使用して、本核酸断片の全部または一部をDNAハイブリダイズプローブとして使用して、あらゆる所望の細菌からのライブラリーをスクリーニングして直接単離できる(C40カロテノイドを生成する細菌が好ましい)。本核酸配列に基づく特定のオリゴヌクレオチドプローブは、技術分野で知られている方法によってデザインして合成できる(マニアティス(Maniatis)、前出)。さらに当業者に知られている方法(例えばランダムプライマーDNA標識、ニックトランスレーション、または末端標識技術)によって、配列全体を直接使用してDNAプローブを合成し、または利用できる生体外(in vitro)転写システムを使用してRNAプローブを合成できる。さらに特定のプライマーをデザインして使用し、本配列の一部(または全長)を増幅できる。得られる増幅生成物を増幅反応中に直接標識し、または増幅反応後に標識して、プローブとして使用し、適切なストリンジェンシー条件下で完全長DNA断片を単離できる。
【0132】
典型的にPCRタイプ増幅技術ではプライマーは異なる配列を有し、互いに相補的でない。所望の試験条件次第で、プライマーの配列は、標的核酸の効率的かつ忠実な複製を提供するようにデザインされるべきである。PCRプライマーデザインの方法は一般的であり、技術分野で周知である。テイン(Thein)およびウォーレス(Wallace)著、「遺伝性疾患診断における特定のハイブリダイズプローブとしてのオリゴヌクレオチドの使用(The use of oligonucleotide as specific hybridization probes in the Diagnosis of Genetic Disorders)」、ヒト遺伝病:実際的アプローチ(Human Genetic Diseases:A Practical Approach)中、K.E.デービス(Davis)編、(1986)、33〜50ページ、IRL:ヘルンドン(Herndon)バージニア(VA);およびリュクリック(Rychlik)W.「分子生物学的方法(Methods in Molecular Biology)」第15巻、31〜39ページ、PCRプロトコル:現行の方法と応用(PCR Protocols:Current Methods and Applications)、ホワイト(White),B.A.編(1993)、ヒューマニア(Humania):ニュージャージー州トトワ(Totowa、NJ)。
【0133】
概してポリメラーゼ連鎖反応プロトコルにおいて、本配列の2本の短い断片を使用して、DNAまたはRNAから相同的遺伝子をコードするより長い核酸断片を増幅しても良い。またポリメラーゼ連鎖反応は、1つのプライマーの配列が本核酸断片から誘導され、その他のプライマーの配列が微生物の遺伝子をコードするmRNA前駆物質の3’末端へのポリアデニル酸のトラクトの存在を利用する、クローン化された核酸断片のライブラリーにおいて実施しても良い。
【0134】
代案としては、第2のプライマー配列は、クローン化ベクターから誘導される配列に基づいても良い。例えば当業者は、RACEプロトコル(フローマン(Frohman)ら
、Proc.Natl.Acad.Sci.USA 85:8998(1988))に従って、PCRを使用して、転写物の一点と3’または5’末端との間の領域のコピーを増幅し、cDNAを作り出すことができる。3’および5’方向に向けたプライマーは、本配列からデザインできる。メリーランド州ゲーサーズバーグのBRL(BRL、Gaithersburg、MD)から商業的に入手できる3’RACEまたは5’RACEシステム系を使用して、特定の3’または5’cDNA断片を単離できる(オハラ(Ohara)ら、Proc.Natl.Acad.Sci.USA 86:5673(1989);ロー(Loh)ら、Science 243:217(1989))。
【0135】
代案としては、相同体の同定のために、ハイブリダイズ試薬としてC40カロテノイド生合成経路の本配列を用いても良い。核酸ハイブリダイズ試験の基本的構成要素には、プローブ、対象の遺伝子または遺伝子断片を含有することが疑われるサンプル、および特定のハイブリダイズ法が含まれる。本発明のプローブは典型的に、検出する核酸配列に相補的である一本鎖核酸配列である。プローブは、検出する核酸配列と「ハイブリダイズ可能」である。プローブの長さは、5個の塩基から数万個の塩基の間で変動しても良く、実施する特定の試験に左右される。典型的に、約15個の塩基から約30個の塩基のプローブ長が適切である。プローブ分子の一部のみが、検出する核酸配列に相補的である必要がある。さらにプローブと標的配列との間の相補性は完璧でなくても良い。ハイブリダイズは不完全に相補的である分子間でも生じ、その結果、ハイブリッドした領域内の特定の塩基の一部は、適切な相補的塩基と対合形成しない。
【0136】
ハイブリダイズ方法は良く定義されている。典型的には、プローブおよびサンプルは、核酸ハイブリダイズを可能にする条件下で混合されなくてはならない。これは適切な濃度および温度条件下において、無機または有機塩存在下で、プローブとサンプルを接触させることを伴う。プローブとサンプル核酸の間であらゆる可能なハイブリダイズが起きるように、プローブおよびサンプル核酸は、十分長い時間接触しなくてはならない。混合物中のプローブまたは標的濃度が、ハイブリダイズが生じるのに必要な時間を決定する。プローブまたは標的濃度が高いほど、必要なハイブリダイズインキュベーション時間は短くなる。場合により、カオトロピック剤を添加しても良い。カオトロピック剤は、ヌクレアーゼ活性を阻害することによって核酸を安定化させる。さらにカオトロピック剤は、室温において短いオリゴヌクレオチドプローブの高感応性でストリンジェントなハイブリダイズを可能にする(ヴァン・ネス(Van Ness)およびチェン(Chen)Nucl.Acids Res.19:5143〜5151(1991))。適切なカオトロピック剤としては、特に塩化グアニジニウム、グアニジニウムチオシアネート、ナトリウムチオシアネート、リチウムテトラクロロ酢酸、過塩素酸ナトリウム、ルビジウムテトラクロロ酢酸、ヨウ化カリウム、およびセシウムトリフルオロ酢酸が挙げられる。典型的にカオトロピック剤は、約3Mの最終濃度で存在する。所望するならば、ハイブリダイズ混合物にホルムアミドを典型的に30〜50%(v/v)で添加できる。
【0137】
様々なハイブリダイズ溶液を用いることができる。典型的にこれらは、約20〜60%、好ましくは30%容積の極性有機溶剤を含んでなる。一般的なハイブリダイズ溶液では、約30〜50%v/vのホルムアミド、約0.15〜1Mの塩化ナトリウム、(例えばクエン酸ナトリウム、トリス−HCl、PIPESまたはHEPESなどの)約0.05〜0.1Mの緩衝液(pH範囲約6〜9)、(例えばドデシル硫酸ナトリウムなどの)約0.05〜0.2%の界面活性剤、または0.5〜20mMのEDTA、ファーマシア(Pharmacia Inc.)からのFICOLL(約300〜500キロダルトン)、ポリビニルピロリドン(約250〜500キロダルトン)、および血清アルブミンを用いる。また典型的なハイブリダイズ溶液には、約0.1〜5mg/mLの未標識のキャリア核酸、(例えば仔ウシ胸腺またはサケ精子DNA、または酵母RNAなどの)断片化核DNA、および場合により約0.5〜2%重量/容積のグリシンも含まれる。(例えばポ
リエチレングリコールなどの)多様な極性水溶性剤または膨潤剤をはじめとする容積排除剤、(例えばポリアクリレートまたはポリメチルアクリレートなどの)アニオン性ポリマー、および(例えば硫酸デキストランなどの)アニオン性糖ポリマーなどその他の添加剤を含めても良い。
【0138】
核酸ハイブリダイズは多様なアッセイ型式に適合できる。最も適切なものの1つは、サンドイッチアッセイ型式である。サンドイッチアッセイは、特に非変性条件下でのハイブリダイズに適合できる。サンドイッチ−タイプアッセイの主要構成要素は固形担体である。固形担体はそれに吸着され、あるいはそれと共有結合的に結合する、未標識で配列の一部分と相補的である固定核酸プローブを有する。
【0139】
本ヌクレオチドおよび推定されるアミノ酸配列の利用可能性は、DNA発現ライブラリーの免疫学的スクリーニングを容易にする。本アミノ酸配列の一部を代表する合成ペプチドを合成しても良い。これらのペプチドを用いて、動物を免疫し、アミノ酸配列を含んでなるペプチドまたはタンパク質に対して特異性があるポリクローナルまたはモノクローナル抗体を生成できる。次にこれらの抗体を使用して、DNA発現ライブラリーをスクリーニングし、対象の完全長DNAクローンを単離する(ラーナー(Lerner)R.A.、Adv.Immunol.36:1(1984)、マニアティス(Maniatis)、前出)。
【0140】
遺伝子構成
パントエア(Pantoea)属内の様々な種からのidiおよびcrtE、crtX、crtY、crtI、crtB、およびcrtZをコードする多様な遺伝子配列が入手できるが、配列番号18(8814bp)として開示される本核酸断片が、ユニークな遺伝子構成crtE−idi−crtY−crtI−crtB−crtZを有するように見える。
・crtE(配列番号1)は、ヌクレオチド2400〜3305に位置し、直接的方向に翻訳される。
・idi(配列番号3)は、ヌクレオチド3343〜4392に位置し、配向方向に翻訳される。
・crtY(配列番号5)は、ヌクレオチド4424〜5593に位置し、配向方向に翻訳される。
・crtI(配列番号7)は、ヌクレオチド5590〜7068に位置し、配向方向に翻訳される。
・crtB(配列番号9)は、ヌクレオチド7068〜7994に位置し、配向方向に翻訳される。
・crtZ(配列番号11)は、ヌクレオチド7909〜8466に位置し、crtE−idi−crtY−crtI−crtBと反対方向に翻訳される。
【0141】
最も「一般的な」遺伝子構成は、P.アナナティス(P.ananatis)(ジェンバンク(登録商標)登録番号D90087)、P.ステワルティ(P.stewartii)(ジェンバンク登録番号AY166713)、およびパントエア・アグロメランスpv.ミレチア(Pantoea agglomerans pv.milletiae)(ジェンバンク(登録商標)登録番号AB076662)で観察されるものであり、カロテン産生性クラスターはcrtEXYIBZ(「crtE−crtX−crtY−crtI−crtB−crtZ」とも記述される)を含んでなる(図5)。P.アグロメランス(P.agglomerans)EHO−10(ジェンバンク(登録商標)登録番号M87280)は、crtE−仮説タンパク質−crtX−crtY−crtI−crtB−crtZのカロテン産生性クラスターを含んでなると注釈される。しかしながら、本願出願人らによる「仮説タンパク質」のバイオインフォマティクス分析は、真のP.アグロメ
ランス(P.agglomerans)EHO−10がcrtE−idi−crtX−crtY−crtI−crtB−crtZを含んでなるとみなされるべきだと判断した。
【0142】
同様に、より遠縁の生物体からのその他のカロテン産生性遺伝子クラスターは、crtE−idi−crtY−crtI−crtB−crtZのユニークな遺伝子構成を含有しない。例えばブラジ根粒菌ダイズ根粒菌属(Bradyrhizobium)ORS278のカンタキサンチン遺伝子クラスターはcrtE−crtY−crtI−crtB−crtWであり、アグロバクテリウム・オーランティアカム(Agrobacterium
aurantiacum)からのアスタキサンチン遺伝子クラスターはcrtW−crtZ−crtY−crtI−crtBであり、フラボバクテリウム(Flavobacterium)sp.R1534からのゼアキサンチン遺伝子クラスターはcrtZ−crtY−crtI−crtB−crtEである(ジロー(Giraud)ら、米国特許仮出願第2003/0087337 A1号)。
【0143】
ここで開示される遺伝子構成は、代謝の遺伝子操作中に、大腸菌(E.coli)およびその他の細菌において工業的に価値あるカロテノイドの生成を最大化するのに有用ないくつかのユニークな利点を与えるかもしれない。第一にイソペンテニルピロリン酸イソメラーゼをコードするidiが、カロテノイド生成(カジワラ(Kajiwara)ら、前出;ワング(Wang)ら、前出;アルブレヒト(Albrecht)ら、前出)に劇的に影響することが実証されているので、またこの遺伝子がここで述べられるカロテン産生性−crtY−crtI−crtB−crtZクラスター中に直接組み込まれるので、オペロンの発現が、下流カロテノイド生合成経路への増大するイソプレノイド流束をもたらすことができ、それによって増大するカロテノイド生成および力価が帰結する。第二に、本カロテン産生性crtE−idi−crtY−crtI−crtB−crtZクラスター中のcrtX遺伝子の不在による利点があるかもしれない。具体的には、CrtXによるゼアキサンチンからゼアキサンチン−β−グルコシドへの転換が排除されることで、ここで述べるカロテン産生性クラスターによってコードされる反応の生成物としてのゼアキサンチンの高レベルの蓄積が可能になる。クラスターから不要なcrtX遺伝子を除去する余計な努力が必要でないため、発現クラスター中のcrtXの不在はまた、β−カロテンの合成にも有益かもしれない。
【0144】
したがって、本発明は、
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、カロテノイド生合成経路酵素をコードする核酸分子
を含んでなる形質転換宿主細胞を提供するステップと、
ここで、該分子の遺伝子構成はcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子は記載順にクラスター化し、crtZの転写はcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起き、
(b)適切な生育条件下で(a)の形質転換宿主細胞を有効量の発酵性炭素基質で生育させる、カロテノイド化合物の製造方法を提供する。
【0145】
微生物における組換え発現
本配列の遺伝子および遺伝子産物は、異種宿主細胞中、特に微生物細胞宿主中で生成しても良い。組換え微生物宿主中の発現は、これまで宿主を使用して可能でなかった新しい生成物合成のために、様々な経路中間体を発現させ、および/または宿主中に既存の経路を調節するのに有用かもしれない。
【0146】
適切な上流イソプレン経路遺伝子をコードする遺伝子、および本発明の下流カロテノイ
ド生合成経路遺伝子の(場合によりその他のcrt遺伝子との)様々な組み合わせを適切な微生物宿主に導入する方法は、一般的である。当業者には明らかなように、特定のカロテノイド生成物の生成のために、宿主生物体への導入が必要とされる特定の機能性は、宿主細胞(およびそのイソプレノイド化合物の天然産生)、基質の利用可能性、および所望の最終生成物に左右される。
【0147】
本crt遺伝子がカロテノイド生成において効果的であるためには、宿主細胞が細胞内に適切なレベルのFPPを有することが必要であることが理解される。FPPは、外来性に提供されても良く、または天然または導入された遺伝的経路のどちらかによって、細胞内に内在性に生成されても良い。したがって特定の宿主細胞が適切なレベルのFPPを生成する遺伝的機構を有さない場合、上流イソプレノイド経路のあらゆる必要な遺伝子を得て、これらの遺伝子を宿主内に遺伝子操作して、下流経路を通じたカロテノイド生合成のための出発原料としてFPPを生成することは、十分に当業者の理解の範囲内であると考察される。IPPをFPPの前駆物質として、良く知られている酢酸/メバロン酸経路を通じて合成しても良い。代案としては、最近の研究は、メバロン酸依存経路が全ての生物体で作動しているわけではないことを実証している。IPP生合成のための代案のメバロン酸非依存経路が、細菌および緑藻類および高等植物で特性決定されている(ホルバック(Horbach)ら、FEMS Microbiol.Lett.111:135〜140(1993);ローマー(Rohmer)ら、Biochem.295:517〜524(1993);シュベンダー(Schwender)ら、Biochem.316:73〜80(1996);アイゼンライヒ(Eisenreich)ら、Proc.Natl.Acad.Sci.USA93:6431〜6436(1996))。
【0148】
例えば、1つもしくはそれ以上の本下流C40カロテノイド生合成経路crtEYIBZ配列をコードするキメラ遺伝子の導入が、選択された宿主微生物におけるカロテノイド化合物の生成をもたらすことが期待される。適切な遺伝的形質転換システムによって、多様な非カロテン産生性宿主を遺伝子操作することが可能であろう。これは例えばエルウィニア・ヘルビコラ(Erwinia herbicola)crt遺伝子を使用して、宿主の大腸菌(E.coli)、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)、サッカロミセス・セレビジエ(Saccharomyces cerevisiae)、ピヒア・パストリス(Pichia pastoris)(酵母)、アスペルギルス・ニドランス(Aspergillus nidulans)(菌・カビ類)、ロドバクター・スフェロイデス(Rhodobacter
sphaeroides)、および高等植物(米国特許第5,656,472号明細書)中で、様々なカロテノイドを生成して示されている。したっがってここで前述したように、アンテラキサンチン、アドニルビン、アドニキサンチン、アスタキサンチン、カンタキサンチン、カプソルブリン、β−クリプトキサンチン、α−カロテン、β−カロテン、ε−カロテン、エキネノン、3−ヒドロキシエキネノン、3’−ヒドロキシエキネノン、γ−カロテン、4−ケト−γ−カロテン、ζ−カロテン、α−クリプトキサンチン、デオキシフレキシキサンチン、ジアトキサンチン、7,8−ジデヒドロアスタキサンチン、フコキサンチン、フコキサンチノール、イソレニエラテン、ラクツカキサンチン、ルテイン、リコピン、ミキソバクトン、ネオキサンチン、ニューロスポレン、ヒドロキシニューロスポレン、ペリジニン、フィトエン、ロドピン、ロドピングルコシド、4−ケト−ルビキサンチン、シホナキサンチン、スフェロイデン、スフェロイデノン、スピリロキサンチン、4−ケト−トルレン、3−ヒドロキシ−4−ケト−トルレン、ウリオリド、ウリオリドアセタート、ビオラキサンチン、ゼアキサンチン−β−ジグルコシド、およびゼアキサンチンを全て、(例えば)以下のcrt酵素機能性の様々な組み合わせを導入することで、ここの技術を使用して微生物宿主中で生成しても良い。CrtE、CrtX、CrtY、CrtI、CrtB、CrtZ、CrtW、CrtO、CrtA、CrtC、CrtD、CrtF、およびCrtU。したがってFPPからのフィトエンの形成は、CrtEおよ
びCrtBと、crtE、crtB、およびcrtIをはじめとするFPPからのリコピン合成に必要なカロテノイド特異的遺伝子と、crtE、crtB、crtI、およびcrtYをはじめとするFPPからのβ−カロテン生成に必要な遺伝子を必要とする。crt遺伝子間の関係のこの理解に基づいて、あらゆる所望のカロテノイド生成物の発現に適した、微生物宿主細胞およびcrt遺伝子を選択することが可能であろう。類似の様式で、例えばidiなどの上流イソプレン経路遺伝子の上方制御によって、発現を増幅しても良い。
【0149】
外来性タンパクの高レベル発現を導く制御配列を含有する微生物発現システムおよび発現ベクターは、当業者には周知である。これらのいずれかを使用してキメラ遺伝子を構築し、本配列の遺伝子産物を生成できる。次に形質転換を通じて、これらキメラ遺伝子を適切な微生物に導入し、酵素の高レベル発現を提供できる。
【0150】
適切な宿主細胞の形質転換のために有用なベクターまたはカセットは、技術分野で周知である。典型的にベクターまたはカセットは、関連性のある遺伝子の転写および翻訳を導く配列、選択可能マーカー、および自律的複製または染色体組み込みができるようにする配列を含有する。適切なベクターは、転写イニシエーション制御を含む遺伝子の5’領域、および転写終結を制御するDNA断片の3’領域を含んでなる。双方の制御領域が、形質転換宿主細胞に相同的な遺伝子から誘導されることが最も好ましいが、このような制御領域は、必ずしも産生宿主として選択された特定種に固有の遺伝子から誘導されなくて良いものと理解される。
【0151】
所望の宿主細胞において本ORFの発現を駆動させるのに有用なイニシエーション調節領域、またはプロモーターは多数有り、当業者には良く知られている。CYC1、HIS3、GAL1、GAL10、ADH1、PGK、PHO5、GAPDH、ADC1、TRP1、URA3、LEU2、ENO、TPI(例えばサッカロミセス(Saccharomyces)における発現に有用)、AOX1(例えばピヒア(Pichia)における発現に有用)、およびlac、ara、tet、trp、lPL、lPR、T7、tac、およびtrc(例えば大腸菌(Escherichia coli)における発現に有用)ならびに例えばバチルス(Bacillus)における発現に有用なamy、apr、nprプロモーターおよび様々なファージプロモーターをはじめとするが、これに限定されるものではない、実質的にこれらの遺伝子を駆動できるあらゆるプロモーターが、本発明において適切である。さらにデオキシ−キシルロースリン酸シンターゼまたはメタノールデヒドロゲナーゼオペロンプロモーター(スプリンガー(Springer)ら、FEMS Microbiol Lett 160:119〜124(1998))、ポリヒドロキシアルカン酸合成のためのプロモーター(フェルナー(Foellner)ら、Appl.Microbiol.Biotechnol.40:284〜291(1993))、メチロトローフ内の天然プラスミドから同定されるプロモーター(欧州特許第296484号明細書)、Plac(トヤマ(Toyama)ら、Microbiology 143:595〜602(1997);欧州特許第62971号明細書)、Ptrc(ブロシウス(Brosius)ら、Gene 27:161〜172(1984))、メタノトローフから同定されたプロモーター(PCT/US03/33698号明細書)、および抗生物質抵抗性に関連したプロモーター[例えばカナマイシン(スプリンガー(Springer)ら、FEMS Microbiol Lett160:119〜124(1998);ウエダ(Ueda)ら、Appl.Environ.Microbiol.57:924〜926(1991))またはテトラサイクリン(米国特許第4,824,786号明細書)]が、C1代謝個体における発現に適している。
【0152】
RBSがベクターによって提供されない場合、発現する遺伝子上流に人工リボソーム結合部位(「RBS」)を含めることが必要である。これは、単一プロモーターが、第1、
第2、第3、およびその他遺伝子群の発現を推進する場合に、発現するオペロンの第2、第3、およびその他の遺伝子のために必要とされることが多い。特定の宿主生物体におけるRBSの好ましい配列を判定する方法は当業者には良く知られており、この合成部位の製造手段についても同様である。
【0153】
終結制御領域はまた、好ましい宿主に固有の様々な遺伝子から誘導されても良い。場合により、終結部位は必要ないかもしれないが、含まれることが最も好ましい。
【0154】
遺伝子をクローン化ベクターに単に挿入するだけでは、それが必要なレベルで成功裏に発現することは確証されない。高発現率の必要性に答えて、転写、翻訳、タンパク質安定性、酸素限界、および宿主細胞からの分泌の側面を制御するいくつかの異なる遺伝的要素を操作することで、多くの特殊化した発現ベクターが作り出されている。より具体的には、遺伝子発現を制御するように操作される分子の特徴として以下が挙げられる。1.)関連転写プロモーターおよびターミネーター配列の性質、2.)リボソーム結合部位の強度、3.)クローンされた遺伝子のコピー数、および遺伝子がプラスミドにあるかまたは宿主細胞のゲノム中に組み込まれているかどうか、4.)合成された外来タンパク質の最終細胞内位置、5.)宿主生物体中の翻訳効率、6.)宿主細胞内のクローン化遺伝子タンパク質の本質的な安定性、および7.)頻度が宿主細胞の好むコドン使用頻度に近づくような、クローン化遺伝子内のコドン使用。これらの各タイプの修正は、C40カロテノイドの発現をさらに最適化する手段として、本発明中に包含される。
【0155】
最後に、C40カロテノイドの蓄積を促進するには、エネルギーまたは炭素のシンクとして機能するかもしれない、標的経路中または競合経路中の特定の遺伝子の発現を低下させまたは排除することが必要かもしれない。代案としては、所望のカロテノイド中間体の上流の様々な遺伝子を過剰発現させ、生成を促進させることが有用かもしれない。上述の目的で遺伝的経路を操作する方法は、技術分野で一般的であり良く知られている。
【0156】
例えばひとたび重要な遺伝的経路が同定および配列決定されると、特定の遺伝子を上方制御して経路の産出量を増大できる。例えばpBR322などのマルチコピー型プラスミド上で、標的遺伝子の追加的コピーを宿主細胞に導入しても良い。代案としては、非天然プロモーターの調節の下におかれるように標的遺伝子を変性しても良い。経路が、細胞サイクル中または発酵生産中の特定点で作動することが所望される場合、制御されたまたは誘導可能プロモーターを使用して、標的遺伝子の天然プロモーターを置き換えても良い。同様に場合によっては、天然または内在性プロモーターを変性させて、遺伝子発現を増大させても良い。例えば内在性プロモーターは、生体内で(in vivo)突然変異、欠失、および/または置換によって変性できる(米国特許第5,565,350号明細書、ザーリン(Zarling)らのPCT/US93/03868号明細書を参照されたい)。
【0157】
代案としては、中断させる遺伝子配列が知られている場合、遺伝子下方制御の最も効果的な方法の1つは、外来DNAを構造的な遺伝子に挿入して転写を中断させる、標的を定めた遺伝子中断である。これは、中断させる遺伝子の一部に対して高度な相同性を有する配列で両脇を挟まれた、挿入するDNA(遺伝子マーカーであることが多い)を含んでなる遺伝子カセットの製造によってもたらされる。カセットの宿主細胞への導入は、細胞の天然DNA複製機序を通じた、構造的遺伝子への外来DNAの挿入をもたらす(例えばハミルトン(Hamilton)ら、J.Bacteriol.171:4617〜4622(1989);バルバス(Balbas)ら、Genes、136:211〜213(1993);ゲルデナー(Gueldener)ら、Nucleic Acid Res.24:2519〜2524(1996);およびスミス(Smith)ら、Methods Mol.Cell.Biol.5:270〜277(1996)参照)。
【0158】
アンチセンス技術は、標的遺伝子配列が知られている場合に遺伝子を下方制御する別の方法である。これを達成するために、所望の遺伝子からの核酸セグメントをクローン化して、RNAのアンチセンス鎖が転写されるようにプロモーターに作動的に結合する。次にこのコンストラクトを宿主細胞に導入し、RNAのアンチセンス鎖を生成する。アンチセンスRNAは、対象のタンパク質をコードするmRNAの蓄積を防止することで、遺伝子発現を阻害する。当業者は、特定の遺伝子の発現を低下させるために、特別な考察がアンチセンス技術の使用に関わることを理解する。例えば、アンチセンス遺伝子の適切な発現レベルは、当業者に知られている異なる調節因子を使用した、異なるキメラ遺伝子の使用を必要とするかもしれない。
【0159】
配列が知られている場合、標的を定めた遺伝子中断およびアンチセンス技術が遺伝子を下方制御する効果的手段を提供するが、配列ベースではないその他のより特異性が低い方法が開発されている。例えば細胞をUV放射線暴露して、次に所望の表現型についてスクリーンしても良い。変異体を作り出すために、化学薬品による変異誘発もまた効果的であり、一般に使用される物質としては、非複製DNAに影響する化学物質(例えばHNO2およびNH2OH)、ならびに複製DNAに影響する薬剤(例えばフレームシフト変異を引き起こすことで注目に値するアクリジン染料)が挙げられる。放射線または化学薬品を使用して変異体を作り出す特定の方法は、技術分野で良く文書化されている。例えばトーマスD.ブロック(Thomas D.Brock)著「バイオテクノロジー:工業微生物学テキストブック(Biotechnology:A Textbook of Industrial Microbiology)」第二版(1989)Sinauer Associates:マサチューセッツ州サンダーランド(Sunderland、MA)、またはデシュパンデ,ムカンド(Deshpande,Mukund)V.、Appl.Biochem.Biotechnol.、36:227〜234(1992)を参照されたい。
【0160】
別の非特異的遺伝子中断方法は、転移因子またはトランスポゾンの使用である。トランスポゾンは、DNAに無作為に挿入される遺伝的因子であるが、後で配列に基づいて検索して、挿入がどこで起きたのか判定できる。生体内(in vivo)および生体外(in vitro)転位法の双方が知られている。どちらの方法にもトランスポザーゼ酵素と組み合わされた転移因子の使用が関与する。転移因子またはトランスポゾンがトランスポザーゼ存在下で核酸断片に接触すると、転移因子が核酸断片中に無作為に挿入される。中断された遺伝子は転移因子配列に基づいて同定されても良いので、技術は、ランダム変異誘発のため、そして遺伝子単離のために有用である。生体外(in vitro)転位のためのキットは市販される(例えばニュージャージー州ブランチバーグのパーキン・エルマー・アプライド・バイオシステムズ(Perkin Elmer Applied Biosystems(Branchburg、NJ))から入手できる酵母Ty1因子に基づく、プライマー・アイランド転位キット、マサチューセッツ州ビバリーのニュー・イングランド・バイオ・ラブズ(New England Biolabs(Beverly、MA))から入手できる細菌性トランスポゾンTn7に基づくゲノム・プライミング・システム、およびウィスコンシン州マディソンのエピセンター・テクノロジーズ(Epicentre Technologies(Madison、WI))から入手できる、Tn5細菌性転移因子に基づくEZ::TNトランスポゾン挿入システムを参照されたい)。
【0161】
本発明の文脈で、上述のいずれかの一方法によって、カロテノイド生合成経路の発現を調節することが有用かもしれない。例えば本発明は、カロテノイド生合成経路中の酵素をコードするいくつかの単離された遺伝子、およびC40カロテノイドの生成をもたらす方法を提供する。したがってここでcrtE、idi、crtY、crtI、crtB、お
よびcrtZ遺伝子の様々な組み合わせを過剰発現して、C40カロテノイドの増大する生成を促進するのに加えて、三炭素化合物(ピルビン酸およびD−グリセルアルデヒド3−リン酸)の最初の縮合を上方制御して、(dxs遺伝子によって媒介される)五炭素化合物D−1−デオキシキシルロース−5−リン酸の生成を増大させることもまた有用かもしれない。これは下流カロテノイド生合成経路に入る炭素の流束を増大させ、C40カロテノイドの増大する生成を可能にする。代案としては(またはそれに加えて)、微生物宿主がこれらのタイプの化合物を合成できれば、crtM/crtN遺伝子をノックアウトして、C30カロテノイドの合成をもたらすることが望ましかもしれない。または天然機能性crtE、crtX、crtY、crtI、crtB、およびcrtZ遺伝子を有するシステムでは、上述のいずれかの一方法によって下流遺伝子(例えばcrtZまたはcrtX)を中断させることで、β−カロテンまたはゼアキサンチンの蓄積をもたらしても良い。
【0162】
好ましい微生物宿主
本遺伝子およびカロテノイド生合成経路の核酸断片の発現のために好ましい異種宿主細胞は、菌・カビ類または細菌系統群中に広く見られ、広範な温度、pH価、および溶剤耐性で生育する微生物宿主である。例えばあらゆる細菌、酵母、および糸状菌類が、本核酸断片の発現のための適切な宿主であることが考察された。転写、翻訳およびタンパク質生合成器官は、それぞれの細胞供給原料に関係なく同一であるので、細胞バイオマスを生成するのに使用される炭素供給材料に関係なく、機能性遺伝子が発現される。大規模微生物生育および機能遺伝子発現は、広範な単純または複合炭水化物、有機酸およびアルコール、および/または飽和炭化水素(例えば光合成または化学的自己栄養宿主の場合は、メタンまたは二酸化炭素などの)を利用しても良い。しかし機能遺伝子は、窒素、亜リン酸、イオウ、酸素、炭素、または小型無機イオンをはじめとするあらゆる微量栄養素の形態および量をはじめとする特定の生育条件によって、阻止または抑圧され制御されても良い。さらに機能遺伝子の制御は、培養に添加される典型的に栄養素またはエネルギー源とは見なされない、特定の制御分子の存在または不在によって達成されても良い。生育速度もまた、遺伝子発現における重要な制御因子であっても良い。
【0163】
適切な宿主株の例としては、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、サッカロミセス(Saccharomyces)、ピヒア(Pichia)、カンジダ(Candida)、ハンゼヌラ(Hansenula)、ヤロウィア(Yarrowia)、ロドスポリジウム(Rhodosporidium)、リポミセス(Lipomyces)、サルモネラ(Salmonella)、バチルス(Bacillus)、アシネトバクター(Acinetobacter)、ザイモモナス(Zymomonas)、アグロバクテリウム(Agrobacterium)、フラボバクテリウム(Flavobacterium)、ロドバクター(Rhodobacter)、ロドコッカス(Rhodococcus)、ストレプトマイセス(Streptomyces)、ブレビバクテリウム(Brevibacterium)、コリネバクテリア(Corynebacteria)、マイコバクテリウム(Mycobacterium)、エシェリヒア(Escherichia)、パントエア(Pantoea)、シュードモナス(Pseudomonas)、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、メチロサイナス(Methylosinus)、メチロマイクロビウム(Methylomicrobium)、メチロシスティス(Methylocystis)、アルカリゲネス(Alcaligenes)、シネコシスティス(Synechocystis)、スネコッカス(Synechococcus)、アナベナ(Anabaena)、チオバチルス(Thiobacillus)、メタノバクテリウム(Methanobacterium)、クレブシエラ(Klebsiella)、メチロフィラス(Methylophilus)、メチロバチルス(Methylobacillus)
、メチロバクテリウム(Methylobacterium)、ヒフォミクロビウム(Hyphomicrobium)、キサントバクター(Xanthobacter)、パラコッカス(Paracoccus)、ノカルジア(Nocardia)、アルスロバクター(Arthrobacter)、ロドシュードモナス(Rhodopseudomonas)、トルロプシス(Torulopsis)、ロドトルラ(Rhodotorula)、およびファフィア(Phaffia)が挙げられるが、これに限定されるものではない。
【0164】
微生物宿主としてのメチロトローフおよびメチロモナス(Methylomonas)sp.16a
組換え微生物源からいくつかのカロテノイドが生成されている[例えばリコピン(lycoene)生成のための大腸菌(E.coli)およびカンジダ・ユチリス(Candia utilis)(ファーマー(Farmer)W.R.およびリヤオ(Liao)J.C.、Biotechnol.Prog.17:57〜61(2001);ワング(Wang)C.ら、Biotechnol Prog.16:922〜926(2000);ミサワ(Misawa)N.およびシマダ(Shimada)H.、J.Biotechnol.59:169〜181(1998);シマダ(Shimada)ら、Appl.Environm.Microbiol.64:2676〜2680(1998));β−カロテン生成のための大腸菌(E.coli)、カンジダ・ユチリス(Candia utilis)、およびパフィア・ロドジマ(Pfaffia rhodozyma)(アルブレヒト(Albrecht)ら、Biotechnol.Lett.21:791〜795(1999);ミウラ(Miura)Y.ら、Appl.Environm.Microbiol.64:1226〜1229(1998);米国特許第5,691,190号明細書);ゼアキサンチン生成のための大腸菌(E.coli)およびカンジダ・ユチリス(Candia utilis)(アルブレヒト(Albrecht)ら、前出;ミウラ(Miura)Y.ら、前出);アスタキサンチン生成のための大腸菌(E.coli)およびパフィア・ロドジマ(Pfaffia rhodozyma)(米国特許第5,466,599号明細書、米国特許第6,015,684号明細書、米国特許第5,182,208号明細書、米国特許第5,972,642号明細書;米国特許第5,656,472号明細書、米国特許第5,545,816号明細書、米国特許第5,530,189号明細書、米国特許第5,530,188号明細書、米国特許第5,429,939号明細書、および米国特許第6,124,113号明細書もまた参照されたい)]が、異なるcrt遺伝子の様々な組み合わせを使用したこれらのカロテノイド生成法には、低い収率と比較的高価な供給材料への依存という欠点がある。したがって微生物宿主中で安価な供給材料から、高収率のカロテノイドを製造する方法を同定することが望ましい。
【0165】
単一エネルギー源として一炭素基質を利用する、いくつかの微生物がある。このような微生物はここで「C1代謝個体」と称される。これらの生物体は、単一エネルギーおよびバイオマス源として炭素−炭素結合が欠如している炭素基質を使用する能力によって特徴付けられる。これらの炭素基質としては、メタン、メタノール、ホルメート、ホルムアルデヒド、ギ酸、メチル化アミン(例えば、モノ−、ジ−、およびトリ−メチルアミン)、メチル化チオール、二酸化炭素、およびその他の様々なあらゆる炭素−炭素結合が欠如している還元炭素化合物が挙げられるが、これに限定されるものではない。
【0166】
全てのC1代謝微生物は、概してメチロトローフとして分類される。メチロトローフは、炭素−炭素結合を含有しない有機化合物を酸化できるあらゆる生物体として定義されても良い。しかし通性メチロトローフ、絶対メチロトローフ、絶対メタノトローフは、全てメチロトローフの様々なサブセットである。具体的には次のようである。
・通性メチロトローフは、炭素−炭素結合を含有しない有機化合物を酸化する能力を有
するが、エネルギーおよびバイオマスのために、糖および複合炭水化物などのその他の炭素基質を使用しても良い。通性メチロトローフ細菌は多くの環境中に見られるが、最も一般には土壌、ごみ埋め立て、および廃棄物処理場から単離される。多くの通性メチロトローフは、プロテオバクテリア(Proteobacteria)のβおよびγ亜群のメンバーである(ハンソン(Hanson)ら、C1化合物上での微生物の生育(Microb.Growth C1 Compounds)[国際シンポジウム]第7回(1993)、p285〜302、マレルJ.コリン(Murrell J.Collin)およびドンP.ケリー(Don P.Kelly)編、インターセプト(Intercept):英国アンドーバー(Andover、UK);マディガン(Madigan)ら、ブロックの微生物生物学(Brock Biology of Microorganisms)、第8版、プレンティスホール(Prentice Hall):ニュージャージー州アッパー・サドル・リバー(Upper Saddle River、NJ)(1997))。
・絶対メチロトローフは、エネルギー生成が、炭素−炭素結合を含有しない有機化合物の使用に限定された生物体である。
・絶対メタノトローフは、メタンを酸化する特徴的能力を有する絶対メチロトローフである。
【0167】
さらに一炭素基質を利用する能力は細菌に限定されず、酵母および菌・カビ類にも及ぶ。いくつかの酵母属は、エネルギー源としてより複雑な材料に加えて一炭素基質を使用できる(すなわちメチロトローフ酵母)。
【0168】
数多くのこれらのメチロトローフ生物体が知られているが、これらの微生物のわずかなものだけが材料合成のための工業プロセスにおいて成功裏に利用されている。そして一炭素基質は対費用効果の高いエネルギー源であるが、これらの微生物の遺伝子操作における困難さ、ならびにそれらの遺伝的機構に関する情報不足が、主に天然生成物合成のためのそれらの使用を制限している。
【0169】
これらの困難さにもかかわらず、多くのメタノトローフはこれらの生物体が色素を合成できるようにする固有のイソプレノイド経路を含有し、様々な非内在性イソプレノイド化合物を生成するために、これらの微生物を遺伝子操作する構想の可能性を提供する。メタノトローフは一炭素基質(すなわちメタンまたはメタノール)をエネルギー源として使用できるので、これらの生物体においてカロテノイドを安価に生成することが可能であろう。メタノトローフがβ−カロテン生成のために遺伝子操作される実施例の1つは、国際公開第02/18617号パンフレットで述べられる。
【0170】
本発明では、一炭素基質を唯一のエネルギー源として使用できる微生物における、カロテノイド化合物生合成に関与する遺伝子発現のための方法が提供される。宿主微生物は、カロテノイドのための代謝前駆物質として、ファルネシルピロリン酸(FPP)を合成する能力を有するあらゆるC1代謝個体であっても良い。より具体的には、本発明で適切な通性メチロトローフ細菌としては、メチロフィラス(Methylophilus)、メチロバチルス(Methylobacillus)、メチロバクテリウム(Methylobacterium)、ヒフォミクロビウム(Hyphomicrobium)、キサントバクター(Xanthobacter)、バチルス(Bacillus)、パラコッカス(Paracoccus)、ノカルジア(Nocardia)、アルスロバクター(Arthrobacter)、ロドシュードモナス(Rhodopseudomonas)、およびシュードモナス(Pseudomonas)が挙げられるが、これに限定されるものではない。本発明で有用な具体的なメチロトローフ酵母としては、カンジダ(Candida)、ハンゼヌラ(Hansenula)、ピヒア(Pichia)、トルロプシス(Torulopsis)、およびロドトルラ(Rhodotorula)が挙げら
れるが、これに限定されるものではない。そして例示的なメタノトローフとしては、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、メチロサイナス(Methylosinus)、メチロシクティス(Methylocyctis)、メチロマイクロビウム(Methylomicrobium)、およびメタノモナス(Methanomonas)属が挙げられるが、これに限定されるものではない。
【0171】
本発明で特に興味深いのは、エネルギー的に好ましい炭素流経路を有する高生育絶対メタノトローフである。例えば出願人は、それを炭素流束操作のために特に有用なものにする、いくつかの経路特性を有するメタノトローフの特定の株を見いだした。この株はメチロモナス(Methylomonas)16a(ATCC PTA2402)(米国特許第6,689,601号明細書)として知られており、この特定の株およびその他の近縁メチロトローフは、C40カロテノイド生成に有用な本発明の遺伝子産物発現のための好ましい微生物宿主である(国際公開第02/18617号パンフレット)。
【0172】
メチロモナス(Methylomonas)sp.16aは、C30カロテノイドを天然に生成する。オドム(Odom)らは、メチロモナス(Methylomonas)16aにおけるC40カロテノイド遺伝子の発現が、C30およびC40カロテノイド混合物を生成することを報告している(国際公開第02/18617号パンフレット)。crtN1、ald、crtN2、およびcrtN3遺伝子をはじめとする(がこれに限定されるものではない)、この株中のC30カロテノイド生成に関与する遺伝子のいくつかが同定されている。ald遺伝子の中断、またはcrtN1/ald/crtN2遺伝子クラスターの発現を推進するプロモーターの中断は、C40カロテノイド生成により適した様々な非着色変異体(「白色変異体」)を作り出す(米国仮出願第60/527083号を本願明細書に援用する)。例えば非着色メチロモナス(Methylomonas)sp.16a株MWM1000は、aldおよびcrtN1遺伝子を中断することで作り出された。
【0173】
メチロモナス(Methylomonas)sp.16a株は、炭素利用経路にいくつかの異常を含有する。例えばゲノム配列データに基づいて、株は2つのヘキソース代謝経路のための遺伝子を含有することが示される。エントナー−ドゥドロフ経路(ケト−デオキシホスホグルコン酸アルドラーゼ酵素を用いる)が株中に存在する。これが絶対メタノトローフにおいて稼働している経路であることは、概して広く認められている。しかしエムデン−マイヤーホフ経路もまた存在する(フルクトースビスリン酸アルドラーゼ酵素を用いる)。この経路が絶対メタノトローフ中に存在しないか、または稼働していないことは良く知られている。エネルギー的には後者の経路が最も好ましく、生物学的に有用なエネルギーのより高い収率を可能にして、究極的にメチロモナス(Methylomonas)16aにおいて、細胞集団生成およびその他の細胞集団依存生成物のより高い収率が得られる。16a株におけるこの経路の活性は、マイクロアレイデータおよびATP還元を測定する生化学的証拠を通じて確認されている。16a株は、エムデン−マイヤーホフおよびエントナー−ドゥドロフ経路酵素の双方を有することが示されているが、データは、エムデン−マイヤーホフ経路酵素がエントナー−ドゥドロフ経路酵素よりも強力に発現することをを示唆する。この結果は意外であり、メタノトローフ細菌解糖代謝に関する既存の信念に反する。出願人は、例えばメチロモナス・クララ(Methylomonas
clara)およびメチロサイナス・スポリウム(Methylosinus sporium)をはじめとする、この特徴を有する別のメタノトローフ細菌を見いだした。この活性は、ほとんどの細菌システム中の酵素の典型的なホスホリル供与体であるATPに対して酵素活性が欠如しているために、メタノトローフにおいて見いだされないままであったように思われる。
【0174】
16a株中のエムデン−マイヤーホフ経路の特に新しく有用な特徴は、重要なホスホフルクトキナーゼステップがATP−依存性でなく、ピロリン酸依存性であることである。この特徴は、ATPの代わりにピロリン酸を使用することで、経路のエネルギー収率を増大させる。
【0175】
メタノトローフ細菌では、リブロース一リン酸経路またはRuMP回路として知られている、反応の循環セットを通じてメタンが生分子に転換される。この経路は3つの相を含んでなり、各相は一連の酵素的ステップである。第1のステップは、ヘキソースまたは六炭糖を形成する、ペントース中へのC−1(ホルムアルデヒド)の「固定」または組み込みである。これは五炭糖(ペントース)とホルムアルデヒドとの縮合反応を通じて起き、ヘキスロース一リン酸シンターゼによって触媒される。第2段階は「開裂」と称され、2個の三炭素分子へのヘキソースの分裂をもたらす。これらの三炭素分子の1つはRuMP経路を通じてリサイクルされ、他の三炭素断片は細胞の生育に用いられる。
【0176】
メタノトローフおよびメチロトローフにおいて、RuMP経路は3つの変種の1つとして生じても良い。しかしこれらの変種の2つのみ、FBP/TA(フルクトースビスホスホターゼ/トランスアルドラーゼ)経路またはKDPG/TA(ケトデオキシホスホグルコン酸/トランスアルドラーゼ)経路が一般に見られる(ディクフイゼン(Dijkhuizen)L.およびドゥブリエ(Devries)G.E.「好気性メタノール利用グラム陰性およびグラム陽性細菌の生理学と生化学(The Physiology and biochemistry of aerobic methanol−utilizing gram negative and gram positive bacteria)」「メタンおよびメタノール利用細菌(Methane and Methanol Utilizers)」中、コリン・マレル(Colin Murrell)およびハワード・ダルトン(Howard Dalton)編より、Plenum:ニューヨーク、1992)。
【0177】
メチロモナス(Methylomonas)sp.16a株は、「開裂」ステップの扱いにおいてユニークであり、重要な中間体としてフルクトースビスリン酸を経由して、この転換を行う遺伝子が見いだされた。フルクトースビスリン酸アルドラーゼおよびトランスアルドラーゼのための遺伝子は、1片のDNA上に一緒にクラスター化して見いだされている。次にケトデオキシホスホグルコン酸中間体が関与するその他の変種のための遺伝子もまた、一緒にクラスター化して見いだされている。入手できる文献は、これらの生物体(絶対メチロトローフおよびメタノトローフ)がKDPG経路のみに依存し、FBP依存固定経路が通性メチロトローフによって用いられることを教示する(ディクフイゼン(Dijkhuizen)ら、前出)。したがって後者の観察は予期されるが、前者はそうでない。絶対メタン利用細菌におけるFBP遺伝子の発見は意外であり、また有用性も示唆する。FBP経路は、KDPG経路で利用されるよりも多くのエネルギー(ATP)が利用されるとという事実のために、宿主微生物にとってエネルギー的に好ましい。したがってFBP経路を用いる生物体は、KDPG経路を用いるものに比べて、エネルギー上の利点および生育上の利点を有するかもしれない。この利点はまた、菌株中のエネルギー要求生成経路にとって有用かもしれない。この経路を使用することで、メタン利用細菌は、単細胞タンパク質、またはRuMP経路を通る炭素の流れから誘導されるあらゆるその他の生成物(例えばカロテノイド)のための生成基盤として、その他のメタン利用生物体に比べて利点を有するかもしれない。
【0178】
したがって本発明は、
(a)メタンおよびメタノールよりなる群から選択されるC1炭素基質で生育し、
(b)ピロリン酸依存性ホスホフルクトキナーゼ酵素をコードする遺伝子を含んでなる、機能的エムデンマイヤーホフ炭素経路を含んでなる、
高生育でエネルギー的に好ましいメチロモナス(Methylomonas)株におけるカロテノイド化合物の製造方法を提供する。
【0179】
C1代謝細菌の形質転換
C1代謝細菌形質の転換技術は十分開発されいないが、当業者に良く知られているその他の細菌のために用いられる一般方法を応用しても良い。メチロバクテリウム・エキストロクエン(Methylobacterium extorquens)AM1(トヤマ(Toyama)H.、ら、FEMS Microbiol.Lett.166:1〜7(1998))、メチロフィラス・メチロトロファス(Methylophilus methylotrophus)AS1(キム(Kim)C.S.およびT.K.ウッド(Wood)、Appl.Microbiol.Biotechnol.48:105〜108(1997))、およびメチロバチルス(Methylobacillus)sp.株12S(ヨシダ(Yoshida)T.ら、Biotechnol.Lett.、23:787〜791(2001))の形質転換のために、電気穿孔が成功裏に使用されている。しかし当業者には良く知られているように、1つの特定のC1代謝利用生物体から別の生物体への具体的電気穿孔パラメーターの推定は、困難かもしれない。
【0180】
供与細胞と受容細胞の直接接触に依存する細菌接合は、C1代謝細菌中への遺伝子の転位のために、より容易に受け入れられることが多い。簡単に述べると、この細菌接合プロセスは、「供与」細胞と「受容」細胞を互いに密接に接触させて一緒に混合することを伴う。接合は、新たに合成された供与DNAの受容細胞中への直接転位を伴う、「供与」細菌と「受容」細菌間の細胞質連絡の形成によって起きる。技術分野で良く知られているように、接合の受容体は、供与細菌からの水平転位を通じてDNAを受容できるあらゆる細胞として定義される。接合転位中の供与体は、接合プラスミド、接合トランスポゾン、または可動性プラスミドを含有する細菌である。供与体プラスミドの物理的転位は、下で述べられる2つの様式の1つによって起きることができる。
1.場合によっては、供与体と受容体のみが接合にとって必要とされる。これは転位されるプラスミドが接合性かつ可動性である自己伝播性のプラスミドの場合(すなわちtra−遺伝子と、Mobタンパク質をコードする遺伝子の双方を保有する)に起きる。一般にプロセスは、1.)oriT中の特定部位で二本鎖プラスミドDNAにニックが入れられるステップと、2.)poreまたはpilus構造を通じて、一本鎖DNAが受容体に放出されるステップと、3.)DNAリラクセース酵素が二本鎖DNAをoriTで開裂して結合し、5’末端を放出する(中間体構造としてリラクソゾームを形成する)ステップと、4.)引き続いて、補助タンパク質の複合体がoriTでアセンブルして、DNA転位プロセスを容易にするステップとを伴う。
2.代案としては、供与体プラスミドの受容体への転位のために「三親交雑」接合が必要とされる。このタイプの接合には、供与細胞、受容細胞、および「ヘルパー」プラスミドが関与する。供与細胞は、可動性プラスミドまたは接合トランスポゾンを保有する。可動性ベクターは、ニッケースをコードする遺伝子であるoriTを含有し、Mobタンパク質をコードする遺伝子を有する。しかしMobタンパク質は、単独ではゲノムの転位達成するのに十分でない。したがって可動性プラスミドは、ヘルパープラスミド(供与体内または「ヘルパー」細胞内に位置する)によって適切な接合システムが提供されない限り、それら自身の転位を促進できない。プラスミドは、poreまたはpilusの形成に関与する転位タンパク質(Tra)をコードするので、接合プラスミドは噛合対の形成およびDNA転位のために必要とされる。
【0181】
C1代謝細菌が関与する成功裏の接合の例としては、ストリアー(Stolyar)ら(Mikrobiologiya 64(5):686−691(1995));モトヤマ(Motoyama)ら(Appl.Micro.Biotech.42(1):67〜72(1994));ロイド(Lloyd)ら(Archives of Micro
biology 171(6):364−370(1999));およびオドム(Odom)ら(国際公開第02/18617号パンフレット)の研究が挙げられる。
【0182】
カロテノイドの生体内(in vitro)生物転換
代案としては、生体内(in vitro)で本願明細書の生物転換を行うことが可能である。CrtE、CrtY、CrtI、CrtB、およびCrtZのための基質が、宿主細胞によって内在性に合成されない場合、基質を外来性に添加することが可能である。この実施態様では、適切なカロテノイド基質を中性洗剤(例えばDMSO)で可溶化してもまたはリン脂質ベシクルと混合しても良い。細胞中への輸送を助けるために、宿主細胞を場合によりトルエンなどの適切な溶剤で透過化処理しても良い。このタイプのカロテノイド基質の生体外(in−vitro)生物転換法は、技術分野に基礎がある(例えばハンドル(Hundle)B.S.ら、FEBS、315:329〜334(1993);およびブラムリー(Bramley)、P.M.ら、Phytochemistry、26:1935〜1939(1987)を参照されたい)。
【0183】
組換え微生物を使用した工業生産
本タンパク質の商業的生産が所望される場合、多様な培養方法を応用しても良い。例えば、組換え微生物宿主から過剰発現する特定の遺伝子産物の大規模生産は、バッチまたは連続の培養方法の双方で生産しても良い。
【0184】
古典的なバッチ培養法は閉鎖システムであり、そこでは培養液の組成が発酵の最初に設定され、培養プロセス中の人為的変化を受けない。したがって培養プロセス開始時に培養液に所望の生物体または生物体群を接種し、生育または代謝活動を生じさせながらシステムには何も添加しない。しかし典型的には「バッチ」培養は、炭素源の添加に関するバッチであり、pHおよび酸素濃度などの因子の調節が試みられることが多い。バッチシステムでは、システムの代謝産物および生物質量組成は、培養が終結する時点まで常に変化する。バッチ培養内で細胞は、静的な遅滞期から高い対数増殖期へ、そして最後に成長率が減退または停止する静止期へ調節される。処置を施さない場合、静止期にある細胞はやがて死滅する。システムによっては対数期にある細胞が、最終生成物または中間体の生産の大部分を担うことが多い。その他のシステムでは、静止または対数期後生成を得ることができる。
【0185】
標準バッチシステムのバリエーションが、流加バッチシステムである。流加バッチ培養プロセスも本発明において適切であり、培養の進行と共に基質が段階的に添加されること以外は、典型的なバッチシステムを含んでなる。流加バッチシステムは、異化代謝産物抑制が細胞の代謝を阻害する傾向があって、培養液中に限定量の基質を有することが望ましい場合に有用である。流加バッチシステム中の実際の基質濃度の測定は困難であるので、pH、溶存酸素、およびCO2などの排ガス分圧などの測定可能因子の変化に基づいて推定される。バッチおよび流加バッチ培養法は、技術分野で一般的であり周知であって、実例はブロック(Brock)(前出)およびデシュパンデ(Deshpande)(前出)にある。
【0186】
本タンパク質の商業的生成は、連続培養によって達成しても良い。連続の培養は開放システムであり、規定の培地をバイオリアクターに連続的に添加して、等量の慣熟培地を工程から同時に除去する。連続培養は、概して細胞を恒常的な高い液相密度に維持し、そこでは細胞が主に対数増殖期にある。代案としては連続培養を固定細胞で実施しても良く、そこでは炭素および養素が連続的に添加され、価値ある生成物、副産物または老廃物は細胞集団から連続的に除去される。細胞固定化は、天然および/または合成材料から構成される広範囲の固体担体を使用して実施しても良い。
【0187】
連続または半連続培養は、細胞生育または最終生成物濃度に影響する1つの因子またはあらゆるいくつかの因子の調節を可能にする。例えば一方法では、炭素源または窒素レベルなどの制限的栄養物質を固定された割合に維持し、その他の全パラメーターの調節ができるようにする。別のシステムでは、培地濁度によって測定される細胞濃度を一定に保ちながら、生育に影響するいくつかの因子を連続的に変化させることができる。連続システムは定常状態生育条件を維持することを目指すので、培地が抜き取られることによる細胞損失は、培養中の細胞生育率に対してバランスが取れていなくてはならない。連続的培養プロセスのために栄養素および成長因子を調節する方法、ならびに生成物形成速度を最大化する技術は工業微生物学の技術分野で良く知られており、多様な方法が上記のブロック(Brock)で詳述される。
【0188】
本発明における発酵培地は、適切な炭素基質を含有しなくてはならない。適切な基質としては、単糖類(例えばグルコースおよびフルクトース)、二糖類(例えばラクトースまたはスクロース)、多糖類(例えばデンプンまたはセルロースまたはそれらの混合物)、再生可能な供給材料(例えば乳清透過液、コーンスティープリーカー、甜菜モラセス、および大麦の麦芽)からの未精製混合物が挙げられるが、これに限定されるものではない。さらに炭素基質は、重要な生化学的中間体への代謝転換が実証されている、二酸化炭素、メタンまたはメタノールなどの一炭素基質であっても良い。一または二炭素基質に加えて、メチロトローフ生物体はまた、メチルアミン、グルコサミン、および代謝活性のための多様なアミノ酸などのいくつかのその他の炭素含有化合物を利用することが知られている。例えば、メチロトローフ酵母は、メチルアミンからの炭素を利用してトレハロースまたはグリセロールを形成することが知られている(ベリオン(Bellion)ら、C1化合物上での微生物の生育(Microb.Growth C1 Compounds)[国際シンポジウム]第7回(1993)、415〜32、マレルJ.コリン(Murrell J.Collin)およびドンP.ケリー(Don P.Kelly)編、インターセプト:英国アンドーバー)。同様に、様々なカンジダ(Candida)種が、アラニンまたはオレイン酸を代謝する(サルター(Sulter)ら、Arch.Microbiol.153:485〜489(1990))。したがって本発明で用いられる炭素源は多種多様な炭素含有基質を包含しても良く、生物体の選択によってのみ制限されることが考察された。
【0189】
植物における組換え生成
植物および藻類もまた、カロテノイド化合物を製造することが知られている。本発明のcrtE、idi、crtY、crtI、crtB、およびcrtZ核酸断片を使用して、微生物タンパク質を発現する能力を有する遺伝子導入植物を作り出しても良い。好ましい植物宿主は、本タンパク質の高生成レベルを支えるいずれかの品種であることができる。適切な緑色植物としては、ダイズ、アブラナ(Brassica napus、B.campestris)、ヒマワリ(Helianthus annus)、綿(Gossypium hirsutum)、トウモロコシ、タバコ(Nicotiana tabacum)、アルファルファ(Medicago sativa)、小麦(Triticum種)、大麦(Hordeum vulgare)、オート麦(Avena sativa、L)、モロコシ(Sorghum bicolor)、イネ(Oryza sativa)、シロイヌナズナ、アブラナ科の野菜(ブロッコリー、カリフラワー、キャベツ、アメリカボウフウなど)、メロン、ニンジン、セロリ、パセリ、トマト、ジャガイモ、オランダイチゴ、落花生、ブドウ、草の種、テンサイ、サトウキビ、マメ、エンドウ豆、ライ麦、亜麻、堅木、軟木、および飼料としての草が挙げられるが、これに限定されるものではない。藻類のとしては、スピルリナ(Spirulina)、ヘモタコックス(Haemotacoccus)、およびデュナリエラ(Dunalliela)などの種商業的に重要な宿主が挙げられるが、これに限定されるものではない。
【0190】
好ましいカロテノイド化合物の過剰発現は、最初に本発明のキメラ遺伝子を構築することによって達成されても良く、その中でコード領域は、所望の発生段階で所望の組織中の遺伝子の発現を導くプロモーターに作動的に結合される。便宜上、キメラ遺伝子は、同一遺伝子に由来するプロモーター配列および翻訳リーダー配列を含んでなっても良い。転写終止シグナルをコードする3’非コード配列もまた、提供されなくてはならない。本キメラ遺伝子はまた、遺伝子発現を容易にするために1つもしくはそれ以上のイントロンも含んでなる。
【0191】
コード領域の発現を誘導できるあらゆるプロモーターとあらゆるターミネーターのあらゆる組み合わせをキメラ遺伝子配列で使用しても良い。プロモーターおよびターミネーターのいくつかの適切な例としては、ノパリンシンターゼ(nos)、オクトピンシンターゼ(ocs)、およびカリフラワーモザイクウィルス(CaMV)遺伝子からのものが挙げられる。使用しても良い効率的な植物プロモーターの1タイプは、高等植物プロモーターである。本発明の遺伝子配列と機能的に連結するこのようなプロモーターは、本遺伝子産物の発現を促進できなくてはならない。本発明で使用しても良い高等植物プロモーターとしては、例えば1.)ダイズからのリブロース−1,5−ビスリン酸カルボキシラーゼの小型サブユニット(ss)のプロモーター(ベリー−ロウ(Berry−Lowe)ら、J.Molecular and App.Gen.、1:483〜498(1982))、および2.)クロロフィルa/b結合タンパク質のプロモーターが挙げられる。これらの2つのプロモーターは、植物細胞において光誘導されることが知られている(例えば「植物の遺伝子操作、農業における展望(Genetic Engineering of Plants,an Agricultural Perspective)」A.キャッシュモア(Cashmore)編、Plenum:ニューヨーク(1983)、p29〜38;コルッジ(Coruzzi)G.ら、J.Biol.l Chem.、258:1399(1983);およびダンスミア(Dunsmuir)P.ら、J.Mol.Appl.Genet.、2:285(1983)参照)。
【0192】
次に本キメラ遺伝子を含んでなるプラスミドベクターを構築できる。プラスミドベクターの選択は、宿主植物を形質転換するのに使用される方法に左右される。当業者は、キメラ遺伝子を含有する宿主細胞を成功裏に形質転換し、選択して増殖するためにプラスミドベクター上に存在しなくてはならない遺伝的要素を良く知っている。当業者はまた、異なる独立形質転換イベントが、異なるレベルおよびパターンの発現をもたらすことを認識するので(ジョーンズ(Jones)ら、EMBO J.4:2411〜2418(1985);デ・アルメイダ(De Almeida)ら、Mol.Gen.Genetics
218:78〜86(1989))、所望の発現レベルおよびパターンを示す系統を得るためには、複数イベントをスクリーニングしなくてはならない。このようなスクリーニングは、DNAブロットのサザン分析(サザン(Southern)、J.Mol.Biol.98:503(1975))、mRNA発現のノーザン分析(クロツェック(Kroczek)、J.Chromatogr.Biomed.Appl.、618(1〜2):133〜145(1993))、タンパク質発現のウエスタン分析、または表現型分析によって達成しても良い。
【0193】
用途によっては、本タンパク質を異なる細胞区画に方向付けることが有用であろう。したがって適切な細胞内標的配列を添加することで、および/または以下のような既存の標的配列を除去することで、酵素をコードするコード配列を変更して、上述のキメラ遺伝子をさらに補完しても良いことが想定された。1.)トランジット配列(キーグストラ(Keegstra)K.、Cell 56:247〜253(1989))、2.)シグナル配列、または3.)小胞体局在化(クリスピールス(Chrispeels)J.J.、Ann.Rev.Plant Phys.Plant Mol.Biol.42:21〜53(1991))または核局在化シグナル(ライケル(Raikhel)、N.、P
lant Phys.100:1627〜1632(1992))をコードする配列。引用した参考文献はこれらの各例を示すが、リストは網羅的でなく、有用なさらに多くの標的シグナルが、将来発見されるかもしれない。
【0194】
タンパク質操作
本crtE、idi、crtY、crtI、crtB、および/またはcrtZヌクレオチドを使用して、向上されたまたは改変された活性を有する遺伝子産物を製造できるかもしれないことが考察された。1.)誤りがちなPCR(メルニコフ(Melnikov)ら、Nucleic Acids Research、27(4):1056〜1062(1999年2月15日))、2.)部位特異的変異誘発(クームズ(Coombs)ら、Proteins(1998)、p259〜311、1プレート(アンジェレッティ,ルース・ホーグ(Angeletti,Ruth Hogue)編、アカデミック:カリフォルニア州サンディエゴ(San Diego、CA))、および3.)「遺伝子シャフリング」(本願明細書に援用する米国特許第5,605,793号明細書、米国特許第5,811,238号明細書、米国特許第5,830,721号明細書、および米国特許第5,837,458号明細書)をはじめとするが、これに限定されるものではない、天然遺伝子配列を変異させて活性が改変または向上された遺伝子産物を生成する様々な方法が知られている。
【0195】
遺伝子シャッフリング法は、その容易な実行、および高率な変異誘発と容易なスクリーニングのために特に魅力的である。遺伝子シャフリングのプロセスは、対象の遺伝子と類似したまたは違っている追加的なDNA領域集団存在下で、対象の遺伝子を特定サイズの断片に制限エンドヌクレアーゼ開裂することを伴う。次に断片のプールを変性させ再アニールして、変異遺伝子を作り出す。次に変異した遺伝子を変更した活性についてスクリーニングする。
【0196】
本発明の本微生物の配列を変異させて、この方法によって変更されまたは増強される活性についてスクリーニングしても良い。配列は二本鎖であるべきで、50bp〜10kB範囲の様々な長さであることができる。配列は技術分野で良く知られている制限エンドヌクレアーゼを使用して、約10bp〜1000bpの範囲の断片に無作為に消化されても良い(マニアティス(Maniatis)、同上)。本微生物の配列に加えて、微生物の配列の全部または部分とハイブリダイズ可能な断片集団を添加しても良い。同様に、本配列とハイブリダイズ可能でない断片集団を添加しても良い。典型的にこれらの追加的断片集団は、総核酸と比較して重量で約10〜20倍過剰に添加される。一般にこのプロセスに従えば、混合物中の異なる特定の核酸断片の数は約100〜約1000個になる。無作為核酸断片の混合集団は、変性されて一本鎖核酸断片を形成し、次に再アニールされる。その他の一本鎖核酸断片との相同性領域を有する一本鎖核酸断片のみが再アニールする。無作為核酸断片は、加熱によって変性されても良い。当業者は、二本鎖核酸を完全に変性するのに必要な条件を判定できる。好ましくは温度は、約80℃〜100℃である。核酸断片を冷却によって再アニールしても良い。好ましくは温度は約20℃〜75℃である。再生は、ポリエチレングリコール(「PEG」)または塩の添加によって加速できる。適切な塩濃度は、0mM〜200mMの範囲であっても良い。次にアニールされた核酸断片は、核酸ポリメラーゼおよびdNTP(すなわちdATP、dCTP、dGTPおよびdTTP)存在下でインキュベートされる。核酸ポリメラーゼは、クレノウ断片、Taqポリメラーゼまたは技術分野で知られているあらゆるその他のDNAポリメラーゼであっても良い。アニールに先だって、アニールと同時に、またはアニール後に、ポリメラーゼを無作為核酸断片に添加しても良い。変性、再生、およびポリメラーゼ存在下でのインキュベーションのサイクルは、所望の回数反復される。好ましくはサイクルは約2〜50回反復され、より好ましくは手順は10〜40回反復される。得られる核酸は約50bp〜約100kBの範囲のより大きな二本鎖ポリヌクレオチドであり、標準クローン化および発
現プロトコルによって、発現および変更された活性についてスクリーニングしても良い(マニアティス(Manatis)、同上)。
【0197】
さらにハイブリッドタンパク質は、遺伝子シャフリング(エクソンシャフリング)法を使用した、機能性領域の融合によってアセンブルできる(ニクソン(Nixon)ら、Proc.Natl.Acad.Sci.USA、94:1069〜1073(1997))。本遺伝子の機能性領域は、その他の遺伝子の機能性領域と組み合わせて、所望の触媒機能を有する新しい酵素作り出すことができる。PCRオーバーラップ伸長法を使用してハイブリッド酵素を構築し、当業者に良く知られている技術使用して、様々な発現ベクター中にクローンしても良い。
【実施例】
【0198】
以下の実施例で本発明をさらに明らかにする。これらの実施例は、本発明の好ましい実施態様を示しながら、例証のみのために提供されるものと理解される。上記考察およびこれらの実施例から、当業者は本発明の本質的特質を把握でき、その範囲と精神を逸脱することなく、本発明の様々な変化と修正を行って、様々な利用法および条件に適合させることができる。
【0199】
一般方法
実施例で使用される標準組換えDNAおよび分子クローン化技術は技術分野で良く知られており、マニアティス(Maniatis)(前出)、シルハビー(Silhavy)ら(前出)、およびオースベルら(Ausubel)(前出)によって述べられている。
【0200】
細菌培養の維持および生育に適した材料および方法は、技術分野で良く知られている。以下の実施例で使用するのに適した技術は、下に述べられる。「一般微生物学方法マニュアル(Manual of Methods for General Bacteriology)」、フィリップス・ゲアハルト(Phillipp Gerhardt)、R.G.E.マリー(R.G.E.Murray)、ラルフN.コスティロウ(Ralph N.Costilow)、ユージーンW.ネスター(Eugene W.Nester)、ウィリスA.ウッド(Willis A.Wood)、ノエルR.クリーグ(Noel R.Krieg)、およびG.ブリッグス・フィリップス(G.Briggs Phillips)編、米国微生物学会、ワシントンD.C.(Washington,D.C.)(1994)またはブロック(Brock)ら(前出)。細菌細胞の生育および維持のために使用される全ての試薬、制限酵素および材料は、特に断りのない限り、ウィスコンシン州ミルウォーキーのアルドリッチ・ケミカルズ(Aldrich Chemicals(Milwaukee、WI))、ミシガン州デトロイトのディフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))、メリーランド州ゲーサーズバーグのギブコ/BRL(GIBCO/BRL(Gaithersburg、MD))、またはミズーリ州セントルイスのシグマケミカル(Sigma Chemical Company(St.Louis、MO))から得た。
【0201】
配列データは、ベクターと挿入特異性プライマーの組み合わせを使用して、染料ターミネーター技術(米国特許第5,366,860号明細書、欧州特許第272,007号明細書)を使用して、ABI自動シーケンサー上で生成した。ミシガン州アナーバーのジーン・コーズ社(Gene Codes Corp.(Ann Arbor、MI))からのシーケンチャー(Sequencher)(商標)バージョン4.0.5中で配列編集およびアセンブリーを実施した。全配列は、双方向で少なくとも2回のカバレッジを示す。遺伝子配列の操作は、メリーランド州ベセズダのインフォマックス(InforMax,Inc.(Bethesda、MD))からのベクターNTIプログラムを使用して達成した。ベクターNTIにおけるデフォルト値を使用して、対合比較を実施した。米国バ
イオテクノロジカル情報センター(NCBI)で設定されたデフォルト値を使用して、BLAST分析を実施した。
【0202】
略語の意味は以下の通り。「sec」は秒を意味し、「min」は分を意味し、「h」は時間を意味し、「d」は日を意味し、「μL」はマイクロリットルを意味し、「mL」はミリリットルを意味し、「L」はリットルを意味し、「μM」はマイクロモル(micromolar)を意味し、「mM」はミリモル(millimolar)を意味し、「M」はモル(molar)を意味し、「mmol」はミリモル(millimole)を意味し、「μmol」マイクロモル(micromole)を意味し、「g」はグラムを意味し、「μg」はマイクログラムを意味し、「ng」はナノグラムを意味し、「U」は単位を意味し、「bp」は塩基対を意味し、「kB」はキロベースを意味する。
【0203】
実施例1
カロテノイド産生株パントエア・アグロメランス(Pantoea agglomerans)DC404の単離
本実施例は、腸内細菌科に属する黄色素細菌株パントエア・アグロメランス(Pantoea agglomerans)DC404株の単離および同定について述べる。この生物体中で生成された天然カロテノイドの分析は、様々なゼアキサンチン前駆物質およびゼアキサンチン誘導体に加えて、ゼアキサンチンの生成を証明する。
【0204】
株単離および分類:新らしいカロテノイド産生細菌株を単離するために、環境サンプルの収集物から着色微生物を単離した。デラウェア州ウィルミントンの家庭菜園土壌を採取して、ルリア・ブロス(LB)に再懸濁した。10μLのループを満たす再懸濁液をLBプレート上に画線して、プレートを30℃でインキュベートした。多様なコロニーの外見を有する着色細菌を選んで、LBプレート上に均一に2回画線して、30℃でインキュベートした。これらのコロニーから、淡黄色で滑らかな半透明コロニーを形成したものを「DC404株」と命名した。
【0205】
タイプDC404株で16S rRNA遺伝子配列決定を実施した。具体的には、プライマーHK12(配列番号13)およびJCR14(配列番号14)を使用して、株の16S rRNA遺伝子をPCRによって増幅した。製造業者(キアジェン(Qiagen))の説明書に従って、QIAquick PCR精製キットを使用して増幅した16S
rRNA遺伝子を精製し、自動ABIシーケンサーで配列決定した。配列決定反応をプライマーHK12、JCR14、およびJCR15(配列番号15)で開始した。アセンブルしたDC404の1324bpの16S rRNA遺伝子配列(配列番号16)をジェンバンク(登録商標)に対するBLASTN検索のためのクエリー配列として使用した(アルトシュル(Altschul)ら、Nucleic Acids Res.25:3389〜3402(1997))。
【0206】
BLAST分析からは、DC404株が腸内細菌科に属することが示された。その16S rRNA遺伝子配列はこの科内のいくつかの異なる種との配列類似性を示し、トップヒット(96%同一性)はネズミチフス菌(Salmonella typhimurium)であった。上で述べた分析に従って、DC404株の16S rRNA遺伝子配列との96%同一性を有するその他の種としては、クレブシエラ・オキシトカ(Klebsiella oxytoca)、エンテロバクター・クロアカ(Enterobacter cloacae)、シトロバクター・アマロナチカス(Citrobacter amalonaticus)、エルウィニア・アミロボラ(Erwinia amylovora)、およびパントエア・アグロメランス(Pantoea agglomerans)が挙げられる。しかし比較的低いヒットの相同性(96%のみの同一性)に一致して、メリーランド州コッキーズヴィルのベクトン・ディキンソン(Becton Dick
inson(Cockeysville、MD))からのBBLエンテロチューブTM IIを使用した生化学的試験は、DC404がサルモネラ(Salmonella)種でないことを示唆する。
【0207】
外部会社であるデラウェア州ニューアークのマイクロビアルID(Microbial
ID,Inc.(Newark、DE))によるその脂肪酸プロフィールに従って、DC404をさらに特性決定した。ガスクロマトグラフィーによる脂肪酸プロファイリングは、DC404株がパントエア・アグロメランス(Pantoea agglomerans)に最も近縁であることを示唆した(類似度指数は0.557であった)。類似度指数は、未知のサンプルの脂肪酸組成が、そのマッチとしてリストされたライブラリーエントリを作成するのに使用した株の平均脂肪酸組成と比較して、どれだけ近いかを表す数値である。未知のサンプルの脂肪酸組成と、ライブラリーエントリの平均値との正確な一致は、類似度指数1.000をもたらす。0.400〜0.600の類似度指数は種の一致であるかもしれず、異型株を示す。DC404株の16S rRNA分析はまた、トップBLASTヒット(96%の同一性を有する)の1つとして、パントエア・アグロメランス(Pantoea agglomerans)を同定したので、パントエア・アグロメランス(Pantoea agglomerans)の異型株としてDC404株を命名した。
【0208】
DC404のカロテノイド分析:DC404中の黄色色素を抽出してHPLCで分析した。株を200mLのLB中で30℃において2日間生育させ、次に細胞を4000gで30minの遠心分離によって収集した。細胞ペレットを10mLのアセトンで抽出した。溶剤を窒素下で乾燥させ、カロテノイドを0.5mLのアセトンに再懸濁した。抽出物をミシガン州アナーバーノのパル・コーポレーション(Pall Corporation(Ann Arbor、MI))からのアクロディスク(Acrodisc)CR25mmシリンジフィルターで濾過して、次にカリフォルニア州フォスター・シティのアジレント(Agilent、(Foster City、CA))からのアジレント(Agilent)シリーズ1100 LC/MSD SIを使用して、0.1mLの10%アセトン+90%HPLC分析用アセトニトリル中で濃縮した。
【0209】
サンプル(20μL)を150mm×4.6mmのZORBAX C18(3.5μm粒子)カラム(アジレントテクノロジーズ(Agilent Technologies,Inc.))に装填した。カラム温度を40℃に保った。流速は1mL/分であり、溶剤ランニングプログラムは次のとおりであった。
・0〜2分:95%緩衝液Aおよび5%緩衝液B、
・2〜10分:95%緩衝液Aおよび5%緩衝液Bから60%緩衝液Aおよび40%緩衝液Bへの直線的濃度勾配、
・10〜12分:60%緩衝液Aおよび40%緩衝液Bから50%緩衝液Aおよび50%緩衝液Bへの直線的濃度勾配、
・12〜18分:50%緩衝液Aおよび50%緩衝液B、
・18〜20分:95%緩衝液Aおよび5%緩衝液B。
【0210】
緩衝液Aは95%アセトニトリルおよび5%dH2Oであり、緩衝液Bは100%テトラデヒドロフランであった。
【0211】
HPLC分析(図3)は、標品との比較で、DC404株がゼアキサンチン(6.28minピーク)、β−クリプトキサンチン(10.23minピーク)およびβ−カロテン(13.02minピーク)を生成したことを示唆した。具体的には、ゼアキサンチンおよびβ−クリプトキサンチン標品は、スイス国ルプシンゲンのカロテナチュール(CaroteNature(Lupsingen、Switzerland))から購入し、
β−カロテン標品は、ミズーリ州セントルイスのシグマ(Sigma(St.Louis、MO))から購入した。MS分析からゼアキサンチンピークの分子量が569であり、β−クリプトキサンチンピークの分子量が553であり、β−カロテンピークの分子量がが537であることを確認した。3.22minに溶出するピークは、その分子量731が示唆するように、ゼアキサンチンモノグルコシドである可能性が高い。
【0212】
実施例2
パントエア・アグロメランス(Pantoea agglomerans)DC404着色コスミドクローンの同定
実施例2は、パントエア・アグロメランス(Pantoea agglomerans)DC404からのゲノムDNAの約40kBの断片を発現できる、大腸菌(E.coli)コスミドクローンの構造について述べる。この形質転換体は、主にゼアキサンチンを生成した。大腸菌(E.coli)コスミドクローン中にゼアキサンチングルコシドは検出されなかった。
【0213】
染色体DNAの調製:DC404株を25mLのLB培地内で30℃で一晩通気して生育させた。細菌性細胞を4,000gで10min遠心分離した。細胞ペレットを5mLの50mMトリス−10mM EDTA(pH8)中に穏やかに再懸濁して、リゾチームを最終濃度2mg/mLに添加した。懸濁液を37℃で1hインキュベートした。次にドデシル硫酸ナトリウムを最終濃度1%に添加して、タンパク質分解酵素Kを100μg/mLで添加した。懸濁液を55℃で2hインキュベートした。懸濁液は透明になり、等容積のフェノール:クロロホルム:イソアミルアルコール(25:24:1)で2回、クロロホルム:イソアミルアルコール(24:1)で1回透明な溶解産物を抽出した。4,000rpmで20minの遠心処理後、水相を注意深く除去して新しい試験管に移した。2容積のエタノールを添加して、密封したガラス・パスツール・ピペットにDNAを穏やかに巻き付けた。DNAを70%エタノールを含有する試験管に浸した。風乾後、DNAをRNaseA(100μg/mL)と共に400μLのTE(10mMトリス−1mM
EDTA、pH8)に再懸濁し、4℃に保管した。DNAの濃度および純度を分光光度法によりOD260/OD280で測定した。
【0214】
コスミドライブラリーの構築:ウィスコンシン州マディソンのエピセンター・テクノロジーズ(Epicentre Technologies(Madison、WI))からのpWEBコスミドクローン化キットを使用して、製造業者の説明書に従って、DC404のコスミドライブラリーを構築した。シリンジニードルを通過させて、ゲノムDNAを剪断した。剪断したDNAを末端修復して、40kBの標準との比較によって低融点アガロース上でサイズ選択した。大きさがほぼ40kBのDNA断片を精製して、ブラントエンドクローン化レディーのpWEBコスミドベクター中にライゲートした。超高効率MaxPlaxラムダ・パッケージング・エキストラクツ(Lambda Packaging Extracts)を使用して、ライブラリーをパッケージし、EPI100大腸菌(E.coli)細胞上に播種した。コスミドライブラリークローンから2個の黄色コロニーが同定された。2個のクローンからのコスミドDNAは、類似した制限酵素消化パターンを有したので、1個のクローン上でさらなる分析を実施した。
【0215】
黄色コスミドクローンのカロテノイド分析:実施例1で述べられるようにしてLC−MSによって、コスミドpWEB−404を含有する大腸菌(E.coli)EPI100中のカロテノイドを分析した。HPLCの結果を図4に示す。そのUVスペクトル、分子量、および標品との比較に基づいて、6.27minのピークはゼアキサンチンと同定された。顕著な量のβ−カロテンまたはβ−クリプトキサンチン中間体のいずれも蓄積せず、ゼアキサンチングルコシドは生成されなかった。DC404(図3)中で観察された顕著なゼアキサンチンモノグルコシドピークは、この大腸菌(E.coli)コスミドクロ
ーン中には存在しなかった。分子重量がゼアキサンチンモノグルコシド(731ダルトン)と一致しなかったので、3.56minの小さなピークはゼアキサンチンモノグルコシドの異性体ではなかった。コスミドpWEB−404を含有する大腸菌(E.coli)内のゼアキサンチングルコシドの不在は、ゼアキサンチングルコシル転移酵素をコードするcrtX遺伝子が、コスミドpWEB−404中に存在しないかもしれないことを示唆する。
【0216】
実施例3
カロテノイド生合成遺伝子の同定
この実施例は、コスミドpWEB−404中のパントエア・アグロメランス(Pantoea agglomerans)DC404 crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子の同定について述べ、その他の知られているパントエア(Panteoa)crt遺伝子に関わるこれら遺伝子の関連性の比較を提供する。その他のパントエア(Panteoa)中の下流カロテノイド生合成経路の遺伝子構成とは対照的に、crtX遺伝子は、ここで述べるP.アグロメランス(P.agglomerans)DC404からのコスミド上に遺伝子クラスターの一部として存在しなかった。
【0217】
HPLC分析は、コスミドpWEB−404が、ゼアキサンチンの合成のための遺伝子を含有するに違いないことを示唆した。カロテノイド合成遺伝子を配列決定するために、ウィスコンシン州マディソンのエピセンター(Epicentre(Madison、WI))からのEZ::TN<TET−1>キットを使用して製造業者の説明書に従って、コスミドDNA pWEB−404に生体外(in vitro)遺伝子転移を行った。TET−1 FP−1フォーワードプライマー(配列番号17)を使用して、トランスポゾン末端から、200個のテトラサイクリン抵抗性トランスポゾンインサートを配列決定した。ミシガン州アナーバーのジーン・コーズ社(Gene Codes Corp.(Ann Arbor、MI))からのシーケンチャー(Sequencher)プログラムによって、配列アセンブリーを実施した。DC404からのカロテノイド生合成経路の6個の遺伝子を含有する、8814bpのcontig(配列番号18)をアセンブルした(図5)。
【0218】
BLAST「nr」データベース(全ての非重複性ジェンバンク(登録商標)CDS翻訳、三次元構造ブルックヘブンタンパク質データバンクから誘導される配列、SWISS−PROTタンパク質配列データベース、EMBL、およびDDBJデータベースを含んでなる)に含有される配列との類似性に対するBLAST(基礎的局在性整列化検索ツール(Basic Locl Alignment Search Tool)アルトシュール(Altschul)ら、前出)検索を行って、crtE、idi、crtY、crtI、crtB、およびcrtZをコードする遺伝子を同定した。米国バイオテクノロジー情報センター(NCBI)が提供するBLASTNアルゴリズムを使用して、「nr」データベースに含有されるあらゆる公的に入手できるDNA配列との類似性について、各配列を分析した。DNA配列を全ての読み取り枠で翻訳し、NCBIが提供するBLASTXアルゴリズムを使用して、「nr」データベースに含有されるあらゆる公的に入手できるタンパク質配列との類似性について比較した(ギッシュ(Gish)W.およびステート(States)D.J.、Nature Genetics 3:266〜272(1993))。
【0219】
BLASTNnrまたはBLASTXnrアルゴリズムのどちらかを使用して、全ての比較を行った。BLAST比較の結果を表2に示し、各遺伝子が最も類似する配列を要約する。表2は、期待値に報告される値を有するBLASTXnrアルゴリズムに基づくデータを示す。期待値は、このサイズのデータベースの検索において絶対的に偶然に期待される、規定のスコアにおけるマッチ数を特定して、マッチの統計学的有意さを推定する。
【0220】
ベクターNTIプログラム(前出)を使用して、ヌクレオチドおよびアミノ酸配列もまた、その他のパントエア(Pantoea)株からのものと比較した。表3に対合比較の同一性を要約する。
【0221】
【表3】
【0222】
【表4】
【0223】
【表5】
【0224】
様々なパントエア(Pantoea)種中のcrt遺伝子クラスター遺伝子構成の比較からは、遺伝子idiおよびcrtXの存在に関する興味深い違い(図5)もまた明らかにされた。イソペンテニルピロリン酸イソメラーゼをコードする遺伝子idiは、上流イソプレノイド経路に関与し、例えばパントエア・アナナティス(Pantoea ananatis)(ジェンバンク(登録商標)登録番号D90087)およびパントエア・ステワルティ(Pantoea stewartii)(ジェンバンク(登録商標)登録番号AY1667131)中のcrtEXYIBZクラスターの遺伝子構成のように、通常、crt遺伝子クラスター内には存在しない。しかしときどき、パントエア・アグロメランス(Pantoea agglomerans)EHO−10(ジェンバンク(登録商標)登録番号M87280)で見られるように、idiがcrt遺伝子オペロンの一部であることが見いだされる。本願明細書の出願人が行ったBLAST分析に従って、P.アグロメランス(P.agglomerans)EHO−10(ジェンバンク(登録商標)登録番号M87280)のcrtE−仮説タンパク質−crtX−crtY−crtI−crtB−crtZのカロテン産生性クラスター内に含有される「仮説タンパク質」は、crtE−idi−crtX−crtY−crtI−crtB−crtZを含んでなると見なされるべきである。同様にidiはまた、パントエア・アグロメランス(Pantoea agglomerans)DC404のcrt遺伝子クラスター中にも存在し、crtE遺伝子の下流である。しかし表3に示すように、DC404株のidiは、EHO−10のidiと68%のみの同一性を有する。
【0225】
crtX遺伝子に関してDC404株は、ここで述べるカロテン産生性遺伝子クラスター内にこの遺伝子が不在である点が、その他のパントエア(Pantoea)種と比べてユニークである(図5)。さらにコスミドクローンはゼアキサンチンのみを生成し、ゼアキサンチングルコシドを生成しないので、DC404からのcrt遺伝子クラスターを含有する大腸菌(E.coli)コスミドクローンpWEB−404はcrtX遺伝子を含有しない。ここで述べる遺伝子クラスター中のcrtX遺伝子の欠如にも関わらず、DC404はゼアキサンチンモノグルコシドを主に生成するので、crtX遺伝子を有さねばならない。crtX遺伝子はDC404染色体中の別の場所に位置し、crt遺伝子クラスターとは連結していないと結論された。DC404株中のcrtXの配列は、今回は同定されなかった。
【0226】
実施例4
メチロモナス(Methylomonas)sp.16aにおけるパントエア・アグロメランス(Pantoea agglomerans)DC404のcrtEidiYIB遺伝子クラスターの発現
以下の実施例は、パントエア・アグロメランス(Pantoea agglomerans)DC404(実施例3)からのcrtEidiYIB遺伝子を含んでなるcrt遺伝子クラスターをメチロモナス(Methylomonas)16a(ATCC PTA2402)中に導入して、β−カロテンなどの望ましい40−炭素カロテノイドを合成できるようにすることについて述べる。
【0227】
最初に、プライマーpWEB404F:5’−GAATTCACTAGTCGAGACGCCGGGTACCAACCAT−3’(配列番号19)およびpWEB404R:5’−GAATTCTAGCGCGGGCGCTGCCAGA−3’(配列番号20)を使用して、PCRによって、crtEidiYIB遺伝子を含有するDC404からの断片を増幅した。以下の熱サイクル条件で、カリフォルニア州ラホーヤのストラタジーン(Stratagene、La Jolla、CA)からのPfu Turboポリメラーゼと共に、コスミドDNA pWEB−404をテンプレートとして使用した。92℃(5min);94℃(1min)、60℃(1min)、72℃(9min)を25サイクル;および72℃(10min)。ゲル電気泳動法に続いて、およそ5.6kBの単一生成物が観察された。パーキン・エルマー(Perkin Elmer)からのTaqポリメラーゼを10分間の72℃の反応で使用して、インビトロゲン(Invitrogen)からのpTrcHis2−TOPO中へのTOPOクローン化のために、追加的な3’アデノシンヌクレオチドを断片に付加した。大腸菌(E.coli)TOP10細胞中への形質転換に続いて、いくつかのコロニーの色が明るい黄色に見え、それらがカロテノイド化合物を生成していることが示唆された。次に遺伝子クラスターをフロリダ州マルコ・アイランドのモビテック(MoBiTec,LLC(Marco Island、FL))からの広範宿主ベクターpBHR1中にサブクローン化し、ウィスコンシン州ミドルタウンのルシジェン(Lucigen(Middletown、WI))からの大腸菌(E.coli)10G細胞中に電気穿孔した。50μg/mLのカナマイシンを含有するLB培地上で、生成したプラスミドpDCQ330を含有する形質転換体を選択した。
【0228】
三親交雑接合噛合によって、プラスミドpDCQ330をメチロモナス(Methylomonas)16a中に転位した。pRK2013(ATCC番号37159)を含有する大腸菌(E.coli)ヘルパー株、およびpDCQ330を含有する大腸菌(E.coli)10G供与株をカナマイシン(50μg/mL)を含有するLB培地中で一晩生育させてLB中で3回洗浄し、元の培養容積のほぼ60倍の濃度に相当する容積のLB中に再懸濁した。
【0229】
メチロモナス(Methylomonas)16a株MWM1000(Δald/crtN1)は、天然C30カロテノイドの合成を中断させるald/crtN1遺伝子の単一クロスオーバーノックアウトを含有した(米国特許出願第60/527083号)。国際公開第02/18617号パンフレットで述べられる一般条件を使用して、この(Δald/crtN1)株を受容体として生育させた。簡単に述べると、少なくとも8:1の気体/液体比を使用して(すなわち160mL総容積中の20mLの硝酸塩液体「BTZ−3」培地)、30℃で絶えず振盪しながら、イリノイ州ウィートンのウィートン・サイエンティフィック(Wheaton Scientific、(Wheaton、IL))からの血清ストッパーしたウィートンボトル内で、メチロモナス(Methylomonas)16a株MWM1000を生育させた。
【0230】
ここで「合成培地」または「BTZ−3」培地とも称される硝酸塩液体培地は、下に示す溶液1と混合された様々な塩(表4および5)から構成され、または規定の硝酸塩が15mMの塩化アンモニウムで置き換えられている。溶液1は、100倍に濃縮された微量元素原液のための組成物を提供する。
【0231】
【表6】
【0232】
【表7】
【0233】
培養のための標準気相は、空気中に25%のメタンを含有する。これらの条件下で、MWM1000受容体をBTZ−3培地中で48h培養してBTZ−3中で3回洗浄し、元の培養容積の150倍の濃度に相当する容積のBTZ−3に再懸濁した。
【0234】
次に0.5%(w/v)酵母抽出物を含有するBTZ−3寒天プレートの表面で、供与体、ヘルパー、および受容細胞のペーストをそれぞれ1:1:2の比率で合わせた。プレートを25%メタン内で16〜72h、30℃に維持して接合を生じさせ、その後細胞ペーストを収集してBTZ−3に再懸濁した。カナマイシン(50μg/mL)を含有するBTZ−3寒天に希釈物を播種して、25%メタン内において30℃で1週間まで培養した。黄色の接合完了体をカナマイシン(50μg/mL)含有BTZ−3寒天上に画線した。
【0235】
カロテノイド組成物の分析のために、カナマイシン(50μg/mL)を含有する25mLのBTZ−3中で接合完了体を培養し、唯一の炭素源として25%のメタン内で30℃で3〜4日間インキュベートした。細胞を遠心分離によって収集し、−20℃で凍結した。解凍後ペレットを抽出し、実施例1で述べるようにして、HPLCによってカロテノイド含量を分析した。
【0236】
pDCQ330を含有するメチロモナス(Methylomonas)16a MWM1000からの抽出物のHPLC分析は、β−カロテンのほぼ排他的名生成を示した(図6)。14minピークの滞留時間、UVスペクトル、および分子重量は、シグマ(Sigma)からのβ−カロテン標品に一致する。これによって、このメタノトローフ宿主内でのDC404からのcrtEidiYIB遺伝子クラスターを利用したC40カロテノイドの合成が確証された。
【図面の簡単な説明】
【0237】
【図1】上流イソプレノイドおよび下流カロテノイド生合成経路を示す。
【図2】CrtE、CrtX、CrtY、CrtI、CrtB、およびCrtZによって触媒される具体的な化学転換を図示する、下流C40カロテノイド生合成経路の一部を示す。
【図3】パントエア・アグロメランス(Pantoea agglomerans)DC404株内に含有されるカロテノイドのHPLC分析の結果を示す。
【図4】コスミドpWEB−404を含んでなる形質転換大腸菌(E.coli)内に含有されるカロテノイドのHPLC分析の結果を示す。
【図5】パントエア・アナナティス(Pantoea ananatis)(ジェンバンク(登録商標)登録番号D90087)、パントエア・ステワルティ(Pantoea stewartii(ジェンバンク(登録商標)登録番号AY166713)、パントエア・アグロメランス(Pantoea agglomerans)EHO−10(ジェンバンク(登録商標)登録番号M87280)、およびパントエア・アグロメランス(Pantoea agglomerans)DC404のカロテノイド遺伝子クラスターの組成の比較を示す。
【図6】pDCQ330を含有するメチロモナス(Methylomonas)16a CrtN1−株からのカロテノイドのHPLC分析を示す。
【関連出願との関係】
【0001】
本願明細書は、2003年6月12日に出願された米国仮願第60/477,874号、および、2003年12月3日に出願された米国仮出願第60/527,083号の優先権の利益を主張する。
【技術分野】
【0002】
本発明は、分子生物学および微生物学の分野に関する。より具体的には、本発明は微生物のカロテノイド化合物産生に有用な酵素(例えばリコピン、β−カロテン、およびゼアキサンチン)をコードする、・アグロメランス(Pantoea agglomerans)から単離される核酸断片に関連する。
【背景技術】
【0003】
カロテノイドは、最も広く分布して構造的に多様性のあるクラスの天然色素の1つを代表し、淡黄色からオレンジから深紅色の色素色を生じる。カロテン産生性組織の顕著な例としては、ニンジン、トマト、赤ピーマン、およびラッパズイセンおよびマリーゴールドの花弁が挙げられる。カロテノイドは、全ての光合成生物体、ならびにいくつかの細菌および菌・カビ類によって合成される。これらの色素は、光合成、栄養、および光酸化損傷に対する保護おいて重要な機能を有する。例えば動物はカロテノイドを合成する能力を有さず、食物源を通じてこれらの栄養的に重要な化合物を得なくてはならない。
【0004】
自然界における600を越える異なるカロテノイドの存在にもかかわらず、工業的には、食物色素、動物飼料、医薬品、および化粧品のためにわずかなカロテノイドが使用されるのみである。これは生産の困難さが主因である。現在、工業目的で使用されるほとんどのカロテノイドは化学合成によって生産されるが、これらの化合物は化学的に製造することが非常に困難である(非特許文献1)。天然カロテノイドは、植物材料の抽出または微生物合成のいずれかによって得ることができる。しかし商業的なカロテノイド生産にはわずかな植物のみが広く使用され、これらの植物におけるカロテノイド合成の生産性は比較的低い。その結果、これらの植物から生成されるカロテノイドは非常に高価である。生合成の生産能力を増大させる一方法は、組換えDNA技術を応用することである(非特許文献2でレビューされている)。したがって非カロテン産生性細菌および酵母中でカロテノイドを生成することにより、最も適切で効率的な産生生物体の質、量、および選択のコントロールを可能にすることが望ましい。後者は、消費者にとって商業的生産経済(ひいては入手可能性)のために、特に重要である。
【0005】
構造的に最も一般的なカロテノイドは、炭素40個(C40)のテルペノイドである。しかし炭素原子30個のみのカロテノイド(C30、ジアポカロテノイド)が、いくつかの種で検出される。これらの各タイプのカロテノイドの生合成は、イソプレン生合成経路、およびその五炭素ユニバーサルイソプレン構成単位であるイソペンテニルピロリン酸(IPP)に由来する。この生合成経路は、2つの部分に分割できる。1)ファルネシルピロリン酸(FPP)の形成をもたらす上流イソプレン経路、および2)FPPを長鎖C30およびC40カロテン産生性化合物に転換する様々なcrt遺伝子を含んでなる下流カロテノイド生合成経路。この経路の双方の部分を図1に示す。
【0006】
典型的にフィトエンの形成が、C40カロテノイドの生合成にユニークな第1のステップを代表する(図1および2)。フィトエンそれ自体は無色のカロテノイドであり、イソペンテニルピロリン酸イソメラーゼ(遺伝子idiによってコードされる)によるIPPからジメチルアリルピロリン酸(DMAPP)への異性化を通じて生じる。この反応に、
一続きの3プレニル転移酵素反応が続き、そこではゲラニルピロリン酸(GPP)、ファルネシルピロリン酸(FPP)、およびゲラニルゲラニルピロリン酸(GGPP)が形成される。GGPP合成酵素をコードする遺伝子crtEが、この後者の反応を司る。最後に2個のGGPP分子が縮合して、フィトエン(PPPP)を形成する。この反応はフィトエンシンターゼ(遺伝子crtBによってコードされる)によって触媒される。
【0007】
リコピンはフィトエンから生成される「着色」カロテノイドである。リコピンは特徴的な赤色を熟したトマトに与え、食物色素として多くの用途を有する。それはまた、いくつかの細菌、菌・カビ類、および緑色植物におけるその他のカロテノイドの生合成の中間体でもある。リコピンは、遺伝子crtI(フィトエンデサチュラーゼをコードする)によって触媒される、8個の水素原子の除去による4つの逐次脱水素化反応を通じて、フィトエンから生合成的に調製される。この反応における中間物(Imtermediaries)は、フィトフルエン、ζ−カロテン、およびニューロスポレンである。
【0008】
リコピンシクラーゼ(CrtY)は、リコピンをβ−カロテンに転換する。β−カロテンは、カラースペクトルが黄色からオレンジの範囲である典型的なカロテンである。これは、マーガリンおよびバターの着色料として、ビタミンA生成源として、そして最近は特定種類の癌に対して防止効果の可能性がある化合物としての用途がある。
【0009】
β−カロテンは、β−カロテン水酸化酵素活性(crtZ遺伝子によってコードされる)によってもたらされる水酸化反応を通じて、ゼアキサンチンに転換する。例えばこれはトウモロコシの種子中に存在する黄色色素である。ゼアキサンチンは雌鶏または錦鯉の飼料内に含有され、それらの着色の重要な色素源である。
【0010】
フィトエン、リコピン、β−カロテン、およびゼアキサンチンの生成をつかさどるカロテノイド生合成遺伝子および酵素に加えて、(i)水素付加、(ii)脱水素化、(iii)環化、(iv)酸化、(v)エステル化/グリコシル化、またはこれらのプロセスのあらゆる組み合わせによって、C40化合物の分子内転換を可能にして、多数のその他の官能性付与カロテノイド化合物を生成する、その他の様々なcrt遺伝子が知られている。
【0011】
腸内細菌科内の細菌の多くは自然に着色しており、これらの生物体がカロテノイドを生成する能力が示される。さらにC40カロテノイド生合成は、かつて全てエルウィニア(Erwinia)属に所属するとされていた、細菌の大まかなグループに分類されていた生物体の小さなグループであるパントエア(Pantoea)属において、特に良く研究されている[エルウィニア(Erwinia)、ペクトバクテリウム(Pectobacterium)、ブレンネリア(Brenneria)gen.nov.、およびパントエア(Pantoea)を含んでなる4つの系統学的群に、かつての大きなエルウィニア(Erwinia)属が再分類された詳細については、非特許文献3を参照されたい]。例えば非特許文献4、非特許文献5などのいくつかのレビューが、カロテノイド色素生合成の遺伝学を考察している。crtEXYIBZをコードする遺伝子配列は、パントエア・アグロメランス(Pantoea agglomerans)(かつてE.ヘルビコラ(E.herbicola)EHO−10(ATCC番号39368)として知られていた)、P.アナナティス(P.ananatis)(かつてE.ウレドボラ(E.uredovora)20D3(ATCC番号19321)として知られていた)、P.ステワルティ(P.stewartii)(かつてE.ステワルティ(E.stewartii)(ATCC番号8200)として知られていた)、およびP.アグロメランスpv.ミレチア(P.agglomerans pv.milletiae)について入手できる(特許文献1、特許文献2、特許文献3、特許文献4、特許文献5、特許文献6、ジェンバンク(登録商標)登録番号M87280、D90087、AY166713、AB07
6662もまたそれぞれ参照されたい)。しかし既存の文献は、これらの詳しく特性決定されたパントエア(Pantoea)種にあまり関連しない生物体におけるcrtEXYIBZをコードする遺伝子配列、およびこれらの配列の遺伝子構成に関する限定的情報のみを提供する。
【0012】
したがって解決すべき問題は、パントエア・アグロメランス(Pantoea agglomerans)(ATCC番号39368)、P.アナナティス(P.ananatis)(ATCC番号19321)、P.ステワルティ(P.stewartii)(ATCC番号8200)、およびP.アグロメランスpv.ミレチア(P.agglomerans pv.milletiae)とより遠縁の生物体から、カロテノイド生合成酵素の全てまたは一部をコードする核酸配列を同定して、カロテノイド生合成経路のより良い理解のための研究を容易にし、これらの経路の操作のための遺伝的ツールを提供して、適切な遺伝子を適切な宿主中に導入し発現することで、カロテノイドを大量に合成する手段を提供することである。これは合成方法に勝るカロテノイド生成をもたらすであろう。
【0013】
本出願人は、パントエア・アグロメランス(Pantoea agglomerans)DC404株と称される黄色着色素細菌から、CrtE、Idi、CrtY、CrtI、CrtB、およびCrtZ酵素をコードするカロテノイド生合成経路中の6つのユニークな読み取り枠(ORF)を単離することで、既述の問題を解決した。またこれらの配列の遺伝子構成は新しい。
【0014】
【特許文献1】米国特許第5,656,472号明細書
【特許文献2】米国特許第5,545,816号明細書
【特許文献3】米国特許第5,530,189号明細書
【特許文献4】米国特許第5,530,188号明細書
【特許文献5】米国特許第5,429,939号明細書
【特許文献6】国際公開第02/079395A2号パンフレット
【非特許文献1】ネリス(Nelis)およびレーンハー(Leenheer)、Appl.Bacteriol.70:181〜191(1991)
【非特許文献2】ミサワ(Misawa)およびシマダ(Shimada)、J.Biotech.59:169〜181(1998)
【非特許文献3】ハウベン(Hauben)ら、Syst.Appl.Microbiol.21(3):384〜397(1998年8月)
【非特許文献4】G.アームストロング(Armstrong)、J.Bact.176:4795〜4802(1994)
【非特許文献5】G.アームストロング(Armstrong)、Annu.Rev.Microbiol.51:629−659(1997)
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明は、リコペン、β−カロテン、およびゼアキサンチンをはじめとする、様々なカロテノイドの合成に関与することが実証されたDC404株から単離される、6つの遺伝子を提供する。遺伝子は同一オペロン上にクラスター化され、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子を含む。crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子のDNA配列は、ORF1〜6および配列番号1、3、5、7、9、および11にそれぞれ対応する。
【0016】
したがって本発明は、カロテノイド生合成経路酵素をコードする単離された核酸分子を提供し、分子の遺伝子構成はcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子は
記載順にクラスター化され、crtZの転写はcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起きる。
【0017】
好ましい実施態様では、crtE−idi−crtY−crtI−crtB−crtZ遺伝子クラスターのカロテノイド生合成経路酵素の少なくとも1つは、配列番号2、4、6、8、10、および12よりなる群から選択されるアミノ酸配列によってコードされる。そしてさらに好ましい実施態様では、単離された酸分子は配列番号18に記載の配列を有するポリヌクレオチドを含んでなる。
【0018】
同様に本発明は、crtE−idi−crtY−crtI−crtB−crtZを含んでなる配列番号18で示される単離された核酸分子、または配列番号18と少なくとも95%の同一性を有する遺伝子または単離された核酸分子を提供し、単離された核酸分子は、全てのポリペプチドcrtE、idi、crtY、crtI、crtB、およびcrtZをコードする。
【0019】
さらに本発明は、
(a)配列番号2、4、6、8、10、および12よりなる群から選択されるアミノ酸配列をコードする単離された核酸分子、
(b)0.1×SSC、0.1%SDSで65℃、および2×SSC、0.1%SDSで洗浄後、0.1×SSC、0.1%SDSというハイブリダイゼーション条件下で(a)とハイブリダイズする単離された核酸分子、および
(c)(a)または(b)と相補的である単離された核酸分子
よりなる群から選択される、カロテノイド生合成経路酵素をコードする単離された核酸分子を提供する。
【0020】
本発明はさらに、細菌、酵母、糸状菌類、藻類、および植物における遺伝子の遺伝的発現に適した制御領域を含んでなる、本遺伝子および遺伝的キメラによってコードされるポリペプチド、ならびに該物質を含んでなる形質転換宿主を提供する。
【0021】
本発明は、
(a)本核酸分子ゲノムのライブラリーを探索し、
(b)本核酸分子とハイブリダイズするDNAクローンを同定し、
(c)ステップ(b)で同定されたクローンを含んでなるゲノム断片を配列決定することを含んでなる、カロテノイド生合成経路酵素をコードする核酸分子を得る方法を提供し、配列決定されたゲノム断片は、カロテノイド生合成酵素をコードする。
【0022】
同様に本発明は、
(a)本核酸配列の一部に対応する少なくとも1つのオリゴヌクレオチドプライマーを合成し、
(b)ステップ(a)のオリゴヌクレオチドプライマーを使用して、クローン化ベクター中に存在するインサートを増幅しすること
を含んでなる、カロテノイド生合成経路酵素をコードする核酸分子を得る方法を提供し、増幅されたインサートは、カロテノイド生合成経路酵素をコードするアミノ酸配列の一部をコードする。
【0023】
好ましい実施態様では、本発明は、
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、カロテノイド生合成経路酵素をコードする核酸分子
を含んでなる形質転換宿主細胞を提供し、
ここで、該分子の遺伝子構成はcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子は記載順にクラスター化し、crtZの転写はcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起き、
(b)適切な生育条件下でステップ(a)の宿主細胞を有効量の発酵性炭素基質に接触させることでカロテノイド化合物が生成することを含んでなるカロテノイド化合物の製造方法を提供する。
【0024】
同様に本発明は、カロテノイド化合物の生成に適した制御領域を含んでなる、本カロテノイド酵素をコードする一組の核酸分子および遺伝子キメラをさらに提供する。
【0025】
具体的な好ましい実施態様では、本発明は例えばメチロモナス(Methylomonas)16a(ATCCの称号PTA2402)のような高生育メタノトローフ細菌株などのC1代謝宿主におけるカロテノイド化合物の製造方法を提供し、C1代謝宿主は、
(a)メタンおよびメタノールよりなる群から選択されるC1炭素基質で生育し、
(b)機能性的エムデン−マイヤーホフ炭素経路を含んでなり、前記経路はピロリン酸依存性ホスホフルクトキナーゼ酵素をコードする遺伝子を含んでなる。
【0026】
さらに本発明は、生物体においてカロテノイド生合成が改変されるように、生物体中で配列番号1、3、5、7、9、および11よりなる群から選択される少なくとも1つのカロテノイド遺伝子を過剰発現するステップを含んでなる、生物体中でカロテノイド生合成を制御する方法を提供する。
【0027】
代案の実施態様では、本発明は、
(i)a)配列番号1、3、5、7、9、および11よりなる群から選択されるカロテノイド生合成経路酵素をコードする単離された核酸分子、
b)ステップ(a)の前記単離された核酸分子とハイブリダイズする第1のヌクレオチド断片集団、および
c)ステップ(a)の前記単離された核酸分子とハイブリッドしない第2のヌクレオチド断片集団
を含んでなるヌクレオチド配列の混合物を制限エンドヌクレアーゼで消化して、
制限酵素消化断片の混合物を生成させ、
(ii)前記制限酵素消化断片の混合物を変性し、
(iii)ステップ(ii)の変性した前記制限酵素消化断片の混合物をポリメラーゼと共にインキュベートし、
(iv)ステップ(ii)および(iii)を繰り返して、改変された生物学的活性を有するタンパク質をコードする突然変異カロテノイド遺伝子を生成させること
を含んでなる方法によって生成された、改変された生物学的活性を有するカロテノイド生合成経路酵素をコードする突然変異遺伝子を提供する。
【0028】
別の実施態様では、本発明は、配列番号16に記載の16s rDNA配列を含んでなるパントエア・アグロメランス(Pantoea agglomerans)DC404株を提供する。
【0029】
配列説明および生物学的寄託
本願明細書の一部を形成する以下の詳細な説明および添付の配列説明によって、本発明をより完全に理解できるであろう。
【0030】
以下の配列は、37C.F.R.1.821〜1.825(「ヌクレオチド配列および/またはアミノ酸配列開示を含む特許出願の要件−配列規則(Requirements
for Patent Applications Containing Nucleotide Sequences and/or Amino Acid Sequence Disclosures−the Sequence Rules)」)を満たし、世界知的所有権機関(WIPO)標準ST.25(1998)およびEPOおよびPCTの配列表要件(規則5.2および49.5(aの2)、および実施細則第208号および附属書C)に一致する。ヌクレオチドおよびアミノ酸配列データのために使用される記号および型式は、37C.F.R.§1.822で述べられる規則に従う。
【0031】
配列番号1〜12は、表1で同定される完全長の遺伝子またはタンパク質である。
【0032】
【表1】
【0033】
配列番号13〜15、および17は、プライマーHK12、JCR14、JCR15、およびTET−1FP−1をそれぞれコードするヌクレオチド配列である。
【0034】
配列番号16は、DC404株の16S rRNA遺伝子配列を提供する。
【0035】
配列番号18は、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子をコードするDC404株からのDNAの8,814bp断片のヌクレオチド配列である。
【0036】
配列番号19は、プライマーpWEB404Fのヌクレオチド配列である。
【0037】
配列番号20は、プライマーpWEB404Rのヌクレオチド配列である。
【0038】
出願人らは、「特許手続上の微生物の寄託の国際的承認に関するブダペスト条約」の条項に従って、以下の生物学的寄託を行った。
【0039】
【表2】
【0040】
ここでの用法では、「ATCC」は、米国VA20110−2209マナッサスのユニバーシティ・ブールヴァード10801のATCCに所在する米国微生物系統保存機関国際寄託局(American Type Culture Collection International Depository Authority)を指す。「国際寄託名」は、ATCCに寄託された培養物の登録番号である。
【0041】
列挙した寄託株は、表示された国際受託機関に少なくとも30年間保存され、それを開示する特許の付与時に一般に公開される。寄託株の利用可能性は、政府の行動によって付与された特許権を失墜させて主題発明を実施する認可とはみなされない。
【発明を実施するための最良の形態】
【0042】
本発明の遺伝子およびそれらの発現生成物は、様々なカロテノイド化合物を生成する能力を有する組換え生物体の作成のために有用である。CrtE、Idi、CrtY、CrtI、CrtB、およびCrtZをコードする核酸断片がパントエア・アグロメランス(Pantoea agglomerans)DC404株から単離され、当業者に良く知られているBLASTおよびFASTAアルゴリズムを使用して、ヌクレオチドおよびタンパク質配列を含む公共データベースとの比較によって同定されている。本発明の遺伝子および遺伝子産物は、カロテノイド化合物の改良または操作のために、多様なやり方で使用しても良い。これらの遺伝子を含んでなる遺伝子クラスターのユニークな遺伝子構成の結果として、さらなる利点があるかもしれない。
【0043】
カロテノイド化合物は化学的に作ることが非常に困難なので、これらの化合物の微生物による生成には一般的な実用的用途がある。(ネリス(Nelis)およびリーンハー(Leenheer)、前出)。ほとんどのカロテノイドは濃色を有し、天然色素または着色剤と見なすことができる。さらに多くのカロテノイドは強力な抗酸化特性を有するので、食餌中にこれらの化合物を含めることは健康的であると考えられる。良く知られている例はβ−カロテン、カンタキサンチン、およびアスタキサンチンである。さらにカロテノイドは、水産養殖で必要とされる要素である。カロテノイド着色はサケおよびエビの生物体の価値に対して極めて重要であるので、これらの水産養殖は本発明の特に有用な用途である(シャヒディ(Shahidi)F.、およびブラウン(Brown)J.A.、Critical reviews in Food Science、38(1):1〜67(1998))。最後にカロテノイドは、ステロイド、香料および香水、および可能な電気光学用途がある化合物の合成において中間体としての用途を有する。
【0044】
下の開示は、パントエア・アグロメランス(Pantoea agglomerans)DC404株からのカロテノイド合成遺伝子の単離、遺伝子工学によるこれらの遺伝子の修飾、および大腸菌(E.coli)、細菌、酵母、菌・カビ類、および高等植物中でのクローン化および発現に適した適合性プラスミド中へのそれらの挿入の詳細な説明を提供する。
【0045】
定義
本開示では、いくつかの用語および略語が使用される。以下の定義が提供される。
【0046】
「読み取り枠」はORFと略記される。
【0047】
「ポリメラーゼ連鎖反応」はPCRと略記される。
【0048】
「高速液体クロマトグラフィー」はHPLCと略記される。
【0049】
用語「イソプレノイド化合物」とは、形式的にイソプレン(2−メチルブタ−1,3−ジエン、CH2=C(CH3)CH=CH2)から誘導される化合物を指し、その骨格は分子中の反復として認識できる。これらの化合物は、イソペンテニルピロリン酸(IPP)から開始して、イソプレノイド経路を通じて生合成的に生成されてイソプレン単位の頭尾縮合によって形成され、例えば長さが炭素5、10、15、20、30、または40個であっても良い分子をもたらす。
【0050】
「カロテノイド生合成経路」という用語は、図1で示すように、本発明の上流イソプレノイド経路、および/または下流カロテノイド生合成経路のメンバーを含んでなる遺伝子を指す。
【0051】
「上流イソプレノイド経路」および「上流経路」という用語は区別なく使用され、ピルビン酸およびグリセルアルデヒド−3−リン酸からファルネシルピロリン酸(FPP)への転換に関与する酵素を指す。これらの酵素としては、「dxs」遺伝子(1−デオキシキシルロース−5−リン酸シンターゼをコードする)、「dxr」遺伝子(1−デオキシキシルロース−5−リン酸レダクトイソメラーゼをコードする)、「ispD」遺伝子(2C−メチル−D−エリスリトールシチジル転移酵素をコードする、ygbPとしても知られている)、「ispE」遺伝子(4−ジホスホシチジル−2−C−メチルエリスリトールキナーゼをコードする、ychBとしても知られている)、「ispF」遺伝子(2C−メチル−D−エリスリトール2,4−シクロ二リン酸シンターゼをコードする、ygbBとしても知られている)、「pyrG」遺伝子(CTPシンターゼをコードする)、ジメチルアリル二リン酸の形成に関与する「lytB」遺伝子、2−C−メチル−D−エリスリトール4−リン酸の合成に関与する「gcpE」遺伝子、「idi」遺伝子(IPPからジメチルアリルピロリン酸への分子内転換を司る)、およびイソプレノイド経路中の「ispA」遺伝子(ゲラニル転移酵素またはファルネシル二リン酸シンターゼをコードする)が挙げられるが、これに限定されるものではない。
【0052】
「Idi」という用語は、idi遺伝子によってコードされるイソペンテニル二リン酸イソメラーゼ(E.C.5.3.3.2)を指す。代表的なidi遺伝子は、配列番号3として提供される。
【0053】
用語「下流カロテノイド生合成経路」および「下流経路」は区別なく使用され、FPPを一連のカロテノイドに転換する酵素を指す。これらとしては(合成がC30カロテノイド生合成にユニークな第1のステップを代表する)ジアポフィトエン、または(合成がC40カロテノイド生合成にユニークな第1のステップを代表する)フィトエンのいずれかの即時の合成に関与する、遺伝子および遺伝子産物が挙げられる。様々なC30〜C40カロテノイド生成をもたらす全ての引き続く反応は、下流カロテノイド生合成経路に含まれる。これらの遺伝子および遺伝子産物には、crtM、crtN、crtN2、crtE、crtX、crtY、crtI、crtB、crtZ、crtW、crtO、crtA、crtC、crtD、crtF、およびcrtUをはじめとするが、これに限定されるものではない「crt」遺伝子の全てが含まれる。最後に「下流カロテノイド生合成酵素」という用語は、CrtM、CrtN、CrtN2、CrtE、CrtX、CrtY、
CrtI、CrtB、CrtZ、CrtW、CrtO、CrtA、CrtC、CrtD、CrtF、およびCrtUをはじめとするが、これに限定されるものではない、本下流経路中のあらゆる酵素を指す包括的用語である。
【0054】
本願明細書では、「カロテノイド化合物」という用語は、イソプレンから形式的に誘導される接合ポリエン炭素骨格を有する炭化水素のクラスとして定義される。このクラスの分子は、トリテルペン(C30ジアポカロテノイド)およびテトラテルペン(C40カロテノイド)およびそれらの酸素化誘導体から構成される。これらの分子は典型的に強力な光吸収特性を有し、長さがC200を超える範囲であっても良い。例えば長さがC35、C50、C60、C70、およびC80であるその他の「カロテノイド化合物」も知られている。
【0055】
「C30ジアポカロテノイド」は、2個の中央メチル基が1,6−位の関係にあり、残りの非末端メチル基が1,5−位の関係にあるように、イソプレノイド単位の配置が分子の中央で逆転するような様式で結合する6個のイソプレノイド単位からなる。全てのC30カロテノイドは、(i)水素付加、(ii)脱水素化、(iii)環化、(iv)酸化、(v)エステル化/グリコシル化、またはこれらのプロセスのいずれかの組み合わせによって、接合二重結合の長い中央鎖を有する非環式C30H42構造から形式的に誘導されても良い。
【0056】
「テトラテルペン」または「C40カロテノイド」は、2個の中央メチル基が1,6−位の関係にあり、残りの非末端メチル基が1,5−位の関係にあるように、イソプレノイド単位の配置が分子の中央で逆転するような様式で結合した8個のイソプレノイド単位からなる。全てのC40カロテノイドは、(i)水素付加、(ii)脱水素化、(iii)環化、(iv)酸化、(v)エステル化/グリコシル化、またはこれらのプロセスのいずれかの組み合わせによって、接合二重結合の長い中央鎖を有する非環式C40H56構造(下の式I)から形式的に誘導されても良い。このクラスはまた、炭素骨格(式I)の再配置、またはこの構造の一部の(形式的)除去から生じる特定の化合物も含む。
【0057】
【化1】
【0058】
便宜上、カロテノイドの式は、簡単な表記法で書かれることが多い(下の式IA)。
【0059】
【化2】
【0060】
式中、破線はイソプレノイド単位中の形式的区分を示す。
【0061】
「官能性付与された」または「官能性付与」という用語は、あらゆるカロテノイド主鎖
部分の(i)水素付加、(ii)脱水素化、(iii)環化、(iv)酸化、または(v)エステル化/グリコシル化を指す。この主鎖は、接合二重結合の長い中央鎖として定義される。官能性付与はまた、上のプロセスのあらゆる組み合わせによって生じても良い。
【0062】
「CrtE」という用語は、crtE遺伝子によってコードされ、トランス−トランス−ファルネシル二リン酸とイソペンテニル二リン酸をピロリン酸とゲラニルゲラニル二リン酸に転換する、ゲラニルゲラニルピロリン酸合成酵素(synthase)を指す。代表的なcrtE遺伝子は、配列番号1として提供される。
【0063】
「CrtY」という用語は、crtY遺伝子によってコードされ、リコペンをβ−カロテンに転換するリコペンシクラーゼ酵素を指す。代表的なcrtY遺伝子は、配列番号5として提供される。
【0064】
「CrtI」という用語は、crtI遺伝子によってコードされるフィトエンデサチュラーゼ酵素を指す。CrtIは4個の二重結合の導入により、フィトフルエン、β−カロテン、およびニューロスポレンの中間物を通じて、フィトエンをリコペンに転換する。代表的なcrtI遺伝子は、配列番号7として提供される。
【0065】
「CrtB」という用語は、プレフィトエン二リン酸からフィトエンへの反応を触媒するcrtB遺伝子によってコードされるフィトエンシンターゼ酵素を指す。代表的なcrtB遺伝子は、配列番号9として提供される。
【0066】
「CrtZ」という用語は、β−カロテンからゼアキサンチンへの水酸化反応を触媒するcrtZ遺伝子によってコードされる、β−カロテン水酸化酵素を指す。代表的なcrtZ遺伝子は、配列番号11として提供される。
【0067】
「CrtX」という用語は、crtX遺伝子によってコードされ、ゼアキサンチンをゼアキサンチン−β−ジグルコシドに転換する、ゼアキサンチングルコシル転移酵素を指す。
【0068】
「遺伝子構成」という用語は、ORFが単一核酸断片上に組織化される順序と方向を指す。
【0069】
本願明細書中では、3つの異なるDNAクラスターの遺伝子構成について述べられ、それぞれは下のように定義される。
1.「crtE−idi−crtY−crtI−crtB−crtZ」または「crtE−idi−crtYIBZ」という用語は、以下の遺伝子構成を有する分子を指す。crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子は述べられる順にクラスター化され、crtZの転写はcrtE、idi、crtY、crtI、およびcrtBと反対の方向で起きる。
2.「crtE−crtX−crtY−crtI−crtB−crtZ」または「crtEXYIBZ」という用語は、以下の遺伝子構成を有する分子を指す。crtE、crtX、crtY、crtI、crtB、およびcrtZ遺伝子は述べられる順にクラスター化され、crtZの転写はcrtE、crtX、crtY、crtI、およびcrtBと反対の方向で起きる。
3.「crtE−idi−crtX−crtY−crtI−crtB−crtZ」または「crtE−idi−crtXYIBZ」という用語は、以下の遺伝子構成を有する分子を指す。crtE、idi、crtX、crtY、crtI、crtB、およびcrtZ遺伝子は述べられる順にクラスター化され、crtZの転写はcrtE、idi、crtX、crtY、crtI、およびcrtBと反対の方向で起きる。
【0070】
「エムデン−マイヤーホフ経路」という用語は、グルコースおよびフルクトースなどのヘキソースをグリセルアルデヒド3−リン酸、ジヒドロキシアセトンリン酸、ホスホエノールピルビン酸、およびピルビン酸などの重要な細胞性三炭素中間体に転換するための一連の生化学的反応を指す。これらの反応は、ATPの形態の生化学的に有用なエネルギーの正味を生じて典型的に進行する。エムデン−マイヤーホフ経路にユニークな重要な酵素は、ホスホフルクトキナーゼおよびフルクトース1,6−ビスリン酸アルドラーゼである。
【0071】
「エントナー−ドゥドロフ経路」という用語は、生化学的に有用なエネルギーの正味の生成なしに、グルコースまたはフルクトースなどのヘキソースを重要な三炭素細胞性中間体ピルビン酸およびグリセルアルデヒド3−リン酸に転換するための一連の生化学的反応を指す。エントナー−ドゥドロフ経路にユニークな重要な酵素は、6−ホスホグルコン酸デヒドラターゼおよびケトデオキシホスホ−グルコン酸アルドラーゼである。
【0072】
「C1炭素基質」または「一炭素基質」という用語は、炭素−炭素結合が欠如するあらゆる炭素含有分子を指す。例はメタン、メタノール、ホルムアルデヒド、ギ酸、ホルマート、メチル化アミン(例えばモノ−、ジ−、およびトリ−メチルアミン)、メチル化チオール、および二酸化炭素である。
【0073】
「C1代謝個体」という用語は、そのエネルギーおよびバイオマスの唯一の供給源として一炭素基質を使用する能力を有する微生物を指す。C1代謝個体は、典型的にメチロトローフおよび/またはメタノトローフである。
【0074】
「メチロトローフ」という用語は、炭素−炭素結合を含有しない有機化合物を酸化できる生物体を意味する。メチロトローフがCH4を酸化できる場合、メチロトローフはまたメタノトローフでもある。
【0075】
「メタノトローフ」または「メタノトローフ細菌」という用語は、その主要な炭素およびエネルギー供給源としてメタンを利用できる原核生物を意味する。メタンから二酸化炭素への完全な酸化は、好気性分解経路によって起きる。本発明で有用なメタノトローフの典型的な例としては、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、およびメチロサイナス(Methylosinus)属が挙げられる(がこれに限定されるものではない)。
【0076】
「高生育メタノトローフ細菌株」という用語は、唯一の炭素およびエネルギー源としてメタンまたはメタノールで生育できる、機能的エムデンマイヤーホフ炭素流経路を有する細菌を指し、高速の生育および代謝されるC1基質1グラム当たりの高い細胞質量生成が得られる。ここで述べられる特定の「高生育メタノトローフ細菌株」は、「メチロモナス(Methylomonas)16a」、「16a」または「メチロモナス(Methylomonas)sp.16a」と称され、これらの用語は区別なく使用されてメチロモナス(Methylomonas)sp.16a(ATCC PTA−2402)株を指す(米国特許第6,689,601号明細書)。
【0077】
メチロモナス(Methylomonas)中の「crt遺伝子クラスター」という用語は、メチロモナス(Methylomonas)sp.16aの天然C30カロテノイド生合成経路において活性であるcrtN1、ald、およびcrtN2を含んでなる読み取り枠を指す。
【0078】
「CrtN1」という用語は、crtN1遺伝子によってコードされ、メチロモナス(Methylomonas)sp.16aの天然カロテノイド生合成経路において活性である酵素を指す。この遺伝子は、メチロモナス(Methylomonas)においてcrt遺伝子クラスター上に位置する第1の遺伝子である。
【0079】
「ALD」という用語は、ald遺伝子によってコードされ、メチロモナス(Methylomonas)sp.16aの天然カロテノイド生合成経路において活性である酵素を指す。この遺伝子は、メチロモナス(Methylomonas)においてcrt遺伝子クラスター上に位置する第2の遺伝子である。
【0080】
「CrtN3」という用語は、crtN3遺伝子によってコードされ、メチロモナス(Methylomonas)sp.16aの天然カロテノイド生合成に影響する酵素を指す。この遺伝子はcrt遺伝子クラスター内に位置せず、代わりにこの遺伝子はメチロモナス(Methylomonas)ゲノムの異なる遺伝子座内に存在する(国際公開第02/18617号パンフレット)。
【0081】
「色素なし」または「白色変異体」または「非着色株」という用語は、天然ピンク色素(例えばC30カロテノイド)を生成しないメチロモナス(Methylomonas)sp.16a細菌を指す。したがって細菌細胞の色はピンクとは対照的に、白色に見える。メチロモナス(Methylomonas)sp.16a白色変異体は、天然C30カロテノイド遺伝子の全てまたは一部を消去して遺伝子操作された。例えばメチロモナス(Methylomonas)sp.16a中のald/crtN1遺伝子または天然crt遺伝子クラスターを駆動するプロモーターのどちらかの中断は、C40カロテノイド生成によりふさわしい非着色(「白色」)変異体を作り出す(国際公開第02/18617号パンフレット)。
【0082】
「メチロモナス(Methylomonas)sp.16aMWM1000」または「MWM1100」という用語は、メチロモナス(Methylomonas)sp.16aに天然であるaldおよびcrtN1遺伝子の一部を除去して作り出された、非着色メタノトローフ細菌株を指す(国際公開第02/18617号パンフレット)。欠失は、MWM1000におけるC30カロテノイドの生成を中断させた。ald/crtN1欠失は「Δald/crtN1」と表される。
【0083】
ここでの用法では、「単離された核酸断片」は、場合により合成、非天然または修飾ヌクレオチド塩基を含有する一本鎖または二本鎖であるRNAまたはDNAのポリマーである。DNAポリマーの形態の単離された核酸断片は、1つまたはそれ以上のcDNA、ゲノムDNAまたは合成DNAの断片を含んでなっても良い。
【0084】
核酸分子は、適切な温度および溶液イオン強度条件下で、核酸分子の一本鎖形態がその他の核酸分子とアニールできる場合、cDNA、ゲノムDNA、またはRNAなどの別の核酸分子と「ハイブリダイズ可能」である。ハイブリダイズおよび洗浄条件については良く知られており、サムブルック(Sambrook)、J.、フリッチュ(Fritsch)、E.F.およびマニアティス(Maniatis)、T.「分子クローン化:実験室マニュアル(Molecular Cloning:A Laboratory Manual)」、第二版、コールド・スプリング・ハーバー・ラボラトリー(Cold Spring Harbor Laboratory)、ニューヨーク州コールド・スプリング・ハーバー(Cold Spring Harbor、NY)(1989)の特に第11章およびその表11.1で例証される。(以下「マニアティス(Maniatis)」)。温度およびイオン強度条件が、ハイブリダイズの「ストリンジェンシー」を定める。ストリンジェンシー条件は、(遠縁の生物からの相同的配列などの)中程度に類似の断
片をスクリーニングするため、そして(機能性酵素を複製する近縁の生物からの遺伝子などの)高度に類似した断片をスクリーニングするために調節できる。ハイブリダイズ後の洗浄が、ストリンジェンシー条件を決定する。1つの好ましい条件の組は、室温において6×SSC、0.5%SDSで15分間に始まり、次に45℃において2×SSC、0.5%SDSで30分間を反復し、次に50℃において0.2×SSC、0.5%SDSを30分間を2回反復する、一連の洗浄を使用する。より好ましいストリンジェンシー条件の組はより高い温度を使用し、そこでは洗浄は、最後の0.2×SSC、0.5%SDS中での2回の30分間の洗浄の温度を60℃に増大させること以外は上述したのと同一である。別の好ましい高度にストリンジェンシー条件の組は、65℃において0.1×SSC、0.1%SDS中での2回の最終洗浄を使用する。さらに別のストリンジェントな条件の組は、例えば0.1×SSC、0.1%SDSで65℃のハイブリダイズ、および2×SSC、0.1%SDSで洗浄後、0.1×SSC、0.1%SDSを含む。
【0085】
ハイブリダイズのストリンジェンシー次第で塩基間のミスマッチは可能であるが、ハイブリダイズは、2つの核酸が相補的配列を含有することを要求する。核酸がハイブリダイズする適切なストリンジェンシーは、技術分野で良く知られた変数である核酸の長さおよび相補性の程度に左右される。2つのヌクレオチド配列間の類似性または相同性の程度が高い程、これらの配列を有する核酸ハイブリッドのTm値は大きくなる。核酸ハイブリダイズの相対安定性(より高いTmに対応する)は、次の順で低下する。RNA:RNA、DNA:RNA、DNA:DNA。長さが100ヌクレオチドを超えるハイブリッドでは、Tmを計算する式が導かれている(サムブルック(Sambrook)ら、前述9.50〜9.51参照)。より短かい核酸、すなわちオリゴヌクレオチドによるハイブリダイズのためにはミスマッチの配置がより重要になり、オリゴヌクレオチドの長さがその特異性を決定する(サムブルック(Sambrook))ら、前述11.7〜11.8参照)。一実施態様では、ハイブリダイズ可能な核酸の長さは少なくともヌクレオチド約10個である。ハイブリダイズ可能な核酸の好ましい最小の長さは少なくともヌクレオチド約15個、より好ましくは少なくともヌクレオチド約20個、そして最も好ましくは長さが少なくともヌクレオチド約30個である。さらに当業者は、温度および洗浄液の塩濃度が、プローブの長さなどの要因次第で必要に応じて調節できることを認識する。
【0086】
アミノ酸またはヌクレオチド配列の「かなりの部分」とは、当業者による配列の手動評価によって、あるいはBLAST(「基礎的局在性整列化検索ツール(Basic Local Alignment Search Tool)」アルトシュル(Altschul)S.F.ら、J.Mol.Biol.215:403〜410(1993))などのアルゴリズムを使用したコンピュータ支援配列比較および同定によって、遺伝子またはポリペプチドの推定上の同定を得るのに十分なポリペプチドまたは遺伝子のヌクレオチド配列のアミノ酸配列を含んでなる部分である。推定的にポリペプチドまたは核酸配列が既知のタンパク質または遺伝子に相同的であると同定するためには、概して10個以上の隣接するアミノ酸または30個以上のヌクレオチド配列が必要である。さらにヌクレオチド配列に関して、20〜30個の隣接するヌクレオチドを含んでなる遺伝子特異的オリゴヌクレオチドプローブを配列依存遺伝子同定法(例えばサザンハイブリダイズ)および単離(例えば細菌コロニーまたはバクテリオファージ・プラークの原位置(in situ)ハイブリダイズ)において使用しても良い。さらにプライマーを含んでなる特定の核酸断片を得るために、塩基12〜15個の短いオリゴヌクレオチドをPCRで増幅プライマーとして使用しても良い。したがってヌクレオチド配列の「かなりの部分」は、配列を含んでなる核酸断片を特異的に同定および/または単離できるようにする十分な配列を含んでなる。本願明細書は、1つまたはそれ以上の特定の微生物タンパク質をコードする部分的または完全アミノ酸およびヌクレオチド配列を教示する。当業者はここで報告される配列の恩恵を被り、当業者に知られている目的のために、今や開示された配列の全てまたはかなりの部分を使用できる。したがって本発明は、添付の配列表で報告される完全な配列、
ならびに上で定義される配列のかなりの部分を含んでなる。
【0087】
「相補的」と言う用語は、互いにハイブリダイズできるヌクレオチド塩基間の関係について述べるために使用される。例えばDNAについて、アデノシンはチミンに相補的であり、シトシンはグアニンに相補的である。したがって本発明は添付の配列表で報告されるような完全な配列、ならびに実質的に類似した核酸配列に相補的である単離された核酸断片も含む。
【0088】
技術分野で既知の「パーセント同一性」と言う用語は、配列を比較して判定される2つ以上のポリペプチド配列または2つ以上のポリヌクレオチド配列の関係である。技術分野において「同一性」は、場合によってはこのような配列ストリング間の整合によって判定されるような、ポリペプチドまたはポリヌクレオチド配列間の配列関連性の程度も意味する。「同一性」および「類似性」は、1)「計算分子生物学(Computational Molecular Biology)(レスク(Lesk)A.M.編)、オックスフォードユニバーシティ(Oxford University)、ニューヨーク(NY)(1988)、2)「バイオコンピューティング:情報科学およびゲノムプロジェクト(Biocomputing:Informatics and Genome Projects)」(スミス(Smith)D.W.編)、アカデミック(Academic)、ニューヨーク(1993)、3)「配列データのコンピュータ分析(Computer Analysis of Sequence Data)」、第一部、(グリフィン(Griffin)A.M.、およびグリフィン(Griffin)H.G.編)、ヒュマーナ(Humana)、ニュージャージー(NJ)(1994)、4)「分子生物学における配列分析(Sequence Analysis in Molecular Biology)」(フォン・ハインェ(von Heinje)G.編)、アカデミック(1987)、5)「配列分析入門(Sequence Analysis Primer)」(グリブスコフ(Gribskov)M.およびデュヴルー(Devereux)J.編)、ストックトン(Stockton)、ニューヨーク(1991)で述べられたものをはじめとするがこれに限定されるものではない既知の方法によって容易に計算できる。同一性を判定する好ましい方法は、試験される配列間に最良の整合を与えるように設計される。同一性および類似性を判定する方法は、一般に入手できるコンピュータプログラムで体系化される。配列整合および同一性百分率の計算は、ウィスコンシンマディソンのDNASTAR(DNASTAR Inc.(Madison、WI))からのLASERGENEバイオインフォマティクス計算スイートのMegalignプログラムを使用して実施しても良い。Clustal整合法(ヒギンズ(Higgins)およびシャープ(Sharp)、CABIOS.5:151〜153(1989))を使用して、デフォルトのパラメータ(GAP PENALTY=10、GAP LENGTH PENALTY=10)で、配列の複数の整合を実施する。Clustal法を使用した対整合のデフォルトのパラメータは、KTUPLE 1、GAP PENALTY=3、WINDOW=5、およびDIAGONALS SAVED=5である。
【0089】
適切な核酸断片(本発明の単離ポリヌクレオチド)は、ここで報告するアミノ酸配列と少なくとも約70%同一の、好ましくは少なくとも約75%同一の、そしてより好ましくは少なくとも約80%同一のポリペプチドをコードする。好ましい核酸断片は、ここで報告するアミノ酸配列と約85%同一のアミノ酸配列をコードする。より好ましい核酸断片は、ここで報告するアミノ酸配列と少なくとも約90%同一のアミノ酸配列をコードする。最も好ましい核酸断片は、ここで報告するアミノ酸配列と少なくとも約95%同一のアミノ酸配列をコードする。適切な核酸断片は上の相同性を有するだけでなく、典型的に少なくとも50個のアミノ酸、好ましくは少なくとも100個のアミノ酸、より好ましくは少なくとも150個のアミノ酸、さらにより好ましくは少なくとも200個のアミノ酸、そして最も好ましくは少なくとも250個のアミノ酸を有するポリペプチドをコードする
。
【0090】
「コドン縮重」とは、コードされるポリペプチドのアミノ酸配列に影響することなく、ヌクレオチド配列の変動を可能にする遺伝コードにおける性質を指す。したがって本発明は、配列番号2、4、6、8、10、および12に記載される、本微生物のポリペプチドをコードするアミノ酸配列の全てまたはかなりの部分をコードするあらゆる核酸断片に関する。当業者は、任意のアミノ酸を特定化するためのヌクレオチドコドンの利用において、特定の宿主細胞によって示される「コドンバイアス」を十分承知している。したがって宿主細胞中における改善された発現のために遺伝子を合成する場合、コドン使用頻度が、宿主細胞の好ましいコドン使用頻度に近くなるようように、遺伝子をデザインすることが望ましい。
【0091】
「合成遺伝子」は、当業者に知られた手順を使用して化学的に合成されるオリゴヌクレオチド構成単位から構築できる。これらの構成単位をライゲートしアニールして遺伝子セグメントを形成し、次にそれを酵素的にアセンブルして遺伝子全体を構成しても良い。DNA配列に関連して「化学的に合成された」とは、構成要素ヌクレオチドが生体内で(in vitro)アセンブルされたことを意味する。DNAの手動化学合成は確立した手順を使用して達成されても良く、あるいはいくつかの市販の機器の1つを使用して自動化学合成を実施できる。したがって遺伝子をヌクレオチド配列の最適化に基づいて、最適な遺伝子発現のために調整し、宿主細胞のコドンバイアスを反映させることができる。当業者は、コドン利用が宿主によって好まれるコドンに偏っている場合の遺伝子発現成功の見込みの真価を認める。好ましいコドンの判定は、配列情報が利用できる宿主細胞から誘導された遺伝子の調査に基づくことができる。
【0092】
「遺伝子」とは、コード配列に先行する制御配列(5’非コード配列)およびその後ろの制御配列(3’非コード配列)を含めた特異性タンパクを発現する核酸断片を指す。「天然遺伝子」とは、それ自体の制御配列を有して自然界に見いだされる遺伝子を指す。「キメラ遺伝子」とは、自然界には一緒に見られない制御およびコード配列を含んでなる天然遺伝子でないあらゆる遺伝子を指す。したがってキメラ遺伝子は、異なる供給源から誘導される制御配列およびコード配列、あるいは同一供給源から誘導されるが、自然界に見られるのとは異なるやり方で配列された制御配列およびコード配列を含んでなっても良い。「内在性遺伝子」とは、生物のゲノム中の自然な部位にある天然遺伝子を指す。「外来性」遺伝子とは、常態では宿主生物に見られないが、遺伝子移入によって宿主生物中に導入される遺伝子を指す。外来遺伝子は、非天然生物に挿入された天然遺伝子、またはキメラ遺伝子を含んでなることができる。「導入遺伝子」とは、形質転換手順によってゲノム中に導入された遺伝子である。
【0093】
「コード配列」とは、特定のアミノ酸配列をコードするDNA配列を指す。「適切な調節配列」とは、コード配列の上流(5’非コード配列)、配列内、または下流(3’非コード配列)に位置して、転写、RNAプロセシングまたは安定性、または関連コード配列の翻訳に影響を及ぼすヌクレオチド配列を指す。調節配列は、プロモーター、翻訳リーダー配列、イントロン、ポリアデニル化認識配列、RNAプロセッシング部位、エフェクター結合部位、およびステム−ループ構造を含んでも良い。
【0094】
「プロモーター」とは、コード配列または機能性RNAの発現を調節できるDNA配列を指す。概してコード配列は、プロモーター配列に対して3’に位置する。プロモーターは、そっくりそのまま天然遺伝子から誘導されても良く、あるいは自然界に見られる異なるプロモーターから誘導される異なる要素からなっても良く、あるいは合成DNAセグメントを含んでなってさえ良い。異なるプロモーターは、異なる組織または細胞タイプ中で、あるいは異なる発達段階において、あるいは異なる環境または生理学的条件に呼応して
、遺伝子の発現を導いても良いことが当業者には理解される。ほとんどの細胞タイプ中でほとんどの場合に遺伝子の発現を引き起こすプロモーターは、一般に「構造プロモーター」と称される。ほとんどの場合、制御配列のはっきりした境界は完全に画定されていないので、異なる長さのDNA断片が同一のプロモーター活性を有しても良いこともさらに認識される。
【0095】
「3’非コード配列」は、コード配列の下流に位置するDNA配列を指し、mRNAプロセッシングまたは遺伝子発現に影響できる調節シグナルをコードするポリアデニル化認識配列(常態では真核細胞に限られる)およびその他の配列を含む。ポリアデニル化シグナルは、通常、mRNA前駆物質の3’末端へのポリアデニル酸トラクトの付加に影響することで特徴付られる。
【0096】
「RNA転写物」とは、DNA配列のRNAポリメラーゼ触媒転写から得られる生成物を指す。RNA転写物がDNA配列の完全な相補的コピーである場合、それは一次転写物と称され、あるいはそれは一次転写物の転写後プロセッシングから誘導されるRNA配列であるかもしれず、成熟RNAと称される。「メッセンジャーRNA」または「mRNA」とはイントロンがなく、細胞によってタンパク質に翻訳されることができるRNAを指す。「cDNA」とは、mRNAに対して相補的であり、それから誘導される二重鎖DNAを指す。「センスRNA」とは、mRNAを含み、細胞によってタンパク質に翻訳されることができるRNA転写物を指す。「アンチセンスRNA」とは、標的一次転写物またはmRNAの全部または一部に相補的であり、標的遺伝子の発現をブロックするRNA転写物を指す(米国特許第5,107,065号明細書、国際公開第99/28508号パンフレット)。アンチセンスRNAの相補性は、特定の遺伝子転写物のあらゆる部分、すなわち5’非コード配列、3’非コード配列、またはコード配列にあっても良い。「機能RNA」とは、翻訳されないがそれでもなお細胞過程に影響するアンチセンスRNA、リボザイムRNA、またはその他のRNAを指す。
【0097】
「作動的に結合した」と言う用語は、1つの機能が他方の機能によって影響される、単一核酸断片上の核酸配列のつながりを指す。例えばプロモーターはコード配列の発現に影響できる場合、そのコード配列と作動的に結合する(すなわちコード配列がプロモーターの転写調節の下にある)。コード配列は、センスまたはアンチセンスオリエンテーションで制御配列に作動的に結合できる。
【0098】
「発現」と言う用語は、ここでの用法では、本発明の核酸断片から誘導されるセンス(mRNA)またはアンチセンスRNAの転写および安定した蓄積を指す。発現はまた、mRNAのポリペプチドへの翻訳を指しても良い。
【0099】
「成熟」タンパク質とは、翻訳後処理されたポリペプチド、すなわちそれから一次翻訳生成物中に存在するあらゆるプレ−またはプロペプチドが除去されたものを指す。「前駆」タンパク質とは、mRNAの翻訳の一次生成物、すなわちプレ−またはプロペプチドがなおも存在するものを指す。プレ−またはプロペプチドは、細胞内局在化シグナルであっても良い(がそれに限定されない)。
【0100】
「シグナルペプチド」という用語は、分泌された成熟タンパク質に先行するアミノ末端ポリペプチドを指す。シグナルペプチドは成熟タンパク質から切断されるので、その中には存在しない。シグナルペプチドは、分泌されたタンパク質を細胞膜を越えて方向付け、転位置する機能を有する。シグナルペプチドはまた、シグナルタンパク質とも称される。
【0101】
「接合」とは、1つの細菌細胞(すなわち「供与体」)から別の(すなわち「受容体」)細菌細胞への一方向性のDNA転位(例えば細菌プラスミドからの)が起きる、特定の
タイプの形質転換を意味する。プロセスは直接的な細胞と細胞の接触を伴う。別の細菌細胞(すなわち「ヘルパー」)が存在して接合を容易にすることもある。
【0102】
「形質転換」とは、遺伝的に安定した遺伝形質をもたらす、宿主生物体中のゲノムへの核酸断片の転移を指す。形質転換核酸断片を含有する宿主生物体は、「遺伝子導入」、「組換え」または「形質転換」生物体と称される。
【0103】
「プラスミド」、「ベクター」、および「カセット」と言う用語は、細胞の中心的代謝の一部ではない遺伝子を運ぶことが多く、通常環状二本鎖DNA断片の形態である染色体外因子を指す。このような因子は、あらゆる供給源から誘導される一本鎖または二本鎖DNAまたはRNAの配列、ゲノム一体化配列、直鎖または環状のファージまたはヌクレオチド配列を自律的に複製するかもしれず、そこではいくつかのヌクレオチド配列が独自の構成に連結または組み換えされ、それは選択された遺伝子産物のために、適切な3’非翻訳配列と共にプロモーター断片およびDNA配列を細胞中に導入することができる。「形質転換カセット」とは、外来性遺伝子を含有し、外来遺伝子に加えて特定の宿主細胞の形質転換を容易にする因子を有する特定のベクターを指す。「発現カセット」とは、外来性遺伝子を含有し、外来性遺伝子に加えて外来性宿主におけるその遺伝子の促進された発現を可能にする因子を有する特定のベクターを指す。
【0104】
「改変された生物学的活性」と言う用語は、アッセイ方法によって測定できる、ヌクレオチド配列によってコードされるタンパク質に関連した活性を指し、活性は、天然配列に関連した活性を超え、またはそれ未満のいずれかである。「増強された生物学的活性」とは、天然配列に関連したものを超える改変された活性を指す。「低下した生物学的活性」とは、天然配列に関連したもの未満の改変された活性を指す。
【0105】
「配列分析ソフトウェア」と言う用語は、ヌクレオチドまたはアミノ酸配列の分析のために有用なあらゆるコンピュータアルゴリズムまたはソフトウェアプログラムを指す。「配列分析ソフトウェア」は、市販のものでも、あるいは独立して開発されても良い。典型的な配列分析ソフトウェアとしては、1.)ウィスコンシン州マディソンのジェネティック・コンピュータ・グループからのGCGパッケージプログラム(Wisconsin Package Version 9.0、Genetics Computer Group(GCG),Madison,WI)、2.)BLASTP、BLASTN、BLASTX(アルシュール(Altschul)ら著、J.Mol.Biol.215:403〜410(1990))、および3.)ウィスコンシン州マディソンのDNASTAR(DNASTAR,Inc.Madison、WI)からのDNASTAR、および4.)スミス−ウォーターマン・アルゴリズムを組み入れたFASTAプログラム(W.R.ピアースン(Pearson)著、Comput.Methods Genome Res.[Proc.Int.Symp.](1994)、1992年会議、111〜20、編集者:スハイ,サンドル(Suhai,Sandor)、Plenum、ニューヨーク州ニューヨーク(New York、NY))、および5.)メリーランド州ベセズダのインフォマックス(InforMax,Inc.(Bethesda、MD))からのベクターNTIプログラムが挙げられるが、これに限定されるものではない。本願明細書の文脈内では、配列分析ソフトウェアを分析のために使用する場合、分析結果は特に断りのない限り、言及されるプログラムの「デフォルト値」に基づくものと理解される。ここでの用法では、「デフォルト値」とは、最初に初期化されたときにソフトウェアに元々ロードされた、あらゆる値またはパラメータの組(製造業者によって設定される)を意味する。
【0106】
ここで使用される標準組換えDNAおよび分子クローン化技術は技術分野で良く知られており、マニアティス(Maniatis)(上述);シルハビー(Silhavy)T
.J.、ベンナン(Bennan)M.L.およびエンクイスト(Enquist)L.W.「遺伝子融合実験(Experiments with Gene Fusions)」コールド・スプリング・ハーバー・ラボラトリー:ニューヨーク州コールド・スプリング・ハーバー(1984);およびオースベル(Ausubel)、F.M.ら「分子生物学現代プロトコル(Current Protocols in Molecular Biology)」、グリーン・パブリッシング・アソシエーツ(Greene Publishing Assoc.)およびワイリー−インターサイエンス(Wiley−Interscience)による出版(1987)で述べられている。
【0107】
カロテノイド生成に関与する遺伝子
カロテノイド化合物の生合成に関与する酵素経路は、ピルビン酸およびグリセルアルデヒド−3−リン酸からファルネシルピロリン酸への転換を提供する上流イソプレノイド経路、およびジアポフィトエンまたはフィトエンいずれかの合成と、全ての引き続いて生成されるカロテノイドを提供する下流カロテノイド生合成経路の2つの部分に都合良く分けることができる(図1)。上流経路は多くの微生物に遍在性であり、これらの場合、所望のカロテノイド生合成のための下流経路を含んでなる遺伝子を導入することだけが必要かもしれない。2つの経路間の区分は、ファルネシルピロリン酸塩(FPP)の合成に関わる。FPPが天然に存在する場合、下流カロテノイド生合成経路の要素のみが必要である。しかしカロテノイド生成において下流経路カロテノイド遺伝子が効果的であるためには、宿主細胞が細胞内に適切なレベルのFPPを有することが必要なことが理解される。FPP合成が宿主細胞によって提供されない場合、FPPの生成に必要な遺伝子を導入することが必要である。これらの各経路について下で詳細に述べる。
【0108】
上流イソプレノイド経路
イソプレノイド生合成は、一般的なC5イソプレンサブユニットであるイソペンテニルピロリン酸塩(IPP)を生じる2つの経路のどちらかを通じて起きる。最初に良く知られている酢酸/メバロン酸経路を通じてIPPを合成しても良い。しかし最近の研究は、メバロン酸依存経路が全ての生物体で作動しているわけではないことを実証している。IPP生合成のための代案のメバロン酸非依存経路が、細菌および緑藻類および高等植物で特性決定されている(ホルバック(Horbach)ら、FEMS Microbiol.Lett.111:135〜140(1993);ローマー(Rohmer)ら、Biochem.295:517〜524(1993);シュベンダー(Schwender)ら、Biochem.316:73〜80(1996);アイゼンライヒ(Eisenreich)ら、Proc.Natl.Acad.Sci.USA 93:6431〜6436(1996))。
【0109】
メバロン酸非依存イソプレノイド経路中の多くのステップが知られている(図1)。例えばIPPの生成をもたらす代案の経路の最初のステップは、コール(Cole)ら(Nature 393:537〜544(1998))によって結核菌(Mycobacterium tuberculosis)において研究されている。経路の第1のステップは、D−1−デオキシキシルロース−5−リン酸として知られている五炭素化合物を生じる、2つの三炭素分子(ピルビン酸およびD−グリセルアルデヒド3−リン酸)の縮合を伴う。この反応はdxs遺伝子によってコードされるDxs酵素によって起きる。次にD−1−デオキシキシルロース−5−リン酸の異性化および還元から、2−C−メチル−D−エリスリトール−4−リン酸が生じる。異性化および還元プロセスに関与する酵素の1つは、遺伝子dxrによってコードされるD−1−デオキシキシルロース−5−リン酸レダクトイソメラーゼ(Dxr)である。2−C−メチル−D−エリスリトール−4−リン酸を引き続いて、注釈なしの遺伝子ygbP(コール(Cole)ら、前出)でコードされる酵素によって、CTP−依存反応において4−ジホスホシチジル−2C−メチル−D−エリスリトールに転換する。しかし最近ygbP遺伝子は、isp遺伝子クラスター
の一部としてispDと改名された(SwissProtein登録番号Q46893)。
【0110】
次にychB遺伝子によってコードされる酵素によって、4−ジホスホシチジル−2C−メチル−D−エリスリトールの第2位のヒドロキシ基をATP−依存反応においてリン酸化できる。この生成物は4−ジホスホシチジル−2C−メチル−D−エリスリトールをリン酸化して、4−ジホスホシチジル−2C−メチル−D−エリスリトール2−リン酸が得られる。ychB遺伝子もまたisp遺伝子クラスターの一部としてispEと改名された(SwissProtein登録番号P24209)。最後にygbB遺伝子の生成物が、CTP−依存様式で、4−ジホスホシチジル−2C−メチル−D−エリスリトール2−リン酸を2C−メチル−D−エリスリトール2,4−シクロ二リン酸に転換する。この遺伝子もまた最近改名されて、isp遺伝子クラスターに属する。具体的には、ygbB遺伝子の新しい名称はispF(SwissProtein登録番号P36663)である。pyrG遺伝子の生成物は、CTPシンターゼとしてこれらの反応において重要である。
【0111】
lytBおよびgcpE(およびおそらくはその他の)遺伝子によってコードされる酵素はイソペンテニルピロリン酸(IPP)およびジメチルアリルピロリン酸(DMAPP)の形成をもたらす反応に関与すると考えられる。IPPは、idi遺伝子によってコードされるイソペンテニル二リン酸イソメラーゼ(または「IPPイソメラーゼ」)を通じてDMAPPに異性化しても良いが、この酵素は生存には必須ではなく、2−C−メチル−D−エリスリトール4−リン酸(MEP)経路を使用するいくつかの細菌では不在でも良い。最近の証拠はMEP経路がIPPの前に分岐して、lytB遺伝子産物を通じてIPPとDMAPPを別個に生成することを示唆する。lytBノックアウト変異は、IPPおよびDMAPPの双方を補った培養液中以外では、大腸菌(E.coli)に対して致死性である。
【0112】
FPPの合成は、IPPのジメチルアリルピロリン酸(DMAPP)への異性化を通じて起きる。この反応に、ispAによって触媒される2つのプレニル転移酵素反応手順が続き、ゲラニルピロリン酸(GPP、炭素数10の分子)およびファルネシルピロリン酸(FPP、炭素数15の分子)の生成をそれぞれもたらす。
【0113】
下流カロテノイド生合成経路
上流イソプレノイド経路と下流カロテノイド経路の間の区分は、幾分主観的である。FPP合成はカロテン産生性および非カロテン産生性細菌の双方で一般的であるので、出願人は、C30ジアポカロテノイドまたはC40カロテノイドのどちらかの形成をもたらす分岐する2経路の化合物へのファルネシルピロリン酸(FPP)の転換を開始するための下流カロテノイド生合成経路中の第1のステップを考察する。
【0114】
C40経路中では、生合成経路の第1のステップは、IPPの添加によって、ファルネシルピロリン酸(FPP)をゲラニルゲラニルピロリン酸(GGPP)として知られている炭素数20の分子に転換するプレニル転移酵素反応で始まる。GGPP合成酵素をコードする遺伝子crtE(EC2.5.1.29)は、このプレニル転移酵素反応を司る。次に2個のGGPP分子の縮合反応が起きて、下流カロテノイド生合成経路の第1の炭素数40の分子であるフィトエン((7,8,11,12,7’,8’,11’,12’−ω−オクタヒドロ−ω,ω−カロテン、またはPPPP)を形成する。この酵素的反応はCrtB(フィトエンシンターゼ、EC2.5.1.−)によって触媒される。
【0115】
引き続く水素付加、脱水素、環化、酸化、またはこれらのプロセスのあらゆる組み合わせによって、化合物フィトエンからC40カロテノイドのスペクトルを生成した。例えば
「赤色」着色スペクトルを与えるリコピンは、遺伝子crtI(フィトエンデサチュラーゼをコードする)によって触媒される、8個の水素原子の除去による4つの逐次脱水素化反応を通じて、フィトエンから生成される(図2参照)。リコピンシクラーゼ(CrtY)は、リコピンをβ−カロテン(β,β−カロテン)に転換する。β−カロテンは、β−カロテン水酸化酵素(crtZ遺伝子によってコードされる)活性に起因する水酸化反応を通じて、ゼアキサンチン((3R,3’R)−β,β−カロテン−3,3’−ジオール)に転換される。
【0116】
フィトエン、リコペン、β−カロテン、およびゼアキサンチンを作り出す組み合わせで使用できるcrtE、crtY、crtI、crtB、およびcrtZに加えて、直鎖C40化合物の分子内転換を可能にし、多数のその他の官能性付与カロテノイド化合物を生成する様々なその他のcrt遺伝子が知られている。当業者は、公的に入手できる文献(例えばジェンバンク(登録商標))、特許文献、およびカロテノイドを生成する能力を有する微生物の実験的分析に従って、様々なその他のcrt遺伝子同定できるであろう。
例えば、
・ゼアキサンチンは、crtX遺伝子(EC2.4.1.−、例えばジェンバンク(登録商標)登録番号D90087、M87280、M90698)によってコードされるゼアキサンチングルコシル転移酵素によってゼアンキサンチン−β−グルコシドに転換できる。
・β−カロテンは、crtW(例えばジェンバンク(登録商標)登録番号AF218415、D45881、D58420、D58422、X86782、Y15112)、crtO(例えばジェンバンク(登録商標)登録番号X86782、Y15112)またはbktによってコードされるβ−カロテンケトラーゼによって、カンタキサンチンに転換できる。エキネノンはこの反応における中間体である。
・カンタキサンチンは、crtZ遺伝子によってコードされるβ−カロテン水酸化酵素によって、アスタキサンチンに転換できる。アドニルビンはこの反応における中間体である。
・ゼアキサンチンは、crtW、crtO、またはbktによってコードされるβ−カロテンケトラーゼによって、アスタキサンチンに転換できる。アドニキサンチンはこの反応における中間体である。
・スフェロイデンは、crtA(例えばジェンバンク(登録商標)登録番号AJ010302、Z11165、X52291)によってコードされるスフェロイデンモノオキシゲナーゼによって、スフェロイデノンに転換できる。
crtC(例えばジェンバンク(登録商標)登録番号AB034704、AF195122、AJ010302、AF287480、U73944、X52291、Z11165、Z21955)、crtD(例えばジェンバンク(登録商標)登録番号AJ010302、X63204、U73944、X52291、Z11165)、およびcrtF(例えばジェンバンク(登録商標)登録番号AB034704、AF288602、AJ010302、X52291、Z11165)遺伝子によってそれぞれコードされる、ヒドロキシニューロスポレンシンターゼ、メトキシニューロスポレンデサチュラーゼ、およびヒドロキシニューロスポレン−O−メチル転移酵素の逐次作用によって、ニューロスポレンはスフェロイデンに転換でき、リコペンはスピリロキサンチンに転換できる。
・β−カロテンは、crtU(例えばジェンバンク(登録商標)登録番号AF047490、AF121947、AF139916、AF195507、AF272737、AF372617、AJ133724、AJ224683、D26095、U38550、X89897、Y15115)によってコードされる、β−カロテンデサチュラーゼによってイソレニエラテンに転換できる。
【0117】
これらの例は制限を意図せず、このC40下流カロテノイド生合成経路内には、その他の多くのカロテノイド遺伝子および生成物が存在する。したがって宿主生物体中で十分な
FPP源が利用できれば、本発明の方法を使用して、場合により植物、動物、および/または細菌源から単離されたあらゆるその他の知られているcrt遺伝子に加えて、ここに示すcrtE、crtY、crtI、crtB、およびcrtZ遺伝子の様々な組み合わせを使用することで、無数の異なるカロテノイドおよびカロテノイド誘導体が作成できる。
【0118】
本発明の有用な生成物は、アンテラキサンチン、アドニルビン、アドニキサンチン、アスタキサンチン、カンタキサンチン、カプソルブリン、β−クリプトキサンチン、α−カロテン、β−カロテン、ε−カロテン、エキネノン、3−ヒドロキシエキネノン、3’−ヒドロキシエキネノン、α−カロテン、4−ケト−γ−カロテン、ζ−カロテン、α−クリプトキサンチン、デオキシフレキシキサンチン、ジアトキサンチン、7,8−ジデヒドロアスタキサンチン、フコキサンチン、フコキサンチノール、イソレニエラテン、ラクツカキサンチン、ルテイン、リコピン、ミキソバクトン、ネオキサンチン、ニューロスポレン、ヒドロキシニューロスポレン、ペリジニン、フィトエン、ロドピン、ロドピングルコシド、4−ケト−ルビキサンチン、シホナキサンチン、スフェロイデン、スフェロイデノン、スピリロキサンチン、4−ケト−トルレン、3−ヒドロキシ−4−ケト−トルレン、ウリオリド、ウリオリドアセタート、ビオラキサンチン、ゼアキサンチン−β−ジグルコシド、およびゼアキサンチンをはじめとするが、これに限定されるものではない、ここで定義されるあらゆるカロテノイド化合物を含むことが構想された。さらに本発明は、ヒドロキシ−、メトキシ−、オキソ−、エポキシ−、カルボキシ−、またはアルデヒド酸官能基、グリコシドエステル、またはスルフェートを作り出す、これらの分子の誘導化を包含する。
【0119】
上流イソプレノイド経路と下流カロテノイド生合成経路との相互作用
全体的なイソプレノイド生合成を促進させることによって、カロテノイド生成を促進する多様な研究が試みられている。特にidiの上方制御は、カロテノイド生成に劇的に影響することが実証されている。例えばカジワラ(Kajiwara)ら(Biochem.J.324:421〜426(1997))は、「IPPイソメラーゼは、原核生物大腸菌(E.coli)のイソプレノイド生合成において影響力が大きいステップを形成し、代謝遺伝子操作による工業的に有用なイソプレノイドの効率的生成の可能性がある」ことを最初に実証した。具体的には、外来性に発現されるIPPイソメラーゼが、エルウィニア(Erwinia)カロテノイド生合成遺伝子クラスターを含んでなる大腸菌(E.coli)において、対照と比べて3.6〜4.5倍高いレベルのリコピン生成を可能にし、同様に1.5〜2.7倍高いレベルのβ−カロテンおよび1.7〜2.1倍高いレベルのフィトエンが生成された。
【0120】
ワング(Wang)らの引き続く研究(Biotech.Bioengineering 62(2):235〜241(1999))は、大腸菌(E.coli)idi遺伝子、アーキオグロバス・フルギダス(Archaeoglobus fulgidus)gps遺伝子、およびアグロバクテリウム・オーランチアカム(Agrobacterium aurantiacum)crtBIYZW遺伝子クラスターで形質転換した大腸菌(E.coli)において、50倍高いアスタキサンチン生成をもたらした。GGPP合成における最後のステップが、カロテノイド生成における第1の速度制御ステップであり、第2の速度制御ステップはIPP異性化であると結論された。そして、最後にアルブレヒト(Albrecht)ら(Biotech.Letters 21:791〜795(1999))は、大腸菌(E.coli)における内在性dxsおよびdxr遺伝子および外来性idi遺伝子(ファフィア・ロドジマ(Phaffia rhodozyma)からの)の過剰発現が、カロテン産生を3.5倍まで刺激できることを見いだした。
【0121】
したがって大腸菌(E.coli)およびその他の細菌における、工業的に価値あるカ
ロテノイド生成の最大化に向けた代謝操作法は、上流イソプレノイド経路中の流束および律速ステップ、ならびに下流カロテノイド生合成経路における発現レベルを注意深く考慮すべきである。上流イソプレノイド経路中の律速遺伝子(例えばidi)の過剰発現は、カロテン産生を劇的に増大できる。
【0122】
P.アグロメランス(P.agglomerans)DC404株のカロテノイド生合成遺伝子および酵素の配列同定
C40カロテノイド生合成経路に関与する遺伝子産物をコードする多様なヌクレオチド配列が、DC404株から単離されている。例えばORFの1および3〜6は、下流カロテノイド生合成経路中のcrtE、Y、I、B、およびZ遺伝子をコードし(図1および2参照)、それらの酵素生成物は、着色カロテノイド、リコペン、β−カロテン、およびゼアキサンチンの生成をもたらす。ORF2は、上流イソプレノイド経路においてidi遺伝子をコードする。これらの6個のORFは、crtE−idi−crtY−crtI−crtB−crtZの遺伝子構成を有する1つの核酸断片(配列番号18)上に構成される。crtE−idi−crtY−crtI−crtB遺伝子は、オペロン内で作動的に結合されているように見え、他方crtZ遺伝子は、反対方向に転写される。
【0123】
DC404株から単離された遺伝子の組全体(crtE−idi−crtY−crtI−crtB−crtZ)は、ここで1つの配列(配列番号18)内に開示される。カロテノイド化合物の生成ために、この遺伝子クラスターをベクターに入れて、微生物宿主中で発現させる。当業者はコードされた全てのポリペプチドが発現して酵素的に活性ならば、その有用性に影響することなく、このような遺伝子クラスターに少数の核酸置換、付加、および欠失(特定の宿主細胞発現のための好ましいコドンの置換などの)があっても良いことを認識するであろう。したがってcrtE−idi−crtY−crtI−crtB−crtZを含んでなる配列番号18で示される単離された核酸分子、配列番号18と少なくとも95%の同一性を有する遺伝子または単離された核酸分子を提供することは本発明の範囲内であり、単離された核酸分子は、crtE、idi、crtY、crtI、crtB、およびcrtZの全てのポリペプチドをコードする。
【0124】
スミス−ウォーターマン位置合わせアルゴリズム(W.R.ピアソン(Pearson)著、Comput.Methods Genome Res.[Proc.Int.Symp.](1994)、1992年会議、111〜20、編集者:スハイ,サンドル(Suhai,Sandor)、Plenum、New York、NY)を使用して、crtEヌクレオチド塩基および推定されるアミノ酸配列(ORF1)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸301個の長さにわたり、ここで報告するCrtEのアミノ酸配列と約65%同一であることが明らかにされる。より好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtEコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtEの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0125】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、idiヌクレオチド塩基および推定されるアミノ酸配列(ORF2)と、公共データベースとの比較から、最も類似した既知の配列が、アミノ酸349個の長さにわたり、ここで報告するIdiのアミノ酸配列と約67%同一であることが明らかにされる。より好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいidiコード核酸配列は、活性タンパク質をコードするもので、ここで報告するidiの核酸配列と少なくとも約70%〜80
%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0126】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、crtYヌクレオチド塩基および推定されるアミノ酸配列(ORF3)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸389個の長さにわたり、ここで報告するCrtYのアミノ酸配列と約61%同一であることが明らかにされる。より好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtYコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtYの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0127】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、crtIヌクレオチド塩基および推定されるアミノ酸配列(ORF4)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸492の長さにわたり、ここで報告するCrtIのアミノ酸配列と約82%同一であることが明らかにされる。好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtIコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtIの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0128】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、crtBヌクレオチド塩基および推定されるアミノ酸配列(ORF5)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸308個の長さにわたり、ここで報告するCrtBのアミノ酸配列と約67%同一であることが明らかにされる。より好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtBコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtBの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0129】
スミス−ウォーターマンのアライメントアルゴリズム(W.R.ピアースン(Pearson)、前出)を使用して、crtZヌクレオチド塩基および推定されるアミノ酸配列(ORF6)と公共データベースとの比較から、最も類似した既知の配列が、アミノ酸185個の長さにわたり、ここで報告するCrtZのアミノ酸配列と約73%同一であることが明らかにされる。好ましいアミノ酸断片は、ここでの配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。同様に本ORFに対応する好ましいcrtZコード核酸配列は、活性タンパク質をコードするもので、ここで報告するcrtZの核酸配列と少なくとも約70%〜80%同一であり、85%〜90%同一の配列が特に適切であり、約95%同一の配列が最も好ましい。
【0130】
相同体の単離
本発明のC40カロテノイド生合成経路の各核酸断片を使用して、同一またはその他の微生物の(または植物)種から相同タンパク質をコードする遺伝子を単離しても良い。配
列依存プロトコルを使用した相同遺伝子の単離は、技術分野で良く知られている。配列依存プロトコルの例としては、以下が挙げられるが、これに限定されるものではない。1.)核酸ハイブリダイズ法、2.)様々な核酸増幅技術の使用によって例証されるようなDNAおよびRNA増幅法[例えばポリメラーゼ連鎖反応(PCR)、ミュリス(Mullis)ら、米国特許第4,683,202号明細書;リガーゼ連鎖反応(LCR)、タボール(Tabor)S.ら、Proc.Natl.Acad.Sci.USA82:1074(1985);または鎖置換増幅(SDA)、ウォーカー(Walker)ら、Proc.Natl.Acad.Sci.USA、89:392(1992)]、および3.)相補によるライブラリー構築およびスクリーニング法。
【0131】
例えば、ここで述べるようなC40カロテノイド生合成経路のものと類似したタンパク質またはポリペプチドをコードする遺伝子は、当業者に良く知られている方法を使用して、本核酸断片の全部または一部をDNAハイブリダイズプローブとして使用して、あらゆる所望の細菌からのライブラリーをスクリーニングして直接単離できる(C40カロテノイドを生成する細菌が好ましい)。本核酸配列に基づく特定のオリゴヌクレオチドプローブは、技術分野で知られている方法によってデザインして合成できる(マニアティス(Maniatis)、前出)。さらに当業者に知られている方法(例えばランダムプライマーDNA標識、ニックトランスレーション、または末端標識技術)によって、配列全体を直接使用してDNAプローブを合成し、または利用できる生体外(in vitro)転写システムを使用してRNAプローブを合成できる。さらに特定のプライマーをデザインして使用し、本配列の一部(または全長)を増幅できる。得られる増幅生成物を増幅反応中に直接標識し、または増幅反応後に標識して、プローブとして使用し、適切なストリンジェンシー条件下で完全長DNA断片を単離できる。
【0132】
典型的にPCRタイプ増幅技術ではプライマーは異なる配列を有し、互いに相補的でない。所望の試験条件次第で、プライマーの配列は、標的核酸の効率的かつ忠実な複製を提供するようにデザインされるべきである。PCRプライマーデザインの方法は一般的であり、技術分野で周知である。テイン(Thein)およびウォーレス(Wallace)著、「遺伝性疾患診断における特定のハイブリダイズプローブとしてのオリゴヌクレオチドの使用(The use of oligonucleotide as specific hybridization probes in the Diagnosis of Genetic Disorders)」、ヒト遺伝病:実際的アプローチ(Human Genetic Diseases:A Practical Approach)中、K.E.デービス(Davis)編、(1986)、33〜50ページ、IRL:ヘルンドン(Herndon)バージニア(VA);およびリュクリック(Rychlik)W.「分子生物学的方法(Methods in Molecular Biology)」第15巻、31〜39ページ、PCRプロトコル:現行の方法と応用(PCR Protocols:Current Methods and Applications)、ホワイト(White),B.A.編(1993)、ヒューマニア(Humania):ニュージャージー州トトワ(Totowa、NJ)。
【0133】
概してポリメラーゼ連鎖反応プロトコルにおいて、本配列の2本の短い断片を使用して、DNAまたはRNAから相同的遺伝子をコードするより長い核酸断片を増幅しても良い。またポリメラーゼ連鎖反応は、1つのプライマーの配列が本核酸断片から誘導され、その他のプライマーの配列が微生物の遺伝子をコードするmRNA前駆物質の3’末端へのポリアデニル酸のトラクトの存在を利用する、クローン化された核酸断片のライブラリーにおいて実施しても良い。
【0134】
代案としては、第2のプライマー配列は、クローン化ベクターから誘導される配列に基づいても良い。例えば当業者は、RACEプロトコル(フローマン(Frohman)ら
、Proc.Natl.Acad.Sci.USA 85:8998(1988))に従って、PCRを使用して、転写物の一点と3’または5’末端との間の領域のコピーを増幅し、cDNAを作り出すことができる。3’および5’方向に向けたプライマーは、本配列からデザインできる。メリーランド州ゲーサーズバーグのBRL(BRL、Gaithersburg、MD)から商業的に入手できる3’RACEまたは5’RACEシステム系を使用して、特定の3’または5’cDNA断片を単離できる(オハラ(Ohara)ら、Proc.Natl.Acad.Sci.USA 86:5673(1989);ロー(Loh)ら、Science 243:217(1989))。
【0135】
代案としては、相同体の同定のために、ハイブリダイズ試薬としてC40カロテノイド生合成経路の本配列を用いても良い。核酸ハイブリダイズ試験の基本的構成要素には、プローブ、対象の遺伝子または遺伝子断片を含有することが疑われるサンプル、および特定のハイブリダイズ法が含まれる。本発明のプローブは典型的に、検出する核酸配列に相補的である一本鎖核酸配列である。プローブは、検出する核酸配列と「ハイブリダイズ可能」である。プローブの長さは、5個の塩基から数万個の塩基の間で変動しても良く、実施する特定の試験に左右される。典型的に、約15個の塩基から約30個の塩基のプローブ長が適切である。プローブ分子の一部のみが、検出する核酸配列に相補的である必要がある。さらにプローブと標的配列との間の相補性は完璧でなくても良い。ハイブリダイズは不完全に相補的である分子間でも生じ、その結果、ハイブリッドした領域内の特定の塩基の一部は、適切な相補的塩基と対合形成しない。
【0136】
ハイブリダイズ方法は良く定義されている。典型的には、プローブおよびサンプルは、核酸ハイブリダイズを可能にする条件下で混合されなくてはならない。これは適切な濃度および温度条件下において、無機または有機塩存在下で、プローブとサンプルを接触させることを伴う。プローブとサンプル核酸の間であらゆる可能なハイブリダイズが起きるように、プローブおよびサンプル核酸は、十分長い時間接触しなくてはならない。混合物中のプローブまたは標的濃度が、ハイブリダイズが生じるのに必要な時間を決定する。プローブまたは標的濃度が高いほど、必要なハイブリダイズインキュベーション時間は短くなる。場合により、カオトロピック剤を添加しても良い。カオトロピック剤は、ヌクレアーゼ活性を阻害することによって核酸を安定化させる。さらにカオトロピック剤は、室温において短いオリゴヌクレオチドプローブの高感応性でストリンジェントなハイブリダイズを可能にする(ヴァン・ネス(Van Ness)およびチェン(Chen)Nucl.Acids Res.19:5143〜5151(1991))。適切なカオトロピック剤としては、特に塩化グアニジニウム、グアニジニウムチオシアネート、ナトリウムチオシアネート、リチウムテトラクロロ酢酸、過塩素酸ナトリウム、ルビジウムテトラクロロ酢酸、ヨウ化カリウム、およびセシウムトリフルオロ酢酸が挙げられる。典型的にカオトロピック剤は、約3Mの最終濃度で存在する。所望するならば、ハイブリダイズ混合物にホルムアミドを典型的に30〜50%(v/v)で添加できる。
【0137】
様々なハイブリダイズ溶液を用いることができる。典型的にこれらは、約20〜60%、好ましくは30%容積の極性有機溶剤を含んでなる。一般的なハイブリダイズ溶液では、約30〜50%v/vのホルムアミド、約0.15〜1Mの塩化ナトリウム、(例えばクエン酸ナトリウム、トリス−HCl、PIPESまたはHEPESなどの)約0.05〜0.1Mの緩衝液(pH範囲約6〜9)、(例えばドデシル硫酸ナトリウムなどの)約0.05〜0.2%の界面活性剤、または0.5〜20mMのEDTA、ファーマシア(Pharmacia Inc.)からのFICOLL(約300〜500キロダルトン)、ポリビニルピロリドン(約250〜500キロダルトン)、および血清アルブミンを用いる。また典型的なハイブリダイズ溶液には、約0.1〜5mg/mLの未標識のキャリア核酸、(例えば仔ウシ胸腺またはサケ精子DNA、または酵母RNAなどの)断片化核DNA、および場合により約0.5〜2%重量/容積のグリシンも含まれる。(例えばポ
リエチレングリコールなどの)多様な極性水溶性剤または膨潤剤をはじめとする容積排除剤、(例えばポリアクリレートまたはポリメチルアクリレートなどの)アニオン性ポリマー、および(例えば硫酸デキストランなどの)アニオン性糖ポリマーなどその他の添加剤を含めても良い。
【0138】
核酸ハイブリダイズは多様なアッセイ型式に適合できる。最も適切なものの1つは、サンドイッチアッセイ型式である。サンドイッチアッセイは、特に非変性条件下でのハイブリダイズに適合できる。サンドイッチ−タイプアッセイの主要構成要素は固形担体である。固形担体はそれに吸着され、あるいはそれと共有結合的に結合する、未標識で配列の一部分と相補的である固定核酸プローブを有する。
【0139】
本ヌクレオチドおよび推定されるアミノ酸配列の利用可能性は、DNA発現ライブラリーの免疫学的スクリーニングを容易にする。本アミノ酸配列の一部を代表する合成ペプチドを合成しても良い。これらのペプチドを用いて、動物を免疫し、アミノ酸配列を含んでなるペプチドまたはタンパク質に対して特異性があるポリクローナルまたはモノクローナル抗体を生成できる。次にこれらの抗体を使用して、DNA発現ライブラリーをスクリーニングし、対象の完全長DNAクローンを単離する(ラーナー(Lerner)R.A.、Adv.Immunol.36:1(1984)、マニアティス(Maniatis)、前出)。
【0140】
遺伝子構成
パントエア(Pantoea)属内の様々な種からのidiおよびcrtE、crtX、crtY、crtI、crtB、およびcrtZをコードする多様な遺伝子配列が入手できるが、配列番号18(8814bp)として開示される本核酸断片が、ユニークな遺伝子構成crtE−idi−crtY−crtI−crtB−crtZを有するように見える。
・crtE(配列番号1)は、ヌクレオチド2400〜3305に位置し、直接的方向に翻訳される。
・idi(配列番号3)は、ヌクレオチド3343〜4392に位置し、配向方向に翻訳される。
・crtY(配列番号5)は、ヌクレオチド4424〜5593に位置し、配向方向に翻訳される。
・crtI(配列番号7)は、ヌクレオチド5590〜7068に位置し、配向方向に翻訳される。
・crtB(配列番号9)は、ヌクレオチド7068〜7994に位置し、配向方向に翻訳される。
・crtZ(配列番号11)は、ヌクレオチド7909〜8466に位置し、crtE−idi−crtY−crtI−crtBと反対方向に翻訳される。
【0141】
最も「一般的な」遺伝子構成は、P.アナナティス(P.ananatis)(ジェンバンク(登録商標)登録番号D90087)、P.ステワルティ(P.stewartii)(ジェンバンク登録番号AY166713)、およびパントエア・アグロメランスpv.ミレチア(Pantoea agglomerans pv.milletiae)(ジェンバンク(登録商標)登録番号AB076662)で観察されるものであり、カロテン産生性クラスターはcrtEXYIBZ(「crtE−crtX−crtY−crtI−crtB−crtZ」とも記述される)を含んでなる(図5)。P.アグロメランス(P.agglomerans)EHO−10(ジェンバンク(登録商標)登録番号M87280)は、crtE−仮説タンパク質−crtX−crtY−crtI−crtB−crtZのカロテン産生性クラスターを含んでなると注釈される。しかしながら、本願出願人らによる「仮説タンパク質」のバイオインフォマティクス分析は、真のP.アグロメ
ランス(P.agglomerans)EHO−10がcrtE−idi−crtX−crtY−crtI−crtB−crtZを含んでなるとみなされるべきだと判断した。
【0142】
同様に、より遠縁の生物体からのその他のカロテン産生性遺伝子クラスターは、crtE−idi−crtY−crtI−crtB−crtZのユニークな遺伝子構成を含有しない。例えばブラジ根粒菌ダイズ根粒菌属(Bradyrhizobium)ORS278のカンタキサンチン遺伝子クラスターはcrtE−crtY−crtI−crtB−crtWであり、アグロバクテリウム・オーランティアカム(Agrobacterium
aurantiacum)からのアスタキサンチン遺伝子クラスターはcrtW−crtZ−crtY−crtI−crtBであり、フラボバクテリウム(Flavobacterium)sp.R1534からのゼアキサンチン遺伝子クラスターはcrtZ−crtY−crtI−crtB−crtEである(ジロー(Giraud)ら、米国特許仮出願第2003/0087337 A1号)。
【0143】
ここで開示される遺伝子構成は、代謝の遺伝子操作中に、大腸菌(E.coli)およびその他の細菌において工業的に価値あるカロテノイドの生成を最大化するのに有用ないくつかのユニークな利点を与えるかもしれない。第一にイソペンテニルピロリン酸イソメラーゼをコードするidiが、カロテノイド生成(カジワラ(Kajiwara)ら、前出;ワング(Wang)ら、前出;アルブレヒト(Albrecht)ら、前出)に劇的に影響することが実証されているので、またこの遺伝子がここで述べられるカロテン産生性−crtY−crtI−crtB−crtZクラスター中に直接組み込まれるので、オペロンの発現が、下流カロテノイド生合成経路への増大するイソプレノイド流束をもたらすことができ、それによって増大するカロテノイド生成および力価が帰結する。第二に、本カロテン産生性crtE−idi−crtY−crtI−crtB−crtZクラスター中のcrtX遺伝子の不在による利点があるかもしれない。具体的には、CrtXによるゼアキサンチンからゼアキサンチン−β−グルコシドへの転換が排除されることで、ここで述べるカロテン産生性クラスターによってコードされる反応の生成物としてのゼアキサンチンの高レベルの蓄積が可能になる。クラスターから不要なcrtX遺伝子を除去する余計な努力が必要でないため、発現クラスター中のcrtXの不在はまた、β−カロテンの合成にも有益かもしれない。
【0144】
したがって、本発明は、
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、カロテノイド生合成経路酵素をコードする核酸分子
を含んでなる形質転換宿主細胞を提供するステップと、
ここで、該分子の遺伝子構成はcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子は記載順にクラスター化し、crtZの転写はcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起き、
(b)適切な生育条件下で(a)の形質転換宿主細胞を有効量の発酵性炭素基質で生育させる、カロテノイド化合物の製造方法を提供する。
【0145】
微生物における組換え発現
本配列の遺伝子および遺伝子産物は、異種宿主細胞中、特に微生物細胞宿主中で生成しても良い。組換え微生物宿主中の発現は、これまで宿主を使用して可能でなかった新しい生成物合成のために、様々な経路中間体を発現させ、および/または宿主中に既存の経路を調節するのに有用かもしれない。
【0146】
適切な上流イソプレン経路遺伝子をコードする遺伝子、および本発明の下流カロテノイ
ド生合成経路遺伝子の(場合によりその他のcrt遺伝子との)様々な組み合わせを適切な微生物宿主に導入する方法は、一般的である。当業者には明らかなように、特定のカロテノイド生成物の生成のために、宿主生物体への導入が必要とされる特定の機能性は、宿主細胞(およびそのイソプレノイド化合物の天然産生)、基質の利用可能性、および所望の最終生成物に左右される。
【0147】
本crt遺伝子がカロテノイド生成において効果的であるためには、宿主細胞が細胞内に適切なレベルのFPPを有することが必要であることが理解される。FPPは、外来性に提供されても良く、または天然または導入された遺伝的経路のどちらかによって、細胞内に内在性に生成されても良い。したがって特定の宿主細胞が適切なレベルのFPPを生成する遺伝的機構を有さない場合、上流イソプレノイド経路のあらゆる必要な遺伝子を得て、これらの遺伝子を宿主内に遺伝子操作して、下流経路を通じたカロテノイド生合成のための出発原料としてFPPを生成することは、十分に当業者の理解の範囲内であると考察される。IPPをFPPの前駆物質として、良く知られている酢酸/メバロン酸経路を通じて合成しても良い。代案としては、最近の研究は、メバロン酸依存経路が全ての生物体で作動しているわけではないことを実証している。IPP生合成のための代案のメバロン酸非依存経路が、細菌および緑藻類および高等植物で特性決定されている(ホルバック(Horbach)ら、FEMS Microbiol.Lett.111:135〜140(1993);ローマー(Rohmer)ら、Biochem.295:517〜524(1993);シュベンダー(Schwender)ら、Biochem.316:73〜80(1996);アイゼンライヒ(Eisenreich)ら、Proc.Natl.Acad.Sci.USA93:6431〜6436(1996))。
【0148】
例えば、1つもしくはそれ以上の本下流C40カロテノイド生合成経路crtEYIBZ配列をコードするキメラ遺伝子の導入が、選択された宿主微生物におけるカロテノイド化合物の生成をもたらすことが期待される。適切な遺伝的形質転換システムによって、多様な非カロテン産生性宿主を遺伝子操作することが可能であろう。これは例えばエルウィニア・ヘルビコラ(Erwinia herbicola)crt遺伝子を使用して、宿主の大腸菌(E.coli)、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)、サッカロミセス・セレビジエ(Saccharomyces cerevisiae)、ピヒア・パストリス(Pichia pastoris)(酵母)、アスペルギルス・ニドランス(Aspergillus nidulans)(菌・カビ類)、ロドバクター・スフェロイデス(Rhodobacter
sphaeroides)、および高等植物(米国特許第5,656,472号明細書)中で、様々なカロテノイドを生成して示されている。したっがってここで前述したように、アンテラキサンチン、アドニルビン、アドニキサンチン、アスタキサンチン、カンタキサンチン、カプソルブリン、β−クリプトキサンチン、α−カロテン、β−カロテン、ε−カロテン、エキネノン、3−ヒドロキシエキネノン、3’−ヒドロキシエキネノン、γ−カロテン、4−ケト−γ−カロテン、ζ−カロテン、α−クリプトキサンチン、デオキシフレキシキサンチン、ジアトキサンチン、7,8−ジデヒドロアスタキサンチン、フコキサンチン、フコキサンチノール、イソレニエラテン、ラクツカキサンチン、ルテイン、リコピン、ミキソバクトン、ネオキサンチン、ニューロスポレン、ヒドロキシニューロスポレン、ペリジニン、フィトエン、ロドピン、ロドピングルコシド、4−ケト−ルビキサンチン、シホナキサンチン、スフェロイデン、スフェロイデノン、スピリロキサンチン、4−ケト−トルレン、3−ヒドロキシ−4−ケト−トルレン、ウリオリド、ウリオリドアセタート、ビオラキサンチン、ゼアキサンチン−β−ジグルコシド、およびゼアキサンチンを全て、(例えば)以下のcrt酵素機能性の様々な組み合わせを導入することで、ここの技術を使用して微生物宿主中で生成しても良い。CrtE、CrtX、CrtY、CrtI、CrtB、CrtZ、CrtW、CrtO、CrtA、CrtC、CrtD、CrtF、およびCrtU。したがってFPPからのフィトエンの形成は、CrtEおよ
びCrtBと、crtE、crtB、およびcrtIをはじめとするFPPからのリコピン合成に必要なカロテノイド特異的遺伝子と、crtE、crtB、crtI、およびcrtYをはじめとするFPPからのβ−カロテン生成に必要な遺伝子を必要とする。crt遺伝子間の関係のこの理解に基づいて、あらゆる所望のカロテノイド生成物の発現に適した、微生物宿主細胞およびcrt遺伝子を選択することが可能であろう。類似の様式で、例えばidiなどの上流イソプレン経路遺伝子の上方制御によって、発現を増幅しても良い。
【0149】
外来性タンパクの高レベル発現を導く制御配列を含有する微生物発現システムおよび発現ベクターは、当業者には周知である。これらのいずれかを使用してキメラ遺伝子を構築し、本配列の遺伝子産物を生成できる。次に形質転換を通じて、これらキメラ遺伝子を適切な微生物に導入し、酵素の高レベル発現を提供できる。
【0150】
適切な宿主細胞の形質転換のために有用なベクターまたはカセットは、技術分野で周知である。典型的にベクターまたはカセットは、関連性のある遺伝子の転写および翻訳を導く配列、選択可能マーカー、および自律的複製または染色体組み込みができるようにする配列を含有する。適切なベクターは、転写イニシエーション制御を含む遺伝子の5’領域、および転写終結を制御するDNA断片の3’領域を含んでなる。双方の制御領域が、形質転換宿主細胞に相同的な遺伝子から誘導されることが最も好ましいが、このような制御領域は、必ずしも産生宿主として選択された特定種に固有の遺伝子から誘導されなくて良いものと理解される。
【0151】
所望の宿主細胞において本ORFの発現を駆動させるのに有用なイニシエーション調節領域、またはプロモーターは多数有り、当業者には良く知られている。CYC1、HIS3、GAL1、GAL10、ADH1、PGK、PHO5、GAPDH、ADC1、TRP1、URA3、LEU2、ENO、TPI(例えばサッカロミセス(Saccharomyces)における発現に有用)、AOX1(例えばピヒア(Pichia)における発現に有用)、およびlac、ara、tet、trp、lPL、lPR、T7、tac、およびtrc(例えば大腸菌(Escherichia coli)における発現に有用)ならびに例えばバチルス(Bacillus)における発現に有用なamy、apr、nprプロモーターおよび様々なファージプロモーターをはじめとするが、これに限定されるものではない、実質的にこれらの遺伝子を駆動できるあらゆるプロモーターが、本発明において適切である。さらにデオキシ−キシルロースリン酸シンターゼまたはメタノールデヒドロゲナーゼオペロンプロモーター(スプリンガー(Springer)ら、FEMS Microbiol Lett 160:119〜124(1998))、ポリヒドロキシアルカン酸合成のためのプロモーター(フェルナー(Foellner)ら、Appl.Microbiol.Biotechnol.40:284〜291(1993))、メチロトローフ内の天然プラスミドから同定されるプロモーター(欧州特許第296484号明細書)、Plac(トヤマ(Toyama)ら、Microbiology 143:595〜602(1997);欧州特許第62971号明細書)、Ptrc(ブロシウス(Brosius)ら、Gene 27:161〜172(1984))、メタノトローフから同定されたプロモーター(PCT/US03/33698号明細書)、および抗生物質抵抗性に関連したプロモーター[例えばカナマイシン(スプリンガー(Springer)ら、FEMS Microbiol Lett160:119〜124(1998);ウエダ(Ueda)ら、Appl.Environ.Microbiol.57:924〜926(1991))またはテトラサイクリン(米国特許第4,824,786号明細書)]が、C1代謝個体における発現に適している。
【0152】
RBSがベクターによって提供されない場合、発現する遺伝子上流に人工リボソーム結合部位(「RBS」)を含めることが必要である。これは、単一プロモーターが、第1、
第2、第3、およびその他遺伝子群の発現を推進する場合に、発現するオペロンの第2、第3、およびその他の遺伝子のために必要とされることが多い。特定の宿主生物体におけるRBSの好ましい配列を判定する方法は当業者には良く知られており、この合成部位の製造手段についても同様である。
【0153】
終結制御領域はまた、好ましい宿主に固有の様々な遺伝子から誘導されても良い。場合により、終結部位は必要ないかもしれないが、含まれることが最も好ましい。
【0154】
遺伝子をクローン化ベクターに単に挿入するだけでは、それが必要なレベルで成功裏に発現することは確証されない。高発現率の必要性に答えて、転写、翻訳、タンパク質安定性、酸素限界、および宿主細胞からの分泌の側面を制御するいくつかの異なる遺伝的要素を操作することで、多くの特殊化した発現ベクターが作り出されている。より具体的には、遺伝子発現を制御するように操作される分子の特徴として以下が挙げられる。1.)関連転写プロモーターおよびターミネーター配列の性質、2.)リボソーム結合部位の強度、3.)クローンされた遺伝子のコピー数、および遺伝子がプラスミドにあるかまたは宿主細胞のゲノム中に組み込まれているかどうか、4.)合成された外来タンパク質の最終細胞内位置、5.)宿主生物体中の翻訳効率、6.)宿主細胞内のクローン化遺伝子タンパク質の本質的な安定性、および7.)頻度が宿主細胞の好むコドン使用頻度に近づくような、クローン化遺伝子内のコドン使用。これらの各タイプの修正は、C40カロテノイドの発現をさらに最適化する手段として、本発明中に包含される。
【0155】
最後に、C40カロテノイドの蓄積を促進するには、エネルギーまたは炭素のシンクとして機能するかもしれない、標的経路中または競合経路中の特定の遺伝子の発現を低下させまたは排除することが必要かもしれない。代案としては、所望のカロテノイド中間体の上流の様々な遺伝子を過剰発現させ、生成を促進させることが有用かもしれない。上述の目的で遺伝的経路を操作する方法は、技術分野で一般的であり良く知られている。
【0156】
例えばひとたび重要な遺伝的経路が同定および配列決定されると、特定の遺伝子を上方制御して経路の産出量を増大できる。例えばpBR322などのマルチコピー型プラスミド上で、標的遺伝子の追加的コピーを宿主細胞に導入しても良い。代案としては、非天然プロモーターの調節の下におかれるように標的遺伝子を変性しても良い。経路が、細胞サイクル中または発酵生産中の特定点で作動することが所望される場合、制御されたまたは誘導可能プロモーターを使用して、標的遺伝子の天然プロモーターを置き換えても良い。同様に場合によっては、天然または内在性プロモーターを変性させて、遺伝子発現を増大させても良い。例えば内在性プロモーターは、生体内で(in vivo)突然変異、欠失、および/または置換によって変性できる(米国特許第5,565,350号明細書、ザーリン(Zarling)らのPCT/US93/03868号明細書を参照されたい)。
【0157】
代案としては、中断させる遺伝子配列が知られている場合、遺伝子下方制御の最も効果的な方法の1つは、外来DNAを構造的な遺伝子に挿入して転写を中断させる、標的を定めた遺伝子中断である。これは、中断させる遺伝子の一部に対して高度な相同性を有する配列で両脇を挟まれた、挿入するDNA(遺伝子マーカーであることが多い)を含んでなる遺伝子カセットの製造によってもたらされる。カセットの宿主細胞への導入は、細胞の天然DNA複製機序を通じた、構造的遺伝子への外来DNAの挿入をもたらす(例えばハミルトン(Hamilton)ら、J.Bacteriol.171:4617〜4622(1989);バルバス(Balbas)ら、Genes、136:211〜213(1993);ゲルデナー(Gueldener)ら、Nucleic Acid Res.24:2519〜2524(1996);およびスミス(Smith)ら、Methods Mol.Cell.Biol.5:270〜277(1996)参照)。
【0158】
アンチセンス技術は、標的遺伝子配列が知られている場合に遺伝子を下方制御する別の方法である。これを達成するために、所望の遺伝子からの核酸セグメントをクローン化して、RNAのアンチセンス鎖が転写されるようにプロモーターに作動的に結合する。次にこのコンストラクトを宿主細胞に導入し、RNAのアンチセンス鎖を生成する。アンチセンスRNAは、対象のタンパク質をコードするmRNAの蓄積を防止することで、遺伝子発現を阻害する。当業者は、特定の遺伝子の発現を低下させるために、特別な考察がアンチセンス技術の使用に関わることを理解する。例えば、アンチセンス遺伝子の適切な発現レベルは、当業者に知られている異なる調節因子を使用した、異なるキメラ遺伝子の使用を必要とするかもしれない。
【0159】
配列が知られている場合、標的を定めた遺伝子中断およびアンチセンス技術が遺伝子を下方制御する効果的手段を提供するが、配列ベースではないその他のより特異性が低い方法が開発されている。例えば細胞をUV放射線暴露して、次に所望の表現型についてスクリーンしても良い。変異体を作り出すために、化学薬品による変異誘発もまた効果的であり、一般に使用される物質としては、非複製DNAに影響する化学物質(例えばHNO2およびNH2OH)、ならびに複製DNAに影響する薬剤(例えばフレームシフト変異を引き起こすことで注目に値するアクリジン染料)が挙げられる。放射線または化学薬品を使用して変異体を作り出す特定の方法は、技術分野で良く文書化されている。例えばトーマスD.ブロック(Thomas D.Brock)著「バイオテクノロジー:工業微生物学テキストブック(Biotechnology:A Textbook of Industrial Microbiology)」第二版(1989)Sinauer Associates:マサチューセッツ州サンダーランド(Sunderland、MA)、またはデシュパンデ,ムカンド(Deshpande,Mukund)V.、Appl.Biochem.Biotechnol.、36:227〜234(1992)を参照されたい。
【0160】
別の非特異的遺伝子中断方法は、転移因子またはトランスポゾンの使用である。トランスポゾンは、DNAに無作為に挿入される遺伝的因子であるが、後で配列に基づいて検索して、挿入がどこで起きたのか判定できる。生体内(in vivo)および生体外(in vitro)転位法の双方が知られている。どちらの方法にもトランスポザーゼ酵素と組み合わされた転移因子の使用が関与する。転移因子またはトランスポゾンがトランスポザーゼ存在下で核酸断片に接触すると、転移因子が核酸断片中に無作為に挿入される。中断された遺伝子は転移因子配列に基づいて同定されても良いので、技術は、ランダム変異誘発のため、そして遺伝子単離のために有用である。生体外(in vitro)転位のためのキットは市販される(例えばニュージャージー州ブランチバーグのパーキン・エルマー・アプライド・バイオシステムズ(Perkin Elmer Applied Biosystems(Branchburg、NJ))から入手できる酵母Ty1因子に基づく、プライマー・アイランド転位キット、マサチューセッツ州ビバリーのニュー・イングランド・バイオ・ラブズ(New England Biolabs(Beverly、MA))から入手できる細菌性トランスポゾンTn7に基づくゲノム・プライミング・システム、およびウィスコンシン州マディソンのエピセンター・テクノロジーズ(Epicentre Technologies(Madison、WI))から入手できる、Tn5細菌性転移因子に基づくEZ::TNトランスポゾン挿入システムを参照されたい)。
【0161】
本発明の文脈で、上述のいずれかの一方法によって、カロテノイド生合成経路の発現を調節することが有用かもしれない。例えば本発明は、カロテノイド生合成経路中の酵素をコードするいくつかの単離された遺伝子、およびC40カロテノイドの生成をもたらす方法を提供する。したがってここでcrtE、idi、crtY、crtI、crtB、お
よびcrtZ遺伝子の様々な組み合わせを過剰発現して、C40カロテノイドの増大する生成を促進するのに加えて、三炭素化合物(ピルビン酸およびD−グリセルアルデヒド3−リン酸)の最初の縮合を上方制御して、(dxs遺伝子によって媒介される)五炭素化合物D−1−デオキシキシルロース−5−リン酸の生成を増大させることもまた有用かもしれない。これは下流カロテノイド生合成経路に入る炭素の流束を増大させ、C40カロテノイドの増大する生成を可能にする。代案としては(またはそれに加えて)、微生物宿主がこれらのタイプの化合物を合成できれば、crtM/crtN遺伝子をノックアウトして、C30カロテノイドの合成をもたらすることが望ましかもしれない。または天然機能性crtE、crtX、crtY、crtI、crtB、およびcrtZ遺伝子を有するシステムでは、上述のいずれかの一方法によって下流遺伝子(例えばcrtZまたはcrtX)を中断させることで、β−カロテンまたはゼアキサンチンの蓄積をもたらしても良い。
【0162】
好ましい微生物宿主
本遺伝子およびカロテノイド生合成経路の核酸断片の発現のために好ましい異種宿主細胞は、菌・カビ類または細菌系統群中に広く見られ、広範な温度、pH価、および溶剤耐性で生育する微生物宿主である。例えばあらゆる細菌、酵母、および糸状菌類が、本核酸断片の発現のための適切な宿主であることが考察された。転写、翻訳およびタンパク質生合成器官は、それぞれの細胞供給原料に関係なく同一であるので、細胞バイオマスを生成するのに使用される炭素供給材料に関係なく、機能性遺伝子が発現される。大規模微生物生育および機能遺伝子発現は、広範な単純または複合炭水化物、有機酸およびアルコール、および/または飽和炭化水素(例えば光合成または化学的自己栄養宿主の場合は、メタンまたは二酸化炭素などの)を利用しても良い。しかし機能遺伝子は、窒素、亜リン酸、イオウ、酸素、炭素、または小型無機イオンをはじめとするあらゆる微量栄養素の形態および量をはじめとする特定の生育条件によって、阻止または抑圧され制御されても良い。さらに機能遺伝子の制御は、培養に添加される典型的に栄養素またはエネルギー源とは見なされない、特定の制御分子の存在または不在によって達成されても良い。生育速度もまた、遺伝子発現における重要な制御因子であっても良い。
【0163】
適切な宿主株の例としては、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、サッカロミセス(Saccharomyces)、ピヒア(Pichia)、カンジダ(Candida)、ハンゼヌラ(Hansenula)、ヤロウィア(Yarrowia)、ロドスポリジウム(Rhodosporidium)、リポミセス(Lipomyces)、サルモネラ(Salmonella)、バチルス(Bacillus)、アシネトバクター(Acinetobacter)、ザイモモナス(Zymomonas)、アグロバクテリウム(Agrobacterium)、フラボバクテリウム(Flavobacterium)、ロドバクター(Rhodobacter)、ロドコッカス(Rhodococcus)、ストレプトマイセス(Streptomyces)、ブレビバクテリウム(Brevibacterium)、コリネバクテリア(Corynebacteria)、マイコバクテリウム(Mycobacterium)、エシェリヒア(Escherichia)、パントエア(Pantoea)、シュードモナス(Pseudomonas)、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、メチロサイナス(Methylosinus)、メチロマイクロビウム(Methylomicrobium)、メチロシスティス(Methylocystis)、アルカリゲネス(Alcaligenes)、シネコシスティス(Synechocystis)、スネコッカス(Synechococcus)、アナベナ(Anabaena)、チオバチルス(Thiobacillus)、メタノバクテリウム(Methanobacterium)、クレブシエラ(Klebsiella)、メチロフィラス(Methylophilus)、メチロバチルス(Methylobacillus)
、メチロバクテリウム(Methylobacterium)、ヒフォミクロビウム(Hyphomicrobium)、キサントバクター(Xanthobacter)、パラコッカス(Paracoccus)、ノカルジア(Nocardia)、アルスロバクター(Arthrobacter)、ロドシュードモナス(Rhodopseudomonas)、トルロプシス(Torulopsis)、ロドトルラ(Rhodotorula)、およびファフィア(Phaffia)が挙げられるが、これに限定されるものではない。
【0164】
微生物宿主としてのメチロトローフおよびメチロモナス(Methylomonas)sp.16a
組換え微生物源からいくつかのカロテノイドが生成されている[例えばリコピン(lycoene)生成のための大腸菌(E.coli)およびカンジダ・ユチリス(Candia utilis)(ファーマー(Farmer)W.R.およびリヤオ(Liao)J.C.、Biotechnol.Prog.17:57〜61(2001);ワング(Wang)C.ら、Biotechnol Prog.16:922〜926(2000);ミサワ(Misawa)N.およびシマダ(Shimada)H.、J.Biotechnol.59:169〜181(1998);シマダ(Shimada)ら、Appl.Environm.Microbiol.64:2676〜2680(1998));β−カロテン生成のための大腸菌(E.coli)、カンジダ・ユチリス(Candia utilis)、およびパフィア・ロドジマ(Pfaffia rhodozyma)(アルブレヒト(Albrecht)ら、Biotechnol.Lett.21:791〜795(1999);ミウラ(Miura)Y.ら、Appl.Environm.Microbiol.64:1226〜1229(1998);米国特許第5,691,190号明細書);ゼアキサンチン生成のための大腸菌(E.coli)およびカンジダ・ユチリス(Candia utilis)(アルブレヒト(Albrecht)ら、前出;ミウラ(Miura)Y.ら、前出);アスタキサンチン生成のための大腸菌(E.coli)およびパフィア・ロドジマ(Pfaffia rhodozyma)(米国特許第5,466,599号明細書、米国特許第6,015,684号明細書、米国特許第5,182,208号明細書、米国特許第5,972,642号明細書;米国特許第5,656,472号明細書、米国特許第5,545,816号明細書、米国特許第5,530,189号明細書、米国特許第5,530,188号明細書、米国特許第5,429,939号明細書、および米国特許第6,124,113号明細書もまた参照されたい)]が、異なるcrt遺伝子の様々な組み合わせを使用したこれらのカロテノイド生成法には、低い収率と比較的高価な供給材料への依存という欠点がある。したがって微生物宿主中で安価な供給材料から、高収率のカロテノイドを製造する方法を同定することが望ましい。
【0165】
単一エネルギー源として一炭素基質を利用する、いくつかの微生物がある。このような微生物はここで「C1代謝個体」と称される。これらの生物体は、単一エネルギーおよびバイオマス源として炭素−炭素結合が欠如している炭素基質を使用する能力によって特徴付けられる。これらの炭素基質としては、メタン、メタノール、ホルメート、ホルムアルデヒド、ギ酸、メチル化アミン(例えば、モノ−、ジ−、およびトリ−メチルアミン)、メチル化チオール、二酸化炭素、およびその他の様々なあらゆる炭素−炭素結合が欠如している還元炭素化合物が挙げられるが、これに限定されるものではない。
【0166】
全てのC1代謝微生物は、概してメチロトローフとして分類される。メチロトローフは、炭素−炭素結合を含有しない有機化合物を酸化できるあらゆる生物体として定義されても良い。しかし通性メチロトローフ、絶対メチロトローフ、絶対メタノトローフは、全てメチロトローフの様々なサブセットである。具体的には次のようである。
・通性メチロトローフは、炭素−炭素結合を含有しない有機化合物を酸化する能力を有
するが、エネルギーおよびバイオマスのために、糖および複合炭水化物などのその他の炭素基質を使用しても良い。通性メチロトローフ細菌は多くの環境中に見られるが、最も一般には土壌、ごみ埋め立て、および廃棄物処理場から単離される。多くの通性メチロトローフは、プロテオバクテリア(Proteobacteria)のβおよびγ亜群のメンバーである(ハンソン(Hanson)ら、C1化合物上での微生物の生育(Microb.Growth C1 Compounds)[国際シンポジウム]第7回(1993)、p285〜302、マレルJ.コリン(Murrell J.Collin)およびドンP.ケリー(Don P.Kelly)編、インターセプト(Intercept):英国アンドーバー(Andover、UK);マディガン(Madigan)ら、ブロックの微生物生物学(Brock Biology of Microorganisms)、第8版、プレンティスホール(Prentice Hall):ニュージャージー州アッパー・サドル・リバー(Upper Saddle River、NJ)(1997))。
・絶対メチロトローフは、エネルギー生成が、炭素−炭素結合を含有しない有機化合物の使用に限定された生物体である。
・絶対メタノトローフは、メタンを酸化する特徴的能力を有する絶対メチロトローフである。
【0167】
さらに一炭素基質を利用する能力は細菌に限定されず、酵母および菌・カビ類にも及ぶ。いくつかの酵母属は、エネルギー源としてより複雑な材料に加えて一炭素基質を使用できる(すなわちメチロトローフ酵母)。
【0168】
数多くのこれらのメチロトローフ生物体が知られているが、これらの微生物のわずかなものだけが材料合成のための工業プロセスにおいて成功裏に利用されている。そして一炭素基質は対費用効果の高いエネルギー源であるが、これらの微生物の遺伝子操作における困難さ、ならびにそれらの遺伝的機構に関する情報不足が、主に天然生成物合成のためのそれらの使用を制限している。
【0169】
これらの困難さにもかかわらず、多くのメタノトローフはこれらの生物体が色素を合成できるようにする固有のイソプレノイド経路を含有し、様々な非内在性イソプレノイド化合物を生成するために、これらの微生物を遺伝子操作する構想の可能性を提供する。メタノトローフは一炭素基質(すなわちメタンまたはメタノール)をエネルギー源として使用できるので、これらの生物体においてカロテノイドを安価に生成することが可能であろう。メタノトローフがβ−カロテン生成のために遺伝子操作される実施例の1つは、国際公開第02/18617号パンフレットで述べられる。
【0170】
本発明では、一炭素基質を唯一のエネルギー源として使用できる微生物における、カロテノイド化合物生合成に関与する遺伝子発現のための方法が提供される。宿主微生物は、カロテノイドのための代謝前駆物質として、ファルネシルピロリン酸(FPP)を合成する能力を有するあらゆるC1代謝個体であっても良い。より具体的には、本発明で適切な通性メチロトローフ細菌としては、メチロフィラス(Methylophilus)、メチロバチルス(Methylobacillus)、メチロバクテリウム(Methylobacterium)、ヒフォミクロビウム(Hyphomicrobium)、キサントバクター(Xanthobacter)、バチルス(Bacillus)、パラコッカス(Paracoccus)、ノカルジア(Nocardia)、アルスロバクター(Arthrobacter)、ロドシュードモナス(Rhodopseudomonas)、およびシュードモナス(Pseudomonas)が挙げられるが、これに限定されるものではない。本発明で有用な具体的なメチロトローフ酵母としては、カンジダ(Candida)、ハンゼヌラ(Hansenula)、ピヒア(Pichia)、トルロプシス(Torulopsis)、およびロドトルラ(Rhodotorula)が挙げら
れるが、これに限定されるものではない。そして例示的なメタノトローフとしては、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、メチロサイナス(Methylosinus)、メチロシクティス(Methylocyctis)、メチロマイクロビウム(Methylomicrobium)、およびメタノモナス(Methanomonas)属が挙げられるが、これに限定されるものではない。
【0171】
本発明で特に興味深いのは、エネルギー的に好ましい炭素流経路を有する高生育絶対メタノトローフである。例えば出願人は、それを炭素流束操作のために特に有用なものにする、いくつかの経路特性を有するメタノトローフの特定の株を見いだした。この株はメチロモナス(Methylomonas)16a(ATCC PTA2402)(米国特許第6,689,601号明細書)として知られており、この特定の株およびその他の近縁メチロトローフは、C40カロテノイド生成に有用な本発明の遺伝子産物発現のための好ましい微生物宿主である(国際公開第02/18617号パンフレット)。
【0172】
メチロモナス(Methylomonas)sp.16aは、C30カロテノイドを天然に生成する。オドム(Odom)らは、メチロモナス(Methylomonas)16aにおけるC40カロテノイド遺伝子の発現が、C30およびC40カロテノイド混合物を生成することを報告している(国際公開第02/18617号パンフレット)。crtN1、ald、crtN2、およびcrtN3遺伝子をはじめとする(がこれに限定されるものではない)、この株中のC30カロテノイド生成に関与する遺伝子のいくつかが同定されている。ald遺伝子の中断、またはcrtN1/ald/crtN2遺伝子クラスターの発現を推進するプロモーターの中断は、C40カロテノイド生成により適した様々な非着色変異体(「白色変異体」)を作り出す(米国仮出願第60/527083号を本願明細書に援用する)。例えば非着色メチロモナス(Methylomonas)sp.16a株MWM1000は、aldおよびcrtN1遺伝子を中断することで作り出された。
【0173】
メチロモナス(Methylomonas)sp.16a株は、炭素利用経路にいくつかの異常を含有する。例えばゲノム配列データに基づいて、株は2つのヘキソース代謝経路のための遺伝子を含有することが示される。エントナー−ドゥドロフ経路(ケト−デオキシホスホグルコン酸アルドラーゼ酵素を用いる)が株中に存在する。これが絶対メタノトローフにおいて稼働している経路であることは、概して広く認められている。しかしエムデン−マイヤーホフ経路もまた存在する(フルクトースビスリン酸アルドラーゼ酵素を用いる)。この経路が絶対メタノトローフ中に存在しないか、または稼働していないことは良く知られている。エネルギー的には後者の経路が最も好ましく、生物学的に有用なエネルギーのより高い収率を可能にして、究極的にメチロモナス(Methylomonas)16aにおいて、細胞集団生成およびその他の細胞集団依存生成物のより高い収率が得られる。16a株におけるこの経路の活性は、マイクロアレイデータおよびATP還元を測定する生化学的証拠を通じて確認されている。16a株は、エムデン−マイヤーホフおよびエントナー−ドゥドロフ経路酵素の双方を有することが示されているが、データは、エムデン−マイヤーホフ経路酵素がエントナー−ドゥドロフ経路酵素よりも強力に発現することをを示唆する。この結果は意外であり、メタノトローフ細菌解糖代謝に関する既存の信念に反する。出願人は、例えばメチロモナス・クララ(Methylomonas
clara)およびメチロサイナス・スポリウム(Methylosinus sporium)をはじめとする、この特徴を有する別のメタノトローフ細菌を見いだした。この活性は、ほとんどの細菌システム中の酵素の典型的なホスホリル供与体であるATPに対して酵素活性が欠如しているために、メタノトローフにおいて見いだされないままであったように思われる。
【0174】
16a株中のエムデン−マイヤーホフ経路の特に新しく有用な特徴は、重要なホスホフルクトキナーゼステップがATP−依存性でなく、ピロリン酸依存性であることである。この特徴は、ATPの代わりにピロリン酸を使用することで、経路のエネルギー収率を増大させる。
【0175】
メタノトローフ細菌では、リブロース一リン酸経路またはRuMP回路として知られている、反応の循環セットを通じてメタンが生分子に転換される。この経路は3つの相を含んでなり、各相は一連の酵素的ステップである。第1のステップは、ヘキソースまたは六炭糖を形成する、ペントース中へのC−1(ホルムアルデヒド)の「固定」または組み込みである。これは五炭糖(ペントース)とホルムアルデヒドとの縮合反応を通じて起き、ヘキスロース一リン酸シンターゼによって触媒される。第2段階は「開裂」と称され、2個の三炭素分子へのヘキソースの分裂をもたらす。これらの三炭素分子の1つはRuMP経路を通じてリサイクルされ、他の三炭素断片は細胞の生育に用いられる。
【0176】
メタノトローフおよびメチロトローフにおいて、RuMP経路は3つの変種の1つとして生じても良い。しかしこれらの変種の2つのみ、FBP/TA(フルクトースビスホスホターゼ/トランスアルドラーゼ)経路またはKDPG/TA(ケトデオキシホスホグルコン酸/トランスアルドラーゼ)経路が一般に見られる(ディクフイゼン(Dijkhuizen)L.およびドゥブリエ(Devries)G.E.「好気性メタノール利用グラム陰性およびグラム陽性細菌の生理学と生化学(The Physiology and biochemistry of aerobic methanol−utilizing gram negative and gram positive bacteria)」「メタンおよびメタノール利用細菌(Methane and Methanol Utilizers)」中、コリン・マレル(Colin Murrell)およびハワード・ダルトン(Howard Dalton)編より、Plenum:ニューヨーク、1992)。
【0177】
メチロモナス(Methylomonas)sp.16a株は、「開裂」ステップの扱いにおいてユニークであり、重要な中間体としてフルクトースビスリン酸を経由して、この転換を行う遺伝子が見いだされた。フルクトースビスリン酸アルドラーゼおよびトランスアルドラーゼのための遺伝子は、1片のDNA上に一緒にクラスター化して見いだされている。次にケトデオキシホスホグルコン酸中間体が関与するその他の変種のための遺伝子もまた、一緒にクラスター化して見いだされている。入手できる文献は、これらの生物体(絶対メチロトローフおよびメタノトローフ)がKDPG経路のみに依存し、FBP依存固定経路が通性メチロトローフによって用いられることを教示する(ディクフイゼン(Dijkhuizen)ら、前出)。したがって後者の観察は予期されるが、前者はそうでない。絶対メタン利用細菌におけるFBP遺伝子の発見は意外であり、また有用性も示唆する。FBP経路は、KDPG経路で利用されるよりも多くのエネルギー(ATP)が利用されるとという事実のために、宿主微生物にとってエネルギー的に好ましい。したがってFBP経路を用いる生物体は、KDPG経路を用いるものに比べて、エネルギー上の利点および生育上の利点を有するかもしれない。この利点はまた、菌株中のエネルギー要求生成経路にとって有用かもしれない。この経路を使用することで、メタン利用細菌は、単細胞タンパク質、またはRuMP経路を通る炭素の流れから誘導されるあらゆるその他の生成物(例えばカロテノイド)のための生成基盤として、その他のメタン利用生物体に比べて利点を有するかもしれない。
【0178】
したがって本発明は、
(a)メタンおよびメタノールよりなる群から選択されるC1炭素基質で生育し、
(b)ピロリン酸依存性ホスホフルクトキナーゼ酵素をコードする遺伝子を含んでなる、機能的エムデンマイヤーホフ炭素経路を含んでなる、
高生育でエネルギー的に好ましいメチロモナス(Methylomonas)株におけるカロテノイド化合物の製造方法を提供する。
【0179】
C1代謝細菌の形質転換
C1代謝細菌形質の転換技術は十分開発されいないが、当業者に良く知られているその他の細菌のために用いられる一般方法を応用しても良い。メチロバクテリウム・エキストロクエン(Methylobacterium extorquens)AM1(トヤマ(Toyama)H.、ら、FEMS Microbiol.Lett.166:1〜7(1998))、メチロフィラス・メチロトロファス(Methylophilus methylotrophus)AS1(キム(Kim)C.S.およびT.K.ウッド(Wood)、Appl.Microbiol.Biotechnol.48:105〜108(1997))、およびメチロバチルス(Methylobacillus)sp.株12S(ヨシダ(Yoshida)T.ら、Biotechnol.Lett.、23:787〜791(2001))の形質転換のために、電気穿孔が成功裏に使用されている。しかし当業者には良く知られているように、1つの特定のC1代謝利用生物体から別の生物体への具体的電気穿孔パラメーターの推定は、困難かもしれない。
【0180】
供与細胞と受容細胞の直接接触に依存する細菌接合は、C1代謝細菌中への遺伝子の転位のために、より容易に受け入れられることが多い。簡単に述べると、この細菌接合プロセスは、「供与」細胞と「受容」細胞を互いに密接に接触させて一緒に混合することを伴う。接合は、新たに合成された供与DNAの受容細胞中への直接転位を伴う、「供与」細菌と「受容」細菌間の細胞質連絡の形成によって起きる。技術分野で良く知られているように、接合の受容体は、供与細菌からの水平転位を通じてDNAを受容できるあらゆる細胞として定義される。接合転位中の供与体は、接合プラスミド、接合トランスポゾン、または可動性プラスミドを含有する細菌である。供与体プラスミドの物理的転位は、下で述べられる2つの様式の1つによって起きることができる。
1.場合によっては、供与体と受容体のみが接合にとって必要とされる。これは転位されるプラスミドが接合性かつ可動性である自己伝播性のプラスミドの場合(すなわちtra−遺伝子と、Mobタンパク質をコードする遺伝子の双方を保有する)に起きる。一般にプロセスは、1.)oriT中の特定部位で二本鎖プラスミドDNAにニックが入れられるステップと、2.)poreまたはpilus構造を通じて、一本鎖DNAが受容体に放出されるステップと、3.)DNAリラクセース酵素が二本鎖DNAをoriTで開裂して結合し、5’末端を放出する(中間体構造としてリラクソゾームを形成する)ステップと、4.)引き続いて、補助タンパク質の複合体がoriTでアセンブルして、DNA転位プロセスを容易にするステップとを伴う。
2.代案としては、供与体プラスミドの受容体への転位のために「三親交雑」接合が必要とされる。このタイプの接合には、供与細胞、受容細胞、および「ヘルパー」プラスミドが関与する。供与細胞は、可動性プラスミドまたは接合トランスポゾンを保有する。可動性ベクターは、ニッケースをコードする遺伝子であるoriTを含有し、Mobタンパク質をコードする遺伝子を有する。しかしMobタンパク質は、単独ではゲノムの転位達成するのに十分でない。したがって可動性プラスミドは、ヘルパープラスミド(供与体内または「ヘルパー」細胞内に位置する)によって適切な接合システムが提供されない限り、それら自身の転位を促進できない。プラスミドは、poreまたはpilusの形成に関与する転位タンパク質(Tra)をコードするので、接合プラスミドは噛合対の形成およびDNA転位のために必要とされる。
【0181】
C1代謝細菌が関与する成功裏の接合の例としては、ストリアー(Stolyar)ら(Mikrobiologiya 64(5):686−691(1995));モトヤマ(Motoyama)ら(Appl.Micro.Biotech.42(1):67〜72(1994));ロイド(Lloyd)ら(Archives of Micro
biology 171(6):364−370(1999));およびオドム(Odom)ら(国際公開第02/18617号パンフレット)の研究が挙げられる。
【0182】
カロテノイドの生体内(in vitro)生物転換
代案としては、生体内(in vitro)で本願明細書の生物転換を行うことが可能である。CrtE、CrtY、CrtI、CrtB、およびCrtZのための基質が、宿主細胞によって内在性に合成されない場合、基質を外来性に添加することが可能である。この実施態様では、適切なカロテノイド基質を中性洗剤(例えばDMSO)で可溶化してもまたはリン脂質ベシクルと混合しても良い。細胞中への輸送を助けるために、宿主細胞を場合によりトルエンなどの適切な溶剤で透過化処理しても良い。このタイプのカロテノイド基質の生体外(in−vitro)生物転換法は、技術分野に基礎がある(例えばハンドル(Hundle)B.S.ら、FEBS、315:329〜334(1993);およびブラムリー(Bramley)、P.M.ら、Phytochemistry、26:1935〜1939(1987)を参照されたい)。
【0183】
組換え微生物を使用した工業生産
本タンパク質の商業的生産が所望される場合、多様な培養方法を応用しても良い。例えば、組換え微生物宿主から過剰発現する特定の遺伝子産物の大規模生産は、バッチまたは連続の培養方法の双方で生産しても良い。
【0184】
古典的なバッチ培養法は閉鎖システムであり、そこでは培養液の組成が発酵の最初に設定され、培養プロセス中の人為的変化を受けない。したがって培養プロセス開始時に培養液に所望の生物体または生物体群を接種し、生育または代謝活動を生じさせながらシステムには何も添加しない。しかし典型的には「バッチ」培養は、炭素源の添加に関するバッチであり、pHおよび酸素濃度などの因子の調節が試みられることが多い。バッチシステムでは、システムの代謝産物および生物質量組成は、培養が終結する時点まで常に変化する。バッチ培養内で細胞は、静的な遅滞期から高い対数増殖期へ、そして最後に成長率が減退または停止する静止期へ調節される。処置を施さない場合、静止期にある細胞はやがて死滅する。システムによっては対数期にある細胞が、最終生成物または中間体の生産の大部分を担うことが多い。その他のシステムでは、静止または対数期後生成を得ることができる。
【0185】
標準バッチシステムのバリエーションが、流加バッチシステムである。流加バッチ培養プロセスも本発明において適切であり、培養の進行と共に基質が段階的に添加されること以外は、典型的なバッチシステムを含んでなる。流加バッチシステムは、異化代謝産物抑制が細胞の代謝を阻害する傾向があって、培養液中に限定量の基質を有することが望ましい場合に有用である。流加バッチシステム中の実際の基質濃度の測定は困難であるので、pH、溶存酸素、およびCO2などの排ガス分圧などの測定可能因子の変化に基づいて推定される。バッチおよび流加バッチ培養法は、技術分野で一般的であり周知であって、実例はブロック(Brock)(前出)およびデシュパンデ(Deshpande)(前出)にある。
【0186】
本タンパク質の商業的生成は、連続培養によって達成しても良い。連続の培養は開放システムであり、規定の培地をバイオリアクターに連続的に添加して、等量の慣熟培地を工程から同時に除去する。連続培養は、概して細胞を恒常的な高い液相密度に維持し、そこでは細胞が主に対数増殖期にある。代案としては連続培養を固定細胞で実施しても良く、そこでは炭素および養素が連続的に添加され、価値ある生成物、副産物または老廃物は細胞集団から連続的に除去される。細胞固定化は、天然および/または合成材料から構成される広範囲の固体担体を使用して実施しても良い。
【0187】
連続または半連続培養は、細胞生育または最終生成物濃度に影響する1つの因子またはあらゆるいくつかの因子の調節を可能にする。例えば一方法では、炭素源または窒素レベルなどの制限的栄養物質を固定された割合に維持し、その他の全パラメーターの調節ができるようにする。別のシステムでは、培地濁度によって測定される細胞濃度を一定に保ちながら、生育に影響するいくつかの因子を連続的に変化させることができる。連続システムは定常状態生育条件を維持することを目指すので、培地が抜き取られることによる細胞損失は、培養中の細胞生育率に対してバランスが取れていなくてはならない。連続的培養プロセスのために栄養素および成長因子を調節する方法、ならびに生成物形成速度を最大化する技術は工業微生物学の技術分野で良く知られており、多様な方法が上記のブロック(Brock)で詳述される。
【0188】
本発明における発酵培地は、適切な炭素基質を含有しなくてはならない。適切な基質としては、単糖類(例えばグルコースおよびフルクトース)、二糖類(例えばラクトースまたはスクロース)、多糖類(例えばデンプンまたはセルロースまたはそれらの混合物)、再生可能な供給材料(例えば乳清透過液、コーンスティープリーカー、甜菜モラセス、および大麦の麦芽)からの未精製混合物が挙げられるが、これに限定されるものではない。さらに炭素基質は、重要な生化学的中間体への代謝転換が実証されている、二酸化炭素、メタンまたはメタノールなどの一炭素基質であっても良い。一または二炭素基質に加えて、メチロトローフ生物体はまた、メチルアミン、グルコサミン、および代謝活性のための多様なアミノ酸などのいくつかのその他の炭素含有化合物を利用することが知られている。例えば、メチロトローフ酵母は、メチルアミンからの炭素を利用してトレハロースまたはグリセロールを形成することが知られている(ベリオン(Bellion)ら、C1化合物上での微生物の生育(Microb.Growth C1 Compounds)[国際シンポジウム]第7回(1993)、415〜32、マレルJ.コリン(Murrell J.Collin)およびドンP.ケリー(Don P.Kelly)編、インターセプト:英国アンドーバー)。同様に、様々なカンジダ(Candida)種が、アラニンまたはオレイン酸を代謝する(サルター(Sulter)ら、Arch.Microbiol.153:485〜489(1990))。したがって本発明で用いられる炭素源は多種多様な炭素含有基質を包含しても良く、生物体の選択によってのみ制限されることが考察された。
【0189】
植物における組換え生成
植物および藻類もまた、カロテノイド化合物を製造することが知られている。本発明のcrtE、idi、crtY、crtI、crtB、およびcrtZ核酸断片を使用して、微生物タンパク質を発現する能力を有する遺伝子導入植物を作り出しても良い。好ましい植物宿主は、本タンパク質の高生成レベルを支えるいずれかの品種であることができる。適切な緑色植物としては、ダイズ、アブラナ(Brassica napus、B.campestris)、ヒマワリ(Helianthus annus)、綿(Gossypium hirsutum)、トウモロコシ、タバコ(Nicotiana tabacum)、アルファルファ(Medicago sativa)、小麦(Triticum種)、大麦(Hordeum vulgare)、オート麦(Avena sativa、L)、モロコシ(Sorghum bicolor)、イネ(Oryza sativa)、シロイヌナズナ、アブラナ科の野菜(ブロッコリー、カリフラワー、キャベツ、アメリカボウフウなど)、メロン、ニンジン、セロリ、パセリ、トマト、ジャガイモ、オランダイチゴ、落花生、ブドウ、草の種、テンサイ、サトウキビ、マメ、エンドウ豆、ライ麦、亜麻、堅木、軟木、および飼料としての草が挙げられるが、これに限定されるものではない。藻類のとしては、スピルリナ(Spirulina)、ヘモタコックス(Haemotacoccus)、およびデュナリエラ(Dunalliela)などの種商業的に重要な宿主が挙げられるが、これに限定されるものではない。
【0190】
好ましいカロテノイド化合物の過剰発現は、最初に本発明のキメラ遺伝子を構築することによって達成されても良く、その中でコード領域は、所望の発生段階で所望の組織中の遺伝子の発現を導くプロモーターに作動的に結合される。便宜上、キメラ遺伝子は、同一遺伝子に由来するプロモーター配列および翻訳リーダー配列を含んでなっても良い。転写終止シグナルをコードする3’非コード配列もまた、提供されなくてはならない。本キメラ遺伝子はまた、遺伝子発現を容易にするために1つもしくはそれ以上のイントロンも含んでなる。
【0191】
コード領域の発現を誘導できるあらゆるプロモーターとあらゆるターミネーターのあらゆる組み合わせをキメラ遺伝子配列で使用しても良い。プロモーターおよびターミネーターのいくつかの適切な例としては、ノパリンシンターゼ(nos)、オクトピンシンターゼ(ocs)、およびカリフラワーモザイクウィルス(CaMV)遺伝子からのものが挙げられる。使用しても良い効率的な植物プロモーターの1タイプは、高等植物プロモーターである。本発明の遺伝子配列と機能的に連結するこのようなプロモーターは、本遺伝子産物の発現を促進できなくてはならない。本発明で使用しても良い高等植物プロモーターとしては、例えば1.)ダイズからのリブロース−1,5−ビスリン酸カルボキシラーゼの小型サブユニット(ss)のプロモーター(ベリー−ロウ(Berry−Lowe)ら、J.Molecular and App.Gen.、1:483〜498(1982))、および2.)クロロフィルa/b結合タンパク質のプロモーターが挙げられる。これらの2つのプロモーターは、植物細胞において光誘導されることが知られている(例えば「植物の遺伝子操作、農業における展望(Genetic Engineering of Plants,an Agricultural Perspective)」A.キャッシュモア(Cashmore)編、Plenum:ニューヨーク(1983)、p29〜38;コルッジ(Coruzzi)G.ら、J.Biol.l Chem.、258:1399(1983);およびダンスミア(Dunsmuir)P.ら、J.Mol.Appl.Genet.、2:285(1983)参照)。
【0192】
次に本キメラ遺伝子を含んでなるプラスミドベクターを構築できる。プラスミドベクターの選択は、宿主植物を形質転換するのに使用される方法に左右される。当業者は、キメラ遺伝子を含有する宿主細胞を成功裏に形質転換し、選択して増殖するためにプラスミドベクター上に存在しなくてはならない遺伝的要素を良く知っている。当業者はまた、異なる独立形質転換イベントが、異なるレベルおよびパターンの発現をもたらすことを認識するので(ジョーンズ(Jones)ら、EMBO J.4:2411〜2418(1985);デ・アルメイダ(De Almeida)ら、Mol.Gen.Genetics
218:78〜86(1989))、所望の発現レベルおよびパターンを示す系統を得るためには、複数イベントをスクリーニングしなくてはならない。このようなスクリーニングは、DNAブロットのサザン分析(サザン(Southern)、J.Mol.Biol.98:503(1975))、mRNA発現のノーザン分析(クロツェック(Kroczek)、J.Chromatogr.Biomed.Appl.、618(1〜2):133〜145(1993))、タンパク質発現のウエスタン分析、または表現型分析によって達成しても良い。
【0193】
用途によっては、本タンパク質を異なる細胞区画に方向付けることが有用であろう。したがって適切な細胞内標的配列を添加することで、および/または以下のような既存の標的配列を除去することで、酵素をコードするコード配列を変更して、上述のキメラ遺伝子をさらに補完しても良いことが想定された。1.)トランジット配列(キーグストラ(Keegstra)K.、Cell 56:247〜253(1989))、2.)シグナル配列、または3.)小胞体局在化(クリスピールス(Chrispeels)J.J.、Ann.Rev.Plant Phys.Plant Mol.Biol.42:21〜53(1991))または核局在化シグナル(ライケル(Raikhel)、N.、P
lant Phys.100:1627〜1632(1992))をコードする配列。引用した参考文献はこれらの各例を示すが、リストは網羅的でなく、有用なさらに多くの標的シグナルが、将来発見されるかもしれない。
【0194】
タンパク質操作
本crtE、idi、crtY、crtI、crtB、および/またはcrtZヌクレオチドを使用して、向上されたまたは改変された活性を有する遺伝子産物を製造できるかもしれないことが考察された。1.)誤りがちなPCR(メルニコフ(Melnikov)ら、Nucleic Acids Research、27(4):1056〜1062(1999年2月15日))、2.)部位特異的変異誘発(クームズ(Coombs)ら、Proteins(1998)、p259〜311、1プレート(アンジェレッティ,ルース・ホーグ(Angeletti,Ruth Hogue)編、アカデミック:カリフォルニア州サンディエゴ(San Diego、CA))、および3.)「遺伝子シャフリング」(本願明細書に援用する米国特許第5,605,793号明細書、米国特許第5,811,238号明細書、米国特許第5,830,721号明細書、および米国特許第5,837,458号明細書)をはじめとするが、これに限定されるものではない、天然遺伝子配列を変異させて活性が改変または向上された遺伝子産物を生成する様々な方法が知られている。
【0195】
遺伝子シャッフリング法は、その容易な実行、および高率な変異誘発と容易なスクリーニングのために特に魅力的である。遺伝子シャフリングのプロセスは、対象の遺伝子と類似したまたは違っている追加的なDNA領域集団存在下で、対象の遺伝子を特定サイズの断片に制限エンドヌクレアーゼ開裂することを伴う。次に断片のプールを変性させ再アニールして、変異遺伝子を作り出す。次に変異した遺伝子を変更した活性についてスクリーニングする。
【0196】
本発明の本微生物の配列を変異させて、この方法によって変更されまたは増強される活性についてスクリーニングしても良い。配列は二本鎖であるべきで、50bp〜10kB範囲の様々な長さであることができる。配列は技術分野で良く知られている制限エンドヌクレアーゼを使用して、約10bp〜1000bpの範囲の断片に無作為に消化されても良い(マニアティス(Maniatis)、同上)。本微生物の配列に加えて、微生物の配列の全部または部分とハイブリダイズ可能な断片集団を添加しても良い。同様に、本配列とハイブリダイズ可能でない断片集団を添加しても良い。典型的にこれらの追加的断片集団は、総核酸と比較して重量で約10〜20倍過剰に添加される。一般にこのプロセスに従えば、混合物中の異なる特定の核酸断片の数は約100〜約1000個になる。無作為核酸断片の混合集団は、変性されて一本鎖核酸断片を形成し、次に再アニールされる。その他の一本鎖核酸断片との相同性領域を有する一本鎖核酸断片のみが再アニールする。無作為核酸断片は、加熱によって変性されても良い。当業者は、二本鎖核酸を完全に変性するのに必要な条件を判定できる。好ましくは温度は、約80℃〜100℃である。核酸断片を冷却によって再アニールしても良い。好ましくは温度は約20℃〜75℃である。再生は、ポリエチレングリコール(「PEG」)または塩の添加によって加速できる。適切な塩濃度は、0mM〜200mMの範囲であっても良い。次にアニールされた核酸断片は、核酸ポリメラーゼおよびdNTP(すなわちdATP、dCTP、dGTPおよびdTTP)存在下でインキュベートされる。核酸ポリメラーゼは、クレノウ断片、Taqポリメラーゼまたは技術分野で知られているあらゆるその他のDNAポリメラーゼであっても良い。アニールに先だって、アニールと同時に、またはアニール後に、ポリメラーゼを無作為核酸断片に添加しても良い。変性、再生、およびポリメラーゼ存在下でのインキュベーションのサイクルは、所望の回数反復される。好ましくはサイクルは約2〜50回反復され、より好ましくは手順は10〜40回反復される。得られる核酸は約50bp〜約100kBの範囲のより大きな二本鎖ポリヌクレオチドであり、標準クローン化および発
現プロトコルによって、発現および変更された活性についてスクリーニングしても良い(マニアティス(Manatis)、同上)。
【0197】
さらにハイブリッドタンパク質は、遺伝子シャフリング(エクソンシャフリング)法を使用した、機能性領域の融合によってアセンブルできる(ニクソン(Nixon)ら、Proc.Natl.Acad.Sci.USA、94:1069〜1073(1997))。本遺伝子の機能性領域は、その他の遺伝子の機能性領域と組み合わせて、所望の触媒機能を有する新しい酵素作り出すことができる。PCRオーバーラップ伸長法を使用してハイブリッド酵素を構築し、当業者に良く知られている技術使用して、様々な発現ベクター中にクローンしても良い。
【実施例】
【0198】
以下の実施例で本発明をさらに明らかにする。これらの実施例は、本発明の好ましい実施態様を示しながら、例証のみのために提供されるものと理解される。上記考察およびこれらの実施例から、当業者は本発明の本質的特質を把握でき、その範囲と精神を逸脱することなく、本発明の様々な変化と修正を行って、様々な利用法および条件に適合させることができる。
【0199】
一般方法
実施例で使用される標準組換えDNAおよび分子クローン化技術は技術分野で良く知られており、マニアティス(Maniatis)(前出)、シルハビー(Silhavy)ら(前出)、およびオースベルら(Ausubel)(前出)によって述べられている。
【0200】
細菌培養の維持および生育に適した材料および方法は、技術分野で良く知られている。以下の実施例で使用するのに適した技術は、下に述べられる。「一般微生物学方法マニュアル(Manual of Methods for General Bacteriology)」、フィリップス・ゲアハルト(Phillipp Gerhardt)、R.G.E.マリー(R.G.E.Murray)、ラルフN.コスティロウ(Ralph N.Costilow)、ユージーンW.ネスター(Eugene W.Nester)、ウィリスA.ウッド(Willis A.Wood)、ノエルR.クリーグ(Noel R.Krieg)、およびG.ブリッグス・フィリップス(G.Briggs Phillips)編、米国微生物学会、ワシントンD.C.(Washington,D.C.)(1994)またはブロック(Brock)ら(前出)。細菌細胞の生育および維持のために使用される全ての試薬、制限酵素および材料は、特に断りのない限り、ウィスコンシン州ミルウォーキーのアルドリッチ・ケミカルズ(Aldrich Chemicals(Milwaukee、WI))、ミシガン州デトロイトのディフコ・ラボラトリーズ(DIFCO Laboratories(Detroit、MI))、メリーランド州ゲーサーズバーグのギブコ/BRL(GIBCO/BRL(Gaithersburg、MD))、またはミズーリ州セントルイスのシグマケミカル(Sigma Chemical Company(St.Louis、MO))から得た。
【0201】
配列データは、ベクターと挿入特異性プライマーの組み合わせを使用して、染料ターミネーター技術(米国特許第5,366,860号明細書、欧州特許第272,007号明細書)を使用して、ABI自動シーケンサー上で生成した。ミシガン州アナーバーのジーン・コーズ社(Gene Codes Corp.(Ann Arbor、MI))からのシーケンチャー(Sequencher)(商標)バージョン4.0.5中で配列編集およびアセンブリーを実施した。全配列は、双方向で少なくとも2回のカバレッジを示す。遺伝子配列の操作は、メリーランド州ベセズダのインフォマックス(InforMax,Inc.(Bethesda、MD))からのベクターNTIプログラムを使用して達成した。ベクターNTIにおけるデフォルト値を使用して、対合比較を実施した。米国バ
イオテクノロジカル情報センター(NCBI)で設定されたデフォルト値を使用して、BLAST分析を実施した。
【0202】
略語の意味は以下の通り。「sec」は秒を意味し、「min」は分を意味し、「h」は時間を意味し、「d」は日を意味し、「μL」はマイクロリットルを意味し、「mL」はミリリットルを意味し、「L」はリットルを意味し、「μM」はマイクロモル(micromolar)を意味し、「mM」はミリモル(millimolar)を意味し、「M」はモル(molar)を意味し、「mmol」はミリモル(millimole)を意味し、「μmol」マイクロモル(micromole)を意味し、「g」はグラムを意味し、「μg」はマイクログラムを意味し、「ng」はナノグラムを意味し、「U」は単位を意味し、「bp」は塩基対を意味し、「kB」はキロベースを意味する。
【0203】
実施例1
カロテノイド産生株パントエア・アグロメランス(Pantoea agglomerans)DC404の単離
本実施例は、腸内細菌科に属する黄色素細菌株パントエア・アグロメランス(Pantoea agglomerans)DC404株の単離および同定について述べる。この生物体中で生成された天然カロテノイドの分析は、様々なゼアキサンチン前駆物質およびゼアキサンチン誘導体に加えて、ゼアキサンチンの生成を証明する。
【0204】
株単離および分類:新らしいカロテノイド産生細菌株を単離するために、環境サンプルの収集物から着色微生物を単離した。デラウェア州ウィルミントンの家庭菜園土壌を採取して、ルリア・ブロス(LB)に再懸濁した。10μLのループを満たす再懸濁液をLBプレート上に画線して、プレートを30℃でインキュベートした。多様なコロニーの外見を有する着色細菌を選んで、LBプレート上に均一に2回画線して、30℃でインキュベートした。これらのコロニーから、淡黄色で滑らかな半透明コロニーを形成したものを「DC404株」と命名した。
【0205】
タイプDC404株で16S rRNA遺伝子配列決定を実施した。具体的には、プライマーHK12(配列番号13)およびJCR14(配列番号14)を使用して、株の16S rRNA遺伝子をPCRによって増幅した。製造業者(キアジェン(Qiagen))の説明書に従って、QIAquick PCR精製キットを使用して増幅した16S
rRNA遺伝子を精製し、自動ABIシーケンサーで配列決定した。配列決定反応をプライマーHK12、JCR14、およびJCR15(配列番号15)で開始した。アセンブルしたDC404の1324bpの16S rRNA遺伝子配列(配列番号16)をジェンバンク(登録商標)に対するBLASTN検索のためのクエリー配列として使用した(アルトシュル(Altschul)ら、Nucleic Acids Res.25:3389〜3402(1997))。
【0206】
BLAST分析からは、DC404株が腸内細菌科に属することが示された。その16S rRNA遺伝子配列はこの科内のいくつかの異なる種との配列類似性を示し、トップヒット(96%同一性)はネズミチフス菌(Salmonella typhimurium)であった。上で述べた分析に従って、DC404株の16S rRNA遺伝子配列との96%同一性を有するその他の種としては、クレブシエラ・オキシトカ(Klebsiella oxytoca)、エンテロバクター・クロアカ(Enterobacter cloacae)、シトロバクター・アマロナチカス(Citrobacter amalonaticus)、エルウィニア・アミロボラ(Erwinia amylovora)、およびパントエア・アグロメランス(Pantoea agglomerans)が挙げられる。しかし比較的低いヒットの相同性(96%のみの同一性)に一致して、メリーランド州コッキーズヴィルのベクトン・ディキンソン(Becton Dick
inson(Cockeysville、MD))からのBBLエンテロチューブTM IIを使用した生化学的試験は、DC404がサルモネラ(Salmonella)種でないことを示唆する。
【0207】
外部会社であるデラウェア州ニューアークのマイクロビアルID(Microbial
ID,Inc.(Newark、DE))によるその脂肪酸プロフィールに従って、DC404をさらに特性決定した。ガスクロマトグラフィーによる脂肪酸プロファイリングは、DC404株がパントエア・アグロメランス(Pantoea agglomerans)に最も近縁であることを示唆した(類似度指数は0.557であった)。類似度指数は、未知のサンプルの脂肪酸組成が、そのマッチとしてリストされたライブラリーエントリを作成するのに使用した株の平均脂肪酸組成と比較して、どれだけ近いかを表す数値である。未知のサンプルの脂肪酸組成と、ライブラリーエントリの平均値との正確な一致は、類似度指数1.000をもたらす。0.400〜0.600の類似度指数は種の一致であるかもしれず、異型株を示す。DC404株の16S rRNA分析はまた、トップBLASTヒット(96%の同一性を有する)の1つとして、パントエア・アグロメランス(Pantoea agglomerans)を同定したので、パントエア・アグロメランス(Pantoea agglomerans)の異型株としてDC404株を命名した。
【0208】
DC404のカロテノイド分析:DC404中の黄色色素を抽出してHPLCで分析した。株を200mLのLB中で30℃において2日間生育させ、次に細胞を4000gで30minの遠心分離によって収集した。細胞ペレットを10mLのアセトンで抽出した。溶剤を窒素下で乾燥させ、カロテノイドを0.5mLのアセトンに再懸濁した。抽出物をミシガン州アナーバーノのパル・コーポレーション(Pall Corporation(Ann Arbor、MI))からのアクロディスク(Acrodisc)CR25mmシリンジフィルターで濾過して、次にカリフォルニア州フォスター・シティのアジレント(Agilent、(Foster City、CA))からのアジレント(Agilent)シリーズ1100 LC/MSD SIを使用して、0.1mLの10%アセトン+90%HPLC分析用アセトニトリル中で濃縮した。
【0209】
サンプル(20μL)を150mm×4.6mmのZORBAX C18(3.5μm粒子)カラム(アジレントテクノロジーズ(Agilent Technologies,Inc.))に装填した。カラム温度を40℃に保った。流速は1mL/分であり、溶剤ランニングプログラムは次のとおりであった。
・0〜2分:95%緩衝液Aおよび5%緩衝液B、
・2〜10分:95%緩衝液Aおよび5%緩衝液Bから60%緩衝液Aおよび40%緩衝液Bへの直線的濃度勾配、
・10〜12分:60%緩衝液Aおよび40%緩衝液Bから50%緩衝液Aおよび50%緩衝液Bへの直線的濃度勾配、
・12〜18分:50%緩衝液Aおよび50%緩衝液B、
・18〜20分:95%緩衝液Aおよび5%緩衝液B。
【0210】
緩衝液Aは95%アセトニトリルおよび5%dH2Oであり、緩衝液Bは100%テトラデヒドロフランであった。
【0211】
HPLC分析(図3)は、標品との比較で、DC404株がゼアキサンチン(6.28minピーク)、β−クリプトキサンチン(10.23minピーク)およびβ−カロテン(13.02minピーク)を生成したことを示唆した。具体的には、ゼアキサンチンおよびβ−クリプトキサンチン標品は、スイス国ルプシンゲンのカロテナチュール(CaroteNature(Lupsingen、Switzerland))から購入し、
β−カロテン標品は、ミズーリ州セントルイスのシグマ(Sigma(St.Louis、MO))から購入した。MS分析からゼアキサンチンピークの分子量が569であり、β−クリプトキサンチンピークの分子量が553であり、β−カロテンピークの分子量がが537であることを確認した。3.22minに溶出するピークは、その分子量731が示唆するように、ゼアキサンチンモノグルコシドである可能性が高い。
【0212】
実施例2
パントエア・アグロメランス(Pantoea agglomerans)DC404着色コスミドクローンの同定
実施例2は、パントエア・アグロメランス(Pantoea agglomerans)DC404からのゲノムDNAの約40kBの断片を発現できる、大腸菌(E.coli)コスミドクローンの構造について述べる。この形質転換体は、主にゼアキサンチンを生成した。大腸菌(E.coli)コスミドクローン中にゼアキサンチングルコシドは検出されなかった。
【0213】
染色体DNAの調製:DC404株を25mLのLB培地内で30℃で一晩通気して生育させた。細菌性細胞を4,000gで10min遠心分離した。細胞ペレットを5mLの50mMトリス−10mM EDTA(pH8)中に穏やかに再懸濁して、リゾチームを最終濃度2mg/mLに添加した。懸濁液を37℃で1hインキュベートした。次にドデシル硫酸ナトリウムを最終濃度1%に添加して、タンパク質分解酵素Kを100μg/mLで添加した。懸濁液を55℃で2hインキュベートした。懸濁液は透明になり、等容積のフェノール:クロロホルム:イソアミルアルコール(25:24:1)で2回、クロロホルム:イソアミルアルコール(24:1)で1回透明な溶解産物を抽出した。4,000rpmで20minの遠心処理後、水相を注意深く除去して新しい試験管に移した。2容積のエタノールを添加して、密封したガラス・パスツール・ピペットにDNAを穏やかに巻き付けた。DNAを70%エタノールを含有する試験管に浸した。風乾後、DNAをRNaseA(100μg/mL)と共に400μLのTE(10mMトリス−1mM
EDTA、pH8)に再懸濁し、4℃に保管した。DNAの濃度および純度を分光光度法によりOD260/OD280で測定した。
【0214】
コスミドライブラリーの構築:ウィスコンシン州マディソンのエピセンター・テクノロジーズ(Epicentre Technologies(Madison、WI))からのpWEBコスミドクローン化キットを使用して、製造業者の説明書に従って、DC404のコスミドライブラリーを構築した。シリンジニードルを通過させて、ゲノムDNAを剪断した。剪断したDNAを末端修復して、40kBの標準との比較によって低融点アガロース上でサイズ選択した。大きさがほぼ40kBのDNA断片を精製して、ブラントエンドクローン化レディーのpWEBコスミドベクター中にライゲートした。超高効率MaxPlaxラムダ・パッケージング・エキストラクツ(Lambda Packaging Extracts)を使用して、ライブラリーをパッケージし、EPI100大腸菌(E.coli)細胞上に播種した。コスミドライブラリークローンから2個の黄色コロニーが同定された。2個のクローンからのコスミドDNAは、類似した制限酵素消化パターンを有したので、1個のクローン上でさらなる分析を実施した。
【0215】
黄色コスミドクローンのカロテノイド分析:実施例1で述べられるようにしてLC−MSによって、コスミドpWEB−404を含有する大腸菌(E.coli)EPI100中のカロテノイドを分析した。HPLCの結果を図4に示す。そのUVスペクトル、分子量、および標品との比較に基づいて、6.27minのピークはゼアキサンチンと同定された。顕著な量のβ−カロテンまたはβ−クリプトキサンチン中間体のいずれも蓄積せず、ゼアキサンチングルコシドは生成されなかった。DC404(図3)中で観察された顕著なゼアキサンチンモノグルコシドピークは、この大腸菌(E.coli)コスミドクロ
ーン中には存在しなかった。分子重量がゼアキサンチンモノグルコシド(731ダルトン)と一致しなかったので、3.56minの小さなピークはゼアキサンチンモノグルコシドの異性体ではなかった。コスミドpWEB−404を含有する大腸菌(E.coli)内のゼアキサンチングルコシドの不在は、ゼアキサンチングルコシル転移酵素をコードするcrtX遺伝子が、コスミドpWEB−404中に存在しないかもしれないことを示唆する。
【0216】
実施例3
カロテノイド生合成遺伝子の同定
この実施例は、コスミドpWEB−404中のパントエア・アグロメランス(Pantoea agglomerans)DC404 crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子の同定について述べ、その他の知られているパントエア(Panteoa)crt遺伝子に関わるこれら遺伝子の関連性の比較を提供する。その他のパントエア(Panteoa)中の下流カロテノイド生合成経路の遺伝子構成とは対照的に、crtX遺伝子は、ここで述べるP.アグロメランス(P.agglomerans)DC404からのコスミド上に遺伝子クラスターの一部として存在しなかった。
【0217】
HPLC分析は、コスミドpWEB−404が、ゼアキサンチンの合成のための遺伝子を含有するに違いないことを示唆した。カロテノイド合成遺伝子を配列決定するために、ウィスコンシン州マディソンのエピセンター(Epicentre(Madison、WI))からのEZ::TN<TET−1>キットを使用して製造業者の説明書に従って、コスミドDNA pWEB−404に生体外(in vitro)遺伝子転移を行った。TET−1 FP−1フォーワードプライマー(配列番号17)を使用して、トランスポゾン末端から、200個のテトラサイクリン抵抗性トランスポゾンインサートを配列決定した。ミシガン州アナーバーのジーン・コーズ社(Gene Codes Corp.(Ann Arbor、MI))からのシーケンチャー(Sequencher)プログラムによって、配列アセンブリーを実施した。DC404からのカロテノイド生合成経路の6個の遺伝子を含有する、8814bpのcontig(配列番号18)をアセンブルした(図5)。
【0218】
BLAST「nr」データベース(全ての非重複性ジェンバンク(登録商標)CDS翻訳、三次元構造ブルックヘブンタンパク質データバンクから誘導される配列、SWISS−PROTタンパク質配列データベース、EMBL、およびDDBJデータベースを含んでなる)に含有される配列との類似性に対するBLAST(基礎的局在性整列化検索ツール(Basic Locl Alignment Search Tool)アルトシュール(Altschul)ら、前出)検索を行って、crtE、idi、crtY、crtI、crtB、およびcrtZをコードする遺伝子を同定した。米国バイオテクノロジー情報センター(NCBI)が提供するBLASTNアルゴリズムを使用して、「nr」データベースに含有されるあらゆる公的に入手できるDNA配列との類似性について、各配列を分析した。DNA配列を全ての読み取り枠で翻訳し、NCBIが提供するBLASTXアルゴリズムを使用して、「nr」データベースに含有されるあらゆる公的に入手できるタンパク質配列との類似性について比較した(ギッシュ(Gish)W.およびステート(States)D.J.、Nature Genetics 3:266〜272(1993))。
【0219】
BLASTNnrまたはBLASTXnrアルゴリズムのどちらかを使用して、全ての比較を行った。BLAST比較の結果を表2に示し、各遺伝子が最も類似する配列を要約する。表2は、期待値に報告される値を有するBLASTXnrアルゴリズムに基づくデータを示す。期待値は、このサイズのデータベースの検索において絶対的に偶然に期待される、規定のスコアにおけるマッチ数を特定して、マッチの統計学的有意さを推定する。
【0220】
ベクターNTIプログラム(前出)を使用して、ヌクレオチドおよびアミノ酸配列もまた、その他のパントエア(Pantoea)株からのものと比較した。表3に対合比較の同一性を要約する。
【0221】
【表3】
【0222】
【表4】
【0223】
【表5】
【0224】
様々なパントエア(Pantoea)種中のcrt遺伝子クラスター遺伝子構成の比較からは、遺伝子idiおよびcrtXの存在に関する興味深い違い(図5)もまた明らかにされた。イソペンテニルピロリン酸イソメラーゼをコードする遺伝子idiは、上流イソプレノイド経路に関与し、例えばパントエア・アナナティス(Pantoea ananatis)(ジェンバンク(登録商標)登録番号D90087)およびパントエア・ステワルティ(Pantoea stewartii)(ジェンバンク(登録商標)登録番号AY1667131)中のcrtEXYIBZクラスターの遺伝子構成のように、通常、crt遺伝子クラスター内には存在しない。しかしときどき、パントエア・アグロメランス(Pantoea agglomerans)EHO−10(ジェンバンク(登録商標)登録番号M87280)で見られるように、idiがcrt遺伝子オペロンの一部であることが見いだされる。本願明細書の出願人が行ったBLAST分析に従って、P.アグロメランス(P.agglomerans)EHO−10(ジェンバンク(登録商標)登録番号M87280)のcrtE−仮説タンパク質−crtX−crtY−crtI−crtB−crtZのカロテン産生性クラスター内に含有される「仮説タンパク質」は、crtE−idi−crtX−crtY−crtI−crtB−crtZを含んでなると見なされるべきである。同様にidiはまた、パントエア・アグロメランス(Pantoea agglomerans)DC404のcrt遺伝子クラスター中にも存在し、crtE遺伝子の下流である。しかし表3に示すように、DC404株のidiは、EHO−10のidiと68%のみの同一性を有する。
【0225】
crtX遺伝子に関してDC404株は、ここで述べるカロテン産生性遺伝子クラスター内にこの遺伝子が不在である点が、その他のパントエア(Pantoea)種と比べてユニークである(図5)。さらにコスミドクローンはゼアキサンチンのみを生成し、ゼアキサンチングルコシドを生成しないので、DC404からのcrt遺伝子クラスターを含有する大腸菌(E.coli)コスミドクローンpWEB−404はcrtX遺伝子を含有しない。ここで述べる遺伝子クラスター中のcrtX遺伝子の欠如にも関わらず、DC404はゼアキサンチンモノグルコシドを主に生成するので、crtX遺伝子を有さねばならない。crtX遺伝子はDC404染色体中の別の場所に位置し、crt遺伝子クラスターとは連結していないと結論された。DC404株中のcrtXの配列は、今回は同定されなかった。
【0226】
実施例4
メチロモナス(Methylomonas)sp.16aにおけるパントエア・アグロメランス(Pantoea agglomerans)DC404のcrtEidiYIB遺伝子クラスターの発現
以下の実施例は、パントエア・アグロメランス(Pantoea agglomerans)DC404(実施例3)からのcrtEidiYIB遺伝子を含んでなるcrt遺伝子クラスターをメチロモナス(Methylomonas)16a(ATCC PTA2402)中に導入して、β−カロテンなどの望ましい40−炭素カロテノイドを合成できるようにすることについて述べる。
【0227】
最初に、プライマーpWEB404F:5’−GAATTCACTAGTCGAGACGCCGGGTACCAACCAT−3’(配列番号19)およびpWEB404R:5’−GAATTCTAGCGCGGGCGCTGCCAGA−3’(配列番号20)を使用して、PCRによって、crtEidiYIB遺伝子を含有するDC404からの断片を増幅した。以下の熱サイクル条件で、カリフォルニア州ラホーヤのストラタジーン(Stratagene、La Jolla、CA)からのPfu Turboポリメラーゼと共に、コスミドDNA pWEB−404をテンプレートとして使用した。92℃(5min);94℃(1min)、60℃(1min)、72℃(9min)を25サイクル;および72℃(10min)。ゲル電気泳動法に続いて、およそ5.6kBの単一生成物が観察された。パーキン・エルマー(Perkin Elmer)からのTaqポリメラーゼを10分間の72℃の反応で使用して、インビトロゲン(Invitrogen)からのpTrcHis2−TOPO中へのTOPOクローン化のために、追加的な3’アデノシンヌクレオチドを断片に付加した。大腸菌(E.coli)TOP10細胞中への形質転換に続いて、いくつかのコロニーの色が明るい黄色に見え、それらがカロテノイド化合物を生成していることが示唆された。次に遺伝子クラスターをフロリダ州マルコ・アイランドのモビテック(MoBiTec,LLC(Marco Island、FL))からの広範宿主ベクターpBHR1中にサブクローン化し、ウィスコンシン州ミドルタウンのルシジェン(Lucigen(Middletown、WI))からの大腸菌(E.coli)10G細胞中に電気穿孔した。50μg/mLのカナマイシンを含有するLB培地上で、生成したプラスミドpDCQ330を含有する形質転換体を選択した。
【0228】
三親交雑接合噛合によって、プラスミドpDCQ330をメチロモナス(Methylomonas)16a中に転位した。pRK2013(ATCC番号37159)を含有する大腸菌(E.coli)ヘルパー株、およびpDCQ330を含有する大腸菌(E.coli)10G供与株をカナマイシン(50μg/mL)を含有するLB培地中で一晩生育させてLB中で3回洗浄し、元の培養容積のほぼ60倍の濃度に相当する容積のLB中に再懸濁した。
【0229】
メチロモナス(Methylomonas)16a株MWM1000(Δald/crtN1)は、天然C30カロテノイドの合成を中断させるald/crtN1遺伝子の単一クロスオーバーノックアウトを含有した(米国特許出願第60/527083号)。国際公開第02/18617号パンフレットで述べられる一般条件を使用して、この(Δald/crtN1)株を受容体として生育させた。簡単に述べると、少なくとも8:1の気体/液体比を使用して(すなわち160mL総容積中の20mLの硝酸塩液体「BTZ−3」培地)、30℃で絶えず振盪しながら、イリノイ州ウィートンのウィートン・サイエンティフィック(Wheaton Scientific、(Wheaton、IL))からの血清ストッパーしたウィートンボトル内で、メチロモナス(Methylomonas)16a株MWM1000を生育させた。
【0230】
ここで「合成培地」または「BTZ−3」培地とも称される硝酸塩液体培地は、下に示す溶液1と混合された様々な塩(表4および5)から構成され、または規定の硝酸塩が15mMの塩化アンモニウムで置き換えられている。溶液1は、100倍に濃縮された微量元素原液のための組成物を提供する。
【0231】
【表6】
【0232】
【表7】
【0233】
培養のための標準気相は、空気中に25%のメタンを含有する。これらの条件下で、MWM1000受容体をBTZ−3培地中で48h培養してBTZ−3中で3回洗浄し、元の培養容積の150倍の濃度に相当する容積のBTZ−3に再懸濁した。
【0234】
次に0.5%(w/v)酵母抽出物を含有するBTZ−3寒天プレートの表面で、供与体、ヘルパー、および受容細胞のペーストをそれぞれ1:1:2の比率で合わせた。プレートを25%メタン内で16〜72h、30℃に維持して接合を生じさせ、その後細胞ペーストを収集してBTZ−3に再懸濁した。カナマイシン(50μg/mL)を含有するBTZ−3寒天に希釈物を播種して、25%メタン内において30℃で1週間まで培養した。黄色の接合完了体をカナマイシン(50μg/mL)含有BTZ−3寒天上に画線した。
【0235】
カロテノイド組成物の分析のために、カナマイシン(50μg/mL)を含有する25mLのBTZ−3中で接合完了体を培養し、唯一の炭素源として25%のメタン内で30℃で3〜4日間インキュベートした。細胞を遠心分離によって収集し、−20℃で凍結した。解凍後ペレットを抽出し、実施例1で述べるようにして、HPLCによってカロテノイド含量を分析した。
【0236】
pDCQ330を含有するメチロモナス(Methylomonas)16a MWM1000からの抽出物のHPLC分析は、β−カロテンのほぼ排他的名生成を示した(図6)。14minピークの滞留時間、UVスペクトル、および分子重量は、シグマ(Sigma)からのβ−カロテン標品に一致する。これによって、このメタノトローフ宿主内でのDC404からのcrtEidiYIB遺伝子クラスターを利用したC40カロテノイドの合成が確証された。
【図面の簡単な説明】
【0237】
【図1】上流イソプレノイドおよび下流カロテノイド生合成経路を示す。
【図2】CrtE、CrtX、CrtY、CrtI、CrtB、およびCrtZによって触媒される具体的な化学転換を図示する、下流C40カロテノイド生合成経路の一部を示す。
【図3】パントエア・アグロメランス(Pantoea agglomerans)DC404株内に含有されるカロテノイドのHPLC分析の結果を示す。
【図4】コスミドpWEB−404を含んでなる形質転換大腸菌(E.coli)内に含有されるカロテノイドのHPLC分析の結果を示す。
【図5】パントエア・アナナティス(Pantoea ananatis)(ジェンバンク(登録商標)登録番号D90087)、パントエア・ステワルティ(Pantoea stewartii(ジェンバンク(登録商標)登録番号AY166713)、パントエア・アグロメランス(Pantoea agglomerans)EHO−10(ジェンバンク(登録商標)登録番号M87280)、およびパントエア・アグロメランス(Pantoea agglomerans)DC404のカロテノイド遺伝子クラスターの組成の比較を示す。
【図6】pDCQ330を含有するメチロモナス(Methylomonas)16a CrtN1−株からのカロテノイドのHPLC分析を示す。
【特許請求の範囲】
【請求項1】
ポリヌクレオチドを含んでなる、カロテノイド生合成経路酵素をコードする単離された核酸分子であって、ポリヌクレオチドの遺伝子構成がcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子が記載順にクラスター化されており、crtZの転写がcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起きる、単離された核酸分子。
【請求項2】
カロテノイド生合成経路酵素の少なくとも1つが、配列番号2、4、6、8、10、および12よりなる群から選択されるアミノ酸配列によってコードされる、請求項1に記載の単離された核酸分子。
【請求項3】
配列番号18に記載の配列を有する、請求項1に記載の単離された酸分子。
【請求項4】
(a)配列番号2、4、6、8、10、および12よりなる群から選択されるアミノ酸配列をコードする単離された核酸分子、
(b)0.1×SSC、0.1%SDSで65℃、および2×SSC、0.1%SDSで洗浄後、0.1×SSC、0.1%SDSというハイブリダイゼーション条件下で(a)とハイブリダイズする単離された核酸分子、および
(c)(a)または(b)と相補的である単離された核酸分子
よりなる群から選択される、カロテノイド生合成経路酵素をコードする単離された核酸分子。
【請求項5】
配列番号1、3、5、7、9、および11よりなる群から選択される、請求項4に記載の単離された核酸分子。
【請求項6】
crtE−idi−crtY−crtI−crtB−crtZ、遺伝子を含んでなる、配列番号18に記載の単離された核酸分子、または配列番号18と少なくとも95%の同一性を有する単離された核酸分子であって、ポリペプチドcrtE、idi、crtY、crtI、crtB、およびcrtZの全てをコードする単離された核酸分子。
【請求項7】
パントエア・アグロメランス(Pantoea agglomerans)DC404株から単離される、請求項1または4に記載の単離された核酸断片。
【請求項8】
請求項4に記載の単離された核酸分子によってコードされる、ポリペプチド。
【請求項9】
配列番号2、4、6、8、10、および12よりなる群から選択される、請求項8に記載のポリペプチド。
【請求項10】
配列番号2に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも70%の同一性を有する少なくとも301アミノ酸のゲラニルゲラニルピロリン酸合成酵素をコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項11】
配列番号4に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも70%の同一性を有する少なくとも349アミノ酸のイソペンテニルピロリン酸イソメラーゼ酵素をコードする第1のヌクレオチド配列を含んでなる単離された核酸分子、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項12】
配列番号6に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも70%の同一性を有する少なくとも389アミノ酸のリコペンシクラーゼをコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項13】
配列番号8に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも82%の同一性を有する少なくとも492アミノ酸のフィトエンデサチュラーゼをコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項14】
配列番号10に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも70%の同一性を有する少なくとも308アミノ酸のフィトエンシンターゼ酵素をコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項15】
配列番号12に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも73%の同一性を有する少なくとも185アミノ酸のβ−カロテン水酸化酵素をコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項16】
適切な制御配列に作動的に結合された、請求項4または10〜15のいずれか一項に記載の単離された核酸分子を含んでなるキメラ遺伝子。
【請求項17】
請求項6に記載の単離された核酸分子を含んでなるベクター。
【請求項18】
請求項16に記載のキメラ遺伝子を含んでなる形質転換宿主細胞。
【請求項19】
請求項6に記載の単離された核酸分子を含んでなる形質転換宿主。
【請求項20】
宿主細胞が、細菌、酵母、糸状菌類、藻類、および緑色植物よりなる群から選択される、請求項18または19に記載の形質転換宿主細胞。
【請求項21】
宿主細胞が、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、サッカロミセス(Saccharomyces)、ピヒア(Pichia)、カンジダ(Candida)、ハンゼヌラ(Hansenula)、ヤロウィア(Yarrowia)、ロドスポリジウム(Rhodosporidium)、リポミセス(Lipomyces)、サルモネラ(Salmonella)、バチルス(Bacillus)、アシネトバクター(Acinetobacter)、ザイモモナス(Zymomonas)、アグロバクテリウム(Agrobacterium)、フラボバクテリウム(Flavobacterium)、ロドバクター(Rhodobacter)、ロドコッカス(Rhodococcus)、ストレプトマイセス(Streptomyces)、ブレビバクテリウム(Brevibacterium)、コリネバクテリア(Corynebacteria)、マイコバクテリウム(Mycobacterium)、エシ
ェリヒア(Escherichia)、パントエア(Pantoea)、シュードモナス(Pseudomonas)、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、メチロサイナス(Methylosinus)、メチロマイクロビウム(Methylomicrobium)、メティロシスティス(Methylocystis)、アルカリゲネス(Alcaligenes)、シネコシスティス(Synechocystis)、シネココックス(Synechococcus)、アナベナ(Anabaena)、チオバチルス(Thiobacillus)、メタノバクテリウム(Methanobacterium)、クレブシエラ(Klebsiella)、メチロフィラス(Methylophilus)、メチロバチルス(Methylobacillus)、メチロバクテリウム(Methylobacterium)、ヒフォミクロビウム(Hyphomicrobium)、キサントバクター(Xanthobacter)、パラコッカス(Paracoccus)、ノカルジア(Nocardia)、アルスロバクター(Arthrobacter)、ロドシュードモナス(Rhodopseudomonas)、トルロプシス(Torulopsis)、ロドトルラ(Rhodotorula)、およびファフィア(Phaffia)よりなる群から選択される、請求項18または19に記載の形質転換宿主細胞。
【請求項22】
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、カロテノイド生合成経路酵素をコードする核酸分子
を含んでなる形質転換宿主細胞を提供し、
ここで、該分子の遺伝子構成がcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子が記載順にクラスター化されており、crtZの転写がcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起き、
(b)適切な生育条件下でステップ(a)の宿主細胞を有効量の発酵性炭素基質に接触させ、これによってカロテノイド化合物が生成することを含んでなる、
カロテノイド化合物の製造方法。
【請求項23】
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、配列番号2、4、6、8、10、および12よりなる群から選択される酵素をコードする一組の核酸分子
を含んでなる形質転換宿主細胞を提供し、
(b)適切な生育条件下でステップ(a)の宿主細胞を有効量の発酵性炭素基質に接触させ、これによってカロテノイド化合物が生成すること
を含んでなるカロテノイド化合物の製造方法。
【請求項24】
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、請求項6に記載の単離された核酸分子を含んでなる形質転換宿主細胞を提供し、
(b)適切な生育条件下でステップ(a)の宿主細胞を有効量の発酵性炭素基質に接触させ、これによってカロテノイド化合物が生成すること
を含んでなるカロテノイド化合物の製造方法。
【請求項25】
形質転換宿主細胞が、C1代謝宿主、細菌、酵母、糸状菌類、藻類、および緑色植物よりなる群から選択される、請求項22〜24のいずれか一項に記載の方法。
【請求項26】
C1代謝宿主がメタノトローフであり、発酵性炭素基質が、メタン、メタノール、ホルムアルデヒド、ギ酸、メチル化アミン、メチル化チオール、および二酸化炭素よりなる群
から選択される、請求項25に記載の方法。
【請求項27】
C1代謝宿主が、
(a)メタンおよびメタノールよりなる群から選択されるC1炭素基質で生育し、
(b)機能的エムデンマイヤーホフ炭素経路を含んでなり、前記経路がピロリン酸依存性ホスホフルクトキナーゼ酵素をコードする遺伝子を含んでなる、
請求項26に記載の方法。
【請求項28】
C1代謝宿主細胞が、高生育メタノトローフ細菌株であり、メチロモナス(Methylomonas)16aとして知られており、ATCC称号PTA 2402を有する、請求項27に記載の方法。
【請求項29】
形質転換宿主細胞が、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、サッカロミセス(Saccharomyces)、ピヒア(Pichia)、カンジダ(Candida)、ハンゼヌラ(Hansenula)、ヤロウィア(Yarrowia)、ロドスポリジウム(Rhodosporidium)、リポミセス(Lipomyces)、サルモネラ(Salmonella)、バチルス(Bacillus)、アシネトバクター(Acinetobacter)、ザイモモナス(Zymomonas)、アグロバクテリウム(Agrobacterium)、フラボバクテリウム(Flavobacterium)、ロドバクター(Rhodobacter)、ロドコッカス(Rhodococcus)、ストレプトマイセス(Streptomyces)、ブレビバクテリウム(Brevibacterium)、コリネバクテリア(Corynebacteria)、マイコバクテリウム(Mycobacterium)、エシェリヒア(Escherichia)、パントエア(Pantoea)、シュードモナス(Pseudomonas)、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、メチロサイナス(Methylosinus)、メチロマイクロビウム(Methylomicrobium)、メティロシスティス(Methylocystis)、アルカリゲネス(Alcaligenes)、シネコシスティス(Synechocystis)、シネココックス(Synechococcus)、アナベナ(Anabaena)、チオバチルス(Thiobacillus)、メタノバクテリウム(Methanobacterium)、クレブシエラ(Klebsiella)、メチロフィラス(Methylophilus)、メチロバチルス(Bacillus)、メチロバクテリウム(Methylobacterium)、ヒフォミクロビウム(Hyphomicrobium)、キサントバクター(Xanthobacter)、パラコッカス(Paracoccus)、ノカルジア(Nocardia)、アルスロバクター(Arthrobacter)、ロドシュードモナス(Rhodopseudomonas)、トルロプシス(Torulopsis)、ロドトルラ(Rhodotorula)、およびファフィア(Phaffia)よりなる群から選択される、請求項25に記載の方法。
【請求項30】
生成されたカロテノイド化合物が、アンテラキサンチン、アドニルビン、アドニキサンチン、アスタキサンチン、カンタキサンチン、カプソルブリン、β−クリプトキサンチン、α−カロテン、β−カロテン、ε−カロテン、エキネノン、3−ヒドロキシエキネノン、3’−ヒドロキシエキネノン、γ−カロテン、4−ケト−γ−カロテン、ζ−カロテン、α−クリプトキサンチン、デオキシフレキシキサンチン、ジアトキサンチン、7,8−ジデヒドロアスタキサンチン、フコキサンチン、フコキサンチノール、イソレニエラテン、ラクツカキサンチン、ルテイン、リコペン、ミキソバクトン、ネオキサンチン、ニューロスポレン、ヒドロキシニューロスポレン、ペリジニン、フィトエン、ロドピン、ロドピングルコシド、4−ケト−ルビキサンチン、シホナキサンチン、スフェロイデン、スフェロイデノン、スピリロキサンチン、4−ケト−トルレン、3−ヒドロキシ−4−ケト−ト
ルレン、ウリオリド、ウリオリドアセタート、ビオラキサンチン、ゼアキサンチン−β−ジグルコシド、およびゼアキサンチンよりなる群から選択される、請求項22〜24のいずれか一項に記載の方法。
【請求項31】
カロテノイド生合成が生物体において改変されるように、生物体において配列番号1、3、5、7、9、11、および18よりなる群から選択される、少なくとも1つのカロテノイド生合成経路遺伝子を過剰発現することを含んでなる、生物体においてカロテノイド生合成を制御する方法。
【請求項32】
前記カロテノイド遺伝子がマルチコピープラスミドで過剰発現される、請求項31に記載の方法。
【請求項33】
前記カロテノイド遺伝子が誘導可能プロモーターまたは調節プロモーターに作動的に結合する、請求項31に記載の方法。
【請求項34】
前記カロテノイド遺伝子がアンチセンス方向で発現される、請求項31に記載の方法。
【請求項35】
前記カロテノイド遺伝子がコード領域への外来性DNAの挿入によって中断される、請求項31に記載の方法。
【請求項36】
配列番号16で示される16s rDNA配列を含んでなるDC404株。
【請求項1】
ポリヌクレオチドを含んでなる、カロテノイド生合成経路酵素をコードする単離された核酸分子であって、ポリヌクレオチドの遺伝子構成がcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子が記載順にクラスター化されており、crtZの転写がcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起きる、単離された核酸分子。
【請求項2】
カロテノイド生合成経路酵素の少なくとも1つが、配列番号2、4、6、8、10、および12よりなる群から選択されるアミノ酸配列によってコードされる、請求項1に記載の単離された核酸分子。
【請求項3】
配列番号18に記載の配列を有する、請求項1に記載の単離された酸分子。
【請求項4】
(a)配列番号2、4、6、8、10、および12よりなる群から選択されるアミノ酸配列をコードする単離された核酸分子、
(b)0.1×SSC、0.1%SDSで65℃、および2×SSC、0.1%SDSで洗浄後、0.1×SSC、0.1%SDSというハイブリダイゼーション条件下で(a)とハイブリダイズする単離された核酸分子、および
(c)(a)または(b)と相補的である単離された核酸分子
よりなる群から選択される、カロテノイド生合成経路酵素をコードする単離された核酸分子。
【請求項5】
配列番号1、3、5、7、9、および11よりなる群から選択される、請求項4に記載の単離された核酸分子。
【請求項6】
crtE−idi−crtY−crtI−crtB−crtZ、遺伝子を含んでなる、配列番号18に記載の単離された核酸分子、または配列番号18と少なくとも95%の同一性を有する単離された核酸分子であって、ポリペプチドcrtE、idi、crtY、crtI、crtB、およびcrtZの全てをコードする単離された核酸分子。
【請求項7】
パントエア・アグロメランス(Pantoea agglomerans)DC404株から単離される、請求項1または4に記載の単離された核酸断片。
【請求項8】
請求項4に記載の単離された核酸分子によってコードされる、ポリペプチド。
【請求項9】
配列番号2、4、6、8、10、および12よりなる群から選択される、請求項8に記載のポリペプチド。
【請求項10】
配列番号2に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも70%の同一性を有する少なくとも301アミノ酸のゲラニルゲラニルピロリン酸合成酵素をコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項11】
配列番号4に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも70%の同一性を有する少なくとも349アミノ酸のイソペンテニルピロリン酸イソメラーゼ酵素をコードする第1のヌクレオチド配列を含んでなる単離された核酸分子、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項12】
配列番号6に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも70%の同一性を有する少なくとも389アミノ酸のリコペンシクラーゼをコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項13】
配列番号8に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも82%の同一性を有する少なくとも492アミノ酸のフィトエンデサチュラーゼをコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項14】
配列番号10に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも70%の同一性を有する少なくとも308アミノ酸のフィトエンシンターゼ酵素をコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項15】
配列番号12に記載の配列を有するポリペプチドと比較したときに、スミス−ウォーターマンのアライメント法に基づいて、少なくとも73%の同一性を有する少なくとも185アミノ酸のβ−カロテン水酸化酵素をコードする第1のヌクレオチド配列、
または第1のヌクレオチド配列の相補体を含んでなる第2のヌクレオチド配列
を含んでなる、単離された核酸分子。
【請求項16】
適切な制御配列に作動的に結合された、請求項4または10〜15のいずれか一項に記載の単離された核酸分子を含んでなるキメラ遺伝子。
【請求項17】
請求項6に記載の単離された核酸分子を含んでなるベクター。
【請求項18】
請求項16に記載のキメラ遺伝子を含んでなる形質転換宿主細胞。
【請求項19】
請求項6に記載の単離された核酸分子を含んでなる形質転換宿主。
【請求項20】
宿主細胞が、細菌、酵母、糸状菌類、藻類、および緑色植物よりなる群から選択される、請求項18または19に記載の形質転換宿主細胞。
【請求項21】
宿主細胞が、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、サッカロミセス(Saccharomyces)、ピヒア(Pichia)、カンジダ(Candida)、ハンゼヌラ(Hansenula)、ヤロウィア(Yarrowia)、ロドスポリジウム(Rhodosporidium)、リポミセス(Lipomyces)、サルモネラ(Salmonella)、バチルス(Bacillus)、アシネトバクター(Acinetobacter)、ザイモモナス(Zymomonas)、アグロバクテリウム(Agrobacterium)、フラボバクテリウム(Flavobacterium)、ロドバクター(Rhodobacter)、ロドコッカス(Rhodococcus)、ストレプトマイセス(Streptomyces)、ブレビバクテリウム(Brevibacterium)、コリネバクテリア(Corynebacteria)、マイコバクテリウム(Mycobacterium)、エシ
ェリヒア(Escherichia)、パントエア(Pantoea)、シュードモナス(Pseudomonas)、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、メチロサイナス(Methylosinus)、メチロマイクロビウム(Methylomicrobium)、メティロシスティス(Methylocystis)、アルカリゲネス(Alcaligenes)、シネコシスティス(Synechocystis)、シネココックス(Synechococcus)、アナベナ(Anabaena)、チオバチルス(Thiobacillus)、メタノバクテリウム(Methanobacterium)、クレブシエラ(Klebsiella)、メチロフィラス(Methylophilus)、メチロバチルス(Methylobacillus)、メチロバクテリウム(Methylobacterium)、ヒフォミクロビウム(Hyphomicrobium)、キサントバクター(Xanthobacter)、パラコッカス(Paracoccus)、ノカルジア(Nocardia)、アルスロバクター(Arthrobacter)、ロドシュードモナス(Rhodopseudomonas)、トルロプシス(Torulopsis)、ロドトルラ(Rhodotorula)、およびファフィア(Phaffia)よりなる群から選択される、請求項18または19に記載の形質転換宿主細胞。
【請求項22】
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、カロテノイド生合成経路酵素をコードする核酸分子
を含んでなる形質転換宿主細胞を提供し、
ここで、該分子の遺伝子構成がcrtE−idi−crtY−crtI−crtB−crtZであり、crtE、idi、crtY、crtI、crtB、およびcrtZ遺伝子が記載順にクラスター化されており、crtZの転写がcrtE、idi、crtY、crtI、およびcrtBとは反対方向に起き、
(b)適切な生育条件下でステップ(a)の宿主細胞を有効量の発酵性炭素基質に接触させ、これによってカロテノイド化合物が生成することを含んでなる、
カロテノイド化合物の製造方法。
【請求項23】
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、配列番号2、4、6、8、10、および12よりなる群から選択される酵素をコードする一組の核酸分子
を含んでなる形質転換宿主細胞を提供し、
(b)適切な生育条件下でステップ(a)の宿主細胞を有効量の発酵性炭素基質に接触させ、これによってカロテノイド化合物が生成すること
を含んでなるカロテノイド化合物の製造方法。
【請求項24】
(a)(i)適切なレベルのファルネシルピロリン酸、および
(ii)適切な制御配列の制御下にある、請求項6に記載の単離された核酸分子を含んでなる形質転換宿主細胞を提供し、
(b)適切な生育条件下でステップ(a)の宿主細胞を有効量の発酵性炭素基質に接触させ、これによってカロテノイド化合物が生成すること
を含んでなるカロテノイド化合物の製造方法。
【請求項25】
形質転換宿主細胞が、C1代謝宿主、細菌、酵母、糸状菌類、藻類、および緑色植物よりなる群から選択される、請求項22〜24のいずれか一項に記載の方法。
【請求項26】
C1代謝宿主がメタノトローフであり、発酵性炭素基質が、メタン、メタノール、ホルムアルデヒド、ギ酸、メチル化アミン、メチル化チオール、および二酸化炭素よりなる群
から選択される、請求項25に記載の方法。
【請求項27】
C1代謝宿主が、
(a)メタンおよびメタノールよりなる群から選択されるC1炭素基質で生育し、
(b)機能的エムデンマイヤーホフ炭素経路を含んでなり、前記経路がピロリン酸依存性ホスホフルクトキナーゼ酵素をコードする遺伝子を含んでなる、
請求項26に記載の方法。
【請求項28】
C1代謝宿主細胞が、高生育メタノトローフ細菌株であり、メチロモナス(Methylomonas)16aとして知られており、ATCC称号PTA 2402を有する、請求項27に記載の方法。
【請求項29】
形質転換宿主細胞が、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、サッカロミセス(Saccharomyces)、ピヒア(Pichia)、カンジダ(Candida)、ハンゼヌラ(Hansenula)、ヤロウィア(Yarrowia)、ロドスポリジウム(Rhodosporidium)、リポミセス(Lipomyces)、サルモネラ(Salmonella)、バチルス(Bacillus)、アシネトバクター(Acinetobacter)、ザイモモナス(Zymomonas)、アグロバクテリウム(Agrobacterium)、フラボバクテリウム(Flavobacterium)、ロドバクター(Rhodobacter)、ロドコッカス(Rhodococcus)、ストレプトマイセス(Streptomyces)、ブレビバクテリウム(Brevibacterium)、コリネバクテリア(Corynebacteria)、マイコバクテリウム(Mycobacterium)、エシェリヒア(Escherichia)、パントエア(Pantoea)、シュードモナス(Pseudomonas)、メチロモナス(Methylomonas)、メチロバクター(Methylobacter)、メチロコックス(Methylococcus)、メチロサイナス(Methylosinus)、メチロマイクロビウム(Methylomicrobium)、メティロシスティス(Methylocystis)、アルカリゲネス(Alcaligenes)、シネコシスティス(Synechocystis)、シネココックス(Synechococcus)、アナベナ(Anabaena)、チオバチルス(Thiobacillus)、メタノバクテリウム(Methanobacterium)、クレブシエラ(Klebsiella)、メチロフィラス(Methylophilus)、メチロバチルス(Bacillus)、メチロバクテリウム(Methylobacterium)、ヒフォミクロビウム(Hyphomicrobium)、キサントバクター(Xanthobacter)、パラコッカス(Paracoccus)、ノカルジア(Nocardia)、アルスロバクター(Arthrobacter)、ロドシュードモナス(Rhodopseudomonas)、トルロプシス(Torulopsis)、ロドトルラ(Rhodotorula)、およびファフィア(Phaffia)よりなる群から選択される、請求項25に記載の方法。
【請求項30】
生成されたカロテノイド化合物が、アンテラキサンチン、アドニルビン、アドニキサンチン、アスタキサンチン、カンタキサンチン、カプソルブリン、β−クリプトキサンチン、α−カロテン、β−カロテン、ε−カロテン、エキネノン、3−ヒドロキシエキネノン、3’−ヒドロキシエキネノン、γ−カロテン、4−ケト−γ−カロテン、ζ−カロテン、α−クリプトキサンチン、デオキシフレキシキサンチン、ジアトキサンチン、7,8−ジデヒドロアスタキサンチン、フコキサンチン、フコキサンチノール、イソレニエラテン、ラクツカキサンチン、ルテイン、リコペン、ミキソバクトン、ネオキサンチン、ニューロスポレン、ヒドロキシニューロスポレン、ペリジニン、フィトエン、ロドピン、ロドピングルコシド、4−ケト−ルビキサンチン、シホナキサンチン、スフェロイデン、スフェロイデノン、スピリロキサンチン、4−ケト−トルレン、3−ヒドロキシ−4−ケト−ト
ルレン、ウリオリド、ウリオリドアセタート、ビオラキサンチン、ゼアキサンチン−β−ジグルコシド、およびゼアキサンチンよりなる群から選択される、請求項22〜24のいずれか一項に記載の方法。
【請求項31】
カロテノイド生合成が生物体において改変されるように、生物体において配列番号1、3、5、7、9、11、および18よりなる群から選択される、少なくとも1つのカロテノイド生合成経路遺伝子を過剰発現することを含んでなる、生物体においてカロテノイド生合成を制御する方法。
【請求項32】
前記カロテノイド遺伝子がマルチコピープラスミドで過剰発現される、請求項31に記載の方法。
【請求項33】
前記カロテノイド遺伝子が誘導可能プロモーターまたは調節プロモーターに作動的に結合する、請求項31に記載の方法。
【請求項34】
前記カロテノイド遺伝子がアンチセンス方向で発現される、請求項31に記載の方法。
【請求項35】
前記カロテノイド遺伝子がコード領域への外来性DNAの挿入によって中断される、請求項31に記載の方法。
【請求項36】
配列番号16で示される16s rDNA配列を含んでなるDC404株。
【図1】

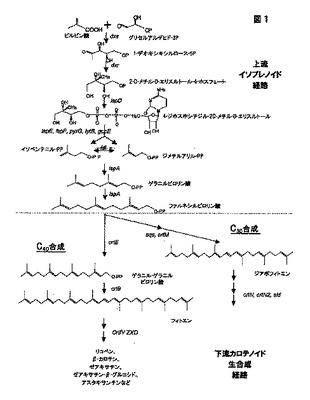
【図2】
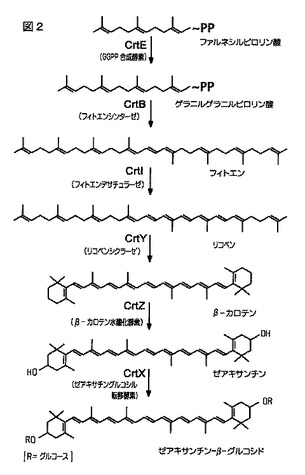
【図3】

【図4】

【図5】
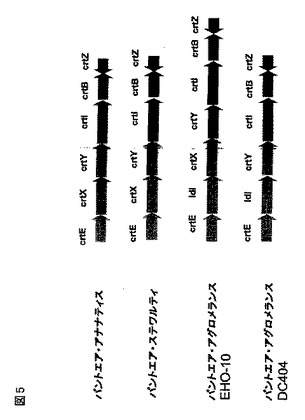
【図6】
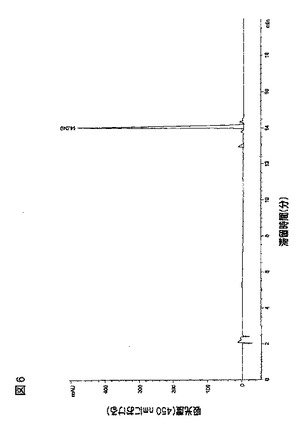
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2007−516697(P2007−516697A)
【公表日】平成19年6月28日(2007.6.28)
【国際特許分類】
【出願番号】特願2006−533795(P2006−533795)
【出願日】平成16年6月10日(2004.6.10)
【国際出願番号】PCT/US2004/019038
【国際公開番号】WO2005/044975
【国際公開日】平成17年5月19日(2005.5.19)
【出願人】(390023674)イー・アイ・デュポン・ドウ・ヌムール・アンド・カンパニー (2,692)
【氏名又は名称原語表記】E.I.DU PONT DE NEMOURS AND COMPANY
【Fターム(参考)】
【公表日】平成19年6月28日(2007.6.28)
【国際特許分類】
【出願日】平成16年6月10日(2004.6.10)
【国際出願番号】PCT/US2004/019038
【国際公開番号】WO2005/044975
【国際公開日】平成17年5月19日(2005.5.19)
【出願人】(390023674)イー・アイ・デュポン・ドウ・ヌムール・アンド・カンパニー (2,692)
【氏名又は名称原語表記】E.I.DU PONT DE NEMOURS AND COMPANY
【Fターム(参考)】
[ Back to top ]