
ガンの予防におけるヒドロキシマタイレシノール
【課題】ヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防。
【解決手段】ヒドロキシマタイレシノールを含む医薬品、食品添加物および食品。ヒドロキシマタイレシノールによる、ヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防、ならびにヒドロキシマタイレシノールによる、ヒトの血清中のエンテロラクトンまたはヒドロキシマタイレシノールのほかの代謝産物の濃度の増加により、ヒトのガン、ある種の非ガン性ホルモン依存性疾患の予防をもたらす。
【解決手段】ヒドロキシマタイレシノールを含む医薬品、食品添加物および食品。ヒドロキシマタイレシノールによる、ヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防、ならびにヒドロキシマタイレシノールによる、ヒトの血清中のエンテロラクトンまたはヒドロキシマタイレシノールのほかの代謝産物の濃度の増加により、ヒトのガン、ある種の非ガン性ホルモン依存性疾患の予防をもたらす。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒドロキシマタイレシノールをヒトに投与することによる、ヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防方法に関する。本発明はさらに、ヒドロキシマタイレシノールをヒトに投与することによるヒトの血清中のエンテロラクトンまたはヒドロキシマタイレシノールのほかの代謝産物の濃度を増加させる方法であって、それによってヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防をもたらす方法に関する。また、本発明はヒドロキシマタイレシノールを含有する医薬品、食品添加物および食品に関する。
【背景技術】
【0002】
本発明の背景、およびとくに実用に関してさらに詳細に述べた例を説明するため、ここで用いた刊行物などを参考文献として組み入れる。
【0003】
リグナン類は2,3−ジベンジルブタン骨格を有するフェノール系化合物の一群である。これらは、前駆体とよばれる桂皮酸、コーヒー酸、フェルラ酸、クマリン酸およびガリウム酸などの単量体単位のカップリングによって形成される(エアおよびロイク、1990年)。リグナンは植物に広く分布しており、さまざまな部位(根、葉、幹、種子、果実)に見られるが、少量である。多くの供給源(種子や果実)において、リグナンは植物の繊維成分と結びついたグリコシド共役体として検出される。哺乳類のリグナン前駆体の最も一般的な供給源食物は未精製の穀物である。食用植物のなかでリグナンの濃度が最も高いのは亜麻仁(flaxseed)であり、次に未精製の穀物、とくにライ麦があげられる。表1にさまざまな植物性食品から生成される哺乳類のリグナンを示す。
【0004】
針葉樹にも多量のリグナンが見られる。リグナンの型は種によって異なり、リグナンの量は樹木の部位によって相違する。トウヒ(Picea abies)の心材に見られる典型的なリグナンには、ヒドロキシマタイレシノール(HMR)、α−コニデンドリン(conidendrin)、コニデンドリン酸、マタイレシノール、イソラリシレシノール(isolariciresinol)、セコイソラリシレシノール(secoisolariciresinol)、リオヴァイル(liovile)、ピセアレシノ−ル(picearesinol)、ラリシレシノールおよびピノレシノールがある(エクマン、1979年)。トウヒ中のリグナンのうち、最も豊富に含まれる単一成分はHMRであり、全リグナンの約60パーセントを占め、主に非共役遊離形で存在する。太い根におけるリグナン濃度は2〜3パーセントである。大量のリグナンは枝の心材(5〜10パーセント)やねじれた部分(twists)の心材に含まれ、とくに木節ではリグナンの量は10パーセントより多い(エクマン、1976年および1979年)。これらの濃度は、リグナン高含有材料として知られる、亜麻を挽いて粉末にしたものと比べ約100倍である。
【0005】
ヒドロキシマタイレシノールの化学構造は
【化1】

である。
【0006】
リグナンはたとえば、圧縮材の繊維から単離することができる。これらの繊維は幹や、紙の品質を悪化させる木節(大き過ぎる木材チップの画分)の圧縮材から生じる(エクマン、1976年)。
【0007】
マタイレシノールやセコイソラリシレシノールなどの植物性リグナンは、腸内微生物(gut microflora)によって、哺乳類リグナン、つまりそれぞれエンテロラクトンおよびエンテロジオールに変換される(アクセルソンら、1982年)。それらは腸肝循環し、グルクロニド共役体として尿に排出される(アクセルソンおよびセッチェル、1982年)。リグナンの化学的な予防作用の実験的な証拠として、脂肪分の多い食品にリグナン高含有亜麻仁粉末(5〜10%)または亜麻仁リグナン(セコイソラリシレシノール−ジグリコシド(SDG))を加えると、ラットにおいて抗エストロゲン感受性DMBA誘発性乳ガンの進行が抑制された(セレイノおよびトンプソン、1991年、1992年、トンプソンら1996年a、1996年b)。それらにより、上皮細胞の増殖、核異常、腫瘍の成長および新しい腫瘍の発生が減少した。また、大量のリグナンの摂取が実験的前立腺および結腸ガンを防ぐ可能性もある。食用ライ麦(リグナンを含む)によって、ラットの移植されたダニングR3327前立腺ガンの成長が初期段階で抑制された(ツァンら、1997年;ランドストロームら、1998年)。明らかな腫瘍を有する動物の割合、腫瘍の大きさ、および成長率はかなり低かった。さらに、亜麻仁またはSDGを加えることによって、ラットの結腸において化学的に誘導された異常陰窩の形成が抑制された(セレイノおよびトンプソン、1992年。ジェナブおよびトンプソン、1996年)。よって、抗腫瘍作用は、弱いエストロゲン−抗エストロゲン様特性および/またはほかのメカニズムによって起こる可能性があるが、それら特性やメカニズムはあまり解明されていない。
【0008】
乳ガンの女性では、エンテロラクトンの尿への排出および血清中濃度が低く(イングラムら、1997年;フルテンら、1998年)、それはリグナンが化学的予防性であることを示している。哺乳類のリグナン(エンテロラクトンおよびエンテロジオール)は、エストロゲンと構造が似ているので、ホルモン性のガン、つまり乳ガンなどを緩和する、と仮定されてきた。エンテロラクトンはMCF−7細胞において弱いエストロゲン様の潜在力を有するが(モサビおよびアドラクルツ、1992年)、マウスの子宮重量においてはエストロゲン様の応答は見られなかった(セッチェルら、1981年)。エストロゲン様活性の兆候として、ラットに妊娠および授乳期間中SDGを与えることによって、離乳時には子宮重量が増加したが、後期にはその効果ははっきりしなかった(タウら、1998年)。また、予想される抗腫瘍効果は、リグナンの抗エストロゲン様作用と結びついている(ウォーターズおよびノーラー、1982年)。哺乳類リグナン、つまりエンテロラクトンによるアロマターゼの阻害は、リグナン高含有植物性食品の消費により乳ガンなどのエストロゲン依存性疾病を低減するメカニズムを提示していることが考えられる(アドラクルツら、1993年;ワンら1994年)。リグナンの潜在的な抗酸化活性もまた、ガンの進行を予防するリグナン作用のメカニズムを示していることが考えられる。さらに、哺乳類のリグナンは、テストステロンの、強い細胞内アンドロゲンである5α−ジヒドロテストステロン(DHT)への変換をヒトにおいて可能な濃度で阻害することを示した(エバンズら、1995年)。DHT濃度の低下は、前立腺ガン(PC)や良性前立腺過形成(BPH)の危険性を緩和するであろう。
【0009】
エンテロラクトンの前駆体としてのリグナンはまた、下部尿道症(LUTS)および女性化乳房を軽減することができる。我々は、動物実験の結果に基づき、膀胱頸部共同運動障害あるいは外括約筋偽共同運動障害として顕われる尿道共同運動障害における筋機能障害の進行に、エストロゲンが重要な役割を果たすことを示唆した(シュトレングら、考察は未発表)。そのような神経筋の変化は、アロマターゼ阻害剤(MPV−2213ad)によって少なくとも部分的に元に戻るが、それはエストロゲンの役割を表している。さらに、女性化乳房はエストロゲンとの接触やアンドロゲンよりエストロゲンが多く存在する場合などに誘発される。女性化乳房はアロマターゼ阻害剤によってうまく治療できる。5α−還元酵素および/またはアロマターゼを阻害するリグナンの能力は、潜在的な抗酸化活性と合わせて、男性においてホルモン性疾病が進行する際のリグナンによる予防作用のメカニズムを示している可能性がある。
【0010】
リグナンに潜在するヒトへの効果に関するデータはない。ヒトにおけるリグナンの作用に関する最近の学説は、亜麻仁粉(つまりリグナン)を加えた食品がもたらす効果の研究から導き出されてきた。女性の食事に亜麻仁粉を含ませた場合、月経周期に変化が現れた(フィップスら、1993年)。正常な月経周期の被験者らでは、常食に加えて1日10gの亜麻仁粉を摂取した場合、黄体期の平均長さがより長くなり、黄体期の血清中の17β−エストラジオールに対するプロゲステロンの比率がより高くなった(フィップスら、1993年)。エストロンまたは17β−エストラジオールの濃度に関しては、亜麻摂取グループと対照グループのあいだに大きな相違点はなかった。また、閉経後の女性の血清中エストロゲン濃度に関しても、亜麻摂取グループと対照グループのあいだに大きな相違点はなかった(ブレジンスキーら、1997年)。亜麻仁粉の追加によって血清中のSHBG(エストラジオールを強力に結合するタンパク質)の濃度が増加した。これは、肝臓組織で見られる典型的なエストロゲン様効果である。一方、SHBGの濃度の増加により、内因性エストロゲンのバイオアべイラビリティは低下する。健康な青年の場合、短期間(6週間)の食事への亜麻仁の添加(マフィンに1日10g)は、テストステロンの血漿中濃度に目立った影響を示さず、男性におけるエストロゲン作用(estrogenicity)の欠如を示している(シュルツら、1991年)。同時にこれらの研究は、リグナンは弱いホルモン(エストロゲン様および抗エストロゲン様)効果を有するが、リグナンの作用機構はそのホルモン効果によってすべて説明され得るわけではない、ということを示す。
【0011】
結論として、以前は動物実験や臨床実験で使用するのに充分な量の単離哺乳類リグナンを入手することはできず、リグナン摂取量を増加させる唯一の可能性は、繊維質の多い亜麻仁などの食品の消費を増加させることであった。効率よくエンテロラクトンに変換し、大量に生成され単離されうるHMRなどのリグナンは、ガン、他のホルモン性疾患および心疾患を化学的に予防するための医薬品および機能性食品などの食品の開発において貴重なものである。
【発明の概要】
【0012】
ある側面において、本発明は有効量のヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体をヒトに投与することからなる、ヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防方法に関する。
【0013】
さらなる側面において、本発明は、有効量のヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体をヒトに投与することからなる、ヒトの血清中のエンテロラクトンまたはヒドロキシマタイレシノールのほかの代謝産物の濃度を増加させる方法であって、それによってヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防をもたらす方法に関する。
【0014】
第3の側面において、本発明は、有効量のヒドロキシマタイレシノールまたは幾何異性体もしくは立体異性体と、薬学的に許容しうる担体とからなる医薬品に関する。
【0015】
第4の側面において、本発明は、ヒドロキシマタイレシノールまたはその幾何異性体もしくは立体異性体を濃縮した液体または固体材料からなる、食品添加物用製品に関する。
【0016】
第5の側面において、本発明は、有効量のヒドロキシマタイレシノールまたはその幾何異性体もしくは立体異性体からなる食品に関する。
【0017】
もう1つの側面において、本発明は、前記有効量のヒドロキシマタイレシノール、その幾何異性体または立体異性体を食品に添加することからなる食品の安定性を高める方法に関する。
【図面の簡単な説明】
【0018】
【図1】図1はJEG−3細胞におけるリグナンによるアロマターゼの濃度相関阻害を示す。
【図2】図2はHMRの存在下および非存在下におけるMCF−7細胞の増殖を示す。
【図3】図3はHMRまたはアロマターゼ阻害剤で処理した未成熟ラットの子宮の湿重量を示す。
【図4】図4はメスのラットにおけるDMBA誘発性乳腺腫瘍に対するHMRの抗腫瘍活性を示す。
【図5】図5は種々の投与量のHMRで処理したラットの、エンテロラクトンの尿への排出を示す。
【発明を実施するための形態】
【0019】
本発明はヒドロキシマタイレシノール(HMR)を食品に添加し、また医薬品として用いることによってヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患を予防する、リグナン、つまりHMRの用途に関する。驚いたことに、HMRはインビボでエンテロラクトンへ代謝されるが、これがリグナンの抗腫瘍特性の原因の一つであると考えられている。インビトロでのHMRの抗酸化活性は大きく、この特性は、HMRが、体内において有害な遊離酸素種に対する保護効果を通して心疾患をも防ぐことができるということを示している。また、本発明は食品の安定性を向上する(つまり栄養価の損失や食品の風味の低下につながる、脂質および色素の酸化とビタミンの喪失とを抑制する)ための食品添加物としてのHMRの用途に関する。
【0020】
本発明の方法は、乳ガン、前立腺ガンおよび結腸ガン、下部尿路症、尿道共同運動障害、膀胱不安定性、膀胱排出口閉塞、良性前立腺過形成および男性の女性化乳房などの非ガン性ホルモン依存性疾患、および血清中の酸化LDLに起因する心疾患の予防に特に効果的である。
【0021】
本発明による医薬品は経口処方であることが好ましい。活性化合物(HMR)の必要量は、予防すべき特定の症状によって異なるが、標準的な投与量は大人で1日約10〜100mgである。
【0022】
本発明の食品添加物において、ヒドロキシマタイレシノールで強化される材料は、HMRとの混合に適し、HMRの特性を損なわない、食用で中毒性のない固体または液体の材料であればどのようなものでもよい。この材料の役割は主に、HMRの確実な投与を容易にすることである。好ましい濃度は、たとえば強化された材料100g中にHMR100mg〜1gである。
【0023】
本発明の食品とは、とくに機能性食品、栄養補助食品、栄養剤、治療用食品、栄養補給食品、健康食品、遺伝子操作食品(designer food)または任意の食品である。食品中のHMRの好ましい濃度は、たとえば食品100gに対してHMR1〜20mgである。
【0024】
本発明の機能性食品はたとえば、バター、マーガリン、ビスケット、パン、ケーキ、アメ、菓子類、ヨーグルトもしくは他の発酵乳製品、またはムースリなどのシリアルの形が可能である。
【0025】
ヒドロキシマタイレシノールの添加は、栄養価の損失と食品の風味の低下につながる、脂質、ビタミンおよび色素の酸化を抑制することを目的として、食品の安定性を向上する場合にとくに有用である。このためにHMRの好ましい濃度は、たとえば約0.1%である。
【0026】
本発明で使用するHMRは、圧縮材の大き過ぎるチップ画分(oversize chip fraction)(枝、ねじれ部分および木節を含む)から単離する。HMRはガンや心疾患などの疾病の予防に用いる。
【0027】
HMRの特性を7つの異なる分析方法によって検討した。
1.インビトロにおける抗酸化力の測定
2.JEG−3細胞におけるアロマターゼ阻害力の測定
3.MCF−7培養細胞におけるエストロゲン様および抗エストロゲン様活性の測定
4.子宮成長の生物学的検定によるエストロゲン様および抗エストロゲン様活性の評価
5.オスの成体ラットにおけるエストロゲン様および抗エストロゲン様活性の測定
6.ラットDMBA誘発性乳ガンモデルにおける抗腫瘍活性の調査
7.種々の量のHMRを投与した後のラットの尿に含まれる代謝産物の分析
【0028】
以前は、HMRが樹木のリグナン成分であることから、その特徴はあまり注目されておらず、生物学的試験を行なうのに充分な量のHMRを単離し、精製することは不可能であった。しかし、トウヒ(spruce)の様々な部位におけるHMRの分布が知られ(エクマン、1976年および1979年)、リグナン、とくにHMRの詳細な研究の機会が得られた。
【0029】
HMRの投与量と尿中のエンテロラクトンの量との間に線形相関が見られた。エンテロラクトンは広く知られた哺乳類のリグナンであり、マタイレジノールから腸内細菌によって、またはエンテロジオールの酸化によって形成される(アクセルソンおよびセッチェル、1981年;アクセルソンら、1982年)。わずかな量の代謝されなかったHMRとほかの代謝産物(エンテロジオールと7−ヒドロキシエンテロラクトン)とが尿中に見られた。それらの量は、毎日のHMRの投与量を増やしても変化しなかった。これらの発見は、HMRが代謝されエンテロラクトンとなり、さらにジメチル化およびジヒドロキシル化を経てHMRから生じたエンテロラクトンは、エンテロジオールには変換しないということを示唆している。HMRの構造に基づいて、7−ヒドロキシエンテロラクトンが主なHMRの代謝産物であると考えられたが、それは違っていた。このヒドロキシル基は代謝の際に脱離する。HMRの代謝は、SDGの代謝とは異なる。SDGは代謝によってエンテロジオールになり、その一部は酸化されエンテロラクトンになる(リカードら、1996年、ランペら、1994年)。HMRはしたがって、エンテロラクトンの直接の前駆体として、SDGよりも有利である。
【0030】
HMRはラットの子宮または男性において、たとえあるとしても弱いエストロゲン様活性しか示さない。HMRはMCF−7細胞において、弱く有意性のないエストロゲン様活性しか示さない。また、HMRに関して抗エストロゲン活性は見られなかった。それゆえ、図2に示すように、HMRがラットのDMBA誘発性腫瘍モデルにおいて非常に有意な抗腫瘍活性を示したことは驚きである。HMRの活性はHMR自身あるいはエンテロラクトンが原因であると考えられる。しかし、ラットにDMBA処理ののち、2種類の投与量(3および15mg/kg)でHMRを与えた場合、HMRの化学的予防作用には投与量との因果関係はなかった。よって、抗腫瘍効果を得るためには、HMRは必ずしもエンテロラクトンに変換される必要はなく、またこれらのリグナンの投与量はより少量でも最大の化学的予防効果を得るのに充分である。
【0031】
表2および3に示すように、HMRは非常に有効な抗酸化剤である。HMRは、既知の最も強力な、脂質過酸化反応の阻害剤のひとつであり、LDLの酸化を防ぐ優れた阻害剤である。LDLの酸化の阻害はヒトにおいて非常に重要であるとされている。なぜなら、酸化LDLの血清中濃度が、アテローム性動脈硬化症といった心疾患を予測する最良のものの一つとされているからである。HMRは食品の安定性を向上する(つまり栄養価の損失と食品の風味の低下につながるビタミン、脂質および色素の酸化を抑制する)食品添加物となりうる。なぜなら、HMRは、食品安定性の向上のために通常用いられる、よく知られた抗酸化剤、すなわちブチル化ヒドロキシアニソール(BHA)およびブチル化ヒドロキシトルエン(BHT)などよりもさらに優れたスーパーオキシドアニオンの捕捉剤およびペルオキシラジカルの捕捉剤であるからである。
【0032】
実験
薬品
リグナンのエストロゲン様作用、抗エストロゲン様作用、芳香族化(aromatization)を阻害する能力そして抗酸化特性を調べるため、様々なリグナンをインビトロで試験した。試験用化合物は以下から入手した。エンテロジオールおよびエンテロラクトンは、イギリス、ロンドンのプランテック社(Plantech)より購入した。2つの7−OHエナンチオマーを有する7−ヒドロキシエンテロラクトンはフィンランド、ヘルシンキ大学応用化学研究室、クリスティナ・ワララ教授(Dr. Kristina Waehaelae)のご好意により提供された。
【0033】
樹木からのHMRの抽出
エクマン(1976年)およびエクマン(1979年)に記載の方法によって、HMR抽出物をノルウェイ・トウヒ(Picea Abies)より単離した。簡単言うと、凍結乾燥粉末心材をヘキサンでソックスレー抽出し、無極性、親油性の抽出物を除去した。その木材試料をアセトン/水(9:1 v/v)を用いて、同じ装置でさらに抽出し、粗リグナンを得た。ヒドロキシマタイレシノール(HMR)およびその異性体を単離し、さらに精製するため、XAD樹脂で再度クロマトグラフィーを行なった。
【0034】
インビトロにおける抗酸化力の測定
リグナンの抗酸化力を4つの異なる方法で評価した。1)脂質過酸化の阻害、2)低比重リポタンパク(LDL)酸化の阻害、3)スーパーオキシド アニオンの捕捉および4)ぺルオキシラジカルの捕捉の分析である。
【0035】
脂質過酸化の阻害は、インビトロでのラット肝臓ミクロソームにおいてtert−ブチルハイドロパーオキサイド誘発脂質過酸化(t−BuOOH−LP)を阻害するリグナンの能力により評価した(アホトゥパら、1997年)。t−BuOOH−LPに対する試験は次のように行なった。緩衝液0.8ml(50mM 炭酸ナトリウム、pH10.2、0.1mM EDTA)を照度計のキュベットにピペットで注入した。最終濃度が1.5mg タンパク質/ml の希釈肝臓ミクロソーム20μlを添加し、ついで、ルミノール6ml(0.5mg/ml)および試験用薬品(test chemicals)を加えた。エタノールまたはジメチルスルホキシド(インキュベーション体積の2%)で希釈した少量の被験化合物をインキュベーション混合物に加え、ビヒクル(vehicle)(エタノールまたはジメチルスルホキシド)の場合と脂質過酸化力を比較した。0.9mMのt−BuOOH 0.05mlを用いて33℃で反応を開始した。化学発光を1分ごとに約45分間測定し、曲線下部の面積(積分)を求めた。化学発光の測定は分析専用ソフトを用いてパーソナルコンピュータに接続したバイオ−オービット(Bio-Orbit)1251照度計(バイオ−オービット社製、ツルク、フィンランド)を用いて行なった。
【0036】
LDLの酸化の阻害は、アホトゥパら(1996年)に記載の方法によって評価した。簡単に言うと、緩衝化ヘパリンを用いて沈殿(precipitation)によりLDLを単離した。リン酸緩衝液中で再度懸濁した後、20mMのCuCl2を添加し、混合液を37℃で3時間インキュベートした。こののちクロロフォルム−メタノールによりLDL脂質を抽出し、窒素下で乾燥し、シクロヘキサンに再び溶解させ、分光光度法を用い、234nmで分析した。吸光度の強度はLDLの酸化を示す。さまざまな化合物のLDL酸化防止力を調べるため、CuCl2添加前のインキュベーション混合物にそれらの化合物を加えた。分析中に被験化合物の妨げとなりうるものは、インキュベーション期間の前後に234nmでの吸収を測定することによって取り除いた。初期濃度(0.1mM)で抗酸化効果を示した化合物に関し、IC−50値(被験化合物によってLDLの酸化が50%阻害された濃度)を求めた。
【0037】
スーパーオキシドアニオン捕捉方法は、キサンチン−キサンチンオキシダーゼ系により調製した条件下で生成されるスーパーオキシドアニオン、および生成した反応性酸素種の照度計による検出にもとづいた(アホトゥパら、1997年)。被験化合物が化学発光を減少させる能力を評価した。IC−50濃度(50%の化学発光が抑制された濃度)を求めた。
【0038】
ペルオキシラジカル捕捉の分析は、2,2'−アゾビス(2−アミジノプロパン)HClの熱分解によるペルオキシラジカルの発生、および化学発光によるその検出にもとづいた(アホトゥパら、1997年)。その結果を化学量論的な因子、つまり被験化合物1モルにつき何モルのペルオキシラジカルが捕捉されうるか、として求めた。
【0039】
JEG−3細胞におけるアロマターゼ阻害力の測定
HMRおよび構造的に関連するリグナン(エンテロラクトン、エンテロジオールおよび7−ヒドロキシエンテロラクトン)の効果を、ヒト絨毛癌細胞株であるJEG−3細胞における3H−アンドロステンジオンからの3H−17β−エストラジオールの生成にもとづいて調べた。JEG−3絨毛癌細胞は、インビトロでアロマターゼ阻害作用を調査し得る有効なアロマターゼ・モデルである(クレッケルら、1991年)。ウシ胎仔血清(FCS)10%含有DMEM中に細胞を保持した。3H−アンドロスト−4−エン,3,17−ジオン(0.5nM)50μl、非標識アンドロステンジオン(0.5nM)50μl、被験化合物(10mM)100μlおよび細胞懸濁液800μl(細胞100万個)をインキュベーション混合物とした。4時間のインキュベーションののち、非標識の担体(アンドロステンジオン、テストステロン、17β−エストラジオールおよびエストロン)を加えた。ジクロロメタン3.0mlを用いてそれらステロイドを2回抽出した。以前に述べられたように、HPLCを用いて放射性標識3H−17β−エストラジオールの分離と定量を行なった(マケラら、1995年)。カラム系は保護カラム、続いてC18 150×3.9mmID分析カラム(テクノパック10C18HPLCテクノロジー;ウェリントンハウス、チェシャー州、イギリス)から構成された。移動相はアセトニトリル/水(35/65)であり、流速は1.2ml/分であった。放射性代謝物質のインライン検出のため、HPLCカラムの溶離液を液体蛍光体(liquid scintillant)と連続的に混合し、インライン放射能検出器を用いてモニターした。
【0040】
MCF−7培養細胞におけるエストロゲン様および抗エストロゲン様活性の測定
MCF−7細胞株(ヒト乳ガン細胞)保存培養物(stock cultures)は、5%FCS、ペニシリン100 U/ml、ストレプトマイシン100μg/ml、インシュリン10μg/mlおよび17β−エストラジオール1nMを加えたフェノールレッドを含まないRPMI培地中、T−75細胞培養びん内で培養した。培地は1週間に3回新しいものと交換した。保存培養物をトリプシン処理によって取り出し、フェノールレッドを含まないベルセン溶液(versene solution)10ml中に懸濁し、800rpmで5分間遠心分離した。5%デキストラン木炭除去(dextran charcoal stripped)FSC(dcFSC)を加えたRPMI培地中に細胞ペレットを慎重に再懸濁し、6穴プレートに50000細胞個/3.0ml溶媒/ウェルの割合で播種した。培養2日目に培地を交換し、被験化合物を加えた。リグナン化合物のエストロゲン様効果を調べるため、リグナン化合物をエタノールに希釈し、細胞培養物に加え、最終濃度を1.0Mとした。各増殖検定において、1.0nM17β−エストラジオールのエタノール溶液をエストロゲン様反応に関する陽性対照として使用した。同量のエタノールを対照ウェルに加えた。また、抗エストロゲン様効果を調べるため、17β−エストラジオールとリグナンとの両溶液を培養細胞に加えた。被験化合物の存在下において5日〜7日間細胞を培養し、培地は2日おきに交換した。細胞増殖は、コールター計数器を用いて溶出した核を数えることによって定量化した。
【0041】
未成熟ラットの子宮向性(uterotropic)試験におけるエストロゲン様および抗エストロゲン様活性の評価
3日であった処理時間を7日に変えた以外は、以前に記述された方法(ジョーダンら、1977年)にもとづき、未成熟ラットの子宮向性検定を行ない、HMRのエストロゲン様活性を評価した。処理時間を延長したのは、被験化合物のエストロゲン性が弱いと予測されるからである。エストラジオールの生合成を防ぐアロマターゼ阻害剤(MPV−2213ad)による未成熟ラットの処置は、非エストロゲン刺激子宮に対応する実験方法上の対照として使用される。
【0042】
オスの成体ラットにおけるエストロゲン様および抗エストロゲン様活性の評価
正常および低アンドロゲンのノーブル(Noble)系オスラット(6〜9月齢)において、HMRのエストロゲン(抗アンドロゲン)様および抗エストロゲン様効果をそれぞれ調べた。オス生殖器系の構造および機能の変化を伴う慢性の低アンドロゲン状態を新生児期のエストロゲン化により誘発した(ジエチルスチルベストロール10.0μg/体重kg(菜種油に溶解)を生後1〜5日間皮下注射)。これらの変化は、MPV−2213adを一日に10〜30mg/体重kg投与することによるアロマターゼ阻害剤処理によって部分的に回複することが知られている(シュトレングら、考察は未発表)。
【0043】
動物に大豆を含まない基礎食(SDS、ウィンザム、エセックス州、イギリス)を与えた。水は自由に飲ませた。正常な動物および低アンドロゲンの動物12匹に、菜種油に溶解して一日量HMR50mg/体重kgをチューブにより補給した。両方の動物モデルのほかの12匹に対し、プラセボ処理として菜種油のみをチューブにより補給した。4週間の処置ののち、動物を殺し、精巣および付属する性腺(腹側前立腺、精嚢および凝固腺)の重量を測定した。血清および精巣テストステロン、ならびに下垂体および血清黄体ホルモン(LH)のレベルを免疫学的検定法によって測定した(ハービストら、1993年)。
【0044】
ラットDMBA誘発性乳ガンモデルにおける抗腫瘍活性の調査
ラットの乳ガンにおけるHMRの抗腫瘍活性を、以前に述べられた方法(カンガスら、1986年)により調査した。生後50日のメスのスプレーグ−ダウレイ(Sprague-Dawley)系ラットにDMBA(ジメチルベンツ[a]アントラセン(dimethylbentz[a]anthracene))12.0mgをチューブにより補給した。約6週間後には明らかな腫瘍が見られ、そののち腫瘍の幅(w)および長さ(l)を1週間に1度測定し、式V=(πw2l)/12にしたがって腫瘍の大きさを求めた。また、1週間に1度ラットの体重を測った。実験開始時に各グループの腫瘍の総数が同じになるように、ラットを3グループに分けた。(1)対照グループ8匹、(2)HMR3.0mg/kg7匹、および(3)HMR15.0mg/kg7匹であり、実験終了前に1匹を殺さなければならなかった。
【0045】
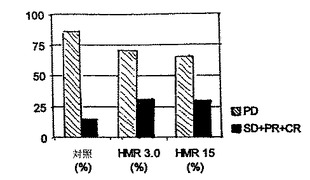
DMBA誘導の9週間後、つまり明白な腫瘍が見られるようになって3週間後のころから、HMRの経口投与を開始し、7.5週間にわたって毎日投与した。実験終了時、腫瘍を成長パターンにより各群、1.成長腫瘍(PD=進行性疾患);2.非成長、安定腫瘍(SD=安定性疾患、腫瘍の大きさに変化なし、または75%に満たない退化が見られる);3.退化腫瘍(PR=部分的な応答、つまり75%を超える腫瘍体積の退化が見られる);4.消失腫瘍(CR=完全な応答、明らかな腫瘍は見られない)に分類した。
【0046】
種々の量のHMRを投与した後のラットの尿に含まれる代謝産物の分析
オスのスプレーグ−ダウレイラット(4月齢)10匹を用いてHMRの代謝をインビボで調査した。代謝調査のあいだ、ラットを2匹1組、12時間の明暗周期で収容し、水は自由に飲ませ、大豆抜きの基礎食(SDS、ウィンザム、エセックス州、イギリス)を与えた。
【0047】
10%エタノール−PEG溶液に溶解したHMRを3、15、25および50mg/体重kgの投与量で1日1回2日間ラットにチューブにより補給した。2回目のチューブによる補給後、0.56Mアスコルビン酸120μl、および防腐剤として0.15Mナトリウムアジド120μlが入った回収器内の代謝物保持器に24時間分の尿を集めた。遠心分離した尿の容量を測定し、−20℃で保管した。前処理として、解凍した尿3.0mlに0.2M酢酸緩衝液750μl(pH4.0±0.1)を加えた。Sep−Pak C18カラム(シリカをベースにした樹脂100mg/カラム)を尿の抽出に用いた。水3.0ml、メタノール3.0mlおよび酢酸緩衝液3.0mlを用いてカラムをあらかじめ調整した。尿をカラムに通し、酢酸緩衝液3.0mlで洗浄したのち、メタノール3.0mlを用いてポリフェノール類(polyphenolics)を溶出した。溶出物を窒素下45℃のウォーターバスで蒸発乾固し、乾燥した残渣を0.2M酢酸緩衝液3.0ml中に再溶解した。エスカルゴ(Helix pomatia)酵素混合物30μlを加え、溶液を37℃でインキュベートし、グルクロニドおよび硫酸エステルをともに加水分解した。フラボン原液(EtOH中100μg/ml)300μlを加水分解した試料に加え、C−18カラムを用いて試料を抽出し、前記のように蒸発乾固し、GC−MSによる分析まで−20℃で保管した。
【0048】
蒸発させた尿試料をピリジンに溶解し、BSTFA:TMCS(10:1)のシリル化試薬を加えシリル化した。シリル化した試料のGC−MS分析は、HP6890−5973GC−MS装置を用いて行なった。GCカラムはHP−1架橋メチルポリシロキサンカラム(15m×直径0.25mm、膜厚0.25μm)を使用し、ヘリウムをキャリヤーガスとして流速1ml/分で用いた。GC炉の温度は昇温速度8℃/分で60℃から290℃に設定し、GC注入器はスプリット率1:15のスプリットモードに設定した。注入器の温度は250℃であった。化合物の同定は質量スペクトルに基づいて行なった。定量計算は内標準に相対的な対象化合物の無修正のピーク面積に基づいて行なった。
【0049】
結果
インビトロでの抗酸化作用の評価
本実験において、HMRはほかのリグナンまたはフラボノイドより脂質過酸化阻害作用が大きかった(表2)。よく知られた抗酸化剤、たとえば水溶性ビタミンE誘導体であるTROLEXや、BHAおよびBHTを、脂質過酸化阻害力、LDL酸化阻害力およびスーパーオキシドならびにペルオキシラジカル捕捉力においてHMRと比較した(表3)。全体として、HMRは最も強力な抗酸化剤であり、すべての検定においてBHAやBHTより効果が大きく、脂質過酸化阻害検定以外のすべての実験においてTROLEXより強力であった。脂質過酸化防止実験では、両化合物はほぼ同じように活性であった。
【0050】
JEG−3細胞におけるアロマターゼ阻害力
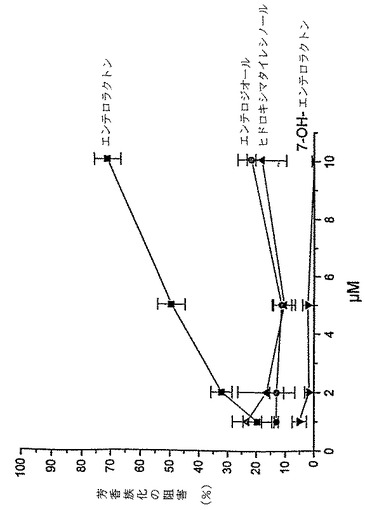
JEG−3細胞における3H−アンドロステンジオンからの3H−17β−エストラジオールの生成の阻害について、様々なHMR濃度で実験した。HMRの阻害力をエンテロラクトン、7−ヒドロキシエンテロラクトンおよびエンテロジオールと比較した。エンテロラクトンは1.0〜10.0μMの濃度範囲内で投与量に依存した芳香族化阻害を生じた。さらに、エンテロジオールには阻害作用はなく、阻害作用にはラクトン環が不可欠であることが示された。7−ヒドロキシエンテロラクトンおよびヒドロキシマタイレシノールには阻害効果はなく(図1)、アロマターゼ阻害におけるリグナン分子内の水酸基の数と位置の重要性を示唆している。
【0051】
MCF−7培養細胞におけるエストロゲン様および抗エストロゲン様活性
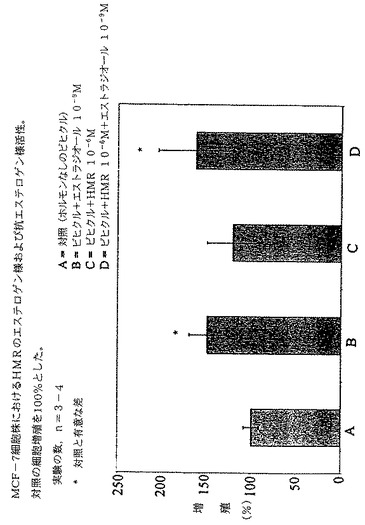
MCF−7細胞増殖において、図2に示すようにHMRは非常に弱い、統計的に有意とは言えないエストロゲン様および抗エストロゲン様活性しか有さなかった。
【0052】
未成熟ラットの子宮向性試験におけるエストロゲン様および抗エストロゲン様活性の評価
図3は未成熟ラットの子宮の成長に対するHMRの作用を例示している。HMRは、未成熟ラットの子宮重量の増加に有意なエストロゲン様効果を示さなかった。HMRにより、子宮重量が減少することもなく、抗エストロゲン様効果もないことが示された。アロマターゼ阻害剤は、予想通り子宮重量の増加を抑制し、アロマターゼ阻害剤の測定の方法は適切であったことが示された。
【0053】
オスの成体ラットにおけるエストロゲン様および抗エストロゲン様活性の評価
HMR処理の4週間後、対照および低アンドロゲングループでは、付属生殖腺および精巣の重量において有意な変化は見られなかった(表4)。テストステロンやLHの濃度にも有意な変化は見られなかった(表5)。これらの結果は、HMRは男性において完全なエストロゲン作用薬ではない、ということを示している。なぜなら、HMRは視床下部−脳下垂体−生殖腺−軸に対して典型的なエストロゲン様作用(LHおよびアンドロゲン分泌の抑制)を及ぼさないからである。また、オスのラットにおける新生児期のエストロゲン化により誘発された変化は、HMRにより元に戻ることはないので、HMRは抗エストロゲンでもない。
【0054】
ラットDMBA誘発性乳ガンモデルにおける抗腫瘍活性の調査
進行腫瘍(PD)と安定腫瘍(SD)、退化腫瘍(PR)および消失腫瘍(CR)の数を表4に示す。HMRの抗腫瘍効果は統計的に非常に有意であることがわかった。本モデルでは、抗腫瘍作用の明確な投与量依存性はなかった。HMRの抗酸化特性および腫瘍成長退化特性はともに、インビボでの抗腫瘍活性と結び付いている、という可能性がある。HMRのインビボでの抗腫瘍活性のメカニズムは依然知られていない。
【0055】
種々の量のHMRを投与した後のラットの尿に含まれる代謝産物の分析
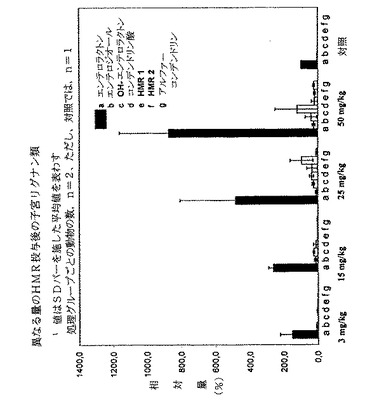
図5は、ラットにおけるHMRの主な分泌代謝物が、生物学的活性化合物であると考えられる、エンテロラクトンであることを示している。これはHMRの化学構造を考慮すると、ヒドロキシエンテロラクトンが主な代謝物であると考えられるので、驚くべきことである。HMRのエンテロラクトンへの代謝は、ラットの肝臓よりむしろ腸内細菌叢(bacterial intestinal flora)によって触媒され得る。
【0056】
結論
ヒドロキシマタイレシノール(HMR)は、DMBA誘発性乳ガンモデルにおいて、他の物質に変化せずに、および/またはエンテロラクトンに変化したのちに抗腫瘍活性を有する。それゆえ、HMRは乳ガン(BC)、前立腺ガン(PC)、結腸癌または良性前立腺過形成(BPH)を患う恐れのあるヒトにおいて、有益な効果をもたらす可能性がある。HMRは代謝によってエンテロラクトンとなり、インビトロにおける芳香族化を阻害する。HMRはアロマターゼ阻害剤の前駆体として下部尿路症(LUTS)、膀胱不安定性、膀胱排出口閉塞、尿道共同運動障害および女性化乳房の進行を抑制し得る。また、HMRには強い抗酸化活性があり、それゆえに食品添加物(抗酸化剤)として用いることができる。医薬品や栄養補助食品(dietary supplement)としてのHMRは、ヒトに対する有益な心血管効果を与え得る。新しい画期的な機能性食品、栄養補給食品、健康食品、治療用食品、遺伝子操作食品、あるいは新規な食品を製造するために、HMRを添加することは実行性のあることである。
【0057】
本発明の方法は、様々な形の実施態様によって具体的に表され、ここに記載したものはわずか一部であるということは充分理解されるであろう。ほかの実施態様が考えられ、それらが本発明の主旨に沿っていることは当業者に明らかであろう。このように、記載した実施態様は例示的なものであり、限定的なものと解釈されるべきではない。
【0058】
【表1】
【0059】
【表2】
【0060】
【表3】
【0061】
【表4】
【0062】
【表5】
【0063】
[参考文献]
Adlercreutz H, Bannwart C, Waehaelae K, Maekelae T, Brunow G, Hase T, Arosemena PJ, Kellis JT, and Vickery LE: Inhibition of human aromatase by mammalian lignans and isoflavonoid phytoestrogens. J Steroid Biochem Mol Biol, 44: 147-153, 1993.
Ahotupa M, Ruutu M, and Maentylae E: Simple methods of quantifying oxidation products and antioxidant potential of low density lipoproteins. Clin Biochem, 29: 139-144, 1996.
Ahotupa M, Maentylae E, and Kangas L: Antioxidant properties of the triphenylethylene antiestrogen drug toremifene. Naunyn-Schmiedeberg's Arch Pharmacol, 356: 297-302, 1997.
Axelson M and Setchell KDR: The excretion of lignans in rats-evidence for an intestinal bacterial source for this new group of compounds. FEBS lett, 123: 337-342, 1981.
Axelson M, Sjoevall J, Gustafsson BE and Setchell KDR: Origin of lignans in mammals and identification of a precursor from plants. Nature, 298: 659-660, 1982.
Ayres D, and Loike, J. Lignans: Chemical, bilogical and clinical properties. Cambridge university press, 1990.
Brzezinski A, Adlercreutz H, Shaoul R, Roesler A, Shmueli A, Tanos V and Schenker JG: Short-term effects of phytoestrogen-rich diet on postmenopausal women. Menopause (The Journal of the North American Menopause Society), 4: 89-94, 1997.
Ekman R: Analysis of lignans in Norway spruce by combined gas chromatography-mass spectrometry. Holzforschung, 30:79-85, 1976.
Ekman R: Distribution of lignans in Norway spruce. Acta Academiae Aboensis, Ser B, 39:1-6, 1979.
Evans BA, Griffiths K and Morton MS. Inhibition of 5α-reductase in genital skin fibroblasts and prostate tissue by dietary lignans and isoflavonoids. J Endocrinol, 147: 295-302, 1995.
Haavisto A-M, Petterson K, Bergendahl M, Perheentupa A, Roser JF, and Huhtaniemi I A. Supersensitive immunofluorometric assay for rat luteinizing hormone. Endocrinology, 132: 1687-1691, 1993.
Hulten K, Adlercreutz H, Winkvist A, Lenner P, Hallmans G and Ågren Å. Low levels of phyto-estrogens in blood as risk factor for breast cancer. In: COST 916 Workshop ‘Phyto-oestrogens: exposure, bioavailability, health benefits and safety concerns', 1998.
Ingram D, Sanders K. Kolybaba M and Lopez D. Case-control study of phyto-oestrogens and breast cancer. Lancet, Oct 4;350(9083): 990-994,1997.
Jenab M and Thompson LU. The influence of flaxseed and lignans on colon carcinogenesis and beta-glucuronidase activity. Carcinogenesis, Jun;17(6):1343-1348, 1996.
Jordan VC, Collins MM, Rowsby L, Prestwich G: A monohydroxylated metabolite of tamoxifen with potent antiestrogenic activity. J Endocrinol, 75: 305-316, 1997.
Kangas L, Nieminen A-L, Blanco G, Groenroos M, Kallio S, Karjalainen A, Perilae M, Soedervall M and Toivola R: A new triphenylethylene compound, Fc-1157a. II Antitumor effects. Cancer Chemother Pharmacol, 17:109-113, 1986.
Lampe JW, Martini MC, Kurzer MS, Adlercreutz H and Slavin JL: Urinary lignan and isoflavonoid excretion in premenopausal women consuming flaxseed powder. Am J Clin Nutr, 60:122-8, 1994.
Landstrom M, Zhang JX, Hallmans G, Aman P, Bergh A, Damber JE, Mazur W, Waehaelae K and Adlercreutz H. Inhibitory effects of soy and rye diets on the development of Dunning R3327 prostateadenocarcinoma in rats. Prostate, Aug 1; 36(3): 151-161, 1998
Mousavi Y and Adlercreutz H: Enterolactone and estradiol inhibit each other's proliferative effect on MCF-7 breast cancer cells in culture. J Steroid Biochem Mol Biol, 41: 615-619, 1992.
Mattinen J, Sjoeholm R and Ekman R. NMR-spectroscopic study of hydroxymatairesinol, the major lignan in Norway spruce (Picea abies) heartwood. ACH models in chemistry, 135(4):583-590, 1998.
Maekelae S, Poutanen M, Lehtimaeki J, Kostian M-L, Santti R and Vihko R. Estrogen-specific 17β-hydroxysteroid oxidoreductase type 1 (E.C.1.1.1.62) as a possible target for the action of phytoestrogens. P.S.E.B.M., 208:51-59, 1995.
Phipps WR, Martini MC, Lampe JW, Slavin JL and Kurzer MS. Effect of flax seed ingestion on the menstrual cycle. J Clin Endocrinol Metab, 77(5):1215-1219, 1993.
Rickard SE, Orcheson LJ, Seidl MM, Luyengi L, Fong HHS and Thompson LU: Dose-dependent production of mammalian lignans in rats and in vitro from the purified precursor secoisolariciresinol diglycoside in flaxseed. J Nutr, 126: 2012-2019, 1996.
Shultz TD, Bonorden WR and Seaman WR. Effect of short-time flaxseed consumption on lignan and sex hormone metabolism in men. Nutrition Research, 11:1089-110, 1991.
Serraino M and Thompson LU: The effect of flaxseed supplementation on early risk markers for mammary carcinogenesis. Cnacer Letters, 60: 135-142, 1991.
Sarraino M and Thompson LU: The effect of flaxseed supplementation on the initiation and promotional stages of mammary tumorigenesis. Nutr Cancer, 17:153-159, 1992.
Setchell KDR, Borriello SP, Gordon H, Lawson AM, Harkness R and Morgan DML. Lignan formation in man - microbaial involvement and possible roles in relation to cancer. Lancet, 4:4-7, 1981.
Streng T, Talo A and Santti R. Unpublished observations.
Thompson LU, Robb P, Serraino M and Cheung F. Mammalian lignan production from various foods. Nutr Cancer, 16: 43-52, 1991.
Thompson LU, Seidl, MM, Rickard SE, Orcheson, LJ, and Fong HHS: Antitumorigenic effect of a mammalian lignan precursors from flaxseed. Nutr Cancer, 26: 159-165, 1996a.
Thompson LU, Rickard SE, Orcheson LJ and Seidl MM: Flaxseed and its lignan and oil components reduce mammary tumor growth at a late stage of carcinogenesis. Carcinogenesis, 17: 1373-1376, 1996b.
Tou JCL, Chen J and Thompson U. Flaxseed and its lignan precursor, secoisolariciresinol diglycoside, affect pregnancy outcome and reproductive development in rats. J Nutr, 128: 1861-1868, 1998.
Wang C, Maekelae T, Hase T, Adlercreutz H and Kurzer MS: Lignans and flavonoids inhibit aromatase enzyme in human adipocytes. J Steroid Biochem Molec Biol, 50: 205-212, 1994.
Waters AP and Knowler JT. Effect of a lignan (HPMF) on RNA synthesis in the rat uterus. J Reprod. Fert, 66:379-381, 1982.
Zhang J-X, Hallmans G, Landstroem M, Bergh A, Damber J-E, Åman P and Adlercreutz H. soy and rye diets inhibit the development of Dunning R3327 prostatic adenocarcinoma in rats. Cancer Letters, 114: 313-314, 1997.
【技術分野】
【0001】
本発明は、ヒドロキシマタイレシノールをヒトに投与することによる、ヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防方法に関する。本発明はさらに、ヒドロキシマタイレシノールをヒトに投与することによるヒトの血清中のエンテロラクトンまたはヒドロキシマタイレシノールのほかの代謝産物の濃度を増加させる方法であって、それによってヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防をもたらす方法に関する。また、本発明はヒドロキシマタイレシノールを含有する医薬品、食品添加物および食品に関する。
【背景技術】
【0002】
本発明の背景、およびとくに実用に関してさらに詳細に述べた例を説明するため、ここで用いた刊行物などを参考文献として組み入れる。
【0003】
リグナン類は2,3−ジベンジルブタン骨格を有するフェノール系化合物の一群である。これらは、前駆体とよばれる桂皮酸、コーヒー酸、フェルラ酸、クマリン酸およびガリウム酸などの単量体単位のカップリングによって形成される(エアおよびロイク、1990年)。リグナンは植物に広く分布しており、さまざまな部位(根、葉、幹、種子、果実)に見られるが、少量である。多くの供給源(種子や果実)において、リグナンは植物の繊維成分と結びついたグリコシド共役体として検出される。哺乳類のリグナン前駆体の最も一般的な供給源食物は未精製の穀物である。食用植物のなかでリグナンの濃度が最も高いのは亜麻仁(flaxseed)であり、次に未精製の穀物、とくにライ麦があげられる。表1にさまざまな植物性食品から生成される哺乳類のリグナンを示す。
【0004】
針葉樹にも多量のリグナンが見られる。リグナンの型は種によって異なり、リグナンの量は樹木の部位によって相違する。トウヒ(Picea abies)の心材に見られる典型的なリグナンには、ヒドロキシマタイレシノール(HMR)、α−コニデンドリン(conidendrin)、コニデンドリン酸、マタイレシノール、イソラリシレシノール(isolariciresinol)、セコイソラリシレシノール(secoisolariciresinol)、リオヴァイル(liovile)、ピセアレシノ−ル(picearesinol)、ラリシレシノールおよびピノレシノールがある(エクマン、1979年)。トウヒ中のリグナンのうち、最も豊富に含まれる単一成分はHMRであり、全リグナンの約60パーセントを占め、主に非共役遊離形で存在する。太い根におけるリグナン濃度は2〜3パーセントである。大量のリグナンは枝の心材(5〜10パーセント)やねじれた部分(twists)の心材に含まれ、とくに木節ではリグナンの量は10パーセントより多い(エクマン、1976年および1979年)。これらの濃度は、リグナン高含有材料として知られる、亜麻を挽いて粉末にしたものと比べ約100倍である。
【0005】
ヒドロキシマタイレシノールの化学構造は
【化1】
である。
【0006】
リグナンはたとえば、圧縮材の繊維から単離することができる。これらの繊維は幹や、紙の品質を悪化させる木節(大き過ぎる木材チップの画分)の圧縮材から生じる(エクマン、1976年)。
【0007】
マタイレシノールやセコイソラリシレシノールなどの植物性リグナンは、腸内微生物(gut microflora)によって、哺乳類リグナン、つまりそれぞれエンテロラクトンおよびエンテロジオールに変換される(アクセルソンら、1982年)。それらは腸肝循環し、グルクロニド共役体として尿に排出される(アクセルソンおよびセッチェル、1982年)。リグナンの化学的な予防作用の実験的な証拠として、脂肪分の多い食品にリグナン高含有亜麻仁粉末(5〜10%)または亜麻仁リグナン(セコイソラリシレシノール−ジグリコシド(SDG))を加えると、ラットにおいて抗エストロゲン感受性DMBA誘発性乳ガンの進行が抑制された(セレイノおよびトンプソン、1991年、1992年、トンプソンら1996年a、1996年b)。それらにより、上皮細胞の増殖、核異常、腫瘍の成長および新しい腫瘍の発生が減少した。また、大量のリグナンの摂取が実験的前立腺および結腸ガンを防ぐ可能性もある。食用ライ麦(リグナンを含む)によって、ラットの移植されたダニングR3327前立腺ガンの成長が初期段階で抑制された(ツァンら、1997年;ランドストロームら、1998年)。明らかな腫瘍を有する動物の割合、腫瘍の大きさ、および成長率はかなり低かった。さらに、亜麻仁またはSDGを加えることによって、ラットの結腸において化学的に誘導された異常陰窩の形成が抑制された(セレイノおよびトンプソン、1992年。ジェナブおよびトンプソン、1996年)。よって、抗腫瘍作用は、弱いエストロゲン−抗エストロゲン様特性および/またはほかのメカニズムによって起こる可能性があるが、それら特性やメカニズムはあまり解明されていない。
【0008】
乳ガンの女性では、エンテロラクトンの尿への排出および血清中濃度が低く(イングラムら、1997年;フルテンら、1998年)、それはリグナンが化学的予防性であることを示している。哺乳類のリグナン(エンテロラクトンおよびエンテロジオール)は、エストロゲンと構造が似ているので、ホルモン性のガン、つまり乳ガンなどを緩和する、と仮定されてきた。エンテロラクトンはMCF−7細胞において弱いエストロゲン様の潜在力を有するが(モサビおよびアドラクルツ、1992年)、マウスの子宮重量においてはエストロゲン様の応答は見られなかった(セッチェルら、1981年)。エストロゲン様活性の兆候として、ラットに妊娠および授乳期間中SDGを与えることによって、離乳時には子宮重量が増加したが、後期にはその効果ははっきりしなかった(タウら、1998年)。また、予想される抗腫瘍効果は、リグナンの抗エストロゲン様作用と結びついている(ウォーターズおよびノーラー、1982年)。哺乳類リグナン、つまりエンテロラクトンによるアロマターゼの阻害は、リグナン高含有植物性食品の消費により乳ガンなどのエストロゲン依存性疾病を低減するメカニズムを提示していることが考えられる(アドラクルツら、1993年;ワンら1994年)。リグナンの潜在的な抗酸化活性もまた、ガンの進行を予防するリグナン作用のメカニズムを示していることが考えられる。さらに、哺乳類のリグナンは、テストステロンの、強い細胞内アンドロゲンである5α−ジヒドロテストステロン(DHT)への変換をヒトにおいて可能な濃度で阻害することを示した(エバンズら、1995年)。DHT濃度の低下は、前立腺ガン(PC)や良性前立腺過形成(BPH)の危険性を緩和するであろう。
【0009】
エンテロラクトンの前駆体としてのリグナンはまた、下部尿道症(LUTS)および女性化乳房を軽減することができる。我々は、動物実験の結果に基づき、膀胱頸部共同運動障害あるいは外括約筋偽共同運動障害として顕われる尿道共同運動障害における筋機能障害の進行に、エストロゲンが重要な役割を果たすことを示唆した(シュトレングら、考察は未発表)。そのような神経筋の変化は、アロマターゼ阻害剤(MPV−2213ad)によって少なくとも部分的に元に戻るが、それはエストロゲンの役割を表している。さらに、女性化乳房はエストロゲンとの接触やアンドロゲンよりエストロゲンが多く存在する場合などに誘発される。女性化乳房はアロマターゼ阻害剤によってうまく治療できる。5α−還元酵素および/またはアロマターゼを阻害するリグナンの能力は、潜在的な抗酸化活性と合わせて、男性においてホルモン性疾病が進行する際のリグナンによる予防作用のメカニズムを示している可能性がある。
【0010】
リグナンに潜在するヒトへの効果に関するデータはない。ヒトにおけるリグナンの作用に関する最近の学説は、亜麻仁粉(つまりリグナン)を加えた食品がもたらす効果の研究から導き出されてきた。女性の食事に亜麻仁粉を含ませた場合、月経周期に変化が現れた(フィップスら、1993年)。正常な月経周期の被験者らでは、常食に加えて1日10gの亜麻仁粉を摂取した場合、黄体期の平均長さがより長くなり、黄体期の血清中の17β−エストラジオールに対するプロゲステロンの比率がより高くなった(フィップスら、1993年)。エストロンまたは17β−エストラジオールの濃度に関しては、亜麻摂取グループと対照グループのあいだに大きな相違点はなかった。また、閉経後の女性の血清中エストロゲン濃度に関しても、亜麻摂取グループと対照グループのあいだに大きな相違点はなかった(ブレジンスキーら、1997年)。亜麻仁粉の追加によって血清中のSHBG(エストラジオールを強力に結合するタンパク質)の濃度が増加した。これは、肝臓組織で見られる典型的なエストロゲン様効果である。一方、SHBGの濃度の増加により、内因性エストロゲンのバイオアべイラビリティは低下する。健康な青年の場合、短期間(6週間)の食事への亜麻仁の添加(マフィンに1日10g)は、テストステロンの血漿中濃度に目立った影響を示さず、男性におけるエストロゲン作用(estrogenicity)の欠如を示している(シュルツら、1991年)。同時にこれらの研究は、リグナンは弱いホルモン(エストロゲン様および抗エストロゲン様)効果を有するが、リグナンの作用機構はそのホルモン効果によってすべて説明され得るわけではない、ということを示す。
【0011】
結論として、以前は動物実験や臨床実験で使用するのに充分な量の単離哺乳類リグナンを入手することはできず、リグナン摂取量を増加させる唯一の可能性は、繊維質の多い亜麻仁などの食品の消費を増加させることであった。効率よくエンテロラクトンに変換し、大量に生成され単離されうるHMRなどのリグナンは、ガン、他のホルモン性疾患および心疾患を化学的に予防するための医薬品および機能性食品などの食品の開発において貴重なものである。
【発明の概要】
【0012】
ある側面において、本発明は有効量のヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体をヒトに投与することからなる、ヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防方法に関する。
【0013】
さらなる側面において、本発明は、有効量のヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体をヒトに投与することからなる、ヒトの血清中のエンテロラクトンまたはヒドロキシマタイレシノールのほかの代謝産物の濃度を増加させる方法であって、それによってヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患の予防をもたらす方法に関する。
【0014】
第3の側面において、本発明は、有効量のヒドロキシマタイレシノールまたは幾何異性体もしくは立体異性体と、薬学的に許容しうる担体とからなる医薬品に関する。
【0015】
第4の側面において、本発明は、ヒドロキシマタイレシノールまたはその幾何異性体もしくは立体異性体を濃縮した液体または固体材料からなる、食品添加物用製品に関する。
【0016】
第5の側面において、本発明は、有効量のヒドロキシマタイレシノールまたはその幾何異性体もしくは立体異性体からなる食品に関する。
【0017】
もう1つの側面において、本発明は、前記有効量のヒドロキシマタイレシノール、その幾何異性体または立体異性体を食品に添加することからなる食品の安定性を高める方法に関する。
【図面の簡単な説明】
【0018】
【図1】図1はJEG−3細胞におけるリグナンによるアロマターゼの濃度相関阻害を示す。
【図2】図2はHMRの存在下および非存在下におけるMCF−7細胞の増殖を示す。
【図3】図3はHMRまたはアロマターゼ阻害剤で処理した未成熟ラットの子宮の湿重量を示す。
【図4】図4はメスのラットにおけるDMBA誘発性乳腺腫瘍に対するHMRの抗腫瘍活性を示す。
【図5】図5は種々の投与量のHMRで処理したラットの、エンテロラクトンの尿への排出を示す。
【発明を実施するための形態】
【0019】
本発明はヒドロキシマタイレシノール(HMR)を食品に添加し、また医薬品として用いることによってヒトのガン、ある種の非ガン性ホルモン依存性疾患および/または心疾患を予防する、リグナン、つまりHMRの用途に関する。驚いたことに、HMRはインビボでエンテロラクトンへ代謝されるが、これがリグナンの抗腫瘍特性の原因の一つであると考えられている。インビトロでのHMRの抗酸化活性は大きく、この特性は、HMRが、体内において有害な遊離酸素種に対する保護効果を通して心疾患をも防ぐことができるということを示している。また、本発明は食品の安定性を向上する(つまり栄養価の損失や食品の風味の低下につながる、脂質および色素の酸化とビタミンの喪失とを抑制する)ための食品添加物としてのHMRの用途に関する。
【0020】
本発明の方法は、乳ガン、前立腺ガンおよび結腸ガン、下部尿路症、尿道共同運動障害、膀胱不安定性、膀胱排出口閉塞、良性前立腺過形成および男性の女性化乳房などの非ガン性ホルモン依存性疾患、および血清中の酸化LDLに起因する心疾患の予防に特に効果的である。
【0021】
本発明による医薬品は経口処方であることが好ましい。活性化合物(HMR)の必要量は、予防すべき特定の症状によって異なるが、標準的な投与量は大人で1日約10〜100mgである。
【0022】
本発明の食品添加物において、ヒドロキシマタイレシノールで強化される材料は、HMRとの混合に適し、HMRの特性を損なわない、食用で中毒性のない固体または液体の材料であればどのようなものでもよい。この材料の役割は主に、HMRの確実な投与を容易にすることである。好ましい濃度は、たとえば強化された材料100g中にHMR100mg〜1gである。
【0023】
本発明の食品とは、とくに機能性食品、栄養補助食品、栄養剤、治療用食品、栄養補給食品、健康食品、遺伝子操作食品(designer food)または任意の食品である。食品中のHMRの好ましい濃度は、たとえば食品100gに対してHMR1〜20mgである。
【0024】
本発明の機能性食品はたとえば、バター、マーガリン、ビスケット、パン、ケーキ、アメ、菓子類、ヨーグルトもしくは他の発酵乳製品、またはムースリなどのシリアルの形が可能である。
【0025】
ヒドロキシマタイレシノールの添加は、栄養価の損失と食品の風味の低下につながる、脂質、ビタミンおよび色素の酸化を抑制することを目的として、食品の安定性を向上する場合にとくに有用である。このためにHMRの好ましい濃度は、たとえば約0.1%である。
【0026】
本発明で使用するHMRは、圧縮材の大き過ぎるチップ画分(oversize chip fraction)(枝、ねじれ部分および木節を含む)から単離する。HMRはガンや心疾患などの疾病の予防に用いる。
【0027】
HMRの特性を7つの異なる分析方法によって検討した。
1.インビトロにおける抗酸化力の測定
2.JEG−3細胞におけるアロマターゼ阻害力の測定
3.MCF−7培養細胞におけるエストロゲン様および抗エストロゲン様活性の測定
4.子宮成長の生物学的検定によるエストロゲン様および抗エストロゲン様活性の評価
5.オスの成体ラットにおけるエストロゲン様および抗エストロゲン様活性の測定
6.ラットDMBA誘発性乳ガンモデルにおける抗腫瘍活性の調査
7.種々の量のHMRを投与した後のラットの尿に含まれる代謝産物の分析
【0028】
以前は、HMRが樹木のリグナン成分であることから、その特徴はあまり注目されておらず、生物学的試験を行なうのに充分な量のHMRを単離し、精製することは不可能であった。しかし、トウヒ(spruce)の様々な部位におけるHMRの分布が知られ(エクマン、1976年および1979年)、リグナン、とくにHMRの詳細な研究の機会が得られた。
【0029】
HMRの投与量と尿中のエンテロラクトンの量との間に線形相関が見られた。エンテロラクトンは広く知られた哺乳類のリグナンであり、マタイレジノールから腸内細菌によって、またはエンテロジオールの酸化によって形成される(アクセルソンおよびセッチェル、1981年;アクセルソンら、1982年)。わずかな量の代謝されなかったHMRとほかの代謝産物(エンテロジオールと7−ヒドロキシエンテロラクトン)とが尿中に見られた。それらの量は、毎日のHMRの投与量を増やしても変化しなかった。これらの発見は、HMRが代謝されエンテロラクトンとなり、さらにジメチル化およびジヒドロキシル化を経てHMRから生じたエンテロラクトンは、エンテロジオールには変換しないということを示唆している。HMRの構造に基づいて、7−ヒドロキシエンテロラクトンが主なHMRの代謝産物であると考えられたが、それは違っていた。このヒドロキシル基は代謝の際に脱離する。HMRの代謝は、SDGの代謝とは異なる。SDGは代謝によってエンテロジオールになり、その一部は酸化されエンテロラクトンになる(リカードら、1996年、ランペら、1994年)。HMRはしたがって、エンテロラクトンの直接の前駆体として、SDGよりも有利である。
【0030】
HMRはラットの子宮または男性において、たとえあるとしても弱いエストロゲン様活性しか示さない。HMRはMCF−7細胞において、弱く有意性のないエストロゲン様活性しか示さない。また、HMRに関して抗エストロゲン活性は見られなかった。それゆえ、図2に示すように、HMRがラットのDMBA誘発性腫瘍モデルにおいて非常に有意な抗腫瘍活性を示したことは驚きである。HMRの活性はHMR自身あるいはエンテロラクトンが原因であると考えられる。しかし、ラットにDMBA処理ののち、2種類の投与量(3および15mg/kg)でHMRを与えた場合、HMRの化学的予防作用には投与量との因果関係はなかった。よって、抗腫瘍効果を得るためには、HMRは必ずしもエンテロラクトンに変換される必要はなく、またこれらのリグナンの投与量はより少量でも最大の化学的予防効果を得るのに充分である。
【0031】
表2および3に示すように、HMRは非常に有効な抗酸化剤である。HMRは、既知の最も強力な、脂質過酸化反応の阻害剤のひとつであり、LDLの酸化を防ぐ優れた阻害剤である。LDLの酸化の阻害はヒトにおいて非常に重要であるとされている。なぜなら、酸化LDLの血清中濃度が、アテローム性動脈硬化症といった心疾患を予測する最良のものの一つとされているからである。HMRは食品の安定性を向上する(つまり栄養価の損失と食品の風味の低下につながるビタミン、脂質および色素の酸化を抑制する)食品添加物となりうる。なぜなら、HMRは、食品安定性の向上のために通常用いられる、よく知られた抗酸化剤、すなわちブチル化ヒドロキシアニソール(BHA)およびブチル化ヒドロキシトルエン(BHT)などよりもさらに優れたスーパーオキシドアニオンの捕捉剤およびペルオキシラジカルの捕捉剤であるからである。
【0032】
実験
薬品
リグナンのエストロゲン様作用、抗エストロゲン様作用、芳香族化(aromatization)を阻害する能力そして抗酸化特性を調べるため、様々なリグナンをインビトロで試験した。試験用化合物は以下から入手した。エンテロジオールおよびエンテロラクトンは、イギリス、ロンドンのプランテック社(Plantech)より購入した。2つの7−OHエナンチオマーを有する7−ヒドロキシエンテロラクトンはフィンランド、ヘルシンキ大学応用化学研究室、クリスティナ・ワララ教授(Dr. Kristina Waehaelae)のご好意により提供された。
【0033】
樹木からのHMRの抽出
エクマン(1976年)およびエクマン(1979年)に記載の方法によって、HMR抽出物をノルウェイ・トウヒ(Picea Abies)より単離した。簡単言うと、凍結乾燥粉末心材をヘキサンでソックスレー抽出し、無極性、親油性の抽出物を除去した。その木材試料をアセトン/水(9:1 v/v)を用いて、同じ装置でさらに抽出し、粗リグナンを得た。ヒドロキシマタイレシノール(HMR)およびその異性体を単離し、さらに精製するため、XAD樹脂で再度クロマトグラフィーを行なった。
【0034】
インビトロにおける抗酸化力の測定
リグナンの抗酸化力を4つの異なる方法で評価した。1)脂質過酸化の阻害、2)低比重リポタンパク(LDL)酸化の阻害、3)スーパーオキシド アニオンの捕捉および4)ぺルオキシラジカルの捕捉の分析である。
【0035】
脂質過酸化の阻害は、インビトロでのラット肝臓ミクロソームにおいてtert−ブチルハイドロパーオキサイド誘発脂質過酸化(t−BuOOH−LP)を阻害するリグナンの能力により評価した(アホトゥパら、1997年)。t−BuOOH−LPに対する試験は次のように行なった。緩衝液0.8ml(50mM 炭酸ナトリウム、pH10.2、0.1mM EDTA)を照度計のキュベットにピペットで注入した。最終濃度が1.5mg タンパク質/ml の希釈肝臓ミクロソーム20μlを添加し、ついで、ルミノール6ml(0.5mg/ml)および試験用薬品(test chemicals)を加えた。エタノールまたはジメチルスルホキシド(インキュベーション体積の2%)で希釈した少量の被験化合物をインキュベーション混合物に加え、ビヒクル(vehicle)(エタノールまたはジメチルスルホキシド)の場合と脂質過酸化力を比較した。0.9mMのt−BuOOH 0.05mlを用いて33℃で反応を開始した。化学発光を1分ごとに約45分間測定し、曲線下部の面積(積分)を求めた。化学発光の測定は分析専用ソフトを用いてパーソナルコンピュータに接続したバイオ−オービット(Bio-Orbit)1251照度計(バイオ−オービット社製、ツルク、フィンランド)を用いて行なった。
【0036】
LDLの酸化の阻害は、アホトゥパら(1996年)に記載の方法によって評価した。簡単に言うと、緩衝化ヘパリンを用いて沈殿(precipitation)によりLDLを単離した。リン酸緩衝液中で再度懸濁した後、20mMのCuCl2を添加し、混合液を37℃で3時間インキュベートした。こののちクロロフォルム−メタノールによりLDL脂質を抽出し、窒素下で乾燥し、シクロヘキサンに再び溶解させ、分光光度法を用い、234nmで分析した。吸光度の強度はLDLの酸化を示す。さまざまな化合物のLDL酸化防止力を調べるため、CuCl2添加前のインキュベーション混合物にそれらの化合物を加えた。分析中に被験化合物の妨げとなりうるものは、インキュベーション期間の前後に234nmでの吸収を測定することによって取り除いた。初期濃度(0.1mM)で抗酸化効果を示した化合物に関し、IC−50値(被験化合物によってLDLの酸化が50%阻害された濃度)を求めた。
【0037】
スーパーオキシドアニオン捕捉方法は、キサンチン−キサンチンオキシダーゼ系により調製した条件下で生成されるスーパーオキシドアニオン、および生成した反応性酸素種の照度計による検出にもとづいた(アホトゥパら、1997年)。被験化合物が化学発光を減少させる能力を評価した。IC−50濃度(50%の化学発光が抑制された濃度)を求めた。
【0038】
ペルオキシラジカル捕捉の分析は、2,2'−アゾビス(2−アミジノプロパン)HClの熱分解によるペルオキシラジカルの発生、および化学発光によるその検出にもとづいた(アホトゥパら、1997年)。その結果を化学量論的な因子、つまり被験化合物1モルにつき何モルのペルオキシラジカルが捕捉されうるか、として求めた。
【0039】
JEG−3細胞におけるアロマターゼ阻害力の測定
HMRおよび構造的に関連するリグナン(エンテロラクトン、エンテロジオールおよび7−ヒドロキシエンテロラクトン)の効果を、ヒト絨毛癌細胞株であるJEG−3細胞における3H−アンドロステンジオンからの3H−17β−エストラジオールの生成にもとづいて調べた。JEG−3絨毛癌細胞は、インビトロでアロマターゼ阻害作用を調査し得る有効なアロマターゼ・モデルである(クレッケルら、1991年)。ウシ胎仔血清(FCS)10%含有DMEM中に細胞を保持した。3H−アンドロスト−4−エン,3,17−ジオン(0.5nM)50μl、非標識アンドロステンジオン(0.5nM)50μl、被験化合物(10mM)100μlおよび細胞懸濁液800μl(細胞100万個)をインキュベーション混合物とした。4時間のインキュベーションののち、非標識の担体(アンドロステンジオン、テストステロン、17β−エストラジオールおよびエストロン)を加えた。ジクロロメタン3.0mlを用いてそれらステロイドを2回抽出した。以前に述べられたように、HPLCを用いて放射性標識3H−17β−エストラジオールの分離と定量を行なった(マケラら、1995年)。カラム系は保護カラム、続いてC18 150×3.9mmID分析カラム(テクノパック10C18HPLCテクノロジー;ウェリントンハウス、チェシャー州、イギリス)から構成された。移動相はアセトニトリル/水(35/65)であり、流速は1.2ml/分であった。放射性代謝物質のインライン検出のため、HPLCカラムの溶離液を液体蛍光体(liquid scintillant)と連続的に混合し、インライン放射能検出器を用いてモニターした。
【0040】
MCF−7培養細胞におけるエストロゲン様および抗エストロゲン様活性の測定
MCF−7細胞株(ヒト乳ガン細胞)保存培養物(stock cultures)は、5%FCS、ペニシリン100 U/ml、ストレプトマイシン100μg/ml、インシュリン10μg/mlおよび17β−エストラジオール1nMを加えたフェノールレッドを含まないRPMI培地中、T−75細胞培養びん内で培養した。培地は1週間に3回新しいものと交換した。保存培養物をトリプシン処理によって取り出し、フェノールレッドを含まないベルセン溶液(versene solution)10ml中に懸濁し、800rpmで5分間遠心分離した。5%デキストラン木炭除去(dextran charcoal stripped)FSC(dcFSC)を加えたRPMI培地中に細胞ペレットを慎重に再懸濁し、6穴プレートに50000細胞個/3.0ml溶媒/ウェルの割合で播種した。培養2日目に培地を交換し、被験化合物を加えた。リグナン化合物のエストロゲン様効果を調べるため、リグナン化合物をエタノールに希釈し、細胞培養物に加え、最終濃度を1.0Mとした。各増殖検定において、1.0nM17β−エストラジオールのエタノール溶液をエストロゲン様反応に関する陽性対照として使用した。同量のエタノールを対照ウェルに加えた。また、抗エストロゲン様効果を調べるため、17β−エストラジオールとリグナンとの両溶液を培養細胞に加えた。被験化合物の存在下において5日〜7日間細胞を培養し、培地は2日おきに交換した。細胞増殖は、コールター計数器を用いて溶出した核を数えることによって定量化した。
【0041】
未成熟ラットの子宮向性(uterotropic)試験におけるエストロゲン様および抗エストロゲン様活性の評価
3日であった処理時間を7日に変えた以外は、以前に記述された方法(ジョーダンら、1977年)にもとづき、未成熟ラットの子宮向性検定を行ない、HMRのエストロゲン様活性を評価した。処理時間を延長したのは、被験化合物のエストロゲン性が弱いと予測されるからである。エストラジオールの生合成を防ぐアロマターゼ阻害剤(MPV−2213ad)による未成熟ラットの処置は、非エストロゲン刺激子宮に対応する実験方法上の対照として使用される。
【0042】
オスの成体ラットにおけるエストロゲン様および抗エストロゲン様活性の評価
正常および低アンドロゲンのノーブル(Noble)系オスラット(6〜9月齢)において、HMRのエストロゲン(抗アンドロゲン)様および抗エストロゲン様効果をそれぞれ調べた。オス生殖器系の構造および機能の変化を伴う慢性の低アンドロゲン状態を新生児期のエストロゲン化により誘発した(ジエチルスチルベストロール10.0μg/体重kg(菜種油に溶解)を生後1〜5日間皮下注射)。これらの変化は、MPV−2213adを一日に10〜30mg/体重kg投与することによるアロマターゼ阻害剤処理によって部分的に回複することが知られている(シュトレングら、考察は未発表)。
【0043】
動物に大豆を含まない基礎食(SDS、ウィンザム、エセックス州、イギリス)を与えた。水は自由に飲ませた。正常な動物および低アンドロゲンの動物12匹に、菜種油に溶解して一日量HMR50mg/体重kgをチューブにより補給した。両方の動物モデルのほかの12匹に対し、プラセボ処理として菜種油のみをチューブにより補給した。4週間の処置ののち、動物を殺し、精巣および付属する性腺(腹側前立腺、精嚢および凝固腺)の重量を測定した。血清および精巣テストステロン、ならびに下垂体および血清黄体ホルモン(LH)のレベルを免疫学的検定法によって測定した(ハービストら、1993年)。
【0044】
ラットDMBA誘発性乳ガンモデルにおける抗腫瘍活性の調査
ラットの乳ガンにおけるHMRの抗腫瘍活性を、以前に述べられた方法(カンガスら、1986年)により調査した。生後50日のメスのスプレーグ−ダウレイ(Sprague-Dawley)系ラットにDMBA(ジメチルベンツ[a]アントラセン(dimethylbentz[a]anthracene))12.0mgをチューブにより補給した。約6週間後には明らかな腫瘍が見られ、そののち腫瘍の幅(w)および長さ(l)を1週間に1度測定し、式V=(πw2l)/12にしたがって腫瘍の大きさを求めた。また、1週間に1度ラットの体重を測った。実験開始時に各グループの腫瘍の総数が同じになるように、ラットを3グループに分けた。(1)対照グループ8匹、(2)HMR3.0mg/kg7匹、および(3)HMR15.0mg/kg7匹であり、実験終了前に1匹を殺さなければならなかった。
【0045】
DMBA誘導の9週間後、つまり明白な腫瘍が見られるようになって3週間後のころから、HMRの経口投与を開始し、7.5週間にわたって毎日投与した。実験終了時、腫瘍を成長パターンにより各群、1.成長腫瘍(PD=進行性疾患);2.非成長、安定腫瘍(SD=安定性疾患、腫瘍の大きさに変化なし、または75%に満たない退化が見られる);3.退化腫瘍(PR=部分的な応答、つまり75%を超える腫瘍体積の退化が見られる);4.消失腫瘍(CR=完全な応答、明らかな腫瘍は見られない)に分類した。
【0046】
種々の量のHMRを投与した後のラットの尿に含まれる代謝産物の分析
オスのスプレーグ−ダウレイラット(4月齢)10匹を用いてHMRの代謝をインビボで調査した。代謝調査のあいだ、ラットを2匹1組、12時間の明暗周期で収容し、水は自由に飲ませ、大豆抜きの基礎食(SDS、ウィンザム、エセックス州、イギリス)を与えた。
【0047】
10%エタノール−PEG溶液に溶解したHMRを3、15、25および50mg/体重kgの投与量で1日1回2日間ラットにチューブにより補給した。2回目のチューブによる補給後、0.56Mアスコルビン酸120μl、および防腐剤として0.15Mナトリウムアジド120μlが入った回収器内の代謝物保持器に24時間分の尿を集めた。遠心分離した尿の容量を測定し、−20℃で保管した。前処理として、解凍した尿3.0mlに0.2M酢酸緩衝液750μl(pH4.0±0.1)を加えた。Sep−Pak C18カラム(シリカをベースにした樹脂100mg/カラム)を尿の抽出に用いた。水3.0ml、メタノール3.0mlおよび酢酸緩衝液3.0mlを用いてカラムをあらかじめ調整した。尿をカラムに通し、酢酸緩衝液3.0mlで洗浄したのち、メタノール3.0mlを用いてポリフェノール類(polyphenolics)を溶出した。溶出物を窒素下45℃のウォーターバスで蒸発乾固し、乾燥した残渣を0.2M酢酸緩衝液3.0ml中に再溶解した。エスカルゴ(Helix pomatia)酵素混合物30μlを加え、溶液を37℃でインキュベートし、グルクロニドおよび硫酸エステルをともに加水分解した。フラボン原液(EtOH中100μg/ml)300μlを加水分解した試料に加え、C−18カラムを用いて試料を抽出し、前記のように蒸発乾固し、GC−MSによる分析まで−20℃で保管した。
【0048】
蒸発させた尿試料をピリジンに溶解し、BSTFA:TMCS(10:1)のシリル化試薬を加えシリル化した。シリル化した試料のGC−MS分析は、HP6890−5973GC−MS装置を用いて行なった。GCカラムはHP−1架橋メチルポリシロキサンカラム(15m×直径0.25mm、膜厚0.25μm)を使用し、ヘリウムをキャリヤーガスとして流速1ml/分で用いた。GC炉の温度は昇温速度8℃/分で60℃から290℃に設定し、GC注入器はスプリット率1:15のスプリットモードに設定した。注入器の温度は250℃であった。化合物の同定は質量スペクトルに基づいて行なった。定量計算は内標準に相対的な対象化合物の無修正のピーク面積に基づいて行なった。
【0049】
結果
インビトロでの抗酸化作用の評価
本実験において、HMRはほかのリグナンまたはフラボノイドより脂質過酸化阻害作用が大きかった(表2)。よく知られた抗酸化剤、たとえば水溶性ビタミンE誘導体であるTROLEXや、BHAおよびBHTを、脂質過酸化阻害力、LDL酸化阻害力およびスーパーオキシドならびにペルオキシラジカル捕捉力においてHMRと比較した(表3)。全体として、HMRは最も強力な抗酸化剤であり、すべての検定においてBHAやBHTより効果が大きく、脂質過酸化阻害検定以外のすべての実験においてTROLEXより強力であった。脂質過酸化防止実験では、両化合物はほぼ同じように活性であった。
【0050】
JEG−3細胞におけるアロマターゼ阻害力
JEG−3細胞における3H−アンドロステンジオンからの3H−17β−エストラジオールの生成の阻害について、様々なHMR濃度で実験した。HMRの阻害力をエンテロラクトン、7−ヒドロキシエンテロラクトンおよびエンテロジオールと比較した。エンテロラクトンは1.0〜10.0μMの濃度範囲内で投与量に依存した芳香族化阻害を生じた。さらに、エンテロジオールには阻害作用はなく、阻害作用にはラクトン環が不可欠であることが示された。7−ヒドロキシエンテロラクトンおよびヒドロキシマタイレシノールには阻害効果はなく(図1)、アロマターゼ阻害におけるリグナン分子内の水酸基の数と位置の重要性を示唆している。
【0051】
MCF−7培養細胞におけるエストロゲン様および抗エストロゲン様活性
MCF−7細胞増殖において、図2に示すようにHMRは非常に弱い、統計的に有意とは言えないエストロゲン様および抗エストロゲン様活性しか有さなかった。
【0052】
未成熟ラットの子宮向性試験におけるエストロゲン様および抗エストロゲン様活性の評価
図3は未成熟ラットの子宮の成長に対するHMRの作用を例示している。HMRは、未成熟ラットの子宮重量の増加に有意なエストロゲン様効果を示さなかった。HMRにより、子宮重量が減少することもなく、抗エストロゲン様効果もないことが示された。アロマターゼ阻害剤は、予想通り子宮重量の増加を抑制し、アロマターゼ阻害剤の測定の方法は適切であったことが示された。
【0053】
オスの成体ラットにおけるエストロゲン様および抗エストロゲン様活性の評価
HMR処理の4週間後、対照および低アンドロゲングループでは、付属生殖腺および精巣の重量において有意な変化は見られなかった(表4)。テストステロンやLHの濃度にも有意な変化は見られなかった(表5)。これらの結果は、HMRは男性において完全なエストロゲン作用薬ではない、ということを示している。なぜなら、HMRは視床下部−脳下垂体−生殖腺−軸に対して典型的なエストロゲン様作用(LHおよびアンドロゲン分泌の抑制)を及ぼさないからである。また、オスのラットにおける新生児期のエストロゲン化により誘発された変化は、HMRにより元に戻ることはないので、HMRは抗エストロゲンでもない。
【0054】
ラットDMBA誘発性乳ガンモデルにおける抗腫瘍活性の調査
進行腫瘍(PD)と安定腫瘍(SD)、退化腫瘍(PR)および消失腫瘍(CR)の数を表4に示す。HMRの抗腫瘍効果は統計的に非常に有意であることがわかった。本モデルでは、抗腫瘍作用の明確な投与量依存性はなかった。HMRの抗酸化特性および腫瘍成長退化特性はともに、インビボでの抗腫瘍活性と結び付いている、という可能性がある。HMRのインビボでの抗腫瘍活性のメカニズムは依然知られていない。
【0055】
種々の量のHMRを投与した後のラットの尿に含まれる代謝産物の分析
図5は、ラットにおけるHMRの主な分泌代謝物が、生物学的活性化合物であると考えられる、エンテロラクトンであることを示している。これはHMRの化学構造を考慮すると、ヒドロキシエンテロラクトンが主な代謝物であると考えられるので、驚くべきことである。HMRのエンテロラクトンへの代謝は、ラットの肝臓よりむしろ腸内細菌叢(bacterial intestinal flora)によって触媒され得る。
【0056】
結論
ヒドロキシマタイレシノール(HMR)は、DMBA誘発性乳ガンモデルにおいて、他の物質に変化せずに、および/またはエンテロラクトンに変化したのちに抗腫瘍活性を有する。それゆえ、HMRは乳ガン(BC)、前立腺ガン(PC)、結腸癌または良性前立腺過形成(BPH)を患う恐れのあるヒトにおいて、有益な効果をもたらす可能性がある。HMRは代謝によってエンテロラクトンとなり、インビトロにおける芳香族化を阻害する。HMRはアロマターゼ阻害剤の前駆体として下部尿路症(LUTS)、膀胱不安定性、膀胱排出口閉塞、尿道共同運動障害および女性化乳房の進行を抑制し得る。また、HMRには強い抗酸化活性があり、それゆえに食品添加物(抗酸化剤)として用いることができる。医薬品や栄養補助食品(dietary supplement)としてのHMRは、ヒトに対する有益な心血管効果を与え得る。新しい画期的な機能性食品、栄養補給食品、健康食品、治療用食品、遺伝子操作食品、あるいは新規な食品を製造するために、HMRを添加することは実行性のあることである。
【0057】
本発明の方法は、様々な形の実施態様によって具体的に表され、ここに記載したものはわずか一部であるということは充分理解されるであろう。ほかの実施態様が考えられ、それらが本発明の主旨に沿っていることは当業者に明らかであろう。このように、記載した実施態様は例示的なものであり、限定的なものと解釈されるべきではない。
【0058】
【表1】
【0059】
【表2】
【0060】
【表3】
【0061】
【表4】
【0062】
【表5】
【0063】
[参考文献]
Adlercreutz H, Bannwart C, Waehaelae K, Maekelae T, Brunow G, Hase T, Arosemena PJ, Kellis JT, and Vickery LE: Inhibition of human aromatase by mammalian lignans and isoflavonoid phytoestrogens. J Steroid Biochem Mol Biol, 44: 147-153, 1993.
Ahotupa M, Ruutu M, and Maentylae E: Simple methods of quantifying oxidation products and antioxidant potential of low density lipoproteins. Clin Biochem, 29: 139-144, 1996.
Ahotupa M, Maentylae E, and Kangas L: Antioxidant properties of the triphenylethylene antiestrogen drug toremifene. Naunyn-Schmiedeberg's Arch Pharmacol, 356: 297-302, 1997.
Axelson M and Setchell KDR: The excretion of lignans in rats-evidence for an intestinal bacterial source for this new group of compounds. FEBS lett, 123: 337-342, 1981.
Axelson M, Sjoevall J, Gustafsson BE and Setchell KDR: Origin of lignans in mammals and identification of a precursor from plants. Nature, 298: 659-660, 1982.
Ayres D, and Loike, J. Lignans: Chemical, bilogical and clinical properties. Cambridge university press, 1990.
Brzezinski A, Adlercreutz H, Shaoul R, Roesler A, Shmueli A, Tanos V and Schenker JG: Short-term effects of phytoestrogen-rich diet on postmenopausal women. Menopause (The Journal of the North American Menopause Society), 4: 89-94, 1997.
Ekman R: Analysis of lignans in Norway spruce by combined gas chromatography-mass spectrometry. Holzforschung, 30:79-85, 1976.
Ekman R: Distribution of lignans in Norway spruce. Acta Academiae Aboensis, Ser B, 39:1-6, 1979.
Evans BA, Griffiths K and Morton MS. Inhibition of 5α-reductase in genital skin fibroblasts and prostate tissue by dietary lignans and isoflavonoids. J Endocrinol, 147: 295-302, 1995.
Haavisto A-M, Petterson K, Bergendahl M, Perheentupa A, Roser JF, and Huhtaniemi I A. Supersensitive immunofluorometric assay for rat luteinizing hormone. Endocrinology, 132: 1687-1691, 1993.
Hulten K, Adlercreutz H, Winkvist A, Lenner P, Hallmans G and Ågren Å. Low levels of phyto-estrogens in blood as risk factor for breast cancer. In: COST 916 Workshop ‘Phyto-oestrogens: exposure, bioavailability, health benefits and safety concerns', 1998.
Ingram D, Sanders K. Kolybaba M and Lopez D. Case-control study of phyto-oestrogens and breast cancer. Lancet, Oct 4;350(9083): 990-994,1997.
Jenab M and Thompson LU. The influence of flaxseed and lignans on colon carcinogenesis and beta-glucuronidase activity. Carcinogenesis, Jun;17(6):1343-1348, 1996.
Jordan VC, Collins MM, Rowsby L, Prestwich G: A monohydroxylated metabolite of tamoxifen with potent antiestrogenic activity. J Endocrinol, 75: 305-316, 1997.
Kangas L, Nieminen A-L, Blanco G, Groenroos M, Kallio S, Karjalainen A, Perilae M, Soedervall M and Toivola R: A new triphenylethylene compound, Fc-1157a. II Antitumor effects. Cancer Chemother Pharmacol, 17:109-113, 1986.
Lampe JW, Martini MC, Kurzer MS, Adlercreutz H and Slavin JL: Urinary lignan and isoflavonoid excretion in premenopausal women consuming flaxseed powder. Am J Clin Nutr, 60:122-8, 1994.
Landstrom M, Zhang JX, Hallmans G, Aman P, Bergh A, Damber JE, Mazur W, Waehaelae K and Adlercreutz H. Inhibitory effects of soy and rye diets on the development of Dunning R3327 prostateadenocarcinoma in rats. Prostate, Aug 1; 36(3): 151-161, 1998
Mousavi Y and Adlercreutz H: Enterolactone and estradiol inhibit each other's proliferative effect on MCF-7 breast cancer cells in culture. J Steroid Biochem Mol Biol, 41: 615-619, 1992.
Mattinen J, Sjoeholm R and Ekman R. NMR-spectroscopic study of hydroxymatairesinol, the major lignan in Norway spruce (Picea abies) heartwood. ACH models in chemistry, 135(4):583-590, 1998.
Maekelae S, Poutanen M, Lehtimaeki J, Kostian M-L, Santti R and Vihko R. Estrogen-specific 17β-hydroxysteroid oxidoreductase type 1 (E.C.1.1.1.62) as a possible target for the action of phytoestrogens. P.S.E.B.M., 208:51-59, 1995.
Phipps WR, Martini MC, Lampe JW, Slavin JL and Kurzer MS. Effect of flax seed ingestion on the menstrual cycle. J Clin Endocrinol Metab, 77(5):1215-1219, 1993.
Rickard SE, Orcheson LJ, Seidl MM, Luyengi L, Fong HHS and Thompson LU: Dose-dependent production of mammalian lignans in rats and in vitro from the purified precursor secoisolariciresinol diglycoside in flaxseed. J Nutr, 126: 2012-2019, 1996.
Shultz TD, Bonorden WR and Seaman WR. Effect of short-time flaxseed consumption on lignan and sex hormone metabolism in men. Nutrition Research, 11:1089-110, 1991.
Serraino M and Thompson LU: The effect of flaxseed supplementation on early risk markers for mammary carcinogenesis. Cnacer Letters, 60: 135-142, 1991.
Sarraino M and Thompson LU: The effect of flaxseed supplementation on the initiation and promotional stages of mammary tumorigenesis. Nutr Cancer, 17:153-159, 1992.
Setchell KDR, Borriello SP, Gordon H, Lawson AM, Harkness R and Morgan DML. Lignan formation in man - microbaial involvement and possible roles in relation to cancer. Lancet, 4:4-7, 1981.
Streng T, Talo A and Santti R. Unpublished observations.
Thompson LU, Robb P, Serraino M and Cheung F. Mammalian lignan production from various foods. Nutr Cancer, 16: 43-52, 1991.
Thompson LU, Seidl, MM, Rickard SE, Orcheson, LJ, and Fong HHS: Antitumorigenic effect of a mammalian lignan precursors from flaxseed. Nutr Cancer, 26: 159-165, 1996a.
Thompson LU, Rickard SE, Orcheson LJ and Seidl MM: Flaxseed and its lignan and oil components reduce mammary tumor growth at a late stage of carcinogenesis. Carcinogenesis, 17: 1373-1376, 1996b.
Tou JCL, Chen J and Thompson U. Flaxseed and its lignan precursor, secoisolariciresinol diglycoside, affect pregnancy outcome and reproductive development in rats. J Nutr, 128: 1861-1868, 1998.
Wang C, Maekelae T, Hase T, Adlercreutz H and Kurzer MS: Lignans and flavonoids inhibit aromatase enzyme in human adipocytes. J Steroid Biochem Molec Biol, 50: 205-212, 1994.
Waters AP and Knowler JT. Effect of a lignan (HPMF) on RNA synthesis in the rat uterus. J Reprod. Fert, 66:379-381, 1982.
Zhang J-X, Hallmans G, Landstroem M, Bergh A, Damber J-E, Åman P and Adlercreutz H. soy and rye diets inhibit the development of Dunning R3327 prostatic adenocarcinoma in rats. Cancer Letters, 114: 313-314, 1997.
【特許請求の範囲】
【請求項1】
ヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体を濃縮した液体または固体材料からなり、食品添加物として用いられる製品。
【請求項2】
前記食品が機能性食品、栄養補助食品、栄養剤、治療用食品、栄養補給食品、健康食品、遺伝子操作食品または任意の食品からなる群から選択される請求項1記載の製品。
【請求項3】
前記食品がバター、マーガリン、ビスケット、パン、ケーキ、アメ、菓子類、ヨーグルトもしくは他の発酵乳製品、またはムースリなどのシリアルの形の機能性食品である請求項1記載の製品。
【請求項4】
有効量のヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体からなる食品。
【請求項5】
前記食品が機能性食品、栄養補助食品、栄養剤、治療用食品、栄養補給食品、健康食品、遺伝子操作食品または任意の食品からなる群から選択される請求項4記載の食品。
【請求項6】
前記食品がバター、マーガリン、ビスケット、パン、ケーキ、アメ、菓子類、ヨーグルトもしくは他の発酵乳製品、またはムースリなどのシリアルの形の機能性食品である請求項5記載の食品。
【請求項7】
前記有効量のヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体を食品に添加することからなる食品の安定性を高める方法。
【請求項8】
食品の栄養価を損ない、味の低下をもたらす脂質、ビタミンおよび色素の酸化を抑えることによって食品の安定性を高めることを特徴とする請求項7記載の方法。
【請求項1】
ヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体を濃縮した液体または固体材料からなり、食品添加物として用いられる製品。
【請求項2】
前記食品が機能性食品、栄養補助食品、栄養剤、治療用食品、栄養補給食品、健康食品、遺伝子操作食品または任意の食品からなる群から選択される請求項1記載の製品。
【請求項3】
前記食品がバター、マーガリン、ビスケット、パン、ケーキ、アメ、菓子類、ヨーグルトもしくは他の発酵乳製品、またはムースリなどのシリアルの形の機能性食品である請求項1記載の製品。
【請求項4】
有効量のヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体からなる食品。
【請求項5】
前記食品が機能性食品、栄養補助食品、栄養剤、治療用食品、栄養補給食品、健康食品、遺伝子操作食品または任意の食品からなる群から選択される請求項4記載の食品。
【請求項6】
前記食品がバター、マーガリン、ビスケット、パン、ケーキ、アメ、菓子類、ヨーグルトもしくは他の発酵乳製品、またはムースリなどのシリアルの形の機能性食品である請求項5記載の食品。
【請求項7】
前記有効量のヒドロキシマタイレシノール、またはその幾何異性体もしくは立体異性体を食品に添加することからなる食品の安定性を高める方法。
【請求項8】
食品の栄養価を損ない、味の低下をもたらす脂質、ビタミンおよび色素の酸化を抑えることによって食品の安定性を高めることを特徴とする請求項7記載の方法。
【図1】

【図2】
【図3】

【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2011−52024(P2011−52024A)
【公開日】平成23年3月17日(2011.3.17)
【国際特許分類】
【出願番号】特願2010−280712(P2010−280712)
【出願日】平成22年12月16日(2010.12.16)
【分割の表示】特願2000−609455(P2000−609455)の分割
【原出願日】平成12年3月9日(2000.3.9)
【出願人】(301062123)ホルモス ヌトラセウティカル オサケ ユキチュア リミテッド (1)
【Fターム(参考)】
【公開日】平成23年3月17日(2011.3.17)
【国際特許分類】
【出願日】平成22年12月16日(2010.12.16)
【分割の表示】特願2000−609455(P2000−609455)の分割
【原出願日】平成12年3月9日(2000.3.9)
【出願人】(301062123)ホルモス ヌトラセウティカル オサケ ユキチュア リミテッド (1)
【Fターム(参考)】
[ Back to top ]