
ガン細胞の老化、アポトーシス誘導剤
【課題】本発明の課題は、CARFの機能をさらに詳細に検討し、その解明をとおして新たな抗ガン剤の提供。
【解決手段】CARFの発現抑制剤あるいは不活性化剤を抗ガン剤として使用する。これらCARFの発現抑制剤あるいは不活性化剤は、ガン細胞に対する老化、アポトーシス誘導剤として機能する。CARF抑制剤としてはこれを標的とするsiRNAあるいはその発現ベクターである。またCARFの不活性化剤としてはこれに対する抗体であることを特徴とする。
【解決手段】CARFの発現抑制剤あるいは不活性化剤を抗ガン剤として使用する。これらCARFの発現抑制剤あるいは不活性化剤は、ガン細胞に対する老化、アポトーシス誘導剤として機能する。CARF抑制剤としてはこれを標的とするsiRNAあるいはその発現ベクターである。またCARFの不活性化剤としてはこれに対する抗体であることを特徴とする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はCARFの発現抑制剤または不活性化剤を利用したガン細胞の老化およ微びアポトーシス誘導剤に関する。
【背景技術】
【0002】
染色体9p21に位置するINK4a遺伝子座はヒトのガン細胞で高頻度に変異が起きている部位である。この遺伝子座は、p16INK4aとARF(alternative reading frame protein)の構造的に全く異なった2つのタンパク質因子をコードしている。この10年間で、この2つのタンパク質因子が主要なガン抑制因子であることを示す、細胞老化、細胞分裂制御、ガン由来細胞に関する多くの証拠が蓄積してきた(非特許文献1-3参照)。このガン抑制因子の機能的欠損は、様々ながんにおいて遺伝子欠損、変異、メチル化によるサイレンシングなどの異なる機構によって引き起こされていることや、p16INK4aによる細胞分裂抑制はpRBのリン酸化の抑制が原因であることも報告されている(非特許文献4参照)。一方、ARFによる細胞分裂の抑制は、p53アンタゴニストであるHDM2(human double minute-2 oncoprotein)を阻害することで、p53機能を促進している(非特許文献5-7参照)。p53とRbを介するがん抑制経路の制御は、細胞老化やがん化の中心であり、その調節因子の知見や分子解析は非常に重要である。
ARF-p53経路の制御を理解するために、我々はp19ARF(マウスARF)に結合するパートナー因子を酵母インターラクティブスクリーンにより探索し、新規のタンパク質CARF(collaborator of ARF)を単離した。我々はCARFに関して既に次のことを明らかにしている(非特許文献8−10)。
(i)CARFはセリンを多く含む核タンパク質である。
(ii)ヒト4p35染色体およびマウス第8染色体に座位する。
(iii)ヒト-マウス間のホモロジーは84.2%である。
(iv)ヒトおよびマウスのARFに結合する、
(v)ARF-CARF複合体は各周辺の領域に局在する。
(vi)ARF-CARF複合体はp53機能を活性化する。
(vii)CARFはARFが無くてもp53と結合でき、HDM2からの分解を防ぐ。
【非特許文献1】Jacobs, J. J., Kieboom, K., Marino, S.,DePinho, R. A., and van Lohuizen, M. (1999) Nature397, 164-168
【非特許文献2】Vogt, M., Haggblom, C., Yeargin,J., Christiansen-Weber, T., and Haas, M. (1998) Cell Growth Differ 9,139-146
【非特許文献3】Wei, W., Hemmer, R. M., and Sedivy,J. M. (2001) Mol Cell Biol 21, 6748-6757.
【非特許文献4】Serrano, M., Lee, H., Chin, L.,Cordon-Cardo, C., Beach, D., and DePinho, R. A. (1996) Cell 85, 27-37
【非特許文献5】Weber,J. D., Jeffers, J. R., Rehg, J. E., Randle, D. H., Lozano, G., Roussel, M. F.,Sherr, C. J., and Zambetti, G. P. (2000) GenesDev 14, 2358-2365.
【非特許文献6】Lloyd,A. C. (2000) Nat Cell Biol 2, E48-50.
【非特許文献7】Zhang, Y., Xiong, Y., and Yarbrough,W. G. (1998) Cell 92, 725-734
【非特許文献8】Wadhwa,R., Sugihara, T., Hasan, M. K., Duncan, E. L., Taira, K., and Kaul, S. C.(2003) Exp Gerontol 38, 245-252
【非特許文献9】Hasan, M. K., Yaguchi, T.,Sugihara, T., Kumar, P. K., Taira, K., Reddel, R. R., Kaul, S. C., and Wadhwa,R. (2002) J Biol Chem 277, 37765-37770
【非特許文献10】Hasan, M. K., Yaguchi, T., Minoda,Y., Hirano, T., Taira, K., Wadhwa, R., and Kaul, S. C. (2004) Biochem J 380, 605-610
【発明の開示】
【発明が解決しようとする課題】
【0003】
本発明の課題は、上記CARFの機能をさらに詳細に検討し、その解明をとおして新たな薬剤を開発する点にある。
【課題を解決するための手段】
【0004】
本発明者等は、鋭意研究の結果、CARF機能について、それがARF-p53-HDM2経路における重要な制御因子であり、細胞分裂およびDNA障害応答において中心的に関与しているとことを示す次の新たな知見を得た。
(i)CARFはHDM2に結合する。
(ii)CARFはHDM2依存的プロテアソーム経路で分解される。
(iii)CARFはHDM2の転写抑制因子として振る舞う。
(iv)CARFは老化細胞で発現上昇し、一方でそれを大量発現させるとヒト正常細胞が早熟性老化細胞になる。
(v)CARFはG2期に発現がピークとなり、細胞周期の調節に関与している。
(vi)CARFは細胞のDNA障害応答の中心に関与する。
(vii)ヒト形質転換細胞におけるCARFの過剰発現はG2増殖抑制を引き起こし、siRNAによるサイレンシングはアポトーシスを促進する。
そして、これらの知見をふまえ、ガン細胞に対するCARFの作用をさらに検討した。その結果、CARFの阻害は、ガン細胞の老化、アポトーシスを誘導するという全く意外な知見を得て本発明を完成するに至った。
すなわち、本発明は以下のとおりのものである。
(1)CARFの発現抑制剤または不活性化剤を有効成分として含有することを特徴とする、抗ガン剤。
(2)抗ガン剤が、ガン細胞の老化あるいはアポトーシス誘導剤であることを特徴とする、上記(1)に記載の抗ガン剤。
(3)CARFの発現抑制剤がCARFを標的とするsiRNAあるいはsiRNA発現ベクターであることを特徴とする、上記(1)または(2)に記載の抗ガン剤。
(4)siRNAが、配列番号5に示されるセンス鎖と配列番号6に示されるアンチセンス鎖の2本鎖オリゴヌクレオチであることを特徴とする上記(3)に記載の抗ガン剤。
(5)CARFの不活性化剤が、CARFに対する抗体であることを特徴とする上記(4)に記載の抗ガン剤。
(6)CARFに対する抗体がモノクローナル抗体である上記(5)に記載の抗ガン剤。
【発明を実施するための最良の形態】
【0005】
上記CARFの全長遺伝子の塩基配列は配列番号1に示され、CARFタンパク質のアミノ酸配列は配列番号2に示される。
本発明は、CARFの作用を阻害する薬剤を使用してガン細胞の老化、あるいはアポトーシスを誘導するものであり、これにより抗ガン作用を発揮させるものである。
CARFの作用を阻害する薬剤としては、CARFの発現抑制剤あるいは不活性化剤が挙げられ、CARFの発現抑制剤の具体例は、例えば、CARFを標的とする、siRNA、リボザイム、又はアンチセンスオリゴヌクレオチドである。
CARFを標的とするsiRNAは、基本的には互いに相補の2本鎖RNAからなるオリゴヌクレオチドであり、このsiRNAのうちの一方(センス鎖)は、標的となるCARF遺伝子(配列番号1)の構造遺伝子部分に対応するmRNA中の塩基配列における、任意の配列、例えば19ヌクレオチドからなる領域を含み、この領域の選定においては、GC含量が例えば30〜55%になる域を選定する。また、siRNAのうちの他方(アンチセンス鎖)は上記センス鎖と相補の塩基配列を有し、このセンスあるいはアンチセンス鎖は、その3‘末端に0〜3塩基の付加配列を有してよい。なお、後記する実施例において使用したsiRNAは、CARF遺伝子の168〜286番目の塩基配列に基づき設計したものであるが、本発明は特にこれに限定されない。
上記のように設計したセンス及びアンチセンスRNAは、例えば、それぞれ常法により化学合成し、合成したRNAを、溶液中で加温することによりアニーリングさせて2本鎖とし,siRNAとすることができる。このようなsiRNAは、例えば、リポソーム等を用いて、生体内に導入することが出来る。
また、他の方法として、上記センスおよびアンチセンスRNAに対応するDNAをそれぞれ合成し、さらに、センス鎖DNA−リンカー―アンチセンス鎖DNAとなるように各DNAを連結し、この連結されたDNAを発現ベクターに導入して、抗ガン剤としてもよい。このようにして得られた組換えベクターは細胞内において、組み込まれたDNAが転写され、低分子二重鎖「ヘアピン」型RNA(shRNA)が発現し、このRNA は細胞内酵素により切断されsiRNAを産生する。
使用するベクターとしては、例えば、非ウイルスベクター、アデノウイルスベクター、レンチウイルスベクター等のベクターが挙げられる。
【0006】
一方、上記センス鎖とアンチセンス鎖DNAを、それぞれ異なる発現ベクターに導入し、細胞内で別々に発現させて、細胞内においてアニールさせsiRNAを生成させるようにしてもよい。これらの発現ベクターを用いる場合には、生体内あるいは細胞内で持続的にsiRNAを産生させることが可能であり、より好ましい。
本発明において用いるアンチセンスオリゴヌクレオチドは、細胞内においてCARF遺伝子の発現に基づき産生されるmRNAの発現を抑制するRNA分子であり、CARFに対応するmRNA中の塩基配列に相補の塩基配列を有し、15〜30の塩基長を有する。アンチセンスオリゴヌクレオチドの構造としては、天然型のホスホジエステル型オリゴマーも用いることが出来るが、ヌクレアーゼによる切断を回避するため、ホスホロチオエート型、ホスホロジチオエート型、ホスホロアミデート型、メチルホスホネート型、メチルホスホノチオエート型の各オリゴマーを用いてもよく、さらに、塩基部分、リボース部分を化学修飾してもよい。これらのアンチセンスオリゴヌクレオチドの設計、製法は常法に従う。
また、本発明において用いるリボザイムは、細胞内においてCARF遺伝子の発現に基づき産生されるmRNAを認識し、該mRNAを切断するRNA分子であり、CARFに対応するmRNA中の塩基配列に相補の塩基配列部分とループ構造部分を有する。このようなリボザイムは、CARFに対応するmRNA中の塩基配列に基づき設計し、常法により合成する。
また、上記天然型のアンチセンスオリゴヌクレオチドあるいはリボザイムをガン治療用に用いる場合は、これらをコードするDNAを上記遺伝子治療用ベクターに挿入し、生体内に導入し、生体内で上記アンチセンスRNA、あるいはリボザイムを持続的に発現させることが好ましい。
【0007】
一方、本願発明において使用する、CARFの不活性剤としては、例えばCARFに対する抗体が挙げられる。抗体は、ポリクローナル抗体及びモノクローナル抗体のいずれでもよい。
ポリクローナル抗体は、CARFを、マウス、ウサギ、ラットまたはヤギ等の動物に免役した後、該動物の血清を採取し、好ましくは、さらに単離精製して得られる。単離精製は、遠心分離、透析、塩析、DEAE−セルロース、プロテインAアガロース等によるクロマトグラフ等の常法のタンパク質の単離精製手段を適宜組み合わせて行えばよい。
また、モノクローナル抗体は、いわゆるハイブリドーマ法により得られる。すなわち、CARFを上記マウス等の動物に免役した後、脾臓細胞を取り出し、該細胞とミエローマ細胞とを細胞融合し、得られた融合細胞をスクリーニングし、さらにクローニングを繰り返して、CARFに対するモノクローナル抗体産生細胞を得る。この細胞を培地に培養することにより、CARFに対するモノクローナル抗体を得ることができる。
上記ポリクローナル抗体及びモノクローナル抗体の製造において、動物に免役する場合においては、フロインドアジュバンドなどのアジュバンドも用いることが出来る。
【実施例】
【0008】
以下に、本発明の実施例を示すが、本発明はこれら実施例により限定されるものではない。
【0009】
実施例1
1)CARF siRNAの作製
CARFに対する、21塩基のsiRNA(配列番号1の遺伝子における168〜286番目塩基配列に対応)をホスホロアミダイトを用いて化学的に合成した。合成RNAは、脱保護し、ゲル抽出して精製した。コントロールに用いたsiRNAおよび標的siRNAの配列は以下に示される。
コントロール用
5’-AAGACCGAGUCCAUGAGGCUT-3’(配列番号3)
5’-GCCUCAUGGACUCGGUCUUUT-3’(配列番号4)
CARF標的用
5’-CGGAGUACCUGAGCCAGAAUT-3’(配列番号5)
5’-UUCUGGCUCAGGUACUCCGUT-3’ (配列番号6)
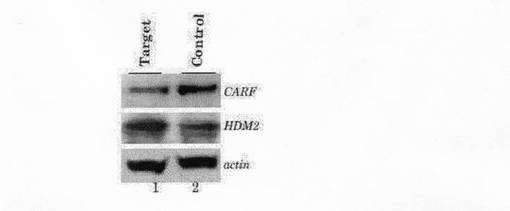
siRNAのアニーリングでは、20uMの各RNA鎖をアニーリング緩衝液(100 mM 酢酸カリウム, 30 mM HEPES-KOH at pH 7.4, 2 mM 酢酸マグネシウム)中で、90℃で1分間の後、37℃に1時間かけて冷却した。二本鎖siRNAのトランスフェクションは、オリゴフェクタミン試薬(インビトロジェン社)を用いて行った。20uMのsiRNAを1-5ulを用い、12ウェル細胞にトランスフェクションし、トランスフェクションから24-48時間後、抗CARF抗体を用いたウェスタンブロッティングや免疫染色で実験を行った。結果を図7に示す。これによれば、上記siRNAが、CARFの発現を特異的に抑制していることが明らかである。
【0010】
2)CARFsiRNAによるガン細胞のアポトーシス誘導
アポトーシスを検出するため、培養ウェル毎に、TUNEL染色を行った。この方法は、アポトーシスに伴うゲノムDNA鎖の切断を、切断後の3’-OH基に蛍光標識することによって検出するものである。
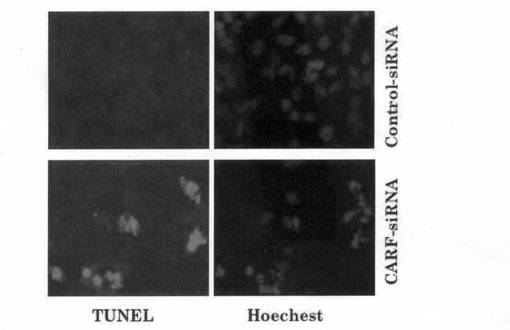
上記1)で得られた、CARF特異的二本鎖siRNA及びCARF発現に影響を与えないコントロール二本鎖siRNAを、上記1)と同様の方法により、それぞれHela細胞に導入した。ここで使用した、CARF培養ウエル毎にsiRNA導入後の細胞を DeadEndTM Fluorometric TUNEL System (Promega社)を用いてTUNEL染色し、アポトーシスを検出した。図14に示されているように、CARF特異的siRNAによるCARF発現の抑制によって、アポトーシスに至った細胞が緑色蛍光を示している。コントロールsiRNAの場合には、CARFの発現に変化は無く、アポトーシスを起こしている細胞も見られなかった。細胞の核は、DNA結合試薬(Hoechest)を用いて染色し、青色に観察される。
【0011】
実施例2
ヒト精巣細胞由来のcDNAを用い、以下のプライマーを使用してPCRを行った。
5’-GGATCCATGGCGCAGGAGGTG-3’(配列番号7)
5’-GTCGACTAGTAATTCTTGAGGA-3’(配列番号8)
サブクローニング後に、PCRで増幅されたDNA領域をBamHIおよびSalIで切断した。切断後のDNA断片を大腸菌発現ベクターpQE30(Qiagen社)のSalIおよびBamHIサイトを用いてクローニングした。HisタグはpQE30ベクターに搭載されている。cDNA配列は、シークエンシングによって確認した。
【0012】
2)ヒトCARFタンパク質の発現
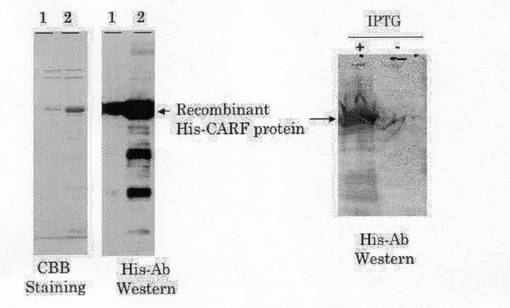
上記で得られた全長CARF cDNA(配列番号1)を大腸菌内発現用ベクター pQE30 (Qiagen)上のSalIとBamHIサイトに組込んだ後、M15大腸菌株に導入し、大腸菌をOD580=0.6になるまで培養し、イソプロピル-1-チオ-β-D-ガラクトピラノシド(IPTG) (0.2 mM)で37℃ 5時間処理で発現誘導した。そこで発現させたHis標識付き組み換えタンパク質は、Ni-NTAアガロース(Qiagen)と結合させることにより単離した。得られたタンパク質の純度および大きさは、ドデシル硫酸ナトリウム・ポリアクリルアミドゲル電気泳動(SDS-PAGE)後、クーマシーブリリアントブルー(CBB)染色により解析した。また、His標識に対する抗体を用いたウェスタンブロッティングにおいても確認した(図3)。図中、レーン1は 、レーン2は別の日に行った実験のサンプルから得られた。左側のパネルがCBB染色で、右側のパネルがウェスタンブロットの結果を示している。
【0013】
3)抗体の調製(図16参照)
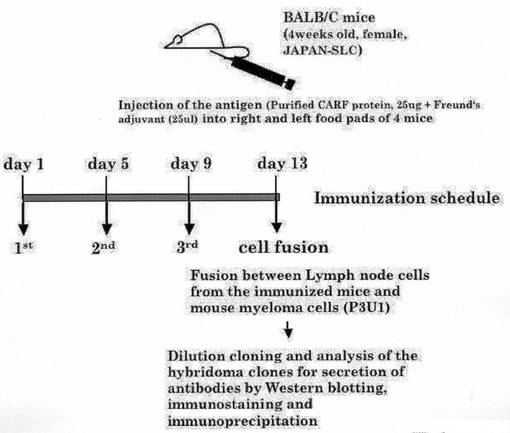
上記2)で得られた精製CARFタンパク質(配列番号2)は、同量のフロイドアジュバントと混合し、抗体産生の抗原として用いた。モノクローナル抗体作製のために、抗原は、4週令Balb/cマウスの足蹠に注入した。マウスは4日間隔で3回、免疫化を行い、13日目に屠殺した。免疫化したマウスから得たリンパ節細胞は、マウスミエローマ細胞(P3U1)と融合させた。ハイブリドーマクローンの希釈クローニングを行い、各クローンの抗体分泌量はウェスタンブロッティング、免疫染色法により検査した。
一方、ポリクローナル抗体のために、50 ugの抗原をウサギの背中の皮下部位に複数回、10日間隔で2回注入し、その後、100 ugの蛋白質を追加免疫のために注射した。追加免疫の10日後、ウサギから採血し、その血清は、ウェスタンブロッティング、免疫染色、免疫沈降解析により抗CARF抗体の検査を行った。
【0014】
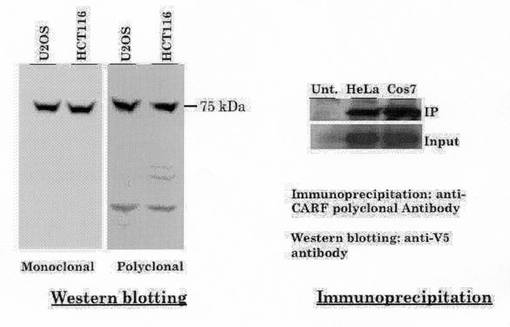
図17左側は、ウェスタンブロッティングを行った結果を示す。10ugのタンパク質を含む細胞抽出液をSDS-PAGE後、PVDF膜にトランスファーし、抗CARF抗体を用いて検出した。ホースラディッシュペルオキシダーゼ(HRP)標識した二次抗体を用い、ECL検出キット(Amersham社)を用いて検出した。モノクローナル抗体においては75kDa付近に唯一のバンドがみられた。また、ポリクローナル抗体についても75kDa付近にほとんど唯一のバンドがみられた。
図17右側は、免疫沈降法を行った結果を示す図である。V5タグ付きCARFタンパク質を細胞で発現させ、抗CARF抗体を用いて免疫沈降(IP)させ、沈降したタンパク質を抗V5タグ抗体を用いてウェスタンブロットすると、V5タグ付きCARFが検出された。これによっても唯一のタンパク質が検出されている。Inputレーンは、HeLa細胞およびCos7細胞におけるV5タグ付きCARFタンパク質を示している。これらの結果は、抗CARF抗体が、CARF抗体に特異的である事を示している。
【0015】
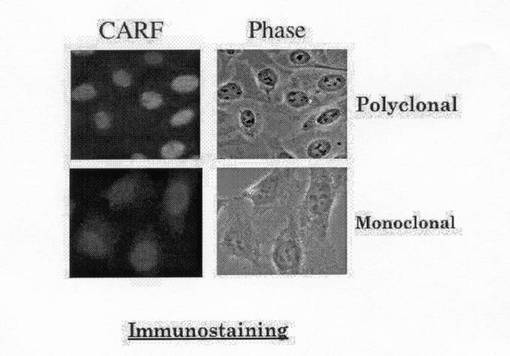
また、図18は、抗CARF抗体を用いたCARFタンパク質の免疫染色を示している。抗CARFモノクローナル抗体および抗CARFポリクローナル抗体において、核への染色が観察された。これは、V5タグ付きCARFタンパク質を細胞に発現させた場合の、抗V5抗体の局在と同等である。これらの結果から、両抗CARF抗体は、CARF特異的であり、CARFタンパク質を不活性化する。
【図面の簡単な説明】
【0016】
【図1】本発明の上記siRNAがCARFの発現を特異的に抑制すること示す、ウェスタンブロッティング及び免疫染色の写真である。
【図2】本発明のsiRNAがHela細胞のアポトーシスを誘導することを示す、TUNEL染色の写真である。
【図3】実施例2,2)で得られたCARFタンパク質をCBB染色及びウエスタンブロッティングで解析した結果を示す写真である。
【図4】本発明の抗CARF抗体を調製するための方法の概要を示すフローチャート図である。
【図5】本発明のポリクローナル抗体及びモノクローナル抗体のCARFに対する特異性を、ウエスタンブロッティングび免疫沈降解析で試験した結果を示す写真である。
【図6】本発明のポリクローナル抗体及びモノクローナル抗体を使用したCARFの免疫染色の結果を示す図である。
【技術分野】
【0001】
本発明はCARFの発現抑制剤または不活性化剤を利用したガン細胞の老化およ微びアポトーシス誘導剤に関する。
【背景技術】
【0002】
染色体9p21に位置するINK4a遺伝子座はヒトのガン細胞で高頻度に変異が起きている部位である。この遺伝子座は、p16INK4aとARF(alternative reading frame protein)の構造的に全く異なった2つのタンパク質因子をコードしている。この10年間で、この2つのタンパク質因子が主要なガン抑制因子であることを示す、細胞老化、細胞分裂制御、ガン由来細胞に関する多くの証拠が蓄積してきた(非特許文献1-3参照)。このガン抑制因子の機能的欠損は、様々ながんにおいて遺伝子欠損、変異、メチル化によるサイレンシングなどの異なる機構によって引き起こされていることや、p16INK4aによる細胞分裂抑制はpRBのリン酸化の抑制が原因であることも報告されている(非特許文献4参照)。一方、ARFによる細胞分裂の抑制は、p53アンタゴニストであるHDM2(human double minute-2 oncoprotein)を阻害することで、p53機能を促進している(非特許文献5-7参照)。p53とRbを介するがん抑制経路の制御は、細胞老化やがん化の中心であり、その調節因子の知見や分子解析は非常に重要である。
ARF-p53経路の制御を理解するために、我々はp19ARF(マウスARF)に結合するパートナー因子を酵母インターラクティブスクリーンにより探索し、新規のタンパク質CARF(collaborator of ARF)を単離した。我々はCARFに関して既に次のことを明らかにしている(非特許文献8−10)。
(i)CARFはセリンを多く含む核タンパク質である。
(ii)ヒト4p35染色体およびマウス第8染色体に座位する。
(iii)ヒト-マウス間のホモロジーは84.2%である。
(iv)ヒトおよびマウスのARFに結合する、
(v)ARF-CARF複合体は各周辺の領域に局在する。
(vi)ARF-CARF複合体はp53機能を活性化する。
(vii)CARFはARFが無くてもp53と結合でき、HDM2からの分解を防ぐ。
【非特許文献1】Jacobs, J. J., Kieboom, K., Marino, S.,DePinho, R. A., and van Lohuizen, M. (1999) Nature397, 164-168
【非特許文献2】Vogt, M., Haggblom, C., Yeargin,J., Christiansen-Weber, T., and Haas, M. (1998) Cell Growth Differ 9,139-146
【非特許文献3】Wei, W., Hemmer, R. M., and Sedivy,J. M. (2001) Mol Cell Biol 21, 6748-6757.
【非特許文献4】Serrano, M., Lee, H., Chin, L.,Cordon-Cardo, C., Beach, D., and DePinho, R. A. (1996) Cell 85, 27-37
【非特許文献5】Weber,J. D., Jeffers, J. R., Rehg, J. E., Randle, D. H., Lozano, G., Roussel, M. F.,Sherr, C. J., and Zambetti, G. P. (2000) GenesDev 14, 2358-2365.
【非特許文献6】Lloyd,A. C. (2000) Nat Cell Biol 2, E48-50.
【非特許文献7】Zhang, Y., Xiong, Y., and Yarbrough,W. G. (1998) Cell 92, 725-734
【非特許文献8】Wadhwa,R., Sugihara, T., Hasan, M. K., Duncan, E. L., Taira, K., and Kaul, S. C.(2003) Exp Gerontol 38, 245-252
【非特許文献9】Hasan, M. K., Yaguchi, T.,Sugihara, T., Kumar, P. K., Taira, K., Reddel, R. R., Kaul, S. C., and Wadhwa,R. (2002) J Biol Chem 277, 37765-37770
【非特許文献10】Hasan, M. K., Yaguchi, T., Minoda,Y., Hirano, T., Taira, K., Wadhwa, R., and Kaul, S. C. (2004) Biochem J 380, 605-610
【発明の開示】
【発明が解決しようとする課題】
【0003】
本発明の課題は、上記CARFの機能をさらに詳細に検討し、その解明をとおして新たな薬剤を開発する点にある。
【課題を解決するための手段】
【0004】
本発明者等は、鋭意研究の結果、CARF機能について、それがARF-p53-HDM2経路における重要な制御因子であり、細胞分裂およびDNA障害応答において中心的に関与しているとことを示す次の新たな知見を得た。
(i)CARFはHDM2に結合する。
(ii)CARFはHDM2依存的プロテアソーム経路で分解される。
(iii)CARFはHDM2の転写抑制因子として振る舞う。
(iv)CARFは老化細胞で発現上昇し、一方でそれを大量発現させるとヒト正常細胞が早熟性老化細胞になる。
(v)CARFはG2期に発現がピークとなり、細胞周期の調節に関与している。
(vi)CARFは細胞のDNA障害応答の中心に関与する。
(vii)ヒト形質転換細胞におけるCARFの過剰発現はG2増殖抑制を引き起こし、siRNAによるサイレンシングはアポトーシスを促進する。
そして、これらの知見をふまえ、ガン細胞に対するCARFの作用をさらに検討した。その結果、CARFの阻害は、ガン細胞の老化、アポトーシスを誘導するという全く意外な知見を得て本発明を完成するに至った。
すなわち、本発明は以下のとおりのものである。
(1)CARFの発現抑制剤または不活性化剤を有効成分として含有することを特徴とする、抗ガン剤。
(2)抗ガン剤が、ガン細胞の老化あるいはアポトーシス誘導剤であることを特徴とする、上記(1)に記載の抗ガン剤。
(3)CARFの発現抑制剤がCARFを標的とするsiRNAあるいはsiRNA発現ベクターであることを特徴とする、上記(1)または(2)に記載の抗ガン剤。
(4)siRNAが、配列番号5に示されるセンス鎖と配列番号6に示されるアンチセンス鎖の2本鎖オリゴヌクレオチであることを特徴とする上記(3)に記載の抗ガン剤。
(5)CARFの不活性化剤が、CARFに対する抗体であることを特徴とする上記(4)に記載の抗ガン剤。
(6)CARFに対する抗体がモノクローナル抗体である上記(5)に記載の抗ガン剤。
【発明を実施するための最良の形態】
【0005】
上記CARFの全長遺伝子の塩基配列は配列番号1に示され、CARFタンパク質のアミノ酸配列は配列番号2に示される。
本発明は、CARFの作用を阻害する薬剤を使用してガン細胞の老化、あるいはアポトーシスを誘導するものであり、これにより抗ガン作用を発揮させるものである。
CARFの作用を阻害する薬剤としては、CARFの発現抑制剤あるいは不活性化剤が挙げられ、CARFの発現抑制剤の具体例は、例えば、CARFを標的とする、siRNA、リボザイム、又はアンチセンスオリゴヌクレオチドである。
CARFを標的とするsiRNAは、基本的には互いに相補の2本鎖RNAからなるオリゴヌクレオチドであり、このsiRNAのうちの一方(センス鎖)は、標的となるCARF遺伝子(配列番号1)の構造遺伝子部分に対応するmRNA中の塩基配列における、任意の配列、例えば19ヌクレオチドからなる領域を含み、この領域の選定においては、GC含量が例えば30〜55%になる域を選定する。また、siRNAのうちの他方(アンチセンス鎖)は上記センス鎖と相補の塩基配列を有し、このセンスあるいはアンチセンス鎖は、その3‘末端に0〜3塩基の付加配列を有してよい。なお、後記する実施例において使用したsiRNAは、CARF遺伝子の168〜286番目の塩基配列に基づき設計したものであるが、本発明は特にこれに限定されない。
上記のように設計したセンス及びアンチセンスRNAは、例えば、それぞれ常法により化学合成し、合成したRNAを、溶液中で加温することによりアニーリングさせて2本鎖とし,siRNAとすることができる。このようなsiRNAは、例えば、リポソーム等を用いて、生体内に導入することが出来る。
また、他の方法として、上記センスおよびアンチセンスRNAに対応するDNAをそれぞれ合成し、さらに、センス鎖DNA−リンカー―アンチセンス鎖DNAとなるように各DNAを連結し、この連結されたDNAを発現ベクターに導入して、抗ガン剤としてもよい。このようにして得られた組換えベクターは細胞内において、組み込まれたDNAが転写され、低分子二重鎖「ヘアピン」型RNA(shRNA)が発現し、このRNA は細胞内酵素により切断されsiRNAを産生する。
使用するベクターとしては、例えば、非ウイルスベクター、アデノウイルスベクター、レンチウイルスベクター等のベクターが挙げられる。
【0006】
一方、上記センス鎖とアンチセンス鎖DNAを、それぞれ異なる発現ベクターに導入し、細胞内で別々に発現させて、細胞内においてアニールさせsiRNAを生成させるようにしてもよい。これらの発現ベクターを用いる場合には、生体内あるいは細胞内で持続的にsiRNAを産生させることが可能であり、より好ましい。
本発明において用いるアンチセンスオリゴヌクレオチドは、細胞内においてCARF遺伝子の発現に基づき産生されるmRNAの発現を抑制するRNA分子であり、CARFに対応するmRNA中の塩基配列に相補の塩基配列を有し、15〜30の塩基長を有する。アンチセンスオリゴヌクレオチドの構造としては、天然型のホスホジエステル型オリゴマーも用いることが出来るが、ヌクレアーゼによる切断を回避するため、ホスホロチオエート型、ホスホロジチオエート型、ホスホロアミデート型、メチルホスホネート型、メチルホスホノチオエート型の各オリゴマーを用いてもよく、さらに、塩基部分、リボース部分を化学修飾してもよい。これらのアンチセンスオリゴヌクレオチドの設計、製法は常法に従う。
また、本発明において用いるリボザイムは、細胞内においてCARF遺伝子の発現に基づき産生されるmRNAを認識し、該mRNAを切断するRNA分子であり、CARFに対応するmRNA中の塩基配列に相補の塩基配列部分とループ構造部分を有する。このようなリボザイムは、CARFに対応するmRNA中の塩基配列に基づき設計し、常法により合成する。
また、上記天然型のアンチセンスオリゴヌクレオチドあるいはリボザイムをガン治療用に用いる場合は、これらをコードするDNAを上記遺伝子治療用ベクターに挿入し、生体内に導入し、生体内で上記アンチセンスRNA、あるいはリボザイムを持続的に発現させることが好ましい。
【0007】
一方、本願発明において使用する、CARFの不活性剤としては、例えばCARFに対する抗体が挙げられる。抗体は、ポリクローナル抗体及びモノクローナル抗体のいずれでもよい。
ポリクローナル抗体は、CARFを、マウス、ウサギ、ラットまたはヤギ等の動物に免役した後、該動物の血清を採取し、好ましくは、さらに単離精製して得られる。単離精製は、遠心分離、透析、塩析、DEAE−セルロース、プロテインAアガロース等によるクロマトグラフ等の常法のタンパク質の単離精製手段を適宜組み合わせて行えばよい。
また、モノクローナル抗体は、いわゆるハイブリドーマ法により得られる。すなわち、CARFを上記マウス等の動物に免役した後、脾臓細胞を取り出し、該細胞とミエローマ細胞とを細胞融合し、得られた融合細胞をスクリーニングし、さらにクローニングを繰り返して、CARFに対するモノクローナル抗体産生細胞を得る。この細胞を培地に培養することにより、CARFに対するモノクローナル抗体を得ることができる。
上記ポリクローナル抗体及びモノクローナル抗体の製造において、動物に免役する場合においては、フロインドアジュバンドなどのアジュバンドも用いることが出来る。
【実施例】
【0008】
以下に、本発明の実施例を示すが、本発明はこれら実施例により限定されるものではない。
【0009】
実施例1
1)CARF siRNAの作製
CARFに対する、21塩基のsiRNA(配列番号1の遺伝子における168〜286番目塩基配列に対応)をホスホロアミダイトを用いて化学的に合成した。合成RNAは、脱保護し、ゲル抽出して精製した。コントロールに用いたsiRNAおよび標的siRNAの配列は以下に示される。
コントロール用
5’-AAGACCGAGUCCAUGAGGCUT-3’(配列番号3)
5’-GCCUCAUGGACUCGGUCUUUT-3’(配列番号4)
CARF標的用
5’-CGGAGUACCUGAGCCAGAAUT-3’(配列番号5)
5’-UUCUGGCUCAGGUACUCCGUT-3’ (配列番号6)
siRNAのアニーリングでは、20uMの各RNA鎖をアニーリング緩衝液(100 mM 酢酸カリウム, 30 mM HEPES-KOH at pH 7.4, 2 mM 酢酸マグネシウム)中で、90℃で1分間の後、37℃に1時間かけて冷却した。二本鎖siRNAのトランスフェクションは、オリゴフェクタミン試薬(インビトロジェン社)を用いて行った。20uMのsiRNAを1-5ulを用い、12ウェル細胞にトランスフェクションし、トランスフェクションから24-48時間後、抗CARF抗体を用いたウェスタンブロッティングや免疫染色で実験を行った。結果を図7に示す。これによれば、上記siRNAが、CARFの発現を特異的に抑制していることが明らかである。
【0010】
2)CARFsiRNAによるガン細胞のアポトーシス誘導
アポトーシスを検出するため、培養ウェル毎に、TUNEL染色を行った。この方法は、アポトーシスに伴うゲノムDNA鎖の切断を、切断後の3’-OH基に蛍光標識することによって検出するものである。
上記1)で得られた、CARF特異的二本鎖siRNA及びCARF発現に影響を与えないコントロール二本鎖siRNAを、上記1)と同様の方法により、それぞれHela細胞に導入した。ここで使用した、CARF培養ウエル毎にsiRNA導入後の細胞を DeadEndTM Fluorometric TUNEL System (Promega社)を用いてTUNEL染色し、アポトーシスを検出した。図14に示されているように、CARF特異的siRNAによるCARF発現の抑制によって、アポトーシスに至った細胞が緑色蛍光を示している。コントロールsiRNAの場合には、CARFの発現に変化は無く、アポトーシスを起こしている細胞も見られなかった。細胞の核は、DNA結合試薬(Hoechest)を用いて染色し、青色に観察される。
【0011】
実施例2
ヒト精巣細胞由来のcDNAを用い、以下のプライマーを使用してPCRを行った。
5’-GGATCCATGGCGCAGGAGGTG-3’(配列番号7)
5’-GTCGACTAGTAATTCTTGAGGA-3’(配列番号8)
サブクローニング後に、PCRで増幅されたDNA領域をBamHIおよびSalIで切断した。切断後のDNA断片を大腸菌発現ベクターpQE30(Qiagen社)のSalIおよびBamHIサイトを用いてクローニングした。HisタグはpQE30ベクターに搭載されている。cDNA配列は、シークエンシングによって確認した。
【0012】
2)ヒトCARFタンパク質の発現
上記で得られた全長CARF cDNA(配列番号1)を大腸菌内発現用ベクター pQE30 (Qiagen)上のSalIとBamHIサイトに組込んだ後、M15大腸菌株に導入し、大腸菌をOD580=0.6になるまで培養し、イソプロピル-1-チオ-β-D-ガラクトピラノシド(IPTG) (0.2 mM)で37℃ 5時間処理で発現誘導した。そこで発現させたHis標識付き組み換えタンパク質は、Ni-NTAアガロース(Qiagen)と結合させることにより単離した。得られたタンパク質の純度および大きさは、ドデシル硫酸ナトリウム・ポリアクリルアミドゲル電気泳動(SDS-PAGE)後、クーマシーブリリアントブルー(CBB)染色により解析した。また、His標識に対する抗体を用いたウェスタンブロッティングにおいても確認した(図3)。図中、レーン1は 、レーン2は別の日に行った実験のサンプルから得られた。左側のパネルがCBB染色で、右側のパネルがウェスタンブロットの結果を示している。
【0013】
3)抗体の調製(図16参照)
上記2)で得られた精製CARFタンパク質(配列番号2)は、同量のフロイドアジュバントと混合し、抗体産生の抗原として用いた。モノクローナル抗体作製のために、抗原は、4週令Balb/cマウスの足蹠に注入した。マウスは4日間隔で3回、免疫化を行い、13日目に屠殺した。免疫化したマウスから得たリンパ節細胞は、マウスミエローマ細胞(P3U1)と融合させた。ハイブリドーマクローンの希釈クローニングを行い、各クローンの抗体分泌量はウェスタンブロッティング、免疫染色法により検査した。
一方、ポリクローナル抗体のために、50 ugの抗原をウサギの背中の皮下部位に複数回、10日間隔で2回注入し、その後、100 ugの蛋白質を追加免疫のために注射した。追加免疫の10日後、ウサギから採血し、その血清は、ウェスタンブロッティング、免疫染色、免疫沈降解析により抗CARF抗体の検査を行った。
【0014】
図17左側は、ウェスタンブロッティングを行った結果を示す。10ugのタンパク質を含む細胞抽出液をSDS-PAGE後、PVDF膜にトランスファーし、抗CARF抗体を用いて検出した。ホースラディッシュペルオキシダーゼ(HRP)標識した二次抗体を用い、ECL検出キット(Amersham社)を用いて検出した。モノクローナル抗体においては75kDa付近に唯一のバンドがみられた。また、ポリクローナル抗体についても75kDa付近にほとんど唯一のバンドがみられた。
図17右側は、免疫沈降法を行った結果を示す図である。V5タグ付きCARFタンパク質を細胞で発現させ、抗CARF抗体を用いて免疫沈降(IP)させ、沈降したタンパク質を抗V5タグ抗体を用いてウェスタンブロットすると、V5タグ付きCARFが検出された。これによっても唯一のタンパク質が検出されている。Inputレーンは、HeLa細胞およびCos7細胞におけるV5タグ付きCARFタンパク質を示している。これらの結果は、抗CARF抗体が、CARF抗体に特異的である事を示している。
【0015】
また、図18は、抗CARF抗体を用いたCARFタンパク質の免疫染色を示している。抗CARFモノクローナル抗体および抗CARFポリクローナル抗体において、核への染色が観察された。これは、V5タグ付きCARFタンパク質を細胞に発現させた場合の、抗V5抗体の局在と同等である。これらの結果から、両抗CARF抗体は、CARF特異的であり、CARFタンパク質を不活性化する。
【図面の簡単な説明】
【0016】
【図1】本発明の上記siRNAがCARFの発現を特異的に抑制すること示す、ウェスタンブロッティング及び免疫染色の写真である。
【図2】本発明のsiRNAがHela細胞のアポトーシスを誘導することを示す、TUNEL染色の写真である。
【図3】実施例2,2)で得られたCARFタンパク質をCBB染色及びウエスタンブロッティングで解析した結果を示す写真である。
【図4】本発明の抗CARF抗体を調製するための方法の概要を示すフローチャート図である。
【図5】本発明のポリクローナル抗体及びモノクローナル抗体のCARFに対する特異性を、ウエスタンブロッティングび免疫沈降解析で試験した結果を示す写真である。
【図6】本発明のポリクローナル抗体及びモノクローナル抗体を使用したCARFの免疫染色の結果を示す図である。
【特許請求の範囲】
【請求項1】
CARFの発現抑制剤または不活性化剤を有効成分として含有することを特徴とする、抗ガン剤。
【請求項2】
抗ガン剤が、ガン細胞の老化あるいはアポトーシス誘導剤であることを特徴とする、請求項1に記載の抗ガン剤。
【請求項3】
CARFの発現抑制剤がCARFを標的とするsiRNAあるいはsiRNA発現ベクターであることを特徴とする、請求項1または2に記載の抗ガン剤。
【請求項4】
siRNAが、配列番号5に示されるセンス鎖と配列番号6に示されるアンチセンス鎖の2本鎖オリゴヌクレオチであることを特徴とする請求項3に記載の抗ガン剤。
【請求項5】
CARFの不活性化剤が、CARFに対する抗体であることを特徴とする請求項4に記載の抗ガン剤。
【請求項6】
CARFに対する抗体がモノクローナル抗体である請求項5に記載の抗ガン剤。
【請求項1】
CARFの発現抑制剤または不活性化剤を有効成分として含有することを特徴とする、抗ガン剤。
【請求項2】
抗ガン剤が、ガン細胞の老化あるいはアポトーシス誘導剤であることを特徴とする、請求項1に記載の抗ガン剤。
【請求項3】
CARFの発現抑制剤がCARFを標的とするsiRNAあるいはsiRNA発現ベクターであることを特徴とする、請求項1または2に記載の抗ガン剤。
【請求項4】
siRNAが、配列番号5に示されるセンス鎖と配列番号6に示されるアンチセンス鎖の2本鎖オリゴヌクレオチであることを特徴とする請求項3に記載の抗ガン剤。
【請求項5】
CARFの不活性化剤が、CARFに対する抗体であることを特徴とする請求項4に記載の抗ガン剤。
【請求項6】
CARFに対する抗体がモノクローナル抗体である請求項5に記載の抗ガン剤。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2008−24606(P2008−24606A)
【公開日】平成20年2月7日(2008.2.7)
【国際特許分類】
【出願番号】特願2006−196050(P2006−196050)
【出願日】平成18年7月18日(2006.7.18)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成20年2月7日(2008.2.7)
【国際特許分類】
【出願日】平成18年7月18日(2006.7.18)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]