
キスペプチン拮抗薬及びその使用
【課題】本発明は、個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療用の薬剤の製造におけるキスペプチンの拮抗薬の使用に関する。本発明はまた、個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療において有用である、キスペプチンの拮抗薬として作用する可能性がある、定義された特定のペプチド分子を提供する。さらに、本発明は、キスペプチンの拮抗薬および/または定義されたペプチドを同定し、かつ/または使用する方法、ならびにその医薬組成物を提供する。
【解決手段】個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療用の薬剤の製造におけるキスペプチンの拮抗薬の使用。
【解決手段】個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療用の薬剤の製造におけるキスペプチンの拮抗薬の使用。
Notice: Undefined index: DEJ in /mnt/www/gzt_disp.php on line 298
【特許請求の範囲】
【請求項1】
個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療用の薬剤の製造におけるキスペプチンの拮抗薬の使用。
【請求項2】
個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療における使用のためのキスペプチンの拮抗薬。
【請求項3】
患者におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療のための、あるいは患者におけるキスペプチン活性により誘発され、または悪化した状態の治療用の薬剤の製造における、以下の配列
X1-G/W-X2-R/(D)R-X3
[ここで、
X1は、FまたはAまたは任意のD-アミノ酸残基であり、
X2は、LまたはAまたは任意のD-アミノ酸残基であり、
X3は、FまたはWであり、
ペプチド分子のC末端アミノ酸残基が当残基上の電荷を除去する基z、
またはそのフラグメントもしくは変異体を含み、
ペプチド配列が
F-G-L-R-F、
F-G-L-R-W、または
F-G-(D)F-R-Fでない]
を含むペプチド分子の使用。
【請求項4】
前記ペプチド配列が
F-G-A-R-W、
F-G-L-(D)R-W、
F-G-(D)L-R-W、または
(D)F-G-L-R-W
でない、請求項3に記載の使用。
【請求項5】
X1が(D)Fである、請求項3または4に記載の使用。
【請求項6】
X2が(D)F、(D)Lおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項3から5のいずれかに記載の使用。
【請求項7】
前記ペプチド配列が
(D)F-W-L-R-W、または
F-G-(D)W-R-F
からなる群から選択される、請求項3から6のいずれかに記載の使用。
【請求項8】
N末端残基が当残基上の電荷を除去する基yを含む、請求項3から7のいずれかに記載の使用。
【請求項9】
X1が当残基上の電荷を除去する基yを含む、請求項8に記載の使用。
【請求項10】
基yがアセチル基、トリフルオロアセチル基、環化アミノ酸、またはN末端における電荷を欠いた合成アミノ酸からなる群から選択される、請求項8または9に記載の使用。
【請求項11】
X3が当残基上の電荷を除去する基zを含む、請求項3から10のいずれかに記載の使用。
【請求項12】
前記配列が
I) ac.F - G - (D)F - R - W.z;
II) ac.F - G - (D)L - R - W.z;
III) ac.F - G - L - (D)R - W.z;
IV) ac.F - G - A - R - W.z;
V) ac.A - G - L - R - W.z;
VI) ac.(D)F - W - L - R - W.z; または
VII) ac.F - G - (D)W - R - F.z.
からなる群から選択される、請求項3から11のいずれかに記載の使用。
【請求項13】
患者におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療のための、あるいは患者におけるキスペプチン活性により誘発され、または悪化した状態の治療用の薬剤の製造における、以下の配列
XA-XB-XC-N-XD-XE-G-XF-R-F
[ここで、
XAは、Yまたは任意のD-アミノ酸残基であり、
XBは、Nまたは任意のD-アミノ酸残基であり、
XCは、Wまたは任意のD-アミノ酸残基であり、
XDは、GまたはSまたは任意のD-アミノ酸残基であり、
XEは、Fまたは(D)Wまたは(D)Lであり、
XFは、WまたはLまたは任意のD-アミノ酸残基であり、
ペプチド分子のC末端アミノ酸残基が当残基上の電荷を除去する基z、
またはそのフラグメントもしくは変異体を含み、
ペプチド配列が
Y - N - W - N - S - F - G - L - R - F;
(D)Y - (D)N - W - N - S - F - G - W - R - F;
(D)Y - (D)N - W - N - G - F - G - W - R - F;
(D)Y - (D)N - W - N - S - F - G - (D)W - R - F; または
(D)Y - (D)N - W - N - G - F - G - (D)W - R - F.
でない]
を含むペプチド分子の使用。
【請求項14】
XAが(D)Fおよび(D)Yおよび(D)Aからなる群から選択されるD-アミノ酸残基である、請求項13に記載の使用。
【請求項15】
XBが(D)Aおよび(D)Nからなる群から選択されるD-アミノ酸残基である、請求項13または14に記載の使用。
【請求項16】
XAまたはXBの1つが(D)Yである場合、他は(D)Nでない、請求項13から15のいずれかに記載の使用。
【請求項17】
XAおよびXBの両方がD-アミノ酸残基でない、請求項13から16のいずれかに記載の使用。
【請求項18】
XFが(D)Wである場合、XAは(D)Fである、請求項13から17のいずれかに記載の使用。
【請求項19】
XCが(D)Aおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項13から18のいずれかに記載の使用。
【請求項20】
XDが(D)Aおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項13から19のいずれかに記載の使用。
【請求項21】
XDがSである場合、XFは(D)Wであり、かつ/またはXAは(D)Yでない、請求項13から20のいずれかに記載の使用。
【請求項22】
XDがSである場合、XFは(D)Wであり、かつ/またはXAは(D)Aである、請求項13から20のいずれかに記載の使用。
【請求項23】
XFが(D)Lおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項13から22のいずれかに記載の使用。
【請求項24】
XEおよびXFの両方が(D)Wである場合、XAは(D)Yでない、請求項13から23のいずれかに記載の使用。
【請求項25】
XEおよびXFの両方が(D)Wである場合、XAは(D)Aである、請求項13から23のいずれかに記載の使用。
【請求項26】
N末端残基が当残基上の電荷を除去する基yを含む、請求項13から25のいずれかに記載の使用。
【請求項27】
X1が当残基上の電荷を除去する基yを含む、請求項26に記載の使用。
【請求項28】
基yがアセチル基、トリフルオロアセチル基、環化アミノ酸、またはN末端における電荷を欠いた合成アミノ酸からなる群から選択される、請求項26または27に記載の使用。
【請求項29】
ペプチド分子のC末端F残基が当残基上の電荷を除去する基zを含む、請求項13から28のいずれかに記載の使用。
【請求項30】
前記配列が
a) Y - N - W - N - G - F - G - L - R - F.z;
b) Y - N - W - N - G - F - G - (D)L - R - F.z;
c) Y - N - W - N - G - F - G - (D)W - R - F.z;
d) Y - N - W - N - G - (D)W - G - L - R - F.z;
e) ac.Y - N - W - N - G - F - G - (D)W - R - F.z;
f) ac.Y - N - W - N - (D)W - F - G - (D)W - R - F.z;
g) ac.(D)Y - N - W - N - G - F - G - (D)W - R - F.z;
h) ac.Y - N - (D)W - N - G - F - G - (D)W - R - F.z;
i) ac.Y - (D)N - W - N - G - F - G - (D)W - R - F.z;
j) ac.Y - N - W - N - (D)A - F - G - (D)W - R - F.z;
k) ac.(D)A - N - W - N - G - F - G - (D)W - R - F.z;
l) ac.Y - N - (D)A - N - G - F - G - (D)W - R - F.z;
m) ac.Y - (D)A - W - N - G - F - G - (D)W - R - F.z;
n) ac.(D)W - N - W - N - G - F - G - (D)W - R - F.z;
o) ac.(D)F - N - W - N - G - F - G - (D)W - R - F.z;
p) ac.(D)Y - N - W - N - G - (D)W - G - (D)W - R - F.z;
q) ac.(D)A - N - W - N - G - (D)W - G - (D)W - R - F.z;
r) ac.(D)A - N - W - N - S - F - G - (D)W - R - F.z;
s) ac.(D)A - N - W - N - G - F - G - W - R - F.z;
t) ac.(D)A - N - W - N - (D)S - F - G - (D)W - R - F.z; または
u) ac.(D)A - N - W - N - G - F - G - (D)L - R - F.z;
からなる群から選択される、請求項13から29のいずれかに記載の使用。
【請求項31】
個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態は、増殖性障害、子宮内膜症、子宮線維症、思春期早発症、子癇前症、子宮内胎児発育遅延(IUGR)、子宮外妊娠、月経痛、高血圧、冠動脈心疾患、中枢神経系(CNS)、膵臓および/または免疫系の病変、排卵の抑制または阻害、受胎能、生殖組織の化学療法誘発性および/または放射線療法誘発性損傷、創傷治癒の抑制または阻害、成長ホルモン産生の抑制または阻害を含む群から選択される、請求項1から30のいずれかに記載の使用。
【請求項32】
増殖性障害が良性前立腺過形成、がん、生殖組織のがん、婦人科がん、前立腺がん、乳がん、卵巣がん、子宮がん、子宮頚がん、子宮内膜がん、黒色腫、膵臓がん、胃がんを含む群から選択される、請求項31に記載の使用。
【請求項33】
キスペプチンの拮抗薬またはキスペプチンの拮抗薬であるそのフラグメントもしくは変異体である、請求項3から32のいずれかに定義されているペプチド分子。
【請求項34】
医療用の請求項1または2に定義されている拮抗薬あるいは請求項3から33のいずれか一項に定義されているペプチド分子。
【請求項35】
キスペプチンの拮抗薬である、請求項34に記載のペプチド分子。
【請求項36】
以下の配列
X1-G/W-X2-R/(D)R-X3
を含むペプチド分子
[ここで、
X1は、FまたはAまたは任意のD-アミノ酸残基であり、
X2は、LまたはAまたは任意のD-アミノ酸残基であり、
X3は、FまたはWであり、
ペプチド分子のC末端アミノ酸残基が当残基上の電荷を除去する基z、
またはそのフラグメントもしくは変異体を含み、
ペプチド配列が
F - G - L - R - F;
F - G - L - R - W;
F - G - (D)F - R - F;
F - G - A - R - W;
F - G - L - (D)R - W;
F - G - (D)L - R - W;
A - G - L - R - W; または
(D)F - G - L - R - W.
でない]。
【請求項37】
X1が(D)Fである、請求項36に記載のペプチド分子。
【請求項38】
X2が(D)F、(D)Lおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項36または37に記載のペプチド分子。
【請求項39】
ペプチド配列が
(D)F-W-L-R-W、または
F-G-(D)W-R-F
からなる群から選択される、請求項36から38のいずれかに記載のペプチド分子。
【請求項40】
N末端残基が当残基上の電荷を除去する基yを含む、請求項36から39のいずれかに記載のペプチド分子。
【請求項41】
X1が当残基上の電荷を除去する基yを含む、請求項40に記載のペプチド分子。
【請求項42】
基yがアセチル基、トリフルオロアセチル基、環化アミノ酸、またはN末端における電荷を欠いた合成アミノ酸から選択される、請求項40または41に記載のペプチド分子。
【請求項43】
X3が当残基上の電荷を除去する基zを含む、請求項36から42のいずれかに記載のペプチド分子。
【請求項44】
ペプチド配列が
I) ac.F - G - (D)F - R - W.z;
II) ac.F - G - (D)L - R - W.z;
III) ac.F - G - L - (D)R - W.z;
IV) ac.F - G - A - R - W.z;
V) ac.A - G - L - R - W.z
VI) ac.(D)F - W - L - R - W.z; または
VII) ac.F - G - (D)W - R - F.z.
からなる群から選択される、請求項36から43のいずれかに記載のペプチド分子。
【請求項45】
以下の配列
XA-XB-XC-N-XD-XE-G-XF-R-F
を含むペプチド分子
[ここで、
XAは、Yまたは任意のD-アミノ酸残基であり、
XBは、Nまたは任意のD-アミノ酸残基であり、
XCは、Wまたは任意のD-アミノ酸残基であり、
XDは、GまたはSまたは任意のD-アミノ酸残基であり、
XEは、Fまたは(D)Wまたは(D)Lであり、
XFは、WまたはLまたは任意のD-アミノ酸残基であり、
ペプチド分子のC末端アミノ酸残基が当残基上の電荷を除去する基z、
またはそのフラグメントもしくは変異体を含み、
ペプチド配列が
Y - N - W - N - S - F - G - L - R - F;
(D)Y - (D)N - W - N - S - F - G - W - R - F;
(D)Y - (D)N - W - N - G - F - G - W - R - F;
(D)Y - (D)N - W - N - S - F - G - (D)W - R - F; または
(D)Y - (D)N - W - N - G - F - G - (D)W - R - F.
でない]。
【請求項46】
XAが(D)Fおよび(D)Yおよび(D)Aからなる群から選択されるD-アミノ酸残基である、請求項45に記載のペプチド分子。
【請求項47】
XBが(D)Aおよび(D)Nからなる群から選択されるD-アミノ酸残基である、請求項45または46に記載のペプチド分子。
【請求項48】
XAまたはXBの1つが(D)Yである場合、他は(D)Nでない、請求項45から47のいずれかに記載のペプチド分子。
【請求項49】
XAおよびXBの両方がD-アミノ酸残基でない、請求項45から48のいずれかに記載のペプチド分子。
【請求項50】
XFが(D)Wである場合、XAは(D)Fである、請求項45から49のいずれかに記載の使用。
【請求項51】
XCが(D)Aおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項45から50のいずれかに記載のペプチド分子。
【請求項52】
XDが(D)Aおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項45から51のいずれかに記載のペプチド分子。
【請求項53】
XDがSである場合、XFは(D)Wであり、かつ/またはXAは(D)Yでない、請求項45から52のいずれかに記載の使用。
【請求項54】
XDがSである場合、XFは(D)Wであり、かつ/またはXAは(D)Aである、請求項45から52のいずれかに記載の使用。
【請求項55】
XFが(D)Lおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項45から54のいずれかに記載のペプチド分子。
【請求項56】
XEおよびXFの両方が(D)Wである場合、XAは(D)Yでない、請求項45から55のいずれかに記載の使用。
【請求項57】
XEおよびXFの両方が(D)Wである場合、XAは(D)Aである、請求項45から55のいずれかに記載の使用。
【請求項58】
N末端残基が当残基上の電荷を除去する基yを含む、請求項45から57のいずれかに記載のペプチド分子。
【請求項59】
X1が当残基上の電荷を除去する基yを含む、請求項58に記載のペプチド分子。
【請求項60】
基yがアセチル基、トリフルオロアセチル基、環化アミノ酸、またはN末端における電荷を欠いた合成アミノ酸からなる群から選択される、請求項58または59に記載のペプチド分子。
【請求項61】
ペプチド分子のC末端F残基が当残基上の電荷を除去する基zを含む、請求項58から60のいずれかに記載のペプチド分子。
【請求項62】
ペプチド配列が
a) Y - N - W - N - G - F - G - L - R - F.z;
b) Y - N - W - N - G - F - G - (D)L - R - F.z;
c) Y - N - W - N - G - F - G - (D)W - R - F.z;
d) Y - N - W - N - G - (D)W - G - L - R - F.z;
e) ac.Y - N - W - N - G - F - G - (D)W - R - F.z;
f) ac.Y - N - W - N - (D)W - F - G - (D)W - R - F.z;
g) ac.(D)Y - N - W - N - G - F - G - (D)W - R - F.z;
h) ac.Y - N - (D)W - N - G - F - G - (D)W - R - F.z;
i) ac.Y - (D)N - W - N - G - F - G - (D)W - R - F.z;
j) ac.Y - N - W - N - (D)A - F - G - (D)W - R - F.z;
k) ac.(D)A - N - W - N - G - F - G - (D)W - R - F.z;
l) ac.Y - N - (D)A - N - G - F - G - (D)W - R - F.z;
m) ac.Y - (D)A - W - N - G - F - G - (D)W - R - F.z;
n) ac.(D)W - N - W - N - G - F - G - (D)W - R - F.z;
o) ac.(D)F - N - W - N - G - F - G - (D)W - R - F.z;
p) ac.(D)Y - N - W - N - G - (D)W - G - (D)W - R - F.z;
q) ac.(D)A - N - W - N - G - (D)W - G - (D)W - R - F.z;
r) ac.(D)A - N - W - N - S - F - G - (D)W - R - F.z;
s) ac.(D)A - N - W - N - G - F - G - W - R - F.z;
t) ac.(D)A - N - W - N - (D)S - F - G - (D)W - R - F.z; または
u) ac.(D)A - N - W - N - G - F - G - (D)L - R - F.z.
からなる群から選択される、請求項58から61のいずれかに記載のペプチド分子。
【請求項63】
zがNH2またはN-プロピルアミドまたはN-エチルアミド(NHEt)またはN-メチルアミドまたはN-ブチルアミドである、請求項3から62のいずれかに定義されているペプチド分子。
【請求項64】
ペプチドがN末端における配列R-R-M-K-W-K-K-Yをさらに含む、請求項3から63のいずれかに定義されているペプチド分子。
【請求項65】
ペプチド配列が
R-R-M-K-W-K-K-Y-(D)A-N-W-N-G-F-G-(D)W-R-F.zまたは
ac.R-R-M-K-W-K-K-Y-(D)A-N-W-N-G-F-G-(D)W-R-F.z
からなる群から選択される、請求項64に定義されているペプチド分子。
【請求項66】
有効な量の請求項1もしくは2に定義されている拮抗薬または請求項63から65のいずれかに記載のペプチド分子および薬学的に許容できる賦形剤もしくは希釈剤を含む医薬組成物。
【請求項67】
請求項66に記載の医薬組成物、または有効な量の請求項1もしくは2に定義されているキスペプチンの拮抗薬、または有効な量の請求項63から65のいずれかに記載のペプチド分子を個体に投与する段階を含む、前記個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態を治療する方法。
【請求項68】
個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態が増殖性障害、子宮内膜症、子宮線維症、思春期早発症、子癇前症、子宮内胎児発育遅延(IUGR)、子宮外妊娠、月経痛、高血圧、冠動脈心疾患、中枢神経系(CNS)、膵臓および/または免疫系の病変、排卵の抑制または阻害、受胎能、生殖組織の化学療法誘発性または放射線療法誘発性損傷、創傷治癒の抑制または阻害、成長ホルモン産生の抑制または阻害を含む群から選択される、請求項67に記載の方法。
【請求項69】
増殖性障害が良性前立腺過形成、がん、生殖組織のがん、婦人科がん、前立腺がん、乳がん、卵巣がん、子宮がん、子宮頚がん、子宮内膜がん、黒色腫、膵臓がん、胃がんを含む群から選択される、請求項68に記載の方法。
【請求項70】
i)試験する化合物を準備する段階、
ii)(i)における化合物がキスペプチン受容体に結合する能力を測定する段階、
iii)(i)における化合物が細胞内のイノシトールリン酸産生のキスペプチン媒介性刺激に拮抗する能力を測定する段階、および
iv)キスペプチン受容体に結合することができ、細胞内のイノシトールリン酸産生のキスペプチン媒介性刺激に拮抗することができる場合に、化合物をキスペプチンの拮抗薬として特定する段階
を含むキスペプチンの拮抗薬として同定する方法。
【請求項71】
実質的に本明細書に記載の通りである拮抗薬またはペプチド分子または医薬組成物。
【請求項72】
実質的に本明細書に記載の通りである使用または方法。
【請求項1】
個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療用の薬剤の製造におけるキスペプチンの拮抗薬の使用。
【請求項2】
個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療における使用のためのキスペプチンの拮抗薬。
【請求項3】
患者におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療のための、あるいは患者におけるキスペプチン活性により誘発され、または悪化した状態の治療用の薬剤の製造における、以下の配列
X1-G/W-X2-R/(D)R-X3
[ここで、
X1は、FまたはAまたは任意のD-アミノ酸残基であり、
X2は、LまたはAまたは任意のD-アミノ酸残基であり、
X3は、FまたはWであり、
ペプチド分子のC末端アミノ酸残基が当残基上の電荷を除去する基z、
またはそのフラグメントもしくは変異体を含み、
ペプチド配列が
F-G-L-R-F、
F-G-L-R-W、または
F-G-(D)F-R-Fでない]
を含むペプチド分子の使用。
【請求項4】
前記ペプチド配列が
F-G-A-R-W、
F-G-L-(D)R-W、
F-G-(D)L-R-W、または
(D)F-G-L-R-W
でない、請求項3に記載の使用。
【請求項5】
X1が(D)Fである、請求項3または4に記載の使用。
【請求項6】
X2が(D)F、(D)Lおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項3から5のいずれかに記載の使用。
【請求項7】
前記ペプチド配列が
(D)F-W-L-R-W、または
F-G-(D)W-R-F
からなる群から選択される、請求項3から6のいずれかに記載の使用。
【請求項8】
N末端残基が当残基上の電荷を除去する基yを含む、請求項3から7のいずれかに記載の使用。
【請求項9】
X1が当残基上の電荷を除去する基yを含む、請求項8に記載の使用。
【請求項10】
基yがアセチル基、トリフルオロアセチル基、環化アミノ酸、またはN末端における電荷を欠いた合成アミノ酸からなる群から選択される、請求項8または9に記載の使用。
【請求項11】
X3が当残基上の電荷を除去する基zを含む、請求項3から10のいずれかに記載の使用。
【請求項12】
前記配列が
I) ac.F - G - (D)F - R - W.z;
II) ac.F - G - (D)L - R - W.z;
III) ac.F - G - L - (D)R - W.z;
IV) ac.F - G - A - R - W.z;
V) ac.A - G - L - R - W.z;
VI) ac.(D)F - W - L - R - W.z; または
VII) ac.F - G - (D)W - R - F.z.
からなる群から選択される、請求項3から11のいずれかに記載の使用。
【請求項13】
患者におけるキスペプチン活性により誘発され、かつ/または悪化した状態の治療のための、あるいは患者におけるキスペプチン活性により誘発され、または悪化した状態の治療用の薬剤の製造における、以下の配列
XA-XB-XC-N-XD-XE-G-XF-R-F
[ここで、
XAは、Yまたは任意のD-アミノ酸残基であり、
XBは、Nまたは任意のD-アミノ酸残基であり、
XCは、Wまたは任意のD-アミノ酸残基であり、
XDは、GまたはSまたは任意のD-アミノ酸残基であり、
XEは、Fまたは(D)Wまたは(D)Lであり、
XFは、WまたはLまたは任意のD-アミノ酸残基であり、
ペプチド分子のC末端アミノ酸残基が当残基上の電荷を除去する基z、
またはそのフラグメントもしくは変異体を含み、
ペプチド配列が
Y - N - W - N - S - F - G - L - R - F;
(D)Y - (D)N - W - N - S - F - G - W - R - F;
(D)Y - (D)N - W - N - G - F - G - W - R - F;
(D)Y - (D)N - W - N - S - F - G - (D)W - R - F; または
(D)Y - (D)N - W - N - G - F - G - (D)W - R - F.
でない]
を含むペプチド分子の使用。
【請求項14】
XAが(D)Fおよび(D)Yおよび(D)Aからなる群から選択されるD-アミノ酸残基である、請求項13に記載の使用。
【請求項15】
XBが(D)Aおよび(D)Nからなる群から選択されるD-アミノ酸残基である、請求項13または14に記載の使用。
【請求項16】
XAまたはXBの1つが(D)Yである場合、他は(D)Nでない、請求項13から15のいずれかに記載の使用。
【請求項17】
XAおよびXBの両方がD-アミノ酸残基でない、請求項13から16のいずれかに記載の使用。
【請求項18】
XFが(D)Wである場合、XAは(D)Fである、請求項13から17のいずれかに記載の使用。
【請求項19】
XCが(D)Aおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項13から18のいずれかに記載の使用。
【請求項20】
XDが(D)Aおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項13から19のいずれかに記載の使用。
【請求項21】
XDがSである場合、XFは(D)Wであり、かつ/またはXAは(D)Yでない、請求項13から20のいずれかに記載の使用。
【請求項22】
XDがSである場合、XFは(D)Wであり、かつ/またはXAは(D)Aである、請求項13から20のいずれかに記載の使用。
【請求項23】
XFが(D)Lおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項13から22のいずれかに記載の使用。
【請求項24】
XEおよびXFの両方が(D)Wである場合、XAは(D)Yでない、請求項13から23のいずれかに記載の使用。
【請求項25】
XEおよびXFの両方が(D)Wである場合、XAは(D)Aである、請求項13から23のいずれかに記載の使用。
【請求項26】
N末端残基が当残基上の電荷を除去する基yを含む、請求項13から25のいずれかに記載の使用。
【請求項27】
X1が当残基上の電荷を除去する基yを含む、請求項26に記載の使用。
【請求項28】
基yがアセチル基、トリフルオロアセチル基、環化アミノ酸、またはN末端における電荷を欠いた合成アミノ酸からなる群から選択される、請求項26または27に記載の使用。
【請求項29】
ペプチド分子のC末端F残基が当残基上の電荷を除去する基zを含む、請求項13から28のいずれかに記載の使用。
【請求項30】
前記配列が
a) Y - N - W - N - G - F - G - L - R - F.z;
b) Y - N - W - N - G - F - G - (D)L - R - F.z;
c) Y - N - W - N - G - F - G - (D)W - R - F.z;
d) Y - N - W - N - G - (D)W - G - L - R - F.z;
e) ac.Y - N - W - N - G - F - G - (D)W - R - F.z;
f) ac.Y - N - W - N - (D)W - F - G - (D)W - R - F.z;
g) ac.(D)Y - N - W - N - G - F - G - (D)W - R - F.z;
h) ac.Y - N - (D)W - N - G - F - G - (D)W - R - F.z;
i) ac.Y - (D)N - W - N - G - F - G - (D)W - R - F.z;
j) ac.Y - N - W - N - (D)A - F - G - (D)W - R - F.z;
k) ac.(D)A - N - W - N - G - F - G - (D)W - R - F.z;
l) ac.Y - N - (D)A - N - G - F - G - (D)W - R - F.z;
m) ac.Y - (D)A - W - N - G - F - G - (D)W - R - F.z;
n) ac.(D)W - N - W - N - G - F - G - (D)W - R - F.z;
o) ac.(D)F - N - W - N - G - F - G - (D)W - R - F.z;
p) ac.(D)Y - N - W - N - G - (D)W - G - (D)W - R - F.z;
q) ac.(D)A - N - W - N - G - (D)W - G - (D)W - R - F.z;
r) ac.(D)A - N - W - N - S - F - G - (D)W - R - F.z;
s) ac.(D)A - N - W - N - G - F - G - W - R - F.z;
t) ac.(D)A - N - W - N - (D)S - F - G - (D)W - R - F.z; または
u) ac.(D)A - N - W - N - G - F - G - (D)L - R - F.z;
からなる群から選択される、請求項13から29のいずれかに記載の使用。
【請求項31】
個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態は、増殖性障害、子宮内膜症、子宮線維症、思春期早発症、子癇前症、子宮内胎児発育遅延(IUGR)、子宮外妊娠、月経痛、高血圧、冠動脈心疾患、中枢神経系(CNS)、膵臓および/または免疫系の病変、排卵の抑制または阻害、受胎能、生殖組織の化学療法誘発性および/または放射線療法誘発性損傷、創傷治癒の抑制または阻害、成長ホルモン産生の抑制または阻害を含む群から選択される、請求項1から30のいずれかに記載の使用。
【請求項32】
増殖性障害が良性前立腺過形成、がん、生殖組織のがん、婦人科がん、前立腺がん、乳がん、卵巣がん、子宮がん、子宮頚がん、子宮内膜がん、黒色腫、膵臓がん、胃がんを含む群から選択される、請求項31に記載の使用。
【請求項33】
キスペプチンの拮抗薬またはキスペプチンの拮抗薬であるそのフラグメントもしくは変異体である、請求項3から32のいずれかに定義されているペプチド分子。
【請求項34】
医療用の請求項1または2に定義されている拮抗薬あるいは請求項3から33のいずれか一項に定義されているペプチド分子。
【請求項35】
キスペプチンの拮抗薬である、請求項34に記載のペプチド分子。
【請求項36】
以下の配列
X1-G/W-X2-R/(D)R-X3
を含むペプチド分子
[ここで、
X1は、FまたはAまたは任意のD-アミノ酸残基であり、
X2は、LまたはAまたは任意のD-アミノ酸残基であり、
X3は、FまたはWであり、
ペプチド分子のC末端アミノ酸残基が当残基上の電荷を除去する基z、
またはそのフラグメントもしくは変異体を含み、
ペプチド配列が
F - G - L - R - F;
F - G - L - R - W;
F - G - (D)F - R - F;
F - G - A - R - W;
F - G - L - (D)R - W;
F - G - (D)L - R - W;
A - G - L - R - W; または
(D)F - G - L - R - W.
でない]。
【請求項37】
X1が(D)Fである、請求項36に記載のペプチド分子。
【請求項38】
X2が(D)F、(D)Lおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項36または37に記載のペプチド分子。
【請求項39】
ペプチド配列が
(D)F-W-L-R-W、または
F-G-(D)W-R-F
からなる群から選択される、請求項36から38のいずれかに記載のペプチド分子。
【請求項40】
N末端残基が当残基上の電荷を除去する基yを含む、請求項36から39のいずれかに記載のペプチド分子。
【請求項41】
X1が当残基上の電荷を除去する基yを含む、請求項40に記載のペプチド分子。
【請求項42】
基yがアセチル基、トリフルオロアセチル基、環化アミノ酸、またはN末端における電荷を欠いた合成アミノ酸から選択される、請求項40または41に記載のペプチド分子。
【請求項43】
X3が当残基上の電荷を除去する基zを含む、請求項36から42のいずれかに記載のペプチド分子。
【請求項44】
ペプチド配列が
I) ac.F - G - (D)F - R - W.z;
II) ac.F - G - (D)L - R - W.z;
III) ac.F - G - L - (D)R - W.z;
IV) ac.F - G - A - R - W.z;
V) ac.A - G - L - R - W.z
VI) ac.(D)F - W - L - R - W.z; または
VII) ac.F - G - (D)W - R - F.z.
からなる群から選択される、請求項36から43のいずれかに記載のペプチド分子。
【請求項45】
以下の配列
XA-XB-XC-N-XD-XE-G-XF-R-F
を含むペプチド分子
[ここで、
XAは、Yまたは任意のD-アミノ酸残基であり、
XBは、Nまたは任意のD-アミノ酸残基であり、
XCは、Wまたは任意のD-アミノ酸残基であり、
XDは、GまたはSまたは任意のD-アミノ酸残基であり、
XEは、Fまたは(D)Wまたは(D)Lであり、
XFは、WまたはLまたは任意のD-アミノ酸残基であり、
ペプチド分子のC末端アミノ酸残基が当残基上の電荷を除去する基z、
またはそのフラグメントもしくは変異体を含み、
ペプチド配列が
Y - N - W - N - S - F - G - L - R - F;
(D)Y - (D)N - W - N - S - F - G - W - R - F;
(D)Y - (D)N - W - N - G - F - G - W - R - F;
(D)Y - (D)N - W - N - S - F - G - (D)W - R - F; または
(D)Y - (D)N - W - N - G - F - G - (D)W - R - F.
でない]。
【請求項46】
XAが(D)Fおよび(D)Yおよび(D)Aからなる群から選択されるD-アミノ酸残基である、請求項45に記載のペプチド分子。
【請求項47】
XBが(D)Aおよび(D)Nからなる群から選択されるD-アミノ酸残基である、請求項45または46に記載のペプチド分子。
【請求項48】
XAまたはXBの1つが(D)Yである場合、他は(D)Nでない、請求項45から47のいずれかに記載のペプチド分子。
【請求項49】
XAおよびXBの両方がD-アミノ酸残基でない、請求項45から48のいずれかに記載のペプチド分子。
【請求項50】
XFが(D)Wである場合、XAは(D)Fである、請求項45から49のいずれかに記載の使用。
【請求項51】
XCが(D)Aおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項45から50のいずれかに記載のペプチド分子。
【請求項52】
XDが(D)Aおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項45から51のいずれかに記載のペプチド分子。
【請求項53】
XDがSである場合、XFは(D)Wであり、かつ/またはXAは(D)Yでない、請求項45から52のいずれかに記載の使用。
【請求項54】
XDがSである場合、XFは(D)Wであり、かつ/またはXAは(D)Aである、請求項45から52のいずれかに記載の使用。
【請求項55】
XFが(D)Lおよび(D)Wからなる群から選択されるD-アミノ酸残基である、請求項45から54のいずれかに記載のペプチド分子。
【請求項56】
XEおよびXFの両方が(D)Wである場合、XAは(D)Yでない、請求項45から55のいずれかに記載の使用。
【請求項57】
XEおよびXFの両方が(D)Wである場合、XAは(D)Aである、請求項45から55のいずれかに記載の使用。
【請求項58】
N末端残基が当残基上の電荷を除去する基yを含む、請求項45から57のいずれかに記載のペプチド分子。
【請求項59】
X1が当残基上の電荷を除去する基yを含む、請求項58に記載のペプチド分子。
【請求項60】
基yがアセチル基、トリフルオロアセチル基、環化アミノ酸、またはN末端における電荷を欠いた合成アミノ酸からなる群から選択される、請求項58または59に記載のペプチド分子。
【請求項61】
ペプチド分子のC末端F残基が当残基上の電荷を除去する基zを含む、請求項58から60のいずれかに記載のペプチド分子。
【請求項62】
ペプチド配列が
a) Y - N - W - N - G - F - G - L - R - F.z;
b) Y - N - W - N - G - F - G - (D)L - R - F.z;
c) Y - N - W - N - G - F - G - (D)W - R - F.z;
d) Y - N - W - N - G - (D)W - G - L - R - F.z;
e) ac.Y - N - W - N - G - F - G - (D)W - R - F.z;
f) ac.Y - N - W - N - (D)W - F - G - (D)W - R - F.z;
g) ac.(D)Y - N - W - N - G - F - G - (D)W - R - F.z;
h) ac.Y - N - (D)W - N - G - F - G - (D)W - R - F.z;
i) ac.Y - (D)N - W - N - G - F - G - (D)W - R - F.z;
j) ac.Y - N - W - N - (D)A - F - G - (D)W - R - F.z;
k) ac.(D)A - N - W - N - G - F - G - (D)W - R - F.z;
l) ac.Y - N - (D)A - N - G - F - G - (D)W - R - F.z;
m) ac.Y - (D)A - W - N - G - F - G - (D)W - R - F.z;
n) ac.(D)W - N - W - N - G - F - G - (D)W - R - F.z;
o) ac.(D)F - N - W - N - G - F - G - (D)W - R - F.z;
p) ac.(D)Y - N - W - N - G - (D)W - G - (D)W - R - F.z;
q) ac.(D)A - N - W - N - G - (D)W - G - (D)W - R - F.z;
r) ac.(D)A - N - W - N - S - F - G - (D)W - R - F.z;
s) ac.(D)A - N - W - N - G - F - G - W - R - F.z;
t) ac.(D)A - N - W - N - (D)S - F - G - (D)W - R - F.z; または
u) ac.(D)A - N - W - N - G - F - G - (D)L - R - F.z.
からなる群から選択される、請求項58から61のいずれかに記載のペプチド分子。
【請求項63】
zがNH2またはN-プロピルアミドまたはN-エチルアミド(NHEt)またはN-メチルアミドまたはN-ブチルアミドである、請求項3から62のいずれかに定義されているペプチド分子。
【請求項64】
ペプチドがN末端における配列R-R-M-K-W-K-K-Yをさらに含む、請求項3から63のいずれかに定義されているペプチド分子。
【請求項65】
ペプチド配列が
R-R-M-K-W-K-K-Y-(D)A-N-W-N-G-F-G-(D)W-R-F.zまたは
ac.R-R-M-K-W-K-K-Y-(D)A-N-W-N-G-F-G-(D)W-R-F.z
からなる群から選択される、請求項64に定義されているペプチド分子。
【請求項66】
有効な量の請求項1もしくは2に定義されている拮抗薬または請求項63から65のいずれかに記載のペプチド分子および薬学的に許容できる賦形剤もしくは希釈剤を含む医薬組成物。
【請求項67】
請求項66に記載の医薬組成物、または有効な量の請求項1もしくは2に定義されているキスペプチンの拮抗薬、または有効な量の請求項63から65のいずれかに記載のペプチド分子を個体に投与する段階を含む、前記個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態を治療する方法。
【請求項68】
個体におけるキスペプチン活性により誘発され、かつ/または悪化した状態が増殖性障害、子宮内膜症、子宮線維症、思春期早発症、子癇前症、子宮内胎児発育遅延(IUGR)、子宮外妊娠、月経痛、高血圧、冠動脈心疾患、中枢神経系(CNS)、膵臓および/または免疫系の病変、排卵の抑制または阻害、受胎能、生殖組織の化学療法誘発性または放射線療法誘発性損傷、創傷治癒の抑制または阻害、成長ホルモン産生の抑制または阻害を含む群から選択される、請求項67に記載の方法。
【請求項69】
増殖性障害が良性前立腺過形成、がん、生殖組織のがん、婦人科がん、前立腺がん、乳がん、卵巣がん、子宮がん、子宮頚がん、子宮内膜がん、黒色腫、膵臓がん、胃がんを含む群から選択される、請求項68に記載の方法。
【請求項70】
i)試験する化合物を準備する段階、
ii)(i)における化合物がキスペプチン受容体に結合する能力を測定する段階、
iii)(i)における化合物が細胞内のイノシトールリン酸産生のキスペプチン媒介性刺激に拮抗する能力を測定する段階、および
iv)キスペプチン受容体に結合することができ、細胞内のイノシトールリン酸産生のキスペプチン媒介性刺激に拮抗することができる場合に、化合物をキスペプチンの拮抗薬として特定する段階
を含むキスペプチンの拮抗薬として同定する方法。
【請求項71】
実質的に本明細書に記載の通りである拮抗薬またはペプチド分子または医薬組成物。
【請求項72】
実質的に本明細書に記載の通りである使用または方法。
【図1−1】



【図1A】

【図1B】
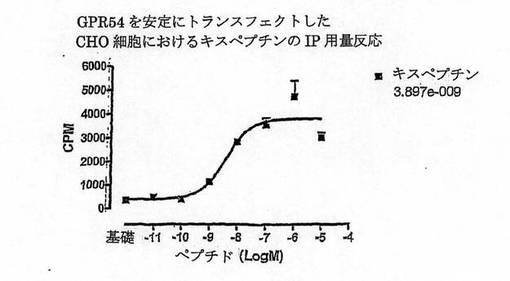
【図2A】
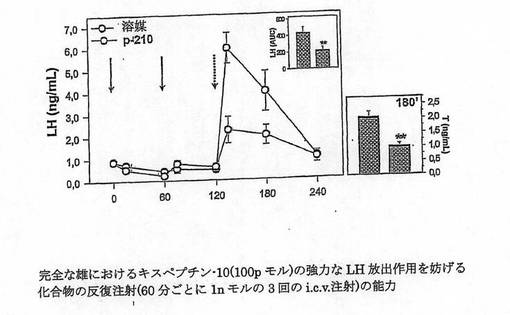
【図2B】
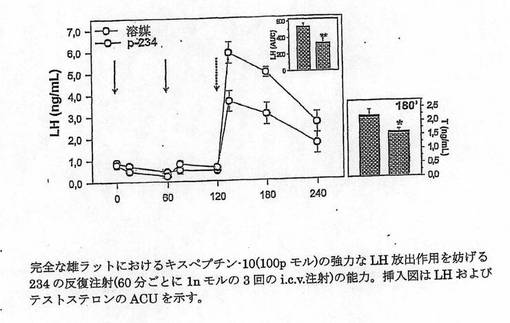
【図2C】
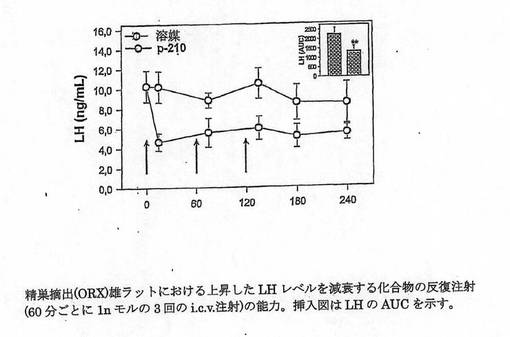
【図3】
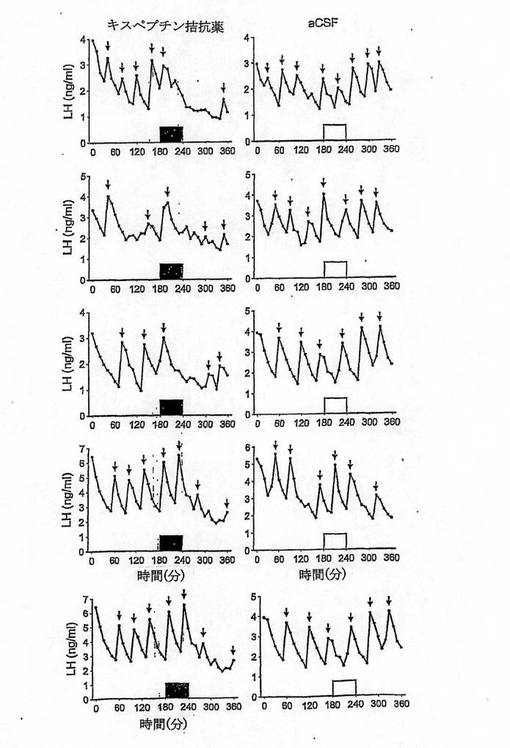
【図4A】
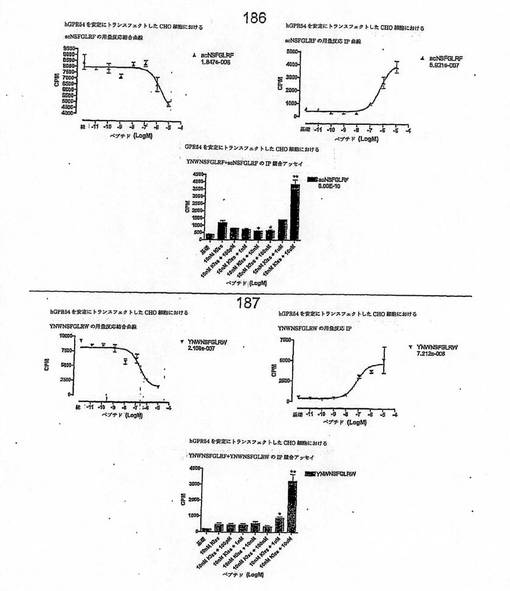
【図4B】
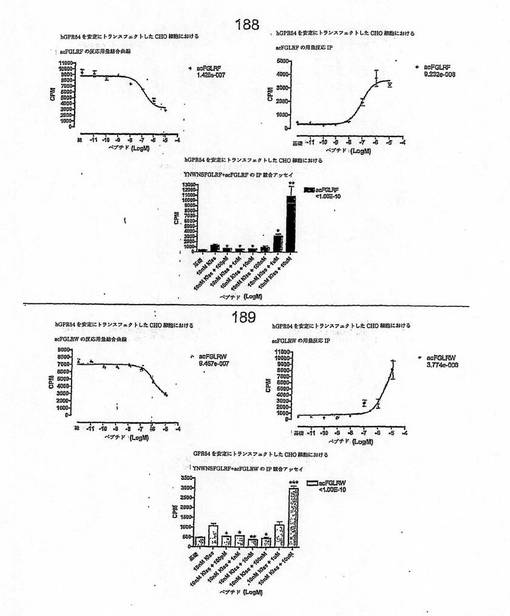
【図4C】
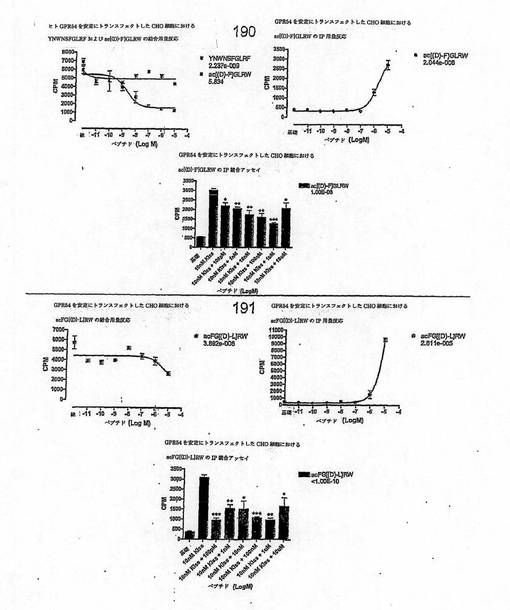
【図4D】
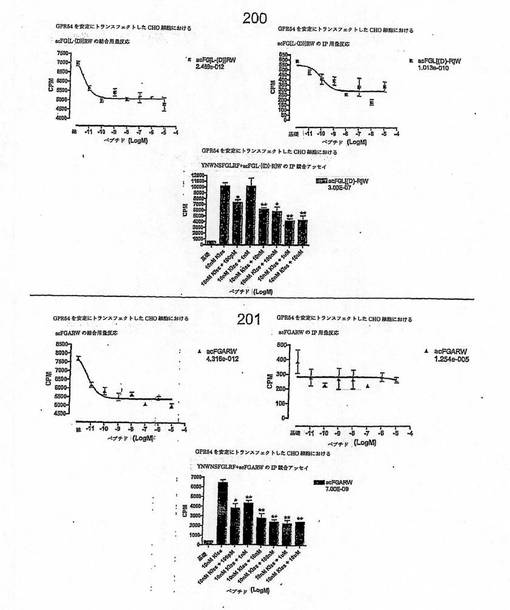
【図4E】
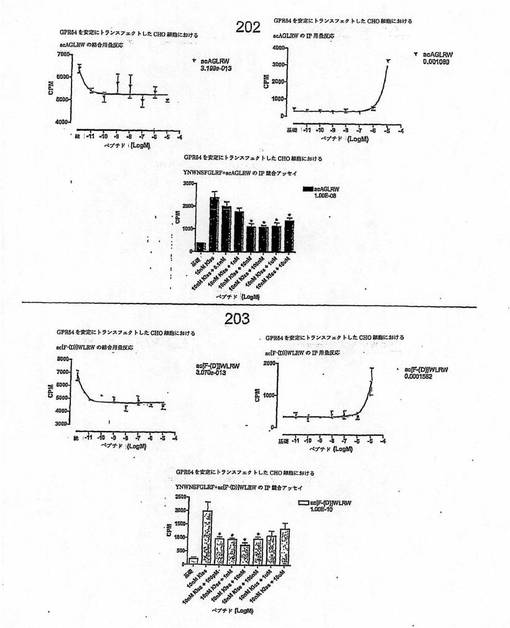
【図4F】
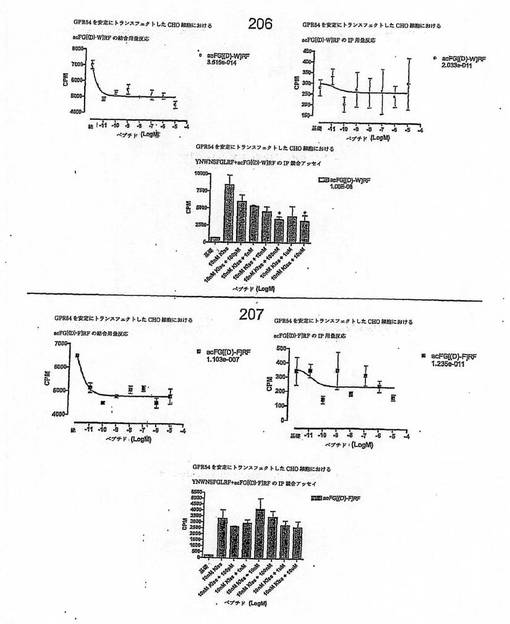
【図4G】
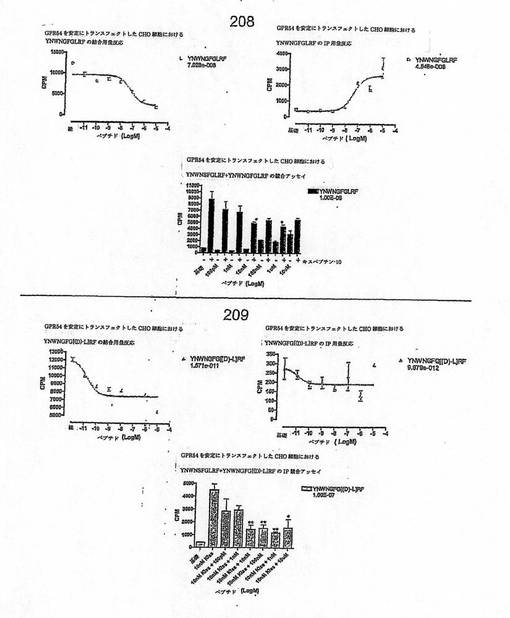
【図4H】

【図4I】
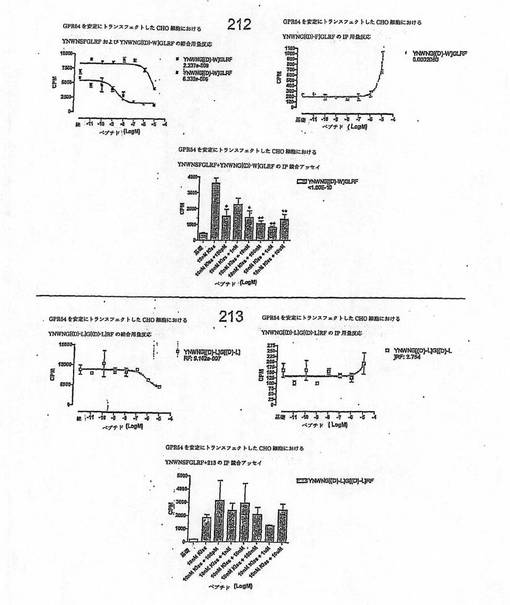
【図4J】
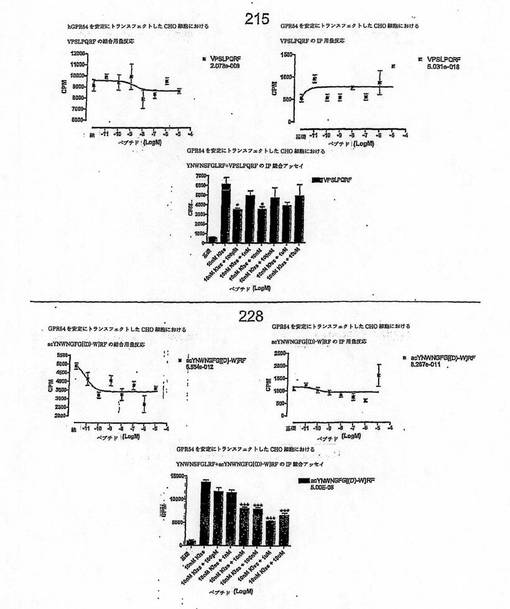
【図4K】
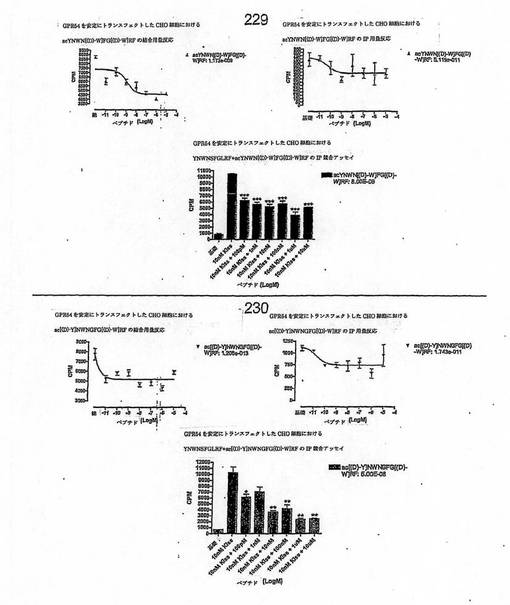
【図4L】
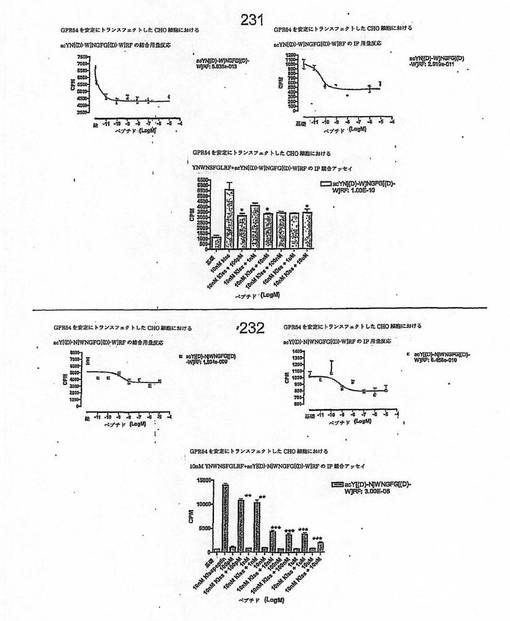
【図4M】

【図4N】
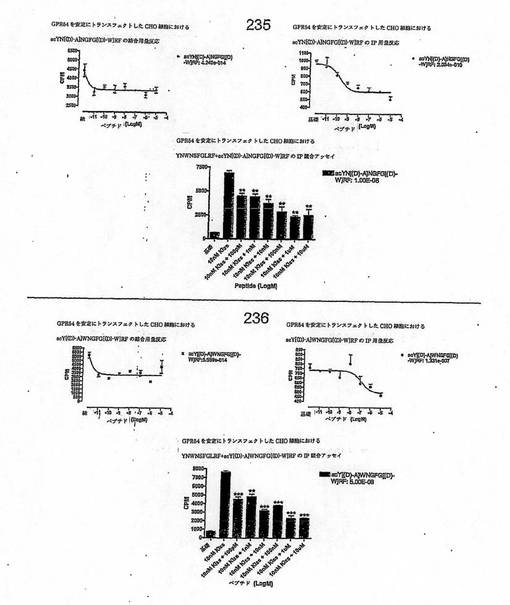
【図4O】
【図4P】

【図4Q】
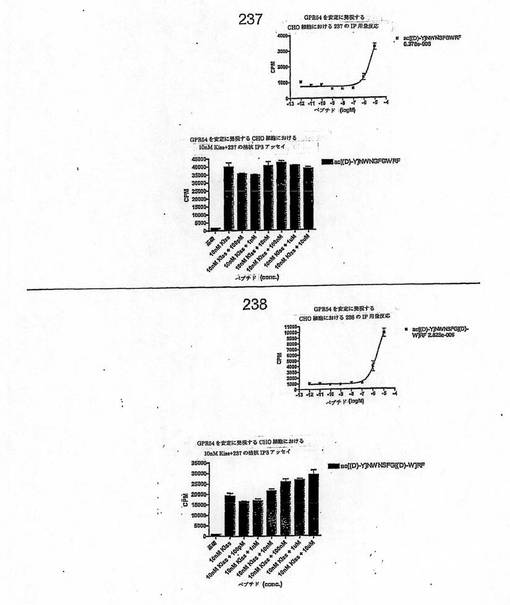
【図4R】
【図4S】
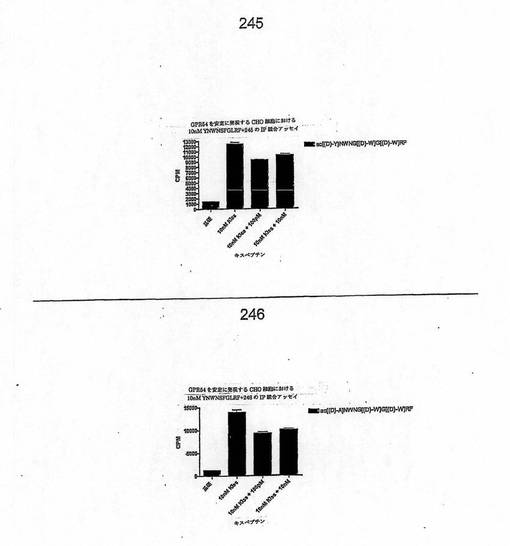
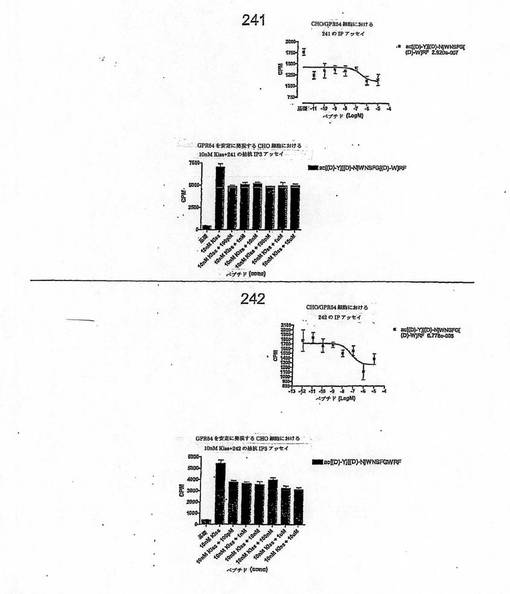
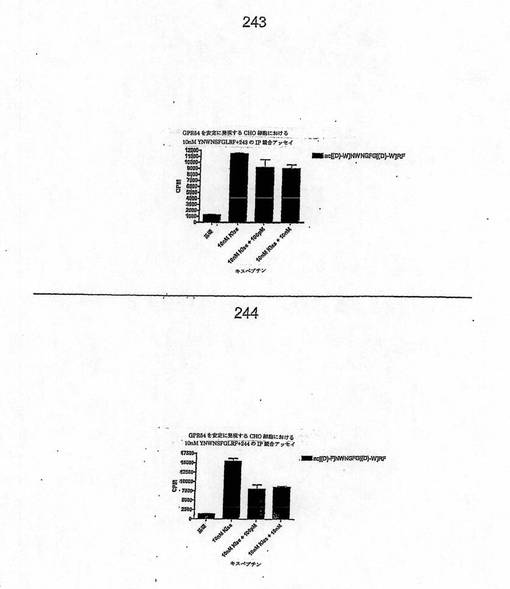
【図4T】
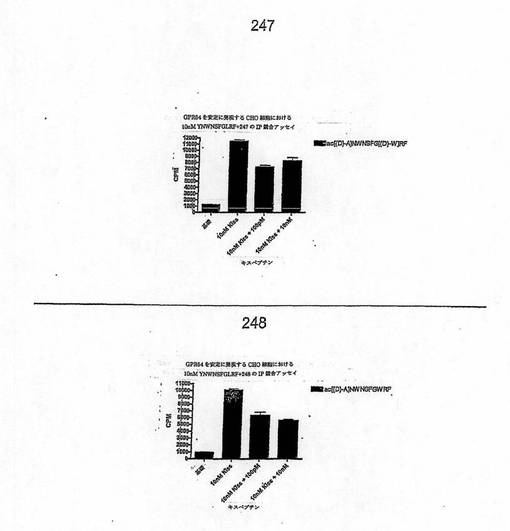
【図5】
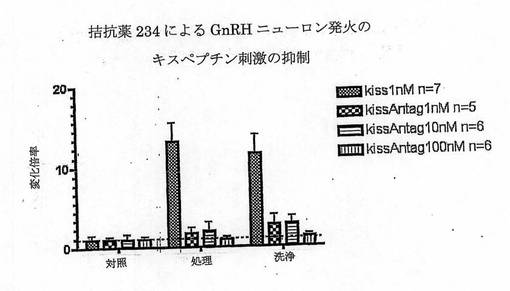
【図6】
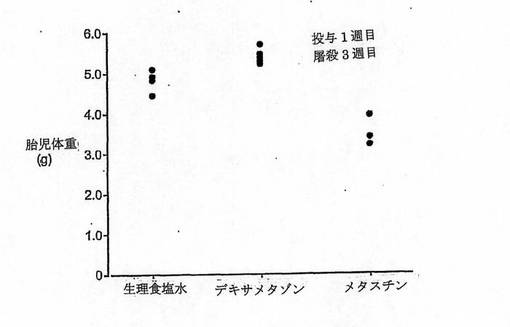
【図7】

【図8】
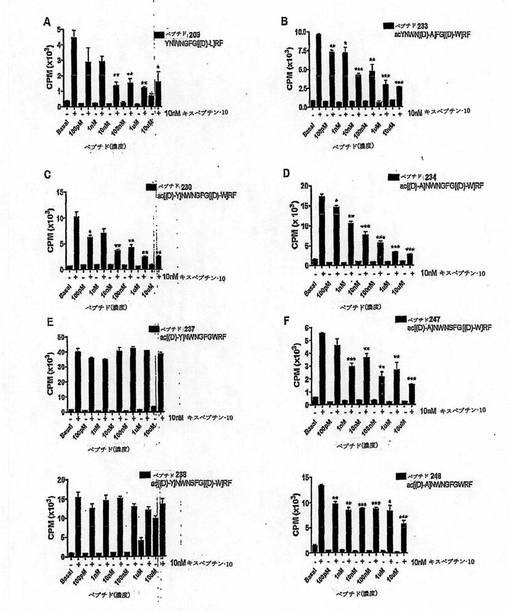
【図9】
【図10】
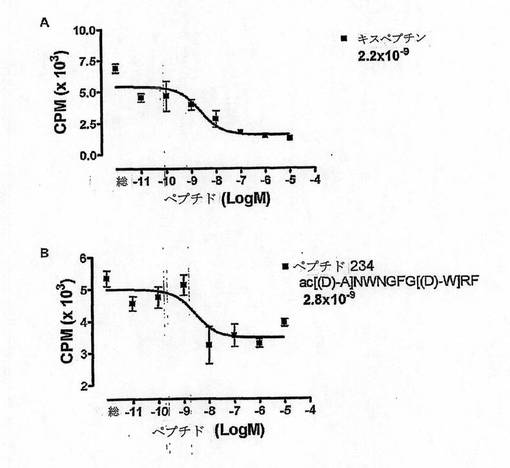
【図11】

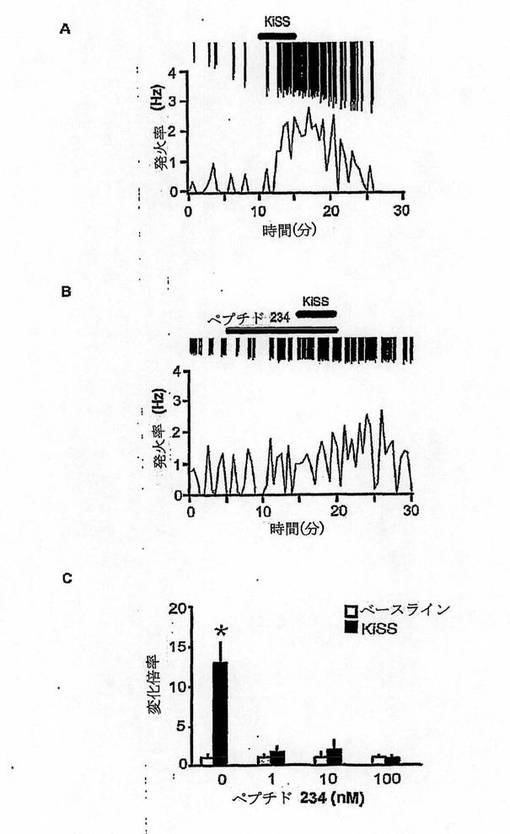
【図12】
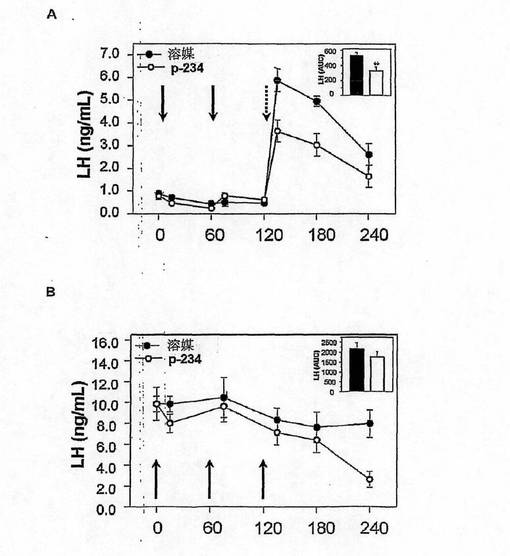
【図13】
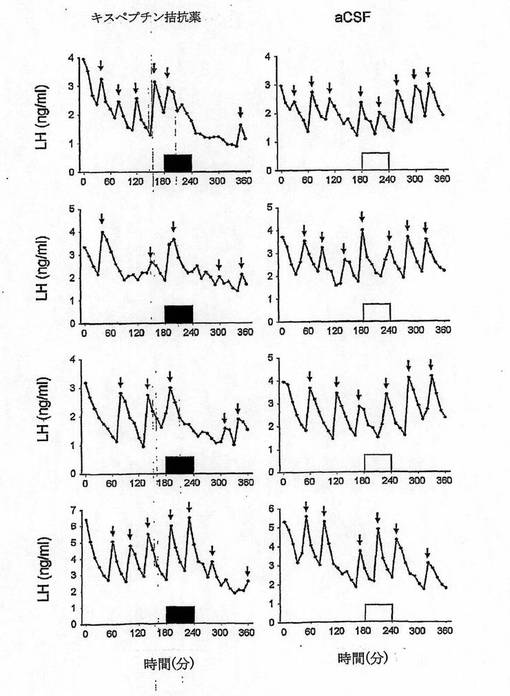
【図14−1】
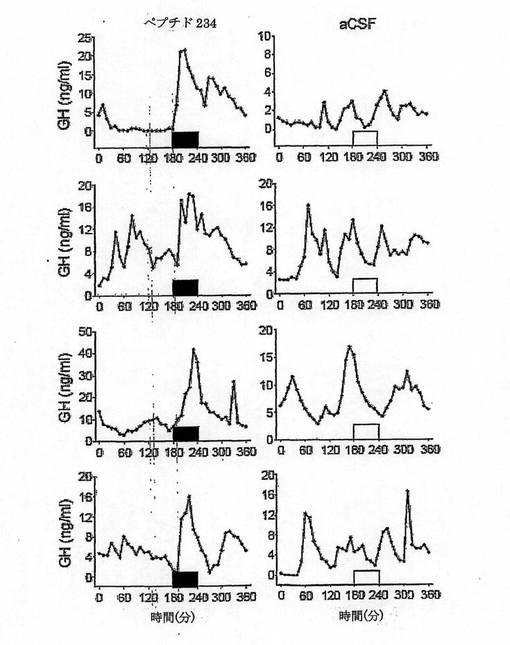
【図14−2】
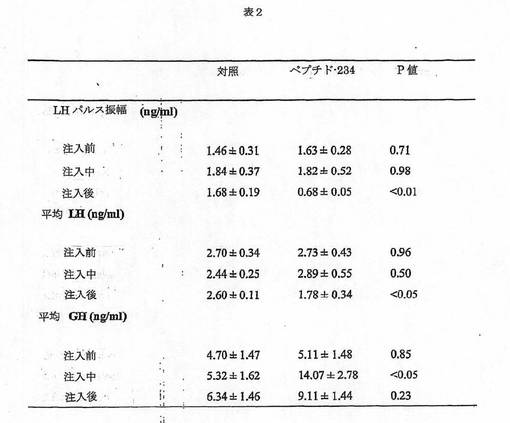
【図15】
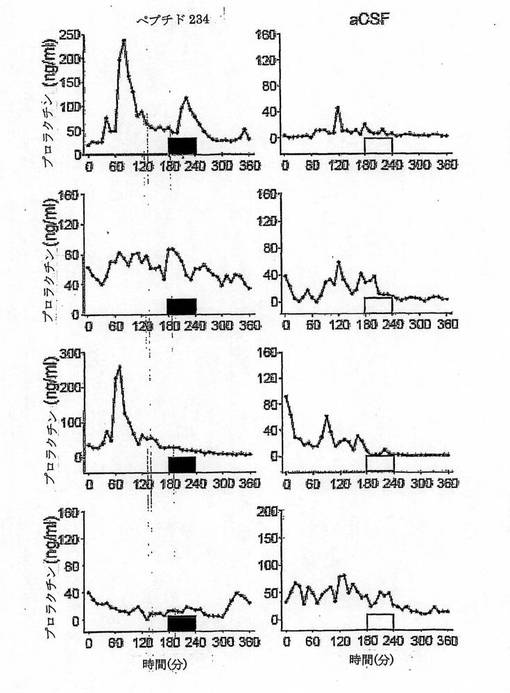
【図16】
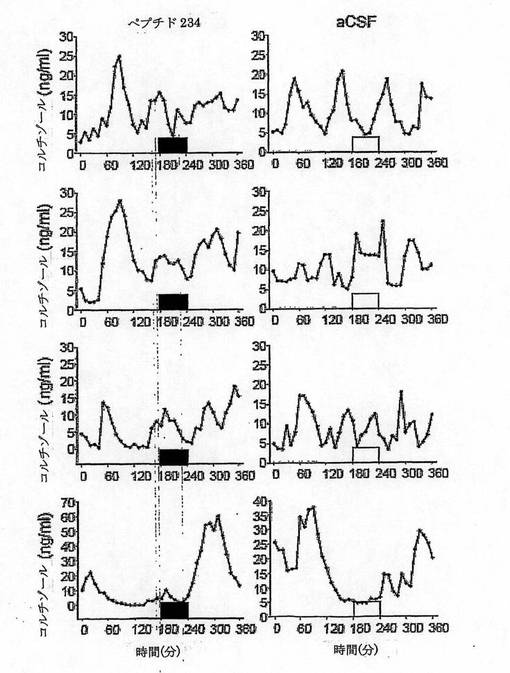
【図17】
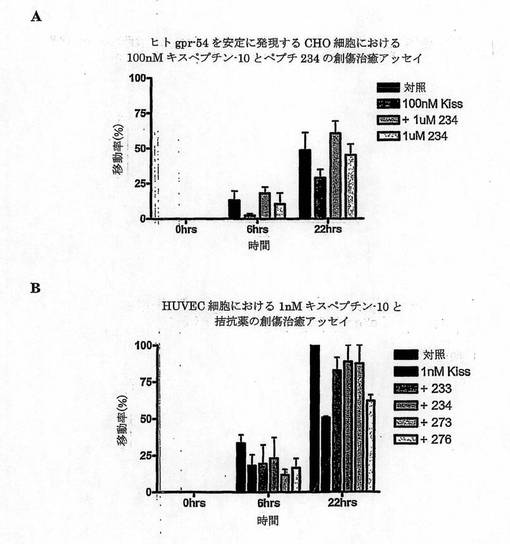
【図1A】
【図1B】
【図2A】
【図2B】
【図2C】
【図3】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図4G】
【図4H】
【図4I】
【図4J】
【図4K】
【図4L】
【図4M】
【図4N】
【図4O】
【図4P】
【図4Q】
【図4R】
【図4S】
【図4T】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14−1】
【図14−2】
【図15】
【図16】
【図17】
【公開番号】特開2013−107896(P2013−107896A)
【公開日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願番号】特願2013−26361(P2013−26361)
【出願日】平成25年2月14日(2013.2.14)
【分割の表示】特願2010−528479(P2010−528479)の分割
【原出願日】平成20年10月8日(2008.10.8)
【出願人】(597166578)メディカル リサーチ カウンシル (60)
【Fターム(参考)】
【公開日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願日】平成25年2月14日(2013.2.14)
【分割の表示】特願2010−528479(P2010−528479)の分割
【原出願日】平成20年10月8日(2008.10.8)
【出願人】(597166578)メディカル リサーチ カウンシル (60)
【Fターム(参考)】
[ Back to top ]