
キナーゼインヒビターを含む培養培地およびその使用
【課題】多くの継代に亘って該幹細胞の自己再生を支持し得る、多能性幹細胞に適した培養方法および培養培地を提供すること。
【解決手段】MEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニストを含む、培養培地。
【解決手段】MEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニストを含む、培養培地。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、多能性幹細胞における自己再生表現型の維持に関する。提供される方法および組成物は、多能性幹細胞、例えば、胚性幹(ES)細胞、特に、ラット、マウス、ウシ、ヒツジ、ブタおよびヒトを含む哺乳類の幹細胞の培養および単離に適する。特に、本発明は、ラット、マウスおよびヒトの多能性細胞の自己再生培養物、ならびにそのための方法および組成物に関する。
【背景技術】
【0002】
血清および白血病抑制因子(LIF)を含む培地の存在下でのインビトロ多能性幹細胞培養物の樹立および維持は、周知である(Smithら、(1998)Nature 336: 688-90)。このような方法は、「許容的」なマウスの系統から多くの継代に亘って多能性胚性幹(ES)細胞を維持するために用いられてきた。多能性幹細胞培養物の維持および自己再生は、フィーダー細胞またはその抽出物(通常はマウス線維芽細胞)の存在下で幹細胞が培養される場合にさらに支持される。このような条件下では、培養物中で多くの継代に亘ってヒトES細胞を多能性状態に維持し得る。
【0003】
多くの場合において、ES細胞は、血清または血清抽出物を含む(したがって、非限定である)培地を用いる、またはヒトES細胞を維持するために用いられる線維芽細胞フィーダー細胞のような他の細胞の存在を必要とする細胞培養条件を用いる場合のみ、維持され得るか、あるいは最もよく維持される。しかし、何らかの不確定の構成成分は、培地中のものであろうとまたは例えばフィーダー細胞によって産生されるものであろうと、ES細胞の増殖および分化についての研究を潜在的に妨害し、または邪魔する。これは、ES細胞およびその子孫の治療および他の適用のための良好な製造実施の発展を妨げる。いくつかの限定されたES細胞培地が知られているが、代替のおよび好ましくは改良された限定培地が必要とされる。
【0004】
本出願人による先の出願であるWO−A−03/095628およびより後の未公開出願において、(1)gp130のアゴニスト(例えば、LIF)および(2)TGF−βスーパーファミリーのアゴニスト(例えば、BMP4)またはIdシグナリング経路のアゴニストを含む無血清培地中でES細胞のような多能性幹細胞を培養することは、複数の継代に亘って幹細胞の自己再生を促進するために用いられる。gp130シグナリングの存在下では、TGF−βスーパーファミリーまたはIdシグナリング経路のアゴニストは、驚くべきことに、分化前のシグナルよりもむしろ自己再生の刺激を提供した。それにもかかわらず、多能性細胞を自己再生状態に維持する効率、およびフィーダー細胞から離してまたはフィーダー条件培地から離して多能性細胞を移すための培地は、常に改良されることが望まれる。
【0005】
Sato Nら、Nat. Med. 2004年1月 10(1) 55-63頁は、血清を含む培地中でのマウスおよびヒトES細胞に対する、グリコーゲン合成酵素キナーゼ3(GSK3)インヒビターである6−ブロモインディルビン−3’−オキシムの効果を記載している。しかし、これらの効果は非常に短い時間枠に亘ってのみ観察され、あまりにも短いため確かな結論を引き出し得なかった。そして、その研究に用いられた非限定培地中の未知の因子の影響は重要であり得る。本発明の発明者らは、その結果を反復することを試みたがこれに失敗し、そして実際にはそれらの記載と反対の効果を見出した。
【0006】
ES細胞の培養培地の調製のためには、個々の培地構成成分をできる限り純粋な形態で提供することが望まれる。しかし、ほとんどの培地構成成分はサイトカインであり、その純度は、それらを細胞系で製造し、次に生産ブロスから可能性のある汚染物質を除去する必要によって損なわれる。いくつかのサイトカインに伴う別の問題は、それらが効果的および無毒性である濃度範囲が狭いことである。より広い範囲の濃度を有しおよび/またはより高い濃度でより毒性が低い培地構成成分は、非常に有用である。サイトカインはまた、貯蔵において安定性が限定され得、そしてより安定な培地構成成分が求められる。
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の目的は、当該技術分野における問題を克服しまたは少なくとも改善すること、好ましくは代替の、より好ましくは改良された、多くの継代に亘って該幹細胞の自己再生を支持し得る、多能性幹細胞に適した培養方法および培養培地を提供することである。本発明のさらなる目的は、細胞の分化が制御される様式で誘導され得るまで多能性幹細胞培養物のインビトロでの維持を可能にする代替の培養系を提供することである。本発明のよりさらなる目的は、多能性幹細胞の誘導および単離を増強し、かつES細胞の単離に対して抵抗性の生物からまたは多能性幹細胞がまだ単離されていない生物からのそれらの誘導および単離を促進する方法および組成物を提供することである。
【課題を解決するための手段】
【0008】
本発明によれば、ES細胞のような多能性幹細胞は、MEKインヒビターおよびGSK3インヒビターを含む、またはMEKインヒビターおよびFGFレセプターのアンタゴニストを含む、(好ましくは無血清の)培地中で培養される。好ましくは、培地は、MEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニスト(例えば、小分子GSK3インヒビターおよび小分子MEKインヒビターおよび小分子FGFRアンタゴニスト)を含む。それによって、幹細胞の複数の継代に亘る自己再生が促進される。したがって、多能性細胞でのGSK3およびMEKの阻害、MEKおよびFGFレセプターシグナリングの阻害、またはGSK3、MEKおよびFGFレセプターシグナリングの阻害は、自己再生の刺激を提供する。
【0009】
本発明は、多くの適用を有する。GSK3およびMEKの阻害の組み合わせ、MEKおよびFGFRの阻害の組み合わせ、またはGSK3、MEKおよびFGFRの阻害の組み合わせが、多能性細胞、特にES細胞を増殖させるため、およびそれらがフィーダー上で誘導または増殖されていた場合、多能性細胞、特にES細胞を、フィーダー細胞またはフィーダー細胞の層(しばしばフィーダーまたはフィーダー細胞として言及される)を伴わずに増殖するように適応させるために用いられ得る。培養物中で幹細胞を増殖させる方法は、GSK3インヒビターおよびMEKインヒビターの存在下、MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下、または好ましくはGSK3インヒビター、MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下で細胞を培養する工程を含む。1以上のGSK3インヒビターおよびMEKインヒビター、1以上のMEKインヒビターおよびFGFRアンタゴニスト、ならびに必要に応じて1以上のMEKインヒビター、GSK3インヒビターおよびFGFRアンタゴニストを含む培養培地が調製され得る。ES細胞は、GSK3インヒビターおよびMEKインヒビターを用いて、MEKインヒビターおよびFGFRアンタゴニストを用いて、またはGSK3インヒビター、MEKインヒビターおよびFGFRアンタゴニストを用いて誘導され得る。
【0010】
本発明の第1の局面によれば、多能性細胞でのGSK3およびMEKの阻害、好ましくはGSK3およびMEKおよびFGFレセプターすべての阻害が、細胞の自己再生を促進するために用いられる。
【図面の簡単な説明】
【0011】
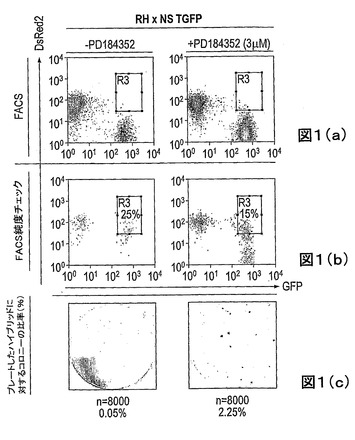
【図1A−C】多能性ES−NSハイブリッドコロニーの形成におけるPD184352の効果の分析に関し、RH×NS TGFP融合の赤色蛍光および緑色蛍光についてのFACS分析:(A)PEG処理の24時間後の融合混合物;(B)AでゲートをかけたFACS分類されたハイブリッドの純度チェック;(C)Aで分類されたハイブリッドをプレートして形成されたコロニーをプレートしたハイブリッドあたりのコロニーの比率(%)を示す。
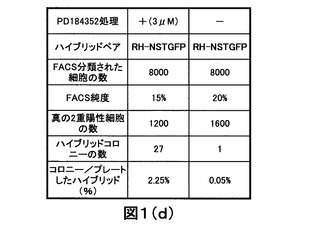
【図1D】多能性ES−NSハイブリッドコロニーの形成におけるPD184352の効果の分析のデータの概要を示す。
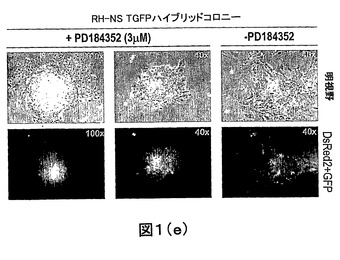
【図1E】多能性ES−NSハイブリッドコロニーの形成におけるPD184352の効果の分析のハイブリッドコロニー形態の例を示す。
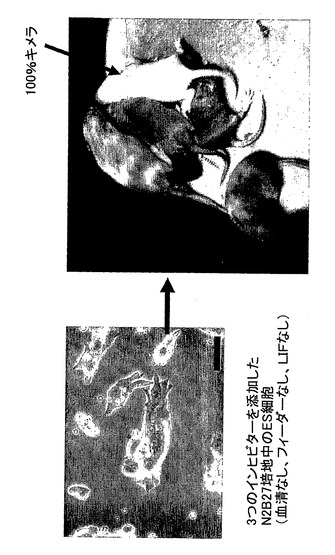
【図2】本発明にしたがって誘導および維持されたマウスのES細胞、ならびにこれらのES細胞による高効率なキメラ発生を示す。
【図3】本発明にしたがって増殖した4継代マウスES細胞を示す。
【図4】本発明にしたがって増殖したマウスES細胞がOct4陽性であることを示す。
【発明を実施するための形態】
【0012】
多能性細胞とは、胚性幹(ES)細胞と呼ばれるものを含むが、これに限定されない。多能性細胞(ES細胞を含む)に特有の性質としては、発生の多能性段階に関わる多重遺伝子の発現、供給源動物に存在するあらゆる組織タイプを代表する細胞に分化する能力、キメラを生じさせる能力、および、特にキメラの生殖系列を生じさせる能力が挙げられる。例えば、ES細胞のような真性多能性細胞は、多能性関連遺伝子Nanog、Oct4、FGF4、Sox−2およびアルカリフォスファターゼのすべてではないとしても多くを発現することが期待される。特に、Nanog、Oct4およびSox−2の発現は、細胞がES細胞である決定的な最初の徴候を提供すると広く考えられている。キメラにおける生殖系列遺伝および3つの一次胚葉(すなわち、内胚葉、中胚葉および外胚葉)のすべてから分化した細胞を含む奇形種または奇形癌を生じる能力もまた、細胞がES細胞であることの決定的な徴候として広く考えられている。
【0013】
GSK3阻害とは、1以上のGSK3酵素の阻害をいう。したがって、GSK3インヒビターはGSK3酵素ファミリーの1つのメンバー、いくつかのメンバー、またはすべてのメンバーを阻害し得る。GSK3酵素ファミリーは周知であり、GSK3−αおよびGSK3−βが挙げられるが、これらに限定されない。多くの変異型が記載されている(例えば、Schafferら; Gene 2003; 302(1-2): 73-81参照)。特定の実施態様ではGSK3−βが阻害される。GSK3−αインヒビターもまた適切であり、そして一般に本発明において使用されるインヒビターは両方ともを阻害する。広い範囲のGSK3インヒビターが公知である(例証として、インヒビターCHIR98014、CHIR99021、AR−AO144−18、TDZD−8、SB216763およびSB415286)。他のインヒビターが公知であり、そして本発明において有用である。さらに、GSK3−βの活性部位の構造は特徴づけられており、特異的なインヒビターおよび非特異的なインヒビターと相互作用する重要残基が同定されている(Bertrandら;J Mol Biol.2003;333(2):393-407)。この構造特性によって、さらなるGSKインヒビターが容易に同定され得る。
【0014】
ある実施態様のインヒビターは、GSK3−βおよびGSK3−αに特異的であり、実質的にerk2を阻害せず、そして実質的にcdc2を阻害しない。インヒビターは、IC50値の比として測定した場合、マウスerk2および/またはヒトcdc2よりもヒトGSK3に対して、好ましくは少なくとも100倍、より好ましくは少なくとも200倍、非常に好ましくは少なくとも400倍の選択性を有する。ここで、GSK3 IC50値とは、ヒトGSK3−βおよびGSK3−αについての平均値をいう。CHIR99021およびCHIR98014について良好な結果が得られ、これらは両方ともGSK3に特異的である。GSK3インヒビターの例は、Bennett Cら、J. Biol. Chem., 277巻, 34号, 2002年8月23日, 30998-31004頁およびRing DBら、Diabetes, 52巻, 2003年3月, 588-595頁に記載されている。CHIR99021の使用に適した濃度は、0.01〜100、好ましくは0.1〜20、より好ましくは0.3〜10μMの範囲にある。
【0015】
GSK3阻害はまた、RNA干渉(RNAi)を用いて好都合に達成し得る。代表的に、GSK3遺伝子のすべてまたは一部に対して相補的である2本鎖RNA分子が多能性細胞に導入され、したがって、GSK3をコードするmRNA分子の特異的な分解を促進する。この転写後機構は、目的のGSK3遺伝子の発現を低減または停止させることとなる。RNAiを用いてGSK3阻害を達成するための適切な技術およびプロトコールは公知である。
【0016】
本明細書中においてMEKインヒビターとは、一般的なMEKインヒビターをいう。したがって、MEKインヒビターとは、MEK1、MEK2およびMEK3を含むタンパク質キナーゼのMEKファミリーメンバーの任意のインヒビターをいう。また、MEK1インヒビター、MEK2インヒビターおよびMEK3インヒビターをいう。MEKインヒビターは、MEKキナーゼファミリーの1つのメンバー、いくつかのメンバーまたはすべてのメンバーを阻害し得る。適切なMEKインヒビターの例としては、当該技術分野で既に公知であり、MEK1インヒビターであるPD184352およびPD98059、MEK1およびMEK2のインヒビターであるU0126およびSL327、ならびに、Daviesら(2000)(Davies SP, Reddy H, Caivano M, Cohen P. Specificity and mechanism of action of some commonly used protein kinase inhibitors. Biochem J. 351, 95-105)に記載されたものが挙げられるが、これらに限定されない。特に、PD184352は、他の公知のMEKインヒビターと比較した場合に、高い特異性および効力を有することが見出されている。他のMEKインヒビターおよびMEKインヒビターのクラスは、Zhangら(2000)Bioorganic & Medicinal Chemistry Letters; 10:2825-2828に記載されている。
【0017】
MEKキナーゼの阻害はまた、RNA干渉(RNAi)を用いて好都合に達成し得る。代表的に、MEK遺伝子のすべてまたは一部に対して相補的である2本鎖RNA分子が多能性細胞に導入され、したがって、MEKをコードするmRNA分子の特異的な分解を促進する。この転写後機構は、目的のMEK遺伝子の発現を低減または停止させることとなる。RNAiを用いてMEK阻害を達成するための適切な技術およびプロトコールは公知である。
【0018】
キナーゼインヒビター(GSK3インヒビターおよびMEKインヒビターを含む)を同定するための多くのアッセイが、公知である。例えば、Daviesら(2000)には、ペプチド基質および放射性標識ATPの存在下でキナーゼをインキュベートするキナーゼアッセイが記載されている。キナーゼによる基質のリン酸化は、基質への標識の取り込みを生じる。各反応のアリコートを、リン酸セルロース紙上に固定化し、そしてリン酸中で洗浄し、遊離ATPを除去する。次いでインキュベーション後の基質の活性を測定し、そしてキナーゼ活性の指標が提供される。キナーゼインヒビター候補の存在下および不在下における相対的なキナーゼ活性は、そのようなアッセイを用いることで容易に決定し得る。Downeyら(1996)J Biol Chem.;271(35):21005-21011にもまた、キナーゼインヒビターの同定に用いられ得るキナーゼ活性についてのアッセイが記載されている。
【0019】
線維芽細胞増殖因子(FGF)レセプター(FGFR)のアンタゴニストとは、代表的にFGFR1および/またはFGFR2を阻害する、FGFレセプターのポリペプチドまたは小分子または他のアンタゴニストをいう。したがって、FGFレセプターアンタゴニストは、FGFレセプターファミリーの1つのメンバー、いくつかのメンバー、またはすべてのメンバーのアンタゴニストであり得、FGFR1、FGFR2、FGFR3およびFGFR4が挙げられるがこれらに限定されない。FGFレセプターファミリーのメンバーは、代表的に、3つの免疫グロブリン様ドメインを含み、そして酸性アミノ酸領域(酸性ボックス)を提示し、この領域は、FGFファミリーメンバーのFGFレセプターへの結合に関与し得る。ある場合には、2つの免疫グロブリン様ドメインのみを含む分子もまた、FGFレセプターとして機能し得る。多くのFGFRアンタゴニストが公知であり、SU5402およびPD173074が挙げられるがこれらに限定されない。多くのFGFRアンタゴニストが公知である(例えば、SU5402およびPD173074)。SU5402の適切な濃度はμMの範囲であり、例えば0.1〜20μM、好ましくは0.5〜10μM、特に1〜5μMの範囲である。発明者らは、PD173074がSU5402の代わりに用いられ得ること、そしてPD173074は約100倍低い濃度で十分に効果的であり、このことはFGFレセプターに対して、より高い親和性を有することと一致することを見出した。したがって、PD173074の適切な濃度は、1〜200nMの範囲、好ましくは5〜100nM、特に10〜50nMの範囲である。ドミナントネガティブ変異体FGFレセプターの導入遺伝子発現によってFGFレセプターシグナリングを阻害することもまた公知である。しかし、本発明の実施態様では、遺伝子導入ベースの拮抗ではなく、小分子アンタゴニストの使用が好ましい。
【0020】
FGFレセプターのアンタゴニストを同定するための適切なアッセイは公知である。例えば、FGFレセプター経由のシグナリングがレポーター遺伝子の発現を活性化する細胞系が、可能性のあるアンタゴニストの活性を評価するために用いられ得る。
【0021】
有利なことに、GSK3インヒビターと組み合わせた、および好ましくはFGFレセプターのアンタゴニストもまた組み合わせたMEKインヒビターの使用は、ES細胞の増殖を改善することが見出された。
【0022】
好ましい実施態様では、約0.1μMと約25μMとの間のMEKインヒビターが用いられる。さらに好ましくは、約0.1μMと約5μMとの間のMEKインヒビターが用いられ、より好ましくは、0.2μMから2μMまでである。
【0023】
本発明による特に好ましい培地は、0.8μMのPD184352、3μMのCHIR99021および/または3μMのSU5402を含む。特に好ましい培地は、0.8μMのPD184352、3μMのCHIR99021および3μMのSU5402を含み、好ましくはN2B27培地中である。SU5402の濃度は、異なる多能性細胞系に合うように最適化され得、代表的には1〜5μMの範囲(例えば、2μM)である。
【0024】
以下の実施例において、発明者らは、自己再生を促進するために、MEKインヒビターとともに、および特定の実施例ではFGFレセプターのアンタゴニストともともにGSK3インヒビターの存在下でマウスES細胞を培養した。他の特定の実施例において、培養物中でマウス多能性細胞の自己再生を促進する方法は、GSK3およびMEKを阻害する工程、またはGSK3、MEKおよびFGFレセプターを阻害する工程を含む。
【0025】
必要に応じて、gp130下流シグナリングの活性化は、GSK3およびMEKを阻害することによって自己再生の促進をさらに増強するためにもまた用いられ得る。gp130下流シグナリングの活性化分子は、時に、gp130アクチベーターまたはgp130アゴニストとも言われる。1以上のgp130下流シグナリング経路の活性化は、gp130を介して作用するサイトカイン、例えばLIFレセプターのサイトカインまたは他のアゴニストの使用によって達成され得る。gp130を介して作用し得、したがってgp130シグナル伝達を活性化し得るサイトカインとしては、LIF、毛様体神経栄養因子(CNTF)、カルジオトロフィン、オンコスタチンM、IL−6+sIL−6レセプター、ハイパーIL−6およびIL−11が挙げられるが、これらに限定されない。適切なサイトカインには、gp130に結合し得および/またはgp130を介してシグナリングを活性化し得る模倣物、融合タンパク質またはキメラが含まれる。血清存在下でgp130を介して作用するサイトカインの役割は十分に確立されているが、血清不在下で未分化細胞を維持するそれらのサイトカインの能力は限られる。
【0026】
本発明の利点は、GSK3インヒビター、MEKインヒビターおよび必要に応じてFGFレセプターのアンタゴニストの存在下では、多能性細胞が限定培地中で増殖し得ることである。GSK3インヒビター、MEKインヒビターおよびFGFレセプターのアンタゴニストの組み合わせを用いることによる特別な利点は、この培地がインスリン、N2B27またはgp130アゴニスト(例えば、LIF)などの他の増殖因子を含有する必要がないことである。したがって、本発明は、血清、血清抽出物、フィーダー細胞およびフィーダー細胞抽出物を含まない培地中でのES細胞の代替のおよび/または改良された培養を可能にする。
【0027】
マウス(Bradleyら、(1984) Nature 309: 255-56)、アメリカミンク(Mol Reprod Dev (1992) 12月; 33(4): 418-31)、ブタおよびヒツジ(J Reprod Fertil Suppl (1991); 43: 255-60)、ハムスター(Dev Biol (1988) 5月; 127(1): 224-7)およびウシ(Roux Arch Dev Biol (1992); 201: 134-141)を含む多くの哺乳類供給源から、目的の胚性幹細胞が報告されている。本明細書の特定の実施例は、マウスおよびヒトのES細胞を用い、そして一次派生物由来のラット細胞もまた用いる。本発明の方法および組成物が、したがって、霊長類(特にヒト)、齧歯類(特にマウスおよびラット)、および鳥類の多能性幹細胞、特にES細胞を含む他の哺乳類の多能性細胞培養物の培養への適応に適切であることが理解される。
【0028】
本発明の第2の局面は、自己再生を促進するための多能性細胞、特にES細胞の培養方法を提供し、該方法は:
(1)GSK3のインヒビター;および
(2)MEKのインヒビター、
を含む培地中で該細胞を維持する工程を含む。
【0029】
好ましくは、該方法は:
(1)GSK3のインヒビター;
(2)MEKのインヒビター;および
(3)FGFレセプターのアンタゴニスト、
を含む培地中で該細胞を維持する工程を含む。
【0030】
本発明の方法は、血清を含まないおよび血清抽出物を含まない培地中で、多能性細胞を増殖(ES細胞の増殖を含む)させるために一般的に用いられ得、これらの細胞は、血清または血清抽出物の存在下で予め継代されている。好ましくは、このような方法はまた、フィーダー細胞および/またはフィーダー細胞抽出物の不在下で行われる。例えば、以下の工程を含むES細胞の培養が行われ得る:
必要に応じてフィーダー上で、培養物中でES細胞を多能性状態に維持する工程;
少なくとも1回該ES細胞を継代する工程;
該培地から血清または血清抽出物を除去しそして(存在する場合)該フィーダーを除去して、該培地がフィーダー、血清および血清抽出物を含まないようにする工程;および
続いて、GSK3のインヒビター、MEKインヒビターおよび必要に応じてFGFRアンタゴニストの存在下でES細胞を多能性状態に維持する工程。
【0031】
さらに、必要に応じて、細胞は、MEKインヒビター、GSK3インヒビターおよびgp130下流シグナリング経路のアクチベーターの存在下で多能性状態に維持され得る。
【0032】
本発明は、ES細胞のトランスフェクトされた集団を得る方法もまた提供し、該方法は:
選択マーカーをコードする構築物でES細胞をトランスフェクトする工程;
該ES細胞をプレートする工程;
MEKインヒビター、GSK3インヒビターおよび必要に応じてFGFRアンタゴニストの存在下で該ES細胞を培養する工程;および
該選択マーカーを発現する細胞を選択する工程、
を含む。
【0033】
さらに、必要に応じて、細胞は、MEKインヒビター、GSK3インヒビターおよびgp130下流シグナリング経路のアクチベーターの存在下で培養される。
【0034】
選択マーカーは、例えばEP−A−0695351に記載のように抗生物質耐性、細胞表面マーカーまたは他の選択マーカーをコードし得、そして好ましくは所望の細胞で選択マーカーを優先的に発現するプロモーターに作動可能に連結した選択マーカーをコードするヌクレオチド配列を含む。
【0035】
さらなる実施態様において、本発明は、多能性(特にES)細胞の培養方法を提供し、該方法は、培養容器(例えばプレート上の個々のウェル)へ個々の細胞を移す工程、および、GSK3インヒビター、MEKインヒビターおよび必要に応じてFGFRアンタゴニストの存在下で該細胞を培養する工程を含み、それにより、すべての細胞が単一細胞の子孫である多能性(特にES)細胞のクローン集団を得る。必要に応じて、細胞はまた、gp130下流シグナリング経路のアクチベーターの存在下で培養され得る。
【0036】
一旦、ES細胞の安定で均一な培養物が得られると、培養条件は、外胚葉、中胚葉または内胚葉の細胞の運命から選択される1以上の細胞タイプへ細胞の分化を方向づけるように変更され得る。サイトカインおよびシグナリング因子の添加または除去は、特定の分化した細胞集団の高い効率での誘導を可能にし得る。非神経外胚葉の運命へのES細胞の分化は、gp130を介して作用するサイトカイン、MEKインヒビターおよびGSK3インヒビターの存在下でES細胞を維持すること、ならびに次いで、サイトカインを除去する一方、GSK3インヒビターおよびMEKインヒビターを維持すること、および/または分化を方向づけ得るさらなるシグナリング分子を添加することによって達成され得る。あるいは、細胞は、MEKインヒビターおよびGSK3インヒビターの存在下で維持され得、次いで、該インヒビターの1つまたは両方を除去すること、および/または分化を方向づけ得るシグナリング分子を添加することによって分化を方向づけ得る。上記の方法はすべて、必要に応じて、プロセスの産物である分化した細胞を得る工程および/または単離する工程を含む。
【0037】
本発明のさらなる局面は、細胞培養培地を提供する。ある培地は、多能性(特にES)細胞の自己再生のためであり、該培地は、GSK3のインヒビター、MEKのインヒビターおよび必要に応じてFGFRアンタゴニストを含む。該培地は、必要に応じてgp130下流シグナリング経路のアクチベーターもまた含み得る。本発明の別の培地は、幹細胞培養培地であり、GSK3のインヒビター、MEKインヒビターおよび必要に応じてFGFRアンタゴニストを含む。すべての培地は、好ましくはさらに基本培地を含む。いくつかの好ましい実施態様では、すべての培地はgp130のアゴニストを含まず、したがって、好ましくはLIFを含まない。
【0038】
本発明は、血清および血清抽出物を含まない培地を提供する。1つのこのような培地は:
基本培地;
MEKインヒビター;
GSK3インヒビター;および
鉄トランスポーター;
を含み、ここで、該培地は、必要に応じて、血清および血清抽出物を含まない。
【0039】
培地は、好ましくはFGFRアンタゴニストもまた含む。培地は、必要に応じてgp130下流シグナリング経路のアクチベーターもまた含み得る。
【0040】
多能性幹細胞(特にラットまたはマウス細胞)に好適な培地は、血清およびgp130アゴニストを含まなくてよく、そしてMEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニストを含む。培地構成成分の置換は、本明細書に記載のように行われ得る。
【0041】
基本培地は、細胞のために炭素および/またはビタミンおよび/またはミネラルの必須供給源を供給する培地である。基本培地は、一般的にタンパク質を含まず、そして細胞の自己再生をそれ自体で支持し得ない。鉄トランスポーターは、鉄の供給源を提供し、または培養培地から鉄を取り込む能力を提供する。適切な鉄トランスポーターとしては、トランスフェリンおよびアポトランスフェリンが挙げられる。培地は、1以上のインスリンまたはインスリン様増殖因子、およびアルブミン(好ましくは組換え体)またはアルブミン代替物をさらに含み、そしてフィーダー細胞およびフィーダー細胞抽出物を含まないことが好ましい。培地は、アポトーシスのインヒビターまたは培養物中の多能性細胞の維持を促進する任意の他の構成成分もまた含み得る。
【0042】
本発明の特定の培地は、さらなる基本培地を含んでまたは含まずに、MEKインヒビター、GSK3インヒビター、インスリン、アルブミンおよびトランスフェリンを含む。この培地では、LIFが、必要に応じて含まれ得、そしてgp130シグナリングの他のアクチベーターによって置換され得る。しかし、好ましい培地は、gp130レセプター結合サイトカインであるLIFを含み、その適切な濃度は、一般的には10U/mlと1000U/mlとの間、より好ましくは50U/mlと500U/mlとの間、さらにより好ましくは100U/mlの範囲にある。GSK3およびMEKインヒビターは、好ましくは本明細書により詳細に記載の通りである。
【0043】
本発明は、胚盤胞から多能性細胞を誘導する方法をさらに提供し、該方法は:
(1)胚盤胞を得る工程;
(2)MEKインヒビター、GSK3インヒビターおよび必要に応じてFGFレセプターのアンタゴニストの存在下で該胚盤胞を培養して、内部細胞塊を得る工程;
(3)該内部細胞塊を解離する工程;
(4)該解離した内部細胞塊から1または複数の細胞を単離する工程;および
(5)該単離した1または複数の細胞を、MEKインヒビター、GSK3インヒビターおよび必要に応じてFGFレセプターのアンタゴニストの存在下で培養する工程、
を含む。
【0044】
必要に応じて、該単離した1または複数の細胞は、MEKインヒビター、GSK3インヒビターおよびgp130下流シグナリングのアクチベーターの存在下で培養される。FGFレセプターのアンタゴニストもまた存在させ得る。
【0045】
好ましくは、前記方法は、胚盤胞をLIF中で、より好ましくは2〜4日の期間に亘って培養する工程を含む。単離した1または複数の細胞は、好ましくは無血清培地中で培養される。代表的には、これらの細胞は、凝集塊として再プレートされる。胚盤胞はまた、好ましくは、無血清培地中で、必要に応じてBMPレセプターのアゴニストの不在下で、培養される。
【0046】
本発明によれば、細胞の培養は、接着培養で行われることがさらに好ましく、これは培養基材上に細胞接着タンパク質を含むことによって促進され得る。本発明によれば、単層培養で多能性細胞を培養することもまた好ましいが、必要に応じて、細胞を懸濁培養でまたはプレ細胞凝集塊として増殖させる;細胞をビーズ上であるいは他の適切な足場(例えば膜または他の3次元構造物)上でもまた増殖させ得る。
【0047】
本発明の実施例において用いられる培養培地は、好ましくは血清アルブミンもまた含む。これは、精製された形態または好ましくは組換え体の形態で用いられ得、そして組換え体の形態の場合には、これはサイトカインなどの可能性のある混入因子の不在という利点を有する。培養培地は、血清アルブミンを含む必要はなく、そしてこの構成成分は除外され得、あるいは別のバルクタンパク質またはWilesらに記載の合成ポリマー(ポリビニルアルコール)に置換され得る。
【0048】
本発明の特に好ましい培地は、完全に限定されるものである。この培地は、不確定のいかなる構成成分、すなわち、その内容物が知られていない構成成分または特定されない不確定因子または変動因子を含み得る構成成分も含まない。完全限定培地を用いることの利点は、多能性細胞の培養および引き続く操作のための効率的および一貫したプロトコールが誘導され得ることである。さらに、細胞を多能性状態に維持することが、より高い効率およびより大きい予測性で達成可能であること、および限定培地を用いて培養される細胞で分化が誘導される場合、分化シグナルへの応答が、非限定培地が用いられる場合より均一であることがわかる。
【0049】
本発明はまた、培養培地の調製を目的とする、培養培地の添加物として用いられ得る濃縮物、および構成成分のキットを提供し、得られる培地は、本発明と一致する。本発明の1つのキットは、第1の容器および第2の容器を含み、第1の容器はMEKインヒビターを含み、そして第2の容器はGSK3インヒビターを含む。好ましくは、キットは、FGFレセプターのアンタゴニストを含む第3の容器を含む。キットはまた、必要に応じて、gp130下流シグナリングのアクチベーターを含むさらなる容器を含み得る。キットは、好ましくは、各容器の内容物が本発明の培養培地を得るように培養培地に添加され得るように処方される。キットは、好ましくは、それらのそれぞれの構成成分の濃縮されたストック溶液を含む。
【0050】
本発明の方法はまた、分化した細胞を得る方法を含み、該方法は、記載のように多能性細胞を培養する工程、および細胞が分化することを可能にするまたは引き起こす工程を含む。ここで、細胞は、多能性幹細胞を含む他の細胞タイプと比較して所望の分化した細胞で差次的発現が可能である選択マーカーを含み、これによって、選択マーカーの差次的発現は、所望の分化した細胞の優先的な単離および/または生存および/または分裂をもたらす。選択マーカーは、所望の分化した細胞で発現し得るが他の細胞タイプでは発現しないか、あるいは所望の分化した細胞と他の細胞タイプとで発現レベルが異なり得、それによって選択マーカーの発現についての選択を可能にする。分化した細胞は、組織幹細胞または組織前駆細胞であり得、そして最終分化した細胞であり得る。
【0051】
一般的に、本発明はまた、本明細書に記載の本発明の方法のいずれかに従うことにより得られる細胞にも及ぶ。本発明の細胞は、創薬のためのアッセイに用いられ得る。本発明の細胞はまた、細胞治療のために用いられ得、したがって、本発明の方法は、MEKの阻害、GSK3の阻害および必要に応じてFGFシグナリングの拮抗の組み合わせを用いて多能性細胞を誘導および/または維持する工程、そこから細胞治療のための細胞を誘導する工程および細胞治療でそれらの細胞を用いる工程を含む。必要に応じて、この組み合わせは、gp130下流シグナリングのアクチベーターの不在下で用いられる。
【0052】
本発明のさらなる局面は、多能性細胞の自己再生を促進するための、MEKおよびFGFレセプターの阻害の使用、必要に応じてGSK3の阻害との組み合わせでの使用に関する。発明者らは、MEKインヒビターとFGFレセプターのアンタゴニストとの組み合わせが、添加サイトカインまたは増殖因子の不在下で無血清培地中で多能性細胞の増殖を支持する場合に効果的であることを見出した。
【0053】
したがって、本発明のさらなる局面は、MEKインヒビターおよびFGFレセプターのアンタゴニストを含む培養培地を提供する。MEKインヒビターおよびFGFレセプターアンタゴニストは、本発明の他の局面に関連して記載される通りである。同様に、培養培地は、本発明の他の局面に関連して本明細書に記載されるさらなる構成成分または因子をさらに含み得る。
【0054】
本発明のさらに他の局面は、多能性細胞のための培養培地の製造におけるMEKインヒビターおよびFGFレセプターのアンタゴニストの使用を提供する。
【0055】
本発明は、多能性細胞を培養する方法、および多能性細胞のトランスフェクトされた集団を得る方法もまた提供し、これらは、本発明の他の局面について記載のように好都合に実施され得る。したがって、本発明のさらなる局面は、自己再生を促進するような多能性細胞の培養方法を提供し、該方法は、MEKインヒビターおよびFGFレセプターのアンタゴニストを含む培地中で細胞を維持する工程を含む。
【0056】
本発明の関連する局面は、多能性細胞の培養方法を提供し、該方法は以下の工程を含む:
必要に応じてフィーダー上で、培養物中でES細胞を多能性状態に維持する工程;
少なくとも1回該ES細胞を継代する工程;
培地から(存在する場合)血清または血清抽出物を除去しそして(存在する場合)該フィーダーを除去して、該培地がフィーダー、血清および血清抽出物を含まないようにする工程;および
続いて、MEKインヒビターおよびFGFレセプターのインヒビターの存在下でES細胞を多能性状態に維持する工程。
【0057】
本発明のさらなる局面は、ES細胞のトランスフェクトされた集団を得る方法を提供し、該方法は:
選択マーカーをコードする構築物でES細胞をトランスフェクトする工程;
該ES細胞をプレートする工程;
MEKインヒビターおよびFGFレセプターアンタゴニストの存在下で該ES細胞を培養する工程、および
該選択マーカーを発現する細胞を選択する工程、
を含む。
【0058】
また、血清および血清抽出物を含まない細胞培養培地が提供され、該培地は:
基本培地;
MEKインヒビター;
FGFレセプターのアンタゴニスト;および
鉄トランスポーター、
を含む。
【0059】
MEKインヒビターとFGFレセプターのアンタゴニストとの組み合わせは、新たな多能性細胞系の誘導にもまた有用である。したがって、本発明のさらなる局面は、胚盤胞から多能性細胞を誘導する方法を提供し、該方法は:
(1)胚盤胞を得る工程;
(2)MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下で該胚盤胞を培養して、内部細胞塊を得る工程;
(3)該内部細胞塊を解離する工程;
(4)該解離した内部細胞塊から1または複数の細胞を単離する工程;および
(5)該単離した1または複数の細胞を、MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下で培養する工程、
を含む。
【0060】
本発明はまた、第1の容器および第2の容器を含むキットを含み、第1の容器はMEKインヒビターを含み、そして第2の容器はFGFレセプターのアンタゴニストを含む。キットはまた、本明細書に記載される他の容器および/または構成成分を含み得る。
【0061】
本発明のさらなる局面は、多能性幹細胞、特にNanogを発現する多能性幹細胞の自己再生の促進において、MEKインヒビターおよびFGFレセプターのアンタゴニストの使用を提供する。関連する局面は、幹細胞集団を増殖させる方法を提供し、該方法は、MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下で幹細胞を培養する工程を含む。
【0062】
本発明の多くの利点は、上記の通りでありまたは明らかである。比較的無毒性および細胞透過性である細胞培養構成成分が同定され得る。本発明の特定の実施態様において用いられるMEKインヒビター、GSK3インヒビターおよびFGFRアンタゴニストは、特に、例えばタンパク質サイトカインの精製と比較して、容易に精製され得る。組換えタンパク質は、作成が高価であり得、そして小分子の培地構成成分は、より安価に生産され、かつ貯蔵がより安定であり得、より広い有効濃度範囲を伴い得る。
【0063】
以下に記載の特定の実施態様は、無血清の完全限定培地でCHIR99021、PD184352および必要に応じてSU5402の組み合わせを用い、そしてマウスES細胞の自己再生の改善を示し、ほとんど分化を示さなかった。BMPの存在下でES細胞を培養する場合、いくつかの神経発生があることが時々報告される。これは、本発明の実施例においては見られなかった。
【0064】
本発明は、ここで、特定の実施例においてさらに説明され、図によって示される。
【0065】
図1は、多能性ES−NSハイブリッドコロニーの形成におけるPD184352の効果の分析結果を示す。(A−C)RH×NS TGFP融合の赤色蛍光および緑色蛍光についてのFACS分析。(A)PEG処理の24時間後の融合混合物;(B)AでゲートをかけたFACS分類されたハイブリッドの純度チェック。(C)Aで分類されたハイブリッドをプレートして形成されたコロニーをプレートしたハイブリッドあたりのコロニーの比率(%)としてスコア付けた。これらのスコアは、FACS分類された細胞の純度を考慮する。(D)データの概要。(E)ハイブリッドコロニー形態の例。
【0066】
図2は、本発明にしたがって誘導および維持されたマウスのES細胞、ならびにこれらのES細胞による高効率なキメラ発生を示す。
【0067】
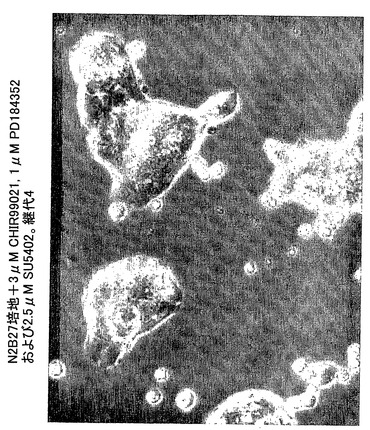
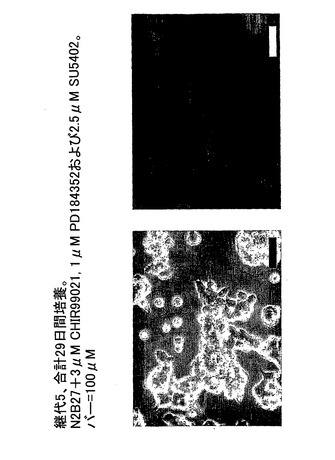
図3は、本発明にしたがって増殖した4継代マウスES細胞を示す。
【0068】
図4は、本発明にしたがって増殖したマウスES細胞がOct4陽性であることを示す。
【0069】
実施例において、用語2i培地または2iは、MEKインヒビターおよびFGFレセプターのアンタゴニストを含む培地を示すために用いられる。用語3i培地または3iは、MEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニストを含む培地を示すために用いられる。
【実施例】
【0070】
GSK−3βインヒビター、MEKインヒビター、培養培地、およびES細胞の自己再生
マウスおよびヒトES細胞を、特に明記のない限りN2B27培地を用いて、そしてGSK−3βインヒビターであるCHIR99021、AR−AO144−18、SB216763およびSB415286、ならびにMEKインヒビターであるPD184352の存在下または不在下で、種々の条件下で増殖させた。
【0071】
N2B27培地の調製:
N2 100×ストック溶液。10mlについて:1mlのインスリン(最終濃度2.5mg/ml)を1mlのアポトランスフェリン(最終濃度10mg/ml)、0.67mlのBSA(最終濃度5mg/ml)、33μlのプロゲステロン(最終濃度2μg/ml)、100μlのプトレシン(最終濃度1.6mg/ml)、10μlの亜セレン酸ナトリウム(最終濃度3μM)および7.187mlのDMEM/F12と混合する。4℃で貯蔵し、そして1カ月以内に使用する。
【0072】
DMEM/F12−N2培地:100mlのDMEM/F12に対して、1mlのN2 100×ストック溶液を添加する。DMEM/F12培地中のN2の各構成成分の最終濃度は:インスリン、25μg/ml;アポトランスフェリン、100μg/ml;プロゲステロン、6ng/ml;プトレシン、16μg/ml;亜セレン酸ナトリウム、30nM;BSA 50μg/mlである。4℃で貯蔵し、そして1カ月以内に使用する。
【0073】
Neurobasal/B27培地:100mlのNeurobasalTM培地に対して、2mlのB27および0.5〜1mlの200mM L−グルタミンを添加する。4℃で貯蔵し、そして1カ月以内に使用する。
【0074】
N2B27培地:DMEM/F12−N2培地をNeurobasal/B27培地と1:1の比で混合する。0.1Mストックから0.1mMの最終濃度になるまでβ−メルカプトエタノールを添加する。4℃で貯蔵し、そして1カ月以内に使用する。
【0075】
実施例1
無血清培地中では、MEKインヒビター+GSK−3βインヒビターは、(1)N2B27培地、および(2)完全限定培地(DMEM/F12−N2)の両方で、マウスES細胞の自己再生を維持するのに十分であった(データは示さず)。ES細胞の自己再生は、MEKインヒビター、GSK−3βインヒビターおよびLIFを含む培地中でさらに改善された(データは示さず)。
【0076】
実施例2
PD184352(MEKのインヒビター)がES細胞中のNanogのレベルを増加させることが示された(データは示さず)。さらに、PD184352で処理されたNanog−/−ES細胞がES細胞の自己再生の増強を示さなくなることが示された。実際、これらの細胞は分化した(データは示さず)。これにより、PD184352によるES自己再生表現型の増強がNanogによって仲介されることが立証された。
【0077】
再プログラミングにおけるPD184352の効果を、細胞融合の前後におけるNS細胞の多分化能への変換を決定することによってもまた研究した。
【0078】
dsRed蛍光タンパク質およびハイグロマイシン耐性を構成的に発現するRH ES細胞を、IRESを経てピューロマイシン耐性に連結した融合タンパク質TauGFPを発現する胎児由来神経幹細胞(NS TGFP)に融合した。融合の一方には、RH細胞を、融合の前後3日間、3μMのPD184352で処理した。コントロールには、PD184352を添加しなかった。融合24時間後、処理した初代ハイブリッドおよび未処理の初代ハイブリッドを分類し、次いでプレートした(図1A−C)。3日後、ハイグロマイシンおよびピューロマイシン選択をES培地に添加した。dsRed2およびGFP蛍光の発現およびES細胞形態の提示についてコロニーのスコアを付けた(図1Dおよび図1E)。結果は、PD184352がES−NSハイブリッドコロニーの形成を45倍増強することを示した。興味深いことに、PD184352で処理したRH細胞中のプレートしたハイブリッドあたりのハイブリッドコロニー形成の比率(%)は、Nanogを過剰発現させているES細胞と比較してちょうど2倍低かった(2.25%対4%)。この結果は、PD184352がES細胞の自己再生を増強するだけでなく、細胞融合前後における再プログラミングもまた増強することを示している。この効果は、処理したRH細胞におけるNanogのレベルの増加によって仲介されていると思われる。したがって、Nanogが内生的に発現する場合、MEKインヒビターがNanogをアップレギュレートするために用いられ得、そして再プログラミングの増強などの付随効果を達成し得る。
【0079】
実施例3
ヒトES細胞を、GSK−3インヒビターであるCHIR99021およびMEKインヒビターであるPD184352を追加した培地で培養した。
【0080】
培養培地へのLIFの添加は、細胞の増殖をさらに改善した(データは示さず)。
【0081】
実施例4
マウスES細胞を、GSK−3インヒビターであるCHIR99021およびMEKインヒビターであるPD184352を追加した培地で培養した。
【0082】
培養培地へのLIFの添加は、細胞の増殖をさらに改善した(データは示さず)。
【0083】
実施例5
マウスおよびヒトES細胞を、CHIR99021、PD184352およびSU5402を含む培地で増殖させ、該培地は以下のように調製した:
【0084】
【表1】

【0085】
培地
DMEM/F12−N2培地の調製
100mlのDMEM/F12(Gibco 42400-010)に対して、1mlのN2 100×ストック溶液を添加する。DMEM/F12培地中のN2の各構成成分の最終濃度は:
インスリン 25μg/ml
プトレシン 16μg/ml
トランスフェリン 100μg/ml
亜セレン酸ナトリウム 30nM
プロゲステロン 6ng/ml
BSA 50μg/ml
である。
【0086】
Neurobasal/B27の調製:
100mlのNeurobasal培地(Gibco 21103-049)に対して、2mlのB27(Gibco 17504-044)および1〜2M L−グルタミン(TC保存 1:100)を添加する。
【0087】
N2B27培地の調製:
DMEM/F12−N2培地をNeurobasal/B27培地と1:1の比で混合する。この培地を、すべての化合物の希釈および細胞の増殖に用いた。
【0088】
この培地は、ヒトES細胞の維持、ならびに129系統マウス由来のES細胞の誘導および維持に用い、そして非許容的なマウス系統であるCBAおよびC56/BL6由来のES細胞の誘導にもまた用いた。
【0089】
実施例6
マウスES細胞を、FGFレセプターのインヒビターおよびMEKインヒビターの存在下で培養した。選択的薬理学的インヒビターSU5402およびPD184352を、それぞれFGFレセプターチロシンキナーゼの阻害およびMEK1/2を経たErk1/2の活性化の阻害のために用いた。発明者らは、どちらのインヒビターの添加も、BMP4を供給することなく、LIFを含むN2B27培地中で強固にES細胞を増殖させるに十分であることを見出した(データは示さず)。未分化の培養物は、これらの条件において、多能性マーカーOct4、NanogおよびRex1の発現を保持しながら継続的に継代され得る。LIF+BMPで維持された培養物中よりもId遺伝子の発現がはるかに低いにも関わらず、神経への運命づけは生じていない。
【0090】
LIFを添加しないN2B27培地にプレートされたES細胞(通常、効率的な神経への運命づけを引き出す条件)は、SU5402またはPD184352のいずれが添加された場合も、数日間に亘ってOct4陽性およびSox1陰性のままである(データは示さず)。しかし、これらの細胞は、継代した後、変わらず分化および/または死滅する。毒性副作用の可能性を低減するために、発明者らは、2.5倍低い用量を用いて、2つのインヒビターを同時に併用した。0.8μMのPD184352+2μMのSU5402を含むN2B27中では、最初にいくらかの分化が観察されるが、継代後はES細胞が生き残って増殖する(データは示さず)。この2インヒビター(2i)条件下では、LIFの存在下よりも生存率はより低く、集団倍加時間はより遅いが、分化は効果的に抑制される。この発見は、ES細胞の自己再生のための最少要件が、細胞増殖および生存率を損なわせることを避けながら、FGFレセプターおよびErkシグナリングから生じる分化シグナルをそらすことであり得る旨示唆している。
【0091】
実施例7
発明者らは、2i培地中におけるES細胞の増殖の減少は、pErkの下流のRskによる阻害的リン酸化の解放の結果生じるグリコーゲン合成酵素キナーゼ3(GSK−3)活性の増加によるものであり得ると結論づけた。CHIR99021は、GSK−3の十分に特徴づけられた高選択性小分子インヒビターであり、これはGSK−3活性を完全にブロックする濃度でサイクリン依存性キナーゼ(CDK)と交差反応しない。発明者らは、血清存在下の培養物に添加した場合、CHIR99021(3μM)は、LIFの存在下であっても、実際に分化を促進することを見出した。無血清のN2B27培地中では、分化応答は減少し、そしていくつかのコロニーは、数日間、形態的に未分化であるように見える。しかし、継代した後、未分化細胞は生き残らない。同様の結果が、広く用いられる他の2つのGSK−3インヒビターであるSB216763およびSB415286でも、(どちらもES細胞にいくぶん有毒のようであったが)得られた。
【0092】
しかしながら、CHIR99021を2iと併用した場合、分化応答は完全に失われる。さらに、CHIR99021は、ES細胞がLIF+血清/BMP中または2i中で代表的に見られる平坦な単層よりもむしろ密集した3次元のコロニーとして増殖するように、2iへの応答を変化させた。3つのインヒビター(3i)中での分化は無視できる程度であり、そしてES細胞は急速に増殖した。最も重要なことに、継代した後、未分化コロニーが高効率で増殖した。2つの独立した親ES細胞系E14Tg2aおよびCGR8の派生体は、3i中で強固な長期増殖を示し、かつ明白な分化はほとんどあるいは全くなかった(データは示さず)。これらは、Oct4、NanogおよびRex1を発現し、系統運命づけマーカーであるGata4、Sox1またはbrachyuryの発現は、それほど示さない(データは示さず)。バルク培養では、ES細胞は、3i中でLIF+BMP中に匹敵する倍加率で増殖し、Oct4−GFP陽性の未分化細胞の比率は、依然として90%を超える。
【0093】
したがって、3i培地は、血清または添加サイトカインの不在下で、分化することなくES細胞を培養するために用いられ得る。
【0094】
実施例8
培養処方物が十分にES細胞の自己再生を持続させるかに関する厳密な試験は、個々の細胞による未分化コロニーの形成である。単一細胞の堆積の後、N2B27+3i中のコロニー形成率は25%(98/384)であり、LIF+BMP(11%、23/192)よりも高い(データは示さず)。これらのコロニーはOct4−GFPを発現し、そして未分化ES細胞として継代可能である。したがって、MEKインヒビター、FGFレセプターのインヒビターおよびGSK3インヒビターを含む培地は、単一細胞に由来する未分化ES細胞コロニーの形成を持続させることが可能である。
【0095】
実施例9
発明者らは、3iが、胚から直接新たなES細胞を誘導する十分な能力を有したのか、あるいは確立した系の適応を反映したのかを検討した。許容的な129系統からの胚盤胞を、ゼラチンコートされたプラスチック上のN2B27+3i中に直接プレートし、5日間培養した。続く内部細胞塊の解離および再プレートの後、12個の胚のうち7個からES細胞コロニーが得られた。これらのうち3個を増殖させ、胚盤胞に注入した。すべてにおいて、キメリズムおよび生殖系列遺伝が高い比率で得られた(表2)。続いて、発明者らは、C57BL/6および非許容的なCBAおよびMF1系統から複数のES細胞を誘導し、3iが胚盤葉上層細胞からES細胞への移行を容易にすることを示した。発明者らは、3iが、発生能を選択または損失させることなく、外因性のLIFおよびBMP/血清の要求からES細胞を解放すると結論付ける。
【0096】
【表2】
【0097】
したがって、ES細胞は、GSK3インヒビターおよびMEKインヒビターの組み合わせ、MEKインヒビターおよびFGFレセプターのアンタゴニストの組み合わせ、または必要に応じてGSK3インヒビター、MEKインヒビターおよびFGFレセプターのアンタゴニストの組み合わせで維持され、そして本発明はまた、培養方法およびそのための培地を提供する。
【技術分野】
【0001】
本発明は、多能性幹細胞における自己再生表現型の維持に関する。提供される方法および組成物は、多能性幹細胞、例えば、胚性幹(ES)細胞、特に、ラット、マウス、ウシ、ヒツジ、ブタおよびヒトを含む哺乳類の幹細胞の培養および単離に適する。特に、本発明は、ラット、マウスおよびヒトの多能性細胞の自己再生培養物、ならびにそのための方法および組成物に関する。
【背景技術】
【0002】
血清および白血病抑制因子(LIF)を含む培地の存在下でのインビトロ多能性幹細胞培養物の樹立および維持は、周知である(Smithら、(1998)Nature 336: 688-90)。このような方法は、「許容的」なマウスの系統から多くの継代に亘って多能性胚性幹(ES)細胞を維持するために用いられてきた。多能性幹細胞培養物の維持および自己再生は、フィーダー細胞またはその抽出物(通常はマウス線維芽細胞)の存在下で幹細胞が培養される場合にさらに支持される。このような条件下では、培養物中で多くの継代に亘ってヒトES細胞を多能性状態に維持し得る。
【0003】
多くの場合において、ES細胞は、血清または血清抽出物を含む(したがって、非限定である)培地を用いる、またはヒトES細胞を維持するために用いられる線維芽細胞フィーダー細胞のような他の細胞の存在を必要とする細胞培養条件を用いる場合のみ、維持され得るか、あるいは最もよく維持される。しかし、何らかの不確定の構成成分は、培地中のものであろうとまたは例えばフィーダー細胞によって産生されるものであろうと、ES細胞の増殖および分化についての研究を潜在的に妨害し、または邪魔する。これは、ES細胞およびその子孫の治療および他の適用のための良好な製造実施の発展を妨げる。いくつかの限定されたES細胞培地が知られているが、代替のおよび好ましくは改良された限定培地が必要とされる。
【0004】
本出願人による先の出願であるWO−A−03/095628およびより後の未公開出願において、(1)gp130のアゴニスト(例えば、LIF)および(2)TGF−βスーパーファミリーのアゴニスト(例えば、BMP4)またはIdシグナリング経路のアゴニストを含む無血清培地中でES細胞のような多能性幹細胞を培養することは、複数の継代に亘って幹細胞の自己再生を促進するために用いられる。gp130シグナリングの存在下では、TGF−βスーパーファミリーまたはIdシグナリング経路のアゴニストは、驚くべきことに、分化前のシグナルよりもむしろ自己再生の刺激を提供した。それにもかかわらず、多能性細胞を自己再生状態に維持する効率、およびフィーダー細胞から離してまたはフィーダー条件培地から離して多能性細胞を移すための培地は、常に改良されることが望まれる。
【0005】
Sato Nら、Nat. Med. 2004年1月 10(1) 55-63頁は、血清を含む培地中でのマウスおよびヒトES細胞に対する、グリコーゲン合成酵素キナーゼ3(GSK3)インヒビターである6−ブロモインディルビン−3’−オキシムの効果を記載している。しかし、これらの効果は非常に短い時間枠に亘ってのみ観察され、あまりにも短いため確かな結論を引き出し得なかった。そして、その研究に用いられた非限定培地中の未知の因子の影響は重要であり得る。本発明の発明者らは、その結果を反復することを試みたがこれに失敗し、そして実際にはそれらの記載と反対の効果を見出した。
【0006】
ES細胞の培養培地の調製のためには、個々の培地構成成分をできる限り純粋な形態で提供することが望まれる。しかし、ほとんどの培地構成成分はサイトカインであり、その純度は、それらを細胞系で製造し、次に生産ブロスから可能性のある汚染物質を除去する必要によって損なわれる。いくつかのサイトカインに伴う別の問題は、それらが効果的および無毒性である濃度範囲が狭いことである。より広い範囲の濃度を有しおよび/またはより高い濃度でより毒性が低い培地構成成分は、非常に有用である。サイトカインはまた、貯蔵において安定性が限定され得、そしてより安定な培地構成成分が求められる。
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の目的は、当該技術分野における問題を克服しまたは少なくとも改善すること、好ましくは代替の、より好ましくは改良された、多くの継代に亘って該幹細胞の自己再生を支持し得る、多能性幹細胞に適した培養方法および培養培地を提供することである。本発明のさらなる目的は、細胞の分化が制御される様式で誘導され得るまで多能性幹細胞培養物のインビトロでの維持を可能にする代替の培養系を提供することである。本発明のよりさらなる目的は、多能性幹細胞の誘導および単離を増強し、かつES細胞の単離に対して抵抗性の生物からまたは多能性幹細胞がまだ単離されていない生物からのそれらの誘導および単離を促進する方法および組成物を提供することである。
【課題を解決するための手段】
【0008】
本発明によれば、ES細胞のような多能性幹細胞は、MEKインヒビターおよびGSK3インヒビターを含む、またはMEKインヒビターおよびFGFレセプターのアンタゴニストを含む、(好ましくは無血清の)培地中で培養される。好ましくは、培地は、MEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニスト(例えば、小分子GSK3インヒビターおよび小分子MEKインヒビターおよび小分子FGFRアンタゴニスト)を含む。それによって、幹細胞の複数の継代に亘る自己再生が促進される。したがって、多能性細胞でのGSK3およびMEKの阻害、MEKおよびFGFレセプターシグナリングの阻害、またはGSK3、MEKおよびFGFレセプターシグナリングの阻害は、自己再生の刺激を提供する。
【0009】
本発明は、多くの適用を有する。GSK3およびMEKの阻害の組み合わせ、MEKおよびFGFRの阻害の組み合わせ、またはGSK3、MEKおよびFGFRの阻害の組み合わせが、多能性細胞、特にES細胞を増殖させるため、およびそれらがフィーダー上で誘導または増殖されていた場合、多能性細胞、特にES細胞を、フィーダー細胞またはフィーダー細胞の層(しばしばフィーダーまたはフィーダー細胞として言及される)を伴わずに増殖するように適応させるために用いられ得る。培養物中で幹細胞を増殖させる方法は、GSK3インヒビターおよびMEKインヒビターの存在下、MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下、または好ましくはGSK3インヒビター、MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下で細胞を培養する工程を含む。1以上のGSK3インヒビターおよびMEKインヒビター、1以上のMEKインヒビターおよびFGFRアンタゴニスト、ならびに必要に応じて1以上のMEKインヒビター、GSK3インヒビターおよびFGFRアンタゴニストを含む培養培地が調製され得る。ES細胞は、GSK3インヒビターおよびMEKインヒビターを用いて、MEKインヒビターおよびFGFRアンタゴニストを用いて、またはGSK3インヒビター、MEKインヒビターおよびFGFRアンタゴニストを用いて誘導され得る。
【0010】
本発明の第1の局面によれば、多能性細胞でのGSK3およびMEKの阻害、好ましくはGSK3およびMEKおよびFGFレセプターすべての阻害が、細胞の自己再生を促進するために用いられる。
【図面の簡単な説明】
【0011】
【図1A−C】多能性ES−NSハイブリッドコロニーの形成におけるPD184352の効果の分析に関し、RH×NS TGFP融合の赤色蛍光および緑色蛍光についてのFACS分析:(A)PEG処理の24時間後の融合混合物;(B)AでゲートをかけたFACS分類されたハイブリッドの純度チェック;(C)Aで分類されたハイブリッドをプレートして形成されたコロニーをプレートしたハイブリッドあたりのコロニーの比率(%)を示す。
【図1D】多能性ES−NSハイブリッドコロニーの形成におけるPD184352の効果の分析のデータの概要を示す。
【図1E】多能性ES−NSハイブリッドコロニーの形成におけるPD184352の効果の分析のハイブリッドコロニー形態の例を示す。
【図2】本発明にしたがって誘導および維持されたマウスのES細胞、ならびにこれらのES細胞による高効率なキメラ発生を示す。
【図3】本発明にしたがって増殖した4継代マウスES細胞を示す。
【図4】本発明にしたがって増殖したマウスES細胞がOct4陽性であることを示す。
【発明を実施するための形態】
【0012】
多能性細胞とは、胚性幹(ES)細胞と呼ばれるものを含むが、これに限定されない。多能性細胞(ES細胞を含む)に特有の性質としては、発生の多能性段階に関わる多重遺伝子の発現、供給源動物に存在するあらゆる組織タイプを代表する細胞に分化する能力、キメラを生じさせる能力、および、特にキメラの生殖系列を生じさせる能力が挙げられる。例えば、ES細胞のような真性多能性細胞は、多能性関連遺伝子Nanog、Oct4、FGF4、Sox−2およびアルカリフォスファターゼのすべてではないとしても多くを発現することが期待される。特に、Nanog、Oct4およびSox−2の発現は、細胞がES細胞である決定的な最初の徴候を提供すると広く考えられている。キメラにおける生殖系列遺伝および3つの一次胚葉(すなわち、内胚葉、中胚葉および外胚葉)のすべてから分化した細胞を含む奇形種または奇形癌を生じる能力もまた、細胞がES細胞であることの決定的な徴候として広く考えられている。
【0013】
GSK3阻害とは、1以上のGSK3酵素の阻害をいう。したがって、GSK3インヒビターはGSK3酵素ファミリーの1つのメンバー、いくつかのメンバー、またはすべてのメンバーを阻害し得る。GSK3酵素ファミリーは周知であり、GSK3−αおよびGSK3−βが挙げられるが、これらに限定されない。多くの変異型が記載されている(例えば、Schafferら; Gene 2003; 302(1-2): 73-81参照)。特定の実施態様ではGSK3−βが阻害される。GSK3−αインヒビターもまた適切であり、そして一般に本発明において使用されるインヒビターは両方ともを阻害する。広い範囲のGSK3インヒビターが公知である(例証として、インヒビターCHIR98014、CHIR99021、AR−AO144−18、TDZD−8、SB216763およびSB415286)。他のインヒビターが公知であり、そして本発明において有用である。さらに、GSK3−βの活性部位の構造は特徴づけられており、特異的なインヒビターおよび非特異的なインヒビターと相互作用する重要残基が同定されている(Bertrandら;J Mol Biol.2003;333(2):393-407)。この構造特性によって、さらなるGSKインヒビターが容易に同定され得る。
【0014】
ある実施態様のインヒビターは、GSK3−βおよびGSK3−αに特異的であり、実質的にerk2を阻害せず、そして実質的にcdc2を阻害しない。インヒビターは、IC50値の比として測定した場合、マウスerk2および/またはヒトcdc2よりもヒトGSK3に対して、好ましくは少なくとも100倍、より好ましくは少なくとも200倍、非常に好ましくは少なくとも400倍の選択性を有する。ここで、GSK3 IC50値とは、ヒトGSK3−βおよびGSK3−αについての平均値をいう。CHIR99021およびCHIR98014について良好な結果が得られ、これらは両方ともGSK3に特異的である。GSK3インヒビターの例は、Bennett Cら、J. Biol. Chem., 277巻, 34号, 2002年8月23日, 30998-31004頁およびRing DBら、Diabetes, 52巻, 2003年3月, 588-595頁に記載されている。CHIR99021の使用に適した濃度は、0.01〜100、好ましくは0.1〜20、より好ましくは0.3〜10μMの範囲にある。
【0015】
GSK3阻害はまた、RNA干渉(RNAi)を用いて好都合に達成し得る。代表的に、GSK3遺伝子のすべてまたは一部に対して相補的である2本鎖RNA分子が多能性細胞に導入され、したがって、GSK3をコードするmRNA分子の特異的な分解を促進する。この転写後機構は、目的のGSK3遺伝子の発現を低減または停止させることとなる。RNAiを用いてGSK3阻害を達成するための適切な技術およびプロトコールは公知である。
【0016】
本明細書中においてMEKインヒビターとは、一般的なMEKインヒビターをいう。したがって、MEKインヒビターとは、MEK1、MEK2およびMEK3を含むタンパク質キナーゼのMEKファミリーメンバーの任意のインヒビターをいう。また、MEK1インヒビター、MEK2インヒビターおよびMEK3インヒビターをいう。MEKインヒビターは、MEKキナーゼファミリーの1つのメンバー、いくつかのメンバーまたはすべてのメンバーを阻害し得る。適切なMEKインヒビターの例としては、当該技術分野で既に公知であり、MEK1インヒビターであるPD184352およびPD98059、MEK1およびMEK2のインヒビターであるU0126およびSL327、ならびに、Daviesら(2000)(Davies SP, Reddy H, Caivano M, Cohen P. Specificity and mechanism of action of some commonly used protein kinase inhibitors. Biochem J. 351, 95-105)に記載されたものが挙げられるが、これらに限定されない。特に、PD184352は、他の公知のMEKインヒビターと比較した場合に、高い特異性および効力を有することが見出されている。他のMEKインヒビターおよびMEKインヒビターのクラスは、Zhangら(2000)Bioorganic & Medicinal Chemistry Letters; 10:2825-2828に記載されている。
【0017】
MEKキナーゼの阻害はまた、RNA干渉(RNAi)を用いて好都合に達成し得る。代表的に、MEK遺伝子のすべてまたは一部に対して相補的である2本鎖RNA分子が多能性細胞に導入され、したがって、MEKをコードするmRNA分子の特異的な分解を促進する。この転写後機構は、目的のMEK遺伝子の発現を低減または停止させることとなる。RNAiを用いてMEK阻害を達成するための適切な技術およびプロトコールは公知である。
【0018】
キナーゼインヒビター(GSK3インヒビターおよびMEKインヒビターを含む)を同定するための多くのアッセイが、公知である。例えば、Daviesら(2000)には、ペプチド基質および放射性標識ATPの存在下でキナーゼをインキュベートするキナーゼアッセイが記載されている。キナーゼによる基質のリン酸化は、基質への標識の取り込みを生じる。各反応のアリコートを、リン酸セルロース紙上に固定化し、そしてリン酸中で洗浄し、遊離ATPを除去する。次いでインキュベーション後の基質の活性を測定し、そしてキナーゼ活性の指標が提供される。キナーゼインヒビター候補の存在下および不在下における相対的なキナーゼ活性は、そのようなアッセイを用いることで容易に決定し得る。Downeyら(1996)J Biol Chem.;271(35):21005-21011にもまた、キナーゼインヒビターの同定に用いられ得るキナーゼ活性についてのアッセイが記載されている。
【0019】
線維芽細胞増殖因子(FGF)レセプター(FGFR)のアンタゴニストとは、代表的にFGFR1および/またはFGFR2を阻害する、FGFレセプターのポリペプチドまたは小分子または他のアンタゴニストをいう。したがって、FGFレセプターアンタゴニストは、FGFレセプターファミリーの1つのメンバー、いくつかのメンバー、またはすべてのメンバーのアンタゴニストであり得、FGFR1、FGFR2、FGFR3およびFGFR4が挙げられるがこれらに限定されない。FGFレセプターファミリーのメンバーは、代表的に、3つの免疫グロブリン様ドメインを含み、そして酸性アミノ酸領域(酸性ボックス)を提示し、この領域は、FGFファミリーメンバーのFGFレセプターへの結合に関与し得る。ある場合には、2つの免疫グロブリン様ドメインのみを含む分子もまた、FGFレセプターとして機能し得る。多くのFGFRアンタゴニストが公知であり、SU5402およびPD173074が挙げられるがこれらに限定されない。多くのFGFRアンタゴニストが公知である(例えば、SU5402およびPD173074)。SU5402の適切な濃度はμMの範囲であり、例えば0.1〜20μM、好ましくは0.5〜10μM、特に1〜5μMの範囲である。発明者らは、PD173074がSU5402の代わりに用いられ得ること、そしてPD173074は約100倍低い濃度で十分に効果的であり、このことはFGFレセプターに対して、より高い親和性を有することと一致することを見出した。したがって、PD173074の適切な濃度は、1〜200nMの範囲、好ましくは5〜100nM、特に10〜50nMの範囲である。ドミナントネガティブ変異体FGFレセプターの導入遺伝子発現によってFGFレセプターシグナリングを阻害することもまた公知である。しかし、本発明の実施態様では、遺伝子導入ベースの拮抗ではなく、小分子アンタゴニストの使用が好ましい。
【0020】
FGFレセプターのアンタゴニストを同定するための適切なアッセイは公知である。例えば、FGFレセプター経由のシグナリングがレポーター遺伝子の発現を活性化する細胞系が、可能性のあるアンタゴニストの活性を評価するために用いられ得る。
【0021】
有利なことに、GSK3インヒビターと組み合わせた、および好ましくはFGFレセプターのアンタゴニストもまた組み合わせたMEKインヒビターの使用は、ES細胞の増殖を改善することが見出された。
【0022】
好ましい実施態様では、約0.1μMと約25μMとの間のMEKインヒビターが用いられる。さらに好ましくは、約0.1μMと約5μMとの間のMEKインヒビターが用いられ、より好ましくは、0.2μMから2μMまでである。
【0023】
本発明による特に好ましい培地は、0.8μMのPD184352、3μMのCHIR99021および/または3μMのSU5402を含む。特に好ましい培地は、0.8μMのPD184352、3μMのCHIR99021および3μMのSU5402を含み、好ましくはN2B27培地中である。SU5402の濃度は、異なる多能性細胞系に合うように最適化され得、代表的には1〜5μMの範囲(例えば、2μM)である。
【0024】
以下の実施例において、発明者らは、自己再生を促進するために、MEKインヒビターとともに、および特定の実施例ではFGFレセプターのアンタゴニストともともにGSK3インヒビターの存在下でマウスES細胞を培養した。他の特定の実施例において、培養物中でマウス多能性細胞の自己再生を促進する方法は、GSK3およびMEKを阻害する工程、またはGSK3、MEKおよびFGFレセプターを阻害する工程を含む。
【0025】
必要に応じて、gp130下流シグナリングの活性化は、GSK3およびMEKを阻害することによって自己再生の促進をさらに増強するためにもまた用いられ得る。gp130下流シグナリングの活性化分子は、時に、gp130アクチベーターまたはgp130アゴニストとも言われる。1以上のgp130下流シグナリング経路の活性化は、gp130を介して作用するサイトカイン、例えばLIFレセプターのサイトカインまたは他のアゴニストの使用によって達成され得る。gp130を介して作用し得、したがってgp130シグナル伝達を活性化し得るサイトカインとしては、LIF、毛様体神経栄養因子(CNTF)、カルジオトロフィン、オンコスタチンM、IL−6+sIL−6レセプター、ハイパーIL−6およびIL−11が挙げられるが、これらに限定されない。適切なサイトカインには、gp130に結合し得および/またはgp130を介してシグナリングを活性化し得る模倣物、融合タンパク質またはキメラが含まれる。血清存在下でgp130を介して作用するサイトカインの役割は十分に確立されているが、血清不在下で未分化細胞を維持するそれらのサイトカインの能力は限られる。
【0026】
本発明の利点は、GSK3インヒビター、MEKインヒビターおよび必要に応じてFGFレセプターのアンタゴニストの存在下では、多能性細胞が限定培地中で増殖し得ることである。GSK3インヒビター、MEKインヒビターおよびFGFレセプターのアンタゴニストの組み合わせを用いることによる特別な利点は、この培地がインスリン、N2B27またはgp130アゴニスト(例えば、LIF)などの他の増殖因子を含有する必要がないことである。したがって、本発明は、血清、血清抽出物、フィーダー細胞およびフィーダー細胞抽出物を含まない培地中でのES細胞の代替のおよび/または改良された培養を可能にする。
【0027】
マウス(Bradleyら、(1984) Nature 309: 255-56)、アメリカミンク(Mol Reprod Dev (1992) 12月; 33(4): 418-31)、ブタおよびヒツジ(J Reprod Fertil Suppl (1991); 43: 255-60)、ハムスター(Dev Biol (1988) 5月; 127(1): 224-7)およびウシ(Roux Arch Dev Biol (1992); 201: 134-141)を含む多くの哺乳類供給源から、目的の胚性幹細胞が報告されている。本明細書の特定の実施例は、マウスおよびヒトのES細胞を用い、そして一次派生物由来のラット細胞もまた用いる。本発明の方法および組成物が、したがって、霊長類(特にヒト)、齧歯類(特にマウスおよびラット)、および鳥類の多能性幹細胞、特にES細胞を含む他の哺乳類の多能性細胞培養物の培養への適応に適切であることが理解される。
【0028】
本発明の第2の局面は、自己再生を促進するための多能性細胞、特にES細胞の培養方法を提供し、該方法は:
(1)GSK3のインヒビター;および
(2)MEKのインヒビター、
を含む培地中で該細胞を維持する工程を含む。
【0029】
好ましくは、該方法は:
(1)GSK3のインヒビター;
(2)MEKのインヒビター;および
(3)FGFレセプターのアンタゴニスト、
を含む培地中で該細胞を維持する工程を含む。
【0030】
本発明の方法は、血清を含まないおよび血清抽出物を含まない培地中で、多能性細胞を増殖(ES細胞の増殖を含む)させるために一般的に用いられ得、これらの細胞は、血清または血清抽出物の存在下で予め継代されている。好ましくは、このような方法はまた、フィーダー細胞および/またはフィーダー細胞抽出物の不在下で行われる。例えば、以下の工程を含むES細胞の培養が行われ得る:
必要に応じてフィーダー上で、培養物中でES細胞を多能性状態に維持する工程;
少なくとも1回該ES細胞を継代する工程;
該培地から血清または血清抽出物を除去しそして(存在する場合)該フィーダーを除去して、該培地がフィーダー、血清および血清抽出物を含まないようにする工程;および
続いて、GSK3のインヒビター、MEKインヒビターおよび必要に応じてFGFRアンタゴニストの存在下でES細胞を多能性状態に維持する工程。
【0031】
さらに、必要に応じて、細胞は、MEKインヒビター、GSK3インヒビターおよびgp130下流シグナリング経路のアクチベーターの存在下で多能性状態に維持され得る。
【0032】
本発明は、ES細胞のトランスフェクトされた集団を得る方法もまた提供し、該方法は:
選択マーカーをコードする構築物でES細胞をトランスフェクトする工程;
該ES細胞をプレートする工程;
MEKインヒビター、GSK3インヒビターおよび必要に応じてFGFRアンタゴニストの存在下で該ES細胞を培養する工程;および
該選択マーカーを発現する細胞を選択する工程、
を含む。
【0033】
さらに、必要に応じて、細胞は、MEKインヒビター、GSK3インヒビターおよびgp130下流シグナリング経路のアクチベーターの存在下で培養される。
【0034】
選択マーカーは、例えばEP−A−0695351に記載のように抗生物質耐性、細胞表面マーカーまたは他の選択マーカーをコードし得、そして好ましくは所望の細胞で選択マーカーを優先的に発現するプロモーターに作動可能に連結した選択マーカーをコードするヌクレオチド配列を含む。
【0035】
さらなる実施態様において、本発明は、多能性(特にES)細胞の培養方法を提供し、該方法は、培養容器(例えばプレート上の個々のウェル)へ個々の細胞を移す工程、および、GSK3インヒビター、MEKインヒビターおよび必要に応じてFGFRアンタゴニストの存在下で該細胞を培養する工程を含み、それにより、すべての細胞が単一細胞の子孫である多能性(特にES)細胞のクローン集団を得る。必要に応じて、細胞はまた、gp130下流シグナリング経路のアクチベーターの存在下で培養され得る。
【0036】
一旦、ES細胞の安定で均一な培養物が得られると、培養条件は、外胚葉、中胚葉または内胚葉の細胞の運命から選択される1以上の細胞タイプへ細胞の分化を方向づけるように変更され得る。サイトカインおよびシグナリング因子の添加または除去は、特定の分化した細胞集団の高い効率での誘導を可能にし得る。非神経外胚葉の運命へのES細胞の分化は、gp130を介して作用するサイトカイン、MEKインヒビターおよびGSK3インヒビターの存在下でES細胞を維持すること、ならびに次いで、サイトカインを除去する一方、GSK3インヒビターおよびMEKインヒビターを維持すること、および/または分化を方向づけ得るさらなるシグナリング分子を添加することによって達成され得る。あるいは、細胞は、MEKインヒビターおよびGSK3インヒビターの存在下で維持され得、次いで、該インヒビターの1つまたは両方を除去すること、および/または分化を方向づけ得るシグナリング分子を添加することによって分化を方向づけ得る。上記の方法はすべて、必要に応じて、プロセスの産物である分化した細胞を得る工程および/または単離する工程を含む。
【0037】
本発明のさらなる局面は、細胞培養培地を提供する。ある培地は、多能性(特にES)細胞の自己再生のためであり、該培地は、GSK3のインヒビター、MEKのインヒビターおよび必要に応じてFGFRアンタゴニストを含む。該培地は、必要に応じてgp130下流シグナリング経路のアクチベーターもまた含み得る。本発明の別の培地は、幹細胞培養培地であり、GSK3のインヒビター、MEKインヒビターおよび必要に応じてFGFRアンタゴニストを含む。すべての培地は、好ましくはさらに基本培地を含む。いくつかの好ましい実施態様では、すべての培地はgp130のアゴニストを含まず、したがって、好ましくはLIFを含まない。
【0038】
本発明は、血清および血清抽出物を含まない培地を提供する。1つのこのような培地は:
基本培地;
MEKインヒビター;
GSK3インヒビター;および
鉄トランスポーター;
を含み、ここで、該培地は、必要に応じて、血清および血清抽出物を含まない。
【0039】
培地は、好ましくはFGFRアンタゴニストもまた含む。培地は、必要に応じてgp130下流シグナリング経路のアクチベーターもまた含み得る。
【0040】
多能性幹細胞(特にラットまたはマウス細胞)に好適な培地は、血清およびgp130アゴニストを含まなくてよく、そしてMEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニストを含む。培地構成成分の置換は、本明細書に記載のように行われ得る。
【0041】
基本培地は、細胞のために炭素および/またはビタミンおよび/またはミネラルの必須供給源を供給する培地である。基本培地は、一般的にタンパク質を含まず、そして細胞の自己再生をそれ自体で支持し得ない。鉄トランスポーターは、鉄の供給源を提供し、または培養培地から鉄を取り込む能力を提供する。適切な鉄トランスポーターとしては、トランスフェリンおよびアポトランスフェリンが挙げられる。培地は、1以上のインスリンまたはインスリン様増殖因子、およびアルブミン(好ましくは組換え体)またはアルブミン代替物をさらに含み、そしてフィーダー細胞およびフィーダー細胞抽出物を含まないことが好ましい。培地は、アポトーシスのインヒビターまたは培養物中の多能性細胞の維持を促進する任意の他の構成成分もまた含み得る。
【0042】
本発明の特定の培地は、さらなる基本培地を含んでまたは含まずに、MEKインヒビター、GSK3インヒビター、インスリン、アルブミンおよびトランスフェリンを含む。この培地では、LIFが、必要に応じて含まれ得、そしてgp130シグナリングの他のアクチベーターによって置換され得る。しかし、好ましい培地は、gp130レセプター結合サイトカインであるLIFを含み、その適切な濃度は、一般的には10U/mlと1000U/mlとの間、より好ましくは50U/mlと500U/mlとの間、さらにより好ましくは100U/mlの範囲にある。GSK3およびMEKインヒビターは、好ましくは本明細書により詳細に記載の通りである。
【0043】
本発明は、胚盤胞から多能性細胞を誘導する方法をさらに提供し、該方法は:
(1)胚盤胞を得る工程;
(2)MEKインヒビター、GSK3インヒビターおよび必要に応じてFGFレセプターのアンタゴニストの存在下で該胚盤胞を培養して、内部細胞塊を得る工程;
(3)該内部細胞塊を解離する工程;
(4)該解離した内部細胞塊から1または複数の細胞を単離する工程;および
(5)該単離した1または複数の細胞を、MEKインヒビター、GSK3インヒビターおよび必要に応じてFGFレセプターのアンタゴニストの存在下で培養する工程、
を含む。
【0044】
必要に応じて、該単離した1または複数の細胞は、MEKインヒビター、GSK3インヒビターおよびgp130下流シグナリングのアクチベーターの存在下で培養される。FGFレセプターのアンタゴニストもまた存在させ得る。
【0045】
好ましくは、前記方法は、胚盤胞をLIF中で、より好ましくは2〜4日の期間に亘って培養する工程を含む。単離した1または複数の細胞は、好ましくは無血清培地中で培養される。代表的には、これらの細胞は、凝集塊として再プレートされる。胚盤胞はまた、好ましくは、無血清培地中で、必要に応じてBMPレセプターのアゴニストの不在下で、培養される。
【0046】
本発明によれば、細胞の培養は、接着培養で行われることがさらに好ましく、これは培養基材上に細胞接着タンパク質を含むことによって促進され得る。本発明によれば、単層培養で多能性細胞を培養することもまた好ましいが、必要に応じて、細胞を懸濁培養でまたはプレ細胞凝集塊として増殖させる;細胞をビーズ上であるいは他の適切な足場(例えば膜または他の3次元構造物)上でもまた増殖させ得る。
【0047】
本発明の実施例において用いられる培養培地は、好ましくは血清アルブミンもまた含む。これは、精製された形態または好ましくは組換え体の形態で用いられ得、そして組換え体の形態の場合には、これはサイトカインなどの可能性のある混入因子の不在という利点を有する。培養培地は、血清アルブミンを含む必要はなく、そしてこの構成成分は除外され得、あるいは別のバルクタンパク質またはWilesらに記載の合成ポリマー(ポリビニルアルコール)に置換され得る。
【0048】
本発明の特に好ましい培地は、完全に限定されるものである。この培地は、不確定のいかなる構成成分、すなわち、その内容物が知られていない構成成分または特定されない不確定因子または変動因子を含み得る構成成分も含まない。完全限定培地を用いることの利点は、多能性細胞の培養および引き続く操作のための効率的および一貫したプロトコールが誘導され得ることである。さらに、細胞を多能性状態に維持することが、より高い効率およびより大きい予測性で達成可能であること、および限定培地を用いて培養される細胞で分化が誘導される場合、分化シグナルへの応答が、非限定培地が用いられる場合より均一であることがわかる。
【0049】
本発明はまた、培養培地の調製を目的とする、培養培地の添加物として用いられ得る濃縮物、および構成成分のキットを提供し、得られる培地は、本発明と一致する。本発明の1つのキットは、第1の容器および第2の容器を含み、第1の容器はMEKインヒビターを含み、そして第2の容器はGSK3インヒビターを含む。好ましくは、キットは、FGFレセプターのアンタゴニストを含む第3の容器を含む。キットはまた、必要に応じて、gp130下流シグナリングのアクチベーターを含むさらなる容器を含み得る。キットは、好ましくは、各容器の内容物が本発明の培養培地を得るように培養培地に添加され得るように処方される。キットは、好ましくは、それらのそれぞれの構成成分の濃縮されたストック溶液を含む。
【0050】
本発明の方法はまた、分化した細胞を得る方法を含み、該方法は、記載のように多能性細胞を培養する工程、および細胞が分化することを可能にするまたは引き起こす工程を含む。ここで、細胞は、多能性幹細胞を含む他の細胞タイプと比較して所望の分化した細胞で差次的発現が可能である選択マーカーを含み、これによって、選択マーカーの差次的発現は、所望の分化した細胞の優先的な単離および/または生存および/または分裂をもたらす。選択マーカーは、所望の分化した細胞で発現し得るが他の細胞タイプでは発現しないか、あるいは所望の分化した細胞と他の細胞タイプとで発現レベルが異なり得、それによって選択マーカーの発現についての選択を可能にする。分化した細胞は、組織幹細胞または組織前駆細胞であり得、そして最終分化した細胞であり得る。
【0051】
一般的に、本発明はまた、本明細書に記載の本発明の方法のいずれかに従うことにより得られる細胞にも及ぶ。本発明の細胞は、創薬のためのアッセイに用いられ得る。本発明の細胞はまた、細胞治療のために用いられ得、したがって、本発明の方法は、MEKの阻害、GSK3の阻害および必要に応じてFGFシグナリングの拮抗の組み合わせを用いて多能性細胞を誘導および/または維持する工程、そこから細胞治療のための細胞を誘導する工程および細胞治療でそれらの細胞を用いる工程を含む。必要に応じて、この組み合わせは、gp130下流シグナリングのアクチベーターの不在下で用いられる。
【0052】
本発明のさらなる局面は、多能性細胞の自己再生を促進するための、MEKおよびFGFレセプターの阻害の使用、必要に応じてGSK3の阻害との組み合わせでの使用に関する。発明者らは、MEKインヒビターとFGFレセプターのアンタゴニストとの組み合わせが、添加サイトカインまたは増殖因子の不在下で無血清培地中で多能性細胞の増殖を支持する場合に効果的であることを見出した。
【0053】
したがって、本発明のさらなる局面は、MEKインヒビターおよびFGFレセプターのアンタゴニストを含む培養培地を提供する。MEKインヒビターおよびFGFレセプターアンタゴニストは、本発明の他の局面に関連して記載される通りである。同様に、培養培地は、本発明の他の局面に関連して本明細書に記載されるさらなる構成成分または因子をさらに含み得る。
【0054】
本発明のさらに他の局面は、多能性細胞のための培養培地の製造におけるMEKインヒビターおよびFGFレセプターのアンタゴニストの使用を提供する。
【0055】
本発明は、多能性細胞を培養する方法、および多能性細胞のトランスフェクトされた集団を得る方法もまた提供し、これらは、本発明の他の局面について記載のように好都合に実施され得る。したがって、本発明のさらなる局面は、自己再生を促進するような多能性細胞の培養方法を提供し、該方法は、MEKインヒビターおよびFGFレセプターのアンタゴニストを含む培地中で細胞を維持する工程を含む。
【0056】
本発明の関連する局面は、多能性細胞の培養方法を提供し、該方法は以下の工程を含む:
必要に応じてフィーダー上で、培養物中でES細胞を多能性状態に維持する工程;
少なくとも1回該ES細胞を継代する工程;
培地から(存在する場合)血清または血清抽出物を除去しそして(存在する場合)該フィーダーを除去して、該培地がフィーダー、血清および血清抽出物を含まないようにする工程;および
続いて、MEKインヒビターおよびFGFレセプターのインヒビターの存在下でES細胞を多能性状態に維持する工程。
【0057】
本発明のさらなる局面は、ES細胞のトランスフェクトされた集団を得る方法を提供し、該方法は:
選択マーカーをコードする構築物でES細胞をトランスフェクトする工程;
該ES細胞をプレートする工程;
MEKインヒビターおよびFGFレセプターアンタゴニストの存在下で該ES細胞を培養する工程、および
該選択マーカーを発現する細胞を選択する工程、
を含む。
【0058】
また、血清および血清抽出物を含まない細胞培養培地が提供され、該培地は:
基本培地;
MEKインヒビター;
FGFレセプターのアンタゴニスト;および
鉄トランスポーター、
を含む。
【0059】
MEKインヒビターとFGFレセプターのアンタゴニストとの組み合わせは、新たな多能性細胞系の誘導にもまた有用である。したがって、本発明のさらなる局面は、胚盤胞から多能性細胞を誘導する方法を提供し、該方法は:
(1)胚盤胞を得る工程;
(2)MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下で該胚盤胞を培養して、内部細胞塊を得る工程;
(3)該内部細胞塊を解離する工程;
(4)該解離した内部細胞塊から1または複数の細胞を単離する工程;および
(5)該単離した1または複数の細胞を、MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下で培養する工程、
を含む。
【0060】
本発明はまた、第1の容器および第2の容器を含むキットを含み、第1の容器はMEKインヒビターを含み、そして第2の容器はFGFレセプターのアンタゴニストを含む。キットはまた、本明細書に記載される他の容器および/または構成成分を含み得る。
【0061】
本発明のさらなる局面は、多能性幹細胞、特にNanogを発現する多能性幹細胞の自己再生の促進において、MEKインヒビターおよびFGFレセプターのアンタゴニストの使用を提供する。関連する局面は、幹細胞集団を増殖させる方法を提供し、該方法は、MEKインヒビターおよびFGFレセプターのアンタゴニストの存在下で幹細胞を培養する工程を含む。
【0062】
本発明の多くの利点は、上記の通りでありまたは明らかである。比較的無毒性および細胞透過性である細胞培養構成成分が同定され得る。本発明の特定の実施態様において用いられるMEKインヒビター、GSK3インヒビターおよびFGFRアンタゴニストは、特に、例えばタンパク質サイトカインの精製と比較して、容易に精製され得る。組換えタンパク質は、作成が高価であり得、そして小分子の培地構成成分は、より安価に生産され、かつ貯蔵がより安定であり得、より広い有効濃度範囲を伴い得る。
【0063】
以下に記載の特定の実施態様は、無血清の完全限定培地でCHIR99021、PD184352および必要に応じてSU5402の組み合わせを用い、そしてマウスES細胞の自己再生の改善を示し、ほとんど分化を示さなかった。BMPの存在下でES細胞を培養する場合、いくつかの神経発生があることが時々報告される。これは、本発明の実施例においては見られなかった。
【0064】
本発明は、ここで、特定の実施例においてさらに説明され、図によって示される。
【0065】
図1は、多能性ES−NSハイブリッドコロニーの形成におけるPD184352の効果の分析結果を示す。(A−C)RH×NS TGFP融合の赤色蛍光および緑色蛍光についてのFACS分析。(A)PEG処理の24時間後の融合混合物;(B)AでゲートをかけたFACS分類されたハイブリッドの純度チェック。(C)Aで分類されたハイブリッドをプレートして形成されたコロニーをプレートしたハイブリッドあたりのコロニーの比率(%)としてスコア付けた。これらのスコアは、FACS分類された細胞の純度を考慮する。(D)データの概要。(E)ハイブリッドコロニー形態の例。
【0066】
図2は、本発明にしたがって誘導および維持されたマウスのES細胞、ならびにこれらのES細胞による高効率なキメラ発生を示す。
【0067】
図3は、本発明にしたがって増殖した4継代マウスES細胞を示す。
【0068】
図4は、本発明にしたがって増殖したマウスES細胞がOct4陽性であることを示す。
【0069】
実施例において、用語2i培地または2iは、MEKインヒビターおよびFGFレセプターのアンタゴニストを含む培地を示すために用いられる。用語3i培地または3iは、MEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニストを含む培地を示すために用いられる。
【実施例】
【0070】
GSK−3βインヒビター、MEKインヒビター、培養培地、およびES細胞の自己再生
マウスおよびヒトES細胞を、特に明記のない限りN2B27培地を用いて、そしてGSK−3βインヒビターであるCHIR99021、AR−AO144−18、SB216763およびSB415286、ならびにMEKインヒビターであるPD184352の存在下または不在下で、種々の条件下で増殖させた。
【0071】
N2B27培地の調製:
N2 100×ストック溶液。10mlについて:1mlのインスリン(最終濃度2.5mg/ml)を1mlのアポトランスフェリン(最終濃度10mg/ml)、0.67mlのBSA(最終濃度5mg/ml)、33μlのプロゲステロン(最終濃度2μg/ml)、100μlのプトレシン(最終濃度1.6mg/ml)、10μlの亜セレン酸ナトリウム(最終濃度3μM)および7.187mlのDMEM/F12と混合する。4℃で貯蔵し、そして1カ月以内に使用する。
【0072】
DMEM/F12−N2培地:100mlのDMEM/F12に対して、1mlのN2 100×ストック溶液を添加する。DMEM/F12培地中のN2の各構成成分の最終濃度は:インスリン、25μg/ml;アポトランスフェリン、100μg/ml;プロゲステロン、6ng/ml;プトレシン、16μg/ml;亜セレン酸ナトリウム、30nM;BSA 50μg/mlである。4℃で貯蔵し、そして1カ月以内に使用する。
【0073】
Neurobasal/B27培地:100mlのNeurobasalTM培地に対して、2mlのB27および0.5〜1mlの200mM L−グルタミンを添加する。4℃で貯蔵し、そして1カ月以内に使用する。
【0074】
N2B27培地:DMEM/F12−N2培地をNeurobasal/B27培地と1:1の比で混合する。0.1Mストックから0.1mMの最終濃度になるまでβ−メルカプトエタノールを添加する。4℃で貯蔵し、そして1カ月以内に使用する。
【0075】
実施例1
無血清培地中では、MEKインヒビター+GSK−3βインヒビターは、(1)N2B27培地、および(2)完全限定培地(DMEM/F12−N2)の両方で、マウスES細胞の自己再生を維持するのに十分であった(データは示さず)。ES細胞の自己再生は、MEKインヒビター、GSK−3βインヒビターおよびLIFを含む培地中でさらに改善された(データは示さず)。
【0076】
実施例2
PD184352(MEKのインヒビター)がES細胞中のNanogのレベルを増加させることが示された(データは示さず)。さらに、PD184352で処理されたNanog−/−ES細胞がES細胞の自己再生の増強を示さなくなることが示された。実際、これらの細胞は分化した(データは示さず)。これにより、PD184352によるES自己再生表現型の増強がNanogによって仲介されることが立証された。
【0077】
再プログラミングにおけるPD184352の効果を、細胞融合の前後におけるNS細胞の多分化能への変換を決定することによってもまた研究した。
【0078】
dsRed蛍光タンパク質およびハイグロマイシン耐性を構成的に発現するRH ES細胞を、IRESを経てピューロマイシン耐性に連結した融合タンパク質TauGFPを発現する胎児由来神経幹細胞(NS TGFP)に融合した。融合の一方には、RH細胞を、融合の前後3日間、3μMのPD184352で処理した。コントロールには、PD184352を添加しなかった。融合24時間後、処理した初代ハイブリッドおよび未処理の初代ハイブリッドを分類し、次いでプレートした(図1A−C)。3日後、ハイグロマイシンおよびピューロマイシン選択をES培地に添加した。dsRed2およびGFP蛍光の発現およびES細胞形態の提示についてコロニーのスコアを付けた(図1Dおよび図1E)。結果は、PD184352がES−NSハイブリッドコロニーの形成を45倍増強することを示した。興味深いことに、PD184352で処理したRH細胞中のプレートしたハイブリッドあたりのハイブリッドコロニー形成の比率(%)は、Nanogを過剰発現させているES細胞と比較してちょうど2倍低かった(2.25%対4%)。この結果は、PD184352がES細胞の自己再生を増強するだけでなく、細胞融合前後における再プログラミングもまた増強することを示している。この効果は、処理したRH細胞におけるNanogのレベルの増加によって仲介されていると思われる。したがって、Nanogが内生的に発現する場合、MEKインヒビターがNanogをアップレギュレートするために用いられ得、そして再プログラミングの増強などの付随効果を達成し得る。
【0079】
実施例3
ヒトES細胞を、GSK−3インヒビターであるCHIR99021およびMEKインヒビターであるPD184352を追加した培地で培養した。
【0080】
培養培地へのLIFの添加は、細胞の増殖をさらに改善した(データは示さず)。
【0081】
実施例4
マウスES細胞を、GSK−3インヒビターであるCHIR99021およびMEKインヒビターであるPD184352を追加した培地で培養した。
【0082】
培養培地へのLIFの添加は、細胞の増殖をさらに改善した(データは示さず)。
【0083】
実施例5
マウスおよびヒトES細胞を、CHIR99021、PD184352およびSU5402を含む培地で増殖させ、該培地は以下のように調製した:
【0084】
【表1】
【0085】
培地
DMEM/F12−N2培地の調製
100mlのDMEM/F12(Gibco 42400-010)に対して、1mlのN2 100×ストック溶液を添加する。DMEM/F12培地中のN2の各構成成分の最終濃度は:
インスリン 25μg/ml
プトレシン 16μg/ml
トランスフェリン 100μg/ml
亜セレン酸ナトリウム 30nM
プロゲステロン 6ng/ml
BSA 50μg/ml
である。
【0086】
Neurobasal/B27の調製:
100mlのNeurobasal培地(Gibco 21103-049)に対して、2mlのB27(Gibco 17504-044)および1〜2M L−グルタミン(TC保存 1:100)を添加する。
【0087】
N2B27培地の調製:
DMEM/F12−N2培地をNeurobasal/B27培地と1:1の比で混合する。この培地を、すべての化合物の希釈および細胞の増殖に用いた。
【0088】
この培地は、ヒトES細胞の維持、ならびに129系統マウス由来のES細胞の誘導および維持に用い、そして非許容的なマウス系統であるCBAおよびC56/BL6由来のES細胞の誘導にもまた用いた。
【0089】
実施例6
マウスES細胞を、FGFレセプターのインヒビターおよびMEKインヒビターの存在下で培養した。選択的薬理学的インヒビターSU5402およびPD184352を、それぞれFGFレセプターチロシンキナーゼの阻害およびMEK1/2を経たErk1/2の活性化の阻害のために用いた。発明者らは、どちらのインヒビターの添加も、BMP4を供給することなく、LIFを含むN2B27培地中で強固にES細胞を増殖させるに十分であることを見出した(データは示さず)。未分化の培養物は、これらの条件において、多能性マーカーOct4、NanogおよびRex1の発現を保持しながら継続的に継代され得る。LIF+BMPで維持された培養物中よりもId遺伝子の発現がはるかに低いにも関わらず、神経への運命づけは生じていない。
【0090】
LIFを添加しないN2B27培地にプレートされたES細胞(通常、効率的な神経への運命づけを引き出す条件)は、SU5402またはPD184352のいずれが添加された場合も、数日間に亘ってOct4陽性およびSox1陰性のままである(データは示さず)。しかし、これらの細胞は、継代した後、変わらず分化および/または死滅する。毒性副作用の可能性を低減するために、発明者らは、2.5倍低い用量を用いて、2つのインヒビターを同時に併用した。0.8μMのPD184352+2μMのSU5402を含むN2B27中では、最初にいくらかの分化が観察されるが、継代後はES細胞が生き残って増殖する(データは示さず)。この2インヒビター(2i)条件下では、LIFの存在下よりも生存率はより低く、集団倍加時間はより遅いが、分化は効果的に抑制される。この発見は、ES細胞の自己再生のための最少要件が、細胞増殖および生存率を損なわせることを避けながら、FGFレセプターおよびErkシグナリングから生じる分化シグナルをそらすことであり得る旨示唆している。
【0091】
実施例7
発明者らは、2i培地中におけるES細胞の増殖の減少は、pErkの下流のRskによる阻害的リン酸化の解放の結果生じるグリコーゲン合成酵素キナーゼ3(GSK−3)活性の増加によるものであり得ると結論づけた。CHIR99021は、GSK−3の十分に特徴づけられた高選択性小分子インヒビターであり、これはGSK−3活性を完全にブロックする濃度でサイクリン依存性キナーゼ(CDK)と交差反応しない。発明者らは、血清存在下の培養物に添加した場合、CHIR99021(3μM)は、LIFの存在下であっても、実際に分化を促進することを見出した。無血清のN2B27培地中では、分化応答は減少し、そしていくつかのコロニーは、数日間、形態的に未分化であるように見える。しかし、継代した後、未分化細胞は生き残らない。同様の結果が、広く用いられる他の2つのGSK−3インヒビターであるSB216763およびSB415286でも、(どちらもES細胞にいくぶん有毒のようであったが)得られた。
【0092】
しかしながら、CHIR99021を2iと併用した場合、分化応答は完全に失われる。さらに、CHIR99021は、ES細胞がLIF+血清/BMP中または2i中で代表的に見られる平坦な単層よりもむしろ密集した3次元のコロニーとして増殖するように、2iへの応答を変化させた。3つのインヒビター(3i)中での分化は無視できる程度であり、そしてES細胞は急速に増殖した。最も重要なことに、継代した後、未分化コロニーが高効率で増殖した。2つの独立した親ES細胞系E14Tg2aおよびCGR8の派生体は、3i中で強固な長期増殖を示し、かつ明白な分化はほとんどあるいは全くなかった(データは示さず)。これらは、Oct4、NanogおよびRex1を発現し、系統運命づけマーカーであるGata4、Sox1またはbrachyuryの発現は、それほど示さない(データは示さず)。バルク培養では、ES細胞は、3i中でLIF+BMP中に匹敵する倍加率で増殖し、Oct4−GFP陽性の未分化細胞の比率は、依然として90%を超える。
【0093】
したがって、3i培地は、血清または添加サイトカインの不在下で、分化することなくES細胞を培養するために用いられ得る。
【0094】
実施例8
培養処方物が十分にES細胞の自己再生を持続させるかに関する厳密な試験は、個々の細胞による未分化コロニーの形成である。単一細胞の堆積の後、N2B27+3i中のコロニー形成率は25%(98/384)であり、LIF+BMP(11%、23/192)よりも高い(データは示さず)。これらのコロニーはOct4−GFPを発現し、そして未分化ES細胞として継代可能である。したがって、MEKインヒビター、FGFレセプターのインヒビターおよびGSK3インヒビターを含む培地は、単一細胞に由来する未分化ES細胞コロニーの形成を持続させることが可能である。
【0095】
実施例9
発明者らは、3iが、胚から直接新たなES細胞を誘導する十分な能力を有したのか、あるいは確立した系の適応を反映したのかを検討した。許容的な129系統からの胚盤胞を、ゼラチンコートされたプラスチック上のN2B27+3i中に直接プレートし、5日間培養した。続く内部細胞塊の解離および再プレートの後、12個の胚のうち7個からES細胞コロニーが得られた。これらのうち3個を増殖させ、胚盤胞に注入した。すべてにおいて、キメリズムおよび生殖系列遺伝が高い比率で得られた(表2)。続いて、発明者らは、C57BL/6および非許容的なCBAおよびMF1系統から複数のES細胞を誘導し、3iが胚盤葉上層細胞からES細胞への移行を容易にすることを示した。発明者らは、3iが、発生能を選択または損失させることなく、外因性のLIFおよびBMP/血清の要求からES細胞を解放すると結論付ける。
【0096】
【表2】
【0097】
したがって、ES細胞は、GSK3インヒビターおよびMEKインヒビターの組み合わせ、MEKインヒビターおよびFGFレセプターのアンタゴニストの組み合わせ、または必要に応じてGSK3インヒビター、MEKインヒビターおよびFGFレセプターのアンタゴニストの組み合わせで維持され、そして本発明はまた、培養方法およびそのための培地を提供する。
【特許請求の範囲】
【請求項1】
MEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニストを含む、培養培地。
【請求項1】
MEKインヒビター、GSK3インヒビターおよびFGFレセプターのアンタゴニストを含む、培養培地。
【図1A−C】

【図1D】
【図1E】
【図2】
【図3】
【図4】
【図1D】
【図1E】
【図2】
【図3】
【図4】
【公開番号】特開2013−66495(P2013−66495A)
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2013−11454(P2013−11454)
【出願日】平成25年1月24日(2013.1.24)
【分割の表示】特願2009−502219(P2009−502219)の分割
【原出願日】平成19年3月30日(2007.3.30)
【出願人】(500219618)ザ・ユニバーシティ・コート・オブ・ザ・ユニバーシティ・オブ・エディンバラ (21)
【氏名又は名称原語表記】The University Court of the University of Edinburgh
【Fターム(参考)】
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願日】平成25年1月24日(2013.1.24)
【分割の表示】特願2009−502219(P2009−502219)の分割
【原出願日】平成19年3月30日(2007.3.30)
【出願人】(500219618)ザ・ユニバーシティ・コート・オブ・ザ・ユニバーシティ・オブ・エディンバラ (21)
【氏名又は名称原語表記】The University Court of the University of Edinburgh
【Fターム(参考)】
[ Back to top ]