
クラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニスト
本発明は、ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1及びインテグリンβ2からなる群から選ばれるタンパク質の膜貫通ドメインに由来するアミノ酸配列を含み、少なくとも1のGxxxGモチーフを含み、最終的に異種配列へと融合されるクラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニスト、当該ペプチドアンタゴニストをコードする核酸、当該ペプチドアンタゴニスト又はそれをコードする核酸を含む医薬組成物、及びその使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、クラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニスト、及び当該アンタゴニストの使用に関する。
【0002】
その構造的及び栄養的役割とは別に、細胞外マトリクス(ECM)は、細胞間連絡の理想的な環境を規定し、そして全ての細胞挙動、例えば増殖、移動、分化、又はアポトーシスを決定する。これらの過程を制御する分子メカニズムは、次第に理解されてきている。神経系では、拡散又は膜貫通シグナルについての3の主要なファミリー(ネトリン、セマフォリン、及びエフリン)は、胚発生のあいだのその機能を有する(TESSIER-LAVIGNE及びGOODMAN, Science, vol.274, p: 1123-33, 1996)。その中で、セマフォリンは、その構造特異性に従って8のクラスに分けられる25超のメンバーから構成されるファミリーを規定し(KOLODKINら、Cell, vol.75, p: 1389-99, 1993)、そしてそれらは、分泌型または膜貫通型セマフォリンのいずれかに分類できる。分泌型セマフォリンはクラスII(無脊椎動物)、III(脊椎動物)、及びV(ウイルス)であり、一方他のクラス(I、IV、及びVI〜VIII)は膜貫通型である。
【0003】
過去5年の間に、生理的及び病理的状態における軸索ガイダンス、細胞移動、細胞分化からアポトーシスに至るセマフォリンの様々な機能の伝達を可能にする伝達経路を解明するために幾つもの研究が設計されてきた。この機能的な多様性は、アクチン細胞骨格リモデリングを導く複数の細胞内経路を選択的動員及び活性化することによりシグナルの統合を調節する高度に動的な受容体複合体を形成するために生じると現在考えられている(CASTELLANI及びROUGON, Curr. Opin. Neurobiol. , vol.12, p:532-41, 2002)。これらの全ては、12〜16個のシステインを有する約500のアミノ酸からなる「semaドメイン」と呼ばれる共通ドメインであって、各セマフォリンの結合特異性を与える共通ドメインを有する(RAPER, Curr. Opin. Neurobiol., vol.10, p: 88-94, 2000)。これらの様々なセマフォリンの中で、クラスIIIセマフォリンは、神経細胞成長円錐の崩壊を誘導しうる。クラスIIIセマフォリンが当初、コラプシンと名づけられた所以である(LUOら、Cell, vol.75, p: 217-227, 1993)。他の残りのファミリーにその名称を与える分子であるsema3Aは、最もよく研究されており、そして全ての場合において、知覚神経及び脊椎運動神経から皮質の錐体神経に至るまで、軸索の忌避因子として記載されてきた(MUELLER, Annu. Rev. Neurosci., vol.22, p: 351-388, 1999)。特に、このセマフォリンは、同じ細胞において2の異なる効果を発揮することができる。これは、皮質ニューロンにおいて示されており、皮質ニューロンでは、sema3Aは、軸索の忌避因子として作用し、かつ樹状突起の化学誘引物質でもある(POLLEUXら、Nature, vol.404, p:567-73, 2000; BAGNARDら、Development, vol.125(24)、p: 5043-53, 1998)。この現象を説明するために、2個の細胞の極において、区別される伝達を確保するメカニズが存在することを考慮する必要がある。区別される伝達の原理だけではなく、分子ヒエラルキーを制御するメカニズムを理解すること、並びに環境変化に応答する細胞挙動の多様性を確保する超分子構造(supra-molecular structures)の形成を解明することが必要とされる。
【0004】
従って、ニューロピリンファミリーの2の既知のメンバー、ニューロピリン-1(NRP1)及びニューロピリン-2(NRP2)の、クラスIIIセマフォリンの伝達カスケードに関与する受容体複合体のリガンド結合サブユニットとしての役割を近年の研究は示した(総説として、Bagnard D. (編) ニューロピリン: from nervous system to vascular and tumor biology. Landes Bioscience- Kluwer Academic/Plenum Publishers Hardbound, ISBN 0-306-47416-6、Advance in Experimental Medicine and Biology Vol.515, p: 140, 2002を参照のこと)。NRP1及びNRP2は、(i)二量化に重要である細胞外部分(RENZIら、J. Neurosci., vol. 19, p:7870-7880, 1999)、膜貫通部分、及び約40のアミノ酸の短い細胞質ドメインを有する1回膜貫通タンパク質である。
【0005】
興味深いことに、NRP1及びNRP2は、伝達能力を持たない短い細胞内ドメインを有する。この所見についての分子的説明は、ニューロピリンがプレキシンファミリーに属する受容体との複合体を形成するということ、並びにプレキシンがニューロピリン/プレキシン複合体の伝達要素であることが見出された際に与えられた(RHOMら、Mech. Dev. , vol.93, p: 95-104, 2000; TAMAGONEら、Cell, vol.99, p:71-80, 1999)。最終的に、クラスIIIセマフォリンによるシグナル伝達は、ニューロピリンとプレキシンとのあいだの複合体形成に依存する。
【0006】
それにもかかわらず、プレキシンとの複合体は、ニューロピリンにより形成される複合体の唯一のタイプではない。
【0007】
ニューロピリンが、アドヘシン分子L1-CAM及びNr-CAMと安定な複合体を形成できるということ(CASTELLANIら、Neuron, vol.27, p: 237-249, 2000)、並びにL1の細胞外ドメインにおける突然変異又は遺伝子標的化マウスにおけるL1の完全な欠失が、sema3Aシグナルの途絶をもたらし、ガイダンスエラーが生じるということが見出された。
【0008】
その結果、チロシン・キナーゼ受容体は、ニューロピリンが関連するシグナル伝達に役割を果たしうる。こうして、DEV神経外胚葉細胞の移動が、Sema3Aにより停止され、そしてNRP1及びVEGFR-1の存在が、当該停止に必要とされるということが観察された(Bagnardら、J Neurosci. , vol.21, p:3332-41, 2001)。この相互作用は、Sema3Aを用いた血管新生のin vitroモデルにおいて、VEGFによる芽形成の阻害を説明する(MIAOら、J. Cell. Biol., vol.146, p:233-242, 1999)。ニューロピリンが、VEGFR-2と(SOKERら、Cell, vol.92, p:735- 45, 1998)、並びにMET(WINBERG et al. , Neuron, vol.32, p:53- 62, 2001)と複合体を形成することも発見された。
【0009】
近年、ニューロピリンが、インテグリンと複合体を形成し、そして当該複合体が、軸索の伸張を促進することができるということが示された(PASTERKAMPら、Nature, vol.424, p:398-405, 2003)。
【0010】
その結果、近年分かったことであるが、上記研究は、神経変性疾患及び癌の潜在的な標的としてクラスIIIセマフォリン/ニューロピリン複合体を同定することに寄与した(GUTTMANN-RAVIVら、Cancer Letter, 2006; CHEDOTALら、Cell Death and Differentiation, 2005を参照のこと)。この文脈では、複合体形成を妨げる薬剤は、明らかに治療潜在性を有し、及び/又は有用な生物学的ツールであろう。
【0011】
このように、GARETHら(Journal of Neurochemistry, vol.92, p: 1180-1190, 2005)は、Sema3A/NRP1複合体のペプチドアンタゴニストを設計するためにアルゴリズムを用いた。この著者は、Sema3A/NRP1二量化に関与するSema3A Igドメインと、受容体の側面二量化を媒介するが、リガンド結合を媒介することはないNRP1 MAMドメインとのあいだにおけるアンタゴニスト・ペプチドを同定した。それにもかかわらず、膜貫通ドメインに位置しないこれらのアンタゴニストは、1μMを超えるIC50を有し、当該濃度は、かなりの濃度であるので、治療においてこのようなアンタゴニストを使用することができない。
【0012】
こうして、治療に使用できるクラスIIIセマフォリン/ニューロピリン複合体の新たなアンタゴニストについて、当該技術分野において認識されている変わらぬ必要性が存在する。
【発明の開示】
【0013】
本発明の目的は、高い活性を有する新たなアンタゴニストを提供することにより当該必要性を満たすことである。
【0014】
本発明者は、NRP1の膜貫通ドメインに相当するペプチドが、10-11M未満のIC50でSema3Aにより誘導される皮質軸索崩壊を阻害することができるということを思いがけずに示した。このペプチドは、2の連続したGxxxG二量化モチーフ(ここで、xは任意のアミノ酸を表す)を含み、当該GxxxGモチーフは、グリコフォリンAにおいて最初に同定された(SENESら、J. MoI. Biol., vol.296, p:921-36, 2000)。このようなモチーフは、erbB受容体(MENDROLAら、J. Biol. Chem. , vol.277, p.4704-12, 2002)のTMセグメントの二量化において働いていると示された。一般的に、小さいアミノ酸-xxx-小さいアミノ酸からなる任意のモチーフ(ここで、小さいアミノ酸のモチーフ定義は、当業者に周知である)は、同等の性質を有する。
【0015】
NRP1のダブルGxxxGモチーフは、高度に保存されており、そしてNRP2のダブルGxxxGモチーフと強いホモロジーを示す(図1)。最終的に、このようなモチーフは、プレキシン・ファミリーのメンバー、Nr-及びL1-CAM、並びにインテグリンを含むニューロピリンの複数のパートナーの膜貫通ドメインにおいて見つかる。全体として、本発明者により得られる結果により、これらのタンパク質の膜貫通ドメインが、セマフォリンシグナル伝達を保証する複合体の形成及び調節において重要な役割を果たすということが示唆される。
【0016】
結果として、1の態様では、本発明は、ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1及びインテグリンβ2からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも1のGxxxGモチーフを含むアミノ酸配列を含み、場合により異種配列に融合されるクラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニストに関する。
【0017】
本明細書に使用される場合、「異種配列」は、ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1又はインテグリンβ2に由来しない任意のアミノ酸配列に関する。当該異種配列は、例えば、特定の細胞内への配置を可能にするか、又は本発明のペプチドアンタゴニストの優れた精製収率(例えばHisタグ)を可能にすることができる。
【0018】
本明細書に使用される場合、「クラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニスト」という用語は、当該複合体形成を妨げ、そして最終的にこのような複合体のシグナル伝達を妨げる合成又は組換えポリペプチドに関する。結果として、本発明のペプチドアンタゴニストは、完全なニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1、及びインテグリンβ2タンパク質を含まない。
【0019】
本明細書に使用されるとき「膜貫通ドメイン」は、細胞膜を横断するペプチドドメインに相当する。当該ドメインは疎水性であり、そしてα-へリックス構造を有する。当業者は、その一般的知識に従って、当該タンパク質のドメインを簡単に同定できる。一例として、タンパク質ドメインの親水性は、Kyte & Doolittle法により決定することができ、そしてα-へリックス構造を形成するタンパク質ドメインの可能性は、Chou&Fasman法により決定することができる。このような方法は、以下のアドレス: http://www.expasy.org/tools/protscale.htmlで利用できる。
【0020】
ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1及びインテグリンβ2の膜貫通ドメインのアミノ酸配列はよく保存されており、そしてタンパク質の完全なアミノ酸配列から簡単に同定することができる。この方法は、当業者に周知である。一例として、ニューロピリン-1アミノ酸配列であって、ムス・ムスクルス(Mus musculus)(P97333)、ホモ・サピエンス(014786)、ラッタス・ノルベギクス(Rattus norvegicus)(Q9QWJ9)、ゼブラフィッシュ(Zebrafish)(Q8QFX6)、及びガルス・ガルス(Gallus gallus)(P79795)に由来する配列;ニューロピリン-2アミノ酸配列であって、ホモ・サピエンス(060462)、ムス・ムスクルス(035375)、ラッタス・ノベルギクス(NP_110496)及びガルス・ガルス(NP_989615)に由来する配列;プレキシンA-1アミノ酸配列であって、ホモ・サピエンス(NP_115618、Q9UIW2)及びムス・ムスクルス(NP_032907、P70206)に由来する配列;プレキシンA-2アミノ酸配列であって、ホモ・サピエンス(CAI40198、Q5JRL6)及びムス・ムスクルス(NP_032908、P70207)に由来する配列;プレキシンA3アミノ酸配列であって、ホモ・サピエンス(NP_032907、P51805)、ゼノパス・トロピカリス(Xenopus tropicalis)(CAI40198)及びムス・ムスクルス(NP_032909、P70208)に由来する配列;プレキシンA4アミノ酸配列であって、ムス・ムスクルス(NP_786926、Q80UG2)、ホモ・サピエンス(Q9HCM2)及びダニオ・レリオ(Danio rerio)(NP_001004495)に由来する配列;Nr-CAMアミノ酸配列であって、ムス・ムスクルス(Q810U4)、ラッタス・ノベルギクス(P97686)、ホモ・サピエンス(Q92823)及びガルス・ガルス(P35331)に由来する配列;L1-CAMアミノ酸配列であって、ホモ・サピエンス(P32004)、タキフグ・ルブリペス(Takifugu rubripes)(Q98902)、ムス・ムスクルス(P11627)及びラッタス・ノベルギクス(Q05695)に由来する配列;インテグリンβ1アミノ酸配列であって、ムス・ムスクルス(P09055)、ホモ・サピエンス(P05556)、フェリス・カツス(Felis catus)(P53713)、ラッタス・ノベルギクス(P49134)、ゼノパス・ラエビス(Xenopus laevis)(P12606)及びガルス・ガルス(P07228)に由来する配列;インテグリンβ2アミノ酸配列であって、ムス・ムスクルス(P11835)、ホモ・サピエンス(P05107)、スス・スクロファ(Sus scrofa)(P53714)、ボス・タウルス(Bos taurus)(P32592)及びシグモドン・ヒスピヅス(Sigmodon hispidus)(AAL38579)に由来する配列を引用することができる。
【0021】
図1は、膜貫通ドメインであって、以下のマウスの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号4:LLTLPAIISIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号5:LTLPAMVGLAAGGGLLLLAITVVLVAY)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCF)、
L1−CAM(配列番号8:GWFIAFVSAIILLLLILLILCFI)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び
インテグリンβ2(配列番号10:VAAIVGGTVVGVVLIGVLLLVIW)
で表される膜貫通ドメインを示す。「GxxxGモチーフ」という用語は、SENESら(上記、2000)に同定されるモチーフに関し、これは図1に示される(下線)。潜在的な「GxxxG」モチーフも示される(点線)。
【0022】
これらの膜貫通ドメインの保存は、図2に明らかに目立っており、図2は、膜貫通ドメインであって、
以下のヒトの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン−2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1−CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
についての同じ膜貫通ドメインを示す。
【0023】
この保存の別の例として、以下の種に由来するニューロピリン-1膜貫通ドメイン:
ガルス・ガルス(配列番号14: ILITIIAMSALGVLLGAICGVVL)、及び
ゼブラフィッシュ(配列番号15:ILITIIAMSALGVFLGAICGVVL)、並びに
ガルス・ガルス由来のニューロピリン-2膜貫通ドメイン(配列番号16:ILVTIIAMSSLGVLLGATCAGLLLY)
を引用することができる。これらは、ヒトニューロピリン-1およびニューロピリン-2膜貫通ドメインとそれぞれ90%超の同一性を有する。
【0024】
好ましい実施態様によると、本発明は、膜貫通ドメインであって、以下のヒトの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン−2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1−CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも1のGxxxGモチーフを含むアミノ酸配列を含み、場合により異種配列に融合されるクラスIIIセマフォリン/ニューロピリン複合体のペプチド性アンタゴニストに関する。
【0025】
別の好ましい実施態様に従って、クラスIIIセマフォリン/ニューロピリン複合体のペプチド性アンタゴニストは、以下の:
ニューロピリン-1(配列番号1、ILITIIAMSALGVLLGAVCGVVL)又は
ニューロピリン-2(配列番号2、ILITIIAMSSLGVLLGATCAGLLLY)膜貫通ドメインに由来するアミノ酸配列を含み、場合により異種配列に融合される。
【0026】
さらに別の好ましい実施態様では、クラスIIIセマフォリン/ニューロピリン複合体のペプチド性アンタゴニストは、少なくとも2個のGxxxGモチーフ、好ましくは少なくとも2個の連続GxxxGモチーフを含むタンパク質の膜貫通ドメインに由来し、そして、以下のヒトの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL),
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
の膜貫通ドメインからなる群から選択されるアミノ酸配列を含み、場合により異種配列に融合される。
【0027】
好ましくは、本発明のペプチドアンタゴニストは、ヒトニューロピリン-1膜貫通ドメイン(配列番号1、ILITIIAMSALGVLLGAVCGVVL)に由来するアミノ酸配列を含む。
【0028】
有利な態様では、上記タンパク質のうちの1つの膜貫通ドメインに由来するアミノ酸配列は、10超のアミノ酸長であり、好ましくは14超のアミノ酸長であり、一例として18超のアミノ酸長、そしてより好ましくは22超のアミノ酸長である。
【0029】
有利な態様では、ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1およびインテグリンβ2からなる群から選ばれるタンパク質のうちの1つに由来するアミノ酸配列は、150未満のアミノ酸長であり、好ましくは100アミノ酸未満のアミノ酸であり、より好ましくは50未満のアミノ酸長である。
【0030】
好ましい実施態様では、ニューロピリン-1及びニューロピリン-2に由来するペプチドアンタゴニストは、クラスIIIセマフォリン二量化に関与するその細胞外ドメインを含まない。これらのドメインは、当業者に周知であり、そしてNEUFELDら(TCM, vol.12(1), p: 13-19、2002)及びBAGNARD (2002, 上記)に記載されている。例えば、これらのドメインは、NRP1及びNRP2のaドメイン(CUBドメイン、セマフォリン結合を保証するa1及びa2ドメインとも呼ばれている)、bドメイン(凝集因子V/VIIIに対するホモログドメイン;b1ドメインとb2ドメインに分けられる。b1は、VEGFイソ型の結合に関与する)、cドメイン(MAMドメイン、NRP1の二量化に関与する)を含む。
【0031】
有利な態様では、当該ペプチドアンタゴニストは、以下の膜貫通ドメインであって:
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr-CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1-CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び、
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
の膜貫通ドメインからなる群から選ばれるアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される。
【0032】
第二の好ましい実施態様では、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、インテグリンβ1、インテグリンβ2、Nr-CAM及びL1-CAMは、シグナル伝達経路に関与するその細胞内ドメインを含まない。これらのドメインは、当業者に周知であり、そしてBAGNARD(上記、2002)に記載され、そして例えば非限定的に、Sex-プレキシンドメイン、PH1A及びPH2Aドメイン又はPRB(プレキシンRac結合ドメイン)ドメインを含む。
【0033】
有利な態様では、当該ペプチドアンタゴニストは、以下の:ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)及びニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)膜貫通ドメインからなる群から選択されるアミノ酸配列又はこの誘導体からなり、場合により異種配列に融合される。好ましくは、当該ペプチド性アンタゴニストは、ニューロピリン-1膜貫通ドメイン(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、又はその誘導体からなり、場合により異種配列に融合される。
【0034】
膜貫通ドメインであって、以下のヒトの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr-CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1-CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)又は
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
の膜貫通ドメイン「に由来する」又は「の誘導体である」アミノ酸配列は、当該膜貫通ドメイン又はその断片と60%長の同一性、例えば70%超又は80%超、好ましくは85%超、最も好ましくは90%超、及び有利な実施態様では95%超の同一性を有するアミノ酸配列に関する。
【0035】
本明細書に使用される場合に、「膜貫通ドメインの断片」は、10超のアミノ酸長、好ましくは14超のアミノ酸長、一例として18超のアミノ酸長、そしてより好ましくは22超のアミノ酸長であるポリペプチドに関する。
【0036】
上記膜貫通ドメインと、本発明のペプチドアンタゴニストのアミノ酸配列とのあいだの同一性の差異は、ペプチドアンタゴニストの膜貫通ドメインアミノ酸配列におけるアミノ酸置換から生じる。
【0037】
好ましくは、これらの膜貫通ドメインにおける置換アミノ酸は、中性及び/又は疎水性アミノ酸であり、そして最も好ましくは疎水性アミノ酸である。このような中性及び疎水性アミノ酸は、当業者に周知である。
【0038】
具体的な実施態様に関し、当該膜貫通ドメイン・アミノ酸配列は、当該膜貫通ドメイン又はその断片との100%の同一性を有する。
【0039】
第二態様では、本発明は、上記ペプチドアンタゴニストをコードする核酸に関する。
【0040】
当該核酸は、RNA又はDNA、好ましくはDNAに対応する。
【0041】
好ましい実施態様では、ペプチドアンタゴニストをコードする核酸は、当該核酸は、原核又は真核細胞、好ましくは真核細胞内で核酸発現を仕向ける遺伝子発現配列に作用可能なように結合されている。「遺伝子発現配列」は、任意の調節性ヌクレオチド配列であり、例えば、それが作用可能なように結合されたペプチドアンタゴニストの核酸の効率的な転写及び翻訳を促進するところのプロモーター配列又はプロモーターエンハンサーの組合せ、並びに原形質膜へペプチドアンタゴニストを適切に標的化することを保証する任意のシグナル配列である。遺伝子発現配列は、例えば、哺乳動物又はウイルスプロモーター、例えば構成的又は誘導性プロモーターであってもよい。構成的哺乳動物プロモーターは、非限定的に、以下の遺伝子:ヒポキサンチン、ホスホリボシル・トランスフェラーゼ(HPTR)、アデノシン・デアミナーゼ、ピルビン酸キナーゼ、βーアクチンのプロモーター、筋肉クレアチン・キナーゼプロモーター、ヒト伸張因子プロモーター及び他の構成的プロモーターを含む。真核細胞において構成的に機能する代表的なウイルスプロモーターとして、例えば、シミアン・ウイルス由来のプロモーター(例えば、SV40)、パピローマウイルス、アデノウイルス、ヒト免疫不全ウイルス(HIV)、サイトメガロウイルス(CMV)、ラウス・サルコーマ・ウイルス(RSV)、B型肝炎ウイルス(HBV)、モロニー白血病ウイルス及び他のレトロウイルスの末端反復(LTR)、並びに単純ヘルペスウイルスのチミジンキナーゼプロモーターが挙げられる。他の構成的プロモーターは、当業者に知られている。本発明の遺伝子発現配列として有用であるプロモーターとして、誘導性プロモーターが挙げられる。誘導性プロモーターは、誘導試薬の存在下で発現される。例えば、メタロチオネインプロモーターは、特定の金属イオンの存在下において転写及び翻訳を促進するように誘導される。他の誘導性プロモーターは、当業者に知られている。
【0042】
一般的に、遺伝子発現配列として、必要に応じて、転写及び翻訳の開始に関して5'非転写及び5'非翻訳配列、例えばTATAボックス、キャップ配列、CAAT配列などをそれぞれ含む。特に、このような5'非転写配列として、作用可能なように結合された抗原核酸を転写調節するためのプロモーター配列を含むプロモーター領域が挙げられる。当該遺伝子発現配列として、エンハンサー配列又は上記上流アクチベーター配列が挙げられる。
【0043】
本明細書に使用される場合に、ペプチドアンタゴニスト核酸配列及び遺伝子発現配列は、遺伝子発現配列の影響下又は調節下で、ペプチドアンタゴニストをコードする配列の発現、又は転写及び/又は翻訳を実行するような方法で共有結合された場合に、「作用可能なように結合される」と呼ばれる。2個のDNA配列は、5'遺伝子発現配列におけるプロモーターの導入が、ペプチドアンタゴニスト配列の転写をもたらす場合、及び2個のDNA配列のあいだの結合性質が、(1)フレームシフト突然変異の導入をもたらさないか、(2)ペプチドアンタゴニスト配列の転写を仕向けるプロモーター領域の能力と干渉しないか、又は(3)対応RNA転写産物がタンパク質へと翻訳される能力と干渉しない場合に作用可能なように結合されるといわれる。こうして、遺伝子発現配列が、抗原核酸配列の効果的な転写を可能にし、その結果得られた転写産物が所望のタンパク質又はポリペプチドに翻訳される場合に、当該遺伝子発現配列は、ペプチドアンタゴニスト核酸配列に作用可能なように結合される。
【0044】
ペプチドアンタゴニスト核酸は、in vivoで単独で、又はベクターと組み合わせてデリバリーされてもよい。この最も広い意味では、「ベクター」は、ペプチドアンタゴニスト核酸を細胞、並びに好ましくはニューロピリン発現細胞へと輸送することを促進できる任意のビヒクルである。好ましくは、ベクターは、ベクターの不存在下でもたらす分解の程度に比べて低い分解性で細胞に核酸を輸送することができる。ベクターは、場合により、ニューロピリン発現細胞において、ペプチドアンタゴニスト核酸の発現を高める上記遺伝子発現配列を含む。一般的に、本発明に有用であるベクターとして、非限定的に、ペプチドアンタゴニスト核酸配列を挿入又は取込むことにより操作されたプラスミド、ファージミド、ウイルス、ウイルス又は細菌起源に由来する他のビヒクルを含む。ウイルスベクターは、好ましいタイプのベクターであり、そして非限定的に、以下のウイルス:レトロウイルス、例えばモロニーマウス白血病ウイルス、ハーベイマウス肉腫ウイルス、マウス乳癌ウイルス、及びラウス肉腫ウイルス;アデノウイルス、アデノ随伴ウイルス;SV40型ウイルス;ポリオーマウイルス;エプステイン-バーウイルス;パピローマ・ウイルス;ヘルペスウイルス;ワクシニア・ウイルス;ポリオウイルス;及びRNAウイルス、例えばレトロウイルス由来の核酸配列を含む。名前が付けられていないが当該技術分野に知られている他のベクターを簡単に使用することができる。
【0045】
好ましいウイルス・ベクターは、非必須遺伝子が、目的の遺伝子で置換されている非細胞変性真核ウイルスに基く。非細胞変性ウイルスとして、レトロウイルス(例えば、レンチウイルス)が挙げられ、レトロウイルスの生活環は、ゲノム・ウイルスRNAをDNAに逆転写し、続いて宿主細胞DNAへとプロウイルス組込みすることに関する。レトロウイルスは、ヒト遺伝子治療のツールとして承認された。最も有用なものは、複製-欠損型である(つまり、所望のタンパク質の合成を仕向けることができるが、感染性粒子を製造することができない)レトロウイルスである。このような遺伝子的に変更されたレトロウイルス発現ベクターは、in vivoにおいて遺伝子を高効率で形質導入することについて一般的に有用性を有する。複製欠損レトロウイルスを産生するための標準的なプロトコル(外来遺伝子物質をプラスミドへと取込み、パッケージング細胞株にプラスミドをトランスフェクションし、パッケージング細胞株により組換えレトロウイルスを産生し、組織培養培地からウイルス粒子を回収し、そして標的細胞をウイルス粒子で感染させることを含む)が、KRIEGLER("A Laboratory Manual," W. H. Freeman CO., New York, 1990)及びMURRY ("Methods in Molecular Biology," vol.7, Humana Press, Inc., Cliffton, N.J., 1991)に与えられる。
【0046】
特定の適用についての好ましいウイルスは、遺伝子治療においてヒトへの使用がすでに承認された二本鎖DNAウイルスであるアデノウイルス及びアデノ随伴ウイルスである。アデノ随伴ウイルスは、複製欠損性であるように遺伝子操作され、そして、広範囲の細胞型及び種へと感染することができる。熱、及び脂質溶媒安定性;肝細胞を含む様々な細胞系統の細胞における高い形質導入効率;並びに重複感染阻害を欠如していること、そうして多重の形質導入を可能にすることといった利点をアデノ随伴ウイルスはさらに有する。報告された様に、アデノ随伴ウイルスは、部位特異的な様式でヒト細胞DNAへと組み込むことができ、それにより、挿入突然変異誘導の可能性を少なくし、そしてレトロウイルス感染の挿入遺伝子発現特徴の変動を少なくする。さらに、野生型アデノ随伴ウイルス感染は、選択圧の不存在下で100回超の継代のあいだ組織培養において継続されており、アデノ随伴ウイルスのゲノム組込みが比較的安定な事象であることを示唆する。アデノ随伴ウイルスは、染色体外の様式で機能することもできる。
【0047】
他のベクターとしてプラスミドベクターが挙げられる。プラスミドベクターは、当該技術分野においてかなり記載されており、そして当業者に周知である。例えば、SANBROOKら、"Molecular Cloning: A Laboratory Manual," 第二版, Cold Spring Harbor Laboratory Press, 1989を参照のこと。ここ数年のあいだに、プラスミドベクターは、in vivoにおいて抗原をコードする遺伝子を細胞へとデリバリーするDNAワクチンとして使用されてきた。プラスミドベクターは、DNAワクチンについて特に利点がある。なぜなら、プラスミドベクターは、多くのウイルスベクターが有するのと同じ安全性についての懸念を有さないからである。しかしながら、宿主細胞に適合するプロモーターを有するプラスミドは、プラスミド内に作用可能なようにコードされた遺伝子からペプチドを発現することができる。いくつかの一般的に使用されるプラスミドとして、pBR322、pUC18、pUC19、pRC/CMV、SV40、及びpBlueScriptが挙げられる。他のプラスミドは、当業者に周知である。さらに、制限酵素及びライゲーション反応を使用してDNAの特定の断片を取り除き、そして加えて、プラスミドをカスタム設計してもよい。プラスミドは、様々な非経口、粘膜、及び局所経路によりデリバリーされうる。例えば、DNAプラスミドは、筋肉内、皮内、皮下、又は他の経路により注射できる。DNAプラスミドは、鼻腔内スプレー又はドロップ、直腸用坐剤、及び経口により投与されてもよい。DNAプラスミドは、遺伝子銃を用いて、表皮又は粘膜表面に投与されてもよい。当該プラスミドは、水溶液中に与えられてもよいし、金粒子上に乾燥されてもよいし、又は、リポソーム、デンドリマー、渦巻き型化(cochleate)、及びマイクロカプセル化を含む他のDNAデリバリーシステムに関して与えられてもよい。
【0048】
核酸ベクターは、細菌及び哺乳動物細胞の両方で活性である選択的マーカーを含むことができる。
【0049】
1の特異的実施態様では、本発明の核酸ベクターは、「裸のDNA」様であるプラスミド、コスミド、又はファージミドに相当する。このような裸のDNAは、非脂質カチオン性ポリマー(WU及びWU、J. Biol. Chem., vol.263, p: 14621-4, 1988)又はリポソーム(BRIGHMANら, Am. J. Med. Sci. vol.298, p: 278-81, 1989)を伴って、細胞取込みを高める複合体を形成した。
【0050】
第二の具体的な実施態様では、核酸ベクターは、in vivo遺伝子治療プロトコルに適用されたウイルスベクターである。適切なウイルスベクターの例として、EP 0871459号、EP 0386882号及びEP 1222300号に記載されるレトロウイルスベクター、並びにUS 2004/ 265273号及びUS 6,638,502号に記載されるアデノウイルスベクターが挙げられる。この場合、ウイルスの内在化は、細胞表面受容体とウイルスエンベロープとの特異的相互作用、続いてウイルス/受容体複合体の受容体媒介性のエンドサイトーシスを介して生じる。
【0051】
第三の態様では、本発明は、上で記載されるペプチドアンタゴニスト、それをコードする核酸、又は当該核酸を含むベクター、医薬として許容されるビヒクルと最終的に関連する核酸ベクターに関する。
【0052】
医薬として許容されるビヒクルの一例として、組成物は、エマルジョン、マイクロエマルジョン、水中油型エマルジョン、無水脂質及び水中油型乳濁液、他のタイプのエマルジョンを含むこともある。当該組成物はまた、1以上の添加物(例えば、希釈剤、賦形剤、安定剤、保存剤)も含む。一般的に、Ullmann's Encyclopedia of Industrial Chemistry、第6版(共編)、1989-1998、Marcel Dekker);及びPharmaceutical Dosage Forms and Drug Delivery Systems (ANSELら、1994、WILLIAMS & WILKINS)を参照のこと。
【0053】
有利な態様では、当該組成物は、10-12M超、より好ましくは10-11M超、そして最も好ましくは10-10M超の濃度を含む。
【0054】
ペプチドアンタゴニスト、核酸又は核酸ベクターは、緩衝液又は水中に溶解されてもよく、又はエマルジョン及びマイクロエマルジョン中に取込まれてもよい。適切な緩衝液として、非限定的に、Ca2+/Mg2+フリーのリン酸緩衝生理食塩水(PBS)、リン酸緩衝生理食塩水(PBS)、通常の生理食塩水(水中の150mM・NaCl)、Tris緩衝液及び界面活性剤が挙げられる。
【0055】
加水分解及び変性を含む、ペプチド不安定性又は変性についての多くの原因が存在する。疎水性相互作用は、分子の集合(つまり、凝集)を引き起こしうる。この結果は、Treg応答の導入の低減を引き起こしうる。このような問題を低減又は予防するために安定剤が加えられてもよい。
【0056】
安定剤として、シクロデキストリン及びその誘導体が挙げられる(例えば、米国特許第5,730,969号を参照のこと)。適切な保存剤、例えばスクロース、マンニトール、ソルビトール、トレハロース、デキストラン、及びグリセリンは、最終的な製剤を安定化するために加えることができる。イオン性及び非イオン性界面活性剤、D-グルコース、D-ガラクトース、D-キシロース、Dガラクツロン酸、トレハロース、デキストラン、ヒドロキシエチル・スターチ、及びそれらの混合物から選ばれる安定剤は、製剤に加えられてもよい。アルカリ金属塩または塩化マグネシウムの添加は、ペプチドを安定化しうる。ペプチドは、デキストラン、硫酸コンドロイチン、スターチ、グリコーゲン、デキストリン、及びアルギン酸塩からなる群から選ばれる多糖と接触させることにより安定化されてもよい。加えられうる他の糖としては、単糖、二糖、糖アルコール、およびそれらの混合物(たとえば、グルコース、マンノース、ガラクトース、フルクトース、スクロース、マルトース、ラクトース、マンニトール、ザイリトール(xylitol)が挙げられる。ポリオールは、ペプチドを安定化しうるし、そして水混和性であるか、又は水溶性である。適切なポリオールは、ポリヒドロキシアルコール、単糖及び二糖、例えば、マンニトール、グリコール(glycrol)、エチレングリコール、プロピレン・グリコール、トリメチルグリコール、ビニル・ピロリドン、グルコース、フルクトース、アラビノース、マンノース、マルトース、スクロース、及びそのポリマーであってもよい。様々な賦形剤は、ペプチドを安定化し、例えば、血清アルブミン、アミノ酸、ヘパリン、脂肪酸、及びリン脂質、界面活性剤、金属、ポリオール、還元剤、金属キレート剤、ポリビニル・ピロリドン、加水分解性ゼラチン、及び硫酸アンモニウムを含む。
【0057】
第四の実施態様では、本発明は、クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関する疾患を患う患者の予防又は治療処置の方法であって、上記組成物を当該患者に投与するステップを含む、前記方法に関する。
【0058】
本明細書に使用される場合、「対象」という用語は、哺乳動物、例えばげっ歯類、ネコ、イヌ、及び霊長類などを指す。当該対象は、ウシ、ブタ、ウマ、ネコ、イヌ、及び最も好ましくはヒトである。
【0059】
クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関連する疾患は、当業者により簡単に決定できる。この様な疾患の一例として、神経変性疾患(例えば、アルツハイマー病、パーキンソン病、中枢神経系病変、病変に伴う脱髄)、癌(例えば、肺癌、乳癌、及び中皮癌、癌腫、神経膠腫)、及び異常血管新生に関与する全ての疾患を挙げることができる。
【0060】
有利な態様では、当該組成物の投与は、10-12M超、好ましくは10-11M超、そして最も好ましくは10-10M超のペプチドアンタゴニストの濃度に相当する。
【0061】
第5態様では、本発明は、クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関する疾患を患う対象の予防又は治療用の医薬の製造のための、上記ペプチドアンタゴニスト、これらをコードする核酸、又は当該核酸を含む核酸ベクターの使用に関する。
【0062】
クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関連する疾患は、当業者のうちの一人により簡単に決定することができる。この様な疾患の一例として、神経変性疾患(例えば、アルツハイマー病、パーキンソン病、中枢神経系病変、病変に伴う脱髄)、癌(例えば、肺癌、乳癌、及び中皮癌、癌腫、神経膠腫)、及び異常血管新生に関与する全ての疾患を挙げることができる。
【0063】
好ましい実施態様では、本発明は、アルツハイマー病、パーキンソン病、中枢神経系病変、及び病変に付随する脱髄を含む群から選ばれる神経変性疾患を患う対象の予防又は治療用の医薬の製造のための、上記ペプチドアンタゴニスト、これらをコードする核酸、又は当該核酸を含む核酸ベクターの使用に関する。
【0064】
第二の好ましい実施態様では、本発明は、肺癌、乳癌、中皮癌、癌腫、及び神経膠腫を含む群から選ばれる癌を患う患者の予防又は治療用の医薬の製造のための、上記ペプチドアンタゴニスト、それをコードする核酸、又は当該核酸を含む拡散ベクターの使用に関する。
【0065】
第三の好ましい実施態様では、本発明は、異常血管新生に関与する疾患を患う対象の予防又は治療のための医薬の製造のための、上記ペプチドアンタゴニスト、それをコードする核酸、又は当該核酸を含む核酸ベクターの使用に関する。
【0066】
有利な実施態様では、当該医薬は、10-12M超、好ましくは10-11M超、そして最も好ましくは10-10M超のペプチドアンタゴニスト濃度の放出を許容する。
【0067】
本発明は、以下の実施例によりさらに例示される。当該実施例は、その範囲を制限することを意図しない。
【実施例】
【0068】
1) NRP1受容体の膜貫通ドメインは、二量化能力を有する
RUSS及びENGELMAN(Proc. Natl. Acad. Sex. USA, vol. 96, p: 863-8, 1999)により記載されるToxCatシステムから得られたToxLucシステムは、NRP1膜貫通ドメイン媒介性の二量化を調査するために使用された。このシステムは、E.coli細胞内膜中での膜貫通へリックス-へリックス・オリゴマー化を計測することを可能にする。NRP1の膜貫通ドメイン(配列番号1、ILITIIAMSALGVLLGAVCGVVL)の二量化能力は、EGF受容体(配列番号17、SIATGMVGALLLLLVVALGIGLFM)、Erb-2タンパク質(配列番号18、SIISAVVGILLVVVLGVVFGILI)及びグリコプロテインA(配列番号19、ITLIIFGVMAGVIGTILLISYGI)のうちの膜貫通ドメインの二量化能力と比較される。
【0069】
特異的融合タンパク質をコードする幾つかの構築体を製造した。これらの融合タンパク質は、NRP1、EGF受容体、Erb2受容体、及びグリコフォリンAそれぞれの膜貫通ドメインに融合されたToxR(二量化依存性転写活性化因子)のN末端DNA結合ドメイン、並びにモノマーのペリプラズム膜アンカー(マルトース結合タンパク質:MBP)を含んだ。
【0070】
ToxRに繋げられ、そしてマルトース結合タンパク質(MBP)に繋げられたキメラタンパク質として、目的のTM配列を細菌DH5a(MM39)中で発現した。TMドメイン媒介性オリゴマー化は、当該システムのオリジナル・バージョンのクロラムフェニコールアセチルトランスフェラーゼ(CAT)をコードする受容体遺伝子のToxRにより活性化される発現をもたらす。便宜上、発明者は、慣用される分子生物学的方法を用いて、ルシフェラーゼの遺伝子により初期のCAT遺伝子を置き換えた。ニューロピリン、EGF受容体、及び野生型erbB2に対応する合成TM配列を、NheI/DpnII断片として新たなプラスミドにクローニングした。グリコプロテインAから得られるTM配列、又はそのG83I突然変異配列(RUSS 及びENGELMAN, Proc. Natl. Acad. Sci. U S A.vol.96(3), p: 863-8、1999) を有するキメラを対照として用いた。Rocheアッセイキットを製品説明書に従って用い、そしてBerthold Microlumat Plate Luminometerを用いて、ルシフェラーゼアッセイを行った。
【0071】
異なる構築物について結果を図3Aに示す。
【0072】
NRP1の膜貫通ドメインを含む構築物で形質転換された細菌は、Erb-2又はEGFRの膜貫通ドメインを含む構築物で形質転換された細菌のルシフェラーゼ発光より有意に高いルシフェラーゼ発光を示し(Erb-2:4.7倍、EGFR:6.1倍)、そしてグリコフォリン-Aの膜貫通ドメインを含む細菌のルシフェラーゼ発光より少し高かった(1.2倍)。興味深いことに、発明者らは、グリコフォリン-AのRMN構造を下にSwissPdbViewerソフトウェアを用いて得られたTM-NRP1の3次元モデルにより、NRP1のTMドメインの二量化能力を確認した。この論理的アプローチは、必要最低限ではあるが、二量化形成を支援するヘリックス間相互作用を示すTM-NRP1の空間的にコンパクトな構成の存在を確認した(図3B)。
【0073】
結論として、NRP1の膜貫通ドメインは、GpA膜貫通ドメインの二量化能力よりも強い効力で二量化を誘導することができる。
【0074】
2) NRP1の膜貫通ドメインのペプチドが、Sema3Aにより引き起こされる皮質軸索崩壊を阻害する
Sema3Aの軸索成長の阻害性性質が、成長円錐の崩壊を誘導する能力に関連すると示唆されている。Sema3A機能性質へのpTM-NRP1ペプチドの効果を調査するために、皮質ニューロンの成長を、Sema3の存在下又は不存在下で、そしてpTM-NRP1又はpTM-NRP1mut(3個のG→V突然変異を有するNRP1 TM)ペプチドの存在下又は不存在下で分析した。
【0075】
ラミニン/ポリ-L-リジン基質を、980μlのGeyの平衡塩類溶液(GBSS、Sigma)を10μlのラミニン(1mg/ml、Sigma)及び10μlポリ-L-リジン(10mg/ml、Sigma)に加えることにより製造した。滅菌カバースリップを、大ディッシュにおき、そして基質を加えた(100μl)。次に、各カバースリップを第二カバースリップで覆うことにより「サンドウィッチ」を製造した。5%CO2を含む空気の下、37℃で少なくとも30分間インキュベートした後に、「サンドウィッチ」を開き、そして各カバースリップを脱イオン水で洗浄した。乾燥させたカバースリップ上で組織片を培養した。
【0076】
E15マウス胚(E1を膣栓の確認による胚発達の初日として定義する)から調製された大脳新皮質を、組織破砕ディスク上に移した。最初の切断後に、ディスクを90°回転させることにより、組織を200×200μmに切断した。皮質キューブをスパチュラを用いて回収し、そしてぺトリ皿の培養倍地中に入れた。基質付のカバースリップを小ペトリ皿(50mmの直径、FALCON)に置いた。750μlの培養培地を加えた後に、カバースリップをインキュベーター内に少なくとも10分間静置すべきである。解剖顕微鏡を用いて、40〜50の皮質組織片を20μlの培養培地中に集め、そして注意深くカバースリップ上に置いた。室温で15分後、多くの組織片を基質へと接着させた。2250μlの培養培地をゆっくり各ディッシュに加えた。次に、組織片培養物を5%CO2を含む空気中において37℃で維持した。
【0077】
18〜24時間後に培養物中で放射状の伸張が見られ、そしてその結果個々の繊維及び成長円錐を分析した。成長円錐について試験された以下の生成物:
・Sema3Aを安定的に発現するHEK293細胞の条件培地から調製された精製Sema3A(100ng/ml)(ここで、抗-Flagシステム(Sigma)を用いることにより精製を行った。)
・pTM-NRP1(10-8、10-9、10-10、及び10-11M)
・10-8MのmpTM-NRP1
・10-8MのpTM-ErbB2w(配列番号18)
を直接培養培地に2時間加えた。
【0078】
インキュベート後に、4%ホルムアルデヒドを15分間培養培地(v/v)中に直接加えた。次に溶液を取り除き、そして4%ホルムアルデヒドに15分間置き換えた。
【0079】
図4Aは、Sema3Aにより引き起こされる皮質軸索崩壊について濃度を増加させたpTM-NRP1ペプチド(10-8、10-9、10-10、及び10-11M)の効果を示す。
【0080】
図4Bは、Sema3Aにより引き起こされる皮質軸索崩壊について、pTM-NRP1又はpTM-NRP1mutペプチド(10-8M)の効果を示す。
【0081】
50%超の崩壊皮質軸索がSema3A処理細胞で観察される一方、対照条件では、10%未満の皮質軸索しか崩壊形態を示さなかったということが結果により示される(図4A)。さらに、高い濃度の野生型pTM-NRP1ペプチドの添加は、約10-11MのIC50で用量依存的な様式で、皮質軸索についてのSema3Aの崩壊効果を抑制した。
【0082】
対照的に、pTM-NRP1mutペプチドの添加は、Sema3Aにより引き起こされる皮質軸索崩壊を阻害しない(図4B)。
【0083】
ペプチド希釈緩衝液を用いそしてGxxxGモチーフを含むErbB2ペプチドを用いた対照実験は、Sema3Aの崩壊効果について影響を示さない。
【0084】
NRP1の膜貫通ドメインを模倣する合成ペプチドの添加が、皮質軸索へのSema3Aの効果を損なわせ、皮質軸索崩壊を阻害するということがこれらの結果により示される。さらに、この生物学的活性は、GxxxGモチーフと関連し、そしてpTM-NRP1ペプチドに特有である。
【0085】
3) NRP1受容体の膜貫通ドメインペプチドは、Sema3Aにより引き起こされるCOS細胞崩壊を阻害する
COS細胞は、セマフォリン受容体を発現しておらず、そしてその結果これらのガイダンスシグナルにもともと感受性ではない。それにもかかわらず、COS細胞にNRP1及びプレキシン-A1を人工的に発現させることは、Sema3Aの細胞崩壊を引き起こさせる。pTM-NRP1ペプチドのSema3A機能性質についての効果を調査するために、NRP1及びプレキシン-A1を発現するCOS細胞の形を、Sema3A及びpTM-NRP1又はpTM-NRP1mutペプチドの有無について分析した。
【0086】
6ウェル-プレート中で、製品説明書に従いリポフェクタミン2000(invitrogen)を用いて、NRP1及びプレキシン-A1をコードする配列を含むpBK-CMVプラスミド(STRATAGENE)1μgをCOS-1細胞にトランスフェクションした。安定的にトランスフェクションされたCOS-1細胞を0.7%ジェネティシンを用いて選別した。安定的にトランスフェクションされたCOS-1細胞は、前もってポリ-L-リジン-皮膜されたガラスカバースリップを用いて12ウェルプレート上で培養した。細胞を、pTM-NRP1又はpTM-NRP1mutペプチド(10-9M)と1時間37℃でインキュベートした。培養培地を取り除き、そしてSema3A(100μl/ml、D-MEM)を発現する構築物で安定的にトランスフェクションされるか又はされていないHEK細胞の条件培地(conditioned medium)に37℃で4時間置換した。最終的に、細胞を2%ホルムアルデヒドで30分間固定し、続いて4%で15分間固定した。試験された各条件について、約400個の細胞を分析した。
【0087】
pTM-NRP1又はpTM-NRP1mutを伴って又は伴わずにSema3Aの存在下又は不存在下でNRP1及びプレキシン-A1を発現するCOS-1細胞の形態を図5Aは示す。図5Bは、Sema3Aにより引き起こされる細胞崩壊についてのpTM-NRP1又はpTM-NRP1mutペプチドの効果を示す(*:p<0.001)。
【0088】
50%超の崩壊細胞がSema3A処理細胞で観察される一方、対照条件では10%未満の細胞しか崩壊形態を示さなかったことが結果により示される(図5B)。さらに、10-9MのpTM-NRP1ペプチドは、Sema3AのCOS-1細胞を崩壊させる効果を完全に損なわせた。対照的に変異GxxxGモチーフを有するpTM-NRP1mutペプチドの10-9Mを加えることが、Sema3Aにより引き起こされる細胞崩壊を阻害することはなかった。
【0089】
これらの結果により、NRP1の膜貫通ドメインを模倣する合成ペプチドを加えることは、そのGxxxGモチーフを有するSema3Aの細胞崩壊効果を損なわせるということが示された。
【0090】
pTM-NRP1がSema3Aシグナルを遮断するメカニズムを解明するために、発明者らは結合アッセイを行った。
【0091】
NRP1発現COS細胞を、AP-Sema3A、つまりSema3Aと分泌型アルカリホスファターゼの融合タンパク質(BAGNARDら、1998)とインキュベートした。
【0092】
野生型COS細胞又はNRP1-発現COS細胞(COS-NRP1)をポリ-L-リジン-皮膜ガラスカバースリップ上で12ウェルプレート上で培養した(0.005mg/ml)。無血清培地中のpTM-NRP1又はpTM-NRP1mut(10-9M)で37℃にて1時間インキュベートした後に、AP-Sema3Aを安定的に発現するHEK細胞から得たアルカリ・ホスファターゼ結合Sema3Aを含む条件培地で90分間置換した(AP-Sema3A;Bagnardら、1998)。セマフォリンを伴わない条件培地(トランスフェクションされていないHEKから得られる)は、対照として役に立った。PBSで3回細胞を洗浄し、そして4%ホルムアルデヒド中で固定して、新たなディッシュに移した。PBS中で3回洗浄後、プレートを50分65℃で暖めた。続いて細胞を1mlのアルカリホスフェート基質(NBT/BCIP、Sigma)と暗所でインキュベートした。45分後、基質を取り除き、そしてガラスカバースリップを洗浄した。慣用される顕微鏡を用いて写真をとり、そしてAxioVision LE Zeissソフトウェアで分析した。試験された各条件につき、約60個の細胞を分析して、光学密度の関数として結合レベルを測定した。Studentのt検定を用いることにより統計分析を行った。
【0093】
図6Aは、pTM-NRP1又はpTM-NRP1mutを伴うか又は伴わずにAP-Semaとインキュベートした後に、野生型COS細胞(対照)又はNRP1発現COS細胞(COS-NRP1)を示す。
【0094】
図6Bは、以前に試験された条件について、細胞あたりの光学密度の平均強度を示す。
【0095】
結果により、AP-Sema3Aの結合が、pTM-NRP1の添加により有意に低減されることが示される(図6A及びB)。驚くべきことに、pTM-NRP1mutの添加は、COS細胞に結合するAP-Sema3Aをブロックしなかった。
【0096】
これらの結果により、NRP1TMのGxxxGxxxGドメインが、Sema3A結合とそれに続く阻害効果を引き起こすのに必須でありそうであると示される。
【0097】
4) NRP1のTMドメインの突然変異が、受容体機能を損なわせる
GxxxGxxxGモチーフの重要な役割を確認するために、突然変異ペプチドとして、全長NRP1のTMドメインに突然変異を導入して、3個全てのグリシン残基をバリンに置き換えた(NRP1mut)。
【0098】
膜貫通領域中に3個の(G→V)変異を有するNRP1タンパク質をコードするプラスミドを、前に記載される様にCOS細胞にトランスフェクションした。次に前と同様に、NRP1の当該突然変異形を発現するCOS細胞中で結合実験を行った。
【0099】
図7Aは、pTM-NRP1を伴って又は伴わずに、AP-Sema3Aとインキュベートした後における野生型COS細胞(対照)、NRP1発現COS細胞(COS-NRP1)、又は3個の(G→V)変異を有するNRP1を発現するCOS細胞(COS-NRP1mut)を示す。
【0100】
図7Bは、前に試験された条件についての細胞あたりの光学密度の平均強度を示す。
【0101】
野生型NRP1を発現するCOS細胞においてかなりの結合が観察された一方、NPR1mutを発現する細胞においてあるとしてもかなり低いAP-Sema3Aの結合しか検出されなかったということが結果により示される(図7A)。結合のかなりの低下は、pTM-NRP1の存在下で得られた結合低下に類似している。驚くべきことに、COS細胞中で当該NRP1突然変異(NRP1mut)をPlexA1と一緒に発現させた場合、Sema3Aは、もはや細胞崩壊を誘導できなかった(図7B)。このことにより、機能的なSema3A受容体の形成についてのNRP1TMドメインのGxxxGxxxGの重要性がさらに確認された。
【0102】
5) TM-NRP1を模倣する合成ペプチドは、セマフォリン受容体複合体の形成を変更する
受容体複合体形成の観点で、TM-NRP1の生物化学的な帰結をさらに調査するために、発明者らは、PC12神経細胞モデルにおいて複合体形成を分析した。興味深いことに、Sema3Aは、NGFから独立した経路を介してこれらの細胞において神経突起の成長を促進することが示された(Schwambornら、J.Biol.Chem., Vol.279(30), P:30923-6, 2004)。
【0103】
PC12(ATCC:CRL-1721)を、4.5gグルコース/L(Gibco)、5%FVS、10%ウマ血清、グルタミン580mg/l及び抗生物質を含むD-MEM培地中で増殖させた。機能アッセイについて、前もってポリ-L-リジン皮膜されたガラスカバースリップを備える12ウェルプレート上でPC12を培養した。PC12細胞を、pTM-NRP1ペプチド(10-9M)を伴って又は伴わずに37℃で1時間インキュベートした。培養培地を取り除き、そしてNGF-含有(100ng/ml;GIBCO)無血清培地に置換し、又はSema3Aを安定的に発現するHEK293細胞から得られた、又はトランスフェクションされていない細胞から得られた条件培地により、12時間37℃で置換された(対照、詳細についてはBAGNARDら、1998を参照のこと)。
【0104】
細胞を2%ホルムアルデヒドで30分間固定し、続いて4%ホルムアルデヒドで15分間固定した。試験された各条件につき、約400個の細胞を分析して、神経突起の伸張を評価した(統計分析を、χ2検定を用いて行った)。
【0105】
図8は、NGF(100ng/ml)、Sema3A、pTM-NRP1(10-9M)を伴って又は伴わずにインキュベートした後におけるPC12細胞の分化アッセイの結果を示す。
図8Bは、各条件について分化した細胞の割合を示す。
【0106】
NGF又はSema3Aの添加が、相乗効果を与えることなくPC12細胞の分化を誘導したということがこの結果により示される(図8A及び8B)。さらに、実験がpTM-NRP1の存在下で行われる場合、NGF効果が維持されている一方、Sema3A誘導性神経突起成長の促進が有意に低下した。これは、ペプチドの添加が、他のシグナル経路に影響することなくSema3A依存性経路の活性化を特異的に遮断するということを示す。
【0107】
6) NRP1受容体の膜貫通ドメインのペプチドが、リガンドSema3Aのその受容体NRP1への結合を拮抗作用する
NRP1-クラスIIIセマフォリンの結合におけるNRP1膜貫通ドメインの役割を調査するため、神経膠腫細胞上にある受容体NRP1へのリガンドSema3Aの結合を、NRP1の膜貫通ドメインペプチド (pTM-NRP1ペプチド;配列番号1)又は変異ペプチド(mpTM-NRP1;配列番号20、ILITIIAMSALVVLLVAVCVVVLYRKR)の存在下又は不存在下で計測した。 これらのpTM-NRP1ペプチドは、自動化ペプチド合成機(Fmoc化学反応、Applied System)により合成した。ペプチド純度を、RP-HPLC(BECKMAN)により90%超として見積もった。
【0108】
セマフォリン受容体を発現するラットC6神経膠腫細胞は、AP-Sema3A、つまり分泌型のSema3Aのアルカリホスファターゼバージョン(ADAMSら、1997, Bagnardら、1998)の結合能力を決定するために使用された。これらの細胞は、10%ウシ胎児血清(PERBIO)、グルタミン0.5mM(GIBCO)、及び抗生物質:100U/mlペニシリン及び100μg/mlストレプトマイシンを含むMEM培地(GIBCO)中で増殖させ、そしてプレーティングした。
【0109】
C6細胞を、96ウェルプレート中で培養し、そして新たに希釈されたpTM-NRP1又はpTM-NRP1mutペプチド(10-12M〜10-10M)を伴って又は伴わずに1時間37℃でインキュベートした。次に培養培地を、AP-Sema3Aを発現する構築物で安定的にトランスフェクションされたHEK細胞の条件培地に置換した(ADAMSら, EMBO J., Vol.16(20), p:6077-86, 1997;BAGNARDら、1998)。非トランスフェクション細胞であるHEKから得た条件培地を内部対照として用いた。pTM-NRP1ペプチドは、自動化ペプチド合成(Fmoc化学、APPLIED SYSTEM)により合成され、そして質量分析器により分析された。ペプチド純度を、RP-HPLC(BECKMAN)により90%超として見積もり、細胞をPBSで洗浄し、そして50μlのアルカリホスファターゼ発光基質(AMERSHAM)とインキュベートした。製品説明書に従って、MICROLUMAT PLUSシステム(BERTHOLD TECHNOLOGIES)を用いて15分後に発光を読み込んだ。各ペプチドについて実験を4回行った。
【0110】
pTM-NRP1ペプチド(黒色棒)又は変異ペプチド(灰色棒)の存在下におけるAP-Sema3Aのその受容体への結合についての結果を図9に示す(*:p<0.005;**:p<0.01、スチューデントt検定)。
【0111】
AP-Sema3AのC6細胞上におけるその受容体NRP1への結合が、野生型pTM-NRP1ペプチドの濃度を増加させて加えることにより、用量依存的な様式で妨げられた。野生型pTMNRP1ペプチドの10-10Mの存在下で、AP-Sema3AのNRP1への結合がペプチドの不存在下の場合に比べて約50%に低下した。野生型pTM-NRP1ペプチドとは対照的に、pTM-NRP1mutペプチドの添加が、AP-Sema3AのNRP1への結合を妨げられなかった。
【0112】
結果として、Sema3Aのその受容体NRP1への結合に、NRP1膜貫通ドメインが関与し、そしてダブルGxxxGモチーフの完全性を必要とする。
【0113】
7) NRP1受容体の膜貫通ドメインペプチドは、セマフォリン受容体複合体の形成を変更する
NRP1受容体複合体の形成において、NRP1受容体の膜貫通ドメインの役割をさらに調査するために、NRP1複合体の形成を、NRP1の膜貫通ドメインペプチド(pTM-NRP1ペプチド)の存在下又は不存在下で、そしてリガンドSema3Aの存在下又は不存在下で、NRP1及びプレキシン-A1を発現するC6細胞上で測定された。
【0114】
NRP1及びプレキシン-A1を発現するC6細胞を、pTM-NRP1ペプチド(10-9M)と1時間インキュベートするか又はインキュベートしなかった。次に培養培地を、Sema3Aを発現する構築物で安定してトランスフェクションされるか又はされていないHEK細胞の培地と取り替えられた。コンフルエントC6を10mM・EDTAを用いて回収し、そして遠心した。ペレットをPBSで洗浄し、そして次に0.1%SDS、1mM・EDTA、1%NP-40、0.5%DOC、2mM・バナジウム塩、及びSDSを伴わないプロテアーゼ阻害剤(PIERCE)を含む溶解緩衝液(Tris-HCl/NaCl;50/150;pH8.0)中に希釈した。4℃で1時間可溶化した後に、ビシンコニン酸法によりタンパク質量を見積もった(BCAタンパク質アッセイ、PIERCE)。
【0115】
スクロース密度勾配沈降実験は、25%、17%、10%、及び3%のスクロースを含む段階的勾配に基いた。溶液を、Hepes/NaCl緩衝液(30/30、pH7.6、0.12%トリトン)及び1Mスクロースを伴うHepes/NaCl緩衝液から作成された。これらの溶液は、線形勾配を形成するために連続して充填された(LERAYら、Arch Biochem Biophys., 1992)。
【0116】
細胞ライセートを超遠心管中の勾配の上に配置し、そしてTL-100超遠心機(BECKMAN)を用いて1時間100000gで遠心し、そして底から分画を回収した(13滴/分画)。
【0117】
Laemmli法に従って、等体積の充填緩衝液をサンプル(62.5mM・Tris-HCl、pH6.8、10%グリセロール、2%SDS、DTT、ブロモフェノールブルー)に加え、そしてこれらを10分間ボイルした。次にサンプルをアクリルアミドゲル(5〜20%)上で一定電圧及び温度で、適切な緩衝液(0.025M・Tris、0.192Mグリシン、pH8.3、0.01%SDS)中でSDS-PAGEにかけた。次にタンパク質を、4℃で、20%エタノール、0.025M・Tris、0.192MグリシンpH8.3、及び0.01%SDSを含む緩衝液中で3時間メタノール活性化ポリビニリデンジフロリド(PVDF)膜に転写した。最後に、PVDF膜を1時間PBS/BSA5%でブロッキングした。
【0118】
次に膜を2時間ポリクローナル抗NRP1(ONCOGENE)で2時間、1:1000の希釈率でインキュベートした。膜をPBS/0.2%TWEEN20中で3回洗浄し、そして二次抗体(A/Gタンパク質、PIERCE、1:100000又はホースラディッシュ・ペルオキシダーゼ結合抗ラビットIgG、AMERSHAM、1:500)とインキュベートした。次に、免疫反応性を、製品説明書に従って、高められた化学発光ウエスタンブロット検出システム(PIERCE)を用いて検出した。
【0119】
結果を図10に示す。各分画におけるNRP1の割合を、明らかにされた総NRP1から計算した。この図では、プレキシン-A1を含むオリゴマーを含むスクロース勾配の重分画は、黒色棒グラフに相当し、NRP1ダイマーを含む中間分画は灰色棒グラフに相当し、そして軽い分画は、ほぼNRP1モノマーから構成され、白色棒グラフに相当する。
【0120】
リガンドSema3Aの不存在下では、NRP1がスクロース勾配の中間分画において主に検出されたということが結果により示される。こうして、NRP1二量体は、そのリガンドSema3Aの不存在下ではオリゴマー形態として主に存在した。
【0121】
対照的に、Sema3Aの存在下では、NRP1受容体は主に重分画中にプレキシン-A1を含むオリゴマー形態として存在した。
【0122】
Sema3Aの存在下におけるpTM-NRP1ペプチドの添加は、NRP1の分布を変更させ、NRP1モノマーの移動レベルに相当する軽い分画中で主に検出された。こうして、NRP1のオリゴマー化は、pTM-NRP1ペプチドの存在により阻害された。それゆえ、NRP1の膜貫通ドメインは、NRP1受容体複合体の形成に関与する。結果として、pTM-NRP1ペプチドの存在下における図3において観察されたSema3A結合の低下は、NRP1オリゴマー化の阻害に関連することもありうる。
【0123】
8) 腫瘍細胞移動のあいだにおけるpTM-NRP1-依存性のNRP1の不活性化の機能的意味
ヒト神経膠腫の優れたモデルであるラットC6神経膠腫細胞株(DAI及びHOLLAND、Biochim. Biophys. Acta, vol.1551, p: M19-27, 2001)は、発明者らのペプチド戦略(pTM-NRP1)を用いることによるNRP1の阻害が、細胞移動及び播種をどれだけ妨げるかを調べるために使用された。
【0124】
C6細胞(ATCC CCL-107)を、PKH26(Sigma)を用いて染色した。注入前に培養培地(5000u/mlペニシリン、5mg/mlストレプトマイシン、200mM・L-グルタミン、及び10%ウシ胎児血清から構成される)中で、氷上で少なくとも2時間、細胞をペプチド(pTM-NRP1 10-8M又は変異pTM-NRP1 10-8M)とインキュベートした。106個の細胞の注射を、以下の座標:ブレグマに対して前後+1.6mm;L、+2mm;皮質表面に対してH、+5mm(antero-posterior, +1.6mm relative to Bregma; L, +2mm; H, +5mm relative to the cortical surface)に従って定位フレームを用いて行った。
【0125】
全ての注射を左線条体で行った。
【0126】
8日の生存期間の後に、動物(4匹のラットからなる3群)を、ペントバルビタールの腹腔内注射により屠殺し、次に100mlのPBSで前もって洗浄し、続いて500mlの2%ホルムアルデヒドで経心臓かん流を行った。脳を4℃で2時間、後固定(post-fixed)し、そして矢状断面(70μm)をビブラトームで調製した。
【0127】
1群の切片を、顕微鏡観察用にPBS-グリセロール(v/v)にマウントし、そして別の群をCD34の免疫染色用に処理した。切片を最初に5%ウシ通常血清を含むPBS中で15分間室温でインキュベートして、非特異的結合部位をブロッキングした。2回目のインキュベートを1時間室温で行いそして次にマウス抗CD34(1:200)を用いて4℃で一晩行った。切片を5分間PBS中で6回洗浄し、そして次にAlexa488に結合されたヤギ抗マウス抗体(1:500;INTERCHIM)と3時間室温でインキュベートした。切片を5分間で6回PBSで洗浄し、そして最終的にPBS-グリセロール(v/v)にマウントし、顕微鏡分析を行った。
【0128】
図11は、pTM-NRP1又はpTM-NRP1mutペプチドを伴うか又は伴わずにC6細胞を脳注射した後のマウス脳切片を示す。腫瘍、皮質、線状体、脳梁(cc)、海馬(Hp)及び側脳室が、顕微鏡写真に写っている。
【0129】
図12は、pTM-NRP1又はpTMNRP1mutを伴って又は伴わずにC6細胞を脳に注射した後において、CD34を免疫染色した後のマウス脳切片を示す。
【0130】
対照条件において、腫瘍が線状体において発達し、そして脳梁及び皮質板に到達したということが結果により示される(図11、n=4)。驚くべきことに、細胞を注射前に、pTM-NRP1で処理した場合に、発明者らは、8日目において腫瘍サイズのかなりの低下を観察した(n=4)。予期される様に、変異pTM-NRP1で処理されたC6細胞は、処理されていない細胞で観察された腫瘍と同様の腫瘍を誘導した(n=4)。こうして、pTM-NRp1の添加は、C6神経膠腫の発達を阻害する。
【0131】
pTM-NRP1の存在下における腫瘍サイズの低下は、CD34、つまり血管新生のマーカー、への免疫反応性のかなりの低減を伴った(図12)。これにより、pTM-NRP1が、VEGFシグナルをブロッキングすることにより抗腫瘍効果を発揮することが示唆された。
【0132】
9) pTM-NRP1が、in vitroにおいてVEGFシグナル伝達を拮抗作用できる
NRP1は、VEGFの受容体である(NEUFELDら、Adv. Exp. Med. Biol., vol.515, p: 81-90, 2002)。その結果、本発明者は、pTM-NRP1がC6細胞においてVEGFシグナル伝達を拮抗作用できるということを確かめた。この目的を達成するために、前に記載されたように調製されたC6腫瘍細胞凝集体(BAGNARDら、1998;及びNASARREら、Neoplasia、Vol.7, p180-189, 2005)を3Dマトリックス(凝固血漿)中で増殖させ、そしてVEGF165で処理した。
【0133】
代表的な腫瘍細胞が、VEGF165(50ng/ml)、pTM-NRP1又はpTMNRP1mut(10-8M)の添加を伴って又は伴わずに凝集することを図13が示す。
【0134】
50ng/mlVEGF165の添加が、C6細胞の凝集体からの移動及び移動鎖(migration chains)の形成を誘導した(図13)。驚くべきことに、pTM-NRP1の添加は、VEGF165依存性C6細胞移動を抑制した。変異型pTM-NRP1の添加は、VEGF165-誘導性C6細胞移動を妨げることができなかった。これらの結果により、pTM-NRP1が、C6細胞におけるVEGF165シグナル伝達を遮断することができるということが示唆される。
【0135】
最後に、これらの結果により、pTM-NRP1が、腫瘍形成の点でNRP1シグナル伝達を阻害するために使用できるということが示唆される。これはVEGF依存性メカニズムを通した腫瘍細胞の移動及び生存におけるNRP1の役割に関する。pTM-NRP1を用いてNRP1を遮断することが、その生存、増殖、及び/又は播種がNRP1依存性のシグナル伝達カスケードを必要とする全ての腫瘍に対して、治療成果を有するということが本発明者らにより提案される。
【図面の簡単な説明】
【0136】
【図1】図1は、膜貫通ドメインであって、以下のマウスの: ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、 ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、 プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、 プレキシンA2(配列番号4:LLTLPAIISIAAGGSLLLIIVIIVLIAY)、 プレキシンA3(配列番号5:LTLPAMVGLAAGGGLLLLAITVVLVAY)、 プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、 Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCF)、 L1−CAM(配列番号8:GWFIAFVSAIILLLLILLILCFI)、 インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び インテグリンβ2(配列番号10:VAAIVGGTVVGVVLIGVLLLVIW) で表される膜貫通ドメインを示す。
【図2】図2は、以下のヒトの: ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、 ニューロピリン−2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、 プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、 プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、 プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、 プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、 Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、 L1−CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、 インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW) についての同じ膜貫通ドメインを示す。
【図3】図3Aは、異なる構築物について結果を示す。
【図4】図4Aは、Sema3Aにより引き起こされる皮質軸索崩壊について濃度を増加させたpTM-NRP1ペプチド(10-8、10-9、10-10、及び10-11M)の効果を示す。図4Bは、Sema3Aにより引き起こされる皮質軸索崩壊について、pTM-NRP1又はpTM-NRP1mutペプチド(10-8M)の効果を示す。
【図5】図5Aは、pTM-NRP1又はpTM-NRP1mutを伴って又は伴わずにSema3Aの存在下又は不存在下でNRP1及びプレキシン-A1を発現するCOS-1細胞の形態を示す。図5Bは、Sema3Aにより引き起こされる細胞崩壊についてのpTM-NRP1又はpTM-NRP1mutペプチドの効果を示す(*:p<0.001)。
【図6】図6Aは、pTM-NRP1又はpTM-NRP1mutを伴うか又は伴わずにAP-Semaとインキュベートした後の野生型COS細胞(対照)又はNRP1発現COS細胞(COS-NRP1)を示す。図6Bは、以前に試験された条件について、細胞あたりの光学密度の平均強度を示す。
【図7】図7Aは、pTM-NRP1を伴って又は伴わずに、AP-Sema3Aとインキュベートした後における野生型COS細胞(対照)、NRP1発現COS細胞(COS-NRP1)、又は3個の(G→V)変異を有するNRP1を発現するCOS細胞(COS-NRP1mut)を示す。図7Bは、前に試験された条件についての細胞あたりの光学密度の平均強度を示す。
【図8】図8は、NGF(100ng/ml)、Sema3A、pTM-NRP1(10-9M)を伴って又は伴わずにインキュベートした後におけるPC12細胞の分化アッセイの結果を示す。図8Bは、各条件について分化した細胞の割合を示す。
【図9】図9は、pTM-NRP1ペプチド(黒色棒)又は変異ペプチド(灰色棒)の存在下におけるAP-Sema3Aのその受容体への結合についての結果を示す(*:p<0.005;**:p<0.01、スチューデントt検定)。
【図10】図10は、プレキシン-A1を含むオリゴマーを含むスクロース勾配の重分画に相当する黒色棒グラフ、NRP1ダイマーを含む中間分画に相当する灰色棒グラフ、そしてほぼNRP1モノマーから構成される軽い分画に相当する白色棒グラフを示す。
【図11】図11は、pTM-NRP1又はpTM-NRP1mutペプチドを伴うか又は伴わずにC6細胞を脳注射した後のマウス脳切片を示す。
【図12】図12は、pTM-NRP1又はpTMNRP1mutを伴って又は伴わずにC6細胞を脳に注射した後において、CD34を免疫染色した後のマウス脳切片を示す。
【図13】図13は、代表的な腫瘍細胞が、VEGF165(50ng/ml)、pTM-NRP1又はpTMNRP1mut(10-8M)の添加を伴って又は伴わずに凝集することを示す。
【技術分野】
【0001】
本発明は、クラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニスト、及び当該アンタゴニストの使用に関する。
【0002】
その構造的及び栄養的役割とは別に、細胞外マトリクス(ECM)は、細胞間連絡の理想的な環境を規定し、そして全ての細胞挙動、例えば増殖、移動、分化、又はアポトーシスを決定する。これらの過程を制御する分子メカニズムは、次第に理解されてきている。神経系では、拡散又は膜貫通シグナルについての3の主要なファミリー(ネトリン、セマフォリン、及びエフリン)は、胚発生のあいだのその機能を有する(TESSIER-LAVIGNE及びGOODMAN, Science, vol.274, p: 1123-33, 1996)。その中で、セマフォリンは、その構造特異性に従って8のクラスに分けられる25超のメンバーから構成されるファミリーを規定し(KOLODKINら、Cell, vol.75, p: 1389-99, 1993)、そしてそれらは、分泌型または膜貫通型セマフォリンのいずれかに分類できる。分泌型セマフォリンはクラスII(無脊椎動物)、III(脊椎動物)、及びV(ウイルス)であり、一方他のクラス(I、IV、及びVI〜VIII)は膜貫通型である。
【0003】
過去5年の間に、生理的及び病理的状態における軸索ガイダンス、細胞移動、細胞分化からアポトーシスに至るセマフォリンの様々な機能の伝達を可能にする伝達経路を解明するために幾つもの研究が設計されてきた。この機能的な多様性は、アクチン細胞骨格リモデリングを導く複数の細胞内経路を選択的動員及び活性化することによりシグナルの統合を調節する高度に動的な受容体複合体を形成するために生じると現在考えられている(CASTELLANI及びROUGON, Curr. Opin. Neurobiol. , vol.12, p:532-41, 2002)。これらの全ては、12〜16個のシステインを有する約500のアミノ酸からなる「semaドメイン」と呼ばれる共通ドメインであって、各セマフォリンの結合特異性を与える共通ドメインを有する(RAPER, Curr. Opin. Neurobiol., vol.10, p: 88-94, 2000)。これらの様々なセマフォリンの中で、クラスIIIセマフォリンは、神経細胞成長円錐の崩壊を誘導しうる。クラスIIIセマフォリンが当初、コラプシンと名づけられた所以である(LUOら、Cell, vol.75, p: 217-227, 1993)。他の残りのファミリーにその名称を与える分子であるsema3Aは、最もよく研究されており、そして全ての場合において、知覚神経及び脊椎運動神経から皮質の錐体神経に至るまで、軸索の忌避因子として記載されてきた(MUELLER, Annu. Rev. Neurosci., vol.22, p: 351-388, 1999)。特に、このセマフォリンは、同じ細胞において2の異なる効果を発揮することができる。これは、皮質ニューロンにおいて示されており、皮質ニューロンでは、sema3Aは、軸索の忌避因子として作用し、かつ樹状突起の化学誘引物質でもある(POLLEUXら、Nature, vol.404, p:567-73, 2000; BAGNARDら、Development, vol.125(24)、p: 5043-53, 1998)。この現象を説明するために、2個の細胞の極において、区別される伝達を確保するメカニズが存在することを考慮する必要がある。区別される伝達の原理だけではなく、分子ヒエラルキーを制御するメカニズムを理解すること、並びに環境変化に応答する細胞挙動の多様性を確保する超分子構造(supra-molecular structures)の形成を解明することが必要とされる。
【0004】
従って、ニューロピリンファミリーの2の既知のメンバー、ニューロピリン-1(NRP1)及びニューロピリン-2(NRP2)の、クラスIIIセマフォリンの伝達カスケードに関与する受容体複合体のリガンド結合サブユニットとしての役割を近年の研究は示した(総説として、Bagnard D. (編) ニューロピリン: from nervous system to vascular and tumor biology. Landes Bioscience- Kluwer Academic/Plenum Publishers Hardbound, ISBN 0-306-47416-6、Advance in Experimental Medicine and Biology Vol.515, p: 140, 2002を参照のこと)。NRP1及びNRP2は、(i)二量化に重要である細胞外部分(RENZIら、J. Neurosci., vol. 19, p:7870-7880, 1999)、膜貫通部分、及び約40のアミノ酸の短い細胞質ドメインを有する1回膜貫通タンパク質である。
【0005】
興味深いことに、NRP1及びNRP2は、伝達能力を持たない短い細胞内ドメインを有する。この所見についての分子的説明は、ニューロピリンがプレキシンファミリーに属する受容体との複合体を形成するということ、並びにプレキシンがニューロピリン/プレキシン複合体の伝達要素であることが見出された際に与えられた(RHOMら、Mech. Dev. , vol.93, p: 95-104, 2000; TAMAGONEら、Cell, vol.99, p:71-80, 1999)。最終的に、クラスIIIセマフォリンによるシグナル伝達は、ニューロピリンとプレキシンとのあいだの複合体形成に依存する。
【0006】
それにもかかわらず、プレキシンとの複合体は、ニューロピリンにより形成される複合体の唯一のタイプではない。
【0007】
ニューロピリンが、アドヘシン分子L1-CAM及びNr-CAMと安定な複合体を形成できるということ(CASTELLANIら、Neuron, vol.27, p: 237-249, 2000)、並びにL1の細胞外ドメインにおける突然変異又は遺伝子標的化マウスにおけるL1の完全な欠失が、sema3Aシグナルの途絶をもたらし、ガイダンスエラーが生じるということが見出された。
【0008】
その結果、チロシン・キナーゼ受容体は、ニューロピリンが関連するシグナル伝達に役割を果たしうる。こうして、DEV神経外胚葉細胞の移動が、Sema3Aにより停止され、そしてNRP1及びVEGFR-1の存在が、当該停止に必要とされるということが観察された(Bagnardら、J Neurosci. , vol.21, p:3332-41, 2001)。この相互作用は、Sema3Aを用いた血管新生のin vitroモデルにおいて、VEGFによる芽形成の阻害を説明する(MIAOら、J. Cell. Biol., vol.146, p:233-242, 1999)。ニューロピリンが、VEGFR-2と(SOKERら、Cell, vol.92, p:735- 45, 1998)、並びにMET(WINBERG et al. , Neuron, vol.32, p:53- 62, 2001)と複合体を形成することも発見された。
【0009】
近年、ニューロピリンが、インテグリンと複合体を形成し、そして当該複合体が、軸索の伸張を促進することができるということが示された(PASTERKAMPら、Nature, vol.424, p:398-405, 2003)。
【0010】
その結果、近年分かったことであるが、上記研究は、神経変性疾患及び癌の潜在的な標的としてクラスIIIセマフォリン/ニューロピリン複合体を同定することに寄与した(GUTTMANN-RAVIVら、Cancer Letter, 2006; CHEDOTALら、Cell Death and Differentiation, 2005を参照のこと)。この文脈では、複合体形成を妨げる薬剤は、明らかに治療潜在性を有し、及び/又は有用な生物学的ツールであろう。
【0011】
このように、GARETHら(Journal of Neurochemistry, vol.92, p: 1180-1190, 2005)は、Sema3A/NRP1複合体のペプチドアンタゴニストを設計するためにアルゴリズムを用いた。この著者は、Sema3A/NRP1二量化に関与するSema3A Igドメインと、受容体の側面二量化を媒介するが、リガンド結合を媒介することはないNRP1 MAMドメインとのあいだにおけるアンタゴニスト・ペプチドを同定した。それにもかかわらず、膜貫通ドメインに位置しないこれらのアンタゴニストは、1μMを超えるIC50を有し、当該濃度は、かなりの濃度であるので、治療においてこのようなアンタゴニストを使用することができない。
【0012】
こうして、治療に使用できるクラスIIIセマフォリン/ニューロピリン複合体の新たなアンタゴニストについて、当該技術分野において認識されている変わらぬ必要性が存在する。
【発明の開示】
【0013】
本発明の目的は、高い活性を有する新たなアンタゴニストを提供することにより当該必要性を満たすことである。
【0014】
本発明者は、NRP1の膜貫通ドメインに相当するペプチドが、10-11M未満のIC50でSema3Aにより誘導される皮質軸索崩壊を阻害することができるということを思いがけずに示した。このペプチドは、2の連続したGxxxG二量化モチーフ(ここで、xは任意のアミノ酸を表す)を含み、当該GxxxGモチーフは、グリコフォリンAにおいて最初に同定された(SENESら、J. MoI. Biol., vol.296, p:921-36, 2000)。このようなモチーフは、erbB受容体(MENDROLAら、J. Biol. Chem. , vol.277, p.4704-12, 2002)のTMセグメントの二量化において働いていると示された。一般的に、小さいアミノ酸-xxx-小さいアミノ酸からなる任意のモチーフ(ここで、小さいアミノ酸のモチーフ定義は、当業者に周知である)は、同等の性質を有する。
【0015】
NRP1のダブルGxxxGモチーフは、高度に保存されており、そしてNRP2のダブルGxxxGモチーフと強いホモロジーを示す(図1)。最終的に、このようなモチーフは、プレキシン・ファミリーのメンバー、Nr-及びL1-CAM、並びにインテグリンを含むニューロピリンの複数のパートナーの膜貫通ドメインにおいて見つかる。全体として、本発明者により得られる結果により、これらのタンパク質の膜貫通ドメインが、セマフォリンシグナル伝達を保証する複合体の形成及び調節において重要な役割を果たすということが示唆される。
【0016】
結果として、1の態様では、本発明は、ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1及びインテグリンβ2からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも1のGxxxGモチーフを含むアミノ酸配列を含み、場合により異種配列に融合されるクラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニストに関する。
【0017】
本明細書に使用される場合、「異種配列」は、ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1又はインテグリンβ2に由来しない任意のアミノ酸配列に関する。当該異種配列は、例えば、特定の細胞内への配置を可能にするか、又は本発明のペプチドアンタゴニストの優れた精製収率(例えばHisタグ)を可能にすることができる。
【0018】
本明細書に使用される場合、「クラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニスト」という用語は、当該複合体形成を妨げ、そして最終的にこのような複合体のシグナル伝達を妨げる合成又は組換えポリペプチドに関する。結果として、本発明のペプチドアンタゴニストは、完全なニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1、及びインテグリンβ2タンパク質を含まない。
【0019】
本明細書に使用されるとき「膜貫通ドメイン」は、細胞膜を横断するペプチドドメインに相当する。当該ドメインは疎水性であり、そしてα-へリックス構造を有する。当業者は、その一般的知識に従って、当該タンパク質のドメインを簡単に同定できる。一例として、タンパク質ドメインの親水性は、Kyte & Doolittle法により決定することができ、そしてα-へリックス構造を形成するタンパク質ドメインの可能性は、Chou&Fasman法により決定することができる。このような方法は、以下のアドレス: http://www.expasy.org/tools/protscale.htmlで利用できる。
【0020】
ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1及びインテグリンβ2の膜貫通ドメインのアミノ酸配列はよく保存されており、そしてタンパク質の完全なアミノ酸配列から簡単に同定することができる。この方法は、当業者に周知である。一例として、ニューロピリン-1アミノ酸配列であって、ムス・ムスクルス(Mus musculus)(P97333)、ホモ・サピエンス(014786)、ラッタス・ノルベギクス(Rattus norvegicus)(Q9QWJ9)、ゼブラフィッシュ(Zebrafish)(Q8QFX6)、及びガルス・ガルス(Gallus gallus)(P79795)に由来する配列;ニューロピリン-2アミノ酸配列であって、ホモ・サピエンス(060462)、ムス・ムスクルス(035375)、ラッタス・ノベルギクス(NP_110496)及びガルス・ガルス(NP_989615)に由来する配列;プレキシンA-1アミノ酸配列であって、ホモ・サピエンス(NP_115618、Q9UIW2)及びムス・ムスクルス(NP_032907、P70206)に由来する配列;プレキシンA-2アミノ酸配列であって、ホモ・サピエンス(CAI40198、Q5JRL6)及びムス・ムスクルス(NP_032908、P70207)に由来する配列;プレキシンA3アミノ酸配列であって、ホモ・サピエンス(NP_032907、P51805)、ゼノパス・トロピカリス(Xenopus tropicalis)(CAI40198)及びムス・ムスクルス(NP_032909、P70208)に由来する配列;プレキシンA4アミノ酸配列であって、ムス・ムスクルス(NP_786926、Q80UG2)、ホモ・サピエンス(Q9HCM2)及びダニオ・レリオ(Danio rerio)(NP_001004495)に由来する配列;Nr-CAMアミノ酸配列であって、ムス・ムスクルス(Q810U4)、ラッタス・ノベルギクス(P97686)、ホモ・サピエンス(Q92823)及びガルス・ガルス(P35331)に由来する配列;L1-CAMアミノ酸配列であって、ホモ・サピエンス(P32004)、タキフグ・ルブリペス(Takifugu rubripes)(Q98902)、ムス・ムスクルス(P11627)及びラッタス・ノベルギクス(Q05695)に由来する配列;インテグリンβ1アミノ酸配列であって、ムス・ムスクルス(P09055)、ホモ・サピエンス(P05556)、フェリス・カツス(Felis catus)(P53713)、ラッタス・ノベルギクス(P49134)、ゼノパス・ラエビス(Xenopus laevis)(P12606)及びガルス・ガルス(P07228)に由来する配列;インテグリンβ2アミノ酸配列であって、ムス・ムスクルス(P11835)、ホモ・サピエンス(P05107)、スス・スクロファ(Sus scrofa)(P53714)、ボス・タウルス(Bos taurus)(P32592)及びシグモドン・ヒスピヅス(Sigmodon hispidus)(AAL38579)に由来する配列を引用することができる。
【0021】
図1は、膜貫通ドメインであって、以下のマウスの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号4:LLTLPAIISIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号5:LTLPAMVGLAAGGGLLLLAITVVLVAY)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCF)、
L1−CAM(配列番号8:GWFIAFVSAIILLLLILLILCFI)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び
インテグリンβ2(配列番号10:VAAIVGGTVVGVVLIGVLLLVIW)
で表される膜貫通ドメインを示す。「GxxxGモチーフ」という用語は、SENESら(上記、2000)に同定されるモチーフに関し、これは図1に示される(下線)。潜在的な「GxxxG」モチーフも示される(点線)。
【0022】
これらの膜貫通ドメインの保存は、図2に明らかに目立っており、図2は、膜貫通ドメインであって、
以下のヒトの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン−2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1−CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
についての同じ膜貫通ドメインを示す。
【0023】
この保存の別の例として、以下の種に由来するニューロピリン-1膜貫通ドメイン:
ガルス・ガルス(配列番号14: ILITIIAMSALGVLLGAICGVVL)、及び
ゼブラフィッシュ(配列番号15:ILITIIAMSALGVFLGAICGVVL)、並びに
ガルス・ガルス由来のニューロピリン-2膜貫通ドメイン(配列番号16:ILVTIIAMSSLGVLLGATCAGLLLY)
を引用することができる。これらは、ヒトニューロピリン-1およびニューロピリン-2膜貫通ドメインとそれぞれ90%超の同一性を有する。
【0024】
好ましい実施態様によると、本発明は、膜貫通ドメインであって、以下のヒトの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン−2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1−CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも1のGxxxGモチーフを含むアミノ酸配列を含み、場合により異種配列に融合されるクラスIIIセマフォリン/ニューロピリン複合体のペプチド性アンタゴニストに関する。
【0025】
別の好ましい実施態様に従って、クラスIIIセマフォリン/ニューロピリン複合体のペプチド性アンタゴニストは、以下の:
ニューロピリン-1(配列番号1、ILITIIAMSALGVLLGAVCGVVL)又は
ニューロピリン-2(配列番号2、ILITIIAMSSLGVLLGATCAGLLLY)膜貫通ドメインに由来するアミノ酸配列を含み、場合により異種配列に融合される。
【0026】
さらに別の好ましい実施態様では、クラスIIIセマフォリン/ニューロピリン複合体のペプチド性アンタゴニストは、少なくとも2個のGxxxGモチーフ、好ましくは少なくとも2個の連続GxxxGモチーフを含むタンパク質の膜貫通ドメインに由来し、そして、以下のヒトの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL),
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
の膜貫通ドメインからなる群から選択されるアミノ酸配列を含み、場合により異種配列に融合される。
【0027】
好ましくは、本発明のペプチドアンタゴニストは、ヒトニューロピリン-1膜貫通ドメイン(配列番号1、ILITIIAMSALGVLLGAVCGVVL)に由来するアミノ酸配列を含む。
【0028】
有利な態様では、上記タンパク質のうちの1つの膜貫通ドメインに由来するアミノ酸配列は、10超のアミノ酸長であり、好ましくは14超のアミノ酸長であり、一例として18超のアミノ酸長、そしてより好ましくは22超のアミノ酸長である。
【0029】
有利な態様では、ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1およびインテグリンβ2からなる群から選ばれるタンパク質のうちの1つに由来するアミノ酸配列は、150未満のアミノ酸長であり、好ましくは100アミノ酸未満のアミノ酸であり、より好ましくは50未満のアミノ酸長である。
【0030】
好ましい実施態様では、ニューロピリン-1及びニューロピリン-2に由来するペプチドアンタゴニストは、クラスIIIセマフォリン二量化に関与するその細胞外ドメインを含まない。これらのドメインは、当業者に周知であり、そしてNEUFELDら(TCM, vol.12(1), p: 13-19、2002)及びBAGNARD (2002, 上記)に記載されている。例えば、これらのドメインは、NRP1及びNRP2のaドメイン(CUBドメイン、セマフォリン結合を保証するa1及びa2ドメインとも呼ばれている)、bドメイン(凝集因子V/VIIIに対するホモログドメイン;b1ドメインとb2ドメインに分けられる。b1は、VEGFイソ型の結合に関与する)、cドメイン(MAMドメイン、NRP1の二量化に関与する)を含む。
【0031】
有利な態様では、当該ペプチドアンタゴニストは、以下の膜貫通ドメインであって:
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr-CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1-CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び、
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
の膜貫通ドメインからなる群から選ばれるアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される。
【0032】
第二の好ましい実施態様では、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、インテグリンβ1、インテグリンβ2、Nr-CAM及びL1-CAMは、シグナル伝達経路に関与するその細胞内ドメインを含まない。これらのドメインは、当業者に周知であり、そしてBAGNARD(上記、2002)に記載され、そして例えば非限定的に、Sex-プレキシンドメイン、PH1A及びPH2Aドメイン又はPRB(プレキシンRac結合ドメイン)ドメインを含む。
【0033】
有利な態様では、当該ペプチドアンタゴニストは、以下の:ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)及びニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)膜貫通ドメインからなる群から選択されるアミノ酸配列又はこの誘導体からなり、場合により異種配列に融合される。好ましくは、当該ペプチド性アンタゴニストは、ニューロピリン-1膜貫通ドメイン(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、又はその誘導体からなり、場合により異種配列に融合される。
【0034】
膜貫通ドメインであって、以下のヒトの:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr-CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1-CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)又は
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
の膜貫通ドメイン「に由来する」又は「の誘導体である」アミノ酸配列は、当該膜貫通ドメイン又はその断片と60%長の同一性、例えば70%超又は80%超、好ましくは85%超、最も好ましくは90%超、及び有利な実施態様では95%超の同一性を有するアミノ酸配列に関する。
【0035】
本明細書に使用される場合に、「膜貫通ドメインの断片」は、10超のアミノ酸長、好ましくは14超のアミノ酸長、一例として18超のアミノ酸長、そしてより好ましくは22超のアミノ酸長であるポリペプチドに関する。
【0036】
上記膜貫通ドメインと、本発明のペプチドアンタゴニストのアミノ酸配列とのあいだの同一性の差異は、ペプチドアンタゴニストの膜貫通ドメインアミノ酸配列におけるアミノ酸置換から生じる。
【0037】
好ましくは、これらの膜貫通ドメインにおける置換アミノ酸は、中性及び/又は疎水性アミノ酸であり、そして最も好ましくは疎水性アミノ酸である。このような中性及び疎水性アミノ酸は、当業者に周知である。
【0038】
具体的な実施態様に関し、当該膜貫通ドメイン・アミノ酸配列は、当該膜貫通ドメイン又はその断片との100%の同一性を有する。
【0039】
第二態様では、本発明は、上記ペプチドアンタゴニストをコードする核酸に関する。
【0040】
当該核酸は、RNA又はDNA、好ましくはDNAに対応する。
【0041】
好ましい実施態様では、ペプチドアンタゴニストをコードする核酸は、当該核酸は、原核又は真核細胞、好ましくは真核細胞内で核酸発現を仕向ける遺伝子発現配列に作用可能なように結合されている。「遺伝子発現配列」は、任意の調節性ヌクレオチド配列であり、例えば、それが作用可能なように結合されたペプチドアンタゴニストの核酸の効率的な転写及び翻訳を促進するところのプロモーター配列又はプロモーターエンハンサーの組合せ、並びに原形質膜へペプチドアンタゴニストを適切に標的化することを保証する任意のシグナル配列である。遺伝子発現配列は、例えば、哺乳動物又はウイルスプロモーター、例えば構成的又は誘導性プロモーターであってもよい。構成的哺乳動物プロモーターは、非限定的に、以下の遺伝子:ヒポキサンチン、ホスホリボシル・トランスフェラーゼ(HPTR)、アデノシン・デアミナーゼ、ピルビン酸キナーゼ、βーアクチンのプロモーター、筋肉クレアチン・キナーゼプロモーター、ヒト伸張因子プロモーター及び他の構成的プロモーターを含む。真核細胞において構成的に機能する代表的なウイルスプロモーターとして、例えば、シミアン・ウイルス由来のプロモーター(例えば、SV40)、パピローマウイルス、アデノウイルス、ヒト免疫不全ウイルス(HIV)、サイトメガロウイルス(CMV)、ラウス・サルコーマ・ウイルス(RSV)、B型肝炎ウイルス(HBV)、モロニー白血病ウイルス及び他のレトロウイルスの末端反復(LTR)、並びに単純ヘルペスウイルスのチミジンキナーゼプロモーターが挙げられる。他の構成的プロモーターは、当業者に知られている。本発明の遺伝子発現配列として有用であるプロモーターとして、誘導性プロモーターが挙げられる。誘導性プロモーターは、誘導試薬の存在下で発現される。例えば、メタロチオネインプロモーターは、特定の金属イオンの存在下において転写及び翻訳を促進するように誘導される。他の誘導性プロモーターは、当業者に知られている。
【0042】
一般的に、遺伝子発現配列として、必要に応じて、転写及び翻訳の開始に関して5'非転写及び5'非翻訳配列、例えばTATAボックス、キャップ配列、CAAT配列などをそれぞれ含む。特に、このような5'非転写配列として、作用可能なように結合された抗原核酸を転写調節するためのプロモーター配列を含むプロモーター領域が挙げられる。当該遺伝子発現配列として、エンハンサー配列又は上記上流アクチベーター配列が挙げられる。
【0043】
本明細書に使用される場合に、ペプチドアンタゴニスト核酸配列及び遺伝子発現配列は、遺伝子発現配列の影響下又は調節下で、ペプチドアンタゴニストをコードする配列の発現、又は転写及び/又は翻訳を実行するような方法で共有結合された場合に、「作用可能なように結合される」と呼ばれる。2個のDNA配列は、5'遺伝子発現配列におけるプロモーターの導入が、ペプチドアンタゴニスト配列の転写をもたらす場合、及び2個のDNA配列のあいだの結合性質が、(1)フレームシフト突然変異の導入をもたらさないか、(2)ペプチドアンタゴニスト配列の転写を仕向けるプロモーター領域の能力と干渉しないか、又は(3)対応RNA転写産物がタンパク質へと翻訳される能力と干渉しない場合に作用可能なように結合されるといわれる。こうして、遺伝子発現配列が、抗原核酸配列の効果的な転写を可能にし、その結果得られた転写産物が所望のタンパク質又はポリペプチドに翻訳される場合に、当該遺伝子発現配列は、ペプチドアンタゴニスト核酸配列に作用可能なように結合される。
【0044】
ペプチドアンタゴニスト核酸は、in vivoで単独で、又はベクターと組み合わせてデリバリーされてもよい。この最も広い意味では、「ベクター」は、ペプチドアンタゴニスト核酸を細胞、並びに好ましくはニューロピリン発現細胞へと輸送することを促進できる任意のビヒクルである。好ましくは、ベクターは、ベクターの不存在下でもたらす分解の程度に比べて低い分解性で細胞に核酸を輸送することができる。ベクターは、場合により、ニューロピリン発現細胞において、ペプチドアンタゴニスト核酸の発現を高める上記遺伝子発現配列を含む。一般的に、本発明に有用であるベクターとして、非限定的に、ペプチドアンタゴニスト核酸配列を挿入又は取込むことにより操作されたプラスミド、ファージミド、ウイルス、ウイルス又は細菌起源に由来する他のビヒクルを含む。ウイルスベクターは、好ましいタイプのベクターであり、そして非限定的に、以下のウイルス:レトロウイルス、例えばモロニーマウス白血病ウイルス、ハーベイマウス肉腫ウイルス、マウス乳癌ウイルス、及びラウス肉腫ウイルス;アデノウイルス、アデノ随伴ウイルス;SV40型ウイルス;ポリオーマウイルス;エプステイン-バーウイルス;パピローマ・ウイルス;ヘルペスウイルス;ワクシニア・ウイルス;ポリオウイルス;及びRNAウイルス、例えばレトロウイルス由来の核酸配列を含む。名前が付けられていないが当該技術分野に知られている他のベクターを簡単に使用することができる。
【0045】
好ましいウイルス・ベクターは、非必須遺伝子が、目的の遺伝子で置換されている非細胞変性真核ウイルスに基く。非細胞変性ウイルスとして、レトロウイルス(例えば、レンチウイルス)が挙げられ、レトロウイルスの生活環は、ゲノム・ウイルスRNAをDNAに逆転写し、続いて宿主細胞DNAへとプロウイルス組込みすることに関する。レトロウイルスは、ヒト遺伝子治療のツールとして承認された。最も有用なものは、複製-欠損型である(つまり、所望のタンパク質の合成を仕向けることができるが、感染性粒子を製造することができない)レトロウイルスである。このような遺伝子的に変更されたレトロウイルス発現ベクターは、in vivoにおいて遺伝子を高効率で形質導入することについて一般的に有用性を有する。複製欠損レトロウイルスを産生するための標準的なプロトコル(外来遺伝子物質をプラスミドへと取込み、パッケージング細胞株にプラスミドをトランスフェクションし、パッケージング細胞株により組換えレトロウイルスを産生し、組織培養培地からウイルス粒子を回収し、そして標的細胞をウイルス粒子で感染させることを含む)が、KRIEGLER("A Laboratory Manual," W. H. Freeman CO., New York, 1990)及びMURRY ("Methods in Molecular Biology," vol.7, Humana Press, Inc., Cliffton, N.J., 1991)に与えられる。
【0046】
特定の適用についての好ましいウイルスは、遺伝子治療においてヒトへの使用がすでに承認された二本鎖DNAウイルスであるアデノウイルス及びアデノ随伴ウイルスである。アデノ随伴ウイルスは、複製欠損性であるように遺伝子操作され、そして、広範囲の細胞型及び種へと感染することができる。熱、及び脂質溶媒安定性;肝細胞を含む様々な細胞系統の細胞における高い形質導入効率;並びに重複感染阻害を欠如していること、そうして多重の形質導入を可能にすることといった利点をアデノ随伴ウイルスはさらに有する。報告された様に、アデノ随伴ウイルスは、部位特異的な様式でヒト細胞DNAへと組み込むことができ、それにより、挿入突然変異誘導の可能性を少なくし、そしてレトロウイルス感染の挿入遺伝子発現特徴の変動を少なくする。さらに、野生型アデノ随伴ウイルス感染は、選択圧の不存在下で100回超の継代のあいだ組織培養において継続されており、アデノ随伴ウイルスのゲノム組込みが比較的安定な事象であることを示唆する。アデノ随伴ウイルスは、染色体外の様式で機能することもできる。
【0047】
他のベクターとしてプラスミドベクターが挙げられる。プラスミドベクターは、当該技術分野においてかなり記載されており、そして当業者に周知である。例えば、SANBROOKら、"Molecular Cloning: A Laboratory Manual," 第二版, Cold Spring Harbor Laboratory Press, 1989を参照のこと。ここ数年のあいだに、プラスミドベクターは、in vivoにおいて抗原をコードする遺伝子を細胞へとデリバリーするDNAワクチンとして使用されてきた。プラスミドベクターは、DNAワクチンについて特に利点がある。なぜなら、プラスミドベクターは、多くのウイルスベクターが有するのと同じ安全性についての懸念を有さないからである。しかしながら、宿主細胞に適合するプロモーターを有するプラスミドは、プラスミド内に作用可能なようにコードされた遺伝子からペプチドを発現することができる。いくつかの一般的に使用されるプラスミドとして、pBR322、pUC18、pUC19、pRC/CMV、SV40、及びpBlueScriptが挙げられる。他のプラスミドは、当業者に周知である。さらに、制限酵素及びライゲーション反応を使用してDNAの特定の断片を取り除き、そして加えて、プラスミドをカスタム設計してもよい。プラスミドは、様々な非経口、粘膜、及び局所経路によりデリバリーされうる。例えば、DNAプラスミドは、筋肉内、皮内、皮下、又は他の経路により注射できる。DNAプラスミドは、鼻腔内スプレー又はドロップ、直腸用坐剤、及び経口により投与されてもよい。DNAプラスミドは、遺伝子銃を用いて、表皮又は粘膜表面に投与されてもよい。当該プラスミドは、水溶液中に与えられてもよいし、金粒子上に乾燥されてもよいし、又は、リポソーム、デンドリマー、渦巻き型化(cochleate)、及びマイクロカプセル化を含む他のDNAデリバリーシステムに関して与えられてもよい。
【0048】
核酸ベクターは、細菌及び哺乳動物細胞の両方で活性である選択的マーカーを含むことができる。
【0049】
1の特異的実施態様では、本発明の核酸ベクターは、「裸のDNA」様であるプラスミド、コスミド、又はファージミドに相当する。このような裸のDNAは、非脂質カチオン性ポリマー(WU及びWU、J. Biol. Chem., vol.263, p: 14621-4, 1988)又はリポソーム(BRIGHMANら, Am. J. Med. Sci. vol.298, p: 278-81, 1989)を伴って、細胞取込みを高める複合体を形成した。
【0050】
第二の具体的な実施態様では、核酸ベクターは、in vivo遺伝子治療プロトコルに適用されたウイルスベクターである。適切なウイルスベクターの例として、EP 0871459号、EP 0386882号及びEP 1222300号に記載されるレトロウイルスベクター、並びにUS 2004/ 265273号及びUS 6,638,502号に記載されるアデノウイルスベクターが挙げられる。この場合、ウイルスの内在化は、細胞表面受容体とウイルスエンベロープとの特異的相互作用、続いてウイルス/受容体複合体の受容体媒介性のエンドサイトーシスを介して生じる。
【0051】
第三の態様では、本発明は、上で記載されるペプチドアンタゴニスト、それをコードする核酸、又は当該核酸を含むベクター、医薬として許容されるビヒクルと最終的に関連する核酸ベクターに関する。
【0052】
医薬として許容されるビヒクルの一例として、組成物は、エマルジョン、マイクロエマルジョン、水中油型エマルジョン、無水脂質及び水中油型乳濁液、他のタイプのエマルジョンを含むこともある。当該組成物はまた、1以上の添加物(例えば、希釈剤、賦形剤、安定剤、保存剤)も含む。一般的に、Ullmann's Encyclopedia of Industrial Chemistry、第6版(共編)、1989-1998、Marcel Dekker);及びPharmaceutical Dosage Forms and Drug Delivery Systems (ANSELら、1994、WILLIAMS & WILKINS)を参照のこと。
【0053】
有利な態様では、当該組成物は、10-12M超、より好ましくは10-11M超、そして最も好ましくは10-10M超の濃度を含む。
【0054】
ペプチドアンタゴニスト、核酸又は核酸ベクターは、緩衝液又は水中に溶解されてもよく、又はエマルジョン及びマイクロエマルジョン中に取込まれてもよい。適切な緩衝液として、非限定的に、Ca2+/Mg2+フリーのリン酸緩衝生理食塩水(PBS)、リン酸緩衝生理食塩水(PBS)、通常の生理食塩水(水中の150mM・NaCl)、Tris緩衝液及び界面活性剤が挙げられる。
【0055】
加水分解及び変性を含む、ペプチド不安定性又は変性についての多くの原因が存在する。疎水性相互作用は、分子の集合(つまり、凝集)を引き起こしうる。この結果は、Treg応答の導入の低減を引き起こしうる。このような問題を低減又は予防するために安定剤が加えられてもよい。
【0056】
安定剤として、シクロデキストリン及びその誘導体が挙げられる(例えば、米国特許第5,730,969号を参照のこと)。適切な保存剤、例えばスクロース、マンニトール、ソルビトール、トレハロース、デキストラン、及びグリセリンは、最終的な製剤を安定化するために加えることができる。イオン性及び非イオン性界面活性剤、D-グルコース、D-ガラクトース、D-キシロース、Dガラクツロン酸、トレハロース、デキストラン、ヒドロキシエチル・スターチ、及びそれらの混合物から選ばれる安定剤は、製剤に加えられてもよい。アルカリ金属塩または塩化マグネシウムの添加は、ペプチドを安定化しうる。ペプチドは、デキストラン、硫酸コンドロイチン、スターチ、グリコーゲン、デキストリン、及びアルギン酸塩からなる群から選ばれる多糖と接触させることにより安定化されてもよい。加えられうる他の糖としては、単糖、二糖、糖アルコール、およびそれらの混合物(たとえば、グルコース、マンノース、ガラクトース、フルクトース、スクロース、マルトース、ラクトース、マンニトール、ザイリトール(xylitol)が挙げられる。ポリオールは、ペプチドを安定化しうるし、そして水混和性であるか、又は水溶性である。適切なポリオールは、ポリヒドロキシアルコール、単糖及び二糖、例えば、マンニトール、グリコール(glycrol)、エチレングリコール、プロピレン・グリコール、トリメチルグリコール、ビニル・ピロリドン、グルコース、フルクトース、アラビノース、マンノース、マルトース、スクロース、及びそのポリマーであってもよい。様々な賦形剤は、ペプチドを安定化し、例えば、血清アルブミン、アミノ酸、ヘパリン、脂肪酸、及びリン脂質、界面活性剤、金属、ポリオール、還元剤、金属キレート剤、ポリビニル・ピロリドン、加水分解性ゼラチン、及び硫酸アンモニウムを含む。
【0057】
第四の実施態様では、本発明は、クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関する疾患を患う患者の予防又は治療処置の方法であって、上記組成物を当該患者に投与するステップを含む、前記方法に関する。
【0058】
本明細書に使用される場合、「対象」という用語は、哺乳動物、例えばげっ歯類、ネコ、イヌ、及び霊長類などを指す。当該対象は、ウシ、ブタ、ウマ、ネコ、イヌ、及び最も好ましくはヒトである。
【0059】
クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関連する疾患は、当業者により簡単に決定できる。この様な疾患の一例として、神経変性疾患(例えば、アルツハイマー病、パーキンソン病、中枢神経系病変、病変に伴う脱髄)、癌(例えば、肺癌、乳癌、及び中皮癌、癌腫、神経膠腫)、及び異常血管新生に関与する全ての疾患を挙げることができる。
【0060】
有利な態様では、当該組成物の投与は、10-12M超、好ましくは10-11M超、そして最も好ましくは10-10M超のペプチドアンタゴニストの濃度に相当する。
【0061】
第5態様では、本発明は、クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関する疾患を患う対象の予防又は治療用の医薬の製造のための、上記ペプチドアンタゴニスト、これらをコードする核酸、又は当該核酸を含む核酸ベクターの使用に関する。
【0062】
クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関連する疾患は、当業者のうちの一人により簡単に決定することができる。この様な疾患の一例として、神経変性疾患(例えば、アルツハイマー病、パーキンソン病、中枢神経系病変、病変に伴う脱髄)、癌(例えば、肺癌、乳癌、及び中皮癌、癌腫、神経膠腫)、及び異常血管新生に関与する全ての疾患を挙げることができる。
【0063】
好ましい実施態様では、本発明は、アルツハイマー病、パーキンソン病、中枢神経系病変、及び病変に付随する脱髄を含む群から選ばれる神経変性疾患を患う対象の予防又は治療用の医薬の製造のための、上記ペプチドアンタゴニスト、これらをコードする核酸、又は当該核酸を含む核酸ベクターの使用に関する。
【0064】
第二の好ましい実施態様では、本発明は、肺癌、乳癌、中皮癌、癌腫、及び神経膠腫を含む群から選ばれる癌を患う患者の予防又は治療用の医薬の製造のための、上記ペプチドアンタゴニスト、それをコードする核酸、又は当該核酸を含む拡散ベクターの使用に関する。
【0065】
第三の好ましい実施態様では、本発明は、異常血管新生に関与する疾患を患う対象の予防又は治療のための医薬の製造のための、上記ペプチドアンタゴニスト、それをコードする核酸、又は当該核酸を含む核酸ベクターの使用に関する。
【0066】
有利な実施態様では、当該医薬は、10-12M超、好ましくは10-11M超、そして最も好ましくは10-10M超のペプチドアンタゴニスト濃度の放出を許容する。
【0067】
本発明は、以下の実施例によりさらに例示される。当該実施例は、その範囲を制限することを意図しない。
【実施例】
【0068】
1) NRP1受容体の膜貫通ドメインは、二量化能力を有する
RUSS及びENGELMAN(Proc. Natl. Acad. Sex. USA, vol. 96, p: 863-8, 1999)により記載されるToxCatシステムから得られたToxLucシステムは、NRP1膜貫通ドメイン媒介性の二量化を調査するために使用された。このシステムは、E.coli細胞内膜中での膜貫通へリックス-へリックス・オリゴマー化を計測することを可能にする。NRP1の膜貫通ドメイン(配列番号1、ILITIIAMSALGVLLGAVCGVVL)の二量化能力は、EGF受容体(配列番号17、SIATGMVGALLLLLVVALGIGLFM)、Erb-2タンパク質(配列番号18、SIISAVVGILLVVVLGVVFGILI)及びグリコプロテインA(配列番号19、ITLIIFGVMAGVIGTILLISYGI)のうちの膜貫通ドメインの二量化能力と比較される。
【0069】
特異的融合タンパク質をコードする幾つかの構築体を製造した。これらの融合タンパク質は、NRP1、EGF受容体、Erb2受容体、及びグリコフォリンAそれぞれの膜貫通ドメインに融合されたToxR(二量化依存性転写活性化因子)のN末端DNA結合ドメイン、並びにモノマーのペリプラズム膜アンカー(マルトース結合タンパク質:MBP)を含んだ。
【0070】
ToxRに繋げられ、そしてマルトース結合タンパク質(MBP)に繋げられたキメラタンパク質として、目的のTM配列を細菌DH5a(MM39)中で発現した。TMドメイン媒介性オリゴマー化は、当該システムのオリジナル・バージョンのクロラムフェニコールアセチルトランスフェラーゼ(CAT)をコードする受容体遺伝子のToxRにより活性化される発現をもたらす。便宜上、発明者は、慣用される分子生物学的方法を用いて、ルシフェラーゼの遺伝子により初期のCAT遺伝子を置き換えた。ニューロピリン、EGF受容体、及び野生型erbB2に対応する合成TM配列を、NheI/DpnII断片として新たなプラスミドにクローニングした。グリコプロテインAから得られるTM配列、又はそのG83I突然変異配列(RUSS 及びENGELMAN, Proc. Natl. Acad. Sci. U S A.vol.96(3), p: 863-8、1999) を有するキメラを対照として用いた。Rocheアッセイキットを製品説明書に従って用い、そしてBerthold Microlumat Plate Luminometerを用いて、ルシフェラーゼアッセイを行った。
【0071】
異なる構築物について結果を図3Aに示す。
【0072】
NRP1の膜貫通ドメインを含む構築物で形質転換された細菌は、Erb-2又はEGFRの膜貫通ドメインを含む構築物で形質転換された細菌のルシフェラーゼ発光より有意に高いルシフェラーゼ発光を示し(Erb-2:4.7倍、EGFR:6.1倍)、そしてグリコフォリン-Aの膜貫通ドメインを含む細菌のルシフェラーゼ発光より少し高かった(1.2倍)。興味深いことに、発明者らは、グリコフォリン-AのRMN構造を下にSwissPdbViewerソフトウェアを用いて得られたTM-NRP1の3次元モデルにより、NRP1のTMドメインの二量化能力を確認した。この論理的アプローチは、必要最低限ではあるが、二量化形成を支援するヘリックス間相互作用を示すTM-NRP1の空間的にコンパクトな構成の存在を確認した(図3B)。
【0073】
結論として、NRP1の膜貫通ドメインは、GpA膜貫通ドメインの二量化能力よりも強い効力で二量化を誘導することができる。
【0074】
2) NRP1の膜貫通ドメインのペプチドが、Sema3Aにより引き起こされる皮質軸索崩壊を阻害する
Sema3Aの軸索成長の阻害性性質が、成長円錐の崩壊を誘導する能力に関連すると示唆されている。Sema3A機能性質へのpTM-NRP1ペプチドの効果を調査するために、皮質ニューロンの成長を、Sema3の存在下又は不存在下で、そしてpTM-NRP1又はpTM-NRP1mut(3個のG→V突然変異を有するNRP1 TM)ペプチドの存在下又は不存在下で分析した。
【0075】
ラミニン/ポリ-L-リジン基質を、980μlのGeyの平衡塩類溶液(GBSS、Sigma)を10μlのラミニン(1mg/ml、Sigma)及び10μlポリ-L-リジン(10mg/ml、Sigma)に加えることにより製造した。滅菌カバースリップを、大ディッシュにおき、そして基質を加えた(100μl)。次に、各カバースリップを第二カバースリップで覆うことにより「サンドウィッチ」を製造した。5%CO2を含む空気の下、37℃で少なくとも30分間インキュベートした後に、「サンドウィッチ」を開き、そして各カバースリップを脱イオン水で洗浄した。乾燥させたカバースリップ上で組織片を培養した。
【0076】
E15マウス胚(E1を膣栓の確認による胚発達の初日として定義する)から調製された大脳新皮質を、組織破砕ディスク上に移した。最初の切断後に、ディスクを90°回転させることにより、組織を200×200μmに切断した。皮質キューブをスパチュラを用いて回収し、そしてぺトリ皿の培養倍地中に入れた。基質付のカバースリップを小ペトリ皿(50mmの直径、FALCON)に置いた。750μlの培養培地を加えた後に、カバースリップをインキュベーター内に少なくとも10分間静置すべきである。解剖顕微鏡を用いて、40〜50の皮質組織片を20μlの培養培地中に集め、そして注意深くカバースリップ上に置いた。室温で15分後、多くの組織片を基質へと接着させた。2250μlの培養培地をゆっくり各ディッシュに加えた。次に、組織片培養物を5%CO2を含む空気中において37℃で維持した。
【0077】
18〜24時間後に培養物中で放射状の伸張が見られ、そしてその結果個々の繊維及び成長円錐を分析した。成長円錐について試験された以下の生成物:
・Sema3Aを安定的に発現するHEK293細胞の条件培地から調製された精製Sema3A(100ng/ml)(ここで、抗-Flagシステム(Sigma)を用いることにより精製を行った。)
・pTM-NRP1(10-8、10-9、10-10、及び10-11M)
・10-8MのmpTM-NRP1
・10-8MのpTM-ErbB2w(配列番号18)
を直接培養培地に2時間加えた。
【0078】
インキュベート後に、4%ホルムアルデヒドを15分間培養培地(v/v)中に直接加えた。次に溶液を取り除き、そして4%ホルムアルデヒドに15分間置き換えた。
【0079】
図4Aは、Sema3Aにより引き起こされる皮質軸索崩壊について濃度を増加させたpTM-NRP1ペプチド(10-8、10-9、10-10、及び10-11M)の効果を示す。
【0080】
図4Bは、Sema3Aにより引き起こされる皮質軸索崩壊について、pTM-NRP1又はpTM-NRP1mutペプチド(10-8M)の効果を示す。
【0081】
50%超の崩壊皮質軸索がSema3A処理細胞で観察される一方、対照条件では、10%未満の皮質軸索しか崩壊形態を示さなかったということが結果により示される(図4A)。さらに、高い濃度の野生型pTM-NRP1ペプチドの添加は、約10-11MのIC50で用量依存的な様式で、皮質軸索についてのSema3Aの崩壊効果を抑制した。
【0082】
対照的に、pTM-NRP1mutペプチドの添加は、Sema3Aにより引き起こされる皮質軸索崩壊を阻害しない(図4B)。
【0083】
ペプチド希釈緩衝液を用いそしてGxxxGモチーフを含むErbB2ペプチドを用いた対照実験は、Sema3Aの崩壊効果について影響を示さない。
【0084】
NRP1の膜貫通ドメインを模倣する合成ペプチドの添加が、皮質軸索へのSema3Aの効果を損なわせ、皮質軸索崩壊を阻害するということがこれらの結果により示される。さらに、この生物学的活性は、GxxxGモチーフと関連し、そしてpTM-NRP1ペプチドに特有である。
【0085】
3) NRP1受容体の膜貫通ドメインペプチドは、Sema3Aにより引き起こされるCOS細胞崩壊を阻害する
COS細胞は、セマフォリン受容体を発現しておらず、そしてその結果これらのガイダンスシグナルにもともと感受性ではない。それにもかかわらず、COS細胞にNRP1及びプレキシン-A1を人工的に発現させることは、Sema3Aの細胞崩壊を引き起こさせる。pTM-NRP1ペプチドのSema3A機能性質についての効果を調査するために、NRP1及びプレキシン-A1を発現するCOS細胞の形を、Sema3A及びpTM-NRP1又はpTM-NRP1mutペプチドの有無について分析した。
【0086】
6ウェル-プレート中で、製品説明書に従いリポフェクタミン2000(invitrogen)を用いて、NRP1及びプレキシン-A1をコードする配列を含むpBK-CMVプラスミド(STRATAGENE)1μgをCOS-1細胞にトランスフェクションした。安定的にトランスフェクションされたCOS-1細胞を0.7%ジェネティシンを用いて選別した。安定的にトランスフェクションされたCOS-1細胞は、前もってポリ-L-リジン-皮膜されたガラスカバースリップを用いて12ウェルプレート上で培養した。細胞を、pTM-NRP1又はpTM-NRP1mutペプチド(10-9M)と1時間37℃でインキュベートした。培養培地を取り除き、そしてSema3A(100μl/ml、D-MEM)を発現する構築物で安定的にトランスフェクションされるか又はされていないHEK細胞の条件培地(conditioned medium)に37℃で4時間置換した。最終的に、細胞を2%ホルムアルデヒドで30分間固定し、続いて4%で15分間固定した。試験された各条件について、約400個の細胞を分析した。
【0087】
pTM-NRP1又はpTM-NRP1mutを伴って又は伴わずにSema3Aの存在下又は不存在下でNRP1及びプレキシン-A1を発現するCOS-1細胞の形態を図5Aは示す。図5Bは、Sema3Aにより引き起こされる細胞崩壊についてのpTM-NRP1又はpTM-NRP1mutペプチドの効果を示す(*:p<0.001)。
【0088】
50%超の崩壊細胞がSema3A処理細胞で観察される一方、対照条件では10%未満の細胞しか崩壊形態を示さなかったことが結果により示される(図5B)。さらに、10-9MのpTM-NRP1ペプチドは、Sema3AのCOS-1細胞を崩壊させる効果を完全に損なわせた。対照的に変異GxxxGモチーフを有するpTM-NRP1mutペプチドの10-9Mを加えることが、Sema3Aにより引き起こされる細胞崩壊を阻害することはなかった。
【0089】
これらの結果により、NRP1の膜貫通ドメインを模倣する合成ペプチドを加えることは、そのGxxxGモチーフを有するSema3Aの細胞崩壊効果を損なわせるということが示された。
【0090】
pTM-NRP1がSema3Aシグナルを遮断するメカニズムを解明するために、発明者らは結合アッセイを行った。
【0091】
NRP1発現COS細胞を、AP-Sema3A、つまりSema3Aと分泌型アルカリホスファターゼの融合タンパク質(BAGNARDら、1998)とインキュベートした。
【0092】
野生型COS細胞又はNRP1-発現COS細胞(COS-NRP1)をポリ-L-リジン-皮膜ガラスカバースリップ上で12ウェルプレート上で培養した(0.005mg/ml)。無血清培地中のpTM-NRP1又はpTM-NRP1mut(10-9M)で37℃にて1時間インキュベートした後に、AP-Sema3Aを安定的に発現するHEK細胞から得たアルカリ・ホスファターゼ結合Sema3Aを含む条件培地で90分間置換した(AP-Sema3A;Bagnardら、1998)。セマフォリンを伴わない条件培地(トランスフェクションされていないHEKから得られる)は、対照として役に立った。PBSで3回細胞を洗浄し、そして4%ホルムアルデヒド中で固定して、新たなディッシュに移した。PBS中で3回洗浄後、プレートを50分65℃で暖めた。続いて細胞を1mlのアルカリホスフェート基質(NBT/BCIP、Sigma)と暗所でインキュベートした。45分後、基質を取り除き、そしてガラスカバースリップを洗浄した。慣用される顕微鏡を用いて写真をとり、そしてAxioVision LE Zeissソフトウェアで分析した。試験された各条件につき、約60個の細胞を分析して、光学密度の関数として結合レベルを測定した。Studentのt検定を用いることにより統計分析を行った。
【0093】
図6Aは、pTM-NRP1又はpTM-NRP1mutを伴うか又は伴わずにAP-Semaとインキュベートした後に、野生型COS細胞(対照)又はNRP1発現COS細胞(COS-NRP1)を示す。
【0094】
図6Bは、以前に試験された条件について、細胞あたりの光学密度の平均強度を示す。
【0095】
結果により、AP-Sema3Aの結合が、pTM-NRP1の添加により有意に低減されることが示される(図6A及びB)。驚くべきことに、pTM-NRP1mutの添加は、COS細胞に結合するAP-Sema3Aをブロックしなかった。
【0096】
これらの結果により、NRP1TMのGxxxGxxxGドメインが、Sema3A結合とそれに続く阻害効果を引き起こすのに必須でありそうであると示される。
【0097】
4) NRP1のTMドメインの突然変異が、受容体機能を損なわせる
GxxxGxxxGモチーフの重要な役割を確認するために、突然変異ペプチドとして、全長NRP1のTMドメインに突然変異を導入して、3個全てのグリシン残基をバリンに置き換えた(NRP1mut)。
【0098】
膜貫通領域中に3個の(G→V)変異を有するNRP1タンパク質をコードするプラスミドを、前に記載される様にCOS細胞にトランスフェクションした。次に前と同様に、NRP1の当該突然変異形を発現するCOS細胞中で結合実験を行った。
【0099】
図7Aは、pTM-NRP1を伴って又は伴わずに、AP-Sema3Aとインキュベートした後における野生型COS細胞(対照)、NRP1発現COS細胞(COS-NRP1)、又は3個の(G→V)変異を有するNRP1を発現するCOS細胞(COS-NRP1mut)を示す。
【0100】
図7Bは、前に試験された条件についての細胞あたりの光学密度の平均強度を示す。
【0101】
野生型NRP1を発現するCOS細胞においてかなりの結合が観察された一方、NPR1mutを発現する細胞においてあるとしてもかなり低いAP-Sema3Aの結合しか検出されなかったということが結果により示される(図7A)。結合のかなりの低下は、pTM-NRP1の存在下で得られた結合低下に類似している。驚くべきことに、COS細胞中で当該NRP1突然変異(NRP1mut)をPlexA1と一緒に発現させた場合、Sema3Aは、もはや細胞崩壊を誘導できなかった(図7B)。このことにより、機能的なSema3A受容体の形成についてのNRP1TMドメインのGxxxGxxxGの重要性がさらに確認された。
【0102】
5) TM-NRP1を模倣する合成ペプチドは、セマフォリン受容体複合体の形成を変更する
受容体複合体形成の観点で、TM-NRP1の生物化学的な帰結をさらに調査するために、発明者らは、PC12神経細胞モデルにおいて複合体形成を分析した。興味深いことに、Sema3Aは、NGFから独立した経路を介してこれらの細胞において神経突起の成長を促進することが示された(Schwambornら、J.Biol.Chem., Vol.279(30), P:30923-6, 2004)。
【0103】
PC12(ATCC:CRL-1721)を、4.5gグルコース/L(Gibco)、5%FVS、10%ウマ血清、グルタミン580mg/l及び抗生物質を含むD-MEM培地中で増殖させた。機能アッセイについて、前もってポリ-L-リジン皮膜されたガラスカバースリップを備える12ウェルプレート上でPC12を培養した。PC12細胞を、pTM-NRP1ペプチド(10-9M)を伴って又は伴わずに37℃で1時間インキュベートした。培養培地を取り除き、そしてNGF-含有(100ng/ml;GIBCO)無血清培地に置換し、又はSema3Aを安定的に発現するHEK293細胞から得られた、又はトランスフェクションされていない細胞から得られた条件培地により、12時間37℃で置換された(対照、詳細についてはBAGNARDら、1998を参照のこと)。
【0104】
細胞を2%ホルムアルデヒドで30分間固定し、続いて4%ホルムアルデヒドで15分間固定した。試験された各条件につき、約400個の細胞を分析して、神経突起の伸張を評価した(統計分析を、χ2検定を用いて行った)。
【0105】
図8は、NGF(100ng/ml)、Sema3A、pTM-NRP1(10-9M)を伴って又は伴わずにインキュベートした後におけるPC12細胞の分化アッセイの結果を示す。
図8Bは、各条件について分化した細胞の割合を示す。
【0106】
NGF又はSema3Aの添加が、相乗効果を与えることなくPC12細胞の分化を誘導したということがこの結果により示される(図8A及び8B)。さらに、実験がpTM-NRP1の存在下で行われる場合、NGF効果が維持されている一方、Sema3A誘導性神経突起成長の促進が有意に低下した。これは、ペプチドの添加が、他のシグナル経路に影響することなくSema3A依存性経路の活性化を特異的に遮断するということを示す。
【0107】
6) NRP1受容体の膜貫通ドメインのペプチドが、リガンドSema3Aのその受容体NRP1への結合を拮抗作用する
NRP1-クラスIIIセマフォリンの結合におけるNRP1膜貫通ドメインの役割を調査するため、神経膠腫細胞上にある受容体NRP1へのリガンドSema3Aの結合を、NRP1の膜貫通ドメインペプチド (pTM-NRP1ペプチド;配列番号1)又は変異ペプチド(mpTM-NRP1;配列番号20、ILITIIAMSALVVLLVAVCVVVLYRKR)の存在下又は不存在下で計測した。 これらのpTM-NRP1ペプチドは、自動化ペプチド合成機(Fmoc化学反応、Applied System)により合成した。ペプチド純度を、RP-HPLC(BECKMAN)により90%超として見積もった。
【0108】
セマフォリン受容体を発現するラットC6神経膠腫細胞は、AP-Sema3A、つまり分泌型のSema3Aのアルカリホスファターゼバージョン(ADAMSら、1997, Bagnardら、1998)の結合能力を決定するために使用された。これらの細胞は、10%ウシ胎児血清(PERBIO)、グルタミン0.5mM(GIBCO)、及び抗生物質:100U/mlペニシリン及び100μg/mlストレプトマイシンを含むMEM培地(GIBCO)中で増殖させ、そしてプレーティングした。
【0109】
C6細胞を、96ウェルプレート中で培養し、そして新たに希釈されたpTM-NRP1又はpTM-NRP1mutペプチド(10-12M〜10-10M)を伴って又は伴わずに1時間37℃でインキュベートした。次に培養培地を、AP-Sema3Aを発現する構築物で安定的にトランスフェクションされたHEK細胞の条件培地に置換した(ADAMSら, EMBO J., Vol.16(20), p:6077-86, 1997;BAGNARDら、1998)。非トランスフェクション細胞であるHEKから得た条件培地を内部対照として用いた。pTM-NRP1ペプチドは、自動化ペプチド合成(Fmoc化学、APPLIED SYSTEM)により合成され、そして質量分析器により分析された。ペプチド純度を、RP-HPLC(BECKMAN)により90%超として見積もり、細胞をPBSで洗浄し、そして50μlのアルカリホスファターゼ発光基質(AMERSHAM)とインキュベートした。製品説明書に従って、MICROLUMAT PLUSシステム(BERTHOLD TECHNOLOGIES)を用いて15分後に発光を読み込んだ。各ペプチドについて実験を4回行った。
【0110】
pTM-NRP1ペプチド(黒色棒)又は変異ペプチド(灰色棒)の存在下におけるAP-Sema3Aのその受容体への結合についての結果を図9に示す(*:p<0.005;**:p<0.01、スチューデントt検定)。
【0111】
AP-Sema3AのC6細胞上におけるその受容体NRP1への結合が、野生型pTM-NRP1ペプチドの濃度を増加させて加えることにより、用量依存的な様式で妨げられた。野生型pTMNRP1ペプチドの10-10Mの存在下で、AP-Sema3AのNRP1への結合がペプチドの不存在下の場合に比べて約50%に低下した。野生型pTM-NRP1ペプチドとは対照的に、pTM-NRP1mutペプチドの添加が、AP-Sema3AのNRP1への結合を妨げられなかった。
【0112】
結果として、Sema3Aのその受容体NRP1への結合に、NRP1膜貫通ドメインが関与し、そしてダブルGxxxGモチーフの完全性を必要とする。
【0113】
7) NRP1受容体の膜貫通ドメインペプチドは、セマフォリン受容体複合体の形成を変更する
NRP1受容体複合体の形成において、NRP1受容体の膜貫通ドメインの役割をさらに調査するために、NRP1複合体の形成を、NRP1の膜貫通ドメインペプチド(pTM-NRP1ペプチド)の存在下又は不存在下で、そしてリガンドSema3Aの存在下又は不存在下で、NRP1及びプレキシン-A1を発現するC6細胞上で測定された。
【0114】
NRP1及びプレキシン-A1を発現するC6細胞を、pTM-NRP1ペプチド(10-9M)と1時間インキュベートするか又はインキュベートしなかった。次に培養培地を、Sema3Aを発現する構築物で安定してトランスフェクションされるか又はされていないHEK細胞の培地と取り替えられた。コンフルエントC6を10mM・EDTAを用いて回収し、そして遠心した。ペレットをPBSで洗浄し、そして次に0.1%SDS、1mM・EDTA、1%NP-40、0.5%DOC、2mM・バナジウム塩、及びSDSを伴わないプロテアーゼ阻害剤(PIERCE)を含む溶解緩衝液(Tris-HCl/NaCl;50/150;pH8.0)中に希釈した。4℃で1時間可溶化した後に、ビシンコニン酸法によりタンパク質量を見積もった(BCAタンパク質アッセイ、PIERCE)。
【0115】
スクロース密度勾配沈降実験は、25%、17%、10%、及び3%のスクロースを含む段階的勾配に基いた。溶液を、Hepes/NaCl緩衝液(30/30、pH7.6、0.12%トリトン)及び1Mスクロースを伴うHepes/NaCl緩衝液から作成された。これらの溶液は、線形勾配を形成するために連続して充填された(LERAYら、Arch Biochem Biophys., 1992)。
【0116】
細胞ライセートを超遠心管中の勾配の上に配置し、そしてTL-100超遠心機(BECKMAN)を用いて1時間100000gで遠心し、そして底から分画を回収した(13滴/分画)。
【0117】
Laemmli法に従って、等体積の充填緩衝液をサンプル(62.5mM・Tris-HCl、pH6.8、10%グリセロール、2%SDS、DTT、ブロモフェノールブルー)に加え、そしてこれらを10分間ボイルした。次にサンプルをアクリルアミドゲル(5〜20%)上で一定電圧及び温度で、適切な緩衝液(0.025M・Tris、0.192Mグリシン、pH8.3、0.01%SDS)中でSDS-PAGEにかけた。次にタンパク質を、4℃で、20%エタノール、0.025M・Tris、0.192MグリシンpH8.3、及び0.01%SDSを含む緩衝液中で3時間メタノール活性化ポリビニリデンジフロリド(PVDF)膜に転写した。最後に、PVDF膜を1時間PBS/BSA5%でブロッキングした。
【0118】
次に膜を2時間ポリクローナル抗NRP1(ONCOGENE)で2時間、1:1000の希釈率でインキュベートした。膜をPBS/0.2%TWEEN20中で3回洗浄し、そして二次抗体(A/Gタンパク質、PIERCE、1:100000又はホースラディッシュ・ペルオキシダーゼ結合抗ラビットIgG、AMERSHAM、1:500)とインキュベートした。次に、免疫反応性を、製品説明書に従って、高められた化学発光ウエスタンブロット検出システム(PIERCE)を用いて検出した。
【0119】
結果を図10に示す。各分画におけるNRP1の割合を、明らかにされた総NRP1から計算した。この図では、プレキシン-A1を含むオリゴマーを含むスクロース勾配の重分画は、黒色棒グラフに相当し、NRP1ダイマーを含む中間分画は灰色棒グラフに相当し、そして軽い分画は、ほぼNRP1モノマーから構成され、白色棒グラフに相当する。
【0120】
リガンドSema3Aの不存在下では、NRP1がスクロース勾配の中間分画において主に検出されたということが結果により示される。こうして、NRP1二量体は、そのリガンドSema3Aの不存在下ではオリゴマー形態として主に存在した。
【0121】
対照的に、Sema3Aの存在下では、NRP1受容体は主に重分画中にプレキシン-A1を含むオリゴマー形態として存在した。
【0122】
Sema3Aの存在下におけるpTM-NRP1ペプチドの添加は、NRP1の分布を変更させ、NRP1モノマーの移動レベルに相当する軽い分画中で主に検出された。こうして、NRP1のオリゴマー化は、pTM-NRP1ペプチドの存在により阻害された。それゆえ、NRP1の膜貫通ドメインは、NRP1受容体複合体の形成に関与する。結果として、pTM-NRP1ペプチドの存在下における図3において観察されたSema3A結合の低下は、NRP1オリゴマー化の阻害に関連することもありうる。
【0123】
8) 腫瘍細胞移動のあいだにおけるpTM-NRP1-依存性のNRP1の不活性化の機能的意味
ヒト神経膠腫の優れたモデルであるラットC6神経膠腫細胞株(DAI及びHOLLAND、Biochim. Biophys. Acta, vol.1551, p: M19-27, 2001)は、発明者らのペプチド戦略(pTM-NRP1)を用いることによるNRP1の阻害が、細胞移動及び播種をどれだけ妨げるかを調べるために使用された。
【0124】
C6細胞(ATCC CCL-107)を、PKH26(Sigma)を用いて染色した。注入前に培養培地(5000u/mlペニシリン、5mg/mlストレプトマイシン、200mM・L-グルタミン、及び10%ウシ胎児血清から構成される)中で、氷上で少なくとも2時間、細胞をペプチド(pTM-NRP1 10-8M又は変異pTM-NRP1 10-8M)とインキュベートした。106個の細胞の注射を、以下の座標:ブレグマに対して前後+1.6mm;L、+2mm;皮質表面に対してH、+5mm(antero-posterior, +1.6mm relative to Bregma; L, +2mm; H, +5mm relative to the cortical surface)に従って定位フレームを用いて行った。
【0125】
全ての注射を左線条体で行った。
【0126】
8日の生存期間の後に、動物(4匹のラットからなる3群)を、ペントバルビタールの腹腔内注射により屠殺し、次に100mlのPBSで前もって洗浄し、続いて500mlの2%ホルムアルデヒドで経心臓かん流を行った。脳を4℃で2時間、後固定(post-fixed)し、そして矢状断面(70μm)をビブラトームで調製した。
【0127】
1群の切片を、顕微鏡観察用にPBS-グリセロール(v/v)にマウントし、そして別の群をCD34の免疫染色用に処理した。切片を最初に5%ウシ通常血清を含むPBS中で15分間室温でインキュベートして、非特異的結合部位をブロッキングした。2回目のインキュベートを1時間室温で行いそして次にマウス抗CD34(1:200)を用いて4℃で一晩行った。切片を5分間PBS中で6回洗浄し、そして次にAlexa488に結合されたヤギ抗マウス抗体(1:500;INTERCHIM)と3時間室温でインキュベートした。切片を5分間で6回PBSで洗浄し、そして最終的にPBS-グリセロール(v/v)にマウントし、顕微鏡分析を行った。
【0128】
図11は、pTM-NRP1又はpTM-NRP1mutペプチドを伴うか又は伴わずにC6細胞を脳注射した後のマウス脳切片を示す。腫瘍、皮質、線状体、脳梁(cc)、海馬(Hp)及び側脳室が、顕微鏡写真に写っている。
【0129】
図12は、pTM-NRP1又はpTMNRP1mutを伴って又は伴わずにC6細胞を脳に注射した後において、CD34を免疫染色した後のマウス脳切片を示す。
【0130】
対照条件において、腫瘍が線状体において発達し、そして脳梁及び皮質板に到達したということが結果により示される(図11、n=4)。驚くべきことに、細胞を注射前に、pTM-NRP1で処理した場合に、発明者らは、8日目において腫瘍サイズのかなりの低下を観察した(n=4)。予期される様に、変異pTM-NRP1で処理されたC6細胞は、処理されていない細胞で観察された腫瘍と同様の腫瘍を誘導した(n=4)。こうして、pTM-NRp1の添加は、C6神経膠腫の発達を阻害する。
【0131】
pTM-NRP1の存在下における腫瘍サイズの低下は、CD34、つまり血管新生のマーカー、への免疫反応性のかなりの低減を伴った(図12)。これにより、pTM-NRP1が、VEGFシグナルをブロッキングすることにより抗腫瘍効果を発揮することが示唆された。
【0132】
9) pTM-NRP1が、in vitroにおいてVEGFシグナル伝達を拮抗作用できる
NRP1は、VEGFの受容体である(NEUFELDら、Adv. Exp. Med. Biol., vol.515, p: 81-90, 2002)。その結果、本発明者は、pTM-NRP1がC6細胞においてVEGFシグナル伝達を拮抗作用できるということを確かめた。この目的を達成するために、前に記載されたように調製されたC6腫瘍細胞凝集体(BAGNARDら、1998;及びNASARREら、Neoplasia、Vol.7, p180-189, 2005)を3Dマトリックス(凝固血漿)中で増殖させ、そしてVEGF165で処理した。
【0133】
代表的な腫瘍細胞が、VEGF165(50ng/ml)、pTM-NRP1又はpTMNRP1mut(10-8M)の添加を伴って又は伴わずに凝集することを図13が示す。
【0134】
50ng/mlVEGF165の添加が、C6細胞の凝集体からの移動及び移動鎖(migration chains)の形成を誘導した(図13)。驚くべきことに、pTM-NRP1の添加は、VEGF165依存性C6細胞移動を抑制した。変異型pTM-NRP1の添加は、VEGF165-誘導性C6細胞移動を妨げることができなかった。これらの結果により、pTM-NRP1が、C6細胞におけるVEGF165シグナル伝達を遮断することができるということが示唆される。
【0135】
最後に、これらの結果により、pTM-NRP1が、腫瘍形成の点でNRP1シグナル伝達を阻害するために使用できるということが示唆される。これはVEGF依存性メカニズムを通した腫瘍細胞の移動及び生存におけるNRP1の役割に関する。pTM-NRP1を用いてNRP1を遮断することが、その生存、増殖、及び/又は播種がNRP1依存性のシグナル伝達カスケードを必要とする全ての腫瘍に対して、治療成果を有するということが本発明者らにより提案される。
【図面の簡単な説明】
【0136】
【図1】図1は、膜貫通ドメインであって、以下のマウスの: ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、 ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、 プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、 プレキシンA2(配列番号4:LLTLPAIISIAAGGSLLLIIVIIVLIAY)、 プレキシンA3(配列番号5:LTLPAMVGLAAGGGLLLLAITVVLVAY)、 プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、 Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCF)、 L1−CAM(配列番号8:GWFIAFVSAIILLLLILLILCFI)、 インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び インテグリンβ2(配列番号10:VAAIVGGTVVGVVLIGVLLLVIW) で表される膜貫通ドメインを示す。
【図2】図2は、以下のヒトの: ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、 ニューロピリン−2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、 プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、 プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、 プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、 プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、 Nr−CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、 L1−CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、 インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW) についての同じ膜貫通ドメインを示す。
【図3】図3Aは、異なる構築物について結果を示す。
【図4】図4Aは、Sema3Aにより引き起こされる皮質軸索崩壊について濃度を増加させたpTM-NRP1ペプチド(10-8、10-9、10-10、及び10-11M)の効果を示す。図4Bは、Sema3Aにより引き起こされる皮質軸索崩壊について、pTM-NRP1又はpTM-NRP1mutペプチド(10-8M)の効果を示す。
【図5】図5Aは、pTM-NRP1又はpTM-NRP1mutを伴って又は伴わずにSema3Aの存在下又は不存在下でNRP1及びプレキシン-A1を発現するCOS-1細胞の形態を示す。図5Bは、Sema3Aにより引き起こされる細胞崩壊についてのpTM-NRP1又はpTM-NRP1mutペプチドの効果を示す(*:p<0.001)。
【図6】図6Aは、pTM-NRP1又はpTM-NRP1mutを伴うか又は伴わずにAP-Semaとインキュベートした後の野生型COS細胞(対照)又はNRP1発現COS細胞(COS-NRP1)を示す。図6Bは、以前に試験された条件について、細胞あたりの光学密度の平均強度を示す。
【図7】図7Aは、pTM-NRP1を伴って又は伴わずに、AP-Sema3Aとインキュベートした後における野生型COS細胞(対照)、NRP1発現COS細胞(COS-NRP1)、又は3個の(G→V)変異を有するNRP1を発現するCOS細胞(COS-NRP1mut)を示す。図7Bは、前に試験された条件についての細胞あたりの光学密度の平均強度を示す。
【図8】図8は、NGF(100ng/ml)、Sema3A、pTM-NRP1(10-9M)を伴って又は伴わずにインキュベートした後におけるPC12細胞の分化アッセイの結果を示す。図8Bは、各条件について分化した細胞の割合を示す。
【図9】図9は、pTM-NRP1ペプチド(黒色棒)又は変異ペプチド(灰色棒)の存在下におけるAP-Sema3Aのその受容体への結合についての結果を示す(*:p<0.005;**:p<0.01、スチューデントt検定)。
【図10】図10は、プレキシン-A1を含むオリゴマーを含むスクロース勾配の重分画に相当する黒色棒グラフ、NRP1ダイマーを含む中間分画に相当する灰色棒グラフ、そしてほぼNRP1モノマーから構成される軽い分画に相当する白色棒グラフを示す。
【図11】図11は、pTM-NRP1又はpTM-NRP1mutペプチドを伴うか又は伴わずにC6細胞を脳注射した後のマウス脳切片を示す。
【図12】図12は、pTM-NRP1又はpTMNRP1mutを伴って又は伴わずにC6細胞を脳に注射した後において、CD34を免疫染色した後のマウス脳切片を示す。
【図13】図13は、代表的な腫瘍細胞が、VEGF165(50ng/ml)、pTM-NRP1又はpTMNRP1mut(10-8M)の添加を伴って又は伴わずに凝集することを示す。
【特許請求の範囲】
【請求項1】
クラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニストであって、ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1及びインテグリンβ2からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも1のGxxxGモチーフを含むアミノ酸配列を含み、場合により異種配列に融合される、前記ペプチドアンタゴニスト。
【請求項2】
以下の:
ヒトニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr-CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1-CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも1のGxxxGモチーフを含むアミノ酸配列を含み、場合により異種配列に融合される、請求項1に記載のペプチドアンタゴニスト。
【請求項3】
ヒトニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)又はニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)の膜貫通ドメイン、好ましくはヒトニューロピリン-1膜貫通ドメイン(配列番号1:ILITIIAMSALGVLLGAVCGVVL)に由来するアミノ酸配列を含み、場合により異種配列に融合される、請求項2に記載のペプチドアンタゴニスト。
【請求項4】
以下の:
ヒトのニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)の膜貫通ドメインを含む群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも2のGxxxGモチーフを含むアミノ酸を含み、場合により異種配列に融合される、請求項2に記載されるペプチドアンタゴニスト。
【請求項5】
ヒトニューロピリン1膜貫通ドメイン(配列番号1、ILITIIAMSALGVLLGAVCGVVL)に由来するアミノ酸配列を含み、場合により異種配列に融合される、請求項3又は4に記載のペプチドアンタゴニスト。
【請求項6】
前記膜貫通ドメインに由来するアミノ酸配列が、10超のアミノ酸長であり、好ましくは14超のアミノ酸長である、請求項1〜5のいずれか一項に記載のペプチドアンタゴニスト。
【請求項7】
ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、Ll-CAM、インテグリンβ1及びインテグリンβ2からなる群から選ばれるタンパク質に由来するアミノ酸配列が、150未満のアミノ酸長、好ましくは100超のアミノ酸長である、請求項1〜6のいずれか一項に記載のペプチドアンタゴニスト。
【請求項8】
前記ペプチドアンタゴニストが、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、インテグリンβ1、インテグリンβ2、Nr-CAM及びL1-CAMからなる群から選ばれるタンパク質の膜貫通ドメインに由来し、ここで当該ペプチドアンタゴニストが、当該タンパク質のシグナル伝達経路と関連する細胞内ドメインを含まない、請求項1〜7のいずれか一項に記載のペプチドアンタゴニスト。
【請求項9】
前記ペプチドアンタゴニストが、ニューロピリン-1及びニューロピリン-2からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして当該ペプチドアンタゴニストが、当該タンパク質のクラスIIIセマフォリン誘導体と関連する細胞外ドメインを含まない、請求項1〜8のいずれか一項に記載のペプチドアンタゴニスト。
【請求項10】
以下の:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr-CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1-CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)の膜貫通ドメインからなる群から選ばれるアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される、請求項8に記載のペプチドアンタゴニスト。
【請求項11】
以下の:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)及び
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)の膜貫通ドメインからなる群から選ばれるアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される、請求項10に記載のペプチドアンタゴニスト。
【請求項12】
以下の:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)の膜貫通ドメインからなる群から選ばれるアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される、請求項10に記載のペプチドアンタゴニスト。
【請求項13】
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)のアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される、請求項11又は12に記載のペプチドアンタゴニスト。
【請求項14】
前記アミノ酸配列が、前記膜貫通ドメイン又はその断片と60%超の同一性、好ましくは70%超の同一性を有する、請求項1又は13に記載のペプチドアンタゴニスト。
【請求項15】
前記アミノ酸配列が、前記膜貫通ドメイン又はその断片と85%超、又は90%超の同一性を有する、請求項14に記載のペプチドアンタゴニスト。
【請求項16】
請求項1〜15のいずれか一項に記載のペプチドアンタゴニストをコードする核酸。
【請求項17】
請求項16に記載の核酸を含む核酸ベクター。
【請求項18】
最終的に医薬として許容されるビヒクルと一緒にされ、請求項1〜15のいずれか一項に記載のペプチドアンタゴニスト、請求項16に記載の核酸、又は請求項17に記載の核酸ベクターを含む、医薬組成物。
【請求項19】
前記組成物が、10-12M超、好ましくは10-11M超の濃度でペプチドアンタゴニストを含むことを特徴とする、請求項18に記載の組成物。
【請求項20】
クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路と関連する疾患を患う対象の予防又は治療のための医薬の製造における、請求項1〜15のいずれか一項に記載されるペプチドアンタゴニスト、請求項16に記載される核酸、又は請求項17に記載される核酸ベクターの使用。
【請求項21】
クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関連する疾患が、神経変性疾患、癌、及び異常血管新生に関連する全ての疾患を含む群から選ばれることを特徴とする、請求項20に記載の使用。
【請求項22】
前記医薬が、10-12M超、好ましくは10-11M超のペプチドアンタゴニストの濃度の放出を可能にする、請求項20又は21のいずれか一項に記載の使用。
【請求項1】
クラスIIIセマフォリン/ニューロピリン複合体のペプチドアンタゴニストであって、ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、L1-CAM、インテグリンβ1及びインテグリンβ2からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも1のGxxxGモチーフを含むアミノ酸配列を含み、場合により異種配列に融合される、前記ペプチドアンタゴニスト。
【請求項2】
以下の:
ヒトニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr-CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1-CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)
からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも1のGxxxGモチーフを含むアミノ酸配列を含み、場合により異種配列に融合される、請求項1に記載のペプチドアンタゴニスト。
【請求項3】
ヒトニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)又はニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)の膜貫通ドメイン、好ましくはヒトニューロピリン-1膜貫通ドメイン(配列番号1:ILITIIAMSALGVLLGAVCGVVL)に由来するアミノ酸配列を含み、場合により異種配列に融合される、請求項2に記載のペプチドアンタゴニスト。
【請求項4】
以下の:
ヒトのニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)、及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)の膜貫通ドメインを含む群から選ばれるタンパク質の膜貫通ドメインに由来し、そして少なくとも2のGxxxGモチーフを含むアミノ酸を含み、場合により異種配列に融合される、請求項2に記載されるペプチドアンタゴニスト。
【請求項5】
ヒトニューロピリン1膜貫通ドメイン(配列番号1、ILITIIAMSALGVLLGAVCGVVL)に由来するアミノ酸配列を含み、場合により異種配列に融合される、請求項3又は4に記載のペプチドアンタゴニスト。
【請求項6】
前記膜貫通ドメインに由来するアミノ酸配列が、10超のアミノ酸長であり、好ましくは14超のアミノ酸長である、請求項1〜5のいずれか一項に記載のペプチドアンタゴニスト。
【請求項7】
ニューロピリン-1、ニューロピリン-2、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、Nr-CAM、Ll-CAM、インテグリンβ1及びインテグリンβ2からなる群から選ばれるタンパク質に由来するアミノ酸配列が、150未満のアミノ酸長、好ましくは100超のアミノ酸長である、請求項1〜6のいずれか一項に記載のペプチドアンタゴニスト。
【請求項8】
前記ペプチドアンタゴニストが、プレキシン-A1、プレキシン-A2、プレキシン-A3、プレキシン-A4、インテグリンβ1、インテグリンβ2、Nr-CAM及びL1-CAMからなる群から選ばれるタンパク質の膜貫通ドメインに由来し、ここで当該ペプチドアンタゴニストが、当該タンパク質のシグナル伝達経路と関連する細胞内ドメインを含まない、請求項1〜7のいずれか一項に記載のペプチドアンタゴニスト。
【請求項9】
前記ペプチドアンタゴニストが、ニューロピリン-1及びニューロピリン-2からなる群から選ばれるタンパク質の膜貫通ドメインに由来し、そして当該ペプチドアンタゴニストが、当該タンパク質のクラスIIIセマフォリン誘導体と関連する細胞外ドメインを含まない、請求項1〜8のいずれか一項に記載のペプチドアンタゴニスト。
【請求項10】
以下の:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)、
プレキシンA1(配列番号3:LLTLPAIVGIGGGGGLLLLVIVAVLIA)、
プレキシンA2(配列番号11:LLTLPAIVSIAAGGSLLLIIVIIVLIAY)、
プレキシンA3(配列番号12:LTLPAMMGLAAGGGLLLLAITAVLVA)、
プレキシンA4(配列番号6:LSLPAIVSIAVAGGLLIIFIVAVLIA)、
Nr-CAM(配列番号7:GWFIGLMCAVALLILILLIVCFI)、
L1-CAM(配列番号13:GWFIGFVSAIILLLLVLLIL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)の膜貫通ドメインからなる群から選ばれるアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される、請求項8に記載のペプチドアンタゴニスト。
【請求項11】
以下の:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)及び
ニューロピリン-2(配列番号2:ILITIIAMSSLGVLLGATCAGLLLY)の膜貫通ドメインからなる群から選ばれるアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される、請求項10に記載のペプチドアンタゴニスト。
【請求項12】
以下の:
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)、
インテグリンβ1(配列番号9:IIPIVAGVVAGIVLIGLALLLIW)及び
インテグリンβ2(配列番号14:IAAIVGGTVAGIVLIGILLLVIW)の膜貫通ドメインからなる群から選ばれるアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される、請求項10に記載のペプチドアンタゴニスト。
【請求項13】
ニューロピリン-1(配列番号1:ILITIIAMSALGVLLGAVCGVVL)のアミノ酸配列又はその誘導体からなり、場合により異種配列に融合される、請求項11又は12に記載のペプチドアンタゴニスト。
【請求項14】
前記アミノ酸配列が、前記膜貫通ドメイン又はその断片と60%超の同一性、好ましくは70%超の同一性を有する、請求項1又は13に記載のペプチドアンタゴニスト。
【請求項15】
前記アミノ酸配列が、前記膜貫通ドメイン又はその断片と85%超、又は90%超の同一性を有する、請求項14に記載のペプチドアンタゴニスト。
【請求項16】
請求項1〜15のいずれか一項に記載のペプチドアンタゴニストをコードする核酸。
【請求項17】
請求項16に記載の核酸を含む核酸ベクター。
【請求項18】
最終的に医薬として許容されるビヒクルと一緒にされ、請求項1〜15のいずれか一項に記載のペプチドアンタゴニスト、請求項16に記載の核酸、又は請求項17に記載の核酸ベクターを含む、医薬組成物。
【請求項19】
前記組成物が、10-12M超、好ましくは10-11M超の濃度でペプチドアンタゴニストを含むことを特徴とする、請求項18に記載の組成物。
【請求項20】
クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路と関連する疾患を患う対象の予防又は治療のための医薬の製造における、請求項1〜15のいずれか一項に記載されるペプチドアンタゴニスト、請求項16に記載される核酸、又は請求項17に記載される核酸ベクターの使用。
【請求項21】
クラスIIIセマフォリン/ニューロピリン複合体シグナル伝達経路に関連する疾患が、神経変性疾患、癌、及び異常血管新生に関連する全ての疾患を含む群から選ばれることを特徴とする、請求項20に記載の使用。
【請求項22】
前記医薬が、10-12M超、好ましくは10-11M超のペプチドアンタゴニストの濃度の放出を可能にする、請求項20又は21のいずれか一項に記載の使用。
【図1】



【図2】

【図3】
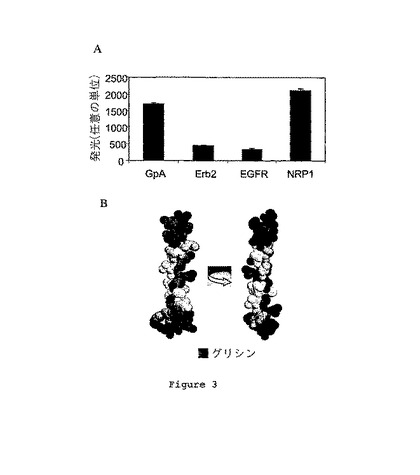
【図4】
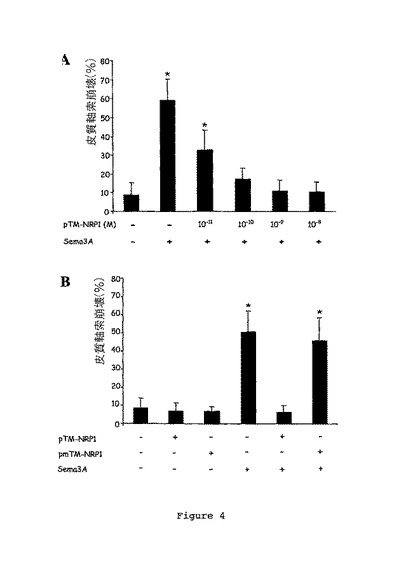
【図5】
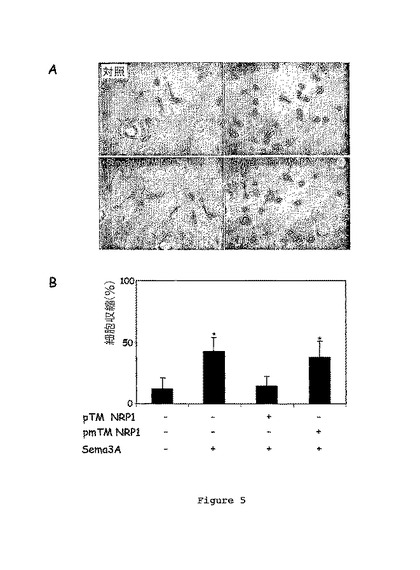
【図6】
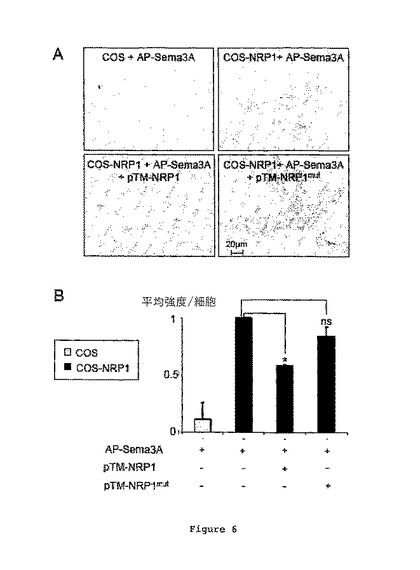
【図7】
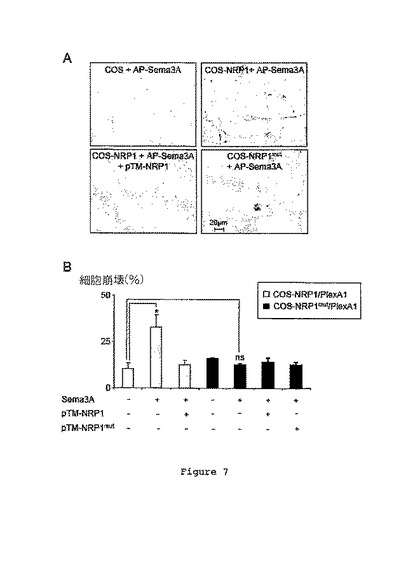
【図8】
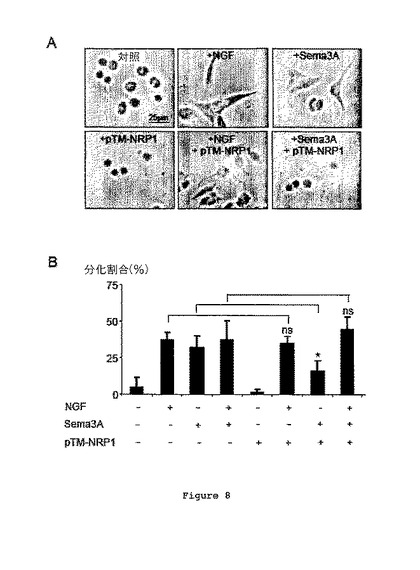
【図9】
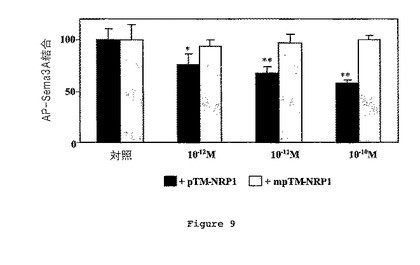
【図10】
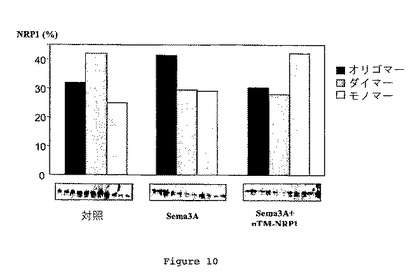
【図11】
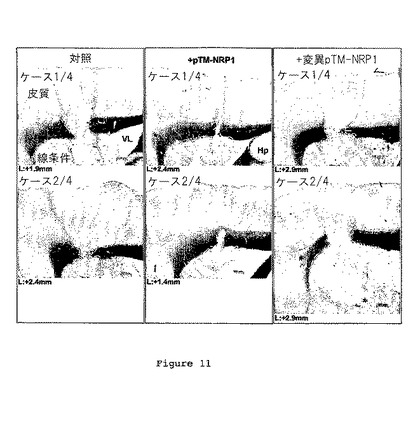
【図12】
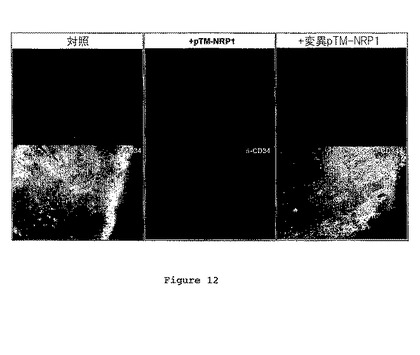
【図13】
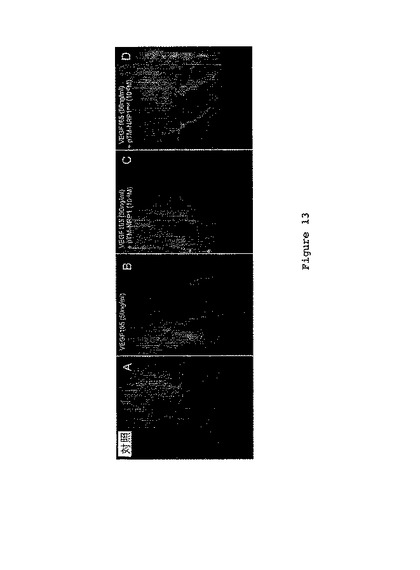
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2008−543347(P2008−543347A)
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願番号】特願2008−519016(P2008−519016)
【出願日】平成18年6月28日(2006.6.28)
【国際出願番号】PCT/IB2006/002334
【国際公開番号】WO2007/000672
【国際公開日】平成19年1月4日(2007.1.4)
【出願人】(500248467)アンスティテュ ナシオナル ドゥ ラ サントゥ エ ドゥ ラ ルシェルシェ メディカル(イーエヌエスエーエールエム) (19)
【Fターム(参考)】
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願日】平成18年6月28日(2006.6.28)
【国際出願番号】PCT/IB2006/002334
【国際公開番号】WO2007/000672
【国際公開日】平成19年1月4日(2007.1.4)
【出願人】(500248467)アンスティテュ ナシオナル ドゥ ラ サントゥ エ ドゥ ラ ルシェルシェ メディカル(イーエヌエスエーエールエム) (19)
【Fターム(参考)】
[ Back to top ]