
クロクマの副甲状腺ホルモン及びクロクマの副甲状腺ホルモンを使用する方法
クロクマの副甲状腺ホルモン(PTH)及びその機能的フラグメントが提供される。さらにまた、cAMPを骨形成細胞で増加させるか、アポトーシスを骨形成細胞で減少させるか、Baxタンパク質対Bcl-2タンパク質の発現レベル比率を骨形成細胞で減少させるか、骨基質タンパク質、転写活性化因子又は転写調節因子の1つ又は2つ以上の発現レベルを骨形成細胞で増加させるか、骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、もしくは骨折の発生率を低下させるか、又は前記の任意の組合せをもたらすためにクロクマのPTH及び機能的フラグメントを使用する方法が提供され、さらにクロクマの副甲状腺ホルモン(PTH)及びその機能的フラグメントに対抗する抗体もまた提供される。

【発明の詳細な説明】
【技術分野】
【0001】
連邦政府後援研究に関する記載
本発明は、米国立衛生研究所(NIAMS AR050420及びNIDDK DK078407)及び米国科学財団(IBN-0343515)により提供された合衆国政府の支援を受けて達成された。合衆国政府は本発明に関して一定の権利を有する。
【背景技術】
【0002】
骨減少疾患は、これまでのところ約4千4百万のアメリカ人の健康を脅かしている(前記には骨粗鬆症を有する1千万人及び低骨量を示し骨粗鬆症発生リスクを有する3千4百万人が含まれる)。骨粗鬆症のアメリカ人の数は2020年までに膨れ上がると予想される。結果として、極めて多くの個体が低骨量による骨折のリスクを有する。50歳を超える白人女性の約40%及び白人男性の13%が、一生の間に股関節部、脊椎又は前腕骨折のリスクを有する。骨粗鬆症関連骨折に付随するコストは2002年には約10億ドルであり、さらに増加し続けると予想される。原発性(年齢性)骨粗鬆症に加えて、不使用骨粗鬆症が、特に卒中又は脊髄損傷により長期的に固定される患者について重要な臨床的課題である。骨折率は、脊髄損傷後の最初の年に健康なコントロールと比較して倍増し、卒中の開始後もまた健康なコントロールと比較して上昇する。不使用は、主として骨格の負荷の低下が不均衡な骨のリモデリングを引き起こす(前記は骨減少をもたらす)という理由から骨折率を増加させる。
【発明の概要】
【0003】
ある実施態様では、本発明は、配列番号:2の連続する少なくとも10アミノ酸残基を含む単離ポリペプチドを提供し、ここで前記ポリペプチドは配列番号:2のアミノ酸残基41及び52の少なくとも1つを含む。ある実施態様では、配列番号:2のアミノ酸残基1−84を含む単離ポリペプチドが提供される。そのようなポリペプチドをコードするポリヌクレオチドもまた提供される。本発明はまた配列番号:1を含む単離ポリヌクレオチドを提供する。
ある実施態様では、本発明はcAMPレベルを骨形成細胞で増加させる方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程はcAMPレベルを骨形成細胞で増加させる。
【0004】
ある実施態様では、本発明はアポトーシスを骨形成細胞で減少させる方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程はアポトーシスを骨形成細胞で減少させる。
ある実施態様では、本発明はBaxタンパク質対Bcl-2タンパク質の発現レベル比率を骨形成細胞で低下させる方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程はBaxタンパク質対Bcl-2タンパク質の発現レベル比率を骨形成細胞で低下させる。
【0005】
ある実施態様では、本発明は、骨基質タンパク質、転写活性化因子又は転写調節因子の発現レベルを骨形成細胞で増加させる方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程は、骨基質タンパク質、転写活性化因子又は転写調節因子の発現レベルを骨形成細胞で増加させる。
ある実施態様では、本発明は、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、もしくは骨折の発生率を減少させるか、又は前記の任意の組合せをもたらす方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程は、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、又は骨折の発生率を減少させる。
【0006】
ある実施態様では、本発明は、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、もしくは骨折の発生率を減少させるか、又は前記の任意の組合せをもたらす方法を提供する。前記方法は、クマの副甲状腺ホルモン又はその機能的フラグメントを含むポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程は、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、又は骨折の発生率を減少させる。
ある実施態様では、本発明は配列番号:2のアミノ酸残基1−34から成る単離ポリペプチドを提供する。ある実施態様では、本発明は配列番号:2のアミノ酸残基1−36から成る単離ポリペプチドを提供する。ある実施態様では、本発明は配列番号:2のアミノ酸残基1−84から成る単離ポリペプチドを提供する。
【図面の簡単な説明】
【0007】
【図1】極限強度、骨強度の測定値、及び灰分比、骨塩含有量の測定値を示す(前記は両方ともクロクマで年齢に合わせて増加する)。
【図2】血清オステオカルシンレベルは、クロクマの冬眠前、冬眠時及び冬眠後プールサンプルで血清副甲状腺ホルモン(PTH)と正の相関性を有することを示す(p=0.0007、n=27)。
【図3】3ヶ月の不使用期間中の標準化した血清吸収マーカー(ICTP)及び血清形成マーカー(PICP及びオステオカルシン)濃度を示す。
【図4】骨芽細胞によって放出されるPGE2の量は、冬眠後に収集した血清で細胞を処理したときに最大であることを示す。
【図5】他の公知のPTH配列と対比した成熟クロクマPTHタンパク質配列を示す。
【図6】ヒト及びクロクマの両PTH 1-34がオステオカルシンをアップレギュレートすることを示す(n=2)。
【図7】アポトーシス関連遺伝子発現に対するヒト及びクロクマのPTH 1-34の作用を示す(n=4)。
【図8】ラット脛骨近位部(左)、骨端軟骨基部から2mm遠位で始まる皮質及び骨梁骨(trabecular bone)の1.6mm解析領域(上右)及び骨梁解析領域(下右)のuCT画像を示す。
【図9】9A−9Fは、模擬卵巣摘出術(OVX)を実施(図9A)又は実際のOVXを実施しその後PTH処置を施さずに6週間(図9B)もしくは14週間(図9C)回復させるか、又はクロクマPTH 1-34の3μg/kg(図9D)、10μg/kg(図9E)もしくは30μg/kg(図9F)による8週間の処置を含む14週間の回復期間後のラットの骨のμCT画像を示す。
【図10】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1-34で処置したラットの骨について総体積の比(BV/TV)としての骨体積のグラフを示す。
【図11】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1-34で処置したラットの骨について骨塩の見掛けの密度のグラフを示す。
【図12】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について骨梁支柱骨(trabecular strut)数のグラフを示す。
【図13】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1-34で処置したラットの骨について骨梁の厚さのグラフを示す。
【図14】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について骨梁骨の鉱化のグラフを示す。
【図15】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について脛骨皮質体積のグラフを示す。
【図16】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について脛骨皮質多孔度のグラフを示す。
【図17】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について大腿骨皮質の強度のグラフを示す。
【図18】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの血清カルシウムレベルのグラフを示す。
【図19】0−100nMのヒト又はクマPTHペプチドで前処理し、続いて血清枯渇に付したMC3T3-S4細胞の相対的カスパーゼ-3活性を示す。標準誤差バーとともにLS平均が提示されている。異なる文字を有するバーは互いに有意に相違する(p<0.005)。
【図20】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの大腿骨の究極負荷を示す。
【図21】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの大腿骨の標準化究極負荷を示す。
【図22】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの骨体積比を示す。
【図23】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの骨梁数を示す。
【図24】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの骨梁の厚さを示す。
【図25】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの骨梁の分離を示す。
【図26】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの見掛けの骨塩密度を示す。
【図27】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの実質的骨塩密度を示す。
【図28】冬眠前、冬眠時及び冬眠後のクマ血清中で培養した細胞のカスパーゼ3/7活性を示す。
【図29】ベヒクル又は種々のPTHペプチドで処理したマウスの大腿骨の究極負荷を示す。
【発明を実施するための形態】
【0008】
ヒト及び他の大半の哺乳動物では、例えば加齢及び長期未使用のような因子は、骨粗鬆症及び骨折リスクの増加をもたらし得る。脊髄損傷による不使用は、特に脛骨及び大腿骨における骨塩密度を顕著に低下させ、さらに大腿骨骨幹の断面慣性モーメントを顕著に低下させる。したがって、骨の曲げ強さは脊髄損傷によって低下し、骨折リスクが増加する。卒中による不使用もまた骨折リスクを増加させる。さらにまた、骨の機械的負荷の低下は、骨形成の持続的低下に加えて迅速な骨吸収増加のために急激な骨減少を引き起こし得る。不使用により誘発される骨リモデリングの変化は皮質内多孔度を増加させ、長骨骨幹の断面特性及び機械的特性を低下させる。負荷低下はまた、骨梁骨の量及び微細構造を大いに低下させる。
【0009】
骨に対する不使用の有害な影響は再可動期間中も持続し得る。ある骨は再可動時に回復し得るが、回復は遅くかつしばしば不完全である。例えば、病臥中の骨減少速度は、再可動時の骨増加よりも3倍以上速く、宇宙飛行時の骨減少の回復は5年後さえも完全ではないことがある。不使用により誘発された骨の変化を再開した活動によって完全に復帰させ得る場合、この再可動期間の長さはしばしば固定期間の2から3倍である。骨に対する機械的負荷を低下させる多くの状況下で、骨形成は低下し、及び/又は骨吸収は増加する。しかしながら、イヌの前肢の固定中に吸収及び形成の両方が増加するが、それでもなおそのような事例において顕著な骨減少が存在する。同様に、脊髄損傷を有する患者で大腿骨の代謝回転が生じ、これは骨減少及び骨折発生率の増加をもたらす。
【0010】
これとは対照的に、クロクマは加齢(図1)によって、又はより重要なことには長期の不使用(冬眠時に生じる)によっても顕著な骨減少に悩まされることはない。冬眠するクロクマは、北方領域では長さがほぼ等しい固定期と活動期を有する。骨代謝の血清マーカーに関するデータ(下記参照)は、クマでは不使用時に吸収と形成の両方が増加し吸収と形成の間のタイムラグ(すなわち復帰期間)は通常のとおりであり、形成の増加は吸収の増加と共役し均衡を保ったままであることを示唆している。クロクマの腸骨稜の生検から得られた組織学的データもまた非活動時の吸収及び形成の増加を示している。しかしながら、クマは、冬眠中に骨梁骨の体積、骨塩密度及び骨塩含有量が低下しないという点で唯一のものである。さらにまた、クロクマでは皮質強度及び灰分比が年齢とともに増加し、多孔度は毎年の不使用期間にもかかわらず低下する。皮質の多孔度は、活動中のハイイログマよりも冬眠中のハイイログマで顕著に低く、さらに大腿骨断面の幾何学及び強度は冬眠によって影響を受けない。
【0011】
クマは多くの固有の生物学的メカニズムを進化させて、食物の無い長い固定化期間を生き残ってきた。これらのメカニズムには、カルシウム及び骨異化作用の他の成分のリサイクルが含まれるように思われる。なぜならば、クマは冬眠中に骨の代謝回転を増加させるが、老廃物を排出しないからである。ヒトでは、病臥により誘発される不使用骨粗鬆症は、主として対応する形成増加を伴わない吸収の増加によって引き起こされる。これは、高カルシウム血症及び尿と糞便のカルシウムによって生じる負のカルシウム均衡をもたらす。クマは冬眠中に排尿も排便もしないので、吸収によって骨から放出されたカルシウムの大半は、骨芽細胞による骨形成を介して骨にリサイクルされるということはありそうなことである。イオン化カルシウムが冬眠時に約23%増加することが見出され、これらはおそらく吸収と形成の間のタイムラグのためであろう。逆説的に、クロクマのPTHレベルはイオン化カルシウムレベルが最高のときにもっとも高い(実施例2、表1)。総合すれば、これらの発見は、クマは骨粗鬆症を回避するために生物学的メカニズムを進化させてきたことを示唆している。
【0012】
大半の動物で不使用時に骨形成が骨吸収と共役しないメカニズムは不明であるが、おそらく機械的及び生化学的因子の両方が必要とされるだろう。機械的緊張の欠如は、骨細胞のアポトーシスの開始及び同時に生じる骨芽細胞活性の低下によって吸収の増加をもたらし得る。例えばヒトPTHのようなホルモンは骨細胞を機械的刺激に対して感受性にし、さらに機械的負荷により相乗的に骨形成を増加させる。ヒトに1日1回投与されたヒトPTHは骨量を増加させ骨折発生率を低下させる。したがって、クロクマでは、循環PTHは低レベルの機械的刺激(おそらく冬眠用器官の震え又は整復による)に対して骨細胞を感受性にして、不使用時の骨形成の維持を助長する。PTHはまた、骨芽細胞の分化を刺激し骨芽細胞のアポトーシスを抑制することによってクロクマで骨形成の維持を助長し得る。
【0013】
PTHは血中カルシウムレベルの主要調節因子であり、したがってクロクマの不使用時の恒常的血清カルシウムレベルの維持に重要な役割を果たす。血清PTHレベルは、活動時及び冬眠時のクロクマにおいて骨形成マーカーオステオカルシンと正の相関性を示し(図2)、オステオカルシン及びPTHは両方とも冬眠中に増加する。さらにまた、クロクマのPTH濃度は、イオン化カルシウム濃度が最高のときにもっとも高い。冬眠中に骨吸収は増加するが総血清カルシウム(tCa)は無変化のままであるので、おそらくPTHのレベル増加は腎臓のカルシウム再吸収の増加を引き起こし、骨への塩のリサイクルを促進し骨形成の増加を均衡させる。これによって、骨塩密度(“BMD”)及び皮質多孔度のような骨稜及び皮質の特性で観察された保存がもたらされる。冬眠中に骨吸収は増加するが、クマは老廃物を冬眠中に排泄しないという事実にもかかわらず血中カルシウム濃度は一定のままである。冬眠中に骨吸収によって遊離したカルシウムはリサイクルされ、骨形成と骨吸収との均衡した共役の維持により骨に戻され得る。このことは冬眠中のクロクマではPTHは同化作用を有するという考えを支持し、冬眠中に均衡のとれた骨リモデリングを維持するクマの特異な能力の説明を提供する。PTHの同化作用は、冬眠からの覚醒に続いて身体活動が開始するときにクロクマで強化され得る。機械的負荷及びヒトPTHは相乗的に作用し、ラットの骨形成をin vivoで、さらに生化学的シグナリングをin vitroで増加させることが以前に示された。春の再可動時に、クロクマの骨形成は血清オステオカルシンが示すように冬眠前のレベルよりも高いまま維持される。
【0014】
クロクマ(ウルスス・アメリカヌス(Ursus americanus))の副甲状腺ホルモン(PTH)をコードするポリヌクレオチドの配列(配列番号:1)が、成熟84アミノ酸PTHタンパク質のポリペプチド配列(配列番号:2)と同様に見出された。さらにまた、完全長PTHタンパク質(配列番号:4)をコードするcDNA(配列番号:3)の配列も決定された(前記完全長PTHタンパク質は25アミノ酸のシグナルペプチド(配列番号:4のアミノ酸残基1−25)及び6アミノ酸のプロペプチド(配列番号:4のアミノ酸残基26−31)を含む)。前記成熟クロクマPTHタンパク質は他の公知のPTHタンパク質とは相違する(図5)。ヒトPTHと比較して、クロクマPTHは、完全長の成熟PTHポリペプチドの合計84アミノ酸残基のうち9アミノ酸残基が異なっている。さらに本明細書にはクロクマPTH及びその機能的フラグメントの種々の使用方法が記載されている。具体的には、クロクマPTHに特異的な抗体の開発のために、配列番号:2の連続する少なくとも10アミノ酸を含み、かつアミノ酸残基41又は52の少なくとも1つを含有するポリペプチドサブフラグメントを使用できることが考えられる。これらの抗体を用いてクロクマPTHを例えばELISAアッセイで定量できる。
【0015】
骨形成細胞表面のPTHレセプターは、細胞内のサイクリックアデノシン一リン酸(cAMP)依存第二メッセンジャーシグナリング経路と共役する。これらのシグナリング経路は順繰りに、骨形成に必要な遺伝子(例えばI型コラーゲン、オステオネクチン及びオステオポンチンをコードするもの)の発現増加をもたらす。cAMP/タンパク質キナーゼA経路は骨形成における組織学的指標及び血清指標の大多数のPTH誘発増加を起こすことができるので、結果としてcAMP応答増加がより強い骨形成をもたらすことができる。あるPTHタンパク質の配列で比較的少数のアミノ酸置換が、天然の形態と比較してより強いサイクリックアデノシン一リン酸(cAMP)産生を刺激できる。例えば、卵巣摘出ラットは、ラットPTH1-34よりもウシPTH1-34の1日25μgの注射に対して25%高い骨形成応答を示した(ラットPTH1-34はウシPTH1-34と比較して5アミノ酸の配列相違を有する。ウシPTH1-34の注射は、処置時に37%高い骨体積比の増加をもたらす。
【0016】
したがって、クロクマPTHのアミノ酸置換によって、ヒトPTHよりも強いcAMP産生が骨形成細胞で惹起されることはありそうなことである。一般的に、PTHは、例えば骨芽細胞のアポトーシス低下、Runx2媒介骨芽細胞分化の増加、SOST系の骨細胞における負のフィードバックのダウンレギュレーション、及び骨基質タンパク質のmRNAの生成増加(いずれもcAMP媒介経路)によってより強い骨形成応答を引き起こす。本発明のメカニズムを理解することが必要というわけではないが、クロクマPTHはおそらく他のPTH形よりも骨形成性が高いと考えられ、これは、クロクマが不使用中に均衡をもった骨リモデリングを維持する固有の能力をもつ理由を説明する。本発明のある実施態様では、骨形成細胞をクロクマPTH又はその機能的フラグメントと接触させる工程は骨形成細胞でcAMPレベルを増加させる。本発明のある実施態様では、配列番号:2のアミノ酸残基1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
【0017】
本明細書で用いられる、PTHポリペプチドと“細胞を接触させる工程”は、in vitro実験の場合には培養溶液にポリペプチドを添加する工程、又はポリペプチド治療薬に適した投与方法を用いて被験者にポリペプチドを投与する工程を含む。“細胞を接触させる工程”はまた、被験者で所望のポリペプチドを合成及び放出させるために、ある発現系で前記ポリペプチドをコードする外因性ポリヌクレオチドを被験者に導入する工程を含む。本明細書で用いられる、“骨形成細胞”には骨芽細胞、骨細胞、骨内皮細胞(bone lining cell)、軟骨芽細胞及び軟骨細胞が含まれるが、ただしこれらに限定されない。適切には骨形成細胞は被験者の内部に存在し得る。
骨形成細胞は規則的に代謝回転を生じ、細胞死の大半はプログラムされた細胞死(又はアポトーシス)による。この規則的な代謝回転速度が存在する場合、骨形成細胞のアポトーシスを低下させるいずれのメカニズムも骨形成細胞数の増加をもたらし、これはおそらく骨の生長を促進させるであろう。したがって、本発明のある実施態様では、骨形成細胞をクロクマPTH又はその機能的フラグメントと接触させる工程は骨形成細胞のアポトーシスを低下させる。本発明のある実施態様では、配列番号:2のアミノ酸1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
【0018】
さらにまた、内因性クロクマPTHのより大きなC-末端フラグメントのいくつかはCPTHR(C-末端PTHレセプター)との結合を介してシーズン性骨リモデリングプロセスで役割を演じる可能性がある。具体的には、クマPTHのC-末端フラグメントは、骨芽細胞形成を妨げることによって及びおそらくは成熟骨細胞活性に影響を与えることによって(前記は通常的には不使用のような吸収性刺激に応答して発生するであろう)、PTH1-84及び1-34の血中カルシウム上昇作用に拮抗し得る(Divieti, P. et al., 2002, Endocrinology 143(1): 171-6)。これは、冬眠中のクマが恒常的カルシウムレベルを維持するために役立ち得る。したがって、本発明のある実施態様では、配列番号:2のアミノ酸残基11−84を含むポリペプチド及びアミノ酸残基7−84を含むポリペプチドから選択される少なくとも1つのポリペプチドと骨細胞及び骨芽細胞を接触させることができる。
タンパク質Baxはアポトーシスを促進するがBcl-2タンパク質は細胞をアポトーシスから保護し、Bax対Bcl-2の発現比率の低下は個々の細胞集団におけるアポトーシスの低下を示唆する。したがって、本発明のある実施態様では、クロクマPTH又はその機能的フラグメントと骨形成細胞を接触させる工程は、Bcl-2タンパク質の発現レベルに対するBaxタンパク質の発現レベルの比率を骨形成細胞で低下させる。本発明のある実施態様では、配列番号:2のアミノ酸残基1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
【0019】
実施例11は、クロクマPTH1-34は培養細胞でBax/Bcl-2発現比率を低下させるが、ヒトPTH1-34はBax/Bcl-2発現比率を増加させることを示す(図7)。したがって、クロクマPTH1-34はヒトPTH1-34よりもアポトーシスの防止により有効であるように思われる。理論に拘束されないが、この相違はヒトPTH1-34とクロクマPTH1-34間での2つのアミノ酸が異なることの結果であるかもしれない。これらのデータは、クマPTHはヒトPTHよりも同化作用が強いことを提唱している。なぜならば、骨芽細胞のアポトーシスの低下はPTH処理によって誘発される骨形成応答に寄与し得るからである。
クロクマPTH又はその機能的フラグメントと骨形成細胞を接触させる工程はまた、骨基質タンパク質、転写活性化因子又は転写調節因子の発現レベルを骨形成細胞で増加させる。本発明のある実施態様では、転写活性化因子はRunx2である。本発明のある実施態様では、転写調節因子はc-fosである。例示的な骨基質タンパク質にはオステオカルシン、オステオポンチン及びコラーゲンI型が含まれるが、ただしこれらに限定されない。本発明のある実施態様では、配列番号:2のアミノ酸残基1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
【0020】
外因性ヒトPTHを用いてヒトの閉経後骨粗鬆症及び加齢性骨粗鬆症が治療されているが、前記PTHは理想的な治療薬ではない。現在のところ組換えヒトPTH1-34(LY333334, Eli Lilly, Indianapolis IN)のみが臨床使用のために承認されており、組換えヒトPTH1-84の1つの型(ALX1-11, NPS Pharmaceuticals, Parsippany, NJ)のみが米国食品医薬品局の承認が考慮されている。LY333334及びALX1-11はin vivoで同じ規模の骨形成を刺激するが、それらの生物学的作用は同一ではない。例えば、PTH1-34はプロコラーゲン-1 mRNAの産生をダウンレギュレートするが、PTH1-84はダウンレギュレートしない(Nasu et al., 1998, Endocr J, 45, 229-34)。さらにまた、ヒトPTHのC-末端部分は、成熟ホルモンから切り離されたとき重要な生物学的機能(例えば骨吸収の阻害)を有する。
LY333334又はALX1-11の長期使用はラットで骨肉腫を生じるが、予備的結果によれば、ヒトPTH1-84はヒトPTH1-34よりも発癌率が低いことが示唆され、これはおそらく、外因性ヒトPTH1-84のC-末端フラグメント(末梢性タンパク分解性プロセッシングにより生じる)はC-末端PTHレセプター(CPTHR)と結合し、骨細胞のアポトーシスを増加させるためであろう。したがって、同化作用は同等であるが、ヒトPTH1-84はひとPTH1-34と比較して優れた骨粗鬆症治療法であり得る。しかしながら、ヒトPTH1-84は失われた骨を完全には回復させることはできない。すなわち、男性及び女性は加齢性骨粗鬆症のために皮質及び海綿質の20‐30%を失うが、ALX1-11をその推奨治療期間中ずっと使用しても8%しか回復させ得ないと言われている。したがって、より強力な骨形成能を有する骨粗鬆症治療が臨床現場で希求されている。
【0021】
ある実施態様では、本発明は、クロクマPTH又はその機能的フラグメントの有効量をその必要がある被験者に投与する工程を含む、骨粗鬆症の治療方法を提供する。種々の実施態様で、骨粗鬆症は、原発性又は加齢性骨粗鬆症及び/又は不使用骨粗鬆症である。本発明のある実施態様では、被験者の骨形成細胞をクロクマPTH又はその機能的フラグメントと接触させる工程は、被験者で骨塩密度を増加させるか、骨量を増加させるか、骨減少を低下させるか、又は骨折の発生率を低下させる。本発明のある実施態様では、配列番号:2のアミノ酸残基1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
適切には、被験者の骨形成細胞をクロクマPTH又はその機能的フラグメントと接触させる工程は、少なくとも約5%又は少なくとも約10%骨塩密度を増加させるか、骨量を増加させるか、骨減少を低下させるか、又は骨折の発生率を低下させる。骨塩密度の増加、骨量の増加、骨減少の低下又は骨折の発生率の低下は、少なくとも約15%、少なくとも約30%、少なくとも約50%、少なくとも約75%、又は少なくとも約90%であり得る。骨塩密度の増加、骨量の増加、骨減少の低下又は骨折の発生率の低下は、治療前及び治療後に当業者に公知の技術により所望の特徴を同一患者で測定することによって決定される。例えば、骨塩密度は、脊髄、手首、腕又は脚内の骨の二重エネルギーX線(DEXA)又はCTスキャンの撮影を含む方法によって決定できる。
【0022】
被験者は適切には哺乳動物で、ヒト、ウマ、イヌ、ネコ、マウス、クマ、ウシ、ブタ又はシカが含まれるが、ただしこれらに限定されない。ある実施態様では、被験者は閉経後の女性である。被験者は骨粗鬆症を有していても又は骨粗鬆症発症のリスクがあってもよい。骨粗鬆症発症のリスク因子には以下が含まれる:50歳以降の骨折の病歴;現時点で低骨量であること;1親等親族の骨折の病歴;女性であること;痩せているか及び/又は骨格が小さいこと;高齢;骨粗鬆症の家族歴;閉経(特に早発性又は外科手術により誘発されたもの)の結果としてのエストロゲン欠乏;月経期間の異常停止(無月経);神経性食欲不振;終生のカルシウム摂取不良;ビタミンD欠乏;一定の薬物の使用(コルチコステロイド、化学療法、抗痙攣薬及び他のもの);一定の慢性的症状、例えば腸のカルシウム吸収を低下させるもの(例えばクローン病)の存在;男性の低テストステロンレベル;非活動的なライフスタイル;現時点で喫煙していること;アルコールの過剰摂取;及び白人又はアジア人であること(ただしアフリカ系アメリカ人及びヒスパニック系も同様に有意なリスクを有する)。さらにまた、女性は閉経後5年から7年でその骨量の20%までを喪失し、いっそう骨粗鬆症に感受性となり得る。
【0023】
クロクマPTH又はその機能的フラグメントはまた、骨粗鬆症と戦うため又は骨粗鬆症発症リスクを有する被験者で骨粗鬆症を予防するための、防止薬(回復薬ではなく)もしくは予防薬、手段として有用である。クマは不使用の間均衡のとれた骨のリモデリングを維持する唯一の動物のようであるので、クロクマPTH又はその機能的フラグメントはまた、例えば宇宙飛行中の宇宙飛行士及び脊髄損傷後の患者で発生する骨格負荷の低下時の骨減少の防止に有用である。
クロクマPTH又はその機能的フラグメントは、カルシウム及び/又はビタミンDと一緒に投与できる。適切には、“ビタミンD”は完全なビタミンDクラスの化合物を指す。クロクマPTH又はその機能的フラグメントはまた、他の同化作用促進又は抗吸収活性物質と一緒に投与できる。適切な同化作用促進活性物質には、ヒトPTHの種々のフラグメント(例えば1-34及び1-84)、フルオリド、GH、インスリン様成長因子I、スタチン及びPYK2キナーゼ阻害剤が含まれる。適切な抗吸収活性物質には、エストロゲン、選択性エストロゲンレセプター調節因子、カルシトニン及びビスホスホネートが含まれるが、ただしこれらに限定されない。ある実施態様では、同化作用を有するクロクマPTHの機能的フラグメント(例えばクロクマPTH1-34)が抗吸収活性物質と一緒に投与される。ある実施態様では、クロクマPTHの種々の機能的フラグメントは互いに組み合わせて投与できる。種々の機能的フラグメントは同時に又は適切な任意の順序で投与できる。
【0024】
カルシウム、ビタミンD、同化作用促進活性物質及び/又は抗吸収活性物質は、クロクマPTH又はその機能的フラグメントと同時に投与するか、又はクロクマPTH又はその機能的フラグメントの前もしくは後に投与できる。カルシウム、ビタミンD、同化作用促進活性物質及び/又は抗吸収活性物質は別々の組成物に存在しても、又はクロクマPTH又はその機能的フラグメントと同じ医薬組成物に存在してもよい。
クロクマPTHもしくはその機能的フラグメントの投与又はクロクマPTHもしくはその機能的フラグメントを含む組成物の投与は、任意の適切な技術によって実施できる。クロクマPTH又はその機能的フラグメントは、例えば経口、経鼻、経直腸及び非経口ルートの投与を含む任意の適切なルートによって投与できる。本明細書で用いられる、非経口という用語には、例えば注射による皮下、皮内、静脈内、筋肉内、腹腔内、クモ膜下投与が含まれるが、ただしこれらに限定されない。上記で考察するように、ポリペプチドの投与には、被験者で外因性ポリヌクレオチドを発現できるようにプロモーターと作動可能に連結した外因性ポリヌクレオチドの投与が含まれる。ポリペプチドの投与にはまた、前記ポリペプチドをコードするポリヌクレオチドを含むウイルスベクターの投与が含まれる。適切には、ウイルスベクターはアデノウイルスベクターである。
【0025】
クロクマPTHもしくはその機能的フラグメント又はクロクマPTHもしくはその機能的フラグメントを含む組成物は、継続的に又は当業者が容易に決定できる明確な間隔をおいて投与することができる。通常の技量を有する臨床医は、被験者に投与されるべきクロクマPTH又はその機能的フラグメントの適切な量を決定できる。
任意の個々の被験者に対する有効用量は多様な因子に左右される。前記因子には、治療される疾患及び疾患の重症度;用いられる個々の化合物の活性;用いられる個々の組成物;被験者の年齢、体重、一般的健康状態、性別及び養生食;投与ルート;用いられるクロクマTPH又はその機能的フラグメントの排出速度又は不活化速度;治療期間;クロクマTPH又はその機能的フラグメントと一緒に用いられる他の医薬;及び医療分野で周知の同様な因子が含まれる。例えば、所望の効果を達成するために必要な用量よりも低いレベルの用量から開始し、所望の効果が達成されるまで投薬量を徐々に増加させることは十分に当業者の水準の範囲内である。
適切には、ある実施態様におけるクロクマTPH又はその機能的フラグメントの投薬量は、0.10μg/kg/日から40μg/kg/日の範囲である。ある実施態様では、投薬量は5μg/kg/日から20μg/kg/日の範囲である。ある実施態様では、投薬量は10μg/kg/日である。ある実施態様では、投薬量は被験者当たり10μg/日から400μg/日の範囲である。ある実施態様では、投薬量は被験者当たり20μg/日から40μg/日の範囲である。ある実施態様では、投薬量は被験者当たり30μg/日である。
【0026】
併用投与のためには、当業者は、クロクマTPH又はその機能的フラグメント及び追加される活性物質(例えばカルシウム、ビタミンD、同化作用促進薬又は抗吸収薬)の両方の適切な投薬量を決定できよう。上記で考察したように、任意の個々の被験者に有効な用量は多様な因子に左右される。ある実施態様では、抗吸収薬の投薬量は1日当たり約5から約100mgであり、カルシウムの投薬量は1日当たり約500から約1500mgであり、ビタミンDの投薬量は1日当たり約500から約1500mgであり、同化作用促進薬の投薬量は1日当たり約0.1mgから約0.5mg又は約10μg/日から400μg/日である。
ある実施態様では、被験者はヒトである。適切には、1日当たりの投薬量がある実施態様では1週間、ある実施態様では1ヶ月、ある実施態様では3カ月間、ある実施態様では6ヶ月間、ある実施態様では1年間、ある実施態様では1年と半年間、ある実施態様では2年間、さらにある実施態様では3年間与えられる。
【0027】
所望の場合は、1日の有効用量を投与目的にふさわしい複数回の用量に分割できる。したがって、シングルドース組成物は1日の用量を構成する量又はそれを複数回に分けた部分を含むことができる。所望の場合は、適切なデリバリー装置に1日の有効量を2日以上のために、例えば7日、14日、21日、28日などのために装填し、このデリバリー装置を用いて、所望の1日1回用量又は1日複数回用量を所望の合計日数の間繰り返して投与する。理解されるところであるが、医療の実施に関する基準及び個々の被験者の症状によって決定されるように、当業者は有効用量及び同時投与治療方式を容易に最適化するであろう。
本発明の方法で有用なクロクマPTH又はその機能的フラグメントを含む組成物は、医薬として有用な組成物の公知の製造方法にしたがって処方することができる。処方は、周知であり当業者が容易に入手できる多数の文献に詳細に記載されている。例えば以下の文献には本開示方法で用いることができる処方が記載されている(Remington's Pharmaceutical Science, E. W. Martin)。一般的には、組成物は、クロクマPTH又はその機能的フラグメントの有効量を適切な担体と一緒にし、組成物の有効な投与を促進することができるように処方されるであろう。
【0028】
本方法で用いられる組成物はまた多様な形態で存在し得る。これら形態には例えば固体、半固体、及び液体投薬形(例えば錠剤、ピル、散剤、液状溶液又は懸濁物、座薬、注射可能及び輸液可能溶液、並びにスプレー)が含まれる。形態は意図する投与態様及び治療薬の適用態様に左右されるであろう。組成物はまた適切には、医薬的に許容できる当業者に公知の通常の賦形剤を含む。賦形剤の例には、注射用の水、エタノール、ジメチルスルホキシド、グリセロール、アルミナ、デンプン、氷酢酸、酢酸ナトリウム、マンニトール、メタクレゾール、組成物のpHを適切な値に調整するための塩酸及び/又は水酸化ナトリウム、並びに等価の或いは適切な担体及び希釈剤が含まれる。所望の適用においてそのような調剤を投与するために、医薬組成物は、担体又は賦形剤を含む全組成物重量を基準にして、総重量が約0.1%から99%、適切には約1から15%の1つ又は2つ以上の本発明のポリペプチドを含むであろう。
【0029】
本明細書で用いられるように、“単離された”核酸分子、ポリヌクレオチド、ポリペプチドなどは、場合によって、天然の状態以外で見出される夾雑物(例えばポリヌクレオチド、ポリペプチドなどの他の種)から少なくとも部分的に精製された成分を指す。単離された核酸、ポリヌクレオチド又はポリペプチドは、前記が本来結合している細胞成分の約50%未満、適切には約75%未満、もっとも適切には約90%未満を含むことができる。残余の細胞成分から(例えばゲル上で)十分にかつ容易に区別できるようにPCRを用いて増幅したポリヌクレオチドは“単離されている”とみなされる。本発明の核酸分子、ポリヌクレオチド及びポリペプチドは“実質的に純粋”であり得る。すなわち、当業界で公知の精製技術を用いて達成可能な最高の純度を有する。
本明細書で用いられるように、“機能的フラグメント”は、より大きなポリペプチド又はポリヌクレオチドの領域又は部分である、あるポリペプチド又はポリヌクレオチドの任意の領域又は部分である。前記領域又は部分はより大きなポリペプチド又はポリヌクレオチドに起因する活性又は機能を有する。例えば、ヒトPTHの機能的フラグメントはヒトPTHの1−34領域である。クロクマPTHの機能的フラグメントには1−34及び1−36が含まれるが、ただしこれらに限定されない。
【0030】
本明細書及び添付の特許請求の範囲で用いられるように、単数形の“a”、“an”、及び“the”には、内容が明瞭にそうでないことを示していないかぎり複数の対応形が含まれる。さらにまた、“or”という語は、内容が明瞭にそうでないことを示していないかぎり一般的には“及び/又は”を含む意味で用いられる。全ての刊行物、特許及び特許出願は、あたかも個々の各刊行物又は特許出願が本明細書に具体的にかつ個々に参照によって含まれるかのように、同じ程度にその全体が全ての目的のために参照により明白に本明細書に含まれる。本開示と参照により含まれる特許、刊行物及び参考文献との間に矛盾がある場合には、本開示が優先する。
本明細書に列挙した数字によるいずれの範囲も、小さい方の値から大きい方の値までの全ての値を含むこともまた特に理解されよう。すなわち、列挙された最小値と最大値の間の数値の可能な全ての組合せが本出願に明瞭に記載されていると解されるべきである。例えば、濃度範囲が1%から50%と記載されている場合は、例えば2%から40%、10%から30%又は1%から3%などの値を本明細書に明瞭に列挙しようとするものである。濃度範囲が“少なくとも5%”である場合には、それより上の全てのパーセンテージ値(100%を含む)もまた明瞭に列挙しようとするものである。これらは特に意図するものの例にすぎない。
以下の実施例は本発明の更なる理解を助けるために提供される。用いられた具体的な材料、方法及び条件は本発明の例示を意図するものであり、本発明の範囲を限定するものではない。
【実施例1】
【0031】
実施例1:クロクマPTH1-84の配列決定
ゲノムDNA抽出
捕獲された雌のクロクマから血液を収集し4℃で保存した。ゲノミックプレップ血液DNA単離キット(GenomicPrep Blood DNA Isolation Kit)(Amersham Biosciences, Piscataway, NJ)を製造業者の指示に従って用い、全血サンプルからゲノムDNAを2週間以内に抽出した。
PCRクローニング及び配列決定
クロクマゲノムDNAをPTHのPCR増幅に用いた。GenBankで入手可能な8つの完全長哺乳動物PTH配列(以下を含む:ウシ(ボス・タウルス(Bos taurus)AAA30749)、ネコ(フェリス・カツス(Felis catus)Q9GL67)、イヌ(カニス・ファミリアリス(Canis familiaris)P52212)、ヒト(ホモ・サピエンス(Homo sapiens)NP_000306)、マカク(マカカ・ファシクラリス(Macaca fascicularis)Q9XT35)、マウス(ムス・ムスクラス(Mus musculus)NP_065648)、ブタ(スス・スクロファ(Sus scrofa)NP_999566)、及びラット(ラトツス・ノルヴェギクス(Rattus norvegicus)NP_058740)のアラインメントを基にして設計したコンセンサスプライマーを用いた。PCR増幅は、20μLの反応体積中で10−15ngのゲノムDNA、100μMのdNTP、各々0.2μMのプライマー及び1ユニットのREDTaq(Sigma, St. Louis, MO)を用いて実施した。PCR生成物は、ウルトラクリーンゲルスピンキット(UltraClean GelSpin Kit)(MoBio Carlsbad, CA)を用いてゲル精製し、さらにTAクローニングキット(Invitrogen, Carlsbad, CA)を用いてpCRIIベクターでクローニングした。DNA配列決定は、DTCSクイックスタート(Quick Startt)キット及びCEQ8000ジネティックアナリシスキット(Beckman Coulter, Fullerton, CA)を製造業者の指示にしたがって用いて実施した。
配列の解析
BlastX(Altschul et al., 1997; Nucleic Acids Res., 25, 3389-402)を用いてGenBankのタンパク質データベースでヌクレオチド配列を検索し、PTHとしてのそれらの推定的同一性を確認した。ClustalWバージョン1.82(Chenna et al., 2003; Nucleic Acids Res., 31, 3497-500)によって多重配列アラインメントを実施した。系統解析は、分子進化遺伝学解析(Molecular Evolutionary Genetics Analysis(MEGA))パッケージ、バージョン3.0(Kumar et al., 2004; Brief Bioinform., 5, 150-63)で実行される近隣結合法(NJ)を用い、アラインメントギャップの操作には、距離計算のためのポアソン修正モデルを用いペア毎に欠失オプションを実施した。
PTHのクローニング及び配列解析
クロクマゲノムDNA由来のPTHのPCR使用クローニングのために種々のプライマーの組合せを用いた。予備的クローンの配列決定結果に基づいて、開始コドンと一致する遺伝子特異的センスプライマーを設計し、停止コドンを含む縮退アンチセンスプライマーと一緒に用いてPTHの完全なコード領域を増幅した。配列確認用クローンを作製するために、停止コドンをカバーする第二のアンチセンスプライマーを設計した。
配列アッセンブリーによって、115アミノ酸の前駆体PTHタンパク質(25アミノ酸のシグナルペプチド及び6アミノ酸のプロペプチドを含む)が明らかになった。推定される成熟タンパク質は84アミノ酸であり、計算分子量9.471ダルトン及びpl 8.1を有する。クロクマPTHは他の哺乳動物のPTHと84−95%の配列類似性を共有し、イヌのPTHともっとも類似する(91%同一性、95%類似性)(図5)。興味深いことに、成熟ホルモンの2つのアミノ酸残基、41及び52はクロクマPTHに固有である。
【実施例2】
【0032】
実施例2:冬眠時の骨吸収及び骨形成マーカーレベル
血清サンプル
捕獲熊研究施設に収容されている5頭のクロクマ(ウルスス・アメリカヌス)から血液を収集した。全てのクマ管理プロトコル(#98-069-F&WS)がヴァージニアポリテクニックインスチチュート(Virginia Polytechnic Institute)及び州立大学の動物管理委員会 (State University Animal Care Committee)によって承認された。ケタミン(100mg/mL)とキシラジン(100mg/mL)の2:1の混合物でクマを麻酔した(投薬量は体重45.5kg当たり前記混合物1ccであった)。体温は冬期収集時には4℃から6℃低く、クマは冬眠状態であることが確認された。冬眠巣穴には尿も糞もなかった。いずれの取扱い手順中にもストレスを示す行動は観察されなかった。クマを麻酔している間に血液を大腿静脈から採集し、サンプルは氷詰め冷却容器で研究室に搬送した。研究室に戻ったら直ちに血液を遠心し、血清を単離しこの血清を華氏-20°で凍結した。血液サンプルは10月の第一日から5月の最終日まで10日毎に各クマから収集した。冬眠は1月の初めに開始し、4月初めに終了した。したがって、収集日は冬眠前活動期、不使用冬眠期及び冬眠後活動的再可動期を包含していた。
クロクマオステオカルシン精製及びRIA手順
クロクマの皮質を砕いて小断片にし、ヘキサン3部及びイソプロパノール2部の混合物で脱脂して凍結乾燥した。前記乾燥骨を液体窒素下で微細粉末に粉砕し、文献(Hauschka et al. (1989), Physiol. Rev., 69, 990-1047)に記載されたようにオステオカルシンを可溶化した。得られたEDTA抽出物から、Colomboらの方法(1993, J. Bone Miner. Res., 8, 733-43)を改変してオステオカルシンを精製した。簡単に記せば、粗EDTA溶液を2倍に希釈し、10gのセプラライト(Sepralyte)C18粒子(Analytichem International, Harbor City, CA)を含むバルクカラムを通過させた。前記カラムは先にメタノールで活性化し、水に0.1%のトリフルオロ酢酸(0.1% TFA)で平衡化してあった。UV吸収が基準値に下がるまで、0.1% TFAによる十分な洗浄に続いて30%メタノール/0.1% TFAで洗浄した。オステオカルシンは、80%メタノール/0.1% TFAで溶出させた。メタノールを送風下で蒸発させ、残留溶液を凍結乾燥した。得られた乾燥タンパク質を0.05Mトリス緩衝液(pH8.0)に懸濁し、先にサンプル緩衝液で平衡化した5mLのバイオラドエコノ-Q(Biorad Econo-Q)カラムに適用した。前記カラムを、0.5Mトリス(pH8.0)中の0.1から0.6MのNaCl勾配で展開した。オステオカルシンは対称性ピーク(カラムから溶出する最後のピーク)として溶出した。このピークがオステオカルシンそのものであることは、分画のアリコットをジアゾベンゼンスルホン酸と反応させ、オステオカルシン含有分画でピーク高に一致する強度の桃色を得ることによって立証した。C18及びEcono-Qカラムはともに新しいもので、他の種に由来するタンパク質に曝露されたことはなかった。他の種に関する以前の経験から、最終オステオカルシンピークは純度が99%を超えると推定された。最終溶出物中のクロクマオステオカルシンの濃度はBCA試薬(Pierce Chemical, Rockford, IL)を用いて決定した。
生化学的解析
PTH、25-OH D、レプチン及びオステオカルシン(骨形成マーカー)について、RIA及びELISAにより血清をアッセイした。
高度に精製したクロクマオステオカルシン及びクロクマ血清を放射能免疫アッセイによって調べた。抗体はモルモット抗ラットオステオカルシンであり、トレーサーは125I標識ラットオステオカルシンであった。ラットオステオカルシン標準物(Biomedical Technologies, Inc, Stoughton, MA)及び精製クロクマオステオカルシンの両方の用量希釈をアッセイに加えた。各アッセイ試験管当たり10μLのクロクマ血清アリコットをデュープリケートでアッセイし、全てのサンプルを同時にアッセイした。前記デュープリケートは5%未満で変動した。
不使用時における形成マーカー及び吸収マーカーの両方の変化を観察するために、5頭のクロクマのオステオカルシン平均値を冬眠期間中の各時点について計算した。これらの値を冬眠期間中の最大オステオカルシン値によって標準化した。同様な計算をPICP(骨形成マーカー)及びICPT(骨吸収マーカー)の測定についても実施した。吸収及び形成マーカーの標準化値を同じグラフにプロットし、不使用時の骨吸収及び骨形成における変化の一次的及び相対的規模を判定した。
上記のようにして入手した冬眠中のクマの血清サンプルを用い、イオン化カルシウム濃度をイオン選択電極(Bayer Rapidlab 865, Leverkusen, Germany)により測定した。
上記のようにして入手した血清サンプルを用い、PTHをELISAキット(Immutopics International, San Clemente, CA)によりアッセイした(変動のアッセイ内係数は4.7%であった)。25-OH DをELISAキット(ALPCO Diagnostics, Windham, NH)でアッセイした(変動のアッセイ内係数は5%であった)。レプチンはRIA(Linco, St. Charles, MO)で測定した(変動のアッセイ内係数は3.4%であった)。IGF-Iは酸エタノール抽出RIA(Nichols Institute Diagnostics, San Juan Capistrano, CA)で測定した(変動のアッセイ内係数は4.3%であった)。血清オステオカルシンは上記のようにRIAにより測定した。全ての血清代謝物について、平均値(あるシーズン内の全てのクマ及び全ての時点について)を各シーズン(冬眠前、冬眠時及び冬眠後)について計算し、ANOVAによって比較した。ANOVAは、フィッシャーのPLSD検定を用いて多重平均比較のために追跡した。自然対数変換を用いてオステオカルシン、PTH、25-OH D及びIGF-Iの自由度の非不変性を修正し、ANOVAを有効にした。線形回帰を用いて、オステオカルシンとホルモンとの間の相関性を判定した。いくつかの血清サンプルの体積は全アッセイを実施するためには不十分であった(各アッセイのサンプルサイズは結果とともに示されている)。
【0033】
結果
骨吸収マーカー(ICPT)は冬眠の開始直後に増加し始めた(図3)。各データの点は5頭のクマの平均値である。10−20日後に、骨形成マーカー(オステオカルシン及びPICP)もまた増加し、冬眠期間中吸収の増加と共役し続けるようであった。これは、吸収と形成との間の1−2週間の組織学的“逆転”期間と一致する。これらのリモデリングマーカーは冬眠期間中ずっと吸収及び形成の増加傾向を示し、形成は吸収との共役及び均衡を維持するようにみえた。平均オステオカルシンレベルは、冬眠前と比較して冬眠中及び冬眠後に高かった(p<0.0001)(表1)。
冬眠時イオン化カルシウムレベルは冬眠前レベルより有意に高かった(p=0.0062)。冬眠から覚醒した後の再可動時に、イオン化カルシウムレベルは冬眠時レベルと比較して有意には増加しなかったが(p=0.37)、しかしながら冬眠前レベルより高いままであった(p=0.015)。
オステオカルシンはPTHと正の相関性を有したが(図2)、25-OH D、レプチン又はIGF-1とは相関性を示さなかった。PTHは、冬眠前(p=0.006)及び冬眠(p=0.014)シーズンよりも冬眠後シーズンで有意に高かった。冬眠前に対して冬眠時のPTH増加は有意ではなかった(p=0.35)。25-OHビタミンDはシーズン性変動を示さなかった(p=0.64)。
血清レプチンは冬眠前と比較して冬眠時に変化しなかったが、冬眠後の再可動中に有意に低かった(p<0.004)(表1)。IGF-1は冬眠前と比較して冬眠時に有意に低下し(p<0.0001)、再可動時にその最高値に達した(表1)。
表1:平均血清代謝物濃度
【0034】
【0035】
平均値は太字で、標準偏差は括弧内に、サンプルサイズは鉤括弧内に示されている。ある代謝物について、同じ上付き文字を有する値は有意(p<0.05)には相違しない。25-OH Dは有意なシーズン性相違を示さなかった。
【実施例3】
【0036】
実施例3:MC-3T3骨芽細胞によるPGE2放出はクマ血清のシーズン性変動によって影響を受ける
骨芽細胞代謝に対するクマ血清のシーズン性変動の影響を判定するために、MC-3T3細胞をクマ血清で処理し、プロスタグランジンE2(PGE2)の放出を定量した。10%ウシ胎児血清(Hyclone, Logan, UT)及び1%ペニシリン-ストレプトマイシン溶液補充アルファ最小必須培養液(Invitrogen, Carlsbad, CA)でMC-3T3細胞を5% CO2下にて37℃で24時間増殖させた。培養液を吸引し、冬眠前、冬眠時及び冬眠後に収集したクマ血清を10%含有する新しい培養液で置き換えた。細胞をさらに24時間増殖させ、続いて培養液を収集し、PGE2解析のために-20℃で凍結した。EDTA中の0.25% トリプシンを用いて細胞を培養皿から取り出し、遠心によって沈殿させ、トリパンブルーと血球計算盤を用いて定量した。
PGE2レベルはBiotrakTM PGE2競合酵素免疫アッセイ(Amersham Biosciences, Piscataway, NJ)を用いて決定した。アッセイは、全ての試験培養液サンプルの50μLを用いデュープリケートで実施した。反応は終末点決定前に1Mの硫酸を用いて停止させ、450nmでマイクロプレートリーダー(VERSAmax, Molecular Devices Corporation, Sunnyvale, CA)を用いて読み取った。前記デュープリケートの光学密度を非特異的結合について修正して平均を算出し、標準曲線と比較して各ウェルのPGE2量を決定した。これらの値を総培養液体積に対して修正し、サンプル中の細胞数によって標準化した。ANOVAを用いて3種の血清グループ間で標準化PGE2を比較した。
クマ血清を用いてin vivoで処理した骨芽細胞系細胞により放出されるPGE2の量は、冬眠前血清(p=0.058)及び冬眠時血清(p=0.014)と比較して冬眠後血清による処理で高かった(図4)。冬眠時血清で処理した細胞のPGE2放出は、冬眠前血清と比較して有意には相違しなかった(p=0.48)。PGE2放出のシーズン性変化は、血清IGF-Iのシーズン性変化と同様な傾向を示した。P-値は冬眠後の値と比較される。冬眠前及び冬眠時の値は互いに相違しなかった(p=0.48)。
【実施例4】
【0037】
実施例4:クマ血清中での培養は冬眠時のBax対Bcl-2の遺伝子発現比を低下させる
上記に記載したように2004から2005年の間に血液サンプルを4頭の雌のクロクマから取り出した。サンプル日付は冬眠前活動期、冬眠不使用期及び冬眠後再可動期を包含する。MC-3T3骨芽細胞系細胞を10%クマ血清含有培養液で24時間培養し、その後バイオラドアクアピュア(BioRad AquaPure)RNA単離キット(#732-6370, Bio-Rad Laboratories, Hercules, CA)を用いて全RNAを単離した。cDNAを作製するために、スーパースクリプトII(Superscript II)逆転写酵素(Invitrogen, Carlsbad, CA)及び0.5μgのオリゴ(dT)12-18プライマーを用い、グラディエントサーモサイクラー(Mastercycler gradient, Eppendorf, Westbury, NY)で以下のように逆転写を実施した:42℃で20分、50℃で10分及び42℃で1時間。プロアポトーシスタンパク質Bax及び抗アポトーシスタンパク質Bcl-2のためのプライマーはプライマークエスト(PrimerQuest)ソフトウェア(Integrated DNA Technologies, Coralville, IA)及びNCBI遺伝子バンク配列を用いて設計した。半定量的PCRをレッドタック(RedTaq)及び以下から成るプロトコルを用いて実施した:94℃で2分、94℃30秒、69.5℃30秒及び72℃1分のサイクル、並びに最終伸長72℃で5分。バンド強度は、イメージJ(ImageJ)ソフトウェアパッケージ(National Institutes of Health, Bethesda, MD)を用いて定量し、3種のハウスキーピング遺伝子(Gapdh、β-アクチン、シクロフィリン)の発現に対して標準化した。
フィッシャーの被保護最小有意差(Fisher's Protected Least Significant Difference)(PLSD)ポストホック検定によるANOVAを用い、3シーズン(冬眠前、冬眠時、冬眠後)についてBax対Bcl-2比を比較した。統計的有意差は得られなかったが(p=0.300)、Bax/Bcl-2比は冬眠前と比較して冬眠中は約42%低下した。統計的有意の欠如はサンプルサイズが小さいことがおそらく関係した(各シーズンについてn=2)。これらのデータは、冬眠中のクマの血清は骨芽細胞のアポトーシスを低下させる生物学的分子を含むことを提唱している。内因性PTH及び骨形成マーカーオステオカルシンは両方とも冬眠時に増加するので(Donahue et al., 2006; J. Exp. Biol., 209, 1630-8)、内因性クマPTHは冬眠時に骨芽細胞のアポトーシスの低下を引き起こし、これは続いて骨形成を増加させるということはありそうである。
【実施例5】
【0038】
実施例5:骨細胞株のcAMPに対するクロクマとヒトのPTH1-84又はそのサブフラグメントの影響の比較
完全長の組換えクロクマPTH(残基1−84)を生成し、骨細胞株(MC-3T3骨芽細胞系細胞及びMLO-Y4骨細胞系細胞)のサイクリックアデノシン一リン酸(cAMP)濃度レベルに対するその影響を精査し、組換えヒトPTH1-84を用いて得られた結果と比較する。クロクマ及びヒトのPTHのサブフラグメント(完全長(1−84)成熟タンパク質のアミノ酸残基1−34、1−36、7−84、11−84及び41−52を含むサブフラグメント)を用いて同等な実験を実施する。いくつかの実験のために、クロクマ及びヒトPTHポリペプチドを固相法により合成する。
多様な形態の組換えクロクマ及びヒトPTHポリペプチドの骨形成細胞中のcAMPレベルに対する影響を決定するために、培養骨細胞(MC-3T3及びMLO-Y4)をヒト又はクロクマPTHの完全長(すなわちアミノ酸残基1−84)ポリペプチド又は上記に列挙したサブフラグメントの1つと10又は30分接触させる。細胞を前記PTHポリペプチドと接触させた後、細胞中のcAMP濃度を下記でさらに説明する競合結合アッセイを用いて測定する。
組換えポリペプチドを用いる実験の全てのために、凍結乾燥ペプチドを1mMの酢酸中で100μMのストック濃度に再構成し、使用前に10μMの作業用ストック濃度に希釈する。
細胞培養
MC-3T3サブクローン14細胞(ATCC, CRL-2594)及びMLO-Y4(以下から入手:L.F. Bonewald, University of Missouri, Kansas City, MO)は、アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%血清(MC-3T3:10%ウシ胎児血清(FBS)、MLO-Y4:5%FBS及び5%ウシ血清)中で5% CO2下で37℃にて維持される。ここに記載した全ての手順が、各アッセイの全ての処理の組合せについてn=6でそれぞれ別個の細胞培養について繰り返される。
細胞内cAMPに対するPTH処理の影響
6ウェルプレートにMC-3T3及びMLO-Y4細胞を適切な密度で播種する(MC-3T3:50,000細胞/cm2、MLO-Y4:15,000細胞/cm2)。細胞を一晩培養して最適な集合状態にする。続いて培養液を吸引し、10%血清+ベヒクル(1mM酢酸)又は10%血清+100nMのPTH(ヒト又はクマ1-84又はそのサブフラグメント)のどちらかを含む培養液と交換する。これらの条件下で細胞を10又は30分培養する(Carter, P. H. et al., 1999, J Biol. Chem. 274(45), 31955-60; Chen, X. et al., 2002, Am. J. Physiol. Cell Physiol. 283(5), C1432-40; Schiller, P. C. et al., 1999, J. Bone Miner. Res. 14(9), 1504-12)。培養後、細胞をトリプシン処理して遠心し、溶解緩衝液に再懸濁する。前記懸濁物を10分間インキュベートして遠心し、細胞屑を分離する。前記細胞溶解物の上清を(2倍希釈に続いて)競合結合アッセイ(サイクリックAMPアッセイ#KGE002, R&D Systems, Minneapolis, MN)によりcAMP濃度についてアッセイする。
試験したポリペプチドの各々について、クロクマPTH系ポリペプチドに応答して細胞性cAMPレベルの増加が認められる。
【実施例6】
【0039】
実施例6:骨細胞株のアポトーシスに対するクロクマとヒトのPTH1-84又はそのサブフラグメントの影響の比較
完全長の組換えクロクマPTH(残基1−84)を生成し、骨細胞株(MC-3T3骨芽細胞系細胞及びMLO-Y4骨細胞系細胞)のアポトーシスに対するその影響を精査し、組換えヒトPTH1-84を用いて得られた結果と比較する。クロクマ及びヒトのPTHのサブフラグメント(完全長(1−84)成熟タンパク質のアミノ酸残基1−34、1−36、7−84、11−84及び41−52を含むサブフラグメント)を用いて同等な実験を実施する。いくつかの実験のために、クロクマ及びヒトPTHポリペプチドを固相法により合成する。
クロクマ及びヒトPTHの骨芽細胞及び骨細胞(プロアポトーシス状態下にある)のアポトーシスを防止する相対的能力を決定するために、細胞をヒト又はクロクマPTH1-84又は上記に列挙したサブフラグメントの1つと一緒に1時間インキュベートする。その後、細胞をデキサメタゾンで6時間処理しアポトーシスを誘発する。アポトーシスは下記でさらに説明するELISAで定量する。
組換えポリペプチドを用いる実験の全てについて、凍結乾燥ペプチドを1mMの酢酸中で100μMのストック濃度に再構成し、使用前に10μMの作業用ストック濃度に希釈する。
さらに追加の実験を0.1%又は10%FBSを用いMC-3T3細胞で実施する。通常量(10%)未満(0.1%)のFBSを用いた追加の実験を実施し、その結果を解析して、通常の血清レベル又は低い血清レベルで実施した実験間で有意に異なる応答が存在するか否かを決定する。本実施例で報告する実験では、結果は使用するFBS量によって影響を受けない。アポトーシス予防実験は、試験したポリペプチドの各々がMC-3T3細胞のアポトーシスを低下又は防止することを示している。
細胞培養
MC-3T3サブクローン14細胞(ATCC, CRL-2594)及びMLO-Y4(以下から入手:L.F. Bonewald, University of Missouri, Kansas City, MO)は、アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%血清(MC-3T3:10%ウシ胎児血清(FBS)、MLO-Y4:5%FBS及び5%ウシ血清)中で5% CO2下にて37℃で維持される。ここに記載した全ての手順が、各アッセイの全ての処理の組合せについてn=6でそれぞれ別個の細胞培養について繰り返される。
アポトーシスに対するPTH処理の影響
6ウェルプレートにMC-3T3を50,000細胞/cm2で播種し、さらにMLO-Y4細胞を15,000細胞/cm2で播種し、細胞を一晩培養して最適な集合状態にする。培養液を吸引し、10%血清+ベヒクル(1mM酢酸)又は10%血清+100nMのPTH(ヒト又はクマ1-84又はそのサブフラグメント)のどちらかを含む培養液と交換する。1時間インキュベートした後(Jilka et al., 1999; J. Clin. Invest., 104, 439-46)、10μMのデキサメタゾン又はそのベヒクル(DMSO)を各ウェルに添加し、細胞を6時間インキュベートする(Bellido, T. et al., 2003, J. Biol. Chem. 278(50), 50259-72.; Jilka et al., 1999, J. Clin. Invest., 104, 439-46)。PTHによるアポトーシスの抑制は自己限定性であるので(Bellido et al., 2003)、アポトーシス誘発中はPTHポリペプチド及びベヒクルはそのままにしておく。6時間後、細胞をトリプシン処理して遠心し、再懸濁して血球計算盤を用いて数える。前記懸濁物から50,000細胞を取り出し溶解緩衝液に入れる。前記溶解物の上清を(遠心した後)解析のために取り出し-20℃で保存する。
アポトーシスはELISA(Cell Death Detection ELISA, #1544675, Roche Applied Science, Indianapolis, IN)により前記溶解物上清から定量する。このアッセイは、細胞溶解物の細胞質分画中の断片化された細胞DNA由来のモノ及びオリゴヌクレオソームを検出し、したがって初期及び中期アポトーシスの良好な測定値を提供する。簡単に記せば、サンプルを緩衝溶液で希釈し、抗ヒストンマウスモノクローナル(クローンH11-4)抗体で被覆したマイクロプレートのウェルに加える。ベヒクル処理細胞の溶解物上清は陰性コントロールとして供する。ペルオキシダーゼ結合抗DNAマウスモノクローナル(クローンMCA-33)抗体を添加した後、光学密度を測定し、各サンプルのアポトーシス量をその対応する陰性コントロールに比して決定する。全てのサンプルをデュープリケートでアッセイする。
試験したポリペプチドの各々が細胞(プロアポトーシス状態下にある)のアポトーシスを低下させた。
【実施例7】
【0040】
実施例7:骨細胞株における遺伝子発現に対するクロクマとヒトのPTH1-84又はそのサブフラグメントの影響の比較
完全長の組換えクロクマPTH(残基1−84)を生成し、骨細胞株(MC-3T3骨芽細胞系細胞及びMLO-Y4骨細胞系細胞)の遺伝子発現レベルに対するその影響を精査し、組換えヒトPTH1-84を用いて得られた結果と比較する。クロクマ及びヒトのPTHのサブフラグメント(完全長(1−84)成熟タンパク質のアミノ酸残基1−34、1−36、7−84、11−84及び41−52を含むサブフラグメント)を用いて同等な実験を実施する。いくつかの実験のために、クロクマ及びヒトPTHポリペプチドを固相法により合成する。
骨基質の調節、転写調節、抗アポトーシス(Bcl-2)遺伝子及びプロアポトーシス遺伝子Baxに対するクロクマ及びヒトPTHの影響を決定するために、細胞をヒト又はクマPTH1-84又はサブフラグメントと1又は3時間培養する。遺伝子発現はリアルタイムPCRにより定量する。
組換えポリペプチドを用いる実験の全てについて、凍結乾燥ペプチドを1mMの酢酸中で100μMのストック濃度に再構成し、使用前に10μMの作業用ストック濃度に希釈する。
さらに追加の実験を0.1%又は10%FBSを用いMC-3T3細胞で実施する。通常量(10%)未満(0.1%)のFBSを用いた追加の実験を実施し、その結果を解析して、通常の血清レベル又は低い血清レベルで実施した実験間で有意に異なる応答が存在するか否かを決定する。本実施例で報告する実験では、結果は使用するFBS量によって影響を受けない。リアルタイムPCRを用いて、ポリペプチド添加後1時間及び3時間の時点の遺伝子発現レベルを判定し、クロクマPTH1-34はMC-3T3細胞で遺伝子発現をアップレギュレートすることを示す。
細胞培養
MC-3T3サブクローン14細胞(ATCC, CRL-2594)及びMLO-Y4(以下から入手:L.F. Bonewald, University of Missouri, Kansas City, MO)は、アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%血清(MC-3T3:10%ウシ胎児血清(FBS)、MLO-Y4:5%FBS及び5%ウシ血清)中で5% CO2下にて37℃で維持する。ここに記載した全ての手順が、各アッセイの全ての処理の組合せについてn=6でそれぞれ別個の細胞培養について繰り返される。
遺伝子発現に対するPTH処理の影響
6ウェルプレートにMC-3T3を50,000細胞/cm2の密度で播種し、さらにMLO-Y4細胞を15,000細胞/cm2の密度で播種し、細胞を一晩培養して最適な集合状態にする。培養液を吸引し、10%血清+ベヒクル(1mM酢酸)又は10%血清+100nMのPTH(ヒト又はクマ1-84又は上記に列挙したサブフラグメント)のどちらかを含む培養液と交換する。細胞をこれらの条件下で1又は3時間培養する(前記時間はc-fos及びオステオカルシンのPTH誘発アップレギュレーションと一致する)(Jiang et al., 2004, J. Biol. Chem., 279, 5329-37; Chen et al., 2002)。SV全RNA単離系(SV Total RNA Isolation System, Promega, Madison, WI)を用いて全RNAを単離する。
cDNAを生成するための逆転写は、スーパースクリプトII(Superscript II)逆転写酵素(Invitrogen, Carlsbad, CA)及び0.5μgのオリゴ(dT)12-18プライマーを用い、グラディエントサーモサイクラー(Mastercycler gradient, Eppendorf, Westbury, NY)で以下のように実施する:42℃で20分、50℃で10分及び42℃で1時間。問題の全ての遺伝子(オステオカルシン、オステオポンチン、I型コラーゲン、c-fos、Runx2、Bax、Bcl-2、SOST)及びハウスキーピング遺伝子(Gapdh、β-アクチン、シクロフィリン)ためのプライマーは、プライマークエスト(PrimerQuest)ソフトウェア(Integrated DNA Technologies, Coralville, IA)及びNCBI遺伝子バンク配列を用いて設計し、PCR条件はMC-3T3及びMLO-Y4細胞由来のRNAを用いて最適化する。リアルタイムPCRは、Mx3000PリアルタイムPCR系(Stratagene, La Jolla, CA)を用いて実施する。プロトコルは、95℃で10分のホットスタート、続いて95℃30秒(変性)、69℃1分(アニーリング)及び72℃1分(伸長)の40サイクルを含む。このプロトコルの例外はc-fosであり、前記は66℃のアニーリング温度を有する。それぞれ25μLの反応物は以下を含む:1x AbsoluteTM qPCR SYBR(商標)グリーンミックス(ABgene, Rochester, NY)、0.1 μMのフォワード及びリバースプライマー、並びに2.5 ngの全RNA相当cDNA鋳型。遺伝子発現は、3種のハウスキーピング遺伝子の幾何平均に対して標準化した相対標準曲線法を用いて決定する。全てのサンプルをデュープリケートで測定し、10%を超える変動係数(CV)を有するサンプルはいずれも解析する。
前記ポリペプチドは、骨基質、転写調節及び転写活性化因子遺伝子のアップレギュレーション、並びにBax/Bcl-2発現比の低下を引き起こす。
【実施例8】
【0041】
実施例8:異なるシーズンのクロクマ血清の骨細胞アポトーシス及び遺伝子発現に対する影響の比較、並びにPTH及びオステオカルシンの血清レベルとの相関性
ヴァージニアテックセンターフォーベアリサーチ(Virginia Tech Center for Bear Research)に2004年から2005年に収容された少なくとも3頭の別個の雌のクロクマ(ウルスス・アメリカヌス)から血液サンプルを収集する。さらに別のクロクマからその後の年に血清を収集する。ヴァージニアポリテクニックインスティチュート(Virginia Polytechnic Institute)及び州立大学の動物管理委員会 ( State University Animal Care Committee)は全てのクマ管理プロトコル(#98-069-F&WS)を承認した。ケタミン(100mg/mL)とキシラジン(100mg/mL)の2:1の混合物でクマを麻酔する(投薬量は体重45.5kg当たり前記混合物1ccである)。クマを麻酔している間に血液サンプルを大腿静脈から採集し、サンプルは氷詰め冷却容器で研究室に搬送する。研究室に戻されたら直ちに血液を遠心して血清を単離し、この血清を-20℃で凍結する。血液サンプルは10月の初めから5月の終わりまで10日毎に各クマから収集する。冬眠は1月の初めに始まり、4月初めに終わる。したがって、収集日は冬眠前活動期、不使用冬眠期及び活動的な冬眠後再可動期を包含している。
クマ血清の10μLアリコットをオステオカルシン濃度について放射能免疫アッセイによりデュープリケートでアッセイする(Patterson-Allen et al., 1982; Anal. Biochem., 120, 1-7)。このアッセイは以前にクマについて有効性が立証された(Donahue et al., 2006; J. Exp. Biol., 209, 1630-8)。抗体はモルモット抗ラットオステオカルシンであり、トレーサーは125I標識ラットオステオカルシンである。クマ血清の100μLアリコットをPTH濃度について(Donahue et al., 2006; J. Exp. Biol., 209, 1630-8)デュープリケートでELISA(Porcine Intact PTH ELISA Kit, #60-3305, Immutopics, Inc., San Clemente, CA)によりアッセイする。このアッセイはPTHの39−84領域と結合し、PTH濃度を比色により提示するためにPTHの13−34領域を必要とする。したがって、前記アッセイは、C-末端サブフラグメント7−84及び11−84と同様に無傷の(1−84)PTHについて良好な測定を提供する。このELISAはクマPTHと交差反応し(Donahue et al., 2006; J. Exp. Biol., 209, 1630-8)、さらにヒトPTHとは100%の交差反応性を有することが示された。このアッセイがクロクマについて有効であることを立証するために、10nMの組換えクロクマ又はヒトPTH1-84を含む培養液サンプルをデュープリケートでアッセイする。PTHサンプルの既知濃度をアッセイの標準曲線から決定した測定濃度と比較する。これらのサンプルから決定される交差反応性におけるいっさいの潜在的な差を、クロクマ血清サンプルの内因性クロクマPTH濃度についての修正として用いる。
培養骨タイプ細胞によるクロクマPTHアポトーシス及び遺伝子発現細胞培養実験のために上記に記載した方法が繰り返されるが、ただし100nM組換えPTH含有培養液を10%クロクマ血清(冬眠前、冬眠時又は冬眠後の期間に由来)含有培養液に置き換える。血清体積は上記に記載したPTH ELISAの後で計算する。
PTHは冬眠前血清よりも冬眠時及び冬眠後血清に多いので、冬眠時及び冬眠後シーズン血清は、冬眠前血清と比較してより強いアポトーシス予防を引き起こす。内因性血清PTH濃度は、より高い血清PTHレベルはより低いアポトーシス率と一致するという点でアポトーシスレベルと反比例する。すなわち、血清PTH濃度はアポトーシスレベルと負の相関性を有する。
【実施例9】
【0042】
実施例9:クマPTHのin vivo試験
クロクマPTH、完全長(1-84)又はそのいくつかの機能的サブフラグメント(1-34、1-36、7-84、11-84、41-52)の1つを骨細胞の同化作用促進刺激についてin vivoで試験し、対応するヒトPTH又はサブフラグメントと比較する。PTHポリペプチドの各々を合成し、皮下注射のために医薬的に適切な担体に懸濁する。クロクマ又はヒト由来の完全長PTH又はその機能的フラグメントを、体重1kg当たり40μgの用量で毎日7週間投与する。クロクマPTH又はその機能的フラグメントは、骨強度、骨量及び骨塩含有量において対応するヒトPTHポリペプチドよりも大きな増加を引き起こす。
【実施例10】
【0043】
実施例10:骨減少を低下させるか又は妨げる予防薬としてのクマPTHの使用
骨粗鬆症は、後肢懸濁物によって(骨粗鬆症の不使用モデル)及び卵巣摘出によって(骨粗鬆症の閉経後モデル)マウスで誘発される。後肢懸濁物の開始後又は卵巣摘出完了後、マウスに通常の用量のクロクマPTH又はその機能的フラグメントを与える。クロクマPTH又はその機能的フラグメントで処置されたマウスは骨減少が未処置マウスより少ない。
【実施例11】
【0044】
実施例11:クマ及びヒトPTH1-34はともにオステオカルシンの遺伝子発現をアップレギュレートするが、クマPTH1-34のみがBax/Bcl-2の発現レベル比を低下させる
MC-3T3細胞をベヒクル又は100nMの合成クマもしくはヒトPTH1-34中で3又は6時間インキュベートした(n=2又は4)。全RNAを単離し、逆転写によりcDNAを生成する。骨基質タンパク質のI型コラーゲン及びオステオカルシン、プロアポトーシスタンパク質Bax、抗アポトーシスタンパク質Bcl-2、並びにハウスキーピング遺伝子のGapdh、β-アクチン及びシクロフィリンのためのプライマーをプライマークエストソフトウェア(Integrated DNA Technologies, Coralville, IA)を用いて設計した。リアルタイムPCRは、Mx3000PリアルタイムPCR系(Stratagene, La Jolla, CA)を用いて実施した。全てのサンプルをデュープリケートで測定した。遺伝子発現は、3種のハウスキーピング遺伝子(Gapdh、β-アクチン及びシクロフィリン)の幾何平均に対して標準化した相対標準曲線法を用いて決定した。アポトーシス関連遺伝子はBax/Bcl-2の発現比として解析した(なぜならばこの比の低下はin vitroでのアポトーシスの低下と密接に関係するからである)。
ヒト又はクマPTH1-34中での培養は、ベヒクルコントロールと比較してI型コラーゲンの発現には影響を与えなかったが、オステオカルシンの発現を実質的にアップレギュレートした(図6)。ヒトPTHとクマPTHとの間には有意な相違は存在しなかった(p>0.09)。
クマPTH中での3時間の培養はBac/Bcl-2の発現比を低下させアポトーシスの低下が推量されたが、ヒトPTH中での3時間の培養は前記の比を増加させアポトーシスの増加が推量された(図7)。クマPTHとヒトPTHとの間の相違は統計的に有意であった(p=0.047)。
【実施例12】
【0045】
実施例12:in vivoラットOVX用量応答実験
90匹の3ヶ月齢雌Sprague-Dawleyラットを入手した。これらラットの80匹に卵巣摘出(OVX)を実施し、10匹に模擬OVX(卵巣は前記動物に残留)を施した。前記OVX動物を更なる実験のために無作為に10匹ずつのグループに割り当てた。
実際の又は模擬OVX術に続いて、全動物を6週間処置せず骨を減少させた。6週間の骨減少期間の後、動物のグループに6週間又は8週間の間(1週間に5日間、毎日)ベヒクル(食塩水)又はクマもしくはヒトPTH1-34の3、10又は30μg/kgを注射した。ヒトに対する従来のPTH1-34の用量は約0.3μg/kgで、一方5μg/kgの用量はラットでは発癌性であった(Tashjian et al., J. Bone Mineral Res. (2008) 23(6): 803-811)。模擬OVXグループ及び実際のOVXラットの1つのグループを6週間で安楽死させ、一方残余の動物はOVX術の後14週で安楽死させた。更なる解析のために安楽死させた動物からサンプルを入手した。
コンピュータ支援マイクロ断層撮影法(μCT)を用いて骨の三次元画像を入手した。続いて前記三次元画像を用いて下記に説明するように骨パラメーターを定量した。測定は、成長板から2mmの位置で始まる1.6mmの厚さで実施した(図8)。骨サンプルは、術後6週間で安楽死させた模擬OVX(図9A、“模擬OVX6週”)及び実際のOVX(図9B、“OVX6週”)とともに術後14週間で安楽死させた実際のOVXラットグループ(図9C、“OVX14週”)から入手し解析した。前記OVXの14グループのラットは、6週間の骨減少期間に続いてベヒクル(食塩水)の注射を8週間与えられ、並行してPTH1-34の注射も8週間与えられた(図9D、9E及び9F)。図9D、9E及び9Fに示すサンプルは、6週間の骨減少期間の後、クロクマPTH1-34(“bbPTH”)を8週間それぞれ3、10又は30μg/kg注射されたラットから入手された。
画像から分かるように、OVX外科手術後6週間(図9B)及び14週間(図9C)のラットの骨は、模擬OVX(図9A)及びbbPTH処理ラット(図9D−9F)の骨と比較して顕著な量の骨減少を示した。クロクマPTH1-34で処置したラットは、ベヒクル(食塩水)のみの注射を与えられたOVXラット(図9C)と比較して、各投薬レベルにおいて用量依存性の骨量増加(図9D−9F)を示した。
模擬OVX、OVX及びPTH処置ラット間の骨パラメーターの相違を定量した。図10は、種々の処理グループについて総体積の比(BV/TV)としての骨体積を示す。卵巣存在下の骨体積比の基準線レベル(“模擬基準線”淡色ひし形)及び卵巣摘出後6週間でPTH処置の非存在下の基準線レベル(“OVX基準線”濃色ひし形)は、“6週”と標識した列の左側に示されている。OVXラットの14週間のベヒクル(食塩水)単独処置(“OVXベヒクル”、三角)は骨体積比の更なる下降をもたらした。OVXのヒト(“hPTH1-34”、四角)又はクロクマ(“bbPTH1-34”、丸)PTH1-34処置は、用量依存態様で骨体積比を増加させた。最高用量の30μg/kgでは、ヒトPTHによる処置は、クマPTH又はベヒクル(食塩水)単独による処置よりも有意に(p<0.05)高い骨体積をもたらした。ヒトPTHによる処置は、試験した3レベルの全てでベヒクル(食塩水)単独処置よりも有意に(p<0.05)高い骨体積比をもたらし、一方、クマPTHは、10μg/kg及び30μg/kgでベヒクル(食塩水)単独処置よりも有意に(高い骨体積比をもたらしたp<0.05)。
同様に、見掛けの骨塩密度をサンプルの各々について決定し(図11)、1 cm3当たりのヒドロキシアパタイトのmg(“App.Dn(mgHA/ccm)”)として示されている。見掛けの骨塩密度は、ベヒクル(食塩水)単独処置OVXラットの骨の場合よりも3、10又は30μg/kgのhPTH又はbbPTHで処置されたOVXラットの骨について有意に高かった(p<0.05)。30μg/kgのhPTHで処置されたOVXラットの見掛けの骨塩密度は、30μg/kgのbbPTHで処置されたOVXラットよりも有意に高かった(p<0.05)。
骨梁支柱骨数及び厚さもまた、模擬OVX、PTH処置無しOVX、及びPTH処置OVXラットの骨について決定した(図12)。記号は、模擬OVXラット(“模擬基準線”、淡色ひし形)、ベヒクル(食塩水)単独処置及び6週安楽死のOVXラット(“OVX基準線”、濃色ひし形)、ベヒクル(食塩水)単独8週間処置及び14週安楽死のOVXラット(“OVX基準線”、濃色ひし形)、及び3、10又は30μg/kgヒト(“hTPH1-34”、四角)又はクマ(“bbPTH1-34”、丸)PTH1-34処置OVXラットについて1ミリメートル当たりの骨梁数(“Tb.N(mm^-1)”)を示す(図12)。
骨梁支柱骨の平均の厚さもまた決定した(図13)。クマPTH又はヒトPTHによる処置は、ベヒクル処置コントロールと比較して、骨梁支柱骨の平均の厚さ(“Tb.Th(mm)”)を有意に増加させた(p<0.05)。
クロクマTPH1-34は、ヒトPTH1-34と比較して用量増加による骨梁骨のより強い鉱化傾向を示した(“M.Dn(mgHA/ccm)”;図14)。OVXラットのクマ(“bbTPH1-34”、丸)又はヒト(“hTPH1-34”、四角)PTH1-34による処置は、ベヒクル(食塩水)単独処置OVXラットよりも有意に(p<0.05)高い骨梁骨鉱化をもたらした。
ヒトPTH1-34(“hTPH”、四角)は、脛骨皮質体積(“BV(mm^2)”、図15)の増加でクマPTH1-34(“bPTH”、丸)よりも優れている。しかしながら全体的には、骨皮質領域の変化は骨の骨梁(bone trabeculae)で観察される変化と比較して軽度であった。
OVXラットの骨の脛骨皮質多孔度は、中等度用量(10μg/kg)及び高用量(30μg/kg)のクマ(“bbTPH1-34”、丸)又はヒト(“hTPH1-34”、四角)PTH1-34でラットを処置したとき、ベヒクル(食塩水)単独処置OVXラットの骨と比較して有意に(p<0.05)低下するように見えた(図16)。しかしながら高用量(30μg/kg)のヒトPTH1-34又はクマPTH1-34は、ベヒクル処置コントロールと比較して有意な(p<0.05)大腿骨皮質強度の増加をもたらした(図17)。
クマPTH1-34がヒトPTH1-34を超えて提供し得るさらに別の利点はより低い血清カルシウムである。高カルシウム血症はヒトでのPTH処置における副作用である。本実験では、血清カルシウムレベルは、ヒトPTH1-34と比較してクマPTH1-34処置ラットで低いように見えるが、ただし結果はp<0.05レベルで有意ではない。しかしながら、統計解析は、サンプルサイズが2倍であればクロクマPTH1-34処置ラットで血清カルシウム濃度は有意に低い(p<0.05)であろうと予想する。したがって、クロクマPTH1-34処置は前記副作用発生率を低下させ得る。
これらの実験は、クロクマPTH1-34は骨梁骨形成の劇的な増加をもたらすという点でヒトPTH1-34と等しく有効であることを示している。ヒトPTH1-34は、最高用量(30μg/kg)でいくつかのパラメーターについてクマPTH1-34よりも有効であることが観察されたが、これはヒトに投与される用量よりも100倍高い。しかしながら、より低くかつ臨床的に対応する用量で、クマPTH1-34は、骨折蓋然性の低下と関係する重要なパラメーターを増加させるという点でヒトPTH1-34と同じように有効であった。
【実施例13】
【0046】
実施例13:クマPTH1-34及びクマPTH1-84は培養マウス骨芽細胞でカスパーゼ-3活性を低下させる
細胞培養
MC-3T3サブクローン4細胞(ATCC, CRL-2594)は標準培養液(89%アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%FBS)中で5% CO2下にて37℃で維持した。細胞は30継代を超えて用いられることはなかった。
血清枯渇により誘発されたアポトーシスのPTHによる減弱
MC3T3細胞を96ウェルプレートに20,000細胞/ウェルの密度で播種し、一晩定着させた。続いて培養液を吸引し、10%FBS及びベヒクル(PBS)又はPTH(10−100nMのどちらかを含む培養液100μLで1時間置き換えた。その後、培養液を吸引し細胞をPBSで洗浄して残留血清を除去した。アポトーシス誘発のために、細胞をコントロール培養液又は無血清培養液で6時間インキュベートした。交換培養液は、89%アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%FBS(コントロール培養液)又は0%FBS(枯渇培養液)のどちらかを含んでいた。枯渇後、培養液を吸引し、細胞をPBSで洗浄し、50μLの新しいPBSを各ウェルに添加した。カスパーゼ3/7活性はDEVD基質の切断によって決定した(カスパーゼ-Glo 3/7アッセイ、#G8093, Promega Corporation)。既知濃度の組換えヒトカスパーゼ-3(#SE-169, BIOMOL Research Laboratories Inc.)を用いて作成した標準曲線に測定したルミネセンス値を適合させた。各96ウェルプレートにつき5ウェルを各処置で用い、手順を最小限10プレートで繰り返した。
統計解析
フィッシャーのPLSDとともにANOVAを用い、反復実験によってブロック化しながら、ペプチド、ベヒクル及びコントロール処置サンプル間でカスパーゼ3/7活性化の減弱を比較した。カスパーゼ3/7活性は、各反復実験内でコントロール培養液(非枯渇細胞)のカスパーゼ3/7レベルに対して標準化し、共分散因子として濃度を処理しながら共分散解析(ANCOVA)を用いてペプチドの用量応答反応態様を比較した。
血清枯渇は、この実験の非枯渇細胞と比較してベヒクル及びPTH前処理サンプルで、カスパーゼ3/7活性(アポトーシスの初期インジケーター)を有意に(p<0.0001)増加させた(図19)。全てのPTH前処理はベヒクル処理細胞と比較してカスパーゼ3/7活性を有意に(p<0.0001)低下させた。カスパーゼ3/7活性のPTH誘発低下は用量依存態様を示さなかったので(p=0.890)、全ての濃度をグループ内比較のためにグループ内でプールした。4ペプチドグループのうちで、bbPTH1-34は血清枯渇誘発アポトーシスの最大の減弱を示した。bbPTH1-34グループの平均カスパーゼ3/7活性は、hPTH1-34グループ(p=0.006)及びいずれのPTH1-84(p≦0.018)よりも有意に低かった(図19)。対照的に、bbPTH1-84とhPTH1-84グループとの間のカスパーゼ3/7活性の相違は有意に近く(p=0.090)、hPTH1-84がより低レベルのアポトーシスを示した(図19)。
【実施例14】
【0047】
実施例14:雄Swiss-WebsterマウスにおけるクマPTH1-34及びクマPTH1-84のin vivo作用
動物
全ての取扱い方法及び実験は、ミシガン工科大学(Michigan Technological University)院内動物管理使用委員会によって承認された。雄のスイス-ウェブスター(Swiss-Webster)マウスを繁殖業者(Harlan, Indianapolis, IN)から入手した。全ての動物を12時間照明/12時間消灯サイクルで維持し、水及び0.95%カルシウム含有餌(Purina Rodent Diet #5001)に自由に接近させた。
第一の実験では、11週齢マウスに毎日(5日/週)2.5nmol/kgのPTH1-34(hPTH1-34又はbbPTH1-34)又は酸性食塩水ベヒクル(0.15M NaCl+0.001N HCl)の皮下注射を8週間実施した(n=10マウス/グループ)。第二の実験では、12週齢マウスに毎日(5日/週)2.5nmol/kgのPTH1-84(hPTH1-84又はbbPTH1-84)又は酸性食塩水ベヒクルの皮下注射を8週間実施した(n=10マウス/グループ)。全てのPTH及びベヒクル処理動物に10mg/kgの投薬量でカルセインの皮下注射を致死12日及び3日前に実施し、骨表面の鉱化を標識した。最後の処置注射から24時間後に、マウスを二酸化炭素で窒息死させた。右大腿骨を10%の中性緩衝ホルマリンで固定した。左大腿骨は0.15M食塩水を浸み込ませたガーゼで包み-20℃で保存した。左脛骨は70%エタノール中にて4℃で保存した。
近位脛骨の骨梁骨の構造及び鉱化
コンピュータ支援微小断層撮影を用いて、左近位脛骨の骨幹端(成長板から0.7mm遠位)における骨梁骨の構造及び鉱化を評価した。実験1の脛骨をμCT35スキャナー(Scanco Medical AG, Basserdorf, Switzerland)により3.5μmの(高)解像でスキャンした。実験2の脛骨はμCT40スキャナー(Scanco Medical AG, Basserdorf, Switzerland)により7μmの(高)解像でスキャンした。骨は70%エタノール中でスキャンした。骨梁骨の体積比(BV/TV、%)、骨梁数(Tb.N, mm-1)、骨梁の厚さ(Tb.Th, mm)、骨梁の分離(Tb.Sp, mm)、骨梁の見掛けの骨塩密度(App.Mn.Dn, mgHA/cm3)及び骨梁組織の骨塩密度(Mat.Mn.Dn, mgHA/cm3)を、製造業者のソフトウェアを用いてコンピュータで計算した。
大腿骨皮質の機械的特性
インストロン(Instron)機械的検査系(Model #8872, Canton, MA)により1mm/分の速度にて3点曲げで左大腿骨に不履行まで負荷をかけた。前方表面が加圧されるように骨の方向を定めた。下方検査固定装置支持部は差し渡し10mm離し、全ての固定装置の接触点を丸めて(半径=1mm)、骨に局所的応力が集中するのを避けた。究極負荷は試験から得られた負荷-ひずみデータを用いて計算した。
皮質骨塩含有量、幾何学的特性及び動的な組織形態計測
曲げ試験に続いて、左大腿骨骨幹を窯炉で完全燃焼させて、骨塩含有量(灰分比)を決定した。右大腿骨骨幹をメチルメタクリレートに包埋し、ダイヤモンド刀で輪切りにして骨幹中央断面を露出させた。デジタルカメラ(Spot Insight QE, Diagnostic Instruments Inc., Sterling Heights, Michigan)を用いて骨幹中央断面画像を捕捉し、シオン画像解析ソフトウェア(Scion Corporation, Frederick, Maryland)を用いて各サンプルについて骨膜面積(Ps.Ar)、皮質面積(Ct.Ar)、骨内膜面積(Es.Ar)を計算した。皮質の厚さ(Ct.Th)は、画像解析ソフトウェア(Bioquant Osteo, Nashville, TN)を用い各解剖学的四分円について及び完全断面について0.1mmの増加幅で計算した。右大腿骨の骨幹中央から得た薄い切片を厚さ50μmにすり潰し、ガラススライドにマウントした。カルセイン標識画像を100X倍率で捕捉し、各大腿骨の骨膜及び骨内膜の骨塩付加成長速度(それぞれPs.MAR及びEs.MAR)並びに表面鉱化(Ps.MS/BS及びEs.MS/MS)を定量した(Bioquant)。
in vivo実験統計解析
動物サイズにおける潜在的多様性を示すために、全ての骨の幾何学的特性、構造的特性及び骨塩特性を体重によって標準化した(31−33)。コンピュータ支援微小断層撮影法によって定量した骨梁骨の特性を、各実験(PTH1-34又はPTH1-84)においてベヒクル、クマPTH及びヒトPTH処置マウス間でANOVAによって比較した。異なるμCT装置で解析を実施したので、PTH1-34及びPTH1-84間ではμCTパラメーターの直接比較は実施しなかった。他の全ての骨特性については、6つの実験グループの全て(PTH1-34及びPTH1-84の両方について)をANOVA及びフィッシャーのPLSDにより比較した。全ての比較についてp<0.05を有意とした。
結果
近位脛骨における骨梁骨の構造及び鉱化:
骨梁骨のμCTのデータは表2及び3に提示されている。PTH1-34実験で実験グループ間に有意差はなかったが、骨梁骨の骨塩密度はbbPTH1-34で処置したマウスで最も低い傾向があった(表2)。対照的に、骨体積比は、hPTH1-84と比較してbbPTH1-84で処置したマウスの近位脛骨でより高かった(表3)。骨体積比に対するbbPTH1-84の影響とhPTH1-84の影響との間における相違は有意で(p=0.013)、bbPTHがより有効な治療方法であった。bbPTH1-84処置マウスにおける骨体積比の増加は骨梁の厚さによって影響を受けたであろう。なぜならば、骨梁の厚さはhPTH1-84で処置したマウスと比較してbbPTH1-84で処置したマウスで大きい傾向があったからである(p=0.089)。骨梁数及び骨梁分離は、PTH1-84実験のグループ間で有意な相違はなかった(表3)。骨梁の見掛けの骨塩密度は、他の処置と比べてbbPTH1-84処置マウスで最も高く、bbPTH1-84の影響ととhPTH1-84の影響との間の相違は統計的に有意であった(p=0.019)(表3)。骨梁の実質的骨塩密度はbbPTH1-84処置マウスで最も高い傾向があった(表3)。
【0048】
表1:PTH1-34実験のマウス近位脛骨における骨梁骨の構造的特性及び骨塩密度
標準偏差は括弧内に示されている。
略語の説明は本文を参照されたい。
【0049】
表2:PTH1-84実験のマウス近位脛骨における骨梁骨の構造的特性及び骨塩密度
標準偏差は括弧内に示されている。
異なる上付き記号はグループ間の有意な相違を示す(p<0.05)。
略語の説明は本文を参照されたい。
【0050】
大腿骨の皮質の機械的特性及び灰分比:
大腿骨の究極負荷はペプチド処置によって増加した(ベヒクル処置マウスと比較して+3%から+15%)(図29)。骨強度の増加はhPTH1-84と比較してbbPTH1-84処置マウスで有意に高かったが(p=0.032)、bbPTH1-34及びhPTH1-34処置マウス間では相違はなかった(p=0.819)。骨塩含有量(灰分比)は処置グループ間で相違はなかった(p=0.180)(データは示されていない)。
大腿骨の皮質の幾何学的特性:
骨の骨膜面積は、PTH1-34実験のベヒクル処置マウスでもっとも高く、PTH1-84実験のベヒクル処置マウスで最も低かった(表3)。bbPTH1-84処置マウスはベヒクル処置マウスと比較して骨膜面積について有意に大きな値を有したが(ただしhTPH1-84ではそうではない)、両PTH-1-34ペプチド処置マウスはベヒクル処置マウスと比較して骨膜面積についてより低い値を示した(表3)。皮質面積も骨内膜面積も処置グループ間で相違はなかった(p>0.136)。皮質の厚さはPTH1-34実験と比較してPTH1-84実験のマウスで大きかったが、処置グループ間で他の相違は存在しなかった(表3)。
【0051】
表3:PTH処置マウスの大腿骨骨幹中央の皮質の幾何学的特性
標準偏差は括弧内に示されている。
異なる上付き記号はグループ間の有意な相違を示す(p<0.05)。
略語の説明は本文を参照されたい。
【0052】
大腿骨における皮質の動的な組織形態計測:
処置グループ間で骨膜もしくは骨内膜の表面鉱化又は骨塩付加成長速度に有意な相違はなかった(p>0.236)(表4)。
【0053】
表4:PTH処置マウス大腿骨骨幹中央の皮質の動的な組織形態計測
標準偏差は括弧内に示されている。
【0054】
結論
これらのデータは、bbPTH1-34はin vitroでhPTH1-34及びhPTH1-84より抗アポトーシス活性が強いことを示唆しているが、bbPTH1-84はマウスで骨体積及び強度をhPTHペプチドよりも大きく増加させる。このことは、bbPTHは骨粗鬆症に対する新規な同化作用性治療薬として潜在能力を有することを提唱している。
これらの実験でhPTHペプチドはベヒクル処置と比較して骨梁骨の体積の増加をもたらさなかった(表1及び2)。これはおそらく比較的低い投薬量がマウスに投与されたからであろう。マウスにおけるPTHの同化作用に関する短期的in vivo実験では、短期間での骨形成及び骨量の大きな増加をもたらすために典型的には10nmol/kg以上の投薬量を用いる。以前の実験では、低いペプチド投薬量では短期間の骨形成性は低く、すなわち1日1nmol/kgのhPTH1-34の6週間注射は、雄のマウスの近位脛骨における骨梁の厚さ(Tb.Th)又は骨塩付加成長速度(MAR)に影響を及ぼさなかったが、より高モル濃度のhPTHの投薬(〜10nmol/kg)はこれらの特性を高めた(以下を参照されたい:Sakai et al. (1999) J Bone Miner Res 14(10):1691-9)。我々の実験では2.5 nmol/kgの投薬量を用いた。なぜならば、前記の値は、マウスで骨芽細胞数を増加させ、骨梁骨の二次性海綿質(trabecular bone secondary spongiosa)の骨芽細胞のアポトーシスを低下させることが示された最小の値であり(以下を参照されたい:Bellido et al. (2003) J Biol Chem 278(50):50259-72)、さらに他の短期in vivo実験で用いられるPTH投薬量と比較して臨床投薬量(20μg/日、70 kgの患者について約0.07 nmol/kg)により近いからである。hPTHペプチドは2.5 nmol/kgでは骨梁骨の質量を増加させなかったが、同じ投薬量のbbPTH1-84で処置したマウスは、hPTH1-84と比較して骨梁骨の特性(表2)及び皮質強度(図29)で改善を示した。骨梁の厚さはbbPTH1-84処置マウスで上昇傾向を示した(表2)。重要なことには、骨梁骨の体積比及び見掛けの骨塩密度はbbPTH1-84によってのみベヒクル処置よりも有意に上昇した(表2)。調べたペプチドアナローグの大半が大腿骨の究極負荷を増加させたが、ベヒクル処置を超える増加はbbPTH1-84についてのみ統計的に有意であった(図29)。総合すれば、これらの発見は、bbPTHはhPTHよりも低い用量で有効な骨形成療法である得るという可能性を高める。
本実験で用いた全てのPTHペプチドが、in vitro血清枯渇によって誘発したアポトーシスシグナリングを減弱させた(図19)。bbPTH1-34は、ヒトPTHよりも強く前骨芽細胞系細胞でアポトーシスシグナリングを低下させた(図19)。最近、我々は予備的実験においてヒト胎児骨芽細胞(hFOB)で同様な結果を見出した。24時間の血清枯渇はhFOBで細胞死(トリパンブルー染色により確認)を増加させたが、100nMのPTH1-34又は1-84による枯渇前1時間のhFOBの前処理はパーセント細胞死を低下させた(ベヒクル処置と比較した細胞死の百分率変化は以下の通りであった:hPTH1-34は-18%、bbPTH1-34は-25%、hTPH1-84は-17%、bbPTH1-84は-21%;p<0.0001、データは示されていない)。bbPTH1-34の抗アポトーシス作用の強化は、bbPTH1-34はin vivoでもっとも強力な骨形成薬剤であり得るであろうという期待をもたらす。しかしながら、bbPTH1-84はbbPTH1-34及びhPTH1-34よりもin vitroで抗アポトーシス活性が低いけれども(図19)、bbPTH1-84はin vivo実験で皮質及び骨梁骨の体積でもっとも大きい増加を引き起こした(表1、表2)。
in vitro実験とin vivo実験の不一致は現時点では説明がつかない。骨芽細胞のアポトーシスの低下は骨梁骨に対するhPTHの同化作用の支配的プロセスであるが、他の生物学的メカニズムがbbPTHの骨体積増加能力を調節することもできる。例えば、骨芽細胞分化の促進(骨芽細胞のアポトーシスの防止ではなく)が主要手段であり、それによってhPTHは骨膜の皮質形成を促進する。この考え方に一致して、大腿骨強度はbbPTH1-84処置マウスで最も高い(図29)。bbPTH1-84はMC3T3細胞で試験した他のペプチドアナローグよりも抗アポトーシス活性は低かったが、脛骨近位の骨体積比もまたbbPTH1-84処置マウスで最も高かった(表1)。hPTHの1‐34フラグメントは完全長のhPTHペプチドの同化作用を再現するために十分であるが、bbPTH1-84はbbPTH1-34及び両hPTHペプチドと比較してin vivoで優れた性能を発揮したという事実は、bbPTHのC末端部分におけるアミノ酸変化はレセプター結合及びレセプター活性化活動に必要とされ得ることを提唱している。
【実施例15】
【0055】
実施例15:卵巣摘出ラットに対するヒトPTH1-84及びクマPTH1-84のin vivoでの作用
繁殖業者の施設で30匹の2.5ヶ月齢雌Sprague-Dawleyラットの卵巣が摘出された(OVX)。更なる実験のためにこのOVXラットを1グループ10匹の3つのグループに無作為に割り当てた。
OVX術の6週間後に、これらラットの皮下にベヒクル(食塩水)又は2.5nmol/kgのクマもしくはヒトPTH1-84を8週間注射した(5日/週で毎日)。最後の注射の24時間後に動物を安楽死させた。前記安楽死させた動物から更なる解析のためにサンプルを入手した。
大腿骨曲げ強度及び近位脛骨の骨梁構造を実施例12に記載したように調べた。
本実験では、大腿骨の究極負荷は、ベヒクル処置と比較してbbPTH1-84処置により有意に増加したが(p=0.013)、ヒトPTH1-84では有意ではなかった(p=0.463)。これらの発見は、bbPTH1-84は、ラットのOVX誘発骨減少の回復にヒトPTH1-84より有効であることを示唆している。しかしながらPTH処置は大腿骨の標準化究極負荷に有意な影響を与えなかった(図21)。この測定は、非標準化負荷よりもより生理学的な骨強度測定である。すなわち、より大きなサンプルサイズがより大きな統計力のために提供されるならばbbPTH1-84はヒトPTH1-84より有意に強力であろう。
骨体積比はPTH処置グループで増加した(図22)。骨梁数はPTH処置によって影響を受けなかったが(図23)、ただし骨梁の厚さはPTH処置グループで増加した(図24)。骨梁分離は処置グループと未処置グループとの間で違いはなかった(図25)。見掛けの骨塩密度はPTH処置グループで増加したが(図26)、実質的骨塩密度は処置グループと未処置グループとの間で違いはなかった(図27)。要約すれば、これらのデータは、ヒト及びクロクマPTH1-84は等しく骨梁骨の体積を増加させるが(図26)、bbPTH1-84はhPTH1-84よりも皮質強度をより大きく増加させることを提唱している(図24及び25)。これらの発見は、bbPTH1-84はヒトに有効な骨粗鬆症薬である可能性を提唱している。
【実施例16】
【0056】
実施例16:冬眠クマ血清はin vitroでカスパーゼ活性を低下させる
11月の初めから5月の初めまで10日毎に6頭のクマから血清を収集した(合計して各クマから6つの活動的冬眠前サンプル(11月−12月)、9つの冬眠時サンプル(1月−3月)、及び4つの活動的冬眠後サンプル(4月−5月))。クマ血清が、アポトーシス刺激に対する骨芽細胞応答に対してシーズン性影響を有するか否かを決定するために、白色壁を有する組織培養プレートにMC3T3-E1前骨芽細胞を各ウェル当たり10,000細胞で、アルファ改変最小必須培養液(MEM(-))に10%ウシ胎児血清を添加した培養液に播種し、一晩定着させた。続いて培養液を吸引し、24時間処置のために2%のクマシーズン血清を含むMEM(-)に置き換えた。再び培養液を吸引し無血清MEM(-)に置き換え、6時間アポトーシスを誘発した。終了時点で培養液を吸引し、50μLのPBS及び50μLのルミネセンスDEVD基質(Caspase-glo 3/7, Promega, Madison, WI)に置き換えた。マイクロプレートリーダー(Synergy HT Multi-Detection Microplate Reader)(Bio-Tek, Winooski VT)でルミネセンスを定量した。図28に示すように、冬眠血清で培養した細胞のカスパーゼ3/7活性は活動期血清で培養した細胞よりも低かった(p<0.0001)。これらの発見は、循環因子(例えばホルモン)のシーズン性変化は、骨芽細胞に抗アポトーシス作用を冬眠時に提供することを示唆している。
【0057】
【0058】
【技術分野】
【0001】
連邦政府後援研究に関する記載
本発明は、米国立衛生研究所(NIAMS AR050420及びNIDDK DK078407)及び米国科学財団(IBN-0343515)により提供された合衆国政府の支援を受けて達成された。合衆国政府は本発明に関して一定の権利を有する。
【背景技術】
【0002】
骨減少疾患は、これまでのところ約4千4百万のアメリカ人の健康を脅かしている(前記には骨粗鬆症を有する1千万人及び低骨量を示し骨粗鬆症発生リスクを有する3千4百万人が含まれる)。骨粗鬆症のアメリカ人の数は2020年までに膨れ上がると予想される。結果として、極めて多くの個体が低骨量による骨折のリスクを有する。50歳を超える白人女性の約40%及び白人男性の13%が、一生の間に股関節部、脊椎又は前腕骨折のリスクを有する。骨粗鬆症関連骨折に付随するコストは2002年には約10億ドルであり、さらに増加し続けると予想される。原発性(年齢性)骨粗鬆症に加えて、不使用骨粗鬆症が、特に卒中又は脊髄損傷により長期的に固定される患者について重要な臨床的課題である。骨折率は、脊髄損傷後の最初の年に健康なコントロールと比較して倍増し、卒中の開始後もまた健康なコントロールと比較して上昇する。不使用は、主として骨格の負荷の低下が不均衡な骨のリモデリングを引き起こす(前記は骨減少をもたらす)という理由から骨折率を増加させる。
【発明の概要】
【0003】
ある実施態様では、本発明は、配列番号:2の連続する少なくとも10アミノ酸残基を含む単離ポリペプチドを提供し、ここで前記ポリペプチドは配列番号:2のアミノ酸残基41及び52の少なくとも1つを含む。ある実施態様では、配列番号:2のアミノ酸残基1−84を含む単離ポリペプチドが提供される。そのようなポリペプチドをコードするポリヌクレオチドもまた提供される。本発明はまた配列番号:1を含む単離ポリヌクレオチドを提供する。
ある実施態様では、本発明はcAMPレベルを骨形成細胞で増加させる方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程はcAMPレベルを骨形成細胞で増加させる。
【0004】
ある実施態様では、本発明はアポトーシスを骨形成細胞で減少させる方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程はアポトーシスを骨形成細胞で減少させる。
ある実施態様では、本発明はBaxタンパク質対Bcl-2タンパク質の発現レベル比率を骨形成細胞で低下させる方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程はBaxタンパク質対Bcl-2タンパク質の発現レベル比率を骨形成細胞で低下させる。
【0005】
ある実施態様では、本発明は、骨基質タンパク質、転写活性化因子又は転写調節因子の発現レベルを骨形成細胞で増加させる方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程は、骨基質タンパク質、転写活性化因子又は転写調節因子の発現レベルを骨形成細胞で増加させる。
ある実施態様では、本発明は、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、もしくは骨折の発生率を減少させるか、又は前記の任意の組合せをもたらす方法を提供する。前記方法は、配列番号:2のアミノ酸残基1−34を含むポリペプチド、配列番号:2のアミノ酸残基1−36を含むポリペプチド、配列番号:2のアミノ酸残基1−84を含むポリペプチドから選択される少なくとも1つのポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程は、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、又は骨折の発生率を減少させる。
【0006】
ある実施態様では、本発明は、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、もしくは骨折の発生率を減少させるか、又は前記の任意の組合せをもたらす方法を提供する。前記方法は、クマの副甲状腺ホルモン又はその機能的フラグメントを含むポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含み、ここで、前記ポリペプチドと前記骨形成細胞を接触させる工程は、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、又は骨折の発生率を減少させる。
ある実施態様では、本発明は配列番号:2のアミノ酸残基1−34から成る単離ポリペプチドを提供する。ある実施態様では、本発明は配列番号:2のアミノ酸残基1−36から成る単離ポリペプチドを提供する。ある実施態様では、本発明は配列番号:2のアミノ酸残基1−84から成る単離ポリペプチドを提供する。
【図面の簡単な説明】
【0007】
【図1】極限強度、骨強度の測定値、及び灰分比、骨塩含有量の測定値を示す(前記は両方ともクロクマで年齢に合わせて増加する)。
【図2】血清オステオカルシンレベルは、クロクマの冬眠前、冬眠時及び冬眠後プールサンプルで血清副甲状腺ホルモン(PTH)と正の相関性を有することを示す(p=0.0007、n=27)。
【図3】3ヶ月の不使用期間中の標準化した血清吸収マーカー(ICTP)及び血清形成マーカー(PICP及びオステオカルシン)濃度を示す。
【図4】骨芽細胞によって放出されるPGE2の量は、冬眠後に収集した血清で細胞を処理したときに最大であることを示す。
【図5】他の公知のPTH配列と対比した成熟クロクマPTHタンパク質配列を示す。
【図6】ヒト及びクロクマの両PTH 1-34がオステオカルシンをアップレギュレートすることを示す(n=2)。
【図7】アポトーシス関連遺伝子発現に対するヒト及びクロクマのPTH 1-34の作用を示す(n=4)。
【図8】ラット脛骨近位部(左)、骨端軟骨基部から2mm遠位で始まる皮質及び骨梁骨(trabecular bone)の1.6mm解析領域(上右)及び骨梁解析領域(下右)のuCT画像を示す。
【図9】9A−9Fは、模擬卵巣摘出術(OVX)を実施(図9A)又は実際のOVXを実施しその後PTH処置を施さずに6週間(図9B)もしくは14週間(図9C)回復させるか、又はクロクマPTH 1-34の3μg/kg(図9D)、10μg/kg(図9E)もしくは30μg/kg(図9F)による8週間の処置を含む14週間の回復期間後のラットの骨のμCT画像を示す。
【図10】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1-34で処置したラットの骨について総体積の比(BV/TV)としての骨体積のグラフを示す。
【図11】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1-34で処置したラットの骨について骨塩の見掛けの密度のグラフを示す。
【図12】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について骨梁支柱骨(trabecular strut)数のグラフを示す。
【図13】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1-34で処置したラットの骨について骨梁の厚さのグラフを示す。
【図14】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について骨梁骨の鉱化のグラフを示す。
【図15】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について脛骨皮質体積のグラフを示す。
【図16】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について脛骨皮質多孔度のグラフを示す。
【図17】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの骨について大腿骨皮質の強度のグラフを示す。
【図18】模擬又は実際のOVX処置に続いてベヒクル(食塩水)又はヒトもしくはクマPTH 1−34で処置したラットの血清カルシウムレベルのグラフを示す。
【図19】0−100nMのヒト又はクマPTHペプチドで前処理し、続いて血清枯渇に付したMC3T3-S4細胞の相対的カスパーゼ-3活性を示す。標準誤差バーとともにLS平均が提示されている。異なる文字を有するバーは互いに有意に相違する(p<0.005)。
【図20】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの大腿骨の究極負荷を示す。
【図21】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの大腿骨の標準化究極負荷を示す。
【図22】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの骨体積比を示す。
【図23】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの骨梁数を示す。
【図24】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの骨梁の厚さを示す。
【図25】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの骨梁の分離を示す。
【図26】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの見掛けの骨塩密度を示す。
【図27】ベヒクル(V)、2.5nmol/kg hPTH 1-84又は2.5nmol/kg bbPTH 1-84で処理したOVXラットの実質的骨塩密度を示す。
【図28】冬眠前、冬眠時及び冬眠後のクマ血清中で培養した細胞のカスパーゼ3/7活性を示す。
【図29】ベヒクル又は種々のPTHペプチドで処理したマウスの大腿骨の究極負荷を示す。
【発明を実施するための形態】
【0008】
ヒト及び他の大半の哺乳動物では、例えば加齢及び長期未使用のような因子は、骨粗鬆症及び骨折リスクの増加をもたらし得る。脊髄損傷による不使用は、特に脛骨及び大腿骨における骨塩密度を顕著に低下させ、さらに大腿骨骨幹の断面慣性モーメントを顕著に低下させる。したがって、骨の曲げ強さは脊髄損傷によって低下し、骨折リスクが増加する。卒中による不使用もまた骨折リスクを増加させる。さらにまた、骨の機械的負荷の低下は、骨形成の持続的低下に加えて迅速な骨吸収増加のために急激な骨減少を引き起こし得る。不使用により誘発される骨リモデリングの変化は皮質内多孔度を増加させ、長骨骨幹の断面特性及び機械的特性を低下させる。負荷低下はまた、骨梁骨の量及び微細構造を大いに低下させる。
【0009】
骨に対する不使用の有害な影響は再可動期間中も持続し得る。ある骨は再可動時に回復し得るが、回復は遅くかつしばしば不完全である。例えば、病臥中の骨減少速度は、再可動時の骨増加よりも3倍以上速く、宇宙飛行時の骨減少の回復は5年後さえも完全ではないことがある。不使用により誘発された骨の変化を再開した活動によって完全に復帰させ得る場合、この再可動期間の長さはしばしば固定期間の2から3倍である。骨に対する機械的負荷を低下させる多くの状況下で、骨形成は低下し、及び/又は骨吸収は増加する。しかしながら、イヌの前肢の固定中に吸収及び形成の両方が増加するが、それでもなおそのような事例において顕著な骨減少が存在する。同様に、脊髄損傷を有する患者で大腿骨の代謝回転が生じ、これは骨減少及び骨折発生率の増加をもたらす。
【0010】
これとは対照的に、クロクマは加齢(図1)によって、又はより重要なことには長期の不使用(冬眠時に生じる)によっても顕著な骨減少に悩まされることはない。冬眠するクロクマは、北方領域では長さがほぼ等しい固定期と活動期を有する。骨代謝の血清マーカーに関するデータ(下記参照)は、クマでは不使用時に吸収と形成の両方が増加し吸収と形成の間のタイムラグ(すなわち復帰期間)は通常のとおりであり、形成の増加は吸収の増加と共役し均衡を保ったままであることを示唆している。クロクマの腸骨稜の生検から得られた組織学的データもまた非活動時の吸収及び形成の増加を示している。しかしながら、クマは、冬眠中に骨梁骨の体積、骨塩密度及び骨塩含有量が低下しないという点で唯一のものである。さらにまた、クロクマでは皮質強度及び灰分比が年齢とともに増加し、多孔度は毎年の不使用期間にもかかわらず低下する。皮質の多孔度は、活動中のハイイログマよりも冬眠中のハイイログマで顕著に低く、さらに大腿骨断面の幾何学及び強度は冬眠によって影響を受けない。
【0011】
クマは多くの固有の生物学的メカニズムを進化させて、食物の無い長い固定化期間を生き残ってきた。これらのメカニズムには、カルシウム及び骨異化作用の他の成分のリサイクルが含まれるように思われる。なぜならば、クマは冬眠中に骨の代謝回転を増加させるが、老廃物を排出しないからである。ヒトでは、病臥により誘発される不使用骨粗鬆症は、主として対応する形成増加を伴わない吸収の増加によって引き起こされる。これは、高カルシウム血症及び尿と糞便のカルシウムによって生じる負のカルシウム均衡をもたらす。クマは冬眠中に排尿も排便もしないので、吸収によって骨から放出されたカルシウムの大半は、骨芽細胞による骨形成を介して骨にリサイクルされるということはありそうなことである。イオン化カルシウムが冬眠時に約23%増加することが見出され、これらはおそらく吸収と形成の間のタイムラグのためであろう。逆説的に、クロクマのPTHレベルはイオン化カルシウムレベルが最高のときにもっとも高い(実施例2、表1)。総合すれば、これらの発見は、クマは骨粗鬆症を回避するために生物学的メカニズムを進化させてきたことを示唆している。
【0012】
大半の動物で不使用時に骨形成が骨吸収と共役しないメカニズムは不明であるが、おそらく機械的及び生化学的因子の両方が必要とされるだろう。機械的緊張の欠如は、骨細胞のアポトーシスの開始及び同時に生じる骨芽細胞活性の低下によって吸収の増加をもたらし得る。例えばヒトPTHのようなホルモンは骨細胞を機械的刺激に対して感受性にし、さらに機械的負荷により相乗的に骨形成を増加させる。ヒトに1日1回投与されたヒトPTHは骨量を増加させ骨折発生率を低下させる。したがって、クロクマでは、循環PTHは低レベルの機械的刺激(おそらく冬眠用器官の震え又は整復による)に対して骨細胞を感受性にして、不使用時の骨形成の維持を助長する。PTHはまた、骨芽細胞の分化を刺激し骨芽細胞のアポトーシスを抑制することによってクロクマで骨形成の維持を助長し得る。
【0013】
PTHは血中カルシウムレベルの主要調節因子であり、したがってクロクマの不使用時の恒常的血清カルシウムレベルの維持に重要な役割を果たす。血清PTHレベルは、活動時及び冬眠時のクロクマにおいて骨形成マーカーオステオカルシンと正の相関性を示し(図2)、オステオカルシン及びPTHは両方とも冬眠中に増加する。さらにまた、クロクマのPTH濃度は、イオン化カルシウム濃度が最高のときにもっとも高い。冬眠中に骨吸収は増加するが総血清カルシウム(tCa)は無変化のままであるので、おそらくPTHのレベル増加は腎臓のカルシウム再吸収の増加を引き起こし、骨への塩のリサイクルを促進し骨形成の増加を均衡させる。これによって、骨塩密度(“BMD”)及び皮質多孔度のような骨稜及び皮質の特性で観察された保存がもたらされる。冬眠中に骨吸収は増加するが、クマは老廃物を冬眠中に排泄しないという事実にもかかわらず血中カルシウム濃度は一定のままである。冬眠中に骨吸収によって遊離したカルシウムはリサイクルされ、骨形成と骨吸収との均衡した共役の維持により骨に戻され得る。このことは冬眠中のクロクマではPTHは同化作用を有するという考えを支持し、冬眠中に均衡のとれた骨リモデリングを維持するクマの特異な能力の説明を提供する。PTHの同化作用は、冬眠からの覚醒に続いて身体活動が開始するときにクロクマで強化され得る。機械的負荷及びヒトPTHは相乗的に作用し、ラットの骨形成をin vivoで、さらに生化学的シグナリングをin vitroで増加させることが以前に示された。春の再可動時に、クロクマの骨形成は血清オステオカルシンが示すように冬眠前のレベルよりも高いまま維持される。
【0014】
クロクマ(ウルスス・アメリカヌス(Ursus americanus))の副甲状腺ホルモン(PTH)をコードするポリヌクレオチドの配列(配列番号:1)が、成熟84アミノ酸PTHタンパク質のポリペプチド配列(配列番号:2)と同様に見出された。さらにまた、完全長PTHタンパク質(配列番号:4)をコードするcDNA(配列番号:3)の配列も決定された(前記完全長PTHタンパク質は25アミノ酸のシグナルペプチド(配列番号:4のアミノ酸残基1−25)及び6アミノ酸のプロペプチド(配列番号:4のアミノ酸残基26−31)を含む)。前記成熟クロクマPTHタンパク質は他の公知のPTHタンパク質とは相違する(図5)。ヒトPTHと比較して、クロクマPTHは、完全長の成熟PTHポリペプチドの合計84アミノ酸残基のうち9アミノ酸残基が異なっている。さらに本明細書にはクロクマPTH及びその機能的フラグメントの種々の使用方法が記載されている。具体的には、クロクマPTHに特異的な抗体の開発のために、配列番号:2の連続する少なくとも10アミノ酸を含み、かつアミノ酸残基41又は52の少なくとも1つを含有するポリペプチドサブフラグメントを使用できることが考えられる。これらの抗体を用いてクロクマPTHを例えばELISAアッセイで定量できる。
【0015】
骨形成細胞表面のPTHレセプターは、細胞内のサイクリックアデノシン一リン酸(cAMP)依存第二メッセンジャーシグナリング経路と共役する。これらのシグナリング経路は順繰りに、骨形成に必要な遺伝子(例えばI型コラーゲン、オステオネクチン及びオステオポンチンをコードするもの)の発現増加をもたらす。cAMP/タンパク質キナーゼA経路は骨形成における組織学的指標及び血清指標の大多数のPTH誘発増加を起こすことができるので、結果としてcAMP応答増加がより強い骨形成をもたらすことができる。あるPTHタンパク質の配列で比較的少数のアミノ酸置換が、天然の形態と比較してより強いサイクリックアデノシン一リン酸(cAMP)産生を刺激できる。例えば、卵巣摘出ラットは、ラットPTH1-34よりもウシPTH1-34の1日25μgの注射に対して25%高い骨形成応答を示した(ラットPTH1-34はウシPTH1-34と比較して5アミノ酸の配列相違を有する。ウシPTH1-34の注射は、処置時に37%高い骨体積比の増加をもたらす。
【0016】
したがって、クロクマPTHのアミノ酸置換によって、ヒトPTHよりも強いcAMP産生が骨形成細胞で惹起されることはありそうなことである。一般的に、PTHは、例えば骨芽細胞のアポトーシス低下、Runx2媒介骨芽細胞分化の増加、SOST系の骨細胞における負のフィードバックのダウンレギュレーション、及び骨基質タンパク質のmRNAの生成増加(いずれもcAMP媒介経路)によってより強い骨形成応答を引き起こす。本発明のメカニズムを理解することが必要というわけではないが、クロクマPTHはおそらく他のPTH形よりも骨形成性が高いと考えられ、これは、クロクマが不使用中に均衡をもった骨リモデリングを維持する固有の能力をもつ理由を説明する。本発明のある実施態様では、骨形成細胞をクロクマPTH又はその機能的フラグメントと接触させる工程は骨形成細胞でcAMPレベルを増加させる。本発明のある実施態様では、配列番号:2のアミノ酸残基1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
【0017】
本明細書で用いられる、PTHポリペプチドと“細胞を接触させる工程”は、in vitro実験の場合には培養溶液にポリペプチドを添加する工程、又はポリペプチド治療薬に適した投与方法を用いて被験者にポリペプチドを投与する工程を含む。“細胞を接触させる工程”はまた、被験者で所望のポリペプチドを合成及び放出させるために、ある発現系で前記ポリペプチドをコードする外因性ポリヌクレオチドを被験者に導入する工程を含む。本明細書で用いられる、“骨形成細胞”には骨芽細胞、骨細胞、骨内皮細胞(bone lining cell)、軟骨芽細胞及び軟骨細胞が含まれるが、ただしこれらに限定されない。適切には骨形成細胞は被験者の内部に存在し得る。
骨形成細胞は規則的に代謝回転を生じ、細胞死の大半はプログラムされた細胞死(又はアポトーシス)による。この規則的な代謝回転速度が存在する場合、骨形成細胞のアポトーシスを低下させるいずれのメカニズムも骨形成細胞数の増加をもたらし、これはおそらく骨の生長を促進させるであろう。したがって、本発明のある実施態様では、骨形成細胞をクロクマPTH又はその機能的フラグメントと接触させる工程は骨形成細胞のアポトーシスを低下させる。本発明のある実施態様では、配列番号:2のアミノ酸1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
【0018】
さらにまた、内因性クロクマPTHのより大きなC-末端フラグメントのいくつかはCPTHR(C-末端PTHレセプター)との結合を介してシーズン性骨リモデリングプロセスで役割を演じる可能性がある。具体的には、クマPTHのC-末端フラグメントは、骨芽細胞形成を妨げることによって及びおそらくは成熟骨細胞活性に影響を与えることによって(前記は通常的には不使用のような吸収性刺激に応答して発生するであろう)、PTH1-84及び1-34の血中カルシウム上昇作用に拮抗し得る(Divieti, P. et al., 2002, Endocrinology 143(1): 171-6)。これは、冬眠中のクマが恒常的カルシウムレベルを維持するために役立ち得る。したがって、本発明のある実施態様では、配列番号:2のアミノ酸残基11−84を含むポリペプチド及びアミノ酸残基7−84を含むポリペプチドから選択される少なくとも1つのポリペプチドと骨細胞及び骨芽細胞を接触させることができる。
タンパク質Baxはアポトーシスを促進するがBcl-2タンパク質は細胞をアポトーシスから保護し、Bax対Bcl-2の発現比率の低下は個々の細胞集団におけるアポトーシスの低下を示唆する。したがって、本発明のある実施態様では、クロクマPTH又はその機能的フラグメントと骨形成細胞を接触させる工程は、Bcl-2タンパク質の発現レベルに対するBaxタンパク質の発現レベルの比率を骨形成細胞で低下させる。本発明のある実施態様では、配列番号:2のアミノ酸残基1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
【0019】
実施例11は、クロクマPTH1-34は培養細胞でBax/Bcl-2発現比率を低下させるが、ヒトPTH1-34はBax/Bcl-2発現比率を増加させることを示す(図7)。したがって、クロクマPTH1-34はヒトPTH1-34よりもアポトーシスの防止により有効であるように思われる。理論に拘束されないが、この相違はヒトPTH1-34とクロクマPTH1-34間での2つのアミノ酸が異なることの結果であるかもしれない。これらのデータは、クマPTHはヒトPTHよりも同化作用が強いことを提唱している。なぜならば、骨芽細胞のアポトーシスの低下はPTH処理によって誘発される骨形成応答に寄与し得るからである。
クロクマPTH又はその機能的フラグメントと骨形成細胞を接触させる工程はまた、骨基質タンパク質、転写活性化因子又は転写調節因子の発現レベルを骨形成細胞で増加させる。本発明のある実施態様では、転写活性化因子はRunx2である。本発明のある実施態様では、転写調節因子はc-fosである。例示的な骨基質タンパク質にはオステオカルシン、オステオポンチン及びコラーゲンI型が含まれるが、ただしこれらに限定されない。本発明のある実施態様では、配列番号:2のアミノ酸残基1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
【0020】
外因性ヒトPTHを用いてヒトの閉経後骨粗鬆症及び加齢性骨粗鬆症が治療されているが、前記PTHは理想的な治療薬ではない。現在のところ組換えヒトPTH1-34(LY333334, Eli Lilly, Indianapolis IN)のみが臨床使用のために承認されており、組換えヒトPTH1-84の1つの型(ALX1-11, NPS Pharmaceuticals, Parsippany, NJ)のみが米国食品医薬品局の承認が考慮されている。LY333334及びALX1-11はin vivoで同じ規模の骨形成を刺激するが、それらの生物学的作用は同一ではない。例えば、PTH1-34はプロコラーゲン-1 mRNAの産生をダウンレギュレートするが、PTH1-84はダウンレギュレートしない(Nasu et al., 1998, Endocr J, 45, 229-34)。さらにまた、ヒトPTHのC-末端部分は、成熟ホルモンから切り離されたとき重要な生物学的機能(例えば骨吸収の阻害)を有する。
LY333334又はALX1-11の長期使用はラットで骨肉腫を生じるが、予備的結果によれば、ヒトPTH1-84はヒトPTH1-34よりも発癌率が低いことが示唆され、これはおそらく、外因性ヒトPTH1-84のC-末端フラグメント(末梢性タンパク分解性プロセッシングにより生じる)はC-末端PTHレセプター(CPTHR)と結合し、骨細胞のアポトーシスを増加させるためであろう。したがって、同化作用は同等であるが、ヒトPTH1-84はひとPTH1-34と比較して優れた骨粗鬆症治療法であり得る。しかしながら、ヒトPTH1-84は失われた骨を完全には回復させることはできない。すなわち、男性及び女性は加齢性骨粗鬆症のために皮質及び海綿質の20‐30%を失うが、ALX1-11をその推奨治療期間中ずっと使用しても8%しか回復させ得ないと言われている。したがって、より強力な骨形成能を有する骨粗鬆症治療が臨床現場で希求されている。
【0021】
ある実施態様では、本発明は、クロクマPTH又はその機能的フラグメントの有効量をその必要がある被験者に投与する工程を含む、骨粗鬆症の治療方法を提供する。種々の実施態様で、骨粗鬆症は、原発性又は加齢性骨粗鬆症及び/又は不使用骨粗鬆症である。本発明のある実施態様では、被験者の骨形成細胞をクロクマPTH又はその機能的フラグメントと接触させる工程は、被験者で骨塩密度を増加させるか、骨量を増加させるか、骨減少を低下させるか、又は骨折の発生率を低下させる。本発明のある実施態様では、配列番号:2のアミノ酸残基1−34又は1−36を含むポリペプチドと骨形成細胞を接触させる。本発明のある実施態様では、配列番号:2を含むポリペプチドと骨形成細胞を接触させる。
適切には、被験者の骨形成細胞をクロクマPTH又はその機能的フラグメントと接触させる工程は、少なくとも約5%又は少なくとも約10%骨塩密度を増加させるか、骨量を増加させるか、骨減少を低下させるか、又は骨折の発生率を低下させる。骨塩密度の増加、骨量の増加、骨減少の低下又は骨折の発生率の低下は、少なくとも約15%、少なくとも約30%、少なくとも約50%、少なくとも約75%、又は少なくとも約90%であり得る。骨塩密度の増加、骨量の増加、骨減少の低下又は骨折の発生率の低下は、治療前及び治療後に当業者に公知の技術により所望の特徴を同一患者で測定することによって決定される。例えば、骨塩密度は、脊髄、手首、腕又は脚内の骨の二重エネルギーX線(DEXA)又はCTスキャンの撮影を含む方法によって決定できる。
【0022】
被験者は適切には哺乳動物で、ヒト、ウマ、イヌ、ネコ、マウス、クマ、ウシ、ブタ又はシカが含まれるが、ただしこれらに限定されない。ある実施態様では、被験者は閉経後の女性である。被験者は骨粗鬆症を有していても又は骨粗鬆症発症のリスクがあってもよい。骨粗鬆症発症のリスク因子には以下が含まれる:50歳以降の骨折の病歴;現時点で低骨量であること;1親等親族の骨折の病歴;女性であること;痩せているか及び/又は骨格が小さいこと;高齢;骨粗鬆症の家族歴;閉経(特に早発性又は外科手術により誘発されたもの)の結果としてのエストロゲン欠乏;月経期間の異常停止(無月経);神経性食欲不振;終生のカルシウム摂取不良;ビタミンD欠乏;一定の薬物の使用(コルチコステロイド、化学療法、抗痙攣薬及び他のもの);一定の慢性的症状、例えば腸のカルシウム吸収を低下させるもの(例えばクローン病)の存在;男性の低テストステロンレベル;非活動的なライフスタイル;現時点で喫煙していること;アルコールの過剰摂取;及び白人又はアジア人であること(ただしアフリカ系アメリカ人及びヒスパニック系も同様に有意なリスクを有する)。さらにまた、女性は閉経後5年から7年でその骨量の20%までを喪失し、いっそう骨粗鬆症に感受性となり得る。
【0023】
クロクマPTH又はその機能的フラグメントはまた、骨粗鬆症と戦うため又は骨粗鬆症発症リスクを有する被験者で骨粗鬆症を予防するための、防止薬(回復薬ではなく)もしくは予防薬、手段として有用である。クマは不使用の間均衡のとれた骨のリモデリングを維持する唯一の動物のようであるので、クロクマPTH又はその機能的フラグメントはまた、例えば宇宙飛行中の宇宙飛行士及び脊髄損傷後の患者で発生する骨格負荷の低下時の骨減少の防止に有用である。
クロクマPTH又はその機能的フラグメントは、カルシウム及び/又はビタミンDと一緒に投与できる。適切には、“ビタミンD”は完全なビタミンDクラスの化合物を指す。クロクマPTH又はその機能的フラグメントはまた、他の同化作用促進又は抗吸収活性物質と一緒に投与できる。適切な同化作用促進活性物質には、ヒトPTHの種々のフラグメント(例えば1-34及び1-84)、フルオリド、GH、インスリン様成長因子I、スタチン及びPYK2キナーゼ阻害剤が含まれる。適切な抗吸収活性物質には、エストロゲン、選択性エストロゲンレセプター調節因子、カルシトニン及びビスホスホネートが含まれるが、ただしこれらに限定されない。ある実施態様では、同化作用を有するクロクマPTHの機能的フラグメント(例えばクロクマPTH1-34)が抗吸収活性物質と一緒に投与される。ある実施態様では、クロクマPTHの種々の機能的フラグメントは互いに組み合わせて投与できる。種々の機能的フラグメントは同時に又は適切な任意の順序で投与できる。
【0024】
カルシウム、ビタミンD、同化作用促進活性物質及び/又は抗吸収活性物質は、クロクマPTH又はその機能的フラグメントと同時に投与するか、又はクロクマPTH又はその機能的フラグメントの前もしくは後に投与できる。カルシウム、ビタミンD、同化作用促進活性物質及び/又は抗吸収活性物質は別々の組成物に存在しても、又はクロクマPTH又はその機能的フラグメントと同じ医薬組成物に存在してもよい。
クロクマPTHもしくはその機能的フラグメントの投与又はクロクマPTHもしくはその機能的フラグメントを含む組成物の投与は、任意の適切な技術によって実施できる。クロクマPTH又はその機能的フラグメントは、例えば経口、経鼻、経直腸及び非経口ルートの投与を含む任意の適切なルートによって投与できる。本明細書で用いられる、非経口という用語には、例えば注射による皮下、皮内、静脈内、筋肉内、腹腔内、クモ膜下投与が含まれるが、ただしこれらに限定されない。上記で考察するように、ポリペプチドの投与には、被験者で外因性ポリヌクレオチドを発現できるようにプロモーターと作動可能に連結した外因性ポリヌクレオチドの投与が含まれる。ポリペプチドの投与にはまた、前記ポリペプチドをコードするポリヌクレオチドを含むウイルスベクターの投与が含まれる。適切には、ウイルスベクターはアデノウイルスベクターである。
【0025】
クロクマPTHもしくはその機能的フラグメント又はクロクマPTHもしくはその機能的フラグメントを含む組成物は、継続的に又は当業者が容易に決定できる明確な間隔をおいて投与することができる。通常の技量を有する臨床医は、被験者に投与されるべきクロクマPTH又はその機能的フラグメントの適切な量を決定できる。
任意の個々の被験者に対する有効用量は多様な因子に左右される。前記因子には、治療される疾患及び疾患の重症度;用いられる個々の化合物の活性;用いられる個々の組成物;被験者の年齢、体重、一般的健康状態、性別及び養生食;投与ルート;用いられるクロクマTPH又はその機能的フラグメントの排出速度又は不活化速度;治療期間;クロクマTPH又はその機能的フラグメントと一緒に用いられる他の医薬;及び医療分野で周知の同様な因子が含まれる。例えば、所望の効果を達成するために必要な用量よりも低いレベルの用量から開始し、所望の効果が達成されるまで投薬量を徐々に増加させることは十分に当業者の水準の範囲内である。
適切には、ある実施態様におけるクロクマTPH又はその機能的フラグメントの投薬量は、0.10μg/kg/日から40μg/kg/日の範囲である。ある実施態様では、投薬量は5μg/kg/日から20μg/kg/日の範囲である。ある実施態様では、投薬量は10μg/kg/日である。ある実施態様では、投薬量は被験者当たり10μg/日から400μg/日の範囲である。ある実施態様では、投薬量は被験者当たり20μg/日から40μg/日の範囲である。ある実施態様では、投薬量は被験者当たり30μg/日である。
【0026】
併用投与のためには、当業者は、クロクマTPH又はその機能的フラグメント及び追加される活性物質(例えばカルシウム、ビタミンD、同化作用促進薬又は抗吸収薬)の両方の適切な投薬量を決定できよう。上記で考察したように、任意の個々の被験者に有効な用量は多様な因子に左右される。ある実施態様では、抗吸収薬の投薬量は1日当たり約5から約100mgであり、カルシウムの投薬量は1日当たり約500から約1500mgであり、ビタミンDの投薬量は1日当たり約500から約1500mgであり、同化作用促進薬の投薬量は1日当たり約0.1mgから約0.5mg又は約10μg/日から400μg/日である。
ある実施態様では、被験者はヒトである。適切には、1日当たりの投薬量がある実施態様では1週間、ある実施態様では1ヶ月、ある実施態様では3カ月間、ある実施態様では6ヶ月間、ある実施態様では1年間、ある実施態様では1年と半年間、ある実施態様では2年間、さらにある実施態様では3年間与えられる。
【0027】
所望の場合は、1日の有効用量を投与目的にふさわしい複数回の用量に分割できる。したがって、シングルドース組成物は1日の用量を構成する量又はそれを複数回に分けた部分を含むことができる。所望の場合は、適切なデリバリー装置に1日の有効量を2日以上のために、例えば7日、14日、21日、28日などのために装填し、このデリバリー装置を用いて、所望の1日1回用量又は1日複数回用量を所望の合計日数の間繰り返して投与する。理解されるところであるが、医療の実施に関する基準及び個々の被験者の症状によって決定されるように、当業者は有効用量及び同時投与治療方式を容易に最適化するであろう。
本発明の方法で有用なクロクマPTH又はその機能的フラグメントを含む組成物は、医薬として有用な組成物の公知の製造方法にしたがって処方することができる。処方は、周知であり当業者が容易に入手できる多数の文献に詳細に記載されている。例えば以下の文献には本開示方法で用いることができる処方が記載されている(Remington's Pharmaceutical Science, E. W. Martin)。一般的には、組成物は、クロクマPTH又はその機能的フラグメントの有効量を適切な担体と一緒にし、組成物の有効な投与を促進することができるように処方されるであろう。
【0028】
本方法で用いられる組成物はまた多様な形態で存在し得る。これら形態には例えば固体、半固体、及び液体投薬形(例えば錠剤、ピル、散剤、液状溶液又は懸濁物、座薬、注射可能及び輸液可能溶液、並びにスプレー)が含まれる。形態は意図する投与態様及び治療薬の適用態様に左右されるであろう。組成物はまた適切には、医薬的に許容できる当業者に公知の通常の賦形剤を含む。賦形剤の例には、注射用の水、エタノール、ジメチルスルホキシド、グリセロール、アルミナ、デンプン、氷酢酸、酢酸ナトリウム、マンニトール、メタクレゾール、組成物のpHを適切な値に調整するための塩酸及び/又は水酸化ナトリウム、並びに等価の或いは適切な担体及び希釈剤が含まれる。所望の適用においてそのような調剤を投与するために、医薬組成物は、担体又は賦形剤を含む全組成物重量を基準にして、総重量が約0.1%から99%、適切には約1から15%の1つ又は2つ以上の本発明のポリペプチドを含むであろう。
【0029】
本明細書で用いられるように、“単離された”核酸分子、ポリヌクレオチド、ポリペプチドなどは、場合によって、天然の状態以外で見出される夾雑物(例えばポリヌクレオチド、ポリペプチドなどの他の種)から少なくとも部分的に精製された成分を指す。単離された核酸、ポリヌクレオチド又はポリペプチドは、前記が本来結合している細胞成分の約50%未満、適切には約75%未満、もっとも適切には約90%未満を含むことができる。残余の細胞成分から(例えばゲル上で)十分にかつ容易に区別できるようにPCRを用いて増幅したポリヌクレオチドは“単離されている”とみなされる。本発明の核酸分子、ポリヌクレオチド及びポリペプチドは“実質的に純粋”であり得る。すなわち、当業界で公知の精製技術を用いて達成可能な最高の純度を有する。
本明細書で用いられるように、“機能的フラグメント”は、より大きなポリペプチド又はポリヌクレオチドの領域又は部分である、あるポリペプチド又はポリヌクレオチドの任意の領域又は部分である。前記領域又は部分はより大きなポリペプチド又はポリヌクレオチドに起因する活性又は機能を有する。例えば、ヒトPTHの機能的フラグメントはヒトPTHの1−34領域である。クロクマPTHの機能的フラグメントには1−34及び1−36が含まれるが、ただしこれらに限定されない。
【0030】
本明細書及び添付の特許請求の範囲で用いられるように、単数形の“a”、“an”、及び“the”には、内容が明瞭にそうでないことを示していないかぎり複数の対応形が含まれる。さらにまた、“or”という語は、内容が明瞭にそうでないことを示していないかぎり一般的には“及び/又は”を含む意味で用いられる。全ての刊行物、特許及び特許出願は、あたかも個々の各刊行物又は特許出願が本明細書に具体的にかつ個々に参照によって含まれるかのように、同じ程度にその全体が全ての目的のために参照により明白に本明細書に含まれる。本開示と参照により含まれる特許、刊行物及び参考文献との間に矛盾がある場合には、本開示が優先する。
本明細書に列挙した数字によるいずれの範囲も、小さい方の値から大きい方の値までの全ての値を含むこともまた特に理解されよう。すなわち、列挙された最小値と最大値の間の数値の可能な全ての組合せが本出願に明瞭に記載されていると解されるべきである。例えば、濃度範囲が1%から50%と記載されている場合は、例えば2%から40%、10%から30%又は1%から3%などの値を本明細書に明瞭に列挙しようとするものである。濃度範囲が“少なくとも5%”である場合には、それより上の全てのパーセンテージ値(100%を含む)もまた明瞭に列挙しようとするものである。これらは特に意図するものの例にすぎない。
以下の実施例は本発明の更なる理解を助けるために提供される。用いられた具体的な材料、方法及び条件は本発明の例示を意図するものであり、本発明の範囲を限定するものではない。
【実施例1】
【0031】
実施例1:クロクマPTH1-84の配列決定
ゲノムDNA抽出
捕獲された雌のクロクマから血液を収集し4℃で保存した。ゲノミックプレップ血液DNA単離キット(GenomicPrep Blood DNA Isolation Kit)(Amersham Biosciences, Piscataway, NJ)を製造業者の指示に従って用い、全血サンプルからゲノムDNAを2週間以内に抽出した。
PCRクローニング及び配列決定
クロクマゲノムDNAをPTHのPCR増幅に用いた。GenBankで入手可能な8つの完全長哺乳動物PTH配列(以下を含む:ウシ(ボス・タウルス(Bos taurus)AAA30749)、ネコ(フェリス・カツス(Felis catus)Q9GL67)、イヌ(カニス・ファミリアリス(Canis familiaris)P52212)、ヒト(ホモ・サピエンス(Homo sapiens)NP_000306)、マカク(マカカ・ファシクラリス(Macaca fascicularis)Q9XT35)、マウス(ムス・ムスクラス(Mus musculus)NP_065648)、ブタ(スス・スクロファ(Sus scrofa)NP_999566)、及びラット(ラトツス・ノルヴェギクス(Rattus norvegicus)NP_058740)のアラインメントを基にして設計したコンセンサスプライマーを用いた。PCR増幅は、20μLの反応体積中で10−15ngのゲノムDNA、100μMのdNTP、各々0.2μMのプライマー及び1ユニットのREDTaq(Sigma, St. Louis, MO)を用いて実施した。PCR生成物は、ウルトラクリーンゲルスピンキット(UltraClean GelSpin Kit)(MoBio Carlsbad, CA)を用いてゲル精製し、さらにTAクローニングキット(Invitrogen, Carlsbad, CA)を用いてpCRIIベクターでクローニングした。DNA配列決定は、DTCSクイックスタート(Quick Startt)キット及びCEQ8000ジネティックアナリシスキット(Beckman Coulter, Fullerton, CA)を製造業者の指示にしたがって用いて実施した。
配列の解析
BlastX(Altschul et al., 1997; Nucleic Acids Res., 25, 3389-402)を用いてGenBankのタンパク質データベースでヌクレオチド配列を検索し、PTHとしてのそれらの推定的同一性を確認した。ClustalWバージョン1.82(Chenna et al., 2003; Nucleic Acids Res., 31, 3497-500)によって多重配列アラインメントを実施した。系統解析は、分子進化遺伝学解析(Molecular Evolutionary Genetics Analysis(MEGA))パッケージ、バージョン3.0(Kumar et al., 2004; Brief Bioinform., 5, 150-63)で実行される近隣結合法(NJ)を用い、アラインメントギャップの操作には、距離計算のためのポアソン修正モデルを用いペア毎に欠失オプションを実施した。
PTHのクローニング及び配列解析
クロクマゲノムDNA由来のPTHのPCR使用クローニングのために種々のプライマーの組合せを用いた。予備的クローンの配列決定結果に基づいて、開始コドンと一致する遺伝子特異的センスプライマーを設計し、停止コドンを含む縮退アンチセンスプライマーと一緒に用いてPTHの完全なコード領域を増幅した。配列確認用クローンを作製するために、停止コドンをカバーする第二のアンチセンスプライマーを設計した。
配列アッセンブリーによって、115アミノ酸の前駆体PTHタンパク質(25アミノ酸のシグナルペプチド及び6アミノ酸のプロペプチドを含む)が明らかになった。推定される成熟タンパク質は84アミノ酸であり、計算分子量9.471ダルトン及びpl 8.1を有する。クロクマPTHは他の哺乳動物のPTHと84−95%の配列類似性を共有し、イヌのPTHともっとも類似する(91%同一性、95%類似性)(図5)。興味深いことに、成熟ホルモンの2つのアミノ酸残基、41及び52はクロクマPTHに固有である。
【実施例2】
【0032】
実施例2:冬眠時の骨吸収及び骨形成マーカーレベル
血清サンプル
捕獲熊研究施設に収容されている5頭のクロクマ(ウルスス・アメリカヌス)から血液を収集した。全てのクマ管理プロトコル(#98-069-F&WS)がヴァージニアポリテクニックインスチチュート(Virginia Polytechnic Institute)及び州立大学の動物管理委員会 (State University Animal Care Committee)によって承認された。ケタミン(100mg/mL)とキシラジン(100mg/mL)の2:1の混合物でクマを麻酔した(投薬量は体重45.5kg当たり前記混合物1ccであった)。体温は冬期収集時には4℃から6℃低く、クマは冬眠状態であることが確認された。冬眠巣穴には尿も糞もなかった。いずれの取扱い手順中にもストレスを示す行動は観察されなかった。クマを麻酔している間に血液を大腿静脈から採集し、サンプルは氷詰め冷却容器で研究室に搬送した。研究室に戻ったら直ちに血液を遠心し、血清を単離しこの血清を華氏-20°で凍結した。血液サンプルは10月の第一日から5月の最終日まで10日毎に各クマから収集した。冬眠は1月の初めに開始し、4月初めに終了した。したがって、収集日は冬眠前活動期、不使用冬眠期及び冬眠後活動的再可動期を包含していた。
クロクマオステオカルシン精製及びRIA手順
クロクマの皮質を砕いて小断片にし、ヘキサン3部及びイソプロパノール2部の混合物で脱脂して凍結乾燥した。前記乾燥骨を液体窒素下で微細粉末に粉砕し、文献(Hauschka et al. (1989), Physiol. Rev., 69, 990-1047)に記載されたようにオステオカルシンを可溶化した。得られたEDTA抽出物から、Colomboらの方法(1993, J. Bone Miner. Res., 8, 733-43)を改変してオステオカルシンを精製した。簡単に記せば、粗EDTA溶液を2倍に希釈し、10gのセプラライト(Sepralyte)C18粒子(Analytichem International, Harbor City, CA)を含むバルクカラムを通過させた。前記カラムは先にメタノールで活性化し、水に0.1%のトリフルオロ酢酸(0.1% TFA)で平衡化してあった。UV吸収が基準値に下がるまで、0.1% TFAによる十分な洗浄に続いて30%メタノール/0.1% TFAで洗浄した。オステオカルシンは、80%メタノール/0.1% TFAで溶出させた。メタノールを送風下で蒸発させ、残留溶液を凍結乾燥した。得られた乾燥タンパク質を0.05Mトリス緩衝液(pH8.0)に懸濁し、先にサンプル緩衝液で平衡化した5mLのバイオラドエコノ-Q(Biorad Econo-Q)カラムに適用した。前記カラムを、0.5Mトリス(pH8.0)中の0.1から0.6MのNaCl勾配で展開した。オステオカルシンは対称性ピーク(カラムから溶出する最後のピーク)として溶出した。このピークがオステオカルシンそのものであることは、分画のアリコットをジアゾベンゼンスルホン酸と反応させ、オステオカルシン含有分画でピーク高に一致する強度の桃色を得ることによって立証した。C18及びEcono-Qカラムはともに新しいもので、他の種に由来するタンパク質に曝露されたことはなかった。他の種に関する以前の経験から、最終オステオカルシンピークは純度が99%を超えると推定された。最終溶出物中のクロクマオステオカルシンの濃度はBCA試薬(Pierce Chemical, Rockford, IL)を用いて決定した。
生化学的解析
PTH、25-OH D、レプチン及びオステオカルシン(骨形成マーカー)について、RIA及びELISAにより血清をアッセイした。
高度に精製したクロクマオステオカルシン及びクロクマ血清を放射能免疫アッセイによって調べた。抗体はモルモット抗ラットオステオカルシンであり、トレーサーは125I標識ラットオステオカルシンであった。ラットオステオカルシン標準物(Biomedical Technologies, Inc, Stoughton, MA)及び精製クロクマオステオカルシンの両方の用量希釈をアッセイに加えた。各アッセイ試験管当たり10μLのクロクマ血清アリコットをデュープリケートでアッセイし、全てのサンプルを同時にアッセイした。前記デュープリケートは5%未満で変動した。
不使用時における形成マーカー及び吸収マーカーの両方の変化を観察するために、5頭のクロクマのオステオカルシン平均値を冬眠期間中の各時点について計算した。これらの値を冬眠期間中の最大オステオカルシン値によって標準化した。同様な計算をPICP(骨形成マーカー)及びICPT(骨吸収マーカー)の測定についても実施した。吸収及び形成マーカーの標準化値を同じグラフにプロットし、不使用時の骨吸収及び骨形成における変化の一次的及び相対的規模を判定した。
上記のようにして入手した冬眠中のクマの血清サンプルを用い、イオン化カルシウム濃度をイオン選択電極(Bayer Rapidlab 865, Leverkusen, Germany)により測定した。
上記のようにして入手した血清サンプルを用い、PTHをELISAキット(Immutopics International, San Clemente, CA)によりアッセイした(変動のアッセイ内係数は4.7%であった)。25-OH DをELISAキット(ALPCO Diagnostics, Windham, NH)でアッセイした(変動のアッセイ内係数は5%であった)。レプチンはRIA(Linco, St. Charles, MO)で測定した(変動のアッセイ内係数は3.4%であった)。IGF-Iは酸エタノール抽出RIA(Nichols Institute Diagnostics, San Juan Capistrano, CA)で測定した(変動のアッセイ内係数は4.3%であった)。血清オステオカルシンは上記のようにRIAにより測定した。全ての血清代謝物について、平均値(あるシーズン内の全てのクマ及び全ての時点について)を各シーズン(冬眠前、冬眠時及び冬眠後)について計算し、ANOVAによって比較した。ANOVAは、フィッシャーのPLSD検定を用いて多重平均比較のために追跡した。自然対数変換を用いてオステオカルシン、PTH、25-OH D及びIGF-Iの自由度の非不変性を修正し、ANOVAを有効にした。線形回帰を用いて、オステオカルシンとホルモンとの間の相関性を判定した。いくつかの血清サンプルの体積は全アッセイを実施するためには不十分であった(各アッセイのサンプルサイズは結果とともに示されている)。
【0033】
結果
骨吸収マーカー(ICPT)は冬眠の開始直後に増加し始めた(図3)。各データの点は5頭のクマの平均値である。10−20日後に、骨形成マーカー(オステオカルシン及びPICP)もまた増加し、冬眠期間中吸収の増加と共役し続けるようであった。これは、吸収と形成との間の1−2週間の組織学的“逆転”期間と一致する。これらのリモデリングマーカーは冬眠期間中ずっと吸収及び形成の増加傾向を示し、形成は吸収との共役及び均衡を維持するようにみえた。平均オステオカルシンレベルは、冬眠前と比較して冬眠中及び冬眠後に高かった(p<0.0001)(表1)。
冬眠時イオン化カルシウムレベルは冬眠前レベルより有意に高かった(p=0.0062)。冬眠から覚醒した後の再可動時に、イオン化カルシウムレベルは冬眠時レベルと比較して有意には増加しなかったが(p=0.37)、しかしながら冬眠前レベルより高いままであった(p=0.015)。
オステオカルシンはPTHと正の相関性を有したが(図2)、25-OH D、レプチン又はIGF-1とは相関性を示さなかった。PTHは、冬眠前(p=0.006)及び冬眠(p=0.014)シーズンよりも冬眠後シーズンで有意に高かった。冬眠前に対して冬眠時のPTH増加は有意ではなかった(p=0.35)。25-OHビタミンDはシーズン性変動を示さなかった(p=0.64)。
血清レプチンは冬眠前と比較して冬眠時に変化しなかったが、冬眠後の再可動中に有意に低かった(p<0.004)(表1)。IGF-1は冬眠前と比較して冬眠時に有意に低下し(p<0.0001)、再可動時にその最高値に達した(表1)。
表1:平均血清代謝物濃度
【0034】
【0035】
平均値は太字で、標準偏差は括弧内に、サンプルサイズは鉤括弧内に示されている。ある代謝物について、同じ上付き文字を有する値は有意(p<0.05)には相違しない。25-OH Dは有意なシーズン性相違を示さなかった。
【実施例3】
【0036】
実施例3:MC-3T3骨芽細胞によるPGE2放出はクマ血清のシーズン性変動によって影響を受ける
骨芽細胞代謝に対するクマ血清のシーズン性変動の影響を判定するために、MC-3T3細胞をクマ血清で処理し、プロスタグランジンE2(PGE2)の放出を定量した。10%ウシ胎児血清(Hyclone, Logan, UT)及び1%ペニシリン-ストレプトマイシン溶液補充アルファ最小必須培養液(Invitrogen, Carlsbad, CA)でMC-3T3細胞を5% CO2下にて37℃で24時間増殖させた。培養液を吸引し、冬眠前、冬眠時及び冬眠後に収集したクマ血清を10%含有する新しい培養液で置き換えた。細胞をさらに24時間増殖させ、続いて培養液を収集し、PGE2解析のために-20℃で凍結した。EDTA中の0.25% トリプシンを用いて細胞を培養皿から取り出し、遠心によって沈殿させ、トリパンブルーと血球計算盤を用いて定量した。
PGE2レベルはBiotrakTM PGE2競合酵素免疫アッセイ(Amersham Biosciences, Piscataway, NJ)を用いて決定した。アッセイは、全ての試験培養液サンプルの50μLを用いデュープリケートで実施した。反応は終末点決定前に1Mの硫酸を用いて停止させ、450nmでマイクロプレートリーダー(VERSAmax, Molecular Devices Corporation, Sunnyvale, CA)を用いて読み取った。前記デュープリケートの光学密度を非特異的結合について修正して平均を算出し、標準曲線と比較して各ウェルのPGE2量を決定した。これらの値を総培養液体積に対して修正し、サンプル中の細胞数によって標準化した。ANOVAを用いて3種の血清グループ間で標準化PGE2を比較した。
クマ血清を用いてin vivoで処理した骨芽細胞系細胞により放出されるPGE2の量は、冬眠前血清(p=0.058)及び冬眠時血清(p=0.014)と比較して冬眠後血清による処理で高かった(図4)。冬眠時血清で処理した細胞のPGE2放出は、冬眠前血清と比較して有意には相違しなかった(p=0.48)。PGE2放出のシーズン性変化は、血清IGF-Iのシーズン性変化と同様な傾向を示した。P-値は冬眠後の値と比較される。冬眠前及び冬眠時の値は互いに相違しなかった(p=0.48)。
【実施例4】
【0037】
実施例4:クマ血清中での培養は冬眠時のBax対Bcl-2の遺伝子発現比を低下させる
上記に記載したように2004から2005年の間に血液サンプルを4頭の雌のクロクマから取り出した。サンプル日付は冬眠前活動期、冬眠不使用期及び冬眠後再可動期を包含する。MC-3T3骨芽細胞系細胞を10%クマ血清含有培養液で24時間培養し、その後バイオラドアクアピュア(BioRad AquaPure)RNA単離キット(#732-6370, Bio-Rad Laboratories, Hercules, CA)を用いて全RNAを単離した。cDNAを作製するために、スーパースクリプトII(Superscript II)逆転写酵素(Invitrogen, Carlsbad, CA)及び0.5μgのオリゴ(dT)12-18プライマーを用い、グラディエントサーモサイクラー(Mastercycler gradient, Eppendorf, Westbury, NY)で以下のように逆転写を実施した:42℃で20分、50℃で10分及び42℃で1時間。プロアポトーシスタンパク質Bax及び抗アポトーシスタンパク質Bcl-2のためのプライマーはプライマークエスト(PrimerQuest)ソフトウェア(Integrated DNA Technologies, Coralville, IA)及びNCBI遺伝子バンク配列を用いて設計した。半定量的PCRをレッドタック(RedTaq)及び以下から成るプロトコルを用いて実施した:94℃で2分、94℃30秒、69.5℃30秒及び72℃1分のサイクル、並びに最終伸長72℃で5分。バンド強度は、イメージJ(ImageJ)ソフトウェアパッケージ(National Institutes of Health, Bethesda, MD)を用いて定量し、3種のハウスキーピング遺伝子(Gapdh、β-アクチン、シクロフィリン)の発現に対して標準化した。
フィッシャーの被保護最小有意差(Fisher's Protected Least Significant Difference)(PLSD)ポストホック検定によるANOVAを用い、3シーズン(冬眠前、冬眠時、冬眠後)についてBax対Bcl-2比を比較した。統計的有意差は得られなかったが(p=0.300)、Bax/Bcl-2比は冬眠前と比較して冬眠中は約42%低下した。統計的有意の欠如はサンプルサイズが小さいことがおそらく関係した(各シーズンについてn=2)。これらのデータは、冬眠中のクマの血清は骨芽細胞のアポトーシスを低下させる生物学的分子を含むことを提唱している。内因性PTH及び骨形成マーカーオステオカルシンは両方とも冬眠時に増加するので(Donahue et al., 2006; J. Exp. Biol., 209, 1630-8)、内因性クマPTHは冬眠時に骨芽細胞のアポトーシスの低下を引き起こし、これは続いて骨形成を増加させるということはありそうである。
【実施例5】
【0038】
実施例5:骨細胞株のcAMPに対するクロクマとヒトのPTH1-84又はそのサブフラグメントの影響の比較
完全長の組換えクロクマPTH(残基1−84)を生成し、骨細胞株(MC-3T3骨芽細胞系細胞及びMLO-Y4骨細胞系細胞)のサイクリックアデノシン一リン酸(cAMP)濃度レベルに対するその影響を精査し、組換えヒトPTH1-84を用いて得られた結果と比較する。クロクマ及びヒトのPTHのサブフラグメント(完全長(1−84)成熟タンパク質のアミノ酸残基1−34、1−36、7−84、11−84及び41−52を含むサブフラグメント)を用いて同等な実験を実施する。いくつかの実験のために、クロクマ及びヒトPTHポリペプチドを固相法により合成する。
多様な形態の組換えクロクマ及びヒトPTHポリペプチドの骨形成細胞中のcAMPレベルに対する影響を決定するために、培養骨細胞(MC-3T3及びMLO-Y4)をヒト又はクロクマPTHの完全長(すなわちアミノ酸残基1−84)ポリペプチド又は上記に列挙したサブフラグメントの1つと10又は30分接触させる。細胞を前記PTHポリペプチドと接触させた後、細胞中のcAMP濃度を下記でさらに説明する競合結合アッセイを用いて測定する。
組換えポリペプチドを用いる実験の全てのために、凍結乾燥ペプチドを1mMの酢酸中で100μMのストック濃度に再構成し、使用前に10μMの作業用ストック濃度に希釈する。
細胞培養
MC-3T3サブクローン14細胞(ATCC, CRL-2594)及びMLO-Y4(以下から入手:L.F. Bonewald, University of Missouri, Kansas City, MO)は、アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%血清(MC-3T3:10%ウシ胎児血清(FBS)、MLO-Y4:5%FBS及び5%ウシ血清)中で5% CO2下で37℃にて維持される。ここに記載した全ての手順が、各アッセイの全ての処理の組合せについてn=6でそれぞれ別個の細胞培養について繰り返される。
細胞内cAMPに対するPTH処理の影響
6ウェルプレートにMC-3T3及びMLO-Y4細胞を適切な密度で播種する(MC-3T3:50,000細胞/cm2、MLO-Y4:15,000細胞/cm2)。細胞を一晩培養して最適な集合状態にする。続いて培養液を吸引し、10%血清+ベヒクル(1mM酢酸)又は10%血清+100nMのPTH(ヒト又はクマ1-84又はそのサブフラグメント)のどちらかを含む培養液と交換する。これらの条件下で細胞を10又は30分培養する(Carter, P. H. et al., 1999, J Biol. Chem. 274(45), 31955-60; Chen, X. et al., 2002, Am. J. Physiol. Cell Physiol. 283(5), C1432-40; Schiller, P. C. et al., 1999, J. Bone Miner. Res. 14(9), 1504-12)。培養後、細胞をトリプシン処理して遠心し、溶解緩衝液に再懸濁する。前記懸濁物を10分間インキュベートして遠心し、細胞屑を分離する。前記細胞溶解物の上清を(2倍希釈に続いて)競合結合アッセイ(サイクリックAMPアッセイ#KGE002, R&D Systems, Minneapolis, MN)によりcAMP濃度についてアッセイする。
試験したポリペプチドの各々について、クロクマPTH系ポリペプチドに応答して細胞性cAMPレベルの増加が認められる。
【実施例6】
【0039】
実施例6:骨細胞株のアポトーシスに対するクロクマとヒトのPTH1-84又はそのサブフラグメントの影響の比較
完全長の組換えクロクマPTH(残基1−84)を生成し、骨細胞株(MC-3T3骨芽細胞系細胞及びMLO-Y4骨細胞系細胞)のアポトーシスに対するその影響を精査し、組換えヒトPTH1-84を用いて得られた結果と比較する。クロクマ及びヒトのPTHのサブフラグメント(完全長(1−84)成熟タンパク質のアミノ酸残基1−34、1−36、7−84、11−84及び41−52を含むサブフラグメント)を用いて同等な実験を実施する。いくつかの実験のために、クロクマ及びヒトPTHポリペプチドを固相法により合成する。
クロクマ及びヒトPTHの骨芽細胞及び骨細胞(プロアポトーシス状態下にある)のアポトーシスを防止する相対的能力を決定するために、細胞をヒト又はクロクマPTH1-84又は上記に列挙したサブフラグメントの1つと一緒に1時間インキュベートする。その後、細胞をデキサメタゾンで6時間処理しアポトーシスを誘発する。アポトーシスは下記でさらに説明するELISAで定量する。
組換えポリペプチドを用いる実験の全てについて、凍結乾燥ペプチドを1mMの酢酸中で100μMのストック濃度に再構成し、使用前に10μMの作業用ストック濃度に希釈する。
さらに追加の実験を0.1%又は10%FBSを用いMC-3T3細胞で実施する。通常量(10%)未満(0.1%)のFBSを用いた追加の実験を実施し、その結果を解析して、通常の血清レベル又は低い血清レベルで実施した実験間で有意に異なる応答が存在するか否かを決定する。本実施例で報告する実験では、結果は使用するFBS量によって影響を受けない。アポトーシス予防実験は、試験したポリペプチドの各々がMC-3T3細胞のアポトーシスを低下又は防止することを示している。
細胞培養
MC-3T3サブクローン14細胞(ATCC, CRL-2594)及びMLO-Y4(以下から入手:L.F. Bonewald, University of Missouri, Kansas City, MO)は、アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%血清(MC-3T3:10%ウシ胎児血清(FBS)、MLO-Y4:5%FBS及び5%ウシ血清)中で5% CO2下にて37℃で維持される。ここに記載した全ての手順が、各アッセイの全ての処理の組合せについてn=6でそれぞれ別個の細胞培養について繰り返される。
アポトーシスに対するPTH処理の影響
6ウェルプレートにMC-3T3を50,000細胞/cm2で播種し、さらにMLO-Y4細胞を15,000細胞/cm2で播種し、細胞を一晩培養して最適な集合状態にする。培養液を吸引し、10%血清+ベヒクル(1mM酢酸)又は10%血清+100nMのPTH(ヒト又はクマ1-84又はそのサブフラグメント)のどちらかを含む培養液と交換する。1時間インキュベートした後(Jilka et al., 1999; J. Clin. Invest., 104, 439-46)、10μMのデキサメタゾン又はそのベヒクル(DMSO)を各ウェルに添加し、細胞を6時間インキュベートする(Bellido, T. et al., 2003, J. Biol. Chem. 278(50), 50259-72.; Jilka et al., 1999, J. Clin. Invest., 104, 439-46)。PTHによるアポトーシスの抑制は自己限定性であるので(Bellido et al., 2003)、アポトーシス誘発中はPTHポリペプチド及びベヒクルはそのままにしておく。6時間後、細胞をトリプシン処理して遠心し、再懸濁して血球計算盤を用いて数える。前記懸濁物から50,000細胞を取り出し溶解緩衝液に入れる。前記溶解物の上清を(遠心した後)解析のために取り出し-20℃で保存する。
アポトーシスはELISA(Cell Death Detection ELISA, #1544675, Roche Applied Science, Indianapolis, IN)により前記溶解物上清から定量する。このアッセイは、細胞溶解物の細胞質分画中の断片化された細胞DNA由来のモノ及びオリゴヌクレオソームを検出し、したがって初期及び中期アポトーシスの良好な測定値を提供する。簡単に記せば、サンプルを緩衝溶液で希釈し、抗ヒストンマウスモノクローナル(クローンH11-4)抗体で被覆したマイクロプレートのウェルに加える。ベヒクル処理細胞の溶解物上清は陰性コントロールとして供する。ペルオキシダーゼ結合抗DNAマウスモノクローナル(クローンMCA-33)抗体を添加した後、光学密度を測定し、各サンプルのアポトーシス量をその対応する陰性コントロールに比して決定する。全てのサンプルをデュープリケートでアッセイする。
試験したポリペプチドの各々が細胞(プロアポトーシス状態下にある)のアポトーシスを低下させた。
【実施例7】
【0040】
実施例7:骨細胞株における遺伝子発現に対するクロクマとヒトのPTH1-84又はそのサブフラグメントの影響の比較
完全長の組換えクロクマPTH(残基1−84)を生成し、骨細胞株(MC-3T3骨芽細胞系細胞及びMLO-Y4骨細胞系細胞)の遺伝子発現レベルに対するその影響を精査し、組換えヒトPTH1-84を用いて得られた結果と比較する。クロクマ及びヒトのPTHのサブフラグメント(完全長(1−84)成熟タンパク質のアミノ酸残基1−34、1−36、7−84、11−84及び41−52を含むサブフラグメント)を用いて同等な実験を実施する。いくつかの実験のために、クロクマ及びヒトPTHポリペプチドを固相法により合成する。
骨基質の調節、転写調節、抗アポトーシス(Bcl-2)遺伝子及びプロアポトーシス遺伝子Baxに対するクロクマ及びヒトPTHの影響を決定するために、細胞をヒト又はクマPTH1-84又はサブフラグメントと1又は3時間培養する。遺伝子発現はリアルタイムPCRにより定量する。
組換えポリペプチドを用いる実験の全てについて、凍結乾燥ペプチドを1mMの酢酸中で100μMのストック濃度に再構成し、使用前に10μMの作業用ストック濃度に希釈する。
さらに追加の実験を0.1%又は10%FBSを用いMC-3T3細胞で実施する。通常量(10%)未満(0.1%)のFBSを用いた追加の実験を実施し、その結果を解析して、通常の血清レベル又は低い血清レベルで実施した実験間で有意に異なる応答が存在するか否かを決定する。本実施例で報告する実験では、結果は使用するFBS量によって影響を受けない。リアルタイムPCRを用いて、ポリペプチド添加後1時間及び3時間の時点の遺伝子発現レベルを判定し、クロクマPTH1-34はMC-3T3細胞で遺伝子発現をアップレギュレートすることを示す。
細胞培養
MC-3T3サブクローン14細胞(ATCC, CRL-2594)及びMLO-Y4(以下から入手:L.F. Bonewald, University of Missouri, Kansas City, MO)は、アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%血清(MC-3T3:10%ウシ胎児血清(FBS)、MLO-Y4:5%FBS及び5%ウシ血清)中で5% CO2下にて37℃で維持する。ここに記載した全ての手順が、各アッセイの全ての処理の組合せについてn=6でそれぞれ別個の細胞培養について繰り返される。
遺伝子発現に対するPTH処理の影響
6ウェルプレートにMC-3T3を50,000細胞/cm2の密度で播種し、さらにMLO-Y4細胞を15,000細胞/cm2の密度で播種し、細胞を一晩培養して最適な集合状態にする。培養液を吸引し、10%血清+ベヒクル(1mM酢酸)又は10%血清+100nMのPTH(ヒト又はクマ1-84又は上記に列挙したサブフラグメント)のどちらかを含む培養液と交換する。細胞をこれらの条件下で1又は3時間培養する(前記時間はc-fos及びオステオカルシンのPTH誘発アップレギュレーションと一致する)(Jiang et al., 2004, J. Biol. Chem., 279, 5329-37; Chen et al., 2002)。SV全RNA単離系(SV Total RNA Isolation System, Promega, Madison, WI)を用いて全RNAを単離する。
cDNAを生成するための逆転写は、スーパースクリプトII(Superscript II)逆転写酵素(Invitrogen, Carlsbad, CA)及び0.5μgのオリゴ(dT)12-18プライマーを用い、グラディエントサーモサイクラー(Mastercycler gradient, Eppendorf, Westbury, NY)で以下のように実施する:42℃で20分、50℃で10分及び42℃で1時間。問題の全ての遺伝子(オステオカルシン、オステオポンチン、I型コラーゲン、c-fos、Runx2、Bax、Bcl-2、SOST)及びハウスキーピング遺伝子(Gapdh、β-アクチン、シクロフィリン)ためのプライマーは、プライマークエスト(PrimerQuest)ソフトウェア(Integrated DNA Technologies, Coralville, IA)及びNCBI遺伝子バンク配列を用いて設計し、PCR条件はMC-3T3及びMLO-Y4細胞由来のRNAを用いて最適化する。リアルタイムPCRは、Mx3000PリアルタイムPCR系(Stratagene, La Jolla, CA)を用いて実施する。プロトコルは、95℃で10分のホットスタート、続いて95℃30秒(変性)、69℃1分(アニーリング)及び72℃1分(伸長)の40サイクルを含む。このプロトコルの例外はc-fosであり、前記は66℃のアニーリング温度を有する。それぞれ25μLの反応物は以下を含む:1x AbsoluteTM qPCR SYBR(商標)グリーンミックス(ABgene, Rochester, NY)、0.1 μMのフォワード及びリバースプライマー、並びに2.5 ngの全RNA相当cDNA鋳型。遺伝子発現は、3種のハウスキーピング遺伝子の幾何平均に対して標準化した相対標準曲線法を用いて決定する。全てのサンプルをデュープリケートで測定し、10%を超える変動係数(CV)を有するサンプルはいずれも解析する。
前記ポリペプチドは、骨基質、転写調節及び転写活性化因子遺伝子のアップレギュレーション、並びにBax/Bcl-2発現比の低下を引き起こす。
【実施例8】
【0041】
実施例8:異なるシーズンのクロクマ血清の骨細胞アポトーシス及び遺伝子発現に対する影響の比較、並びにPTH及びオステオカルシンの血清レベルとの相関性
ヴァージニアテックセンターフォーベアリサーチ(Virginia Tech Center for Bear Research)に2004年から2005年に収容された少なくとも3頭の別個の雌のクロクマ(ウルスス・アメリカヌス)から血液サンプルを収集する。さらに別のクロクマからその後の年に血清を収集する。ヴァージニアポリテクニックインスティチュート(Virginia Polytechnic Institute)及び州立大学の動物管理委員会 ( State University Animal Care Committee)は全てのクマ管理プロトコル(#98-069-F&WS)を承認した。ケタミン(100mg/mL)とキシラジン(100mg/mL)の2:1の混合物でクマを麻酔する(投薬量は体重45.5kg当たり前記混合物1ccである)。クマを麻酔している間に血液サンプルを大腿静脈から採集し、サンプルは氷詰め冷却容器で研究室に搬送する。研究室に戻されたら直ちに血液を遠心して血清を単離し、この血清を-20℃で凍結する。血液サンプルは10月の初めから5月の終わりまで10日毎に各クマから収集する。冬眠は1月の初めに始まり、4月初めに終わる。したがって、収集日は冬眠前活動期、不使用冬眠期及び活動的な冬眠後再可動期を包含している。
クマ血清の10μLアリコットをオステオカルシン濃度について放射能免疫アッセイによりデュープリケートでアッセイする(Patterson-Allen et al., 1982; Anal. Biochem., 120, 1-7)。このアッセイは以前にクマについて有効性が立証された(Donahue et al., 2006; J. Exp. Biol., 209, 1630-8)。抗体はモルモット抗ラットオステオカルシンであり、トレーサーは125I標識ラットオステオカルシンである。クマ血清の100μLアリコットをPTH濃度について(Donahue et al., 2006; J. Exp. Biol., 209, 1630-8)デュープリケートでELISA(Porcine Intact PTH ELISA Kit, #60-3305, Immutopics, Inc., San Clemente, CA)によりアッセイする。このアッセイはPTHの39−84領域と結合し、PTH濃度を比色により提示するためにPTHの13−34領域を必要とする。したがって、前記アッセイは、C-末端サブフラグメント7−84及び11−84と同様に無傷の(1−84)PTHについて良好な測定を提供する。このELISAはクマPTHと交差反応し(Donahue et al., 2006; J. Exp. Biol., 209, 1630-8)、さらにヒトPTHとは100%の交差反応性を有することが示された。このアッセイがクロクマについて有効であることを立証するために、10nMの組換えクロクマ又はヒトPTH1-84を含む培養液サンプルをデュープリケートでアッセイする。PTHサンプルの既知濃度をアッセイの標準曲線から決定した測定濃度と比較する。これらのサンプルから決定される交差反応性におけるいっさいの潜在的な差を、クロクマ血清サンプルの内因性クロクマPTH濃度についての修正として用いる。
培養骨タイプ細胞によるクロクマPTHアポトーシス及び遺伝子発現細胞培養実験のために上記に記載した方法が繰り返されるが、ただし100nM組換えPTH含有培養液を10%クロクマ血清(冬眠前、冬眠時又は冬眠後の期間に由来)含有培養液に置き換える。血清体積は上記に記載したPTH ELISAの後で計算する。
PTHは冬眠前血清よりも冬眠時及び冬眠後血清に多いので、冬眠時及び冬眠後シーズン血清は、冬眠前血清と比較してより強いアポトーシス予防を引き起こす。内因性血清PTH濃度は、より高い血清PTHレベルはより低いアポトーシス率と一致するという点でアポトーシスレベルと反比例する。すなわち、血清PTH濃度はアポトーシスレベルと負の相関性を有する。
【実施例9】
【0042】
実施例9:クマPTHのin vivo試験
クロクマPTH、完全長(1-84)又はそのいくつかの機能的サブフラグメント(1-34、1-36、7-84、11-84、41-52)の1つを骨細胞の同化作用促進刺激についてin vivoで試験し、対応するヒトPTH又はサブフラグメントと比較する。PTHポリペプチドの各々を合成し、皮下注射のために医薬的に適切な担体に懸濁する。クロクマ又はヒト由来の完全長PTH又はその機能的フラグメントを、体重1kg当たり40μgの用量で毎日7週間投与する。クロクマPTH又はその機能的フラグメントは、骨強度、骨量及び骨塩含有量において対応するヒトPTHポリペプチドよりも大きな増加を引き起こす。
【実施例10】
【0043】
実施例10:骨減少を低下させるか又は妨げる予防薬としてのクマPTHの使用
骨粗鬆症は、後肢懸濁物によって(骨粗鬆症の不使用モデル)及び卵巣摘出によって(骨粗鬆症の閉経後モデル)マウスで誘発される。後肢懸濁物の開始後又は卵巣摘出完了後、マウスに通常の用量のクロクマPTH又はその機能的フラグメントを与える。クロクマPTH又はその機能的フラグメントで処置されたマウスは骨減少が未処置マウスより少ない。
【実施例11】
【0044】
実施例11:クマ及びヒトPTH1-34はともにオステオカルシンの遺伝子発現をアップレギュレートするが、クマPTH1-34のみがBax/Bcl-2の発現レベル比を低下させる
MC-3T3細胞をベヒクル又は100nMの合成クマもしくはヒトPTH1-34中で3又は6時間インキュベートした(n=2又は4)。全RNAを単離し、逆転写によりcDNAを生成する。骨基質タンパク質のI型コラーゲン及びオステオカルシン、プロアポトーシスタンパク質Bax、抗アポトーシスタンパク質Bcl-2、並びにハウスキーピング遺伝子のGapdh、β-アクチン及びシクロフィリンのためのプライマーをプライマークエストソフトウェア(Integrated DNA Technologies, Coralville, IA)を用いて設計した。リアルタイムPCRは、Mx3000PリアルタイムPCR系(Stratagene, La Jolla, CA)を用いて実施した。全てのサンプルをデュープリケートで測定した。遺伝子発現は、3種のハウスキーピング遺伝子(Gapdh、β-アクチン及びシクロフィリン)の幾何平均に対して標準化した相対標準曲線法を用いて決定した。アポトーシス関連遺伝子はBax/Bcl-2の発現比として解析した(なぜならばこの比の低下はin vitroでのアポトーシスの低下と密接に関係するからである)。
ヒト又はクマPTH1-34中での培養は、ベヒクルコントロールと比較してI型コラーゲンの発現には影響を与えなかったが、オステオカルシンの発現を実質的にアップレギュレートした(図6)。ヒトPTHとクマPTHとの間には有意な相違は存在しなかった(p>0.09)。
クマPTH中での3時間の培養はBac/Bcl-2の発現比を低下させアポトーシスの低下が推量されたが、ヒトPTH中での3時間の培養は前記の比を増加させアポトーシスの増加が推量された(図7)。クマPTHとヒトPTHとの間の相違は統計的に有意であった(p=0.047)。
【実施例12】
【0045】
実施例12:in vivoラットOVX用量応答実験
90匹の3ヶ月齢雌Sprague-Dawleyラットを入手した。これらラットの80匹に卵巣摘出(OVX)を実施し、10匹に模擬OVX(卵巣は前記動物に残留)を施した。前記OVX動物を更なる実験のために無作為に10匹ずつのグループに割り当てた。
実際の又は模擬OVX術に続いて、全動物を6週間処置せず骨を減少させた。6週間の骨減少期間の後、動物のグループに6週間又は8週間の間(1週間に5日間、毎日)ベヒクル(食塩水)又はクマもしくはヒトPTH1-34の3、10又は30μg/kgを注射した。ヒトに対する従来のPTH1-34の用量は約0.3μg/kgで、一方5μg/kgの用量はラットでは発癌性であった(Tashjian et al., J. Bone Mineral Res. (2008) 23(6): 803-811)。模擬OVXグループ及び実際のOVXラットの1つのグループを6週間で安楽死させ、一方残余の動物はOVX術の後14週で安楽死させた。更なる解析のために安楽死させた動物からサンプルを入手した。
コンピュータ支援マイクロ断層撮影法(μCT)を用いて骨の三次元画像を入手した。続いて前記三次元画像を用いて下記に説明するように骨パラメーターを定量した。測定は、成長板から2mmの位置で始まる1.6mmの厚さで実施した(図8)。骨サンプルは、術後6週間で安楽死させた模擬OVX(図9A、“模擬OVX6週”)及び実際のOVX(図9B、“OVX6週”)とともに術後14週間で安楽死させた実際のOVXラットグループ(図9C、“OVX14週”)から入手し解析した。前記OVXの14グループのラットは、6週間の骨減少期間に続いてベヒクル(食塩水)の注射を8週間与えられ、並行してPTH1-34の注射も8週間与えられた(図9D、9E及び9F)。図9D、9E及び9Fに示すサンプルは、6週間の骨減少期間の後、クロクマPTH1-34(“bbPTH”)を8週間それぞれ3、10又は30μg/kg注射されたラットから入手された。
画像から分かるように、OVX外科手術後6週間(図9B)及び14週間(図9C)のラットの骨は、模擬OVX(図9A)及びbbPTH処理ラット(図9D−9F)の骨と比較して顕著な量の骨減少を示した。クロクマPTH1-34で処置したラットは、ベヒクル(食塩水)のみの注射を与えられたOVXラット(図9C)と比較して、各投薬レベルにおいて用量依存性の骨量増加(図9D−9F)を示した。
模擬OVX、OVX及びPTH処置ラット間の骨パラメーターの相違を定量した。図10は、種々の処理グループについて総体積の比(BV/TV)としての骨体積を示す。卵巣存在下の骨体積比の基準線レベル(“模擬基準線”淡色ひし形)及び卵巣摘出後6週間でPTH処置の非存在下の基準線レベル(“OVX基準線”濃色ひし形)は、“6週”と標識した列の左側に示されている。OVXラットの14週間のベヒクル(食塩水)単独処置(“OVXベヒクル”、三角)は骨体積比の更なる下降をもたらした。OVXのヒト(“hPTH1-34”、四角)又はクロクマ(“bbPTH1-34”、丸)PTH1-34処置は、用量依存態様で骨体積比を増加させた。最高用量の30μg/kgでは、ヒトPTHによる処置は、クマPTH又はベヒクル(食塩水)単独による処置よりも有意に(p<0.05)高い骨体積をもたらした。ヒトPTHによる処置は、試験した3レベルの全てでベヒクル(食塩水)単独処置よりも有意に(p<0.05)高い骨体積比をもたらし、一方、クマPTHは、10μg/kg及び30μg/kgでベヒクル(食塩水)単独処置よりも有意に(高い骨体積比をもたらしたp<0.05)。
同様に、見掛けの骨塩密度をサンプルの各々について決定し(図11)、1 cm3当たりのヒドロキシアパタイトのmg(“App.Dn(mgHA/ccm)”)として示されている。見掛けの骨塩密度は、ベヒクル(食塩水)単独処置OVXラットの骨の場合よりも3、10又は30μg/kgのhPTH又はbbPTHで処置されたOVXラットの骨について有意に高かった(p<0.05)。30μg/kgのhPTHで処置されたOVXラットの見掛けの骨塩密度は、30μg/kgのbbPTHで処置されたOVXラットよりも有意に高かった(p<0.05)。
骨梁支柱骨数及び厚さもまた、模擬OVX、PTH処置無しOVX、及びPTH処置OVXラットの骨について決定した(図12)。記号は、模擬OVXラット(“模擬基準線”、淡色ひし形)、ベヒクル(食塩水)単独処置及び6週安楽死のOVXラット(“OVX基準線”、濃色ひし形)、ベヒクル(食塩水)単独8週間処置及び14週安楽死のOVXラット(“OVX基準線”、濃色ひし形)、及び3、10又は30μg/kgヒト(“hTPH1-34”、四角)又はクマ(“bbPTH1-34”、丸)PTH1-34処置OVXラットについて1ミリメートル当たりの骨梁数(“Tb.N(mm^-1)”)を示す(図12)。
骨梁支柱骨の平均の厚さもまた決定した(図13)。クマPTH又はヒトPTHによる処置は、ベヒクル処置コントロールと比較して、骨梁支柱骨の平均の厚さ(“Tb.Th(mm)”)を有意に増加させた(p<0.05)。
クロクマTPH1-34は、ヒトPTH1-34と比較して用量増加による骨梁骨のより強い鉱化傾向を示した(“M.Dn(mgHA/ccm)”;図14)。OVXラットのクマ(“bbTPH1-34”、丸)又はヒト(“hTPH1-34”、四角)PTH1-34による処置は、ベヒクル(食塩水)単独処置OVXラットよりも有意に(p<0.05)高い骨梁骨鉱化をもたらした。
ヒトPTH1-34(“hTPH”、四角)は、脛骨皮質体積(“BV(mm^2)”、図15)の増加でクマPTH1-34(“bPTH”、丸)よりも優れている。しかしながら全体的には、骨皮質領域の変化は骨の骨梁(bone trabeculae)で観察される変化と比較して軽度であった。
OVXラットの骨の脛骨皮質多孔度は、中等度用量(10μg/kg)及び高用量(30μg/kg)のクマ(“bbTPH1-34”、丸)又はヒト(“hTPH1-34”、四角)PTH1-34でラットを処置したとき、ベヒクル(食塩水)単独処置OVXラットの骨と比較して有意に(p<0.05)低下するように見えた(図16)。しかしながら高用量(30μg/kg)のヒトPTH1-34又はクマPTH1-34は、ベヒクル処置コントロールと比較して有意な(p<0.05)大腿骨皮質強度の増加をもたらした(図17)。
クマPTH1-34がヒトPTH1-34を超えて提供し得るさらに別の利点はより低い血清カルシウムである。高カルシウム血症はヒトでのPTH処置における副作用である。本実験では、血清カルシウムレベルは、ヒトPTH1-34と比較してクマPTH1-34処置ラットで低いように見えるが、ただし結果はp<0.05レベルで有意ではない。しかしながら、統計解析は、サンプルサイズが2倍であればクロクマPTH1-34処置ラットで血清カルシウム濃度は有意に低い(p<0.05)であろうと予想する。したがって、クロクマPTH1-34処置は前記副作用発生率を低下させ得る。
これらの実験は、クロクマPTH1-34は骨梁骨形成の劇的な増加をもたらすという点でヒトPTH1-34と等しく有効であることを示している。ヒトPTH1-34は、最高用量(30μg/kg)でいくつかのパラメーターについてクマPTH1-34よりも有効であることが観察されたが、これはヒトに投与される用量よりも100倍高い。しかしながら、より低くかつ臨床的に対応する用量で、クマPTH1-34は、骨折蓋然性の低下と関係する重要なパラメーターを増加させるという点でヒトPTH1-34と同じように有効であった。
【実施例13】
【0046】
実施例13:クマPTH1-34及びクマPTH1-84は培養マウス骨芽細胞でカスパーゼ-3活性を低下させる
細胞培養
MC-3T3サブクローン4細胞(ATCC, CRL-2594)は標準培養液(89%アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%FBS)中で5% CO2下にて37℃で維持した。細胞は30継代を超えて用いられることはなかった。
血清枯渇により誘発されたアポトーシスのPTHによる減弱
MC3T3細胞を96ウェルプレートに20,000細胞/ウェルの密度で播種し、一晩定着させた。続いて培養液を吸引し、10%FBS及びベヒクル(PBS)又はPTH(10−100nMのどちらかを含む培養液100μLで1時間置き換えた。その後、培養液を吸引し細胞をPBSで洗浄して残留血清を除去した。アポトーシス誘発のために、細胞をコントロール培養液又は無血清培養液で6時間インキュベートした。交換培養液は、89%アルファ最小必須培養液、1%ペニシリン-ストレプトマイシン及び10%FBS(コントロール培養液)又は0%FBS(枯渇培養液)のどちらかを含んでいた。枯渇後、培養液を吸引し、細胞をPBSで洗浄し、50μLの新しいPBSを各ウェルに添加した。カスパーゼ3/7活性はDEVD基質の切断によって決定した(カスパーゼ-Glo 3/7アッセイ、#G8093, Promega Corporation)。既知濃度の組換えヒトカスパーゼ-3(#SE-169, BIOMOL Research Laboratories Inc.)を用いて作成した標準曲線に測定したルミネセンス値を適合させた。各96ウェルプレートにつき5ウェルを各処置で用い、手順を最小限10プレートで繰り返した。
統計解析
フィッシャーのPLSDとともにANOVAを用い、反復実験によってブロック化しながら、ペプチド、ベヒクル及びコントロール処置サンプル間でカスパーゼ3/7活性化の減弱を比較した。カスパーゼ3/7活性は、各反復実験内でコントロール培養液(非枯渇細胞)のカスパーゼ3/7レベルに対して標準化し、共分散因子として濃度を処理しながら共分散解析(ANCOVA)を用いてペプチドの用量応答反応態様を比較した。
血清枯渇は、この実験の非枯渇細胞と比較してベヒクル及びPTH前処理サンプルで、カスパーゼ3/7活性(アポトーシスの初期インジケーター)を有意に(p<0.0001)増加させた(図19)。全てのPTH前処理はベヒクル処理細胞と比較してカスパーゼ3/7活性を有意に(p<0.0001)低下させた。カスパーゼ3/7活性のPTH誘発低下は用量依存態様を示さなかったので(p=0.890)、全ての濃度をグループ内比較のためにグループ内でプールした。4ペプチドグループのうちで、bbPTH1-34は血清枯渇誘発アポトーシスの最大の減弱を示した。bbPTH1-34グループの平均カスパーゼ3/7活性は、hPTH1-34グループ(p=0.006)及びいずれのPTH1-84(p≦0.018)よりも有意に低かった(図19)。対照的に、bbPTH1-84とhPTH1-84グループとの間のカスパーゼ3/7活性の相違は有意に近く(p=0.090)、hPTH1-84がより低レベルのアポトーシスを示した(図19)。
【実施例14】
【0047】
実施例14:雄Swiss-WebsterマウスにおけるクマPTH1-34及びクマPTH1-84のin vivo作用
動物
全ての取扱い方法及び実験は、ミシガン工科大学(Michigan Technological University)院内動物管理使用委員会によって承認された。雄のスイス-ウェブスター(Swiss-Webster)マウスを繁殖業者(Harlan, Indianapolis, IN)から入手した。全ての動物を12時間照明/12時間消灯サイクルで維持し、水及び0.95%カルシウム含有餌(Purina Rodent Diet #5001)に自由に接近させた。
第一の実験では、11週齢マウスに毎日(5日/週)2.5nmol/kgのPTH1-34(hPTH1-34又はbbPTH1-34)又は酸性食塩水ベヒクル(0.15M NaCl+0.001N HCl)の皮下注射を8週間実施した(n=10マウス/グループ)。第二の実験では、12週齢マウスに毎日(5日/週)2.5nmol/kgのPTH1-84(hPTH1-84又はbbPTH1-84)又は酸性食塩水ベヒクルの皮下注射を8週間実施した(n=10マウス/グループ)。全てのPTH及びベヒクル処理動物に10mg/kgの投薬量でカルセインの皮下注射を致死12日及び3日前に実施し、骨表面の鉱化を標識した。最後の処置注射から24時間後に、マウスを二酸化炭素で窒息死させた。右大腿骨を10%の中性緩衝ホルマリンで固定した。左大腿骨は0.15M食塩水を浸み込ませたガーゼで包み-20℃で保存した。左脛骨は70%エタノール中にて4℃で保存した。
近位脛骨の骨梁骨の構造及び鉱化
コンピュータ支援微小断層撮影を用いて、左近位脛骨の骨幹端(成長板から0.7mm遠位)における骨梁骨の構造及び鉱化を評価した。実験1の脛骨をμCT35スキャナー(Scanco Medical AG, Basserdorf, Switzerland)により3.5μmの(高)解像でスキャンした。実験2の脛骨はμCT40スキャナー(Scanco Medical AG, Basserdorf, Switzerland)により7μmの(高)解像でスキャンした。骨は70%エタノール中でスキャンした。骨梁骨の体積比(BV/TV、%)、骨梁数(Tb.N, mm-1)、骨梁の厚さ(Tb.Th, mm)、骨梁の分離(Tb.Sp, mm)、骨梁の見掛けの骨塩密度(App.Mn.Dn, mgHA/cm3)及び骨梁組織の骨塩密度(Mat.Mn.Dn, mgHA/cm3)を、製造業者のソフトウェアを用いてコンピュータで計算した。
大腿骨皮質の機械的特性
インストロン(Instron)機械的検査系(Model #8872, Canton, MA)により1mm/分の速度にて3点曲げで左大腿骨に不履行まで負荷をかけた。前方表面が加圧されるように骨の方向を定めた。下方検査固定装置支持部は差し渡し10mm離し、全ての固定装置の接触点を丸めて(半径=1mm)、骨に局所的応力が集中するのを避けた。究極負荷は試験から得られた負荷-ひずみデータを用いて計算した。
皮質骨塩含有量、幾何学的特性及び動的な組織形態計測
曲げ試験に続いて、左大腿骨骨幹を窯炉で完全燃焼させて、骨塩含有量(灰分比)を決定した。右大腿骨骨幹をメチルメタクリレートに包埋し、ダイヤモンド刀で輪切りにして骨幹中央断面を露出させた。デジタルカメラ(Spot Insight QE, Diagnostic Instruments Inc., Sterling Heights, Michigan)を用いて骨幹中央断面画像を捕捉し、シオン画像解析ソフトウェア(Scion Corporation, Frederick, Maryland)を用いて各サンプルについて骨膜面積(Ps.Ar)、皮質面積(Ct.Ar)、骨内膜面積(Es.Ar)を計算した。皮質の厚さ(Ct.Th)は、画像解析ソフトウェア(Bioquant Osteo, Nashville, TN)を用い各解剖学的四分円について及び完全断面について0.1mmの増加幅で計算した。右大腿骨の骨幹中央から得た薄い切片を厚さ50μmにすり潰し、ガラススライドにマウントした。カルセイン標識画像を100X倍率で捕捉し、各大腿骨の骨膜及び骨内膜の骨塩付加成長速度(それぞれPs.MAR及びEs.MAR)並びに表面鉱化(Ps.MS/BS及びEs.MS/MS)を定量した(Bioquant)。
in vivo実験統計解析
動物サイズにおける潜在的多様性を示すために、全ての骨の幾何学的特性、構造的特性及び骨塩特性を体重によって標準化した(31−33)。コンピュータ支援微小断層撮影法によって定量した骨梁骨の特性を、各実験(PTH1-34又はPTH1-84)においてベヒクル、クマPTH及びヒトPTH処置マウス間でANOVAによって比較した。異なるμCT装置で解析を実施したので、PTH1-34及びPTH1-84間ではμCTパラメーターの直接比較は実施しなかった。他の全ての骨特性については、6つの実験グループの全て(PTH1-34及びPTH1-84の両方について)をANOVA及びフィッシャーのPLSDにより比較した。全ての比較についてp<0.05を有意とした。
結果
近位脛骨における骨梁骨の構造及び鉱化:
骨梁骨のμCTのデータは表2及び3に提示されている。PTH1-34実験で実験グループ間に有意差はなかったが、骨梁骨の骨塩密度はbbPTH1-34で処置したマウスで最も低い傾向があった(表2)。対照的に、骨体積比は、hPTH1-84と比較してbbPTH1-84で処置したマウスの近位脛骨でより高かった(表3)。骨体積比に対するbbPTH1-84の影響とhPTH1-84の影響との間における相違は有意で(p=0.013)、bbPTHがより有効な治療方法であった。bbPTH1-84処置マウスにおける骨体積比の増加は骨梁の厚さによって影響を受けたであろう。なぜならば、骨梁の厚さはhPTH1-84で処置したマウスと比較してbbPTH1-84で処置したマウスで大きい傾向があったからである(p=0.089)。骨梁数及び骨梁分離は、PTH1-84実験のグループ間で有意な相違はなかった(表3)。骨梁の見掛けの骨塩密度は、他の処置と比べてbbPTH1-84処置マウスで最も高く、bbPTH1-84の影響ととhPTH1-84の影響との間の相違は統計的に有意であった(p=0.019)(表3)。骨梁の実質的骨塩密度はbbPTH1-84処置マウスで最も高い傾向があった(表3)。
【0048】
表1:PTH1-34実験のマウス近位脛骨における骨梁骨の構造的特性及び骨塩密度
標準偏差は括弧内に示されている。
略語の説明は本文を参照されたい。
【0049】
表2:PTH1-84実験のマウス近位脛骨における骨梁骨の構造的特性及び骨塩密度
標準偏差は括弧内に示されている。
異なる上付き記号はグループ間の有意な相違を示す(p<0.05)。
略語の説明は本文を参照されたい。
【0050】
大腿骨の皮質の機械的特性及び灰分比:
大腿骨の究極負荷はペプチド処置によって増加した(ベヒクル処置マウスと比較して+3%から+15%)(図29)。骨強度の増加はhPTH1-84と比較してbbPTH1-84処置マウスで有意に高かったが(p=0.032)、bbPTH1-34及びhPTH1-34処置マウス間では相違はなかった(p=0.819)。骨塩含有量(灰分比)は処置グループ間で相違はなかった(p=0.180)(データは示されていない)。
大腿骨の皮質の幾何学的特性:
骨の骨膜面積は、PTH1-34実験のベヒクル処置マウスでもっとも高く、PTH1-84実験のベヒクル処置マウスで最も低かった(表3)。bbPTH1-84処置マウスはベヒクル処置マウスと比較して骨膜面積について有意に大きな値を有したが(ただしhTPH1-84ではそうではない)、両PTH-1-34ペプチド処置マウスはベヒクル処置マウスと比較して骨膜面積についてより低い値を示した(表3)。皮質面積も骨内膜面積も処置グループ間で相違はなかった(p>0.136)。皮質の厚さはPTH1-34実験と比較してPTH1-84実験のマウスで大きかったが、処置グループ間で他の相違は存在しなかった(表3)。
【0051】
表3:PTH処置マウスの大腿骨骨幹中央の皮質の幾何学的特性
標準偏差は括弧内に示されている。
異なる上付き記号はグループ間の有意な相違を示す(p<0.05)。
略語の説明は本文を参照されたい。
【0052】
大腿骨における皮質の動的な組織形態計測:
処置グループ間で骨膜もしくは骨内膜の表面鉱化又は骨塩付加成長速度に有意な相違はなかった(p>0.236)(表4)。
【0053】
表4:PTH処置マウス大腿骨骨幹中央の皮質の動的な組織形態計測
標準偏差は括弧内に示されている。
【0054】
結論
これらのデータは、bbPTH1-34はin vitroでhPTH1-34及びhPTH1-84より抗アポトーシス活性が強いことを示唆しているが、bbPTH1-84はマウスで骨体積及び強度をhPTHペプチドよりも大きく増加させる。このことは、bbPTHは骨粗鬆症に対する新規な同化作用性治療薬として潜在能力を有することを提唱している。
これらの実験でhPTHペプチドはベヒクル処置と比較して骨梁骨の体積の増加をもたらさなかった(表1及び2)。これはおそらく比較的低い投薬量がマウスに投与されたからであろう。マウスにおけるPTHの同化作用に関する短期的in vivo実験では、短期間での骨形成及び骨量の大きな増加をもたらすために典型的には10nmol/kg以上の投薬量を用いる。以前の実験では、低いペプチド投薬量では短期間の骨形成性は低く、すなわち1日1nmol/kgのhPTH1-34の6週間注射は、雄のマウスの近位脛骨における骨梁の厚さ(Tb.Th)又は骨塩付加成長速度(MAR)に影響を及ぼさなかったが、より高モル濃度のhPTHの投薬(〜10nmol/kg)はこれらの特性を高めた(以下を参照されたい:Sakai et al. (1999) J Bone Miner Res 14(10):1691-9)。我々の実験では2.5 nmol/kgの投薬量を用いた。なぜならば、前記の値は、マウスで骨芽細胞数を増加させ、骨梁骨の二次性海綿質(trabecular bone secondary spongiosa)の骨芽細胞のアポトーシスを低下させることが示された最小の値であり(以下を参照されたい:Bellido et al. (2003) J Biol Chem 278(50):50259-72)、さらに他の短期in vivo実験で用いられるPTH投薬量と比較して臨床投薬量(20μg/日、70 kgの患者について約0.07 nmol/kg)により近いからである。hPTHペプチドは2.5 nmol/kgでは骨梁骨の質量を増加させなかったが、同じ投薬量のbbPTH1-84で処置したマウスは、hPTH1-84と比較して骨梁骨の特性(表2)及び皮質強度(図29)で改善を示した。骨梁の厚さはbbPTH1-84処置マウスで上昇傾向を示した(表2)。重要なことには、骨梁骨の体積比及び見掛けの骨塩密度はbbPTH1-84によってのみベヒクル処置よりも有意に上昇した(表2)。調べたペプチドアナローグの大半が大腿骨の究極負荷を増加させたが、ベヒクル処置を超える増加はbbPTH1-84についてのみ統計的に有意であった(図29)。総合すれば、これらの発見は、bbPTHはhPTHよりも低い用量で有効な骨形成療法である得るという可能性を高める。
本実験で用いた全てのPTHペプチドが、in vitro血清枯渇によって誘発したアポトーシスシグナリングを減弱させた(図19)。bbPTH1-34は、ヒトPTHよりも強く前骨芽細胞系細胞でアポトーシスシグナリングを低下させた(図19)。最近、我々は予備的実験においてヒト胎児骨芽細胞(hFOB)で同様な結果を見出した。24時間の血清枯渇はhFOBで細胞死(トリパンブルー染色により確認)を増加させたが、100nMのPTH1-34又は1-84による枯渇前1時間のhFOBの前処理はパーセント細胞死を低下させた(ベヒクル処置と比較した細胞死の百分率変化は以下の通りであった:hPTH1-34は-18%、bbPTH1-34は-25%、hTPH1-84は-17%、bbPTH1-84は-21%;p<0.0001、データは示されていない)。bbPTH1-34の抗アポトーシス作用の強化は、bbPTH1-34はin vivoでもっとも強力な骨形成薬剤であり得るであろうという期待をもたらす。しかしながら、bbPTH1-84はbbPTH1-34及びhPTH1-34よりもin vitroで抗アポトーシス活性が低いけれども(図19)、bbPTH1-84はin vivo実験で皮質及び骨梁骨の体積でもっとも大きい増加を引き起こした(表1、表2)。
in vitro実験とin vivo実験の不一致は現時点では説明がつかない。骨芽細胞のアポトーシスの低下は骨梁骨に対するhPTHの同化作用の支配的プロセスであるが、他の生物学的メカニズムがbbPTHの骨体積増加能力を調節することもできる。例えば、骨芽細胞分化の促進(骨芽細胞のアポトーシスの防止ではなく)が主要手段であり、それによってhPTHは骨膜の皮質形成を促進する。この考え方に一致して、大腿骨強度はbbPTH1-84処置マウスで最も高い(図29)。bbPTH1-84はMC3T3細胞で試験した他のペプチドアナローグよりも抗アポトーシス活性は低かったが、脛骨近位の骨体積比もまたbbPTH1-84処置マウスで最も高かった(表1)。hPTHの1‐34フラグメントは完全長のhPTHペプチドの同化作用を再現するために十分であるが、bbPTH1-84はbbPTH1-34及び両hPTHペプチドと比較してin vivoで優れた性能を発揮したという事実は、bbPTHのC末端部分におけるアミノ酸変化はレセプター結合及びレセプター活性化活動に必要とされ得ることを提唱している。
【実施例15】
【0055】
実施例15:卵巣摘出ラットに対するヒトPTH1-84及びクマPTH1-84のin vivoでの作用
繁殖業者の施設で30匹の2.5ヶ月齢雌Sprague-Dawleyラットの卵巣が摘出された(OVX)。更なる実験のためにこのOVXラットを1グループ10匹の3つのグループに無作為に割り当てた。
OVX術の6週間後に、これらラットの皮下にベヒクル(食塩水)又は2.5nmol/kgのクマもしくはヒトPTH1-84を8週間注射した(5日/週で毎日)。最後の注射の24時間後に動物を安楽死させた。前記安楽死させた動物から更なる解析のためにサンプルを入手した。
大腿骨曲げ強度及び近位脛骨の骨梁構造を実施例12に記載したように調べた。
本実験では、大腿骨の究極負荷は、ベヒクル処置と比較してbbPTH1-84処置により有意に増加したが(p=0.013)、ヒトPTH1-84では有意ではなかった(p=0.463)。これらの発見は、bbPTH1-84は、ラットのOVX誘発骨減少の回復にヒトPTH1-84より有効であることを示唆している。しかしながらPTH処置は大腿骨の標準化究極負荷に有意な影響を与えなかった(図21)。この測定は、非標準化負荷よりもより生理学的な骨強度測定である。すなわち、より大きなサンプルサイズがより大きな統計力のために提供されるならばbbPTH1-84はヒトPTH1-84より有意に強力であろう。
骨体積比はPTH処置グループで増加した(図22)。骨梁数はPTH処置によって影響を受けなかったが(図23)、ただし骨梁の厚さはPTH処置グループで増加した(図24)。骨梁分離は処置グループと未処置グループとの間で違いはなかった(図25)。見掛けの骨塩密度はPTH処置グループで増加したが(図26)、実質的骨塩密度は処置グループと未処置グループとの間で違いはなかった(図27)。要約すれば、これらのデータは、ヒト及びクロクマPTH1-84は等しく骨梁骨の体積を増加させるが(図26)、bbPTH1-84はhPTH1-84よりも皮質強度をより大きく増加させることを提唱している(図24及び25)。これらの発見は、bbPTH1-84はヒトに有効な骨粗鬆症薬である可能性を提唱している。
【実施例16】
【0056】
実施例16:冬眠クマ血清はin vitroでカスパーゼ活性を低下させる
11月の初めから5月の初めまで10日毎に6頭のクマから血清を収集した(合計して各クマから6つの活動的冬眠前サンプル(11月−12月)、9つの冬眠時サンプル(1月−3月)、及び4つの活動的冬眠後サンプル(4月−5月))。クマ血清が、アポトーシス刺激に対する骨芽細胞応答に対してシーズン性影響を有するか否かを決定するために、白色壁を有する組織培養プレートにMC3T3-E1前骨芽細胞を各ウェル当たり10,000細胞で、アルファ改変最小必須培養液(MEM(-))に10%ウシ胎児血清を添加した培養液に播種し、一晩定着させた。続いて培養液を吸引し、24時間処置のために2%のクマシーズン血清を含むMEM(-)に置き換えた。再び培養液を吸引し無血清MEM(-)に置き換え、6時間アポトーシスを誘発した。終了時点で培養液を吸引し、50μLのPBS及び50μLのルミネセンスDEVD基質(Caspase-glo 3/7, Promega, Madison, WI)に置き換えた。マイクロプレートリーダー(Synergy HT Multi-Detection Microplate Reader)(Bio-Tek, Winooski VT)でルミネセンスを定量した。図28に示すように、冬眠血清で培養した細胞のカスパーゼ3/7活性は活動期血清で培養した細胞よりも低かった(p<0.0001)。これらの発見は、循環因子(例えばホルモン)のシーズン性変化は、骨芽細胞に抗アポトーシス作用を冬眠時に提供することを示唆している。
【0057】
【0058】
【特許請求の範囲】
【請求項1】
配列番号:2の連続する少なくとも10アミノ酸残基を含む単離ポリペプチドであって、前記ポリペプチドが配列番号:2のアミノ酸残基41及び52の少なくとも1つを含む、前記単離ポリペプチド。
【請求項2】
ポリペプチドが配列番号:2を含む、請求項1に記載のポリペプチド。
【請求項3】
請求項1に記載のポリペプチドをコードする単離ポリヌクレオチド。
【請求項4】
ポリペプチドが配列番号:2を含む、請求項3に記載の単離ポリヌクレオチド。
【請求項5】
ポリヌクレオチドが配列番号:1を含む、請求項4に記載の単離ポリヌクレオチド。
【請求項6】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と骨形成細胞を接触させる工程を含む、cAMPレベルを骨形成細胞で増加させる方法。
【請求項7】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項6に記載の方法。
【請求項8】
ポリペプチドが配列番号:2を含む、請求項6に記載の方法。
【請求項9】
骨形成細胞が被験者中に存在する、請求項6に記載の方法。
【請求項10】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と骨形成細胞を接触させる工程を含む、アポトーシスを骨形成細胞において減少させる方法。
【請求項11】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項10に記載の方法。
【請求項12】
ポリペプチドが配列番号:2を含む、請求項10に記載の方法。
【請求項13】
骨形成細胞が被験者中に存在する、請求項10に記載の方法。
【請求項14】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と骨形成細胞を接触させる工程を含む、Baxタンパク質対Bcl-2タンパク質の発現レベル比率を骨形成細胞において低下させる方法。
【請求項15】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項14に記載の方法。
【請求項16】
ポリペプチドが配列番号:2を含む、請求項14に記載の方法。
【請求項17】
骨形成細胞が被験者中に存在する、請求項14に記載の方法。
【請求項18】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と骨形成細胞を接触させる工程を含む、骨基質タンパク質、転写活性化因子又は転写調節因子の1つ又は2つ以上の発現レベルを骨形成細胞において増加させる方法。
【請求項19】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項18に記載の方法。
【請求項20】
ポリペプチドが配列番号:2を含む、請求項18に記載の方法。
【請求項21】
骨形成細胞が被験者中に存在する、請求項18に記載の方法。
【請求項22】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含む、被験者において骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、もしくは骨折の発生率を減少させるか、又は前記の任意の組合せをもたらす方法。
【請求項23】
被験者が骨粗鬆症に罹患した閉経後の女性である、請求項22に記載の方法。
【請求項24】
被験者にさらにビタミンD及びカルシウムを投与する工程を含む、請求項22に記載の方法。
【請求項25】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項22に記載の方法。
【請求項26】
ポリペプチドが配列番号:2を含む、請求項22に記載の方法。
【請求項27】
クマの副甲状腺ホルモン又はその機能的フラグメントを含むポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含む、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、もしくは骨折の発生率を減少させるか、又は前記の任意の組合せをもたらす方法。
【請求項28】
請求項1に記載のポリペプチドに対抗する抗体。
【請求項29】
配列番号:2のアミノ酸残基1−34から成る単離ポリペプチド。
【請求項30】
配列番号:2のアミノ酸残基1−36から成る単離ポリペプチド。
【請求項31】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含む、被験者で骨体積を増加させるか、骨塩の見掛け密度を増加させるか、骨梁支柱骨の数を増加させるか、骨梁支柱骨の厚さを増加させるか、骨梁骨の鉱化を増加させるか、皮質体積を増加させるか、皮質の多孔度を低下させるか、もしくは血清カルシウムを低下させるか、又は前記の任意の組合せをもたらす方法。
【請求項32】
被験者が、骨粗鬆症を罹患する閉経後の女性である、請求項31に記載の方法。
【請求項33】
被験者にさらにビタミンD及びカルシウムを投与する工程を含む、請求項31に記載の方法。
【請求項34】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項31に記載の方法。
【請求項35】
ポリペプチドが配列番号:2を含む、請求項31に記載の方法。
【請求項1】
配列番号:2の連続する少なくとも10アミノ酸残基を含む単離ポリペプチドであって、前記ポリペプチドが配列番号:2のアミノ酸残基41及び52の少なくとも1つを含む、前記単離ポリペプチド。
【請求項2】
ポリペプチドが配列番号:2を含む、請求項1に記載のポリペプチド。
【請求項3】
請求項1に記載のポリペプチドをコードする単離ポリヌクレオチド。
【請求項4】
ポリペプチドが配列番号:2を含む、請求項3に記載の単離ポリヌクレオチド。
【請求項5】
ポリヌクレオチドが配列番号:1を含む、請求項4に記載の単離ポリヌクレオチド。
【請求項6】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と骨形成細胞を接触させる工程を含む、cAMPレベルを骨形成細胞で増加させる方法。
【請求項7】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項6に記載の方法。
【請求項8】
ポリペプチドが配列番号:2を含む、請求項6に記載の方法。
【請求項9】
骨形成細胞が被験者中に存在する、請求項6に記載の方法。
【請求項10】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と骨形成細胞を接触させる工程を含む、アポトーシスを骨形成細胞において減少させる方法。
【請求項11】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項10に記載の方法。
【請求項12】
ポリペプチドが配列番号:2を含む、請求項10に記載の方法。
【請求項13】
骨形成細胞が被験者中に存在する、請求項10に記載の方法。
【請求項14】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と骨形成細胞を接触させる工程を含む、Baxタンパク質対Bcl-2タンパク質の発現レベル比率を骨形成細胞において低下させる方法。
【請求項15】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項14に記載の方法。
【請求項16】
ポリペプチドが配列番号:2を含む、請求項14に記載の方法。
【請求項17】
骨形成細胞が被験者中に存在する、請求項14に記載の方法。
【請求項18】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と骨形成細胞を接触させる工程を含む、骨基質タンパク質、転写活性化因子又は転写調節因子の1つ又は2つ以上の発現レベルを骨形成細胞において増加させる方法。
【請求項19】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項18に記載の方法。
【請求項20】
ポリペプチドが配列番号:2を含む、請求項18に記載の方法。
【請求項21】
骨形成細胞が被験者中に存在する、請求項18に記載の方法。
【請求項22】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含む、被験者において骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、もしくは骨折の発生率を減少させるか、又は前記の任意の組合せをもたらす方法。
【請求項23】
被験者が骨粗鬆症に罹患した閉経後の女性である、請求項22に記載の方法。
【請求項24】
被験者にさらにビタミンD及びカルシウムを投与する工程を含む、請求項22に記載の方法。
【請求項25】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項22に記載の方法。
【請求項26】
ポリペプチドが配列番号:2を含む、請求項22に記載の方法。
【請求項27】
クマの副甲状腺ホルモン又はその機能的フラグメントを含むポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含む、被験者で骨塩密度を高めるか、骨量を増加させるか、骨減少を低下させるか、もしくは骨折の発生率を減少させるか、又は前記の任意の組合せをもたらす方法。
【請求項28】
請求項1に記載のポリペプチドに対抗する抗体。
【請求項29】
配列番号:2のアミノ酸残基1−34から成る単離ポリペプチド。
【請求項30】
配列番号:2のアミノ酸残基1−36から成る単離ポリペプチド。
【請求項31】
配列番号:2のアミノ酸残基1−34を含むポリペプチドの有効量と被験者の骨形成細胞を接触させる工程を含む、被験者で骨体積を増加させるか、骨塩の見掛け密度を増加させるか、骨梁支柱骨の数を増加させるか、骨梁支柱骨の厚さを増加させるか、骨梁骨の鉱化を増加させるか、皮質体積を増加させるか、皮質の多孔度を低下させるか、もしくは血清カルシウムを低下させるか、又は前記の任意の組合せをもたらす方法。
【請求項32】
被験者が、骨粗鬆症を罹患する閉経後の女性である、請求項31に記載の方法。
【請求項33】
被験者にさらにビタミンD及びカルシウムを投与する工程を含む、請求項31に記載の方法。
【請求項34】
ポリペプチドが配列番号:2のアミノ酸残基1−36を含む、請求項31に記載の方法。
【請求項35】
ポリペプチドが配列番号:2を含む、請求項31に記載の方法。
【図1】

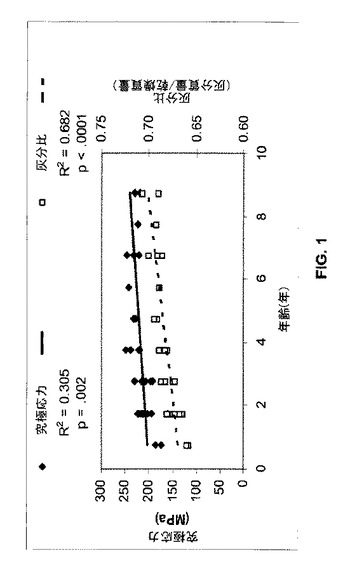
【図2】
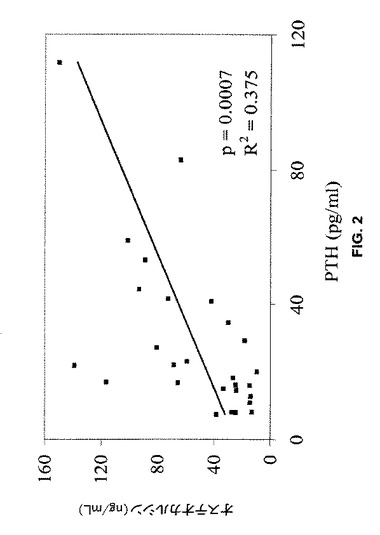
【図3】
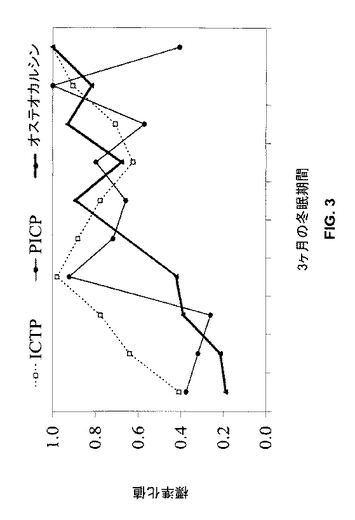
【図4】
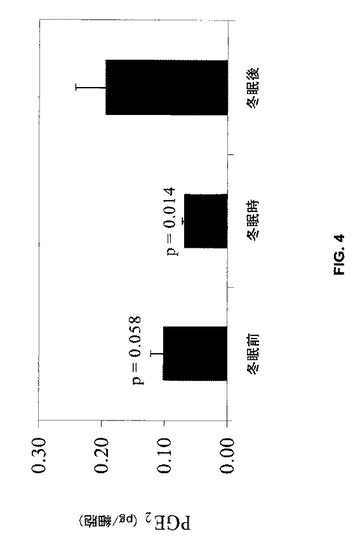
【図5】
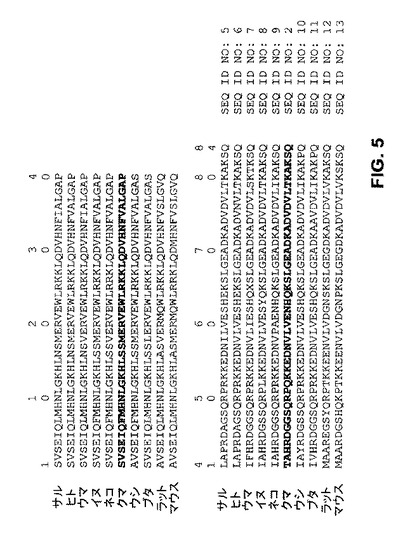
【図6】
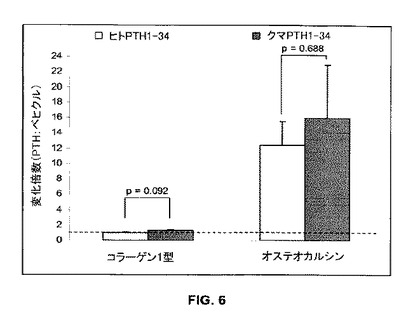
【図7】
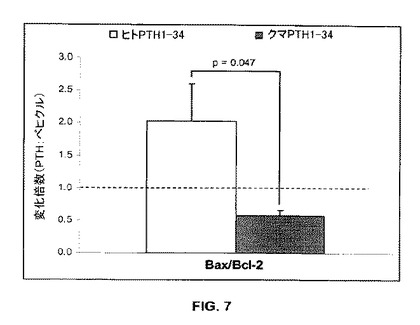
【図8】
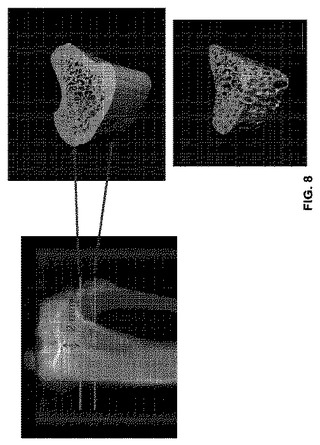
【図9A】

【図9B】

【図9C】

【図9D】

【図9E】

【図9F】

【図10】
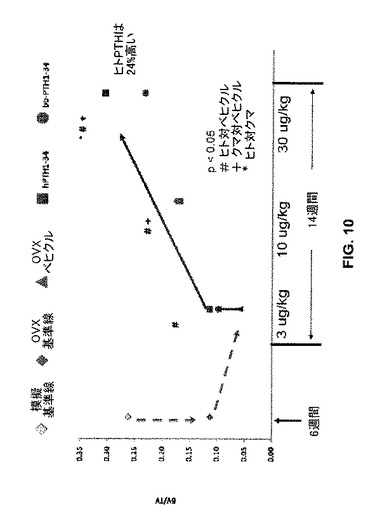
【図11】
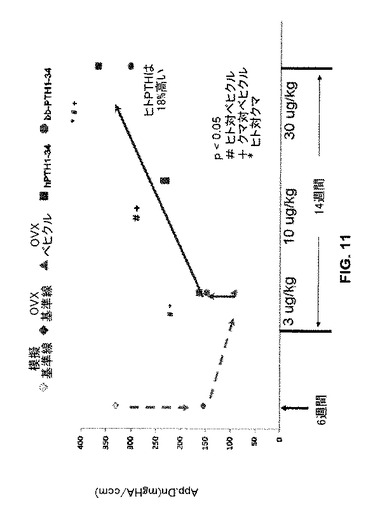
【図12】
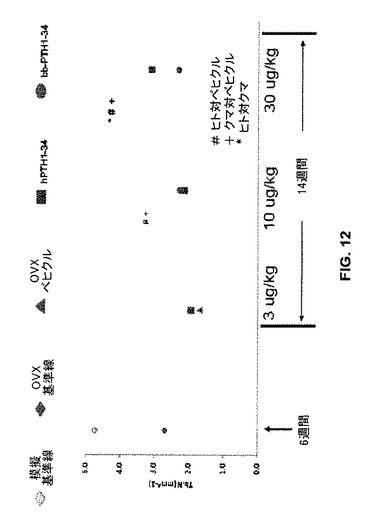
【図13】
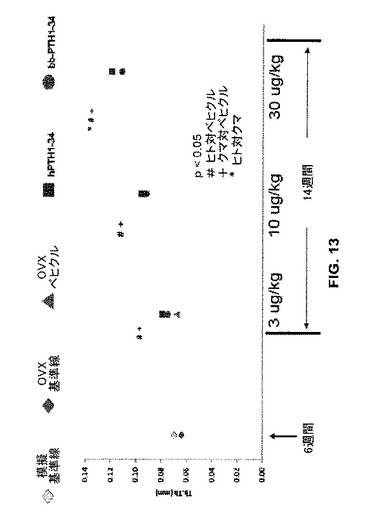
【図14】
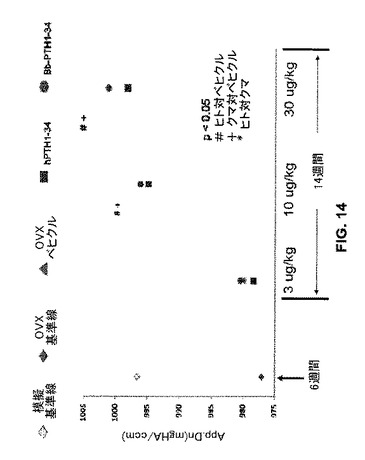
【図15】
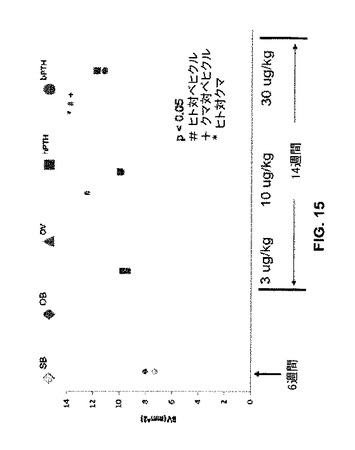
【図16】
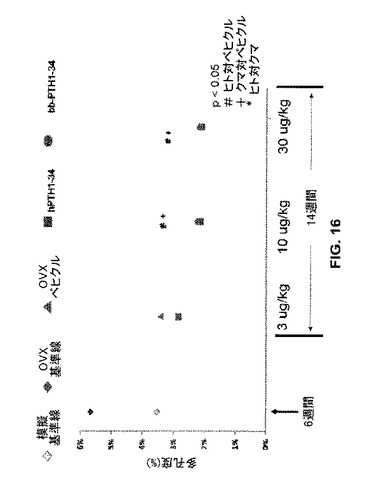
【図17】

【図18】
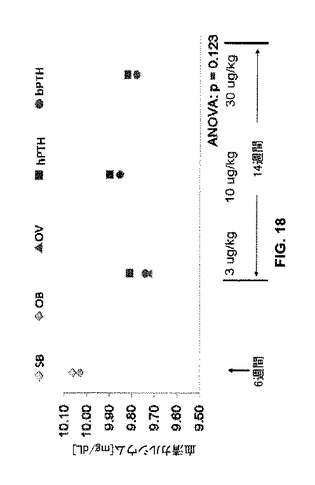
【図19】
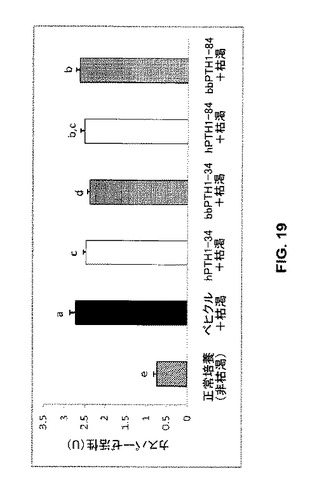
【図20】
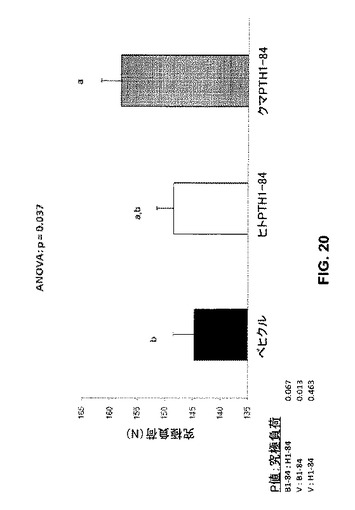
【図21】
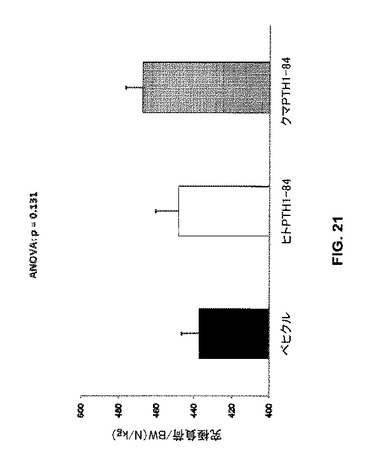
【図22】
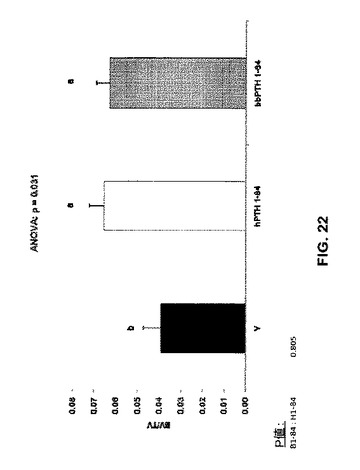
【図23】
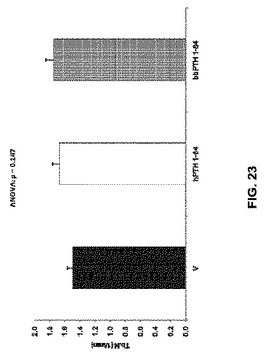
【図24】
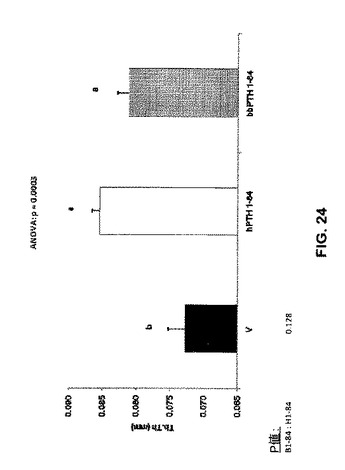
【図25】
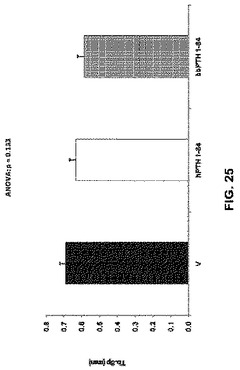
【図26】
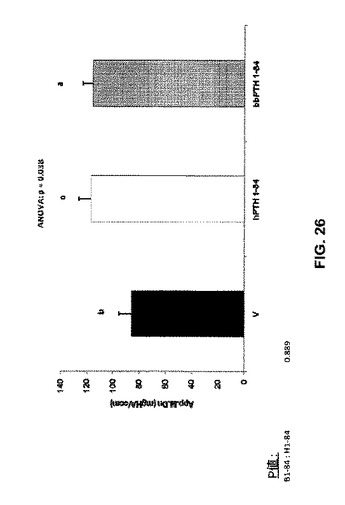
【図27】
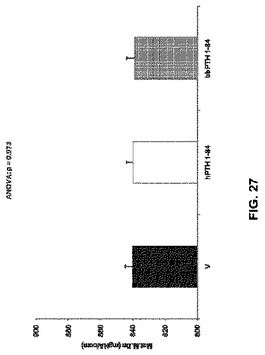
【図28】
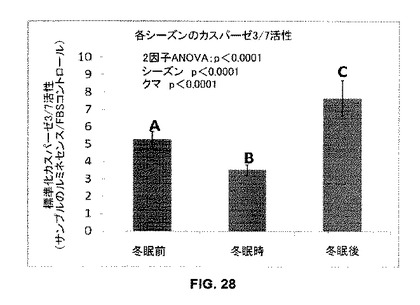
【図29】
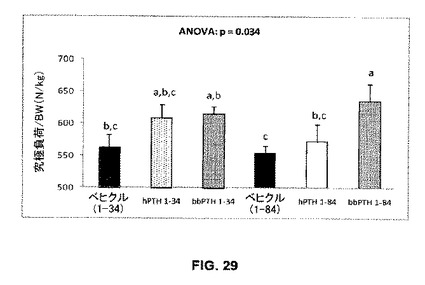
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図9C】
【図9D】
【図9E】
【図9F】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【公表番号】特表2013−512688(P2013−512688A)
【公表日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2012−543070(P2012−543070)
【出願日】平成21年12月7日(2009.12.7)
【国際出願番号】PCT/US2009/066974
【国際公開番号】WO2011/071480
【国際公開日】平成23年6月16日(2011.6.16)
【出願人】(508275168)ミシガン テクノロジカル ユニバーシティ (2)
【Fターム(参考)】
【公表日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願日】平成21年12月7日(2009.12.7)
【国際出願番号】PCT/US2009/066974
【国際公開番号】WO2011/071480
【国際公開日】平成23年6月16日(2011.6.16)
【出願人】(508275168)ミシガン テクノロジカル ユニバーシティ (2)
【Fターム(参考)】
[ Back to top ]