
クロストリジウム・サーモセラム由来のタンパク質複合体を構築するためのタンパク質及びその利用
【課題】酵母などの糖鎖修飾が予想される真核微生物で生産させるのに適しており、しかも、コヘシン−ドッケリン結合の結合能力に優れる、ドッケリンを有するタンパク質及びその利用の提供。
【解決手段】クロストリジウム・サーモセラム由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体を構築するためのタンパク質として、クロストリジウム・サーモセラム由来のタイプIドッケリン中のコヘシン結合能に関連した配列において、糖鎖修飾予測部位に相当する部位に相当する部位のアミノ酸がアスパラギン酸である、少なくとも一つのドッケリン特異的配列を含むドッケリンを有する、タンパク質を用いる。
【解決手段】クロストリジウム・サーモセラム由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体を構築するためのタンパク質として、クロストリジウム・サーモセラム由来のタイプIドッケリン中のコヘシン結合能に関連した配列において、糖鎖修飾予測部位に相当する部位に相当する部位のアミノ酸がアスパラギン酸である、少なくとも一つのドッケリン特異的配列を含むドッケリンを有する、タンパク質を用いる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、クロストリジウム・サーモセラム由来のタンパク質複合体を構築するためのタンパク質及びその利用に関する。
【背景技術】
【0002】
近年、有限である石油資源を代替するものとして、植物の光合成作用に由来するバイオマス資源への期待が高まってきており、バイオマスをエネルギーや各種材料に利用するための各種の試みがなされている。バイオマスを、エネルギー源やその他の原料として有効利用するためには、バイオマスを動物や微生物が容易に利用可能な炭素源に糖化することが必要である。
【0003】
典型的なバイオマスであるセルロースやヘミセルロースを利用するには、これらを糖化(分解)する優れたセルラーゼが必要である。こうしたセルラーゼ源として、一部の細菌が生産するセルロソームが着目されている。セルロソームは、細菌の細胞表層に形成されるタンパク質の複合体であって、セルラーゼが結合する骨格タンパク質(スキャホールディンタンパク質ともいう。)とセルラーゼとを備えている。スキャホールディンタンパク質は、コヘシンという部位を有しており、セルラーゼは、このコヘシンに対して、自身が備えるドッケリンを介して結合することが知られている。セルロソームは、細菌細胞表層に多種のセルラーゼを高密度でかつ大量に提供することができる。
【0004】
近年、遺伝子工学的に人工的なセルロソームを構築する検討がなされている。ここで、セルロソームの構築に関連して、セルロソームの構築する基礎となるコヘシンとドッケリンとの結合性(コヘシン−ドッケリン結合性)について種々の検討がなされている。例えば、クロストリジウム・サーモセラム(Clostridium thermocellum)のドッケリンのアミノ酸を数残基削除またはアラニンスキャンし、コヘシンとの結合性を評価して結合能に重要な残基を考察したものがある(非特許文献1)。この文献によれば、大腸菌で生産したドッケリンは、そのアミノ酸配列中のアスパラギンをアラニンに置換した際に、コヘシンとの結合量を70%程度は維持するが、構造安定化に寄与するカルシウムイオンとの相互作用が弱くなることが報告されている。また、C. thermocellumのドッケリン中の2つのへリックスを構成する2つの繰り返し配列のうち、一方の繰り返し配列のアミノ酸配列中にあるST(セリン−スレオニン)をAA(アラニン−アラニン)に置換することで結合能を損失させると、もう一方のへリックスでコヘシンと結合することも報告されている(非特許文献2)。さらに、コヘシンに関するものであるが、C. thermocellumのコヘシンのアミノ酸を数残基置換し、ドッケリンとの結合能を評価し、あるスレオニンをロイシンに置換する事で、C. thermocellum由来のドッケリンとの結合能を失い、本来は相互作用しないクロストリジウム・セルロリティカム(Clostridium cellulolyticum)由来のドッケリンと結合するようになることが報告されている(非特許文献3)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】A. KARPOL et al., Biochem. J 410, 331-338(2008)
【非特許文献2】A. L. Carvalho et al., PNAS 104(9), 3089-3094(2007)
【非特許文献3】A. Mechaly et al., J. Biol. Chem. 276(13), 9883-9888(2001)
【発明の概要】
【発明が解決しようとする課題】
【0006】
人工的なセルロソームの構築にあたっては、酵母などでセルラーゼを大量に生産し分泌させることが好ましいと考えられる。また、酵母等の真核微生物の細胞表層にセルロソームを構築できれば、セルロソームによって分解したグルコースを直接酵母の炭素源として、種々の有用物質を効率的に生産することができる。しかしながら、細菌等の原核生物に由来する異種タンパク質を酵母等の真核生物で生産させた場合、巨大糖鎖修飾がタンパク質間の相互作用に影響を与える可能性がある。
【0007】
ところで、以上の報告によれば、ドッケリンドメインのアミノ酸置換によりコヘシン−ドッケリン結合の結合能が低下したり(非特許文献1,2)、結合特異性が変化したり(非特許文献3)するなどの影響を与えることが理解される。しかしながら、コヘシン−ドッケリン結合の結合能力の向上については報告されていない。また、こうした報告は、いずれも、タンパク質への糖鎖修飾がなされない大腸菌で生産したコヘシン及びドッケリンについてなされているに過ぎない。したがって、糖鎖修飾が生じる酵母などの真核微生物における、ドッケリンドメインのアミノ酸置換のコヘシン−ドッケリン結合への影響は全く報告されていないのが現状である。
【0008】
そこで、本明細書の開示は、クロストリジウム・サーモセラム由来のタンパク質複合体を酵母などの糖鎖修飾が予想される真核微生物で生産させるのに適しており、しかも、コヘシン−ドッケリン結合の結合能力に優れる、ドッケリンを有するタンパク質及びその利用を提供することを一つの目的とする。
【課題を解決するための手段】
【0009】
本発明者らが、DDBJ(http://www.ddbj.nig.ac.jp/index-j.html)により、C.thermocellumのドッケリンの検索を行い、C. thermocellumのゲノム上で帰属されている72種類のドッケリンを見出し、さらに、UniProt(http://www.uniprot.org/)等を用いて、そのドッケリン中において、コヘシン−ドッケリン結合に関連すると思われる特異的な配列を見出し、これらの特異的配列につきマルチプルアラインメント等を実施して解析した。この結果、これらの142種類の特異的配列の類似性は90%を超えていた。このため、これらの特異的配列は、いずれもコヘシンとの結合能を有していると考えられた。
【0010】
また、発明者らは、これらの特異的配列のうち、約80%の113種類の関連配列には糖鎖修飾予測部位を有するが、残りの29種類には糖鎖修飾部位を有していないことを見出した。そして、本発明者らは、C. thermocellum由来のドッケリンの骨格タンパク質結合領域付近に存在する2箇所の糖鎖修飾予測部位をターゲットとして、当該部位のアスパラギンをアラニンまたはアスパラギン酸に置換してみた。ドッケリンのアスパラギンをアラニンに置換することで糖鎖修飾を解除したが、コヘシンとの結合能が向上しなかった。アスパラギンをアラニンに置換したドッケリンは酵母で生産すると安定な構造を取れずコヘシンと結合できなかった可能性がある。一方、ターゲット部位のアスパラギンをアスパラギン酸に置換したドッケリンを酵母で生産すると、コヘシン−ドッケリン間の結合量が増加し、結果として、酵母の糖化能力を向上させることができることがわかった。
【0011】
以上のことから、本来的に糖鎖修飾予測部位を有していないか、あるいは当該予測部位を有しているときには、その予測部位のアスパラギンをアスパラギン酸に置換して糖鎖修飾を解除することで、コヘシン−ドッケリン結合の結合能力を向上し、糖鎖修飾可能性のある真核微生物でも糖化能力を向上できるという知見を得た。本明細書の開示によれば、以下の手段が提供される。
【0012】
本明細書の開示によれば、C. thermocellum由来のタイプIコヘシンを有する骨格を利用したタンパク質複合体を構築するためのタンパク質であって、C. thermocellum由来のタイプIドッケリン中のコヘシン結合能に関連したドッケリン特異的配列であって、以下の(a)及び(b):
(a)内在性のN型糖鎖修飾予測部位を有しない
(b)内在性のN型糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されている
のいずれかを充足する少なくとも一つのドッケリン特異的配列を含むドッケリンを有するタンパク質が提供される。
【0013】
前記(a)を充足する前記ドッケリン特異的配列は、前記内在性のN型糖鎖修飾予測部位がアスパラギン酸であってもよい。
【0014】
また、本明細書に開示されるタンパク質は、セルロース分解促進活性を有していてもよく、さらに、前記セルロース分解促進活性はセルラーゼ活性であってもよい。また、セルロース分解促進活性は、クロストリジウム・サーモセラム由来のアミノ酸配列によって付与されていてもよい。
【0015】
本明細書の開示によれば、細胞表層にクロストリジウム・サーモセラム由来の骨格タンパク質を利用したタンパク質複合体を備える真核微生物であって、クロストリジウム・サーモセラム由来の骨格タンパク質と、前記骨格タンパク質に結合する本明細書に開示される前記タンパク質と、を備える、真核微生物が提供される。
【0016】
本明細書の開示によれば、有用物質の生産方法であって、本明細書に開示される前記真核微生物であって、前記ドッケリンタンパク質がセルロース分解促進活性を有する真核微生物を用いて、セルロース含有材料を炭素源として発酵する工程、を用いて、セルロース含有材料を糖化し、発酵する工程、を備える、方法が提供される。
【図面の簡単な説明】
【0017】
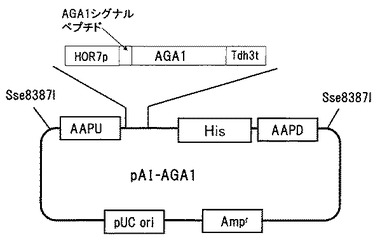
【図1】実施例1で作製したpAI-AGA1ベクターを示す図である。
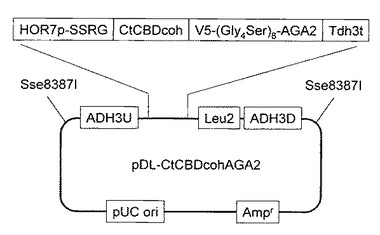
【図2】実施例2で作製したLeu2マーカーおよびADH3相同領域を持つ、pDL-CtCBDCohAGA2ベクターを示す図である。
【図3】Cel48Sドッケリン遺伝子において、そのアミノ酸配列の18番目と50番目のアスパラギンをアラニンに置換したアミノ酸配列及びその遺伝子配列を示す図である。
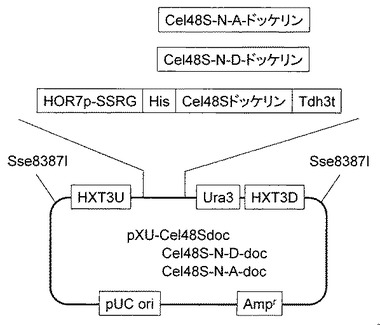
【図4】実施例3で作製したpXU-Cel48Sdocベクター、pXU-Cel48S-N-A-docベクター、およびpXU-Cel48S-N-D-docベクターを示す図である。
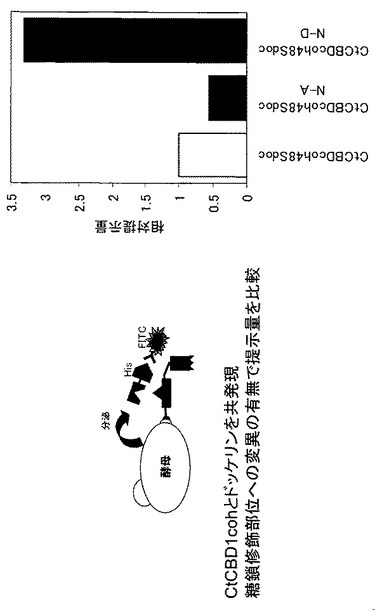
【図5】アミノ酸置換したCel48Sドッケリンを含むタンパク質複合体表層提示酵母におけるドッケリン提示量を示す図である。
【図6】Xyn10Cドッケリン遺伝子において、そのアミノ酸配列の18番目と54番目のアスパラギンをアラニンに置換したアミノ酸配列及びその遺伝子配列を示す図である。
【図7】実施例5で作製したpXU-Xyn10CdocベクターとpXU-Xyn10C-N-D-docベクターを示す図である。
【図8】アミノ酸置換したXyn10Cドッケリンを含むタンパク質複合体表層提示酵母におけるドッケリン提示量を示す図である。
【図9】実施例7で作製したpXU-Cel8A-Cel48SdocとpXU-Cel8A-Cel48S-N-D-docベクターを示す図である。
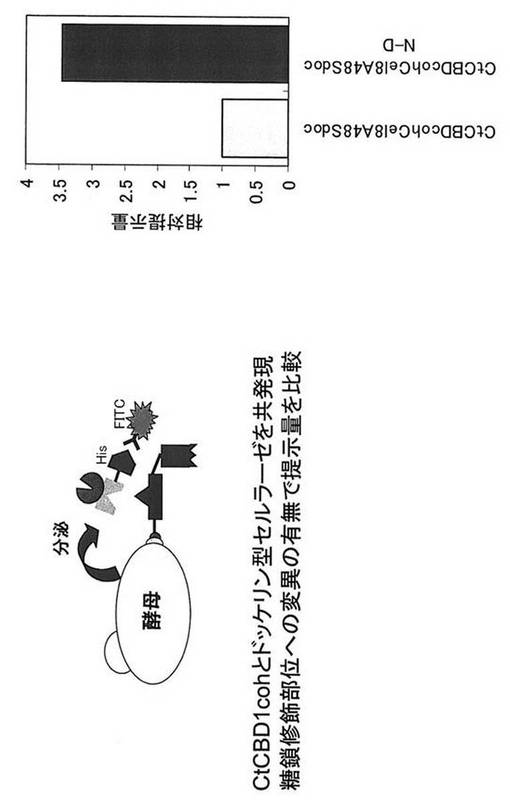
【図10】アミノ酸置換したドッケリン型セルラーゼを含むタンパク質複合体表層提示酵母におけるセルラーゼ提示量を示す図である。
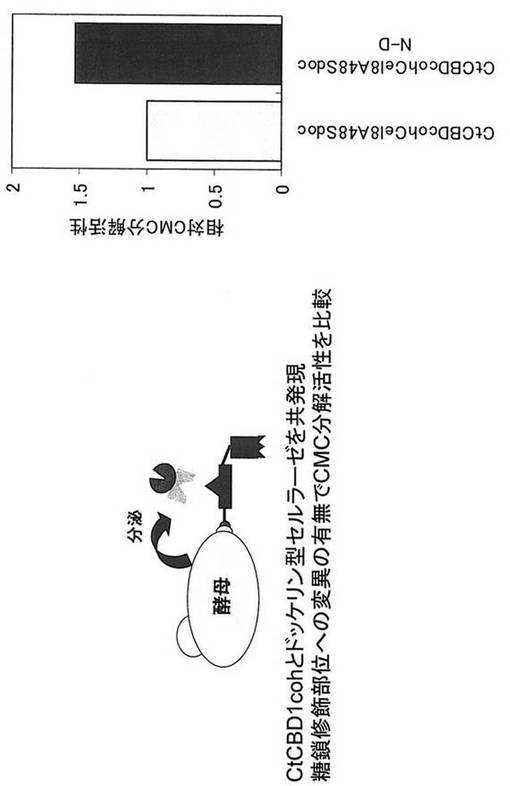
【図11】アミノ酸置換したドッケリン型セルラーゼを含むタンパク質複合体表層提示酵母のCMC分解活性を示す図である。
【発明を実施するための形態】
【0018】
本明細書の開示は、C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体を構築するためのタンパク質、当該タンパク質を含むタンパク質複合体を備える真核微生物及び当該真核微生物を用いる有用物質の生産方法に関する。
【0019】
本明細書に開示されるタンパク質は、C. thermocellum由来のタイプIドッケリン中のコヘシン結合能に関連した配列であって、内在性のN型糖鎖修飾予測部位を有しない又は内在性のN型糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されている、少なくとも一つのドッケリン特異的配列を有している。このため、本明細書に開示されるタンパク質は、酵母などの糖鎖修飾が予想される真核微生物で生産させたときであっても、糖鎖修飾が解除される。この結果、本明細書に開示されるタンパク質は、C. thermocellum由来の骨格タンパク質のタイプIコヘシンへの結合能に優れており、当該タンパク質の結合量及び/又は密度に優れたタンパク質複合体を構築できるものとなっている。
【0020】
また、本明細書に開示される真核微生物は、本明細書に開示されるタンパク質の集積量及び/又は集積密度に優れるタンパク質複合体を細胞表層に備えることができる。このため、本タンパク質の有する機能が増強された真核微生物を得ることができる。本タンパク質は、真核微生物で生産した場合においても、コヘシンへの結合能が優れているため、本明細書に開示される真核微生物が、前記骨格タンパク質及び前記タンパク質は、前記真核微生物が自己生産するものであってもよい。また、前記真核微生物は酵母であってもよい。
【0021】
さらに、本明細書に開示される有用物質の生産方法は、本明細書に開示される真核微生物であって、前記タンパク質がセルロース分解促進活性を有するタンパク質である真核微生物を用いてセルロース含有材料を炭素源として発酵する工程、を備える。本明細書に開示される真核微生物は、セルロース分解促進活性が増強されているため、セルロース含有材料を炭素源として効率的に発酵できる。
【0022】
(C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体を構築するためのタンパク質)
本明細書に開示されるタンパク質は、C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体の構築に特に適したタンパク質である。本タンパク質は、C. thermocellum由来のタイプIドッケリン中のコヘシン結合能に関連した配列であって、(a)内在性のN型糖鎖修飾予測部位を有しない、(b)内在性のN型糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されている、のいずれかを充足する少なくとも一つのドッケリン特異的配列を含むドッケリンを有することができる。
【0023】
C. thermocellumは、セルロソーム生産微生物として知られている。また、C. thermocellumは、セルラーゼ活性を有し、タイプIドッケリンを含むタンパク質を生産している。DDBJ(http://www.ddbj.nig.ac.jp/index-j.html)によるC. thermocellumのゲノムの検索結果に基づけば、C. thermocellumのタイプIドッケリンのアミノ酸配列として、以下の表1の配列番号1〜72に示す72種類のアミノ酸配列が挙げられる。表1において示すlocus(配列名)は、各ドッケリンの名称である。したがって、表2〜表21のlocus欄に示すドッケリン名で特定されるアミノ酸配列は、表1に記載の同一の名称のドッケリンのアミノ酸配列に由来している。
【0024】
【表1】

【0025】
さらに、これらの72種類のタイプIドッケリン中には、類似したアミノ酸配列であってコヘシン結合能に関連したドッケリン特異的配列を見出すことができる。ドッケリン特異的配列は、24アミノ酸からなる天然由来のコンセンサス配列(関連配列)からなることができる。72種類のタイプIドッケリンに内在するこの関連配列として、以下の表2に示す配列番号73〜214の合計142種類のアミノ酸配列が挙げられる。なお、これらのアミノ酸配列は、UniProt(http://www.uniprot.org/)、InterPro(http://www.ebi.ac.uk/interpro/)及びPfam(http://pfam.sanger.ac.uk)などのデータベースにより取得することができる。表2における1st欄にはN末端側の関連配列が記載され、2nd欄にはC末端側の関連配列が記載されている。
【0026】
【表2】
【0027】
表1に示すドッケリンについて相同性検索を行うと、これらのアミノ酸配列間では「相同性」が90%以上を超えることがない一方、表2に示す関連配列間で「類似性」は90%以上である。以上のことから、表1に挙げられたドッケリンは、いずれも、同等の機能を発揮していると推論できる。したがって、表2に示す関連配列が当該機能を担っていることが推論できる。
【0028】
表1に示すドッケリン、すなわち、表2に示す関連配列におけるN型糖鎖修飾予測部位は、酵母などの真核微生物でN型糖鎖修飾を受けるコンセンサス配列であるN−X−T又はN−X−S(Nはアスパラギン、Xはプロリン以外のアミノ酸、Tはスレオニン、Sはセリン)におけるNの位置となることが知られている(A. Herscovics et al., The FASEB Journal (6): 540-550(1993))。ドッケリン又はその関連配列におけるN−X−T/Sは、既に説明したデータベース等を適宜利用することなどにより検出できる。また、ドッケリン又はその関連配列におけるN型糖鎖修飾予測部位に相当する部位は、そのアミノ酸配列中の上記コンセンサス配列を備えてはいないが、Nに相当しうる部位をいう。N型糖鎖修飾予測部位に相当する部位は、当該部位を含む可能性のあるアミノ酸配列を他の公知のドッケリン又はその関連配列のアミノ酸配列とマルチプルアラインメントなどを利用して比較することにより、検出することができる。ドッケリンにおけるN型糖鎖修飾予測部位及び当該部位に相当する部位は、典型的には、関連配列を構成するアミノ酸配列を24個以下程度のアミノ酸からなるアミノ酸配列とするとき、そのN末端側から、通常、9個目がN型糖鎖修飾予測部位又は当該部位に相当する部位となっている。
【0029】
本タンパク質は、N型糖鎖修飾予測部位を有していないドッケリン特異的配列を少なくとも一つ含むドッケリンを有することが好ましい。そして、N型糖鎖修飾予測部位に相当する部位のアミノ酸がアスパラギン酸(D)であるドッケリン特異的配列を少なくとも一つ有することが好ましい。N型糖鎖修飾部位を有しないこと及び/又は糖鎖修飾予測部位に相当する部位がアスパラギン酸であることで、酵母などの真核微生物によるN型糖鎖修飾が解除されるものと考えられる。糖鎖修飾予測部位に相当する部位がアスパラギン酸であるドッケリン特異的配列は、本来的にドッケリンに内在するものであってもよいし、N型糖鎖修飾部位を有していて、そのアスパラギン(N)がアスパラギン酸(D)に置換されたものであってもよい。
【0030】
こうしたドッケリン特異的配列の一つの態様として、表1に開示されるドッケリン及び表2に開示されるこれらのドッケリンにおける関連配列が内在性のN型糖鎖修飾予測部位を有するとき、そのアスパラギンがアスパラギン酸に置換されているドッケリン特異的配列が挙げられる。本タンパク質は、こうしたドッケリン特異的配列を少なくとも一つ含んだドッケリンを有していればよい。こうしたNからDへの置換対象部位を有する関連配列としては、以下の113種類の関連配列が挙げられる。したがって、好ましいドッケリン特異的配列は、以下の関連配列において(N−X−T/S)のNがDに置換された配列で表される。
【0031】
【表3】
【0032】
本タンパク質は、こうしたドッケリン特異的配列を1又は2つ含むドッケリンを備えることができるが、典型的には、関連配列中の糖鎖修飾予測部位においてアスパラギンがアスパラギン酸に置換されていることが好ましい。こうしたドッケリンとしては、以下の表に示すドッケリンが挙げられる。これらの表においては、ドッケリンが有する関連配列で特徴付けて示している。したがって、好ましいドッケリンは、以下の表において各ドッケリンが有する1又は2つの関連配列において(N−X−T/S)のNがDに置換されたドッケリン特異的配列を有することができる。
【0033】
【表4】
【0034】
表4に示すドッケリンは、ドッケリン中に二つの関連配列を有し、それぞれの関連配列がN型糖鎖修飾予測部位を有している。こうしたドッケリンにあっては、二つの関連配列のいずれか又は双方のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで、好ましいドッケリンを得ることができる。
【0035】
【表5】
【0036】
表5に示すドッケリンは、ドッケリン中に1又は2つの関連配列を有し、N末端側の関連配列にN型糖鎖修飾予測部位を有している。こうしたドッケリンにあっては、この関連配列のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで、好ましいドッケリンを得ることができる。
【0037】
【表6】
【0038】
表6に示すドッケリンは、ドッケリン中に二つの関連配列を有し、C末端側の関連配列にN型糖鎖修飾予測部位を有している。こうしたドッケリンにあっては、この関連配列のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換してドッケリン特異的配列とすることで、好ましいドッケリンを得ることができる。
【0039】
本タンパク質において好ましいドッケリンを含むドッケリン特異的配列は、既に文献等においてコヘシンとの結合能が確認されている以下の表に示すC. thermocellumのタイプIドッケリンとの関係で選択される。以下の表においては、ドッケリンは、二つの関連配列で特徴付けて示している。これらのドッケリンに含まれるいずれかの関連配列とのアミノ酸配列の類似性が90%以上の関連配列を有するとき、この関連配列中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列を含む好ましいドッケリンを得ることができる。
【0040】
【表7】
【0041】
【表8】
【0042】
表8に示すドッケリンは、N末端側に一つの関連配列を有し、この関連配列が既にコヘシン結合能が確認された表7に示す8種のドッケリン中のいずれかの関連配列とのアミノ酸配列の類似性が90%以上である。この関連配列において、N型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで好ましいドッケリンを得ることができる。
【0043】
【表9】
【0044】
表9に示すドッケリンは、いずれも2つの関連配列を有し、C末端側の関連配列が既にコヘシン結合能が確認された表7に示す8種のドッケリン中のいずれかの関連配列とのアミノ酸配列の類似性が90%以上である。この関連配列において、N型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで好ましいドッケリンを得ることができる。
【0045】
【表10A】
【表10B】
【0046】
表10に示すドッケリンは、いずれも2つの関連配列を有し、二つの関連配列が既にコヘシン結合能が確認された表7に示す8種のドッケリン中のいずれかの関連配列とのアミノ酸配列の類似性が90%以上である。これらの関連配列のいずれか又は双方において、N型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで好ましいドッケリンを得ることができる。
【0047】
さらに、ドッケリンが、表7に示すコヘシン結合能を有する既知のいずれかのドッケリンのアミノ酸配列とアミノ酸配列の類似性が90%以上であるとき、このドッケリン中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで好ましいドッケリンを得ることができる。
【0048】
【表11】
【0049】
表11に示すドッケリンは、そのアミノ酸配列が、表7に示す8種のドッケリン中のいずれかのアミノ酸配列との類似性が90%以上である。このアミノ酸配列において、少なくとも一つのN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換することで好ましいドッケリンを得ることができる。
【0050】
さらにまた、ドッケリンが、表7に示すコヘシン結合能を有する既知のいずれかのドッケリンの関連配列のアミノ酸配列との相同性が90%以上である関連配列を有するとき、このドッケリン中のその関連配列中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換することで好ましいドッケリンを得ることができる。また、前記既知のドッケリンのアミノ酸配列と90%以上の相同性を有するドッケリン中のN型糖鎖修飾予測部位も好ましい置換対象である。こうしたドッケリンは、表7に示すドッケリンが該当するのみである。これらのドッケリンにおける関連配列中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換することで好ましいドッケリンを得ることができる。
【0051】
ドッケリン特異的配列の他の一つの態様は、表1に開示されるドッケリン及び表2に開示されるこれらのドッケリンにおける関連配列において内在性のN型糖鎖修飾予測部位を有しないドッケリン特異的配列が挙げられる。本タンパク質は、こうしたドッケリン特異的配列を少なくとも一つ含むドッケリンを有していればよい。この種のドッケリン特異的配列となる関連配列としては、以下の29種類の関連配列が挙げられる。
【0052】
【表12】
【0053】
本タンパク質は、表12に示すドッケリン特異的配列を1又は2つ含むドッケリンを備えていることができるが、典型的には、上記ドッケリン特異的配列を本来的に有するドッケリンであることが好ましい。こうしたドッケリンとしては、以下の表に示すドッケリンが挙げられる。これらの表においては、ドッケリンは、ドッケリンが有する関連配列で特徴付けて示している。
こうしたドッケリン特異的配列において、N型糖鎖修飾予測部位に相当する部位のアミノ酸がアスパラギン酸であることが好ましい。こうしたドッケリン特異的配列を1又は2つ有するドッケリンが好ましい。
【0054】
【表13】
【0055】
表13に示すドッケリンは、ドッケリン中に二つの関連配列を有し、いずれの関連配列にもN型糖鎖修飾予測部位を有していない。
【0056】
本タンパク質において好ましいドッケリンが含む内在性のN型糖鎖修飾予測部位を有しないドッケリン特異的配列は、既に文献等においてコヘシンとの結合能が確認されている表7に示すC. thermocellumのタイプIドッケリンとの関係で選択される。これらのドッケリンに含まれるいずれかの関連配列とのアミノ酸配列の類似性が90%以上の関連配列を有するとき、この関連配列中のN型糖鎖修飾予測部位に相当する部位がアスパラギン酸である天然型ドッケリン特異的配列であれば好ましいドッケリンとして用いることができる。
【0057】
【表14】
【0058】
表14に示すドッケリンは、2つの関連配列を有し、これらの関連配列がいずれも既にコヘシン結合能が確認された表7に示す8種のドッケリン中のいずれかの関連配列とのアミノ酸配列の類似性が90%以上である。そして、これらの関連配列のいずれか又は双方が、天然型ドッケリン特異的配列である。なお、天然型ドッケリン特異的配列でない関連配列中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換してもよい。
【0059】
さらに、ドッケリンが、前記既知のいずれかのドッケリンのアミノ酸配列とアミノ酸配列の類似性が90%以上であるとき、このドッケリンに内在する関連配列中のN型糖鎖修飾予測部位に相当する部位がアスパラギン酸である天然型ドッケリン特異的配列を少なくとも一つ有していれば、好ましいドッケリンとして用いることができる。
【0060】
【表15】
【0061】
表15に示すドッケリンは、そのアミノ酸配列が、表7に示す8種のドッケリン中のいずれかのアミノ酸配列との類似性が90%以上である。このアミノ酸配列において、少なくとも一つのN型糖鎖修飾予測部位に相当する部位がアスパラギン酸である天然型ドッケリン特異的配列を有することで好ましいドッケリンとして用いることができる。なお、N型糖鎖修飾予測部位がアスパラギンである他の関連配列を有する場合には、当該アスパラギンをアスパラギン酸に置換してもよい。
【0062】
本タンパク質は、ドッケリン以外に活性部位を備えることができる。活性部位の種類は用途に応じて適宜決定される。本タンパク質は、ドッケリンと活性部位とを適宜組み合わせた人工的なタンパク質であってもよい。また、例えば、セルロソームの構成タンパク質であるセルラーゼであって、ドッケリンを本来的に有するセルラーゼをそのままあるいは適宜改変して用いることもできる。
【0063】
本タンパク質は、バイオマスに由来するセルロース含有材料の糖化利用に際しては、例えば、セルロース分解促進活性を有することができる。すなわち、、セルロース分解促進活性部位を備えることができる。セルロース分解促進活性とは、セルラーゼ活性、セルロース結合活性、セルロース緩和活性などが挙げられる。
【0064】
セルラーゼ活性部位は、公知のセルラーゼにおける活性部位を適宜利用できる。セルラーゼとしては、エンドグルカナーゼ(EC 3.2.1.74)、セロビオヒドロラーゼ(EC 3.2.1.91)及びβ−グルコシダーゼ(EC23.2.4.1、EC 3.2.1.21)が挙げられる。なお、セルラーゼは、そのアミノ酸配列の類似性に基づきGHF(Glycoside Hydrolase family)(http://www.cazy.org/fam/acc.gh.html)の13(5,6,7,8,9,10,12,44,45,48,51,61,74)のファミリーに分類されている。異なるファミリーに分類される同種又は異種のセルラーゼを組み合わせてもよい。
【0065】
セルラーゼとしては、特に限定しないが、それ自体活性の高いセルラーゼであることが好ましい。このようなセルラーゼとしては、ファネロケーテ(Phanerochaete)属菌、Trichoderma reeseiなどのトリコデルマ属(Trichoderma)菌、フザリウム属(Fusarium)菌、トレメテス属(Tremetes)菌、ペニシリウム属(Penicillium)菌、フミコーラ属(Humicola)菌、アクレモニウム属(Acremonium)菌、アスペルギルス属(Aspergillus)菌等の糸状菌の他に、クロストリジウム属(Clostridium)菌、シュードモナス属(Pseudomonas)菌、セルロモナス属(Cellulomonas)菌、ルミノコッカス属(Ruminococcus)菌、バチルス属(Bacillus)菌等の細菌、スルフォロバス属(Sulfolobus)菌等の始原菌、さらにストレプトマイセス属(Streptomyces)菌、サーモアクチノマイセス属(Thermoactinomyces)菌などの放射菌由来のセルラーゼが挙げられる。なお、こうしたセルラーゼ又はその活性部位は、人工的に改変されていてもよい。
【0066】
本タンパク質は、C. thermocellum由来のタンパク質でもあるため、セルロース分解促進活性は、クロストリジウム・サーモセラム由来のアミノ酸配列によって付与されていることが好ましい。
【0067】
本タンパク質は、バイオマスの有効利用を考慮したとき、ヘミセルラーゼ活性部位を備えていてもよい。さらに、リグニンペルオキシダーゼ、マンガンペルオキシダーゼ及びラッカーゼなどのリグニン分解酵素が挙げられる。また、例えば、セルロース緩和タンパク質であるスウォレニンやエクスパンシン、セルロソームやセルラーゼの構成部分であるセルロース結合ドメイン(タンパク質)が挙げられる。また、キシラナーゼやヘミセルラーゼ等のその他のバイオマス分解酵素も挙げられる。これらのタンパク質は、いずれもセルロースへのセルラーゼのアクセシビリティを向上させることができる。
【0068】
本タンパク質は、真核微生物において細胞外分泌性を備えていることが好ましい。すなわち、真核微生物において分泌タンパク質として生産されるタンパク質であることが好ましい。セルラーゼなどの酵素は、本来的に細胞外分泌のためのシグナルを有していることが多い。ドッケリンタンパク質に細胞外分泌性を付与するには、公知の分泌シグナルを用いることができる。分泌シグナルは、用いる真核微生物の種類に応じて適宜選択される。分泌シグナル等については後段にて説明する。
【0069】
当業者であればこのような本タンパク質を適当な宿主微生物で遺伝子組換え等により生産させることができるほか、化学合成によっても得ることができる。
【0070】
以上説明した、本タンパク質は、特定のドッケリンを有していることから、C. thermocellum由来のタイプIコヘシンに対する結合性が向上しており、こうしたコヘシンを有する骨格タンパク質上への集積量や集積密度を高めることができる。
【0071】
(C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質)
本タンパク質は、C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質との複合体の構築のためのタンパク質として好適である。C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質は、C. thermocellum由来の1又は2以上のタイプIコヘシンをを備えることができる。コヘシンは、セルロソーム生産微生物の形成するセルロソームにおけるタイプI等の骨格タンパク質に備えられる触媒活性のあるセルラーゼ等を非共有結合で結合するドメインとして知られている(粟冠ら、蛋白質核酸酵素、Vol.44、No.10(1999)、p41-p50、Demain, A. L., et al., Microbiol Mol. Biol Rev., 69(1), 124-54(2005), Doi, R. H., et al., J. Bacterol., 185(20), 5907-5914(2003)等)。本タンパク質を結合するためのC. thermocellum由来の骨格タンパク質としては、タイプI骨格タンパク質上のタイプIコヘシンドメインを少なくとも有している。そのほか、タイプII骨格タンパク質上のタイプIIコヘシンドメイン及びタイプIII骨格タンパク質上のタイプIIIコヘシンドメインを備えていてもよい。こうした各種タイプのコヘシンドメインとしては、各種セルロソーム生産微生物において多数その配列が決定されている。これらの各種のタイプのコヘシンのアミノ酸配列及びDNA配列は、NCBIのHP(http://www.ncbi.nlm.nih.gov/)等を介してアクセス可能な各種のタンパク質データベースやDNA配列のデータベースにより容易に取得することができる。
【0072】
C. thermocellum由来のコヘシンを有する骨格タンパク質は、C. thermocellum由来のタイプIコヘシンを有していればよく、C. thermocellum由来の骨格タンパク質自体でなくともよく、人工タンパク質であってもよい。また、骨格タンパク質は、C. thermocellum由来の天然のタイプIコヘシン又はその結合性を有する限りそのコヘシンのアミノ酸配列において1又は2以上の変異(付加、挿入、欠失及び置換)を導入した改変コヘシンを備えていてもよい。コヘシンタンパク質は、こうしたコヘシン等を適当なインターバルを置いて複数個備えることもできる。コヘシンタンパク質の全体のアミノ酸配列や、コヘシン間のアミノ酸配列の種類や有無は、タイプI骨格タンパク質のアミノ酸配列及び当該アミノ酸配列に適宜変異を導入した配列を利用できる。
【0073】
骨格タンパク質は、タイプI〜IIIから選択される骨格タンパク質のセルロース結合ドメイン(CBD)を有していてもよい。CBDは、各種骨格タンパク質において基質であるセルロースに結合するドメインとして知られている(前述粟冠ら)。セルロース結合ドメインは、1個又は2個以上有していてもよい。各種のセルロソーム生産微生物のセルロソームにおけるCBDのアミノ酸配列及びDNA配列の多くが決定されている。これらの各種のCBDのアミノ酸配列及びDNA配列は、NCBIのHP(http://www.ncbi.nlm.nih.gov/)等を介してアクセス可能な各種のタンパク質データベースやDNA配列のデータベースにより容易に取得することができる。
【0074】
骨格タンパク質は、真核微生物において細胞外分泌性又は細胞表層提示性を備えていることが好ましい。すなわち、真核微生物において分泌タンパク質として生産されるタンパク質であることが好ましいほか、真核微生物の細胞表層に提示されるタンパク質であることが好ましい。コヘシンタンパク質に細胞外分泌性又は細胞表層提示性を付与するには、公知の分泌シグナルや表層提示用のシステムを用いることができる。
【0075】
当業者であればこのような各種ドメインを適宜有する骨格タンパク質を適当な宿主微生物で遺伝子組換え等により生産させることができる。こうした各種骨格タンパク質のコヘシンドメインを有するコヘシンタンパク質は、化学合成によっても得ることができる。
【0076】
以上説明した、本タンパク質は、特定のドッケリンを有していることから、C. thermocellum由来のタイプIコヘシンに対する結合性が向上しており、こうしたコヘシンを有する骨格タンパク質上への集積量や集積密度を高めることができる。
【0077】
(タンパク質複合体)
本明細書の開示によれば、C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質と、この骨格タンパク質に結合された本タンパク質と、を備えるタンパク質複合体も提供される。このタンパク質複合体は、本タンパク質の集積量及び/又は集積密度が高くなっているため、本タンパク質の活性が増強されたものとなっている。
【0078】
(タンパク質複合体を細胞表層に備える真核微生物)
本明細書に開示される真核微生物は、本明細書に開示されるタンパク質複合体を細胞表層に備えている。本真核微生物は、タンパク質複合体を構成する骨格タンパク質と本タンパク質とを細胞外から供給して自己集合的に細胞表層にて構築されたものであってもよいが、好ましくは、これらのタンパク質を自己生産する。本タンパク質は、真核微生物において生産してもその糖鎖修飾系による糖鎖修飾を解除又は抑制して、対コヘシン結合性が向上しているからである。
【0079】
本タンパク質がセルラーゼ活性等のセルロース分解促進活性を有するとき、真核微生物の細胞表層にセルラーゼなどのセルロース分解促進活性を有するタンパク質を集積させたタンパク質複合体を構築することができる。こうした真核微生物は、細胞表層でセルロース含有材料を分解糖化して得られるグルコースを炭素源として利用できる。
【0080】
こうしたタンパク質をコードするコード化DNAは、宿主となる真核微生物内において当該タンパク質を発現可能に保持されていればよく、その保持形態は特に限定されない。例えば、真核微生物で作動可能なプロモーターの制御下に連結されるとともに適切なターミネーターをその下流に有した状態で保持されている。プロモーターは、構成的プロモーターであっても誘導的プロモーターであってもよい。このような状態のDNAは、宿主染色体内に組み込まれた形態であってもよいし、宿主核内に保持される2μプラスミドや核外に保持されるプラスミドのような形態であってもよい。一般には、こうした外来DNAの導入に伴って、宿主において利用可能な選択マーカー遺伝子も同時に保持されている。
【0081】
真核微生物において生産するドッケリンタンパク質やコヘシンタンパク質に細胞外分泌性や細胞表層提示性を付与することが好ましい。なかでも、本タンパク質には細胞外分泌性を付与することが好ましく、骨格タンパク質には、さらに細胞外に分泌させて細胞表層に提示させる細胞表層提示性を付与することが好ましい。細胞外分泌性を付与するには、分泌シグナルを付与することができる。分泌シグナルとしては、例えば、Rhizopus oryzaeやC. albicansのグルコアミラーゼ遺伝子の分泌シグナル、酵母インベルターゼリーダー、α因子リーダーなどが挙げられる。また、凝集性タンパク質あるいはその一部を用いることでタンパク質を真核微生物の表層に提示した状態に分泌させることができる。例えば、凝集性タンパク質であるα−アグルチニンをコードするSAG1遺伝子の5’領域の320アミノ酸残基からなるペプチドがある。また、所望のタンパク質を細胞表層に提示するためのポリペプチドや手法は、WO01/79483号公報や、特開2003−235579号公報、WO2002/042483号パンフレット、WO2003/016525号パンフレット、特開2006−136223号公報、藤田らの文献(藤田ら,2004. Appl Environ Microbiol 70:1207-1212および藤田ら, 2002. Appl Environ Microbiol 68:5136-5141.)、村井ら, 1998. Appl Environ Microbiol 64:4857-4861.に開示されている。
【0082】
真核微生物としては、特に限定されないで、例えば、公知の各種酵母を利用できる。後述するエタノール発酵等を考慮すると、サッカロマイセス・セレビジエ(Saccharomyces cerevisiae)等のサッカロマイセス属の酵母、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)等のシゾサッカロマイセス属の酵母、キャンディダ・シェハーテ(Candida shehatae)等のキャンディダ属の酵母、ピヒア・スティピティス(Pichia stipitis)等のピヒア属の酵母、ハンセヌラ(Hansenula)属の酵母、トリコスポロン(Trichosporon)属の酵母、ブレタノマイセス(Brettanomyces)属の酵母、パチソレン(Pachysolen)属の酵母、ヤマダジマ(Yamadazyma)属の酵母、クルイベロマイセス・マーキシアヌス(Kluyveromyces marxianus)、クルイベロマイセス・ラクティス(Kluveromyces lactis)等のクルイベロマイセス属の酵母が挙げられる。なかでも、工業的利用性等の観点からサッカロマイセス属酵母が好ましい。なかでも、サッカロマイセス・セレビジエが好ましい。
【0083】
外来タンパク質を発現する真核微生物は、モレキュラークローニング第3版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載されている方法に準じて作製することができる。本タンパク質や骨格タンパク質を真核微生物において発現させるためのベクター及びその構築方法も、同様に、当業者において周知である。なお、ベクターの形態は、使用形態に応じて様々な形態を採ることができる。例えば、DNA断片の形態を採ることができるほか、2マイクロプラスミドなどの適当な酵母用ベクターの形態を採ることもできる。このようなベクターで真核微生物を形質転換することによって本明細書に開示される真核微生物を得ることができる。形質転換にあたり、従来公知の各種方法、例えば、トランスフォーメーション法や、トランスフェクション法、接合法、プロトプラスト法、エレクトロポレーション法、リポフェクション法、酢酸リチウム法等を用いることができる。
【0084】
(有用物質の生産方法)
本明細書に開示される有用物質の生産方法は、本明細書に開示される真核微生物であって、本タンパク質がセルロース分解促進活性を有する真核微生物を用いて、セルロース含有材料を炭素源として発酵する工程、を用いて、セルロース含有材料を糖化し、発酵する工程、
を備えることができる。この方法によれば、この真核微生物を用いてセルロース含有材料を直接分解糖化し、グルコース等として利用できることになる。前記発酵工程の実施により、用いた真核微生物が有している有用物質生産能力に応じて有用物質が生産される。
【0085】
有用物質は、真核微生物がグルコースなどを発酵することにより得る生産物であり、真核微生物の種類によっても異なるし、発酵条件によっても異なる。有用物質としては特に限定しないが、酵母やその他の真核微生物がグルコースを利用して生産可能なものであればよい。有用物質は、酵母などの真核微生物におけるグルコースからの代謝系の1種又は2種以上の酵素を遺伝子組換えにより置換、追加等して合成できるようになった本来の代謝物でない化合物であってもよい。有用物質としては、例えば、エタノールなどのほか、C3〜C5の低級アルコール、乳酸などの有機酸の他、イソプレノド合成経路の追加によるファインケミカル(コエンザイムQ10、ビタミン及びその原料等)、解糖系の改変によるグリセリン、プラスチック・化成品原料など、バイオリファイナリー技術が対象とする材料が挙げられる。有用物質の生産工程終了後、培養液から有用物質含有画分を回収する工程、さらにこれを精製又は濃縮する工程を実施することもできる。回収工程や精製等の工程は有用物質の種類等に応じて適宜選択される。
【0086】
真核微生物が細胞表層にタンパク質複合体として保持する本タンパク質は、2種類以上のセルロース分解促進活性を有していることが好ましい。セルラーゼとしては、例えば、エンドグルカナーゼとセロビオヒドロラーゼ等の活性をそれぞれ有する2種類以上を用いることが好ましい。
【0087】
なお、セルロース含有材料は、D−グルコースがβ−1,4結合でグリコシド結合したβ−グルカンであるセルロースを含有する材料である。セルロース含有材料としては、セルロースを含有していればよく、どのような由来や形態であってもよい。したがって、セルロース系材料としては、例えば、リグノセルロース系材料、結晶性セルロース材料、可溶性セルロース材料(非晶性セルロース材料)、不溶性セルロース材料などの各種セルロース系材料等が含まれる。リグノセルロース系材料としては、例えば、木本植物の木質部や葉部及び草本植物の葉、茎、根等においてリグニン等を複合した状態のリグノセルロース系材料が挙げられる。こうしたリグノセルロース系材料としては、例えば、稲ワラ、麦ワラ、トウモロコシの茎葉、バガス等の農業廃棄物、収集された木、枝、枯葉等又はこれらを解繊して得られるチップ、おがくず、チップなどの製材工場廃材、間伐材や被害木などの林地残材、建設廃材等の廃棄物であってもよい。結晶性セルロース系材料及び不溶性セルロース系材料としては、リグノセルロース系材料からリグニン等を分離後の結晶性セルロース及び不溶性セルロースを含む結晶性又は不溶性セルロース系材料が挙げられる。セルロース材料としては、また、使用済み紙製容器、古紙、使用済みの衣服などの使用済み繊維製品、パルプ廃液を由来としてもよい。
【0088】
セルロース含有材料は、セルラーゼと接触させるのに先立ってセルラーゼによる分解を容易化するために適当な前処理等がなされていてもよい。例えば、硫酸、塩酸、リン酸、硝酸などの無機酸による酸性条件下、セルロースを部分加水分解することにより、セルロースを非晶質化あるいは低分子化できる。この他、超臨界水、アルカリ、加圧熱水などの処理によってもセルロースの非晶質化又は低分子化を行うことができる。
【0089】
セルロース含有材料は、グルコースがβ-1,4-グリコシド結合により重合した重合体及びその誘導体を含んでいる。グルコースの重合度は特に限定しない。また、誘導体としては、カルボキシメチル化、アルデヒド化、若しくはエステル化などの誘導体が挙げられる。セルロース は、結晶性セルロースであってもよいし、非結晶性セルロースであってもよい。
【0090】
本明細書において同一性又は類似性とは、当該技術分野で知られているとおり、配列を比較することにより決定される、2以上のタンパク質あるいは2以上のポリヌクレオチドの間の関係である。当該技術で“同一性 ”とは、タンパク質またはポリヌクレオチド配列の間のアラインメントによって、あるいは場合によっては、一続きのそのような配列間のアラインメントによって決定されるような、タンパク質またはポリヌクレオチド配列の間の配列不変性の程度を意味する。また、類似性とは、タンパク質またはポリヌクレオチド配列の間のアラインメントによって、あるいは場合によっては、一続きの部分的な配列間のアラインメントによって決定されるような、タンパク質またはポリヌクレオチド配列の間の相関性の程度を意味する。より具体的には、配列の同一性と保存性(配列中の特定アミノ酸又は配列における物理化学特性を維持する置換)によって決定される。なお、類似性は、後述するBLASTの配列相同性検索結果においてSimilarity と称される。同一性及び類似性を決定する方法は、対比する配列間で最も長くアラインメントするように設計される方法であることが好ましい。同一性及び類似性を決定するための方法は、公衆に利用可能なプログラムとして提供されている。例えば、AltschulらによるBLAST (Basic Local Alignment Search Tool) プログラム(たとえば、Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ., J. Mol. Biol., 215: p403-410 (1990), Altschyl SF, Madden TL, Schaffer AA, Zhang J, Miller W, Lipman DJ., Nucleic Acids Res. 25: p3389-3402 (1997))を利用し決定することができる。BLASTのようなソフトウェアを用いる場合の条件は、特に限定するものではないが、デフォルト値を用いるのが好ましい。
【実施例】
【0091】
以下、本発明を、実施例を挙げて具体的に説明するが、本発明はこれらの実施例に限定されるものではない。なお、以下に述べる遺伝子組換え操作はMolecular Cloning: A Laboratory Manual (T. Maniatis, et al., Cold Spring Harbor Laboratory) に従い行った。
【実施例1】
【0092】
PCR法により常法に従い増幅後クローニングしたaga1遺伝子の上流にAAP1相同領域とHOR7プロモーター、下流にTdh3ターミネーターとHis3マーカーおよびAAP1相同領域を持つ、pAI-AGA1ベクター(図1)を作製した。このベクターを用いて酵母S.cerevisiae BY4741に形質転換を行い、aga1を細胞表層に大量提示する酵母BY-AGA1を取得した。
【実施例2】
【0093】
C.thermocellumのゲノムから、CBD-コヘシンをPCR法により常法に従い増幅後クローニングした(配列番号215)。取得した遺伝子の上流にADH3相同領域とHOR7プロモーター、下流にV5-tag、aga2、Tdh3ターミネーター、Leu2マーカーおよびADH3相同領域を持つ、pDL-CtCBDCohAGA2ベクターを作製した(図2)。作製したベクターを実施例1で取得した酵母BY-AGA1に導入して、C.thermocellum 由来コヘシンを細胞表層に提示する酵母CtCBDcohを取得した。
【実施例3】
【0094】
C.thermocellum のゲノムからCel48Sドッケリン遺伝子をPCR法により常法に従い増幅後クローニングした(配列番号216)。取得したCel48Sドッケリン遺伝子を鋳型として48Sdock-N18A-Fw と48Sdock-N50A-Rvの2種類のプライマー(配列番号217,218)を利用して 18番目と50番目のアスパラギンをアラニンに置換した遺伝子を取得した。同様にCel48Sドッケリン遺伝子を鋳型として48Sdock-N18D-Fw と48Sdock-N50D-Rvの2種類のプライマー(配列番号219、220)を利用して 18番目と50番目のアスパラギンをアスパラギン酸に置換した遺伝子を取得した(図3)。取得したそれぞれの遺伝子の上流にHXT3相同領域、HOR7プロモーターおよびHis-tag、下流にTdh3ターミネーター、Ura3マーカーおよびHXT3相同領域を持つ、pXU-Cel48Sdocベクター、pXU-Cel48S-N-A-docベクター、およびpXU-Cel48S-N-D-docベクターを作成した(図4)。取得したベクターを実施例2で取得した酵母CtCBDcohにそれぞれ導入することで、C.thermocellum 由来コヘシンを細胞表層に提示し、ドッケリンまたはアミノ酸置換したドッケリンを同時生産するCtCBDcoh48SdocとCtCBDcoh48SdocN-A 、およびCtCBDcoh48SdocN-Dを取得した。
【実施例4】
【0095】
実施例3で取得した3種類の酵母CtCBDcoh48SdocとCtCBDcoh48SdocN-A 、およびCtCBDcoh48SdocN-Dを、それぞれYP+2%グルコース培地で30℃、24時間培養し、OD600=0.5、62.5μl相当量集菌し、PBS溶液で洗浄を行い、PBS + 1mg/ml BSA + anti-His-FITC溶液と混合して4℃、30分間反応し、PBS溶液で2回洗浄後、Flow Cytometryで酵母細胞表層上のドッケリン提示量を評価した。アスパラギンをアラニンに置換する事でCel48Sドッケリンの提示量は約半分に低下した。一方、アスパラギンをアスパラギン酸に置換する事でCel48Sドッケリンの提示量は3.3倍に向上した(図5)。
【実施例5】
【0096】
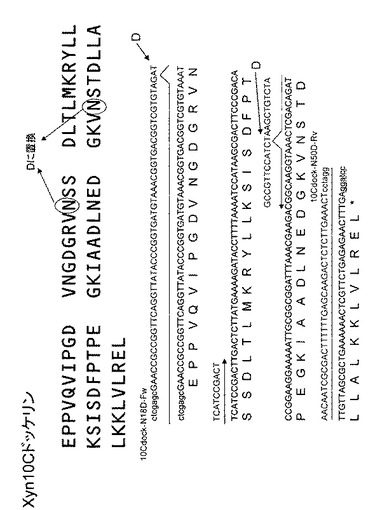
C.thermocellum のゲノムからXyn10Cドッケリン遺伝子をPCR法により常法に従い増幅後クローニングした(配列番号221)。取得したXyn10Cドッケリン遺伝子を鋳型として10Cdock-N18D-Fw と10Cdock-N50D-Rvの2種類のプライマー(配列番号222、223)を利用して 18番目と54番目のアスパラギンをアスパラギン酸に置換した遺伝子を取得した(図6)。取得したそれぞれの遺伝子の上流にHXT3相同領域、HOR7プロモーターおよびHis-tag、下流にTdh3ターミネーター、Ura3マーカーおよびHXT3相同領域を持つ、pXU-Xyn10CdocベクターとpXU-Xyn10C-N-D-docベクターを作成した(図7)。取得したベクターを実施例2で取得した酵母CtCBDcohにそれぞれ導入することで、C.thermocellum 由来コヘシンを細胞表層に提示し、ドッケリンまたはアミノ酸置換したドッケリンを同時生産するCtCBDcoh10CdocとCtCBDcoh10CdocN-Dを取得した。
【実施例6】
【0097】
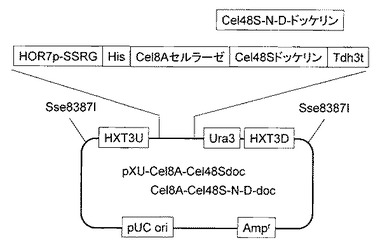
実施例5で取得した2種類の酵母CtCBDcoh10CdocとCtCBDcoh10CdocN-Dを、それぞれYP+2%グルコース培地で30℃、24時間培養し、OD600=0.5、62.5μl相当量集菌し、PBS溶液で洗浄を行い、PBS + 1mg/ml BSA + anti-His-FITC溶液と混合して4℃、30分間反応し、PBS溶液で2回洗浄後、Flow Cytometryで酵母細胞表層上のドッケリン提示量を評価した。Xyn10Cドッケリンでもアスパラギンをアスパラギン酸に置換する事でドッケリンの提示量が1.8倍に提示量が向上した(図8)。今回ターゲットとしたドッケリン内2箇所のアスパラギンはC. thermocellumのゲノム上で帰属されている142種類のドッケリンのうち82%で保存されている。2種類のドッケリンCel48SとXyn10Cでアミノ酸置換が同様の効果を示した事から、C. thermocellumが持つほとんどの酵素群に対して利用可能であると考えられる。
【実施例7】
【0098】
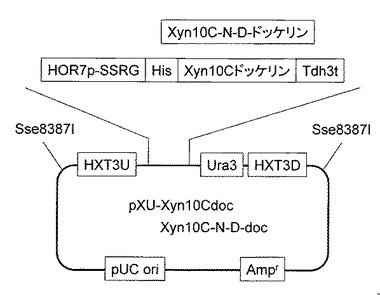
C.thermocellumのゲノムからCel8Aセルラーゼ遺伝子をPCR法により常法に従い増幅後クローニングした(配列番号224)。取得した遺伝子を、実施例3で取得したCel48Sドッケリン遺伝子、またはCel48Sドッケリンの18番目と50番目のアスパラギンをアスパラギン酸に置換した遺伝子とそれぞれ接合し、それぞれの遺伝子の上流にHXT3相同領域、HOR7プロモーターおよびHis-tag、下流にTdh3ターミネーター、Ura3マーカーおよびHXT3相同領域を持つ、pXU-Cel8A-Cel48SdocとpXU-Cel8A-Cel48S-N-D-docベクターを作製した(図9)。取得したベクターを実施例2で取得した酵母CtCBDcohにそれぞれ導入することで、C.thermocellum 由来コヘシンを細胞表層に提示し、ドッケリン型セルラーゼまたはアミノ酸を置換したドッケリン型セルラーゼを同時生産するCtCBDcohCel8A48SdocとCtCBDcohCel8A48SdocN-Dを取得した。
【実施例8】
【0099】
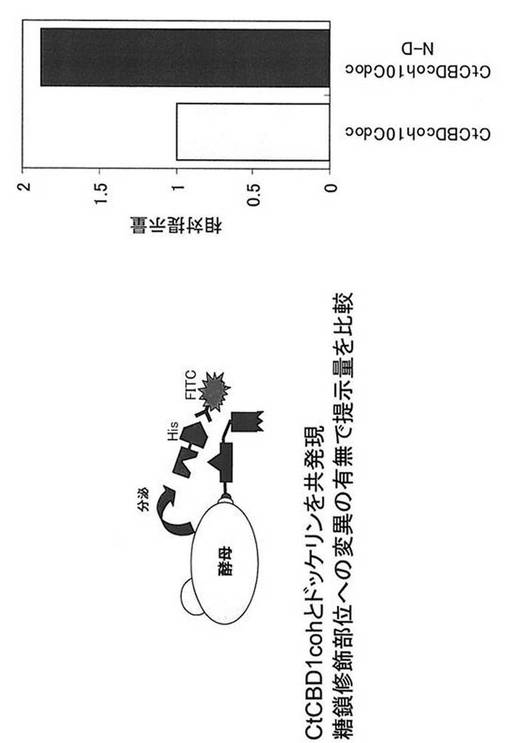
実施例7で取得した2種類の酵母CtCBDcohCel8A48SdocとCtCBDcohCel8A48SdocN-DをYP+2%グルコース培地で30℃、24時間培養し、OD600=0.5、62.5μl相当量集菌し、PBS溶液で1回洗浄を行い、PBS + 1mg/ml BSA + anti-His-FITC溶液と混合して4℃、30分間反応し、PBS溶液で2回洗浄後、Flow Cytometryで酵母細胞表層上のCelA提示量を評価した。アミノ酸を置換する事でCelA提示量の向上が認められた(図10)。
【実施例9】
【0100】
実施例7で取得した2種類の酵母CtCBDcohCel8A48SdocとCtCBDcohCel8A48SdocN-DをYP+2%グルコース培地で30℃、24時間培養し、OD600=1、1ml相当量集菌し、50mM 酢酸緩衝液 pH6.0溶液で洗浄後、1% CMC, 20mM 酢酸緩衝液pH6.0溶液に混合し、40℃で2時間CMC分解反応を行った。アミノ酸を置換する事でCMC分解活性の向上が認められ(図11)、酵母の糖化能力を向上させることが示された。
【配列表フリーテキスト】
【0101】
配列番号217,218,219,220,222,223:プライマー
【技術分野】
【0001】
本発明は、クロストリジウム・サーモセラム由来のタンパク質複合体を構築するためのタンパク質及びその利用に関する。
【背景技術】
【0002】
近年、有限である石油資源を代替するものとして、植物の光合成作用に由来するバイオマス資源への期待が高まってきており、バイオマスをエネルギーや各種材料に利用するための各種の試みがなされている。バイオマスを、エネルギー源やその他の原料として有効利用するためには、バイオマスを動物や微生物が容易に利用可能な炭素源に糖化することが必要である。
【0003】
典型的なバイオマスであるセルロースやヘミセルロースを利用するには、これらを糖化(分解)する優れたセルラーゼが必要である。こうしたセルラーゼ源として、一部の細菌が生産するセルロソームが着目されている。セルロソームは、細菌の細胞表層に形成されるタンパク質の複合体であって、セルラーゼが結合する骨格タンパク質(スキャホールディンタンパク質ともいう。)とセルラーゼとを備えている。スキャホールディンタンパク質は、コヘシンという部位を有しており、セルラーゼは、このコヘシンに対して、自身が備えるドッケリンを介して結合することが知られている。セルロソームは、細菌細胞表層に多種のセルラーゼを高密度でかつ大量に提供することができる。
【0004】
近年、遺伝子工学的に人工的なセルロソームを構築する検討がなされている。ここで、セルロソームの構築に関連して、セルロソームの構築する基礎となるコヘシンとドッケリンとの結合性(コヘシン−ドッケリン結合性)について種々の検討がなされている。例えば、クロストリジウム・サーモセラム(Clostridium thermocellum)のドッケリンのアミノ酸を数残基削除またはアラニンスキャンし、コヘシンとの結合性を評価して結合能に重要な残基を考察したものがある(非特許文献1)。この文献によれば、大腸菌で生産したドッケリンは、そのアミノ酸配列中のアスパラギンをアラニンに置換した際に、コヘシンとの結合量を70%程度は維持するが、構造安定化に寄与するカルシウムイオンとの相互作用が弱くなることが報告されている。また、C. thermocellumのドッケリン中の2つのへリックスを構成する2つの繰り返し配列のうち、一方の繰り返し配列のアミノ酸配列中にあるST(セリン−スレオニン)をAA(アラニン−アラニン)に置換することで結合能を損失させると、もう一方のへリックスでコヘシンと結合することも報告されている(非特許文献2)。さらに、コヘシンに関するものであるが、C. thermocellumのコヘシンのアミノ酸を数残基置換し、ドッケリンとの結合能を評価し、あるスレオニンをロイシンに置換する事で、C. thermocellum由来のドッケリンとの結合能を失い、本来は相互作用しないクロストリジウム・セルロリティカム(Clostridium cellulolyticum)由来のドッケリンと結合するようになることが報告されている(非特許文献3)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】A. KARPOL et al., Biochem. J 410, 331-338(2008)
【非特許文献2】A. L. Carvalho et al., PNAS 104(9), 3089-3094(2007)
【非特許文献3】A. Mechaly et al., J. Biol. Chem. 276(13), 9883-9888(2001)
【発明の概要】
【発明が解決しようとする課題】
【0006】
人工的なセルロソームの構築にあたっては、酵母などでセルラーゼを大量に生産し分泌させることが好ましいと考えられる。また、酵母等の真核微生物の細胞表層にセルロソームを構築できれば、セルロソームによって分解したグルコースを直接酵母の炭素源として、種々の有用物質を効率的に生産することができる。しかしながら、細菌等の原核生物に由来する異種タンパク質を酵母等の真核生物で生産させた場合、巨大糖鎖修飾がタンパク質間の相互作用に影響を与える可能性がある。
【0007】
ところで、以上の報告によれば、ドッケリンドメインのアミノ酸置換によりコヘシン−ドッケリン結合の結合能が低下したり(非特許文献1,2)、結合特異性が変化したり(非特許文献3)するなどの影響を与えることが理解される。しかしながら、コヘシン−ドッケリン結合の結合能力の向上については報告されていない。また、こうした報告は、いずれも、タンパク質への糖鎖修飾がなされない大腸菌で生産したコヘシン及びドッケリンについてなされているに過ぎない。したがって、糖鎖修飾が生じる酵母などの真核微生物における、ドッケリンドメインのアミノ酸置換のコヘシン−ドッケリン結合への影響は全く報告されていないのが現状である。
【0008】
そこで、本明細書の開示は、クロストリジウム・サーモセラム由来のタンパク質複合体を酵母などの糖鎖修飾が予想される真核微生物で生産させるのに適しており、しかも、コヘシン−ドッケリン結合の結合能力に優れる、ドッケリンを有するタンパク質及びその利用を提供することを一つの目的とする。
【課題を解決するための手段】
【0009】
本発明者らが、DDBJ(http://www.ddbj.nig.ac.jp/index-j.html)により、C.thermocellumのドッケリンの検索を行い、C. thermocellumのゲノム上で帰属されている72種類のドッケリンを見出し、さらに、UniProt(http://www.uniprot.org/)等を用いて、そのドッケリン中において、コヘシン−ドッケリン結合に関連すると思われる特異的な配列を見出し、これらの特異的配列につきマルチプルアラインメント等を実施して解析した。この結果、これらの142種類の特異的配列の類似性は90%を超えていた。このため、これらの特異的配列は、いずれもコヘシンとの結合能を有していると考えられた。
【0010】
また、発明者らは、これらの特異的配列のうち、約80%の113種類の関連配列には糖鎖修飾予測部位を有するが、残りの29種類には糖鎖修飾部位を有していないことを見出した。そして、本発明者らは、C. thermocellum由来のドッケリンの骨格タンパク質結合領域付近に存在する2箇所の糖鎖修飾予測部位をターゲットとして、当該部位のアスパラギンをアラニンまたはアスパラギン酸に置換してみた。ドッケリンのアスパラギンをアラニンに置換することで糖鎖修飾を解除したが、コヘシンとの結合能が向上しなかった。アスパラギンをアラニンに置換したドッケリンは酵母で生産すると安定な構造を取れずコヘシンと結合できなかった可能性がある。一方、ターゲット部位のアスパラギンをアスパラギン酸に置換したドッケリンを酵母で生産すると、コヘシン−ドッケリン間の結合量が増加し、結果として、酵母の糖化能力を向上させることができることがわかった。
【0011】
以上のことから、本来的に糖鎖修飾予測部位を有していないか、あるいは当該予測部位を有しているときには、その予測部位のアスパラギンをアスパラギン酸に置換して糖鎖修飾を解除することで、コヘシン−ドッケリン結合の結合能力を向上し、糖鎖修飾可能性のある真核微生物でも糖化能力を向上できるという知見を得た。本明細書の開示によれば、以下の手段が提供される。
【0012】
本明細書の開示によれば、C. thermocellum由来のタイプIコヘシンを有する骨格を利用したタンパク質複合体を構築するためのタンパク質であって、C. thermocellum由来のタイプIドッケリン中のコヘシン結合能に関連したドッケリン特異的配列であって、以下の(a)及び(b):
(a)内在性のN型糖鎖修飾予測部位を有しない
(b)内在性のN型糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されている
のいずれかを充足する少なくとも一つのドッケリン特異的配列を含むドッケリンを有するタンパク質が提供される。
【0013】
前記(a)を充足する前記ドッケリン特異的配列は、前記内在性のN型糖鎖修飾予測部位がアスパラギン酸であってもよい。
【0014】
また、本明細書に開示されるタンパク質は、セルロース分解促進活性を有していてもよく、さらに、前記セルロース分解促進活性はセルラーゼ活性であってもよい。また、セルロース分解促進活性は、クロストリジウム・サーモセラム由来のアミノ酸配列によって付与されていてもよい。
【0015】
本明細書の開示によれば、細胞表層にクロストリジウム・サーモセラム由来の骨格タンパク質を利用したタンパク質複合体を備える真核微生物であって、クロストリジウム・サーモセラム由来の骨格タンパク質と、前記骨格タンパク質に結合する本明細書に開示される前記タンパク質と、を備える、真核微生物が提供される。
【0016】
本明細書の開示によれば、有用物質の生産方法であって、本明細書に開示される前記真核微生物であって、前記ドッケリンタンパク質がセルロース分解促進活性を有する真核微生物を用いて、セルロース含有材料を炭素源として発酵する工程、を用いて、セルロース含有材料を糖化し、発酵する工程、を備える、方法が提供される。
【図面の簡単な説明】
【0017】
【図1】実施例1で作製したpAI-AGA1ベクターを示す図である。
【図2】実施例2で作製したLeu2マーカーおよびADH3相同領域を持つ、pDL-CtCBDCohAGA2ベクターを示す図である。
【図3】Cel48Sドッケリン遺伝子において、そのアミノ酸配列の18番目と50番目のアスパラギンをアラニンに置換したアミノ酸配列及びその遺伝子配列を示す図である。
【図4】実施例3で作製したpXU-Cel48Sdocベクター、pXU-Cel48S-N-A-docベクター、およびpXU-Cel48S-N-D-docベクターを示す図である。
【図5】アミノ酸置換したCel48Sドッケリンを含むタンパク質複合体表層提示酵母におけるドッケリン提示量を示す図である。
【図6】Xyn10Cドッケリン遺伝子において、そのアミノ酸配列の18番目と54番目のアスパラギンをアラニンに置換したアミノ酸配列及びその遺伝子配列を示す図である。
【図7】実施例5で作製したpXU-Xyn10CdocベクターとpXU-Xyn10C-N-D-docベクターを示す図である。
【図8】アミノ酸置換したXyn10Cドッケリンを含むタンパク質複合体表層提示酵母におけるドッケリン提示量を示す図である。
【図9】実施例7で作製したpXU-Cel8A-Cel48SdocとpXU-Cel8A-Cel48S-N-D-docベクターを示す図である。
【図10】アミノ酸置換したドッケリン型セルラーゼを含むタンパク質複合体表層提示酵母におけるセルラーゼ提示量を示す図である。
【図11】アミノ酸置換したドッケリン型セルラーゼを含むタンパク質複合体表層提示酵母のCMC分解活性を示す図である。
【発明を実施するための形態】
【0018】
本明細書の開示は、C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体を構築するためのタンパク質、当該タンパク質を含むタンパク質複合体を備える真核微生物及び当該真核微生物を用いる有用物質の生産方法に関する。
【0019】
本明細書に開示されるタンパク質は、C. thermocellum由来のタイプIドッケリン中のコヘシン結合能に関連した配列であって、内在性のN型糖鎖修飾予測部位を有しない又は内在性のN型糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されている、少なくとも一つのドッケリン特異的配列を有している。このため、本明細書に開示されるタンパク質は、酵母などの糖鎖修飾が予想される真核微生物で生産させたときであっても、糖鎖修飾が解除される。この結果、本明細書に開示されるタンパク質は、C. thermocellum由来の骨格タンパク質のタイプIコヘシンへの結合能に優れており、当該タンパク質の結合量及び/又は密度に優れたタンパク質複合体を構築できるものとなっている。
【0020】
また、本明細書に開示される真核微生物は、本明細書に開示されるタンパク質の集積量及び/又は集積密度に優れるタンパク質複合体を細胞表層に備えることができる。このため、本タンパク質の有する機能が増強された真核微生物を得ることができる。本タンパク質は、真核微生物で生産した場合においても、コヘシンへの結合能が優れているため、本明細書に開示される真核微生物が、前記骨格タンパク質及び前記タンパク質は、前記真核微生物が自己生産するものであってもよい。また、前記真核微生物は酵母であってもよい。
【0021】
さらに、本明細書に開示される有用物質の生産方法は、本明細書に開示される真核微生物であって、前記タンパク質がセルロース分解促進活性を有するタンパク質である真核微生物を用いてセルロース含有材料を炭素源として発酵する工程、を備える。本明細書に開示される真核微生物は、セルロース分解促進活性が増強されているため、セルロース含有材料を炭素源として効率的に発酵できる。
【0022】
(C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体を構築するためのタンパク質)
本明細書に開示されるタンパク質は、C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体の構築に特に適したタンパク質である。本タンパク質は、C. thermocellum由来のタイプIドッケリン中のコヘシン結合能に関連した配列であって、(a)内在性のN型糖鎖修飾予測部位を有しない、(b)内在性のN型糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されている、のいずれかを充足する少なくとも一つのドッケリン特異的配列を含むドッケリンを有することができる。
【0023】
C. thermocellumは、セルロソーム生産微生物として知られている。また、C. thermocellumは、セルラーゼ活性を有し、タイプIドッケリンを含むタンパク質を生産している。DDBJ(http://www.ddbj.nig.ac.jp/index-j.html)によるC. thermocellumのゲノムの検索結果に基づけば、C. thermocellumのタイプIドッケリンのアミノ酸配列として、以下の表1の配列番号1〜72に示す72種類のアミノ酸配列が挙げられる。表1において示すlocus(配列名)は、各ドッケリンの名称である。したがって、表2〜表21のlocus欄に示すドッケリン名で特定されるアミノ酸配列は、表1に記載の同一の名称のドッケリンのアミノ酸配列に由来している。
【0024】
【表1】
【0025】
さらに、これらの72種類のタイプIドッケリン中には、類似したアミノ酸配列であってコヘシン結合能に関連したドッケリン特異的配列を見出すことができる。ドッケリン特異的配列は、24アミノ酸からなる天然由来のコンセンサス配列(関連配列)からなることができる。72種類のタイプIドッケリンに内在するこの関連配列として、以下の表2に示す配列番号73〜214の合計142種類のアミノ酸配列が挙げられる。なお、これらのアミノ酸配列は、UniProt(http://www.uniprot.org/)、InterPro(http://www.ebi.ac.uk/interpro/)及びPfam(http://pfam.sanger.ac.uk)などのデータベースにより取得することができる。表2における1st欄にはN末端側の関連配列が記載され、2nd欄にはC末端側の関連配列が記載されている。
【0026】
【表2】
【0027】
表1に示すドッケリンについて相同性検索を行うと、これらのアミノ酸配列間では「相同性」が90%以上を超えることがない一方、表2に示す関連配列間で「類似性」は90%以上である。以上のことから、表1に挙げられたドッケリンは、いずれも、同等の機能を発揮していると推論できる。したがって、表2に示す関連配列が当該機能を担っていることが推論できる。
【0028】
表1に示すドッケリン、すなわち、表2に示す関連配列におけるN型糖鎖修飾予測部位は、酵母などの真核微生物でN型糖鎖修飾を受けるコンセンサス配列であるN−X−T又はN−X−S(Nはアスパラギン、Xはプロリン以外のアミノ酸、Tはスレオニン、Sはセリン)におけるNの位置となることが知られている(A. Herscovics et al., The FASEB Journal (6): 540-550(1993))。ドッケリン又はその関連配列におけるN−X−T/Sは、既に説明したデータベース等を適宜利用することなどにより検出できる。また、ドッケリン又はその関連配列におけるN型糖鎖修飾予測部位に相当する部位は、そのアミノ酸配列中の上記コンセンサス配列を備えてはいないが、Nに相当しうる部位をいう。N型糖鎖修飾予測部位に相当する部位は、当該部位を含む可能性のあるアミノ酸配列を他の公知のドッケリン又はその関連配列のアミノ酸配列とマルチプルアラインメントなどを利用して比較することにより、検出することができる。ドッケリンにおけるN型糖鎖修飾予測部位及び当該部位に相当する部位は、典型的には、関連配列を構成するアミノ酸配列を24個以下程度のアミノ酸からなるアミノ酸配列とするとき、そのN末端側から、通常、9個目がN型糖鎖修飾予測部位又は当該部位に相当する部位となっている。
【0029】
本タンパク質は、N型糖鎖修飾予測部位を有していないドッケリン特異的配列を少なくとも一つ含むドッケリンを有することが好ましい。そして、N型糖鎖修飾予測部位に相当する部位のアミノ酸がアスパラギン酸(D)であるドッケリン特異的配列を少なくとも一つ有することが好ましい。N型糖鎖修飾部位を有しないこと及び/又は糖鎖修飾予測部位に相当する部位がアスパラギン酸であることで、酵母などの真核微生物によるN型糖鎖修飾が解除されるものと考えられる。糖鎖修飾予測部位に相当する部位がアスパラギン酸であるドッケリン特異的配列は、本来的にドッケリンに内在するものであってもよいし、N型糖鎖修飾部位を有していて、そのアスパラギン(N)がアスパラギン酸(D)に置換されたものであってもよい。
【0030】
こうしたドッケリン特異的配列の一つの態様として、表1に開示されるドッケリン及び表2に開示されるこれらのドッケリンにおける関連配列が内在性のN型糖鎖修飾予測部位を有するとき、そのアスパラギンがアスパラギン酸に置換されているドッケリン特異的配列が挙げられる。本タンパク質は、こうしたドッケリン特異的配列を少なくとも一つ含んだドッケリンを有していればよい。こうしたNからDへの置換対象部位を有する関連配列としては、以下の113種類の関連配列が挙げられる。したがって、好ましいドッケリン特異的配列は、以下の関連配列において(N−X−T/S)のNがDに置換された配列で表される。
【0031】
【表3】
【0032】
本タンパク質は、こうしたドッケリン特異的配列を1又は2つ含むドッケリンを備えることができるが、典型的には、関連配列中の糖鎖修飾予測部位においてアスパラギンがアスパラギン酸に置換されていることが好ましい。こうしたドッケリンとしては、以下の表に示すドッケリンが挙げられる。これらの表においては、ドッケリンが有する関連配列で特徴付けて示している。したがって、好ましいドッケリンは、以下の表において各ドッケリンが有する1又は2つの関連配列において(N−X−T/S)のNがDに置換されたドッケリン特異的配列を有することができる。
【0033】
【表4】
【0034】
表4に示すドッケリンは、ドッケリン中に二つの関連配列を有し、それぞれの関連配列がN型糖鎖修飾予測部位を有している。こうしたドッケリンにあっては、二つの関連配列のいずれか又は双方のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで、好ましいドッケリンを得ることができる。
【0035】
【表5】
【0036】
表5に示すドッケリンは、ドッケリン中に1又は2つの関連配列を有し、N末端側の関連配列にN型糖鎖修飾予測部位を有している。こうしたドッケリンにあっては、この関連配列のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで、好ましいドッケリンを得ることができる。
【0037】
【表6】
【0038】
表6に示すドッケリンは、ドッケリン中に二つの関連配列を有し、C末端側の関連配列にN型糖鎖修飾予測部位を有している。こうしたドッケリンにあっては、この関連配列のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換してドッケリン特異的配列とすることで、好ましいドッケリンを得ることができる。
【0039】
本タンパク質において好ましいドッケリンを含むドッケリン特異的配列は、既に文献等においてコヘシンとの結合能が確認されている以下の表に示すC. thermocellumのタイプIドッケリンとの関係で選択される。以下の表においては、ドッケリンは、二つの関連配列で特徴付けて示している。これらのドッケリンに含まれるいずれかの関連配列とのアミノ酸配列の類似性が90%以上の関連配列を有するとき、この関連配列中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列を含む好ましいドッケリンを得ることができる。
【0040】
【表7】
【0041】
【表8】
【0042】
表8に示すドッケリンは、N末端側に一つの関連配列を有し、この関連配列が既にコヘシン結合能が確認された表7に示す8種のドッケリン中のいずれかの関連配列とのアミノ酸配列の類似性が90%以上である。この関連配列において、N型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで好ましいドッケリンを得ることができる。
【0043】
【表9】
【0044】
表9に示すドッケリンは、いずれも2つの関連配列を有し、C末端側の関連配列が既にコヘシン結合能が確認された表7に示す8種のドッケリン中のいずれかの関連配列とのアミノ酸配列の類似性が90%以上である。この関連配列において、N型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで好ましいドッケリンを得ることができる。
【0045】
【表10A】
【表10B】
【0046】
表10に示すドッケリンは、いずれも2つの関連配列を有し、二つの関連配列が既にコヘシン結合能が確認された表7に示す8種のドッケリン中のいずれかの関連配列とのアミノ酸配列の類似性が90%以上である。これらの関連配列のいずれか又は双方において、N型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで好ましいドッケリンを得ることができる。
【0047】
さらに、ドッケリンが、表7に示すコヘシン結合能を有する既知のいずれかのドッケリンのアミノ酸配列とアミノ酸配列の類似性が90%以上であるとき、このドッケリン中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換したドッケリン特異的配列とすることで好ましいドッケリンを得ることができる。
【0048】
【表11】
【0049】
表11に示すドッケリンは、そのアミノ酸配列が、表7に示す8種のドッケリン中のいずれかのアミノ酸配列との類似性が90%以上である。このアミノ酸配列において、少なくとも一つのN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換することで好ましいドッケリンを得ることができる。
【0050】
さらにまた、ドッケリンが、表7に示すコヘシン結合能を有する既知のいずれかのドッケリンの関連配列のアミノ酸配列との相同性が90%以上である関連配列を有するとき、このドッケリン中のその関連配列中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換することで好ましいドッケリンを得ることができる。また、前記既知のドッケリンのアミノ酸配列と90%以上の相同性を有するドッケリン中のN型糖鎖修飾予測部位も好ましい置換対象である。こうしたドッケリンは、表7に示すドッケリンが該当するのみである。これらのドッケリンにおける関連配列中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換することで好ましいドッケリンを得ることができる。
【0051】
ドッケリン特異的配列の他の一つの態様は、表1に開示されるドッケリン及び表2に開示されるこれらのドッケリンにおける関連配列において内在性のN型糖鎖修飾予測部位を有しないドッケリン特異的配列が挙げられる。本タンパク質は、こうしたドッケリン特異的配列を少なくとも一つ含むドッケリンを有していればよい。この種のドッケリン特異的配列となる関連配列としては、以下の29種類の関連配列が挙げられる。
【0052】
【表12】
【0053】
本タンパク質は、表12に示すドッケリン特異的配列を1又は2つ含むドッケリンを備えていることができるが、典型的には、上記ドッケリン特異的配列を本来的に有するドッケリンであることが好ましい。こうしたドッケリンとしては、以下の表に示すドッケリンが挙げられる。これらの表においては、ドッケリンは、ドッケリンが有する関連配列で特徴付けて示している。
こうしたドッケリン特異的配列において、N型糖鎖修飾予測部位に相当する部位のアミノ酸がアスパラギン酸であることが好ましい。こうしたドッケリン特異的配列を1又は2つ有するドッケリンが好ましい。
【0054】
【表13】
【0055】
表13に示すドッケリンは、ドッケリン中に二つの関連配列を有し、いずれの関連配列にもN型糖鎖修飾予測部位を有していない。
【0056】
本タンパク質において好ましいドッケリンが含む内在性のN型糖鎖修飾予測部位を有しないドッケリン特異的配列は、既に文献等においてコヘシンとの結合能が確認されている表7に示すC. thermocellumのタイプIドッケリンとの関係で選択される。これらのドッケリンに含まれるいずれかの関連配列とのアミノ酸配列の類似性が90%以上の関連配列を有するとき、この関連配列中のN型糖鎖修飾予測部位に相当する部位がアスパラギン酸である天然型ドッケリン特異的配列であれば好ましいドッケリンとして用いることができる。
【0057】
【表14】
【0058】
表14に示すドッケリンは、2つの関連配列を有し、これらの関連配列がいずれも既にコヘシン結合能が確認された表7に示す8種のドッケリン中のいずれかの関連配列とのアミノ酸配列の類似性が90%以上である。そして、これらの関連配列のいずれか又は双方が、天然型ドッケリン特異的配列である。なお、天然型ドッケリン特異的配列でない関連配列中のN型糖鎖修飾予測部位のアスパラギンをアスパラギン酸に置換してもよい。
【0059】
さらに、ドッケリンが、前記既知のいずれかのドッケリンのアミノ酸配列とアミノ酸配列の類似性が90%以上であるとき、このドッケリンに内在する関連配列中のN型糖鎖修飾予測部位に相当する部位がアスパラギン酸である天然型ドッケリン特異的配列を少なくとも一つ有していれば、好ましいドッケリンとして用いることができる。
【0060】
【表15】
【0061】
表15に示すドッケリンは、そのアミノ酸配列が、表7に示す8種のドッケリン中のいずれかのアミノ酸配列との類似性が90%以上である。このアミノ酸配列において、少なくとも一つのN型糖鎖修飾予測部位に相当する部位がアスパラギン酸である天然型ドッケリン特異的配列を有することで好ましいドッケリンとして用いることができる。なお、N型糖鎖修飾予測部位がアスパラギンである他の関連配列を有する場合には、当該アスパラギンをアスパラギン酸に置換してもよい。
【0062】
本タンパク質は、ドッケリン以外に活性部位を備えることができる。活性部位の種類は用途に応じて適宜決定される。本タンパク質は、ドッケリンと活性部位とを適宜組み合わせた人工的なタンパク質であってもよい。また、例えば、セルロソームの構成タンパク質であるセルラーゼであって、ドッケリンを本来的に有するセルラーゼをそのままあるいは適宜改変して用いることもできる。
【0063】
本タンパク質は、バイオマスに由来するセルロース含有材料の糖化利用に際しては、例えば、セルロース分解促進活性を有することができる。すなわち、、セルロース分解促進活性部位を備えることができる。セルロース分解促進活性とは、セルラーゼ活性、セルロース結合活性、セルロース緩和活性などが挙げられる。
【0064】
セルラーゼ活性部位は、公知のセルラーゼにおける活性部位を適宜利用できる。セルラーゼとしては、エンドグルカナーゼ(EC 3.2.1.74)、セロビオヒドロラーゼ(EC 3.2.1.91)及びβ−グルコシダーゼ(EC23.2.4.1、EC 3.2.1.21)が挙げられる。なお、セルラーゼは、そのアミノ酸配列の類似性に基づきGHF(Glycoside Hydrolase family)(http://www.cazy.org/fam/acc.gh.html)の13(5,6,7,8,9,10,12,44,45,48,51,61,74)のファミリーに分類されている。異なるファミリーに分類される同種又は異種のセルラーゼを組み合わせてもよい。
【0065】
セルラーゼとしては、特に限定しないが、それ自体活性の高いセルラーゼであることが好ましい。このようなセルラーゼとしては、ファネロケーテ(Phanerochaete)属菌、Trichoderma reeseiなどのトリコデルマ属(Trichoderma)菌、フザリウム属(Fusarium)菌、トレメテス属(Tremetes)菌、ペニシリウム属(Penicillium)菌、フミコーラ属(Humicola)菌、アクレモニウム属(Acremonium)菌、アスペルギルス属(Aspergillus)菌等の糸状菌の他に、クロストリジウム属(Clostridium)菌、シュードモナス属(Pseudomonas)菌、セルロモナス属(Cellulomonas)菌、ルミノコッカス属(Ruminococcus)菌、バチルス属(Bacillus)菌等の細菌、スルフォロバス属(Sulfolobus)菌等の始原菌、さらにストレプトマイセス属(Streptomyces)菌、サーモアクチノマイセス属(Thermoactinomyces)菌などの放射菌由来のセルラーゼが挙げられる。なお、こうしたセルラーゼ又はその活性部位は、人工的に改変されていてもよい。
【0066】
本タンパク質は、C. thermocellum由来のタンパク質でもあるため、セルロース分解促進活性は、クロストリジウム・サーモセラム由来のアミノ酸配列によって付与されていることが好ましい。
【0067】
本タンパク質は、バイオマスの有効利用を考慮したとき、ヘミセルラーゼ活性部位を備えていてもよい。さらに、リグニンペルオキシダーゼ、マンガンペルオキシダーゼ及びラッカーゼなどのリグニン分解酵素が挙げられる。また、例えば、セルロース緩和タンパク質であるスウォレニンやエクスパンシン、セルロソームやセルラーゼの構成部分であるセルロース結合ドメイン(タンパク質)が挙げられる。また、キシラナーゼやヘミセルラーゼ等のその他のバイオマス分解酵素も挙げられる。これらのタンパク質は、いずれもセルロースへのセルラーゼのアクセシビリティを向上させることができる。
【0068】
本タンパク質は、真核微生物において細胞外分泌性を備えていることが好ましい。すなわち、真核微生物において分泌タンパク質として生産されるタンパク質であることが好ましい。セルラーゼなどの酵素は、本来的に細胞外分泌のためのシグナルを有していることが多い。ドッケリンタンパク質に細胞外分泌性を付与するには、公知の分泌シグナルを用いることができる。分泌シグナルは、用いる真核微生物の種類に応じて適宜選択される。分泌シグナル等については後段にて説明する。
【0069】
当業者であればこのような本タンパク質を適当な宿主微生物で遺伝子組換え等により生産させることができるほか、化学合成によっても得ることができる。
【0070】
以上説明した、本タンパク質は、特定のドッケリンを有していることから、C. thermocellum由来のタイプIコヘシンに対する結合性が向上しており、こうしたコヘシンを有する骨格タンパク質上への集積量や集積密度を高めることができる。
【0071】
(C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質)
本タンパク質は、C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質との複合体の構築のためのタンパク質として好適である。C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質は、C. thermocellum由来の1又は2以上のタイプIコヘシンをを備えることができる。コヘシンは、セルロソーム生産微生物の形成するセルロソームにおけるタイプI等の骨格タンパク質に備えられる触媒活性のあるセルラーゼ等を非共有結合で結合するドメインとして知られている(粟冠ら、蛋白質核酸酵素、Vol.44、No.10(1999)、p41-p50、Demain, A. L., et al., Microbiol Mol. Biol Rev., 69(1), 124-54(2005), Doi, R. H., et al., J. Bacterol., 185(20), 5907-5914(2003)等)。本タンパク質を結合するためのC. thermocellum由来の骨格タンパク質としては、タイプI骨格タンパク質上のタイプIコヘシンドメインを少なくとも有している。そのほか、タイプII骨格タンパク質上のタイプIIコヘシンドメイン及びタイプIII骨格タンパク質上のタイプIIIコヘシンドメインを備えていてもよい。こうした各種タイプのコヘシンドメインとしては、各種セルロソーム生産微生物において多数その配列が決定されている。これらの各種のタイプのコヘシンのアミノ酸配列及びDNA配列は、NCBIのHP(http://www.ncbi.nlm.nih.gov/)等を介してアクセス可能な各種のタンパク質データベースやDNA配列のデータベースにより容易に取得することができる。
【0072】
C. thermocellum由来のコヘシンを有する骨格タンパク質は、C. thermocellum由来のタイプIコヘシンを有していればよく、C. thermocellum由来の骨格タンパク質自体でなくともよく、人工タンパク質であってもよい。また、骨格タンパク質は、C. thermocellum由来の天然のタイプIコヘシン又はその結合性を有する限りそのコヘシンのアミノ酸配列において1又は2以上の変異(付加、挿入、欠失及び置換)を導入した改変コヘシンを備えていてもよい。コヘシンタンパク質は、こうしたコヘシン等を適当なインターバルを置いて複数個備えることもできる。コヘシンタンパク質の全体のアミノ酸配列や、コヘシン間のアミノ酸配列の種類や有無は、タイプI骨格タンパク質のアミノ酸配列及び当該アミノ酸配列に適宜変異を導入した配列を利用できる。
【0073】
骨格タンパク質は、タイプI〜IIIから選択される骨格タンパク質のセルロース結合ドメイン(CBD)を有していてもよい。CBDは、各種骨格タンパク質において基質であるセルロースに結合するドメインとして知られている(前述粟冠ら)。セルロース結合ドメインは、1個又は2個以上有していてもよい。各種のセルロソーム生産微生物のセルロソームにおけるCBDのアミノ酸配列及びDNA配列の多くが決定されている。これらの各種のCBDのアミノ酸配列及びDNA配列は、NCBIのHP(http://www.ncbi.nlm.nih.gov/)等を介してアクセス可能な各種のタンパク質データベースやDNA配列のデータベースにより容易に取得することができる。
【0074】
骨格タンパク質は、真核微生物において細胞外分泌性又は細胞表層提示性を備えていることが好ましい。すなわち、真核微生物において分泌タンパク質として生産されるタンパク質であることが好ましいほか、真核微生物の細胞表層に提示されるタンパク質であることが好ましい。コヘシンタンパク質に細胞外分泌性又は細胞表層提示性を付与するには、公知の分泌シグナルや表層提示用のシステムを用いることができる。
【0075】
当業者であればこのような各種ドメインを適宜有する骨格タンパク質を適当な宿主微生物で遺伝子組換え等により生産させることができる。こうした各種骨格タンパク質のコヘシンドメインを有するコヘシンタンパク質は、化学合成によっても得ることができる。
【0076】
以上説明した、本タンパク質は、特定のドッケリンを有していることから、C. thermocellum由来のタイプIコヘシンに対する結合性が向上しており、こうしたコヘシンを有する骨格タンパク質上への集積量や集積密度を高めることができる。
【0077】
(タンパク質複合体)
本明細書の開示によれば、C. thermocellum由来のタイプIコヘシンを有する骨格タンパク質と、この骨格タンパク質に結合された本タンパク質と、を備えるタンパク質複合体も提供される。このタンパク質複合体は、本タンパク質の集積量及び/又は集積密度が高くなっているため、本タンパク質の活性が増強されたものとなっている。
【0078】
(タンパク質複合体を細胞表層に備える真核微生物)
本明細書に開示される真核微生物は、本明細書に開示されるタンパク質複合体を細胞表層に備えている。本真核微生物は、タンパク質複合体を構成する骨格タンパク質と本タンパク質とを細胞外から供給して自己集合的に細胞表層にて構築されたものであってもよいが、好ましくは、これらのタンパク質を自己生産する。本タンパク質は、真核微生物において生産してもその糖鎖修飾系による糖鎖修飾を解除又は抑制して、対コヘシン結合性が向上しているからである。
【0079】
本タンパク質がセルラーゼ活性等のセルロース分解促進活性を有するとき、真核微生物の細胞表層にセルラーゼなどのセルロース分解促進活性を有するタンパク質を集積させたタンパク質複合体を構築することができる。こうした真核微生物は、細胞表層でセルロース含有材料を分解糖化して得られるグルコースを炭素源として利用できる。
【0080】
こうしたタンパク質をコードするコード化DNAは、宿主となる真核微生物内において当該タンパク質を発現可能に保持されていればよく、その保持形態は特に限定されない。例えば、真核微生物で作動可能なプロモーターの制御下に連結されるとともに適切なターミネーターをその下流に有した状態で保持されている。プロモーターは、構成的プロモーターであっても誘導的プロモーターであってもよい。このような状態のDNAは、宿主染色体内に組み込まれた形態であってもよいし、宿主核内に保持される2μプラスミドや核外に保持されるプラスミドのような形態であってもよい。一般には、こうした外来DNAの導入に伴って、宿主において利用可能な選択マーカー遺伝子も同時に保持されている。
【0081】
真核微生物において生産するドッケリンタンパク質やコヘシンタンパク質に細胞外分泌性や細胞表層提示性を付与することが好ましい。なかでも、本タンパク質には細胞外分泌性を付与することが好ましく、骨格タンパク質には、さらに細胞外に分泌させて細胞表層に提示させる細胞表層提示性を付与することが好ましい。細胞外分泌性を付与するには、分泌シグナルを付与することができる。分泌シグナルとしては、例えば、Rhizopus oryzaeやC. albicansのグルコアミラーゼ遺伝子の分泌シグナル、酵母インベルターゼリーダー、α因子リーダーなどが挙げられる。また、凝集性タンパク質あるいはその一部を用いることでタンパク質を真核微生物の表層に提示した状態に分泌させることができる。例えば、凝集性タンパク質であるα−アグルチニンをコードするSAG1遺伝子の5’領域の320アミノ酸残基からなるペプチドがある。また、所望のタンパク質を細胞表層に提示するためのポリペプチドや手法は、WO01/79483号公報や、特開2003−235579号公報、WO2002/042483号パンフレット、WO2003/016525号パンフレット、特開2006−136223号公報、藤田らの文献(藤田ら,2004. Appl Environ Microbiol 70:1207-1212および藤田ら, 2002. Appl Environ Microbiol 68:5136-5141.)、村井ら, 1998. Appl Environ Microbiol 64:4857-4861.に開示されている。
【0082】
真核微生物としては、特に限定されないで、例えば、公知の各種酵母を利用できる。後述するエタノール発酵等を考慮すると、サッカロマイセス・セレビジエ(Saccharomyces cerevisiae)等のサッカロマイセス属の酵母、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)等のシゾサッカロマイセス属の酵母、キャンディダ・シェハーテ(Candida shehatae)等のキャンディダ属の酵母、ピヒア・スティピティス(Pichia stipitis)等のピヒア属の酵母、ハンセヌラ(Hansenula)属の酵母、トリコスポロン(Trichosporon)属の酵母、ブレタノマイセス(Brettanomyces)属の酵母、パチソレン(Pachysolen)属の酵母、ヤマダジマ(Yamadazyma)属の酵母、クルイベロマイセス・マーキシアヌス(Kluyveromyces marxianus)、クルイベロマイセス・ラクティス(Kluveromyces lactis)等のクルイベロマイセス属の酵母が挙げられる。なかでも、工業的利用性等の観点からサッカロマイセス属酵母が好ましい。なかでも、サッカロマイセス・セレビジエが好ましい。
【0083】
外来タンパク質を発現する真核微生物は、モレキュラークローニング第3版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載されている方法に準じて作製することができる。本タンパク質や骨格タンパク質を真核微生物において発現させるためのベクター及びその構築方法も、同様に、当業者において周知である。なお、ベクターの形態は、使用形態に応じて様々な形態を採ることができる。例えば、DNA断片の形態を採ることができるほか、2マイクロプラスミドなどの適当な酵母用ベクターの形態を採ることもできる。このようなベクターで真核微生物を形質転換することによって本明細書に開示される真核微生物を得ることができる。形質転換にあたり、従来公知の各種方法、例えば、トランスフォーメーション法や、トランスフェクション法、接合法、プロトプラスト法、エレクトロポレーション法、リポフェクション法、酢酸リチウム法等を用いることができる。
【0084】
(有用物質の生産方法)
本明細書に開示される有用物質の生産方法は、本明細書に開示される真核微生物であって、本タンパク質がセルロース分解促進活性を有する真核微生物を用いて、セルロース含有材料を炭素源として発酵する工程、を用いて、セルロース含有材料を糖化し、発酵する工程、
を備えることができる。この方法によれば、この真核微生物を用いてセルロース含有材料を直接分解糖化し、グルコース等として利用できることになる。前記発酵工程の実施により、用いた真核微生物が有している有用物質生産能力に応じて有用物質が生産される。
【0085】
有用物質は、真核微生物がグルコースなどを発酵することにより得る生産物であり、真核微生物の種類によっても異なるし、発酵条件によっても異なる。有用物質としては特に限定しないが、酵母やその他の真核微生物がグルコースを利用して生産可能なものであればよい。有用物質は、酵母などの真核微生物におけるグルコースからの代謝系の1種又は2種以上の酵素を遺伝子組換えにより置換、追加等して合成できるようになった本来の代謝物でない化合物であってもよい。有用物質としては、例えば、エタノールなどのほか、C3〜C5の低級アルコール、乳酸などの有機酸の他、イソプレノド合成経路の追加によるファインケミカル(コエンザイムQ10、ビタミン及びその原料等)、解糖系の改変によるグリセリン、プラスチック・化成品原料など、バイオリファイナリー技術が対象とする材料が挙げられる。有用物質の生産工程終了後、培養液から有用物質含有画分を回収する工程、さらにこれを精製又は濃縮する工程を実施することもできる。回収工程や精製等の工程は有用物質の種類等に応じて適宜選択される。
【0086】
真核微生物が細胞表層にタンパク質複合体として保持する本タンパク質は、2種類以上のセルロース分解促進活性を有していることが好ましい。セルラーゼとしては、例えば、エンドグルカナーゼとセロビオヒドロラーゼ等の活性をそれぞれ有する2種類以上を用いることが好ましい。
【0087】
なお、セルロース含有材料は、D−グルコースがβ−1,4結合でグリコシド結合したβ−グルカンであるセルロースを含有する材料である。セルロース含有材料としては、セルロースを含有していればよく、どのような由来や形態であってもよい。したがって、セルロース系材料としては、例えば、リグノセルロース系材料、結晶性セルロース材料、可溶性セルロース材料(非晶性セルロース材料)、不溶性セルロース材料などの各種セルロース系材料等が含まれる。リグノセルロース系材料としては、例えば、木本植物の木質部や葉部及び草本植物の葉、茎、根等においてリグニン等を複合した状態のリグノセルロース系材料が挙げられる。こうしたリグノセルロース系材料としては、例えば、稲ワラ、麦ワラ、トウモロコシの茎葉、バガス等の農業廃棄物、収集された木、枝、枯葉等又はこれらを解繊して得られるチップ、おがくず、チップなどの製材工場廃材、間伐材や被害木などの林地残材、建設廃材等の廃棄物であってもよい。結晶性セルロース系材料及び不溶性セルロース系材料としては、リグノセルロース系材料からリグニン等を分離後の結晶性セルロース及び不溶性セルロースを含む結晶性又は不溶性セルロース系材料が挙げられる。セルロース材料としては、また、使用済み紙製容器、古紙、使用済みの衣服などの使用済み繊維製品、パルプ廃液を由来としてもよい。
【0088】
セルロース含有材料は、セルラーゼと接触させるのに先立ってセルラーゼによる分解を容易化するために適当な前処理等がなされていてもよい。例えば、硫酸、塩酸、リン酸、硝酸などの無機酸による酸性条件下、セルロースを部分加水分解することにより、セルロースを非晶質化あるいは低分子化できる。この他、超臨界水、アルカリ、加圧熱水などの処理によってもセルロースの非晶質化又は低分子化を行うことができる。
【0089】
セルロース含有材料は、グルコースがβ-1,4-グリコシド結合により重合した重合体及びその誘導体を含んでいる。グルコースの重合度は特に限定しない。また、誘導体としては、カルボキシメチル化、アルデヒド化、若しくはエステル化などの誘導体が挙げられる。セルロース は、結晶性セルロースであってもよいし、非結晶性セルロースであってもよい。
【0090】
本明細書において同一性又は類似性とは、当該技術分野で知られているとおり、配列を比較することにより決定される、2以上のタンパク質あるいは2以上のポリヌクレオチドの間の関係である。当該技術で“同一性 ”とは、タンパク質またはポリヌクレオチド配列の間のアラインメントによって、あるいは場合によっては、一続きのそのような配列間のアラインメントによって決定されるような、タンパク質またはポリヌクレオチド配列の間の配列不変性の程度を意味する。また、類似性とは、タンパク質またはポリヌクレオチド配列の間のアラインメントによって、あるいは場合によっては、一続きの部分的な配列間のアラインメントによって決定されるような、タンパク質またはポリヌクレオチド配列の間の相関性の程度を意味する。より具体的には、配列の同一性と保存性(配列中の特定アミノ酸又は配列における物理化学特性を維持する置換)によって決定される。なお、類似性は、後述するBLASTの配列相同性検索結果においてSimilarity と称される。同一性及び類似性を決定する方法は、対比する配列間で最も長くアラインメントするように設計される方法であることが好ましい。同一性及び類似性を決定するための方法は、公衆に利用可能なプログラムとして提供されている。例えば、AltschulらによるBLAST (Basic Local Alignment Search Tool) プログラム(たとえば、Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ., J. Mol. Biol., 215: p403-410 (1990), Altschyl SF, Madden TL, Schaffer AA, Zhang J, Miller W, Lipman DJ., Nucleic Acids Res. 25: p3389-3402 (1997))を利用し決定することができる。BLASTのようなソフトウェアを用いる場合の条件は、特に限定するものではないが、デフォルト値を用いるのが好ましい。
【実施例】
【0091】
以下、本発明を、実施例を挙げて具体的に説明するが、本発明はこれらの実施例に限定されるものではない。なお、以下に述べる遺伝子組換え操作はMolecular Cloning: A Laboratory Manual (T. Maniatis, et al., Cold Spring Harbor Laboratory) に従い行った。
【実施例1】
【0092】
PCR法により常法に従い増幅後クローニングしたaga1遺伝子の上流にAAP1相同領域とHOR7プロモーター、下流にTdh3ターミネーターとHis3マーカーおよびAAP1相同領域を持つ、pAI-AGA1ベクター(図1)を作製した。このベクターを用いて酵母S.cerevisiae BY4741に形質転換を行い、aga1を細胞表層に大量提示する酵母BY-AGA1を取得した。
【実施例2】
【0093】
C.thermocellumのゲノムから、CBD-コヘシンをPCR法により常法に従い増幅後クローニングした(配列番号215)。取得した遺伝子の上流にADH3相同領域とHOR7プロモーター、下流にV5-tag、aga2、Tdh3ターミネーター、Leu2マーカーおよびADH3相同領域を持つ、pDL-CtCBDCohAGA2ベクターを作製した(図2)。作製したベクターを実施例1で取得した酵母BY-AGA1に導入して、C.thermocellum 由来コヘシンを細胞表層に提示する酵母CtCBDcohを取得した。
【実施例3】
【0094】
C.thermocellum のゲノムからCel48Sドッケリン遺伝子をPCR法により常法に従い増幅後クローニングした(配列番号216)。取得したCel48Sドッケリン遺伝子を鋳型として48Sdock-N18A-Fw と48Sdock-N50A-Rvの2種類のプライマー(配列番号217,218)を利用して 18番目と50番目のアスパラギンをアラニンに置換した遺伝子を取得した。同様にCel48Sドッケリン遺伝子を鋳型として48Sdock-N18D-Fw と48Sdock-N50D-Rvの2種類のプライマー(配列番号219、220)を利用して 18番目と50番目のアスパラギンをアスパラギン酸に置換した遺伝子を取得した(図3)。取得したそれぞれの遺伝子の上流にHXT3相同領域、HOR7プロモーターおよびHis-tag、下流にTdh3ターミネーター、Ura3マーカーおよびHXT3相同領域を持つ、pXU-Cel48Sdocベクター、pXU-Cel48S-N-A-docベクター、およびpXU-Cel48S-N-D-docベクターを作成した(図4)。取得したベクターを実施例2で取得した酵母CtCBDcohにそれぞれ導入することで、C.thermocellum 由来コヘシンを細胞表層に提示し、ドッケリンまたはアミノ酸置換したドッケリンを同時生産するCtCBDcoh48SdocとCtCBDcoh48SdocN-A 、およびCtCBDcoh48SdocN-Dを取得した。
【実施例4】
【0095】
実施例3で取得した3種類の酵母CtCBDcoh48SdocとCtCBDcoh48SdocN-A 、およびCtCBDcoh48SdocN-Dを、それぞれYP+2%グルコース培地で30℃、24時間培養し、OD600=0.5、62.5μl相当量集菌し、PBS溶液で洗浄を行い、PBS + 1mg/ml BSA + anti-His-FITC溶液と混合して4℃、30分間反応し、PBS溶液で2回洗浄後、Flow Cytometryで酵母細胞表層上のドッケリン提示量を評価した。アスパラギンをアラニンに置換する事でCel48Sドッケリンの提示量は約半分に低下した。一方、アスパラギンをアスパラギン酸に置換する事でCel48Sドッケリンの提示量は3.3倍に向上した(図5)。
【実施例5】
【0096】
C.thermocellum のゲノムからXyn10Cドッケリン遺伝子をPCR法により常法に従い増幅後クローニングした(配列番号221)。取得したXyn10Cドッケリン遺伝子を鋳型として10Cdock-N18D-Fw と10Cdock-N50D-Rvの2種類のプライマー(配列番号222、223)を利用して 18番目と54番目のアスパラギンをアスパラギン酸に置換した遺伝子を取得した(図6)。取得したそれぞれの遺伝子の上流にHXT3相同領域、HOR7プロモーターおよびHis-tag、下流にTdh3ターミネーター、Ura3マーカーおよびHXT3相同領域を持つ、pXU-Xyn10CdocベクターとpXU-Xyn10C-N-D-docベクターを作成した(図7)。取得したベクターを実施例2で取得した酵母CtCBDcohにそれぞれ導入することで、C.thermocellum 由来コヘシンを細胞表層に提示し、ドッケリンまたはアミノ酸置換したドッケリンを同時生産するCtCBDcoh10CdocとCtCBDcoh10CdocN-Dを取得した。
【実施例6】
【0097】
実施例5で取得した2種類の酵母CtCBDcoh10CdocとCtCBDcoh10CdocN-Dを、それぞれYP+2%グルコース培地で30℃、24時間培養し、OD600=0.5、62.5μl相当量集菌し、PBS溶液で洗浄を行い、PBS + 1mg/ml BSA + anti-His-FITC溶液と混合して4℃、30分間反応し、PBS溶液で2回洗浄後、Flow Cytometryで酵母細胞表層上のドッケリン提示量を評価した。Xyn10Cドッケリンでもアスパラギンをアスパラギン酸に置換する事でドッケリンの提示量が1.8倍に提示量が向上した(図8)。今回ターゲットとしたドッケリン内2箇所のアスパラギンはC. thermocellumのゲノム上で帰属されている142種類のドッケリンのうち82%で保存されている。2種類のドッケリンCel48SとXyn10Cでアミノ酸置換が同様の効果を示した事から、C. thermocellumが持つほとんどの酵素群に対して利用可能であると考えられる。
【実施例7】
【0098】
C.thermocellumのゲノムからCel8Aセルラーゼ遺伝子をPCR法により常法に従い増幅後クローニングした(配列番号224)。取得した遺伝子を、実施例3で取得したCel48Sドッケリン遺伝子、またはCel48Sドッケリンの18番目と50番目のアスパラギンをアスパラギン酸に置換した遺伝子とそれぞれ接合し、それぞれの遺伝子の上流にHXT3相同領域、HOR7プロモーターおよびHis-tag、下流にTdh3ターミネーター、Ura3マーカーおよびHXT3相同領域を持つ、pXU-Cel8A-Cel48SdocとpXU-Cel8A-Cel48S-N-D-docベクターを作製した(図9)。取得したベクターを実施例2で取得した酵母CtCBDcohにそれぞれ導入することで、C.thermocellum 由来コヘシンを細胞表層に提示し、ドッケリン型セルラーゼまたはアミノ酸を置換したドッケリン型セルラーゼを同時生産するCtCBDcohCel8A48SdocとCtCBDcohCel8A48SdocN-Dを取得した。
【実施例8】
【0099】
実施例7で取得した2種類の酵母CtCBDcohCel8A48SdocとCtCBDcohCel8A48SdocN-DをYP+2%グルコース培地で30℃、24時間培養し、OD600=0.5、62.5μl相当量集菌し、PBS溶液で1回洗浄を行い、PBS + 1mg/ml BSA + anti-His-FITC溶液と混合して4℃、30分間反応し、PBS溶液で2回洗浄後、Flow Cytometryで酵母細胞表層上のCelA提示量を評価した。アミノ酸を置換する事でCelA提示量の向上が認められた(図10)。
【実施例9】
【0100】
実施例7で取得した2種類の酵母CtCBDcohCel8A48SdocとCtCBDcohCel8A48SdocN-DをYP+2%グルコース培地で30℃、24時間培養し、OD600=1、1ml相当量集菌し、50mM 酢酸緩衝液 pH6.0溶液で洗浄後、1% CMC, 20mM 酢酸緩衝液pH6.0溶液に混合し、40℃で2時間CMC分解反応を行った。アミノ酸を置換する事でCMC分解活性の向上が認められ(図11)、酵母の糖化能力を向上させることが示された。
【配列表フリーテキスト】
【0101】
配列番号217,218,219,220,222,223:プライマー
【特許請求の範囲】
【請求項1】
C. thermocellum由来のタイプIコヘシンを有する骨格を利用したタンパク質複合体を構築するためのタンパク質であって、C. thermocellum由来のタイプIドッケリン中のコヘシン結合能に関連したドッケリン特異的配列であって、以下の(a)及び(b):
(a)内在性のN型糖鎖修飾予測部位を有しない
(b)内在性のN型糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されている
のいずれかを充足する少なくとも一つのドッケリン特異的配列を含むドッケリンを有するタンパク質。
【請求項2】
前記(b)を充足するドッケリン特異的配列は、以下の表から選択されるアミノ酸配列の前記N糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されたアミノ酸配列である、請求項1に記載のタンパク質。
【表16】
【請求項3】
前記ドッケリンは、以下の表に記載の1又は2つのアミノ酸配列からなるドッケリン特異的配列によって特徴付けられるとともに、前記(b)を充足する前記ドッケリン特異的配列を少なくとも一つ含む、請求項1又は2に記載のタンパク質。
【表17A】
【表17B】
【表17C】
【請求項4】
前記ドッケリンは、以下の表から選択されるアミノ酸配列によって特徴付けられるとともに、前記(b)を充足する前記ドッケリン特異的配列を少なくとも一つ含む、請求項1〜3のいずれかに記載のタンパク質。
【表18】
【請求項5】
前記(a)を充足するドッケリン特異的配列は、N型糖鎖修飾予測部位に相当する部位のアミノ酸がアスパラギン酸である、請求項1〜4のいずれかに記載のタンパク質。
【請求項6】
前記(a)を充足するドッケリン特異的配列は、以下の表から選択されるアミノ酸配列である、請求項1〜5のいずれかに記載のタンパク質。
【表19】
【請求項7】
前記ドッケリンは、以下の表に記載の2つのアミノ酸配列からなるドッケリン特異的配列によって特徴付けられるとともに、前記(a)を充足するドッケリン特異的配列を少なくとも一つ含む、請求項5又は6に記載のタンパク質。
【表20】
【請求項8】
前記ドッケリンは、以下の表から選択されるアミノ酸配列によって特徴付けられるとともに、前記(a)を充足するドッケリン特異的配列を少なくとも一つ含む、請求項5〜7のいずれかに記載のタンパク質。
【表21】
【請求項9】
セルロース分解促進活性を有する、請求項1〜8のいずれかに記載のタンパク質。
【請求項10】
前記セルロース分解促進活性はセルラーゼ活性である、請求項9に記載のタンパク質。
【請求項11】
前記セルロース分解促進活性は、クロストリジウム・サーモセラム由来のアミノ酸配列によって付与されている、請求項9又は10に記載のタンパク質。
【請求項12】
細胞表層にクロストリジウム・サーモセラム由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体を備える真核微生物であって、
クロストリジウム・サーモセラム由来のタイプIコヘシンを有する骨格タンパク質と、
前記骨格タンパク質に結合する請求項1〜11のいずれかに記載のタンパク質と、
を備える、真核微生物。
【請求項13】
前記骨格タンパク質及び前記タンパク質は、前記真核微生物が自己生産する、請求項12に記載の真核微生物。
【請求項14】
前記真核微生物は酵母である、請求項12又は13に記載の真核微生物。
【請求項15】
有用物質の生産方法であって、
請求項12〜14のいずれかに記載の真核微生物であって、前記タンパク質がセルロース分解促進活性を有する真核微生物を用いて、セルロース含有材料を炭素源として発酵する工程、を用いて、セルロース含有材料を糖化し、発酵する工程、
を備える、方法。
【請求項1】
C. thermocellum由来のタイプIコヘシンを有する骨格を利用したタンパク質複合体を構築するためのタンパク質であって、C. thermocellum由来のタイプIドッケリン中のコヘシン結合能に関連したドッケリン特異的配列であって、以下の(a)及び(b):
(a)内在性のN型糖鎖修飾予測部位を有しない
(b)内在性のN型糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されている
のいずれかを充足する少なくとも一つのドッケリン特異的配列を含むドッケリンを有するタンパク質。
【請求項2】
前記(b)を充足するドッケリン特異的配列は、以下の表から選択されるアミノ酸配列の前記N糖鎖修飾予測部位のアスパラギンがアスパラギン酸に置換されたアミノ酸配列である、請求項1に記載のタンパク質。
【表16】
【請求項3】
前記ドッケリンは、以下の表に記載の1又は2つのアミノ酸配列からなるドッケリン特異的配列によって特徴付けられるとともに、前記(b)を充足する前記ドッケリン特異的配列を少なくとも一つ含む、請求項1又は2に記載のタンパク質。
【表17A】
【表17B】
【表17C】
【請求項4】
前記ドッケリンは、以下の表から選択されるアミノ酸配列によって特徴付けられるとともに、前記(b)を充足する前記ドッケリン特異的配列を少なくとも一つ含む、請求項1〜3のいずれかに記載のタンパク質。
【表18】
【請求項5】
前記(a)を充足するドッケリン特異的配列は、N型糖鎖修飾予測部位に相当する部位のアミノ酸がアスパラギン酸である、請求項1〜4のいずれかに記載のタンパク質。
【請求項6】
前記(a)を充足するドッケリン特異的配列は、以下の表から選択されるアミノ酸配列である、請求項1〜5のいずれかに記載のタンパク質。
【表19】
【請求項7】
前記ドッケリンは、以下の表に記載の2つのアミノ酸配列からなるドッケリン特異的配列によって特徴付けられるとともに、前記(a)を充足するドッケリン特異的配列を少なくとも一つ含む、請求項5又は6に記載のタンパク質。
【表20】
【請求項8】
前記ドッケリンは、以下の表から選択されるアミノ酸配列によって特徴付けられるとともに、前記(a)を充足するドッケリン特異的配列を少なくとも一つ含む、請求項5〜7のいずれかに記載のタンパク質。
【表21】
【請求項9】
セルロース分解促進活性を有する、請求項1〜8のいずれかに記載のタンパク質。
【請求項10】
前記セルロース分解促進活性はセルラーゼ活性である、請求項9に記載のタンパク質。
【請求項11】
前記セルロース分解促進活性は、クロストリジウム・サーモセラム由来のアミノ酸配列によって付与されている、請求項9又は10に記載のタンパク質。
【請求項12】
細胞表層にクロストリジウム・サーモセラム由来のタイプIコヘシンを有する骨格タンパク質を利用したタンパク質複合体を備える真核微生物であって、
クロストリジウム・サーモセラム由来のタイプIコヘシンを有する骨格タンパク質と、
前記骨格タンパク質に結合する請求項1〜11のいずれかに記載のタンパク質と、
を備える、真核微生物。
【請求項13】
前記骨格タンパク質及び前記タンパク質は、前記真核微生物が自己生産する、請求項12に記載の真核微生物。
【請求項14】
前記真核微生物は酵母である、請求項12又は13に記載の真核微生物。
【請求項15】
有用物質の生産方法であって、
請求項12〜14のいずれかに記載の真核微生物であって、前記タンパク質がセルロース分解促進活性を有する真核微生物を用いて、セルロース含有材料を炭素源として発酵する工程、を用いて、セルロース含有材料を糖化し、発酵する工程、
を備える、方法。
【図1】

【図2】
【図3】

【図4】
【図5】
【図6】
【図7】
【図9】
【図8】
【図10】
【図11】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図9】
【図8】
【図10】
【図11】
【公開番号】特開2011−219399(P2011−219399A)
【公開日】平成23年11月4日(2011.11.4)
【国際特許分類】
【出願番号】特願2010−88952(P2010−88952)
【出願日】平成22年4月7日(2010.4.7)
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【Fターム(参考)】
【公開日】平成23年11月4日(2011.11.4)
【国際特許分類】
【出願日】平成22年4月7日(2010.4.7)
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【Fターム(参考)】
[ Back to top ]