
クロマチン構造の検出方法
ゲノムDNAにおけるDNA修飾剤のアクセス可能性を決定するための方法および組成物を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
関連特許出願の相互参照
本出願は、2008年12月2日に提出された米国仮特許出願第61/119,280号の優先権の恩典を主張し、それはあらゆる目的で参照により本明細書に組み入れられる。
【背景技術】
【0002】
発明の背景
細胞内のほとんどのDNAは、一群のヒストンタンパク質の周りにパッケージングされ、ヌクレオソームとして知られる構造をとる。このヌクレオソームDNAは、DNAを緻密に凝縮させるらせん状構造へとさらにパッケージングされうる。この緻密なパッケージングは、転写因子および転写機構へのDNAのアクセスを制限しうる。このような様式でパッケージングされたゲノムDNAは時にクロマチンと称される。
【0003】
クロマチンは、DNAが疎にパッケージングされ、アクセス可能(accessible)であり、常にではないが一般に転写能力があるユークロマチンと、DNAが緻密にパッケージングされ、アクセス不能(inaccessible)であり、常にではないが一般的には転写的にサイレントであるヘテロクロマチンという、2つの主なグループに分類される。
【0004】
これらの2つのクロマチン状態間の推移を制御するものはエピジェネティクス(epigenetics)である。エピジェネティックなイベントには主にDNAメチル化およびヒストン修飾の2つがある。これらのイベントは、DNAがどのようにパッケージングされるか、およびDNAが転写に関して活性であるかそれともサイレントであるかに影響を及ぼす。
【発明の概要】
【0005】
発明の概略
本発明は、染色体DNAを分析するための方法であって、DNA修飾剤に対する染色体上のDNA領域のアクセス可能性(accessibility)を決定し、任意で、該アクセス可能性をクロマチン構造と関連づける段階を非限定的に含む方法を提供する。いくつかの態様において、本方法は以下の段階を含む:
a.以下を同時に行う段階:
i.細胞の細胞膜を透過処理するかまたは崩壊させる段階;および
ii.該細胞を、DNA切断剤またはDNA修飾剤と、該剤が該細胞内のゲノムDNAを切断または修飾するような条件下で接触させる段階であって、ゲノムDNAの異なる領域が該剤によって異なる程度で切断または修飾され、それにより、切断されたDNA領域および無傷のDNA領域、または修飾されたDNA領域および修飾されていないDNA領域が生成される、段階;ならびに
b.少なくとも1つの、無傷の、もしくは修飾されていない、もしくは修飾されたDNA領域の物理的特徴もしくは量を検出するか、または少なくとも1つの、無傷の、もしくは修飾されていない、もしくは修飾されたDNA領域のクローニング、単離、もしくはヌクレオチド配列決定を行う段階。
【0006】
いくつかの態様において、本方法はさらに、前記量を前記細胞における前記DNA領域のクロマチン構造と関連づける段階を含む。
【0007】
いくつかの態様において、本方法は、少なくとも第1の無傷の、または修飾されていない、または修飾された染色体DNA領域、および第2の無傷の、または修飾されていない、または修飾された染色体DNA領域の物理的特徴または量を、検出する段階;ならびに第1および第2のDNA領域の物理的特徴または量を比較する段階を含む。
【0008】
いくつかの態様において、本方法は、第1の染色体DNA領域および第2の染色体DNA領域の無傷のコピーの数を定量し、それにより、DNA切断剤に対する第1および第2のDNA領域の相対的なアクセス可能性を評価する段階を含む。
【0009】
いくつかの態様においては、ゲノムDNAを、接触させる段階の後でかつ検出する段階の前に、単離する。
【0010】
いくつかの態様においては、細胞を人工の培養表面に直接的または間接的に付着させながら、細胞を透過処理して、DNA切断剤またはDNA修飾剤と接触させる。
【0011】
いくつかの態様において、透過処理する段階は、細胞を、細胞膜を透過性にする剤と接触させることを含む。いくつかの態様において、細胞膜を透過性にする剤はリゾ脂質(lysolipid)である。いくつかの態様において、リゾ脂質はリゾホスファチジルコリンである。
【0012】
いくつかの態様において、透過処理するかまたは崩壊させる段階は、非イオン性界面活性剤を用いて細胞膜を崩壊させることを含む。いくつかの態様において、非イオン性界面活性剤は、NP40、Tween20およびTriton X-100の群から選択される。
【0013】
いくつかの態様においては、細胞をDNA切断剤と接触させ、該DNA切断剤は酵素である。
【0014】
いくつかの態様においては、細胞をDNA修飾剤と接触させ、該DNA修飾剤はメチルトランスフェラーゼおよびDNA修飾性化学物質からなる群より選択される。いくつかの態様において、DNA切断剤はDNA切断酵素であり;ゲノムDNA中の修飾部は切断部位であり;かつ、検出する段階は切断後のDNA領域の無傷のコピーの量を定量することを含む。いくつかの態様において、検出する段階は定量的増幅を含む。いくつかの態様において、定量的増幅は、定量的ポリメラーゼ連鎖反応(任意でリアルタイム性)および定量的ライゲーション媒介ポリメラーゼ連鎖反応(任意でリアルタイム性)からなる群より選択される。
【0015】
いくつかの態様において、DNA切断剤は、DNアーゼおよび制限酵素から選択される。いくつかの態様において、DNA修飾剤はメチルトランスフェラーゼであり;修飾部は、細胞のネイティブなメチル化酵素によってはメチル化されないであろうヌクレオチド配列中のヌクレオチド上のメチル基であり;かつ、検出する段階は、修飾部の有無を検出する方法を含む。いくつかの態様において、メチルトランスフェラーゼはDAMメチルトランスフェラーゼであり、定量する段階は、修飾されたDNAを、DAMメチルトランスフェラーゼによってメチル化されたDNA配列を認識する制限酵素を用いて切断することを含む。いくつかの態様において、メチルトランスフェラーゼはDNA中のシトシンにメチル部分を付加し、検出する段階は、修飾されたDNAをメチル化特異的制限酵素を用いて切断すること、および/または修飾されたDNAをバイサルファイト(bisulfite)で処理することを含む。
【0016】
いくつかの態様において、DNA修飾剤はクロマチン構造に対して立体障害を有する分子であり、ここで該分子はDNA修飾性化学物質と連結されている。いくつかの態様において、DNA修飾性化学物質は、硫酸ジメチルおよびヒドラジンからなる群より選択される。
【0017】
いくつかの態様において、検出する段階は、標的DNA領域の少なくとも一部分および対照DNA領域の少なくとも一部分を定量することを含み、ここで対照DNA領域は、以下のいずれかである配列を含む:
i.ある動物の本質的にすべての細胞においてアクセス可能である;または
ii.ある動物のほとんどの細胞においてアクセス不能である;または
iii.細胞型もしくは生育環境に応じてさまざまなアクセス可能性を有する。
【0018】
いくつかの態様において、対照DNA領域は、以下のそれぞれを用いて定量的に増幅される:
DNA修飾剤の潜在的な修飾部位を含まない対照DNA領域の一部分の増幅をプライミングするプライマー;および
DNA修飾剤の潜在的な修飾部位を少なくとも1つ含む対照DNA領域の一部分の増幅をプライミングするプライマー。
【0019】
いくつかの態様において、物理的特徴はDNAメチル化である。
【0020】
いくつかの態様において、段階a.は前記DNAをDNA切断剤と接触させることを含み、段階b.は無傷のDNAをバイサルファイトと接触させることを含む。
【0021】
いくつかの態様において、本方法はさらに、バイサルファイトで処理したDNAの融解温度を決定し、該融解温度をDNAメチル化の有無または程度と関連づける段階を含む。
【0022】
いくつかの態様において、段階bは、以下のもののライブラリーを調製することを含む:無傷のDNA領域;または修飾されたDNA領域;または修飾されていないDNA領域。
【0023】
いくつかの態様において、段階bは、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域を、ポリヌクレオチドのライブラリーと、該DNA領域と該ライブラリーの1つまたは複数のメンバーとのハイブリダイゼーションを可能にする条件下で接触させること、および該DNA領域と該1つまたは複数のメンバーとのハイブリダイゼーションを検出することを含む。いくつかの態様において、ライブラリーはマイクロアレイ上に構成されている。
【0024】
いくつかの態様において、段階bは、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域を増幅することを含む。
【0025】
いくつかの態様において、段階bは、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域のヌクレオチド配列決定を行うことを含む。
【0026】
本発明はまた、DNA修飾剤に対する染色体上の遺伝子座のアクセス可能性を決定するためのキットも提供する。いくつかの態様において、本キットは、細胞膜の透過処理剤または崩壊剤;および制限酵素、DNアーゼまたは他のDNA修飾剤を含む。
【0027】
いくつかの態様において、細胞膜の透過処理剤または崩壊剤と、制限酵素またはDNアーゼとは、同じ容器内で同じ緩衝液中にある。
【0028】
いくつかの態様において、細胞膜の透過処理剤または崩壊剤と、制限酵素またはDNアーゼとは、別々の容器内にある。
【0029】
いくつかの態様において、本キットはさらにバイサルファイトを含む。
【0030】
いくつかの態様において、本キットはさらに、真核生物のゲノムDNAのある領域の増幅のためのプライマー対を含む。
【0031】
いくつかの態様において、本キットはさらに以下のものを含む:
a.DNA修飾剤の潜在的な修飾部位を含まない対照DNA領域の一部分の増幅をプライミングするプライマー;および/または
b.DNA修飾剤の潜在的な修飾部位を少なくとも1つ含む対照DNA領域の一部分の増幅をプライミングするプライマー。
【0032】
いくつかの態様において、本キットは以下のものを含む:
a.修飾剤に対してアクセス不能である対照DNA領域の一部分の増幅をプライミングするプライマー;および/または
b.修飾剤に対してアクセス可能である対照DNA領域の一部分の増幅をプライミングするプライマー。
【0033】
本発明のその他の態様は、本明細書の残りの部分を読めば明らかになるであろう。
【0034】
定義
細胞膜を「透過処理すること」とは、本明細書で用いる場合、修飾剤が細胞内に入ることを可能にするように細胞膜の完全性を低減させることを指す。透過処理された細胞膜を有する細胞は一般に細胞膜を保ち、そのため細胞の構造は実質的に無傷のままに保たれる。対照的に、細胞膜を「崩壊させること」とは、本明細書で用いる場合、細胞の構造が無傷のままでは保たれないように細胞膜の完全性を低減させることを指す。例えば、細胞膜を非イオン性界面活性剤と接触させることは、細胞膜を除去および/または溶解させ、それにより、少なくともいくらかの染色体構造を保っているゲノムDNAに対する修飾剤のアクセスを可能にする。
【0035】
「DNA修飾剤」とは、本明細書で用いる場合、DNAを検出可能な様式に改変する分子のことを指す。例示的な修飾には、DNAのニッキングもしくは切断、または導入もしくはDNAからの化学部分の除去が含まれる。DNA切断を引き起こさないDNA修飾剤には、DNAメチラーゼが非限定的に含まれる。
【0036】
「DNA領域」とは、本明細書で用いる場合、ゲノムDNA内部の関心対象の標的配列のことを指す。DNA領域は、関心対象であって用いるDNA修飾剤がアクセス可能である、任意の長さでありうる。いくつかの態様において、DNA領域は単一塩基対を含みうるが、ゲノムDNA内部の短い配列セグメント(例えば、2〜100、2〜500、50〜500bp)またはより長いセグメント(例えば、100〜10,000、100〜1000または1000〜5000bp)でもありうる。DNA領域内のDNAの量は、時には、PCR反応において増幅させようとする配列の量によって決まる。例えば、標準的なPCR反応は一般に約35〜5000塩基対を増幅することができる。
【0037】
異なる「程度」の修飾とは、複数の試料間での、または1つもしくは複数の試料中の2つもしくはそれ以上のDNA領域間での、1つまたは複数のDNA領域の修飾されたコピー数(実数または相対数)が異なることを指す。例えば、2つのDNA領域(便宜上「領域A」および「領域B」と呼ぶ)の100個のコピーがそれぞれ、ある細胞内の染色体DNA上に存在するならば、異なる程度での修飾の一例は、領域Aの10個のコピーが修飾され、一方、領域Bの70個のコピーが修飾された場合であると考えられる。
【0038】
「オリゴヌクレオチド」または「ポリヌクレオチド」または「核酸」という用語は、リボース核酸(RNA)もしくはデオキシリボース核酸(DNA)の重合体またはそれらの類似体に対応しうる、単量体の重合体のことを互換的に指す。これには、RNAおよびDNA、さらにはそれらの修飾形態、ペプチド核酸(PNA)、ロックド核酸(locked nucleic acid, LNA(商標))といったヌクレオチドの重合体が含まれる。ある用途において、核酸は、複数の種類の単量体、例えば、RNAサブユニットおよびDNAサブユニットの両方を含む重合体でありうる。
【0039】
核酸は典型的には一本鎖または二本鎖であり、一般にホスホジエステル結合を含むが、場合によっては、本明細書で概説するように、代替的な骨格を有しうる核酸類似体が含められ、これには例えば、ホスホルアミド(Beaucage et al. (1993) Tetrahedron 49(10):1925およびその中の参考文献;Letsinger (1970) J. Org. Chem. 35:3800;Sprinzl et al. (1977) Eur. J. Biochem. 81:579;Letsinger et al. (1986) Nucl. Acids Res. 14: 3487;Sawai et al. (1984) Chem. Lett. 805;Letsinger et al. (1988) J. Am. Chem. Soc. 110:4470;およびPauwels et al. (1986) Chemica Scripta 26:1419)、ホスホロチオエート(Mag et al. (1991) Nucleic Acids Res. 19:1437および米国特許第5,644,048号)、ホスホロジチオエート(Briu et al. (1989) J. Am. Chem. Soc. 111:2321)、O-メチルホスホロアミダイト結合(O-methylphophoroamidite linkage)(Eckstein, Oligonucleotides and Analogues: A Practical Approach, Oxford University Press (1992))、ならびにペプチド核酸骨格および結合(Eghoim (1992) J. Am. Chem. Soc. 114:1895;Meier et al. (1992) Chem. Int. Ed. Engl. 31:1008;Nielsen (1993) Nature 365:566;およびCarlsson et al. (1996) Nature 380:207)が非限定的に含まれ、これらの参考文献はそれぞれ参照により組み入れられる。他の類似体核酸には、正に荷電した骨格(Denpcy et al. (1995) Proc. Natl. Acad. Sci. USA 92:6097);非イオン性骨格(米国特許第5,386,023号、第5,637,684号、第5,602,240号、第5,216,141号および第4,469,863号;Angew (1991) Chem. Intl. Ed. English 30: 423;Letsinger et al. (1988) J. Am. Chem. Soc. 110:4470;Letsinger et al. (1994) Nucleoside & Nucleotide 13:1597;Chapters 2 and 3, ASC Symposium Series 580, "Carbohydrate Modifications in Antisense Research", Ed. Y. S. Sanghvi and P. Dan Cook;Mesmaeker et al. (1994) Bioorganic & Medicinal Chem. Lett. 4: 395;Jeffs et al. (1994) J. Biomolecular NMR 34:17;Tetrahedron Lett. 37:743 (1996))、および、米国特許第5,235,033号および第5,034,506号ならびにChapters 6 and 7, ASC Symposium Series 580, Carbohydrate Modifications in Antisense Research, Ed. Y. S. Sanghvi and P. Dan Cookに記載されたものを含む非リボース骨格を有するものが含まれ、これらの参考文献はそれぞれ参照により組み入れられる。1つまたは複数の炭素環式糖を含む核酸も、核酸の定義の範囲に含まれる(Jenkins et al. (1995) Chem. Soc. Rev. pp169-176、これは参照により組み入れられる)。いくつかの核酸類似体は、例えば、Rawls, C & E News Jun. 2, 1997 page 35にも記載されており、これは参照により組み入れられる。リボース-リン酸骨格のこれらの修飾は、標識部分などの追加部分の付加を容易にするため、または生理的環境におけるそのような分子の安定性および半減期を変更するために行ってもよい。
【0040】
核酸中に典型的に認められる、天然に存在する複素環式塩基(例えば、アデニン、グアニン、チミン、シトシンおよびウラシル)に加えて、核酸類似体にはまた、天然に存在しない複素環式塩基または他の修飾塩基も含まれ、その多くは本明細書中に記載されているか、そうでなければ参照されている。特に、天然に存在しない多くの塩基は、例えば、Seela et al. (1991) Helv. Chim. Acta 74:1790, Grein et al. (1994) Bioorg. Med. Chem. Lett. 4:971-976、およびSeela et al. (1999) Helv. Chim. Acta 82:1640にさらに記載されており、これらはそれぞれ参照により組み入れられる。さらに例を挙げるならば、融解温度(Tm)変更因子として作用するヌクレオチド中に用いられるある種の塩基が任意で含められる。例えば、これらのいくつかには、7-デアザプリン(例えば、7-デアザグアニン、7-デアザアデニンなど)、ピラゾロ[3,4-d]ピリミジン、プロピニル-dN(例えば、プロピニル-dU、プロピニル-dCなど)などが含まれる。例えば、参照により組み入れられる、Seelaに対して1999年11月23日に発行された「7-デアザ-2'-デオキシグアノシンヌクレオチドの合成」と題する米国特許第5,990,303号を参照のこと。他の代表的な複素環式塩基には、例えば、ヒポキサンチン、イノシン、キサンチン;2-アミノプリン、2,6-ジアミノプリン、2-アミノ-6-クロロプリン、ヒポキサンチン、イノシンおよびキサンチンの8-アザ誘導体;アデニン、グアニン、2-アミノプリン、2,6-ジアミノプリン、2-アミノ-6-クロロプリン、ヒポキサンチン、イノシンおよびキサンチンの7-デアザ-8-アザ誘導体;6-アザシトシン;5-フルオロシトシン;5-クロロシトシン;5-ヨードシトシン;5-ブロモシトシン;5-メチルシトシン;5-プロピニルシトシン;5-ブロモビニルウラシル;5-フルオロウラシル;5-クロロウラシル;5-ヨードウラシル;5-ブロモウラシル;5-トリフルオロメチルウラシル;5-メトキシメチルウラシル;5-エチニルウラシル;5-プロピニルウラシルなどが含まれる。
【0041】
DNA修飾剤に対するDNA領域の「アクセス可能性」とは、本明細書で用いる場合、細胞の染色体内の特定のDNA領域が、特定のDNA修飾剤によって接触されて修飾される能力のことを指す。本発明の範囲を限定することは意図しないが、DNA領域を含む特定のクロマチン構造は、DNA修飾剤が特定のDNA領域を修飾する能力に影響を及ぼすと考えられている。例えば、DNA領域がヒストンタンパク質の周りに巻き付けられていることもあり、さらに、関心対象のDNA領域に対するDNA修飾剤のアクセスを防止または抑制するさらなるヌクレオソーム構造を有することもある。
【0042】
「II-S型制限酵素」は、当技術分野におけるその通常の意味で用いられ、DNAにおける特定の認識配列を認識し、続いてそのDNA分子をその認識配列の外側で切断する制限酵素のことを指す。例示的なII-S型制限酵素には、MnlI、FokIおよびAlwIが非限定的に含まれる。
【図面の簡単な説明】
【0043】
【図1】クロマチンの概略図を図示している。真核生物DNAは、DNAが疎にパッケージングされ、アクセス可能であり、転写能力があるユークロマチンと、DNAが緻密にパッケージングされ、アクセス不能であり、転写的にサイレントであるヘテロクロマチンという、2つの一般的状態に分類することができる。エピジェネティクスが、これらの2つの状態間の推移を制御する。本明細書に記載されたアッセイは、クロマチン構造、エピジェネティックなイベントの機能的帰結を評価することができる。
【図2】アッセイの概略図を図示している。培地を吸引し、透過処理/消化用緩衝液を添加する。ヌクレアーゼが細胞内に拡散し、核の中に入って、アクセス可能なクロマチンを消化するが、アクセス不能なクロマチン(図の下方の太線として表されている)は消化されない。
【図3】例示的な作業の流れの概略図を図示している。
【図4】Mnl 1をプローブとして用いた場合に生成されるデータを評価するための分析アプローチを図示している。各アッセイでは2つのDNA試料を分析し、一方のDNA試料はヌクレアーゼを含まない緩衝液で処理した細胞から単離した対照であり、もう一方の試料は緩衝液およびヌクレアーゼで処理した細胞から単離する。両方の試料を、トータルプライマーのセットおよび無傷プライマーのセットを用いるリアルタイムPCRによって分析する。(A)ΔCt(切断なし)は、対照DNA試料のデータを用いてトータルCtから無傷Ctを差し引くことによって算出される。(B)ΔCt(切断あり)は、ヌクレアーゼ処理したDNA試料のデータを用いてトータルCtから無傷Ctを差し引くことによって算出される。アクセス可能な遺伝子は、MnlI部位で切断され、有する無傷DNAがより少なく、ΔCt(切断あり)がより高く、それは消化の程度に比例し、クロマチンのアクセス可能性がより大きいことを反映する。アクセス不能な、または「ロックダウン(locked-down)された」DNAの量は、式2∧(ΔCt(切断なし)−ΔCt(切断あり))によって算出される。
【図5】分析した標的遺伝子の略図である。グレーの領域は転写される領域を表している。Mnl 1部位およびTATAボックスの位置を指し示している。GAPDHはほとんどの細胞株で発現されるハウスキーピング遺伝子であり、アクセス可能なクロマチン構造内にあると予想される。ヘモグロビンβ鎖(HBB)は、ほとんどの細胞でサイレント化しており、アクセス不能なクロマチン構造にあると予想される。グルタチオン-s-トランスフェラーゼπ(GSTP1)は、前立腺癌ではエピジェネティックに不活性化されており、非癌性RWPE-1前立腺細胞では発現されるが、癌性LNCaP前立腺細胞ではサイレント化している。
【図6】分析した標的遺伝子、およびプライマーセットの位置を図示している。遺伝子はすべて、どのMnlI I部位も横切らず、すべてのDNA鎖を増幅すると考えられるトータルプライマーのセット、さらには2つ〜4つのMnlI部位を横切り、これらの部位で切断されないDNAのみを増幅すると考えられる無傷プライマーのセットを用いて分析した。
【図7】アガロースゲル電気泳動を用いたgDNA完全性の分析の概要を提示している。Hela細胞を、ヌクレアーゼ(M、MnlI;D、DNアーゼI)を含まない透過処理/消化用緩衝液、リゾレシチンを含まない透過処理/消化用緩衝液、またはヌクレアーゼおよびリゾレシチンの両方を含む透過処理/消化用緩衝液で処理した。gDNAを単離して、アガロースゲル電気泳動によって分析した。これらの結果は、有意なgDNA消化のためには、ヌクレアーゼおよびリゾレシチンの両方が必要なことを指し示している。
【図8】MnlIで処理したHela細胞におけるHBBに関して、蛍光をサイクル数の関数として図示している。Hela細胞は前述の通りに処理した。gDNAを単離して、HBB遺伝子を増幅するプライマーを用いて分析した。HBB無傷プライマーのセットを用いた場合、完全な反応物(+、x)のCt値は対照反応物と同程度である。このことは、HBBがHela細胞においてアクセス不能なクロマチン立体配置内にあることを意味する。
【図9】MnlIで処理したHela細胞におけるGAPDHに関して、蛍光をサイクル数の関数として図示している。Hela細胞を前述の通りに処理し、gDNAを単離して、GAPDH遺伝子を増幅するプライマーを用いて分析した。GAPDH無傷プライマーのセットを用いた場合、完全な反応物(+、x)のCt値は対照反応物よりも高い(曲線が右方移動している)。このことは、GAPDHがHela細胞においてアクセス可能なクロマチン立体配置内にあることを意味する。
【図10】MnlIで処理したHela細胞に関して、蛍光をサイクル数の関数として図示している。Hela細胞を前述の通りに処理し、gDNAを単離して、GSTP1遺伝子を増幅するプライマーを用いて分析した。GSTP1無傷プライマーのセットを用いた場合、完全な反応物(+、x)のCt値は対照反応物よりも高い(曲線が右方移動している)。このことは、GSTP1がHela細胞においてアクセス可能なクロマチン立体配置内にあることを意味する。
【図11】3つのDNA領域(HBB、GAPDHおよびGSTP1)のデータを表形式で図示しており、アウトプットは全DNAに占めるアクセス不能であったパーセンテージとして算出している。
【図12】MnlIで処理した前立腺細胞におけるHBBに関して、蛍光をサイクル数の関数として図示している。いずれの細胞株においても、HBB無傷プライマーのセットを用いた+酵素反応のCt値(x)は、HBBトータルプライマーのセットを用いた+酵素反応(o)と同程度である。このことは、HBBがRWPE-1細胞およびLNCaP細胞においてアクセス不能なクロマチン立体配置内にあることを意味する。
【図13】MnlIで処理した前立腺細胞におけるGAPDHに関して、蛍光をサイクル数の関数として図示している。いずれの細胞株においても、GAPDH無傷プライマーのセットを用いた+酵素反応のCt値(x)は、GAPDHトータルプライマーのセットを用いた+酵素反応(o)よりも高い(曲線は右方移動している)。このことは、GAPDHがRWPE-1細胞およびLNCaP細胞においてアクセス可能なクロマチン立体配置内にあることを意味する。
【図14】MnlIで処理した前立腺細胞におけるGSTP1に関して、蛍光をサイクル数の関数として図示している。RWPE-1細胞において、GSTP1無傷プライマーのセットを用いた+酵素反応のCt値(x)は、GSTP1トータルプライマーのセットを用いた+酵素反応(o)よりも高い(曲線は右方移動している)。このことは、GSTP1がRWPE-1細胞においてアクセス可能なクロマチン立体配置内にあることを意味する。対照的に、LNCaP細胞では、GSTP1無傷プライマーのセットを用いた+酵素反応のCt値(x)は、GSTP1トータルプライマーのセットを用いた+酵素反応(o)と同程度である。このことは、GSTP1がLNCaP細胞においてアクセス不能なクロマチン立体配置内にあることを意味する。
【図15】前立腺細胞からのデータの概要を図示している。アクセス不能なDNAのパーセンテージは、材料および方法の項に記載した通りに算出した。これらの結果は、いずれの細胞株においても、HBBはアクセス不能なクロマチン構造内にあり、GAPDHはアクセス可能なクロマチン構造内にあることを指し示している。GSTP1はRWPE-1細胞においてはアクセス可能なクロマチン内にあり、LNCaP細胞においてはアクセス不能なクロマチン構造内にある。
【図16】DNアーゼIをクロマチン構造プローブとして用いた場合に生成されたデータを評価するための分析アプローチを図示している。各アッセイで2つのDNA試料を分析し、一方のDNA試料はヌクレアーゼを含まない緩衝液で処理した細胞から単離した対照であり、もう一方の試料は緩衝液およびヌクレアーゼで処理した細胞から単離する。両方の試料を、アクセス不能な参照遺伝子(HBB)を増幅するプライマーセットを用いるリアルタイムPCRによって分析する。ΔCt(参照)はアクセス不能性クロマチンのヌクレアーゼ消化の程度を表し、+ヌクレアーゼの曲線に伴うCtからヌクレアーゼなしの曲線に伴うCtを差し引くことによって算出される。ΔCt(標的)は標的クロマチン領域のヌクレアーゼ消化の程度を表し、+ヌクレアーゼの曲線に伴うCtからヌクレアーゼなしの曲線に伴うCtを差し引くことによって算出される。標的遺伝子に伴うアクセス不能な、すなわち「ロックダウンされた」DNAの量は、式2∧(ΔCt(参照)−ΔCt(標的))によって算出される。
【図17】分析した標的遺伝子、およびプライマーセットの位置の概略図である。グレーの領域は転写される領域を表している。矢印付きの黒の領域は、各プライマーの位置および方向を表している。
【図18】DNアーゼIで処理した細胞株におけるHBBに関して、蛍光をサイクル数の関数として図示している。すべての細胞株において、+ヌクレアーゼ反応物のCt値(グレーの線)は、ヌクレアーゼなしの反応物(黒の線)と同程度である。このことは、HBBがすべての細胞株においてアクセス不能なクロマチン立体配置内にあることを意味する。
【図19】DNアーゼIで処理した細胞株におけるGAPDHに関して、蛍光をサイクル数の関数として図示している。すべての細胞株において、+ヌクレアーゼ反応物のCt値(グレーの線)は、ヌクレアーゼなしの反応物のCt値(黒の線)よりも有意に高い。このことはGAPDHがすべての細胞株においてアクセス可能なクロマチン立体配置内にあることを意味する。
【図20】DNアーゼIで処理した細胞株におけるGSTP1に関して、蛍光をサイクル数の関数として図示している。本発明者らは、ΔCt(GSTP1)値において細胞株依存的な差を確認している。Hela細胞およびHCT15細胞ではΔCt(GSTP1)が大きく、このことはGSTP1がこれらの細胞株においてアクセス可能な立体配置内にあることを指し示している。LNCaP細胞ではΔCt(GSTP1)が小さく、このことはGSTP1がアクセス不能なクロマチン内にあることを指し示している。PC3細胞ではΔCt(GSTP1)が中間値を有し、このことはGSTP1が部分的にアクセス可能なクロマチン構造内にあることを指し示している。これらの結果は、種々の細胞株において検出されたGSTP1 mRNA発現のレベルと非常によく合致する(図21参照)。
【図21】DNアーゼIをクロマチン構造プローブとして用いた実験によるデータの概要を図示している。アクセス不能なDNAのパーセンテージは、図16で説明した通りに算出した。GAPDHおよびGSTP1のRNA発現の相対的レベルは、種々の細胞株から単離したmRNAをqRT-PCRによって分析することによって算出した。これらの結果は、標的遺伝子のクロマチン構造が標的遺伝子の発現レベルとよく相関することを指し示している。このことは、記載したアッセイが、遺伝子調節に対するエピジェネティックな影響を評価するための有用なツールであることを意味する。
【発明を実施するための形態】
【0044】
詳細な説明
I.序論
本発明は、細胞膜を透過処理するかまたは崩壊させ、細胞内のゲノムDNAを修飾して、続いてさまざまな遺伝子座における修飾の程度を定量することによって、クロマチン構造の分析を可能にする。特定の遺伝子座での修飾の程度は、修飾剤に対する染色体のその部分のアクセス可能性を反映し、それ故にクロマチンの状態を反映する。
【0045】
本発明の1つの利点は、例えば、細胞を、透過処理剤とDNA修飾剤または切断剤とを含む1つの緩衝液と接触させることによって、透過処理および細胞内の無傷のクロマチンの修飾を同時に行うことができるという発見である。これは、結果の極めて迅速な生成を可能にする。さらに、この方法は、従前のいくつかのクロマチン分析方法で必要とされたような、核の単離といった煩雑で、場合によってはアーチファクトを誘発する可能性もある段階をなくす。
【0046】
II.一般的方法
本発明の方法は、細胞の透過処理および細胞とDNA修飾剤との接触を同時に、細胞内のゲノムDNAが修飾剤に対して(クロマチン構造の違いが原因で)さまざまなアクセス可能性を有するような条件下で行い、続いて、DNA領域における修飾の量を定量することを伴う。DNAのさまざまなアクセス可能性は、ゲノムDNAのヌクレオソーム構造を反映する可能性がある。例えば、いくつかの態様において、DNA修飾剤に対してよりアクセス可能であるDNA領域は、より「粗」なクロマチン構造内にある可能性が高い。
【0047】
種々の真核生物細胞を本発明に用いることができる。いくつかの態様において、細胞は、ヒトの、または非ヒトの哺乳動物細胞を非限定的に含む動物細胞である。非ヒト哺乳動物細胞には、霊長動物細胞、マウス細胞、ラット細胞、ブタ細胞およびウシ細胞が非限定的に含まれる。いくつかの態様において、細胞は植物細胞である。細胞は、例えば、培養初代細胞、不死化培養細胞であってもよく、または生検試料もしくは組織試料に由来し、任意で培養した上でアッセイする前に分裂するように刺激したものでもよい。培養細胞は、透過処理および/またはDNA修飾の段階の前および/または最中に、浮遊していてもよく、または付着していてもよい。細胞が動物組織、生検試料などに由来してもよい。例えば、細胞は腫瘍生検試料に由来しうる。
【0048】
本方法は、あるDNA領域に対するアクセス可能性を、その同じ領域からの転写と関連づけることを含みうる。いくつかの態様においては、アクセス可能性と遺伝子発現との間の相関関係を決定するために実験を行い、その後に特定のDNA領域に対するDNA修飾剤のアクセス可能性を用いて、そのDNA領域からの転写を予測することができる。いくつかの態様においては、DNA領域からの転写、およびDNA修飾剤に対するその領域のアクセス可能性を、両方とも決定する。転写を測定するための多岐にわたる方法が公知であり、それにはノーザンブロット法およびRT-PCRが非限定的に含まれる。
【0049】
いくつかの態様においては、ある領域のDNAメチル化状態を、DNA修飾剤に対するDNA領域のアクセス可能性と関連づけることができる。いくつかの態様においては、アクセス可能性とその領域におけるDNAメチル化との相関関係を決定するために実験を行い、その後に特定のDNA領域に対するDNA修飾剤のアクセス可能性を利用して、そのDNA領域からのDNAメチル化を予測することができる。いくつかの態様においては、DNA領域のメチル化、およびDNA修飾剤に対するその領域のアクセス可能性を、両方とも決定する。DNAメチル化を測定するための多岐にわたる方法が公知であり、それには、バイサルファイトの使用(例えば、配列決定において、および/またはメチル化感受性制限酵素との組み合わせで(例えば、Eads et al., Nucleic Acids Research 28(8): E32 (2002)を参照)および高分解能融解アッセイ(HRM)(例えば、Wodjacz et al, Nucleic Acids Research 35(6):e41 (2007)を参照)が非限定的に含まれる。
【0050】
本発明は、透過処理/DNA修飾後の、細胞のゲノム中の第1のDNA領域と第2のDNA領域の量または他の物理的特徴の比較を提供する。代替的に、または加えて、2つの異なる細胞における第1のDNA領域の量または他の物理的特徴を比較することもできる。例えば、2つの細胞は、罹患しているおよび健常な細胞または組織、異なる細胞型、異なる発生段階(幹細胞または始原細胞を非限定的に含む)などであってよい。このようにして、本発明の方法を用いることにより、細胞間のクロマチン構造の違いを検出すること、および/または1つの細胞内の2つもしくはそれ以上のDNA領域(例えば、遺伝子)間の相対的なクロマチン構造を決定することができる。加えて、同じ細胞または異なる細胞における特定の領域のクロマチン構造に対する薬物刺激、化学刺激または環境的刺激の影響を決定することもできる。
【0051】
III.細胞の透過処理および崩壊
細胞膜は、当技術分野において公知の任意のやり方で透過処理すること、または崩壊させることができる。本明細書で説明しているように、本方法は、DNAの単離の前にゲノムDNAを接触させることを伴い、それ故に、細胞膜の透過処理または崩壊の方法は、ヌクレオソームまたはクロマチンの構造が破壊されるように細胞のゲノムDNAの構造を崩壊させることはないと考えられる。
【0052】
いくつかの態様においては、細胞膜を、細胞膜を透過性にするかまたは崩壊させる剤と接触させる。リゾ脂質は、細胞膜を透過性にする例示的なクラスの剤である。例示的なリゾ脂質には、リゾホスファチジルコリン(当技術分野においてリゾレシチンとしても公知である)またはモノパルミトイルホスファチジルコリンが非限定的に含まれる。種々のリゾ脂質が、例えば、WO/2003/052095号にも記載されている。
【0053】
非イオン性界面活性剤は、細胞膜を崩壊させる例示的なクラスの剤である。例示的な非イオン性界面活性剤には、NP40、Tween 20およびTriton X-100が非限定的に含まれる。
【0054】
本発明の1つの利点は、透過処理剤とDNA切断剤またはDNA修飾剤との同時送達である。したがって、いくつかの態様においては、両方の剤を含む緩衝液を細胞と接触させる。緩衝液は、細胞クロマチンの構造を維持させながら両方の剤の活性を維持させるのに適合しているべきである。
【0055】
または、エレクトロポレーションまたは微粒子銃法を、DNA修飾剤が細胞内に導入され、従ってゲノムDNAと接触しうるように、細胞膜を透過処理するために用いることもできる。多岐にわたるエレクトロポレーション法が周知であり、本明細書に記載されたDNA修飾剤の送達用に適合させることができる。例示的なエレクトロポレーション法には、WO/2000/062855号に記載されたものが非限定的に含まれる。微粒子銃法には、米国特許第5,179,022号に記載されたものが非限定的に含まれる。
【0056】
IV.DNA修飾剤
透過処理の後に、または透過処理と同時に(例えば、エレクトロポレーション中、または透過処理剤とのインキュベーション中に)、DNA修飾剤を導入し、その結果、該剤がゲノムDNAと接触し、それによってDNAに修飾を導入する。多岐にわたるDNA修飾剤を本発明に従って用いることができる。
【0057】
いくつかの態様においては、透過処理剤の除去後に、任意で緩衝液の変更とともに、DNA修飾剤を、透過処理を行った細胞と接触させる。または、いくつかの好ましい態様においては、1つまたは複数の中間段階を省いて(例えば、緩衝液の変更、細胞の洗浄などを省いて)、DNA修飾剤をゲノムDNAと接触させる。上述した通り、この後者のアプローチは、手間のかかる作業および必要な時間の量を減らすために好都合であり得、さらに、アッセイにおける誤差および混入をもたらす可能性のある原因も減らす。
【0058】
用いるDNA修飾剤の量、ならびにDNA修飾剤との反応時間は、用いる剤に依存すると考えられる。当業者は、用いる剤に応じて条件をどのように調整すべきかを認識しているであろう。一般に、DNA修飾段階の条件は、「完全な」消化が達成されないように調整する。したがって、例えば、いくつかの態様において、修飾段階の条件は、陽性対照‐すなわち、修飾がアクセス可能であって、生じる対照‐が、100%未満ではあるが高いレベル、例えば、80〜95%、80〜99%、85〜95%、90〜98%などで生じるように設定される。
【0059】
A.制限酵素
いくつかの態様において、DNA修飾剤は制限酵素である。したがって、これらの態様において、ゲノムDNAに導入される修飾は、配列特異的な一本鎖(例えば、ニック)または二本鎖切断イベントである。多岐にわたる制限酵素が公知であり、本発明に用いることができる。
【0060】
あらゆる種類の制限酵素を用いることができる。I型酵素はDNAを、それらの認識配列から離れたところでランダムに切断する。II型酵素はDNAを、それらの認識配列の付近または内部の定まった位置で切断する。いくつかのII型酵素はDNAを、それらの認識配列の内部で切断する。II-S型酵素は、それらの認識配列の外側を片側で切断する。より正確には「IV型」と呼ばれる、第3の主な種類のII型酵素は、それらの認識配列の外側を切断する。例えば、連続配列を認識するもの(例えば、AcuI:CTGAAG)は、片側のみを切断する;不連続配列を認識するもの(例えば、BcgI:CGANNNNNNTGC)は両側を切断し、その認識配列を含む小さな断片を遊離させる。III型はそれらの認識配列の外側を切断し、切断を遂行するためには、そのような配列が反対向きで2つ同一のDNA分子内に必要である。
【0061】
本発明の方法は、あらゆる種類の制限酵素または他のDNA切断酵素と用いるために適合させることができる。いくつかの態様において、酵素は、認識配列の比較的付近(例えば、5、10または20塩基対以内)を切断する1つまたは複数のものである。そのような酵素は、切断を達成するためにアクセス可能でなければならないDNAの範囲が認識配列それ自体よりも長く、それ故に「緻密」なクロマチン構造内にないDNAをより広範な範囲で含みうるため、クロマチン構造をアッセイする上で特に有用な可能性がある。配列特異的な制限酵素は、改善された定量的結果を与える可能性があり、これは一部には、本明細書に(例えば、実施例に)記載したように、同じDNA領域に基づく対照を設計しうるためである。したがって、例えば、配列非特異的エンドヌクレアーゼ(「DNアーゼ」)による消化と比較して、総コピー数および消化されたコピー数をより正確に決定することができる。DNアーゼIとは異なり、II-S型制限酵素による切断は、関心対象のDNA領域に対するヒストンの結合に対して高感度でありうる。認識配列の外側を切断する例示的な酵素には、例えば、II-S型酵素、III型酵素およびIV型酵素が含まれる。II-S型制限酵素には、MnlI、FokIおよびAlwIが非限定的に含まれる。
【0062】
いくつかの態様においては、複数(例えば、2つ、3つ、4つなど)の制限酵素を用いる。酵素の組み合わせは、すべてが1つの型由来である酵素の組み合わせを含んでもよく、または異なる型の混合物であってもよい。
【0063】
本明細書に記載した通りに、無傷のまたは切断されたDNAを、その後に別々に検出して定量し、DNA領域の無傷のおよび/または切断されたコピーの数を決定する。
【0064】
いくつかの態様においては、透過処理剤または膜崩壊剤を、制限酵素の前に添加する。いくつかの態様においては、制限酵素と透過処理剤または崩壊剤とを同時に添加する(例えば、適切な緩衝液中に、またはそれとともに)。仮に両方の剤を最初に同時に細胞と接触させない場合でも、透過処理は持続的過程であるため、透過処理とDNA修飾剤への接触とを同時に行うことを依然として達成することができる。したがって、例えば、透過処理剤の添加のすぐ後に(透過処理が実質的に完了する前に)DNA修飾剤を添加することは、透過処理および細胞をDNA修飾剤と接触させることを「同時に」行うことと見なすことができる。「同時に」とは、透過処理および修飾剤の添加の間に中間操作(緩衝液の変更、遠心処理などを非限定的に含む)を行わないことを意味する。
【0065】
いくつかの態様においては、0.5%リゾレシチン(w/v)、50mM NaCl、10mM Tris-HCl pH 7.4、10mM MgCl2、1mM DTT、100ug/ml BSAおよび0〜500単位/mlのMnlI(または他の制限酵素)を用いる。いくつかの態様においては、0.25%リゾレシチン(w/v)、50mM NaCl、10mM Tris-HCl pH 7.4、10mM MgCl2、1mM DTT、100ug/ml BSAおよび0〜500単位/mlのMnlI(または他の制限酵素)を用いる。いくつかの態様においては、0.75%リゾレシチン(w/v)、50mM NaCl、10mM Tris-HCl pH 7.4、10mM MgCl2、1mM DTT、100ug/ml BSAおよび0〜500単位/mlのMnlI(または他の制限酵素)を用いる。いくつかの態様においては、1%リゾレシチン(w/v)、50mM NaCl、10mM Tris-HCl pH 7.4、10mM MgCl2、1mM DTT、100ug/ml BSAおよび0〜800単位/mlのMnlI(または他の制限酵素)を用いる。
【0066】
透過処理および消化の後に、任意で同時に溶解/停止用緩衝液および/または温度上昇を加えることにより、消化を任意で停止させて、細胞を溶解する。例示的な溶解/停止用緩衝液には、反応を停止させて細胞を溶解するのに十分なキレート剤および界面活性剤が含まれうる。例えば、いくつかの態様において、溶解/停止用緩衝液は、100mM Tris-HCl pH 8、100mM NaCl、100mM EDTA、5% SDS(w/v)および3mg/mlのプロテイナーゼKを含む。いくつかの態様において、溶解/停止用緩衝液は、100mM Tris-HCl pH 8、100mM NaCl、100mM EDTA、1% SDS(w/v)および3mg/mlのプロテイナーゼKを含む。いくつかの態様において、溶解/停止用緩衝液は、200mM Tris-HCl pH 8、100mM NaCl、500mM EDTA、5% SDS(w/v)および5mg/mlのプロテイナーゼKを含む。
【0067】
B.DNアーゼ
いくつかの態様においては、DNAに配列非特異的な様式で切れ目またはニックを入れる酵素をDNA修飾剤として用いる。したがって、いくつかの態様において、DNA修飾剤は配列非特異的エンドヌクレアーゼ(本明細書では「DNアーゼ」とも称する)である。
【0068】
あらゆる配列非特異的エンドヌクレアーゼ(例えば、DNアーゼI、II、III、IV、V、VI、VIIのいずれか)を、本発明に従って用いることができる。例えば、DNアーゼIを非限定的に含む、あらゆるDNアーゼを用いることができる。用いるDNアーゼには、天然に存在するDNアーゼ、さらには改変DNアーゼが含まれうる。改変DNアーゼの一例はTURBO DNアーゼ(Ambion)であり、これは「活性亢進」および塩耐性を可能にする突然変異を含む。例示的なDNアーゼには、ウシ膵臓DNアーゼI(例えば、New England Biolabsから入手可能)が非限定的に含まれる。
【0069】
本明細書に記載した通りに、無傷のDNAをその後に別々に検出して定量し、DNA領域の無傷のおよび/または切断されたコピーの数を定量することができる。
【0070】
いくつかの態様においては、透過処理剤または膜崩壊剤を、DNアーゼの前に添加する。いくつかの態様においては、DNアーゼと透過処理剤または崩壊剤とを同時に添加する(例えば、適切な緩衝液とともに)。いくつかの態様において、透過処理/消化用緩衝液は、0.25%リゾレシチン(w/v)、10mM Tris-HCl pH 7.4、2.5mM MgCl2、0.5mM CaCl2および0〜200単位/mlのDNアーゼIを含む。いくつかの態様において、透過処理/消化用緩衝液は、0.5%リゾレシチン(w/v)、10mM Tris-HCl pH 7.4、2.5mM MgCl2、0.5mM CaCl2および0〜200単位/mlのDNアーゼIを含む。いくつかの態様において、透過処理/消化用緩衝液は、0.75%リゾレシチン(w/v)、10mM Tris-HCl pH 7.4、2.5mM MgCl2、0.5mM CaCl2および0〜500単位/mlのDNアーゼIを含む。いくつかの態様において、透過処理/消化用緩衝液は、0.25%リゾレシチン(w/v)、10mM Tris-HCl pH 7.4、2.5mM MgCl2、0.5mM CaCl2および0〜500単位/mlのDNアーゼIを含む。透過処理および溶解は、例えば、制限酵素に関して上述した通りに停止させることができる。
【0071】
他の箇所で考察しているように、DNアーゼまたは他の一般的なDNA切断剤の使用を、一方が標的であり、もう一方が、試験条件のいずれにおいても概ね常にアクセス可能であるかまたは概ね常にアクセス不能であるDNA領域である、少なくとも2つの異なるDNA領域間の切断の程度をモニターすることによって向上させることができる。そのような遺伝子の例は本明細書中の他の箇所で考察されており、公知であるかまたは同定することができる。例えば、「ハウスキーピング」遺伝子を範囲に含むDNA領域は概ね常にアクセス可能である。続いて、対照と比較した残りの標的の相対量を用いて、標的DNA領域での相対的なクロマチン構造を決定することができる。
【0072】
C.メチルトランスフェラーゼ
本発明のいくつかの態様において、DNA修飾剤はDNAに対する共有結合修飾を生成させる。例えば、いくつかの態様において、本発明のDNA修飾剤はメチルトランスフェラーゼである。
【0073】
種々のメチルトランスフェラーゼが当技術分野において公知であり、本発明に用いることができる。いくつかの態様において、用いるメチルトランスフェラーゼはDNA中のアデノシンに対してメチル部分を付加する。そのようなメチルトランスフェラーゼの例には、DAMメチルトランスフェラーゼが非限定的に含まれる。アデノシンは真核生物細胞ではメチル化されないため、特定のDNA領域におけるメチル化アデノシンの存在は、DAMメチルトランスフェラーゼ(または類似の活性を有する他のメチルトランスフェラーゼ)がDNA領域にアクセス可能であったことを指し示している。アデノシンのメチル化は、例えば、認識配列がメチル化アデノシンを含む制限酵素を用いて検出することができる。そのような酵素の一例には、DpnIが非限定的に含まれる。続いて、制限酵素による切断を、本明細書に記載した通りに(例えば、無傷のDNAは増幅されるが切断されたDNAはそうでないように‐またはLM-PCRを用いて、切断されたDNAは増幅するが無傷のDNAは増幅しないように)定量する。
【0074】
いくつかの態様において、メチルトランスフェラーゼはGC配列中のシトシンをメチル化する。そのようなメチルトランスフェラーゼの例には、MCviPIが非限定的に含まれる。例えば、Xu et al., Nuc. Acids Res. 26(17): 3961-3966 (1998)を参照のこと。GC配列は真核生物細胞ではメチル化されないため、特定のDNA領域におけるメチル化GC配列の存在は、DNA修飾剤(すなわち、GC配列中のシトシンをメチル化するメチルトランスフェラーゼ)がDNA領域にアクセス可能であったことを指し示している。メチル化GC配列は、任意のさまざまな手法を用いて同定することができる。いくつかの態様において、メチル化GC配列を検出するための方法は、バイサルファイト変換を含む。バイサルファイト変換は、DNAを、非メチル化シトシンをウラシルに変換するために十分な量のバイサルファイトと接触させることを含む。メチル化シトシンは変換されない。このため、GC配列を含むDNA領域を、GC配列中のシトシンをメチル化するメチルトランスフェラーゼと接触させ、単離して、続いてバイサルファイトと接触させることができる。GC配列中のCがメチル化されていなければ、そのCはU(またはその後に増幅された場合はT)に変換されると考えられるが、一方、メチル化CはCのままであると考えられる。ヌクレオチド配列決定、ならびにプライマー伸長またはプライマーに基づく増幅および/またはメチル化感受性制限消化を伴う方法を非限定的に含む、任意のさまざまな方法を用いて、バイサルファイトにより変換されたCの有無を検出することができる(例えば、MSnuPE、MSPまたはメチルライト(Methlyllight)、高分解能融解分析;ピロシークエンシングなど)。例えば、Fraga, et al., Biotechniques 33:632, 634, and 636-649 (2002);El-Maarri 0 Adv Exp Med Biol 544:197-204 (2003);Laird, Nat Rev Cancer 3:253266 (2003);およびCallinan, Hum Mol Genet 15 Spec No 1:R95-101 (2006)を参照のこと。
【0075】
いくつかの態様において、メチルトランスフェラーゼは、CG(「CpG」としても知られる)配列中のシトシンをメチル化する。そのようなメチルトランスフェラーゼの例には、M.SssIが非限定的に含まれる。そのようなメチルトランスフェラーゼの使用は一般に、典型的にはメチル化されないDNA領域に対する使用に限定される。これは、CG配列が真核生物細胞では内因性にメチル化され、それ故に、メチル化が稀であるようなDNA領域を除き、CG配列が内因性メチルトランスフェラーゼではなく修飾剤によってメチル化されると想定することは一般に不可能であるためである。GC配列の場合と同様に、CG配列のメチル化は、バイサルファイト変換を伴う方法を含む、任意のさまざまな方法によって検出することができる。
【0076】
D.化学物質
いくつかの態様において、DNA修飾剤はDNA修飾性化学物質である。大部分のDNA修飾性化学物質はクロマチンと比較して相対的に小さいため、融合パートナーを伴わないDNA修飾性化学物質の使用は、異なるDNA領域のアクセス可能性の程度に仮に違いがあるにしてもわずかであると考えられることから、状況によっては有効でない。このため、いくつかの態様において、DNA修飾剤は、DNA修飾性化学物質と連結された立体障害を有する分子を含む。立体障害を有する分子は、クロマチン構造に応じてDNA修飾剤のアクセス可能性に違いをもたらす任意のタンパク質または他の分子でありうる。これは例えば、結果を、本明細書に記載したDNアーゼまたは制限酵素を用いた結果と比較することによって検討することができる。
【0077】
いくつかの態様において、立体障害を有する分子は、少なくとも5、7、10または15kDのサイズであると考えられる。おそらく当業者は、立体障害を備える分子としてポリペプチドを用いることが好都合であると判断するであろう。DNA修飾剤がDNAを修飾する能力を著しく妨害することのない任意のポリペプチドを用いることができる。いくつかの態様において、ポリペプチドは、以下でさらに詳細に考察している二本鎖配列非特異的な核酸結合ドメインである。
【0078】
本発明のDNA修飾性化学物質は、立体障害を有する分子と直接連結させること、またはリンカーを介して連結させることができる。種々のホモおよびヘテロ二官能性リンカーが公知であり、この目的に用いることができる。
【0079】
例示的なDNA修飾性化学物質には、ヒドラジン(および、例えば、Mathison et al., Toxicology and Applied Pharmacology 127(1):91-98 (1994)に記載されているようなその誘導体)および硫酸ジメチルが非限定的に含まれる。いくつかの態様において、ヒドラジンはDNA中のグアノシンにメチル基を導入するか、他の様式でDNAを損傷させる。いくつかの態様において、硫酸ジメチルはグアニンをメチル化するか、または、グアニン中に存在するイミダゾール環を開裂させることにより、DNA中のグアニンの塩基特異的切断をもたらす。
【0080】
DNA修飾性化学物質による修飾の検出は、生じたDNA修飾の種類に依存すると考えられる。いくつかの態様において、硫酸ジメチルまたはヒドラジンによる修飾を検出するために、DNAをピペリジンにより高温(90℃)で処理する。DNAはDNA修飾部位で切断を受け、その切断を、本明細書に記載したヌクレアーゼ切断を検出するのと同じやり方で検出することができる。
【0081】
E.DNA修飾剤を改良するためのDNA結合ドメイン
いくつかの態様においては、本発明のDNA修飾剤または切断剤を、二本鎖配列非特異的な核酸結合ドメイン(例えば、DNA結合ドメイン)と融合させるかまたは他の様式で連結させる。DNA修飾剤がポリペプチドである場合には、二本鎖配列非特異的な核酸結合ドメインを、例えば、DNA修飾剤とのタンパク質融合物として、組換えDNA技術を介して合成することができる。二本鎖配列非特異的な核酸結合ドメインは、二本鎖核酸と配列非依存的な様式で結合するタンパク質またはタンパク質の定まった領域であり、すなわち結合は、特定の配列に対する全体的な選好性を示さない。いくつかの態様において、二本鎖核酸結合タンパク質は、二本鎖核酸に対して一本鎖核酸よりも10倍またはそれ以上高い親和性を示す。二本鎖核酸結合タンパク質は、本発明のいくつかの態様において耐熱性である。そのようなタンパク質の例には、古細菌小型塩基性DNA結合タンパク質Sac7dおよびSso7d(例えば、Choli et al., Biochimica et Biophysica Acta 950:193-203, 1988;Baumann et al., Structural Biol. 1:808-819, 1994;およびGao et al, Nature Struc. Biol. 5:782-786, 1998を参照)、古細菌HMf様タンパク質(例えば、Stanch et al., J. Molec. Biol. 255:187-203, 1996;Sandman et al., Gene 150:207-208, 1994を参照)、およびPCNA相同体(例えば、Cann et al., J. Bacteriology 181:6591-6599, 1999;Shamoo and Steitz, Cell:99, 155-166, 1999;De Felice et al., J. Molec. Biol. 291, 47-57, 1999;およびZhang et al., Biochemistry 34:10703-10712, 1995を参照)が非限定的に含まれる。DNA結合ドメインに関するそのほかの情報については、欧州特許第1283875B1も参照のこと。
【0082】
Sso7dおよびSac7d
Sso7dおよびSac7dは、それぞれ超好熱性古細菌スルホロブス・ソルファタリカス(Sulfolobus solfataricus)およびS.アシドカルダリウス(S. acidocaldarius)に由来する小型(約7,000kd MW)の塩基性染色体タンパク質である。これらのタンパク質はリジンに富み、高い耐熱性、酸安定性および化学安定性を有する。それらはDNAと配列非依存的な様式で結合し、結合すると、DNAのTMをいくつかの条件下では最大40℃上昇させる(McAfee et al., Biochemistry 34:10063-10077, 1995)。これらのタンパク質およびそれらの相同体は、典型的には、高温でゲノムDNAを安定化させることに関与すると考えられている。
【0083】
HMF様タンパク質
HMf様タンパク質は、DNAと直接相互作用すると考えられている真核生物H4ヒストンとアミノ酸配列および構造の両方の点で相同性のある古細菌ヒストンである。HMfファミリーのタンパク質は溶液中で安定な二量体を形成し、いくつかのHMf相同体が耐熱性種(例えば、メタノテルムス・フェルビダス(Methanothermus fervidus)およびパイロコッカス属GB-3a菌株)から同定されている。HMfファミリーのタンパク質は、Taq DNAポリメラーゼまたは固有の処理能力が低い任意のDNA修飾酵素と結合させると、その酵素がDNA基質に沿ってスライドする能力を強化し、それ故にその処理能力を高める。例えば、二量体HMf様タンパク質を、Taq DNAポリメラーゼのN末端に、例えば化学修飾を介して共有結合で連結させ、それによりそのポリメラーゼの処理能力を改善することができる。
【0084】
当業者は、当技術分野において公知であるその他の二本鎖配列非特異的な核酸結合ドメインを認知していると考えられ、それらを本明細書に記載したように用いることもできる。
【0085】
F.DNA修飾段階後のDNAの単離
いくつかの態様においては、DNA修飾/切断の段階の後に、利用可能な任意の方法に従ってゲノムDNAを細胞から単離する。その後の定量段階のために許容しうる純度のDNAがもたらされる限り、本質的にあらゆるDNA精製手順を用いることができる。例えば、標準的な細胞溶解試薬を、細胞を溶解するために用いることができる。任意でプロテアーゼ(プロテイナーゼKを非限定的に含む)を用いることができる。DNAは、当技術分野において公知である通りに混合物から単離することができる。いくつかの態様においては、フェノール/クロロホルム抽出を用い、DNAをその後に(例えば、エタノールにより)沈殿させて、精製する。いくつかの態様においては、必要に応じて(例えば、RNアーゼにより、またはDNA精製カラムにより)RNAを除去または分解する。
【0086】
任意で、ゲノムDNAを、中間精製段階を省いて、細胞溶解物から直接的に増幅するかまたは他の様式で検出する。
【0087】
いくつかの態様においては、無傷の、修飾された、または修飾されていないDNAを単離して、ライブラリーへとクローニングする。場合によっては、1つまたは複数の特定の無傷の、修飾された、または修飾されていない配列を単離および/またはクローニングする。または、無傷の、修飾された、または修飾されていないDNA領域を有する試料を用いて、そのような領域に関して濃縮されたライブラリーを調製する。DNA切断剤との接触後に無傷のDNAは、その剤に対して、よりアクセス可能性が低かったDNAを表す。同様に、DNA修飾剤との接触後に修飾されていないDNAは、よりアクセス可能性の低いDNAを表す。その反対に、修飾されたDNAは、修飾剤に対して、よりアクセス可能性が高かったDNAを表す。上記の態様のいくつかにおいては、クローニングの前に、無傷のDNAを切断されたDNAから精製し(例えば、分離し)、かつ/または修飾されたDNAを修飾されていないDNAから精製し、それにより、1つのクラスのDNAに関してのクローニングプールを濃縮する。修飾された/修飾されていないDNAに関する濃縮は、修飾の性質に応じて異なると考えられる。いくつかの態様においては、修飾された(または修飾されていないDNA)と特異的に結合する親和性薬剤を用いて、修飾されたDNAを修飾されていないDNAから分離する。
【0088】
いくつかの態様においては、サブトラクティブライブラリーを作製する。例えば、本発明の方法において無傷の、修飾された、または修飾されていない罹患細胞DNA領域に関して濃縮されたライブラリーを作製し、その後に健常細胞由来の対応するライブラリーを用いてサブトラクションして、それにより、両方とも無傷であるか、修飾されているか、または修飾されておらず、かつ特定の疾患に対して特異的である、差次的なDNA配列ライブラリーを作製することができる。癌細胞を非限定的に含む、任意の罹患細胞を用いることができる。代替的なサブトラクティブ戦略を、例えば、異なる細胞型、細胞時期、薬物治療などの間で用いることもできる。
【0089】
G.DNAの物理的特徴の検出
細胞とDNA修飾剤またはDNA切断剤との接触の後に、DNAの任意のさまざまな物理的特徴を検出することができる。物理的特徴には、DNAメチル化、融解温度、GC含有量、ヌクレオチド配列、およびポリヌクレオチドとハイブリダイズする能力が非限定的に含まれる。そのような特徴を検出するための種々の方法が公知であり、用いることができる。いくつかの態様において、DNA修飾/切断の段階の後に決定される物理的特徴は、DNAフットプリンティング(例えば、DNAの特定の領域に対する1つまたは複数の特定のタンパク質の能力)を決定することを含まない。例えば、1つの非限定的な態様において、例えばqPCRを用いる無傷のDNAの定量は、DNAフットプリント法を含まない。
【0090】
いくつかの態様において、物理的特徴はDNAメチル化である。例えば、相対的にアクセス可能なDNAがDNA切断剤によって切断されれば、残りの無傷のDNA(アクセス可能性がより低いDNAに相当する)を単離し、続いてメチル化状態に関して分析することができる。非常にさまざまなDNAメチル化検出方法が公知である。いくつかの態様においては、DNA修飾剤または切断剤との接触後に、DNAをバイサルファイトと接触させ、それにより、DNA中のメチル化されていないシトシンをウラシルに変換する。続いて、特定のDNA領域のメチル化を、本明細書で考察したものを含む、任意のさまざまなメチル化検出方法によって決定することができる。いくつかの態様において、高分解能融解アッセイ(HRM)を用いて、バイサルファイト変換後のメチル化状態を検出する。この方法では、バイサルファイト変換後にDNA領域を増幅し、その結果得られたアンプリコンの融解温度を決定する。融解温度はシトシンがバイサルファイトによって変換されたか否か(およびその後に増幅反応において「T」としてコピーされたか否か)に応じて異なると考えられるため、アンプリコンの融解温度をメチル化含有量と相関させることができる。
【0091】
V.標的DNA領域
DNA領域は、ゲノムDNA内部の関心対象の標的配列である。細胞のゲノムDNA中の任意のDNA配列を、本明細書に記載した通りに、DNA修飾剤のアクセス可能性に関して評価することができる。例えば、複数の異なる細胞型において、処理していない細胞と薬物刺激、化学刺激もしくは環境的刺激に曝露された細胞との間で、または正常組織と罹患組織との間で異なるアクセス可能性を呈する、関心対象のDNA領域を同定するために、DNA領域をスクリーニングする。したがって、いくつかの態様において、本発明の方法は、アクセス可能性の変化が疾患(またはその欠如)のマーカーとして作用するDNA領域を同定するために用いられる。例示的な疾患には、癌が非限定的に含まれる。癌でない細胞と比較して癌細胞において、変化したDNAメチル化および/またはクロマチン構造を有する、数多くの遺伝子が記載されている。
【0092】
いくつかの態様において、DNA領域は、特定の細胞の罹患状態または発生状態に応じて、アクセス可能性に差異を伴うことが判明している。これらの態様において、本発明の方法は、診断ツールまたは予後予測ツールとして用いることができる。本発明の方法を用いて診断または予後予測がひとたび確定されれば、その診断または予後予測を考慮して、治療レジメンを設定すること、または既存の治療レジメンを変更することができる。例えば、本発明の方法に従った癌細胞の検出は、癌細胞が検出された個体に対する化学療法薬および/または放射線の投与につながりうる。
【0093】
種々のDNA領域を、研究目的で、および/または試薬が予想通りに機能することを確認するための対照DNA領域として、検出することができる。例えば、いくつかの態様においては、ある動物の本質的にすべての細胞においてアクセス可能であるDNA領域をアッセイする。そのようなDNA領域は、例えば、アクセス可能性に関する陽性対照として有用である。そのようなDNA領域は、例えば、構成的であるかまたはほぼ構成的である遺伝子の内部または近傍に見いだすことができる。そのような遺伝子には、一般に「ハウスキーピング」遺伝子と呼ばれるもの、すなわち、基本的な生物機能を維持するためにその発現が必要とされる遺伝子が含まれる。そのような遺伝子の例には、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ(GAPDH)およびβアクチン(ACTB)が非限定的に含まれる。DNA領域には、任意でプロモーターの少なくとも一部分を含む、そのような遺伝子の全体または一部分が含まれうる。
【0094】
いくつかの態様において、DNA領域は、ある動物のほとんどの細胞においてアクセス不能であるDNAの少なくとも一部分を含む。そのようなDNA領域は、例えば、アクセス可能性に関する陰性対照として有用である。この文脈における「アクセス不能な」とは、そのコピーが、DNA領域のコピーの20%前後を上回らない割合でしか修飾されないDNA領域のことを指す。そのような遺伝子配列の例には、「異質染色質」性として一般に認識されているものが含まれ、極めて特定の細胞型のみで発現される(例えば、組織特異的または臓器特異的な様式で発現される)遺伝子が含まれる。(特定の細胞型を除いて)一般にアクセス不能である例示的な遺伝子には、ヘモグロビン-β鎖(HBB)および免疫グロブリン軽鎖κ(IGK)が非限定的に含まれる。
【0095】
いくつかの態様において、DNA領域は、細胞の罹患状態に応じて異なるアクセス可能性を有するか、またはさもなければ細胞の型または生育環境に応じてさまざまなアクセス可能性を有する、遺伝子配列である。例えば、ある種の遺伝子は、癌でない細胞では一般にアクセス不能であるが、癌細胞ではアクセス可能である。さまざまなアクセス可能性を有する遺伝子には、例えば、グルタチオン-s-トランスフェラーゼπ(GSTP1)が含まれる。
【0096】
いくつかの態様において、本発明のDNA領域は、以下の遺伝子の1つまたは複数からの遺伝子配列(例えば、プロモーター配列)から選択される:1型カドヘリン1(E-カドヘリン)、シトクロムP450-1A1(CYP1A1)、Ras関連ドメインファミリー1A(RASSF1A)、p15、p16、細胞死関連プロテインキナーゼ1(DAPK)、腺腫性結腸ポリポーシス(APC)、メチルグアニン-DNAメチルトランスフェラーゼ(MGMT)、乳癌1遺伝子(BRCA1)およびhMLH。
【0097】
いくつかの態様において、DNA領域は、例えば、異なる細胞型の間、異なる条件の間、正常細胞と罹患細胞との間などで異なるアクセス可能性を有する領域を同定するために、ランダムに選択される。
【0098】
VI.標的遺伝子座のコピーの定量
DNA修飾を定量するための方法は、ゲノムDNAに導入されるDNA修飾の種類に依存すると考えられる。例えば、二本鎖DNA切断イベント(例えば、制限酵素もしくはDNアーゼによって導入されるような、または、メチルトランスフェラーゼ処理後もしくは本明細書に記載されたDNA修飾性化学物質による修飾後に、例えばメチル化感受性もしくはメチル化依存性制限酵素によって続いて導入されるような修飾)を、関心対象のDNA領域を含むアンプリコンを生成するように設計された増幅反応を用いて好都合に検出することができる。配列特異的制限酵素を用いた場合のような定まった部位での切断イベントの場合には、潜在的な切断部位にまたがるアンプリコンが生成されるようにプライマーを設計する。無傷のDNAのみが増幅されると考えられる。全DNAの量も判明しているならば、切断されたDNAの量を、全DNAと無傷のDNAとの間の差として計算することができる。DNAの総量は、当技術分野において公知である任意のDNA定量法に従って決定することができる。いくつかの態様において、全DNAの量は、修飾とは無関係にDNAを増幅するプライマーのセットを設計することにより、好都合に決定することができる。これは例えば、同じ遺伝子領域内または別のDNA領域内にある、潜在的な切断部位をまたがないプライマーを設計することによって達成することができる。DNアーゼIなどの配列非特異的ヌクレアーゼを用いる場合のような不定部位での切断イベントの場合には、アクセス不能な参照遺伝子の使用を内部対照として組み入れるべきである。
【0099】
以下により詳細に考察するように、定量的増幅(例えばリアルタイムPCRを含む)方法は、DNA領域の無傷のコピーの量の決定を可能にし、さまざまな対照とともに用いる場合、細胞におけるコピーの総数と比較して無傷のDNAの相対量を決定するために用いることができる。従って、DNA領域の修飾されたコピーまたは修飾されていないコピーの実数または相対数(例えば、コピーの総数に対して相対的、または第2のDNA領域の修飾されたコピーまたは修飾されていないコピーの数に対して相対的)を、算出することができる。
【0100】
本発明のいくつかの態様において、DNA領域の修飾されたコピーの数は直接的に決定される。例えば、特定の付着末端または平滑末端の存在時にのみ起こると考えられる特定のライゲーションイベントを検出することなどにより、制限酵素切断を検出して定量することもできる。例えば、制限酵素によって生成される付着末端に対して相補的な付着末端を含む核酸アダプターを、切断されたゲノムDNAとライゲートさせることができる。続いて、(例えば、定量的増幅方法により)ライゲーションイベントの数を検出して定量することができる。
【0101】
いくつかの態様においては、ライゲーション媒介PCR(LM-PCR)を用いて、DNA領域の切断されたコピーの数を定量する。LM-PCRの方法は当技術分野において公知であり、Pfeifer et al., Science 246: 810-813 (1989)において最初に記載されている。LM-PCRは、必要に応じて、定量的結果のためにリアルタイムで行うことができる。
【0102】
定量的増幅方法(例えば、定量的PCRまたは定量的線形増幅)は、核酸テンプレートを増幅し、増幅されたDNAの量を直接的または間接的に(例えば、Ct値を決定して)決定し、続いて増幅サイクル数に基づいて最初のテンプレートの量を算出することを伴う。反応を用いたDNA遺伝子座の増幅は周知である(米国特許第4,683,195号および第4,683,202号;PCR PROTOCOLS: A GUIDE TO METHODS AND APPLICATIONS (Innis et al., eds, 1990)を参照)。典型的には、PCRを用いてDNAテンプレートを増幅する。しかし、代替的な増幅方法も記載されており、それらの代替的な方法がそれらの方法が切断されたDNAを増幅するよりも高い程度で無傷のDNAを増幅する限り、それを用いることもできる。定量的増幅の方法は、例えば、米国特許第6,180,349号;第6,033,854号;および第5,972,602号、さらには例えば、Gibson et al., Genome Research 6:9951001 (1996);DeGraves, et al., Biotechniques 34(1):106-10, 112-5 (2003);Deiman B, et al., Mol Biotechnol. 20(2):163-79 (2002)に記載されている。増幅は「リアルタイム」でモニターすることができる。
【0103】
いくつかの態様において、定量的増幅は、増幅(例えば、PCR)反応のサイクルにおけるテンプレートのコピーを表すシグナル(例えば、プローブの蛍光)のモニタリングに基づく。PCRの初期サイクルでは、形成されるアンプリコンの量が、アッセイから測定可能であるシグナル出力を支持しないため、非常に弱いシグナルしか観察されない。初期サイクルの後には、形成されるアンプリコンの量が増加するため、シグナル強度は測定可能なレベルまで増大し、PCRが非対数期に入る後期のサイクルではプラトーに達する。シグナル強度対サイクル数のプロットを通じて、測定可能なシグナルがPCR反応から得られる特定のサイクルを推定し、それを用いてPCRの開始前の標的の量を逆算することができる。この方法によって決定される特定のサイクル数は、典型的には、サイクル閾値(Ct)と呼ばれる。例示的な方法は、例えば、Heid et al. Genome Methods 6:986-94 (1996)に、加水分解プローブに関連して記載されている。
【0104】
増幅産物の検出のための1つの方法は、5'-3'エキソヌクレアーゼ「加水分解」PCRアッセイ(TaqMan(商標)アッセイとも呼ばれる)である(米国特許第5,210,015号および第5,487,972号;Holland et al., PNAS USA 88: 7276-7280 (1991);Lee et al., Nucleic Acids Res. 21: 3761-3766 (1993))。このアッセイは、増幅反応の間に、二重標識された蛍光発生プローブ(「TaqMan(商標)」プローブ)のハイブリダイゼーションおよび切断によって特定のPCR産物の蓄積を検出する。蛍光発生プローブは、蛍光レポーター色素およびクエンチャー色素の両方で標識されたオリゴヌクレオチドからなる。PCRの間に、このプローブは、増幅されるセグメントとそれがハイブリダイズした場合に、しかもその場合にのみ、DNAポリメラーゼの5'-エキソヌクレアーゼ活性によって切断される。プローブの切断は、レポーター色素の蛍光強度の増加を生じさせる。
【0105】
エネルギー転移の使用に依拠して増幅産物を検出するもう1つの方法は、Tyagi and Kramer, Nature Biotech. 14:303-309 (1996)に記載された「ビーコンプローブ」法であり、これは米国特許第5,119,801号および第5,312,728号の主題でもある。この方法は、ヘアピン構造を形成することができるオリゴヌクレオチドハイブリダイゼーションプローブを用いる。ハイブリダイゼーションプローブの一方の末端(5'末端または3'末端かのいずれか)にはドナーフルオロフォアがあり、もう一方の末端にはアクセプター部分がある。TyagiとKramerの方法の場合には、このアクセプター部分はクエンチャーであり、すなわち、アクセプターはドナーによって放出されたエネルギーを吸収するが、その後に自ら蛍光を発することはない。したがって、ビーコンがオープン構造である場合はドナーフルオロフォアの蛍光を検出しうるが、ビーコンがヘアピン(閉鎖)構造である場合はドナーフルオロフォアの蛍光は消光する。PCRに用いる場合、PCR産物の一方の鎖とハイブリダイズする分子ビーコンプローブはオープン構造であり、蛍光が検出されるが、ハイブリダイズしないままのものは蛍光を発しないと考えられる(Tyagi and Kramer, Nature Biotechnol. 14: 303-306 (1996))。その結果、蛍光の量はPCR産物の量が増加するにつれて増加し、それ故にPCRの進行の尺度として用いることができる。当業者は、その他の定量的増幅法も利用可能であることを認識しているであろう。
【0106】
核酸の定量的増幅を行うためのさまざまな他の手法も公知である。例えば、いくつかの方法は、オリゴヌクレオチドが標的核酸とハイブリダイズした場合に蛍光の変化が生じるように構築されている1つまたは複数のプローブオリゴヌクレオチドを用いる。例えば、そのような1つの方法は、2つのオリゴプローブをアンプリコンとアニーリングさせる、蛍光共鳴エネルギー転移(FRET)を利用する二重フルオロフォアアプローチ、例えば、LightCycler(商標)ハイブリダイゼーションプローブである。オリゴヌクレオチドは、効率的なエネルギー転移に適合する距離で隔てられたフルオロフォアと頭-尾方向でハイブリダイズするように設計される。核酸と結合するかまたは伸長産物に組み入れられた場合にシグナルを発するように構築された標識オリゴヌクレオチドの他の例には、以下が含まれる:Scorpions(商標)プローブ(例えば、Whitcombe et al., Nature Biotechnology 17:804-807, 1999および米国特許第6,326,145号)、Sunrise(商標)(またはAmplifluor(商標))プローブ(例えば、Nazarenko et al., Nuc. Acids Res. 25:2516-2521, 1997および米国特許第6,117,635号)、および、クエンチャーがなくてもシグナル低下をもたらす二次構造を形成し、かつ標的とハイブリダイズした場合に増大したシグナルを発するプローブ(例えば、Lux probes(商標))。
【0107】
他の態様においては、二本鎖DNA中に挿入された時にシグナルを生じるインターカレート剤を用いうる。例示的な剤には、SYBR GREEN(商標)、SYBR GOLD(商標)およびEVAGREEN(商標)が含まれる。これらの剤はテンプレート特異的ではないため、シグナルはテンプレート特異的増幅に基づいて生成されると想定される。テンプレート配列の融解点は一般に、例えばプライマー-ダイマーなどよりもはるかに高いと考えられるため、このことはテンプレートの関数としてシグナルをモニターすることによって確認することができる。
【0108】
いくつかの態様において、DNA領域の量は、試料中のヌクレオチド配列のコピーを決定し、続いて試料中の同じ配列を有するコピーの相対数または絶対数を決定することによって決定される。
【0109】
本発明の方法による修飾された(または修飾されていない)DNA領域の定量は、いくつかの態様において、DNA領域の修飾されたコピーまたは修飾されていないコピーの、その同じ領域のコピーの総数と比較した相対量(例えば、比またはパーセンテージなどの標準化された値)を決定することによって、さらに改良することができる。いくつかの態様において、1つのDNA領域の修飾されたコピーまたは修飾されていないコピーの相対量を、第2の(またはそれ以上の)DNA領域の修飾されたコピーまたは修飾されていないコピーの数と比較する。いくつかの態様において、2つまたはそれ以上のDNA領域の間で比較する場合には、各DNA領域の修飾されたコピーまたは修飾されていないコピーの相対量を、まず、DNA領域のコピーの総数に対して標準化する。または、同じ試料から得られる場合には、いくつかの態様において、各DNA領域のコピーの総数はおおよそ同じであると仮定し、そのため2つまたはそれ以上のDNA領域の間で比較する場合に、各値をコピーの総数に対してまず標準化することなしに、修飾されたコピーまたは修飾されていないコピーの相対量(例えば、比またはパーセンテージ)を各DNA領域間で決定することもできる。
【0110】
いくつかの態様においては、修飾されたコピーまたは修飾されていないコピーの実際の量または相対量(例えば、全DNAとの対比)を、対照値と比較する。対照値は、例えば、特定のDNA領域に対するアクセス可能性が特定の値を上回るかそれ未満であるかを知りたい場合に、好都合に用いることができる。例えば、特定のDNA領域が正常細胞では典型的にはアクセス可能であるが罹患細胞ではアクセス不能である(またはその反対の)状況では、修飾されたコピーまたは修飾されていないコピーの実数または相対数を、単に、(例えば、20%超またはそれ未満が修飾されているかまたは修飾されていない、80%超またはそれ未満が修飾されているかまたは修飾されていない、などの)対照値と比較してもよい。または、対照値が、対照DNA領域に関する過去データまたは予想されるデータであってもよい。これらの場合には、対照DNA領域の実際の量または相対量を(任意で数回)決定し、その結果得られたデータを用いて、関心対象のDNA領域に関して決定された修飾されたコピーまたは修飾されていないコピーの実数または相対数と比較することのできる対照値を作成することができる。
【0111】
本明細書に記載された方法のための計算は、コンピュータベースの計算およびツールを含みうる。これらのツールは、従来設計の汎用コンピュータシステム(本明細書では「ホストコンピュータ」と称する)によって実行可能なコンピュータプログラムの形態で提供されることが好都合である。ホストコンピュータは数多くのさまざまなハードウェア構成部分を用いて構成されてよく、さまざまな寸法および方式で製造することができる(例えば、デスクトップPC、ラップトップ、タブレットPC、ハンディ型コンピュータ、サーバー、ワークステーション、大型汎用コンピュータ)。モニター、キーボード、ディスクドライブ、CDおよび/またはDVDドライブなどの標準的な構成部品が含まれてもよい。ホストコンピュータをネットワークに結びつける場合、接続は任意の適した伝達媒体(例えば、有線媒体、光媒体および/または無線媒体)ならびに任意の適した通信プロトコル(例えば、TCP/IP)を介して提供されうる;ホストコンピュータが適したネットワーク用ハードウェア(例えば、モデム、イーサネットカード、WiFiカード)を含んでもよい。ホストコンピュータは、UNIX、Linux、Microsoft Windows、MacOSまたは任意の他のオペレーティングシステムを含む、さまざまなオペレーティングシステムの任意のものを実行してよい。
【0112】
本発明の実行局面に関するコンピュータコードは、PERL、C、C++、Java、JavaScript、VBScript、AWK、または、ホストコンピュータ上で実行されうるか、もしくはホストコンピュータ上で実行されるようにコンパイルされうる、任意の他のスクリプト言語またはプログラム言語を含むさまざまな言語で記述されうる。また、コードを、アセンブリ言語またはマシン言語のような低水準言語で記述するかまたは分散させてもよい。
【0113】
ホストコンピュータシステムは、ユーザーがツールの動作を制御するインターフェースを好都合に提供する。本明細書に記載された例において、ソフトウェアツールはスクリプト(例えば、PERLを用いる)として実行され、その遂行はユーザーにより、LinuxまたはUNIXなどのオペレーティングシステムの標準的なコマンドラインインターフェースから開始される。当業者は、コマンドをオペレーティングシステムに対して適宜適合させうることを認識しているであろう。他の態様において、ユーザーがポインティングデバイスを用いて動作を制御することを可能にするグラフィカルユーザーインターフェースが提供されてもよい。したがって、本発明は、いかなる特定のユーザーインターフェースにも限定されない。
【0114】
本発明のさまざまな特徴を組み入れたスクリプトまたはプログラムを、記憶および/または伝達のために、さまざまなコンピュータ可読媒体上に符号化して記録してもよい。適した媒体の例には、磁気ディスクまたはテープ、コンパクトディスク(CD)またはDVD(デジタル多目的ディスク)などの光学記憶媒体、フラッシュメモリー、ならびにインターネットを含む種々のプロトコルに準拠する有線、光および/または無線ネットワークを介した伝達用に適合化されたキャリア信号が含まれる。
【0115】
VII.反応混合物
本発明はまた、本明細書に記載された試薬の1つまたは複数を、任意で(クロマチン状態を決定しようとする)真核細胞とともに含む、反応混合物も提供する。いくつかの態様において、反応混合物は、例えば、DNA修飾剤(例えば、制限酵素、DNアーゼ、メチルトランスフェラーゼまたはDNA修飾性化学物質)および細胞透過処理剤および/または細胞崩壊剤、ならびに真核細胞を含む。また、本明細書に記載された他の試薬(バイサルファイトを非限定的に含む)を本発明の反応混合物中に含めることもできる。
【0116】
VIII.キット
本発明はまた、本発明のアクセス可能性アッセイを行うためのキットも提供する。キットは任意で、書面による使用説明書、または電子的な使用説明書(例えば、CD-ROMまたはDVD上)も含みうる。本発明のキットは、例えば、DNA修飾剤、ならびに細胞透過処理剤および/または細胞崩壊剤を含みうる。DNA修飾剤には、例えば、制限酵素、DNアーゼ、メチルトランスフェラーゼまたはDNA修飾性化学物質を含む、本明細書中に詳細に記載されたものが含まれうる。いくつかの態様において、DNA修飾剤は、MnlI、FokIおよびAlwIを非限定的に含むII-S型制限酵素である。本発明のキットは、透過処理剤およびDNA修飾剤を、同じバイアル/容器内(およびそれ故に同じ緩衝液中)に含む。または、透過処理剤およびDNA修飾剤が別々のバイアル/容器内にあってもよい。
【0117】
本発明のキットはまた、1つまたは複数の対照細胞および/または核酸も含みうる。例示的な対照核酸には、例えば、ある動物の本質的にすべての細胞においてアクセス可能である(例えば、ハウスキーピング遺伝子配列またはそのプロモーター)か、またはある動物のほとんどの細胞においてアクセス不能であるかのいずれかである遺伝子配列を含むものが含まれる。いくつかの態様において、本キットは、そのような遺伝子配列を増幅するためのプライマーの1つまたは複数のセットを(遺伝子配列または細胞がキットに実際に含まれるか否かにかかわらず)含む。例えば、いくつかの態様において、本キットは、DNA修飾剤ならびに細胞透過処理剤および/または細胞崩壊剤、ならびに対照DNA領域を増幅するための1つまたは複数のプライマーセット、ならびに任意で、第2のDNA領域、例えば標的DNA領域を増幅するための1つまたは複数のプライマーセットを含む。DNA修飾剤が制限酵素である態様においては、1つのDNA領域当たりに、制限酵素の潜在的な切断部位を少なくとも1つ(例えば、1つ、2つ、3つ、4つなど)含むDNA領域の一部分を増幅するための(例えば、修飾されていないコピーの数を計算するために有用な)一つのプライマーセット、および、潜在的な切断部位を含まないDNA領域の一部分を増幅するための(例えば、コピーの総数を計算するために有用な)少なくとも第2のプライマーセットという、少なくとも2つのプライマーセットを含めることができる。
【0118】
いくつかの態様において、本発明のキットは、以下のもののうち1つまたは複数を含む:
(i)細胞膜の透過処理剤または崩壊剤;
(ii)制限酵素、DNアーゼまたは他のDNA修飾剤;
(iii)修飾剤によるさらなる修飾を防止することのできる「停止」用溶液;
(iv)核酸の抽出および/または精製のための材料(例えば、ゲノムDNAの精製のため、および/または「停止」用溶液の成分などの非DNA成分の除去のためのスピンカラム);
(vi)DNAのPCR/qPCR増幅のための試薬、任意で、テンプレートおよび/またはポリメラーゼのほかにPCRまたはqPCRのために必要なすべての成分を含む1つの混合物;
(vii)特定の標的遺伝子のPCR/qPCR増幅のためのプライマーセット。
【実施例】
【0119】
以下の実施例は、特許請求する本発明を限定するためではなく、それを例示するために提供される。
【0120】
実施例1:一般的アプローチ
本アッセイは、DNA修飾剤の、細胞内のクロマチンの異なる部分に対するアクセス可能性の違いを利用する。図2に図示されているように、DNA修飾剤は、クロマチンのある部分に、他の部分よりも容易にアクセスすることができる。図3は、アッセイの例示的な作業の流れを図示している。付着細胞を出発材料として24ウェルプレート中で増殖させた。この態様においては、2つのウェルを各実験に対して用いる:一方のウェルはヌクレアーゼを含まない透過処理用緩衝液で処理する;もう一方のウェルはヌクレアーゼを含む透過処理用緩衝液で処理する。細胞からリアルタイムPCRに至るまでの過程全体には約3時間がかかり、その結果は細胞採取の当日に得ることができる。
【0121】
図4は、MnlIをクロマチン構造プローブとして用いた場合の、細胞から単離したDNAの分析に関する概略図を図示している。この場合におけるアッセイは、「ヌクレアーゼなし」の対照と同時並行して行う。2つのプライマーセットを各遺伝子に対して用いる。一方のプライマーセットはMnlI消化部位にまたがらないが、もう一方のプライマーセットは少なくとも1つのMnlI部位にまたがる。ヌクレアーゼを含まない透過処理用試薬で処理した細胞は、両方のプライマーセットから完全な増幅(すなわち、すべてのコピーが増幅される)をもたらし、それ故に図4の左のグラフに図示されているように同程度のCt値を有するはずである。デルタCt値(トータルCtから無傷Ctを差し引いたもの)は、プライマー特性に基づく、切断されなかったDNAにおけるこれらのプライマーセット間のCt値の差を反映している。
【0122】
対照的に、MnlIで処理した細胞では、アクセス可能な遺伝子はMnlI部位で切断されたはずである。MnlI部位にまたがらないプライマーセットは、依然としてすべてのDNA分子を増幅すると考えられ(コピーのトータル数の尺度として機能する)、従ってCtはDNAの全コピーの数量を反映する。(少なくとも1つのMnlI部位にまたがる)もう一方のプライマーセットは、切断されなかった配列、すなわち無傷の配列のみを増幅しうると考えられる。消化の程度に応じて、Ct値は推移し、消化の量に比例して大きくなると考えられる。続いて、トータルCtから無傷Ctを差し引くことによってデルタCtを算出することができ、もしDNAがアクセス可能であれば、これは負の数字をもたらすと考えられる。アクセス不能であるMnlI部位のパーセンテージは、図4に示された式によって算出される。
【0123】
さまざまな種類のDNA領域に対するこの手法の適用は、図5〜6に図示されている。図5は、GAPDH、HBBおよびGSTP1遺伝子の少なくとも一部を示している。MnlI、TATAボックス、プロモーター領域およびエクソンが示されている。GAPDHは陽性対照として用いた。GAPDHはほとんどの細胞株で発現されるハウスキーピング遺伝子であり、アクセス可能であるはずである。ヘモグロビンβ鎖遺伝子は陰性対照として選択した。ヘモグロビンはほとんどの細胞で発現されず、エピジェネティックにサイレント化されており、アクセス不能なクロマチン構造内にあると予想される。最後に、前立腺癌においてエピジェネティックに不活性化されるグルタチオン-s-トランスフェラーゼπすなわちGSTP1も分析した。図6は、増幅のための標的とすることのできる領域を示している。各遺伝子について、どのMnlI部位にもまたがらず、すべてのDNA鎖を増幅すると考えられるトータルプライマーのセットを用いる。少なくとも1つ(例えば、2つ〜4つ)のMnlI部位にまたがり、これらの部位で切断されないDNA鎖のみを増幅すると考えられる「無傷」プライマーのセットも用いる。
【0124】
実施例2:Hela細胞においてMnlIを用いたデータ
この実施例は、以上に概説した実験アプローチを用いて生成されたデータの結果を示している。
【0125】
材料および方法
化学物質 L-α-リゾホスファチジルコリン(リゾレシチン)はSigma-Aldrichから購入した。MnlI、DNアーゼI、BSAおよびプロテイナーゼKは、New England Biolabsから購入した。RNアーゼAはQiagenから購入した。組織培養プレートはVWRから購入した。iQ SYBRはBio-Radから入手した。
【0126】
細胞の処理 24ウェルプレート中で増殖させた細胞を、それらが集密度90%に達した時点で処理した。培地を吸引し、100ulの透過処理/消化用緩衝液を細胞の上から穏やかに重ねた。MnlIで処理した細胞に関して、透過処理/消化用緩衝液は、リゾレシチン、NaCl、Tris-HCl、MgCl2、DTT、BSAおよびMnlIからなっていた。DNアーゼIで処理した細胞に関して、透過処理/消化用緩衝液は、リゾレシチン、Tris-HCl、MgCl2、CaCl2およびDNアーゼIからなっていた。透過処理した細胞を、続いて37℃で1時間インキュベートした。インキュベーション後に、25ulの溶解/停止用溶液(100mM Tris-HCl pH 7.4、100mM NaCl、100mM EDTA、5% N-ラウロイルサルコシン(w/v)、80ug/mlのRNアーゼAおよび3mg/mlのプロテイナーゼK)を透過処理/消化用緩衝液に添加し、細胞溶解物を37℃で10分間インキュベートした。
【0127】
DNAの単離 細胞溶解物を収集し、市販の核酸精製キット(Aurum, Bio-Rad)を標準的な手順に従って用いてDNAを単離した。DNAを100ul中に溶出させ、DNAの濃度を、NanoDrop 1000分光光度計(Thermo Scientific)を用いて決定した。続いて試料を1μl当たり1〜10ngのDNAという濃度に希釈した。
【0128】
分析した遺伝子 本発明者らは、3つのヒト遺伝子のプロモーター領域を分析した:グリセルアルデヒド-3-リン酸デヒドロゲナーゼ(GAPDH)、ヘモグロビンβ(HBB)およびグルタチオン-s-トランスフェラーゼπ(GSTP1)。GAPDHはハウスキーピング遺伝子である;これはほとんどの細胞で発現され、アクセス可能なクロマチン構造を有すると予想される。HBBはほとんどの細胞で発現されず、アクセス不能なクロマチン構造を有すると予想される。GSTP1は前立腺癌においてエピジェネティックに不活性化されている。非癌性前立腺細胞株であるRWPE1ではGSTP1はアクセス可能なクロマチン構造内にあって、発現され、癌性前立腺細胞株LNCaPではGSTP1はアクセス不能なクロマチン構造内にあり、発現されない(Okino, S.T., et al., Chromatin changes on the GSTP1 promoter associated with its inactivation in prostate cancer. Mol Carcinog, 2007. 46(10): P. 839-46)。GSTP1 mRNAはHela細胞およびHCT15細胞株でも高発現される;PC3細胞株ではGSTP1 mRNAはHela細胞およびHCT15細胞で見出されるレベルのおよそ半分のレベルで発現される。
【0129】
リアルタイムPCR MnlIで処理した細胞から単離したDNA試料を、Chromo4連続蛍光検出器(MJ Research)を取り付けたPTC-200サーマルサイクラーにおいて、各プライマーセットを用いて分析した。各反応物は容積20ulとし、50ngのDNA、500nMの各プライマーおよび10ulのiQ SYBR green supermix(Bio-Rad)を含めた。PCR条件は、96℃ 10分間;96℃ 30秒間および68℃ 1分間を40サイクル;72℃ 5分間;0.2℃間隔で各段階で5秒間保つ72℃から98℃までの融解曲線、とした。DNアーゼIで処理した細胞から単離したDNA試料を、CFX96リアルタイムPCR検出システム(Bio-Rad)において分析した。各反応物は容積20ulとし、5ngのDNA、500nMの各プライマーおよび10ulのiTaq Fast SYBR green supermix with ROX(Bio-Rad)を含めた。PCR条件は、95℃ 5分間;95℃ 30秒間および72℃ 1分間を40サイクル;72℃ 5分間;0.2℃間隔で各段階で5秒間保つ72℃から95℃までの融解曲線、とした。
【0130】
データ分析 アクセス不能なクロマチンのレベルを、MnlIをヌクレアーゼプローブとして用いて決定するために、本発明者らは以下の計算を行った。ΔCt(切断なし)は、MnlIを含まない消化/透過処理用緩衝液で処理した細胞から単離したDNAを分析した場合の(無傷Ct−トータルCt)として算出した。ΔCt(切断あり)は、MnlIで処理した細胞から単離したDNAを分析した場合の(無傷Ct−トータルCt)として算出した。アクセス不能なクロマチンのレベルは、2∧(ΔCt(切断なし)−ΔCt(切断あり))によって算出した(図4参照)。
【0131】
DNアーゼIはアクセス可能なクロマチンにおいてトータルおよび無傷の増幅領域の両方の内部を切断するため、GAPDHおよびGSTP1に関するアクセス不能なクロマチンのレベルをDNアーゼIを用いて決定するために、本発明者らは、本発明者らがMnlIを分析する場合に用いたのと同じ計算を用いることはできなかった。本発明者らはHBBプロモーターがDNアーゼI消化に対する抵抗性が高いことを見いだし、本発明者らは、アクセス不能性クロマチンの低い消化レベルを反映するHBB ΔCtを内部参照標準として用いた(図16に図示されている)。ΔCt(参照)は、(HBBの+ヌクレアーゼCt−HBBのヌクレアーゼなしCt)として算出した。ΔCt(標的)は、GAPDH/GSTP1の+ヌクレアーゼCt−GAPDH/GSTP1のヌクレアーゼなしCtとして算出した。GAPDHまたはGSTP1標的遺伝子のアクセス不能なクロマチンのレベルは、2∧(ΔCt(参照)-ΔCt(標的))として算出した。
【0132】
結果
Hela細胞を、ヌクレアーゼを含まない消化用緩衝液、リゾレシチン(細胞透過処理用試薬)およびヌクレアーゼを含まない消化用緩衝液、またはリゾレシチンおよびヌクレアーゼ(MnlI[M]またはDNアーゼI[D])を含む完全な消化用緩衝液のいずれかで処理した。
【0133】
続いて、ゲノムDNA(gDNA)を細胞から単離し、アガロースゲル電気泳動に供した(図7参照)。このデータから、gDNAを消化するためにはリゾレシチンおよびヌクレアーゼが必要であることが示された。リゾレシチンまたはヌクレアーゼのいずれかを欠く反応物ではごくわずかなgDNA消化が検出された。このことは、リゾレシチンによって透過処理した細胞では、内因性ヌクレアーゼによるクロマチンの消化は有意でないこと、および消化用緩衝液中に存在するヌクレアーゼはリゾレシチンの非存在下では細胞内に入らないことを示している。
【0134】
その後に、MnlIで処理した細胞から単離したDNA試料を、図6に示したプライマーセットを用いるリアルタイムPCRによって分析した。図8に示されているように、陰性対照反応ではヘモグロビンβ鎖遺伝子(これはHela細胞ではサイレント化している)を分析した。トータルプライマーのセットおよび無傷プライマーのセットに関して、Ct値はほぼ同じであった。このことは、この遺伝子領域がHela細胞においてアクセス不能なクロマチン構造内にあったことを示している。
【0135】
図9は、陽性対照であるGAPDHに関する結果を示している。GAPDHに関するCt値は、陰性対照においてHBBに関して得られたCt値とほぼ同じであったが、「無傷」プライマーセットを用いた増幅は右方に移動した。このことは、「トータル」DNAよりも無傷が少ないことを示している。このことは、MnlIがGAPDHプロモーター内の自身の認識部位にアクセスしてそれを消化したことを意味する。このため本発明者らは、GAPDHプロモーターはHela細胞においてアクセス可能なクロマチン内にあったと推論している。
【0136】
図10は、GSTπの分析による結果を示している。GSTπは前立腺癌では不活性化されており、Hela細胞では発現されている。完全な反応物のCt値は、無傷プライマーセットの反応物では右方に移動したが、トータルプライマーセットの反応物ではそうではなかった。このことは、Hela細胞ではGSTπがアクセス可能なクロマチン構造内にあることを示している。
【0137】
図11は、上記のデータを表にまとめたものである。算出された値は、「トータル」プライマーセットによって決定されたコピーの総数と比較した、無傷(それ故にヌクレアーゼに対してアクセス不能性)であったコピーのパーセンテージである。
【0138】
実施例3:前立腺癌細胞においてMnlIを用いたデータ
上記の方法を、アクセス可能性がGSTP1の発現と関係しているか否かを判断するために種々の細胞種に対しても適用した。非癌性ヒト前立腺に由来し、GSTP1が高発現されるRWPE-1細胞;および癌性ヒト前立腺に由来し、GSTP1がエピジェネティックな修飾によってサイレント化しているLNCaP細胞という、2つの細胞型を用いた。
【0139】
これらの細胞の分析方法は、本質的には上記の通りとした。図12は、対照反応(すなわち、ヘモグロビン遺伝子について)を示している。いずれの細胞株においても、酵素を含まなかった試料におけるデルタCtは、酵素を含むものとほぼ同じである。すなわち、デルタCtは約0であった。このことはこの遺伝子が両方の細胞株においてアクセス不能であったことを示している。
【0140】
図13は、GAPDH遺伝子を分析した陽性対照反応の結果を示している。酵素を伴わない反応に関して、トータルおよび無傷Ctはほぼ同じであった。しかし、酵素を含む反応については、無傷Ctは右方に移動した。このことは無傷のDNAがより少なく、それ故にGAPDH遺伝子がアクセス可能なクロマチン構造内にあったことを示している。
【0141】
図14は、GSTπ遺伝子の分析によるデータを示している。非癌性RWPE-1細胞株では、無傷Ctが酵素の存在下で右方に移動した。対照的に、癌性LNCaP細胞株では、無傷Ctは検知しうる程度には変化しなかった。このことは、GSTP1がRWPE-1細胞ではアクセス可能なクロマチン内にあり、LNCaP細胞ではアクセス不能なクロマチン内にあったことを示している。
【0142】
これらの実験を4回繰り返した。結果の概要は図15に示されており、この場合もアクセス不能であった(または「ロックされた(locked)」)コピーのパーセンテージを算出している。
【0143】
実施例4:DNアーゼIを用いたデータ
この実施例は、MnlIではなくDNアーゼIをヌクレアーゼとした方法の使用によるデータを示す。この実施例において、本発明者らは、4つのヒト癌細胞株:Hela(子宮頸癌)、PC3(前立腺癌)、LNCaP(前立腺癌)およびHCT15(結腸癌)を分析した。本発明者らはまた、本発明者らがMnlIの例で分析したのと同じ遺伝子も分析した:HBB、GAPDHおよびGSTP1。
【0144】
DNアーゼIはDNAを定まった部位では切断しないため、本発明者らは、本発明者らがMnlIをクロマチン構造プローブとして用いた場合と同じデータ分析戦略を用いることはできなかった。その代わりに、本発明者らは、エピジェネティックにサイレント化される参照遺伝子(HBB)を、アクセス不能性クロマチンの消化の程度を反映する内部対照として利用した。この戦略を用いると、特定の標的遺伝子のクロマチン構造を、図16に詳細に示した通りに決定することができる。図17は、分析した遺伝子、およびクロマチン構造を評価するために用いたプライマーの位置の概略描写である。
【0145】
図18に示されているように、DNアーゼIはすべての細胞株において、HBBを検知しうる程度には切断しなかった(ヌクレアーゼなしの曲線と+ヌクレアーゼの曲線はほぼ重なる)。この結果は、HBBがすべての細胞株でアクセス不能なクロマチン立体配置内にあることを示しており、参照遺伝子対照としてのその有用性を実証している。HBB mRNAはこれらの細胞株のいずれにおいても発現されないため(非提示データ)、これらの結果は予想されたことでもある。
【0146】
GAPDH遺伝子に関する本発明者らの分析を図19に示している。すべての細胞株において、本発明者らは、+ヌクレアーゼの曲線がヌクレアーゼなしの曲線に対して有意に右方移動していることを確認している。このことは、GAPDHがすべての細胞株においてアクセス可能なクロマチン構造内にあることを意味する。これらの結果は、GAPDHがすべてではないにしてもほとんどの細胞株で発現される(非提示データ)「ハウスキーピング」遺伝子であることから、予想されたことである。
【0147】
GSTP1 mRNAは、分析した細胞株においてさまざまなレベルで発現される。Hela細胞およびHCT15細胞は多くの量のGSTP1 mRNAを発現し、PC3細胞は中程度の量(Hela細胞およびHCT15細胞で見出される量の約半分)を発現し、LNCaP細胞はGSTP1 mRNAを検知しうる程度では発現しない(非提示データ)。GSTP1クロマチン構造に関する本発明者らの分析を図20に示している。Hela細胞およびHCT15細胞では、本発明者らは+ヌクレアーゼの曲線がヌクレアーゼなしの曲線に対して有意に右方移動していることを確認しており、このことはGSTP1がアクセス可能なクロマチン構造内にあることを意味する。PC3細胞では、本発明者らは+ヌクレアーゼの曲線がヌクレアーゼなしの曲線に対して中程度に右方移動していることを確認しており、このことはGSTP1が部分的にアクセス可能なクロマチン構造内にあることを意味する。最後に、LNCaP細胞では、本発明者らは+ヌクレアーゼの曲線およびヌクレアーゼなしの曲線がほぼ重なることを確認している。このことはGSTP1がLNCaP細胞においてアクセス不能なクロマチン立体配置内にあることを意味する。重要なこととして、本発明者らは、分析したすべての細胞株において、GSTP1クロマチン構造とGSTP1 mRNA発現との間に優れた相関関係を確認している。
【0148】
図21は、結果の概要を示している。アクセス不能なDNAのパーセンテージを図16に記載した通りに算出した。それを「ロックされている」クロマチンとして示している。GAPDHおよびGSTP1のRNA発現の相対的レベルは、種々の細胞株から単離したmRNAをqRT-PCRにより標準的な手順によって分析することによって算出した。それらの結果は、標的遺伝子のクロマチン構造が標的遺伝子の発現レベルと良く相関していることを示している。このことは、記載したアッセイが、遺伝子発現に対するエピジェネティックな影響を評価するための有用なツールであることを意味する。
【0149】
概要
本発明者らは、培養細胞におけるクロマチン構造を評価することのできる迅速かつ高感度なアッセイを開発した。本発明者らのアッセイには、他のクロマチン構造アッセイを上回るいくつかの利点がある(Okino, S.T., et al., Mol Carcinog 46(10):839-46 (2007);Okino, S.T. and J.P. Whitlock, Jr., Mol Cell Biol 15(7): p. 3714-21 (1995);Rao, S., E. Procko, and M.F. Shannon, Jimmunol 167(8):4494-503 (2001);Kilgore, J.A., et al., Methods 41(3):320-32 (2007))。最も重要なこととして、本発明者らの手順では、本発明者らは、透過処理した細胞において、クロマチンを、1つの段階でインサイチューで消化する。他の手順は核の単離を必要とし、それはより多くの手間のかかる作業、より多くの時間、および出発材料としてより多くの細胞を必要とする。加えて、本発明者らのプライマーセットの設計は、クロマチンのアクセス可能性に関してより正確で、内部対照を有する評価をもたらす。
【0150】
実施例5:DNAメチル化を評価するための変法
本発明のもう1つの用途は、アクセス不能なクロマチン領域のDNAメチル化状態を決定することである。DNAメチル化とは、5-メチルシトシンを生成させる、シトシンの5位での天然のメチル化のことを指す。DNAメチル化は、遺伝子活性およびクロマチン構造の調節と関連づけられている;異常なDNAメチル化パターンは、ヒトの病態および特定の罹患状態と関連性がある。この変法では、前述の手順を、PCR増幅の段階を含まないが、その手前まで行う。続いて、精製されたゲノムDNAを、メチル化されていないシトシンがウラシルに変換され、5-メチルシトシンは変化しないように、バイサルファイトによって標準的な手順を用いて改変する。続いて、特定のDNA領域を標的とするプライマーセットを用いて、ヌクレアーゼで処理した細胞から精製された、バイサルファイトで改変されたDNAを増幅する。こうすることにより、元々アクセス不能なクロマチン立体配置内にあり、それ故にDNA修飾剤によって切断されなかったDNAを増幅することができる。続いて、増幅されたDNAを、融解曲線分析、高分解能融解分析、バイサルファイト-DNA配列決定、デジタルPCR、メチル化特異的PCR、ピロシークエンシングなどの標準的な手順によって、DNAメチル化に関して分析する。このような分析は、アクセス不能なクロマチン領域のDNAメチル化状態を決定することができ、エピジェネティックな遺伝子不活性化と関連性のあるDNAメチル化の特定のパターンに関する有意義な情報を提供することができる。
【0151】
本明細書に記載された例および態様が例示のみを目的としていること、ならびにそれらを考慮してさまざまな改変および変更が当業者には想起されると考えられ、それらが本出願の趣旨および範囲ならびに添付の特許請求の範囲に含まれることは、理解されるであろう。本明細書中に引用された刊行物、特許および特許出願はすべて、それらの全体が、あらゆる目的で参照により組み入れられる。
【技術分野】
【0001】
関連特許出願の相互参照
本出願は、2008年12月2日に提出された米国仮特許出願第61/119,280号の優先権の恩典を主張し、それはあらゆる目的で参照により本明細書に組み入れられる。
【背景技術】
【0002】
発明の背景
細胞内のほとんどのDNAは、一群のヒストンタンパク質の周りにパッケージングされ、ヌクレオソームとして知られる構造をとる。このヌクレオソームDNAは、DNAを緻密に凝縮させるらせん状構造へとさらにパッケージングされうる。この緻密なパッケージングは、転写因子および転写機構へのDNAのアクセスを制限しうる。このような様式でパッケージングされたゲノムDNAは時にクロマチンと称される。
【0003】
クロマチンは、DNAが疎にパッケージングされ、アクセス可能(accessible)であり、常にではないが一般に転写能力があるユークロマチンと、DNAが緻密にパッケージングされ、アクセス不能(inaccessible)であり、常にではないが一般的には転写的にサイレントであるヘテロクロマチンという、2つの主なグループに分類される。
【0004】
これらの2つのクロマチン状態間の推移を制御するものはエピジェネティクス(epigenetics)である。エピジェネティックなイベントには主にDNAメチル化およびヒストン修飾の2つがある。これらのイベントは、DNAがどのようにパッケージングされるか、およびDNAが転写に関して活性であるかそれともサイレントであるかに影響を及ぼす。
【発明の概要】
【0005】
発明の概略
本発明は、染色体DNAを分析するための方法であって、DNA修飾剤に対する染色体上のDNA領域のアクセス可能性(accessibility)を決定し、任意で、該アクセス可能性をクロマチン構造と関連づける段階を非限定的に含む方法を提供する。いくつかの態様において、本方法は以下の段階を含む:
a.以下を同時に行う段階:
i.細胞の細胞膜を透過処理するかまたは崩壊させる段階;および
ii.該細胞を、DNA切断剤またはDNA修飾剤と、該剤が該細胞内のゲノムDNAを切断または修飾するような条件下で接触させる段階であって、ゲノムDNAの異なる領域が該剤によって異なる程度で切断または修飾され、それにより、切断されたDNA領域および無傷のDNA領域、または修飾されたDNA領域および修飾されていないDNA領域が生成される、段階;ならびに
b.少なくとも1つの、無傷の、もしくは修飾されていない、もしくは修飾されたDNA領域の物理的特徴もしくは量を検出するか、または少なくとも1つの、無傷の、もしくは修飾されていない、もしくは修飾されたDNA領域のクローニング、単離、もしくはヌクレオチド配列決定を行う段階。
【0006】
いくつかの態様において、本方法はさらに、前記量を前記細胞における前記DNA領域のクロマチン構造と関連づける段階を含む。
【0007】
いくつかの態様において、本方法は、少なくとも第1の無傷の、または修飾されていない、または修飾された染色体DNA領域、および第2の無傷の、または修飾されていない、または修飾された染色体DNA領域の物理的特徴または量を、検出する段階;ならびに第1および第2のDNA領域の物理的特徴または量を比較する段階を含む。
【0008】
いくつかの態様において、本方法は、第1の染色体DNA領域および第2の染色体DNA領域の無傷のコピーの数を定量し、それにより、DNA切断剤に対する第1および第2のDNA領域の相対的なアクセス可能性を評価する段階を含む。
【0009】
いくつかの態様においては、ゲノムDNAを、接触させる段階の後でかつ検出する段階の前に、単離する。
【0010】
いくつかの態様においては、細胞を人工の培養表面に直接的または間接的に付着させながら、細胞を透過処理して、DNA切断剤またはDNA修飾剤と接触させる。
【0011】
いくつかの態様において、透過処理する段階は、細胞を、細胞膜を透過性にする剤と接触させることを含む。いくつかの態様において、細胞膜を透過性にする剤はリゾ脂質(lysolipid)である。いくつかの態様において、リゾ脂質はリゾホスファチジルコリンである。
【0012】
いくつかの態様において、透過処理するかまたは崩壊させる段階は、非イオン性界面活性剤を用いて細胞膜を崩壊させることを含む。いくつかの態様において、非イオン性界面活性剤は、NP40、Tween20およびTriton X-100の群から選択される。
【0013】
いくつかの態様においては、細胞をDNA切断剤と接触させ、該DNA切断剤は酵素である。
【0014】
いくつかの態様においては、細胞をDNA修飾剤と接触させ、該DNA修飾剤はメチルトランスフェラーゼおよびDNA修飾性化学物質からなる群より選択される。いくつかの態様において、DNA切断剤はDNA切断酵素であり;ゲノムDNA中の修飾部は切断部位であり;かつ、検出する段階は切断後のDNA領域の無傷のコピーの量を定量することを含む。いくつかの態様において、検出する段階は定量的増幅を含む。いくつかの態様において、定量的増幅は、定量的ポリメラーゼ連鎖反応(任意でリアルタイム性)および定量的ライゲーション媒介ポリメラーゼ連鎖反応(任意でリアルタイム性)からなる群より選択される。
【0015】
いくつかの態様において、DNA切断剤は、DNアーゼおよび制限酵素から選択される。いくつかの態様において、DNA修飾剤はメチルトランスフェラーゼであり;修飾部は、細胞のネイティブなメチル化酵素によってはメチル化されないであろうヌクレオチド配列中のヌクレオチド上のメチル基であり;かつ、検出する段階は、修飾部の有無を検出する方法を含む。いくつかの態様において、メチルトランスフェラーゼはDAMメチルトランスフェラーゼであり、定量する段階は、修飾されたDNAを、DAMメチルトランスフェラーゼによってメチル化されたDNA配列を認識する制限酵素を用いて切断することを含む。いくつかの態様において、メチルトランスフェラーゼはDNA中のシトシンにメチル部分を付加し、検出する段階は、修飾されたDNAをメチル化特異的制限酵素を用いて切断すること、および/または修飾されたDNAをバイサルファイト(bisulfite)で処理することを含む。
【0016】
いくつかの態様において、DNA修飾剤はクロマチン構造に対して立体障害を有する分子であり、ここで該分子はDNA修飾性化学物質と連結されている。いくつかの態様において、DNA修飾性化学物質は、硫酸ジメチルおよびヒドラジンからなる群より選択される。
【0017】
いくつかの態様において、検出する段階は、標的DNA領域の少なくとも一部分および対照DNA領域の少なくとも一部分を定量することを含み、ここで対照DNA領域は、以下のいずれかである配列を含む:
i.ある動物の本質的にすべての細胞においてアクセス可能である;または
ii.ある動物のほとんどの細胞においてアクセス不能である;または
iii.細胞型もしくは生育環境に応じてさまざまなアクセス可能性を有する。
【0018】
いくつかの態様において、対照DNA領域は、以下のそれぞれを用いて定量的に増幅される:
DNA修飾剤の潜在的な修飾部位を含まない対照DNA領域の一部分の増幅をプライミングするプライマー;および
DNA修飾剤の潜在的な修飾部位を少なくとも1つ含む対照DNA領域の一部分の増幅をプライミングするプライマー。
【0019】
いくつかの態様において、物理的特徴はDNAメチル化である。
【0020】
いくつかの態様において、段階a.は前記DNAをDNA切断剤と接触させることを含み、段階b.は無傷のDNAをバイサルファイトと接触させることを含む。
【0021】
いくつかの態様において、本方法はさらに、バイサルファイトで処理したDNAの融解温度を決定し、該融解温度をDNAメチル化の有無または程度と関連づける段階を含む。
【0022】
いくつかの態様において、段階bは、以下のもののライブラリーを調製することを含む:無傷のDNA領域;または修飾されたDNA領域;または修飾されていないDNA領域。
【0023】
いくつかの態様において、段階bは、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域を、ポリヌクレオチドのライブラリーと、該DNA領域と該ライブラリーの1つまたは複数のメンバーとのハイブリダイゼーションを可能にする条件下で接触させること、および該DNA領域と該1つまたは複数のメンバーとのハイブリダイゼーションを検出することを含む。いくつかの態様において、ライブラリーはマイクロアレイ上に構成されている。
【0024】
いくつかの態様において、段階bは、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域を増幅することを含む。
【0025】
いくつかの態様において、段階bは、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域のヌクレオチド配列決定を行うことを含む。
【0026】
本発明はまた、DNA修飾剤に対する染色体上の遺伝子座のアクセス可能性を決定するためのキットも提供する。いくつかの態様において、本キットは、細胞膜の透過処理剤または崩壊剤;および制限酵素、DNアーゼまたは他のDNA修飾剤を含む。
【0027】
いくつかの態様において、細胞膜の透過処理剤または崩壊剤と、制限酵素またはDNアーゼとは、同じ容器内で同じ緩衝液中にある。
【0028】
いくつかの態様において、細胞膜の透過処理剤または崩壊剤と、制限酵素またはDNアーゼとは、別々の容器内にある。
【0029】
いくつかの態様において、本キットはさらにバイサルファイトを含む。
【0030】
いくつかの態様において、本キットはさらに、真核生物のゲノムDNAのある領域の増幅のためのプライマー対を含む。
【0031】
いくつかの態様において、本キットはさらに以下のものを含む:
a.DNA修飾剤の潜在的な修飾部位を含まない対照DNA領域の一部分の増幅をプライミングするプライマー;および/または
b.DNA修飾剤の潜在的な修飾部位を少なくとも1つ含む対照DNA領域の一部分の増幅をプライミングするプライマー。
【0032】
いくつかの態様において、本キットは以下のものを含む:
a.修飾剤に対してアクセス不能である対照DNA領域の一部分の増幅をプライミングするプライマー;および/または
b.修飾剤に対してアクセス可能である対照DNA領域の一部分の増幅をプライミングするプライマー。
【0033】
本発明のその他の態様は、本明細書の残りの部分を読めば明らかになるであろう。
【0034】
定義
細胞膜を「透過処理すること」とは、本明細書で用いる場合、修飾剤が細胞内に入ることを可能にするように細胞膜の完全性を低減させることを指す。透過処理された細胞膜を有する細胞は一般に細胞膜を保ち、そのため細胞の構造は実質的に無傷のままに保たれる。対照的に、細胞膜を「崩壊させること」とは、本明細書で用いる場合、細胞の構造が無傷のままでは保たれないように細胞膜の完全性を低減させることを指す。例えば、細胞膜を非イオン性界面活性剤と接触させることは、細胞膜を除去および/または溶解させ、それにより、少なくともいくらかの染色体構造を保っているゲノムDNAに対する修飾剤のアクセスを可能にする。
【0035】
「DNA修飾剤」とは、本明細書で用いる場合、DNAを検出可能な様式に改変する分子のことを指す。例示的な修飾には、DNAのニッキングもしくは切断、または導入もしくはDNAからの化学部分の除去が含まれる。DNA切断を引き起こさないDNA修飾剤には、DNAメチラーゼが非限定的に含まれる。
【0036】
「DNA領域」とは、本明細書で用いる場合、ゲノムDNA内部の関心対象の標的配列のことを指す。DNA領域は、関心対象であって用いるDNA修飾剤がアクセス可能である、任意の長さでありうる。いくつかの態様において、DNA領域は単一塩基対を含みうるが、ゲノムDNA内部の短い配列セグメント(例えば、2〜100、2〜500、50〜500bp)またはより長いセグメント(例えば、100〜10,000、100〜1000または1000〜5000bp)でもありうる。DNA領域内のDNAの量は、時には、PCR反応において増幅させようとする配列の量によって決まる。例えば、標準的なPCR反応は一般に約35〜5000塩基対を増幅することができる。
【0037】
異なる「程度」の修飾とは、複数の試料間での、または1つもしくは複数の試料中の2つもしくはそれ以上のDNA領域間での、1つまたは複数のDNA領域の修飾されたコピー数(実数または相対数)が異なることを指す。例えば、2つのDNA領域(便宜上「領域A」および「領域B」と呼ぶ)の100個のコピーがそれぞれ、ある細胞内の染色体DNA上に存在するならば、異なる程度での修飾の一例は、領域Aの10個のコピーが修飾され、一方、領域Bの70個のコピーが修飾された場合であると考えられる。
【0038】
「オリゴヌクレオチド」または「ポリヌクレオチド」または「核酸」という用語は、リボース核酸(RNA)もしくはデオキシリボース核酸(DNA)の重合体またはそれらの類似体に対応しうる、単量体の重合体のことを互換的に指す。これには、RNAおよびDNA、さらにはそれらの修飾形態、ペプチド核酸(PNA)、ロックド核酸(locked nucleic acid, LNA(商標))といったヌクレオチドの重合体が含まれる。ある用途において、核酸は、複数の種類の単量体、例えば、RNAサブユニットおよびDNAサブユニットの両方を含む重合体でありうる。
【0039】
核酸は典型的には一本鎖または二本鎖であり、一般にホスホジエステル結合を含むが、場合によっては、本明細書で概説するように、代替的な骨格を有しうる核酸類似体が含められ、これには例えば、ホスホルアミド(Beaucage et al. (1993) Tetrahedron 49(10):1925およびその中の参考文献;Letsinger (1970) J. Org. Chem. 35:3800;Sprinzl et al. (1977) Eur. J. Biochem. 81:579;Letsinger et al. (1986) Nucl. Acids Res. 14: 3487;Sawai et al. (1984) Chem. Lett. 805;Letsinger et al. (1988) J. Am. Chem. Soc. 110:4470;およびPauwels et al. (1986) Chemica Scripta 26:1419)、ホスホロチオエート(Mag et al. (1991) Nucleic Acids Res. 19:1437および米国特許第5,644,048号)、ホスホロジチオエート(Briu et al. (1989) J. Am. Chem. Soc. 111:2321)、O-メチルホスホロアミダイト結合(O-methylphophoroamidite linkage)(Eckstein, Oligonucleotides and Analogues: A Practical Approach, Oxford University Press (1992))、ならびにペプチド核酸骨格および結合(Eghoim (1992) J. Am. Chem. Soc. 114:1895;Meier et al. (1992) Chem. Int. Ed. Engl. 31:1008;Nielsen (1993) Nature 365:566;およびCarlsson et al. (1996) Nature 380:207)が非限定的に含まれ、これらの参考文献はそれぞれ参照により組み入れられる。他の類似体核酸には、正に荷電した骨格(Denpcy et al. (1995) Proc. Natl. Acad. Sci. USA 92:6097);非イオン性骨格(米国特許第5,386,023号、第5,637,684号、第5,602,240号、第5,216,141号および第4,469,863号;Angew (1991) Chem. Intl. Ed. English 30: 423;Letsinger et al. (1988) J. Am. Chem. Soc. 110:4470;Letsinger et al. (1994) Nucleoside & Nucleotide 13:1597;Chapters 2 and 3, ASC Symposium Series 580, "Carbohydrate Modifications in Antisense Research", Ed. Y. S. Sanghvi and P. Dan Cook;Mesmaeker et al. (1994) Bioorganic & Medicinal Chem. Lett. 4: 395;Jeffs et al. (1994) J. Biomolecular NMR 34:17;Tetrahedron Lett. 37:743 (1996))、および、米国特許第5,235,033号および第5,034,506号ならびにChapters 6 and 7, ASC Symposium Series 580, Carbohydrate Modifications in Antisense Research, Ed. Y. S. Sanghvi and P. Dan Cookに記載されたものを含む非リボース骨格を有するものが含まれ、これらの参考文献はそれぞれ参照により組み入れられる。1つまたは複数の炭素環式糖を含む核酸も、核酸の定義の範囲に含まれる(Jenkins et al. (1995) Chem. Soc. Rev. pp169-176、これは参照により組み入れられる)。いくつかの核酸類似体は、例えば、Rawls, C & E News Jun. 2, 1997 page 35にも記載されており、これは参照により組み入れられる。リボース-リン酸骨格のこれらの修飾は、標識部分などの追加部分の付加を容易にするため、または生理的環境におけるそのような分子の安定性および半減期を変更するために行ってもよい。
【0040】
核酸中に典型的に認められる、天然に存在する複素環式塩基(例えば、アデニン、グアニン、チミン、シトシンおよびウラシル)に加えて、核酸類似体にはまた、天然に存在しない複素環式塩基または他の修飾塩基も含まれ、その多くは本明細書中に記載されているか、そうでなければ参照されている。特に、天然に存在しない多くの塩基は、例えば、Seela et al. (1991) Helv. Chim. Acta 74:1790, Grein et al. (1994) Bioorg. Med. Chem. Lett. 4:971-976、およびSeela et al. (1999) Helv. Chim. Acta 82:1640にさらに記載されており、これらはそれぞれ参照により組み入れられる。さらに例を挙げるならば、融解温度(Tm)変更因子として作用するヌクレオチド中に用いられるある種の塩基が任意で含められる。例えば、これらのいくつかには、7-デアザプリン(例えば、7-デアザグアニン、7-デアザアデニンなど)、ピラゾロ[3,4-d]ピリミジン、プロピニル-dN(例えば、プロピニル-dU、プロピニル-dCなど)などが含まれる。例えば、参照により組み入れられる、Seelaに対して1999年11月23日に発行された「7-デアザ-2'-デオキシグアノシンヌクレオチドの合成」と題する米国特許第5,990,303号を参照のこと。他の代表的な複素環式塩基には、例えば、ヒポキサンチン、イノシン、キサンチン;2-アミノプリン、2,6-ジアミノプリン、2-アミノ-6-クロロプリン、ヒポキサンチン、イノシンおよびキサンチンの8-アザ誘導体;アデニン、グアニン、2-アミノプリン、2,6-ジアミノプリン、2-アミノ-6-クロロプリン、ヒポキサンチン、イノシンおよびキサンチンの7-デアザ-8-アザ誘導体;6-アザシトシン;5-フルオロシトシン;5-クロロシトシン;5-ヨードシトシン;5-ブロモシトシン;5-メチルシトシン;5-プロピニルシトシン;5-ブロモビニルウラシル;5-フルオロウラシル;5-クロロウラシル;5-ヨードウラシル;5-ブロモウラシル;5-トリフルオロメチルウラシル;5-メトキシメチルウラシル;5-エチニルウラシル;5-プロピニルウラシルなどが含まれる。
【0041】
DNA修飾剤に対するDNA領域の「アクセス可能性」とは、本明細書で用いる場合、細胞の染色体内の特定のDNA領域が、特定のDNA修飾剤によって接触されて修飾される能力のことを指す。本発明の範囲を限定することは意図しないが、DNA領域を含む特定のクロマチン構造は、DNA修飾剤が特定のDNA領域を修飾する能力に影響を及ぼすと考えられている。例えば、DNA領域がヒストンタンパク質の周りに巻き付けられていることもあり、さらに、関心対象のDNA領域に対するDNA修飾剤のアクセスを防止または抑制するさらなるヌクレオソーム構造を有することもある。
【0042】
「II-S型制限酵素」は、当技術分野におけるその通常の意味で用いられ、DNAにおける特定の認識配列を認識し、続いてそのDNA分子をその認識配列の外側で切断する制限酵素のことを指す。例示的なII-S型制限酵素には、MnlI、FokIおよびAlwIが非限定的に含まれる。
【図面の簡単な説明】
【0043】
【図1】クロマチンの概略図を図示している。真核生物DNAは、DNAが疎にパッケージングされ、アクセス可能であり、転写能力があるユークロマチンと、DNAが緻密にパッケージングされ、アクセス不能であり、転写的にサイレントであるヘテロクロマチンという、2つの一般的状態に分類することができる。エピジェネティクスが、これらの2つの状態間の推移を制御する。本明細書に記載されたアッセイは、クロマチン構造、エピジェネティックなイベントの機能的帰結を評価することができる。
【図2】アッセイの概略図を図示している。培地を吸引し、透過処理/消化用緩衝液を添加する。ヌクレアーゼが細胞内に拡散し、核の中に入って、アクセス可能なクロマチンを消化するが、アクセス不能なクロマチン(図の下方の太線として表されている)は消化されない。
【図3】例示的な作業の流れの概略図を図示している。
【図4】Mnl 1をプローブとして用いた場合に生成されるデータを評価するための分析アプローチを図示している。各アッセイでは2つのDNA試料を分析し、一方のDNA試料はヌクレアーゼを含まない緩衝液で処理した細胞から単離した対照であり、もう一方の試料は緩衝液およびヌクレアーゼで処理した細胞から単離する。両方の試料を、トータルプライマーのセットおよび無傷プライマーのセットを用いるリアルタイムPCRによって分析する。(A)ΔCt(切断なし)は、対照DNA試料のデータを用いてトータルCtから無傷Ctを差し引くことによって算出される。(B)ΔCt(切断あり)は、ヌクレアーゼ処理したDNA試料のデータを用いてトータルCtから無傷Ctを差し引くことによって算出される。アクセス可能な遺伝子は、MnlI部位で切断され、有する無傷DNAがより少なく、ΔCt(切断あり)がより高く、それは消化の程度に比例し、クロマチンのアクセス可能性がより大きいことを反映する。アクセス不能な、または「ロックダウン(locked-down)された」DNAの量は、式2∧(ΔCt(切断なし)−ΔCt(切断あり))によって算出される。
【図5】分析した標的遺伝子の略図である。グレーの領域は転写される領域を表している。Mnl 1部位およびTATAボックスの位置を指し示している。GAPDHはほとんどの細胞株で発現されるハウスキーピング遺伝子であり、アクセス可能なクロマチン構造内にあると予想される。ヘモグロビンβ鎖(HBB)は、ほとんどの細胞でサイレント化しており、アクセス不能なクロマチン構造にあると予想される。グルタチオン-s-トランスフェラーゼπ(GSTP1)は、前立腺癌ではエピジェネティックに不活性化されており、非癌性RWPE-1前立腺細胞では発現されるが、癌性LNCaP前立腺細胞ではサイレント化している。
【図6】分析した標的遺伝子、およびプライマーセットの位置を図示している。遺伝子はすべて、どのMnlI I部位も横切らず、すべてのDNA鎖を増幅すると考えられるトータルプライマーのセット、さらには2つ〜4つのMnlI部位を横切り、これらの部位で切断されないDNAのみを増幅すると考えられる無傷プライマーのセットを用いて分析した。
【図7】アガロースゲル電気泳動を用いたgDNA完全性の分析の概要を提示している。Hela細胞を、ヌクレアーゼ(M、MnlI;D、DNアーゼI)を含まない透過処理/消化用緩衝液、リゾレシチンを含まない透過処理/消化用緩衝液、またはヌクレアーゼおよびリゾレシチンの両方を含む透過処理/消化用緩衝液で処理した。gDNAを単離して、アガロースゲル電気泳動によって分析した。これらの結果は、有意なgDNA消化のためには、ヌクレアーゼおよびリゾレシチンの両方が必要なことを指し示している。
【図8】MnlIで処理したHela細胞におけるHBBに関して、蛍光をサイクル数の関数として図示している。Hela細胞は前述の通りに処理した。gDNAを単離して、HBB遺伝子を増幅するプライマーを用いて分析した。HBB無傷プライマーのセットを用いた場合、完全な反応物(+、x)のCt値は対照反応物と同程度である。このことは、HBBがHela細胞においてアクセス不能なクロマチン立体配置内にあることを意味する。
【図9】MnlIで処理したHela細胞におけるGAPDHに関して、蛍光をサイクル数の関数として図示している。Hela細胞を前述の通りに処理し、gDNAを単離して、GAPDH遺伝子を増幅するプライマーを用いて分析した。GAPDH無傷プライマーのセットを用いた場合、完全な反応物(+、x)のCt値は対照反応物よりも高い(曲線が右方移動している)。このことは、GAPDHがHela細胞においてアクセス可能なクロマチン立体配置内にあることを意味する。
【図10】MnlIで処理したHela細胞に関して、蛍光をサイクル数の関数として図示している。Hela細胞を前述の通りに処理し、gDNAを単離して、GSTP1遺伝子を増幅するプライマーを用いて分析した。GSTP1無傷プライマーのセットを用いた場合、完全な反応物(+、x)のCt値は対照反応物よりも高い(曲線が右方移動している)。このことは、GSTP1がHela細胞においてアクセス可能なクロマチン立体配置内にあることを意味する。
【図11】3つのDNA領域(HBB、GAPDHおよびGSTP1)のデータを表形式で図示しており、アウトプットは全DNAに占めるアクセス不能であったパーセンテージとして算出している。
【図12】MnlIで処理した前立腺細胞におけるHBBに関して、蛍光をサイクル数の関数として図示している。いずれの細胞株においても、HBB無傷プライマーのセットを用いた+酵素反応のCt値(x)は、HBBトータルプライマーのセットを用いた+酵素反応(o)と同程度である。このことは、HBBがRWPE-1細胞およびLNCaP細胞においてアクセス不能なクロマチン立体配置内にあることを意味する。
【図13】MnlIで処理した前立腺細胞におけるGAPDHに関して、蛍光をサイクル数の関数として図示している。いずれの細胞株においても、GAPDH無傷プライマーのセットを用いた+酵素反応のCt値(x)は、GAPDHトータルプライマーのセットを用いた+酵素反応(o)よりも高い(曲線は右方移動している)。このことは、GAPDHがRWPE-1細胞およびLNCaP細胞においてアクセス可能なクロマチン立体配置内にあることを意味する。
【図14】MnlIで処理した前立腺細胞におけるGSTP1に関して、蛍光をサイクル数の関数として図示している。RWPE-1細胞において、GSTP1無傷プライマーのセットを用いた+酵素反応のCt値(x)は、GSTP1トータルプライマーのセットを用いた+酵素反応(o)よりも高い(曲線は右方移動している)。このことは、GSTP1がRWPE-1細胞においてアクセス可能なクロマチン立体配置内にあることを意味する。対照的に、LNCaP細胞では、GSTP1無傷プライマーのセットを用いた+酵素反応のCt値(x)は、GSTP1トータルプライマーのセットを用いた+酵素反応(o)と同程度である。このことは、GSTP1がLNCaP細胞においてアクセス不能なクロマチン立体配置内にあることを意味する。
【図15】前立腺細胞からのデータの概要を図示している。アクセス不能なDNAのパーセンテージは、材料および方法の項に記載した通りに算出した。これらの結果は、いずれの細胞株においても、HBBはアクセス不能なクロマチン構造内にあり、GAPDHはアクセス可能なクロマチン構造内にあることを指し示している。GSTP1はRWPE-1細胞においてはアクセス可能なクロマチン内にあり、LNCaP細胞においてはアクセス不能なクロマチン構造内にある。
【図16】DNアーゼIをクロマチン構造プローブとして用いた場合に生成されたデータを評価するための分析アプローチを図示している。各アッセイで2つのDNA試料を分析し、一方のDNA試料はヌクレアーゼを含まない緩衝液で処理した細胞から単離した対照であり、もう一方の試料は緩衝液およびヌクレアーゼで処理した細胞から単離する。両方の試料を、アクセス不能な参照遺伝子(HBB)を増幅するプライマーセットを用いるリアルタイムPCRによって分析する。ΔCt(参照)はアクセス不能性クロマチンのヌクレアーゼ消化の程度を表し、+ヌクレアーゼの曲線に伴うCtからヌクレアーゼなしの曲線に伴うCtを差し引くことによって算出される。ΔCt(標的)は標的クロマチン領域のヌクレアーゼ消化の程度を表し、+ヌクレアーゼの曲線に伴うCtからヌクレアーゼなしの曲線に伴うCtを差し引くことによって算出される。標的遺伝子に伴うアクセス不能な、すなわち「ロックダウンされた」DNAの量は、式2∧(ΔCt(参照)−ΔCt(標的))によって算出される。
【図17】分析した標的遺伝子、およびプライマーセットの位置の概略図である。グレーの領域は転写される領域を表している。矢印付きの黒の領域は、各プライマーの位置および方向を表している。
【図18】DNアーゼIで処理した細胞株におけるHBBに関して、蛍光をサイクル数の関数として図示している。すべての細胞株において、+ヌクレアーゼ反応物のCt値(グレーの線)は、ヌクレアーゼなしの反応物(黒の線)と同程度である。このことは、HBBがすべての細胞株においてアクセス不能なクロマチン立体配置内にあることを意味する。
【図19】DNアーゼIで処理した細胞株におけるGAPDHに関して、蛍光をサイクル数の関数として図示している。すべての細胞株において、+ヌクレアーゼ反応物のCt値(グレーの線)は、ヌクレアーゼなしの反応物のCt値(黒の線)よりも有意に高い。このことはGAPDHがすべての細胞株においてアクセス可能なクロマチン立体配置内にあることを意味する。
【図20】DNアーゼIで処理した細胞株におけるGSTP1に関して、蛍光をサイクル数の関数として図示している。本発明者らは、ΔCt(GSTP1)値において細胞株依存的な差を確認している。Hela細胞およびHCT15細胞ではΔCt(GSTP1)が大きく、このことはGSTP1がこれらの細胞株においてアクセス可能な立体配置内にあることを指し示している。LNCaP細胞ではΔCt(GSTP1)が小さく、このことはGSTP1がアクセス不能なクロマチン内にあることを指し示している。PC3細胞ではΔCt(GSTP1)が中間値を有し、このことはGSTP1が部分的にアクセス可能なクロマチン構造内にあることを指し示している。これらの結果は、種々の細胞株において検出されたGSTP1 mRNA発現のレベルと非常によく合致する(図21参照)。
【図21】DNアーゼIをクロマチン構造プローブとして用いた実験によるデータの概要を図示している。アクセス不能なDNAのパーセンテージは、図16で説明した通りに算出した。GAPDHおよびGSTP1のRNA発現の相対的レベルは、種々の細胞株から単離したmRNAをqRT-PCRによって分析することによって算出した。これらの結果は、標的遺伝子のクロマチン構造が標的遺伝子の発現レベルとよく相関することを指し示している。このことは、記載したアッセイが、遺伝子調節に対するエピジェネティックな影響を評価するための有用なツールであることを意味する。
【発明を実施するための形態】
【0044】
詳細な説明
I.序論
本発明は、細胞膜を透過処理するかまたは崩壊させ、細胞内のゲノムDNAを修飾して、続いてさまざまな遺伝子座における修飾の程度を定量することによって、クロマチン構造の分析を可能にする。特定の遺伝子座での修飾の程度は、修飾剤に対する染色体のその部分のアクセス可能性を反映し、それ故にクロマチンの状態を反映する。
【0045】
本発明の1つの利点は、例えば、細胞を、透過処理剤とDNA修飾剤または切断剤とを含む1つの緩衝液と接触させることによって、透過処理および細胞内の無傷のクロマチンの修飾を同時に行うことができるという発見である。これは、結果の極めて迅速な生成を可能にする。さらに、この方法は、従前のいくつかのクロマチン分析方法で必要とされたような、核の単離といった煩雑で、場合によってはアーチファクトを誘発する可能性もある段階をなくす。
【0046】
II.一般的方法
本発明の方法は、細胞の透過処理および細胞とDNA修飾剤との接触を同時に、細胞内のゲノムDNAが修飾剤に対して(クロマチン構造の違いが原因で)さまざまなアクセス可能性を有するような条件下で行い、続いて、DNA領域における修飾の量を定量することを伴う。DNAのさまざまなアクセス可能性は、ゲノムDNAのヌクレオソーム構造を反映する可能性がある。例えば、いくつかの態様において、DNA修飾剤に対してよりアクセス可能であるDNA領域は、より「粗」なクロマチン構造内にある可能性が高い。
【0047】
種々の真核生物細胞を本発明に用いることができる。いくつかの態様において、細胞は、ヒトの、または非ヒトの哺乳動物細胞を非限定的に含む動物細胞である。非ヒト哺乳動物細胞には、霊長動物細胞、マウス細胞、ラット細胞、ブタ細胞およびウシ細胞が非限定的に含まれる。いくつかの態様において、細胞は植物細胞である。細胞は、例えば、培養初代細胞、不死化培養細胞であってもよく、または生検試料もしくは組織試料に由来し、任意で培養した上でアッセイする前に分裂するように刺激したものでもよい。培養細胞は、透過処理および/またはDNA修飾の段階の前および/または最中に、浮遊していてもよく、または付着していてもよい。細胞が動物組織、生検試料などに由来してもよい。例えば、細胞は腫瘍生検試料に由来しうる。
【0048】
本方法は、あるDNA領域に対するアクセス可能性を、その同じ領域からの転写と関連づけることを含みうる。いくつかの態様においては、アクセス可能性と遺伝子発現との間の相関関係を決定するために実験を行い、その後に特定のDNA領域に対するDNA修飾剤のアクセス可能性を用いて、そのDNA領域からの転写を予測することができる。いくつかの態様においては、DNA領域からの転写、およびDNA修飾剤に対するその領域のアクセス可能性を、両方とも決定する。転写を測定するための多岐にわたる方法が公知であり、それにはノーザンブロット法およびRT-PCRが非限定的に含まれる。
【0049】
いくつかの態様においては、ある領域のDNAメチル化状態を、DNA修飾剤に対するDNA領域のアクセス可能性と関連づけることができる。いくつかの態様においては、アクセス可能性とその領域におけるDNAメチル化との相関関係を決定するために実験を行い、その後に特定のDNA領域に対するDNA修飾剤のアクセス可能性を利用して、そのDNA領域からのDNAメチル化を予測することができる。いくつかの態様においては、DNA領域のメチル化、およびDNA修飾剤に対するその領域のアクセス可能性を、両方とも決定する。DNAメチル化を測定するための多岐にわたる方法が公知であり、それには、バイサルファイトの使用(例えば、配列決定において、および/またはメチル化感受性制限酵素との組み合わせで(例えば、Eads et al., Nucleic Acids Research 28(8): E32 (2002)を参照)および高分解能融解アッセイ(HRM)(例えば、Wodjacz et al, Nucleic Acids Research 35(6):e41 (2007)を参照)が非限定的に含まれる。
【0050】
本発明は、透過処理/DNA修飾後の、細胞のゲノム中の第1のDNA領域と第2のDNA領域の量または他の物理的特徴の比較を提供する。代替的に、または加えて、2つの異なる細胞における第1のDNA領域の量または他の物理的特徴を比較することもできる。例えば、2つの細胞は、罹患しているおよび健常な細胞または組織、異なる細胞型、異なる発生段階(幹細胞または始原細胞を非限定的に含む)などであってよい。このようにして、本発明の方法を用いることにより、細胞間のクロマチン構造の違いを検出すること、および/または1つの細胞内の2つもしくはそれ以上のDNA領域(例えば、遺伝子)間の相対的なクロマチン構造を決定することができる。加えて、同じ細胞または異なる細胞における特定の領域のクロマチン構造に対する薬物刺激、化学刺激または環境的刺激の影響を決定することもできる。
【0051】
III.細胞の透過処理および崩壊
細胞膜は、当技術分野において公知の任意のやり方で透過処理すること、または崩壊させることができる。本明細書で説明しているように、本方法は、DNAの単離の前にゲノムDNAを接触させることを伴い、それ故に、細胞膜の透過処理または崩壊の方法は、ヌクレオソームまたはクロマチンの構造が破壊されるように細胞のゲノムDNAの構造を崩壊させることはないと考えられる。
【0052】
いくつかの態様においては、細胞膜を、細胞膜を透過性にするかまたは崩壊させる剤と接触させる。リゾ脂質は、細胞膜を透過性にする例示的なクラスの剤である。例示的なリゾ脂質には、リゾホスファチジルコリン(当技術分野においてリゾレシチンとしても公知である)またはモノパルミトイルホスファチジルコリンが非限定的に含まれる。種々のリゾ脂質が、例えば、WO/2003/052095号にも記載されている。
【0053】
非イオン性界面活性剤は、細胞膜を崩壊させる例示的なクラスの剤である。例示的な非イオン性界面活性剤には、NP40、Tween 20およびTriton X-100が非限定的に含まれる。
【0054】
本発明の1つの利点は、透過処理剤とDNA切断剤またはDNA修飾剤との同時送達である。したがって、いくつかの態様においては、両方の剤を含む緩衝液を細胞と接触させる。緩衝液は、細胞クロマチンの構造を維持させながら両方の剤の活性を維持させるのに適合しているべきである。
【0055】
または、エレクトロポレーションまたは微粒子銃法を、DNA修飾剤が細胞内に導入され、従ってゲノムDNAと接触しうるように、細胞膜を透過処理するために用いることもできる。多岐にわたるエレクトロポレーション法が周知であり、本明細書に記載されたDNA修飾剤の送達用に適合させることができる。例示的なエレクトロポレーション法には、WO/2000/062855号に記載されたものが非限定的に含まれる。微粒子銃法には、米国特許第5,179,022号に記載されたものが非限定的に含まれる。
【0056】
IV.DNA修飾剤
透過処理の後に、または透過処理と同時に(例えば、エレクトロポレーション中、または透過処理剤とのインキュベーション中に)、DNA修飾剤を導入し、その結果、該剤がゲノムDNAと接触し、それによってDNAに修飾を導入する。多岐にわたるDNA修飾剤を本発明に従って用いることができる。
【0057】
いくつかの態様においては、透過処理剤の除去後に、任意で緩衝液の変更とともに、DNA修飾剤を、透過処理を行った細胞と接触させる。または、いくつかの好ましい態様においては、1つまたは複数の中間段階を省いて(例えば、緩衝液の変更、細胞の洗浄などを省いて)、DNA修飾剤をゲノムDNAと接触させる。上述した通り、この後者のアプローチは、手間のかかる作業および必要な時間の量を減らすために好都合であり得、さらに、アッセイにおける誤差および混入をもたらす可能性のある原因も減らす。
【0058】
用いるDNA修飾剤の量、ならびにDNA修飾剤との反応時間は、用いる剤に依存すると考えられる。当業者は、用いる剤に応じて条件をどのように調整すべきかを認識しているであろう。一般に、DNA修飾段階の条件は、「完全な」消化が達成されないように調整する。したがって、例えば、いくつかの態様において、修飾段階の条件は、陽性対照‐すなわち、修飾がアクセス可能であって、生じる対照‐が、100%未満ではあるが高いレベル、例えば、80〜95%、80〜99%、85〜95%、90〜98%などで生じるように設定される。
【0059】
A.制限酵素
いくつかの態様において、DNA修飾剤は制限酵素である。したがって、これらの態様において、ゲノムDNAに導入される修飾は、配列特異的な一本鎖(例えば、ニック)または二本鎖切断イベントである。多岐にわたる制限酵素が公知であり、本発明に用いることができる。
【0060】
あらゆる種類の制限酵素を用いることができる。I型酵素はDNAを、それらの認識配列から離れたところでランダムに切断する。II型酵素はDNAを、それらの認識配列の付近または内部の定まった位置で切断する。いくつかのII型酵素はDNAを、それらの認識配列の内部で切断する。II-S型酵素は、それらの認識配列の外側を片側で切断する。より正確には「IV型」と呼ばれる、第3の主な種類のII型酵素は、それらの認識配列の外側を切断する。例えば、連続配列を認識するもの(例えば、AcuI:CTGAAG)は、片側のみを切断する;不連続配列を認識するもの(例えば、BcgI:CGANNNNNNTGC)は両側を切断し、その認識配列を含む小さな断片を遊離させる。III型はそれらの認識配列の外側を切断し、切断を遂行するためには、そのような配列が反対向きで2つ同一のDNA分子内に必要である。
【0061】
本発明の方法は、あらゆる種類の制限酵素または他のDNA切断酵素と用いるために適合させることができる。いくつかの態様において、酵素は、認識配列の比較的付近(例えば、5、10または20塩基対以内)を切断する1つまたは複数のものである。そのような酵素は、切断を達成するためにアクセス可能でなければならないDNAの範囲が認識配列それ自体よりも長く、それ故に「緻密」なクロマチン構造内にないDNAをより広範な範囲で含みうるため、クロマチン構造をアッセイする上で特に有用な可能性がある。配列特異的な制限酵素は、改善された定量的結果を与える可能性があり、これは一部には、本明細書に(例えば、実施例に)記載したように、同じDNA領域に基づく対照を設計しうるためである。したがって、例えば、配列非特異的エンドヌクレアーゼ(「DNアーゼ」)による消化と比較して、総コピー数および消化されたコピー数をより正確に決定することができる。DNアーゼIとは異なり、II-S型制限酵素による切断は、関心対象のDNA領域に対するヒストンの結合に対して高感度でありうる。認識配列の外側を切断する例示的な酵素には、例えば、II-S型酵素、III型酵素およびIV型酵素が含まれる。II-S型制限酵素には、MnlI、FokIおよびAlwIが非限定的に含まれる。
【0062】
いくつかの態様においては、複数(例えば、2つ、3つ、4つなど)の制限酵素を用いる。酵素の組み合わせは、すべてが1つの型由来である酵素の組み合わせを含んでもよく、または異なる型の混合物であってもよい。
【0063】
本明細書に記載した通りに、無傷のまたは切断されたDNAを、その後に別々に検出して定量し、DNA領域の無傷のおよび/または切断されたコピーの数を決定する。
【0064】
いくつかの態様においては、透過処理剤または膜崩壊剤を、制限酵素の前に添加する。いくつかの態様においては、制限酵素と透過処理剤または崩壊剤とを同時に添加する(例えば、適切な緩衝液中に、またはそれとともに)。仮に両方の剤を最初に同時に細胞と接触させない場合でも、透過処理は持続的過程であるため、透過処理とDNA修飾剤への接触とを同時に行うことを依然として達成することができる。したがって、例えば、透過処理剤の添加のすぐ後に(透過処理が実質的に完了する前に)DNA修飾剤を添加することは、透過処理および細胞をDNA修飾剤と接触させることを「同時に」行うことと見なすことができる。「同時に」とは、透過処理および修飾剤の添加の間に中間操作(緩衝液の変更、遠心処理などを非限定的に含む)を行わないことを意味する。
【0065】
いくつかの態様においては、0.5%リゾレシチン(w/v)、50mM NaCl、10mM Tris-HCl pH 7.4、10mM MgCl2、1mM DTT、100ug/ml BSAおよび0〜500単位/mlのMnlI(または他の制限酵素)を用いる。いくつかの態様においては、0.25%リゾレシチン(w/v)、50mM NaCl、10mM Tris-HCl pH 7.4、10mM MgCl2、1mM DTT、100ug/ml BSAおよび0〜500単位/mlのMnlI(または他の制限酵素)を用いる。いくつかの態様においては、0.75%リゾレシチン(w/v)、50mM NaCl、10mM Tris-HCl pH 7.4、10mM MgCl2、1mM DTT、100ug/ml BSAおよび0〜500単位/mlのMnlI(または他の制限酵素)を用いる。いくつかの態様においては、1%リゾレシチン(w/v)、50mM NaCl、10mM Tris-HCl pH 7.4、10mM MgCl2、1mM DTT、100ug/ml BSAおよび0〜800単位/mlのMnlI(または他の制限酵素)を用いる。
【0066】
透過処理および消化の後に、任意で同時に溶解/停止用緩衝液および/または温度上昇を加えることにより、消化を任意で停止させて、細胞を溶解する。例示的な溶解/停止用緩衝液には、反応を停止させて細胞を溶解するのに十分なキレート剤および界面活性剤が含まれうる。例えば、いくつかの態様において、溶解/停止用緩衝液は、100mM Tris-HCl pH 8、100mM NaCl、100mM EDTA、5% SDS(w/v)および3mg/mlのプロテイナーゼKを含む。いくつかの態様において、溶解/停止用緩衝液は、100mM Tris-HCl pH 8、100mM NaCl、100mM EDTA、1% SDS(w/v)および3mg/mlのプロテイナーゼKを含む。いくつかの態様において、溶解/停止用緩衝液は、200mM Tris-HCl pH 8、100mM NaCl、500mM EDTA、5% SDS(w/v)および5mg/mlのプロテイナーゼKを含む。
【0067】
B.DNアーゼ
いくつかの態様においては、DNAに配列非特異的な様式で切れ目またはニックを入れる酵素をDNA修飾剤として用いる。したがって、いくつかの態様において、DNA修飾剤は配列非特異的エンドヌクレアーゼ(本明細書では「DNアーゼ」とも称する)である。
【0068】
あらゆる配列非特異的エンドヌクレアーゼ(例えば、DNアーゼI、II、III、IV、V、VI、VIIのいずれか)を、本発明に従って用いることができる。例えば、DNアーゼIを非限定的に含む、あらゆるDNアーゼを用いることができる。用いるDNアーゼには、天然に存在するDNアーゼ、さらには改変DNアーゼが含まれうる。改変DNアーゼの一例はTURBO DNアーゼ(Ambion)であり、これは「活性亢進」および塩耐性を可能にする突然変異を含む。例示的なDNアーゼには、ウシ膵臓DNアーゼI(例えば、New England Biolabsから入手可能)が非限定的に含まれる。
【0069】
本明細書に記載した通りに、無傷のDNAをその後に別々に検出して定量し、DNA領域の無傷のおよび/または切断されたコピーの数を定量することができる。
【0070】
いくつかの態様においては、透過処理剤または膜崩壊剤を、DNアーゼの前に添加する。いくつかの態様においては、DNアーゼと透過処理剤または崩壊剤とを同時に添加する(例えば、適切な緩衝液とともに)。いくつかの態様において、透過処理/消化用緩衝液は、0.25%リゾレシチン(w/v)、10mM Tris-HCl pH 7.4、2.5mM MgCl2、0.5mM CaCl2および0〜200単位/mlのDNアーゼIを含む。いくつかの態様において、透過処理/消化用緩衝液は、0.5%リゾレシチン(w/v)、10mM Tris-HCl pH 7.4、2.5mM MgCl2、0.5mM CaCl2および0〜200単位/mlのDNアーゼIを含む。いくつかの態様において、透過処理/消化用緩衝液は、0.75%リゾレシチン(w/v)、10mM Tris-HCl pH 7.4、2.5mM MgCl2、0.5mM CaCl2および0〜500単位/mlのDNアーゼIを含む。いくつかの態様において、透過処理/消化用緩衝液は、0.25%リゾレシチン(w/v)、10mM Tris-HCl pH 7.4、2.5mM MgCl2、0.5mM CaCl2および0〜500単位/mlのDNアーゼIを含む。透過処理および溶解は、例えば、制限酵素に関して上述した通りに停止させることができる。
【0071】
他の箇所で考察しているように、DNアーゼまたは他の一般的なDNA切断剤の使用を、一方が標的であり、もう一方が、試験条件のいずれにおいても概ね常にアクセス可能であるかまたは概ね常にアクセス不能であるDNA領域である、少なくとも2つの異なるDNA領域間の切断の程度をモニターすることによって向上させることができる。そのような遺伝子の例は本明細書中の他の箇所で考察されており、公知であるかまたは同定することができる。例えば、「ハウスキーピング」遺伝子を範囲に含むDNA領域は概ね常にアクセス可能である。続いて、対照と比較した残りの標的の相対量を用いて、標的DNA領域での相対的なクロマチン構造を決定することができる。
【0072】
C.メチルトランスフェラーゼ
本発明のいくつかの態様において、DNA修飾剤はDNAに対する共有結合修飾を生成させる。例えば、いくつかの態様において、本発明のDNA修飾剤はメチルトランスフェラーゼである。
【0073】
種々のメチルトランスフェラーゼが当技術分野において公知であり、本発明に用いることができる。いくつかの態様において、用いるメチルトランスフェラーゼはDNA中のアデノシンに対してメチル部分を付加する。そのようなメチルトランスフェラーゼの例には、DAMメチルトランスフェラーゼが非限定的に含まれる。アデノシンは真核生物細胞ではメチル化されないため、特定のDNA領域におけるメチル化アデノシンの存在は、DAMメチルトランスフェラーゼ(または類似の活性を有する他のメチルトランスフェラーゼ)がDNA領域にアクセス可能であったことを指し示している。アデノシンのメチル化は、例えば、認識配列がメチル化アデノシンを含む制限酵素を用いて検出することができる。そのような酵素の一例には、DpnIが非限定的に含まれる。続いて、制限酵素による切断を、本明細書に記載した通りに(例えば、無傷のDNAは増幅されるが切断されたDNAはそうでないように‐またはLM-PCRを用いて、切断されたDNAは増幅するが無傷のDNAは増幅しないように)定量する。
【0074】
いくつかの態様において、メチルトランスフェラーゼはGC配列中のシトシンをメチル化する。そのようなメチルトランスフェラーゼの例には、MCviPIが非限定的に含まれる。例えば、Xu et al., Nuc. Acids Res. 26(17): 3961-3966 (1998)を参照のこと。GC配列は真核生物細胞ではメチル化されないため、特定のDNA領域におけるメチル化GC配列の存在は、DNA修飾剤(すなわち、GC配列中のシトシンをメチル化するメチルトランスフェラーゼ)がDNA領域にアクセス可能であったことを指し示している。メチル化GC配列は、任意のさまざまな手法を用いて同定することができる。いくつかの態様において、メチル化GC配列を検出するための方法は、バイサルファイト変換を含む。バイサルファイト変換は、DNAを、非メチル化シトシンをウラシルに変換するために十分な量のバイサルファイトと接触させることを含む。メチル化シトシンは変換されない。このため、GC配列を含むDNA領域を、GC配列中のシトシンをメチル化するメチルトランスフェラーゼと接触させ、単離して、続いてバイサルファイトと接触させることができる。GC配列中のCがメチル化されていなければ、そのCはU(またはその後に増幅された場合はT)に変換されると考えられるが、一方、メチル化CはCのままであると考えられる。ヌクレオチド配列決定、ならびにプライマー伸長またはプライマーに基づく増幅および/またはメチル化感受性制限消化を伴う方法を非限定的に含む、任意のさまざまな方法を用いて、バイサルファイトにより変換されたCの有無を検出することができる(例えば、MSnuPE、MSPまたはメチルライト(Methlyllight)、高分解能融解分析;ピロシークエンシングなど)。例えば、Fraga, et al., Biotechniques 33:632, 634, and 636-649 (2002);El-Maarri 0 Adv Exp Med Biol 544:197-204 (2003);Laird, Nat Rev Cancer 3:253266 (2003);およびCallinan, Hum Mol Genet 15 Spec No 1:R95-101 (2006)を参照のこと。
【0075】
いくつかの態様において、メチルトランスフェラーゼは、CG(「CpG」としても知られる)配列中のシトシンをメチル化する。そのようなメチルトランスフェラーゼの例には、M.SssIが非限定的に含まれる。そのようなメチルトランスフェラーゼの使用は一般に、典型的にはメチル化されないDNA領域に対する使用に限定される。これは、CG配列が真核生物細胞では内因性にメチル化され、それ故に、メチル化が稀であるようなDNA領域を除き、CG配列が内因性メチルトランスフェラーゼではなく修飾剤によってメチル化されると想定することは一般に不可能であるためである。GC配列の場合と同様に、CG配列のメチル化は、バイサルファイト変換を伴う方法を含む、任意のさまざまな方法によって検出することができる。
【0076】
D.化学物質
いくつかの態様において、DNA修飾剤はDNA修飾性化学物質である。大部分のDNA修飾性化学物質はクロマチンと比較して相対的に小さいため、融合パートナーを伴わないDNA修飾性化学物質の使用は、異なるDNA領域のアクセス可能性の程度に仮に違いがあるにしてもわずかであると考えられることから、状況によっては有効でない。このため、いくつかの態様において、DNA修飾剤は、DNA修飾性化学物質と連結された立体障害を有する分子を含む。立体障害を有する分子は、クロマチン構造に応じてDNA修飾剤のアクセス可能性に違いをもたらす任意のタンパク質または他の分子でありうる。これは例えば、結果を、本明細書に記載したDNアーゼまたは制限酵素を用いた結果と比較することによって検討することができる。
【0077】
いくつかの態様において、立体障害を有する分子は、少なくとも5、7、10または15kDのサイズであると考えられる。おそらく当業者は、立体障害を備える分子としてポリペプチドを用いることが好都合であると判断するであろう。DNA修飾剤がDNAを修飾する能力を著しく妨害することのない任意のポリペプチドを用いることができる。いくつかの態様において、ポリペプチドは、以下でさらに詳細に考察している二本鎖配列非特異的な核酸結合ドメインである。
【0078】
本発明のDNA修飾性化学物質は、立体障害を有する分子と直接連結させること、またはリンカーを介して連結させることができる。種々のホモおよびヘテロ二官能性リンカーが公知であり、この目的に用いることができる。
【0079】
例示的なDNA修飾性化学物質には、ヒドラジン(および、例えば、Mathison et al., Toxicology and Applied Pharmacology 127(1):91-98 (1994)に記載されているようなその誘導体)および硫酸ジメチルが非限定的に含まれる。いくつかの態様において、ヒドラジンはDNA中のグアノシンにメチル基を導入するか、他の様式でDNAを損傷させる。いくつかの態様において、硫酸ジメチルはグアニンをメチル化するか、または、グアニン中に存在するイミダゾール環を開裂させることにより、DNA中のグアニンの塩基特異的切断をもたらす。
【0080】
DNA修飾性化学物質による修飾の検出は、生じたDNA修飾の種類に依存すると考えられる。いくつかの態様において、硫酸ジメチルまたはヒドラジンによる修飾を検出するために、DNAをピペリジンにより高温(90℃)で処理する。DNAはDNA修飾部位で切断を受け、その切断を、本明細書に記載したヌクレアーゼ切断を検出するのと同じやり方で検出することができる。
【0081】
E.DNA修飾剤を改良するためのDNA結合ドメイン
いくつかの態様においては、本発明のDNA修飾剤または切断剤を、二本鎖配列非特異的な核酸結合ドメイン(例えば、DNA結合ドメイン)と融合させるかまたは他の様式で連結させる。DNA修飾剤がポリペプチドである場合には、二本鎖配列非特異的な核酸結合ドメインを、例えば、DNA修飾剤とのタンパク質融合物として、組換えDNA技術を介して合成することができる。二本鎖配列非特異的な核酸結合ドメインは、二本鎖核酸と配列非依存的な様式で結合するタンパク質またはタンパク質の定まった領域であり、すなわち結合は、特定の配列に対する全体的な選好性を示さない。いくつかの態様において、二本鎖核酸結合タンパク質は、二本鎖核酸に対して一本鎖核酸よりも10倍またはそれ以上高い親和性を示す。二本鎖核酸結合タンパク質は、本発明のいくつかの態様において耐熱性である。そのようなタンパク質の例には、古細菌小型塩基性DNA結合タンパク質Sac7dおよびSso7d(例えば、Choli et al., Biochimica et Biophysica Acta 950:193-203, 1988;Baumann et al., Structural Biol. 1:808-819, 1994;およびGao et al, Nature Struc. Biol. 5:782-786, 1998を参照)、古細菌HMf様タンパク質(例えば、Stanch et al., J. Molec. Biol. 255:187-203, 1996;Sandman et al., Gene 150:207-208, 1994を参照)、およびPCNA相同体(例えば、Cann et al., J. Bacteriology 181:6591-6599, 1999;Shamoo and Steitz, Cell:99, 155-166, 1999;De Felice et al., J. Molec. Biol. 291, 47-57, 1999;およびZhang et al., Biochemistry 34:10703-10712, 1995を参照)が非限定的に含まれる。DNA結合ドメインに関するそのほかの情報については、欧州特許第1283875B1も参照のこと。
【0082】
Sso7dおよびSac7d
Sso7dおよびSac7dは、それぞれ超好熱性古細菌スルホロブス・ソルファタリカス(Sulfolobus solfataricus)およびS.アシドカルダリウス(S. acidocaldarius)に由来する小型(約7,000kd MW)の塩基性染色体タンパク質である。これらのタンパク質はリジンに富み、高い耐熱性、酸安定性および化学安定性を有する。それらはDNAと配列非依存的な様式で結合し、結合すると、DNAのTMをいくつかの条件下では最大40℃上昇させる(McAfee et al., Biochemistry 34:10063-10077, 1995)。これらのタンパク質およびそれらの相同体は、典型的には、高温でゲノムDNAを安定化させることに関与すると考えられている。
【0083】
HMF様タンパク質
HMf様タンパク質は、DNAと直接相互作用すると考えられている真核生物H4ヒストンとアミノ酸配列および構造の両方の点で相同性のある古細菌ヒストンである。HMfファミリーのタンパク質は溶液中で安定な二量体を形成し、いくつかのHMf相同体が耐熱性種(例えば、メタノテルムス・フェルビダス(Methanothermus fervidus)およびパイロコッカス属GB-3a菌株)から同定されている。HMfファミリーのタンパク質は、Taq DNAポリメラーゼまたは固有の処理能力が低い任意のDNA修飾酵素と結合させると、その酵素がDNA基質に沿ってスライドする能力を強化し、それ故にその処理能力を高める。例えば、二量体HMf様タンパク質を、Taq DNAポリメラーゼのN末端に、例えば化学修飾を介して共有結合で連結させ、それによりそのポリメラーゼの処理能力を改善することができる。
【0084】
当業者は、当技術分野において公知であるその他の二本鎖配列非特異的な核酸結合ドメインを認知していると考えられ、それらを本明細書に記載したように用いることもできる。
【0085】
F.DNA修飾段階後のDNAの単離
いくつかの態様においては、DNA修飾/切断の段階の後に、利用可能な任意の方法に従ってゲノムDNAを細胞から単離する。その後の定量段階のために許容しうる純度のDNAがもたらされる限り、本質的にあらゆるDNA精製手順を用いることができる。例えば、標準的な細胞溶解試薬を、細胞を溶解するために用いることができる。任意でプロテアーゼ(プロテイナーゼKを非限定的に含む)を用いることができる。DNAは、当技術分野において公知である通りに混合物から単離することができる。いくつかの態様においては、フェノール/クロロホルム抽出を用い、DNAをその後に(例えば、エタノールにより)沈殿させて、精製する。いくつかの態様においては、必要に応じて(例えば、RNアーゼにより、またはDNA精製カラムにより)RNAを除去または分解する。
【0086】
任意で、ゲノムDNAを、中間精製段階を省いて、細胞溶解物から直接的に増幅するかまたは他の様式で検出する。
【0087】
いくつかの態様においては、無傷の、修飾された、または修飾されていないDNAを単離して、ライブラリーへとクローニングする。場合によっては、1つまたは複数の特定の無傷の、修飾された、または修飾されていない配列を単離および/またはクローニングする。または、無傷の、修飾された、または修飾されていないDNA領域を有する試料を用いて、そのような領域に関して濃縮されたライブラリーを調製する。DNA切断剤との接触後に無傷のDNAは、その剤に対して、よりアクセス可能性が低かったDNAを表す。同様に、DNA修飾剤との接触後に修飾されていないDNAは、よりアクセス可能性の低いDNAを表す。その反対に、修飾されたDNAは、修飾剤に対して、よりアクセス可能性が高かったDNAを表す。上記の態様のいくつかにおいては、クローニングの前に、無傷のDNAを切断されたDNAから精製し(例えば、分離し)、かつ/または修飾されたDNAを修飾されていないDNAから精製し、それにより、1つのクラスのDNAに関してのクローニングプールを濃縮する。修飾された/修飾されていないDNAに関する濃縮は、修飾の性質に応じて異なると考えられる。いくつかの態様においては、修飾された(または修飾されていないDNA)と特異的に結合する親和性薬剤を用いて、修飾されたDNAを修飾されていないDNAから分離する。
【0088】
いくつかの態様においては、サブトラクティブライブラリーを作製する。例えば、本発明の方法において無傷の、修飾された、または修飾されていない罹患細胞DNA領域に関して濃縮されたライブラリーを作製し、その後に健常細胞由来の対応するライブラリーを用いてサブトラクションして、それにより、両方とも無傷であるか、修飾されているか、または修飾されておらず、かつ特定の疾患に対して特異的である、差次的なDNA配列ライブラリーを作製することができる。癌細胞を非限定的に含む、任意の罹患細胞を用いることができる。代替的なサブトラクティブ戦略を、例えば、異なる細胞型、細胞時期、薬物治療などの間で用いることもできる。
【0089】
G.DNAの物理的特徴の検出
細胞とDNA修飾剤またはDNA切断剤との接触の後に、DNAの任意のさまざまな物理的特徴を検出することができる。物理的特徴には、DNAメチル化、融解温度、GC含有量、ヌクレオチド配列、およびポリヌクレオチドとハイブリダイズする能力が非限定的に含まれる。そのような特徴を検出するための種々の方法が公知であり、用いることができる。いくつかの態様において、DNA修飾/切断の段階の後に決定される物理的特徴は、DNAフットプリンティング(例えば、DNAの特定の領域に対する1つまたは複数の特定のタンパク質の能力)を決定することを含まない。例えば、1つの非限定的な態様において、例えばqPCRを用いる無傷のDNAの定量は、DNAフットプリント法を含まない。
【0090】
いくつかの態様において、物理的特徴はDNAメチル化である。例えば、相対的にアクセス可能なDNAがDNA切断剤によって切断されれば、残りの無傷のDNA(アクセス可能性がより低いDNAに相当する)を単離し、続いてメチル化状態に関して分析することができる。非常にさまざまなDNAメチル化検出方法が公知である。いくつかの態様においては、DNA修飾剤または切断剤との接触後に、DNAをバイサルファイトと接触させ、それにより、DNA中のメチル化されていないシトシンをウラシルに変換する。続いて、特定のDNA領域のメチル化を、本明細書で考察したものを含む、任意のさまざまなメチル化検出方法によって決定することができる。いくつかの態様において、高分解能融解アッセイ(HRM)を用いて、バイサルファイト変換後のメチル化状態を検出する。この方法では、バイサルファイト変換後にDNA領域を増幅し、その結果得られたアンプリコンの融解温度を決定する。融解温度はシトシンがバイサルファイトによって変換されたか否か(およびその後に増幅反応において「T」としてコピーされたか否か)に応じて異なると考えられるため、アンプリコンの融解温度をメチル化含有量と相関させることができる。
【0091】
V.標的DNA領域
DNA領域は、ゲノムDNA内部の関心対象の標的配列である。細胞のゲノムDNA中の任意のDNA配列を、本明細書に記載した通りに、DNA修飾剤のアクセス可能性に関して評価することができる。例えば、複数の異なる細胞型において、処理していない細胞と薬物刺激、化学刺激もしくは環境的刺激に曝露された細胞との間で、または正常組織と罹患組織との間で異なるアクセス可能性を呈する、関心対象のDNA領域を同定するために、DNA領域をスクリーニングする。したがって、いくつかの態様において、本発明の方法は、アクセス可能性の変化が疾患(またはその欠如)のマーカーとして作用するDNA領域を同定するために用いられる。例示的な疾患には、癌が非限定的に含まれる。癌でない細胞と比較して癌細胞において、変化したDNAメチル化および/またはクロマチン構造を有する、数多くの遺伝子が記載されている。
【0092】
いくつかの態様において、DNA領域は、特定の細胞の罹患状態または発生状態に応じて、アクセス可能性に差異を伴うことが判明している。これらの態様において、本発明の方法は、診断ツールまたは予後予測ツールとして用いることができる。本発明の方法を用いて診断または予後予測がひとたび確定されれば、その診断または予後予測を考慮して、治療レジメンを設定すること、または既存の治療レジメンを変更することができる。例えば、本発明の方法に従った癌細胞の検出は、癌細胞が検出された個体に対する化学療法薬および/または放射線の投与につながりうる。
【0093】
種々のDNA領域を、研究目的で、および/または試薬が予想通りに機能することを確認するための対照DNA領域として、検出することができる。例えば、いくつかの態様においては、ある動物の本質的にすべての細胞においてアクセス可能であるDNA領域をアッセイする。そのようなDNA領域は、例えば、アクセス可能性に関する陽性対照として有用である。そのようなDNA領域は、例えば、構成的であるかまたはほぼ構成的である遺伝子の内部または近傍に見いだすことができる。そのような遺伝子には、一般に「ハウスキーピング」遺伝子と呼ばれるもの、すなわち、基本的な生物機能を維持するためにその発現が必要とされる遺伝子が含まれる。そのような遺伝子の例には、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ(GAPDH)およびβアクチン(ACTB)が非限定的に含まれる。DNA領域には、任意でプロモーターの少なくとも一部分を含む、そのような遺伝子の全体または一部分が含まれうる。
【0094】
いくつかの態様において、DNA領域は、ある動物のほとんどの細胞においてアクセス不能であるDNAの少なくとも一部分を含む。そのようなDNA領域は、例えば、アクセス可能性に関する陰性対照として有用である。この文脈における「アクセス不能な」とは、そのコピーが、DNA領域のコピーの20%前後を上回らない割合でしか修飾されないDNA領域のことを指す。そのような遺伝子配列の例には、「異質染色質」性として一般に認識されているものが含まれ、極めて特定の細胞型のみで発現される(例えば、組織特異的または臓器特異的な様式で発現される)遺伝子が含まれる。(特定の細胞型を除いて)一般にアクセス不能である例示的な遺伝子には、ヘモグロビン-β鎖(HBB)および免疫グロブリン軽鎖κ(IGK)が非限定的に含まれる。
【0095】
いくつかの態様において、DNA領域は、細胞の罹患状態に応じて異なるアクセス可能性を有するか、またはさもなければ細胞の型または生育環境に応じてさまざまなアクセス可能性を有する、遺伝子配列である。例えば、ある種の遺伝子は、癌でない細胞では一般にアクセス不能であるが、癌細胞ではアクセス可能である。さまざまなアクセス可能性を有する遺伝子には、例えば、グルタチオン-s-トランスフェラーゼπ(GSTP1)が含まれる。
【0096】
いくつかの態様において、本発明のDNA領域は、以下の遺伝子の1つまたは複数からの遺伝子配列(例えば、プロモーター配列)から選択される:1型カドヘリン1(E-カドヘリン)、シトクロムP450-1A1(CYP1A1)、Ras関連ドメインファミリー1A(RASSF1A)、p15、p16、細胞死関連プロテインキナーゼ1(DAPK)、腺腫性結腸ポリポーシス(APC)、メチルグアニン-DNAメチルトランスフェラーゼ(MGMT)、乳癌1遺伝子(BRCA1)およびhMLH。
【0097】
いくつかの態様において、DNA領域は、例えば、異なる細胞型の間、異なる条件の間、正常細胞と罹患細胞との間などで異なるアクセス可能性を有する領域を同定するために、ランダムに選択される。
【0098】
VI.標的遺伝子座のコピーの定量
DNA修飾を定量するための方法は、ゲノムDNAに導入されるDNA修飾の種類に依存すると考えられる。例えば、二本鎖DNA切断イベント(例えば、制限酵素もしくはDNアーゼによって導入されるような、または、メチルトランスフェラーゼ処理後もしくは本明細書に記載されたDNA修飾性化学物質による修飾後に、例えばメチル化感受性もしくはメチル化依存性制限酵素によって続いて導入されるような修飾)を、関心対象のDNA領域を含むアンプリコンを生成するように設計された増幅反応を用いて好都合に検出することができる。配列特異的制限酵素を用いた場合のような定まった部位での切断イベントの場合には、潜在的な切断部位にまたがるアンプリコンが生成されるようにプライマーを設計する。無傷のDNAのみが増幅されると考えられる。全DNAの量も判明しているならば、切断されたDNAの量を、全DNAと無傷のDNAとの間の差として計算することができる。DNAの総量は、当技術分野において公知である任意のDNA定量法に従って決定することができる。いくつかの態様において、全DNAの量は、修飾とは無関係にDNAを増幅するプライマーのセットを設計することにより、好都合に決定することができる。これは例えば、同じ遺伝子領域内または別のDNA領域内にある、潜在的な切断部位をまたがないプライマーを設計することによって達成することができる。DNアーゼIなどの配列非特異的ヌクレアーゼを用いる場合のような不定部位での切断イベントの場合には、アクセス不能な参照遺伝子の使用を内部対照として組み入れるべきである。
【0099】
以下により詳細に考察するように、定量的増幅(例えばリアルタイムPCRを含む)方法は、DNA領域の無傷のコピーの量の決定を可能にし、さまざまな対照とともに用いる場合、細胞におけるコピーの総数と比較して無傷のDNAの相対量を決定するために用いることができる。従って、DNA領域の修飾されたコピーまたは修飾されていないコピーの実数または相対数(例えば、コピーの総数に対して相対的、または第2のDNA領域の修飾されたコピーまたは修飾されていないコピーの数に対して相対的)を、算出することができる。
【0100】
本発明のいくつかの態様において、DNA領域の修飾されたコピーの数は直接的に決定される。例えば、特定の付着末端または平滑末端の存在時にのみ起こると考えられる特定のライゲーションイベントを検出することなどにより、制限酵素切断を検出して定量することもできる。例えば、制限酵素によって生成される付着末端に対して相補的な付着末端を含む核酸アダプターを、切断されたゲノムDNAとライゲートさせることができる。続いて、(例えば、定量的増幅方法により)ライゲーションイベントの数を検出して定量することができる。
【0101】
いくつかの態様においては、ライゲーション媒介PCR(LM-PCR)を用いて、DNA領域の切断されたコピーの数を定量する。LM-PCRの方法は当技術分野において公知であり、Pfeifer et al., Science 246: 810-813 (1989)において最初に記載されている。LM-PCRは、必要に応じて、定量的結果のためにリアルタイムで行うことができる。
【0102】
定量的増幅方法(例えば、定量的PCRまたは定量的線形増幅)は、核酸テンプレートを増幅し、増幅されたDNAの量を直接的または間接的に(例えば、Ct値を決定して)決定し、続いて増幅サイクル数に基づいて最初のテンプレートの量を算出することを伴う。反応を用いたDNA遺伝子座の増幅は周知である(米国特許第4,683,195号および第4,683,202号;PCR PROTOCOLS: A GUIDE TO METHODS AND APPLICATIONS (Innis et al., eds, 1990)を参照)。典型的には、PCRを用いてDNAテンプレートを増幅する。しかし、代替的な増幅方法も記載されており、それらの代替的な方法がそれらの方法が切断されたDNAを増幅するよりも高い程度で無傷のDNAを増幅する限り、それを用いることもできる。定量的増幅の方法は、例えば、米国特許第6,180,349号;第6,033,854号;および第5,972,602号、さらには例えば、Gibson et al., Genome Research 6:9951001 (1996);DeGraves, et al., Biotechniques 34(1):106-10, 112-5 (2003);Deiman B, et al., Mol Biotechnol. 20(2):163-79 (2002)に記載されている。増幅は「リアルタイム」でモニターすることができる。
【0103】
いくつかの態様において、定量的増幅は、増幅(例えば、PCR)反応のサイクルにおけるテンプレートのコピーを表すシグナル(例えば、プローブの蛍光)のモニタリングに基づく。PCRの初期サイクルでは、形成されるアンプリコンの量が、アッセイから測定可能であるシグナル出力を支持しないため、非常に弱いシグナルしか観察されない。初期サイクルの後には、形成されるアンプリコンの量が増加するため、シグナル強度は測定可能なレベルまで増大し、PCRが非対数期に入る後期のサイクルではプラトーに達する。シグナル強度対サイクル数のプロットを通じて、測定可能なシグナルがPCR反応から得られる特定のサイクルを推定し、それを用いてPCRの開始前の標的の量を逆算することができる。この方法によって決定される特定のサイクル数は、典型的には、サイクル閾値(Ct)と呼ばれる。例示的な方法は、例えば、Heid et al. Genome Methods 6:986-94 (1996)に、加水分解プローブに関連して記載されている。
【0104】
増幅産物の検出のための1つの方法は、5'-3'エキソヌクレアーゼ「加水分解」PCRアッセイ(TaqMan(商標)アッセイとも呼ばれる)である(米国特許第5,210,015号および第5,487,972号;Holland et al., PNAS USA 88: 7276-7280 (1991);Lee et al., Nucleic Acids Res. 21: 3761-3766 (1993))。このアッセイは、増幅反応の間に、二重標識された蛍光発生プローブ(「TaqMan(商標)」プローブ)のハイブリダイゼーションおよび切断によって特定のPCR産物の蓄積を検出する。蛍光発生プローブは、蛍光レポーター色素およびクエンチャー色素の両方で標識されたオリゴヌクレオチドからなる。PCRの間に、このプローブは、増幅されるセグメントとそれがハイブリダイズした場合に、しかもその場合にのみ、DNAポリメラーゼの5'-エキソヌクレアーゼ活性によって切断される。プローブの切断は、レポーター色素の蛍光強度の増加を生じさせる。
【0105】
エネルギー転移の使用に依拠して増幅産物を検出するもう1つの方法は、Tyagi and Kramer, Nature Biotech. 14:303-309 (1996)に記載された「ビーコンプローブ」法であり、これは米国特許第5,119,801号および第5,312,728号の主題でもある。この方法は、ヘアピン構造を形成することができるオリゴヌクレオチドハイブリダイゼーションプローブを用いる。ハイブリダイゼーションプローブの一方の末端(5'末端または3'末端かのいずれか)にはドナーフルオロフォアがあり、もう一方の末端にはアクセプター部分がある。TyagiとKramerの方法の場合には、このアクセプター部分はクエンチャーであり、すなわち、アクセプターはドナーによって放出されたエネルギーを吸収するが、その後に自ら蛍光を発することはない。したがって、ビーコンがオープン構造である場合はドナーフルオロフォアの蛍光を検出しうるが、ビーコンがヘアピン(閉鎖)構造である場合はドナーフルオロフォアの蛍光は消光する。PCRに用いる場合、PCR産物の一方の鎖とハイブリダイズする分子ビーコンプローブはオープン構造であり、蛍光が検出されるが、ハイブリダイズしないままのものは蛍光を発しないと考えられる(Tyagi and Kramer, Nature Biotechnol. 14: 303-306 (1996))。その結果、蛍光の量はPCR産物の量が増加するにつれて増加し、それ故にPCRの進行の尺度として用いることができる。当業者は、その他の定量的増幅法も利用可能であることを認識しているであろう。
【0106】
核酸の定量的増幅を行うためのさまざまな他の手法も公知である。例えば、いくつかの方法は、オリゴヌクレオチドが標的核酸とハイブリダイズした場合に蛍光の変化が生じるように構築されている1つまたは複数のプローブオリゴヌクレオチドを用いる。例えば、そのような1つの方法は、2つのオリゴプローブをアンプリコンとアニーリングさせる、蛍光共鳴エネルギー転移(FRET)を利用する二重フルオロフォアアプローチ、例えば、LightCycler(商標)ハイブリダイゼーションプローブである。オリゴヌクレオチドは、効率的なエネルギー転移に適合する距離で隔てられたフルオロフォアと頭-尾方向でハイブリダイズするように設計される。核酸と結合するかまたは伸長産物に組み入れられた場合にシグナルを発するように構築された標識オリゴヌクレオチドの他の例には、以下が含まれる:Scorpions(商標)プローブ(例えば、Whitcombe et al., Nature Biotechnology 17:804-807, 1999および米国特許第6,326,145号)、Sunrise(商標)(またはAmplifluor(商標))プローブ(例えば、Nazarenko et al., Nuc. Acids Res. 25:2516-2521, 1997および米国特許第6,117,635号)、および、クエンチャーがなくてもシグナル低下をもたらす二次構造を形成し、かつ標的とハイブリダイズした場合に増大したシグナルを発するプローブ(例えば、Lux probes(商標))。
【0107】
他の態様においては、二本鎖DNA中に挿入された時にシグナルを生じるインターカレート剤を用いうる。例示的な剤には、SYBR GREEN(商標)、SYBR GOLD(商標)およびEVAGREEN(商標)が含まれる。これらの剤はテンプレート特異的ではないため、シグナルはテンプレート特異的増幅に基づいて生成されると想定される。テンプレート配列の融解点は一般に、例えばプライマー-ダイマーなどよりもはるかに高いと考えられるため、このことはテンプレートの関数としてシグナルをモニターすることによって確認することができる。
【0108】
いくつかの態様において、DNA領域の量は、試料中のヌクレオチド配列のコピーを決定し、続いて試料中の同じ配列を有するコピーの相対数または絶対数を決定することによって決定される。
【0109】
本発明の方法による修飾された(または修飾されていない)DNA領域の定量は、いくつかの態様において、DNA領域の修飾されたコピーまたは修飾されていないコピーの、その同じ領域のコピーの総数と比較した相対量(例えば、比またはパーセンテージなどの標準化された値)を決定することによって、さらに改良することができる。いくつかの態様において、1つのDNA領域の修飾されたコピーまたは修飾されていないコピーの相対量を、第2の(またはそれ以上の)DNA領域の修飾されたコピーまたは修飾されていないコピーの数と比較する。いくつかの態様において、2つまたはそれ以上のDNA領域の間で比較する場合には、各DNA領域の修飾されたコピーまたは修飾されていないコピーの相対量を、まず、DNA領域のコピーの総数に対して標準化する。または、同じ試料から得られる場合には、いくつかの態様において、各DNA領域のコピーの総数はおおよそ同じであると仮定し、そのため2つまたはそれ以上のDNA領域の間で比較する場合に、各値をコピーの総数に対してまず標準化することなしに、修飾されたコピーまたは修飾されていないコピーの相対量(例えば、比またはパーセンテージ)を各DNA領域間で決定することもできる。
【0110】
いくつかの態様においては、修飾されたコピーまたは修飾されていないコピーの実際の量または相対量(例えば、全DNAとの対比)を、対照値と比較する。対照値は、例えば、特定のDNA領域に対するアクセス可能性が特定の値を上回るかそれ未満であるかを知りたい場合に、好都合に用いることができる。例えば、特定のDNA領域が正常細胞では典型的にはアクセス可能であるが罹患細胞ではアクセス不能である(またはその反対の)状況では、修飾されたコピーまたは修飾されていないコピーの実数または相対数を、単に、(例えば、20%超またはそれ未満が修飾されているかまたは修飾されていない、80%超またはそれ未満が修飾されているかまたは修飾されていない、などの)対照値と比較してもよい。または、対照値が、対照DNA領域に関する過去データまたは予想されるデータであってもよい。これらの場合には、対照DNA領域の実際の量または相対量を(任意で数回)決定し、その結果得られたデータを用いて、関心対象のDNA領域に関して決定された修飾されたコピーまたは修飾されていないコピーの実数または相対数と比較することのできる対照値を作成することができる。
【0111】
本明細書に記載された方法のための計算は、コンピュータベースの計算およびツールを含みうる。これらのツールは、従来設計の汎用コンピュータシステム(本明細書では「ホストコンピュータ」と称する)によって実行可能なコンピュータプログラムの形態で提供されることが好都合である。ホストコンピュータは数多くのさまざまなハードウェア構成部分を用いて構成されてよく、さまざまな寸法および方式で製造することができる(例えば、デスクトップPC、ラップトップ、タブレットPC、ハンディ型コンピュータ、サーバー、ワークステーション、大型汎用コンピュータ)。モニター、キーボード、ディスクドライブ、CDおよび/またはDVDドライブなどの標準的な構成部品が含まれてもよい。ホストコンピュータをネットワークに結びつける場合、接続は任意の適した伝達媒体(例えば、有線媒体、光媒体および/または無線媒体)ならびに任意の適した通信プロトコル(例えば、TCP/IP)を介して提供されうる;ホストコンピュータが適したネットワーク用ハードウェア(例えば、モデム、イーサネットカード、WiFiカード)を含んでもよい。ホストコンピュータは、UNIX、Linux、Microsoft Windows、MacOSまたは任意の他のオペレーティングシステムを含む、さまざまなオペレーティングシステムの任意のものを実行してよい。
【0112】
本発明の実行局面に関するコンピュータコードは、PERL、C、C++、Java、JavaScript、VBScript、AWK、または、ホストコンピュータ上で実行されうるか、もしくはホストコンピュータ上で実行されるようにコンパイルされうる、任意の他のスクリプト言語またはプログラム言語を含むさまざまな言語で記述されうる。また、コードを、アセンブリ言語またはマシン言語のような低水準言語で記述するかまたは分散させてもよい。
【0113】
ホストコンピュータシステムは、ユーザーがツールの動作を制御するインターフェースを好都合に提供する。本明細書に記載された例において、ソフトウェアツールはスクリプト(例えば、PERLを用いる)として実行され、その遂行はユーザーにより、LinuxまたはUNIXなどのオペレーティングシステムの標準的なコマンドラインインターフェースから開始される。当業者は、コマンドをオペレーティングシステムに対して適宜適合させうることを認識しているであろう。他の態様において、ユーザーがポインティングデバイスを用いて動作を制御することを可能にするグラフィカルユーザーインターフェースが提供されてもよい。したがって、本発明は、いかなる特定のユーザーインターフェースにも限定されない。
【0114】
本発明のさまざまな特徴を組み入れたスクリプトまたはプログラムを、記憶および/または伝達のために、さまざまなコンピュータ可読媒体上に符号化して記録してもよい。適した媒体の例には、磁気ディスクまたはテープ、コンパクトディスク(CD)またはDVD(デジタル多目的ディスク)などの光学記憶媒体、フラッシュメモリー、ならびにインターネットを含む種々のプロトコルに準拠する有線、光および/または無線ネットワークを介した伝達用に適合化されたキャリア信号が含まれる。
【0115】
VII.反応混合物
本発明はまた、本明細書に記載された試薬の1つまたは複数を、任意で(クロマチン状態を決定しようとする)真核細胞とともに含む、反応混合物も提供する。いくつかの態様において、反応混合物は、例えば、DNA修飾剤(例えば、制限酵素、DNアーゼ、メチルトランスフェラーゼまたはDNA修飾性化学物質)および細胞透過処理剤および/または細胞崩壊剤、ならびに真核細胞を含む。また、本明細書に記載された他の試薬(バイサルファイトを非限定的に含む)を本発明の反応混合物中に含めることもできる。
【0116】
VIII.キット
本発明はまた、本発明のアクセス可能性アッセイを行うためのキットも提供する。キットは任意で、書面による使用説明書、または電子的な使用説明書(例えば、CD-ROMまたはDVD上)も含みうる。本発明のキットは、例えば、DNA修飾剤、ならびに細胞透過処理剤および/または細胞崩壊剤を含みうる。DNA修飾剤には、例えば、制限酵素、DNアーゼ、メチルトランスフェラーゼまたはDNA修飾性化学物質を含む、本明細書中に詳細に記載されたものが含まれうる。いくつかの態様において、DNA修飾剤は、MnlI、FokIおよびAlwIを非限定的に含むII-S型制限酵素である。本発明のキットは、透過処理剤およびDNA修飾剤を、同じバイアル/容器内(およびそれ故に同じ緩衝液中)に含む。または、透過処理剤およびDNA修飾剤が別々のバイアル/容器内にあってもよい。
【0117】
本発明のキットはまた、1つまたは複数の対照細胞および/または核酸も含みうる。例示的な対照核酸には、例えば、ある動物の本質的にすべての細胞においてアクセス可能である(例えば、ハウスキーピング遺伝子配列またはそのプロモーター)か、またはある動物のほとんどの細胞においてアクセス不能であるかのいずれかである遺伝子配列を含むものが含まれる。いくつかの態様において、本キットは、そのような遺伝子配列を増幅するためのプライマーの1つまたは複数のセットを(遺伝子配列または細胞がキットに実際に含まれるか否かにかかわらず)含む。例えば、いくつかの態様において、本キットは、DNA修飾剤ならびに細胞透過処理剤および/または細胞崩壊剤、ならびに対照DNA領域を増幅するための1つまたは複数のプライマーセット、ならびに任意で、第2のDNA領域、例えば標的DNA領域を増幅するための1つまたは複数のプライマーセットを含む。DNA修飾剤が制限酵素である態様においては、1つのDNA領域当たりに、制限酵素の潜在的な切断部位を少なくとも1つ(例えば、1つ、2つ、3つ、4つなど)含むDNA領域の一部分を増幅するための(例えば、修飾されていないコピーの数を計算するために有用な)一つのプライマーセット、および、潜在的な切断部位を含まないDNA領域の一部分を増幅するための(例えば、コピーの総数を計算するために有用な)少なくとも第2のプライマーセットという、少なくとも2つのプライマーセットを含めることができる。
【0118】
いくつかの態様において、本発明のキットは、以下のもののうち1つまたは複数を含む:
(i)細胞膜の透過処理剤または崩壊剤;
(ii)制限酵素、DNアーゼまたは他のDNA修飾剤;
(iii)修飾剤によるさらなる修飾を防止することのできる「停止」用溶液;
(iv)核酸の抽出および/または精製のための材料(例えば、ゲノムDNAの精製のため、および/または「停止」用溶液の成分などの非DNA成分の除去のためのスピンカラム);
(vi)DNAのPCR/qPCR増幅のための試薬、任意で、テンプレートおよび/またはポリメラーゼのほかにPCRまたはqPCRのために必要なすべての成分を含む1つの混合物;
(vii)特定の標的遺伝子のPCR/qPCR増幅のためのプライマーセット。
【実施例】
【0119】
以下の実施例は、特許請求する本発明を限定するためではなく、それを例示するために提供される。
【0120】
実施例1:一般的アプローチ
本アッセイは、DNA修飾剤の、細胞内のクロマチンの異なる部分に対するアクセス可能性の違いを利用する。図2に図示されているように、DNA修飾剤は、クロマチンのある部分に、他の部分よりも容易にアクセスすることができる。図3は、アッセイの例示的な作業の流れを図示している。付着細胞を出発材料として24ウェルプレート中で増殖させた。この態様においては、2つのウェルを各実験に対して用いる:一方のウェルはヌクレアーゼを含まない透過処理用緩衝液で処理する;もう一方のウェルはヌクレアーゼを含む透過処理用緩衝液で処理する。細胞からリアルタイムPCRに至るまでの過程全体には約3時間がかかり、その結果は細胞採取の当日に得ることができる。
【0121】
図4は、MnlIをクロマチン構造プローブとして用いた場合の、細胞から単離したDNAの分析に関する概略図を図示している。この場合におけるアッセイは、「ヌクレアーゼなし」の対照と同時並行して行う。2つのプライマーセットを各遺伝子に対して用いる。一方のプライマーセットはMnlI消化部位にまたがらないが、もう一方のプライマーセットは少なくとも1つのMnlI部位にまたがる。ヌクレアーゼを含まない透過処理用試薬で処理した細胞は、両方のプライマーセットから完全な増幅(すなわち、すべてのコピーが増幅される)をもたらし、それ故に図4の左のグラフに図示されているように同程度のCt値を有するはずである。デルタCt値(トータルCtから無傷Ctを差し引いたもの)は、プライマー特性に基づく、切断されなかったDNAにおけるこれらのプライマーセット間のCt値の差を反映している。
【0122】
対照的に、MnlIで処理した細胞では、アクセス可能な遺伝子はMnlI部位で切断されたはずである。MnlI部位にまたがらないプライマーセットは、依然としてすべてのDNA分子を増幅すると考えられ(コピーのトータル数の尺度として機能する)、従ってCtはDNAの全コピーの数量を反映する。(少なくとも1つのMnlI部位にまたがる)もう一方のプライマーセットは、切断されなかった配列、すなわち無傷の配列のみを増幅しうると考えられる。消化の程度に応じて、Ct値は推移し、消化の量に比例して大きくなると考えられる。続いて、トータルCtから無傷Ctを差し引くことによってデルタCtを算出することができ、もしDNAがアクセス可能であれば、これは負の数字をもたらすと考えられる。アクセス不能であるMnlI部位のパーセンテージは、図4に示された式によって算出される。
【0123】
さまざまな種類のDNA領域に対するこの手法の適用は、図5〜6に図示されている。図5は、GAPDH、HBBおよびGSTP1遺伝子の少なくとも一部を示している。MnlI、TATAボックス、プロモーター領域およびエクソンが示されている。GAPDHは陽性対照として用いた。GAPDHはほとんどの細胞株で発現されるハウスキーピング遺伝子であり、アクセス可能であるはずである。ヘモグロビンβ鎖遺伝子は陰性対照として選択した。ヘモグロビンはほとんどの細胞で発現されず、エピジェネティックにサイレント化されており、アクセス不能なクロマチン構造内にあると予想される。最後に、前立腺癌においてエピジェネティックに不活性化されるグルタチオン-s-トランスフェラーゼπすなわちGSTP1も分析した。図6は、増幅のための標的とすることのできる領域を示している。各遺伝子について、どのMnlI部位にもまたがらず、すべてのDNA鎖を増幅すると考えられるトータルプライマーのセットを用いる。少なくとも1つ(例えば、2つ〜4つ)のMnlI部位にまたがり、これらの部位で切断されないDNA鎖のみを増幅すると考えられる「無傷」プライマーのセットも用いる。
【0124】
実施例2:Hela細胞においてMnlIを用いたデータ
この実施例は、以上に概説した実験アプローチを用いて生成されたデータの結果を示している。
【0125】
材料および方法
化学物質 L-α-リゾホスファチジルコリン(リゾレシチン)はSigma-Aldrichから購入した。MnlI、DNアーゼI、BSAおよびプロテイナーゼKは、New England Biolabsから購入した。RNアーゼAはQiagenから購入した。組織培養プレートはVWRから購入した。iQ SYBRはBio-Radから入手した。
【0126】
細胞の処理 24ウェルプレート中で増殖させた細胞を、それらが集密度90%に達した時点で処理した。培地を吸引し、100ulの透過処理/消化用緩衝液を細胞の上から穏やかに重ねた。MnlIで処理した細胞に関して、透過処理/消化用緩衝液は、リゾレシチン、NaCl、Tris-HCl、MgCl2、DTT、BSAおよびMnlIからなっていた。DNアーゼIで処理した細胞に関して、透過処理/消化用緩衝液は、リゾレシチン、Tris-HCl、MgCl2、CaCl2およびDNアーゼIからなっていた。透過処理した細胞を、続いて37℃で1時間インキュベートした。インキュベーション後に、25ulの溶解/停止用溶液(100mM Tris-HCl pH 7.4、100mM NaCl、100mM EDTA、5% N-ラウロイルサルコシン(w/v)、80ug/mlのRNアーゼAおよび3mg/mlのプロテイナーゼK)を透過処理/消化用緩衝液に添加し、細胞溶解物を37℃で10分間インキュベートした。
【0127】
DNAの単離 細胞溶解物を収集し、市販の核酸精製キット(Aurum, Bio-Rad)を標準的な手順に従って用いてDNAを単離した。DNAを100ul中に溶出させ、DNAの濃度を、NanoDrop 1000分光光度計(Thermo Scientific)を用いて決定した。続いて試料を1μl当たり1〜10ngのDNAという濃度に希釈した。
【0128】
分析した遺伝子 本発明者らは、3つのヒト遺伝子のプロモーター領域を分析した:グリセルアルデヒド-3-リン酸デヒドロゲナーゼ(GAPDH)、ヘモグロビンβ(HBB)およびグルタチオン-s-トランスフェラーゼπ(GSTP1)。GAPDHはハウスキーピング遺伝子である;これはほとんどの細胞で発現され、アクセス可能なクロマチン構造を有すると予想される。HBBはほとんどの細胞で発現されず、アクセス不能なクロマチン構造を有すると予想される。GSTP1は前立腺癌においてエピジェネティックに不活性化されている。非癌性前立腺細胞株であるRWPE1ではGSTP1はアクセス可能なクロマチン構造内にあって、発現され、癌性前立腺細胞株LNCaPではGSTP1はアクセス不能なクロマチン構造内にあり、発現されない(Okino, S.T., et al., Chromatin changes on the GSTP1 promoter associated with its inactivation in prostate cancer. Mol Carcinog, 2007. 46(10): P. 839-46)。GSTP1 mRNAはHela細胞およびHCT15細胞株でも高発現される;PC3細胞株ではGSTP1 mRNAはHela細胞およびHCT15細胞で見出されるレベルのおよそ半分のレベルで発現される。
【0129】
リアルタイムPCR MnlIで処理した細胞から単離したDNA試料を、Chromo4連続蛍光検出器(MJ Research)を取り付けたPTC-200サーマルサイクラーにおいて、各プライマーセットを用いて分析した。各反応物は容積20ulとし、50ngのDNA、500nMの各プライマーおよび10ulのiQ SYBR green supermix(Bio-Rad)を含めた。PCR条件は、96℃ 10分間;96℃ 30秒間および68℃ 1分間を40サイクル;72℃ 5分間;0.2℃間隔で各段階で5秒間保つ72℃から98℃までの融解曲線、とした。DNアーゼIで処理した細胞から単離したDNA試料を、CFX96リアルタイムPCR検出システム(Bio-Rad)において分析した。各反応物は容積20ulとし、5ngのDNA、500nMの各プライマーおよび10ulのiTaq Fast SYBR green supermix with ROX(Bio-Rad)を含めた。PCR条件は、95℃ 5分間;95℃ 30秒間および72℃ 1分間を40サイクル;72℃ 5分間;0.2℃間隔で各段階で5秒間保つ72℃から95℃までの融解曲線、とした。
【0130】
データ分析 アクセス不能なクロマチンのレベルを、MnlIをヌクレアーゼプローブとして用いて決定するために、本発明者らは以下の計算を行った。ΔCt(切断なし)は、MnlIを含まない消化/透過処理用緩衝液で処理した細胞から単離したDNAを分析した場合の(無傷Ct−トータルCt)として算出した。ΔCt(切断あり)は、MnlIで処理した細胞から単離したDNAを分析した場合の(無傷Ct−トータルCt)として算出した。アクセス不能なクロマチンのレベルは、2∧(ΔCt(切断なし)−ΔCt(切断あり))によって算出した(図4参照)。
【0131】
DNアーゼIはアクセス可能なクロマチンにおいてトータルおよび無傷の増幅領域の両方の内部を切断するため、GAPDHおよびGSTP1に関するアクセス不能なクロマチンのレベルをDNアーゼIを用いて決定するために、本発明者らは、本発明者らがMnlIを分析する場合に用いたのと同じ計算を用いることはできなかった。本発明者らはHBBプロモーターがDNアーゼI消化に対する抵抗性が高いことを見いだし、本発明者らは、アクセス不能性クロマチンの低い消化レベルを反映するHBB ΔCtを内部参照標準として用いた(図16に図示されている)。ΔCt(参照)は、(HBBの+ヌクレアーゼCt−HBBのヌクレアーゼなしCt)として算出した。ΔCt(標的)は、GAPDH/GSTP1の+ヌクレアーゼCt−GAPDH/GSTP1のヌクレアーゼなしCtとして算出した。GAPDHまたはGSTP1標的遺伝子のアクセス不能なクロマチンのレベルは、2∧(ΔCt(参照)-ΔCt(標的))として算出した。
【0132】
結果
Hela細胞を、ヌクレアーゼを含まない消化用緩衝液、リゾレシチン(細胞透過処理用試薬)およびヌクレアーゼを含まない消化用緩衝液、またはリゾレシチンおよびヌクレアーゼ(MnlI[M]またはDNアーゼI[D])を含む完全な消化用緩衝液のいずれかで処理した。
【0133】
続いて、ゲノムDNA(gDNA)を細胞から単離し、アガロースゲル電気泳動に供した(図7参照)。このデータから、gDNAを消化するためにはリゾレシチンおよびヌクレアーゼが必要であることが示された。リゾレシチンまたはヌクレアーゼのいずれかを欠く反応物ではごくわずかなgDNA消化が検出された。このことは、リゾレシチンによって透過処理した細胞では、内因性ヌクレアーゼによるクロマチンの消化は有意でないこと、および消化用緩衝液中に存在するヌクレアーゼはリゾレシチンの非存在下では細胞内に入らないことを示している。
【0134】
その後に、MnlIで処理した細胞から単離したDNA試料を、図6に示したプライマーセットを用いるリアルタイムPCRによって分析した。図8に示されているように、陰性対照反応ではヘモグロビンβ鎖遺伝子(これはHela細胞ではサイレント化している)を分析した。トータルプライマーのセットおよび無傷プライマーのセットに関して、Ct値はほぼ同じであった。このことは、この遺伝子領域がHela細胞においてアクセス不能なクロマチン構造内にあったことを示している。
【0135】
図9は、陽性対照であるGAPDHに関する結果を示している。GAPDHに関するCt値は、陰性対照においてHBBに関して得られたCt値とほぼ同じであったが、「無傷」プライマーセットを用いた増幅は右方に移動した。このことは、「トータル」DNAよりも無傷が少ないことを示している。このことは、MnlIがGAPDHプロモーター内の自身の認識部位にアクセスしてそれを消化したことを意味する。このため本発明者らは、GAPDHプロモーターはHela細胞においてアクセス可能なクロマチン内にあったと推論している。
【0136】
図10は、GSTπの分析による結果を示している。GSTπは前立腺癌では不活性化されており、Hela細胞では発現されている。完全な反応物のCt値は、無傷プライマーセットの反応物では右方に移動したが、トータルプライマーセットの反応物ではそうではなかった。このことは、Hela細胞ではGSTπがアクセス可能なクロマチン構造内にあることを示している。
【0137】
図11は、上記のデータを表にまとめたものである。算出された値は、「トータル」プライマーセットによって決定されたコピーの総数と比較した、無傷(それ故にヌクレアーゼに対してアクセス不能性)であったコピーのパーセンテージである。
【0138】
実施例3:前立腺癌細胞においてMnlIを用いたデータ
上記の方法を、アクセス可能性がGSTP1の発現と関係しているか否かを判断するために種々の細胞種に対しても適用した。非癌性ヒト前立腺に由来し、GSTP1が高発現されるRWPE-1細胞;および癌性ヒト前立腺に由来し、GSTP1がエピジェネティックな修飾によってサイレント化しているLNCaP細胞という、2つの細胞型を用いた。
【0139】
これらの細胞の分析方法は、本質的には上記の通りとした。図12は、対照反応(すなわち、ヘモグロビン遺伝子について)を示している。いずれの細胞株においても、酵素を含まなかった試料におけるデルタCtは、酵素を含むものとほぼ同じである。すなわち、デルタCtは約0であった。このことはこの遺伝子が両方の細胞株においてアクセス不能であったことを示している。
【0140】
図13は、GAPDH遺伝子を分析した陽性対照反応の結果を示している。酵素を伴わない反応に関して、トータルおよび無傷Ctはほぼ同じであった。しかし、酵素を含む反応については、無傷Ctは右方に移動した。このことは無傷のDNAがより少なく、それ故にGAPDH遺伝子がアクセス可能なクロマチン構造内にあったことを示している。
【0141】
図14は、GSTπ遺伝子の分析によるデータを示している。非癌性RWPE-1細胞株では、無傷Ctが酵素の存在下で右方に移動した。対照的に、癌性LNCaP細胞株では、無傷Ctは検知しうる程度には変化しなかった。このことは、GSTP1がRWPE-1細胞ではアクセス可能なクロマチン内にあり、LNCaP細胞ではアクセス不能なクロマチン内にあったことを示している。
【0142】
これらの実験を4回繰り返した。結果の概要は図15に示されており、この場合もアクセス不能であった(または「ロックされた(locked)」)コピーのパーセンテージを算出している。
【0143】
実施例4:DNアーゼIを用いたデータ
この実施例は、MnlIではなくDNアーゼIをヌクレアーゼとした方法の使用によるデータを示す。この実施例において、本発明者らは、4つのヒト癌細胞株:Hela(子宮頸癌)、PC3(前立腺癌)、LNCaP(前立腺癌)およびHCT15(結腸癌)を分析した。本発明者らはまた、本発明者らがMnlIの例で分析したのと同じ遺伝子も分析した:HBB、GAPDHおよびGSTP1。
【0144】
DNアーゼIはDNAを定まった部位では切断しないため、本発明者らは、本発明者らがMnlIをクロマチン構造プローブとして用いた場合と同じデータ分析戦略を用いることはできなかった。その代わりに、本発明者らは、エピジェネティックにサイレント化される参照遺伝子(HBB)を、アクセス不能性クロマチンの消化の程度を反映する内部対照として利用した。この戦略を用いると、特定の標的遺伝子のクロマチン構造を、図16に詳細に示した通りに決定することができる。図17は、分析した遺伝子、およびクロマチン構造を評価するために用いたプライマーの位置の概略描写である。
【0145】
図18に示されているように、DNアーゼIはすべての細胞株において、HBBを検知しうる程度には切断しなかった(ヌクレアーゼなしの曲線と+ヌクレアーゼの曲線はほぼ重なる)。この結果は、HBBがすべての細胞株でアクセス不能なクロマチン立体配置内にあることを示しており、参照遺伝子対照としてのその有用性を実証している。HBB mRNAはこれらの細胞株のいずれにおいても発現されないため(非提示データ)、これらの結果は予想されたことでもある。
【0146】
GAPDH遺伝子に関する本発明者らの分析を図19に示している。すべての細胞株において、本発明者らは、+ヌクレアーゼの曲線がヌクレアーゼなしの曲線に対して有意に右方移動していることを確認している。このことは、GAPDHがすべての細胞株においてアクセス可能なクロマチン構造内にあることを意味する。これらの結果は、GAPDHがすべてではないにしてもほとんどの細胞株で発現される(非提示データ)「ハウスキーピング」遺伝子であることから、予想されたことである。
【0147】
GSTP1 mRNAは、分析した細胞株においてさまざまなレベルで発現される。Hela細胞およびHCT15細胞は多くの量のGSTP1 mRNAを発現し、PC3細胞は中程度の量(Hela細胞およびHCT15細胞で見出される量の約半分)を発現し、LNCaP細胞はGSTP1 mRNAを検知しうる程度では発現しない(非提示データ)。GSTP1クロマチン構造に関する本発明者らの分析を図20に示している。Hela細胞およびHCT15細胞では、本発明者らは+ヌクレアーゼの曲線がヌクレアーゼなしの曲線に対して有意に右方移動していることを確認しており、このことはGSTP1がアクセス可能なクロマチン構造内にあることを意味する。PC3細胞では、本発明者らは+ヌクレアーゼの曲線がヌクレアーゼなしの曲線に対して中程度に右方移動していることを確認しており、このことはGSTP1が部分的にアクセス可能なクロマチン構造内にあることを意味する。最後に、LNCaP細胞では、本発明者らは+ヌクレアーゼの曲線およびヌクレアーゼなしの曲線がほぼ重なることを確認している。このことはGSTP1がLNCaP細胞においてアクセス不能なクロマチン立体配置内にあることを意味する。重要なこととして、本発明者らは、分析したすべての細胞株において、GSTP1クロマチン構造とGSTP1 mRNA発現との間に優れた相関関係を確認している。
【0148】
図21は、結果の概要を示している。アクセス不能なDNAのパーセンテージを図16に記載した通りに算出した。それを「ロックされている」クロマチンとして示している。GAPDHおよびGSTP1のRNA発現の相対的レベルは、種々の細胞株から単離したmRNAをqRT-PCRにより標準的な手順によって分析することによって算出した。それらの結果は、標的遺伝子のクロマチン構造が標的遺伝子の発現レベルと良く相関していることを示している。このことは、記載したアッセイが、遺伝子発現に対するエピジェネティックな影響を評価するための有用なツールであることを意味する。
【0149】
概要
本発明者らは、培養細胞におけるクロマチン構造を評価することのできる迅速かつ高感度なアッセイを開発した。本発明者らのアッセイには、他のクロマチン構造アッセイを上回るいくつかの利点がある(Okino, S.T., et al., Mol Carcinog 46(10):839-46 (2007);Okino, S.T. and J.P. Whitlock, Jr., Mol Cell Biol 15(7): p. 3714-21 (1995);Rao, S., E. Procko, and M.F. Shannon, Jimmunol 167(8):4494-503 (2001);Kilgore, J.A., et al., Methods 41(3):320-32 (2007))。最も重要なこととして、本発明者らの手順では、本発明者らは、透過処理した細胞において、クロマチンを、1つの段階でインサイチューで消化する。他の手順は核の単離を必要とし、それはより多くの手間のかかる作業、より多くの時間、および出発材料としてより多くの細胞を必要とする。加えて、本発明者らのプライマーセットの設計は、クロマチンのアクセス可能性に関してより正確で、内部対照を有する評価をもたらす。
【0150】
実施例5:DNAメチル化を評価するための変法
本発明のもう1つの用途は、アクセス不能なクロマチン領域のDNAメチル化状態を決定することである。DNAメチル化とは、5-メチルシトシンを生成させる、シトシンの5位での天然のメチル化のことを指す。DNAメチル化は、遺伝子活性およびクロマチン構造の調節と関連づけられている;異常なDNAメチル化パターンは、ヒトの病態および特定の罹患状態と関連性がある。この変法では、前述の手順を、PCR増幅の段階を含まないが、その手前まで行う。続いて、精製されたゲノムDNAを、メチル化されていないシトシンがウラシルに変換され、5-メチルシトシンは変化しないように、バイサルファイトによって標準的な手順を用いて改変する。続いて、特定のDNA領域を標的とするプライマーセットを用いて、ヌクレアーゼで処理した細胞から精製された、バイサルファイトで改変されたDNAを増幅する。こうすることにより、元々アクセス不能なクロマチン立体配置内にあり、それ故にDNA修飾剤によって切断されなかったDNAを増幅することができる。続いて、増幅されたDNAを、融解曲線分析、高分解能融解分析、バイサルファイト-DNA配列決定、デジタルPCR、メチル化特異的PCR、ピロシークエンシングなどの標準的な手順によって、DNAメチル化に関して分析する。このような分析は、アクセス不能なクロマチン領域のDNAメチル化状態を決定することができ、エピジェネティックな遺伝子不活性化と関連性のあるDNAメチル化の特定のパターンに関する有意義な情報を提供することができる。
【0151】
本明細書に記載された例および態様が例示のみを目的としていること、ならびにそれらを考慮してさまざまな改変および変更が当業者には想起されると考えられ、それらが本出願の趣旨および範囲ならびに添付の特許請求の範囲に含まれることは、理解されるであろう。本明細書中に引用された刊行物、特許および特許出願はすべて、それらの全体が、あらゆる目的で参照により組み入れられる。
【特許請求の範囲】
【請求項1】
以下の段階を含む、染色体DNAを分析するための方法:
a.以下を同時に行う段階:
i.細胞の細胞膜を透過処理するかまたは崩壊させる段階;および
ii.該細胞を、DNA切断剤またはDNA修飾剤と、該剤が該細胞内のゲノムDNAを切断または修飾するような条件下で接触させる段階であって、ゲノムDNAの異なる領域が該剤によって異なる程度で切断または修飾され、それにより、切断されたDNA領域および無傷のDNA領域、または修飾されたDNA領域および修飾されていないDNA領域が生成される、段階;ならびに
b.少なくとも1つの、無傷の、もしくは修飾されていない、もしくは修飾されたDNA領域の量を検出するか、または少なくとも1つの、無傷の、もしくは修飾されていない、もしくは修飾されたDNA領域のクローニング、単離、もしくはヌクレオチド配列決定を行う段階。
【請求項2】
少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域の前記量を、前記細胞における該DNA領域のクロマチン構造と関連づける段階を含む、請求項1記載の方法。
【請求項3】
少なくとも第1の無傷の、または修飾されていない、または修飾された染色体DNA領域、および第2の無傷の、または修飾されていない、または修飾された染色体DNA領域の量を、検出する段階;ならびに
第1および第2のDNA領域の該量を比較する段階
を含む、請求項1記載の方法。
【請求項4】
第1の染色体DNA領域および第2の染色体DNA領域の無傷のコピーの数を定量し、それにより、DNA切断剤に対する第1および第2のDNA領域の相対的なアクセス可能性(accessibility)を評価する段階を含む、請求項1記載の方法。
【請求項5】
接触させる段階の後でかつ検出する段階の前に、前記ゲノムDNAを単離する、請求項1記載の方法。
【請求項6】
前記細胞を人工の培養表面に直接的または間接的に付着させながら、前記細胞を透過処理して、DNA切断剤またはDNA修飾剤と接触させる、請求項1記載の方法。
【請求項7】
透過処理する段階が、前記細胞を、細胞膜を透過性にする剤と接触させることを含む、請求項1記載の方法。
【請求項8】
細胞膜を透過性にする前記剤がリゾ脂質(lysolipid)である、請求項7記載の方法。
【請求項9】
リゾ脂質がリゾホスファチジルコリンである、請求項8記載の方法。
【請求項10】
透過処理するかまたは崩壊させる段階が、非イオン性界面活性剤を用いて細胞膜を崩壊させることを含む、請求項1記載の方法。
【請求項11】
前記非イオン性界面活性剤が、NP40、Tween20およびTriton X-100の群より選択される、請求項10記載の方法。
【請求項12】
前記細胞をDNA切断剤と接触させ、かつ該DNA切断剤が酵素である、請求項1記載の方法。
【請求項13】
前記細胞をDNA修飾剤と接触させ、かつ該DNA修飾剤がメチルトランスフェラーゼおよびDNA修飾性化学物質からなる群より選択される、請求項1記載の方法。
【請求項14】
DNA切断剤がDNA切断酵素であり;
ゲノムDNA中の修飾部が、DNAが切断されている部位であり;かつ
検出する段階が、切断後のDNA領域の無傷のコピーの量を定量することを含む、請求項12記載の方法。
【請求項15】
検出する段階が定量的増幅を含む、請求項14記載の方法。
【請求項16】
定量的増幅が、定量的ポリメラーゼ連鎖反応および定量的ライゲーション媒介ポリメラーゼ連鎖反応からなる群より選択される、請求項15記載の方法。
【請求項17】
DNA切断剤がDNアーゼおよび制限酵素から選択される、請求項12記載の方法。
【請求項18】
DNA修飾剤がメチルトランスフェラーゼであり;
修飾部が、細胞のネイティブなメチル化酵素によってはメチル化されないヌクレオチド配列中のヌクレオチド上のメチル基であり;かつ
検出する段階が、該修飾部の有無を検出する方法を含む、請求項13記載の方法。
【請求項19】
メチルトランスフェラーゼがDAMメチルトランスフェラーゼであり、かつ定量する段階が、修飾されたDNAを、DAMメチルトランスフェラーゼによってメチル化されたDNA配列を認識する制限酵素を用いて切断することを含む、請求項18記載の方法。
【請求項20】
メチルトランスフェラーゼがDNA中のシトシンにメチル部分を付加し、かつ検出する段階が、修飾されたDNAをメチル化特異的制限酵素を用いて切断すること、および/または修飾されたDNAをバイサルファイト(bisulfite)で処理することを含む、請求項18記載の方法。
【請求項21】
DNA修飾剤が、クロマチン構造に対して立体障害を有する分子であり、該分子がDNA修飾性化学物質と連結されている、請求項1記載の方法。
【請求項22】
DNA修飾性化学物質が、硫酸ジメチルおよびヒドラジンからなる群より選択される、請求項21記載の方法。
【請求項23】
検出する段階が、標的DNA領域の少なくとも一部分および対照DNA領域の少なくとも一部分を定量することを含み、対照DNA領域が、
i.ある動物の本質的にすべての細胞においてアクセス可能であるか;または
ii.ある動物のほとんどの細胞においてアクセス不能であるかの
いずれかである配列を含む、請求項1記載の方法。
【請求項24】
対照DNA領域が、
DNA修飾剤の潜在的な修飾部位を含まない対照DNA領域の一部分の増幅をプライミングするプライマー;および
DNA修飾剤の潜在的な修飾部位を少なくとも1つ含む対照DNA領域の一部分の増幅をプライミングするプライマー
のそれぞれを用いて定量的に増幅される、請求項1記載の方法。
【請求項25】
前記DNA領域のDNAメチル化を決定する、請求項1記載の方法。
【請求項26】
段階a.が、前記DNAをDNA切断剤と接触させることを含み、かつ段階b.が、前記無傷のDNAをバイサルファイトと接触させることを含む、請求項25記載の方法。
【請求項27】
バイサルファイトで処理したDNAの融解温度を決定し、該融解温度をDNAメチル化の有無または程度と関連づける段階をさらに含む、請求項26記載の方法。
【請求項28】
段階bが、
無傷のDNA領域;または
修飾されたDNA領域;または
修飾されていないDNA領域
のライブラリーを調製することを含む、請求項1記載の方法。
【請求項29】
段階bが、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域を、ポリヌクレオチドのライブラリーと、該DNA領域と該ライブラリーの1つまたは複数のメンバーとのハイブリダイゼーションを可能にする条件下で接触させること、および該DNA領域と該1つまたは複数のメンバーとのハイブリダイゼーションを検出することを含む、請求項1記載の方法。
【請求項30】
前記ライブラリーがマイクロアレイ上に構成されている、請求項29記載の方法。
【請求項31】
段階bが、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域を増幅することを含む、請求項1記載の方法。
【請求項32】
段階bが、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域のヌクレオチド配列決定を行うことを含む、請求項1記載の方法。
【請求項33】
以下を含む、DNA修飾剤に対する染色体上の遺伝子座のアクセス可能性を決定するためのキット:
細胞膜の透過処理剤または崩壊剤;
制限酵素またはDNアーゼ。
【請求項34】
細胞膜の透過処理剤または崩壊剤と、制限酵素またはDNアーゼとが、同じ容器内で同じ緩衝液中にある、請求項33記載のキット。
【請求項35】
細胞膜の透過処理剤または崩壊剤と、制限酵素またはDNアーゼとが、別々の容器内にある、請求項33記載のキット。
【請求項36】
バイサルファイトをさらに含む、請求項33記載のキット。
【請求項37】
真核生物のゲノムDNAのある領域の増幅のためのプライマー対をさらに含む、請求項33記載のキット。
【請求項38】
a.DNA修飾剤の潜在的な修飾部位を含まない対照DNA領域の一部分の増幅をプライミングするプライマー;および/または
b.DNA修飾剤の潜在的な修飾部位を少なくとも1つ含む対照DNA領域の一部分の増幅をプライミングするプライマー
を含む、請求項33記載のキット。
【請求項39】
a.修飾剤に対してアクセス不能である対照DNA領域の一部分の増幅をプライミングするプライマー;および/または
b.修飾剤に対してアクセス可能である対照DNA領域の一部分の増幅をプライミングするプライマー
を含む、請求項33記載のキット。
【請求項1】
以下の段階を含む、染色体DNAを分析するための方法:
a.以下を同時に行う段階:
i.細胞の細胞膜を透過処理するかまたは崩壊させる段階;および
ii.該細胞を、DNA切断剤またはDNA修飾剤と、該剤が該細胞内のゲノムDNAを切断または修飾するような条件下で接触させる段階であって、ゲノムDNAの異なる領域が該剤によって異なる程度で切断または修飾され、それにより、切断されたDNA領域および無傷のDNA領域、または修飾されたDNA領域および修飾されていないDNA領域が生成される、段階;ならびに
b.少なくとも1つの、無傷の、もしくは修飾されていない、もしくは修飾されたDNA領域の量を検出するか、または少なくとも1つの、無傷の、もしくは修飾されていない、もしくは修飾されたDNA領域のクローニング、単離、もしくはヌクレオチド配列決定を行う段階。
【請求項2】
少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域の前記量を、前記細胞における該DNA領域のクロマチン構造と関連づける段階を含む、請求項1記載の方法。
【請求項3】
少なくとも第1の無傷の、または修飾されていない、または修飾された染色体DNA領域、および第2の無傷の、または修飾されていない、または修飾された染色体DNA領域の量を、検出する段階;ならびに
第1および第2のDNA領域の該量を比較する段階
を含む、請求項1記載の方法。
【請求項4】
第1の染色体DNA領域および第2の染色体DNA領域の無傷のコピーの数を定量し、それにより、DNA切断剤に対する第1および第2のDNA領域の相対的なアクセス可能性(accessibility)を評価する段階を含む、請求項1記載の方法。
【請求項5】
接触させる段階の後でかつ検出する段階の前に、前記ゲノムDNAを単離する、請求項1記載の方法。
【請求項6】
前記細胞を人工の培養表面に直接的または間接的に付着させながら、前記細胞を透過処理して、DNA切断剤またはDNA修飾剤と接触させる、請求項1記載の方法。
【請求項7】
透過処理する段階が、前記細胞を、細胞膜を透過性にする剤と接触させることを含む、請求項1記載の方法。
【請求項8】
細胞膜を透過性にする前記剤がリゾ脂質(lysolipid)である、請求項7記載の方法。
【請求項9】
リゾ脂質がリゾホスファチジルコリンである、請求項8記載の方法。
【請求項10】
透過処理するかまたは崩壊させる段階が、非イオン性界面活性剤を用いて細胞膜を崩壊させることを含む、請求項1記載の方法。
【請求項11】
前記非イオン性界面活性剤が、NP40、Tween20およびTriton X-100の群より選択される、請求項10記載の方法。
【請求項12】
前記細胞をDNA切断剤と接触させ、かつ該DNA切断剤が酵素である、請求項1記載の方法。
【請求項13】
前記細胞をDNA修飾剤と接触させ、かつ該DNA修飾剤がメチルトランスフェラーゼおよびDNA修飾性化学物質からなる群より選択される、請求項1記載の方法。
【請求項14】
DNA切断剤がDNA切断酵素であり;
ゲノムDNA中の修飾部が、DNAが切断されている部位であり;かつ
検出する段階が、切断後のDNA領域の無傷のコピーの量を定量することを含む、請求項12記載の方法。
【請求項15】
検出する段階が定量的増幅を含む、請求項14記載の方法。
【請求項16】
定量的増幅が、定量的ポリメラーゼ連鎖反応および定量的ライゲーション媒介ポリメラーゼ連鎖反応からなる群より選択される、請求項15記載の方法。
【請求項17】
DNA切断剤がDNアーゼおよび制限酵素から選択される、請求項12記載の方法。
【請求項18】
DNA修飾剤がメチルトランスフェラーゼであり;
修飾部が、細胞のネイティブなメチル化酵素によってはメチル化されないヌクレオチド配列中のヌクレオチド上のメチル基であり;かつ
検出する段階が、該修飾部の有無を検出する方法を含む、請求項13記載の方法。
【請求項19】
メチルトランスフェラーゼがDAMメチルトランスフェラーゼであり、かつ定量する段階が、修飾されたDNAを、DAMメチルトランスフェラーゼによってメチル化されたDNA配列を認識する制限酵素を用いて切断することを含む、請求項18記載の方法。
【請求項20】
メチルトランスフェラーゼがDNA中のシトシンにメチル部分を付加し、かつ検出する段階が、修飾されたDNAをメチル化特異的制限酵素を用いて切断すること、および/または修飾されたDNAをバイサルファイト(bisulfite)で処理することを含む、請求項18記載の方法。
【請求項21】
DNA修飾剤が、クロマチン構造に対して立体障害を有する分子であり、該分子がDNA修飾性化学物質と連結されている、請求項1記載の方法。
【請求項22】
DNA修飾性化学物質が、硫酸ジメチルおよびヒドラジンからなる群より選択される、請求項21記載の方法。
【請求項23】
検出する段階が、標的DNA領域の少なくとも一部分および対照DNA領域の少なくとも一部分を定量することを含み、対照DNA領域が、
i.ある動物の本質的にすべての細胞においてアクセス可能であるか;または
ii.ある動物のほとんどの細胞においてアクセス不能であるかの
いずれかである配列を含む、請求項1記載の方法。
【請求項24】
対照DNA領域が、
DNA修飾剤の潜在的な修飾部位を含まない対照DNA領域の一部分の増幅をプライミングするプライマー;および
DNA修飾剤の潜在的な修飾部位を少なくとも1つ含む対照DNA領域の一部分の増幅をプライミングするプライマー
のそれぞれを用いて定量的に増幅される、請求項1記載の方法。
【請求項25】
前記DNA領域のDNAメチル化を決定する、請求項1記載の方法。
【請求項26】
段階a.が、前記DNAをDNA切断剤と接触させることを含み、かつ段階b.が、前記無傷のDNAをバイサルファイトと接触させることを含む、請求項25記載の方法。
【請求項27】
バイサルファイトで処理したDNAの融解温度を決定し、該融解温度をDNAメチル化の有無または程度と関連づける段階をさらに含む、請求項26記載の方法。
【請求項28】
段階bが、
無傷のDNA領域;または
修飾されたDNA領域;または
修飾されていないDNA領域
のライブラリーを調製することを含む、請求項1記載の方法。
【請求項29】
段階bが、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域を、ポリヌクレオチドのライブラリーと、該DNA領域と該ライブラリーの1つまたは複数のメンバーとのハイブリダイゼーションを可能にする条件下で接触させること、および該DNA領域と該1つまたは複数のメンバーとのハイブリダイゼーションを検出することを含む、請求項1記載の方法。
【請求項30】
前記ライブラリーがマイクロアレイ上に構成されている、請求項29記載の方法。
【請求項31】
段階bが、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域を増幅することを含む、請求項1記載の方法。
【請求項32】
段階bが、少なくとも1つの、無傷の、または修飾されていない、または修飾されたDNA領域のヌクレオチド配列決定を行うことを含む、請求項1記載の方法。
【請求項33】
以下を含む、DNA修飾剤に対する染色体上の遺伝子座のアクセス可能性を決定するためのキット:
細胞膜の透過処理剤または崩壊剤;
制限酵素またはDNアーゼ。
【請求項34】
細胞膜の透過処理剤または崩壊剤と、制限酵素またはDNアーゼとが、同じ容器内で同じ緩衝液中にある、請求項33記載のキット。
【請求項35】
細胞膜の透過処理剤または崩壊剤と、制限酵素またはDNアーゼとが、別々の容器内にある、請求項33記載のキット。
【請求項36】
バイサルファイトをさらに含む、請求項33記載のキット。
【請求項37】
真核生物のゲノムDNAのある領域の増幅のためのプライマー対をさらに含む、請求項33記載のキット。
【請求項38】
a.DNA修飾剤の潜在的な修飾部位を含まない対照DNA領域の一部分の増幅をプライミングするプライマー;および/または
b.DNA修飾剤の潜在的な修飾部位を少なくとも1つ含む対照DNA領域の一部分の増幅をプライミングするプライマー
を含む、請求項33記載のキット。
【請求項39】
a.修飾剤に対してアクセス不能である対照DNA領域の一部分の増幅をプライミングするプライマー;および/または
b.修飾剤に対してアクセス可能である対照DNA領域の一部分の増幅をプライミングするプライマー
を含む、請求項33記載のキット。
【図1】

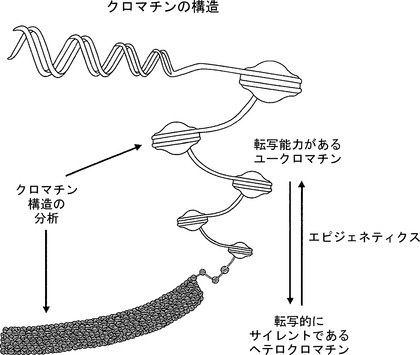
【図2】
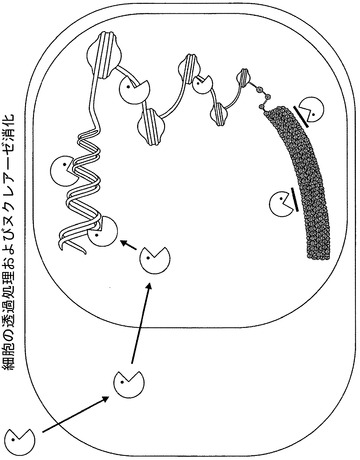
【図3】
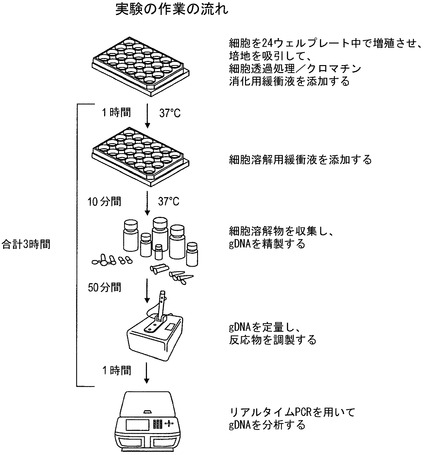
【図4】
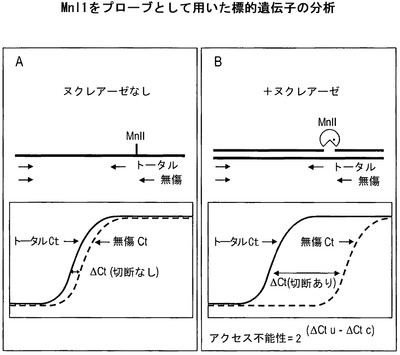
【図5】
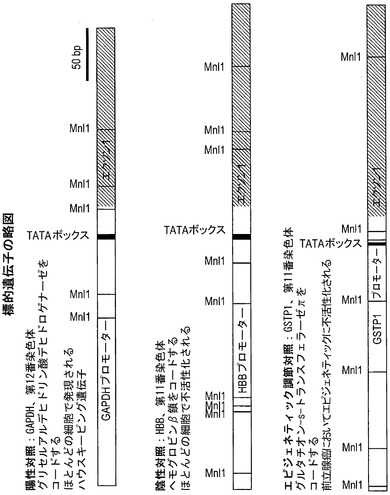
【図6】
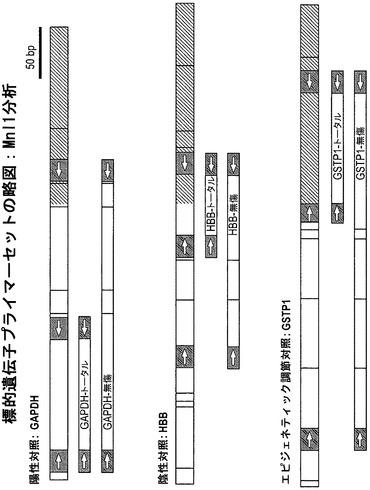
【図7】
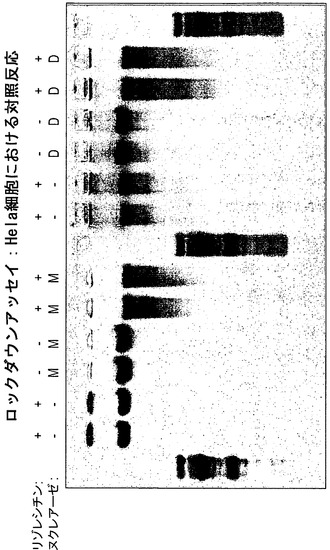
【図8】
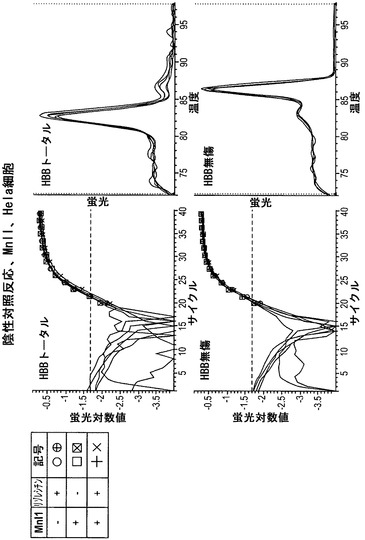
【図9】
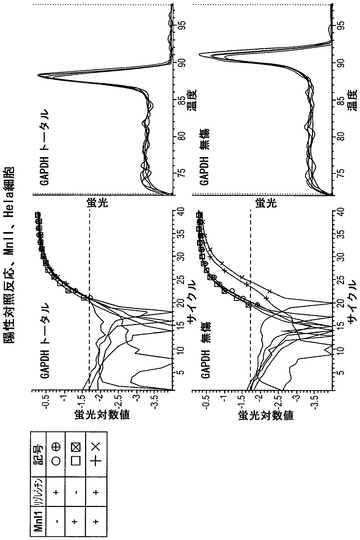
【図10】
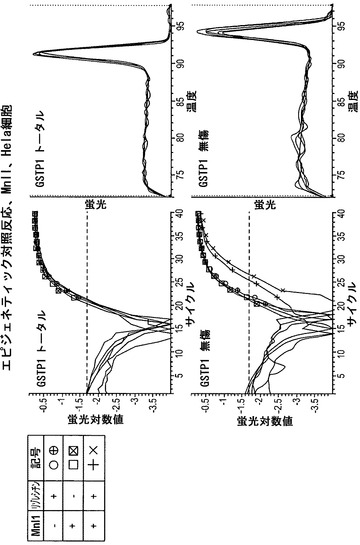
【図11】

【図12】
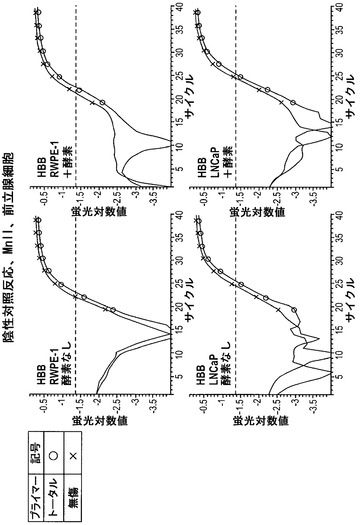
【図13】
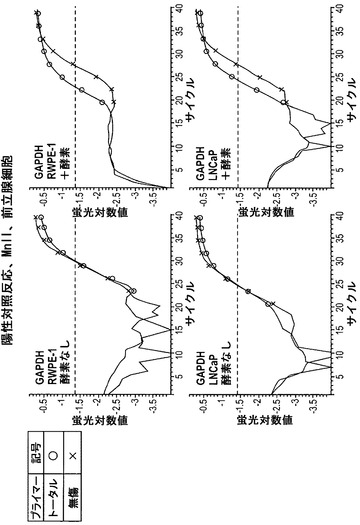
【図14】
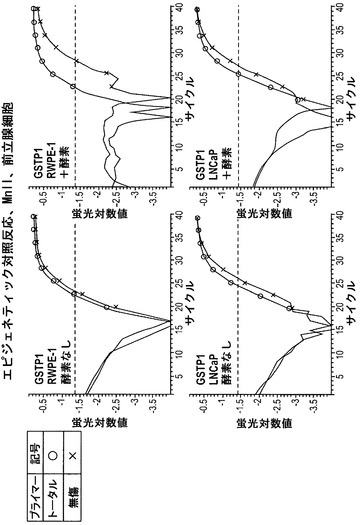
【図15】

【図16】
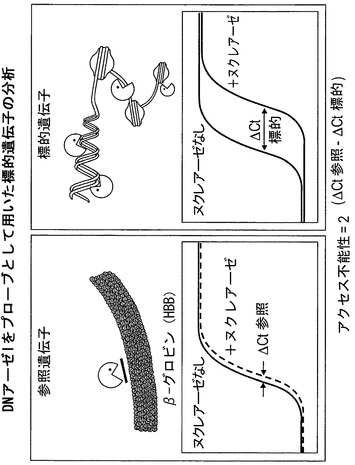
【図17】

【図18】
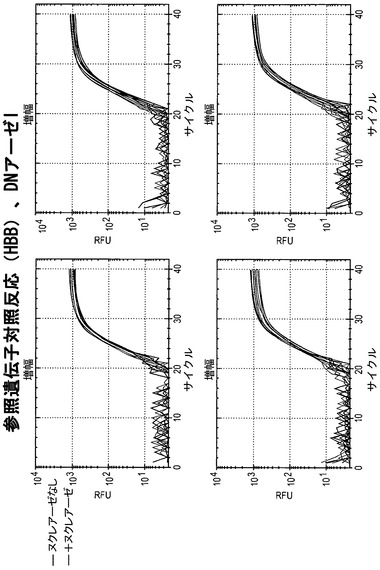
【図19】
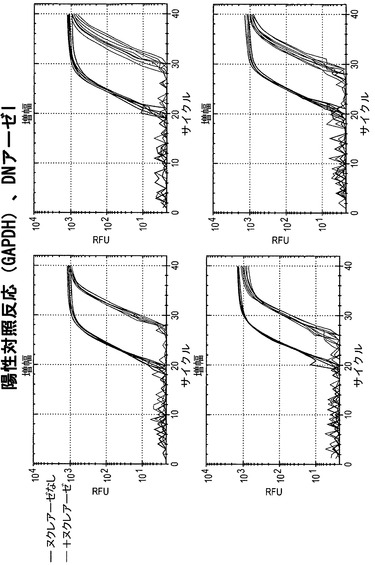
【図20】
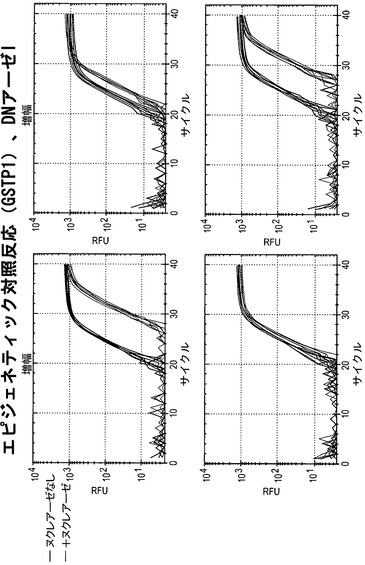
【図21】
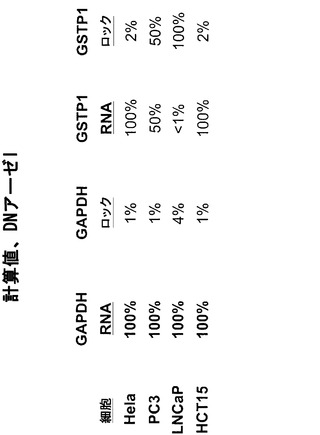
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【公表番号】特表2012−510294(P2012−510294A)
【公表日】平成24年5月10日(2012.5.10)
【国際特許分類】
【出願番号】特願2011−539555(P2011−539555)
【出願日】平成21年11月13日(2009.11.13)
【国際出願番号】PCT/US2009/064425
【国際公開番号】WO2010/065266
【国際公開日】平成22年6月10日(2010.6.10)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.イーサネット
2.UNIX
3.Linux
4.WINDOWS
5.JAVA
6.JAVASCRIPT
【出願人】(507190880)バイオ−ラッド ラボラトリーズ インコーポレーティッド (25)
【Fターム(参考)】
【公表日】平成24年5月10日(2012.5.10)
【国際特許分類】
【出願日】平成21年11月13日(2009.11.13)
【国際出願番号】PCT/US2009/064425
【国際公開番号】WO2010/065266
【国際公開日】平成22年6月10日(2010.6.10)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.イーサネット
2.UNIX
3.Linux
4.WINDOWS
5.JAVA
6.JAVASCRIPT
【出願人】(507190880)バイオ−ラッド ラボラトリーズ インコーポレーティッド (25)
【Fターム(参考)】
[ Back to top ]