
クローン原性内皮前駆細胞の単離、増殖および使用
内皮性コロニー形成細胞(EPC)の階層は、哺乳動物の臍帯血、臍静脈、および大動脈から同定された。高増殖能内皮性コロニー形成細胞(HPP−ECFC)と名付けられた新規に単離された細胞が、単離され、そして特徴付けられた。臍帯血に由来する内皮細胞か、またはHUVECおよびHAECに由来する内皮細胞の増殖能およびクローン原性能を試験する単一細胞アッセイが開発された。EPCは、脈管壁中に常在することが判明している。ヒト臍帯血に由来する高増殖能内皮性コロニー形成細胞(HPP−ECPCS)に由来する細胞の支持細胞層の使用は、再増殖性の造血幹細胞および造血前駆細胞の成長および生存を刺激する。成長および生存の刺激は、インビトロ培養における前駆細胞の数の増加、およびNOD/SCID免疫不全マウス移植系におけるヒト細胞移植のレベルの増加によって決定される。

【発明の詳細な説明】
【背景技術】
【0001】
(発明の背景)
内皮細胞および造血細胞を生じる幹細胞分化に関するモデルは、幹細胞およびその子孫の臨床的価値のため、関心のあるものである。幹細胞および前駆細胞の特徴は、それらの増殖する能力および機能的子孫を生じる能力であり、そして前駆細胞は、そのクローン原性能によって同定される。以前に使用されていた方法は、前駆体を同定するために、一種類の内皮細胞が単離および特徴付けされていたことを保証しない。
【0002】
ほとんどの哺乳動物において、正常な、成熟動脈、静脈、および毛細管におけるインビボでの内皮細胞増殖は、存在していたとしても、非常に遅いことが報告されている。いくつかの実験動物(例えば、ブタおよびイヌ)において、放射標識研究は、0.6%〜3.0%の内皮細胞が、特定の脈管の限局された範囲に限られた分裂細胞によって毎日ターンオーバーすることを実証した。これらの分裂内皮細胞が独特であり、そして他の成熟した内皮に欠けている増殖能を有するかどうかは、未確定のままである。
【0003】
好対照なことに、インビトロにおいて、ヒト脈管由来または動物脈管由来である内皮細胞のプレーティングは、最初に、活発な内皮細胞増殖を伴う。例えば、臍静脈内皮細胞(HUVEC)およびウシ大動脈内皮細胞(BAEC)は、内皮細胞機能のインビトロ分析のための、2つの一般に研究されるモデルである。HUVEC細胞およびBAEC細胞の両方は、培養の初期においては良く増殖するが、経時的に細胞分裂は弱々しくなり、そして細胞は老化して15継代〜20継代の後、分裂できなくなる。脈管由来である各々の内皮細胞が同様の増殖能を有するのか否か、または細胞の一部のみが増殖し得るのか否かは、不明である。
【0004】
脈管新生(新脈管形成)は、既存の脈管からの新規の脈管の形成のプロセスである;これは、成体被験体において、新規の脈管を生じると報告されているプロセスである。最近、骨髄由来循環内皮前駆細胞(EPC)が記載され、そして、これらの細胞はまた、少なくともいくつかの実験的マウスの虚血モデルまたは腫瘍モデルにおいて、新規の脈管の形成に役割を果たすことが報告されている。相反する証拠は、発生の間の骨髄由来(EPC)が正常動脈、静脈、および毛細管の内皮の内張り(lining)に寄与せず、そして新脈管形成において軽微な役割のみを果たすことを示す。循環EPCと正常脈管に存在する増殖能を有する上記内皮細胞との間の関係は、不明である。
【0005】
脈管新生治療のためのEPCの使用、または患者の心臓血管疾患の危険および進行を評価するためのバイオマーカーとしてのEPCの使用を支持する新たな証拠は、蓄積し、そして興味をかきたてている。しかしながら、EPCの一様な定義は無く、このことは、これらの研究の解釈を問題のあるものとし、そして臨床使用のために適切な細胞型の再現を妨げる。(例えば、造血、腸、神経細胞の)幹細胞および前駆細胞の特徴は、それらの増殖する能力、および機能的な子孫を生じる能力であるが、EPCは、選択された細胞表面抗原の発現によって第一に規定される。分子の細胞表面発現に唯一依存することは、その発現が上記細胞の生理学的状態によって変化し得るので、問題となり得る。個々の内皮細胞またはEPCにおける増殖能(内因性応答)を評価するためのアッセイは報告されておらず、したがって、比較分析は利用不可能である。
【0006】
以前の研究は、「内皮前駆細胞」と名付けられた細胞の集団が、細胞表面抗原CD34を発現する分類された細胞か、または単核細胞のいずれかを、規定された培養条件において培養することによって、ヒト臍帯血からか、または成体末梢血から単離され得ることを報告した。
【0007】
造血前駆細胞および内皮前駆細胞は、卵黄嚢および胚の発生において、多くの細胞表面マーカーを共有し、そして多数の遺伝子の遺伝子破壊が、造血細胞および内皮細胞の両方の発達に影響する。したがって、これらの系統は、共通の前駆体(血管芽細胞)から起こると仮定される。造血細胞の発達における幹細胞および前駆細胞の階層が、報告された。上記階層内の造血前駆細胞は、それらのクローン原性および増殖能によって同定される。遺伝的研究が、内皮細胞の起源が造血細胞の発達に密接に関連することを明白に示しているにもかかわらず、増殖能の違いに基づいて幹細胞および前駆細胞の同様の階層を支持する証拠は確立されていない。すなわち、造血細胞系に類似する個々の細胞のクローン原性および増殖能によって区別され得るEPCの階層は、報告されていない。
【0008】
造血幹細胞および前駆細胞(HSC/Ps)の両方は、成体の末梢血と比較して、臍帯において豊富である。現在、臍帯血は、種々の血液学的な障害および悪性疾患を有する患者の移植のための、造血幹細胞の代替的な資源として使用されている。
【0009】
毎年、何千もの患者が造血幹細胞(HSC)の移植を必要としている。この患者のほぼ2/3が、移植のためのヒト白血球抗原(HLA)適合性一致を見出せ得ずにいる。これは、特に、多くの民族集団および代表的でない少数派(under−represented minority)に関してあてはまる。白人患者の1/3のみが、適切に一致した同胞の移植片(最低限の対宿主性移植片病(GVHD)合併症を伴う、最も適合性のある供給源)を見出す。
【0010】
ヒト臍帯血は、臨床移植のためのHSCの代替的な供給源であることが知られている。ドナー臍帯血が、レシピエントに対して完全に主要組織適合性が一致するか、または不一致かにより、種々の先天性または後天性の血液学的障害のための処置として、臍帯血細胞は、調整された(conditioned)宿主に移植され、そして再増殖する。たとえこの臍帯血移植片が上記レシピエントにとって2つ以上の座で不一致であったとしても、GVHDの発生率および重症度は、同様の不一致の成体の骨髄移植片または動員(mobilized)末梢血移植片の移植に関して観察されるGVHDの発生率および重症度と比較して、有意に小さい。
【発明の開示】
【発明が解決しようとする課題】
【0011】
移植に関する臍帯血のより広範な使用に対する制限するものとしては、限られた数のHSCおよび前駆細胞のみしか移植片中に存在しないという事実が挙げられる。ほとんどの患者が、一致した同胞のドナーを有さないので、ほとんどの臍帯血移植片は、不一致のレシピエントへと移植される。多くの研究が、移植片中の臍帯血の用量は、上記移植片が血縁関係の無いドナーに由来する場合、患者の生存に対して重要であることを報告する。移植に関連する死亡率は、1.7×105個のCD34+細胞/kgの臍帯血移植片を受けたレシピエントにおいて20%、これに対して移植片中のより少量のCD34+細胞を受けたレシピエントにおいて75%と報告されている。エキソビボで臍帯血HSCを効率的に増殖させ、移植片中の細胞数を増加させるための方法を発見することは、臨床的移植のための大きな進歩であり、そして大きな商業的市場を有する。
【0012】
臍帯血HSC増殖のためのアプローチは、目覚しいものではない。ほとんどの研究において、臍帯血単核細胞または単離されたCD34+細胞に対する種々の成長因子の添加は、総細胞数における増加、コロニー形成単位細胞(CFC)数における増加、および免疫不全(NOD/SCID)マウスまたはヤギ胎仔における短期間の前駆細胞移植における増加と、相関している。しかしながら、NOD/SCIDマウスにおけるSCID再増殖細胞(SRC)頻度、またはヤギ胎仔における長期間の移植によって測定されるようなHSCの数の増加については、ほとんどのアプローチが効果的でない。ヒト患者における増殖された臍帯血HSCの使用の結果は、失望すべきものであった。
【課題を解決するための手段】
【0013】
(発明の要旨)
単一細胞コロニーアッセイは、末梢血および臍帯血から、ならびに臍帯または成体血管から単離された内皮細胞から単離された、哺乳動物の内皮前駆細胞(EPC)の間での新規の階層を説明するために開発された。ヒト、ウシ、ブタおよびラットの生物学的試料に由来する前駆細胞の異なる集団は、クローン原性および増殖能に基づいて同定された。
【0014】
内皮前駆細胞(EPC)は、成体末梢血および臍帯血から単離され、そしてエキソビボで指数関数的に増殖された。対照的に、ヒト臍静脈内皮細胞(HUVEC)または血管壁に由来するヒト大動脈内皮細胞(HAEC)は、広義には、分化した成熟内皮細胞(EC)としてみなされ、そしてそれらは、EPC研究のための「コントロール」として利用される。しかしながら、成体および臍帯血由来のEPCと同様に、血管壁由来であるHUVECおよびHAECは、インビトロにおいて少なくとも40集団倍加の間継代され得る。EPCを、それらの増殖能およびクローン原性能に基づいて識別する、新規の単一細胞析出(deposition)アッセイの利用は、HUVECまたはHAEC中に、EPCが常在することを見出した。単一細胞クローン原性アッセイは、EPCの増殖能およびクローン原性能に基づいて、EPCの新規の階層を明確にするために開発された。EPCの完全な階層は、血管壁由来のHUVECおよびHAECにおいて同定され、そしてそれらのクローン原性能および増殖能によって識別された。EPCの多様性がヒト脈管において存在し、そして、脈管の一体の維持におけるEPCの起源および機能の両方を決定するための、概念的な枠組みを提供する。したがって、EPCは、臨床的使用(例えば、移植片)のために、末梢血またはヒト脈管の生検のいずれかから容易に得られる。
【0015】
単離された内皮コロニー形成細胞は、以下の特徴を有する:
(a)内皮細胞の特有の細胞表面抗原(例えば、CD31、CD105、CD146、およびCD144)を発現する;
(b)造血細胞に特有の細胞表面抗原(例えば、CD45、およびCD14)を発現しない;
(c)アセチル化LDLを取り込む;および
(d)マトリゲル(matrigel)において、毛細管様の管を形成する。
【0016】
HPP−ECFCとして分類される、単離された細胞はまた、
(a)単一細胞からプレートされた場合、少なくとも約2000細胞の、少なくとも二次コロニーへと再プレートする;
(b)高増殖性を示す;
(c)単一細胞から増殖する;および
(d)高レベルのテロメラーゼ(HeLa細胞によって発現されるテロメラーゼレベルの、少なくとも34%)を発現する。HPP−ECFCはまた、>0.8である、高い核 対 細胞質比を示し、細胞直径は、<22ミクロンであり、そして少なくとも107個の子孫が、単一細胞に由来する。
【0017】
内皮コロニー形成細胞を単離するための方法は、以下の工程:
(a)細胞外マトリックスタンパク質でコートされた支持体上で、生物学的試料からの細胞を培養する工程;
(b)上記支持体に接着する細胞を選択し、そして再プレート可能なコロニーを形成する工程;および
(c)上記コロニーから単一細胞を選択する工程
を包含する。
【0018】
上記生物学的試料は、哺乳動物の臍帯血、または血管であり得る。ヒト供給源、ウシ供給源、ブタ供給源、およびラット供給源が、適切である。内皮細胞の型に関する単一細胞アッセイは、以下の工程:
(a)特定の分類方法を使用し、生物学的試料の細胞を分類する工程;
(b)規定された条件下で、細胞外マトリックスタンパク質上で、この単一分類の細胞を培養する工程;および
(c)特定のコロニーのサイズ、形態、および増殖能を列挙し、上記内皮細胞の型(例えば、HPPP−ECFC)を決定する工程
を包含する。
【0019】
HPP−ECFCを富化する方法は、以下の工程:
(a)特定の分類方法を使用し、生物学的試料の細胞を分類する工程;および
(b)規定された条件下で、細胞外マトリックスタンパク質上で単一分類の細胞を培養する工程
を包含する。
【0020】
エキソビボにおいて造血幹細胞を増殖させるための方法は、以下の工程:
(a)コラーゲンコートされた固体支持体上で、HPP−ECFC細胞を培養する工程;および
(b)HPP−ECFC細胞とともに共培養することによって、造血幹細胞(HSC)を増殖させる工程であって、ここで、上記HPP−ECFCはヒト臍帯血細胞由来であり、かつ上記HSC細胞はヒト骨髄由来である、工程
を包含する。
【0021】
哺乳動物中の移植片における、造血幹細胞の百分率を改善するための方法は、以下の工程:
(a)ヒト骨髄細胞を臍帯血HPP−ECFCとともに共培養し、生成物を形成させる工程;および
(b)適切な量の上記生成物を、上記哺乳動物中に移植する工程であって、ここで、上記細胞は、ヒト骨髄由来であるCD45+細胞であり、かつ上記哺乳動物は、NOD−SCIDマウスである、工程
を包含する。
【0022】
自己の臍帯血もしくは血縁関係の無い臍帯血、動員成体末梢血、または骨髄由来HSCとの共培養における、臍帯血の高増殖能内皮コロニー形成細胞(HPP−ECFC)は、HSC細胞の数を増加させ、そして結果としてHSCの増加、およびHSC再増殖活性の増大をもたらし、レシピエント被験体におけるより高いレベルの移植につながる。
【0023】
ヒト臍帯血からの高増殖能内皮コロニー形成細胞(HPP−ECFC)由来の細胞の支持細胞層(feeder layer)の使用は、再増殖性の造血幹細胞および造血前駆細胞の成長および生存を刺激する。成長および生存の刺激は、インビトロ培養中の前駆細胞の数の増加、およびNOD/SCID免疫不全マウス移植系におけるヒト細胞移植レベルの増加によって決定された。
【発明を実施するための最良の形態】
【0024】
(発明の詳細な説明)
高増殖能内皮コロニー形成細胞(HPP−ECFC)と名付けられた新規の内皮細胞前駆体は、成体血液EPCにおける20〜30倍加と比較して、高増殖能(100集団倍加まで)を示す。HPP−ECFCは、臍帯血のみからではなく、臍帯および成体血管からも単離される。HPP−ECFC細胞は、単一細胞レベルで再プレートされ得、そして細胞の大多数は、少なくとも二次HPP−ECFCの再生を伴って増幅する(図7)。予想外なことに、HPP−ECFCコロニー形成は、臍帯血においては存在するが、成体末梢血においては存在しない(図5d)。さらに、単層のHPP−ECFC由来の臍帯血内皮細胞は、成体LPP−ECFCと比較(エキソビボ培養の同じ期間の間)して、集団倍加時間(PDT)において2.5倍の減少を示し、そして累積的集団倍加レベル(CPDL)において少なくとも2倍の増加を示した。異なった方法論を利用して臍帯血から単離された内皮前駆細胞の他の集団とは対照的に、臍帯血HPP−ECFC子孫は、内皮細胞抗原を均一に発現し、かつ造血特異的細胞抗原を発現しない。したがって、HPP−ECFCは、ヒト臍帯血において富化されており、成体末梢血においては見出されなかった。HPP−ECFCはまた、哺乳動物の血管(例えば、臍静脈管およびヒト大動脈管)由来である内皮細胞において見出された(図11)。これらのHPP−ECFCは、プレートした新鮮な臍帯血単核細胞の培養物において、10日間以内に出現するのに対し、成体血液のLPP−ECFCでは、プレート後14日間前に、ほとんど出現しない。
【0025】
さらに、臍帯血コロニーは、成体コロニーと比較して、一貫してより大きく出現する(図1)。成体内皮細胞コロニーと臍帯血内皮細胞コロニーとの間には、そのサイズ、頻度、および出現の時期において、明瞭な差異が存在する(図1、図3、図10)。これらの知見は、臍帯血EPCは、HPP−ECFC、LLP−ECFC、およびクラスターからなるのに対し、成体血液EPCは、LPP−ECFCおよびクラスターからなることを示す。
【0026】
HPP−ECFC、LPP−ECFC、クラスター、および成熟内皮細胞の完全な階層は、生きている哺乳動物のドナー中の任意の血管から単離され得る。
【0027】
EPCは、造血細胞前駆体の規定のために利用された技術と同様の用語法を使用して、同定され得る(図7)。HPP−ECFCは、二次HPP−ECFCへの再プレートに加えて、内皮前駆体の全てのその後の段階を生じる。単一細胞から生じる低い増殖能の内皮コロニー形成細胞(LPP−ECFC)は、コロニーを形成し、それは50個より多くの細胞を含むが、再プレートにおいて、二次LPP−ECFCコロニーさえも形成しない。それらは、内皮細胞クラスター(50個未満の細胞)を生じる。後に、内皮細胞クラスターは、単一細胞から生じ得るが、50個未満の細胞を含み得、かつコロニーまたはクラスターへと再プレートし得ない。
【0028】
造血幹細胞および造血前駆細胞は、成体末梢血と比較して、臍帯血において富化されている。1つの興味深い知見は、成体末梢血と比較して、EPCはまた、臍帯血において富化されている。さらに、臍帯血由来のEPCは高レベルのテロメラーゼ活性を含み、このことは、これらの細胞が、細胞老化の明白な徴候を伴わずに、少なくとも100集団倍加の間増殖し得ることを説明し得る。単一細胞のレベルで、いくつかの臍帯血EPC由来の細胞は、107倍〜1012倍増殖し得る。本発明者らの知る限りでは、他の一次哺乳動物内皮前駆細胞または成熟細胞は、同様の成長特性能力またはクローン原性能力を有すると同定されていない。
【0029】
(ヒト臍帯血から単離された内皮細胞コロニー(EPC)の特徴付け)
細胞は、培養物中で増殖した同時に、内皮細胞表現型を維持していた。前記した臍帯血から単離された内皮前駆細胞に対して、本発明は、ヒト臍帯血から単離した細胞(すなわち、臍帯血HPP−ECFC)および前駆体が、単一細胞を用いて開始した場合でさえ、少なくとも100集団倍加の間培養され得、そして指数関数的に増殖し得ることに関する。さらに、これらの細胞は、造血細胞特異的表面抗原を発現せず(CD45およびCD14もまたほとんど発現しない)、そしてメチセルロース(methycellulose)アッセイにおいて造血細胞コロニーを形成しない。さらに、上記HPP−ECFC前駆体は、マトリゲル中で迅速に脈管を形成し、IL−1刺激またはTNF−α刺激のいずれかに応答してVCAM−1を上方制御し、そしてそれらの内皮細胞アイデンティティ(identity)を確認する内皮細胞特異的抗原を発現する。これらの細胞は、高増殖能内皮コロニー形成細胞(HPP−ECFC)と命名された。
【0030】
臍帯血HPP−ECFCは、成体血液(これは、LPP−ECFC、クラスター、および成熟内皮細胞からなる)と比較して、より大きな複製動態を示す。内皮前駆体は、AC133、CD34、およびFlk1を発現すると報告される一方で、HPP−ECFC子孫およびEOC子孫は、AC133抗原、CD34抗原、およびFlk1抗原を発現する細胞の同様の頻度を示す。したがって、これらの細胞表面マーカーは、異なる増殖能を有する細胞を区別し得ない。
【0031】
内皮増殖細胞は、成体末梢血から単離したMNCの培養の2週間〜4週間後に出現し、そして内皮増殖細胞は、それらのインビトロにおける指数関数的な成長によって特徴付けられる(しかしながら、EOCは、より低いレベルのテロメラーゼ示し、二次LPP−ECFCへと再プレートされず、そしてTPP−ECFCよりずっと前に複製的老化に達する)。対照的に、ヒト臍帯血MNC由来のHPP−ECFCは、I型コラーゲンでコートされた組織培養プレート上の完全EGM−2培地中において、臍帯血MNCの培養の5日後〜10日後に出現した。分散した接着細胞コロニーが出現し、そして「丸石」形態の内皮細胞を示した(図1)。この形態および上記コロニーの出現は、前述した成体末梢血由来のEOCコロニーに関する形態および出現と似通っているが、区別されるものであり、かつ接着循環内皮細胞またはマクロファージと明白に区別される。上記HPP−ECFCコロニーは大きくて、そして小さくて円い細胞、長くて薄い細胞、および大きくて平らな円い細胞の混合物からなる一方で、上記EOCは、ほぼ均一に長くて薄い細胞から構成される。このコロニー由来の細胞は継代培養され、そしてこれらのコロニー由来の増殖した細胞は、免疫表現型決定(immunophenotyping)、機能的試験、および成長動態の測定のために使用された。最初の継代の後、上記細胞は、「丸石」形態を有する紡錘体形状の細胞の単層を形成した。免疫表現型決定は、上記細胞が、内皮細胞表面抗原(CD31、CD141、CD105、CD146、CD144、vWF、およびflk−1)を一様に発現することを明らかにした(図2a、図2b)。上記細胞は、造血細胞表面特有の抗原(CD45およびCD14)を発現せず、このことは、上記単層が、造血細胞を混入していないことを確証した(図1、図3、図10)。
【0032】
上記細胞が、アセチル化低密度リポタンパク質(Ac−LDL)または(Dil−AC−LDL)を取り込んだことで、接着コロニーに由来する上記単層が内皮細胞であったことを確認した。これらの細胞機能は、内皮細胞に特有である。上記接着コロニーから継代培養された細胞は、Ac−LDLを均一に取り込み、異なる数の細胞を播種した後マトリゲル中で脈管を形成し、そしてrhTNF−α刺激またはrhIL−1刺激の両方に応答してVCAM−1を上方制御した(図2c〜図2e)。
【0033】
臍帯血HPP−ECFCおよび子孫の成長動態は、時間の関数として測定された。特筆すべきことに、臍帯血HPP−ECFCおよび子孫は、培養において、少なくとも100集団倍加の間、老化の徴候を伴わずに指数関数的に増殖し得、そして細胞数は、培養物中で、100日間の期間にわたって1020倍に増加した。臍帯血内皮細胞の代表的な成長曲線は、インビトロにおける、これらの細胞の増殖能を例証する。したがって、免疫表現型決定、機能的試験、および成長動態の分析に基づいて、内皮細胞(HPP−ECFCと命名される)のコロニーは、臍帯血MNCから独自に培養され得、そして指数関数的に増殖可能な内皮細胞の集密的単層になるまで継代され得ることが示される。
【0034】
(HPP−ECFCコロニーはヒト臍帯血中に存在するが、成体末梢血中には存在しない)
50mL〜100mLの末梢血が、健康な成体ドナーまたは正常期胎児の臍帯から集められ、そしてMNCが単離された。細胞は、細胞外成熟分子でコートされた組織培養プレート上で、完全EGM−2培地中に播種され、そして次の1週間〜6週間にわたってコロニー形成が観察された。等容積の血液あたりのコロニーの数は、成体末梢血と比較して、臍帯血において15倍増加していた(表I、図1a、図1b)。同数の臍帯MNCおよび成体MNCがプレートされた場合、コロニー形成における類似の相違がまた認められた。成体EOCコロニーは、代表的には、培養の開始の2週間後〜4週間後に形成されるが、臍帯血HPP−ECFCコロニーは、5日間〜10日間以内に出現する。最終的に、臍帯血および成体末梢血に由来する内皮細胞からフローサイトメトリーによって単離された細胞の免疫表現型決定は、コロニー細胞が、内皮細胞表面抗原(CD31、CD105、CD146、CD144、vWF、およびflk−1)を一様に発現し、そして造血細胞表面抗原(CD45およびCD14)を発現しないことを明らかにした。このことは、それらの内皮細胞アイデンティティを確証した。上記成体EOCの免疫表現型決定は、以前に公表された研究と一致した。したがって、内皮コロニー形成細胞HPP−ECFCは、臍帯血中に存在し、成体末梢血EOC(これは、LPP−ECFCを表す)と比較して異なる細胞型を表す。
【0035】
(臍帯血HPP−ECFCの増殖速度は、成体血液EOCよりも速い)
成体末梢血と比較して、臍帯血由来の細胞のコロニー形成の出現の頻度および時期における相違を考慮するとき、1つの疑問は、成体血および臍帯血の増殖動態における相違が存在するのかしないのか、であった。HPP−ECFCの初期継代単層は、臍帯血から樹立され、そしてEOCは成体末梢血から樹立され、そして細胞は、I型コラーゲンコートされたプレート上で完全EGM−2培地中で培養された。長期培養における集団倍加時間(PDT)および累積集団倍加レベル(CPDL)を決定するために、投入細胞数を計数した(これらは、臍帯血および成体血液由来の細胞の増殖動態を、定量化および比較するために使用される測定である)。上記PDTおよびCPDLを正確に定量化するために、細胞を、少なくとも10継代の間培養した。異なるドナーに由来する複数の細胞株の試験の結果は、成体EOCコントロールと比較して、臍帯血HPP−ECFCのPDTにおいて2.5倍の減少が存在したことを示した。さらに、PDTにおける減少と一致して、臍帯血細胞の培養は、成体EOCの連続的な継代と比較して、CPDLにおいて有意な増加を示した。したがって、臍帯細胞および成体細胞の両方は、培養において増殖し得るが、臍帯血HPP−ECFCおよび子孫の増殖能は、成体EOCと比較してより大きい。臍帯血由来のHPP−ECFCはまた、成体血液EOCと比較して、単一細胞レベルでより大きい増殖能を示す。
【0036】
臍帯血HPP−ECFC由来の内皮細胞または成体EOCの、単一細胞レベルでの、個々の増殖能力およびクローン原性能力が決定された。新規の実験方法が、単一臍帯血HPP−ECFC由来の内皮細胞および成体EOCの増殖能力およびクローン原性能力を定量化するために設計された。
【0037】
初期継代臍帯血HPP−ECFC由来または成体EOC由来の内皮細胞の子孫は、緑色蛍光タンパク質(GFP)をコードするレトロウイルスを用いて最初に形質導入され、そしてGFPの発現に関して選択された。臍帯内皮細胞および成体内皮細胞の両方の形質導入効率は、95%より高かった。その後の選択は、1つのGFPを発現するHPP−ECFC由来内皮細胞または成体EOC由来の内皮細胞は、(>直径100ミクロンおよび<9ポンド/平方インチのシーズフロー圧(sheath flow pressure)を有するソートノズル(sorting nozzle)を使用して)蛍光サイトメトリー分類によって、I型コラーゲンでコートされ、そして200μlのEGM−2培地で満たされた96ウェル組織培養プレートの1つのウェル中にプレートされた。直後に、個々のウェルは試験され、1つの内皮細胞のみが各ウェル中に置かれたことを確実にされた。次いで、内皮細胞は14日間培養され、そしてその培地の2分の1は、4日間毎に新鮮なEGM−2培地を用いて交換された。14日間の終わりに、GFPを発現する内皮細胞の数が計算された。
【0038】
少なくとも1回の細胞分裂を起こしている単一細胞の数は、成体EOC由来の内皮細胞と比較して、臍帯血HPP−ECFC由来の内皮細胞について有意に多かった。14日間の終了の後、各々のウェル中の細胞の数のスコアリングにおいて、単一臍帯血HPP−ECFC由来の内皮細胞は、成体EOC由来の内皮細胞と比較して、より多く分裂し、そしてより大きなコロニーを生成したことが明らかであった。成体EOC由来の内皮細胞と比較した、単一臍帯血HPP−ECFC由来の内皮細胞の、分裂およびコロニーを形成する能力における差異に起因して、各ウェル中の(少なくとも1回の細胞分裂を示した)細胞の数が計数された。ほとんどの単一成体EOC由来の(分裂した)内皮細胞は、2個と50個の間の細胞のクラスターを生成したが、一部は、500細胞までの二次コロニーを生じた。しかし、ただ1つの>2000細胞のコロニーが、任意の単一に分類された成体EOC由来の内皮細胞から生じた。しかしながら、60%を超える臍帯血HPP−ECFC由来の(分裂した)内皮細胞は、少なくとも2000細胞からなる、良好に限局性の二次コロニーを形成し、そして多数の単一に分類された細胞が、>10,000細胞からなるコロニーを生じた(本発明者らの知る限りでは、成体EOC由来の内皮細胞は、このようなコロニーを生成することは決して無かった)。
【0039】
これらの細胞がさらなるコロニーを形成し得るかどうかを決定するために、単一成体EOC由来の内皮細胞または単一臍帯血HPP−ECFC由来の内皮細胞のいずれかに由来する二次細胞コロニーが、連続的に再プレートされた。複数の独立した実験において、単一成体EOC由来の内皮細胞に由来する二次コロニーは、24ウェルまたは6ウェルのI型コラーゲンコートされた組織培養プレート中に再プレートされた後は、三次コロニーを生じることは決して無い。プレートされた単一細胞は静止状態のまま残り、かつ増殖しなかった。しかしながら、単一臍帯血HPP−ECFC由来の内皮細胞(2000細胞より多くを生じた)に由来する二次コロニーのほとんどは、同じ実験条件下において再プレートされ、三次内皮コロニーを形成し得た。単一一次臍帯血HPP−ECFC由来の内皮細胞は、二次コロニーを生成し得、これは、引き続いて連続的に継代され、107個〜1012個の内皮細胞を生成し得る。
【0040】
造血高増殖能コロニー形成細胞(HPP−CFC;インビトロにおけるクローン原性アッセイにおいて培養され得る、最も初期の多分化能の造血前駆体)に対する、臍帯血由来の内皮コロニー形成細胞の独特かつ新規に同定された集団の類似性を考慮して、これらの細胞は、「高増殖能内皮コロニー形成細胞(HPP−ECFC)」と名付けられる。要約すれば、これらの細胞は、以下の点で前記の成体EOCと異なる:
(1)成体EOCと同一の実験条件下において培養した場合、HPP−ECFCは、より高い増殖動態を有する、
(2)プレートされた成体末梢MNC由来の成体EOCと比較して、HPP−ECFCは、臍帯血MNCからの培養において、より早い時点で出現する、
(3)成体EOCと比較して、HPP−ECFCは、単一細胞レベルにおいてより高いクローン原性能を有する、
(4)HPP−ECFCは、連続的に再プレートされ、少なくとも二次HPP−ECFCコロニーを形成し得る一方で/他方では、成体EOCは、この潜在力を示さず、かつHPP−ECFCは高いレベルのテロメラーゼを示す。
【0041】
(EPC由来の臍帯血内皮細胞および成体内皮細胞の増殖動態)
異なる系統の前駆細胞は、それらのクローン原性能および増殖能によって規定および区別される。臍帯血のコロニー形成および成体EPCのコロニー形成における相違に起因して、EPC由来の臍帯血内皮細胞および成体内皮細胞の増殖動態が比較された。最初に、この細胞が二次コロニーを形成し、集密まで成長するか否かを試験するために、限界細胞希釈で、臍帯血内皮細胞コロニーおよび生体内皮細胞コロニーに由来する細胞がプレートされた。興味深いことに、成体EPCコロニーおよび臍帯血EPCコロニーの両方に由来する細胞子孫は、集密にまで成長する前に、種々のサイズの二次細胞コロニーを形成した(図3)。しかしながら、臍帯血EPC由来の細胞子孫に由来するコロニーは、成体のコロニーと比較して、一貫してより大きく、そしてより小さい細胞を含んだ(図3)。
【0042】
細胞単層は、EPC由来の臍帯血内皮細胞および生体内皮細胞の増殖能を決定するために連続的に継代された。特筆すべきことに、臍帯血EPC由来の細胞は、明白な老化の徴候を伴わずに、少なくとも100集団倍加の間増殖し得た。対照的に、成体EPC由来の細胞は、20〜30集団倍加の間のみ継代され得た(図4a)。臍帯血EPC由来細胞および成体EPC由来細胞の増殖動態を定量化および比較するために、培養において規定された期間(60日間)の間に、集団倍加時間(PDT)および累積集団倍加レベル(CPDL)が計算された。成体EPC由来の細胞と比較して、臍帯血EPC由来の細胞の、PDTにおいて2.5倍の減少、およびCDPLにおいて1.5倍の増加があった(図4b、図4c)。成体EPCのPDTおよびCDPLは、健康な成体ドナーから単離したEPC由来の細胞の増殖動態を試験した、2つの最近の報告と同様であった。
【0043】
rhVEGF刺激またはrhbFGF刺激(これらは、2つの内皮細胞分裂促進因子)のいずれかに応答する、臍帯血EPC由来の細胞および成体EPC由来の細胞の増殖が比較された。臍帯血EPC由来の細胞および成体EPC由来の細胞は、血清飢餓にされ、次いで、rhVEGFまたはrhbFGFの存在下かまた非存在下において培養された。細胞は16時間培養され、そしてDNA合成を測定するために収集する前にトリチウム化チミジンでパルス(pulse)された。臍帯血EPC由来の細胞は、成体EPC由来の細胞と比較して、rhVEGF刺激またはrhbFGF刺激のいずれかに応答して、より大きなDNA合成を示した(図4d)。まとめると(collectively)、これらの結果は、短期間アッセイおよび長期間アッセイの両方において、臍帯血EPC由来の細胞の増殖速度および増殖能力は、成体EPC由来の細胞よりも大きいことを示す。さらに、臍帯血EPC由来の内皮細胞および成体EPC由来の内皮細胞は、限界希釈でプレートされた場合、種々のサイズおよび形態の別個の細胞コロニーを形成する。
【0044】
(EPCコロニー由来の単一臍帯血内皮細胞および成体内皮細胞のクローン原性能および増殖能の定量化)
臍帯血EPCコロニーおよび成体EPCコロニーは、異なる増殖能およびクローン原性能を有する細胞を生じる。しかしながら、前駆細胞のクローン原性能についての厳密な試験は、単一細胞が、他の細胞の非存在下において分裂し、そしてコロニーを形成するか否かを決定するものである。したがって、EPCコロニー由来の単一臍帯血内皮細胞および単一生体内皮細胞の増殖能およびクローン原性能を定量化するために、アッセイが開発された。
【0045】
最初のEPCコロニーに由来する臍帯血内皮細胞および成体内皮細胞は、EGFPをコードするレトロウイルスを用いて形質導入され、そしてEGFP発現について選択される。選択に続き、1つのEGFPを発現する内皮細胞が、I型コラーゲンでコートされた96ウェル組織培養プレートの1ウェル中にFACSによってプレートされ、そして完全EGM−2培地で満たされた。内皮細胞は培養され、そして14日目の終わりにEGFPを発現する内皮細胞の数が本明細書中に記載されるように計数された。この方法は、図5aに図示される。
【0046】
特筆すべきことに、少なくとも1回の細胞分裂を起こした単一細胞の百分率は、臍帯血内皮細胞について、成体細胞と比較して5倍増加した(図5b)。さらに、単一臍帯血内皮細胞に由来する細胞子孫の平均数は、個々の成体細胞に由来する細胞の数と比較して100倍大きかった(図5c)。分裂した単一成体内皮細胞の80%超は、2〜50細胞の数の範囲におよぶ細胞の小さなコロニーまたはクラスターを生じた(図5d)。しかしながら、いくつかの単一成体内皮細胞は、500個を超える細胞を含むコロニーを形成しなかった(図5d)。対照的に、少なくとも60%の単一でプレートされた分裂した臍帯血内皮細胞は、14日間の培養期間において、2,000個と10,000個の間の細胞を含む良好に限局したコロニーを形成する(図5d)。単一臍帯血細胞または単一成体細胞に由来する細胞の、種々の内皮細胞のコロニーおよびクラスターのサイズおよび形態の顕微鏡写真は、図5eに示される。これらの単一細胞の研究は、臍帯のEPCおよび成体のEPCの異なる型が存在することを示し(これは、それらの増殖能およびクローン原性能によって区別され得る)、そしてEPCは、造血前駆細胞の階層と類似した増殖能の階層を示す。
【0047】
(単一臍帯血内皮細胞の細胞子孫は、連続的に再プレートされ得、そして長期間培養において指数関数的に増殖され得る)
造血細胞系において、最も増殖性の細胞型は、高増殖能コロニー形成細胞(HPP−ECFC)と名付けられた。このHPP−ECFCは、大きな細胞コロニーを形成するその能力によって規定され、連続的なプレーティングにおいて、少なくとも二次コロニーを形成する潜在性を有する個々の細胞を生じる。単一のプレートされた臍帯血EPC由来細胞または成体EPC由来細胞に由来するクローン性子孫は、トリプシン処理され、再プレートされ、そして24ウェル組織培養プレート中で7日間培養された。1000個を超える単一成体EPC由来の細胞のクローン性子孫を、24ウェルプレート中にプレーティングした後、14日間の培養の後、上記ウェル中で二次コロニー1個のみが検出された(図6a)。対照的に、単一でプレートされた臍帯血EPC由来の細胞のクローン性子孫の約2分の1(421のうちの205)は、24ウェルプレート中で、二次コロニーを形成したか、または集密まで急速に成長した(図6a)。単一臍帯血内皮細胞の上記子孫に由来する二次内皮細胞コロニーまたは集密細胞単層の代表的な顕微鏡写真は、図6bに示される。5日以内に集密まで急速に成長したこれらのウェル中では、二次コロニーは検出されなかったので、限界希釈分析を、この集密単層上で実施した。この単層由来のプレートされた単一細胞のうちの少なくとも9パーセントは、100細胞より多くを含む内皮細胞コロニーを形成した。この結果は、臍帯血EPCに由来する個々の細胞が、二次コロニーを形成し得ることを実証した。
【0048】
単一でプレートされた臍帯血EPC由来内皮細胞の、長期増殖能が試験された。単一臍帯血内皮細胞に由来する二次コロニーまたは集密細胞単層は、組織培養プレートを徐々に大きくして連続的に継代された。もともと3つの異なる臍帯血ドナーに由来する、11の単一内皮細胞の細胞子孫が試験された。長期間の培養において、単一臍帯血内皮細胞は、少なくとも107細胞を生じた(図6c)。試験された上記11の単一臍帯血内皮細胞の平均CPDLは、30.8であった。したがって、臍帯血中の高増殖性EPCの集団は、二次コロニーおよび三次コロニーを形成する。
【0049】
(EPC由来の臍帯血内皮細胞は、高レベルのテロメラーゼ活性を含む)
臍帯血EPCに由来する内皮細胞は、ヘーフリック限度を超えて、少なくとも100集団倍加の間、連続的に継代される(図4a)。唯一他に報告されている同様の成長動態を有する一次内皮細胞は、テロメラーゼを過剰発現するように遺伝的に操作された内皮細胞のみである。したがって、臍帯血EPC由来の細胞および成体EPC由来の細胞において、テロメラーゼ活性を、それらの成長動態における相違についての可能性のある分子的説明として測定した。初期継代臍帯血EPC由来の子孫および後期継代臍帯血EPC由来の子孫の両方は、成体EPC由来の細胞と比較して、有意に上昇したレベルのテロメラーゼ活性を示し、テロメラーゼを過剰発現する上記の一次内皮細胞株を連想させる(図6d〜図6e)。したがって、臍帯血EPC由来の細胞は、大きな増殖能と一致して、培養における連続的な継代に伴って高レベルのテロメラーゼ活性(同数のHeLa細胞のテロメラーゼ活性の34±10%)を保持する。
【0050】
(エキソビボにおける、再増殖性の幹細胞および前駆細胞の増殖)
自己の臍帯血もしくは血縁関係の無い臍帯血、動員成体末梢血、または骨髄由来のHSCとともに共培養される、臍帯血の高増殖能内皮コロニー形成細胞(HPP−ECFC)は、HSC細胞の数を拡大させ、そして結果として、レシピエント被験体における、より高いレベルの移植につながるHSCの増加、およびHSC再増殖活性の増加をもたらす。
【0051】
臍帯血由来のHPP−ECFCとヒトHSCの共培養は、造血前駆細胞数を増加させ、そしてNOD/SCIDマウスにおけるヒト造血細胞の移植(インビボにおけるヒトHSC機能の測定のためのアッセイ)を増強した。
【0052】
ヒト臍帯血CD34+CD38−細胞または動員成体末梢血CD34+CD38−細胞(HSC活性に富む)と、7日間まで(サイトカイン添加有りで)共培養されたヒト臍帯血HPP−ECFC由来の内皮細胞は、結果として、準致死的照射されたNOD/SCIDマウスにおいて、>100倍ヒトCD45+細胞移植を増強する(図12)。
【0053】
上記HPP−ECFCの収集、単離、および増殖の方法、ならびに上記HPP−ECFCとヒト幹細胞との共培養のための特定の方法は、新規である。HPP−ECFCは、任意の臍帯血試料から収集され得、増殖され、凍結され、そして保存される。これらの細胞は、次いで、ヒト臍帯血細胞試料、骨髄由来細胞試料、または動員成体末梢血幹細胞試料および動員成体末梢血前駆細胞試料を増殖させるために、解凍され、増殖され、そして共培養において使用され得る。この増殖生成物は、次いで、(規制当局の承認後に)移植目的のために使用され得る。
【実施例】
【0054】
(材料と方法)
(成体末梢血試料および臍帯血試料)
健康なヒトボランティア(22歳と50歳との間の年齢の男性および女性)から、新鮮な血液試料(50ml〜100ml)を、静脈穿刺によって収集し、そしてクエン酸リン酸デキストロース(citrate phosphate dextrose)溶液中で抗凝血化した。健康な新生児(38週在胎齢〜40週在胎齢、男児および女児)からのヒト臍帯血試料(20ml〜70ml)を、抗血液凝固剤としてクエン酸リン酸デキストロース溶液を含む滅菌シリンジ中で収集した。分娩および出産の前に、全ての母親から書面によるインフォームドコンセントを得た。Indiana University School of Medicineの施設内倫理委員会は、全てのプロトコールを承認した。
【0055】
(バフィコート細胞の調製)
ヒト単核細胞(MNC)を、成体末梢血または臍帯血のいずれかから得た。手短に言えば、20ml〜100mlの新鮮な血液を、ハンクス平衡塩溶液(HBSS)(Invitrogen、Grand Island、NY)で1対1に希釈し、そして等容積のFicoll−Paque(Amersham Biosciences)(フィコール密度勾配物質)上にオーバーレイした。細胞を、室温で、1800rpm(740×g)で30分間遠心した。MNCを、単離し、そして10%〜20%ウシ胎仔血清(Hyclone、Logan、UT)、2%ペニシリン/ストレプトマイシン(Invitrogen)、および0.25μg/mlのアンホテリシンB(Invitrogen)を補充したEBM−2培地(Cambrex、Walkersville、MD)(完全EGM−2培地)を用いて3回洗浄した。
【0056】
(内皮成長細胞の培養および定量分析)
バフィコートMNCを、最初に、10%ウシ胎仔血清、2%ペニシリン/ストレプトマイシン、および0.25μg/mlのアンホテリシンBを補充した、12mlのEGM−2培地(Cambrex)(完全EGM−2培地)中に再懸濁した。次いで、前もって細胞外マトリックスタンパク質(例えば、I型ラット尾部コラーゲン(BD Biosciences)、ビトロネクチン、フィブロネクチン、10型コラーゲン、ポリリジン)でコートした6ウェル組織培養プレート(BD Biosciences、Bedford MA)の3つの別々のウェル上に、4mlの細胞を播種した。このプレートを、加湿インキュベーター中で、37℃、5% CO2でインキュベートした。24時間の培養後、非接着細胞および残屑を慎重に吸引し、そして残存する接着細胞を、2mlのEGM−2培地で1回洗浄した。洗浄後、各ウェルに4mlのEGM−2培地を添加した。培養の7日目までEGM−2培地を毎日交換し、次いで最初の継代まで隔日で交換した。
【0057】
細胞のコロニーは、培養の5日目と22日目との間に最初に出現し、そして、良好に限局された丸石の様に見える細胞の単層として同定された(図1c)。倒立顕微鏡を使用して、40×拡大率で、目視検査によってコロニーを数えた。
【0058】
継代のために、上記のオリジナルのコラーゲンコートした組織培養プレートから、0.05%トリプシン−0.53mM EDTA(Invitrogen)を使用して細胞を取り出し、10mlのEGM−2培地中に再懸濁し、そしてI型ラット尾部コラーゲンでコートされた75cm2組織培養フラスコ上にプレートした。内皮細胞の単層を、続いて、90%〜100%の集密に達した後に継代した。
【0059】
(HUVECおよびHAECの培養)
動脈管または静脈管から内皮細胞を直接単離するために、2つのアプローチを使用した。第1の手段において、切断された脈管の一端中に20G平滑末端針を挿入し、そして滅菌生理食塩水を使用して脈管含有物(血液細胞を有する血漿)を反対の末端に流し出した。次いで、上記脈管(長さ3cm〜5cm)の各末端を分離するために、脈管鉗子(vascular clamp)を適用した。23G針によって、脈管壁を介してハンクス平衡塩溶液(HBSS)中の0.1%コラゲナーゼの溶液を注入し、そしてこの脈管セグメントを、37℃で5分間インキュベートした。次いで、この脈管の一端から上記脈管鉗子を取り除き、そして細胞解離緩衝液(Gibco)の注入(上記脈管の「開放」末端の反対の、上記脈管の遠位末端を通して注入した)によって上記内皮細胞を放出した。この脈管セグメントを、最小限の(a minimum of)10mLの細胞解離緩衝液を通して注入した。この懸濁された細胞を、350×gで遠心分離し、そして10% FBSを含むEBM−2培地中で洗浄し、計数し、そしてトリパンブルー排除を使用して生存度を調べた。
【0060】
第2のアプローチは、直径のより大きな脈管(>1cm)について最も適切である。この脈管を、全長に沿って切開し、そして内皮管腔が露出するように開いた。残存するあらゆる血液細胞および血漿を、HBSSで洗い流した。この内皮を、ラバーポリスマン(rubber policemen)を用いたフィルムスクレーピング(firm scraping)によって、一方の端から端まで動かして取り出した。上記ラバーポリスマンに付着している細胞を、10% FBSを含むEBM−2の溶液中でこのラバーポリスマンを回旋することによって、(細胞外マトリックスタンパク質で予めコートした)6cm組織培養ウェル中に洗い落とした。毎日、目視検査を実施しながら細胞を培養した。内皮のコロニーは、3日間〜10日間のうちに出現する。この接着内皮コロニーを、トリプシン−EDTAによって取り出し、そして細胞外マトリックスタンパク質でコートしたT 25フラスコに移した。3回継代され、凍結保存されたヒト臍静脈内皮細胞(HUVEC)およびヒト大動脈内皮細胞(HAEC)を、Cambrexから得た。継代のために、I型ラット尾部コラーゲンでプレコートされた75cm2組織培養フラスコの中の完全EGM−2培地中に細胞を播種した。
【0061】
(EPCの成長動態および複製能の評価)
第一継代時の細胞を、トリパンブルー排除アッセイ(Sigma、St.Louis、MO)よって数えた。次いで、細胞の単層を90%集密まで成長させ、そして継代した。各継代において、成長動態曲線、集団倍加時間(PDT)、および累積集団倍加レベル(CPDL)の計算のために、細胞を数えた。
【0062】
継代の間に生じた集団倍加(PD)数を、以下の方程式に従って計算した:
PD=log2(CH/CS)
ここで、CHは、収集のときの生存細胞の数であり、そしてCSは、播種した細胞の数である。全ての以前のPDの和は、各継代におけるCPDLを決定した。その継代に関するPDの数で割った細胞の播種と収集との間の時間間隔を使用することによって、PDTを導き出した。
【0063】
(マトリゲルアッセイおよびアセチル化低密度リポタンパク質(Ac−LDLまたはDil−Ac−LDL)の取り込み)
マトリゲルアッセイを実施した。手短に言うと、初期継代(2代〜3代)の、HPP−ECFC由来の内皮細胞またはEPC由来の内皮細胞を、前もって30μlのマトリゲル(BD Biosciences)でコートした96ウェル組織培養プレート上に、ウェルあたり5000〜20,000細胞の細胞密度で播種した。キャピラリー様の管の形成について、細胞を2時間毎に観察した。
【0064】
接着したHPP−ECFCおよび子孫またはEPCおよび子孫の、Ac−LDLまたはDil−Ac−LDLを取り込む能力を評価するために、6ウェルのI型ラット尾部コラーゲンコートされた組織培養プレート中で培養された細胞の培地中に、10μg/mlのAc−LDL(Biomedical Technologies Inc.、Stoughton、MA)を添加した。37℃で、30分間または4時間、細胞をインキュベートし、次いで1.5μg/mlのDAPI(Sigma)で染色されたリン酸緩衝生理食塩水(PBS)を用いて3回洗浄し、そしてAc−LDLまたはDil−Ac−LDLの取り込みを、蛍光顕微鏡を使用することによって試験した。
【0065】
(蛍光サイトメトリーによる内皮細胞の免疫表現型決定)
初期継代(1代〜2代または3代〜4代)の、HPP−ECFCおよび子孫、またはEPCおよび子孫(5×105個)を、100μlのPBSおよび2%FBS中で、以下に概説されるように、異なる濃度の一次抗体またはアイソタイプコントロール抗体とともに4℃で30分間〜60分間インキュベートした。2% FBSを含むPBSで、細胞を3回洗浄し、そして蛍光活性細胞ソーティング(fluorescence activated cell sorting)(FACS(著作権))(Becton Dickinson、San Diego、CA)によって分析した。フルオレセインイソチオシアネート(FITC)(BD Pharmingen、San Diego、CA)に結合したヒトCD31に対して直接的に結合した一次マウスモノクローナル抗体を1:20希釈で、アロフィコシアニン(APC)(BD Pharmingen)に結合したヒトCD34を1:25希釈で、FITC(BD Pharmingen)に結合したヒトCD14を1:10希釈で、FITC(BD Pharmingen)に結合したヒトCD45を1:10希釈で、APC(BD Pharmingen)に結合したヒトCD117を1:100希釈で、フィコエリトリン(PE)(BD Pharmingen)に結合したCD146を1:10希釈で、PE(Miltenyi Biotec、Auburn、CA)に結合したヒトAC133を1:5希釈で、FITC(Cymbus Biotechnology、Chandlers Ford、UK)に結合したヒトCD141を1:10希釈で、Alexa Fluor 647(Alexa Fluor 647モノクローナル抗体標識キット、Molecular Probes、Eugene、OR)に結合したヒトCD105(BD Pharmingen)を1:100希釈で、およびAlexa Fluor 647に結合したヒトCD144を1:100希釈で使用した。
【0066】
細胞アゴニストによる活性化後、脈管細胞接着分子(VCAM−1)の細胞表面発現について試験するために、血清飢餓内皮細胞を、10ng/mlの組み換えヒトインターロイキン−1(IL−1)(Peprotech、Rocky Hill、NJ)か、または10ng/mlの組み換えヒト腫瘍壊死因子−α(TNF−α)(Peprotech)のいずれかを用いて、37℃で4時間刺激した。刺激の後、FITC(BD Pharmingen)に結合したヒトVCAM−1に対する一次抗体を1:20希釈で利用して、VCAM−1の細胞表面発現を試験した。免疫表現型決定(immunopherotyping)およびUCAM−1発現に対する全てのアイソタイプコントロールに関して、以下の抗体を使用した:FITC(BD Pharmingen)に結合したマウスIgG2a,κ、FITC(BD Pharmingen)に結合したマウスIgG1,κ、PE(BD Pharmingen)に結合したマウスIgG1,κ、およびAPC(BD Pharmingen)に結合したマウスIgG1,κ。
【0067】
フォン・ビルブラント因子(vWF)およびflk−1の細胞表面発現の検出のために、室温で10分間、アセトン中で細胞を固定し、PBSで2回洗浄し、そしてPBS、3%脱脂粉乳、および0.1% Triton X−100(Sigma)を用いて、30分間ブロックおよび浸透させた。本発明者らは、2μg/mlのヒトvWFに対する一次抗体(Dako、Carpenteria、CA)、および2μg/mlのヒトflk−1に対するビオチン化一次抗体(Sigma)を、1:20希釈で使用した。vWFに対して使用された二次抗体は、1:100希釈でのFITC(BD Pharmingen)に結合したヤギ抗ウサギ抗体であり、そしてflk−1に対して使用された二次抗体は、1:100希釈でのAPC(BD Pharmingen)に結合したストレプトアビジン(strepavidin)であった。vWFに対するアイソタイプコントロールについては、本発明者らは、ウサギIg一次抗体(Dako)を1:100希釈で、1:100希釈でのFITC(BD Pharmingen)に結合した抗ウサギIg二次抗体とともに使用した。flk−1に対するアイソタイプコントロールついては、本発明者らは、ビオチン化マウスIgG1,κ(BD Pharmingen)一次抗体を1:100希釈で、1:100希釈でのストレプトアビジン(strepavidin)APC二次抗体(BD Pharmingen)ともに使用した。
【0068】
(テロメラーゼ活性アッセイ)
テロメラーゼ活性の検出のために、TRAP−ezeテロメラーゼ検出キット(Oncor、Gaithersburg、MD)の形式で、テロメア反復増幅プロトコール(TRAP)を利用した。手短に言うと、培養した1000個のHPP−ECFCコロニーまたはEPCコロニーを、濾紙上に吸着させ、そしてTRAPアッセイ緩衝液中に溶解した。この溶解した材料をPCR増幅に供し、そしてそのPCR生成物(6bp増分のラダー)を非変性ポリアクリルアミドゲル上で電気泳動し、そしてDNA染色または32Pを用いた放射標識によって可視化した。PCR生成物を、そのままでか、または1/10希釈でか、もしくは1/100希釈でロードし、そして染色強度のレベルをHELA細胞株(1000細胞)ポジティブコントロールと比較した。
【0069】
(チミジン取り込みアッセイ)
内皮コロニー由来の内皮細胞を、成長因子を欠乏させ、そして5% FBSを補充したEBM−2培地中で24時間培養した。次に、3×104個の細胞を、I型コラーゲンで予めコートした6ウェル組織培養皿の各ウェル中にプレートし、1% FBSを補充したEBM−2培地中で16時間培養した。次いで、細胞を、血清を含まないEBM−2培地中でさらに8時間培養し、静止状態を確実にした。10% FBSを補充したEBM−2培地中で、25ng/mlの組み換えヒト脈管内皮成長因子(rhVEGF)(Peprotech)、25ng/mlの組み換えヒト塩基性線維芽細胞成長因子(rhbFGF)(Peprotech)、または成長因子無し(図示されるように)で、37℃、5% CO2の加湿インキュベーター中で細胞を刺激した。成長因子またはFBSを含まないEGM−2培地中で、いくつかの細胞を培養した。細胞を16時間培養し、そして収集の5時間前に、1μCiのトリチウム化チミジン(Perkin Elmer Life Sciences Products、Boston、MA)を添加した。細胞を、0.1N水酸化ナトリウムで1時間溶解した。溶解物を5mlの液体シンチラント(liquid scintilant)(Fisher Scientific、St.Louis、MO)中に収集し、そしてβ放射を測定した。アッセイを、三つ組み(triplicate)で行った。
【0070】
(GALV偽型MFG−EGFPの生成)
MFG−EGFPレトロウイルスベクターは、モロニーマウス白血病ウイルスの長い末端反復配列(LTR)の制御の下で増強された緑色蛍光タンパク質(EGFP)を発現する。そしてこれは、Pollokら(2001)によって以前に記載されている。GALV偽型ベクターの生成のための、アンホトロピック(amphotrophic)なMFG−EGFPクローンからの上清を使用し、PG13パッケージング株(American Type Culture Collection(ATCC)、Manassas、VA)を感染させ、そして感染した細胞を、単一細胞クローニングによって単離した。個々のクローンを、5×105個のヒト赤白血球病細胞(HEL)(ATCC)を感染させ、そして上清の終点希釈後48時間の百分率EGFP発現を決定することによって、力価についてスクリーニングした。MFG−EGFPクローン5は、0.5×106感染単位/ml〜1×106感染単位/mlの力価を有し、そしてこれを実験に使用した。
【0071】
(内皮細胞のレトロウイルス形質導入)
初期継代(1代〜2代)内皮コロニー由来の内皮細胞を、等量の出発の力価のMGF−EGFP上清を用いて形質導入した。6ウェル非組織培養プレートを、5μg/cm2フィブロネクチンCH−296(Takara Shuzo、Otsu、Japan)を用いて、室温で2時間かまたは4℃で一晩コートした。プレートをPBSで1回洗浄し、そして形質導入のために内皮細胞を5×104細胞/cm2でプレートした。完全EGM−2培地で1:1に希釈したレトロウイルス上清を用いて細胞を、一晩の培養の間完全EGM−2培地の交換を行い、連続2日間4時間感染させた。第2ラウンドの感染の後、細胞を収集し、計数し、そして蛍光サイトメトリーによってEGFP発現について分析した。
【0072】
(単一細胞アッセイ)
上記MFG−EGFPレトロウイルスで形質導入した初期継代(1代〜4代)内皮コロニー由来の内皮細胞を、蛍光サイトメトリーによって、EGFP発現について分類した。FACS Vantage Sorter(Becton Dickenson)(≦9ポンド/平方インチのシーズフロー圧で≧100ミクロンのソートノズル)を使用し、200μlの完全EGM−2培地を含んだI型コラーゲンで予めコートした96ウェル平底組織培養プレートの各ウェル中に、EGFPを発現する1つの単一内皮細胞を置いた。50×拡大率の蛍光顕微鏡下で個々のウェルを検査し、1個の細胞のみが各ウェル中に置かれていることを確実にした。加湿インキュベーター中で、37℃、5%CO2で細胞を培養した。培地を、100μl取り出し、それを100μlの新鮮な完全EGM−2培地で置き換えることによって、4日間毎に交換した。第14日目に、プレートした1個の細胞からの内皮細胞の成長について、各ウェルを検査した。単一内皮細胞の分裂の頻度を定量化するために、100×拡大率の蛍光顕微鏡を用いて2個以上の内皮細胞を有する細胞の数を計数した。ウェルあたりの細胞の数を数えるために、100×拡大率の蛍光顕微鏡を用いた目視検査によって、上記細胞を計数するか(ウェルあたり50細胞未満)、または上記細胞をトリプシン処理し、そしてトリパンブルー排除アッセイを利用して血球計を用いてそれらを計数した(ウェルあたり50細胞超)。
【0073】
単一細胞に由来する内皮細胞の長期間増殖能および長期間複製能を、決定した。培養の開始の14日後、50細胞より多くを含む個々のウェルを、トリプシン処理し、500μlの完全EGM−2培地中に収集し、そしてI型コラーゲンでコートした24ウェル組織培養皿に継代培養した。この細胞の継代培養の4日後、その培地を吸引し、そして500μlの新鮮な完全EGM−2培地で置き換えた。7日目に、50×拡大率の蛍光顕微鏡を用いた目視検査によって、コロニー成長または細胞集密についてウェルを検査した。次いで、細胞をトリプシン処理し、計数し、そしてI型コラーゲンで予めコートした6ウェル組織培養プレート中に継代培養した。6ウェルプレートにおける培養の7日後、本明細書中で開示される条件下で、長期間培養について、集密な細胞単層を含んだ10ウェル〜12ウェルを選択した。各試料について、PDTおよびCPDLを計算した。
【0074】
(CDマーカー)
CD14(リポ多糖レセプター)
CD31(血小板内皮細胞接着分子)
CD34(シアロムチン)
CD45(共通白血球抗原)
CD105(エンドグリン)
CD117(c−Kitレセプター)
CD133(プロミニン1)
CD141(トロンボモジュリン)
CD144(脈管内皮カドヘリン)
CD146(内皮関連抗原、S−エンド−1)
flk−1(胎児肝臓キナーゼ−1、脈管内皮成長因子2に対するレセプター)。
【0075】
(EPCの共焦点画像化)
10% FBSを添加したEBM−2培地を使用し、T75フラスコ中で4日間、3代〜5代継代したEPCを成長させた。細胞が集密に達した場合、培地を吸引し、上記フラスコに5mlの滅菌PBSを添加し、次いで吸引し、トリプシン−EDTAを添加し、そしてこのフラスコを37℃で5分間インキュベートした。このトリプシンをクエンチするために、5mLの10% FBSを含むEBM−2培地を添加し、そして遊離したEPCを350×Gで10分間遠心分離した。ペレット化した細胞をPBSで洗浄し、次いで10% FBSを含むEBM−2培地中に再懸濁した。
【0076】
ガラス製チャンバースライド(4チャンバー構造;Corning)を、4℃で一晩、細胞外マトリックスタンパク質(例えば、1型コラーゲンもしくは4型コラーゲン、フィブロネクチン、またはビトロネクチン)でコートし、次いで翌朝に(in the morning)滅菌PBSで洗浄した。このPBSを吸引し、そして10% FBSを含むEBM−2培地中の細胞を、チャンバーあたり50細胞で添加し、そして5%CO2中で、37℃で7日間インキュベートした。
【0077】
EPCを含むスライドをPBSで2回洗浄し、そしてアセトン中で10分間室温で細胞を固定し、PBSで2回洗浄し、そしてPBS、3%脱脂粉乳、および0.1% Triton X100を用いて、30分間、ブロックおよび浸透させた。この細胞の細胞膜を強調するために、核染色のための1.5mg/ml DAPIとともに、フィコエリトリン(PE)に結合したCD146に対する一次抗体(1□g/mL)を、上記固定した細胞に添加した。30分間のインキュベーション後、PBS中で細胞を2回洗浄し、そしてZeiss 510共焦点顕微鏡を使用して蛍光について検査した。7×拡大率で倍率を保持したまま、40×水浸対物レンズを介して、紫外レーザー(351/364nm励起)およびヘリウム−ネオンレーザー(543nm励起)を使用して上記DAPIおよびPE−標識した細胞を励起した。画像を一平面中に取り込み、そして提示のための単色画像として表示した。NIH Imageソフトウェアを使用し、種々のEPCコロニー型に由来する細胞の、核の直径および細胞質の直径を定量化した。
【0078】
(表1)
成体末梢血単核細胞および臍帯血単核細胞から単離した内皮前駆細胞コロニーの、出現の数および時期の一覧表。
【0079】
【表1】
。
【0080】
(HPP−ECFCおよびCD34+細胞の共培養は、NOD−SCID再増殖細胞を増殖させる)
NOD/SCIDマウスにおける骨髄(marrow)再増殖細胞(SRC)を有する(harbor)ことが以前に示された、ヒトCD34+骨髄細胞を単離した。代表的には、5%〜50%のヒトCD45+キメラ現象のレベルを達成するために、0.5×106個〜1.0×106個のヒト骨髄CD34+細胞を、NOD/SCIDマウス中に注入する。最初に、ヒト骨髄からの収集の日に、9×103個のCD34+細胞のみをコントロールとしてNOD−SCIDマウス中に注入した。SCF、G−CSF、TPO、およびFlt−3の存在下で、9×103個のCD34+細胞を7日間培養した。これらは、エキソビボにおいてHSCを最大に増殖させるために現在使用されている成長因子である。成長因子の不在下で、臍帯血HPP−ECFC由来の子孫の単層とともに9×103個のCD34+細胞を7日間共培養した。培養の7日後、この培養したCD34細胞をNOD−SCIDマウス中に注入し、そしてヒト細胞の存在下で、移植の4週間後、移植されたマウスの末梢血を試験した。CD34+細胞の成長因子との7日間の共培養は、ヒト骨髄から単離した直後に注入したCD34+と比較して、移植の8週間後のNOD−SCIDマウス中で検出されたヒト細胞の百分率を10倍増大させた(図12)。以前の研究と比較して、非常に限られた数の細胞の注入にもかかわらず、臍帯血HPP−ECFC由来の細胞とのCD34+細胞の共培養は、移植の8週間後のNOD−SCIDマウスにおいて検出されたヒト細胞の百分率を260倍増大させた(図12)。ヒト骨髄系統およびリンパ系統の両方が移植の8週間後に検出された。このことは、造血系の多系統の再構成が、臍帯血HPP−ECFCとのCD34+細胞の共培養によって達成されたことを示した。
【0081】
HPP−ECFC由来の細胞の集密な単層の2つのT75フラスコを用いて開始して、カルシウムまたはマグネシウムを欠いたハンクス平衡塩溶液(HBBS)で最初に細胞を洗浄し、次いで各フラスコに1分間1.5mLのトリプシンEDTA(Gibco)を添加した。次に、10%ウシ胎仔血清(FBS)(Hyclone)を含む8.5mLの内皮基礎培地2(EBM2)(Cambrex)を添加し、そして懸濁した細胞を収集し、そして血球計上でトリパンブルー排除を介して計数した。
【0082】
コラーゲン1で予めコートした6ウェル組織培養プレート(BD Biosciences)上に、HPP−ECFC由来の細胞を、3×105細胞/ウェルでプレートした。10% FBSを補充した内皮成長培地2(EGM2)(Cambrex)を用いて細胞を培養し、そして一晩培養した。翌朝、その集密細胞単層をEBM2+10% FBSで2回洗浄し、次いで、蛍光活性化細胞分類によって収集し、4mLのEBM2+10% FBS+ヒト巨核球成長由来因子(MGDF)(100ng/mL)、顆粒球コロニー刺激因子(G−CSF)(100ng/mL)、および幹細胞因子(SCF)(100ng/mL)、ならびにflt−3リガンド(100ng/mL)中に再懸濁した9,000個のCD34+CD38dimLin−(CD4、CD8、CD11b、CD14、CD24、CD31、CD33、およびグリコホリンA)成体ヒト骨髄由来細胞とともに共培養した。37℃、5% CO2加湿インキュベーター中で7日間、障害無しに細胞を培養した。いくつかの培養物において、EBM2+10% FBS中でHPP−ECFCとともに、そして成長因子無添加で、CD34+細胞を共培養した。
【0083】
7日間の共培養の後、5mLのピペットを使用して非接着細胞および培地(4mL)を吸引した。2mLリン酸緩衝生理食塩水(PBS)で1回ウェルを洗浄し、そして最初の吸引物にそのPBSとともに非接着細胞を添加した。同じウェルに対して、1mLの細胞分離緩衝液(Gibco)を室温で4分間添加し、次いで、吸引および上記最初の吸引物への添加の前に、この細胞分離緩衝液およびバラバラになった細胞をそのウェル中で穏徐に滴定した。最後に、上記HPP−ECFC単層を2mLのPBSで最後に1回洗浄し、そして上記最初の吸引物中に、わずかな細胞を含むこの溶液を添加した。培地および細胞の最終容積は、9mLであった。
【0084】
この細胞懸濁物を、室温で1500rpm(514×g)で10分間遠心分離した。溶液を取り除き、細胞ペレットを機械的に取り出し(dislodge)、次いで1/2mL〜1mLのEBM2+10% FBS中に再懸濁した。血球計上において、トリパンブルー中で細胞を計数した。回収した細胞を、前駆体アッセイにおいてプレートするか、またはNOD/SCIDマウス中に静脈内注入した。
【0085】
上記に概説された方法は、ヒト移植のための移植片を提供するために改変され得る。この場合には、HPP−ECFC子孫は、臍帯血HPP−ECFCの存在下で、大数の(自己または同種異系のヒト臍帯血由来、動員末梢血由来、または骨髄由来の)CD34+造血幹細胞が増殖し得るように、T75フラスコ中または灌流チャンバー系にプレートされる。細胞に直接触れることなくドナーCD34+細胞がHPP−ECFC子孫とともに培養され得るような系が使用され、したがって、上記ドナーCD34+細胞は増殖され得、回収され得、そして臍帯血HPP−ECFC子孫の「混入」したドナー細胞無しで、ヒト患者中に移植され得る。
【0086】
(文献)
以下の文献は、開示される方法および組成物が本発明の実施において使用される程度に、参考として本発明の範囲中に援用される。
【0087】
【表2】
【0088】
【表3】
【0089】
【表4】
【0090】
【表5】
【0091】
【表6】
【図面の簡単な説明】
【0092】
【図1−1】成体末梢血および臍帯血に由来する、内皮前駆細胞コロニーの単離。図1aは、成体末梢血(AB)および臍帯血(CB)20mlあたりの単離されたEPCコロニーの数。結果は、成体ドナーについての18の独立した実験および臍帯血についての13の独立した実験についての、EPCコロニーの平均数±SEMの平均数を示す。*対応のあるスチューデントt検定によって、P<0.0001。図1bは、等容積の成体末梢血(AB)および臍帯血(CB)からの培養開始後の、初期EPCコロニー出現の時間である。結果は、成体ドナーについての18の独立した実験および臍帯血についての13の独立した実験についての、初期EPCコロニー出現の前の平均日数±SEMの前の平均日数を示す。*対応のあるスチューデントt検定によって、P<0.0001。
【図1−2】成体末梢血および臍帯血に由来する、内皮前駆細胞コロニーの単離。図1cは、成体血液に由来する内皮コロニーの、代表的な低分解能顕微鏡写真および高分解能顕微鏡写真である。図1dは、CBに由来する内皮細胞コロニーの、代表的な低分解能顕微鏡写真および高分解能顕微鏡写真である。図1eは、低分解能拡大率および高分解能拡大率での、成体内皮細胞コロニーに由来する内皮細胞単層である。図1fは、低拡大率および高拡大率での、CB内皮細胞コロニーに由来する、内皮細胞単層である。写真中のスケールバーは、100μm(c、d、e、fの左側カラム)、および10μm(c、d、e、fの右側カラム)を表す。
【図2】成体EPC由来および臍帯血EPC由来の内皮細胞の、表現型分析および機能分析。図2a〜図2bは、蛍光サイトメトリーによる、成体EPCコロニー(図2a)または臍帯血EPCコロニー(図2b)のいずれかに由来する細胞単層の免疫表現型決定である。臍帯血EPC由来の細胞および成体EPC由来の細胞の両方は、CD31、CD141、CD105、CD146、CD144、vWF、およびFlk−1を発現するが、CD45およびCD14を発現しない。いくつかの臍帯血細胞および成体細胞は、CD34、CD133、およびCD117を発現する。示されているものは、同様の結果を有する、異なる成体細胞単層を利用した18の独立した実験および異なる臍帯血細胞単層を使用した13の独立した実験からの、代表的なデータである。アイソタイプコントロールは、試験した各表面抗原についての各ヒストグラム上にグレーでオーバーレイされる。(図2c)成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞は、DiI−Ac−LDLを取り込む(50×拡大率)。DiI−Ac−LDLを取り込み(赤色)、かつDAPIで染色された(青色)、成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞についての代表的な写真が示される。示されているものは、同様の結果を有する、異なる成体細胞単層を利用した18の独立した実験および異なる臍帯血細胞単層を使用した13の独立した実験からの、代表的なデータである。写真中のスケールバーは、100μmを表す。(図2d)毛細管様の構造の形成のためにマトリゲル中にプレートされた、増強された緑色蛍光タンパク質(EGFP)を発現するように遺伝的に操作された成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞(50×拡大率)。示されているものは、同様の結果を有する、異なる成体細胞単層を利用した18の独立した実験および異なる臍帯血細胞単層を使用した13の独立した実験からの、代表的なデータである。写真中のスケールバーは、100μmを表す。(図2e)成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞は、rhTNF−αか、またはrhIL−1のいずれかに応答して、脈管細胞接着分子(VCAM−1)の細胞表面発現を上方制御する。示されているものは、同様の結果を有する、異なる成体細胞単層を利用した18の独立した実験および異なる臍帯血細胞単層を使用した13の独立した実験からの、代表的なデータである。VCAM−1に関するアイソタイプコントロールは、各ヒストグラム上にグレーでオーバーレイされる。
【図3】成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞を、低い細胞密度で再プレートした7日後に形成された、形態学的に明確に異なる二次コロニーの代表的な写真。
【図4】臍帯血内皮前駆細胞コロニーおよび成体内皮前駆細胞コロニーに由来する内皮細胞子孫の成長動態。(図4a)単核細胞から収集された、成体血液(AB)EPC由来の内皮細胞および臍帯血(CB)EPC由来の内皮細胞のエキソビボ増殖。黒色の四角(box)は、各継代における総細胞数を表す。各継代において、細胞は上で示した内皮細胞表面抗原(図2a〜図2b)を均一に発現したが、造血細胞特異的抗原(CD45およびCD14)を発現しなかった。臍帯血EPC由来の内皮細胞および成体EPC由来の内皮細胞についての代表的な成長曲線を示す。異なるドナーに由来する、11の他の臍帯血内皮細胞単層および成体内皮細胞単層は、同様の成長動態を示した。(図4b〜図4c)60日間の培養の間の、臍帯血(CB)EPC由来の内皮細胞および成体血液(AB)EPC由来の内皮細胞の集団倍加時間(PDT)および累積集団倍加レベル(CPDL)。結果は、6つの独立した実験のPDTの平均数±SEMの平均数およびCPDLの平均数±SEMの平均数を表す。*対応のあるスチューデントt検定によって、P<0.01。(図4d)臍帯血(CB)EPC由来の内皮細胞および成体血液(AB)EPC由来の内皮細胞のDNA合成。初期継代(1代〜2代)の臍帯血EPC由来の内皮細胞は、10% FBS、rhVEGF、およびrhbFGFに応答して、成体細胞と比較して増大したDNA合成を示す。結果は、異なるドナーに由来する内皮細胞を利用した4つの独立した実験の平均を表す。*対応のあるスチューデントt検定によって、P<0.01。
【図5】EPCコロニーに由来する、単一臍帯血内皮細胞および単一成体内皮細胞のクローン原性能および増殖能の定量化。(図5a)成体EPCコロニーか、または臍帯EPCコロニーのいずれかに由来する内皮細胞を使用した単一細胞アッセイの概略図。(図5b)培養の14日後、少なくとも1回の細胞分裂を起こした成体血液(AB)EPC由来の内皮細胞または臍帯血(CB)EPC由来の内皮細胞の百分率。結果は、異なるドナーに由来する単一内皮細胞を使用した5つの独立した実験の平均を表す。*対応のあるスチューデントt検定によって、P<0.01。(図5c)培養の14日後の、単一成体血液(AB)EPC由来の内皮細胞または単一臍帯血(CB)EPC由来の内皮細胞に由来する細胞子孫の平均数。結果は、異なるドナーに由来する単一内皮細胞を使用した5つの独立した実験の平均を表す。*対応のあるスチューデントt検定によって、P<0.01。(図5d)定量可能な範囲を示す細胞数を有するコロニーを生じる、分裂する単一細胞の百分率(HPP、LPP、クラスター)。(図5e)単一臍帯血EPC由来の内皮細胞または単一成体EPC由来の内皮細胞に由来する、種々の内皮細胞クラスター(<50細胞)、LPP(約51細胞〜2000細胞)、およびHPP(約2000細胞〜>10,000細胞)の代表的な写真(50×拡大率)。結果は、異なるドナーに由来する細胞を利用した4つの他の独立した実験を表す。写真中のスケールバーは、100μmを表す。LPP−ECFC(51細胞〜2000細胞)およびHPP−ECFC(2000細胞>10,000細胞)の範囲は、概算値である。
【図6】単一臍帯血EPC由来の内皮細胞または単一成体EPC由来の内皮細胞に由来する細胞子孫の、再プレート能および長期間培養。(図6a)24ウェル組織培養プレートにおいて、培養の7日後に二次コロニーを形成するか、または細胞集密まで急速に成長する単一臍帯血(CB)EPC由来の内皮細胞または単一成体(AB)EPC由来の内皮細胞に由来する細胞子孫の百分率。結果は、4人の異なるドナーに由来する細胞を使用した4つの独立した実験の平均±SEMを表す。*対応のあるスチューデントt検定によって、P<0.01。(図6b)培養の7日後における、24ウェルプレート中の、単一のプレートされた臍帯血由来の内皮細胞の細胞子孫に由来する、二次内皮細胞コロニーまたは集密細胞単層の代表的な写真(50×拡大率)。写真中のスケールバーは、100μmを表す。(図6c)長期間培養における、11の単一のプレートされた、3人の異なる臍帯血ドナーから単離された内皮細胞の細胞子孫の成長動態。黒色の四角は、各継代における細胞の総数を表す。(図6d)異なるドナーから単離された、1000個の初期継代の成体血EPC由来の内皮細胞(レーン1〜レーン4)および臍帯血EPC由来の内皮細胞(レーン5〜レーン8)のテロメラーゼ活性。15のCDPLで、成体細胞および臍帯細胞を試験した。Pは、ポジティブコントロールとして使用した1000個のHeLa細胞におけるテロメラーゼ活性を示し、そしてNは、ネガティブコントロールを示す。成体試料におけるテロメラーゼ活性の平均レベルは、HeLa細胞のテロメラーゼ活性の4±4%であり、そして臍帯血試料のテロメラーゼ活性の平均レベルは、HeLa細胞のテロメラーゼ活性の34±10%であった。(図6e)初期継代および後期継代の成体EPC由来の内皮細胞(a)および臍帯血EPC由来の内皮細胞(c)のテロメラーゼ活性の比較。PDは、試験した細胞の累積集団倍加レベルを示す。Pは、ポジティブコントロールとして使用したHeLa細胞におけるテロメラーゼ活性を示す。Nは、ネガティブコントロールを示す。3人の異なるドナーに由来する初期継代および後期継代の臍帯血EPC由来の内皮細胞および成体EPC由来の内皮細胞を利用した3つの他の実験は、同様の結果を示した。
【図7】前駆細胞の別個の集団の増殖能およびクローン原性能に基づく、内皮前駆細胞階層のモデル。高増殖能内皮コロニー形成細胞(HPP−ECFC)は、再プレートによって少なくとも二次コロニーを形成する、1つの細胞から生じる大きなコロニーである。HPP−ECFCは、全てのその後の段階の内皮前駆体を生じ、さらに二次HPP−ECFCへと再プレートされる。単一細胞から生じた低増殖能内皮コロニー形成細胞(LPP−ECFC)は、50細胞より多くを含むコロニーを形成するが、再プレートによってLPP−ECFCの二次コロニーを形成しない。内皮細胞クラスター(EC−クラスター)は、1つの細胞から生じ得るが、50細胞未満しか含まない。これは、HPP−ECFCコロニーおよびLPP−ECFCコロニーにおいて見出されるより小さな細胞と比較して、代表的にはより大きい(図10を参照のこと)。成熟した末期分化内皮細胞は、分裂しない。
【図8】臍帯血EPC、HUVEC、およびHAEC由来の内皮細胞の免疫表現型分析。(図8a〜図8b)臍帯血EPC(a)、臍静脈(b)、またはヒト大動脈(c)に由来する細胞単層の、蛍光サイトメトリーによる免疫表現形決定。臍帯血EPC由来のEC、HUVEC、およびHAECは、CD31、CD141、CD105、CD146、CD144、vWF、およびFlk−1を発現するが、CD45およびCD14を発現しない。5つの異なる臍帯血EC単層、5つの異なるHUVEC試料、および5つの異なるHAEC試料を用いた5つの独立した実験からの代表的なデータは、同様の結果を示した。アイソタイプコントロールは、試験した各表面抗原について、各ヒストグラムに白色でオーバーレイされる。
【図9】臍帯血EPCコロニー、HUVEC、およびHAECに由来する単一内皮細胞のクローン原性能および増殖能の定量化。(図9a)培養の14日後に、少なくとも1回の細胞分裂を起こす単一臍帯血(CB)EPC由来のEC、HUVEC、またはHAECの百分率。結果は、異なるドナーに由来する単一ECを使用した5つの独立した実験の平均を表す。(図9b)個々のウェル中において分裂する単一細胞の百分率(図5を参照のこと)。培養の14日後に、個々のウェル中において(分類されたような)細胞のコロニーを生じる、単一CB EPC由来のEC、HUVEC、またはHAECの百分率。*単一CB由来のEC 対 単一HUVECまたは単一HAECのいずれかの比較のための、対応のあるスチューデントt検定によって、P<0.01。(図9c)単一臍帯血EPC由来のEC、HUVEC、またはHAECに由来する、種々のECクラスター(<50細胞)またはコロニー(>50細胞)の代表的な写真(50×拡大率)。結果は、異なるドナーからの細胞を利用した4つの他の独立した実験の代表的なものである。写真中のスケールバーは、100μmを表す。(図9d)24ウェル組織培養プレートにおいて、培養の7日後に、二次コロニーを形成するかまたは細胞集密まで急速に成長した、単一臍帯血EPC由来のEC、HUVEC、またはHAECに由来する細胞子孫の百分率。結果は、4人の異なるドナーに由来する細胞を使用した、4つの独立した実験の平均±SEMを表す。*単一CB由来のEC 対 単一HUVECまたは単一HAECのいずれかの比較のための、対応のあるスチューデントt検定によって、P<0.01。(図9e)培養の7日後における24ウェルプレート中の、単一のプレートされた臍帯血EPC由来のEC、HUVEC、またはHAECの細胞子孫に由来する二次ECコロニーまたは集密細胞単層の代表的な写真(50×拡大率)。写真中のスケールバーは、100μmを表す。(図9f)HUVECに由来するHPP−ECFCコロニー(HPP)およびLPP−ECFCコロニー(LPP)におけるテロメラーゼ活性。Pos.は、ポジティブコントロールとして使用したHeLa細胞におけるテロメラーゼ活性を示し、そしてNeg.は、ネガティブコントロールを示す。結果は、4つの他の独立した実験の代表的なものである。臍帯血ECおよび臍帯血HAECから単離された、HPP−ECFCコロニーとLPP−ECFCコロニーとの間において、テロメラーゼ活性における、同様の相違を観察した。
【図10】HPP−ECFCの単色画像(図10a)、LPP−ECFCの単色画像(図10b)、内皮クラスターの単色画像(図10c)、および成熟した分化内皮細胞の単色画像(図10d)。HPP−ECFCは、最小の細胞質拡散(cytoplasmic spreading)(直径は12ミクロン〜22ミクロンで変化)を有し、>0.8の核 対 細胞質比を有する小さな細胞(核直径8ミクロン〜10ミクロン)である。LPP−ECFCは、サイズに関してより不均一(heterogenous)であるが、HPP−ECFCと比較して、より大きい。LPP−ECFC核は、10.5ミクロン〜12.5ミクロンでサイズが変化し、そして>0.4の比を有するが、<0.5であるより大きな細胞質拡散(25ミクロン〜60ミクロンで変化)を有する。内皮クラスターは、13.0ミクロン〜16.5ミクロンで変化する核を有する、ほぼ成熟した内皮細胞であり、そして>0.2であるが、<0.3である核 対 細胞質比を有する、65ミクロン〜80ミクロンで変化する細胞質直径を有する。成熟した分化内皮細胞は、大きくて非常に良好に拡散した細胞であり、17.0ミクロン〜22.0ミクロンの範囲の核直径および85ミクロン〜105ミクロンの細胞質直径を有し、そして内皮クラスターと同様の核 対 細胞質比を有する。したがって、HPP−ECFCは、任意の他のEPCよりも非常に明確に小さく、そして成熟内皮細胞よりもかなり小さい。
【図11】明確に分類されたサイズのコロニーを生じる、分裂した単一のプレートされたウシ、ブタ、およびラットの大動脈内皮細胞の百分率。
【図12】(A)単離の同日に注入したヒト骨髄由来のCD34+細胞の移植;(B)最大刺激濃度のG−CSF、TPO、SCF、またはFlt−3を用いた7日間の培養;または(C)臍帯血HPP−ECFCとの7日間の共培養(ここで、同期(gated)細胞は、ヒトCD45抗原についてポジティブに染色されている)の、8週間後のNOD−SCIDマウスの末梢血において、フローサイトメトリーによって検出されたヒト細胞(CD45ポジティブ)のキメラ現象の百分率;グラフは、同様の結果を有する3つの独立した実験の代表的なものである。
【背景技術】
【0001】
(発明の背景)
内皮細胞および造血細胞を生じる幹細胞分化に関するモデルは、幹細胞およびその子孫の臨床的価値のため、関心のあるものである。幹細胞および前駆細胞の特徴は、それらの増殖する能力および機能的子孫を生じる能力であり、そして前駆細胞は、そのクローン原性能によって同定される。以前に使用されていた方法は、前駆体を同定するために、一種類の内皮細胞が単離および特徴付けされていたことを保証しない。
【0002】
ほとんどの哺乳動物において、正常な、成熟動脈、静脈、および毛細管におけるインビボでの内皮細胞増殖は、存在していたとしても、非常に遅いことが報告されている。いくつかの実験動物(例えば、ブタおよびイヌ)において、放射標識研究は、0.6%〜3.0%の内皮細胞が、特定の脈管の限局された範囲に限られた分裂細胞によって毎日ターンオーバーすることを実証した。これらの分裂内皮細胞が独特であり、そして他の成熟した内皮に欠けている増殖能を有するかどうかは、未確定のままである。
【0003】
好対照なことに、インビトロにおいて、ヒト脈管由来または動物脈管由来である内皮細胞のプレーティングは、最初に、活発な内皮細胞増殖を伴う。例えば、臍静脈内皮細胞(HUVEC)およびウシ大動脈内皮細胞(BAEC)は、内皮細胞機能のインビトロ分析のための、2つの一般に研究されるモデルである。HUVEC細胞およびBAEC細胞の両方は、培養の初期においては良く増殖するが、経時的に細胞分裂は弱々しくなり、そして細胞は老化して15継代〜20継代の後、分裂できなくなる。脈管由来である各々の内皮細胞が同様の増殖能を有するのか否か、または細胞の一部のみが増殖し得るのか否かは、不明である。
【0004】
脈管新生(新脈管形成)は、既存の脈管からの新規の脈管の形成のプロセスである;これは、成体被験体において、新規の脈管を生じると報告されているプロセスである。最近、骨髄由来循環内皮前駆細胞(EPC)が記載され、そして、これらの細胞はまた、少なくともいくつかの実験的マウスの虚血モデルまたは腫瘍モデルにおいて、新規の脈管の形成に役割を果たすことが報告されている。相反する証拠は、発生の間の骨髄由来(EPC)が正常動脈、静脈、および毛細管の内皮の内張り(lining)に寄与せず、そして新脈管形成において軽微な役割のみを果たすことを示す。循環EPCと正常脈管に存在する増殖能を有する上記内皮細胞との間の関係は、不明である。
【0005】
脈管新生治療のためのEPCの使用、または患者の心臓血管疾患の危険および進行を評価するためのバイオマーカーとしてのEPCの使用を支持する新たな証拠は、蓄積し、そして興味をかきたてている。しかしながら、EPCの一様な定義は無く、このことは、これらの研究の解釈を問題のあるものとし、そして臨床使用のために適切な細胞型の再現を妨げる。(例えば、造血、腸、神経細胞の)幹細胞および前駆細胞の特徴は、それらの増殖する能力、および機能的な子孫を生じる能力であるが、EPCは、選択された細胞表面抗原の発現によって第一に規定される。分子の細胞表面発現に唯一依存することは、その発現が上記細胞の生理学的状態によって変化し得るので、問題となり得る。個々の内皮細胞またはEPCにおける増殖能(内因性応答)を評価するためのアッセイは報告されておらず、したがって、比較分析は利用不可能である。
【0006】
以前の研究は、「内皮前駆細胞」と名付けられた細胞の集団が、細胞表面抗原CD34を発現する分類された細胞か、または単核細胞のいずれかを、規定された培養条件において培養することによって、ヒト臍帯血からか、または成体末梢血から単離され得ることを報告した。
【0007】
造血前駆細胞および内皮前駆細胞は、卵黄嚢および胚の発生において、多くの細胞表面マーカーを共有し、そして多数の遺伝子の遺伝子破壊が、造血細胞および内皮細胞の両方の発達に影響する。したがって、これらの系統は、共通の前駆体(血管芽細胞)から起こると仮定される。造血細胞の発達における幹細胞および前駆細胞の階層が、報告された。上記階層内の造血前駆細胞は、それらのクローン原性および増殖能によって同定される。遺伝的研究が、内皮細胞の起源が造血細胞の発達に密接に関連することを明白に示しているにもかかわらず、増殖能の違いに基づいて幹細胞および前駆細胞の同様の階層を支持する証拠は確立されていない。すなわち、造血細胞系に類似する個々の細胞のクローン原性および増殖能によって区別され得るEPCの階層は、報告されていない。
【0008】
造血幹細胞および前駆細胞(HSC/Ps)の両方は、成体の末梢血と比較して、臍帯において豊富である。現在、臍帯血は、種々の血液学的な障害および悪性疾患を有する患者の移植のための、造血幹細胞の代替的な資源として使用されている。
【0009】
毎年、何千もの患者が造血幹細胞(HSC)の移植を必要としている。この患者のほぼ2/3が、移植のためのヒト白血球抗原(HLA)適合性一致を見出せ得ずにいる。これは、特に、多くの民族集団および代表的でない少数派(under−represented minority)に関してあてはまる。白人患者の1/3のみが、適切に一致した同胞の移植片(最低限の対宿主性移植片病(GVHD)合併症を伴う、最も適合性のある供給源)を見出す。
【0010】
ヒト臍帯血は、臨床移植のためのHSCの代替的な供給源であることが知られている。ドナー臍帯血が、レシピエントに対して完全に主要組織適合性が一致するか、または不一致かにより、種々の先天性または後天性の血液学的障害のための処置として、臍帯血細胞は、調整された(conditioned)宿主に移植され、そして再増殖する。たとえこの臍帯血移植片が上記レシピエントにとって2つ以上の座で不一致であったとしても、GVHDの発生率および重症度は、同様の不一致の成体の骨髄移植片または動員(mobilized)末梢血移植片の移植に関して観察されるGVHDの発生率および重症度と比較して、有意に小さい。
【発明の開示】
【発明が解決しようとする課題】
【0011】
移植に関する臍帯血のより広範な使用に対する制限するものとしては、限られた数のHSCおよび前駆細胞のみしか移植片中に存在しないという事実が挙げられる。ほとんどの患者が、一致した同胞のドナーを有さないので、ほとんどの臍帯血移植片は、不一致のレシピエントへと移植される。多くの研究が、移植片中の臍帯血の用量は、上記移植片が血縁関係の無いドナーに由来する場合、患者の生存に対して重要であることを報告する。移植に関連する死亡率は、1.7×105個のCD34+細胞/kgの臍帯血移植片を受けたレシピエントにおいて20%、これに対して移植片中のより少量のCD34+細胞を受けたレシピエントにおいて75%と報告されている。エキソビボで臍帯血HSCを効率的に増殖させ、移植片中の細胞数を増加させるための方法を発見することは、臨床的移植のための大きな進歩であり、そして大きな商業的市場を有する。
【0012】
臍帯血HSC増殖のためのアプローチは、目覚しいものではない。ほとんどの研究において、臍帯血単核細胞または単離されたCD34+細胞に対する種々の成長因子の添加は、総細胞数における増加、コロニー形成単位細胞(CFC)数における増加、および免疫不全(NOD/SCID)マウスまたはヤギ胎仔における短期間の前駆細胞移植における増加と、相関している。しかしながら、NOD/SCIDマウスにおけるSCID再増殖細胞(SRC)頻度、またはヤギ胎仔における長期間の移植によって測定されるようなHSCの数の増加については、ほとんどのアプローチが効果的でない。ヒト患者における増殖された臍帯血HSCの使用の結果は、失望すべきものであった。
【課題を解決するための手段】
【0013】
(発明の要旨)
単一細胞コロニーアッセイは、末梢血および臍帯血から、ならびに臍帯または成体血管から単離された内皮細胞から単離された、哺乳動物の内皮前駆細胞(EPC)の間での新規の階層を説明するために開発された。ヒト、ウシ、ブタおよびラットの生物学的試料に由来する前駆細胞の異なる集団は、クローン原性および増殖能に基づいて同定された。
【0014】
内皮前駆細胞(EPC)は、成体末梢血および臍帯血から単離され、そしてエキソビボで指数関数的に増殖された。対照的に、ヒト臍静脈内皮細胞(HUVEC)または血管壁に由来するヒト大動脈内皮細胞(HAEC)は、広義には、分化した成熟内皮細胞(EC)としてみなされ、そしてそれらは、EPC研究のための「コントロール」として利用される。しかしながら、成体および臍帯血由来のEPCと同様に、血管壁由来であるHUVECおよびHAECは、インビトロにおいて少なくとも40集団倍加の間継代され得る。EPCを、それらの増殖能およびクローン原性能に基づいて識別する、新規の単一細胞析出(deposition)アッセイの利用は、HUVECまたはHAEC中に、EPCが常在することを見出した。単一細胞クローン原性アッセイは、EPCの増殖能およびクローン原性能に基づいて、EPCの新規の階層を明確にするために開発された。EPCの完全な階層は、血管壁由来のHUVECおよびHAECにおいて同定され、そしてそれらのクローン原性能および増殖能によって識別された。EPCの多様性がヒト脈管において存在し、そして、脈管の一体の維持におけるEPCの起源および機能の両方を決定するための、概念的な枠組みを提供する。したがって、EPCは、臨床的使用(例えば、移植片)のために、末梢血またはヒト脈管の生検のいずれかから容易に得られる。
【0015】
単離された内皮コロニー形成細胞は、以下の特徴を有する:
(a)内皮細胞の特有の細胞表面抗原(例えば、CD31、CD105、CD146、およびCD144)を発現する;
(b)造血細胞に特有の細胞表面抗原(例えば、CD45、およびCD14)を発現しない;
(c)アセチル化LDLを取り込む;および
(d)マトリゲル(matrigel)において、毛細管様の管を形成する。
【0016】
HPP−ECFCとして分類される、単離された細胞はまた、
(a)単一細胞からプレートされた場合、少なくとも約2000細胞の、少なくとも二次コロニーへと再プレートする;
(b)高増殖性を示す;
(c)単一細胞から増殖する;および
(d)高レベルのテロメラーゼ(HeLa細胞によって発現されるテロメラーゼレベルの、少なくとも34%)を発現する。HPP−ECFCはまた、>0.8である、高い核 対 細胞質比を示し、細胞直径は、<22ミクロンであり、そして少なくとも107個の子孫が、単一細胞に由来する。
【0017】
内皮コロニー形成細胞を単離するための方法は、以下の工程:
(a)細胞外マトリックスタンパク質でコートされた支持体上で、生物学的試料からの細胞を培養する工程;
(b)上記支持体に接着する細胞を選択し、そして再プレート可能なコロニーを形成する工程;および
(c)上記コロニーから単一細胞を選択する工程
を包含する。
【0018】
上記生物学的試料は、哺乳動物の臍帯血、または血管であり得る。ヒト供給源、ウシ供給源、ブタ供給源、およびラット供給源が、適切である。内皮細胞の型に関する単一細胞アッセイは、以下の工程:
(a)特定の分類方法を使用し、生物学的試料の細胞を分類する工程;
(b)規定された条件下で、細胞外マトリックスタンパク質上で、この単一分類の細胞を培養する工程;および
(c)特定のコロニーのサイズ、形態、および増殖能を列挙し、上記内皮細胞の型(例えば、HPPP−ECFC)を決定する工程
を包含する。
【0019】
HPP−ECFCを富化する方法は、以下の工程:
(a)特定の分類方法を使用し、生物学的試料の細胞を分類する工程;および
(b)規定された条件下で、細胞外マトリックスタンパク質上で単一分類の細胞を培養する工程
を包含する。
【0020】
エキソビボにおいて造血幹細胞を増殖させるための方法は、以下の工程:
(a)コラーゲンコートされた固体支持体上で、HPP−ECFC細胞を培養する工程;および
(b)HPP−ECFC細胞とともに共培養することによって、造血幹細胞(HSC)を増殖させる工程であって、ここで、上記HPP−ECFCはヒト臍帯血細胞由来であり、かつ上記HSC細胞はヒト骨髄由来である、工程
を包含する。
【0021】
哺乳動物中の移植片における、造血幹細胞の百分率を改善するための方法は、以下の工程:
(a)ヒト骨髄細胞を臍帯血HPP−ECFCとともに共培養し、生成物を形成させる工程;および
(b)適切な量の上記生成物を、上記哺乳動物中に移植する工程であって、ここで、上記細胞は、ヒト骨髄由来であるCD45+細胞であり、かつ上記哺乳動物は、NOD−SCIDマウスである、工程
を包含する。
【0022】
自己の臍帯血もしくは血縁関係の無い臍帯血、動員成体末梢血、または骨髄由来HSCとの共培養における、臍帯血の高増殖能内皮コロニー形成細胞(HPP−ECFC)は、HSC細胞の数を増加させ、そして結果としてHSCの増加、およびHSC再増殖活性の増大をもたらし、レシピエント被験体におけるより高いレベルの移植につながる。
【0023】
ヒト臍帯血からの高増殖能内皮コロニー形成細胞(HPP−ECFC)由来の細胞の支持細胞層(feeder layer)の使用は、再増殖性の造血幹細胞および造血前駆細胞の成長および生存を刺激する。成長および生存の刺激は、インビトロ培養中の前駆細胞の数の増加、およびNOD/SCID免疫不全マウス移植系におけるヒト細胞移植レベルの増加によって決定された。
【発明を実施するための最良の形態】
【0024】
(発明の詳細な説明)
高増殖能内皮コロニー形成細胞(HPP−ECFC)と名付けられた新規の内皮細胞前駆体は、成体血液EPCにおける20〜30倍加と比較して、高増殖能(100集団倍加まで)を示す。HPP−ECFCは、臍帯血のみからではなく、臍帯および成体血管からも単離される。HPP−ECFC細胞は、単一細胞レベルで再プレートされ得、そして細胞の大多数は、少なくとも二次HPP−ECFCの再生を伴って増幅する(図7)。予想外なことに、HPP−ECFCコロニー形成は、臍帯血においては存在するが、成体末梢血においては存在しない(図5d)。さらに、単層のHPP−ECFC由来の臍帯血内皮細胞は、成体LPP−ECFCと比較(エキソビボ培養の同じ期間の間)して、集団倍加時間(PDT)において2.5倍の減少を示し、そして累積的集団倍加レベル(CPDL)において少なくとも2倍の増加を示した。異なった方法論を利用して臍帯血から単離された内皮前駆細胞の他の集団とは対照的に、臍帯血HPP−ECFC子孫は、内皮細胞抗原を均一に発現し、かつ造血特異的細胞抗原を発現しない。したがって、HPP−ECFCは、ヒト臍帯血において富化されており、成体末梢血においては見出されなかった。HPP−ECFCはまた、哺乳動物の血管(例えば、臍静脈管およびヒト大動脈管)由来である内皮細胞において見出された(図11)。これらのHPP−ECFCは、プレートした新鮮な臍帯血単核細胞の培養物において、10日間以内に出現するのに対し、成体血液のLPP−ECFCでは、プレート後14日間前に、ほとんど出現しない。
【0025】
さらに、臍帯血コロニーは、成体コロニーと比較して、一貫してより大きく出現する(図1)。成体内皮細胞コロニーと臍帯血内皮細胞コロニーとの間には、そのサイズ、頻度、および出現の時期において、明瞭な差異が存在する(図1、図3、図10)。これらの知見は、臍帯血EPCは、HPP−ECFC、LLP−ECFC、およびクラスターからなるのに対し、成体血液EPCは、LPP−ECFCおよびクラスターからなることを示す。
【0026】
HPP−ECFC、LPP−ECFC、クラスター、および成熟内皮細胞の完全な階層は、生きている哺乳動物のドナー中の任意の血管から単離され得る。
【0027】
EPCは、造血細胞前駆体の規定のために利用された技術と同様の用語法を使用して、同定され得る(図7)。HPP−ECFCは、二次HPP−ECFCへの再プレートに加えて、内皮前駆体の全てのその後の段階を生じる。単一細胞から生じる低い増殖能の内皮コロニー形成細胞(LPP−ECFC)は、コロニーを形成し、それは50個より多くの細胞を含むが、再プレートにおいて、二次LPP−ECFCコロニーさえも形成しない。それらは、内皮細胞クラスター(50個未満の細胞)を生じる。後に、内皮細胞クラスターは、単一細胞から生じ得るが、50個未満の細胞を含み得、かつコロニーまたはクラスターへと再プレートし得ない。
【0028】
造血幹細胞および造血前駆細胞は、成体末梢血と比較して、臍帯血において富化されている。1つの興味深い知見は、成体末梢血と比較して、EPCはまた、臍帯血において富化されている。さらに、臍帯血由来のEPCは高レベルのテロメラーゼ活性を含み、このことは、これらの細胞が、細胞老化の明白な徴候を伴わずに、少なくとも100集団倍加の間増殖し得ることを説明し得る。単一細胞のレベルで、いくつかの臍帯血EPC由来の細胞は、107倍〜1012倍増殖し得る。本発明者らの知る限りでは、他の一次哺乳動物内皮前駆細胞または成熟細胞は、同様の成長特性能力またはクローン原性能力を有すると同定されていない。
【0029】
(ヒト臍帯血から単離された内皮細胞コロニー(EPC)の特徴付け)
細胞は、培養物中で増殖した同時に、内皮細胞表現型を維持していた。前記した臍帯血から単離された内皮前駆細胞に対して、本発明は、ヒト臍帯血から単離した細胞(すなわち、臍帯血HPP−ECFC)および前駆体が、単一細胞を用いて開始した場合でさえ、少なくとも100集団倍加の間培養され得、そして指数関数的に増殖し得ることに関する。さらに、これらの細胞は、造血細胞特異的表面抗原を発現せず(CD45およびCD14もまたほとんど発現しない)、そしてメチセルロース(methycellulose)アッセイにおいて造血細胞コロニーを形成しない。さらに、上記HPP−ECFC前駆体は、マトリゲル中で迅速に脈管を形成し、IL−1刺激またはTNF−α刺激のいずれかに応答してVCAM−1を上方制御し、そしてそれらの内皮細胞アイデンティティ(identity)を確認する内皮細胞特異的抗原を発現する。これらの細胞は、高増殖能内皮コロニー形成細胞(HPP−ECFC)と命名された。
【0030】
臍帯血HPP−ECFCは、成体血液(これは、LPP−ECFC、クラスター、および成熟内皮細胞からなる)と比較して、より大きな複製動態を示す。内皮前駆体は、AC133、CD34、およびFlk1を発現すると報告される一方で、HPP−ECFC子孫およびEOC子孫は、AC133抗原、CD34抗原、およびFlk1抗原を発現する細胞の同様の頻度を示す。したがって、これらの細胞表面マーカーは、異なる増殖能を有する細胞を区別し得ない。
【0031】
内皮増殖細胞は、成体末梢血から単離したMNCの培養の2週間〜4週間後に出現し、そして内皮増殖細胞は、それらのインビトロにおける指数関数的な成長によって特徴付けられる(しかしながら、EOCは、より低いレベルのテロメラーゼ示し、二次LPP−ECFCへと再プレートされず、そしてTPP−ECFCよりずっと前に複製的老化に達する)。対照的に、ヒト臍帯血MNC由来のHPP−ECFCは、I型コラーゲンでコートされた組織培養プレート上の完全EGM−2培地中において、臍帯血MNCの培養の5日後〜10日後に出現した。分散した接着細胞コロニーが出現し、そして「丸石」形態の内皮細胞を示した(図1)。この形態および上記コロニーの出現は、前述した成体末梢血由来のEOCコロニーに関する形態および出現と似通っているが、区別されるものであり、かつ接着循環内皮細胞またはマクロファージと明白に区別される。上記HPP−ECFCコロニーは大きくて、そして小さくて円い細胞、長くて薄い細胞、および大きくて平らな円い細胞の混合物からなる一方で、上記EOCは、ほぼ均一に長くて薄い細胞から構成される。このコロニー由来の細胞は継代培養され、そしてこれらのコロニー由来の増殖した細胞は、免疫表現型決定(immunophenotyping)、機能的試験、および成長動態の測定のために使用された。最初の継代の後、上記細胞は、「丸石」形態を有する紡錘体形状の細胞の単層を形成した。免疫表現型決定は、上記細胞が、内皮細胞表面抗原(CD31、CD141、CD105、CD146、CD144、vWF、およびflk−1)を一様に発現することを明らかにした(図2a、図2b)。上記細胞は、造血細胞表面特有の抗原(CD45およびCD14)を発現せず、このことは、上記単層が、造血細胞を混入していないことを確証した(図1、図3、図10)。
【0032】
上記細胞が、アセチル化低密度リポタンパク質(Ac−LDL)または(Dil−AC−LDL)を取り込んだことで、接着コロニーに由来する上記単層が内皮細胞であったことを確認した。これらの細胞機能は、内皮細胞に特有である。上記接着コロニーから継代培養された細胞は、Ac−LDLを均一に取り込み、異なる数の細胞を播種した後マトリゲル中で脈管を形成し、そしてrhTNF−α刺激またはrhIL−1刺激の両方に応答してVCAM−1を上方制御した(図2c〜図2e)。
【0033】
臍帯血HPP−ECFCおよび子孫の成長動態は、時間の関数として測定された。特筆すべきことに、臍帯血HPP−ECFCおよび子孫は、培養において、少なくとも100集団倍加の間、老化の徴候を伴わずに指数関数的に増殖し得、そして細胞数は、培養物中で、100日間の期間にわたって1020倍に増加した。臍帯血内皮細胞の代表的な成長曲線は、インビトロにおける、これらの細胞の増殖能を例証する。したがって、免疫表現型決定、機能的試験、および成長動態の分析に基づいて、内皮細胞(HPP−ECFCと命名される)のコロニーは、臍帯血MNCから独自に培養され得、そして指数関数的に増殖可能な内皮細胞の集密的単層になるまで継代され得ることが示される。
【0034】
(HPP−ECFCコロニーはヒト臍帯血中に存在するが、成体末梢血中には存在しない)
50mL〜100mLの末梢血が、健康な成体ドナーまたは正常期胎児の臍帯から集められ、そしてMNCが単離された。細胞は、細胞外成熟分子でコートされた組織培養プレート上で、完全EGM−2培地中に播種され、そして次の1週間〜6週間にわたってコロニー形成が観察された。等容積の血液あたりのコロニーの数は、成体末梢血と比較して、臍帯血において15倍増加していた(表I、図1a、図1b)。同数の臍帯MNCおよび成体MNCがプレートされた場合、コロニー形成における類似の相違がまた認められた。成体EOCコロニーは、代表的には、培養の開始の2週間後〜4週間後に形成されるが、臍帯血HPP−ECFCコロニーは、5日間〜10日間以内に出現する。最終的に、臍帯血および成体末梢血に由来する内皮細胞からフローサイトメトリーによって単離された細胞の免疫表現型決定は、コロニー細胞が、内皮細胞表面抗原(CD31、CD105、CD146、CD144、vWF、およびflk−1)を一様に発現し、そして造血細胞表面抗原(CD45およびCD14)を発現しないことを明らかにした。このことは、それらの内皮細胞アイデンティティを確証した。上記成体EOCの免疫表現型決定は、以前に公表された研究と一致した。したがって、内皮コロニー形成細胞HPP−ECFCは、臍帯血中に存在し、成体末梢血EOC(これは、LPP−ECFCを表す)と比較して異なる細胞型を表す。
【0035】
(臍帯血HPP−ECFCの増殖速度は、成体血液EOCよりも速い)
成体末梢血と比較して、臍帯血由来の細胞のコロニー形成の出現の頻度および時期における相違を考慮するとき、1つの疑問は、成体血および臍帯血の増殖動態における相違が存在するのかしないのか、であった。HPP−ECFCの初期継代単層は、臍帯血から樹立され、そしてEOCは成体末梢血から樹立され、そして細胞は、I型コラーゲンコートされたプレート上で完全EGM−2培地中で培養された。長期培養における集団倍加時間(PDT)および累積集団倍加レベル(CPDL)を決定するために、投入細胞数を計数した(これらは、臍帯血および成体血液由来の細胞の増殖動態を、定量化および比較するために使用される測定である)。上記PDTおよびCPDLを正確に定量化するために、細胞を、少なくとも10継代の間培養した。異なるドナーに由来する複数の細胞株の試験の結果は、成体EOCコントロールと比較して、臍帯血HPP−ECFCのPDTにおいて2.5倍の減少が存在したことを示した。さらに、PDTにおける減少と一致して、臍帯血細胞の培養は、成体EOCの連続的な継代と比較して、CPDLにおいて有意な増加を示した。したがって、臍帯細胞および成体細胞の両方は、培養において増殖し得るが、臍帯血HPP−ECFCおよび子孫の増殖能は、成体EOCと比較してより大きい。臍帯血由来のHPP−ECFCはまた、成体血液EOCと比較して、単一細胞レベルでより大きい増殖能を示す。
【0036】
臍帯血HPP−ECFC由来の内皮細胞または成体EOCの、単一細胞レベルでの、個々の増殖能力およびクローン原性能力が決定された。新規の実験方法が、単一臍帯血HPP−ECFC由来の内皮細胞および成体EOCの増殖能力およびクローン原性能力を定量化するために設計された。
【0037】
初期継代臍帯血HPP−ECFC由来または成体EOC由来の内皮細胞の子孫は、緑色蛍光タンパク質(GFP)をコードするレトロウイルスを用いて最初に形質導入され、そしてGFPの発現に関して選択された。臍帯内皮細胞および成体内皮細胞の両方の形質導入効率は、95%より高かった。その後の選択は、1つのGFPを発現するHPP−ECFC由来内皮細胞または成体EOC由来の内皮細胞は、(>直径100ミクロンおよび<9ポンド/平方インチのシーズフロー圧(sheath flow pressure)を有するソートノズル(sorting nozzle)を使用して)蛍光サイトメトリー分類によって、I型コラーゲンでコートされ、そして200μlのEGM−2培地で満たされた96ウェル組織培養プレートの1つのウェル中にプレートされた。直後に、個々のウェルは試験され、1つの内皮細胞のみが各ウェル中に置かれたことを確実にされた。次いで、内皮細胞は14日間培養され、そしてその培地の2分の1は、4日間毎に新鮮なEGM−2培地を用いて交換された。14日間の終わりに、GFPを発現する内皮細胞の数が計算された。
【0038】
少なくとも1回の細胞分裂を起こしている単一細胞の数は、成体EOC由来の内皮細胞と比較して、臍帯血HPP−ECFC由来の内皮細胞について有意に多かった。14日間の終了の後、各々のウェル中の細胞の数のスコアリングにおいて、単一臍帯血HPP−ECFC由来の内皮細胞は、成体EOC由来の内皮細胞と比較して、より多く分裂し、そしてより大きなコロニーを生成したことが明らかであった。成体EOC由来の内皮細胞と比較した、単一臍帯血HPP−ECFC由来の内皮細胞の、分裂およびコロニーを形成する能力における差異に起因して、各ウェル中の(少なくとも1回の細胞分裂を示した)細胞の数が計数された。ほとんどの単一成体EOC由来の(分裂した)内皮細胞は、2個と50個の間の細胞のクラスターを生成したが、一部は、500細胞までの二次コロニーを生じた。しかし、ただ1つの>2000細胞のコロニーが、任意の単一に分類された成体EOC由来の内皮細胞から生じた。しかしながら、60%を超える臍帯血HPP−ECFC由来の(分裂した)内皮細胞は、少なくとも2000細胞からなる、良好に限局性の二次コロニーを形成し、そして多数の単一に分類された細胞が、>10,000細胞からなるコロニーを生じた(本発明者らの知る限りでは、成体EOC由来の内皮細胞は、このようなコロニーを生成することは決して無かった)。
【0039】
これらの細胞がさらなるコロニーを形成し得るかどうかを決定するために、単一成体EOC由来の内皮細胞または単一臍帯血HPP−ECFC由来の内皮細胞のいずれかに由来する二次細胞コロニーが、連続的に再プレートされた。複数の独立した実験において、単一成体EOC由来の内皮細胞に由来する二次コロニーは、24ウェルまたは6ウェルのI型コラーゲンコートされた組織培養プレート中に再プレートされた後は、三次コロニーを生じることは決して無い。プレートされた単一細胞は静止状態のまま残り、かつ増殖しなかった。しかしながら、単一臍帯血HPP−ECFC由来の内皮細胞(2000細胞より多くを生じた)に由来する二次コロニーのほとんどは、同じ実験条件下において再プレートされ、三次内皮コロニーを形成し得た。単一一次臍帯血HPP−ECFC由来の内皮細胞は、二次コロニーを生成し得、これは、引き続いて連続的に継代され、107個〜1012個の内皮細胞を生成し得る。
【0040】
造血高増殖能コロニー形成細胞(HPP−CFC;インビトロにおけるクローン原性アッセイにおいて培養され得る、最も初期の多分化能の造血前駆体)に対する、臍帯血由来の内皮コロニー形成細胞の独特かつ新規に同定された集団の類似性を考慮して、これらの細胞は、「高増殖能内皮コロニー形成細胞(HPP−ECFC)」と名付けられる。要約すれば、これらの細胞は、以下の点で前記の成体EOCと異なる:
(1)成体EOCと同一の実験条件下において培養した場合、HPP−ECFCは、より高い増殖動態を有する、
(2)プレートされた成体末梢MNC由来の成体EOCと比較して、HPP−ECFCは、臍帯血MNCからの培養において、より早い時点で出現する、
(3)成体EOCと比較して、HPP−ECFCは、単一細胞レベルにおいてより高いクローン原性能を有する、
(4)HPP−ECFCは、連続的に再プレートされ、少なくとも二次HPP−ECFCコロニーを形成し得る一方で/他方では、成体EOCは、この潜在力を示さず、かつHPP−ECFCは高いレベルのテロメラーゼを示す。
【0041】
(EPC由来の臍帯血内皮細胞および成体内皮細胞の増殖動態)
異なる系統の前駆細胞は、それらのクローン原性能および増殖能によって規定および区別される。臍帯血のコロニー形成および成体EPCのコロニー形成における相違に起因して、EPC由来の臍帯血内皮細胞および成体内皮細胞の増殖動態が比較された。最初に、この細胞が二次コロニーを形成し、集密まで成長するか否かを試験するために、限界細胞希釈で、臍帯血内皮細胞コロニーおよび生体内皮細胞コロニーに由来する細胞がプレートされた。興味深いことに、成体EPCコロニーおよび臍帯血EPCコロニーの両方に由来する細胞子孫は、集密にまで成長する前に、種々のサイズの二次細胞コロニーを形成した(図3)。しかしながら、臍帯血EPC由来の細胞子孫に由来するコロニーは、成体のコロニーと比較して、一貫してより大きく、そしてより小さい細胞を含んだ(図3)。
【0042】
細胞単層は、EPC由来の臍帯血内皮細胞および生体内皮細胞の増殖能を決定するために連続的に継代された。特筆すべきことに、臍帯血EPC由来の細胞は、明白な老化の徴候を伴わずに、少なくとも100集団倍加の間増殖し得た。対照的に、成体EPC由来の細胞は、20〜30集団倍加の間のみ継代され得た(図4a)。臍帯血EPC由来細胞および成体EPC由来細胞の増殖動態を定量化および比較するために、培養において規定された期間(60日間)の間に、集団倍加時間(PDT)および累積集団倍加レベル(CPDL)が計算された。成体EPC由来の細胞と比較して、臍帯血EPC由来の細胞の、PDTにおいて2.5倍の減少、およびCDPLにおいて1.5倍の増加があった(図4b、図4c)。成体EPCのPDTおよびCDPLは、健康な成体ドナーから単離したEPC由来の細胞の増殖動態を試験した、2つの最近の報告と同様であった。
【0043】
rhVEGF刺激またはrhbFGF刺激(これらは、2つの内皮細胞分裂促進因子)のいずれかに応答する、臍帯血EPC由来の細胞および成体EPC由来の細胞の増殖が比較された。臍帯血EPC由来の細胞および成体EPC由来の細胞は、血清飢餓にされ、次いで、rhVEGFまたはrhbFGFの存在下かまた非存在下において培養された。細胞は16時間培養され、そしてDNA合成を測定するために収集する前にトリチウム化チミジンでパルス(pulse)された。臍帯血EPC由来の細胞は、成体EPC由来の細胞と比較して、rhVEGF刺激またはrhbFGF刺激のいずれかに応答して、より大きなDNA合成を示した(図4d)。まとめると(collectively)、これらの結果は、短期間アッセイおよび長期間アッセイの両方において、臍帯血EPC由来の細胞の増殖速度および増殖能力は、成体EPC由来の細胞よりも大きいことを示す。さらに、臍帯血EPC由来の内皮細胞および成体EPC由来の内皮細胞は、限界希釈でプレートされた場合、種々のサイズおよび形態の別個の細胞コロニーを形成する。
【0044】
(EPCコロニー由来の単一臍帯血内皮細胞および成体内皮細胞のクローン原性能および増殖能の定量化)
臍帯血EPCコロニーおよび成体EPCコロニーは、異なる増殖能およびクローン原性能を有する細胞を生じる。しかしながら、前駆細胞のクローン原性能についての厳密な試験は、単一細胞が、他の細胞の非存在下において分裂し、そしてコロニーを形成するか否かを決定するものである。したがって、EPCコロニー由来の単一臍帯血内皮細胞および単一生体内皮細胞の増殖能およびクローン原性能を定量化するために、アッセイが開発された。
【0045】
最初のEPCコロニーに由来する臍帯血内皮細胞および成体内皮細胞は、EGFPをコードするレトロウイルスを用いて形質導入され、そしてEGFP発現について選択される。選択に続き、1つのEGFPを発現する内皮細胞が、I型コラーゲンでコートされた96ウェル組織培養プレートの1ウェル中にFACSによってプレートされ、そして完全EGM−2培地で満たされた。内皮細胞は培養され、そして14日目の終わりにEGFPを発現する内皮細胞の数が本明細書中に記載されるように計数された。この方法は、図5aに図示される。
【0046】
特筆すべきことに、少なくとも1回の細胞分裂を起こした単一細胞の百分率は、臍帯血内皮細胞について、成体細胞と比較して5倍増加した(図5b)。さらに、単一臍帯血内皮細胞に由来する細胞子孫の平均数は、個々の成体細胞に由来する細胞の数と比較して100倍大きかった(図5c)。分裂した単一成体内皮細胞の80%超は、2〜50細胞の数の範囲におよぶ細胞の小さなコロニーまたはクラスターを生じた(図5d)。しかしながら、いくつかの単一成体内皮細胞は、500個を超える細胞を含むコロニーを形成しなかった(図5d)。対照的に、少なくとも60%の単一でプレートされた分裂した臍帯血内皮細胞は、14日間の培養期間において、2,000個と10,000個の間の細胞を含む良好に限局したコロニーを形成する(図5d)。単一臍帯血細胞または単一成体細胞に由来する細胞の、種々の内皮細胞のコロニーおよびクラスターのサイズおよび形態の顕微鏡写真は、図5eに示される。これらの単一細胞の研究は、臍帯のEPCおよび成体のEPCの異なる型が存在することを示し(これは、それらの増殖能およびクローン原性能によって区別され得る)、そしてEPCは、造血前駆細胞の階層と類似した増殖能の階層を示す。
【0047】
(単一臍帯血内皮細胞の細胞子孫は、連続的に再プレートされ得、そして長期間培養において指数関数的に増殖され得る)
造血細胞系において、最も増殖性の細胞型は、高増殖能コロニー形成細胞(HPP−ECFC)と名付けられた。このHPP−ECFCは、大きな細胞コロニーを形成するその能力によって規定され、連続的なプレーティングにおいて、少なくとも二次コロニーを形成する潜在性を有する個々の細胞を生じる。単一のプレートされた臍帯血EPC由来細胞または成体EPC由来細胞に由来するクローン性子孫は、トリプシン処理され、再プレートされ、そして24ウェル組織培養プレート中で7日間培養された。1000個を超える単一成体EPC由来の細胞のクローン性子孫を、24ウェルプレート中にプレーティングした後、14日間の培養の後、上記ウェル中で二次コロニー1個のみが検出された(図6a)。対照的に、単一でプレートされた臍帯血EPC由来の細胞のクローン性子孫の約2分の1(421のうちの205)は、24ウェルプレート中で、二次コロニーを形成したか、または集密まで急速に成長した(図6a)。単一臍帯血内皮細胞の上記子孫に由来する二次内皮細胞コロニーまたは集密細胞単層の代表的な顕微鏡写真は、図6bに示される。5日以内に集密まで急速に成長したこれらのウェル中では、二次コロニーは検出されなかったので、限界希釈分析を、この集密単層上で実施した。この単層由来のプレートされた単一細胞のうちの少なくとも9パーセントは、100細胞より多くを含む内皮細胞コロニーを形成した。この結果は、臍帯血EPCに由来する個々の細胞が、二次コロニーを形成し得ることを実証した。
【0048】
単一でプレートされた臍帯血EPC由来内皮細胞の、長期増殖能が試験された。単一臍帯血内皮細胞に由来する二次コロニーまたは集密細胞単層は、組織培養プレートを徐々に大きくして連続的に継代された。もともと3つの異なる臍帯血ドナーに由来する、11の単一内皮細胞の細胞子孫が試験された。長期間の培養において、単一臍帯血内皮細胞は、少なくとも107細胞を生じた(図6c)。試験された上記11の単一臍帯血内皮細胞の平均CPDLは、30.8であった。したがって、臍帯血中の高増殖性EPCの集団は、二次コロニーおよび三次コロニーを形成する。
【0049】
(EPC由来の臍帯血内皮細胞は、高レベルのテロメラーゼ活性を含む)
臍帯血EPCに由来する内皮細胞は、ヘーフリック限度を超えて、少なくとも100集団倍加の間、連続的に継代される(図4a)。唯一他に報告されている同様の成長動態を有する一次内皮細胞は、テロメラーゼを過剰発現するように遺伝的に操作された内皮細胞のみである。したがって、臍帯血EPC由来の細胞および成体EPC由来の細胞において、テロメラーゼ活性を、それらの成長動態における相違についての可能性のある分子的説明として測定した。初期継代臍帯血EPC由来の子孫および後期継代臍帯血EPC由来の子孫の両方は、成体EPC由来の細胞と比較して、有意に上昇したレベルのテロメラーゼ活性を示し、テロメラーゼを過剰発現する上記の一次内皮細胞株を連想させる(図6d〜図6e)。したがって、臍帯血EPC由来の細胞は、大きな増殖能と一致して、培養における連続的な継代に伴って高レベルのテロメラーゼ活性(同数のHeLa細胞のテロメラーゼ活性の34±10%)を保持する。
【0050】
(エキソビボにおける、再増殖性の幹細胞および前駆細胞の増殖)
自己の臍帯血もしくは血縁関係の無い臍帯血、動員成体末梢血、または骨髄由来のHSCとともに共培養される、臍帯血の高増殖能内皮コロニー形成細胞(HPP−ECFC)は、HSC細胞の数を拡大させ、そして結果として、レシピエント被験体における、より高いレベルの移植につながるHSCの増加、およびHSC再増殖活性の増加をもたらす。
【0051】
臍帯血由来のHPP−ECFCとヒトHSCの共培養は、造血前駆細胞数を増加させ、そしてNOD/SCIDマウスにおけるヒト造血細胞の移植(インビボにおけるヒトHSC機能の測定のためのアッセイ)を増強した。
【0052】
ヒト臍帯血CD34+CD38−細胞または動員成体末梢血CD34+CD38−細胞(HSC活性に富む)と、7日間まで(サイトカイン添加有りで)共培養されたヒト臍帯血HPP−ECFC由来の内皮細胞は、結果として、準致死的照射されたNOD/SCIDマウスにおいて、>100倍ヒトCD45+細胞移植を増強する(図12)。
【0053】
上記HPP−ECFCの収集、単離、および増殖の方法、ならびに上記HPP−ECFCとヒト幹細胞との共培養のための特定の方法は、新規である。HPP−ECFCは、任意の臍帯血試料から収集され得、増殖され、凍結され、そして保存される。これらの細胞は、次いで、ヒト臍帯血細胞試料、骨髄由来細胞試料、または動員成体末梢血幹細胞試料および動員成体末梢血前駆細胞試料を増殖させるために、解凍され、増殖され、そして共培養において使用され得る。この増殖生成物は、次いで、(規制当局の承認後に)移植目的のために使用され得る。
【実施例】
【0054】
(材料と方法)
(成体末梢血試料および臍帯血試料)
健康なヒトボランティア(22歳と50歳との間の年齢の男性および女性)から、新鮮な血液試料(50ml〜100ml)を、静脈穿刺によって収集し、そしてクエン酸リン酸デキストロース(citrate phosphate dextrose)溶液中で抗凝血化した。健康な新生児(38週在胎齢〜40週在胎齢、男児および女児)からのヒト臍帯血試料(20ml〜70ml)を、抗血液凝固剤としてクエン酸リン酸デキストロース溶液を含む滅菌シリンジ中で収集した。分娩および出産の前に、全ての母親から書面によるインフォームドコンセントを得た。Indiana University School of Medicineの施設内倫理委員会は、全てのプロトコールを承認した。
【0055】
(バフィコート細胞の調製)
ヒト単核細胞(MNC)を、成体末梢血または臍帯血のいずれかから得た。手短に言えば、20ml〜100mlの新鮮な血液を、ハンクス平衡塩溶液(HBSS)(Invitrogen、Grand Island、NY)で1対1に希釈し、そして等容積のFicoll−Paque(Amersham Biosciences)(フィコール密度勾配物質)上にオーバーレイした。細胞を、室温で、1800rpm(740×g)で30分間遠心した。MNCを、単離し、そして10%〜20%ウシ胎仔血清(Hyclone、Logan、UT)、2%ペニシリン/ストレプトマイシン(Invitrogen)、および0.25μg/mlのアンホテリシンB(Invitrogen)を補充したEBM−2培地(Cambrex、Walkersville、MD)(完全EGM−2培地)を用いて3回洗浄した。
【0056】
(内皮成長細胞の培養および定量分析)
バフィコートMNCを、最初に、10%ウシ胎仔血清、2%ペニシリン/ストレプトマイシン、および0.25μg/mlのアンホテリシンBを補充した、12mlのEGM−2培地(Cambrex)(完全EGM−2培地)中に再懸濁した。次いで、前もって細胞外マトリックスタンパク質(例えば、I型ラット尾部コラーゲン(BD Biosciences)、ビトロネクチン、フィブロネクチン、10型コラーゲン、ポリリジン)でコートした6ウェル組織培養プレート(BD Biosciences、Bedford MA)の3つの別々のウェル上に、4mlの細胞を播種した。このプレートを、加湿インキュベーター中で、37℃、5% CO2でインキュベートした。24時間の培養後、非接着細胞および残屑を慎重に吸引し、そして残存する接着細胞を、2mlのEGM−2培地で1回洗浄した。洗浄後、各ウェルに4mlのEGM−2培地を添加した。培養の7日目までEGM−2培地を毎日交換し、次いで最初の継代まで隔日で交換した。
【0057】
細胞のコロニーは、培養の5日目と22日目との間に最初に出現し、そして、良好に限局された丸石の様に見える細胞の単層として同定された(図1c)。倒立顕微鏡を使用して、40×拡大率で、目視検査によってコロニーを数えた。
【0058】
継代のために、上記のオリジナルのコラーゲンコートした組織培養プレートから、0.05%トリプシン−0.53mM EDTA(Invitrogen)を使用して細胞を取り出し、10mlのEGM−2培地中に再懸濁し、そしてI型ラット尾部コラーゲンでコートされた75cm2組織培養フラスコ上にプレートした。内皮細胞の単層を、続いて、90%〜100%の集密に達した後に継代した。
【0059】
(HUVECおよびHAECの培養)
動脈管または静脈管から内皮細胞を直接単離するために、2つのアプローチを使用した。第1の手段において、切断された脈管の一端中に20G平滑末端針を挿入し、そして滅菌生理食塩水を使用して脈管含有物(血液細胞を有する血漿)を反対の末端に流し出した。次いで、上記脈管(長さ3cm〜5cm)の各末端を分離するために、脈管鉗子(vascular clamp)を適用した。23G針によって、脈管壁を介してハンクス平衡塩溶液(HBSS)中の0.1%コラゲナーゼの溶液を注入し、そしてこの脈管セグメントを、37℃で5分間インキュベートした。次いで、この脈管の一端から上記脈管鉗子を取り除き、そして細胞解離緩衝液(Gibco)の注入(上記脈管の「開放」末端の反対の、上記脈管の遠位末端を通して注入した)によって上記内皮細胞を放出した。この脈管セグメントを、最小限の(a minimum of)10mLの細胞解離緩衝液を通して注入した。この懸濁された細胞を、350×gで遠心分離し、そして10% FBSを含むEBM−2培地中で洗浄し、計数し、そしてトリパンブルー排除を使用して生存度を調べた。
【0060】
第2のアプローチは、直径のより大きな脈管(>1cm)について最も適切である。この脈管を、全長に沿って切開し、そして内皮管腔が露出するように開いた。残存するあらゆる血液細胞および血漿を、HBSSで洗い流した。この内皮を、ラバーポリスマン(rubber policemen)を用いたフィルムスクレーピング(firm scraping)によって、一方の端から端まで動かして取り出した。上記ラバーポリスマンに付着している細胞を、10% FBSを含むEBM−2の溶液中でこのラバーポリスマンを回旋することによって、(細胞外マトリックスタンパク質で予めコートした)6cm組織培養ウェル中に洗い落とした。毎日、目視検査を実施しながら細胞を培養した。内皮のコロニーは、3日間〜10日間のうちに出現する。この接着内皮コロニーを、トリプシン−EDTAによって取り出し、そして細胞外マトリックスタンパク質でコートしたT 25フラスコに移した。3回継代され、凍結保存されたヒト臍静脈内皮細胞(HUVEC)およびヒト大動脈内皮細胞(HAEC)を、Cambrexから得た。継代のために、I型ラット尾部コラーゲンでプレコートされた75cm2組織培養フラスコの中の完全EGM−2培地中に細胞を播種した。
【0061】
(EPCの成長動態および複製能の評価)
第一継代時の細胞を、トリパンブルー排除アッセイ(Sigma、St.Louis、MO)よって数えた。次いで、細胞の単層を90%集密まで成長させ、そして継代した。各継代において、成長動態曲線、集団倍加時間(PDT)、および累積集団倍加レベル(CPDL)の計算のために、細胞を数えた。
【0062】
継代の間に生じた集団倍加(PD)数を、以下の方程式に従って計算した:
PD=log2(CH/CS)
ここで、CHは、収集のときの生存細胞の数であり、そしてCSは、播種した細胞の数である。全ての以前のPDの和は、各継代におけるCPDLを決定した。その継代に関するPDの数で割った細胞の播種と収集との間の時間間隔を使用することによって、PDTを導き出した。
【0063】
(マトリゲルアッセイおよびアセチル化低密度リポタンパク質(Ac−LDLまたはDil−Ac−LDL)の取り込み)
マトリゲルアッセイを実施した。手短に言うと、初期継代(2代〜3代)の、HPP−ECFC由来の内皮細胞またはEPC由来の内皮細胞を、前もって30μlのマトリゲル(BD Biosciences)でコートした96ウェル組織培養プレート上に、ウェルあたり5000〜20,000細胞の細胞密度で播種した。キャピラリー様の管の形成について、細胞を2時間毎に観察した。
【0064】
接着したHPP−ECFCおよび子孫またはEPCおよび子孫の、Ac−LDLまたはDil−Ac−LDLを取り込む能力を評価するために、6ウェルのI型ラット尾部コラーゲンコートされた組織培養プレート中で培養された細胞の培地中に、10μg/mlのAc−LDL(Biomedical Technologies Inc.、Stoughton、MA)を添加した。37℃で、30分間または4時間、細胞をインキュベートし、次いで1.5μg/mlのDAPI(Sigma)で染色されたリン酸緩衝生理食塩水(PBS)を用いて3回洗浄し、そしてAc−LDLまたはDil−Ac−LDLの取り込みを、蛍光顕微鏡を使用することによって試験した。
【0065】
(蛍光サイトメトリーによる内皮細胞の免疫表現型決定)
初期継代(1代〜2代または3代〜4代)の、HPP−ECFCおよび子孫、またはEPCおよび子孫(5×105個)を、100μlのPBSおよび2%FBS中で、以下に概説されるように、異なる濃度の一次抗体またはアイソタイプコントロール抗体とともに4℃で30分間〜60分間インキュベートした。2% FBSを含むPBSで、細胞を3回洗浄し、そして蛍光活性細胞ソーティング(fluorescence activated cell sorting)(FACS(著作権))(Becton Dickinson、San Diego、CA)によって分析した。フルオレセインイソチオシアネート(FITC)(BD Pharmingen、San Diego、CA)に結合したヒトCD31に対して直接的に結合した一次マウスモノクローナル抗体を1:20希釈で、アロフィコシアニン(APC)(BD Pharmingen)に結合したヒトCD34を1:25希釈で、FITC(BD Pharmingen)に結合したヒトCD14を1:10希釈で、FITC(BD Pharmingen)に結合したヒトCD45を1:10希釈で、APC(BD Pharmingen)に結合したヒトCD117を1:100希釈で、フィコエリトリン(PE)(BD Pharmingen)に結合したCD146を1:10希釈で、PE(Miltenyi Biotec、Auburn、CA)に結合したヒトAC133を1:5希釈で、FITC(Cymbus Biotechnology、Chandlers Ford、UK)に結合したヒトCD141を1:10希釈で、Alexa Fluor 647(Alexa Fluor 647モノクローナル抗体標識キット、Molecular Probes、Eugene、OR)に結合したヒトCD105(BD Pharmingen)を1:100希釈で、およびAlexa Fluor 647に結合したヒトCD144を1:100希釈で使用した。
【0066】
細胞アゴニストによる活性化後、脈管細胞接着分子(VCAM−1)の細胞表面発現について試験するために、血清飢餓内皮細胞を、10ng/mlの組み換えヒトインターロイキン−1(IL−1)(Peprotech、Rocky Hill、NJ)か、または10ng/mlの組み換えヒト腫瘍壊死因子−α(TNF−α)(Peprotech)のいずれかを用いて、37℃で4時間刺激した。刺激の後、FITC(BD Pharmingen)に結合したヒトVCAM−1に対する一次抗体を1:20希釈で利用して、VCAM−1の細胞表面発現を試験した。免疫表現型決定(immunopherotyping)およびUCAM−1発現に対する全てのアイソタイプコントロールに関して、以下の抗体を使用した:FITC(BD Pharmingen)に結合したマウスIgG2a,κ、FITC(BD Pharmingen)に結合したマウスIgG1,κ、PE(BD Pharmingen)に結合したマウスIgG1,κ、およびAPC(BD Pharmingen)に結合したマウスIgG1,κ。
【0067】
フォン・ビルブラント因子(vWF)およびflk−1の細胞表面発現の検出のために、室温で10分間、アセトン中で細胞を固定し、PBSで2回洗浄し、そしてPBS、3%脱脂粉乳、および0.1% Triton X−100(Sigma)を用いて、30分間ブロックおよび浸透させた。本発明者らは、2μg/mlのヒトvWFに対する一次抗体(Dako、Carpenteria、CA)、および2μg/mlのヒトflk−1に対するビオチン化一次抗体(Sigma)を、1:20希釈で使用した。vWFに対して使用された二次抗体は、1:100希釈でのFITC(BD Pharmingen)に結合したヤギ抗ウサギ抗体であり、そしてflk−1に対して使用された二次抗体は、1:100希釈でのAPC(BD Pharmingen)に結合したストレプトアビジン(strepavidin)であった。vWFに対するアイソタイプコントロールについては、本発明者らは、ウサギIg一次抗体(Dako)を1:100希釈で、1:100希釈でのFITC(BD Pharmingen)に結合した抗ウサギIg二次抗体とともに使用した。flk−1に対するアイソタイプコントロールついては、本発明者らは、ビオチン化マウスIgG1,κ(BD Pharmingen)一次抗体を1:100希釈で、1:100希釈でのストレプトアビジン(strepavidin)APC二次抗体(BD Pharmingen)ともに使用した。
【0068】
(テロメラーゼ活性アッセイ)
テロメラーゼ活性の検出のために、TRAP−ezeテロメラーゼ検出キット(Oncor、Gaithersburg、MD)の形式で、テロメア反復増幅プロトコール(TRAP)を利用した。手短に言うと、培養した1000個のHPP−ECFCコロニーまたはEPCコロニーを、濾紙上に吸着させ、そしてTRAPアッセイ緩衝液中に溶解した。この溶解した材料をPCR増幅に供し、そしてそのPCR生成物(6bp増分のラダー)を非変性ポリアクリルアミドゲル上で電気泳動し、そしてDNA染色または32Pを用いた放射標識によって可視化した。PCR生成物を、そのままでか、または1/10希釈でか、もしくは1/100希釈でロードし、そして染色強度のレベルをHELA細胞株(1000細胞)ポジティブコントロールと比較した。
【0069】
(チミジン取り込みアッセイ)
内皮コロニー由来の内皮細胞を、成長因子を欠乏させ、そして5% FBSを補充したEBM−2培地中で24時間培養した。次に、3×104個の細胞を、I型コラーゲンで予めコートした6ウェル組織培養皿の各ウェル中にプレートし、1% FBSを補充したEBM−2培地中で16時間培養した。次いで、細胞を、血清を含まないEBM−2培地中でさらに8時間培養し、静止状態を確実にした。10% FBSを補充したEBM−2培地中で、25ng/mlの組み換えヒト脈管内皮成長因子(rhVEGF)(Peprotech)、25ng/mlの組み換えヒト塩基性線維芽細胞成長因子(rhbFGF)(Peprotech)、または成長因子無し(図示されるように)で、37℃、5% CO2の加湿インキュベーター中で細胞を刺激した。成長因子またはFBSを含まないEGM−2培地中で、いくつかの細胞を培養した。細胞を16時間培養し、そして収集の5時間前に、1μCiのトリチウム化チミジン(Perkin Elmer Life Sciences Products、Boston、MA)を添加した。細胞を、0.1N水酸化ナトリウムで1時間溶解した。溶解物を5mlの液体シンチラント(liquid scintilant)(Fisher Scientific、St.Louis、MO)中に収集し、そしてβ放射を測定した。アッセイを、三つ組み(triplicate)で行った。
【0070】
(GALV偽型MFG−EGFPの生成)
MFG−EGFPレトロウイルスベクターは、モロニーマウス白血病ウイルスの長い末端反復配列(LTR)の制御の下で増強された緑色蛍光タンパク質(EGFP)を発現する。そしてこれは、Pollokら(2001)によって以前に記載されている。GALV偽型ベクターの生成のための、アンホトロピック(amphotrophic)なMFG−EGFPクローンからの上清を使用し、PG13パッケージング株(American Type Culture Collection(ATCC)、Manassas、VA)を感染させ、そして感染した細胞を、単一細胞クローニングによって単離した。個々のクローンを、5×105個のヒト赤白血球病細胞(HEL)(ATCC)を感染させ、そして上清の終点希釈後48時間の百分率EGFP発現を決定することによって、力価についてスクリーニングした。MFG−EGFPクローン5は、0.5×106感染単位/ml〜1×106感染単位/mlの力価を有し、そしてこれを実験に使用した。
【0071】
(内皮細胞のレトロウイルス形質導入)
初期継代(1代〜2代)内皮コロニー由来の内皮細胞を、等量の出発の力価のMGF−EGFP上清を用いて形質導入した。6ウェル非組織培養プレートを、5μg/cm2フィブロネクチンCH−296(Takara Shuzo、Otsu、Japan)を用いて、室温で2時間かまたは4℃で一晩コートした。プレートをPBSで1回洗浄し、そして形質導入のために内皮細胞を5×104細胞/cm2でプレートした。完全EGM−2培地で1:1に希釈したレトロウイルス上清を用いて細胞を、一晩の培養の間完全EGM−2培地の交換を行い、連続2日間4時間感染させた。第2ラウンドの感染の後、細胞を収集し、計数し、そして蛍光サイトメトリーによってEGFP発現について分析した。
【0072】
(単一細胞アッセイ)
上記MFG−EGFPレトロウイルスで形質導入した初期継代(1代〜4代)内皮コロニー由来の内皮細胞を、蛍光サイトメトリーによって、EGFP発現について分類した。FACS Vantage Sorter(Becton Dickenson)(≦9ポンド/平方インチのシーズフロー圧で≧100ミクロンのソートノズル)を使用し、200μlの完全EGM−2培地を含んだI型コラーゲンで予めコートした96ウェル平底組織培養プレートの各ウェル中に、EGFPを発現する1つの単一内皮細胞を置いた。50×拡大率の蛍光顕微鏡下で個々のウェルを検査し、1個の細胞のみが各ウェル中に置かれていることを確実にした。加湿インキュベーター中で、37℃、5%CO2で細胞を培養した。培地を、100μl取り出し、それを100μlの新鮮な完全EGM−2培地で置き換えることによって、4日間毎に交換した。第14日目に、プレートした1個の細胞からの内皮細胞の成長について、各ウェルを検査した。単一内皮細胞の分裂の頻度を定量化するために、100×拡大率の蛍光顕微鏡を用いて2個以上の内皮細胞を有する細胞の数を計数した。ウェルあたりの細胞の数を数えるために、100×拡大率の蛍光顕微鏡を用いた目視検査によって、上記細胞を計数するか(ウェルあたり50細胞未満)、または上記細胞をトリプシン処理し、そしてトリパンブルー排除アッセイを利用して血球計を用いてそれらを計数した(ウェルあたり50細胞超)。
【0073】
単一細胞に由来する内皮細胞の長期間増殖能および長期間複製能を、決定した。培養の開始の14日後、50細胞より多くを含む個々のウェルを、トリプシン処理し、500μlの完全EGM−2培地中に収集し、そしてI型コラーゲンでコートした24ウェル組織培養皿に継代培養した。この細胞の継代培養の4日後、その培地を吸引し、そして500μlの新鮮な完全EGM−2培地で置き換えた。7日目に、50×拡大率の蛍光顕微鏡を用いた目視検査によって、コロニー成長または細胞集密についてウェルを検査した。次いで、細胞をトリプシン処理し、計数し、そしてI型コラーゲンで予めコートした6ウェル組織培養プレート中に継代培養した。6ウェルプレートにおける培養の7日後、本明細書中で開示される条件下で、長期間培養について、集密な細胞単層を含んだ10ウェル〜12ウェルを選択した。各試料について、PDTおよびCPDLを計算した。
【0074】
(CDマーカー)
CD14(リポ多糖レセプター)
CD31(血小板内皮細胞接着分子)
CD34(シアロムチン)
CD45(共通白血球抗原)
CD105(エンドグリン)
CD117(c−Kitレセプター)
CD133(プロミニン1)
CD141(トロンボモジュリン)
CD144(脈管内皮カドヘリン)
CD146(内皮関連抗原、S−エンド−1)
flk−1(胎児肝臓キナーゼ−1、脈管内皮成長因子2に対するレセプター)。
【0075】
(EPCの共焦点画像化)
10% FBSを添加したEBM−2培地を使用し、T75フラスコ中で4日間、3代〜5代継代したEPCを成長させた。細胞が集密に達した場合、培地を吸引し、上記フラスコに5mlの滅菌PBSを添加し、次いで吸引し、トリプシン−EDTAを添加し、そしてこのフラスコを37℃で5分間インキュベートした。このトリプシンをクエンチするために、5mLの10% FBSを含むEBM−2培地を添加し、そして遊離したEPCを350×Gで10分間遠心分離した。ペレット化した細胞をPBSで洗浄し、次いで10% FBSを含むEBM−2培地中に再懸濁した。
【0076】
ガラス製チャンバースライド(4チャンバー構造;Corning)を、4℃で一晩、細胞外マトリックスタンパク質(例えば、1型コラーゲンもしくは4型コラーゲン、フィブロネクチン、またはビトロネクチン)でコートし、次いで翌朝に(in the morning)滅菌PBSで洗浄した。このPBSを吸引し、そして10% FBSを含むEBM−2培地中の細胞を、チャンバーあたり50細胞で添加し、そして5%CO2中で、37℃で7日間インキュベートした。
【0077】
EPCを含むスライドをPBSで2回洗浄し、そしてアセトン中で10分間室温で細胞を固定し、PBSで2回洗浄し、そしてPBS、3%脱脂粉乳、および0.1% Triton X100を用いて、30分間、ブロックおよび浸透させた。この細胞の細胞膜を強調するために、核染色のための1.5mg/ml DAPIとともに、フィコエリトリン(PE)に結合したCD146に対する一次抗体(1□g/mL)を、上記固定した細胞に添加した。30分間のインキュベーション後、PBS中で細胞を2回洗浄し、そしてZeiss 510共焦点顕微鏡を使用して蛍光について検査した。7×拡大率で倍率を保持したまま、40×水浸対物レンズを介して、紫外レーザー(351/364nm励起)およびヘリウム−ネオンレーザー(543nm励起)を使用して上記DAPIおよびPE−標識した細胞を励起した。画像を一平面中に取り込み、そして提示のための単色画像として表示した。NIH Imageソフトウェアを使用し、種々のEPCコロニー型に由来する細胞の、核の直径および細胞質の直径を定量化した。
【0078】
(表1)
成体末梢血単核細胞および臍帯血単核細胞から単離した内皮前駆細胞コロニーの、出現の数および時期の一覧表。
【0079】
【表1】
。
【0080】
(HPP−ECFCおよびCD34+細胞の共培養は、NOD−SCID再増殖細胞を増殖させる)
NOD/SCIDマウスにおける骨髄(marrow)再増殖細胞(SRC)を有する(harbor)ことが以前に示された、ヒトCD34+骨髄細胞を単離した。代表的には、5%〜50%のヒトCD45+キメラ現象のレベルを達成するために、0.5×106個〜1.0×106個のヒト骨髄CD34+細胞を、NOD/SCIDマウス中に注入する。最初に、ヒト骨髄からの収集の日に、9×103個のCD34+細胞のみをコントロールとしてNOD−SCIDマウス中に注入した。SCF、G−CSF、TPO、およびFlt−3の存在下で、9×103個のCD34+細胞を7日間培養した。これらは、エキソビボにおいてHSCを最大に増殖させるために現在使用されている成長因子である。成長因子の不在下で、臍帯血HPP−ECFC由来の子孫の単層とともに9×103個のCD34+細胞を7日間共培養した。培養の7日後、この培養したCD34細胞をNOD−SCIDマウス中に注入し、そしてヒト細胞の存在下で、移植の4週間後、移植されたマウスの末梢血を試験した。CD34+細胞の成長因子との7日間の共培養は、ヒト骨髄から単離した直後に注入したCD34+と比較して、移植の8週間後のNOD−SCIDマウス中で検出されたヒト細胞の百分率を10倍増大させた(図12)。以前の研究と比較して、非常に限られた数の細胞の注入にもかかわらず、臍帯血HPP−ECFC由来の細胞とのCD34+細胞の共培養は、移植の8週間後のNOD−SCIDマウスにおいて検出されたヒト細胞の百分率を260倍増大させた(図12)。ヒト骨髄系統およびリンパ系統の両方が移植の8週間後に検出された。このことは、造血系の多系統の再構成が、臍帯血HPP−ECFCとのCD34+細胞の共培養によって達成されたことを示した。
【0081】
HPP−ECFC由来の細胞の集密な単層の2つのT75フラスコを用いて開始して、カルシウムまたはマグネシウムを欠いたハンクス平衡塩溶液(HBBS)で最初に細胞を洗浄し、次いで各フラスコに1分間1.5mLのトリプシンEDTA(Gibco)を添加した。次に、10%ウシ胎仔血清(FBS)(Hyclone)を含む8.5mLの内皮基礎培地2(EBM2)(Cambrex)を添加し、そして懸濁した細胞を収集し、そして血球計上でトリパンブルー排除を介して計数した。
【0082】
コラーゲン1で予めコートした6ウェル組織培養プレート(BD Biosciences)上に、HPP−ECFC由来の細胞を、3×105細胞/ウェルでプレートした。10% FBSを補充した内皮成長培地2(EGM2)(Cambrex)を用いて細胞を培養し、そして一晩培養した。翌朝、その集密細胞単層をEBM2+10% FBSで2回洗浄し、次いで、蛍光活性化細胞分類によって収集し、4mLのEBM2+10% FBS+ヒト巨核球成長由来因子(MGDF)(100ng/mL)、顆粒球コロニー刺激因子(G−CSF)(100ng/mL)、および幹細胞因子(SCF)(100ng/mL)、ならびにflt−3リガンド(100ng/mL)中に再懸濁した9,000個のCD34+CD38dimLin−(CD4、CD8、CD11b、CD14、CD24、CD31、CD33、およびグリコホリンA)成体ヒト骨髄由来細胞とともに共培養した。37℃、5% CO2加湿インキュベーター中で7日間、障害無しに細胞を培養した。いくつかの培養物において、EBM2+10% FBS中でHPP−ECFCとともに、そして成長因子無添加で、CD34+細胞を共培養した。
【0083】
7日間の共培養の後、5mLのピペットを使用して非接着細胞および培地(4mL)を吸引した。2mLリン酸緩衝生理食塩水(PBS)で1回ウェルを洗浄し、そして最初の吸引物にそのPBSとともに非接着細胞を添加した。同じウェルに対して、1mLの細胞分離緩衝液(Gibco)を室温で4分間添加し、次いで、吸引および上記最初の吸引物への添加の前に、この細胞分離緩衝液およびバラバラになった細胞をそのウェル中で穏徐に滴定した。最後に、上記HPP−ECFC単層を2mLのPBSで最後に1回洗浄し、そして上記最初の吸引物中に、わずかな細胞を含むこの溶液を添加した。培地および細胞の最終容積は、9mLであった。
【0084】
この細胞懸濁物を、室温で1500rpm(514×g)で10分間遠心分離した。溶液を取り除き、細胞ペレットを機械的に取り出し(dislodge)、次いで1/2mL〜1mLのEBM2+10% FBS中に再懸濁した。血球計上において、トリパンブルー中で細胞を計数した。回収した細胞を、前駆体アッセイにおいてプレートするか、またはNOD/SCIDマウス中に静脈内注入した。
【0085】
上記に概説された方法は、ヒト移植のための移植片を提供するために改変され得る。この場合には、HPP−ECFC子孫は、臍帯血HPP−ECFCの存在下で、大数の(自己または同種異系のヒト臍帯血由来、動員末梢血由来、または骨髄由来の)CD34+造血幹細胞が増殖し得るように、T75フラスコ中または灌流チャンバー系にプレートされる。細胞に直接触れることなくドナーCD34+細胞がHPP−ECFC子孫とともに培養され得るような系が使用され、したがって、上記ドナーCD34+細胞は増殖され得、回収され得、そして臍帯血HPP−ECFC子孫の「混入」したドナー細胞無しで、ヒト患者中に移植され得る。
【0086】
(文献)
以下の文献は、開示される方法および組成物が本発明の実施において使用される程度に、参考として本発明の範囲中に援用される。
【0087】
【表2】
【0088】
【表3】
【0089】
【表4】
【0090】
【表5】
【0091】
【表6】
【図面の簡単な説明】
【0092】
【図1−1】成体末梢血および臍帯血に由来する、内皮前駆細胞コロニーの単離。図1aは、成体末梢血(AB)および臍帯血(CB)20mlあたりの単離されたEPCコロニーの数。結果は、成体ドナーについての18の独立した実験および臍帯血についての13の独立した実験についての、EPCコロニーの平均数±SEMの平均数を示す。*対応のあるスチューデントt検定によって、P<0.0001。図1bは、等容積の成体末梢血(AB)および臍帯血(CB)からの培養開始後の、初期EPCコロニー出現の時間である。結果は、成体ドナーについての18の独立した実験および臍帯血についての13の独立した実験についての、初期EPCコロニー出現の前の平均日数±SEMの前の平均日数を示す。*対応のあるスチューデントt検定によって、P<0.0001。
【図1−2】成体末梢血および臍帯血に由来する、内皮前駆細胞コロニーの単離。図1cは、成体血液に由来する内皮コロニーの、代表的な低分解能顕微鏡写真および高分解能顕微鏡写真である。図1dは、CBに由来する内皮細胞コロニーの、代表的な低分解能顕微鏡写真および高分解能顕微鏡写真である。図1eは、低分解能拡大率および高分解能拡大率での、成体内皮細胞コロニーに由来する内皮細胞単層である。図1fは、低拡大率および高拡大率での、CB内皮細胞コロニーに由来する、内皮細胞単層である。写真中のスケールバーは、100μm(c、d、e、fの左側カラム)、および10μm(c、d、e、fの右側カラム)を表す。
【図2】成体EPC由来および臍帯血EPC由来の内皮細胞の、表現型分析および機能分析。図2a〜図2bは、蛍光サイトメトリーによる、成体EPCコロニー(図2a)または臍帯血EPCコロニー(図2b)のいずれかに由来する細胞単層の免疫表現型決定である。臍帯血EPC由来の細胞および成体EPC由来の細胞の両方は、CD31、CD141、CD105、CD146、CD144、vWF、およびFlk−1を発現するが、CD45およびCD14を発現しない。いくつかの臍帯血細胞および成体細胞は、CD34、CD133、およびCD117を発現する。示されているものは、同様の結果を有する、異なる成体細胞単層を利用した18の独立した実験および異なる臍帯血細胞単層を使用した13の独立した実験からの、代表的なデータである。アイソタイプコントロールは、試験した各表面抗原についての各ヒストグラム上にグレーでオーバーレイされる。(図2c)成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞は、DiI−Ac−LDLを取り込む(50×拡大率)。DiI−Ac−LDLを取り込み(赤色)、かつDAPIで染色された(青色)、成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞についての代表的な写真が示される。示されているものは、同様の結果を有する、異なる成体細胞単層を利用した18の独立した実験および異なる臍帯血細胞単層を使用した13の独立した実験からの、代表的なデータである。写真中のスケールバーは、100μmを表す。(図2d)毛細管様の構造の形成のためにマトリゲル中にプレートされた、増強された緑色蛍光タンパク質(EGFP)を発現するように遺伝的に操作された成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞(50×拡大率)。示されているものは、同様の結果を有する、異なる成体細胞単層を利用した18の独立した実験および異なる臍帯血細胞単層を使用した13の独立した実験からの、代表的なデータである。写真中のスケールバーは、100μmを表す。(図2e)成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞は、rhTNF−αか、またはrhIL−1のいずれかに応答して、脈管細胞接着分子(VCAM−1)の細胞表面発現を上方制御する。示されているものは、同様の結果を有する、異なる成体細胞単層を利用した18の独立した実験および異なる臍帯血細胞単層を使用した13の独立した実験からの、代表的なデータである。VCAM−1に関するアイソタイプコントロールは、各ヒストグラム上にグレーでオーバーレイされる。
【図3】成体EPC由来の内皮細胞および臍帯血EPC由来の内皮細胞を、低い細胞密度で再プレートした7日後に形成された、形態学的に明確に異なる二次コロニーの代表的な写真。
【図4】臍帯血内皮前駆細胞コロニーおよび成体内皮前駆細胞コロニーに由来する内皮細胞子孫の成長動態。(図4a)単核細胞から収集された、成体血液(AB)EPC由来の内皮細胞および臍帯血(CB)EPC由来の内皮細胞のエキソビボ増殖。黒色の四角(box)は、各継代における総細胞数を表す。各継代において、細胞は上で示した内皮細胞表面抗原(図2a〜図2b)を均一に発現したが、造血細胞特異的抗原(CD45およびCD14)を発現しなかった。臍帯血EPC由来の内皮細胞および成体EPC由来の内皮細胞についての代表的な成長曲線を示す。異なるドナーに由来する、11の他の臍帯血内皮細胞単層および成体内皮細胞単層は、同様の成長動態を示した。(図4b〜図4c)60日間の培養の間の、臍帯血(CB)EPC由来の内皮細胞および成体血液(AB)EPC由来の内皮細胞の集団倍加時間(PDT)および累積集団倍加レベル(CPDL)。結果は、6つの独立した実験のPDTの平均数±SEMの平均数およびCPDLの平均数±SEMの平均数を表す。*対応のあるスチューデントt検定によって、P<0.01。(図4d)臍帯血(CB)EPC由来の内皮細胞および成体血液(AB)EPC由来の内皮細胞のDNA合成。初期継代(1代〜2代)の臍帯血EPC由来の内皮細胞は、10% FBS、rhVEGF、およびrhbFGFに応答して、成体細胞と比較して増大したDNA合成を示す。結果は、異なるドナーに由来する内皮細胞を利用した4つの独立した実験の平均を表す。*対応のあるスチューデントt検定によって、P<0.01。
【図5】EPCコロニーに由来する、単一臍帯血内皮細胞および単一成体内皮細胞のクローン原性能および増殖能の定量化。(図5a)成体EPCコロニーか、または臍帯EPCコロニーのいずれかに由来する内皮細胞を使用した単一細胞アッセイの概略図。(図5b)培養の14日後、少なくとも1回の細胞分裂を起こした成体血液(AB)EPC由来の内皮細胞または臍帯血(CB)EPC由来の内皮細胞の百分率。結果は、異なるドナーに由来する単一内皮細胞を使用した5つの独立した実験の平均を表す。*対応のあるスチューデントt検定によって、P<0.01。(図5c)培養の14日後の、単一成体血液(AB)EPC由来の内皮細胞または単一臍帯血(CB)EPC由来の内皮細胞に由来する細胞子孫の平均数。結果は、異なるドナーに由来する単一内皮細胞を使用した5つの独立した実験の平均を表す。*対応のあるスチューデントt検定によって、P<0.01。(図5d)定量可能な範囲を示す細胞数を有するコロニーを生じる、分裂する単一細胞の百分率(HPP、LPP、クラスター)。(図5e)単一臍帯血EPC由来の内皮細胞または単一成体EPC由来の内皮細胞に由来する、種々の内皮細胞クラスター(<50細胞)、LPP(約51細胞〜2000細胞)、およびHPP(約2000細胞〜>10,000細胞)の代表的な写真(50×拡大率)。結果は、異なるドナーに由来する細胞を利用した4つの他の独立した実験を表す。写真中のスケールバーは、100μmを表す。LPP−ECFC(51細胞〜2000細胞)およびHPP−ECFC(2000細胞>10,000細胞)の範囲は、概算値である。
【図6】単一臍帯血EPC由来の内皮細胞または単一成体EPC由来の内皮細胞に由来する細胞子孫の、再プレート能および長期間培養。(図6a)24ウェル組織培養プレートにおいて、培養の7日後に二次コロニーを形成するか、または細胞集密まで急速に成長する単一臍帯血(CB)EPC由来の内皮細胞または単一成体(AB)EPC由来の内皮細胞に由来する細胞子孫の百分率。結果は、4人の異なるドナーに由来する細胞を使用した4つの独立した実験の平均±SEMを表す。*対応のあるスチューデントt検定によって、P<0.01。(図6b)培養の7日後における、24ウェルプレート中の、単一のプレートされた臍帯血由来の内皮細胞の細胞子孫に由来する、二次内皮細胞コロニーまたは集密細胞単層の代表的な写真(50×拡大率)。写真中のスケールバーは、100μmを表す。(図6c)長期間培養における、11の単一のプレートされた、3人の異なる臍帯血ドナーから単離された内皮細胞の細胞子孫の成長動態。黒色の四角は、各継代における細胞の総数を表す。(図6d)異なるドナーから単離された、1000個の初期継代の成体血EPC由来の内皮細胞(レーン1〜レーン4)および臍帯血EPC由来の内皮細胞(レーン5〜レーン8)のテロメラーゼ活性。15のCDPLで、成体細胞および臍帯細胞を試験した。Pは、ポジティブコントロールとして使用した1000個のHeLa細胞におけるテロメラーゼ活性を示し、そしてNは、ネガティブコントロールを示す。成体試料におけるテロメラーゼ活性の平均レベルは、HeLa細胞のテロメラーゼ活性の4±4%であり、そして臍帯血試料のテロメラーゼ活性の平均レベルは、HeLa細胞のテロメラーゼ活性の34±10%であった。(図6e)初期継代および後期継代の成体EPC由来の内皮細胞(a)および臍帯血EPC由来の内皮細胞(c)のテロメラーゼ活性の比較。PDは、試験した細胞の累積集団倍加レベルを示す。Pは、ポジティブコントロールとして使用したHeLa細胞におけるテロメラーゼ活性を示す。Nは、ネガティブコントロールを示す。3人の異なるドナーに由来する初期継代および後期継代の臍帯血EPC由来の内皮細胞および成体EPC由来の内皮細胞を利用した3つの他の実験は、同様の結果を示した。
【図7】前駆細胞の別個の集団の増殖能およびクローン原性能に基づく、内皮前駆細胞階層のモデル。高増殖能内皮コロニー形成細胞(HPP−ECFC)は、再プレートによって少なくとも二次コロニーを形成する、1つの細胞から生じる大きなコロニーである。HPP−ECFCは、全てのその後の段階の内皮前駆体を生じ、さらに二次HPP−ECFCへと再プレートされる。単一細胞から生じた低増殖能内皮コロニー形成細胞(LPP−ECFC)は、50細胞より多くを含むコロニーを形成するが、再プレートによってLPP−ECFCの二次コロニーを形成しない。内皮細胞クラスター(EC−クラスター)は、1つの細胞から生じ得るが、50細胞未満しか含まない。これは、HPP−ECFCコロニーおよびLPP−ECFCコロニーにおいて見出されるより小さな細胞と比較して、代表的にはより大きい(図10を参照のこと)。成熟した末期分化内皮細胞は、分裂しない。
【図8】臍帯血EPC、HUVEC、およびHAEC由来の内皮細胞の免疫表現型分析。(図8a〜図8b)臍帯血EPC(a)、臍静脈(b)、またはヒト大動脈(c)に由来する細胞単層の、蛍光サイトメトリーによる免疫表現形決定。臍帯血EPC由来のEC、HUVEC、およびHAECは、CD31、CD141、CD105、CD146、CD144、vWF、およびFlk−1を発現するが、CD45およびCD14を発現しない。5つの異なる臍帯血EC単層、5つの異なるHUVEC試料、および5つの異なるHAEC試料を用いた5つの独立した実験からの代表的なデータは、同様の結果を示した。アイソタイプコントロールは、試験した各表面抗原について、各ヒストグラムに白色でオーバーレイされる。
【図9】臍帯血EPCコロニー、HUVEC、およびHAECに由来する単一内皮細胞のクローン原性能および増殖能の定量化。(図9a)培養の14日後に、少なくとも1回の細胞分裂を起こす単一臍帯血(CB)EPC由来のEC、HUVEC、またはHAECの百分率。結果は、異なるドナーに由来する単一ECを使用した5つの独立した実験の平均を表す。(図9b)個々のウェル中において分裂する単一細胞の百分率(図5を参照のこと)。培養の14日後に、個々のウェル中において(分類されたような)細胞のコロニーを生じる、単一CB EPC由来のEC、HUVEC、またはHAECの百分率。*単一CB由来のEC 対 単一HUVECまたは単一HAECのいずれかの比較のための、対応のあるスチューデントt検定によって、P<0.01。(図9c)単一臍帯血EPC由来のEC、HUVEC、またはHAECに由来する、種々のECクラスター(<50細胞)またはコロニー(>50細胞)の代表的な写真(50×拡大率)。結果は、異なるドナーからの細胞を利用した4つの他の独立した実験の代表的なものである。写真中のスケールバーは、100μmを表す。(図9d)24ウェル組織培養プレートにおいて、培養の7日後に、二次コロニーを形成するかまたは細胞集密まで急速に成長した、単一臍帯血EPC由来のEC、HUVEC、またはHAECに由来する細胞子孫の百分率。結果は、4人の異なるドナーに由来する細胞を使用した、4つの独立した実験の平均±SEMを表す。*単一CB由来のEC 対 単一HUVECまたは単一HAECのいずれかの比較のための、対応のあるスチューデントt検定によって、P<0.01。(図9e)培養の7日後における24ウェルプレート中の、単一のプレートされた臍帯血EPC由来のEC、HUVEC、またはHAECの細胞子孫に由来する二次ECコロニーまたは集密細胞単層の代表的な写真(50×拡大率)。写真中のスケールバーは、100μmを表す。(図9f)HUVECに由来するHPP−ECFCコロニー(HPP)およびLPP−ECFCコロニー(LPP)におけるテロメラーゼ活性。Pos.は、ポジティブコントロールとして使用したHeLa細胞におけるテロメラーゼ活性を示し、そしてNeg.は、ネガティブコントロールを示す。結果は、4つの他の独立した実験の代表的なものである。臍帯血ECおよび臍帯血HAECから単離された、HPP−ECFCコロニーとLPP−ECFCコロニーとの間において、テロメラーゼ活性における、同様の相違を観察した。
【図10】HPP−ECFCの単色画像(図10a)、LPP−ECFCの単色画像(図10b)、内皮クラスターの単色画像(図10c)、および成熟した分化内皮細胞の単色画像(図10d)。HPP−ECFCは、最小の細胞質拡散(cytoplasmic spreading)(直径は12ミクロン〜22ミクロンで変化)を有し、>0.8の核 対 細胞質比を有する小さな細胞(核直径8ミクロン〜10ミクロン)である。LPP−ECFCは、サイズに関してより不均一(heterogenous)であるが、HPP−ECFCと比較して、より大きい。LPP−ECFC核は、10.5ミクロン〜12.5ミクロンでサイズが変化し、そして>0.4の比を有するが、<0.5であるより大きな細胞質拡散(25ミクロン〜60ミクロンで変化)を有する。内皮クラスターは、13.0ミクロン〜16.5ミクロンで変化する核を有する、ほぼ成熟した内皮細胞であり、そして>0.2であるが、<0.3である核 対 細胞質比を有する、65ミクロン〜80ミクロンで変化する細胞質直径を有する。成熟した分化内皮細胞は、大きくて非常に良好に拡散した細胞であり、17.0ミクロン〜22.0ミクロンの範囲の核直径および85ミクロン〜105ミクロンの細胞質直径を有し、そして内皮クラスターと同様の核 対 細胞質比を有する。したがって、HPP−ECFCは、任意の他のEPCよりも非常に明確に小さく、そして成熟内皮細胞よりもかなり小さい。
【図11】明確に分類されたサイズのコロニーを生じる、分裂した単一のプレートされたウシ、ブタ、およびラットの大動脈内皮細胞の百分率。
【図12】(A)単離の同日に注入したヒト骨髄由来のCD34+細胞の移植;(B)最大刺激濃度のG−CSF、TPO、SCF、またはFlt−3を用いた7日間の培養;または(C)臍帯血HPP−ECFCとの7日間の共培養(ここで、同期(gated)細胞は、ヒトCD45抗原についてポジティブに染色されている)の、8週間後のNOD−SCIDマウスの末梢血において、フローサイトメトリーによって検出されたヒト細胞(CD45ポジティブ)のキメラ現象の百分率;グラフは、同様の結果を有する3つの独立した実験の代表的なものである。
【特許請求の範囲】
【請求項1】
単離された内皮性コロニー形成細胞であって、以下の特徴:
(a)内皮細胞に特有の細胞表面抗原を発現する;
(b)造血細胞に特有の細胞表面抗原を発現しない;
(c)アセチル化LDLを取り込む;および
(d)マトリゲルにおいて、毛細管様の管を形成する、
を有する、単離された内皮性コロニー形成細胞。
【請求項2】
請求項1に記載の単離された細胞であって、ここで、前記内皮細胞に特有の細胞表面抗原は、CD31、CD105、CD144、およびCD146である、細胞。
【請求項3】
請求項1に記載の単離された細胞であって、ここで、前記造血細胞に特有の細胞表面抗原は、CD14およびCD45である、細胞。
【請求項4】
請求項1に記載の単離された細胞であって、以下:
(a)単一細胞からプレートされた場合、少なくとも約2000個の細胞の、少なくとも二次コロニーへと再プレートする;
(b)高増殖性を示す;
(c)単一細胞から増殖する;および
(d)高レベルのテロメラーゼを発現する、
としてさらに定義される、細胞。
【請求項5】
請求項4に記載の細胞であって、ここで、高レベルのテロメラーゼ発現は、HeLa細胞によって発現されるテロメラーゼ発現の、少なくとも約34%である、細胞。
【請求項6】
請求項4に記載の単離された細胞であって、>0.8である、高い核 対 細胞質比を示すものとしてさらに定義される、細胞。
【請求項7】
請求項4に記載の方法であって、少なくとも107個の子孫が単一細胞に由来する、方法。
【請求項8】
請求項4に記載の単離された細胞であって、<22ミクロンの細胞直径を有するものとしてさらに定義される、細胞。
【請求項9】
請求項1に記載の細胞を単離するための方法であって、該方法は、以下の工程:
(a)細胞外マトリックスタンパク質でコートされた支持体上で、生物学的試料に由来する細胞を培養する工程;
(b)該支持体に接着し、かつ再プレート可能なコロニーを形成する細胞を選択する工程;および
(c)該コロニーから単一細胞を選択する工程
を包含する、方法。
【請求項10】
請求項9に記載の方法であって、ここで、前記生物学的試料は、哺乳動物の血液である、方法。
【請求項11】
請求項9に記載の方法であって、ここで、前記生物学的試料は、哺乳動物の血管である、方法。
【請求項12】
請求項9に記載の方法によって単離される、細胞。
【請求項13】
内皮細胞の型に関する単一細胞アッセイであって、以下の工程:
(a)特定の分類方法を使用し、生物学的試料の細胞を分類する工程;
(b)規定された条件下で、細胞外マトリックスタンパク質上で該単一分類の細胞を培養する工程;および
(c)特定のコロニーのサイズ、形態、および増殖能を列挙し、該内皮細胞の型を決定する工程
を包含する、単一細胞アッセイ。
【請求項14】
請求項13に記載の単一細胞アッセイであって、ここで、前記内皮細胞の型は、HPP−ECFCを含む、単一細胞アッセイ。
【請求項15】
請求項12に記載の細胞を富化するための方法であって、該方法は、以下の工程:
(a)特定の分類方法を使用し、生物学的試料の細胞を分類する工程;および
(b)規定された条件下で、細胞外マトリックスタンパク質上で単一分類の細胞を培養する工程
を包含する、方法。
【請求項16】
エキソビボにおいて造血幹細胞を増殖させるための方法であって、該方法は、以下の工程:
(a)コラーゲンコートされた固体支持体上で、HPP−ECFC細胞を培養する工程;および
(b)HPP−ECFC細胞とともに共培養することによって、造血幹細胞(HSC)を増殖させる工程
を包含する、方法。
【請求項17】
請求項16に記載の方法であって、ここで、前記HPP−ECFCは、ヒト臍帯血細胞由来であり、そして前記HSC細胞は、ヒト骨髄由来である、方法。
【請求項18】
哺乳動物中の移植片における、造血幹細胞の百分率を改善するための方法であって、該方法は、以下の工程:
(a)ヒト骨髄細胞を臍帯血HPP−ECFCとともに共培養し、生成物を形成させる工程;および
(b)適切な量の該生成物を、該哺乳動物中に移植する工程
を包含する、方法。
【請求項19】
請求項18に記載の方法であって、ここで、前記細胞は、ヒト骨髄由来であるCD45+細胞であり、そして前記哺乳動物は、NOD−SCIDマウスである、方法。
【請求項1】
単離された内皮性コロニー形成細胞であって、以下の特徴:
(a)内皮細胞に特有の細胞表面抗原を発現する;
(b)造血細胞に特有の細胞表面抗原を発現しない;
(c)アセチル化LDLを取り込む;および
(d)マトリゲルにおいて、毛細管様の管を形成する、
を有する、単離された内皮性コロニー形成細胞。
【請求項2】
請求項1に記載の単離された細胞であって、ここで、前記内皮細胞に特有の細胞表面抗原は、CD31、CD105、CD144、およびCD146である、細胞。
【請求項3】
請求項1に記載の単離された細胞であって、ここで、前記造血細胞に特有の細胞表面抗原は、CD14およびCD45である、細胞。
【請求項4】
請求項1に記載の単離された細胞であって、以下:
(a)単一細胞からプレートされた場合、少なくとも約2000個の細胞の、少なくとも二次コロニーへと再プレートする;
(b)高増殖性を示す;
(c)単一細胞から増殖する;および
(d)高レベルのテロメラーゼを発現する、
としてさらに定義される、細胞。
【請求項5】
請求項4に記載の細胞であって、ここで、高レベルのテロメラーゼ発現は、HeLa細胞によって発現されるテロメラーゼ発現の、少なくとも約34%である、細胞。
【請求項6】
請求項4に記載の単離された細胞であって、>0.8である、高い核 対 細胞質比を示すものとしてさらに定義される、細胞。
【請求項7】
請求項4に記載の方法であって、少なくとも107個の子孫が単一細胞に由来する、方法。
【請求項8】
請求項4に記載の単離された細胞であって、<22ミクロンの細胞直径を有するものとしてさらに定義される、細胞。
【請求項9】
請求項1に記載の細胞を単離するための方法であって、該方法は、以下の工程:
(a)細胞外マトリックスタンパク質でコートされた支持体上で、生物学的試料に由来する細胞を培養する工程;
(b)該支持体に接着し、かつ再プレート可能なコロニーを形成する細胞を選択する工程;および
(c)該コロニーから単一細胞を選択する工程
を包含する、方法。
【請求項10】
請求項9に記載の方法であって、ここで、前記生物学的試料は、哺乳動物の血液である、方法。
【請求項11】
請求項9に記載の方法であって、ここで、前記生物学的試料は、哺乳動物の血管である、方法。
【請求項12】
請求項9に記載の方法によって単離される、細胞。
【請求項13】
内皮細胞の型に関する単一細胞アッセイであって、以下の工程:
(a)特定の分類方法を使用し、生物学的試料の細胞を分類する工程;
(b)規定された条件下で、細胞外マトリックスタンパク質上で該単一分類の細胞を培養する工程;および
(c)特定のコロニーのサイズ、形態、および増殖能を列挙し、該内皮細胞の型を決定する工程
を包含する、単一細胞アッセイ。
【請求項14】
請求項13に記載の単一細胞アッセイであって、ここで、前記内皮細胞の型は、HPP−ECFCを含む、単一細胞アッセイ。
【請求項15】
請求項12に記載の細胞を富化するための方法であって、該方法は、以下の工程:
(a)特定の分類方法を使用し、生物学的試料の細胞を分類する工程;および
(b)規定された条件下で、細胞外マトリックスタンパク質上で単一分類の細胞を培養する工程
を包含する、方法。
【請求項16】
エキソビボにおいて造血幹細胞を増殖させるための方法であって、該方法は、以下の工程:
(a)コラーゲンコートされた固体支持体上で、HPP−ECFC細胞を培養する工程;および
(b)HPP−ECFC細胞とともに共培養することによって、造血幹細胞(HSC)を増殖させる工程
を包含する、方法。
【請求項17】
請求項16に記載の方法であって、ここで、前記HPP−ECFCは、ヒト臍帯血細胞由来であり、そして前記HSC細胞は、ヒト骨髄由来である、方法。
【請求項18】
哺乳動物中の移植片における、造血幹細胞の百分率を改善するための方法であって、該方法は、以下の工程:
(a)ヒト骨髄細胞を臍帯血HPP−ECFCとともに共培養し、生成物を形成させる工程;および
(b)適切な量の該生成物を、該哺乳動物中に移植する工程
を包含する、方法。
【請求項19】
請求項18に記載の方法であって、ここで、前記細胞は、ヒト骨髄由来であるCD45+細胞であり、そして前記哺乳動物は、NOD−SCIDマウスである、方法。
【図1−1】

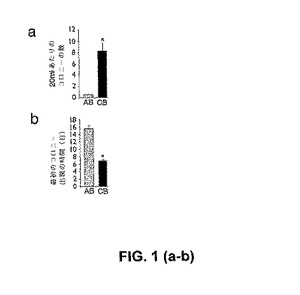
【図1−2】
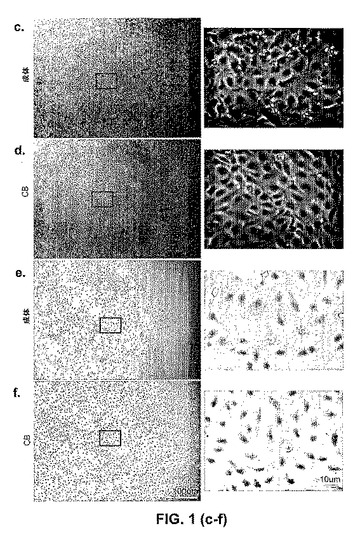
【図2】
【図3】

【図4】
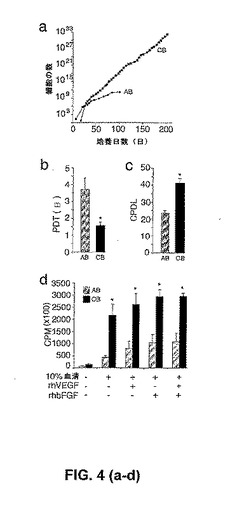
【図5】
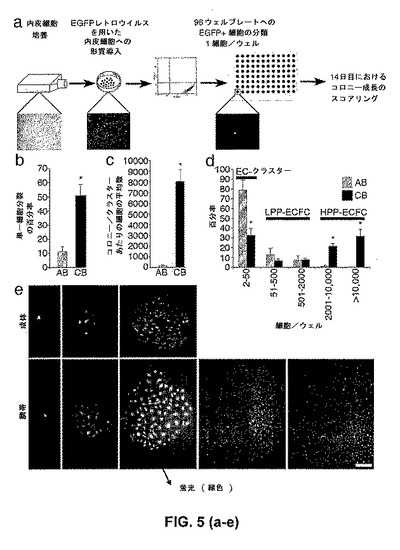
【図6】
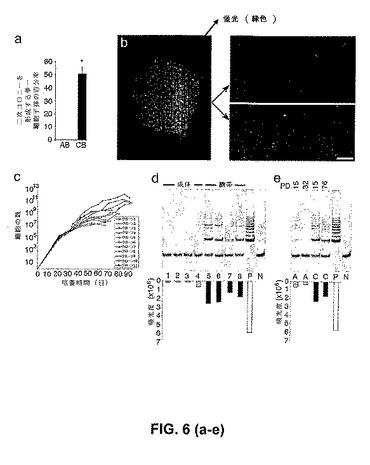
【図7】
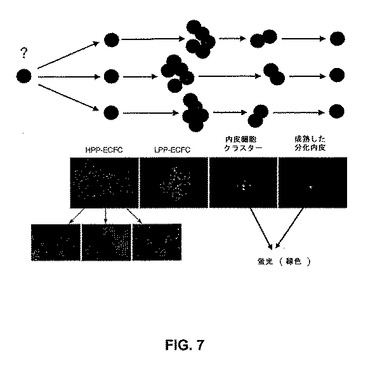
【図8】
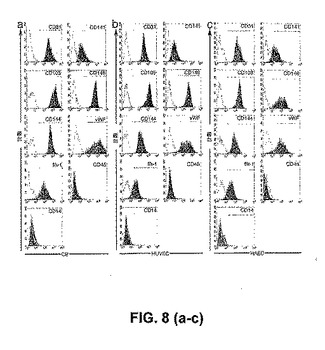
【図9】
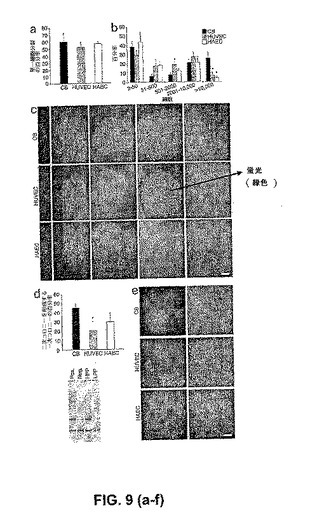
【図10(a−b)】
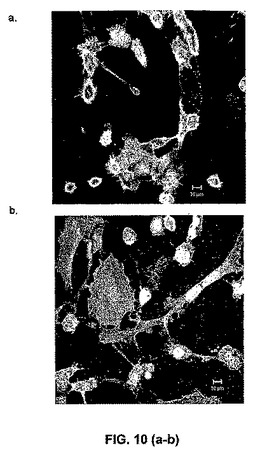
【図10(c−d)】
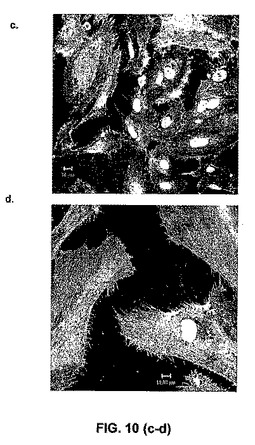
【図11】
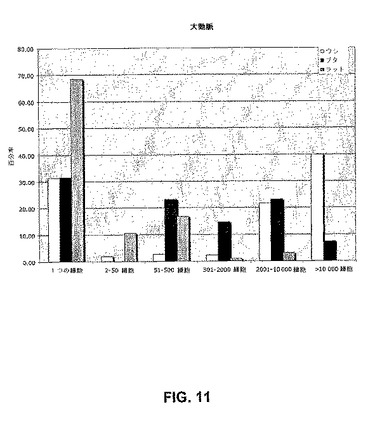
【図12】
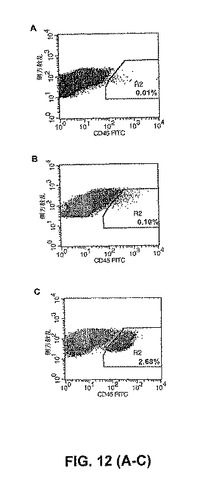
【図1−2】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10(a−b)】
【図10(c−d)】
【図11】
【図12】
【公表番号】特表2007−521817(P2007−521817A)
【公表日】平成19年8月9日(2007.8.9)
【国際特許分類】
【出願番号】特願2006−552372(P2006−552372)
【出願日】平成17年2月9日(2005.2.9)
【国際出願番号】PCT/US2005/004267
【国際公開番号】WO2005/078073
【国際公開日】平成17年8月25日(2005.8.25)
【出願人】(506265705)インディアナ ユニバーシティ リサーチ アンド テクノロジー コーポレイション (4)
【Fターム(参考)】
【公表日】平成19年8月9日(2007.8.9)
【国際特許分類】
【出願日】平成17年2月9日(2005.2.9)
【国際出願番号】PCT/US2005/004267
【国際公開番号】WO2005/078073
【国際公開日】平成17年8月25日(2005.8.25)
【出願人】(506265705)インディアナ ユニバーシティ リサーチ アンド テクノロジー コーポレイション (4)
【Fターム(参考)】
[ Back to top ]