
グリカン特異的分析ツール
触媒的に不活性であり、かつその基質親和性が少なくとも1.2倍に高められた突然変異した糖質プロセシング酵素であるレクテンズ分子を提供する。さらに、そのようなレクテンズを作成するための方法および使用方法も提供する。レクテンズアプローチに従ったそのほかの突然変異したタンパク質もさらに提供する。

【発明の詳細な説明】
【技術分野】
【0001】
連邦政府による資金提供を受けた研究の記載
本発明の開発中に行われた研究の一部は、米国国立衛生研究所(National Institutes of Health)によって授与されたR41GM086991号の下で米国政府基金を利用した。したがって、米国政府は本発明において一定の権利を有する。
【0002】
発明の分野
本発明は、グリカン特異的分析ツール、それらの使用方法、およびグリカン特異的分析ツールを作成するための工程を対象とする。他の分析ツールも本明細書においてさらに提供する。
【背景技術】
【0003】
グリカンは、脂質およびタンパク質と結びついた状態で認められることの多い複合糖質である。複合糖質はタンパク質および細胞の表面に存在することから、それらは往々にして、生物認識プロセスにおける機能的位置を占めている。オリゴ糖および多糖の複雑な形状、機能性および動的特性は、これらの分子が生体情報のエンコーダーとして分子間相互作用において機能することを可能にする。
【0004】
糖質認識は正常な生物発生の不可欠な部分であるが、先天性免疫系によっても、宿主生物が病原体の表面に提示された糖質に基づいて外来性病原体を同定することを可能にするように使用されうる。逆に、多くの細菌性およびウイルス性病原体はまず、宿主細胞表面の糖質と特異的に結合することによって宿主組織に付着する。このため、糖質に基づく宿主-病原体相互作用を妨害することのできる、または抗細菌ワクチンとして機能しうる治療薬を開発することには関心が寄せられている。また、異常グリコシル化はある種の癌および他の疾患のマーカーであり、そのためにそれらは診断的および治療的な用途の標的ともなっている。例えば、O結合型N-アセチルグルコサミンによる細胞内タンパク質の修飾状態(O-GlcNAc化)は、疾患、特に2型糖尿病によって引き起こされる変化の重要なバイオマーカーである。
【0005】
生物発生および疾患におけるグリカンの重要性にもかかわらず、現在のところ十分なグリカン特異的分析ツールはなく、このことが疾患の診断および治療における異常グリコシル化の活用を遅らせてきた。例えば、細胞へのO-GlcNAc取り入れおよびその後のタンパク質の提示のモニタリングのための現行の方法は、N-アジドアセチルグルコサミン(GlcNAz)などの標識試薬の外因性取り込みに基づいている。残念ながら、この方法は、単離された組織試料またはタンパク質試料におけるO-GlcNAcの分析には適用できない。グライコミクス/プロテオミクス解析に適用しうる1つの代替的なO-GlcNAc標識アプローチは化学酵素的タグ標識を用いる。この方法の重大な限界は、それが他のGlcNAcが末端にある複合グリカンも標識することである。このため、異常グリコシル化を探索しようとして生体試料を調べるために用いることのできる、明確な糖質特異性を備えた分析ツールは依然として必要とされている。
【0006】
現在のところ、グリカン特異的分析の用途に用いられている主なタイプの生体分子は、糖結合タンパク質(レクチン)および抗体の2つである。これらのタイプの試薬のいずれにも伴う主な欠点は、糖質とタンパク質との間の特徴的に弱い相互作用であり、その解離定数は典型的には、レクチンについてはミリモルからマイクロモルの範囲、抗体についてはマイクロモルからナノモルの範囲である。さらに、抗体を用いる際の大きな問題は、糖質が免疫原としては極めて劣ることである。それらは一般にT細胞依存性応答を生じさせることができず、そのためほとんどの場合はIgMクラス抗体を産生させるが、これは分析的および診断的な用途には不都合である。重鎖および軽鎖の可変ドメインからなる単鎖キメラ(scFv)は、不安定性を来す恐れがある。さらに、抗体を利用するグリカン特異的分析の手法には、抗体の選択性がコンテキスト依存性であることに起因する欠点もある。または、レクチンは、その特異性の幅広さから、分析用途のためのそれらの使用には限界がある。このため、糖質配列に対する十分な特異性を有し、なおかつ広範囲にわたるグリカン内の配列を認識することのできる分析用試薬を開発することには依然として需要が存在する。
【発明の概要】
【0007】
本発明は、その触媒活性を消失させながらもその基質特異性は維持するように突然変異させた糖質プロセシング酵素を含む、レクテンズ(lectenz)を提供する。ある態様において、本発明のレクテンズは、グリカンに対して、野生型酵素のKmよりも高い親和性を有する。また、本発明のレクテンズが、著しく低下したkoff速度を有してもよい。他の態様において、本発明は、触媒的に不活性な糖質プロセシング酵素を含むレクテンズであって、その不活性酵素が野生型残基とは異なる1つまたは複数のアミノ酸残基を含み、前記残基が、結合した基質に近接するが、気相(ΔEMM)相互作用エネルギーに対する寄与が約|0.5kcal/mol|未満である残基、全(ΔG)相互作用エネルギーに対する寄与が約|0.5kcal/mol|未満である残基、または結合相互作用エネルギーに対して不利に寄与する任意の残基、およびそれらの組み合わせからなるリストから選択される、レクテンズを提示する。基質に近接する残基は、本明細書では一般に基質の5Å以内にあると考えるが、それよりも遠い場合も近い場合も考えうる。
【0008】
別の局面において、本発明は、レクテンズを作製するための方法を提供する。いくつかの態様において、本方法は:(a)糖質プロセシング酵素の配列を、突然変異した場合に糖質プロセシング酵素のグリカンに対する親和性または酵素-グリカン複合体の安定性に影響を及ぼしうると考えられる1つまたは複数のアミノ酸残基に関して解析する段階;(b)糖質プロセシング酵素が段階(a)で同定された少なくとも1つの突然変異したアミノ酸を有する、酵素-グリカン複合体の結合エネルギーを予測するための計算シミュレーションを行う段階;(c)段階(a)および(b)で同定された突然変異を含む糖質プロセシング酵素を、それらが複合体を形成する能力に関して検査する段階;ならびに(d)グリカンに対する結合親和性がWT酵素のそれよりも高い、段階(c)による突然変異体を同定する段階、を含む。
【0009】
本発明の別の局面は、レクテンズをグリカン特異的分析の用途に用いる方法を提供する。ある態様において、本発明のレクテンズは親和性試薬として、または組織染色のための媒体として用いることができる。他の態様において、レクテンズは、生体試料を特定のグリコフォームに関して濃縮するために用いることができる。さらに他の態様において、レクテンズには、糖タンパク質上の特異的なグリコシル化部位を決定するという用途もある。本発明の他の局面は、活性治療薬の標的化送達のための媒体としてのレクテンズの使用を伴う。
【0010】
ある態様において、糖質プロセシング酵素はグリコシダーゼ酵素である。他の態様において、糖質プロセシング酵素はグリコシルトランスフェラーゼ酵素である。他の態様において、糖質プロセシング酵素は多糖リアーゼ酵素である。他の態様において、糖質プロセシング酵素はスルファターゼ酵素である。他の態様において、糖質プロセシング酵素はスルホトランスフェラーゼ酵素である。他の態様において、糖質プロセシング酵素はリガーゼ酵素である。さらなる態様において、糖質プロセシング酵素はアミダーゼ酵素である。さらに別の態様において、糖質プロセシング酵素はエピメラーゼ酵素である。
【図面の簡単な説明】
【0011】
【図1】糖質プロセシング酵素(この例ではノイラミニダーゼ)とその糖質結合性レクテンズ類似体との関係を描写している。
【図2】レクテンズの親和性を最適化するための、計算による方法とインビトロディスプレイライブラリー法を組み合わせるプロトコールを描写している。
【図3】PNGアーゼF(PNGase F)複合体におけるCα位置のRMSDを描写している。
【図4】キトビオースのPNGアーゼFとの結合に関する水素結合相互作用の配置図を描写している。
【図5】左の画像では、PNGアーゼFの結合部位における二糖リガンドの4.5Å以内にある残基(ダークグレー)を描写している。右の画像では、溶媒接触可能表面を、結合のために最も重要であると同定された残基に印を付した上で描写している。
【図6】速度論的なオン速度およびオフ速度の変動を突然変異および温度(10℃および25℃)の関数として指し示しているSPRセンサーグラム(sensogram)を描写している。
【図7】酵母におけるAga2タンパク質と融合させたタンパク質ディスプレイライブラリーを描写している。c-mycタグ標識タンパク質に対する蛍光標識抗原の結合の検出を図示している。
【図8】高親和性クローンの選択を指し示す、フローサイトメトリーを介した細胞選別を描写している。
【図9】不活性酵素と比べた予備的レクテンズの親和性の強化を描写している。
【0012】
本出願の図版および図面が必ずしも一定の縮尺で描かれてはいないこと、ならびにこれらの図面および図版は本発明を単に例示しているに過ぎず、それを限定するものではないことは理解されるであろう。
【発明を実施するための形態】
【0013】
発明の詳細な説明
以下の記載では、本発明の十分な理解が得られるように、説明の目的で、具体的な数字、パラメーターおよび試薬を記述している。しかし、本発明を、これらの具体的な細目がなくても実施しうることは明らかであろう。場合によっては、本発明が分かりにくくならないように、周知の特徴を省いてもよく、または単純化してもよい。
【0014】
記載された態様、および本明細書における「1つの(one)態様」、「1つの(an)態様」、「1つの例示的態様」などへの言及は、記載された態様が特定の特徴、構造または特質を含みうるものの、あらゆる態様が必ずしもその特定の特徴、構造または特質を含まなくてもよいことを指し示している。その上、そのような語句は、必ずしも、同じ態様について言及しているわけでもない。さらに、特定の特徴、構造または特質を1つの態様に関連して記載している場合には、明示的に記載されているか否かにかかわらず、他の態様に関連してそのような特徴、構造または特質を生じさせることが当業者の知識の範囲内にあることは理解されるであろう。
【0015】
I.定義
本明細書で用いる場合、糖質プロセシング酵素とは、糖質含有基質に対して作用する任意の酵素を指して用いられる用語である。その例には、グリコシダーゼ、グリコシルトランスフェラーゼが含まれるが、それらには限定されない。
【0016】
本明細書で用いる場合、「グリコシダーゼ」という用語は、グリコシド結合の加水分解を触媒する酵素を指して用いられる。「グリコシド結合」という用語は、糖質分子を別の糖質分子に連結させるか、または糖質分子をタンパク質分子に連結させるか、または糖質分子を脂質分子に連結させる、あるタイプの官能基のことを指す。「糖質」という用語は、一般式Cm(H2O)nの有機化合物を指すものとする。本発明において、「糖質」、「複合糖質」および「グリカン」という用語は互換的に用いられる。
【0017】
「触媒的に不活性な突然変異体」または「突然変異体」または「不活性酵素」という用語は互換的に用いられ、その触媒活性の少なくとも95%を喪失していて、かつ触媒的に活性な酵素とは異なるアミノ酸組成を有する酵素のことを指す。別に記述する場合を除き、触媒的に不活性な突然変異体による化学結合切断の速度は、最大でも、あらゆる触媒が存在しない同一の条件下で測定される結合切断速度の5%を上回らない。「触媒的に活性な酵素」とは、化学結合の加水分解を触媒しうるタンパク質を指すものとする。「野生型(WT)酵素」とは、生物において天然に存在するのと同じ遺伝子配列を有し、ヒト介入によって改変されていない遺伝子によってコードされる酵素のことを指す。当然ながら、野生型酵素の天然に存在する多型形態がこの定義に含まれることは理解されるであろう。さらに、それ以外の点ではタンパク質の天然の開始コドンも終止コドンも変化させない、タンパク質の精製または単離に用いられるタグまたは他の修飾などの修飾が、本発明におけるWT酵素の定義の範囲に含まれることも理解されるであろう。本明細書で用いる場合、「リガンド」および「基質」という用語は互換的に用いられ、WT酵素または突然変異体酵素が結合することのできる分子のことを指す。
【0018】
本発明のレクテンズは、グリカンに対して、野生型酵素のKmよりも高い親和性を有する。Kmの意味を理解するためには、酵素作用のモデルを用いる必要があるであろう。最も単純なモデルはミカエリス・メンテンの古典的モデルであり、これは多くの種類の酵素に有用であることが証明されている(式1)。
【0019】
基質(S)は、第1の反応において酵素(E)と可逆的に結合する。ほとんどの場合には、この段階を測定することはできない。測定されるのは、第2の反応によって生み出される生成物(P)である。ミカエリス・メンテン定数(Km)は式2で定義される。
【0020】
Kmが、酵素と基質との間の結合の強さの尺度となる結合定数ではないことに注意されたい。その値は、酵素に対する基質の親和性だけでなく、酵素と結合した基質が生成物に変換される速度も含む。k2がk-1よりもはるかに小さい場合にのみ、Kmは結合親和性に等しいと考えられる。野生型酵素の状況ではWT酵素がリガンドに作用するため、親和性の値を直接測定することは難しく、この理由から不活性酵素のKdを野生型酵素のKmと比較することが好都合であることが理解されるであろう。本明細書で用いる場合、「親和性」という用語は、2つの分子間の引力のことを意味する。レクテンズの親和性は、通常はWT酵素のKmと対比して測定されるが、そのリガンドに対する解離定数Kdの、WT酵素の不活性形態に対する相対的な低下という観点で表現することもできる。解離定数Kdは、複合体がその成分へと解離する性向の尺度となる平衡定数である。一般的反応:
があるとし、式中、iEおよびSはそれぞれ不活性酵素および基質を示し、iESは不活性-酵素-基質複合体である。この場合、対応する解離定数Kdは:
として定義され、式中、[iE]および[S]はそれぞれ遊離した不活性-酵素および基質の濃度を示し、[iES]は不活性-酵素-基質複合体の濃度である。したがって、ある態様において、レクテンズの親和性の増大は、レクテンズの親和性を、親和性を最適化する別の突然変異を受けていない触媒的に不活性な形態のWT酵素と比較することによって測定される。本発明において、タンパク質のそのリガンドに対する親和性は、解離定数(Kd)または会合定数(Ka)として表現することができる。
【0021】
「ΔG」という表現は、結合のギブス自由エネルギーのことを指す。ギブス自由エネルギーは、複合体を1つにまとめる力に抗しながら、複合体を、それ以上の分離に必要な余分な仕事量を無視しうる十分な距離まで隔てられた構成要素部分に分解させる際に行わなければならない仕事量を表す熱力学的ポテンシャルである。「ΔEMM」という表現は、気相における分子力学自由エネルギーのことを指す。「特異性」または「酵素特異性」という用語は互換的に用いられ、酵素が、種々のリガンドの集団から、特異的な分子構造を含むリガンドを認識して選択する能力のことを指す。「非特異的」結合または相互作用という用語は、個々の分子または残基のいかなる特異的認識にも識別にも基づかない、分子または残基の間での弱い相互作用イベントのことを指す。
【0022】
「分子動力学(MD)」という用語は、公知の物理的過程の近似によって原子および分子をある期間にわたって相互作用させて粒子の運動の概観を得る、コンピュータシミュレーションの一形態を指すものとする。古典的MDシミュレーションは、古典的力場から導き出されるエネルギーおよび力を用いるニュートンの運動方程式によって規定される。古典的力場は、分子または分子の凝集物における原子位置を分子または凝集物のポテンシャルエネルギーと関係づける数学モデルである。「エヴァルト処理(Ewald treatment)」または「エヴァルト和」という用語は、本明細書で用いる場合、周期構造系(例えば、結晶)、特に静電エネルギーの相互作用エネルギーを計算するための方法を記述している。「ベルレ(Verlet)アルゴリズム」または「ベルレ積分」という用語は、ニュートンの運動方程式を積分するために用いられる数値的方法のことを意味する。
【0023】
II.レクテンズ
本発明の1つの局面はレクテンズを提供する。本発明のレクテンズは、所与のグリカンに対して野生型酵素と実質的に同じ特異性を有し、かつグリカンに対する親和性がWT酵素と比較して高くなっている、糖質プロセシング酵素の触媒的に不活性な突然変異体を含む。本明細書で用いる場合、「実質的に同じ」という用語は、野生型酵素の少なくとも60%であるグリコシダーゼ突然変異体の特異性を記述するものとする。いくつかの態様において、突然変異体の特異性は、WT酵素の少なくとも70%である。少なくとも1つの態様において、突然変異したグリコシダーゼはその基質に対して、同じ基質に対する野生型酵素に比して少なくとも85%特異的である。他の態様において、突然変異したグリコシダーゼは、同じ基質に対する野生型酵素に比して少なくとも95%特異的である。
【0024】
遺伝子操作されたレクテンズに関して、活性部位残基の突然変異は、酵素を不活性化するのみならず、親和性を強化する可能性のある経路も与える(図1)。いくつかの態様において、グリカンに対するレクテンズの親和性は、野生型酵素のそれよりも高くなる。ある態様において、レクテンズのその基質に対する親和性は、解離定数Kdの観点で表現することができる(式3および4)。解離定数が小さいほど、レクテンズはより強固に基質と結合している。本発明のいくつかの態様において、グリカンに対するレクテンズの解離定数(Kd)は、WT糖質プロセシング酵素のKmより少なくとも約1.2倍〜約1,000倍小さい。他の態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKmより少なくとも約2倍小さい。ある態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKmより少なくとも約10倍小さい。ある態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKmより少なくとも約10,000倍小さい。さらなる態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKmより少なくとも約100,000倍小さい。
【0025】
また、本発明のレクテンズの親和性向上を、WT酵素の不活性突然変異体のそれと対比したKdの低下という観点で表現しうることも理解されるであろう。すなわち、グリカンに対するレクテンズのKdは、不活性なWT糖質プロセシング酵素のそれより少なくとも約1.2倍〜約1,000倍小さい。他の態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKdより少なくとも約2倍小さい。ある態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKdより少なくとも約10倍小さい。ある態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKdより少なくとも約10,000倍小さい。さらなる態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKdより少なくとも約100,000倍小さい。
【0026】
本発明のレクテンズは、いかなる特定の糖質プロセシング酵素にも限定されない。そうではなくて、本発明は、糖質を特異的に認識することのできる、任意のグリコシダーゼ酵素またはグリコシルトランスフェラーゼ酵素、タンパク質またはポリペプチドに対して幅広く適用可能である。本発明に適したグリコシダーゼの例には、ラクターゼ、アミラーゼ、キチナーゼ、スクラーゼ、マルターゼ、ノイラミニダーゼ、インベルターゼ、ヒアルロニダーゼおよびリゾチームが非限定的に含まれる。酵素委員会(Enzyme Commission)(EC)番号3.2.-.-、式中「-」は数字である、によって分類されているグリコシダーゼが本明細書に含まれることは理解されるであろう。本発明のグリコシダーゼは反転型グリコシダーゼでも保持型グリコシダーゼでもありうる。1つの態様において、本発明のレクテンズは、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から単離されたPNGアーゼFから調製される。別の態様において、レクテンズは、バクテロイデス・テタイオタオミクロン(Bacteroides thetaiotaomicron)から単離されたβ-O-GlcNAcアーゼに関して決定されているWT配列を用いて、組換えβ-O-GlcNAcアーゼから調製される。さらに別の態様においては、ウェルシュ菌(Clostridium perfringens)由来のノイラミニダーゼを、レクテンズを調製するために用いる。グリコシダーゼのほかに、本発明に用いるために適した糖質プロセシング酵素には、EC番号2.4.-.-として指定されたものを含むグリコシルフェラーゼ、およびEC番号4.2.2.-として指定されたものを含む多糖(polysacharide)リアーゼが含まれる。他の糖質プロセシング酵素には、糖質エステラーゼ、スルファターゼ、スルホトランスフェラーゼ、または糖質基質に対して作用する他の任意の酵素が含まれる。本発明の触媒的に不活性な糖質プロセシング酵素は、原核生物または真核生物、ならびに他のものから単離された糖質プロセシング酵素から調製することができる。
【0027】
ある態様において、本発明のレクテンズは高い耐熱性を有すると考えられる。高い耐熱性とは、レクテンズがその三次構造を、生理的に許容される緩衝液中で少なくとも40℃の温度で少なくとも30分間にわたって保つことを意味する。生理的に許容される緩衝液とは一般に、0〜1%のNaCl濃度および0〜20mMのグルコース濃度、pH 6〜8の0.01〜0.2Mリン酸緩衝液のことを指す。ある態様において、レクテンズは正しくフォールディングした状態を、生理的に許容される緩衝液中で少なくとも60℃の温度で少なくとも30分間保つ。他の態様において、レクテンズはその三次構造を、生理的に許容される緩衝液中で少なくとも80℃の温度で少なくとも30分間保つ。必要であれば、本発明のレクテンズは、好熱性生物または超好熱性生物から単離されたグリコシダーゼから調製することができる。本明細書における適した糖質プロセシング酵素を単離しうる好熱性生物および超好熱性生物の例には、サーマス・サーモフィルス(Thermus thermophilus)、スピロヘータ・アメリカナ(Spirochaeta americana)、パイロコッカス・フリオサス(Pyrococcus furiosus)、メタノパイラス・カンドレリ(Methanopyrus kandleri)、パイロロバス・フマリ(Pyrolobus fumarii)、ジオサーモバクテリウム・フェリレデュセンス(Geothermobacterium ferrireducens)およびアーケオグロブス・フルギダス(Archaeoglobus fulgidus)が非限定的に含まれる。
【0028】
いくつかの態様において、本発明のレクテンズは長い速度論的オフ速度(kinetic off-rate)によって特徴づけられる。速度論的オフ速度は、解離速度定数(koff)、またはリガンドがタンパク質から解離する速度(式5参照)によって測定される。一般に、基質-不活性酵素複合体の形成は、以下の速度論的機序によって記述しうる:
式中、iEおよびSはそれぞれ不活性酵素および基質を示し、iESは不活性酵素-基質複合体であり、konおよびkoffはそれぞれ複合体形成および解離に関する速度定数である。少なくともいくつかの態様において、本発明のレクテンズはそのリガンドから、WT酵素のそれよりも少なくとも約2倍の遅さの速度で解離すると考えられる。1つの態様において、レクテンズ-グリカン複合体解離の速度は、WT-グリカン複合体のそれよりも少なくとも5倍の遅さである。さらに別の態様において、レクテンズ-グリカン複合体の解離の速度は、WTのそれよりも少なくとも10倍の遅さである。また、比較をレクテンズのオフ速度と不活性化されたWT酵素との間で行うこともできる。少なくともいくつかの態様において、本発明のレクテンズはそのリガンドから、不活性WT酵素のそれよりも少なくとも約2倍の遅さの速度で解離すると考えられる。1つの態様において、レクテンズ-グリカン複合体解離の速度は、不活性WT酵素とグリカンとの複合体のそれよりも少なくとも5倍の遅さである。さらに別の態様において、レクテンズ-グリカン複合体の解離の速度は、不活性WT酵素のそれよりも少なくとも10倍の遅さである。
【0029】
ある態様において、本発明の触媒的に不活性な突然変異体は、WT残基と異なっていて、かつ、酵素-基質複合体中で基質に近接するが、全(ΔG)相互作用エネルギーに対する寄与が約-0.5kcal/molを上回る残基、または結合相互作用エネルギーに対して不利に寄与する任意の残基、およびそれらの組み合わせから選択される、1つまたは複数のアミノ酸残基を有する。基質に近接するが強い相互作用を形成しないこの残基セットを、ぬるい(tepid)残基セットと命名してもよいであろう。ある態様において、突然変異体グリコシダーゼは、ぬるい残基セット中に2つまたはそれ以上の突然変異を有する。他の態様において、突然変異体グリコシダーゼは、これらの位置に3つもしくはそれ以上、4つもしくはそれ以上、またはさらには5つもしくはそれ以上の突然変異を有する。
【0030】
III.レクテンズを作製するためのコンピュータ支援による方法
本発明の別の局面は、糖質プロセシング酵素の触媒的に不活性な突然変異体を含むレクテンズを作製するためのコンピュータ支援による方法であって、
(a)糖質プロセシング酵素の配列を、突然変異した場合に酵素を不活性化しうると考えられる1つまたは複数のアミノ酸残基に関して解析する段階;
(b)WT酵素-グリカン複合体の、または糖質プロセシング酵素が段階(a)で同定された少なくとも1つの突然変異したアミノ酸を有する複合体の結合エネルギーを予測するために計算シミュレーションを行う段階;
(c)残基を、それらの予測される相互作用エネルギーに基づいて、2つの群に、すなわち、酵素の特異性を規定するのに必須である第1の残基群、および基質に近接するが特異性を規定するのに必須ではない第2の残基群(この第2のセットを本明細書では、ぬるい残基と称する)に細分類する段階;
(d)段階(a)、(b)および(c)で同定された突然変異を含む糖質プロセシング酵素を、それらが酵素-グリカン複合体を形成する能力に関して検査する段階;ならびに
(e)グリカンに対する結合親和性がWTグリコシダーゼのそれよりも少なくとも1.2倍高い、段階(d)による突然変異体を同定する段階、
を含む方法を提供する。
【0031】
図2中の資料形態に略図として示したこの態様は、本発明のレクテンズを得るための1つの態様のみを表しているに過ぎないものと理解されたい。本明細書においてさらに詳細に記述している図2の諸段階を、本発明のレクテンズを得るために、本明細書において説明した通りに改変することができる。具体的には、例えば、図2中の特定の段階が、本明細書中の他の箇所でさらに詳細に説明されている「必須な残基を同定する」段階のための「計算によるアラニンスキャニング」といった特定の手法について言及している点で、それは本発明の単なる1つの態様に過ぎず、同じ段階を行うための他のアプローチも存在する。
【0032】
本発明のある態様は、レクテンズへの変換のために適している糖質プロセシング酵素の同定を伴う。レクテンズの生成は特定の糖質プロセシング酵素には限定されないが、その活性部位残基が公知である、かつ/または触媒的に不活性な突然変異体が記載されている、糖質プロセシング酵素を選択することが好ましいであろう。触媒残基の同定は、多くの糖質プロセシング酵素に関して部位指定突然変異誘発を用いて行われており、多くの場合には酵素の速度論的動態の測定によって確かめられている。しかし、不活性酵素(第1世代レクテンズ)は、多くの場合、レクチンと同等な親和性を有する。
【0033】
一般に、コンピュータ支援による突然変異誘発手法を実施する前には、関心対象のリガンドに対する触媒的に不活性な糖質プロセシング酵素の結合親和性を予測することが好ましい。糖質プロセシング酵素のそのリガンドに対する親和性を特徴づけるためには、いくつかの周知の手法がある。1つのそのような手法は、AMBER/GLYCAMタンパク質/糖質力場を使用する分子動力学(MD)手法である。
【0034】
エネルギー解析の前に、MDシミュレーションの安定性およびコンフォメーション平衡化のレベルを決定するために、Cα原子の位置の根平均二乗差(root mean squared difference)(RMSD)をシミュレーション時間の関数として求めることができる。そのようなデータに基づき、平均RMSDが安定であるか否かおよび約0Å〜4Åの範囲内にあるか否かを判定することができる。また、コンフォメーション平衡に達するために必要な時間を、このアプローチを通じて判別することも可能である。平衡化していないデータセットの部分はその後の解析に当たって省くことが一般に好ましい。5ナノ秒のデータを収集し、システムが平衡化するまでに約1ナノ秒を費やした1つの態様では、最初の1ナノ秒のデータを省いた。上記の5ナノ秒/1ナノ秒の態様は例示に過ぎず、本発明を限定するものではないことは理解されるであろう。
【0035】
結合部位におけるリガンド安定性は、グリカンと糖質プロセシング酵素との間の分子間水素結合を評価することによって評価することができる。水素結合およびそれらの占有率に関する平均値を収集することができ、可能であれば、結晶学的に決定された値に沿ってそれらを収集する。結合部位におけるリガンドの位置のRMSDのモニタリングにより、結合部位におけるリガンド安定性を決定することも可能である。MDシミュレーションが安定であって、かつリガンドとタンパク質との間の実験的相互作用を再現しうることが確かめられたならば、そのシステムをその後の解析に使用することができる。典型的なシミュレーションは、圧力および温度が一定(NPT)の条件下で、または体積および温度が一定(NVT)の条件下で行うことができる。これらのシミュレーションは、AMBERのSANDERモジュール、およびTIP3P水モデルを用いて行うことができる。ある態様においては、タンパク質力場パラメーターをParm99セットから採用し、糖質パラメーターをGLYCAM06から採用する。また、非周期的境界条件下で陰溶媒モデルを用いるシミュレーションを行うこともできる。
【0036】
AMBERは、構造に基づくリガンド設計、ならびにあらゆる複合分子系における構造および自由エネルギーの理解のための、シミュレーションに基づく方法を提供する分子モデリングおよびシミュレーション用のパッケージである。AMBERはUniversity of California、San Francisco校で開発され、そこから入手可能である。AMBERの内部の他のモジュールを用いて、これらのMDシミュレーションを行うことができる。実際には、MDシミュレーションを行うためのAMBER以外のプログラムも存在する。そのようなプログラムも本発明に適用可能である。
【0037】
ある態様において、グリカン-タンパク質複合体の初期座標は、可能であれば、不活性酵素-基質、活性酵素-阻害薬または酵素-生成物複合体からの結晶学的データから選択することができる。相同性モデリングまたは比較モデリングによって作成されたモデルのような、タンパク質の理論モデルを用いることもできる。遊離した酵素の構造のみが入手可能である場合には、AutoDockまたは別の等価なプログラムを用いて共複合体(co-complex)を予測することができる。AutoDockは、3D構造が判明している受容体に低分子がどのように結合するかを予測するために設計された自動ドッキングツール・スイートである。他のドッキングプログラムも存在し、それらも本発明に適用可能であると考えられる。
【0038】
ある態様においては、ヒスチジンプロトン付加の状態を、可能であれば分子内水素結合から推測することができ、そうでなければヒスチジンをNεで中性でプロトン付加されているとして扱うことができる。複合体上の任意の実効電荷は、適した数の対イオン(Cl-またはNa+)の付加によって中和することができる。典型的には、オリゴ糖-タンパク質複合体は、周期的立方体内で例えばほぼ10,000個のTIP3P水分子によって溶媒和されると考えられ、この枠の端と溶質の最も近い原子との間の最小距離は10Åである。周期的境界条件は、直接空間カットオフ距離を12Åとする長距離静電作用に関するエヴァルト処理と合わせて適用することができる。これらのパラメーターが本発明を限定するものでないことは理解されるであろう。実際には、TIP3Pは計算化学のために用いられる古典的水モデルの1つに過ぎないことが理解されるであろう。TIP4P、TIP5P、SPC、BNSなどの他の水モデルを本発明に用いることもできる。また、水を、誘電定数、距離依存的誘電定数、一般化ボルンモデルなどの陰溶媒和モデルを用いて、またはポワソン・ボルツマン近似によって近似することもできる。
【0039】
ある態様においては、初期コンフィギュレーションを、SANDERモジュールを用いてエネルギー極小化することができる。1つの態様において、初期コンフィギュレーションは、5,000サイクルの最急降下およびSANDERモジュールを用いた25,000サイクルの共役勾配エネルギー極小化を含む。続いてシステム全体を、加熱および続いて冷却することにより、シミュレーションアニーリングを行う。ある態様において、シミュレーションアニーリングは、50psで5Kから300Kにし、続いて5Kでの冷却をさらに50ps行うことを含む。初期原子速度はボルツマン分布から指定することができ、一般に5Kとする。生成動態段階の前に、システム全体を150psで再び5Kから300Kに加熱することによって熱平衡化することができる。2fsの時間段階を用い、ベルレアルゴリズムを用いて運動方程式を積分することができる。水素を含む結合は、SHAKEアルゴリズムを用いてそれらの平衡長に拘束することができる。これらのパラメーターは例示に過ぎず、本発明を限定するものではないことは理解されるであろう。
【0040】
本発明の方法は、いかなる特定のリガンドにも限定されない。本発明に適しているリガンドには、任意の天然もしくは合成脂質またはそれらの誘導体が含まれる。適したリガンドの例には、ラクトース、スクロース、マルトース、トレハロース、セロビオース、キトビオース、N結合型オリゴ糖、O結合型オリゴ糖、オリゴ糖、単糖、末端分枝型および非分枝型のα-(2,3)およびα-(2,6)-Neu5Ac、高マンノースN-グリカン上のα-(1-2)-man、糖タンパク質および糖脂質上のα-Gal、グリコサミノグリカン(ヘパリン、ヘパラン、コンドロイチン、ヒアルロン酸およびそれらの硫酸化類似体)、糖タンパク質および糖脂質上のβ-N-およびβ-またはα-O-GlcNAc、糖タンパク質および糖脂質上のβ-Gal、N-グリカン上のα-1,2/3/6 Man、N結合型またはO結合型グリカン上のα-Fucが非限定的に含まれる。
【0041】
A.糖質プロセシング酵素-グリカン複合体の安定性に影響を及ぼす可能性のある残基を解析するための、コンピュータ支援による方法
レクテンズへの変換のための糖質プロセシング酵素がひとたび選択されれば、その配列を、突然変異した場合に酵素-グリカン複合体の親和性または安定性または特異性に影響を及ぼす可能性のあるアミノ酸残基に関して解析することができる。
【0042】
いくつかの態様において、配列解析は、計算による突然変異誘発によって行うことができる。1つの態様においては、タンパク質安定性および/またはリガンド親和性に影響を及ぼすキー(key)タンパク質残基を、計算による飽和突然変異誘発実験を用いて同定することができる。計算による飽和突然変異誘発はアミノ酸配列レベルで実施され、これは、1つのアミノ酸側鎖の別のものによる置き換えに続いて、基質と酵素との間の相互作用の親和性または安定性または特異性に対する置き換えの影響について計算解析を行うことを伴う。他の態様において、標的化またはランダム計算による突然変異誘発を行うことができる。他の態様において、酵素内の各アミノ酸によって酵素-グリカン複合体の安定性または特異性に加えられる寄与を、グリカン-酵素複合体に関して直接計算することもできる。
【0043】
ある態様において、タンパク質安定性またはリガンド親和性またはリガンド特異性に影響を及ぼす「ホットスポット」キータンパク質残基は、計算によるアラニンスキャニング突然変異誘発(ASM)を用いて同定される。ASMは、個々の残基のアラニンによる逐次的置き換えによって行うことができる。ASMは、タンパク質の機能、安定性および形状に関与する残基を同定することができる。各アラニン置換により、タンパク質の機能性に対する個々のアミノ酸の寄与を検討する。この手法の全般的な概観は、Kollman, P.A., et al., Calculating Structures and Freeエネルギーof Complex Molecules: Combining Molecular Mechanics and 連続 Models. Acc. Chem. Res., 2000. 33(12): p.889-97およびArakat, N., et al., Exploiting Elements of Transcriptional Machinery to Enhance Protein Stability JMB, 2007. 366(103-116)に提示されている。1つの典型的な例では、WT酵素または酵素-基質複合体のMDシミュレーションを行い、データを収集する。その後に、すべて(または選択したもののみ)の残基の側鎖をCβ位置まで切断して短縮し、その結果、それらをアラニンに変換させる。不足している水素原子を加えて完全なアラニンを形成させる。この手順を、MDシミュレーションで収集した構造のすべてについて繰り返す。アラニンに変換させたところで、酵素または複合体の安定性に対する突然変異の影響をMDデータから計算することができる。これらの影響は、複合体もしくはタンパク質の構造の変化をモニターすることによって、またはタンパク質もしくは複合体に関連したエネルギーの変化をモニターすることによって決定される。また、アラニン突然変異を導入した後に別個のMDシミュレーションを行うこともできる。アラニンスキャニング突然変異誘発は、当技術分野において公知の唯一のスキャニング突然変異誘発法ではない。このため、この手法の考察は例示に過ぎず、本発明を限定するものではない。
【0044】
さらに他の態様において、キー残基を、野生型複合体におけるそれらのエネルギー寄与の大きさによって、または結合したリガンドに対するそれらの近接性によって同定することもできる。1つのそのような態様においては、水素結合、ファンデルワールス接触、および/または水媒介接触によって基質と直接相互作用する、同定された「ホットスポット」キー残基を含む、アミノ酸残基のサブセットを作り出すことができる。非水素(または重)原子との間に約3.2Åを上回らずに位置する任意の残基を、ファンデルワールス接触によって、または水素結合によって基質と直接相互作用するとみなすことができる。非水素原子との間に約4.5Åを上回らずに位置する任意の残基を、水媒介接触によって、または非特異的静電相互作用によって基質と相互作用するとみなしてよいであろう。他の態様において、サブセットは、気相(ΔEMM)または全(ΔG)相互作用エネルギーのいずれかに対する寄与が少なくとも約-0.5kcal/mol未満であるとして同定された任意の残基を含むと考えられる。
【0045】
または、ホットスポットを、特異的に有利な突然変異を予測するため、またはポテンシャル的に有利な突然変異のクラス(中性、荷電性、疎水性など)を同定する(クラスフォーカシング)ための、さらなる理論的解析に供することもできる。Hayes, R.J., et al., Combining computational and experimental screening for rapid optimization of protein properties. Proc Natl Acad Sci U S A, 2002. 99(25): p.15926-31は、クラスフォーカシングの手法を記載している。
【0046】
いくつかの態様においては、レクテンズ特異性が劣化する恐れのある不適切な機会を避けることが好ましいであろう。これらの態様においては、基質との相互作用に直接関与する残基を、突然変異誘発のために最初は選択しないことにする。他の態様においては、非特異的に相互作用するが、リガンド結合に対して有意に有利に寄与する残基(相互作用エネルギーの算出により同定)も、初期の突然変異誘発試験から除外するものとする。続いて、残りのすべての残基、特に結合に対して不利に寄与する任意のものを、1回目の突然変異誘発のために選考する。いくつかの態様においては、以前に除外したサブセットのメンバーを、初回の実験的および理論的な突然変異誘発を完了した後に、突然変異誘発に供することができる。
【0047】
B.糖質プロセシング酵素-グリカン複合体の結合エネルギーを予測するためのコンピュータ支援による計算シミュレーション法
いくつかの態様においては、上記の計算による突然変異誘発法のいずれかによって行った理論的突然変異の、糖質プロセシング酵素-グリカン複合体の結合エネルギーに対する影響を予測することが重要と考えられる。ある態様において、結合エネルギーは、熱力学的積分(TI)としても知られる自由エネルギー摂動法を用いて算出することができる。TIを用いることで、キー構造モイエティーの結合に対するエネルギー寄与を定量することができる。Straatsma, Holonomic Constraint Contributions to Free Energy Differences from Thermodynamic Integration Molecular Dynamics Simulations. Chem. Phys. Lett., 1992. 196: p.297-302, Zacharias et al., Inversion of Receptor Binding Preferences by Mutagenesis: Free Energy Thermodynamic Integration Studies of Sugar Binding to L-Arabinose Binding Proteins. Biochemistry, 1993. 32: p.7428-7434、およびChipot and Kollman, Alternative Approaches to Potential of Mean Force Calculations: Free Energy Perturbation versus Thermodynamic Integration Case Study of Some Representative Nonpolar Interactions. J Comput Chem, 1996. 17(9): p.1112-1131は、TIの方法論を記載している。TIは一般に、きわめて類似したリガンドに対する相対的結合エネルギーを検討する目的に限定されるが、キー構造モイエティーの結合に対するエネルギー寄与を定量することもできる。TIシミュレーションの典型的な一例では、自由エネルギーを初期残基を一連の漸進的段階を通じて最終的なものに変換する理論的プロセスを計算し、その間に各状態からの寄与率を変化させるという改変MD条件下でシミュレーションを行う。この非物質的プロセスは、各状態に関するエネルギー関数を数学的に混合することによって行われ、これは時に計算アルケミーと呼ばれることもある。
【0048】
さらに他の態様において、結合の全自由エネルギー(ΔG)は、基質とタンパク質(反応物)との間の相互作用エネルギーの直接分解によって算出することができる。ΔGの直接算出は、分子力学(MM)エネルギー推定値と、脱溶媒和自由エネルギーを捕らえるためのポワソン・ボルツマン(PB)または一般化ボルン(GB)などの連続溶媒モデルとを組み合わせる。これらの算出は一般に、コンフォメーションエントロピーによる追加の寄与を別に計算することを必要とする。
【0049】
限定的ではないが一例を挙げると、1つの典型的なMM-GB/PB算出では、MD軌道から抽出したそれぞれの構造「スナップショット」について、自由エネルギーをタンパク質(ΔGタンパク質)、リガンド(ΔGリガンド)および複合体(ΔG複合体)に関して計算する。関心対象の酵素によっては、データの初期部分を切り捨ててシステムを平衡化させる。例えば5nsの軌道では最初の1nsを切り捨てることができる。これらのモデルでは、データのスナップショットを設定間隔で収集する。単なる例示に過ぎないが、残りの4nsから、分子力学(MM)結合エネルギー解析のために2,000個のスナップショットを(2ps間隔で)選択することができる。続いて、結合自由エネルギー(ΔG)を減算によって計算することができる(式6参照)。軌道全体にわたって平均することにより、最終的な平均相互作用エネルギー(<ΔG結合>)が得られる。
<ΔG>=<ΔG複合体>-<ΔGタンパク質>-<ΔGリガンド>[6]
式中、平均はMDスナップショット全体にわたるものである。
【0050】
構成要素の自由エネルギーは、エネルギーを3つのカテゴリー、すなわち分子力学(静電およびファンデルワールス)エネルギー、溶媒和エネルギーおよびエントロピーエネルギー(式7参照)に分けることによって計算することができる。
<ΔG>=<ΔEMM>−T<ΔSMM>+<ΔG溶媒和>[7]
【0051】
解析の前に、水分子を溶媒和軌道から除去することができる。続いて、溶媒和によるエネルギー寄与を、その相対的速さの点から大型のタンパク質-糖質複合体への適用に非常に適している一般化ボルン(GB)陰溶媒和モデルの適用によって入手することができる。MM-GBSAの結果は、より厳密なMM-PBSA解析(ポワソン-ボルツマン陰溶媒近似に基づく)によるものに十分に匹敵する。GB近似はまた、計算によるアラニンスキャニングにおいても有効に働くことが最近示されている。少なくとも1つの態様において、糖質-タンパク質相互作用エネルギーを計算するためのGB法は、Tsui and Case, Theory and Applications of the Generalized Born Solvation Model in Macromolecular Simulations. Biopolymers, 2001. 56: p.275-291におけるGBパラメーター表示を使用する。
【0052】
ある態様において、エントロピーへの振動、並進および回転の寄与はエネルギーが極小化された座標の通常の様式解析から導き出すことができ、一方、コンフォメーションエントロピーは該当する内部座標の共分散行列の解析から推算する。Karplus and Kushick, Method for Estimating the Configurational Entropy of Macromolecules. Macromol., 1981. 14: p.325-332を参照。糖質の場合には、グリコシド間(inter-glycosidic)ねじれ角に関連したコンフォメーションエントロピーに焦点を絞ることが特に好都合かつ適切である。主として束縛回転に起因するコンフォメーションエントロピーの変化を、各オリゴ糖の遊離型および結合型における骨格ねじれ角の動きから推算することができる。結合状態および遊離状態におけるねじれ角に関する共分散行列の行列式から、相対コンフォメーションエントロピーを導き出すことができる。
【0053】
いくつかの態様においては、結合エネルギーを、古典的力学力場を用いて算出する。一般に、かかわる分子に関係する原子間特性を力場としてパラメーター表示する。AMBER力場を用いるためには、力場のパラメーターに関する値(例えば、力の定数、平衡結合長および結合角、電荷を入力する)。これらのパラメーターセットはかなり多く存在し、AMBERソフトウエアのユーザーマニュアルに詳細に記載されている。各パラメーターは名称を持ち、ある特定のタイプの分子に関するパラメーターを与える。
【0054】
1つの態様においては、GLYCAM/AMBER糖質力場を用いて結合解析を実施する。GLYCAM06パラメーターはさまざまな生体分子力場とともに用いることができる。GLYCAM06と適合性のある力場の例には、AMBER、CHARMM、NWCHEMなどが非限定的に含まれる。ある態様においては、GLYCAMパラメーターを、タンパク質に関するAMBERパラメーターによって補強することができる。GLYCAM06はいかなるデフォールトパラメーターも汎用パラメーターも使用しておらず、何らかの特定のクラスの生体分子にはもはや限定されないが、低分子力場の趣旨には十分に拡張可能である。GLYCAM06パラメーターは、例えば、Kirschner et al., GLYCAM06: A Generalizable Biomolecular Force Field. Carbohydrates. J. Comput. Chem., 2007. Early View (DOI 10.1002/jcc.20820)に記載されている。
【0055】
ある補足的な態様においては、計算シミュレーションを、コンフォメーションサンプリングを達成するために行う。そのような手法には、分子動力学シミュレーション、モンテカルロシミュレーション、または側鎖回転異性体の検索が含まれる。
【0056】
C.糖質プロセシング酵素突然変異体の発現および検査
本明細書に記載した計算による突然変異誘発および分子シミュレーションの方法を用いて、予測された望ましいリガンド結合特質を有する糖質プロセシング酵素突然変異体が同定されれば、親和性および複合体安定性の予測を、実験的突然変異誘発を用いて確かめることができる。いくつかの態様においては、関心対象の糖質プロセシング酵素のコード配列を、適した種から単離されたゲノムDNAから増幅し、適したベクター中にサブクローニングする。遺伝子クローニングおよびタンパク質過剰発現の定型的な方法は記載されている。ゲノムDNA由来の関心対象の糖質プロセシング酵素のコード配列を、選択した種から単離して、任意の適したベクター中にサブクローニングすることができる。いくつかの態様においては、適したアフィニティータグを備えた関心対象の糖質プロセシング酵素を発現するように、ベクターを遺伝子操作することができる。タンパク質のタグ標識は、アフィニティークロマトグラフィー手法を用いたその精製を容易にすると考えられる。1つの態様において、糖質プロセシング酵素に、ヘキサヒスチジンタグによるタグ標識を行うことができる。別の態様においては、糖質プロセシング酵素を、抗原ペプチドタグを含むように遺伝子操作することができる。適したベクターの例には、pOPH6、pETおよびpBADが非限定的に含まれる。pOPH6は、選択した大腸菌株に発現のために導入して形質転換を行わせることができる。本発明は、タンパク質の過剰発現に関して、特定の大腸菌株には限定されない。適した株の例にはDH5αが含まれる。
【0057】
一晩培養した細胞培養物(5〜10ml)を、十分な量の炭素源、ミネラル、イオン、抗生物質および他の試薬を含む適した量の栄養培地(例えば、ルリア・ベルターニ培地)中に接種することができる。一般に、これらのバッチ生産は小規模、すなわち100〜200mlであるが、より大容積のバッチを調製することもできる。抗生物質の選択は、大腸菌株およびクローニングされたベクターの遺伝子操作された耐性に依存すると考えられる。例えば、DH5α中にクローニングされたpOPH6ベクターの場合には、80〜120μg/mlアンピシリン、0.8〜1.5%v/vグリセロール、80〜150mMリン酸カリウム(pH 7.0)および0.2〜1.5mMイソプロピルβ-D-1-チオガラクトピラノシド(IPTG)を含むルリア-ベルターニ培地を用いうるであろう。培地を、導入から4〜25時間後に収集することができる。
【0058】
いくつかの態様においては、過剰発現された酵素を単離することができる。種々の方法をタンパク質精製のために利用することができる。1つの態様においては、過剰発現されたグリコシダーゼを含む清澄化された培地を、適したアフィニティーカラムに通すことができる。ヘキサヒスチジンアフィニティータグが付加されたタンパクを、ニッケルまたはコバルトとともに固定化された樹脂を用いて精製することができる。例えば、清澄化された培地を、適した量の塩およびイミダゾールを含む冷ローディング緩衝液で希釈し、Hi-Trap IMACカラム(Amersham Pharmacia Biotech)に2ml/分で通すことができる。抗原タグ標識タンパク質は、それを抗原特異的抗体とともに固定化された樹脂に通すことによって精製することができる。組換え酵素を、適したゲル濾過カラムを用いて脱塩処理することできる。
【0059】
ある態様においては、実験的突然変異誘発を、確立されたプロトコールに従った部位指定突然変異誘発を用いて実施する。一例を挙げると、部位指定突然変異誘発を、適したグリコシダーゼの遺伝子を含むベクター(例えば、pOPH6)上で、Stratagene(La Jolla, CA)製のQuikChange(商標)突然変異誘発キットを用いて行う。それぞれの突然変異体に対するセンスプライマーおよびアンチセンスプライマーは、遺伝子の配列を用いて設計することができ、自動DNA合成装置(Integrated DNA Technologies, Inc.)によって合成することができる。約10〜約20ngのプラスミドおよび約5〜約20ピコモルのセンスプライマーおよびアンチセンスプライマーを、製造元のプロトコールに従ったポリメラーゼ連鎖反応(PCR)混合物に添加することができる。PCRは、熱サイクル制御ユニット(MJ Research, Cambridge, MA)上で行うことができる。DNAテンプレートは、製造元のプロトコールに従った適したエンドヌクレアーゼの添加によって消化することができる。いくつかの態様においては、所望の突然変異のみが生成されたことを確認するために、それぞれの突然変異体の全コード領域を完全にシークエンシングする。いくつかの態様においては、確認された突然変異体DNAをテンプレートとして用いることで、単一のアミノ酸突然変異に用いたのと同じ手順を用いて複数の突然変異を作り出すことができる。
【0060】
アミノ酸突然変異体が、計算による飽和突然変異誘発のための方法を介して同定されたところで、突然変異体タンパク質のライブラリーを、望ましい結合特質を持つ突然変異体に関してスクリーニングすることができる。タンパク質-リガンド相互作用のハイスループットスクリーニングのために用いられる数多くの技術が、当技術分野において利用可能である。本発明に適したそのような技術の例には、ツーハイブリッドシステム、mRNAディスプレイ、ファージディスプレイ、酵母ディスプレイ、リボソームディスプレイおよび細菌ディスプレイが非限定的に含まれる。本発明のアプローチは、ライブラリーにおいてランダム化すべきであるタンパク質中の部位を同定するための、さらなる主観的な方策である。したがって、本発明の計算解析を組み合わせることにより、フォーカストバイオコンビナトリアルライブラリーを設計し、続いて構築することができる。それらの設計および構築によるそのようなライブラリーは、ライブラリースクリーニングのためのはるかに効率的なアプローチを提供する。
【0061】
いくつかの態様において、タンパク質-リガンド相互作用のハイスループットスクリーニングは、突然変異誘発ディスプレイライブラリーを作り出すことによって行うことができる。1つのそのようなライブラリーシステムは、GENEARTによって合成することができる。1つの態様においては、そのライブラリーをファージ上にディスプレイすることができる。ファージディスプレイライブラリーは、当技術分野において十分に確立されたプロトコールを用いて構築することができる。例を挙げると、関心対象のタンパク質またはペプチドをコードするDNAライブラリーを、M13繊維状ファージのpIII遺伝子またはpVIII遺伝子中にライゲートする。続いて、ファージ遺伝子およびインサートDNAハイブリッドを、TG1またはXL1-Blue大腸菌などの大腸菌細胞に導入して形質転換を行わせる。「ファージミド」ベクターを用いる場合には(簡略化されたディスプレイ構築物ベクター)、ファージ粒子は、それらが、ファージDNAのパッケージング、およびマイナー(pIII)またはメジャー(pVIII)コートタンパク質のいずれかの上での成熟ビリオンとそれらの外殻の一部としての関連タンパク質断片との集合を可能にするヘルパーファージに感染するまでは、大腸菌細胞から放出されないと考えられる。pIII遺伝子またはpVIII遺伝子中への多くのさまざまなDNA断片の取り入れによってライブラリーが生成され、それから関心対象のメンバーを単離することができる。該当するDNA標的またはタンパク質標的をウェルの表面に固定化することにより、そのような標的の1つと結合するタンパク質をその表面にディスプレイするファージは、洗浄によっても保持されるが、他のものは除去されると考えられる。保持されたものを溶出させて、より多くのファージ(ヘルパーファージによる細菌の感染により)を産生させるため、およびそのようにして該当する(すなわち、結合性)ファージが濃縮されたファージ混合物を生産するために用いることができる。これらの段階の反復サイクリングを、望ましくない材料を除去することによる金の試料の濃縮に準拠して「パニング」と称する。
【0062】
他の態様においては、突然変異誘発ディスプレイライブラリーを酵母上にディスプレイさせることができる。酵母ディスプレイでは、関心対象のタンパク質をAga2pタンパク質との融合物として酵母の表面にディスプレイさせる。Aga2pタンパク質は天然において酵母によって用いられ、酵母細胞接合時の細胞-細胞接触を媒介する。このため、Aga2pを介したタンパク質のディスプレイは、そのタンパク質を細胞表面から突出させ、酵母細胞壁上の他の分子との相互作用の可能性を最小限に抑える。Aga2遺伝子、HA(血球凝集素)タグ、ディスプレイされるタンパク質およびc-mycタグの融合物としての発現ベクターを図示している図7を参照されたい。酵母ディスプレイライブラリーは、当技術分野において十分に確立されたプロトコールを用いて構築することができる。いくつかの態様においては、関心対象の遺伝子を、AGA2遺伝子とインフレームになるように、選択したベクター中にクローニングすることができる。適したベクターの例には、pYD1ベクター(Invitrogen)およびpPNL6(Pacific Northwest National Laboratory)が非限定的に含まれる。その結果得られた構築物を、続いて、染色体に組み込まれたAGA1遺伝子を含む、適した酵母株(例えば、EBY100 S.セレビシエ(S. cerevisiae))に導入して形質転換を行わせる。ベクター由来のAga2融合タンパク質およびEBY100宿主株中のAga1タンパク質はいずれも、ガラクトースの非存在下ではクローニングされたタンパク質の検出可能な発現を許容しない厳密に調節されるプロモーターであるGAL1プロモーターによって調節される。ガラクトースを用いた導入に応じて、Aga1タンパク質およびAga2融合タンパク質が分泌経路内で会合し、クローニングされた突然変異体が細胞表面にディスプレイされる。
【0063】
突然変異した糖質プロセシング酵素を細胞表面にディスプレイする突然変異誘発ライブラリーがひとたび構築されれば、望ましい結合特性および複合体形成特性を有する突然変異体を同定するためにそれをスクリーニングすることができる。選択したリガンドと高親和性複合体を形成しうる突然変異体を同定するために用いられるアッセイシステムの基本原理は、ディスプレイライブラリーおよびリガンドを含む反応混合物を、2つの試薬が相互作用して結合し、それによって複合体を形成するのに十分な条件および時間の下で調製することを伴う。続いて、結合パートナー間の複合体の形成を捕らえる。反応が完了した後に、反応しなかった構成要素は除去されるが(例えば、洗浄により)、形成された複合体は細胞表面に保持されると考えられる。細胞表面に係留された複合体の検出は、さまざまなやり方で達成することができる。いくつかの態様においては、リガンドを直接的または間接的に前標識することができる。リガンドを標識する場合、細胞表面に固定化された標識の検出により、複合体が形成されたことが指し示される。結合パートナーを前標識しない場合には、間接的標識を用いて、表面に係留された複合体を検出することができる。分子の標識は周知であり、例えば、タンパク質、核酸、糖質、カルボン酸のビオチン化のための、アミン反応性物質およびチオール反応性物質を含む、多数のビオチン化剤が公知である。ビオチン化された基質は、アビジンまたはストレプトアビジンを介してビオチン化構成要素と結びつけることができる。
【0064】
いくつかの態様においては、ディスプレイライブラリーのサイズを、ビオチン化N結合型糖ペプチドと低親和性ないし高親和性で結合する酵母を含むように濃縮することができる。低親和性相互作用を直接測定することは難しい。ビオチン化N結合型糖ペプチドを、ストレプトアビジンでコーティングされた常磁性ビーズ(例えば、Invitrogen)の表面に固定化することにより、ライブラリーを、ビーズ表面上の標的と結合するタンパク質をディスプレイする酵母に関して濃縮することができる。N結合型糖ペプチドでコーティングされた常磁性ビーズによって捕らえられた酵母を磁石で単離し、結合しなかった酵母を洗い流し、パニングの工程を繰り返す。いくつかの態様において、ライブラリーを、ライブラリーの初期サイズおよびパニングの回数に応じて細胞106〜108個に減少させることができる。少なくとも1つの態様においては、ライブラリーを約1×107個に減少させる。
【0065】
ある態様においては、高親和性突然変異体の初期スクリーニングを実施することができる。これらの態様において、スクリーニングはフローサイトメトリーによって行うことができる。スクリーニングは、特定のビオチン化糖ペプチドと結合する突然変異体のみを選択することによって行うことができる。一例を挙げると、図9に詳しく示しているように、C末端c-mycタグを有するAga2p融合タンパク質を発現する酵母を抗c-myc mAbとともにインキュベートし、その後に完全長グリコシダーゼクローンを発現した酵母を検出するために蛍光二次mAbを添加する。ある態様において、ビオチン化糖ペプチドと結合する突然変異体の検出は、ストレプトアビジンの添加によって達成することができる。1つの態様において、ストレプトアビジンは、適した蛍光標識(例えば、PE-Cy5)で標識することができる。ストレプトアビジン-PE-Cy5を用いて、糖ペプチドと結合する酵素クローンを蛍光標識することができる。糖ペプチドに対して高い親和性を有するc-mycを発現する細胞のみを選別することができる(図8)。いくつかの態様において、個々の酵母クローン上にディスプレイされた選択したグリコシダーゼ突然変異体の解離定数を決定することができる。少なくとも1つの態様において、解離定数はフローサイトメトリーによって決定することができる。
【0066】
ある態様においては、予備的スクリーニングアッセイにおいて同定された高親和性結合物を、さらなる結合試験のために発現させて精製することができる。生成されたレクテンズの結合親和性は、当技術分野において公知の種々の手法(例えば、フィルター結合アッセイ、電気泳動移動度シフトアッセイ(EMSA)、表面プラズモン共鳴(SPR)など)によって分析することができる。1つの態様においては、結合定数を、Biacore装置を用いるSPRによって測定する。SPRを用いて複合体の結合定数を評価するために用いられる1つの典型的なアッセイは、関心対象のレクテンズを、適したカップリング方法(例えば、アミンカップリング方法)によって20〜30℃でSPRチップ表面に固定化し、モック誘導体化したフローセルを参照表面とすることを含む。結合分析は、ランニング緩衝液の連続フロー(10〜50μl/min)を用いてさまざまな温度で行うことができる。ランニング緩衝液は、15〜25mM Tris-HCl(pH 7.5)、100〜200mM NaCl、0.5〜1.5mM EDTA、0.0025〜0.0075% P20界面活性剤でありうる。リガンドは、適した濃度範囲を得るための各々のランニング緩衝液中での系列希釈によって調製することができる。リガンドの結合は、レクテンズの低密度固定化表面上で一連の濃度(0.625〜10μM)で分析することができる。最大平衡センサーグラム値を用いて、飽和結合曲線をプロットし、平衡解離定数(Kd)の値を直接計算することができる。
【0067】
本発明の1つの局面において、レクテンズを作製するための方法は、酵素-基質または酵素阻害薬複合体に関する結晶学的データを計算による突然変異誘発のための基盤として用いる段階をさらに含む。一般に、レクテンズの基質との結晶化および結晶成長パラメーターの最適化は、当技術分野において記載されている方法および手順によって行うことができる。結晶化のために適した条件は、個々の事例ごとに決定されるであろう。結晶を回折に関して検査し、最も高い分解能で回折する結晶をデータ収集のために用いることができる。ある態様においては、分子置き換えを、複合体の構造を解明するために用いることが考えられる。これらの態様においては、生成されたレクテンズ-グリカン複合体のX線データを用いて、MDシミュレーションを検証すること、および/または新たなシミュレーションを開始することができる。
【0068】
本発明の全体を通して、グリカンプロセシング酵素から導き出されたレクテンズに関して例示されているものの、本発明が任意の酵素-基質複合体に対して広く適用可能であることは理解されるであろう。したがって、以下の実施例に限定されることなく、本発明の範囲をさらに例示することのみを目的として、このレクテンズアプローチを、プロテアーゼ、リパーゼ、キナーゼ(kinease)、ホスファターゼ、ヒドロラーゼ、イソメラーゼなどの酵素を、酵素基質に対する特異性を維持している受容体タンパク質に変換するために用いることができる。
【0069】
さらに、本発明は糖質プロセシング酵素に限定されない。これは糖質結合タンパク質にもさらに適用可能である。実際に、これはタンパク質-リガンド相互作用全般に適用可能である。
【0070】
V.使用方法
本発明の別の局面は、本明細書に記載されたレクテンズの使用方法を提供する。本明細書に記載されたレクテンズの非常にさまざまな潜在的な用途が、当業者には直ちに明らかであろう。以下は、そのような試薬の潜在的な有用性を説明している態様のうちごく一部を挙げたものである。
【0071】
ある態様において、本発明のレクテンズは、グリカン特異的分析ツールにおける用途のために用いることができる。本発明のレクテンズに基づくグリカン特異的分析ツールは、環境的、発酵、食品および医学の領域を含む多くの領域において検出方法として用いうる可能性があり、ヒトまたは動物におけるインビボまたはインビトロ検知にも用いうると考えられる。
【0072】
いくつかの態様において、本明細書に記載した規定された糖質特異性を有するレクテンズは、異常グリコシル化を探索しようとして生体試料を調べるために用いることができる。生体試料の例には、任意の生体液、組織または臓器が非限定的に含まれる。生体液の例には、血液、尿、血清、唾液、脳脊髄液および精液が非限定的に含まれる。他の態様において、本発明のレクテンズは、糖質に基づく生体液中の標的分析物レベルの検出のために用いることができる。標的分析物の例には、N結合型またはO結合型グリカン、グリコサミノグリカン(ヘパリンを含む)などの内因性に認められる分子、植物多糖、糖質に基づく薬物などの外因性に消費される種、および、その表面が特異な複合グリカンでコーティングされていることの多い病原体が非限定的に含まれる。他の態様において、本明細書に記載されたレクテンズは、医薬品探索、および、グリカンに基づく新たな化合物の生物活性の評価における用途もある。
【0073】
いくつかの具体的な態様において、本明細書に記載されたレクテンズは、異常グリコシル化を呈する疾患を診断および/または治療するために用いることができる。1つの態様において、本発明のレクテンズは、糖タンパク質、糖脂質、および/または種々の糖質エピトープを含むある特定の腫瘍抗原を検出するために用いることができる。数多くのこれらの腫瘍抗原が、新生物性病態においてアップレギュレートされることが見いだされている。新生物性障害の発症および進行を知らせることのできる、ならびに本発明のレクテンズによって検出することのできる腫瘍抗原の例には、結腸直腸癌、胃癌、膵癌、肺癌および乳癌ならびに発生過程にある胎児と関連性のある糖タンパク質である癌胎児性抗原(CEA);膵癌の患者で認められる糖脂質中に存在する糖質抗原19-9(CA 19-9)またはシアル酸付加ルイスA抗原;ならびに乳癌と関連性のある糖質抗原15-3(CA15-3)が非限定的に含まれる。
【0074】
抗原の存在が癌性細胞への形質転換を必ずしも指し示すわけではないが、細胞内におけるその局在は、CEAの場合のように、その指標となる。この理由から、選択性が高くかつ高親和性の分析ツールが必要とされている。診断検査は現在、糖タンパク質のペプチド部分または糖脂質の糖部分に対して産生されることの多い抗体に依拠しているが、しかし、その厳密なエピトープは今現在、明らかにされつつある途上に過ぎない。グリカンの特徴が決定されている例では、複数のグリコフォームが往々にして存在する(例えば、CEA)。これらのグリコフォームを識別しうる試薬がないことから、現時点では、グリコシル化のわずかな差異が病態、癌のタイプまたは組織内局在と相関する程度を決定することは不可能である。現在、これらの課題は、主として単離された糖タンパク質のMS分析によって対処することができるが、それらは複数のグリコフォームの混合物として検討される。典型的には、行われる唯一のレベルのグリコフォーム-フォーカシングは、レクチン(コンカナバリンA(Con A))アフィニティークロマトグラフィーを用いる高マンノース含有グリカンの濃縮である。より効率的な実験室分析および定型的な臨床診断手法は、グリコフォーム特異的試薬がないために依然として著しく制約されている。
【0075】
本発明のレクテンズは、生体試料中の任意の所与の糖タンパク質中に存在する各グリコフォームの相対的存在量を定量するために特に有用である。本明細書で用いる場合、「グリコフォーム」という用語は、特定のタイプの糖タンパク質が結びついているタンパク質のタイプのことを指す。2つのタンパク質は、それらが同じ糖タンパク質を保有しているならば同じグリコフォームのものと考えられる。いくつかの態様において、本発明のレクテンズは、生体試料を特定のグリコフォームに関して濃縮するために用いることができる。他の態様において、本明細書に記載された方法によって生成されたレクテンズは、グリカンが結びつくタンパク質表面上の特異的グリコシル化部位を同定するために用いることができる。これらの態様において、特定のオリゴ糖に対して特異的なレクテンズは、任意の糖タンパク質のタンパク質分解消化物から完全な糖ペプチドを分離するために用いられると考えられる。例えば、PNGアーゼ-Fにより導き出されたレクテンズは、N結合型糖ペプチドを、糖タンパク質の典型的なプロテアーゼ消化によって生じるであろう他の糖ペプチドまたはペプチドから分離するために用いることができる。関心対象の分析物における試料の濃縮は、糖ペプチド画分のさらなる特徴決定において大きな力添えとなる。特に、濃縮はペプチド配列およびグリカン構造の同定を容易にし、それは完全なタンパク質内部のグリコシル化部位の同定およびグリコシル化部位に存在する特定のグリカンの特徴決定を可能にする。
【0076】
他の態様において、本発明のレクテンズは、生体液、組織、臓器または生細胞におけるタンパク質の特異的グリカン修飾のモニタリングにおける用途があると考えられる。本発明の方法によって遺伝子操作されたレクテンズはタンパク質の実体には依存しないと考えられ、すなわちそれらはコンテキスト非依存性であると考えられ、かつ所与のグリカンを含む任意のタンパク質を認識しうると考えられ、そのため所与のグリカン修飾の検出のために極めて有用であると考えられる。
【0077】
さらに他の態様において、本発明のレクテンズは、細胞または組織をインビトロまたはインビボで染色するために用いることができる。
【0078】
他の態様において、レクテンズを、糖タンパク質中の特定の糖タンパク質またはグリコシル化部位に対して特異的であるように開発することができる。そのようなレクテンズは、製薬産業または研究産業で用いるための組換え糖タンパク質の生産過程で生じる可能性のある、混合物中の特定の糖タンパク質をモニターするために用いうると考えられる。
【0079】
前記の諸態様において、レクテンズに着色剤または色素によるタグ付加を施して、関心対象の細胞または組織または糖タンパク質または糖ペプチドまたはオリゴ糖または多糖を含む生体試料に適用することができる。
【0080】
ある態様において、本発明のレクテンズは治療薬として用いることができる。これらの態様において、特定のレクテンズの設計は、ヒト相同体が存在するグリコシダーゼに基づくことができる。これにより、そのようなレクテンズに免疫反応性がないことが保証されると考えられる。ある態様において、本発明のレクテンズを、活性治療薬の送達用に改変することができる。本発明のレクテンズは明確に定められたグリカン特異性を有するため、治療薬の送達を、特定のグリカンをディスプレイするような細胞、組織または臓器のみに標的化することができる。部位特異的送達のために用いうる治療薬の例には、さまざまな化学療法薬、抗生物質および抗イルス薬、毒素、放射性同位体、サイトカインなどが非限定的に含まれる。
【0081】
ある態様において、本発明のレクテンズは、例えばアフィニティークロマトグラフィーを含むアフィニティー分離のための試薬として用いることができる。アフィニティークロマトグラフィーは、生化学的混合物を、レクテンズとグリカンとの間のそれのような高度に特異的な生物的相互作用に基づいて分離する方法である。本発明は、いかなる特定の設計にもクロマトグラフィー系にも限定されない。一般に、レクテンズは固体支持体と共有結合性に結びついているか、または他の様式で固定化されていると考えられ、固定相を構成すると考えられる。ある態様において、レクテンズにより誘導体化された固定相をカラムクロマトグラフィーに用いることができる。これらの態様においては、固体固定相の粒子を用いて、管の内容積全体を満たすことができる(充填カラム)。または、固相粒子を管の内壁上またはそれに沿って集中させ、管の中間部分で、生体試料の通路を制限せずに開かれたままにすること(すなわち、移動相)もできる(中空カラム)。他の態様においては、レクテンズにより誘導体化された固定相をバッチクロマトグラフィーのために用いることができる。これらの態様においては、固定相を容器に添加して、生体試料と混合することができる。前記の例は一般にアフィニティークロマトグラフィーに対象を絞っているが、これらの原理が他のアフィニティー精製プロトコールにも容易に適用されることは理解されるであろう。
【実施例】
【0082】
実施例1
レクテンズへの変換のための標的酵素
糖質-バイオセンサー(レクテンズ)への変換のための標的酵素
表1に提示しているのは、レクテンズとしての再設計に供することのできる3種の初期グリコシダーゼである。レクテンズ1は、グライコミクス解析のすべての局面において広範な用途があると考えられる。レクテンズ2は糖尿病におけるグリカンの分析をさらに進めるために極めて重要であると考えられ、レクテンズ3はヒトおよびトリのインフルエンザ受容体を比較して特徴づける上で有用であると考えられる。
【0083】
(表1)糖質-バイオセンサー(レクテンズ)への変換のための初期標的酵素
【0084】
レクテンズ1:N結合型グリカンの濃縮のためのグライコミクス親和性試薬(コアにフコースを含まないN結合型オリゴ糖に対して特異的)
多くの用途において、特に臨床的な組織および液体の分析を伴うものにおいて、疾患の進行に応じたタンパク質および糖タンパク質の発現の変化を同定することには大きな関心が寄せられている。特定の分子の発現の変化を病態と高度に相関づけることが可能であれば、それらを利用して、有望な診断薬、ならびにおそらくは、罹患した細胞および組織を画像化するための試薬を開発することができるであろう。これらの分析のすべての基礎にある難しさは、これらの混合物があまりにも複雑であるため、試料中に存在する個々の種の大半のものの、質量分析を用いるその後の同定および精製を可能にする分離方法を開発することが極めて難易度が高いことに理由がある。さらに、説得力のある証拠により、タンパク質上に発現されるグリカンの変化は疾患に関する補足的なマーカーとして役立てうることが実証されている。このため、病態との相関づけのための同定および精製を行えるように、糖タンパク質からタンパク質を、糖ペプチドからペプチドを迅速に分離することを可能にすることが非常に求められている。例えば、血清において、構成要素の半分はグリコシル化されていないと推定されており、中でもアルブミンは最も群を抜いて存在量が多い。これらのタンパク質を糖タンパク質から除去することができれば、複雑性は2分の1に減る。
【0085】
さらに、グライコミクスシークエンシングは、特定のN結合型グリカン構造を各ペプチド上の各グリコシル化部位にマッピングする目的で、トリプシンにより生成される糖ペプチドの解析に対象を絞っている。このタイプの分析では、タンパク質分解消化によって生成される種の大半は圧倒的にグリコシル化されていないペプチドである。これらを関心対象の糖ペプチドと分離することができれば、部位特異的グリカンの情報を得るための解析を大きく簡略化することができると考えられる。スカフォールド用のPNGアーゼFの選択は、この酵素がさらなる解析のためにN-グリカンを糖タンパク質および糖ペプチドから取り出すために広く用いられているという事実に基づく。これは活性が強く、ペプチドがコアに脊椎動物では発現されないα-1-3フコシル化を含まない限り、その作用は普遍的である。さらに、これは脊椎動物のようにα-1-6フコシル化を発現する種に対して完全な活性を有する。PNGアーゼFレクテンズは、抽出物、液体から、さらには質量分析による詳細な構造解析のために、精製された糖タンパク質から、糖タンパク質および糖ペプチドを分離するために、極めて有用であると考えられる。この試薬は、プロテオミクス/グライコミクス解析において、干し草山(haystack)(タンパク質分解ペプチド)から針(糖ペプチド)を抽出するために用いうると考えられる。
【0086】
初期PNGアーゼレクテンズのスカフォールドは、フラボバクテリウム・メニンゴセプチカムによって産生される酵素から生成させることができ、これは基質との複合体における結晶構造が報告されている。結晶構造の存在はレクテンズ-グリカン親和性の計算による最適化にとって大きな意味がある。加えて、予備的な点突然変異誘発試験により、PNGアーゼFを完全に不活性にする単一の突然変異(D60N)も同定されている。
【0087】
レクテンズ2:糖尿病関連タンパク質高GlcNac化に対するバイオセンサー(β-O-GlcNAcに対して特異的)
中性の核細胞質ヘキソサミニダーゼは1975年に最初に記載され、1994年にO-GlcNAcアーゼとして特徴づけられた。これがクローニングされたのは最近になってであり(CCRCのDr. L. Wellsによる)、その結果、核細胞質性の中性β-N-アセチルグルコサミニダーゼ(O-GlcNAcアーゼ、OGA、EC 3.2.1.52)と同定された。ヘキソサミニダーゼAまたはBとは異なり、OGAはサイトゾルに局在していて核ではその度合いは少なく、至適pHは中性であり、かつ、GalNAcを触媒しない上にそれによって阻害もされない。アポトーシスにおけるO-GlcNAcアーゼの役割はまだ解明されていない;しかし、この酵素のポリペプチドの中央部付近での切断は酵素活性に影響を及ぼさず、このことはN末端の「ヒアルロニダーゼ様」ドメインがあれば活性にとって十分であることを示唆する。ごく最近、ウェルシュ菌由来のO-GlcNAcアーゼの結晶構造が、遊離状態および阻害薬と複合体を形成したものの両方について報告された。それらはヒトタンパク質と高い相同性を有しており、計算による検討のための優れた基盤をもたらす。
【0088】
レクテンズ3:ヒトインフルエンザ受容体α-2,6結合型ノイラミン酸に対するバイオセンサー(α-Neu5Ac-(2,6)-β-Galに対して特異的)
ヒト細胞表面糖質(ノイラミン酸またはシアル酸)は、インフルエンザウイルスの血球凝集素タンパク質の標的となる。血球凝集素は、細胞表面のシアル酸受容体と結合することにより、ウイルスの宿主細胞との接着およびその中への侵入を媒介する。ヒトインフルエンザウイルスは、ヒト上気道の上皮細胞の表面に認められる主要なタイプであるα-2,6結合によってガラクトースと連結したシアル酸と選好的に結合する。トリウイルスは、トリ腸上皮の表面に認められ、ヒト気道ではその程度がより少ないα-2,3結合と結合する傾向にある。これらからみて、この受容体は宿主特異性の鍵となる決定要因であり、ポテンシャルインフルエンザの予防および治療を可能にする可能性のある重要な標的である。本発明者らの初期レクテンズスカフォールドは、UGAの研究者らによってP.マルトシダ(P. multocida)から単離され、クローニングされて大腸菌において発現されている2,6-シアリダーゼnanBになるであろう(Dr. M. Lee, J Bacteriol. 2000 December; 182(24): 6874-6883)。これは現在までのところ、ヒト2-6結合に対する顕著な選好性を有する唯一のノイラミニダーゼである。この酵素の3D構造(Q27701)は、PDBテンプレート2SLI(同じファミリーに属するヒドロラーゼ)から比較モデリングによって作成され、Modbaseデータベースに寄託されており、これは計算解析のための基盤として役立つと考えられる。推定された活性部位に、AutoDockをGLYCAMパラメーターによって用いてリガンドをドッキングさせることができる。同時に、構造の特徴決定を、タンパク質結晶学、NMR分光法および部位指定突然変異誘発を用いて実験的に行うことができる。
【0089】
PNGアーゼF-キトビオース複合体のMDシミュレーション
PNGアーゼF-キトビオース複合体の5nsの完全溶媒和MDシミュレーションを、nPT条件(300K、1atm)で、複合体に関するAMBER/GLYCAMタンパク質/糖質力場および実験的X線構造を使用して行った。エネルギー解析の前に、シミュレーションの安定性およびコンフォメーション平衡化のレベルを明らかにするために、Cα原子の位置の根平均二乗差(RMSD)をシミュレーション時間の関数として決定した(図3)。そのデータに基づき、平均RMSD(1.5A)が妥当であることが明らかになった;しかし、シミュレーションが平衡化に至るのは遅かった。それ故に、データの最初の1nsはその後の解析には含めなかった。
【0090】
キトビオースとタンパク質との間の分子間水素結合をモニターすることにより、結合部位におけるリガンド安定性を評価した(図4)。水素結合に関する平均値およびそれらの占有率を表2に、結晶学的に決定した値とともに提示している。
【0091】
(表2)キトビオースおよびPNGアーゼFとの間で計算した、選択した水素結合の長さおよび占有率
a 標準的な3.3Åカットオフを用いて決定した占有率
b Kuhn et al., Crystal-Structure of Peptide-N-4-(N-Acetyl-Beta-D-Glucosaminyl) Asparagine Amidase-F at 2.2-Angstrom Resolution. Biochemistry, 1994. 33(39): p. 11699-11706.
【0092】
MDシミュレーションが安定であり、かつリガンドとタンパク質との間の実験的相互作用を再現しうることを確かめた上で、続いてそれをその後の結合エネルギーおよびアラニンスキャニング解析に使用した。
【0093】
相互作用エネルギー解析によるホットスポットおよびキー残基の同定
本発明者らは、AMBERに組み込まれているような一般化ボルン(GB)連続溶媒モデルを、タンパク質-リガンド相互作用のために開発された溶媒パラメーターとともに用いた。本発明者らは以前、これらの溶媒パラメーターが、糖質-抗体および糖質-レクチンのMM-GB結合エネルギー予測によく機能することを報告している。結合エネルギーに対する寄与を、PNGアーゼF中の313アミノ酸に関して計算した。相互作用エネルギーを、リガンドの4.5Å以内にあるすべての残基(接触区域内)、ならびに、全MM(ファンデルワールスおよび静電)相互作用エネルギー(ΔEMM)または全結合自由エネルギー(ΔG結合)に対する寄与が少なくとも-0.5kcal/mol未満であった他の任意のものについて表3に提示している。また、基質結合に対して不利に寄与した任意の残基、例えばD60およびE206なども含めている。
【0094】
全相互作用エネルギー(-13.7kcal/mol)は、同程度のサイズのガレクチン-1-LacNAc複合体に関して計算したデータと同等であり、ガレクチン-1の場合と同じように、コンフォメーションエントロピーおよびコンフィギュレーションエントロピーを省略しているために、実験から得られた親和性を過大評価している。にもかかわらず、残基ごとの相互作用エネルギーを用いて、キーアミノ酸残基を同定することができる。D60およびE206は明らかな例外であったが、残基ごとの正味結合エネルギーの大半は有利であった(図5)。D60は最も主要な触媒残基であり、一方、E206およびE118は反応中間体の安定化のために重要であると考えられる。したがって、これらの2つの残基が、加水分解反応の二糖生成物にとって最も不安定化させるものであることは意義深いものの、おそらく驚くには当たらない。
【0095】
不利な可能性のある相互作用は、負の溶媒和自由エネルギーΔGGBによって相殺される不利なΔEMMを有するD57によっても示されている。ガレクチン-1-LacNAc複合体について観察されているように、準エンタルピー寄与(ΔEMM)およびエントロピー関連脱溶媒和ΔGGBの項は多くの場合、大きさが同程度であって符号が逆であることから、溶媒和モデルの選択を決定的なものにする。一般に、理論的計算の精度に限定して依拠するのではなく、鍵になる可能性のあるそのような部位すべてをディスプレイライブラリーに含める。
【0096】
図5は、左の画像では:PNGアーゼFの結合部位において二糖リガンド(赤)の4.5Å以内にある残基を表している。右の画像では:溶媒接触可能表面を、結合のために最も重要であると同定された残基に印を付した上で表している。
【0097】
表4の中のデータとの直接比較が、2つの類似した糖質-タンパク質複合体(ガレクチン-1-LacNAcおよびCon A-トリマンノシド)に関して可能であり、これらはいずれもAMBER/GLYCAM力場およびGB溶媒和近似を使用している。表3では、ガレクチン-1およびCon AのMM-GB解析の場合と同じように、分子力学エネルギー(-33.8kcal/mol)の大半は静電相互作用(-20.8)から生じている。しかし、ファンデルワールス寄与および静電寄与(それぞれ-12.9kcal/molおよび-20.8kcal/mol)はいずれも、ガレクチン-1と結合した類縁の二糖について観察されたものよりも小さく(ファンデルワールスについては-17〜-24kcal/mol、静電については-30.8〜-67.5kcal/mol)、このことはPNGアーゼFにおいて側鎖最適化を介した親和性強化の余地があることを強く示唆する。さらに、静電寄与が脱溶媒和自由エネルギーによってほぼ打ち消されることも見てとることができる。この現象は以前の研究のいずれにおいても観察されており、エントロピー-エンタルピー補償の現れである可能性がある。このため、GB計算の誤差の打ち消しにより、利点を得ることができる。
【0098】
表3におけるエネルギーに基づき、公知の不活性D60A突然変異体を作製し、エネルギーを再計算した。結合エネルギーはD60A突然変異(全ΔEMM=-35.5、ΔGGB=19.6およびΔG結合=-15.9、表5を参照のこと)では顕著に改善しており、親和性の正味の利得はおよそ2kcal/molであった。
【0099】
(表3)基質であるキトビオース(β-GlcNAc-(1,4)-β-GlcNAc-OH)と結合した野生型PNGアーゼFに関する結合自由エネルギーに対する残基寄与(kcal/mol)
【0100】
(表4)キトビオースと結合したPNGアーゼFのアラニンスキャニングおよび静電スキャニングによって同定された有利な突然変異体に関する全相互作用エネルギー(kcal/mol)
【0101】
引き続き、可能性のあるさらなるキー残基を探すためにD60A突然変異体に対してアラニンスキャニングを行った(表5)。ある程度の親和性の強化(-0.5kcal/mol)を伴う1つの二重突然変異体が後に同定され(D60A/E206A)、一方、5つの残基はリガンド結合のために決定的であることが確認された(W251、W191、W120、W59およびR61)。
【0102】
(表5)キトビオースと結合したPNGアーゼFのD60N突然変異体に関してアラニンスキャニングから予測された相対的a相互作用エネルギー(kcal/mol)
a 野生型配列(D60)との対比。 b 二重突然変異体は初期突然変異体(D60A)との対比。
【0103】
側鎖リパッキング実験を最初に行うのではなく、不活性突然変異体を、有利な静電相互作用につながると考えられる位置に関してスキャニングすることを行った。このスキャニングを、直接接触区域における残基のすべてにわたり、理論的に正および負のプローブ残基(実験的設計を参照)について行った(表6)。
【0104】
荷電残基に対する突然変異のうち、全結合エネルギーの強化につながると予想されたものはなかったが、いくつかの可能性のある突然変異が分子力学相互作用の改善につながることが示唆された。すなわち、残基E206、S155、E118およびY62をそれぞれArgおよびLysに突然変異させ、エネルギーを再計算することができる。その結果得られた結合自由エネルギーを用いて、クローニングおよび過剰発現を行うための何らかの特異的な点突然変異体を選択することができる。このことは、糖質-タンパク質相互作用が水素結合のネットワークの複雑なネットワークによって特徴づけられること、およびそのネットワークの摂動が有利であることは稀であると考えられることからみて、おそらく驚くには当たらない。この場合も、いずれも酵素機構における関与が示されているE206およびE118が、親和性最適化のための可能性のあるキー残基として同定されていることは意義深い。また、負に荷電した残基に対する突然変異のうち、親和性の改善につながると予測されたものがなかったことも、ディスプレイライブラリーの設計のためには注目すべきである。
【0105】
(表6)静電アラニンスキャニングにより予測された、キー残基に関するD60A突然変異体に対して相対的な相互作用エネルギー
【0106】
計算による親和性データに基づき、クローニング用にいくつかの突然変異体を選択し、大腸菌において過剰発現させた。実験による親和性分析の結果を、以下の項に提示する。
【0107】
PNGアーゼFレクテンズに関する実験から得られた結合親和性の測定
表7に提示しているのは、主として高マンノースオリゴ糖によって占有される単一のN-グリコシル化部位を含む変性RNアーゼBと、PNGアーゼFの突然変異体との間の相互作用に関して、表面プラズモン共鳴(SPR)を用いて測定した解離定数である。
【0108】
(表7)変性糖タンパク質RNアーゼBとPNGアーゼFのレクテンズ突然変異体との間の相互作用に関して測定した解離定数
a Km。 b 報告されている不活性突然変異体。 c 野生型との対比。
【0109】
計算によって予測されたように、E206A突然変異体およびD60A突然変異体はいずれも結合を顕著に強化する。また、静電スキャニングから示唆されたように、E206での正電荷(KまたはR)は少しではあるが親和性のさらなる増大をもたらす。現時点で、側鎖リパッキング実験または飽和突然変異誘発の恩典を受けていない第1世代レクテンズはマイクロモルの親和性を有し、これは2つの点突然変異だけでマイクロモルレベルに達した。これらの予備的突然変異が、高マンノースオリゴ糖に対してレクチンCon Aによって示されたものとほぼ等しいまで(Kdがほぼ1×10-6Mに等しい)に、PNGアーゼレクテンズの親和性を強化したことは注目に値する。
【0110】
セクションBにおいて述べたように、高親和性はバイオセンサーにとって唯一の望ましい特性である。高親和性は試薬をアフィニティークロマトグラフィーに使用することを可能にすると考えられる。しかし、バイオセンサーを組織染色などの用途に首尾良く用いようとするならば、遅いオフ速度(koff)を達成することも重要である。
【0111】
SPRは、koffの変動を評価するための好都合な方法を提供する。図6の検討は、突然変異体D60AとE206Aとの間で有意に異なる速度論的挙動を指し示している。いずれの突然変異体もKdについて同程度の値を有するが(表7)、D60Aは速いオン速度およびオフ速度を示した(10℃および25℃の両方)が、一方、E206Aはいずれの温度でも大幅に低いオフ速度を提示する。このことは、結合の速度論的動態を調整して、ある範囲にわたる特性を達成しうると考えられることから、極めて意義深い特徴である。
【0112】
さらなる親和性強化
計算によるデータに基づき、以下の7つの残基:D57、Y62、E118、S155、I156、G192およびE206を含む不活性D60A突然変異体に関するフォーカスト酵母ディスプレイライブラリーを開発した。これらの7つの位置を20種のアミノ酸すべてについてランダム化し、その結果、アミノ酸レベルで207(ほぼ109に等しい)種のクローンという理論的多様性を得た。
【0113】
加えて、D60A突然変異体を使用して、同じ残基Y62、E118、I156、S155、G192、E206およびD57のサブセットに対して計算による側鎖リパッキング実験を行うこともできる。計算による突然変異誘発をキー残基の同じセットに対して行うことにより、本発明者らは、計算解析が最適化された突然変異誘発データを再現する範囲を見いだすことができると予想している。計算方法(陰溶媒和モデルなど)の変更の実施を、その精度を高めるために行うことができる。
【0114】
実施例2
レクテンズの指向性進化
PNGアーゼF酵素の不活性D60A突然変異体を基にして、DNAライブラリーを作り出した。計算解析により同定された残基D57、Y62、E118、S155、I156、G192およびE206を、20種のアミノ酸すべてをコードするようにDNAレベルで改変した。このライブラリーを酵母ディスプレイベクターpPNL6中にクローニングし、酵母に導入して形質転換を行った。
【0115】
磁気ビーズ上に捕捉したdRNアーゼBに対してこのライブラリーのパニングを2ラウンド行い、続いて3ラウンド目にはフローサイトメトリーによってc-myc陽性酵母を選別した。この3ラウンドを1回繰り返し、合計6ラウンドとした。表8は、第6ラウンドからの18種のクローンのDNAのシークエンシングによる酵母クローンの濃縮を示している。
【0116】
(表8)第6ラウンドからのクローンの濃縮
【0117】
クローンR6.1.13を競合アッセイを用いる機能分析のために選択し、細菌において発現させて精製した。このアッセイでは、R6.1.13の1μM溶液のうち50μLを、dRNアーゼBビーズとともにプレインキュベートした。同様に、不活性酵素D60A突然変異体の1μM溶液のうち50μLをdRNアーゼBビーズとともにプレインキュベートした。プレインキュベートした各溶液に対して、Con Aレクチン(DyLight 488で蛍光標識)を最終濃度100nMとなるように添加した。標識したCon Aも、対照としてdRNアーゼBの存在下または非存在下で最終濃度100nMとなるようにビーズに添加した。ビーズの蛍光をフローサイトメトリーによって測定した。結合したCon Aの画分を、dRNアーゼBの存在下および非存在下におけるビーズとともにビーズの蛍光に対して標準化した(図9参照)。
【0118】
クローンR6.1.13タンパク質は、不活性酵素D60A突然変異体と比較して、dRNアーゼビーズに対するCon A結合の阻害のおよそ36%の増大を示し、このことは親和性強化を指し示している。このクローンは、クローン濃縮が3/18と少しであったことによって指し示されるように完全には最適化されておらず(表8)、そのため、さらに濃縮ラウンドを重ねることにより、さらなる親和性向上を得ることができる。
【技術分野】
【0001】
連邦政府による資金提供を受けた研究の記載
本発明の開発中に行われた研究の一部は、米国国立衛生研究所(National Institutes of Health)によって授与されたR41GM086991号の下で米国政府基金を利用した。したがって、米国政府は本発明において一定の権利を有する。
【0002】
発明の分野
本発明は、グリカン特異的分析ツール、それらの使用方法、およびグリカン特異的分析ツールを作成するための工程を対象とする。他の分析ツールも本明細書においてさらに提供する。
【背景技術】
【0003】
グリカンは、脂質およびタンパク質と結びついた状態で認められることの多い複合糖質である。複合糖質はタンパク質および細胞の表面に存在することから、それらは往々にして、生物認識プロセスにおける機能的位置を占めている。オリゴ糖および多糖の複雑な形状、機能性および動的特性は、これらの分子が生体情報のエンコーダーとして分子間相互作用において機能することを可能にする。
【0004】
糖質認識は正常な生物発生の不可欠な部分であるが、先天性免疫系によっても、宿主生物が病原体の表面に提示された糖質に基づいて外来性病原体を同定することを可能にするように使用されうる。逆に、多くの細菌性およびウイルス性病原体はまず、宿主細胞表面の糖質と特異的に結合することによって宿主組織に付着する。このため、糖質に基づく宿主-病原体相互作用を妨害することのできる、または抗細菌ワクチンとして機能しうる治療薬を開発することには関心が寄せられている。また、異常グリコシル化はある種の癌および他の疾患のマーカーであり、そのためにそれらは診断的および治療的な用途の標的ともなっている。例えば、O結合型N-アセチルグルコサミンによる細胞内タンパク質の修飾状態(O-GlcNAc化)は、疾患、特に2型糖尿病によって引き起こされる変化の重要なバイオマーカーである。
【0005】
生物発生および疾患におけるグリカンの重要性にもかかわらず、現在のところ十分なグリカン特異的分析ツールはなく、このことが疾患の診断および治療における異常グリコシル化の活用を遅らせてきた。例えば、細胞へのO-GlcNAc取り入れおよびその後のタンパク質の提示のモニタリングのための現行の方法は、N-アジドアセチルグルコサミン(GlcNAz)などの標識試薬の外因性取り込みに基づいている。残念ながら、この方法は、単離された組織試料またはタンパク質試料におけるO-GlcNAcの分析には適用できない。グライコミクス/プロテオミクス解析に適用しうる1つの代替的なO-GlcNAc標識アプローチは化学酵素的タグ標識を用いる。この方法の重大な限界は、それが他のGlcNAcが末端にある複合グリカンも標識することである。このため、異常グリコシル化を探索しようとして生体試料を調べるために用いることのできる、明確な糖質特異性を備えた分析ツールは依然として必要とされている。
【0006】
現在のところ、グリカン特異的分析の用途に用いられている主なタイプの生体分子は、糖結合タンパク質(レクチン)および抗体の2つである。これらのタイプの試薬のいずれにも伴う主な欠点は、糖質とタンパク質との間の特徴的に弱い相互作用であり、その解離定数は典型的には、レクチンについてはミリモルからマイクロモルの範囲、抗体についてはマイクロモルからナノモルの範囲である。さらに、抗体を用いる際の大きな問題は、糖質が免疫原としては極めて劣ることである。それらは一般にT細胞依存性応答を生じさせることができず、そのためほとんどの場合はIgMクラス抗体を産生させるが、これは分析的および診断的な用途には不都合である。重鎖および軽鎖の可変ドメインからなる単鎖キメラ(scFv)は、不安定性を来す恐れがある。さらに、抗体を利用するグリカン特異的分析の手法には、抗体の選択性がコンテキスト依存性であることに起因する欠点もある。または、レクチンは、その特異性の幅広さから、分析用途のためのそれらの使用には限界がある。このため、糖質配列に対する十分な特異性を有し、なおかつ広範囲にわたるグリカン内の配列を認識することのできる分析用試薬を開発することには依然として需要が存在する。
【発明の概要】
【0007】
本発明は、その触媒活性を消失させながらもその基質特異性は維持するように突然変異させた糖質プロセシング酵素を含む、レクテンズ(lectenz)を提供する。ある態様において、本発明のレクテンズは、グリカンに対して、野生型酵素のKmよりも高い親和性を有する。また、本発明のレクテンズが、著しく低下したkoff速度を有してもよい。他の態様において、本発明は、触媒的に不活性な糖質プロセシング酵素を含むレクテンズであって、その不活性酵素が野生型残基とは異なる1つまたは複数のアミノ酸残基を含み、前記残基が、結合した基質に近接するが、気相(ΔEMM)相互作用エネルギーに対する寄与が約|0.5kcal/mol|未満である残基、全(ΔG)相互作用エネルギーに対する寄与が約|0.5kcal/mol|未満である残基、または結合相互作用エネルギーに対して不利に寄与する任意の残基、およびそれらの組み合わせからなるリストから選択される、レクテンズを提示する。基質に近接する残基は、本明細書では一般に基質の5Å以内にあると考えるが、それよりも遠い場合も近い場合も考えうる。
【0008】
別の局面において、本発明は、レクテンズを作製するための方法を提供する。いくつかの態様において、本方法は:(a)糖質プロセシング酵素の配列を、突然変異した場合に糖質プロセシング酵素のグリカンに対する親和性または酵素-グリカン複合体の安定性に影響を及ぼしうると考えられる1つまたは複数のアミノ酸残基に関して解析する段階;(b)糖質プロセシング酵素が段階(a)で同定された少なくとも1つの突然変異したアミノ酸を有する、酵素-グリカン複合体の結合エネルギーを予測するための計算シミュレーションを行う段階;(c)段階(a)および(b)で同定された突然変異を含む糖質プロセシング酵素を、それらが複合体を形成する能力に関して検査する段階;ならびに(d)グリカンに対する結合親和性がWT酵素のそれよりも高い、段階(c)による突然変異体を同定する段階、を含む。
【0009】
本発明の別の局面は、レクテンズをグリカン特異的分析の用途に用いる方法を提供する。ある態様において、本発明のレクテンズは親和性試薬として、または組織染色のための媒体として用いることができる。他の態様において、レクテンズは、生体試料を特定のグリコフォームに関して濃縮するために用いることができる。さらに他の態様において、レクテンズには、糖タンパク質上の特異的なグリコシル化部位を決定するという用途もある。本発明の他の局面は、活性治療薬の標的化送達のための媒体としてのレクテンズの使用を伴う。
【0010】
ある態様において、糖質プロセシング酵素はグリコシダーゼ酵素である。他の態様において、糖質プロセシング酵素はグリコシルトランスフェラーゼ酵素である。他の態様において、糖質プロセシング酵素は多糖リアーゼ酵素である。他の態様において、糖質プロセシング酵素はスルファターゼ酵素である。他の態様において、糖質プロセシング酵素はスルホトランスフェラーゼ酵素である。他の態様において、糖質プロセシング酵素はリガーゼ酵素である。さらなる態様において、糖質プロセシング酵素はアミダーゼ酵素である。さらに別の態様において、糖質プロセシング酵素はエピメラーゼ酵素である。
【図面の簡単な説明】
【0011】
【図1】糖質プロセシング酵素(この例ではノイラミニダーゼ)とその糖質結合性レクテンズ類似体との関係を描写している。
【図2】レクテンズの親和性を最適化するための、計算による方法とインビトロディスプレイライブラリー法を組み合わせるプロトコールを描写している。
【図3】PNGアーゼF(PNGase F)複合体におけるCα位置のRMSDを描写している。
【図4】キトビオースのPNGアーゼFとの結合に関する水素結合相互作用の配置図を描写している。
【図5】左の画像では、PNGアーゼFの結合部位における二糖リガンドの4.5Å以内にある残基(ダークグレー)を描写している。右の画像では、溶媒接触可能表面を、結合のために最も重要であると同定された残基に印を付した上で描写している。
【図6】速度論的なオン速度およびオフ速度の変動を突然変異および温度(10℃および25℃)の関数として指し示しているSPRセンサーグラム(sensogram)を描写している。
【図7】酵母におけるAga2タンパク質と融合させたタンパク質ディスプレイライブラリーを描写している。c-mycタグ標識タンパク質に対する蛍光標識抗原の結合の検出を図示している。
【図8】高親和性クローンの選択を指し示す、フローサイトメトリーを介した細胞選別を描写している。
【図9】不活性酵素と比べた予備的レクテンズの親和性の強化を描写している。
【0012】
本出願の図版および図面が必ずしも一定の縮尺で描かれてはいないこと、ならびにこれらの図面および図版は本発明を単に例示しているに過ぎず、それを限定するものではないことは理解されるであろう。
【発明を実施するための形態】
【0013】
発明の詳細な説明
以下の記載では、本発明の十分な理解が得られるように、説明の目的で、具体的な数字、パラメーターおよび試薬を記述している。しかし、本発明を、これらの具体的な細目がなくても実施しうることは明らかであろう。場合によっては、本発明が分かりにくくならないように、周知の特徴を省いてもよく、または単純化してもよい。
【0014】
記載された態様、および本明細書における「1つの(one)態様」、「1つの(an)態様」、「1つの例示的態様」などへの言及は、記載された態様が特定の特徴、構造または特質を含みうるものの、あらゆる態様が必ずしもその特定の特徴、構造または特質を含まなくてもよいことを指し示している。その上、そのような語句は、必ずしも、同じ態様について言及しているわけでもない。さらに、特定の特徴、構造または特質を1つの態様に関連して記載している場合には、明示的に記載されているか否かにかかわらず、他の態様に関連してそのような特徴、構造または特質を生じさせることが当業者の知識の範囲内にあることは理解されるであろう。
【0015】
I.定義
本明細書で用いる場合、糖質プロセシング酵素とは、糖質含有基質に対して作用する任意の酵素を指して用いられる用語である。その例には、グリコシダーゼ、グリコシルトランスフェラーゼが含まれるが、それらには限定されない。
【0016】
本明細書で用いる場合、「グリコシダーゼ」という用語は、グリコシド結合の加水分解を触媒する酵素を指して用いられる。「グリコシド結合」という用語は、糖質分子を別の糖質分子に連結させるか、または糖質分子をタンパク質分子に連結させるか、または糖質分子を脂質分子に連結させる、あるタイプの官能基のことを指す。「糖質」という用語は、一般式Cm(H2O)nの有機化合物を指すものとする。本発明において、「糖質」、「複合糖質」および「グリカン」という用語は互換的に用いられる。
【0017】
「触媒的に不活性な突然変異体」または「突然変異体」または「不活性酵素」という用語は互換的に用いられ、その触媒活性の少なくとも95%を喪失していて、かつ触媒的に活性な酵素とは異なるアミノ酸組成を有する酵素のことを指す。別に記述する場合を除き、触媒的に不活性な突然変異体による化学結合切断の速度は、最大でも、あらゆる触媒が存在しない同一の条件下で測定される結合切断速度の5%を上回らない。「触媒的に活性な酵素」とは、化学結合の加水分解を触媒しうるタンパク質を指すものとする。「野生型(WT)酵素」とは、生物において天然に存在するのと同じ遺伝子配列を有し、ヒト介入によって改変されていない遺伝子によってコードされる酵素のことを指す。当然ながら、野生型酵素の天然に存在する多型形態がこの定義に含まれることは理解されるであろう。さらに、それ以外の点ではタンパク質の天然の開始コドンも終止コドンも変化させない、タンパク質の精製または単離に用いられるタグまたは他の修飾などの修飾が、本発明におけるWT酵素の定義の範囲に含まれることも理解されるであろう。本明細書で用いる場合、「リガンド」および「基質」という用語は互換的に用いられ、WT酵素または突然変異体酵素が結合することのできる分子のことを指す。
【0018】
本発明のレクテンズは、グリカンに対して、野生型酵素のKmよりも高い親和性を有する。Kmの意味を理解するためには、酵素作用のモデルを用いる必要があるであろう。最も単純なモデルはミカエリス・メンテンの古典的モデルであり、これは多くの種類の酵素に有用であることが証明されている(式1)。
【0019】
基質(S)は、第1の反応において酵素(E)と可逆的に結合する。ほとんどの場合には、この段階を測定することはできない。測定されるのは、第2の反応によって生み出される生成物(P)である。ミカエリス・メンテン定数(Km)は式2で定義される。
【0020】
Kmが、酵素と基質との間の結合の強さの尺度となる結合定数ではないことに注意されたい。その値は、酵素に対する基質の親和性だけでなく、酵素と結合した基質が生成物に変換される速度も含む。k2がk-1よりもはるかに小さい場合にのみ、Kmは結合親和性に等しいと考えられる。野生型酵素の状況ではWT酵素がリガンドに作用するため、親和性の値を直接測定することは難しく、この理由から不活性酵素のKdを野生型酵素のKmと比較することが好都合であることが理解されるであろう。本明細書で用いる場合、「親和性」という用語は、2つの分子間の引力のことを意味する。レクテンズの親和性は、通常はWT酵素のKmと対比して測定されるが、そのリガンドに対する解離定数Kdの、WT酵素の不活性形態に対する相対的な低下という観点で表現することもできる。解離定数Kdは、複合体がその成分へと解離する性向の尺度となる平衡定数である。一般的反応:
があるとし、式中、iEおよびSはそれぞれ不活性酵素および基質を示し、iESは不活性-酵素-基質複合体である。この場合、対応する解離定数Kdは:
として定義され、式中、[iE]および[S]はそれぞれ遊離した不活性-酵素および基質の濃度を示し、[iES]は不活性-酵素-基質複合体の濃度である。したがって、ある態様において、レクテンズの親和性の増大は、レクテンズの親和性を、親和性を最適化する別の突然変異を受けていない触媒的に不活性な形態のWT酵素と比較することによって測定される。本発明において、タンパク質のそのリガンドに対する親和性は、解離定数(Kd)または会合定数(Ka)として表現することができる。
【0021】
「ΔG」という表現は、結合のギブス自由エネルギーのことを指す。ギブス自由エネルギーは、複合体を1つにまとめる力に抗しながら、複合体を、それ以上の分離に必要な余分な仕事量を無視しうる十分な距離まで隔てられた構成要素部分に分解させる際に行わなければならない仕事量を表す熱力学的ポテンシャルである。「ΔEMM」という表現は、気相における分子力学自由エネルギーのことを指す。「特異性」または「酵素特異性」という用語は互換的に用いられ、酵素が、種々のリガンドの集団から、特異的な分子構造を含むリガンドを認識して選択する能力のことを指す。「非特異的」結合または相互作用という用語は、個々の分子または残基のいかなる特異的認識にも識別にも基づかない、分子または残基の間での弱い相互作用イベントのことを指す。
【0022】
「分子動力学(MD)」という用語は、公知の物理的過程の近似によって原子および分子をある期間にわたって相互作用させて粒子の運動の概観を得る、コンピュータシミュレーションの一形態を指すものとする。古典的MDシミュレーションは、古典的力場から導き出されるエネルギーおよび力を用いるニュートンの運動方程式によって規定される。古典的力場は、分子または分子の凝集物における原子位置を分子または凝集物のポテンシャルエネルギーと関係づける数学モデルである。「エヴァルト処理(Ewald treatment)」または「エヴァルト和」という用語は、本明細書で用いる場合、周期構造系(例えば、結晶)、特に静電エネルギーの相互作用エネルギーを計算するための方法を記述している。「ベルレ(Verlet)アルゴリズム」または「ベルレ積分」という用語は、ニュートンの運動方程式を積分するために用いられる数値的方法のことを意味する。
【0023】
II.レクテンズ
本発明の1つの局面はレクテンズを提供する。本発明のレクテンズは、所与のグリカンに対して野生型酵素と実質的に同じ特異性を有し、かつグリカンに対する親和性がWT酵素と比較して高くなっている、糖質プロセシング酵素の触媒的に不活性な突然変異体を含む。本明細書で用いる場合、「実質的に同じ」という用語は、野生型酵素の少なくとも60%であるグリコシダーゼ突然変異体の特異性を記述するものとする。いくつかの態様において、突然変異体の特異性は、WT酵素の少なくとも70%である。少なくとも1つの態様において、突然変異したグリコシダーゼはその基質に対して、同じ基質に対する野生型酵素に比して少なくとも85%特異的である。他の態様において、突然変異したグリコシダーゼは、同じ基質に対する野生型酵素に比して少なくとも95%特異的である。
【0024】
遺伝子操作されたレクテンズに関して、活性部位残基の突然変異は、酵素を不活性化するのみならず、親和性を強化する可能性のある経路も与える(図1)。いくつかの態様において、グリカンに対するレクテンズの親和性は、野生型酵素のそれよりも高くなる。ある態様において、レクテンズのその基質に対する親和性は、解離定数Kdの観点で表現することができる(式3および4)。解離定数が小さいほど、レクテンズはより強固に基質と結合している。本発明のいくつかの態様において、グリカンに対するレクテンズの解離定数(Kd)は、WT糖質プロセシング酵素のKmより少なくとも約1.2倍〜約1,000倍小さい。他の態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKmより少なくとも約2倍小さい。ある態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKmより少なくとも約10倍小さい。ある態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKmより少なくとも約10,000倍小さい。さらなる態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKmより少なくとも約100,000倍小さい。
【0025】
また、本発明のレクテンズの親和性向上を、WT酵素の不活性突然変異体のそれと対比したKdの低下という観点で表現しうることも理解されるであろう。すなわち、グリカンに対するレクテンズのKdは、不活性なWT糖質プロセシング酵素のそれより少なくとも約1.2倍〜約1,000倍小さい。他の態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKdより少なくとも約2倍小さい。ある態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKdより少なくとも約10倍小さい。ある態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKdより少なくとも約10,000倍小さい。さらなる態様において、グリカンに対するレクテンズの解離定数は、WT糖質プロセシング酵素のKdより少なくとも約100,000倍小さい。
【0026】
本発明のレクテンズは、いかなる特定の糖質プロセシング酵素にも限定されない。そうではなくて、本発明は、糖質を特異的に認識することのできる、任意のグリコシダーゼ酵素またはグリコシルトランスフェラーゼ酵素、タンパク質またはポリペプチドに対して幅広く適用可能である。本発明に適したグリコシダーゼの例には、ラクターゼ、アミラーゼ、キチナーゼ、スクラーゼ、マルターゼ、ノイラミニダーゼ、インベルターゼ、ヒアルロニダーゼおよびリゾチームが非限定的に含まれる。酵素委員会(Enzyme Commission)(EC)番号3.2.-.-、式中「-」は数字である、によって分類されているグリコシダーゼが本明細書に含まれることは理解されるであろう。本発明のグリコシダーゼは反転型グリコシダーゼでも保持型グリコシダーゼでもありうる。1つの態様において、本発明のレクテンズは、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から単離されたPNGアーゼFから調製される。別の態様において、レクテンズは、バクテロイデス・テタイオタオミクロン(Bacteroides thetaiotaomicron)から単離されたβ-O-GlcNAcアーゼに関して決定されているWT配列を用いて、組換えβ-O-GlcNAcアーゼから調製される。さらに別の態様においては、ウェルシュ菌(Clostridium perfringens)由来のノイラミニダーゼを、レクテンズを調製するために用いる。グリコシダーゼのほかに、本発明に用いるために適した糖質プロセシング酵素には、EC番号2.4.-.-として指定されたものを含むグリコシルフェラーゼ、およびEC番号4.2.2.-として指定されたものを含む多糖(polysacharide)リアーゼが含まれる。他の糖質プロセシング酵素には、糖質エステラーゼ、スルファターゼ、スルホトランスフェラーゼ、または糖質基質に対して作用する他の任意の酵素が含まれる。本発明の触媒的に不活性な糖質プロセシング酵素は、原核生物または真核生物、ならびに他のものから単離された糖質プロセシング酵素から調製することができる。
【0027】
ある態様において、本発明のレクテンズは高い耐熱性を有すると考えられる。高い耐熱性とは、レクテンズがその三次構造を、生理的に許容される緩衝液中で少なくとも40℃の温度で少なくとも30分間にわたって保つことを意味する。生理的に許容される緩衝液とは一般に、0〜1%のNaCl濃度および0〜20mMのグルコース濃度、pH 6〜8の0.01〜0.2Mリン酸緩衝液のことを指す。ある態様において、レクテンズは正しくフォールディングした状態を、生理的に許容される緩衝液中で少なくとも60℃の温度で少なくとも30分間保つ。他の態様において、レクテンズはその三次構造を、生理的に許容される緩衝液中で少なくとも80℃の温度で少なくとも30分間保つ。必要であれば、本発明のレクテンズは、好熱性生物または超好熱性生物から単離されたグリコシダーゼから調製することができる。本明細書における適した糖質プロセシング酵素を単離しうる好熱性生物および超好熱性生物の例には、サーマス・サーモフィルス(Thermus thermophilus)、スピロヘータ・アメリカナ(Spirochaeta americana)、パイロコッカス・フリオサス(Pyrococcus furiosus)、メタノパイラス・カンドレリ(Methanopyrus kandleri)、パイロロバス・フマリ(Pyrolobus fumarii)、ジオサーモバクテリウム・フェリレデュセンス(Geothermobacterium ferrireducens)およびアーケオグロブス・フルギダス(Archaeoglobus fulgidus)が非限定的に含まれる。
【0028】
いくつかの態様において、本発明のレクテンズは長い速度論的オフ速度(kinetic off-rate)によって特徴づけられる。速度論的オフ速度は、解離速度定数(koff)、またはリガンドがタンパク質から解離する速度(式5参照)によって測定される。一般に、基質-不活性酵素複合体の形成は、以下の速度論的機序によって記述しうる:
式中、iEおよびSはそれぞれ不活性酵素および基質を示し、iESは不活性酵素-基質複合体であり、konおよびkoffはそれぞれ複合体形成および解離に関する速度定数である。少なくともいくつかの態様において、本発明のレクテンズはそのリガンドから、WT酵素のそれよりも少なくとも約2倍の遅さの速度で解離すると考えられる。1つの態様において、レクテンズ-グリカン複合体解離の速度は、WT-グリカン複合体のそれよりも少なくとも5倍の遅さである。さらに別の態様において、レクテンズ-グリカン複合体の解離の速度は、WTのそれよりも少なくとも10倍の遅さである。また、比較をレクテンズのオフ速度と不活性化されたWT酵素との間で行うこともできる。少なくともいくつかの態様において、本発明のレクテンズはそのリガンドから、不活性WT酵素のそれよりも少なくとも約2倍の遅さの速度で解離すると考えられる。1つの態様において、レクテンズ-グリカン複合体解離の速度は、不活性WT酵素とグリカンとの複合体のそれよりも少なくとも5倍の遅さである。さらに別の態様において、レクテンズ-グリカン複合体の解離の速度は、不活性WT酵素のそれよりも少なくとも10倍の遅さである。
【0029】
ある態様において、本発明の触媒的に不活性な突然変異体は、WT残基と異なっていて、かつ、酵素-基質複合体中で基質に近接するが、全(ΔG)相互作用エネルギーに対する寄与が約-0.5kcal/molを上回る残基、または結合相互作用エネルギーに対して不利に寄与する任意の残基、およびそれらの組み合わせから選択される、1つまたは複数のアミノ酸残基を有する。基質に近接するが強い相互作用を形成しないこの残基セットを、ぬるい(tepid)残基セットと命名してもよいであろう。ある態様において、突然変異体グリコシダーゼは、ぬるい残基セット中に2つまたはそれ以上の突然変異を有する。他の態様において、突然変異体グリコシダーゼは、これらの位置に3つもしくはそれ以上、4つもしくはそれ以上、またはさらには5つもしくはそれ以上の突然変異を有する。
【0030】
III.レクテンズを作製するためのコンピュータ支援による方法
本発明の別の局面は、糖質プロセシング酵素の触媒的に不活性な突然変異体を含むレクテンズを作製するためのコンピュータ支援による方法であって、
(a)糖質プロセシング酵素の配列を、突然変異した場合に酵素を不活性化しうると考えられる1つまたは複数のアミノ酸残基に関して解析する段階;
(b)WT酵素-グリカン複合体の、または糖質プロセシング酵素が段階(a)で同定された少なくとも1つの突然変異したアミノ酸を有する複合体の結合エネルギーを予測するために計算シミュレーションを行う段階;
(c)残基を、それらの予測される相互作用エネルギーに基づいて、2つの群に、すなわち、酵素の特異性を規定するのに必須である第1の残基群、および基質に近接するが特異性を規定するのに必須ではない第2の残基群(この第2のセットを本明細書では、ぬるい残基と称する)に細分類する段階;
(d)段階(a)、(b)および(c)で同定された突然変異を含む糖質プロセシング酵素を、それらが酵素-グリカン複合体を形成する能力に関して検査する段階;ならびに
(e)グリカンに対する結合親和性がWTグリコシダーゼのそれよりも少なくとも1.2倍高い、段階(d)による突然変異体を同定する段階、
を含む方法を提供する。
【0031】
図2中の資料形態に略図として示したこの態様は、本発明のレクテンズを得るための1つの態様のみを表しているに過ぎないものと理解されたい。本明細書においてさらに詳細に記述している図2の諸段階を、本発明のレクテンズを得るために、本明細書において説明した通りに改変することができる。具体的には、例えば、図2中の特定の段階が、本明細書中の他の箇所でさらに詳細に説明されている「必須な残基を同定する」段階のための「計算によるアラニンスキャニング」といった特定の手法について言及している点で、それは本発明の単なる1つの態様に過ぎず、同じ段階を行うための他のアプローチも存在する。
【0032】
本発明のある態様は、レクテンズへの変換のために適している糖質プロセシング酵素の同定を伴う。レクテンズの生成は特定の糖質プロセシング酵素には限定されないが、その活性部位残基が公知である、かつ/または触媒的に不活性な突然変異体が記載されている、糖質プロセシング酵素を選択することが好ましいであろう。触媒残基の同定は、多くの糖質プロセシング酵素に関して部位指定突然変異誘発を用いて行われており、多くの場合には酵素の速度論的動態の測定によって確かめられている。しかし、不活性酵素(第1世代レクテンズ)は、多くの場合、レクチンと同等な親和性を有する。
【0033】
一般に、コンピュータ支援による突然変異誘発手法を実施する前には、関心対象のリガンドに対する触媒的に不活性な糖質プロセシング酵素の結合親和性を予測することが好ましい。糖質プロセシング酵素のそのリガンドに対する親和性を特徴づけるためには、いくつかの周知の手法がある。1つのそのような手法は、AMBER/GLYCAMタンパク質/糖質力場を使用する分子動力学(MD)手法である。
【0034】
エネルギー解析の前に、MDシミュレーションの安定性およびコンフォメーション平衡化のレベルを決定するために、Cα原子の位置の根平均二乗差(root mean squared difference)(RMSD)をシミュレーション時間の関数として求めることができる。そのようなデータに基づき、平均RMSDが安定であるか否かおよび約0Å〜4Åの範囲内にあるか否かを判定することができる。また、コンフォメーション平衡に達するために必要な時間を、このアプローチを通じて判別することも可能である。平衡化していないデータセットの部分はその後の解析に当たって省くことが一般に好ましい。5ナノ秒のデータを収集し、システムが平衡化するまでに約1ナノ秒を費やした1つの態様では、最初の1ナノ秒のデータを省いた。上記の5ナノ秒/1ナノ秒の態様は例示に過ぎず、本発明を限定するものではないことは理解されるであろう。
【0035】
結合部位におけるリガンド安定性は、グリカンと糖質プロセシング酵素との間の分子間水素結合を評価することによって評価することができる。水素結合およびそれらの占有率に関する平均値を収集することができ、可能であれば、結晶学的に決定された値に沿ってそれらを収集する。結合部位におけるリガンドの位置のRMSDのモニタリングにより、結合部位におけるリガンド安定性を決定することも可能である。MDシミュレーションが安定であって、かつリガンドとタンパク質との間の実験的相互作用を再現しうることが確かめられたならば、そのシステムをその後の解析に使用することができる。典型的なシミュレーションは、圧力および温度が一定(NPT)の条件下で、または体積および温度が一定(NVT)の条件下で行うことができる。これらのシミュレーションは、AMBERのSANDERモジュール、およびTIP3P水モデルを用いて行うことができる。ある態様においては、タンパク質力場パラメーターをParm99セットから採用し、糖質パラメーターをGLYCAM06から採用する。また、非周期的境界条件下で陰溶媒モデルを用いるシミュレーションを行うこともできる。
【0036】
AMBERは、構造に基づくリガンド設計、ならびにあらゆる複合分子系における構造および自由エネルギーの理解のための、シミュレーションに基づく方法を提供する分子モデリングおよびシミュレーション用のパッケージである。AMBERはUniversity of California、San Francisco校で開発され、そこから入手可能である。AMBERの内部の他のモジュールを用いて、これらのMDシミュレーションを行うことができる。実際には、MDシミュレーションを行うためのAMBER以外のプログラムも存在する。そのようなプログラムも本発明に適用可能である。
【0037】
ある態様において、グリカン-タンパク質複合体の初期座標は、可能であれば、不活性酵素-基質、活性酵素-阻害薬または酵素-生成物複合体からの結晶学的データから選択することができる。相同性モデリングまたは比較モデリングによって作成されたモデルのような、タンパク質の理論モデルを用いることもできる。遊離した酵素の構造のみが入手可能である場合には、AutoDockまたは別の等価なプログラムを用いて共複合体(co-complex)を予測することができる。AutoDockは、3D構造が判明している受容体に低分子がどのように結合するかを予測するために設計された自動ドッキングツール・スイートである。他のドッキングプログラムも存在し、それらも本発明に適用可能であると考えられる。
【0038】
ある態様においては、ヒスチジンプロトン付加の状態を、可能であれば分子内水素結合から推測することができ、そうでなければヒスチジンをNεで中性でプロトン付加されているとして扱うことができる。複合体上の任意の実効電荷は、適した数の対イオン(Cl-またはNa+)の付加によって中和することができる。典型的には、オリゴ糖-タンパク質複合体は、周期的立方体内で例えばほぼ10,000個のTIP3P水分子によって溶媒和されると考えられ、この枠の端と溶質の最も近い原子との間の最小距離は10Åである。周期的境界条件は、直接空間カットオフ距離を12Åとする長距離静電作用に関するエヴァルト処理と合わせて適用することができる。これらのパラメーターが本発明を限定するものでないことは理解されるであろう。実際には、TIP3Pは計算化学のために用いられる古典的水モデルの1つに過ぎないことが理解されるであろう。TIP4P、TIP5P、SPC、BNSなどの他の水モデルを本発明に用いることもできる。また、水を、誘電定数、距離依存的誘電定数、一般化ボルンモデルなどの陰溶媒和モデルを用いて、またはポワソン・ボルツマン近似によって近似することもできる。
【0039】
ある態様においては、初期コンフィギュレーションを、SANDERモジュールを用いてエネルギー極小化することができる。1つの態様において、初期コンフィギュレーションは、5,000サイクルの最急降下およびSANDERモジュールを用いた25,000サイクルの共役勾配エネルギー極小化を含む。続いてシステム全体を、加熱および続いて冷却することにより、シミュレーションアニーリングを行う。ある態様において、シミュレーションアニーリングは、50psで5Kから300Kにし、続いて5Kでの冷却をさらに50ps行うことを含む。初期原子速度はボルツマン分布から指定することができ、一般に5Kとする。生成動態段階の前に、システム全体を150psで再び5Kから300Kに加熱することによって熱平衡化することができる。2fsの時間段階を用い、ベルレアルゴリズムを用いて運動方程式を積分することができる。水素を含む結合は、SHAKEアルゴリズムを用いてそれらの平衡長に拘束することができる。これらのパラメーターは例示に過ぎず、本発明を限定するものではないことは理解されるであろう。
【0040】
本発明の方法は、いかなる特定のリガンドにも限定されない。本発明に適しているリガンドには、任意の天然もしくは合成脂質またはそれらの誘導体が含まれる。適したリガンドの例には、ラクトース、スクロース、マルトース、トレハロース、セロビオース、キトビオース、N結合型オリゴ糖、O結合型オリゴ糖、オリゴ糖、単糖、末端分枝型および非分枝型のα-(2,3)およびα-(2,6)-Neu5Ac、高マンノースN-グリカン上のα-(1-2)-man、糖タンパク質および糖脂質上のα-Gal、グリコサミノグリカン(ヘパリン、ヘパラン、コンドロイチン、ヒアルロン酸およびそれらの硫酸化類似体)、糖タンパク質および糖脂質上のβ-N-およびβ-またはα-O-GlcNAc、糖タンパク質および糖脂質上のβ-Gal、N-グリカン上のα-1,2/3/6 Man、N結合型またはO結合型グリカン上のα-Fucが非限定的に含まれる。
【0041】
A.糖質プロセシング酵素-グリカン複合体の安定性に影響を及ぼす可能性のある残基を解析するための、コンピュータ支援による方法
レクテンズへの変換のための糖質プロセシング酵素がひとたび選択されれば、その配列を、突然変異した場合に酵素-グリカン複合体の親和性または安定性または特異性に影響を及ぼす可能性のあるアミノ酸残基に関して解析することができる。
【0042】
いくつかの態様において、配列解析は、計算による突然変異誘発によって行うことができる。1つの態様においては、タンパク質安定性および/またはリガンド親和性に影響を及ぼすキー(key)タンパク質残基を、計算による飽和突然変異誘発実験を用いて同定することができる。計算による飽和突然変異誘発はアミノ酸配列レベルで実施され、これは、1つのアミノ酸側鎖の別のものによる置き換えに続いて、基質と酵素との間の相互作用の親和性または安定性または特異性に対する置き換えの影響について計算解析を行うことを伴う。他の態様において、標的化またはランダム計算による突然変異誘発を行うことができる。他の態様において、酵素内の各アミノ酸によって酵素-グリカン複合体の安定性または特異性に加えられる寄与を、グリカン-酵素複合体に関して直接計算することもできる。
【0043】
ある態様において、タンパク質安定性またはリガンド親和性またはリガンド特異性に影響を及ぼす「ホットスポット」キータンパク質残基は、計算によるアラニンスキャニング突然変異誘発(ASM)を用いて同定される。ASMは、個々の残基のアラニンによる逐次的置き換えによって行うことができる。ASMは、タンパク質の機能、安定性および形状に関与する残基を同定することができる。各アラニン置換により、タンパク質の機能性に対する個々のアミノ酸の寄与を検討する。この手法の全般的な概観は、Kollman, P.A., et al., Calculating Structures and Freeエネルギーof Complex Molecules: Combining Molecular Mechanics and 連続 Models. Acc. Chem. Res., 2000. 33(12): p.889-97およびArakat, N., et al., Exploiting Elements of Transcriptional Machinery to Enhance Protein Stability JMB, 2007. 366(103-116)に提示されている。1つの典型的な例では、WT酵素または酵素-基質複合体のMDシミュレーションを行い、データを収集する。その後に、すべて(または選択したもののみ)の残基の側鎖をCβ位置まで切断して短縮し、その結果、それらをアラニンに変換させる。不足している水素原子を加えて完全なアラニンを形成させる。この手順を、MDシミュレーションで収集した構造のすべてについて繰り返す。アラニンに変換させたところで、酵素または複合体の安定性に対する突然変異の影響をMDデータから計算することができる。これらの影響は、複合体もしくはタンパク質の構造の変化をモニターすることによって、またはタンパク質もしくは複合体に関連したエネルギーの変化をモニターすることによって決定される。また、アラニン突然変異を導入した後に別個のMDシミュレーションを行うこともできる。アラニンスキャニング突然変異誘発は、当技術分野において公知の唯一のスキャニング突然変異誘発法ではない。このため、この手法の考察は例示に過ぎず、本発明を限定するものではない。
【0044】
さらに他の態様において、キー残基を、野生型複合体におけるそれらのエネルギー寄与の大きさによって、または結合したリガンドに対するそれらの近接性によって同定することもできる。1つのそのような態様においては、水素結合、ファンデルワールス接触、および/または水媒介接触によって基質と直接相互作用する、同定された「ホットスポット」キー残基を含む、アミノ酸残基のサブセットを作り出すことができる。非水素(または重)原子との間に約3.2Åを上回らずに位置する任意の残基を、ファンデルワールス接触によって、または水素結合によって基質と直接相互作用するとみなすことができる。非水素原子との間に約4.5Åを上回らずに位置する任意の残基を、水媒介接触によって、または非特異的静電相互作用によって基質と相互作用するとみなしてよいであろう。他の態様において、サブセットは、気相(ΔEMM)または全(ΔG)相互作用エネルギーのいずれかに対する寄与が少なくとも約-0.5kcal/mol未満であるとして同定された任意の残基を含むと考えられる。
【0045】
または、ホットスポットを、特異的に有利な突然変異を予測するため、またはポテンシャル的に有利な突然変異のクラス(中性、荷電性、疎水性など)を同定する(クラスフォーカシング)ための、さらなる理論的解析に供することもできる。Hayes, R.J., et al., Combining computational and experimental screening for rapid optimization of protein properties. Proc Natl Acad Sci U S A, 2002. 99(25): p.15926-31は、クラスフォーカシングの手法を記載している。
【0046】
いくつかの態様においては、レクテンズ特異性が劣化する恐れのある不適切な機会を避けることが好ましいであろう。これらの態様においては、基質との相互作用に直接関与する残基を、突然変異誘発のために最初は選択しないことにする。他の態様においては、非特異的に相互作用するが、リガンド結合に対して有意に有利に寄与する残基(相互作用エネルギーの算出により同定)も、初期の突然変異誘発試験から除外するものとする。続いて、残りのすべての残基、特に結合に対して不利に寄与する任意のものを、1回目の突然変異誘発のために選考する。いくつかの態様においては、以前に除外したサブセットのメンバーを、初回の実験的および理論的な突然変異誘発を完了した後に、突然変異誘発に供することができる。
【0047】
B.糖質プロセシング酵素-グリカン複合体の結合エネルギーを予測するためのコンピュータ支援による計算シミュレーション法
いくつかの態様においては、上記の計算による突然変異誘発法のいずれかによって行った理論的突然変異の、糖質プロセシング酵素-グリカン複合体の結合エネルギーに対する影響を予測することが重要と考えられる。ある態様において、結合エネルギーは、熱力学的積分(TI)としても知られる自由エネルギー摂動法を用いて算出することができる。TIを用いることで、キー構造モイエティーの結合に対するエネルギー寄与を定量することができる。Straatsma, Holonomic Constraint Contributions to Free Energy Differences from Thermodynamic Integration Molecular Dynamics Simulations. Chem. Phys. Lett., 1992. 196: p.297-302, Zacharias et al., Inversion of Receptor Binding Preferences by Mutagenesis: Free Energy Thermodynamic Integration Studies of Sugar Binding to L-Arabinose Binding Proteins. Biochemistry, 1993. 32: p.7428-7434、およびChipot and Kollman, Alternative Approaches to Potential of Mean Force Calculations: Free Energy Perturbation versus Thermodynamic Integration Case Study of Some Representative Nonpolar Interactions. J Comput Chem, 1996. 17(9): p.1112-1131は、TIの方法論を記載している。TIは一般に、きわめて類似したリガンドに対する相対的結合エネルギーを検討する目的に限定されるが、キー構造モイエティーの結合に対するエネルギー寄与を定量することもできる。TIシミュレーションの典型的な一例では、自由エネルギーを初期残基を一連の漸進的段階を通じて最終的なものに変換する理論的プロセスを計算し、その間に各状態からの寄与率を変化させるという改変MD条件下でシミュレーションを行う。この非物質的プロセスは、各状態に関するエネルギー関数を数学的に混合することによって行われ、これは時に計算アルケミーと呼ばれることもある。
【0048】
さらに他の態様において、結合の全自由エネルギー(ΔG)は、基質とタンパク質(反応物)との間の相互作用エネルギーの直接分解によって算出することができる。ΔGの直接算出は、分子力学(MM)エネルギー推定値と、脱溶媒和自由エネルギーを捕らえるためのポワソン・ボルツマン(PB)または一般化ボルン(GB)などの連続溶媒モデルとを組み合わせる。これらの算出は一般に、コンフォメーションエントロピーによる追加の寄与を別に計算することを必要とする。
【0049】
限定的ではないが一例を挙げると、1つの典型的なMM-GB/PB算出では、MD軌道から抽出したそれぞれの構造「スナップショット」について、自由エネルギーをタンパク質(ΔGタンパク質)、リガンド(ΔGリガンド)および複合体(ΔG複合体)に関して計算する。関心対象の酵素によっては、データの初期部分を切り捨ててシステムを平衡化させる。例えば5nsの軌道では最初の1nsを切り捨てることができる。これらのモデルでは、データのスナップショットを設定間隔で収集する。単なる例示に過ぎないが、残りの4nsから、分子力学(MM)結合エネルギー解析のために2,000個のスナップショットを(2ps間隔で)選択することができる。続いて、結合自由エネルギー(ΔG)を減算によって計算することができる(式6参照)。軌道全体にわたって平均することにより、最終的な平均相互作用エネルギー(<ΔG結合>)が得られる。
<ΔG>=<ΔG複合体>-<ΔGタンパク質>-<ΔGリガンド>[6]
式中、平均はMDスナップショット全体にわたるものである。
【0050】
構成要素の自由エネルギーは、エネルギーを3つのカテゴリー、すなわち分子力学(静電およびファンデルワールス)エネルギー、溶媒和エネルギーおよびエントロピーエネルギー(式7参照)に分けることによって計算することができる。
<ΔG>=<ΔEMM>−T<ΔSMM>+<ΔG溶媒和>[7]
【0051】
解析の前に、水分子を溶媒和軌道から除去することができる。続いて、溶媒和によるエネルギー寄与を、その相対的速さの点から大型のタンパク質-糖質複合体への適用に非常に適している一般化ボルン(GB)陰溶媒和モデルの適用によって入手することができる。MM-GBSAの結果は、より厳密なMM-PBSA解析(ポワソン-ボルツマン陰溶媒近似に基づく)によるものに十分に匹敵する。GB近似はまた、計算によるアラニンスキャニングにおいても有効に働くことが最近示されている。少なくとも1つの態様において、糖質-タンパク質相互作用エネルギーを計算するためのGB法は、Tsui and Case, Theory and Applications of the Generalized Born Solvation Model in Macromolecular Simulations. Biopolymers, 2001. 56: p.275-291におけるGBパラメーター表示を使用する。
【0052】
ある態様において、エントロピーへの振動、並進および回転の寄与はエネルギーが極小化された座標の通常の様式解析から導き出すことができ、一方、コンフォメーションエントロピーは該当する内部座標の共分散行列の解析から推算する。Karplus and Kushick, Method for Estimating the Configurational Entropy of Macromolecules. Macromol., 1981. 14: p.325-332を参照。糖質の場合には、グリコシド間(inter-glycosidic)ねじれ角に関連したコンフォメーションエントロピーに焦点を絞ることが特に好都合かつ適切である。主として束縛回転に起因するコンフォメーションエントロピーの変化を、各オリゴ糖の遊離型および結合型における骨格ねじれ角の動きから推算することができる。結合状態および遊離状態におけるねじれ角に関する共分散行列の行列式から、相対コンフォメーションエントロピーを導き出すことができる。
【0053】
いくつかの態様においては、結合エネルギーを、古典的力学力場を用いて算出する。一般に、かかわる分子に関係する原子間特性を力場としてパラメーター表示する。AMBER力場を用いるためには、力場のパラメーターに関する値(例えば、力の定数、平衡結合長および結合角、電荷を入力する)。これらのパラメーターセットはかなり多く存在し、AMBERソフトウエアのユーザーマニュアルに詳細に記載されている。各パラメーターは名称を持ち、ある特定のタイプの分子に関するパラメーターを与える。
【0054】
1つの態様においては、GLYCAM/AMBER糖質力場を用いて結合解析を実施する。GLYCAM06パラメーターはさまざまな生体分子力場とともに用いることができる。GLYCAM06と適合性のある力場の例には、AMBER、CHARMM、NWCHEMなどが非限定的に含まれる。ある態様においては、GLYCAMパラメーターを、タンパク質に関するAMBERパラメーターによって補強することができる。GLYCAM06はいかなるデフォールトパラメーターも汎用パラメーターも使用しておらず、何らかの特定のクラスの生体分子にはもはや限定されないが、低分子力場の趣旨には十分に拡張可能である。GLYCAM06パラメーターは、例えば、Kirschner et al., GLYCAM06: A Generalizable Biomolecular Force Field. Carbohydrates. J. Comput. Chem., 2007. Early View (DOI 10.1002/jcc.20820)に記載されている。
【0055】
ある補足的な態様においては、計算シミュレーションを、コンフォメーションサンプリングを達成するために行う。そのような手法には、分子動力学シミュレーション、モンテカルロシミュレーション、または側鎖回転異性体の検索が含まれる。
【0056】
C.糖質プロセシング酵素突然変異体の発現および検査
本明細書に記載した計算による突然変異誘発および分子シミュレーションの方法を用いて、予測された望ましいリガンド結合特質を有する糖質プロセシング酵素突然変異体が同定されれば、親和性および複合体安定性の予測を、実験的突然変異誘発を用いて確かめることができる。いくつかの態様においては、関心対象の糖質プロセシング酵素のコード配列を、適した種から単離されたゲノムDNAから増幅し、適したベクター中にサブクローニングする。遺伝子クローニングおよびタンパク質過剰発現の定型的な方法は記載されている。ゲノムDNA由来の関心対象の糖質プロセシング酵素のコード配列を、選択した種から単離して、任意の適したベクター中にサブクローニングすることができる。いくつかの態様においては、適したアフィニティータグを備えた関心対象の糖質プロセシング酵素を発現するように、ベクターを遺伝子操作することができる。タンパク質のタグ標識は、アフィニティークロマトグラフィー手法を用いたその精製を容易にすると考えられる。1つの態様において、糖質プロセシング酵素に、ヘキサヒスチジンタグによるタグ標識を行うことができる。別の態様においては、糖質プロセシング酵素を、抗原ペプチドタグを含むように遺伝子操作することができる。適したベクターの例には、pOPH6、pETおよびpBADが非限定的に含まれる。pOPH6は、選択した大腸菌株に発現のために導入して形質転換を行わせることができる。本発明は、タンパク質の過剰発現に関して、特定の大腸菌株には限定されない。適した株の例にはDH5αが含まれる。
【0057】
一晩培養した細胞培養物(5〜10ml)を、十分な量の炭素源、ミネラル、イオン、抗生物質および他の試薬を含む適した量の栄養培地(例えば、ルリア・ベルターニ培地)中に接種することができる。一般に、これらのバッチ生産は小規模、すなわち100〜200mlであるが、より大容積のバッチを調製することもできる。抗生物質の選択は、大腸菌株およびクローニングされたベクターの遺伝子操作された耐性に依存すると考えられる。例えば、DH5α中にクローニングされたpOPH6ベクターの場合には、80〜120μg/mlアンピシリン、0.8〜1.5%v/vグリセロール、80〜150mMリン酸カリウム(pH 7.0)および0.2〜1.5mMイソプロピルβ-D-1-チオガラクトピラノシド(IPTG)を含むルリア-ベルターニ培地を用いうるであろう。培地を、導入から4〜25時間後に収集することができる。
【0058】
いくつかの態様においては、過剰発現された酵素を単離することができる。種々の方法をタンパク質精製のために利用することができる。1つの態様においては、過剰発現されたグリコシダーゼを含む清澄化された培地を、適したアフィニティーカラムに通すことができる。ヘキサヒスチジンアフィニティータグが付加されたタンパクを、ニッケルまたはコバルトとともに固定化された樹脂を用いて精製することができる。例えば、清澄化された培地を、適した量の塩およびイミダゾールを含む冷ローディング緩衝液で希釈し、Hi-Trap IMACカラム(Amersham Pharmacia Biotech)に2ml/分で通すことができる。抗原タグ標識タンパク質は、それを抗原特異的抗体とともに固定化された樹脂に通すことによって精製することができる。組換え酵素を、適したゲル濾過カラムを用いて脱塩処理することできる。
【0059】
ある態様においては、実験的突然変異誘発を、確立されたプロトコールに従った部位指定突然変異誘発を用いて実施する。一例を挙げると、部位指定突然変異誘発を、適したグリコシダーゼの遺伝子を含むベクター(例えば、pOPH6)上で、Stratagene(La Jolla, CA)製のQuikChange(商標)突然変異誘発キットを用いて行う。それぞれの突然変異体に対するセンスプライマーおよびアンチセンスプライマーは、遺伝子の配列を用いて設計することができ、自動DNA合成装置(Integrated DNA Technologies, Inc.)によって合成することができる。約10〜約20ngのプラスミドおよび約5〜約20ピコモルのセンスプライマーおよびアンチセンスプライマーを、製造元のプロトコールに従ったポリメラーゼ連鎖反応(PCR)混合物に添加することができる。PCRは、熱サイクル制御ユニット(MJ Research, Cambridge, MA)上で行うことができる。DNAテンプレートは、製造元のプロトコールに従った適したエンドヌクレアーゼの添加によって消化することができる。いくつかの態様においては、所望の突然変異のみが生成されたことを確認するために、それぞれの突然変異体の全コード領域を完全にシークエンシングする。いくつかの態様においては、確認された突然変異体DNAをテンプレートとして用いることで、単一のアミノ酸突然変異に用いたのと同じ手順を用いて複数の突然変異を作り出すことができる。
【0060】
アミノ酸突然変異体が、計算による飽和突然変異誘発のための方法を介して同定されたところで、突然変異体タンパク質のライブラリーを、望ましい結合特質を持つ突然変異体に関してスクリーニングすることができる。タンパク質-リガンド相互作用のハイスループットスクリーニングのために用いられる数多くの技術が、当技術分野において利用可能である。本発明に適したそのような技術の例には、ツーハイブリッドシステム、mRNAディスプレイ、ファージディスプレイ、酵母ディスプレイ、リボソームディスプレイおよび細菌ディスプレイが非限定的に含まれる。本発明のアプローチは、ライブラリーにおいてランダム化すべきであるタンパク質中の部位を同定するための、さらなる主観的な方策である。したがって、本発明の計算解析を組み合わせることにより、フォーカストバイオコンビナトリアルライブラリーを設計し、続いて構築することができる。それらの設計および構築によるそのようなライブラリーは、ライブラリースクリーニングのためのはるかに効率的なアプローチを提供する。
【0061】
いくつかの態様において、タンパク質-リガンド相互作用のハイスループットスクリーニングは、突然変異誘発ディスプレイライブラリーを作り出すことによって行うことができる。1つのそのようなライブラリーシステムは、GENEARTによって合成することができる。1つの態様においては、そのライブラリーをファージ上にディスプレイすることができる。ファージディスプレイライブラリーは、当技術分野において十分に確立されたプロトコールを用いて構築することができる。例を挙げると、関心対象のタンパク質またはペプチドをコードするDNAライブラリーを、M13繊維状ファージのpIII遺伝子またはpVIII遺伝子中にライゲートする。続いて、ファージ遺伝子およびインサートDNAハイブリッドを、TG1またはXL1-Blue大腸菌などの大腸菌細胞に導入して形質転換を行わせる。「ファージミド」ベクターを用いる場合には(簡略化されたディスプレイ構築物ベクター)、ファージ粒子は、それらが、ファージDNAのパッケージング、およびマイナー(pIII)またはメジャー(pVIII)コートタンパク質のいずれかの上での成熟ビリオンとそれらの外殻の一部としての関連タンパク質断片との集合を可能にするヘルパーファージに感染するまでは、大腸菌細胞から放出されないと考えられる。pIII遺伝子またはpVIII遺伝子中への多くのさまざまなDNA断片の取り入れによってライブラリーが生成され、それから関心対象のメンバーを単離することができる。該当するDNA標的またはタンパク質標的をウェルの表面に固定化することにより、そのような標的の1つと結合するタンパク質をその表面にディスプレイするファージは、洗浄によっても保持されるが、他のものは除去されると考えられる。保持されたものを溶出させて、より多くのファージ(ヘルパーファージによる細菌の感染により)を産生させるため、およびそのようにして該当する(すなわち、結合性)ファージが濃縮されたファージ混合物を生産するために用いることができる。これらの段階の反復サイクリングを、望ましくない材料を除去することによる金の試料の濃縮に準拠して「パニング」と称する。
【0062】
他の態様においては、突然変異誘発ディスプレイライブラリーを酵母上にディスプレイさせることができる。酵母ディスプレイでは、関心対象のタンパク質をAga2pタンパク質との融合物として酵母の表面にディスプレイさせる。Aga2pタンパク質は天然において酵母によって用いられ、酵母細胞接合時の細胞-細胞接触を媒介する。このため、Aga2pを介したタンパク質のディスプレイは、そのタンパク質を細胞表面から突出させ、酵母細胞壁上の他の分子との相互作用の可能性を最小限に抑える。Aga2遺伝子、HA(血球凝集素)タグ、ディスプレイされるタンパク質およびc-mycタグの融合物としての発現ベクターを図示している図7を参照されたい。酵母ディスプレイライブラリーは、当技術分野において十分に確立されたプロトコールを用いて構築することができる。いくつかの態様においては、関心対象の遺伝子を、AGA2遺伝子とインフレームになるように、選択したベクター中にクローニングすることができる。適したベクターの例には、pYD1ベクター(Invitrogen)およびpPNL6(Pacific Northwest National Laboratory)が非限定的に含まれる。その結果得られた構築物を、続いて、染色体に組み込まれたAGA1遺伝子を含む、適した酵母株(例えば、EBY100 S.セレビシエ(S. cerevisiae))に導入して形質転換を行わせる。ベクター由来のAga2融合タンパク質およびEBY100宿主株中のAga1タンパク質はいずれも、ガラクトースの非存在下ではクローニングされたタンパク質の検出可能な発現を許容しない厳密に調節されるプロモーターであるGAL1プロモーターによって調節される。ガラクトースを用いた導入に応じて、Aga1タンパク質およびAga2融合タンパク質が分泌経路内で会合し、クローニングされた突然変異体が細胞表面にディスプレイされる。
【0063】
突然変異した糖質プロセシング酵素を細胞表面にディスプレイする突然変異誘発ライブラリーがひとたび構築されれば、望ましい結合特性および複合体形成特性を有する突然変異体を同定するためにそれをスクリーニングすることができる。選択したリガンドと高親和性複合体を形成しうる突然変異体を同定するために用いられるアッセイシステムの基本原理は、ディスプレイライブラリーおよびリガンドを含む反応混合物を、2つの試薬が相互作用して結合し、それによって複合体を形成するのに十分な条件および時間の下で調製することを伴う。続いて、結合パートナー間の複合体の形成を捕らえる。反応が完了した後に、反応しなかった構成要素は除去されるが(例えば、洗浄により)、形成された複合体は細胞表面に保持されると考えられる。細胞表面に係留された複合体の検出は、さまざまなやり方で達成することができる。いくつかの態様においては、リガンドを直接的または間接的に前標識することができる。リガンドを標識する場合、細胞表面に固定化された標識の検出により、複合体が形成されたことが指し示される。結合パートナーを前標識しない場合には、間接的標識を用いて、表面に係留された複合体を検出することができる。分子の標識は周知であり、例えば、タンパク質、核酸、糖質、カルボン酸のビオチン化のための、アミン反応性物質およびチオール反応性物質を含む、多数のビオチン化剤が公知である。ビオチン化された基質は、アビジンまたはストレプトアビジンを介してビオチン化構成要素と結びつけることができる。
【0064】
いくつかの態様においては、ディスプレイライブラリーのサイズを、ビオチン化N結合型糖ペプチドと低親和性ないし高親和性で結合する酵母を含むように濃縮することができる。低親和性相互作用を直接測定することは難しい。ビオチン化N結合型糖ペプチドを、ストレプトアビジンでコーティングされた常磁性ビーズ(例えば、Invitrogen)の表面に固定化することにより、ライブラリーを、ビーズ表面上の標的と結合するタンパク質をディスプレイする酵母に関して濃縮することができる。N結合型糖ペプチドでコーティングされた常磁性ビーズによって捕らえられた酵母を磁石で単離し、結合しなかった酵母を洗い流し、パニングの工程を繰り返す。いくつかの態様において、ライブラリーを、ライブラリーの初期サイズおよびパニングの回数に応じて細胞106〜108個に減少させることができる。少なくとも1つの態様においては、ライブラリーを約1×107個に減少させる。
【0065】
ある態様においては、高親和性突然変異体の初期スクリーニングを実施することができる。これらの態様において、スクリーニングはフローサイトメトリーによって行うことができる。スクリーニングは、特定のビオチン化糖ペプチドと結合する突然変異体のみを選択することによって行うことができる。一例を挙げると、図9に詳しく示しているように、C末端c-mycタグを有するAga2p融合タンパク質を発現する酵母を抗c-myc mAbとともにインキュベートし、その後に完全長グリコシダーゼクローンを発現した酵母を検出するために蛍光二次mAbを添加する。ある態様において、ビオチン化糖ペプチドと結合する突然変異体の検出は、ストレプトアビジンの添加によって達成することができる。1つの態様において、ストレプトアビジンは、適した蛍光標識(例えば、PE-Cy5)で標識することができる。ストレプトアビジン-PE-Cy5を用いて、糖ペプチドと結合する酵素クローンを蛍光標識することができる。糖ペプチドに対して高い親和性を有するc-mycを発現する細胞のみを選別することができる(図8)。いくつかの態様において、個々の酵母クローン上にディスプレイされた選択したグリコシダーゼ突然変異体の解離定数を決定することができる。少なくとも1つの態様において、解離定数はフローサイトメトリーによって決定することができる。
【0066】
ある態様においては、予備的スクリーニングアッセイにおいて同定された高親和性結合物を、さらなる結合試験のために発現させて精製することができる。生成されたレクテンズの結合親和性は、当技術分野において公知の種々の手法(例えば、フィルター結合アッセイ、電気泳動移動度シフトアッセイ(EMSA)、表面プラズモン共鳴(SPR)など)によって分析することができる。1つの態様においては、結合定数を、Biacore装置を用いるSPRによって測定する。SPRを用いて複合体の結合定数を評価するために用いられる1つの典型的なアッセイは、関心対象のレクテンズを、適したカップリング方法(例えば、アミンカップリング方法)によって20〜30℃でSPRチップ表面に固定化し、モック誘導体化したフローセルを参照表面とすることを含む。結合分析は、ランニング緩衝液の連続フロー(10〜50μl/min)を用いてさまざまな温度で行うことができる。ランニング緩衝液は、15〜25mM Tris-HCl(pH 7.5)、100〜200mM NaCl、0.5〜1.5mM EDTA、0.0025〜0.0075% P20界面活性剤でありうる。リガンドは、適した濃度範囲を得るための各々のランニング緩衝液中での系列希釈によって調製することができる。リガンドの結合は、レクテンズの低密度固定化表面上で一連の濃度(0.625〜10μM)で分析することができる。最大平衡センサーグラム値を用いて、飽和結合曲線をプロットし、平衡解離定数(Kd)の値を直接計算することができる。
【0067】
本発明の1つの局面において、レクテンズを作製するための方法は、酵素-基質または酵素阻害薬複合体に関する結晶学的データを計算による突然変異誘発のための基盤として用いる段階をさらに含む。一般に、レクテンズの基質との結晶化および結晶成長パラメーターの最適化は、当技術分野において記載されている方法および手順によって行うことができる。結晶化のために適した条件は、個々の事例ごとに決定されるであろう。結晶を回折に関して検査し、最も高い分解能で回折する結晶をデータ収集のために用いることができる。ある態様においては、分子置き換えを、複合体の構造を解明するために用いることが考えられる。これらの態様においては、生成されたレクテンズ-グリカン複合体のX線データを用いて、MDシミュレーションを検証すること、および/または新たなシミュレーションを開始することができる。
【0068】
本発明の全体を通して、グリカンプロセシング酵素から導き出されたレクテンズに関して例示されているものの、本発明が任意の酵素-基質複合体に対して広く適用可能であることは理解されるであろう。したがって、以下の実施例に限定されることなく、本発明の範囲をさらに例示することのみを目的として、このレクテンズアプローチを、プロテアーゼ、リパーゼ、キナーゼ(kinease)、ホスファターゼ、ヒドロラーゼ、イソメラーゼなどの酵素を、酵素基質に対する特異性を維持している受容体タンパク質に変換するために用いることができる。
【0069】
さらに、本発明は糖質プロセシング酵素に限定されない。これは糖質結合タンパク質にもさらに適用可能である。実際に、これはタンパク質-リガンド相互作用全般に適用可能である。
【0070】
V.使用方法
本発明の別の局面は、本明細書に記載されたレクテンズの使用方法を提供する。本明細書に記載されたレクテンズの非常にさまざまな潜在的な用途が、当業者には直ちに明らかであろう。以下は、そのような試薬の潜在的な有用性を説明している態様のうちごく一部を挙げたものである。
【0071】
ある態様において、本発明のレクテンズは、グリカン特異的分析ツールにおける用途のために用いることができる。本発明のレクテンズに基づくグリカン特異的分析ツールは、環境的、発酵、食品および医学の領域を含む多くの領域において検出方法として用いうる可能性があり、ヒトまたは動物におけるインビボまたはインビトロ検知にも用いうると考えられる。
【0072】
いくつかの態様において、本明細書に記載した規定された糖質特異性を有するレクテンズは、異常グリコシル化を探索しようとして生体試料を調べるために用いることができる。生体試料の例には、任意の生体液、組織または臓器が非限定的に含まれる。生体液の例には、血液、尿、血清、唾液、脳脊髄液および精液が非限定的に含まれる。他の態様において、本発明のレクテンズは、糖質に基づく生体液中の標的分析物レベルの検出のために用いることができる。標的分析物の例には、N結合型またはO結合型グリカン、グリコサミノグリカン(ヘパリンを含む)などの内因性に認められる分子、植物多糖、糖質に基づく薬物などの外因性に消費される種、および、その表面が特異な複合グリカンでコーティングされていることの多い病原体が非限定的に含まれる。他の態様において、本明細書に記載されたレクテンズは、医薬品探索、および、グリカンに基づく新たな化合物の生物活性の評価における用途もある。
【0073】
いくつかの具体的な態様において、本明細書に記載されたレクテンズは、異常グリコシル化を呈する疾患を診断および/または治療するために用いることができる。1つの態様において、本発明のレクテンズは、糖タンパク質、糖脂質、および/または種々の糖質エピトープを含むある特定の腫瘍抗原を検出するために用いることができる。数多くのこれらの腫瘍抗原が、新生物性病態においてアップレギュレートされることが見いだされている。新生物性障害の発症および進行を知らせることのできる、ならびに本発明のレクテンズによって検出することのできる腫瘍抗原の例には、結腸直腸癌、胃癌、膵癌、肺癌および乳癌ならびに発生過程にある胎児と関連性のある糖タンパク質である癌胎児性抗原(CEA);膵癌の患者で認められる糖脂質中に存在する糖質抗原19-9(CA 19-9)またはシアル酸付加ルイスA抗原;ならびに乳癌と関連性のある糖質抗原15-3(CA15-3)が非限定的に含まれる。
【0074】
抗原の存在が癌性細胞への形質転換を必ずしも指し示すわけではないが、細胞内におけるその局在は、CEAの場合のように、その指標となる。この理由から、選択性が高くかつ高親和性の分析ツールが必要とされている。診断検査は現在、糖タンパク質のペプチド部分または糖脂質の糖部分に対して産生されることの多い抗体に依拠しているが、しかし、その厳密なエピトープは今現在、明らかにされつつある途上に過ぎない。グリカンの特徴が決定されている例では、複数のグリコフォームが往々にして存在する(例えば、CEA)。これらのグリコフォームを識別しうる試薬がないことから、現時点では、グリコシル化のわずかな差異が病態、癌のタイプまたは組織内局在と相関する程度を決定することは不可能である。現在、これらの課題は、主として単離された糖タンパク質のMS分析によって対処することができるが、それらは複数のグリコフォームの混合物として検討される。典型的には、行われる唯一のレベルのグリコフォーム-フォーカシングは、レクチン(コンカナバリンA(Con A))アフィニティークロマトグラフィーを用いる高マンノース含有グリカンの濃縮である。より効率的な実験室分析および定型的な臨床診断手法は、グリコフォーム特異的試薬がないために依然として著しく制約されている。
【0075】
本発明のレクテンズは、生体試料中の任意の所与の糖タンパク質中に存在する各グリコフォームの相対的存在量を定量するために特に有用である。本明細書で用いる場合、「グリコフォーム」という用語は、特定のタイプの糖タンパク質が結びついているタンパク質のタイプのことを指す。2つのタンパク質は、それらが同じ糖タンパク質を保有しているならば同じグリコフォームのものと考えられる。いくつかの態様において、本発明のレクテンズは、生体試料を特定のグリコフォームに関して濃縮するために用いることができる。他の態様において、本明細書に記載された方法によって生成されたレクテンズは、グリカンが結びつくタンパク質表面上の特異的グリコシル化部位を同定するために用いることができる。これらの態様において、特定のオリゴ糖に対して特異的なレクテンズは、任意の糖タンパク質のタンパク質分解消化物から完全な糖ペプチドを分離するために用いられると考えられる。例えば、PNGアーゼ-Fにより導き出されたレクテンズは、N結合型糖ペプチドを、糖タンパク質の典型的なプロテアーゼ消化によって生じるであろう他の糖ペプチドまたはペプチドから分離するために用いることができる。関心対象の分析物における試料の濃縮は、糖ペプチド画分のさらなる特徴決定において大きな力添えとなる。特に、濃縮はペプチド配列およびグリカン構造の同定を容易にし、それは完全なタンパク質内部のグリコシル化部位の同定およびグリコシル化部位に存在する特定のグリカンの特徴決定を可能にする。
【0076】
他の態様において、本発明のレクテンズは、生体液、組織、臓器または生細胞におけるタンパク質の特異的グリカン修飾のモニタリングにおける用途があると考えられる。本発明の方法によって遺伝子操作されたレクテンズはタンパク質の実体には依存しないと考えられ、すなわちそれらはコンテキスト非依存性であると考えられ、かつ所与のグリカンを含む任意のタンパク質を認識しうると考えられ、そのため所与のグリカン修飾の検出のために極めて有用であると考えられる。
【0077】
さらに他の態様において、本発明のレクテンズは、細胞または組織をインビトロまたはインビボで染色するために用いることができる。
【0078】
他の態様において、レクテンズを、糖タンパク質中の特定の糖タンパク質またはグリコシル化部位に対して特異的であるように開発することができる。そのようなレクテンズは、製薬産業または研究産業で用いるための組換え糖タンパク質の生産過程で生じる可能性のある、混合物中の特定の糖タンパク質をモニターするために用いうると考えられる。
【0079】
前記の諸態様において、レクテンズに着色剤または色素によるタグ付加を施して、関心対象の細胞または組織または糖タンパク質または糖ペプチドまたはオリゴ糖または多糖を含む生体試料に適用することができる。
【0080】
ある態様において、本発明のレクテンズは治療薬として用いることができる。これらの態様において、特定のレクテンズの設計は、ヒト相同体が存在するグリコシダーゼに基づくことができる。これにより、そのようなレクテンズに免疫反応性がないことが保証されると考えられる。ある態様において、本発明のレクテンズを、活性治療薬の送達用に改変することができる。本発明のレクテンズは明確に定められたグリカン特異性を有するため、治療薬の送達を、特定のグリカンをディスプレイするような細胞、組織または臓器のみに標的化することができる。部位特異的送達のために用いうる治療薬の例には、さまざまな化学療法薬、抗生物質および抗イルス薬、毒素、放射性同位体、サイトカインなどが非限定的に含まれる。
【0081】
ある態様において、本発明のレクテンズは、例えばアフィニティークロマトグラフィーを含むアフィニティー分離のための試薬として用いることができる。アフィニティークロマトグラフィーは、生化学的混合物を、レクテンズとグリカンとの間のそれのような高度に特異的な生物的相互作用に基づいて分離する方法である。本発明は、いかなる特定の設計にもクロマトグラフィー系にも限定されない。一般に、レクテンズは固体支持体と共有結合性に結びついているか、または他の様式で固定化されていると考えられ、固定相を構成すると考えられる。ある態様において、レクテンズにより誘導体化された固定相をカラムクロマトグラフィーに用いることができる。これらの態様においては、固体固定相の粒子を用いて、管の内容積全体を満たすことができる(充填カラム)。または、固相粒子を管の内壁上またはそれに沿って集中させ、管の中間部分で、生体試料の通路を制限せずに開かれたままにすること(すなわち、移動相)もできる(中空カラム)。他の態様においては、レクテンズにより誘導体化された固定相をバッチクロマトグラフィーのために用いることができる。これらの態様においては、固定相を容器に添加して、生体試料と混合することができる。前記の例は一般にアフィニティークロマトグラフィーに対象を絞っているが、これらの原理が他のアフィニティー精製プロトコールにも容易に適用されることは理解されるであろう。
【実施例】
【0082】
実施例1
レクテンズへの変換のための標的酵素
糖質-バイオセンサー(レクテンズ)への変換のための標的酵素
表1に提示しているのは、レクテンズとしての再設計に供することのできる3種の初期グリコシダーゼである。レクテンズ1は、グライコミクス解析のすべての局面において広範な用途があると考えられる。レクテンズ2は糖尿病におけるグリカンの分析をさらに進めるために極めて重要であると考えられ、レクテンズ3はヒトおよびトリのインフルエンザ受容体を比較して特徴づける上で有用であると考えられる。
【0083】
(表1)糖質-バイオセンサー(レクテンズ)への変換のための初期標的酵素
【0084】
レクテンズ1:N結合型グリカンの濃縮のためのグライコミクス親和性試薬(コアにフコースを含まないN結合型オリゴ糖に対して特異的)
多くの用途において、特に臨床的な組織および液体の分析を伴うものにおいて、疾患の進行に応じたタンパク質および糖タンパク質の発現の変化を同定することには大きな関心が寄せられている。特定の分子の発現の変化を病態と高度に相関づけることが可能であれば、それらを利用して、有望な診断薬、ならびにおそらくは、罹患した細胞および組織を画像化するための試薬を開発することができるであろう。これらの分析のすべての基礎にある難しさは、これらの混合物があまりにも複雑であるため、試料中に存在する個々の種の大半のものの、質量分析を用いるその後の同定および精製を可能にする分離方法を開発することが極めて難易度が高いことに理由がある。さらに、説得力のある証拠により、タンパク質上に発現されるグリカンの変化は疾患に関する補足的なマーカーとして役立てうることが実証されている。このため、病態との相関づけのための同定および精製を行えるように、糖タンパク質からタンパク質を、糖ペプチドからペプチドを迅速に分離することを可能にすることが非常に求められている。例えば、血清において、構成要素の半分はグリコシル化されていないと推定されており、中でもアルブミンは最も群を抜いて存在量が多い。これらのタンパク質を糖タンパク質から除去することができれば、複雑性は2分の1に減る。
【0085】
さらに、グライコミクスシークエンシングは、特定のN結合型グリカン構造を各ペプチド上の各グリコシル化部位にマッピングする目的で、トリプシンにより生成される糖ペプチドの解析に対象を絞っている。このタイプの分析では、タンパク質分解消化によって生成される種の大半は圧倒的にグリコシル化されていないペプチドである。これらを関心対象の糖ペプチドと分離することができれば、部位特異的グリカンの情報を得るための解析を大きく簡略化することができると考えられる。スカフォールド用のPNGアーゼFの選択は、この酵素がさらなる解析のためにN-グリカンを糖タンパク質および糖ペプチドから取り出すために広く用いられているという事実に基づく。これは活性が強く、ペプチドがコアに脊椎動物では発現されないα-1-3フコシル化を含まない限り、その作用は普遍的である。さらに、これは脊椎動物のようにα-1-6フコシル化を発現する種に対して完全な活性を有する。PNGアーゼFレクテンズは、抽出物、液体から、さらには質量分析による詳細な構造解析のために、精製された糖タンパク質から、糖タンパク質および糖ペプチドを分離するために、極めて有用であると考えられる。この試薬は、プロテオミクス/グライコミクス解析において、干し草山(haystack)(タンパク質分解ペプチド)から針(糖ペプチド)を抽出するために用いうると考えられる。
【0086】
初期PNGアーゼレクテンズのスカフォールドは、フラボバクテリウム・メニンゴセプチカムによって産生される酵素から生成させることができ、これは基質との複合体における結晶構造が報告されている。結晶構造の存在はレクテンズ-グリカン親和性の計算による最適化にとって大きな意味がある。加えて、予備的な点突然変異誘発試験により、PNGアーゼFを完全に不活性にする単一の突然変異(D60N)も同定されている。
【0087】
レクテンズ2:糖尿病関連タンパク質高GlcNac化に対するバイオセンサー(β-O-GlcNAcに対して特異的)
中性の核細胞質ヘキソサミニダーゼは1975年に最初に記載され、1994年にO-GlcNAcアーゼとして特徴づけられた。これがクローニングされたのは最近になってであり(CCRCのDr. L. Wellsによる)、その結果、核細胞質性の中性β-N-アセチルグルコサミニダーゼ(O-GlcNAcアーゼ、OGA、EC 3.2.1.52)と同定された。ヘキソサミニダーゼAまたはBとは異なり、OGAはサイトゾルに局在していて核ではその度合いは少なく、至適pHは中性であり、かつ、GalNAcを触媒しない上にそれによって阻害もされない。アポトーシスにおけるO-GlcNAcアーゼの役割はまだ解明されていない;しかし、この酵素のポリペプチドの中央部付近での切断は酵素活性に影響を及ぼさず、このことはN末端の「ヒアルロニダーゼ様」ドメインがあれば活性にとって十分であることを示唆する。ごく最近、ウェルシュ菌由来のO-GlcNAcアーゼの結晶構造が、遊離状態および阻害薬と複合体を形成したものの両方について報告された。それらはヒトタンパク質と高い相同性を有しており、計算による検討のための優れた基盤をもたらす。
【0088】
レクテンズ3:ヒトインフルエンザ受容体α-2,6結合型ノイラミン酸に対するバイオセンサー(α-Neu5Ac-(2,6)-β-Galに対して特異的)
ヒト細胞表面糖質(ノイラミン酸またはシアル酸)は、インフルエンザウイルスの血球凝集素タンパク質の標的となる。血球凝集素は、細胞表面のシアル酸受容体と結合することにより、ウイルスの宿主細胞との接着およびその中への侵入を媒介する。ヒトインフルエンザウイルスは、ヒト上気道の上皮細胞の表面に認められる主要なタイプであるα-2,6結合によってガラクトースと連結したシアル酸と選好的に結合する。トリウイルスは、トリ腸上皮の表面に認められ、ヒト気道ではその程度がより少ないα-2,3結合と結合する傾向にある。これらからみて、この受容体は宿主特異性の鍵となる決定要因であり、ポテンシャルインフルエンザの予防および治療を可能にする可能性のある重要な標的である。本発明者らの初期レクテンズスカフォールドは、UGAの研究者らによってP.マルトシダ(P. multocida)から単離され、クローニングされて大腸菌において発現されている2,6-シアリダーゼnanBになるであろう(Dr. M. Lee, J Bacteriol. 2000 December; 182(24): 6874-6883)。これは現在までのところ、ヒト2-6結合に対する顕著な選好性を有する唯一のノイラミニダーゼである。この酵素の3D構造(Q27701)は、PDBテンプレート2SLI(同じファミリーに属するヒドロラーゼ)から比較モデリングによって作成され、Modbaseデータベースに寄託されており、これは計算解析のための基盤として役立つと考えられる。推定された活性部位に、AutoDockをGLYCAMパラメーターによって用いてリガンドをドッキングさせることができる。同時に、構造の特徴決定を、タンパク質結晶学、NMR分光法および部位指定突然変異誘発を用いて実験的に行うことができる。
【0089】
PNGアーゼF-キトビオース複合体のMDシミュレーション
PNGアーゼF-キトビオース複合体の5nsの完全溶媒和MDシミュレーションを、nPT条件(300K、1atm)で、複合体に関するAMBER/GLYCAMタンパク質/糖質力場および実験的X線構造を使用して行った。エネルギー解析の前に、シミュレーションの安定性およびコンフォメーション平衡化のレベルを明らかにするために、Cα原子の位置の根平均二乗差(RMSD)をシミュレーション時間の関数として決定した(図3)。そのデータに基づき、平均RMSD(1.5A)が妥当であることが明らかになった;しかし、シミュレーションが平衡化に至るのは遅かった。それ故に、データの最初の1nsはその後の解析には含めなかった。
【0090】
キトビオースとタンパク質との間の分子間水素結合をモニターすることにより、結合部位におけるリガンド安定性を評価した(図4)。水素結合に関する平均値およびそれらの占有率を表2に、結晶学的に決定した値とともに提示している。
【0091】
(表2)キトビオースおよびPNGアーゼFとの間で計算した、選択した水素結合の長さおよび占有率
a 標準的な3.3Åカットオフを用いて決定した占有率
b Kuhn et al., Crystal-Structure of Peptide-N-4-(N-Acetyl-Beta-D-Glucosaminyl) Asparagine Amidase-F at 2.2-Angstrom Resolution. Biochemistry, 1994. 33(39): p. 11699-11706.
【0092】
MDシミュレーションが安定であり、かつリガンドとタンパク質との間の実験的相互作用を再現しうることを確かめた上で、続いてそれをその後の結合エネルギーおよびアラニンスキャニング解析に使用した。
【0093】
相互作用エネルギー解析によるホットスポットおよびキー残基の同定
本発明者らは、AMBERに組み込まれているような一般化ボルン(GB)連続溶媒モデルを、タンパク質-リガンド相互作用のために開発された溶媒パラメーターとともに用いた。本発明者らは以前、これらの溶媒パラメーターが、糖質-抗体および糖質-レクチンのMM-GB結合エネルギー予測によく機能することを報告している。結合エネルギーに対する寄与を、PNGアーゼF中の313アミノ酸に関して計算した。相互作用エネルギーを、リガンドの4.5Å以内にあるすべての残基(接触区域内)、ならびに、全MM(ファンデルワールスおよび静電)相互作用エネルギー(ΔEMM)または全結合自由エネルギー(ΔG結合)に対する寄与が少なくとも-0.5kcal/mol未満であった他の任意のものについて表3に提示している。また、基質結合に対して不利に寄与した任意の残基、例えばD60およびE206なども含めている。
【0094】
全相互作用エネルギー(-13.7kcal/mol)は、同程度のサイズのガレクチン-1-LacNAc複合体に関して計算したデータと同等であり、ガレクチン-1の場合と同じように、コンフォメーションエントロピーおよびコンフィギュレーションエントロピーを省略しているために、実験から得られた親和性を過大評価している。にもかかわらず、残基ごとの相互作用エネルギーを用いて、キーアミノ酸残基を同定することができる。D60およびE206は明らかな例外であったが、残基ごとの正味結合エネルギーの大半は有利であった(図5)。D60は最も主要な触媒残基であり、一方、E206およびE118は反応中間体の安定化のために重要であると考えられる。したがって、これらの2つの残基が、加水分解反応の二糖生成物にとって最も不安定化させるものであることは意義深いものの、おそらく驚くには当たらない。
【0095】
不利な可能性のある相互作用は、負の溶媒和自由エネルギーΔGGBによって相殺される不利なΔEMMを有するD57によっても示されている。ガレクチン-1-LacNAc複合体について観察されているように、準エンタルピー寄与(ΔEMM)およびエントロピー関連脱溶媒和ΔGGBの項は多くの場合、大きさが同程度であって符号が逆であることから、溶媒和モデルの選択を決定的なものにする。一般に、理論的計算の精度に限定して依拠するのではなく、鍵になる可能性のあるそのような部位すべてをディスプレイライブラリーに含める。
【0096】
図5は、左の画像では:PNGアーゼFの結合部位において二糖リガンド(赤)の4.5Å以内にある残基を表している。右の画像では:溶媒接触可能表面を、結合のために最も重要であると同定された残基に印を付した上で表している。
【0097】
表4の中のデータとの直接比較が、2つの類似した糖質-タンパク質複合体(ガレクチン-1-LacNAcおよびCon A-トリマンノシド)に関して可能であり、これらはいずれもAMBER/GLYCAM力場およびGB溶媒和近似を使用している。表3では、ガレクチン-1およびCon AのMM-GB解析の場合と同じように、分子力学エネルギー(-33.8kcal/mol)の大半は静電相互作用(-20.8)から生じている。しかし、ファンデルワールス寄与および静電寄与(それぞれ-12.9kcal/molおよび-20.8kcal/mol)はいずれも、ガレクチン-1と結合した類縁の二糖について観察されたものよりも小さく(ファンデルワールスについては-17〜-24kcal/mol、静電については-30.8〜-67.5kcal/mol)、このことはPNGアーゼFにおいて側鎖最適化を介した親和性強化の余地があることを強く示唆する。さらに、静電寄与が脱溶媒和自由エネルギーによってほぼ打ち消されることも見てとることができる。この現象は以前の研究のいずれにおいても観察されており、エントロピー-エンタルピー補償の現れである可能性がある。このため、GB計算の誤差の打ち消しにより、利点を得ることができる。
【0098】
表3におけるエネルギーに基づき、公知の不活性D60A突然変異体を作製し、エネルギーを再計算した。結合エネルギーはD60A突然変異(全ΔEMM=-35.5、ΔGGB=19.6およびΔG結合=-15.9、表5を参照のこと)では顕著に改善しており、親和性の正味の利得はおよそ2kcal/molであった。
【0099】
(表3)基質であるキトビオース(β-GlcNAc-(1,4)-β-GlcNAc-OH)と結合した野生型PNGアーゼFに関する結合自由エネルギーに対する残基寄与(kcal/mol)
【0100】
(表4)キトビオースと結合したPNGアーゼFのアラニンスキャニングおよび静電スキャニングによって同定された有利な突然変異体に関する全相互作用エネルギー(kcal/mol)
【0101】
引き続き、可能性のあるさらなるキー残基を探すためにD60A突然変異体に対してアラニンスキャニングを行った(表5)。ある程度の親和性の強化(-0.5kcal/mol)を伴う1つの二重突然変異体が後に同定され(D60A/E206A)、一方、5つの残基はリガンド結合のために決定的であることが確認された(W251、W191、W120、W59およびR61)。
【0102】
(表5)キトビオースと結合したPNGアーゼFのD60N突然変異体に関してアラニンスキャニングから予測された相対的a相互作用エネルギー(kcal/mol)
a 野生型配列(D60)との対比。 b 二重突然変異体は初期突然変異体(D60A)との対比。
【0103】
側鎖リパッキング実験を最初に行うのではなく、不活性突然変異体を、有利な静電相互作用につながると考えられる位置に関してスキャニングすることを行った。このスキャニングを、直接接触区域における残基のすべてにわたり、理論的に正および負のプローブ残基(実験的設計を参照)について行った(表6)。
【0104】
荷電残基に対する突然変異のうち、全結合エネルギーの強化につながると予想されたものはなかったが、いくつかの可能性のある突然変異が分子力学相互作用の改善につながることが示唆された。すなわち、残基E206、S155、E118およびY62をそれぞれArgおよびLysに突然変異させ、エネルギーを再計算することができる。その結果得られた結合自由エネルギーを用いて、クローニングおよび過剰発現を行うための何らかの特異的な点突然変異体を選択することができる。このことは、糖質-タンパク質相互作用が水素結合のネットワークの複雑なネットワークによって特徴づけられること、およびそのネットワークの摂動が有利であることは稀であると考えられることからみて、おそらく驚くには当たらない。この場合も、いずれも酵素機構における関与が示されているE206およびE118が、親和性最適化のための可能性のあるキー残基として同定されていることは意義深い。また、負に荷電した残基に対する突然変異のうち、親和性の改善につながると予測されたものがなかったことも、ディスプレイライブラリーの設計のためには注目すべきである。
【0105】
(表6)静電アラニンスキャニングにより予測された、キー残基に関するD60A突然変異体に対して相対的な相互作用エネルギー
【0106】
計算による親和性データに基づき、クローニング用にいくつかの突然変異体を選択し、大腸菌において過剰発現させた。実験による親和性分析の結果を、以下の項に提示する。
【0107】
PNGアーゼFレクテンズに関する実験から得られた結合親和性の測定
表7に提示しているのは、主として高マンノースオリゴ糖によって占有される単一のN-グリコシル化部位を含む変性RNアーゼBと、PNGアーゼFの突然変異体との間の相互作用に関して、表面プラズモン共鳴(SPR)を用いて測定した解離定数である。
【0108】
(表7)変性糖タンパク質RNアーゼBとPNGアーゼFのレクテンズ突然変異体との間の相互作用に関して測定した解離定数
a Km。 b 報告されている不活性突然変異体。 c 野生型との対比。
【0109】
計算によって予測されたように、E206A突然変異体およびD60A突然変異体はいずれも結合を顕著に強化する。また、静電スキャニングから示唆されたように、E206での正電荷(KまたはR)は少しではあるが親和性のさらなる増大をもたらす。現時点で、側鎖リパッキング実験または飽和突然変異誘発の恩典を受けていない第1世代レクテンズはマイクロモルの親和性を有し、これは2つの点突然変異だけでマイクロモルレベルに達した。これらの予備的突然変異が、高マンノースオリゴ糖に対してレクチンCon Aによって示されたものとほぼ等しいまで(Kdがほぼ1×10-6Mに等しい)に、PNGアーゼレクテンズの親和性を強化したことは注目に値する。
【0110】
セクションBにおいて述べたように、高親和性はバイオセンサーにとって唯一の望ましい特性である。高親和性は試薬をアフィニティークロマトグラフィーに使用することを可能にすると考えられる。しかし、バイオセンサーを組織染色などの用途に首尾良く用いようとするならば、遅いオフ速度(koff)を達成することも重要である。
【0111】
SPRは、koffの変動を評価するための好都合な方法を提供する。図6の検討は、突然変異体D60AとE206Aとの間で有意に異なる速度論的挙動を指し示している。いずれの突然変異体もKdについて同程度の値を有するが(表7)、D60Aは速いオン速度およびオフ速度を示した(10℃および25℃の両方)が、一方、E206Aはいずれの温度でも大幅に低いオフ速度を提示する。このことは、結合の速度論的動態を調整して、ある範囲にわたる特性を達成しうると考えられることから、極めて意義深い特徴である。
【0112】
さらなる親和性強化
計算によるデータに基づき、以下の7つの残基:D57、Y62、E118、S155、I156、G192およびE206を含む不活性D60A突然変異体に関するフォーカスト酵母ディスプレイライブラリーを開発した。これらの7つの位置を20種のアミノ酸すべてについてランダム化し、その結果、アミノ酸レベルで207(ほぼ109に等しい)種のクローンという理論的多様性を得た。
【0113】
加えて、D60A突然変異体を使用して、同じ残基Y62、E118、I156、S155、G192、E206およびD57のサブセットに対して計算による側鎖リパッキング実験を行うこともできる。計算による突然変異誘発をキー残基の同じセットに対して行うことにより、本発明者らは、計算解析が最適化された突然変異誘発データを再現する範囲を見いだすことができると予想している。計算方法(陰溶媒和モデルなど)の変更の実施を、その精度を高めるために行うことができる。
【0114】
実施例2
レクテンズの指向性進化
PNGアーゼF酵素の不活性D60A突然変異体を基にして、DNAライブラリーを作り出した。計算解析により同定された残基D57、Y62、E118、S155、I156、G192およびE206を、20種のアミノ酸すべてをコードするようにDNAレベルで改変した。このライブラリーを酵母ディスプレイベクターpPNL6中にクローニングし、酵母に導入して形質転換を行った。
【0115】
磁気ビーズ上に捕捉したdRNアーゼBに対してこのライブラリーのパニングを2ラウンド行い、続いて3ラウンド目にはフローサイトメトリーによってc-myc陽性酵母を選別した。この3ラウンドを1回繰り返し、合計6ラウンドとした。表8は、第6ラウンドからの18種のクローンのDNAのシークエンシングによる酵母クローンの濃縮を示している。
【0116】
(表8)第6ラウンドからのクローンの濃縮
【0117】
クローンR6.1.13を競合アッセイを用いる機能分析のために選択し、細菌において発現させて精製した。このアッセイでは、R6.1.13の1μM溶液のうち50μLを、dRNアーゼBビーズとともにプレインキュベートした。同様に、不活性酵素D60A突然変異体の1μM溶液のうち50μLをdRNアーゼBビーズとともにプレインキュベートした。プレインキュベートした各溶液に対して、Con Aレクチン(DyLight 488で蛍光標識)を最終濃度100nMとなるように添加した。標識したCon Aも、対照としてdRNアーゼBの存在下または非存在下で最終濃度100nMとなるようにビーズに添加した。ビーズの蛍光をフローサイトメトリーによって測定した。結合したCon Aの画分を、dRNアーゼBの存在下および非存在下におけるビーズとともにビーズの蛍光に対して標準化した(図9参照)。
【0118】
クローンR6.1.13タンパク質は、不活性酵素D60A突然変異体と比較して、dRNアーゼビーズに対するCon A結合の阻害のおよそ36%の増大を示し、このことは親和性強化を指し示している。このクローンは、クローン濃縮が3/18と少しであったことによって指し示されるように完全には最適化されておらず(表8)、そのため、さらに濃縮ラウンドを重ねることにより、さらなる親和性向上を得ることができる。
【特許請求の範囲】
【請求項1】
糖質プロセシング酵素を含むレクテンズ(lectenz)であって、該酵素が、
(i)該酵素の触媒活性を消失させ;かつ
(ii)不活性化された酵素のその基質に対する親和性を野生型酵素と比較して高める
1つまたは複数の突然変異を有する、レクテンズ。
【請求項2】
突然変異した糖質プロセシング酵素が、酵素の特異性の変化を実質的に全く有しない、請求項1記載のレクテンズ。
【請求項3】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約1.2倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項4】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約2倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項5】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約10倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項6】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約100倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項7】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約1,000倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項8】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約10,000倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項9】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約100,000倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項10】
糖質プロセシング酵素が、グリコシダーゼ、グリコシルトランスフェラーゼ、多糖リアーゼ、糖質エステラーゼ、スルファターゼ、スルホトランスフェラーゼ、リガーゼ、エピメラーゼ、および糖質基質に対して作用する他の任意の酵素からなる群より選択される、請求項1〜9のいずれか一項に記載のレクテンズ。
【請求項11】
糖質プロセシング酵素が原核生物由来の遺伝子によってコードされる、請求項1〜10のいずれか一項に記載のレクテンズ。
【請求項12】
糖質プロセシング酵素が真核生物由来の遺伝子によってコードされる、請求項1〜10のいずれか一項に記載のレクテンズ。
【請求項13】
糖質プロセシング酵素が反転型グリコシダーゼである、請求項1〜12のいずれか一項に記載のレクテンズ。
【請求項14】
糖質プロセシング酵素が保持型グリコシダーゼである、請求項1〜12のいずれか一項に記載のレクテンズ。
【請求項15】
糖質プロセシング酵素がPNGアーゼF(PNGase F)である、請求項1〜14のいずれか一項に記載のレクテンズ。
【請求項16】
PNGアーゼFが、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)由来の遺伝子によってコードされる、請求項15記載のレクテンズ。
【請求項17】
糖質プロセシング酵素がβ-O-GlcNAcアーゼ(β-O-GlcNAcase)である、請求項1〜12のいずれか一項に記載のレクテンズ。
【請求項18】
β-O-GlcNAcアーゼが、B.テタイオタオミクロン(B. thetaiotaomicron)由来の遺伝子によってコードされる、請求項17記載のレクテンズ。
【請求項19】
糖質プロセシング酵素がノイラミニダーゼである、請求項1〜12のいずれか一項に記載のレクテンズ。
【請求項20】
ノイラミニダーゼが、ウェルシュ菌(Clostridium perfringens)由来の遺伝子によってコードされる、請求項19記載のレクテンズ。
【請求項21】
1つまたは複数の突然変異がさらに、酵素の速度論的オフ速度(kinetic off-rate)を、野生型酵素と比較して少なくとも1.2倍低下させる、請求項1〜20のいずれか一項に記載のレクテンズ。
【請求項22】
触媒的に不活性な糖質プロセシング酵素を含むレクテンズであって、該酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約1.2倍高く、かつ該酵素が、該酵素の特異性の低下を実質的に全く有しない、レクテンズ。
【請求項23】
1つまたは複数の残基が水素結合によって基質と相互作用することができる、請求項22記載のレクテンズ。
【請求項24】
1つまたは複数の残基が基質とファンデルワールス接触を形成することができる、請求項22〜23のいずれか一項に記載のレクテンズ。
【請求項25】
1つまたは複数の残基が水媒介接触(water mediated contact)によって基質と相互作用することができる、請求項23〜23のいずれか一項に記載のレクテンズ。
【請求項26】
1つまたは複数の残基が、非特異的な非結合性相互作用によって基質と相互作用することができる、請求項22〜25のいずれか一項に記載のレクテンズ。
【請求項27】
1つまたは複数の残基の、気相(ΔEMM)相互作用エネルギーに対する寄与が、少なくとも約-0.5kcal/mol未満である、請求項22〜26のいずれか一項に記載のレクテンズ。
【請求項28】
1つまたは複数の残基の、全(ΔG)相互作用エネルギーに対する寄与が、少なくとも約-0.5kcal/mol未満である、請求項22〜27のいずれか一項に記載のレクテンズ。
【請求項29】
糖質プロセシング酵素を含むレクテンズを作製するための方法であって、該酵素が、
(i)該酵素の触媒活性を消失させ;かつ
(ii)酵素のその基質に対する親和性を野生型酵素と比較して高める
1つまたは複数の突然変異を有しており、
該方法が、
(a)糖質プロセシング酵素の配列を、突然変異した場合に該酵素を不活性化しうる1つまたは複数のアミノ酸残基に関して解析する段階;
(b)段階(a)で同定された少なくとも1つの突然変異したアミノ酸を有するグリコシダーゼとグリカンとの複合体の結合エネルギーを予測するための計算シミュレーションを行う段階;
(c)残基を、それらの予測される相互作用エネルギーに基づいて、酵素の特異性を規定するのに必須である第1の残基群、および基質に近接するが特異性を規定するのに必須ではない第2の残基群という2つの群に細分類する段階;
(d)段階(a)および(b)および(c)で同定された突然変異を含むグリコシダーゼを、それらがグリコシダーゼ-グリカン複合体を形成する能力に関して検査する段階;ならびに
(e)グリカンに対する結合親和性がWTグリコシダーゼのそれよりも少なくとも1.2倍高い、段階(d)による突然変異体を同定する段階、
を含む、方法。
【請求項30】
突然変異した糖質プロセシング酵素が、酵素の特異性の変化を実質的に全く有しない、請求項29記載の方法。
【請求項31】
糖質プロセシング酵素の配列を、計算によるアラニンスキャニング突然変異誘発を用いて解析する、請求項29〜30のいずれか一項に記載の方法。
【請求項32】
糖質プロセシング酵素の配列を、ΔGの直接算出を用いて解析する、請求項29〜31のいずれか一項に記載の方法。
【請求項33】
糖質プロセシング酵素の配列を、熱力学的積分算出を用いて解析する、請求項29〜32のいずれか一項に記載の方法。
【請求項34】
コンフォメーションサンプリングを実行するために計算シミュレーションを行う、請求項29〜33のいずれか一項に記載の方法。
【請求項35】
コンフォメーションサンプリングが、分子動力学シミュレーション、モンテカルロシミュレーション、または側鎖回転異性体の検索を含む、請求項29〜34のいずれか一項に記載の方法。
【請求項36】
計算による力場(computational force field)を用いてシミュレーションを行う、請求項29〜35のいずれか一項に記載の方法。
【請求項37】
糖質プロセシング酵素を含むレクテンズに生体試料を触れさせる段階を含む、生体試料を調べる方法であって、該酵素が、
(i)酵素の触媒活性を消失させ;かつ
(ii)酵素のその基質に対する親和性を野生型酵素と比較して高める
1つまたは複数の突然変異を有する、方法。
【請求項38】
酵素の特異性の変化を実質的に全く有しない突然変異した糖質プロセシング酵素をさらに含む、請求項37記載の方法。
【請求項39】
異常グリコシル化を呈する疾患を診断または治療する段階をさらに含む、請求項37〜38のいずれか一項に記載の方法。
【請求項40】
糖質、糖タンパク質、糖ペプチドまたは他の糖質含有分子の、親和性に基づく精製、濃縮またはモニタリングをさらに含む、請求項37〜38のいずれか一項に記載の方法。
【請求項41】
糖質結合タンパク質を含む新規生体分子を作製するための方法であって、該タンパク質が、
(i)タンパク質のその基質に対する親和性を野生型タンパク質と比較して高め;
(iii)タンパク質の特異性の変化を実質的に全く有しない
1つまたは複数の突然変異を有しており;
該方法が、
(a)糖質-タンパク質複合体の結合エネルギーを予測するために計算シミュレーションを行う段階であって、該タンパク質が;
(b)残基を、それらの予測される相互作用エネルギーに基づいて、タンパク質の特異性を規定するのに必須である第1の残基群、および糖質リガンドに近接するが特異性を規定するのに必須ではない第2の残基群という2つの群に細分類する段階;
(c)段階(a)および(b)で同定された突然変異を含むタンパク質を、それらがタンパク質-グリカン複合体を形成する能力に関して検査する段階;ならびに
(d)グリカンに対する結合親和性がWTタンパク質のそれよりも少なくとも1.2倍高い、段階(c)による突然変異体を同定する段階、
を含む、方法。
【請求項42】
フォーカストバイオコンビナトリアルライブラリーを作製するために用いられる、請求41記載の方法。
【請求項43】
フォーカストバイオコンビナトリアルライブラリーを作製するために用いられる、請求項29記載の方法。
【請求項1】
糖質プロセシング酵素を含むレクテンズ(lectenz)であって、該酵素が、
(i)該酵素の触媒活性を消失させ;かつ
(ii)不活性化された酵素のその基質に対する親和性を野生型酵素と比較して高める
1つまたは複数の突然変異を有する、レクテンズ。
【請求項2】
突然変異した糖質プロセシング酵素が、酵素の特異性の変化を実質的に全く有しない、請求項1記載のレクテンズ。
【請求項3】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約1.2倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項4】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約2倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項5】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約10倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項6】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約100倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項7】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約1,000倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項8】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約10,000倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項9】
酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約100,000倍高い、請求項1〜2のいずれか一項に記載のレクテンズ。
【請求項10】
糖質プロセシング酵素が、グリコシダーゼ、グリコシルトランスフェラーゼ、多糖リアーゼ、糖質エステラーゼ、スルファターゼ、スルホトランスフェラーゼ、リガーゼ、エピメラーゼ、および糖質基質に対して作用する他の任意の酵素からなる群より選択される、請求項1〜9のいずれか一項に記載のレクテンズ。
【請求項11】
糖質プロセシング酵素が原核生物由来の遺伝子によってコードされる、請求項1〜10のいずれか一項に記載のレクテンズ。
【請求項12】
糖質プロセシング酵素が真核生物由来の遺伝子によってコードされる、請求項1〜10のいずれか一項に記載のレクテンズ。
【請求項13】
糖質プロセシング酵素が反転型グリコシダーゼである、請求項1〜12のいずれか一項に記載のレクテンズ。
【請求項14】
糖質プロセシング酵素が保持型グリコシダーゼである、請求項1〜12のいずれか一項に記載のレクテンズ。
【請求項15】
糖質プロセシング酵素がPNGアーゼF(PNGase F)である、請求項1〜14のいずれか一項に記載のレクテンズ。
【請求項16】
PNGアーゼFが、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)由来の遺伝子によってコードされる、請求項15記載のレクテンズ。
【請求項17】
糖質プロセシング酵素がβ-O-GlcNAcアーゼ(β-O-GlcNAcase)である、請求項1〜12のいずれか一項に記載のレクテンズ。
【請求項18】
β-O-GlcNAcアーゼが、B.テタイオタオミクロン(B. thetaiotaomicron)由来の遺伝子によってコードされる、請求項17記載のレクテンズ。
【請求項19】
糖質プロセシング酵素がノイラミニダーゼである、請求項1〜12のいずれか一項に記載のレクテンズ。
【請求項20】
ノイラミニダーゼが、ウェルシュ菌(Clostridium perfringens)由来の遺伝子によってコードされる、請求項19記載のレクテンズ。
【請求項21】
1つまたは複数の突然変異がさらに、酵素の速度論的オフ速度(kinetic off-rate)を、野生型酵素と比較して少なくとも1.2倍低下させる、請求項1〜20のいずれか一項に記載のレクテンズ。
【請求項22】
触媒的に不活性な糖質プロセシング酵素を含むレクテンズであって、該酵素のその基質に対する親和性が、野生型酵素と比較して少なくとも約1.2倍高く、かつ該酵素が、該酵素の特異性の低下を実質的に全く有しない、レクテンズ。
【請求項23】
1つまたは複数の残基が水素結合によって基質と相互作用することができる、請求項22記載のレクテンズ。
【請求項24】
1つまたは複数の残基が基質とファンデルワールス接触を形成することができる、請求項22〜23のいずれか一項に記載のレクテンズ。
【請求項25】
1つまたは複数の残基が水媒介接触(water mediated contact)によって基質と相互作用することができる、請求項23〜23のいずれか一項に記載のレクテンズ。
【請求項26】
1つまたは複数の残基が、非特異的な非結合性相互作用によって基質と相互作用することができる、請求項22〜25のいずれか一項に記載のレクテンズ。
【請求項27】
1つまたは複数の残基の、気相(ΔEMM)相互作用エネルギーに対する寄与が、少なくとも約-0.5kcal/mol未満である、請求項22〜26のいずれか一項に記載のレクテンズ。
【請求項28】
1つまたは複数の残基の、全(ΔG)相互作用エネルギーに対する寄与が、少なくとも約-0.5kcal/mol未満である、請求項22〜27のいずれか一項に記載のレクテンズ。
【請求項29】
糖質プロセシング酵素を含むレクテンズを作製するための方法であって、該酵素が、
(i)該酵素の触媒活性を消失させ;かつ
(ii)酵素のその基質に対する親和性を野生型酵素と比較して高める
1つまたは複数の突然変異を有しており、
該方法が、
(a)糖質プロセシング酵素の配列を、突然変異した場合に該酵素を不活性化しうる1つまたは複数のアミノ酸残基に関して解析する段階;
(b)段階(a)で同定された少なくとも1つの突然変異したアミノ酸を有するグリコシダーゼとグリカンとの複合体の結合エネルギーを予測するための計算シミュレーションを行う段階;
(c)残基を、それらの予測される相互作用エネルギーに基づいて、酵素の特異性を規定するのに必須である第1の残基群、および基質に近接するが特異性を規定するのに必須ではない第2の残基群という2つの群に細分類する段階;
(d)段階(a)および(b)および(c)で同定された突然変異を含むグリコシダーゼを、それらがグリコシダーゼ-グリカン複合体を形成する能力に関して検査する段階;ならびに
(e)グリカンに対する結合親和性がWTグリコシダーゼのそれよりも少なくとも1.2倍高い、段階(d)による突然変異体を同定する段階、
を含む、方法。
【請求項30】
突然変異した糖質プロセシング酵素が、酵素の特異性の変化を実質的に全く有しない、請求項29記載の方法。
【請求項31】
糖質プロセシング酵素の配列を、計算によるアラニンスキャニング突然変異誘発を用いて解析する、請求項29〜30のいずれか一項に記載の方法。
【請求項32】
糖質プロセシング酵素の配列を、ΔGの直接算出を用いて解析する、請求項29〜31のいずれか一項に記載の方法。
【請求項33】
糖質プロセシング酵素の配列を、熱力学的積分算出を用いて解析する、請求項29〜32のいずれか一項に記載の方法。
【請求項34】
コンフォメーションサンプリングを実行するために計算シミュレーションを行う、請求項29〜33のいずれか一項に記載の方法。
【請求項35】
コンフォメーションサンプリングが、分子動力学シミュレーション、モンテカルロシミュレーション、または側鎖回転異性体の検索を含む、請求項29〜34のいずれか一項に記載の方法。
【請求項36】
計算による力場(computational force field)を用いてシミュレーションを行う、請求項29〜35のいずれか一項に記載の方法。
【請求項37】
糖質プロセシング酵素を含むレクテンズに生体試料を触れさせる段階を含む、生体試料を調べる方法であって、該酵素が、
(i)酵素の触媒活性を消失させ;かつ
(ii)酵素のその基質に対する親和性を野生型酵素と比較して高める
1つまたは複数の突然変異を有する、方法。
【請求項38】
酵素の特異性の変化を実質的に全く有しない突然変異した糖質プロセシング酵素をさらに含む、請求項37記載の方法。
【請求項39】
異常グリコシル化を呈する疾患を診断または治療する段階をさらに含む、請求項37〜38のいずれか一項に記載の方法。
【請求項40】
糖質、糖タンパク質、糖ペプチドまたは他の糖質含有分子の、親和性に基づく精製、濃縮またはモニタリングをさらに含む、請求項37〜38のいずれか一項に記載の方法。
【請求項41】
糖質結合タンパク質を含む新規生体分子を作製するための方法であって、該タンパク質が、
(i)タンパク質のその基質に対する親和性を野生型タンパク質と比較して高め;
(iii)タンパク質の特異性の変化を実質的に全く有しない
1つまたは複数の突然変異を有しており;
該方法が、
(a)糖質-タンパク質複合体の結合エネルギーを予測するために計算シミュレーションを行う段階であって、該タンパク質が;
(b)残基を、それらの予測される相互作用エネルギーに基づいて、タンパク質の特異性を規定するのに必須である第1の残基群、および糖質リガンドに近接するが特異性を規定するのに必須ではない第2の残基群という2つの群に細分類する段階;
(c)段階(a)および(b)で同定された突然変異を含むタンパク質を、それらがタンパク質-グリカン複合体を形成する能力に関して検査する段階;ならびに
(d)グリカンに対する結合親和性がWTタンパク質のそれよりも少なくとも1.2倍高い、段階(c)による突然変異体を同定する段階、
を含む、方法。
【請求項42】
フォーカストバイオコンビナトリアルライブラリーを作製するために用いられる、請求41記載の方法。
【請求項43】
フォーカストバイオコンビナトリアルライブラリーを作製するために用いられる、請求項29記載の方法。
【図1】

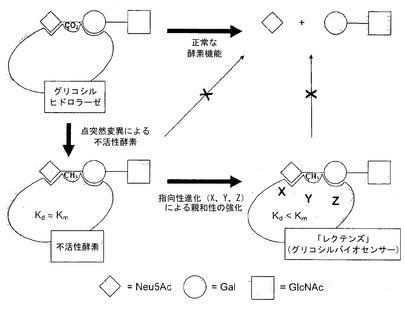
【図2】
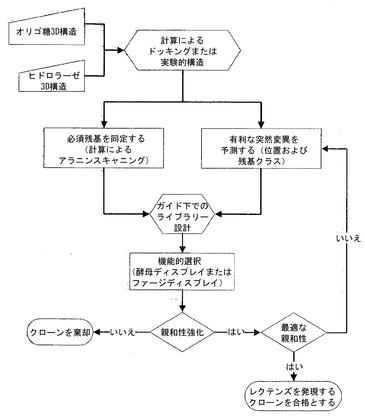
【図3】
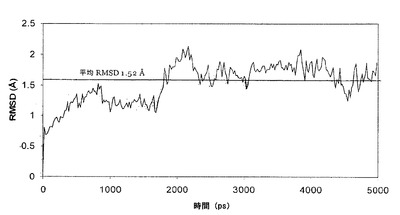
【図4】
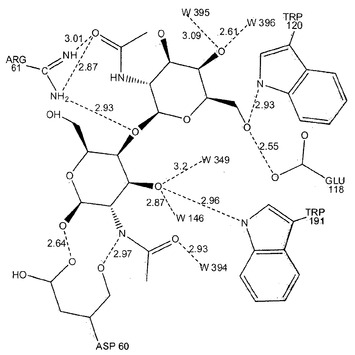
【図5】
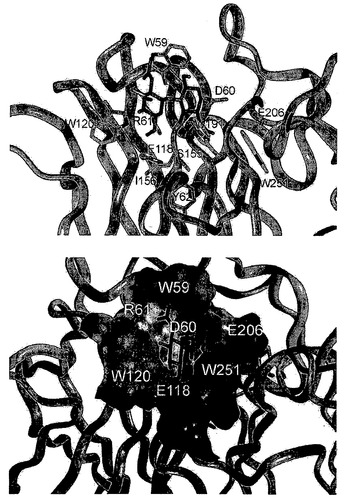
【図6】
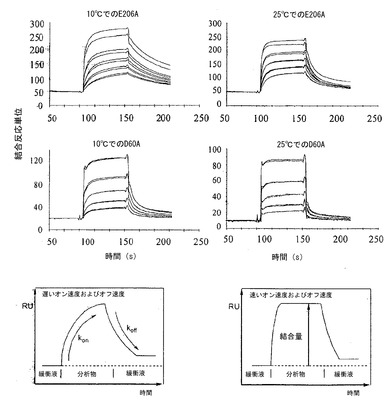
【図7】
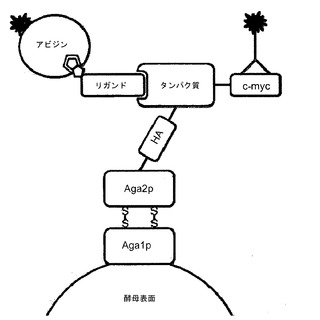
【図8】
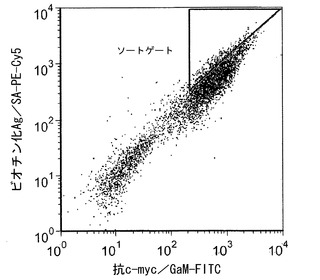
【図9】
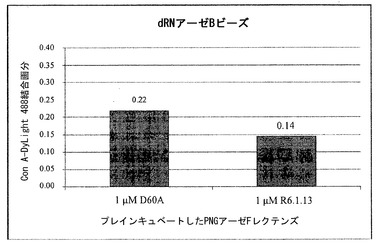
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2012−511333(P2012−511333A)
【公表日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願番号】特願2011−540910(P2011−540910)
【出願日】平成21年12月10日(2009.12.10)
【国際出願番号】PCT/US2009/067582
【国際公開番号】WO2010/068817
【国際公開日】平成22年6月17日(2010.6.17)
【出願人】(511140356)ユニバーシティ オブ ジョージア リサーチ ファウンデーション インコーポレイテッド (1)
【出願人】(511140367)グリコセンサーズ アンド ダイアグノスティクス リミテッド ライアビリティ カンパニー (1)
【Fターム(参考)】
【公表日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願日】平成21年12月10日(2009.12.10)
【国際出願番号】PCT/US2009/067582
【国際公開番号】WO2010/068817
【国際公開日】平成22年6月17日(2010.6.17)
【出願人】(511140356)ユニバーシティ オブ ジョージア リサーチ ファウンデーション インコーポレイテッド (1)
【出願人】(511140367)グリコセンサーズ アンド ダイアグノスティクス リミテッド ライアビリティ カンパニー (1)
【Fターム(参考)】
[ Back to top ]